ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
Данная заявка испрашивает приоритет согласно предварительной заявке на патент США № 62/617157, поданной 12 января 2018 года, которая настоящим включена посредством ссылки во всей своей полноте.
ОПИСАНИЕ ТЕКСТОВОГО ФАЙЛА, ПОДАННОГО В ЭЛЕКТРОННОМ ВИДЕ
Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и настоящим включен посредством ссылки во всей своей полноте. Копия перечня последовательностей в машиночитаемой форме, которая была создана 20 декабря 2018 года, называется A-2189-WO-PCT_SeqList_ST25 и имеет размер 490 килобайт.
ОБЛАСТЬ ТЕХНИКИ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к области биофармацевтических препаратов. В частности, настоящее изобретение относится к антителам, которые специфически связываются с рецептором I типа гипофизарного полипептида человека, активирующего аденилатциклазу (PAC1), и эффективно ингибируют его биологическую активность. Настоящее изобретение также относится к фармацевтическим композициям, содержащим антитела, а также к способам получения и применения таких антител.
ПРЕДПОСЫЛКИ СОЗДАНИЯ ИЗОБРЕТЕНИЯ
Мигрени представляют собой эпизодические головные боли, которые могут включать значительную боль, часто сопровождаются тошнотой, рвотой и чрезвычайной чувствительностью к свету (фотофобией) и звуку (фонофобией), и иногда им предшествуют сенсорные предупреждающие симптомы или признаки (ауры). Мигрень представляет собой широко распространенное во всем мире заболевание: примерно 12% европейской популяции и 18% женщин, 6% мужчин в Соединенных Штатах страдают от приступов мигрени (Lipton et al, Neurology, Vol. 68:343-349, 2007; Lipton et al., Headache, Vol. 41:646-657, 2001). Исследование по оценке распространенности мигрени в Соединенных Штатах показало, что почти у половины популяции пациентов с мигренью было три или более мигреней в месяц (Lipton et al, Neurology, Vol. 68:343-349, 2007). Кроме того, мигрени связаны с рядом психиатрических и медицинских сопутствующих заболеваний, таких как депрессия и сосудистые нарушения (Buse et al., J. Neurol. Neurosurg. Psychiatry, Vol. 81:428-432, 2010; Bigal et al., Neurology, Vol. 72:1864-1871, 2009). Большинство современных способов терапии мигрени либо плохо переносятся, либо являются неэффективными (Loder et al., Headache, Vol. 52:930-945, 2012; Lipton et al, 2001); таким образом, мигрень остается неудовлетворенной медицинской потребностью.
Основной компонент патогенеза мигрени включает активацию тригеминоваскулярной системы. Высвобождение тригеминального и парасимпатического нейротрансмиттеров из периваскулярных нервных волокон (Sánchez-del-Rio and Reuter, Curr. Opin. Neurol., Vol. 17(3):289-93, 2004) приводит к вазодилатации черепных кровеносных сосудов и, как предполагается, это связано с возникновением мигренозных головных болей (Edvinsson, Cephalagia, Vol. 33(13): 1070-1072, 2013; Goadsby et al., New Engl J Med., Vol. 346(4):257-270, 2002).
Гипофизарные активирующие аденилатциклазу полипептиды (PACAP) представляют собой пептиды из 38 аминокислот (PACAP38) или 27 аминокислот (PACAP27), которые впервые были выделены из овечьего гипоталамического экстракта на основании их способности стимулировать образование циклического AMP (cAMP) в клетках передней доли гипофиза (Miyata et al., Biochem Biophys Res Commun., Vol. 164:567-574, 1989; Miyata et al., Biochem Biophys Res Commun., Vol.170:643-648, 1990). PACAP принадлежит к суперсемейству VIP/секретин/глюкагон. Последовательность PACAP27 соответствует 27 N-концевым аминокислотам PACAP38 и характеризуется 68% идентичностью с вазоактивным интестинальным полипептидом (VIP) (Pantaloni et al., J. Biol. Chem., Vol. 271: 22146-22151, 1996; Pisegna and Wank, Proc. Natl. Acad. Sci. USA, Vol. 90: 6345-49, 1993; Campbell and Scanes, Growth Regul., Vol. 2:175-191, 1992). Основной формой пептида PACAP в организме человека является PACAP38, и фармакология PACAP38, как было показано, не отличается от фармакологии PACAP27. Сообщалось о трех рецепторах PACAP: один рецептор, который связывает PACAP с высокой аффинностью и характеризуется гораздо более низкой аффинностью к VIP (рецептор PAC1), и два рецептора, которые одинаково хорошо распознают PACAP и VIP (рецепторы VPAC1 и VPAC2) (Vaudry et al., Pharmacol Rev., Vol. 61:283-357, 2009).
Экспериментальные модели мигрени человека, применяющие PACAP в качестве провокационного вещества для индукции мигренеподобных головных болей, поддерживают подход к антагонизму сигнального пути PACAP/PAC1 в качестве лечения для профилактики мигрени. Уровень PACAP38 повышается в плазме крови во время спонтанных приступов мигрени у пациентов с мигренью, и эти повышенные уровни PACAP38 могут быть нормализованы с помощью суматриптана, терапевтического средства при острой мигрени (Tuka et al., Cephalalgia, Vol. 33: 1085-1095, 2013; Zagami et al., Ann. Clin. Transl. Neurol., Vol.1: 1036-1040, 2014). Инфузия PACAP38 вызывает головные боли у здоровых субъектов и мигренеподобные головные боли у пациентов с мигренью (Schytz et al., Brain, Vol. 132:16-25, 2009; Amin et al., Brain, Vol. 137: 779-794, 2014; Guo et al., Cephalalgia, Vol. 37:125-135, 2017). Однако в той же модели VIP не вызывает мигренеподобных головных болей у пациентов с мигренью (Rahmann et al., Cephalalgia, Vol. 28:226-236, 2008). Отсутствие индукции мигренеподобной головной боли от инфузии VIP предполагает, что влияние пептида PACAP38 опосредовано рецептором PAC1, а не рецепторами VPAC1 или VPAC2, так как VIP характеризуется гораздо более высокой аффинностью к последним двум рецепторам. Это мнение также подтверждается исследованиями на животных, в которых антагонисты рецептора PAC1 ингибируют ноцицептивную активность нейронов в тригеминоцервикальном комплексе в модели мигрени in vivo (Akerman et al., Sci. Transl. Med., Vol. 7: 308ra157, 2015; Hoffmann et al., Cephalalgia, Vol. 37 (1S): 3, Abstract OC-BA-004, 2017). Взятые вместе, эти данные предполагают, что фармакологические средства, которые ингибируют PACAP-активацию рецептора PAC1, обладают потенциалом к лечению мигрени.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение частично основано на разработке и получении антител с высокой аффинностью, которые специфически связываются с PAC1 человека и эффективно ингибируют его. Антитела по настоящему изобретению обладают повышенной ингибирующей активностью против PAC1 человека по сравнению с ранее описанными антителами к PAC1 при значениях IC50 в пикомолярном диапазоне. Выделенные антитела и их антигенсвязывающие фрагменты можно применять для ингибирования, подавления или модулирования биологической активности PAC1 человека, включая ингибирование или снижение индуцированной PACAP активации PAC1, подавление или снижение вазодилатации, а также снижение выраженности или лечение симптомов мигрени и других сосудистых головных болей.
В некоторых вариантах осуществления выделенные антитела и их антигенсвязывающие фрагменты специфически связываются с PAC1 человека по эпитопу, который содержит одну или несколько аминокислот, выбранных из Asp59, Asn60, Ile61, Arg116, Asn117, Thr119, Glu120, Asp121, Gly122, Trp123, Ser124, Glu125, Pro126, Phe127, Pro128, His129, Tyr130, Phe131, Asp132, и Gly135 под SEQ ID NO: 1. В определенных вариантах осуществления эпитоп содержит по меньшей мере аминокислоты Asn60, Ile61, Glu120 и Asp121 PAC1 человека. В этих и других вариантах осуществления антитела и антигенсвязывающие фрагменты по настоящему изобретению содержат специфические аминокислоты в конкретных положениях в пределах вариабельных областей легкой цепи и тяжелой цепи, которые взаимодействуют с этими остатками эпитопа. Например, в одном варианте осуществления антитела или антигенсвязывающие фрагменты содержат вариабельную область легкой цепи, в которой аминокислота в положении 29 в соответствии с нумерацией AHo представляет собой основную аминокислоту (например, аргинин или лизин), которая взаимодействует с аминокислотами Glu120 или Asp121 PAC1 человека. В другом варианте осуществления антитела или антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, в которой аминокислота в положении 61 в соответствии с нумерацией AHo представляет собой гидрофобную, основную или нейтральную гидрофильную аминокислоту (например, изолейцин, валин, лейцин, глутамин, аспарагин, аргинин или лизин), которая взаимодействует с аминокислотами Asn60 или Ile61 PAC1 человека. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, в которой аминокислота в положении 66 в соответствии с нумерацией AHo представляет собой основную или нейтральную гидрофильную аминокислоту (например, глутамин, аспарагин, аргинин или лизин), которая взаимодействует с аминокислотами Asn60 или Ile61 PAC1 человека.
Антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению способны ингибировать индуцированную лигандом активацию рецептора PAC1. Например, в некоторых вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты ингибируют индуцированную PACAP активацию PAC1 человека при IC50, составляющей менее 500 пМ, как измерено с помощью клеточного анализа cAMP. В других вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты ингибируют индуцированную PACAP активацию PAC1 человека при IC50, составляющей менее 300 пМ, как измерено с помощью клеточного анализа cAMP. В определенных вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей от приблизительно 50 пМ до приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP.
В некоторых вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению перекрестно реагируют с рецепторами PAC1 из других видов. В одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты специфически связываются с рецептором PAC1 яванского макака и ингибируют его индуцированную PACAP активацию. В таком варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты способны ингибировать индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей от приблизительно 0,1 нМ до приблизительно 1 нМ или от приблизительно 50 пМ до приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP. В другом варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты специфически связываются с рецептором PAC1 крысы и ингибируют его индуцированную PACAP активацию. Антитела к PAC1 или антигенсвязывающие фрагменты способны ингибировать индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее 10 нМ, например, при IC50, составляющей от приблизительно 0,1 нМ до приблизительно 10 нМ или от приблизительно 100 пМ до приблизительно 2 нМ, как измерено с помощью клеточного анализа cAMP.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую определяющие комплементарность области CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую определяющие комплементарность области CDRH1, CDRH2 и CDRH3. Вариабельные области легкой цепи и тяжелой цепи или CDR могут происходить из любого из антител к PAC1, описанных в данном документе. Например, в некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 5-16; CDRL2, содержащую последовательность под SEQ ID NO: 26; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 36-38; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 88-96; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 106-166; и CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 171-177. В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 17-25; CDRL2, содержащую последовательность, выбранную из SEQ ID NO: 27-35; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 39-51; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 97-105; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 167-170; CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 178-190.
В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична или на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 54-66 и SEQ ID NO: 68-87. В этих и других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична или на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 191-312. В одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат вариабельную область легкой цепи, содержащую последовательность, выбранную из SEQ ID NO: 54-66 и вариабельную область тяжелой цепи, содержащую последовательность, выбранную из SEQ ID NO: 191-295. В другом варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат вариабельную область легкой цепи, содержащую последовательность, выбранную из SEQ ID NO: 68-87 и вариабельную область тяжелой цепи, содержащую последовательность, выбранную из SEQ ID NO: 296-312.
В любом из вариантов осуществления, описанных в данном документе, включая варианты осуществления, описанные выше, антитело к PAC1 или антигенсвязывающий фрагмент по настоящему изобретению представляет собой моноклональное антитело или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления моноклональное антитело или его антигенсвязывающий фрагмент представляют собой химерное антитело или его антигенсвязывающий фрагмент. В других вариантах осуществления моноклональное антитело или его антигенсвязывающий фрагмент представляют собой гуманизированное антитело или его антигенсвязывающий фрагмент. В еще одних вариантах осуществления моноклональное антитело или его антигенсвязывающий фрагмент представляют собой полностью человеческое антитело или его антигенсвязывающий фрагмент. Моноклональное антитело может быть любого изотипа, такого как IgG1, IgG2, IgG3 или IgG4 человека. В одном конкретном варианте осуществления моноклональное антитело представляет собой антитело IgG1 человека. В другом конкретном варианте осуществления моноклональное антитело представляет собой антитело IgG2 человека. Моноклональное антитело может содержать легкую цепь, которая содержит константную область каппа-цепи человека. В некоторых вариантах осуществления константная область каппа-цепи человека содержит последовательность под SEQ ID NO: 318 или под SEQ ID NO: 319. Таким образом, антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут содержать легкую цепь, которая содержит любую из последовательностей вариабельных областей легкой цепи, перечисленных в таблице 1A, слитую с константной областью каппа-цепи человека, содержащей последовательность под SEQ ID NO: 318 или SEQ ID NO: 319. В других вариантах осуществления моноклональное антитело может содержать легкую цепь, которая содержит константную область лямбда-цепи человека. В определенных вариантах осуществления константная область лямбда-цепи человека содержит последовательность под SEQ ID NO: 315. Таким образом, в некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут содержать легкую цепь, которая содержит любую из последовательностей вариабельных областей легкой цепи, перечисленных в таблице 1A, слитую с константной областью лямбда-цепи человека, содержащей последовательность под SEQ ID NO: 315.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут содержать одну или несколько модификаций, которые влияют на гликозилирование антитела или антигенсвязывающего фрагмента. В некоторых вариантах осуществления антитело или антигенсвязывающий фрагмент содержит одну или несколько мутаций для снижения или устранения гликозилирования. В таких вариантах осуществления агликозилированное антитело может содержать в своей тяжелой цепи мутацию по аминокислотному положению N297 (согласно схеме нумерации EU), такую как мутация N297G. Агликозилированное антитело может содержать дополнительные мутации для стабилизации структуры антитела. Такие мутации могут включать пары замен цистеина, такие как A287C и L306C, V259C и L306C, R292C и V302C, а также V323C и I332C (аминокислотные положения согласно схеме нумерации EU). В одном варианте осуществления агликозилированное антитело содержит мутации R292C и V302C (согласно схеме нумерации EU) в своей тяжелой цепи. В определенных вариантах осуществления агликозилированное антитело к PAC1 содержит константную область тяжелой цепи, содержащую аминокислотную последовательность под SEQ ID NO: 324 или SEQ ID NO: 325.
Настоящее изобретение также включает выделенные полинуклеотиды и векторы экспрессии, кодирующие антитела к PAC1 и антигенсвязывающие фрагменты, описанные в данном документе, а также клетки-хозяева, такие как клетки CHO, содержащие кодирующие полинуклеотиды и векторы экспрессии. В определенных вариантах осуществления настоящее изобретение включает способы получения антител к PAC1 и антигенсвязывающих фрагментов, описанных в данном документе. В одном варианте осуществления способ включает культивирование клетки-хозяина, содержащей вектор экспрессии, кодирующий антитело к PAC1 или антигенсвязывающий фрагмент, в условиях, которые обеспечивают экспрессию антитела или антигенсвязывающего фрагмента, и выделение антитела или антигенсвязывающего фрагмента из культуральной среды или клетки-хозяина.
Антитела к PAC1 или антигенсвязывающие фрагменты, описанные в данном документе, можно применять для производства фармацевтической композиции или лекарственного препарата для лечения или предупреждения состояний, связанных с биологической активностью PAC1, таких как головная боль, мигрень, кластерная головная боль и вазомоторные симптомы. Таким образом, в настоящем изобретении также предусмотрена фармацевтическая композиция, содержащая антитело к PAC1 или антигенсвязывающий фрагмент, описанный в данном документе, и фармацевтически приемлемое вспомогательное вещество. Фармацевтические композиции можно применять в любом из способов, описанных в данном документе.
В определенных вариантах осуществления в настоящем изобретении предусмотрены способы лечения или предупреждения состояния, связанного с головной болью, у пациента, нуждающегося в этом, включающие введение пациенту эффективного количества антитела к PAC1 или антигенсвязывающего фрагмента, описанного в данном документе. В некоторых вариантах осуществления состояние, связанное с головной болью, подлежащее лечению или предупреждению с помощью способов по настоящему изобретению, представляет собой мигрень. Мигрень может представлять собой эпизодическую мигрень или хроническую мигрень. В других вариантах осуществления состояние, связанное с головной болью, подлежащее лечению или предупреждению с помощью способов по настоящему изобретению, представляет собой кластерную головную боль. В определенных вариантах осуществления в способах предусмотрено профилактическое лечение этих состояний.
В некоторых вариантах осуществления способов по настоящему изобретению способы включают введение пациенту второго терапевтического средства от головной боли в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению. Второе терапевтическое средство от головной боли может представлять собой терапевтическое средство от острой головной боли, такое как агонист рецепторов серотонина (например, агонист рецептора серотонина 5HT1B, 5HT1D и/или 5HT1F). В некоторых вариантах осуществления терапевтическое средство от острой головной боли представляет собой триптан, эрготамин, нестероидное противовоспалительное лекарственное средство или опиоид. В других вариантах осуществления второе терапевтическое средство от головной боли, подлежащее введению в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой профилактическое терапевтическое средство от головной боли, такое как противоэпилептическое средство, бета-блокатор, антидепрессант или онаботулотоксин А. Второе терапевтическое средство от головной боли можно вводить пациенту до, после или одновременно с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению.
В определенных вариантах осуществления способов по настоящему изобретению способы включают введение пациенту антагониста пути CGRP в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению. Антагонист пути CGRP может являться антагонистом рецептора CGRP человека, таким как антитело, которое специфически связывается с рецептором CGRP человека. В одном варианте осуществления антагонист пути CGRP, вводимый в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой эренумаб. В других вариантах осуществления антагонист пути CGRP может являться антагонистом лиганда CGRP, таким как антитело, которое специфически связывается с α-CGRP и/или β-CGRP человека. В одном таком варианте осуществления антагонист пути CGRP, вводимый в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой фреманезумаб. В другом таком варианте осуществления антагонист пути CGRP, вводимый в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой галканезумаб. В еще одном таком варианте осуществления антагонист пути CGRP, вводимый в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой эптинезумаб.
Настоящее изобретение также включает способы подавления вазодилатации у пациента, нуждающегося в этом. В одном варианте осуществления способ включает введение пациенту эффективного количества любого из антител к PAC1 или антигенсвязывающих фрагментов, описанных в данном документе. В некоторых вариантах осуществления пациент, нуждающийся в лечении, характеризуется состоянием, связанным с головной болью, таким как мигрень или кластерная головная боль. В других вариантах осуществления пациент, нуждающийся в лечении, характеризуется вазомоторными симптомами (например, приливами, покраснением лица, потливостью и ночными потливостями), такими как симптомы, связанные с менопаузой.
Конкретно предусмотрено применение антител к PAC1 или антигенсвязывающих фрагментов в любом из способов, раскрытых в данном документе, или для получения лекарственных препаратов для введения в соответствии с любым из способов, раскрытых в данном документе. Например, настоящее изобретение включает антитело к PAC1 или антигенсвязывающий фрагмент для применения в способе лечения или предупреждения состояния, связанного с головной болью, у пациента, нуждающегося в этом. Состояние, связанное с головной болью, включает мигрень (например, эпизодическую и хроническую мигрень) и кластерную головную боль. В некоторых вариантах осуществления в настоящем изобретении предусмотрено антитело к PAC1 или антигенсвязывающий фрагмент для применения в способе подавления вазодилатации у пациента, нуждающегося в этом. В таких вариантах осуществления у пациента может быть диагностировано состояние, связанное с головной болью, или он может характеризоваться состоянием, связанным с головной болью. В других вариантах осуществления в настоящем изобретении предусмотрено антитело к PAC1 или антигенсвязывающий фрагмент для применения в способе подавления активации рецептора PAC1 человека у пациента, характеризующегося состоянием, связанным с головной болью. Состояние, связанное с головной болью, может представлять собой мигрень (например, эпизодическую или хроническую мигрень) или кластерную головную боль.
Настоящее изобретение также включает применение антитела к PAC1 или антигенсвязывающего фрагмента для получения лекарственного препарата для лечения или предупреждения состояния, связанного с головной болью, у пациента, нуждающегося в этом. Состояние, связанное с головной болью, включает мигрень (например, эпизодическую и хроническую мигрень) и кластерную головную боль. В определенных вариантах осуществления настоящее изобретение охватывает применение антитела к PAC1 или антигенсвязывающего фрагмента для получения лекарственного препарата для подавления вазодилатации у пациента, нуждающегося в этом. В таких вариантах осуществления у пациента может быть диагностировано состояние, связанное с головной болью, или он может характеризоваться состоянием, связанным с головной болью. В других вариантах осуществления настоящее изобретение включает применение антитела к PAC1 или антигенсвязывающего фрагмента для получения лекарственного препарата для подавления активации рецептора PAC1 человека у пациента, характеризующегося состоянием, связанным с головной болью. Состояние, связанное с головной болью, может представлять собой мигрень (например, эпизодическую или хроническую мигрень) или кластерную головную боль.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Фигура 1A представляет собой вид спереди кристаллической структуры комплекса между внеклеточным доменом (ECD) PAC1 человека и Fab-фрагментом антитела 29G4v9. VL=вариабельная область легкой цепи; CL=константная область легкой цепи; VH=вариабельная область тяжелой цепи; и CH1=константная область тяжелой цепи CH1.
Фигура 1B представляет собой вид сбоку кристаллической структуры комплекса между ECD PAC1 человека и Fab-фрагментом антитела 29G4v9. Вид отображает структуру, показанную на фиг. 1А, повернутую на 90° влево. Данный вид демонстрирует, что вариабельная область легкой цепи (VL) расположена позади вариабельной области тяжелой цепи (VH), а константная область легкой цепи (CL) расположена позади константной области тяжелой цепи CH1.
Фигура 2A представляет собой расширенный вид участка контакта между ECD PAC1 человека и CDR1 легкой цепи (аминокислоты 24-34 из SEQ ID NO: 3) Fab 29G4v9. Зона 1 содержит остатки Glu120 и Asp121 ECD PAC1 человека (SEQ ID NO: 1), тогда как зона 3 содержит остаток Phe127 ECD PAC1 человека.
Фигура 2B представляет собой расширенный вид участка контакта между ECD PAC1 человека и остатком Arg93 в CDR3 легкой цепи из Fab 29G4v9.
Фигура 3A представляет собой расширенный вид участка контакта между ECD PAC1 человека и аминокислотами Arg31 и Phe32 CDR1 тяжелой цепи из Fab 29G4v9. Две аминокислоты CDR1 тяжелой цепи лежат по обе стороны от остатка Phe131 ECD PAC1 человека.
Фигура 3B представляет собой расширенный вид участка контакта между областью ECD PAC1 человека, содержащей аминокислотные остатки Asn60 и Ile61 (относительно SEQ ID NO: 1), и аминокислотами Tyr53, Asp54 и Gly56 CDR2 тяжелой цепи из Fab 29G4v9.
Фигура 3C представляет собой расширенный вид участка контакта между ECD PAC1 человека и аминокислотами Val102, Leu103 и Thr104 CDR3 тяжелой цепи из Fab 29G4v9.
Фигура 4 представляет собой схему процесса отбора мутантных вариантов с улучшенным связыванием из библиотеки мутантных вариантов Fab антител на основе дрожжевого дисплея.
Фигура 5 представляет собой схему основывающегося на скорости диссоциации процесса отбора мутантных вариантов с высокой аффинностью связывания из библиотеки на основе дрожжевого дисплея.
На Фигуре 6 показан ингибирующий эффект антител к PAC1 при индуцированном максадиланом увеличении притока крови к коже у крыс. Одно из семи антител (420653, 420845, 420943, 421873, 420889 (PL-50347), 421091 (PL-50350) и 421051 (PL-50351)) вводили крысам внутривенно в дозе 0,1 мг/кг или 0,3 мг/кг за двадцать четыре часа до стимуляции 10 нг максадилана (внутрикожная инъекция). Приток крови к коже оценивали через 30 минут после стимуляции максадиланом посредством лазерной доплеровской визуализации. *p<0,05, **p<0,01 по сравнению с группой, получавшей носитель, с помощью однофакторного ANOVA с последующим MCT Даннетта.
На фигуре 7A показан дозозависимый эффект антитела 420653 к PAC1 при индуцированном максадиланом увеличении притока крови к коже у крыс. Антитело вводили крысам внутривенно в одной из семи доз, находящихся в диапазоне 0,01-3 мг/кг, за двадцать четыре часа до стимуляции 10 нг максадилана (внутрикожная инъекция). Приток крови к коже оценивали через 30 минут после стимуляции максадиланом посредством лазерной доплеровской визуализации. *p<0,05, ****p<0,0001 по сравнению с группой, получавшей носитель, с помощью однофакторного ANOVA с последующим MCT Даннетта.
На фигуре 7B показан дозозависимый эффект антитела 420845 к PAC1 при индуцированном максадиланом увеличении притока крови к коже у крыс. Антитело вводили крысам внутривенно в одной из пяти доз, находящихся в диапазоне 0,01-3 мг/кг, за двадцать четыре часа до стимуляции 10 нг максадилана (внутрикожная инъекция). Приток крови к коже оценивали через 30 минут после стимуляции максадиланом посредством лазерной доплеровской визуализации. ****p<0,0001 по сравнению с группой, получавшей носитель, с помощью однофакторного ANOVA с последующим MCT Даннетта.
На фигуре 7C показан дозозависимый эффект антитела 420943 к PAC1 при индуцированном максадиланом увеличении притока крови к коже у крыс. Антитело вводили крысам внутривенно в одной из шести доз, находящихся в диапазоне 0,1-30 мг/кг, за двадцать четыре часа до стимуляции 10 нг максадилана (внутрикожная инъекция). Приток крови к коже оценивали через 30 минут после стимуляции максадиланом посредством лазерной доплеровской визуализации. ****p<0,0001 по сравнению с группой, получавшей носитель, с помощью однофакторного ANOVA с последующим MCT Даннетта.
На фигуре 7D показан дозозависимый эффект антитела 421873 к PAC1 при индуцированном максадиланом увеличении притока крови к коже у крыс. Антитело вводили крысам внутривенно в одной из пяти доз, находящихся в диапазоне 0,3-30 мг/кг, за двадцать четыре часа до стимуляции 10 нг максадилана (внутрикожная инъекция). Приток крови к коже оценивали через 30 минут после стимуляции максадиланом посредством лазерной доплеровской визуализации. *p<0,05, ****p<0,0001 по сравнению с группой, получавшей носитель, с помощью однофакторного ANOVA с последующим MCT Даннетта.
Фигура 8A представляет собой профиль зависимости концентрации в сыворотке крови от времени для антител 420653, 420845, 420943 и 421873 к PAC1 после введения однократной внутривенной дозы 1 мг/кг у крыс.
Фигура 8B представляет собой профиль зависимости концентрации в сыворотке крови от времени для антител 420653, 420845, 420943 и 421873 к PAC1 после введения однократной внутривенной дозы 5 мг/кг у крыс.
Фигура 8C представляет собой профиль зависимости концентрации в сыворотке крови от времени для антител 420653, 420845, 420943 и 421873 к PAC1 после введения однократной внутривенной дозы 25 мг/кг у крыс.
Фигура 9 представляет собой профиль зависимости концентрации в сыворотке крови от времени для антител 420653, 420845, 420943, 421873 и 29G4v22 к PAC1 после введения однократной внутривенной дозы 10 мг/кг у яванских макаков.
На фигурах 10A и 10B во временной динамике показаны ингибирующие эффекты антител 420653 и 29G4v22 к PAC1 на индуцированное максадиланом увеличение притока крови к коже у яванских макаков. Двадцать четыре яванских макака получили однократную внутривенную болюсную инъекцию антитела 420653 в дозе 0,1 мг/кг или 3 мг/кг или антитела 29G4v22 в дозе 10 мг/кг в день 0. В дни 2, 4, 7, 10, 14, 21, 28 и 36 после введения доз антител измеряли ответы после введения максадилана. В каждом случае приток крови к коже измеряли посредством лазерной доплеровской визуализации через 30 минут после внутрикожной инъекции 1 нг максадилана. Все данные выражены в виде среднего значения ± стандартная ошибка среднего значения. На фигуре 10A показано изменение в % от исходного уровня притока крови к коже, индуцированное внутрикожной инъекцией максадилана в указанные дни после обработки антителами. *p<0,05, **p<0,01, ***p<0,001 по сравнению с днем 0 в той же группе обработки с помощью однофакторного ANOVA с последующим применением критерия множественных сравнений Бонферрони. Восемь животных в группе за исключением дня 36, когда четыре животных тестировали с применением антитела 420653 в дозе 3 мг/кг, как обозначено символом "^". На фигуре 10B показан ингибирующий эффект обработки антителами на индуцированное максадиланом увеличение притока крови к коже, выраженный в виде ингибирования в %. #p<0,05, ##p<0,01, ###p<0,001 сравнение между антителом 420653 при 3 мг/кг и антителом 29G4v22 при 10 мг/кг в каждый момент времени с помощью двухвыборочного непарного t-теста.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к выделенным антителам и их антигенсвязывающим фрагментам, которые специфически связываются с рецептором I типа гипофизарного полипептида, активирующего аденилатциклазу (PAC1), в частности PAC1 человека. Антитела по настоящему изобретению обладают повышенной ингибирующей активностью против PAC1 человека по сравнению с ранее описанными антагонистическими антителами к PAC1. Антитела по настоящему изобретению также перекрестно реагируют с рецептором PAC1 крысы и рецептором PAC1 яванского макака, что позволяет проводить доклиническую оценку in vivo антител у этих видов.
PAC1 человека представляет собой белок из 468 аминокислот (эталонная последовательность NCBI NP_001109.2), кодируемый геном ADCYAP1R1 на хромосоме 7. Рецептор PAC1 человека представляет собой сопряженный с G-белком рецептор, который положительно связан с аденилатциклазой. Активация рецептора PAC1 человека его эндогенными лигандами (например, PACAP38 или PACAP27) приводит к повышению уровня внутриклеточного циклического AMP (cAMP). Аминокислотная последовательность PAC1 человека представлена ниже под SEQ ID NO: 1.
1 MAGVVHVSLA ALLLLPMAPA MHSDCIFKKE QAMCLEKIQR ANELMGFNDS SPGCPGMWDN
61 ITCWKPAHVG EMVLVSCPEL FRIFNPDQVW ETETIGESDF GDSNSLDLSD MGVVSRNCTE
121 DGWSEPFPHY FDACGFDEYE SETGDQDYYY LSVKALYTVG YSTSLVTLTT AMVILCRFRK
181 LHCTRNFIHM NLFVSFMLRA ISVFIKDWIL YAEQDSNHCF ISTVECKAVM VFFHYCVVSN
241 YFWLFIEGLY LFTLLVETFF PERRYFYWYT IIGWGTPTVC VTVWATLRLY FDDTGCWDMN
301 DSTALWWVIK GPVVGSIMVN FVLFIGIIVI LVQKLQSPDM GGNESSIYLR LARSTLLLIP
361 LFGIHYTVFA FSPENVSKRE RLVFELGLGS FQGFVVAVLY CFLNGEVQAE IKRKWRSWKV
421 NRYFAVDFKH RHPSLASSGV NGGTQLSILS KSSSQIRMSG LPADNLAT
Аминокислоты 1-23 белка PAC1 человека (SEQ ID NO: 1) составляют сигнальный пептид, который обычно удален из зрелого белка. Зрелый белок PAC1 человека имеет основную структуру сопряженного с G-белком рецептора, состоящего из семи трансмембранных доменов, внеклеточного домена, состоящего из N-концевой области и трех внеклеточных петель, трех внутриклеточных петель и С-концевого цитоплазматического домена. N-концевой внеклеточный домен примерно находится в области, состоящей из аминокислот 24-153 из SEQ ID NO: 1, и первый из семи трансмембранных доменов начинается с аминокислоты 154 из SEQ ID NO: 1. С-концевой цитоплазматический домен примерно расположен в области, состоящей из аминокислот 397-468 из SEQ ID NO: 1. См. Blechman and Levkowitz, Front. Endocrinol., Vol. 4 (55): 1-19, 2013 для определения расположения доменов в пределах аминокислотной последовательности. Термины "PAC1 человека", "рецептор PAC1 человека", "hPAC1" и "рецептор hPAC1" применяются взаимозаменяемо и могут относиться к полипептиду под SEQ ID NO: 1, полипептиду под SEQ ID NO: 1 минус сигнальный пептид (аминокислоты 1-23), аллельным вариантам PAC1 человека или сплайс-вариантам PAC1 человека.
В настоящем изобретении предусмотрены антитела, которые специфически связываются с PAC1 человека. "Антитело" представляет собой белок, который содержит антигенсвязывающий фрагмент, который специфически связывается с антигеном, и остов или часть каркаса, что позволяет антигенсвязывающему фрагменту принимать конформацию, которая способствует связыванию антитела с антигеном. Применяемый в данном документе термин "антитело" обычно относится к тетрамерному белку-иммуноглобулину, содержащему два полипептида легкой цепи (приблизительно 25 кДа каждый) и два полипептида тяжелой цепи (приблизительно 50-70 кДа каждый). Термин "легкая цепь" или "легкая цепь иммуноглобулина" относится к полипептиду, содержащему от аминоконца к карбоксильному концу одну вариабельную область легкой цепи (VL) иммуноглобулина и один константный домен легкой цепи (CL) иммуноглобулина. Константный домен легкой цепи (CL) иммуноглобулина может представлять собой константный домен каппа-цепи (κ) человека или константный домен лямбда-цепи (λ) человека. Термин "тяжелая цепь" или "тяжелая цепь иммуноглобулина" относится к полипептиду, содержащему от аминоконца к карбоксильному концу одну вариабельную область тяжелой цепи иммуноглобулина (VH), константный домен 1 тяжелой цепи иммуноглобулина (CH1), шарнирную область иммуноглобулина, константный домен 2 тяжелой цепи иммуноглобулина (CH2), константный домен 3 тяжелой цепи иммуноглобулина (CH3) и необязательно константный домен 4 тяжелой цепи иммуноглобулина (CH4). Тяжелые цепи классифицируют как мю- (μ), дельта- (Δ), гамма- (γ), альфа- (α) и эпсилон-(ε) цепи, и они определяют изотип антитела как IgM, IgD, IgG, IgA и IgE соответственно. Классы антител IgG и IgA дополнительно делят на подклассы, а именно IgG1, IgG2, IgG3 и IgG4, а также IgA1 и IgA2 соответственно. Тяжелые цепи в антителах IgG, IgA и IgD содержат три константных домена (СН1, СН2 и СН3), тогда как тяжелые цепи в антителах IgM и IgE содержат четыре константных домена (СН1, СН2, СН3 и СН4). Константные домены тяжелой цепи иммуноглобулина могут происходить из иммуноглобулина любого изотипа, включая подтипы. Цепи антитела связаны друг с другом посредством межполипептидных дисульфидных связей между CL-доменом и CH1-доменом (т. е. между легкой и тяжелой цепями) и между шарнирными областями двух тяжелых цепей антитела.
Настоящее изобретение также включает антигенсвязывающие фрагменты антител к PAC1, описанных в данном документе. "Антигенсвязывающий фрагмент", применяемый в данном документе взаимозаменяемо со "связывающим фрагментом" или "фрагментом", представляет собой часть антитела, в которой отсутствуют по меньшей мере некоторые аминокислоты, присутствующие в полноразмерной тяжелой цепи и/или легкой цепи, но которая все еще способна специфически связываться с антигеном. Антигенсвязывающий фрагмент включает без ограничения одноцепочечный вариабельный фрагмент (scFv), нанотело (например, VH-домен тяжелой цепи верблюжьих антител; VHH-фрагмент , см. Cortez-Retamozo et al., Cancer Research, Vol. 64:2853-57, 2004), Fab-фрагмент, Fab'-фрагмент, F(ab')2-фрагмент, Fv-фрагмент, Fd-фрагмент и фрагмент определяющей комплементарность области (CDR), и может быть получен из любого источника, представляющего собой млекопитающее, такое как человек, мышь, крыса, кролик или верблюд. Антигенсвязывающие фрагменты могут конкурировать за связывание целевого антигена с интактным антителом, и фрагменты могут быть получены посредством модификации интактных антител (например, посредством ферментативного или химического расщепления) или синтеза de novo с применением технологий рекомбинантной ДНК или синтеза пептидов. В некоторых вариантах осуществления антигенсвязывающий фрагмент содержит по меньшей мере одну CDR из антитела, которое связывается с антигеном, например, CDR3 тяжелой цепи из антитела, которое связывается с антигеном. В других вариантах осуществления антигенсвязывающий фрагмент содержит все три CDR из тяжелой цепи антитела, которое связывается с антигеном, или все три CDR из легкой цепи антитела, которое связывается с антигеном. В еще других вариантах осуществления антигенсвязывающий фрагмент содержит все шесть CDR из антитела, которое связывается с антигеном (три из тяжелой цепи и три из легкой цепи).
Термин "выделенная молекула" (где молекула представляет собой, например, полипептид, полинуклеотид, антитело или антигенсвязывающий фрагмент) означает молекулу, которая в силу своего происхождения или источника получения (1) не связана со связанными естественным путем компонентами, которые сопровождают ее в ее природном состоянии, (2) практически не содержит других молекул того же вида, (3) экспрессируется клеткой другого вида или (4) не встречается в природе. Таким образом, молекула, которая синтезирована химическим путем или экспрессируется в клеточной системе, отличной от клетки, из которой она происходит естественным путем, будет считаться "выделенной" из ее связанных естественным путем компонентов. Молекулу также можно сделать практически свободной от связанных естественным путем компонентов посредством выделения с применением методик очистки, хорошо известных из уровня техники. Чистоту или гомогенность молекулы можно оценивать с помощью ряда средств, хорошо известных из уровня техники. Например, чистоту образца полипептида можно оценивать с помощью электрофореза в полиакриламидном геле и окрашивания геля для визуализации полипептида с применением методик, хорошо известных из уровня техники. Для определенных целей более высокую разрешающую способность можно обеспечивать посредством применения HPLC или других средств для очистки, хорошо известных из уровня техники.
В определенных вариантах осуществления настоящего изобретения антитела или их антигенсвязывающие фрагменты специфически связываются с PAC1 человека. Антитело или его антигенсвязывающий фрагмент "специфически связывается" с целевым антигеном, если он обладает значительно более высокой аффинностью связывания и, следовательно, способен отличать этот антиген по сравнению с его аффинностью к другим неродственным белкам в аналогичных условиях анализа связывания. Антитела или антигенсвязывающие фрагменты, которые специфически связывают антиген, могут характеризоваться равновесной константой диссоциации (KD), составляющей ≤ 1×10-6 M. Антитело или связывающий фрагмент специфически связывает антиген с "высокой аффинностью", если KD составляет ≤ 1×10-8 M. В одном варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 5×10-9 M. В другом варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 1×10-9 M. В еще одном варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 5×10-10 M. В другом варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 1×10-10 M. В определенных вариантах осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 5×10-11 M. В других вариантах осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 1×10-11 M. В одном конкретном варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 5×10-12 M. В другом конкретном варианте осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека с KD, составляющей ≤ 1×10-12 M.
Аффинность определяют с применением различных методик, примером которых является анализ аффинности ELISA. В различных вариантах осуществления аффинность определяют с помощью анализа поверхностного плазмонного резонанса (например, анализ на основе BIAcore®). С применением этой методологии можно измерить константу скорости ассоциации (ka в M-1с-1) и константу скорости диссоциации (kd в с-1). Равновесную константу диссоциации (KD в M) можно затем рассчитать из отношения кинетических констант скорости (kd/ka). В некоторых вариантах осуществления аффинность определяют посредством кинетического способа, такого как анализ кинетического исключения (KinExA), как описано в Rathanaswami et al. Analytical Biochemistry, Vol. 373:52-60, 2008. С помощью анализа KinExA можно измерять равновесную константу диссоциации (KD в M) и константу скорости ассоциации (ka в M-1с-1). Константа скорости диссоциации (kd в с-1) может быть вычислена из этих значений (KD x ka). В других вариантах осуществления аффинность определяют с помощью способа интерферометрии биослоя, такого как описанный в Kumaraswamy et al., Methods Mol. Biol., Vol. 1278:165-82, 2015 и используемый в системах Octet® (Pall ForteBio). Кинетические константы (ka и kd) и константы аффинности (KD) могут быть рассчитаны в режиме реального времени с помощью способа интерферометрии биослоя. В некоторых вариантах осуществления антитела или связывающие фрагменты, описанные в данном документе, проявляют требуемые характеристики, такие как авидность связывания, измеренную с помощью kd (константы скорости диссоциации) для PAC1 человека, составляющей приблизительно 10-2, 10-3, 10-4, 10-5, 10-6, 10-7, 10-8, 10-9, 10-10 s-1 или ниже (более низкие значения указывают на более высокую авидность связывания) и/или аффинность связывания, измеренную с помощью KD (равновесной константы диссоциации) для PAC1 человека, составляющей приблизительно 10-8, 10-9, 10-10, 10-11, 10-12 M или ниже (более низкие значения указывают на более высокую аффинность связывания).
Предпочтительно, антитела или связывающие фрагменты по настоящему изобретению не связываются в значительной степени или перекрестно не реагируют с другими представителями семейства рецепторов секретина/глюкагона, такими как рецептор VPAC1 человека (эталонная последовательность в NCBI NP_004615.2) и рецептор VPAC2 человека (эталонная последовательность в NCBI NP_003373.2). Как применяется в настоящем документе, антитело или связывающий фрагмент "не связывается в значительной степени" с целевым антигеном, если он обладает аффинностью связывания с этим антигеном, которая сравнима с его аффиностью к другим неродственным белкам в аналогичных условиях анализа связывания. Антитела или связывающие фрагменты, которые не связываются в значительной степени с целевым антигеном, могут также включать такие белки, которые не генерируют статистически отличающийся от отрицательного контроля сигнал при анализе аффинности, такой как описанный в данном документе для целевого антигена. В качестве примера антитело, которое генерирует значение сигнала при анализе на основе ELISA или BIAcore® для определения связывания с VPAC1 человека, которое статистически не отличается от значения сигнала, полученного с отрицательным контролем (например, буферным раствором без антитела), будет считаться не связывающимся в значительной степени с VPAC1 человека. Антитела или связывающие фрагменты, которые не связывают в значительной степени антиген, могут характеризоваться равновесной константой диссоциации (KD) для этого антигена, составляющей более 1×10-6 M, более 1×10-5 M, более 1×10-4 M или более 1×10-3 M. Таким образом, в определенных вариантах осуществления антитела и связывающие фрагменты по настоящему изобретению селективно связываются с PAC1 человека относительно VPAC1 человека и VPAC2 человека. Другими словами, антитела и связывающие фрагменты по настоящему изобретению не связываются в значительной степени с VPAC1 человека или VPAC2 человека.
Антитела и антигенсвязывающие фрагменты по настоящему изобретению способны ингибировать, подавлять или модулировать один или несколько видов биологической активности рецептора PAC1 человека. Биологические активности рецептора PAC1 человека включают без ограничения индукцию путей передачи сигнала, опосредованной рецептором PACAP, индукцию вазодилатации и подавление вазоконстрикции. В некоторых вариантах осуществления антитела или связывающие фрагменты по настоящему изобретению ингибируют связывание PACAP (например, PACAP38 или PACAP27) с рецептором PAC1 человека. "Ингибирование связывания" происходит, когда избыток антител или связывающих фрагментов уменьшает количество рецептора PAC1 человека, связанного с PACAP, или наоборот, например, на по меньшей мере приблизительно 40%, приблизительно 50%, приблизительно 60%, приблизительно 70% приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95%, приблизительно 97%, приблизительно 99% или более, например, согласно измерению связывания с помощью анализа конкурентного связывания in vitro. Константы ингибирования (Ki), которые указывают на то, насколько эффективны антитела или антигенсвязывающие фрагменты по настоящему изобретению при предотвращении связывания PACAP с рецептором PAC1 человека, можно рассчитать из таких анализов конкурентного связывания. В качестве примера, радиоактивный изотоп (например, 125I) присоединяют к лиганду рецептора (например, PACAP38) и с помощью анализа измеряют степень связывания помеченного радиоактивной меткой лиганда с рецептором PAC1 человека при увеличивающихся концентрациях антитела к PAC1 или его связывающего фрагмента. Значение Ki можно рассчитать с применением уравнения Ki=IC50/(1+([L]/Kd)), где [L] представляет собой концентрацию применяемого радиолиганда (например, PACAP38, помеченного с помощью 125I), и Kd представляет собой константу диссоциации радиолиганда. См. например, Keen M, MacDermot J (1993) Analysis of receptors by radioligand binding. В: Wharton J, Polak JM (eds) Receptor autoradiography, principles and practice. Oxford University Press, Oxford. Чем ниже значение Ki для антагониста, тем более эффективным является антагонист. В некоторых вариантах осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению конкурируют за связывание с рецептором PAC1 человека с помеченным радиоактивной меткой лигандом PACAP с Ki, составляющей ≤ 1 нМ. В других вариантах осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению конкурируют за связывание с рецептором PAC1 человека с помеченным радиоактивной меткой лигандом PACAP с Ki, составляющей ≤ 500 пМ. В еще одних вариантах осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению конкурируют за связывание с рецептором PAC1 человека с помеченным радиоактивной меткой лигандом PACAP с Ki, составляющей ≤ 200 пМ. В других определенных вариантах осуществления антитела или их антигенсвязывающие фрагменты по настоящему изобретению конкурируют за связывание с рецептором PAC1 человека с помеченным радиоактивной меткой лигандом PACAP с Ki, составляющей ≤ 100 пМ.
В определенных вариантах осуществления антитела и антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную лигандом активацию рецептора PAC1 человека. Лиганд может представлять собой эндогенный лиганд рецептора, такой как PACAP38 или PACAP27, или лиганд может являться другом известным агонистом рецептора, таким как максадилан. Максадилан представляет собой пептид из 65 аминокислот, первоначально выделенный из москита, который обладает исключительной селективностью в отношении PAC1 по сравнению с VPAC1 или VPAC2 и, таким образом, может применяться в качестве селективного в отношении PAC1 агониста (Lerner et al., J Biol Chem., Vol. 266(17):11234-11236, 1991; Lerner et al., Peptides, Vol. 28(9): 1651-1654, 2007). Различные анализы для оценки активации рецепторов PAC1 известны из уровня техники и включают анализы на основе клеток, с помощью которых измеряют индуцированные лигандом мобилизацию кальция и образование cAMP. Иллюстративный клеточный анализ cAMP описан в примере 3. Другие подходящие анализы активации рецептора PAC1 описаны в Dickson et al., Ann. N. Y. Acad. Sci., Vol. 1070:239-42, 2006; Bourgault et al., J. Med. Chem., Vol. 52: 3308-3316, 2009; публикации патента США № 2011/0229423, все из которых настоящим включены посредством ссылки во всей своей полноте.
Ингибирующую активность антител или антигенсвязывающих фрагментов в отношении активации рецептора PAC1 можно количественно определить посредством расчета IC50 при любом функциональном анализе рецептора, таком как анализы, описанные выше. "IC50" представляет собой дозу/концентрацию, необходимую для достижения 50% ингибирования биологической или биохимической функции. В случае радиоактивных лигандов IC50 представляет собой концентрацию конкурирующего лиганда, при которой вытесняется 50% специфического связывания радиолиганда. IC50 любого конкретного вещества или антагониста можно определить посредством построения кривой доза-ответ и определения влияния различных концентраций лекарственного средства или антагониста на обратимую активность агониста в конкретном функциональном анализе. Значения IC50 можно вычислить для данного антагониста или лекарственного средства посредством определения концентрации, необходимой для ингибирования половины максимального биологического ответа агониста. Таким образом, значение IC50 для любого антитела к PAC1 или его связывающего фрагмента по настоящему изобретению можно вычислить посредством определения концентрации антитела или связывающего фрагмента, необходимой для ингибирования половины максимального биологического ответа лиганда (например, PACAP27, PACAP38 или максадилана) при активации рецептора PAC1 человека в любом функциональном анализе, таком как анализ cAMP, описанный в примерах. Под антителом к PAC1 или его связывающим фрагментом, который ингибирует индуцированную лигандом (например, индуцированную PACAP) активацию рецептора PAC1, понимается нейтрализующее или антагонистическое антитело или связывающий фрагмент рецептора PAC1.
В определенных вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP (индуцированную PACAP38 или PACAP27) активацию рецептора PAC1 человека. Например, антитела или антигенсвязывающие фрагменты могут ингибировать индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей менее приблизительно 10 нМ, менее приблизительно 8 нМ, менее приблизительно 5 нМ, менее приблизительно 3 нМ, менее приблизительно 1 нМ, менее приблизительно 800 пМ, менее приблизительно 500 пМ, менее приблизительно 400 пМ, менее приблизительно 300 пМ, менее приблизительно 200 пМ или менее приблизительно 100 пМ, как измерено с помощью анализа мобилизации кальция на основе клеток или анализа cAMP. В одном конкретном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей менее приблизительно 5 нМ, как измерено с помощью клеточного анализа cAMP. В другом конкретном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей менее приблизительно 1 нМ, как измерено с помощью клеточного анализа cAMP. В еще одном конкретном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей менее приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP. В другом варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей менее приблизительно 300 пМ, как измерено с помощью клеточного анализа cAMP. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей от приблизительно 0,1 нМ до приблизительно 1 нМ, как измерено с помощью клеточного анализа cAMP. В других вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека при IC50, составляющей от приблизительно 50 пМ до приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP.
В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению связываются с рецепторами PAC1 из других видов и ингибируют их. Например, антитела или антигенсвязывающие фрагменты связываются с рецептором PAC1 яванского макака и ингибируют его (эталонная последовательность в NCBI XP_015303041.1). В таких вариантах осуществления антитела или антигенсвязывающие фрагменты ингибируют индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей менее приблизительно 10 нМ, менее приблизительно 8 нМ, менее приблизительно 5 нМ, менее приблизительно 3 нМ, менее приблизительно 1 нМ, менее приблизительно 800 пМ, менее приблизительно 500 пМ, менее приблизительно 400 пМ, менее приблизительно 300 пМ, менее приблизительно 200 пМ или менее приблизительно 100 пМ, как измерено с помощью анализа мобилизации кальция на основе клеток или анализа cAMP. В одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей менее приблизительно 1 нМ, как измерено с помощью клеточного анализа cAMP. В другом варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей менее приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей от приблизительно 0,1 нМ до приблизительно 1 нМ, как измерено с помощью клеточного анализа cAMP. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 яванского макака при IC50, составляющей от приблизительно 50 пМ до приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP.
В определенных вариантах осуществления антитела или антигенсвязывающие фрагменты связываются с рецептором PAC1 крысы и ингибируют его (эталонная последовательность в NCBI NP_598195.1). В этих вариантах осуществления антитела или антигенсвязывающие фрагменты могут ингибировать индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее приблизительно 10 нМ, менее приблизительно 8 нМ, менее приблизительно 5 нМ, менее приблизительно 3 нМ, менее приблизительно 1 нМ, менее приблизительно 800 пМ, менее приблизительно 500 пМ, менее приблизительно 400 пМ, менее приблизительно 300 пМ, менее приблизительно 200 пМ или менее приблизительно 100 пМ, как измерено с помощью анализа мобилизации кальция на основе клеток или анализа cAMP. В одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее приблизительно 10 нМ, как измерено с помощью клеточного анализа cAMP. В другом варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее приблизительно 5 нМ, как измерено с помощью клеточного анализа cAMP. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее приблизительно 500 пМ, как измерено с помощью клеточного анализа cAMP. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей менее приблизительно 300 пМ, как измерено с помощью клеточного анализа cAMP. В некоторых вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей от приблизительно 0,1 нМ до приблизительно 10 нМ, как измерено с помощью клеточного анализа cAMP. В еще одном варианте осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 крысы при IC50, составляющей от приблизительно 100 пМ до приблизительно 2 нМ, как измерено с помощью клеточного анализа cAMP. В некоторых вариантах осуществления, в которых антитела или связывающие фрагменты по настоящему изобретению перекрестно реагируют с рецепторами PAC1 из других видов, антитела или связывающие фрагменты способны ингибировать индуцированную PACAP активацию рецептора PAC1 со сравнимыми уровнями эффективности. Например, антитело или связывающий фрагмент по настоящему изобретению способен ингибировать индуцированную PACAP активацию рецептора PAC1 человека при IC50, аналогичной IC50 для антитела или связывающего фрагмента для ингибирования индуцированной PACAP активации рецептора PAC1 яванского макака или крысы. Перекрестная реактивность с рецепторами PAC1 других видов антител или связывающих фрагментов по настоящему изобретению может являться преимущественной, поскольку антитела или связывающие фрагменты можно оценивать на дополнительных доклинических животных моделях в отношении терапевтической эффективности.
Как правило, антитела или антигенсвязывающие фрагменты по настоящему изобретению не ингибируют в значительной степени индуцированную лигандом активацию (например, индуцированную PACAP38, PACAP27 или VIP) рецептора VPAC1 человека или рецептора VPAC2. Как применяется в данном документе, антитело или антигенсвязывающий фрагмент "в значительной степени не будет ингибировать" активацию рецептора или связывание лиганда с его рецептором, если нет статистической разницы между индуцированной лигандом активацией рецептора или связыванием лиганда с рецептором в присутствии или в отсутствие антитела или антигенсвязывающего фрагмента. Например, если количество cAMP, продуцирование которого индуцировано с помощью PACAP или VIP в клетках, экспрессирующих рецептор VPAC1 человека в присутствии антитела или связывающего фрагмента, статистически не отличается от количества, продуцируемого в отсутствие антитела или связывающего фрагмента, тогда будет считаться, что антитело или связывающий фрагмент в значительной степени не ингибирует индуцированную PACAP/VIP активацию рецептора VPAC1 человека. Подобным образом, если количество PACAP или VIP, связанного с рецептором VPAC1 человека в присутствии избытка антитела или связывающего фрагмента, статистически не отличается от количества PACAP или VIP, связанного с рецептором в отсутствие антитела или связывающего фрагмента, то будет считаться, что антитело или связывающий фрагмент в значительной степени не ингибирует связывание PACAP или VIP с рецептором VPAC1 человека. В определенных вариантах осуществления антитела или связывающие фрагменты по настоящему изобретению ингибируют индуцированную PACAP активацию рецептора PAC1 человека, но не ингибируют в значительной степени индуцированную PACAP активацию рецептора VPAC1 человека или рецептора VPAC2 человека. Таким образом, антитела и связывающие фрагменты по настоящему изобретению селективно ингибируют рецептор PAC1 человека по сравнению с рецептором VPAC1 человека и рецептором VPAC2 человека.
Антитела и связывающие фрагменты по настоящему изобретению способны в некоторых вариантах осуществления связываться с конкретной областью или эпитопом PAC1 человека. Применяемый в данном документе термин "эпитоп" относится к любой детерминанте, способной специфически связываться антителом или его фрагментом. Эпитоп представляет собой область антигена, которая связывается антителом или связывающим фрагментом или взаимодействует с антителом или связывающим фрагментом, который нацеливается на этот антиген, и когда антиген представляет собой белок, содержит специфические аминокислоты, которые непосредственно контактируют или взаимодействуют с антителом или связывающим фрагментом. Эпитоп может быть образован как смежными аминокислотами, так и несмежными аминокислотами, расположенными рядом из-за сворачивания белка в третичную структуру. "Линейный эпитоп" представляет собой эпитоп, где первичная аминокислотная последовательность образует распознаваемый эпитоп. Линейный эпитоп обычно содержит по меньшей мере 3 или 4 аминокислоты и более часто по меньшей мере 5, по меньшей мере 6 или по меньшей мере 7 аминокислот, например, от приблизительно 8 до приблизительно 10 аминокислот в уникальной последовательности. "Конформационный эпитоп", в отличие от линейного эпитопа, представляет собой группу аминокислот, которые не являются смежными (например, аминокислотные остатки в полипептиде, которые не являются смежными в первичной последовательности полипептида, но являются таковыми в контексте третичной и четвертичной структуры полипептида, расположены достаточно близко друг к другу, чтобы быть связанными антителом или его связывающим фрагментом). Эпитопные детерминанты могут содержать химически активные поверхностные группы молекул, такие как аминокислоты, боковые цепи сахаров, фосфорильные или сульфонильные группы, и могут иметь специфические характеристики трехмерной структуры и/или специфические характеристики заряда. Как правило, антитела или связывающие фрагменты, специфические в отношении конкретной целевой молекулы, будут предпочтительно распознавать эпитоп на целевой молекуле в сложной смеси из белков и/или макромолекул.
В определенных вариантах осуществления антитела или антигенсвязывающие фрагменты по настоящему изобретению связываются с PAC1 человека по эпитопу в пределах N-концевого внеклеточного домена (ECD) PAC1 человека (SEQ ID NO: 4). В связанных вариантах осуществления антитела или антигенсвязывающие фрагменты связываются с PAC1 человека по эпитопу в пределах аминокислот 24-153 SEQ ID NO: 1. В других связанных вариантах осуществления антитела или антигенсвязывающие фрагменты связываются с PAC1 человека по эпитопу в пределах аминокислот 50-140 SEQ ID NO: 1. Как описано в примере 1, кристаллическая структура комплекса N-концевого ECD PAC1 человека и Fab-области нейтрализующего антитела к PAC1 позволила выявить ключевые аминокислоты в пределах ECD PAC1 человека, которые содержали участок контакта для связывания с Fab к PAC1. Эти аминокислоты основного участка контакта, все из которых содержат по меньшей мере один атом, не представляющий собой водород, на расстоянии 5 Å или меньше от атома, не представляющего собой водород, в Fab включают Asp59, Asn60, Ile61, Arg116, Asn117, Thr119, Asp121, Gly122, Trp123, Ser124, Glu125, Pro126, Phe127, Pro128, His129, Tyr130, Phe131, Asp132 и Gly135 (номера аминокислотных положений относительно SEQ ID NO: 1). Таким образом, в некоторых вариантах осуществления антитела или связывающие фрагменты по настоящему изобретению связываются с PAC1 человека по эпитопу, содержащему одну или несколько аминокислот, выбранных из Asp59, Asn60, Ile61, Arg116, Asn117, Thr119, Glu120, Asp121, Gly122, Trp123, Ser124, Glu125, Pro126, Phe127, Pro128, His129, Tyr130, Phe131, Asp132 и Gly135 из SEQ ID NO: 1. В других вариантах осуществления антитела или связывающие фрагменты связываются с PAC1 человека по эпитопу, содержащему по меньшей мере аминокислоты Asn60, Ile61, Glu120 и Asp121 из SEQ ID NO: 1. В еще одних вариантах осуществления антитела или связывающие фрагменты связываются с PAC1 человека по эпитопу, содержащему по меньшей мере аминокислоты Asn60, Ile61, Glu120, Asp121, Phe127 и Phe131 из SEQ ID NO: 1. В других определенных вариантах осуществления антитела или связывающие фрагменты связываются с PAC1 человека по эпитопу, содержащему все из аминокислот, выбранных из Asp59, Asn60, Ile61, Arg116, Asn117, Thr119, Glu120, Asp121, Gly122, Trp123, Ser124, Glu125, Pro126, Phe127, Pro128, His129, Tyr130, Phe131, Asp132 и Gly135 из SEQ ID NO: 1.
Кристаллическая структура комплекса Fab-ECD PAC1 человека, описанная в примере 1, также позволила выявить важные остатки в CDR тяжелой и легкой цепей Fab, которые взаимодействовали с аминокислотами в ECD PAC1 человека, тем самым позволив идентифицировать ключевые аминокислоты в паратопе антитела. "Паратоп" представляет собой область антитела, которая распознает и связывается с целевым антигеном. Остатки паратопа включают Gln27, Gly30, Arg31 и Ser32 в вариабельной области легкой цепи (SEQ ID NO: 3) и Arg31, Phe32, Tyr53, Asp54, Gly56 в вариабельной области тяжелой цепи (SEQ ID NO: 2). Конкретные мутации нескольких этих остатков в паратопе были разработаны для улучшения взаимодействия с остатками основного участка контакта (т.е. остатками в эпитопе) в ECD PAC1 человека, что приводит к повышению аффинности связывания и ингибирующей активности по сравнению с исходным антителом (см. примеры 1-3). Gln27, Gly30, Arg31 и Ser32 в вариабельной области легкой цепи под SEQ ID NO: 3 или под SEQ ID NO: 52 соответствуют аминокислотным положениям 29, 32, 39 и 40 согласно нумерации AHo соответственно, и Arg31, Phe32, Tyr53, Asp54, Gly56 в вариабельной области тяжелой цепи под SEQ ID NO: 2 или под SEQ ID NO: 191 соответствуют аминокислотным положениям 33, 39, 60, 61 и 66 согласно нумерации AHo соответственно. Схема нумерации AHo представляет собой схему нумерации на основе структуры, которая вводит гэпы в CDR-области для минимизации отклонения от средней структуры выравненных доменов (Honegger and Pluckthun, J. Mol. Biol. 309(3):657-670; 2001). В схеме нумерации AHo структурно эквивалентные положения в различных антителах будут иметь одинаковый номер остатка.
В некоторых вариантах осуществления в настоящем изобретении предусмотрено антитело или его антигенсвязывающий фрагмент, которые специфически связываются с PAC1 человека (SEQ ID NO: 1), где антитело или его антигенсвязывающий фрагмент содержат: (i) вариабельную область легкой цепи, в которой аминокислота в положении 29 в соответствии с нумерацией AHo представляет собой основную аминокислоту, которая взаимодействует с аминокислотами Glu120 или Asp121 PAC1 человека, и (ii) вариабельную область тяжелой цепи, в которой аминокислота в положении 61 в соответствии с нумерацией AHo представляет собой гидрофобную, основную или нейтральную гидрофильную аминокислоту, которая взаимодействует с аминокислотами Asn60 или Ile61 PAC1 человека. Как применяется в данном документе, указано, что одна аминокислота "взаимодействует" с другой аминокислотой, когда один или несколько атомов в одной аминокислоте образуют нековалентные связи с одним или несколькими атомами в другой аминокислоте, например, посредством ван-дер-ваальсовых, гидрофобных или электростатических сил. Основные аминокислоты включают аргинин, лизин и гистидин, тогда как нейтральные гидрофильные аминокислоты включают аспарагин, глутамин, серин и треонин. Гидрофобные аминокислоты включают фенилаланин, триптофан, тирозин, аланин, изолейцин, лейцин и валин. В некоторых вариантах осуществления аминокислота в положении 29 в соответствии с нумерацией AHo в вариабельной области легкой цепи представляет собой лизин или аргинин. В одном конкретном варианте осуществления аминокислота в положении 29 в соответствии с нумерацией AHo в вариабельной области легкой цепи представляет собой лизин. В связанных вариантах осуществления аминокислота в положении 61 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой изолейцин, лейцин, валин, глутамин, аспарагин, аргинин или лизин. В одном варианте осуществления аминокислота в положении 61 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой изолейцин. В другом варианте осуществления аминокислота в положении 61 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой глутамин или аспарагин. В еще одном варианте осуществления аминокислота в положении 61 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой аргинин.
В некоторых вариантах осуществления аминокислота в положении 66 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи антитела или его антигенсвязывающего фрагмента представляет собой основную или нейтральную гидрофильную аминокислоту, которая взаимодействует с аминокислотами Asn60 или Ile61 PAC1 человека (SEQ ID NO: 1). Аминокислота в положении 66 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи может представлять собой глутамин, аспарагин, аргинин или лизин. В одном варианте осуществления аминокислота в положении 66 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой аргинин. В другом варианте осуществления аминокислота в положении 66 в соответствии с нумерацией AHo в вариабельной области тяжелой цепи представляет собой аспарагин.
Было обнаружено, что взаимодействия паратопа и эпитопа, описанные выше, коррелируют с улучшениями ингибирующей активности при значениях IC50 в пикомолярном диапазоне. См. пример 3. Таким образом, антитела или их антигенсвязывающие фрагменты, содержащие вышеуказанные аминокислоты в положениях 29 в вариабельной области легкой цепи и положениях 61 и/или 66 в вариабельной области тяжелой цепи в соответствии с нумерацией AHo, и взаимодействующие с указанными остатками в рецепторе PAC1 человека (Glu120, Asp121, Asn60 и/или Ile61 из SEQ ID NO: 1), как ожидается, будут обладать усиленной ингибирующей активностью, например, ингибируют индуцированную PACAP активацию PAC1 человека при IC50, составляющей менее 500 пМ, как измерено с помощью клеточного анализа cAMP. В некоторых вариантах осуществления такие антитела или антигенсвязывающие фрагменты способны подавлять индуцированную PACAP активацию PAC1 человека при IC50, составляющей менее 300 пМ, как измерено с помощью клеточного анализа cAMP. В этих и других вариантах осуществления антитела или их антигенсвязывающие фрагменты могут содержать вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности под SEQ ID NO: 52, и вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности под SEQ ID NO: 191.
Антитела или антигенсвязывающие фрагменты по настоящему изобретению могут содержать одну или несколько определяющих комплементарность областей (CDR) из вариабельных областей легкой и тяжелой цепей антител, которые специфически связываются с PAC1 человека, как описано в данном документе. Термин "CDR" относится к определяющей комплементарность области (также называемой "минимальными единицами распознавания" или "гипервариабельной областью") в пределах вариабельных последовательностей антител. Существуют три CDR вариабельной области тяжелой цепи (CDRH1, CDRH2 и CDRH3) и три CDR вариабельной области легкой цепи (CDRL1, CDRL2 и CDRL3). Применяемый в данном документе термин "CDR-область" относится к группе из трех CDR, которые встречаются в одной вариабельной области (т. е. три CDR легкой цепи или три CDR тяжелой цепи). CDR в каждой из двух цепей обычно выравнены по каркасным областям (FR) с образованием структуры, которая специфически связывается с конкретным эпитопом или доменом на целевом белке (например, PAC1 человека). Как правило, от N-конца к С-концу встречающиеся в природе вариабельные области как легкой, так и тяжелой цепей соответствуют следующему порядку этих элементов: FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Была разработана система нумерации для присвоения номеров аминокислотам, которые занимают положения в каждом из этих доменов. Эта система нумерации определена в Kabat Sequences of Proteins of Immunological Interest (1987 and 1991, NIH, Bethesda, MD) или Chothia & Lesk, 1987, J. Mol. Biol. 196:901-917; Chothia et al., 1989, Nature 342:878-883. С помощью этой системы можно идентифицировать определяющие комплементарность области (CDR) и каркасные области (FR) данного антитела. Другие системы нумерации аминокислот в цепях иммуноглобулина включают IMGT® (Международная информационная система ImMunoGeneTics; Lefranc et al., Dev. Comp. Immunol. 29:185-203; 2005) и AHo (Honegger and Pluckthun, J. Mol. Biol. 309(3):657-670; 2001).
В определенных вариантах осуществления антитела к PAC1 или их антигенсвязывающие фрагменты по настоящему изобретению содержат по меньшей мере одну вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и по меньшей мере одну вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, из любого из антител к PAC1, описанных в данном документе. Вариабельные области легкой цепи и тяжелой цепи, а также связанные CDR иллюстративных антител к PAC1 человека приведены ниже в таблицах 1A и 1B соответственно.
(SEQ ID NO: 52)
(SEQ ID NO: 26)
(SEQ ID NO: 53)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 56)
(SEQ ID NO: 26)
(SEQ ID NO: 57)
(SEQ ID NO: 26)
(SEQ ID NO: 58)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 59)
(SEQ ID NO: 26)
(SEQ ID NO: 60)
(SEQ ID NO: 26)
(SEQ ID NO: 61)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 54)
(SEQ ID NO: 26)
(SEQ ID NO: 56)
(SEQ ID NO: 26)
(SEQ ID NO: 57)
(SEQ ID NO: 26)
(SEQ ID NO: 58)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 56)
(SEQ ID NO: 26)
(SEQ ID NO: 57)
(SEQ ID NO: 26)
(SEQ ID NO: 58)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 56)
(SEQ ID NO: 26)
(SEQ ID NO: 57)
(SEQ ID NO: 26)
(SEQ ID NO: 58)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 56)
(SEQ ID NO: 26)
(SEQ ID NO: 57)
(SEQ ID NO: 26)
(SEQ ID NO: 58)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 55)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 63)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 64)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 62)
(SEQ ID NO: 26)
(SEQ ID NO: 65)
(SEQ ID NO: 26)
(SEQ ID NO: 65)
(SEQ ID NO: 26)
(SEQ ID NO: 65)
(SEQ ID NO: 26)
(SEQ ID NO: 66)
(SEQ ID NO: 26)
(SEQ ID NO: 67)
(SEQ ID NO: 27)
(SEQ ID NO: 68)
(SEQ ID NO: 28)
(SEQ ID NO: 69)
(SEQ ID NO: 29)
(SEQ ID NO: 70)
(SEQ ID NO: 28)
(SEQ ID NO: 71)
(SEQ ID NO: 30)
(SEQ ID NO: 72)
(SEQ ID NO: 30)
(SEQ ID NO: 73)
(SEQ ID NO: 28)
(SEQ ID NO: 74)
(SEQ ID NO: 31)
(SEQ ID NO: 75)
(SEQ ID NO: 32)
(SEQ ID NO: 76)
(SEQ ID NO: 28)
(SEQ ID NO: 77)
(SEQ ID NO: 28)
(SEQ ID NO: 78)
(SEQ ID NO: 31)
(SEQ ID NO: 79)
(SEQ ID NO: 31)
(SEQ ID NO: 80)
(SEQ ID NO: 28)
(SEQ ID NO: 81)
(SEQ ID NO: 33)
(SEQ ID NO: 79)
(SEQ ID NO: 31)
(SEQ ID NO: 82)
(SEQ ID NO: 34)
(SEQ ID NO: 83)
(SEQ ID NO: 31)
(SEQ ID NO: 84)
(SEQ ID NO: 31)
(SEQ ID NO: 85)
(SEQ ID NO: 31)
(SEQ ID NO: 86)
(SEQ ID NO: 35)
(SEQ ID NO: 87)
(SEQ ID NO: 27)
(SEQ ID NO: 78)
(SEQ ID NO: 31)
(SEQ ID NO: 88)
(SEQ ID NO: 132)
(SEQ ID NO: 91)
(SEQ ID NO: 94)
Антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут содержать одну или несколько CDR легкой цепи (т. е. CDRL) и/или CDR тяжелой цепи (т. е. CDRH), представленных в таблицах 1A и 1B соответственно. В некоторых вариантах осуществления антитела к PAC1 или их связывающие фрагменты получают из антитела 29G4v9, 29G4v10 или 29G4v22 (т. е. содержат одну или несколько замен в одной или нескольких CDRL и/или CDRH антитела 29G4v9, 29G4v10 или 29G4v22). Например, в определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат одну или несколько CDR легкой цепи, выбранных из (i) CDRL1, выбранной из SEQ ID NO: 5-16, (ii) CDRL2 под SEQ ID NO: 26 и (iii) CDRL3, выбранной из SEQ ID NO: 36-38, и (iv) CDRL под (i), (ii) и (iii), которая содержит одну или несколько, например, одну, две, три, четыре или более аминокислотных замен (например, консервативных аминокислотных замен), делеций или вставок не более пяти, четырех, трех, двух или одной аминокислоты. В этих и других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат одну или несколько CDR тяжелой цепи, выбранных из (i) CDRH1, выбранной из SEQ ID NO: 88-96, (ii) CDRH2, выбранной из SEQ ID NO: 106-166 и (iii) CDRH3, выбранной из SEQ ID NO: 171-177, и (iv) CDRH под (i), (ii) и (iii), которая содержит одну или несколько, например, одну, две, три, четыре или более аминокислотных замен (например, консервативных аминокислотных замен), делеций или вставок не более пяти, четырех, трех, двух или одной аминокислоты аминокислоты.
В других вариантах осуществления антитела к PAC1 или их связывающие фрагменты получают из антитела 19H8 (т. е. содержат одну или несколько замен в одной или нескольких CDRL и/или CDRH антитела 19H8). В таких вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат одну или несколько CDR легкой цепи, выбранных из (i) CDRL1, выбранной из SEQ ID NO: 17-25, (ii) CDRL2, выбранной из SEQ ID NO: 27-35 и (iii) CDRL3, выбранной из SEQ ID NO: 39-51, и (iv) CDRL под (i), (ii) и (iii), которая содержит одну или несколько, например, одну, две, три, четыре или более аминокислотных замен (например, консервативных аминокислотных замен), делеций или вставок не более пяти, четырех, трех, двух или одной аминокислоты. В этих и других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты содержат одну или несколько CDR тяжелой цепи, выбранных из (i) CDRH1, выбранной из SEQ ID NO: 97-105, (ii) CDRH2, выбранной из SEQ ID NO: 167-170, и (iii) CDRH3, выбранной из SEQ ID NO: 178-190, и (iv) CDRH под (i), (ii) и (iii), которая содержит одну или несколько, например, одну, две, три, четыре или более аминокислотных замен (например, консервативных аминокислотных замен), делеций или вставок не более пяти, четырех, трех, двух или одной аминокислоты аминокислоты.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты могут содержать 1, 2, 3, 4, 5 или 6 вариантных форм CDR, перечисленных в таблицах 1A и 1B, причем каждая характеризуется по меньшей мере 80%, 85%, 90% или 95% идентичностью последовательности с последовательностью CDR, перечисленной в таблицах 1A и 1B. В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты включают 1, 2, 3, 4, 5 или 6 CDR, перечисленных в таблицах 1A и 1B, каждая из которых отличается не более чем на 1, 2, 3, 4 или 5 аминокислот из CDR, перечисленных в этих таблицах. В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 5-16, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRL2, содержащую последовательность под SEQ ID NO: 26 или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 36-38, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 88-96, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 106-166, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; и CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 171-177, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены. В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 5-16; CDRL2, содержащую последовательность под SEQ ID NO: 26; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 36-38; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 88-96; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 106-166; и CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 171-177.
В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 17-25, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRL2, содержащую последовательность, выбранную из SEQ ID NO: 27-35, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 39-51, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 97-105, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 167-170, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены; и CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 178-190, или ее вариант, содержащий одну, две, три или четыре аминокислотные замены. В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат CDRL1, содержащую последовательность, выбранную из SEQ ID NO: 17-25; CDRL2, содержащую последовательность, выбранную из SEQ ID NO: 27-35; CDRL3, содержащую последовательность, выбранную из SEQ ID NO: 39-51; CDRH1, содержащую последовательность, выбранную из SEQ ID NO: 97-105; CDRH2, содержащую последовательность, выбранную из SEQ ID NO: 167-170; и CDRH3, содержащую последовательность, выбранную из SEQ ID NO: 178-190.
В конкретных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, где: (a) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно; (b) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно; (c) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 12, 26 и 36 соответственно; или (d) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 13, 26 и 36 соответственно.
В других конкретных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где: (a) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 171 соответственно; (b) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 116 и 171 соответственно; (c) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 134 и 171 соответственно; (d) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 92, 145 и 174 соответственно; (e) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 172 соответственно; (f) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 128 и 172 соответственно; (g) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 153 и 171 соответственно; (i) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 154 и 171 соответственно; или (j) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 155 и 171 соответственно.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где:
(a) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 171 соответственно;
(b) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 116 и 171 соответственно;
(c) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 134 и 171 соответственно;
(d) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 92, 145 и 174 соответственно;
(e) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 172 соответственно;
(f) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 128 и 172 соответственно;
(g) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 134 и 171 соответственно;
(h) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 12, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 153 и 171 соответственно;
(i) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 12, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 154 и 171 соответственно; или
(j) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 13, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 155 и 171 соответственно.
В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 171 соответственно. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 116 и 171 соответственно. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 134 и 171 соответственно. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 5, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 92, 145 и 174 соответственно. В одном конкретном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 7, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 108 и 172 соответственно. В другом конкретном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 12, 26 и 36 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 88, 153 и 171 соответственно.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, где: (a) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 25, 31 и 42 соответственно; (b) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 20, 30 и 44 соответственно; (c) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 33 и 42 соответственно; (d) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 23, 31 и 50 соответственно; (e) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 31 и 44 соответственно; (f) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 27 и 39 соответственно; (g) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 18, 31 и 46 соответственно; (h) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 28 и 40 соответственно; (i) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 18, 30 и 43 соответственно; или (j) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 22, 28 и 49 соответственно.
В определенных других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где: (a) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 190 соответственно; (b) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно; (c) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 99, 168 и 187 соответственно; (d) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 97, 167 и 178 соответственно; (e) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 189 соответственно; (f) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 179 соответственно; или (g) CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 102, 169 и 182 соответственно.
В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где:
(a) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 25, 31 и 42 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 190 соответственно;
(b) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 20, 30 и 44 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно;
(c) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 33 и 42 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 99, 168 и 187 соответственно;
(d) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 23, 31 и 50 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 97, 167 и 178 соответственно;
(e) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 31 и 44 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 189 соответственно;
(f) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 27 и 39 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно;
(g) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 18, 31 и 46 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 179 соответственно;
(h) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 28 и 40 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 179 соответственно;
(i) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 18, 30 и 43 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно; или
(j) CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 22, 28 и 49 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 102, 169 и 182 соответственно.
В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 25, 31 и 42 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 190 соответственно. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 20, 30 и 44 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 33 и 42 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 99, 168 и 187 соответственно. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 23, 31 и 50 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 97, 167 и 178 соответственно. В одном конкретном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 31 и 44 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 98, 168 и 189 соответственно. В другом конкретном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую CDRH1, CDRH2 и CDRH3, где CDRL1, CDRL2 и CDRL3 имеют последовательность под SEQ ID NO: 17, 27 и 39 соответственно, и CDRH1, CDRH2 и CDRH3 имеют последовательность под SEQ ID NO: 100, 168 и 182 соответственно.
В определенных вариантах осуществления антитела и антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область тяжелой цепи (VH) иммуноглобулина и вариабельную область легкой цепи (VL) иммуноглобулина из антитела, которое специфически связывается с PAC1 человека, такого как антитела, описанные в данном документе. Термин "вариабельная область", применяемый в данном документе взаимозаменяемо с термином "вариабельный домен" (вариабельная область легкой цепи (VL), вариабельная область тяжелой цепи (VH)), относится к области в каждой из легкой и тяжелой цепей иммуноглобулина, которая участвует непосредственно в связывании антитела с антигеном. Как обсуждалось выше, области вариабельной легкой и тяжелой цепей имеют одинаковую общую структуру, и каждая область содержит четыре каркасных (FR) области, последовательности которых являются в значительной степени консервативными, соединенными тремя CDR. Каркасные области принимают конформацию бета-листа, и CDR могут образовывать петли, соединяющие структуру бета-листа. CDR в каждой цепи удерживаются в своей трехмерной структуре каркасными областями и вместе с CDR из другой цепи образуют антигенсвязывающий сайт.
Таким образом, в некоторых вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению могут содержать вариабельную область легкой цепи, выбранную из LV-01 - LV-15, как показано в таблице 1A, и/или вариабельную область тяжелой цепи, выбранную из HV-01 - HV-105, как показано в таблице 1B, и связывающие фрагменты, производные и варианты этих вариабельных областей легкой цепи и тяжелой цепи. В других вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению могут содержать вариабельную область легкой цепи, выбранную из LV-16 - LV-36, как показано в таблице 1A, и/или вариабельную область тяжелой цепи, выбранную из HV-106 - HV-122, как показано в таблице 1B, и связывающие фрагменты, производные и варианты этих вариабельных областей легкой цепи и тяжелой цепи.
Каждая из вариабельных областей легкой цепи, перечисленных в таблице 1A, может быть объединена с любой из вариабельных областей тяжелой цепи, перечисленных в таблице 1B, для образования антитела к PAC1 или его антигенсвязывающего фрагмента по настоящему изобретению. Примеры таких комбинаций включают без ограничения: (i) LV-03 и любую из HV-03 - HV-26, HV-32 - HV-47 и HV-57 - HV-62; (ii) LV-04 и любую из HV-11 - HV-91; (iii) LV-05 и любую из HV-01, HV-03, HV-07, HV-09 и HV-10; (iv) LV-06 и любую из HV-01, HV-03, HV-07, HV-09 и HV-10; (v) LV-07 и любую из HV-01, HV-03, HV-07, HV-09 и HV-10; (vi) LV-08 и HV-01; (vii) LV-09 и HV-01; (viii) LV-10 и HV-01; (ix) LV-11 и любую из HV-92, HV-93, HV-95 - HV-97 и HV-99 - HV-101; (x) LV-12 и HV-94; (xi) LV-13 и HV-98; (xii) LV-14 и любую из HV-102 - HV-104; (xiii) LV-15 и HV-105; (xiv) LV-17 и HV-107; (xv) LV-18 и HV-108; (xvi) LV-19 и HV-109; (xvii) LV-20 и HV-110; (xviii) LV-21 и HV-110; (xix) LV-22 и HV-111; (xx) LV-23 и HV-107; (xxi) LV-24 и HV-112; (xxii) LV-25 и HV-113; (xxiii) LV-26 и HV-114; (xxiv) LV-27 и HV-106 или HV-110; (xxv) LV-28 и HV-115 или HV-118; (xxvi) LV-29 и HV-116; (xxvii) LV-30 и HV-117; (xxviii) LV-31 и HV-119; (xxix) LV-32 и HV-120; (xxx) LV-33 и HV-121; (xxxi) LV-34 и HV-122; (xxxii) LV-35 и HV-110; и (xxxiii) LV-36 и HV-110.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-03 (SEQ ID NO: 54), и вариабельную область тяжелой цепи, содержащую последовательность HV-04 (SEQ ID NO: 194). В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-03 (SEQ ID NO: 54), и вариабельную область тяжелой цепи, содержащую последовательность HV-13 (SEQ ID NO: 203). В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-04 (SEQ ID NO: 55), и вариабельную область тяжелой цепи, содержащую последовательность HV-38 (SEQ ID NO: 228). В еще других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-04 (SEQ ID NO: 55), и вариабельную область тяжелой цепи, содержащую последовательность HV-84 (SEQ ID NO: 274). В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-03 (SEQ ID NO: 54), и вариабельную область тяжелой цепи, содержащую последовательность HV-24 (SEQ ID NO: 214). В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-04 (SEQ ID NO: 55), и вариабельную область тяжелой цепи, содержащую последовательность HV-50 (SEQ ID NO: 240). В одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-03 (SEQ ID NO: 54), и вариабельную область тяжелой цепи, содержащую последовательность HV-38 (SEQ ID NO: 228). В другом варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-11 (SEQ ID NO: 62), и вариабельную область тяжелой цепи, содержащую последовательность HV-92 (SEQ ID NO: 282). В еще одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-11 (SEQ ID NO: 62), и вариабельную область тяжелой цепи, содержащую последовательность HV-93 (SEQ ID NO: 283). В еще одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-12 (SEQ ID NO: 63), и вариабельную область тяжелой цепи, содержащую последовательность HV-94 (SEQ ID NO: 284).
В определенных других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-34 (SEQ ID NO: 85), и вариабельную область тяжелой цепи, содержащую последовательность HV-122 (SEQ ID NO: 312). В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-21 (SEQ ID NO: 72), и вариабельную область тяжелой цепи содержащую последовательность HV-110 (SEQ ID NO: 300). В других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-30 (SEQ ID NO: 81), и вариабельную область тяжелой цепи, содержащую последовательность HV-117 (SEQ ID NO: 307). В еще других вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-27 (SEQ ID NO: 78), и вариабельную область тяжелой цепи, содержащую последовательность HV-106 (SEQ ID NO: 296). В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-32 (SEQ ID NO: 83), и вариабельную область тяжелой цепи, содержащую последовательность HV-120 (SEQ ID NO: 310). В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-36 (SEQ ID NO: 87), и вариабельную область тяжелой цепи, содержащую последовательность HV-110 (SEQ ID NO: 300). В одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-23 (SEQ ID NO: 74), и вариабельную область тяжелой цепи, содержащую последовательность HV-107 (SEQ ID NO: 297). В другом варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-17 (SEQ ID NO: 68), и вариабельную область тяжелой цепи, содержащую последовательность HV-107 (SEQ ID NO: 297). В еще одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-20 (SEQ ID NO: 71), и вариабельную область тяжелой цепи, содержащую последовательность HV-110 (SEQ ID NO: 300). В еще одном варианте осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению содержат вариабельную область легкой цепи, содержащую последовательность LV-26 (SEQ ID NO: 77), и вариабельную область тяжелой цепи, содержащую последовательность HV-114 (SEQ ID NO: 304).
В некоторых вариантах осуществления антитела к PAC1 или их антигенсвязывающие фрагменты содержат вариабельную область легкой цепи, содержащую последовательность смежных аминокислот, которая отличается от последовательности вариабельной области легкой цепи в таблице 1A, т. е. VL, выбранную из LV-01 - LV-36, только по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотным остаткам, где каждое такое различие последовательностей независимо представляет собой либо делецию, вставку, либо замену одной аминокислоты, причем делеции, вставки и/или замены приводят к изменениям не более 15 аминокислот относительно вышеуказанных последовательностей вариабельного домена. Вариабельная область легкой цепи в некоторых антителах к PAC1 и связывающих фрагментах содержит последовательность аминокислот, которая характеризуется по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичностью последовательности с аминокислотными последовательностями под SEQ ID NO: 52-87 (т. е. вариабельными областями легкой цепи в таблице 1A).
В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности, выбранной из SEQ ID NO: 54-66. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 54-66. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, выбранную из SEQ ID NO: 54-66. В некоторых вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 54. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 55. В еще одних вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 62. В еще других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 63.
В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности, выбранной из SEQ ID NO: 68-87. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, которая на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 68-87. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность, выбранную из SEQ ID NO: 68-87. В определенных вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 68. В некоторых вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 71. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 72. В еще одних вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 74. В еще других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 77. В определенных вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 78. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 81. В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 83. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 85. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 87.
В этих и других вариантах осуществления антитела к PAC1 и их антигенсвязывающие фрагменты содержат вариабельную область тяжелой цепи, содержащую последовательность смежных аминокислот, которая отличается от последовательности вариабельной области тяжелой цепи в таблице 1B, т. е. VH, выбранную из HV-01 - HV-122, только по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотным остаткам, где каждое такое различие последовательностей независимо представляет собой либо делецию, вставку, либо замену одной аминокислоты, причем делеции, вставки и/или замены приводят к изменениям не более 15 аминокислот относительно вышеуказанных последовательностей вариабельного домена. Вариабельная область тяжелой цепи в некоторых антителах к PAC1 и антигенсвязывающих фрагментах содержит последовательность аминокислот, которая характеризуется по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичностью последовательности с аминокислотными последовательностями под SEQ ID NO: 191-312 (т. е. вариабельными областями тяжелой цепи в таблице 1B).
В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности, выбранной из SEQ ID NO: 191-295. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 191-295. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, выбранную из SEQ ID NO: 191-295. В некоторых вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 194. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 203. В еще одних вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 214. В еще других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 228. В определенных вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 240. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 274. В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 282. В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 283. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 284.
В другом варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 90% идентична последовательности, выбранной из SEQ ID NO: 296-312. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, которая на по меньшей мере 95% идентична последовательности, выбранной из SEQ ID NO: 296-312. В еще одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность, выбранную из SEQ ID NO: 296-312. В некоторых вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 296. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 297. В еще одних вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 300. В еще других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 304. В определенных вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 307. В других вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 310. В одном варианте осуществления антитело к PAC1 или его антигенсвязывающий фрагмент содержат вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 312.
Применяемый в данном документе термин "идентичность" относится к взаимосвязи между последовательностями двух или более полипептидных молекул или двух или более молекул нуклеиновой кислоты, как определено при выравнивании и сравнении последовательностей. Применяемый в данном документе термин "процент идентичности" означает процент идентичных остатков аминокислот или нуклеотидов в сравниваемых молекулах и рассчитывается на основе размера наименьшей из сравниваемых молекул. Для этих расчетов гэпы в выравниваниях (если таковые имеются) должны быть учтены с помощью определенной математической модели или компьютерной программы (т. е. "алгоритма"). Способы, которые можно применять для расчета идентичности выравниваемых нуклеиновых кислот или полипептидов, включают способы, описанные в Computational Molecular Biology, (Lesk, A. M., ed.), 1988, New York: Oxford University Press; Biocomputing Informatics and Genome Projects, (Smith, D. W., ed.), 1993, New York: Academic Press; Computer Analysis of Sequence Data, Part I, (Griffin, A. M., and Griffin, H. G., eds.), 1994, New Jersey: Humana Press; von Heinje, G., 1987, Sequence Analysis in Molecular Biology, New York: Academic Press; Sequence Analysis Primer, (Gribskov, M. and Devereux, J., eds.), 1991, New York: M. Stockton Press и Carillo et al., 1988, SIAM J. Applied Math. 48:1073. Например, идентичность последовательности может быть определена с помощью стандартных способов, которые обычно применяют для сравнения сходства в положении аминокислот двух полипептидов. С применением компьютерной программы, такой как BLAST или FASTA, две полипептидные или две полинуклеотидные последовательности выравнивают для оптимального соответствия их соответствующих остатков (либо по всей длине одной или обеих последовательностей, либо по предварительно определенной части одной или обеих последовательностей). В программах предусмотрен штраф за открытие по умолчанию и штраф за гэп по умолчанию, и матрицу оценки, такую как PAM 250 (Dayhoff et al., в Atlas of Protein Sequence and Structure, vol. 5, supp. 3, 1978) или BLOSUM62 (Henikoff et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:10915-10919), можно применять совместно с компьютерной программой. Например, процент идентичности затем может быть рассчитан как общее количество идентичных совпадений, умноженное на 100, а затем разделенное на сумму длины более длинной последовательности в пределах совпадающего промежутка и количества промежутков, введенных в более длинные последовательности с целью выравнивания двух последовательностей. При расчете процента идентичности сравниваемые последовательности выравнивают способом, который дает наибольшее совпадение между последовательностями.
Программный пакет GCG представляет собой компьютерную программу, которую можно применять для определения процента идентичности, при этом в пакет входит GAP (Devereux et al., 1984, Nucl. Acid Res. 12:387; Genetics Computer Group, Университет штата Висконсин, Мэдисон, WI). Компьютерный алгоритм GAP применяют для выравнивания двух полипептидов или двух полинуклеотидов, для которых должен быть определен процент идентичности последовательностей. Последовательности выравнивают для оптимального совпадения их соответствующих аминокислот или нуклеотидов ("совпадающий промежуток", определяемый алгоритмом). Штраф за открытие гэпа (который рассчитывается как 3x средняя диагональ, где "средняя диагональ" представляет собой среднее значение диагонали применяемой матрицы сравнения; "диагональ" представляет собой балл или число, присвоенное каждому полному аминокислотному совпадению в соответствии с определенной матрицей сравнения) и штраф за продление гэпа (который, как правило, составляет 1/10 долю от штрафа за открытие гэпа), а также матрицу сравнения, такую как PAM 250 или BLOSUM 62, применяют вместе с алгоритмом. В определенных вариантах осуществления в этом алгоритме также применяется стандартная матрица сравнения (см. Dayhoff et al., (1978) Atlas of Protein Sequence and Structure 5:345-352 для матрицы сравнения PAM 250; Henikoff et al., 1992, Proc. Natl. Acad. Sci. U.S.A. 89:10915-10919 для матрицы сравнения BLOSUM 62).
Рекомендуемые параметры для определения процента идентичности полипептидов или нуклеотидных последовательностей с применением программы GAP включают следующие:
Алгоритм: Needleman et al. 1970, J. Mol. Biol. 48:443-453;
Матрица сравнения: BLOSUM 62 из Henikoff et al., 1992, см. выше;
Штраф за гэп: 12 (но без штрафа за концевые гэпы)
Штраф за длину гэпа: 4
Пороговое значение степени сходства: 0
Применение определенных схем выравнивания для выравнивания двух аминокислотных последовательностей может приводить к совпадению только короткой области двух последовательностей, и эта небольшая выровненная область может характеризоваться очень высокой степенью идентичности последовательностей, даже если отсутствует значительная взаимосвязь между двумя полноразмерными последовательностями. Соответственно, выбранный способ выравнивания (программа GAP) может быть, если требуется, скорректирован для обеспечения выравнивания, которое охватывает по меньшей мере 50 смежных аминокислот целевого полипептида.
Антитела к PAC1 по настоящему изобретению могут содержать любую константную область иммуноглобулина. Термин "константная область", применяемый в данном документе взаимозаменяемо с термином "константный домен", относится ко всем доменам антитела, отличным от вариабельной области. Константная область не участвует непосредственно в связывании антигена, но проявляет различные эффекторные функции. Как описано выше, антитела делятся на конкретные изотипы (IgA, IgD, IgE, IgG и IgM) и подтипы (IgG1, IgG2, IgG3, IgG4, IgA1 IgA2) в зависимости от аминокислотной последовательности константной области их тяжелых цепей. Константная область легкой цепи может представлять собой, например, константную область легкой цепи каппа- или лямбда-типа, например, константную область легкой цепи каппа- или лямбда-типа человека, которые обнаружены во всех пяти изотипах антител. Примеры последовательностей константной области легкой цепи иммуноглобулина человека показаны в следующей таблице.
Константная область тяжелой цепи антител к PAC1 по настоящему изобретению может представлять собой, например, константную область тяжелой цепи альфа-, дельта-, эпсилон-, гамма- или мю-типа, например, константную область тяжелой цепи альфа-, дельта-, эпсилон-, гамма- или мю-типа человека. В некоторых вариантах осуществления антитела к PAC1 содержат константную область тяжелой цепи из иммуноглобулина IgG1, IgG2, IgG3 или IgG4, такого как иммуноглобулин IgG1, IgG2, IgG3 или IgG4 человека. В одном варианте осуществления антитело к PAC1 содержит константную область тяжелой цепи из иммуноглобулина IgG1 человека. В таких вариантах осуществления константная область иммуноглобулина IgG1 человека может содержать одну или несколько мутаций для предотвращения гликозилирования антитела, как описано более подробно в данном документе. В другом варианте осуществления антитело к PAC1 содержит константную область тяжелой цепи из иммуноглобулина IgG2 человека. В еще одном варианте осуществления антитело к PAC1 содержит константную область тяжелой цепи из иммуноглобулина IgG4 человека. Примеры последовательностей константной области тяжелой цепи IgG1, IgG2 и IgG4 человека показаны ниже в таблице 3.
Каждая из вариабельных областей легкой цепи, раскрытых в таблице 1A, и каждая из вариабельных областей тяжелой цепи, раскрытых в таблице 1B, могут быть присоединены к вышеуказанным константным областям легкой цепи (таблица 2) и константным областям тяжелой цепи (таблица 3) с образованием полной легкой и тяжелой цепей антитела соответственно. Кроме того, каждая из образованных таким образом последовательностей тяжелых и легких цепей может быть объединена с другой с образованием полной структуры антитела. Следует понимать, что вариабельные области тяжелой цепи и легкой цепи, предусмотренные в данном документе, также могут быть присоединены к другим константным доменам, содержащим последовательности, отличные от иллюстративных последовательностей, перечисленных выше.
Антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут представлять собой любые из антител к PAC1 или антигенсвязывающих фрагментов, раскрытых в данном документе. Например, в определенных вариантах осуществления антитело к PAC1 представляет собой антитело к PAC1, выбранное из любых из антител, перечисленных в таблицах 9, 10, 12, 13, 14, 19 и 20. В некоторых вариантах осуществления антитела к PAC1 содержат вариабельную область легкой цепи и/или вариабельную область тяжелой цепи, содержащую одну или несколько аминокислотных замен, указанных в таблицах 9, 10, 12, 13, 19 или 20. Например, в одном варианте осуществления антитело к PAC1 содержит вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 52 с мутацией в одном или нескольких из аминокислотных положений 27, 28, 30, 31, 32 и/или 93. В определенных вариантах осуществления мутация выбрана из Q27K, Q27R, Q27H, S28A, G30M, G30W, R31Q, R31L, R31H, R31W, R31Y, S32A, S32N, S32L, R93F, R93M, R93Y или их комбинаций. В этих и других вариантах осуществления антитело к PAC1 содержит вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 191 с мутацией в одном или нескольких из аминокислотных положений 31, 32, 49, 52, 53, 54, 56, 57, 62, 70, 102, 103 и/или 104. В некоторых вариантах осуществления мутация выбрана из R31F, R31H, R31Y, R31M, R31K, F32Y, A49G, S52N, S52T, Y53F, Y53H, Y53S, D54I, D54L, D54N, D54R, D54Q, D54Y, D54F, D54M, D54S, D54T, G56Q, G56N, G56R, G56H, G56A, G56S, G56T, G56D, N57A, N57F, N57G, N57S, N57Q, N57L, N57T, E62R, I70V, I70L, I70M, V102P, V102L, V102I, V102F, V102M, L103M, T104S или их комбинаций.
В другом варианте осуществления антитело к PAC1 содержит вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 67 с мутацией в одном или нескольких из аминокислотных положений 28, 30, 31, 34, 49, 50, 51, 52, 53, 91, 92, 93, 94 и/или 96. Мутация может быть выбрана из S28Y, S28T, S28K, S28P, S28Q, S28R, S28M, S30A, S30V, R31Q, N34V, N34S, Y49F, A50V, A50G, A51G, A51S, S52Q, S52H, S52N, S52Y, S52R, S52L, S53I, S53Y, S53N, S53M, S53H, S53R, S91A, Y92I, S93G, S93Q, S93I, S93N, S93M, P94E, P94M, P94N, P94Q, P94A, P94T, P94I, F96Y или их комбинаций. В этих и других вариантах осуществления антитело к PAC1 содержит вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 296 с мутацией в одном или нескольких из аминокислотных положений 31, 32, 33, 57, 58, 60, 103, 105, 106, 107 и/или 110. В некоторых вариантах осуществления мутация выбрана из S31N, N32R, N32K, N32H, S33L, S33Q, S33Y, S33V, S33D, S57G, K58Q, S60K, T103M, T103Q, T103R, T103V, K105N, K105E, K105D, K105I, K105A, K105S, K105Q, Q106G, Q106E, L107D, L107N, L110M, L110F или их комбинаций.
В определенных вариантах осуществления антитело к PAC1 или антигенсвязывающий фрагмент по настоящему изобретению выбран из антител 420653, 420845, 420943, 421873, 420889, 421091, 421051, 480711, 480706, 480713, 448730, 448195, 448788, 448901, 448689, 448202, 452128, 448924, 3574 и 3575 или их антигенсвязывающих фрагментов, вариабельная область и последовательности CDR которых приведены в таблицах 1A и 1B. В некоторых вариантах осуществления антитело к PAC1 представляет собой антитело, выбранное из антител 420653, 420845, 420943, 421873, 420889, 421091, 421051, 480711, 480706 и 480713. В других вариантах осуществления антитело к PAC1 представляет собой антитело, выбранное из антител 448730, 448195, 448788, 448901, 448689, 448202, 452128, 448924, 3574 и 3575. Последовательности полноразмерных легких цепей и полноразмерных тяжелых цепей этих иллюстративных антител к PAC1 человека приведены ниже в таблицах 4A и 4B соответственно.
caccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 542)
caccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 547)
caccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 550)
caccatctgtcttcatcttcccgccatctgatgagcagttgaaatctggaactgcctctgttgtgtgcctgctgaataacttctatcccagagaggccaaagtacagtggaaggtggataacgccctccaatcgggtaactcccaggagagtgtcacagagcaggacagcaaggacagcacctacagcctcagcagcaccctgacgctgagcaaagcagactacgagaaacacaaagtctacgcctgcgaagtcacccatcagggcctgagctcgcccgtcacaaagagcttcaacaggggagagtgt (SEQ ID NO: 542)
TCCAGCCTGGGAGGTCCCTGCGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGATTTGCCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATCATATGATGGAGGAAATAAATACTATGCAGAGTCCGTGAAGGGCCGGTTCACCATCTCCAGAGACAATTCCAAGAACACCCTGTATCTGCAAATGAACAGCCTGAGAGCTGAGGACACGGCTCTGTTTTACTGTGCGAGAGGATACGATGTTTTGACTGGTTACCCCGACTACTGGGGCCAGGGAACCCTGGTCACCGTCTCTAGTGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGTGCGAGGAGCAGTACGGCAGCACGTACCGTTGCGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTGTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA (SEQ ID NO: 552)
TCCAGCCTGGGAGGTCCCTGCGACTCTCCTGTGCAGCCTCTGGATTCACCTTCAGTAGATTTGCCATGCACTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGGTGTTATCAACTATCGTGGACATGGTAAATACTATGCAGAGTCCGTGAAGGGCCGGTTCACCGTGTCCAGAGACAATTCCAAGAACACCCTGTATCTGCAAATGAACAGCCTGAGAGCTGAGGACACGGCTCTGTTTTACTGTGCGAGAGGATACGATGTTTTGACTGGTTACCCCGACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCTAGTGCCTCCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGTGCGAGGAGCAGTACGGCAGCACGTACCGTTGCGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTGTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGC
GACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTATAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAA (SEQ ID NO: 560)
RGMWNQNWFLDHWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPCEEQYGSTYRCVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK (SEQ ID NO: 536)
ccatcggtcttccccctggcaccctcctccaagagcacctctgggggcacagcggccctgggctgcctggtcaaggactacttccccgaaccggtgacggtgtcgtggaactcaggcgccctgaccagcggcgtgcacaccttcccggctgtcctacagtcctcaggactctactccctcagcagcgtggtgaccgtgccctccagcagcttgggcacccagacctacatctgcaacgtgaatcacaagcccagcaacaccaaggtggacaagaaagttgagcccaaatcttgtgacaaaactcacacatgcccaccgtgcccagcacctgaactcctggggggaccgtcagtcttcctcttccccccaaaacccaaggacaccctcatgatctcccggacccctgaggtcacatgcgtggtggtggacgtgagccacgaagaccctgaggtcaagttcaactggtacgtggacggcgtggaggtgcataatgccaagacaaagccgtgcgaggagcagtacggcagcacgtaccgttgcgtcagcgtcctcaccgtcctgcaccaggactggctgaatggcaaggagtacaagtgcaaggtgtccaacaaagccctcccagcccccatcgagaaaaccatctccaaagccaaagggcagccccgagaaccacaggtgtacaccctgcccccatcccgggaggagatgaccaagaaccaggtcagcctgacctgcctggtcaaaggcttctatcccagcgacatcgccgtggagtgggagagcaatgggcagccggagaacaactacaagaccacgcctcccgtgctggactccgacggctccttcttcctctatagcaagctcaccgtggacaagagcaggtggcagcaggggaacgtcttctcatgctccgtgatgcatgaggctctgcacaaccactacacgcagaagagcctctccctgtctccgggtaaa (SEQ ID NO: 565)
В определенных вариантах осуществления антитела к PAC1 по настоящему изобретению могут содержать легкую цепь, выбранную из LC-01 - LC-05, как показано в таблице 4A, и/или тяжелую цепь, выбранную из HC-01 - HC-11, как показано в таблице 4B, и связывающие фрагменты, производные и варианты этих легких цепей и тяжелых цепей. В других вариантах осуществления антитела к PAC1 по настоящему изобретению могут содержать легкую цепь, выбранную из LC-06 - LC-15, как показано в таблице 4A, и/или тяжелую цепь, выбранную из HC-12 - HC-18, как показано в таблице 4B, и связывающие фрагменты, производные и варианты этих легких цепей и тяжелых цепей.
Каждая из легких цепей, перечисленных в таблице 4A, может быть объединена с любой из тяжелых цепей, перечисленных в таблице 4B, с образованием антитела к PAC1 или его антигенсвязывающего фрагмента по настоящему изобретению. Например, в определенных вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-03 (SEQ ID NO: 506), и тяжелую цепь, содержащую последовательность HC-03 (SEQ ID NO: 521). В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-03 (SEQ ID NO: 506), и тяжелую цепь, содержащую последовательность HC-04 (SEQ ID NO: 522). В других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-01 (SEQ ID NO: 504), и тяжелую цепь, содержащую последовательность HC-05 (SEQ ID NO: 523). В еще других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-01 (SEQ ID NO: 504), и тяжелую цепь, содержащую последовательность HC-06 (SEQ ID NO: 524). В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-03 (SEQ ID NO: 506), и тяжелую цепь, содержащую последовательность HC-07 (SEQ ID NO: 525). В определенных вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-01 (SEQ ID NO: 504), и тяжелую цепь, содержащую последовательность HC-08 (SEQ ID NO: 526). В одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-03 (SEQ ID NO: 506), и тяжелую цепь, содержащую последовательность HC-05 (SEQ ID NO: 523). В другом варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-04 (SEQ ID NO: 507), и тяжелую цепь, содержащую последовательность HC-09 (SEQ ID NO: 527). В еще одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-04 (SEQ ID NO: 507), и тяжелую цепь, содержащую последовательность HC-10 (SEQ ID NO: 528). В еще одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-05 (SEQ ID NO: 508), и тяжелую цепь, содержащую последовательность HC-11 (SEQ ID NO: 529).
В определенных других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-07 (SEQ ID NO: 510), и тяжелую цепь, содержащую последовательность HC-13 (SEQ ID NO: 531). В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-08 (SEQ ID NO: 511), и тяжелую цепь, содержащую последовательность HC-14 (SEQ ID NO: 532). В других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-09 (SEQ ID NO: 512), и тяжелую цепь, содержащую последовательность HC-15 (SEQ ID NO: 533). В еще других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-10 (SEQ ID NO: 513), и тяжелую цепь, содержащую последовательность HC-16 (SEQ ID NO: 534). В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-11 (SEQ ID NO: 514), и тяжелую цепь, содержащую последовательность HC-17 (SEQ ID NO: 535). В определенных вариантах осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-12 (SEQ ID NO: 515), и тяжелую цепь, содержащую последовательность HC-17 (SEQ ID NO: 535). В одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-13 (SEQ ID NO: 516), и тяжелую цепь, содержащую последовательность HC-14 (SEQ ID NO: 532). В другом варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-14 (SEQ ID NO: 517), и тяжелую цепь, содержащую последовательность HC-18 (SEQ ID NO: 536). В еще одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-06 (SEQ ID NO: 509), и тяжелую цепь, содержащую последовательность HC-14 (SEQ ID NO: 532). В еще одном варианте осуществления антитела к PAC1 по настоящему изобретению содержат легкую цепь, содержащую последовательность LC-15 (SEQ ID NO: 518), и тяжелую цепь, содержащую последовательность HC-12 (SEQ ID NO: 530).
В некоторых вариантах осуществления антитела к PAC1 содержат легкую цепь, содержащую последовательность смежных аминокислот, которая отличается от последовательности легкой цепи в таблице 4A, т. е. легкую цепь, выбранную из LC-01 - LC-15, только по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотным остаткам, где каждое такое различие последовательностей независимо представляет собой либо делецию, вставку, либо замену одной аминокислоты, причем делеции, вставки и/или замены приводят к изменениям не более чем 15 аминокислот относительно вышеуказанных последовательностей легкой цепи. Легкая цепь в некоторых антителах к PAC1 содержит последовательность аминокислот, которая характеризуется по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичностью последовательности с аминокислотными последовательностями под SEQ ID NO: 504-518 (т. е. легкими цепями в таблице 4A).
В этих и других вариантах осуществления антитела к PAC1 содержат тяжелую цепь, содержащую последовательность смежных аминокислот, которая отличается от последовательности тяжелой цепи в таблице 4B, т. е. тяжелую цепь, выбранную из HC-01 - HC-18, только по 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 аминокислотным остаткам, где каждое такое различие последовательностей независимо представляет собой либо делецию, вставку, либо замену одной аминокислоты, причем делеции, вставки и/или замены приводят к изменениям не более чем 15 аминокислот относительно вышеуказанных последовательностей тяжелой цепи. Тяжелая цепь в некоторых антителах к PAC1 содержит последовательность аминокислот, которая характеризуется по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 97% или по меньшей мере 99% идентичностью последовательности с аминокислотными последовательностями под SEQ ID NO: 519-536 (т. е. тяжелыми цепями в таблице 4B).
Антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению могут представлять собой моноклональные антитела, поликлональные антитела, рекомбинантные антитела, антитела человека, гуманизированные антитела, химерные антитела, или мультиспецифические антитела, или их антигенсвязывающие фрагменты. В определенных вариантах осуществления антитело к PAC1 представляет собой моноклональное антитело. В таких вариантах осуществления антитело к PAC1 может представлять собой химерное антитело, гуманизированное антитело или полностью человеческое антитело, содержащее константный домен иммуноглобулина человека. В этих и других вариантах осуществления антитело к PAC1 представляет собой антитело IgG1, IgG2, IgG3 или IgG4 человека. Таким образом, антитело к PAC1 может в некоторых вариантах осуществления содержать константный домен IgG1, IgG2, IgG3 или IgG4 человека. В одном варианте осуществления антитело к PAC1 представляет собой моноклональное антитело IgG1 человека. В другом варианте осуществления антитело к PAC1 представляет собой моноклональное антитело IgG2 человека. В еще одном варианте осуществления антитело к PAC1 представляет собой моноклональное антитело IgG4 человека.
Применяемый в данном документе термин "моноклональное антитело" (или "mAb") относится к антителу, полученному из популяции практически гомогенных антител, то есть отдельные антитела, составляющие популяцию, являются идентичными за исключением возможно встречающихся в природе мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифическими и направлены против отдельного антигенного сайта или эпитопа, в отличие от препаратов на основе поликлональных антител, которые обычно включают различные антитела, направленные против различных эпитопов. Моноклональные антитела можно получать с применением любой методики, известной из уровня техники, например, посредством иммортализации клеток селезенки, собранных у животного после завершения схемы иммунизации. Клетки селезенки могут быть иммортализованы с применением любой методики, известной из уровня техники, например, посредством их слияния с клетками миеломы с получением гибридом. См. например, Antibodies; Harlow and Lane, Cold Spring Harbor Laboratory Press, 1-е издание, например, от 1988 года, или 2-е издание, например, от 2014 года. Клетки миеломы для применения в процедурах слияния для получения гибридом предпочтительно представляют собой клетки, которые не продуцируют антитела, обладают высокой эффективностью слияния и характеризуются дефицитами ферментов, что делает их неспособными к росту в определенных селективных средах, которые поддерживают рост только требуемых слитых клеток (гибридом). Примеры подходящих линий клеток для применения для слияний с клетками мыши включают без ограничения Sp-20, P3-X63/Ag8, P3-X63-Ag8.653, NS1/1.Ag 4 1, Sp210-Ag14, FO, NSO/U, MPC-11, MPC11-X45-GTG 1.7 и S194/5XXO Bul. Примеры подходящих линий клеток, применяемых для слияний с клетками крысы, включают без ограничения R210.RCY3, Y3-Ag 1.2.3, IR983F и 4B210. Другие линии клеток, применяемые для слияний клеток, представляют собой U-266, GM1500-GRG2, LICR-LON-HMy2 и UC729-6.
В некоторых случаях линию клеток гибридомы получают посредством иммунизации животного (например, кролика, крысы, мыши или трансгенного животного, имеющего последовательности иммуноглобулина человека) иммуногеном PAC1 (см. например, WO 2014/144632); сбора клеток селезенки у иммунизированного животного; слияния собранных клеток селезенки с линией клеток миеломы с получением тем самым клеток гибридомы; получения линий клеток гибридомы из клеток гибридомы и идентификации линии клеток гибридомы, которая продуцирует антитело, которое связывается с PAC1. Другим применяемым способом получения моноклональных антител является способ SLAM, описанный в Babcook et al., Proc. Natl. Acad. Sci. USA, Vol. 93: 7843-7848, 1996.
Моноклональные антитела, секретируемые линией клеток гибридомы, могут быть очищены с применением любой методики, известной из уровня техники, такой как протеин А-сефароза, гидроксиапатитная хроматография, гель-электрофорез, диализ или аффинная хроматография. Супернатанты гибридом или mAb можно дополнительно подвергать скринингу для идентификации mAb с определенными свойствами, такими как способность связывать PAC1 (например, PAC1 человека, PAC1 яванского макака или PAC1 крысы); перекрестная реактивность с другими представителями семейства PAC1 (например, VPAC1 человека или VPAC2 человека); способность блокировать или препятствовать связыванию лиганда PACAP с PAC1 или способность функционально блокировать индуцированную PACAP активацию рецептора PAC1, например, с применением анализа cAMP, как описано в данном документе.
В некоторых вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению представляют собой химерные или гуманизированные антитела или их антигенсвязывающие фрагменты на основе последовательностей CDR и вариабельных областей антител, описанных в данном документе. Химерное антитело представляет собой антитело, состоящее из сегментов белка из различных антител, которые ковалентно соединены с образованием функциональных легких или тяжелых цепей иммуноглобулина или их связывающих фрагментов. Как правило, часть тяжелой цепи и/или легкой цепи идентична или гомологична соответствующей последовательности в антителах, полученных из определенного вида или принадлежащих к определенному классу или подклассу антител, тогда как остальная(остальные) часть(части) цепи(цепей) идентична(идентичны) или гомологична(гомологичны) соответствующей последовательности в антителах, полученных из другого вида или принадлежащих к другому классу или подклассу антител. Для способов, относящихся к химерным антителам, см. например, патент США № 4816567 и Morrison et al., 1985, Proc. Natl. Acad. Sci. USA 81:6851-6855, каждый из которых настоящим включен посредством ссылки во всей своей полноте.
Как правило, целью получения химерного антитела является создание химеры, в которой количество аминокислот из предполагаемых видов является максимальным. Одним из примеров является "CDR-привитое" антитело, в котором антитело содержит одну или несколько CDR из конкретного вида или принадлежащих определенному классу или подклассу антител, тогда как остальная(остальные) часть(части) цепи(цепей) антитела идентична(идентичны) или гомологична(гомологичны) соответствующей последовательности в антителах, полученных из другого вида или принадлежащих другому классу или подклассу антител. Прививание CDR описано, например, в патенте США № 6180370, № 5693762, № 5693761, № 5585089 и № 5530101. Для применения на людях вариабельную область или отобранные CDR из антитела грызуна или кролика зачастую прививают на антитело человека, заменяя встречающиеся в природе вариабельные области или CDR антитела человека.
Одним из применимых типов химерного антитела является "гуманизированное" антитело. Как правило, гуманизированное антитело получают из моноклонального антитела, первоначально происходящего из животного, отличного от человека, такого как грызун или кролик. Определенные аминокислотные остатки в этом моноклональном антителе, как правило, из частей антитела, которые не распознают антиген, модифицируют таким образом, чтобы они были гомологичными соответствующим остаткам в антителе человека соответствующего изотипа. Гуманизацию можно осуществлять, например, с применением различных способов посредством замены по меньшей мере части вариабельной области грызуна или кролика на соответствующие области антитела человека (см. например, патент США № 5585089 и № 5693762; Jones et al., 1986, Nature 321:522-525; Riechmann et al., 1988, Nature 332:323-27 и Verhoeyen et al., 1988, Science 239:1534-1536).
В одном аспекте CDR вариабельных областей легкой и тяжелой цепей антител, предусмотренных в данном документе (см. таблицы 1A и 1B), прививают на каркасные области (FR) из антител из одного и того же или различных филогенетических видов. Например, CDR вариабельных областей тяжелой и легкой цепей, перечисленных в таблицах 1A и 1B, могут быть привиты к консенсусным FR человека. Для создания консенсусных FR человека FR из нескольких аминокислотных последовательностей тяжелой цепи или легкой цепи человека могут быть выровнены для идентификации консенсусной аминокислотной последовательности. В качестве альтернативы, привитые вариабельные области из одной тяжелой или легкой цепи можно применять с константной областью, которая отличается от константной области этой конкретной тяжелой или легкой цепи, раскрытой в данном документе. В других вариантах осуществления привитые вариабельные области являются частью одноцепочечного Fv-антитела.
В определенных вариантах осуществления антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению представляют собой полностью человеческие антитела или их антигенсвязывающие фрагменты. Полностью человеческие антитела, которые специфически связываются с PAC1 человека, можно получать с применением иммуногенов или их фрагментов, описанных в WO 2014/144632, таких как полипептиды, состоящие из последовательности под SEQ ID NO: 1 или SEQ ID NO: 4. "Полностью человеческое антитело" представляет собой антитело, которое содержит вариабельные и константные области, полученные из последовательностей иммуноглобулинов зародышевой линии человека или указывающие на них. Одним конкретным способом, предусмотренным для осуществления получения полностью человеческих антител, является "гуманизация" гуморальной иммунной системы мыши. Введение локусов иммуноглобулинов (Ig) человека в геном мышей, у которых были инактивированы эндогенные гены Ig, является одним из способов получения полностью человеческих моноклональных антител (mAb) у мыши - животного, которое можно иммунизировать любым требуемым антигеном. Применение полностью человеческих антител может минимизировать иммуногенные и аллергические ответы, которые иногда могут быть вызваны введением мышиных или полученных из мыши mAb людям в качестве терапевтических средств.
Полностью человеческие антитела можно получать посредством иммунизации трансгенных животных (обычно мышей), которые способны продуцировать репертуар антител человека в отсутствие продуцирования эндогенных иммуноглобулинов. Применяемые для этой цели антигены, как правило, содержат шесть или более смежных аминокислот и необязательно конъюгированы с носителем, таким как гаптен. См. например, Jakobovits et al., 1993, Proc. Natl. Acad. Sci. USA 90:2551-2555; Jakobovits et al., 1993, Nature 362:255-258 и Bruggermann et al., 1993, Year in Immunol. 7:33. В одном примере такого способа трансгенных животных получают посредством инактивации эндогенных локусов иммуноглобулина мыши, кодирующих тяжелые и легкие цепи иммуноглобулина мыши, и вставки в геном мыши больших фрагментов геномной ДНК человека, содержащей локусы, которые кодируют белки тяжелых и легких цепей человека. Частично модифицированных животных, которые имеют неполный набор локусов иммуноглобулинов человека, подвергают кроссбридингу с получением животного, обладающего всеми требуемыми модификациями иммунной системы. В случае введения иммуногена эти трансгенные животные продуцируют антитела, которые являются иммуноспецифическими в отношении иммуногена, однако имеют скорее человеческие, а не мышиные аминокислотные последовательности, в том числе вариабельные области. Для дополнительной информации по таким способам см. например, WO96/33735 и WO94/02602. Дополнительные способы, относящиеся к применению трансгенных мышей для получения антител человека, описаны в патенте США № 5545807; № 6713610; № 6673986; № 6162963; № 5939598; № 5545807; № 6300129; № 6255458; № 5877397; № 5874299 и № 5545806; в публикациях согласно PCT WO 91/10741, WO 90/04036, WO 94/02602, WO 96/30498, WO 98/24893, а также в EP 546073B1 и EP 546073A1.
Описанные выше трансгенные мыши, обозначенные как мыши "HuMab", содержат минилокус гена иммуноглобулина человека, который кодирует нереаранжированные последовательности тяжелых (мю- и гамма-) цепей и легкой каппа-цепи иммуноглобулина человека вместе с целевыми мутациями, которые инактивируют локусы эндогенных мю- и каппа-цепей (Lonberg et al., 1994, Nature 368:856-859). Соответственно, мыши проявляют сниженную экспрессию IgM или каппа-белков мыши, и в ответ на иммунизацию введенные трансгены тяжелой и легкой цепей человека претерпевают переключение класса и соматическую мутацию с образованием высокоаффинных моноклональных антител IgG-каппа человека (Lonberg and Huszar, 1995, Intern. Rev. Immunol. 13: 65-93; Harding and Lonberg, 1995, Ann. N.Y Acad. Sci. 764:536-546). Получение мышей HuMab подробно описано в Taylor et al., 1992, Nucleic Acids Research 20:6287-6295; Chen et al., 1993, International Immunology 5:647-656; Tuaillon et al., 1994, J. Immunol. 152:2912-2920; Lonberg et al., 1994, Nature 368:856-859; Lonberg, 1994, Handbook of Exp. Pharmacology 113:49-101; Taylor et al., 1994, International Immunology 6:579-591; Lonberg and Huszar, 1995, Intern. Rev. Immunol. 13:65-93; Harding and Lonberg, 1995, Ann. N.Y Acad. Sci. 764:536-546; Fishwild et al., 1996, Nature Biotechnology 14:845-851; указанные выше ссылки настоящим включены посредством ссылки во всей своей полноте. См. дополнительно патент США № 5545806; № 5569825; № 5625126; № 5633425; № 5789650; № 5877397; № 5661016; № 5814318; № 5874299; и № 5770429; а также патент США № 5545807; международную публикацию №№ WO 93/1227; WO 92/22646; и WO 92/03918, раскрытия всех из которых настоящим включены посредством ссылки во всей своей полноте. Технологии, применяемые для получения антител человека в этих трансгенных мышах, раскрыты также в WO 98/24893 и Mendez et al., 1997, Nature Genetics 15:146-156, которые настоящим включены посредством ссылки. Например, линии трансгенных мышей HCo7 и HCo12 можно применять для получения полностью человеческих антител к PAC1. Одной конкретной линией трансгенных мышей, подходящей для получения полностью человеческих антител к PAC1, является линия трансгенных мышей XenoMouse®, описанная в патентах США №№ 6114598; 6162963; 6833268; 7049426; 7064244; Green et al., 1994, Nature Genetics 7:13-21; Mendez et al., 1997, Nature Genetics 15:146-156; Green and Jakobovitis, 1998, J. Ex. Med, 188:483-495; Green, 1999, Journal of Immunological Methods 231:11-23; Kellerman and Green, 2002, Current Opinion in Biotechnology 13, 593-597, все из которых настоящим включены посредством ссылки во всей своей полноте.
Антитела человеческого происхождения также можно получать с применением методик фагового дисплея. Фаговый дисплей описан, например, в Dower et al., WO 91/17271, McCafferty et al., WO 92/01047 и Caton and Koprowski, Proc. Natl. Acad. Sci. USA, 87:6450-6454 (1990), каждый из которых включен в данный документ посредством ссылки во всей своей полноте. Антитела, полученные с помощью технологии с применением фагов, как правило, получают в виде антигенсвязывающих фрагментов, например, Fv- или Fab-фрагментов у бактерий и, таким образом, они лишены эффекторных функций. Эффекторные функции можно вводить посредством одной из двух стратегий: Фрагменты можно конструировать либо в виде полных антител для экспрессии в клетках млекопитающих, либо в виде фрагментов биспецифических антител со вторым сайтом связывания, способным запускать эффекторную функцию, если это требуется. Как правило, Fd-фрагмент (VH-CH1) и легкую цепь (VL-CL) антител отдельно клонируют с помощью ПЦР и случайным образом рекомбинируют в комбинаторных библиотеках для фагового дисплея, которые затем могут быть отобраны для связывания с конкретным антигеном. Фрагменты антител экспрессируются на поверхности фага, и отбор Fv или Fab (а следовательно фага, содержащего ДНК, кодирующую фрагмент антитела) посредством связывания антигена осуществляется посредством нескольких раундов связывания антигена и повторной амплификации, - процедуры, называемой пэннингом. Фрагменты антител, специфические в отношении антигена, обогащаются и, наконец, выделяются. Методики фагового дисплея также можно применять в подходе для гуманизации моноклональных антител грызунов, называемом "управляемым отбором" (см. Jespers, L. S. et al., 1994, Bio/Technology 12, 899-903). Для этого Fd-фрагмент моноклонального антитела мыши может быть отображен в комбинации с библиотекой легких цепей человека, и полученная в результате библиотека гибридных Fab может быть затем отобрана с помощью антигена. Таким образом, Fd-фрагмент мыши обеспечивает шаблон для управления отбором. Затем отобранные легкие цепи человека объединяют с библиотекой Fd-фрагментов человека. Отбор полученной библиотеки приводит к получению полностью человеческого Fab.
Как только клетки, продуцирующие антитела к PAC1 в соответствии с настоящим изобретением, были получены с применением любой из описанных выше иммунизации и других методик, гены специфических антител могут быть клонированы посредством выделения и амплификации с них ДНК или mRNA в соответствии со стандартными процедурами, описанными в данном документе. Полученные из них антитела можно секвенировать, и идентифицированными CDR и ДНК, кодирующей CDR, можно манипулировать, как описано в данном документе, для получения других антител к PAC1 или антигенсвязывающих фрагментов в соответствии с настоящим изобретением.
В определенных вариантах осуществления антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению могут содержать одну или несколько мутаций или модификаций в константной области. Например, константные области тяжелой цепи или Fc-области антител к PAC1 могут содержать одну или несколько аминокислотных замен, которые оказывают влияние на гликозилирование, эффекторную функцию и/или связывание антитела Fcγ-рецептором. Термин "Fc-область" относится к С-концевой области тяжелой цепи иммуноглобулина, которая может быть получена посредством расщепления интактного антитела с помощью папаина. Fc-область иммуноглобулина обычно содержит два константных домена, домен CH2 и домен CH3, и необязательно содержит домен CH4. В определенных вариантах осуществления Fc-область представляет собой Fc-область из иммуноглобулина IgG1, IgG2, IgG3 или IgG4. В некоторых вариантах осуществления Fc-область содержит домены CH2 и CH3 из иммуноглобулина IgG1 человека или IgG2 человека. Fc-область может сохранять эффекторную функцию, такую как связывание C1q, комплементзависимая цитотоксичность (CDC), связывание с рецептором Fc, антителозависимая клеточно-опосредованная цитотоксичность (ADCC) и фагоцитоз. В других вариантах осуществления Fc-область может быть модифицирована для снижения или устранения эффекторной функции, как описано более подробно ниже.
Если не указано иное посредством ссылки на конкретную последовательность, то в настоящей заявке и формуле изобретения нумерация аминокислотных остатков в тяжелой цепи или легкой цепи иммуноглобулина соответствует нумерации AHo, как описано в Honegger and Pluckthun, J. Mol. Biol. 309(3):657-670; 2001, или нумерации EU, как описано в Edelman et al., Proc. Natl. Acad. USA, Vol. 63: 78-85 (1969). Применяемая в данном документе схема нумерации AHo обычно применяется при ссылке на положение аминокислоты в вариабельных областях, тогда как схема нумерации EU обычно применяется при ссылке на положение аминокислоты в константной области иммуноглобулина.
Замена аминокислоты в аминокислотной последовательности в данном документе, как правило, обозначается однобуквенным сокращением аминокислотного остатка в конкретном положении, за которым следует числовое положение аминокислоты относительно исходной представляющей интерес последовательности, за которой следует однобуквенное сокращение для замененного аминокислотного остатка. Например, "T30D" означает замену остатка треонина на остаток аспарагиновой кислоты по аминокислотному положению 30 относительно исходной представляющей интерес последовательности. В другом примере "S218G" означает замену остатка серина на остаток глицина по аминокислотному положению 218 относительно исходной представляющей интерес аминокислотной последовательности.
Одна из функций Fc-области иммуноглобулина заключается в том, чтобы сообщать иммунной системе в случае, если иммуноглобулин связывается со своей мишенью. Это обычно называют "эффекторной функцией". Взаимодействие приводит к антителозависимой клеточной цитотоксичности (ADCC), антителозависимому клеточному фагоцитозу (ADCP) и/или комплементзависимой цитотоксичности (CDC). ADCC и ADCP опосредуются связыванием Fc-области с рецепторами Fc на поверхности клеток иммунной системы. CDC опосредуется связыванием Fc-области с белками системы комплемента, например, C1q. В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат одну или несколько аминокислотных замен в Fc-области для усиления эффекторной функции, в том числе активности ADCC, активности CDC, активности ADCP и/или клиренса или увеличения периода полужизни антитела. Иллюстративные аминокислотные замены (в соответствии со схемой нумерации EU), которые могут усиливать эффекторную функцию, включают без ограничения E233L, L234I, L234Y, L235S, G236A, S239D, F243L, F243V, P247I, D280H, K290S, K290E, K290N, K290Y, R292P, E294L, Y296W, S298A, S298D, S298V, S298G, S298T, T299A, Y300L, V305I, Q311M, K326A, K326E, K326W, A330S, A330L, A330M, A330F, I332E, D333A, E333S, E333A, K334A, K334V, A339D, A339Q, P396L или комбинации любых из вышеперечисленных.
В других вариантах осуществления антитела к PAC1 по настоящему изобретению содержат одну или несколько аминокислотных замен в Fc-области для снижения эффекторной функции. Иллюстративные аминокислотные замены (в соответствии со схемой нумерации EU), которые могут снижать эффекторную функцию, включают без ограничения C220S, C226S, C229S, E233P, L234A, L234V, V234A, L234F, L235A, L235E, G237A, P238S, S267E, H268Q, N297A, N297G, V309L, E318A, L328F, A330S, A331S, P331S или комбинации любых из вышеперечисленных.
Гликозилирование может способствовать эффекторной функции антител, в частности антител IgG1. Таким образом, в некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению могут содержать одну или несколько аминокислотных замен, которые влияют на уровень или тип гликозилирования антител. Гликозилирование полипептидов обычно является либо N-связанным, либо O-связанным. N-связанное относится к присоединению углеводного фрагмента к боковой цепи остатка аспарагина. Трипептидные последовательности аспарагин-Х-серин и аспарагин-Х-треонин, где Х представляет собой любую аминокислоту за исключением пролина, представляют собой последовательности распознавания для ферментативного присоединения углеводного фрагмента к боковой цепи аспарагина. Таким образом, присутствие любой из этих трипептидных последовательностей в полипептиде создает потенциальный сайт гликозилирования. O-связанное гликозилирование относится к присоединению одного из сахаров, представляющих собой N-ацетилгалактозамин, галактозу или ксилозу, к гидроксиаминокислоте, наиболее часто серину или треонину, хотя также можно применять 5-гидроксипролин или 5-гидроксилизин.
В определенных вариантах осуществления гликозилирование описанных в данном документе антител к PAC1 усиливается посредством добавления одного или нескольких сайтов гликозилирования, например, к Fc-области антитела. Добавление сайтов гликозилирования к антителу можно удобно осуществлять посредством изменения аминокислотной последовательности таким образом, чтобы она содержала одну или несколько описанных выше трипептидных последовательностей (для сайтов N-связанного гликозилирования). Изменение также можно осуществлять посредством замены или добавления одного или нескольких остатков серина или треонина к исходной последовательности (для сайтов O-связанного гликозилирования). Для удобства аминокислотная последовательность антитела может быть изменена посредством изменений на уровне ДНК, в частности, посредством мутации ДНК, кодирующей целевой полипептид, в предварительно выбранных основаниях таким образом, что образуются кодоны, которые будут транслироваться в требуемые аминокислоты.
Настоящее изобретение также охватывает получение молекул антител с измененной структурой углеводов, что приводит к изменению эффекторной активности, включая антитела с отсутствующим или сниженным уровнем фукозилирования, которые проявляют улучшенную активность ADCC. Из уровня техники известны различные способы снижения или устранения фукозилирования. Например, эффекторная активность ADCC опосредуется связыванием молекулы антитела с рецептором FcγRIII, который, как было показано, зависит от углеводной структуры N-связанного гликозилирования остатка N297 домена CH2. Нефукозилированные антитела связывают этот рецептор с повышенной аффинностью и запускают эффекторные функции, опосредованные FcγRIII, более эффективно, чем нативные фукозилированные антитела. Например, рекомбинантное получение нефукозилированного антитела в клетках линии СНО, в которых фермент альфа-1,6-фукозилтрансфераза был нокаутирован, приводит к получению антитела со 100-кратно повышенной активностью ADCC (см. Yamane-Ohnuki et al., Biotechnol Bioeng. 87(5):614-22, 2004). Подобные эффекты могут быть достигнуты посредством снижения активности фермента альфа-1,6-фукозилтрансферазы или других ферментов в пути фукозилирования, например, посредством обработки с помощью siRNA или антисмысловой РНК, конструирования линий клеток для нокаута фермента(ов) или культивирования с селективными ингибиторами гликозилирования (см. Rothman et al., Mol Immunol. 26 (12): 1113-23, 1989). Некоторые штаммы клеток-хозяев, например, Lec13 или линия клеток гибридомы YB2/0 крысы, естественным путем продуцируют антитела с более низкими уровнями фукозилирования (см. Shields et al., J Biol Chem. 277(30):26733-40, 2002 и Shinkawa et al., J Biol Chem. 278(5):3466-73, 2003). Также было установлено, что повышение уровня разветвленного углевода, например, посредством рекомбинантного продуцирования антитела в клетках, которые сверхэкспрессируют фермент GnTIII, также повышает активность ADCC (см. Umana et al., Nat Biotechnol. 17(2):176-80, 1999).
В других вариантах осуществления гликозилирование описанных в данном документе антител к PAC1 снижается или устраняется посредством удаления одного или нескольких сайтов гликозилирования, например, из Fc-области антитела. В некоторых вариантах осуществления антитело к PAC1 представляет собой агликозилированное моноклональное антитело человека, например, агликозилированное моноклональное антитело IgG1 человека. Аминокислотные замены, которые устраняют или модифицируют сайты N-связанного гликозилирования, могут снижать или устранять N-связанное гликозилирование антитела. В определенных вариантах осуществления антитела к PAC1, описанные в данном документе, содержат в своей тяжелой цепи мутацию в положении N297 (в соответствии со схемой нумерации EU), такую как N297Q, N297A или N297G. В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат Fc-область из антитела IgG1 человека с мутацией в положении N297. В одном конкретном варианте осуществления антитела к PAC1 по настоящему изобретению содержат Fc-область из антитела IgG1 человека с мутацией N297G. Например, в некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат константную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 324.
Для улучшения стабильности молекул, содержащих мутацию N297, Fc-область антител к PAC1 может быть дополнительно сконструирована. Например, в некоторых вариантах осуществления одна или несколько аминокислот в Fc-области заменены на цистеин для содействия образованию дисульфидной связи в димерном состоянии. Таким образом, остатки, соответствующие V259, A287, R292, V302, L306, V323 или I332 (в соответствии со схемой нумерации EU) Fc-области IgG1, могут быть заменены на цистеин. Предпочтительно, конкретные пары остатков заменены на цистеин таким образом, что они предпочтительно образуют дисульфидную связь друг с другом, таким образом ограничивая или предотвращая скремблирование дисульфидных связей. Предпочтительные пары включают без ограничения A287C и L306C, V259C и L306C, R292C и V302C, а также V323C и I332C. В определенных вариантах осуществления антитела к PAC1,описанные в данном документе, содержат Fc-область из антитела IgG1 человека с мутациями R292C и V302C. В таких вариантах осуществления Fc-область также может содержать мутацию N297, такую как мутация N297G. В некоторых вариантах осуществления антитела к PAC1 по настоящему изобретению содержат константную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 325.
Также могут быть желательны модификации антител к PAC1 по настоящему изобретению для увеличения периода полужизни в сыворотке крови, например, посредством включения или добавления эпитопа, связывающего рецептор реутилизации (например, посредством мутации в соответствующей области или посредством включения эпитопа в пептидную метку, которая затем сливается с антителом на любом конце или в середине, например, посредством синтеза ДНК или пептида; см., например WO96/32478), или добавления молекул, таких как PEG или другие водорастворимые полимеры, включая полисахаридные полимеры. Эпитоп, связывающий рецептор реутилизации, предпочтительно состоит из области, где любые один или несколько аминокислотных остатков из одной или двух петель Fc-области переносятся в аналогичное положение антитела. Еще более предпочтительно, переносятся три или более остатков из одной или двух петель Fc-области. Еще более предпочтительно, эпитоп взят из домена СН2 Fc-области (например, Fc-области IgG) и перенесен в область СН1, СН3 или VH или в более чем одну такую область антитела. В качестве альтернативы, эпитоп взят из домена СН2 Fc-области и перенесен в область CL или область VL, либо в обе эти области антитела. См. международные заявки WO 97/34631 и WO 96/32478 для описания вариантов Fc и их взаимодействия с рецептором реутилизации.
Настоящее изобретение включает один или несколько выделенных полинуклеотидов или выделенных нуклеиновых кислот, кодирующих антитела к PAC1 или антигенсвязывающие фрагменты, описанные в данном документе. Кроме того, настоящее изобретение охватывает векторы, содержащие нуклеиновые кислоты, клетки-хозяева или линии клеток, содержащие нуклеиновые кислоты, и способы получения антител к PAC1 и антигенсвязывающих фрагментов по настоящему изобретению. Нуклеиновые кислоты содержат, например, полинуклеотиды, которые кодируют все или часть антитела или антигенсвязывающего фрагмента, например, одну или обе цепи антитела по настоящему изобретению, или его фрагмент, производное или вариант, полинуклеотиды, достаточные для применения в качестве гибридизационных зондов, ПЦР-праймеры или праймеры для секвенирования для идентификации, анализа, мутации или амплификации полинуклеотида, кодирующего полипептид, антисмысловые олигонуклеотиды для ингибирования экспрессии полинуклеотида и последовательности, комплементарные вышеуказанным последовательностям. Нуклеиновые кислоты могут быть любой длины, подходящей для требуемого применения или функции, и могут содержать одну или несколько дополнительных последовательностей, например, регуляторных последовательностей, и/или быть частью большей нуклеиновой кислоты, например, вектора. Молекулы нуклеиновой кислоты по настоящему изобретению включают ДНК и РНК как в одноцепочечной, так и в двухцепочечной форме, а также соответствующие комплементарные последовательности. ДНК включает, например, cDNA, геномную ДНК, химически синтезированную ДНК, ДНК, амплифицированную с помощью ПЦР, и их комбинации. Молекулы нуклеиновой кислоты по настоящему изобретению включают полноразмерные гены или молекулы cDNA, а также комбинацию их фрагментов. Нуклеиновые кислоты по настоящему изобретению можно получать из человеческих источников, а также видов, отличных от человека.
Соответствующие аминокислотные последовательности из иммуноглобулина или его области (например, вариабельной области, Fc-области и т. д.) или представляющего интерес полипептида могут быть определены посредством прямого секвенирования белка, и подходящие кодирующие нуклеотидные последовательности могут быть сконструированы в соответствии с универсальной таблицей кодонов. В качестве альтернативы, геномную ДНК или cDNA, кодирующую моноклональные антитела или их связывающие фрагменты по настоящему изобретению, можно выделять и секвенировать из клеток, продуцирующих такие антитела, с помощью общепринятых процедур (например, с помощью олигонуклеотидных зондов, которые способны специфически связываться с генами, кодирующими тяжелую и легкую цепи моноклональных антител).
Термин "выделенная нуклеиновая кислота", который применяется в данном документе взаимозаменяемо с термином "выделенный полинуклеотид", представляет собой нуклеиновую кислоту, которая была отделена от смежных генетических последовательностей, присутствующих в геноме организма, из которого нуклеиновая кислота была выделена, в случае нуклеиновых кислот, выделенных из встречающихся в природе источников. В случае нуклеиновых кислот, синтезированных ферментативно с матрицы или химически, таких как ПЦР-продукты, молекулы cDNA или, например, олигонуклеотиды, понятно, что нуклеиновые кислоты, образующиеся в результате таких процессов, представляют собой выделенные нуклеиновые кислоты. Выделенная молекула нуклеиновой кислоты относится к молекуле нуклеиновой кислоты в виде отдельного фрагмента или в качестве компонента более крупной конструкции на основе нуклеиновой кислоты. В одном предпочтительном варианте осуществления нуклеиновые кислоты практически свободны от загрязняющего эндогенного материала. Молекулу нуклеиновой кислоты предпочтительно получают из ДНК или РНК, выделенной по меньшей мере один раз, в практически чистой форме и в количестве или концентрации, позволяющих идентифицировать, манипулировать и восстанавливать составляющие ее нуклеотидные последовательности с помощью стандартных биохимических способов (таких как способы, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989)). Такие последовательности предпочтительно предусмотрены и/или сконструированы в виде открытой рамки считывания, не прерываемой внутренними нетранслируемыми последовательностями или интронами, которые обычно присутствуют в эукариотических генах. Последовательности нетранслируемой ДНК могут быть представлены 5'- или 3'-концами открытой рамки считывания, где то же самое не мешает манипуляции или экспрессии кодирующей области. Если не указано иное, то левый конец любой обсуждаемой в данном документе одноцепочечной полинуклеотидной последовательности является 5'-концом; левое направление двухцепочечных полинуклеотидных последовательностей называется 5'-направлением. Направление от 5'- к 3'-концу, в котором происходит наращивание образующихся РНК-транскриптов, называется направлением транскрипции; области последовательности цепи ДНК, содержащие такую же последовательность, что и РНК-транскрипт, которые расположены в 5'-направлении относительно 5'-конца РНК-транскрипта, называются "вышележащими последовательностями"; области последовательности цепи ДНК, содержащие такую же последовательность, что и РНК-транскрипт, которые расположены в 3'-направлении относительно 3'-конца РНК-транскрипта, называются "нижележащими последовательностями".
Настоящее изобретение также включает нуклеиновые кислоты, которые гибридизуются в умеренно жестких условиях и, более предпочтительно, в очень жестких условиях с нуклеиновыми кислотами, кодирующими полипептиды, описанные в данном документе. Основные параметры, влияющие на выбор условий гибридизации, и руководство для разработки подходящих условий приведены в Sambrook, Fritsch, and Maniatis (1989, Molecular Cloning: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y., главы 9 и 11, и Current Protocols in Molecular Biology, 1995, Ausubel et al., eds., John Wiley & Sons, Inc., разделы 2.10 и 6.3-6.4), и могут быть легко определены специалистами в данной области техники исходя, например, из длины и/или нуклеотидного состава ДНК. Один из способов достижения умеренно жестких условий включает применение раствора для предварительной промывки, содержащего 5 x SSC, 0,5% SDS, 1,0 мМ EDTA (pH 8,0), буфер для гибридизации с приблизительно 50% формамида, 6 x SSC и температуры гибридизации, составляющей приблизительно 55°C (или других подобных растворов для гибридизации, таких как раствор, содержащий приблизительно 50% формамида, с температурой гибридизации, составляющей приблизительно 42°C), и условиями промывки, составляющими приблизительно 60°C, в 0,5 x SSC, 0,1% SDS. Как правило, очень жесткие условия определяются как условия гибридизации, указанные выше, но с промывкой при примерно 68°С, 0,2 x SSC, 0,1% SDS. SSPE (1 x SSPE представляет собой 0,15 М NaCl, 10 мМ NaH2PO4 и 1,25 мМ EDTA, рН 7,4) можно заменить на SSC (1 x SSC представляет собой 0,15 М NaCl и 15 мМ цитрата натрия) в буферах для гибридизации и промывки; промывки осуществляют в течение 15 минут после завершения гибридизации. Следует понимать, что температуру промывки и концентрацию промывочной соли можно корректировать по мере необходимости для достижения требуемой степени жесткости с применением основных принципов, с помощью которых регулируют условия реакции гибридизации и стабильность дуплекса, как известно специалистам в данной области техники и дополнительно описано ниже (см. например, Sambrook et al., 1989).
При гибридизации нуклеиновой кислоты с целевой нуклеиновой кислотой с неизвестной последовательностью предполагают, что длина гибрида равна длине гибридизующейся нуклеиновой кислоты. При гибридизации нуклеиновых кислот с известной последовательностью длину гибрида можно определить посредством выравнивания последовательностей нуклеиновых кислот и идентификации области или областей оптимальной комплементарности последовательностей. Температура гибридизации для гибридов, предполагаемая длина которых составляет меньше чем 50 пар оснований, должна быть на 5-10°C ниже температуры плавления (Tm) гибрида, где Tm определяют в соответствии со следующими уравнениями. Для гибридов длиной меньше чем 18 пар оснований Tm (°C) = 2 (количество оснований A+T) + 4(количество оснований G+C). Для гибридов длиной более 18 пар оснований Tm (°C) = 81,5+16,6 (log10 [Na+]) + 0,41 (% G+C) - (600/N), где N представляет собой количество оснований в гибриде, и [Na+] представляет собой концентрацию ионов натрия в буфере для гибридизации ([Na+] для 1 x SSC=0,165M). Предпочтительно, каждая такая гибридизующаяся нуклеиновая кислота имеет длину, которая составляет по меньшей мере 15 нуклеотидов (или более предпочтительно по меньшей мере 18 нуклеотидов, или по меньшей мере 20 нуклеотидов, или по меньшей мере 25 нуклеотидов, или по меньшей мере 30 нуклеотидов, или по меньшей мере 40 нуклеотидов, или наиболее предпочтительно по меньшей мере 50 нуклеотидов) или по меньшей мере 25% (более предпочтительно по меньшей мере 50%, или по меньшей мере 60%, или по меньшей мере 70% и наиболее предпочтительно по меньшей мере 80%) длины нуклеиновой кислоты по настоящему изобретению, с которой она гибридизуется, и характеризуется по меньшей мере 60% идентичностью последовательности (более предпочтительно по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 81%, по меньшей мере 82%, по меньшей мере 83%, по меньшей мере 84%, по меньшей мере 85%, по меньшей мере 86%, по меньшей мере 87%, по меньшей мере 88%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99%, и наиболее предпочтительно по меньшей мере 99,5%) с нуклеиновой кислотой по настоящему изобретению, с которой она гибридизуется, где идентичность последовательности определяют посредством сравнения последовательностей гибридизующихся нуклеиновых кислот, если они выровнены, чтобы максимизировать перекрывание и идентичность при минимизации гэпов в последовательности, как более подробно описано выше.
Варианты антител к PAC1 и антигенсвязывающих фрагментов, описанных в данном документе, можно получать посредством сайт-специфического мутагенеза нуклеотидов в ДНК, кодирующей полипептид, с применением кассетного или ПЦР-мутагенеза или других методик, хорошо известных из уровня техники, таких как методики, описанные в примере 3, для получения ДНК, кодирующей вариант, и последующей экспрессии рекомбинантной ДНК в клеточной культуре, как описано в данном документе. Однако антитела или антигенсвязывающие фрагменты, содержащие вариантные CDR, имеющие не более приблизительно 100-150 остатков, можно получать посредством синтеза in vitro с применением известных методик. Варианты обычно проявляют ту же качественную биологическую активность, что и встречающийся в природе аналог, например, связывание с антигеном. Такие варианты включают, например, делеции и/или вставки, и/или замены остатков в пределах аминокислотных последовательностей антител. Для получения конечной конструкции выполняют любую комбинацию из делеции, вставки и замены при условии, что конечная конструкция обладает требуемыми характеристиками. Изменения аминокислот также могут изменять посттрансляционные процессы, которым подвергается антитело, такие как изменение количества или положения сайтов гликозилирования. В определенных вариантах осуществления варианты антитела получают с целью модификации тех аминокислотных остатков, которые непосредственно участвуют в связывании эпитопа. В других вариантах осуществления модификация остатков, которые непосредственно не участвуют в связывании эпитопа, или остатков, не участвующих каким-либо образом в связывании эпитопа, является желательной для целей, обсуждаемых в данном документе. Предполагается мутагенез в пределах любой из CDR-областей и/или каркасных областей. Специалист в данной области техники может применять методики ковариационного анализа для конструирования полезных модификаций в аминокислотной последовательности антитела. См. например, Choulier, et al., Proteins 41:475-484, 2000; Demarest et al., J. Mol. Biol. 335:41-48, 2004; Hugo et al., Protein Engineering 16(5):381-86, 2003; Aurora et al., публикацию патента США № 2008/0318207 A1; Glaser et al., публикацию патента США № 2009/0048122 A1; Urech et al., WO 2008/110348 A1; Borras et al., WO 2009/000099 A2. Такие модификации, определенные с помощью ковариационного анализа, могут улучшать характеристики эффективности, фармакокинетики, фармакодинамики и/или технологичности антитела.
В таблицах 4A и 4B показаны иллюстративные последовательности нуклеиновых кислот, кодирующие соответственно полные легкую и тяжелую цепи антител к PAC1, описанных в данном документе. В таблицах 5A и 5B показаны иллюстративные последовательности нуклеиновых кислот, кодирующие соответственно вариабельные области легкой и тяжелой цепей антител к PAC1, описанных в данном документе. Полинуклеотиды, кодирующие вариабельные области антител к PAC1, можно применять, необязательно с нуклеиновыми кислотами, кодирующими константные области легкой и тяжелой цепей, перечисленные в таблицах 2 и 3 соответственно, для конструирования антител и антигенсвязывающих фрагментов по настоящему изобретению.
iPS:420665; iPS:420672;
iPS:420679; iPS:420686; iPS:420837; iPS:420841;
iPS:420845; iPS:420849;
iPS:420853; iPS:420857;
iPS:420861; iPS:420865;
iPS:420869; iPS:420873;
iPS:420877; iPS:420881;
iPS:420885; iPS:420889;
iPS:420893; iPS:420897; iPS:421027; iPS:421031;
iPS:421035; iPS:421039;
iPS:421043; iPS:421047;
iPS:421051; iPS:421055;
iPS:421059; iPS:421063;
iPS:421067; iPS:421071;
iPS:421075; iPS:421079;
iPS:421083; iPS:421087; iPS:421147; iPS:421207;
iPS:421211; iPS:421215;
iPS:421219; iPS:421223
iPS:420704; iPS:420711;
iPS:420718; iPS:420725;
iPS:420732; iPS:420739;
iPS:420746; iPS:420753;
iPS:420760; iPS:420767;
iPS:420774; iPS:420781;
iPS:420788; iPS:420795; iPS:420802; iPS:420809;
iPS:420816; iPS:420823;
iPS:420830; iPS:420901;
iPS:420908; iPS:420915;
iPS:420922; iPS:420929;
iPS:420936; iPS:420943;
iPS:420950; iPS:420957;
iPS:420964; iPS:420971;
iPS:420978; iPS:420985;
iPS:420992; iPS:420999;
iPS:421006; iPS:421013; iPS:421020; iPS:421091;
iPS:421098; iPS:421105;
iPS:421112; iPS:421119;
iPS:421126; iPS:421133;
iPS:421140; iPS:421163;
iPS:391578; iPS:421170;
iPS:421176; iPS:421182; iPS:421239; iPS:421246;
iPS:421253; iPS:421260;
iPS:421267; iPS:421286;
iPS:421293; iPS:421300;
iPS:421307; iPS:421326;
iPS:421333; iPS:421340;
iPS:421347; iPS:421354; iPS:421373; iPS:421380;
iPS:421387; iPS:421394;
iPS:421855; iPS:421861;
iPS:421867; iPS:421873;
iPS:421879; iPS:421885;
iPS:421891; iPS:421897;
iPS:421903; iPS:421909;
iPS:421915
iPS:421282; iPS:421322;
iPS:421369
iPS:480704; iPS:480710
iPS:480717
iPS:391478; iPS:421157; iPS:421189; iPS:421195;
iPS:421201
iPS:421231; iPS:421235
iPS:421278; iPS:421282
iPS:421318; iPS:421322
iPS:421365; iPS:421369
AGTCCGTGAAGGGCCGGTTCACCGTGTCCAGAGACAATTCCAAGAACACCCTGTATCTGCAAATGAACAGCCTGAGAGCTGAGGACACGGCTCTGTTTTACTGTGCGAGAGGATACGATGTTTTGACTGGTTACCCCGACTACTGGGGCCAGGGAACCCTGGTCACCGTGTCTAGT
Выделенные нуклеиновые кислоты, кодирующие антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению, могут содержать нуклеотидную последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична любой из нуклеотидных последовательностей, перечисленных в таблицах 4A, 4B, 5A и 5B. В некоторых вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область легкой цепи антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 328-342. В других вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область легкой цепи антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 343-363. В определенных вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область легкой цепи антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 328-342. В определенных других вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область легкой цепи антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 343-363. В связанных вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 364-468. В других связанных вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 469-485. В некоторых вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 364-468. В других вариантах осуществления выделенная нуклеиновая кислота, кодирующая вариабельную область тяжелой цепи антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 469-485.
В некоторых вариантах осуществления выделенная нуклеиновая кислота, кодирующая легкую цепь антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 537-541. В других вариантах осуществления выделенная нуклеиновая кислота, кодирующая легкую цепь антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 542-551. В определенных вариантах осуществления выделенная нуклеиновая кислота, кодирующая легкую цепь антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 537-541. В определенных других вариантах осуществления выделенная нуклеиновая кислота, кодирующая легкую цепь антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 542-551. В связанных вариантах осуществления выделенная нуклеиновая кислота, кодирующая тяжелую цепь антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 552-562. В других связанных вариантах осуществления выделенная нуклеиновая кислота, кодирующая тяжелую цепь антитела к PAC1, содержит последовательность, которая на по меньшей мере 80% идентична, на по меньшей мере 90% идентична, на по меньшей мере 95% идентична или на по меньшей мере 98% идентична последовательности, выбранной из SEQ ID NO: 563-569. В некоторых вариантах осуществления выделенная нуклеиновая кислота, кодирующая тяжелую цепь антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 552-562. В других вариантах осуществления выделенная нуклеиновая кислота, кодирующая тяжелую цепь антитела к PAC1, содержит последовательность, выбранную из SEQ ID NO: 563-569.
Последовательности нуклеиновых кислот, предусмотренные в таблицах 4A, 4B, 5A и 5B, являются только иллюстративными. Как будет понятно специалистам в данной области техники, из-за вырожденности генетического кода может быть получено чрезвычайно большое количество нуклеиновых кислот, все из которых кодируют CDR, вариабельные области и тяжелые и легкие цепи или другие компоненты антител и антигенсвязывающих фрагментов, описанных в данном документе. Таким образом, идентифицировав конкретную аминокислотную последовательность, специалисты в данной области техники могут получить любое количество различных нуклеиновых кислот с помощью простой модификации последовательности одного или нескольких кодонов таким путем, который не приводит к изменению аминокислотной последовательности кодируемого белка.
Настоящее изобретение также включает векторы, содержащие одну или несколько нуклеиновых кислот, кодирующих один или несколько компонентов антител или антигенсвязывающих фрагментов по настоящему изобретению (например, вариабельные области, легкие цепи и тяжелые цепи). Термин "вектор" означает любую молекулу или единицу (например, нуклеиновую кислоту, плазмиду, бактериофаг или вирус), применяемую для переноса кодирующей белок информации в клетку-хозяина. Примеры векторов включают без ограничения плазмиды, вирусные векторы, неэписомные векторы млекопитающих и векторы экспрессии, например, рекомбинантные векторы экспрессии. Применяемый в данном документе термин "вектор экспрессии" или "конструкция для экспрессии" относится к рекомбинантной молекуле ДНК, содержащей требуемую кодирующую последовательность и соответствующие регуляторные последовательности нуклеиновой кислоты, необходимые для экспрессии функционально связанной кодирующей последовательности в конкретной клетке-хозяине. Вектор экспрессии может включать без ограничения последовательности, которые оказывают влияние на транскрипцию, трансляцию или регулируют их и при наличии интронов оказывают влияние на сплайсинг РНК кодирующей области, функционально связанной с ними. Последовательности нуклеиновых кислот, необходимые для экспрессии в прокариотах, включают промотор, необязательно последовательность оператора, сайт связывания рибосомы и, возможно, другие последовательности. Известно, что эукариотические клетки используют промоторы, энхансеры и сигналы терминации и полиаденилирования.
Если требуется, секреторная сигнальная пептидная последовательность также может необязательно кодироваться вектором экспрессии, функционально связанным с представляющей интерес кодирующей последовательностью таким образом, что экспрессируемый полипептид может секретироваться рекомбинантной клеткой-хозяином для более легкого выделения представляющего интерес полипептида из клетки. Например, в некоторых вариантах осуществления сигнальные пептидные последовательности могут быть присоединены к аминоконцу/слиты с аминоконцом любой из полипептидных последовательностей вариабельных областей, перечисленных в таблицах 1A и 1B, или любой из полных полипептидных последовательностей цепей, перечисленных в таблицах 4A и 4B. В определенных вариантах осуществления сигнальный пептид, имеющий аминокислотную последовательность MDMRVPAQLLGLLLLWLRGARC (SEQ ID NO: 486), слит с аминоконцом любой из полипептидных последовательностей вариабельных областей из таблиц 1A и 1B или полных полипептидных последовательностей цепей из таблиц 4A и 4B. В других вариантах осуществления сигнальный пептид, имеющий аминокислотную последовательность MAWALLLLTLLTQGTGSWA (SEQ ID NO: 487), слит с аминоконцом любой из полипептидных последовательностей вариабельных областей из таблиц 1A и 1B или полных полипептидных последовательностей цепей из таблиц 4A и 4B. В еще других вариантах осуществления сигнальный пептид, имеющий аминокислотную последовательность MTCSPLLLTLLIHCTGSWA (SEQ ID NO: 488), слит с аминоконцом любой из полипептидных последовательностей вариабельных областей из таблиц 1A и 1B или полных полипептидных последовательностей цепей из таблиц 4A и 4B. Другие подходящие сигнальные пептидные последовательности, которые могут быть слиты с аминоконцом полипептидных последовательностей вариабельных областей или полных полипептидных последовательностей цепей, описанных в данном документе, включают: MEAPAQLLFLLLLWLPDTTG (SEQ ID NO: 489), MEWTWRVLFLVAAATGAHS (SEQ ID NO: 490), METPAQLLFLLLLWLPDTTG (SEQ ID NO: 491), METPAQLLFLLLLWLPDTTG (SEQ ID NO: 492), MKHLWFFLLLVAAPRWVLS (SEQ ID NO: 493), MEWSWVFLFFLSVTTGVHS (SEQ ID NO: 494), MDIRAPTQLLGLLLLWLPGAKC (SEQ ID NO: 495), MDIRAPTQLLGLLLLWLPGARC (SEQ ID NO: 496), MDTRAPTQLLGLLLLWLPGATF (SEQ ID NO: 497), MDTRAPTQLLGLLLLWLPGARC (SEQ ID NO: 498), METGLRWLLLVAVLKGVQC (SEQ ID NO: 499), METGLRWLLLVAVLKGVQCQE (SEQ ID NO: 500) и MDMRAPTQLLGLLLLWLPGARC (SEQ ID NO: 501). Другие сигнальные или секреторные пептиды известны специалистам в данной области техники и могут быть слиты с любой из полипептидных цепей вариабельных областей, перечисленных в таблицах 1A и 1B, или полных полипептидных последовательностей цепей из таблиц 4A и 4B, например, для облегчения или оптимизации экспрессии в конкретных в клетках-хозяевах.
Как правило, векторы экспрессии, применяемые в клетках-хозяевах для получения антител к PAC1 и антигенсвязывающих фрагментов по настоящему изобретению, будут содержать последовательности для поддержания плазмиды и для клонирования и экспрессии экзогенных нуклеотидных последовательностей, кодирующих компоненты антител и антигенсвязывающих фрагментов. Такие последовательности, обобщенно называемые "фланкирующими последовательностями", в определенных вариантах осуществления, как правило, будут содержать одну или несколько из следующих нуклеотидных последовательностей: промотор, одну или несколько энхансерных последовательностей, точку начала репликации, последовательность терминации транскрипции, полную последовательность интрона, содержащую донорный и акцепторный сайты сплайсинга, последовательность, кодирующую лидерную последовательность для секреции полипептида, сайт связывания рибосомы, последовательность полиаденилирования, полилинкерную область для вставки нуклеиновой кислоты, кодирующей подлежащий экспрессии полипептид, и элемент селектируемого маркера. Каждая из этих последовательностей рассматривается ниже.
Необязательно, вектор может содержать последовательность, кодирующую "метку", т. е., молекулу олигонуклеотида, расположенную на 5′- или 3′-конце последовательности, кодирующей полипептид; олигонуклеотидную последовательность метки, кодирующую polyHis (например, hexaHis), FLAG®, HA (гемагглютинин вируса гриппа), myc или другую молекулу метки, для которой существуют коммерчески доступные антитела. Как правило, эта метка слита с полипептидом при экспрессии полипептида, и она может служить в качестве средства для аффинной очистки или выявления полипептида из клетки-хозяина. Аффинную очистку можно осуществлять, например, с помощью колоночной хроматографии с применением антител к метке в качестве аффинной матрицы. Необязательно, впоследствии метка может быть удалена из очищенного полипептида с помощью различных способов, таких как применение определенных пептидаз для расщепления.
Фланкирующие последовательности могут быть гомологичными (т. е. из того же вида и/или штамма, что и клетка-хозяин), гетерологичными (т. е. из вида, отличного от вида или штамма клетки-хозяина), гибридными (т. е. комбинацией фланкирующих последовательностей из более чем одного источника), синтетическими или нативными. Таким образом, источником фланкирующей последовательности может быть любой прокариотический или эукариотический организм, любой позвоночный или беспозвоночный организм или любое растение при условии, что фланкирующая последовательность является функциональной и может быть активирована посредством механизма клетки-хозяина.
Фланкирующие последовательности, применяемые в векторах по настоящему изобретению, можно получать любым из нескольких способов, хорошо известных из уровня техники. Как правило, применяемые в данном документе фланкирующие последовательности будут предварительно идентифицированы с помощью картирования и/или расщепления рестрикционными эндонуклеазами и, следовательно, могут быть выделены из соответствующего источника, представляющего собой ткань, с применением подходящих рестрикционных эндонуклеаз. В некоторых случаях может быть известна полная нуклеотидная последовательность фланкирующей последовательности. В данном случае фланкирующая последовательность может быть синтезирована с помощью рутинных способов синтеза или клонирования нуклеиновых кислот.
Вне зависимости от того, известна вся или только часть фланкирующей последовательности, ее можно получить с помощью полимеразной цепной реакции (ПЦР) и/или посредством скрининга геномной библиотеки с подходящим зондом, таким как олигонуклеотид и/или фрагмент фланкирующей последовательности из того же или другого вида. Если фланкирующая последовательность неизвестна, то фрагмент ДНК, содержащий фланкирующую последовательность, можно выделить из более крупного участка ДНК, который может содержать, например, кодирующую последовательность и даже другой ген или гены. Выделение можно осуществлять посредством расщепления рестрикционными эндонуклеазами для получения соответствующего фрагмента ДНК с последующим выделением с помощью очистки в агарозном геле, колоночной хроматографии Qiagen® (Чатсворт, Калифорния) или других способов, известных специалисту в данной области техники. Выбор подходящих ферментов для осуществления этой цели будет совершенно очевиден для специалиста в данной области техники.
Как правило, точка начала репликации является частью коммерчески приобретенных прокариотических векторов экспрессии, и точка начала репликации способствует амплификации вектора в клетке-хозяине. Если выбранный вектор не содержит точку начала репликации, то ее можно химически синтезировать на основе известной последовательности и лигировать с вектором. Например, точка начала репликации из плазмиды pBR322 (New England Biolabs, Беверли, Массачусетс) подходит для большинства грамотрицательных бактерий, и различные источники вирусного происхождения (например, SV40, вирус полиомы, аденовирус, вирус везикулярного стоматита (VSV) или папилломавирусы, такие как HPV или BPV) подходят для клонирования векторов в клетках млекопитающих. В целом, компонент точки начала репликации не требуется для векторов экспрессии млекопитающих (например, точка начала репликации SV40 часто используется только потому, что она также содержит промотор ранних генов вируса).
Как правило, последовательность терминации транскрипции расположена в 3'-направлении относительно конца кодирующей полипептид области и служит для терминации транскрипции. Обычно последовательность терминации транскрипции в прокариотических клетках представляет собой G-C-богатый фрагмент, за которым следует последовательность поли-Т. Хотя последовательность легко клонировать из библиотеки или даже коммерчески приобрести в качестве части вектора, ее также можно легко синтезировать с применением известных способов синтеза нуклеиновых кислот.
Ген селектируемого маркера кодирует белок, необходимый для выживания и роста клетки-хозяина, растущей в селективной культуральной среде. Типичные гены селективных маркеров кодируют белки, которые (a) придают устойчивость к антибиотикам или другим токсинам, например, ампициллину, тетрациклину или канамицину в случае прокариотических клеток-хозяев; (b) восполняют ауксотрофные дефициты в клетке; или (c) обеспечивают необходимые питательные вещества, недоступные из комплексной или определенной среды. Конкретные селектируемые маркеры представляют собой ген устойчивости к канамицину, ген устойчивости к ампициллину и ген устойчивости к тетрациклину. Преимущественно, можно также применять ген устойчивости к неомицину для отбора как в прокариотических, так и в эукариотических клетках-хозяевах.
Для амплификации гена, который будет экспрессироваться, можно применять другие селектируемые гены. Амплификация представляет собой процесс, при котором гены, необходимые для продуцирования белка, необходимого для роста или выживания клеток, повторяются в тандеме в пределах хромосом последующих поколений рекомбинантных клеток. Примеры подходящих селектируемых маркеров для клеток млекопитающих включают дигидрофолатредуктазу (DHFR) и гены тимидинкиназы, не содержащие промоторов. Трансформанты клеток млекопитающих помещают в условия давления отбора, в которых только лишь трансформанты адаптированы для выживания вследствие присутствия в векторе селектируемого гена. Давление отбора устанавливают посредством культивирования трансформированных клеток в условиях, при которых концентрация средства для отбора в среде последовательно увеличивается, приводя тем самым к амплификации как селектируемого гена, так и ДНК, которая кодирует другой ген, такой как один или несколько компонентов антител или антигенсвязывающих фрагментов, описанных в данном документе. В результате с амплифицированной ДНК синтезируются повышенные количества полипептида.
Сайт связывания рибосомы обычно необходим для инициации трансляции mRNA и характеризуется наличием последовательности Шайна-Дальгарно (прокариоты) или последовательности Козак (эукариоты). Как правило, этот элемент расположен в направлении 3'-конца относительно промотора и в направлении 5'-конца относительно кодирующей последовательности полипептида, подлежащего экспрессии. В определенных вариантах осуществления одна или несколько кодирующих областей могут быть функционально связаны с внутренним сайтом связывания рибосомы (IRES), что позволяет осуществлять трансляцию двух открытых рамок считывания с одного РНК-транскрипта.
В некоторых случаях, например, если в системе экспрессии эукариотической клетки-хозяина требуется гликозилирование, можно манипулировать с различными пре- или пропоследовательностями для улучшения гликозилирования или выхода. Например, можно изменять сайт расщепления пептидазой конкретного сигнального пептида или добавлять пропоследовательности, которые также могут оказывать влияние на гликозилирование. Конечный белковый продукт может содержать в положении -1 (по отношению к первой аминокислоте зрелого белка) одну или несколько дополнительных аминокислот, связанных с экспрессией, которые могли быть не полностью удалены. Например, конечный белковый продукт может содержать один или несколько аминокислотных остатков, находящихся в сайте расщепления пептидазой, присоединенных к аминоконцу. В качестве альтернативы, применение некоторых сайтов расщепления ферментами может приводить к получению слегка усеченной формы требуемого полипептида, если фермент осуществляет разрезание в такой области в пределах зрелого полипептида.
Векторы экспрессии и векторы для клонирования по настоящему изобретению будут, как правило, содержать промотор, который распознается организмом-хозяином и функционально связан с молекулой, кодирующей полипептид. Применяемый в данном документе термин "функционально связанный" относится к связыванию двух или более последовательностей нуклеиновых кислот таким образом, что образуется молекула нуклеиновой кислоты, способная направлять транскрипцию данного гена и/или синтез требуемой молекулы белка. Например, регуляторная последовательность в векторе, которая "функционально связана" с кодирующей белок последовательностью, лигируется с ней таким образом, что экспрессия кодирующей белок последовательности достигается в условиях, совместимых с транскрипционной активностью регуляторных последовательностей. Более конкретно, последовательность промотора и/или энхансера, включая любую комбинацию цис-действующих элементов регуляции транскрипции, функционально связана с кодирующей последовательностью, если она стимулирует или модулирует транскрипцию кодирующей последовательности в соответствующей клетке-хозяине или другой системе экспрессии.
Промоторы представляют собой нетранскрибируемые последовательности, расположенные выше (т. е. в 5'-направлении) стартового кодона структурного гена (как правило, в пределах приблизительно 100-1000 п. о.), которые регулируют транскрипцию структурного гена. Обычно промоторы группируют в один из двух классов: индуцибельные промоторы и конститутивные промоторы. Индуцибельные промоторы инициируют повышенные уровни транскрипции с ДНК, находящейся под их контролем, в ответ на некоторое изменение в условиях культивирования, такое как наличие или отсутствие питательного вещества или изменение температуры. С другой стороны, конститутивные промоторы приводят к постоянной транскрипции гена, с которым они функционально связаны, то есть они осуществляют незначительную регуляцию экспрессии гена или такая регуляция отсутствует. Хорошо известно большое количество промоторов, распознаваемых различными потенциальными клетками-хозяевами. Подходящий промотор функционально связан с ДНК, кодирующей, например, тяжелую цепь, легкую цепь или другой компонент антител и антигенсвязывающих фрагментов по настоящему изобретению с помощью удаления промотора из исходной ДНК посредством расщепления рестрикционным ферментом и вставки требуемой последовательности промотора в вектор.
Также из уровня техники хорошо известны промоторы, подходящие для применения с дрожжевыми клетками-хозяевами. Дрожжевые энхансеры предпочтительно применяют с дрожжевыми промоторами. Подходящие промоторы для применения с клетками-хозяевами млекопитающих хорошо известны и включают без ограничения промоторы, полученные из геномов вирусов, таких как вирус полиомы, вирус оспы кур, аденовирус (например, серотипы 2, 8 или 9 аденовируса), вирус папилломы крупного рогатого скота, вирус саркомы птиц, цитомегаловирус, ретровирусы, вирус гепатита В и вирус обезьян 40 (SV40). Другие подходящие промоторы млекопитающих включают гетерологичные промоторы млекопитающих, например, промоторы генов, кодирующих белки теплового шока, и промотор актина.
Дополнительные промоторы, которые могут представлять интерес, включают без ограничения: промотор ранних генов SV40 (Benoist and Chambon, 1981, Nature 290:304-310); промотор CMV (Thornsen et al., 1984, Proc. Natl. Acad. U.S.A. 81:659-663); промотор, содержащийся в 3'-концевом длинном повторе вируса саркомы Рауса (Yamamoto et al., 1980, Cell 22:787-797); промотор тимидинкиназы вируса герпеса (Wagner et al., 1981, Proc. Natl. Acad. Sci. U.S.A. 78: 1444-1445); промоторные и регуляторные последовательности из гена металлотионина (Prinster et al., 1982, Nature 296:39-42); и прокариотические промоторы, такие как промотор бета-лактамазы (Villa-Kamaroff et al., 1978, Proc. Natl. Acad. Sci. U.S.A. 75:3727-3731); или промотор tac (DeBoer et al., 1983, Proc. Natl. Acad. Sci. U.S.A. 80:21-25). Также представляют интерес следующие транскрипционные регуляторные области животных, которые проявляют тканевую специфичность и применялись в трансгенных животных: регуляторная область гена эластазы I, которая активна в ацинарных клетках поджелудочной железы (Swift et al., 1984, Cell 38:639-646; Ornitz et al., 1986, Cold Spring Harbor Symp. Quant. Biol. 50:399-409; MacDonald, 1987, Hepatology 7:425-515); регуляторная область гена инсулина, которая активна в бета-клетках поджелудочной железы (Hanahan, 1985, Nature 315: 115-122); регуляторная область гена иммуноглобулина, которая активна в лимфоидных клетках (Grosschedl et al., 1984, Cell 38:647-658; Adames et al., 1985, Nature 318:533-538; Alexander et al., 1987, Mol. Cell. Biol. 7: 1436-1444); регуляторная область вируса опухоли молочной железы мыши, которая активна в клетках яичка, молочной железы, лимфоидных и тучных клетках (Leder et al., 1986, Cell 45:485-495); регуляторная область гена альбумина, которая активна в печени (Pinkert et al., 1987, Genes and Devel. 1:268-276); регуляторная область гена альфа-фетопротеина, которая активна в печени (Krumlauf et al., 1985, Mol. Cell. Biol. 5: 1639-1648; Hammer et al., 1987, Science 253:53-58); регуляторная область гена альфа-1-антитрипсина, которая активна в печени (Kelsey et al., 1987, Genes and Devel. 1: 161-171); регуляторная область гена бета-глобина, которая активна в миелоидных клетках (Mogram et al., 1985, Nature 315:338-340; Kollias et al., 1986, Cell 46:89-94); регуляторная область гена основного белка миелина, которая активна в олигодендроцитах в головном мозге (Readhead et al., 1987, Cell 48:703-712); регуляторная область гена легкой цепи 2 миозина, которая активна в скелетных мышцах (Sani, 1985, Nature 314:283-286); и регуляторная область гена гонадотропного рилизинг-гормона, которая активна в гипоталамусе (Mason et al., 1986, Science 234: 1372-1378).
Последовательность энхансера может быть вставлена в вектор для усиления транскрипции ДНК, кодирующей компонент антител или антигенсвязывающих фрагментов (например, легкой цепи, тяжелой цепи или вариабельных областей) высших эукариот. Энхансеры представляют собой цис-действующие элементы ДНК длиной обычно приблизительно 10-300 п. о., которые воздействуют на промотор для усиления транскрипции. Энхансеры являются относительно независимыми от ориентации и положения, они были обнаружены в положениях как 5', так и 3' относительно транскрипционной единицы. Известно несколько энхансерных последовательностей, доступных из генов млекопитающих (например, генов глобина, эластазы, альбумина, альфа-фетопротеина и инсулина). Как правило, обычно применяют энхансер из вируса. Известные из уровня техники энхансер SV40, энхансер промотора ранних генов цитомегаловируса, энхансер вируса полиомы и энхансеры аденовируса представляют собой иллюстративные энхансерные элементы для активации эукариотических промоторов. Хотя энхансер может находиться в векторе как с 5'-, так и с 3'-конца относительно кодирующей последовательности, как правило, он расположен в 5'-сайте относительно промотора. Последовательность, кодирующая соответствующую нативную или гетерологичную сигнальную последовательность (лидерную последовательность или сигнальный пептид), может быть встроена в вектор экспрессии для стимуляции внеклеточной секреции антитела или антигенсвязывающего фрагмента, как описано выше. Выбор сигнального пептида или лидерной последовательности зависит от типа клеток-хозяев, в которых должно продуцироваться антитело или антигенсвязывающий фрагмент, и гетерологичная сигнальная последовательность может заменять нативную сигнальную последовательность. Примеры сигнальных пептидов описаны выше. Другие сигнальные пептиды, которые являются функциональными в клетках-хозяевах млекопитающих, включают сигнальную последовательность интерлейкина-7 (IL-7), описанную в патенте США № 4965195; сигнальную последовательность рецептора интерлейкина-2, описанную в Cosman et al.,1984, Nature 312:768; сигнальный пептид рецептора интерлейкина-4, описанный в европейском патенте № 0367566; сигнальный пептид рецептора интерлейкина-1 I типа, описанный в патенте США № 4968607; и сигнальный пептид рецептора интерлейкина-1 II типа, описанный в европейском патенте № 0460846.
Предусмотренные векторы экспрессии могут быть сконструированы на основе начального вектора, такого как коммерчески доступный вектор. Такие векторы могут содержать все необходимые фланкирующие последовательности или не содержать их. Если одна или несколько из описанных в данном документе фланкирующих последовательностей еще не присутствуют в векторе, то их можно получить отдельно и лигировать с вектором. Способы, применяемые для получения каждой из фланкирующих последовательностей, хорошо известны специалисту в данной области техники. Векторы экспрессии могут быть введены в клетки-хозяева для получения таким образом белков, включая антитела и антигенсвязывающие фрагменты, кодируемые нуклеиновыми кислотами, описанными в данном документе.
В определенных вариантах осуществления нуклеиновые кислоты, кодирующие различные компоненты антител к PAC1 или антигенсвязывающих фрагментов по настоящему изобретению, могут быть вставлены в один и тот же вектор экспрессии. Например, нуклеиновая кислота, кодирующая легкую цепь или вариабельную область антитела к PAC1, может быть клонирована в том же векторе, что и нуклеиновая кислота, кодирующая тяжелую цепь или вариабельную область антитела к PAC1. В таких вариантах осуществления две нуклеиновые кислоты могут быть разделены внутренним сайтом посадки рибосомы (IRES) и находиться под контролем одного промотора таким образом, что легкая цепь и тяжелая цепь экспрессируются с одного и того же mRNA-транскрипта. В качестве альтернативы, две нуклеиновые кислоты могут находиться под контролем двух отдельных промоторов таким образом, что легкая цепь и тяжелая цепь экспрессируются с двух отдельных mRNA-транскриптов. В некоторых вариантах осуществления нуклеиновую кислоту, кодирующую легкую цепь или вариабельную область антитела к PAC1, клонируют в одном векторе экспрессии, а нуклеиновую кислоту, кодирующую тяжелую цепь или вариабельную область антитела к PAC1, клонируют во втором векторе экспрессии. В таких вариантах осуществления клетка-хозяин может быть котрансфицирована двумя векторами экспрессии для получения полных антител или антигенсвязывающих фрагментов по настоящему изобретению.
После того как вектор был сконструирован, и одна или несколько молекул нуклеиновой кислоты, кодирующих компоненты антител и антигенсвязывающих фрагментов, описанных в данном документе, были вставлены в соответствующий(соответствующие) сайт(сайты) вектора или векторов, готовый(готовые) вектор(векторы) может(могут) быть вставлен(вставлены) в подходящую клетку-хозяина для амплификации и/или экспрессии полипептида. Таким образом, настоящее изобретение охватывает выделенную клетку-хозяина или линию клеток, содержащую один или несколько векторов экспрессии, кодирующих компоненты антител к PAC1 или антигенсвязывающих фрагментов, описанных в данном документе. Применяемый в данном документе термин "клетка-хозяин" относится к клетке, которая была трансформирована или способна трансформироваться нуклеиновой кислотой и вследствие этого экспрессирует представляющий интерес ген. Данный термин включает потомство родительской клетки вне зависимости от того, идентично или нет это потомство по морфологии или по генетической структуре исходной родительской клетке, до тех пор, пока присутствует представляющий интерес ген. Клетка-хозяин, которая содержит выделенную нуклеиновую кислоту по настоящему изобретению, предпочтительно функционально связанную с по меньшей мере одной последовательностью для регуляции экспрессии (например, промотором или энхансером), представляет собой "рекомбинантную клетку-хозяина".
Трансформацию вектором экспрессии для антитела к PAC1 или антигенсвязывающего фрагмента выбранной клетки-хозяина можно осуществлять с помощью хорошо известных способов, включая трансфекцию, инфекцию, совместное осаждение с фосфатом кальция, электропорацию, микроинъекцию, липофекцию, трансфекцию, опосредованную DEAE-декстраном, или другие известные методики. Выбранный способ частично будет зависеть от типа применяемой клетки-хозяина. Эти способы и другие подходящие способы хорошо известны специалисту в данной области техники и приведены, например, в Sambrook et al., 2001.
При культивировании в соответствующих условиях клетка-хозяин синтезирует антитело или антигенсвязывающий фрагмент, который впоследствии может быть собран из культуральной среды (если клетка-хозяин секретирует его в среду) или непосредственно из клетки-хозяина, продуцирующей его (если он не секретируется). Выбор подходящей клетки-хозяина будет зависеть от различных факторов, таких как требуемые уровни экспрессии, модификации полипептидов, которые требуются или необходимы для активности (такие как гликозилирование или фосфорилирование), и легкость фолдинга в биологически активную молекулу.
Иллюстративные клетки-хозяева включают прокариотические, дрожжевые клетки или клетки высших эукариот. Прокариотические клетки-хозяева включают эубактерии, такие как грамотрицательные или грамположительные организмы, например, Enterobacteriaceae, такие как Escherichia, например, E. coli, Enterobacter, Erwinia, Klebsiella, Proteus, Salmonella, например, Salmonella typhimurium, Serratia, например, Serratia marcescans и Shigella, а также Bacillus, например, B. subtilis и B. licheniformis, Pseudomonas и Streptomyces. Эукариотические микроорганизмы, такие как мицелиальные грибы или дрожжи, являются подходящими хозяевами для клонирования или экспрессии рекомбинантных полипептидов. Saccharomyces cerevisiae или обыкновенные пекарские дрожжи наиболее часто применяются среди низших эукариотических микроорганизмов-хозяев. Однако ряд других родов, видов и штаммов являются общедоступными и применимыми в данном документе, такие как Pichia, например, P. pastoris, Schizosaccharomyces pombe; Kluyveromyces, Yarrowia; Candida; Trichoderma reesia; Neurospora crassa; Schwanniomyces, например, Schwanniomyces occidentalis; и мицелиальные грибы, такие как, например, Neurospora, Penicillium, Tolypocladium и хозяева Aspergillus, такие как A. nidulans и A. niger.
Клетки-хозяева для экспрессии гликозилированных антител и антигенсвязывающих фрагментов могут быть получены из многоклеточных организмов. Примеры клеток беспозвоночных включают клетки растений и насекомых. Были идентифицированы многочисленные бакуловирусные штаммы и варианты и соответствующие пермиссивные клетки-хозяева насекомых из таких хозяев, как Spodoptera frugiperda (гусеница), Aedes aegypti (комар), Aedes albopictus (комар), Drosophila melanogaster (плодовая мушка) и Bombyx mori. Общедоступно множество вирусных штаммов для трансфекции таких клеток, например, вариант L-1 Autographa californica NPV и штамм Bm-5 Bombyx mori NPV.
Клетки-хозяева позвоночных также являются подходящими хозяевами, и рекомбинантное получение антител и антигенсвязывающих фрагментов из таких клеток стало рутинной процедурой. Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны из уровня техники и включают без ограничения иммортализованные линии клеток, доступные из Американской коллекции типовых культур (АТСС), включая без ограничения клетки яичника китайского хомяка (СНО), включая клетки линии CHOK1 (ATCC CCL61), DXB-11, DG-44 и клетки яичника китайского хомяка/-DHFR (CHO, Urlaub et al, Proc. Natl. Acad. Sci. USA 77: 4216, 1980); линию клеток CV1 почки обезьяны, трансформированную с помощью SV40 (COS-7, ATCC CRL 1651); эмбриональную линию клеток почки человека (клетки линии 293 или 293, субклонированные для роста в суспензионной культуре (Graham et al, J. Gen Virol. 36: 59, 1977); клетки почки детеныша хомяка (BHK, ATCC CCL 10); клетки Сертоли мыши (TM4, Mather, Biol. Reprod. 23: 243-251, 1980); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1587); клетки карциномы шейки матки человека (HELA, ATCC CCL 2); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крысы линии buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки гепатомы человека (Hep G2, HB 8065); опухоль молочной железы мыши (MMT 060562, ATCC CCL51); клетки линии TRI (Mather et al., Annals N.Y. Acad. Sci. 383: 44-68, 1982); клетки линии MRC 5 или клетки линии FS4; клетки миеломы млекопитающих и ряд других линий клеток. В определенных вариантах осуществления линии клеток могут быть выбраны посредством определения того, какие линии клеток обладают высокими уровнями экспрессии и конститутивно продуцируют антитела и антигенсвязывающие фрагменты со свойствами, заключающимися в связывании PAC1. В другом варианте осуществления может быть выбрана линия клеток из линии дифференцировки B-клеток, которая не продуцирует свое собственное антитело, но обладает способностью продуцировать и секретировать гетерологичное антитело. Клетки линии СНО являются предпочтительными клетками-хозяевами в некоторых вариантах осуществления для экспрессии антител к PAC1 и антигенсвязывающих фрагментов по настоящему изобретению.
Клетки-хозяева трансформируют или трансфицируют с помощью описанных выше нуклеиновых кислот или векторов для получения антител к PAC1 или антигенсвязывающих фрагментов и культивируют в общепринятых питательных средах, модифицированных в соответствии с необходимостью индукции промоторов, отбора трансформантов или амплификации генов, кодирующих требуемые последовательности. Кроме того, новые векторы и трансфицированные линии клеток с несколькими копиями транскрипционных единиц, разделенных селективным маркером, особенно применимы для экспрессии антител и антигенсвязывающих фрагментов. Таким образом, в настоящем изобретении также предусмотрен способ получения антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе, включающий культивирование клетки-хозяина, содержащей один или несколько векторов экспрессии, описанных в данном документе, в культуральной среде в условиях, обеспечивающих экспрессию антитела или антигенсвязывающего фрагмента, кодируемого одним или несколькими векторами экспрессии; и выделение антитела или антигенсвязывающего фрагмента из культуральной среды или клетки-хозяина.
Клетки-хозяева, применяемые для получения антител или антигенсвязывающих фрагментов по настоящему изобретению, можно культивировать в различных средах. Коммерчески доступные среды, такие как Ham's F10 (Sigma), минимальная питательная среда (MEM, Sigma), RPMI-1640 (Sigma) и модифицированная по Дульбекко среда Игла (DMEM, Sigma), подходят для культивирования клеток-хозяев. Кроме того, любую из сред, описанных в Ham et al., Meth. Enz. 58: 44, 1979; Barnes et al., Anal. Biochem. 102: 255, 1980; патентах США №№ 4767704; 4657866; 4927762; 4560655; или 5122469; WO90103430; WO 87/00195; или патенте США № 30985 можно применять в качестве культуральной среды для клеток-хозяев. Любую из этих сред можно, при необходимости, дополнить гормонами и/или другими факторами роста (такими как инсулин, трансферрин или эпидермальный фактор роста), солями (такими как хлорид натрия, кальций, магний и фосфат), буферами (такими как HEPES), нуклеотидами (такими как аденозин и тимидин), антибиотиками (такими как лекарственное средство Gentamycin™), микроэлементами (определяемыми как неорганические соединения, обычно присутствующие в конечных концентрациях в микромолярном диапазоне) и глюкозой или эквивалентным источником энергии. Любые другие необходимые добавки также могут быть включены в соответствующих концентрациях, которые должны быть известны специалистам в данной области техники. Условия культивирования, такие как температура, рН и т. п. являются такими, которые ранее применялись для клетки-хозяина, выбранной для экспрессии, и будут очевидны специалисту в данной области техники.
В ходе культивирования клеток-хозяев антитело или антигенсвязывающий фрагмент можно получать внутриклеточно, в периплазматическом пространстве, или он может секретироваться непосредственно в среду. Если антитело или антигенсвязывающий фрагмент получают внутриклеточно, то на первой стадии дебрис в форме частиц либо клетки-хозяева или лизированные фрагменты удаляют, например, посредством центрифугирования или ультрафильтрации. Антитело или антигенсвязывающий белок можно очищать с применением, например, гидроксиапатитной хроматографии, катионо- или анионообменной хроматографии или предпочтительно аффинной хроматографии с применением представляющего(представляющих) интерес антигена(антигенов) или белка А или белка G в качестве аффинного лиганда. Белок A можно применять для очистки белков, которые содержат полипептиды на основе тяжелых цепей γ1, γ2 или γ4 человека (Lindmark et al., J. Immunol. Meth. 62: 1-13, 1983). Белок G рекомендуется для всех изотипов мыши и для γ3 человека (Guss et al., EMBO J. 5: 1567-1575, 1986). Матрица, к которой присоединяется аффинный лиганд, наиболее часто является агарозой, но также доступны другие матрицы. Механически стабильные матрицы, такие как стекло с контролируемым размером пор или поли(стиролдивинил)бензол, обеспечивают более высокие скорости потока и меньшее время обработки, чем те, которые можно достигнуть с применением агарозы. Если белок содержит домен CH3, для очистки можно применять смолу Bakerbond ABXTM (J. T. Baker, Phillipsburg, N.J.). Другие методики очистки белков, такие как осаждение этанолом, обращенно-фазовая HPLC, хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония также возможны в зависимости от конкретного антитела или антигенсвязывающего фрагмента, который необходимо выделить.
В определенных вариантах осуществления в настоящем изобретении предусмотрена композиция (например, фармацевтическая композиция), содержащая одно или множество антител к PAC1 или антигенсвязывающих фрагментов по настоящему изобретению (например, моноклональных антител к PAC1 или их связывающих фрагментов) вместе с фармацевтически приемлемыми разбавителями, носителями, вспомогательными веществами, солюбилизаторами, эмульгаторами, консервантами и/или адъювантами. Фармацевтические композиции можно применять в любом из способов, описанных в данном документе. Фармацевтические композиции по настоящему изобретению включают без ограничения жидкие, замороженные и лиофилизированные композиции. "Фармацевтически приемлемый" относится к молекулам, соединениям и композициям, которые нетоксичны для людей-реципиентов в используемых дозах и концентрациях и/или не вызывают аллергических или нежелательных реакций при введении людям. В некоторых вариантах осуществления фармацевтическая композиция может содержать материалы для составления, предназначенные для модификации, поддержания или сохранения, например, рН, осмолярности, вязкости, прозрачности, цвета, изотоничности, запаха, стерильности, стабильности, скорости растворения или высвобождения, адсорбции или проникающей способности композиции. В таких вариантах осуществления подходящие материалы для составления включают без ограничения аминокислоты (такие как глицин, глутамин, аспарагин, аргинин или лизин); противомикробные вещества; антиоксиданты (такие как аскорбиновая кислота, сульфит натрия или гидросульфит натрия); буферы (такие как боратный, бикарбонатный, Tris-HCl, цитратные, фосфатные буферы или буферы на основе других органических кислот); объемообразующие средства (такие как маннит или глицин); хелатирующие средства (такие как этилендиаминтетрауксусная кислота (EDTA)); комплексообразующие средства (такие как кофеин, поливинилпирролидон, бета-циклодекстрин или гидроксипропил-бета-циклодекстрин); наполнители; моносахариды; дисахариды и другие углеводы (такие как глюкоза, манноза или декстрины); белки (такие как сывороточный альбумин, желатин или иммуноглобулины); средства для окрашивания и разбавления, эмульгирующие средства, гидрофильные полимеры (такие как поливинилпирролидон); низкомолекулярные полипептиды; солеобразующие противоионы (такие как ион натрия); консерванты (такие как хлорид бензалкония, бензойная кислота, салициловая кислота, тимеросал, фенетиловый спирт, метилпарабен, пропилпарабен, хлоргексидин, сорбиновая кислота или пероксид водорода); растворители (такие как глицерин, пропиленгликоль или полиэтиленгликоль); сахарные спирты (такие как маннит или сорбит); суспендирующие средства; поверхностно-активные вещества или смачивающие средства (такие как плюроники, PEG, сорбитановые сложные эфиры, полисорбаты, такие как полисорбат 20, полисорбат 80, тритон, трометамин, лецитин, холестерин, тилоксапал); средства, повышающие стабильность (такие как сахароза или сорбит); средства, повышающие тоничность (такие как галогениды щелочных металлов, предпочтительно хлорид натрия или калия, маннит, сорбит); среды-носители для доставки; разбавители; вспомогательные вещества и/или фармацевтические адъюванты. Способы и подходящие материалы для составления молекул для терапевтического применения известны в фармацевтических дисциплинах и описаны, например, в REMINGTON's PHARMACEUTICAL SCIENCES, 18th Edition, (A.R. Genrmo, ed.), 1990, Mack Publishing Company.
В некоторых вариантах осуществления фармацевтическая композиция по настоящему изобретению содержит стандартный фармацевтический носитель, такой как стерильный забуференный фосфатом физиологический раствор, бактериостатическая вода и т. п. Можно применять различные водные носители, например, воду, забуференную воду, 0,4% физиологический раствор, 0,3% глицин и т. п., и они могут включать другие белки для повышения стабильности, такие как альбумин, липопротеин, глобулин и т. д., подвергнутые небольшим химическим модификациям или т. п.
Иллюстративные концентрации антител или антигенсвязывающих фрагментов в составе могут находиться в диапазоне от приблизительно 0,1 мг/мл до приблизительно 200 мг/мл или от приблизительно 0,1 мг/мл до приблизительно 50 мг/мл, или от приблизительно 0,5 мг/мл до приблизительно 25 мг/мл или, в качестве альтернативы, от приблизительно 2 мг/мл до приблизительно 10 мг/мл. Водный состав антитела или антигенсвязывающего фрагмента можно получать в буферном растворе с доведенным pH, например, при рН в диапазоне от приблизительно 4,5 до приблизительно 6,5 или от приблизительно 4,8 до приблизительно 5,5 или, в качестве альтернативы, приблизительно 5,0. Примеры буферов, подходящих для рН в этом диапазоне, включают ацетатные (например, ацетат натрия), сукцинатные (такие как сукцинат натрия), глюконатные, гистидиновые, цитратные и буферы на основе других органических кислот. Концентрация буфера может составлять от приблизительно 1 мМ до приблизительно 200 мМ или от приблизительно 10 мМ до приблизительно 60 мМ, в зависимости, например, от буфера и требуемой изотоничности состава.
В состав может быть включено средство для регуляции тоничности, которое также способно стабилизировать антитело или антигенсвязывающий фрагмент. Иллюстративные средства для регуляции тоничности включают полиолы, такие как маннит, сахароза или трегалоза. Предпочтительно, водная композиция является изотонической, хотя подходящими могут быть гипертонические или гипотонические растворы. Иллюстративные концентрации полиола в составе могут находиться в диапазоне от приблизительно 1% до приблизительно 15% вес/объем.
Поверхностно-активное вещество также можно добавлять к составу для снижения агрегации антитела или антигенсвязывающего фрагмента в составе и/или минимизации образования частиц в составе, и/или снижения адсорбции. Иллюстративные поверхностно-активные вещества включают неионные поверхностно-активные вещества, такие как полисорбаты (например, полисорбат 20 или полисорбат 80) или полоксамеры (например, полоксамер 188). Иллюстративные концентрации поверхностно-активного вещества могут находиться в диапазоне от приблизительно 0,001% до приблизительно 0,5% или от приблизительно 0,005% до приблизительно 0,2% или, в качестве альтернативы, от приблизительно 0,004% до приблизительно 0,01% вес/объем.
В одном варианте осуществления состав содержит идентифицированные выше средства (т. е. антитело или антигенсвязывающий фрагмент, буфер, полиол и поверхностно-активное вещество) и фактически не содержит одного или нескольких консервантов, таких как бензиловый спирт, фенол, м-крезол, хлорбутанол и хлорид бензетония. В другом варианте осуществления в состав может быть включен консервант, например, в концентрациях, находящихся в диапазоне от приблизительно 0,1% до приблизительно 2% или, в качестве альтернативы, от приблизительно 0,5% до приблизительно 1%. Один или несколько других фармацевтически приемлемых носителей, вспомогательных веществ или стабилизаторов, таких как те, которые описаны в REMINGTON'S PHARMACEUTICAL SCIENCES, 18th Edition, (AR Genrmo, ed.), 1990, Mack Publishing Company, могут быть включены в состав при условии, что они не оказывают неблагоприятного воздействия на требуемые характеристики состава.
Терапевтические составы антитела или антигенсвязывающего фрагмента получают для хранения посредством смешивания антитела или антигенсвязывающего фрагмента, характеризующегося требуемой степенью чистоты, с необязательными физиологически приемлемыми носителями, вспомогательными веществами или стабилизаторами (REMINGTON'S PHARMACEUTICAL SCIENCES, 18th Edition, (A.R. Genrmo, ed.), 1990, Mack Publishing Company) в виде лиофилизированных составов или водных растворов. Приемлемые носители, вспомогательные вещества или стабилизаторы являются нетоксичными для реципиентов в используемых дозах и концентрациях и включают описанные выше приемлемые носители, вспомогательные вещества или стабилизаторы, такие как буферы (например, фосфатный, цитратный и буферы на основе других органических кислот); антиоксиданты (например, аскорбиновая кислота и метионин); консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт, алкилпарабены, такие как метил- или пропилпарабен; катехин; резорцин; циклогексанол; 3-пентанол и м-крезол); низкомолекулярные (например, с меньше чем приблизительно 10 остатками) полипептиды; белки (такие как сывороточный альбумин, желатин или иммуноглобулины); гидрофильные полимеры (например, поливинилпирролидон); аминокислоты (например, глицин, глутамин, аспарагин, гистидин, аргинин или лизин); моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу, мальтозу или декстрины; хелатирующие средства, такие как EDTA; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как ионы натрия; комплексы с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные вещества, такие как полисорбаты (например, полисорбат 20 или полисорбат 80) или полоксамеры (например, полоксамер 188), или полиэтиленгликоль (PEG).
В одном варианте осуществления подходящий состав по настоящему изобретению содержит изотонический буфер, такой как фосфатный, ацетатный или Трис-буфер, в комбинации со средством для регуляции тоничности, таким как полиол, сорбит, сахароза или хлорид натрия, который регулирует тоничность и стабилизирует. Одним примером такого средства для регуляции тоничности является 5% сорбит или сахароза. Кроме того, состав может необязательно включать поверхностно-активное вещество в количестве от 0,01% до 0,02% вес/объем, например, для предотвращения агрегации или улучшения стабильности. Значение pH состава может находиться в диапазоне 4,5-6,5 или 4,5-5,5. Другие иллюстративные описания фармацевтических составов для антител и антигенсвязывающих фрагментов можно найти в публикации патента США № 2003/0113316 и патенте США № 6171586, каждый из которых настоящим включен посредством ссылки во всей своей полноте.
Составы, применяемые для введения in vivo, должны быть стерильными. Композиции по настоящему изобретению можно стерилизовать с помощью общепринятых, хорошо известных методик стерилизации. Например, стерилизация легко осуществляется посредством фильтрации через стерильные фильтрующие мембраны. Полученные растворы могут быть упакованы для применения или отфильтрованы в асептических условиях и лиофилизированы, причем лиофилизированный препарат объединяют со стерильным раствором перед введением.
Способ сублимационного высушивания часто используется для стабилизации полипептидов для длительного хранения, особенно если полипептид относительно нестабилен в жидких композициях. Цикл лиофилизации обычно состоит из трех стадий: замораживание, первичное высушивание и вторичное высушивание (см. Williams and Polli, Journal of Parenteral Science and Technology, Volume 38, Number 2, pages 48-59, 1984). На стадии замораживания раствор охлаждают до тех пор, пока он не будет надлежащим образом заморожен. Основная часть воды в растворе на этой стадии образует лед. Лед сублимирует на стадии первичного высушивания, которую осуществляют посредством снижения давления в камере до значения ниже давления пара льда с применением вакуума. Наконец, сорбированную или связанную воду удаляют на стадии вторичного высушивания при пониженном давлении в камере и повышенной температуре хранения. С помощью данного способа получают материал, известный как лиофилизированная масса. После этого масса может быть восстановлена перед применением. Стандартная практика восстановления лиофилизированного материала заключается в добавлении объема чистой воды (обычно эквивалентного объему, удаленному во время лиофилизации), хотя для получения фармацевтических препаратов для парентерального введения иногда применяют разбавленные растворы антибактериальных средств (см. Chen, Drug Development and Industrial Pharmacy, Volume 18: 1311-1354, 1992).
Было отмечено, что в некоторых случаях вспомогательные вещества действуют как стабилизаторы для продуктов, подвергнутых сублимационному высушиванию (см. Carpenter et al., Volume 74: 225-239, 1991). Например, известные вспомогательные вещества включают полиолы (включая маннит, сорбит и глицерин); сахара (включая глюкозу и сахарозу) и аминокислоты (включая аланин, глицин и глутаминовую кислоту). Кроме того, полиолы и сахара также часто применяют для защиты полипептидов от замораживания и повреждения, вызванного высушиванием, и для повышения стабильности при хранении в высушенном состоянии. В целом, сахара, в частности дисахариды, эффективны как в способе сублимационного высушивания, так и при хранении. Другие классы молекул, включая моно- и дисахариды и полимеры, такие как PVP, также были описаны как стабилизаторы лиофилизированных продуктов.
Для инъекции фармацевтический состав и/или лекарственный препарат могут быть представлены в виде порошка, подходящего для восстановления соответствующим раствором, как описано выше. Их примеры включают без ограничения высушенные посредством сублимационного высушивания, высушенные посредством ротационного высушивания или высушенные посредством высушивания распылением порошки, аморфные порошки, гранулы, осадки или частицы. Для инъекции составы могут необязательно содержать стабилизаторы, модификаторы рН, поверхностно-активные вещества, модификаторы биодоступности и их комбинации.
Можно получать препараты с замедленным высвобождением. Подходящие примеры препаратов с замедленным высвобождением включают полупроницаемые матрицы из твердых гидрофобных полимеров, содержащие антитело или антигенсвязывающий фрагмент, причем эти матрицы имеют форму формованных изделий, например, пленок или микрокапсулы. Примеры матриц с замедленным высвобождением включают сложные полиэфиры, гидрогели (например, поли(2-гидроксиэтилметакрилат) или поли(виниловый спирт)), полилактиды (патент США № 3773919), сополимеры L-глутаминовой кислоты и этил-L-глутамата, неразлагаемый этиленвинилацетат, разлагаемые сополимеры молочной кислоты и гликолевой кислоты, такие как Lupron Depot™ (инъецируемые микросферы, состоящие из сополимера молочной кислоты и гликолевой кислоты и ацетата лейпролида) и поли-D-(-)-3-гидроксимасляную кислоту. Хотя полимеры, такие как этиленвинилацетат и сополимер молочной кислоты и гликолевой кислоты обеспечивают возможность высвобождения молекул в течение более 100 дней, определенные гидрогели высвобождают белки в течение более коротких периодов времени. Если инкапсулированные полипептиды остаются в организме в течение длительного времени, они могут денатурировать или агрегировать в результате воздействия влаги при 37°C, что приводит к потере биологической активности и возможным изменениям в иммуногенности. Для стабилизации могут быть разработаны рациональные стратегии в зависимости от используемого механизма. Например, если обнаружено, что механизм агрегации заключается в образовании межмолекулярных S-S-связей посредством тиодисульфидного обмена, то стабилизация может быть достигнута посредством модификации сульфгидрильных остатков, лиофилизации из кислых растворов, контроля содержания влаги, применения соответствующих добавок и разработки специфических полимерных матричных композиций.
Составы по настоящему изобретению могут быть разработаны для кратковременного, быстрого высвобождения, длительного действия или замедленного высвобождения, как описано в данном документе. Таким образом, фармацевтические составы также могут быть составлены для контролируемого высвобождения или для медленного высвобождения.
Конкретные дозы могут быть скорректированы в зависимости от заболевания, нарушения или состояния, подлежащего лечению (например, эпизодическая мигрень, хроническая мигрень или кластерная головная боль), возраста, веса тела, общего состояния здоровья, пола и рациона питания субъекта, интервалов между введением доз, путей введения, скорости выведения и комбинаций лекарственных средств.
Антитела к PAC1 или антигенсвязывающие фрагменты по настоящему изобретению можно вводить любыми подходящими способами, включая парентеральное, подкожное, внутривенное, интраперитонеальное, внутрилегочное и интраназальное и, если требуется для местного лечения, внутриочаговое введение. Парентеральное введение включает внутривенное, внутриартериальное, интраперитонеальное, внутримышечное, внутрикожное или подкожное введение. Кроме того, антитело или антигенсвязывающий фрагмент подходящим образом вводят путем импульсной инфузии, в частности при снижающихся дозах антитела или антигенсвязывающего фрагмента. Предпочтительно, дозу вводят путем инъекций, наиболее предпочтительно внутривенных или подкожных инъекций, частично в зависимости от того, является ли введение краткосрочным или хроническим. Предусматриваются другие способы введения, включая местное, в частности трансдермальное, трансмукозальное, ректальное, пероральное или местное введение, например, через катетер, расположенный рядом с нужным участком. Антитело или антигенсвязывающий фрагмент по настоящему изобретению можно вводить в физиологическом растворе в дозе, находящейся в диапазоне 0,01-100 мг/кг с частотой от ежедневной до еженедельной или ежемесячной.
Антитела к PAC1 и антигенсвязывающие фрагменты, описанные в данном документе, применимы для лечения или снижения выраженности состояния, связанного с биологической активностью рецептора PAC1 у пациента, нуждающегося в этом. Таким образом, в данном документе раскрыты антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению для применения в способах лечения. Термин "пациент" включает пациентов-людей и применяется взаимозаменяемо с термином "субъект". Применяемый в данном документе термин "осуществление лечения" или "лечение" представляет собой вмешательство, выполненное с целью предупреждения развития или изменения патологии нарушения. Соответственно "лечение" относится как к терапевтическому лечению, так и к профилактическим или превентивным мерам. К нуждающимся в лечении относятся те, у кого уже диагностировано нарушение или те, кто страдает нарушением или состоянием, а также те, у которых необходимо предупредить нарушение или состояние. Термин "лечение" включает любой признак успеха в снижении выраженности повреждения, патологии или состояния, включая любой объективный или субъективный параметр, такой как смягчение боли, ремиссия, ослабление симптомов или способствование лучшей переносимости повреждения, патологии или состояния пациента, замедление темпа дегенерации или ухудшения, способствование менее изнурительному протеканию конечной стадии дегенерации или улучшение физического или психического здоровья пациента. Лечение или снижение выраженности симптомов может быть основано на объективных или субъективных параметрах, включая результаты физического осмотра, самостоятельного предоставления сведений пациентом, психоневрологических обследований и/или психиатрической оценки.
Соответственно, в некоторых вариантах осуществления в настоящем изобретении предусматривается способ лечения или предупреждения состояния, связанного с биологической активностью рецептора PAC1 (например, состояния, связанного с индуцированной PACAP активацией рецептора PAC1) у пациента, нуждающегося в этом, включающий введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе. Сигнальный путь PACAP/PAC1 задействован в различных физиологических процессах, включая сердечно-сосудистую функцию, метаболическую и эндокринную функцию, воспаление, реакцию на стресс, регуляцию сосудистого тонуса и регуляцию автономной нервной системы, в частности баланса между симпатической и парасимпатической системами. См, например, Tanida et al., Regulatory Peptides, Vol. 161: 73-80, 2010; Moody et al., Curr. Opin. Endocrinol. Diabetes Obes., Vol. 18: 61-67, 2011; и Hashimoto et al., Current Pharmaceutical Design, Vol. 17: 985-989, 2011. Состояния, связанные с аберрантной или чрезмерной активацией сигнального пути PACAP/PAC1, включают без ограничения состояния, связанные с головной болью, такие как мигрень, кластерная головная боль, головная боль напряженного типа, гемиплегическая мигрень и ретинальная мигрень; состояния воспаления кожи; хроническую боль, такую как невропатическая боль; тревожные расстройства; синдром раздраженного кишечника; и вазомоторные симптомы, такие как приливы, покраснение лица, потливость и ночные потливости. Таким образом, антитела к PAC1 и их антигенсвязывающие фрагменты по настоящему изобретению можно вводить пациентам для предупреждения , снижения выраженности или лечения любого из этих состояний или нарушений или других состояний, связанных с аберрантной или избыточной биологической активностью рецептора PAC1. В определенных вариантах осуществления в настоящем изобретении предусмотрены способы лечения или предупреждения состояния, связанного с головной болью (например, эпизодической мигрени, хронической мигрени, кластерной головной боли, головной боли напряженного типа, гемиплегической мигрени и ретинальной мигрени), у пациента, нуждающегося в этом, включающие введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, как описано в данном документе. В некоторых вариантах осуществления в настоящем изобретении предусмотрен способ ингибирования активации рецептора PAC1 человека у пациента, характеризующегося состоянием, связанным с головной болью, включающий введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе. В одном варианте осуществления пациент характеризуется состоянием мигренозной головной боли, таким как эпизодическая мигрень или хроническая мигрень. В другом варианте осуществления пациент характеризуется состоянием кластерной головной боли.
Как правило, "эффективное количество" представляет собой количество, достаточное для уменьшения тяжести и/или частоты симптомов, устранения симптомов и/или основной причины, предупреждения возникновения симптомов и/или их основной причины и/или улучшения или устранения повреждения, которое является результатом конкретного состояния или связано с ним. В некоторых вариантах осуществления эффективное количество представляет собой терапевтически эффективное количество или профилактически эффективное количество. "Терапевтически эффективное количество" представляет собой количество, достаточное для лечения болезненного состояния или симптома(симптомов), в частности состояния или симптома(симптомов), связанного с болезненным состоянием, или иного предупреждения, препятствования, замедления или обращения прогрессирования болезненного состояния или любого другого нежелательного симптома, связанного с заболеванием в какой бы то ни было форме (т. е. которое обеспечивает "терапевтическую эффективность"). "Профилактически эффективное количество" представляет собой количество антитела или антигенсвязывающего фрагмента, которое при введении субъекту будет оказывать желаемый профилактический эффект, например, предупреждение или задержку возникновения (или повторного появления) состояния или снижение вероятности возникновения (или повторного появления) состояния. Полный терапевтический или профилактический эффект не обязательно возникает в результате введения одной дозы и может возникнуть только после введения серии доз. Таким образом, терапевтически или профилактически эффективное количество может быть введено за одно или несколько введений.
В определенных вариантах осуществления в настоящем изобретении предусматриваются способы подавления вазодилатации у пациента, нуждающегося в этом, включающие введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе. Лиганды рецептора PAC1, такие как PACAP38 и VIP, являются сильными вазодилататорами, и блокирование связывания этих лигандов с рецептором PAC1 может подавлять вазодилатацию и снижать выраженность состояний, связанных с аберрантной или избыточной вазодилатацией, таких как состояния, связанные с головной болью, приливы и покраснение. В одном варианте осуществления пациент характеризуется состоянием, связанным с головной болью, таким как мигрень или кластерная головная боль. В другом варианте осуществления пациент характеризуется вазомоторными симптомами (например, приливами, покраснением лица, потливостью и ночными потливостями). В соответствующем варианте осуществления пациент характеризуется вазомоторными симптомами, связанными с менопаузой.
В некоторых вариантах осуществления способов по настоящему изобретению состояние, связанное с головной болью, подлежащее лечению, предупреждению или снижению выраженности представляет собой мигрень. Таким образом, настоящее изобретение включает способ лечения, предупреждения или снижения выраженности мигрени у пациента, нуждающегося в этом, включающий введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе. Мигренозные головные боли представляют собой рецидивирующие головные боли продолжительностью от приблизительно 4 до приблизительно 72 часов, которые характеризуются односторонней пульсирующей болью, и/или болью со степенью выраженности от умеренной до тяжелой, и/или болью, которая усугубляется при физической активности. Мигренозные головные боли часто сопровождаются тошнотой, рвотой и/или чувствительностью к свету (фотофобией), звуку (фонофобией) или запаху. У некоторых пациентов началу мигренозной головной боли предшествует аура. Аура, как правило, является визуальным, сенсорным, языковым или моторным нарушением, которое сигнализирует о скором наступлении головной боли. Способы, описанные в данном документе, позволяют предупреждать, лечить или снижать выраженность одного или нескольких симптомов мигренозных головных болей с аурой либо без нее у пациентов-людей.
PACAP38 посредством активации своих рецепторов индуцирует вазодилатацию, в частности вазодилатацию сосудистой системы твердой мозговой оболочки (Schytz et al., Neurotherapeutics, Vol. 7(2):191-196, 2010). В частности, сигнальный каскад рецептора PACAP38/PAC1 задействован в патофизиологии мигрени (Amin et al., Brain, Vol. 137: 779-794, 2014). Инфузия PACAP38, который характеризуется более высокой аффинностью к рецептору PAC1, чем к рецепторам VPAC1 и VPAC2, вызывает мигренеподобную головную боль у пациентов с мигренью (Schytz et al., Brain 132:16-25, 2009; Amin et al., Brain, Vol. 137: 779-794, 2014; Guo et al., Cephalalgia, Vol. 37:125-135, 2017). Кроме того, уровни PACAP38 повышаются во внутричерепном кровообращении у пациентов, испытывающих приступ мигрени, и уровни PACAP38 снижаются после лечения симптомов мигрени с помощью триптанов (Tuka et al., Cephalalgia, Vol. 33, 1085-1095, 2013; Zagami et al., Ann. Clin. Transl. Neurol., Vol. 1: 1036-1040, 2014). Эти сообщения предполагают, что эндогенное высвобождение PACAP38 является важным триггером мигренозных головных болей, и его эффекты в основном опосредованы активацией рецептора PAC1.
В некоторых вариантах осуществления пациенты, подлежащие лечению в соответствии со способами по настоящему изобретению, характеризуются эпизодической мигренью, страдают от нее или имеют диагноз эпизодической мигрени. Диагноз "эпизодическая мигрень" ставится, когда пациенты с мигренью в анамнезе (например, по меньшей мере пять приступов мигрени в течение жизни) проводят 14 или меньше дней с наличием мигренозной головной боли в месяц. "День с мигренозной головной болью" включает любой календарный день, в течение которого пациент испытывает начало, продолжение или рецидив "мигренозной головной боли" с аурой продолжительностью более 30 минут или без нее. "Мигренозная головная боль" представляет собой головную боль, связанную с тошнотой или рвотой или чувствительностью к свету или звуку и/или головной болью, характеризующейся по меньшей мере двумя из следующих болевых признаков: односторонней болью, пульсирующей болью, интенсивностью боли от средней до сильной или болью, усугубляющейся при физической активности. В определенных вариантах осуществления пациенты, характеризующиеся эпизодической мигренью, страдающие от нее или имеющие диагноз эпизодической мигрени, проводят по меньшей мере четыре, но менее 15 дней с наличием мигренозной головной боли в месяц в среднем. В связанных вариантах осуществления пациенты, характеризующиеся эпизодической мигренью, страдающие от нее или имеющие диагноз эпизодической мигрени, проводят менее 15 дней с головной болью в месяц в среднем. Применяемый в данном документе термин "день с головной болью" представляет собой любой календарный день, когда пациент испытывает мигренозную головную боль, как определено в данном документе, или любую головную боль, которая длится более 30 минут или требует острого лечения головной боли.
В определенных вариантах осуществления пациенты, подлежащие лечению в соответствии со способами по настоящему изобретению, характеризуются хронической мигренью, страдают от нее или имеют диагноз хронической мигрени. Диагноз "хроническая мигрень" ставится, когда пациенты с мигренью (т. е. пациенты с по меньшей мере пятью приступами мигренозной головной боли в течение жизни) проводят 15 или больше дней с головной болью в месяц и по меньшей мере 8 дней с наличием головной боли, которые являются днями с мигренозной головной болью. В некоторых вариантах осуществления пациенты, характеризующиеся хронической мигренью, страдающие от нее или имеющие диагноз хронической мигрени, проводят 15 или больше дней с мигренозной головной болью в месяц в среднем. В определенных вариантах осуществления способов, описанных в данном документе, введение антитела к PAC1 или антигенсвязывающего фрагмента по настоящему изобретению предупреждает, снижает или задерживает прогрессирование мигрени от эпизодической мигрени до хронической мигрени у пациента.
В некоторых вариантах осуществления в настоящем изобретении предусматривается способ лечения, предупреждения или снижения выраженности кластерной головной боли у пациента, нуждающегося в этом, включающий введение пациенту эффективного количества антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе. Кластерная головная боль представляет собой состояние, которое включает, как наиболее характерную черту, повторяющиеся сильные головные боли на одной стороне головы, обычно вокруг глаза (см. Nesbitt et al., BMJ, Vol. 344:e2407, 2012). Кластерные головные боли часто возникают периодически: спонтанные ремиссии прерывают активные периоды боли. Кластерные головные боли часто сопровождаются черепными вегетативными симптомами, такими как слезотечение, заложенность носа, птоз, сужение зрачка, покраснение лица, потливость и припухлость вокруг глаза, часто ограниченные на стороне головы с болью. Средний возраст начала возникновения кластерной головной боли составляет ~ 30-50 лет. Она более распространена у мужчин при соотношении мужчин и женщин от 2,5:1 до 3,5:1. Для лечения кластерной головной боли использовали стимуляцию крылонебного ганглия (SPG). Система нейростимуляции, которая обеспечивает низкоуровневую (но с высокой частотой, с физиологической блокировкой) электростимуляцию SPG, продемонстрировала эффективность в ослабления острой изнурительной боли, относящейся к кластерной головной боли, в недавнем клиническом испытании (см. Schoenen J, et al., Cephalalgia, Vol. 33(10):816-30, 2013). С учетом этого наблюдения и поскольку PACAP является одним из основных нейротрансмиттеров в SPG, ожидается, что подавление передачи сигналов PACAP/PAC1 антителом к PAC1 или его антигенсвязывающим фрагментом, описанным в данном документе, должно иметь эффективность при лечении кластерной головной боли у людей.
Другие состояния, связанные с сигнальным путем PACAP/PAC1, которые можно лечить в соответствии со способами по настоящему изобретению, включают без ограничения воспалительные состояния кожи, как например розацеа (см. публикацию патента США № 20110229423), синдромы хронической боли, как например невропатическая боль (см. Jongsma et al., Neuroreport, Vol. 12: 2215-2219, 2001; Hashimoto et al., Annals of the New York Academy of Sciences, Vol. 1070: 75-89, 2006), головные боли напряженного типа, гемиплегическая мигрень, ретинальная мигрень, тревожные расстройства, как например посттравматическое стрессовое расстройство (см. Hammack and May, Biol. Psychiatry, Vol.78(3):167-177, 2015), синдром раздраженного кишечника и вазомоторные симптомы (например, приливы, покраснение лица, потливость и ночная потливость), как например симптомы, связанные с менопаузой. В одном варианте осуществления состояние, подлежащее лечению с помощью введения антитела к PAC1 или его антигенсвязывающего фрагмента по настоящему изобретению, представляет собой хроническую боль. В другом варианте осуществления состояние, подлежащее лечению с помощью введения антитела к PAC1 или его антигенсвязывающего фрагмента по настоящему изобретению, представляет собой невропатическую боль.
В любом из способов, описанных в данном документе, лечение может включать профилактическое лечение. Профилактическое лечение относится к лечению, разработанному для проведения до наступления состояния или приступа (например, до приступа мигрени или начала эпизода кластерной головной боли) для снижения частоты, тяжести и/или продолжительности симптомов (например, мигрени или кластерных головных болей) у больного.
В некоторых вариантах осуществления способы по настоящему изобретению для лечения или предупреждения состояния, связанного с головной болью, у пациента включают введение пациенту антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе, в комбинации с одним или несколькими средствами, подходящими для острого или профилактического лечения мигренозной головной боли или другого расстройства с головной болью, описанного в данном документе. Применяемый в данном документе термин "комбинированная терапия" охватывает введение двух соединений (например, антитела к PAC1 и дополнительного средства) последовательным образом (т. е. каждое соединение вводится в разное время в любом порядке), а также введение двух соединений по сути одновременно. По сути одновременное введение включает параллельное введение и может выполняться путем введения одного состава, содержащего оба соединения (например, одного состава, содержащего фиксированное соотношение обоих соединений, или предварительно заполненного шприца с фиксированным соотношением каждого соединения), или параллельного введения отдельных составов, содержащих каждое из соединений. Таким образом, в определенных вариантах осуществления способы по настоящему изобретению включают введение антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе, со вторым терапевтическим средством от головной боли.
В определенных вариантах осуществления второе терапевтическое средство от головной боли может представлять собой терапевтическое средство от острой головной боли, используемое для острого лечения головной боли или мигрени. В некоторых вариантах осуществления терапевтическое средство от острой головной боли представляет собой агонист рецептора (5-гидрокситриптамин; 5-HT) серотонина, например агонист рецептора 5HT1 серотонина. Терапевтическое средство от острой головной боли может представлять собой агонист рецепторов 5HT1B, 5HT1D и/или 5HT1F серотонина. Такие агонисты серотониновых рецепторов включают без ограничения триптаны (например, алмотриптан, фроватриптан, ризатриптан, суматриптан, наратриптан, элетриптан и золмитриптан), эрготамины (например, дигидроэрготамин и тартрат эрготамина) и агонисты 5HT1F-селективных рецепторов серотонина, например ласмидитан. Другие подходящие терапевтические средства от острой головной боли включают нестероидные противовоспалительные лекарственные средства (например, ацетилсалициловая кислота, ибупрофен, напроксен, индометацин и диклофенак) и опиоиды (например, кодеин, морфин, гидрокодон, фентанил, меперидин и оксикодон). В одном варианте осуществления терапевтическое средство от острой головной боли, которое вводят в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой триптан. В другом варианте осуществления терапевтическое средство от острой головной боли, которое вводят в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой эрготамин. В еще одном варианте осуществления терапевтическое средство от острой головной боли, которое вводят в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой нестероидное противовоспалительное лекарственное средство. В еще одном варианте осуществления терапевтическое средство от острой головной боли, которое вводят в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению, представляет собой опиоид.
В некоторых вариантах осуществления второе терапевтическое средство от головной боли представляет собой профилактическое терапевтическое средство от головной боли, используемое для профилактического лечения головных болей или мигрени. В одном варианте осуществления профилактическим терапевтическим средством от головной боли является противоэпилептическое средство, такое как дивальпроекс, вальпроат натрия, вальпроевая кислота, топирамат или габапентин. В другом варианте осуществления профилактическое терапевтическое средство от головной боли представляет собой бета-блокатор, такой как пропранолол, тимолол, атенолол, метопролол или надолол. В еще одном варианте осуществления профилактическое терапевтическое средство от головной боли представляет собой антидепрессант, такой как трициклический антидепрессант (например, амитриптилин, нортриптилин, доксепин и флуоксетин). В еще другом варианте осуществления профилактическое терапевтическое средство от головной боли представляет собой онаботулотоксин А.
В определенных вариантах осуществления способы настоящего изобретения включают введение антитела к PAC1 или его антигенсвязывающего фрагмента, описанного в данном документе, с антагонистом сигнального пути кальцитонин-ген-связанного пептида (CGRP) (т. е. подавляет активацию или передачу сигналов рецептора CGRP лигандом CGRP). Например, антитело к PAC1 или антигенсвязывающий фрагмент по настоящему изобретению можно вводить в комбинации с антагонистом пути CGRP для лечения или предупреждения состояния, связанного с головной болью (например, мигрени или кластерной головной боли), у пациента, нуждающегося в этом. В некоторых вариантах осуществления антагонист пути CGRP представляет собой антагонист рецептора CGRP человека. Антагонисты рецептора CGRP включают низкомолекулярные ингибиторы рецептора CGRP, такие как описанные в публикации патента США № 20060142273 и патентах США №№ 7842808; 7772244; 7754732; 7569578; 8685965; 8569291; 8377955; 8372859; 8143266; 7947677 и 7625901, все из которых включены в данный документ посредством ссылки во всей своей полноте. Антагонисты рецептора CGRP могут также включать пептидные антагонисты рецептора, такие как описанные в патенте США № 8168592, который включен в данный документ посредством ссылки во всей своей полноте. В определенных вариантах осуществления антагонист рецептора CGRP, подлежащий введению с антителами к PAC1 и антигенсвязывающими фрагментами по настоящему изобретению, представляет собой моноклональное антитело, которое специфически связывается с рецептором CGRP человека, как например антитела, описанные в патенте США № 9102731 и публикации патента США № 20160311913, все из которых включены в данный документ посредством ссылки во всей своей полноте. В одном конкретном варианте осуществления способов по настоящему изобретению антитело к PAC1 или его антигенсвязывающий фрагмент, описанный в данном документе, вводят в комбинации с моноклональным антителом к рецептору CGRP, содержащим вариабельную область легкой цепи, содержащую последовательность под SEQ ID NO: 502 (последовательность, приведенную ниже), и вариабельную область тяжелой цепи, содержащую последовательность под SEQ ID NO: 503 (последовательность, приведенную ниже), для лечения или предупреждения состояния, связанного с головной болью (например мигрени или кластерной головной боли), у пациента. В другом конкретном варианте осуществления способов по настоящему изобретению моноклональное антитело к рецептору CGRP, которое вводят в комбинации с антителом к PAC1 или антигенсвязывающим фрагментом по настоящему изобретению для лечения или предупреждения состояния, связанного с головной болью (например, мигрени или кластерной головной боли), у пациента, представляет собой эренумаб.
Последовательность вариабельной области легкой цепи иллюстративного моноклонального антитела к рецептору CGRP:
QSVLTQPPSV SAAPGQKVTI SCSGSSSNIG NNYVSWYQQL PGTAPKLLIY DNNKRPSGIP DRFSGSKSGT STTLGITGLQ TGDEADYYCG TWDSRLSAVV FGGGTKLTVL (SEQ ID NO: 502).
Последовательность вариабельной области тяжелой цепи иллюстративного моноклонального антитела к рецептору CGRP:
QVQLVESGGG VVQPGRSLRL SCAASGFTFS SFGMHWVRQA PGKGLEWVAV ISFDGSIKYS VDSVKGRFTI SRDNSKNTLF LQMNSLRAED TAVYYCARDR LNYYDSSGYY HYKYYGMAVW GQGTTVTVSS (SEQ ID NO: 503).
В некоторых вариантах осуществления антагонист пути CGRP, подлежащий введению с антителами к PAC1 или антигенсвязывающими фрагментами по настоящему изобретению для лечения или предупреждения состояния, связанного с головной болью (например, мигрени или кластерной головной боли), у пациента, представляет собой антагонист лиганда CGRP. Антагонист лиганда CGRP может являться "ловушкой", или растворимым рецептором CGRP, или другим белком, который связывается с лигандом CGRP, такой как антитело к лиганду CGRP. Антитела к лиганду CGRP известны в уровне техники и описаны, например, в WO 2007/054809; WO 2007/076336; WO 2011/156324 и WO 2012/162243, все из которых включены в данный документ посредством ссылки во всей своей полноте. В определенных вариантах осуществления антагонист лиганда CGRP, подлежащий введению с антителами к PAC1 и антигенсвязывающими фрагментами по настоящему изобретению, представляет собой моноклональное антитело, которое специфически связывается с α-CGRP человека и/или β-CGRP человека. В одном варианте осуществления антитело к лиганду CGRP представляет собой фреманезумаб. В другом варианте осуществления антитело к лиганду CGRP представляет собой галканезумаб. В еще одном варианте осуществления антитело к лиганду CGRP представляет собой эптинезумаб.
Настоящее изобретение также включает применение антитела к PAC1 и антигенсвязывающих фрагментов в любом из способов, раскрытых в данном документе. Например, в определенных вариантах осуществления в настоящем изобретении предусматривается антитело к PAC1 или его антигенсвязывающий фрагмент, описанный в данном документе, для использования в способе лечения или предупреждения состояния, связанного с головной болью, у пациента, нуждающегося в этом. В некоторых таких вариантах осуществления состояние, связанное с головной болью, представляет собой мигрень. Мигрень может представлять собой эпизодическую мигрень или хроническую мигрень. В других вариантах осуществления состояние, связанное с головной болью, представляет собой кластерную головную боль. В некоторых вариантах осуществления способ лечения или предупреждения состояния, связанного с головной болью, включает введение второго терапевтического средства от головной боли в комбинации с антителом к PAC1 или его антигенсвязывающим фрагментом. В одном варианте осуществления второе терапевтическое средство от головной боли представляет собой терапевтическое средство от острой головной боли, такое как агонист серотониновых рецепторов 5HT1B, 5HT1D и/или 5HT1F (например, триптан или эрготамин), нестероидное противовоспалительное лекарственное средство или опиоид. В другом варианте осуществления второе терапевтическое средство от головной боли представляет собой профилактическое терапевтическое средство от головной боли, такое как противоэпилептическое средство, бета-блокатор, антидепрессант, онаботулотоксин А или антагонист пути CGRP. В некоторых вариантах осуществления антагонист пути CGRP представляет собой антагонист рецептора CGRP человека. В одном конкретном варианте осуществления антагонист рецептора CGRP человека представляет собой моноклональное антитело, которое специфически связывается с рецептором CGRP человека, такое как эренумаб. В других вариантах осуществления антагонист пути CGRP представляет собой антагонист лиганда CGRP, такой как моноклональное антитело, которое специфически связывается с α-CGRP человека и/или β-CGRP человека. В определенных вариантах осуществления антитело к лиганду CGRP представляет собой фреманезумаб, галканезумаб или эптинезумаб.
В некоторых вариантах осуществления в настоящем изобретении предусматривается антитело к PAC1 или антигенсвязывающий фрагмент, описанный в данном документе, для применения в способе подавления вазодилатации у пациента, нуждающегося в этом. В таких вариантах осуществления пациент может быть диагностирован или иметь состояние, связанное с головной болью, такое как мигрень (например, эпизодическая или хроническая мигрень) или кластерная головная боль. В других вариантах осуществления в настоящем изобретении предусматривается антитело к PAC1 или антигенсвязывающий фрагмент, описанный в данном документе, для применения в способе подавления активации рецептора PAC1 человека у пациента, характеризующегося состоянием, связанным с головной болью. Состояние, связанное с головной болью, может представлять собой мигрень (например, эпизодическую или хроническую мигрень) или кластерную головную боль.
Конкретно предусмотрено применение антител к PAC1 или их антигенсвязывающих фрагментов для получения лекарственных препаратов для введения в соответствии с любым из способов, раскрытых в данном документе. Например, в некоторых вариантах осуществления настоящее изобретение охватывает применение антитела к PAC1 или антигенсвязывающего фрагмента, описанного в данном документе, для получения лекарственного препарата для лечения или предупреждения состояния, связанного с головной болью, у пациента, нуждающегося в этом. В некоторых таких вариантах осуществления состояние, связанное с головной болью, представляет собой мигрень. Мигрень может представлять собой эпизодическую мигрень или хроническую мигрень. В других вариантах осуществления состояние, связанное с головной болью, представляет собой кластерную головную боль. В определенных вариантах осуществления антитело к PAC1 или его антигенсвязывающий фрагмент составлены для введения в составе со вторым терапевтическим средством от головной боли. В одном варианте осуществления второе терапевтическое средство от головной боли представляет собой терапевтическое средство от острой головной боли, такое как агонист серотониновых рецепторов 5HT1B, 5HT1D и/или 5HT1F (например, триптан или эрготамин), нестероидное противовоспалительное лекарственное средство или опиоид. В другом варианте осуществления второе терапевтическое средство от головной боли представляет собой профилактическое терапевтическое средство от головной боли, такое как противоэпилептическое средство, бета-блокатор, антидепрессант, онаботулотоксин А или антагонист пути CGRP. В некоторых вариантах осуществления антагонист пути CGRP представляет собой антагонист рецептора CGRP человека. В одном конкретном варианте осуществления антагонист рецептора CGRP человека представляет собой моноклональное антитело, которое специфически связывается с рецептором CGRP человека, такое как эренумаб. В других вариантах осуществления антагонист пути CGRP представляет собой антагонист лиганда CGRP, такой как моноклональное антитело, которое специфически связывается с α-CGRP человека и/или β-CGRP человека. В определенных вариантах осуществления антитело к лиганду CGRP представляет собой фреманезумаб, галканезумаб или эптинезумаб.
В некоторых вариантах осуществления настоящее изобретение включает применение антитела к PAC1 или антигенсвязывающего фрагмента, описанного в данном документе, для получения лекарственного препарата для подавления вазодилатации у пациента, нуждающегося в этом. В таких вариантах осуществления пациент может быть диагностирован или иметь состояние, связанное с головной болью, такое как мигрень (например, эпизодическая или хроническая мигрень) или кластерная головная боль. В других вариантах осуществления настоящее изобретение включает применение антитела к PAC1 или антигенсвязывающего фрагмента, описанного в данном документе, для получения лекарственного препарата для подавления активации рецептора PAC1 человека у пациента, характеризующегося состоянием, связанным с головной болью. Состояние, связанное с головной болью, может представлять собой мигрень (например, эпизодическую или хроническую мигрень) или кластерную головную боль.
Антитела к PAC1 и антигенсвязывающие фрагменты по настоящему изобретению также применимы для выявления PAC1 человека в биологических образцах и идентификации клеток или тканей, которые экспрессируют PAC1 человека. Например, антитела к PAC1 и антигенсвязывающие фрагменты можно использовать в диагностических анализах, например иммуноанализах для обнаружения и/или количественного определения PAC1, экспрессируемого в ткани или клетке. Кроме того, антитела к PAC1 и антигенсвязывающие фрагменты, описанные в данном документе, можно использовать для подавления формирования PAC1 комплекса с PACAP, таким образом модулируя биологическую активность PAC1 в клетке или ткани. Такая биологическая активность включает повышение внутриклеточного уровня cAMP и вазодилатацию.
Антитела к PAC1 и антигенсвязывающие фрагменты, описанные в данном документе, можно использовать для диагностических целей для выявления, диагностики или мониторинга заболеваний и/или состояний, связанных с PAC1, включающих мигрень, кластерную головную боль и тревожные расстройства, такие как посттравматическое стрессовое расстройство. Также предусмотрены способы выявления присутствия PAC1 в образце с применением классических иммуногистологических способов, известных специалистам в данной области техники (например, Tijssen, 1993, Practice and Theory of Enzyme Immunoassays, Vol 15 (Eds R.H. Burdon and P.H. van Knippenberg, Elsevier, Amsterdam); Zola, 1987, Monoclonal Antibodies: A Manual of Techniques, pp. 147-158 (CRC Press, Inc.); Jalkanen et al., 1985, J. Cell. Biol. 101:976-985; Jalkanen et al., 1987, J. Cell Biol. 105:3087-3096). Примеры способов, применимых для выявления наличия PAC1, включают иммуноанализы, такие как твердофазный иммуноферментный анализ (ELISA) и радиоиммуноанализ (RIA), с применением антител к PAC1 и антигенсвязывающих фрагментов, описанных в данном документе. Выявление PAC1 можно осуществлять in vivo или in vitro.
Для диагностических областей применения антитело к PAC1 или антигенсвязывающий фрагмент можно подвергать мечению с помощью выявляемой группы мечения. Подходящие группы мечения включают без ограничения радиоизотопы или радионуклиды (например, 3H, 14C, 15N, 35S, 90Y, 99Tc, 111In, 125I, 131I), флуоресцентные группы (например, FITC, родамин, люминофоры на основе комплексов лантанидов), ферментные группы (например, пероксидазу хрена, β-галактозидазу, люциферазу, щелочную фосфатазу), хемилюминесцентные группы, биотинильные группы или заданные полипептидные эпитопы, распознаваемые вторичным репортером (например, парные последовательности лейциновых застежек, сайты связывания для вторичных антител, домены связывания металлов, эпитопные метки). В некоторых вариантах осуществления группу мечения присоединяют к антителу или антигенсвязывающему фрагменту с помощью спейсерных плеч различной длины для уменьшения потенциального стерического затруднения. Различные способы мечения белков известны из уровня техники и могут быть применены.
В другом варианте осуществления антитела к PAC1 и антигенсвязывающие фрагменты, описанные в данном документе, можно использовать для идентификации клетки или клеток, которые экспрессируют PAC1. В конкретном варианте осуществления в антитело или антигенсвязывающий фрагмент вводят метку в форме группы мечения и обнаруживают связывание меченого антитела или антигенсвязывающего белка с PAC1. Антитела или антигенсвязывающие фрагменты также можно применять в анализах иммунопреципитации в биологических образцах. В дополнительном конкретном варианте осуществления связывания антитела или антигенсвязывающего фрагмента с PAC1 обнаруживают in vivo. В дополнительном конкретном варианте осуществления антитело или антигенсвязывающий фрагмент выделяют и проводят измерения с применением методов, известных в данной области техники. См., например, Harlow and Lane, 1988, Antibodies: A Laboratory Manual, New York: Cold Spring Harbor (ред.1991 и периодические дополнения); John E. Coligan, ed., 1993, Current Protocols In Immunology New York: John Wiley & Sons.
Следующие примеры, в том числе проведенные эксперименты и достигнутые результаты, предоставлены только для иллюстративных целей и не должны рассматриваться как ограничивающие объем прилагаемой формулы изобретения.
ПРИМЕРЫ
Пример 1 Управляемое кристаллической структурой конструирование антител к PAC1 человека
Определяли кристаллическую структуру комплекса между внеклеточным доменом (ECD) PAC1 человека и Fab-фрагментом нейтрализующего антитела к PAC1 человека (29G4v9). ECD PAC1 человека и Fab к PAC1 очищали по отдельности, и затем обеспечивали образование их комплекса друг с другом при молярном соотношении 1:1. Образец последовательно прогоняли через колонку для гель-фильтрации, уравновешенную в 20 мМ TRIS с pH 7,5, 50 мМ NaCl, 5 мМ EDTA, и концентрировали до 35 мг/мл, и фильтровали.
Последовательности для тяжелой цепи (содержащей вариабельную область (VH), константную область CH1, верхнюю шарнирную область и сайт расщепления для каспазы III) и легкой цепи (содержащей вариабельную область (VL) и константную область CL) Fab-фрагмента перечислены ниже. Последовательность конструкции на основе ECD PAC1 человека представлена ниже, и она содержала аминокислоты 26-143 PAC1 человека (SEQ ID NO: 1) без области между аминокислотами 89-109.
Аминокислотная последовательность тяжелой цепи Fab 29G4v9, состоящей из VH-области (аминокислоты 1-120), CH1-области (аминокислоты 121-218), верхней шарнирной области (аминокислоты 219-221) и сайта расщепления для каспазы III (222-226):
QVQLVESGGG VVQPGRSLRL SCAASGFTFS RFAMHWVRQA PGKGLEWVAV ISYDGGNKYY AESVKGRFTI SRDNSKNTLY LQMNSLRAED TALFYCARGY DVLTGYPDYW GQGTLVTVSS ASTKGPSVFP LAPCSRSTSE STAALGCLVK DYFPEPVTVS WNSGALTSGV HTFPAVLQSS GLYSLSSVVT VPSSNFGTQT YTCNVDHKPS NTKVDKTVER KGDEVD (SEQ ID NO: 2).
Аминокислотная последовательность легкой цепи Fab 29G4v9, состоящей из VL-области (аминокислоты 1-108) и CL-области (аминокислоты 109-214):
DIQLTQSPSF LSASVGDRVT ITCRASQSIG RSLHWYQQKP GKAPKLLIKY ASQSLSGVPS RFSGSGSGTE FTLTISSLQP EDFATYYCHQ SSRLPFTFGP GTKVDIKRTV AAPSVFIFPP SDEQLKSGTA SVVCLLNNFY PREAKVQWKV DNALQSGNSQ ESVTEQDSKD STYSLSSTLT LSKADYEKHK VYACEVTHQG LSSPVTKSFN RGEC (SEQ ID NO: 3).
Аминокислотная последовательность конструкции на основе ECD PAC1 человека:
GSMAHSDGIF KKEQAMCLEK IQRANELMGF NDSSPGCPGM WDNITCWKPA HVGEMVLVSC PELFRIFNPD QDMGVVSRNC TEDGWSEPFP HYFDACGFDE YESET (SEQ ID NO: 4).
Сначала очищенные ECD PAC1 человека и Fab к PAC1 сокристаллизировали с применением метода диффузии пара в варианте с сидячей каплей с помощью коммерчески доступных наборов для скрининга. Условия кристаллизирования (набор № 61 комплекта Qiagen MPD (134561)) дополнительно расширяли с применением метода диффузии пара в варианте с висячей каплей. Белок и буфер для кристаллизации (0,1 М лимонной кислоты с pH 4,0, 40% MPD) смешивали 1:1 в висячих каплях объемом 1 мкл над резервуаром с раствором буфера для кристаллизации при 4°C. Стержневидные кристаллы образовывались за несколько недель.
Кристалл уравновешивали в буфере для кристаллизации в качестве криопротектора и замораживали в жидком азоте для транспортировки в Национальную лабораторию Лоуренса в Беркли для сбора данных. Набор данных собирали на базе лаборатории Advanced Light Source с помощью источника синхротронного излучения Beamline 5.0.2 на CCD-детекторе ADSC-Q315r (λ=1,000Å). Данные интегрировали и масштабировали с применением HKL2000 (Otwinowski and Minor, Methods Enzymology, Vol. 276, 307-326, 1997), и они были полными на 99,8% до 2,00 Å, при этом значение Rслияния составляло 0,081 (полные на 98,0% при последнем слое 2,07-2,00 Å с I/σ=2,35). Кристаллы принадлежат к орторомбической пространственной группе P21212 с параметрами элементарной ячейки a=65,2 Å, b=177,9 Å, c=53,8 Å, α=90°, β=90°, γ=90°. Кристаллическую структуру определяли посредством молекулярного замещения с применением PhaserMR (Winn et al., Acta Crystallogr., Sect. D: Biol. Crystallogr., Vol. 67: 235-242, 2011). Специально изготовленную Fab-структуру применяли в качестве первой модели поиска для определения компонента, представляющего собой Fab к PAC1. В последующем молекулярном замещении применяли PDBID: 2JOD (Sun et al., Proc.Natl.Acad.Sci.USA, Vol. 104: 7875, 2007), структуру ECD PAC1 человека, полученную с помощью ЯМР, в качестве исходной модели для определения компонента, представляющего собой ECD PAC1 человека. В асимметричной единице содержится одна молекула ECD и одна молекула Fab. Структуру уточняли с применением как Refmac5 (Winn et al., Acta Crystallogr., Sect. D: Biol. Crystallogr., Vol. 67: 235-242, 2011; Murshudov et al., Acta Crystallogr., Sect. D: Biol. Crystallogr., Vol. 67: 355-367, 2011), так и Phenix.refine (Adams et al., Acta Crystallogr., Sect. D: Biol. Crystallogr., Vol. 66: 213-221, 2010), и построение модели проводили с применением программы графического представления Coot (Emsley and Cowtan, Acta Crystallogr., Sect. D: Biol. Crystallogr., Vol. 2004, 60: 2126-2132, 2004).
Структуру комплекса ECD PAC1 человека : Fab к PAC1 уточняли до 2,00 Å, при этом значение R-фактора составляло 20% и значение Rв свободной форме составляло 23%. Вид спереди и вид сбоку структуры комплекса показаны на фигурах 1A и 1B соответственно. Взаимодействие между Fab 29G4v9 и ECD PAC1 состоит из гидрофобных, электростатических взаимодействий и взаимодействий посредством водородных связей. Взаимодействие между Fab 29G4v9 и ECD PAC1 характеризуется значениями площади утопленной поверхности (1613 Å2) и комплементарности формы (0,695), которые являются типичными для взаимодействий антитело-антиген. Все аминокислоты в ECD PAC1, которые содержали по меньшей мере один атом, отличный от атома водорода, на расстоянии 5 Å или меньше от атома, отличного от атома водорода, в Fab 29G4v9, определяли в качестве аминокислот основного участка контакта в ECD PAC1. Расстояния для атомов рассчитывали с помощью программы PyMOL (DeLano, W.L. The PyMOL Molecular Graphics System. (Palo Alto, 2002)). Аминокислоты основного участка контакта в ECD PAC1 включают Asp59, Asn60, Ile61, Arg116, Asn117, Thr119, Asp121, Gly122, Trp123, Ser124, Glu125, Pro126, Phe127, Pro128, His129, Tyr130, Phe131, Asp132 и Gly135, при этом номера аминокислотных положений указаны относительно SEQ ID NO: 1.
Участок контакта между Fab 29G4v9 и ECD PAC1 в кристаллической структуре анализировали, чтобы идентифицировать области, где взаимодействия между двумя молекулами являлись субоптимальными. На основе структурного анализа в вариабельных областях легкой и тяжелой цепи конструировали мутации аминокислот для усиления взаимодействия в таких областях между Fab и ECD PAC1, чтобы обеспечить улучшение в отношении аффинности связывания и/или ингибирующей удельной активности антитела. В частности, в результате анализа взаимодействия между легкой цепью Fab и ECD PAC1 выявили четыре области, где мутации в легкой цепи Fab могут, возможно, обеспечить улучшение в отношении взаимодействий с PAC1. В случае зоны 1, области, содержащей аминокислоты Glu120 и Asp121 ECD PAC1, была предложена мутация с заменой Gln27 в CDR1 легкой цепи (SEQ ID NO: 3) на лизин, тирозин или аргинин для обеспечения лучшей обусловленной зарядами комплементарности или потенциала связывания посредством атомов водорода с аминокислотами Glu120 и Asp121 PAC1 (фигура 2A). Зона 2 представляет собой область с положительным электростатическим потенциалом, и была предложена мутация с заменой Ser28 в CDR1 легкой цепи (SEQ ID NO: 3) на глутамат для обеспечения лучшей обусловленной зарядами комплементарности (фигура 2A). Зона 3 представляет собой гидрофобную область, содержащую остаток Phe127 ECD PAC1, и она обладает потенциалом связывания посредством атомов водорода. Gly30, Arg31 и Ser32 в CDR1 легкой цепи находятся несколько вдали от зоны 3 (фигура 2A). Таким образом, было предложено несколько мутаций в этих трех сайтах, кратко описанных в таблице 6 ниже, для обеспечения улучшения в отношении гидрофобных взаимодействий или взаимодействий, обусловленных связыванием посредством атомов водорода, между этими тремя остатками и этой зоной ECD PAC1. Arg93 в CDR3 легкой цепи сидит в кармане из остатков с отрицательным электростатическим потенциалом PAC1 (зона 4; фигура 2B). Однако, вследствие геометрии, Arg93 не образует каких-либо непосредственных водородных связей с остатками PAC1. Были предложены мутации с заменой Arg93 в CDR3 легкой цепи на глутамин, лизин, гистидин или аспарагин для обеспечения альтернативной обусловленной зарядами комплементарности или потенциала связывания посредством атомов водорода с остатками в ECD PAC1 (фигура 2B).
В результате анализа взаимодействия между тяжелой цепью Fab 29G4v9 и ECD PAC1 выявили три основные области, где мутации в тяжелой цепи Fab могут обеспечить улучшение в отношении взаимодействий с ECD PAC1. Как показано на фигуре 3A, зона 5 охватывает аминокислоту Phe131 PAC1, и ее можно поделить на две подзоны, которые лежат по обе стороны от Phe131. Arg31 и Phe32 в CDR1 тяжелой цепи лежат в этих подзонах, и мутации в этих сайтах были предположены для обеспечения улучшения в отношении гидрофобных взаимодействий с остатком Phe131 PAC1 или обеспечения альтернативных взаимодействий на основе обусловленной зарядами комплементарности. Перечень мутаций см. в таблице 6. В случае зоны 6, которая содержит остатки Asn60 и Ile61 ECD PAC1 (относительно SEQ ID NO: 1), мутации остатков Tyr53, Asp54 и Gly56 CDR2 тяжелой цепи были предложены для обеспечения улучшения в отношении гидрофобных взаимодействий или обеспечения альтернативного связывания посредством атомов водорода с остатками Asn60 и Ile61 PAC1 (фигура 3B). В случае зоны 7, области с гидрофобными остатками и некоторым отрицательным электростатическим потенциалом, мутации остатков Val102, Leu103 и Thr104 CDR3 тяжелой цепи, представленные в таблице 6, были предложены для обеспечения улучшения в отношении гидрофобных взаимодействий или обеспечения альтернативных взаимодействий, обусловленных связыванием посредством атомов водорода (фигура 3C).
Сводные данные по конкретным мутациям, предложенным для обеспечения улучшения в отношении взаимодействия между антителом к PAC1 и рецептором PAC1 человека, исходя из анализа кристаллической структуры комплекса Fab/ECD PAC1, представлены в таблице 6 ниже.
В дополнение к мутациям, разработку которых осуществляли с помощью анализа взаимодействующих аминокислот между Fab к PAC1 и ECD PAC1 человека, как описано выше, также осуществляли анализ созревания аффинности in silico с применением кристаллической структуры, чтобы идентифицировать дополнительные мутации для обеспечения улучшения в отношении аффинности связывания и/или ингибирующей удельной активности антитела к PAC1. Аминокислотные остатки в Fab 29G4v9, вовлеченные в связывание с ECD PAC1 человека, идентифицировали путем визуального изучения кристаллической структуры комплекса, описанного выше. Остатки из этого участка контакта антитела были выбраны для виртуального внесения мутаций по всем аминокислотам, кроме цистеина. Влияние мутации на взаимодействие, представляющее собой связывание антитела/ECD PAC1, оценивали путем расчета изменения значения свободной энергии связывания (ΔΔGсвязывания) при мутации конкретного остатка с применением программного обеспечения для молекулярного моделирования Discovery Studio от Biovia. Отрицательное значение ΔΔGсвязывания указывает на то, что мутация приводит к более сильному связыванию с ECD PAC1 по сравнению с исходной молекулой. С помощью этих расчетов идентифицировали примерно 65 мутаций, которые приводили к отрицательному значению ΔΔGсвязывания. Эти 65 мутаций далее сократили до 50 (18 в легкой цепи и 32 в тяжелой цепи) на основе визуального изучения и анализа "смоделированной" структуры таких вариантов с ECD PAC1. Сводные данные по этим 50 мутациям, для которых было предсказано обеспечение повышения аффинности связывания антитела к PAC1 на основе расчетов свободной энергии связывания, представлены в таблице 7 ниже.
По сравнению со структурным анализом, подход in silico привел к идентификации дополнительных мутаций, как предусматривающих другие аминокислоты, так и другие положения в легкой и/или тяжелой цепи.
Мутации, описанные в данном примере, вводили в антитело к PAC1 путем его рекомбинантного получения и испытывали в отношении способности подавлять индуцированную посредством PACAP активацию PAC1 человека в клеточном анализе in vitro, как описано в примере 3 в данном документе.
Пример 2. Созревание аффинности антитела к PAC1 человека 29G4v10 в дрожжевом дисплее
Один высокоэффективный подход для обеспечения созревания аффинности предусматривает конструирование библиотек на основе дрожжевого дисплея для мутантных вариантов Fab антитела и селекцию молекул, характеризующихся улучшенным связыванием, посредством активируемой флуоресценцией сортировки клеток (FACS) (фигура 4). Для получения антител к PAC1 с улучшенной ингибирующей удельной активностью для антитела 29G4v10 (VH-область под SEQ ID NO: 191; VL-область под SEQ ID NO: 52) обеспечивали созревание аффинности с помощью FACS библиотеки Fab на основе дрожжевого дисплея. Управление разработкой библиотек осуществляли на основе определенной структуры сокристалла, описанной в примере 1, комплекса между ECD PAC1 человека и Fab 29G4v9, CDR которого практически идентичны таковым из 29G4v10. С помощью структурного анализа изначально идентифицировали шесть и семь положений для мутагенеза в CDR в пределах легкой цепи (LC) и тяжелой цепи (HC) соответственно и были предложены стратегии усиления аффинности. Варианты с точечными мутациями были разработаны для обеспечения улучшения в отношении гидрофобных взаимодействий и обусловленной зарядами комплементарности и для обеспечения альтернативных взаимодействий, представляющих собой водородные связи, по всему участку контакта. См. пример 1. С этих вариантов с точечными мутациями начинали инженерный подход, который включал повторяющиеся раунды продуцирования, определения характеристик и целесообразного комбинирования полезных мутаций. Целью разработки дрожжевых библиотек являлось дополнение линейного подхода, описанного в примере 1, посредством более полного изучения комбинаций мутаций между 13 положениями и оптимизации стратегии диверсификации по каждому выбранному положению.
За счет получения отдельных библиотек LC и HC теоретические значения разнообразия поддерживали на уровне удобного в управлении размера, составляющего < 107. Библиотеку LC разрабатывали для применения насыщающего мутагенеза с применением MIX19 по пяти из шести выбранных положений (Gln27, Gly30, Arg31, Ser32 и Arg93 из SEQ ID NO: 52). MIX19 представляет собой смесь тримеров амидофосфитов (кодон), кодирующих все аминокислоты, кроме цистеина. В оставшемся положении легкой цепи (Ser28 из SEQ ID NO: 52) использовали мутацию с заменой на серин, глутамат, аланин или стоп-кодон. Для изучения всех семи положений в HC (Arg31, Phe32, Tyr53, Gly56, Val102, Leu103 и Thr104 из SEQ ID NO: 191) в одной библиотеке авторы настоящего изобретения разработали специальную смесь из девяти кодонов (MIX9) для диверсификации по пяти из семи положений CDR тяжелой цепи (Arg31, Phe32, Tyr53, Val102 и Leu103 из SEQ ID NO: 191). Специальная MIX9 содержала гидрофобные аминокислоты, основные аминокислоты и доноры и акцепторы водородной связи (например, F, L, Y, M, Q, H, K, R и S). Важно, что эта специальная MIX9 не содержала цистеина, аспарагина и триптофана, для сведения к минимуму внесения отрицательно влияющих элементов в последовательность, которые могут создавать риски в отношении последующей технологичности. Для остальных двух положений в тяжелой цепи осуществляли мутацию с заменой на глицин, аланин, серин и треонин (Gly56 и Thr104 из SEQ ID NO: 191). В целом, для этой первой кампании были разработаны библиотека mutHC с теоретическим значением разнообразия, составляющим 8,5×106, и библиотека mutLC с теоретическим значением разнообразия, составляющим 9,9×106, и запас выборок сконструированных библиотек превышал теоретические значения разнообразия в >9 раз.
Две сконструированные библиотеки Fab, mutHC и mutLC, обогащали в отношении связывания с ECD PAC1 человека с применением FACS, повышая жесткость с каждым последующим раундом путем снижения концентрации ECD, применяемой для связывания (раунд 1: 30 нМ ECD PAC1; раунд 2: 0,67 нМ ECD PAC1 и раунд 3: 0,2 нМ ECD PAC1). Идентично обрабатываемый образец дрожжей с Fab 29G4v10 в рабочем порядке применяли для осуществления гейтирования конкретно клонов дрожжей, демонстрирующих улучшенное связывание относительно исходного Fab. Нормализованное значение соотношения связывание/дисплей для каждого клона рассчитывали путем деления медианного значения флуоресцентного сигнала связывания для каждого клона на медианное значение флуоресцентного сигнала дисплея для каждого клона. К раунду 3 накопление молекул, характеризующихся улучшенным связыванием, происходило в библиотеке mutHC, но не в библиотеке mutLC. В результате укороченного скрининга ~200 клонов дрожжей из пула раунда 3 mutHC получали 11 молекул, характеризующихся умеренно улучшенным связыванием (таблица 8). Варианты с улучшенной аффинностью получали рекомбинантным путем и оценивали в отношении функциональной активности in vitro, как описано в примере 3. По счастливой случайности, сайт изомеризации аспартата в пределах исходного Fab 29G4v10, являющийся элементом, потенциально отрицательно влияющим на технологичность, был удален во всех этих 11 уникальных мутантных вариантах.
сигнал дисплея (B/D)
D54K;
G56S
D54S;
G56S
G56A
T104S
F32Y
D54K;
G56A
F32Y
G56A
F32Y
D54Y;
G56S
D54F
F32Y
D54M;
G56T
D54R;
G56T
T104S
Для получения дополнительных улучшений в отношении аффинности также конструировали библиотеку молекул с перетасованными цепями, в которой обеспечивали объединение мутаций, накопленных в отсортированных пулах mutHC и mutLC. Для обеспечения улучшения в отношении проведения различия между основными связывающими клонами реализовали стратегию отбора и скрининга, основывающуюся на скорости диссоциации при связывании (фигура 5). Сначала клетки дрожжей, экспонирующие мутантные варианты Fab, насыщали биотинилированным ECD PAC1 человека и тщательно промывали, после чего подвергали продолжительной инкубации в буфере, содержащем большой избыток немеченого ECD PAC1 человека (не более 24 часов). Инкубация с немеченым ECD PAC1 человека, которая происходила при температурах, находящихся в диапазоне от 25°C до 37°C, обеспечивала необратимость событий диссоциации от клеток. Клетки, сохраняющие наибольший уровень связывания с биотинилированным PAC1, тем самым демонстрирующие наименьшую скорость диссоциации, выделяли с помощью FACS после окрашивания с помощью флюоресцирующего конъюгата стрептавидина (фигура 5). Клоны дрожжей, экспонирующие Fab, демонстрирующие наименьшие скорости диссоциации, характеризовались наивысшим уровнем флуоресценции.
При основывающейся на скорости диссоциации сортировке библиотеки молекул с перетасованными цепями идентично обрабатываемый образец дрожжей с Fab 29G4v10 использовали, чтобы конкретно выделять клетки с большим уровнем связывания биотинилированного PAC1 после конкуренции с немеченым PAC1 в течение ночи. Из двух пулов, собранных при разных значениях жесткости гейтирования, ~600 отдельных клонов дрожжей подвергали скринингу с применением анализа скорости диссоциации при связывании и идентифицировали более 190 клонов с более высоким уровнем остающегося связывания с PAC1, чем в случае исходного Fab 29G4v10. Специфичность связывания этих перспективных клонов оценивали путем подтверждения того, что отсутствует связывание клонов с ECD неродственных рецепторов (белка запрограммированной гибели клеток 1 (PD1) и рецептора желудочного ингибиторного полипептида (GIPR)). Также во время скрининга удаляли мутантные варианты, содержащие дополнительные сайты с аномалиями цистеина, сайты N-связанного гликозилирования, изомеризации аспартата, дезамидирования аспарагина и окисления триптофана относительно первоначальной последовательности 29G4v10. Основные 30 мутантных вариантов с изменениями в отношении связывания показаны в таблице 9 с изменениями последовательности в CDR от исходного антитела 29G4v10 и уровнем связывания ECD PAC1 человека в процентах после анализа скорости диссоциации. Более высокий уровень ассоциации в процентах для связывания ECD PAC1 указывает на то, что мутантные варианты Fab характеризуются меньшей скоростью диссоциации.
F32Y
G56S
R31Q
F32Y
D54F;
G56S
R31Q
F32Y
G56S
R31L;
S32A
F32Y
D54Y;
G56T
R31Q
D54S;
G56A
T104S
R31Q
R31W
R31W
R31L;
S32A
F32Y
T104S
G30W;
R31K
F32Y
D54R;
G56S
S28E;
G30M;
R31S;
S32L;
L33H;
ΔH341
S28A;
G30S;
R31N;
S32L;
L33H;
ΔH341
F32Y
D54M;
G56T
S28A;
G30W;
R31H;
S32N
F32Y
G56S
R31Q
F32Y
G56A
R31W
F32Y
D54R;
G56S
R31Q
F32Y
G56A
R31Q
F32Y
D54S;
G56S
L103M
R31W
G56A
T104S
S28A;
R31F
R31W
F32Y
D54S
T104S
R31M
F32Y
T104S
S28A;
G30F;
R31G;
S32N
D54S;
G56A
T104S
S28A;
G30W;
R31H;
S32N
F32Y
D54R;
G56S
S28A;
G30Y;
R31H;
S32N
D54H;
G56T
L103M;
T104S
R31W
S28A;
G30F;
R31G;
S32N
G56S
S28A;
G30W;
R31H;
S32N
D54Q;
G56S
R31W
D54R;
G56T
R31Q
F32Y
G56A
R31Q
F32Y
D54H
R31Q
Для достижения дополнительных улучшений в отношении аффинности разрабатывали совокупность библиотек второго поколения. В случае библиотек первого поколения, описанных выше, для обеспечения диверсификации фокусировались на подсовокупности положений CDR исходного антитела 29G4v10, которые находились в пределах 4,5Å от ECD PAC1 в кристаллической структуре (пример 1). В случае библиотек второго поколения изучали мутагенез в областях, которые по большей части не были затронуты в библиотеках первого поколения, таких как CDR2 и CDR3 легкой цепи. В случае нескольких положений, уже изученных на библиотеках первого поколения, для обеспечения диверсификации в пределах ограниченной совокупности нейтральных/полезных мутаций, которые возникли после трех раундов сортировки, применяли данные по тенденциям секвенирования. Наконец, некоторые утопленные остатки каркаса, которые могут влиять на конформации CDR, являлись мишенью ограниченной диверсификации, при этом в стратегии внесения мутаций предпочтение отдавали аминокислотам, часто встречающимся в том же положении в других зародышевых линиях VH3 и VK3 человека.
Всего четыре библиотеки Fab второго поколения были разработаны таким образом, чтобы мишенями диверсификации являлись 24 остатка CDR тяжелой цепи (аминокислоты 27, 29, 31-34, 49, 52-57, 70, 72, 77, 79, 98, 100-104 и 106 из SEQ ID NO: 191) и 17 остатков CDR легкой цепи (аминокислоты 27, 28, 30-32, 34, 46, 49-54, 92-94 и 96 из SEQ ID NO: 52). Ограниченное разнообразие по многим положениям, являющимся мишенями, позволяло принимать к рассмотрению большее количество положений CDR в пределах одной библиотеки и осуществлять совместную оптимизацию HCDR1 с HCDR3 и LCDR1 с LCDR3. Четыре библиотеки, которые были сконструированы, обеспечивали 8-24-кратное перекрытие теоретических значений разнообразия, находящихся в диапазоне от 7×106 до 1×107.
В четырех сконструированных библиотеках Fab обеспечивали накопление в отношении связывания с ECD PAC1 человека с применением FACS, как описано выше. К раунду 3 только в случае библиотек mutHCDR2 и mutLCDR1-LCDR3 получали пулы с уровнем связывания, эквивалентным таковому исходного антитела 29G4v10 или превосходящим его. Следовательно, эти два пула переносили на следующий этап для получения библиотеки молекул с перетасованными цепями mutH2/mutL1L3 для обеспечения дополнительных улучшений в отношении аффинности. Для обеспечения накопления мутантных вариантов, характеризующихся наивысшей аффинностью, применяли 2B10, наиболее эффективный клон дрожжей из библиотек первого поколения (таблица 9), для установления более жестких параметров гейтирования при сортировании. После конкуренции на основе скорости диссоциации в течение ночи с немеченым PAC1 при 30°C или 37°C, можно было отсортировать клоны дрожжей со значительно сниженными значениями скорости диссоциации по сравнению с клоном 2B10. В результате ограниченного скрининга ~200 клонов выявили, что ~100 клонов характеризовались меньшими скоростями диссоциации, чем 2B10, однако большинство из перспективных связывающих молекул содержали в пределах HCDR2 элемент, обуславливающий негативную предрасположенность к потенциальному дезамидированию аспарагина. Неспецифическое связывание с ECD PD1 или GIPR было минимальным для всех клонов, подвергнутых скринингу. В результате применения жестких фильтров по связыванию и последовательности получили десять основных мутантных вариантов со значительно улучшенным уровнем связывания PAC1 по сравнению с таковым клона 2B10 (уровень ассоциации в процентах после конкуренции при 37°C в течение ночи был >2 раза выше) и без каких-либо элементов последовательности, потенциально обуславливающих негативную предрасположенность (таблица 10). Изменения последовательности от исходного антитела 29G4v10 для этих основных десяти мутантных вариантов и уровень связывания ECD PAC1 человека в процентах после анализа скорости диссоциации при 30°C и 37°C показаны в таблице 10. Уровень ассоциации в процентах рассчитывали в виде нормализованного значения уровня связывания после анализа скорости диссоциации, деленного на нормализованное значение связывания без анализа скорости диссоциации. Более высокий уровень ассоциации в процентах для связывания ECD PAC1 указывает на то, что мутантные варианты Fab характеризуются меньшей скоростью диссоциации.
G56A;
N57F
R31W
D54R;
G56H;
N57G
R31W
D54T;
N57A
S28A;
R31W
D54F;
G56D;
N57A
R31W
Y53F;
D54Q;
G56T;
N57T
R31W
G56D;
N57L
R31W
D54S;
N57S
R31W
G56A;
N57S
R31W
G56A;
N57Q
R31W
G56Q;
N57F
R31Y
F32Y
G56S
R31L;
S32A
Как показано в таблице 10, все из этих главных десяти мутантных вариантов с изменениями в отношении связывания содержали мутацию Q27K в CDRL1, и все кроме одного содержали мутацию R31W в CDRL1. Все мутантные варианты содержали мутации по аминокислотным положениям D54 и N57 в CDRH2, и большинство также содержали мутацию по аминокислотному положению G56 в CDRH2. Консервативные мутации по аминокислотным положениям 49 и 70 в каркасной области 2 (FR2) и каркасной области 3 (FR3) тяжелой цепи соответственно также наблюдали во многих из этих десяти мутантных вариантов.
В конечном итоге на основе структуры ECD PAC1 в комплексе с Fab 29G4v9, близкородственном варианте нейтрализующих антител к PAC1 29G4v22 и 29G4v10, комбинаторные библиотеки мутантных вариантов 29G4v10 были разработаны и отсортированы в отношении улучшенного связывания с ECD PAC1. Чтобы выделить молекулы, характеризующиеся наиболее улучшенным связыванием, накопленные мутации объединяли путем конструирования библиотек молекул с перетасованными CDR и/или молекул с перетасованными цепями для осуществления отбора при более жестких условиях. В результате скринингов отдельных клонов дрожжей с Fab после сортировки получали молекулы, характеризующиеся улучшенным связыванием с ECD PAC1 человека, со значительно меньшими скоростями диссоциации при связывании по сравнению с таковыми для исходного антитела 29G4v10 и минимальным уровнем неспецифического связывания. Подсовокупность вариантов с улучшенной аффинностью получали рекомбинантным путем и оценивали в отношении функциональной активности in vitro, как описано в примере 3.
Пример 3. Функциональная активность in vitro вариантов антитела к PAC1 человека
Чтобы оценить эффект мутаций в вариабельных областях тяжелой и легкой цепи, идентифицированных с помощью анализа структуры сокристалла (пример 1) или библиотек на основе дрожжевого дисплея (пример 2), в отношении ингибирующей удельной активности антитела к PAC1, варианты получали с помощью способов рекомбинантной экспрессии в виде полных двухвалентных моноклональных антител и/или в виде одновалентных продуктов слияния Fab-Fc (например, Fab-области, слитой с димерной Fc-областью IgG) и оценивали в клеточном анализе cAMP, как описано более подробно ниже. Легкие цепи моноклональных антител и Fab-фрагментов содержали вариабельную область легкой цепи из указанного варианта антитела, слитую с константной областью легкой цепи каппа человека, содержащей последовательность под SEQ ID NO: 318 или SEQ ID NO: 319. Тяжелые цепи моноклональных антител и продуктов слияния Fab-Fc содержали вариабельную область тяжелой цепи из указанного варианта антитела, слитую с агликозилированной стабилизированной дисульфидными связями константной областью IgG1z человека, содержащей последовательность под SEQ ID NO: 325.
Последовательности вариантов антитела к PAC1 получали с помощью сайт-направленного мутагенеза (SDM) или с помощью сборки Golden Gate (GGA) в тех случаях, где SDM не приносил ожидаемого результата. При сайт-направленном мутагенезе использовали парные праймеры для мутагенеза, которые фланкировали сайт мутации. Реакции для осуществления ПЦР полного вектора проводили с применением матриц, представляющих собой двухнитевую плазмидную ДНК. Праймеры для всех требующихся в конкретном клоне мутаций объединяли в мастер-микс праймеров, при этом за отдельную реакцию происходило включение от одной до нескольких мутаций. После амплификации матричную плазмидную ДНК удаляли путем расщепления с помощью DpnI, эндонуклеазы, которая предпочтительно вносит разрывы в метилированную ДНК. Затем продуктом SDM трансформировали компетентные клетки для роста и скрининга с помощью секвенирования. Реакции для осуществления SDM проводили с применением набора QuikChange Lightning Multi Site-Directed Mutagenesis Kit (Agilent), следуя указаниям производителя.
Если SDM не приносил ожидаемого результата, использовали альтернативную стратегию клонирования. Вкратце, при GGA для разрезания и бесшовного лигирования друг с другом нескольких фрагментов ДНК использовали ферменты рестрикции II типа и ДНК-лигазу T4 (Engler et al., PLOS One, Vol. 3(11): e3647, 2008). В этом примере в состав нескольких фрагментов ДНК входили (i) синтетическая последовательность нуклеиновой кислоты (gBlock, Integrated DNA Technologies, Коралвилл, Айова), кодирующая консенсусную последовательность Kozak, последовательность сигнального пептида и последовательность вариабельной области антитела; (ii) фрагмент константного домена антитела, высвобожденный из вектора для экспрессии частей целевой молекулы; и (iii) остов вектора экспрессии. Реакционные смеси для GGA состояли из 10 нг gBlock, 10 нг вектора для экспрессии части целевой молекулы, 10 нг вектора экспрессии, 1 мкл 10x реакционного буфера Fast Digest+0,5 мМ АТФ (Thermo Fisher, Уолтем, Массачусетс), 0,5 мкл Esp3I FastDigest (Thermo Fisher, Уолтем, Массачусетс), 1 мкл ДНК-лигазы T4 (5 МЕ/мкл, Thermo Fisher, Уолтем, Массачусетс) и воды для доведения до 10 мкл. Реакции проводили в ходе 15 циклов, состоящих из стадии расщепления при 37°C продолжительностью 2 минуты и стадии лигирования при 16°C продолжительностью 3 минуты. После 15 циклов следовала конечная стадия расщепления при 37°C продолжительностью 5 минут и стадия инактивации фермента при 80°C продолжительностью 5 минут.
После клонирования полипептиды антитела к PAC1, из которых первые 22 аминокислоты представляли собой сигнальный пептид VK1 (MDMRVPAQLLGLLLLWLRGARC; SEQ ID NO: 486), получали с помощью транзиентной трансфекции клеток HEK 293 соответствующими cDNA. Клетки при концентрации 1,5×106 клеток/мл трансфицировали с помощью 0,5 мг/л ДНК (0,5 мг/л PAC1 в векторе pTT5) или (0,1 мг/л PAC1 в векторе pTT5 с 0,4 мг/л пустого вектора pTT5) (Durocher et al., NRCC, Nucleic Acids. Res., Vol.30: e9, 2002) с 4 мл PEI/мг ДНК в среде F17 (Thermo Fisher). Через 1 час после трансфекции к культурам добавляли дрожжевой экстракт и глюкозу, и затем клетки выращивали в суспензии с применением среды для экспрессии F17 с добавлением 0,1% коллифора, 6 мМ L-глутамина и 50 мкг/мл генетицина в течение 6 дней, после чего кондиционированные среды собирали для очистки.
Варианты антитела к PAC1 очищали от кондиционированных сред с применением аффинной хроматографии с белком A (MabSelect SuRe, GE Healthcare Life Sciences, Литл Чалфонт, Бакингемшир, Великобритания) с последующей катионообменной хроматографией (колонки SP Sepharose High Performance (SP HP) (GE Healthcare Life Sciences). Концентрацию белка каждого очищенного пула определяли по поглощению в УФ-диапазоне при 280 нм (A280) с применением NanoDrop 2000 (Thermo Fisher Scientific, Рокфорд, Иллинойс, США). Очищенные пулы диализировали против 2 л 10 мМ ацетата натрия, 9% сахарозы, pH 5,2 (A52Su) с применением колб для диализа Slide-A-Lyzer с MWCO 20 кДа (Thermo Fisher Scientific) в течение 2 часов при 4°C с осторожным перемешиванием на плите магнитной мешалки. Использованный диализат декантировали, добавляли 2 л свежего A52Su, и диализ продолжали в течение ночи. После диализа образцы концентрировали с применением ультрафильтрационных концентраторов с MWCO 30 кДа (Thermo Fisher Scientific), которые концентрировали при 2000 x g посредством бакет-ротора, пока концентрация каждого образца не составляла примерно 40 мг/мл исходя из измерения A280. Конечные продукты анализировали в отношении чистоты главного пика с применением микрокапиллярного электрофореза посредством системы Caliper LabChip GXII (PerkinElmer, Уолтем, Массачусетс, США) и эксклюзионной хроматографии с применением колонки ACQUITY UPLC Protein BEH SEC, 200 Å, 4,6×300 мм (Waters Corporation, Милфорд, Массачусетс, США). Содержание эндотоксинов измеряли с применением Endosafe-MCS (Charles River, Уилмингтон, Массачусетс, США) и картриджей для PTS 0,05 ЕЭ/мл (Charles River).
Функциональную активность очищенных моноклональных антител или слитых белков Fab-Fc оценивали с применением клеточного анализа активности рецептора PAC1 на основе содержания cAMP. Как PACAP38, так и PACAP27 являются агонистами рецептора PAC1, активация которого приводит в результате к повышению внутриклеточного содержания cAMP. В анализе использовали линию клеток, происходящую из нейробластомы человека (SH-SY5Y; Biedler JL et al., Cancer Res., Vol. 38: 3751-3757, 1978), полученную из ATCC (номер по ATCC CRL-2266; "клетки CRL-2266"). Клетки CRL-2266 экспрессируют рецептор PAC1 человека эндогенно (Monaghan et al., J Neurochem., Vol. 104(1): 74-88, 2008). Кроме того, линию клеток CHO, стабильно экспрессирующую рецептор PAC1 крысы (№ доступа в GenBank NM_133511.2) или рецептор PAC1 яванского макака (эталонная последовательность в NCBI XP_015303041.1), применяли вместо клеток CRL2266 для анализов, чтобы оценить наличие межвидовой перекрестной реактивности у антител или продуктов слияния Fab-Fc к PAC1 по рецепторам PAC1 крысы и яванского макака. Для измерения содержания cAMP применяли набор для анализа cAMP LANCE Ultra (PerkinElmer, Бостон, Массачусетс).
В день анализа замороженные клетки CRL-2266 размораживали при 37°C и один раз промывали с помощью буфера для анализа. 10 мкл клеточной суспензии, содержащей 2000 клеток, добавляли в 96-луночные белые микропланшеты с половинным объемом лунок. После добавления 5 мкл варианта моноклонального антитела или слитого белка Fab-Fc к PAC1 (кривая зависимости "доза-эффект", построенная по 10 точкам: концентрация находится в диапазоне от 1 мкМ до 0,5 фМ) смесь инкубировали в течение 30 мин. при комнатной температуре. Затем в качестве агониста добавляли 5 мкл PACAP38 человека (конечная концентрация 10 пМ), и смесь дополнительно инкубировали в течение 15 мин. при комнатной температуре. После стимуляции с помощью PACAP38 человека добавляли 20 мкл смеси для выявления и инкубировали в течение 45 минут при комнатной температуре. Планшеты считывали на приборе EnVision (PerkinElmer, Бостон, Массачусетс) при длине волны излучения 665 нм. Данные обрабатывали и анализировали с помощью Prizm (GraphPad Software Inc.) с отображением POC (процент контроля, где контроль определен как активность агониста, применяемого в анализе) в виде функции от испытуемой концентрации антагониста (например, варианта антитела или слитого белка Fab-Fc к PAC1) и аппроксимировали с помощью стандартных кривых нелинейной регрессии с получением значений IC50. POC рассчитывали следующим образом:
 Одновалентные слитые белки Fab-Fc получали с помощью мутаций, кратко описанных в таблицах 6 и 7, и испытывали в отношении функциональной активности в клеточном анализе cAMP, описанном выше. Мутантные варианты слитых белков Fab-Fc разделяли на две отдельные группы: мутации, которые обеспечивают улучшение ингибирующей удельной активности в отношении рецептора PAC1 человека по сравнению с исходной молекулой, и мутации, определенные как нейтральные (таблица 11). Мутации определяли как нейтральные, если мутация характеризовалась средней удельной активностью, являющейся менее чем в 1,5 раза слабее, чем таковая для исходной молекулы, и по меньшей мере одним значением удельной активности, являющимся выше, чем у исходной молекулы в одном и том же цикле.
Одновалентные слитые белки Fab-Fc получали с помощью мутаций, кратко описанных в таблицах 6 и 7, и испытывали в отношении функциональной активности в клеточном анализе cAMP, описанном выше. Мутантные варианты слитых белков Fab-Fc разделяли на две отдельные группы: мутации, которые обеспечивают улучшение ингибирующей удельной активности в отношении рецептора PAC1 человека по сравнению с исходной молекулой, и мутации, определенные как нейтральные (таблица 11). Мутации определяли как нейтральные, если мутация характеризовалась средней удельной активностью, являющейся менее чем в 1,5 раза слабее, чем таковая для исходной молекулы, и по меньшей мере одним значением удельной активности, являющимся выше, чем у исходной молекулы в одном и том же цикле.
Мутации, которые обеспечивали повышение ингибирующей удельной активности в отношении рецептора PAC1 человека, лежали в трех различных областях трехмерного пространства на поверхности антитела. Второй раунд вариантов антитела и слитых белков Fab-Fc получали путем объединения мутаций в этих трех областях для потенциального обеспечения дополнительного повышения удельной активности. Кроме того, включали некоторые из нейтральных мутаций, поскольку существовала вероятность, что они не будут оказывать существенного влияния на связывание, однако могут обеспечивать механизмы для модифицирования биофизических характеристик антитела. Вариабельные области, содержащие требующиеся мутации, включали в двухвалентное моноклональное антитело, содержащее агликозилированную Fc-область IgG1 человека и/или одновалентный слитый белок Fab-Fc. Fc-область агликозилированного IgG1-антитела человека содержала последовательность Fc-области IgG1z человека с мутациями N297G, R292C и V302C согласно нумерации EU (SEQ ID NO: 325). Варианты антитела и слитые белки Fab-Fc с комбинациями мутаций оценивали в отношении функциональной активности в клеточном анализе cAMP. Результаты показаны в таблице 12 ниже.
(двухвалентное связывание мишени)
(одновалентное связывание мишени)
(в планшете)1
(в планшете)1
(0,09)
(0,13)
(0,23)
(0,05)
(0,28)
(0,19)
(0,50)
(0,50)
(0,97)
(0,22)
(0,31)
(0,39)
(0,19)
G56R
(0,02)
(0,17)
G56R
(0,06)
(0,48)
G56R
(0,05)
(0,20)
G56R
(0,29)
(0,13)
G56N
(0,21)
(0,63)
G56N
(0,04)
(0,58)
G56N
(0,18)
(0,32)
G56N
(0,13)
(0,34)
E62R
(0,04)
(0,13)
E62R
(0,05)
(0,11)
E62R
(0,06)
(0,85)
E62R
(0,54)
(0,30)
(0,20)
(0,05)
(0,43)
(0,17)
(0,83)
(0,09)
(1,91)
E62R
(0,05)
(0,01)
E62R
(0,20)
(0,28)
(0,07)
(0,43)
(0,42)
(0,42)
(0,64)
(0,16)
G56R
(0,10)
(0,18)
G56R
(0,00)
(0,10)
G56R
(0,04)
(0,14)
G56R
(0,04)
(0,74)
G56N
(0,06)
(0,73)
G56N
(0,13)
(0,27)
G56N
(0,01)
(0,37)
G56N
(0,03)
(1,03)
E62R
(0,06)
E62R
(0,40)
E62R
(0,51)
E62R
(0,27)
(0,01)
(0,04)
(0,01)
(0,39)
(0,08)
(0,33)
(0,18)
(0,38)
G56R;
E62R
(0,61)
G56R;
E62R
(1,11)
G56R;
E62R
(0,20)
G56R;
E62R
(0,21)
G56N;
E62R
(0,00)
(0,27)
G56N;
E62R
(0,01)
(0,54)
G56N;
E62R
(0,01)
(1,42)
G56N;
E62R
(0,06)
(1,38)
G56R
(0,01)
(1,54)
G56R
(0,09)
(1,29)
G56R
(0,00)
(1,74)
G56R
(0,01)
(1,26)
G56N
(0,32)
G56N
(0,07)
(0,84)
G56N
(0,18)
(1,89)
G56N
(0,17)
(1,13)
E62R
(0,03)
(0,61)
E62R
(0,01)
(0,53)
G56R;
E62R
(0,41)
G56R;
E62R
(0,31)
G56R;
E62R
(0,29)
G56R;
E62R
(2,17)
G56N;
E62R
(0,53)
G56N;
E62R
(0,00)
G56N;
E62R
(0,01)
G56N;
E62R
(0,04)
G56R
(0,04)
G56R
(0,01)
G56R
(0,04)
G56R
(0,14)
G56N
G56N
(0,07)
G56N
(0,29)
G56N
(0,05)
G56R;
E62R
(0,00)
G56R;
E62R
(0,01)
G56R;
E62R
(0,04)
G56R;
E62R
G56N;
E62R
G56N;
E62R
G56N;
E62R
G56N;
E62R
G56N;
E62R
(0,37)
G30W
S32N
(0,09)
(0,56)
D54N
G56A
G56R
E62R
E62R
(0,36)
(0,13)
2Это значение представляет собой кратное увеличение IC50 по сравнению со средним значением IC50 для исходной молекулы 29G4v10, представленной в формате либо mAb, либо слитого белка Fab-Fc.
ND=не определено
Наибольшие улучшения удельной активности наблюдали, когда антитела содержали мутацию в положении Q27 (Q27K) в вариабельной области легкой цепи и мутацию в положении D54 (D54I, D54Q или D54N) и/или положении G56 (G56R или G56N) в вариабельной области тяжелой цепи. Q27 в вариабельной области легкой цепи под SEQ ID NO: 52 соответствует аминокислотному положению 29 в нумерации согласно AHo. D54 и G56 в вариабельной области тяжелой цепи под SEQ ID NO: 191 соответствуют аминокислотным положениям 61 и 66 в нумерации согласно AHo соответственно. Основная аминокислота, такая как лизин или аргинин, в положении Q27 в вариабельной области легкой цепи предположительно обеспечивает улучшенную обусловленную зарядами комплементарность с кислотными аминокислотами Glu120 и Asp121 в ECD PAC1 (см. зону 1 на фигуре 2A). Гидрофобные остатки (например, изолейцин) или нейтральные гидрофильные остатки (например, глутамин или аспарагин) в положении D54 в вариабельной области тяжелой цепи и основные остатки (например, аргинин) или нейтральные гидрофильные остатки (например, аспарагин) в положении G56 в вариабельной области тяжелой цепи по всей видимости обеспечивают улучшение в отношении гидрофобного взаимодействия или связывания посредством атомов водорода с аминокислотными остатками Asn60 и Ile61 в ECD PAC1 (см. зону 6 на фигуре 3B).
Основные одиннадцать мутантных вариантов из скрининга библиотеки MutHC (таблица 8) были представлены в формате агликозилированных моноклональных антител IgG1 и одновалентных молекул Fab-Fc, как описано выше, для продуцирования и испытания функциональности в анализе содержания cAMP (таблица 13). Умеренно улучшенное связывание на дрожжах в случае этой совокупности вариантов привело к улучшению функции блокирования рецептора PAC1 в случае четырех из 11 mAb и семи из 11 Fab-Fc. Тем не менее, в виде антитела мутантный вариант iPS:421873 показал приблизительно 4-кратное улучшение функции блокирования PAC1 по сравнению с исходным антителом 29G4v10. Градации активности мутантных вариантов по большей части согласовались между двумя форматами, при этом для Fab-Fc была последовательно показана более слабая функция (более высокие значения IC50), чем в случае mAb, предположительно из-за потери авидности.
Таблица 13. Ингибирующая удельная активность in vitro вариантов из скрининга библиотеки MutHC
(двухвалентное связывание мишени)
(одновалентное связывание мишени)
(в планшете)1
(в планшете)1
F32Y
D54K
G56S
D54S;
G56S
G56A
T104S
F32Y
D54K;
G56A
F32Y
G56A
F32Y
D54Y;
G56S
D54F
F32Y
D54M;
G56T
D54R;
G56T
T104S
1Это значение представляет собой кратное увеличение IC50 по сравнению со значением IC50 для исходной молекулы 29G4v10, представленной в формате либо mAb, либо слитого белка Fab-Fc, которую анализировали параллельно с каждым вариантом молекулы.
2Это значение представляет собой кратное увеличение IC50 по сравнению со средним значением IC50 для исходной молекулы 29G4v10, представленной в формате либо mAb, либо слитого белка Fab-Fc.
Подсовокупность мутантных вариантов 29G4v10 из библиотек молекул с перетасованными цепями (таблицы 9 и 10) преобразовывали в агликозилированные моноклональные антитела IgG1 и испытывали в отношении функциональной активности в клеточном анализе cAMP. Результаты показаны в таблице 14 ниже.
Таблица 14. Ингибирующая удельная активность in vitro вариантов из библиотек молекул с перетасованными цепями на основе дрожжевого дисплея
129G4v22 представляет собой еще одно нейтрализующее антитело к PAC1, которое характеризуется значительным структурным сходством с 29G4v10. Последовательности VL и VH для 29G4v22 представлены под SEQ ID NO: 53 и 192 соответственно.
2Ранее зафиксированное значение, полученное из предыдущего анализа и включенное для целей сравнения.
При представлении в формате моноклонального антитела (mAb) мутантные варианты 29G4v10 из библиотеки молекул с перетасованными цепями второго поколения (таблица 10) являлись более сильными антагонистами PAC1, чем мутантные варианты из библиотеки молекул с перетасованными цепями первого поколения (таблица 9), свидетельствуя о том, что усиленная функция блокирования PAC1 коррелирует с улучшенным связыванием и меньшими скоростями диссоциации при связывании, в частности. Однако, неожиданно, большинство мутантных вариантов в виде mAb являлись менее активными, чем антитело iPS:421873, мутантный вариант с наибольшей удельной активностью из скрининга библиотеки MutHC (таблица 13), несмотря на их предположительно усиленное связывание с ECD PAC1 человека. Тем не менее, мутантный вариант 30_D05 демонстрировал приблизительно 10-кратное и 67-кратное улучшение функции по сравнению с исходным антителом 29G4v10 и контрольным антителом 29G4v22 соответственно. Одним общим признаком, присущим mAb с высокой удельной активностью, S8_30_D05 и iPS:421873, является мутация D54R в пределах HCDR2, которая отсутствует во всех остальных мутантных вариантах mAb с меньшей удельной активностью. Интересно, что все варианты с созревшей аффинностью демонстрировали перекрестную реактивность с рецептором PAC1 крысы, в отличие от исходного антитела 29G4v10 и контрольного антитела 29G4v22 (таблица 14).
Пример 4. Функциональная активность in vivo вариантов антитела к PAC1 человека
Максадилан является пептидом с вазодилатирующим действием и агонистом рецептора PAC1. При внутрикожном введении максадилан вызывает локальное повышение притока крови к коже, величину которого можно измерить с помощью лазерной доплеровской визуализации. Подавление этого эффекта с помощью антагонистов PAC1 (например, антител к PAC1) может служить в качестве трансляционной фармакодинамической модели антагонизма в отношении биологической активности PAC1. Подсовокупность вариантов антител к PAC1, которые демонстрировали улучшенную ингибирующую удельную активность in vitro при представлении в формате моноклональных антител из примера 3, испытывали в отношении эффективности ингибирования активации рецептора PAC1 in vivo путем применения модели притока крови к коже у крысы.
Фармакодинамическая модель in vivo
Не использовавшиеся ранее в опытах самцы крыс Sprague Dawley в возрасте 8-12 недель на момент исследования приобретали у Charles River Laboratories. Все процедуры в данном примере проводили в соответствии с Законом о благополучии животных, Руководством по содержанию и использованию лабораторных животных и требованиями Управления по обеспечению благосостояния лабораторных животных. Животных содержали группой в нестерильной проветриваемой микроизоляторной системе для содержания в помещении, аккредитованном Ассоциацией по аттестации и аккредитации содержания лабораторных животных Amgen (AAALAC). Животные имели свободный доступ к гранулированному корму (Harlan Teklad 2020X, Индианаполис, Индиана) и воде (полученной на месте с помощью обратного осмоса) через автоматическую поилку.
Подсовокупность антител к PAC1 испытывали на фармакодинамической (PD) модели индуцированного посредством максадилана увеличения притока крови к коже (MIIBF) у крысы с помощью лазерной доплеровской визуализации. Свежий раствор максадилана для введения в виде доз (Bachem, H6734.0500) получали ежедневно путем разбавления исходного раствора максадилана (0,5 мг/мл) в 1X фосфатно-солевом буферном растворе (PBS) до конечной концентрации 0,5 мкг/мл. Все антитела к PAC1 (Ab) получали в растворе с 10 мМ ацетата натрия, 9% сахарозы, pH 5,2 (A52Su), при разных концентрациях, в зависимости от дозы, требующейся для эксперимента, и вводили посредством одной болюсной i.v.-инъекции за один день до измерения притока крови к коже (DBF) с помощью лазерной доплеровской визуализации.
Устройство для лазерной доплеровской визуализации (LDI-2, Moor Instruments, Ltd, Уилмингтон, Делавэр) применяли для измерения DBF на выбритой части кожи живота мыши с применением лазерного луча малой мощности с длиной волны 633 нм, генерируемого в трубке гелий-неонового газового лазера. Разрешающая способность измерительного прибора составляла 0,2-2 мм, при этом расстояние сканирования между отверстием прибора и поверхностью ткани составляло 30 см. DBF измеряли и выражали в виде либо % изменения от исходного уровня [100 x (отдельное значение плотности потока после воздействия максадиланом - отдельное значение плотности потока на исходном уровне)/отдельное значение плотности потока на исходном уровне], либо % ингибирования DBF [среднее значение % изменения от BL для среды-носителя - отдельное значение % изменения от BL у крысы, обработанной антителом)/среднее значение % изменения от BL для среды-носителя] для количественного определения амплитуды обусловленного антителами эффекта.
В день испытания после осуществления наркоза с помощью пропофола брюшную область крысы выбривали и каждое животное помещали в положение лежа на спине на подушку с циркулирующей теплой водой, выполненную с возможностью контроля температуры, для поддержания стабильной температуры тела во время исследования. После стабилизационного периода продолжительностью 10-15 минут на живот крысы помещали резиновое О-образное кольцо (внутренний диаметр 0,925 см, O-Rings West, Сиэтл, Вашингтон) (не размещая его непосредственно на видимом кровеносном сосуде). После помещения О-образного кольца на выбранной области осуществляли измерение исходного уровня (BL) DBF. После сканирования для определения BL вводили максадилан, представляющий собой агонист PAC1 (20 мкл раствора с концентрацией 0,5 мкг/мл), путем внутрикожной инъекции в центр О-образного кольца. DBF измеряли через 30 мин. после инъекции максадилана или через 24 ± 1,5 часа после обработки с помощью антитела. С помощью O-кольца определяют область, представляющую интерес, в которой анализировали DBF.
Все результаты измерения DBF выражали в виде среднего значения ± SEM. Для оценки статистической значимости эффектов Ab к PAC1 относительно таковых при обработке средой-носителем применяли однофакторный ANOVA с последующим применением критерия множественных сравнений Даннетта (MCT). Для определения значимости между двумя группами применяли p-значение, составляющее < 0,05.
Скрининговая оценка с введением одной внутривенной (i.v.) дозы
Крыс предварительно обрабатывали одним из 7 разных антител к PAC1 (420653, 420845, 420943, 421873, 420889 (PL-50347), 421091 (PL-50350) и 421051 (PL-50351)) за 24 часа до стимуляции максадиланом (20 мкл раствора с концентрацией 0,5 мкг/мл) при дозе 0,1 мг/кг или 0,3 мг/кг, что приводило в результате к снижению значения MIIBF по сравнению с таковым в случае группы, обрабатываемой средой-носителем (A52Su). В момент времени 30 мин. после обработки максадиланом имело место статистически значимое подавление MIIBF при 0,3 мг/кг по сравнению с таковым в случае группы среды-носителя для пяти из семи испытуемых антител (фигура 6). Значения конечной концентрации в сыворотке крови в момент времени 24 ± 1,5 часа для шести из семи антител перечислены в таблице 15 ниже.
Таблица 15. Конечная концентрация в сыворотке крови в случае исследования (скринингового) с одной инъекцией
Оценка зависимости "доза-ответ"
Крыс предварительно обрабатывали с помощью 4 разных антител к PAC1 (420653, 420845, 420943 и 421873) за 24 часа до стимуляции максадиланом (20 мкл раствора с концентрацией 0,5 мкг/мл) при дозе, находящийся в диапазоне от 0,01 мг/кг до 30 мг/кг. Дозозависимое снижение MIIBF по сравнению с группой, обрабатываемой средой-носителем, наблюдали в случае каждого из четырех антител (фигуры 7A-7D). Ab 420653 оказывало значительный эффект при дозе всего 0,06 мг/кг со значениями ингибирующего эффекта, составляющими 44%, 68%, 86%, 95% и 101% при 0,06, 0,1, 0,3, 1 и 3 мг/кг соответственно (фигура 7A). Ab 420845 оказывало значительный эффект при 0,3, 1 и 3 мг/кг со значением ингибирования, составляющим 79% 102% и 107% соответственно (фигура 7B). Ab 420943 и 421873 характеризовались немного меньшей удельной активностью, чем Ab 420845 и Ab 420653, но все же оказывали значительный ингибирующий эффект при 1 мг/кг. % ингибирования DBF в случае Ab 420943 составлял 56%, 46%, 52% и 81% при 1, 3, 10, 30 мг/кг соответственно (фигура 7C). % ингибирования DBF в случае Ab 421873 составлял 34%, 55%, 72% и 100% при 1, 3, 10 и 30 мг/кг соответственно (фигура 7D).
Результаты экспериментов, описанных в данном примере, показывают, что антитела, которые эффективно ингибируют индуцированную лигандом активацию рецептора PAC1 in vitro, также ингибируют активацию рецептора PAC1 in vivo, согласно оценке с помощью анализа притока крови к коже, модели опосредованного PAC1 вазодилатирующего действия.
Пример 5. Фармакокинетические характеристики вариантов антитела к PAC1 человека
Предварительные фармакокинетические (PK) исследования четырех антител к PAC1 (420653, 420845, 420943 и 421873) проводили на не использовавшихся ранее в опытах самцах крыс Sprague Dawley и не использовавшихся ранее в опытах самцах яванских макак. Исходное антитело 29G4v10 или структурно родственное антитело 29G4v22 (VL, содержащая SEQ ID NO: 53, и VH, содержащая SEQ ID NO: 192) оценивали в качестве контроля. Испытуемые антитела вводили в виде доз исследуемым животным путем внутривенного болюсного введения. Образцы крови собирали в указанные моменты времени после введения дозы и обрабатывали с получением сыворотки крови. Все препараты сыворотки хранили при температуре примерно -70°C (±10°C) до передачи для следующего анализа.
Для измерения количества испытуемого антитела в образцах сыворотки крови от крыс и яванских макак после введения доз разрабатывали колориметрический твердофазный иммуноферментный анализ (ELISA) с применением мышиных моноклональных антител (mAb), направленных против Fc IgG человека (Amgen, Inc., Калифорния, США). Планшеты для микротитрования покрывали мышиным mAb к Fc человека при концентрации 2 мкг/мл. Покрытые планшеты для микротитрования блокировали с помощью I-block (Applied Biosystems, Калифорния, США). Стандартные образцы для количественного определения (STD) и образцы для контроля качества (QC) получали путем добавления известного количества испытуемого антитела в 100% сыворотку крови от изучаемых видов. STD, QC, холостой образец и образцы для исследования разбавляли 1:30 в буфере для анализа (1X PBS с 1 M NaCl, 1% BSA и 0,5% Tween 20). Разбавленные STD, QC, холостой образец и образцы для исследования инкубировали в покрытых планшетах для микротитрования в течение 1 ч. при 25°C без встряхивания. После стадии промывки в планшеты для микротитрования добавляли мышиное mAb к Fc человека, конъюгированное с пероксидазой хрена (HRP), при 30 нг/мл в буфере для анализа и инкубировали в течение 1 ч. при 25°C без встряхивания. После конечной стадии промывки в планшеты для микротитрования добавляли раствор тетраметилбензидина (TMB) в качестве субстрата для пероксидазы (KPL Inc., MD, США). В присутствии HRP TMB вызывал появление колориметрического сигнала, который был пропорциональным количеству связанного Fc человека, присутствующего в STD, QC и образцах для исследования. Продолжительность проявления цвета зависела от количества аналита, и его останавливали путем добавления 2 н. серной кислоты. Оптическую плотность (OD) измеряли при 450 нм по отношению к 650 нм с применением устройства для считывания микротитрационных планшетов SpectraMax 340PC (Molecular Devices, Калифорния, США) и программного обеспечения SoftMax Pro. Регрессионный анализ данных анализа осуществляли с применением логистической (авто-оценка) регрессионной модели c весовым коэффициентом 1/y. Динамический диапазон анализа составлял от 20 нг/мл до 2000 нг/мл.
При PK-исследованиях как на крысах, так и на яванских макаках проводили некомпартментный анализ по отдельным данным концентрации в сыворотке в номинальный момент времени с применением Phoenix® WinNonlin® (версия 6.4; Certara, Нью-Джерси, США). Отдельные значения концентрации менее нижнего предела количественного определения (LLOQ, 20 нг/мл) сообщали как "ниже предела количественного определения" (BQL) и принимали равными нулю для расчета сводной статистики. Средние значения концентрации менее LLOQ не сообщали и не наносили на график. Все значения концентрации менее LLOQ исключали из некомпартментного анализа. Для PK-анализа применяли номинальные значения дозы и номинальные значения времени отбора образцов. Оценивали следующие PK-параметры.
• Значение исходной концентрации (C0) после внутривенного введения оценивали с помощью экстраполяция назад к нулевому моменту времени с применением первых 2 наблюдаемых уменьшающихся значений концентрации.
• Площадь под кривой зависимости концентрации от времени в диапазоне от нулевого момента времени до бесконечности (AUCбескон.) рассчитывали с помощью линейного метода трапеций.
• Период полувыведения в конечной фазе (t1/2,z) рассчитывали как ln2/λz. λz представляет собой константу скорости первого порядка лекарственного средства, относящуюся к конечной части кривой.
• Системное выведение (CL) рассчитывали как CL=Доза/AUCбескон. после внутривенного введения.
• Объем распределения в равновесном состоянии (Vss) оценивали как Vss=CL x MRTбескон. (среднее время удержания от нулевого момента времени до бесконечности).
Сводные данные по ингибирующей удельной активности in vitro четырех антител, а также исходного антитела 29G4v10 и контрольного антитела 29G4v22 к PAC1 человека, PAC1 крысы и PAC1 яв. мак. представлены в таблице 16 ниже. Антитела оценивали в отношении способности ингибировать индуцированную лигандом (PACAP38 или максадиланом) активацию рецептора PAC1 из разных видов с применением анализа содержания cAMP, описанного в примере 3.
Таблица 16. Сводные данные по ингибирующей удельной активности in vitro для вариантов антитела к PAC1
(n=3)
(n=3)
(n=3)
(n=3)
(n=3)
(n=2)
Крысам внутривенно вводили одно из четырех антител при дозе 1 мг/кг, 5 мг/кг и 25 мг/кг. Образцы крови собирали в различные моменты времени после введения дозы, и концентрацию антитела измеряли в образцах сыворотки крови в каждый из моментов времени с применением анализа ELISA, описанного выше. Сводные данные по PK-параметрам для исследования на крысах представлены в таблице 17 ниже, и профили концентрации в сыворотке крови в зависимости от времени для каждой из доз показаны на фигурах 8A-8C.
Таблица 17. Сводные данные по PK-параметрам для вариантов антитела к PAC1 у крыс
кг)
(мкг/
мл)
мл)
(мл/
кг)
ч./
кг)
(ч.)
Яванским макакам внутривенно вводили одно из четырех антител при дозе 10 мг/кг. Образцы крови собирали в различные моменты времени после введения дозы, и концентрацию антитела измеряли в образцах сыворотки крови в каждый из моментов времени с применением анализа ELISA, описанного выше. Сводные данные по PK-параметрам для исследования на яванских макаках представлены в таблице 18 ниже, и профили концентрации в сыворотке крови в зависимости от времени показаны на фигуре 9. PK-профиль антитела 29G4v22 показан для сравнения.
Таблица 18. Сводные данные по PK-параметрам для вариантов антитела к PAC1 у яванских макаков
(мкг/
мл)
мл)
(мл/кг)
кг)
(ч.)
При этих дозах все четыре из вариантов антитела к PAC1 характеризовались более продолжительным временем полужизни в сыворотке крови, чем контрольное антитело 29G4v22. Интересно, что хотя все четыре варианта антитела к PAC1 демонстрировали большую ингибирующую удельную активность in vitro в отношении рецептора PAC1 по сравнению с контрольным антителом 29G4v22 (таблица 16), PK-профиль для некоторых из вариантов был менее предпочтительным у яванских макаков. Например, Ab 420845 характеризовалось более быстрой скоростью выведения и более низкой общей экспозицией по сравнению с контрольным антителом 29G4v22. Однако, Ab 420653 характеризовалось PK-профилем, сопоставимым с таковым в случае контрольного антитела 29G4v22 (фигура 9), свидетельствуя о том, что Ab 420653 можно вводить при более низкой дозе при той же частоте введения дозы с достижением аналогичного фармакологического эффекта.
Пример 6. Обеспечение созревания аффинности в дрожжевом дисплее антитела к PAC1 человека 19H8
Для получения дополнительных антител к PAC1 с улучшенной удельной ингибирующей активностью для антитела 19H8 (VH-область под SEQ ID NO: 296; VL-область под SEQ ID NO: 67) обеспечивали созревание аффинности с помощью FACS библиотеки Fab на основе дрожжевого дисплея с применением способов, описанных в примере 2. Антитело 19H8 структурно отличается от антител 29G4v9, 29G4v10 и 29G4v22, однако также демонстрирует очень сильную нейтрализующую активность в отношении рецептора PAC1 человека.
Поскольку изначально для комплекса ECD PAC1-Fab 19H8 не была доступна информация по кристаллической структуре, была получена модель с применением гомолога, чтобы идентифицировать остатки, находящиеся на поверхности, в пределах каждой петли CDR. Поскольку ожидается, что ECD PAC1 образует непосредственные контакты с остатками CDR, находящимися на поверхности, было высказано предположение, что изменение природы этих контактов или создание новых контактов посредством комплексного мутагенеза может привести к улучшению связывания антитела с ECD PAC1. Чтобы ограничить теоретические значения разнообразия каждой библиотеки до удобных в управлении 106-107, на один CDR идентифицировали не более пяти положений для осуществления насыщающего мутагенеза с применением MIX19 и на один CDR конструировали по одной отдельной библиотеке. Из-за опасений относительно правильности смоделированных конформаций петли CDRH3 большинство остатков, подверженных воздействию растворителя, в пределах данной петли не возможно идентифицировать с достаточной степенью точности. Поэтому была рассмотрена возможность осуществления диверсификации по восьми из 11 остатков CDR в пределах CDRH3, за исключением остатков в начале и в конце петли. Чтобы сократить число положений для осуществления насыщающего мутагенеза до пяти, исключали два остатка триптофана и один остаток фенилаланина в пределах CDRH3, поскольку ароматические остатки часто являются критически важными посредниками в белок-белковых взаимодействиях. Всего было разработано шесть библиотек Fab с отдельными CDR, чтобы мишенями диверсификации являлись 16 остатков CDR тяжелой цепи и 15 остатков CDR легкой цепи, и запас выборок сконструированных библиотек превышал теоретические значения разнообразия в 4,5-46 раз. Сводные данные по положениям, являющимся мишенями в случае каждой библиотеки CDR, представлены ниже.
Библиотеки CDR тяжелой цепи (аминокислотные положения указаны относительно SEQ ID NO: 296):
библиотека CDRH1: Asp27 (расположен рядом с CDRH1 в FR1), Ser31, Asn32, Ser33 и Thr35;
библиотека CDRH2: Tyr54, Tyr55, Ser57, Lys58, Ser60 и His62;
библиотека CDRH3: Thr103, Lys105, Gln106, Leu107 и Leu110.
Библиотеки CDR легкой цепи (аминокислотные положения указаны относительно SEQ ID NO: 67):
библиотека CDRL1: Ser28, Ser30, Arg31, Tyr32 и Asn34;
библиотека CDRL2: Tyr49 (расположен рядом с CDRL2 в FR2), Ala50, Ala51, Ser52 и Ser53;
библиотека CDRL3: Ser91, Tyr92, Ser93, Pro94 и Phe96.
В шести библиотеках Fab с отдельными CDR обеспечивали накопление в отношении связывания с ECD PAC1 человека с применением FACS, повышая жесткость с каждым последующим раундом путем снижения концентрации ECD PAC1, применяемой для связывания (раунд 1: 30 нМ ECD PAC1; раунд 2: 0,67 нМ ECD PAC1 и раунд 3: 0,2 нМ ECD PAC1). Для обеспечения дополнительных улучшений в отношении аффинности также были сконструированы две библиотеки Fab с перетасованными CDR, в которых обеспечили объединение мутаций, накопленных в библиотеках с отдельными CDR (одна для тяжелой цепи и одна для легкой цепи), и конечная библиотека молекул с перетасованными цепями, в которой обеспечили объединение мутаций, накопленных в каждой библиотеке молекул с перетасованными CDR. Библиотеки молекул с перетасованными CDR и молекул с перетасованными цепями подвергали процедурам отбора в отношении связывания ECD PAC1 при более жестких условиях с применением способа отбора на основе скорости диссоциации при связывании, описанного в примере 2 и изображенного на фигуре 5. Сортировка библиотеки молекул с перетасованными цепями, основывающаяся на конечной скорости диссоциации, предполагала, что большинство из клонов дрожжей в этом пуле были значительно улучшены в отношении связывания ECD PAC1 по сравнению с исходным антителом 19H8.
Примерно 800 отдельных клонов дрожжей подвергли скринингу в отношении улучшенного связывания с ECD PAC1 человека. Мутантные последовательности с элементами последовательности, обуславливающих негативную предрасположенность (например, сайты с аномалиями цистеина, сайты N-связанного гликозилирования, сайты изомеризации аспартата, дезамидирования аспарагина и окисления триптофана) удаляли. Основные ~200 уникальные связывающие молекулы были отобраны для использования во вторичном скрининге, где их ранжировали по скорости диссоциации при связывании и оценивали в отношении неспецифического связывания. Во вторичном скрининге >80% клонов характеризовались меньшими скоростями диссоциации при связывании, чем исходное антитело 19H8, что было измерено по более высокому уровню ассоциации в процентах биотинилированного ECD PAC1 человека после конкуренции в течение ночи с немеченым ECD PAC1 при 30°C. В измерениях представлены нижние пределы улучшения скорости диссоциации, поскольку биотинилированный ECD PAC1 полностью диссоциировал из исходного 19H8 уже после одного часа конкуренции при 30°C. Поскольку ни один из подвергнутых скринингу клонов не демонстрировал связывание с ECD неродственных рецепторов PD1 и GIPR, чтобы дополнительно уменьшить пул клонов, применяли дополнительные фильтры для последовательностей для удаления мутантных вариантов, CDR которых содержали сайты расщепления фурином, дополнительные остатки триптофана и большее количество ковариационных нарушений, чем исходное антитело 19H8. Для ранжирования клонов применяли дополнительный анализ связывания с применением биотинилированного ECD VPAC2 человека для определения степени неспецифического связывания с VPAC2 человека - рецептором, структурно родственным рецептору PAC1 человека. Основные 20 мутантных вариантов, демонстрирующие наиболее низкие скорости диссоциации при связывании с ECD PAC1 человека и наименьшую величину связывания VPAC2 человека среди клонов дрожжей, которые были включены во вторичный скрининг, перечислены в таблица 19 ниже. Более высокий уровень ассоциации в процентах для связывания ECD PAC1 указывает на то, что мутантные варианты Fab характеризуются меньшей скоростью диссоциации.
Таблица 19. Основные молекулы, характеризующиеся улучшенным связыванием, из библиотек на основе дрожжевого дисплея для исходного антитела 19H8
N32R;
S33L
S60K
L107D
A51G;
S52Q; S53R
Y92I;
S93G;
P94M; F96Y
N32R;
S33L
S60K
L107D
Y92I;
S93Q;
P94E; F96Y
S33Q
S60K
L107D
A51G;
S52Q; S53R
Y92I;
S93I;
P94N; F96Y
S33Q
L107N
Y92I; S93N; P94Q; F96Y
S33Q
L107N
N34V
N32K;
S33Q
K58Q;
S60K
Q106G;
L107D;
L110M
A51G;
S52Q; S53R
Y92I;
S93Q;
P94Q; F96Y
N32R;
S33L
S60K
L107D
S52Y; S53N
Y92I;
S93M;
P94A; F96Y
N32K;
S33Q
S60K
L107D
R31Q
S52R; S53Y
Y92I;
S93Q;
P94N; F96Y
S33Y
S60K
Q106G;
L107D;
L110M
S52L; S53Y
Y92I;
S93Q;
P94T; F96Y
S33Y
K58Q; S60K
L107N
A51G;
S52Q; S53R
Y92I; P94I; F96Y
N32K;
S33Q
S60K
L107N
S52Y; S53N
Y92I;
S93I;
P94Q; F96Y
N32K;
S33Q
S60K
Q106G;
L107D
S52Y; S53N
Y92I;
S93Q;
P94N; F96Y
S33V
S60K
Q106E;
L107D
A51G;
S52Q; S53R
S93Q;
P94T; F96Y
S33Q
S60K
Q106G;
L107N
S52R; S53I
Y92I;
S93I;
P94N; F96Y
N32H;
S33Q
S60K
L107D
S52Y; S53N
Y92I;
S93Q;
P94N; F96Y
S33D
S60K
Q106E;
L107D
A51G;
S53I
Y92I;
S93Q;
P94N; F96Y
N32R;
S33L
S60K
Q106G;
L107D;
L110F
S52Y; S53N
S93Q;
P94Q; F96Y
N32R;
S33L
S60K
L107D
S53R
S93Q;
P94N; F96Y
S33L
Q106G; L107N
Y92I; S93I; P94N; F96Y
N32K;
S33Q
S60K
L107N
S52R;
S53N
S93Q;
P94T; F96Y
Варианты с улучшенной аффинностью получали с помощью способов рекомбинантной экспрессии в виде полных двухвалентных моноклональных антител и/или в виде одновалентных продуктов слияния Fab-Fc (например, Fab-область, слитая с димерной Fc-областью IgG) и оценивали в отношении функциональной активности in vitro в клеточном анализе cAMP, описанном в примере 3. Результаты функционального анализа показаны в таблице 20 ниже.
Таблица 20. Ингибирующая удельная активность in vitro вариантов антитела 19H8 и слитых белков Fab-Fc
(двухвалентное связывание мишени)
(одновалентное связывание мишени)
N32R;
S33L
S60K
L107D
A51G;
S52Q; S53R
Y92I;
S93G;
P94M; F96Y
N32R;
S33L
S60K
L107D
Y92I;
S93Q;
P94E; F96Y
S33Q
S60K
L107D
A51G;
S52Q; S53R
Y92I;
S93I;
P94N; F96Y
S33Q
L107N
Y92I; S93N; P94Q; F96Y
S33Q
L107N
N34V
N32K;
S33Q
K58Q;
S60K
Q106G;
L107D;
L110M
A51G;
S52Q; S53R
Y92I;
S93Q;
P94Q; F96Y
N32R;
S33L
S60K
L107D
S52Y; S53N
Y92I;
S93M;
P94A; F96Y
N32K;
S33Q
S60K
L107D
R31Q
S52R; S53Y
Y92I;
S93Q;
P94N; F96Y
S60K
L107D
A51G;
S52Q; S53R
Y92I;
S93V;
P94Q; F96Y
S33Y
K58Q; S60K
L107N
A51G;
S52Q; S53R
Y92I; P94I; F96Y
N32K;
S33Q
S60K
L107N
S52Y; S53N
Y92I;
S93I;
P94Q; F96Y
N32K;
S33Q
S60K
Q106G;
L107D
S52Y; S53N
Y92I;
S93Q;
P94N; F96Y
S33V
S60K
Q106E;
L107D
A51G;
S52Q; S53R
S93Q;
P94T; F96Y
S33Q
S60K
Q106G;
L107N
S52R; S53I
Y92I;
S93I;
P94N; F96Y
N32H;
S33Q
S60K
L107D
S52Y; S53N
Y92I;
S93Q;
P94N; F96Y
S33D
S60K
Q106E;
L107D
A51G;
S53I
Y92I;
S93Q;
P94N; F96Y
N32R;
S33L
S60K
Q106G;
L107D;
L110F
S52Y; S53N
S93Q;
P94Q; F96Y
N32K;
S33Q
S57R;
K58T;
S60K
K105S;
Q106E;
L107D
Y92I;
S93Q;
P94Q; F96Y
S33L
Q106G; L107N
Y92I; S93I; P94N; F96Y
N32K;
S33Q
S60K
L107N
S52R;
S53N
S93Q;
P94T; F96Y
N32K;
S33Q
S60K
L107N
Y92I; S93I; P94Q; F96Y
1Ранее зафиксированное значение, полученное из предыдущего анализа и включенное для целей сравнения.
При представлении в формате слитых белков Fab-Fc варианты 19H8 демонстрировали повышение удельной ингибирующей активности в 2-58 раз по сравнению с исходным слитым белок Fab-Fc 19H8. Многие из вариантов 19H8 характеризовались более высокой удельной активностью, чем варианты 29G4v10 при представлении в формате слитых белков Fab-Fc (сравните результаты в таблице 20 с результатами в таблице 12 для слитых белков Fab-Fc). При представлении в формате двухвалентных моноклональных антител варианты 19H8 также демонстрировали сильную нейтрализующую активность в отношении PAC1 человека, при этом значения IC50 находились в диапазоне одноразрядных наномолярных или пикомолярных значений.
Пример 7. Функциональная активность in vivo варианта антитела к PAC1 человека у яванских макаков
Чтобы оценить уровень занятости мишени in vivo антителом к PAC1 420653, оценивали способность антитела ингибировать индуцированное максадиланом повышение притока крови к коже у яванских макаков. Максадилан представляет собой селективный агонист рецептора PAC1 и может активировать рецептор у грызунов, яванских макаков и людей. Как описано в примере 4, внутрикожное введение максадилана вызывает локальное повышение притока крови к коже, которое можно измерить с помощью лазерной доплеровской визуализации. Подавление этого эффекта антителом к PAC1 может служить в качестве трансляционной фармакодинамической модели антагонизма в отношении биологической активности PAC1.
Для данного исследования применяли самцов яванского макака в возрасте 5-8 лет. Свежий раствор максадилана для введения в виде доз (Bachem, H6734.0500) получали ежедневно путем разбавления исходного раствора максадилана (0,5 мг/мл) в 1X фосфатно-солевом буферном растворе (PBS). Антитело к PAC1 420653 или 29G4v22, которое испытывали в качестве контроля, получали в растворе с 10 мМ ацетата натрия, 9% сахарозы, 0,01% Tween-80, pH 5,2 (A52SuT) при разных концентрациях, в зависимости от дозы, требующейся для эксперимента, и вводили посредством внутривенной (i.v.) инфузии.
Устройство для лазерной доплеровской визуализации (LDI2-IR, Moor Instruments, Ltd, Уилмингтон, Делавэр) применяли для измерения притока крови к коже (DBF) на выбритой части кожи либо вентральной области предплечья, либо медиальной области бедра с применением лазерного луча малой мощности с длиной волны 633 нм, генерируемого в трубке гелий-неонового газового лазера. В прицельном пучке излучения с длиной волны в инфракрасном диапазоне объединяли с таковыми в видимом красном диапазоне, чтобы придать более высокое значения весового коэффициента притоку крови в более глубоких слоях кожи (0,6-1 мм). Падающий свет рассеивался неподвижной тканью и движущейся кровью. Движущаяся кровь в микроциркуляторной части сосудистого русла обусловливала доплеровский сдвиг для света. Свет, характеризующийся наличием сдвига, от движущейся крови и свет без сдвига от ткани затем направлялся на 2 квадратичных детектора. Выявленные флуктуации интенсивности затем подергали обработке с получением параметров потока (пропорциональных притоку крови в ткани) и концентрации (пропорциональных концентрации движущихся клеток крови). Разрешающая способность измерительного прибора составляла 0,2-2 мм, при этом расстояние сканирования между отверстием прибора и поверхностью ткани составляло 20-100 см.
DBF измеряли в виде плотности потока (относительные единицы) и выражали в виде % изменения от исходного уровня через 30 мин. после введения максадилана [100 x (отдельное значение плотности потока после воздействия максадиланом - отдельное значение плотности потока на исходном уровне)/отдельное значение плотности потока на исходном уровне]. Единицы измерения плотности потока от двух отдельных O-колец усредняли для каждого сеанса испытания. Ингибирующий эффект антител к PAC1 в отношении индуцированного максадиланом повышения притока крови (MIIBF), определенный у отдельных животных, выражали в виде % ингибирования [100 x (среднее значение % изменения от исходного уровня в день 0 - отдельное значение % изменения от исходного уровня у животного, обработанного антителом)/среднее значение % изменения от исходного уровня в день 0].
После осуществления наркоза и стабилизации жизненных показателей в течение 15-20 минут резиновые О-образные кольца (внутренний диаметр 0,925 см, O-Rings West, Сиэтл, Вашингтон) помещали на расстоянии приблизительно 0,6-1 см друг от друга на предварительно выбритой коже вентральной области предплечья или медиальной области бедра, не размещая их непосредственно на видимом кровеносном сосуде. Конечность, используемая для каждой конкретной сессии испытания, являлась предварительно определенной, таким образом, чтобы в разные дни испытания в ходе исследования использовались разные конечности. После помещения O-колец на кожу осуществляли измерение исходного уровня. После сканирования для определения исходного уровня в центр O-колец осуществляли внутрикожную инъекцию раствора 1 нг максадилана в 20 мкл среды-носителя (PBS). O-кольца служили в качестве представляющей интерес области при анализе данных.
До осуществления введений антител к PAC1 в день 0 получали данные динамики MIIBF. Доза максадилана 1 нг была выбрана на основе результатов предыдущего анализа зависимости "доза-ответ" для максадилана у яванских макаков. После измерения исходного уровня DBF до обработки максадиланом динамику ответа в виде DBF на внутрикожную инъекцию максадилана оценивали путем получения сканограмм лазерной доплеровской визуализации через 5, 10, 15, 20, 25 и 30 минут после применения максадилана. Чтобы идентифицировать особей, отвечающих на воздействие максадиланом, и исключить особей, не отвечающих на воздействие максадиланом, из включения в исследование, реализовали процедуру предварительного скрининга до введения антитела. Животных считали отвечающими на воздействие максадиланом и включали в исследование, если у них имелось изменение притока DBF ≥ 60 единиц измерения плотности потока (среднее значение DBF через 30 минут после воздействия максадиланом - среднее значение на исходном уровне).
Двадцать четыре яванских макака, идентифицированные как отвечающие на воздействие максадиланом, получали либо антитело 420653 при дозе 0,1 мг/кг или 3 мг/кг, либо антитело 29G4v22 при дозе 10 мг/кг в день 0. Антитела вводили посредством i.v.-инфузии в подкожную вену при скорости 1 мл/мин. с помощью шприцевого инфузионного насоса. Измерения DBF после воздействия максадиланом осуществляли в дни 2, 4, 7, 10, 14, 21, 28 и 36, используя разные конечности. В каждом случае DBF измеряли с помощью лазерной доплеровской визуализации в течение 30 минут после внутрикожной инъекции 1 нг максадилана. Образцы цельной крови для фармакокинетического (PK) анализа собирали в нескольких моментах времени, в том числе до введения дозы антитела, через 30 мин., 1, 2, 4, 7, 10, 14, 21, 28, 35 или 36 и 42 дня после обработки антителом.
Как показано на фигуре 10A, антитело 420653 при 3 мг/кг, но не при 0,1 мг/кг в значительной степени предотвращало MIIBF в день 2, день 7, день 14 и день 21 (после обработки антителом) по сравнению с днем 0 (до обработки антителом) (n=8 животных/группа; p-значение с поправкой Бонферрони=0,0087). Антитело 29G4v22 при 10 мг/кг также в значительной степени предотвращало MIIBF в день 2 и день 7 (n=8 животных/группа) (фигура 10A). Максимальный ингибирующий эффект антитела 420653 при 3 мг/кг наблюдали в день 2, при этом уровень ингибирования составлял 96%, при этом при 0,1 мг/кг антитело 420653 вызывало максимальный уровень ингибирования, составляющий 39%, в день 4, который не являлся статистически значимым (фигура 10B). Антитело 29G4v22 при 10 мг/кг вызывало максимальный уровень ингибирования, составляющий 63%, в день 7 (фигура 10B). Сравнение антитела 420653 (3 мг/кг) и антитела 29G4v22 (10 мг/кг) в одних и тех же моментах времени показало значительные отличия в день 2, день 7, день 14 и день 21 (фигура 10B).
Концентрации антитела в сыворотке в случае антитела 420653 и антитела 29G4v22 в дни, когда осуществляли измерения DBF, показаны ниже в таблице 21. Нижний предел количественного определения (LLOQ) или ниже уровня количественного определения (BQL) составлял 10 нг/мл в случае антитела 420653 и 50 нг/мл в случае антитела 29G4v22.
Таблица 21. Концентрации антител 420653 и 29G4v22 в сыворотке у яванских макаков после обработки с помощью антитела
*Меньшее количество точек данных PK обусловлено тем, что для некоторых образцов получили LLOQ/BQL.
В общем, эти результаты демонстрируют, что антитело 420653 в значительной степени ослабляло MIIBF у яванских макаков при введении при дозе 3 мг/кг. Ингибирующий эффект антитела 420653 при 3 мг/кг в отношении MIIBF являлся более устойчивым, чем таковой антитела 29G4v22 при более высокой дозе, составляющей 10 мг/кг.
Все публикации, патенты и патентные заявки, обсуждаемые и цитируемые в данном документе, включены в данный документ посредством ссылки в полном их объеме. Необходимо понимать, что раскрытое изобретение не ограничивается конкретной описанной методологией, протоколами и материалами, поскольку они могут варьироваться. Кроме того, необходимо понимать, что используемая в данном документе терминология предназначена только для описания конкретных вариантов осуществления изобретения и не предназначена для ограничения объема прилагаемой формулы изобретения.
Специалистам в данной области техники будет понятно, или они способны определить с использованием только обычного экспериментирования, наличие многих эквивалентов конкретных вариантов осуществления настоящего изобретения, описанного в данном документе. Подразумевается, что такие эквиваленты охвачены приведенной ниже формулой изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ЭМДЖЕН ИНК.
<120> АНТИТЕЛА К PAC1 И ВАРИАНТЫ ИХ ПРИМЕНЕНИЯ
<130> A-2189-WO-PCT
<150> 62/617,157
<151> 2018-01-12
<160> 569
<170> PatentIn версия 3.5
<210> 1
<211> 468
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> PAC1 человека
<400> 1
Met Ala Gly Val Val His Val Ser Leu Ala Ala Leu Leu Leu Leu Pro
1. 5 10 15
Met Ala Pro Ala Met His Ser Asp Cys Ile Phe Lys Lys Glu Gln Ala
20 25 30
Met Cys Leu Glu Lys Ile Gln Arg Ala Asn Glu Leu Met Gly Phe Asn
35 40 45
Asp Ser Ser Pro Gly Cys Pro Gly Met Trp Asp Asn Ile Thr Cys Trp
50 55 60
Lys Pro Ala His Val Gly Glu Met Val Leu Val Ser Cys Pro Glu Leu
65 70 75 80
Phe Arg Ile Phe Asn Pro Asp Gln Val Trp Glu Thr Glu Thr Ile Gly
85 90 95
Glu Ser Asp Phe Gly Asp Ser Asn Ser Leu Asp Leu Ser Asp Met Gly
100 105 110
Val Val Ser Arg Asn Cys Thr Glu Asp Gly Trp Ser Glu Pro Phe Pro
115 120 125
His Tyr Phe Asp Ala Cys Gly Phe Asp Glu Tyr Glu Ser Glu Thr Gly
130 135 140
Asp Gln Asp Tyr Tyr Tyr Leu Ser Val Lys Ala Leu Tyr Thr Val Gly
145 150 155 160
Tyr Ser Thr Ser Leu Val Thr Leu Thr Thr Ala Met Val Ile Leu Cys
165 170 175
Arg Phe Arg Lys Leu His Cys Thr Arg Asn Phe Ile His Met Asn Leu
180 185 190
Phe Val Ser Phe Met Leu Arg Ala Ile Ser Val Phe Ile Lys Asp Trp
195 200 205
Ile Leu Tyr Ala Glu Gln Asp Ser Asn His Cys Phe Ile Ser Thr Val
210 215 220
Glu Cys Lys Ala Val Met Val Phe Phe His Tyr Cys Val Val Ser Asn
225 230 235 240
Tyr Phe Trp Leu Phe Ile Glu Gly Leu Tyr Leu Phe Thr Leu Leu Val
245 250 255
Glu Thr Phe Phe Pro Glu Arg Arg Tyr Phe Tyr Trp Tyr Thr Ile Ile
260 265 270
Gly Trp Gly Thr Pro Thr Val Cys Val Thr Val Trp Ala Thr Leu Arg
275 280 285
Leu Tyr Phe Asp Asp Thr Gly Cys Trp Asp Met Asn Asp Ser Thr Ala
290 295 300
Leu Trp Trp Val Ile Lys Gly Pro Val Val Gly Ser Ile Met Val Asn
305 310 315 320
Phe Val Leu Phe Ile Gly Ile Ile Val Ile Leu Val Gln Lys Leu Gln
325 330 335
Ser Pro Asp Met Gly Gly Asn Glu Ser Ser Ile Tyr Leu Arg Leu Ala
340 345 350
Arg Ser Thr Leu Leu Leu Ile Pro Leu Phe Gly Ile His Tyr Thr Val
355 360 365
Phe Ala Phe Ser Pro Glu Asn Val Ser Lys Arg Glu Arg Leu Val Phe
370 375 380
Glu Leu Gly Leu Gly Ser Phe Gln Gly Phe Val Val Ala Val Leu Tyr
385 390 395 400
Cys Phe Leu Asn Gly Glu Val Gln Ala Glu Ile Lys Arg Lys Trp Arg
405 410 415
Ser Trp Lys Val Asn Arg Tyr Phe Ala Val Asp Phe Lys His Arg His
420 425 430
Pro Ser Leu Ala Ser Ser Gly Val Asn Gly Gly Thr Gln Leu Ser Ile
435 440 445
Leu Ser Lys Ser Ser Ser Gln Ile Arg Met Ser Gly Leu Pro Ala Asp
450 455 460
Asn Leu Ala Thr
465
<210> 2
<211> 226
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ECD PAC1 человека
<400> 2
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Asn Phe Gly Thr Gln Thr Tyr Thr Cys Asn Val Asp His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Thr Val Glu Arg Lys Gly Asp Glu
210 215 220
Val Asp
225
<210> 3
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ECD PAC1 человека
<400> 3
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 4
<211> 105
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> ECD PAC1 человека
<400> 4
Gly Ser Met Ala His Ser Asp Gly Ile Phe Lys Lys Glu Gln Ala Met
1. 5 10 15
Cys Leu Glu Lys Ile Gln Arg Ala Asn Glu Leu Met Gly Phe Asn Asp
20 25 30
Ser Ser Pro Gly Cys Pro Gly Met Trp Asp Asn Ile Thr Cys Trp Lys
35 40 45
Pro Ala His Val Gly Glu Met Val Leu Val Ser Cys Pro Glu Leu Phe
50 55 60
Arg Ile Phe Asn Pro Asp Gln Asp Met Gly Val Val Ser Arg Asn Cys
65 70 75 80
Thr Glu Asp Gly Trp Ser Glu Pro Phe Pro His Tyr Phe Asp Ala Cys
85 90 95
Gly Phe Asp Glu Tyr Glu Ser Glu Thr
100 105
<210> 5
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 5
Arg Ala Ser Gln Ser Val Gly Arg Ser Leu His
1. 5 10
<210> 6
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 6
Arg Ala Ser Gln Ser Ile Gly Arg Ser Leu His
1. 5 10
<210> 7
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 7
Arg Ala Ser Lys Ser Val Gly Arg Ser Leu His
1. 5 10
<210> 8
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 8
Arg Ala Ser Gln Ser Val Trp Arg Ser Leu His
1. 5 10
<210> 9
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 9
Arg Ala Ser Gln Ser Val Gly Arg Asn Leu His
1. 5 10
<210> 10
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 10
Arg Ala Ser Lys Ser Val Trp Arg Ser Leu His
1. 5 10
<210> 11
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 11
Arg Ala Ser Lys Ser Val Gly Arg Asn Leu His
1. 5 10
<210> 12
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 12
Arg Ala Ser Lys Ser Val Gly Trp Ser Leu His
1. 5 10
<210> 13
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 13
Arg Ala Ser Lys Ser Val Gly Tyr Ser Leu His
1. 5 10
<210> 14
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 14
Arg Ala Ser Lys Ala Val Gly Trp Ser Leu His
1. 5 10
<210> 15
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 15
Arg Ala Ser Lys Ser Val Gly Gln Ser Leu His
1. 5 10
<210> 16
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 16
Arg Ala Ser Arg Ser Val Gly Leu Ala Leu His
1. 5 10
<210> 17
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 17
Arg Ala Ser Gln Ser Ile Ser Arg Tyr Leu Asn
1. 5 10
<210> 18
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 18
Arg Ala Ser Gln Tyr Ile Val Arg Tyr Leu Asn
1. 5 10
<210> 19
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 19
Arg Ala Ser Gln Thr Ile Val Arg Tyr Leu Asn
1. 5 10
<210> 20
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 20
Arg Ala Ser Gln Lys Ile Ala Arg Tyr Leu Val
1. 5 10
<210> 21
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 21
Arg Ala Ser Gln Pro Ile Ala Gln Tyr Leu Asn
1. 5 10
<210> 22
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 22
Arg Ala Ser Gln Pro Ile Ser Arg Tyr Leu Ser
1. 5 10
<210> 23
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 23
Arg Ala Ser Gln Gln Ile Ala Arg Tyr Leu Asn
1. 5 10
<210> 24
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 24
Arg Ala Ser Gln Arg Ile Ala Arg Tyr Leu Asn
1. 5 10
<210> 25
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 25
Arg Ala Ser Gln Met Ile Ala Arg Tyr Leu Asn
1. 5 10
<210> 26
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 26
Tyr Ala Ser Gln Ser Leu Ser
1. 5
<210> 27
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 27
Ala Ala Ser Ser Leu Gln Ser
1. 5
<210> 28
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 28
Ala Gly Gln Arg Leu Gln Ser
1. 5
<210> 29
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 29
Ala Ala His His Leu Gln Ser
1. 5
<210> 30
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 30
Ala Ala Asn Met Leu Gln Ser
1. 5
<210> 31
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 31
Ala Ser Tyr Asn Leu Gln Ser
1. 5
<210> 32
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 32
Ala Gly Arg Tyr Leu Gln Ser
1. 5
<210> 33
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 33
Ala Gly Arg Ile Leu Gln Ser
1. 5
<210> 34
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 34
Ala Gly Ser Ile Leu Gln Ser
1. 5
<210> 35
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 35
Gly Ala Arg Asn Leu Gln Ser
1. 5
<210> 36
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 36
His Gln Ser Ser Arg Leu Pro Phe Thr
1. 5
<210> 37
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 37
His Gln Ser Ser Met Leu Pro Phe Thr
1. 5
<210> 38
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 38
His Gln Ser Ser Phe Leu Pro Phe Thr
1. 5
<210> 39
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 39
Gln Gln Ser Tyr Ser Pro Pro Phe Thr
1. 5
<210> 40
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 40
Gln Gln Ala Ile Gly Met Pro Tyr Thr
1. 5
<210> 41
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 41
Gln Gln Ala Ile Gln Glu Pro Tyr Thr
1. 5
<210> 42
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 42
Gln Gln Ala Ile Ile Asn Pro Tyr Thr
1. 5
<210> 43
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 43
Gln Gln Ala Ile Asn Gln Pro Tyr Thr
1. 5
<210> 44
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 44
Gln Gln Ser Ile Gln Gln Pro Tyr Thr
1. 5
<210> 45
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 45
Gln Gln Ala Ile Gln Gln Pro Tyr Thr
1. 5
<210> 46
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 46
Gln Gln Ala Ile Met Ala Pro Tyr Thr
1. 5
<210> 47
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 47
Gln Gln Ala Ile Gln Asn Pro Tyr Thr
1. 5
<210> 48
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 48
Gln Gln Ala Ile Val Gln Pro Tyr Thr
1. 5
<210> 49
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 49
Gln Gln Ala Ile Ser Ile Pro Tyr Thr
1. 5
<210> 50
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 50
Gln Gln Ala Ile Ile Gln Pro Tyr Thr
1. 5
<210> 51
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 51
Gln Gln Ser Ile Gln Thr Pro Tyr Thr
1. 5
<210> 52
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 52
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 53
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 53
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 54
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 54
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 55
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 55
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 56
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 56
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Trp Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 57
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 57
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Arg Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 58
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 58
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Met Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 59
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 59
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Trp Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 60
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 60
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Arg Asn
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 61
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 61
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Met Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 62
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 62
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Trp Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 63
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 63
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Tyr Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 64
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 64
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ala Val Gly Trp Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 65
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 65
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Gln Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 66
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 66
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Arg Ser Val Gly Leu Ala
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Phe Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 67
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 67
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Tyr Ser Pro Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 68
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 68
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gly Met Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 69
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 69
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala His His Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Glu Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 70
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 70
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 71
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 71
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Asn Met Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Asn Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 72
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 72
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Lys Ile Ala Arg Tyr
20 25 30
Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Asn Met Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 73
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 73
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg
100 105
<210> 74
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 74
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Met Ala Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 75
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 75
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Pro Ile Ala Gln Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Gly Arg Tyr Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 76
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 76
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Val Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 77
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 77
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Pro Ile Ser Arg Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ser Ile Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 78
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 78
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Gln Ile Ala Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 79
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 79
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 80
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 80
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Thr Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 81
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 81
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Gly Arg Ile Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 82
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 82
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Arg Ile Ala Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Ser Ile Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 83
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 83
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 84
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 84
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 85
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 85
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Met Ile Ala Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 86
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 86
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Arg Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Thr Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 87
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 87
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Tyr Ser Pro Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 88
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 88
Arg Phe Ala Met His
1. 5
<210> 89
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 89
His Phe Ala Met His
1. 5
<210> 90
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 90
Arg Tyr Ala Met His
1. 5
<210> 91
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 91
Lys Tyr Ala Met His
1. 5
<210> 92
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 92
Lys Phe Ala Met His
1. 5
<210> 93
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 93
His Tyr Ala Met His
1. 5
<210> 94
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 94
Tyr Phe Ala Met His
1. 5
<210> 95
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 95
Phe Tyr Ala Met His
1. 5
<210> 96
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 96
Tyr Tyr Ala Met His
1. 5
<210> 97
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 97
Ser Asn Ser Ala Thr Trp Asn
1. 5
<210> 98
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 98
Asn Arg Leu Ala Thr Trp Asn
1. 5
<210> 99
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 99
Ser Arg Gln Ala Thr Trp Asn
1. 5
<210> 100
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 100
Asn Lys Gln Ala Thr Trp Asn
1. 5
<210> 101
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 101
Ser Asn His Ala Thr Trp Asn
1. 5
<210> 102
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 102
Ser Arg Tyr Ala Thr Trp Asn
1. 5
<210> 103
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 103
Ser His Val Ala Thr Trp Asn
1. 5
<210> 104
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 104
Asn His Gln Ala Thr Trp Asn
1. 5
<210> 105
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 105
Ser Arg Asp Ala Thr Trp Asn
1. 5
<210> 106
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 106
Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 107
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 107
Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 108
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 108
Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 109
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 109
Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 110
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 110
Val Ile Ser Tyr Tyr Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 111
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 111
Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 112
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 112
Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 113
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 113
Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 114
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 114
Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 115
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 115
Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 116
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 116
Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 117
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 117
Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 118
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 118
Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 119
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 119
Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 120
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 120
Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 121
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 121
Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 122
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 122
Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 123
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 123
Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 124
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 124
Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 125
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 125
Val Ile Ser Tyr Tyr Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 126
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 126
Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 127
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 127
Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 128
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 128
Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 129
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 129
Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 130
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 130
Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 131
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 131
Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 132
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 132
Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 133
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 133
Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 134
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 134
Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 135
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 135
Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 136
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 136
Val Ile Ser Phe Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 137
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 137
Val Ile Ser Tyr Asp Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 138
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 138
Val Ile Ser Phe Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 139
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 139
Val Ile Ser Tyr Asn Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 140
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 140
Val Ile Ser Phe Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 141
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 141
Val Ile Ser Phe Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 142
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 142
Val Ile Ser Tyr Asp Gly Ala Asn Lys Tyr Tyr Ala Arg Ser Val Lys
1. 5 10 15
Gly
<210> 143
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 143
Val Ile Ser Phe Lys Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 144
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 144
Val Ile Ser Phe Ser Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 145
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 145
Val Ile Ser Tyr Arg Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 146
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 146
Val Ile Ser Tyr Ser Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 147
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 147
Val Ile Ser Phe Lys Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 148
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 148
Val Ile Ser Tyr Arg Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 149
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 149
Val Ile Ser Phe Tyr Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 150
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 150
Val Ile Ser Phe Phe Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 151
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 151
Val Ile Ser Phe Met Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 152
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 152
Val Ile Ser His Arg Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 153
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 153
Val Ile Asn Tyr Arg Gly His Gly Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 154
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 154
Val Ile Ser Phe Ser Gly Gly Ser Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 155
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 155
Val Ile Ser Tyr Thr Gly Gln Phe Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 156
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 156
Val Ile Ser Tyr Thr Gly Ala Gln Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 157
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 157
Val Ile Ser Tyr Ser Gly Ala Ser Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 158
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 158
Val Ile Ser Tyr Ser Gly Ala Phe Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 159
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 159
Val Ile Thr Tyr Thr Gly Gly Ala Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 160
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 160
Val Ile Asn Phe Gln Gly Thr Thr Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 161
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 161
Val Ile Ser Tyr Ser Gly Asp Leu Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 162
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 162
Val Ile Asn Tyr Phe Gly Asp Ala Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 163
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 163
Val Ile Ser Ser Phe Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 164
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 164
Val Ile Ser Tyr Ser Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 165
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 165
Val Ile Ser His Tyr Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 166
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 166
Val Ile Ser Tyr Gln Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val Lys
1. 5 10 15
Gly
<210> 167
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 167
Arg Thr Tyr Tyr Arg Ser Lys Trp Ser Asn His Tyr Ala Val Ser Val
1. 5 10 15
Lys Ser
<210> 168
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 168
Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala Val Ser Val
1. 5 10 15
Lys Ser
<210> 169
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 169
Arg Thr Tyr Tyr Arg Gly Gln Trp Lys Asn His Tyr Ala Val Ser Val
1. 5 10 15
Lys Ser
<210> 170
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 170
Arg Thr Tyr Phe Arg Arg Thr Trp Lys Asn His Tyr Ala Val Ser Val
1. 5 10 15
Lys Ser
<210> 171
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 171
Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 172
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 172
Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 173
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 173
Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 174
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 174
Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 175
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 175
Gly Tyr Asp Met Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 176
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 176
Gly Tyr Asp Leu Leu Ser Gly Tyr Pro Asp Tyr
1. 5 10
<210> 177
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 177
Gly Tyr Asp Pro Leu Thr Gly Tyr Pro Asp Tyr
1. 5 10
<210> 178
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 178
Gly Thr Trp Lys Gln Leu Trp Phe Leu Asp His
1. 5 10
<210> 179
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 179
Gly Thr Trp Asn Gln Asp Trp Phe Leu Asp His
1. 5 10
<210> 180
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 180
Gly Thr Trp Asp Gln Asp Trp Phe Leu Asp His
1. 5 10
<210> 181
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 181
Gly Thr Trp Glu Gln Asp Trp Phe Leu Asp His
1. 5 10
<210> 182
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 182
Gly Met Trp Asn Gln Asn Trp Phe Leu Asp His
1. 5 10
<210> 183
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 183
Gly Thr Trp Ile Gly Asp Trp Phe Met Asp His
1. 5 10
<210> 184
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 184
Gly Thr Trp Ile Gln Asp Trp Phe Leu Asp His
1. 5 10
<210> 185
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 185
Gly Met Trp Ser Gly Asp Trp Phe Leu Asp His
1. 5 10
<210> 186
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 186
Gly Met Trp Ser Glu Asp Trp Phe Leu Asp His
1. 5 10
<210> 187
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 187
Gly Met Trp Gln Gly Asn Trp Phe Leu Asp His
1. 5 10
<210> 188
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 188
Gly Gln Trp Asn Glu Asp Trp Phe Leu Asp His
1. 5 10
<210> 189
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 189
Gly Arg Trp Glu Gly Asp Trp Phe Phe Asp His
1. 5 10
<210> 190
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 190
Gly Val Trp Ile Gly Asn Trp Phe Leu Asp His
1. 5 10
<210> 191
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 191
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 192
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 192
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Leu Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 193
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 193
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 194
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 194
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 195
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 195
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 196
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 196
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 197
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 197
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 198
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 198
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 199
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 199
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 200
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 200
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 201
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 201
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 202
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 202
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 203
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 203
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 204
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 204
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 205
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 205
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 206
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 206
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 207
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 207
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 208
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 208
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 209
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 209
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 210
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 210
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 211
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 211
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 212
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 212
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 213
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 213
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 214
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 214
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 215
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 215
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 216
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 216
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 217
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 217
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 218
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 218
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 219
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 219
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 220
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 220
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 221
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 221
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 222
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 222
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 223
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 223
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 224
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 224
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 225
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 225
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 226
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 226
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 227
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 227
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 228
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 228
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 229
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 229
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 230
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 230
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 231
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 231
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 232
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 232
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 233
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 233
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 234
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 234
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 235
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 235
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 236
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 236
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 237
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 237
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 238
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 238
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 239
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 239
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 240
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 240
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 241
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 241
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 242
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 242
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 243
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 243
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 244
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 244
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 245
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 245
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 246
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 246
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 247
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 247
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Tyr Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 248
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 248
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 249
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 249
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 250
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 250
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 251
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 251
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 252
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 252
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 253
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 253
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 254
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 254
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 255
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 255
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 256
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 256
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 257
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 257
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 258
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 258
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 259
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 259
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 260
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 260
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 261
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 261
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 262
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 262
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 263
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 263
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 264
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 264
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 265
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 265
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ala Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 266
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 266
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 267
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 267
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 268
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 268
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 269
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 269
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 270
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 270
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 271
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 271
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Lys Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Lys Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 272
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 272
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 273
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 273
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Ser Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Met Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 274
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 274
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Lys Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Arg Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 275
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 275
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ser Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Ser Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 276
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 276
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Lys Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 277
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 277
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Arg Gly Ala Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 278
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 278
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Tyr Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 279
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 279
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Phe Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 280
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 280
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Met Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Phe Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 281
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 281
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser His Arg Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Ser Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 282
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 282
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Asn Tyr Arg Gly His Gly Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 283
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 283
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Phe Ser Gly Gly Ser Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Leu Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 284
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 284
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Tyr Thr Gly Gln Phe Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 285
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 285
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Tyr Thr Gly Ala Gln Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Met Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 286
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 286
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Tyr Ser Gly Ala Ser Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Met Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 287
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 287
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ser Gly Ala Phe Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 288
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 288
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Thr Tyr Thr Gly Gly Ala Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 289
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 289
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Asn Phe Gln Gly Thr Thr Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 290
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 290
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Tyr Ser Gly Asp Leu Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 291
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 291
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Asn Tyr Phe Gly Asp Ala Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 292
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 292
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Phe Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Ser Phe Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 293
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 293
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ser Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 294
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 294
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Tyr Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser His Tyr Gly Thr Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Pro Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 295
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 295
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser His Tyr
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Ser Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 296
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 296
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
Ser Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Ser Lys Trp Ser Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Lys Gln Leu Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 297
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 297
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Asn Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 298
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 298
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Asp Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 299
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 299
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Glu Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 300
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 300
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Asn Gln Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 301
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 301
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Gln Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Ile Gly Asp Trp Phe Met Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 302
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 302
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Ile Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 303
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 303
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
His Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Asp Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 304
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 304
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Tyr Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Gln Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Asn Gln Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 305
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 305
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Ser Gly Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 306
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 306
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser His
20 25 30
Val Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Ser Glu Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 307
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 307
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Gln Gly Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 308
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 308
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn His
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Ile Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 309
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 309
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Asp Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Gln Trp Asn Glu Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 310
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 310
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Arg Trp Glu Gly Asp Trp Phe Phe Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 311
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 311
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Phe Arg Arg Thr Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Ser Glu Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 312
<211> 123
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 312
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Val Trp Ile Gly Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 313
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 313
Gly Gln Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 314
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 314
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 315
<211> 105
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 315
Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu
1. 5 10 15
Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe
20 25 30
Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val
35 40 45
Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys
50 55 60
Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser
65 70 75 80
His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu
85 90 95
Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 316
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 316
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
35 40 45
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Lys Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
100 105
<210> 317
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 317
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
1. 5 10 15
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Val Ser Asp
20 25 30
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro
35 40 45
Val Lys Val Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn
50 55 60
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
65 70 75 80
Ser His Arg Ser Tyr Ser Cys Arg Val Thr His Glu Gly Ser Thr Val
85 90 95
Glu Lys Thr Val Ala Pro Ala Glu Cys Ser
100 105
<210> 318
<211> 106
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 318
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
1. 5 10 15
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
20 25 30
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
35 40 45
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
50 55 60
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
65 70 75 80
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
85 90 95
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 319
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 319
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1. 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 320
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 320
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 321
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 321
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 322
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 322
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 323
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 323
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 324
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 324
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Gly Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 325
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 325
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu
165 170 175
Glu Gln Tyr Gly Ser Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 326
<211> 326
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 326
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1. 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Asn Phe Gly Thr Gln Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Thr Val Glu Arg Lys Cys Cys Val Glu Cys Pro Pro Cys Pro Ala Pro
100 105 110
Pro Val Ala Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
115 120 125
Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
130 135 140
Val Ser His Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
145 150 155 160
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
165 170 175
Ser Thr Phe Arg Val Val Ser Val Leu Thr Val Val His Gln Asp Trp
180 185 190
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
195 200 205
Ala Pro Ile Glu Lys Thr Ile Ser Lys Thr Lys Gly Gln Pro Arg Glu
210 215 220
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn
225 230 235 240
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
245 250 255
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
260 265 270
Thr Pro Pro Met Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
275 280 285
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
290 295 300
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
305 310 315 320
Ser Leu Ser Pro Gly Lys
325
<210> 327
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 327
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu
165 170 175
Glu Gln Tyr Gly Ser Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 328
<211> 326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 328
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgtac 326
<210> 329
<211> 326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 329
gatatccagc tcactcaatc gccatcattt ctctccgctt cggtaggcga ccgggtcacg 60
atcacatgca gggcgtcgca aagcattggg aggtcgttgc attggtatca gcagaaaccc 120
ggaaaggccc cgaaacttct gatcaaatac gcatcacaaa gcttgagcgg tgtgccgtcg 180
cgcttctccg gttccggaag cggaacggaa ttcacgctta caatctcctc actgcagccc 240
gaggatttcg cgacctatta ctgtcaccag tcatccagac tcccgtttac ttttggccct 300
gggaccaagg tggacattaa gcgtac 326
<210> 330
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 330
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 331
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 331
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 332
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 332
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtctgg cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 333
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 333
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtcgga cgaaacttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 334
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 334
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgatgt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 335
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 335
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa gtccgtctgg cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 336
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 336
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa gtccgtcgga cgaaacttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 337
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 337
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgatgt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa g 321
<210> 338
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 338
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa atccgtcggg tggagcttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgta 325
<210> 339
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 339
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa atccgtcggg tacagcttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgta 325
<210> 340
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 340
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa agccgtcggg tggagcttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgta 325
<210> 341
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 341
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa atcagtcggt cagtctttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgcgtt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgta 325
<210> 342
<211> 325
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 342
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatcccg ttcagtcggt ctggctttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgttct tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgta 325
<210> 343
<211> 326
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 343
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctatgct gcatccagtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag agttacagtc ccccattcac tttcggccct 300
gggaccaaag tggatatcaa acgtac 326
<210> 344
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 344
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcggta tgccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 345
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 345
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct gctcatcatt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccagg aaccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 346
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 346
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gactattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatca acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 347
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 347
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct gctaacatgt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcaacc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 348
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 348
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gaaaattgct cgttacttag tttggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct gctaacatgt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 349
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 349
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa acgt 324
<210> 350
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 350
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatgg ctccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 351
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 351
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gcctattgct cagtacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct ggtcgttact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccaga acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 352
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 352
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcgttc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 353
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 353
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gccgatttct cgttacttat cttggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatctcta tcccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 354
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 354
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gcagattgct cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatcc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 355
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 355
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccaga acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 356
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 356
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gactattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccaga ctccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 357
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 357
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct ggtcgtatct tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatca acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 358
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 358
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gcgtattgct cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggttctatct tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccaga acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 359
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 359
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 360
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 360
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 361
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 361
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gatgattgct cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatca acccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 362
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 362
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacggt gctcgtaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccaga ctccatacac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 363
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 363
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctatgct gcatccagtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag agttacagtc ccccattcac tttcggccct 300
gggaccaaag tggatatcaa a 321
<210> 364
<211> 362
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 364
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctcctca 360
gc 362
<210> 365
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 365
caagttcagt tggtggagtc tggagccgaa gtagtaaagc caggagcttc agtgaaagtc 60
tcttgtaaag caagtggatt cacgtttagc cgctttgcca tgcattgggt gcggcaagct 120
cccggtcagg ggttggagtg gatgggagtt attagctatg acgggggcaa taagtactac 180
gccgagtctg ttaagggtcg ggtcacaatg acacgggaca cctcaaccag tacactctat 240
atggaactgt ctagcctgag atccgaggac accgctgtgt attattgcgc tagggggtac 300
gatgtattga cgggttatcc tgattactgg gggcagggga cactcgtaac cgtctctagt 360
<210> 366
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 366
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 367
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 367
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 368
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 368
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 369
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 369
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 370
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 370
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 371
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 371
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 372
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 372
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 373
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 373
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 374
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 374
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 375
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 375
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 376
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 376
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 377
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 377
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 378
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 378
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 379
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 379
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 380
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 380
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 381
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 381
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 382
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 382
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 383
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 383
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 384
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 384
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 385
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 385
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 386
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 386
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 387
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 387
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 388
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 388
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 389
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 389
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 390
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 390
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 391
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 391
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 392
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 392
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 393
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 393
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 394
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 394
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 395
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 395
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 396
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 396
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 397
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 397
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 398
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 398
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 399
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 399
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 400
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 400
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 401
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 401
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 402
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 402
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 403
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 403
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 404
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 404
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 405
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 405
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 406
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 406
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 407
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 407
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 408
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 408
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 409
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 409
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 410
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 410
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaaacaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 411
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 411
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 412
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 412
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 413
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 413
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 414
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 414
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 415
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 415
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 416
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 416
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 417
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 417
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 418
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 418
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 419
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 419
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 420
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 420
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatt acggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 421
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 421
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 422
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 422
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 423
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 423
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattcg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 424
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 424
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 425
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 425
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 426
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 426
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 427
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 427
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 428
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 428
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattca acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 429
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 429
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 430
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 430
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 431
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 431
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 432
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 432
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 433
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 433
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattcg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 434
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 434
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 435
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 435
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 436
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 436
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 437
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 437
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattcg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 438
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 438
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggagccaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 439
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 439
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 440
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 440
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 441
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 441
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 442
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 442
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattcg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 443
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 443
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 444
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 444
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt aagtacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattca agggaagcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 445
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 445
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 446
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 446
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattca gcggaagcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 447
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 447
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt aagtttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc gcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 448
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 448
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata gcggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga gcggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 449
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 449
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattca agggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 450
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 450
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc gcggagccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 451
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 451
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattct acggaagcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 452
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 452
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattct tcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 453
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 453
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcattca tgggaaccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatttcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 454
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 454
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt tactttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcacacc gcggaaccaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga gcggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
<210> 455
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 455
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atcaactatc gtggacatgg taaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 456
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 456
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atatcttttt ctggaggttc taaatactat 180
gcagagtccg tgaagggccg gttcaccttg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 457
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 457
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atctcttata ctggacagtt caaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 458
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 458
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atatcttata ctggagctca gaaatactat 180
gcagagtccg tgaagggccg gttcaccatg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 459
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 459
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atatcttatt ctggagcttc taaatactat 180
gcagagtccg tgaagggccg gttcaccatg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 460
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 460
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggctgtt atatcttatt ctggagcttt caaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 461
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 461
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt ataacttata ctggaggtgc taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 462
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 462
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atcaactttc agggaactac taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 463
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 463
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atatcttatt ctggagatct gaaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 464
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 464
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atcaactatt tcggagacgc taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 465
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 465
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgagactc 60
tcctgtgcag cctctggatt caccttcagt ttctacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atctcatctt tcggaagtaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgctga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 466
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 466
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt tactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atctcatact ctggaagtaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgctga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 467
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 467
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt tactacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atctcacatt acggaactaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatcctctga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 468
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 468
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt cattacgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atctcatacc agggaagtaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgctga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
<210> 469
<211> 370
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 469
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct agcaacagtg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat attacaggtc caagtggtct 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaacgt ggaaacagct atggttcctt gaccactggg gccagggaac cctggtcacc 360
gtctctagtg 370
<210> 470
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 470
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt ggaaccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 471
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 471
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt gggaccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 472
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 472
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgtcagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt gggaacagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 473
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 473
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggaaccagaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 474
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 474
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg tcagtggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt ggatcggtga ctggttcatg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 475
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 475
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt ggatccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 476
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 476
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctaaccatg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt gggaccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 477
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 477
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgttacg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg tcagtggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggaaccagaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 478
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 478
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggtctggtga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 479
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 479
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcatgttg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggtctgaaga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 480
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 480
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgtcagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggcagggtaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 481
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 481
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccatcagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt ggatccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 482
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 482
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgtgacg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggacagt ggaacgaaga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 483
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 483
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggacgtt gggaaggtga ctggttcttc gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 484
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 484
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat acttcaggcg tacttggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggtctgaaga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 485
<211> 369
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 485
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggagttt ggatcggtaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtctctagt 369
<210> 486
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 486
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Arg Gly Ala Arg Cys
20
<210> 487
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 487
Met Ala Trp Ala Leu Leu Leu Leu Thr Leu Leu Thr Gln Gly Thr Gly
1. 5 10 15
Ser Trp Ala
<210> 488
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 488
Met Thr Cys Ser Pro Leu Leu Leu Thr Leu Leu Ile His Cys Thr Gly
1. 5 10 15
Ser Trp Ala
<210> 489
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 489
Met Glu Ala Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1. 5 10 15
Asp Thr Thr Gly
20
<210> 490
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 490
Met Glu Trp Thr Trp Arg Val Leu Phe Leu Val Ala Ala Ala Thr Gly
1. 5 10 15
Ala His Ser
<210> 491
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 491
Met Glu Thr Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1. 5 10 15
Asp Thr Thr Gly
20
<210> 492
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 492
Met Glu Thr Pro Ala Gln Leu Leu Phe Leu Leu Leu Leu Trp Leu Pro
1. 5 10 15
Asp Thr Thr Gly
20
<210> 493
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 493
Met Lys His Leu Trp Phe Phe Leu Leu Leu Val Ala Ala Pro Arg Trp
1. 5 10 15
Val Leu Ser
<210> 494
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 494
Met Glu Trp Ser Trp Val Phe Leu Phe Phe Leu Ser Val Thr Thr Gly
1. 5 10 15
Val His Ser
<210> 495
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 495
Met Asp Ile Arg Ala Pro Thr Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Pro Gly Ala Lys Cys
20
<210> 496
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 496
Met Asp Ile Arg Ala Pro Thr Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Pro Gly Ala Arg Cys
20
<210> 497
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 497
Met Asp Thr Arg Ala Pro Thr Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Pro Gly Ala Thr Phe
20
<210> 498
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 498
Met Asp Thr Arg Ala Pro Thr Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Pro Gly Ala Arg Cys
20
<210> 499
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 499
Met Glu Thr Gly Leu Arg Trp Leu Leu Leu Val Ala Val Leu Lys Gly
1. 5 10 15
Val Gln Cys
<210> 500
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 500
Met Glu Thr Gly Leu Arg Trp Leu Leu Leu Val Ala Val Leu Lys Gly
1. 5 10 15
Val Gln Cys Gln Glu
20
<210> 501
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 501
Met Asp Met Arg Ala Pro Thr Gln Leu Leu Gly Leu Leu Leu Leu Trp
1. 5 10 15
Leu Pro Gly Ala Arg Cys
20
<210> 502
<211> 110
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 502
Gln Ser Val Leu Thr Gln Pro Pro Ser Val Ser Ala Ala Pro Gly Gln
1. 5 10 15
Lys Val Thr Ile Ser Cys Ser Gly Ser Ser Ser Asn Ile Gly Asn Asn
20 25 30
Tyr Val Ser Trp Tyr Gln Gln Leu Pro Gly Thr Ala Pro Lys Leu Leu
35 40 45
Ile Tyr Asp Asn Asn Lys Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Lys Ser Gly Thr Ser Thr Thr Leu Gly Ile Thr Gly Leu Gln
65 70 75 80
Thr Gly Asp Glu Ala Asp Tyr Tyr Cys Gly Thr Trp Asp Ser Arg Leu
85 90 95
Ser Ala Val Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105 110
<210> 503
<211> 130
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 503
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Phe
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Phe Asp Gly Ser Ile Lys Tyr Ser Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Arg Leu Asn Tyr Tyr Asp Ser Ser Gly Tyr Tyr His Tyr
100 105 110
Lys Tyr Tyr Gly Met Ala Val Trp Gly Gln Gly Thr Thr Val Thr Val
115 120 125
Ser Ser
130
<210> 504
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 504
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 505
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 505
Asp Ile Gln Leu Thr Gln Ser Pro Ser Phe Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 506
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 506
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Arg Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 507
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 507
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1. 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Ser Val Gly Trp Ser
20 25 30
Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Lys Tyr Ala Ser Gln Ser Leu Ser Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys His Gln Ser Ser Arg Leu Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 508
<211> 142
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 508
Leu Thr Ile Ser Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys
1. 5 10 15
His Gln Ser Ser Arg Leu Pro Phe Thr Phe Gly Pro Gly Thr Lys Val
20 25 30
Asp Ile Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro
35 40 45
Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu
50 55 60
Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn
65 70 75 80
Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser
85 90 95
Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala
100 105 110
Asp Tyr Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly
115 120 125
Leu Ser Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
130 135 140
<210> 509
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 509
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Tyr Ser Pro Pro Phe
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 510
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 510
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Met Ile Ala Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 511
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 511
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Lys Ile Ala Arg Tyr
20 25 30
Leu Val Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Asn Met Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 512
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 512
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Gly Arg Ile Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Asn Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 513
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 513
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Ile Gln Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 514
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 514
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Met Ala Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 515
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 515
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Gly Met Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 516
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 516
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Tyr Ile Val Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Asn Met Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Asn Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 517
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 517
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Pro Ile Ser Arg Tyr
20 25 30
Leu Ser Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Phe Ala Gly Gln Arg Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ser Ile Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 518
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 518
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Ile Thr Ile Thr Cys Arg Ala Ser Gln Gln Ile Ala Arg Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ser Tyr Asn Leu Gln Ser Gly Ile Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Asn Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ala Ile Ile Gln Pro Tyr
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 519
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 519
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 520
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 520
Gln Val Gln Leu Val Glu Ser Gly Ala Glu Val Val Lys Pro Gly Ala
1. 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Val Ile Ser Tyr Asp Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Leu Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 521
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 521
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 522
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 522
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Arg Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 523
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 523
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Gln Gly Asn Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 524
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 524
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Lys Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Arg Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Leu Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 525
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 525
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Ile Gly Gly Asn Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 526
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 526
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asn Gly Arg Asn Lys Tyr Tyr Ala Arg Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Ile Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 527
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 527
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Asn Tyr Arg Gly His Gly Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 528
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 528
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Phe Ser Gly Gly Ser Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Leu Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 529
<211> 450
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 529
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Phe
20 25 30
Ala Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Val Ile Ser Tyr Thr Gly Gln Phe Lys Tyr Tyr Ala Glu Ser Val
50 55 60
Lys Gly Arg Phe Thr Val Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Leu Phe Tyr Cys
85 90 95
Ala Arg Gly Tyr Asp Val Leu Thr Gly Tyr Pro Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser Thr Tyr Arg
290 295 300
Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 530
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 530
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Asn
20 25 30
Ser Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Ser Lys Trp Ser Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Lys Gln Leu Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 531
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 531
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Val Trp Ile Gly Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 532
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 532
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Lys
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Asn Gln Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 533
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 533
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Gln Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Gln Gly Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 534
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 534
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Arg Trp Glu Gly Asp Trp Phe Phe Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 535
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 535
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Asn Arg
20 25 30
Leu Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Lys Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Thr Trp Asn Gln Asp Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 536
<211> 453
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 536
Gln Val Gln Leu Gln Gln Ser Gly Pro Gly Leu Val Lys Pro Ser Gln
1. 5 10 15
Thr Leu Ser Leu Thr Cys Ala Ile Ser Gly Asp Ser Val Ser Ser Arg
20 25 30
Tyr Ala Thr Trp Asn Trp Ile Arg Gln Ser Pro Ser Arg Gly Leu Glu
35 40 45
Trp Leu Gly Arg Thr Tyr Tyr Arg Gly Gln Trp Lys Asn His Tyr Ala
50 55 60
Val Ser Val Lys Ser Arg Ile Thr Ile Asn Pro Asp Thr Ser Lys Ser
65 70 75 80
Gln Phe Ser Leu Gln Leu Asn Ser Val Thr Pro Glu Asp Thr Ala Val
85 90 95
Tyr Tyr Cys Ala Arg Gly Met Trp Asn Gln Asn Trp Phe Leu Asp His
100 105 110
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Cys Glu Glu Gln Tyr Gly Ser
290 295 300
Thr Tyr Arg Cys Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 537
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 537
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccca gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 538
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 538
gatatccagc tcactcaatc gccatcattt ctctccgctt cggtaggcga ccgggtcacg 60
atcacatgca gggcgtcgca aagcattggg aggtcgttgc attggtatca gcagaaaccc 120
ggaaaggccc cgaaacttct gatcaaatac gcatcacaaa gcttgagcgg tgtgccgtcg 180
cgcttctccg gttccggaag cggaacggaa ttcacgctta caatctcctc actgcagccc 240
gaggatttcg cgacctatta ctgtcaccag tcatccagac tcccgtttac ttttggccct 300
gggaccaagg tggacattaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 539
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 539
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa gtccgtcgga cgatcattgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 540
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 540
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa atccgtcggg tggagcttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 541
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 541
gagatcgtac ttactcagtc acccgccaca ttgtccctga gcccgggtga acgggcgacc 60
ctcagctgcc gagcatccaa atccgtcggg tacagcttgc actggtacca acaaaaaccg 120
ggccaggccc ccagacttct gatcaagtat gcgtcacaga gcttgtcggg tattcccgct 180
cgcttttcgg ggtcgggatc cgggacagat ttcacgctca caatctcctc gctggaaccc 240
gaggacttcg cggtctacta ttgtcatcag tcatcgaggt tgcctttcac gtttggacca 300
gggaccaagg tggacattaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 542
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 542
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctatgct gcatccagtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag agttacagtc ccccattcac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 543
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 543
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gatgattgct cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatca acccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 544
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 544
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gaaaattgct cgttacttag tttggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct gctaacatgt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 545
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 545
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct ggtcgtatct tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatca acccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 546
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 546
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag tctatccagc agccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 547
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 547
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatgg ctccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 548
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 548
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gagcattagc aggtatttaa attggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcggta tgccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 549
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 549
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gtacattgtt cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct gctaacatgt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcaacc agccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 550
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 550
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gccgatttct cgttacttat cttggtatca acagaaacca 120
gggaaagccc ctaaactcct gatcttcgct ggtcagcgtt tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatctcta tcccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 551
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 551
gacatccaga tgacccagtc tccatcctcc ctgtctgcat ctgtaggaga cagaatcacc 60
atcacttgcc gggcaagtca gcagattgct cgttacttaa actggtatca acagaaacca 120
gggaaagccc ctaaactcct gatctacgct tcttacaact tgcaaagtgg gatcccatca 180
aggttcagcg gcagtggatc tgggacagat ttcactctca ccatcaacag tctgcaacct 240
gaagattttg caacttactt ctgtcaacag gctatcatcc agccatacac tttcggccct 300
gggaccaaag tggatatcaa acgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gt 642
<210> 552
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 552
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatg atggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 553
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 553
caagttcagt tggtggagtc tggagccgaa gtagtaaagc caggagcttc agtgaaagtc 60
tcttgtaaag caagtggatt cacgtttagc cgctttgcca tgcattgggt gcggcaagct 120
cccggtcagg ggttggagtg gatgggagtt attagctatg acgggggcaa taagtactac 180
gccgagtctg ttaagggtcg ggtcacaatg acacgggaca cctcaaccag tacactctat 240
atggaactgt ctagcctgag atccgaggac accgctgtgt attattgcgc tagggggtac 300
gatgtattga cgggttatcc tgattactgg gggcagggga cactcgtaac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacggc 900
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 554
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 554
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 555
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 555
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggacgcaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 556
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 556
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc agggaaacaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 557
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 557
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt aagtttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatatc gcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatctgttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 558
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 558
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata tcggaggaaa taaatactat 180
gcagagtccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 559
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 559
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtggcagtt atatcatata acggacgcaa taaatactat 180
gcacgctccg tgaagggccg gttcaccatc tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatatcttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtctctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 560
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 560
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atcaactatc gtggacatgg taaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 561
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 561
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atatcttttt ctggaggttc taaatactat 180
gcagagtccg tgaagggccg gttcaccttg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 562
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 562
caggtgcagc tggtggagtc tgggggaggc gtggtccagc ctgggaggtc cctgcgactc 60
tcctgtgcag cctctggatt caccttcagt agatttgcca tgcactgggt ccgccaggct 120
ccaggcaagg ggctggagtg ggtgggtgtt atctcttata ctggacagtt caaatactat 180
gcagagtccg tgaagggccg gttcaccgtg tccagagaca attccaagaa caccctgtat 240
ctgcaaatga acagcctgag agctgaggac acggctctgt tttactgtgc gagaggatac 300
gatgttttga ctggttaccc cgactactgg ggccagggaa ccctggtcac cgtgtctagt 360
gcctccacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 420
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 480
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 540
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 600
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 660
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 720
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 780
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 840
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgtgcgagga gcagtacggc 900
agcacgtacc gttgcgtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 960
gagtacaagt gcaaggtgtc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 1020
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 1080
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 1140
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 1200
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 1260
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 1320
cagaagagcc tctccctgtc tccgggtaaa 1350
<210> 563
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 563
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct agcaacagtg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat attacaggtc caagtggtct 180
aatcattatg cagtatctgt gaaaagtcga atcaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaacgt ggaaacagct atggttcctt gaccactggg gccagggaac cctggtcacc 360
gtgtctagtg cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggcaaa 1359
<210> 564
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 564
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggagttt ggatcggtaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<210> 565
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 565
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aacaaacagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggaaccagaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<210> 566
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 566
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgtcagg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggcagggtaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<210> 567
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 567
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggacgtt gggaaggtga ctggttcttc gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<210> 568
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 568
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct aaccgtctgg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg taaatggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaactt ggaaccagga ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<210> 569
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид
<400> 569
caggtacagc tgcagcagtc aggtccagga ctggtgaagc cctcgcagac cctctcactc 60
acctgtgcca tctccgggga cagtgtctct tctcgttacg ctacttggaa ctggatcagg 120
cagtccccat cgagaggcct tgagtggctg ggaaggacat actacagggg tcagtggaaa 180
aatcattatg cagtatctgt gaaaagtcga ataaccatca accccgacac gtccaagagc 240
cagttctccc tgcagctgaa ctctgtgact cccgaggaca cggctgtgta ttactgtgca 300
agaggaatgt ggaaccagaa ctggttcctg gaccactggg gccagggaac cctggtcacc 360
gtgtcctcag cctccaccaa gggcccatcg gtcttccccc tggcaccctc ctccaagagc 420
acctctgggg gcacagcggc cctgggctgc ctggtcaagg actacttccc cgaaccggtg 480
acggtgtcgt ggaactcagg cgccctgacc agcggcgtgc acaccttccc ggctgtccta 540
cagtcctcag gactctactc cctcagcagc gtggtgaccg tgccctccag cagcttgggc 600
acccagacct acatctgcaa cgtgaatcac aagcccagca acaccaaggt ggacaagaaa 660
gttgagccca aatcttgtga caaaactcac acatgcccac cgtgcccagc acctgaactc 720
ctggggggac cgtcagtctt cctcttcccc ccaaaaccca aggacaccct catgatctcc 780
cggacccctg aggtcacatg cgtggtggtg gacgtgagcc acgaagaccc tgaggtcaag 840
ttcaactggt acgtggacgg cgtggaggtg cataatgcca agacaaagcc gtgcgaggag 900
cagtacggca gcacgtaccg ttgcgtcagc gtcctcaccg tcctgcacca ggactggctg 960
aatggcaagg agtacaagtg caaggtgtcc aacaaagccc tcccagcccc catcgagaaa 1020
accatctcca aagccaaagg gcagccccga gaaccacagg tgtacaccct gcccccatcc 1080
cgggaggaga tgaccaagaa ccaggtcagc ctgacctgcc tggtcaaagg cttctatccc 1140
agcgacatcg ccgtggagtg ggagagcaat gggcagccgg agaacaacta caagaccacg 1200
cctcccgtgc tggactccga cggctccttc ttcctctata gcaagctcac cgtggacaag 1260
agcaggtggc agcaggggaa cgtcttctca tgctccgtga tgcatgaggc tctgcacaac 1320
cactacacgc agaagagcct ctccctgtct ccgggtaaa 1359
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИТЕЛА К CD40L И СПОСОБЫ ЛЕЧЕНИЯ СВЯЗАННЫХ С CD40L ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2016 |
|
RU2727646C2 |
| АНТИТЕЛА, ИНДУЦИРУЮЩИЕ ИММУННУЮ ТОЛЕРАНТНОСТЬ, ИНДУЦИРОВАННЫЕ ЛИМФОЦИТЫ И ТЕРАПЕВТИЧЕСКИЙ АГЕНТ/СПОСОБ КЛЕТОЧНОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ИНДУЦИРОВАННЫХ ЛИМФОЦИТОВ | 2019 |
|
RU2816592C2 |
| АНТИТЕЛО К EPHA4 | 2016 |
|
RU2719158C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| CD47-АНТИГЕНСВЯЗЫВАЮЩАЯ ЕДИНИЦА И ЕЕ ПРИМЕНЕНИЕ | 2018 |
|
RU2780859C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| Композиция для предупреждения и лечения заболевания кожи, содержащая вещество, специфично связывающееся с пептидом, имеющим происхождение из виментина | 2018 |
|
RU2751486C1 |
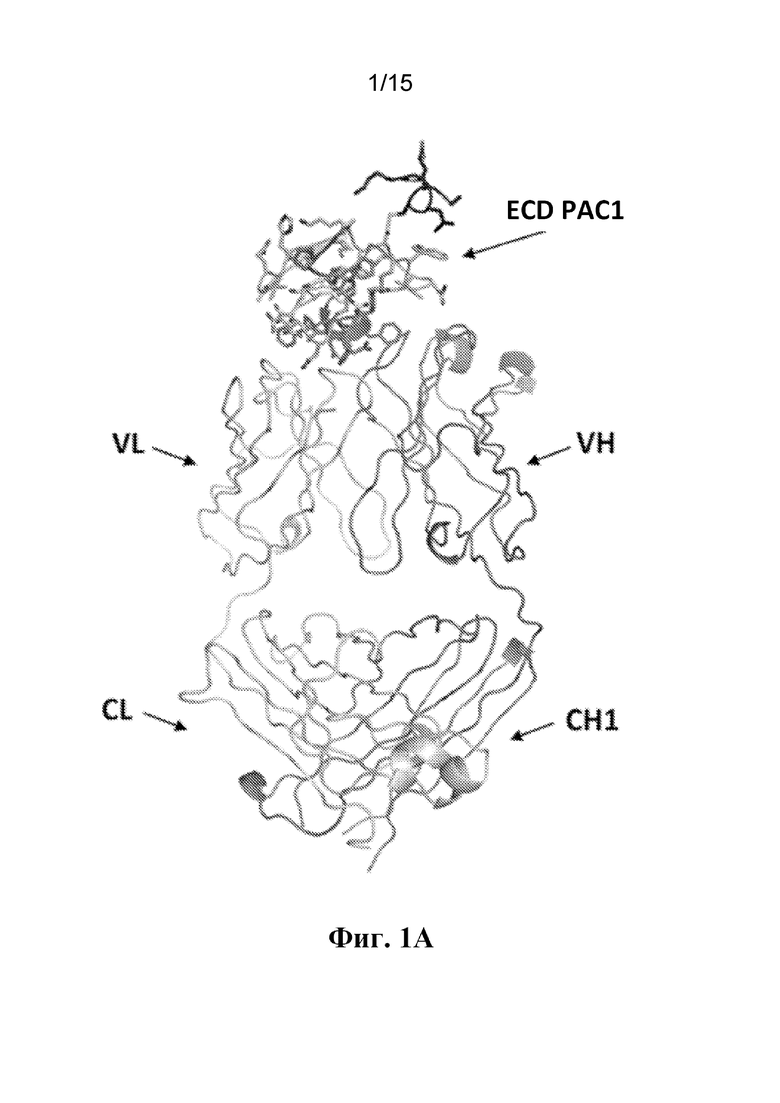
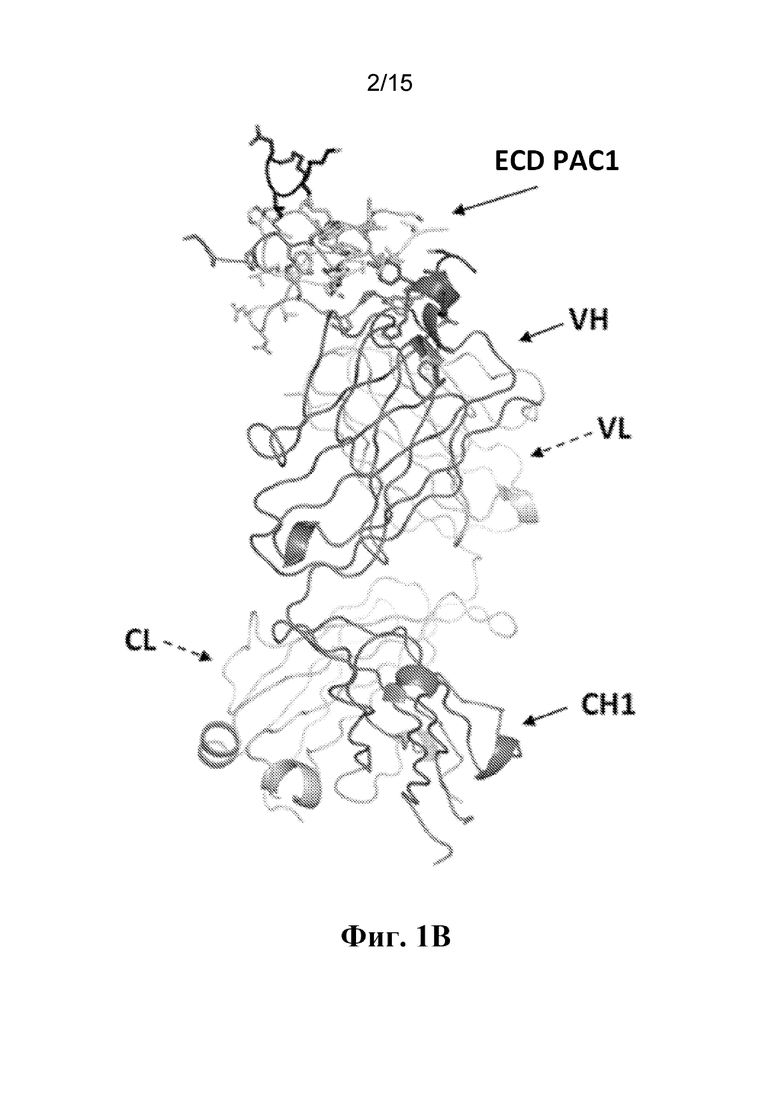
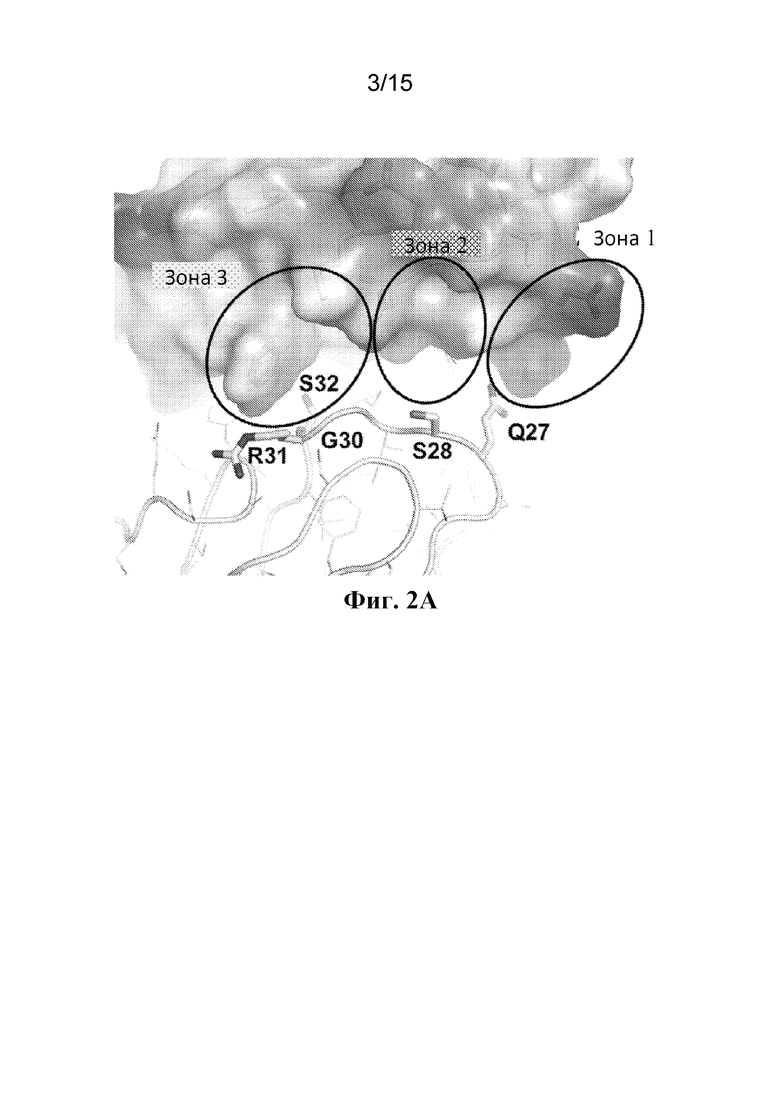
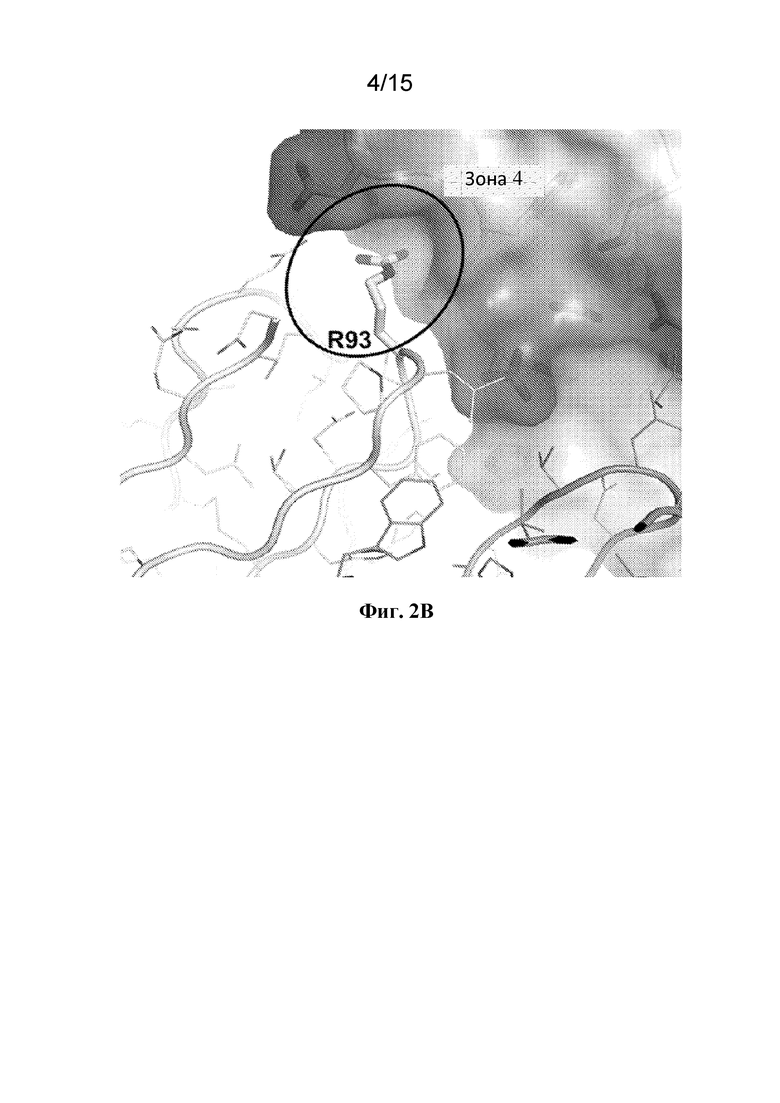
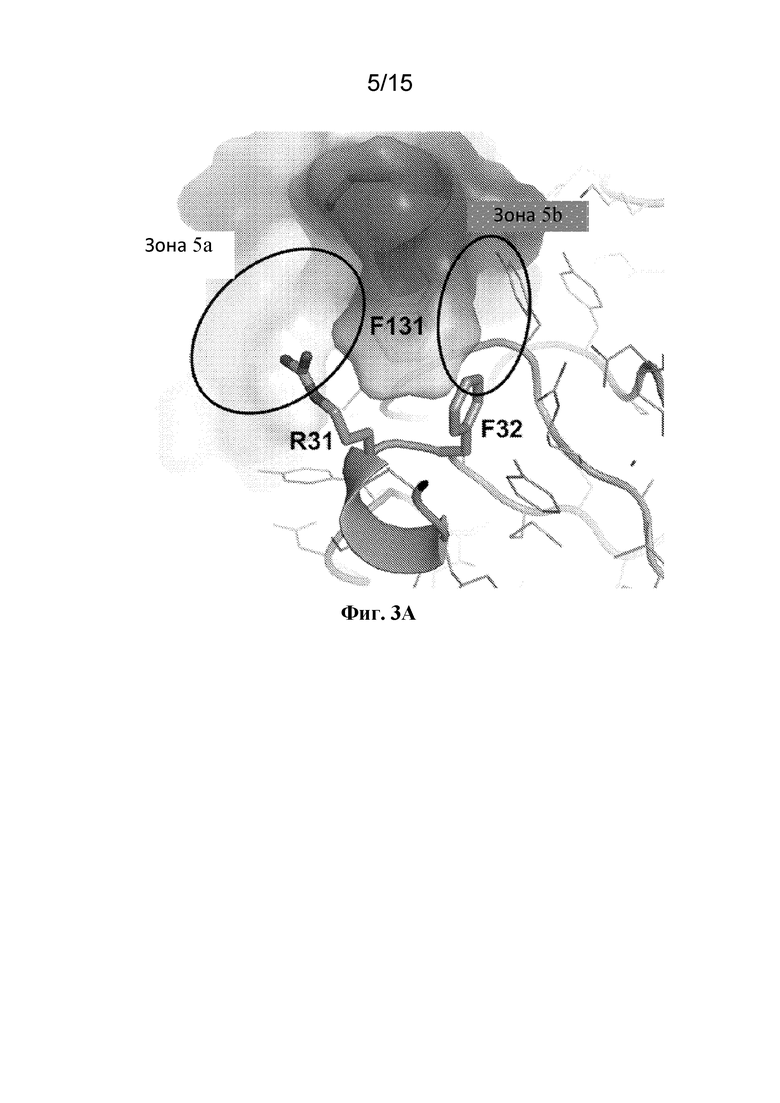
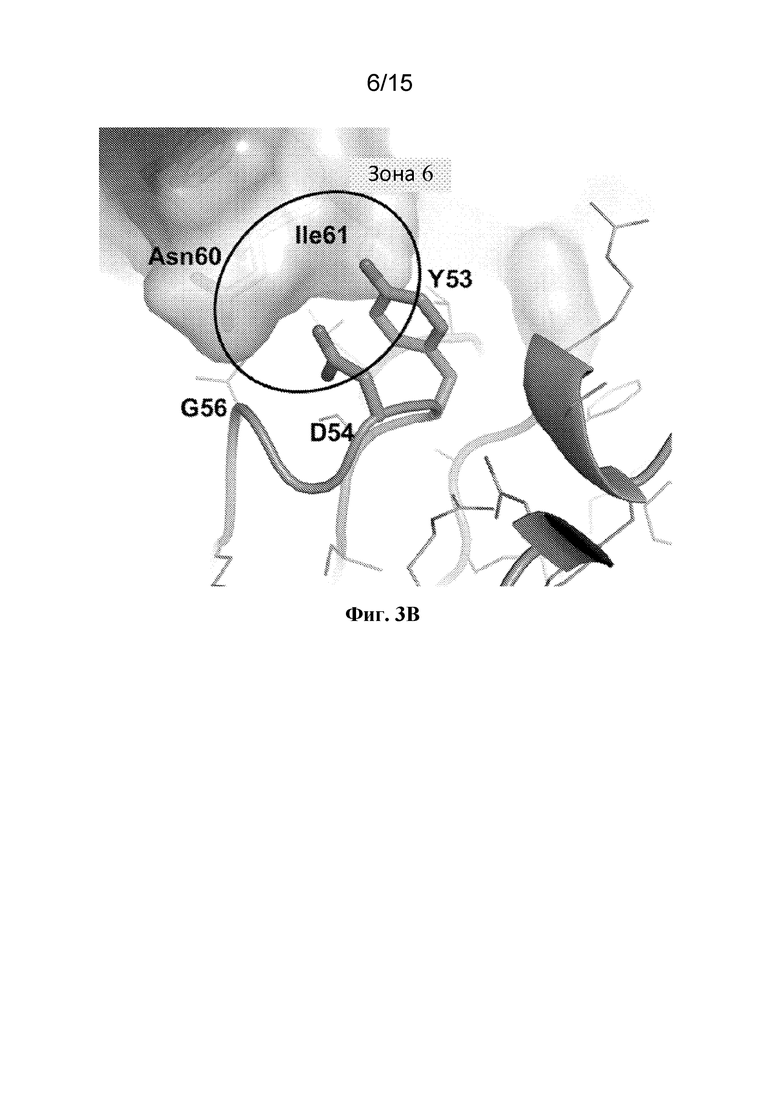
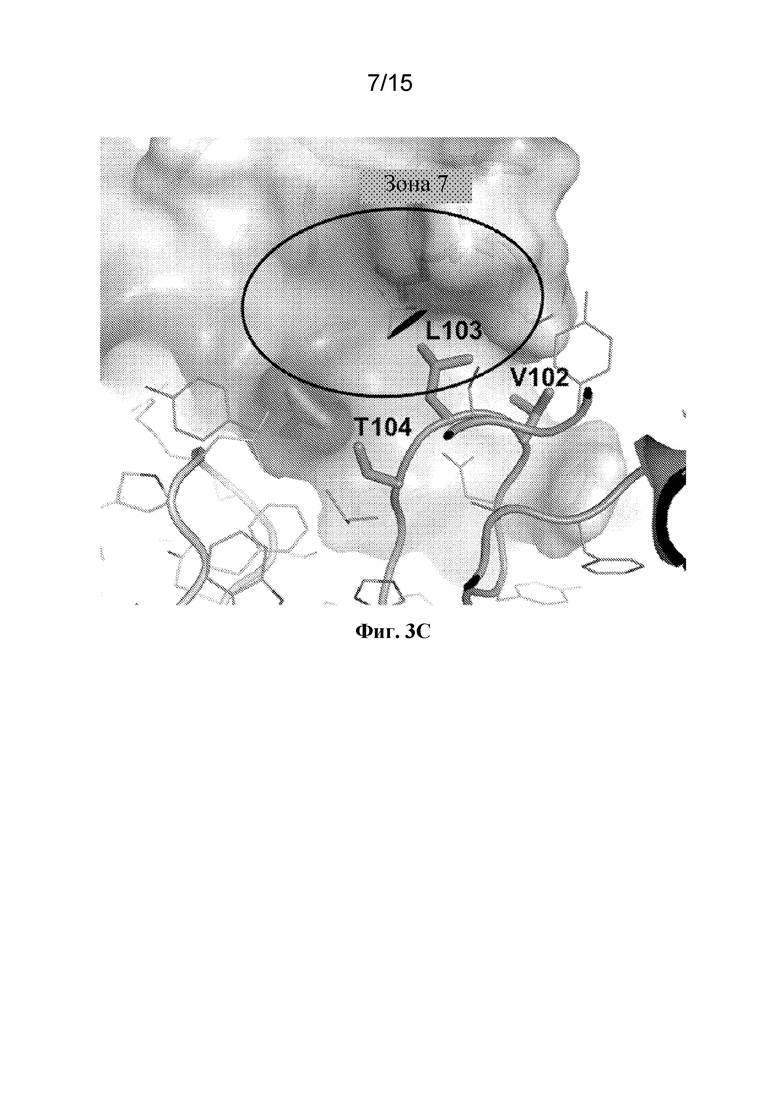
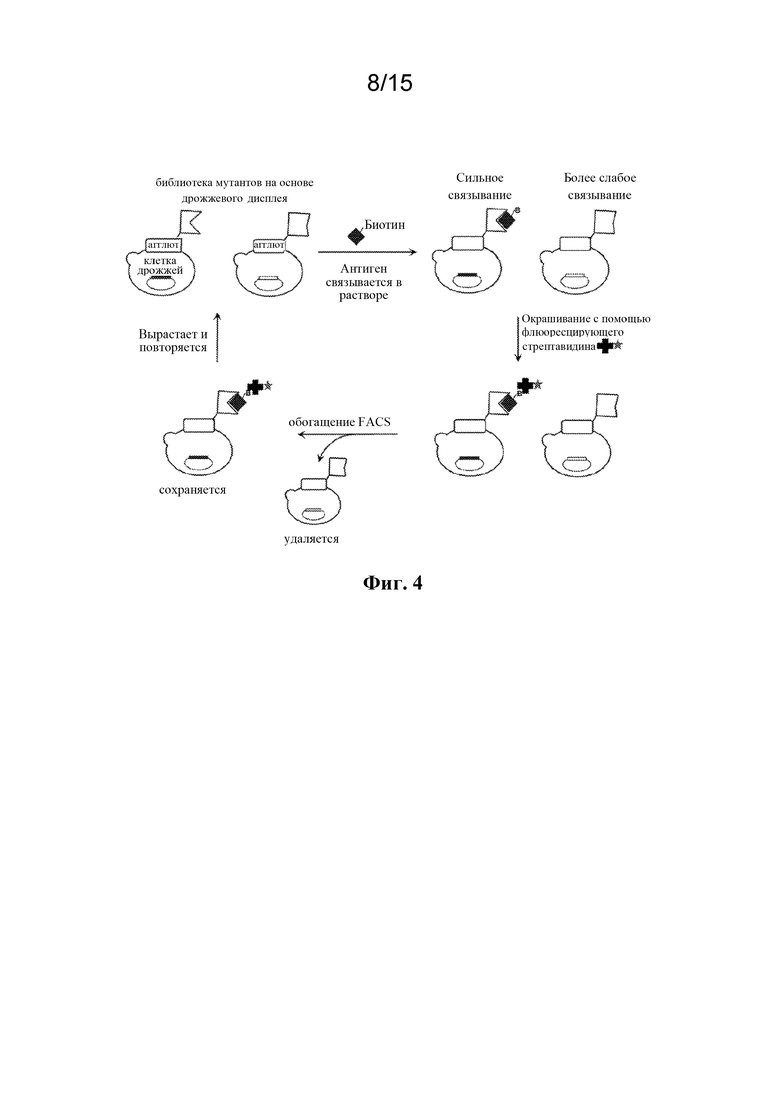
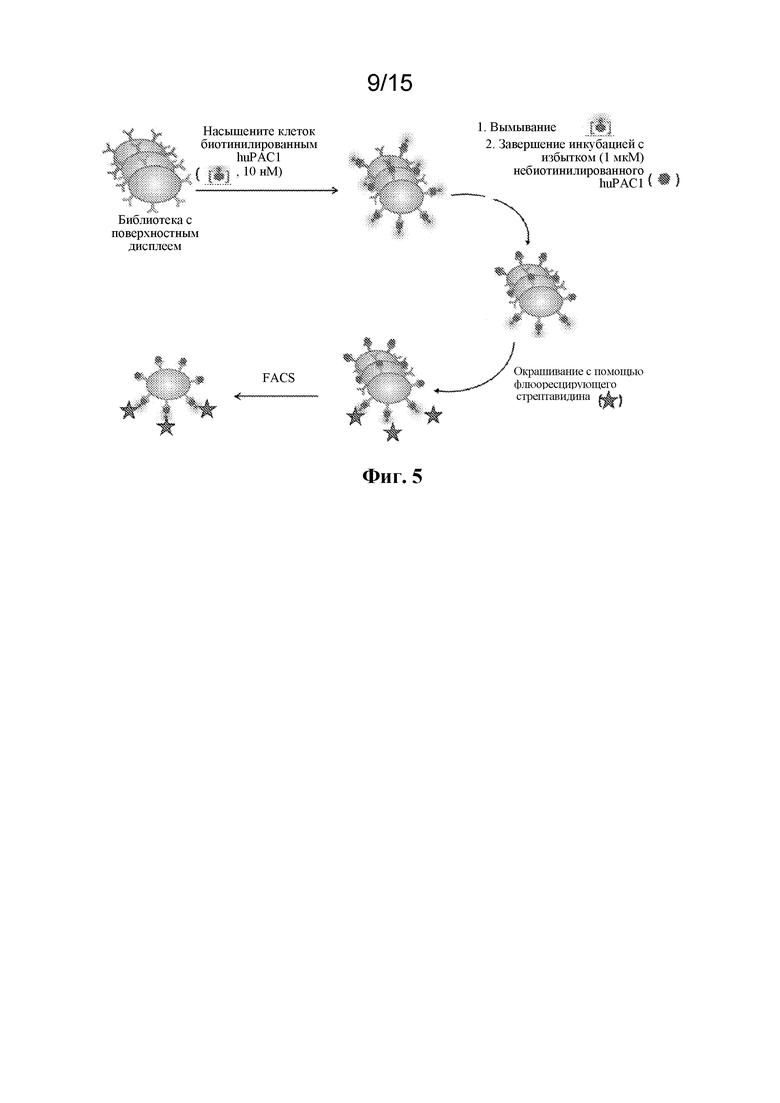
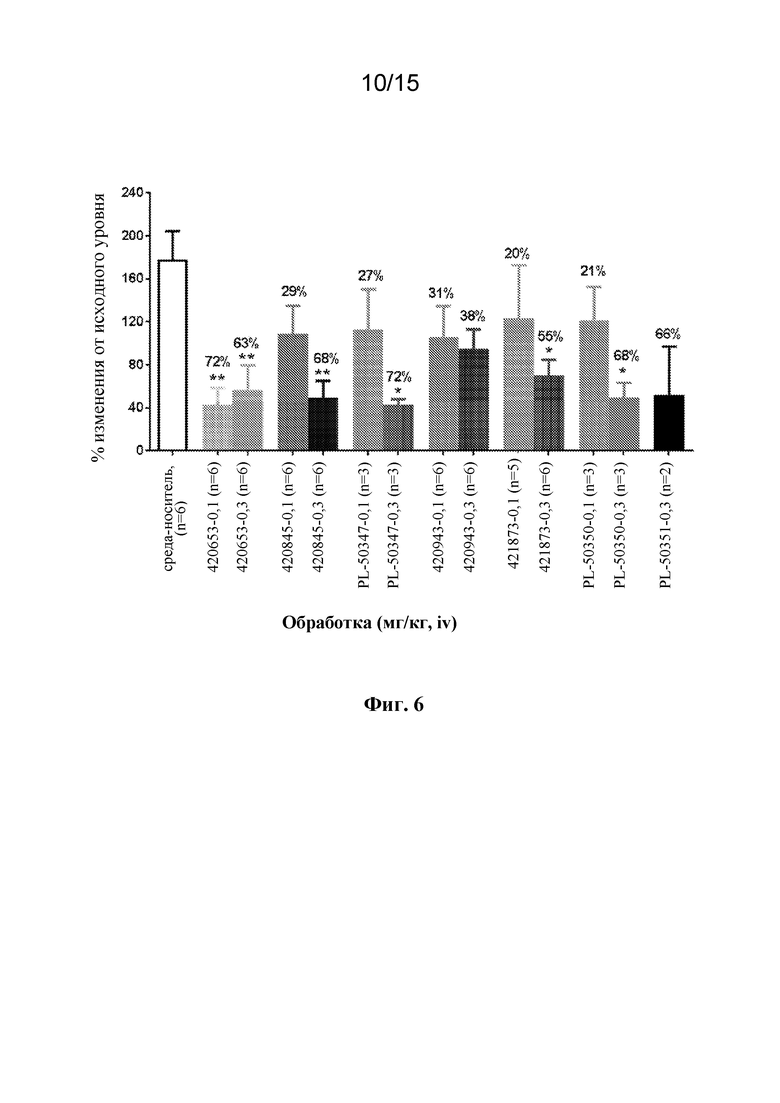
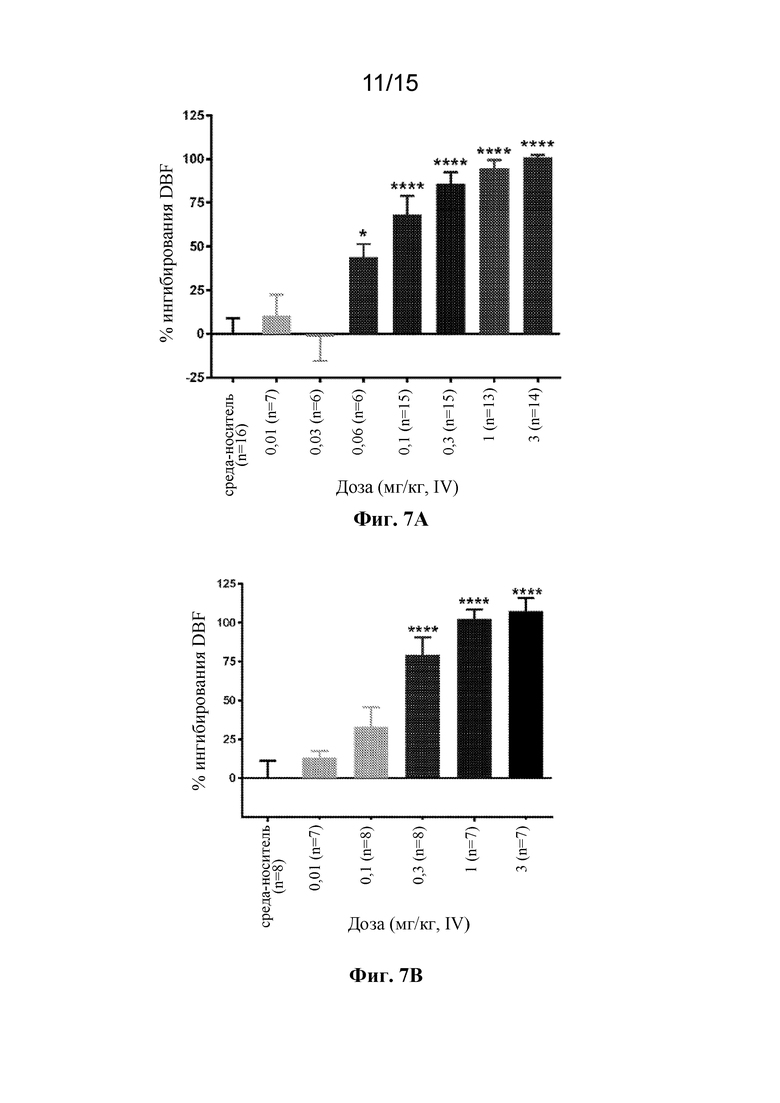
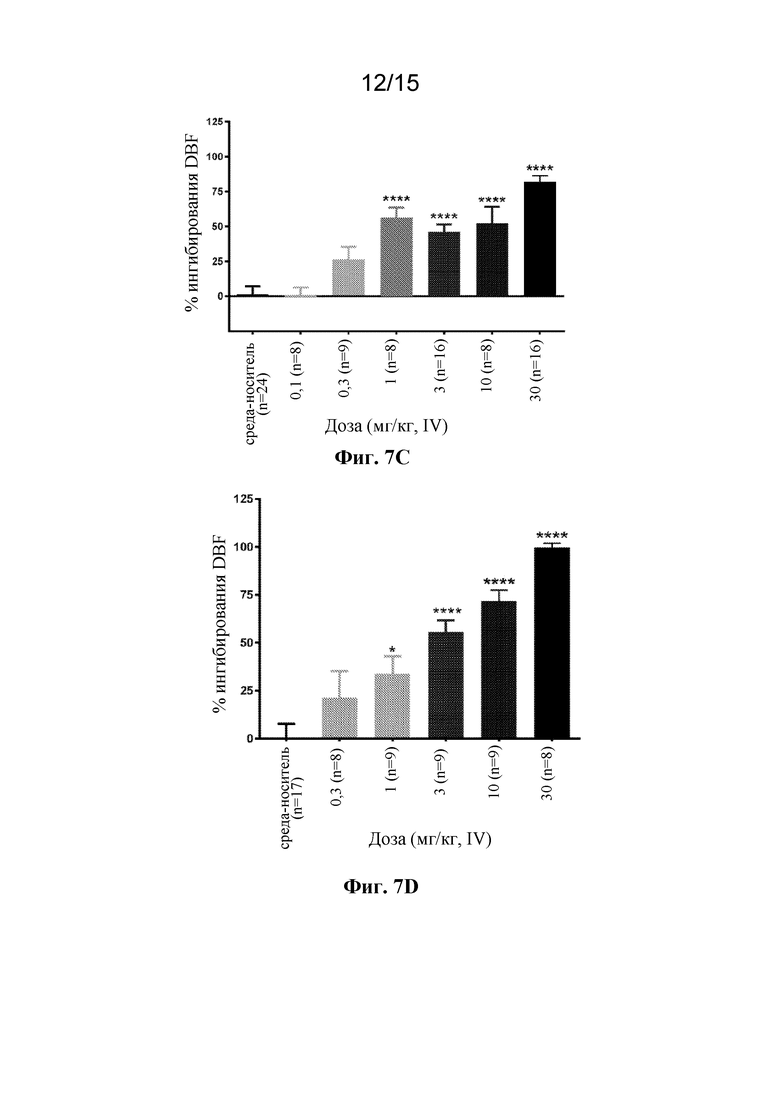
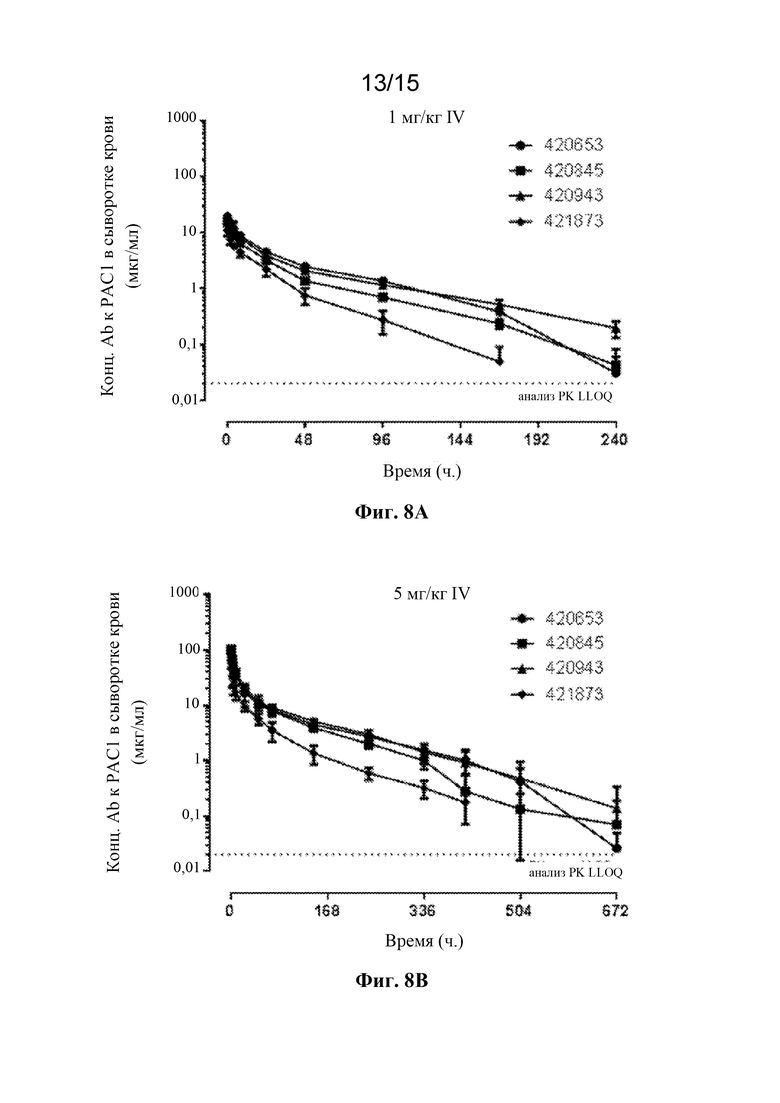
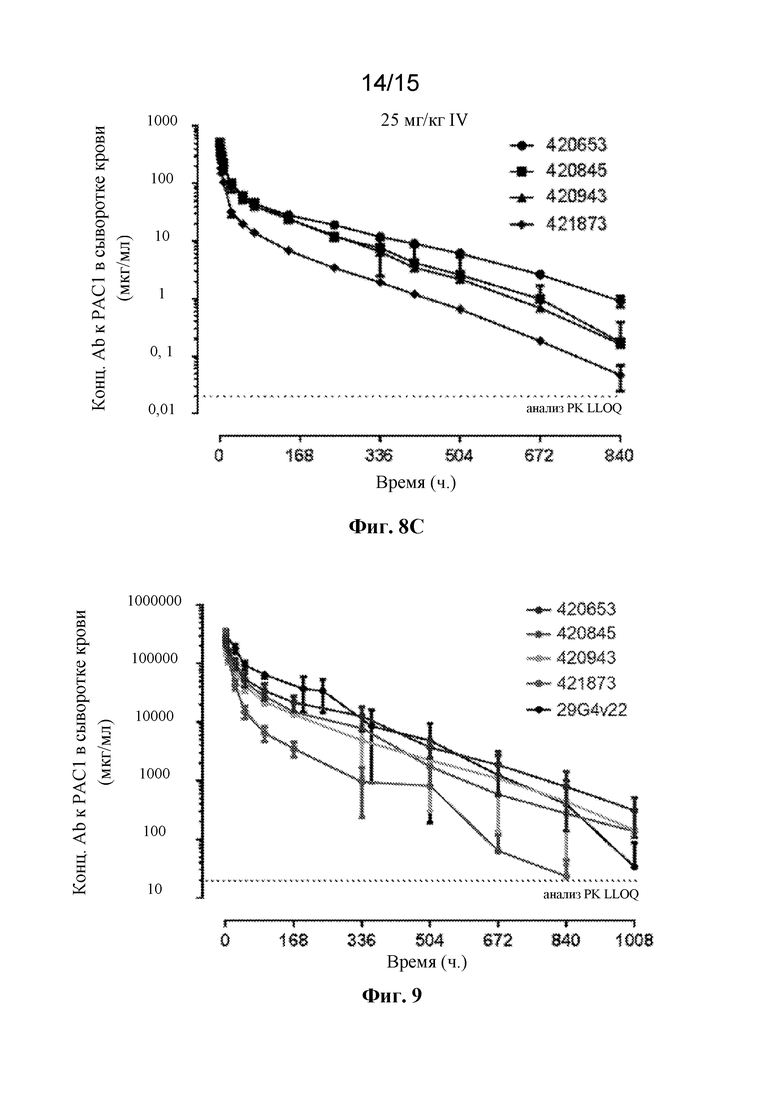
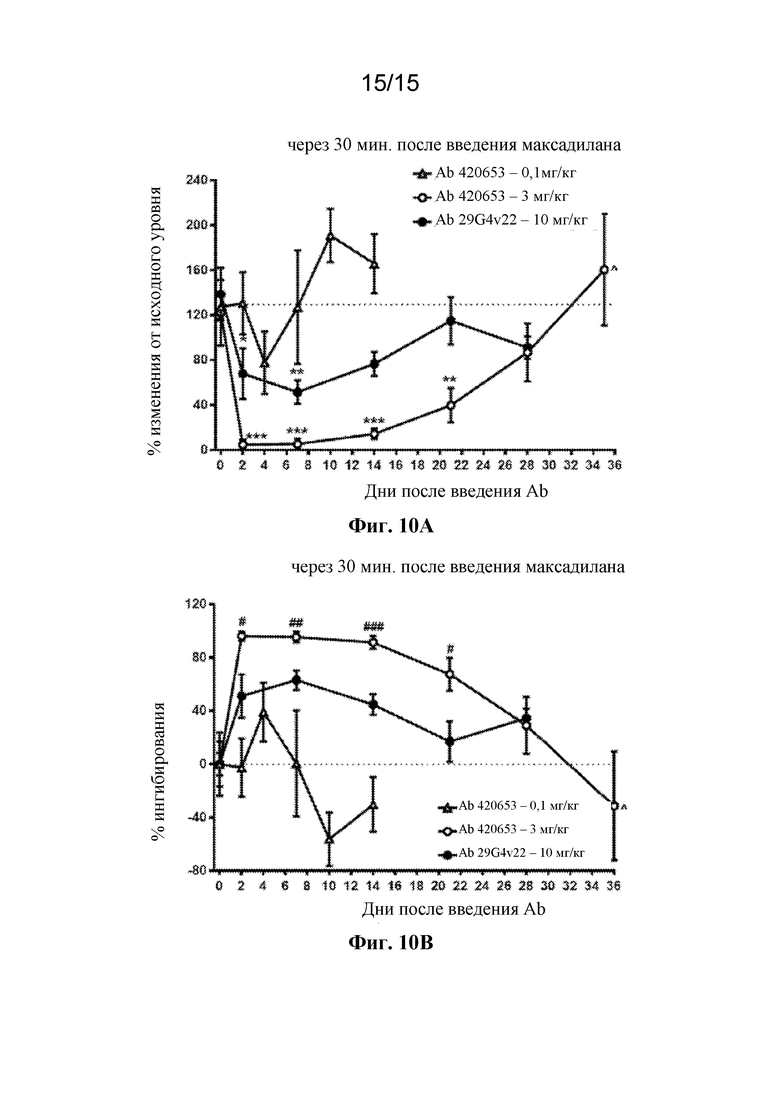
Изобретение относится к области биотехнологии. Предложены нейтрализующие антитела к рецептору I типа гипофизарного полипептида человека, активирующего аденилатциклазу (PAC1), а также фармацевтические композиции, содержащие такие антитела. Также предложены полинуклеотиды, векторы экспрессии и клетки-хозяева для получения антител. Кроме того, описаны способы лечения или предупреждения состояний, связанных с головной болью, таких как мигрень и кластерная головная боль, с применением указанных нейтрализующих антител. Изобретение обеспечивает повышенную ингибирующую активность против PAC1 человека. 9 н. и 35 з.п. ф-лы, 20 ил., 24 табл., 7 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфически связывается с рецептором I типа гипофизарного полипептида человека, активирующего аденилатциклазу (PAC1) человека,
где антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую определяющие комплементарность области CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую определяющие комплементарность области CDRH1, CDRH2 и CDRH3, где
(a) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:7, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 108 и 171 соответственно;
(b) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:7, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 116 и 171 соответственно;
(c) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:5, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 134 и 171 соответственно;
(d) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:5, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:92, 145 и 174 соответственно;
(e) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:7, 26 и 36 соответственн, и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 108 и 172 соответственно;
(f) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:5, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 128 и 172 соответственно;
(g) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:7, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 134 и 171 соответственно;
(h) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:12, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 153 и 171 соответственно;
(i) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:12, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 154 и 171 соответственно; или
(j) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:13, 26 и 36 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:88, 155 и 171 соответственно.
2. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1, где вариабельная область легкой цепи содержит
(i) последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из SEQ ID NO:54-66,
(ii) последовательность, которая по меньшей мере на 95% идентична последовательности, выбранной из SEQ ID NO:54-66, или
(iii) последовательность, выбранную из SEQ ID NO:54-66.
3. Выделенное антитело или его антигенсвязывающий фрагмент по п. 1 или 2, где вариабельная область тяжелой цепи содержит
(i) последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из SEQ ID NO:191-295,
(ii) последовательность, которая по меньшей мере на 95% идентична последовательности, выбранной из SEQ ID NO:191-295, или
(iii) последовательность, выбранную из SEQ ID NO:191-295.
4. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-3, где
(a) вариабельная область легкой цепи содержит последовательность SEQ ID NO:54, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:194;
(b) вариабельная область легкой цепи содержит последовательность SEQ ID NO:54, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:203;
(c) вариабельная область легкой цепи содержит последовательность SEQ ID NO:55, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:228;
(d) вариабельная область легкой цепи содержит последовательность SEQ ID NO:55, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:274;
(e) вариабельная область легкой цепи содержит последовательность SEQ ID NO:54, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:214;
(f) вариабельная область легкой цепи содержит последовательность SEQ ID NO:55, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:240;
(g) вариабельная область легкой цепи содержит последовательность SEQ ID NO:54, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:228;
(h) вариабельная область легкой цепи содержит последовательность SEQ ID NO:62, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:282;
(i) вариабельная область легкой цепи содержит последовательность SEQ ID NO:62, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:283; или
(j) вариабельная область легкой цепи содержит последовательность SEQ ID NO:63, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:284.
5. Выделенное антитело или его антигенсвязывающий фрагмент, которое специфически связывается с PAC1 человека,
где антитело или его антигенсвязывающий фрагмент содержит вариабельную область легкой цепи, содержащую определяющие комплементарность области CDRL1, CDRL2 и CDRL3, и вариабельную область тяжелой цепи, содержащую определяющие комплементарность области CDRH1, CDRH2 и CDRH3, где
(a) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:25, 31 и 42 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:98, 168 и 190 соответственно;
(b) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:20, 30 и 44 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:100, 168 и 182 соответственно;
(c) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:17, 33 и 42 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:99, 168 и 187 соответственно;
(d) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:23, 31 и 50 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:97, 167 и 178 соответственно;
(e) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:17, 31 и 44 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:98, 168 и 189 соответственно;
(f) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:17, 27 и 39 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:100, 168 и 182 соответственно; или
(g) CDRL1, CDRL2 и CDRL3 имеют последовательности SEQ ID NO:18, 31 и 46 соответственно и CDRH1, CDRH2 и CDRH3 имеют последовательности SEQ ID NO:98, 168 и 179 соответственно.
6. Выделенное антитело или его антигенсвязывающий фрагмент по п. 5, где вариабельная область легкой цепи содержит
(i) последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из SEQ ID NO:68-87,
(ii) последовательность, которая по меньшей мере на 95% идентична последовательности, выбранной из SEQ ID NO:68-87, или
(iii) последовательность, выбранную из SEQ ID NO:68-87.
7. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 5 или 6, где вариабельная область тяжелой цепи содержит
(i) последовательность, которая по меньшей мере на 90% идентична последовательности, выбранной из SEQ ID NO:296-312,
(ii) последовательность, которая по меньшей мере на 95% идентична последовательности, выбранной из SEQ ID NO:296-312, или
(iii) последовательность, выбранную из SEQ ID NO:296-312.
8. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 5-7, где
(a) вариабельная область легкой цепи содержит последовательность SEQ ID NO:85, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:312;
(b) вариабельная область легкой цепи содержит последовательность SEQ ID NO:72, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:300;
(c) вариабельная область легкой цепи содержит последовательность SEQ ID NO:81, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:307;
(d) вариабельная область легкой цепи содержит последовательность SEQ ID NO:78, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:296;
(e) вариабельная область легкой цепи содержит последовательность SEQ ID NO:83, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:310;
(f) вариабельная область легкой цепи содержит последовательность SEQ ID NO:87, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:300; или
(g) вариабельная область легкой цепи содержит последовательность SEQ ID NO:74, и вариабельная область тяжелой цепи содержит последовательность SEQ ID NO:297.
9. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-8, где антитело или его антигенсвязывающий фрагмент представляет собой моноклональное антитело или его антигенсвязывающий фрагмент.
10. Выделенное антитело или его антигенсвязывающий фрагмент по п. 9, где моноклональное антитело или его антигенсвязывающий фрагмент представляет собой гуманизированное антитело или его антигенсвязывающий фрагмент или полностью человеческое антитело или его антигенсвязывающий фрагмент.
11. Выделенное антитело или его антигенсвязывающий фрагмент по п. 9 или 10, где моноклональное антитело содержит легкую цепь, которая содержит константную область каппа-цепи человека.
12. Выделенное антитело или его антигенсвязывающий фрагмент по п. 11, где константная область каппа-цепи человека содержит последовательность SEQ ID NO:318 или SEQ ID NO:319.
13. Выделенное антитело или его антигенсвязывающий фрагмент по п. 9 или 10, где моноклональное антитело содержит легкую цепь, которая содержит константную область лямбда-цепи человека.
14. Выделенное антитело или его антигенсвязывающий фрагмент по п. 13, где константная область лямбда-цепи человека содержит последовательность SEQ ID NO:315.
15. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 9-14, где моноклональное антитело представляет собой антитело IgG1, IgG2, IgG3 или IgG4 человека.
16. Выделенное антитело или его антигенсвязывающий фрагмент по п. 15, где моноклональное антитело представляет собой антитело IgG1 человека.
17. Выделенное антитело или его антигенсвязывающий фрагмент по п. 16, где моноклональное антитело представляет собой агликозилированное антитело IgG1 человека.
18. Выделенное антитело или его антигенсвязывающий фрагмент по п. 17, где моноклональное антитело содержит в своей тяжелой цепи мутацию в аминокислотном положении N297 согласно нумерации EU.
19. Выделенное антитело или его антигенсвязывающий фрагмент по п. 18, где мутация представляет собой N297G.
20. Выделенное антитело или его антигенсвязывающий фрагмент по п. 18 или 19, где моноклональное антитело дополнительно содержит в своей тяжелой цепи мутации R292C и V302C согласно нумерации EU.
21. Выделенное антитело или его антигенсвязывающий фрагмент по п. 17, где моноклональное антитело содержит константную область тяжелой цепи, которая содержит последовательность SEQ ID NO:324 или SEQ ID NO:325.
22. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-21, где антитело или его антигенсвязывающий фрагмент ингибирует индуцированную PACAP активацию PAC1 крысы при IC50, составляющей менее 10 нМ, как измерено с помощью клеточного анализа cAMP.
23. Выделенное антитело, которое специфически связывается с PAC1 человека, где антитело содержит легкую цепь и тяжелую цепь, где
(a) легкая цепь содержит последовательность SEQ ID NO:506, и тяжелая цепь содержит последовательность SEQ ID NO:521;
(b) легкая цепь содержит последовательность SEQ ID NO:506, и тяжелая цепь содержит последовательность SEQ ID NO:522;
(c) легкая цепь содержит последовательность SEQ ID NO:504, и тяжелая цепь содержит последовательность SEQ ID NO:523;
(d) легкая цепь содержит последовательность SEQ ID NO:504, и тяжелая цепь содержит последовательность SEQ ID NO:524;
(e) легкая цепь содержит последовательность SEQ ID NO:506, и тяжелая цепь содержит последовательность SEQ ID NO:525;
(f) легкая цепь содержит последовательность SEQ ID NO:504, и тяжелая цепь содержит последовательность SEQ ID NO:526;
(g) легкая цепь содержит последовательность SEQ ID NO:506, и тяжелая цепь содержит последовательность SEQ ID NO:523;
(h) легкая цепь содержит последовательность SEQ ID NO:507, и тяжелая цепь содержит последовательность SEQ ID NO:527;
(i) легкая цепь содержит последовательность SEQ ID NO:507, и тяжелая цепь содержит последовательность SEQ ID NO:528;
(j) легкая цепь содержит последовательность SEQ ID NO:508, и тяжелая цепь содержит последовательность SEQ ID NO:529;
(k) легкая цепь содержит последовательность SEQ ID NO:510, и тяжелая цепь содержит последовательность SEQ ID NO:531;
(l) легкая цепь содержит последовательность SEQ ID NO:511, и тяжелая цепь содержит последовательность SEQ ID NO:532;
(m) легкая цепь содержит последовательность SEQ ID NO:512, и тяжелая цепь содержит последовательность SEQ ID NO:533;
(n) легкая цепь содержит последовательность SEQ ID NO:513, и тяжелая цепь содержит последовательность SEQ ID NO:534;
(o) легкая цепь содержит последовательность SEQ ID NO:514, и тяжелая цепь содержит последовательность SEQ ID NO:535;
(p) легкая цепь содержит последовательность SEQ ID NO:509, и тяжелая цепь содержит последовательность SEQ ID NO:532; или
(q) легкая цепь содержит последовательность SEQ ID NO:518, и тяжелая цепь содержит последовательность SEQ ID NO:530.
24. Фармацевтическая композиция для лечения или профилактики состояния, связанного с головной болью, содержащая выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-23 и фармацевтически приемлемое вспомогательное вещество.
25. Выделенный полинуклеотид, который кодирует выделенное антитело или его антигенсвязывающий фрагмент по любому из пп. 1-23.
26. Вектор экспрессии, содержащий полинуклеотид по п. 25.
27. Клетка-хозяин для получения антитела или антигенсвязывающего фрагмента, которое специфически связывается с PAC1 человека, содержащая вектор экспрессии по п. 26.
28. Способ получения антитела или его антигенсвязывающего фрагмента, которое специфически связывается с PAC1 человека, где способ включает культивирование клетки-хозяина по п. 27 в условиях, которые обеспечивают экспрессию антитела или его антигенсвязывающего фрагмента, и выделение антитела или его антигенсвязывающего фрагмента из культуральной среды или клетки-хозяина.
29. Способ лечения или профилактики состояния, связанного с головной болью, у пациента, где способ включает введение пациенту эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп. 1-23.
30. Способ по п. 29, где состояние, связанное с головной болью, представляет собой мигрень.
31. Способ по п. 30, где мигрень представляет собой эпизодическую мигрень.
32. Способ по п. 30, где мигрень представляет собой хроническую мигрень.
33. Способ по п. 29, где состояние, связанное с головной болью, представляет собой кластерную головную боль.
34. Способ по любому из пп. 29-33, где антитело или его антигенсвязывающий фрагмент вводят пациенту в качестве профилактического лечения.
35. Способ по любому из пп. 29-34, дополнительно включающий введение пациенту второго терапевтического средства от головной боли.
36. Способ по п. 35, где второе терапевтическое средство от головной боли представляет собой терапевтическое средство от острой головной боли.
37. Способ по п. 36, где терапевтическое средство от острой головной боли представляет собой агонист рецепторов серотонина 5HT1B, 5HT1D и/или 5HT1F.
38. Способ по п. 37, где агонист рецептора серотонина представляет собой триптан или эрготамин.
39. Способ по п. 36, где терапевтическое средство от острой головной боли представляет собой нестероидное противовоспалительное лекарственное средство или опиоид.
40. Способ по п. 35, где второе терапевтическое средство от головной боли представляет собой профилактическое терапевтическое средство от головной боли.
41. Способ по п. 40, где профилактическое терапевтическое средство от головной боли представляет собой противоэпилептическое средство, бета-блокатор, антидепрессант, онаботулотоксин А или антагонист пути CGRP.
42. Способ по п. 41, где антагонист пути CGRP представляет собой антагонист рецептора CGRP человека.
43. Способ по п. 42, где антагонист рецептора CGRP человека представляет собой моноклональное антитело, которое специфически связывается с рецептором CGRP человека.
44. Способ по п. 43, где моноклональное антитело к рецептору CGRP представляет собой эренумаб.
| US 9365653 B2, 14.06.2016 | |||
| US 8741291 B2, 03.06.2014 | |||
| JACOBSEN F.W | |||
| et al., Engineering an IgG Scaffold Lacking Effector Function with Optimized Developability, THE JOURNAL OF BIOLOGICAL CHEMISTRY, 2017, Vol | |||
| ПРИСПОСОБЛЕНИЕ ДЛЯ УСТРАНЕНИЯ СКОЛЬЖЕНИЯ КОЛЕС АВТОМОБИЛЕЙ | 1920 |
|
SU292A1 |
| Кипятильник для воды | 1921 |
|
SU5A1 |
| STEPHEN D | |||
| SILBERSTEIN, Current management: migraine headache, CNS Spectrums, 2017, vol.22, pp.4-12 | |||
| WO | |||

