Изобретение относится к медицине, а именно к биохимии, фармакологии и клинической фармакологии, и может быть использовано для изучения механизмов дифференцировки миобластов и оценки влияния на нее фармакологических веществ.
Линия клеток C2C12 представляет собой мышиные миобластные клетки, происходящие из сателлитных клеток, обладающие способностью спонтанно дифференцироваться в мышечные волокна после удаления сыворотки [D. Yaffe, O. Saxel. Serial passaging and differentiation of myogenic cells isolated from dystrophic mouse muscle. Nature. Vol. 270. P. 725 – 727. 1977; H.M. Blau, C.P. Chiu, C. Webster. Cytoplasmic activation of human nuclear genes in stable heterocaryons. Cell. Vol. 32, P. 1171-1180. 1983]. При этом дифференцируется около 50% клеток, а остальные миобласты остаются в недифференцированном состоянии [N. Yoshida, S. Yoshida, K. Koishi, K. Masuda, Y. Nabeshima. Cell heterogeneity upon myogenic differentiation: down-regulation of MyoD and Myf-5 generates 'reserve cells. J. Cell Sci. Vol. 111. P. 769-779. 1998].
Данная клеточная линия является классической моделью для изучения механизмов дифференцировки миобластов и оценки влияния на нее фармакологических веществ. Известно, что флаваноид (-)-Epicatechin индуцируeт миогенную дифференцировку клеток линии С2С12 за счет индукции прегнан Х рецептора [M. Ortiz-Flores et al. PXR is a target of (-)-epicatechin in skeletal muscle. Heliyon. Vol. 6, N 10, e05357 (2020); doi: 10.1016/j.heliyon.2020.e05357].
Известо, что альфа-магостин активирует миогенную дифференцировку миобластов [M. Lin, S. Zhou, K. Sakamoto. Alpha Mangostin promotes myogenic differentiation of C2C12 mouse myoblast cells. Biochemical and Biophysical Research Communications
Vol. 528, Is.1, 2020, P. 193-198. DOI: 10.1016 / j.bbrc.2020.04.128].
Известно, что эпикатехин вызывает MyoD-зависимую дифференцировку миобластов и миогенную конверсию фибробластов [Lee S.J. et al. Epicatechin elicits MyoD-dependent myoblast differentiation and myogenic conversion of fibroblasts. Plos One. 2017. DOI: 10.1371 / journal.pone.0175271].
Техническим результатом изобретения является разработка простого, эффективного способа активации миогенной дифференцировки миобластов клеточной линии C2C12.
Для достижения технического результата выполнено следующее исследование.
Эксперимент был проведен in vitro на линии клеток мышиных миобластов С2С12, предоставленной Институтом биологии гена (Москва). Клетки культивировали при 37ºС и 5% содержании СО2 в инкубаторе WS-189C (World Science, Корея) в Дульбекко модифицированной среде Игла (DMEM) с высоким содержанием глюкозы (4500 мг/л) (Sigma-Aldrich, Германия), содержащей L-глутамин (4 мМ) (Sigma-Aldrich, Германия), 10% эмбриональной бычьей сыворотки (Sigma-Aldrich, Германия), 100 ЕД/мл и 100 мкг/мл пенициллина и стрептомицина (Sigma-Aldrich, Германия) соответственно. После достижения 70−90% конфлюентности клетки снимали с флакона добавлением раствора трипсин-ЭДТА (0,25% трипсина и 0,2% ЭДТА, Sigma-Aldrich, Германия), высевали в 6-луночные планшеты (Сorning, США).
Были сформированы следующие экспериментальные группы:
- клетки до дифференцировки – инкубация 7 дней с питательной средой, содержащей 10% эмбриональной бычьей сыворотки (n=3);
- индукция дифференцировки миобластов – клетки инкубировали 7 дней с дифференцировочной питательной средой, содержащей 2% лошадиной сыворотки (Sigma-Aldrich, Германия), L-глутамин (4 мМ), 100 ЕД/мл и 100 мкг/мл пенициллина и стрептомицина (n=3) [J. Sin et al. Mitophagy is required for mitochondrial biogenesis and myogenic differentiation of C2C12 myoblasts. Autophagy. Vol.12 (2), P. 369-380. 2016; doi: 10.1080/15548627.2015.1115172];
- индукция дифференцировки миобластов с добавлением этилметилгидроксипиридина сукцината (ЭМГПС) (Фармасофт, Россия) – клетки инкубировали 7 дней в дифференцировочной питательной среде, содержащей этилметилгидроксипиридина сукцинат в концентрации 10 мкМ в течение 7 дней (n=3).
Визуализацию клеток выполняли с помощью инвертированного микроскопа Olympus CKX-53 («Olympus», Япония).
После окончания экспозиции, клетки снимали с лунок раствором трипсин-ЭДТА (Sigma-Aldrich, США), трижды промывали раствором фосфатного буфера (BioRad, США) и лизировали в NP40 Cell Lysis Buffer Thermo (Thermo Fisher Scientific, США) c добавлением смеси ингибиторов протеиназ (Sigma-Aldrich, США) в течение 30 минут при +4°С и постоянном перемешивании из расчета 107 клеток на 100 мкл буфера. Полученный лизат центрифугировали при 13000 об/мин в течение 10 минут (AvantiJXN-3, BeckmanCoulter, США).
Цитоплазматические белки (30 мкг) подвергали электрофорезу с использованием TGX Stain-Free FastCast Acrylamide Kit (Bio-Rad, США) в буферной системе Laemmli (BioRad,США). Перед загрузкой образцы обрабатывались в соответствии с протоколом Bio-Rad, их смешивали с буфером для образцов Laemmli (Bio-Rad, США), содержащем 2,5% β-меркаптоэтанола (Bio-Rad, США) в соотношении 1:3, инкубировали 10 мин при температуре 70°C. Гели прогоняли при 100 В в течение 90 мин.
Для определения относительного количества миозина, α-актина, Myod, миогенина, методом вестерн-блот использовали первичные мышиные моноклональные антитела (sc-377460 MyoD (G-1) Monoclonal Antibody, Santa cruz biotechnology,inc, США; sc-12732 myogenin (F5D) Monoclonal Antibody, Santa cruz biotechnology,inc, США; sc-53092, MYH1/2/3 (N3.36) Monoclonal Antibody, Santa cruz biotechnology,inc, США; sc-53142, Smooth Muscle Actin (B4), Monoclonal Antibody, Santa cruz biotechnology,inc, США) в концентрации 1:200. Визуализацию первичных антител осуществляли с использованием вторичных кроличьих антител (Rabbit anti-Mouse IgG (H+L) Secondary Antibody, HRP, Invitrogen) в разведении 1:4000.
Белки визуализировали хемилюминесценцией с помощью Chemi Doc XRS+ (Bio-Rad, США). Интенсивность полученных полос (бэндов) анализировали денситометрически с помощью программного обеспечения ImageLab (Bio-Rad, США).
Количество белков оценивали относительно содержания белка домашнего хозяйства GAPDH (первичные GAPDH Loading Control Monoclonal Antibody (GA1R), DyLight 68, Invitrogen, США, разведение 1:1000, вторичные антитела - вторичные кроличьи антитела к первичным антителам GAPDH - Rabbitanti-Mouse IgG (H+L) Secondary Antibody, HRP, Invitrogen, США, разведение 1:4000).
Полученные результаты анализировали с помощью программ «Stat Soft Statistica 13.0» (США, номер лицензии JPZ811I521319AR25ACD-W) и Microsoft Excel for MAC ver. 16.24 (ID02984-001-000001). Результаты представлены в виде M±SD. Для оценки статистической значимости различий использовали дисперсионный анализ (ANOVA), множественные сравнения выполняли с помощью критерия Ньюмена-Кейлса. Статистически значимыми считали различия при p<0,05.
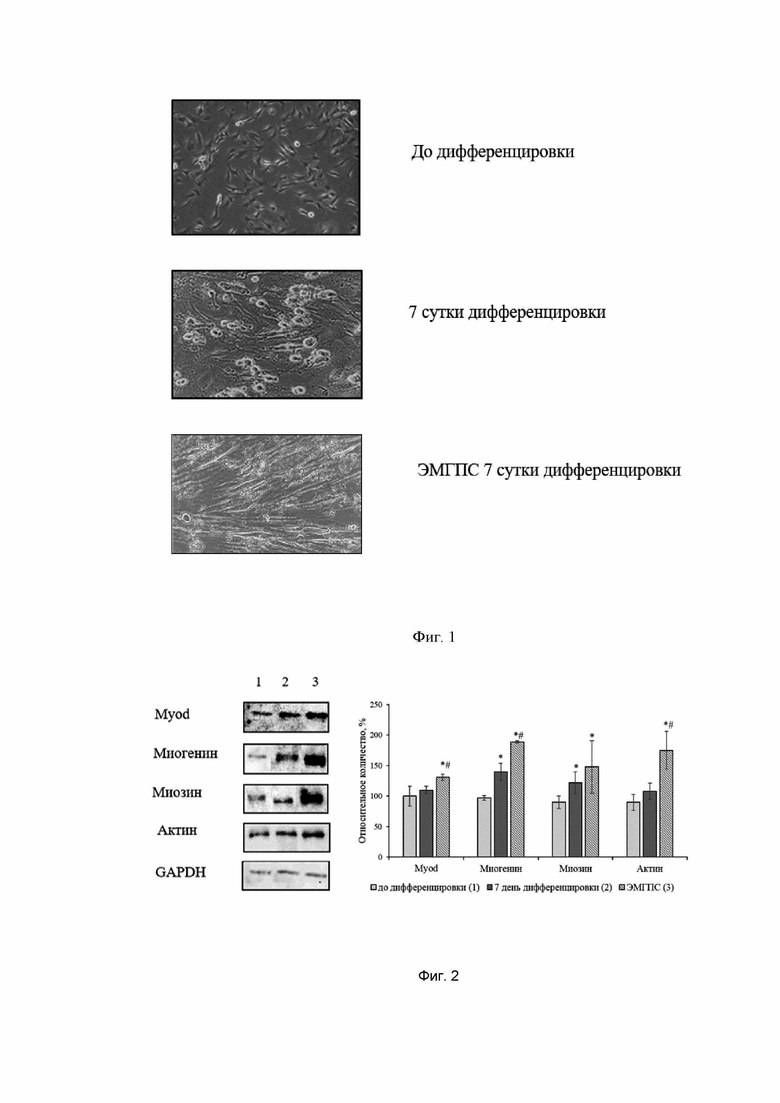
Смена питательной среды, содержащей 10% эмбриональной бычьей сыворотки, на среду с содержанием 2% лошадиной сыворотки запускало процесс дифференцировки миобластов. Клетки меняли свою форму: из отростчатых или веретенообразных они превращались в более вытянутые. В дальнейшем отдельные миобласты сливались и формировали миотубы - веретенообразные структуры длиной 100-600 мкм, шириной 30-50 мкм с несколькими ядрами. Максимально выраженные изменения наблюдались на 7 день (фиг. 1).
Добавление этилметилгидроксипиридина сукцината усиливало процесс дифференцировки миобластов клеточной линии С2С12. Большее количество клеток меняли форму и сливались в миотубы.
Данные морфологические изменения сопровождались следующей динамикой в уровне белков клеток (фиг. 2). Количество миозина в клетках линии С2С12 на 7 день эксперимента, при использовании обычной дифференцировочной среды и среды с добавлением этилметилгидроксипиридина сукцината статистически значимо увеличивалось по сравнению с показателями клеток до дифференцировки на 35,6% (р=0,004) и 64,4% (р=0,001) соответственно. В группах с тестируемыми веществами отличий от клеток на 7 день при использовании обычной дифференцировочной среды отмечено не было.
Уровень α-актина статистически значимо не отличался в клетках до и после дифференцировки. В то же время при использовании этилметилгидроксипиридина сукцината отмечалось увеличение содержания α-актина на 94,4 % (р=0,02) по сравнению с клетками до дифференцировки и на 62% (р=0,02) по сравнению с клетками на 7 день дифференцировки.
Содержание Myod (Myogenic determination gene number 1), который является главным регулятором миогенеза, в клетках линии С2С12 до дифференцировки и на 7 день дифференцировки не отличался. Однако, внесение в дифференцировочную питательную среду этилметилгидроксипиридина сукината приводило к увеличению количества Myod на 31,1 % (р=0,016) по сравнению с клетками до дифференцировки, а также на 19,1% (р=0,002) по сравнению с клетками на 7 день дифференцировки.
Количество миогенина, другого регуляторного белка, статистически значимо возрастало в клетках на 7 день при обычных условиях дифференцировки на 44,3% (р=0,02), а при добавлении этилметигидроксипиридина сукцината в питательную среду на 94,8% (р=0,007) по сравнению с клетками линии С2С12 до дифференцировки. Этилметилгидроксипиридина сукцинат также стимулировал увеличение количества миогенина по сравнению с клетками на 7 день в обычных условиях дифференцировки на 35% (р=0,03).
Краткое описание чертежей
Фиг. 1. Влияние этилметилгидроксипиридина сукцината 10 мкМ на дифференцировку клеток линии С2С12, х200. Фазово-контрастная микроскопия.
Фиг. 2. Относительное количество Myod, миогенина, миозина, актина в клетках линии С2С12 при воздействии этилметилгидроксипиридина сукцината в концентрации 10 мкМ в течение 7 дней. Примечание: статистически значимые отличия * - p<0,05 по сравнению с клетками до дифференцировки; # - p<0,05 по сравнению с клетками на 7 день дифференцировки
Таким образом, показано, что инкубирование клеточной линии С2С12 в дифференцировочной питательной среде с добавлением этилметилгидроксипиридина сукцината до достижения концентрации в среде 10 мкМ в течение 7 дней ускоряет процесс миогенной дифференцировки миобластов, о чем свидетельствуют морфологические изменения клеток, а также повышение уровня специфических мышечных белков α-актина, миозина и факторов миогенной дифференцировки – MyoD и миогенина.
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ стимуляции миогенеза | 2021 |
|
RU2776045C1 |
| КОМПОЗИЦИИ ИЗ КУЛЬТИВИРУЕМОГО МЯСА | 2018 |
|
RU2778255C2 |
| СПОСОБ ПОВЫШЕНИЯ КОЛИЧЕСТВА БЕЛКА РЕЗИСТЕНТНОСТИ РАКА МОЛОЧНОЙ ЖЕЛЕЗЫ В ЭКСПЕРИМЕНТЕ НА КЛЕТОЧНОЙ ЛИНИИ ГЕПАТОЦЕЛЛЮЛЯРНОЙ КАРЦИНОМЫ ЧЕЛОВЕКА | 2023 |
|
RU2803336C1 |
| Способы получения происходящих из мышц клеток | 2018 |
|
RU2796929C2 |
| Генетическая конструкция и способ экспрессии ERVW-1 и MYMK | 2025 |
|
RU2839754C1 |
| КОМПОЗИЦИИ ГУМАНИЗИРОВАННЫХ СЛИТЫХ БЕЛКОВ NOTCH И СПОСОБЫ ЛЕЧЕНИЯ | 2008 |
|
RU2532830C2 |
| СПОСОБ ПОЛУЧЕНИЯ ПОПУЛЯЦИИ СТРОМАЛЬНЫХ КЛЕТОК ЖИРОВОЙ ТКАНИ, ИНДУЦИРОВАННЫХ К НЕЙРАЛЬНОЙ ДИФФЕРЕНЦИРОВКЕ | 2008 |
|
RU2396345C1 |
| СПОСОБ ПОЛУЧЕНИЯ МИОБЛАСТОВ, ИСПОЛЬЗОВАНИЕ БИОПТАТА ДЕСНЫ, ПРЕПАРАТ МИОБЛАСТОВ ДЛЯ ЛЕЧЕНИЯ ПАТОЛОГИЙ МЫШЕЧНОЙ ТКАНИ И СПОСОБ ЕГО ПОЛУЧЕНИЯ | 2014 |
|
RU2576842C2 |
| Штамм гибридных культивируемых клеток Homo sapiens/Mus musculus 1B9C7 - продуцент человеческих моноклональных антител, специфичных к протеолитическому домену ботулинического токсина типа A | 2021 |
|
RU2783897C1 |
| ШТАММ ГИБРИДНЫХ КЛЕТОК ЖИВОТНЫХ Mus musculus 3F11 - ПРОДУЦЕНТ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ, СПЕЦИФИЧНЫХ К БОТУЛИНИЧЕСКОМУ ТОКСИНУ ТИПА B | 2014 |
|
RU2566553C1 |
Настоящее изобретение относится к клеточной биологии и медицине, в частности к способу активации миогенной дифференцировки миобластов линии клеток C2C12. Для осуществления способа указанные клетки инкубируют в течение 7 дней с дифференцировочной питательной средой, содержащей 2% лошадиной сыворотки, L-глутамин в концентрации 4 мМ, пенициллин в концентрации 100 ЕД/мл и стрептомицин в концентрации 100 мкг/мл с добавлением в среду этилметилгидроксипиридина сукцината до достижения концентрации 10 мкМ. Настоящее изобретение позволяет повысить эффективность и упростить способ активации миогенной дифференцировки миобластов клеточной линии C2C12, и может быть использовано для изучения механизмов дифференцировки миобластов и оценки влияния на нее фармакологических веществ. 2 ил.
Способ активации миогенной дифференцировки миобластов линии клеток C2C12, включающий инкубирование клеток в течение 7 дней с дифференцировочной питательной средой, содержащей 2% лошадиной сыворотки, L-глутамин в концентрации 4 мМ, пенициллин в концентрации 100 ЕД/мл и стрептомицин в концентрации 100 мкг/мл, отличающийся добавлением в среду этилметилгидроксипиридина сукцината до достижения концентрации 10 мкМ.
| BLAND CHRISTOPHER S | |||
| et al., Global regulation of alternative splicing during myogenic differentiation, Nucleic Acids Research, 2010, Vol | |||
| Способ сужения чугунных изделий | 1922 |
|
SU38A1 |
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| Устройство для электрической колокольной сигнализации на одно путных железных дорогах | 1927 |
|
SU7651A1 |
| ARNESON-WISSINK PAIGE C | |||
| et al., The wasting-associated metabolite succinate disrupts myogenesis and impairs skeletal muscle regeneration, JCSM Rapid | |||

