ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к составу ДНК, лиофилизированному из композиции, содержащей плазмидную ДНК, соль и углевода, где указанная плазмидная ДНК содержит ген HGF или его вариант.
ПРЕДПОСЫЛКИ К СОЗДАНИЮ ИЗОБРЕТЕНИЯ
Лиофилизация является, зачастую, предпочтительной технологией приготовления лекарственных веществ, поскольку в лиофилизированном состоянии повышается долгосрочная стабильность многих веществ. Однако для плазмидной ДНК лиофилизированные составы не являются составами выбора. В большинстве клинических испытаний, использующих в качестве вектора доставки голую (некомплексированная плазмида) ДНК, предпочтительным составом был жидкий состав.
Несмотря на то что лиофилизированная плазмидная ДНК может быть предпочтительной формой для хранения, считалось, что лиофилизированные составы плазмидной ДНК обуславливали уменьшение эффективности экспрессии гена. Лиофилизация является причиной удаления области гидратации вокруг молекулы. Для ДНК выяснено, что имеется приблизительно 20 молекул воды на нуклеотидную пару, наиболее прочно связанных с ДНК, которые никогда не образуют структуру, подобную льду, при низкотемпературном охлаждении. При дегидратации ДНК с помощью гигроскопичных солей при 0% влажности остается только пять или шесть молекул воды. Таким образом, лиофилизация может повысить стабильность ДНК при долгосрочном хранении, но также может служить причиной некоторого повреждения во время изначального процесса лиофилизации потенциально из-за изменений во вторичной структуре ДНК или концентрации реакционноспособных элементов, таких как загрязняющие металлы. Таким образом, грубая структурная перестройка плазмиды может являться потенциальным механизмом уменьшения эффективности экспрессии гена лиофилизированной плазмидной ДНК.
В работе Poxon et al, Pharmaceutical Development and Technology 5:115-122 (2000), авторы продемонстрировали, что лиофилизация плазмидной ДНК (pRL-CMV) приводит к статистически значимой потере эффективности трансфекции. Биофункциональный анализ, оценивающий трансфекционную активность, продемонстрировал потерю более чем 75% активности плазмидной ДНК после лиофилизации по сравнению с контрольной плазмидой, которая осталась в растворе. Несмотря на то что Poxon et al использовали углеводы для улучшения in vitro сниженной трансфекционной активности нетерапевтической плазмиды pRL-CMV, экспрессирующей люциферазу Renilla, хранящейся в ЭДТА буфере, Poxon et al не рассматривали in vivo применения лиофилизированных препаратов голой ДНК для лечения или профилактики заболевания.
В силу вышеизложенного в уровне техники существует необходимость в стабильном лиофилизированном составе, который не сказывается на эффективности экспрессии гена. Настоящее изобретение обеспечивает лиофилизированный состав плазмидной ДНК, который не только сохраняет биологическую активность экспрессируемого гена, но, в некоторых случаях, способен усилить биологическую активность.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к лиофилизированному ДНК-составу. В одном аспекте данного изобретения ДНК-состав, перед лиофилизацией, содержит плазмидную ДНК, соль и углевод; и данная плазмидная ДНК содержит ген HGF или его вариант. В другом аспекте данного изобретения данный ДНК-состав является лиофилизированным. В другом аспекте данного изобретения лиофилизированный ДНК-состав является восстановленным.
В одном варианте осуществления углевод в ДНК-составе по настоящему изобретению представляет собой моно-, олиго- или полисахарид, такой как сахароза, глюкоза, лактоза, трегалоза, арабиноза, пентоза, рибоза, ксилоза, галактоза, гексоза, идоза, манноза, талоза, гептоза, фруктоза, глюконовая кислота, сорбитол, маннитол, метил-α-глюкопиранозид, мальтоза, изоаскорбиновая кислота, аскорбиновая кислота, лактон, сорбоза, глюкаровая кислота, эритроза, треоза, аллоза, альтроза, гулоза, эритрулоза, рибулоза, ксилулоза, псикоза, тагатоза, глюкуроновая кислота, галактуроновая кислота, маннактуроновая кислота, глюкозамин, галактозамин, нейраминовая кислота, арабинаны, фруктаны, фуканы, галактаны, галактуронаны, глюканы, маннаны, ксиланы, леван, фукоидан, каррагинан, галактокаролаза, пектины, пектиновые кислоты, амилоза, пуллулан, гликоген, амилопектин, целлюлоза, декстран, циклодекстрин, пустулан, хитин, агароза, кератин, хондроитин, дерматан, гиалуроновая кислота, альгиновая кислота, ксаниановая камедь или крахмал.
В некоторых вариантах осуществления данного изобретения углеводом является сахароза или маннитол.
В другом варианте осуществления углевод в ДНК-составе по настоящему изобретению присутствует в количестве, выбранном из группы, состоящей из количества от приблизительно 0,05% до приблизительно 30%, от приблизительно 0,1% до приблизительно 15%, от приблизительно 0,2% до приблизительно 10%, от приблизительно 0,5% до 5%, от приблизительно 0,75% до 3%, от приблизительно 0,8% до 2% и от приблизительно 0,8% до 1,5%. В определенных вариантах осуществления данный углевод представляет собой сахарозу или маннитол. В некоторых других вариантах осуществления углевод в ДНК-составе находится в количестве приблизительно 1,1 %.
В другом варианте осуществления соль ДНК-состава выбрана из группы, состоящей из NaCl или KCl. В дополнительных вариантах осуществления данная соль в ДНК-составе присутствует в количестве, выбранном из группы, состоящей из количества приблизительно от 0,01% до 10%, приблизительно от 0,1% до 5%, приблизительно от 0,1% до 4%, приблизительно от 0,5% до 2%, приблизительно от 0,8% до 1,5%, приблизительно от 0,8% до 1,2% вес/объем. В некоторых вариантах осуществления данная соль в ДНК-составе присутствует в количестве приблизительно 0,9% вес/объем.
В другом варианте осуществления плазмидная ДНК по данному изобретению содержит ген HGF или его вариант. В некоторых вариантах осуществления ген HGF представляет собой ген HGF млекопитающего или его вариант. В дополнительных вариантах осуществления ген HGF представляет собой ген человека или его вариант. В некоторых аспектах данного изобретения ген HGF представляет собой гибридный ген HGF, например, гибридный ген HGF, содержащий кДНК HGF и природный или чужеродный интрон, или его фрагмент, например, природный интрон 4, или его фрагмент, гена HGF человека. В конкретных вариантах осуществления гибридный ген HGF содержит HGF-X2 (SEQ ID NO: 13), HGF-X3 (SEQ ID NO: 14), HGF- X6 (SEQ ID NO: 8), HGF-X7 (SEQ ID NO: 9) или HGF-X8 (SEQ ID NO: 10). В дополнительных вариантах осуществления плазмидная ДНК, содержащая гибридный ген HGF, выбрана из группы, состоящей из: pCK-HGF-X2, pCK-HGF-X3, pCK-HGF-X6, pCK-HGF-X7, pCK-HGF-X8, pCP-HGF-X2, pCP-HGF-X3, pCP-HGF-X6, pCP-HGF-X7 и pCP-HGF-X8, где HGF-X2, HGF-X3, HGF-X6, HGF-X7 и HGF-X8 соответствуют последовательностям SEQ ID NOs: 13-14 и 8-10 соответственно.
Лиофилизированные составы ДНК поддерживают или усиливают экспрессию плазмидной ДНК. В некоторых аспектах лиофилизированный состав ДНК обеспечивает усиленную биологическую активность экспрессируемого белка. В некоторых других аспектах данного изобретения усиленная экспрессия плазмидной ДНК, или увеличенная биологическая активность экспрессируемого белка, обусловлена присутствием углевода в составе. В некоторых вариантах осуществления этот углевод представляет собой сахарозу или маннитол.
Данное изобретение также обеспечивает восстановленный состав лиофилизированной плазмидной ДНК. В некоторых вариантах осуществления лиофилизированная ДНК восстановлена в фармацевтически приемлемом растворе. В дополнительных вариантах осуществления фармацевтически приемлемый раствор выбран из группы, состоящей из воды, PBS, TE, Tris буфера и физиологического раствора.
В другом варианте осуществления плазмидная ДНК в восстановленном лиофилизированном составе присутствует в конечной концентрации приблизительно 1 нг/мл, приблизительно 5 нг/мл, приблизительно 10 нг/мл, приблизительно 50 нг/мл, приблизительно 100 нг/мл, приблизительно 250 нг/мл, приблизительно 500 нг/мл, приблизительно 1 мкг/мл, приблизительно 5 мкг/мл, приблизительно 10 мкг/мл, приблизительно 50 мкг/мл, приблизительно 100 мкг/мл, приблизительно 200 мкг/мл, приблизительно 300 мкг/мл, приблизительно 400 мкг/мл, приблизительно 500 мкг/мл, приблизительно 600 мкг/мл, приблизительно 700 мкг/мл, приблизительно 800 мкг/мл, приблизительно 900 мкг/мл, приблизительно 1 мг/мл, приблизительно 2 мг/мл, приблизительно 2,5 мг/мл, приблизительно 3 мг/мл, приблизительно 3,5 мг/мл, приблизительно 4 мг/мл, приблизительно 4,5 мг/мл, приблизительно 5 мг/мл, приблизительно 5,5 мг/мл, приблизительно 6 мг/мл, приблизительно 7 мг/мл, приблизительно 8 мг/мл, приблизительно 9 мг/мл, приблизительно 10 мг/мл, приблизительно 20 мг/мл или приблизительно 30 мг/мл. В другом варианте осуществления конечная концентрация плазмидной ДНК в восстановленном лиофилизированном составе составляет приблизительно от 1 нг/мл до приблизительно 30 мг/мл. В некоторых аспектах конечная концентрация плазмидной ДНК в восстановленном лиофилизированном составе составляет от приблизительно 100 мкг/мл до приблизительно 2,5 мг/мл. В дополнительных аспектах конечная концентрация плазмидной ДНК в восстановленном лиофилизированном составе составляет от приблизительно 500 мкг/мл до приблизительно 1 мг/мл.
Настоящее изобретение также относится к способу лечения или профилактики ишемической болезни или заболевания печени у человека, включающему введение композиции, восстановленной из лиофилизированного состава ДНК гепатоцитарного фактора роста (HGF), где ДНК-состав содержит плазмидную ДНК, соль и углевод; и где данная плазмидная ДНК содержит ген HGF или его вариант. В некоторых аспектах композицию, восстановленную из лиофилизированного состава ДНК HGF, вводят прямой инъекцией.
Настоящее изобретение, кроме того, относится к способу получения лиофилизированного состава ДНК HGF, включающему: (a) получение ДНК-состава, содержащего плазмидную ДНК, соль и углевод, где плазмидная ДНК содержит ген HGF, или его вариант; и (b) лиофилизацию ДНК-состава.
Этапы лиофилизации могут включать подвергание ДНК-состава по данному изобретению процессу заморозки при минусовых температурах (например, -10°С до -50°С), и затем подвергание одному или нескольким циклам высушивания, которые содержат постепенное нагревание ДНК-состава до температуры приблизительно от 20°С до температуры приблизительно равной или ниже 30°С, при которых происходит лиофилизация в течение времени от приблизительно 50 до приблизительно 100 часов. В дополнительном аспекте данного изобретения способ лиофилизации включает: (a) получение водосодержащего ДНК-состава, содержащего плазмидную ДНК, соль и углевод, где плазмидная ДНК содержит ген HGF, или его вариант; (b) охлаждение раствора ДНК-состава до температуры от приблизительно -10°С до приблизительно -50°С, до тех пор, пока не замерзнет; (c) высушивание ДНК-состава путем нагревания до температуры от приблизительно 20°С до приблизительно 30°С; и (d) восстановление лиофилизированного ДНК-состава, имеющего влагосодержание от приблизительно 0,1 процента по весу до приблизительно 5 процентов по весу, исходя из общей массы восстановленного ДНК-состава.
В некоторых вариантах осуществления ДНК-состав лиофилизируют при условиях, включающих (a) от приблизительно 30 до приблизительно 50 часов при температуре выше или равной приблизительно -50°С и ниже, чем приблизительно 0°С, и (b) приблизительно от 20 часов до приблизительно 50 часов при температуре от выше, или равной, приблизительно 0°С до ниже, или равной, приблизительно 30°С, постепенно, где самая низкая (a) температура составляет от приблизительно -50°C до приблизительно -30°C, и самая высокая (b) температура составляет между приблизительно 20°С и 30°С. В одном аспекте ДНК-состав лиофилизирован при условиях -50°C в течение 4 часов, -40°C в течение 12 часов, -30°C в течение 6 часов, -20°C в течение 6 часов, -10°C в течение 6 часов, 0°C в течение 6 часов, 10°C в течение 6 часов и 30° C в течение 24 часов, постепенно. В другом аспекте ДНК-состав лиофилизирован в условиях 5°C в течение 1 минуты, -50°C в течение 2 часов, -40°C в течение 6 часов, -35°C в течение 3 часов, -30°C в течение 6 часов, -25°C в течение 3 часов, -20°C в течение 3 часов, -15°C в течение 3 часов, -10°C в течение 6 часов, -5°C в течение 3 часов, 0°C в течение 6 часов, и 30°C в течение 17 часов, постепенно. В другом аспекте ДНК-состав лиофилизирован в условиях 5°C в течение 1 минуты, -10°C в течение 1 минуты, -20°C в течение 1 минуты, -30°C в течение 1 минуты, -50°C в течение 1 минуты, -50°C в течение 2 часов, -45°C в течение 6 часов, -40°C в течение 3 часов, -35°C в течение 6 часов, -30°C в течение 3 часов, -25°C в течение 6 часов, -20°C в течение 3 часов, -15°C в течение 6 часов, -10°C в течение 3 часов, -5°C в течение 6 часов, 0°C в течение 12 часов, 10°C в течение 3 часов, 20°C в течение 6 часов и 30°C в течение 29 часов, постепенно.
Данное изобретение дополнительно относится к лиофилизированному составу нуклеиновых кислот или восстановленному лиофилизированному составу нуклеиновых кислот, как вышеупомянуто, где нуклеиновая кислота представляет собой РНК, кодирующую HGF, или его вариант.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
Рассмотренные выше и другие объекты и характеристики настоящего изобретения станут очевидны из последующего описания данного изобретения в сочетании с прилагаемыми чертежами, на которых:
на фиг.1 представлена гистограмма сравнения экспрессии HGF in vitro среди различных составов. Уровни экспрессии HGF были измерены, используя ELISA в супернатантах культуры, выделенных из клеток 293Т, трансфицированных лиофилизированной плазмидной ДНК pCK-HGF-X7, находящейся в смеси с 0,9% NaCl, в конечной концентрации ДНК 0,5 мг/мл, с сахарозой 0,25% (столбик 3), 1,1% (столбик 4), 5% (столбик 5), 10% (столбик 6) или 20% (столбик 7) или с маннитолом 1,2% (столбик 8), 4,85% (столбик 9) или 10% (столбик 10). Контрольные реакции с отрицательным контролем (столбик 1) и нелиофилизированной ДНК (столбик 2) использовали для сравнения.
На фиг.2 представлена гистограмма сравнения экспрессии HGF in vivo между нелиофилизированной и лиофилизированной pCK-HGF-X7. Мышам в переднюю большеберцовую мышцу делали инъекцию 100 мкг нелиофилизированной pCK-HGF-X7, содержащей 0,9% NaCl (NL-HGF-X7), или pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl (L-HGF-X7). После безболезненного умерщвления мышей в День 7 уровни экспрессии HGF определяли посредством ELISA лизатов мышечной ткани. Уровни экспрессии HGF представлены для отрицательного контроля (столбик 1), нелиофилизированной pCK-HGF-X7, содержащей 0,9% NaCl (NL-HGF-X7; столбик 2), и для pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl (L-HGF-X7; столбик 3).
На фиг.3 представлена схема эксперимента с использованием модели ишемической болезни сердца у свиньи. NL-HGF-X7 представляет собой нелиофилизированную pCK-HGF-X7, содержащую 0,9% NaCl. L-HGF-X7 представляет собой pCK-HGF-X7, лиофилизированную с 1,1% сахарозой и 0,9% NaCl.
На фиг.4 представлена гистограмма, описывающая действие нелиофилизированной и лиофилизированной pCK-HGF-X7 на перфузию миокарда. Демонстрируется процент положительной динамики перфузии миокарда по сравнению с исходным уровнем при использовании модели ишемической болезни сердца у свиньи. Результаты представлены для свиней, которым сделали инъекцию только плазмиды (pCK; столбик 1), нелиофилизированной pCK-HGF-X7, содержащей 0,9% NaCl (NL-HGF-X7; столбик 2), и pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl (L-HGF-X7; столбик 3).
На фиг.5 представлена гистограмма, демонстрирующая воздействие нелиофилизированной и лиофилизированной pCK-HGF-X7 на утолщение стенки. Демонстрируется процент положительной динамики утолщения стенки в пограничной зоне ишемии левого желудочка, куда была сделана инъекция, по сравнению с исходным уровнем при использовании модели ишемической болезни сердца у свиньи. Результаты представлены для свиней, которым сделали инъекцию только плазмиды (pCK; столбик 1), нелиофилизированной pCK-HGF-X7, содержащей 0,9% NaCl (NL-HGF-X7; столбик 2), и pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl (L-HGF-X7; столбик 3).
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения
Термин «ДНК», или «нуклеиновая кислота», или «фрагмент нуклеиновой кислоты» относится к любому одному или нескольким участкам нуклеиновой кислоты, например, фрагментам ДНК или РНК, находящимся в полинуклеотиде или конструкции. Нуклеиновая кислота, или ее фрагмент, может быть представлена линейной (например, мРНК) или кольцевой (например, плазмида) формой, а также двухнитевой или однонитевой формами. Под «изолированной» нуклеиновой кислотой или полинуклеотидом подразумевается молекула нуклеиновой кислоты, ДНК или РНК, которая была выделена из ее естественного окружения. Например, рекомбинантный полинуклеотид, содержащийся в векторе, рассматривается как изолированный в целях настоящего изобретения. Дополнительные примеры изолированного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах, или очищенные (частично или в большей степени) полинуклеотиды в растворе. Изолированные молекулы РНК включают in vivo или in vitro транскрипты РНК полинуклеотидов по настоящему изобретению. Изолированные полинуклеотиды или нуклеиновые кислоты, в соответствии с настоящим изобретением, дополнительно включают такие молекулы, полученные синтетическим путем.
В настоящем документе «кодирующая область» представляет собой часть нуклеиновой кислоты, которая содержит в себе кодоны, транслируемые в аминокислоты. Несмотря на то что «стоп-кодон» (TAG, TGA, или TAA) не транслируется в аминокислоту, его можно рассматривать как часть кодирующей области, но любые фланкирующие последовательности, например, промотры, участки связывания рибосомы, терминаторы транскрипции и тому подобное, не являются частью кодирующей области. Две или несколько нуклеиновых кислот или фрагментов нуклеиновой кислоты по настоящему изобретению могут присутствовать в одной полинуклеотидной конструкции, например, в одной плазмиде, или в отдельных полинуклеотидных конструкциях, например, отдельных (различных) плазмидах. Кроме того, любая нуклеиновая кислота, или фрагмент нуклеиновой кислоты, может кодировать единственный HGF полипептид или фрагмент, производное, или его вариант, например, или может кодировать более чем один полипептид, например, нуклеиновая кислота может кодировать два или несколько полипептидов. Дополнительно, нуклеиновая кислота может включать регуляторный элемент, такой как промотор, участок связывания рибосом, или терминатор транскрипции, или может кодировать гетерогенные кодирующие области, слитые с кодирующей последовательностью HGF, например, специализированные элементы или мотивы, такие как секреторный сигнальный пептид или гетерогенный функциональный домен.
В случае с ДНК полинуклеотид, содержащий нуклеиновую кислоту, которая кодирует полипептид, обычно также включает промотор и/или другие контрольные элементы транскрипции или трансляции, функционально связанные с фрагментом нуклеиновой кислоты, кодирующим данный полипептид. Функциональная связь представляет собой связь фрагмента нуклеиновой кислоты, кодирующего генный продукт, например, полипептида, с одной или несколькими регуляторными последовательностями, таким образом, чтобы поместить экспрессию данного генного продукта под влияние или контроль данной регуляторной последовательности (последовательностей).
ДНК полинуклеотид по настоящему изобретению может представлять собой кольцевую или линеаризированную плазмиду или вектор, или другую линейную ДНК, которая может также быть неинфекционной и неинтегрированной (а именно не интегрируется в геном клеток позвоночного). Линеаризированная плазмида представляет собой плазмиду, которая ранее была кольцевой, но была линеаризирована, например, посредством расщепления эндонуклеазой рестрикции. Используемые в настоящем описании термины плазмида и вектор могут использоваться взаимозаменяемо.
Термин «лиофилизированная ДНК» относится к любой ДНК, которая получена в сухом виде посредством быстрого замораживания и обезвоживания, в замороженном состоянии в условиях глубокого вакуума. «Лиофилизация» или «лиофильная сушка» относится к процессу замораживания и высушивания раствора. Лиофилизированная ДНК зачастую получена готовой для использования посредством добавления стерильной дистиллированной воды.
«Вектор» относится к любому носителю для клонирования и/или переноса нуклеиновой кислоты в клетку-хозяина. Вектор может представлять собой репликон, к которому другой сегмент ДНК может быть присоединен, чтобы осуществить репликацию данного присоединенного сегмента. «Репликон» относится к любому генетическому элементу (например, плазмида, фаг, космида, хромосома, вирус), который действует как автономная единица репликации ДНК in vivo, а именно способна к репликации под собственным контролем. Термин «вектор» включает носители для встраивания данной нуклеиновой кислоты в клетку in vitro, ex vivo или in vivo. Большое число векторов, известных в уровне техники, можно использовать для манипуляций с нуклеиновыми кислотами, инкорпорации реакционных элементов промоторов в гены и тому подобное. Возможные векторы включают, например, плазмиды, такие как pBR322 или плазмидные производные pUC, или вектор Bluescript. Например, вставка фрагментов ДНК, соответствующих реакционным элементам и промоторам, в подходящий вектор может быть осуществлена посредством лигирования соответствующих фрагментов ДНК в выбранный вектор, который имеет комплиментарные «липкие» концы. Альтернативно, данные концы молекул ДНК могут быть ферментативно модифицированы, или любой сайт может быть получен посредством лигирующих нуклеотидных последовательностей (линкеры) в концах ДНК. Такие векторы могут быть сконструированы таким образом, чтобы содержать селектируемые маркеры генов, которые обеспечены для выбора клеток. Такие маркеры делают возможной идентификацию и/или выбор клеток-хозяев, которые экспрессируют белки, кодируемые данным маркером.
Дополнительные векторы включают липоплексы (катионный комплекс липосома-ДНК), полиплексы (катионный комплекс полимер-ДНК) и комплексы белок-ДНК. Кроме нуклеиновой кислоты вектор может также содержать в себе одну или несколько регуляторных областей, и/или селектируемые маркеры, пригодные для отбора, оценки и контроля результатов переноса нуклеиновой кислоты (перенос в какую ткань, длительность экспрессии и тому подобное).
Термин «плазмида» относится к экстрахромосомному элементу, зачастую несущему ген, который не является частью центрального метаболизма данной клетки, и обычно находится в виде кольцевых двухнитевых молекул ДНК. Такие элементы могут представлять собой автономно реплицирующиеся последовательности, последовательности, интегрирующиеся в геном, фаговые или нуклеотидные последовательности, линейные, кольцевые или сверхспиральные одно- или двухнитевых ДНК или РНК, полученных из любого источника, в которых некоторое количество нуклеотидных последовательностей соединено или рекомбинировано в уникальную конструкцию, которая обеспечивает введение в клетку промоторного фрагмента и последовательности ДНК для выбранного генного продукта наряду с подходящей 3'-нетранслируемой последовательностью. Используемый в настоящем документе термин «плазмида» относится к конструкции, состоящей из генетического материала (например, нуклеиновых кислот). Как правило, плазмида содержит участок начала репликации, который является функциональным в бактериальных клетках-хозяевах, например, Escherichia coli, и селектируемые маркеры для выявления бактериальных клеток-хозяев, содержащих данную плазмиду.
Плазмиды по настоящему изобретению могут включать генетические элементы, как описано здесь, организованные таким образом, что встроенные кодирующие последовательности могут быть транскрибированы и транслированы в эукариотических клетках. В некоторых вариантах осуществления, описанных здесь, плазмида представляет собой закрытую кольцевую молекулу ДНК.
Термин «экспрессия» относится к биологическому производству продукта, кодируемого кодирующей последовательностью. В большинстве случаев ДНК-последовательность, в том числе кодирующая последовательность, транскрибируется в форму матричной РНК (мРНК). Эта матричная РНК затем транслируется в форму полипептидного продукта, который обладает соответствующей биологической активностью. Также процесс экспрессии может включать дополнительные этапы преобразования продукта транскрипции РНК, такие как сплайсинг, для удаления интронов, и/или посттрансляционный процессинг полипептидного продукта.
Термин «экспрессирующий вектор» относится к вектору, плазмиде или носителю, сконструированному для обеспечения экспрессии встроенной нуклеотидной последовательности, после трансформации в клетку-хозяина. Клонируемый ген, а именно встроенная последовательность нуклеиновой кислоты, например, ген HGF, или его вариант, обычно помещается под контроль контрольных элементов, таких как промотор, минимальный промотор, энхансер или тому подобное. Области контроля инициации, или промоторы, пригодные для запуска экспрессии нуклеиновой кислоты в требуемой клетке-хозяине, многочисленны и хорошо известны специалисту в уровне техники. Фактически любой промотор, способный запускать экспрессию этих генов, может быть использован в экспрессирующих векторах, в том числе, но не ограничиваясь ими, вирусные промоторы, бактериальные промоторы, промоторы животного происхожления, промоторы млекопитающих, синтетические промоторы, конститутивные промоторы, тканеспецифические промоторы, промоторы, связанные с заболеванием или патогенезом, промоторы, специфические для дифференциации, индуцибельные промоторы, промоторы, регулируемые светом; в том числе, но не ограничиваясь ими, область раннего промотора SV40 (SV40), промотор, содержащий 3' длинный концевой повтор (LTR) вируса саркомы Рауса (RSV), E1A или главный поздний промотор (MLP) аденовируса (Ad), предранний промотор цитомегаловируса человека (HCMV), промотор тимидинкиназы (ТК) вируса простого герпеса (HSV), промотор IE1 бакуловируса, промотор фактора элонгации 1 альфа (EF1), промотор глицеральдегид-3-фосфат дегидрогеназы (GAPDH), промотор фосфоглицераткиназы (PGK), промотор убихитина С (Ubc), промотор альбумина, регуляторные последовательности промотора металлотионеина-L и области транскрипционного контроля мыши, широкораспространенные промоторы (HPRT, виментин, β-актин, тубулин и тому подобное), промоторы промежуточных филаментов (десмин, нейрофиламенты, кератин, GFAP и тому подобное), промоторы терапевтических генов (MDR, CFTR или типа фактора VIII и тому подобное), промоторы, связанные с заболеванием или патогенезом, и промоторы, которые демонстрируют тканеспецифичность и используются у трансгенных животных, такие как контрольная область гена эластазы I, который активен в ацинарных клетках поджелудочной железы; контрольная область гена инсулина, активного в бета-клетках поджелудочной железы, контрольная область гена иммуноглобулина, активного в лимфоидных клетках, контрольная область вируса опухоли молочной железы мышей, активного в тестикулярных клетках, клетках молочной железы, лимфоцитах и мастоцитах; контрольные области Apo AI и Apo AII гена альбумина, активного в печени, контрольная область гена альфа-фетопротеина, активного в печени, контрольная область гена альфа 1-антитрипсина, активного в печени, контрольная область гена бета-глобина, активного в миелоидных клетках, контрольная область гена основного миелинового белка, активного в клетках олигодендроглии головного мозга, контрольная область гена легкой цепи-2 миозина, активного в скелетной мускулатуре, и контрольная область гена
гонадотропин-высвобождающего гормона, активного в гипоталамусе, промотор пируваткиназы, промотор виллина, промотор интестинального белка, связывающего жирные кислоты, промотор β-актина гладкомышечной клетки и тому подобное. Кроме того, эти экспрессирующие последовательности могут быть модифицированы посредством добавления энхансера или регуляторных последовательностей и тому подобного. Неограничивающие примеры экспрессирующих векторов по данному изобретению включают в себя pCK (Lee et al, Biochem. Biophys. Res. Commun. 272:230 (2000); WO 2000/040737) и pCP (pCDNA3.1, Invitrogen, USA).
«Конструкция» в настоящем документе в широком смысле означает композицию, которая не встречается в природе. Конструкция может быть получена посредством синтетических технологий, например посредством получения рекомбинантной ДНК и экспрессии или методов химического синтеза для нуклеиновых или аминокислот. Конструкция также может быть произведена посредством добавления или присоединения одного вещества к другому таким образом, что полученный результат не встречается в природе в такой форме.
«Ген» означает полинуклеотид, содержащий нуклеотиды, который кодирует функциональную молекулу, в том числе функциональные молекулы, получаемые в результате только транскрипции (например, биоактивные виды РНК), или в результате транскрипции и трансляции (например, полипептид). Термин «ген» включает в себя нуклеиновые кислоты кДНК и геномную ДНК. «Ген» также относится к фрагменту нуклеиновой кислоты, который экспрессирует специфическую РНК, белок или полипептид, в том числе регуляторные последовательности, предшествующие (5' некодирующие последовательности) и последующие (3' некодирующие последовательности) данной кодирующей последовательности. «Нативный ген» означает ген, который в природе обнаружен со своими собственными регуляторными последовательностями. «Химерный ген» означает любой ген, который не является нативным геном, включающий в себя регуляторные и/или кодирующие последовательности, которые в природе вместе не встречаются. Соответственно, химерный ген может содержать в себе регуляторные последовательности и кодирующие последовательности, которые получены из различных источников, или регуляторные последовательности и кодирующие последовательности, полученные из одного и того же источника, но расположенные в порядке, отличном от такового, обнаруженного в природе. Химерный ген может содержать кодирующие последовательности, полученные из различных источников, и/или регуляторные последовательности, полученные из различных источников. «Эндогенный ген» означает нативный ген в его естественном расположении в геноме организма. «Чужеродный» ген или «гетерогенный» ген означает ген, который в нормальных условиях не обнаружен в организме-хозяине, но который встроен в данный организм-хозяина посредством генного переноса. Чужеродные гены могут содержать в себе нативные гены, встроенные в ненативный организм, или химерные гены. «Трансген» означает ген, который был встроен в данную клетку посредством процедуры генного переноса.
«Гетерогенная ДНК» относится к ДНК, которая по природе не содержится в данной клетке, или хромосомном участке данной клетки. Гетерогенная ДНК может содержать в себе ген, чужеродный данной клетке.
Выражения «выделенный» или «биологически чистый» относятся к веществу, которое в большей степени или преимущественно свободно от компонентов, которые обычно сопутствуют данному веществу в природных условиях. Таким образом, изолированные пептиды в соответствии с данным изобретением предпочтительно не содержат в себе веществ, обычно связанных с данными пептидами в естественных условиях.
Лиофилизированные ДНК-составы
ДНК-состав по изобретению, перед лиофилизацией, в своем составе имеет некоторые наполнители, в том числе углевод и соль.
Как описано в настоящем документе, стабильность лиофилизированного ДНК-состава для применения в качестве диагностического или терапевтического агента может быть увеличена посредством составления, до лиофилизиции, смеси данной ДНК с водным раствором, содержащим в себе стабилизирующее количество углевода.
Углевод в ДНК-составе по изобретению представляет собой моно-, олиго- или полисахарид, такой как сахароза, глюкоза, лактоза, трегалоза, арабиноза, пентоза, рибоза, ксилоза, галактоза, гексоза, идоза, манноза, талоза, гептоза, фруктоза, глюконовая кислота, сорбитол, маннитол, метил α-глюкопиранозид, мальтоза, изоаскорбиновая кислота, аскорбиновая кислота, лактон, сорбоза, глюкаровая кислота, эритроза, треоза, аллоза, альтроза, гулоза, эритрулоза, рибулоза, ксилулоза, псикоза, тагатоза, глюкуроновая кислота, галактуроновая кислота, маннуроновая кислота, глюкозамин, галактозамин, нейраминовая кислота, арабинаны, фруктаны, фуканы, галактаны, галактуронаны, глюканы, маннаны, ксиланы, леван, фукоидан, каррагинан, галактокаролоза, пектины, пектиновые кислоты, амилоза, пуллулан, гликоген, амилопектин, целлюлоза, декстран, циклодекстрин, пустулан, хитин, агароза, кератин, хондроитин, дерматан, гиалуроновая кислота, альгиновая кислота, ксантановая камедь или крахмал.
В одном аспекте данный углевод представляет собой маннитол или сахарозу.
Данный углеводный раствор перед лиофилизацией может представлять собой раствор углевода только в воде или может включать в себя буфер. Примеры таких буферов включают в себя PBS, HEPES, TRIS или TRIS/EDTA. Как правило, данный углеводный раствор объединен с данной ДНК до конечной концентрации сахарозы от приблизительно 0,05% до приблизительно 30%, обычно от приблизительно 0,1% до приблизительно 15% сахарозы, например от 0,2% до приблизительно 5%, 10% или 15% сахарозы, предпочтительно в диапазоне между приблизительно 0,5% и 10% сахарозы, от 1% до 5% сахарозы, от 1% до 3% сахарозы и наиболее предпочтительно приблизительно 1,1% сахарозы.
Соль в данной ДНК-композиции по данному изобретению представляет собой NaCl или KCl. В некоторых аспектах данная соль представляет собой NaCl. В дополнительных аспектах данная соль в данном ДНК-составе присутствует в количестве, выбранном из группы, включающей количества приблизительно от 0,001% до приблизительно 10%, приблизительно между 0,1% и 5%, приблизительно между 0,1% и 4%, приблизительно между 0,5% и 2%, приблизительно между 0,8% и 1,5%, приблизительно между 0,8% и 1,2% вес/объем. В некоторых вариантах осуществления данная соль в данном ДНК-составе присутствует в количестве приблизительно 0,9% вес/объем.
В ДНК-составе по данному изобретению конечная концентрация плазмидной ДНК составляет приблизительно от 1 нг/мл 30 нг/мл. Например, состав по настоящему изобретению может иметь конечную концентрацию плазмиды приблизительно 1 нг/мл, приблизительно 5 нг/мл, приблизительно 10 нг/мл, приблизительно 50 нг/мл, приблизительно 100 нг/мл, приблизительно 200 нг/мл, приблизительно 500 нг/мл, приблизительно 1 мкг/мл, приблизительно 5 мкг/мл, приблизительно 10 мкг/мл, приблизительно 50 мкг/мл, приблизительно 100 мкг/мл, приблизительно 200 мкг/мл, приблизительно 400 мкг/мл, приблизительно 500 мкг/мл, приблизительно 600 мкг/мл, приблизительно 800 мкг/мл, приблизительно 1 мг/мл, приблизительно 2 мг/мл, приблизительно 2,5 мг/мл, приблизительно 3 мг/мл, приблизительно 3,5 мг/мл, приблизительно 4 мг/мл, приблизительно 4,5 мг/мл, приблизительно 5 мг/мл, приблизительно 5,5 мг/мл, приблизительно 6 мг/мл, приблизительно 7 мг/мл, приблизительно 8 мг/мл, приблизительно 9 мг/мл, приблизительно 10 мг/мл, приблизительно 20 мг/мл, или приблизительно 30 мг/мл. В некоторых вариантах осуществления данного изобретения конечная концентрация ДНК составляет приблизительно от 100 мкг/мл до приблизительно 2,5 мг/мл. В конкретных вариантах осуществления данного изобретения конечная концентрация данной ДНК составляет приблизительно от 0,5 мг/мл до 1 мг/мл.
ДНК-состав по настоящему изобретению лиофилизирован в стандартных условиях, известных в уровне техники. Способ лиофилизации ДНК-состава по изобретению может включать в себя (a) помещение контейнера, например, флакона, с ДНК-составом, например, ДНК-составом, содержащим плазмидную ДНК, соль и углевод, где плазмидная ДНК содержит ген HGF или его вариант, в лиофилизатор, где лиофилизатор имеет температуру запуска приблизительно от 5°С до приблизительно -50°С; (b) охлаждение данного ДНК-состава до температур ниже нуля (например, от -10° C до -50°C); и (c) основательное высушивание ДНК-состава. Условия лиофилизации, например, температура и продолжительность, ДНК-состава по данному изобретению могут быть подобраны исполнителем, имеющим рядовые навыки в уровне техники, с учетом факторов, которые оказывают влияние на параметры лиофилизации, например, тип используемого агрегата для лиофильной сушки, используемое количество ДНК и размер используемого контейнера.
Контейнер, содержащий лиофилизированный состав ДНК, может быть затем герметически закрыт и может храниться в течение длительного периода времени при различных температурах (например, от комнатной температуры до -180°С, предпочтительно приблизительно от 2-8°С до приблизительно -80°С, более предпочтительно, приблизительно от -20°С до приблизительно -80°С и наиболее предпочтительно, приблизительно -20°С). В некоторых аспектах лиофилизированные ДНК-составы предпочтительно стабильны в диапазоне температур приблизительно от 2-8°С до приблизительно -80°С в течение, по меньшей мере, 6 месяцев без значительной утраты активности. Стабильное хранение состава плазмидной ДНК также может представлять собой хранение плазмидной ДНК в стабильной форме в течение длительных периодов времени перед как таковым использованием для исследования или лечения, основанного на применении плазмиды. Время хранения может быть несколько месяцев, 1 год, 5 лет, 10 лет, 15 лет или вплоть до 20 лет. Предпочтительно, данная композиция стабильна в течение, по меньшей мере, приблизительно 3 лет.
HGF плазмидной ДНК
Настоящее изобретение обеспечивает лиофилизированный ДНК-состав, где ДНК-состав, перед лиофилизацией, содержит плазмидную ДНК, и данная плазмидная ДНК включает ген HGF или его вариант.
Гепатоцитарный фактор роста (HGF) представляет собой гликопротеин, связывающий гепарин, также известный как рассеивающий фактор, или гепатопоэтин-А. Эндогенный ген, кодирующий HGF человека, расположен на хромосоме 7q21.1 и содержит в себе 18 экзонов и 17 интронов, имеющих нуклеотидную последовательность SEQ ID NO: 1 (Seki T., et al, Gene 102:213-219 (1991)). Транскрипт размером около 6 т.п.н. считывается с гена HGF, а затем из него синтезируется полипептидный предшественник HGF, состоящий из 728 аминокислот (SEQ ID NO: 2). В то же самое время посредством альтернативного сплайсинга гена HGF также синтезируется полипептидный предшественник dHGF, состоящий из 723 аминокислот. Биологически неактивные предшественники могут быть превращены в активные формы дисульфидносвязанного гетеродимера под действием протеазы в сыворотке. В данных гетеродимерах альфа-цепь, имеющая высокую молекулярную массу, формирует четыре домена типа «двойная петля» и N-концевую петлю «шпильку» как предактивированная пептидная область плазминогена. Данные домены типа «двойной петли» тройной дисульфидсвязанной петлеобразной структуры состоят приблизительно из 80 аминокислот и могут иметь важную функцию в белок-белковых взаимодействиях. Низкомолекулярная бета-цепь формирует неактивный серин-протеаза подобный домен. dHGF, состоящий из 723 аминокислот, представляет собой полипептид с делецией пяти амонокислот в 1-ом домене типа «двойная петля» альфа-цепи, а именно F, L, P, S и S.
HGF, секретируемый клетками мезодермального происхождения, имеет различные биологические функции, например, 1) вовлечение эпителиальных клеток в тубулярную структуру; 2) стимулирование васкуляризации от эндотелиальных клеток in vitro и in vivo; 3) регенерация печени и почек благодаря своей противоапоптотической активности; 4) органогенез почек, яичников и семенников; 5) контроль остеогенеза; 6) стимуляция роста и дифференциации кроветворных клеток-предшественников эритроидного ряда; и 7) рост нейрональных аксонов (Stella, M.C. and Comoglio, P.M., The International Journal of Biochemistry & Cell Biology 31:1357-1362 (1999)). Основываясь на этих разнообразных функциях, HGF, или ген, кодирующий HGF или его вариант, может быть показан в качестве терапевтического агента для лечения ишемической болезни или заболеваний печени. Фактически, in vivo, HGF может существовать или как HGF, или как dHGF, и по этой причине совместная экспрессия HGF и dHGF является важной для максимального терапевтического эффекта. Гибридный ген HGF, который может одновременно экспрессировать HGF и dHGF с высокой эффективностью для генной терапии, представляет собой вариант HGF, который может быть потенциально предпочтителен для использования в композиции плазмидной ДНК по настоящему изобретению.
Данный гибридный ген HGF ранее был описан в международной заявке № WO 03/078568 и патенте США № 2005/0079581 A1, содержания которых включены здесь посредством ссылки. Данный гибридный ген HGF получен посредством вставки природного или чужеродного интрона между экзонами 4 и 5 в кДНК HGF. Данный гибридный ген HGF имеет более высокую эффективность экспрессии, чем кДНК HGF, и одновременно экспрессирует два гетеротипа HGF и dHGF (делетированный вариант HGF).
Термин «изоформа HGF» относится к любому полипептиду HGF, имеющему аминокислотную последовательность, которая, по меньшей мере, на 80% идентична (например, по меньшей мере, на 90% или 95% идентична) аминокислотной последовательности HGF, естественным образом продуцируемого у животных, в том числе все аллельные варианты. В одном варианте осуществления данный термин означает изоформы, которые, как известно, обладают пролиферативной активностью. Изоформы HGF включают в себя, без ограничения, flHGF, dHGF, NK1, NK2 и NK4, например, соответствующие последовательностям SEQ ID NOs: 2-6, и их варианты (например, варианты NK2, SEQ ID NOs: 11-12).
Термин «flHGF» означает полноразмерный белок HGF животного, например, млекопитающего, например, аминокислоты 1-728 (SEQ ID NO: 2) HGF человека.
Термин «dHGF» означает делетированный вариант белка HGF, продуцируемый в результате альтернативного сплайсинга гена HGF у животного, например, млекопитающего, например, HGF человека, состоящий из 723 аминокислот (SEQ ID NO: 3) с делецией пяти аминокислот в 1-м домене типа «двойной петли» альфа-цепи (F, L, P, S и S) из последовательности полноразмерного HGF.
Термин «NK1» означает изоформу HGF животного, например, млекопитающего, например, человека, состоящую из N-концевой петли «шпильки» и домена типа «двойная петля» 1.
Термин «NK2» означает изоформу HGF животного, например, млекопитающего, например, человека, состоящую из N-концевой петли «шпильки», домена типа «двойной петли» 1 и домена типа «двойной петли» 2.
Термин «NK4» означает изоформу HGF животного, например, млекопитающего, например, человека, состоящую из N-концевой петли «шпильки», домена типа «двойной петли» 1, домена типа «двойной петли» 2, домена типа «двойной петли» 3 и домена типа «двойной петли» 4.
Структура и функция HGF были широко исследованы и специалисту в уровне техники известны аминокислоты в последовательности HGF, которые являются важными для сохранения в значительной степени всей биологической активности данного белка и которые являются предпочтительно не измененными или лишь консервативно изменены в последовательности любого варианта HGF. Смотри, например, Hartmann et al, Proc. Natl. Acad. Sci USA 89:11574 (1992); Lokker et al, EMBO J. 11:2503 (1992), Zhou et al, Structure 6:109 (1998), Ultsch et al, Structure 6:1383 (1998), Shimizu et al, Biochem. Biophys. Res. Commun. 189:1329 (1992), Yoshiyama et al, Biochem. Biophys. Res. Commun. 175:660 (1991), каждая из которых включена здесь посредством ссылки во всей полноте. Например, выяснено, что N-концевая петля «шпилька» и домен типа «двойной петли» 1 необходимы для клеточной пролиферативной активности. Другие аминокислоты, которые не являются критичными для биологической активности, могут быть более свободно делетированы/или замещены. Специалист в уровне техники может получить варианты изоформ HGF, используя рутинные технологии мутагенеза, такие как описаны в указанных выше ссылочных материалах, и идентифицировать варианты, сохраняющие по существу всю биологическую активность изоформы HGF.
Вариант осуществления данного гибридного гена HGF по настоящему изобретению, содержащий в себе природный интрон, состоит из 7113 п.н. и имеет нуклеотидную последовательность SEQ ID NO: 7.
Гибридный ген HGF может включать в себя фрагмент природного интрона, при желании, имеющего небольшую рекомбинантную последовательность, встроенную в него между экзонами 4 и 5 кДНК HGF. Здесь такой гибридный ген HGF, включающий в себя фрагмент природного интрона, обозначен как «HGF-X». Примеры гибридных генов HGF включают в себя HGF-X2 (SEQ ID NO: 13), HGF-X3 (SEQ ID NO: 14), HGF-X6 (SEQ ID NO: 8), HGF-X7 (SEQ ID NO: 9) и HGF-X8 (SEQ ID NO: 10).
Применение и способы лечения
Как описано выше, HGF обладает различными биологическими функциями, и, основываясь на этих различных функциях, HGF, ген, кодирующий HGF, или его вариант, может быть показан в качестве терапевтического агента для лечения ишемической болезни или заболеваний печени. В настоящем изобретении HGF ДНК-состав применяется после восстановления лиофилизированного ДНК-состава.
Термин «восстановленный» или «восстановление» означает восстановление первоначальной формы, например, посредством восстановления влагосодержания, вещества, предварительно видоизмененного в целях консервации и хранения, например, восстановление жидкого состояния состава плазмидной ДНК, который ранее был высушен и сохранен. Лиофилизированная композиция по настоящему изобретению может быть восстановлена в любом водном растворе, с получением стабильного, монодисперсного раствора, пригодного для применения. Такие водные растворы включают в себя, но не ограничены ими: стерильную воду, TE, PBS, буфер Tris или физиологический раствор.
Концентрация восстановленной ДНК в способах данного изобретения подобрана в зависимости от множества факторов, в том числе доставляемого количества состава, возраста и массы тела субъекта, способа доставки и пути введения и иммуногенности доставляемого антигена.
Этот восстановленный лиофилизированный состав ДНК по данному изобретению может вводиться в организм перорально или посредством парентеральных путей введения, например, посредством внутривенной, внутримышечной, интарэндокардиальной, интрамиокардиальной, интраперикардиальной, интравентрикулярной, внутрисуставной, внутрикожной, внутрицеребральной, внутрипочечной, внутрипеченочной, внутриселезеночной, внутрилимфатической, подкожной, внутрь брюшной полости, внутритестикулярной, внутриовариальной, внутриматочной, интрастернальной, внутритрахеальной, внутриплевральной, интраторакальной, интрадуральной, интраспинальной, интрамедуллярной, интрамуральной, intrascorionic и артериальной инъекции или инфузии или местно посредством ректального, интранозального, ингаляционного или внутриглазного пути введения. В некоторых вариантах изобретения способ доставки является внутримышечным, интрамиокардиальным, внутривенным, внутрицеребральным или внутрипочечным.
Следует понимать, что стандартная суточная доза восстановленного лиофилизированного ДНК-состава по настоящему изобретению должна быть определена с учетом различных факторов, включающих в себя состояния, относительно которых проводится лечение, выбранный путь введения, возраст, пол и массу тела конкретного пациента, и тяжесть симптомов у данного пациента, и может вводиться в однократной дозе или дробными дозами. Следовательно, в любом случае, суточная доза не должна рассматриваться как ограничение объема данного изобретения.
Термины «лечить», «лечение» или «курс лечения» при ишемической болезни или заболевании печени в настоящем документе относятся к введению в организм пациента действующего агента, например, HGF, например гибридного HGF, или его варианта, в количестве, достаточном для обеспечения облегчения одного или нескольких симптомов ишемической болезни, или заболевания печени, или профилактики прогрессирования ишемической болезни или заболевания печени.
«Ишемическая болезнь» означает заболевание, связанное с недостаточным снабжением кровью части тела (например, сердца или мозга) в результате затруднения притока артериальной крови (как в результате сужения просвета артерий в результате спазма или в результате заболевания). Примеры ишемических заболеваний включают в себя заболевание коронарных артерий (CAD) и заболевание периферических артерий (PAD).
Термин «заболевание печени» применяется ко многим заболеваниям и нарушениям, которые служат причиной неправильного функционирования или прекращения функционирования печени. HGF является главным агентом, стимулирующим пролиферацию гепатоцитов, и действует сообща с трансформирующим фактором роста-альфа и гепаринсвязывающим эпидермальным фактором роста во время регенерации печени. Кроме того, благодаря противоапоптотическим действиям HGF делает повреждение печени менее тяжелым в экспериментальных моделях скоротечной печеночной недостаточности на животных и уменьшает фиброз печени у животных с циррозом печени. Следовательно, HGF рассматривается не только как индуцирующий регенерацию печени, но также как ингибирующий прогрессирование заболевания и уменьшающий фиброз печени у пациентов, страдающих трудноизлечимыми заболеваниями печени. Что касается лечения заболевания печени, восстановленный лиофилизированный ДНК-состав по данному изобретению можно применять в соответствии с вышеупомянутыми способами доставки. В некоторых вариантах осуществления способ доставки при лечении заболеваний печени будет внутривенным, внутриартериальным или внутрипеченочным.
В некоторых аспектах данного изобретения восстановленный HGF ДНК-состав может содержать в себе две или несколько изоформ HGF. Данные изоформы HGF могут быть предварительно лиофилизированы по отдельности или в одном и том же ДНК-составе. Обе из этих лиофилизированных изоформ, после восстановления, могут быть введены в организм раздельно, или в одно и то же время, то есть введены совместно; могут быть введены или совместно введены раздельные восстановленные композиции плазмидной ДНК для двух или нескольких изоформ HGF, или может быть введена единственная экспрессирующая плазмида, содержащая в себе гены для двух или нескольких изоформ HGF и способная экспрессировать данные гены двух или нескольких изоформ HGF. Например, две изоформы, flHGF и dHGF, могут быть введены в организм с использованием двух отдельных плазмид. Альтернативно, данные две отдельные плазмиды, содержащие в себе гены flHGF и dHGF, могут быть использованы при совместном введении. Наконец, может быть использована единственная экспрессирующая плазмида, содержащая в себе гены и flHGF, и dHGF. В некоторых аспектах данного изобретения flHGF и dHGF в одной и той же экспрессирующей плазмиде закодированы одним и тем же полинуклеотидом или раздельными полинуклеотидами.
Существует целый ряд способов включения более одного полинуклеотида, способного к экспрессии изоформы HGF, в одну и ту же плазмиду. В том числе, например, использование последовательностей участков внутренней посадки рибосомы (IRES), парных промоторов/полигенных экспрессирующих кластеров, и слитых белков. Данные две или несколько изоформ, экспрессируемые одной и той же плазмидой, или двумя раздельными плазмидами, как обсуждалось ранее, выбраны из группы, содержащей flHGF, dHGF, NK1, NK2, и NK4, или выбраны из группы, содержащей последовательности SEQ ID NOs: 2-6. Данные две или несколько изоформ могут также включать дополнительные изоформы HGF, известные специалисту, имеющему обычные навыки в уровне техники.
В некоторых аспектах данного изобретения данная плазмидная ДНК вводится посредством прямой внутриклеточной инъекции и, более предпочтительно, с помощью шприца или катетера. Катетеры использовались для введения рекомбинантных генов in vivo (смотри, например, E.G. Nabel, et al., Proc. Natl. Acad. Sci. USA 89, 5157 (1992); E.G. Nabel, et al, Science 249, 1285 (1990); E.G. Nabel, et al, Science 244, 1342 (1989); E.G. Nabel, et al., J. Clin. Invest. 91, 1822 (1993); G..Plautz, et al., Circ. 83, 578 (1991); E.G. Nabel, et al., Nature (1993) (в печати)). Использование катетера обеспечивает возможность доставки данной плазмидной ДНК в клетки, доступ к которым с помощью шприца затруднен.
Данная плазмидная ДНК может быть введена посредством внутриартериальной или внутривенной инъекции и, более предпочтительно, с помощью шприца или катетера. Например, для доставки плазмидной ДНК в сердце можно использовать бедренную артерию; для доставки плазмидной ДНК в печень можно использовать воротную вену.
Введение данной плазмидной ДНК по данному изобретению также может быть выполнено посредством генного переноса в клетки-мишени, in situ, для оптимизации последующей доставки генов in vivo.
При практическом применении настоящего изобретения будут использовать, если не указано особо, обычные технологические приемы клеточной биологии, клеточной культуры, молекулярной биологии (в том числе ПЦР), вакцинологии, микробиологии, рекомбинантных ДНК и иммунологии, которые входят в уровень техники. Такие технологические приемы подробно объяснены в литературе. Смотри, например, Molecular Cloning A Laboratory Manual, 2nd Ed., Sambrook et al, ed., Cold Spring Harbor Laboratory Press: (1989); DNA Cloning, Volumes I and II (D.N. Glover ed., 1985); Oligonucleotide Synthesis (M.J. Gait ed., 1984); Mullis et al. патент США № 4683195; Nucleic Acid Hybridization (B.D. Hames & S. J. Higgins eds. 1984); Transcription And Translation (B.D. Hames & S. J. Higgins eds. 1984); Culture Of Animal Cells (R.I. Freshney, Alan R. Liss, Inc., 1987); Immobilized Cells And Enzymes (IRL Press, 1986); B. Perbal, A Practical Guide To Molecular Cloning (1984); научный труд, Methods In Enzymology (Academic Press, Inc., N. Y.); Gene Transfer Vectors For Mammalian Cells (J.H. Miller and M.P. Calos eds., 1987, Cold Spring Harbor Laboratory); Methods In Enzymology, Том 154 и 155 (Wu et al. eds.), Immunochemical Methods In Cell And Molecular Biology (Mayer and Walker, eds., Academic Press, London, 1987); и у Ausubel et al., Current Protocols in Molecular Biology, John Wiley and Sons, Baltimore, Maryland (1989). Каждый из данных справочных материалов, процитированных в этом абзаце, включен здесь в полном объеме посредством ссылки.
Следующие примеры приведены здесь только с иллюстративной целью и не предназначены для ограничения объема данного изобретения.
Пример 1: Подготовка плазмиды
В данном эксперименте использовали плазмиду pCK-HGF-X7 (WO 03/078568), которая предназначена для экспрессии белка гепатоцитарного фактора роста (HGF).
E. coli (TOP10, Invitrogen, USA) была трансформирована с pCK-HGF-X7, и одиночная колония была изолирована. Затем данную одиночную колонию культивировали в среде LB, содержащей в себе 30 мкг/мл канамицина. Плазмидную ДНК очищали с помощью набора EndoFree plasmid Giga kit (Qiagen, USA), и ресуспендировали в физиологическом растворе, содержащим 0,9% NaCl, при конечной концентрации ДНК от 1,0 до 2,0 мг/мл.
Пример 2: Лиофилизация
Композиции pCK-HGF-X7 были приготовлены в физиологическом растворе, содержащем 0,9% NaCl, при конечной концентрации ДНК 0,5 мг/мл или 1 мг/мл, с сахарозой (0,25, 1,1, 5, 10 или 20% вес/объем) или маннитолом (1,2, 4,85 или 10% вес/объем). В таблице 1А и 1В представлено процентное содержание сахарозы и маннитола, соответственно, и соответствующие соотношения углевод/ДНК (вес/вес) для протестированных составов pCK-HGF-X7.
к ДНК (вес/вес)
к ДНК (вес/вес)
Затем суспендированную плазмидную ДНК лиофилизировали с использованием устройства Production-Master Freeze Dryer (C&H Cooling & Heating Systems, Korea). Температура была снижена до -50°С в течение 4 часов при 100 мТорр. Затем, температуру подняли до -40°С на 12 часов, -30°C на 6 часов, -20°C на 6 часов, -10°C на 6 часов, 0°C на 6 часов, 10°C на 6 часов и 30°C на 24 часа, постепенно, при 28-29 мТорр. Лиофилизированную плазмидную ДНК хранили при -20°С до момента анализа.
Данную суспендированную плазмидную ДНК также лиофилизировали с использованием устройства Production-Master Freeze Dryer (C&H Cooling & Heating Systems, Korea). Температура была снижена до 5°С на 1 минуту и -50°С в течение 2 часов при 100 мТорр. Затем температуру подняли до -40°С на 6 часов, -35°С на 3 часа, -30°C на 6 часов, -25°С на 3 часа, -20°C на 3 часа, -15°С на 3 часа, -10°C на 6 часов, -5°С на 3 часа, 0°C на 6 часов, и 30°C на 17 часов, постепенно, при 28-29 мТорр. Лиофилизированную плазмидную ДНК хранили при -20°С до момента анализа.
Данную суспендированную плазмидную ДНК также лиофилизировали с использованием устройства Production-Master Freeze Dryer (C&H Cooling & Heating Systems, Korea). Температура была снижена до 5°С на 1 минуту, -10°С на 1 минуту, -20° на 1 минуту, -30°С на 1 минуту, и -50°С на 1 минуту при 150 мТорр. Температуру -50°С поддерживали еще на протяжении 2 часов при 150 мТорр. Затем температуру подняли до -45°С на 6 часов, -40°С на 3 часа, -35°С на 6 часов, -30°C на 3 часа, -25°С на 6 часов, -20°C на 3 часа, -15°С на 6 часов, -10°C на 3 часа, -5°С на 6 часов, 0°C на 12 часов, 10°С на 3 часа, 20°С на 6 часов и 30°C на 29 часов, постепенно, при 30 мТорр. Лиофилизированную плазмидную ДНК хранили при -20°С до момента анализа.
Лиофилизированные составы, полученные, как описано выше, анализировали на предмет эффективности экспрессии гена in vitro в соответствии с методами, описанными в Примере 3. Результаты in vitro для этих композиций были одинаковыми.
Пример 3: Влияние лиофилизации на эффективность экспрессии гена in vitro плазмидной ДНК
1. Материалы и методы
Для оценки воздействия лиофилизации на эффективность экспрессии генов плазмидной ДНК данную лиофилизированную плазмидную ДНК трансфицировали в клетки 293Т и измеряли уровень экспрессии HGF. В качестве контроля нелиофилизированная плазмидная ДНК также была трансфицирована.
Четыре микрограмма pCK-HGF-X7 в различных композициях (как описано в Примере 1) трансфицировали в 1×10 6 293T клеток с помощью FuGENE6 (Roche Diagnostics, Germany) (n=5). Перед траснфекцией 1 мг лиофилизированной плазмидной ДНК был восстановлен в 2 мл воды для инъекций до конечной концентрации 0,5 мг/мл.
Через два дня после трансфекции супернатанты культуры были получены и проанализированы на предмет экспрессии HGF с использованием набора для иммуноферментного анализа HGF человека human HGF ELISA kit (R&D Systems, MN, USA), в соответствии с инструкциями производителя. Результаты ELISA были статистически обработаны с помощью критерия множественного сравнения Даннета, используя программу SPSS (версия 13.0, SPSS. Inc, USA).
2. Результаты и обсуждение
Результаты экспрессии гена HGF представлены на фиг.1. Вопреки опубликованным ранее сообщениям лиофилизация не влияла на in vitro эффективность экспрессии гена плазмидной ДНК. Среди различных композиций уровень HGF, произведенный плазмидой pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl, был значительно выше такового, произведенного нелиофилизированной плазмидой pCK-HGF-X7 (p=0,001) (фиг.1).
Эти результаты свидетельствую о том, что лиофилизация композиции, содержащей в себе 1,1% сахарозы и 0,9% NaCl, была бы более подходящей для pCK-HGF-X7, чем нелиофилизированная композиция.
Пример 4: Сравнительный анализ in vivo экспрессии генов между нелиофилизированной и лиофилизированной pCK-HGF-X7
1. Материалы и методы
Для каждой группы получали по тринадцать пятинедельных мышей BALB/c (самцы, Charles River) и обеспечивали их едой и водой неограниченно. Мышам предоставили отдых в течение 7 дней перед началом эксперимента.
Мышам инъецировали 100 мкг нелиофилизированной pCK-HGF-X7, содержащей в себе 0,9% NaCl (NL-HGF-X7), или лиофилизированной pCK-HGF-X7, содержащей в себе 1,1% сахарозу и 0,9% NaCl (L-HGF-X7), в краниальную большеберцовую мышцу, а затем мышей безболезненно умерщвляли на 7 день после обработки. Перед инъекцией лиофилизированная плазмидная ДНК была восстановлена в воде до конечной концентрации 0,5 мг/мл. Для измерения уровня экспрессии белка HGF мышцы, в которые была произведена инъекция, были собраны, и мышечная ткань была лизирована с помощью 500 мкл буфера для лизиса клеток (50 мМ NaCl, 0,2% додецилфульфат натрия, 0,5% дезоксихолат натрия, 2% IGEPAL CA-630, 25 мМ Tris-HCl, pH7,4, 1 мМ фенилметилсульфонил фторид) в течение 16 часов при 4°С. Лизаты центрифугировали при 12000 об/мин в течение 5 минут, супернатанты собраны и проанализированы на предмет экспрессии HGF с помощью набора для иммуноферментного анализа HGF человека human HGF ELISA kit (R&D Systems).
Результаты ELISA были статистически обработаны с помощью однофакторного дисперсионного анализа ANOVA и последующего теста Тьюки, используя программу SPSS (версия 13.0).
2. Результаты и обсуждение
В среднем 246 нг/мл белка HGF вырабатывалось у животных, которым была введена pCK-HGF-X7, лиофилизированная с 1,1% сахарозы и 0,9% NaCl (L-HGF-X7), в то время как у животных, которым была введена нелиофилизированная pCK-HGF-X7, экспрессировалось только 76 нг/мл HGF (фиг.2). Этот результат свидетельствует о том, что pCK-HGF-X7, лиофилизированная с 1,1% сахарозой и 0,9% NaCl, может экспрессировать белок HGF более эффективно, чем нелиофилизирвоанная pCK-HGF-X7 (p<0,001).
Пример 5: Сравнительный анализ терапевтических эффектов на модель ишемической болезни сердца у свиньи между нелиофилизированной и лиофилизированной pCK-HGF-X7
1. Материалы и методы
(1) Животные
Были получены одиннадцать свиней йоркширской породы (самцы, от 28 до 30 кг, Clinical Research Institute in Seoul National University Hospital), которых обеспечили едой два раза в день и водой без ограничения. Свиньям дали 7 дней отдыха перед экспериментом. Полный план эксперимента представлен на фиг.3.
(2) Создание модели ишемической болезни сердца у свиней
Каждой свинье инъекционно внутримышечно были введены ксилазин (2 мг/кг), кетамин (20 мг/кг) и атропин (0,05 мг/кг). Через двадцать минут в поверхностную бедренную артерию для постоянного контроля кровяного давления был введен стилет-катетер Medicut диаметром 22. Внутривенно был введен тиопентан натрия (10 мг/кг), и осуществлена эндотрахеальная интубация через рототрахеальный путь. Анастезию поддерживали посредством ингаляционного поступления энфлюрана. Во время данной операции поддерживали приточно-вытяжную вентиляцию и содержание кислорода 30%~40%. Проводили непрерывный контроль электрокардиограммы, насыщения кислородом и артериального кровяного давления.
Затем осуществили левостороннюю торакотомию. После открытия перикарда с последующим осмотром левой передней нисходящей коронарной артерии (LAD) внутривенно вводили 2% лидокаин (1 мг/кг) и дистальную одну третью часть LAD лигировали на протяжении 3 минут, максимально оставляя вторую диагональную ветвь. Реперфузию (ишемическое предсостояние) осуществляли в течение 5 минут с использованием 5-0 полипропиленовых нитей, укрепленных небольшим кусочком Нелатона (4 Fr). После этого однократного ишемического предсостояния была лигирована дистальная LAD и депрессия или подъем ST-сегмента был подтвержден на контролируемой электрокардиограмме. Дополнительное введение лидокаина (1 мг/мл) осуществили внутривенно через 15 минут после лигирования, и перикард и раны после торакотомии были закрыты. Единственная 28 Fr плевральная дренажная трубка, подключенная для аспирации стенки, была удалена незамедлительно после достаточного восстановления самостоятельного дыхания с последующим удалением эндотрахеальной трубки.
Все протоколы были утверждены Комитетом по уходу и использованию животных Национального Университета г.Сеула (Seoul National University Animal Care and Use Committee).
(3) Внутримиокардиальное введение плазмид
Через двадцать восемь дней после лигирования коронарной артерии была проведена повторная торакотомия. Общая доза 1 мг лиофилизированной с 1,1% сахарозой и 0,9% NaCl плазмиды pCK-HGF-X7 (L-HGF-X7, n=3), или нелиофилизированной pCK-HGF-X7, содержащей в себе 0,9% NaCl (NL-HGF-X7, n=4), посредством игл для инъекций инсулина калибром 27, была введена в переднелатеральную пограничную зону ишемии, которая расположена между фиброзной зоной инфаркта и макроскопически нормальным миокардом вдоль хода второй диагональной ветви. Всего было пять участков введения. В каждый участок было введено по 0,2 мг плазмидной ДНК и расстояние между участками введения составляло 1,5 см. Данная лиофилизированная плазмидная ДНК перед введением была восстановлена с помощью воды до конечной концентрации 1 мг/мл. В качестве контроля идентичное количество нелиофилизированной pCK, содержащей 0,9% NaCl (n=4), было введено в переднелатеральную пограничную зону ишемии. Места введения инъекции были промаркированы метками шовного материала с использованием металлических колец.
(4) Однофотонная эмиссионная компьютерная томография миокарда
Через двадцать шесть дней после хирургического вызывания инфаркта миокарда была осуществлена 99m Tc-MIBI синхронизированная однофотонная эмиссионная компьютерная томография (SPECT) (Vertex EPIC, ADAC Labs, CA., USA) для определения исходного уровня перед введением плазмид. Синхронизированную SPECT повторно проводили через 28 дней (на день 54 после индукции инфаркта миокарда).
Для сегментарного анализы была выбрана 20-сегментная модель. Шесть сегментов, соответствующих основанию сердца, не учитывались при данном анализе, поскольку эта область может беспрепятственно находиться под влиянием ослабления диафрагмы или некоторых артефактов вокруг сердца; также по причине того, что основание сердца находилось вдалеке от участков дистальной коронарной лигатуры и инъекции плазмиды.
Изображения SPECT, сконструированные посредством синхронизированной электрокардиографии, были проанализированы с помощью программы для автоматического определения количества (AutoQUANT, ADAC Labs, CA., USA), которая, как полагают, устраняет возможные ошибки оценки, обусловленные любыми действиями исполнителя.
Объем сегментальной перфузии был количественно определен посредством измерения поглощения 99m Tc-MIBI и рассчитан как процент максимального потребления. Когда рассчитанная таким образом сегментальная перфузия была менее 70%, этот участок определялся как слабоперфузируемая область и использовался в качестве мишени для доставки плазмиды. Сегменты, остававшиеся хорошо перфузируемыми даже после коронарного лигирования, также были исключены, поскольку они возможно не получают никакой пользы от данного терапевтического ангиогенеза. Утолщение стенки в систолическую фазу было обозначено как процент окончания диастолического утолщения стенки на синхронизированных изображениях.
(5) Статистические вычисления
Данные представлены в виде процента положительной динамики по сравнению с изначальным уровнем. Все данные были проанализированы с использованием SPSS (версия 13.0). Статистический анализ миокардиальной перфузии и сегментарного утолщения стенки был осуществлен с помощью t-критерия Стьюдента для парных выборок.
2. Результаты
Внутри каждой обрабатываемой группы были проведены сравнения изменений в сегментарной перфузии до и после введения плазмидной ДНК. Базовые значения для средней сегментарной перфузии, полученные в день 26 после лигирования LAD, составляли 39,0±14,6, 43,4±13,4 и 36,9±16,3% для групп, получавших pCK, NL-HGF-X7 и L-HGF-X7, соответственно. 99m Tc-MIBI синхронизированная SPECT, проведенная в день 54, показала, что средние значения для сегментарной перфузии в группах, получавших pCK и NL-HGF-X7, были 37,8±13,9% и 44,0±14,5%, соответственно, что не являлость значимым отличием от базовых значений, полученных в день 26 (p=0,320 для pCK и 0,721 для NL-HGF-X7). В отличие от этого среднее значение сегментарной перфузии в группе, получавшей L-HGF-X7, составляло 41,2±17,6%, демонстрируя значительное повышение относительно базового значения (p=0,003). При сравнении между группами абсолютной величины процентного увеличения сегментарной перфузии от базового значения процентное увеличение сегментарной перфузии в группе, получавшей L-HGF-X7, было на 14,74% выше, чем таковое в группе, получавшей pCK (p=0,003), в то время как в группе, получавшей NL-HGF-X7, не наблюдалось значительной разницы с группой, получавшей pCK (p=0,254) (Фиг.4).
В каждой обработанной группе также были сравнены изменения в сегментарном утолщении стенки до и после введения ДНК. В день 26 средние значения сегментарного утолщения стенки составляли 24,7±16,5, 33,4±15,9 и 16,5±15,9% для групп, получавших pCK, NL-HGF-X7 и L-HGF-X7, соответственно, и не наблюдалось никаких значительных различий между группами (p=NS). В день 54, среднее значение сегментарного утолщения стенки для групп, получавших pCK, NL-HGF-X7 и L-HGF-X7, составляло 27,9±18,4, 43,1±11,8 и 30,2±10,7% соответственно. При сравнении между группами абсолютной величины процентного увеличения сегментарного утолщения стенки от базового значения, процентное увеличение в группе, получавшей L-HGF-X7, было 83,54%, что было значительно выше, чем таковое в группе, получавшей NL-HGF-X7 (28,99%) (фиг.5).
Эти результаты свидетельствуют о том, что интрамиокардиальное введение данной лиофилизированной композиции (L-HGF-X7) может более эффективно увеличить региональное кровообращение и утолщение стенки в пограничной области ишемии левого желудочка, куда поставили инъекцию, по сравнению с нелиофилизированной композицией (NL-HGF-X7). Без привязки к теории это происходит, вероятно, в результате ангиогенной и противофиброзной активности экспрессируемого HGF-X7.
Выводы
Сегментарная перфузия и утолщение стенки были значительно увеличены в группе, получавшей лиофилизированную pCK-HGF-X7, по сравнению с таковыми в группах, получавших нелиофилизированные pCK и pCK-HGF-X7.
Эти результаты показывают, что интрамиокардиальное введение pCK-HGF-X7, лиофилизированной с 1,1% сахарозой и 0,9% NaCl, пораженным свиньям, может эффективно и стабильно повышать региональную перфузию и утолщение стенки в ишемическом миокарде по сравнению с введением нелиофилизированной pCK-HGF-X7.
Несмотря на то что данное изобретение было описано в отношении вышеуказанных специфических вариантов осуществления, следует понимать, что специалистом в уровне техники могут быть проведены различные модификации и изменения данного изобретения, которые также находятся в диапазоне данного изобретения, как обозначено в прилагаемых пунктах формулы изобретения.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> VIROMED CO., LTD.
<120> ЛИОФИЛИЗИРОВАННЫЕ ДНК-СОСТАВЫ ДЛЯ УВЕЛИЧЕННОЙ ЭКСПРЕССИИ ПЛАЗМИДНОЙ ДНК
ДНК
<130> PCA810080VML
<150> US 61/043,605
<151> 2008-04-09
<160> 14
<170> KopatentIn 1.71
<210> 1
<211> 2187
<212> ДНК
<213> Homo sapiens
<400> 1
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
agctttttgc cttcgagcta tcggggtaaa gacctacagg aaaactactg tcgaaatcct 540
cgaggggaag aagggggacc ctggtgtttc acaagcaatc cagaggtacg ctacgaagtc 600
tgtgacattc ctcagtgttc agaagttgaa tgcatgacct gcaatgggga gagttatcga 660
ggtctcatgg atcatacaga atcaggcaag atttgtcagc gctgggatca tcagacacca 720
caccggcaca aattcttgcc tgaaagatat cccgacaagg gctttgatga taattattgc 780
cgcaatcccg atggccagcc gaggccatgg tgctatactc ttgaccctca cacccgctgg 840
gagtactgtg caattaaaac atgcgctgac aatactatga atgacactga tgttcctttg 900
gaaacaactg aatgcatcca aggtcaagga gaaggctaca ggggcactgt caataccatt 960
tggaatggaa ttccatgtca gcgttgggat tctcagtatc ctcacgagca tgacatgact 1020
cctgaaaatt tcaagtgcaa ggacctacga gaaaattact gccgaaatcc agatgggtct 1080
gaatcaccct ggtgttttac cactgatcca aacatccgag ttggctactg ctcccaaatt 1140
ccaaactgtg atatgtcaca tggacaagat tgttatcgtg ggaatggcaa aaattatatg 1200
ggcaacttat cccaaacaag atctggacta acatgttcaa tgtgggacaa gaacatggaa 1260
gacttacatc gtcatatctt ctgggaacca gatgcaagta agctgaatga gaattactgc 1320
cgaaatccag atgatgatgc tcatggaccc tggtgctaca cgggaaatcc actcattcct 1380
tgggattatt gccctatttc tcgttgtgaa ggtgatacca cacctacaat agtcaattta 1440
gaccatcccg taatatcttg tgccaaaacg aaacaattgc gagttgtaaa tgggattcca 1500
acacgaacaa acataggatg gatggttagt ttgagataca gaaataaaca tatctgcgga 1560
ggatcattga taaaggagag ttgggttctt actgcacgac agtgtttccc ttctcgagac 1620
ttgaaagatt atgaagcttg gcttggaatt catgatgtcc acggaagagg agatgagaaa 1680
tgcaaacagg ttctcaatgt ttcccagctg gtatatggcc ctgaaggatc agatctggtt 1740
ttaatgaagc ttgccaggcc tgctgtcctg gatgattttg ttagtacgat tgatttacct 1800
aattatggat gcacaattcc tgaaaagacc agttgcagtg tttatggctg gggctacact 1860
ggattgatca actatgatgg cctattacga gtggcacatc tctatataat gggaaatgag 1920
aaatgcagcc agcatcatcg agggaaggtg actctgaatg agtctgaaat atgtgctggg 1980
gctgaaaaga ttggatcagg accatgtgag ggggattatg gtggcccact tgtttgtgag 2040
caacataaaa tgagaatggt tcttggtgtc attgttcctg gtcgtggatg tgccattcca 2100
aatcgtcctg gtatttttgt ccgagtagca tattatgcaa aatggataca caaaattatt 2160
ttaacatata aggtaccaca gtcatag 2187
<210> 2
<211> 728
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu
195 200 205
Val Glu Cys Met Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp
210 215 220
His Thr Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro
225 230 235 240
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp
245 250 255
Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr
260 265 270
Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys
275 280 285
Ala Asp Asn Thr Met Asn Asp Thr Asp Val Pro Leu Glu Thr Thr Glu
290 295 300
Cys Ile Gln Gly Gln Gly Glu Gly Tyr Arg Gly Thr Val Asn Thr Ile
305 310 315 320
Trp Asn Gly Ile Pro Cys Gln Arg Trp Asp Ser Gln Tyr Pro His Glu
325 330 335
His Asp Met Thr Pro Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu Asn
340 345 350
Tyr Cys Arg Asn Pro Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr Thr
355 360 365
Asp Pro Asn Ile Arg Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys Asp
370 375 380
Met Ser His Gly Gln Asp Cys Tyr Arg Gly Asn Gly Lys Asn Tyr Met
385 390 395 400
Gly Asn Leu Ser Gln Thr Arg Ser Gly Leu Thr Cys Ser Met Trp Asp
405 410 415
Lys Asn Met Glu Asp Leu His Arg His Ile Phe Trp Glu Pro Asp Ala
420 425 430
Ser Lys Leu Asn Glu Asn Tyr Cys Arg Asn Pro Asp Asp Asp Ala His
435 440 445
Gly Pro Trp Cys Tyr Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr Cys
450 455 460
Pro Ile Ser Arg Cys Glu Gly Asp Thr Thr Pro Thr Ile Val Asn Leu
465 470 475 480
Asp His Pro Val Ile Ser Cys Ala Lys Thr Lys Gln Leu Arg Val Val
485 490 495
Asn Gly Ile Pro Thr Arg Thr Asn Ile Gly Trp Met Val Ser Leu Arg
500 505 510
Tyr Arg Asn Lys His Ile Cys Gly Gly Ser Leu Ile Lys Glu Ser Trp
515 520 525
Val Leu Thr Ala Arg Gln Cys Phe Pro Ser Arg Asp Leu Lys Asp Tyr
530 535 540
Glu Ala Trp Leu Gly Ile His Asp Val His Gly Arg Gly Asp Glu Lys
545 550 555 560
Cys Lys Gln Val Leu Asn Val Ser Gln Leu Val Tyr Gly Pro Glu Gly
565 570 575
Ser Asp Leu Val Leu Met Lys Leu Ala Arg Pro Ala Val Leu Asp Asp
580 585 590
Phe Val Ser Thr Ile Asp Leu Pro Asn Tyr Gly Cys Thr Ile Pro Glu
595 600 605
Lys Thr Ser Cys Ser Val Tyr Gly Trp Gly Tyr Thr Gly Leu Ile Asn
610 615 620
Tyr Asp Gly Leu Leu Arg Val Ala His Leu Tyr Ile Met Gly Asn Glu
625 630 635 640
Lys Cys Ser Gln His His Arg Gly Lys Val Thr Leu Asn Glu Ser Glu
645 650 655
Ile Cys Ala Gly Ala Glu Lys Ile Gly Ser Gly Pro Cys Glu Gly Asp
660 665 670
Tyr Gly Gly Pro Leu Val Cys Glu Gln His Lys Met Arg Met Val Leu
675 680 685
Gly Val Ile Val Pro Gly Arg Gly Cys Ala Ile Pro Asn Arg Pro Gly
690 695 700
Ile Phe Val Arg Val Ala Tyr Tyr Ala Lys Trp Ile His Lys Ile Ile
705 710 715 720
Leu Thr Tyr Lys Val Pro Gln Ser
725
<210> 3
<211> 723
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr Cys Arg Asn Pro Arg
165 170 175
Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser Asn Pro Glu Val Arg
180 185 190
Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu Val Glu Cys Met Thr
195 200 205
Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp His Thr Glu Ser Gly
210 215 220
Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro His Arg His Lys Phe
225 230 235 240
Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp Asp Asn Tyr Cys Arg
245 250 255
Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr Thr Leu Asp Pro His
260 265 270
Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys Ala Asp Asn Thr Met
275 280 285
Asn Asp Thr Asp Val Pro Leu Glu Thr Thr Glu Cys Ile Gln Gly Gln
290 295 300
Gly Glu Gly Tyr Arg Gly Thr Val Asn Thr Ile Trp Asn Gly Ile Pro
305 310 315 320
Cys Gln Arg Trp Asp Ser Gln Tyr Pro His Glu His Asp Met Thr Pro
325 330 335
Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu Asn Tyr Cys Arg Asn Pro
340 345 350
Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr Thr Asp Pro Asn Ile Arg
355 360 365
Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys Asp Met Ser His Gly Gln
370 375 380
Asp Cys Tyr Arg Gly Asn Gly Lys Asn Tyr Met Gly Asn Leu Ser Gln
385 390 395 400
Thr Arg Ser Gly Leu Thr Cys Ser Met Trp Asp Lys Asn Met Glu Asp
405 410 415
Leu His Arg His Ile Phe Trp Glu Pro Asp Ala Ser Lys Leu Asn Glu
420 425 430
Asn Tyr Cys Arg Asn Pro Asp Asp Asp Ala His Gly Pro Trp Cys Tyr
435 440 445
Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr Cys Pro Ile Ser Arg Cys
450 455 460
Glu Gly Asp Thr Thr Pro Thr Ile Val Asn Leu Asp His Pro Val Ile
465 470 475 480
Ser Cys Ala Lys Thr Lys Gln Leu Arg Val Val Asn Gly Ile Pro Thr
485 490 495
Arg Thr Asn Ile Gly Trp Met Val Ser Leu Arg Tyr Arg Asn Lys His
500 505 510
Ile Cys Gly Gly Ser Leu Ile Lys Glu Ser Trp Val Leu Thr Ala Arg
515 520 525
Gln Cys Phe Pro Ser Arg Asp Leu Lys Asp Tyr Glu Ala Trp Leu Gly
530 535 540
Ile His Asp Val His Gly Arg Gly Asp Glu Lys Cys Lys Gln Val Leu
545 550 555 560
Asn Val Ser Gln Leu Val Tyr Gly Pro Glu Gly Ser Asp Leu Val Leu
565 570 575
Met Lys Leu Ala Arg Pro Ala Val Leu Asp Asp Phe Val Ser Thr Ile
580 585 590
Asp Leu Pro Asn Tyr Gly Cys Thr Ile Pro Glu Lys Thr Ser Cys Ser
595 600 605
Val Tyr Gly Trp Gly Tyr Thr Gly Leu Ile Asn Tyr Asp Gly Leu Leu
610 615 620
Arg Val Ala His Leu Tyr Ile Met Gly Asn Glu Lys Cys Ser Gln His
625 630 635 640
His Arg Gly Lys Val Thr Leu Asn Glu Ser Glu Ile Cys Ala Gly Ala
645 650 655
Glu Lys Ile Gly Ser Gly Pro Cys Glu Gly Asp Tyr Gly Gly Pro Leu
660 665 670
Val Cys Glu Gln His Lys Met Arg Met Val Leu Gly Val Ile Val Pro
675 680 685
Gly Arg Gly Cys Ala Ile Pro Asn Arg Pro Gly Ile Phe Val Arg Val
690 695 700
Ala Tyr Tyr Ala Lys Trp Ile His Lys Ile Ile Leu Thr Tyr Lys Val
705 710 715 720
Pro Gln Ser
<210> 4
<211> 207
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser
195 200 205
<210> 5
<211> 290
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu
195 200 205
Val Glu Cys Met Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp
210 215 220
His Thr Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro
225 230 235 240
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp
245 250 255
Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr
260 265 270
Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys
275 280 285
Glu Thr
290
<210> 6
<211> 470
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu
195 200 205
Val Glu Cys Met Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp
210 215 220
His Thr Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro
225 230 235 240
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp
245 250 255
Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr
260 265 270
Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys
275 280 285
Ala Asp Asn Thr Met Asn Asp Thr Asp Val Pro Leu Glu Thr Thr Glu
290 295 300
Cys Ile Gln Gly Gln Gly Glu Gly Tyr Arg Gly Thr Val Asn Thr Ile
305 310 315 320
Trp Asn Gly Ile Pro Cys Gln Arg Trp Asp Ser Gln Tyr Pro His Glu
325 330 335
His Asp Met Thr Pro Glu Asn Phe Lys Cys Lys Asp Leu Arg Glu Asn
340 345 350
Tyr Cys Arg Asn Pro Asp Gly Ser Glu Ser Pro Trp Cys Phe Thr Thr
355 360 365
Asp Pro Asn Ile Arg Val Gly Tyr Cys Ser Gln Ile Pro Asn Cys Asp
370 375 380
Met Ser His Gly Gln Asp Cys Tyr Arg Gly Asn Gly Lys Asn Tyr Met
385 390 395 400
Gly Asn Leu Ser Gln Thr Arg Ser Gly Leu Thr Cys Ser Met Trp Asp
405 410 415
Lys Asn Met Glu Asp Leu His Arg His Ile Phe Trp Glu Pro Asp Ala
420 425 430
Ser Lys Leu Asn Glu Asn Tyr Cys Arg Asn Pro Asp Asp Asp Ala His
435 440 445
Gly Pro Trp Cys Tyr Thr Gly Asn Pro Leu Ile Pro Trp Asp Tyr Cys
450 455 460
Pro Ile Ser Arg Cys Glu
465 470
<210> 7
<211> 7113
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гибрид синтетического гепатоцитарного фактора роста
<400> 7
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atccagtata ttaataaaat ccctttttgt attcaatgag 2280
ggaaacacat aattttcatc aattagcagc ttattggaat atctgcatga tggtttaaca 2340
cttttaagtg ttgactaaag attaatttta cagaaaatag aaaaagaaat atgtttctgt 2400
ctggaggaat gatttattgt tgacccctaa attgaaatat tttactagtg gcttaatgga 2460
aagatgatga aagatgatga aattaatgta gaagcttaac tagaaaatca ggtgacctga 2520
tatctacatc tgtatccttc attggccacc cagcattcat taatgaatca gatgatggaa 2580
tagatcaagt ttcctaggaa cacagtgaat attaaaagaa aacaaaggga gcctagcacc 2640
tagaagacct agtttatatt tcaaagtata tttggatgta acccaatttt aaacatttcc 2700
tcacttgtct ctcttaaagc cttgccaaca gcaaggacag agaaccaaaa atagtgtata 2760
tatgaataaa tgcttattac agaatctgct gactggcaca tgctttgtgt gtaatgggtt 2820
ctcataaaca cttgttgaat gaacacacat aagtgaaaga gcatggctag gcttcatccc 2880
ttggtcaaat atggggtgct aaagaaaagc aggggaaata cattgggaca ctaacaaaaa 2940
aaaacagtta atttaggtaa aagataaaat acaccacaga atgaagaaaa gagatgaccc 3000
agactgctct ttaaccttca tgtcctagag aggtttttga tatgaattgc attcagaatt 3060
gtggaaagga gcccatcttt tctcttcatt ttgattttat taactccaat gggggaattt 3120
tattcgtgtt ttggccatat ctacttttga tttctacatt attctctctt cctttctacc 3180
tgtatttgtc ctaataaatt gttgacttat taattcacta cttcctcaca gctttttttt 3240
ggctttacaa atccactgga aaggtatatg ggtgtatcac tttgtgtatt tcggtgtgca 3300
tgtgtagagg ggacaaaaat cctctctcaa actataaata ttgagtattt gtgtattgaa 3360
catttgctat aactactagg tttcttaaat aatcttaata tataaaatga tatagaaaaa 3420
gggaaattat agttcgtatt attcatctaa gtgaagagat taaaacccag ggagtaaata 3480
aattgtctaa ggactaaggt tgtatactat ttaggtgata gatatggggc aaccgtatgg 3540
gttttatgat taacaaataa acttctcacc actctaccat atcaactttt ccataaaaga 3600
gagctatagt attctttgct taaataaatt tgattagtgc atgacttctt gaaaacatat 3660
aaagcaaaag tcacatttga ttctatcaga aaagtgagta agccatggcc caaacaaaag 3720
atgcattaaa atattctgga atgatggagc taaaagtaag aaaaatgact ttttaaaaaa 3780
gtttactgtt aggaattgtg aaattatgct gaattttagt tgcattataa tttttgtcag 3840
tcatacggtc tgacaacctg tcttatttct atttccccat atgaggaatg ctagttaagt 3900
atggatatta actattacta cttagatgca ttgaagttgc ataatatgga taatacttca 3960
ctggttccct gaaaatgttt agttagtaat aagtctctta cactatttgt tttgtccaat 4020
aatttatatt ttctgaagac ttaactctag aatacactca tgtcaaaatg aaagaatttc 4080
attgcaaaat attgcttggt acatgacgca tacctgtatt tgttttgtgt cacaacatga 4140
aaaatgatgg tttattagaa gtttcattgg gtaggaaaca catttgaatg gtatttacta 4200
agatactaaa atccttggac ttcactctaa ttttagtgcc atttagaact caaggtctca 4260
gtaaaagtag aaataaagcc tgttaacaaa acacaagctg aatattaaaa atgtaactgg 4320
attttcaaag aaatgtttac tggtattacc tgtagatgta tattctttat tatgatcttt 4380
tgtgtaaagt ctggcagaca aatgcaatat ctaattgttg agtccaatat cacaagcagt 4440
acaaaagtat aaaaaagact tggccttttc taatgtgtta aaatacttta tgctggtaat 4500
aacactaaga gtagggcact agaaatttta agtgaagata atgtgttgca gttactgcac 4560
tcaatggctt actattataa accaaaactg ggatcactaa gctccagtca gtcaaaatga 4620
tcaaaattat tgaagagaat aagcaattct gttctttatt aggacacagt agatacagac 4680
tacaaagtgg agtgtgctta ataagaggta gcatttgtta agtgtcaatt actctattat 4740
cccttggagc ttctcaaaat aaccatataa ggtgtaagat gttaaaggtt atggttacac 4800
tcagtgcaca ggtaagctaa taggctgaga gaagctaaat tacttactgg ggtctcacag 4860
taagaaagtg agctgaagtt tcagcccaga tttaactgga ttctgggctc tttattcatg 4920
ttacttcatg aatctgtttc tcaattgtgc agaaaaaagg gggctattta taagaaaagc 4980
aataaacaaa caagtaatga tctcaaataa gtaatgcaag aaatagtgag atttcaaaat 5040
cagtggcagc gatttctcag ttctgtccta agtggccttg ctcaatcacc tgctatcttt 5100
tagtggagct ttgaaattat gtttcagaca acttcgattc agttctagaa tgtttgactc 5160
agcaaattca caggctcatc tttctaactt gatggtgaat atggaaattc agctaaatgg 5220
atgttaataa aattcaaacg ttttaaggac agatgaaaat gacagaattt taaggtaaaa 5280
tatatgaagg aatataagat aaaggatttt tctaccttca gcaaaaacat acccactaat 5340
tagtaaaatt aataggcaaa aaaaagttgc atgctcttat actgtaatga ttatcatttt 5400
aaaactagct ttttgccttc gagctatcgg ggtaaagacc tacaggaaaa ctactgtcga 5460
aatcctcgag gggaagaagg gggaccctgg tgtttcacaa gcaatccaga ggtacgctac 5520
gaagtctgtg acattcctca gtgttcagaa gttgaatgca tgacctgcaa tggggagagt 5580
tatcgaggtc tcatggatca tacagaatca ggcaagattt gtcagcgctg ggatcatcag 5640
acaccacacc ggcacaaatt cttgcctgaa agatatcccg acaagggctt tgatgataat 5700
tattgccgca atcccgatgg ccagccgagg ccatggtgct atactcttga ccctcacacc 5760
cgctgggagt actgtgcaat taaaacatgc gctgacaata ctatgaatga cactgatgtt 5820
cctttggaaa caactgaatg catccaaggt caaggagaag gctacagggg cactgtcaat 5880
accatttgga atggaattcc atgtcagcgt tgggattctc agtatcctca cgagcatgac 5940
atgactcctg aaaatttcaa gtgcaaggac ctacgagaaa attactgccg aaatccagat 6000
gggtctgaat caccctggtg ttttaccact gatccaaaca tccgagttgg ctactgctcc 6060
caaattccaa actgtgatat gtcacatgga caagattgtt atcgtgggaa tggcaaaaat 6120
tatatgggca acttatccca aacaagatct ggactaacat gttcaatgtg ggacaagaac 6180
atggaagact tacatcgtca tatcttctgg gaaccagatg caagtaagct gaatgagaat 6240
tactgccgaa atccagatga tgatgctcat ggaccctggt gctacacggg aaatccactc 6300
attccttggg attattgccc tatttctcgt tgtgaaggtg ataccacacc tacaatagtc 6360
aatttagacc atcccgtaat atcttgtgcc aaaacgaaac aattgcgagt tgtaaatggg 6420
attccaacac gaacaaacat aggatggatg gttagtttga gatacagaaa taaacatatc 6480
tgcggaggat cattgataaa ggagagttgg gttcttactg cacgacagtg tttcccttct 6540
cgagacttga aagattatga agcttggctt ggaattcatg atgtccacgg aagaggagat 6600
gagaaatgca aacaggttct caatgtttcc cagctggtat atggccctga aggatcagat 6660
ctggttttaa tgaagcttgc caggcctgct gtcctggatg attttgttag tacgattgat 6720
ttacctaatt atggatgcac aattcctgaa aagaccagtt gcagtgttta tggctggggc 6780
tacactggat tgatcaacta tgatggccta ttacgagtgg cacatctcta tataatggga 6840
aatgagaaat gcagccagca tcatcgaggg aaggtgactc tgaatgagtc tgaaatatgt 6900
gctggggctg aaaagattgg atcaggacca tgtgaggggg attatggtgg cccacttgtt 6960
tgtgagcaac ataaaatgag aatggttctt ggtgtcattg ttcctggtcg tggatgtgcc 7020
attccaaatc gtcctggtat ttttgtccga gtagcatatt atgcaaaatg gatacacaaa 7080
attattttaa catataaggt accacagtca tag 7113
<210> 8
<211> 4679
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ген синтетического HGF-X6
<400> 8
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatc ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tcccttcctt tctacctgta tttgtcctaa taaattgttg acttattaat 780
tcactacttc ctcacagctt ttttttggct ttacaaatcc actggaaagg tatatgggtg 840
tatcactttg tgtatttcgg tgtgcatgtg tagaggggac aaaaatcctc tctcaaacta 900
taaatattga gtatttgtgt attgaacatt tgctataact actaggtttc ttaaataatc 960
ttaatatata aaatgatata gaaaaaggga aattatagtt cgtattattc atctaagtga 1020
agagattaaa acccagggag taaataaatt gtctaaggac taaggttgta tactatttag 1080
gtgatagata tggggcaacc gtatgggttt tatgattaac aaataaactt ctcaccactc 1140
taccatatca acttttccat aaaagagagc tatagtattc tttgcttaaa taaatttgat 1200
tagtgcatga cttcttgaaa acatataaag caaaagtcac atttgattct atcagaaaag 1260
tgagtaagcc atggcccaaa caaaagatgc attaaaatat tctggaatga tggagctaaa 1320
agtaagaaaa atgacttttt aaaaaagttt actgttagga attgtgaaat tatgctgaat 1380
tttagttgca ttataatttt tgtcagtcat acggtctgac aacctgtctt atttctattt 1440
ccccatatga ggaatgctag ttaagtatgg atattaacta ttactactta gatgcattga 1500
agttgcataa tatggataat acttcactgg ttccctgaaa atgtttagtt agtaataagt 1560
ctcttacact atttgttttg tccaataatt tatattttct gaagacttaa ctctagaata 1620
cactcatgtc aaaatgaaag aatttcattg caaaatattg cttggtacat gacgcatacc 1680
tgtatttgtt ttgtgtcaca acatgaaaaa tgatggttta ttagaagttt cattgggtag 1740
gaaacacatt tgaatggtat ttactaagat actaaaatcc ttggacttca ctctaatttt 1800
agtgccattt agaactcaag gtctcagtaa aagtagaaat aaagcctgtt aacaaaacac 1860
aagctgaata ttaaaaatgt aactggattt tcaaagaaat gtttactggt attacctgta 1920
gatgtatatt ctttattatg atcttttgtg taaagtctgg cagacaaatg caatatctaa 1980
ttgttgagtc caatatcaca agcagtacaa aagtataaaa aagacttggc cttttctaat 2040
gtgttaaaat actttatgct ggtaataaca ctaagagtag ggcactagaa attttaagtg 2100
aagataatgt gttgcagtta ctgcactcaa tggcttacta ttataaacca aaactgggat 2160
cactaagctc cagtcagtca aaatgatcaa aattattgaa gagaataagc aattctgttc 2220
tttattagga cacagtagat acagactaca aagtggagtg tgcttaataa gaggtagcat 2280
ttgttaagtg tcaattactc tattatccct tggagcttct caaaataacc atataaggtg 2340
taagatgtta aaggttatgg ttacactcag tgcacaggta agctaatagg ctgagagaag 2400
ctaaattact tactggggtc tcacagtaag aaagtgagct gaagtttcag cccagattta 2460
actggattct gggctcttta ttcatgttac ttcatgaatc tgtttctcaa ttgtgcagaa 2520
aaaagggggc tatttataag aaaagcaata aacaaacaag taatgatctc aaataagtaa 2580
tgcaagaaat agtgagattt caaaatcagt ggcagcgatt tctcagttct gtcctaagtg 2640
gccttgctca atcacctgct atcttttagt ggagctttga aattatgttt cagacaactt 2700
cgattcagtt ctagaatgtt tgactcagca aattcacagg ctcatctttc taacttgatg 2760
gtgaatatgg aaattcagct aaatggatgt taataaaatt caaacgtttt aaggacagat 2820
gaaaatgaca gaattttaag gtaaaatata tgaaggaata taagataaag gatttttcta 2880
ccttcagcaa aaacataccc actaattagt aaaattaata ggcaaaaaaa agttgcatgc 2940
tcttatactg taatgattat cattttaaaa ctagcttttt gccttcgagc tatcggggta 3000
aagacctaca ggaaaactac tgtcgaaatc ctcgagggga agaaggggga ccctggtgtt 3060
tcacaagcaa tccagaggta cgctacgaag tctgtgacat tcctcagtgt tcagaagttg 3120
aatgcatgac ctgcaatggg gagagttatc gaggtctcat ggatcataca gaatcaggca 3180
agatttgtca gcgctgggat catcagacac cacaccggca caaattcttg cctgaaagat 3240
atcccgacaa gggctttgat gataattatt gccgcaatcc cgatggccag ccgaggccat 3300
ggtgctatac tcttgaccct cacacccgct gggagtactg tgcaattaaa acatgcgctg 3360
acaatactat gaatgacact gatgttcctt tggaaacaac tgaatgcatc caaggtcaag 3420
gagaaggcta caggggcact gtcaatacca tttggaatgg aattccatgt cagcgttggg 3480
attctcagta tcctcacgag catgacatga ctcctgaaaa tttcaagtgc aaggacctac 3540
gagaaaatta ctgccgaaat ccagatgggt ctgaatcacc ctggtgtttt accactgatc 3600
caaacatccg agttggctac tgctcccaaa ttccaaactg tgatatgtca catggacaag 3660
attgttatcg tgggaatggc aaaaattata tgggcaactt atcccaaaca agatctggac 3720
taacatgttc aatgtgggac aagaacatgg aagacttaca tcgtcatatc ttctgggaac 3780
cagatgcaag taagctgaat gagaattact gccgaaatcc agatgatgat gctcatggac 3840
cctggtgcta cacgggaaat ccactcattc cttgggatta ttgccctatt tctcgttgtg 3900
aaggtgatac cacacctaca atagtcaatt tagaccatcc cgtaatatct tgtgccaaaa 3960
cgaaacaatt gcgagttgta aatgggattc caacacgaac aaacatagga tggatggtta 4020
gtttgagata cagaaataaa catatctgcg gaggatcatt gataaaggag agttgggttc 4080
ttactgcacg acagtgtttc ccttctcgag acttgaaaga ttatgaagct tggcttggaa 4140
ttcatgatgt ccacggaaga ggagatgaga aatgcaaaca ggttctcaat gtttcccagc 4200
tggtatatgg ccctgaagga tcagatctgg ttttaatgaa gcttgccagg cctgctgtcc 4260
tggatgattt tgttagtacg attgatttac ctaattatgg atgcacaatt cctgaaaaga 4320
ccagttgcag tgtttatggc tggggctaca ctggattgat caactatgat ggcctattac 4380
gagtggcaca tctctatata atgggaaatg agaaatgcag ccagcatcat cgagggaagg 4440
tgactctgaa tgagtctgaa atatgtgctg gggctgaaaa gattggatca ggaccatgtg 4500
agggggatta tggtggccca cttgtttgtg agcaacataa aatgagaatg gttcttggtg 4560
tcattgttcc tggtcgtgga tgtgccattc caaatcgtcc tggtattttt gtccgagtag 4620
catattatgc aaaatggata cacaaaatta ttttaacata taaggtacca cagtcatag 4679
<210> 9
<211> 3679
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ген синтетического HGF-X7
<400> 9
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatc ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tcctgggtag gaaacacatt tgaatggtat ttactaagat actaaaatcc 780
ttggacttca ctctaatttt agtgccattt agaactcaag gtctcagtaa aagtagaaat 840
aaagcctgtt aacaaaacac aagctgaata ttaaaaatgt aactggattt tcaaagaaat 900
gtttactggt attacctgta gatgtatatt ctttattatg atcttttgtg taaagtctgg 960
cagacaaatg caatatctaa ttgttgagtc caatatcaca agcagtacaa aagtataaaa 1020
aagacttggc cttttctaat gtgttaaaat actttatgct ggtaataaca ctaagagtag 1080
ggcactagaa attttaagtg aagataatgt gttgcagtta ctgcactcaa tggcttacta 1140
ttataaacca aaactgggat cactaagctc cagtcagtca aaatgatcaa aattattgaa 1200
gagaataagc aattctgttc tttattagga cacagtagat acagactaca aagtggagtg 1260
tgcttaataa gaggtagcat ttgttaagtg tcaattactc tattatccct tggagcttct 1320
caaaataacc atataaggtg taagatgtta aaggttatgg ttacactcag tgcacaggta 1380
agctaatagg ctgagagaag ctaaattact tactggggtc tcacagtaag aaagtgagct 1440
gaagtttcag cccagattta actggattct gggctcttta ttcatgttac ttcatgaatc 1500
tgtttctcaa ttgtgcagaa aaaagggggc tatttataag aaaagcaata aacaaacaag 1560
taatgatctc aaataagtaa tgcaagaaat agtgagattt caaaatcagt ggcagcgatt 1620
tctcagttct gtcctaagtg gccttgctca atcacctgct atcttttagt ggagctttga 1680
aattatgttt cagacaactt cgattcagtt ctagaatgtt tgactcagca aattcacagg 1740
ctcatctttc taacttgatg gtgaatatgg aaattcagct aaatggatgt taataaaatt 1800
caaacgtttt aaggacagat gaaaatgaca gaattttaag gtaaaatata tgaaggaata 1860
taagataaag gatttttcta ccttcagcaa aaacataccc actaattagt aaaattaata 1920
ggcaaaaaaa agttgcatgc tcttatactg taatgattat cattttaaaa ctagcttttt 1980
gccttcgagc tatcggggta aagacctaca ggaaaactac tgtcgaaatc ctcgagggga 2040
agaaggggga ccctggtgtt tcacaagcaa tccagaggta cgctacgaag tctgtgacat 2100
tcctcagtgt tcagaagttg aatgcatgac ctgcaatggg gagagttatc gaggtctcat 2160
ggatcataca gaatcaggca agatttgtca gcgctgggat catcagacac cacaccggca 2220
caaattcttg cctgaaagat atcccgacaa gggctttgat gataattatt gccgcaatcc 2280
cgatggccag ccgaggccat ggtgctatac tcttgaccct cacacccgct gggagtactg 2340
tgcaattaaa acatgcgctg acaatactat gaatgacact gatgttcctt tggaaacaac 2400
tgaatgcatc caaggtcaag gagaaggcta caggggcact gtcaatacca tttggaatgg 2460
aattccatgt cagcgttggg attctcagta tcctcacgag catgacatga ctcctgaaaa 2520
tttcaagtgc aaggacctac gagaaaatta ctgccgaaat ccagatgggt ctgaatcacc 2580
ctggtgtttt accactgatc caaacatccg agttggctac tgctcccaaa ttccaaactg 2640
tgatatgtca catggacaag attgttatcg tgggaatggc aaaaattata tgggcaactt 2700
atcccaaaca agatctggac taacatgttc aatgtgggac aagaacatgg aagacttaca 2760
tcgtcatatc ttctgggaac cagatgcaag taagctgaat gagaattact gccgaaatcc 2820
agatgatgat gctcatggac cctggtgcta cacgggaaat ccactcattc cttgggatta 2880
ttgccctatt tctcgttgtg aaggtgatac cacacctaca atagtcaatt tagaccatcc 2940
cgtaatatct tgtgccaaaa cgaaacaatt gcgagttgta aatgggattc caacacgaac 3000
aaacatagga tggatggtta gtttgagata cagaaataaa catatctgcg gaggatcatt 3060
gataaaggag agttgggttc ttactgcacg acagtgtttc ccttctcgag acttgaaaga 3120
ttatgaagct tggcttggaa ttcatgatgt ccacggaaga ggagatgaga aatgcaaaca 3180
ggttctcaat gtttcccagc tggtatatgg ccctgaagga tcagatctgg ttttaatgaa 3240
gcttgccagg cctgctgtcc tggatgattt tgttagtacg attgatttac ctaattatgg 3300
atgcacaatt cctgaaaaga ccagttgcag tgtttatggc tggggctaca ctggattgat 3360
caactatgat ggcctattac gagtggcaca tctctatata atgggaaatg agaaatgcag 3420
ccagcatcat cgagggaagg tgactctgaa tgagtctgaa atatgtgctg gggctgaaaa 3480
gattggatca ggaccatgtg agggggatta tggtggccca cttgtttgtg agcaacataa 3540
aatgagaatg gttcttggtg tcattgttcc tggtcgtgga tgtgccattc caaatcgtcc 3600
tggtattttt gtccgagtag catattatgc aaaatggata cacaaaatta ttttaacata 3660
taaggtacca cagtcatag 3679
<210> 10
<211> 2729
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Ген синтетического HGF-X8
<400> 10
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatc ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgga tccttatgtt tcagacaact tcgattcagt tctagaatgt ttgactcagc 780
aaattcacag gctcatcttt ctaacttgat ggtgaatatg gaaattcagc taaatggatg 840
ttaataaaat tcaaacgttt taaggacaga tgaaaatgac agaattttaa ggtaaaatat 900
atgaaggaat ataagataaa ggatttttct accttcagca aaaacatacc cactaattag 960
taaaattaat aggcaaaaaa aagttgcatg ctcttatact gtaatgatta tcattttaaa 1020
actagctttt tgccttcgag ctatcggggt aaagacctac aggaaaacta ctgtcgaaat 1080
cctcgagggg aagaaggggg accctggtgt ttcacaagca atccagaggt acgctacgaa 1140
gtctgtgaca ttcctcagtg ttcagaagtt gaatgcatga cctgcaatgg ggagagttat 1200
cgaggtctca tggatcatac agaatcaggc aagatttgtc agcgctggga tcatcagaca 1260
ccacaccggc acaaattctt gcctgaaaga tatcccgaca agggctttga tgataattat 1320
tgccgcaatc ccgatggcca gccgaggcca tggtgctata ctcttgaccc tcacacccgc 1380
tgggagtact gtgcaattaa aacatgcgct gacaatacta tgaatgacac tgatgttcct 1440
ttggaaacaa ctgaatgcat ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc 1500
atttggaatg gaattccatg tcagcgttgg gattctcagt atcctcacga gcatgacatg 1560
actcctgaaa atttcaagtg caaggaccta cgagaaaatt actgccgaaa tccagatggt 1620
ctgaatcacc ctggtgtttt accactgatc caaacatccg agttggctac tgctcccaaa 1680
ttccaaactg tgatatgtca catggacaag attgttatcg tgggaatggc aaaaattata 1740
tgggcaactt atcccaaaca agatctggac taacatgttc aatgtgggac aagaacatgg 1800
aagacttaca tcgtcatatc ttctgggaac cagatgcaag taagctgaat gagaattact 1860
gccgaaatcc agatgatgat gctcatggac cctggtgcta cacgggaaat ccactcattc 1920
cttgggatta ttgccctatt tctcgttgtg aaggtgatac cacacctaca atagtcaatt 1980
tagaccatcc cgtaatatct tgtgccaaaa cgaaacaatt gcgagttgta aatgggattc 2040
caacacgaac aaacatagga tggatggtta gtttgagata cagaaataaa catatctgcg 2100
gaggatcatt gataaaggag agttgggttc ttactgcacg acagtgtttc ccttctcgag 2160
acttgaaaga ttatgaagct tggcttggaa ttcatgatgt ccacggaaga ggagatgaga 2220
aatgcaaaca ggttctcaat gtttcccagc tggtatatgg ccctgaagga tcagatctgg 2280
ttttaatgaa gcttgccagg cctgctgtcc tggatgattt tgttagtacg attgatttac 2340
ctaattatgg atgcacaatt cctgaaaaga ccagttgcag tgtttatggc tggggctaca 2400
ctggattgat caactatgat ggcctattac gagtggcaca tctctatata atgggaaatg 2460
agaaatgcag ccagcatcat cgagggaagg tgactctgaa tgagtctgaa atatgtgctg 2520
gggctgaaaa gattggatca ggaccatgtg agggggatta tggtggccca cttgtttgtg 2580
agcaacataa aatgagaatg gttcttggtg tcattgttcc tggtcgtgga tgtgccattc 2640
caaatcgtcc tggtattttt gtccgagtag catattatgc aaaatggata cacaaaatta 2700
ttttaacata taaggtacca cagtcatag 2729
<210> 11
<211> 285
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr Cys Arg Asn Pro Arg
165 170 175
Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser Asn Pro Glu Val Arg
180 185 190
Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu Val Glu Cys Met Thr
195 200 205
Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp His Thr Glu Ser Gly
210 215 220
Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro His Arg His Lys Phe
225 230 235 240
Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp Asp Asn Tyr Cys Arg
245 250 255
Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr Thr Leu Asp Pro His
260 265 270
Thr Arg Trp Glu Tyr Cys Ala Ile Lys Thr Cys Glu Thr
275 280 285
<210> 12
<211> 296
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Met Trp Val Thr Lys Leu Leu Pro Ala Leu Leu Leu Gln His Val Leu
1 5 10 15
Leu His Leu Leu Leu Leu Pro Ile Ala Ile Pro Tyr Ala Glu Gly Gln
20 25 30
Arg Lys Arg Arg Asn Thr Ile His Glu Phe Lys Lys Ser Ala Lys Thr
35 40 45
Thr Leu Ile Lys Ile Asp Pro Ala Leu Lys Ile Lys Thr Lys Lys Val
50 55 60
Asn Thr Ala Asp Gln Cys Ala Asn Arg Cys Thr Arg Asn Lys Gly Leu
65 70 75 80
Pro Phe Thr Cys Lys Ala Phe Val Phe Asp Lys Ala Arg Lys Gln Cys
85 90 95
Leu Trp Phe Pro Phe Asn Ser Met Ser Ser Gly Val Lys Lys Glu Phe
100 105 110
Gly His Glu Phe Asp Leu Tyr Glu Asn Lys Asp Tyr Ile Arg Asn Cys
115 120 125
Ile Ile Gly Lys Gly Arg Ser Tyr Lys Gly Thr Val Ser Ile Thr Lys
130 135 140
Ser Gly Ile Lys Cys Gln Pro Trp Ser Ser Met Ile Pro His Glu His
145 150 155 160
Ser Phe Leu Pro Ser Ser Tyr Arg Gly Lys Asp Leu Gln Glu Asn Tyr
165 170 175
Cys Arg Asn Pro Arg Gly Glu Glu Gly Gly Pro Trp Cys Phe Thr Ser
180 185 190
Asn Pro Glu Val Arg Tyr Glu Val Cys Asp Ile Pro Gln Cys Ser Glu
195 200 205
Val Glu Cys Met Thr Cys Asn Gly Glu Ser Tyr Arg Gly Leu Met Asp
210 215 220
His Thr Glu Ser Gly Lys Ile Cys Gln Arg Trp Asp His Gln Thr Pro
225 230 235 240
His Arg His Lys Phe Leu Pro Glu Arg Tyr Pro Asp Lys Gly Phe Asp
245 250 255
Asp Asn Tyr Cys Arg Asn Pro Asp Gly Gln Pro Arg Pro Trp Cys Tyr
260 265 270
Thr Leu Asp Pro His Thr Arg Trp Glu Tyr Cys Ala Ile Lys Asn Met
275 280 285
Arg Asp Ile Thr Trp Ala Leu Asn
290 295
<210> 13
<211> 6190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гибрид синтетического гепатоцитарного фактора роста (HGF)-X2
<400> 13
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atcccttcct ttctacctgt atttgtccta ataaattgtt 2280
gacttattaa ttcactactt cctcacagct tttttttggc tttacaaatc cactggaaag 2340
gtatatgggt gtatcacttt gtgtatttcg gtgtgcatgt gtagagggga caaaaatcct 2400
ctctcaaact ataaatattg agtatttgtg tattgaacat ttgctataac tactaggttt 2460
cttaaataat cttaatatat aaaatgatat agaaaaaggg aaattatagt tcgtattatt 2520
catctaagtg aagagattaa aacccaggga gtaaataaat tgtctaagga ctaaggttgt 2580
atactattta ggtgatagat atggggcaac cgtatgggtt ttatgattaa caaataaact 2640
tctcaccact ctaccatatc aacttttcca taaaagagag ctatagtatt ctttgcttaa 2700
ataaatttga ttagtgcatg acttcttgaa aacatataaa gcaaaagtca catttgattc 2760
tatcagaaaa gtgagtaagc catggcccaa acaaaagatg cattaaaata ttctggaatg 2820
atggagctaa aagtaagaaa aatgactttt taaaaaagtt tactgttagg aattgtgaaa 2880
ttatgctgaa ttttagttgc attataattt ttgtcagtca tacggtctga caacctgtct 2940
tatttctatt tccccatatg aggaatgcta gttaagtatg gatattaact attactactt 3000
agatgcattg aagttgcata atatggataa tacttcactg gttccctgaa aatgtttagt 3060
tagtaataag tctcttacac tatttgtttt gtccaataat ttatattttc tgaagactta 3120
actctagaat acactcatgt caaaatgaaa gaatttcatt gcaaaatatt gcttggtaca 3180
tgacgcatac ctgtatttgt tttgtgtcac aacatgaaaa atgatggttt attagaagtt 3240
tcattgggta ggaaacacat ttgaatggta tttactaaga tactaaaatc cttggacttc 3300
actctaattt tagtgccatt tagaactcaa ggtctcagta aaagtagaaa taaagcctgt 3360
taacaaaaca caaactgaat attaaaaatg taactggatt ttcaaagaaa tgtttactgg 3420
tattacctgt agatgtatat tctttattat gatcttttgt gtaaagtctg gcagacaaat 3480
gcaatatcta attgttgagt ccaatatcac aagcagtaca aaagtataaa aaagacttgg 3540
ccttttctaa tgtgttaaaa tactttatgc tggtaataac actaagagta gggcactaga 3600
aattttaagt gaagataatg tgttgcagtt actgcactca atggcttact attataaacc 3660
aaaactggga tcactaagct ccagtcagtc aaaatgatca aaattattga agagaataag 3720
caattctgtt ctttattagg acacagtaga tacagactac aaagtggagt gtgcttaata 3780
agaggtagca tttgttaagt gtcaattact ctattatccc ttggagcttc tcaaaataac 3840
catataaggt gtaagatgtt aaaggttatg gttacactca gtgcacaggt aagctaatag 3900
gctgagagaa gctaaattac ttactggggt ctcacagtaa gaaagtgagc tgaagtttca 3960
gcccagattt aactggattc tgggctcttt attcatgtta cttcatgaat ctgtttctca 4020
attgtgcaga aaaaaggggg ctatttataa gaaaagcaat aaacaaacaa gtaatgatct 4080
caaataagta atgcaagaaa tagtgagatt tcaaaatcag tggcagcgat ttctcagttc 4140
tgtcctaagt ggccttgctc aatcacctgc tatcttttag tggagctttg aaattatgtt 4200
tcagacaact tcgattcagt tctagaatgt ttgactcagc aaattcacag gctcatcttt 4260
ctaacttgat ggtgaatatg gaaattcagc taaatggatg ttaataaaat tcaaacgttt 4320
taaggacaga tggaaatgac agaattttaa ggtaaaatat atgaaggaat ataagataaa 4380
ggatttttct accttcagca aaaacatacc cactaattag taaaattaat aggcgaaaaa 4440
aagttgcatg ctcttatact gtaatgatta tcattttaaa actagctttt tgccttcgag 4500
ctatcggggt aaagacctac aggaaaacta ctgtcgaaat cctcgagggg aagaaggggg 4560
accctggtgt ttcacaagca atccagaggt acgctacgaa gtctgtgaca ttcctcagtg 4620
ttcagaagtt gaatgcatga cctgcaatgg ggagagttat cgaggtctca tggatcatac 4680
agaatcaggc aagatttgtc agcgctggga tcatcagaca ccacaccggc acaaattctt 4740
gcctgaaaga tatcccgaca agggctttga tgataattat tgccgcaatc ccgatggcca 4800
gccgaggcca tggtgctata ctcttgaccc tcacacccgc tgggagtact gtgcaattaa 4860
aacatgcgct gacaatacta tgaatgacac tgatgttcct ttggaaacaa ctgaatgcat 4920
ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc atttggaatg gaattccatg 4980
tcagcgttgg gattctcagt atcctcacga gcatgacatg actcctgaaa atttcaagtg 5040
caaggaccta cgagaaaatt actgccgaaa tccagatggg tctgaatcac cctggtgttt 5100
taccactgat ccaaacatcc gagttggcta ctgctcccaa attccaaact gtgatatgtc 5160
acatggacaa gattgttatc gtgggaatgg caaaaattat atgggcaact tatcccaaac 5220
aagatctgga ctaacatgtt caatgtggga caagaacatg gaagacttac atcgtcatat 5280
cttctgggaa ccagatgcaa gtaagctgaa tgagaattac tgccgaaatc cagatgatga 5340
tgctcatgga ccctggtgct acacgggaaa tccactcatt ccttgggatt attgccctat 5400
ttctcgttgt gaaggtgata ccacacctac aatagtcaat ttagaccatc ccgtaatatc 5460
ttgtgccaaa acgaaacaat tgcgagttgt aaatgggatt ccaacacgaa caaacatagg 5520
atggatggtt agtttgagat acagaaataa acatatctgc ggaggatcat tgataaagga 5580
gagttgggtt cttactgcac gacagtgttt cccttctcga gacttgaaag attatgaagc 5640
ttggcttgga attcatgatg tccacggaag aggagatgag aaatgcaaac aggttctcaa 5700
tgtttcccag ctggtatatg gccctgaagg atcagatctg gttttaatga agcttgccag 5760
gcctgctgtc ctggatgatt ttgttagtac gattgattta cctaattatg gatgcacaat 5820
tcctgaaaag accagttgca gtgtttatgg ctggggctac actggattga tcaactatga 5880
tggcctatta cgagtggcac atctctatat aatgggaaat gagaaatgca gccagcatca 5940
tcgagggaag gtgactctga atgagtctga aatatgtgct ggggctgaaa agattggatc 6000
aggaccatgt gagggggatt atggtggccc acttgtttgt gagcaacata aaatgagaat 6060
ggttcttggt gtcattgttc ctggtcgtgg atgtgccatt ccaaatcgtc ctggtatttt 6120
tgtccgagta gcatattatg caaaatggat acacaaaatt attttaacat ataaggtacc 6180
acagtcatag 6190
<210> 14
<211> 5190
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Гибрид синтетического гепатоцитарного фактора роста (HGF)-X3
<400> 14
atgtgggtga ccaaactcct gccagccctg ctgctgcagc atgtcctcct gcatctcctc 60
ctgctcccca tcgccatccc ctatgcagag ggacaaagga aaagaagaaa tacaattcat 120
gaattcaaaa aatcagcaaa gactacccta atcaaaatag atccagcact gaagataaaa 180
accaaaaaag tgaatactgc agaccaatgt gctaatagat gtactaggaa taaaggactt 240
ccattcactt gcaaggcttt tgtttttgat aaagcaagaa aacaatgcct ctggttcccc 300
ttcaatagca tgtcaagtgg agtgaaaaaa gaatttggcc atgaatttga cctctatgaa 360
aacaaagact acattagaaa ctgcatcatt ggtaaaggac gcagctacaa gggaacagta 420
tctatcacta agagtggcat caaatgtcag ccctggagtt ccatgatacc acacgaacac 480
aggtaagaac agtatgaaga aaagagatga agcctctgtc ttttttacat gttaacagtc 540
tcatattagt ccttcagaat aattctacaa tcctaaaata acttagccaa cttgctgaat 600
tgtattacgg caaggtttat atgaattcat gactgatatt tagcaaatga ttaattaata 660
tgttaataaa atgtagccaa aacaatatct taccttaatg cctcaatttg tagatctcgg 720
tatttgtgaa ataataacgt aaacttcgtt taaaaggatt cttcttcctg tctttgagaa 780
agtacggcac tgtgcagggg gagaggttga ttgtgaaaaa tcagaggtag atgagaatct 840
tactgagggc tgagggttct ttaaccttgg tggatctcaa cattggttgc acattaaaat 900
cacctgctgc aagcccttga cgaatcttac ttagaagatg acaacacaga acaattaaat 960
cagaatctct ggggagaata gggcaccagt attttttgag ctcccaccat gattccaaag 1020
tgcagccaaa tttgagaacc actgctaaaa gctcaagctt cagattgacc agcttttcca 1080
tctcacctat cgcctaaaga ccaaattgga taaatgtgtt cattacgaca gatgggtact 1140
atttaaagat gagtaaacac aatatactta ggctcgtcag actgagagtt ttaatcatca 1200
ctgaggaaaa acatagatat ctaatactga ctggagtatt agtcaaggct tatttcacac 1260
acaattttat cagaaaccaa agtagtttaa aacagctctc cccttattag taatgcattg 1320
gagggtttac tttaccatgt accttgctga gcactgtacc ttgttaatct catttacttg 1380
taatgagaac cacacagcgg gtagttttat tggttctatt ttacctacat gacaaaactg 1440
aagcataaaa acacttagta agttttcagt gtcatgcaca actaggaagt gacatggcca 1500
gaatataagc ccagtcacca tcactctata acctgcgctt ttaacaactt cagggcatga 1560
cacatttggc cggtcagtag aacccatgct gtgatttgtt tttgcagtgg tggtgatgac 1620
tgccttgttg aatccacttt ttattctatt ccattttggg gacacaattc tgcaagatga 1680
ttcttcatta ggaaacagag atgagttatt gaccaacaca gaaagaaaaa gagtttgttg 1740
ctccacactg ggattaaacc tatgatcttg gcctaattaa cactagctag taagtgtcca 1800
agctgatcat ctctacaaca tttcaataac agaaaacaac aattttcaaa attagttact 1860
tacaattatg tagaaatgcc tctaaaacac agtattttcc ttatattaca aaaacaaaaa 1920
ttataattgg ttttgtcctc ttttgagagt ttgcatggtg ttactccctg catagtgaag 1980
aaaacatttt atttaagtag atggatctaa gtttttcatg aacaaaggaa tgacatttga 2040
aatcaatcct accctagtcc aggagaatgc attagattaa cctagtagag gtcttatttc 2100
accctgagtt ttctatgatc gtgattctct gctggaggag taattgtgaa atagatctct 2160
ctgggaactg gcttcctagt ccaatcagct cttttaccaa tgaacacttc cttgtgatat 2220
agatgtttat ggccgagagg atcctgggta ggaaacacat ttgaatggta tttactaaga 2280
tactaaaatc cttggacttc actctaattt tagtgccatt tagaactcaa ggtctcagta 2340
aaagtagaaa taaagcctgt taacaaaaca caaactgaat attaaaaatg taactggatt 2400
ttcaaagaaa tgtttactgg tattacctgt agatgtatat tctttattat gatcttttgt 2460
gtaaagtctg gcagacaaat gcaatatcta attgttgagt ccaatatcac aagcagtaca 2520
aaagtataaa aaagacttgg ccttttctaa tgtgttaaaa tactttatgc tggtaataac 2580
actaagagta gggcactaga aattttaagt gaagataatg tgttgcagtt actgcactca 2640
atggcttact attataaacc aaaactggga tcactaagct ccagtcagtc aaaatgatca 2700
aaattattga agagaataag caattctgtt ctttattagg acacagtaga tacagactac 2760
aaagtggagt gtgcttaata agaggtagca tttgttaagt gtcaattact ctattatccc 2820
ttggagcttc tcaaaataac catataaggt gtaagatgtt aaaggttatg gttacactca 2880
gtgcacaggt aagctaatag gctgagagaa gctaaattac ttactggggt ctcacagtaa 2940
gaaagtgagc tgaagtttca gcccagattt aactggattc tgggctcttt attcatgtta 3000
cttcatgaat ctgtttctca attgtgcaga aaaaaggggg ctatttataa gaaaagcaat 3060
aaacaaacaa gtaatgatct caaataagta atgcaagaaa tagtgagatt tcaaaatcag 3120
tggcagcgat ttctcagttc tgtcctaagt ggccttgctc aatcacctgc tatcttttag 3180
tggagctttg aaattatgtt tcagacaact tcgattcagt tctagaatgt ttgactcagc 3240
aaattcacag gctcatcttt ctaacttgat ggtgaatatg gaaattcagc taaatggatg 3300
ttaataaaat tcaaacgttt taaggacaga tggaaatgac agaattttaa ggtaaaatat 3360
atgaaggaat ataagataaa ggatttttct accttcagca aaaacatacc cactaattag 3420
taaaattaat aggcgaaaaa aagttgcatg ctcttatact gtaatgatta tcattttaaa 3480
actagctttt tgccttcgag ctatcggggt aaagacctac aggaaaacta ctgtcgaaat 3540
cctcgagggg aagaaggggg accctggtgt ttcacaagca atccagaggt acgctacgaa 3600
gtctgtgaca ttcctcagtg ttcagaagtt gaatgcatga cctgcaatgg ggagagttat 3660
cgaggtctca tggatcatac agaatcaggc aagatttgtc agcgctggga tcatcagaca 3720
ccacaccggc acaaattctt gcctgaaaga tatcccgaca agggctttga tgataattat 3780
tgccgcaatc ccgatggcca gccgaggcca tggtgctata ctcttgaccc tcacacccgc 3840
tgggagtact gtgcaattaa aacatgcgct gacaatacta tgaatgacac tgatgttcct 3900
ttggaaacaa ctgaatgcat ccaaggtcaa ggagaaggct acaggggcac tgtcaatacc 3960
atttggaatg gaattccatg tcagcgttgg gattctcagt atcctcacga gcatgacatg 4020
actcctgaaa atttcaagtg caaggaccta cgagaaaatt actgccgaaa tccagatggg 4080
tctgaatcac cctggtgttt taccactgat ccaaacatcc gagttggcta ctgctcccaa 4140
attccaaact gtgatatgtc acatggacaa gattgttatc gtgggaatgg caaaaattat 4200
atgggcaact tatcccaaac aagatctgga ctaacatgtt caatgtggga caagaacatg 4260
gaagacttac atcgtcatat cttctgggaa ccagatgcaa gtaagctgaa tgagaattac 4320
tgccgaaatc cagatgatga tgctcatgga ccctggtgct acacgggaaa tccactcatt 4380
ccttgggatt attgccctat ttctcgttgt gaaggtgata ccacacctac aatagtcaat 4440
ttagaccatc ccgtaatatc ttgtgccaaa acgaaacaat tgcgagttgt aaatgggatt 4500
ccaacacgaa caaacatagg atggatggtt agtttgagat acagaaataa acatatctgc 4560
ggaggatcat tgataaagga gagttgggtt cttactgcac gacagtgttt cccttctcga 4620
gacttgaaag attatgaagc ttggcttgga attcatgatg tccacggaag aggagatgag 4680
aaatgcaaac aggttctcaa tgtttcccag ctggtatatg gccctgaagg atcagatctg 4740
gttttaatga agcttgccag gcctgctgtc ctggatgatt ttgttagtac gattgattta 4800
cctaattatg gatgcacaat tcctgaaaag accagttgca gtgtttatgg ctggggctac 4860
actggattga tcaactatga tggcctatta cgagtggcac atctctatat aatgggaaat 4920
gagaaatgca gccagcatca tcgagggaag gtgactctga atgagtctga aatatgtgct 4980
ggggctgaaa agattggatc aggaccatgt gagggggatt atggtggccc acttgtttgt 5040
gagcaacata aaatgagaat ggttcttggt gtcattgttc ctggtcgtgg atgtgccatt 5100
ccaaatcgtc ctggtatttt tgtccgagta gcatattatg caaaatggat acacaaaatt 5160
attttaacat ataaggtacc acagtcatag 5190
| название | год | авторы | номер документа |
|---|---|---|---|
| ЛЕЧЕНИЕ НЕЙРОПАТИИ ДНК-КОНСТРУКЦИЯМИ, КОДИРУЮЩИМИ IGF-1, И ДНК- КОНСТРУКЦИЯМИ, КОДИРУЮЩИМИ HGF | 2019 |
|
RU2781980C2 |
| ЛИОФИЛИЗИРОВАННЫЕ ФАРМАЦЕВТИЧЕСКИЕ КОМПОЗИЦИИ ДЛЯ ГЕНОТЕРАПИИ ГОЛОЙ ДНК | 2019 |
|
RU2795471C2 |
| МОДИФИЦИРОВАННЫЕ МИТОХОНДРИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2811467C2 |
| АНТИТЕЛО ПРОТИВ LILRB1 И ЕГО ПРИМЕНЕНИЯ | 2021 |
|
RU2813373C1 |
| КОМПОЗИЦИЯ ДЛЯ ОБЛЕГЧЕНИЯ ИЛИ ЛЕЧЕНИЯ БОЛИ | 2017 |
|
RU2725830C1 |
| АНТИ-LILRB1 АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2801535C1 |
| Новый вариант О-сукцинилгомосеринтрансферазы и способ получения О-сукцинилгомосерина с использованием этого варианта | 2018 |
|
RU2747493C1 |
| МУТИРОВАННЫЙ БЕЛОК SPOT И СПОСОБ ПОЛУЧЕНИЯ L-АМИНОКИСЛОТ С ЕГО ПРИМЕНЕНИЕМ | 2022 |
|
RU2837657C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| Новый мутант О-сукцинилгомосеринтрансферазы и способ получения О-сукцинилгомосерина с использованием этого мутанта | 2018 |
|
RU2747494C1 |




























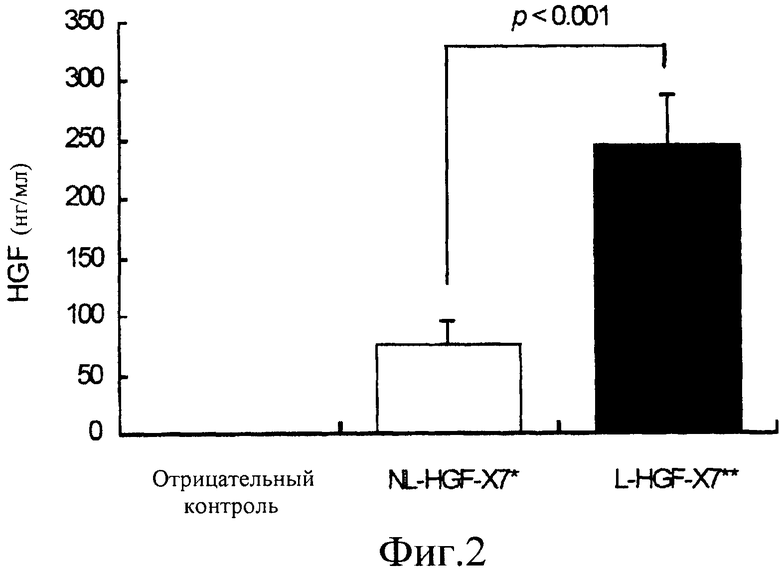
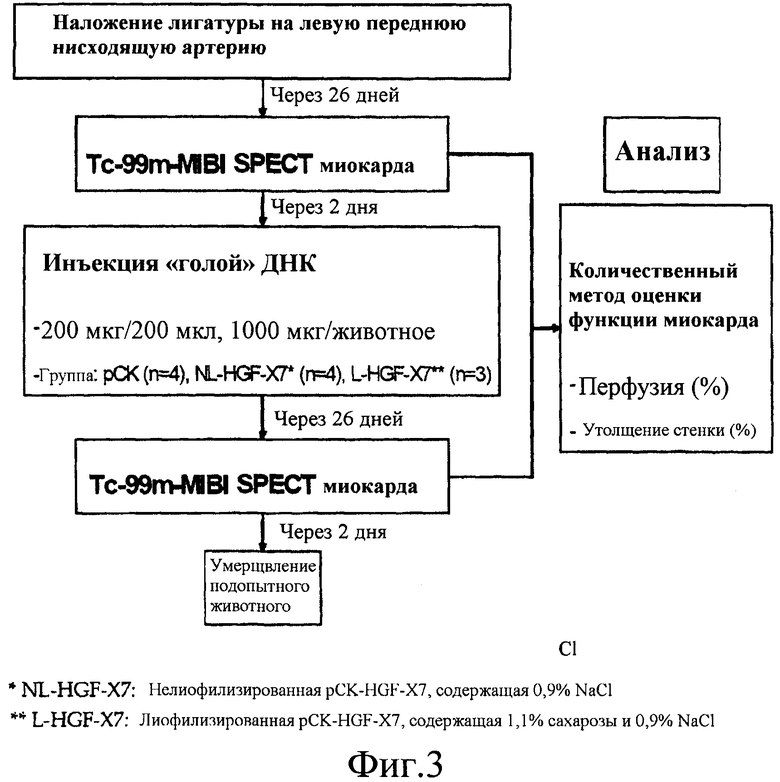
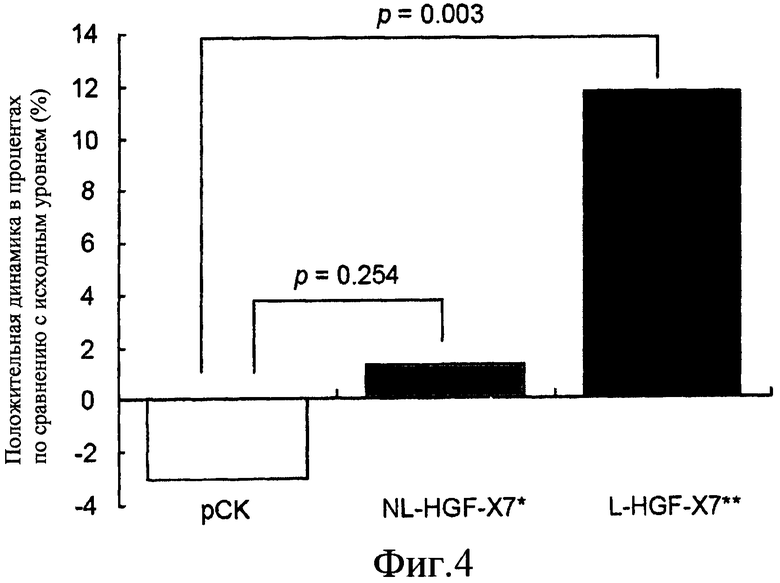
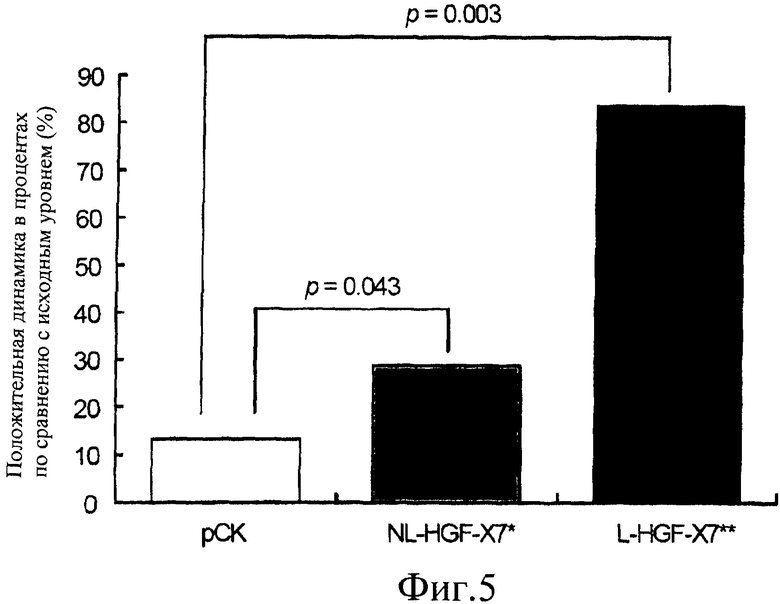
Изобретение относится к области биотехнологии, а именно к лиофилизированному ДНК-составу для применения в способе лечения ишемического заболевания и к способу лечения ишемического заболевания у индивидуума. Лиофилизированный ДНК-состав содержит плазмидную ДНК, соль и углевод, где указанная плазмидная ДНК содержит ген HGF или его вариант и где указанный ген HGF или его вариант выбран из группы, состоящей из flHGF, dHGF, NK1, NK2, NK4 и их смеси. Способ лечения ишемического заболевания у человека или млекопитающего включает введение композиции, восстановленной из лиофилизированного ДНК-состава, путем прямой инъекции. Предложенное изобретение позволяет эффективно лечить ишемическое заболевание у человека или млекопитающего. 2 н. и 12 з.п. ф-лы, 5 ил., 2 табл., 5 пр.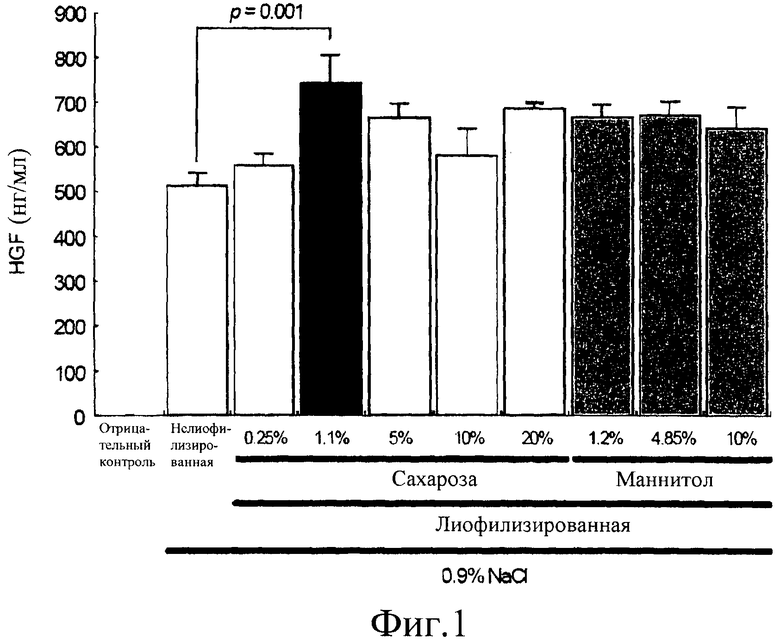
1. Лиофилизированный ДНК-состав для применения в способе лечения ишемического заболевания, который содержит плазмидную ДНК, соль и углевод, где указанная плазмидная ДНК содержит ген HGF или его вариант, и где указанный ген HGF или его вариант выбран из группы, состоящей из flHGF, dHGF, NK1, NK2, NK4 и их смеси, и указанный способ включает введение композиции, восстановленной из указанного лиофилизированного ДНК-состава.
2. ДНК-состав по п.1, где указанный углевод представляет собой моно-, олиго- или полисахарид, выбранный из группы, состоящей из сахарозы, глюкозы, лактозы, трегалозы, арабинозы, пентозы, рибозы, ксилозы, галактозы, гексозы, идозы, маннозы, талозы, гептозы, фруктозы, глюконовой кислоты, сорбитола, маннитола, метил-α-глюкопиранозида, мальтозы, лактона, сорбозы, глюкаровой кислоты, эритрозы, треозы, аллозы, альтрозы, гулозы, эритрулозы, рибулозы, ксилулозы, псикозы, тагатозы, глюкуроновой кислоты, галактуроновой кислоты, маннуроновой кислоты, глюкозамина, галактозамина, нейраминовой кислоты, арабинанов, фруктанов, фуканов, галактанов, галактуронанов, глюканов, маннанов, ксиланов, левана, фукоидана, каррагинана, галактокаролозы, пектинов, пектиновых кислот, амилозы, пуллулана, гликогена, амилопектина, целлюлозы, декстрана, пустулана, хитина, агарозы, кератина, хондроитина, дерматана, гиалуроновой кислоты, альгиновой кислоты, ксантановой камеди, крахмала и их смесей.
3. ДНК-состав по п.1, где указанный углевод присутствует в количестве, выбранном из группы, состоящей из количества между приблизительно 0,05% и приблизительно 30%, между приблизительно 0,1% и приблизительно 15%, между приблизительно 0,2% и приблизительно 10%, между приблизительно 0,5% и 5%, между приблизительно 0,75% и 3%, между приблизительно 0,8% и 2% и между приблизительно 0,8% и 1,5%.
4. ДНК-состав по п.1, где указанный углевод присутствует в количестве от приблизительно 0,1% до приблизительно 10%.
5. ДНК-состав по п.2, где указанный углевод выбран из группы, состоящей из сахарозы, маннитола и их смеси.
6. ДНК-состав по п.1, где указанная соль выбрана из группы, состоящей из NaCl, KCl и их смеси.
7. ДНК-состав по п.1, где указанная соль присутствует в количестве, выбранном из группы, состоящей из количества между приблизительно 0,001% и приблизительно 10%, между приблизительно 0,1% и 5%, между приблизительно 0,5% и 2%, между приблизительно 0,8% и 1,5% и между приблизительно 0,8% и 1,2%.
8. ДНК-состав по п.1, где указанная плазмидная ДНК содержит гибридный ген HGF.
9. ДНК-состав по п.8, где указанный гибридный ген HGF выбран из группы, состоящей из HGF-X2, HGF-X3, HGF-X6, HGF-X7 и HGF-X8.
10. ДНК-состав по п.9, где указанная плазмидная ДНК выбрана из группы, состоящей из: pCK-HGF-X2, pCK-HGF-Х3, pCK-HGF-Х6, pCK-HGF-X7, pCK-HGF-X8, pCP-HGF-X2, pCP-HGF-Х3, pCP-HGF-Х6, pCP-HGF-X7 и pCP-HGF-Х8.
11. ДНК-состав по п.1, где указанная плазмидная ДНК присутствует в концентрации от приблизительно 1 нг/мл до приблизительно 30 мг/мл.
12. Способ лечения ишемического заболевания у индивидуума, включающий введение композиции, восстановленной из лиофилизированного ДНК-состава, путем прямой инъекции, где указанный лиофилизированный ДНК-состав содержит плазмидную ДНК, соль и углевод;
где указанная плазмидная ДНК содержит ген HGF или его вариант;
где указанный ген HGF или его вариант выбран из группы, состоящей из flHGF, dHGF, NK1, NK2, NK4 и их смеси; и
где указанным индивидуумом является человек или млекопитающее.
13. Способ по п.12, где указанная лиофилизированная ДНК восстановлена в фармацевтически приемлемом растворе.
14. Способ по п.13, где указанный фармацевтически приемлемый раствор выбран из группы, состоящей из воды, PBS, ТЕ, буфера Tris, физиологического раствора и их смеси.
| JO J | |||
| ET AL., Liver targeting of plasmid DNA with a cationized pullulan for tumor suppression, J Nanosci Nanotechnol, 2006, v.6, no.9-10, pp.2853-2859 | |||
| MORISHITA R | |||
| ET AL., Therapeutic angiogenesis using hepatocyte growth factor (HGF), Curr Gene Ther, 2004, v.4, no.2, pp.199-206 | |||
| CN 1358543 A, 17.07.2002 | |||
| WO 03078568 A2, 25.09.2003 | |||
| US 2003176347 A, 18.09.2003 | |||
| Высоковакуумное разъемное соединение | 1978 |
|
SU838221A1 |
| СОДЕРЖАЩИЕ ПЭГ КОНЪЮГАТЫ HGF-NK4 | 2002 |
|
RU2293574C2 |

