ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Настоящая заявка относится к области биомедицины и конкретно к мультиспефицическому слитому белку, а также его применению в лечении опухоли и/или аутоиммунного заболевания.
УРОВЕНЬ ТЕХНИКИ
[0002] В настоящее время в области терапии опухолей существует два основных способа - введение таргетированных лекарственных средств и иммунотерапия. Может иметь место взаимодействие между двумя способами терапии, что вызывает более сильный цитотоксический эффект, таким образом, уменьшая опухоль непрерывно и постоянно. Однако взаимодействие между таргетированными лекарственными средствами и иммунотерапией является очень сложным, и на общий противоопухолевый эффект и профиль токсичности комбинированной терапии могут влиять различные факторы, такие как биологический вид, дозы, порядок введения, лекарственные формы и т.п.
[0003] Антитело против белка программируемой гибели 1 (PD-1) представляет собой иммунотерапевтическое средство. PD-1 экспрессируется в активированных T-клетках, B-клетках и миелоидных клетках и имеет два лиганда, т.е. лиганд белка программируемой гибели 1 (PD-L1) и PD-L2. Ингибиторы PD-1 и/или PD-L1 могут специфически связываться с PD-L1 на опухолевых клетках, вызывая противоопухолевый эффект, но частота клинических ответов является низкой, и могут иметь место некоторые побочные эффекты, например, пневмония, колит, гепатит и т.д.
[0004] Белок CD47 представляет собой тип трансмембранного гликопротеина, являющегося членом суперсемейства иммуноглобулинов. В дополнение к экспрессии клетками нормальных тканей, CD47 гиперэкспрессируется многими опухолевыми клетками. CD47 на поверхности опухолевых клеток связывается с SIRPα на поверхности макрофагов, предотвращая фагоцитоз опухолевых клеток макрофагами, что считают одним из механизмов, посредством которого опухоли избегают иммунологического надзора. Блокирование взаимодействия между белком CD47 и SIRPα может ингибировать рост опухоли.
[0005] Однако, существующие реагенты, используемые для блокирования взаимодействия между белком CD47 и SIRPα, имеют ограниченную активность распознавания, их аффинности к CD47 всегда недостаточны, таким образом, они имеют ограниченную способность к ингибированию опухолей. Кроме того, существующие лекарственные средства на основе антител, нацеленные на CD47, могут вызывать побочные эффекты, такие как анемия или тромбоцитопения. Таким образом, необходимо получить эффективное терапевтическое средство, специфически нацеленное и на белок CD47, и на ассоциированные опухолевые антигены.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0006] Настоящее изобретение относится к слитому белку, включающему первый связывающий домен, специфически связывающийся с PD-L1, и второй связывающий домен, специфически связывающийся с белком CD47. Настоящее изобретение также относится к иммуноконъюгату, включающему слитый белок; молекуле нуклеиновой кислоты, кодирующей слитый белок; вектору, композиции и клетке, способным включать и/или экспрессировать слитый белок; и способу получения слитого белка. Слитый белок, иммуноконъюгат, молекула нуклеиновой кислоты, вектор, композиция и клетка по настоящему изобретению имеют одно или более из следующих свойств: 1) способность одновременно специфически связываться с белком CD47 и PD-L1; 2) способность специфически блокировать взаимодействие между белком CD47 и SIRPα; 3) способность специфически блокировать взаимодействие между PD-1 и PD-L1; 4) способность эффективно ингибировать рост и/или пролиферацию опухолей или опухолевых клеток.
[0007] В одном из аспектов настоящее изобретение относится к слитому белку, включающему первый связывающий домен, специфически связывающийся с PD-L1; и второй связывающий домен, специфически связывающийся с белком CD47; где второй связывающий домен содержит мутанта варианта 1 SIRPα человека, мутант содержит замену, делецию или добавление аминокислотного остатка в одном или более положениях от положения 33 до положения 149 по сравнению с последовательностью, приведенной в SEQ ID NO: 29.
[0008] В некоторых вариантах осуществления мутант содержит замены аминокислот в одном или более аминокислотных остатках, выбранных из группы, состоящей из: I61, V63, E77, Q82, K83, E84, V93, D95, L96, K98, N100, R107, G109 и V132.
[0009] В некоторых вариантах осуществления мутант содержит замены аминокислот в аминокислотных остатках, выбранных из группы, состоящей из: (1) I61, V63, E77, E84, V93, L96, K98, N100 и V132; (2) I61, E77, Q82, K83 и E84; (3) I61, V63, K83, E84 и V132; (4) I61, E77, E84, R107 и V132; (5) I61, V63, E77, K83, E84 и N100; (6) I61, E77, Q82, K83, E84 и R107; (7) I61, E77, Q82, E84, V93, L96, N100, R107, G109 и V132; (8) I61, E77, Q82, K83, E84 и V132; (9) I61; (10) I61, D95, L96, G109 и V132; (11) I61, D95, L96, K98, G109 и V132; (12) I61, E77, E84, V93, R107 и V132; (13) E77, L96, N100, G109 и V132; (14) I61, V63, Q82, E84, D95, L96, N100 и V132; (15) I61, E77, Q82, K83, E84, V93, D95, L96, K98, N100 и V132; (16) I61, E77, Q82, K83, E84 и V93; (17) I61, V63, E77, K83, E84, D95, L96, K98 и N100; (18) I61, V63, E77, K83, D95, L96, K98, N100 и G109; (19) I61, E77, Q82, E84, V93, D95, L96, K98 и N100; и (20) I61, V63, E77, Q82 и E84.
[0010] В некоторых вариантах осуществления мутант содержит одну или более замен аминокислот, выбранных из группы, состоящей из: I61L/V/F, V63I, E77I/N/Q/K/H/M/R/N/V/L, Q82S/R/G/N, K83R, E84K/H/D/R/G, V93L/A, D95H/R/E, L96S/T, K98R, N100G/K/D/E, R107N/S, G109R/H и V132L/R/I/S.
[0011] В некоторых вариантах осуществления мутант содержит замены аминокислот, выбранные из группы, состоящей из: (1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132L; (2) I61V, E77N, Q82S, K83R и E84H; (3) I61F, V63I, K83R, E84K и V132I; (4) I61L, E77Q, E84D, R107N и V132I; (5) I61L, V63I, E77K, K83R, E84D и N100G; (6) I61V, E77H, Q82R, K83R, E84H и R107S; (7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R; (8) I61L, E77M, Q82G, K83R, E84D и V132L; (9) I61L; (10) I61F, D95H, L96S, G109H и V132S; (11) I61F, D95H, L96S, K98R, G109H и V132S; (12) I61L, E77Q, E84D, V93A, R107N и V132I; (13) E77K, L96S, N100K, G109H и V132L; (14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I; (15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L; (16) I61V, E77N, Q82S, K83R, E84H и V93A; (17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E; (18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R; (19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и (20) I61L, V63I, E77N, Q82G и E84G.
[0012] В некоторых вариантах осуществления мутант содержит аминокислотную последовательность, приведенную в любой из SEQ ID NO: 30-49.
[0013] В некоторых вариантах осуществления первый связывающий домен содержит антитело или его антигенсвязывающий фрагмент или вариант. В некоторых вариантах осуществления антитело выбрано из группы, состоящей из: моноклонального антитела, одноцепочечного антитела, химерного антитела, гуманизированного антитела и полностью человеческого антитела. В некоторых вариантах осуществления антигенсвязывающий фрагмент выбран из группы, состоящей из: Fab, Fab', F(ab')2, F(ab)2, dAb, выделенной определяющей комплементарность области CDR, Fv и scFv.
[0014] В некоторых вариантах осуществления PD-L1 является PD-L1 человека.
[0015] В некоторых вариантах осуществления антитело содержит тяжелую цепь антитела или его фрагмента, тяжелая цепь антитела или его фрагмента содержит HCDR1-3, и HCDR1 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 4 и SEQ ID NO: 18. В некоторых вариантах осуществления HCDR2 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 5 и SEQ ID NO: 19. В некоторых вариантах осуществления HCDR3 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 6 и SEQ ID NO: 20. В некоторых вариантах осуществления тяжелая цепь антитела или его фрагмента содержит вариабельную область тяжелой цепи VH, и вариабельная область тяжелой цепи VH содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 8 и SEQ ID NO: 22. В некоторых вариантах осуществления тяжелая цепь антитела или его фрагмента содержит константную область тяжелой цепи, и константная область тяжелой цепи содержит IgG. В некоторых вариантах осуществления IgG выбран из группы, состоящей из: IgG1 и IgG4. В некоторых вариантах осуществления тяжелая цепь антитела содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 13 и SEQ ID NO: 27.
[0016] В некоторых вариантах осуществления антитело содержит легкую цепь антитела или его фрагмента, легкая цепь антитела или его фрагмента содержит LCDR1-3, LCDR1 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 15. В некоторых вариантах осуществления LCDR2 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 2 и SEQ ID NO: 16. В некоторых вариантах осуществления LCDR3 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 3 и SEQ ID NO: 17. В некоторых вариантах осуществления легкая цепь антитела или его фрагмента содержит вариабельную область легкой цепи VL, и вариабельная область легкой цепи VL содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 7 и SEQ ID NO: 21. В некоторых вариантах осуществления легкая цепь антитела или его фрагмента содержит константную область легкой цепи, и константная область легкой цепи включает Igκ. В некоторых вариантах осуществления легкая цепь антитела содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 11 и SEQ ID NO: 25.
[0017] В некоторых вариантах осуществления первый связывающий домен находится на N-конце второго связывающего домена. В некоторых вариантах осуществления слитый белок дополнительно содержит линкер, линкер находится на C-конце первого связывающего домена и N-конце второго связывающего домена. В некоторых вариантах осуществления линкер содержит аминокислотную последовательность, приведенную в SEQ ID NO: 52.
[0018] В некоторых вариантах осуществления слитый белок содержит по меньшей мере два вторых связывающих домена. В некоторых вариантах осуществления каждый из вторых связывающих доменов соответствующим образом находится на C-конце первого связывающего домена.
[0019] В другом аспекте настоящее изобретение относится к иммуноконъюгату, содержащему слитый белок.
[0020] В другом аспекте настоящее изобретение относится к одной или более выделенным молекулам нуклеиновой кислоты, кодирующим слитый белок или иммуноконъюгат.
[0021] В другом аспекте настоящее изобретение относится к одному или более векторам, содержащим молекулы нуклеиновой кислоты.
[0022] В другом аспекте настоящее изобретение относится к композиции, содержащей слитый белок, иммуноконъюгат или молекулу нуклеиновой кислоты и, необязательно, фармацевтически приемлемые эксципиенты.
[0023] В другом аспекте настоящее изобретение относится к клетке, содержащей слитый белок, иммуноконъюгат, молекулу нуклеиновой кислоты или вектор.
[0024] В другом аспекте настоящее изобретение относится к способу получения слитого белка, включающему культивирование клетки в условиях, делающих возможной экспрессию слитого белка.
[0025] В другом аспекте настоящее изобретение относится к применению слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки в получении лекарственного средства, где лекарственное средство используют для лечения опухоли.
[0026] В некоторых вариантах осуществления опухоль включает солидную опухоль и несолидную опухоль.
[0027] В некоторых вариантах осуществления солидная опухоль и несолидная опухоль включают множественную миелому, лейкоз, неходжкинскую лимфому, лимфому Ходжкина, нейроглиому, герминому, саркому, мезотелиому, плацентому, злокачественное новообразование головного мозга, злокачественное новообразование костной ткани, рак кожи, рак носоглотки, рак легких, рак ротовой полости, рак пищевода, рак желудка, рак печени, рак поджелудочной железы, рак предстательной железы, рак кишечника, рак молочной железы, рак шейки матки, рак яичников и рак яичка, опухоль лобных пазух, гипофарингеальный рак, ольфакторную нейробластому, рак языка, карциному десен, карциному ампулы фатерова сосочка, рак толстого кишечника, рак прямой кишки, рак почки, карциному мочеточника, рак мочевого пузыря, рак полового члена, карциному фаллопиевых труб, рак века, ретинобластому.
[0028] В другом аспекте настоящее изобретение относится к слитому белку, иммуноконъюгату, молекуле нуклеиновой кислоты, вектору, композиции или клетке, используемым для лечения опухоли.
[0029] В другом аспекте настоящее изобретение относится к способу блокирования взаимодействия между белком PD-L1 и PD-1, включающему введение нуждающемуся в этом индивидууму эффективного количества слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки.
[0030] В другом аспекте настоящее изобретение относится к способу блокирования взаимодействия между белком CD47 и SIRPα, включающему введение нуждающемуся в этом индивидууму эффективного количества слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки.
[0031] В другом аспекте настоящее изобретение относится к способу ингибирования роста и/или пролиферации опухолей или опухолевых клеток, включающему введение нуждающемуся в этом индивидууму эффективного количества слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки.
[0032] В другом аспекте настоящее изобретение относится к способу профилактики или лечения опухолей у индивидуума, включающему введение нуждающемуся в этом индивидууму эффективного количества слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки.
[0033] В другом аспекте настоящее изобретение относится к слитому белку, иммуноконъюгату, молекуле нуклеиновой кислоты, вектору, композиции или клетке, используемой для профилактики или лечения опухолей у индивидуума.
[0034] Другие аспекты и преимущества настоящего изобретения могут быть понятны специалистам в этой области из следующего подробного описания. В следующем подробном описании приведены и описаны лишь примеры вариантов осуществления настоящего изобретения. Как будет понятно специалистам в этой области, содержание настоящей заявки позволяет специалистам в этой области осуществлять изменения в описанных конкретных вариантах осуществления без отклонения от сущности и объема изобретения, к которому относится настоящая заявка. Соответственно, сопутствующие чертежи и описание изобретения представляют собой исключительно примеры, и не предназначены для ограничения изобретения.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0035] Конкретные признаки настоящего изобретения приведены в сопутствующей формуле изобретения. Характеристики и преимущества настоящего изобретения будут более понятны с учетом примеров вариантов осуществления и сопутствующих чертежей, подробно описанных ниже. Сопутствующие чертежи в кратком изложении описаны ниже:
[0036] На фиг. 1 показан пример структуры слитого белка по настоящему изобретению.
[0037] На фиг. 2-3 показана способность слитого белка по настоящему изобретению к связыванию с PD-L1.
[0038] На фиг. 4 показана способность слитого белка по настоящему изобретению к связыванию с CD47.
[0039] На фиг. 5 показана способность слитого белка по настоящему изобретению к одновременному связыванию с PD-L1 и CD47.
[0040] На фиг. 6 показано, что слитый белок по настоящему изобретению конкурентно блокирует связывание между CD47 и его лигандом SIRPα.
[0041] На фиг. 7-8 показано, что слитый белок по настоящему изобретению конкурентно блокирует связывание между PD-1 и PD-L1.
[0042] На фиг. 9-10 показано, что рост объема опухоли в модели лимфомы человека на мышах ингибируют с использованием слитого белка по настоящему изобретению.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0043] Реализация настоящего изобретения будет проиллюстрирована в следующих конкретных вариантах осуществления, и другие преимущества и эффекты настоящего изобретения будут понятны специалисту в этой области из описания изобретения.
[0044] В настоящей заявке термин "слитый белок", как правило, относится к белку, получаемому при слиянии двух или более белков или полипептидов. Слитый белок можно получать искусственно посредством технологии рекомбинантных ДНК. Например, гены или молекулы нуклеиновой кислоты, кодирующие два или более белка или полипептида, можно связывать друг с другом с получением слитого гена или слитой молекулы нуклеиновой кислоты, которая может кодировать слитый белок. Трансляция слитых генов может приводить к продуцированию единого полипептида, который может обладать свойствами по меньшей мере одного или даже каждого из двух или более белков или полипептидов до слияния.
[0045] В настоящем описании термин "специфически связывается", как правило, относится к неслучайной реакции связывания между двумя молекулами, такой как реакция между антителом и антигеном, против которого продуцируется антитело. То, что одно антитело является специфическим для конкретного антигена, означает связывание с антигеном с аффинностью (KD) ≤10-5 M (например, 10-6 M, 10-7 M, 10-8 M, 10-9 M, 10-10 M и т.д.), где термин "KD" относится к отношению скорости диссоциации и скорости связывания (koff/kon), которое можно определять способом, известным специалистам в этой области.
[0046] В настоящей заявке термин "связывающий домен", как правило, означает домен, который может специфически связываться и/или распознавать специфический эпитоп на мишени (например, антигене). В настоящей заявке термин "домен", как правило, относится к практически сферической структурной области, четко отделенной в структуре субъединиц белка. Например, полипептидная цепь, прежде всего, может представлять собой регулярную вторичную структуру, образованную смежными аминокислотными остатками в некоторых областях, затем смежные вторичные структурные фрагменты могут собираться с образованием супервторичной структуры; в связи с этим, полипептидная цепь может сворачиваться в третичную структуру, являющуюся почти сферической. В случае более крупных белковых молекул или субъединиц полипептидная цепь зачастую может представлять собой третичную структуру, образованную в результате ассоциации двух или более относительно независимых региональных структур, которые можно четко различать в пространстве, и такую относительно независимую региональную структуру можно обозначать как домен.
[0047] В настоящей заявке термины "первый", "второй", как в выражениях "первый связывающий домен" и "второй связывающий домен", используют исключительно для различения в рамках описания.
[0048] В настоящей заявке термин "белок CD47", как правило, относится, к интегрин-ассоциированному белку (IAP), являющемуся многопроходным трансмембранным рецептором, принадлежащим к суперсемейству иммуноглобулинов. Например, белок CD47 может связываться с мембранными интегринами, а также с их лигандами тромбоспондином-1 (TSP-1) и сигнальным регуляторным белком альфа (SIRPα). Белок CD47 широко экспрессируется на поверхности мембраны клетки. В настоящей заявке белок CD47 может включать любые варианты, изотипы и видовые гомологи CD47 человека. Аминокислотная последовательность белка CD47 человека указана как CEJ95640.1 в GenBank. Белок CD47 может экспрессироваться естественным образом клетками или экспрессироваться на клетках, трансфицированных с использованием генов CD47.
[0049] В настоящей заявке термин "SIRPα", как правило, относится к регуляторному мембранному гликопротеину из семейства SIRP, который можно использовать в качестве лиганда белка CD47. В настоящей заявке SIRPα может включать SIRPα человека, например, вариант 1 SIRPα и вариант 2 SIRPα. Вариант 2 SIRPα отличается от варианта 1 SIRPα на 13 аминокислот, и его аминокислотная последовательность указана как CAA71403.1 в GenBank. В настоящей заявке термин "вариант 1 SIRPα", как правило, относится к белку SIRPα, аминокислотная последовательность которого указана как NCBI RefSeq NP_542970.1 (остатки 31-504 составляют зрелый тип), аминокислотная последовательность варианта 1 SIRPα приведена в SEQ ID NO: 29.
[0050] Термин "антитело", как правило, относится к белку, содержащему один или более полипептидов, по существу, кодируемых генами иммуноглобулинов или фрагментами генов иммуноглобулинов. Например, гены иммуноглобулинов могут включать гены константной области κ, λ, α, γ, δ, ε и μ, а также многочисленные гены вариабельной области иммуноглобулина. Например, легкую цепь можно классифицировать как κ или λ, что может определять типы иммуноглобулина, соответственно: Igκ и Igλ. Тяжелую цепь можно классифицировать как γ, μ, α, δ или ε, что, в свою очередь, определяет типы иммуноглобулина, соответственно: IgG, IgM, IgA, IgD и IgE. Например, антитело может иметь структурные единицы, содержащие тетрамеры, каждый из тетрамеров может состоять из двух пар одинаковых полипептидных цепей, и каждая пара содержит "легкую цепь" (приблизительно 25 кДа) и "тяжелую цепь" (приблизительно 50-70 кДа), N-конец каждого члена может определять вариабельную область из приблизительно от 100 до 110 или более аминокислот, главным образом, отвечающую за распознавание антигенов. Например, термины "вариабельная область легкой цепи (VL)" и "вариабельная область тяжелой цепи (VH)", как правило, относятся к вариабельной области легкой цепи и тяжелой цепи, соответственно. Антитело может существовать в виде полного иммуноглобулина или в виде множества полностью охарактеризованных фрагментов, получаемых посредством расщепления различными пептидазами или экспрессии de novo.
[0051] В настоящем изобретении термин "антигенсвязывающий фрагмент", как правило, относится к одной или более частям полноразмерного антитела, где части, по существу, сохраняют способность связываться с тем же антигеном (например, PD-L1), что и антитело, и они могут конкурировать с полноразмерным антителом за специфическое связывание с антигеном. В целом, см. Fundamental Immunology, Ch. 7 (Paul, W., ed., Edition 2, Raven Press, N.Y. (1989)), полное содержание которой включено в настоящее описание в качестве ссылки. Антигенсвязывающие фрагменты можно получать с помощью технологии рекомбинантных ДНК или ферментативного или химического расщепления полного антитела. В некоторых случаях, антигенсвязывающие фрагменты включают Fab, Fab', F(ab')2, (Fab)2, Fd, Fv, dAb и определяющую комплементарность область (CDR), одноцепочечное антитело (например, scFv), химерное антитело, диатело и полипептид, содержащий по меньшей мере часть антитела, достаточную для придания полипептиду специфического связывания с антигеном. Для получения антигенсвязывающих фрагментов антитела из указанного антитела и скрининга антигенсвязывающих фрагментов антитела в терминах специфичности тем же образом, что и полные антитела, можно использовать общепринятые технологии, известные специалистам в этой области (например, технологии рекомбинантных ДНК или ферментативного или химического расщепления). Например, для получения F(ab')2антитела в шарнирной области ниже дисульфидной связи можно расщеплять пепсином.
[0052] В настоящей заявке термин "Fab", как правило, относится, к фрагментам антител, состоящим из доменов VL, VH, CL и CH1.
[0053] В настоящей заявке термин "Fab'", как правило, относится к фрагментам антител с несколькими дополнительными остатками на карбокси-конце домена CH1 по сравнению с Fab-фрагментами. Например, Fab' может содержать один или более цистеинов, происходящих из шарнирной области антитела.
[0054] В настоящей заявке термин "F(ab)2", как правило, относится к антигенсвязывающим фрагментам, полученным из спаренных Fab-фрагментов, связанных с цистеином.
[0055] В настоящей заявке термин "dAb-фрагменты", как правило, относится к фрагментам антител, состоящим из доменов VH (Ward et al, Nature 341:544-546 (1989)).
[0056] В настоящей заявке термин "определяющие комплементарность области CDR", как правило, относится к 3 гипервариабельным областям (HVR) вариабельной области легкой цепи (VL) и вариабельной области тяжелой цепи (VH), и гипервариабельные области также известны как определяющие комплементарность области, т.к. эти области могут иметь точную пространственную комплементарность в отношении антигенных детерминант.
[0057] В настоящей заявке термин "Fv-фрагменты", как правило, относится к фрагментам антител, состоящим из одноплечевых доменов VL и VH антитела.
[0058] В настоящей заявке термин "scFv", как правило, относится к молекуле, состоящей из вариабельной области тяжелой цепи и вариабельной области легкой цепи антитела, связанных коротким пептидным линкером, также известной как одноцепочечное антитело.
[0059] В настоящей заявке термин "моноклональное антитело", как правило, относится к группе антител, являющихся, по существу, гомологичными, и различные антитела, содержащиеся в этой группе, могут быть идентичными, за исключением возможного наличия природных мутаций в незначительных количествах. Моноклональные антитела являются высокоспецифическими конкретно в отношении одного антигенного участка. Кроме того, в отличие от препарата поликлональных антител, содержащего разные антитела против разных детерминант (эпитопов), определение "моноклональное" для каждого моноклонального антитела против одной детерминанты на антигене не следует интерпретировать как требование к получению антител любыми конкретными способами. Например, моноклональные антитела можно получать гибридомным способом или в бактериальных, эукариотических животных или растительных клетках способами рекомбинантной ДНК, а также их можно получать из фаговой библиотеки антител, например, с использованием технологии, описанной в Clackson et al., Nature, 352:624-628 (1991) и Marks et al., Mol. Biol., 222:581-597 (1991).
[0060] В настоящей заявке термин "химерное антитело", как правило, относится к такому антителу, где часть каждой аминокислотной последовательности тяжелой цепи или легкой цепи гомологична соответствующей аминокислотной последовательности в антителе конкретного биологического вида или принадлежит к конкретной категории, и остальные сегменты цепи гомологичны соответствующим последовательностям других биологических видов. Например, вариабельные области легкой цепи и тяжелой цепи получают из вариабельных областей антитела одного вида животных (например, мышей, крыс и т.д.), в то время как константная часть гомологична последовательности антитела другого биологического вида (например, человека). Например, в случае получения химерных антител не принадлежащие человеку B-клетки или гибридомные клетки можно использовать для получения вариабельных областей, а константные области, комбинируемые с ними, получают из человека. Вариабельные области обладают преимуществом простого получения, и на их специфичность не влияет источник получения константных областей, комбинируемых с ними. В то же время, т.к. константные области химерных антител можно получать из человека, менее вероятно, что химерные антитела вызовут иммунный ответ во время инъекции, чем при использовании антител с не принадлежащими человеку константными областями.
[0061] В настоящей заявке термин "гуманизированное антитело", как правило, относится к модифицированному антителу, где снижена иммуногенность антитела, иммуноглобулинового связывающего белка и полипептида, полученного из не являющихся человеком видов (например, мышей или крыс), в отношении людей, но они все равно сохраняют антигенсвязывающие свойства исходного антитела. Например, гуманизированное антитело можно получать способами генетической инженерии, и не принадлежащие человеку связывающие домены можно гуманизировать с помощью пересадки CDR (Jones et al., Nature 321:522 (1986)) и ее варианта, включая реконструирование (Verhoeyen, et al., 1988 Science 239:1534-1536; Riechmann, et al., 1988 Nature 332:323-337; Tempest, et al., Bio/Technol 1991 9:266-271), гиперхимеризацию (Queen, et al., 1989 Proc Natl Acad Sci USA 86:10029-10033; Co, et al., 1991, Proc Natl Acad Sci USA 88:2869-2873; Co, et al., 1992 J Immunol 148:1149-1154) и венирование (Mark, et al., "Derivation of therapeutically active humanized and veneered anti-CD18 antibodies" в: Metcalf B W, Dalton B J, eds. Cellular adhesion: molecular definition to therapeutic potential. New York: Plenum Press, 1994: 291-312). В случае других областей, например, шарнирной области и доменах константной области, полученных из не принадлежащих человеку источников, их также можно гуманизировать.
[0062] В настоящей заявке термин "полностью человеческое антитело", как правило, относится к антителу, полученному посредством экспрессии генов, кодирующих антитело человека, в генетически сконструированном животном с делецией генов антител. Например, используя трансгенную или трансхромосомную технологию, все гены, кодирующие антитело человека, можно переносить в генетически сконструированное животное с делецией генов антител таким образом, что животное может экспрессировать антитело человека.
[0063] Белок, полипептид и/или аминокислотную последовательность, указанные в настоящей заявке, также следует понимать как включающие следующий диапазон: варианты или гомологи, имеющие те же или схожие функции, что и белок или полипептид.
[0064] В настоящей заявке вариант может являться белком или полипептидом с заменой, делецией или добавлением одной или более аминокислот в аминокислотной последовательности белка и/или полипептида (например, антитела или его фрагмента, специфически связывающегося с белком PD-L1). Например, функциональный вариант может содержать белок или полипептид с модификациями аминокислот посредством по меньшей мере одной, например 1-30, 1-20 или 1-10, кроме того, например, 1, 2, 3, 4 или 5 замен, делеций и/или инсерций аминокислот. Функциональный вариант может, по существу, сохранять биологические характеристики белка или полипептида до модификации (например, замены, делеции или добавления). Например, функциональный вариант может сохранять по меньшей мере 60%, 70%, 80%, 90% или 100% биологических характеристик белка или полипептида до модификации (например, антигенсвязывающую способность). Например, замена может являться консервативной заменой.
[0065] В настоящей заявке гомолог может являться белком или полипептидом, имеющим по меньшей мере приблизительно 85% (например, по меньшей мере приблизительно 85%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или более) гомологии в отношении аминокислотной последовательности белка и/или полипептида (например, антитела или его фрагмента, специфически связывающегося с белком PD-L1).
[0066] В настоящей заявке термин "гомология", как правило, относится к сравнимости, сходству или соответствию между двумя или более последовательностями. "Процент гомологии последовательности" можно вычислять следующим образом: две сравниваемые последовательности сравнивают в окне сравнения для определения количества положений одинаковых оснований нуклеиновых кислот (например, A, T, C, G, I) или одинаковых аминокислотных остатков (например, Ala, Pro, Ser, Thr, Gly, Val, Leu, Ile, Phe, Tyr, Trp, Lys, Arg, His, Asp, Glu, Asn, Gln, Cys и Met), содержащихся в двух последовательностях, для получения количества совпадающих положений, которое разделяют на общее количество положений в окне сравнения (т.е. размер окна), а затем умножают на 100 для получения процента гомологии последовательности. Сравнение для определения процента гомологии последовательности можно осуществлять разными способами, известными в этой области, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение BLAST, BLAST-2, ALIGN или Megalign (DNASTAR). Специалисты в этой области могут определять параметры, подходящие для выравнивания последовательностей, включая любые алгоритмы, необходимые для осуществления максимального выравнивания в диапазоне сравниваемой полноразмерной последовательности или в целевой области последовательности. Гомологию также можно определять следующими способами: FASTA и BLAST. Описание алгоритма FASTA можно найти в Improved Tool for Comparison of Biological Stquences, W. R. Pearson and D. J. Lipman, Proc. Natl. Acad. Sci., 85: 2444-2448, 1988; и Fast and Sensitive Protein Comparability Search, D. J. Lipman and W. R. Pearson, Science, 227: 1435-1441, 1989. Описание алгоритма BLAST можно найти в A Basic Search Tool for Local Alignment, S. Altschul, W. Gish, W. Miller, E. W. Myers and D. Lipman, Journal of Molecular Biology, 215: 403-410, 1990.
[0067] В настоящей заявке термин "PD-L1", как правило, относится к лиганду белка программируемой гибели-1 (PD-1), также известному как CD274, B7-H или B7H1. PD-1 отрицательно модулирует передачу сигнала T-клеточного антигенного рецептора посредством взаимодействия со специфическим лигандом (PD-L). PD-L1 может являться прогностическим показателем для различных опухолей. В настоящей заявке PD-L1 может являться PD-L1 человека. Gene ID PD-L1 человека в GenBank является 29126.
[0068] В настоящей заявке термин "PD-1", как правило, относится к члену семейства синуклеинов, также известному как NACP, PARK1 или PARK4. В настоящей заявке PD1 может являться PD1 человека. Gene ID PD-L1 человека в GenBank является 6622.
[0069] Как правило, в полипептидной цепи аминогруппа связана с карбоксильной группой так, что образуется единая цепь, но на двух концах белка остаются аминокислотные остатки, не образующие пептидные связи, соответственно, являющиеся концом полипептидной цепи, несущим свободные аминогруппы, и концом полипептидной цепи, несущим карбоксильные группы, соответственно. В настоящей заявке термин "N-конец", как правило, относится к концу полипептидной цепи с аминокислотным остатком, несущим свободные аминогруппы. В настоящей заявке термин "C-конец", как правило, относится к концу полипептидной цепи с аминокислотным остатком, несущим свободные карбоксильные группы.
[0070] В настоящей заявке термин "молекула нуклеиновой кислоты", как правило, относится к нуклеотиду, дезоксирибонуклеотиду, рибонуклеотиду или их аналогу в выделенных формах любой длины, выделенному из природного окружения или синтезированному искусственно.
[0071] В настоящей заявке термин "иммуноконъюгат", как правило, относится к молекуле полипептида с иммунологической функцией, в котором одна или более гетерогенных молекул (включая, в качестве неограничивающих примеров, цитотоксин) конъюгированы. В настоящей заявке термины "конъюгировать", "связывать" и "подвергать слиянию" можно использовать взаимозаменяемо, и они, как правило, относятся к двум или более химическим элементам, последовательностям или компонентам, связанным друг с другом, например, способами, включающими химическую конъюгацию или рекомбинацию. Гетерогенная молекула может являться цитотоксином, химиотерапевтическим средством и т.д. Например, слитый белок по настоящему изобретению можно конъюгировать с одной или более гетерогенными молекулами (например, цитотоксином) для получения иммуноконъюгата.
[0072] В настоящем изобретении термин "вектор" относится к носителю для доставки нуклеиновой кислоты, в котором полинуклеотид, кодирующий некоторый белок, можно встраивать таким образом, что белок может экспрессироваться. Вектор позволяет элементу генетического материала, который он несет, экспрессироваться в клетке-хозяине благодаря трансформации, трансдукции или трансфекции клетки-хозяина. Например, вектор включают: плазмиду; фагмиду; космиду; искусственные хромосомы, такие как искусственная дрожжевая хромосома (YAC), искусственная бактериальная хромосома (BAC) или искусственная хромосома, полученная из P1 (PAC); фаги, такие как фаг λ или фаг M13; и животное вирусы и т.п. Типы вирусов животных, используемых в качестве вектора, включают ретровирус (включая лентивирус), аденовирус, аденоассоциированный вирус, вирус герпеса (например, вирус простого герпеса), поксвирус, бакуловирус, вирус папилломы, паповавирус (например, SV40). Вектор может содержать различные элементы контроля экспрессии, включая промоторную последовательность, последовательность инициации транскрипции, энхансерную последовательность, элемент для селекции и репортерный ген. Кроме того, вектор также может содержать точки начала репликации. Вектор также может содержать компонент, способствующий его проникновению в клетку, такой как, в качестве неограничивающих примеров, вирион, липосома или белковое покрытие.
[0073] В настоящей заявке термин "опухоль", как правило относится, к неоплазиям, образующимся при пролиферации локальных гистоцитов в организмах млекопитающих (например, клетках или их частях) под действием различных онкогенных факторов. В настоящей заявке опухоль может включать солидную опухоль и несолидную опухоль. Солидная опухоль может включать нейроглиому, герминому, саркому, мезотелиому, плацентому, злокачественное новообразование головного мозга, злокачественное новообразование костной ткани, рак кожи, рак носоглотки, рак легких, рак ротовой полости, рак пищевода, рак желудка, рак печени, рак поджелудочной железы, рак предстательной железы, рак кишечника, рак молочной железы, рак шейки матки, рак яичников и рак яичка. В настоящей заявке несолидная опухоль может включать множественную миелому, лейкоз, неходжкинскую лимфому, лимфому Ходжкина.
[0074] В настоящей заявке термин "включает", как правило, означает включение конкретно определенных признаков, не исключающее другие факторы.
[0075] В настоящей заявке термин "приблизительно", как правило, относится к колебаниям в диапазоне на 0,5%-10% выше или ниже определенного значения, например, колебаниям в диапазоне на 0,5%, 1%, 1,5%, 2%, 2,5%, 3%, 3,5%, 4%, 4,5%, 5%, 5,5%, 6%, 6,5%, 7%, 7,5%, 8%, 8,5%, 9%, 9,5% или 10% выше или ниже определенного значения.
[0076] Слитый белок
[0077] В одном из аспектов настоящее изобретение относится к слитому белку, слитый белок может содержать первый связывающий домен и второй связывающий домен. Первый связывающий домен может специфически связываться с PD-L1; второй связывающий домен может специфически связываться с белком CD47, второй связывающий домен может содержать мутанта варианта 1 SIRPα человека, и мутант содержит замену, делецию или добавление аминокислотных остатков в одном или более (например, 1-2, 1-3, 1-4, 1-5, 1-6, 1-7, 1-8, 1-9, 1-10 или более) положениях от положения 33 до положения 149 по сравнению с последовательностью, приведенной в SEQ ID NO: 29. Слитый белок по настоящему изобретению может специфически связываться и с опухолеспецифическим антигеном, и с белком CD47, таким образом, играя роль в лечении опухолей и/или аутоиммунных заболеваний.
[0078] В настоящей заявке термин "первый связывающий домен", как правило, относится к домену, который может специфически связываться с PD-L1. Термин "второй связывающий домен", как правило, относится к домену, который может специфически связываться с белком CD47.
[0079] Второй связывающий домен, специфически связывающийся с CD47
[0080] В настоящей заявке мутант (например, мутант варианта 1 SIRPα человека, специфически связывающийся с белком CD47) содержит замены аминокислот в одном или более (например, 1-2, 1-3, 1-4, 1-5, 1-6, 1-7, 1-8, 1-9, 1-10 или более) аминокислотных остатках, выбранных из группы, состоящей из: I61, V63, E77, Q82, K83, E84, V93, D95, L96, K98, N100, R107, G109 и V132.
[0081] В настоящей заявке положения аминокислотных остатков в замене аминокислоты означают номера остатков, определенных с учетом аминокислотной последовательности, приведенной в SEQ ID NO: 29.
[0082] В настоящей заявке термин "замена аминокислоты Xn" означает, что замена аминокислоты встречается в остатке X в положении n в соответствующей аминокислотной последовательности, приведенной в SEQ ID NO: 29, где n является положительным целым числом, X является сокращенным обозначением любого аминокислотного остатка. Например, термин "замена аминокислоты I61" означает, что замена аминокислоты встречается в остатке I в положении 61 в соответствующей аминокислотной последовательности, приведенной в SEQ ID NO: 29.
[0083] В настоящей заявке замены аминокислот могут являться неконсервативными заменами. Неконсервативные замены могут включать замену аминокислотных остатков в целевых белках или полипептидах в неконсервативной форме, например, замену аминокислотного остатка с некоторым размером боковой цепи или некоторым свойством (например, гидрофильного) другим аминокислотным остатком с другим размером боковой цепи или другим свойством (например, гидрофобным).
[0084] В настоящей заявке замены аминокислот также могут являться консервативными заменами. Консервативные замены может включать замену аминокислотных остатков в целевых белках или полипептидах в консервативной форме, например, замену аминокислотного остатка с некоторым размером боковой цепи или некоторым свойством (например, гидрофильного) другим аминокислотным остатком с таким же или схожим размером боковой цепи или тем же или схожим свойством (например, также гидрофильным). Такие консервативные замены, как правило, не влияют значительно на структуру или функцию образующегося белка. В настоящей заявке вариант аминокислотной последовательности слитого белка или его фрагмента может содержать консервативные аминокислотные замены, не изменяющие значительно структуру или функцию белка (например, мутант варианта 1 SIRPα человека, блокирующий CD47 и специфически связывающийся с белком CD47).
[0085] Например, взаимные замены различных аминокислот в каждой из следующих групп можно считать консервативными заменами в рамках изобретения: группа аминокислот с неполярными боковыми цепями: аланин, валин, лейцин, изолейцин, пролин, фенилаланин, триптофан и метионин; группа незаряженных аминокислот с полярными боковыми цепями: глицин, серин, треонин, цистеин, тирозин, аспарагин и глутамин; группа отрицательно заряженных аминокислот с полярными боковыми цепями: аспарагиновая кислота и глутаминовая кислота; положительно заряженные основные аминокислоты: лизин, аргинин и гистидин; и аминокислоты, несущие фенильные группы: фенилаланин, триптофан и тирозин.
[0086] В настоящей заявке мутант может содержать замены аминокислот в аминокислотных остатках, выбранных из группы, состоящей из: (1) I61, V63, E77, E84, V93, L96, K98, N100 и V132; (2) I61, E77, Q82, K83 и E84; (3) I61, V63, K83, E84 и V132; (4) I61, E77, E84, R107 и V132; (5) I61, V63, E77, K83, E84 и N100; (6) I61, E77, Q82, K83, E84 и R107; (7) I61, E77, Q82, E84, V93, L96, N100, R107, G109 и V132; (8) I61, E77, Q82, K83, E84 и V132; (9) I61; (10) I61, D95, L96, G109 и V132; (11) I61, D95, L96, K98, G109 и V132; (12) I61, E77, E84, V93, R107 и V132; (13) E77, L96, N100, G109 и V132; (14) I61, V63, Q82, E84, D95, L96, N100 и V132; (15) I61, E77, Q82, K83, E84, V93, D95, L96, K98, N100 и V132; (16) I61, E77, Q82, K83, E84 и V93; (17) I61, V63, E77, K83, E84, D95, L96, K98 и N100; (18) I61, V63, E77, K83, D95, L96, K98, N100 и G109; (19) I61, E77, Q82, E84, V93, D95, L96, K98 и N100; и (20) I61, V63, E77, Q82 и E84.
[0087] В настоящей заявке мутант может содержать одну или более (например, 1-2, 1-3, 1-4, 1-5, 1-6, 1-7, 1-8, 1-9, 1-10 или более) замен аминокислот, выбранных из группы, состоящей из: I61L/V/F, V63I, E77I/N/Q/K/H/M/R/N/V/L, Q82S/R/G/N, K83R, E84K/H/D/R/G, V93L/A, D95H/R/E, L96S/T, K98R, N100G/K/D/E, R107N/S, G109R/H и V132L/R/I/S.
[0088] В настоящей заявке замена аминокислоты "XnY/Z" означает, что остаток X в положении n в соответствующей аминокислотной последовательности, приведенной в SEQ ID NO: 29, заменяют аминокислотным остатком Y или аминокислотным остатком Z, где n является положительным целым числом, X, Y и Z являются сокращенными обозначениями любых независимых аминокислотных остатков, и X отличается от Y или Z. Например, замена аминокислоты "I61L/V/F" означает, что остаток I в положении 61 в соответствующей аминокислотной последовательности, приведенной в SEQ ID NO: 29, заменяют аминокислотным остатком L, V или F.
[0089] В настоящей заявке мутант может содержать замены аминокислот, выбранные из группы, состоящей из: (1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132L; (2) I61V, E77N, Q82S, K83R и E84H; (3) I61F, V63I, K83R, E84K и V132I; (4) I61L, E77Q, E84D, R107N и V132I; (5) I61L, V63I, E77K, K83R, E84D и N100G; (6) I61V, E77H, Q82R, K83R, E84H и R107S; (7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R; (8) I61L, E77M, Q82G, K83R, E84D и V132L; (9) I61L; (10) I61F, D95H, L96S, G109H и V132S; (11) I61F, D95H, L96S, K98R, G109H и V132S; (12) I61L, E77Q, E84D, V93A, R107N и V132I; (13) E77K, L96S, N100K, G109H и V132L; (14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I; (15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L; (16) I61V, E77N, Q82S, K83R, E84H и V93A; (17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E; (18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R; (19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и (20) I61L, V63I, E77N, Q82G и E84G.
[0090] В настоящей заявке в контексте варианта 1 SIRPα человека (аминокислотной последовательности, приведенной в SEQ ID NO: 29, т.е. остатков в положениях 33-149 в аминокислотной последовательности SIRPα человека) мутанты варианта 1 SIRPα, содержащие замены аминокислот из указанных выше групп (1)-(20), соответственно, последовательно обозначены как M1, M5, M12, M35, M37, M41, M57, M67, M81, M82, M84, M91, M99, M102, M111, M122, M126, M130, M135 и M145. Мутанты варианта 1 SIRPα могут последовательно содержать аминокислотные последовательности, приведенные в любой из SEQ ID NO: 30-49.
[0091] В некоторых вариантах осуществления мутант варианта 1 SIRPα является M91, и мутант варианта 1 SIRPα содержит аминокислотную последовательность, приведенную в SEQ ID NO: 41.
[0092] Первый связывающий домен, специфически связывающийся с PD-L1
[0093] В настоящей заявке первый связывающий домен может содержать антитело или его антигенсвязывающий фрагмент или вариант. Например, антитело может быть выбрано из группы, состоящей из: моноклонального антитела, одноцепочечного антитела, химерного антитела, гуманизированного антитела и полностью человеческого антитела. Например, антигенсвязывающий фрагмент выбран из группы, состоящей из: Fab, Fab', (Fab')2, F(ab)2, dAb, выделенной определяющей комплементарность области CDR, Fv и scFv.
[0094] Антитело или его антигенсвязывающий фрагмент по настоящему изобретению может приводить к уничтожению опухолевых клеток и/или ингибировать рост опухоли посредством специфического связывания белка PD-L1. Например, опухоль может включать PD-L1-положительную опухоль. Например, PD-L1-положительная опухоль может быть выбрана из группы, состоящей из: рака желудка, рака молочной железы, рака шейки матки, рака легких, опухоли головы и шеи, меланомы, глиомы, лимфоэпителиомы, рака пищевода или колоректального рака. В настоящей заявке антитело и его антигенсвязывающий фрагмент может приводить к уничтожению рака желудка, рака молочной железы, рака шейки матки, рака легких, опухоли головы и шеи, меланомы, глиомы, лимфоэпителиомы, рака пищевода или колоректального рака клетки или ингибировать рост клеток рака желудка, рака молочной железы, рака шейки матки, рака легких, опухоли головы и шеи, меланомы, глиомы, лимфоэпителиомы, рака пищевода или колоректального рака.
[0095] Белок PD-L1 по настоящему изобретению может являться белком PD-L1 человека или его функциональным фрагментом. Например, белок PD-L1 может не являться белком PD-L1 мыши или белком PD-L1 крысы. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент по изобретению, по существу, не связывается с белком PD-L1 мыши или белком PD-L1 крысы.
[0096] Антитело или его антигенсвязывающий фрагмент по изобретению может конкурировать с референсным антителом за связывание с белком PD-L1. Референсное антитело может содержать вариабельную область легкой цепи и вариабельную область тяжелой цепи. Например, вариабельная область легкой цепи референсного антитела может содержать LCDR1-3, LCDR1 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 15; LCDR2 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 2 и SEQ ID NO: 16; и LCDR3 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 3 и SEQ ID NO: 17. Вариабельная область тяжелой цепи референсного антитела может содержать HCDR1-3, HCDR1 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 4 и SEQ ID NO: 18; HCDR2 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 5 и SEQ ID NO: 19; и HCDR3 может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 6 и SEQ ID NO: 20.
[0097] Например, аминокислотная последовательность вариабельной области легкой цепи референсного антитела может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 7 и SEQ ID NO: 21, и аминокислотная последовательность вариабельной области тяжелой цепи референсного антитела может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 8 и SEQ ID NO: 22. Кроме того, например, легкая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 11 и SEQ ID NO: 25; и тяжелая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 13 и SEQ ID NO: 27. Например, легкая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 11, и тяжелая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 13. Например, легкая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 25, и тяжелая цепь референсного антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 27.
[0098] Антитело или его антигенсвязывающий фрагмент по изобретению может содержать легкую цепь антитела или его фрагмента. Например, легкая цепь антитела или его фрагмента может содержать константную область Igκ, например, она может содержать константную область Igκ человека.
[0099] Например, легкая цепь антитела или его фрагмента может содержать LCDR1, и LCDR1 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 1. Легкая цепь антитела или его фрагмента может содержать LCDR2, и LCDR2 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 2. Легкая цепь антитела или его фрагмента может содержать LCDR3, и LCDR3 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 3. Кроме того, например, легкая цепь антитела или его фрагмента может содержать LCDR1, и LCDR1 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 15. Легкая цепь антитела или его фрагмента может содержать LCDR2, и LCDR2 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 16. Легкая цепь антитела или его фрагмента может содержать LCDR3, и LCDR3 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 17.
[0100] Легкая цепь антитела или его фрагмента в настоящей заявке может содержать вариабельную область легкой цепи VL, и аминокислотная последовательность вариабельной области легкой цепи VL может являться SEQ ID NO: 7. В некоторых вариантах осуществления аминокислотная последовательность легкой цепи антитела или его фрагмента может являться SEQ ID NO: 11. Кроме того, например, аминокислотная последовательность вариабельной области легкой цепи VL может являться SEQ ID NO: 21. В некоторых вариантах осуществления аминокислотная последовательность легкой цепи антитела или его фрагмента может являться SEQ ID NO: 25.
[0101] Антитело или его антигенсвязывающий фрагмент по изобретению может содержать тяжелую цепь антитела или его фрагмента. Например, тяжелая цепь антитела или его фрагмента дополнительно содержит константную область человека. При этом константная область человека может включать константную область IgG человека. При этом константная область IgG может включать константную область IgG1 или IgG4 человека.
[0102] Например, тяжелая цепь антитела или его фрагмента может содержать HCDR1, и HCDR1 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 4. Тяжелая цепь антитела или его фрагмента может содержать HCDR2, и HCDR2 может содержать аминокислотную последовательность, приведенную в SEQ ID NO 5. Тяжелая цепь антитела или его фрагмента может содержать HCDR3, и HCDR3 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 6. Кроме того, например, тяжелая цепь антитела или его фрагмента может содержать HCDR1, и HCDR1 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 18. Тяжелая цепь антитела или его фрагмента может содержать HCDR2, и HCDR2 может содержать аминокислотную последовательность, приведенную в SEQ ID NO 19. Тяжелая цепь антитела или его фрагмента может содержать HCDR3, и HCDR3 может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 20.
[0103] Тяжелая цепь антитела или его фрагмента может содержать вариабельную область тяжелой цепи VH, и вариабельная область тяжелой цепи VH может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 8. В некоторых вариантах осуществления тяжелая цепь антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 13. Кроме того, например, вариабельная область тяжелой цепи VH может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 22. В некоторых вариантах осуществления тяжелая цепь антитела может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 27.
[0104] В некоторых вариантах осуществления аминокислотная последовательность легкой цепи антитела или его антигенсвязывающего фрагмента по изобретению содержит SEQ ID NO: 11; и аминокислотная последовательность его тяжелой цепи содержит SEQ ID NO: 13; или аминокислотная последовательность легкой цепи антитела или его антигенсвязывающего фрагмента по изобретению содержит SEQ ID NO: 25; и аминокислотная последовательность его тяжелой цепи содержит SEQ ID NO: 27.
[0105] В некоторых вариантах осуществления аминокислотная последовательность LCDR1 в антителе или его антигенсвязывающем фрагменте по настоящему изобретению может содержать SEQ ID NO: 1; аминокислотная последовательность LCDR2 может содержать SEQ ID NO: 2; аминокислотная последовательность LCDR3 может содержать SEQ ID NO: 3; и аминокислотная последовательность HCDR1 может содержать SEQ ID NO: 4; или аминокислотная последовательность HCDR2 может содержать SEQ ID NO: 5; аминокислотная последовательность HCDR3 может содержать SEQ ID NO: 6. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1201 или антитело, содержащее те же LCDR1-3 и HCDR1-3, что и антитело SG1201. В некоторых вариантах осуществления легкая цепь антитела или его антигенсвязывающий фрагмент по изобретению может содержать вариабельную область легкой цепи, и аминокислотная последовательность вариабельной области легкой цепи может содержать SEQ ID NO: 7; и его тяжелая цепь может содержать вариабельную область тяжелой цепи, и аминокислотная последовательность вариабельной области тяжелой цепи может содержать SEQ ID NO: 8. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1201 или антитело, содержащее ту же вариабельную область легкой цепи и вариабельную область тяжелой цепи, что и антитело SG1201. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент по настоящему изобретению может содержать легкую цепь и тяжелую цепь, аминокислотная последовательность легкой цепи приведена в SEQ ID NO: 11 и аминокислотная последовательность тяжелой цепи приведена в SEQ ID NO: 13. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1201 или содержать те же аминокислотные последовательности легкой цепи и тяжелой цепи, что и антитело SG1201.
[0106] В некоторых вариантах осуществления антитело по настоящему изобретению может являться SG1201. Аминокислотные последовательности LCDR1-3 антитела SG1201 приведены в SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, соответственно; аминокислотная последовательность VL приведена в SEQ ID NO: 7; аминокислотная последовательность легкой цепи приведена в SEQ ID NO: 11; аминокислотные последовательности HCDR1-3 приведены в SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6,соответственно; аминокислотная последовательность VH приведена в SEQ ID NO: 8; и аминокислотная последовательность тяжелой цепи приведена в SEQ ID NO: 13.
[0107] В некоторых вариантах осуществления аминокислотная последовательность LCDR1 в антителе или его антигенсвязывающем фрагменте по настоящему изобретению может содержать SEQ ID NO: 15; аминокислотная последовательность LCDR2 может содержать SEQ ID NO: 16; аминокислотная последовательность LCDR3 может содержать SEQ ID NO: 17; и аминокислотная последовательность HCDR1 может содержать SEQ ID NO: 18; аминокислотная последовательность HCDR2 может содержать SEQ ID NO: 19; аминокислотная последовательность HCDR3 может содержать SEQ ID NO: 20. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1202 или антитело, содержащее те же LCDR1-3 и HCDR1-3, что и антитело SG1202. В некоторых вариантах осуществления легкая цепь антитела или его антигенсвязывающий фрагмент по изобретению может содержать вариабельную область легкой цепи, и аминокислотная последовательность вариабельной области легкой цепи может содержать SEQ ID NO: 21; и его тяжелая цепь может содержать вариабельную область тяжелой цепи, аминокислотная последовательность вариабельной области тяжелой цепи может содержать SEQ ID NO: 22. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1202 или антитело, содержащее ту же вариабельную область легкой цепи и вариабельную область тяжелой цепи, что и антитело SG1202. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент по изобретению может содержать легкую цепь и тяжелую цепь, аминокислотная последовательность легкой цепи приведена в SEQ ID NO: 25, и аминокислотная последовательность тяжелой цепи приведена в SEQ ID NO: 27. Например, антитело или его антигенсвязывающий фрагмент может включать антитело SG1202 или содержать те же аминокислотные последовательности легкой цепи и тяжелой цепи, что и антитело SG1202.
[0108] В некоторых вариантах осуществления антитело по настоящему изобретению может являться SG1202. Аминокислотные последовательности LCDR1-3 антитела SG1202 приведены в SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17, соответственно; аминокислотная последовательность VL приведена в SEQ ID NO: 21; аминокислотные последовательности HCDR1-3 приведены в SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, соответственно; аминокислотная последовательность VH приведена в SEQ ID NO: 22; аминокислотная последовательность легкой цепи приведена в SEQ ID NO: 25; и аминокислотная последовательность тяжелой цепи приведена в SEQ ID NO: 27.
[0109] Антитело или его антигенсвязывающий фрагмент по изобретению также может содержать одну или более случайных мутаций (например, одну или более, например, одну или несколько замен аминокислот) в аминокислотной последовательности легкой цепи и/или тяжелой цепи SG1201 и/или SG1202. Например, антитело или его антигенсвязывающий фрагмент может содержать одну или более случайных мутаций (например, одну или более, например, одну или несколько замен аминокислот) в одном или более положениях каркасных областей L-FR1-4 в вариабельной области легкой цепи SG1201 и/или SG1202, и/или содержать одну или более случайных мутаций (например, одну или более, например, одну или несколько замен аминокислот) в одном или более положениях каркасных областей H-FR1-4 в вариабельной области тяжелой цепи SG1201 и/или SG1202.
[0110] Связь между первым связывающим доменом и вторым связывающим доменом
[0111] В настоящей заявке первый связывающий домен может находиться на N-конце второго связывающего домена. Например, C-конец первого связывающего домена можно связывать с N-концом второго связывающего домена косвенно с помощью линкера. В некоторых случаях C-конец первого связывающего домена также можно связывать с N-концом второго связывающего домена напрямую (например, в рамке считывания).
[0112] В настоящей заявке слитый белок также может содержать линкер, который может находиться на C-конце первого связывающего домена и N-конце второго связывающего домена. Например, в слитом белке C-конец первого связывающего домена можно связывать с N-концом линкера, и C-конец линкера можно связывать с N-концом второго связывающего домена. Например, первый связывающий домен, линкер и второй связывающий домен могут содержаться в слитом белке последовательно от N-конца до C-конца.
[0113] В настоящей заявке линкер может содержать аминокислотную последовательность, приведенную в SEQ ID NO: 52.
[0114] В некоторых случаях слитый белок может содержать по меньшей мере 2 (например, по меньшей мере 2, по меньшей мере 3, по меньшей мере 4, по меньшей мере 5, по меньшей мере 6, по меньшей мере 7, по меньшей мере 8, по меньшей мере 9, по меньшей мере 10 или более) вторых связывающих доменов. В настоящей заявке каждый из вторых связывающих доменов может находиться на C-конце первого связывающего домена, соответственно. В настоящей заявке более двух вторых связывающих доменов можно связывать с C-концом первого связывающего домена прямо или косвенно.
[0115] В настоящей заявке слитый белок может содержать первый связывающий домен, специфически связывающийся с PD-L1, и второй связывающий домен, специфически связывающийся с белком CD47, где второй связывающий домен может содержать мутанта варианта 1 SIRPα человека, C-конец антитела, специфически связывающегося с PD-L1, или его антигенсвязывающего фрагмента или варианта можно прямо или косвенно связывать с N-концом мутанта варианта 1 SIRPα человека. Например, второй связывающий домен может содержать по меньшей мере двух мутантов варианта 1 SIRPα человека, и N-концы двух мутантов варианта 1 SIRPα человека связывают с C-концами антитела, специфически связывающегося с PD-L1, или его антигенсвязывающего фрагмента или варианта, соответственно.
[0116] Например, как показано на фиг. 1, первый связывающий домен слитого белка (SG12473) может содержать SG1201, и его второй связывающий домен может содержать двух мутантов M91 варианта 1 SIRPα, последовательность используемого линкера 1 приведена в SEQ ID NO: 52, N-концы двух M91 связывают с C-концами двух тяжелых цепей SG1201 с помощью линкера 1, соответственно. В слитом белке M91 связывают с C-концом тяжелой цепи SG1201 для получения второй полипептидной цепи, и легкую цепь SG1201 можно обозначать как первую полипептидную цепь. Аминокислотные последовательности второй полипептидной цепи и первой полипептидной цепи SG12473 приведены в SEQ ID NO: 53 и SEQ ID NO: 11, соответственно.
[0117] Например, как показано на фиг. 1, первый связывающий домен слитого белка (SG12474) может содержать SG1202, его второй связывающий домен может содержать двух мутантов M91 варианта 1 SIRPα, последовательность используемого линкера 1 приведена в SEQ ID NO: 52, N-концы двух M91 связывают с C-концами двух тяжелых цепей SG1202 с помощью линкера 1, соответственно. В слитом белке M91 связывают с C-концом тяжелой цепи SG1202 для получения второй полипептидной цепи, и легкую цепь SG1202 можно обозначать как первую полипептидную цепь. Аминокислотные последовательности второй полипептидной цепи и первой полипептидной цепи SG12474 приведены в SEQ ID NO: 54 и SEQ ID NO: 25, соответственно.
[0118] Молекула нуклеиновой кислоты, вектор и клетка, а также способ получения
[0119] В другом аспекте настоящее изобретение относится к одной или более выделенным молекулам нуклеиновой кислоты, кодирующим слитый белок или иммуноконъюгат. Например, каждая молекула нуклеиновой кислоты из одной или более молекул нуклеиновой кислоты может кодировать целое антитело или его антигенсвязывающий фрагмент, а также может кодировать его часть (например, одно или более из HCDR1-3, LCDR1-3, VL, VH, легких цепей или тяжелых цепей).
[0120] Молекула нуклеиновой кислоты по настоящему изобретению может быть выделенной. Например, ее можно получать или синтезировать указанными ниже способами: (i) посредством амплификации in vitro, например, молекулу можно получать посредством амплификации с помощью полимеразной цепной реакции (ПЦР), (ii) молекулу можно получать посредством клонирования с рекомбинацией, (iii) молекулу можно очищать, например, фракционировать посредством ферментативного расщепления и электрофореза в геле, или, (iv) молекулу можно синтезировать, например, посредством химического синтеза. В некоторых вариантах осуществления выделенная нуклеиновая кислота является молекулой нуклеиновой кислоты, полученной посредством технологии рекомбинантных ДНК.
[0121] Способы рекомбинантной ДНК и молекулярного клонирования включают способы, описанные в Sambrook, J., Fritsch, E. F. and Maniatis, T. Molecular Cloning: A Laboratory Manual; Cold Spring Harbor Laboratory Press: Cold Spring Harbor, (1989) (Maniatis) и T. J. Silhavy, M. L. Bennan and L. W. Enquist, Experiments with Gene Fusions, Cold Spring Harbor Laboratory, Cold Spring Harbor, N.Y. (1984), а также Ausubel, F. M. et al, Current Protocols in Molecular biology, pub. by Greene Publishing Assoc. and Wiley-Interscience (1987). В кратком изложении, нуклеиновые кислоты можно получать из фрагментов геномной ДНК, кДНК и РНК, и все из нуклеиновых кислот можно выделять из клеток напрямую или получать посредством рекомбинации различными способами амплификации (включая, в качестве неограничивающих примеров, ПЦР и RT-ПЦР).
[0122] Прямой химический синтез нуклеиновых кислот, как правило, включает последовательное добавление 3'-кэпированных и 5'-кэпированных мономеров нуклеотидов к концевому 5'-гидроксилу растущей полимерной цепи нуклеотида, где каждое добавление происходит посредством нуклеофильной атаки концевого 5'-гидроксила растущей цепи на 3'-положение добавляемых мономеров, и мономеры, как правило, являются фосфорными производными, такими как фосфотриэфир, фосфорамидит и т.д. См., например, Matteuci et al, Tet. Lett. 521:719 (1980); патент США № 4500707 Caruthers et al; и патенты США №№ 5436327 и 5700637 Southern et al. В другом аспекте настоящее изобретение относится к вектору, содержащему выделенный полинуклеотид по настоящему изобретению. Вектор может являться любой линейной нуклеиновой кислотой, плазмидой, фагмидой, космидой, РНК-вектором, вирусным вектором и т.п. Неограничивающие примеры вирусного вектора могут включать ретровирус, аденовирус и аденоассоциированный вирус. В некоторых вариантах осуществления вектор является экспрессирующим вектором, например, вектором фагового дисплея.
[0123] В другом аспекте настоящее изобретение относится к одному или более векторам, включающим молекулы нуклеиновой кислоты. Например, вектор может содержать одну или более молекул нуклеиновой кислоты по настоящему изобретению. Каждый вектор может содержать одну или более молекул нуклеиновой кислоты. Кроме того, вектор может дополнительно содержать другие гены, такие как маркерные гены, делающий возможной селекцию вектора в подходящих клетках-хозяевах и в подходящих условиях. Кроме того, вектор может дополнительно содержать элемент контроля экспрессии, делающий возможной правильную экспрессию кодирующих областей в подходящем хозяине. Такой элемент контроля хорошо известен специалистам в этой области. Например, он может включать промоторы, участки связывания рибосом, энхансеры и другие элементы контроля для регуляции транскрипции генов или трансляции мРНК и т.п. В некоторых вариантах осуществления последовательности контроля экспрессии являются настраиваемыми элементами. Конкретные структуры последовательностей контроля экспрессии могут меняться в зависимости от биологического вида или функций типов клеток, но, как правило, содержат 5'-нетранскрибируемые последовательности и 5'- и 3'-нетранслируемые последовательности, участвующие в инициации транскрипции и трансляции, соответственно, например, TATA-кассеты, кэпированные последовательности, последовательности CAAT и т.д. Например, 5'-нетранскрибируемая последовательность контроля экспрессии может содержать промоторную область, которая может содержать промоторную последовательность для транскрипции и контроля функционально связанных нуклеиновых кислот. Последовательность контроля экспрессии может дополнительно включать энхансерную последовательность или вышележащую активаторную последовательность. В настоящей заявке подходящие промоторы могут включать, например, промоторы полимераз SP6, T3 и T7, промотор U6RNA человека, промотор CMV и искусственный гибридный промотор (например, CMV), в котором некоторую часть промотора можно подвергать слиянию с некоторой частью промотора гена другого клеточного белка (например, GAPDH человека, глицеральдегид-3-фосфатдегидрогеназы), и он может содержать или не содержать дополнительные интроны. Одну или более молекул нуклеиновой кислоты по настоящему изобретению можно функционально связывать с элементами контроля экспрессии.
[0124] Вектор может содержать, например, плазмиду, космиду, вирус, фаг или другие векторы, общеупотребительные, например, в генетической инженерии. В некоторых вариантах осуществления вектор может являться экспрессирующим вектором.
[0125] Вектор также может содержать один или более селективных маркерных гена, которые после экспрессии могут придавать один или более фенотипических признаков, которые можно использовать для селекции или идентификации клеток-хозяев, несущих вектор, другими способами. Неограничивающие примеры подходящих селективных маркеров для эукариот включают дигидрофолатредуктазу и ген резистентности к неомицину.
[0126] В другом аспекте настоящее изобретение относится к клетке, содержащей слитый белок, иммуноконъюгат, молекулу нуклеиновой кислоты или вектор. Клетка может являться клеткой-хозяином. Например, клетка может включать различные типы клеток, как указано ниже: прокариотические клетки, такие как Escherichia coli или Bacillus subtilis, грибковые клетки, такие как дрожжевые клетки или Aspergillus, клетки насекомых, такие как S2 Drosophila клетки или Sf9, или клетки животных, такие как фибробласты, клетки CHO, клетки COS, клетки NSO, клетки HeLa, клетки BHK, клетки HEK 293 или клетки человека.
[0127] Например, вектор можно стабильно или транзиторно встраивать в клетку-хозяина разными известными способами. Например, один из способов включает обработку хлоридом кальция, где вектор встраивают посредством осаждения кальцием. Также можно использовать другие соли схожими способами, например, фосфат кальция. Кроме того, можно использовать электропорацию (т.е. используя электрический ток для повышения проницаемости клеток для нуклеиновых кислот). Другие примеры способов трансформации включают микроинъекцию, опосредованную DEAE-декстраном трансформацию и тепловой шок в присутствии ацетата лития. Для трансфекции клеток-хозяев также можно использовать липидные комплексы, липосомы и дендримеры.
[0128] Если в клетку-хозяина встраивают гетерогенные последовательности, для идентификации клеток-хозяев, в которые можно встраивать вектор, можно использовать различные способы. Один из примеров способов селекции включает субкультивирование отдельной клетки для получения отдельной колонии, а затем тестирование экспрессии желаемого белкового продукта. Для другого способа необходима селекция клетки-хозяина, содержащей гетерогенную последовательность, на основе фенотипических признаков, обеспечиваемых экспрессией селективных маркерных генов, включенных в вектор.
[0129] Например, встраивание различных гетерогенных последовательностей по настоящему изобретению в клетку-хозяина можно подтверждать следующими способами, такими как ПЦР, Саузерн-блоттинг или нозерн-блоттинг. Например, нуклеиновые кислоты можно получать из полученной клетки-хозяина, и конкретные целевые последовательности можно амплифицировать посредством ПЦР с использованием праймеров, специфических для целевых последовательностей. Продукты амплификации подвергают электрофорезу в агарозном геле, электрофорезу в полиакриламидном геле или капиллярному электрофорезу, а затем окрашивают бромистым этидием, раствором SYBR Green или т.п. или подвергают детекции ДНК с помощью УФ. Альтернативно, в реакциях гибридизации можно использовать зонды нуклеиновых кислот, специфические для целевых последовательностей. Экспрессию конкретных последовательностей генов можно определять посредством гибридизации с использованием ПЦР или нозерн-блоттинга или посредством детекции обратной транскрипции соответствующей мРНК с использованием иммунологических анализов антител, реагирующих с кодируемыми продуктами генов. Неограничивающие примеры иммунологических анализов включают ELISA, радиоиммунологический анализ и иммунологический анализ сэндвич-типа.
[0130] Кроме того, встраивание различных гетерогенных последовательностей по настоящему изобретению в клетку-хозяина можно подтверждать с помощью ферментативной активности ферментов (например, ферментативных маркеров), кодируемых гетерогенными последовательностями. Ферменты можно определять различными известными в этой области способами. Как правило, ферментативную активность можно определять по образованию продуктов или превращению субстрата ферментативной реакции. Реакцию можно проводить in vitro или in vivo.
[0131] В другом аспекте настоящее изобретение относится к способу получения слитого белка, который может включать культивирование клетки в условиях, делающих возможной экспрессию слитого белка. Например, можно использовать подходящие среды для культивирования, подходящую температуру и время культивирования, и все эти способы известны специалистам в этой области.
[0132] В некоторых случаях способ может дополнительно включать стадии разделения и/или очистки слитого белка. Например, слитый белок по настоящему изобретению можно очищать и выделять посредством аффинной хроматографии с использованием протеин G-агарозы или протеин A-агарозы или посредством электрофореза в геле и/или высокоэффективной жидкостной хроматографии.
[0133] Иммуноконъюгат, композиция и применение
[0134] В другом аспекте настоящее изобретение относится к иммуноконъюгату, содержащему слитый белок. Например, иммуноконъюгат может являться конъюгатом слитый белок-лекарственное средство (ADC), в котором слитый белок по настоящему изобретению конъюгируют с одним или более терапевтическими средствами, и терапевтические средства включают, в качестве неограничивающих примеров, цитотоксические средства, радиотоксические средства (например, радиоактивные изотопы) и/или иммунные ингибиторы (например, любые средства, уничтожающие клетки посредством ингибирования иммунных ответов) и т.п. В некоторых вариантах осуществления терапевтические средства могут являться средствами, с помощью которых можно лечить опухолеассоциированные заболевания или нарушения.
[0135] Конъюгацию можно осуществлять с помощью пептидного линкера (например, расщепляемого линкера) или другими способами. Например, линкер может являться кислото-неустойчивым линкером, пептидаза-чувствительным линкером, фотолабильным линкером и т.п.
[0136] В другом аспекте настоящее изобретение относится к композиции, содержащей слитый белок, иммуноконъюгат или молекулу нуклеиновой кислоты и, необязательно, фармацевтически приемлемые эксципиенты.
[0137] Например, фармацевтически приемлемые эксципиенты могут включать буферные средства, антиоксиданты, консерванты, низкомолекулярные полипептиды, белки, гидрофильные полимеры, аминокислоты, сахар, хелатирующие средства, противоионы, комплексные соединения с металлами и/или неионные поверхностно-активные вещества и т.п.
[0138] В настоящей заявке композицию можно составлять с использованием фармацевтически приемлемых носителей или дилюентов и любых других известных вспомогательных средств и эксципиентов общепринятыми способами, например, описанными в Remington: The Science and Practice of Pharmacy, Edition 19, Gennaro ed., Mack Publishing Co., Easton, PA, 1995.
[0139] В настоящей заявке композицию можно составлять для перорального введения, внутривенного введения, внутримышечного введения, введения in situ в опухолевые очаги, ингаляции, ректального введения, вагинального введения, трансдермального введения или введения с помощью подкожного резервуара.
[0140] Например, композицию можно использовать для ингибирования роста опухоли. Например, композиция по настоящему изобретению может ингибировать или замедлять развитие или прогрессирование заболеваний, уменьшать размер опухолей (даже, по существу, приводить к элиминации опухолей) и/или облегчать и/или стабилизировать статус заболеваний.
[0141] Например, композиция по настоящему изобретению может представлять собой подходящую форму для перорального введения, такую как таблетки, капсулы, пилюли, порошки, препараты с замедленным высвобождением, растворы, суспензии; или для парентеральной инъекции, такую как стерильные растворы, суспензии или эмульсии; или для локального введения в виде мази или крема; или для ректального введения в виде суппозиториев. Композиция может представлять собой стандартную лекарственную форму, подходящую для однократной дозы с точной дозировкой. Композиция может дополнительно содержать общепринятые носители или эксципиенты для лекарственных средств. Кроме того, композиция может содержать другие лекарственные средства или средства, носители, адъюванты и т.д.
[0142] Композиция по настоящему изобретению может содержать терапевтически эффективное количество слитого белка. Терапевтически эффективное количество представляет собой дозу, необходимую для профилактики и/или лечения (по меньшей мере частичного лечения) заболеваний или нарушений (например, опухолей) и/или любых их осложнений у индивидуума, страдающего или имеющего риск развития этих заболеваний или нарушений. Конкретное количество/концентрацию в дозе можно изменять в зависимости от способа введения и потребности пациента, и его можно определять, например, с учетом размера пациента, вязкости и/или масса тела. Например, подходящая доза может составлять приблизительно 0,1 мг или от 1 мг/кг/сутки до приблизительно 50 мг/кг/сутки; иногда доза может быть более высокой. Следует понимать, что специалист в этой области (например, врач или фармацевт) легко может корректировать эти конкретные дозы с учетом конкретного пациента, препаратов и/или статуса заболевания.
[0143] В настоящей заявке термины "лечение", или "излечение", или "облегчение", или "улучшение" можно использовать в настоящей заявке взаимозаменяемо, и они относятся к способам, которыми можно достигать благоприятных или желаемых результатов (включая, в качестве неограничивающих примеров, терапевтические благоприятные эффекты и/или профилактические благоприятные эффекты). В настоящей заявке терапевтические благоприятные эффекты, как правило, относятся к эрадикации или снижению тяжести основных заболеваний, подвергаемых лечению. Кроме того, терапевтические благоприятные эффекты реализуют посредством эрадикации или снижения тяжести основных заболеваний или уменьшения частоты одного или более физических симптомов, ассоциированных с основными заболеваниями, таким образом, чтобы наблюдать улучшение у индивидуума (хотя индивидуум все равно может страдать основными заболеваниями). Что касается профилактических благоприятных эффектов, композицию можно вводить индивидууму, имеющему риск развития конкретного заболевания, или индивидууму, имеющему один или более физических симптомов указанного заболевания, даже если заболевание может быть не диагностировано.
[0144] В другом аспекте настоящее изобретение относится к применению слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки в получении лекарственного средства, где лекарственное средство можно использовать для лечения опухоли.
[0145] В другом аспекте слитый белок, иммуноконъюгат, молекулу нуклеиновой кислоты, вектор, композицию или клетку по настоящему изобретению можно использовать для лечения опухоли.
[0146] В другом аспекте настоящее изобретение относится к способу лечения опухоли, включающему введение индивидууму слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки по настоящему изобретению.
[0147] В другом аспекте настоящее изобретение относится к способу блокирования взаимодействия между белком CD47 и SIRPα, включающему введение (например, введение нуждающемуся в этом индивидууму или в клетки или биологические образцы) слитого белка или композиции по настоящему изобретению.
[0148] В другом аспекте настоящее изобретение относится к способу блокирования взаимодействия между PD-L1 и PD1, включающему введение (например, введение нуждающемуся в этом индивидууму или в клетки или биологические образцы) слитого белка или композиции по настоящему изобретению.
[0149] В другом аспекте настоящее изобретение относится к способу ингибирования опухоли или роста и/или пролиферации опухолевых клеток, включающему приведение слитого белка или композиции по настоящему изобретению в контакт с опухолью или опухолевыми клетками. Например, приведение в контакт можно осуществлять in vitro.
[0150] В другом аспекте настоящее изобретение относится к способу ингибирования опухоли или роста и/или пролиферации опухолевых клеток, включающему введение нуждающемуся в этом индивидууму эффективного количества слитого белка, иммуноконъюгата, молекулы нуклеиновой кислоты, вектора, композиции или клетки.
[0151] В некоторых вариантах осуществления опухоль может включать солидную опухоль и несолидную опухоль.
[0152] В некоторых вариантах осуществления солидная опухоль и несолидная опухоль может включать множественную миелому, лейкоз, неходжкинскую лимфому, лимфому Ходжкина, нейроглиому, герминому, саркому, мезотелиому, плацентому, злокачественное новообразование головного мозга, злокачественное новообразование костной ткани, рак кожи, рак носоглотки, рак легких, рак ротовой полости, рак пищевода, рак желудка, рак печени, рак поджелудочной железы, рак предстательной железы, рак кишечника, рак молочной железы, рак шейки матки, рак яичников и рак яичка, опухоль лобных пазух, гипофарингеальный рак, ольфакторную нейробластому, рак языка, карциному десен, карциному ампулы фатерова сосочка, рак толстого кишечника, рак прямой кишки, рак почки, карциному мочеточника, рак мочевого пузыря, рак полового члена, карциному фаллопиевых труб, рак века, ретинобластому.
[0153] В другом аспекте настоящее изобретение относится к слитому белку, иммуноконъюгату, молекуле нуклеиновой кислоты, вектору, композиции или клетке, которую можно использовать для лечения опухоли или аутоиммунное заболевание
[0154] В настоящей заявке термин "индивидуум", как правило, относится к человеку или не являющимся человеком животным, включая, в качестве неограничивающих примеров, кошку, собаку, лошадь, свинью, корову, овцу, козу, кролика, мышь, крысу или обезьяну.
[0155] Без связи с какой-либо теорией, приведенные ниже варианты осуществления предназначены исключительно для описания слитого белка, способа получения и применения по настоящему изобретению, а не ограничения объема настоящего изобретения.
[0156] Примеры
[0157] Пример 1. Конструирование слитого белка
[0158] С учетом структуры слитого белка, показанной на фиг. 1, используя антитело против PD-L1 атезолизумаб от Roche (SG1201) и мутанта M91 (SEQ ID: NO 41) варианта 1 SIRPα например, выбранный линкер 1 (SEQ ID: NO 52) используют для последовательного соединения от N-конца к C-концу SG1201, линкера и двух M91, где N-концы двух M91 связывают с C-концом тяжелой цепи SG1201 соответствующим образом для получения слитого белка SG12473.
[0159] Аминокислотные последовательности LCDR1-3 антитела SG1201 приведены в SEQ ID NO: 1, SEQ ID NO: 2 и SEQ ID NO: 3, соответственно; аминокислотная последовательность VL приведена в SEQ ID NO: 7; нуклеотидная последовательность, кодирующая VL, приведена в SEQ ID NO: 9; аминокислотные последовательности HCDR1-3 антитела SG1201 приведены в SEQ ID NO: 4, SEQ ID NO: 5 и SEQ ID NO: 6, соответственно; аминокислотная последовательность VH приведена в SEQ ID NO: 8; нуклеотидная последовательность, кодирующая VH, приведена в SEQ ID NO: 10. Аминокислотная последовательность легкой цепи антитела SG1201 приведена в SEQ ID NO: 11; и нуклеотидная последовательность, кодирующая его легкую цепь, приведена в SEQ ID NO: 12. Аминокислотная последовательность тяжелой цепи антитела SG1201 приведена в SEQ ID NO: 13; и нуклеотидная последовательность, кодирующая его тяжелую цепь, приведена в SEQ ID NO: 14.
[0160] Слитый белок SG12473 состоит из первой полипептидной цепи и второй полипептидной цепи, где первая полипептидная цепь является легкой цепью SG1201, аминокислотная последовательность которой приведена в SEQ ID NO: 11; вторая полипептидная цепь является полипептидной цепью, полученной посредством связывания тяжелой цепи SG1201 с мутацией в Fc-области и M91 с помощью линкера 1, аминокислотная последовательность которой приведена в SEQ ID NO: 53.
[0161] С учетом структуры слитого белка, показанной на фиг. 1, используя антитело против PD-L1 дурвалумаб от AstraZeneca (SG1202) и мутанта M91 (SEQ ID: NO 41) варианта 1 SIRPα, например, выбранный линкер 1 (SEQ ID: NO 52) используют для последовательного соединения от N-конца к C-концу SG1202, линкера и двух M91, где N-концы двух M91 связывают с C-концом тяжелой цепи SG1202 соответствующим образом для получения слитого белка SG12474.
[0162] Аминокислотные последовательности LCDR1-3 антитела SG1202 приведены в SEQ ID NO: 15, SEQ ID NO: 16 и SEQ ID NO: 17, соответственно; аминокислотная последовательность VL приведена в SEQ ID NO: 21; нуклеотидная последовательность, кодирующая VL, приведена в SEQ ID NO: 23; аминокислотные последовательности HCDR1-3 антитела SG1202 приведены в SEQ ID NO: 18, SEQ ID NO: 19 и SEQ ID NO: 20, соответственно; аминокислотная последовательность VH приведена в SEQ ID NO: 22; нуклеотидная последовательность, кодирующая VH, приведена в SEQ ID NO: 24. Аминокислотная последовательность легкой цепи антитела SG1202 приведена в SEQ ID NO: 25; нуклеотидная последовательность, кодирующая его легкую цепь, приведена в SEQ ID NO: 26. Аминокислотная последовательность тяжелой цепи антитела SG1202 приведена в SEQ ID NO: 27; и нуклеотидная последовательность, кодирующая его тяжелую цепь, приведена в SEQ ID NO: 28.
[0163] Слитый белок SG12474 состоит из первой полипептидной цепи и второй полипептидной цепи, где первая полипептидная цепь является легкой цепью SG1202, аминокислотная последовательность которой приведена в SEQ ID NO: 25; вторая полипептидная цепь является полипептидной цепью, полученной посредством связывания между тяжелой цепью SG1202 с мутацией в Fc-области и M91 с помощью линкера 1, аминокислотная последовательность которой приведена в SEQ ID NO: 54.
[0164] При этом аминокислотная последовательность IgG1-Fc приведена в SEQ ID NO: 50; и аминокислотная последовательность Fc с мутацией приведена в SEQ ID NO: 51.
[0165] Слитый белок SS002M91 из мутанта M91 (SEQ ID: NO 41) варианта 1 SIRPα и IgG1-Fc является гомодимером, и аминокислотная последовательность его мономера приведена в SEQ ID NO: 55.
[0166] Пример 2. Детекция активности связывания двух антигенов
[0167] (1) Используя гуманизированное антитело против разных антигенов в качестве контроля, активность связывания между соответствующим бифункциональным белком и родственным антигеном оценивали посредством ELISA.
[0168] PD-L1 (человеческого происхождения, белок PD-L1/B7-H1/CD274 (гистидиновая метка), приобретенным в Sino Biological Co.) покрывали полоски планшетов для ELISA при 4°C в течение ночи; после промывки PBST добавляли 10% эмбриональной телячьей сыворотки и блокировали при 37°C в течение 1 часа; добавляли разные концентрации антитела SG1201 и слитого белка SG12473 или разные концентрации антитела SG1202 и слитого белка SG12474 и проводили реакцию при 37°C в течение 1 часа; после промывки PBST добавляли меченое пероксидазой хрена вторичное антитело козы против IgG человека (HRP-меченое антитело козы против IgG человека, Thermo Fisher Scientific), проводили реакцию при 37°C в течение 30 минут и промывали PBST 5 раз; в каждую лунку добавляли 100 мкл TMB (eBioscience), помещали в темноту при комнатной температуре (20±5°C) на 1-2 мин; затем в каждую лунку добавляли 100 мкл стоп-раствора 2 Н H2SO4 для прекращения реакции субстрата, значения OD считывали при 450 нм с помощью спектрофотометра для чтения микропланшетов и анализировали способность бифункционального белка связываться с соответствующим антигеном-мишенью.
[0169] Результаты приведены на фиг. 2-3. На фиг. 2-3 показано, что, хотя типы антител против PD-L1 в слитом белке SG12473 и слитом белке SG12474 отличались, это не влияло на способности слитого белка SG12473 и слитого белка SG12474 связываться с PD-L1.
[0170] (2) Используя мутанта M91 рецептора CD47 в качестве контроля, оценивали активность связывания между соответствующим бифункциональным белком и CD47 посредством ELISA.
[0171] CD47 (человеческого происхождения, белок CD47 (гистидиновая метка), приобретенным в Sino Biological Co.) покрывали полоски планшетов для ELISA при 4°C в течение ночи; после промывки PBST добавляли 10% эмбриональной телячьей сыворотки и блокировали при 37°C в течение 1 часа; добавляли разные концентрации слитого белка SS002M91, слитого белка SG12473 и слитого белка SG12474 и проводили реакцию при 37°C в течение 1 часа; после промывки PBST добавляли меченое пероксидазой хрена вторичное антитело козы против IgG человека (HRP-меченое антитело козы против IgG человека, Thermo Fisher Scientific), проводили реакцию при 37°C в течение 30 минут и промывали PBST 5 раз; в каждую лунку добавляли 100 мкл TMB (eBioscience), помещали в темноту при комнатной температуре (20±5°C) на 1-2 мин; затем в каждую лунку добавляли 100 мкл стоп-раствора 2 Н H2SO4 для прекращения реакции субстрата, значения OD считывали при 450 нм с помощью спектрофотометра для чтения микропланшетов и анализировали способность слитого белка SG12473 и слитого белка SG12474 связываться с CD47.
[0172] Результаты приведены на фиг. 4. На фиг. 4 показано, что, хотя типы антител против PD-L1 в слитом белке SG12473 и слитом белке SG12474 отличались, это не влияло на способности слитого белка SG12473 и слитого белка SG12474 связываться с CD47.
[0173] Пример 3. Детекция активности одновременного связывания двух антигенов
[0174] Используя гуманизированное антитело против разных антигенов в качестве контроля, оценивали биологическую активность слитого белка SG12473 и слитого белка SG12474 в отношении одновременного связывания двух антигенов посредством ELISA.
[0175] PD-L1 покрывали полоски планшетов для ELISA при 4°C в течение ночи; после промывки PBST добавляли 10% эмбриональной телячьей сыворотки и блокировали при 37°C в течение 1 часа; добавляли разные концентрации антитела SG1201, слитого белка SG12473, антитела SG1202 и слитого белка SG12474 и проводили реакцию при 37°C в течение 1 часа; после промывки PBST добавляли меченый биотином CD47 (биотин-Fc-CD47), проводили реакцию при 37°C в течение 30 минут и промывали PBST 5 раз; добавляли меченый пероксидазой хрена авидин (стрептавидин-HRP, Jiaxuan Bio.), проводили реакцию при 37°C в течение 30 минут и промывали PBST 5 раз; в каждую лунку добавляли 100 мкл TMB (eBioscience), помещали в темноту при комнатной температуре (20±5°C) на 1-2 мин; затем в каждую лунку добавляли 100 мкл стоп-раствора 2 Н H2SO4 для прекращения реакции субстрата, значения OD считывали при 450 нм с помощью спектрофотометра для чтения микропланшетов, и анализировали способности слитого белка SG12473 и слитого белка SG12474 одновременно связываться с PD-L1 и CD47.
[0176] Результаты приведены на фиг. 5. На фиг. 5 показано, что, хотя типы антител против PD-L1 в слитом белке SG12473 и слитом белке SG12474 отличались, это не влияло на способности слитого белка SG12473 и слитого белка SG12474 одновременно связываться с PD-L1 и CD47.
[0177] Пример 4. Анализ активности блокирования взаимодействия CD47/SIRPα
[0178] Используя слитый белок SS002M91 в качестве контроля, оценивали биологическую активность слитых белков SG12473 и SG12474 в отношении блокирования взаимодействия CD47/SIRPα.
[0179] SIRPα-His покрывали планшет для анализа в количестве 1 мкг/мл в течение ночи при 4°C; после промывки PBST добавляли 10% эмбриональной телячьей сыворотки и блокировали при 37°C в течение 1 часа; SS002M91, SG12473, SG12474 градиентно разводили с помощью 10% эмбриональной телячьей крови соответствующим образом, в образцы добавляли биотин-Fc-CD47 до конечной концентрации 2 мкг/мл, и предварительно инкубировали при 37°C в течение 30 мин, как и первичное антитело; после промывки планшета для анализа PBST добавляли первичное антитело и инкубировали при 37°C в течение 1 часа; после 5-кратной промывки PBST добавляли меченый пероксидазой хрена авидин (стрептавидин-HRP, Jiaxuan Bio.) и инкубировали при 37°C в течение 30 минут; после 5-кратной промывки PBST в каждую лунку добавляли 100 мкл TMB (eBioscience) и помещали в темноту при комнатной температуре (20±5°C) на 1-5 мин; затем в каждую лунку добавляли 100 мкл стоп-раствора 2 Н H2SO4 для прекращения реакции субстрата, значения OD считывали при 450 нм с помощью спектрофотометра для чтения микропланшетов и анализировали блокирующие эффекты SS002M91, SG12473, SG12474 в отношении CD47/SIRPα.
[0180] Результаты приведены на фиг. 6. На фиг. 6 показано, что, как и слитый белок SS002M91, слитые белки SG12473, SG12474 могут конкурентно блокировать связывание между CD47 и его лигандом SIRPα. При этом значение IC50 SG12473 составляло 1,26 нМ, значение IC50 SG12474 составляло 0,77 нМ, и значение IC50 SS002M91 составляло 1,16 нМ.
[0181] Пример 5. Анализ активности блокирования взаимодействия PD-1/PD-L1
[0182] Используя SG1201 и SG1202 в качестве контроля, оценивали биологическую активность слитых белков SG12473 и SG12474 в отношении блокирования взаимодействия PD-1/PD-L1.
[0183] PD-L1-Fc покрывали планшет для анализа в количестве 2 мкг/мл в течение ночи при 4°C; после промывки PBST добавляли 10% эмбриональной телячьей сыворотки и блокировали при 37°C в течение 1 часа; SG1201, SG12473, SG1202 и SG12474 градиентно разводили 10% эмбриональной телячьей крови соответствующим образом, в образцы добавляли биотин-Fc-PD1 до конечной концентрации 1 мкг/мл и предварительно инкубировали при 37°C в течение 30 мин, как и первичное антитело; после промывки планшета для анализа PBST добавляли первичное антитело и инкубировали при 37°C в течение 1 часа; после 5-кратной промывки PBST добавляли меченый пероксидазой хрена авидин (стрептавидин-HRP, Jiaxuan Bio.) и инкубировали при 37°C в течение 30 минут; после 5-кратной промывки PBST в каждую лунку добавляли 100 мкл TMB (eBioscience) и помещали в темноту при комнатной температуре (20±5°C) на 1-5 мин; затем в каждую лунку добавляли 100 мкл стоп-раствора 2 Н H2SO4 для прекращения реакции субстрата, значения OD считывали при 450 нм с помощью спектрофотометра для чтения микропланшетов и анализировали блокирующие эффекты SG1201, SG12473, SG1202 и SG12474 в отношении PD-1/PD-L1.
[0184] Как показано на фиг. 7-8, как и SG1201 и SG1202, слитые белки SG12473 и SG12474 могут конкурентно блокировать связывание между PD-1 и PD-L1. При этом значение IC50 SG1201 составляет 11,23 нМ, значение IC50 SG12473 составляет 13,22 нМ, значение IC50 SG1202 составляет 10,89 нМ, и значение IC50 SG12474 составляет 9,12 нМ.
[0185] Пример 6. Слитый белок ингибирует активность опухоли in vivo
[0186] Самкам мышей NCG в модели на животных MiXeno подкожно гетерогенно трансплантировали линию клеток лимфомы человека KARPAS-299 для оценки ингибиторного эффекта слитого белка SG12473 в отношении активности опухоли.
[0187] Для тестирования выбирали самок мышей NCG возрастом 6-8 недель (приобретенных в Jiangsu Jicui Yaokang Biotechnology Co. Ltd.). Мышам подкожно инокулировали клетки KARPAS-299 и периодически отслеживали профили роста опухолей. Когда опухоли достигали среднего размера 35 мм3, мышей случайным образом распределяли по группам по размеру опухоли и массе тела мышей, подлежащих введению. В день рандомизации (приблизительно в день 4) свежеполученные PBMC человека (донора) инокулировали через хвостовую вену для получения гуманизированной модели KARPAS-299 на мышах.
[0188] Выборку разделяли на группу, которой вводили контрольный растворитель, группу SG1201, а также группы низкой, средней и высокой дозы SG12473, при этом в каждой группе было 12 мышей, и для каждых 6 мышей использовали PBMC из одного источника. Лекарственные средства вводили посредством интраперитонеальной инъекции дважды в неделю всего в течение трех недель. В частности,
[0189] Группа 1A и группа 1B: группы, которым вводили контрольный растворитель
[0190] Группа 2A и группа 2B: группы SG1201, SG1201 в дозе 5 мг/кг
[0191] Группа 3A и группа 3B: группы низких доз SG12473, SG12473 в дозе 5 мг/кг
[0192] Группа 4A и группа 4B: группы средних доз SG12473, SG12473 в дозе 10 мг/кг
[0193] Группа 5A и группа 5B: группы высоких доз SG12473, SG12473 в дозе 20 мг/кг.
[0194] Терапевтическую эффективность оценивали по относительному ингибированию роста опухоли (TGI), и безопасность оценивали с учетом изменений массы животных и уровня смертности.
[0195] Результаты свидетельствовали о том, что у мышей, которым вводили PBMC от донора A, наблюдали значимые противоопухолевые ингибиторные эффекты во всех из групп низкой, средней и высокой дозы SG12473. Как видно из результатов, показанных на фиг. 9, TGI в группах низкой, средней и высокой дозы SG12473 после завершения введения составляло 44,71%, 47,16% и 96,49% соответственно; и через 4 дня после завершения введения TGI составляло 43,71%, 47,92% и 95,94%, соответственно, демонстрируя статистически значимые различия относительно групп, которым вводили контрольный растворитель (все значения p составляли <0,01). Не наблюдали значимого противоопухолевого ингибиторного эффекта в группах SG1201.
[0196] У мышей, которым вводили PBMC от донора B наблюдали значимые противоопухолевые ингибиторные эффекты во всех из групп низкой, средней и высокой дозы SG12473. Как видно из результатов, показанных на фиг. 10, в группах низкой, средней и высокой дозы SG12473 после завершения введения составляло 36,96%, 62,06% и 98,90% соответственно; и через 4 дня после завершения введения TGI составляло 33,44%, 57,57% и 98,83% соответственно, демонстрируя статистически значимые различия относительно групп, которым вводили контрольный растворитель (все значения p составляли <0,01). Не наблюдали значимого противоопухолевого ингибиторного эффекта в группах SG1201.
[0197] Не наблюдали гибели животных в каждой из исследуемых групп и очевидной токсичности лекарственного средства, и переносимость в течение лечения была хорошей.
[0198] Изложенное выше подробное описание представлено посредством объяснения и примеров, и не предназначено для ограничения объема формулы изобретения. Различные изменения вариантов осуществления, представленных в настоящей заявке, очевидны специалистам в этой области и входят в объем формулы изобретения и эквивалентов.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> HANGZHOU SUMGEN BIOTECH CO., LTD.
<120> СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ
<130> 0070-PA-017RU
<160> 55
<170> PatentIn version 3.5
<210> 1
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR1 SG1201
<400> 1
Arg Ala Ser Gln Asp Val Ser Thr Ala Val Ala
1 5 10
<210> 2
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR2 SG1201
<400> 2
Ser Ala Ser Phe Leu Tyr Ser
1 5
<210> 3
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR3 SG1201
<400> 3
Gln Gln Tyr Leu Tyr His Pro Ala Thr
1 5
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 SG1201
<400> 4
Asp Ser Trp Ile His
1 5
<210> 5
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR2 SG1201
<400> 5
Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
1 5 10 15
Gly
<210> 6
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR3 SG1201
<400> 6
Arg His Trp Pro Gly Gly Phe Asp Tyr
1 5
<210> 7
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL SG1201
<400> 7
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 8
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH SG1201
<400> 8
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 9
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность VL SG1201
<400> 9
gacatccaga tgacccagag ccctagcagc ctgagcgcca gcgtgggcga cagggtgacc 60
atcacctgca gggccagcca ggacgtgagc accgccgtgg cctggtacca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcctagc 180
aggttcagcg gcagcggcag cggcaccgac ttcaccctga ccatcagcag cctgcagcct 240
gaggacttcg ccacctacta ctgccagcag tacctgtacc accctgccac cttcggccag 300
ggcaccaagg tggagatcaa g 321
<210> 10
<211> 354
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность VH SG1201
<400> 10
gaggtgcagc tggtggagag cggcggcggc ctggtgcagc ctggcggcag cctgaggctg 60
agctgcgccg ccagcggctt caccttcagc gacagctgga tccactgggt gaggcaggcc 120
cctggcaagg gcctggagtg ggtggcctgg atcagccctt acggcggcag cacctactac 180
gccgacagcg tgaagggcag gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgag ggccgaggac accgccgtgt actactgcgc caggaggcac 300
tggcctggcg gcttcgacta ctggggccag ggcaccctgg tgaccgtgag cagc 354
<210> 11
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь SG1201
<400> 11
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Ser Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Leu Tyr His Pro Ala
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 12
<211> 645
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность легкой цепи SG1201
<400> 12
gacatccaga tgacccagag ccctagcagc ctgagcgcca gcgtgggcga cagggtgacc 60
atcacctgca gggccagcca ggacgtgagc accgccgtgg cctggtacca gcagaagcct 120
ggcaaggccc ctaagctgct gatctacagc gccagcttcc tgtacagcgg cgtgcctagc 180
aggttcagcg gcagcggcag cggcaccgac ttcaccctga ccatcagcag cctgcagcct 240
gaggacttcg ccacctacta ctgccagcag tacctgtacc accctgccac cttcggccag 300
ggcaccaagg tggagatcaa gcgtacggtg gctgcaccat ctgtcttcat cttcccgcca 360
tctgatgagc agttgaaatc tggaactgcc tctgttgtgt gcctgctgaa taacttctat 420
cccagagagg ccaaagtaca gtggaaggtg gataacgccc tccaatcggg taactcccag 480
gagagtgtca cagagcagga cagcaaggac agcacctaca gcctcagcag caccctgacg 540
ctgagcaaag cagactacga gaaacacaaa gtctacgcct gcgaagtcac ccatcagggc 600
ctgagctcgc ccgtcacaaa gagcttcaac aggggagagt gttag 645
<210> 13
<211> 448
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь SG1201
<400> 13
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 14
<211> 1344
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность тяжелой цепи SG1201
<400> 14
gaggtgcagc tggtggagag cggcggcggc ctggtgcagc ctggcggcag cctgaggctg 60
agctgcgccg ccagcggctt caccttcagc gacagctgga tccactgggt gaggcaggcc 120
cctggcaagg gcctggagtg ggtggcctgg atcagccctt acggcggcag cacctactac 180
gccgacagcg tgaagggcag gttcaccatc agcgccgaca ccagcaagaa caccgcctac 240
ctgcagatga acagcctgag ggccgaggac accgccgtgt actactgcgc caggaggcac 300
tggcctggcg gcttcgacta ctggggccag ggcaccctgg tgaccgtgag cagcgctagc 360
accaagggcc catcggtctt ccccctggca ccctcctcca agagcacctc tgggggcaca 420
gcggccctgg gctgcctggt caaggactac ttccccgaac cggtgacggt gtcgtggaac 480
tcaggcgccc tgaccagcgg cgtgcacacc ttcccggctg tcctacagtc ctcaggactc 540
tactccctca gcagcgtggt gaccgtgccc tccagcagct tgggcaccca gacctacatc 600
tgcaacgtga atcacaagcc cagcaacacc aaggtggaca agaaagttga gcccaaatct 660
tgtgacaaaa ctcacacatg cccaccgtgc ccagcacctg aactcctggg gggaccgtca 720
gtcttcctct tccccccaaa acccaaggac accctcatga tctcccggac ccctgaggtc 780
acgtgcgtgg tggtggacgt gagccacgaa gaccccgagg tcaagttcaa ctggtacgtg 840
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta cgccagcacg 900
taccgtgtgg tcagcgtcct caccgtcctg caccaggact ggctgaatgg caaggagtac 960
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 1020
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccggga agagatgacc 1080
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 1140
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgctggac 1200
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 1260
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaaccacta cacgcagaag 1320
agcctctccc tgtctccggg taaa 1344
<210> 15
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR1 SG1202
<400> 15
Arg Ala Ser Gln Arg Val Ser Ser Ser Tyr Leu Ala
1 5 10
<210> 16
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR2 SG1202
<400> 16
Asp Ala Ser Ser Arg Ala Thr
1 5
<210> 17
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LCDR3 SG1202
<400> 17
Gln Gln Tyr Gly Ser Leu Pro Trp Thr
1 5
<210> 18
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR1 SG1202
<400> 18
Arg Tyr Trp Met Ser
1 5
<210> 19
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR2 SG1202
<400> 19
Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val Lys
1 5 10 15
Gly
<210> 20
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> HCDR3 SG1202
<400> 20
Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr
1 5 10
<210> 21
<211> 108
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VL SG1202
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 22
<211> 121
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VH SG1202
<400> 22
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 23
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность VL SG1202
<400> 23
gagatcgtgc tgacccagtc ccctggcacc ctgtctctgt ctcccggcga gagagccacc 60
ctgtcttgcc gggcctccca gcgggtgtcc tcctcctacc tggcctggta tcagcagaaa 120
cccggacagg cccctaggct gctgatctac gacgcctcct ccagagccac cggcatccct 180
gaccggttct ccggctctgg ctccggcacc gacttcaccc tgaccatctc ccggctggaa 240
cctgaggact ttgccgtgta ttactgccag cagtacggct ccctgccttg gaccttcggc 300
cagggaacca aggtggagat caaa 324
<210> 24
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность VH SG1202
<400> 24
gaggtgcagc tggtcgagtc tggcggagga ctggtgcagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc cggtactgga tgtcttgggt gcgccaggct 120
cctggcaagg gactggaatg ggtggccaac atcaaacagg atggctctga gaagtactac 180
gtggactccg tgaagggccg gttcaccatc tccagggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgag ggccgaggac accgccgtgt actactgtgc ccgggagggc 300
ggatggttcg gcgagctggc cttcgattac tggggccagg gcaccctggt gacagtgtcc 360
tct 363
<210> 25
<211> 215
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Легкая цепь SG1202
<400> 25
Glu Ile Val Leu Thr Gln Ser Pro Gly Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Arg Val Ser Ser Ser
20 25 30
Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu
35 40 45
Ile Tyr Asp Ala Ser Ser Arg Ala Thr Gly Ile Pro Asp Arg Phe Ser
50 55 60
Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Arg Leu Glu
65 70 75 80
Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Tyr Gly Ser Leu Pro
85 90 95
Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala
100 105 110
Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser
115 120 125
Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu
130 135 140
Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser
145 150 155 160
Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu
165 170 175
Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val
180 185 190
Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys
195 200 205
Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 26
<211> 648
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность легкой цепи SG1202
<400> 26
gagatcgtgc tgacccagtc ccctggcacc ctgtctctgt ctcccggcga gagagccacc 60
ctgtcttgcc gggcctccca gcgggtgtcc tcctcctacc tggcctggta tcagcagaaa 120
cccggacagg cccctaggct gctgatctac gacgcctcct ccagagccac cggcatccct 180
gaccggttct ccggctctgg ctccggcacc gacttcaccc tgaccatctc ccggctggaa 240
cctgaggact ttgccgtgta ttactgccag cagtacggct ccctgccttg gaccttcggc 300
cagggaacca aggtggagat caaacgtacg gtggctgcac catctgtctt catcttcccg 360
ccatctgatg agcagttgaa atctggaact gcctctgttg tgtgcctgct gaataacttc 420
tatcccagag aggccaaagt acagtggaag gtggataacg ccctccaatc gggtaactcc 480
caggagagtg tcacagagca ggacagcaag gacagcacct acagcctcag cagcaccctg 540
acgctgagca aagcagacta cgagaaacac aaagtctacg cctgcgaagt cacccatcag 600
ggcctgagct cgcccgtcac aaagagcttc aacaggggag agtgttag 648
<210> 27
<211> 451
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Тяжелая цепь SG1202
<400> 27
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 28
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность тяжелой цепи SG1202
<400> 28
gaggtgcagc tggtcgagtc tggcggagga ctggtgcagc ctggcggctc cctgagactg 60
tcttgcgccg cctccggctt caccttctcc cggtactgga tgtcttgggt gcgccaggct 120
cctggcaagg gactggaatg ggtggccaac atcaaacagg atggctctga gaagtactac 180
gtggactccg tgaagggccg gttcaccatc tccagggaca acgccaagaa ctccctgtac 240
ctgcagatga actccctgag ggccgaggac accgccgtgt actactgtgc ccgggagggc 300
ggatggttcg gcgagctggc cttcgattac tggggccagg gcaccctggt gacagtgtcc 360
tctgctagca ccaagggccc atcggtcttc cccctggcac cctcctccaa gagcacctct 420
gggggcacag cggccctggg ctgcctggtc aaggactact tccccgaacc ggtgacggtg 480
tcgtggaact caggcgccct gaccagcggc gtgcacacct tcccggctgt cctacagtcc 540
tcaggactct actccctcag cagcgtggtg accgtgccct ccagcagctt gggcacccag 600
acctacatct gcaacgtgaa tcacaagccc agcaacacca aggtggacaa gaaagttgag 660
cccaaatctt gtgacaaaac tcacacatgc ccaccgtgcc cagcacctga actcctgggg 720
ggaccgtcag tcttcctctt ccccccaaaa cccaaggaca ccctcatgat ctcccggacc 780
cctgaggtca cgtgcgtggt ggtggacgtg agccacgaag accccgaggt caagttcaac 840
tggtacgtgg acggcgtgga ggtgcataat gccaagacaa agccgcggga ggagcagtac 900
gccagcacgt accgtgtggt cagcgtcctc accgtcctgc accaggactg gctgaatggc 960
aaggagtaca agtgcaaggt ctccaacaaa gccctcccag cccccatcga gaaaaccatc 1020
tccaaagcca aagggcagcc ccgagaacca caggtgtaca ccctgccccc atcccgggaa 1080
gagatgacca agaaccaggt cagcctgacc tgcctggtca aaggcttcta tcccagcgac 1140
atcgccgtgg agtgggagag caatgggcag ccggagaaca actacaagac cacgcctccc 1200
gtgctggact ccgacggctc cttcttcctc tacagcaagc tcaccgtgga caagagcagg 1260
tggcagcagg ggaacgtctt ctcatgctcc gtgatgcatg aggctctgca caaccactac 1320
acgcagaaga gcctctccct gtctccgggt aaa 1353
<210> 29
<211> 117
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 30
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M1
<400> 30
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gln Lys Lys Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Ser
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 31
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M5
<400> 31
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 32
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M12
<400> 32
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Arg Lys Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 33
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M35
<400> 33
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 34
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M37
<400> 34
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 35
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M41
<400> 35
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg His Leu Ile Tyr
35 40 45
Asn Arg Arg His Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Ser Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 36
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M57
<400> 36
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Ile Leu Ile Tyr
35 40 45
Asn Gly Lys Arg Gly His Phe Pro Arg Val Thr Thr Leu Ser Asp Thr
50 55 60
Thr Lys Arg Gly Asn Met Asp Phe Ser Ile Ser Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Arg Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 37
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M67
<400> 37
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Met Leu Ile Tyr
35 40 45
Asn Gly Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 38
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M81
<400> 38
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 39
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M82
<400> 39
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 40
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M84
<400> 40
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Phe Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser His Ser
50 55 60
Thr Arg Arg Asn Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ser Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 41
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M91
<400> 41
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 42
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M99
<400> 42
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Ile Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Lys Leu Ile Tyr
35 40 45
Asn Gln Lys Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Ser
50 55 60
Thr Lys Arg Lys Asn Met Asp Phe Ser Ile Arg Ile His Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 43
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M102
<400> 43
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Glu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Arg Ser
50 55 60
Thr Lys Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 44
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M111
<400> 44
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Arg Leu Ile Tyr
35 40 45
Asn Asn Arg Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Leu Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 45
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M122
<400> 45
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Ser Arg His Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 46
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M126
<400> 46
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Asp Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Thr
50 55 60
Thr Arg Arg Glu Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 47
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M130
<400> 47
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Val Leu Ile Tyr
35 40 45
Asn Gln Arg Glu Gly His Phe Pro Arg Val Thr Thr Val Ser Glu Ser
50 55 60
Thr Arg Arg Asp Asn Met Asp Phe Ser Ile Arg Ile Arg Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 48
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M135
<400> 48
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Val Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Leu Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Leu Ser Glu Thr
50 55 60
Thr Arg Arg Gly Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 49
<211> 117
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> M145
<400> 49
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Ile Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Asn Leu Ile Tyr
35 40 45
Asn Gly Lys Gly Gly His Phe Pro Arg Val Thr Thr Val Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Arg Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Val Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser
115
<210> 50
<211> 233
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> IgG1-FC
<400> 50
Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 51
<211> 232
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мутантный FC
<400> 51
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 52
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер 1
<400> 52
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 53
<211> 575
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вторая полипептидная цепь SG12473
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Ser
20 25 30
Trp Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Trp Ile Ser Pro Tyr Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg His Trp Pro Gly Gly Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Gln Val Ile Gln
450 455 460
Pro Asp Lys Ser Val Leu Val Ala Ala Gly Glu Thr Ala Thr Leu Arg
465 470 475 480
Cys Thr Ala Thr Ser Leu Leu Pro Val Gly Pro Ile Gln Trp Phe Arg
485 490 495
Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr Asn Gln Lys Asp Gly His
500 505 510
Phe Pro Arg Val Thr Thr Ala Ser Asp Leu Thr Lys Arg Asn Asn Met
515 520 525
Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr Pro Ala Asp Ala Gly Thr
530 535 540
Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser Pro Asp Asp Ile Glu Phe
545 550 555 560
Lys Ser Gly Ala Gly Thr Glu Leu Ser Val Arg Ala Lys Pro Ser
565 570 575
<210> 54
<211> 578
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Вторая полипептидная цепь SG12474
<400> 54
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Gly Trp Phe Gly Glu Leu Ala Phe Asp Tyr Trp Gly
100 105 110
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Glu Leu Gln
450 455 460
Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly Glu Thr Ala
465 470 475 480
Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly Pro Ile Gln
485 490 495
Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr Asn Gln Lys
500 505 510
Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu Thr Lys Arg
515 520 525
Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr Pro Ala Asp
530 535 540
Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser Pro Asp Asp
545 550 555 560
Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val Arg Ala Lys
565 570 575
Pro Ser
<210> 55
<211> 350
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мономер SS002M91
<400> 55
Glu Leu Gln Val Ile Gln Pro Asp Lys Ser Val Leu Val Ala Ala Gly
1 5 10 15
Glu Thr Ala Thr Leu Arg Cys Thr Ala Thr Ser Leu Leu Pro Val Gly
20 25 30
Pro Ile Gln Trp Phe Arg Gly Ala Gly Pro Gly Arg Gln Leu Ile Tyr
35 40 45
Asn Gln Lys Asp Gly His Phe Pro Arg Val Thr Thr Ala Ser Asp Leu
50 55 60
Thr Lys Arg Asn Asn Met Asp Phe Ser Ile Asn Ile Gly Asn Ile Thr
65 70 75 80
Pro Ala Asp Ala Gly Thr Tyr Tyr Cys Val Lys Phe Arg Lys Gly Ser
85 90 95
Pro Asp Asp Ile Glu Phe Lys Ser Gly Ala Gly Thr Glu Leu Ser Val
100 105 110
Arg Ala Lys Pro Ser Leu Glu Pro Lys Ser Cys Asp Lys Thr His Thr
115 120 125
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
130 135 140
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
145 150 155 160
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
165 170 175
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
180 185 190
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
195 200 205
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
210 215 220
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
225 230 235 240
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
245 250 255
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val
260 265 270
Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly
275 280 285
Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
290 295 300
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp
305 310 315 320
Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
325 330 335
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
340 345 350
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИВАЛЕТНЫЕ И ПОЛИСПЕЦИФИЧНЫЕ GITR-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2016 |
|
RU2753439C2 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800923C2 |
| Слитые молекулы, происходящие от Cholix-токсина, для пероральной доставки биологически активных нагрузок | 2015 |
|
RU2723178C2 |
| НОВЫЙ СЛИТЫЙ БЕЛОК, СПЕЦИФИЧНЫЙ ДЛЯ CD137 И PD-L1 | 2019 |
|
RU2818349C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2728861C1 |
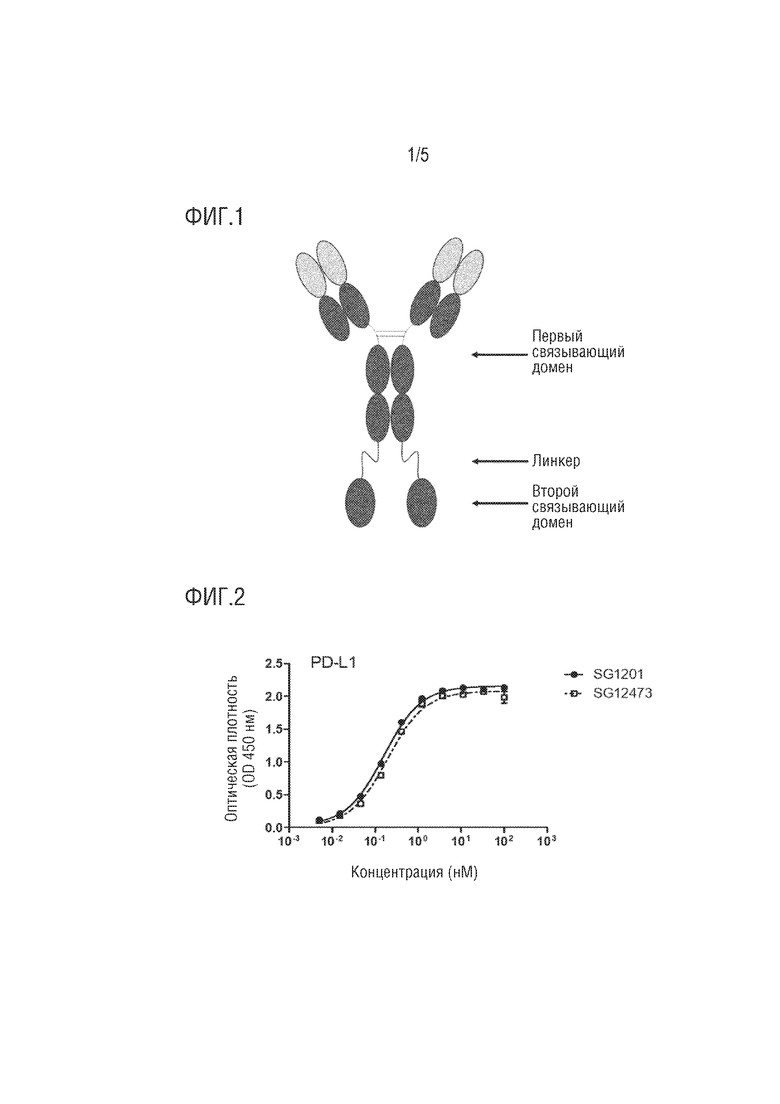




Настоящее изобретение относится к области биотехнологии, конкретно к рекомбинантным слитым белкам, содержащим мутант домена SIRPα человека, и может быть использовано в медицине в лечении опухоли. Сконструирован слитый белок, который содержит первый связывающий домен, специфически связывающийся с PD-L1, связанный прямо или косвенно через линкер со вторым связывающим доменом, специфически связывающимся с белком CD47. При этом второй связывающий домен содержит мутант домена SIRPα человека. Изобретение позволяет получить слитый белок, способный одновременно специфически связываться с белком CD47 и PD-L1, специфически блокировать взаимодействие между белком CD47 и SIRPα, а также между PD-1 и PD-L1 и эффективно ингибировать рост и/или пролиферацию опухолей или опухолевых клеток. 10 н. и 25 з.п. ф-лы, 10 ил., 6 пр.
1. Слитый белок для связывания с белком CD47 и белком PD-L1, содержащий:
первый связывающий домен, специфически связывающийся с PD-L1; и
второй связывающий домен, специфически связывающийся с белком CD47;
где указанный второй связывающий домен содержит мутанта домена SIRPα человека, где указанный домен SIRPα человека содержит аминокислотные остатки в положениях 33-149 варианта 1 SIRPα человека, где указанные аминокислотные остатки в положениях 33-149 варианта 1 SIRPα человека представлены SEQ ID NO: 29, и где указанный мутант содержит аминокислотные замены, выбранные из группы, состоящей из:
(1) I61L, V63I, E77I, E84K, V93L, L96S, K98R, N100G и V132L;
(2) I61V, E77N, Q82S, K83R и E84H;
(3) I61F, V63I, K83R, E84K и V132I;
(4) I61L, E77Q, E84D, R107N и V132I;
(5) I61L, V63I, E77K, K83R, E84D и N100G;
(6) I61V, E77H, Q82R, K83R, E84H и R107S;
(7) I61L, E77I, Q82G, E84R, V93L, L96T, N100G, R107S, G109R и V132R;
(8) I61L, E77M, Q82G, K83R, E84D и V132L;
(9) I61L;
(10) I61F, D95H, L96S, G109H и V132S;
(11) I61F, D95H, L96S, K98R, G109H и V132S;
(12) I61L, E77Q, E84D, V93A, R107N и V132I;
(13) E77K, L96S, N100K, G109H и V132L;
(14) I61L, V63I, Q82G, E84G, D95R, L96S, N100D и V132I;
(15) I61L, E77R, Q82N, K83R, E84G, V93L, D95E, L96T, K98R, N100D и V132L;
(16) I61V, E77N, Q82S, K83R, E84H и V93A;
(17) I61V, V63I, E77V, K83R, E84D, D95E, L96T, K98R и N100E;
(18) I61L, V63I, E77V, K83R, D95E, L96S, K98R, N100D и G109R;
(19) I61V, E77L, Q82G, E84G, V93L, D95E, L96T, K98R и N100G; и
(20) I61L, V63I, E77N, Q82G и E84G,
где первый связывающий домен и второй связывающий домен связаны прямо или косвенно через линкер между ними.
2. Слитый белок по п.1, где указанный мутант содержит аминокислотную последовательность, приведенную в любой из SEQ ID NO: 30-49.
3. Слитый белок по п.1, где указанный первый связывающий домен содержит антитело или его антигенсвязывающий фрагмент.
4. Слитый белок по п.3, где указанное антитело выбрано из группы, состоящей из: моноклонального антитела, одноцепочечного антитела, химерного антитела, гуманизированного антитела и полностью человеческого антитела.
5. Слитый белок по п.3, где указанный антигенсвязывающий фрагмент выбран из группы, состоящей из: Fab, Fab', F(ab)2, dAb, выделенной определяющей комплементарность области CDR, Fv и scFv.
6. Слитый белок по п.1, где указанный PD-L1 является PD-L1 человека.
7. Слитый белок по п.3, где указанное антитело содержит тяжелую цепь антитела или его фрагмента, указанная тяжелая цепь антитела или его фрагмента содержит HCDR1-3, указанная HCDR1 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 4 и SEQ ID NO: 18.
8. Слитый белок по п.7, где указанная HCDR2 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 5 и SEQ ID NO: 19.
9. Слитый белок по любому из пп.7,8, где указанная HCDR3 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 6 и SEQ ID NO: 20.
10. Слитый белок по п.7, где указанная тяжелая цепь антитела или его фрагмента содержит вариабельную область тяжелой цепи VH, и указанная вариабельная область тяжелой цепи VH содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 8 и SEQ ID NO: 22.
11. Слитый белок по п.7, где указанная тяжелая цепь антитела или его фрагмента содержит константную область тяжелой цепи, и указанная константная область тяжелой цепи содержит IgG.
12. Слитый белок по п.11, где указанный IgG выбран из группы, состоящей из: IgG1 и IgG4.
13. Слитый белок по п.7, где указанная тяжелая цепь антитела содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 13 и SEQ ID NO: 27.
14. Слитый белок по п.3, где указанное антитело содержит легкую цепь антитела или его фрагмента, указанная легкая цепь антитела или его фрагмента содержит LCDR1-3, указанная LCDR1 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 1 и SEQ ID NO: 15.
15. Слитый белок по п.14, где указанная LCDR2 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 2 и SEQ ID NO: 16.
16. Слитый белок по любому из пп.14,15, где указанная LCDR3 содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 3 и SEQ ID NO: 17.
17. Слитый белок по п.14, где указанная легкая цепь антитела или его фрагмента содержит вариабельную область легкой цепи VL, и указанная вариабельная область легкой цепи VL содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 7 и SEQ ID NO: 21.
18. Слитый белок по п.14, где указанная легкая цепь антитела или его фрагмента содержит константную область легкой цепи, и константная область легкой цепи содержит Igκ.
19. Слитый белок по п.14, где указанная легкая цепь антитела содержит аминокислотную последовательность, приведенную в любой из последовательностей из группы, состоящей из: SEQ ID NO: 11 и SEQ ID NO: 25.
20. Слитый белок по п.1, где указанный первый связывающий домен находится на N-конце второго связывающего домена.
21. Слитый белок по п.1, где указанный слитый белок дополнительно содержит линкер, линкер находится на C-конце первого связывающего домена и N-конце второго связывающего домена.
22. Слитый белок по п.21, где указанный линкер содержит аминокислотную последовательность, приведенную в SEQ ID NO: 52.
23. Слитый белок по п.1, содержащий по меньшей мере два вторых связывающих домена.
24. Слитый белок по п.23, где каждый из указанных вторых связывающих доменов находится на C-конце первого связывающего домена, соответственно.
25. Иммуноконъюгат для лечения заболевания, содержащий слитый белок по любому из пп.1-24 и одно или более терапевтических средств.
26. Выделенная молекула нуклеиновой кислоты, кодирующая слитый белок по любому из пп.1-24.
27. Экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты по п.26.
28. Композиция для связывания с белком CD47 и белком PD-L1, содержащая эффективное количество слитого белка по любому из пп.1-24 и фармацевтически приемлемый эксципиент.
29. Клетка для экспрессии слитого белка по любому из пп. 1-24, содержащая экспрессирующий вектор по п.27.
30. Способ получения слитого белка по любому из пп.1-24, включающий культивирование клетки по п.29 в условиях, делающих возможной экспрессию указанного слитого белка и выделение указанного слитого белка.
31. Применение слитого белка по любому из пп.1-24, иммуноконъюгата по п.25, молекулы нуклеиновой кислоты по п.26, экспрессирующего вектора по п.27, композиции по п.28 или клетки по п.29 для применения в лечении опухоли.
32. Применение по п.31, где указанная опухоль включает солидную опухоль и несолидную опухоль.
33. Применение по п.32, где указанная солидная опухоль и несолидная опухоль включают множественную миелому, лейкоз, неходжкинскую лимфому, лимфому Ходжкина, нейроглиому, герминому, саркому, мезотелиому, плацентому, злокачественное новообразование головного мозга, злокачественное новообразование костной ткани, рак кожи, рак носоглотки, рак легких, рак ротовой полости, рак пищевода, рак желудка, рак печени, рак поджелудочной железы, рак предстательной железы, рак кишечника, рак молочной железы, рак шейки матки, рак яичников и рак яичка, опухоль лобных пазух, гипофарингеальный рак, ольфакторную нейробластому, рак языка, карциному десен, карциному ампулы фатерова сосочка, рак толстого кишечника, рак прямой кишки, рак почки, карциному мочеточника, рак мочевого пузыря, рак полового члена, карциному фаллопиевых труб, рак века, ретинобластому.
34. Способ in vitro блокирования взаимодействия между белком CD47 и SIRPα, включающий введение эффективного количества слитого белка по любому из пп.1-24, иммуноконъюгата по п.25, молекулы нуклеиновой кислоты по п.26, экспрессирующего вектора по п.27, композиции по п.28 или клетки по п.29.
35. Способ in vitro ингибирования роста и/или пролиферации опухолей или опухолевых клеток, включающий введение эффективного количества слитого белка по любому из пп.1-24, иммуноконъюгата по п.25, молекулы нуклеиновой кислоты по п.26, экспрессирующего вектора по п.27, композиции по п.28 или клетки по п.29.
| WO 2018205936 A1, 15.11.2018 | |||
| BONING LIU et al., Elimination of tumor by CD47/PD-L1 dual-targeting fusion protein that engages innate and adaptive immune responses, MABS, 2017, v | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Аппарат для очищения воды при помощи химических реактивов | 1917 |
|
SU2A1 |
| Способ очищения амида ортотолуолсульфокислоты | 1921 |
|
SU315A1 |
| CN 107459578 A, 12.12.2017 | |||
| CN 109535263 A, 29.03.2019 | |||
| LEE H.T | |||
| et al., Molecular mechanism of PD-1/PD-L1 blockade via anti-PD-L1 antibodies | |||
