Область техники
Настоящее изобретение относится к способу получения активной формы производного аналога инсулина длительного действия с использованием клострипаина и, более конкретно, к способу получения активной формы производного аналога инсулина длительного действия, в котором аминокислота аргинин (Arg) в положении 22 В-цепи инсулина заменена на лизин (Lys), таким образом, что аналог инсулина может быть превращен в активную форму без расщепления В-цепи, даже когда он взаимодействует с клострипаином.
Предшествующий уровень техники
Диабет представляет собой метаболическое заболевание, отличающееся высокими уровнями глюкозы, и развивается в результате сочетания генетических факторов и факторов окружающей среды. Диабет включает диабет 1-го типа, диабет 2-го типа, гестационный диабет и другие состояния, которые вызывают гипергликемию. Диабет подразумевает нарушение метаболизма, при котором поджелудочная железа продуцирует недостаточные количества инсулина или при котором клетки организма человека не способны соответствующим образом отвечать на инсулин, и, таким образом, их способность поглощать глюкозу нарушается. В результате, глюкоза накапливается в крови.
Диабет 1-го типа, также называемый инсулинозависимым сахарным диабетом (IDDM - от англ. insulin-dependent diabetes mellitus) и юношеским диабетом, обусловлен разрушением бета-клеток, приводящим к абсолютному дефициту инсулина. С другой стороны, диабет 2-го типа, известный как инсулиннезависимый сахарный диабет (NIDDM - от англ. non-insulin dependent diabetes mellitus) и приобретенный диабет, ассоциирован с преобладающей инсулинорезистентностью и, таким образом, относительным дефицитом инсулина и/или преобладающим дефектом секреции инсулина с инсулинорезистентностью.
В частности, диабет ассоциирован с разными осложнениями, такими как сердечно-сосудистое заболевание и ретинопатия, и, вследствие этого, представляет собой заболевание, которое становится очень тяжелым, если не оказывается надлежащий контроль течения заболевания, как например, контроль уровня глюкозы в крови. Ожидается, что мировой рынок лекарственных средств против диабета расширится с $ 41,7 миллиарда в 2015 году до $ 66,1 миллиардов к 2022 году, и, как ожидается, станет вторым по величине после рынка противораковых лекарственных средств. Глобально, имеется 422 миллиона взрослых людей с диабетом в 2014 году, что составляет 8,5% всего взрослого населения, почти в два раза больше, по сравнению с 4,5% в 1980 году (ВОЗ (Всемирная организация здравоохранения), 2016 год). Кроме того, общие всемирные расходы на здравоохранение из-за диабета оцениваются на уровне $ 673 миллиардов, и ожидается, что число пациентов с диабетом в возрасте 20-79 лет будет увеличиваться до примерно 620 миллионов к 2040 году.
Наиболее репрезентативные способы лечения диабета включают способ введения инсулина для контроля уровня глюкозы в крови пациента до нормального уровня. Инсулин представляет собой гормон, регулирующий уровень глюкозы в крови, который секретируется в поджелудочной железе человека. Он функционирует, осуществляя перенос избытка глюкозы в крови к клеткам, снабжая клетки энергией и поддерживая уровни глюкозы в крови на нормальных уровнях.
Инсулин подвергается разным посттрансляционным модификациям на протяжении пути образования. Продукция и секреция главным образом независимы; полученный инсулин хранится в ожидании секреции. Как С-пептид, так и зрелый инсулин являются биологически активными.
У млекопитающих инсулин синтезируется в бета-клетках поджелудочной железы. Инсулин состоит из двух полипептидных цепей (А-цепи и В-цепи), которые связаны друг с другом дисульфидными связями. Однако, инсулин сначала синтезируется в виде одного единственного полипептида, называемого препроинсулином, в бета-клетках поджелудочной железы. Препроинсулин содержит сигнальный пептид из 24 аминокислот, который направляет выделяющуюся полипептидную цепь к шероховатому эндоплазматическому ретикулуму. Сигнальный пептид транслоцируется в просвет шероховатого эндоплазматического ретикулума и затем расщепляется, с образованием, вследствие этого, проинсулина. В шероховатом эндоплазматическом ретикулуме проинсулин сворачивается в правильной конформации, и образуется три дисульфидные связи. Через 5-10 минут после его сборки в эндоплазматическом ретикулуме проинсулин транспортируется в транс-сеть Гольджи, где образуются незрелые гранулы.
Проинсулин подвергается созреванию в активный инсулин под действием клеточных эндопептидаз, известных как прогормонконвертазы (РС1 и РС2), а также карбоксипептидазы Е, которая представляет собой экзопротеазу. Эндопептидазы осуществляют расщепление в 2 положениях, высвобождая фрагмент, называемый С-пептидом, и оставляя 2 пептидные цепи, В- и А-цепи, связанные 2 дисульфидными связями. Каждый из сайтов расщепления расположен после пары основных остатков (лизин (Lys)-64 и аргинин (Arg)-65, и аргинин (Arg)-31 и аргинин (Arg)-32). После расщепления С-пептида данные 2 пары основных остатков удаляются под действием карбоксипептидазы. С-пептид расположен в центральной части проинсулина, и первичная последовательность проинсулина находится в следующем порядке «В-С-А» (В-цепь и А-цепь идентифицировали, исходя из массы, и С-пептид был открыт позже).
Полученный зрелый инсулин (активный инсулин) упакован внутри зрелых гранул, ожидая метаболические сигналы (например, лейцин (Leu), аргинин (Arg), глюкоза и манноза) и стимуляцию блуждающего нерва для экзоцитоза из клетки в кровоток.
Для лечения диабета вводят активный инсулин. Технологии получения активного инсулина с использованием технологии рекомбинантных ДНК выглядят следующим образом. Сначала, в Eli Lilly Corp. использовали способ, который включает следующие стадии: экспрессия А-цепи и В-цепи по отдельности с использованием Е.coli, и смешивание А-цепи и В-цепи in vitro с образованием дисульфидных мостиков, со связыванием, таким образом, А- и В-цепей друг с другом дисульфидными связями. Однако, в данном способе имелась проблема, заключающаяся в том, что эффективность продукции являлась низкой. В таком случае, в Eli Lilly Corp. разработали способ, который включает следующие стадии: экспрессия проинсулина; образование дисульфидных связей in vitro; и затем отщепление С-пептида от продукта под действием трипсина и карбоксипептидазы В, с продукцией, таким образом, инсулина.
В Novo Nordisk Corp. разработали способ, который включает следующие стадии: экспрессия минипроинсулина, содержащего В-цепь и А-цепь, связанные двумя основными аминокислотами, в дрожжах; и затем обработка минипроинсулина трипсином с продукцией инсулина. Данный способ имеет преимущества, заключающиеся в том, что дисульфидные связи образуются во время экспрессии и секреции минипроинсулина и что минипроинсулин легко выделяется и очищается, поскольку он секретируется в культуральную среду. Однако, данный способ было сложно применять для крупномасштабного производства, сопоставимо с продукцией с использованием Е. coli.
С тех пор, активно ведется разработка новых способов получения инсулина посредством технологии рекомбинантных ДНК. Hoechst AG разработали способ, который включает следующие стадии: экспрессия нового производного инсулина или препроинсулина в Е. coli; и образование дисульфидных связей in vitro с последующей обработкой лизилэндопептидазой или клострипаином и карбоксипептидазой В, с получением, таким образом, инсулина. В Bio-Technology General Corp. разработали способ, в котором слитый белок, содержащий супероксиддисмутазу (SOD - от англ. superoxide dismutase), связанную с проинсулином, экспрессируется в Е. coli для повышения эффективности экспрессии и эффективности образования дисульфидных связей in vitro. Превращение проинсулина в инсулин проводили под действием трипсина и карбоксипептидазы В. Как описано выше, многие способы получения инсулина посредством технологии рекомбинантных ДНК пробовали и улучшали в отношении эффективности экспрессии, эффективности образования дисульфидных связей и процесса превращения проинсулина в инсулин (KR 10-2001-7013921).
Авторы настоящего изобретения сделали всесторонние попытки разработать способ получения, подходящий для нового производного аналога инсулина длительного действия, разработанного авторами настоящего изобретения, которое имеет увеличенный период полувыведения in vivo, приводя к улучшению длительности действия, по сравнению с нативным инсулином, и, как результат, обнаружили, что применение клострипаина делает возможным эффективное получение нового производного аналога инсулина длительного действия, разработанного авторами настоящего изобретения, таким образом, осуществляя настоящее изобретение.
Раскрытие изобретения
Техническая проблема
Цель настоящего изобретения заключается в предложении способа получения активной формы нового производного аналога инсулина длительного действия.
Техническое решение
Для достижения указанной выше цели, согласно настоящему изобретению предложен способ получения активной формы производного аналога инсулина длительного действия, причем способ включает стадию взаимодействия производного аналога инсулина, которое содержит аналог инсулина, содержащий вариант В-цепи инсулина, представленный аминокислотной последовательностью SEQ ID NO 2, аргинин (Arg) в положении аминокислоты 22 В-цепи инсулина замещен лизином (Lys) в нативном инсулине, и слитый с ним альбумин-связывающий домен, с клострипаином.
Согласно настоящему изобретению также предложено производное аналога инсулина длительного действия, полученное данным способом.
Краткое описание графических материалов
На Фиг. 1 показаны результаты анализа стабильности производного аналога инсулина длительного действия, сконструированного посредством замены аргинина (Arg) в положении 22 В-цепи инсулина лизином (Lys), посредством обработки клострипаином. Масс-спектрометрия показала, что когда в В-цепи инсулина содержался аргинин (Arg) в положении 22, происходило ее расщепление, а когда аргинин (Arg) был заменен лизином (Lys), расщепления В-цепи не происходило.
На Фиг. 2 показаны результаты анализа экспрессии производных аналога инсулина длительного действия 1-10 в рекомбинантных Е.coli посредством SDS-PAGE (от англ. sodium dodecyl sulphate - polyacrylamide gel electrophoresis - электрофорез в полиакриламидном геле в присутствии додецилсульфата натрия). Даже когда одну или более мутаций вводили в последовательность инсулина, сохранялась экспрессия белка в Е.coli.
На Фиг. 3 показаны результаты отслеживания солюбилизации и повторного сворачивания аналога инсулина 4, слитого с альбумин-связывающий доменом (ABD, от англ. - albumin-binding domain), посредством ОФ-ВЭЖХ (Обращенно-фазовая высоко-эффективная жидкостная хроматография). Когда структура белка была несвернута в результате солюбилизации и сопровождалась индукцией повторного сворачивания, наблюдали сдвиг времени удерживания в ОФ-ВЭЖХ вследствие образования трехмерной структуры.
На Фиг. 4 показаны результаты SDS-PAGE, проводимого для анализа эффективности получения активной формы аналога инсулина 4, слитого с ABD, посредством обработки клострипаином в зависимости от добавления ДТТ (Дитиотреитол).
На Фиг. 5 показаны результаты SDS-PAGE, проводимого для анализа эффективности получения активной формы аналога инсулина 4, слитого с ABD, в зависимости от времени обработки клострипаином и СрВ.
На Фиг. 6 показаны результаты SDS-PAGE, проводимого для определения диапазона рН, в котором активную форму аналога инсулина 4, слитого с ABD, получают с высокой эффективностью при одновременной обработке клострипаином и СрВ.
На Фиг. 7 показаны результаты SDS-PAGE, проводимого для анализа эффективности получения активной формы аналога инсулина 4, слитого с ABD, в зависимости от дополнительного добавления клострипаина и ДТТ.
На Фиг. 8 показаны результаты подтверждения эффектов повышения эффективности превращения в активную форму слитого с ABD аналога инсулина 4 и уменьшения примесей в оптимальных условиях ферментативной реакции в соответствии с примером настоящего изобретения (молекулярная масса основной примеси: 10296 Да; молекулярная масса активной формы инсулина: 11238 Да).
На Фиг. 9 показаны результаты SDS-PAGE, проводимого для определения температурного диапазона, в котором активную форму аналога инсулина 4, слитого с ABD, получают с высокой эффективностью при одновременной обработке клострипаином и СрВ.
На Фиг. 10 показаны результаты анализа аналога инсулина 4, слитого с ABD, посредством SDS-PAGE после расщепления под действием клострипаина и СрВ и его очистки.
На Фиг. 11 показаны результаты анализа - эксклюзионной хроматографии, проводимого для исследования того, будет ли образовываться гексамер, когда цинк и фенол добавляют к аналогу инсулина 4, слитому с ABD, в соответствии с примером настоящего изобретения.
На Фиг. 12 показаны результаты оценки способности аналогов инсулина, слитых с ABD, снижать уровень глюкозы в крови в соответствии с примером настоящего изобретения.
Наилучший способ осуществления изобретения
Если не определено иное, все технические и научные термины, используемые в данном документе, имеют то же значение, обычно подразумеваемое одним из средних специалистов в области, к которой принадлежит данное раскрытие. В общем, номенклатура, используемая в данном документе, хорошо известна и обычно используется в данной области.
Аналог инсулина, полученный в результате осуществления замены одной или более аминокислот нативного инсулина, имеет увеличенный период полувыведения in vivo, и, таким образом, может использоваться в качестве основного средства инсулинотерапии. Когда данный аналог инсулина связывается с альбумином в организме человека, его период полувыведения in vivo может дополнительно увеличиваться, таким образом, что аналог инсулина может быть использован в виде композиции для еженедельного введения.
Авторы настоящего изобретения смогли обнаружить, что когда одна или более аминокислот нативного инсулина заменены, период полувыведения in vivo инсулина может быть увеличен, и когда с ним дополнительно слит альбумин-связывающий домен, инсулин может связываться с альбумином in vivo, и, таким образом, его период полувыведения может дополнительно увеличиваться. Однако, когда альбумин-связывающий домен был слит с аналогом инсулина, содержащим одну или более аминокислотных замен, и когда трипсин, используемый в известном уровне техники, использовали в процессе превращения аналога инсулина в активную форму, возникала проблема, заключающаяся в том, что происходило расщепление самого альбумин-связывающего домена, затрудняя получение активной формы, существующей на протяжении длительного периода времени.
Соответственно, в настоящем изобретении были сделаны попытки использовать клострипаин в качестве фермента, который позволяет превращать инсулин в активную форму, не индуцируя расщепление альбумин-связывающего домена. Однако, клострипаин индуцировал расщепление самого инсулина, и, по этой причине, аминокислоты инсулина заменяли, предотвращая, вследствие этого, расщепление самого инсулина.
Таким образом, в одном аспекте настоящее изобретение направлено на способ получения активной формы производного аналога инсулина длительного действия, причем способ включает стадию взаимодействия производного аналога инсулина с клострипаином, в котором аналог инсулина, содержащий вариант В-цепи инсулина, представленный аминокислотной последовательностью SEQ ID NO 2, аргинин (Arg) в положении аминокислоты 22 В-цепи инсулина заменен лизином (Lys) в нативном инсулине, слит с альбумин-связывающим доменом.
В настоящем изобретении фраза «аналог инсулина, содержащий вариант В-цепи инсулина, представленный аминокислотной последовательностью SEQ ID NO 2, аргинин (Arg) в положении аминокислоты 22 В-цепи инсулина заменен лизином (Lys) в нативном инсулине» означает, что аналог инсулина может дополнительно содержать вариант аминокислоты в А-цепи инсулина или В-цепи инсулина помимо замены аргинина (Arg) на лизин (Lys) в положении аминокислоты 22 В-цепи нативного инсулина.
В настоящем изобретении аналог инсулина может дополнительно содержать одну или более аминокислотных замен, выбранных из группы, состоящей из нижеследующего: замена валина (Val) на лейцин (Leu) в положении аминокислоты 3 А-цепи инсулина, представленной аминокислотной последовательностью SEQ ID NO 4; замена треонина (Thr) на аспарагиновую кислоту (Asp) в положении аминокислоты 8 А-цепи инсулина, представленной аминокислотной последовательностью SEQ ID NO 4; замена изолейцина (Не) на лизин (Lys) в положении аминокислоты 10 А-цепи инсулина, представленной аминокислотной последовательностью SEQ ID NO 4; замена тирозина (Туг) на глутаминовую кислоту (Glu) в положении аминокислоты 14 А-цепи инсулина, представленной аминокислотной последовательностью SEQ ID NO 4; замена тирозина (Tyr) на фенилаланин (Phe) в положении аминокислоты 19 А-цепи инсулина, представленной аминокислотной последовательностью SEQ ID NO 4; замена гистидина (His) на треонин (Thr) в положении аминокислоты 5 варианта В-цепи инсулина, представленного аминокислотной последовательностью SEQ ID NO 2; замена серина (Ser) на аспарагиновую кислоту (Asp) в положении аминокислоты 9 варианта В-цепи инсулина, представленного аминокислотной последовательностью SEQ ID NO 2; замена глутаминовой кислоты (Glu) на аланин (Ala) в положении аминокислоты 13 варианта В-цепи инсулина, представленного аминокислотной последовательностью SEQ ID NO 2; замена лейцина (Leu) на глутамин (Gln) в положении аминокислоты 17 варианта В-цепи инсулина, представленного аминокислотной последовательностью SEQ ID NO 2; и замена фенилаланина (Phe) на серии (Ser) в положении аминокислоты 24 варианта В-цепи инсулина, представленного аминокислотной последовательностью SEQ ID NO 2, но не ограничивается ими.
В одном примере настоящего изобретения альбумин-связывающий домен может содержать альбумин-связывающий мотив, представленный следующей аминокислотной последовательностью:
GVSDFYKKLIXaKAKTVEGVEALKXbXcI,
где:
Ха независимо выбран из D и Е,
Xb независимо выбран из D и Е, и
Xc независимо выбран из А и Е.
В одном примере настоящего изобретения Ха представляет собой D, Xb представляет собой D, и Xc представляет собой А.
В одном примере настоящего изобретения альбумин-связывающий домен может содержать следующую аминокислотную последовательность:
LAX3AKX6X7ANX10ELDX14Y-[BM]-LX43X44LP,
где:
[ВМ] представляет собой альбумин-связывающий мотив, как определено в предыдущем абзаце,
Х3 независимо выбран из С, Е, Q и S;
X6 независимо выбран из С, Е и S;
X7 независимо выбран из А и S;
Х10 независимо выбран из A, R и S;
Х14 независимо выбран из А, С, K и S;
Х43 независимо выбран из А и K; и
Х44 независимо выбран из А, Е и S.
Альбумин-связывающий домен может быть представлен аминокислотными последовательностями, выбранными из группы, состоящей из SEQ ID NO: 6-13, но не ограничивается ими. Альбумин-связывающий домен может предпочтительно быть представлен аминокислотной последовательностью SEQ ID NO 6.
В настоящем изобретении предпочтительно, чтобы альбумин-связывающий домен был слит с С-концом А-цепи в инсулине или аналоге инсулина В-цепь - С-цепь - А-цепь в отношении эффективности повторного сворачивания белка, эффективности ферментативной реакции и активности полученного производного инсулина или производных аналога инсулина. В связи с этим, линкер может быть введен между инсулином (или аналогом инсулина) и альбумин-связывающим доменом.
В настоящем изобретении полинуклеотид, кодирующий аналог инсулина, и нуклеотид, кодирующий альбумин-связывающий домен, можно вводить в рекомбинантный вектор таким образом, чтобы аналог инсулина и альбумин-связывающий домен могли экспрессироваться в виде слитого белка.
Функция производного аналога инсулина согласно настоящему изобретению изменяется в зависимости от трехмерной структуры производного. Таким образом, возможно вносить небольшие изменения в аминокислотную последовательность производного аналога инсулина длительного действия согласно настоящему изобретению, не влияя на функцию данного производного. Таким образом, настоящее изобретение включает варианты альбумин-связывающего домена или производного аналога инсулина длительного действия, которые сохраняют свойство связывания альбумина или высокую устойчивость к ферментативному расщеплению. Например, аминокислотные остатки, принадлежащие к конкретной функциональной группе аминокислотных остатков (например, гидрофобность, гидрофильность, полярность и т.д.), могут быть заменены другими аминокислотными остатками, принадлежащими к той же функциональной группе.
В том виде, в котором они используются в данном документе, термины «альбумин-связывающий» и «аффинность связывания в отношении альбумина» относятся к свойству полипептида или белка, которое можно тестировать посредством применения технологии поверхностного плазмонного резонанса, как например, с помощью прибора Biacore. Например, аффинность связывания альбумина можно тестировать в эксперименте, в котором альбумин или его фрагмент иммобилизуют на сенсорном чипе прибора, и образец, содержащий полипептид или белок, подлежащий тестированию, пропускают через чип.
В качестве альтернативы, полипептид или белок, подлежащий тестированию, иммобилизуют на сенсорном чипе прибора, и образец, содержащий альбумин или его фрагмент, пропускают через чип. В связи с этим, альбумин может представлять собой альбумин сыворотки, происходящей из млекопитающего, как например, альбумин сыворотки человека. Специалисты в данной области могут интерпретировать результаты, полученные с помощью таких экспериментов, для установления количественного показателя аффинности связывания полипептида или белка в отношении альбумина. Если количественный показатель является желательным, например, для определения значения Ко для взаимодействия, могут быть также использованы методы поверхностного плазмонного резонанса. Величины связывания могут, например, быть определены на приборе Biacore 2000 (Biacore АВ). Альбумин подходящим образом иммобилизован на измерительном сенсорном чипе, и образцы полипептида или белка, чья аффинность подлежит определению, получают посредством серийного разведения и инъецируют в случайном порядке. Значения KD можно, затем, рассчитывать на основании результатов, используя, например, 1:1 модель связывания Ленгмюра программного обеспечения BIAevaluation 4.1, предоставленного производителем прибора.
В одном воплощении настоящего изобретения альбумин, с которым связывается производное аналога инсулина, выбран из альбумина сыворотки человека, альбумина сыворотки крысы, альбумина сыворотки яванского макака и альбумина сыворотки мыши, но не ограничивается ими.
В одном конкретном воплощении альбумин, с которым связывается производное аналога инсулина, представляет собой альбумин сыворотки человека.
Между тем, описание альбумин-связывающего мотива и альбумин-связывающего домена (или альбумин-связывающего полипептида) истолковывается как включающее содержание, раскрытое в выложенной для всеобщего ознакомления публикации корейского патента №10-2015-0058454, соответствующей WO 2014048977.
Аналог инсулина и альбумин-связывающий домен связаны друг с другом пептидной связью; полипептидным линкером; или непептидильным линкером, выбранным из группы, состоящей из полиэтиленгликоля, полипропиленгликоля, сополимера этиленгликоля и пропиленгликоля, полиоксиэтилированного полиола, поливинилового спирта, полисахаридов, декстрана, простого поливинилэтилового эфира, биоразлагаемых полимеров, жирных кислот, нуклеотидов, липидных полимеров, хитина, гиалуроновой кислоты и их комбинаций, но не ограничиваются ими.
В настоящем изобретении линкер, состоящий из повтора последовательности (GGGGS)n (где n представляет собой целое число, находящееся в интервале от 1 до 6), может быть вставлен между аналогом инсулина и альбумин-связывающим доменом. Это основано на результатах экспериментов, указывающих на то, что когда линкер не был введен, выход повторного сворачивания был низким, а когда два или более повторов последовательности (GGGGS) были введены, отсутствовало значимое различие в выходе повторного сворачивания. Между тем, при увеличении числа (n) повторов данной последовательности, скорость превращения в активную форму инсулина под действием клострипаина повышалась, но фармакокинетика in vivo аналога инсулина не подвергалась значимому влиянию. Принимая во внимание все описанные выше результаты, линкер предпочтительно состоит из 2-4 повторов последовательности GGGGS.
Таким образом, в настоящем изобретении аналог инсулина и альбумин-связывающий домен могут быть связаны друг с другом полипептидным линкером, и полипептидный линкер может содержать (GGGGS)n (где n представляет собой целое число, находящееся в интервале от 1 до 6, но не ограничивается ими). Предпочтительно, полипептидный линкер может быть представлен аминокислотной последовательностью SEQ ID NO 5.
В настоящем изобретении восстанавливающий агент можно добавлять во время реакции с клострипаином. Восстанавливающий агент может быть выбран из группы, состоящей из цистеина, β-меркаптоэтанола, ТСЕР (от англ. Tris(2-carboxyethyl)phosphine hydrochloride - Трис (2-карбоксиэтил) фосфин гидрохлорид), GSH (от англ. glutathione - глутатион) и ДТТ, но не ограничивается ими. В качестве восстанавливающего агента предпочтительно добавляют 0,1-0,5 мМ ДТТ. Более предпочтительно добавляют 0,1-0,4 мМ ДТТ.
ДТТ может быть добавлен только один раз на начальной стадии, на которой производное аналога инсулина взаимодействует с клострипаином, а именно, в момент времени, в который инициируется реакция, и ДТТ может дополнительно быть добавлен 2-5 раз после его добавления в момент времени, в который инициирована реакция. Однако, наиболее предпочтительно ввиду удобства и эффективности способа получения, чтобы ДТТ был добавлен на начальной стадии реакции производного аналога инсулина с клострипаином и дополнительно был добавлен через 3-6 часов после реакции. Клострипаин можно добавлять в концентрации 0,1-5 единиц, предпочтительно 0,5-2 единиц, на мг белка, но не ограничивается ими.
В настоящем изобретении производное аналога инсулина могут дополнительно приводить во взаимодействие с карбоксипептидазой В (СрВ), которую добавляют во время или после реакции с клострипаином. В связи с этим, СрВ можно добавлять вместе с клострипаином при инициации реакции.
В настоящем изобретении клострипаин и/или карбоксипептидаза В (СрВ) приводится(ятся) во взаимодействие в условиях рН 6,0-9,0, предпочтительно 6,5-7,5, и в температурных условиях 4-40°С, предпочтительно, 30-40°С, но не ограничивается ими.
СрВ можно добавлять в концентрации 0,001-1 единиц, предпочтительно 0,001-0,1 единиц, на мг белка, но, не ограничиваясь ими.
В другом аспекте настоящее изобретение направлено на активную форму производного аналога инсулина, полученную описанным выше способом.
В настоящем изобретении производное аналога инсулина можно получать в рекомбинантном микроорганизме, в котором рекомбинантный вектор содержит полинуклеотид, кодирующий производное аналога, посредством лизиса и очистки культивируемого рекомбинантного микроорганизма.
Рекомбинантный вектор согласно настоящему изобретению может быть сконструирован в качестве вектора для общепринятого клонирования или экспрессии, и может быть сконструирован в качестве вектора для применения прокариотической или эукариотической клетки в качестве клетки-хозяина.
В том виде, в котором он используется в данном документе, термин «вектор» относится к рекомбинантному вектору, способному экспрессировать целевой белок в соответствующей клетке-хозяине, который представляет собой генетическую конструкцию, включающую существенные регуляторные факторы, функционально связанные, для обеспечения экспрессии вставки нуклеиновой кислоты. Согласно настоящему изобретению можно получать рекомбинантный вектор, который включает нуклеиновую кислоту, кодирующую производное аналога инсулина, и данное производное аналога инсулина по настоящему изобретению можно получать посредством трансформации или трансфекции клетки-хозяина рекомбинантным вектором.
В настоящем изобретении нуклеиновая кислота, кодирующая аналог инсулина и производное аналога, функционально связана с последовательностью контроля экспрессии нуклеиновой кислоты. В том виде, в котором он используется в данном документе, термин «функционально связан» относится к функциональной связи между последовательностью контроля экспрессии нуклеиновой кислоты (например, промотором, сигнальной последовательностью, сайтом связывания рибосом, последовательностью терминации транскрипции и т.д.) и другой нуклеотидной последовательностью, и, таким образом, регуляторная последовательность может контролировать транскрипцию и/или трансляцию другой нуклеотидной последовательности.
В том виде, в котором он используется в данном документе, термин «промотор» относится к нетранслируемой последовательности нуклеиновой кислоты, расположенной выше кодирующей области, которая включает сайт связывания полимеразы, и обладает активностью инициации транскрипции гена, расположенного ниже промотора, в мРНК, а именно, сайт ДНК, с которым связывается полимераза и инициирует транскрипцию гена, и он расположен в 5' области участка инициации транскрипции мРНК.
Например, когда вектор по настоящему изобретению представляет собой рекомбинантный вектор, и в качестве клетки-хозяина используется прокариотическая клетка, обычно включены сильный промотор, способный стимулировать транскрипцию (такой как промотор tac, промотор lac, промотор lacUV5, промотор lpp, промотор pLλ, промотор pRλ, промотор rac5, промотор amp, промотор recA, промотор SP6, промотор trp, промотор trc, промотор phoA, промотор araBAD, промотор Т5 и промотор Т7), сайт связывания рибосом для инициации трансляции и последовательности терминации транскрипции/трансляции.
Кроме того, вектор, который может быть использован в настоящем изобретении, может быть получен посредством манипуляций с плазмидами (например, pSC101, pGV1106, pACYC177, ColE1, pKT230, рМЕ290, pBR322, pUC8/9, pUC6, pBD9, pHC79, pIJ61, pLAFR1, pHV14, серия pGEX, серия pET, серия pPICZa, pUC19 и т.д.), фагами (например, λgt4λB, λ-Charon, λΔz1, M13 и т.д.) или вирусами (например, SV40 и т.д.), которые обычно используются в данной области, но, не ограничиваясь ими.
Между тем, когда вектор по настоящему изобретению представляет собой рекомбинантный вектор, и эукариотическую клетку используют в качестве клетки-хозяина, можно использовать промоторы, происходящие из геномов клеток млекопитающего (например, промотор металлотионеина), промоторы, происходящие из вирусов млекопитающего (например, поздний промотор аденовируса, 7,5K промотор вируса осповакцины, промотор SV40, промотор цитомегаловируса и промотор tk HSV (от англ. Herpes Simpiex Virus - вирус простого герпеса)), и, в общем, вектор включает полиаденилированную последовательность (например, терминатор гормона роста крупного рогатого скота и полиаденилированную последовательность, происходящую из SV40) в качестве последовательности терминации транскрипции.
Кроме того, рекомбинантный вектор по настоящему изобретению включает ген устойчивости к антибиотику, обычно используемый в данной области в качестве селективного маркера, и может включать, например, гены устойчивости к ампициллину, гентамицину, карбенициллину, хлорамфениколу, стрептомицину, канамицину, генетицину, неомицину и тетрациклину.
Рекомбинантный вектор по настоящему изобретению может дополнительно включать другую последовательность для облегчения очистки собираемых целевых белков, а именно, инсулина и/или его аналога. Последовательность, подлежащая дополнительному включению, может представлять собой последовательность метки для очистки белка, например, глутатион-S-трансферазу (Pharmacia, США), мальтоза-связывающий белок (NEB, США), FLAG (IBI, США), 6-гистидин и т.д., но виды последовательности, необходимой для очистки целевых белков, не ограничиваются ими. Слитые белки, экспрессируемые рекомбинантным вектором, включая указанную выше последовательность метки, могут быть очищены посредством аффинной хроматографии. Например, когда подвергают слиянию глутатион S-трансферазу, можно использовать глутатион, который представляет собой субстрат данного фермента, и когда используется 6-гистидиновая метка, желательный целевой белок может быть с легкостью собран посредством колонки Ni-NTA. Рекомбинантный микроорганизм, трансформированный вектором, может быть конструирован, используя рекомбинантный вектор, содержащий полинуклеотид, кодирующий аналог инсулина и/или производное аналога.
В том виде, в котором он используется в данном документе, термин «трансформация» относится к способу введения ДНК в клетку-хозяина и получения ДНК, реплицируемой в ней, в виде хромосомного фактора или посредством выполнения хромосомной интеграции, которая является явлением искусственного осуществления генетического изменения посредством введения экзогенной ДНК в клетку.
Способ трансформации, используемый в настоящем изобретении, может представлять собой любой способ трансформации, и его можно легко осуществлять в соответствии с общепринятым способом, используемым в данной области. Примеры обычно используемого способа трансформации могут включать способ осаждения с использованием CaCl2, способ Hanahan с улучшенной эффективностью, используя диметилсульфоксид (ДМСО) в качестве восстанавливающего агента в способе осаждения с использованием CaCl2, электропорацию, способ осаждения с использованием CaPO4, способ слияния протопластов, способ перемешивания с использованием карбидокремниевого волокна, способ трансформации, опосредованной агробактериями, способ трансформации с использованием ПЭГ (полиэтиленгликоль), трансформации, опосредованные сульфатом декстрана, липофектамином и сушкой/давлением, и т.д.
Способ трансформации рекомбинантным вектором, включающим нуклеиновую кислоту, кодирующую аналог инсулина и/или производное аналога согласно настоящему изобретению, может не ограничиваться данными способами, но любой способ трансформации или трансфекции, обычно используемый в данной области, можно использовать без ограничения.
Рекомбинантный трансформант по настоящему изобретению может быть получен посредством введения рекомбинантного вектора, включающего нуклеиновую кислоту, кодирующую производное аналога инсулина, в клетку-хозяина. Соответствующий хозяин, подлежащий использованию в настоящем изобретении, может особым образом не ограничиваться при условии, что он может экспрессировать нуклеиновую кислоту по настоящему изобретению. Примеры соответствующего хозяина могут включать бактерии, принадлежащие к роду Escherichia, как например, Е. coli, бактерии, принадлежащие к роду Bacillus, как например, Bacillus subtilis, бактерии, принадлежащие к роду Pseudomonas, как например, Pseudomonas putida, дрожжи, как например Pichia pastoris, Saccharomyces cerevisiae и Schizosaccharomyces pombe, клетку насекомого, как например Spodoptera frugiperda (SF9), и животные клетки, такие как СНО (от англ. - Chinese Hamster Ovary - яичник китайского хомячка), COS и BSC, но не ограничиваются ими.
В том виде, в котором он используется в данном документе, термин «активная форма» означает зрелую форму инсулина, аналога инсулина или производного аналога инсулина, способную регулировать уровни глюкозы в крови in vivo, и относится к инсулину, аналогу инсулина или производному аналога инсулина, которую получают посредством удаления С-пептида из формы проинсулина и которая содержит А-цепь и В-цепь инсулина.
Примеры
Далее в данном документе, настоящее изобретение будет описано более подробно со ссылкой на примеры. Специалисту, обладающему средней квалификацией в данной области, будет очевидно, что данные примеры представлены исключительно в иллюстративных целях, и не должны истолковываться как ограничивающие объем настоящего изобретения.
В приведенных ниже примерах аналог инсулина, слитый с ABD, используют в том же значении, как производное аналога инсулина длительного действия в настоящем изобретении.
Пример 1: Конструирование экспрессионного вектора и штамма с инсулином, слитым с ABD
Человеческий инсулин синтезируется в форме пре-про-инсулина, препоследовательность отщепляется в эндоплазматическом ретикулуме, и проинсулин обрабатывается в аппарате Гольджи и эндоплазматическом ретикулуме, с образованием, таким образом, зрелого инсулина. На основе данного факта, для получения рекомбинантного инсулина способом экспрессии белка проинсулина в Е. coli и затем удаления C-цепи обработкой трипсином, конструировали проинсулин. Для повышения эффективности экспрессии проинсулина в Е. coli и эффективности очистки, метку слияния включали в N-конец, и проводили оптимизацию кодонов.
Число участков инсулина, с которыми может быть слит альбумин-связывающий домен (ABD), теоретически равно четырем. Однако, N-конец А-цепи представляет собой положение, важное для активности инсулина и, вследствие этого, его исключали из положений слияния. Несмотря на то, что N-конец В-цепи важен для образования гексамера инсулина, он был включен в положения слияния, поскольку активность инсулина могла сохраняться. Если ABD слит между В-цепью и С-цепью, он может оказывать действие на сворачивание белка. Таким образом, конструкцию инсулина конструировали либо в порядке В-С-А или порядке А-С-В в качестве структуры кандидата. Наконец, генные структуры для экспрессии следующих трех форм инсулина, слитого с ABD, конструировали следующим образом:
Ndel - Метка слияния - В-цепь - С-цепь - А-цепь - Линкер - ABD - EcoRI,
Ndel - Метка слияния - ABD - Линкер - В-цепь - С-цепь - А-цепь - EcoRI и
Ndel - Метка слияния - А-цепь - С-цепь - В-цепь - линкер - ABD - EcoRI.
В качестве экспрессионного вектора использовали вектор pJ401(DNA 2.0). Вектор расщепляли рестриктазами Ndel и EcoRI, и затем ДНК-фрагменты разделяли посредством электрофореза в 1%-ном агарозном геле. Генетические структуры для экспрессии слитых белков ABD и ДНК-фрагменты, полученные из экспрессионного вектора, как описано выше, лигировали друг с другом, используя ДНК-лигазу Т4, конструируя, таким образом, плазмиды. Далее, каждой из данных плазмид трансформировали Е. coli BL21(DE3) способом на основе хлорида кальция. Отбирали трансформированные штаммы, обладающие устойчивостью к канамицину, и из них выделяли ДНК. То, была ли ДНК правильно вставлена, определяли аналитическим способом на основе расщепления эндонуклеазами рестрикции.
Когда конструировали ген, имеющий следующую структуру: Ndel - метка слияния - ABD - Линкер - В-цепь - С-цепь - А-цепь - EcoRI, его определяли ненадлежащим из-за очень низкого выхода повторного сворачивания белка. Кроме того, когда конструировали ген, имеющий следующую структуру Ndel - метка слияния - А-цепь - С-цепь - В-цепь - Линкер - ABD - EcoRI, слитый с ABD инсулин обладал активностью инсулина, но он демонстрировал низкий выход повторного сворачивания и выход ферментативной обработки, таким образом, он считался неподходящим в качестве способа получения активной формы инсулина. Таким образом, среди данных трех форм инсулина, слитого с ABD, структуру Ndel - метка слияния - В-цепь - С-цепь - А-цепь - линкер - ABD - EcoRI отбирали и использовали в последующем эксперименте.
Между А-цепью и альбумин-связывающим доменом (ABD) вставляли линкер, состоящий из 1-6 повторов последовательности (GGGGS). В данном случае, когда линкер не вводили, выход повторного сворачивания был очень низким, но когда вводили линкер, состоящий из двух или более повторов последовательности (GGGGS)n, не было значимого различия в выходе повторного сворачивания. Между тем, при увеличении n в последовательности (GGGGS)n, скорость превращения в активную форму инсулина под действием клострипаина повышалась, но фармакокинетика in vivo инсулина значимо не подвергалась воздействию. Принимая во внимание все описанные выше результаты, конструировали линкер, состоящий из 2-4 повторов последовательности GGGGS.
Аминокислотные последовательности каждого из фрагментов для инсулина, слитого с ABD, используемого в настоящем изобретении, показаны ниже в Таблице 1.

Пример 2: Конструирование слитых с ABD аналогов инсулина, имеющих модифицированные аминокислотные последовательности инсулина
Для получения инсулина, используя рекомбинантные Е. coli, способ превращения проинсулина в активную форму посредством применения трипсина является необходимым. Однако, трипсин расщепляет двухосновные аминокислоты с высокой эффективностью и также расщепляет единичные аминокислоты, такие как лизин (Lys) или аргинин (Arg), осложняя получение желательной активной формы инсулина. Кроме того, последовательность ABD также включает некоторое количество остатков лизина (Lys) и аргинина (Arg), дополнительно осложняя получение желательного инсулина, слитого с ABD, обладающего активностью, посредством применения трипсина.
По этой причине, клострипаин использовали в качестве фермента, способного заменять трипсин для индукции превращения в активную форму. В данном случае, когда клострипаин взаимодействовал с инсулином, слитым с ABD, происходило расщепление аргинина (Arg) в положении 22 В-цепи инсулина. Для решения данной проблемы аргинин (Arg) в положении 22 В-цепи заменяли лизином (Lys). В данном случае расщепление в положении 22 В-цепи инсулина под действием клострипаина значимо уменьшалось и, таким образом, активная форма слитого с ABD инсулина могла быть эффективно получена (Фиг. 1).
Помимо мутации, как описано выше, дополнительные мутации вводили в инсулин, слитый с ABD, для дополнительного увеличения стабильности и in vivo периода полувыведения инсулина, слитого с ABD. Пять аминокислот в каждой из А-цепи и В-цепи заменяли, и замененные положения показаны ниже в Таблице 2.

Оптимизацию кодонов для экспрессии генов в Е. coli проводили, используя алгоритм GeneArt, и синтезировали гены, имеющие замененные аминокислоты.
Плазмиды, сконструированные как описано выше, вводили в Е. coli BL21 (DE3) таким же образом, как описано в Примере 1, конструируя, таким образом, штаммы Е coli.
Пример 3: Экспрессия аналогов инсулина, слитых с ABD
Для экспрессии аналогов инсулина, слитых с ABD, каждый из рекомбинантных штаммов Е. coli инокулировали в 100 мл среды LB и культивировали при встряхивании при 37°С в течение 16 часов, и культуры использовали в качестве культур для посева. 2 л среды LB добавляли в 7-л ферментер (New Brunswick BioFlo), стерилизовали и затем инокулировали с культурой для посева. Культивирование проводили в условиях температуры 35°С, скорости потока воздуха 3 об/об/мин и скорости перемешивания 1000 об/мин, и рН во время культивирования поддерживали на уровне 6,8 с помощью аммония и фосфорной кислоты. В момент времени, в который источник углерода в среде истощался, начинали подачу и в тот же момент времени экспрессию белка индуцировали IPTG (от англ. isopropylthiogalactoside - изопропилтиогалактозид). Дополнительное культивирование проводили в течение 10 часов после индукции экспрессии, и рекомбинантные штаммы выделяли, используя центрифугу.
Аналоги инсулина, слитые с ABD, экспрессировали в виде телец включения в штамме Е. coli, и даже когда аминокислотные мутации вводили в домены инсулина, уровни экспрессии обнаруживались в векторной системе, используемой в настоящем изобретении (Фиг. 2).
Пример 4: Индукция лизиса клеток и солюбилизации/повторного сворачивания
Каждый из штаммов, экспрессирующих инсулин, слитый с ABD, или аналоги инсулина, слитые с ABD, суспендировали в лизирующем буфере (20 мМ Tris, 10 мМ ЭДТА, 10% сахароза, 0,2 М NaCl, рН 8,0), и клетки лизировали, используя гомогенизатор высокого давления. Лизированные клетки центрифугировали в высокоскоростной центрифуге при 7000 об/мин, и растворимый белок и некоторые обломки клеток удаляли, выделяя, таким образом, осадок, включающий тельце включения. Выделенное тельце включения промывали буфером (содержащим 1% Triton Х-100, 0,2 М NaCl и 1 М мочевину) и затем центрифугировали при 7000 об/мин. Осажденное тельце включения дополнительно два раза промывали дистиллированной водой и затем хранили при -80°С до применения.
Хранящееся в замороженном виде тельце включения растворяли в солюбилизирующем буфере (25 мМ Tris, 8 М мочевина, 30 мМ цистеин-HCl, рН 10,5), и затем разбавляли в буфере для повторного сворачивания (25 мМ Tris-HCl, рН 10,5) и подвергали повторному сворачиванию при 4°С в течение 16 часов. Происходило ли повторное сворачивание, определяли посредством анализа ОФ-ВЭЖХ. Когда солюбилизированный раствор анализировали посредством ОФ-ВЭЖХ, пик белка наблюдался на уровне примерно 20 минут вследствие высокой гидрофобности, но когда происходило повторное сворачивание, наблюдался сдвиг данного пика к 16 минутам (Фиг. 3). Причина этого состоит в том, что когда протекает повторное сворачивание, гидрофобность снижается, по сравнению с гидрофобностью в солюбилизированном растворе. Повторное сворачивание продолжали до тех пор, пока сдвига пика белка больше не наблюдалось во время анализа посредством ОФ-ВЭЖХ.
Пример 5: Превращение в активную форму посредством применения фермента
Для получения инсулина, используя рекомбинантный штамм Е. coli, требуется превращение проинсулина в активную форму посредством применения трипсина. Однако, инсулин, слитый с ABD, или аналог инсулина, слитый с ABD, имеет целый ряд сайтов расщепления трипсином в своей последовательности, и по этой причине клострипаин использовали в качестве фермента, способного заменить трипсин для индукции превращения в активную форму.
Когда индуцировали превращение в активную форму, используя клострипаин, происходило расщепление в положении 22 В-цепи инсулина, осложняя получение желательной формы инсулина. По данной причине, аналог инсулина, слитый с ABD, полученный посредством замены в положении 22 В-цепи аргинина (Arg) на лизин (Lys), использовали для получения активной формы инсулина. Между тем, слитый с ABD аналог инсулина 4 использовали во всех экспериментах, если дополнительно конкретно не определено иное.
Клострипаин содержит цистеин (Cys) в своем активном сайте, и, таким образом, требует восстанавливающих условий для демонстрации ферментативной активности. Кроме того, когда раствор для повторного сворачивания обрабатывают восстанавливающим агентом для сохранения активности клострипаина, дисульфидная связь в области инсулина с высокой долей вероятности будет разорвана. По данной причине, требуется установить условия, которые повышают выход ферментативной реакции без разрыва дисульфидной связи. Таким образом, используя восстанавливающий агент ДТТ, содержащийся в растворе ферментативной реакции, проводят эксперименты по концентрации обработки и дополнительной обработки.
Для превращения аналога инсулина, слитого с ABD, в форме предшественника в активную форму, также дополнительно требуется карбоксипептидаза В (СрВ). СрВ представляет собой фермент, который расщепляет основные аминокислоты на С-конце, и конкретные условия также тестировали. Конкретно, оценивали время обработки клострипаином и СрВ, возможность одновременной обработки, количество ферментативной обработки, температуру ферментативной обработки и соответствующий уровень рН.
Поскольку клострипаин существует в виде проформы, для его активации индуцировали авторасщепление. С этой целью сублимированный клострипаин (Worthington, США) растворяли в дистиллированной воде и затем активировали при 4°С в течение 30 минут после добавления активирующего буфера (500 мМ Tris, 50 мМ ДТТ, 25 мМ CaCl2, рН 7,8). Активированный клострипаин добавляли к повторно свернутому белку в концентрации 0,1-5 единиц на мг белка и позволяли взаимодействовать при 25-40°С в течение 2-8 часов. После реакции с клострипаином СрВ добавляли в концентрации 0,001-1 единица на 1 мг белка и давали прореагировать. Ферментативную реакцию останавливали, снижая рН до 3,5 или меньше посредством применения HCl.
5-1: Оценка эффективности превращения в активную форму после добавления ДТТ
Для исследования условий ферментативной реакции для клострипаина и СрВ ДТТ добавляли к раствору для повторного сворачивания до концентрации 0-0,4 мМ, и затем за продукцией активной формы инсулина под действием клострипаина наблюдали посредством SDS-PAGE. Было показано, что когда добавляли более чем 0,5 мМ ДТТ, дисульфидные связи между А-цепью и В-цепью инсулина были разорваны (данные не показаны).
Между тем, как может быть видно на Фиг. 4, ДТТ не добавляли к раствору ферментативной реакции, скорость превращения аналога инсулина, слитого с ABD, в активную форму была низкой, но при повышении концентрации ДТТ эффективность превращения в активную форму повышалась.
5-2: Оценка эффективности превращения в активную форму инсулина в разные моменты времени добавления СрВ
Для превращения аналога проинсулина, слитого с ABD, в активную форму аналог проинсулина должен быть обработан как клострипаином, так и СрВ. В связи с этим, определяли момент времени добавления СрВ. Для расщепления С-пептида инсулина и метки слияния сначала нужно провести обработку клострипаином, и для удаления аргинина (Arg) из сайта расщепления затем нужно провести обработку СрВ. Однако, для сокращения времени получения, обработку СрВ через 2-4 часа после обработки клострипаином сравнивали с одновременной обработкой клострипаином и СрВ.
В результате, как можно видеть на Фиг. 5, картина ферментативной реакции при одновременной обработке клострипаином и СрВ не отличалась от картины ферментативной реакции в условиях, при которых сначала проводят обработку клострипаином. Таким образом, для улучшения эффективности способа выбирали одновременную обработку клострипаином и СрВ.
5-3: рН условия при одновременной обработке клострипаином и СрВ
Известно, что клострипаин демонстрирует оптимальную активность при рН 7,4-7,8, и СрВ демонстрирует оптимальную активность при рН 9,0. Поскольку оптимальные диапазоны рН двух ферментов отличаются друг от друга, диапазон рН, демонстрирующий наивысшую эффективность превращения в активную форму инсулина, определяли посредством одновременной обработки двумя данными ферментами. Теоретические значения pl всех аналогов инсулина, слитых с ABD, показанных в таблице 2 согласно настоящему изобретению, находились близко к 6, и, таким образом, обращали внимание на осаждение. Соответственно, рН 6,0 или ниже исключали. Кроме того, при рН 9,0 или выше агрегат увеличивался. По данной причине оценку проводили при рН от 6,0 до 9,0.
Одновременная обработка двумя данными ферментами в диапазоне рН 6,5-8,5 демонстрировала высокую эффективность превращения в активную форму инсулина. Кроме того, как показано на Фиг. 6, эффективность превращения в активную форму инсулина была особенно высокой в диапазоне рН 6,5-7,5.
5-4: Оценка эффективности превращения в активную форму инсулина после дополнительного добавления клострипаина и ДТТ
Как показано в Примере 5-1, когда добавляют ДТТ, эффективность превращения в активную форму инсулина повышалась. Однако, период полувыведения ДТТ, как известно, уменьшается быстро при высоких рН и температурах, и, таким образом, полагали, что будет трудно сохранять активность клострипаина на протяжении длительного времени ферментативной реакции. Соответственно, исследовали способ, способный сохранять активность клострипаина на протяжении длительного времени.
В связи с этим, способ, в котором 1 U/мг клострипаина дополнительно добавляют через 4,5 часа после начала ферментативной реакции, оценивали в сравнении со способом, в котором дополнительно добавляют 0,2 мМ ДТТ. В результате, как показано на Фиг. 7, даже когда дополнительно добавляли клострипаин, эффективность превращения в активную форму значимо не повышалась, но когда дополнительно добавляли ДТТ, превращение в активную форму значимо увеличивалось, даже когда клострипаин дополнительно не добавляли. Кроме того, когда и клострипаин, и ДТТ дополнительно добавляли, наблюдали, что количество непревращенной формы было относительно маленьким, и эффективность превращения в активную форму повышалась.
Однако анализ раствора ферментативной реакции посредством масс-спектрометрии указывал на то, что когда добавляли ДТТ, превращение в активную форму инсулина значимо увеличивалось, но когда ДТТ и клострипаин добавляли одновременно, вместо этого индуцировалось неспецифичное расщепление инсулина, приводя к увеличению количества примесей. Таким образом, можно было видеть, что добавление только лишь восстанавливающего агента ДТТ было преимущественным для превращения в активную форму инсулина (Фиг. 8).
5-5: Оценка эффективности превращения в активную форму инсулина при разных температурах
Оценку проводили для определения температурного интервала, в котором клострипаин и СрВ будут демонстрировать оптимальные активности. В данное время, рассматривая технологический процесс в промышленном масштабе, температуру 4°С или ниже и температуру 40°С или выше исключали, и оценку проводили при температуре от 10 до 30°С. В результате, как показано на Фиг. 9, было видно, что при повышении температуры с 10°С до 20°С и 30°С, количество непревращенной формы было меньше, и превращение в активную форму протекало быстрее. Кроме того, было показано, что, когда концентрация ДТТ повышалась, превращение в активную форму протекало быстрее.
Пример 6: Очистка аналога инсулина, слитого с ABD
Образец, ферментативно обработанный клострипаином и СрВ, сначала очищали посредством хроматографии на ионообменной смоле, используя Fractogel® EMD COO- (М) (Merck) в соответствии с инструкцией производителя, и затем дополнительно очищали посредством обращенно-фазовой хроматографии, используя PharmprepO Р100 RP-18e (Merck) в соответствии с инструкцией производителя.
В результате, как может быть видно на Фиг. 10, активная форма беспримесного аналога инсулина, слитого с ABD, могла быть очищена двумя данными способами очистки.
Пример 7: Измерение аффинностей связывания аналогов инсулина, слитых с ABD, в отношении альбумина
Для измерения аффинностей связывания белков - аналогов инсулина, слитых с ABD, в отношении альбумина использовали аналитический метод поверхностный плазмонный резонанс (SPR - от англ. surface plasmon resonance, BIACORE 3000, GE healthcare). Рекомбинантный человеческий сывороточный альбумин иммобилизовали на чипе СМ5 способом связывания аминогрупп, и ABD или аналоги инсулина, слитые с ABD, разведенные до пяти и более концентраций, связывались с ним, и измеряли их аффинности в отношении человеческого сывороточного альбумина.
В результате, как может быть видно ниже в Таблице 3, аффинности аналогов инсулина в отношении человеческого сывороточного альбумина сохранялись на уровнях пМ, даже при том, что они были ниже, чем аффинности самого ABD.
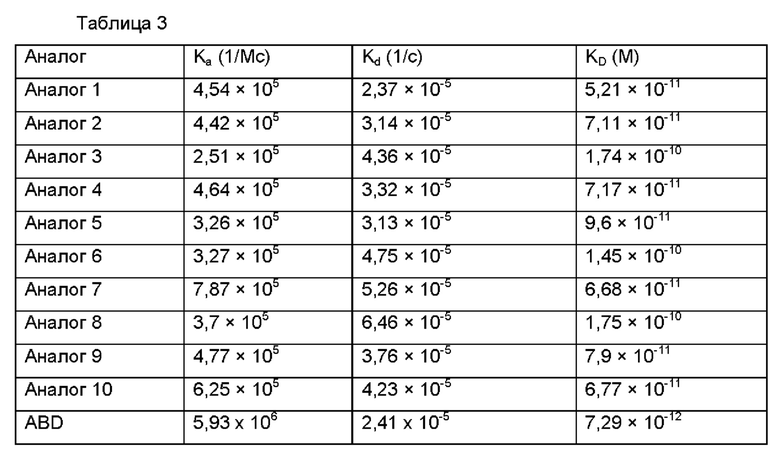
Пример 8: Сравнение аффинностей нативного инсулина и аналогов инсулина, слитых с ABD, в отношении инсулинового рецептора
Для измерения афинностей связывания нативного инсулина и аналогов инсулина, слитых с ABD, в отношении инсулинового рецептора, использовали метод анализа поверхностный плазмонный резонанс (SPR, BIACORE 3000, GE healthcare). Инсулиновый рецептор иммобилизовали на чипе СМ5 способом связывания аминогрупп, и каждый из нативного инсулина и аналогов инсулина, слитых с ABD, разведенных до пяти или более концентраций, связывался с ним, и измеряли их аффинности в отношении инсулинового рецептора.
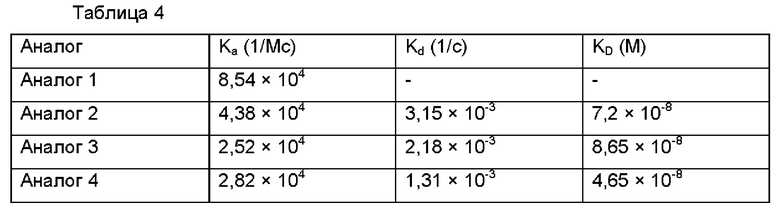
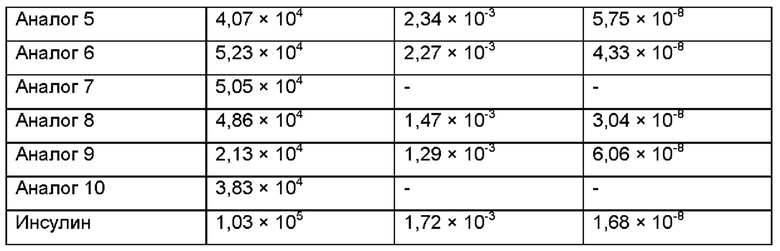
В результате, как можно видеть ниже в Таблице 4, аффинности аналогов инсулина, слитых с ABD, уменьшались, по сравнению с аффинностью нативного инсулина. В частности, аффинность аналога 3 демонстрировала наибольшее уменьшение и падала до уровня примерно 19,4% относительно нативного инсулина.
Эффективности аналогов инсулина, слитых с ABD, оценивали в сравнении с эффективностью инсулина гларгин в моделях диабета 1-го типа, индуцируемого стрептозотоцином. Аналоги 1, 3 и 10 исключали из кандидатов, поскольку они демонстрировали способность снижения уровня глюкозы в крови 50% или меньше, в сравнении с инсулином гларгин.
Пример 9: Синтез гексамера
Инсулин связывается с цинком in vivo, образуя, вследствие этого, стабильную гексамерную структуру. Свойство инсулина образовывать гексамер также используется в разработке препарата и может играть важную роль в увеличении периода полувыведения in vivo инсулина. Соответственно, будут ли аналоги инсулина, слитые с ABD, сохранять свойство образования гексамеров, анализировали посредством эксклюзионной хроматографии. В результате, было показано, что аналог 4 сохранял способность образовывать гексамер посредством добавления цинка и фенола (Фиг. 11). Образование гексамера также наблюдали во всех аналогах, за исключением аналогов 7, 9 и 10. Таким образом, среди аналогов, имеющих последовательности, показанные в Таблице 2, аналоги 7 и 9 дополнительно исключали из кандидатов.
Пример 10: Оценка фармакокинетики in vivo и способности снижения уровня глюкозы в крови аналогов инсулина, слитых с ABD
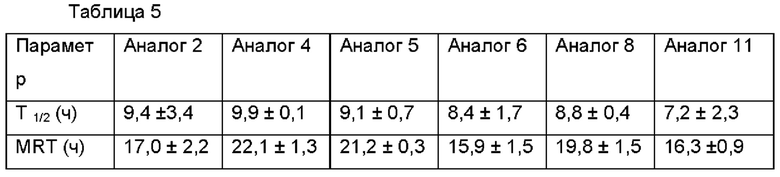
Для оценки in vivo фармакокинетики шести аналогов инсулина, слитых с ABD, каждый из аналогов инсулина вводили подкожно нормальным крысам SD (возраст 6 недель), и затем брали образцы крови в моменты времени 0, 1, 4, 8, 24, 48, 72 и 96 часов. Концентрацию каждого аналога инсулина, слитого с ABD, оставшегося в крови в каждый из моментов времени, измеряли, используя ELISA (от англ. enzyme-linked immunosorbent assay - твердофазный иммуноферментный анализ). Кроме того, используя часть отобранных образцов крови, уровни глюкозы в крови в зависимости от времени измеряли с помощью устройства, контролирующего уровень глюкозы в крови.
В результате, как можно видеть ниже в Таблице 5, аналоги инсулина, слитые с ABD, демонстрировали значимо увеличенные периоды полувыведения, по сравнению с нативным инсулином, как известно, имеющим период полувыведения 5 минут. Аналог 11, полученный посредством замены только в положении 22 В-цепи инсулина, демонстрировал период полувыведения 7,2 часов, и аналог 4, демонстрирующий самое большое увеличение периода полувыведения, по оценке имел период полувыведения 9,9 часов.
Анализировали время, на протяжении которого уровни глюкозы в крови у нормальных животных сохранялись на пониженных уровнях. В результате, было показано, что аналоги 5, 6 и 8 имели увеличенные периоды полувыведения, но поддерживали уровни глюкозы в крови на протяжении короткого периода времени. По сравнению с данными аналогами, аналог 4 обладал наилучшей способностью снижать уровни глюкозы в крови и сохранял свою эффективность на протяжении самого длительного периода времени (Фиг. 12).
Несмотря на то, что настоящее изобретение подробно описано со ссылкой на конкретные признаки, специалистам в данной области будет очевидно, что данное описание предназначено исключительно для предпочтительного воплощения и не ограничивает объем настоящего изобретения. Таким образом, существенный объем настоящего изобретения будет определяться прилагаемой формулой изобретения и ее эквивалентами.
Промышленная применимость
В общепринятом способе превращения проинсулина в активную форму посредством применения трипсина альбумин-связывающий домен нового производного аналога инсулина длительного действия, впервые разработанного авторами настоящего изобретения, также расщепляется, таким образом, осложняя превращение нового производного аналога инсулина длительного действия в активную форму. Для преодоления данной трудности клострипаин использовали для превращения производного аналога инсулина согласно настоящему изобретению в активную форму. Таким образом, способ по настоящему изобретению может эффективно использоваться для получения длительно действующего терапевтического средства для лечения диабета.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> DAEWOONG PHARMACEUTICAL CO., LTD.
<120> СПОСОБ ПОЛУЧЕНИЯ АКТИВНОЙ ФОРМЫ ПРОИЗВОДНОГО АНАЛОГА ИНСУЛИНА
ДЛИТЕЛЬНОГО ДЕЙСТВИЯ С ИСПОЛЬЗОВАНИЕМ КЛОСТРИПАИНА
<130> P17-B267
<160> 13
<170> KoPatentIn 3.0
<210> 1
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> Метка слияния
<400> 1
Met Ala Thr Thr Ser Thr Gly Asn Ser Ala His His His His His His
1 5 10 15
Ser Ser Gly Ser Ala Arg
20
<210> 2
<211> 30
<212> Белок
<213> homo sapiens
<400> 2
Phe Val Asn Gln His Leu Cys Gly Ser His Leu Val Glu Ala Leu Tyr
1 5 10 15
Leu Val Cys Gly Glu Lys Gly Phe Phe Tyr Thr Pro Lys Thr
20 25 30
<210> 3
<211> 35
<212> Белок
<213> homo sapiens
<400> 3
Arg Arg Glu Ala Glu Asp Leu Gln Val Gly Gln Val Glu Leu Gly Gly
1 5 10 15
Gly Pro Gly Ala Gly Ser Leu Gln Pro Leu Ala Leu Glu Gly Ser Leu
20 25 30
Gln Ala Arg
35
<210> 4
<211> 21
<212> Белок
<213> homo sapiens
<400> 4
Gly Ile Val Glu Gln Cys Cys Thr Ser Ile Cys Ser Leu Tyr Gln Leu
1 5 10 15
Glu Asn Tyr Cys Asn
20
<210> 5
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 5
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 6
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид (ABD)
<400> 6
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 7
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 7
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 8
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 8
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Glu Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 9
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 9
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Asp Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Glu Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 10
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 10
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Glu Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 11
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 11
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Glu Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Asp Glu Ile Leu Ala Ala Leu Pro
35 40 45
<210> 12
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 12
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Glu Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Glu Ala Ile Leu Ala Ala Leu Pro
35 40 45
<210> 13
<211> 46
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сконструированный альбумин-связывающий полипептид
<400> 13
Leu Ala Glu Ala Lys Glu Ala Ala Asn Ala Glu Leu Asp Ser Tyr Gly
1 5 10 15
Val Ser Asp Phe Tyr Lys Lys Leu Ile Glu Lys Ala Lys Thr Val Glu
20 25 30
Gly Val Glu Ala Leu Lys Glu Glu Ile Leu Ala Ala Leu Pro
35 40 45
<---
Настоящее изобретение относится к области биотехнологии, конкретно к получению производных инсулина, и может быть использовано в медицине для лечения диабета. Предложенное изобретение позволяет получить активную форму производного аналога инсулина длительного действия, таким образом, что аналог инсулина может быть превращен в активную форму без расщепления В-цепи, даже когда он взаимодействует с клострипаином. В традиционном способе превращения проинсулина в активную форму посредством применения трипсина, альбумин-связывающий домен расщепляется, затрудняя превращение производного аналога инсулина длительного действия в активную форму. Способ получения согласно настоящему изобретению преодолевает данную трудность, и, таким образом, он может эффективно использоваться для получения длительно действующего терапевтического средства для лечения диабета. 2 н. и 12 з.п. ф-лы, 12 ил., 5 табл., 10 пр.
1. Способ получения активной формы производного аналога инсулина длительного действия, включающий
- стадию взаимодействия производного аналога инсулина, в котором альбумин-связывающий домен слит с аналогом инсулина посредством линкера, с клострипаином,
где во время реакции с клострипаином добавляют восстанавливающий агент,
где производное аналога инсулина дополнительно приводят во взаимодействие с карбоксипептидазой В (СрВ), которую добавляют во время или после реакции с клострипаином,
где указанный аналог инсулина содержит А-цепь инсулина или ее вариант, вариант В-цепи инсулина и С-цепь инсулина, где А-цепь инсулина или ее вариант
i) представлены SEQ ID NO 4;
ii) представлены аминокислотной последовательностью, содержащей замену треонина (Thr) на аспарагиновую кислоту (Asp) в положении аминокислоты 8 в SEQ ID NO 4; или
iii) представлены аминокислотной последовательностью, содержащей замену тирозина (Tyr) на глутаминовую кислоту (Glu) в положении аминокислоты 14 в SEQ ID NO 4;
где вариант В-цепи инсулина, представленный аминокислотной последовательностью SEQ ID NO 2, аргинин (Arg) в положении аминокислоты 22 В-цепи инсулина заменен лизином (Lys) в нативном инсулине,
где линкер представляет собой полипептидный линкер (GGGGS)n (где n представляет собой целое число, находящееся в интервале от 1 до 6),
где производное аналога инсулина содержит вариант В-цепи инсулина, С-цепь инсулина, А-цепь инсулина или ее вариант, линкер и альбумин-связывающий домен в этом порядке; и
- стадию очистки активной формы производного аналога инсулина длительного действия.
2. Способ по п. 1, в котором альбумин-связывающий домен содержит альбумин-связывающий мотив, представленный следующей аминокислотной последовательностью:
GVSDFYKKLIXaKAKTVEGVEALKXbXcI,
где:
Xa независимо выбран из D и Е,
Xb независимо выбран из D и Е, и
Xc независимо выбран из А и Е.
3. Способ по п. 2, в котором альбумин-связывающий домен содержит следующую аминокислотную последовательность:
LAX3AKX6X7ANX10ELDX14Y-[BM]-LX43X44LP,
где:
[ВМ] представляет собой альбумин-связывающий мотив, как определено в п. 2,
X3 независимо выбран из С, Е, Q и S;
Х6 независимо выбран из С, Е и S;
Х7 независимо выбран из А и S;
Х10 независимо выбран из A, R и S;
Х14 независимо выбран из А, С, K и S;
Х43 независимо выбран из А и K; и
Х44 независимо выбран из А, Е и S.
4. Способ по п. 3, в котором альбумин-связывающий домен представлен аминокислотными последовательностями, выбранными из группы, состоящей из SEQ ID NO: 6-13.
5. Способ п. 4, в котором альбумин-связывающий домен представлен аминокислотной последовательностью SEQ ID NO 6.
6. Способ по п. 1, в котором аналог инсулина и альбумин-связывающий домен слиты друг с другом посредством пептидной связи; полипептидного линкера; или непептидильного линкера, выбранного из группы, состоящей из полиэтиленгликоля, полипропиленгликоля, сополимера этиленгликоля и пропиленгликоля, полиоксиэтилированного полиола, поливинилового спирта, полисахаридов, декстрана, простого поливинилэтилового эфира, биоразлагаемых полимеров, жирных кислот, нуклеотидов, липидных полимеров, хитина, гиалуроновой кислоты и их комбинаций.
7. Способ по п. 1, в котором полипептидный линкер представлен аминокислотной последовательностью SEQ ID NO 5.
8. Способ по п. 1, в котором восстанавливающий агент выбран из группы, состоящей из цистеина, β-меркаптоэтанола, Трис-(2-карбоксиэтил)-фосфин гидрохлорида (ТСЕР), глутатиона (GSH) и дитиотреитола (ДТТ).
9. Способ по п. 8, в котором добавляют от 0,1 до 0,5 мМ ДТТ в качестве восстанавливающего агента.
10. Способ по п. 9, в котором ДТТ добавляют на начальной стадии, на которой производное аналога инсулина взаимодействует с клострипаином, и дополнительно добавляют через 3-6 часов после реакции.
11. Способ по п. 1, в котором клострипаин и/или карбоксипептидазу В (СрВ) приводят во взаимодействие в условиях рН 6,0-9,0.
12. Способ по п. 11, в котором клострипаин и/или карбоксипептидазу В (СрВ) приводят во взаимодействие в условиях рН 6,5-7,5.
13. Способ по п. 1, в котором клострипаин и/или карбоксипептидазу В (СрВ) приводят во взаимодействие в условиях 4-40°С.
14. Активная форма производного аналога инсулина длительного действия, полученная способом по п. 1.
| KR 20150138101 А, 09.12.2015, KATSOYANNIS P.G | |||
| et al., A synthetic human insulin analogue modified at position B-22 | |||
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |
| Chem | |||
| Soc., Perkin Trans | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Машина для добывания торфа и т.п. | 1922 |
|
SU22A1 |

