1. ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к иммуноглобулинам, связывающим ADAMTS5, и более конкретно, к полипептидам, содержащим или в основном состоящим из одного или более таких иммуноглобулинов (также обозначенным в настоящем описании как «иммуноглобулин(ы) по изобретению» и «полипептиды по изобретению», соответственно). Изобретение также относится к конструкциям, содержащим такие иммуноглобулины, например, одиночные вариабельные домены иммуноглобулинов (ISVD), или полипептиды, так же, как к нуклеиновым кислотам, кодирующим такие иммуноглобулины или полипептиды (также обозначенным в настоящем описании как «нуклеиновая кислота (кислоты) по изобретению»; к способам получения таких иммуноглобулинов, полипептидов и конструкций; к клеткам-хозяевам, экспрессирующим или способным экспрессировать такие иммуноглобулины или полипептиды; к композициям, и в частности, к фармацевтическим композициям, содержащим такие иммуноглобулины, полипептиды, конструкции, нуклеиновые кислоты и/или клетки-хозяева; и к применениям иммуноглобулинов, полипептидов, конструкций, нуклеиновых кислот, клеток-хозяев и/или композиций, в частности, для профилактических и/или терапевтических целей, таких как профилактические и/или терапевтические цели, упомянутые в настоящем описании. Другие аспекты, варианты осуществления, преимущества и применения изобретения станут понятными из дальнейшего описания в настоящем документе.
2. УРОВЕНЬ ТЕХНИКИ ДЛЯ ИЗОБРЕТЕНИЯ
Остеоартрит (OA) является одной из наиболее распространенных причин инвалидности во всем мире. Он поражает 30 миллионов американцев и является наиболее распространенным нарушением суставов. Предполагают, что он поразит более 20 процентов популяции США к 2025 г. Это заболевание не является системным и обычно ограничено несколькими суставами. Однако заболевание может возникать во всех суставах, наиболее часто, в суставах коленей, бедер, рук, плеч и позвоночника. OA характеризуется прогрессирующей эрозией суставного хряща (хряща, покрывающего кости), приводящей к хронической боли и инвалидности. В конечном счете заболевание приводит к полному разрушению суставного хряща, склерозу подлежащей кости, формированию остеофита и т.д., которые все приводят к потере подвижности и боли. Остеоартрит можно определять как разнообразную группу состояний, характеризующуюся комбинацией симптомов, признаков, возникающих в результате дефектов в суставном хряще и изменений в соседних тканях, включая кость, сухожилия и мышцу. Боль является наиболее выраженным симптомом OA и наиболее частой причиной обращения пациентов за медицинской помощью. Не существует излечивающего лечения для OA, т.е. современные способы лечения не ингибируют структурное разрушение сустава с OA. Управление течением заболевания ограничено видами лечения, которые являются в лучшем случае паллиативными и вносят небольшой вклад в борьбу с лежащей в основе причиной прогрессирования заболевания. Несмотря на то, что начало заболевания может являться многофакторным, разрушение хряща, по-видимому, является результатом неконтролируемого протеолитического разрушения внеклеточного матрикса (ECM). Наиболее распространенными компонентами ECM суставного хряща являются коллаген (в первую очередь, коллаген II) и протеогликаны, в основном, аггрекан (Kiani et al., 2002 Cell Research 12:19-32).
Аггрекан является важным для надлежащего функционирования суставного хряща, поскольку он обеспечивает структуру гидратированного геля, придающую хрящу свойства несения нагрузки. Аггрекан представляет собой большую, многомодульную молекулу (2317 аминокислот), экспрессируемую хондроцитами. Его коровый белок состоит из трех глобулярных доменов (G1, G2 и G3) и области большой протяженности между G2 и G3 для присоединения гликозаминогликановой цепи. Эта протяженная область содержит два домена, один замещенный цепями кератансульфата (домен KS) и один с цепями хондроитинсульфата (домен CS). Домен CS имеет 100-150 присоединенных к нему цепей гликозаминогликана (GAG). Аггрекан формирует большие комплексы с гиалуронаном, в котором 50-100 молекул аггрекана взаимодействуют посредством домена G1 и связывают белок с одной молекулой гиалуронана. После поглощения воды (из-за содержания GAG), эти комплексы формируют обратимо деформируемый гель, устойчивый к сжатию. Структура, удержание жидкости и функция суставного хряща связаны с содержанием в матриксе аггрекана, и с количеством хондроитинсульфата, связанного с интактным коровым белком. Структурно, OA характеризуется деградацией аггрекана, прогрессирующим образом высвобождающей домены G3 и G2 (приводящей к «дефляции» хряща) и в конечном счете, к высвобождению домена G1 и деградации коллагена, необратимо разрушающим структуру хряща. Наиболее значимый для патогенеза OA участок расщепления аггрекана локализован в последовательности TEGE373↓374ARGS. Этот участок расщепления расположен в интерглобулярном домене (IGD) аггрекана. Антитела, узнающие неоэпитоп 374ARGS, привели к открытию аггреканазы-1, как доказано, представляющей собой ADAMTS4, и аггреканазы-2, представляющей собой ADAMTS5. Затем показали, что другие родственные ферменты ADAMTS, включая ADAMTS1, -8, -9, -15 и -20, имеют активность аггреканазы. ADAMTS16 и 18 также являются слабыми аггреканазами. ADAMTS и матриксные металлопротеиназы (MMP) разделяют участок связывания с аггреканом, который является очень сходным как по последовательности, так и по общей форме (El Bakali et al., 2014 Future Medicinal Chemistry (Review) 6:1399).
Ферменты ADAMTS (дезинтегрины и металлопротеиназы с тромбоспондиновыми мотивами) представляют собой секретированные, мультидоменные, ассоциированные с матриксом цинковые металлоэндопептидазы, имеющие разнообразные роли в морфогенезе и патофизиологическом ремоделировании ткани, в воспалении и в биологии сосудов (Kelwick et al., 2015 Genome Biology 16:113). Человеческое семейство включает 19 членов, которые можно разделить на подгруппы на основании их известных субстратов. Аггреканазы или протеогликаназы включают ADAMTS1, -4, -5, -8, 9, -15 и -20, которые могут расщеплять связывающие гиалуронан внеклеточные белки хондроитинсульфат-протеогликаны (CSPG), включая аггрекан, версикан, бревикан и нейрокан. Двумя наиболее предпочтительными участками расщепления в бычьем аггрекане являются KEEE1667↓1668GLGS, за которым следует GELE1480↓1481GRGT. Затем расщепление происходит в NITEGE373↓374ARGS в IGD (в котором MMP не расщепляют), высвобождая вышеупомянутый неоэпитоп, и в TAQE1771↓1772AGEG и VSQE1871↓1872LGQR в области CS-2 (Fosang et al., 2008 European Cells and Materials 15:11-26). Эти участки расщепления являются высоко консервативными у человека, быка, мышей и крыс.
Различные цепи доказательств показывают, что ADAMTS5 является принципиальным ферментом, вовлеченным в патогенез остеоартрита. ADAMTS5 является главной аггреканазой, присутствующей в хряще. В эксплантатах и хондроцитах человеческого хряща, нокдаун ADAMTS5 ослаблял разрушение аггрекана, позволяя предполагать, что этот фермент может играть роль в тканях человека. Экспрессию этого фермента усиливают цитокины, такие как интерлейкин-1 и онкостатин-M, провоцирующие разрушение аггрекана в тканях. Образованные посредством ADAMTS5 фрагменты аггрекана детектированы в синовиальной жидкости и сыворотке пациентов с OA (Germaschewski et al., 2014 Osteoarthritis Cartilage 22:690-697).
ADAMTS5 сначала синтезируется как неактивный белок, включающий домен протеазы на N-конце и вспомогательный домен на C-конце. Домен протеазы состоит из сигнального пептида, продомена с последовательностью узнавания фурина и каталитического домена. Продомен расщепляется посредством конвертаз пробелка для получения активных ферментов. ADAMTS5 дополнительно содержит вспомогательные домены, которые активно участвуют в узнавании субстрата и модулируют аффинность протеазы для ее субстрата (субстратов) («экзосайты»). Подобный дезинтегрину домен, центральный подобный тромбоспондину (TS) повтор типа I, богатый цистеином домен, спейсерная область и дополнительный мотив TS ADAMTS5 представляют собой вспомогательные домены с потенциальными функциями экзосайтов. Богатый цистеином домен, по-видимому, является необходимым для связывания и заякоривания ADAMTS5 на гликозаминогликанах. Наибольшая вариабельность среди членов семейства ADAMTS обнаружена в этих вспомогательных доменах (Kelwick et al., 2015 Genome Biology 16:113).
Модифицирующие течение заболевания противоостеоартритные лекарственные средства (DMOAD), которые можно определять как лекарственные средства, ингибирующие структурное прогрессирование заболевания и, в идеале, также улучшающие симптомы и/или функцию, являются широко востребованными. DMOAD, вероятно, будут прописаны на длительные периоды при этом хроническом заболевании стареющей популяции, таким образом, необходимы данные о превосходной безопасности в целевой популяции со множеством сопутствующих заболеваний и потенциальными взаимодействиями между лекарственными средствами.
Несколько фармацевтических компаний разработали низкомолекулярные ингибиторы ADAMTS5. Заявлено, что некоторые из этих соединений являются специфическими для ADAMTS5, в то время как другие оказывают также эффект на другие члены семейства ADAMTS или даже на MMP. Эти случаи перекрестного ингибирования считают ответственными за скелетно-мышечный синдром, побочный эффект, вызванный ингибиторами широкого спектра и включающий артралгию, миалгию, тугоподвижность суставов и тендинит (Santamaria et al., 2015 Biochem J 471:391-401). Ингибитор аггреканазы AGG-523 от Wyeth использовали в 5 клинических исследованиях фазы I у здоровых субъектов и пациентов с OA, но не использовали в дальнейшем. Никакие другие низкомолекулярные ингибиторы ADAMTS не вошли в какую-либо дальнейшую клиническую разработку в качестве потенциального DMOAD (Bondeson et al., 2015 Drug Discovery 10:5-14). Действительно, несмотря на ряд недавних клинических исследований, конкретно исследующих DMOAD, никаких таких лекарственных средств не одобрено до настоящего времени (El Bakali et al., 2014 Future Medicinal Chemistry (Review) 6:1399).
С учетом успеха направленной биологической терапии с использованием антител («Ab»), возник интерес к разработке сходных терапевтических способов для OA. Исследование в Rottapharm моноклонального антитела (mAb) CRB0017, нацеленного против спейсерного домена ADAMTS5, показало, что у мышей, внутрисуставное введение этого mAb значительно предотвращало прогрессирование заболевания зависимым от дозы образом (Chiusaroli et al., 2013 Osteoarthritis Cartilage 21:1807). Не присутствовало ни сравнения с системным введением, ни оценки, до какой степени mAb подтекало из синовиального пространства. В другом исследовании использовали системное введение mAb 12F4 у мышей, при этом показано как структурная модификация заболевания, так и облегчение связанного с болью поведения (Miller et al., 2014 Osteoarthritis Cartilage 22iii, S35). Однако, однократное введение mAb 12F4 у яванского макака вызвало фокальное кровотечение, зависимое от дозы увеличение среднего артериального давления и аномалии сердечной проводимости (более конкретно, повышения ST и желудочковые аритмии по ECG), указывающие на сердечную ишемию, которые продолжались вплоть до 8 месяцев после введения однократной дозы (Larkin et al., 2014 Osteoarthritis Cartilage 22iii, S483). Эти побочные эффекты остановили дальнейшую клиническую разработку mAb 12F4.
В WO2008/074840 от Ablynx NV описано получение нанотел® против членов семейства дезинтегрина и металлопротеиназ (ADAM), включая ADAMTS5.
Терапевтическим вмешательствам в суставы дополнительно препятствует сложность нацеливания лекарственных средств на суставной хрящ. Поскольку суставной хрящ представляет собой лишенную сосудов и лимфы ткань, традиционные способы доставки лекарственного средства (пероральный, внутривенный, внутримышечный) в конечном счете зависят от переноса через синовиальное пространство лекарственных средств из капилляров синовиальной оболочки в хрящ посредством пассивной диффузии. Это делает необходимой разработку внутрисуставной (IA) доставки лекарственных средств.
С другой стороны, IA доставка терапевтических белков ограничена их быстрым выведением из суставной щели и отсутствуем удержания внутри хряща; и ограничением крупными суставами. Время удержания в синовиальном пространстве лекарственного средства в суставе часто составляет менее 24 час (Edwards 2011 Vet J 190:15-21; Larsen et al., 2008 J Pham Sci 97:4622-4654). Из-за быстрого выведения большинства инъецированных IA лекарственных средств, необходимы частые инъекции для поддержания эффективной концентрации (Owen et al., 1994 Br J Clin Pharmacol 38:349-355). Кроме того, IA доставка терапевтических белков не является практически осуществимой для небольших суставов, что препятствует лечению, например, пальцев при OA.
Остается необходимость в эффективных DMOAD.
3. СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Целью настоящего изобретения является предоставление полипептидов против OA с улучшенными профилактическими, терапевтическими и/или фармакологическими свойствами, в дополнение к другим преимущественным свойствам (например, таким как улучшенная простота получения, хорошая стабильность и/или уменьшенная стоимость), по сравнению с аминокислотными последовательностями и антителами предшествующего уровня техники. В частности, целью настоящего изобретения является предоставление полипептидов, ингибирующих ADAMTS, и особенно, ингибирующих ADAMTS5.
ADAM, ADAMTS и MMP разделяют участки связывания с аггреканом, которые являются очень сходными как по последовательности, так и по общей форме. Поскольку различные другие члены семейств ADAM (включая TACE) и ADAMTS имеют разнообразные роли в нормальной физиологии, сильно ассоциированные со многими распространенными патологическими состояниями, включая астму, артрит, злокачественные опухоли, нарушения соединительной ткани или тромботическую тромбоцитопеническую пурпуру (El Bakali et al., выше), авторы настоящего изобретения признают важность сохранения избирательности. Для эффективного прекращения только активности ADAMTS5, нацеливание на каталитический домен, по-видимому, является наилучшим вариантом. Однако, каталитические домены ADAMTS4 и ADAMTS5 разделяют высокую степень сходства последовательности (ср. El Bakali et al., выше). Кроме того, оказалось, что высокая консервативность последовательности каталитического домена между различными видами удерживает от сильного иммунного ответа.
Неожиданно, ISVD по изобретению соответствуют этим кажущимся взаимоисключающими требованиям ингибирования (ферментной) активности ADAMTS5 с одной стороны, с сохранением в то же время избирательности, с другой стороны.
Различные моновалентные ISVD по настоящему изобретению являлись равноактивными с общепринятым двухвалентным антителом mAb 12F4 H4L0 в ферментном анализе AlphaLISA. Кроме того, ISVD по настоящему изобретению являлись также равноактивными с общепринятым двухвалентным антителом mAb12F4 H4L0 при высокой или даже избыточной концентрации субстрата аггрекана, напоминающей концентрацию в суставах. С другой стороны, в анализе бычьего эксплантата ex vivo, даже более близко напоминающем физиологические условия, большинство моновалентных ISVD имели лучшую IC50, чем сравнительное mAb 12F4 H4L0 («12F4») и mAb CRB0017 от Rottapharm. В анализе человеческого эксплантата ex vivo, ISVD по настоящему изобретению также были существенно лучшими, чем общепринятое антитело CRB0017 предшествующего уровня техники, как показано по IC50.
После дополнительной модификации ISVD с учетом различных разнообразных и благоприятных признаков, включая стабильность, аффинность и ингибирующую активность, так же как минимизацию иммуногенности, эти ISVD затем оценивали in vivo.
Для системного введения ISVD по настоящему изобретению показано сильное ингибирование активности аггреканазы in vivo, как оценено у яванского макака. Кроме того, ISVD также являлись безопасными для использования, в отличие от антитела 12F4 предшествующего уровня техники. Также в модели дестабилизации медиального мениска (DMM) in vivo на мышах, для ISVD по настоящему изобретению показано структурное преимущество, вплоть до 50%, как для профилактического, так и для терапевтического лечения посредством системного введения. Раннее лечение с использованием ISVD по настоящему изобретению вызывало зависимое от дозы, значимое и значительное симптоматическое преимущество во время индуцированного рассечением передней крестообразной связки и резекцией медиального мениска (ACLT+tMx) OA у крыс.
ISVD в конечном счете предназначены для ингибирования ADAMTS5 в суставах и таким образом, должны выдерживать условия синовиальной жидкости в суставах, которая содержит различные протеазы. Наряду с благоприятными характеристиками выше, показано, что выделенные ISVD являлись чрезвычайно стабильными в синовиальной жидкости. Предполагают, что эта стабильность позволяет режим менее частого дозирования.
Соответственно, настоящее изобретение относится к полипептиду, содержащему по меньшей мере 1 одиночный вариабельный домен иммуноглобулина (ISVD), связывающий дезинтегрин и металлопротеиназу с тромбоспондиновыми мотивами (ADAMTS), предпочтительно, где указанный ADAMTS выбран из группы, состоящей из ADAMTS1 - ADAMTS19, предпочтительно, ADAMTS5, ADAMTS4, ADAMTS1, ADAMTS8, ADAMTS9 и ADAMTS15, и ADAMTS20, наиболее предпочтительно, ADAMTS5. В одном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, связывающий ADAMTS, предпочтительно, ADAMTS5, не связывается с ADAMTS4, MMP1 или MMP14.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном, состоит из 4 каркасных областей (FR1-FR4, соответственно) и 3 определяющих комплементарность областей (CDR1-CDR3, соответственно), в которых (i) CDR1 выбрана из группы, состоящей из SEQ ID NO: 21, 35, 20, 22, 25, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34; и аминокислотных последовательностей, имеющих 1, 2 или 3 отличие(отличия) аминокислот от SEQ ID NO: 21, 35, 20, 22, 25, 33, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34; (ii) CDR2 выбрана из группы, состоящей из SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и аминокислотных последовательностей, имеющих 1, 2 или 3 отличие(отличия) аминокислот от SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и (iii) CDR3 выбрана из группы, состоящей из SEQ ID NO: SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70; и аминокислотных последовательностей, имеющих 1, 2, 3 или 4 отличие(отличия) аминокислот от SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых (i) CDR1 выбрана из группы, состоящей из (a) SEQ ID NO: 22; и (b) аминокислотной последовательности, имеющей 1, 2, 3, 4, 5 или 6 отличие(отличий) аминокислот от SEQ ID NO: 22, где в положении 2 S заменен на R; в положении 3 A заменен на T; в положении 4 V заменен на F; в положении 6 V заменен на S; в положении 7 N заменен на Y; и/или в положении 10 A заменен на G; (ii) CDR2 представляет собой SEQ ID NO: 36; и (iii) CDR3 представляет собой SEQ ID NO: 54.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых (i) CDR1 представляет собой SEQ ID NO: 33; (ii) CDR2 выбрана из группы, состоящей из (c) SEQ ID NO: 50; и (d) аминокислотной последовательности, имеющей 1, 2, или 3 отличие (отличий) аминокислот от SEQ ID NO: 50, где в положении 8 M заменен на I; в положении 9 P заменен на T; и/или в положении 10 Y заменен на F; и (iii) CDR3 выбрана из группы, состоящей из (e) SEQ ID NO: 68; и (f) аминокислотной последовательности, имеющей 1 или 2 отличие (отличий) аминокислот от SEQ ID NO: 68, где в положении 5 F заменен на L; и/или в положении 11 D заменен на E.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых (i) CDR1 представляет собой SEQ ID NO: 28; (ii) CDR2 выбрана из группы, состоящей из (c) SEQ ID NO: 44; и (d) аминокислотной последовательности, имеющей 1, 2, или 3 отличие (отличий) аминокислот от SEQ ID NO: 44, где в положении 3 S заменен на T; в положении 4 R заменен на W; в положении 8 T заменен на I; и/или в положении 9 T заменен на L; и (iii) CDR3 выбрана из группы, состоящей из (e) SEQ ID NO: 62; и (f) аминокислотной последовательности, имеющей 1 или 2 отличие(отличий) аминокислот от SEQ ID NO: 62, где в положении 1 G заменен на S; и/или в положении 14 D заменен на E.
В предпочтительных вариантах осуществления всех аспектов по изобретению, одиночный вариабельный домен иммуноглобулина (ISVD) по изобретению, предпочтительно, состоит из или в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей CDR1, CDR2 и CDR3, как описано в настоящем описании выше и ниже. Предпочтительные каркасные последовательности описаны, например, в таблице A-2 ниже и могут быть использованы в ISVD по изобретению. Предпочтительно, CDR, описанные в таблице A-2, сочетают с соответствующими каркасными областями одной и той же конструкции ISVD.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD выбран из группы ISVD, где: CDR1 выбрана из группы, состоящей из SEQ ID NO: 21, 35, 20, 22, 25, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34; CDR2 выбрана из группы, состоящей из SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и CDR3 выбрана из группы, состоящей из SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD выбран из группы ISVD, где:
CDR1 представляет собой SEQ ID NO: 21, CDR2 представляет собой SEQ ID NO: 37, и CDR3 представляет собой SEQ ID NO: 55;
CDR1 представляет собой SEQ ID NO: 35, CDR2 представляет собой SEQ ID NO: 53, и CDR3 представляет собой SEQ ID NO: 118;
CDR1 представляет собой SEQ ID NO: 35, CDR2 представляет собой SEQ ID NO: 53, и CDR3 представляет собой SEQ ID NO: 71;
CDR1 представляет собой SEQ ID NO: 20, CDR2 представляет собой SEQ ID NO: 36, и CDR3 представляет собой SEQ ID NO: 54;
CDR1 представляет собой SEQ ID NO: 22, CDR2 представляет собой SEQ ID NO: 36, и CDR3 представляет собой SEQ ID NO: 54;
CDR1 представляет собой SEQ ID NO: 25, CDR2 представляет собой SEQ ID NO: 40, и CDR3 представляет собой SEQ ID NO: 58;
CDR1 представляет собой SEQ ID NO: 33, CDR2 представляет собой SEQ ID NO: 50, и CDR3 представляет собой SEQ ID NO: 68;
CDR1 представляет собой SEQ ID NO: 33, CDR2 представляет собой SEQ ID NO: 51, и CDR3 представляет собой SEQ ID NO: 69;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 44, и CDR3 представляет собой SEQ ID NO: 62;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 45, и CDR3 представляет собой SEQ ID NO: 63;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 43, и CDR3 представляет собой SEQ ID NO: 61;
CDR1 представляет собой SEQ ID NO: 24, CDR2 представляет собой SEQ ID NO: 39, и CDR3 представляет собой SEQ ID NO: 57;
CDR1 представляет собой SEQ ID NO: 23, CDR2 представляет собой SEQ ID NO: 38, и CDR3 представляет собой SEQ ID NO: 56;
CDR1 представляет собой SEQ ID NO: 26, CDR2 представляет собой SEQ ID NO: 41, и CDR3 представляет собой SEQ ID NO: 59;
CDR1 представляет собой SEQ ID NO: 27, CDR2 представляет собой SEQ ID NO: 119, и CDR3 представляет собой SEQ ID NO: 60;
CDR1 представляет собой SEQ ID NO: 27, CDR2 представляет собой SEQ ID NO: 42, и CDR3 представляет собой SEQ ID NO: 60;
CDR1 представляет собой SEQ ID NO: 29, CDR2 представляет собой SEQ ID NO: 46, и CDR3 представляет собой SEQ ID NO: 64;
CDR1 представляет собой SEQ ID NO: 30, CDR2 представляет собой SEQ ID NO: 47, и CDR3 представляет собой SEQ ID NO: 65;
CDR1 представляет собой SEQ ID NO: 31, CDR2 представляет собой SEQ ID NO: 48, и CDR3 представляет собой SEQ ID NO: 66;
CDR1 представляет собой SEQ ID NO: 32, CDR2 представляет собой SEQ ID NO: 49, и CDR3 представляет собой SEQ ID NO: 67; и
CDR1 представляет собой SEQ ID NO: 34, CDR2 представляет собой SEQ ID NO: 52, и CDR3 представляет собой SEQ ID NO: 70.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, в котором CDR1 представляет собой SEQ ID NO: 21, CDR2 представляет собой SEQ ID NO: 37, и CDR3 представляет собой SEQ ID NO: 55.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, в котором указанный ISVD выбран из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 8, 117, 12, 13, 14, 15 и 18.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид связывается с ADAMTS5 с KD между 1E-07 M и 1E-13 M, например, между 1E-08 M и 1E-12 M, предпочтительно, не более 1E-07 M, предпочтительно, ниже, чем 1E-08 M или 1E-09 M, или даже ниже, чем 1E 10 M, например, 5E-11 M, 4E-11 M, 3E 11 M, 2E-11 M, 1,7E-11 M, 1E 11 M или даже 5E-12 M, 4E-12 M, 3E 12 M, 1E-12 M, например, как определено посредством KinExA, или альтернативно, посредством Gyrolab.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует активность ADAMTS5 с IC50 между 1E-07 M и 1E-12 M, например, между 1E-08 M и 1E-11 M, например, как определено посредством анализа FRET для человека или AlphaLISA для человека.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует (ферментную) активность ADAMTS5 с IC50 не более 1E-07 M, предпочтительно, 1E-08 M, 5E-09 M или 4E-9 M, 3E-9 M, 2E-9 M, например, 1E-9 M.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид модулирует ADAMTS5 с EC50 между 1E-07 M и 1E-12 M, например, между 1E-08 M и 1E-11 M, например, как определено посредством ELISA связывания.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид связывается с ADAMTS5 со скоростью диссоциации менее чем 1E-04 с-1, например, как определено посредством SPR.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный ADAMTS5 представляет собой ADAMTS5 человека (SEQ ID NO: 149), ADAMTS5 быка (SEQ ID NO: 150), ADAMTS5 крысы (SEQ ID NO: 151), ADAMTS5 морской свинки (SEQ ID NO: 152), ADAMTS5 мыши (SEQ ID NO: 153) или ADAMTS5 яванского макака, (SEQ ID NO: 154), предпочтительно, ADAMTS5 человека, наиболее предпочтительно SEQ ID NO: 149.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид является антагонистом активности ADAMTS5, например, протеазной активности, такой как расщепление аггрекана, версикана, бревикана, нейрокана, декорина и/или бигликана, предпочтительно, расщепление аггрекана; предпочтительно, является антагонистом аггреканазной активности ADAMTS5.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид блокирует связывание ADAMTS5 с аггреканом по меньшей мере на 20%, например, по меньшей мере на 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или даже больше, например, как определено посредством FRET, AlplaLISA или ELISA.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует протеазную активность ADAMTS5, например, ингибирует протеолиз субстрата, такого как аггрекан, версикан, бревикан, нейрокан, декорин и/или бигликан, предпочтительно, аггрекан.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, где по меньшей мере 1 ISVD специфически связывается с ADAMTS, предпочтительно ADAMTS5, предпочтительно, выбранным из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18, предпочтительно, где указанные по меньшей мере 2 ISVD специфически связываются с ADAMTS, предпочтительно, ADAMTS5.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, содержащему два или более ISVD, которые специфически связываются с ADAMTS5, где (a) по меньшей мере «первый» ISVD специфически связывается с первой антигенной детерминантой, эпитопом, частью, доменом, субъединицей или конформацией ADAMTS5; и где (b) по меньшей мере «второй» ISVD специфически связывается с второй антигенной детерминантой, эпитопом, частью, доменом, субъединицей или конформацией ADAMTS5, отличной от первой антигенной детерминанты, эпитопа, части, домена, субъединицы или конформации, соответственно, предпочтительно где указанный «первый» ISVD, специфически связывающий ADAMTS5, выбран из группы, состоящей из SEQ ID NO: 2, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 8, 117, 12, 13, 14, 15 и 18, предпочтительно, где указанный «второй» ISVD, специфически связывающий ADAMTS5, представляет собой SEQ ID NO: 118 или 19, даже более предпочтительно, указанный полипептид выбран из группы, состоящей из SEQ ID NO: 127 (клон 130 049-093-Alb), SEQ ID NO: 126 (клон 129 2F3-093-Alb), SEQ ID NO: 127 (клон 130 049-093-Alb) и SEQ ID NO: 128 (клон 131 9D3-093-Alb).
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, дополнительно содержащему ISVD, связывающий сывороточный альбумин, предпочтительно где указанный ISVD, связывающий сывороточный альбумин, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых CDR1 представляет собой SEQ ID NO: 146, CDR2 представляет собой SEQ ID NO: 147, и CDR3 представляет собой SEQ ID NO: 148, даже более предпочтительно, где указанный ISVD, связывающий сывороточный альбумин, выбран из группы, состоящей из ALB8 (SEQ ID NO: 131), ALB23 (SEQ ID NO: 132), ALB129 (SEQ ID NO: 133), ALB132 (SEQ ID NO: 134), ALB11 (SEQ ID NO: 135), ALB11 (S112K)-A (SEQ ID NO: 136), ALB82 (SEQ ID NO: 137), ALB82-A (SEQ ID NO: 138), ALB82-AA (SEQ ID NO: 139), ALB82-AAA (SEQ ID NO: 140), ALB82-G (SEQ ID NO: 141), ALB82-GG (SEQ ID NO: 142), ALB82-GGG (SEQ ID NO: 143), ALB92 (SEQ ID NO: 144), и ALB223 (SEQ ID NO: 145), даже более предпочтительно, где указанный полипептид выбран из группы, состоящей из SEQ ID NO: 129 (клон 577 2F3SO-Alb), SEQ ID NO: 130 (клон 579 2F3 SO-093-Alb), SEQ ID NO: 120 (клон 4 2A12-Alb), SEQ ID NO: 121 (клон 5 2D7-Alb), SEQ ID NO: 122 (клон 6 2F3-Alb), SEQ ID NO: 123 (клон 69 049-Alb), SEQ ID NO: 124 (клон 70 9D3-Alb), SEQ ID NO: 125 (клон 71 3B2-Alb), SEQ ID NO: 126 (клон 129 2F3-093-Alb), SEQ ID NO: 127 (клон 130 049-093-Alb), и SEQ ID NO: 128 (клон 131 9D3-093-Alb).
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, дополнительно содержащему по меньшей мере один ISVD, специфически связывающий аггрекан, предпочтительно, выбранный из группы, состоящей из SEQ ID NO: 156 (нанотело 00745 PEA114F08) и SEQ ID NO: 157 (нанотело 00747 PEA604F02).
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, специфически связывающих аггрекан, где указанные по меньшей мере 2 ISVD, специфически связывающие аггрекан, могут являться одинаковыми или различными, предпочтительно где указанные по меньшей мере 2 ISVD, специфически связывающие аггрекан, независимо выбраны из группы, состоящей из SEQ ID NO: 156 и 157, даже более предпочтительно, где указанный ISVD, специфически связывающий аггрекан, специфически связывает аггрекан человека [SEQ ID NO: 155]. Предпочтительно, указанный ISVD, специфически связывающий аггрекан, специфически связывает аггрекан собаки (см. также таблица 2), аггрекан быка, аггрекан крысы; аггрекан свиньи; аггрекан мыши, аггрекан кролика; аггрекан яванского макака и/или аггрекан макака-резуса. Предпочтительно, указанный ISVD, специфически связывающий аггрекан, предпочтительно связывается с хрящевой тканью, такой как хрящ и/или мениск.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид имеет стабильность в течение по меньшей мере 7 суток, например, 14 суток, 21 суток, 1 месяца, 2 месяцев или даже 3 месяцев в синовиальной жидкости (SF) при 37°C.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанные по меньшей мере два ISVD напрямую связаны друг с другом или связаны посредством линкера, предпочтительно, указанный линкер выбран из группы, состоящей из SEQ ID NO: 158 to 174 (т.е. SEQ ID NO: 158, 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173 и 174), предпочтительно, SEQ ID NO: 169.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, дополнительно содержащему C-концевое удлинение, предпочтительно, где указанное C-концевое удлинение представляет собой C-концевое удлинение (X)n, в котором n представляет собой 1-10, предпочтительно 1-5, например, 1, 2, 3, 4 или 5 (и предпочтительно, 1 или 2, например, 1); и каждый X представляет собой (предпочтительно, природный) аминокислотный остаток, который выбран независимо, и предпочтительно, независимо выбран из группы, состоящей из аланина (A), глицина (G), валина (V), лейцина (L) или изолейцина (I).
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид имеет по меньшей мере 80%, 90%, 95% или 100% идентичность последовательности с любой из SEQ ID NO: 1-19 (т.е. SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 и 19), 116-117 или 120-130 (т.е. SEQ ID NO: 120, 121, 122, 123, 124, 125, 126, 127, 128, 129 и 130).
В дополнительном предпочтительном аспекте, изобретение относится к способу лечения и/или предотвращения заболеваний или нарушений у индивидуума, например, тех, в которые вовлечена активность ADAMTS5, включающему введение полипептида по любому из пп. 1-43 указанному индивидууму в количестве, эффективном для лечения или предотвращения симптома указанного заболевания или нарушения, предпочтительно, где указанные заболевания или нарушения выбраны из группы, состоящей из артропатий и хондродистрофий, артритного заболевания, такого как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматического разрыва или открепления, ахондроплазии, реберного хондрита, спондилоэпиметафизарной дисплазии, межпозвонковой грыжи, заболевания дегенерации поясничного диска, дегенеративного заболевания сустава, и рецидивирующих полихондрита, рассекающего остеохондрита и аггреканопатий. Более предпочтительно, указанное заболевание или нарушение представляет собой артритное заболевание и наиболее предпочтительно, остеоартрит.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, для использования в качестве лекарственного средства.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, для использования в лечении или предотвращении симптома ассоциированного с ADAMTS5 заболевания, например, такого как, артропатии и хондродистрофии, артритное заболевание, такое как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматический разрыв или открепление, ахондроплазия, реберный хондрит, спондилоэпиметафизарная дисплазия, межпозвонковая грыжа, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава, и рецидивирующие полихондрит, рассекающий остеохондрит и аггреканопатии. Более предпочтительно, указанное заболевание представляет собой артритное заболевание и наиболее предпочтительно, остеоартрит.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид перекрестно блокирует связывание с ADAMTS5 по меньшей мере одного из полипептидов, представленных любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18, и/или его связывание с ADAMTS5 перекрестно блокировано по меньшей мере одним из полипептидов, представленных любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
В дополнительном предпочтительном аспекте, изобретение относится к полипептиду, который перекрестно блокирует связывание с ADAMTS5 полипептида, представленного любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18, и/или связывание которого с ADAMTS5 перекрестно блокировано по меньшей мере одним из полипептидов, представленных любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18, где указанный полипептид содержит по меньшей мере один VH, VL, dAb, одиночный вариабельный домен иммуноглобулина (ISVD), специфически связывающие ADAMTS5, где связывание с ADAMTS5 модулирует активность ADAMTS5.
Другие аспекты, преимущества, приложения и применения полипептидов и композиций станут понятными из дальнейшего описания в настоящем документе. Некоторые документы процитированы на протяжении текста этого описания. Полное содержание каждого из документов, процитированных в настоящем описании (включая все патенты, патентные заявки, научные публикации, спецификации производителя, инструкции и т.д.), либо выше, либо ниже, таким образом, приведено в качестве ссылки. Ничего в настоящем описании не следует рассматривать как допущение того, что это изобретение не имеет прав задним числом на такое описание предшествующего изобретения.
4. ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: ISVD, связывающие ADAMTS5, не ингибируют активность MMP1 (A) или MMP14 (B).
Фигура 2: ISVD, связывающие ADAMTS5, не ингибируют активность ADAMTS4
Фигура 3: Уровни связывания предсуществующих антител (preAb) с конструкцией нанотела 581 (A) и конструкцией нанотела 579 (B) из 3 групп образцов доноров, полученных от: здоровых субъектов, субъектов с остеоартритом и доноров с остаточным связыванием preAb.
Фигура 4: Конструкция нанотела 581 («C011400581») не связывает ADAMTS1 человека (A) и не связывает ADAMTS4 человека или ADAMTS15 человека (B).
Фигура 5: Активность в системе человеческого эксплантата. Показано накопление GAG в супернатанте через 7 суток.
Фигура 6: A) Зависимость от времени высвобождения GAG в супернатанте в бычьей системе совместной культуры.
B) Расчет площади под кривой (AUC) для высвобождения GAG с течением времени (7-28 суток). Количество повторов n=4. Данные показаны в формате «ящик с усами» от мин. до макс.
C) Данные показывают содержание GAG [нг/мг хряща] после расщепления папаином. Количество повторов n=4. Данные показаны в формате «ящик с усами» от мин. до макс.
Фигура 7: Расчет площади под кривой (AUC) для высвобождения exAGNx1 с течением времени (7-28 суток). Количество повторов n=4. Данные показаны в формате «ящик с усами» от мин. до макс.
Фигура 8: Расчет площади под кривой (AUC) для высвобождения C2M с течением времени (7-28 суток). Количество повторов n=4. Данные показаны в формате «ящик с усами» от мин. до макс.
Фигура 9: Расчет площади под кривой (AUC) для высвобождения C3M с течением времени (7-28 суток). Количество повторов n=4. Данные показаны в формате «ящик с усами» от мин. до макс.
Фигура 10: Ингибирование активности аггреканазы у NHP.
Фигура 11: Ингибирование дегенерации хряща. Медианная сумма дегенерации хряща показана при профилактическом (A) и терапевтическом (B) лечении в модели DMM на мышах.
Фигура 12: Симптоматическое поведение в модели хирургического OA на крысах.
Фигура 13: Ширина медиальной дегенерации хряща большеберцовой кости.
Фигура 14: Эффект нанотела против ADAMTS-5 на вызванную аггреканазой деградацию аггрекана ex vivo в культурах хряща (A, B, C) и совместных культурах хряща и синовиальной мембраны (D, E). Перечисленные концентрации представляют собой концентрацию нанотела. Статистический анализ проводили с использованием обычного однофакторного ANOVA или двухфакторного ANOVA. Статистическую значимость признавали, когда p<0,05.
5. ПОДРОБНОЕ ОПИСАНИЕ
Остается необходимость в безопасных и эффективных лекарственных средствах против OA, в частности, DMOAD. Эти лекарственные средства должны удовлетворять различным и часто противоположным требованиям, особенно при предполагаемом широко применимом формате. Таким образом, формат должен, предпочтительно, являться применимым для широкого диапазона пациентов. Формат должен, предпочтительно, являться безопасным и не должен индуцировать инфекции из-за частого введения. Кроме того, формат должен, предпочтительно, являться дружественным для пациента, например, формат должен позволять удобный режим дозирования и способ введения, например, системное введение. Например, является предпочтительным, чтобы формат не удалялся из кровотока немедленно после введения. Однако продление времени полужизни, предпочтительно, не должно приводить к неспецифической активности и побочным эффектам или ограничивать эффективность.
Настоящее изобретение реализует по меньшей мере одно из этих требований.
На основании не являющихся общепринятыми способов скрининга, характеризации и комбинирования, авторы настоящего изобретения неожиданно обнаружили, что одиночные вариабельные домены иммуноглобулинов (ISVD) действовали необычайно хорошо в экспериментах in vitro и in vivo.
Кроме того, авторам настоящего изобретения удалось переконструировать ISVD, дополнительно превосходящие сравнительные лекарственные средства в облегчении OA. Кроме того, показано также, что ISVD по изобретению являются значительно более безопасными, чем соединения предшествующего уровня техники.
Целью настоящего изобретения является предоставление полипептидов, являющихся антагонистами ADAMTS, в частности ADAMTS5, с улучшенными профилактическими, терапевтическими и/или фармакологическими свойствами, включая лучший профиль безопасности, по сравнению с аминокислотными последовательностями и антителами предшествующего уровня техники.
Соответственно, настоящее изобретение относится к ISVD и полипептидам, которые нацелены против и/или которые могут специфически связывать (как определено в настоящем описании) ADAMTS5.
Соответственно, настоящее изобретение относится к ISVD и полипептидам, которые нацелены против и/или которые могут специфически связывать (как определено в настоящем описании) ADAMTS и модулировать их активность, в частности, к полипептиду, содержащему по меньшей мере один ISVD, специфически связывающий ADAMTS5, где связывание с ADAMTS5 модулирует активность ADAMTS5.
Если не указано или не определено иное, все используемые термины имеют их общепринятое в данной области значение, понятное специалисту в данной области. Приведена ссылка, например, на стандартные руководства, такие как Sambrook et al. (Molecular Cloning: A Laboratory Manual (2nd Ed.) Vols. 1-3, Cold Spring Harbor Laboratory Press, 1989), F. Ausubel et al. (Current protocols in molecular biology, Green Publishing and Wiley Interscience, New York, 1987), Lewin (Genes II, John Wiley & Sons, New York, N.Y., 1985), Old et al. (Principles of Gene Manipulation: An Introduction to Genetic Engineering (2nd edition) University of California Press, Berkeley, CA, 1981); Roitt et al. (Immunology (6th Ed.) Mosby/Elsevier, Edinburgh, 2001), Roitt et al. (Roitt's Essential Immunology (10th Ed.) Blackwell Publishing, UK, 2001), и Janeway et al. (Immunobiology (6th Ed.) Garland Science Publishing/Churchill Livingstone, New York, 2005), так же, как на общий уровень техники, процитированные в настоящем описании.
Если не указано иное, все методы, стадии, способы и манипуляции, не описанные конкретно подробно, могут быть осуществлены и осуществлены способом, известным по существу, как понятно специалисту в данной области. Снова приведена ссылка, например, на стандартные руководства и общий уровень техники, упомянутые в настоящем описании, и на дополнительные ссылки, процитированные в этих документах; так же как, например, на следующие обзоры Presta (Adv. Drug Deliv. Rev. 58 (5-6): 640-56, 2006), Levin and Weiss (Mol. Biosyst. 2(1): 49-57, 2006), Irving et al. (J. Immunol. Methods 248(1-2): 31-45, 2001), Schmitz et al. (Placenta 21 Suppl. A: S106-12, 2000), Gonzales et al. (Tumour Biol. 26(1): 31-43, 2005), в которых описаны способы белковой инженерии, такие как аффинное созревание и другие способы улучшения специфичности и других желательных свойств белков, таких как иммуноглобулины.
Следует отметить, что, в рамках изобретения, формы единственного числа включают ссылки на множественное число, если контекст явно не указывает на иное. Таким образом, например, ссылка на «реагент» включает один или более таких различных реагентов, и ссылка на «способ» включает ссылку на эквивалентные стадии и способы, известные обычным специалистам в данной области, которые могут модифицировать или заменять способы, описанные в настоящем описании.
Если не указано иное, термин «по меньшей мере», предшествующий серии элементов, следует понимать как относящийся к каждому элементу в сериях. Специалисту в данной области известно, или он может определять, с использованием не более, чем общепринятых экспериментов, множество эквивалентов конкретных вариантов осуществления изобретения, описанных в настоящем описании. Такие эквиваленты предназначены для включение в настоящее изобретение.
Термин «и/или» при использовании в любом месте настоящего описания включает значение «и», «или» и «все или любая другая комбинация элементов, связанных указанным термином».
Термин «около» или «приблизительно», в рамках изобретения, означает в переделах 20%, предпочтительно, в переделах 15%, более предпочтительно, в переделах 10%, и наиболее предпочтительно, в переделах 5% от данного значения или диапазона.
На протяжении этого описания и следующей ниже формулы изобретения, если контекст не требует иного, слово «включать», и его варианты, такие как «включает» и «включающий», понимают как подразумевающие включение указанного целого числа или стадии, или группы целых чисел или стадий, но не исключение любого другого целого числа или стадии, или группы целых чисел или стадий. В рамках изобретения, термин «включающий» можно заменять термином «содержащий» или «заключающий», или иногда, при использовании в настоящем описании, термином «имеющий».
Термин «последовательность», в рамках изобретения (например, в терминах, подобных терминам «последовательность иммуноглобулина», «последовательность антитела», «последовательность вариабельного домена», «последовательность VHH» или «белковая последовательность»), следует в общем понимать как включающий соответствующую аминокислотную последовательность, так же как кодирующие ее последовательности нуклеиновых кислот или нуклеотидов, если контекст не требует более ограниченной интерпретации.
Аминокислотные последовательности интерпретируют для обозначения одиночной аминокислоты или неразветвленной последовательности из двух или более аминокислот, в зависимости от контекста. Нуклеотидные последовательности интерпретируют для обозначения неразветвленной последовательности из 3 или более нуклеотидов.
Аминокислоты представляют собой L-аминокислоты, обычно обнаруживаемые в природных белках. Аминокислотные остатки указывают в соответствии со стандартным трехбуквенным или однобуквенным кодом аминокислот. Приведена ссылка, например, на таблицу A-2 на странице 48 WO 08/020079. Аминокислотные последовательности, содержащие D-аминокислоты, не предназначены для включения в это определение. Любую аминокислотную последовательность, содержащую посттрансляционно модифицированные аминокислоты, можно описать как аминокислотную последовательность, первоначально транслированную, с использованием символов, показанных в этой таблице A-2, с положениями модификаций; например, гидроксилирования или гликозилирования, однако, эти модификации не следует явно указывать в аминокислотной последовательности. Любой пептид или белок, который можно экспрессировать в форме последовательности с модифицированными связями, перекрестными сшивками и концевыми кэпами, непептидильными связями и т.д., включен в это определение, все, как известно в данной области.
Термины «белок», «пептид», «белок/пептид» и «полипептид» используют взаимозаменяемо на протяжении этого описания, и каждый имеет одинаковое значение для целей этого описания. Каждый термин относится к органическому соединению, составленному из линейной цепи из двух или более аминокислот. Соединение может иметь десять или более аминокислот; двадцать пять или более аминокислот; пятьдесят или более аминокислот; сто или более аминокислот, двести или более аминокислот и даже триста или более аминокислот. Специалисту в данной области понятно, что полипептиды, как правило, содержат меньше аминокислот, чем белки, хотя не существует принятой в данной области точки отсечения для количества аминокислот, устанавливающей отличия полипептидов и белков; что полипептиды можно получать химическим синтезом или рекомбинантными способами; и что белки, как правило, получают in vitro или in vivo рекомбинантными способами, как известно в данной области. По традиции, амидная связь в первичной структуре полипептидов расположена в том порядке, в котором записаны аминокислоты, в котором амино-конец (N-конец) полипептида всегда находится слева, в то время как кислый конец (C-конец) находится справа.
Последовательность нуклеиновой кислоты или аминокислотную последовательность считают «(находящейся в) (по существу) выделенной (форме)» - например, по сравнению с реакционной средой или культуральной средой, из которой она получена - когда она отделена по меньшей мере от одного другого компонента, с которым она обычно ассоциирована в указанных источнике или среде, такого как другая нуклеиновая кислота, другой белок/полипептид, другой биологический компонент или макромолекула, или по меньшей мере один контаминант, примесь или минорный компонент. В частности, последовательность нуклеиновой кислоты или аминокислотную последовательность считают «(по существу) выделенной», когда она очищена по меньшей мере в 2 раза, в частности, по меньшей мере в 10 раз, более конкретно, по меньшей мере в 100 раз и вплоть до 1000 раз или более. Нуклеиновая кислота или аминокислота, находящаяся «в (по существу) выделенной форме», является, предпочтительно, по существу гомогенной, как определено с использованием подходящего способа, такого как подходящий хроматографический способ, например, электрофорез в полиакриламидном геле.
Когда говорят, что нуклеотидная последовательность или аминокислотная последовательность «содержит» другую нуклеотидную последовательность или аминокислотную последовательность, соответственно, или «в основном состоит из» другой нуклеотидной последовательности или аминокислотной последовательности, это может означать, что последняя нуклеотидная последовательность или аминокислотная последовательность введена в первую упомянутую нуклеотидную последовательность или аминокислотную последовательность, соответственно, но более обычно, это в общем означает, что первая упомянутая нуклеотидная последовательность или аминокислотная последовательность содержит внутри своей последовательности фрагмент из нуклеотидов или аминокислотных остатков, соответственно, имеющий такую же нуклеотидную последовательность или аминокислотную последовательность, соответственно, как последняя последовательность, независимо от того, каким образом первая упомянутая последовательность была фактически образована или получена (что может, например, быть осуществлено любым подходящим способом, описанным в настоящем описании). В качестве неограничивающего примера, когда говорят, что полипептид по изобретению содержит одиночный вариабельный домен иммуноглобулина («ISVD»), это может означать, что указанная последовательность одиночного вариабельного домена иммуноглобулина введена в последовательность полипептида по изобретению, но более обычно, это в общем означает, что полипептид по изобретению содержит внутри своей последовательности последовательность одиночных вариабельных доменов иммуноглобулинов, независимо от того, каким образом указанный полипептид по изобретению образован или получен. Также, когда говорят, что нуклеиновая кислота или нуклеотидная последовательность содержит другую нуклеотидную последовательность, первая упомянутая нуклеиновая кислота или нуклеотидная последовательность, предпочтительно, является такой, что когда она экспрессируется до продукта экспрессии (например, полипептида), аминокислотная последовательность, кодируемая последней нуклеотидной последовательностью, формирует часть указанного продукта экспрессии (иными словами, что последняя нуклеотидная последовательность находится в той же рамке считывания, что и первая упомянутая, более длинная нуклеиновая кислота или нуклеотидная последовательность). Также, когда говорят, что конструкция по изобретению содержит полипептид или ISVD, это может означать, что указанная конструкция по меньшей мере включает указанный полипептид или ISVD, соответственно, но более обычно, это означает, что указанная конструкция включает группы, остатки (например, аминокислотные остатки), фрагменты и/или связывающие единицы, в дополнение к указанному полипептиду или ISVD, независимо от того, каким образом указанный полипептид или ISVD связан с указанными группами, остатками (например, аминокислотными остатками), фрагментами и/или связывающими единицами и независимо от того, каким образом указанная конструкция образована или получена.
Под «по существу состоит из» понимают, что ISVD, используемый по изобретению, либо является точно таким же, как ISVD по изобретению, либо соответствует ISVD по изобретению, имея ограниченное количество аминокислотных остатков, например, 1-20 аминокислотных остатков, например, 1-10 аминокислотных остатков и предпочтительно 1-6 аминокислотных остатков, например, 1, 2, 3, 4, 5 или 6 аминокислотных остатков, добавленных на амино-конце, на карбокси-конце, или как на амино-конце, так и на карбокси-конце ISVD.
Для целей сравнения двух или более нуклеотидных последовательностей, процент «идентичности последовательности» между первой нуклеотидной последовательностью и второй нуклеотидной последовательностью можно рассчитывать посредством деления [количества нуклеотидов в первой нуклеотидной последовательности, которые являются идентичными нуклеотидам в соответствующих положениях во второй нуклеотидной последовательности] на [общее количество нуклеотидов в первой нуклеотидной последовательности] и умножения на [100%], при котором каждую делецию, вставку, замену или добавление нуклеотида во второй нуклеотидной последовательности - по сравнению с первой нуклеотидной последовательностью - рассматривают как отличие в одном нуклеотиде (положении). Альтернативно, степень идентичности последовательности между двумя или более нуклеотидными последовательностями можно рассчитывать с использованием известного компьютерного алгоритма выравнивания последовательностей, такого как NCBI Blast v2.0, с использованием стандартных установок. Некоторые другие способы, компьютерные алгоритмы и условия для определения степени идентичности последовательности, например, описаны в WO 04/037999, EP 0967284, EP 1085089, WO 00/55318, WO 00/78972, WO 98/49185 и GB 2357768. Обычно, для цели определения процента «идентичности последовательности» между двумя нуклеотидными последовательностями, в соответствии со способом расчета, описанным в настоящем описании выше, нуклеотидную последовательность с наибольшим количеством нуклеотидов принимают как «первую» нуклеотидную последовательность, и другую нуклеотидную последовательность принимают как «вторую» нуклеотидную последовательность.
Для целей сравнения двух или более аминокислотных последовательностей, процент «идентичности последовательности» между первой аминокислотной последовательностью и второй аминокислотной последовательностью (также обозначенный в настоящем описании как «идентичность аминокислот») можно рассчитывать посредством деления [количества аминокислотных остатков в первой аминокислотной последовательностью, которые являются идентичными аминокислотным остаткам в соответствующих положениях во второй аминокислотной последовательности] на [общее количество аминокислотных остатков в первой аминокислотной последовательности] и умножения на [100%], при котором каждую делецию, вставку, замену или добавление аминокислотного остатка во второй аминокислотной последовательности - по сравнению с первой аминокислотной последовательностью - рассматривают как отличие в одном аминокислотном остатке (положение), т.е. как «отличие аминокислоты», как определено в настоящем описании. Альтернативно, степень идентичности последовательности между двумя аминокислотными последовательностями можно рассчитывать с использованием известного компьютерного алгоритма, такого как упомянутые выше, для определения степени идентичности последовательностей для нуклеотидных последовательностей, снова с использованием стандартных установок. Обычно, для цели определения процента «идентичности последовательности» между двумя аминокислотными последовательностями, в соответствии со способом расчета, описанным в настоящем описании выше, аминокислотную последовательность с наибольшим количеством аминокислотных остатков принимают как «первую» аминокислотную последовательность, и другую аминокислотную последовательность принимают как «вторую» аминокислотная последовательность.
Также, при определении степени идентичности последовательности между двумя аминокислотными последовательностями, специалист в данной области может принимать во внимание так называемые «консервативные» аминокислотные замены, которые в общем можно описать как аминокислотные замены, в которых аминокислотный остаток заменен на другой аминокислотный остаток сходной химической структуры, и которые оказывают небольшое влияние или по существу не оказывают влияния на функцию, активность или другие биологические свойства полипептида. Такие консервативные аминокислотные замены хорошо известны в данной области, например, из WO 04/037999, GB 335768, WO 98/49185, WO 00/46383 и WO 01/09300; и (предпочтительные) типы и/или комбинации таких замен можно выбирать на основании соответствующих объяснений из WO 04/037999, так же как WO 98/49185, и из дополнительных ссылок, процитированных в этих документах.
Такие консервативные замены предпочтительно представляют собой замены, в которых одна аминокислота из следующих групп (a) - (e) заменена на другой аминокислотный остаток в пределах той же группы: (a) небольшие алифатические, неполярные или слабо полярные остатки: Ala, Ser, Thr, Pro и Gly; (b) полярные, отрицательно заряженные остатки и их (незаряженные) амиды: Asp, Asn, Glu и Gln; (c) полярные, положительно заряженные остатки: His, Arg и Lys; (d) большие алифатические, неполярные остатки: Met, Leu, Ile, Val и Cys; и (e) ароматические остатки: Phe, Tyr и Trp. Особенно предпочтительными консервативными заменами являются следующие: Ala на Gly или на Ser; Arg на Lys; Asn на Gln или на His; Asp на Glu; Cys на Ser; Gln на Asn; Glu на Asp; Gly на Ala или на Pro; His на Asn или на Gln; Ile на Leu или на Val; Leu на Ile или на Val; Lys на Arg, на Gln или на Glu; Met на Leu, на Tyr или на Ile; Phe на Met, на Leu или на Tyr; Ser на Thr; Thr на Ser; Trp на Tyr; Tyr на Trp; и/или Phe на Val, на Ile или на Leu.
Любые аминокислотные замены, применимые к полипептидам, описанным в настоящем описании, могут быть также основаны на анализе частот вариантов аминокислот между гомологичными белками из различных видов, разработанном в Schulz et al. («Principles of Protein Structure», Springer-Verlag, 1978), на анализах структурообразующих потенциалов, разработанном в Chou and Fasman (Biochemistry 13: 211, 1974; Adv. Enzymol., 47: 45-149, 1978), и на анализе паттернов гидрофобности в белках, разработанном в Eisenberg et al. (Proc. Natl. Acad Sci. USA 81: 140-144, 1984), Kyte and Doolittle (J. Molec. Biol. 157: 105-132, 1981), и Goldman et al. (Ann. Rev. Biophys. Chem. 15: 321-353, 1986), полное содержание всех из которых приведено в настоящем описании в качестве ссылки. Информация о первичной, вторичной и третичной структуре нанотел приведена в описании в настоящем документе и на общем уровне техники, процитированном выше. Также, для этой цели, кристаллическая структура домена VHH ламы приведена, например, в Desmyter et al. (Nature Structural Biology, 3: 803, 1996), Spinelli et al. (Natural Structural Biology, 3: 752-757, 1996) и Decanniere et al. (Structure, 7 (4): 361, 1999). Дополнительную информацию о некоторых аминокислотных остатках, которые в общепринятых VH доменах формируют поверхность контакта VH/VL, и потенциальной камелизации в этих положениях можно обнаружить и на предшествующем уровне техники, процитированном выше.
Аминокислотные последовательности и последовательности нуклеиновой кислоты называют «точно такими же», если они имеют 100% идентичность последовательности (как определено в настоящем описании) на протяжении полной длины.
При сравнении двух аминокислотных последовательностей, термин «отличие аминокислот(ы)» относится к вставке, делеции или замене одного аминокислотного остатка в некотором положении первой последовательности, по сравнению со второй последовательностью; понятно, что две аминокислотные последовательности могут содержать одно, два или более таких отличий аминокислот. Более конкретно, в ISVD и/или полипептидах по настоящему изобретению, термин «отличие аминокислот(ы)» относится к вставке, делеции или замене одного аминокислотного остатка в некотором положении последовательности CDR, указанной в b), d) или f), по сравнению с последовательностью CDR, соответственно, из a), c) или e); понятно, что последовательность CDR из b), d) и f) может содержать одно, два, три, четыре или максимум пять таких отличий аминокислот, по сравнению с последовательностью CDR, соответственно, из a), c) или e).
«Отличие аминокислот(ы)» может представлять собой любую одну, две, три, четыре или максимум пять замен, делеций или вставок, или любую их комбинацию, которые либо улучшают свойства связывающей ADAMTS5 молекулы по изобретению, такой как полипептид по изобретению, или которые по меньшей мере не ухудшают слишком сильно желательные свойства или баланс или комбинацию желательных свойств связывающей ADAMTS5 молекулы по изобретению, такой как полипептид по изобретению. В этом отношении, полученная связывающая ADAMTS5 молекула по изобретению, такая как полипептид по изобретению, должна по меньшей мере связывать ADAMTS5 с такой же, приблизительно такой же или более высокой аффинностью, по сравнению с полипептидом, содержащим одну или более последовательностей CDR без одной, двух, трех, четырех или максимум, пяти замен, делеций или вставок. Аффинность можно измерять любым подходящим способом, известным в данной области, но, предпочтительно, ее измеряют способом, как описано в разделе примеры.
В этом отношении, аминокислотная последовательность CDR в соответствии с b), d) и/или f), как указано ниже, может представлять собой аминокислотную последовательность, полученную из аминокислотной последовательности в соответствии с a), c) и/или e), соответственно, посредством аффинного созревания с использованием одного или более способов аффинного созревания, как известно по существу, или как описано в примерах. Например, и в зависимости от организма-хозяина, используемого для экспрессии полипептида по изобретению, такие делеции и/или замены можно конструировать таким способом, что один или более участков посттрансляционной модификации (таких как один или более участков гликозилирования) удаляют, как находится в компетенции специалиста в данной области (ср. примеры).
«Семейство нанотел», «семейство VHH» или «семейство», в рамках изобретения относится к группе последовательностей нанотел и/или VHH, которые имеют идентичную длину (т.е. они имеют одинаковое количество аминокислот в последовательности) и у которых аминокислотная последовательность между положением 8 и положением 106 (в соответствии с нумерацией по Kabat) имеет идентичность аминокислотной последовательности 89% или более.
Термины «эпитоп» и «антигенная детерминанта», которые можно использовать взаимозаменяемо, относятся к части макромолекулы, такой как полипептид или белок, узнаваемой антигенсвязывающими молекулами, такими как иммуноглобулины, общепринятые антитела, одиночные вариабельные домены иммуноглобулинов и/или полипептиды по изобретению, и более конкретно, антигенсвязывающим участком указанных молекул. Эпитопы определяют минимальный участок связывания для иммуноглобулина, и таким образом, представляют собой мишень для специфичности иммуноглобулина.
Часть антигенсвязывающей молекулы (такой как иммуноглобулин, общепринятое антитело, одиночный вариабельный домен иммуноглобулина и/или полипептид по изобретению), узнающую эпитоп, называют «паратопом».
Аминокислотную последовательность (такую как одиночный вариабельный домен иммуноглобулина, антитело, полипептид по изобретению, или в общем антигенсвязывающий белок или полипептид, или его фрагмент), которая может «связываться с» или «специфически связываться с», которая «имеет аффинность для» и/или которая «имеет специфичность для» определенного эпитопа, антигена или белка (или по меньшей мере для одной его части, фрагмента или эпитопа), называют «против» или «нацеленной против» указанного эпитопа, антигена или белка, или она является «связывающей» молекулой по отношению к такому эпитопу, антигену или белку, или ее называют «анти»-эпитоп, «анти»-антиген или «анти»-белок (например, «анти»-ADAMTS5).
Аффинность обозначает силу или стабильность молекулярного взаимодействия. Аффинность обычно приводят как KD, или константу диссоциации, имеющую размерность моль/литр (или M). Аффинность можно также выражать как константу связывания, KA, которая равна 1/KD и имеет размерность (моль/литр)-1 (или M-1). В настоящем описании, стабильность взаимодействия между двумя молекулами может быть в основном выражена в отношении значения KD их взаимодействия; специалисту в данной области понятно, что с учетом отношения KA =1/KD, определение силы молекулярного взаимодействия по его значению KD можно также использовать для расчета соответствующего значения KA. Значение KD характеризует силу молекулярного взаимодействия также в термодинамическом смысле, поскольку оно относится к изменению свободной энергии (DG) связывания посредством хорошо известного соотношения DG=RT.ln(KD) (эквивалентно, DG=-RT.ln(KA)), где R равно газовой постоянной, T равно абсолютной температуре, и ln обозначает натуральный логарифм.
KD для биологических взаимодействий, которые считают значительными (например, специфическими), как правило, лежат в диапазоне от 10-12 M (0,001 нМ) до 10-5 M (10000 нМ). Чем сильнее взаимодействие, тем ниже его KD.
KD можно также выражать как соотношение скорости диссоциации комплекса, обозначенной как koff, к скорости его связывания, обозначенной kon (так что KD =koff/kon и KA=kon/koff). Скорость диссоциации koff имеет размерность с-1 (где с представляет собой обозначение SI для секунды). kon для скорости связывания имеет размерность M-1с-1. Скорость связывания может меняться между 102 M-1с-1 и приблизительно 107 M-1с-1, приближаясь к константе скорости ограниченного диффузией связывания для бимолекулярного взаимодействия. Скорость диссоциации связана с временем полужизни данного молекулярного взаимодействия соотношением t1/2=ln(2)/koff. Скорость диссоциации может меняться между 10-6 с-1 (почти необратимый комплекс с t1/2 несколько суток) и 1 с-1 (t1/2=0,69 с).
Специфическое связывание антигенсвязывающего белка, такого как ISVD, с антигеном или антигенной детерминантой можно определять посредством любого подходящего способа, известного по существу, включая, например, анализы насыщающего связывания и/или анализы конкурентного связывания, такие как радиоиммуноанализы (RIA), ферментные иммуноанализы (EIA) и конкурентные сэндвич-анализы, и их различные варианты, известные по существу в данной области; так же как посредством других способов, упомянутых в настоящем описании.
Аффинность молекулярного взаимодействия между двумя молекулами можно измерять посредством различных способов, известных по существу, таких как хорошо известный биосенсорный способ поверхностного плазмонного резонанса (SPR) (см., например, Ober et al., 2001, Intern. Immunology 13: 1551-1559) где одну молекулу иммобилизуют на биосенсорном чипе, и другую молекулу пропускают над иммобилизованной молекулой в проточных условиях, получая измерения kon, koff, и таким образом, значения KD (или KA). Это можно, например, проводить с использованием хорошо известных устройств BIACORE® (Pharmacia Biosensor AB, Uppsala, Sweden). Анализ кинетического исключения (KINEXA®) (Drake et al., 2004, Analytical Biochemistry 328: 35-43) измеряет события связывания в растворе без мечения партнеров по связыванию и основан на кинетическом исключении диссоциации комплекса. Анализы аффинности в растворе можно также проводить с использованием системы иммуноанализа GYROLAB®, которая предоставляет платформу для автоматизированного биоанализа и быстрый оборот образцов (Fraley et al. 2013, Bioanalysis 5: 1765-74), или ELISA.
Специалисту в данной области понятно также, что измеренная KD может соответствовать кажущейся KD, если способ измерения каким-либо образом влияет на аффинность связывания, присущую подразумеваемым молекулам, например, из-за артефактов, связанных с покрытием биосенсора одной молекулой. Также кажущуюся KD можно измерять, если одна молекула содержит более одного участка узнавания для другой молекулы. В такой ситуации на измеренную аффинность может влиять авидность взаимодействия между двумя молекулами. В частности, точное измерение KD может являться достаточно трудоемким, и как следствие, часто значения кажущейся KD определяют для оценки силы связывания двух молекул. Следует отметить, что при условии, что все измерения проводят согласованным способом (например, сохраняя условия анализа неизменными), измерения кажущейся KD можно использовать в качестве приближения истинной KD и таким образом, в настоящем описании KD и кажущуюся KD следует рассматривать с равной важностью или правомочностью.
Термин «специфичность» относится к количеству различных типов антигенов или антигенных детерминант, с которыми могут связываться конкретные антигенсвязывающая молекула или антигенсвязывающий белок (такие как ISVD или полипептид по изобретению). Специфичность антигенсвязывающего белка можно определять на основании аффинности и/или авидности, например, как описано на страницах 53-56 из WO 08/020079 (содержание которой приведено в настоящем описании в качестве ссылки), в которой также описаны предпочтительные способы измерения связывания между антигенсвязывающей молекулой (такой как полипептид или ISVD по изобретению) и соответствующим антигеном. Как правило, антигенсвязывающие белки (такие как ISVD и/или полипептиды по изобретению) связываются со своим антигеном с константой диссоциации (KD) 10-5-10-12 моль/литр или менее и предпочтительно, 10-7-1012 моль/литр или менее, и более предпочтительно, 10-8-10-12 моль/литр (т.е. с константой связывания (KA) 105-1012 литров/моль или более и предпочтительно, 107-1012 литров/моль или более, и более предпочтительно, 108-1012 литров/моль). Любое значение KD более чем 10-4 моль/литр (или любое значение KA ниже чем 104 литров/моль), как правило, считают указывающим на неспецифическое связывание. Предпочтительно, моновалентный ISVD по изобретению связывается с желательным антигеном с аффинностью менее чем 500 нМ, предпочтительно, менее чем 200 нМ, более предпочтительно, менее чем 10 нМ, например, менее чем 500 нМ, например, такой как между 10 и 5 нМ или менее. Приведена также ссылка на раздел n) на страницах 53-56 из WO 08/020079.
Считают, что ISVD и/или полипептид является «специфическим для» (первой) мишени или антигена, по сравнению с другой (второй) мишенью или антигеном, когда он связывается с первым антигеном с аффинностью (как описано выше, и соответствующим образом выражено как значение KD, значение KA, скорость Koff и/или скорость Kon), которая по меньшей мере в 10 раз, например, по меньшей мере в 100 раз, и предпочтительно, по меньшей мере в 1000 раз или более, лучше чем аффинность, с которой ISVD и/или полипептид связывается с второй мишенью или антигеном. Например, ISVD и/или полипептид может связываться с первой мишенью или антигеном с значением KD, которое по меньшей мере в 10 раз меньше, например, по меньшей мере в 100 раз меньше, и предпочтительно по меньшей мере в 1000 раз меньше или даже менее этого, чем KD, с которой указанный ISVD и/или полипептид связывается со второй мишенью или антигеном. Предпочтительно, когда ISVD и/или полипептид является «специфическим для» первой мишени или антигена, по сравнению с второй мишенью или антигеном, он является нацеленным против (как определено в настоящем описании) указанной первой мишени или антигена, но не нацеленным против указанной второй мишени или антигена.
Специфическое связывание антигенсвязывающего белка с антигеном или антигенной детерминантой можно определять любым подходящим способом, известным по существу, включая, например, анализы насыщающего связывания и/или анализы конкурентного связывания, такие как радиоиммунные анализы (RIA), ферментные иммуноанализы (EIA) и их различные варианты, известные в данной области; так же, как другие способы, упомянутые в настоящем описании.
Предпочтительным способом, который можно использовать для оценки аффинности, является способ 2-стадийного ELISA (твердофазного иммуноферментного анализа) из Friguet et al., 1985 (J. Immunol. Methods 77: 305-19). Этот способ обеспечивает измерение равновесия связывания в фазе раствора и исключает возможные артефакты, связанные с адсорбцией одной из молекул на подложке, такой как пластик. Как понятно специалисту в данной области, константа диссоциации может представлять собой фактическую или кажущуюся константу диссоциации. Способы определения константы диссоциации понятны специалисту в данной области, и включают, например, способы, упомянутые на страницах 53-56 из WO 08/020079.
Наконец, следует отметить, что во многих ситуациях специалист в данной области может считать удобным определение аффинности связывания относительно одной и той же эталонной молекулы. Например, для оценки силы связывания между молекулами A и B, можно, например, использовать эталонную молекулу C, которая, как известно, связывается с B, и которая является подходящим образом меченной с использованием группы флуорофора или хромофора, или другой химической группы, такой как биотин, для простоты детекции в ELISA или FACS (активированной флуоресценцией сортировке клеток) или другом формате (флуорофор для флуоресцентной детекции, хромофор для детекции поглощения света, биотин для опосредованной стрептавидином детекции в ELISA). Как правило, эталонную молекулу C поддерживают при фиксированной концентрации, и концентрацию A меняют для данной концентрации или количества B. В результате, получают значение IC50, соответствующее концентрации A, при которой сигнал, измеряемый для C в отсутствие A, уменьшается вдвое. При условии, что KD эталон.., KD эталонной молекулы, известна, так же как общая концентрация сэталон. эталонной молекулы, кажущуюся KD для взаимодействия A-B можно получить из следующей формулы: KD = IC50/(1+сэталон./KDэталнн.). Следует отметить, что если сэталон. << KDэталон., KD ≈ IC50. При условии, что измерение IC50 проводят постоянным способом (например, сохраняя сэталон. фиксированным) для сравниваемых связывающих молекул, различия в силе или стабильности молекулярного взаимодействия можно оценивать посредством сравнения IC50, и это измерение признают эквивалентным KD или кажущейся KD на протяжении этого текста.
Половина максимальной ингибирующей концентрации (IC50) также может являться показателем эффективности соединения для ингибирования биологической или биохимической функции, например, фармакологического эффекта. Этот количественный показатель показывает, как много полипептида или ISVD (например, нанотела) необходимо для ингибирования данного биологического процесса (или компонента процесса, т.е. фермента, клетки, клеточного рецептора, хемотаксиса, анаплазии, метастазирования, инвазивности и т.д.) наполовину. Иными словами, она составляет половину максимальной (50%) ингибирующей концентрации (IC) вещества (50% IC, или IC50). Значения IC50 можно рассчитывать для данного антагониста, такого как полипептид или ISVD (например, нанотело) по изобретению, посредством определения концентрации, необходимой для ингибирования половины максимального биологического ответа агониста. KD лекарственного средства можно определять посредством построения кривой зависимости ответа от дозы и проверки эффекта различных концентраций антагониста, такого как полипептид или ISVD (например, нанотело) по изобретению, на обращение активности агониста.
Термин «половина максимальной эффективной концентрации» (EC50) относится к концентрации соединения, которая индуцирует половину ответа между фоном и максимумом после указанного времени экспозиции. В настоящем контексте ее используют в качестве показателя активности полипептида, ISVD (например, нанотела). EC50 на градуированной кривой зависимости ответа от дозы представляет концентрацию соединения, когда наблюдают 50% от его максимального эффекта. Концентрацию, предпочтительно, выражают в молярных единицах.
В биологических системах, небольшие изменения концентрации лиганда, как правило, приводят к быстрым изменениям ответа, следующим сигмоидальной функции. Точка перегиба, в которой увеличение ответа с увеличением концентрации лиганда начинает замедляться, представляет собой EC50. Это можно определять математически посредством вывода кривой наилучшего соответствия. Основываться на графике для оценки, является удобным в большинстве случаев. В случае, когда EC50 представлена в разделе примеры, эксперименты были разработаны, чтобы отражать KD настолько точно, насколько возможно. Иными словами, значения EC50 можно тогда рассматривать, как значения KD. Термин «средняя KD» относится к среднему значению KD, полученному по меньшей мере в 1, но предпочтительно, более чем в 1, например, по меньшей мере в 2 экспериментах. Термин «средний» относится к математическому термину «средний» (сумме данных, деленной на количество значений данных).
Это относится также к IC50, которая является показателем соединения применительно к его ингибированию (50% ингибирования). Для анализов конкурентного связывания и анализов функциональных антагонистов, IC50 является наиболее распространенным суммарным показателем кривой зависимости ответа от дозы. Для анализов агонистов/стимуляторов, наиболее распространенным суммарным показателем является EC50.
Константа ингибирования (Ki) представляет собой показатель того, насколько активным является ингибитор; она представляет собой концентрацию, необходимую для получения половины максимального ингибирования. В отличие от IC50, которая может меняться в зависимости от условий эксперимента, Ki является абсолютным значением, и его часто обозначают как константа ингибирования лекарственного средства. Константу ингибирования Ki можно рассчитывать с использованием уравнения Ченга Прусова:
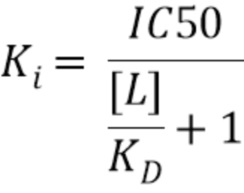 ,
,
в котором [L] представляет собой фиксированную концентрацию лиганда.
Термин «активность» полипептида и/или ISVD по изобретению, в рамках изобретения, является функцией количества полипептида и/или ISVD по изобретению, необходимого для возникновения его специфического эффекта. Он относится к способности указанного полипептида и/или ISVD по изобретению модулировать и/или частично или полностью ингибировать активность ADAMTS5. Более конкретно, он может относиться к способности указанного полипептида и/или ISVD уменьшать или даже полностью ингибировать активность ADAMTS5, как определено в настоящем описании. Таким образом, он может относиться к способности указанного полипептида и/или ISVD ингибировать активность ADAMTS5, например, ферментную активность, такую как протеолиз, например, активность протеазы и/или активность эндопептидазы, так же, как связывание субстрата, включая, но без ограничения, аггрекан, версикан, бревикан, нейрокан, декорин и/или бигликан, предпочтительно, расщепление аггрекана. Указанный полипептид и/или ISVD, предпочтительно, является антагонистом активности аггреканазы ADAMTS5. Активность можно измерять посредством любого подходящего анализа, известного в данной области или описанного в настоящем описании. В рамках изобретения, «активность аггреканазы» определяют как протеолитическое расщепление аггрекана.
«Эффективность» полипептида по изобретению измеряет максимальную силу собственно эффекта, при насыщающих концентрациях полипептид. Эффективность показывает максимальный ответ, достижимый для полипептида по изобретению. Она относится к способности полипептида оказывать желательный (терапевтический) эффект.
В одном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид связывается с ADAMTS5 с KD между 1E-07 M и 1E-13 M, например, между 1E-08 M и 1E-12 M, предпочтительно, не более 1E-07 M, предпочтительно, ниже чем 1E-08 M или 1E-09 M, или даже ниже, чем 1E 10 M, например, 5E-11 M, 4E-11 M, 3E 11 M, 2E-11 M, 1,7E-11 M, 1E 11 M, или даже 5E-12 M, 4E-12 M, 3E12 M, 1E-12 M, например, как определено посредством Gyrolab или KinExA.
В одном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид модулирует ADAMTS5 с EC50 между 1E-07 M и 1E-12 M, например, между 1E-08 M и 1E-11 M, например, как определено посредством ELISA связывания.
В одном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид связывается с ADAMTS5 со скоростью диссоциации менее чем 5E-04 с-1, например, менее чем 1E-04 с-1 или 5E-05 с-1, или даже менее чем 1E-05 с-1, например, как определено посредством SPR.
В одном аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует активность ADAMTS5 с IC50 между 1E-07 M и 1E-12 M, например, между 1E-08 M и 1E-11 M, например, как определено посредством анализа FRET для человека или AlphaLISA для человека.
В одном аспекте изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует ферментную активность ADAMTS5 с IC50 не более 1E-07 M, предпочтительно, 1E-08 M, 5E-09 M, или 4E-9 M, 3E-9 M, 2E-9 M, например, 1E-9 M.
Говорят, что аминокислотная последовательность, такая как ISVD или полипептид, имеет «перекрестную реакционную способность» для двух различных антигенов или антигенных детерминант (например, таких как ADAMTS5 из различных видов млекопитающих, например, таких как ADAMTS5 человека, ADAMTS5 быка, ADAMTS5 крысы, ADAMTS5 морской свинки, ADAMTS5 мыши или ADAMTS5 яванского макака), если она является специфической для (как определено в настоящем описании) этих различных антигенов или антигенных детерминант. Понятно, что ISVD или полипептид можно считать имеющим перекрестную реакционную способность, хотя аффинность связывания для двух различных антигенов может отличаться, например, в 2, 5, 10, 50, 100 раз или даже больше, при условии, что он является специфическим для (как определено в настоящем описании) этих различных антигенов или антигенных детерминант.
ADAMTS5 также является известным как ADAMTS11, ADMP-2 или аггреканаза-2.
Соответствующую структурную информацию для ADAMTS5 можно обнаружить, например, в номерах доступа в UniProt, как описано в таблице 1 ниже (ср. таблицу B).
Таблица 1
«ADAMTS5 человека» относится к ADAMTS5, содержащему аминокислотную последовательность из SEQ ID NO: 149. В одном аспекте, полипептид по изобретению специфически связывается с ADAMTS5 из Human sapiens, Mus musculus, Cavia Porcellus, Bos taurus, Macaca mulatta и/или Rattus norvegicus, предпочтительно, ADAMTS5 человека, предпочтительно, SEQ ID NO: 149.
Термины «(перекрестно) блокировать», «(перекрестно) блокированный», «(перекрестно) блокирующие», «конкурентное связывание», «(перекрестно) конкурировать», «(перекрестно) конкурирующие» и «(перекрестная) конкуренция» используют в настоящем описании взаимозаменяемо для обозначения способности иммуноглобулина, антитела, ISVD, полипептида или другой связывающей молекулы мешать связыванию других иммуноглобулинов, антител, ISVD, полипептидов или связывающих молекул с данной мишенью. Степень, в которой иммуноглобулин, антитело, ISVD, полипептид или другая связывающая молекула способны мешать связыванию другой с мишенью, и таким образом, то, можно ли их назвать перекрестно блокирующими по изобретению, можно определять с использованием анализов конкурентного связывания, общепринятых в данной области, например, таких как скрининг очищенных ISVD против ISVD, экспонированных на фаге, в конкурентном ELISA. Особенно подходящие количественные анализы перекрестного блокирования включают ELISA.
Другие способы определения того, являются ли иммуноглобулин, антитело, ISVD, полипептид или другая связывающая молекула, нацеленные против мишени, (перекрестно) блокирующим, способными (перекрестно) блокировать, конкурентно связывающими или способными (перекрестно) конкурировать, как определено в настоящем описании, можно оценивать в «сэндвич-анализе» на основе SPR, например, таком как описано в разделе примеры. Другие подходящие способы описаны, например, в Xiao-Chi Jia et al. (Journal of Immunological Methods 288: 91-98, 2004), Miller et al. (Journal of Immunological Methods 365: 118-125, 2011).
Соответственно, настоящее изобретение относится к полипептиду, как описано в настоящем описании, такому, как представлено посредством SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18 (ср. таблицу A-1), где указанный полипептид конкурирует с перекрестно блокирующим полипептидом, например, как определено посредством конкурентного ELISA.
Настоящее изобретение относится к способу определения конкурентов, таких как полипептиды, конкурирующих с полипептидом, как описано в настоящем описании, таким, как представлено посредством любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18, где полипептид, как описано в настоящем описании, проявляет конкуренцию или перекрестное блокирование для конкурента, такого как полипептид, применительно к связыванию с ADAMTS5, например, таким как ADAMTS5 человека (SEQ ID NO: 149), где связывание с ADAMTS5 конкурента уменьшено по меньшей мере на 5%, например, на 10%, 20%, 30%, 40%, 50% или даже больше, например, на 80%, 90% или даже 100% (т.е. фактически не поддается детекции в данном анализе) в присутствии полипептида по изобретению, по сравнению со связыванием с ADAMTS5 конкурента в отсутствие полипептида по изобретению. Конкуренцию и перекрестное блокирующие можно определять любыми способами, известными в данной области, например, такими как конкурентный ELISA или анализ FACS. В одном аспекте настоящее изобретение относится к полипептиду по изобретению, где указанный полипептид перекрестно блокирует связывание с ADAMTS5 по меньшей мере одного из полипептидов, представленных посредством SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18, и/или его связывание с ADAMTS5 перекрестно блокировано по меньшей мере одним из полипептидов, представленным посредством SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18.
Настоящее изобретение также относится к конкурентам, конкурирующим с полипептидом, как описано в настоящем описании, таким как SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18, где конкурент проявляет конкуренцию или перекрестное блокирование для полипептида, как описано в настоящем описании, применительно к связыванию с ADAMTS5, где связывание с ADAMTS5 полипептида по изобретению уменьшено по меньшей мере на 5%, например, на 10%, 20%, 30%, 40%, 50% или даже больше, например, на 80%, или даже больше, например, по меньшей мере на 90% или даже 100% (т.е. фактически не поддается детекции в данном анализе) в присутствии указанного конкурента, по сравнению со связыванием с ADAMTS5 полипептида по изобретению в отсутствие указанного конкурента. В одном аспекте настоящее изобретение относится к полипептиду, который перекрестно блокирует связывание с ADAMTS5 полипептида по изобретению, такого как одна из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18, и/или его связывание с ADAMTS5 перекрестно блокировано по меньшей мере одной из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18, предпочтительно, где указанный полипептид содержит по меньшей мере один VH, VL, dAb или ISVD, специфически связывающие ADAMTS5, где связывание с ADAMTS5 модулирует активность ADAMTS5.
«Виды ADAMTS5 активности» и «виды активности ADAMTS5» (эти термины используют в настоящем описании взаимозаменяемо) включают, но без ограничения, виды ферментной активности, такие как протеолиз, например, активность протеазы (также называемую протеазной или пептидазной активностью), и виды активности эндопептидазы, с одной стороны, и виды активности экзосайтов, например, такие как узнавание и/или связывание субстрата, например, посредством подобного дезинтегрину домена, центрального подобного тромбоспондину (TS) повтора типа I, богатого цистеином домена, спейсерной области и/или дополнительных мотивов TS. Виды активности ADAMTS5 включают связывание и/или протеолиз субстратов, таких как связывающие гиалуронан внеклеточные белки хондроитинсульфат-протеогликаны (CSPG), такие как аггрекан, версикан, бревикан, нейрокан, декорин и бигликан. В рамках изобретения, протеолиз представляет собой распад белков на более мелкие полипептиды или аминокислоты посредством гидролиза пептидных связей, которые соединяют аминокислоты вместе в полипептидной цепи.
В контексте настоящего изобретения, «модуляция» или «модулировать», в общем, означает изменять активность ADAMTS5, как измерено с использованием подходящего анализа in vitro, клеточного анализа или анализа in vivo (такого как анализы, упомянутые в настоящем описании). В частности, «модуляция» или «модулировать» может означать либо уменьшение, либо ингибирование активности, или альтернативно, увеличение активности ADAMTS5, как измерено с использованием подходящего анализа in vitro, клеточного анализа или анализа in vivo (например, такого как анализы, упомянутые в настоящем описании), по меньшей мере на 1%, предпочтительно, по меньшей мере на 5%, например, по меньшей мере на 10% или по меньшей мере на 25%, например, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 80% или на 90%, или более, по сравнению с активностью ADAMTS5 в таком же анализе в таких же условиях, но в отсутствие ISVD или полипептида по изобретению.
Соответственно, настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид модулирует активность ADAMTS5, предпочтительно, ингибирует активность ADAMTS5.
Соответственно, настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует протеазную активность ADAMTS5, например, ингибирует протеолиз субстрата, такого как аггрекан, версикан, бревикан, нейрокан, декорин и/или бигликан.
Соответственно, настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид блокирует связывание ADAMTS5 с субстратом, таким как аггрекан, версикан, бревикан, нейрокан, декорин и/или бигликан, где указанный субстрат, предпочтительно, представляет собой аггрекан.
В одном аспекте изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид блокирует связывание ADAMTS5 с аггреканом по меньшей мере на 20%, например, по меньшей мере на 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или даже больше, например, как определено посредством ELISA.
В одном аспекте изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид является антагонистом или ингибирует активность ADAMTS5, такую как (i) протеазная активность, предпочтительно расщепление аггрекана, версикана, бревикана, нейрокана, декорина и/или бигликана, предпочтительно, расщепление аггрекана; предпочтительно, является антагонистом аггреканазной активности ADAMTS5; (ii) связывание субстрата с ADAMTS5, например, с экзосайтом ADAMTS5, например, подобным дезинтегрину доменом, центральным подобным тромбоспондину (TS) повтором типа I, богатым цистеином доменом, спейсерной областью или дополнительным мотивом TS.
Соответственно, настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид ингибирует протеазную активность ADAMTS5, предпочтительно, по меньшей мере на 5%, например, на 10%, 20%, 30%, 40%, 50% или даже больше, например, по меньшей мере на 60%, 70%, 80%, 90%, 95% или даже больше, как определено посредством любого подходящего способа, известного в данной области, например, такого как ферментные анализы ингибирования, или как описано в разделе примеры.
Несмотря на то, что ADAM, ADAMTS и MMP разделяют участок связывания с аггреканом, который является очень сходным как по последовательности, так и по общей форме, например, каталитические домены ADAMTS4 и ADAMTS5 разделяют высокую степень сходства последовательности, авторам настоящего изобретения удалось идентифицировать ISVD, которые были специфическими для мишени, как показано в примерах. Специфичность для мишени также может исключать или по меньшей мере ограничивать скелетно-мышечный синдром, который является побочным эффектом, вызванным ингибиторами широкого спектра.
В одном аспекте изобретение относится к связывающей ADAMTS5 молекуле, такой как ISVD и полипептид по изобретению, где указанная связывающая ADAMTS5 молекула не связывает ADAMTS4, ADAMTS1, ADAMTS15, MMP1 и/или MMP14 (мембранного типа). Предпочтительно, настоящее изобретение относится к полипептиду, как определено в настоящем описании, где указанный ISVD, связывающий ADAMTS5, не связывает ADAMTS4, MMP1 или MMP14.
Если не указано иное, термины «иммуноглобулин» и «последовательность иммуноглобулина» - независимо от того, используют ли их в настоящем описании для обозначения антитела из тяжелых цепей или общепринятого 4-цепочечного антитела - используют в качестве общего термина для включения полноразмерного антитела, его индивидуальных цепей, так же как всех его частей, доменов или фрагментов (включая, но без ограничения, антигенсвязывающие домены или фрагменты, такие как домены VHH или домены VH/VL, соответственно).
Термин «домен» (полипептида или белка), в рамках изобретения, относится к свернутой белковой структуре, имеющей способность сохранять свою третичную структуру независимо от остального белка. Как правило, домены являются ответственными за отдельные функциональные свойства белков, и во многих случаях, могут быть добавлены, удалены или перенесены в другие белки без потери функции остального белка и/или остальных доменов.
Термин «домен иммуноглобулина», в рамках изобретения, относится к глобулярной области цепи антитела (например, такой как цепь общепринятого 4-цепочечного антитела или антитела из тяжелых цепей), или к полипептиду, который в основном состоит из такой глобулярной области. Домены иммуноглобулинов отличаются тем, что они сохраняют характеристики иммуноглобулинового сворачивания молекул антител, которое состоит из двухслойного сэндвича из приблизительно семи антипараллельных бета-цепей, аранжированных в два бета-листа, необязательно, стабилизированные посредством консервативной дисульфидной связи.
Термин «вариабельный домен иммуноглобулина», в рамках изобретения, обозначает домен иммуноглобулина, в основном состоящий из четырех «каркасных областей», обозначенных в данной области и в настоящем описании ниже как «каркасная область 1» или «FR1»; как «каркасная область 2» или «FR2»; как «каркасная область 3» или «FR3»; и как «каркасная область 4» или «FR4», соответственно; где каркасные области прерываются тремя «определяющими комплементарность областями» или «CDR», обозначенными в данной области и в настоящем описании ниже как «определяющая комплементарность область 1» или «CDR1»; как «определяющая комплементарность область 2» или «CDR2»; и как «определяющая комплементарность область 3» или «CDR3», соответственно. Таким образом, общую структуру или последовательность вариабельного домена иммуноглобулина можно указывать следующим образом: FR1 - CDR1 - FR2 - CDR2 - FR3 - CDR3 - FR4. Вариабельный домен(ы) иммуноглобулина придает специфичность антителу для антигена, поскольку несет антигенсвязывающий участок.
Термин «одиночный вариабельный домен иммуноглобулина» (сокращенно обозначенный в настоящем описании как «ISVD» или «ISV»), и взаимозаменяемо используемый с ним «одиночный вариабельный домен», определяет молекулы, где антигенсвязывающий участок присутствует на одиночном домене иммуноглобулина и сформирован посредством одиночного домена иммуноглобулина. Это отличает одиночные вариабельные домены иммуноглобулинов от «общепринятых» иммуноглобулинов или их фрагментов, где два домена иммуноглобулинов, в частности, два вариабельных домена, взаимодействуют с формированием антигенсвязывающего участка. Как правило, в общепринятых иммуноглобулинах, вариабельный домен тяжелой цепи (VH) и вариабельный домен легкой цепи (VL) взаимодействуют с формированием антигенсвязывающего участка. В последнем случае, определяющие комплементарность области (CDR) как VH, так и VL, вносят вклад в антигенсвязывающий участок, т.е. всего 6 CDR вовлечены в формирование антигенсвязывающего участка.
С учетом вышеуказанного определения, антигенсвязывающий домен общепринятого 4-цепочечного антитела (такого как молекула IgG, IgM, IgA, IgD или IgE; известная в данной области) или фрагмента Fab, фрагмента F(ab')2, фрагмента Fv, например, связанного дисульфидной связью фрагмента Fv или scFv, или диатела (все известны в данной области), полученный из такого общепринятого 4-цепочечного антитела, обычно не рассматривают как одиночный вариабельный домен иммуноглобулина, поскольку, в этих случаях, связывание с соответствующим эпитопом антигена в норме не происходит с одним (одиночным) доменом иммуноглобулина, но происходит с парой (ассоциированных) доменов иммуноглобулина, таких как вариабельные домены легких и тяжелых цепей, т.е. с парой VH-VL доменов иммуноглобулина, которые совместно связываются с эпитопом соответствующего антигена.
В отличие от этого, ISVD являются способными специфически связываться с эпитопом антигена без спаривания с дополнительным вариабельным доменом иммуноглобулина. Участок связывания ISVD сформирован одиночным доменом VHH, VH или VL. Таким образом, антигенсвязывающий участок ISVD сформирован не более, чем тремя CDR.
Таким образом, одиночный вариабельный домен может представлять собой последовательность вариабельного домена легкой цепи (например, VL-последовательность) или ее подходящий фрагмент; или последовательность вариабельного домена тяжелой цепи (например, VH-последовательность или VHH-последовательность) или ее подходящий фрагмент; при условии, что он является способным формировать одиночную антигенсвязывающую единицу (т.е. функциональную антигенсвязывающую единицу, которая в основном состоит из одиночного вариабельного домена, так что одиночный антигенсвязывающий домен не обязательно должен взаимодействовать с другим вариабельным доменом для формирования функциональной антигенсвязывающей единицы).
В одном варианте осуществления изобретения, ISVD представляют собой последовательности вариабельного домена тяжелой цепи (например, VH-последовательность); более конкретно, ISVD могут представлять собой последовательности вариабельного домена тяжелой цепи, полученные из общепринятого четырех-цепочечного антитела или последовательности вариабельного домена тяжелой цепи, полученные из антитела из тяжелых цепей.
Например, ISVD может представлять собой (одно-)доменное антитело (или аминокислоту, подходящую для использования для (одно-)доменного антитела), «dAb» или dAb (или аминокислоту, подходящую для использования для dAb) или нанотело (как определено в настоящем описании, и включая, но без ограничения, VHH); другие одиночные вариабельные домены, или любой подходящий фрагмент любого одного из них.
В частности, ISVD может представлять собой нанотело® (как определено в настоящем описании) или его подходящий фрагмент. [Примечание: нанотело®, нанотела® и наноклон® являются зарегистрированными торговыми наименованиями от Ablynx N.V.] Для общего описания нанотел, приведена ссылка на дополнительное описание ниже, так же как на предшествующий уровень техники, процитированный в настоящем описании, например, как описано в WO 08/020079 (страница 16).
«Домены VHH», также известные как VHH, домены VHH, фрагменты VHH антител и антитела VHH, первоначально описаны как антигенсвязывающий (вариабельный) домен иммуноглобулина из «антител из тяжелых цепей» (т.е. из «антител, лишенных легких цепей»; Hamers-Casterman et al. 1993 Nature 363: 446-448). Термин «домен VHH» выбран для установления отличий этих вариабельных доменов от вариабельных доменов тяжелых цепей, присутствующих в общепринятых 4-цепочечных антителах (обозначенных в настоящем описании как «домены VH» или «домены VH») и от вариабельных доменов легких цепей, присутствующих в общепринятых 4-цепочечных антителах (обозначенных в настоящем описании как «домены VL» или «домены VL»). Для дополнительного описания VHH и нанотел, приведена ссылка на обзорную статью от Muyldermans (Reviews in Molecular Biotechnology 74: 277-302, 2001), так же, как на следующие патентные заявки, упомянутые в качестве общего уровня техники: WO 94/04678, WO 95/04079 и WO 96/34103 от Vrije Universiteit Brussel; WO 94/25591, WO 99/37681, WO 00/40968, WO 00/43507, WO 00/65057, WO 01/40310, WO 01/44301, EP 1134231 и WO 02/48193 от Unilever; WO 97/49805, WO 01/21817, WO 03/035694, WO 03/054016 и WO 03/055527 от Vlaams Instituut voor Biotechnologie (VIB); WO 03/050531 от Algonomics N.V. и Ablynx N.V.; WO 01/90190 от National Research Council of Canada; WO 03/025020 (= EP 1433793) от Institute of Antibodies; так же как WO 04/041867, WO 04/041862, WO 04/041865, WO 04/041863, WO 04/062551, WO 05/044858, WO 06/40153, WO 06/079372, WO 06/122786, WO 06/122787 и WO 06/122825, от Ablynx N.V. и дальнейшие опубликованные патентные заявки от Ablynx N.V. Приведена также ссылка на дополнительные литературные источники предшествующего уровня техники, упомянутые в этих заявках, и в частности, на список литературы, упомянутый на страницах 41-43 Международной заявки WO 06/040153, где этот список и ссылки приведены в настоящем описании в качестве ссылки. Как описано в этих ссылках, нанотела (в частности, последовательности VHH и частично гуманизированные нанотела) можно, в частности, характеризовать по присутствию одного или более «характерных остатков» в одной или более из каркасных последовательностей. Дополнительное описание нанотел, включая гуманизацию и/или камелизацию нанотел, так же как другие модификации, части или фрагменты, производные или «слитые нанотела», мультивалентные конструкции (включая некоторые неограничивающие примеры линкерных последовательностей) и различные модификации для увеличения времени полужизни нанотел и их препаратов можно обнаружить, например, в WO 08/101985 и WO 08/142164. Для дополнительного общего описания нанотел, приведена ссылка на предшествующий уровень техники, процитированный в настоящем описании, например, такой как описано в WO 08/020079 (страница 16).
В частности, каркасные последовательности, присутствующие в связывающих ADAMTS5 молекулах по изобретению, таких как ISVD и/или полипептиды по изобретению, могут содержать один или более характерных остатков (например, как описано в WO 08/020079 (таблицы A-3 - A-8)), так что связывающая ADAMTS5 молекула по изобретению представляет собой нанотело. Некоторые предпочтительные, но неограничивающие примеры (подходящих комбинаций) таких каркасных последовательностей станут понятными из дальнейшего описания в настоящем документе (см., например, таблицу A-2). Как правило, нанотела (в частности, последовательности VHH и частично гуманизированные нанотела) можно, в частности, характеризовать по присутствию одного или более «характерных остатков» в одной или более из каркасных последовательностей (как, например, дополнительно описано в WO 08/020079, страница 61, строка 24 - страница 98, строка 3).
Более конкретно, изобретение относится к связывающим ADAMTS5 молекулам, содержащим по меньшей мере один одиночный вариабельный домен иммуноглобулина, представляющий собой аминокислотную последовательность с (общей) структурой
FR1 - CDR1 - FR2 - CDR2 - FR3 - CDR3 - FR4,
в которой FR1 - FR4 относятся к каркасным областям 1-4, соответственно, и в которой CDR1-CDR3 относятся к определяющим комплементарность областям 1-3, соответственно, и которая:
i) имеет по меньшей мере 80%, более предпочтительно 90%, даже более предпочтительно, 95% идентичность аминокислот по меньшей мере с одной из аминокислотных последовательностей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 или 18 (см. таблицу A-1), в которых для целей определения степени идентичности аминокислот, аминокислотные остатки, формирующие последовательности CDR, не учитывают. В этом отношении, приведена также ссылка на таблицу A-2, в которой перечислены последовательности каркасной области 1 (SEQ ID NO: 72-84), последовательности каркасной области 2 (SEQ ID NO: 85-94), последовательности каркасной области 3 (SEQ ID NO: 95-113) и последовательности каркасной области 4 (SEQ ID NO: 114-115) одиночных вариабельных доменов иммуноглобулинов из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18; или
ii) комбинации каркасных последовательностей, как показано в таблице A-2;
и в которых:
iii) предпочтительно один или более из аминокислотных остатков в положениях 11, 37, 44, 45, 47, 83, 84, 103, 104 и 108, в соответствии с нумерацией по Kabat выбраны из характерных остатков, таких как упомянуто в таблице A-3 - таблице A-8 из WO 08/020079.
Связывающие ADAMTS5 молекулы по изобретению, такие как ISVD и/или полипептиды по изобретению, могут также содержать специфические мутации/аминокислотные остатки, описанные в следующих одновременно поданных Предварительных патентных заявках США, всех озаглавленных «Improved immunoglobulin variable domains»: US 61/994552, поданной 16 мая 2014 г.; US 61/014015, поданной 18 июня 2014 г.; US 62/040167, поданной 21 августа 2014 г.; и US 62/047560, поданной 8 сентября 2014 г. (все принадлежат Ablynx N.V.).
В частности, связывающие ADAMTS5 молекулы по изобретению, такие как ISVD и/или полипептиды по изобретению, могут соответствующим образом содержать (i) K или Q в положении 112; или (ii) K или Q в положении 110 в комбинации с V в положении 11; или (iii) T в положении 89; или (iv) L в положении 89 с K или Q в положении 110; или (v) V в положении 11 и L в положении 89; или любую подходящую комбинацию (i) - (v).
Как также описано в указанных одновременно поданных Предварительных патентных заявках США, когда связывающая ADAMTS5 молекула по изобретению, такая как ISVD и/или полипептид по изобретению, содержит мутации в соответствии с одним из (i) - (v) выше (или их подходящую комбинацию):
- аминокислотный остаток в положении 11, предпочтительно, выбран из L, V или K (и наиболее предпочтительно, представляет собой V); и/или
- аминокислотный остаток в положении 14 предпочтительно, соответствующим образом выбран из A или P; и/или
- аминокислотный остаток в положении 41 предпочтительно, соответствующим образом выбран из A или P; и/или
- аминокислотный остаток в положении 89 предпочтительно, соответствующим образом выбран из T, V или L; и/или
- аминокислотный остаток в положении 108 предпочтительно, соответствующим образом выбран из Q или L; и/или
- аминокислотный остаток в положении 110 предпочтительно, соответствующим образом выбран из T, K или Q; и/или
- аминокислотный остаток в положении 112 предпочтительно, соответствующим образом выбран из S, K или Q.
Как упомянуто в указанных одновременно поданных Предварительных патентных заявках США, указанные мутации являются эффективными для предотвращения или уменьшения связывания так называемых «предсуществующих антител» с иммуноглобулинами и соединениями по изобретению. Для этой цели связывающие ADAMTS5 молекулы по изобретению, такие как ISVD и/или полипептиды по изобретению, могут также содержать (необязательно в комбинации с указанными мутациями) C-концевое удлинение (X)n (в котором n представляет собой 1-10, предпочтительно 1-5, например, 1, 2, 3, 4 или 5 (и предпочтительно, 1 или 2, например, 1); и каждый X представляет собой (предпочтительно, природный) аминокислотный остаток, который выбран независимо, и предпочтительно, независимо выбран из группы, состоящей из аланина (A), глицина (G), валина (V), лейцина (L) или изолейцина (I)), см., например, Предварительные патентные заявки США, так же как WO 12/175741. В частности, связывающая ADAMTS5 молекула по изобретению, такая как ISVD и/или полипептид по изобретению, может содержать такое C-концевое удлинение, когда она формирует C-конец содержащего ее белка, полипептида или другого соединения или конструкции (см., например, указанные Предварительные патентные заявки США, так же как WO 12/175741).
Связывающая ADAMTS5 молекула по изобретению может представлять собой иммуноглобулин, такой как одиночный вариабельный домен иммуноглобулина, полученный любым подходящим способом и из любого подходящего источника, и может, например, представлять собой природные последовательности VHH (т.е. из подходящего вида верблюдовых) или синтетические или полусинтетические аминокислотные последовательности, включая, но без ограничения, «гуманизированные» (как определено в настоящем описании) нанотела или последовательности VHH, «камелизированные» (как определено в настоящем описании) последовательности иммуноглобулинов (и в частности, камелизированные последовательности вариабельного домена тяжелой цепи), так же, как нанотела, полученные такими способами, как аффинное созревание (например, начиная с синтетических, случайных или природных последовательностей иммуноглобулинов), прививка CDR, маскировка поверхностных остатков, комбинирование фрагментов, происходящих из различных последовательностей иммуноглобулинов, сборка посредством ПЦР с использованием перекрывающихся праймеров, и сходные способы конструирования последовательностей иммуноглобулинов, хорошо известные специалисту в данной области; или любую подходящую комбинацию любого из вышеуказанного, как далее описано в настоящем описании. Также, когда иммуноглобулин содержит последовательность VHH, указанный иммуноглобулин можно соответствующим образом гуманизировать, как далее описано в настоящем описании, таким образом, чтобы получить один или более дополнительно (частично или полностью) гуманизированных иммуноглобулинов по изобретению. Подобным образом, когда иммуноглобулин содержит синтетическую или полусинтетическую последовательность (такую как частично гуманизированная последовательность), указанный иммуноглобулин можно, необязательно, дополнительно соответствующим образом гуманизировать, снова, как описано в настоящем описании, снова таким образом, чтобы получить один или более дополнительно (частично или полностью) гуманизированных иммуноглобулинов по изобретению.
«Доменные антитела», также известные как «Dab», «Доменные Антитела» и «dAb» (термины «доменные антитела» и «dAb» используют в качестве торговых наименований в группе компаний GlaxoSmithKline), описаны, например, в EP 0368684, Ward et al. (Nature 341: 544-546, 1989), Holt et al. (Tends in Biotechnology 21: 484-490, 2003) и WO 03/002609 так же как, например, в WO 04/068820, WO 06/030220, WO 06/003388 и других опубликованных патентных заявках от Domantis Ltd. Доменные антитела в основном соответствуют доменам VH или VL из не относящихся к верблюдовым млекопитающих, в частности, из человеческих 4-цепочечных антител. Для связывания с эпитопом в форме одиночного антигенсвязывающего домена, т.е. без спаривания с доменом VL или VH, соответственно, необходим специфический отбор по таким свойствам связывания антигена, например, с использованием библиотеки последовательностей человеческих одиночных доменов VH или VL. Доменные антитела имеют, подобно VHH, молекулярную массу от приблизительно 13 до приблизительно 16 кДа и, если происходят из полностью человеческих последовательностей, не требуют гуманизации, например, для терапевтического использования для человека.
Следует также отметить, что, хотя и наименее предпочтительно в контексте настоящего изобретения, поскольку они не происходят из млекопитающих, одиночные вариабельные домены можно получать из конкретных видов акул (например, так называемые «домены IgNAR», см., например, WO 05/18629).
Настоящее изобретение относится, в частности, к ISVD, где указанные ISVD выбраны из группы, состоящей из VHH, гуманизированных VHH и камелизированных VH.
Аминокислотные остатки домена VHH пронумерованы в соответствии с общей нумерацией доменов VH, приведенной в Kabat et al. («Sequence of proteins of immunological interest», US Public Health Services, NIH Bethesda, MD, Publication No. 91), как применимо к доменам VHH из верблюдовых, как показано, например, на фигуре 2 из Riechmann and Muyldermans (J. Immunol. Methods 231: 25-38, 1999), все как известно в данной области. Альтернативные способы нумерации аминокислотных остатков доменов VH, которые также можно использовать аналогичным способом для доменов VHH, известны в данной области. Однако, в настоящем описании, формуле изобретения и фигурах, следуют нумерации в соответствии с Kabat, применимой для доменов VHH, как описано выше, если не указано иное.
Следует отметить, что - как хорошо известно в данной области для доменов VH и для доменов VHH - общее количество аминокислотных остатков в каждой из CDR может меняться и может не соответствовать общему количеству аминокислотных остатков, указанных в нумерации по Kabat (то есть, одно или более положений в соответствии с нумерацией по Kabat может не являться занятым в фактической последовательности, или фактическая последовательность может содержать больше аминокислотных остатков, чем количество, допустимое в нумерации по Kabat). Это означает, что, в общем, нумерация в соответствии с Kabat может соответствовать или может не соответствовать фактической нумерации аминокислотных остатков в фактической последовательности. Общее количество аминокислотных остатков в домене VH и домене VHH обычно может лежать в диапазоне от 110 до 120, часто между 112 и 115. Однако следует отметить, что более короткие и более длинные последовательности также могут являться подходящими для целей, описанных в настоящем описании.
Применительно к CDR, как хорошо известно в данной области, существует множество соглашений для определения и описания CDR из фрагмента VH или VHH, таких как определение Kabat (которое основано на изменчивости последовательности и является наиболее общепринятым) и определение Chothia (которое основано на локализации областей структурных петель). Приведена ссылка, например, на web-сайт http://www.bioinf.org.uk/abs/. Для целей по настоящему описанию и формуле изобретения, CDR наиболее предпочтительно определяют на основании определения Abm (которое основано на программном обеспечении для моделирования антител AbM от Oxford Molecular), поскольку его рассматривают как оптимальный компромисс между определениями Kabat и Chothia (ср. http://www.bioinf.org.uk/abs/) в рамках изобретения, FR1 содержит аминокислотные остатки в положениях 1-25, CDR1 содержит аминокислотные остатки в положениях 26-35, FR2 содержит аминокислоты в положениях 36-49, CDR2 содержит аминокислотные остатки в положениях 50-58, FR3 содержит аминокислотные остатки в положениях 59-94, CDR3 содержит аминокислотные остатки в положениях 95-102, и FR4 содержит аминокислотные остатки в положениях 103-113.
В значении по настоящему изобретению, термин «одиночный вариабельный домен иммуноглобулина» или «одиночный вариабельный домен» содержит полипептиды, происходящие из не относящегося к человеку источника, предпочтительно, из верблюдовых, предпочтительно, из антитела из тяжелых цепей верблюдовых. Они могут являться гуманизированными, как описано в настоящем описании. Кроме того, термин включает полипептиды, происходящие из не относящихся к верблюдовым источников, например, мыши или человека, которые были «камелизированы», как описано в настоящем описании.
Таким образом, ISVD, такие как доменные антитела и нанотела (включая VHH домены), можно подвергать гуманизации. В частности, гуманизированные ISVD, такие как нанотела (включая VHH домены), могут представлять собой ISVD, которые являются такими, как в общем определено в настоящем описании, но в которых присутствует по меньшей мере один аминокислотный остаток (и в частности, по меньшей мере в одном из каркасных остатков) который представляет собой и/или который соответствует гуманизирующей замене (как определено в настоящем описании). Гуманизирующие замены, которые потенциально можно использовать, можно определять посредством сравнения последовательности каркасных областей природной последовательности VHH с соответствующей каркасной последовательностью из одной или более близко родственных человеческих последовательностей VH, после чего одну или более гуманизирующих замен (или их комбинаций), которые потенциально можно использовать, определенные таким образом, можно вводить в указанную последовательность VHH (любым способом, известным по существу, как далее описано в настоящем описании) и полученные гуманизированные последовательности VHH можно тестировать по аффинности для мишени, по стабильности, по простоте и уровню экспрессии, и/или по другим желательным свойствам. Таким образом, посредством ограниченного количества проб и ошибок, специалист в данной области может определять другие подходящие гуманизирующие замены (или их подходящие комбинации), на основании описания в настоящем документе. Также, на основании вышеизложенного, (каркасные области) ISVD, такого как нанотело (включая домены VHH) можно частично гуманизировать или полностью гуманизировать.
Другой особенно предпочтительный класс ISVD по изобретению содержит ISVD с аминокислотной последовательностью, которая соответствует аминокислотной последовательности природного домена VH, но которая была «камелизирована», т.е. посредством замены одного или более аминокислотных остатков в аминокислотной последовательности природного домена VH из общепринятого 4-цепочечного антитела на один или более из аминокислотных остатков, встречающихся в соответствующем положении(положениях) в домене VHH антитела из тяжелых цепей. Это можно осуществлять способом, известным по существу, который понятен специалисту в данной области, например, на основании описания в настоящем документе. Такие «камелизирующие» замены предпочтительно вводят в положениях аминокислот, формирующих поверхность контакта и/или присутствующих на поверхности контакта VH-VL, и/или в так называемых характерных остатках верблюдовых, как определено в настоящем описании (см. также, например, WO 94/04678 и Davies and Riechmann (1994 и 1996)). Предпочтительно, последовательность VH, которую используют в качестве исходного материала или исходной точки для получения или конструирования камелизированных одиночных вариабельных доменов иммуноглобулинов, предпочтительно, представляет собой последовательность VH млекопитающего, более предпочтительно, последовательность VH человека, например, последовательность VH3. Однако, следует отметить, что такие камелизированные одиночные вариабельные домены иммуноглобулинов по изобретению можно получать любым подходящим способом, известным по существу, и таким образом, они не являются строго ограниченными полипептидами, полученными с использованием полипептида, содержащего природный домен VH, в качестве исходного материала. Приведена ссылка на Davies and Riechmann (FEBS 339: 285-290, 1994; Biotechnol. 13: 475-479, 1995; Prot. Eng. 9: 531-537, 1996) и Riechmann and Muyldermans (J. Immunol. Methods 231: 25-38, 1999)
Например, снова как далее описано в настоящем описании, как «гуманизацию», так и «камелизацию», можно осуществлять посредством получения нуклеотидной последовательности, кодирующей природный домен VHH или домен VH, соответственно, и затем изменения, способом, известным по существу, одного или более кодонов в указанной нуклеотидной последовательности таким образом, чтобы новая нуклеотидная последовательность кодировала «гуманизированный» или «камелизированный» ISVD по изобретению, соответственно. Затем эту нуклеиновую кислоту можно экспрессировать способом, известным по существу, таким образом, чтобы получить желательные ISVD по изобретению. Альтернативно, на основании аминокислотной последовательности природного домена VHH или домена VH, соответственно, аминокислотную последовательность желательных гуманизированных или камелизированных ISVD по изобретению, соответственно, можно конструировать и затем синтезировать de novo с использованием способов пептидного синтеза, известных по существу. Также на основании аминокислотной последовательности или нуклеотидной последовательности природного домена VHH или домена VH, соответственно, нуклеотидную последовательность, кодирующую желательные гуманизированные или камелизированные ISVD по изобретению, соответственно, можно конструировать и затем синтезировать de novo с использованием способов синтеза нуклеиновых кислот, известных по существу, после чего полученную таким образом нуклеиновую кислоту можно экспрессировать способом, известным по существу, таким образом, чтобы получать желательные ISVD по изобретению.
ISVD, такие как доменные антитела и нанотела (включая домены VHH и гуманизированные домены VHH), можно также подвергать аффинному созреванию посредством введения одного или более изменений в аминокислотную последовательность одной или более CDR, где изменения приводят к улучшенной аффинности полученного ISVD для соответствующего антигена, по сравнению с соответствующей исходной молекулой. Аффинно зрелые молекулы ISVD по изобретению можно получать способами, известными в данной области, например, как описано в Marks et al. (Biotechnology 10:779-783, 1992), Barbas, et al. (Proc. Nat. Acad. Sci, USA 91: 3809-3813, 1994), Shier et al. (Gene 169: 147-155, 1995), Yelton et al. (Immunol. 155: 1994-2004, 1995), Jackson et al. (J. Immunol. 154: 3310-9, 1995), Hawkins et al. (J. MoI. Biol. 226: 889 896, 1992), Johnson and Hawkins (Affinity maturation of antibodies using phage display, Oxford University Press, 1996).
Способ конструирования/отбора и/или получения полипептида, начиная с ISVD, такого как VH, VL, VHH, доменное антитело или нанотело, также обозначают в настоящем описании как «форматирование» указанного ISVD; и ISVD, составляющий часть полипептида, обозначают как являющийся «форматированным» или находящийся «в формате» указанного полипептида. Примеры способов, которыми ISVD можно форматировать, и примеры таких форматов понятны специалисту в данной области на основании описания в настоящем документе; и такой форматированный одиночный вариабельный домен иммуноглобулина составляет дополнительный аспект изобретения.
Предпочтительные CDR показаны в таблице A-2.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых
(i) CDR1 выбрана из группы, состоящей из SEQ ID NO: 21, 35, 20, 22, 25, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34; и
аминокислотных последовательностей, имеющих 1, 2 или 3 отличие(отличий) аминокислот от SEQ ID NO: 21, 35, 20, 22, 25, 33, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34;
(ii) CDR2 выбрана из группы, состоящей из SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и
аминокислотных последовательностей, имеющих 1, 2 или 3 отличие(отличий) аминокислот от SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и
(iii) CDR3 выбрана из группы, состоящей из SEQ ID NO: SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70; и
аминокислотных последовательностей, имеющих 1, 2, 3 или 4 отличие(отличий) аминокислот от SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых
(i) CDR1 выбрана из группы, состоящей из
(a) SEQ ID NO: 22; и
(b) аминокислотной последовательности, имеющей 1, 2, 3, 4, 5 или 6 отличие(отличий) аминокислот от SEQ ID NO: 22, где
- в положении 2 S заменен на R;
- в положении 3 A заменен на T;
- в положении 4 V заменен на F;
- в положении 6 V заменен на S;
- в положении 7 N заменен на Y; и/или
- в положении 10 A заменен на G;
(ii) CDR2 представляет собой SEQ ID NO: 36; и
(iii) CDR3 представляет собой SEQ ID NO: 54.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых
(i) CDR1 представляет собой SEQ ID NO: 33;
(ii) CDR2 выбрана из группы, состоящей из
(c) SEQ ID NO: 50; и
(d) аминокислотной последовательности, имеющей 1, 2 или 3 отличие(отличий) аминокислот от SEQ ID NO: 50, где
- в положении 8 M заменен на I;
- в положении 9 P заменен на T; и/или
- в положении 10 Y заменен на F; и
(iii) CDR3 выбрана из группы, состоящей из
(e) SEQ ID NO: 68; и
(f) аминокислотной последовательности, имеющей 1 или 2 отличие(отличий) аминокислот от SEQ ID NO: 68, где
- в положении 5 F заменен на L; и/или
- в положении 11 D заменен на E.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD, специфически связывающий ADAMTS5, в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых
(i) CDR1 представляет собой SEQ ID NO: 28;
(ii) CDR2 выбрана из группы, состоящей из
(c) SEQ ID NO: 44; и
(d) аминокислотной последовательности, имеющей 1, 2 или 3 отличие (отличий) аминокислот от SEQ ID NO: 44, где
- в положении 3 S заменен на T;
- в положении 4 R заменен на W;
- в положении 8 T заменен на I; и/или
- в положении 9 T заменен на L; и
(iii) CDR3 выбрана из группы, состоящей из
(e) SEQ ID NO: 62; и
(f) аминокислотной последовательности, имеющей 1 или 2 отличие (отличий) аминокислот от SEQ ID NO: 62, где
- в положении 1 G заменен на S; и/или
- в положении 14 D заменен на E.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD специфически связывается с ADAMTS5 и в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых
- CDR1 выбрана из группы, состоящей из SEQ ID NO: 21, 35, 20, 22, 25, 33, 28, 24, 23, 26, 27, 29, 30, 31, 32 и 34;
- CDR2 выбрана из группы, состоящей из SEQ ID NO: 37, 53, 36, 40, 50, 51, 44, 45, 43, 39, 38, 41, 119, 42, 46, 47, 48, 49 и 52; и
- CDR3 выбрана из группы, состоящей из SEQ ID NO: 55, 118, 71, 54, 58, 68, 69, 62, 63, 61, 57, 56, 59, 60, 64, 65, 66, 67 и 70.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD специфически связывается с ADAMTS5 и в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых указанный ISVD выбран из группы ISVD, где:
CDR1 представляет собой SEQ ID NO: 21, CDR2 представляет собой SEQ ID NO: 37, и CDR3 представляет собой SEQ ID NO: 55;
CDR1 представляет собой SEQ ID NO: 35, CDR2 представляет собой SEQ ID NO: 53, и CDR3 представляет собой SEQ ID NO: 118;
CDR1 представляет собой SEQ ID NO: 35, CDR2 представляет собой SEQ ID NO: 53, и CDR3 представляет собой SEQ ID NO: 71;
CDR1 представляет собой SEQ ID NO: 20, CDR2 представляет собой SEQ ID NO: 36, и CDR3 представляет собой SEQ ID NO: 54;
CDR1 представляет собой SEQ ID NO: 22, CDR2 представляет собой SEQ ID NO: 36, и CDR3 представляет собой SEQ ID NO: 54;
CDR1 представляет собой SEQ ID NO: 25, CDR2 представляет собой SEQ ID NO: 40, и CDR3 представляет собой SEQ ID NO: 58;
CDR1 представляет собой SEQ ID NO: 33, CDR2 представляет собой SEQ ID NO: 50, и CDR3 представляет собой SEQ ID NO: 68;
CDR1 представляет собой SEQ ID NO: 33, CDR2 представляет собой SEQ ID NO: 51, и CDR3 представляет собой SEQ ID NO: 69;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 44, и CDR3 представляет собой SEQ ID NO: 62;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 45, и CDR3 представляет собой SEQ ID NO: 63;
CDR1 представляет собой SEQ ID NO: 28, CDR2 представляет собой SEQ ID NO: 43, и CDR3 представляет собой SEQ ID NO: 61;
CDR1 представляет собой SEQ ID NO: 24, CDR2 представляет собой SEQ ID NO: 39, и CDR3 представляет собой SEQ ID NO: 57;
CDR1 представляет собой SEQ ID NO: 23, CDR2 представляет собой SEQ ID NO: 38, и CDR3 представляет собой SEQ ID NO: 56;
CDR1 представляет собой SEQ ID NO: 26, CDR2 представляет собой SEQ ID NO: 41, и CDR3 представляет собой SEQ ID NO: 59;
CDR1 представляет собой SEQ ID NO: 27, CDR2 представляет собой SEQ ID NO: 119, и CDR3 представляет собой SEQ ID NO: 60;
CDR1 представляет собой SEQ ID NO: 27, CDR2 представляет собой SEQ ID NO: 42, и CDR3 представляет собой SEQ ID NO: 60;
CDR1 представляет собой SEQ ID NO: 29, CDR2 представляет собой SEQ ID NO: 46, и CDR3 представляет собой SEQ ID NO: 64;
CDR1 представляет собой SEQ ID NO: 30, CDR2 представляет собой SEQ ID NO: 47, и CDR3 представляет собой SEQ ID NO: 65;
CDR1 представляет собой SEQ ID NO: 31, CDR2 представляет собой SEQ ID NO: 48, и CDR3 представляет собой SEQ ID NO: 66;
CDR1 представляет собой SEQ ID NO: 32, CDR2 представляет собой SEQ ID NO: 49, и CDR3 представляет собой SEQ ID NO: 67; и
CDR1 представляет собой SEQ ID NO: 34, CDR2 представляет собой SEQ ID NO: 52, и CDR3 представляет собой SEQ ID NO: 70.
В конкретном предпочтительном аспекте, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD специфически связывается с ADAMTS5 и в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых CDR1 представляет собой или содержит SEQ ID NO: 21, CDR2 представляет собой SEQ ID NO: 37, и CDR3 представляет собой SEQ ID NO: 55.
В частности, настоящее изобретение относится к ISVD, как описано в настоящем описании, где указанный ISVD специфически связывается с ADAMTS5 и в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых указанный ISVD выбран из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 8, 117, 12, 13, 14, 15 и 18.
Понятно, что, без ограничения, одиночные вариабельные домены иммуноглобулинов по настоящему изобретению можно использовать в качестве «строительных блоков» для получения полипептида, который может, необязательно, содержать один или более дополнительных одиночных вариабельных доменов иммуноглобулинов, которые могут служить в качестве строительных блоков (т.е. против того же самого или другого эпитопа на ADAMTS5 и/или против одного или более антигенов, белков или мишеней, отличных от ADAMTS5).
Полипептид по изобретению (также обозначенный в настоящем описании как «конструкция нанотела») содержит по меньшей мере один ISVD, связывающий ADAMTS, предпочтительно ADAMTS5, например, два ISVD, связывающие ADAMTS5, и предпочтительно, также ISVD, связывающий альбумин. В полипептиде по изобретению, ISVD могут быть связаны напрямую или связаны посредством линкера. Даже более предпочтительно, полипептид по изобретению содержит C-концевое удлинение. Как подробно описано в настоящем описании, C-концевое удлинение в основном предотвращает/прекращает связывание предсуществующих антител/факторов в большинстве образцов от субъектов/пациентов-людей. C-концевое удлинение присутствует на C-конце последнего аминокислотного остатка (обычно остатка серина) последнего (локализованного наиболее близко к C-концу) ISVD.
Как дополнительно уточнено ниже, ISVD можно получать из домена VHH, VH или VL, однако, ISVD выбирают таким образом, чтобы они не формировали комплементарные пары доменов VH и VL в полипептидах по изобретению. Нанотело, VHH и гуманизированный VHH являются необычными в том смысле, что они происходят из природных антител верблюдовых, не имеющих легких цепей, и действительно, эти домены не могут ассоциировать с легкими цепями верблюдовых для формирования комплементарных пар VHH и VL. Таким образом, полипептиды по настоящему изобретению не содержат комплементарных ISVD и/или не формируют комплементарных пар ISVD, например, таких как комплементарные пары VH/VL.
В общем, полипептиды или конструкции, содержащие или в основном состоящие из одиночного строительного блока, одиночного ISVD или одиночного нанотела, обозначены в настоящем описании как «моновалентные» полипептиды и «моновалентные конструкции», соответственно. Полипептиды или конструкции, содержащие два или более строительных блоков (например, такие как ISVD) также обозначены в настоящем описании как «мультивалентные» полипептиды или конструкции, и строительные блоки/ISVD, присутствующие в таких полипептидах или конструкциях, также обозначены в настоящем описании как находящиеся в «мультивалентном формате». Например, «двухвалентный» полипептид может содержать два ISVD, необязательно, связанные посредством линкерной последовательности, в то время как «трехвалентный» полипептид может содержать три ISVD, необязательно связанные посредством двух линкерных последовательностей; в то время как «тетравалентный» полипептид может содержать четыре ISVD, необязательно связанные посредством трех линкерных последовательностей, и т.д.
В мультивалентном полипептиде, два или более ISVD могут являться одинаковыми или различными, и могут быть направлены против одного и того же антигена или антигенной детерминанты (например, против одной и той же части (частей) или эпитопа (эпитопов), или против различных частей или эпитопов), или могут, альтернативно, быть направлены против различных антигенов или антигенных детерминант; или могут представлять собой любую подходящую комбинацию этого. Полипептиды и конструкции, содержащие по меньшей мере два строительных блока (например, такие как ISVD) в которых по меньшей мере один строительный блок направлен против первого антигена (т.е. ADAMTS5), и по меньшей мере один строительный блок направлен против второго антигена (т.е. отличного от ADAMTS5), можно также обозначать как «мультиспецифические» полипептиды и конструкции, и строительные блоки (например, такие как ISVD), присутствующие в таких полипептидах и конструкциях, также обозначены в настоящем описании как находящиеся в «мультиспецифическом формате». Таким образом, например, «биспецифический» полипептид по изобретению представляет собой полипептид, содержащий по меньшей мере один ISVD, направленный против первого антигена (т.е. ADAMTS5) и по меньшей мере один дополнительный ISVD, направленный против второго антигена (т.е. отличного от ADAMTS5), в то время как «триспецифический» полипептид по изобретению представляет собой полипептид, содержащий по меньшей мере один ISVD, направленный против первого антигена (т.е. ADAMTS5), по меньшей мере один дополнительный ISVD, направленный против второго антигена (т.е. отличного от ADAMTS5) и по меньшей мере один дополнительный ISVD, направленный против третьего антигена (т.е. отличного как от ADAMTS5, так и от второго антигена); и т.д.
В одном аспекте, настоящее изобретение относится к полипептиду, содержащему по меньшей мере 2 ISVD, где по меньшей мере 1 ISVD специфически связывается с ADAMTS, предпочтительно ADAMTS5, более предпочтительно, выбранным из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
В одном аспекте, настоящее изобретение относится к полипептиду, содержащему по меньшей мере 2 ISVD, где указанные по меньшей мере 2 ISVD специфически связываются с ADAMTS, предпочтительно ADAMTS5, более предпочтительно, каждый ISVD из указанных 2 ISVD независимо выбран из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
«Мультипаратопные» полипептиды и «мультипаратопные» конструкции, например, такие как «бипаратопные» полипептиды или конструкции и «трипаратопные» полипептиды или конструкции, содержат или в основном состоят из двух или более строительных блоков, какждый из которых имеет отличный паратоп.
Соответственно, ISVD по изобретению, связывающие ADAMTS5, могут находиться в основном в выделенной форме (как определено в настоящем описании), или они могут формировать часть конструкции или полипептида, которые могут содержать или в основном состоять из одного или более ISVD, связывающих ADAMTS5 и которые могут, необязательно, дополнительно содержать одну или более дополнительных аминокислотных последовательностей (все, необязательно, связанные посредством одного или более подходящих линкеров). Настоящее изобретение относится к полипептиду или конструкции, которые содержат или в основном состоят из по меньшей мере одного ISVD по изобретению, такого как один или более ISVD по изобретению (или их подходящие фрагменты), связывающие ADAMTS5.
Один или более ISVD по изобретению можно использовать в качестве строительных блоков в таких полипептиде или конструкции, таким образом, чтобы получать моновалентные, мультивалентные или мультипаратопные полипептид или конструкцию по изобретению, соответственно, все, как описано в настоящем описании. Настоящее изобретение, таким образом, также относится к полипептиду, представляющему собой моновалентную конструкцию, содержащую или в основном состоящую из одного моновалентного полипептида или ISVD по изобретению.
Настоящее изобретение, таким образом, также относится к полипептиду или конструкции, представляющим собой мультивалентный полипептид или мультивалентную конструкцию, соответственно, например, такие как двухвалентные или трехвалентные полипептид или конструкция, содержащие или в основном состоящие из двух или более ISVD по изобретению (для мультивалентных и мультиспецифических полипептидов, содержащих один или более доменов VHH, и их получения, приведена также ссылка на Conrath et al. (J. Biol. Chem. 276: 7346-7350, 2001), так же как, например, на WO 96/34103, WO 99/23221 и WO 2010/115998).
В одном аспекте, в его самой простой форме, мультивалентный полипептид или конструкция по изобретению представляет собой двухвалентный полипептид или конструкцию по изобретению, содержащие первый ISVD, такой как нанотело, направленное против ADAMTS5, и идентичный второй ISVD, такой как нанотело, направленное против ADAMTS5, где указанный первый и указанный второй ISVD, такие как нанотела, могут, необязательно являться связанными посредством линкерной последовательности (как определено в настоящем описании). В другой форме, мультивалентный полипептид или конструкция по изобретению могут представлять собой трехвалентный полипептид или конструкцию по изобретению, содержащие первый ISVD, такой как нанотело, направленное против ADAMTS5, идентичный второй ISVD, такой как нанотело, направленное против ADAMTS5, и идентичный третий ISVD, такой как нанотело, направленное против ADAMTS5, в которых указанный первый, второй и третий ISVD, такие как нанотела, могут, необязательно являться связанными посредством одной или более, и в частности, двух, линкерных последовательностей. В одном аспекте, изобретение относится к полипептиду или конструкции, содержащим или в основном состоящим из по меньшей мере двух ISVD по изобретению, например, 2, 3 или 4 ISVD (или их подходящих фрагментов), связывающих ADAMTS5. Два или более ISVD могут, необязательно являться связанными посредством одного или более пептидных линкеров.
В другом аспекте, мультивалентный полипептид или конструкция по изобретению могут представлять собой биспецифические полипептид или конструкцию по изобретению, содержащие первый ISVD, такой как нанотело, направленное против ADAMTS5, и второй ISVD, такой как нанотело, направленное против второго антигена, например, такого как аггрекан, в которых указанный первый и второй ISVD, такие как нанотела, могут, необязательно являться связанными посредством линкерной последовательности (как определено в настоящем описании); в то время как мультивалентный полипептид или конструкция по изобретению могут также представлять собой триспецифические полипептид или конструкцию по изобретению, содержащие первый ISVD, такой как нанотело, направленное против ADAMTS5, второй ISVD, такой как нанотело, направленное против второго антигена, например, такого как аггрекан, и третий ISVD, такой как нанотело, направленное против третьего антигена, в которых указанный первый, второй и третий ISVD, такие как нанотела, могут, необязательно являться связанными посредством одной или более, и в частности, двух, линкерных последовательностей.
Изобретение, кроме того, относится к мультивалентному полипептиду, содержащему или (в основном) состоящему из по меньшей мере одного ISVD (или его подходящих фрагментов), связывающего ADAMTS5, предпочтительно, ADAMTS5 человека, и одного дополнительного ISVD, такого как ISVD, связывающий аггрекан.
Особенно предпочтительные двухвалентные, биспецифические полипептиды или конструкции по изобретению представляют собой полипептиды или конструкции, показанные в Примерах, описанных в настоящем описании и в таблице A-1 (ср. SEQ ID NO: 120-130 (т.е. SEQ ID NO: 120, 121, 122, 123, 124, 125, 126, 127, 128, 129 и 130), наиболее предпочтительно, SEQ ID NO: 129 и 130).
В предпочтительном аспекте, полипептид или конструкция по изобретению содержит или в основном состоит из по меньшей мере двух ISVD, где указанные по меньшей мере два ISVD могут являться одинаковыми или различными, но из них по меньшей мере один ISVD направлен против ADAMTS5, предпочтительно, указанный ISVD, связывающий ADAMTS5, выбран из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
Два или более ISVD, присутствующие в мультивалентном полипептиде или конструкции по изобретению, могут состоять из последовательности вариабельного домена легкой цепи (например, VL-последовательности) или из последовательности вариабельного домена тяжелой цепи (например, VH-последовательности); они могут состоять из последовательности вариабельного домена тяжелой цепи, полученной из общепринятого четырех-цепочечного антитела, или из последовательности вариабельного домена тяжелой цепи, полученной из антитела из тяжелых цепей. В предпочтительном аспекте, они состоят из доменного антитела (или аминокислоты, подходящей для использования для доменного антитела), однодоменного антитела (или аминокислоты, подходящей для использования для однодоменного антитела), из «dAb» (или аминокислоты, подходящей для использования для dAb), из нанотела® (включая, но без ограничения, VHH), из гуманизированной последовательности VHH, из гуманизированной последовательности VH; или из последовательности VHH, полученной посредством аффинного созревания. Два или более одиночных вариабельных доменов иммуноглобулинов могут состоять из частично или полностью гуманизированного нанотела, или частично или полностью гуманизированного VHH.
В одном аспекте по изобретению, первый ISVD и второй ISVD, присутствующие в мультипаратопных (предпочтительно бипаратопных или трипаратопных) полипептиде или конструкции по изобретению, не проявляют (перекрестную) конкуренцию друг с другом за связывание с ADAMTS5 и, таким образом, принадлежат к разным семействам. Соответственно, настоящее изобретение относится к мультипаратопным (предпочтительно, бипаратопным) полипептиду или конструкции, содержащим два или более ISVD, где каждый ISVD принадлежит к другому семейству. В одном аспекте, первый ISVD из этих мультипаратопных (предпочтительно бипаратопных) полипептида или конструкции по изобретению не осуществляет перекрестное блокирование связывания с ADAMTS5 второго ISVD из этих мультипаратопных (предпочтительно, бипаратопных) полипептида или конструкции по изобретению, и/или связывание первого ISVD с ADAMTS5 не подвергается перекрестному блокированию вторым ISVD. В другом аспекте, первый ISVD из мультипаратопных (предпочтительно, бипаратопных) полипептида или конструкции по изобретению перекрестно блокирует связывание с ADAMTS5 второго ISVD из этих мультипаратопных (предпочтительно, бипаратопных) полипептида или конструкции по изобретению, и/или связывание первого ISVD его с ADAMTS5 перекрестно блокировано вторым ISVD.
В особенно предпочтительном аспекте, полипептид или конструкция по изобретению содержит или в основном состоит из трех или более ISVD, из которых по меньшей мере два ISVD направлены против ADAMTS5. Следует понимать, что указанные по меньшей мере два ISVD, направленные против ADAMTS5, могут являться одинаковыми или различными, могут быть направлены против одного и того же эпитопа или различных эпитопов ADAMTS5, могут принадлежать к одной и той же эпитопной группе или к различных эпитопным группам, и/или могут связываться с одинаковыми или различными доменами ADAMTS5.
Относительная аффинность может зависеть от локализации ISVD в полипептиде. Следует понимать, что порядок ISVD в полипептиде по изобретению (ориентацию) может выбирать, в соответствии с необходимостью, специалист в данной области. Порядок индивидуальных ISVD, так же как то, содержит ли полипептид линкер, является вопросом выбора дизайна. Некоторые ориентации, с линкерами или без, могут обеспечивать предпочтительные характеристики связывания, по сравнению с другими ориентациями. Например, порядок первого ISVD (например, ISVD 1) и второго ISVD (например, ISVD 2) в полипептиде по изобретению может представлять собой (от N-конца к C-концу): (i) ISVD 1 (например, нанотело 1) - [линкер] - ISVD 2 (например, нанотело 2) - [C-концевое удлинение]; или (ii) ISVD 2 (например, нанотело 2) - [линкер]- ISVD 1 (например, нанотело 1) - [C-концевое удлинение]; (где группы в квадратных скобках, т.е. линкер и C-концевое удлинение, являются необязательными). Все ориентации включены в изобретение. Полипептиды, содержащие ориентацию ISVD, обеспечивающую желательные характеристики связывания, можно легко идентифицировать посредством общепринятого скрининга, например, как проиллюстрировано в разделе примеры. Предпочтительный порядок представляет собой от N-конца к C-концу: ISVD, связывающий ADAMTS5 - [линкер] - ISVD, связывающий Альбумин или аггрекан - [C-концевое удлинение], где группы в квадратных скобках являются необязательными.
В одном аспекте, настоящее изобретение относится к полипептиду, содержащему два или более ISVD, которые специфически связываются с ADAMTS5, где
a) по меньшей мере «первый» ISVD специфически связывается с первой антигенной детерминантой, эпитопом, частью, доменом, субъединицей или конформацией ADAMTS5, предпочтительно, где указанный «первый» ISVD, специфически связывающий ADAMTS5, выбран из группы, состоящей из SEQ ID NO: 2, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 8, 117, 12, 13, 14, 15 и 18; и
b) по меньшей мере «второй» ISVD специфически связывается с второй антигенной детерминантой, эпитопом, частью, доменом, субъединицей или конформацией ADAMTS5, отличной от первой антигенной детерминанты, эпитопа, части, домена, субъединицы или конформации, соответственно, предпочтительно, где указанный «второй» ISVD, специфически связывающий ADAMTS5, представляет собой SEQ ID NO: 116 или 19.
В предпочтительном аспекте, полипептид или конструкция по изобретению содержит или в основном состоит из по меньшей мере двух ISVD, связывающих ADAMTS5, где указанные по меньшей мере два ISVD могут являться одинаковыми или различными, которые независимо выбраны из группы, состоящей из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
В следующем аспекте, изобретение относится к мультипаратопным (предпочтительно, бипаратопным) полипептиду или конструкции, содержащим два или более ISVD, направленные против ADAMTS5, связывающие один и тот же эпитоп(ы), который связываются с любой из SEQ ID NO: 2, 116, 19, 1, 3, 6, 16, 17, 10, 11, 9, 5, 4, 7, 117, 8, 12, 13, 14, 15 и 18.
В следующем аспекте, изобретение относится к полипептиду, как описано в настоящем описании, где указанный полипептид имеет по меньшей мере 80%, 90%, 95% или 100% идентичность последовательности с любой из SEQ ID NO: 1-19 (т.е. SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18 и 19), 116-117 или 120-130 (т.е. SEQ ID NO: 120, 121, 122, 123, 124, 125, 126, 127, 128, 129 и 130).
В одном аспекте, настоящее изобретение относится к полипептиду, как описано в настоящем описании, который выбран из группы, состоящей из SEQ ID NO: 127 (клон 130 049-093-Alb), SEQ ID NO: 126 (клон 129 2F3-093-Alb), SEQ ID NO: 127 (клон 130 049-093-Alb) и SEQ ID NO: 128 (клон 131 9D3-093-Alb).
В данной области существует необходимость в более эффективных видах терапии против нарушений, поражающих хрящ в суставах, таких как остеоартрит. Особенно при системном введении, время удержания большинства лекарственных средств является недостаточным. Авторы настоящего изобретения выдвинули гипотезу, что эффективность терапевтического лекарственного средства, такого как конструкция, полипептид и ISVD по изобретению, можно значительно увеличивать посредством связывания терапевтического лекарственного средства с группой, которая увеличивает время полужизни лекарственного средства и следовательно, увеличивает удержание лекарственного средства, но которая не должна нарушать эффективность указанного терапевтического лекарственного средства.
В конкретном аспекте изобретения, конструкция или полипептид по изобретению могут иметь группу, придающую увеличенное время полужизни, по сравнению с соответствующими конструкцией или полипептидом по изобретению без указанной группы. Некоторые предпочтительные, но неограничивающие примеры таких конструкций и полипептидов по изобретению станут понятными специалисту в данной области, на основании дальнейшего описания в настоящем документе, и например, включают ISVD или полипептиды по изобретению, химически модифицированные для увеличения времени полужизни (например, посредством пегилирования); связывающие ADAMTS5 молекулы по изобретению, такие как ISVD и/или полипептиды по изобретению, содержащие по меньшей мере один дополнительный участок связывания для связывания с сывороточным белком (таким как сывороточный альбумин); или полипептиды по изобретению, содержащие по меньшей мере один ISVD по изобретению, связанный по меньшей мере с одной группой (и в частности, по меньшей мере с одной аминокислотной последовательностью), увеличивающей время полужизни аминокислотной последовательности по изобретению. Примеры конструкций по изобретению, таких как полипептиды по изобретению, содержащих такие продлевающие время полужизни группы или ISVD, станут понятными специалисту в данной области на основании дальнейшего описания в настоящем документе; и, например, включают, без ограничения, полипептиды, в которых один или более ISVD по изобретению соответствующим образом связаны с одним или более сывороточными белками или их фрагментами (такими как (человеческий) сывороточный альбумин или его подходящие фрагменты) или с одной или более связывающими единицами, которые могут связываться с сывороточными белками (например, такие как доменные антитела, одиночные вариабельные домены иммуноглобулинов, подходящие для использования в качестве доменного антитела, однодоменные антитела, одиночные вариабельные домены иммуноглобулинов подходящие для использования в качестве однодоменного антитела, dAb, одиночные вариабельные домены иммуноглобулинов, подходящие для использования в качестве dAb, или нанотела, которые могут связываться с сывороточными белками, такими как сывороточный альбумин (такой как человеческий сывороточный альбумин), сывороточные иммуноглобулины, такие как IgG, или трансферрин; приведена ссылка на дополнительное описание и ссылки, упомянутые в настоящем описании); полипептиды, в которых аминокислотная последовательность по изобретению связана с фрагментом Fc (таким как человеческий Fc) или его соответствующей частью или фрагментом; или полипептиды, в которых один или более одиночных вариабельных доменов иммуноглобулинов по изобретению соответствующим образом связаны с одним или более малыми белками или пептидами, которые могут связываться с сывороточными белками, такие как, без ограничения, белки и пептиды, описанные в WO 91/01743, WO 01/45746, WO 02/076489, WO2008/068280, WO2009/127691 и PCT/EP2011/051559.
В одном аспекте настоящее изобретение относится к конструкции по изобретению или к полипептиду, где указанная конструкция или указанный полипептид дополнительно содержит группу, связывающую сывороточный белок, или сывороточный белок. Предпочтительно, указанная группа, связывающая сывороточный белок, связывает сывороточный альбумин, такой как человеческий сывороточный альбумин.
В одном аспекте, настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему ISVD, связывающий сывороточный альбумин.
В общем, конструкции или полипептиды по изобретению с увеличенным временем полужизни, предпочтительно, имеют время полужизни, по меньшей мере в 1,5 раза, предпочтительно, по меньшей мере в 2 раза, например, по меньшей мере, в 5 раз, например, по меньшей мере в 10 раз или более чем в 20 раз, превышающее собственное время полужизни соответствующих конструкций или полипептидов по изобретению, т.е. без группы, придающей увеличенное время полужизни. Например, конструкции или полипептиды по изобретению с увеличенным временем полужизни могут иметь время полужизни, например, у человека, увеличенное более чем на 1 час, предпочтительно, более чем на 2 часа, более предпочтительно более чем на 6 часов, например, более чем на 12 часов, или даже более чем на 24, 48 или 72 часов, по сравнению с собственно соответствующими конструкциями или полипептидами по изобретению, т.е. без группы, придающей увеличенное время полужизни.
В предпочтительном, но неограничивающем аспекте изобретения, конструкции по изобретению и полипептиды по изобретению, имеют время полужизни в сыворотке например, у человека, увеличенное более чем на 1 час, предпочтительно, более чем на 2 часа, более предпочтительно, более чем на 6 часов, например, более чем на 12 часов, или даже более чем на 24, 48 или 72 часов, по сравнению с собственно соответствующими конструкциями или полипептидами по изобретению, т.е. без группы, придающей увеличенное время полужизни.
В другом предпочтительном, но неограничивающем аспекте изобретения, такие конструкции по изобретению, например, полипептиды по изобретению, имеют время полужизни в сыворотке у человека по меньшей мере приблизительно 12 часов, предпочтительно, по меньшей мере 24 часа, более предпочтительно, по меньшей мере 48 часов, даже более предпочтительно, по меньшей мере 72 часа или более. Например, конструкции или полипептиды по изобретению могут иметь время полужизни по меньшей мере 5 суток (например, приблизительно 5-10 суток), предпочтительно, по меньшей мере 9 суток (например, приблизительно 9-14 суток), более предпочтительно, по меньшей мере приблизительно 10 суток (например, приблизительно 10-15 суток), или по меньшей мере приблизительно 11 суток (например, приблизительно 11-16 суток), более предпочтительно, по меньшей мере приблизительно 12 суток (например, приблизительно 12-18 суток или более), или более чем 14 суток (например, приблизительно 14-19 суток).
В особенно предпочтительном, но неограничивающем аспекте изобретения, изобретение относится к конструкции по изобретению и к полипептиду по изобретению, содержащим, помимо одного или более строительных блоков, связывающих ADAMTS5, по меньшей мере один строительный блок, связывающий сывороточный альбумин, такой как ISVD, связывающий сывороточный альбумин, такой как человеческий сывороточный альбумин, как описано в настоящем описании. Предпочтительно, указанный ISVD, связывающий сывороточный альбумин, содержит или в основном состоит из 4 каркасных областей (FR1 - FR4, соответственно) и 3 определяющих комплементарность областей (CDR1 - CDR3, соответственно), в которых CDR1 представляет собой SFGMS, CDR2 представляет собой SISGSGSDTLYADSVKG, и CDR3 представляет собой GGSLSR. Предпочтительно, указанный ISVD, связывающий человеческий сывороточный альбумин, выбран из группы, состоящей из Alb8, Alb23, Alb129, Alb132, Alb11, Alb11 (S112K)-A, Alb82, Alb82-A, Alb82-AA, Alb82-AAA, Alb82-G, Alb82-GG, Alb82-GGG, Alb92 или Alb223 (ср. таблицу D).
В одном аспекте, настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере один ISVD, связывающий ADAMTS5, и ISVD, связывающий сывороточный альбумин, предпочтительно, выбранный из группы, состоящей из SEQ ID NO: 129 (клон 577 2F3*-Alb), SEQ ID NO: 130 (клон 579 2F3*-093-Alb), SEQ ID NO: 120 (клон 4 2A12-Alb), SEQ ID NO: 121 (клон 5 2D7-Alb), SEQ ID NO: 122 (клон 6 2F3-Alb), SEQ ID NO: 123 (клон 69 049-Alb), SEQ ID NO: 124 (клон 70 9D3-Alb), SEQ ID NO: 125 (клон 71 3B2-Alb), SEQ ID NO: 126 (клон 129 2F3-093-Alb), SEQ ID NO: 127 (клон 130 049-093-Alb) и SEQ ID NO: 128 (клон 131 9D3-093-Alb)(ср. таблицу A-1).
В одном варианте осуществления, настоящее изобретение относится к конструкции по изобретению, такой как полипептид, содержащий группу, связывающую сывороточный белок, где указанная группа, связывающая сывороточный белок, представляет собой полипептид, не основанный на антителе.
В данной области существует необходимость в более эффективных видах терапии против нарушений, поражающих хрящ в суставах, таких как остеоартрит. Даже при внутрисуставном введении, время удержания большинства лекарственных средств для лечения пораженного хряща является недостаточным. Авторы настоящего изобретения выдвинули гипотезу, что эффективность терапевтического лекарственного средства, такого как конструкция, полипептид и ISVD по изобретению, можно модулировать посредством связывания терапевтического лекарственного средства с группой, которая может «заякоривать» лекарственное средство в суставе и следовательно, увеличивать удержание лекарственного средства, но которая не должна нарушать эффективность указанного терапевтического лекарственного средства (эта группа также обозначена в настоящем описании как «заякоривающий в хряще белок» или «CAP»). Эта концепция заякоривания может модулировать не только эффективность лекарственного средства, но также специфичность действия для пораженного заболеванием сустава, посредством уменьшения токсичности и побочных эффектов, таким образом расширяя число возможных применимых лекарственных средств.
Предусматривают, что формат молекулы для клинического применения содержит один или два строительных блока, такие как ISVD, связывающие ADAMTS5, и один или более строительных блоков, например, ISVD, с таким удерживающим механизмом действия, и возможно, дополнительные группы. В одновременно поданной заявке показано, что такие форматы сохраняют как связывание ADAMTS5, так и терапевтический эффект, например, ингибирующую активность, так же как свойства удержания. Один или более строительных блоков, таких как ISVD, с удерживающим механизмом действия, могут представлять собой любой строительный блок, имеющий эффект удержания («строительный блок CAP») при заболеваниях, в которые вовлечен ADAMTS5, таких как артритное заболевание, остеоартрит, спондилоэпиметафизарная дисплазия, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава, ревматоидный артрит, рассекающий остеохондрит, аггреканопатии.
«Строительный блок CAP» используют для направления, заякоривания и/или удержания других, например, терапевтических, строительных блоков, таких как ISVD, связывающие ADAMTS5, в желательном участке, например, таком как сустав, в котором указанный другой, например, терапевтический, строительный блок должен оказывать свой эффект, например, связывание и/или ингибирование ADAMTS5.
Авторы настоящего изобретения выдвинули дополнительную гипотезу, что связывающие аггрекан молекулы, такие как ISVD, связывающие аггрекан, могут потенциально функционировать как такой якорь, хотя аггрекан является сильно гликозилированным и деградированным при различных нарушениях, поражающих хрящ в суставах. Кроме того, с учетом стоимости и интенсивного тестирования в различных моделях на животных, необходимых до того, как лекарственное средство может поступать в клинику, такие связывающие аггрекан молекулы должны, предпочтительно, иметь широкую перекрестную реакционную способность, например, связывающие аггрекан молекулы должны связываться с аггреканом из различных видов.
С использованием различных изобретательских способов иммунизации, скрининга и характеризации, авторам настоящего изобретения удалось идентифицировать различные связывающие аггрекан молекулы с превосходными признаками избирательности, стабильности и специфичности, обеспечивающими продленное удержание и активность в суставе (ср. одновременно поданную заявку).
В одном аспекте, настоящее изобретение относится к способу уменьшения и/или ингибирования оттока композиции, полипептида или конструкции из сустава, где указанный способ включает введение фармацевтически активного количества по меньшей мере одного полипептида по изобретению, конструкции по изобретению или композиции по изобретению нуждающемуся в этом индивидууму.
По настоящему изобретению, термин «уменьшение и/или ингибирование оттока» обозначает уменьшение и/или ингибирование вытекающего потока композиции, полипептида или конструкции изнутри сустава наружу. Предпочтительно, отток уменьшают и/или ингибируют по меньшей мере на 10%, например, по меньшей мере на 20%, 30%, 40% или 50% или даже больше, например, по меньшей мере на 60%, 70%, 80%, 90% или даже 100%, по сравнению с оттоком вышеупомянутых композиции, полипептида или конструкции в суставе в таких же условиях но в отсутствие связывающей аггрекан молекулы по изобретению, например, ISVD, связывающих аггрекан.
Предусматривают, что, помимо заболеваний, в которые вовлечен ADAMTS5, таких как артритное заболевание, остеоартрит, спондилоэпиметафизарная дисплазия, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава, ревматоидный артрит, рассекающий остеохондрит и аггреканопатии, связывающие аггрекан молекулы по изобретению можно также использовать при различных других заболеваниях, поражающих хрящ, таких как артропатии и хондродистрофии, артритное заболевание (такое как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматический разрыв или открепление), ахондроплазия, реберный хондрит, спондилоэпиметафизарная дисплазия, межпозвонковая грыжа, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава и рецидивирующий полихондрит (совместно названных в настоящем описании как «ассоциированные с аггреканом заболевания»).
Указанный строительный блок CAP, например, ISVD, связывающие аггрекан, предпочтительно, связывается с хрящевой тканью, такой как хрящ и/или мениск. В предпочтительном аспекте, строительный блок CAP проявляет перекрестную реакционную способность для других видов и специфически связывает один или более из аггрекана человека (SEQ ID NO: 155), аггрекана собаки, аггрекана быка, аггрекана крысы; аггрекана свиньи; аггрекана мыши, аггрекана кролика; аггрекана яванского макака и/или аггрекана макака-резуса. Соответствующую структурную информацию для аггрекана можно обнаружить, например, в номерах доступа (UniProt) как показано в таблице 2 ниже.
Предпочтительным строительным блоком CAP является ISVD, связывающий аггрекан, предпочтительно, аггрекан человека, предпочтительно, представленный SEQ ID NO: 155, как показано в таблице B.
Таблица 2
Настоящее изобретение, таким образом, относится к полипептиду или конструкции по изобретению, дополнительно содержащим по меньшей мере один строительный блок CAP.
Настоящее изобретение, таким образом, относится к полипептиду или конструкции по изобретению, дополнительно содержащим по меньшей мере один ISVD, специфически связывающий аггрекан, предпочтительно, выбранный из ISVD, представленных SEQ ID NO: 156 и 157.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, специфически связывающих аггрекан.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, специфически связывающих аггрекан, где указанные по меньшей мере 2 ISVD, специфически связывающие аггрекан, могут являться одинаковыми или различными.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, специфически связывающих аггрекан, где указанные по меньшей мере 2 ISVD, специфически связывающие аггрекан, независимо выбраны из группы, состоящей из SEQ ID NO: 156-157.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему по меньшей мере 2 ISVD, специфически связывающих аггрекан, где указанные по меньшей мере 2 ISVD, специфически связывающие аггрекан, представлены SEQ ID NO: 156-157.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, содержащему ISVD, специфически связывающий аггрекан, где указанный ISVD, специфически связывающий аггрекан, специфически связывает аггрекан человека [SEQ ID NO: 155].
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий аггрекан, специфически связывает аггрекан человека (SEQ ID NO: 155), аггрекан собаки, аггрекан быка, аггрекан крысы; аггрекан свиньи; аггрекан мыши, аггрекан кролика; аггрекан яванского макака и/или аггрекан макака-резуса.
В одном аспекте настоящее изобретение относится к полипептиду, как описано в настоящем описании, где указанный ISVD, специфически связывающий аггрекан, предпочтительно, связывается с хрящевой тканью, такой как хрящ и/или мениск.
Следует понимать, что ISVD, полипептид и конструкция по изобретению являются, предпочтительно, стабильными. Стабильность полипептида, конструкции или ISVD по изобретению можно измерять посредством общепринятых анализов, известных специалисту в данной области. Типичные анализы включают (без ограничения) анализы, в которых определяют активность указанных полипептида, конструкции или ISVD, с последующей инкубацией в синовиальной жидкости в течение желательного периода времени, после чего активность определяют снова.
В одном аспекте настоящее изобретение относится к ISVD, полипептиду или конструкции по изобретению, имеющим стабильность по меньшей мере 7 суток, например, по меньшей мере 14 суток, 21 сутки, 1 месяц, 2 месяца или даже 3 месяца в синовиальной жидкости (SF) при 37°C.
Желательную активность терапевтического строительного блока, например, ISVD, связывающего ADAMTS5, в мультивалентных полипептиде или конструкции по изобретению можно измерять посредством общепринятых анализов, известных специалисту в данной области.
В одном аспекте, настоящее изобретение относится к конструкции, как описано в настоящем описании, содержащей по меньшей мере один ISVD или полипептид и одно или более из других групп, остатков, мотивов или связывающих единиц. Одно или более из других групп, остатков, мотивов или связывающих единиц предпочтительно, выбраны из группы, состоящей из молекулы полиэтиленгликоля, сывороточных белков или их фрагментов, связывающих единиц, которые могут связываться с сывороточными белками, фрагмента Fc и малых белков или пептидов, которые могут связываться с сывороточными белками, дополнительных аминокислотных остатков, меток или других функциональных групп, например, токсинов, меток, радиоактивных химических соединений и т.д.
В одном варианте осуществления, как упомянуто ниже, настоящее изобретение относится к конструкции по изобретению, такой как полипептид, содержащей группу, обеспечивающую продление времени полужизни, где указанная группа представляет собой PEG. Таким образом, настоящее изобретение относится также к конструкции или полипептиду по изобретению, содержащим PEG.
Дополнительные аминокислотные остатки могут или могут не изменять, преобразовывать или иным образом влиять на другие (биологические) свойства полипептида по изобретению и могут или могут не добавлять дополнительную функциональность полипептиду по изобретению. Например, такие аминокислотные остатки:
a) могут содержать N-концевой остаток Met, например, в результате экспрессии в гетерологичной клетке-хозяине или организме-хозяине.
b) могут формировать сигнальную последовательность или лидерную последовательность, управляющую секрецией полипептида из клетки-хозяина после синтеза (например, для получения пре-, про- или препро-формы полипептида по изобретению, в зависимости от клетки-хозяина, используемой для экспрессии полипептида по изобретению). Подходящие секреторные лидерные пептиды известны специалисту в данной области, и могут являться такими, как далее описано в настоящем описании. Обычно такая лидерная последовательность может быть связана с N-концом полипептида, хотя изобретение, в его самом широком смысле, не является ограниченным этим;
c) могут формировать «метку», например, аминокислотную последовательность или остаток, который позволяет или облегчают очистку полипептида, например, с использованием аффинных способов, направленных против указанных последовательности или остатка. затем, указанную последовательность или остаток можно удалять (например, посредством химического или ферментного отщепления) для получения полипептида (для этой цели, метка может, необязательно, являться связанной с аминокислотной последовательностью или полипептидной последовательностью посредством расщепляемой линкерной последовательности или может содержать расщепляемый мотив). Некоторыми предпочтительными, но неограничивающими примерами таких остатков являются множественные остатки гистидина, остатки глутатиона и myc-метка, такая как AAAEQKLISEEDLNGAA;
d) могут представлять собой один или более аминокислотных остатков, которые были функционализированы и/или которые могут служить участком для присоединения функциональных групп. Подходящие аминокислотные остатки и функциональные группы известны специалисту в данной области и включают, но без ограничения, аминокислотные остатки и функциональные группы, упомянутые в настоящем описании для производных полипептидов по изобретению.
Настоящее изобретению относится также к конструкциям, содержащим полипептид и/или ISVD по изобретению, которые дополнительно содержат другие функциональные группы, например, токсины, метки, радиоактивные химические соединения и т.д.
Другие группы, остатки, мотивы или связывающие единицы могут, например, представлять собой химические группы, остатки, мотивы, которые сами по себе могут являться или могут не являться биологически и/или фармакологически активными. Например, и без ограничения, такие группы можно связывать с одним или более ISVD или полипептидами по изобретению таким образом, чтобы получить «производное» полипептида или конструкции по изобретению.
Соответственно, изобретение, в его самом широком смысле, относится также к конструкциям и/или полипептидам, являющимся производными конструкций и/или полипептидов по изобретению. Такие производные можно, в общем, получать посредством модификации, и в частности, посредством химической и/или биологической (например, ферментной) модификации, конструкций и/или полипептидов по изобретению и/или одного или более аминокислотных остатков, формирующих полипептид по изобретению.
Примеры таких модификаций, так же, как примеры аминокислотных остатков в последовательностях полипептидов, которые можно модифицировать таким образом (т.е. либо в остове белка, но предпочтительно, на боковой цепи), методы и способы, которые можно использовать для введения таких модификаций, и потенциальные применения и преимущества таких модификаций известны специалисту в данной области (см. также Zangi et al., Nat Biotechnol 31(10):898-907, 2013).
Например, такая модификация может включать введение (например, посредством ковалентного связывания или любым другим подходящим способом) одного или более из (функциональных) групп, остатков или мотивов в или на полипептид по изобретению, и в частности, одного или более из функциональных групп, остатков или мотивов, придающих одно или более желательных свойств или функциональностей конструкции и/или полипептиду по изобретению. Примеры таких функциональных групп известны специалисту в данной области.
Например, такая модификация может включать введение (например, посредством ковалентного связывания или любым другим подходящим способом) одного или более функциональных мотивов, которые увеличивают время полужизни, растворимость и/или абсорбцию конструкции или полипептида по изобретению, которые уменьшают иммуногенность и/или токсичность конструкции или полипептида по изобретению, которые исключают или ослабляют любые нежелательные побочные эффекты конструкции или полипептида по изобретению, и/или которые придают другие обеспечивающие преимущество свойства и/или уменьшают нежелательные свойства конструкции или полипептида по изобретению; или любой комбинации двух или более из вышеуказанного. Примеры таких функциональных мотивов и способов их введения известны специалисту в данной области, и могут, в общем, включать функциональные мотивы и способы, упомянутые в общем уровне техники, процитированном в настоящем описании выше, так же как функциональные мотивы и способы, известные по существу для модификации фармацевтических белков, и в частности, для модификации антител или фрагментов антител (включая ScFv и однодоменные антитела), для которых приведена ссылка, например, на Remington (Pharmaceutical Sciences, 16th ed., Mack Publishing Co., Easton, PA, 1980). Такие функциональные мотивы можно, например, связывать напрямую (например, ковалентно) с полипептидом по изобретению, или необязательно, посредством подходящего линкера или спейсера, снова, как известно специалисту в данной области.
Одним конкретным примером является производное полипептида или конструкции по изобретению, где полипептид или конструкция по изобретению химически модифицированы для увеличения их времени полужизни (например, посредством пегилирования). Это является одним из наиболее широко используемых способов увеличения времени полужизни и/или уменьшения иммуногенности фармацевтических белков и включает присоединение подходящего фармакологически пригодного полимера, такого как поли(этиленгликоль) (PEG) или его производных (таких как метоксиполи(этиленгликоль) или mPEG). В общем, можно использовать любую подходящую форму пегилирования, такую как пегилирование, используемое в данной области для антител и фрагментов антител (включая, но без ограничения, (одно)доменные антитела и ScFv); приведена ссылка, например, на Chapman (Nat. Biotechnol. 54: 531-545, 2002), Veronese and Harris (Adv. Drug Deliv. Rev. 54: 453-456, 2003), Harris and Chess (Nat. Rev. Drug. Discov. 2: 214-221, 2003) и WO 04/060965. Различные реагенты для пегилирования белков также являются коммерчески доступными, например, из Nektar Therapeutics, USA.
Предпочтительно, используют сайт-специфческое пегилирование, в частности, посредством остатка цистеина (см., например, Yang et al. (Protein Engineering 16: 761-770, 2003). Например, для этой цели, PEG можно присоединять к остатку цистеина, встречающемуся в природе в полипептиде по изобретению, конструкцию или полипептид по изобретению можно модифицировать так, чтобы соответствующим образом вводить один или более остатков цистеина для присоединения PEG, или аминокислотную последовательность, содержащую один или более остатков цистеина для присоединения PEG, можно сливать с N- и/или C-концом конструкции или полипептида по изобретению, все с использованием способов белковой инженерии, известных по существу специалисту в данной области.
Предпочтительно, для конструкций или полипептидов по изобретению, используют PEG с молекулярной массой более чем 5000, например, более чем 10000, и менее чем 200000, например, менее чем 100000; например, в диапазоне 20000-80000.
Другая, обычно менее предпочтительная, модификация включает N-связанное или O-связанное гликозилирование, обычно в качестве части котрансляционной и/или посттрансляционной модификации, в зависимости от клетки-хозяина, используемой для экспрессии полипептида по изобретению.
Другая модификация может включать введение одной или более поддающихся детекции меток или других испускающих сигнал групп или мотивов, в зависимости от намеченного использования полипептида или конструкции по изобретению. Подходящие метки и способы их присоединения, использования и детекции известны специалисту в данной области, и например, включают, но без ограничения, флуоресцентные метки (такие как флуоресцеин, изотиоцианат, родамин, фикоэритрин, фикоцианин, аллофикоцианин, o-фтальдегид и флуоресцамин, и флуоресцентные металлы, такие как 152Eu или другие металлы из группы лантанидов), фосфоресцентные метки, хемилюминесцентные метки или биолюминесцентные метки (такие как люминал, изолюминол, ароматический сложный эфир акридиния, имидазол, соли акридиния, сложный эфир оксалата, диоксетан или GFP и его аналоги), радиоактивные изотопы (такие как 3H, 125I, 32P, 35S, 14C, 51Cr, 36Cl, 57Co, 58Co, 59Fe и 75Se), металлы, хелаты металлов или катионов металлов (например, таких как катионы металлов 99mTc, 123I, 111In, 131I, 97Ru, 67Cu, 67Ga и 68Ga или другие металлы или катионы металлов, особенно подходящие для использования в диагностике и визуализации in vivo, in vitro или in situ, такие как (157Gd, 55Mn, 162Dy, 52Cr и 56Fe)), так же как хромофоры и ферменты (такие как малатдегидрогеназа, нуклеаза стафилококка, дельта-V-стероид-изомераза, алкогольдегидрогеназа дрожжей, альфа-глицерофосфатдегидрогеназа, триозофосфатизомераза, биотин-авидин-пероксидаза, пероксидаза хрена, щелочная фосфатаза, аспарагиназа, глюкоза оксидаза, β-галактозидаза, рибонуклеаза, уреаза, каталаза, глюкоза-VI-фосфатдегидрогеназа, глюкоамилаза и ацетилхолинэстераза). Другие пригодные метки известны специалисту в данной области, и например, включают мотивы, которые можно детектировать с использованием спектроскопии NMR или ESR.
Такие меченые полипептиды и конструкции по изобретению можно, например, использовать для анализов in vitro, in vivo или in situ (включая иммуноанализы, известные по существу, такие как ELISA, RIA, EIA и другие «сэндвич-анализы», и т.д.) так же как для целей диагностики и визуализации in vivo, в зависимости от выбора конкретной метки.
Как понятно специалисту в данной области, другая модификация может включать введение хелатообразующей группы, например, для образования хелата одного из металлов или катионов металлов, упомянутых выше. Пригодные хелатообразующие группы, например, включают, без ограничения, диэтилентриаминпентауксусную кислоту (DTPA) или этилендиаминтетрауксусную кислоту (ЭДТА).
Другая модификация может включать введение функциональной группы, представляющей собой одну часть из специфической связывающейся пары, такой как связывающаяся пара биотин-(стрепт)авидин. Такую функциональную группу можно использовать для связывания полипептида по изобретению с другим белком, полипептидом или химическим соединением, связанным с другой половиной связывающейся пары, т.е. посредством формирования связывающейся пары. Например, конструкцию или полипептид по изобретению можно конъюгировать с биотином, и связывать с другим белком, полипептидом, соединением или носителем, конъюгированным с авидином или стрептавидином. Например, такие конъюгированные конструкцию или полипептид по изобретению можно использовать в качестве репортера, например, в диагностической системе, где испускающее поддающийся детекции сигнал средство конъюгировано с авидином или стрептавидином. Такие связывающиеся пары можно, например, также использовать для связывания конструкции или полипептида по изобретению с носителем, включая носители, пригодные для фармацевтических целей. Один неограничивающий пример представляет собой липосомные составы, описанные в Cao and Suresh (Journal of Drug Targeting 8: 257, 2000). Такие связывающиеся пары можно также использовать для связывания терапевтически действующего вещества с полипептидом по изобретению.
Другие потенциальные химические и ферментные модификации известны специалисту в данной области. Такие модификации можно также вводить для исследовательских целей (например, для исследования взаимосвязей функция-активность). Приведена ссылка, например, на Lundblad and Bradshaw (Biotechnol. Appl. Biochem. 26: 143-151, 1997).
Предпочтительно, конструкции, полипептиды и/или производные являются такими, что они связывают ADAMTS5, с аффинностью (соответствующим образом измеренной и/или выраженной как значение KD (истинной или кажущейся), значение KA (истинной или кажущейся), kon скорости или скорости связывания и/или koff или скорости диссоциации, или альтернативно, как значение IC50, как далее описано в настоящем описании), то есть, как определено в настоящем описании (например, как определено для полипептидов по изобретению).
Такие конструкции и/или полипептиды по изобретению и их производные могут также находиться в основном в выделенной форме (как определено в настоящем описании).
В одном аспекте, настоящее изобретение относится к конструкции по изобретению, которая содержит или в основном состоит из ISVD по изобретению или полипептида по изобретению, и которая дополнительно содержит одно или более из других групп, остатков, мотивов или связывающих единиц, которые, необязательно, связаны посредством одного или более пептидных линкеров.
В одном аспекте, настоящее изобретение относится к конструкции по изобретению, в которой одно или более из других групп, остатков, мотивов или связывающих единиц выбраны из группы, состоящей из молекулы полиэтиленгликоля, сывороточных белков или их фрагментов, связывающих единиц, которые могут связываться с сывороточными белками, фрагмента Fc и малых белков или пептидов, которые могут связываться с сывороточными белками.
В конструкциях по изобретению, таких как полипептиды по изобретению, два или более строительных блоков, например, таких как ISVD, и необязательно, одно или более из других групп, лекарственных средств, средств, остатков, мотивов или связывающих единицы могут быть напрямую связаны друг с другом (например, как описано в WO 99/23221) и/или могут быть связаны друг с другом посредством одного или более пригодных спейсеров или линкеров, или любой их комбинации. Пригодные спейсеры или линкеры для использования в мультивалентных и мультиспецифических полипептидов известны специалисту в данной области, и могут в общем представлять собой любой линкер или спейсер, используемый в данной области для связывания аминокислотных последовательностей. Предпочтительно, указанный линкер или спейсер является пригодным для использования в конструирования конструкций, белков или полипептидов, предназначенных для фармацевтического применения.
Например, полипептид по изобретению может, например, представлять собой трехвалентный, триспецифический полипептид, содержащий один строительный блок, такой как ISVD, связывающий ADAMTS5, ISVD, связывающий альбумин, и потенциально, другой строительный блок, такой как третий ISVD, в которых указанный первый, второй и третий строительные блоки, такие как ISVD, могут, необязательно являться связанными посредством одной или более, и в частности 2, линкерных последовательностей. Также настоящее изобретение относится к конструкции или полипептиду по изобретению, содержащим первый ISVD, связывающий ADAMTS5, и, возможно, второй ISVD, связывающий альбумин, и/или, возможно, третий ISVD и/или, возможно, четвертый ISVD, где указанный первый ISVD и/или указанный второй ISVD и/или, возможно, указанный третий ISVD и/или возможно, указанный четвертый ISVD связаны посредством линкеров, в частности, 3 линкеров.
Некоторые особенно предпочтительные линкеры включают линкеры, используемые в данной области для связывания фрагментов антител или доменов антител. Они включают линкеры, упомянутые в общем уровне техники, процитированном выше, так же, как, например, линкеры, используемые в данной области для конструирования диател или фрагментов ScFv (в этом отношении, однако, следует отметить, что, в то время как в диателах и фрагментах ScFv, используемая линкерная последовательность должна иметь длину, степень гибкости и другие свойства, позволяющие сближение соответствующих доменов VH и VL для формирования полного антигенсвязывающего участка, не существует конкретного ограничения длины или гибкости линкера, используемого в полипептиде по изобретению, поскольку каждый ISVD, такой как нанотело, сам формирует полный антигенсвязывающий участок).
Например, линкер может представлять собой подходящую аминокислотную последовательность, и в частности, аминокислотные последовательности из между 1 и 50, предпочтительно, между 1 и 30, например, между 1 и 10 аминокислотных остатков. Некоторые примеры таких аминокислотных последовательностей включают линкеры gly-ser, например, типа (glyxsery)2, такие как (например, (gly4ser)3 или (gly3ser2)3, как описано в WO 99/42077, и линкеры GS30, GS15, GS9 и GS7, описанные в заявках от Ablynx, упомянутых в настоящем описании (см., например, WO 06/040153 и WO 06/122825), так же, как подобные шарнирам области, такие как шарнирные области природной тяжелой цепи антитела или сходные последовательности (такие, как описано в WO 94/04678). Предпочтительные линкеры показаны в таблице C.
Некоторые другие особенно предпочтительные линкеры представляют собой поли-аланин (такой как AAA), так же как линкеры GS30 (см. также SEQ ID NO: 85 в WO 06/122825) и GS9 (см. также SEQ ID NO: 84 в WO 06/122825).
Другие пригодные линкеры, как правило, включают органические соединения или полимеры, в частности, линкеры, пригодные для использования в белках для фармацевтического применения. Например, мотивы поли(этиленгликоля) использовали для связывания доменов антитела, см., например, WO 04/081026.
В объем изобретения включено то, что длина, степень гибкости и/или другие свойства используемых линкера(линкеров) (хотя и не критично, как это обычно бывает для линкеров, используемых в фрагментах ScFv) могут оказывать некоторое влияние на свойства конечной конструкции по изобретению, такой как полипептид по изобретению, включая, но без ограничения, аффинность, специфичность или авидность для хемокина, или для одного или более из других антигенов. На основании описания в настоящем документе, специалист в данной области способен определять оптимальный линкер(ы) для использования в конкретной конструкции по изобретению, такой как полипептид по изобретению, необязательно, после некоторых ограниченных общепринятых экспериментов.
Например, в мультивалентных полипептидах по изобретению, содержащих строительные блоки, ISVD или нанотела, направленные против ADAMTS5 и другой мишени, длина и гибкость линкера, предпочтительно, являются такими, которые позволяют каждому строительному блоку, такому как ISVD, по изобретению, присутствующему в полипептиде, связываться с родственной ему мишенью, например, антигенной детерминантой на каждой из мишеней. Снова, на основании описания в настоящем документе, специалист в данной области может определять оптимальный линкер(ы) для использования в конструкции по изобретению, такой как полипептид по изобретению, необязательно, после некоторых ограниченных общепринятых экспериментов.
В объем изобретения включено также то, что используемые линкер(ы) придают одно или более других обеспечивающих преимущества свойств или функциональностей конструкциям по изобретению, таким как полипептиды по изобретению, и/или предоставляют один или более участков для образования производных и/или для присоединения функциональных групп (например, как описано в настоящем описании для производных ISVD по изобретению). Например, линкеры, содержащие один или более заряженных аминокислотных остатков, могут обеспечивать улучшенные гидрофильные свойства, в то время как линкеры, формирующие или содержащие малые эпитопы или метки, можно использовать для целей детекции, идентификации и/или очистки. Снова, на основании описания в настоящем документе, специалист в данной области способен определять оптимальные линкеры для использования в конкретном полипептиде по изобретению, необязательно, после некоторых ограниченных общепринятых экспериментов.
Наконец, когда два или более линкеров используют в конструкциях, таких как полипептиды по изобретению, эти линкеры могут являться одинаковыми или различными. Снова, на основании описания в настоящем документе, специалист в данной области способен определять оптимальные линкеры для использования в конкретной конструкции или полипептиде по изобретению, необязательно, после некоторых ограниченных общепринятых экспериментов.
Обычно, для простоты экспрессии и продукции, конструкция по изобретению, такая как полипептид по изобретению, представляет собой линейный полипептид. Однако изобретение, в его самом широком смысле, не является ограниченным этим. Например, когда конструкция по изобретению, такая как полипептид по изобретению, содержит три или более строительных блоков, ISVD или нанотел, можно связывать их с использованием линкера с тремя или более «плечами», где каждое «плечо» связано со строительным блоком, ISVD или нанотелом, таким образом, чтобы получить «звездообразную» конструкцию. Является также возможным, хотя обычно менее предпочтительным, использовать кольцевые конструкции.
Соответственно, настоящее изобретение относится к конструкции по изобретению, такой как полипептид по изобретению, где указанные ISVD напрямую связаны друг с другом или связаны посредством линкера.
Соответственно, настоящее изобретение относится к конструкции по изобретению, такой как полипептид по изобретению, где первый ISVD и/или второй ISVD, и/или возможно, ISVD, связывающий сывороточный альбумин, связаны посредством линкера.
Соответственно, настоящее изобретение относится к конструкции по изобретению, такой как полипептид по изобретению, где указанный линкер выбран из группы, состоящей из линкеров из 3A, 5GS, 7GS, 9GS, 10GS, 15GS, 18GS, 20GS, 25GS, 30GS, 35GS, поли-A, 8GS, 40GS, шарнира G1, шарнира 9GS-G1, верхней длинной шарнирной области ламы и шарнира G3, например, таких как представлено в таблице C (SEQ ID NO: 158-174).
Соответственно, настоящее изобретение относится к конструкции по изобретению, такой как полипептид по изобретению, где указанный полипептид выбран из группы, состоящей из SEQ ID NO: 120-130.
Изобретение, кроме того, относится к способам получения конструкций, полипептидов, ISVD, нуклеиновых кислот, клеток-хозяев и композиций, описанных в настоящем описании.
Мультивалентные полипептиды по изобретению можно, как правило, получать способом, который включает по меньшей мере стадию связывания соответствующим образом ISVD и/или моновалентного полипептида по изобретению с одним или более дополнительными ISVD, необязательно, посредством одного или более подходящих линкеров, таким образом, чтобы получить мультивалентный полипептид по изобретению. Полипептиды по изобретению можно также получать способом, который, как правило, включает по меньшей мере стадии получения нуклеиновой кислоты, кодирующей полипептид по изобретению, экспрессии указанной нуклеиновой кислоты соответствующим способом и выделения экспрессированного полипептида по изобретению. Такие способы можно осуществлять образом, известным по существу, известным специалисту в данной области, например, на основании методов и способов, далее описанных в настоящем описании.
Способ получения мультивалентных полипептидов по изобретению может включать по меньшей мере стадии связывания двух или более ISVD по изобретению и например, одного или более линкеров, соответствующим способом. ISVD по изобретению (и линкеры) можно связывать любым способом, как известно в данной области и как далее описано в настоящем описании. Предпочтительные способы включают соединение последовательностей нуклеиновой кислоты, кодирующих ISVD по изобретению (и линкеры) для получения генетической конструкции, экспрессирующей мультивалентный полипептид. Способы соединения аминокислот или нуклеиновых кислот известны специалисту в данной области, и снова приведена ссылка на стандартные руководства, такие как Sambrook et al. и Ausubel et al., упомянутые выше, так же как на примеры ниже.
Соответственно, настоящее изобретение также относится к применению ISVD по изобретению для получения мультивалентного полипептида по изобретению. Способ получения мультивалентного полипептида может включать связывание ISVD по изобретению по меньшей мере с одним дополнительным ISVD по изобретению, необязательно, посредством одного или более линкеров. Затем ISVD по изобретению используют в качестве связывающего домена или строительного блока для предоставления и/или получения мультивалентного полипептида, содержащего 2 (например, в двухвалентном полипептиде), 3 (например, в трехвалентном полипептид), 4 (например, в тетравалентном) или более (например, в мультивалентном полипептиде) строительных блоков. В этом отношении, ISVD по изобретению можно использовать в качестве связывающего домена или связывающей единицы для предоставления и/или получения мультивалентного, например, двухвалентного, трехвалентного или тетравалентного полипептида по изобретению, содержащего 2, 3, 4 или более строительных блоков.
Соответственно, настоящее изобретение также относится к применению полипептида ISVD по изобретению (как описано в настоящем описании) для получения мультивалентного полипептида. Способ получения мультивалентного полипептида может включать связывание ISVD по изобретению по меньшей мере с одним дополнительным ISVD по изобретению, необязательно, посредством одного или более линкеров.
Полипептиды и нуклеиновые кислоты по изобретению можно получать способом, известным по существу, как станет понятно специалисту в данной области из дальнейшего описания в настоящем документе. Например, полипептиды по изобретению можно получать любым способом, известным по существу для получения антител и в частности, для получения фрагментов антител (включая, но без ограничения, (одно)доменные антитела и фрагменты ScFv). Некоторые предпочтительные, но неограничивающие способы получения полипептидов и нуклеиновых кислот, включают методы и способы, описанные в настоящем описании.
Способ получения полипептида по изобретению может включать следующие стадии: экспрессии, в подходящих клетке-хозяине или организме-хозяине (также обозначенным в настоящем описании как «хозяин по изобретению»), или в другой подходящей системе экспрессии, нуклеиновой кислоты, кодирующей указанный полипептид по изобретению (также обозначенной в настоящем описании как «нуклеиновая кислота по изобретению»); необязательно, с последующими выделением и/или очисткой полученного таким образом полипептида по изобретению.
В частности, такой способ может включать стадии: культивирования и/или поддержания хозяина по изобретению в условиях, являющихся такими, что указанный хозяин по изобретению экспрессирует и/или продуцирует по меньшей мере один полипептид по изобретению; необязательно, с последующими выделением и/или очисткой полученного таким образом полипептида по изобретению.
Соответственно, настоящее изобретение также относится к нуклеиновой кислоте или нуклеотидной последовательности, кодирующей полипептид, ISVD или конструкцию по изобретению (также обозначенной как «нуклеиновая кислота по изобретению»).
Нуклеиновая кислота по изобретению может находиться в форме одно- или двухцепочечной ДНК или РНК. В соответствии с одним вариантом осуществления изобретения, нуклеиновая кислота по изобретению находится в по существу выделенной форме, как определено в настоящем описании. Нуклеиновая кислота по изобретению может также находиться в форме, присутствовать в и/или являться частью вектора, например, экспрессирующего вектора, например, такого как плазмида, космида или YAC, которые снова могут находиться в по существу выделенной форме. Соответственно, настоящее изобретение также относится к экспрессирующему вектору, содержащему нуклеиновую кислоту или нуклеотидную последовательность по изобретению.
Нуклеиновые кислоты по изобретению можно подготавливать или получать способом, известным по существу, на основании информации о полипептидах по изобретению, приведенной в настоящем описании, и/или их можно выделять из подходящего природного источника, также, как понятно специалисту в данной области, для получения нуклеиновой кислоты по изобретению, также несколько нуклеотидных последовательностей, таких как по меньшей мере две нуклеиновые кислоты, кодирующие ISVD, по изобретению и например, нуклеиновые кислоты, кодирующие один или более линкеров, можно соединять вместе соответствующим способом. Способы получения нуклеиновых кислот по изобретению известны специалисту в данной области и могут, например, включать, но без ограничения, автоматизированный синтез ДНК; сайт-направленный мутагенез; комбинирование двух или более природных и/или синтетических последовательностей (или двух или более их частей), введение мутаций, приводящих к экспрессии укороченного продукта экспрессии; введение одного или более участков рестрикции (например, для получения кассет и/или областей, которые можно легко расщеплять и/или лигировать с использованием подходящих рестрикционных ферментов), и/или введение мутаций посредством реакции ПЦР с использованием одного или более «несовпадающих» праймеров. Эти и другие способы известны специалисту в данной области, и снова приведена ссылка на стандартные руководства, такие как Sambrook et al. и Ausubel et al., упомянутые выше, так же как на примеры ниже.
В предпочтительном, но неограничивающем варианте осуществления, генетическая конструкция по изобретению содержит
a) по меньшей мере одну нуклеиновую кислоту по изобретению;
b) функционально связанные с ней один или более регуляторных элементов, такие как промотор и, необязательно, подходящий терминатор; и необязательно, также
c) один или более дополнительных элементов генетических конструкций, известных по существу;
в которых термины «регуляторный элемент», «промотор», «терминатор» и «функционально связанный» имеют их обычное в данной области значение.
Генетические конструкции по изобретению можно, в основном, получать посредством связывания, соответствующим образом, нуклеотидной последовательности(последовательностей) по изобретению с одним или более дополнительными элементами, описанными выше, например, с использованием способов, описанных в общих руководствах, таких как Sambrook et al. и Ausubel et al., упомянутых выше.
Нуклеиновые кислоты по изобретению и/или генетические конструкции по изобретению можно использовать для трансформации клетки-хозяина или организма-хозяина, т.е. для экспрессии и/или продукции полипептида по изобретению. Пригодные хозяева или клетки-хозяева известны специалисту в данной области, и могут, например, представлять собой любые пригодные клетку или линию клеток грибов, прокариот или эукариот, или любой пригодный грибковый, прокариотический или (не относящийся к человеку) эукариотический организм, так же как другие клетки-хозяева или (не относящиеся к человеку) хозяева, известные по существу для экспрессии и продукции антител и фрагментов антител (включая, но без ограничения, (одно)доменные антитела и фрагменты ScFv), известные специалисту в данной области. Приведена также ссылка на общий уровень техники, процитированный в настоящем описании выше, так же как например, на WO 94/29457; WO 96/34103; WO 99/42077; Frenken et al. (Res Immunol. 149: 589-99, 1998); Riechmann and Muyldermans (1999), выше; van der Linden (J. Biotechnol. 80: 261-70, 2000); Joosten et al. (Microb. Cell Fact. 2: 1, 2003); Joosten et al. (Appl. Microbiol. Biotechnol. 66: 384-92, 2005); и дополнительные ссылки, процитированные в настоящем описании. Кроме того, полипептиды по изобретению можно также экспрессировать и/или продуцировать в бесклеточных системах экспрессии, и подходящие примеры таких систем известны специалисту в данной области. Подходящие способы трансформации хозяина или клетки-хозяина по изобретению известны специалисту в данной области и могут зависеть от намеченных клетки-хозяина/организма-хозяина и генетической конструкции, подлежащей использованию. Снова приведена ссылка на руководства и патентные заявки, упомянутые выше. Трансформированная клетка-хозяин (которая может находиться в форме стабильной линии клеток) или организмы-хозяева (которые могут находиться в форме стабильной мутантной линии или штамма) составляют дополнительные аспекты настоящего изобретения. Соответственно, настоящее изобретение относится к хозяину или клетке-хозяину, содержащей нуклеиновую кислоту по изобретению или экспрессирующий вектор по изобретению. Предпочтительно, эти клетки-хозяева или организмы-хозяева являются такими, что они экспрессируют или являются (по меньшей мере) способными экспрессировать (например, в подходящих условиях) полипептид по изобретению (и, в случае организма-хозяина: по меньшей мере в одной его клетке, части, ткани или органе). Изобретение также включает следующие поколения, потомство и/или потомков клетки-хозяина или организма-хозяина по изобретению, которые можно, например, получать посредством деления клеток или посредством полового или неполового воспроизведения.
Для продукции/получения экспрессии полипептидов по изобретению, трансформированную клетку-хозяина или трансформированного организма-хозяина можно, как правило, сохранять, поддерживать и/или культивировать в таких условиях, что (желательный) полипептид по изобретению экспрессируется/продуцируется. Подходящие условия известны специалисту в данной области и обычно могут зависеть от используемого клетки-хозяина/организма-хозяина, так же, как от регуляторных элементов, контролирующих экспрессию (соответствующей) нуклеотидной последовательности по изобретению. Снова приведена ссылка на руководства и патентные заявки, упомянутые выше в разделах о генетических конструкциях по изобретению.
Полипептид по изобретению можно затем выделять из клетки-хозяина/организма-хозяина и/или из среды, в которой культивировали указанную клетку-хозяина или организм-хозяина, с использованием способов выделения и/или очистки белка, известных по существу, таких как способы (препаративной) хроматографии и/или электрофореза, способы дифференциальной преципитации, аффинные способы (например, с использованием специфической, отщепляемой аминокислотной последовательности, слитой с полипептидом по изобретению) и/или препаративные иммунологические способы (т.е. с использованием антител против полипептида, подлежащего выделению).
В одном аспекте изобретение относится к способу получения конструкции, полипептида или ISVD по изобретению, включающему по меньшей мере стадии: (a) экспрессии, в подходящих клетке-хозяине или организме-хозяине или в другой подходящей системе экспрессии, последовательности нуклеиновой кислоты по изобретению; необязательно, с последующими (b) выделением и/или очисткой конструкции, полипептида ISVD по изобретению.
В одном аспекте изобретение относится к композиции, содержащей конструкцию, полипептид, ISVD или нуклеиновую кислоту по изобретению.
Как упомянуто выше, остается необходимость в безопасных и эффективных лекарственных средствах против OA. На основании не являющихся общепринятыми способов скрининга, характеризации и комбинирования, авторы настоящего изобретения идентифицировали ISVD, связывающие и ингибирующие ADAMTS5. Эти связывающие ADAMTS5 молекулы имели исключительно хорошее действие в экспериментах in vitro и in vivo. Кроме того, показано также, что ISVD по изобретению являются значительно более эффективными, чем соединения предшествующего уровня техники. Настоящее изобретение, таким образом, относится к ISVD и полипептидам, являющимся антагонистами ADAMTS, в частности, ADAMTS5, с улучшенными профилактическими, терапевтическими и/или фармакологическими свойствами, включая лучший профиль безопасности, по сравнению с аминокислотными последовательностями и антителами предшествующего уровня техники. Кроме того, эти связывающие ADAMTS5 молекулы, при связывании с ISVD, связывающими альбумин, имели увеличенное удержание у субъекта, их можно было вводить системно в то же время с сохранением активности.
В одном аспекте настоящее изобретение относится к способу лечения или предотвращения у индивидуума заболеваний или нарушений, например, тех, в которые вовлечена активность ADAMTS5, включающему введение ISVD или полипептида по изобретению указанному индивидууму в количестве, эффективном для лечения или предотвращения симптома указанного заболевания или нарушения.
В одном аспекте настоящее изобретение относится к композиции по изобретению, ISVD по изобретению, полипептиду по изобретению, и/или конструкции по изобретению для использования в качестве лекарственного средства.
В другом аспекте изобретение относится к применению ISVD, полипептида и/или конструкции по изобретению в получении фармацевтической композиции для предотвращения и/или лечения по меньшей мере одного ассоциированного с ADAMTS5 заболевания; и/или для использования в одном или более из способов лечения, упомянутых в настоящем описании.
Изобретение также относится к применению ISVD, полипептида и/или конструкции по изобретению в получении фармацевтической композиции для предотвращения и/или лечения по меньшей мере одного заболевания или нарушения, которое можно предотвращать и/или лечить посредством модуляции активности ADAMTS, предпочтительно, ADAMTS5, например, ингибирования деградации аггрекана.
Изобретение также относится к применению ISVD, полипептида, соединения и/или конструкции по изобретению в получении фармацевтической композиции для предотвращения и/или лечения по меньшей мере одного заболевания, нарушения или состояния, которое можно предотвращать и/или лечить посредством введения ISVD, полипептида, соединения и/или конструкции по изобретению пациенту.
Изобретение, кроме того, относится к ISVD, полипептиду, соединению и/или конструкции по изобретению или к содержащей их фармацевтической композиции для использования в предотвращении и/или лечении по меньшей мере одного ассоциированного с ADAMTS5 заболевания.
Предусматривают, что связывающие ADAMTS5 молекулы по изобретению можно использовать при различных заболеваниях, поражающих хрящ, таких как артропатии и хондродистрофии, артритное заболевание, такое как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматический разрыв или открепление, ахондроплазия, реберный хондрит, спондилоэпиметафизарная дисплазия, межпозвонковая грыжа, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава и рецидивирующий полихондрит, рассекающий остеохондрит и аггреканопатии, и неалкогольный стеатогепатит (NASH) (совместно обозначенные в настоящем описании как «ассоциированные с ADAMTS5 заболевания»).
В одном аспекте настоящее изобретение относится к композиции, ISVD, полипептиду и/или конструкции по изобретению для использования в лечении или предотвращении симптома ассоциированного с ADAMTS5 заболевания, например, такого как артропатии и хондродистрофии, артритное заболевание, такое как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматический разрыв или открепление, ахондроплазия, реберный хондрит, спондилоэпиметафизарная дисплазия, межпозвонковая грыжа, заболевание дегенерации поясничного диска, дегенеративное заболевание сустава и рецидивирующий полихондрит, рассекающий остеохондрит и аггреканопатии, и NASH.
В одном аспекте настоящее изобретение относится к способу предотвращения или лечения артропатий и хондродистрофий, артритного заболевания, такого как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматического разрыва или открепления, ахондроплазии, реберного хондрита, спондилоэпиметафизарной дисплазии, межпозвонковой грыжи, заболевания дегенерации поясничного диска, дегенеративного заболевания сустава, и рецидивирующего полихондрита и NASH, где указанный способ включает введение, нуждающемуся в этом субъекту, фармацевтически активного количества по меньшей мере композиции, иммуноглобулина, полипептида или конструкции по изобретению нуждающемуся в этом индивидууму.
В одном аспекте настоящее изобретение относится к применению ISVD, полипептида, композиции или конструкции по изобретению, в получении фармацевтической композиции для лечения или предотвращения, например, артропатий и хондродистрофий, артритного заболевания, такого как остеоартрит, ревматоидный артрит, подагрический артрит, псориатический артрит, травматического разрыва или открепления, ахондроплазии, реберного хондрита, спондилоэпиметафизарной дисплазии, межпозвонковой грыжи, заболевания дегенерации поясничного диска, дегенеративного заболевания сустава и рецидивирующих полихондрита, рассекающего остеохондрита и аггреканопатий, и NASH.
Ожидают также, что, посредством связывания с аггреканом, конструкции и/или полипептиды по изобретению могут уменьшать или ингибировать активность члена семейства сериновых протеаз, катепсинов, матриксных металлопротеиназ (MMP), например, такого как MMP20, но также ADAMTS4 (аггреканазы-1) и/или ADAMTS11, для деградации аггрекана.
В контексте настоящего изобретения, термин «предотвращение и/или лечение» не только включает предотвращение и/или лечение заболевания, но также, в общем, включает предотвращение начала заболевания, замедление или обращение прогрессирования заболевания, предотвращение или замедление начала одного или более симптомов, ассоциированных с заболеванием, уменьшение и/или облегчение одного или более симптомов, ассоциированных с заболеванием, уменьшение тяжести и/или длительности заболевания и/или любых симптомов, ассоциированных с ним, и/или предотвращение дальнейшего увеличения тяжести заболевания и/или любых симптомов, ассоциированных с ним, предотвращение, уменьшение или обращение любого физиологического повреждения, вызванного заболеванием, и, в общем, любое фармакологическое действие, обеспечивающее преимущество для пациента, подвергаемого лечению.
Режим дозирования определяют лечащий терапевт и клинические факторы. Как хорошо известно в области медицины, дозирование для любого одного пациента зависит от множества факторов, включая рост, массу, площадь поверхности тела, возраст пациента, конкретное соединение, подлежащее введению, активность используемого полипептида (включая антитела), время и способ введения, общее состояние здоровья и комбинацию с другими видами терапии или лечения. Белковое фармацевтически активное вещество может присутствовать в количестве между 1 г и 100 мг/кг массы тела на дозу; однако, предусмотрены также дозы ниже или выше этого иллюстративного диапазона. Если режим представляет собой непрерывную инфузию, оно может лежать в диапазоне от 1 пг до 100 мг на килограмм массы тела в минуту.
ISVD, полипептид или конструкцию по изобретению можно использовать в концентрации, например, 0,01, 0,1, 0,5, 1, 2, 5, 10, 20 или 50 пг/мл, для ингибирования и/или нейтрализации биологической функции ADAMTS5 по меньшей мере приблизительно на 50%, предпочтительно, на 75%, более предпочтительно, на 90%, 95% или вплоть до 99%, и наиболее предпочтительно, приблизительно на 100% (по существу полностью), как проанализировано способами, хорошо известными в данной области.
Как правило, режим лечения может включать введение одного или более ISVD, полипептидов и/или конструкций по изобретению, или одной или более содержащих их композиций, в одном или более фармацевтически эффективных количествах или дозах. Конкретные количество (количества) или дозы, подлежащие введению, может определять клиницист, снова на основании факторов, процитированных выше. Применимые дозы конструкций, полипептидов, и/или ISVD по изобретению можно определять посредством сравнения их активности in vitro и активности in vivo в моделях на животных. Способы экстраполяции эффективных доз у мышей и других животных, для человека известны в данной области; например, см. US 4938949.
Как правило, в зависимости от конкретного заболевания, нарушения или состояния, подлежащего лечению, активности конкретных ISVD, полипептида и/или конструкции по изобретению, подлежащих использованию, конкретного способа введения и конкретных используемых фармацевтического состава или композиции, клиницист способен определять подходящий режим дозирования.
Количество конструкций, полипептидов и/или ISVD по изобретению, необходимое для использования в лечении, может меняться не только в зависимости от конкретных выбранных иммуноглобулина, полипептида, соединения и/или конструкции, но также от способа введения, характера состояния, подлежащего лечению, и возраста и состояния пациента, и в конечном счете определяется решением лечащего терапевта или клинициста. Также доза конструкций, полипептидов и/или ISVD по изобретению меняется в зависимости от клетки, опухоли, ткани, трансплантата или органа-мишени.
Желательную дозу можно удобным образом представлять в виде однократной дозы или в виде дробных доз, вводимых с соответствующими интервалами, например, в форме двух, трех, четырех или более частей доз в сутки. Собственно часть дозы можно делить дополнительно, например, на несколько дискретных введений со свободными интервалами.
Режим введения может включать длительное, ежесуточное лечение. Под «длительным» понимают длительность по меньшей мере две недели и предпочтительно, несколько недель, месяцев или лет. Необходимые модификации в этом диапазоне дозирования может определять специалист в данной области с использованием только общепринятых экспериментов, принимая во внимание объяснения в настоящем описании. См. Remington's Pharmaceutical Sciences (Martin, E.W., ed. 4), Mack Publishing Co., Easton, PA. Дозу может также корректировать терапевт индивидуума в случае какого-либо осложнения.
Обычно, в вышеуказанном способе, можно использовать ISVD, полипептид и/или конструкцию по изобретению. Однако, в объем изобретения включено использование двух или более ISVD, полипептидов и/или конструкций по изобретению в комбинации.
ISVD, полипептиды и/или конструкции по изобретению можно использовать в комбинации с одним или более дополнительными фармацевтически активными соединениями или компонентами, т.е. в форме комбинированного режима лечения, который может приводить или может не приводить к синергическому эффекту.
Фармацевтическая композиция может также содержать по меньшей мере одно дополнительное действующее вещество, например, одно или более дополнительных антител или их антигенсвязывающих фрагментов, пептидов, белков, нуклеиновых кислот, органических и неорганических молекул.
Снова, клиницист способен выбирать такие дополнительные соединения или компоненты, так же как подходящий комбинированный режим лечения, на основании факторов, процитированных выше, и своей экспертной оценки.
В частности, ISVD, полипептиды и/или конструкции по изобретению можно использовать в комбинации с другими фармацевтически активными соединениями или компонентами, которые используют или можно использовать для предотвращения и/или лечения заболеваний, нарушений и состояний, процитированных в настоящем описании, в результате чего можно получать или можно не получать синергический эффект. Примеры таких соединений и компонентов, также как способов, методов и фармацевтических составов или композиций для их введения понятны клиницисту.
Когда два или более вещества или компонента предназначены для использования в качестве части комбинированного режима лечения, их можно вводить посредством одного и того же способа введения или посредством различных способов введения, по существу в одно и то же время или в различные периоды времени (например, по существу одновременно, последовательно или в соответствии с альтернативным режимом). Когда вещества или компоненты предназначены для введения одновременно, посредством одного и того же способа введения, их можно вводить в форме различных фармацевтических составов или композиций, или части комбинированного фармацевтического состава или композиции, как известно специалисту в данной области.
Также, когда два или более активных вещества или способа предназначены для использования в качестве части комбинированного режима лечения, каждое из веществ или компонентов можно вводить в таком же количестве и в соответствии с таким же режимом, как используют, когда соединение или компонент используют самостоятельно, и такое комбинированное использование может приводить или может не приводить к синергическому эффекту. Однако, когда комбинированное использование двух или более активных веществ или компонентов приводит к синергическому эффекту, может также являться возможным уменьшать количество одного, более одного или всех из веществ или компонентов, подлежащих введению, в то же время все еще достигая желательного терапевтического действия. Это, например, можно использовать для исключения, ограничения или уменьшения любых нежелательных побочных эффектов, ассоциированных с использованием одного или более из веществ или компонентов, когда их используют в обычных количествах, все еще с получением в то же время желательного фармацевтического или терапевтического эффекта.
Эффективность режима лечения, используемого по изобретению, можно определять и/или отслеживать любым способом, известным по существу для рассматриваемого заболевания, нарушения или состояния, как понятно клиницисту. Клиницист способен также, когда это целесообразно, и в зависимости от конкретного случая, изменять или модифицировать конкретный режим лечения, так, чтобы достигать желательного терапевтического эффекта, для исключения, ограничения или уменьшения нежелательных побочных эффектов, и/или для достижения целесообразного равновесия между достижением терапевтического эффекта с одной стороны и исключением, ограничением или уменьшением нежелательных побочных эффектов с другой стороны.
Как правило, режиму лечения можно следовать, пока не будет достигнут желательный терапевтический эффект, и/или так долго, как необходимо поддерживать желательный терапевтический эффект. Снова, это может определять клиницист.
Таким образом, в следующем аспекте, изобретение относится к фармацевтической композиции, содержащей по меньшей мере одну конструкцию по изобретению, по меньшей мере один полипептид по изобретению, по меньшей мере один ISVD по изобретению, или по меньшей мере одну нуклеиновую кислоту по изобретению и по меньшей мере один пригодный носитель, разбавитель или наполнитель (т.е. пригодный для фармацевтического использования), и необязательно, одно или более дополнительных активных веществ. В конкретном аспекте, изобретение относится к фармацевтической композиции, содержащей конструкцию, полипептид, ISVD или нуклеиновую кислоту по изобретению, предпочтительно по меньшей мере одну из SEQ ID NO: OOO, и по меньшей мере один пригодный носитель, разбавитель или наполнитель (т.е. пригодный для фармацевтического использования), и необязательно, одно или более дополнительных активных веществ.
Субъект, подлежащий лечению, может представлять собой любое теплокровное животное, но, в частности, представляет собой млекопитающее, и более конкретно, человека. В ветеринарных применениях, субъект, подлежащий лечению, включает любое животное, которое вывели для коммерческих целей или содержат в качестве домашнего животного. Как известно специалисту в данной области, субъект, подлежащий лечению, может, в частности, представлять собой индивидуума, страдающего от или подверженного риску заболеваний, нарушений и состояний, упомянутых в настоящем описании. Таким образом, в предпочтительном варианте осуществления изобретения, фармацевтические композиции, содержащие полипептид по изобретению, предназначены для использования в медицине или диагностике. Предпочтительно, фармацевтические композиции предназначены для использования в медицине человека, но их можно также использовать для ветеринарных целей.
Снова, в такой фармацевтической композиции, одно или более из иммуноглобулинов, полипептидов, соединений и/или конструкций по изобретению, или кодирующих их нуклеотидов, и/или содержащую их фармацевтическую композицию, можно также соответствующим образом комбинировать с одним или более другими активными компонентами, такими как упомянутые в настоящем описании.
Изобретение также относится к композиции (например, но без ограничения, к фармацевтической композиции или препарату, как далее описано в настоящем описании) для использования, либо in vitro (например, в анализе in vitro или клеточном анализе), либо in vivo (например, в одной клетке или многоклеточном организме, и в частности, у млекопитающего, и более конкретно, у человека, например, у человека, подверженного риску или страдающего от заболевания, нарушения или состояния по изобретению).
Следует понимать, что ссылка на лечение включает как лечение развившихся симптомов, так и профилактическое лечение, если явно не указано иное.
В общем, для фармацевтического использования, конструкции, полипептиды и/или ISVD по изобретению можно составлять в форме фармацевтического препарата или композиции, содержащих по меньшей мере одну конструкцию, полипептид и/или ISVD по изобретению и по меньшей мере один фармацевтически приемлемый носитель, разбавитель или наполнитель и/или адъювант, и необязательно, одно или более из фармацевтически активных полипептидов и/или соединений. В качестве неограничивающих примеров, такой состав может находиться в форме, подходящей для перорального введения, для парентерального введения (например, посредством внутривенной, внутримышечной или подкожной инъекции или внутривенной инфузии), для местного введения, для введения посредством ингаляции, посредством кожного пластыря, посредством имплантата, посредством суппозитория и т.д., где парентеральное введение является предпочтительным. Такие подходящие для введения формы - которые могут являться твердыми, полутвердыми или жидкими, в зависимости от способа введения - так же как способы и носители для использования в их получении, известны специалисту в данной области, и далее описаны в настоящем описании. Такой фармацевтический препарат или композицию в общем обозначают в настоящем описании как «фармацевтическая композиция».
Можно упомянуть иллюстративные эксципиенты, дезинтегрирующие средства, связующие, наполнители и смазывающие средства. Примеры дезинтегрирующих средств включают агар-агар, альгины, карбонат кальция, целлюлозу, коллоидный диоксид кремния, камеди, алюмосиликат магния, метилцеллюлозу и крахмал. Примеры связующих включают микрокристаллическую целлюлозу, гидроксиметилцеллюлозу, гидроксипропилцеллюлозу и поливинилпирролидон. Примеры наполнителей включают карбонат кальция, фосфат кальция, трехосновный сульфат кальция, карбоксиметилцеллюлозу кальция, целлюлозу, декстрин, декстрозу, фруктозу, лактит, лактозу, карбонат магния, оксид магния, мальтит, мальтодекстрины, мальтозу, сорбит, крахмал, сахарозу, сахар и ксилит. Примеры смазывающих средств включают агар, этилолеат, этиллаурат, глицерин, глицерилпальмитостеарат, гидрогенизированное растительное масло, оксид магния, стеараты, маннит, полоксамер, гликоли, бензоат натрия, лаурилсульфат натрия, стеарил натрия, сорбит и тальк. Обычные стабилизаторы, консерванты, увлажняющие средства и эмульгаторы, улучшающие консистенцию средства, улучшающие вкус средства, соли для изменения осмотического давления, буферные вещества, солюбилизаторы, разбавители, смягчающие средства, красители и маскирующие средства, и антиоксиданты следует принимать во внимание в качестве фармацевтических вспомогательных средств.
Пригодные носители включают, но без ограничения, карбонат магния, стеарат магния, тальк, сахар, лактозу, пектин, декстрин, крахмал, желатин, трагакант, метилцеллюлозу, карбоксиметилцеллюлозу натрия, воск с низкой точкой плавления, масло какао, воду, спирты, полиолы, глицерин, растительные масла и т.п.
В общем, конструкции, полипептиды и/или ISVD по изобретению можно составлять и вводить любым подходящим способом, известным по существу. Приведена ссылка, например, на общий уровень техники, процитированный выше (и в частности, на WO 04/041862, WO 04/041863, WO 04/041865, WO 04/041867 и WO 08/020079) так же как на стандартные руководства, такие как Remington's Pharmaceutical Sciences, 18th Ed., Mack Publishing Company, USA (1990), Remington, the Science and Practice of Pharmacy, 21st Edition, Lippincott Williams and Wilkins (2005); или Handbook of Therapeutic Antibodies (S. Dubel, Ed.), Wiley, Weinheim, 2007 (см., например, страницы 252-255).
В конкретном аспекте, изобретение относится к фармацевтической композиции, которая содержит конструкцию, полипептид, ISVD или нуклеиновую кислоту по изобретению, и которая дополнительно содержит по меньшей мере один фармацевтически приемлемый носитель, разбавитель или наполнитель и/или адъювант, и необязательно, содержит одно или более из дополнительных фармацевтически активных полипептидов и/или соединений.
Конструкции, полипептиды, и/или ISVD по изобретению можно составлять и вводить любым способом, известным по существу для общепринятых антител и фрагментов антител (включая ScFv и диатела) и других фармацевтически активных белков. Такие составы и способы их получения известны специалисту в данной области, и например, включают препараты, предпочтительные как пригодные для парентерального введения (например, внутривенного, внутрибрюшинного, подкожного, внутримышечного, внутрипросветного, внутриартериального, интратекального, интраназального или внутрибронхиального введения), а также для местного (т.е. чрескожного или внутрикожного) введения.
Препараты для парентерального введения могут, например, представлять собой стерильные растворы, суспензии, дисперсии или эмульсии, пригодные для инфузии или инъекции. Пригодные носители или разбавители для таких препаратов, например, включают, без ограничения, носители или разбавители, упомянутые на странице 143 WO 08/020079. Обычно, водные растворы или суспензии являются более предпочтительными.
Конструкции, полипептиды, и/или ISVD по изобретению можно также вводить с использованием способов доставки, известных в генотерапии, см., например, Патент США No. 5399346, способы доставки для генотерапии из которого приведены в качестве ссылки. С использованием способа доставки для генотерапии, первичные клетки, трансфицированные геном, кодирующим конструкцию, полипептид и/или ISVD по изобретению, можно, кроме того, трансфицировать с использованием тканеспецифических промоторов для нацеливания на специфические органы, ткани, трансплантаты, опухоли или клетки, и можно, кроме того, трансфицировать с использованием сигнальных и стабилизирующих последовательностей для субклеточно локализованной экспрессии.
В соответствии с дополнительными аспектами изобретения, полипептид по изобретению можно использовать для дополнительных применений in vivo и in vitro. Например, полипептиды по изобретению можно использовать для диагностических целей, например, в анализах, разработанных для детекции и/или количественной оценки присутствия ADAMTS5 и/или для очистки ADAMTS5. Полипептиды можно также тестировать в моделях конкретных заболеваний на животных и для проведения исследований токсикологии, безопасности и дозирования.
Наконец, изобретение относится к набору, содержащему по меньшей мере один полипептид по изобретению, по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую указанные компоненты, вектор или векторную систему по изобретению, и/или клетку-хозяина по изобретению. Предусматривают, что набор можно предлагать в различных формах, например, в форме диагностического набора.
Изобретение в настоящее время дополнительно описано посредством следующих неограничивающих предпочтительных аспектов, примеров и фигур.
Полное содержание всех ссылок (включая литературные источники, выданные патенты, опубликованные патентные заявки и поданные одновременно патентные заявки), процитированные на протяжении этой заявки, таким образом, явно приведено в качестве ссылки, в частности, для объяснений, процитированных в настоящем описании выше.
Последовательности описаны в основном содержании описания и в отдельном списке последовательностей, в соответствии со стандартом WIPO ST.25. SEQ ID, обозначенная конкретным номером, должна быть одинаковой в основном содержании описания и в отдельном списке последовательностей. Например, SEQ ID no.: 1 должна определять одинаковую последовательность в обоих, в основном содержании описания и в отдельном списке последовательностей. В случае несоответствия между определением последовательности в основном содержании описания и в отдельном списке последовательностей (если, например, SEQ ID no.: 1 в основном содержании описания ошибочно соответствует SEQ ID no.: 2 в отдельном списке последовательностей), тогда ссылку на конкретную последовательность в заявке, в частности, конкретных вариантах осуществления, следует понимать как ссылку на последовательность в основном содержании заявки, а не в отдельном списке последовательностей. Иными словами, несоответствие между определением/обозначением последовательности в основном содержании описания и в отдельном списке последовательностей следует разрешать посредством коррекции отдельного списка последовательностей применительно к последовательностям и их обозначению, описанным в основном содержании заявки, которое включает описание, примеры, фигуры и формулу изобретения.
ПРИМЕРЫ
Пример 1. Получение рекомбинантного белка ADAMTS5 из различных видов
Различные форматы рекомбинантного белка ADAMTS5 из быка, крысы, морской свинки, мыши и яванского макака получали в собственной лаборатории посредством системы экспрессии HEK293 Flp-In™ с использованием реагента для трансфекции FuGENE® HD (Promega). Через двое суток после трансфекции, селективную среду, содержащую 100 мкг/мл гигромицина B, добавляли к клеткам для отбора стабильно экспрессирующих клеток.
Стабильно экспрессирующие клетки размножали. Кондиционированную среду, содержащую секретированный ADAMTS5, собирали каждые сутки из клеток, очищали посредством хроматографии на HisTrap, и за этим следовала замена буфера на буфер 50 мМ Hepes, pH 7,5. Чистоту белка подтверждали посредством SDS-PAGE.
Пример 2. Иммунизация лам с использованием белка ADAMTS5, клонирование репертуаров фрагментов антител только из тяжелой цепи и получение фага
2.1 Иммунизации
После одобрения этического комитета отделения ветеринарной медицины (University Ghent, Belgium), 3 лам иммунизировали с использованием рекомбинантного белка ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD).
2.2 Клонирование репертуаров фрагментов антител только из тяжелой цепи и получение фага
После последней инъекции иммуногена, собирали образцы крови. Из этих образцов крови, мононуклеарные клетки периферической крови (PBMC) получали с использованием фиколла-гипака, в соответствии с инструкциями производителя (Amersham Biosciences, Piscataway, NJ, US). Из PBMC выделяли тотальную РНК и использовали в качестве исходного материала для RT-ПЦР для амплификации кодирующих VHH/нанотело фрагментов ДНК, в основном как описано в WO 05/044858. Затем фаг получали в соответствии со стандартными способами (см., например, литературу предшествующего уровня техники и заявки, поданные Ablynx N.V., процитированные в настоящем описании) и хранили после стерилизации фильтрацией при 4°C для дальнейшего использования.
Пример 3. Отбор VHH, специфических для ADAMTS5 человека посредством фагового дисплея
Репертуары VHH, полученные от всех лам и клонированные в фаговых библиотеках, использовали для отбора связывающих ADAMTS5 человека нанотел. Рекомбинантный белок ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD) иммобилизовали при 5 мкг/мл на планшете Maxisorp (Nunc, Wiesbaden, Germany), рядом с отрицательным контролем с 0 мкг/мл антигена. После инкубации с фаговыми библиотеками и интенсивной промывки, связанные фаги элюировали трипсином (1 мг/мл), и использовали для инфекции клеток E. coli. Инфицированные клетки E. coli либо использовали для получения фага для следующего цикла отбора, либо рассевали на чашки с агаром для анализа. Кроме того, синтетические библиотеки использовали в трех последовательных циклах отбора.
Выходы всех циклов отбора анализировали по коэффициенту обогащения, представляющему собой количество фагов, присутствующих в элюате, по сравнению с контролем. Наилучшие условия отбора выбраны для дальнейшего анализа.
Для скрининга выхода отбора по специфическим связывающим членам, отдельные колонии отбирали из чашек с агаром и выращивали в 1 мл 96-луночных планшетах с глубокими лунками. Контролируемую LacZ экспрессию VHH индуцировали посредством добавления IPTG. Периплазматические экстракты получали в соответствии со стандартными способами (см., например, литературу предшествующего уровня техники и заявки, поданные Ablynx N.V., процитированные в настоящем описании.)
Пример 4. Скрининг периплазматических экстрактов по функциональным блокирующим нанотелам
На первой стадии, периплазматические экстракты тестировали по связыванию с рекомбинантным ADAMTS5 человека посредством ELISA связывания. Кратко, рекомбинантным ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD) в концентрации 1 мкг/мл покрывали 384-луночные планшеты MaxiSorp (Nunc, Wiesbaden, Germany). Лунки блокировали раствором казеина (1%). После добавления 10-кратного разведения периплазматических экстрактов, связывание нанотела детектировали с использованием конъюгата антитела мыши против Flag с HRP (Sigma, St. Louis, US) и последующей ферментной реакции в присутствии субстрата esTMB (3,3',5,5'-тетраметилбензидина) (SDT, Brussels, Belgium). Клоны, для которых показаны сигналы ELISA, превышающие сумму средних сигналов для контрольных не имеющих отношения к делу нанотел и 3-кратное стандартное отклонение для контрольных не имеющих отношения к делу нанотел, считали кодирующими положительные связывающие ADAMTS5 человека нанотела.
Для идентификации нанотел, способных предотвращать опосредованное ADAMTS5 расщепление аггрекана, клоны тестировали в основанном на FRET ферментном анализе ADAMTS5 человека, с использованием 50 мМ HEPES (pH 7,5), 100 мМ NaCl, 5 мМ CaCl2*2H2O, 0,1% CHAPS, 5% глицерина в качестве буфера для анализа. Кратко, периплазматические экстракты, содержащие связывающие нанотела против ADAMTS5, инкубировали в 384-луночных планшетах OptiPlate (PerkinElmer, Waltham, MA, US) с 10 мкл 25 мкМ связанного с гасителем флуорогенного человеческого пептидного субстрата (Abz-TEGEARGSVI-Dap(Dnp)-KK-NH2, Anaspec, Serain Belgium, кат. # 60431-1) и 10 мкл 25 нМ ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD). Способность нанотел предотвращать опосредованное ADAMTS5 расщепление мониторировали каждую минуту в течение 2 часов на считывателе для планшетов Tecan Infinite M1000. Данные анализировали с использованием программного обеспечения Graphpad Prism. Из кривой прогрессирования ферментной реакции, определяли начальную скорость превращения для отрицательного контроля (v0), так же как скорость превращения для каждого клона нанотела в планшете (vi) и фоновый сигнал (Sb). С использованием формулы 100*(1 - (vi - Sb) /(v0 - Sb)), рассчитывали процент ингибирования.
Периплазматический экстракт, содержащий не относящееся к делу нанотело, использовали в качестве отрицательного контроля. Периплазматические экстракты, содержащие нанотела против ADAMTS5, которые являлись способными уменьшать сигнал флуоресценции более чем на 50%, по сравнению с сигналом отрицательного контроля, считали ингибирующими нанотелами. Затем определяли последовательность ДНК положительных клонов.
Обобщение данных скрининга периплазматических экстрактов приведено в таблице 4.1. Аминокислотные последовательности нанотел против ADAMTS5 показаны в таблице A-1.
Таблица 4.1 Результаты скрининга периплазматических экстрактов, содержащих нанотела против ADAMTS5
После клонирования и секвенирования, идентифицировали различные семейства, состоящие из клонов с различиями в CDR, но имеющих сходные характеристики связывания и ингибирования.
После клонирования и секвенирования, нанотело 02A12 идентифицировано как член семейства клона 09D03 (ср. таблицы A-1 и A-2). Изменчивость последовательности областей CDR показана в таблицах 4.1A, 4.1B и 4.1C ниже. Аминокислотные последовательности CDR клона 09D03 использовали в качестве эталона, с которым сравнивали CDR членов семейства (CDR1 начинается в положении по Kabat 26, CDR2 начинается в положении по Kabat 50, и CDR3 начинается в положении по Kabat 95).
Таблица 4.1A (CDR1 09D03)
* Вплоть до 6 мутаций CDR1 в одном клоне (SEQ ID NO: 22).
Таблица 4.1B (CDR21 09D03)
* Вплоть до 0 мутаций CDR2 в одном клоне (SEQ ID NO: 36).
Таблица 4.1C (CDR3 09D03)
* Вплоть до 0 мутаций CDR в одном клоне (SEQ ID NO: 54).
После клонирования и секвенирования, клон 09A05 идентифицирован как член семейства клона 03B02 (ср. таблицы A-1 и A-2). Изменчивость последовательности областей CDR показана в таблицах 4.1D, 4.1E и 4.1F ниже. Аминокислотные последовательности CDR клона 03B02 использовали в качестве эталона, с которым сравнивали CDR членов семейства (CDR1 начинается в положении по Kabat 26, CDR1 начинается в положении по Kabat 50, и CDR3 начинается в положении по Kabat 95).
Таблица 4.1D (CDR1 03B02)
* Вплоть до 0 мутаций CDR1 в одном клоне (SEQ ID NO: 33).
Таблица 4.1E (CDR2 03B02)
* Вплоть до 3 мутаций CDR2 в одном клоне (SEQ ID NO: 50).
Таблица 4.1F (CDR3 03B02)
* Вплоть до 2 мутаций CDR в одном клоне (SEQ ID NO: 68).
После клонирования и секвенирования клоны 03D02 и 02C10 идентифицированы как члены семейства клона 03D01 (ср. таблицы A-1 и A-2). Изменчивость последовательности областей CDR показана в таблицах 4.1G, 4.1H и 4.1I ниже. Аминокислотные последовательности CDR клона 03D01 использовали в качестве эталона, с которым сравнивали CDR членов семейства (CDR1 начинается в положении по Kabat 26, CDR2 начинается в положении по Kabat 50, и CDR3 начинается в положении по Kabat 95).
Таблица 4.1G (CDR1 03D01)
* Вплоть до 0 мутаций CDR1 в одном клоне (SEQ ID NO: 28).
Таблица 4.1H (CDR2 03D01)
* Вплоть до 3 мутаций CDR2 в одном клоне (SEQ ID NO: 44).
Таблица 4.1I (CDR3 03D01)
* Вплоть до 1 мутации CDR в одном клоне (SEQ ID NO: 62).
Вышеуказанные результаты показывают, что некоторая изменчивость аминокислот является допустимой, с сохранением в то же время характеристик связывания и ингибирования. Примечательно, что связывание с ADAMTS5 не является прогностическим фактором для ингибирования активности ADAMTS5. Например, клон 3F04 является сильным связывающим членом, но слабым ингибитором.
Пример 5. Характеризация очищенных моновалентных нанотел против ADAMTS5
Клоны, отобранные при скрининге, описанном в примере 4, характеризовали дополнительно. После трансформации E. coli (TG-1) выбранными нанотелами, экспрессию индуцировали добавлением 1 мМ IPTG и позволяли ей продолжаться в течение 4 часов при 37°C. После центрифугирования культур клеток, периплазматические экстракты получали посредством замораживания-размораживания осадков с последующим центрифугированием. Эти экстракты затем использовали в качестве исходного материала для очистки посредством IMAC на колонках HisTrap FF crude 1 мл (GE healthcare, Buckinghamshire, United Kingdom) с последующим обессоливанием посредством центрифугируемых колонок Zeba (Pierce, Rockford, IL, USA), получая по меньшей мере 95% чистоту, как оценено посредством SDS-PAGE.
5.1 Оценка блокирующих ADAMTS5 нанотел в ферментных анализах ADAMTS5
Способность нанотел предотвращать опосредованное ADAMTS5 расщепление аггрекана подтверждали в ферментном анализе на основе FRET, в основном, как описано в примере 4. Данные анализировали с использованием программного обеспечения Graphpad Prism. Из кривой прогрессирования ферментной реакции, определяли начальную скорость превращения (vi), и эти значения наносили на график как функцию концентраций ингибитора, и рассчитывали значения IC50. В сущности, анализ FRET для человека с использованием очищенных нанотел подтвердил результаты, описанные в примере 4 с использованием периплазматических экстрактов. Значения IC50 лежали в диапазоне между 1,8E-09 M и 3,7E-08 M. Примечательно, что хотя общепринятое mAb 12F4 имело несколько лучшую IC50 1,0E-09 M, чем моновалентные нанотела, для всех нанотел показана лучшая эффективность (данные не представлены).
После анализа на основе FRET, проводили анализ AlphaLISA (Perkin Elmer, Waltham, MA, US) на основе ADAMTS5 человека с биотинилированным 43-членным олигопептидом аггрекана в качестве субстрата. После расщепления посредством ADAMTS5 этого субстрата, высвобождается продукт биотинилированный неоэпитоп ARGSV, который можно детектировать посредством донорных бусин стрептавидин-AlphaScreen и антитела против неоэпитопа («ARGSV»), связанного с покрытыми антителами против IgG мыши акцепторными бусинами AlphaLISA, приводящими в результате к образованию сигнала люминесценции AlphaScreen при лазерном возбуждении.
Для определения ингибирующего потенциала нанотел, серийные разведения очищенных нанотел (вместе с положительным контролем и отрицательным контрольным не относящимся к делу нанотелом) инкубировали с ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD) в течение 15 минут при комнатной температуре в 384-луночных планшетах Optiplate (Perkin Elmer, Waltham, MA, US). После добавления субстрата биотинилированного 43-членного олигопептида аггрекана (Biosyntan, Berlin, Germany) и инкубации в течение 3 часов при 37°C, реакцию останавливали. Первую стадию детекции проводили посредством добавления 5 мкл раствора для детекции 1, содержащего антитело против неоэпитопа аггрекана «ARGSV», mouse BC3 mAb (MDBioproducts, Egg, Switzerland) и донорных бусин AlphaScreen со стрептавидином (Perkin Elmer). После 1 час инкубации при комнатной температуре, добавляли 5 мкл второго раствора для детекции, содержащего акцепторные бусины AlphaLISA против антител мыши (Perkin Elmer). Планшеты инкубировали for 2 часов при комнатной температуре в темноте. Измерение проводили посредством считывания планшетов на считывателе для планшетов со множеством меток Envision Multi label Plate reader (Perkin Elmer, Waltham, MA, US).
Для определения способности нанотел предотвращать опосредованное ADAMTS5 расщепление субстрата, уменьшение сигнала анализировали как функцию концентрации нанотела, и рассчитывали значения IC50.
Результаты обобщены в таблице 5.1.
Таблица 5.1 Активность (IC50) и % ингибирования ADAMTS5 для нанотел и эталонных соединений в ферментном анализе AlphaLISA для человека.
В анализе AlphaLISA, по меньшей мере для трех моновалентных нанотел (2F03, 11B06 и 9D03) показаны значения IC50, сходные с общепринятым mAb 12F4, в то время как остальные имели более высокие значения. Однако для всех нанотел показано около 100% ингибирования. Примечательно, что нанотело 3B03, имеющее значение IC50, по меньшей мере в 100 выше, чем mAb 12F04, все еще имеет 100% ингибирования.
5.2 Оценка блокирующих ADAMTS5 нанотел в анализе бычьего эксплантата
Для оценки способности нанотел блокировать деградацию хряща в анализе ex vivo, в котором субстрат присутствует в состоянии, более близком к физиологическому состоянию, по сравнению с биохимическими анализами, проводили анализ бычьего эксплантата. Кратко, получали свежеприготовленные срезы эксплантата бычьего хряща (диаметром 4 мм) из коленных суставов коровы и инкубировали в 96-луночных планшетах в присутствии IL-1α для индукции деградации хряща. В качестве показателя деградации хряща/аггрекана, высвобождение GAG детектировали в супернатанте после 5 суток инкубации (37 °C, 7,5% CO2) посредством метахроматического красителя 1,9-диметилметиленового синего (излучение при 633 нм). Хондроитинсульфат включали в качестве стандарта анализа. Эффективность определяли посредством индуцированных IL-1α контрольных образцов без соединения (0%) и в присутствии MSC2310852A (100% эффект).
Результаты обобщены в таблице 5.2.
Таблица 5.2 Значения IC50 для моновалентных нанотел в анализе бычьего эксплантата.
5.3 Группировка эпитопов
«Сэндвич-анализ» на основе SPR на устройстве Biacore T100 проводили для группировки нанотел против ADAMTS5 в различные эпитопные группы. С этой целью, каждое нанотело против ADAMTS5 иммобилизовали на сенсорном чипе CM5 посредством присоединения по аминогруппе с использованием химических реакций EDC и NHS. После стадии связывания 100 нМ ADAMTS5, инъецировали очищенные нанотела в концентрации 1 мкМ, каждое в одном цикле и без регенерации, с временем поверхностного контакта 120 секунд и скорости потока 45 мкл/минуту. Кривые обрабатывали с использованием программного обеспечения Biacore T100 Evaluation и оценивали по дополнительному связыванию с фиксированным ADAMTS5 (другая группа) или отсутствию связывания (та же группа).
Нанотела можно было разделить на две группы: одна большая группа («группа 1»), которая содержит все ингибирующие нанотела, имеющие сходные или перекрывающиеся эпитопы, и вторая группа, которая содержит связывающее, нефункциональное нанотело 3F04, узнающее другой эпитоп на ADAMTS5 («группа 2»).
В таблице 5.3 обобщены связывающие эпитопы тестированных нанотел.
Таблица 5.3 эпитопные группы нанотел против ADAMTS5
5.4 Перекрестная для видов реакционная способность
Перекрестную для видов реакционную способность сначала оценивали посредством основанного на SPR анализа скорости диссоциации на устройстве Biacore T100. Нанотела тестировали по связыванию с ADAMTS5 человека, яванского макака («cyno»), морской свинки, мыши и быка. С этой целью, рекомбинантный ADAMTS5 иммобилизовали на чипе CM5 посредством присоединения по аминогруппе с использованием EDC и NHS. Очищенные нанотела инъецировали в течение 2 минут в концентрации 100 нМ и позволяли диссоциацию в течение 15 мин при скорости потока 45 мкл/мин. Скорости диссоциации для каждого индивидуального нанотела определяли посредством определения соответствия модели взаимодействия 1:1 (модели Ленгмюра) индивидуальных кривых диссоциации с использованием программного обеспечения BIA Evaluation. В качестве эталона, скорости диссоциации для ADAMTS5 человека определяли в каждом эксперименте.
Результаты обобщены в таблице 5.4A.
Таблица 5.4A Обобщение данных перекрестной для видов реакционной способности моновалентных нанотел против ADAMTS5
Для каждого тестированного нанотела, сходные скорости диссоциации наблюдали для всех тестированных видов.
Кроме того, перекрестную реакционную способность для ADAMTS5 яванского макака и морской свинки исследовали также посредством определения активности с использованием AlphaLISA, в основном, как описано в примере 5.1, но с использованием биотинилированного олигопептидного субстрата 43-аггрекана яванского макака и биотинилированного олигопептидного субстрата 43-аггрекана морской свинки. Для определения способности нанотел предотвращать опосредованное ADAMTS5 расщепление субстрата, анализировали уменьшение сигнала как функцию концентрации нанотела, и рассчитывали значения IC50.
Полученная активность обобщена в таблице 5.4B.
Таблица 5.4B Обобщение данных перекрестной для видов реакционной способности моновалентных нанотел против ADAMTS5
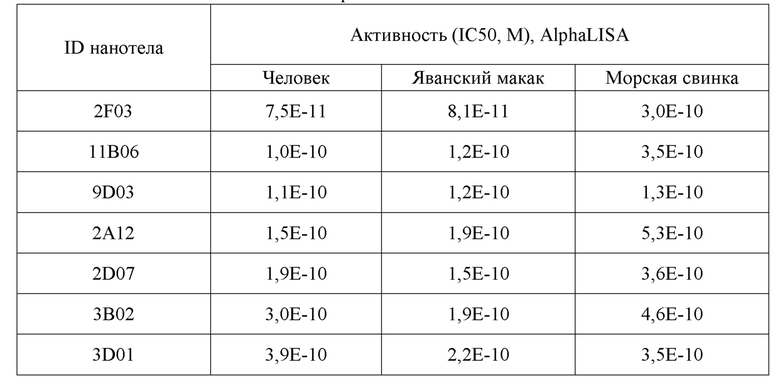
Все нанотела и контрольные соединения блокировали ADAMTS5 человека и яванского макака со сходной активностью. В AlphaLISA для морской свинки, для наиболее активных нанотел показана немного более низкая активность, чем в AlphaLISA для человека, но это, наиболее вероятно, является результатом более низкой чувствительности анализа из-за увеличенной концентрации фермента.
Перекрестную реакционную способность для крысы нанотел 2A12, 2F03, 093 и 049 оценивали посредством определения аффинности на основании SPR с использованием Biacore T100. Для контроля, определяли также аффинность для ADAMTS5 человека. Для этого, ADAMTS5 крысы и человека иммобилизовали на чипе CM5 посредством присоединения по аминогруппе, с использованием химических реакций EDC и NHS. Очищенные нанотела инъецировали в течение 2 минут при различных концентрациях (между 1 и 1000 нМ) и позволяли диссоциацию в течение 25 мин при скорости потока 45 мкл/мин. Кинетические константы рассчитывали по сенсограммам с использованием программного обеспечения BIAEvaluation (для взаимодействия 1:1).
Как представлено в таблице 5.4C, сходную аффинность для ADAMTS5 человека и ADAMTS5 крысы получали для всех тестированных нанотел. Аффинность нанотела 069 с увеличенным временем полужизни (ср. ниже) являлась сходной с аффинностью его моновалентного эквивалента (представляющего собой нанотело 049), что показывает, что добавление ALB11 на C-конце не оказывает эффекта на аффинность. Аффинность бипаратопного нанотела 130 с увеличенным временем полужизни была выше для обоих видов (в 15-раз для ADAMTS5 человека и в 10 раз для ADAMTS5 крысы), по сравнению с аффинностью нанотела 069 с увеличенным временем полужизни (Таблица 5.4C).
Таблица 5.4C Обобщение аффинности для ADAMTS5 человека и ADAMTS5 крысы
5.5 Ингибирование активности MMP-1 и MMP-14
Для подтверждения избирательности нанотел для ADAMTS5, ингибирование активности ферментов MMP-1 и MMP-14 оценивали посредством анализов на основе FRET с использованием соответствующих ферментов. Кратко, активированную MMP-1 или MMP-14 человека инкубировали в течение 30 минут при комнатной температуре с 10 мкл серийных разведений нанотела. После инкубации, добавляли 20 мкл соответственно 5 мкМ или 2,5 мкМ флуорогенного пептидного субстрата (флуорогенного субстрата для MMP Mca-PLGL-Dpa-AR-NH2 (R&D Systems, кат. #ES001)). Способность нанотел предотвращать опосредованное MMP-1 и MMP-14 расщепление мониторировали каждую минуту в течение 2 часов при 37°C на считывателе для планшетов Tecan Infinite M1000.
Полученные кривые титрования показаны на фигуре 1.
В то время как природные ингибиторы TIMP2 и TIMP3 ингибировали активность MMP-1 и MMP-14, ни для одного из тестированных нанотел не показано ингибирования.
5.6 Ингибирование активности ADAMTS4 человека
Для оценки ингибирования ADAMTS4 человека, проводили анализ, сходный с AlphaLISA для ADAMTS5 человека, в основном, как описано в примере 5.1, но с использованием ADAMTS4 человека (R&D Systems, Minneapolis, US; кат. # 4307-AD). Для определения способности нанотел предотвращать опосредованное ADAMTS4 человека расщепление субстрата, анализировали уменьшение сигнала как функцию от концентрации нанотела, и рассчитывали значения IC50.
Результаты показаны на фигуре 2.
В то время как малая молекула MSC2310852A ингибировала активность ADAMTS4 человека, ни для одного из тестированных нанотел или mAb 12F4 не показана ингибирующая активность (см. фигуру 2). Моноклональное антитело 12F4 (H4L0) описано как избирательное по сравнению с ADAMTS4 в WO 2011/002968.
Пример 6. Форматирование нанотел
6.1 Нокаут мотивов N-гликозилирования
Мотивы N-гликозилирования, присутствующие в нанотеле 11B06 (положение N52) и нанотеле 3F04 (положение N110f), подвергали нокауту перед форматированием. Участки рандомизировали посредством библиотеки кодонов NNK. Поскольку эти положения локализованы в CDR, мутация мотива может потенциально влиять на характеристики связывания. Соответственно, проводили скрининг библиотек по возможным заменам, которые не влияют на свойство связывания, посредством анализа скорости диссоциации на основе SPR для ADAMTS5 человека, с использованием устройства Biacore T100, в основном, как описано в примере 5.4.
Неожиданно, замена аминокислоты N52 на серин в нанотеле 11B06 и аминокислоты N100F на глутамин в нанотеле 3F04 не влияли на связывание (данные не представлены).
Таким образом, в качестве строительных блоков для дальнейшего форматирования, нанотела 049 (=11B06 (N52S)) и 093 (3F04 (N100FQ)) использовали для представления нанотел 11B06 и 3F04, соответственно (см. таблицу A-1).
6.2 Получение форматированных нанотел
Для получения конструкций нанотел с увеличенным временем полужизни, которые блокируют активность аггреканазы ADAMTS5 человека, нанотела 2A12, 2D07, 2F03, 049, 9D03 и 3B02 сливали с нанотелом против человеческого сывороточного альбумина (HSA) ALB11 (см. таблицу 6.3). Кроме того, конструкции ALB11 с увеличенным временем полужизни получали также из комбинации двух нанотел для различных связывающих эпитопов (комбинации члена группы 1 и члена группы 2). Конструкции форматированного нанотела экспрессировали в Pichia pastoris, секретировали в культуральную среду и подвергали аффинной очистке на бусинах Poros MabCaptureA с белком A (Applied Biosystems, Bleiswijk, Netherlands кат. # 4374729). Подтверждали целостность конструкций нанотела.
6.3 Активность в AlphaLISA в отсутствие и в присутствии HSA
Конструкции форматированного нанотела тестировали в AlphaLISA, как описано в примере 5.1. Кроме того, влияние связывания HSA на активность нанотела исследовали посредством проведения эксперимента в присутствии избытка HSA (конечная концентрация 4,2 мкМ).
Значения IC50 для всех конструкций нанотел с увеличенным временем полужизни перечислены в таблице 6.3.
Таблица 6.3. Активность конструкций форматированного нанотела (+/- HSA), как определено в AlphaLISA
ND=не определяли ; 093*=3F04 (N100fQ); 0492*=11B06 (N52S).
Результаты показывают, что ни форматирование ALB11, ни связывание HSA не влияет на активность конструкций нанотела.
6.4 Связывание с HSA
Аффинность нанотела ALB11 для HSA определяли посредством SPR на Biacore T100. С этой целью, HSA иммобилизовали на чипе CM5 посредством присоединения по аминогруппе и обрабатывали в основном, как описано в примере 5.3. В качестве эталона, определяли также аффинность моновалентного ALB11, которая составляла 3,2 нМ.
Результаты обобщены в таблице 6.4.
Таблица 6.4 Аффинность конструкций нанотела HLE для HSA
ND=не определяли; *FNb=функциональное нанотело; 0492*=11B06 (N52S); 0933*=3F04 (N100fQ).
Все конструкции нанотела ADAMTS5 с увеличенным временем полужизни имели сходную аффинность (см. таблицу 6.4), которая была ниже, чем аффинность моновалентного ALB11 для HSA.
6.5 Определение аффинности для ADAMTS5 человека с использованием KinExA
Аффинность конструкции форматированного нанотела 069 (049-35GS-ALB11) и конструкции нанотела 130 (049-35GS-093-ALB11) (см. также таблицу 6.5) определяли в растворе с использованием анализа кинетического исключения на устройстве KinExA3000 (Sapidyne, Boise, USA). С этой целью, ADAMTS5 человека связывали с бусинами PMMA в соответствии с инструкциями производителя, подробно описанными в Darling and Brault (2004 Assay Drug Dev Technol. 2:647-57). Фиксированную концентрацию конструкции нанотела (50 нМ конструкции нанотела 069 или 5 нМ конструкции нанотела 130) добавляли к серийным разведениям ADAMTS5 человека в диапазоне от 2 нМ до 0,2 нМ и инкубировали при комнатной температуре в течение 16 часов, пока не достигали равновесия. Затем, смеси инъецировали посредством автодозатора KinExA на колонку, набитую конъюгированными с ADAMTS5 человека бусинами PMMA для связывания свободных конструкций нанотела на бусинах и детекции с использованием меченного Alexa647 HSA. Процент свободной конструкции нанотела наносили на график как функцию титрованного ADAMTS5 человека и подбирали соответствующую кривую с использованием программного обеспечения KinExA Pro v3.2.6.
Результаты показаны в таблице 6.5.
Таблица 6.5 Аффинность 2 конструкций нанотела для связывания с ADAMTS5 человека, как определено посредством KinExA
049*=11B06 (N52S); 0932*=3F04 (N100fQ).
Приблизительно в 20 раз более высокую аффинность наблюдали для конструкции нанотела 130, по сравнению с конструкцией нанотела 069. Эти данные подтверждают авидное связывание на ADAMTS5 человека конструкции нанотела 130.
Пример 7. Оптимизация последовательности (SO) нанотел
Иллюстративные нанотело 2F03, нанотело 049 (11B06 (N52S)) и нанотело 093 (3F04 (N100fQ)) подвергали процессу оптимизации последовательности. Оптимизация последовательности представляет собой способ, в котором последовательность исходного нанотела подвергают мутагенезу, и этот способ включает гуманизацию (i) нанотела и нокаут посттрансляционных модификаций (ii), так же как эпитопов для потенциальных предсуществующих антител (iii). Нанотело ALB11 также подвергали мутации для нокаута эпитопов для потенциальных предсуществующих антител.
(i) для целей гуманизации, последовательность исходного нанотела подвергают мутагенезу для получения последовательности нанотела, которая является более идентичной человеческой зародышевой консенсусной последовательности IGHV3-IGHJ. Специфические аминокислоты в каркасных областях (за исключением так называемых характерных остатков), отличающиеся между нанотелом и человеческим зародышевым консенсусом IGHV3-IGHJ, изменяют до человеческих эквивалентов, таким образом, чтобы сохранять интактными структуру, активность и стабильность белка. Известно, что некоторые характерные остатки являются критическими для стабильности, активности и аффинности нанотела, и таким образом, их не подвергают мутагенезу.
(ii) аминокислоты, которые присутствуют в CDR, и для которых существует экспериментальное доказательство, что они являются чувствительными к посттрансляционным модификациям (PTM), изменяют таким образом, чтобы инактивировать участок PTM, в то же время сохраняя интактными структуру, активность и стабильность белка.
(iii) последовательность нанотела оптимизируют, без влияния на структуру, активность и стабильность белка, для минимизации связывания любых природных предсуществующих антител и уменьшения потенциала стимуляции вызванного лечением иммуногенного ответа.
Для получения оптимизированных по последовательности конструкции форматированного нанотела 581 и конструкции нанотела 579, соответствующие строительные блоки соединяли посредством линкеров 35GS. В результате получены конструкция нанотела 581 (2F3SO-35GS линкер-Alb11) и конструкция нанотела 579 (2F3SO-35GS линкер-093SO-35GS линкер-Alb11), соответственно (ср. таблицу A-1). Конструкции продуцировали в Pichia pastoris в форме лишенных метки белков и очищали посредством аффинной хроматографии с белком A, с последующим обессоливанием. Подтверждали целостность конструкций нанотел.
7.1 Аффинность для ADAMTS5 человека
Аффинность конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) определяли с использованием KinExA, как описано в примере 6.5, с несколькими незначительными адаптациями к описанному способу. Фиксированную концентрацию 20 нМ конструкции нанотела 581 инкубировали вместе с серийными разведениями ADAMTS5 человека (2,2-кратными серийными разведениями в диапазоне между 20 нМ и 0,32 нМ и пустым контролем без ADAMTS5). Для тестирования влияния альбумина, HSA добавляли к этой смеси для предварительной инкубации в избранных экспериментах с ADAMTS5 человека, в концентрации, в 100 раз больше его KD для конструкции нанотела 581. Смеси инкубировали и позволяли достигнуть равновесия в течение 24 часов перед инъекцией посредством автодозатора KinExA на колонку, набитую конъюгированными с ADAMTS5 человека бусинами PMMA. Связанную конструкцию нанотела 581 детектировали с использованием меченного AF647 средства против нанотела, узнающего конструкцию нанотела 581 (полученную в Ablynx).
Результаты представлены в таблице 7.1.
Таблица 7.1 Аффинность конструкции нанотела 581 для ADAMTS5 человека в присутствии и в отсутствие HSA
Результаты показывают, что аффинность конструкции нанотела 581 для ADAMTS5 человека составляет 3,65 нМ без HSA и 4,84 нМ с HSA.
7.2 Аффинность для сывороточного альбумина
Аффинность строительного блока ALB11 в конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) для связывания с сывороточным альбумином (SA) человека, яванского макака, морской свинки, мыши и крысы определяли с использованием SPR, как описано в примере 5.3.
Результаты показаны в таблице 7.2.
Таблица 7.2 Аффинность (KD, нМ) конструкции нанотела 581 для связывания с сывороточным альбумином человека, яванского макака, морской свинки, мыши и крысы
* скорость диссоциации находится вне предела детекции: > 5,0E-01 (1/с).
7.3 Связывание пре-Ab
Проводили скрининг конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) и конструкции нанотела 579 (2F3SO-35GS линкер-093SO-35GS линкер-Alb11) по связыванию предсуществующего антитела (пре-Ab) из 3 соответствующих групп образцов сыворотки доноров (образцов от здоровых субъектов, образцов OA и смещенной группы образцов с известным остаточным связыванием пре-Ab с другими нанотелами), посредством технологии SPR на устройстве ProteOn XPR36. Образцы крови анализировали по связыванию с конструкциями нанотел против ADAMTS5, которые были связаны с HSA. Уровни связывания предсуществующих антител определяли после коррекции сенсограммы по двум эталонам, посредством установки точек регистрации на 125 секунд (5 секунд после окончания связывания). Анализ проводили с использованием ProteOn manager 3.1 (Bio-Rad Laboratories, Inc.).
Результаты показаны на фигуре 3.
Обе конструкции нанотел имели низкие остаточные уровни связывания пре-Ab, где для конструкции нанотела 581 явно показано наименьшее остаточное связывание пре-Ab (ср. фигуру 3.).
7.4 Функциональность в AlphaLISA для человека
AlphaLISA проводили с использованием конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) с использованием ADAMTS5 человека, в основном, как описано в примере 5.1.
Результаты показаны в таблице 7.4.
Таблица 7.4 Активность (нМ) конструкции нанотела 581, как определено посредством AlphaLISA.
Результаты показывают, что активность конструкции нанотела 581 составляет 188 нМ.
7.5 Получение профилей иммуногенности в анализе дендритных клеток - T-клеток
Относительную иммуногенность определяли в анализе пролиферации дендритных клеток - T-клеток. По существу, нанотела тестировали применительно к группе из 50 образцов клеток здоровых доноров, содержащих наиболее распространенные аллели HLA класса II, таким образом, представляющих большинство глобальной популяции. Иммунный ответ оценивали с использованием пролиферации T-клеток в качестве суррогатного маркера образования антител против лекарственного средства на основе антител. Гемоцианин морского блюдечка (KLH) использовали в качестве положительного контроля. Положительный контроль KLH приводил к положительному ответу у всех 50 доноров. Ни один из доноров не тестирован как положительный для конструкции нанотела 581 (2F3SO-35GS линкер-Alb11). Конструкция нанотела 579 (2F3SO-35GS линкер-093SO-35GS линкер-Alb11) приводила к положительному ответу у трех доноров, или 6%. В нулевых условиях (только со средой), у 2 из 50 доноров, или 4%, обнаружен положительный ответ. Эти результаты использовали для установления общего порога ответа более чем у 3 из 50 доноров, соответствующего общему порогу ответа >6%. Общий иммуногенный потенциал тестированных нанотел считали низким, поскольку процент значительных, положительных ответов составлял ≤6% (см. таблицу 7.5).
Таблица 7.5 Результат построения профилей иммуногенности, как определено в анализе DC - T-клеток
7.6 Связывание с ADAMTS1 человека
Связывание с ADAMTS1 человека тестировали в ELISA связывания (см. фигуру 4AA). Кратко, с использованием 18,5 нМ ADAMTS1 человека (R&D Systems, Minneapolis, US; кат. # 2197-AD) и ADAMTS5 человека (R&D Systems, Minneapolis, US; кат. # 2198-AD) покрывали 384-луночные планшеты MaxiSorp (Nunc, Wiesbaden, Germany). Лунки блокировали раствором казеина (1%), и серийные разведения конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) и mAb против ADAMTS1 человека (R&D Systems, Minneapolis, US; кат. # MAB2197) тестировали по связыванию с обоими использованными для покрытия белками. После детекции с использованием конъюгированного с HRP средства против нанотела, узнающего конструкцию нанотела 581 (полученную в Ablynx), и конъюгированного с HRP антитела против антител мыши (Abcam, Cambridge, UK), соответственно, и последующей ферментной реакции в присутствии субстрата esTMB (3,3',5,5'-тетраметилбензидина) (SDT, Brussels, Belgium), измеряли OD450 нм.
Кривую зависимости ответа от дозы наблюдали для конструкции нанотела 581 с использованием ADAMTS5 человека и для mAb против ADAMTS1 человека с использованием ADAMTS1 человека, показывая, что конструкция нанотела 581 не связывается с ADAMTS1 человека.
7.7 Связывание с ADAMTS4 и ADAMTS15 человека
Связывание с ADAMTS4 и ADAMTS15 человека тестировали в ELISA связывания (см. фигуру 4B). Кратко, с использованием 1 мкг/мл ADAMTS4 человека (R&D Systems, Minneapolis, US; кат. # 4307-AD) или ADAMTS15 человека (R&D Systems, Minneapolis, US; кат. # 5149-AD) покрывали в течение ночи при 1 мкг/мл 96-луночные планшеты Maxisorp ELISA (Nunc, Wiesbaden, Germany). После блокирования планшетов с использованием SuperBlock T20 (PBS) (Thermo Scientific Pierce; кат. # 37516) раститровку для кривой зависимости ответа от дозы конструкции нанотела 581 («C011400581») или положительного контрольного mAb против ADAMTS4 (R&D Systems, Minneapolis, US; кат. # MAB4307), или положительного контрольного mAb против ADAMTS15 (R&D Systems, Minneapolis, US; кат. # MAB5149) наносили на покрытый планшет, начиная с 1 мкМ, и инкубировали в течение 1 часа при комнатной температуре. Связанное нанотело C011400581 детектировали с использованием биотинилированного средства против нанотела, узнающего конструкцию нанотела 581 (полученную в Ablynx), в то время как положительные контрольные mAb детектировали с использованием биотинилированного pAb козы против антител мыши (Jackson Immuno Research; кат. # 115-065-062). Для обоих стрептавидин-HRP использовали в качестве вторичного средства для детекции (Thermo Scientific Pierce; кат. # 21126). Затем планшеты визуализировали посредством добавления sTMB) (SDT, Brussels, Belgium). Все разведения и средства для детекции подготавливали в разбавителе для анализа (= PBS+10% Superblock+0,05% Tween20). Реакцию окрашивания останавливали через 2,5 минуты посредством добавления 1M HCl. Оптическую плотность измеряли при 450 нм с использованием 620 нм в качестве контрольной длины волны.
7.8 Перекрестная для видов реакционная способность с использованием определения аффинности KinExA
Аффинность конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) для ADAMTS5 нескольких видов определяли с использованием KinExA, как описано в примере 7.1 Для всех видов, конъюгированные с ADAMTS5 человека бусины PMMA получали и использовали для связывания свободной конструкции нанотела 581, как описано выше. Связанную конструкцию нанотела 581 детектировали с использованием меченного AF647 средства против нанотела, узнающего конструкцию нанотела 581 (полученную в Ablynx). Результаты показаны в таблице 7.7
Таблица 7.7 Аффинность конструкции нанотела 581 для ADAMTS5 яванского макака, крысы, мыши и быка
Пример 8. Активность в анализе человеческого эксплантата
Конструкцию нанотела 581 (2F3SO-35GS линкер-Alb11) оценивали по способности блокировать деградацию хряща в анализе ex vivo, в основном, как описано в примере 5.2, но теперь в анализе человеческого эксплантата. Кратко, получали свежеприготовленные срезы эксплантата человеческого хряща (диаметром 4 мм) из коленных суставов человека и инкубировали в 96-луночных планшетах в присутствии 10 нг/мл IL-1α для индукции деградации хряща). В качестве показателя деградации хряща/аггрекана, высвобождение GAG детектировали в супернатанте после 7 суток инкубации (37°C, 7,5% CO2) посредством метахроматического красителя 1,9-диметилметиленового синего (излучение при 633 нм). хондроитинсульфат включали в качестве стандарта анализа. Эффективность определяли посредством индуцированного IL-1β и OSM контроля без соединения и в присутствии MSC2310852A. CRB0017 представляет собой mAb против ADAMTS5, подлежащее исследованию в клинических исследованиях фазы I (Rottapharm).
Результаты обобщены на фигуре 5 и в таблице 8.
Таблица 8
Для нанотел показана высокая активность и 100% ингибирование деградации хряща (потери GAG) в анализе человеческого эксплантата. Как показано в таблице 8, нанотела являются по меньшей мере в 10 раз лучшими, чем CRB0017, в анализе бычьего эксплантата.
Пример 9. Модуляция C2M, C3M и exAGNX1 в бычьей модели совместной культуры
Исследовали эффект конструкции нанотела 581 («581» на фигурах) на высвобождение и содержание GAG, C2M (маркера деградации коллагена II), C3M (маркера деградации коллагена III) и exAGNx1 (маркера деградации аггрекана, характеризующегося неоэпитопом TEGE) в бычьей системе совместной культуры. В этих системах совместной культуры, эксплантаты хряща из колена быка инкубировали вместе с синовиальной мембраной от того же животного в течение 28 суток. Наивысшее высвобождение GAG детектировали после 7 суток в культуре (фигура 6A). Конструкцию нанотела 581 тестировали в трех концентрациях: 1 нМ, 10 нМ, 100 нМ. Инкубация эксплантатов хряща с синовиальной мембраной приводила к увеличению высвобождения GAG. Конструкция нанотела 581 ингибировала высвобождение GAG (фигура 6B).
В соответствии с высвобождением GAG, анализ содержания оставшегося GAG в эксплантате хряща (после расщепления папаином) выявлял уменьшение содержания GAG после совместной инкубации эксплантатов с синовиальной мембраной, по сравнению с эксплантатами отдельно (фигура 6C).
Параллельно с анализом высвобождения GAG, измеряли высвобождение exAGNx1 в качестве специфического маркера деградации аггрекана. Анализ площади под кривой (AUC) показал сильную индукцию высвобождения exAGNx1, когда эксплантаты хряща и синовиальную мембрану инкубировали вместе. Кроме того, конструкция нанотела 581 ингибировала высвобождение exAGNx1 зависимым от концентрации образом с полным ингибированием при 100 нМ 581 (фигура 7).
Для определения эффекта конструкции нанотела 581 на коллагеновую сеть хряща в системе совместной культуры, определяли C2M, в качестве маркера деградации коллагена II. В отличие от маркеров деградации аггрекановой сети (GAG и exAGNx1), уровень C2M начинал увеличиваться между сутками 14 и 21. Инкубация конструкции нанотела 581 в системе совместной культуры ингибировала высвобождение C2M (фигура 8).
Определяли дополнительный маркер синовиального воспаления, C3M (т.е. маркер деградации коллагена типа III). Анализ зависимости от времени высвобождения C3M показал, что уровень C3M начинал увеличиваться через 14 суток, достигая максимума приблизительно на сутки 21. Обработка конструкцией нанотела 581 ингибировала C3M при всех трех концентрациях (фигура 9).
Пример 10. Ингибирование активности аггреканазы у NHP
Способность конструкции нанотела 581 (2F3SO-35GS линкер-Alb11; «C011400581») ингибировать активность аггреканазы in vivo оценивали в модели на яванском макаке, нечеловекообразном примате (NHP). Кратко, конструкцию нанотела 581 вводили в группах из 3 самцов и 3 самок яванских макаков один раз в неделю посредством подкожного (s.c.) введения при уровнях дозирования 6, 30 или 150 мг/кг в течение 4 недель. Сопутствующую контрольную группу обрабатывали носителем, т.е. 20 мМ гистидином, 8% сахарозой, 0,01% Tween 20 pH 6,0.
Ингибирование активности аггреканазы измеряли в образцах сыворотки, собранных в нескольких временных точках, посредством определения уровня фрагментов деградации аггрекана, характеризующихся присутствием неоэпитопа ARGS в сыворотке. Для всех подвергнутых обработке конструкцией нанотела 581 животных показан сходный профиль, т.е. уровни ARGS уменьшались после первого дозирования и достигали очень низких уровней около или ниже нижнего предела диапазона измерений (LLMR) анализа (0,08 нМ) между 48 часов и 120 часов. Затем, для уровней ARGS показано устойчивое максимальное уменьшение около или ниже LLMR, и они не возвращались к исходным до окончания исследования через 4 недели.
Обобщение результатов показано на фигуре 10.
В заключения, специфические для ADAMTS5 нанотела привлекают мишень после системного введения и активно модулируют уровни неоэпитопа ARGS in vivo при всех тестированных уровнях дозирования. Кроме того, в отличие от mAb предшествующего уровня техники, не наблюдали связанных с тестируемым препаратом патологических аритмий. Кроме того, не было отмечено доказательств какого-либо связанного с тестированием повышения ST у самцов и самок обезьян, подвергнутых лечению нанотелами, при любом из тестированных уровней дозирования.
Пример 11. Ингибирование дегенерации хряща
Для дополнительной оценки способности нанотел и конструкций нанотел ингибировать дегенерацию хряща in vivo, использовали модель DMM (дестабилизации медиального мениска) на мышах. Кратко, медиальный мениск подвергали хирургической дестабилизации. Иллюстративную конструкцию нанотела 581 (2F3SO-35GS линкер-Alb11) либо вводили подкожно за 3 суток до индукции DMM (профилактически), либо вводили через 3 суток после индукции DMM (терапевтически), в различных концентрациях. После 8 недель лечения, животных умерщвляли, и коленные суставы извлекали. Коленные суставы погружали в парафиновые блоки, получали срезы и окрашивали с использованием толуидинового синего. Затем срезы оценивали в баллах по нескольким параметрам, включая суммарную дегенерацию медиального хряща.
Результаты показаны на фигуре 11.
Результаты показывают, что присутствует структурное преимущество вплоть до 50% как для профилактического, так и для терапевтического лечения (s.c.) с использованием нанотел в модели DMM на мышах.
Пример 12. Симптоматическое преимущество в хирургической модели OA на крысах
Для оценки способности нанотел обеспечивать симптоматическое преимущество, использовали хирургическую модель OA на крысах. Кратко, крыс подвергали хирургической обработке ACLt и tMx для индукции OA на сутки 0. В хирургической модели ACLt и tMx (рассечения передней крестообразной связки, расширенного с использованием медиальной менискэктомии), конструкцию нанотела 581 вводили s.c. каждые вторые сутки с суток 3 и далее. В еженедельном режиме, определяли симптоматическое преимущество посредством анализа походки (на подиуме), так же как уменьшения диаметра сустава.
Результаты показаны на фигуре 12.
Остеоартрит индуцировали (на сутки 0) у взрослых самцов крыс (Lister Hooded; средняя масса тела 346±20 г) посредством хирургической операции рассечения передней крестообразной связки (ACLT)+резекции медиального мениска (tMx) на правом коленном суставе. Обработку носителем или тестируемым препаратом (sc) начинали на сутки 3 после хирургической операции и продолжали каждые вторые сутки до суток 42. В контрольной группе здоровых не проводили хирургического вмешательства, но вводили sc носитель в тех же временных точках, что и в группах дозирования. (A) Нарушение походки с течением времени рассчитывали как «% преимущество по сравнению с носителем». Среднее для группы «здоровые+плацебо» = 100% преимущество. Среднее для группы ««ACLT tMx+носитель» = 0% преимущество. (B) Среднее «преимущество по сравнению с носителем» во всех исследованных временных точках после начала обработки. Показано среднее±SEM для 14-15 крыс/группу. *= p<0,05, рассчитано с использованием однофакторного ANOVA и критерия Даннетта.
Ранняя обработка с использованием нанотел вызывала зависимое от дозы, значимое и значительное симптоматическое преимущество при индуцированном ACLT+tMx OA.
Пример 13. Исследования токсичности у яванских макаков
4-недельное исследование субхронической токсичности у яванских макаков проводили для получения информации о системной токсичности, местной переносимости и фармакологической безопасности нанотел. Иллюстративную конструкцию нанотела 581 (2F3SO-35GS линкер-Alb11) вводили s.c. в группах из 3 самцов и 3 самок яванских макаков (Macaca fascicularis) (один раз в неделю посредством подкожного введения в дорсальную область в дозах 6, 30 или 150 мг/кг в течение 4 недель. Сопутствующую контрольную группу подвергали обработке носителем, 20 мМ гистидином, 8% сахарозой, 0,01% Tween 20 pH 6,0.
Связанных с тестируемым препаратом признаков местной непереносимости не отмечено ни на одном из тестированных уровней дозирования. Не отмечено связанных с тестируемым препаратом эффектов на поведение, массу тела, потребление пищи, гематологические и биохимические параметры, типирование лимфоцитов, уровни CRP, параметры анализа мочи, зрительные и слуховые функции, и массу органов ни у одного из животных при любом уровне дозирования (данные не представлены). Ни макроскопическое исследование при некропсии, ни гистопатологическое исследование не выявило никаких локальных или системных изменений органов, связанных с обработкой тестируемым препаратом, ни у одного из животных, исследуемых при любом тестированном уровне дозирования.
Поскольку опубликовано, что для mAb 12F4 показано очаговое эндокардиальное кровоизлияние, зависимое от дозы увеличение кровяного давления без доказательств обратимости, так же как аномалии сердечной проводимости (повышение ST и желудочковые аритмии) (Larkin, et al., The highs and lows of translational drug development: Antibody-mediated inhibition of ADAMTS-5 for osteoarthritis disease modification, OARSI conference 2014: Paris; Renninger et al., Identification of Altered Cardiovascular Function Produced by a Novel Biologic Compound in a Stand Alone Safety Pharmacology Primate Study, in SPS meeting, 2013), обширную неинвазивную телеметрию с использованием системы EMKA проводили для животных.
Не отмечено связанного с тестируемым препаратом влияния на какой-либо из телеметрических параметров (с использованием неинвазивной системы EMKA) у какого-либо из животных на каком-либо уровне дозирования. В частности, не наблюдали патологических аритмий и не присутствовало доказательства связанного с каким-либо тестируемым препаратом повышения ST у самцов и самок обезьян, подвергнутых лечению с использованием нанотел, на каких-либо тестированных уровнях дозирования (данные не представлены).
Эти исследования подтверждают, что специфические для ADAMTS5 нанотела можно считать безопасными.
Пример 14. Исследования in vivo на модели MMT DMOAD у крыс
Чтобы показать эффективность in vivo ингибиторов ADAMTS5, слитых со связывающей CAP молекулой по изобретению, использовали модель хирургически индуцированного разрыва медиального мениска (MMT) на крысах. Кратко, нанотело против ADAMTS5 соединяли со связывающей CAP молекулой (обозначено как конструкция нанотела C010100954 или конструкция нанотела 954). Крыс подвергали операции на одном колене для индукции подобных OA симптомов. Лечение начинали через 3 суток после хирургического вмешательства, посредством IA инъекции. Гистопатологию проводили на сутки 42 после хирургического вмешательства. Промежуточные и конечные образцы сыворотки отбирали для исследовательского анализа биомаркеров. Определяли медиальную и общую значительную ширину дегенерации хряща, так же как процент уменьшения дегенерации хряща. Использовали 20 животных на группу.
Субхрящевой дефект в медиальной большеберцовой кости показан на фигуре 13.
Результаты показывают, что ширину хряща существенно уменьшали посредством конструкции ADAMTS5-CAP через 42 суток, по сравнению с носителем. Эти результаты дополнительно позволяют предполагать, что
(a) группа CAP не оказывает отрицательного влияния на активность нанотела против ADAMTS5;
(b) группа CAP обеспечивает удержание нанотела против ADAMTS5; и
(c) нанотело против ADAMTS5 оказывает положительный эффект на ширину хряща, даже после присоединения группы CAP.
Пример 15.
Способы: Эксплантаты бычьего хряща от четырех животных (BEX), так же как эксплантаты человеческого хряща из восьми замененных хирургически коленных суставов (HEX) и из одного здорового человеческого коленного сустава (hHEX), культивировали в течение вплоть до 21 суток только в среде (w/o), в присутствии провоспалительных цитокинов (онкостатина M [10 нг/мл]+TNFα [20 нг/мл] (O+T)) или O+T с иллюстративной конструкцией нанотела 581 (2F3SO-35GS линкер-Alb11) [1 мкМ - 1 нМ]. Хрящ и синовиальную мембрану от коров (bCC) и из 4 замененных пораженных остеоартритом человеческих коленных суставов (hCC) совместно культивировали ex vivo в течение вплоть до 28 суток только в среде (w/o), с O+T или O+T плюс иллюстративная конструкция нанотела 581 (2F3SO-35GS линкер-Alb11) [1 мкМ - 0,6 нМ]. Хрящ нарезали на эксплантаты равного размера с использованием пробойника для биопсии. Синовиальные мембраны нарезали на эксплантаты равного размера (30 мг [±3 мг]) посредством скальпеля. Метаболическую активность эксплантатов оценивали посредством аламарового синего. Обновление ткани хряща оценивали с использованием твердофазного иммуноферментного анализа (ELISA) для измерения хорошо охарактеризованных биомаркеров деградации (huARGS, exAGNxI, C2M) и формирования (ProC2) в кондиционированной среде. ProC2 и C2M представляют собой метаболиты формирования и деградации коллагена типа II, соответственно, в то время как exAGNxI и huARGS представляют собой метаболиты деградированного посредством ADAMTS-5 аггрекана. Опубликованы средние значения и стандартная ошибка среднего (SEM). Статистический анализ выполняли с использованием однофакторного дисперсионного анализа (ANOVA) с критерием множественного сравнения Данна или двухфакторного ANOVA с критерием множественного сравнения Даннетта, принимая нормальное распределение.
Результаты: Метаболическая активность BEX, HEX и bCC являлась стабильной на протяжении периода культивирования, в то время как метаболическая активность в hCC и hHEX значительно падала от суток 14 в условиях обработки O+T, по сравнению с w/o. В культурах, стимулированных с использованием O+T, метаболиты деградированного посредством ADAMTS-5 аггрекана достигали макисимума в пределах первой недели культивирования, за исключением hHEX, в котором уровни huARGS и exAGNxI увеличивались немного позже. Деградация коллагена типа II, C2M, посредством O+T, достигала макисмума после суток 19. Формирование коллагена типа II, Pro-C2, оставалось относительно стабильным на протяжени культивирования, по сравнению с контролем w/o.
Обработка иллюстративной конструкцией нанотела 581 (2F3SO-35GS линкер-Alb11) в комбинации с O+T зависимым от дозы образом уменьшала уровень huARGS на сутки 5 в BEX (наивысшая доза: 8% из O+T), HEX (наивысшая доза: 40% из O+T), bCC (наивысшая доза: 10% из O+T), hCC (наивысшая доза: 40% из O+T), и hHEX (наивысшая доза: 24% из O+T) (фигура 14). IC50 иллюстративной конструкции нанотела 581 (2F3SO-35GS линкер-Alb11), на основании уменьшения уровня huARGS, лежала в диапазоне от 300 нМ в BEX до <15 нМ в HEX, hHEX, bCC и hCC (фигура 14). Эффект иллюстративной конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) на exAGNxI являлся сходным с huARGS в тестированных культурах. Иллюстративная конструкция нанотела 581 (2F3SO-35GS линкер-Alb11) уменьшала также уровень C2M (маркера деградации коллагена типа II), значимо и зависимым от дозы образом, хотя эффект был меньше, чем для маркеров деградации аггрекана. Для иллюстративной конструкции нанотела 581 (2F3SO-35GS линкер-Alb11) не показано эффекта на метаболит формирования коллагена типа II Pro-C2 в каком-либо из тестированных условий.
Заключения: В этом исследовании, авторы настоящего изобретения показали, что иллюстративная конструкция нанотела 581 (2F3SO-35GS линкер-Alb11) оказывала защитные эффекты на хрящ, благодаря своему зависимому от дозы ингибированию опосредованной ADAMTS-5 деградации аггрекана и опосредованной MMP деградации коллагена типа II в провоспалительных условиях в культурах бычьего и человеческого хряща ex vivo и в совместных культурах хряща и синовиальной мембраны. 2F3SO-35GS линкер-Alb (SEQ ID NO: 129) является одним из предпочтительных вариантов осуществления изобретения.
Пример 16. Конструкции нанотел по настоящему изобретению превосходят нанотела предществующего уровня техники
В WO 2008/074840 описаны различные нанотела против ADAMTS5. В настоящем эксперименте проводили сравнение между 26 нанотелами, описанными в WO2008/074840, и нанотелами по настоящему изобретению, представленными иллюстративным нанотелом 2F3so (2F3*).
Все конструкции тестировали в анализе AlphaLISA, как описано в примере 5.1. Нанотело 2F3so (2F3*) представляет собой связывающий ADAMTS5 моновалентный строительный блок из C011400581 (SEQ ID NO: 129). В качестве отрицательного контроля, использовали не имеющее отношения к делу нанотело IRR00028.
Сначала активность нанотел предшествующего уровня техники определяли в AlphaLISA. Результаты показаны в таблице 16.1.
Таблица 16.1 Активность нанотел предшествующего уровня техники в AlphaLISA
Результаты показывают, что только 7 нанотел предшествующего уровня техники имеют блокирующую активность. Остальные нанотела предшествующего уровня техники не имели активности или усиливали активность.
На второй фазе этого сравнительного эксперимента, блокирующие нанотела сравнивали по отдельности с иллюстративным моновалентным нанотелом 2F3so (2F3*).
Значения IC50 для конструкций нанотел перечислены в таблице 16.2. Кратность различий с иллюстративным моновалентным нанотелом 2F3so (2F3*) также указана для простоты сравнения.
Таблица 16.2 Активность конструкций нанотел предшествующего уровня техники по сравнению с нанотелом 2F3so (2F3*), как определено посредством AlphaLISA
Можно заключить, что конструкции нанотел по настоящему изобретению превосходят нанотела предшествующего уровня техники против ADAMTS5.
Таблица A-1. Наименование и короткое описание («ID»), SEQ ID NO: («SEQ») и аминокислотные последовательности моновалентных и мультивалентных нанотел против ADAMTS5
* SO (оптимизированный по последовательности) вариант.
Таблица A-2. Последовательности CDR и каркасных областей, плюс предпочтительные комбинации, как представлено в формуле I, а именно, FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4 (следующие термины: «ID» относится к данному SEQ ID NO)
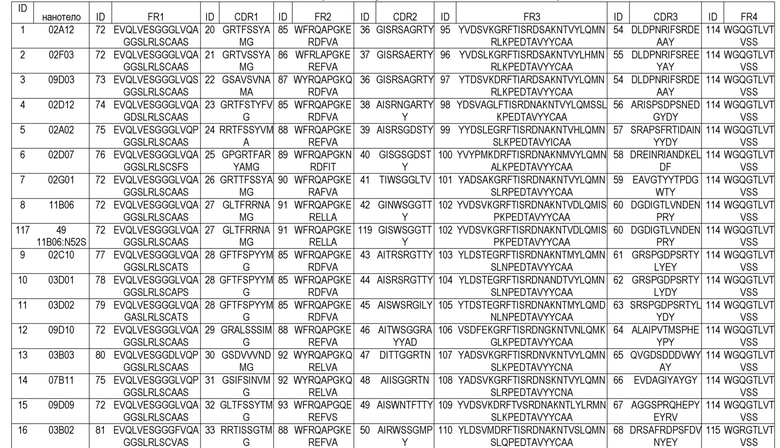
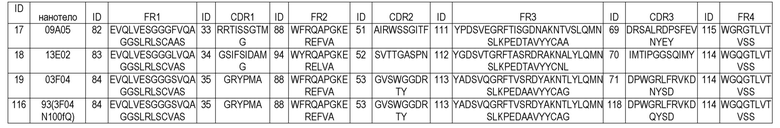
Таблица B. Прочие аминокислотные последовательности: Наименование и короткое описание («ID»), SEQ ID NO: («SEQ») и аминокислотные последовательности («последовательности»)
Таблица C. Различные линкерные последовательности («ID» относится к SEQ ID NO, в рамках изобретения)
Таблица D. Связывающие сывороточный альбумин последовательности ISVD («ID» относится к SEQ ID NO, в рамках изобретения), включая последовательности CDR
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Merck Patent GmbH
Ablynx N.V.
<120> Связывающие ADAMTS иммуноглобулины
<130> MEPAT.0103
<150> EP17174403.0
<151> 2017-06-02
<160> 174
<170> BiSSAP 1.3.6
<210> 1
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 1
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 2
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Val Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Leu Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr Tyr Val Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu His Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 3
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 3
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Ala Val Ser Val Asn
20 25 30
Ala Met Ala Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Asp Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Asp Arg Phe Thr Ile Ala Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 4
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 4
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Asp
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Thr Tyr
20 25 30
Phe Val Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Ala Ile Ser Arg Asn Gly Ala Arg Thr Tyr Tyr Tyr Asp Ser Val
50 55 60
Ala Gly Leu Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Ala Arg Ile Ser Pro Ser Asp Pro Ser Asn Glu Asp Gly Tyr
100 105 110
Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 5
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 5
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Arg Arg Thr Phe Ser Ser Tyr
20 25 30
Val Met Ala Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Ser Arg Ser Gly Asp Ser Thr Tyr Tyr Tyr Asp Ser Leu
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val His
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Ile Cys
85 90 95
Ala Ala Ser Arg Ala Pro Ser Phe Arg Thr Ile Asp Ala Ile Asn Tyr
100 105 110
Tyr Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 6
<211> 126
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Phe Ser Gly Pro Gly Arg Thr Phe Ala
20 25 30
Arg Tyr Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Asn Arg Asp
35 40 45
Phe Ile Thr Gly Ile Ser Gly Ser Gly Asp Ser Thr Tyr Tyr Val Tyr
50 55 60
Pro Met Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Val Tyr Leu Gln Met Asn Ala Leu Lys Pro Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Ala Asp Arg Glu Ile Asn Arg Ile Ala Asn Asp Lys Glu
100 105 110
Leu Asp Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 7
<211> 123
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 7
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Thr Phe Ser Ser
20 25 30
Tyr Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Ala Phe
35 40 45
Val Ala Thr Ile Trp Ser Gly Gly Leu Thr Val Tyr Ala Asp Ser Ala
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Glu Ala Val Gly Thr Tyr Tyr Thr Pro Asp Gly Trp Thr Tyr
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 8
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Arg Arg Asn
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Leu
35 40 45
Ala Gly Ile Asn Trp Ser Gly Gly Thr Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Asp
65 70 75 80
Leu Gln Met Ile Ser Pro Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Gly Asp Ile Gly Thr Leu Val Asn Asp Glu Asn Pro Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 9
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 9
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Tyr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Ala Ile Thr Arg Ser Arg Gly Thr Thr Tyr Tyr Leu Asp Ser Thr
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Asn Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Gly Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Glu
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 10
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Pro Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Tyr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Ala Ile Ser Arg Ser Arg Gly Thr Thr Tyr Tyr Leu Asp Ser Thr
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Asn Asp Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Asn Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Gly Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 11
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 11
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Ala
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Ser Pro Tyr
20 25 30
Tyr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Ala Ile Ser Trp Ser Arg Gly Ile Leu Tyr Tyr Thr Asp Ser Thr
50 55 60
Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Met Tyr
65 70 75 80
Leu Gln Met Asp Asn Leu Asn Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Ser Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 12
<211> 127
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Ala Leu Ser Ser Ser
20 25 30
Ile Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Thr Trp Ser Gly Gly Arg Ala Tyr Tyr Ala Asp Val Ser
50 55 60
Asp Phe Glu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Gly Lys Asn
65 70 75 80
Thr Val Asn Leu Gln Met Lys Gly Leu Lys Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Ala Ala Leu Ala Ile Pro Val Thr Met Ser Pro His
100 105 110
Glu Tyr Pro Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 13
<211> 121
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Asp Val Val Val Asn
20 25 30
Asp Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Asp Ile Thr Thr Gly Gly Arg Thr Asn Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Asn
85 90 95
Ala Gln Val Gly Asp Ser Asp Asp Asp Val Trp Tyr Ala Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 14
<211> 119
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 14
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Ile Phe Ser Ile Asn
20 25 30
Val Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Ala Ile Ile Ser Gly Gly Arg Thr Asn Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Asn
85 90 95
Ala Glu Val Asp Ala Gly Ile Tyr Ala Tyr Gly Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 15
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 15
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Ser Ser Tyr
20 25 30
Thr Met Gly Trp Phe Arg Gln Ala Pro Gly Gln Glu Arg Glu Phe Val
35 40 45
Ser Ala Ile Ser Trp Asn Thr Phe Thr Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Asp Arg Phe Thr Val Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Arg Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Ala Gly Gly Ser Pro Arg Gln His Glu Pro Tyr Glu Tyr Arg
100 105 110
Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 16
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 16
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Phe Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Arg Arg Thr Ile Ser Ser Gly
20 25 30
Thr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Arg Trp Ser Ser Gly Met Pro Tyr Tyr Leu Asp Ser Val
50 55 60
Met Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Gln Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Arg Ser Ala Phe Arg Asp Pro Ser Phe Asp Val Asn Tyr
100 105 110
Glu Tyr Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 17
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Phe Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Arg Arg Thr Ile Ser Ser Gly
20 25 30
Thr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Arg Trp Ser Ser Gly Ile Thr Phe Tyr Pro Asp Ser Val
50 55 60
Glu Gly Arg Phe Thr Ile Ser Gly Asp Asn Ala Lys Asn Thr Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Arg Ser Ala Leu Arg Asp Pro Ser Phe Glu Val Asn Tyr
100 105 110
Glu Tyr Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
115 120 125
<210> 18
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Ser Ile Phe Ser Ile Asp
20 25 30
Ala Met Gly Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Val
35 40 45
Ala Ser Val Thr Thr Gly Ala Ser Pro Asn Tyr Gly Asp Ser Val Thr
50 55 60
Gly Arg Phe Thr Ala Ser Arg Asp Arg Ala Lys Asn Ala Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys Asn
85 90 95
Leu Ile Met Thr Ile Pro Gly Gly Ser Gln Ile Met Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 19
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 19
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Arg Tyr Pro Met Ala Trp
20 25 30
Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser
35 40 45
Trp Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Gln Gly Arg Phe
50 55 60
Thr Val Ser Arg Asp Tyr Ala Lys Asn Thr Leu Tyr Leu Gln Met Asn
65 70 75 80
Ser Leu Lys Pro Glu Asp Ala Ala Val Tyr Tyr Cys Ala Gly Asp Pro
85 90 95
Trp Gly Arg Leu Phe Arg Val Lys Asp Asn Tyr Ser Asp Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 20
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 20
Gly Arg Thr Phe Ser Ser Tyr Ala Met Gly
1 5 10
<210> 21
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 21
Gly Arg Thr Val Ser Ser Tyr Ala Met Gly
1 5 10
<210> 22
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 22
Gly Ser Ala Val Ser Val Asn Ala Met Ala
1 5 10
<210> 23
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 23
Gly Arg Thr Phe Ser Thr Tyr Phe Val Gly
1 5 10
<210> 24
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 24
Arg Arg Thr Phe Ser Ser Tyr Val Met Ala
1 5 10
<210> 25
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 25
Gly Pro Gly Arg Thr Phe Ala Arg Tyr Ala Met Gly
1 5 10
<210> 26
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 26
Gly Arg Thr Thr Phe Ser Ser Tyr Ala Met Gly
1 5 10
<210> 27
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 27
Gly Leu Thr Phe Arg Arg Asn Ala Met Gly
1 5 10
<210> 28
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 28
Gly Phe Thr Phe Ser Pro Tyr Tyr Met Gly
1 5 10
<210> 29
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 29
Gly Arg Ala Leu Ser Ser Ser Ile Met Gly
1 5 10
<210> 30
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 30
Gly Ser Asp Val Val Val Asn Asp Met Gly
1 5 10
<210> 31
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 31
Gly Ser Ile Phe Ser Ile Asn Val Met Gly
1 5 10
<210> 32
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 32
Gly Leu Thr Phe Ser Ser Tyr Thr Met Gly
1 5 10
<210> 33
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 33
Arg Arg Thr Ile Ser Ser Gly Thr Met Gly
1 5 10
<210> 34
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 34
Gly Ser Ile Phe Ser Ile Asp Ala Met Gly
1 5 10
<210> 35
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 35
Gly Arg Tyr Pro Met Ala
1 5
<210> 36
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 36
Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr
1 5 10
<210> 37
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 37
Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr
1 5 10
<210> 38
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 38
Ala Ile Ser Arg Asn Gly Ala Arg Thr Tyr Tyr
1 5 10
<210> 39
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 39
Ala Ile Ser Arg Ser Gly Asp Ser Thr Tyr
1 5 10
<210> 40
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 40
Gly Ile Ser Gly Ser Gly Asp Ser Thr Tyr
1 5 10
<210> 41
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 41
Thr Ile Trp Ser Gly Gly Leu Thr Val
1 5
<210> 42
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 42
Gly Ile Asn Trp Ser Gly Gly Thr Thr Tyr
1 5 10
<210> 43
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 43
Ala Ile Thr Arg Ser Arg Gly Thr Thr Tyr
1 5 10
<210> 44
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 44
Ala Ile Ser Arg Ser Arg Gly Thr Thr Tyr
1 5 10
<210> 45
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 45
Ala Ile Ser Trp Ser Arg Gly Ile Leu Tyr
1 5 10
<210> 46
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 46
Ala Ile Thr Trp Ser Gly Gly Arg Ala Tyr Tyr Ala Asp
1 5 10
<210> 47
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 47
Asp Ile Thr Thr Gly Gly Arg Thr Asn
1 5
<210> 48
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 48
Ala Ile Ile Ser Gly Gly Arg Thr Asn
1 5
<210> 49
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 49
Ala Ile Ser Trp Asn Thr Phe Thr Thr Tyr
1 5 10
<210> 50
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 50
Ala Ile Arg Trp Ser Ser Gly Met Pro Tyr
1 5 10
<210> 51
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 51
Ala Ile Arg Trp Ser Ser Gly Ile Thr Phe
1 5 10
<210> 52
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 52
Ser Val Thr Thr Gly Ala Ser Pro Asn
1 5
<210> 53
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 53
Gly Val Ser Trp Gly Gly Asp Arg Thr Tyr
1 5 10
<210> 54
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 54
Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala Tyr
1 5 10 15
<210> 55
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 55
Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala Tyr
1 5 10 15
<210> 56
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 56
Ala Arg Ile Ser Pro Ser Asp Pro Ser Asn Glu Asp Gly Tyr Asp Tyr
1 5 10 15
<210> 57
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 57
Ser Arg Ala Pro Ser Phe Arg Thr Ile Asp Ala Ile Asn Tyr Tyr Asp
1 5 10 15
Tyr
<210> 58
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 58
Asp Arg Glu Ile Asn Arg Ile Ala Asn Asp Lys Glu Leu Asp Phe
1 5 10 15
<210> 59
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 59
Glu Ala Val Gly Thr Tyr Tyr Thr Pro Asp Gly Trp Thr Tyr
1 5 10
<210> 60
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 60
Asp Gly Asp Ile Gly Thr Leu Val Asn Asp Glu Asn Pro Arg Tyr
1 5 10 15
<210> 61
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 61
Gly Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Glu Tyr
1 5 10 15
<210> 62
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 62
Gly Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Asp Tyr
1 5 10 15
<210> 63
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 63
Ser Arg Ser Pro Gly Asp Pro Ser Arg Thr Tyr Leu Tyr Asp Tyr
1 5 10 15
<210> 64
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 64
Ala Leu Ala Ile Pro Val Thr Met Ser Pro His Glu Tyr Pro Tyr
1 5 10 15
<210> 65
<211> 13
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 65
Gln Val Gly Asp Ser Asp Asp Asp Val Trp Tyr Ala Tyr
1 5 10
<210> 66
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 66
Glu Val Asp Ala Gly Ile Tyr Ala Tyr Gly Tyr
1 5 10
<210> 67
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 67
Ala Gly Gly Ser Pro Arg Gln His Glu Pro Tyr Glu Tyr Arg Val
1 5 10 15
<210> 68
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 68
Asp Arg Ser Ala Phe Arg Asp Pro Ser Phe Asp Val Asn Tyr Glu Tyr
1 5 10 15
<210> 69
<211> 16
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 69
Asp Arg Ser Ala Leu Arg Asp Pro Ser Phe Glu Val Asn Tyr Glu Tyr
1 5 10 15
<210> 70
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 70
Ile Met Thr Ile Pro Gly Gly Ser Gln Ile Met Tyr
1 5 10
<210> 71
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 71
Asp Pro Trp Gly Arg Leu Phe Arg Val Lys Asp Asn Tyr Ser Asp
1 5 10 15
<210> 72
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 72
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 73
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 73
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 74
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 74
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Asp
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 75
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 75
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 76
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 76
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Phe Ser
20 25
<210> 77
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 77
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser
20 25
<210> 78
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 78
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Pro Ser
20 25
<210> 79
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 79
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Ala
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser
20 25
<210> 80
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 80
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 81
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 81
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Phe Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser
20 25
<210> 82
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 82
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Phe Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser
20 25
<210> 83
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser
20 25
<210> 84
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 84
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser
20 25
<210> 85
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 85
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val Ala
1 5 10
<210> 86
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 86
Trp Phe Arg Leu Ala Pro Gly Lys Glu Arg Glu Phe Val Ala
1 5 10
<210> 87
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 87
Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Asp Phe Val Ala
1 5 10
<210> 88
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 88
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala
1 5 10
<210> 89
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 89
Trp Phe Arg Gln Ala Pro Gly Lys Asn Arg Asp Phe Ile Thr
1 5 10
<210> 90
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 90
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Ala Phe Val Ala
1 5 10
<210> 91
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 91
Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Leu Ala
1 5 10
<210> 92
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 92
Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Glu Leu Val Ala
1 5 10
<210> 93
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 93
Trp Phe Arg Gln Ala Pro Gly Gln Glu Arg Glu Phe Val Ser
1 5 10
<210> 94
<211> 14
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 94
Trp Tyr Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Val Ala
1 5 10
<210> 95
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 95
Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala
1 5 10 15
Lys Asn Thr Val Tyr Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 96
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 96
Tyr Val Asp Ser Leu Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala
1 5 10 15
Lys Asn Thr Val Tyr Leu His Met Asn Arg Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 97
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 97
Tyr Thr Asp Ser Val Lys Asp Arg Phe Thr Ile Ala Arg Asp Ser Ala
1 5 10 15
Lys Asn Thr Val Tyr Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 98
<211> 38
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 98
Tyr Asp Ser Val Ala Gly Leu Phe Thr Ile Ser Arg Asp Asn Ala Lys
1 5 10 15
Asn Thr Val Tyr Leu Gln Met Ser Ser Leu Lys Pro Glu Asp Thr Ala
20 25 30
Val Tyr Tyr Cys Ala Ala
35
<210> 99
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 99
Tyr Tyr Asp Ser Leu Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Val His Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Ile Cys Ala Ala
35
<210> 100
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 100
Tyr Val Tyr Pro Met Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Met Val Tyr Leu Gln Met Asn Ala Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 101
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 101
Tyr Ala Asp Ser Ala Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 102
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 102
Tyr Val Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Val Asp Leu Gln Met Ile Ser Pro Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 103
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 103
Tyr Leu Asp Ser Thr Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Met Tyr Leu Gln Met Asn Ser Leu Asn Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 104
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 104
Tyr Leu Asp Ser Thr Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Asn Asp Thr Val Tyr Leu Gln Met Asn Ser Leu Asn Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 105
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 105
Tyr Thr Asp Ser Thr Glu Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Met Tyr Leu Gln Met Asp Asn Leu Asn Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 106
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 106
Val Ser Asp Phe Glu Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Gly
1 5 10 15
Lys Asn Thr Val Asn Leu Gln Met Lys Gly Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 107
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 107
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val
1 5 10 15
Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Asn Ala
35
<210> 108
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 108
Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser
1 5 10 15
Lys Asn Thr Val Tyr Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Asn Ala
35
<210> 109
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 109
Tyr Val Asp Ser Val Lys Asp Arg Phe Thr Val Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Leu Tyr Leu Arg Met Asn Ser Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 110
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 110
Tyr Leu Asp Ser Val Met Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala
1 5 10 15
Lys Asn Thr Val Ser Leu Gln Met Asn Ser Leu Gln Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 111
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 111
Tyr Pro Asp Ser Val Glu Gly Arg Phe Thr Ile Ser Gly Asp Asn Ala
1 5 10 15
Lys Asn Thr Val Ser Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Ala Ala
35
<210> 112
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 112
Tyr Gly Asp Ser Val Thr Gly Arg Phe Thr Ala Ser Arg Asp Arg Ala
1 5 10 15
Lys Asn Ala Leu Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Thr
20 25 30
Ala Val Tyr Tyr Cys Asn Leu
35
<210> 113
<211> 39
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 113
Tyr Ala Asp Ser Val Gln Gly Arg Phe Thr Val Ser Arg Asp Tyr Ala
1 5 10 15
Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser Leu Lys Pro Glu Asp Ala
20 25 30
Ala Val Tyr Tyr Cys Ala Gly
35
<210> 114
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 114
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 115
<211> 11
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 115
Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser
1 5 10
<210> 116
<211> 120
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 116
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Arg Tyr Pro Met Ala Trp
20 25 30
Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser
35 40 45
Trp Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Gln Gly Arg Phe
50 55 60
Thr Val Ser Arg Asp Tyr Ala Lys Asn Thr Leu Tyr Leu Gln Met Asn
65 70 75 80
Ser Leu Lys Pro Glu Asp Ala Ala Val Tyr Tyr Cys Ala Gly Asp Pro
85 90 95
Trp Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 117
<211> 124
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 117
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Arg Arg Asn
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Leu
35 40 45
Ala Gly Ile Ser Trp Ser Gly Gly Thr Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Asp
65 70 75 80
Leu Gln Met Ile Ser Pro Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Gly Asp Ile Gly Thr Leu Val Asn Asp Glu Asn Pro Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 118
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 118
Asp Pro Trp Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp
1 5 10 15
<210> 119
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 119
Gly Ile Ser Trp Ser Gly Gly Thr Thr Tyr
1 5 10
<210> 120
<211> 274
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 120
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Asp Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly
180 185 190
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
195 200 205
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
210 215 220
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu
225 230 235 240
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr
245 250 255
Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val
260 265 270
Ser Ser
<210> 121
<211> 276
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 121
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ser Phe Ser Gly Pro Gly Arg Thr Phe Ala
20 25 30
Arg Tyr Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Asn Arg Asp
35 40 45
Phe Ile Thr Gly Ile Ser Gly Ser Gly Asp Ser Thr Tyr Tyr Val Tyr
50 55 60
Pro Met Lys Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Met
65 70 75 80
Val Tyr Leu Gln Met Asn Ala Leu Lys Pro Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Ala Asp Arg Glu Ile Asn Arg Ile Ala Asn Asp Lys Glu
100 105 110
Leu Asp Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly
115 120 125
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
145 150 155 160
Ser Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly
165 170 175
Asn Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser
180 185 190
Phe Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp
195 200 205
Val Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser
210 215 220
Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu
225 230 235 240
Tyr Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr
245 250 255
Cys Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val
260 265 270
Thr Val Ser Ser
275
<210> 122
<211> 274
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 122
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Val Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Leu Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr Tyr Val Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu His Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly
180 185 190
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
195 200 205
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
210 215 220
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu
225 230 235 240
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr
245 250 255
Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val
260 265 270
Ser Ser
<210> 123
<211> 274
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 123
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Arg Arg Asn
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Leu
35 40 45
Ala Gly Ile Ser Trp Ser Gly Gly Thr Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Asp
65 70 75 80
Leu Gln Met Ile Ser Pro Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Gly Asp Ile Gly Thr Leu Val Asn Asp Glu Asn Pro Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly
180 185 190
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
195 200 205
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
210 215 220
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu
225 230 235 240
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr
245 250 255
Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val
260 265 270
Ser Ser
<210> 124
<211> 274
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 124
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Ala Val Ser Val Asn
20 25 30
Ala Met Ala Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Asp Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Asp Arg Phe Thr Ile Ala Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly
180 185 190
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
195 200 205
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
210 215 220
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu
225 230 235 240
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr
245 250 255
Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val
260 265 270
Ser Ser
<210> 125
<211> 275
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 125
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Phe Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Arg Arg Thr Ile Ser Ser Gly
20 25 30
Thr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Arg Trp Ser Ser Gly Met Pro Tyr Tyr Leu Asp Ser Val
50 55 60
Met Asp Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Ser
65 70 75 80
Leu Gln Met Asn Ser Leu Gln Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Arg Ser Ala Phe Arg Asp Pro Ser Phe Asp Val Asn Tyr
100 105 110
Glu Tyr Trp Gly Arg Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly
115 120 125
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
130 135 140
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
145 150 155 160
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn
165 170 175
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
180 185 190
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
195 200 205
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
210 215 220
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
225 230 235 240
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
245 250 255
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
260 265 270
Val Ser Ser
275
<210> 126
<211> 429
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 126
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Val Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Leu Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr Tyr Val Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu His Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly Ser
165 170 175
Leu Arg Leu Ser Cys Val Ala Ser Gly Arg Tyr Pro Met Ala Trp Phe
180 185 190
Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser Trp
195 200 205
Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Gln Gly Arg Phe Thr
210 215 220
Val Ser Arg Asp Tyr Ala Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
225 230 235 240
Leu Lys Pro Glu Asp Ala Ala Val Tyr Tyr Cys Ala Gly Asp Pro Trp
245 250 255
Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp Trp Gly Gln Gly
260 265 270
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
275 280 285
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
290 295 300
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
305 310 315 320
Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser Leu Arg Leu Ser Cys
325 330 335
Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly Met Ser Trp Val Arg
340 345 350
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser
355 360 365
Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
370 375 380
Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu Gln Met Asn Ser Leu
385 390 395 400
Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr Ile Gly Gly Ser Leu
405 410 415
Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val Ser Ser
420 425
<210> 127
<211> 429
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 127
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ala Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Leu Thr Phe Arg Arg Asn
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Leu Leu
35 40 45
Ala Gly Ile Ser Trp Ser Gly Gly Thr Thr Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Val Asp
65 70 75 80
Leu Gln Met Ile Ser Pro Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Gly Asp Ile Gly Thr Leu Val Asn Asp Glu Asn Pro Arg
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly Ser
165 170 175
Leu Arg Leu Ser Cys Val Ala Ser Gly Arg Tyr Pro Met Ala Trp Phe
180 185 190
Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser Trp
195 200 205
Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Gln Gly Arg Phe Thr
210 215 220
Val Ser Arg Asp Tyr Ala Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
225 230 235 240
Leu Lys Pro Glu Asp Ala Ala Val Tyr Tyr Cys Ala Gly Asp Pro Trp
245 250 255
Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp Trp Gly Gln Gly
260 265 270
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
275 280 285
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
290 295 300
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
305 310 315 320
Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser Leu Arg Leu Ser Cys
325 330 335
Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly Met Ser Trp Val Arg
340 345 350
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser
355 360 365
Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
370 375 380
Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu Gln Met Asn Ser Leu
385 390 395 400
Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr Ile Gly Gly Ser Leu
405 410 415
Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val Ser Ser
420 425
<210> 128
<211> 429
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 128
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Ala Val Ser Val Asn
20 25 30
Ala Met Ala Trp Tyr Arg Gln Ala Pro Gly Lys Gln Arg Asp Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Gly Arg Thr Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Asp Arg Phe Thr Ile Ala Arg Asp Ser Ala Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Arg Leu Lys Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Asp Glu Ala Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Ser Val Gln Ala Gly Gly Ser
165 170 175
Leu Arg Leu Ser Cys Val Ala Ser Gly Arg Tyr Pro Met Ala Trp Phe
180 185 190
Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser Trp
195 200 205
Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Gln Gly Arg Phe Thr
210 215 220
Val Ser Arg Asp Tyr Ala Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
225 230 235 240
Leu Lys Pro Glu Asp Ala Ala Val Tyr Tyr Cys Ala Gly Asp Pro Trp
245 250 255
Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp Trp Gly Gln Gly
260 265 270
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
275 280 285
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
290 295 300
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
305 310 315 320
Ser Gly Gly Gly Leu Val Gln Pro Gly Asn Ser Leu Arg Leu Ser Cys
325 330 335
Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly Met Ser Trp Val Arg
340 345 350
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser
355 360 365
Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
370 375 380
Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu Gln Met Asn Ser Leu
385 390 395 400
Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys Thr Ile Gly Gly Ser Leu
405 410 415
Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val Ser Ser
420 425
<210> 129
<211> 275
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 129
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Val Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr Tyr Val Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly
180 185 190
Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser
195 200 205
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
210 215 220
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu
225 230 235 240
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Thr
245 250 255
Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val
260 265 270
Ser Ser Ala
275
<210> 130
<211> 430
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 130
Asp Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Val Ser Ser Tyr
20 25 30
Ala Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Gly Ile Ser Arg Ser Ala Glu Arg Thr Tyr Tyr Val Asp Ser Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ala Asp Leu Asp Pro Asn Arg Ile Phe Ser Arg Glu Glu Tyr Ala
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly
115 120 125
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
130 135 140
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu
145 150 155 160
Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly Ser
165 170 175
Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Tyr Pro Met Ala Trp Phe
180 185 190
Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val Ala Gly Val Ser Trp
195 200 205
Gly Gly Asp Arg Thr Tyr Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr
210 215 220
Ile Ser Arg Asp Tyr Ser Lys Asn Thr Leu Tyr Leu Gln Met Asn Ser
225 230 235 240
Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Ala Gly Asp Pro Phe
245 250 255
Gly Arg Leu Phe Arg Val Lys Asp Gln Tyr Ser Asp Trp Gly Gln Gly
260 265 270
Thr Leu Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
275 280 285
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
290 295 300
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Val Glu
305 310 315 320
Ser Gly Gly Gly Val Val Gln Pro Gly Asn Ser Leu Arg Leu Ser Cys
325 330 335
Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe Gly Met Ser Trp Val Arg
340 345 350
Gln Ala Pro Gly Lys Gly Leu Glu Trp Val Ser Ser Ile Ser Gly Ser
355 360 365
Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys Gly Arg Phe Thr Ile
370 375 380
Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr Leu Gln Met Asn Ser Leu
385 390 395 400
Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Thr Ile Gly Gly Ser Leu
405 410 415
Ser Arg Ser Ser Gln Gly Thr Leu Val Thr Val Ser Ser Ala
420 425 430
<210> 131
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 131
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 132
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 132
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 133
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 133
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala
115
<210> 134
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 134
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Thr Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala
115
<210> 135
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 135
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 136
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 136
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Lys
100 105 110
Val Ser Ser Ala
115
<210> 137
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 137
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 138
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 138
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala
115
<210> 139
<211> 117
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 139
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ala
115
<210> 140
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 140
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala Ala Ala
115
<210> 141
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 141
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Gly
115
<210> 142
<211> 117
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 142
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Gly Gly
115
<210> 143
<211> 118
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 143
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Asn
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Thr Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Gly Gly Gly
115
<210> 144
<211> 115
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 144
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser
115
<210> 145
<211> 116
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 145
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Phe
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ser Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Thr Ile Gly Gly Ser Leu Ser Arg Ser Ser Gln Gly Thr Leu Val Thr
100 105 110
Val Ser Ser Ala
115
<210> 146
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 146
Ser Phe Gly Met Ser
1 5
<210> 147
<211> 17
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 147
Ser Ile Ser Gly Ser Gly Ser Asp Thr Leu Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 148
<211> 6
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 148
Gly Gly Ser Leu Ser Arg
1 5
<210> 149
<211> 930
<212> PRT
<213> Homo sapiens
<220>
<223> Аминокислотная последовательность
<400> 149
Met Leu Leu Gly Trp Ala Ser Leu Leu Leu Cys Ala Phe Arg Leu Pro
1 5 10 15
Leu Ala Ala Val Gly Pro Ala Ala Thr Pro Ala Gln Asp Lys Ala Gly
20 25 30
Gln Pro Pro Thr Ala Ala Ala Ala Ala Gln Pro Arg Arg Arg Gln Gly
35 40 45
Glu Glu Val Gln Glu Arg Ala Glu Pro Pro Gly His Pro His Pro Leu
50 55 60
Ala Gln Arg Arg Arg Ser Lys Gly Leu Val Gln Asn Ile Asp Gln Leu
65 70 75 80
Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly Gly Arg
85 90 95
Arg Phe Leu Leu Asp Leu Glu Arg Asp Gly Ser Val Gly Ile Ala Gly
100 105 110
Phe Val Pro Ala Gly Gly Gly Thr Ser Ala Pro Trp Arg His Arg Ser
115 120 125
His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser Leu Ala
130 135 140
Val Phe Asp Leu Cys Gly Gly Leu Asp Gly Phe Phe Ala Val Lys His
145 150 155 160
Ala Arg Tyr Thr Leu Lys Pro Leu Leu Arg Gly Pro Trp Ala Glu Glu
165 170 175
Glu Lys Gly Arg Val Tyr Gly Asp Gly Ser Ala Arg Ile Leu His Val
180 185 190
Tyr Thr Arg Glu Gly Phe Ser Phe Glu Ala Leu Pro Pro Arg Ala Ser
195 200 205
Cys Glu Thr Pro Ala Ser Thr Pro Glu Ala His Glu His Ala Pro Ala
210 215 220
His Ser Asn Pro Ser Gly Arg Ala Ala Leu Ala Ser Gln Leu Leu Asp
225 230 235 240
Gln Ser Ala Leu Ser Pro Ala Gly Gly Ser Gly Pro Gln Thr Trp Trp
245 250 255
Arg Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln Val Glu Leu Leu
260 265 270
Leu Val Ala Asp Ala Ser Met Ala Arg Leu Tyr Gly Arg Gly Leu Gln
275 280 285
His Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Arg Leu Tyr Ser His
290 295 300
Ala Ser Ile Glu Asn His Ile Arg Leu Ala Val Val Lys Val Val Val
305 310 315 320
Leu Gly Asp Lys Asp Lys Ser Leu Glu Val Ser Lys Asn Ala Ala Thr
325 330 335
Thr Leu Lys Asn Phe Cys Lys Trp Gln His Gln His Asn Gln Leu Gly
340 345 350
Asp Asp His Glu Glu His Tyr Asp Ala Ala Ile Leu Phe Thr Arg Glu
355 360 365
Asp Leu Cys Gly His His Ser Cys Asp Thr Leu Gly Met Ala Asp Val
370 375 380
Gly Thr Ile Cys Ser Pro Glu Arg Ser Cys Ala Val Ile Glu Asp Asp
385 390 395 400
Gly Leu His Ala Ala Phe Thr Val Ala His Glu Ile Gly His Leu Leu
405 410 415
Gly Leu Ser His Asp Asp Ser Lys Phe Cys Glu Glu Thr Phe Gly Ser
420 425 430
Thr Glu Asp Lys Arg Leu Met Ser Ser Ile Leu Thr Ser Ile Asp Ala
435 440 445
Ser Lys Pro Trp Ser Lys Cys Thr Ser Ala Thr Ile Thr Glu Phe Leu
450 455 460
Asp Asp Gly His Gly Asn Cys Leu Leu Asp Leu Pro Arg Lys Gln Ile
465 470 475 480
Leu Gly Pro Glu Glu Leu Pro Gly Gln Thr Tyr Asp Ala Thr Gln Gln
485 490 495
Cys Asn Leu Thr Phe Gly Pro Glu Tyr Ser Val Cys Pro Gly Met Asp
500 505 510
Val Cys Ala Arg Leu Trp Cys Ala Val Val Arg Gln Gly Gln Met Val
515 520 525
Cys Leu Thr Lys Lys Leu Pro Ala Val Glu Gly Thr Pro Cys Gly Lys
530 535 540
Gly Arg Ile Cys Leu Gln Gly Lys Cys Val Asp Lys Thr Lys Lys Lys
545 550 555 560
Tyr Tyr Ser Thr Ser Ser His Gly Asn Trp Gly Ser Trp Gly Ser Trp
565 570 575
Gly Gln Cys Ser Arg Ser Cys Gly Gly Gly Val Gln Phe Ala Tyr Arg
580 585 590
His Cys Asn Asn Pro Ala Pro Arg Asn Asn Gly Arg Tyr Cys Thr Gly
595 600 605
Lys Arg Ala Ile Tyr Arg Ser Cys Ser Leu Met Pro Cys Pro Pro Asn
610 615 620
Gly Lys Ser Phe Arg His Glu Gln Cys Glu Ala Lys Asn Gly Tyr Gln
625 630 635 640
Ser Asp Ala Lys Gly Val Lys Thr Phe Val Glu Trp Val Pro Lys Tyr
645 650 655
Ala Gly Val Leu Pro Ala Asp Val Cys Lys Leu Thr Cys Arg Ala Lys
660 665 670
Gly Thr Gly Tyr Tyr Val Val Phe Ser Pro Lys Val Thr Asp Gly Thr
675 680 685
Glu Cys Arg Leu Tyr Ser Asn Ser Val Cys Val Arg Gly Lys Cys Val
690 695 700
Arg Thr Gly Cys Asp Gly Ile Ile Gly Ser Lys Leu Gln Tyr Asp Lys
705 710 715 720
Cys Gly Val Cys Gly Gly Asp Asn Ser Ser Cys Thr Lys Ile Val Gly
725 730 735
Thr Phe Asn Lys Lys Ser Lys Gly Tyr Thr Asp Val Val Arg Ile Pro
740 745 750
Glu Gly Ala Thr His Ile Lys Val Arg Gln Phe Lys Ala Lys Asp Gln
755 760 765
Thr Arg Phe Thr Ala Tyr Leu Ala Leu Lys Lys Lys Asn Gly Glu Tyr
770 775 780
Leu Ile Asn Gly Lys Tyr Met Ile Ser Thr Ser Glu Thr Ile Ile Asp
785 790 795 800
Ile Asn Gly Thr Val Met Asn Tyr Ser Gly Trp Ser His Arg Asp Asp
805 810 815
Phe Leu His Gly Met Gly Tyr Ser Ala Thr Lys Glu Ile Leu Ile Val
820 825 830
Gln Ile Leu Ala Thr Asp Pro Thr Lys Pro Leu Asp Val Arg Tyr Ser
835 840 845
Phe Phe Val Pro Lys Lys Ser Thr Pro Lys Val Asn Ser Val Thr Ser
850 855 860
His Gly Ser Asn Lys Val Gly Ser His Thr Ser Gln Pro Gln Trp Val
865 870 875 880
Thr Gly Pro Trp Leu Ala Cys Ser Arg Thr Cys Asp Thr Gly Trp His
885 890 895
Thr Arg Thr Val Gln Cys Gln Asp Gly Asn Arg Lys Leu Ala Lys Gly
900 905 910
Cys Pro Leu Ser Gln Arg Pro Ser Ala Phe Lys Gln Cys Leu Leu Lys
915 920 925
Lys Cys
930
<210> 150
<211> 636
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 150
Met Leu Leu Gly Trp Ala Ala Leu Met Leu Cys Ala Leu Arg Leu Pro
1 5 10 15
Pro Val Ala Ala Gly Pro Thr Ala Ala Pro Ala Gln Asp Lys Ala Gly
20 25 30
Gln Pro Arg Ala Ala Ala Val Ala Ala Ala Ala Gln Pro Arg Gly Arg
35 40 45
Arg Gly Glu Glu Ala Gln Glu Pro Ala Glu Pro Pro Gly His Pro His
50 55 60
Pro Leu Ala Pro Gln Arg Gly Ser Arg Gly Leu Val Gln Asn Ile Asp
65 70 75 80
Gln Leu Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly
85 90 95
Gly Arg Arg Phe Leu Leu Asp Leu Glu Arg Asp Asp Ser Val Gly Ala
100 105 110
Ala Gly Leu Val Pro Ala Gly Gly Gly Pro Asn Ala Thr Arg Arg His
115 120 125
Arg Gly His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser
130 135 140
Leu Ala Val Phe Asp Leu Cys Gly Gly Leu Asp Gly Phe Phe Ala Val
145 150 155 160
Lys Arg Ala Arg Tyr Thr Leu Gln Pro Leu Leu Arg Gly Pro Trp Ala
165 170 175
Glu Ala Glu Gly Asp Ala Arg Val Tyr Gly Asp Glu Ser Ala Arg Ile
180 185 190
Leu His Val Tyr Thr Arg Glu Gly Phe Ser Phe Glu Ala Leu Pro Pro
195 200 205
Arg Thr Ser Cys Glu Thr His Ala Ser Pro Pro Gly Ala Arg Glu Arg
210 215 220
Pro Pro Ala Pro Ser Arg Pro Asp Gly Arg Trp Ala Leu Ala Pro Gln
225 230 235 240
Gln Leu Pro Gly Gln Ser Ala Pro Ser Ser Asp Gly Ser Gln Gly Pro
245 250 255
Arg Thr Trp Trp Arg Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln
260 265 270
Val Glu Leu Leu Leu Val Ala Asp Ala Ser Met Ala Arg Met Tyr Gly
275 280 285
Arg Gly Leu Gln His Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Lys
290 295 300
Leu Tyr Ser His Ala Ser Ile Glu Asn His Ile Arg Leu Val Val Val
305 310 315 320
Lys Val Val Val Leu Gly Asp Lys Asp Lys Ser Leu Glu Val Ser Lys
325 330 335
Asn Ala Ala Thr Thr Leu Lys Asn Phe Cys Lys Trp Gln His Gln His
340 345 350
Asn Gln Leu Gly Asp Asp His Glu Glu His Tyr Asp Ala Ala Ile Leu
355 360 365
Phe Thr Arg Glu Asp Leu Cys Gly His His Ser Cys Asp Thr Leu Gly
370 375 380
Met Ala Asp Val Gly Thr Ile Cys Ser Pro Glu Arg Ser Cys Ala Val
385 390 395 400
Ile Glu Asp Asp Gly Leu His Ala Ala Phe Thr Val Ala His Glu Ile
405 410 415
Gly His Leu Leu Gly Leu Ser His Asp Asp Ser Lys Phe Cys Glu Glu
420 425 430
Asn Phe Gly Ser Thr Glu Asp Lys Arg Leu Met Ser Ser Ile Leu Thr
435 440 445
Ser Ile Asp Ala Ser Lys Pro Trp Ser Lys Cys Thr Ser Ala Thr Ile
450 455 460
Thr Glu Phe Leu Asp Asp Gly His Gly Asn Cys Leu Leu Asp Leu Pro
465 470 475 480
Arg Lys Gln Ile Pro Gly Pro Glu Glu Leu Pro Gly Gln Thr Tyr Asp
485 490 495
Ala Ser Gln Gln Cys Asn Leu Thr Phe Gly Pro Glu Tyr Ser Val Cys
500 505 510
Pro Gly Met Asp Val Cys Ala Arg Leu Trp Cys Ala Val Val Arg Gln
515 520 525
Gly Gln Met Val Cys Leu Thr Lys Lys Leu Pro Ala Val Glu Gly Thr
530 535 540
Pro Cys Gly Lys Gly Arg Ile Cys Leu Gln Gly Lys Cys Val Asp Lys
545 550 555 560
Thr Lys Lys Lys Tyr Tyr Ser Thr Ser Ser His Gly Asn Trp Gly Ser
565 570 575
Trp Gly Ser Trp Gly Gln Cys Ser Arg Ser Cys Gly Gly Gly Val Gln
580 585 590
Phe Ala Tyr Arg His Cys Asn Asn Pro Ala Pro Arg Asn Asn Gly Arg
595 600 605
Tyr Cys Thr Gly Lys Arg Ala Ile Tyr Arg Ser Cys Ser Val Thr Pro
610 615 620
Cys Pro His His His His His His His His His His
625 630 635
<210> 151
<211> 629
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 151
Met Arg Leu Glu Trp Ala Ser Leu Leu Leu Leu Leu Leu Leu Leu Cys
1 5 10 15
Ala Ser Cys Leu Ala Leu Ala Ala Asp Asn Pro Ala Ala Ala Pro Ala
20 25 30
Gln Asp Lys Thr Arg Gln Pro Arg Ala Ala Ala Ala Ala Ala Gln Pro
35 40 45
Asp Gln Arg Gln Trp Glu Glu Thr Gln Glu Arg Gly His Pro Gln Pro
50 55 60
Leu Ala Arg Gln Arg Arg Ser Ser Gly Leu Val Gln Asn Ile Asp Gln
65 70 75 80
Leu Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly Gly
85 90 95
Arg Arg Phe Leu Leu Asp Leu Glu Arg Asp Asp Thr Val Gly Ala Ala
100 105 110
Gly Gly Ile Val Thr Ala Gly Gly Leu Ser Ala Ser Ser Gly His Arg
115 120 125
Gly His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser Leu
130 135 140
Ala Val Phe Asp Leu Cys Gly Gly Leu Asp Gly Phe Phe Ala Val Lys
145 150 155 160
His Ala Arg Tyr Thr Leu Lys Pro Leu Leu Arg Gly Ser Trp Ala Glu
165 170 175
Ser Glu Arg Val Tyr Gly Asp Gly Ser Ser Arg Ile Leu His Val Tyr
180 185 190
Thr Arg Glu Gly Phe Ser Phe Glu Ala Leu Pro Pro Arg Thr Ser Cys
195 200 205
Glu Thr Pro Ala Ser Pro Ser Gly Ala Gln Glu Ser Pro Ser Val His
210 215 220
Ser Ser Ser Arg Arg Arg Thr Glu Leu Ala Pro Gln Leu Leu Asp His
225 230 235 240
Ser Ala Phe Ser Pro Ala Gly Asn Ala Gly Pro Gln Thr Trp Trp Arg
245 250 255
Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln Val Glu Leu Leu Leu
260 265 270
Val Ala Asp Ser Ser Met Ala Lys Met Tyr Gly Arg Gly Leu Gln His
275 280 285
Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Arg Leu Tyr Ser His Ala
290 295 300
Ser Ile Glu Asn His Ile Arg Leu Ala Val Val Lys Val Val Val Leu
305 310 315 320
Thr Asp Lys Ser Leu Glu Val Ser Lys Asn Ala Ala Thr Thr Leu Lys
325 330 335
Asn Phe Cys Lys Trp Gln His Gln His Asn Gln Leu Gly Asp Asp His
340 345 350
Glu Glu His Tyr Asp Ala Ala Ile Leu Phe Thr Arg Glu Asp Leu Cys
355 360 365
Gly His His Ser Cys Asp Thr Leu Gly Met Ala Asp Val Gly Thr Ile
370 375 380
Cys Ser Pro Glu Arg Ser Cys Ala Val Ile Glu Asp Asp Gly Leu His
385 390 395 400
Ala Ala Phe Thr Val Ala His Glu Ile Gly His Leu Leu Gly Leu Ser
405 410 415
His Asp Asp Ser Lys Phe Cys Glu Glu Asn Phe Gly Ser Thr Glu Asp
420 425 430
Lys Arg Leu Met Ser Ser Ile Leu Thr Ser Ile Asp Ala Ser Lys Pro
435 440 445
Trp Ser Lys Cys Thr Ser Ala Thr Ile Thr Glu Phe Leu Asp Asp Gly
450 455 460
His Gly Asn Cys Leu Leu Asp Val Pro Arg Lys Gln Ile Leu Gly Pro
465 470 475 480
Glu Glu Leu Pro Gly Gln Thr Tyr Asp Ala Thr Gln Gln Cys Asn Leu
485 490 495
Thr Phe Gly Pro Glu Tyr Ser Val Cys Pro Gly Met Asp Val Cys Ala
500 505 510
Arg Leu Trp Cys Ala Val Val Arg Gln Gly Gln Met Val Cys Leu Thr
515 520 525
Lys Lys Leu Pro Ala Val Glu Gly Thr Pro Cys Gly Lys Gly Arg Ile
530 535 540
Cys Leu Gln Gly Lys Cys Val Asp Lys Thr Lys Lys Lys Tyr Tyr Ser
545 550 555 560
Thr Ser Ser His Gly Asn Trp Gly Ser Trp Gly Pro Trp Gly Gln Cys
565 570 575
Ser Arg Ser Cys Gly Gly Gly Val Gln Phe Ala Tyr Arg His Cys Asn
580 585 590
Asn Pro Ala Pro Arg Asn Ser Gly Arg Tyr Cys Thr Gly Lys Arg Ala
595 600 605
Ile Tyr Arg Ser Cys Ser Val Ile Pro Cys Pro His His His His His
610 615 620
His His His His His
625
<210> 152
<211> 632
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 152
Met Leu Leu Gly Trp Ala Ser Leu Leu Leu Cys Ala Phe Arg Leu Pro
1 5 10 15
Gln Ala Ala Ala Ser Ala Ala Ala Ala Pro Ala Gln Asp Lys Ala Gly
20 25 30
Gln Pro Arg Ala Ala Ala Ala Ala Pro Gln Pro Arg Arg Arg Gln Gly
35 40 45
Glu His Ala Pro Leu Arg Val Glu Pro Pro Gly His Pro His Ala Leu
50 55 60
Ala Pro Gln Arg Arg Gly Arg Gly Leu Leu Gln Ser Ile Asp Arg Leu
65 70 75 80
Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly Gly Arg
85 90 95
Arg Phe Leu Leu Asp Leu Glu Arg Asp Gly Ser Val Gly Ala Ala Gly
100 105 110
Leu Phe Pro Ala Gly Gly Gly Leu Ser Ala Pro Arg Arg His Arg Ser
115 120 125
His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser Leu Ala
130 135 140
Val Phe Asp Leu Cys Gly Gly Leu Arg Gly Phe Phe Ala Val Lys His
145 150 155 160
Ala Arg Tyr Thr Val Lys Pro Leu Leu Arg Gly Pro Trp Ala Glu Ala
165 170 175
Asp Thr Pro Arg Val Tyr Gly Asp Glu Ser Ala Arg Ile Pro His Val
180 185 190
Tyr Thr Arg Glu Gly Phe Ser Phe Glu Ala Leu Pro Pro Arg Ala Ser
195 200 205
Cys Glu Thr Pro Ala Ser Gln Pro Gly Pro His Glu Arg Pro Pro Ala
210 215 220
His Asn Ser Pro Gly Arg His Ser Thr Val Asp Pro Gln Leu Pro Glu
225 230 235 240
Leu Ser Ala Leu Ser Pro Ala Gly Asp Pro Gly Gln Gln Ile Trp Trp
245 250 255
Arg Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln Val Glu Leu Leu
260 265 270
Leu Val Ala Asp Gly Ser Met Ala Lys Met Tyr Gly Arg Gly Leu Gln
275 280 285
His Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Arg Leu Tyr Ser His
290 295 300
Ala Ser Ile Glu Asn His Ile Arg Leu Ala Val Val Lys Val Val Val
305 310 315 320
Leu Gly Asp Lys Asp Lys Ser Leu Glu Val Ser Lys Asn Ala Ala Thr
325 330 335
Thr Leu Lys Asn Phe Cys Lys Trp Gln His Gln His Asn Gln Leu Gly
340 345 350
Asp Asp His Glu Glu His Tyr Asp Ala Ala Ile Leu Phe Thr Arg Glu
355 360 365
Asp Leu Cys Gly His His Ser Cys Asp Thr Leu Gly Met Ala Asp Val
370 375 380
Gly Thr Ile Cys Ser Pro Glu Arg Ser Cys Ala Val Ile Glu Asp Asp
385 390 395 400
Gly Leu His Ala Ala Phe Thr Val Ala His Glu Ile Gly His Leu Leu
405 410 415
Gly Leu Ser His Asp Asp Ser Lys Phe Cys Glu Glu Asn Phe Gly Leu
420 425 430
Thr Glu Asp Lys Arg Leu Met Ser Ser Ile Leu Thr Ser Ile Asp Ala
435 440 445
Ser Lys Pro Trp Ser Lys Cys Thr Ser Ala Thr Met Thr Glu Phe Leu
450 455 460
Asp Asp Gly His Gly Asn Cys Leu Leu Asp Val Pro Arg Lys Gln Ile
465 470 475 480
Pro Ser Pro Glu Glu Leu Pro Gly Gln Thr Tyr Asp Ala Thr Gln Gln
485 490 495
Cys Asn Leu Thr Phe Gly Pro Glu Tyr Ser Val Cys Pro Gly Met Asp
500 505 510
Val Cys Ala Arg Leu Trp Cys Ala Val Val Arg Gln Gly Gln Met Val
515 520 525
Cys Leu Thr Lys Lys Leu Pro Ala Val Glu Gly Thr Pro Cys Gly Lys
530 535 540
Gly Arg Ile Cys Leu Gln Gly Lys Cys Val Asp Lys Thr Lys Lys Lys
545 550 555 560
Tyr Tyr Ser Thr Ser Ser His Gly Asn Trp Gly Ser Trp Gly Pro Trp
565 570 575
Gly Gln Cys Ser Arg Ser Cys Gly Gly Gly Val Gln Phe Ala Tyr Arg
580 585 590
His Cys Asn Asn Pro Ala Pro Arg Asn Ser Gly Arg Tyr Cys Thr Gly
595 600 605
Lys Arg Ala Ile Tyr Arg Ser Cys Ser Val Thr Pro Cys Pro His His
610 615 620
His His His His His His His His
625 630
<210> 153
<211> 632
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 153
Met Arg Leu Glu Trp Ala Pro Leu Leu Leu Leu Leu Leu Leu Leu Ser
1 5 10 15
Ala Ser Cys Leu Ser Leu Ala Ala Asp Ser Pro Ala Ala Ala Pro Ala
20 25 30
Gln Asp Lys Thr Arg Gln Pro Gln Ala Ala Ala Ala Ala Ala Glu Pro
35 40 45
Asp Gln Pro Gln Gly Glu Glu Thr Arg Glu Arg Gly His Leu Gln Pro
50 55 60
Leu Ala Gly Gln Arg Arg Ser Gly Gly Leu Val Gln Asn Ile Asp Gln
65 70 75 80
Leu Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly Gly
85 90 95
Arg Arg Phe Leu Leu Asp Leu Glu Arg Asp Asp Thr Val Gly Ala Ala
100 105 110
Gly Ser Ile Val Thr Ala Gly Gly Gly Leu Ser Ala Ser Ser Gly His
115 120 125
Arg Gly His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser
130 135 140
Leu Ala Val Phe Asp Leu Cys Gly Gly Leu Asp Gly Phe Phe Ala Val
145 150 155 160
Lys His Ala Arg Tyr Thr Leu Lys Pro Leu Leu Arg Gly Ser Trp Ala
165 170 175
Glu Tyr Glu Arg Ile Tyr Gly Asp Gly Ser Ser Arg Ile Leu His Val
180 185 190
Tyr Asn Arg Glu Gly Phe Ser Phe Glu Ala Leu Pro Pro Arg Ala Ser
195 200 205
Cys Glu Thr Pro Ala Ser Pro Ser Gly Pro Gln Glu Ser Pro Ser Val
210 215 220
His Ser Arg Ser Arg Arg Arg Ser Ala Leu Ala Pro Gln Leu Leu Asp
225 230 235 240
His Ser Ala Phe Ser Pro Ser Gly Asn Ala Gly Pro Gln Thr Trp Trp
245 250 255
Arg Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln Val Glu Leu Leu
260 265 270
Leu Val Ala Asp Ser Ser Met Ala Arg Met Tyr Gly Arg Gly Leu Gln
275 280 285
His Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Arg Leu Tyr Ser His
290 295 300
Ala Ser Ile Glu Asn His Ile Arg Leu Ala Val Val Lys Val Val Val
305 310 315 320
Leu Thr Asp Lys Asp Thr Ser Leu Glu Val Ser Lys Asn Ala Ala Thr
325 330 335
Thr Leu Lys Asn Phe Cys Lys Trp Gln His Gln His Asn Gln Leu Gly
340 345 350
Asp Asp His Glu Glu His Tyr Asp Ala Ala Ile Leu Phe Thr Arg Glu
355 360 365
Asp Leu Cys Gly His His Ser Cys Asp Thr Leu Gly Met Ala Asp Val
370 375 380
Gly Thr Ile Cys Ser Pro Glu Arg Ser Cys Ala Val Ile Glu Asp Asp
385 390 395 400
Gly Leu His Ala Ala Phe Thr Val Ala His Glu Ile Gly His Leu Leu
405 410 415
Gly Leu Ser His Asp Asp Ser Lys Phe Cys Glu Glu Asn Phe Gly Thr
420 425 430
Thr Glu Asp Lys Arg Leu Met Ser Ser Ile Leu Thr Ser Ile Asp Ala
435 440 445
Ser Lys Pro Trp Ser Lys Cys Thr Ser Ala Thr Ile Thr Glu Phe Leu
450 455 460
Asp Asp Gly His Gly Asn Cys Leu Leu Asp Leu Pro Arg Lys Gln Ile
465 470 475 480
Leu Gly Pro Glu Glu Leu Pro Gly Gln Thr Tyr Asp Ala Thr Gln Gln
485 490 495
Cys Asn Leu Thr Phe Gly Pro Glu Tyr Ser Val Cys Pro Gly Met Asp
500 505 510
Val Cys Ala Arg Leu Trp Cys Ala Val Val Arg Gln Gly Gln Met Val
515 520 525
Cys Leu Thr Lys Lys Leu Pro Ala Val Glu Gly Thr Pro Cys Gly Lys
530 535 540
Gly Arg Val Cys Leu Gln Gly Lys Cys Val Asp Lys Thr Lys Lys Lys
545 550 555 560
Tyr Tyr Ser Thr Ser Ser His Gly Asn Trp Gly Ser Trp Gly Pro Trp
565 570 575
Gly Gln Cys Ser Arg Ser Cys Gly Gly Gly Val Gln Phe Ala Tyr Arg
580 585 590
His Cys Asn Asn Pro Ala Pro Arg Asn Ser Gly Arg Tyr Cys Thr Gly
595 600 605
Lys Arg Ala Ile Tyr Arg Ser Cys Ser Val Thr Pro Cys Pro His His
610 615 620
His His His His His His His His
625 630
<210> 154
<211> 632
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 154
Met Leu Leu Gly Trp Ala Ser Leu Leu Leu Cys Ala Phe Arg Leu Pro
1 5 10 15
Leu Ala Ala Ala Gly Pro Ala Ala Ala Pro Ala Gln Asp Lys Ala Gly
20 25 30
Gln Pro Ala Thr Ala Ala Ala Ala Ala Gln Pro Arg Arg Arg Gln Gly
35 40 45
Glu Glu Val Gln Glu Arg Thr Glu Pro Pro Gly His Pro His Pro Leu
50 55 60
Ala Gln Arg Arg Ser Ser Lys Gly Leu Val Gln Asn Ile Asp Gln Leu
65 70 75 80
Tyr Ser Gly Gly Gly Lys Val Gly Tyr Leu Val Tyr Ala Gly Gly Arg
85 90 95
Arg Phe Leu Leu Asp Leu Glu Arg Asp Gly Ser Val Gly Thr Ala Gly
100 105 110
Phe Val Pro Thr Glu Gly Gly Thr Ser Ala Pro Trp Arg His Arg Ser
115 120 125
His Cys Phe Tyr Arg Gly Thr Val Asp Gly Ser Pro Arg Ser Leu Ala
130 135 140
Val Phe Asp Leu Cys Gly Gly Leu Asp Gly Phe Phe Ala Val Lys His
145 150 155 160
Ala Arg Tyr Thr Leu Lys Pro Leu Leu Arg Gly Pro Trp Ala Glu Glu
165 170 175
Glu Thr Arg Arg Val Tyr Gly Asp Gly Ser Ala Arg Ile Leu His Val
180 185 190
Tyr Thr Arg Glu Gly Phe Ser Phe Glu Ala Leu Gln Pro Arg Ala Ser
195 200 205
Cys Glu Thr Pro Ala Ser Thr Pro Glu Pro His Glu Arg Pro Pro Ala
210 215 220
His Ser Asn Pro Gly Gly Arg Ala Ala Leu Ala Ser Gln Leu Leu Asp
225 230 235 240
Gln Ser Ala Val Ser Pro Ala Gly Gly Pro Gly Pro Gln Thr Trp Trp
245 250 255
Arg Arg Arg Arg Arg Ser Ile Ser Arg Ala Arg Gln Val Glu Leu Leu
260 265 270
Leu Val Ala Asp Ala Ser Met Ala Arg Leu Tyr Gly Arg Gly Leu Gln
275 280 285
His Tyr Leu Leu Thr Leu Ala Ser Ile Ala Asn Arg Leu Tyr Ser His
290 295 300
Ala Ser Ile Glu Asn His Ile Arg Leu Ala Val Val Lys Val Val Val
305 310 315 320
Leu Gly Asp Lys Asp Lys Ser Leu Glu Val Ser Lys Asn Ala Ala Thr
325 330 335
Thr Leu Lys Asn Phe Cys Lys Trp Gln His Gln His Asn Gln Leu Gly
340 345 350
Asp Asp His Glu Glu His Tyr Asp Ala Ala Ile Leu Phe Thr Arg Glu
355 360 365
Asp Leu Cys Gly His His Ser Cys Asp Thr Leu Gly Met Ala Asp Val
370 375 380
Gly Thr Ile Cys Ser Pro Glu Arg Ser Cys Ala Val Ile Glu Asp Asp
385 390 395 400
Gly Leu His Ala Ala Phe Thr Val Ala His Glu Ile Gly His Leu Leu
405 410 415
Gly Leu Ser His Asp Asp Ser Lys Phe Cys Glu Glu Thr Phe Gly Ser
420 425 430
Thr Glu Asp Lys Arg Leu Met Ser Ser Ile Leu Thr Ser Ile Asp Ala
435 440 445
Ser Lys Pro Trp Ser Lys Cys Thr Ser Ala Thr Ile Thr Glu Phe Leu
450 455 460
Asp Asp Gly His Gly Asn Cys Leu Leu Asp Gln Pro Arg Lys Gln Ile
465 470 475 480
Leu Gly Pro Glu Glu Leu Pro Gly Gln Thr Tyr Asp Ala Thr Gln Gln
485 490 495
Cys Asn Leu Thr Phe Gly Pro Glu Tyr Ser Val Cys Pro Gly Met Asp
500 505 510
Val Cys Ala Arg Leu Trp Cys Ala Val Val Arg Gln Gly Gln Met Val
515 520 525
Cys Leu Thr Lys Lys Leu Pro Ala Val Glu Gly Thr Pro Cys Gly Lys
530 535 540
Gly Arg Ile Cys Leu Gln Gly Lys Cys Val Asp Lys Thr Lys Lys Lys
545 550 555 560
Tyr Tyr Ser Thr Ser Ser His Gly Asn Trp Gly Ser Trp Gly Ser Trp
565 570 575
Gly Gln Cys Ser Arg Ser Cys Gly Gly Gly Val Gln Phe Ala Tyr Arg
580 585 590
His Cys Asn Asn Pro Ala Pro Arg Asn Asn Gly Arg Tyr Cys Thr Gly
595 600 605
Lys Arg Ala Ile Tyr Arg Ser Cys Gly Leu Met Pro Cys Pro His His
610 615 620
His His His His His His His His
625 630
<210> 155
<211> 2415
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 155
Met Thr Thr Leu Leu Trp Val Phe Val Thr Leu Arg Val Ile Thr Ala
1 5 10 15
Ala Val Thr Val Glu Thr Ser Asp His Asp Asn Ser Leu Ser Val Ser
20 25 30
Ile Pro Gln Pro Ser Pro Leu Arg Val Leu Leu Gly Thr Ser Leu Thr
35 40 45
Ile Pro Cys Tyr Phe Ile Asp Pro Met His Pro Val Thr Thr Ala Pro
50 55 60
Ser Thr Ala Pro Leu Ala Pro Arg Ile Lys Trp Ser Arg Val Ser Lys
65 70 75 80
Glu Lys Glu Val Val Leu Leu Val Ala Thr Glu Gly Arg Val Arg Val
85 90 95
Asn Ser Ala Tyr Gln Asp Lys Val Ser Leu Pro Asn Tyr Pro Ala Ile
100 105 110
Pro Ser Asp Ala Thr Leu Glu Val Gln Ser Leu Arg Ser Asn Asp Ser
115 120 125
Gly Val Tyr Arg Cys Glu Val Met His Gly Ile Glu Asp Ser Glu Ala
130 135 140
Thr Leu Glu Val Val Val Lys Gly Ile Val Phe His Tyr Arg Ala Ile
145 150 155 160
Ser Thr Arg Tyr Thr Leu Asp Phe Asp Arg Ala Gln Arg Ala Cys Leu
165 170 175
Gln Asn Ser Ala Ile Ile Ala Thr Pro Glu Gln Leu Gln Ala Ala Tyr
180 185 190
Glu Asp Gly Phe His Gln Cys Asp Ala Gly Trp Leu Ala Asp Gln Thr
195 200 205
Val Arg Tyr Pro Ile His Thr Pro Arg Glu Gly Cys Tyr Gly Asp Lys
210 215 220
Asp Glu Phe Pro Gly Val Arg Thr Tyr Gly Ile Arg Asp Thr Asn Glu
225 230 235 240
Thr Tyr Asp Val Tyr Cys Phe Ala Glu Glu Met Glu Gly Glu Val Phe
245 250 255
Tyr Ala Thr Ser Pro Glu Lys Phe Thr Phe Gln Glu Ala Ala Asn Glu
260 265 270
Cys Arg Arg Leu Gly Ala Arg Leu Ala Thr Thr Gly His Val Tyr Leu
275 280 285
Ala Trp Gln Ala Gly Met Asp Met Cys Ser Ala Gly Trp Leu Ala Asp
290 295 300
Arg Ser Val Arg Tyr Pro Ile Ser Lys Ala Arg Pro Asn Cys Gly Gly
305 310 315 320
Asn Leu Leu Gly Val Arg Thr Val Tyr Val His Ala Asn Gln Thr Gly
325 330 335
Tyr Pro Asp Pro Ser Ser Arg Tyr Asp Ala Ile Cys Tyr Thr Gly Glu
340 345 350
Asp Phe Val Asp Ile Pro Glu Asn Phe Phe Gly Val Gly Gly Glu Glu
355 360 365
Asp Ile Thr Val Gln Thr Val Thr Trp Pro Asp Met Glu Leu Pro Leu
370 375 380
Pro Arg Asn Ile Thr Glu Gly Glu Ala Arg Gly Ser Val Ile Leu Thr
385 390 395 400
Val Lys Pro Ile Phe Glu Val Ser Pro Ser Pro Leu Glu Pro Glu Glu
405 410 415
Pro Phe Thr Phe Ala Pro Glu Ile Gly Ala Thr Ala Phe Ala Glu Val
420 425 430
Glu Asn Glu Thr Gly Glu Ala Thr Arg Pro Trp Gly Phe Pro Thr Pro
435 440 445
Gly Leu Gly Pro Ala Thr Ala Phe Thr Ser Glu Asp Leu Val Val Gln
450 455 460
Val Thr Ala Val Pro Gly Gln Pro His Leu Pro Gly Gly Val Val Phe
465 470 475 480
His Tyr Arg Pro Gly Pro Thr Arg Tyr Ser Leu Thr Phe Glu Glu Ala
485 490 495
Gln Gln Ala Cys Pro Gly Thr Gly Ala Val Ile Ala Ser Pro Glu Gln
500 505 510
Leu Gln Ala Ala Tyr Glu Ala Gly Tyr Glu Gln Cys Asp Ala Gly Trp
515 520 525
Leu Arg Asp Gln Thr Val Arg Tyr Pro Ile Val Ser Pro Arg Thr Pro
530 535 540
Cys Val Gly Asp Lys Asp Ser Ser Pro Gly Val Arg Thr Tyr Gly Val
545 550 555 560
Arg Pro Ser Thr Glu Thr Tyr Asp Val Tyr Cys Phe Val Asp Arg Leu
565 570 575
Glu Gly Glu Val Phe Phe Ala Thr Arg Leu Glu Gln Phe Thr Phe Gln
580 585 590
Glu Ala Leu Glu Phe Cys Glu Ser His Asn Ala Thr Ala Thr Thr Gly
595 600 605
Gln Leu Tyr Ala Ala Trp Ser Arg Gly Leu Asp Lys Cys Tyr Ala Gly
610 615 620
Trp Leu Ala Asp Gly Ser Leu Arg Tyr Pro Ile Val Thr Pro Arg Pro
625 630 635 640
Ala Cys Gly Gly Asp Lys Pro Gly Val Arg Thr Val Tyr Leu Tyr Pro
645 650 655
Asn Gln Thr Gly Leu Pro Asp Pro Leu Ser Arg His His Ala Phe Cys
660 665 670
Phe Arg Gly Ile Ser Ala Val Pro Ser Pro Gly Glu Glu Glu Gly Gly
675 680 685
Thr Pro Thr Ser Pro Ser Gly Val Glu Glu Trp Ile Val Thr Gln Val
690 695 700
Val Pro Gly Val Ala Ala Val Pro Val Glu Glu Glu Thr Thr Ala Val
705 710 715 720
Pro Ser Gly Glu Thr Thr Ala Ile Leu Glu Phe Thr Thr Glu Pro Glu
725 730 735
Asn Gln Thr Glu Trp Glu Pro Ala Tyr Thr Pro Val Gly Thr Ser Pro
740 745 750
Leu Pro Gly Ile Leu Pro Thr Trp Pro Pro Thr Gly Ala Glu Thr Glu
755 760 765
Glu Ser Thr Glu Gly Pro Ser Ala Thr Glu Val Pro Ser Ala Ser Glu
770 775 780
Glu Pro Ser Pro Ser Glu Val Pro Phe Pro Ser Glu Glu Pro Ser Pro
785 790 795 800
Ser Glu Glu Pro Phe Pro Ser Val Arg Pro Phe Pro Ser Val Glu Leu
805 810 815
Phe Pro Ser Glu Glu Pro Phe Pro Ser Lys Glu Pro Ser Pro Ser Glu
820 825 830
Glu Pro Ser Ala Ser Glu Glu Pro Tyr Thr Pro Ser Pro Pro Glu Pro
835 840 845
Ser Trp Thr Glu Leu Pro Ser Ser Gly Glu Glu Ser Gly Ala Pro Asp
850 855 860
Val Ser Gly Asp Phe Thr Gly Ser Gly Asp Val Ser Gly His Leu Asp
865 870 875 880
Phe Ser Gly Gln Leu Ser Gly Asp Arg Ala Ser Gly Leu Pro Ser Gly
885 890 895
Asp Leu Asp Ser Ser Gly Leu Thr Ser Thr Val Gly Ser Gly Leu Thr
900 905 910
Val Glu Ser Gly Leu Pro Ser Gly Asp Glu Glu Arg Ile Glu Trp Pro
915 920 925
Ser Thr Pro Thr Val Gly Glu Leu Pro Ser Gly Ala Glu Ile Leu Glu
930 935 940
Gly Ser Ala Ser Gly Val Gly Asp Leu Ser Gly Leu Pro Ser Gly Glu
945 950 955 960
Val Leu Glu Thr Ser Ala Ser Gly Val Gly Asp Leu Ser Gly Leu Pro
965 970 975
Ser Gly Glu Val Leu Glu Thr Thr Ala Pro Gly Val Glu Asp Ile Ser
980 985 990
Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Thr Ala Pro Gly Val Glu
995 1000 1005
Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Thr Ala Pro
1010 1015 1020
Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr
1025 1030 1035 1040
Thr Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val
1045 1050 1055
Leu Glu Thr Thr Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser
1060 1065 1070
Gly Glu Val Leu Glu Thr Ala Ala Pro Gly Val Glu Asp Ile Ser Gly
1075 1080 1085
Leu Pro Ser Gly Glu Val Leu Glu Thr Ala Ala Pro Gly Val Glu Asp
1090 1095 1100
Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Ala Ala Pro Gly
1105 1110 1115 1120
Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Ala
1125 1130 1135
Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu
1140 1145 1150
Glu Thr Ala Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly
1155 1160 1165
Glu Val Leu Glu Thr Ala Ala Pro Gly Val Glu Asp Ile Ser Gly Leu
1170 1175 1180
Pro Ser Gly Glu Val Leu Glu Thr Ala Ala Pro Gly Val Glu Asp Ile
1185 1190 1195 1200
Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Ala Ala Pro Gly Val
1205 1210 1215
Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Ala Ala
1220 1225 1230
Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu
1235 1240 1245
Thr Ala Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro Ser Gly Glu
1250 1255 1260
Val Leu Glu Thr Ala Ala Pro Gly Val Glu Asp Ile Ser Gly Leu Pro
1265 1270 1275 1280
Ser Gly Glu Val Leu Glu Thr Thr Ala Pro Gly Val Glu Glu Ile Ser
1285 1290 1295
Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Thr Ala Pro Gly Val Asp
1300 1305 1310
Glu Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr Thr Ala Pro
1315 1320 1325
Gly Val Glu Glu Ile Ser Gly Leu Pro Ser Gly Glu Val Leu Glu Thr
1330 1335 1340
Ser Thr Ser Ala Val Gly Asp Leu Ser Gly Leu Pro Ser Gly Gly Glu
1345 1350 1355 1360
Val Leu Glu Ile Ser Val Ser Gly Val Glu Asp Ile Ser Gly Leu Pro
1365 1370 1375
Ser Gly Glu Val Val Glu Thr Ser Ala Ser Gly Ile Glu Asp Val Ser
1380 1385 1390
Glu Leu Pro Ser Gly Glu Gly Leu Glu Thr Ser Ala Ser Gly Val Glu
1395 1400 1405
Asp Leu Ser Arg Leu Pro Ser Gly Glu Glu Val Leu Glu Ile Ser Ala
1410 1415 1420
Ser Gly Phe Gly Asp Leu Ser Gly Val Pro Ser Gly Gly Glu Gly Leu
1425 1430 1435 1440
Glu Thr Ser Ala Ser Glu Val Gly Thr Asp Leu Ser Gly Leu Pro Ser
1445 1450 1455
Gly Arg Glu Gly Leu Glu Thr Ser Ala Ser Gly Ala Glu Asp Leu Ser
1460 1465 1470
Gly Leu Pro Ser Gly Lys Glu Asp Leu Val Gly Ser Ala Ser Gly Asp
1475 1480 1485
Leu Asp Leu Gly Lys Leu Pro Ser Gly Thr Leu Gly Ser Gly Gln Ala
1490 1495 1500
Pro Glu Thr Ser Gly Leu Pro Ser Gly Phe Ser Gly Glu Tyr Ser Gly
1505 1510 1515 1520
Val Asp Leu Gly Ser Gly Pro Pro Ser Gly Leu Pro Asp Phe Ser Gly
1525 1530 1535
Leu Pro Ser Gly Phe Pro Thr Val Ser Leu Val Asp Ser Thr Leu Val
1540 1545 1550
Glu Val Val Thr Ala Ser Thr Ala Ser Glu Leu Glu Gly Arg Gly Thr
1555 1560 1565
Ile Gly Ile Ser Gly Ala Gly Glu Ile Ser Gly Leu Pro Ser Ser Glu
1570 1575 1580
Leu Asp Ile Ser Gly Arg Ala Ser Gly Leu Pro Ser Gly Thr Glu Leu
1585 1590 1595 1600
Ser Gly Gln Ala Ser Gly Ser Pro Asp Val Ser Gly Glu Ile Pro Gly
1605 1610 1615
Leu Phe Gly Val Ser Gly Gln Pro Ser Gly Phe Pro Asp Thr Ser Gly
1620 1625 1630
Glu Thr Ser Gly Val Thr Glu Leu Ser Gly Leu Ser Ser Gly Gln Pro
1635 1640 1645
Gly Val Ser Gly Glu Ala Ser Gly Val Leu Tyr Gly Thr Ser Gln Pro
1650 1655 1660
Phe Gly Ile Thr Asp Leu Ser Gly Glu Thr Ser Gly Val Pro Asp Leu
1665 1670 1675 1680
Ser Gly Gln Pro Ser Gly Leu Pro Gly Phe Ser Gly Ala Thr Ser Gly
1685 1690 1695
Val Pro Asp Leu Val Ser Gly Thr Thr Ser Gly Ser Gly Glu Ser Ser
1700 1705 1710
Gly Ile Thr Phe Val Asp Thr Ser Leu Val Glu Val Ala Pro Thr Thr
1715 1720 1725
Phe Lys Glu Glu Glu Gly Leu Gly Ser Val Glu Leu Ser Gly Leu Pro
1730 1735 1740
Ser Gly Glu Ala Asp Leu Ser Gly Lys Ser Gly Met Val Asp Val Ser
1745 1750 1755 1760
Gly Gln Phe Ser Gly Thr Val Asp Ser Ser Gly Phe Thr Ser Gln Thr
1765 1770 1775
Pro Glu Phe Ser Gly Leu Pro Ser Gly Ile Ala Glu Val Ser Gly Glu
1780 1785 1790
Ser Ser Arg Ala Glu Ile Gly Ser Ser Leu Pro Ser Gly Ala Tyr Tyr
1795 1800 1805
Gly Ser Gly Thr Pro Ser Ser Phe Pro Thr Val Ser Leu Val Asp Arg
1810 1815 1820
Thr Leu Val Glu Ser Val Thr Gln Ala Pro Thr Ala Gln Glu Ala Gly
1825 1830 1835 1840
Glu Gly Pro Ser Gly Ile Leu Glu Leu Ser Gly Ala His Ser Gly Ala
1845 1850 1855
Pro Asp Met Ser Gly Glu His Ser Gly Phe Leu Asp Leu Ser Gly Leu
1860 1865 1870
Gln Ser Gly Leu Ile Glu Pro Ser Gly Glu Pro Pro Gly Thr Pro Tyr
1875 1880 1885
Phe Ser Gly Asp Phe Ala Ser Thr Thr Asn Val Ser Gly Glu Ser Ser
1890 1895 1900
Val Ala Met Gly Thr Ser Gly Glu Ala Ser Gly Leu Pro Glu Val Thr
1905 1910 1915 1920
Leu Ile Thr Ser Glu Phe Val Glu Gly Val Thr Glu Pro Thr Ile Ser
1925 1930 1935
Gln Glu Leu Gly Gln Arg Pro Pro Val Thr His Thr Pro Gln Leu Phe
1940 1945 1950
Glu Ser Ser Gly Lys Val Ser Thr Ala Gly Asp Ile Ser Gly Ala Thr
1955 1960 1965
Pro Val Leu Pro Gly Ser Gly Val Glu Val Ser Ser Val Pro Glu Ser
1970 1975 1980
Ser Ser Glu Thr Ser Ala Tyr Pro Glu Ala Gly Phe Gly Ala Ser Ala
1985 1990 1995 2000
Ala Pro Glu Ala Ser Arg Glu Asp Ser Gly Ser Pro Asp Leu Ser Glu
2005 2010 2015
Thr Thr Ser Ala Phe His Glu Ala Asn Leu Glu Arg Ser Ser Gly Leu
2020 2025 2030
Gly Val Ser Gly Ser Thr Leu Thr Phe Gln Glu Gly Glu Ala Ser Ala
2035 2040 2045
Ala Pro Glu Val Ser Gly Glu Ser Thr Thr Thr Ser Asp Val Gly Thr
2050 2055 2060
Glu Ala Pro Gly Leu Pro Ser Ala Thr Pro Thr Ala Ser Gly Asp Arg
2065 2070 2075 2080
Thr Glu Ile Ser Gly Asp Leu Ser Gly His Thr Ser Gln Leu Gly Val
2085 2090 2095
Val Ile Ser Thr Ser Ile Pro Glu Ser Glu Trp Thr Gln Gln Thr Gln
2100 2105 2110
Arg Pro Ala Glu Thr His Leu Glu Ile Glu Ser Ser Ser Leu Leu Tyr
2115 2120 2125
Ser Gly Glu Glu Thr His Thr Val Glu Thr Ala Thr Ser Pro Thr Asp
2130 2135 2140
Ala Ser Ile Pro Ala Ser Pro Glu Trp Lys Arg Glu Ser Glu Ser Thr
2145 2150 2155 2160
Ala Ala Ala Pro Ala Arg Ser Cys Ala Glu Glu Pro Cys Gly Ala Gly
2165 2170 2175
Thr Cys Lys Glu Thr Glu Gly His Val Ile Cys Leu Cys Pro Pro Gly
2180 2185 2190
Tyr Thr Gly Glu His Cys Asn Ile Asp Gln Glu Val Cys Glu Glu Gly
2195 2200 2205
Trp Asn Lys Tyr Gln Gly His Cys Tyr Arg His Phe Pro Asp Arg Glu
2210 2215 2220
Thr Trp Val Asp Ala Glu Arg Arg Cys Arg Glu Gln Gln Ser His Leu
2225 2230 2235 2240
Ser Ser Ile Val Thr Pro Glu Glu Gln Glu Phe Val Asn Asn Asn Ala
2245 2250 2255
Gln Asp Tyr Gln Trp Ile Gly Leu Asn Asp Arg Thr Ile Glu Gly Asp
2260 2265 2270
Phe Arg Trp Ser Asp Gly His Pro Met Gln Phe Glu Asn Trp Arg Pro
2275 2280 2285
Asn Gln Pro Asp Asn Phe Phe Ala Ala Gly Glu Asp Cys Val Val Met
2290 2295 2300
Ile Trp His Glu Lys Gly Glu Trp Asn Asp Val Pro Cys Asn Tyr His
2305 2310 2315 2320
Leu Pro Phe Thr Cys Lys Lys Gly Thr Val Ala Cys Gly Glu Pro Pro
2325 2330 2335
Val Val Glu His Ala Arg Thr Phe Gly Gln Lys Lys Asp Arg Tyr Glu
2340 2345 2350
Ile Asn Ser Leu Val Arg Tyr Gln Cys Thr Glu Gly Phe Val Gln Arg
2355 2360 2365
His Met Pro Thr Ile Arg Cys Gln Pro Ser Gly His Trp Glu Glu Pro
2370 2375 2380
Arg Ile Thr Cys Thr Asp Ala Thr Thr Tyr Lys Arg Arg Leu Gln Lys
2385 2390 2395 2400
Arg Ser Ser Arg His Pro Arg Arg Ser Arg Pro Ser Thr Ala His
2405 2410 2415
<210> 156
<211> 122
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 156
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Ser Thr Phe Ile Ile Asn
20 25 30
Val Val Arg Trp Tyr Arg Arg Ala Pro Gly Lys Gln Arg Glu Leu Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Asn Ala Asn Tyr Val Asp Ser Val Arg
50 55 60
Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys Asn
85 90 95
Val Pro Thr Thr His Tyr Gly Gly Val Tyr Tyr Gly Pro Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala
115 120
<210> 157
<211> 125
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 157
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Arg Thr Phe Ser Ser Tyr
20 25 30
Thr Met Gly Trp Phe Arg Gln Ala Pro Gly Lys Glu Arg Glu Phe Val
35 40 45
Ala Ala Ile Ser Trp Ser Gly Gly Arg Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Val Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Pro Glu Asp Thr Ala Leu Tyr Tyr Cys
85 90 95
Ala Ala Tyr Arg Arg Arg Arg Ala Ser Ser Asn Arg Gly Leu Trp Asp
100 105 110
Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala
115 120 125
<210> 158
<400> 158
000
<210> 159
<211> 5
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 159
Gly Gly Gly Gly Ser
1 5
<210> 160
<211> 7
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 160
Ser Gly Gly Ser Gly Gly Ser
1 5
<210> 161
<211> 8
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 161
Gly Gly Gly Gly Gly Gly Gly Ser
1 5
<210> 162
<211> 9
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 162
Gly Gly Gly Gly Ser Gly Gly Gly Ser
1 5
<210> 163
<211> 10
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 163
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 164
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 164
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 165
<211> 18
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 165
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Gly Gly
1 5 10 15
Gly Ser
<210> 166
<211> 20
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 166
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 167
<211> 25
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 167
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 168
<211> 30
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 168
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25 30
<210> 169
<211> 35
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 169
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser
35
<210> 170
<211> 40
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 170
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
20 25 30
Gly Gly Ser Gly Gly Gly Gly Ser
35 40
<210> 171
<211> 15
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 171
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
<210> 172
<211> 24
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 172
Gly Gly Gly Gly Ser Gly Gly Gly Ser Glu Pro Lys Ser Cys Asp Lys
1 5 10 15
Thr His Thr Cys Pro Pro Cys Pro
20
<210> 173
<211> 12
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 173
Glu Pro Lys Thr Pro Lys Pro Gln Pro Ala Ala Ala
1 5 10
<210> 174
<211> 62
<212> PRT
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность
<400> 174
Glu Leu Lys Thr Pro Leu Gly Asp Thr Thr His Thr Cys Pro Arg Cys
1 5 10 15
Pro Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
20 25 30
Glu Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro Glu
35 40 45
Pro Lys Ser Cys Asp Thr Pro Pro Pro Cys Pro Arg Cys Pro
50 55 60
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИПЕПТИДЫ, СВЯЗЫВАЮЩИЕСЯ С ADAMTS5, MMP13 И АГГРЕКАНОМ | 2018 |
|
RU2786659C2 |
| ММР13-СВЯЗЫВАЮЩИЕ ИММУНОГЛОБУЛИНЫ | 2018 |
|
RU2784069C2 |
| УСОВЕРШЕНСТВОВАННЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ СЫВОРОТОЧНЫЙ АЛЬБУМИН | 2018 |
|
RU2797270C2 |
| УСОВЕРШЕНСТВОВАННЫЕ СВЯЗЫВАЮЩИЕСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ ВЕЩЕСТВА | 2018 |
|
RU2789495C2 |
| УСОВЕРШЕНСТВОВАННЫЕ АГЕНТЫ, СВЯЗЫВАЮЩИЕ TNF | 2016 |
|
RU2774823C2 |
| УЛУЧШЕННЫЕ ОДИНОЧНЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНА, СВЯЗЫВАЮЩИЕСЯ С СЫВОРОТОЧНЫМ АЛЬБУМИНОМ | 2017 |
|
RU2765384C2 |
| PD1/CTLA4-СВЯЗЫВАЮЩИЕ ВЕЩЕСТВА | 2016 |
|
RU2755724C2 |
| УЛУЧШЕННЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНА | 2015 |
|
RU2809788C2 |
| УЛУЧШЕННЫЕ ВАРИАБЕЛЬНЫЕ ДОМЕНЫ ИММУНОГЛОБУЛИНА | 2015 |
|
RU2746738C2 |
| ИММУНОГЛОБУЛИНЫ, СВЯЗЫВАЮЩИЕ АГГРЕКАН | 2018 |
|
RU2771818C2 |
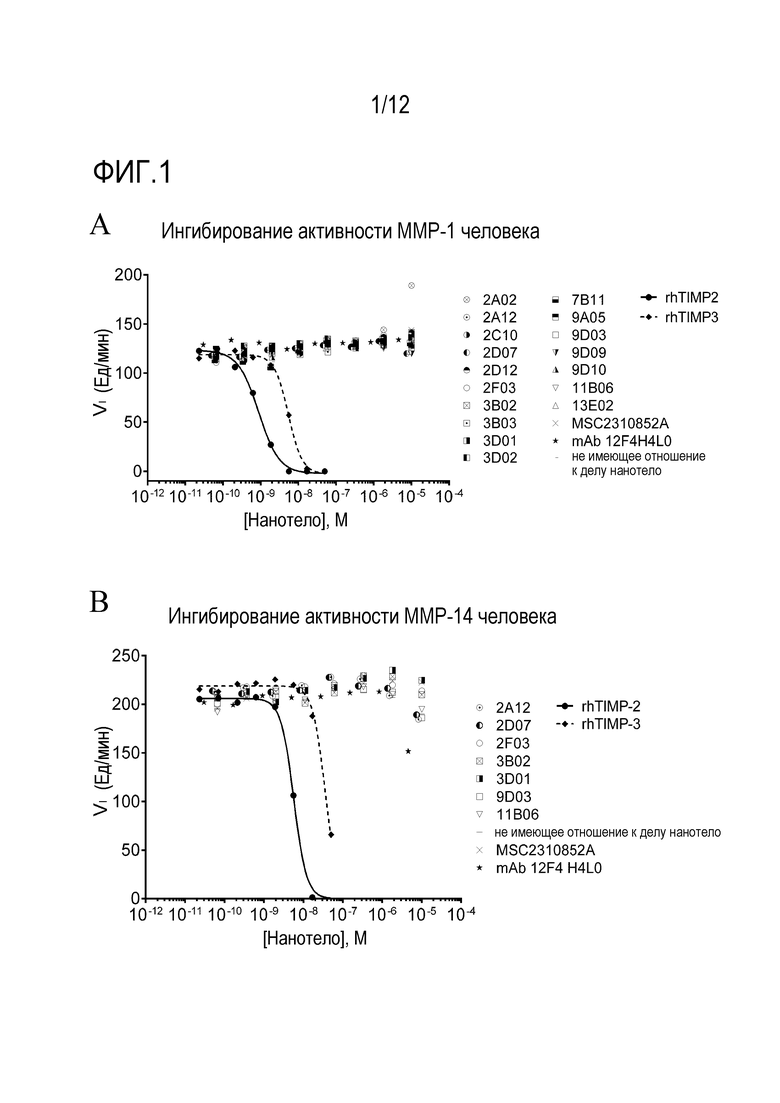

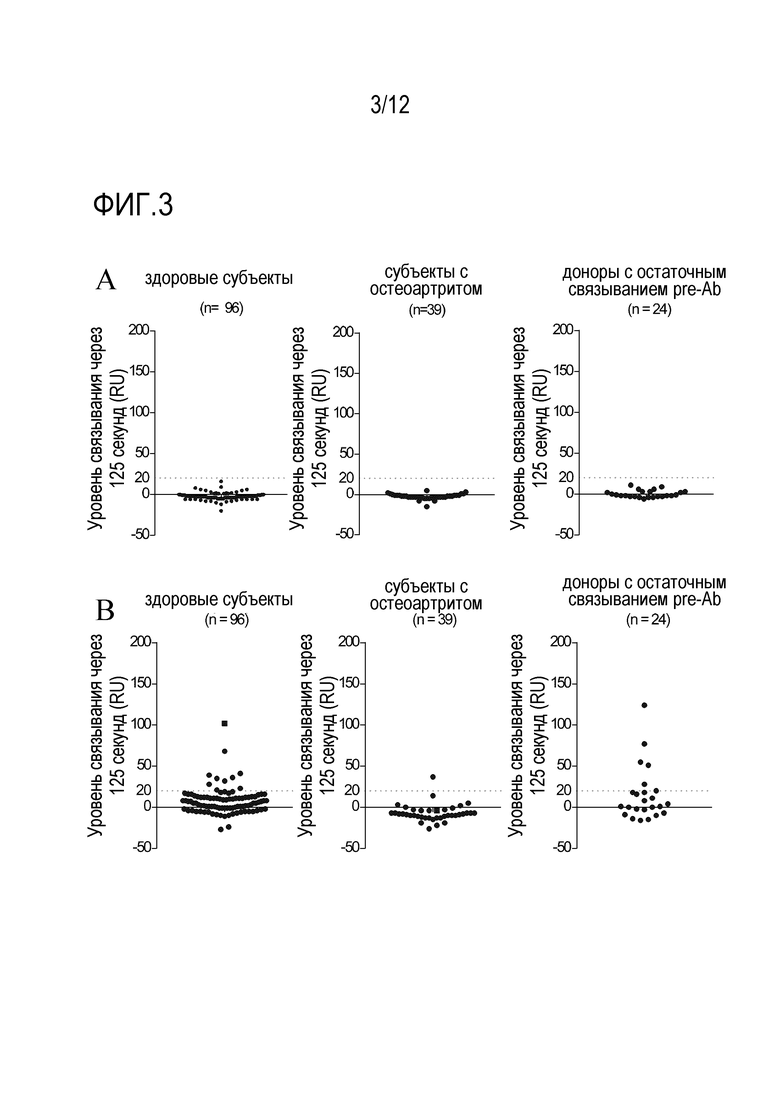
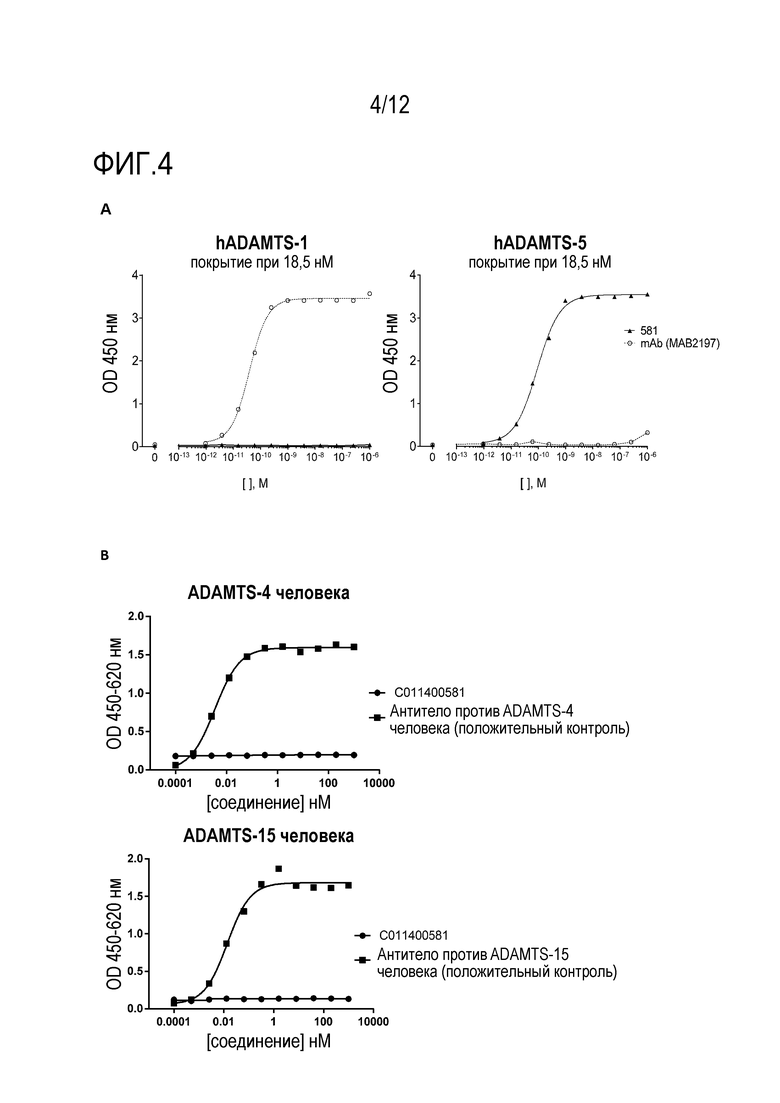
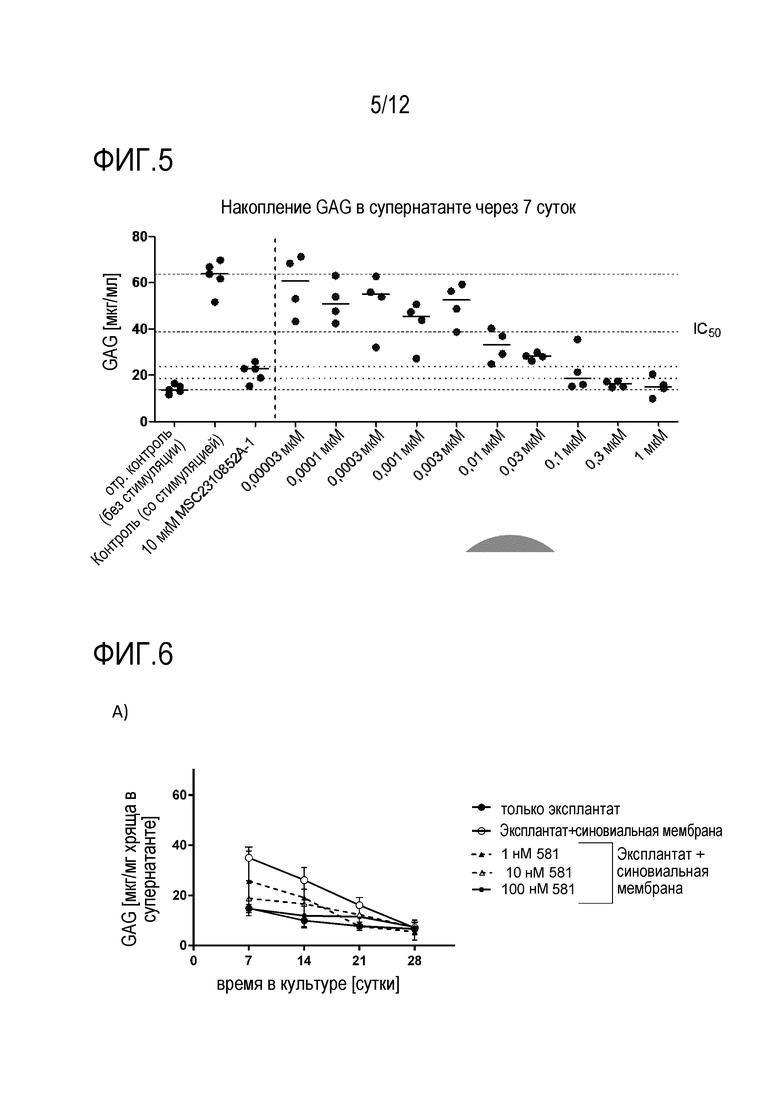
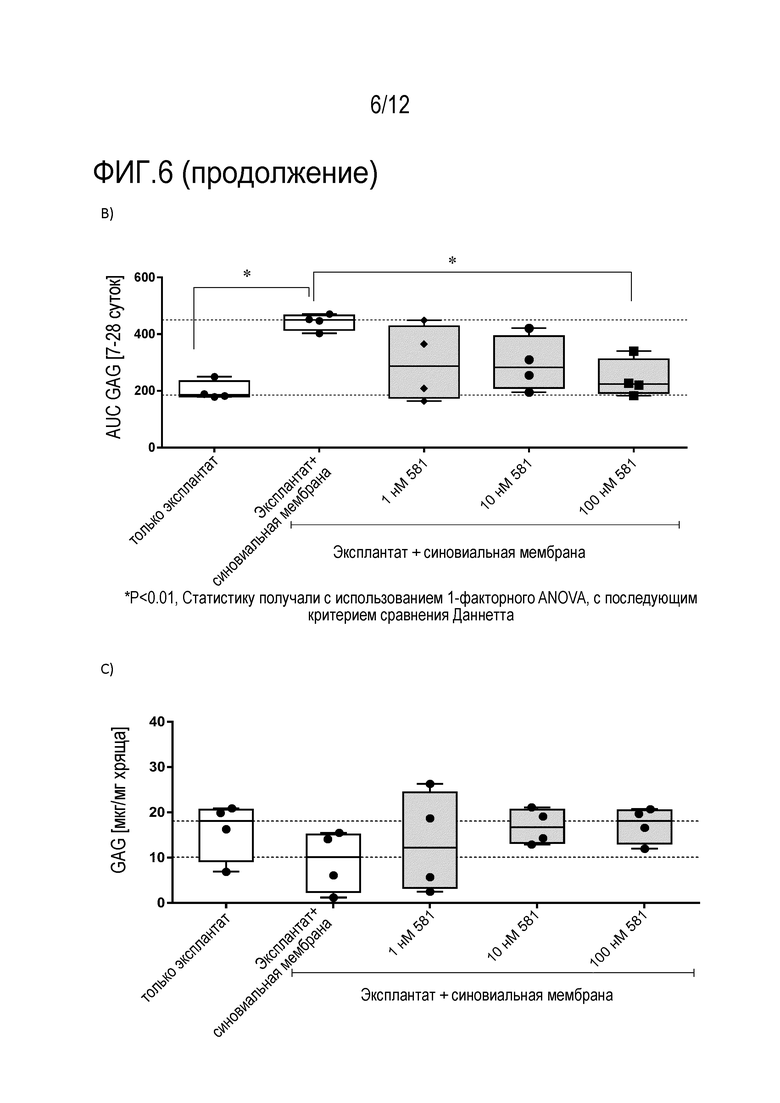
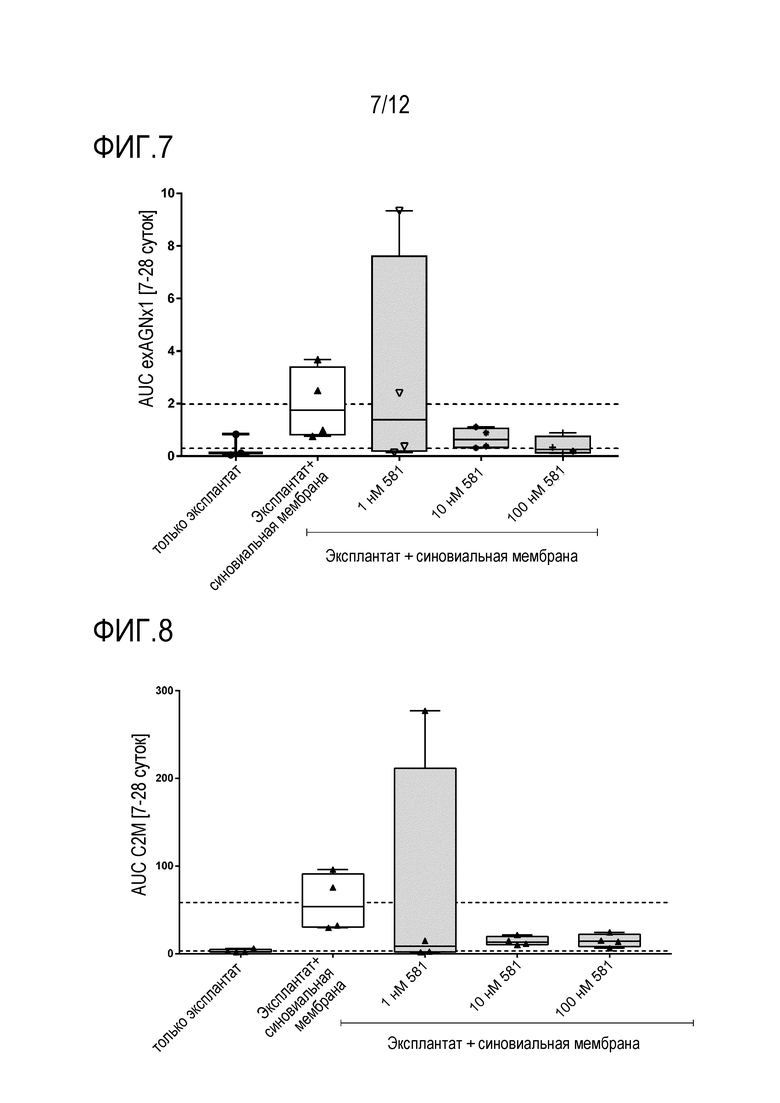
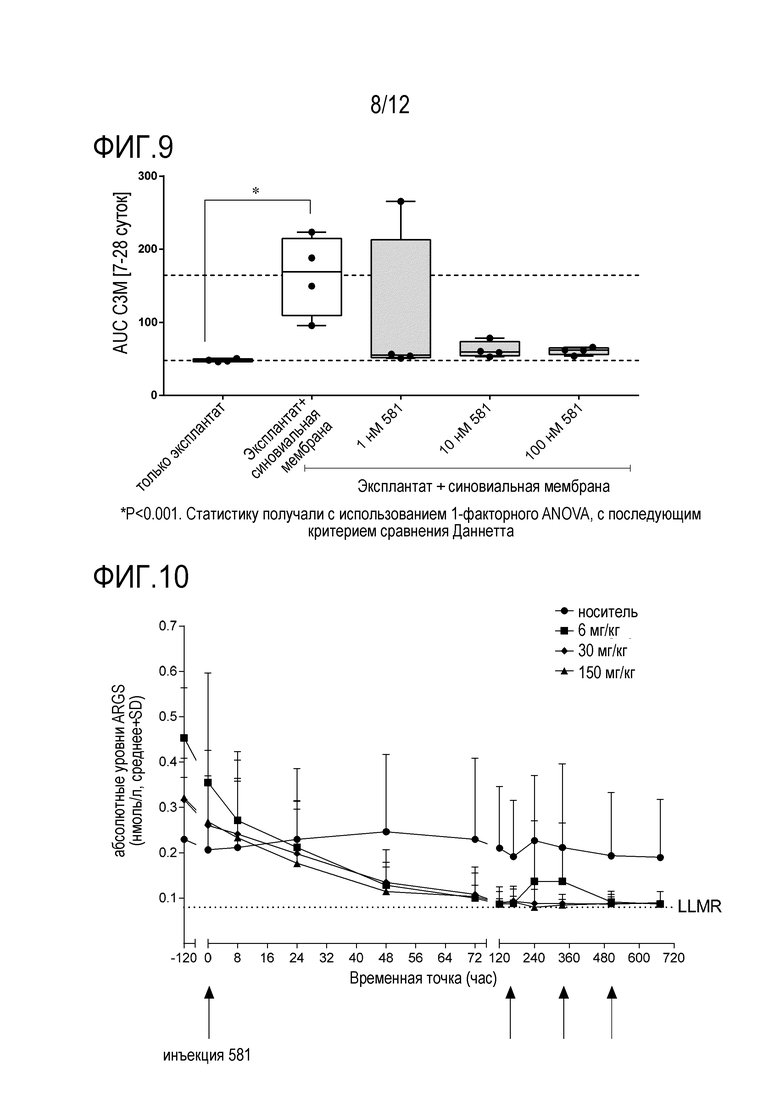
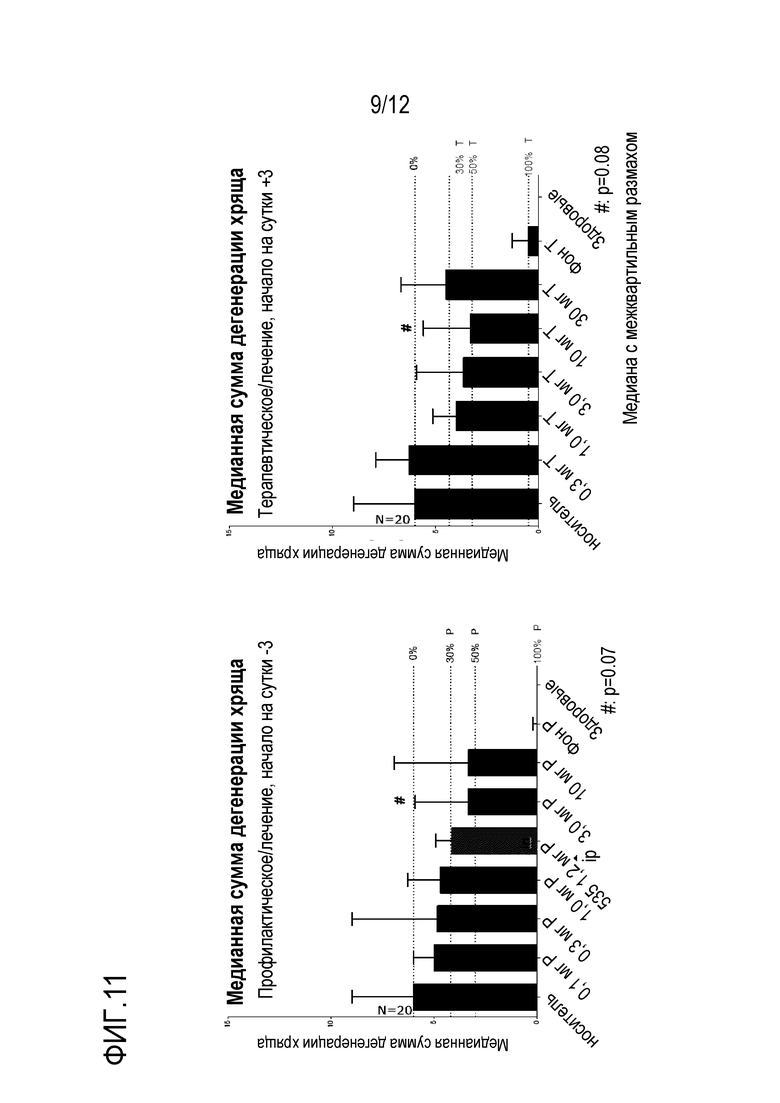
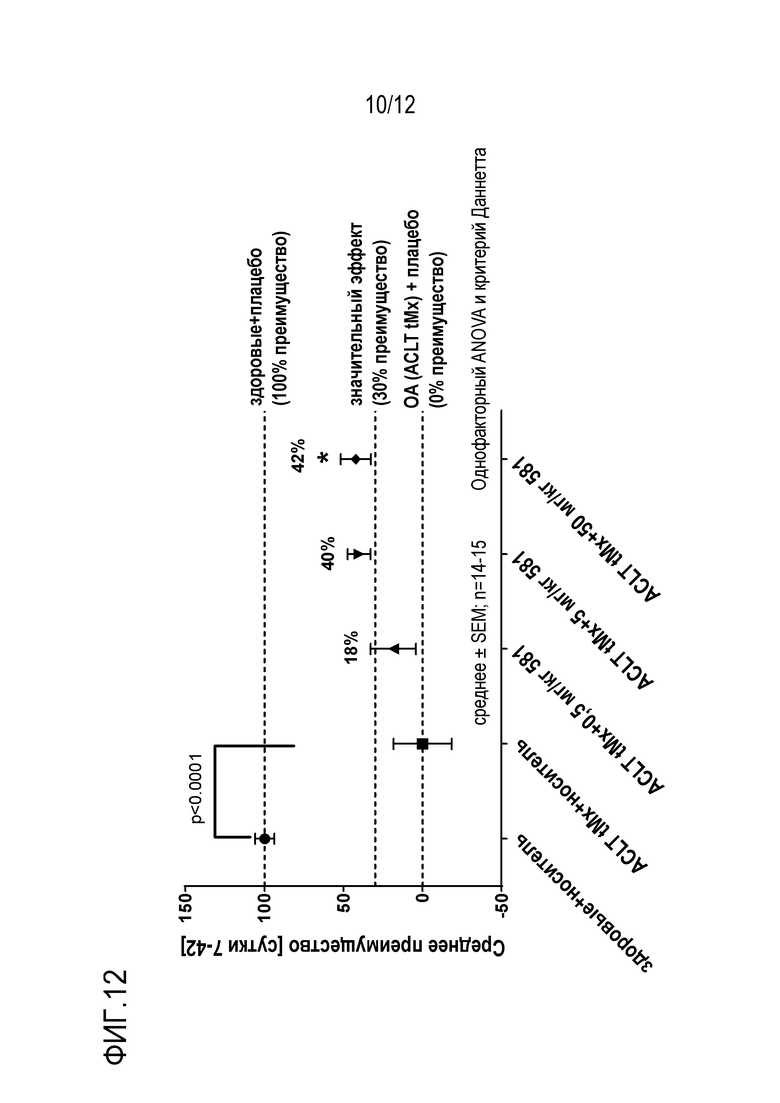
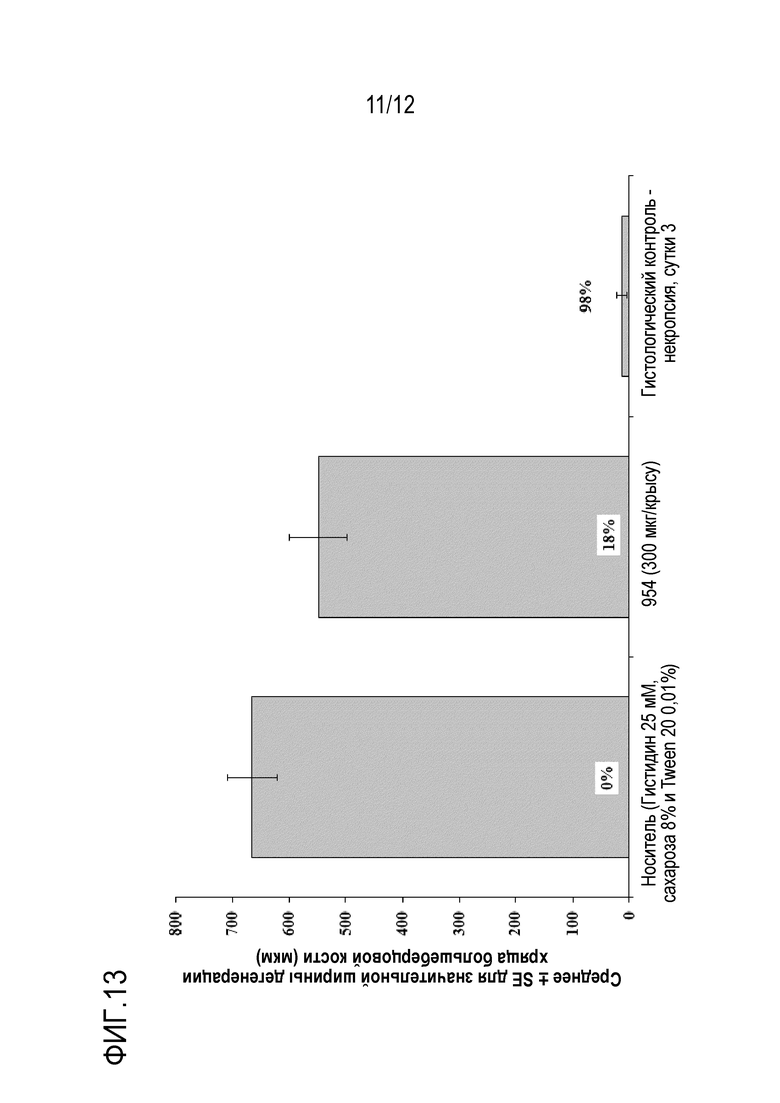
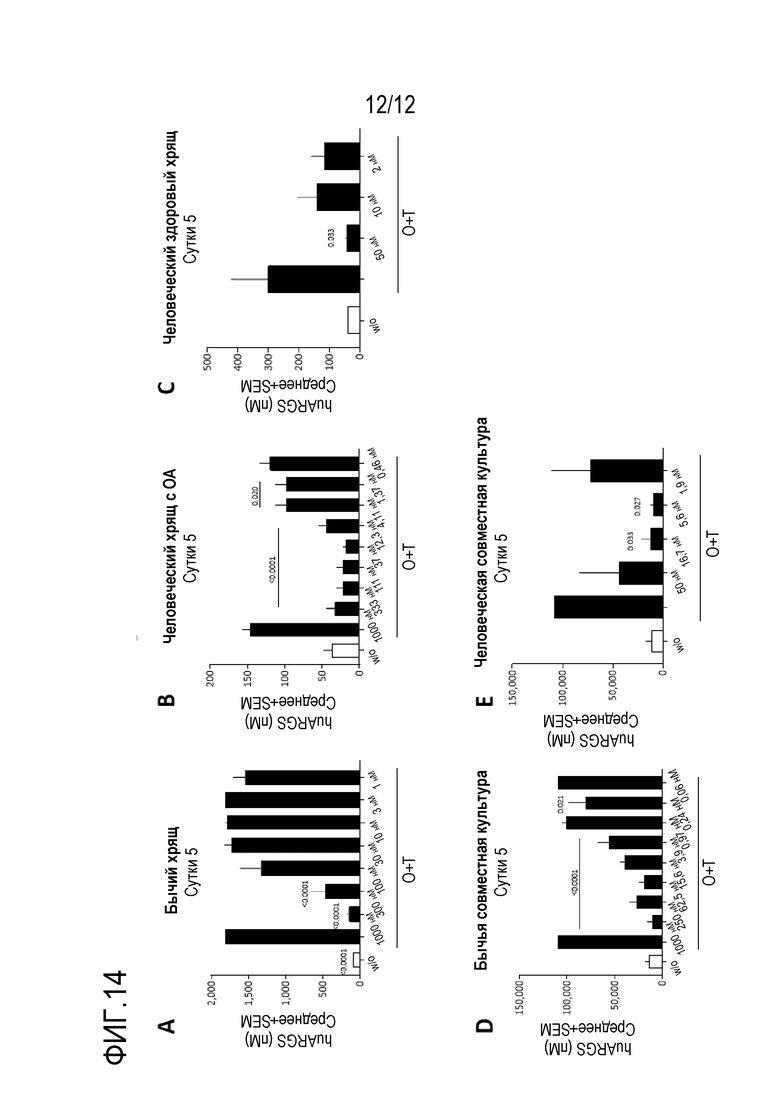
Настоящее изобретение относится к области биотехнологии, конкретно к иммуноглобулинам, связывающим дезинтегрин и металлопротеиназу с тромбоспондиновыми мотивами 5 (ADAMTS5), и может быть использовано в медицине для лечения или предотвращения у индивидуума заболевания или нарушения, в которые вовлечена активность ADAMTS5. Предложен полипептид, связывающий ADAMTS5, содержащий по меньшей мере 1 одиночный вариабельный домен иммуноглобулина (ISVD), связывающий ADAMTS5 и содержащий 3 определяющих комплементарность области: CDR1 с SEQ ID NO: 21, CDR2 с SEQ ID NO: 37 и CDR3 с SEQ ID NO: 55. Изобретение обеспечивает предоставление полипептидов, являющихся антагонистами ADAMTS5 в части ингибирования его ферментной активности с сохранением избирательности, проявляющих улучшенные профилактические, терапевтические и/или фармакологические свойства, включая лучший профиль безопасности, по сравнению с известными антителами к ADAMTS5. 2 н. и 24 з.п. ф-лы, 14 ил., 36 табл., 16 пр.
1. Полипептид, связывающий дезинтегрин и металлопротеиназу с тромбоспондиновыми мотивами 5 (ADAMTS5), содержащий по меньшей мере 1 одиночный вариабельный домен иммуноглобулина (ISVD), связывающий ADAMTS5, где указанный ISVD, связывающий ADAMTS5, содержит 3 определяющих комплементарность области, где CDR1 представляет собой SEQ ID NO: 21, CDR2 представляет собой SEQ ID NO: 37 и CDR3 представляет собой SEQ ID NO: 55.
2. Полипептид по п. 1, где указанный ISVD состоит из или в основном состоит из 4 каркасных областей и указанных 3 определяющих комплементарность областей.
3. Полипептид по п. 1, где указанный полипептид имеет последовательность SEQ ID NO: 129 или 130, или последовательность, имеющую по меньшей мере 95% идентичность последовательности SEQ ID NO: 129 или 130.
4. Полипептид по любому из пп. 1–3, где указанный полипептид связывается с ADAMTS5 с KD менее чем 1E-08 М.
5. Полипептид по любому из пп. 1-3, где указанный полипептид ингибирует ферментную активность ADAMTS5 с IC50 не более 1E–9 M.
6. Полипептид по любому из пп. 1–3, где указанный полипептид модулирует ADAMTS5 с EC50 между 1E–07 M и 1E–12 M.
7. Полипептид по любому из пп. 1–3, где указанный полипептид связывается с ADAMTS5 со скоростью диссоциации менее чем 1E–04 с–1.
8. Полипептид по любому из пп. 1–3, где указанный ADAMTS5 представляет собой ADAMTS5 человека имеет последовательность SEQ ID NO: 149.
9. Полипептид по любому из пп. 1–3, где указанный полипептид является антагонистом аггреканазной активности ADAMTS5.
10. Полипептид по любому из пп. 1–3, где указанный полипептид ингибирует связывание ADAMTS5 с аггреканом по меньшей мере на 20%.
11. Полипептид по любому из пп. 1–3, где указанный полипептид ингибирует протеазную активность ADAMTS5.
12. Полипептид по любому из пп. 1–3, дополнительно содержащий второй ISVD.
13. Полипептид по п. 12, где второй ISVD специфически связывается с ADAMTS5.
14. Полипептид по любому из пп. 1–3, дополнительно содержащий ISVD, связывающийся с сывороточным альбумином.
15. Полипептид по п. 14, где указанный ISVD, связывающий сывороточный альбумин, в основном состоит из 4 каркасных областей и 3 определяющих комплементарность областей, в которых CDR1 имеет последовательность SEQ ID NO: 146, CDR2 имеет последовательность SEQ ID NO: 147, и CDR3 имеет последовательность SEQ ID NO: 148.
16. Полипептид по п. 14, где указанный ISVD, связывающий сывороточный альбумин, выбран из группы, состоящей из ALB8, имеющего последовательность SEQ ID NO: 131, ALB23, имеющего последовательность SEQ ID NO: 132, ALB129, имеющего последовательность SEQ ID NO: 133, ALB132, имеющего последовательность SEQ ID NO: 134, ALB11, имеющего последовательность SEQ ID NO: 135, ALB11, имеющего последовательность (S112K)–A SEQ ID NO: 136, ALB82, имеющего последовательность SEQ ID NO: 137, ALB82–A, имеющего последовательность SEQ ID NO: 138, ALB82–AA, имеющего последовательность SEQ ID NO: 139, ALB82–AAA, имеющего последовательность SEQ ID NO: 140, ALB82–G, имеющего последовательность SEQ ID NO: 141, ALB82–GG, имеющего последовательность SEQ ID NO: 142, ALB82–GGG, имеющего последовательность SEQ ID NO: 143, ALB92, имеющего последовательность SEQ ID NO: 144 и ALB223, имеющего последовательность SEQ ID NO: 145.
17. Полипептид по п. 14, имеющий последовательность, выбранную из группы:
SEQ ID NO: 129 (клон 577 2F3SO–Alb),
SEQ ID NO: 130 (клон 579 2F3SO–093–Alb),
SEQ ID NO: 122 (клон 6 2F3–Alb), и
SEQ ID NO: 126 (клон 129 2F3–093–Alb).
18. Полипептид по п. 12, где ISVD, связывающий ADAMTS5, напрямую или посредством линкера связан со вторым ISVD.
19. Полипептид по п. 18, где указанный линкер имеет любую из последовательностей SEQ ID NO: 158–174.
20. Полипептид по любому из пп. 1–3, где указанный полипептид имеет по меньшей мере 90% идентичность последовательности с любой из SEQ ID NO: 2, 129 или 130.
21. Способ лечения или предотвращения у индивидуума заболевания или нарушения, в которые вовлечена активность ADAMTS5, включающий введение полипептида по любому из пп. 1–20 указанному индивидууму в количестве, эффективном для лечения или предотвращения симптома указанного заболевания или нарушения.
22. Способ по п. 21, где указанное заболевание или нарушение выбрано из группы, состоящей из артропатии, хондродистрофии, артритного заболевания, ахондроплазии, реберного хондрита, спондилоэпиметафизарной дисплазии, межпозвонковой грыжи, заболевания дегенерации поясничного диска, дегенеративного заболевания сустава, рецидивирующего полихондрита, рассекающего остеохондрита и аггреканопатии.
23. Способ по п. 22, где артритное заболевание выбрано из группы, состоящей из остеоартрита, ревматоидного артрита, подагрического артрита, псориатического артрита, травматического разрыва и открепления.
24. Полипептид по любому из пп. 1–20 для применения в качестве лекарственного средства.
25. Полипептид по любому из пп. 1–20 для применения в лечении или предотвращении у индивидуума заболевания или нарушения, в которые вовлечена активность ADAMTS5, где заболевание или нарушение выбрано из артропатии хондродистрофии, артритного заболевания, ахондроплазии, реберного хондрита, спондилоэпиметафизарной дисплазии, межпозвонковой грыжи, заболевания дегенерации поясничного диска, дегенеративного заболевания сустава, рецидивирующего полихондрита, рассекающего остеохондрита и аггреканопатии.
26. Полипептид по п. 25, где артритное заболевание выбрано из группы, состоящей из остеоартрита, ревматоидного артрита, подагрического артрита, псориатического артрита, травматического разрыва и открепления.
| WO 2008074840 A2, 26.06.2008, WO 2015173325 A2, 19.11.2015, WO2011002968 A2, 06.01.2011, SANTAMARIA S | |||
| Кипятильник для воды | 1921 |
|
SU5A1 |
| et al., Strategies for extended serum half-life of protein therapeutics, | |||

