ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к агентам, которые можно применять для лечения Т-клеточной лимфомы или лейкоза.
УРОВЕНЬ ТЕХНИКИ
Лимфоидные злокачественные новообразования можно главным образом разделить на те, которые происходят из Т-клеток, либо из В-клеток. Т-клеточные злокачественные новообразования представляют собой клинически и биологически гетерогенную группу нарушений, которые в совокупности включают 10-20% неходжкинских лимфом и 20% острых лейкозов. Наиболее часто выявляемыми гистологическими подтипами являются периферическая Т-клеточная лимфома, неспецифицированная (ПТКЛН); ангиоиммунобластная Т-клеточная лимфома (АИТЛ) и анапластическая крупноклеточная лимфома (АКЛ). Из всех острых лимфобластных лейкозов (ОЛЛ) около 20% имеют Т-клеточный фенотип.
Эти заболевания обычно ведут себя агрессивно по сравнению, например, с B-клеточными злокачественными новообразованиями, при этом выживаемость в течение 5 лет составляет всего 30%. В случае Т-клеточной лимфомы, данные заболевания ассоциированы с высокой долей пациентов с диссеминированным заболеванием, неблагоприятным значением Международного прогностического индекса (IPI) и распространенностью экстранодальной формы заболевания. Химиотерапия сама по себе обычно не эффективна, и менее 30% пациентов вылечивают с помощью существующих способов лечения.
Кроме того, в отличие от В-клеточных злокачественных новообразований, при которых иммунотерапевтические средства, такие как моноклональное антитело против CD20 ритуксимаб, значительно улучшают результаты, в настоящее время не существует эквивалентно эффективного, минимально токсичного иммунотерапевтического средства для лечения Т-клеточных злокачественных новообразований. Большой проблемой в разработке иммунотерапевтических средств для Т-клеточных нарушений является значительное частичное совпадение экспрессии маркеров клональных и нормальных Т-клеток, при этом не существует одного антигена, который позволял бы однозначно идентифицировать клональные (злокачественные) клетки.
Аналогичная проблема существует при нацеливании на пан-В-клеточный антиген для лечения В-клеточного злокачественного новообразования. Однако в данном случае сопутствующее истощение В-клеточного компонента приводит к относительно небольшой иммуносупрессии, которая легко переносится большинством пациентов. Кроме того, при терапевтических подходах, которые приводят к особенно длительному истощению нормального B-клеточного компонента, его потеря может быть в значительной степени компенсирована путем введения пула молекул иммуноглобулина. Ситуация полностью отличается при нацеливании на Т-клеточные злокачественные новообразования. В этом случае сопутствующее истощение Т-клеточного компонента приводит к тяжелой иммуносупрессии и сильной токсичности. Кроме того, не существует удовлетворительного способа уменьшения последствий потери Т-клеточного компонента.
Токсичность частично иллюстрируют клинические эффекты терапевтического моноклонального антитела Алемтузумаб. Указанный агент лизирует клетки, которые экспрессируют CD52, и обладает некоторой эффективностью при Т-клеточных злокачественных новообразованиях. Полезность этого агента в значительной степени ограничена сильно выраженным клеточным иммунодефицитом, в основном вследствие истощения Т-клеток, сопровождающимся заметно повышенным риском инфекции.
Таким образом, существует потребность в новом способе направленного лечения Т-клеточных злокачественных новообразований, который не связан с вышеуказанными недостатками.
ОПИСАНИЕ ЧЕРТЕЖЕЙ
Фигура 1: Диаграмма комплекса αβ Т-клеточный рецептор/CD3. Т-клеточный рецептор формируется из 6 различных белковых цепей, которые должны собираться в эндоплазматическом ретикулуме, чтобы быть экспрессированными на поверхности клетки. Четыре белка комплекса CD3 (CD3ζ, CD3γ, CD3ε и CD3δ) окружают Т-клеточный рецептор (TCR). Этот TCR придает комплексу специфичность к конкретному антигену и состоит из двух цепей: TCRα и TCRβ. Каждая цепь TCR имеет вариабельный участок, дистальный по отношению к мембране, и константный участок, проксимальный по отношению к мембране. Почти все Т-клеточные лимфомы и многие Т-клеточные лейкозы экспрессируют комплекс TCR/CD3.
Фигура 2: Сегрегация β-константной области Т-клеточного рецептора (TRBC)-1 и TRBC2 во время реаранжировки Т-клеточного рецептора. Каждая бета-цепь TCR формируется в результате геномной рекомбинации отдельных бета-вариабельной (variable; V), разнообразной (diversity; D), соединительной (joining; J) и константной (TRBC) областей. Геном человека содержит два очень похожих и функционально эквивалентных локуса TRBC, известных как TRBC1 и TRBC2. Во время реаранжировки генов происходит рекомбинация TCR J-области либо с TRBC1, либо с TRBC2. Эта реаранжировка является постоянной. Т-клетки экспрессируют множество копий одного TCR на своей поверхности, следовательно, каждая Т-клетка будет экспрессировать TCR, константная область β -цепи которого кодируется либо TRBC1, либо TRBC2.
Фигура 3: Выравнивание человеческих TRBC1 и TRBC2 на уровне аминокислот. Константная цепь TCRβ, кодируемая TRBC1 и TRBC2, различается только отличиями в 4-х аминокислотах: K / N в положении 3 TRBC; N / K в положении 4 TRBC; F / Y в положении 36 TRBC; V / E в положении 135 TRBC.
Фигура 4: Схематические диаграммы, иллюстрирующие различные типы антител, на которые ссылаются при создании гуманизированных связывающих молекул против TRBC1.
Фигура 5: Выбор графта тяжелой и легкой цепи. Были созданы гуманизированные домены VH и VL, включающие CDR из JOVI-1 вместе с различными человеческими каркасными участками. Химерные антитела, содержащие гуманизированные VH с мышиными VL доменами или гуманизированные VL с мышиными VH доменами, были созданы и сравнены с контрольным химерным антителом, имеющим мышиные VH и VL (Jovi-1 химера HC / Jovi-1 химера LC). Гуманизированные антитела также были созданы с помощью комбинаций гуманизированных VH и VL. Все антитела были протестированы на связывание с TRBC1 методом ELISA.
Фигура 6: Связывание TRBC1/TRBC2 обратно мутированными конструкциями. Ряд конструкций обратно мутированных VH был создан на основе каркасного участка H-AF062256, как показано в Таблице 1. Эти домены VH были использованы для создания гуманизированных антител в комбинации с доменом VL, имеющим человеческий каркасный участок 3aaz. Антитела были протестировали на связывание с TRBC1 и TRBC2 с помощью ELISA.
Фигура 7: Стабильность конструкций антител против TRBC1 (Tm°C). Стабильность антител, содержащих обратно мутированные конструкции, показанные в Таблице 1, с гуманизированной легкой цепью 3aaz по сравнению с мышиным антителом JOVI-1.
Фигура 8: Стабильность конструкций антител против TRBC1 (TmºC).
Фигура 9: Схематическая диаграмма, иллюстрирующая гуманизированный химерный антигенный рецептор (CAR) против TRBC1
Фигура 10: CAR-опосредованное специфичное уничтожение TRBC1+ мишеней. Нетрансфицированные Т-клетки или Т-клетки, экспрессирующие мышиный CAR против TRBC1 (положительный контроль), гуманизированный CAR против TRBC1 или CAR против EGFRvIII (отрицательный контроль) были совместно культивированы с клетками-мишенями Raji, экспрессирующими TRBC1 или TRBC2, и уничтожение было проанализировано с использованием проточной цитометрии.
Фигура 11: Пролиферация CAR-экспрессирующих T-клеток после совместного культивирования с клетками-мишенями. Нетрансфицированные Т-клетки, Т-клетки, экспрессирующие мышиный CAR против TRBC1, или Т-клетки, экспрессирующие гуманизированный CAR против TRBC1, были совместно культивированы с клетками-мишенями Raji, экспрессирующими TRBC1 или TRBC2, в течение 72 часов, и была измерена пролиферация Т-клеток.
Фигура 12: Высвобождение цитокинов после совместного культивирования CAR-экспрессирующих Т-клеток и клеток-мишеней. Нетрансфицированные Т-клетки или Т-клетки, экспрессирующие мышиный CAR против TRBC1 (положительный контроль), гуманизированный CAR против TRBC1 или СAR против EGFRvIII (отрицательный контроль) были совместно культивированы с клетками-мишенями Raji, экспрессирующими TRBC1 или TRBC2. Высвобождение IFNγ и IL-2 было измерено через 24 часа.
Фигура 13: Влияние гуманизированного CAR против TRBC1 на рост опухоли в мышиной модели NSG.
Фигура 14: Экспрессия маркеров истощения после совместного культивирования с клетками-мишенями. Активированные РВМС были трансдуцированы CAR на основе мышиного JOVI1 (mJOVI) или CAR на основе гуманизированного Jovi-1 (шарнирный участок H1L), истощены по CD56-экспрессирующим клеткам и совместно культивированы с клетками-мишенями Raji, экспрессирующими TRBC1-TCR. Через 96 часов клетки были окрашены антителами против PD1, против LAG3 и против Tim3, затем проанализированы проточной цитометрией.
КРАТКОЕ ИЗЛОЖЕНИЕ АСПЕКТОВ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения разработали ряд гуманизированных антигенсвязывающих доменов, которые связывают человеческий TRBC1. Антигенсвязывающий домен может быть использован в различных терапевтических форматах, включая химерный антигенный рецептор (CAR), терапевтическое антитело, конъюгат антитело-лекарственное средство (ADC) и биспецифичный рекрутер Т-клеток (BiTE), для истощения популяции злокачественных TRBC1-экспрессирующих Т-клеток у субъекта без влияния на здоровые TRBC2-экспрессирующие Т-клетки.
Таким образом, согласно первому аспекту, в настоящем изобретении предложен антигенсвязывающий домен против TRBC1, который включает:
а) домен VH, имеющий аминокислотную последовательность, выбранную из SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12, SEQ ID No. 13, SEQ ID No. 14, SEQ ID No. 15, SEQ ID No. 16, SEQ ID No. 17 и SEQ ID No. 18; и
б) домен VL, имеющий аминокислотную последовательность, выбранную из SEQ ID No. 19, SEQ ID No. 20, SEQ ID No. 21, SEQ ID No. 22, SEQ ID No. 23, SEQ ID No. 24, SEQ ID No. 25, SEQ ID No. 26, SEQ ID No. 27, SEQ ID No. 28, SEQ ID No. 29, SEQ ID No. 30, SEQ ID No. 31, SEQ ID No. 32, SEQ ID No. 33, SEQ ID No. 34.
Согласно второму аспекту, в настоящем изобретении предложен химерный антигенный рецептор (CAR), который содержит антигенсвязывающий домен против TRBC1 в соответствии с первым аспектом настоящего изобретения.
Согласно третьему аспекту, в настоящем изобретении предложено антитело, которое содержит антигенсвязывающий домен против TRBC1 в соответствии с первым аспектом настоящего изобретения.
Согласно четвертому аспекту, в настоящем изобретении предложен биспецифичный рекрутер Т-клеток (BiTE), который содержит антигенсвязывающий домен против TRBC1 в соответствии с первым аспектом настоящего изобретения.
Согласно пятому аспекту, в настоящем изобретении предложен конъюгат антитело-лекарственное средство, который содержит антигенсвязывающий домен против TRBC1 в соответствии с первым аспектом настоящего изобретения.
Согласно шестому аспекту, в настоящем изобретении предложена последовательность нуклеиновой кислоты, которая кодирует CAR в соответствии со вторым аспектом настоящего изобретения.
Согласно седьмому аспекту, в настоящем изобретении предложен вектор, содержащий последовательность нуклеиновой кислоты в соответствии с шестым аспектом настоящего изобретения.
Согласно восьмому аспекту, в настоящем изобретении предложена клетка, содержащая CAR в соответствии со вторым аспектом настоящего изобретения.
Согласно девятому аспекту, в настоящем изобретении предложен способ получения клетки в соответствии с восьмым аспектом настоящего изобретения, который включает стадию введения в клетку нуклеиновой кислоты в соответствии с шестым аспектом настоящего изобретения или вектора в соответствии с седьмым аспектом настоящего изобретения.
Согласно десятому аспекту, в настоящем изобретении предложена фармацевтическая композиция, которая содержит множество клеток в соответствии с восьмым аспектом настоящего изобретения, антитело в соответствии с третьим аспектом настоящего изобретения, BiTE в соответствии с четвертым аспектом настоящего изобретения или конъюгат антитело-лекарственное средство в соответствии с пятым аспектом настоящего изобретения.
Согласно одиннадцатому аспекту, в настоящем изобретении предложена фармацевтическая композиция в соответствии с десятым аспектом настоящего изобретения для применения в лечении TRBC1-экспрессирующей Т-клеточной лимфомы или лейкоза у субъекта.
Согласно двенадцатому аспекту, в настоящем изобретении предложен способ лечения TRBC1-экспрессирующей Т-клеточной лимфомы или лейкоза у субъекта, который включает стадию введения субъекту фармацевтической композиции в соответствии с десятым аспектом настоящего изобретения.
Согласно тринадцатому аспекту, в настоящем изобретении предложено применение фармацевтической композиции в соответствии с десятым аспектом настоящего изобретения в производстве лекарственного средства для лечения TRBC1-экспрессирующей Т-клеточной лимфомы или лейкоза у субъекта.
TRBC1-экспрессирующая Т-клеточная лимфома или лейкоз могут быть выбраны из следующего: периферическая Т-клеточная лимфома, неспецифицированная (ПТКЛН), ангиоиммунобластная Т-клеточная лимфома (АИТЛ), анапластическая крупноклеточная лимфома (АКЛ), Т-клеточная лимфома, ассоциированная с энтеропатией (ТКЛЭ), гепатолиенальная Т-клеточная лимфома (ГЛТЛ), экстранодальная NK/T-клеточная лимфома, назальный тип, Т-клеточная лимфома кожи, первичная АКЛ кожи, Т-клеточный пролимфоцитарный лейкоз и Т-клеточный острый лимфобластный лейкоз.
ПОДРОБНОЕ ОПИСАНИЕ
В настоящем изобретении предложены агенты, такие как химерные антигенные рецепторы (CAR), которые селективно связывают TRBC1. Такие агенты могут применяться в способах лечения Т-клеточной лимфомы или лейкоза у субъекта. Т-клеточные злокачественные новообразования являются клональными, поэтому они экспрессируют либо TRBC1, либо TRBC2. При введении TCRB1-селективного агента субъекту агент вызывает селективное истощение популяции TRBC1-экспрессирующих злокачественных Т-клеток вместе с TRBC1-экспрессирующими нормальными Т-клетками, но не вызывает истощения популяции TRBC2-экспрессирующих нормальных Т-клеток.
TCR β КОНСТАНТНЫЙ РЕГИОН (TRBC)
Т-клеточный рецептор (TCR) экспрессируется на поверхности Т-лимфоцитов и отвечает за распознавание антигенов, связанных с молекулами главного комплекса гистосовместимости (МНС). Когда TCR взаимодействует с антигенным пептидом и MHC (пептид/MHC), T-лимфоцит активируется посредством ряда биохимических событий, опосредованных ассоциированными ферментами, корецепторами, специализированными адаптерными молекулами и активированными или высвобожденными транскрипционными факторами.
TCR представляет собой связанный дисульфидной связью, заякоренный на мембране гетеродимер, обычно состоящий из высоко вариабельных альфа (α) и бета (β) цепей, экспрессируемых как часть комплекса с инвариантными молекулами цепей CD3. Т-клетки, экспрессирующие этот рецептор, называют α:β (или αβ) Т-клетками (~ 95% общего количества Т-клеток). Меньшая часть Т-клеток экспрессирует альтернативный рецептор, образованный вариабельными гамма (γ) и дельта (δ) цепями, и указанные клетки называются γδ Т-клетками (~ 5% от общего количества Т-клеток).
Каждая α и β цепь состоит из двух внеклеточных доменов: вариабельного (V) участка и константного (C) участка, оба из доменов суперсемейства иммуноглобулинов (IgSF), образующих антипараллельные β-листы. Константный участок расположен проксимально к клеточной мембране, за ним следуют трансмембранный участок и короткий цитоплазматический хвост, в то время как вариабельный участок связывается с комплексом пептид/МНС (см. Фигуру 1). Константный участок TCR состоит из коротких соединительных последовательностей, в которых остаток цистеина образует дисульфидные связи, которые образуют связь между двумя цепями.
Вариабельные домены как α-цепи, так и β-цепи TCR имеют три гипервариабельных или определяющих комплементарность участка (CDR). Вариабельный участок β-цепи также имеет дополнительную область гипервариабельности (HV4), однако она обычно не контактирует с антигеном и поэтому не считается CDR.
TCR также содержит до пяти инвариантных цепей γ, δ, ε (совместно именуемых CD3) и ζ. Субъединицы CD3 и ζ опосредуют сигналинг TCR через специфичные цитоплазматические домены, которые взаимодействуют с молекулами вторичного мессенджера и адаптера после распознавания антигена αβ или γδ. Экспрессии комплекса TCR на клеточной поверхности предшествует парная сборка субъединиц, в которой играют роль трансмембранные и внеклеточные домены TCR α и β и CD3 γ и δ.
Таким образом TCR обычно состоят из комплекса CD3 и α- и β-цепей TCR, которые, в свою очередь, состоят из вариабельных и константных участков (Фигура 1).
Локус (Chr7:q34), который обеспечивает β-константный участок TCR (TRBC), дуплицировался в ходе эволюции с образованием двух почти идентичных и функционально эквивалентных генов: TRBC1 и TRBC2 (Фигура 2), которые отличаются только 4 аминокислотами в зрелом белке, продуцируемом каждым (Фигура 3). Каждый TCR будет содержать взаимоисключающим образом либо TRBC1, либо TRBC2, и, таким образом, каждая αβ T-клетка будет экспрессировать либо TRBC1, либо TRBC2 взаимоисключающим образом.
Несмотря на сходство между последовательностями TRBC1 и TRBC2, их можно различить. Аминокислотные последовательности TRBC1 и TRBC2 могут быть различены in situ на поверхности клетки, например, Т-клетки.
АНТИГЕНСВЯЗЫВАЮЩИЙ ДОМЕН
В рамках настоящего изобретения предложен гуманизированный антигенсвязывающий домен против TRBC1, который имеет вариабельную тяжелую цепь (VH) и вариабельную легкую цепь (VL), которые содержат следующие определяющие комплементарность участки (CDR):
VH CDR1: GYTFTGY (SEQ ID No. 1);
VH CDR2: NPYNDD (SEQ ID No. 2);
VH CDR3: GAGYNFDGAYRFFDF (SEQ ID No. 3);
VL CDR1: RSSQRLVHSNGNTYLH (SEQ ID No. 4);
VL CDR2: RVSNRFP (SEQ ID No. 5); и
VL CDR3: SQSTHVPYT (SEQ ID No. 6).
Антигенсвязывающий домен включает человеческие каркасные участки или человеческие каркасные участки с одной или несколькими мутациями. Например, каркасный участок (участки) может содержать одну или больше замен по сравнению с последовательностью человеческого каркасного участка. Замены могут представлять собой «обратные мутации», когда одна или больше аминокислот замещены эквивалентным остатком из последовательности мышиного антитела. Последовательность вариабельной тяжелой цепи (VH) мышиного антитела показана ниже как SEQ ID No. 7, а последовательность вариабельной легкой цепи (VL) показана как SEQ ID No. 8. В обеих последовательностях последовательности CDR выделены жирным шрифтом и подчеркнуты.
SEQ ID No. 7 - мышиная VH Jovi-1
EVRLQQSGPDLIKPGASVKMSCKASGYTFTGYVMHWVKQRPGQGLEWIGFINPYNDD IQSNERFRGKATLTSDKSSTTAYMELSSLTSEDSAVYYCARGAGYNFDGAYRFFDFWGQGTT LTVSS
SEQ ID No. 8 - мышиная VL Jovi-1
DVVMTQSPLSLPVSLGDQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPKLLIYRVSN RFPGVPDRFSGSGSGTDFTLKISRVEAEDLGIYFCSQSTHVPYTFGGGTKLEIKR
Последовательность гуманизированной VH, содержащая мышиные CDR JOVI-1 SEQ ID No. 1, 2 и 3, может содержать 6 или менее, 5 или менее, 4 или менее, 3 или менее, 2 или 1 мутацию по сравнению с последовательностью человеческого каркасного участка дикого типа.
Последовательность гуманизированной VL, содержащая мышиные CDR JOVI-1 SEQ ID No. 4, 5 и 6, может содержать 6 или менее, 5 или менее, 4 или менее, 3 или менее, 2 или 1 мутацию по сравнению с последовательностью человеческого каркасного участка дикого типа.
Последовательность VH может содержать CDR VH JOVI-1 совместно с человеческим каркасным участком H-AF062256. Эта последовательность показана как SEQ ID No. 9. Последовательности CDR подчеркнуты.
SEQ ID No. 9 - Гуманизированное Jovi-1, каркасный участок H-AF062256
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTRDTSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQ GTMVTVSS
Последовательность VH может содержать CDR VH JOVI-1 совместно с человеческим каркасным участком H-EF177999. Эта последовательность показана как SEQ ID No. 10. Последовательности CDR подчеркнуты.
SEQ ID No. 10 - Гуманизированное Jovi-1, каркасный участок H-EF177999
EVQLVESGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYND DIQSNERFRGRVTMTRDTSTSTVYMELSSLRSEDTAVYYCARGAGYNFDGAYRFFDFWGQGT LVTVSS
Последовательность VH может содержать CDR VH JOVI-1 совместно с человеческим каркасным участком H-KF688165. Эта последовательность показана как SEQ ID No. 11. Последовательности CDR подчеркнуты.
SEQ ID No. 11 - Гуманизированное Jovi-1, каркасный участок H-KF688165
QVQLVQSGAEVKKPGASVKVSCKASEYSFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTRDTSISTAYMEVSSLTSDDAAIYYCARGAGYNFDGAYRFFDFWGQG TLVTVSS
Последовательность VH может содержать последовательность SEQ ID No. 9, 10 или 11, с одной или более мутациями, такими как обратные мутации. Последовательность VH может содержать 6 или менее, 5 или менее, 4 или менее, 3 или менее, 2 или 1 мутацию по сравнению с последовательностью человеческого каркасного участка дикого типа. Например, последовательность VH может содержать последовательность SEQ ID No. 9, с одним из наборов обратных мутаций, показанных в Таблице 1 в Примерах.
Последовательность VH может содержать одну из последовательностей SEQ ID No. 12-18. Последовательности CDR подчеркнуты, а обратные мутации показаны жирным шрифтом.
SEQ ID No. 12 - мутация K73
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTRDKSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQ GTMVTVSS
SEQ ID No. 13 - мутация S71
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTSDTSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQG TMVTVSS
SEQ ID No. 14 - мутации S71, K73
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTSDKSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQ GTMVTVSS
SEQ ID No. 15 - мутация I48
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWIGFINPYND DIQSNERFRGRVTMTRDTSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQGT MVTVSS
SEQ ID No. 16 - мутация I48, K73
QVQLVQSGAEVKKPGASVKVSCKASGYTFTGYVMHWVRQAPGQGLEWIGFINPYND DIQSNERFRGRVTMTRDKSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQG TMVTVSS
SEQ ID No. 17 - мутации M20, S71, K73
QVQLVQSGAEVKKPGASVKMSCKASGYTFTGYVMHWVRQAPGQGLEWMGFINPYN DDIQSNERFRGRVTMTSDKSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQ GTMVTVSS
SEQ ID No. 18 - мутации M20, I48
QVQLVQSGAEVKKPGASVKMSCKASGYTFTGYVMHWVRQAPGQGLEWIGFINPYND DIQSNERFRGRVTMTRDTSISTAYMELSRLRSDDTAVYYCARGAGYNFDGAYRFFDFWGQGT MVTVSS
Последовательность VL может содержать CDR VL JOVI-1 совместно с человеческим каркасным участком 3aaz. Эта последовательность показана как SEQ ID No. 19. Последовательности CDR подчеркнуты.
SEQ ID No. 19
DIVMTQSPLSLPVTPGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGQGTKLEIK
Последовательность VL может содержать последовательность SEQ ID No. 19, с одной или более мутациями, такими как обратные мутации. Последовательность VL может содержать 6 или менее, 5 или менее, 4 или менее, 3 или менее, 2 или 1 мутацию по сравнению с последовательностью человеческого каркасного участка дикого типа. Например, последовательность VL может содержать одну из последовательностей SEQ ID No. 20-34. Последовательности CDR подчеркнуты, а обратные мутации показаны жирным шрифтом.
SEQ ID No. 20
DIVMTQSPLSLPVTLGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 21
DIVMTQSPLSLPVTLGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 22
DIVMTQSPLSLPVTLGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 23
DIVMTQSPLSLPVTLGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 24
DIVMTQSPLSLPVTLGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 25
DIVMTQSPLSLPVTLGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVFYCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 26
DIVMTQSPLSLPVTLGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 27
DIVMTQSPLSLPVTLGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 28
DIVMTQSPLSLPVTPGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 29
DIVMTQSPLSLPVTPGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 30
DIVMTQSPLSLPVTPGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 31
DIVMTQSPLSLPVTPGEQASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 32
DIVMTQSPLSLPVTPGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGGGTKLEIK
SEQ ID No. 33
DIVMTQSPLSLPVTPGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYFCSQSTHVPYTFGQGTKLEIK
SEQ ID No. 34
DIVMTQSPLSLPVTPGEPASISCRSSQRLVHSNGNTYLHWYLQKPGQSPRLLIYRVSNR FPGVPDRFSGSGSGTDFTLKISRVEAEDVGVYYCSQSTHVPYTFGGGTKLEIK
Антигенсвязывающий домен против TRBC1 может содержать:
а) домен VH, который содержит CDR VH JOVI-1 совместно с человеческим каркасным участком H-AF062256 или его вариантом; и
б) домен VH, который содержит CDR VL JOVI-1 совместно с человеческим каркасным участком 3aaz или его вариантом.
Вариант может иметь 5 или менее, 4 или менее, 3 или менее, 2 или 1 мутацию по сравнению с последовательностью человеческого каркасного участка дикого типа.
Домен VH может содержать последовательность SEQ ID No. 9, SEQ ID No. 12, SEQ ID No. 13, SEQ ID No. 14, SEQ ID No. 15, SEQ ID No. 16, SEQ ID No. 17 или SEQ ID No. 18. Домен VL может содержать последовательность SEQ ID No. 19, SEQ ID No. 20, SEQ ID No. 21, SEQ ID No. 22, SEQ ID No. 23, SEQ ID No. 24, SEQ ID No. 25, SEQ ID No. 26, SEQ ID No. 27, SEQ ID No. 28, SEQ ID No. 29, SEQ ID No. 30, SEQ ID No. 31, SEQ ID No. 32, SEQ ID No. 33, SEQ ID No. 34
Антигенсвязывающий домен против TRBC1 может содержать:
а) домен VH, который содержит последовательность SEQ ID No 9; и
б) домен VH, который содержит последовательность SEQ ID No. 19.
АНТИТЕЛО
Антигенсвязывающий домен по первому аспекту настоящего изобретения может представлять собой антитело или его функциональный фрагмент. Антитело может представлять собой терапевтическое антитело, такое как истощающее антитело. Антитело может представлять собой биспецифичное антитело, которое связывает TRBC1 вместе с другим антигеном. Антитело может, например, представлять собой перенаправляющее антитело с двойной аффинностью.
Термин «истощающее антитело» используют в общепринятом смысле для обозначения антитела, которое связывается с антигеном (то есть TRBC1), присутствующим на Т-клетке-мишени, и опосредует гибель Т-клетки-мишени. Следовательно, введение истощающего антитела субъекту приводит к сокращению/снижению количества клеток в организме субъекта, которые экспрессируют целевой антиген.
Используемый в настоящей заявке термин «антитело» обозначает полипептид, имеющий антигенсвязывающий сайт, который содержит по меньшей мере один определяющий комплементарность участок CDR. Антитело может содержать 3 CDR и иметь антигенсвязывающий сайт, который эквивалентен сайту доменного антитела (dAb). Антитело может содержать 6 CDR и иметь антигенсвязывающий сайт, который эквивалентен сайту классической молекулы антитела. Остальная часть полипептида может представлять собой любую последовательность, которая обеспечивает подходящий каркас для антигенсвязывающего сайта и ориентирует его соответствующим образом для его связывания с антигеном. Антитело может представлять собой целую молекулу иммуноглобулина или ее часть, такую как Fab, F(ab)'2, Fv, одноцепочечный Fv (ScFv) фрагмент и гибрид scFv-Fc, или диатело, триатело или нанотело, которая сохраняет антигенную специфичность целого антитела. Антитело может представлять собой бифункциональное антитело. Антитело может представлять собой антитело, отличное от антитела человека, химерное, гуманизированное или полностью человеческое антитело.
КОНЪЮГАТЫ
Антитело может представлять собой конъюгат антитела и другого агента или антитела, например, конъюгат может представлять собой детектируемый компонент или химиотерапевтический компонент.
Детектируемый компонент может представлять собой флуоресцентный фрагмент, например, флуоресцентный пептид. «Флуоресцентный пептид» обозначает полипептид, который после возбуждения излучает свет с детектируемой длиной волны. Примеры флуоресцентных белков включают, но не ограничиваются ими, флуоресцеина изотиоцианат (FITC), фикоэритрин (PE), аллофикоцианин (APC), зеленый флуоресцентный белок (GFP), усиленный GFP, красный флуоресцентный белок (RFP), синий флуоресцентный белок (BFP) и mCherry.
Используемый в настоящей заявке термин «химиотерапевтический компонент» относится к компоненты, который разрушителен для клетки, то есть такой компонент снижает жизнеспособность клетки. Химиотерапевтический компонент может представлять собой цитотоксическое лекарственное средство. Предполагаемый химиотерапевтический агент включает, без ограничения, следующее: алкилирующие агенты, нитрозомочевина, этиленимины/метилмеламин, алкилсульфонаты, антиметаболиты, аналоги пиримидина, эпиподофилотоксины, ферменты, такие как L-аспарагиназа; модификаторы биологического ответа, такие как IFNα, IL-2, G-CSF и GM-CSF; координационные комплексы платины, такие как цисплатин и карбоплатин, антрацендионы, замещенная мочевина, такая как гидроксимочевина, производные метилгидразина, включая N-метилгидразин (MIH) и прокарбазин, адренокортикальные супрессанты, такие как митотан (o, p'-DDD) и аминоглутетимид; гормоны и антагонисты, включая антагонисты адренокортикостероидов, такие как преднизон и его эквиваленты, дексаметазон и аминоглютетимид; прогестины, такие как гидроксипрогестерона капроат, медроксипрогестерона ацетат и мегестрола ацетат; эстрогены, такие как эквиваленты диэтилстилбестрола и этинилэстрадиола; антиэстрогены, такие как тамоксифен; андрогены, включая тестостерона пропионат и флуоксиместерон/эквиваленты; антиандрогены, такие как флутамид, аналоги гонадотропин-рилизинг-гормона и леупролид; и нестероидные антиандрогены, такие как флутамид.
Конъюгат TRBC1-специфичное антитело-лекарственное средство обеспечивает целенаправленную доставку химиотерапевтического компонента в клетки, которые экспрессируют TRBC1.
БИСПЕЦИФИЧНЫЕ РЕКРУТЕРЫ Т-КЛЕТОК
Было разработано большое разнообразие молекул, которые основаны на базовой концепции наличия двух антителоподобных связывающих доменов.
Биспецифичные молекулы, рекрутирующие Т-клетки, представляют собой класс биспецифичных молекул типа антител, которые были разработаны, главным образом, для использования в качестве противораковых лекарственных средств. Они направляют иммунную систему хозяина, точнее цитотоксическую активность Т-клеток, против клетки-мишени, такой как раковая клетка. В этих молекулах один связывающий домен связывается с Т-клеткой через рецептор CD3, а другой - с клетками-мишенями, такими как опухолевая клетка (через специфичную для опухоли молекулу). Поскольку биспецифичная молекула связывает как клетку-мишень, так и Т-клетку, она сближает клетку-мишень с Т-клеткой, так что Т-клетка может оказывать свое воздействие, например, цитотоксическое действие на раковую клетку. Образование комплекса Т-клетка:биспецифичное Ат:раковая клетка индуцирует передачу сигналов в Т-клетке, приводя, например, к высвобождению цитотоксических медиаторов. В идеале агент вызывает желаемую передачу сигналов только в присутствии клетки-мишени, что приводит к селективному уничтожению.
Биспецифичные молекулы, рекрутирующие Т-клетки, были разработаны в ряде различных форматов, но одним из наиболее распространенных является гибридная молекула, состоящая из двух одноцепочечных вариабельных фрагментов (scFv) различных антител. Их иногда называют BiTEs (Биспецифичные рекрутеры Т-клеток).
Таким образом, в настоящем изобретении предложена биспецифичная молекула, которая селективно распознает TRBC1 и способна активировать Т-клетку. Например, агент может представлять собой BiTE. Агент может содержать:
(i) первый домен, который связывает TRBC1, имеющий антигенсвязывающий домен, как определено выше; и
(ii) второй домен, способный активировать Т-клетку.
Биспецифичная молекула может содержать сигнальный пептид, способствующий ее продукции. Сигнальный пептид может вызывать секрецию биспецифичной молекулы клеткой-хозяином, так что биспецифичная молекула может быть собрана из супернатанта клетки-хозяина.
Сигнальный пептид может находиться на аминоконце молекулы. Биспецифичная молекула может иметь общую формулу: Сигнальный пептид - первый домен - второй домен.
Биспецифичная молекула может содержать спейсерную последовательность для соединения первого домена со вторым доменом и пространственного разделения двух доменов.
Спейсерная последовательность может, например, содержать шарнир IgG1 или стебель CD8. Линкер может в качестве альтернативы содержать альтернативную линкерную последовательность, которая имеет такие же характеристики длины и/или расстояния между доменами, что и шарнир IgG1 или стебель CD8.
ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR)
В настоящем изобретении предложен CAR, который селективно распознает TRBC1.
Химерные антигенные рецепторы (CAR), также известные как химерные Т-клеточные рецепторы, искусственные Т-клеточные рецепторы и химерные иммунорецепторы, представляют собой сконструированные рецепторы, которые придают произвольную специфичность иммунной эффекторной клетке. В случае классического CAR специфичность моноклонального антитела придают Т-клетке. Кодирующие CAR нуклеиновые кислоты могут быть перенесены в T-клетки с использованием, например, ретровирусных векторов. Таким образом, может быть сгенерировано большое количество специфичных в отношении рака Т-клеток для адоптивного переноса клеток. Фаза I клинических исследований показывает эффективность этого подхода.
Связывающий антиген-мишень домен CAR обычно объединен через спейсер и трансмембранный домен с эндодоменом, который включает или связывается с внутриклеточным доменом передачи сигналов Т-клеток. Когда CAR связывает антиген-мишень, это приводит к передаче активирующего сигнала в Т-клетку, на которой он экспрессируется.
CAR также может содержать трансмембранный домен, который проходит через мембрану. Он может содержать гидрофобную альфа-спираль. Трансмембранный домен может быть получен из CD28, что обеспечивает хорошую стабильность рецептора.
Эндодомен представляет собой часть CAR, участвующую в передаче сигнала. Эндодомен содержит или ассоциирован с внутриклеточным сигнальным доменом Т-клеток. После распознавания антигена рецепторы кластеризуются, и сигнал передается в клетку. Наиболее часто используемым компонентом передачи сигналов Т-клеток является компонент CD3-дзета, который содержит 3 ITAM. Данное взаимодействие передает сигнал активации к Т-клетке после связывания антигена. CD3-дзета может не обеспечивать полностью компетентный сигнал активации, и могут потребоваться дополнительные костимулирующие сигналы. Например, химерные CD28 и OX40 могут быть использованы с CD3-дзета для передачи пролиферативного сигнала / сигнала выживания, или все три могут быть использованы вместе.
Эндодомен CAR может содержать эндодомен CD28 и OX40, и эндодомен CD3-дзета.
В качестве альтернативы, CAR по второму аспекту настоящего изобретения может не иметь внутриклеточного сигнального домена, но может быть способен связываться с отдельной молекулой, что обеспечивает сигнальную функциональность.
Ранее были описаны системы передачи сигналов CAR, которые включают две части: CAR, который содержит антигенсвязывающий домен и трансмембранный домен; и внутриклеточный компонент передачи сигнала, который содержит внутриклеточный сигнальный домен. Один или несколько костимулирующих доменов могут быть расположены на CAR и/или внутриклеточном компоненте передачи сигнала.
Гетеродимеризация между CAR и компонентом внутриклеточной передачи сигнала создает функциональную систему CAR. Гетеродимеризация может происходить спонтанно, как описано в WO2016/124930; или это может происходить только в присутствии химического индуктора димеризации (CID), как описано в WO2015/150771. В третьей альтернативе гетеродимеризация нарушается присутствием агента, такого как конкретная малая молекула, поэтому передача сигнала, опосредованная CAR, происходит только в отсутствие данного агента. Такая система описана в WO2016/030691.
CAR может содержать сигнальный пептид, так что, когда CAR экспрессируется внутри клетки, такой как T-клетка, образующийся белок направляется в эндоплазматический ретикулум и затем на поверхность клетки, где он экспрессируется.
CAR может содержать спейсерную последовательность для соединения TRBC-связывающего домена с трансмембранным доменом и пространственного отделения TRBC-связывающего домена от мембраны. Гибкий спейсер позволяет TRBC-связывающему домену ориентироваться в разных направлениях, чтобы обеспечить связывание TRBC.
Спейсерная последовательность может, например, содержать Fc-область IgG1, шарнир IgG1 или стебель CD8, или их комбинацию.
НУКЛЕИНОВАЯ КИСЛОТА
В настоящем изобретении также предложена нуклеиновая кислота, кодирующая BiTE или CAR, как определено выше.
Используемые в настоящей заявке термины «полинуклеотид», «нуклеотид» и «нуклеиновая кислота» предназначены для использования в качестве синонимов друг друга.
Специалисту в данной области будет понятно, что многочисленные различные полинуклеотиды и нуклеиновые кислоты могут кодировать один и тот же полипептид в результате вырожденности генетического кода. Кроме того, следует понимать, что квалифицированные специалисты могут, используя обычные методы, делать замены нуклеотидов, которые не влияют на полипептидную последовательность, кодируемую полинуклеотидами, описанными в настоящей заявке, чтобы отразить использование кодонов в любом конкретном организме-хозяине, в котором должны быть экспрессированы полипептиды.
В соответствии с настоящим изобретением нуклеиновые кислоты могут содержать ДНК или РНК. Они могут быть одноцепочечными или двухцепочечными. Они также могут быть полинуклеотидами, которые включают в себя синтетические или модифицированные нуклеотиды. Ряд различных типов модификации олигонуклеотидов известен в данной области. Они включают метилфосфонатные и фосфоротиоатные остовы, добавление акридиновых или полилизиновых цепей на 3' и/или 5' концах молекулы. Для целей применения в соответствии с настоящей заявкой, следует понимать, что полинуклеотиды могут быть модифицированы любым способом, доступным в данной области техники. Такие модификации могут быть выполнены с целью повышения активности in vivo или продолжительности жизни представляющих интерес полинуклеотидов.
Термины «вариант», «гомолог» или «производное» в отношении нуклеотидной последовательности включают любую замену, вариацию, модификацию, замещение, делецию или добавление одной (или более) нуклеиновой кислоты из или в последовательность.
В настоящем изобретении также предложена конструкция нуклеиновой кислоты, которая содержит первую нуклеиновую кислоту, кодирующую CAR, как определено выше; и вторую нуклеиновую кислоту, кодирующую ген самоубийства.
Подходящие гены самоубийства для использования в CAR-экспрессирующей клетке согласно настоящему изобретению включают RQR8, который описан в WO2013/153391; и RapCasp9, который описан в WO2016/135470.
В конструкции нуклеиновой кислоты, описанной выше, первая и вторая последовательности нуклеиновой кислоты могут располагаться в любом порядке.
ВЕКТОР
В настоящем изобретении также предложен вектор или набор векторов, который содержит одну или более последовательность(ей) нуклеиновой кислоты или конструкцию(й) нуклеиновой кислоты согласно настоящему изобретению. Такой вектор может быть использован для введения последовательности(ей) нуклеиновой кислоты или конструкции(й) в клетку-хозяин, например, чтобы она экспрессировала CAR, имеющий антигенсвязывающий домен, в соответствии с первым аспектом настоящего изобретения.
Вектор может, например, представлять собой плазмиду или вирусный вектор, такой как ретровирусный вектор или лентивирусный вектор, или вектор на основе транспозона, или синтетическую мРНК.
Вектор может быть способен трансфицировать или трансдуцировать Т-клетку или NK-клетку.
КЛЕТКА
В настоящем изобретении также предложена клетка, такая как иммунная клетка, содержащая CAR в соответствии с первым аспектом настоящего изобретения.
Клетка может содержать нуклеиновую кислоту, конструкцию нуклеиновой кислоты или вектор в соответствии с настоящим изобретением.
Клетка может представлять собой Т-клетку или естественный киллер (NK).
Т-клетка может представлять собой Т-клетки или Т-лимфоциты, которые представляют собой тип лимфоцитов, которые играют центральную роль в клеточно-опосредованном иммунитете. Их можно отличить от других лимфоцитов, таких как B-клетки и естественные киллеры (NK-клетки), по наличию T-клеточного рецептора (TCR) на поверхности клетки. Существуют различные типы Т-клеток, как показано ниже.
Хелперные Т-хелперные клетки (ТН-клетки) помогают другим лейкоцитам в иммунологических процессах, включая созревание В-клеток в плазматические клетки и В-клетки памяти и активацию цитотоксических Т-клеток и макрофагов. TH-клетки экспрессируют CD4 на своей поверхности. TH-клетки становятся активированными, когда им презентуют пептидные антигены молекулами MHC класса II на поверхности антигенпрезентирующих клеток (APC). Эти клетки могут дифференцироваться в один из нескольких подтипов, включая TH1, TH2, TH3, TH17, Th9 или TFH, которые секретируют разные цитокины для облегчения различных типов иммунных ответов.
Цитолитические Т-клетки (ТC-клетки или CTL) разрушают инфицированные вирусом клетки и опухолевые клетки, а также участвуют в отторжении трансплантата. CTL экспрессируют CD8 на своей поверхности. Эти клетки распознают свои мишени путем связывания с антигеном, ассоциированным с МНС класса I, который присутствует на поверхности всех содержащих ядро клеток. Через IL-10, аденозин и другие молекулы, секретируемые регуляторными Т-клетками, клетки CD8+ могут быть инактивированы до состояния анергии, что предотвращает аутоиммунные заболевания, такие как экспериментальный аутоиммунный энцефаломиелит.
Т-клетки памяти представляют собой подгруппу антиген-специфичных Т-клеток, которые сохраняются в течение длительного времени после устранения инфекции. Данные клетки быстро размножаются до большого количества эффекторных Т-клеток при повторном воздействии когнатного антигена, таким образом обеспечивая иммунную систему «памятью» против прошлых инфекций. Т-клетки памяти включают три подтипа: Т-клетки центральной памяти (клетки TCM) и два типа T-клеток эффекторной памяти (клетки TEM и клетки TEMRA). Клетки памяти могут быть либо CD4+, либо CD8+. Т-клетки памяти обычно экспрессируют белок клеточной поверхности CD45RO.
Регуляторные Т-клетки (Treg клетки), ранее известные как супрессорные Т-клетки, имеют решающее значение для поддержания иммунологической толерантности. Их основная роль заключается в том, чтобы отключить опосредованный Т-клетками иммунитет к концу иммунной реакции и подавить аутореактивные Т-клетки, которые избежали процесса отрицательного отбора в тимусе.
Описаны два основных класса CD4+ Treg клеток - натуральные Treg клетки и адаптивные Treg клетки.
Натуральные Treg клетки (также известные как CD4+CD25+FoxP3+ Treg клетки) возникают в тимусе и связаны с взаимодействиями между развивающимися T-клетками как с миелоидными (CD11c+), так и с плазмоцитоидными (CD123+) дендритными клетками, которые были активированы TSLP. Натуральные Treg клетки можно отличить от других Т-клеток по наличию внутриклеточной молекулы, называемой FoxP3. Мутации гена FOXP3 могут предотвращать развитие регуляторных Т-клеток, вызывая фатальное аутоиммунное заболевание IPEX.
Адаптивные Treg клетки (также известные как клетки Tr1 или клетки Th3) могут возникать во время нормального иммунного ответа.
Клетка может представлять собой естественный киллер (или NK-клетку). NK-клетки являются частью врожденной иммунной системы. NK-клетки обеспечивают быстрые ответы на врожденные сигналы от инфицированных вирусом клеток независимым от МНС образом.
NK-клетки (принадлежащие к группе врожденных лимфоидных клеток) определяют как большие гранулярные лимфоциты (LGL) и представляют собой третий тип клеток, дифференцированных от общего лимфоидного предшественника, генерирующего В- и Т-лимфоциты. Известно, что NK-клетки дифференцируются и созревают в костном мозге, лимфатическом узле, селезенке, миндалинах и тимусе, откуда они затем попадают в кровообращение.
Клетки CAR согласно настоящему изобретению могут быть клетками любого типа, упомянутого выше.
Т- или NK-клетки, экспрессирующие CAR в соответствии с первым аспектом настоящего изобретения, могут быть созданы ex vivo или из собственной периферической крови пациента (от первого лица), либо в процессе трансплантации гемопоэтических стволовых клеток из донорской периферической крови (от второго лица), или периферической крови от несвязанного донора (от третьего лица).
В качестве альтернативы, T- или NK-клетки, экспрессирующие CAR в соответствии с первым аспектом настоящего изобретения, могут быть получены в результате дифференцировки ex vivo индуцибельных клеток-предшественников или эмбриональных клеток-предшественников в T- или NK- клетки. В качестве альтернативы, может быть использована иммортализованная Т-клеточная линия, которая сохраняет свою литическую функцию и может действовать как терапевтическое средство.
Во всех этих вариантах реализации клетки CAR генерируют путем введения ДНК или РНК, кодирующей CAR, одним из многих способов, включая трансдукцию вирусным вектором, трансфекцию ДНК или РНК.
CAR-экспрессирующая клетка согласно настоящему изобретению может представлять собой ex vivo T- или NK-клетку от субъекта. Т- или NK-клетки могут быть из образца мононуклеарных клеток периферической крови (РВМС). Т- или NK-клетки могут быть активированы и/или размножены перед трансдукцией нуклеиновой кислотой, кодирующей CAR, в соответствии с первым аспектом настоящего изобретения, например, путем обработки моноклональным антителом против CD3.
T- или NK-клетки согласно настоящему изобретению могут быть получены посредством:
(i) выделения образца, содержащего Т- или NK-клетки, от субъекта или других источников, перечисленных выше; и
(ii) трансдукции или трансфекции T- или NK-клеток последовательностью (последовательностями) нуклеиновой кислоты, кодирующей CAR согласно настоящему изобретению.
Затем Т- или NK-клетки могут быть очищены, например, отобраны на основе экспрессии антигенсвязывающего домена антигенсвязывающего полипептида.
В настоящем изобретении также предложен набор, который содержит T- или NK-клетку, содержащую CAR, в соответствии с первым аспектом настоящего изобретения.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
В настоящем изобретении также предложена фармацевтическая композиция, содержащая терапевтический компонент, такой как CAR-экспрессирующая клетка, терапевтическое антитело или их конъюгат, или биспецифичный рекрутер T-клеток согласно настоящему изобретению. Фармацевтическая композиция может дополнительно содержать фармацевтически приемлемый носитель, разбавитель или вспомогательное вещество. Фармацевтическая композиция может необязательно содержать один или несколько других фармацевтически активных полипептидов и/или соединений. Такой состав может, например, быть в форме, подходящей для внутривенной инфузии.
Т-КЛЕТОЧНАЯ ЛИМФОМА И/ИЛИ ЛЕЙКОЗ
В настоящем изобретении предложены агенты, клетки и способы лечения Т-клеточной лимфомы и/или лейкоза.
Способ лечения Т-клеточной лимфомы и/или лейкоза относится к терапевтическому применению агента. При этом агент может быть введен субъекту, имеющему заболевание Т-клеточной лимфомой и/или лейкозом, чтобы уменьшить, снизить или улучшить, по меньшей мере, один симптом, связанный с заболеванием, и/или замедлить, снизить или заблокировать прогрессирование заболевания.
Способ согласно настоящему изобретению может быть использован для лечения любой лимфомы и/или лейкоза, связанных с клональной экспансией клетки, экспрессирующей Т-клеточный рецептор (TCR), содержащий TRBC1.
Способ согласно настоящему изобретению можно использовать для лечения Т-клеточной лимфомы, при которой злокачественная Т-клетка экспрессирует TCR, содержащий TRBC1. «Лимфома» используется в настоящей заявке в соответствии со своим стандартным значением для обозначения рака, который обычно развивается в лимфатических узлах, но может также поражать селезенку, костный мозг, кровь и другие органы. Лимфома обычно представлена в виде солидной опухоли из лимфоидных клеток. Первичным симптомом, связанным с лимфомой, является лимфаденопатия, хотя вторичные (B) симптомы могут включать лихорадку, ночные потения, потерю веса, потерю аппетита, усталость, дыхательные расстройства и зуд.
Способ согласно настоящему изобретению может быть использован для лечения Т-клеточного лейкоза, при котором злокачественная Т-клетка экспрессирует TCR, содержащий TRBC1. «Лейкоз» используется в настоящей заявке в соответствии со стандартным значением для обозначения рака крови или костного мозга.
Ниже приведен иллюстративный, неисчерпывающий список заболеваний, которые можно лечить способом согласно настоящему изобретению.
ПЕРИФЕРИЧЕСКАЯ Т-КЛЕТОЧНАЯ ЛИМФОМА
Периферические Т-клеточные лимфомы являются относительно редкими лимфомами и составляют менее 10% всех неходжкинских лимфом (НХЛ). Однако они связаны с агрессивным клиническим течением, а причины и точное клеточное происхождение большинства Т-клеточных лимфом до сих пор недостаточно четко определены.
Лимфома обычно сначала проявляется в виде отека на шее, подмышках или в паху. Дополнительный отек может возникнуть там, где расположены другие лимфатические узлы, например, в селезенке. В целом, увеличенные лимфатические узлы могут вторгаться в пространство кровеносных сосудов, нервов или желудка, что приводит к опухшим рукам и ногам, покалыванию и онемению или ощущению переполнения, соответственно. Симптомы лимфомы также включают неспецифические симптомы, такие как лихорадка, озноб, необъяснимая потеря веса, ночные потения, вялость и зуд.
Классификация ВОЗ использует морфологические и иммунофенотипические признаки в сочетании с клиническими аспектами и в некоторых случаях генетику для определения прогностически и терапевтически значимой категоризации периферических Т-клеточных лимфом (Swerdlow et al.; WHO classification of tumours of haematopoietic and lymphoid tissues. 4th ed.; Lyon: IARC Press; 2008). Анатомическая локализация неопластических Т-клеток частично совпадает с их предполагаемыми нормальными клеточными аналогами и функциями, и как таковые Т-клеточные лимфомы связаны с лимфатическими узлами и периферической кровью. Этот подход позволяет лучше понять некоторые проявления Т-клеточных лимфом, включая их клеточное распределение, некоторые аспекты морфологии и даже соответствующие клинические данные.
Наиболее распространенной из Т-клеточных лимфом является периферическая Т-клеточная лимфома, неспецифицированная (ПТКЛН), составляющая в целом 25%, за которой следует ангиоиммунобластная Т-клеточная лимфома (АИТЛ) (18,5%).
ПЕРИФЕРИЧЕСКАЯ Т-КЛЕТОЧНАЯ ЛИМФОМА, НЕСПЕЦИФИЦИРОВАННАЯ (ПТКЛН)
ПТКЛН включает более 25% всех периферических Т-клеточных лимфом и NK/Т-клеточных лимфом и является наиболее распространенным подтипом. Она определяется по диагнозу исключения, не соответствующему ни одному из конкретных образований зрелых Т-клеточных лимфом, перечисленных в действующем перечне ВОЗ 2008 года. Как таковая, она аналогична диффузной В-крупноклеточной лимфоме, неспецифицированной (ДВКЛ, NOS).
Большинство пациентов представляет собой взрослых со средним возрастом 60 лет и соотношением мужчин и женщин 2:1. Большинство случаев имеют узловое происхождение, однако экстранодальные проявления встречаются примерно у 13% пациентов и чаще всего затрагивают кожу и желудочно-кишечный тракт.
Цитологический спектр очень широк: от полиморфного до мономорфного. Были описаны три морфологически определенных варианта, включая вариант лимфоэпителиоида (Lennert), вариант T-зоны и фолликулярный вариант. Лимфоэпителиоидный вариант ПТКЛ содержит обильные фоновые эпителиоидные гистиоциты и обычно является положительным по CD8. Это было связано с лучшим прогнозом. Фолликулярный вариант ПТКЛН появляется как потенциально отдельное клиникопатологическое образование.
Большинство ПТКЛН имеют фенотип зрелых Т-клеток, и большинство случаев являются CD4-положительными. В 75% случаев наблюдается вариабельная потеря по меньшей мере одного пан-Т-клеточного маркера (CD3, CD2, CD5 или CD7), причем CD7 и CD5 подавляются чаще всего. CD30 и редко CD15 могут быть экспрессированы, причем CD15 является неблагоприятной прогностической характеристикой. Экспрессия CD56, хотя и необычна, также оказывает негативное прогностическое влияние. Дополнительные неблагоприятные патологические прогностические факторы включают индекс пролиферации, превышающий 25% на основе экспрессии KI-67, и присутствие более 70% трансформированных клеток. Иммунофенотипический анализ этих лимфом дал мало понимания их биологии.
АНГИОИММУНОБЛАСТНАЯ Т-КЛЕТОЧНАЯ ЛИМФОМА (АИТЛ)
АИТЛ является системным заболеванием, характеризующимся полиморфным инфильтратом, затрагивающим лимфатические узлы, выраженными венулами с высоким эндотелием (ВЭВ) и периваскулярной экспансией сетей фолликулярных дендритных клеток (ФДК). АИТЛ рассматривается как de-novo T-клеточная лимфома, образованная из фолликулярных хелперных αβ T-клеток (TFH), обычно встречающихся в зародышевых центрах.
АИТЛ является вторым по распространенности заболеванием среди периферических T-клеточных лимфом и NK/T-клеточных лимфом, составляя около 18,5% случаев. Оно возникает у людей от среднего возраста до пожилого, со средним возрастом 65 лет и примерно одинаковой заболеваемостью у мужчин и женщин. Клинически, пациенты обычно имеют позднюю стадию заболевания с генерализованной лимфаденопатией, гепатоспленомегалией и выраженными конституциональными симптомами. Обычно присутствует кожная сыпь с ассоциированным зудом. Часто встречается поликлональная гипергаммаглобулинемия, связанная с аутоиммунными явлениями.
В АИТЛ описаны три различных морфологических паттерна. Раннее поражение АИТЛ (Паттерн I) обычно демонстрирует сохраненную архитектуру с характерными гиперпластическими фолликулами. Распространение опухоли локализуется на периферии фолликулов. В Паттерне II архитектура узлов частично стерта с сохранением нескольких регрессивных фолликулов. Субкапсулярные синусы сохранены и даже расширены. Паракортес содержит разветвляющиеся ВЭВ, и происходит пролиферация ФДК за пределами B-клеточного фолликула. Неопластические клетки имеют размер от небольшого до среднего с минимальной цитологической атипией. Они часто имеют прозрачную или бледную цитоплазму и могут иметь различные T-клеточные мембраны. Полиморфный воспалительный фон обычно понятен.
Хотя AITL представляет собой злокачественное новообразование Т-клеток, наблюдается характерная экспансия В-клеток и плазматических клеток, что, вероятно, отражает функцию неопластических клеток в качестве клеток TFH. Присутствуют как EBV-положительные, так и EBV-негативные B-клетки. Изредка атипичные В-клетки могут напоминать клетки Ходжкина/Рида-Штернберга морфологически и иммунофенотипически, что иногда приводит к проблемам в диагностике этого заболевания. Пролиферация B-клеток при АИТЛ может быть обширной, и у некоторых пациентов развиваются вторичные EBV-положительные диффузные В-крупноклеточные лимфомы (ДВКЛ) или - реже - EBV-негативные B-клеточные опухоли, часто с плазмоцитарной дифференциацией.
Неопластические CD4-положительные Т-клетки АИТЛ демонстрируют сильную экспрессию CD10 и CD279 (PD-1) и являются положительными по CXCL13. CXCL13 приводит к усилению рекрутирования B-клеток в лимфатические узлы посредством адгезии к ВЭВ, активации B-клеток, плазмоцитарной дифференциации и расширению сети ФДК, что способствует морфологическим и клиническим особенностям АИТЛ. Интенсивная экспрессия PD-1 в перифолликулярных опухолевых клетках особенно полезна при различении паттерна I АИТЛ от реактивной фолликулярной и паракортикальной гиперплазии.
Фолликулярный вариант ПТКЛН представляет собой другое заболевание с фенотипом TFH. В отличие от АИТЛ, у него нет выраженных ВЭВ или внефолликулярной экспансии сети ФДК. Неопластические клетки могут образовывать внутрифолликулярные агрегаты, напоминая В-клеточную фолликулярную лимфому, но также могут иметь паттерн межфолликулярного роста или включать расширенные мантийные зоны. Клинически фолликулярный вариант ПТКЛН отличается от АИТЛ, поскольку пациенты чаще оказываются на ранней стадии заболевания с частичным поражением лимфатических узлов и могут не иметь конституциональных симптомов, связанных с АИТЛ.
АНАПЛАСТИЧЕСКАЯ КРУПНОКЛЕТОЧНАЯ ЛИМФОМА (АКЛ)
АКЛ может быть подразделен на АКЛ-«киназа анапластической лимфомы» (ALK)+ или АКЛ-ALK-.
АКЛ-ALK+ является одним из наиболее четко определенных заболеваний среди периферических Т-клеточных лимфом с характерными «отличительными клетками», несущими подковообразные ядра и экспрессирующими ALK и CD30. На его долю приходится около 7% всех периферических Т-клеточных и NK-клеточных лимфом, и оно наиболее распространено в первые три десятилетия жизни. Пациенты часто страдают лимфаденопатией, но поражение экстранодальных участков (кожи, костей, мягких тканей, легких, печени) и В-симптомы являются распространенным явлением.
АКЛ, ALK+ демонстрирует широкий морфологический спектр с 5 различными описанными паттернами, но все варианты содержат некоторые отличительные клетки. Клетки с характерными признаками имеют эксцентричные подковообразные или почковидные ядра и заметную перинуклеарную эозинофильную область Гольджи. Рост опухолевых клеток имеет когезионный характер с предрасположенностью к вовлечению синуса. Меньшие опухолевые клетки преобладают в мелкоклеточном варианте, а в лимфогистиоцитарном варианте обильные гистиоциты маскируют присутствие опухолевых клеток, многие из которых являются небольшими.
По определению, во всех случаях наблюдается ALK и CD30-положительность, причем их экспрессия обычно слабее в более мелких опухолевых клетках. Часто наблюдается потеря пан-Т-клеточных маркеров, в 75% случаев отсутствует экспрессия CD3 на поверхности.
Экспрессия ALK является результатом характерного рекуррентного генетического изменения, заключающегося в перестройке гена ALK на хромосоме 2p23 в один из множества партнерских генов, что приводит к экспрессии химерного белка. Наиболее распространенным партнерским геном, встречающимся в 75% случаев, является ген Нуклеофозмина (NPM1) на хромосоме 5q35, что приводит к t(2;5)(p23;q35). Клеточное распределение ALK в разных вариантах транслокации может варьироваться в зависимости от гена-партнера.
АКЛ-ALK- включена в качестве предварительной категории в классификацию ВОЗ 2008 года. Ее определяют как CD30-положительную Т-клеточную лимфому, которая морфологически неотличима от АКЛ-ALK+ с когезионным характером роста и наличием отличительных клеток, но лишенная экспрессии белка ALK.
Пациенты, как правило, представляют собой взрослых в возрасте от 40 до 65 лет, в отличие от АКЛ-ALK +, которая чаще встречается у детей и молодых взрослых. АКЛ-ALK- может поражать как лимфатические узлы, так и экстранодальные ткани, хотя последнее наблюдается реже, чем в АКЛ-ALK+. Большинство случаев АКЛ-ALK- демонстрируют сглаживание архитектуры лимфатических узлов листами когезионных неопластических клеток с типичными «отличительными» чертами. В отличие от АКЛ-ALK+, мелкоклеточный морфологический вариант не выявляется.
В отличие от своего аналога ALK+, АКЛ-ALK- демонстрирует большее сохранение экспрессии поверхностного Т-клеточного маркера, в то время как экспрессия цитотоксических маркеров и эпителиального мембранного антигена (ЭМА) менее вероятна. Сигнатуры экспрессии генов и рекуррентный хромосомный дисбаланс различны в АКЛ-ALK- и АКЛ-ALK+, подтверждая, что они являются различными заболеваниями на молекулярном и генетическом уровне.
АКЛ-ALK- клинически отличается от АКЛ-ALK+ и ПТКЛН со значительными различиями в прогнозе между этими тремя различными заболеваниями. Общая 5-летняя выживаемость у АКЛ-ALK- составляет 49%, что не так хорошо, как у АКЛ-ALK+ (при 70%), но в то же время значительно лучше, чем у ПТКЛН (32%).
Т-КЛЕТОЧНАЯ ЛИМФОМА, АССОЦИИРОВАННАЯ С ЭНТЕРОПАТИЕЙ (ТКЛЭ)
ТКЛЭ - это агрессивное новообразование, которое, как считается, происходит от интраэпителиальных Т-клеток кишечника. В классификации ВОЗ 2008 года признаны два морфологически, иммуногистохимически и генетически различных типа ТКЛЭ: тип I (представляющий большинство ТКЛЭ) и тип II (составляющий 10-20% случаев).
ТКЛЭ типа I обычно ассоциируется с явной или бессимптомной глютен-чувствительной энтеропатией и чаще встречается у пациентов из Северной Европы в связи с высокой распространенностью целиакии в этой популяции.
Чаще всего поражения ТКЛЭ обнаруживаются в тощей кишке или подвздошной кишке (90% случаев) с редкими проявлениями в двенадцатиперстной кишке, толстой кишке, желудке или в областях за пределами желудочно-кишечного тракта. Кишечные поражения обычно многоочаговые с изъязвлением слизистой оболочки. Клиническое течение ТКЛЭ является агрессивным, большинство пациентов умирает от заболевания или осложнений заболевания в течение 1 года.
Цитологический спектр ТКЛЭ типа I широк, и в некоторых случаях может содержать анапластические клетки. Существует полиморфный воспалительный фон, который в некоторых случаях может скрывать неопластический компонент. Слизистая оболочка кишечника в участках, прилегающих к опухоли, часто демонстрирует признаки целиакии с притуплением ворсинок и увеличением числа интраэпителиальных лимфоцитов (IEL), которые могут представлять собой пораженные клетки-предшественники.
По данным иммуногистохимии, неопластические клетки часто являются CD3+CD4-CD8-CD7+CD5-CD56-βF1+ и содержат цитотоксические белки, связанные с гранулами (TIA-1, гранзим B, перфорин). CD30 частично экспрессирован почти во всех случаях. CD103, который является рецептором хоминга слизистой оболочки, может быть экспрессирован в ТКЛЭ.
ТКЛЭ типа II, также называемая CD56+ мономорфной эпителиотропной Т-клеточной лимфомой кишечника, определяют как опухоль кишечника, состоящую из мономорфных T-клеток малого и среднего размера, которые экспрессируют CD8 и CD56. Часто наблюдают латеральное распространение опухоли в слизистой оболочке и отсутствие воспалительного фона. В большинстве случаев экспрессируется γδ TCR, однако есть случаи, связанные с αβ TCR.
ТКЛЭ типа II имеет более широкое распространение во всем мире, чем ТКЛЭ типа I, и часто наблюдается в азиатских или испаноязычных популяциях, в которых целиакия встречается редко. У лиц европейского происхождения ТКЛЭ II составляет около 20% Т-клеточных лимфом кишечника, с историей целиакии, по крайней мере, в ряде случаев. Клиническое течение является агрессивным.
ГЕПАТОЛИЕНАЛЬНАЯ Т-КЛЕТОЧНАЯ ЛИМФОМА (ГЛТЛ)
ГЛТЛ является агрессивным системным новообразованием, обычно возникающим из цитотоксических γδ Т-клеток врожденной иммунной системы, однако в редких случаях оно также может возникнуть из αβ Т-клеток. Это одна из самых редких Т-клеточных лимфом, которая обычно поражает подростков и молодых взрослых (средний возраст 35 лет) с сильным преобладанием мужчин.
ЭКСТРАНОДАЛЬНАЯ NK/T-КЛЕТОЧНАЯ ЛИМФОМА, НАЗАЛЬНЫЙ ТИП
Экстранодальная NK/T-клеточная лимфома, назальный тип, является агрессивным заболеванием, часто с деструктивными поражениями средней линии и некрозом. Большинство случаев имеют NK-клеточное происхождение, но некоторые случаи происходят от цитотоксических T-клеток. Общепризнано, что данное заболевание ассоциировано с вирусом Эпштейна-Барра (EBV).
Т-КЛЕТОЧНАЯ ЛИМФОМА КОЖИ
Способ согласно настоящему изобретению также может быть использован для лечения Т-клеточной лимфомы кожи.
Т-клеточная лимфома кожи (ТКЛК) характеризуется миграцией злокачественных Т-клеток в кожу, что вызывает появление различных поражений. Эти поражения меняют форму по мере прогрессирования заболевания, обычно начиная с того, что кажется сыпью, и, в конечном итоге, образуя бляшки и опухоли перед метастазированием в другие части тела.
Т-клеточные лимфомы кожи включают те, которые упомянуты в следующем иллюстративном, неисчерпывающем списке; грибовидный микоз, педжетоидный ретикулез, синдром Сезари, гранулематозная вялая кожа, лимфоматоидный папулез, парапсориаз лихеноидный хронический, CD30+ кожная Т-клеточная лимфома, вторичная кожная CD30+ крупноклеточная лимфома, CD30- кожная крупноклеточная Т-клеточная лимфома не грибовидный микоз, плеоморфная Т-клеточная лимфома, лимфома Леннерта, подкожная Т-клеточная лимфома и ангиоцентрическая лимфома.
Признаки и симптомы ТКЛК варьируют в зависимости от конкретного заболевания, из которых два наиболее распространенных типа - грибовидный микоз и синдром Сезари. Классический грибовидный микоз делится на три стадии:
Пятно (атрофическое или неатрофическое): Неспецифический дерматит, пятна на нижней части туловища и ягодиц; минимальный/отсутствующий зуд;
Бляшка: Интенсивно зудящие бляшки, лимфаденопатия; и
Опухоль: Склонна к изъязвлению.
Синдром Сезари определяется эритродермией и лейкозом. Признаки и симптомы включают отечную кожу, лимфаденопатию, ладонный и/или подошвенный гиперкератоз, алопецию, дистрофию ногтей, эктропион и гепатоспленомегалию.
Из всех первичных кожных лимфом 65% относятся к Т-клеточному типу. Наиболее распространенным иммунофенотипом является CD4-положительный. Общей патофизиологии для этих заболеваний не существует, так как термин Т-клеточная лимфома кожи охватывает широкий спектр нарушений.
Первичные этиологические механизмы развития Т-клеточной лимфомы кожи (то есть грибовидного микоза) не выяснены. Грибовидному микозу может предшествовать опосредованная Т-клетками хроническая воспалительная болезнь кожи, которая может иногда прогрессировать до фатальной лимфомы.
ПЕРВИЧНАЯ АКЛ КОЖИ (АКЛК)
АКЛК часто неотличима от АКЛ-ALK- по морфологии. Она определяется как кожная опухоль из крупных клеток с анапластической, плеоморфной или иммунобластной морфологией с более чем 75% клеток, экспрессирующих CD30. Вместе с лимфоматоидным папулезом (LyP) АКЛК относится к спектру первичных CD30-положительных Т-клеточных лимфопролиферативных нарушений, которые как группа составляют вторую наиболее распространенную после грибовидного микоза группу Т-клеточных лимфопролифераций.
Профиль иммуногистохимического окрашивания достаточно схож с АКЛ-ALK- с большей долей случаев, окрашивающих положительно на цитотоксические маркеры. По крайней мере 75% опухолевых клеток должны быть положительными по CD30. CD15 также может быть экспрессирован, и когда происходит вовлечение лимфатического узла, дифференциация с классической лимфомой Ходжкина может быть затруднена. Редкие случаи АКЛ-ALK+ могут иметь локализованные кожные повреждения и напоминать C-ALCL.
Т-КЛЕТОЧНЫЙ ОСТРЫЙ ЛИМФОБЛАСТНЫЙ ЛЕЙКОЗ
Т-клеточный острый лимфобластный лейкоз (Т-ОЛЛ) составляет около 15% и 25% от общего количества ОЛЛ в педиатрической и взрослой когортах соответственно. Пациенты обычно имеют высокое количество лейкоцитов и могут иметь органомегалию, особенно увеличение средостения и вовлечение ЦНС.
Способ согласно настоящему изобретению может быть использован для лечения Т-ОЛЛ, который связан со злокачественной Т-клеткой, которая экспрессирует TCR, включающий TRBC1.
Т-КЛЕТОЧНЫЙ ПРОЛИМФОЦИТАРНЫЙ ЛЕЙКОЗ (Т-ПЛЛ)
Т-клеточный пролимфоцитарный лейкоз (Т-ПЛЛ) представляет собой зрелый Т-клеточный лейкоз с агрессивным поведением и склонностью к поражению крови, костного мозга, лимфатических узлов, печени, селезенки и кожи. Т-ПЛЛ поражает, главным образом, взрослых в возрасте старше 30 лет. Другие названия включают Т-клеточный хронический лимфоцитарный лейкоз, «холмистый» тип Т-клеточного лейкоза и Т-пролимфоцитарный лейкоз/Т-клеточный лимфоцитарный лейкоз.
В периферической крови Т-ПЛЛ состоит из лимфоцитов среднего размера с одиночными ядрышками и базофильной цитоплазмой со случайными пузырьками или выступами. Ядра, как правило, имеют округлую или овальную форму, причем у отдельных пациентов имеются клетки с более нерегулярным ядерным контуром, сходным с церебриформной ядерной формой, наблюдаемой при синдроме Сезари. Мелкоклеточный вариант составляет 20% всех случаев Т-ПЛЛ, а вариант, подобный клеткам Сезари (церебриформный), наблюдается в 5% случаев.
Т-ПЛЛ обладает иммунофенотипом зрелого (посттимического) T-лимфоцита, и неопластические клетки, как правило, положительны по пан-Т-антигенам CD2, CD3 и CD7 и негативны по TdT и CD1a. Иммунофенотип CD4+/CD8- присутствует в 60% случаев, иммунофенотип CD4+/CD8+ присутствует в 25%, а иммунофенотип CD4-/CD8+ присутствует в 15% случаев.
ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ
Способ согласно настоящему изобретению может включать стадию введения агента в форме фармацевтической композиции.
Агент может быть введен с фармацевтически приемлемым носителем, разбавителем, наполнителем или адъювантом. Выбор фармацевтического носителя, вспомогательного вещества или разбавителя может быть сделан с учетом предполагаемого пути введения и стандартной фармацевтической практики. Фармацевтические композиции могут включать в качестве носителя (или в дополнение к нему), вспомогательное вещество или разбавитель, любое подходящее связующее(ие), смазывающее вещество(а), суспендирующий агент(ы), покрывающий агент(ы), солюбилизирующий агент(ы) и другие агенты-носители.
ВВЕДЕНИЕ
Введение агента может быть выполнено с использованием любого из множества путей, которые делают активный ингредиент биодоступным. Например, агент можно вводить пероральным и парентеральным путями, внутрибрюшинно, внутривенно, подкожно, чрескожно, внутримышечно, посредством локальной доставки, например, через катетер или стент.
Как правило, врач определяет фактическую дозировку, которая будет наиболее подходящей для отдельного субъекта, и она будет зависеть от возраста, веса и реакции конкретного пациента. Дозировка такова, что ее достаточно, чтобы уменьшить или истощить количество клональных Т-клеток, экспрессирующих TRBC1.
Далее изобретение будет описано с помощью Примеров, которые предназначены для помощи специалисту в данной области техники при реализации настоящего изобретения и никоим образом не предназначены для ограничения объема настоящего изобретения.
ПРИМЕРЫ
Пример 1 - Выбор графта тяжелой и легкой цепи
Гуманизированные домены VH сконструировали на основе CDR VH JOVI-1 и следующих каркасных участков VH человека: H-AF062256, H-EF177999, H-KF688165. Гуманизированные домены VL сконструировали с человеческим каркасным участком 3aaz.
Антитела получали с использованием или гуманизированного домена VH и мышиного домена VL JOVI-1; мышиного домена VH и гуманизированного домена VL; или гуманизированного домена VH и гуманизированного домена VL (см. Фигуру 4).
Связывание с TRBC1 протестировали с помощью ELISA, и результаты показаны на Фигуре 5.
Было обнаружено, что все комбинации химерной и гуманизированной связующей молекулы способны связывать TRBC1, и связывание было сходным с химерным антителом, имеющим мышиные домены VH и VL.
Пример 2 - Создание и тестирование обратно мутированных конструкций
Была создана серия обратно мутированных связующих молекул для человеческого каркасного участка H-AF062256, как показано в Таблице 1, в которых обратные мутации показаны жирным шрифтом.
Таблица 1
Связующие молекулы имели гуманизированный домен VL, включающий каркасный участок 3aaz.
Связывание с TRBC1 и TRBC2 протестировали с помощью ELISA, и результаты показаны на Фигуре 6. Все конструкции связывали TRBC1, но не TRBC2 и демонстрировали сходную EC50 с химерным антителом, имеющим мышиные домены VH и VL (Jovi-Mu). Это показывает, что после пересадки CDR специфичность и аффинность мышиного антитела сохраняются в гуманизированных антителах.
Пример 3 - Исследование стабильности гуманизированных mAb и scFv
Стабильность связующих молекул, описанных в Примере 2, протестировали методом дифференциальной сканирующей флуориметрии. Белки хранили в PBS в концентрации 150 мкг/мл перед добавлением красителя SYPRO Orange при соотношении белок:краситель 5000:1. Растворы смешивали и помещали в амплификатор для количественной ПЦР и работали в режиме FRET. Растворы выдерживали при 15°С в течение 10 мин, после чего следовало повышение температуры до 95°С с шагом 0,5°С и выдержкой 30 сек на каждом шаге. Показания флуоресценции получали после каждого шага. Для получения значения Tm (константа равновесия; развернутый белок = свернутый белок) первая производная изменения флуоресценции (ΔRFU/Δ°C) была построена в виде графика зависимости от изменения температуры (Δ°C).
Результаты показаны на Фигуре 7. Было обнаружено, что использование человеческого каркасного участка повышает стабильность связующей молекулы в форматах mAb.
Аналогичный эксперимент провели для исследования стабильности эквивалентных связующих молекул в формате scFv. Для этого исследования использовали дифференциальную сканирующую колориметрию. Эксперименты проводили с использованием системы CAP DSC. Белки в буфере для хранения (1 × PBS) помещали в ячейку для образца калориметра, а контрольную ячейку заполняли только буфером для хранения. Ячейки стабилизировали внутри калориметра в течение 1 часа при 25°C, а затем нагревали до конечной температуры 100°C со скоростью 200°C в час. Температура денатурации, Tm, соответствующая максимуму пика перехода, была определена по меньшей мере из двух повторных опытов и варьировала не более чем на 0,25°C.
Результаты сравнения одной связующей молекулы scFv показаны на Фигуре 8. Температура плавления мышиного scFv Jovi-1 была 61°C, тогда как температура плавления гуманизированного scFv (H-AF1, 3aaz) составляла 65°C.
Пример 4 - Получение химерного антигенного рецептора (CAR) с гуманизированным антигенсвязывающим доменом против TRBC1
Был разработан CAR второго поколения, имеющий 41BB и CD3-дзета эндодомен и антигенсвязывающий домен, содержащий гуманизированный scFv JOVI-1 (H-AF1, 3aaz), как схематически показано на Фигуре 9. Первичные человеческие Т-клетки от нормальных доноров были трансдуцированы с помощью ретровирусных векторов, экспрессирующих CAR против TRBC1 или нерелевантный CAR против EGFRvIII в качестве отрицательного контроля. Способность клеток уничтожать клетки-мишени, экспрессирующие TRBC1 или TRBC2, исследовали с помощью проточной цитометрии.
Результаты показаны на Фигуре 10. Т-клетки, экспрессирующие гуманизированный CAR против TRBC1, уничтожали TRBC1-экспрессирующие, но не TRBC2-экспрессирующие клетки-мишени.
Пролиферацию Т-клеток измеряли после 72 часов совместного культивирования с клетками-мишенями, экспрессирующими TRBC1 или TRBC2, и результаты показаны на Фигуре 11. Т-клетки, экспрессирующие либо CAR на основе мышиного Jovi-1, либо гуманизированный CAR, показали увеличение пролиферации, когда их совместно культивировали с TRBC1-экспрессирующими, но не TRBC2-экспрессирующими клетками-мишенями.
Высвобождение цитокинов измеряли после 24 часов совместного культивирования с TRBC1- или TRBC2-экспрессирующими мишенями. Результаты показаны на Фигуре 12. Т-клетки, экспрессирующие либо CAR на основе мышиного Jovi-1, либо гуманизированный CAR, показали увеличение высвобождения IFNγ и IL-2 при совместном культивировании с TRBC1-экспрессирующими, но не TRBC2-экспрессирующими клетками-мишенями.
Пример 5 - Гуманизированный CAR против TRBC1 (αTRBC1) убирает опухоль в мышиной модели NSG
Самкам мышей NSG в возрасте 7-8 недель имплантировали клетки Jurkat, трансдуцированные для экспрессии CD19-Fluc (3 × 106 клеток на одно животное в 0,1 мл PBS). На 7-й день мыши (n=8/ группа) получали внутривенные инфузии 1,0 × 106 Т-клеток с CAR против TRBC1 (aTRBC1) или имитационно трансдуцированных клеток (NT). Рост опухолей проверяли в дни исследования 6, 9, 12 и 15 с помощью биолюминесцентной визуализации (BLI). Кратко, мышам вводили (s.c.) 150 мг/кг D-люциферина за 15 минут до визуализации. Через 10 минут после введения D-люцеферина мышей анестезировали и помещали в камеру для визуализации и визуализировали на предмет люминесценции (вентральный вид и дорсальный вид; до 5 мышей лежали рядом друг с другом в порядке клетки). Продолжительность и биннинг (чувствительность) захвата изображения были получены и обработаны с использованием программного обеспечения Living Image 4.3.1.
Результаты показаны на Фигуре 13. К 12 дню гуманизированные анти-TRBC1 CAR-T-клетки привели к устранению опухоли у всех животных.
Пример 6 - Сравнение влияния мышиных и гуманизированных анти- TRBC1 CAR T-клеток на истощение
Активированные РВМС были трансдуцированы векторами, экспрессирующими CAR на основе мышиного Jovi-1 или CAR на основе гуманизированного Jovi-1, который содержит CDR из JOVI-1 с каркасными участками VH H-AF062256 и каркасными участками VL 3aaz. Клетки собирали через два дня и выдерживали в культуральной среде с 50 Ед/мл IL-2 в течение еще двух дней. Трансдуцированные Т-клетки были очищены от CD56-экспрессирующих клеток с использованием набора для позитивной селекции EasySep CD56. Совместное культивирование TRBC1-TCR-экспрессирующих мишеней Raji с CAR T-клетками проводили следующим образом: клетки-мишени высевали в количестве 50000 клеток на лунку при соотношении эффектор: мишень 1:1 в 96-луночных планшетах с U-образным дном. Через 96 часов совместные культуры собирали и клетки окрашивали антителами против PD1, против LAG3 и против Tim3, затем анализировали проточной цитометрией.
Результаты показаны на Фигуре 14. Т-клетки, экспрессирующие гуманизированный CAR против TRBC1, экспрессировали более низкие уровни всех трех маркеров истощения, чем Т-клетки, экспрессирующие мышиный CAR. Это было верно как для CD4+, так и для CD8+ Т-клеток. Поэтому Т-клетки, экспрессирующие гуманизированный CAR, неожиданно становятся менее истощенными при воздействии на клетки-мишени, чем T-клетки, экспрессирующие CAR с scFv, полученным на основе мышиного антитела Jovi-1.
Истощение Т-клеток - это состояние дисфункции Т-клеток, которое возникает во время многих хронических инфекций и рака. Данное состояние определяется плохой эффекторной функцией. Чтобы CAR-T-клетка была эффективной в уничтожении клеток-мишеней, предпочтительно избегать или снижать скорость истощения Т-клеток.
Все публикации, упомянутые в приведенном выше описании, включены в настоящий документ посредством ссылки. Различные модификации и вариации описанных способов и системы согласно настоящему изобретению будут понятны для специалистов в данной области техники без отступления от объема и сущности настоящего изобретения. Хотя настоящее изобретение было описано в связи с конкретными предпочтительными вариантами осуществления, следует понимать, что заявленное изобретение не должно быть чрезмерно ограничено такими конкретными вариантами осуществления. Действительно, различные модификации описанных способов осуществления настоящего изобретения, которые понятны для специалистов в области молекулярной биологии или в смежных областях, предназначены для включения в объем следующей формулы изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> ОТОЛУС ЛИМИТЕД, GB
<120> АНТИГЕНСВЯЗЫВАЮЩИЕ ДОМЕНЫ ПРОТИВ TRBC1
<130> P112494PCT
<150> GB 1709203.2
<151> 2017-06-09
<160> 36
<170> PatentIn версия 3.5
<210> 1
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> участок определяющий комплементарность (CDR) вариабельной
области тяжелой цепи (VH) 1, VH CDR1
<400> 1
Gly Tyr Thr Phe Thr Gly Tyr
1 5
<210> 2
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH CDR2
<400> 2
Asn Pro Tyr Asn Asp Asp
1 5
<210> 3
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> VH CDR3
<400> 3
Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp Phe
1 5 10 15
<210> 4
<211> 16
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1 вариабельной области легкой цепи (VL)
<400> 4
Arg Ser Ser Gln Arg Leu Val His Ser Asn Gly Asn Thr Tyr Leu His
1 5 10 15
<210> 5
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> VL CDR2
<400> 5
Arg Val Ser Asn Arg Phe Pro
1 5
<210> 6
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> VL CDR3
<400> 6
Ser Gln Ser Thr His Val Pro Tyr Thr
1 5
<210> 7
<211> 124
<212> Белок
<213> Mus musculus
<400> 7
Glu Val Arg Leu Gln Gln Ser Gly Pro Asp Leu Ile Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Lys Ala Thr Leu Thr Ser Asp Lys Ser Ser Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
115 120
<210> 8
<211> 113
<212> Белок
<213> Mus musculus
<400> 8
Asp Val Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Ser Leu Gly
1 5 10 15
Asp Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Lys Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Leu Gly Ile Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg
<210> 9
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированное Jovi-1, каркасный участок H-AF062256
<400> 9
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 10
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированное Jovi-1, каркасный участок H-EF177999
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 11
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Гуманизированное Jovi-1, каркасный участок H-KF688165
<400> 11
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Glu Tyr Ser Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Val Ser Ser Leu Thr Ser Asp Asp Ala Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 12
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной тяжелой цепи (VH) с обратной
мутацией, мутация
K73
<400> 12
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 13
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутация S71
<400> 13
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Ser Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 14
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутации S71, K73
<400> 14
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Ser Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 15
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутация I48
<400> 15
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 16
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутации I48, K73
<400> 16
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 17
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутации M20, S71, K73
<400> 17
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Ser Asp Lys Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 18
<211> 124
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VH с обратной мутацией, мутации M20, I48
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Gly Tyr
20 25 30
Val Met His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Phe Ile Asn Pro Tyr Asn Asp Asp Ile Gln Ser Asn Glu Arg Phe
50 55 60
Arg Gly Arg Val Thr Met Thr Arg Asp Thr Ser Ile Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Arg Leu Arg Ser Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Gly Ala Gly Tyr Asn Phe Asp Gly Ala Tyr Arg Phe Phe Asp
100 105 110
Phe Trp Gly Gln Gly Thr Met Val Thr Val Ser Ser
115 120
<210> 19
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> JOVI-1 VL CDR c человеческим каркасным участком 3aaz
<400> 19
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 20
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность вариабельной области легкой цепи (VL)
с обратными мутациями
<400> 20
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 21
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 21
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 22
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 22
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 23
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 23
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 24
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 24
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 25
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 25
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Phe Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 26
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 26
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 27
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратной мутацией
<400> 27
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Leu Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 28
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 28
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 29
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 29
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 30
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 30
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 31
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратной мутацией
<400> 31
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Gln Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 32
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратными мутациями
<400> 32
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 33
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратной мутацией
<400> 33
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Phe Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 34
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность VL с обратной мутацией
<400> 34
Asp Ile Val Met Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Arg Leu Val His Ser
20 25 30
Asn Gly Asn Thr Tyr Leu His Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Arg Leu Leu Ile Tyr Arg Val Ser Asn Arg Phe Pro Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Ser Gln Ser
85 90 95
Thr His Val Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 35
<211> 175
<212> Белок
<213> Homo sapiens
<400> 35
Asp Leu Asn Lys Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Thr Gly Phe Phe Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys Glu
50 55 60
Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu Arg
65 70 75 80
Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys Gln
85 90 95
Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp Arg
100 105 110
Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg Ala
115 120 125
Asp Cys Gly Phe Thr Ser Val Ser Tyr Gln Gln Gly Val Leu Ser Ala
130 135 140
Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val
145 150 155 160
Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
<210> 36
<211> 175
<212> Белок
<213> Homo sapiens
<400> 36
Asp Leu Lys Asn Val Phe Pro Pro Glu Val Ala Val Phe Glu Pro Ser
1 5 10 15
Glu Ala Glu Ile Ser His Thr Gln Lys Ala Thr Leu Val Cys Leu Ala
20 25 30
Thr Gly Phe Tyr Pro Asp His Val Glu Leu Ser Trp Trp Val Asn Gly
35 40 45
Lys Glu Val His Ser Gly Val Ser Thr Asp Pro Gln Pro Leu Lys Glu
50 55 60
Gln Pro Ala Leu Asn Asp Ser Arg Tyr Cys Leu Ser Ser Arg Leu Arg
65 70 75 80
Val Ser Ala Thr Phe Trp Gln Asn Pro Arg Asn His Phe Arg Cys Gln
85 90 95
Val Gln Phe Tyr Gly Leu Ser Glu Asn Asp Glu Trp Thr Gln Asp Arg
100 105 110
Ala Lys Pro Val Thr Gln Ile Val Ser Ala Glu Ala Trp Gly Arg Ala
115 120 125
Asp Cys Gly Phe Thr Ser Glu Ser Tyr Gln Gln Gly Val Leu Ser Ala
130 135 140
Thr Ile Leu Tyr Glu Ile Leu Leu Gly Lys Ala Thr Leu Tyr Ala Val
145 150 155 160
Leu Val Ser Ala Leu Val Leu Met Ala Met Val Lys Arg Lys Asp
165 170 175
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2830064C2 |
| ХИМЕРНЫЙ АНТИГЕННЫЙ РЕЦЕПТОР (CAR) С АНТИГЕНСВЯЗЫВАЮЩИМИ ДОМЕНАМИ К КОНСТАНТНОЙ ОБЛАСТИ β Т-КЛЕТОЧНОГО РЕЦЕПТОРА | 2015 |
|
RU2744046C2 |
| СВЯЗЫВАЮЩИЙ ДОМЕН | 2019 |
|
RU2829934C2 |
| Т-КЛЕТКИ С ДВУМЯ ХИМЕРНЫМИ АНТИГЕННЫМИ РЕЦЕПТОРАМИ CAR | 2021 |
|
RU2839662C1 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ, НАЦЕЛЕННЫЕ НА FLT3 | 2018 |
|
RU2820859C2 |
| КЛЕТКА | 2015 |
|
RU2768019C2 |
| Химерные антигенные рецепторы, нацеливающиеся на CD70 | 2019 |
|
RU2801824C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ИММУНОТЕРАПИИ ПРОТИВ CD20 | 2017 |
|
RU2782699C2 |
| CD123-СПЕЦИФИЧЕСКИЕ ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2015 |
|
RU2727290C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ КЛЕТОК, ЭКСПРЕССИРУЮЩИХ РЕКОМБИНАНТНЫЙ РЕЦЕПТОР, И РОДСТВЕННЫЕ КОМПОЗИЦИИ | 2019 |
|
RU2835579C2 |
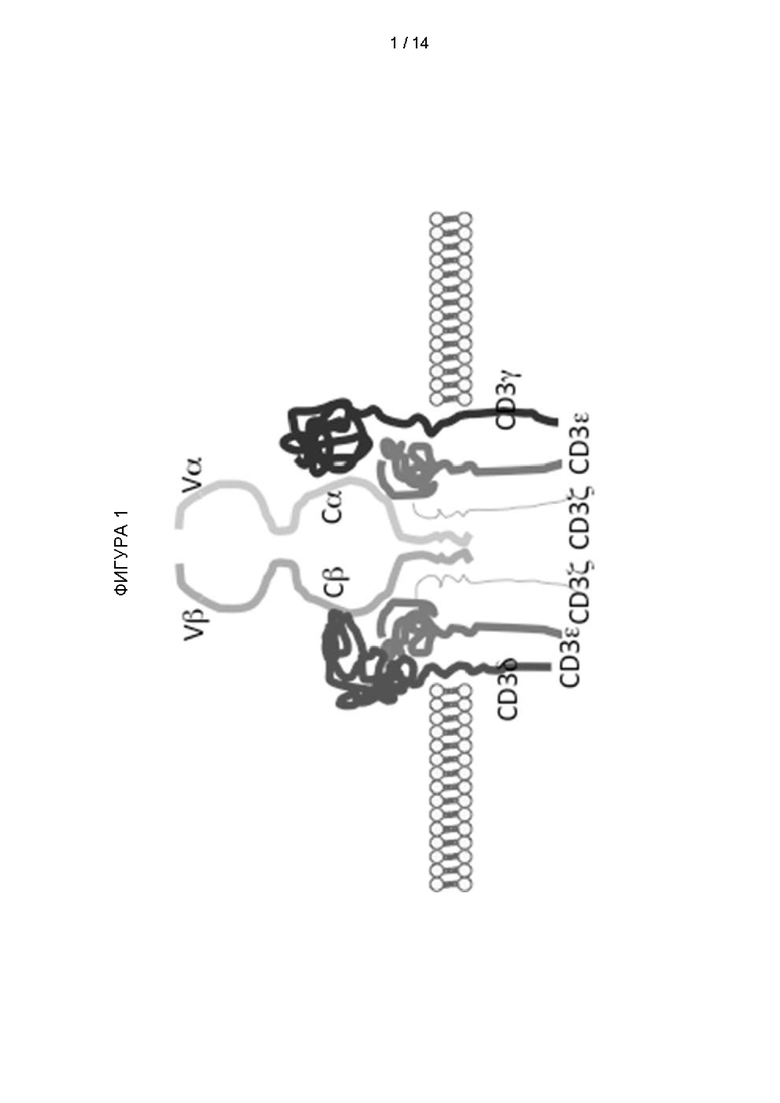
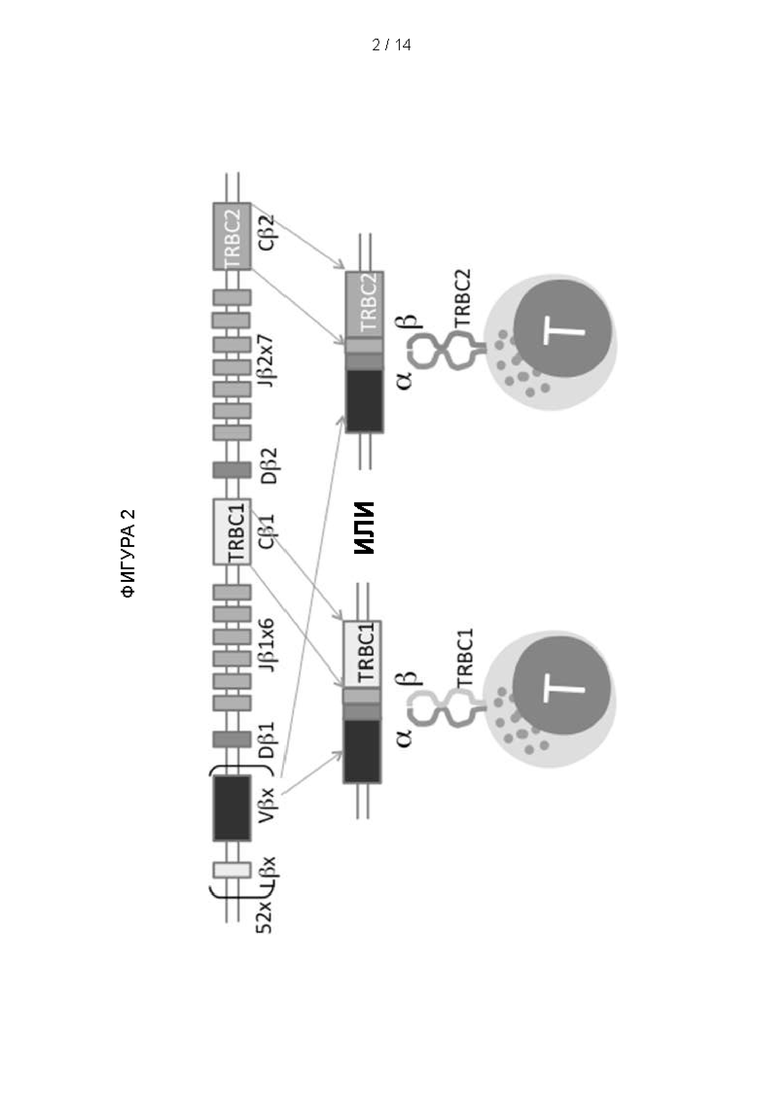
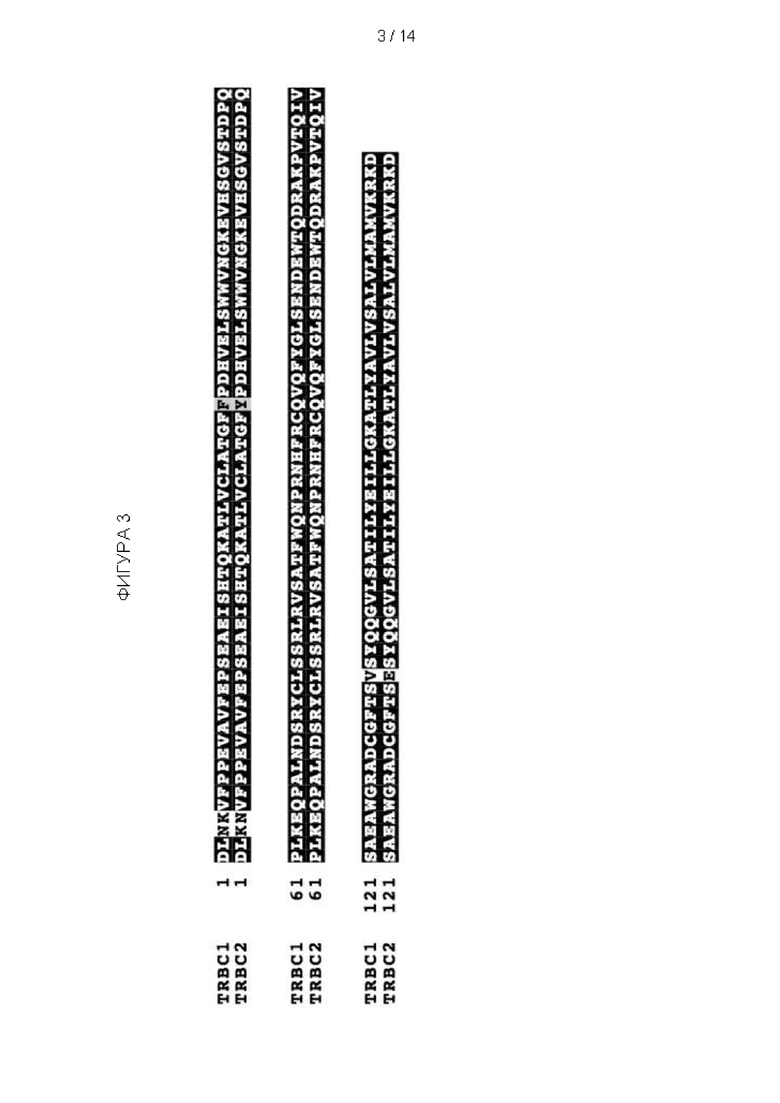
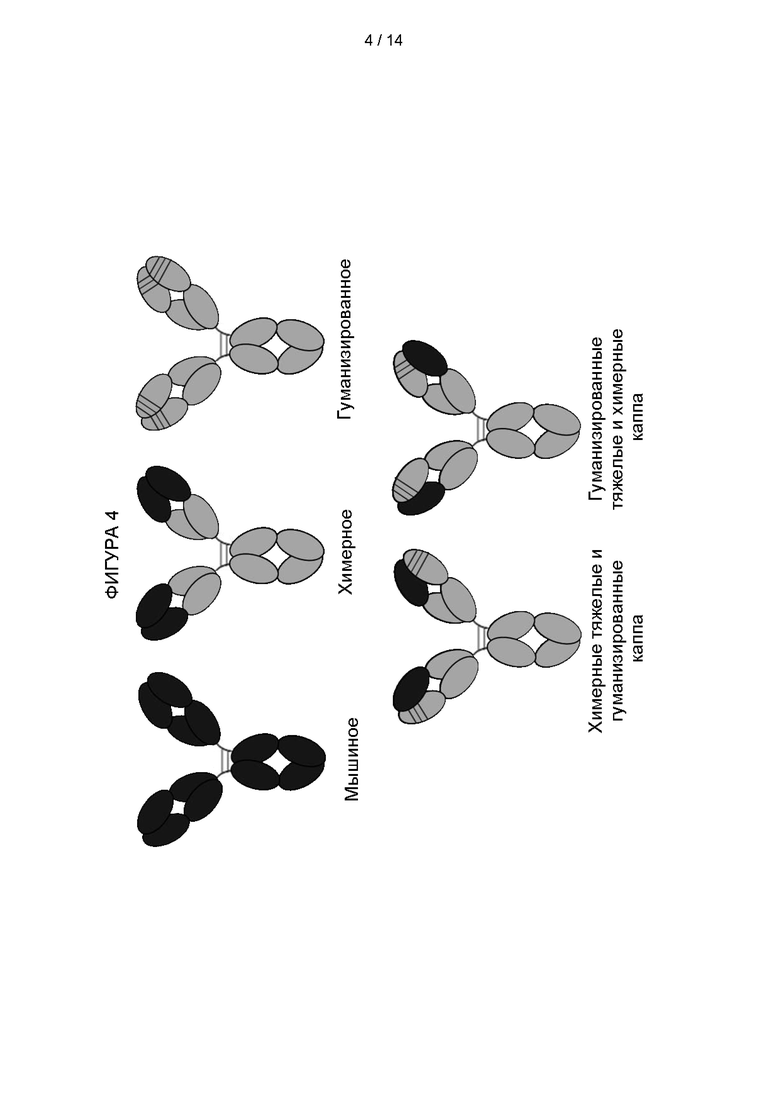
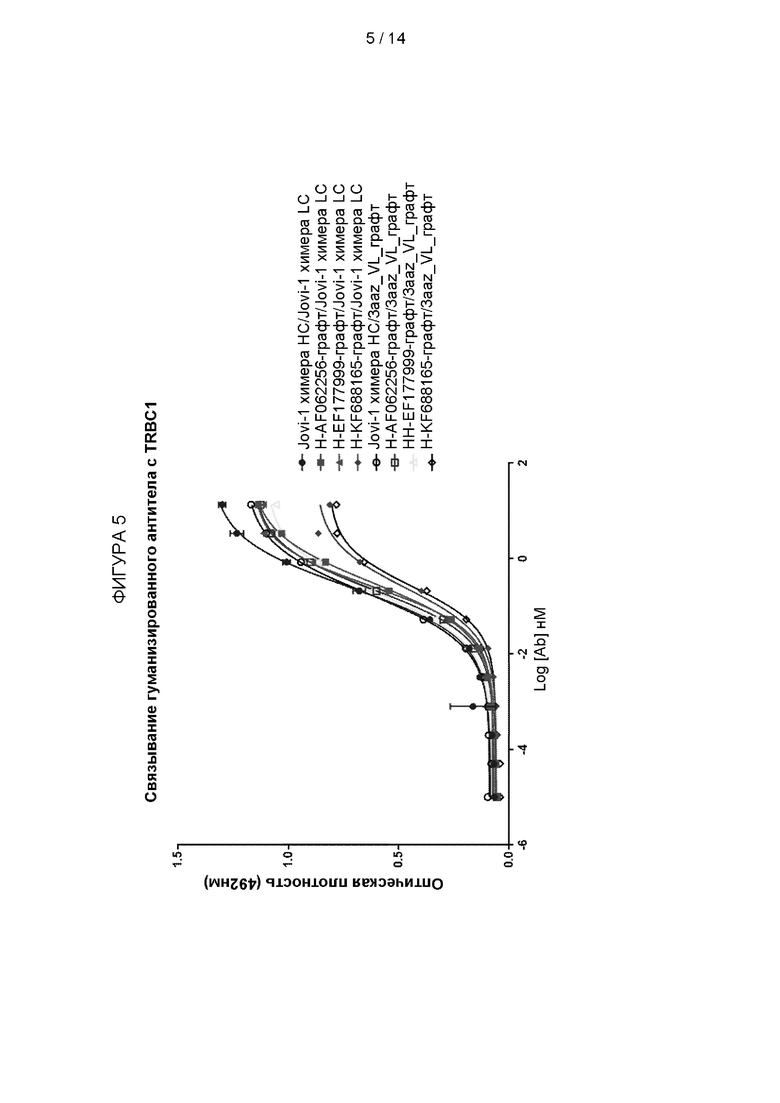
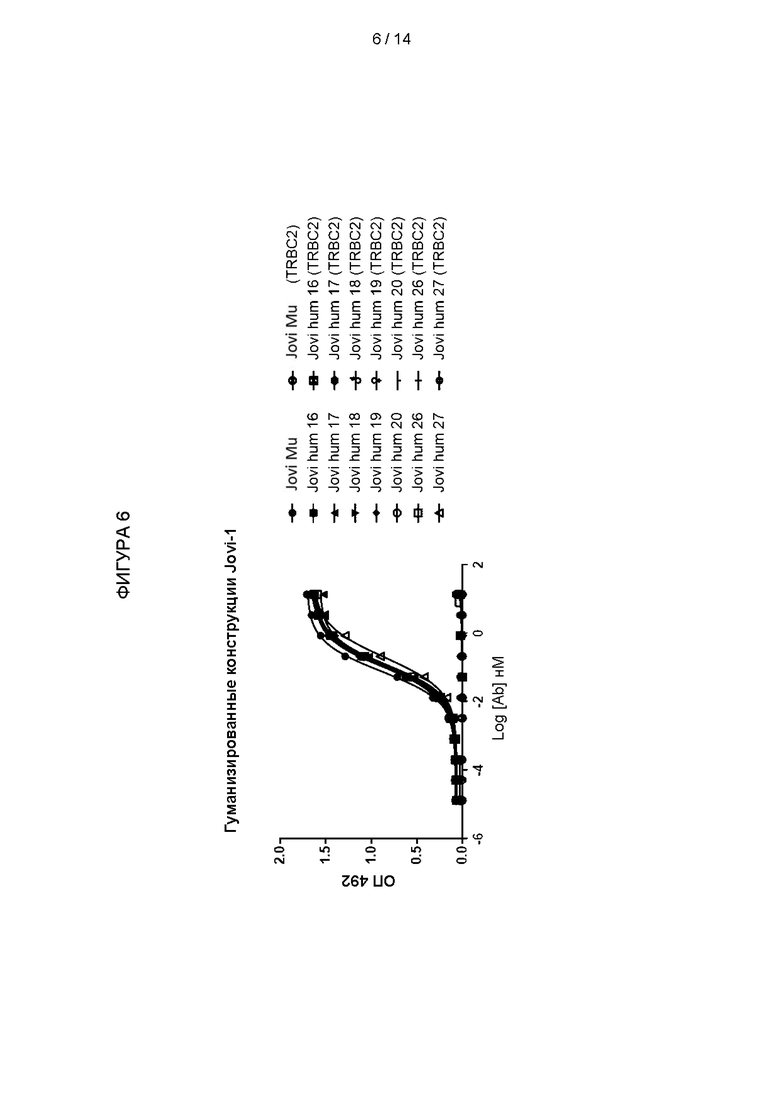
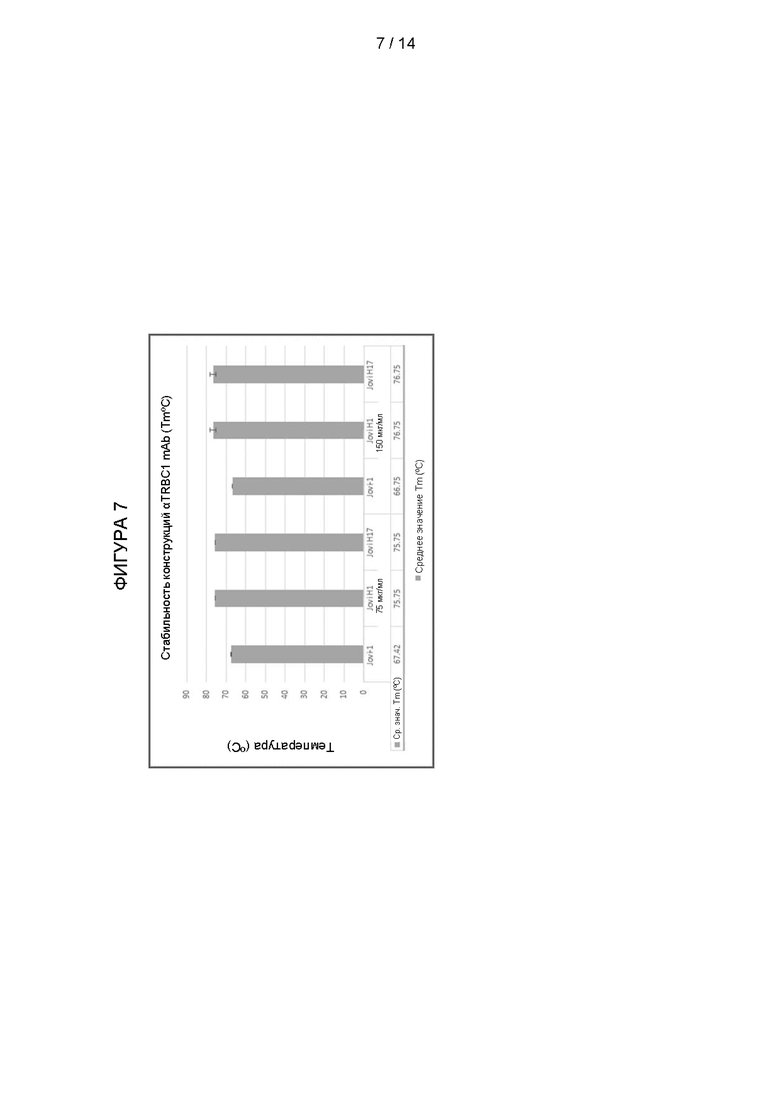
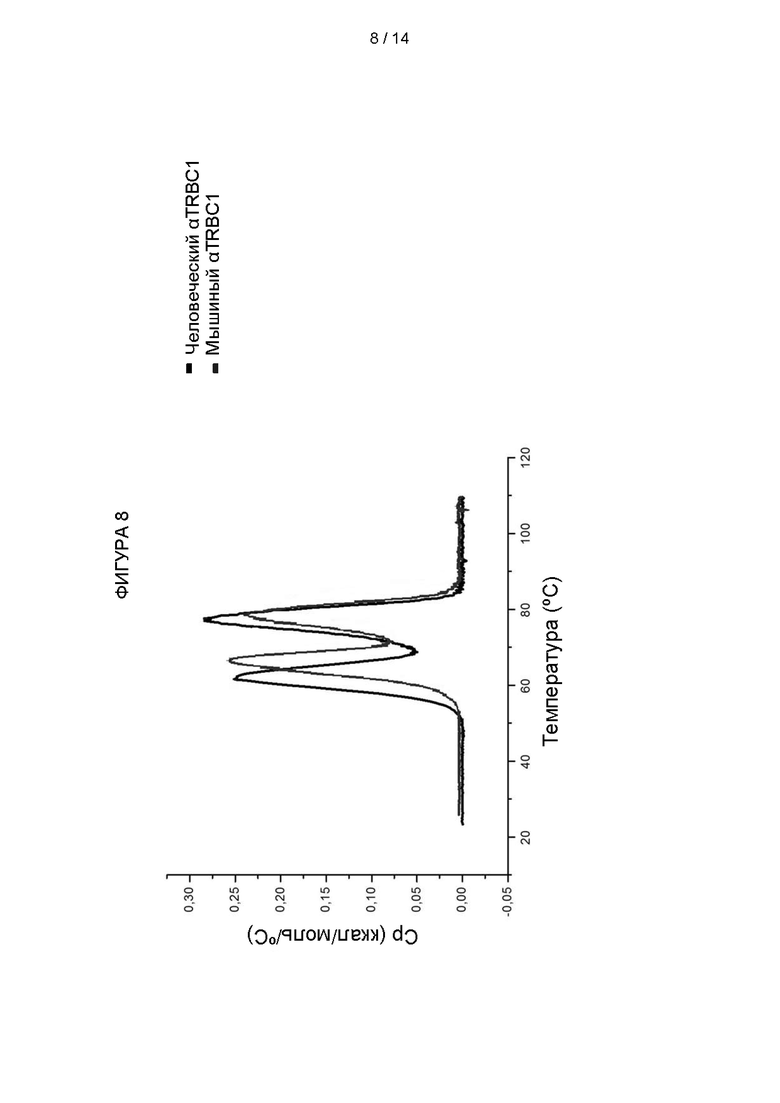
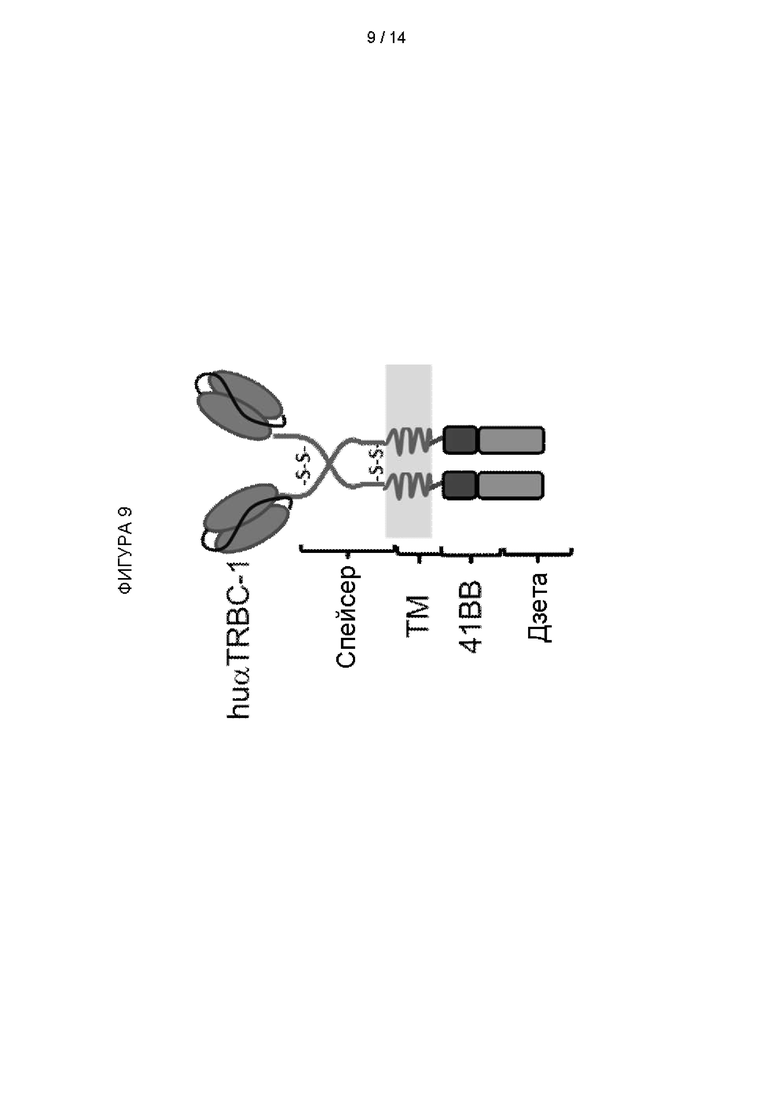
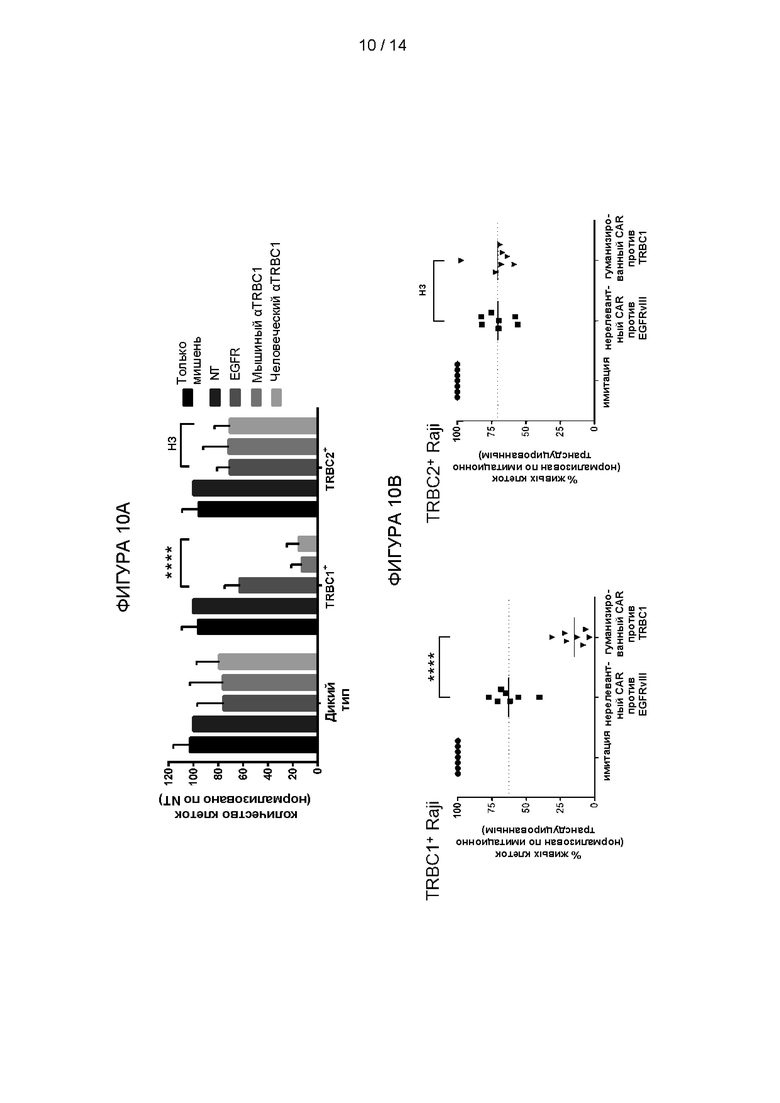

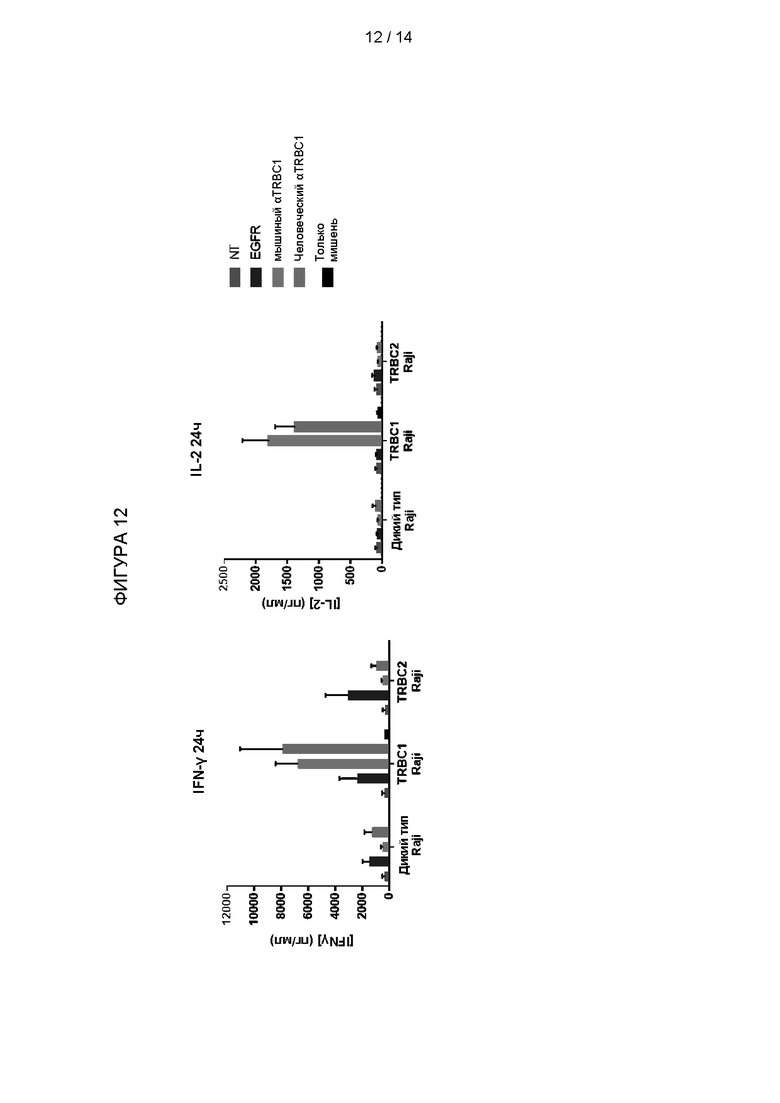
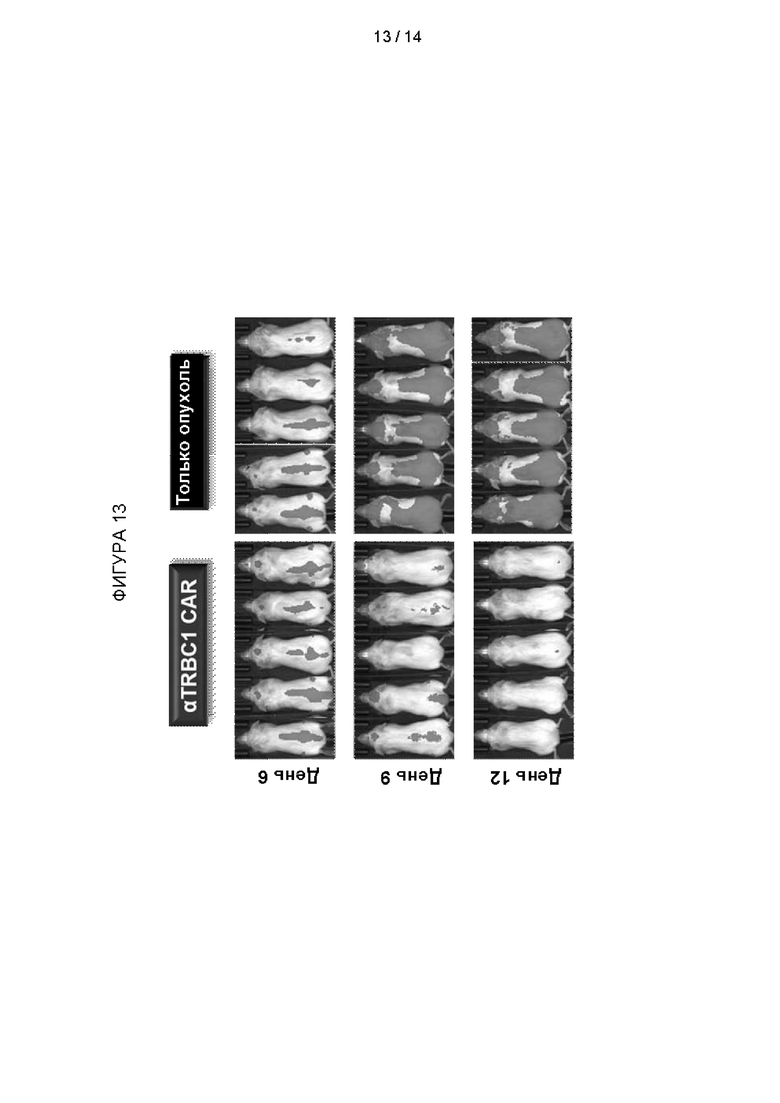
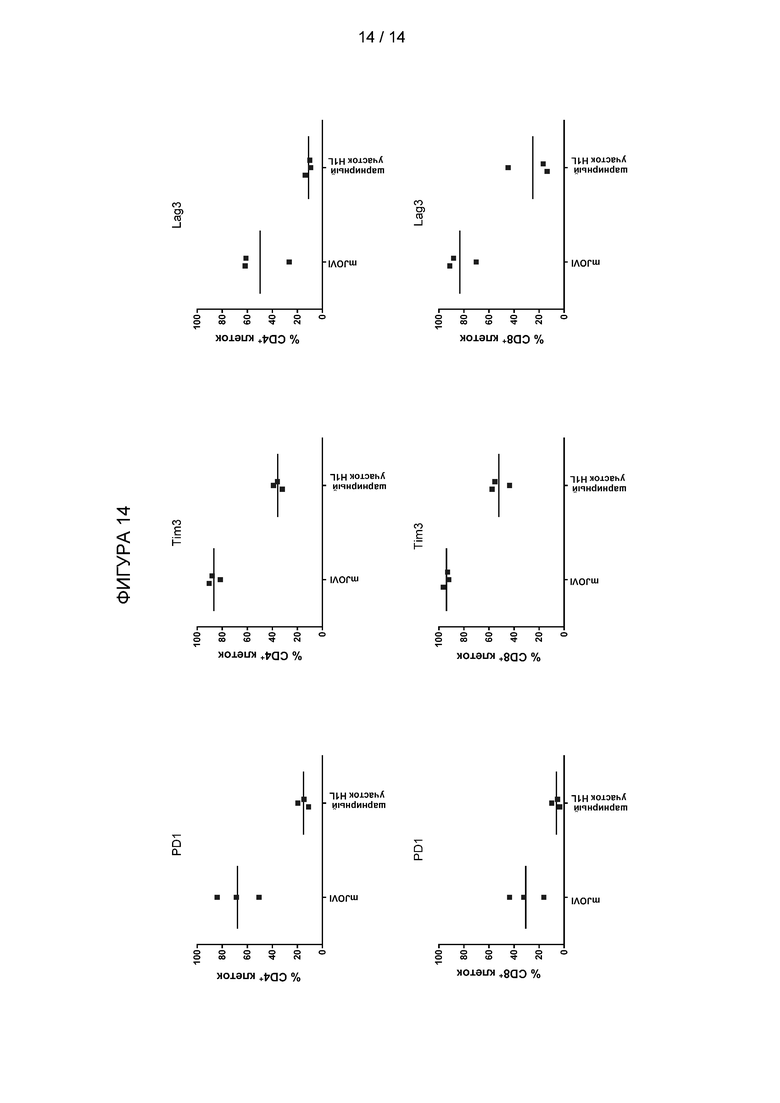
Группа изобретений относится к биотехнологии. Представлены: антигенсвязывающий домен против TRBC1, который охарактеризован последовательностями вариабельных цепей, химерный антигенный рецептор (CAR), способный связывать TRBC1, антитело, способное связывать TRBC1, которое содержит антигенсвязывающий домен против TRBC1, биспецифичный рекрутер T-клеток (BiTE), который способен распознавать TRBC1 и активировать Т-клетку, и конъюгат антитело-лекарственное средство. Также предложены: молекула нуклеиновой кислоты, которая кодирует CAR, вектор для обеспечения экспрессии CAR, цитолитическая клетка и способ ее получения, способ лечения TRBC1-экспрессирующих Т-клеточных лимфомы или лейкоза у субъекта, фармацевтическая композиция для элиминации клеток, которые экспрессируют TRBC1, и её применение при производстве лекарственного средства. Изобретение позволяет использовать его в области терапии рака, в частности для лечения TRBC1-экспрессирующих Т-клеточных лимфомы или лейкоза у субъекта. 12 н. и 4 з.п. ф-лы, 14 ил., 1 табл., 6 пр.
1. Антигенсвязывающий домен против TRBC1, который содержит:
а) домен VH, имеющий аминокислотную последовательность, выбранную из SEQ ID No. 9, SEQ ID No. 10, SEQ ID No. 11, SEQ ID No. 12, SEQ ID No. 13, SEQ ID No. 14, SEQ ID No. 15, SEQ ID No. 16, SEQ ID No. 17 и SEQ ID No. 18; и
б) домен VL, имеющий аминокислотную последовательность, выбранную из SEQ ID No. 19, SEQ ID No. 20, SEQ ID No. 21, SEQ ID No. 22, SEQ ID No. 23, SEQ ID No. 24, SEQ ID No. 25, SEQ ID No. 26, SEQ ID No. 27, SEQ ID No. 28, SEQ ID No. 29, SEQ ID No. 30, SEQ ID No. 31, SEQ ID No. 32, SEQ ID No. 33 и SEQ ID No. 34.
2. Химерный антигенный рецептор (CAR), способный связывать TRBC1, при этом указанный CAR содержит антигенсвязывающий домен против TRBC1 по п. 1.
3. Антитело, способное связывать TRBC1, которое содержит антигенсвязывающий домен против TRBC1 по п. 1.
4. Биспецифичный рекрутер T-клеток (BiTE), который способен распознавать TRBC1 и активировать Т-клетку, при этом указанный BiTE содержит антигенсвязывающий домен против TRBC1 по п. 1.
5. Конъюгат антитело-лекарственное средство, который содержит антигенсвязывающий домен против TRBC1 по п. 1 для доставки указанного лекарственного средства в клетки, которые экспрессируют TRBC1.
6. Молекула нуклеиновой кислоты, которая кодирует CAR по п. 2.
7. Вектор для обеспечения экспрессии CAR по п. 2 в клетке, содержащий молекулу нуклеиновой кислоты по п. 6.
8. Цитолитическая клетка, содержащая CAR по п. 2, где указанная клетка представляет собой Т-клетку или NK-клетку.
9. Способ получения цитолитической клетки в соответствии с п. 8, который включает стадию введения нуклеиновой кислоты по п. 6 или вектора по п. 7 в клетку.
10. Фармацевтическая композиция для элиминации клеток, которые экспрессируют TRBC1, которая содержит в эффективном количестве множество цитолитических клеток по п. 8, антитело по п. 3, BiTE по п. 4 или конъюгат антитело-лекарственное средство по п. 5.
11. Фармацевтическая композиция по п. 10 для применения при лечении TRBC1-экспрессирующих Т-клеточных лимфомы или лейкоза у субъекта.
12. Фармацевтическая композиция по п. 11, отличающаяся тем, что ТRBC1-экспрессирующие Т-клеточные лимфома или лейкоз выбраны из: периферической Т-клеточной лимфомы, неспецифицированной (ПТКЛН); ангиоиммунобластной Т-клеточной лимфомы (АИТЛ), анапластической крупноклеточной лимфомы (АККЛ), Т-клеточной лимфомы, ассоциированной с энтеропатией (ТКЛЭ), гепатолиенальной Т-клеточной лимфомы (ГЛТЛ), экстранодальной NK/T-клеточной лимфомы, назальный тип, Т-клеточной лимфомы кожи, первичной кожной АККЛ, Т-клеточного пролимфоцитарного лейкоза и Т-клеточного острого лимфобластного лейкоза.
13. Способ лечения TRBC1-экспрессирующих Т-клеточных лимфомы или лейкоза у субъекта, который включает стадию введения фармацевтической композиции по п. 10 субъекту.
14. Способ по п. 13, отличающийся тем, что ТRBC1-экспрессирующие Т-клеточные лимфома или лейкоз выбраны из: периферической Т-клеточной лимфомы, неспецифицированной (ПТКЛН); ангиоиммунобластной Т-клеточной лимфомы (АИТЛ), анапластической крупноклеточной лимфомы (АККЛ), Т-клеточной лимфомы, ассоциированной с энтеропатией (ТКЛЭ), гепатолиенальной Т-клеточной лимфомы (ГЛТЛ), экстранодальной NK/T-клеточной лимфомы, назальный тип, Т-клеточной лимфомы кожи, первичной кожной АККЛ, Т-клеточного пролимфоцитарного лейкоза и Т-клеточного острого лимфобластного лейкоза.
15. Применение фармацевтической композиции по п. 10 при производстве лекарственного средства для лечения TRBC1-экспрессирующих Т-клеточных лимфомы или лейкоза у субъекта.
16. Применение по п. 15, отличающееся тем, что ТRBC1-экспрессирующие Т-клеточные лимфома или лейкоз выбраны из: периферической Т-клеточной лимфомы, неспецифицированной (ПТКЛН); ангиоиммунобластной Т-клеточной лимфомы (АИТЛ), анапластической крупноклеточной лимфомы (АККЛ), Т-клеточной лимфомы, ассоциированной с энтеропатией (ТКЛЭ), гепатолиенальной Т-клеточной лимфомы (ГЛТЛ), экстранодальной NK/T-клеточной лимфомы, назальный тип, Т-клеточной лимфомы кожи, первичной кожной АККЛ, Т-клеточного пролимфоцитарного лейкоза и Т-клеточного острого лимфобластного лейкоза.
| WO 2015132598 A1, 11.09.2015 | |||
| WO 03093318 A1, 13.11.2003 | |||
| WO 2013074916 A1, 23.05.2013 | |||
| ОДНОЦЕПОЧЕЧНЫЕ РЕКОМБИНАНТНЫЕ Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2003 |
|
RU2355703C2 |

