1. ОБЛАСТЬ ИЗОБРЕТЕНИЯ
[0001] Изобретение относится к применению уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуумов, имеющих фенилкетонурию (ФКУ), для оптимизации эффективной дозы терапевтического средства против ФКУ. Также изобретение относится к способам лечения индивидуумов, имеющих ФКУ, включающим введение эффективного количества терапевтического средства против ФКУ, где эффективное количество представляет собой количество, которое нормализует уровни аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуума. Также изобретение относится к оптимизированной кодирующей последовательности фенилаланингидроксилазы (PAH), которую можно использовать в таких векторах в качестве векторов на основе рекомбинантного аденоассоциированного вируса (rAAV) для достижения длительной экспрессии PAH - фермента, ответственного за метаболизм фенилаланина - в печени индивидуума. Также изобретение относится к способам лечения с использованием векторов в ген-заместительном подходе.
2. УРОВЕНЬ ТЕХНИКИ
[0002] Фенилкетонурия (ФКУ) представляет собой врожденную ошибку метаболизма аминокислот, которая является результатом снижения активности печеночной фенилаланингидроксилазы (PAH) - фермента, ответственного за метаболизм фенилаланина. Пациенты с мутациями PAH, которые ведут к ФКУ и гиперфенилаланинемии (HPA), имеют повышенные уровни фенилаланина, снижение нейропсихологического функционирования и ослабление когнитивного развития. Для пациентов с тяжелой ФКУ существует возможность необратимой умственной отсталости, если уровни фенилаланина не поддерживать низкими с использованием пищевых ограничений. Неврологические симптомы ФКУ обуславливаются аномальным продуцированием ряда нейротрансмиттеров у индивидуумов, имеющих ФКУ, в результате утраты PAH, которая требуется для конвертирования фенилаланина в метаболит-предшественник, требуемый для синтеза ряда нейротрансмиттеров.
[0003] Современное лечение ФКУ включает пожизненное придерживание диеты, которая имеет низкое содержание аминокислоты фенилаланина. Эту диетотерапию трудно поддерживать, и она не всегда устраняет разрушительные неврологические эффекты, которые могут вызываться повышенными уровнями фенилаланина. Не идеальный контроль рациона в ходе беременности может привести к патологии родов. Кроме того, для пациентов с ФКУ/HPA очень трудно жить нормальной жизнью, соблюдая ограничительную диету, и диетотерапия может быть ассоциирована с дефицитом некоторых питательных веществ, который в некоторых случаях может быть вредоносным для развития головного мозга. Большинство диетических продуктов с низким содержанием фенилаланина имеют настолько неудовлетворительные органолептические свойства, что соблюдение режима их приема при таком лечении нарушается. Таким образом, разработка терапевтического лечения заменила бы или дополнила современное диетическое лечение и предупредила бы неврологические повреждения, которые имеют индивидуумы с ФКУ, в частности, пациенты с наиболее тяжелыми формами заболевания. Однако оптимальным терапевтическим средством против ФКУ было бы средство, которое нормализует уровни аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров, уровни которых изменены в результате недостаточной активности PAH. Таким образом, существует клиническая потребность в терапевтических средствах против ФКУ, которые при доставке в эффективном количестве способны нормализовать уровни конкретных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуумов, имеющих ФКУ.
[0004] Генная терапия обеспечивает возможность излечения через непрерывное эндогенное продуцирование PAH после однократного введения вектора. Она была бы значительным клиническим достижением со значительными предпосылками для других врожденных нарушений, для которых отсутствует эффективное лечение. Аденоассоциированный вирус (AAV) представляет собой небольшой безоболочечный вирус животных с дефектом репликации, который инфицирует людей и некоторые другие виды приматов. Некоторые признаки AAV делают этот вирус привлекательным носителем для доставки терапевтических белков посредством генной терапии, включая, например, то, что AAV известен тем, что он не вызывает заболеваний у человека и индуцирует мягкий иммунный ответ, и что векторы на основе AAV могут инфицировать как делящиеся, так и покоящиеся клетки без встраивания в геном клетки-хозяина. Векторы для генной терапии с использованием AAV успешно применялись в некоторых клинических испытаниях, например, для доставки фактора IX человека (FIX) в печень взрослых людей для лечения гемофилии B.
[0005] Несмотря на его положительные качества, векторы для генной терапии на основе AAV имеют некоторые недостатки. В частности, вместимость векторов AAV для клонирования ограничена вследствие способности упаковывания ДНК вируса. Одноцепочечный ДНК-геном AAV дикого типа имеет размер приблизительно 4,7 тысяч пар оснований (т.п.н.). На практике, в вирусных частицах AAV оказываются полностью упакованными, т.е. являются полноразмерными, геномы AAV размеров вплоть до приблизительно 5,0 т.п.н. Учитывая, что геном нуклеиновой кислоты в векторах AAV должен иметь два инвертированных концевых повтора (ITR) AAV размером приблизительно 145 оснований, способность упаковывания ДНК вектора AAV является такой, что может быть инкапсидировано максимум приблизительно 4,4 т.п.н. кодирующей белок последовательности.
[0006] ФКУ обуславливает несколько новых проблем вследствие определенных молекулярных и биохимических свойств PAH, касающихся размера кДНК PAH и эффективности экспрессии белка PAH, а также уникальных функциональных свойств фермента, таких как клеточная локализация, активность регуляции и потенциал к гетеродимеризации с мутантной PAH. Было предпринято несколько попыток опосредуемой вектором экспрессии PAH на мышах. См., например, Harding et al., Complete correction of hyperphenylalaninemia following liver-directed, recombinant AAV2/8 vector mediated gene therapy in murine phenylketonuria Gene Ther. 2006 Mar; 13(5):457-6, и Viecelli et al, Treatment of Phenylketonuria Using Minicircle-Based Naked-DNA Gene Transfer to Murine Liver Hepatology. 2014 Sep; 60(3): 1035-1043 (также см. WO2018126112). Однако оценка эффективности доставки, иммунной стимуляции, долговременной стабильности экспрессии и безопасности либо отсутствует, либо ее результаты не оптимальны. Таким образом, необходимы более эффективные векторы AAV.hPAH для лечения ФКУ.
3. СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[0007] Варианты осуществления, описанные в настоящем описании, относятся к вектору на основе AAV для генной терапии для доставки функциональной фенилаланингидроксилазы (PAH) человека индивидууму, нуждающемуся в этом.
[0008] В одном аспекте изобретение относится к применению аденоассоциированного вируса (AAV) с дефицитом репликации для доставки гена фенилаланингидроксилазы (PAH) человека в клетки печени пациентов (людей), у которых диагностирована ФКУ. Рекомбинантный вектор на основе AAV (rAAV), используемый для доставки гена hPAH ("rAAV.hPAH" или "AAV-PAH"), должен иметь тропизм к печени (например, rAAV, имеющий капсид AAV5), и трансген hPAH должен контролироваться специфическими для печени элементами контроля экспрессии. В одном варианте осуществления элементы контроля экспрессии включают один или более из следующих: энхансер; промотор; интрон; и сигнал полиA. Такие элементы дополнительно описаны в настоящем описании.
[0009] В одном варианте осуществления трансген hPAH находится в геноме рекомбинантного вектора на основе аденоассоциированного вируса (rAAV), который содержит (a) последовательность 5'-инвертированного концевого повтора (ITR) AAV; (b) промотор; (c) кодон-оптимизированную последовательность, кодирующую фенилаланингидроксилазу человека (hPAH); и (d) AAV 3'-ITR. В конкретном варианте осуществления кодон-оптимизированная последовательность PAH представляет собой SEQ ID NO: 7. Промотор может представлять собой синтетическую промоторную последовательность, содержащую части промотора hAAT, энхансер печеночной области контроля (HCR) и энхансер ApoE. В конкретном варианте осуществления последовательность промотора представляет собой SEQ ID NO: 6. 5'-ITR AAV и/или 3'-ITR AAV могут быть из гетерологичного псевдотипа AAV. В конкретном варианте осуществления последовательности ITR происходят из AAV2. В другом варианте осуществления геном вектора дополнительно содержит сигнальную последовательность полиаденилирования, которая может представлять собой сигнал полиадениллирования бычьего гормона роста (bGH). В другом варианте осуществления геном вектора дополнительно содержит интрон. В некоторых вариантах осуществления интрон представляет собой составную последовательность интрона глобина/AIAT. В конкретном варианте осуществления последовательность интрона представляет собой SEQ ID NO: 14. Геном вектора может иметь размер от приблизительно 4 т.п.н. до приблизительно 5 т.п.н. В конкретном варианте осуществления последовательность генома вектора представляет собой SEQ ID NO: 18.
[0010] В другом аспекте изобретение относится к частице рекомбинантного аденоассоциированного вируса (rAAV), содержащей капсид AAV и геном вектора, как описано в одном или нескольких из вариантов осуществления настоящего изобретения. В одном варианте осуществления капсид AAV представляет собой капсид AAV5. В другом варианте осуществления частица rAAV представляет собой ApoE-HCR-hAAT.cG-AIATI.hPAHcol.bGH. В другом варианте осуществления частица rAAV предоставлена посредством фармацевтической композиции.
[0011] В другом аспекте изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, обладающую по меньшей мере 80% гомологией с нуклеотидной последовательностью SEQ ID NO: 7, и кодирующая функциональную PAH. В одном варианте осуществления функциональная PAH представляет собой PAH человека. В одном варианте осуществления кодирующая последовательность для hPAH является кодон-оптимизированной для экспрессии у человека. Такая последовательность может обладать менее чем 80% идентичностью с нативной кодирующей последовательностью hPAH (SEQ ID NO: 1). В одном варианте осуществления кодирующая последовательность hPAH представляет собой последовательность, показанную в SEQ ID NO: 7. В другом варианте осуществления кодон-оптимизированная нуклеиновая кислота PAH имеет сниженное содержание динуклеотида CpG. В конкретном варианте осуществления количество динуклеотидов CpG составляет менее 25.
[0012] В другом аспекте изобретение относится к фармацевтической композиции, содержащей геном вектора, описанный в настоящем описании, или частицу rAAV, описанную в настоящем описании, и фармацевтически приемлемый носитель. В другом аспекте изобретение относится к иммуногенной композиции, содержащей векторный геном, описанный в настоящем описании, или частицу rAAV, описанную в настоящем описании, и фармацевтически приемлемый носитель. В другом аспекте изобретение относится к вакцине, содержащей векторный геном, описанный в настоящем описании, или частицу rAAV, описанную в настоящем описании, и фармацевтически приемлемый носитель.
[0013] Следующий аспект, описанный в настоящем описании, относится к способу экспрессии белка у индивидуума, включающему введение индивидууму композиции, содержащей частицу rAAV, описанную в настоящем описании, тем самым экспрессируя кодируемый белок PAH в печени индивидуума.
[0014] В другом аспекте изобретение относится к способу лечения пациента, имеющего фенилкетонурию, который включает введение векторного генома rAAV, описанного в настоящем описании, частицы rAAV, описанной в настоящем описании, фармацевтической композиции, описанной в настоящем описании, иммуногенной композиции, описанной в настоящем описании, или вакцины, описанной в настоящем описании. В одном варианте осуществления частицу rAAV доставляют в количестве от приблизительно 1×1012 до приблизительно 1×1015 мкг/кг в водной суспензии. Другой вариант осуществления, предусматриваемый в рамках настоящего изобретения, относится к применению генома вектора rAAV, описанного в настоящем описании, частицы rAAV, описанной в настоящем описании, фармацевтической композиции, описанной в настоящем описании, иммуногенной композиции, описанной в настоящем описании, или вакцины, описанной в настоящем описании, для лечения ФКУ у индивидуума.
[0015] Изобретение относится к способу лечения индивидуума, имеющего фенилкетонурию (ФКУ), включающему стадии введения эффективного количества терапевтического средства против ФКУ индивидууму, где эффективное количество терапевтического средства против ФКУ представляет собой количество, которое изменяет уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров у индивидуума.
[0016] В одном варианте осуществления способ включает стадию измерения уровней одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров в биологическом образце, полученном от индивидуума, после введения эффективного количества терапевтического средства против ФКУ, где эффективное количество терапевтического средства против ФКУ представляет собой количество, которое изменяет уровни одного или более нейротрансмиттеров или метаболитов нейтротрансмиттеров у индивидуума. В другом варианте осуществления способ приводит к смягчению нейрокогнитивного симптома фенилкетонурии (ФКУ) у индивидуума, имеющего ФКУ. В некоторых вариантах осуществления нейрокогнитивные симптомы включают снижение IQ, дефицит внимания и дефицит способности к целенаправленной деятельности, включая стратегическое планирование, тормозной контроль, кратковременную память и когнитивную гибкость. В другом варианте осуществления одна или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, включают фенилэтиламин, фенилэтаноламин, тирамин, дофамин, норадреналин, триптамин, гидрокситриптамин, фенилуксусную кислоту, фенилацетилглутамин, миндальную кислоту, гидроксифенилуксусную кислоту, 3,4-дигидроксифенилуксусную кислоту (DOPAC), 3,4-дигидроминдальную кислоту (DOMA), гомованилиновую кислоту, ванилилминдальную кислоту, дигидроксифенилэтиленгликоль (DOPEG), метилфенилэтиленгликоль (MOPEG), индолуксусную кислоту и гидроксииндолуксусную кислоту.
[0017] В другом варианте осуществления образец, полученный от индивидуума, включает кровь, сыворотку, плазму, цереброспинальную жидкость (CSF) и мочу. В определенном варианте осуществления образец представляет собой CSF. В одном варианте осуществления образец представляет собой плазму.
[0018] В другом варианте осуществления терапевтическое средство против ФКУ включает KUVAN® (сапроптерина дигидрохлорид), PALYNZIQ® (пегвалиаза-pqpz) и/или средство генной терапии против ФКУ. В следующем варианте осуществления терапевтическое средство против ФКУ включает клеточную терапию против ФКУ (см., например, Harding, C., Clin Genet., Aug; 74(2) pages 97-104 (2008)) и/или фармакологический шаперон, который поддерживает стабильность белка PAH, помогает в случае неправильно свернутой PAH и/или повышает ферментативную активность (см. например, Santos-Sierra et al., Human Molecular Genetics, Volume 21, Issue 8, 15 April 2012, Pages 1877-1887; Angel et al., J Clin Invest. 2008 Aug 1; 118(8): 2858-2867).
[0019] В другом варианте осуществления генная терапия против ФКУ включает вектор на основе аденоассоциированного вируса (AAV), который экспрессирует трансген, содержащий фенилаланингидроксилазу (PAH) или пегвалиазу. В другом варианте осуществления эффективное количество средства для генной терапии против ФКУ составляет более 1E12 вг/кг массы тела индивидуума. В другом варианте осуществления эффективное количество средства генной терапии против ФКУ составляет более 1E11 вг/кг массы тела индивидуума. В другом варианте осуществления эффективное количество средства генной терапии против ФКУ превышает 1E10 вг/кг массы тела индивидуума.
[0020] В другом варианте осуществления терапевтическое средство против ФКУ выбрано при условии, что оно не является KUVAN®. В другом варианте осуществления терапевтическое средство против ФКУ выбрано при условии, что оно не является PALYNZIQ® (пегвалиаза-pqpz). В следующих вариантах осуществления терапевтическое средство против ФКУ выбрано при условии, что вектор AAV не является AAV8 или AAVHSC15.
[0021] В другом варианте осуществления уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров измеряют по меньшей мере через один месяц после введения терапевтического средства против ФКУ. В другом варианте осуществления уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров измеряют по меньшей мере через три месяца после введения терапевтического средства против ФКУ. В другом варианте осуществления уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров измеряют по меньшей мере через шесть месяцев после введения терапевтического средства против ФКУ.
[0022] В других вариантах осуществления измененные уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров находятся в пределах 5% от эталонного уровня одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, где эталонный уровень получен в качестве среднего значения уровней одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, полученных для нейротипических индивидуумов (например, по меньшей мере 5, 10, 15 или более нейротипических индивидуумов). В других вариантах осуществления измененные уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров находятся в пределах 10% от эталонного уровня одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, где эталонный уровень получен в качестве среднего значения уровней одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, полученных для нейротипических индивидуумов (например, по меньшей мере 5, 10, 15 или более нейротипических индивидуумов). В следующих вариантах осуществления измененные уровни одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров находятся в пределах 20% от эталонного уровня одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, где эталонный уровень получен в качестве среднего значения уровней одной или более аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров, полученных от нейротипических индивидуумов (например, по меньшей мере 5, 10, 15 или более нейротипических индивидуумов).
[0023] В другом варианте осуществления изобретение относится к способу измерения уровня аминокислот, нейротрансмиттеров или метаболитов нейротрансмиттеров в биологическом образце от индивидуума, включающему стадии получения биологического образца от индивидуума; преципитации образца с холодным ацетонитрилом, содержащим меченный тяжелой меткой внутренний стандарт; осаждения образца центрифугированием; переноса супернатанта в свежий контейнер; добавления карбоната натрия и бензоилхлорида или этилхлорформиата и пиридина в супернатант; добавления муравьиной кислоты в супернатант после реакции ее с карбонатом натрия и бензоилхлоридом или этилхлорформиатом и пиридином; и анализа полученного супернатанта с реагентами посредством LC/MS, тем самым выявляя и определяя уровень нейротрансмиттера или метаболита нейротрансмиттера. В некоторых вариантах осуществления уровни аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров определяют с использованием способа определения результата реакции образца с бензоилхлоридом или этилхлорформиатом и пиридином.
[0024] В рамках изобретения, "вектор AAV" относится к нуклеиновым кислотам, либо одноцепочечным, либо двухцепочечным, имеющим последовательность 5 '-инвертированного концевого повтора (ITR) AAV и последовательность 3'-ITR AAV, фланкирующие кодирующую белок последовательность (в одном варианте осуществления последовательность, кодирующую функциональный терапевтический белок, например PAH), функционально связанную с элементами контроля транскрипции, которые являются гетерологичными для вирусного генома AAV, т.е. одним или более промоторами и/или энхансерами и, необязательно, последовательностью полиаденилирования, и/или одним или более интронами, встроенными между экзонами кодирующей белок последовательности. Одноцепочечный вектор AAV относится к нуклеиновым кислотам, которые присутствуют в геноме вирусной частицы AAV, и могут представлять собой либо смысловую цепь, либо антисмысловую цепь последовательностей нуклеиновых кислот, описанных в настоящем описании. Размер таких одноцепочечных нуклеиновых кислот указывается в виде количества оснований. Двухцепочечный вектор на основе AAV относится к нуклеиновым кислотам, которые присутствуют в ДНК плазмид, например pUC19, или геноме двухцепочечного вируса, например, бакуловируса, используемого для экспрессии или переноса нуклеиновых кислот вектора на основе AAV. Размер таких двухцепочечных нуклеиновых кислот указывается в виде количества пар оснований (п.о.).
[0025] Термин "инвертированный концевой повтор (ITR)", в рамках изобретения, относится к известным в данной области участкам, находящимся на 5'- и 3'-концах генома AAV, которые функционируют в цис-формате в качестве ориджинов репликации ДНК и в качестве сигналов для упаковки вирусного генома. ITR AAV, вместе с кодирующей областью rep AAV, обеспечивает эффективное вырезание и высвобождение, и встраивание нуклеотидной последовательности, находящейся между двумя фланкирующими ITR в геном клетки-хозяина. Последовательности определенных ассоциированных с AAV ITR описаны Yan et al., J. Virol. (2005) vol. 79, pp. 364-379, которая включена в настоящее описание в качестве ссылки в полном объеме. Последовательности ITR, которые применимы в рамках настоящего изобретения, могут представлять собой полноразмерные ITR AAV дикого типа или их фрагменты, которые сохраняют функциональную способность, или могут представлять собой варианты последовательности полноразмерных ITR AAV дикого типа, которые способны функционировать в цис-формате и в качестве ориджинов репликации. ITR AAV, пригодные в рекомбинантных векторах PAH AAV согласно вариантам осуществления, описанным в настоящем описании, могут происходить из любого известного серотипа AAV, и в некоторых вариантах осуществления происходят из серотипа AAV2 или AAV5.
[0026] "Элемент регуляции транскрипции" относится к нуклеотидным последовательностям генов, вовлеченным в регуляцию генетической транскрипции, включающим промотор, положительные элементы ответа, последовательности активаторов и энхансеров для связывания факторов транскрипции для способствования связыванию РНК-полимеразы и стимуляции экспрессии, или последовательности операторов или сайленсеров, с которым связываются белки-репрессоры для блокирования связывания РНК-полимеразы и предотвращения экспрессии. Термин "специфический для печени элемент регуляции транскрипции" относится к регуляторному элементу, который специфически модулирует экспрессию генов в ткани печени. Примеры специфических для печени элементов регуляции транскрипции включают, но не ограничиваются ими, промотор транстиретина мыши (mTTR), эндогенный промотор фактора VIII человека (F8), промотор альфа-1-антитрипсина человека (hAAT) и его активные фрагменты, минимальный промотор альбумина человека и промотор альбумина мыши. Промоторы также могут включать универсальные промоторы, такие как CBA, или вирусные промоторы, такие как CMV. Также предусматриваются энхансеры, происходящие из специфических для печени участков связывания факторов транскрипции, такие как EBP, DBP, HNF1, HNF3, HNF4, HNF6 и Enh1.
[0027] В одном варианте осуществления вектор AAV содержит нуклеиновую кислоту, кодирующую функционально активный белок фенилаланингидроксилазы (PAH). Кодирующая PAH последовательность может представлять собой последовательность дикого типа, кодон-оптимизированную последовательность или вариант (см., например, Fang et al., Gene Ther., vol. 1, pages 247-254 (1994); Eisensmith et al., J. Inherit. Metab. Dis., vol. 19, pages 412-423 (1996); Nagasaki et al., Pediatr. Res., vol. 45, pages 465-473 (1999); Laipis et al., Mol. Ther., vol. 7, pages S391-S392 (2003); Knappskog PM и Martínez A., FEBS Lett., vol. 409, pages 7-11 (1997); Khan et al., J Biol Chem., vol. 12, pages 4359-4367 (2019); Wang GA et al., Proc Natl Acad Sci U S A., vol. 98, pages 1537-42 (2001)).
[0028] В рамках изобретения, PAH дикого типа имеет следующую аминокислотную последовательность:
MSTAVLENPG LGRKLSDFGQ ETSYIEDNCN QNGAISLIFS LKEEVGALAK VLRLFEENDV NLTHIESRPS RLKKDEYEFF THLDKRSLPA LTNIIKILRH DIGATVHELS RDKKKDTVPW FPRTIQELDR FANQILSYGA ELDADHPGFK DPVYRARRKQ FADIAYNYRH GQPIPRVEYM EEEKKTWGTV FKTLKSLYKT HACYEYNHIF PLLEKYCGFH EDNIPQLEDV SQFLQTCTGF RLRPVAGLLS SRDFLGGLAF RVFHCTQYIR HGSKPMYTPE PDICHELLGH VPLFSDRSFA QFSQEIGLAS LGAPDEYIEK LATIYWFTVE FGLCKQGDSI KAYGAGLLSS FGELQYCLSE KPKLLPLELE KTAIQNYTVT EFQPLYYVAE SFNDAKEKVR NFAATIPRPF SVRYDPYTQR IEVLDNTQQL KILADSINSE IGILCSALQK IK (SEQ ID NO:2).
[0029] В других вариантах осуществления рекомбинантный вектор AAV содержит нуклеиновую кислоту, содержащую 5 '-инвертированный концевой повтор (ITR) AAV2 (который может быть модифицированным или может не быть модифицированным, как известно в данной области), специфическую для печени область регуляции транскрипции, кодон-оптимизированную кодирующую область терапевтического белка, необязательно один или более интронов, последовательность полиаденилирования и 3 '-ITR AAV2 (который может быть или может не быть модифицированным, как известно в данной области). В некоторых вариантах осуществления терапевтический белок представляет собой PAH человека или его варианты. В других вариантах осуществления специфическая для печени область регуляции транскрипции содержит укороченную последовательность энхансера ApoE; проксимальный промотор альфа анти-трипсина человека (hAAT) из 186 оснований, включающий 42 оснований 5 '-нетранслируемой области (UTR); один или более энхансеров, выбранных из группы, состоящей из (i) энхансера ApoE/C1 человека из 34 оснований, (ii) дистальной области X промотора AAT человека из 32 оснований, и (iii) 80 дополнительных оснований дистального элемента проксимального промотора AAT человека; и нуклеиновую кислоту, кодирующую PAH человека. В другом варианте осуществления специфическая для печени область регуляции транскрипции содержит энхансерную последовательность α-микроглобулина и проксимальный промотор альфа-антитрипсина человека из 186 оснований (AAT).
[0030] Другие варианты осуществления, описанные в настоящем описании, относятся к векторным конструкциям, кодирующим функциональный полипептид PAH, где конструкции содержат один или более из индивидуальных элементов описанных выше конструкций и их комбинации, в одной или более различных ориентации(ях). Другой вариант осуществления, описанный в настоящем описании, относится к конструкциям в противоположной ориентации. В другом варианте осуществления предусматриваются рекомбинантные вирусные частицы AAV, содержащие описанные в настоящем описании векторы PAH AAV, и их применение для лечения ФКУ у индивидуумов. В одном варианте осуществления индивидуумами являются малолетние индивидуумы.
[0031] Векторы AAV, описанные в настоящем описании, в форме единичной цепи имеют длину менее чем приблизительно 7,0 т.п.н., или имеют длину менее 6,5 т.п.н., или имеют длину менее 6,4 т.п.н., или имеют длину менее 6,3 т.п.н., или имеют длину менее 6,2 т.п.н., или имеют длину менее 6,0 т.п.н., или имеют длину менее 5,8 т.п.н., или имеют длину менее 5,6 т.п.н, или имеют длину менее 5,5 т.п.н., или имеют длину менее 5,4 т.п.н., или имеют длину менее 5,3 т.п.н., или имеют длину менее 5,2 т.п.н., или имеют длину менее 5,0 т.п.н., или имеют длину менее 4,8 т.п.н., или имеют длину менее 4,6 т.п.н., или имеют длину менее 4,5 т.п.н., или имеют длину менее 4,4 т.п.н., или имеют длину менее 4,3 т.п.н., или имеют длину менее 4,2 т.п.н., или имеют длину менее 4,1 т.п.н., или имеют длину менее 4,0 т.п.н., или имеют длину менее 3,9 т.п.н., или имеют длину менее 3,8 т.п.н., или имеют длину менее 3,7 т.п.н., или имеют длину менее 3,6 т.п.н., или имеют длину менее 3,5 т.п.н., или имеют длину менее 3,4 т.п.н., или имеют длину менее 3,3 т.п.н., или имеют длину менее 3,2 т.п.н., или имеют длину менее 3,1 т.п.н., или имеют длину менее 3,0 т.п.н., или имеют длину менее 2,9 т.п.н., или имеют длину менее 2,8 т.п.н., или имеют длину менее 2,7 т.п.н., или имеют длину менее 2,6 т.п.н. Векторы AAV, описанные в настоящем описании, в форме единичной цепи имеют длину в диапазоне от приблизительно 5,0 т.п.н. до приблизительно 6,5 т.п.н., или в диапазоне от приблизительно 4,8 т.п.н. до приблизительно 5,2 т.п.н., или от 4,8 т.п.н. до 5,3 т.п.н., или в диапазоне от приблизительно 4,9 т.п.н. до приблизительно 5,5 т.п.н., или от приблизительно 4,8 т.п.н. до приблизительно 6,0 т.п.н., или от приблизительно 5,0 т.п.н. до 6,2 т.п.н. или от приблизительно 5,1 т.п.н. до приблизительно 6,3 т.п.н., или от приблизительно 5,2 т.п.н. до приблизительно 6,4 т.п.н., или от приблизительно 5,5 т.п.н. до приблизительно 6,5 т.п.н., или в диапазоне от приблизительно 4,0 т.п.н. до приблизительно 5,0 т.п.н., или в диапазоне от приблизительно 3,8 т.п.н. до приблизительно 4,8 т.п.н., или от 3,6 т.п.н. до 4,6 т.п.н., или в диапазоне от приблизительно 3,4 т.п.н. до приблизительно 4,4 т.п.н., или в диапазоне от приблизительно 3,2 т.п.н. до приблизительно 4,2 т.п.н., или в диапазоне от приблизительно 3,0 т.п.н. до 4,0 т.п.н., или в диапазоне от приблизительно 3,0 т.п.н. до приблизительно 4,0 т.п.н., или в диапазоне от приблизительно 2,8 т.п.н. до приблизительно 3,8 т.п.н., или в диапазоне от приблизительно 2,6 т.п.н. до приблизительно 3,6 т.п.н., или в диапазоне от приблизительно 5,0 т.п.н. до приблизительно 4,5 т.п.н., или в диапазоне от приблизительно 4,5 т.п.н. до приблизительно 4,0 т.п.н., или в диапазоне от приблизительно 3,5 т.п.н. до приблизительно 4,0 т.п.н., или в диапазоне от приблизительно 3,0 т.п.н. до приблизительно 3,5 т.п.н., или в диапазоне от 2,5 т.п.н. до 3,0 т.п.н.
[0032] В другом варианте осуществления изобретение относится к способам получения частиц рекомбинантного аденоассоциированного вируса (AAV), содержащих любой из векторов AAV, описанных в настоящем описании. Способы включают стадии культивирования клетки, трансфицированной любым из векторов AAV, описанных в настоящем описании (совместно с различными генами cap и rep AAV), и выделение частиц рекомбинантного терапевтического вируса AAV из супернатанта трансфицированной клетки.
[0033] Клетки, пригодные для продуцирования рекомбинантного AAV, описанного в настоящем описании, представляют собой любые клетки, подверженные бакуловирусной инфекции, включая клетки насекомых, такие как High Five, Sf9, Se301, SeIZD2109, SeUCR1, Sf9, Sf900+, Sf21, BTI-TN-5B1-4, MG-1, Tn368, HzAm1, BM-N, Ha2302, Hz2E5 и Ao38. В другом варианте осуществления можно использовать клетки млекопитающих, такие как HEK293, HeLa, CHO, NSO, SP2/0, PER.C6, Vero, RD, BHK, HT 1080, A549, Cos-7, ARPE-19 и MRC-5.
[0034] Также изобретение относится к рекомбинантной вирусной частице, содержащей любой из векторов AAV, описанных в настоящем описании, или любой вирусной частице, полученной вышеуказанными способами.
[0035] "Вирион AAV", или "вирусная частица AAV", или "векторная частица AAV", или "вирус AAV" относятся к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и инкапсидированного полинуклеотидного вектора AAV, как описано в настоящем описании. Если частица содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от генома AAV дикого типа, такой как трансген, подлежащий доставке в клетку млекопитающего), его, как правило, называют "векторной частицей AAV" или просто "вектором AAV". Таким образом, продуцирование векторных частиц AAV обязательно включает продуцирование вектора AAV, поскольку такой вектор находится в векторной частице AAV.
[0036] В рамках изобретения "терапевтический вирус AAV" относится к вириону AAV, вирусной частице AAV, векторной частице AAV, или вирусу AAV, которые содержат гетерологичный полинуклеотид, который кодирует терапевтический белок.
[0037] В рамках изобретения "терапевтический белок" относится к полипептиду, который обладает биологической активностью, которая заменяет или компенсирует утрату или снижение активности эндогенного белка. Например, функциональная фенилаланингидроксилаза (PAH) является терапевтическим белком против фенилкетонурии (ФКУ).
[0038] В другом варианте осуществления изобретение относится к применению эффективного количества рекомбинантного вируса AAV PAH для получения лекарственного средства для лечения индивидуума, страдающего ФКУ. В одном варианте осуществления индивидуумом, страдающим ФКУ, является человек. В одном варианте осуществления лекарственное средство вводят посредством внутривенного (в/в) введения. В другом варианте осуществления введение лекарственного средства приводит к экспрессии белка PAH в кровотоке индивидуума, достаточной для изменения уровней метаболита нейротрансмиттера или нейротрансмиттера у индивидуума. В некоторых вариантах осуществления лекарственное средство также включает профилактический и/или терапевтический кортикостероид для предупреждения и/или лечения какой-либо гепатотоксичности, ассоциированной с введением вируса AAV PAH. Лекарственное средство, содержащее профилактическое или терапевтическое кортикостероидное средство, может включать по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60 или более мг/в сутки кортикостероида. В некоторых вариантах осуществления лекарственное средство, содержащее профилактический или терапевтический кортикостероид, можно вводить на протяжении непрерывного периода, составляющего по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10 недель или более. В другом варианте осуществления терапия ФКУ, описанная в настоящем описании, необязательно включает добавки с тирозином.
[0039] Другие варианты осуществления станут очевидными специалисту в данной области после прочтения настоящего описания.
4. ОПИСАНИЕ ЧЕРТЕЖЕЙ
[0040] На фиг.1 представлена хроматограмма жидкостной хроматографии - масс-спектрометрии (LC/MS) для происходящих из фенилаланина аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров после реакции с этилхлорформиатом и пиридином.
[0041] На фиг.2A-I представлен набор графиков, сравнивающих уровни различных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров, измеренные в головном мозге мышей дикого типа и мышей Enu2. Как видно на графиках, уровень фенилаланина был повышен у мышей Enu2 по сравнению с мышами дикого типа, в то время как уровень всех других измеренных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров был снижен у мышей Enu2 по сравнению с мышами дикого типа.
[0042] На фиг.3A-I представлен набор графиков, сравнивающих уровни различных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров, измеренные в головном мозге мышей дикого типа и Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH. Доза 2E14 вг/кг AAV-PAH увеличивала уровни измеренных аминокислот (за исключением фенилаланина, уровень которого снижался до уровней дикого типа), нейротрансмиттеров и метаболитов нейротрансмиттеров до уровней, измеренных у мышей дикого типа. Напротив, доза 2E13 вг/кг AAV-PAH не восстанавливала в полной мере уровни измеренных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров.
[0043] На фиг.4 представлен график, демонстрирующий корреляцию между уровнями фенилаланина в головном мозге и плазме. Уровни фенилаланина измеряли как в головном мозге, так и в плазме мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, или 2E13 вг/кг AAV-PAH, или 2E14 вг/кг AAV-PAH. На графике показано, что существует корреляция между уровнями фенилаланина в головном мозге и плазме у каждого протестированного животного независимо от статуса лечения.
[0044] На фиг.5A-D представлен набор графиков, демонстрирующих уровни аминокислот (фенилаланин (A) и тирозин (B)) и метаболитов нейротрансмиттеров (DOPAC (C) и гомованилиновая кислота (D)) в плазме мышей дикого типа и мышей Enu2. Мыши Enu2 имеют более низкие уровни метаболитов нейротрансмиттеров в их плазме по сравнению с мышами дикого типа. Результаты измерения посредством масс-спектрометрии для каждого из анализируемых соединений предоставлено в качестве площади пика, установленной из количества импульсов в минуту (cpm) обнаруженного сигнала.
[0045] На фиг.6A-D представлен набор графиков, демонстрирующих корреляцию каждого из фенилаланина (A), тирозина (B), DOPAC (C) и гомованилиновой кислоты (D) в головном мозге и в плазме мышей дикого типа и мышей Enu2.
[0046] На фиг.7A-I представлен набор графиков, сравнивающих уровни различных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров, измеренные в плазме мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH. Доза 2E14 вг/кг AAV-PAH увеличивала уровни определяемых аминокислот (за исключением фенилаланина, уровень которого снижался до уровней дикого типа), нейротрансмиттеров и метаболитов нейротрансмиттеров относительно уровней, наблюдаемых у мышей дикого типа. Напротив, доза 2E13 вг/кг AAV-PAH не полностью восстанавливала уровни определяемых аминокислот, метаболитов нейротрансмиттеров и нейротрансмиттеров.
[0047] На фиг.8A-C представлен набор графиков, демонстрирующих корреляцию между уровнями в головном мозге и плазме фенилаланина (A), DOPAC (B) и гомованилиновой кислоты (C) у мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH.
[0048] На фиг.9A-D представлен набор графиков, демонстрирующих отрицательную корреляцию между плазмой и головным мозгом уровней фенилаланина и уровней DOPAC (A, B) и гомованилиновой кислоты (C, D) у мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH.
[0049] На фиг.10A-C представлена химическая структура (A) и набор графиков, демонстрирующих уровни дофамина (B) и норадреналина (C) в головном мозге мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH.
[0050] На фиг.11A-C представлена химическая структура (A) и набор графиков, демонстрирующих уровни дофамина (B) и DOPAC (C) в головном мозге мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH.
[0051] На фиг.12A-B представлен набор графиков, демонстрирующих уровни DOPAC (A) и гомованилиновой кислоты (B) в плазме мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH.
[0052] На фиг.13A-D представлена корреляция между уровнями DOPAC (A, B) и гомованилиновой кислоты (C, D) как в головном мозге, так и в плазме, мышей дикого типа, которым вводили носитель, и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH. Эти данные демонстрируют, что уровни в плазме DOPAC и гомованилиновой кислоты могут выступать в качестве суррогатных маркеров для уровней этих метаболитов нейротрансмиттеров в головном мозге.
[0053] На фиг.14A-F представлен набор графиков, демонстрирующих уровни в головном мозге и плазме фенилаланина (A, D), тирозина (B, E) и триптофана (C, F) у мышей Enu2, которым вводили носитель, и у мышей Enu2, которым вводили Peg-PAL (PegPal-10k) в течение трех суток.
[0054] На фиг.15A-C представлен набор графиков, который демонстрирует уровни нейротрансмиттеров дофамина (A), норадреналина (B) и серотонина (C) в головном мозге мышей Enu2, которым вводили носитель, и мышей Enu2, которым вводили Peg-PAL (PegPal-10k) в течение трех суток.
[0055] На фиг.16A-F представлен набор графиков, который демонстрирует уровни DOPAC (A, D), гомованилиновой кислоты (B, E) и 5-гидроксииндолуксусной кислоты (C, F) в головном мозге и плазме мышей Enu2, которым вводили носитель, и мышей Enu2, которым вводили Peg-PAL (PegPal-10k), в течение трех суток.
[0056] На фиг.17A-C представлен набор графиков, который демонстрирует уровни фенилаланина (A), тирозина (B) и триптофана (C) в плазме мышей Enu2 через 72 часа после введения либо носителя, либо однократной дозы PALYNZIQ® (пегвалиаза-pqpz).
[0057] На фиг.18A-F представлен набор графиков, который демонстрирует уровни фенилаланина (A), тирозина (B) и триптофана (C) в головном мозге мышей Enu2 через 72 часа после введения либо носителя, либо однократной дозы PALYNZIQ® (пегвалиаза-pqpz), и корреляцию между указанными маркерами после введения PALYNZIQ® (пегвалиаза-pqpz) у мышей Enu2 (D-F).
[0058] На фиг.19A-H представлен набор графиков, который демонстрирует уровни фенэтиламина (A), дофамина (B), норадреналина (C) и серотонина (D) в плазме и головном мозге мышей Enu2 через 72 часа после введения либо носителя, либо однократной дозы PALYNZIQ® (пегвалиаза-pqpz), и корреляцию между указанными маркерами после введения PALYNZIQ® (пегвалиаза-pqpz) у мышей Enu2 (E-H).
[0059] На фиг.20A-H представлен набор графиков, который демонстрирует уровни фенилацетилглицина (A), гомованилиновой кислоты (B), 3-метокси-4-гидроксифенилгликоля (C) и 5-гидроксииндолуксусной кислоты (D) в плазме мышей Enu2 через 72 часа после введения либо носителя, либо однократной дозы PALYNZIQ® (пегвалиаза-pqpz), и корреляцию между указанными маркерами после введения PALYNZIQ® (пегвалиаза-pqpz) у мышей Enu2 (E-H).
[0060] На фиг.21A-21C представлены уровни гомованилиновой кислоты (HVA) (A), 3-метокси-4-гидроксифенилгликоля (MOPEG) (B) и 5-гидроксииндолуксусной кислоты (5HIAA) (C) в головном мозге мышей Enu2 через 72 часа после введения либо носителя, либо однократной дозы PALYNZIQ® (пегвалиаза-pqpz).
[0061] На фиг.22A-F показано, что дополнение посредством Tyr в комбинации с введением PALYNZIQ® (пегвалиаза-pqpz) повышает уровни норадреналина у мышей PAHEnu2 и мышей PAHWT после введения либо носителя, либо PALYNZIQ® (пегвалиаза-pqpz). Уровни в плазме Tyr (C) и уровни в головном мозге Tyr (D), дофамина (E) и норадреналина (F) определяли у мышей PAHEnu2 и PAHWT. Для определения значимости использовали критерий множественных сравнений Тьюки с внесением поправки в величины p (**** = p<0,0001; *** = p<0,001; ** = p<0,01; *=p<0,05) на A-F. Tyr, тирозин.
[0062] На фиг.23A-T показано, что на уровни в плазме ключевых метаболитов нейротрансмиттеров влияло снижение уровня Phe после PALYNZIQ® (пегвалиаза-pqpz) в течение 12 месяцев. Все данные относятся к образцам, полученным от подгруппы индивидуумов, которые достигли уровней Phe либо <360 мкМ (группа 1), либо >900 мкМ (группа 2). (A-D) Все метаболиты нейротрансмиттеров имели тенденцию к достижению контрольных величин через 12 месяцев. (E-H) Корреляция нейротрансмиттеров метаболитов в плазме и Phe в плазме. Критерий множественных сравнений с внесением поправки в величины p использовали для определения значимости (**** = p<0,0001; *** = p<0,001; ** = p<0,01; *=p<0,05) на A-D. Для E-H, r, значения p и 95% доверительные интервалы вычисляли посредством линейной аппроксимации к логарифмически преобразованным данным. Для E-L включены данные, доступные на исходным уровне, через 6 месяцев и 12 месяцев. Затененные области соответствуют верхнему и нижнему пределу у контрольных индивидуумов. Снижение уровня Phe в плазме у большинства индивидуумов с ФКУ приводит к изменению других метаболитов нейротрансмиттеров в группе >900 мкМ (M-P) с большей тенденцией у индивидуумов с ФКУ в группе с <360 мкМ (Q-T) после лечения PALYNZIQ® (пегвалиаза-pqpz) в течение 12 месяцев. 5-гидроксииндолуксусная кислота (5HIAA); контроль (Ctrl); гомованилиновая кислота (HVA); 3-метокси-4-гидроксифенилгликоль (MOPEG); фенилацетилглицин (PAG); фенилаланин (Phe).
[0063] На фиг.24A-D показано, что подшкала ADHDi для подгруппы индивидуумов с показателями ADHDi ≥9 на исходном уровне обратно коррелировала с повышением MOPEG. Показана корреляция изменения подшкалы ADHDi и изменения HVA в плазме с показателями ADHDi >9 (n=17) (A) или <9 (n=7) (B) на исходном уровне, или MOPEG с ADHDi показателями >9 (C) или <9 (D) на исходном уровне; показатель >9 указывает на симптомы ослабленного внимания. Синими и красными точками указаны индивидуумы из групп <360 и >900 мкМ, соответственно. Величины r и p для каждого набора данных определяли после аппроксимации линейной регрессии к каждому набору данных. 5HIAA, 5-гидроксииндолуксусная кислота; ADHD-RS IV, домен подшкалы ослабления внимания оценочной шкалы синдрома дефицита внимания и гиперактивности; 3-метокси-4-гидроксифенилгликоль (MOPEG); фенилацетилглицин (PAG); фенилаланин (Phe); фенилкетонурия (ФКУ).
[0064] На фиг.25 представлен каскад биосинтеза нейротрансмиттеров. Метаболиты Phe и другие нейротрансмиттеры, происходящие из Tyr и Trp, можно определять в плазме в качестве суррогатных для метаболизма печени. Синим цветом указаны молекулы, образующиеся в плазме, в то время как красным указаны метаболиты, первичным источником происхождения которых является головной мозг или желудочно-кишечный тракт (см., например, Lambert et al., Life Sci., volume 57 pages255-67 (1995); Ruddell et al., J Hepatol., vol. 48 pages 666-675 (2008) (см. фиг.1 и легенду Ruddell)). 5-HIAA, 5-гидроксииндолуксусная кислота; AADC, декарбоксилаза аминокислот; ALDH2, альдегиддегидрогеназа; AR, альдегидредуктаза; COMT, катехо-O-метилтрансфераза; DBH, дофамин бета-гидроксилаза; DOPAC, 3,4-дигидроксифенилксусная кислота; DOPEG, 3,4-дигидроксифенилэтиленгликоль; HVA, гомованилиновая кислота; IAA, индолуксусная кислота; LAT1, большой переносчик 1 нейтральных аминокислот L-типа; MA, миндальная кислота; MAO, моноаминоксидаза; MOPEG, 3-метокси-4-гидроксифенилгликоль; PAA, фенилуксусная кислота; PAG, фенилацетилглицин; PAH, фенилаланингидроксилаза; PEA, фенилэтиламин; TH, тирозингидроксилаза; TPH, триптофангидроксилаза.
[0065] На фиг.26 представлено схематическое представление вектора для генной терапии ApoE-HCR-hAAT.GI.hPAH.bGH (также называемого вектором #1 или вектором WT-hPAH).
[0066] На фиг.27A-B представлено расщепление ферментами рестрикции вектора для генной терапии ApoE-HCR-hAAT.GI.hPAH.bGH, включая схему карты рестрикции (A), и анализ посредством гель-электрофореза расщепленного ферментами рестрикции вектора (B).
[0067] На фиг.28A-B представлен анализ посредством dd-ПЦР вектора для генной терапии ApoE-HCR-hAAT.GI.hPAH.bGH посредством демонстрации результатов анализа с использованием гель-электрофореза реакции dd-ПЦР (A), и схемы вектора для генной терапии с количественным определением амплифицированных областей ДНК (B).
[0068] На фиг.29A-B сравниваются уровни Phe в плазме, измеренные у мышей дикого типа и мышей Enu2, которым вводили носитель, 2E13 вг/кг или 2E14 вг/кг AAV-PAH (A) или 6E13 вг/кг AAV-PAH (B).
[0069] На фиг.30 показано, что у мышей ENU2, которым вводили AAV5-PAH, возрастает скорость повышения массы тела.
[0070] На фиг.31 показано, что цвет шерсти мышей ENU2 возвращается к цвету дикого типа при введении AAV5-PAH.
[0071] На фиг.32A-E показано, что высокая доза AAV-PAH нормализует уровни аминокислот Phe (A) и Tyr (B), и нейротрансмиттеров (C-D) у мышей ENU2 до уровней дикого типа. Также описан химический каскад (E).
[0072] На фиг.33 показано, что у мышей ENU2, которым вводили вектор ApoE-HCR-hAAT.GI.muPAH.bGH, снижались уровни фенилаланина на срок 60 недель.
[0073] На фиг.34 показано минимальное изменение уровней Phe, вызванное повышением уровня белка PAH у мышей дикого типа, которым вводили вектор ApoE-HCR-hAAT.GI.hPAH.bGH или вектор ApoE-HCR-hAAT.GI.muPAH.bGH.
[0074] На фиг.35 представлен вестерн-блоттинг, который демонстрирует увеличенные уровни белка PAH у мышей, которым проводили введение в соответствии с фиг.34, несмотря на минимальное изменение уровня Phe.
[0075] На фиг.36A-B представлено количественное определение окрашивания (A) и окрашивания (B), которое показывает отсутствие значительного окрашивания TUNEL в печени животных, которым вводили AAV5-hPAH в дозе 2E13 или 2E14 вг/кг через 12 недель после введения дозы.
[0076] На фиг.37 представлено схематическое представление вектора ApoE-HCR-hAAT.hPAH-tI2.bGH.
[0077] На фиг.38 представлен график, на котором показано, что уровни Phe в плазме снижались у мышей, которым вводили вектор ApoE-HCR-hAAT.hPAH-tI2.bGH, дозозависимым образом.
[0078] На фиг.39A-D представлены графики, на которых показано, что высокая доза вектора ApoE-HCR-hAAT.hPAH-tI2.bGH корректирует (A-C) уровни аминокислот и (D) уровни нейротрансмиттеров в головном мозге.
[0079] На фиг.40 представлена схема вектора ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH.
[0080] На фиг.41 представлено выравнивание последовательностей ДНК codop-hPAH.
[0081] На фиг.42A-C проанализированы последовательности codop-hPAH с картами, которые сравнивают процентную идентичность последовательностей (A), взаимосвязи согласно кладрограмме (B) и содержание GC (C).
[0082] На фиг.43A-B представлены конструкции вектора AAV5-PAH и структура ДНК.
[0083] На фиг.44A-B представлено сравнение кодон-оптимизированных версий hPAH посредством трансдукции или трансфекции клеток печени HEPG2.
[0084] На фиг.45A-B представлено сравнение кодон-оптимизированных версий hPAH посредством трансдукции AML12.
[0085] На фиг.46A-B показаны уровни Phe у мышей Enu2, которым вводили конструкции AAV-codop PAH и которые имеют различные концентрации фенилаланина в плазме через две недели (A) и на протяжении 6 недель (B).
[0086] На фиг.47 показана корреляция между увеличенным изменением массы тела и снижением процентного изменения Phe, измеренного у мышей ENU2, которым вводили конструкции AAV-codop PAH.
[0087] На фиг.48A-B приведено сравнение V1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH), V1+ интрон LG (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH), Geneius1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH) и вектора (ApoE-HCR-hAAT.cG-AIATI.hPAHco1.bGH), обозначаемого в настоящем описании как "вектор 2", посредством трансфекции HepG2.
[0088] На фиг.49A-C представлена кривая доза-эффект для AAV5-PAH, модифицированного Geneius-1 или Large Intron (A). Варианты AAV5-PAH вводили мышам ENU2 в различных дозах и определяли Phe в плазме через 2 недели (B) и 4 недели (C).
[0089] На фиг.50A-C представлен эффект дозы (A) и распределение PAH (B, C) для конструкций GENEius-1 и Large Intron AAV-PAH.
[0090] На фиг.51A-B представлены эффекты дозы для вариантов AAV5-PAH у самцов и самок мышей ENU2 в течение 5 недель (A). Эффект дозы вариантов AAV5-PAH, которым вводили мышам ENU2 в момент времени 2 недели, исследовали для более определенной идентификации различий в эффективности введения различных доз вектора 2 (B).
[0091] На фиг.52 представлен график уровней фенилаланина у мышей ENU2 при введении возрастающих концентраций вектора 2.
[0092] На фиг.53 приводится сравнение уровней Phe у самцов мышей ENU2 через 2 недели, которым вводили 2e13 вг/кг векторов-кандидатов.
[0093] На фиг.54A-B представлены уровни ALT в плазме у мышей ENU2 в предварительных образцах крови (A) или через 2 недели после введения каждой из конструкций вектора 2, Codop и LgIntron (B).
[0094] На фиг.55 представлено графическое количественное определение окрашивания TUNEL и окрашивания ткани печени у мышей ENU2, которым вводили конструкции AAV-PAH Codop или LgIntron.
[0095] На фиг.56A-B графически представлено количественное определение окрашивания IBA (A) и окрашивания IBA (B) ткани печени у мышей ENU2, которым вводили конструкции AAV-PAH Codop или LgIntron, которое демонстрирует отсутствие статистически значимых отличий в количестве очагов IBA-1(+).
[0096] На фиг.57A-C представлено окрашивание посредством H&E (гематоксилин и эозин) ткани печени у мышей ENU2, которым вводили носитель (A), конструкцию AAV-PAH Codop (B), или конструкцию AAV-PAH LgIntron (C) по сравнению с нормальной тканью мыши дикого типа.
[0097] На фиг.58A-C показано, то уровень phe в плазме мышей ENU2, которым вводили AAV5-muPAH, восстанавливается до уровней WT (A), в то время как уровни ALT остаются теми же между группами (B). У мышей ENU2, которым вводили AAV5-muPAH, изменялся цвет шерсти на темно-коричневый, аналогично WT (C).
[0098] На фиг.59A-B показано, что уровни аминокислот (A) и нейротрансмиттеров (B) в головном мозге восстанавливались до уровней WT у мышей ENU2, которым вводили AAV5-muPAH.
[0099] На фиг.60A-B показано, что у мышей дикого типа, которым вводили AAV5-hPAH, сохранялись нормальные уровни Phe (A) на протяжении исследования, в то время как экспрессия белка PAH возрастала в два раза по сравнению с мышами WT, которым вводили носитель (B).
[00100] На фиг.61A-C представлена ДНК hPAH (A) и белок (B) в окрашенных гепатоцитах у мышей ENU2, которым вводили AAV-hPAH, и количественное определение (C).
[00101] На фиг.62A-C представлена оценка маркеров воспаления после введения AAV5-hPAH мышам WT и ENU2. Количества макрофагов (A) IBA1 и (B) CD68 (активированные M1/M2) измеряли в тканях печени мышей. iNOS (активированные M1), провоспалительный маркер макрофагов продемонстрировало отсутствие повышения между группами (C). Графическое количественное определение окрашивания (D).
[00102] На фиг.63 представлено окрашивание TUNEL ткани печени мышей ENU2 или WT, которым вводили AAV-hPAH. Окрашивание демонстрирует отсутствие значительного повышения апоптотической гибели клеток в группах, в которых проводили генную терапию, по сравнению с контрольными мышами.
[00103] На фиг.64 представлено биораспределение вектора 2, доставляемого при двух различных уровнях дозы яванским макакам посредством в/в инъекции. Результаты представлены в качестве процента геномной ДНК вектора, экстрагированной из печени.
[00104] На фиг.65A-B представлены уровни фенилаланина (A) и тирозина (B) у яванских макаков, которым вводили либо высокую дозу, либо низкую дозу, вектора 2 в течение 30 суток.
[00105] На фиг.66A-C показано, что введение мышам ENU2 вектора 2 приводило к коррекции уровней аминокислот в гомогенатах головного мозга.
[00106] На фиг.67A-D показано, что введение мышам ENU2 вектора 2 приводило к коррекции уровней нейротрансмиттеров в гомогенатах головного мозга.
[00107] На фиг.68A-C показана коррекция метаболитов нейротрансмиттеров, наблюдаемая при использовании высокой дозы вектора 2.
5. ПОДРОБНОЕ ОПИСАНИЕ
[00108] В одном варианте осуществления изобретение относится к применению уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуумов, имеющих фенилкетонурию (ФКУ), для оптимизации эффективной дозы терапевтического средства против ФКУ. В другом варианте осуществления изобретение относится к способам лечения индивидуумов, имеющих ФКУ, включающим введение эффективного количества терапевтического средства против ФКУ, где эффективное количество представляет собой количество, которое нормализует уровни аминокислот, метаболитов нейротрансмиттеров и нейротрансмиттеров у индивидуумов.
[00109] В одном варианте осуществления изобретение относится к векторам AAV, кодирующим функционально активные терапевтические белки (например, полностью упакованные векторы AAV PAH, векторы AAV PAH, и выделенные молекулы нуклеиновых кислот, содержащие нуклеотидную последовательность, обладающую значительной гомологией с нуклеотидной последовательностью SEQ ID NO: 1). В другом варианте осуществления рекомбинантные векторы на основе AAV, кодирующие терапевтические белки, имеют увеличенную экспрессию трансгена, а также увеличенный выход продуцирования вируса AAV и упрощенную очистку. В некоторых вариантах осуществления внесение одного или более интронов в кодирующую терапевтический белок область, оптимизация кодонов и/или перекомпоновка количества и положения энхансеров повышают экспрессию.
[00110] Если не указано иное, технические и научные термины, используемые в настоящем описании, имеют то же значение, которое обычно подразумевает специалист в области, к которой относится настоящее изобретение. См., например, Singleton et al., Dictionary of Microbiology and Molecular Biology 2nd ed., J. Wiley & Sons (New York, N.Y. 1994); Sambrook et al., Molecular Cloning, A Laboratory Manual, Cold Springs Harbor Press (Cold Springs Harbor, N.Y. 1989). Для целей настоящего изобретения ниже определены следующие термины.
[00111] В рамках изобретения, в контексте доставки генов термин "вектор" или "вектор для доставки генов" может относиться к частице, которая выполняет функцию носителя для доставки генов и которая содержит нуклеиновую кислоту (т.е. геном вектора), упакованную, например, в оболочку или капсид. Вектор для доставки генов может представлять собой вирусный ген для доставки генов или невирусный ген для доставки генов. Альтернативно в некоторых контекстах термин "вектор" можно использовать только для обозначения генома вектора. Вирусные векторы, пригодные для применения в рамках настоящего изобретения, могут представлять собой векторы на основе парвовируса, аденовируса, ретровируса, лентивируса или вируса простого герпеса. Парвовирус может представлять собой аденовирус-ассоциированный вирус (AAV).
[00112] В рамках изобретения, "вектор на основе AAV" относится к нуклеиновым кислотам, как одноцепочечным, так и двухцепочечным, имеющим последовательность 5 '-инвертированного концевого повтора (ITR) AAV и 3 '-ITR AAV, фланкирующую кодирующую белок последовательность (в одном варианте осуществления последовательность, кодирующую функциональный терапевтический белок, например, PAH), функционально связанную с элементами регуляции транскрипции, которые являются гетерологичными для генома вируса AAV, т.е. одним или нескольким промоторами и/или энхансерами и необязательно последовательностью полиаденилирования, и/или одним или более интронами, встроенными между экзонами кодирующей белок последовательности. Одноцепочечный вектор на основе AAV относится к нуклеиновым кислотам, которые присутствуют в геноме частицы вируса AAV и могут представлять собой любо смысловую цепь, либо антисмысловую цепь последовательностей нуклеиновых кислот, описанных в настоящем описании. Размер таких одноцепочечных нуклеиновых кислот указывают в количестве оснований. Двухцепочечный вектор AAV относится к нуклеиновым кислотам, которые присутствуют в ДНК плазмид, например, pUC19, или в геном двухцепочечного вируса, например, бакуловируса, используемого для экспрессии или переноса нуклеиновых кислот вектора AAV. Размер таких двухцепочечных нуклеиновых кислот указывают в количестве пар оснований (п.о.).
[00113] Термин "инвертированный концевой повтор (ITR)", в рамках изобретения, относится к известным в данной области областям, встречающимся на 5 '- и 3 '-концах генома AAV, которые функционируют в цис-формате в качестве ориджинов репликации ДНК и в качестве сигналов для упаковывания вирусного генома. ITR AAV, вместе с кодирующей областью rep AAV, обеспечивает эффективное вырезание и высвобождение, и встраивание нуклеотидной последовательности, находящейся между двумя фланкирующими ITR в геном клетки-хозяина. Последовательности определенных ассоциированных с AAV ITR описаны Yan et al., J. Virol. (2005) vol. 79, pp. 364-379, которая включена в настоящее описание в качестве ссылки в полном объеме. Последовательности ITR, которые применимы в рамках настоящего изобретения, могут представлять собой полноразмерные ITR AAV дикого типа или их фрагменты, которые сохраняют функциональную способность, или могут представлять собой варианты последовательности полноразмерных ITR AAV дикого типа, которые способны функционировать в цис-формате и в качестве ориджинов репликации. ITR AAV, пригодные в рекомбинантных векторах PAH AAV согласно вариантам осуществления, описанным в настоящем описании, могут происходить из любого известного серотипа AAV, и в некоторых вариантах осуществления происходят из серотипа AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 или AAV9.
[00114] Термин "последовательности контроля" относится к последовательностям ДНК, необходимым для экспрессии функционально связанной кодирующей последовательности в конкретном организме-хозяине. Последовательности контроля, пригодные, например, для прокариот, включают промотор, необязательно последовательность оператора и участок связывания рибосомы. Известно, что эукариотические клетки используют промоторы, сигналы полиаденилирования и энхансеры.
[00115] "Элемент регуляции транскрипции" относится к нуклеотидным последовательностям гена, вовлеченным в регуляцию генетической транскрипции, включающим промотор, положительные элементы ответа, последовательности активаторов и энхансеров для связывания факторов транскрипции для способствования связыванию РНК-полимеразы и стимуляции экспрессии, или последовательности операторов или сайленсеров, с которым связываются белки-репрессоры для блокирования связывания РНК-полимеразы и предотвращения экспрессии. Термин "специфический для печени элемент регуляции транскрипции" относится к регуляторному элементу, который специфически модулирует экспрессию генов в ткани печени. Примеры специфических для печени элементов регуляции транскрипции включают, но не ограничиваются ими, промотор транстиретина мыши (mTTR), эндогенный промотор фактора VIII человека (F8), промотор альфа-1-антитрипсина человека (hAAT) и его активные фрагменты, минимальный промотор альбумина человека и промотор альбумина мыши. Также предусматриваются энхансеры, происходящие из специфических для печени участков связывания факторов транскрипции, такие как EBP, DBP, HNF1, HNF3, HNF4, HNF6 и Enh1.
[00116] В рамках изобретения, термин "функционально связанный" используют для описания связи между регуляторными элементами и геном или его кодирующей областью. Как правило, экспрессию гена помещают под контроль одного или более регуляторных элементов, например, но не ограничиваясь ими, конститутивных или индуцибельных промоторов, тканеспецифических регуляторных элементов и энхансеров. Ген или кодирующую область называют "функционально связанными" или "эффективно связанными с" или "функционально ассоциированными с" регуляторными элементами, что означает, что этот ген или кодирующая область контролируется или находится под влиянием регуляторного элемента. Например, промотор функционально связан с кодирующей последовательностью, если промотор влияет на транскрипцию или экспрессию кодирующей последовательности.
[00117] В одном варианте осуществления вектор AAV содержит нуклеиновую кислоту, кодирующую функционально активный белок фенилаланингидроксилазы (PAH). Кодирующая PAH последовательность может представлять собой последовательность дикого типа, кодон-оптимизированную последовательность или вариант (см., например, Fang et al., Gene Ther., vol. 1, pages 247-254 (1994); Eisensmith et al., J. Inherit. Metab. Dis., vol. 19, pages 412-423 (1996); Nagasaki et al., Pediatr. Res., vol. 45, pages 465-473 (1999); и Laipis et al., Mol. Ther., vol. 7, pages S391-S392 (2003)).
[00118] В рамках изобретения, PAH дикого типа имеет следующую последовательность нуклеиновой кислоты:
[00119] ATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAA (SEQ ID NO:1)
[00120] В рамках изобретения, PAH дикого типа имеет следующую аминокислотную последовательность:
[00121] MSTAVLENPG LGRKLSDFGQ ETSYIEDNCN QNGAISLIFS LKEEVGALAK VLRLFEENDV NLTHIESRPS RLKKDEYEFF THLDKRSLPA LTNIIKILRH DIGATVHELS RDKKKDTVPW FPRTIQELDR FANQILSYGA ELDADHPGFK DPVYRARRKQ FADIAYNYRH GQPIPRVEYM EEEKKTWGTV FKTLKSLYKT HACYEYNHIF PLLEKYCGFH EDNIPQLEDV SQFLQTCTGF RLRPVAGLLS SRDFLGGLAF RVFHCTQYIR HGSKPMYTPE PDICHELLGH VPLFSDRSFA QFSQEIGLAS LGAPDEYIEK LATIYWFTVE FGLCKQGDSI KAYGAGLLSS FGELQYCLSE KPKLLPLELE KTAIQNYTVT EFQPLYYVAE SFNDAKEKVR NFAATIPRPF SVRYDPYTQR IEVLDNTQQL KILADSINSE IGILCSALQK IK (SEQ ID NO:2).
[00122] Согласно первому аспекту изобретения предусматривается вектор AAV, содержащий нуклеотидную последовательность, обладающую значительной гомологией с нуклеотидной последовательностью SEQ ID NO: 1 и кодирующую функциональный PAH. Термин "значительная гомология" может быть дополнительно определен с использованием термина "процентная (%) гомология". Он подробно обсуждается в настоящем описании далее.
[00123] Термин "выделенный", когда его используют в отношении молекулы нуклеиновой кислоты по настоящему изобретению, как правило, относится к последовательности нуклеиновой кислоты, которая идентифицирована и отделена от по меньшей мере одной посторонней нуклеиновой кислоты, с которой она обычно ассоциирована в ее природном источнике. Выделенная нуклеиновая кислота может присутствовать в форме или в условиях, которые отличаются от формы и условий, в которых она встречается в природе. Таким образом, выделенные молекулы нуклеиновых кислот отличаются от молекул нуклеиновых кислот, существующих в клетках в природе.
[00124] В рамках изобретения, термин "вариант" относится к полинуклеотиду (или полипептиду), имеющему последовательность, по существу сходную с эталонным полинуклеотидом (или полипептидом). Методики внесения нуклеотидных и аминокислотных замен в полинуклеотид, белок или полипептид известны специалисту в данной области (см., например, Sambrook et al. (1989)). В случае полинуклеотида вариант может иметь делеции, замены, вставки одного или более нуклеотидов на 5′-конце, 3′-конце и/или одном или нескольких или внутренних участках по сравнению с эталонным полинуклеотидом. Сходства и/или различия последовательностей между вариантом и эталонным полинуклеотидом могут быть выявлены с использованием общепринятых способов, известных в данной области, например, способов полимеразной цепной реакции (ПЦР) и гибридизации. Варианты полинуклеотидов также включают полинуклеотиды синтетического происхождения, такие как полинуклеотиды, полученные, например, с использованием сайт-направленного мутагенеза. Как правило, вариант полинуклеотида, включая, но не ограничиваясь ими, ДНК, может обладать по меньшей мере приблизительно 50%, приблизительно 55%, приблизительно 60%, приблизительно 65%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или более идентичностью последовательностью с эталонным полинуклеотидом, как определяют посредством программ для выравнивания последовательностей, известных специалистам в данной области. В случае полипептида вариант может иметь делеции, замены, вставки одной или более аминокислот по сравнению с эталонным полипептидом. Сходства и/или различия последовательностей между вариантом и эталонным полипептидом можно выявлять с использованием общепринятых способов, известных в данной области, например, вестерн-блоттинга. Как правило, вариант полипептида может обладать по меньшей мере приблизительно 60%, приблизительно 65%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или более идентичностью последовательности с эталонным полипептидом при определении с использованием программ для выравнивания последовательностей, известных специалистам в данной области.
[00125] Термины "идентичность", "гомология" и их грамматические варианты означают, что две или более из упоминаемых структур являются одинаковыми, когда они являются "выровненными" последовательностями. Таким образом, в качестве примера, когда две полипептидных последовательности идентичны, они имеют одинаковую аминокислотную последовательность, по меньшей мере в пределах упоминаемой области или части. Когда две полинуклеотидных последовательности идентичны, они имеют одинаковую полинуклеотидную последовательность, по меньшей мере в пределах упоминаемой области или части. Идентичность может представлять собой идентичность на протяжении определенного участка (области или домена) последовательности. "Участок" или "область" идентичности относится к частям двух или более упоминаемых структур, которые являются одинаковыми. Таким образом, когда две белковых последовательности или последовательности нуклеиновых кислот являются идентичности на протяжении оного или нескольких участков или областей, они обладают идентичностью в этой области. "Выровненные" последовательности относятся к нескольким полинуклеотидным или белковым (аминокислотным) последовательностям, часто содержащим поправки на отсутствующие или дополнительные основания или аминокислоты (пропуски) по сравнению с эталонной последовательностью. "Значительная гомология" означает, что молекула является структурно или функционально консервативной, так что она имеет или спрогнозировано, что она имеет по меньшей мере частичную структуру или функцию одной или более из структур или функций (например, биологическая функция или активность) эталонной молекулы, или характерной/соответствующей области или части эталонной молекулы, с которой она обладает гомологией.
[00126] "Процентную идентичность (%) последовательностей нуклеиновых кислот" определяют как процент нуклеотидов в последовательности-кандидате, которые являются идентичными с эталонной последовательностью после выравнивания соответствующих последовательностей и внесения пропусков, при необходимости, для достижения максимальной процентной идентичности последовательностей. Выравнивание для целей определения процентной идентичности последовательностей нуклеиновых кислот можно проводить различными способами, известными в данной области, например, с использованием общедоступного компьютерного программного обеспечения, такого как программное обеспечение ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания на протяжении полной длины сравниваемых последовательностей.
[00127] "Процентную (%) идентичность аминокислотных последовательностей" в отношении аминокислотных последовательностей PAH, описанных в настоящем описании, определяют в качестве процента аминокислотных остатков в последовательности-кандидате, которые идентичны аминокислотным остаткам в последовательности полипептида PAH после выравнивания последовательностей и внесения пропусков, если необходимо, для достижения максимальной процентной идентичности последовательностей, и не считая какие-либо консервативные замены частью идентичности последовательностей. Выравнивание для целей определения процентной идентичности аминокислотных последовательностей можно проводить различными способами, известными в данной области, например, с использованием общедоступного программного обеспечения, такого как программное обеспечение ALIGN или Megalign (DNASTAR). Специалисты в данной области могут определить соответствующие параметры для проведения выравнивания, включая любые алгоритмы, необходимые для достижения максимального выравнивания на протяжении всей длины сравниваемых последовательностей.
[00128] "Оптимизация кодонов" или "кодон-оптимизированный" относятся к изменениям, внесенным в нуклеотидную последовательность, так чтобы она с большей вероятностью экспрессировалась на относительно высоком уровне по сравнению с не кодон-оптимизированной последовательностью. Она не изменяет аминокислоты, которые кодирует каждый из кодонов.
[00129] В рамках изобретения, "интрон" в широком значении определяют как последовательность нуклеотидов, которая может быть удалена посредством сплайсинга РНК. "РНК-сплайсинг" означает вырезание интронов из пре-мРНК с образованием зрелой мРНК. Вставку интрона в экспрессированную последовательность можно проводить любым способом, известным в данной области. Единственным ограничением по встраиванию интрона являются ограничения упаковывания вирусных частиц AAV (приблизительно 5 т.п.н.).
[00130] "Вирион AAV", или "вирусная частица AAV", или "векторная частица AAV", или "вирус AAV" относятся к вирусной частице, состоящей по меньшей мере из одного капсидного белка AAV и инкапсидированного полинуклеотидного вектора AAV, как описано в настоящем описании. Если частица содержит гетерологичный полинуклеотид (т.е. полинуклеотид, отличный от генома AAV дикого типа, такой как трансген, подлежащий доставке в клетку млекопитающего), его, как правило, называют "векторной частицей AAV" или просто "вектором AAV". Таким образом, продуцирование векторных частиц AAV обязательно включает продуцирование вектора AAV, поскольку такой вектор находится в векторной частице AAV.
[00131] В рамках изобретения, "терапевтический вирус AAV" относится к вириону AAV, вирусной частице AAV, векторной частице AAV, или вирусу AAV, которые содержат гетерологичный полинуклеотид, который кодирует терапевтический белок. "Вектор на основе AAV", в рамках изобретения, относится к вектору, содержащему один или более представляющих интерес полинуклеотидов (или трансгенов), которые фланкируются последовательностями концевых повторов (ITR) AAV и функционально связаны с одним или более элементами контроля экспрессии. Такие векторы на основе AAV могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они присутствуют в клетке-хозяине, трансфицированной вектором, кодирующим и экспрессирующим продукты генов rep и cap.
[00132] В рамках изобретения "терапевтический белок" относится к полипептиду, который обладает биологической активностью, которая заменяет или компенсирует утрату или снижение активности эндогенного белка. Например, функциональная фенилаланингидроксилаза (PAH) является терапевтическим белком против фенилкетонурии (ФКУ).
[00133] "Нейротрансмиттер", в рамках изобретения, относится к химическому соединению, высвобождающемуся из нервной клетки, которое, таким образом, передает импульс от нервной клетки к другому нерву, мышце, органу или другой ткани. Нейротрансмиттер является передатчиком неврологической информации от одной клетки к другой. В некоторых вариантах осуществления нейротрансмиттеры включают фенэтиламин, тирамин, дофамин, норадреналин, адреналин, триптамин и серотонин. "Метаболит нейротрансмиттера", в рамках изобретения, относится к продуктам после деградации нейротрансмиттеров после одной или двух ферментативных стадий. Неограничивающие примеры метаболитов нейротрансмиттеров включают фенилуксусную кислоту, фенилацетилглицин, фенилацетилглутамин, DOPAC, гомованилиновую кислоту, дигидроксифенилэтиленгликоль (DOPEG), 3-метокси-4-гидроксифенилгликоль (MHPG, MOPEG), индолуксусную кислоту и 5-гидроксииндолуксусную кислоту. Неограничивающие примеры нейротрансмиттеров и метаболитов также представлены на фиг.24.
[00134] "Фенилкетонурия (ФКУ)", в рамках изобретения, относится к наследственному метаболическому заболеванию, вызываемому дефицитом фермента фенилаланингидроксилазы (PAH). Этот приводит к повышенным уровням фенилаланина (Phe) и сниженным уровням нейротрансмиттеров и метаболитов нейротрансмиттеров, которые могут влиять на функцию головного мозга, вызывая тяжелую умственную отсталость, поведенческие расстройства, замедленную речь и судорожные припадки.
[00135] "Лечить" или "лечение", в рамках изобретения, относится к терапевтическому лечению, которое относится к лечению, проводимому у индивидуума, у которого присутствуют признак или симптомы патологии, т.е. ФКУ, для цели уменьшения или устранения этих признаков или симптомов. Эти признаки или симптомы могут быть биохимическими, клеточными, гистологическими, функциональными, субъективными или объективными. "Лечить" или "лечение" относится к снижению или облегчению прогрессирования, тяжести и/или длительности заболевания (или симптома, связанного с ним), ассоциированного с повышенными уровнями фенилаланина (например, ФКУ).
[00136] "Смягчать", в рамках изобретения, относится к действию, уменьшающему тяжесть симптомов, прогрессирование или длительность заболевания.
[00137] В рамках изобретения, "стабильное лечащий" или "стабильное лечение" относится к использованию терапевтического вируса AAV, вводимого индивидууму, где у индивидуума стабильно экспрессируется терапевтический белок, экспрессируемый терапевтическим вирусом AAV. Стабильно экспрессируемый терапевтический белок означает, что белок экспрессируется на протяжении клинически значимого периода времени. "Клинически значимый период времени", в рамках изобретения, означает экспрессию на терапевтически эффективных уровнях в течение периода времени, при котором происходит значительное влияние на качество жизни индивидуума. В некоторых вариантах осуществления значительное влияние на качество жизни демонстрируется отсутствием необходимости во введении альтернативных терапевтических средств внутривенно или подкожно. В некоторых вариантах осуществления клинически значимый период времени представляет собой выражение, означающее по меньшей мере шесть месяцев, по меньшей мере восемь месяцев, по меньшей мере один год, по меньшей мере два года, по меньшей мере три года, по меньшей мере четыре года, по меньшей мере пять лет, по меньшей мере шесть лет, по меньшей мере семь лет, по меньшей мере восемь лет, по меньшей мере девять лет, по меньшей мере десять лет, или всю жизнь индивидуума.
[00138] В рамках изобретения, термин "эффективное количество" относится к количеству, достаточному для обеспечения благоприятных или желательных биологических и/или клинических результатов.
[00139] В рамках изобретения, "индивидуум" относится к животному, которое является объектом лечения, наблюдения или эксперимента. "Животное" включает хладнокровных и теплокровных позвоночных и беспозвоночных, таких как рыбы, моллюски, пресмыкающиеся и, в частности, млекопитающие. "Млекопитающее", в рамках изобретения, относится к индивидууму, принадлежащему классу Mammalia, и включает, но не ограничивается ими, людей, домашних и сельскохозяйственных животных, животных зоопарков, спортивных животных и домашних питомцев. Неограничивающие примеры млекопитающих включают мышей; крыс; кроликов; морских свинок; собак; кошек; овец; коз; коров; лошадей; приматов, таких как макаки, шимпанзе и человекообразные обезьяны, и, в частности, людей. В некоторых вариантах осуществления млекопитающим является человек. Однако в некоторых вариантах осуществления млекопитающим не является человек.
[00140] Как правило, "фармацевтически приемлемый носитель" представляет собой носитель, который не является токсичным или чрезмерно вредоносным для клеток. Иллюстративные фармацевтически приемлемые носители включают стерильную не содержащую пирогенов воду и стерильный не содержащий пирогенов фосфатно-солевой буфер. Фармацевтически приемлемые носители включают физиологически приемлемые носители. Термин "фармацевтически приемлемый носитель" включает любые и все растворители, диспергенты, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие всасывание средства и т.п., которые являются физиологически совместимыми".
[00141] Под "фармацевтически приемлемым" подразумевают материал, который не является биологически или иным образом нежелательным, т.е. материал может быть введен индивидууму без возникновения каких-либо нежелательных биологических эффектов. Таким образом, такую фармацевтическую композицию можно использовать, например, для трансфекции клеток ex vivo или введения вирусной частицы или клетки непосредственно индивидууму.
[00142] Носитель может быть пригоден для парентерального введения, которое включает внутривенное, внутрибрюшинное или внутримышечное введение. Альтернативно носитель может быть пригоден для сублингвального или перорального введения. Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для приготовления стерильных инъекционных растворов или дисперсий непосредственно перед введением. Применение таких сред и средств для фармацевтически активных веществ хорошо известно в данной области. За исключением случаев, когда какая-либо общепринятая среда или средство несовместимы с активным соединением, предусматривается их применение в фармацевтических композициях в рамках настоящего изобретения.
[00143] "Нейротипический" или "нейротипический индивидуум", в рамках изобретения, относится к индивидууму или группе индивидуумов, которые не страдают от и не имеют симптомов каких-либо неврологических или нейрокогнитивных заболеваний или состояний. В некоторых вариантах осуществления нейротипические индивидуумы имеют уровни нейротрансмиттеров и метаболитов нейротрансмиттеров, которые находятся в нормально диапазоне для здорового человека или в нормальном диапазоне при определении из средних уровней для группы здоровых людей. При использовании в отношении животных, таких как мыши, уровни нейротрансмиттеров и метаболитов нейротрансмиттера у нейротипических индивидуумов представляют собой уровни, измеренные у контрольных животных дикого типа.
[00144] KUVAN® (сапроптерин дигидрохлорид) представляет собой первое и единственное одобренное FDA лекарственное средство против ФКУ для снижения уровней Phe в крови у пациентов с гиперфенилаланинемией (HPA) вследствие отвечающей на тетрагидробиоптерин (BH4-) ФКУ. KUVAN® представляет собой фармацевтический состав BH4, натуральный кофактор для фермента PAH, который стимулирует активность остаточного фермента PAH в отношении метаболизма Phe в тирозин. KUVAN® описан в патентах США № 7732599, 8003126, 7566462 и 8178670, все из которых включены в настоящее описание в качестве ссылок в полном объеме.
[00145] PALYNZIQ® (пегвалиаза-pqpz) (пегилированная рекомбинантная фенилаланинаммиаклиаза или "PEG-PAL") представляет собой экспериментальное средство фермент-заместительной терапии для лечения фенилкетонурии (ФКУ). PALYNZIQ® (пегвалиаза-pqpz) описана в патентах США № 7531341, 7534595, 7537923, 7790433, 7560263, 955734, все из которых включены в настоящее описание в качестве ссылок в полном объеме.
[00146] "Терапия ФКУ", в рамках изобретения, относится к любому терапевтическому вмешательству у индивидуума, имеющего ФКУ, которое смягчает симптомы ФКУ или снижает уровни фенилаланина в сыворотке. "Генная терапия ФКУ", в рамках изобретения, относится к любому терапевтическому вмешательству у индивидуума, имеющего ФКУ, которое вовлекает замещение или восстановление активности фенилаланингидроксилазы посредством доставки одной или более молекул нуклеиновых кислот в клетки индивидуума, которые замещают или восстанавливают экспрессию биологически активного фермента фенилаланингидроксилазы. В некоторых вариантах осуществления генная терапия ФКУ относится к генной терапии, вовлекающей вектор на основе аденоассоциированного вируса (AAV), который экспрессирует фенилаланингидроксилазу человека.
[00147] Термин "нейрокогнитивные симптомы", в рамках изобретения, относится к специфическим неврологическим, поведенческим и когнитивным симптомам, ассоциированным с индивидуумами, имеющими фенилкетонурию. В частности, утрата активности фенилаланингидроксилазы приводит к неспособности индивидуумов, имеющих фенилкетонурию, продуцировать достаточные уровни нейротрансмиттеров. Неспособность продуцировать достаточные уровни нейротрансмиттеров прямо приводит к ряду неврологических, когнитивных и поведенческих симптомов. В одном варианте осуществления нейрокогнитивные симптомы включают снижение IQ, дефицит внимания и нарушение способности к целенаправленной деятельности, включая стратегическое планирование, тормозной контроль, кратковременную память и когнитивную гибкость.
[00148] Мыши Enu2 описаны Shedlovsky A, et al. (Mouse models of human phenylketonuria, (1993), Genetics, vol. 134, pages 1205-1210). Эти мыши имеют миссенс-мутацию T835C в экзоне 7 их гена фенилаланингидроксилазы, которая приводит к замене фенилаланина на серин в положении аминокислоты 263 (F263S) фермента. Гомозиготные мыши с мутацией Enu2 имеют тяжелую гиперфенилфенилаланинемию. Они являются гипопигментированными, если только их не поддерживать на рационе с низким содержанием фенилаланина. Самки являются фертильными, но не выкармливают своих детенышей, когда их поддерживают на стандартном рационе для мышей. Цвет шерсти исходного штамма, BTBR +Ttf/tf, является черным и рыжевато-коричневым (at/at). Этот штамм также является гомозиготным по гену tufted (tf/tf), что приводит к различным паттернам линьки шерсти мыши. Эти эффекты, ограниченные шерстью мыши, могут приводить к тому, что мыши выглядят неправильно сформированными.
5.1 ВЕКТОРЫ AAV
[00149] В рамках изобретения, термин "AAV" является стандартным сокращенным обозначением для аденоассоциированного вируса. Аденоассоциированный вирус представляет собой одноцепочечный ДНК-парвовирус который растет только в клетках, которым предоставляются определенные функции путем совместной инфекции вирусом-помощником. В настоящее время существует тринадцать охарактеризованных серотипов AAV. Общая информация и обзор AAV могут быть найдены, например, в Carter, 1989, Handbook of Parvoviruses, Vol. 1, pp. 169-228; и Berns, 1990, Virology, pp. 1743-1764, Raven Press, (New York). Однако является полностью ожидаемым, что эти же принципы будут применимы к дополнительным серотипам AAV, поскольку хорошо известно, что различные серотипы являются довольно близко родственными, как структурно, так и функционально, даже не генетическом уровне (см., например, Blacklowe, 1988, pp. 165-174 Parvoviruses and Human Disease, J. R. Pattison, ed.; и Rose, Comprehensive Virology 3:1-61 (1974)). Например, все серотипы AAV, по-видимому, демонстрируют в высокой степени сходные свойства репликации, опосредуемые гомологичными генами rep; и все содержат три родственных капсидных белка. Степень родственности далее устанавливаются анализом гетеродуплексов, который выявляет обширную перекрестную гибридизацию между серотипами по длине генома; и присутствие аутологичных самогибридизующихся сегментов на концах, которые соответствуют "последовательностям инвертированных концевых повторов" (ITR). Сходные паттерны инфекционности также указывают на то, что репликационные функции каждого серотипа находятся под сходным регуляторным контролем.
[00150] В одном варианте осуществления векторы на основе AAV могут реплицироваться и упаковываться в инфекционные вирусные частицы, когда они присутствуют в клетке-хозяине, трансфицированной вектором, кодирующим и экспрессирующим продукты генов rep и cap.
[00151] В некоторых вариантах осуществления вирусный вектор содержит 5′-инвертированный концевой повтор (ITR) AAV и 3′-ITR AAV, промотор, полинуклеотид, кодирующий PAH, и посттранскрипционный регуляторный элемент, где промотор, полинуклеотид, кодирующий PAH, и посттранскрипционный регуляторный элемент находятся ниже 5′-ITR AAV и выше 3′-ITR AAV. Вирусный вектор можно использовать, например, для достижения высоких уровней PAH у индивидуума для терапевтических целей.
[00152] В некоторых вариантах осуществления рекомбинантный вектор AAV содержит нуклеиновую кислоту, содержащую 5 '-инвертированный концевой повтор (ITR) AAV2 (который может быть модифицированным или может не быть модифицированным, как известно в данной области), специфическую для печени область регуляции транскрипции, кодон-оптимизированную кодирующую область терапевтического белка, необязательно один или более интронов, последовательность полиаденилирования, и 3 '-ITR AAV2 (который может быть модифицированным или может не быть модифицированным, как известно в данной области). В некоторых вариантах осуществления терапевтический белок представляет собой PAH человека или его варианты. В других вариантах осуществления специфическая для печени область регуляции транскрипции содержит укороченную последовательность энхансера ApoE; проксимальный промотор альфа-антитрипсина человека (hAAT) из 186 оснований, включающий 42 оснований 5 '-нетранслируемой области (UTR); один или более энхансеров, выбранных из группы, состоящей из (i) энхансера ApoE/C1 человека из 34 оснований, (ii) дистальной области X промотора AAT человека из 32 оснований, и (iii) 80 дополнительных оснований дистального элемента проксимального промотора AAT человека; и нуклеиновую кислоту, кодирующую PAH человека. В другом варианте осуществления специфическая для печени область регуляции транскрипции содержит энхансерную последовательность α-микроглобулина и проксимальный промотор альфа-антитрипсина человека из 186 оснований (AAT).
[00153] Другие варианты осуществления, описанные в настоящем описании, относятся к векторным конструкциям, кодирующим функциональный полипептид PAH, где конструкции содержат один или более из индивидуальных элементов описанных выше конструкций и их комбинации, в одной или более различных ориентации(ях). Другой вариант осуществления, описанный в настоящем описании, относится к конструкциям в противоположной ориентации. В другом варианте осуществления предусматриваются рекомбинантные вирусные частицы AAV, содержащие описанные в настоящем описании векторы PAH AAV, и их применение для лечения ФКУ у индивидуумов. В одном варианте осуществления индивидуумами являются малолетние индивидуумы.
[00154] Векторы AAV, описанные в настоящем описании, в форме единичной цепи имеют длину менее чем приблизительно 7,0 т.п.н., или имеют длину менее 6,5 т.п.н., или имеют длину менее 6,4 т.п.н., или имеют длину менее 6,3 т.п.н., или имеют длину менее 6,2 т.п.н., или имеют длину менее 6,0 т.п.н., или имеют длину менее 5,8 т.п.н., или имеют длину менее 5,6 т.п.н, или имеют длину менее 5,5 т.п.н., или имеют длину менее 5,4 т.п.н., или имеют длину менее 5,3 т.п.н., или имеют длину менее 5,2 т.п.н. или имеют длину менее 5,0 т.п.н. Векторы AAV, описанные в настоящем описании, в форме единичной цепи имеют длину в диапазоне от приблизительно 5,0 т.п.н. до приблизительно 6,5 т.п.н., или в диапазоне от приблизительно 4,8 т.п.н. до приблизительно 5,2 т.п.н., или от 4,8 т.п.н. до 5,3 т.п.н., или в диапазоне от приблизительно 4,9 т.п.н. до приблизительно 5,5 т.п.н., или от приблизительно 4,8 т.п.н. до приблизительно 6,0 т.п.н., или от приблизительно 5,0 т.п.н. до 6,2 т.п.н. или от приблизительно 5,1 т.п.н. до приблизительно 6,3 т.п.н., или от приблизительно 5,2 т.п.н. до приблизительно 6,4 т.п.н., или от приблизительно 5,5 т.п.н. до приблизительно 6,5 т.п.н.
[00155] Получение вирусного вектора можно проводить с использованием любых подходящих способов генной инженерии, хорошо известных в данной области, включая, но не ограничиваясь ими, стандартные способы расщепления эндонуклеазами рестрикции, лигирование, трансформацию, очистку плазмиды и секвенирования ДНК, например, как описано в Sambrook et al. (Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press, N.Y. (1989)).
[00156] Вирусный вектор может включать последовательности из генома любого известного организма. Последовательности могут быть включены в их нативной форме или могут быть модифицированными каким-либо образом для достижения желаемой активности. Например, последовательности могут содержать инсерции, делеции или замены.
5.1.1 Промотор
[00157] Различные промоторы могут быть функционально связаны с нуклеиновой кислотой, содержащей кодирующую область представляющего интерес белка в вирусном векторе, описанном в настоящем описании. В некоторых вариантах осуществления промотор может контролировать экспрессию представляющего интерес белка в клетке, инфицированной вирусом, происходящим из вирусного вектора, такой как клетка-мишень. Промотор может быть встречающимся в природе или не встречающимся в природе. В некоторых вариантах осуществления промотор представляет собой синтетический промотор. В одном варианте осуществления синтетический промотор содержит последовательности, которые не существуют в природе и которые предназначены для регуляции активности функционально связанного гена. В другом варианте осуществления синтетический промотор содержит фрагменты натуральных для формирования новых участков последовательности ДНК, которые не существуют в природе. Синтетические промоторы, как правило, состоят из регуляторных элементов, промоторов, энхансеров, интронов, доноров и акцепторов сплайсинга, которые предназначены для обеспечения усиленной тканеспецифической экспрессии. Примеры промоторов включают, но не ограничиваются ими, вирусные промоторы, промоторы растений и промоторы млекопитающих. В другом варианте осуществления промотор представляет собой специфический для печени промотор. Конкретные примеры специфических для печени промоторов включают LP1, HLP, HCR-hAAT, ApoE-hAAT, LSP, TBG и TTR. Эти промоторы более подробно описаны в следующих источниках литературы: LP1: Nathwani A. et al. Blood. 2006 April 1; 107(7): 2653-2661; гибридный специфический для печени промотор (HLP): McIntosh J. et al. Blood. 2013 Apr 25; 121(17): 3335-3344; HCR-hAAT: Miao CH et al. Mol Ther. 2000;1: 522-532; ApoE-hAAT: Okuyama T et al. Human Gene Therapy, 7, 637-645 (1996); LSP: Wang L et al. Proc Natl Acad Sci U S A. 1999 March 30; 96(7): 3906-3910, промотор тироксин-связывающего глобулина (TBG): Yan et al., Gene 506:289-294 (2012), и промотор транстиретина (TTR): Costa et al., Mol. Cell. Biol. 8:81-90 (1988).
[00158] В некоторых вариантах осуществления промотор включает промоторный комплекс антитрипсина альфа 1 человека (hAAT). В некоторых вариантах осуществления промотор включает по меньшей мере часть промотора hAAT. Часть промотора hAAT может содержать последовательность нуклеиновой кислоты, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или более идентичностью последовательности с SEQ ID NO: 3.
[00159] В некоторых вариантах осуществления промотор включает специфический для печени энхансер. В некоторых вариантах осуществления промотор включает энхансер области контроля печени (HCR). В некоторых вариантах осуществления промотор включает по меньшей мере часть энхансера HCR. Например, энхансер HCR может включать последовательность нуклеиновой кислоты, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, или более, идентичностью последовательности с SEQ ID NO: 4.
[00160] В некоторых вариантах осуществления промотор включает специфический для печени энхансер аполипопротеина E (ApoE). В некоторых вариантах осуществления промотор включает по меньшей мере часть энхансера ApoE. Например, энхансер ApoE может включать последовательность нуклеиновой кислоты, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, или более, идентичностью последовательности с SEQ ID NO: 4.
[00161] В некоторых вариантах осуществления промотор представляет собой синтетический промотор, содержащий по меньшей мере часть промотора hAAT, по меньшей мере часть энхансера HCR и по меньшей мере часть энхансера ApoE. В некоторых вариантах осуществления промотор может включать последовательность нуклеиновой кислоты, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или более идентичностью последовательности с SEQ ID NO: 6.
[00162] В некоторых вариантах осуществления промотор включает множество копий одного или более энхансеров, идентифицированных выше. В некоторых вариантах осуществления промоторные конструкции включают один или более индивидуальных энхансерных элементов, описанных выше, и их комбинаций, в одной или более различных ориентации(ях).
[00163] В некоторых вариантах осуществления промотор функционально связан с полинуклеотидом, кодирующим один или более представляющих интерес белков. В некоторых вариантах осуществления промотор функционально связан с полинуклеотидом, кодирующим белок PAH.
[00164] Размер промотора может варьироваться. Вследствие ограниченной упаковывающей способности AAV, предпочтительно использовать промотор небольшого размера, но который в то же время позволяет продуцирование на высоком уровне представляющего интерес белка(ов) в клетках-хозяевах. Например, в некоторых вариантах осуществления промотор имеет размет не более приблизительно 1,5 т.п.н., не более приблизительно 1,4 т.п.н., не более приблизительно 1,35 т.п.н., не более приблизительно 1,3 т.п.н., не более приблизительно 1,25 т.п.н., не более приблизительно 1,2 т.п.н., не более приблизительно 1,15 т.п.н., не более приблизительно 1,1 т.п.н., не более приблизительно 1,05 т.п.н., не более приблизительно 1 т.п.н., не более приблизительно 800 пар оснований, не более приблизительно 600 пар оснований, не более приблизительно 400 пар оснований, не более приблизительно 200 пар оснований или не более приблизительно 100 пар оснований.
5.1.2 Регуляторные элементы
[00165] В вирусных векторах можно использовать различные дополнительные регуляторные элементы, например, энхансеры, для дальнейшего повышения уровня экспрессии представляющего интерес белка в клетке-хозяине, сигнал полиаденилирования, последовательность связывания рибосомы и/или консенсусный акцепторный или донорный участок сплайсинга. В некоторых вариантах осуществления регуляторный элемент может способствовать поддержанию рекомбинантной молекулы ДНК внехромосомно в клетке-хозяине и/или повышать эффективность вектора (например, области присоединения к каркасу/матриксу (S/MAR)). Такие регуляторные элементы хорошо известны в данной области.
[00166] Вирусные векторы, описанные в настоящем описании, могут включать регуляторные элементы, такие как область инициации транскрипции и/или область терминации транскрипции. Примеры области терминации транскрипции включают, но не ограничиваются ими, сигнальные последовательности полиаденилирования. Примеры сигнальных последовательностей полиаденилирования включают, но не ограничиваются ими, поли(A) бычьего гормона роста (bGH), поздний поли(A) SV40, поли(A) бета-глобина кролика (rBG), последовательности поли(A) тимидинкиназы (TK), и любые их варианты. В некоторых вариантах осуществления область терминации транскрипции находится ниже посттранскрипционного регуляторного элемента. В некоторых вариантах осуществления область терминации транскрипции представляет собой сигнальную последовательность полиаденилирования. В некоторых вариантах осуществления область терминации транскрипции представляет собой последовательность поли(A) bGH.
[00167] В некоторых вариантах осуществления вирусные векторы могут включать дополнительные последовательности инициации транскрипции и трансляции, и/или дополнительные терминаторы транскрипции и трансляции, которые известны в данной области.
5.1.3 Представляющий интерес белок
[00168] В рамках изобретения, "представляющий интерес белок" может представлять собой любой белок PAH, включая его встречающиеся в природе или не встречающиеся в природе варианты. В некоторых вариантах осуществления полинуклеотид, кодирующий один или более представляющих интерес белков PAH, может быть встроен в вирусные векторы, описанные в настоящем описании, где полинуклеотид функционально связан с промотором. В некоторых случаях промотор может контролировать экспрессию представляющего интерес белка(ов) в клетке-хозяине (например, клетке печени человека).
[00169] В первом аспекте настоящее изобретение относится к выделенной молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, гомологичную нуклеотидной последовательности SEQ ID NO: 1, и кодирующей функциональную фенилаланингидроксилазу дикого типа (PAH) (SEQ ID NO:2).
[00170] Как описано в настоящем описании, нуклеотидная последовательность, кодирующая белок PAH, может быть модифицирована для повышения эффективности экспрессии белка. Способы, которые можно использовать для повышения транскрипции и/или трансляции гена, описанного в настоящем описании, конкретно не ограничены. Например, нуклеотидную последовательность можно модифицировать, чтобы она лучше отражала использование кодонов хозяином для повышения экспрессии гена (например, продуцирования белка) у хозяина (например, млекопитающего). В качестве другого неограничивающего примера модификации, один или более доноров сплайсинга и/или акцепторов сплайсинга в нуклеотидной последовательности представляющего интерес белка модифицируют для уменьшения возможности ненужного сплайсинга. В качестве другого неограничивающего примера модификации, один или более интронов могут быть встроены в или рядом с нуклеотидной последовательностью представляющего интерес белка для оптимизации упаковывания вектора AAV и усиления экспрессии.
[00171] В некоторых вариантах осуществления молекула нуклеиновой кислоты обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или по меньшей мере 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 1. В одном варианте осуществления молекула нуклеиновой кислоты кодирует функциональный белок PAH, т.е. она кодирует PAH, которая при экспрессии обладает функциональностью PAH дикого типа. В некоторых вариантах осуществления молекула нуклеиновой кислоты при экспрессии в подходящей системе (например, клетке-хозяине) продуцирует функциональный белок PAH и на относительно высоком уровне. Поскольку PAH, которая продуцируется, является функциональной, она будет иметь конформацию, которая является такой же, как по меньшей мере у части PAH дикого типа. В некоторых вариантах осуществления функциональный белок PAH, продуцируемый, как описано в настоящем описании, позволяет по меньшей мере некоторое снижение уровней Phe у индивидуума, страдающего от ФКУ.
[00172] В другом варианте осуществления нуклеотидная последовательность, кодирующая функциональную PAH, имеет улучшенное использование кодонов, предпочтительное для клеток человека, по сравнению с встречающейся в природе нуклеотидной последовательностью, кодирующей соответствующую не кодон-оптимизированную последовательность. Адаптивность нуклеотидной последовательности, кодирующей функциональную PAH, к использованию кодонов клетками человека, может быть выражена в качестве индекса адаптации кодонов (CAI). Индекс адаптации кодонов определяют в настоящем описании как меру относительной адаптивности использования кодонов гена в направлении использования кодонов в генах человека с высокой экспрессией. Относительная адаптивность (w) каждого кодона представляет собой соотношение использования каждого кодона и использования наиболее распространенного кодона для той же аминокислоты. CAI определяют как геометрическое среднее значение этих величин относительной адаптивности. Несинонимичные кодоны и кодоны терминации (зависимые от генетического кода) исключаются. Величины CAI находятся в диапазоне от 0 до 1, причем более высокие величины указывают на более высокую долю наиболее распространенных кодонов (см. Sharp and Li, 1987, Nucleic Acid Research 15: 1281-1295; также см.: Kim et al., Gene. 1997, 199:293-301; zur Megede et al., Journal of Virology, 2000, 74: 2628-2635). В некоторых вариантах осуществления молекула нуклеиновой кислоты, кодирующая PAH, имеет CAI по меньшей мере 0,75, 0,80, 0,85, 0,90, 0,95 или 0,99.
[00173] Нуклеотидная последовательность SEQ ID NO:7, также обозначаемая как GENEius 1 (hPAHco1), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH человека, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO:1). Нуклеотидная последовательность SEQ ID NO:7 представляет собой последовательность PAH, кодон-оптимизированную с использованием программного обеспечения для оптимизации кодонов Operon/Eurofins Genomics совместно с уменьшением вручную содержания динуклеотидов CpG и удалением какой-либо дополнительной ORF в смысловом и антисмысловом направлении. Было показано, что содержание динуклеотидоов CpG активирует TLR9 в дендритных клетках, что ведет к потенциальной активации иммунитета и ответам CTL. Доставляемым продуктом авторов изобретения в геноме вектора AAV является оцДНК, что, таким образом, снижает содержание CpG, что может снизить воспаление печени и уровень ALT.
[00174] В одном варианте осуществления молекула нуклеиновой кислоты обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 7. В другом варианте осуществления нуклеотидная последовательность может обладать по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующих положениях) SEQ ID NO: 7. В другом варианте осуществления молекула нуклеиновой кислоты имеет приблизительно 218 изменений кодонов по сравнению с последовательностью, кодирующей PAH дикого типа, и имеет CAI приблизительно 0,96.
[00175] В одном варианте осуществления SEQ ID NO: 7 и последовательности, которые являются сходными с ней, т.е. последовательности, которые обладают относительно высоким уровнем гомологии, демонстрируют неожиданно высокие уровни экспрессии функционального белка. В этом отношении, SEQ ID NO: 7, 8, 9, 10, 11, 12 и 13 представляют собой кодон-оптимизированные последовательности нуклеиновой кислоты PAH, % гомология которых составляет 80,7%, 80,6%, 96,5%, 96,6%, 77,0%, 80,5% и 76,7%, соответственно, по сравнению с SEQ ID NO: 1.
[00176] В одном варианте осуществления молекула нуклеиновой кислоты обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 8. В одном варианте осуществления нуклеотидная последовательность также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, идентичных кодонам (в соответствующих положениях) SEQ ID NO: 8. В другом варианте осуществления молекула нуклеиновой кислоты имеет приблизительно 219 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,96.
[00177] Нуклеотидная последовательность SEQ ID NO: 8, также обозначаемая как GENEius 2 (hPAHco2), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 8 представляет собой последовательность PAH, которая является кодон-оптимизированной с использованием программного обеспечения для оптимизации кодонов Operon/Eurofins Genomics совместно с уменьшением вручную содержанием динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00178] В одном варианте осуществления молекула нуклеиновой кислоты обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 9. В другом варианте осуществления нуклеотидная последовательность также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующем положении) SEQ ID NO: 9. В другом варианте осуществления молекула нуклеиновой кислоты имеет приблизительно 37 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,79.
[00179] Нуклеотидная последовательность SEQ ID NO: 9, также обозначаемая как Nathwani (NW2-Cop, hPAHco3), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 9 представляет собой последовательность PAH человека, кодон-оптимизированную с использованием таблицы использования кодонов, которая использовалась Amit Nathwani для оптимизации кодонов фактора VIII.
[00180] В одном варианте осуществления молекула нуклеиновой кислоты обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 10. В другом варианте осуществления нуклеотидная последовательность также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующем положении) SEQ ID NO: 10. В другом варианте осуществления молекула нуклеиновой кислоты имеет приблизительно 34 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,79.
[00181] Нуклеотидная последовательность SEQ ID NO: 10, также обозначаемая как Nathwani-RCG (NW-RCG, hPAHco4), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 10 представляет собой последовательность PAH человека, кодон-оптимизированную с использованием таблицы использования кодонов, которая использовалась Amit Nathwani для оптимизации кодонов фактора VIII совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00182] В определенном варианте осуществления молекула нуклеиновой кислоты, описанная в настоящем описании, обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 11. В определенном варианте осуществления нуклеотидная последовательность, описанная в настоящем описании, также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующем положении) SEQ ID NO: 11. В другом варианте осуществления изобретение относится к молекула нуклеиновой кислоты, которая имеет приблизительно 228 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,84.
[00183] Нуклеотидная последовательность SEQ ID NO: 11, также обозначаемая как ATUM (DNA2.0-op/D20-P, hPAHco5), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 11 представляет собой последовательность PAH человека, кодон-оптимизированную с использованием алгоритма оптимизации кодонов DNA2.0.
[00184] В определенном варианте осуществления изобретение относится к молекула нуклеиновой кислоты, которая обладает по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологией, или 98% гомологией с нуклеотидной последовательностью SEQ ID NO: 12. В определенном варианте осуществления нуклеотидная последовательность также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующем положении) SEQ ID NO: 12. В другом варианте осуществления изобретение относится к молекула нуклеиновой кислоты, которая имеет приблизительно 228 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,85.
[00185] Нуклеотидная последовательность SEQ ID NO: 12, также обозначаемая как ATUM-RCG (DNA2.0-Cop1, hPAHco6), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 12 представляет собой последовательность PAH человека, кодон-оптимизированную с использованием алгоритма оптимизации кодонов DNA2.0 совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00186] В некоторых вариантах осуществления изобретение относится к молекуле нуклеиновой кислоты, которая имеет по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95% гомологию, или 98% гомологию с нуклеотидной последовательностью SEQ ID NO: 13. В определенном варианте осуществления нуклеотидная последовательность также может иметь по меньшей мере приблизительно 335, по меньшей мере приблизительно 360, по меньшей мере приблизительно 380, по меньшей мере приблизительно 405, по меньшей мере приблизительно 425, по меньшей мере приблизительно 440 из всех кодонов, кодирующих функциональную PAH, которые являются идентичными кодонам (в соответствующем положении) SEQ ID NO: 13. В другом варианте осуществления изобретение относится к молекуле нуклеиновой кислоты, которая имеет приблизительно 238 изменений кодонов по сравнению с кодирующей последовательностью PAH дикого типа, и имеет CAI приблизительно 0,95.
[00187] Нуклеотидная последовательность SEQ ID NO: 13, также обозначаемая как JCAT (hPAHco7), представляет собой кодон-оптимизированную последовательность нуклеиновой кислоты PAH, которая основана на нуклеотидной последовательности PAH человека дикого типа (SEQ ID NO: 1). Нуклеотидная последовательность SEQ ID NO: 13 представляет собой последовательность PAH человека, кодон-оптимизированную с использованием Java Codon Adaptation Tool (www.jcat.de; Grote et al., Nucleic Acids Res. 2005 Jul 1; 33) совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00188] Как правило, оптимизация кодонов не изменяет аминокислоту, которую кодирует каждый кодон. Она просто изменяет нуклеотидную последовательность так, чтобы она с большей вероятностью экспрессировалась на относительно высоком уровне по сравнению с не кодон-оптимизированной последовательностью. Это означает, что нуклеотидные последовательности нуклеиновой кислоты, описанной в настоящем описании, и, например, SEQ ID NO: 7 (или SEQ ID NO: 8-13) могут различаться, но, когда они транслируются, аминокислотная последовательность продуцируемого белка является одинаковой.
[00189] Однако в одном варианте осуществления молекула нуклеиновой кислоты может кодировать белок, имеющий от 0 до 350, от 0 до 300, от 0 до 250, от 0 до 200, от 0 до 150, от 0 до 100, от 0 до 50, от 0 до 30, от 0 до 20, от 0 до 15, от 0 до 10, или от 0 до 5 аминокислотных замен относительно белка, кодируемого нуклеотидной последовательностью SEQ ID NO: 7, SEQ ID NO: 8, SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11, SEQ ID NO: 12 или SEQ ID NO: 13. Если молекула нуклеиновой кислоты кодирует белок, содержащий последовательность, имеющую изменения любой из аминокислот, белок, тем не менее, должен быть функциональным белком. Специалисту в данной области будет понятно, что незначительные изменения могут быть внесены в некоторые из аминокислот белка без неблагоприятного влияния на функцию белка.
[00190] В некоторых вариантах осуществления кодон-оптимизированная молекула нуклеиновой кислоты hPAH имеет содержание динуклеотидов CpG, составляющее менее 25, менее 20, менее 15 или менее 10. В другом варианте осуществления кодон-оптимизированная молекула нуклеиновой кислоты hPAH имеет содержание GC, составляющее менее 65%, менее 60%, или менее 58%.
[00191] Специалист в данной области без труда способен получить молекулу нуклеиновой кислоты, описанную в настоящем описании. Это можно проводить, например, с использованием химического синтеза данной последовательности. Кроме того, специалистам в данной области будут известны подходящие способы определения того, экспрессирует ли нуклеиновая кислота, описанная в настоящем описании, функциональный белок. Например, один подходящий способ in vitro вовлекает встраивание нуклеиновой кислоты в вектор, такой как вектор на основе AAV, трансдукцию клеток-хозяев, таких как клетки 293T или HeLa, вектором, и оценку активности PAH. Альтернативно подходящий способ in vivo вовлекает трансдукцию вектора, содержащего нуклеиновую кислоту, мышам ФКУ и оценку функциональной PAH в плазме мышей. Подходящие способы более подробно описаны ниже.
[00192] В некоторых вариантах осуществления вектор содержит один или более интронов. Интроны могут облегчать процессинг РНК-транскрипта в клетках-хозяевах млекопитающих, повышать экспрессию представляющего интерес белка и/или оптимизировать упаковывание вектора в частицы AAV. Неограничивающими примерами такого интрона являются интрон β-глобина, интрон AIAT и/или интрон hPAH. В некоторых вариантах осуществления интрон представляет собой синтетический интрон. Например, синтетический интрон может включать нуклеотидную последовательность, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99%, или более, идентичностью последовательности с SEQ ID NO: 14. Положение и размер интрона в векторе могут варьироваться. В некоторых вариантах осуществления интрон находится между промотором и последовательностью, кодирующими представляющий интерес белок. В некоторых вариантах осуществления интрон находится в промоторе. В некоторых вариантах осуществления интрон включает энхансерный элемент. В некоторых вариантах осуществления интрон находится в последовательности, кодирующей представляющий интерес белок, предпочтительно между экзонами последовательности, кодирующей представляющий интерес белок. В некоторых вариантах осуществления интрон может содержать весь или часть встречающегося в природе интрона в последовательности, кодирующей представляющий интерес белок. В некоторых вариантах осуществления интрон представляет собой второй интрон PAH. Альтернативно интрон представляет собой укороченную форму 2-го интрона hPAH размером 2116 п.н. В других вариантах осуществления последовательность интрона представляет собой составной интрон бета-глобина/AIAT.
[00193] Включение интронного элемента может повышать экспрессию по сравнению с экспрессией в отсутствие интронного элемента (см. например Kurachi et al., 1995, J Biol Chem. 1995 Mar 10;270(10):5276-81.). Векторы на основе AAV, как правило, вмещают вставки ДНК, имеющие определенный диапазон размера, который обычно составляет от приблизительно 4 т.п.н. до приблизительно 5,2 т.п.н., или несколько больше. Однако не существует минимального размера для упаковывания и небольшие векторные геномы упаковываются очень эффективно. Интроны и фрагменты интронов (например, часть интрона 2 PAH) удовлетворяют этому требованию, также повышая экспрессию. Таким образом, настоящее изобретение не ограничивается включением последовательностей интрона 2 PAH в вектор AAV, и включают другие интроны или другие последовательности ДНК вместо частей интрона 2 PAH. Кроме того, другие 5 '- и 3 '-нетранслируемые области нуклеиновых кислот можно использовать вместо 5 '- и 3 '-нетранслируемых областей, указанных для PAH человека.
[00194] "Часть интрона 2 PAH", в рамках изобретения, означает область интрона 2 PAH, имеющую нуклеотидную длину от приблизительно 0,1 т.п.н. до приблизительно 2,1 т.п.н., которая повышает экспрессию PAH, как правило, приблизительно в 1,5 раза или более на матрице плазмиды или вирусного вектора по сравнению с экспрессией PAH в отсутствии части интрона 2 PAH. Более конкретной частью является часть интрона 2 PAH размером 2116 п.н.
[00195] Полинуклеотиды и полипептиды, включающие модифицированные формы, можно получать с использованием различных способов стандартного клонирования, технологий рекомбинантных ДНК, посредством экспрессии в клетках или трансляции in vitro, и способов химического синтеза, известных специалистам в данной области (Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd edition).
5.1.4 Способы доставки генов
[00196] Также изобретение относится к вектору, содержащему молекулу нуклеиновой кислоты по настоящему изобретению. В одном варианте осуществления вектор может представлять собой вектор для доставки генов. Такой вектор для доставки генов может представлять собой вирусный вектор для доставки гена или не вирусный вектор для доставки генов. Вирусные векторы включают векторы на основе лентивирусов, аденовирусов или вирусов герпеса. Предпочтительно, он представляет собой рекомбинантный аденоассоциированный вирусный (rAAV) вектор. Альтернативно можно использовать невирусные системы, в том числе с использованием голой ДНК (с областями присоединения хроматина или без них) или конъюгированной ДНК, которую вводят в клетки различными способами трансфекции, таких как липиды или электропорация.
[00197] Последовательности неограничивающих примеров векторов на основе AAV предоставлены в SEQ ID NO: 15-23. Например, нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-HCR-hAAT, интрон бета-глобина, кодирующую последовательность дикого типа для PAH человека, и последовательность поли(A) bGH указана в SEQ ID NO: 15 ((вектор 1), ApoE-HCR-hAAT.GI.hPAH.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT, интрон бета-глобина, кодон-оптимизированную последовательность PAH человека (hPAH-Genius1), и последовательность поли(A) bGH, указана в SEQ ID NO: 16 (ApoE-HCR-hAAT.GI.hPAHco1.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT с составным интроном глобина/AIAT, кодирующей последовательностью дикого типа для PAH человека, и последовательностью поли(A) bGH указана в SEQ ID NO: 17 ((V1-LG-Intron), ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT с составным интроном глобина/AIAT, кодон-оптимизированной последовательностью PAH человека, и последовательностью поли(A) bGH указана в SEQ ID NO: 18 ((LGI-hPAH-Geneius1), ApoE-HCR-hAAT.cG-AIATI.hPAHco1.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT, кодирующую последовательность дикого типа для PAH человека с включенным укороченным 2-м интроном, и последовательность поли(A) bGH, указана в SEQ ID NO: 19 ((hPAH+укороченный 2-й интрон), ApoE-HCR-hAAT.hPAH-tI2.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT, интрон бета-глобина, кодон-оптимизированную последовательность PAH человека (hPAH-Genius2), и последовательность поли(A) bGH, указана в SEQ ID NO: 20 ((pFB-ApoE-hAAT-hPAH-Genius2), ApoE-HCR-hAAT.GI.hPAHco2.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор ApoE-hAAT, интрон бета-глобина, кодон-оптимизированную последовательность PAH человека (hPAH-JCAT), и последовательность поли(A) bGH, указана в SEQ ID NO: 21 ((pFB-ApoE-hAAT-hPAH-JCAT), ApoE-HCR-hAAT.GI.hPAHco3.bGH); нуклеотидная последовательность для вектора на основе AAV, включающая промотор TBG с двумя участками макроглобулина, интроном бета-глобина, кодирующей последовательностью дикого типа для PAH человека, и последовательностью поли(A) bGH, указана в SEQ ID NO: 22 ((pFB-hPAHV1-TBG2uGlob), TBG.GI.hPAH.bGH); и нуклеотидная последовательность для вектора на основе AAV, включающая промотор TTR, интрон бета-глобина, кодирующие последовательности дикого типа для PAH человека, и последовательность поли(A) bGH, указана в SEQ ID NO: 23 ((pFB-hPAHV1-TTR), TTR.GI.hPAH.bGH).
[00198] В некоторых вариантах осуществления вектор на основе AAV содержит нуклеотидную последовательность, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или более идентичностью последовательности с SEQ ID NO: 15-23. В некоторых вариантах осуществления вектор на основе AAV содержит нуклеотидную последовательность, обладающую по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или более идентичностью последовательности с SEQ ID NO: 18.
5.1.5 Невирусная доставка генов
[00199] Невирусную доставку генов можно проводить с использованием голой ДНК, которая является наиболее простым способом невирусной трансфекции. Может быть возможным, например, введение нуклеиновой кислоты, описанной в настоящем описании, с использованием голой плазмидной ДНК. Альтернативно, можно использовать способы, такие как электропорация, сонопорация или использование "генной пушки", которая стреляет покрытыми ДНК золотыми частицами в клетки с использованием, например, газа высокого давления или инверсной пушки калибра 0,22 (Helios® Gene Gun System (BIO-RAD)).
[00200] Для повышения доставки нуклеиновой кислоты в клетку может быть необходимой защита ее от повреждения, и ее вхождение в клетку может быть облегчено. Для этого можно использовать липоплексы и полиплексы, которые способны защищать нуклеиновую кислоту от нежелательной деградации в ходе процесса трансфекции.
[00201] Плазмидная ДНК может быть покрыта липидами в организованной структуре, такой как мицелла или липосома. Когда организованная структура находится в комплексе с ДНК, ее называют липоплексом. Для конструирования липоплексов для синтетических векторов можно использовать анионные и нейтральные липиды. В одном варианте осуществления можно использовать катионные липиды вследствие их положительного заряда для уплотнения отрицательно заряженных молекул ДНК, чтобы облегчить инкапсулирование ДНК в липосомы. Для формирования липоплексов может быть необходимым добавление липидов-помощников (обычно электронейтральные липиды, такие как DOPE) к катионным липидам (Dabkowska et al., J R Soc Interface. 2012 Mar 7; 9(68): 548-561).
[00202] В некоторых вариантах осуществления для доставки нуклеиновой кислоты можно использовать комплексы полимеров с ДНК, называемые полиплексами. Большинство полиплексов состоят из катионных полимеров, и их образование регулируется ионными взаимодействиями. Полиплексы, как правило, не могут высвобождать нагруженную на них ДНК в цитоплазму. Таким образом, может быть необходимой котрансфекция с лизирующими эндосомы агентами (для лизиса эндосомы, которая образуется в ходе эндоцитоза - процесса, посредством которого полиплекс входит в клетку), такими как инактивированный аденовирус (Akinc et al., The Journal of Gene Medicine. 7 (5): 657-63).
[00203] В некоторых вариантах осуществления для доставки нуклеиновой кислоты можно использовать гибридные способы, которые сочетают в себе два или более способов. Одним из примеров являются виросомы; в них комбинируются липосомы с инактивированным ВИЧ или вирусом гриппа. В другом варианте осуществления другие способы вовлекают смешение других вирусных векторов с катионными липидами или гибридизацию вирусов, и их можно использовать для доставки нуклеиновой кислоты (Khan, Firdos Alam, Biotechnology Fundamentals, CRC Press, Nov 18, 2015, p. 395).
5.1.6 Вирусные частицы
[00205] В одном варианте осуществления для доставки нуклеиновой кислоты можно использовать подходящий вектор для вирусной доставки генов. В некоторых вариантах осуществления вирусные векторы, пригодные для применения в рамках настоящего изобретения, могут представлять собой парвовирус, аденовирус, ретровирус, лентивирус или вирус простого герпеса. Парвовирус может представлять собой аденовирус-ассоциированный вирус (AAV).
[00206] Таким образом, настоящее изобретение относится к векторам для доставки генов (содержащим нуклеиновую кислоту, описанную в настоящем описании) на основе парвовирусов, в частности, депендовирусов, таких как инфекционный AAV человека или обезьяны, и их компонентов (например, геном парвовируса животных) для применения в качестве векторов для введения и/или экспрессии полипептида фактора VIII в клетке млекопитающего. Таким образом, термин "парвовирусный", в рамках изобретения, охватывает депендовирусы, такие как любой тип AAV.
[00207] Вирусы семейства Parvoviridae представляют собой мелкие ДНК-вирусы животных. Семейство Parvoviridae может быть подразделено на два подсемейства: Parvovirinae, которые инфицируют позвоночных, и Densovirinae, которые инфицируют насекомых. Представителей подсемейства Parvovirinae называют в настоящем описании парвовирусами, и они включают род Dependovirus. Как может быть установлено из названия их рода, представители Dependovirus являются уникальными тем, что им обычно требуется совместная инфекция с вирусом-помощником, таким как аденовирус или вирус герпеса, для продуктивной инфекции в клеточной культуре. Род Dependovirus включает AAV, который обычно инфицирует людей (например, серотипы 1, 2, 3A, 3B, 4, 5 и 6), приматов (например, серотипы 1 и 4), и родственные вирусы, которые инфицируют других теплокровных животных (например, аденоассоциированные вирусы животных подсемейства бычьих, животных семейства собачьих, животных семейства лошадиных, мышей, крыс и животных семейства овечьих) в дополнение к птицам и пресмыкающимся. Дополнительная информация, касающаяся парвовирусов и других представителей Parvoviridae описана в Kenneth I. Berns, "Parvoviridae: The Viruses and Their Replication", Chapter 69, Fields Virology (3d Ed. 1996). Для удобства настоящее изобретение дополнительно проиллюстрировано и описано в настоящем описании применительно к AAV. Однако понятно, что настоящее изобретение не ограничивается AAV, а в равной степени может применяться в отношении других парвовирусов.
[00208] Гены "rep" и "cap" AAV представляют собой гены, кодирующие белки репликации и инкапсидирования, соответственно. Гены rep и cap AAV обнаружены во всех серотипах AAV, исследованных на сегодняшний день, и описаны в настоящем описании и в цитированных ссылках. В AAV дикого типа гены rep и cap обычно встречаются рядом друг с другом в вирусных геномах (т.е. они "сопряжены" в качестве примыкающих или перекрывающихся транскрипционных элементов), и они обычно являются консервативными среди серотипов AAV. Гены rep и cap AAV также индивидуально и в совокупности называют "генами упаковывания AAV". Гены cap AAV для применения в рамках настоящего изобретения кодируют белки Cap, которые способны упаковывать векторы на основе AAV в присутствии rep и функции помощника аденовируса, и способны связывать рецепторы клеток-мишеней. В некоторых вариантах осуществления ген cap AAV кодирует капсидный белок, имеющий аминокислотную последовательность, происходящую из конкретного серотипа AAV.
[00209] Последовательности AAV, используемые для получения AAV, могут происходить из генома любого серотипа AAV. Как правило, серотипы AAV имеют геномные последовательности, обладающие значительной гомологией на уровнях аминокислот и нуклеиновых кислот, обеспечивают сходный набор генетических функций, продуцируют вирионы, которые являются по существу физически и функционально эквивалентными, и реплицируются и собираются посредством практически идентичных механизмов. Для геномной последовательности серотипов AAV и обсуждения геномного сходства см., например, номер доступа GenBank U89790; номер доступа GenBank J01901; номер доступа GenBank AF043303; номер доступа GenBank AF085716; Chiorini et al., J. Vir. (1997) vol. 71, pp. 6823-6833; Srivastava et al., J. Vir. (1983) vol. 45, pp. 555-564; Chiorini et al., J. Vir. (1999) vol. 73, pp. 1309-1319; Rutledge et al., J. Vir. (1998) vol. 72, pp. 309-319; и Wu et al., J. Vir. (2000) vol. 74, pp. 8635-8647).
[00210] Геномная организация всех известных серотипов AAV является в высокой степени сходной. Геном AAV представляет собой линейную одноцепочечную молекулу ДНК, длина которой составляет менее чем приблизительно 5000 нуклеотидов (нт). Инвертированные концевые повторы (ITR) фланкируют уникальные кодирующие нуклеотидные последовательности для неструктурных белков репликации (Rep) и структурных белков (VP). Белки VP формируют капсид. Концевые 145 нт являются самокомплементарными и организованы так, чтобы мог образовываться энергетически стабильный внутримолекулярный дуплекс, формирующий T-образную шпильку. Эти шпилечные структуры функционируют в качестве ориджина для репликации вирусной ДНК, выполняя роль праймеров для клеточного комплекса ДНК-полимеразы. Гены Rep кодируют белки Rep: Rep78, Rep68, Rep52 и Rep40. Rep78 и Rep68 транскрибируются с промотора p5, и Rep 52 и Rep40 транскрибируются с промотора p19. Гены cap кодируют белки VP, VP1, VP2 и VP3. Гены cap транскрибируются с промотора p40. ITR, используемые в векторах согласно настоящим вариантам осуществления, могут соответствовать тому же серотипу, что и ассоциированные гены cap, или могут отличаться. В одном варианте осуществления ITR, используемые в рамках настоящего изобретения, соответствуют серотипу AAV2, и гены cap соответствуют серотипу AAV5.
[00211] Известно, что белки VP AAV определяют тропность к клеткам вириона AAV. Кодирующие белок VP последовательности являются значительно менее консервативными, чем белки и гены Rep среди различных серотипов AAV. Способность последовательностей Rep и ITR перекрестно дополнять соответствующие последовательности других серотипов позволяет получение псевдотипированных частиц AAV, содержащих капсидные белки одного серотипа (например, AAV1, 5 или 8) и последовательности Rep и/или ITR другого серотипа AAV (например, AAV2). Такие псевдотипированные частицы rAAV являются частью настоящего изобретения.
[00212] В одном варианте осуществления последовательности ITR AAV для применения в контексте настоящего изобретения происходят из AAV1, AAV2, AAV4 и/или AAV6. Аналогично, в одном варианте осуществления кодирующие Rep (Rep78 и Rep52) последовательности происходят из AAV1, AAV2, AAV4 и/или AAV6. Однако последовательности, кодирующие капсидные белки VP1, VP2 и VP3 для применения в контексте настоящего изобретения могут быть получены из любого из известных 42 серотипов, например, из AAV1, AAV2, AAV3, AAV4, AAVS, AAV6, AAV7, AAV8 или AAV9 или вновь разработанных AAV-подобных частиц, полученных, например, способами шаффлинга капсида и библиотек капсида AAV.
[00213] Также в контексте настоящего изобретения можно использовать модифицированные последовательности "AAV", например, для получения векторов для генной терапии на основе AAV. Такие модифицированные последовательности, например, включают последовательности, обладающие по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, или более идентичностью нуклеотидной и/или аминокислотной последовательности (например, последовательность, обладающая приблизительно 75-99% идентичностью нуклеотидной последовательности) с ITR, Rep или VP AAV1, AAV2, AAV3, AAV4, AAV5, AAV6, AAV7, AAV8 или AAV9, и их можно использовать вместо последовательностей ITR, Rep или VP AAV дикого типа.
[00214] В некоторых вариантах осуществления последовательность нуклеиновой кислоты, кодирующая капсидный белок AAV, функционально связана с последовательностями контроля экспрессии для экспрессии в конкретном типе клеток, таком как клетки Sf9 или HEK. Для применения вариантов осуществления на практике можно использовать способы экспрессии чужеродных генов в клетках-хозяевах насекомых или клетках-хозяевах млекопитающих, известные специалисту в данной области. Методология молекулярной инженерии и экспрессии полипептидов в клетках насекомых описана, например, в Summers and Smith (1986) A Manual of Methods for Baculovirus Vectors and Insect Culture Procedures, Texas Agricultural Experimental Station Bull. No. 7555, College Station, Tex.; Luckow (1991) In Prokop et al., Cloning and Expression of Heterologous Genes in Insect Cells with Baculovirus Vectors ' Recombinant DNA Technology and Applications, 97-152; King, L. A. and R. D. Possee (1992) The baculovirus expression system, Chapman and Hall, United Kingdom; O 'Reilly, D. R., L. K. Miller, V. A. Luckow (1992) Baculovirus Expressing Vectors: A Laboratory Manual, New York; W.H. Freeman and Richardson, C. D. (1995) Baculovirus Expression Protocols, Methods in Molecular Biology, volume 39; патент США № 4745051; US2003148506 и WO 03/074714, все из которых включены в качестве ссылок в полном объеме. Особенно пригодным промотором для транскрипции нуклеотидной последовательности, кодирующей капсидный белок AAV, является, например, промотор полиэдра. Однако в данной области известны другие промоторы, активные в клетках насекомых, например, промоторы p10, p35 или IE-1, и также предусматриваются другие промоторы, описанные в приведенных выше ссылках.
[00215] Использование клеток насекомых для экспрессии гетерологичных белков подробно описано, также как и способы введения нуклеиновых кислот, таких как векторы, например, совместимые с клетками насекомых векторы, в такие клетки и способы поддержания таких клеток в культуре. (См., например, METHODS IN MOLECULAR BIOLOGY, ed. Richard, Humana Press, N J (1995); O 'Reilly et al., BACULOVIRUS EXPRESSION VECTORS, A LABORATORY MANUAL, Oxford Univ. Press (1994); Samulski et al., J. Vir. (1989) vol. 63, pp.3822-3828; Kajigaya et al., Proc. Nat 'l. Acad. Sci. USA (1991) vol. 88, pp. 4646-4650; Ruffing et al., J. Vir. (1992) vol. 66, pp. 6922-6930; Kirnbauer et al., Vir. (1996) vol. 219, pp. 37-44; Zhao et al., Vir. (2000) vol. 272, pp. 382-393 и патент США № 6204059). В некоторых вариантах осуществления конструкция нуклеиновой кислоты, кодирующая AAV в клетках насекомых, представляет собой совместимый с клеткой насекомых вектор. "Совместимый с клеткой насекомых вектор" или "вектор", в рамках изобретения, относится к молекуле нуклеиновой кислоты, способной к продуктивной трансформации или трансфекции насекомого или клетки насекомого. Иллюстративные биологические векторы включают плазмиды, линейные молекулы нуклеиновых кислот и рекомбинантные вирусы. Можно использовать любой вектор при условии, что он совместим с клетками насекомых. Вектор может встраиваться в геном клеток-насекомых, однако присутствие вектора в клетке насекомого не должно быть постоянным и также включены временные эписомальные векторы. Векторы можно вводить любыми известными способами, например, посредством химической обработки клеток, электропорации или инфекции. В некоторых вариантах осуществления вектор представляет собой бакуловирус, вирусный вектор или плазмиду. В одном варианте осуществления вектор представляет собой бакуловирус, т.е. конструкция представляет собой бакуловирусный вектор. Бакуловирусные векторы и способы их применения описаны в цитированных выше ссылках, касающихся молекулярной инженерии клеток насекомых.
5.2 СПОСОБЫ ПОЛУЧЕНИЯ РЕКОМБИНАНТНЫХ AAV
[00216] Настоящее изобретение относится к материалам и способам для продуцирования рекомбинантных AAV в клетках насекомых или млекопитающих. В некоторых вариантах осуществления вирусная конструкция дополнительно содержит промотор и участок рестрикции ниже промотора, чтобы обеспечить встраивание полинуклеотида, кодирующего один или более представляющих интерес белков, где промотор и участок рестрикции находятся ниже 5 '-ITR AAV и выше 3 '-ITR AAV. В некоторых вариантах осуществления вирусная конструкция, кроме того, содержит посттранскрипционный регуляторный элемент ниже участка рестрикции и выше 3 '-ITR AAV. В некоторых вариантах осуществления вирусная конструкция, кроме того, содержит полинуклеотид, встроенный в участок рестрикции и функционально связанный с промотором, где полинуклеотид содержит кодирующую область представляющего интерес белка. Как будет понятно специалисту в данной области, любой из векторов на основе AAV, описанных в настоящей заявке, можно использовать в способе в качестве вирусной конструкции для получения рекомбинантного AAV.
[00217] В некоторых вариантах осуществления функции помощника предоставляются одной или несколькими плазмидами-помощниками или вирусами-помощниками, содержащими гены-помощники аденовирусов или бакуловирусов. Неограничивающие примеры генов-помощников аденовирусов или бакуловирусов включают, но не ограничиваются ими, E1A, E1B, E2A, E4 и VA, которые могут обеспечить функции помощников для упаковывания AAV.
[00218] Вирусы-помощники AAV известны в данной области и включают, например, вирусы из семейства Adenoviridae и семейства Herpesviridae. Примеры вирусов-помощников AAV включают, но не ограничиваются ими, вирус-помощник SAdV-13 и SAdV-13-подобный вирус-помощник, описанный в публикации США № 20110201088 (содержание которой включено в настоящее описание в качестве ссылки), и векторы-помощники pHELP (Applied Viromics). Квалифицированному специалисту будет понятно, что в рамках настоящего изобретения можно использовать любой вирус-помощник или плазмиду-помощника для AAV, которые могут обеспечить адекватную функцию помощника AAV.
[00219] В некоторых вариантах осуществления в плазмиде присутствуют гены cap AAV. Плазмида, кроме того, может содержать ген rep AAV, который может соответствовать или может не соответствовать тому же серотипу, что и гены cap. В рамках настоящего изобретения для получения рекомбинантного AAV можно использовать гены cap и/или ген rep из любого серотипа AAV (включая, но не ограничиваясь ими, AAV1, AAV2, AAV4, AAV5, AAV6, AAV7, AAV8, AAV9, AAV10, AAV11, AAV12, AAV13 и любые их варианты). В некоторых вариантах осуществления гены cap AAV кодируют капсид из серотипа 1, серотипа 2, серотипа 4, серотипа 5, серотипа 6, серотипа 7, серотипа 8, серотипа 9, серотипа 10, серотипа 11, серотипа 12, серотипа 13 или их вариантов.
[00220] В некоторых вариантах осуществления клетку насекомого или клетку млекопитающего можно трансфицировать плазмидой-помощником или вирусом-помощником, вирусной конструкцией и плазмидой, кодирующей гены cap AAV; и рекомбинантный вирус AAV можно собирать в различные моменты времени после котрансфекции. Например, рекомбинантный вирус AAV можно собирать приблизительно через 12 часов, приблизительно через 24 часа, приблизительно через 36 часов, приблизительно через 48 часов, приблизительно через 72 часа, приблизительно через 96 часов, приблизительно через 120 часов, или в момент времени между любыми из этих двух моментов времени после котрансфекции.
[00221] Рекомбинантный AAV также можно получать с использованием любых общепринятых способов, известных в данной области, пригодных для продуцирования инфекционного рекомбинантного AAV. В некоторых случаях рекомбинантный AAV можно получать с использованием клетки насекомого или клетки млекопитающего, которая стабильно экспрессирует некоторые из необходимых компонентов, для продуцирования частиц AAV. Например, плазмида (или множество плазмид), содержащих гены rep и cap AAV, и селективный маркер, такой как ген резистентности к неомицину, может встраиваться в геном клетки. Затем клетку насекомого или млекопитающего можно совместно инфицировать вирусом-помощником (например, аденовирус или бакуловирус, обеспечивающий функции помощников) и вирусным вектором, содержащим 5 '- и 3 '-ITR AAV (и нуклеотидную последовательность, кодирующую гетерологичный белок, если желательно). Преимуществами этого способа является то, что можно проводить селекцию клеток, и они пригодны для крупномасштабного продуцирования рекомбинантного AAV. В качестве другого неограничивающего примера вместо плазмид можно использовать аденовирус или бакуловирус для введения генов rep и cap в упаковывающие клетки. В качестве другого неограничивающего примера, для продуцирования рекомбинантного AAV, как вирусный вектор, содержащий 5 '- и 3 '-LTR AAV, так и гены rep-cap, могут стабильно встраиваться в ДНК продуцирующих клеток, и функции помощников могут обеспечиваться аденовирусом дикого типа.
[00222] В одном варианте осуществления изобретение относится к способу получения вектора на основе AAV для доставки генов, включающему стадии:
(a) получения клетки насекомого, содержащей одну или более конструкций нуклеиновых кислот, содержащих:
(i) молекулу нуклеиновой кислоты, описанную в настоящем описании, которая фланкируется по меньшей мере одной нуклеотидной последовательностью инвертированного концевого повтора AAV;
(ii) первую экспрессирующую кассету, содержащую нуклеотидную последовательность, кодирующую один или более белков Rep AAV, которые функционально связаны с промотором, который способен контролировать экспрессию белка(ов) Rep в клетке насекомого;
(iii) вторую экспрессирующую кассету, содержащую нуклеотидную последовательность, кодирующую один или более капсидных белков AAV, которые функционально связаны с промотором, который способен контролировать экспрессию капсидного белка(ов) в клетке насекомого;
(iv) и необязательно AAP и MAAP, содержащиеся в мРНК VP2/3,
(b) культивирование клетки насекомого согласно (a) в условиях, способствующих экспрессии Rep и капсидных белков; и, необязательно,
(c) выделение вектора для доставки генов на основе AAV.
[00223] Как правило, также способ получения вектора для доставки генов на основе AAV согласно изобретению включает: введение в клетку, обеспечивающую репликацию AAV, (a) нуклеотидной последовательности, кодирующей матрицу для продуцирования генома вектора по настоящему изобретению (как подробно описано в настоящем описании); (b) нуклеотидных последовательностей, достаточных для репликации матрицы для продуцирования векторного генома (первая экспрессирующая кассета, определенная выше); (c) нуклеотидных последовательностей, достаточных для упаковывания векторного генома в капсид AAV (вторая экспрессирующая кассета, определенная выше), в условиях, достаточных для репликации и упаковывания векторного генома в капсид AAV, в результате чего частицы AAV, содержащие векторный геном, инкапсидированный в капсид AAV, продуцируются в клетке.
[00224] Способ по изобретению может включать стадию аффинной очистки (вирионов, содержащих) рекомбинантного парвовирусного (rAAV) вектора с использованием антитела против AAV, в одном варианте осуществления иммобилизованного антитела. В другом варианте осуществления антитело против AAV представляет собой моноклональное антитело. Одним из антител для применения в рамках настоящего изобретения является одноцепочечное антитело животных семейства верблюжьих или его фрагмент, как получают, например, из верблюдов или лам (см., например, Muyldermans, 2001, Biotechnol. 74: 277-302). Антитело для аффинной очистки rAAV представляет собой антитело, которое специфически связывает эпитоп на капсидном белке AAV, причем в одном варианте осуществления эпитоп представляет собой эпитоп, который присутствует на капсидном белке более чем одного серотипа AAV. Например, антитело может быть индуцировано или отобрано на основе специфического связывания с капсидом AAV2, но в то же время оно может также специфически связываться с капсидами AAV1, AAV3, AAV5, AAV6, AAV8 или AAV9.
5.3 ТИПЫ КЛЕТОК, ИСПОЛЬЗУЕМЫЕ ДЛЯ ПРОДУЦИРОВАНИЯ AAV
[00225] Вирусные частицы, содержащие векторы на основе AAV согласно вариантам осуществления, можно получать с использованием любого типа клеток беспозвоночных, который позволяет продуцирование AAV или биологических продуктов, и который можно поддерживать в культуре. Например, линия клеток насекомых может представлять собой линию из Spodoptera frugiperda, такую как SF9, SF21, SF900+, клеточные линии дрозофил, клеточные линии комаров, например, клеточные линии, происходящие из Aedes albopictus, клеточные линии домашнего тутового шелкопряда, например, клеточные линии Bombyx mori, клеточные линии Trichoplusia ni, такие как клетки High Five, или клеточные линии Lepidoptera, такие как клеточные линии Ascalapha odorata. В одном варианте осуществления клетки насекомых представляют собой клетки из видов насекомых, чувствительных к бакуловирусной инфекции, включая High Five, Sf9, Se301, SeIZD2109, SeUCR1, Sf9, Sf900+, Sf21, BTI-TN-5B1-4, MG-1, Tn368, HzAm1, BM-N, Ha2302, Hz2E5 и Ao38.
[00226] Бакуловирусы представляют собой оболочечные ДНК-вирусы членистоногих, два представителя из которых являются хорошо известными экспрессирующими векторами для продуцирования рекомбинантных белков в клеточных культурах. Бакуловирусы имеют кольцевые двухцепочечные геномы (80-200 т.п.о.), которые могут быть модифицированы способами инженерии для обеспечения доставки крупного геномного содержимого в конкретные клетки. Вирусы, используемые в качестве вектора, как правило, представляют собой мультикапсидный нуклеополиэдровирус Autographa californica (AcMNPV) или нуклеополиэдровирус Bombyx mori (BmNPV) (Kato et al., (2010), Applied Microbiology and Biotechnology, vol. 85, Issue 3, pp 459-470).
[00227] Бакуловирусы часто используют для инфицирования клеток насекомых для экспрессии рекомбинантных белков. В частности, экспрессию гетерологичных генов у насекомых можно проводить, как описано, например, в патенте США № 4745051; EP 127839; EP 155476; Vlak et al., (1988), Journal of General Virology, vol. 68, pp 765-776; Miller et al., (1988), Annual Review of Microbiology, vol. 42, pp 177-179; Carbonell et al., (1998), Gene, vol. 73, Issue 2, pp 409-418; Maeda et al., (1985), Nature, vol. 315, pp 592-594; Lebacq-Veheyden et al., (1988), Molecular and Cellular Biology, vol. 8, no. 8, pp 3129-3135; Smith et al., (1985), PNAS, vol. 82, pp 8404-8408; и Miyajima et al., (1987), Gene, vol. 58, pp 273-281. Многочисленные штаммы и варианты бакуловирусов и соответствующие пермиссивные клетки-хозяева насекомых, которые можно использовать для продуцирования белков, описаны в Luckow et al., (1988), Nature Biotechnology, vol. 6, pp 47-55; Maeda et al., (1985), Nature, vol. 315, pp 592-594; и McKenna et al., (1998), Journal of Invertebrate Pathology, vol. 71, Issue 1, pp 82-90.
[00228] В другом варианте осуществления способы, описанные в настоящем описании, проводят с любым типом клеток млекопитающих, который позволяет репликацию AAV или продуцирование биологических продуктов и который можно поддерживать в культуре. В одном варианте осуществления используемые клетки млекопитающих могут представлять собой клетки HEK293, HeLa, CHO, NSO, SP2/0, PER.C6, Vero, RD, BHK, HT 1080, A549, Cos-7, ARPE-19 и MRC-5.
5.3.1 Организм-хозяин и/или клетки
[00229] В следующем варианте осуществления изобретение относится к хозяину, содержащему вектор, описанный выше. В одном варианте осуществления вектор способен экспрессировать молекулу нуклеиновой кислоты, описанную в настоящем описании, у хозяина. Хозяин может представлять собой любого подходящего хозяина. В другом варианте осуществления терапевтическое средство против ФКУ включает клеточную терапию против ФКУ (см., например, Harding, C., Clin Genet., Aug; 74(2) pages 97-104 (2008).
[00230] В рамках изобретения, термин "хозяин" относится к организмам и/или клеткам, которые содержат молекулу нуклеиновой кислоты или вектор по настоящему изобретению, а также к организмам и/или клеткам, которые являются пригодными для применения для экспрессии рекомбинантного гена или белка. Подразумевается, что настоящее изобретение не ограничивается каким-либо конкретным типом клеток или организмов. Действительно, предусматривается, что в качестве хозяина можно использовать любой подходящий организм и/или клетку. Клетка-хозяин может быть в форме единичной клетки, популяции сходных или различных клеток, например, в форме культуры (такой как жидкая культура или культура на твердом субстрате), организма или его части. В одном варианте осуществления клетка-хозяин может позволить экспрессию молекулы нуклеиновой кислоты, описанной в настоящем описании. Таким образом, клетка-хозяин может представлять собой, например, клетку бактерий, дрожжей, насекомых или млекопитающих.
[00231] В другом варианте осуществления изобретение относится к средству для доставки нуклеиновой кислоты, описанной в настоящем описании, в широкий диапазон клеток, включая делящиеся и не делящиеся клетки. Настоящее изобретение можно использовать для доставки нуклеиновой кислоты, описанной в настоящем описании, в клетку in vitro, например, для получения полипептида, кодируемого такой молекулой нуклеиновой кислоты, in vitro, или для генной терапии ex vivo.
[00232] Молекула нуклеиновой кислоты, вектор, клетки и способы/применение по настоящему изобретению, кроме того, являются пригодными в способе доставки нуклеиновой кислоты, описанной в настоящем описании, хозяину, как правило, хозяину, страдающему от ФКУ.
[00233] Настоящее изобретение может использоваться как для ветеринарных, так и для медицинских применений. Подходящие индивидуумы для способов генной доставки, как описано в настоящем описании, включают как птиц, так и млекопитающих, причем предпочтительными являются млекопитающие. Термин "птица", в рамках изобретения, включает, но не ограничивается ими, кур, уток, гусей, перепелов, индеек и фазанов. Термин "млекопитающее", в рамках изобретения, включает, но не ограничивается ими, людей, животных подсемейства бычьих, животных семейства овечьих, животных семейства козьих, животных семейства лошадиных, животных семейства кошачьих, животных семейства собачьих, зайцеобразных, и т.д. Люди включают новорожденных, младенцев, подростков и взрослых.
5.4 ФАРМАЦЕВТИЧЕСКИЕ СОСТАВЫ
[00234] В одном варианте осуществления изобретение относится к фармацевтической композиции, содержащей нуклеиновую кислоту или вектор по изобретению и фармацевтически приемлемый носитель и/или другое медицинское средство, фармацевтическое средство или адъювант и т.д.
[00235] В других вариантах осуществления изобретение относится к фармацевтическим составам экспрессирующего терапевтический белок вектора на основе AAV/вирионов, пригодные для введения индивидуумам, страдающим от генетического нарушения. В некоторых вариантах осуществления фармацевтические составы, описанные в настоящем описании, представляют собой жидкие составы, которые содержат экспрессирующие рекомбинантный терапевтический белок вирионы AAV, продуцированные из векторов, описанных в настоящем описании, где концентрация рекомбинантных вирионов AAV в составе может широко варьироваться. В некоторых вариантах осуществления концентрация рекомбинантного вириона AAV в составе может находиться в диапазоне от 1E12 вг/мл до 2E16 мг/мл. В одном варианте осуществления концентрация рекомбинантного вириона AAV в составе составляет приблизительно 2E13 вг/мл.
[00236] В других вариантах осуществления фармацевтический состав AAV, описанный в настоящем описании, содержит один или более фармацевтически приемлемых эксципиентов для получения состава с преимущественными свойствами для хранения и/или введения индивидуумам для лечения генетического нарушения. В некоторых вариантах осуществления фармацевтические составы, описанные в настоящем описании, могут храниться при -65°C в течение периода, составляющего по меньшей мере 2 недели, в одном варианте осуществления по меньшей мере 4 недели, в другом варианте осуществления по меньшей мере 6 недель и в другом варианте осуществления по меньшей мере приблизительно 8 недель, без поддающегося обнаружению изменения стабильности. В этом отношении, термин "стабильный" означает, что рекомбинантный вирус AAV, присутствующий в составе, по существу сохраняет его физическую стабильность, химическую стабильность и/или биологическую активность в ходе хранения. В некоторых вариантах осуществления рекомбинантный вирус AAV, присутствующий в фармацевтическом составе, сохраняет по меньшей мере приблизительно 80% его биологической активности у пациента-человека в ходе хранения в течение заданного периода времени при -65°C, в других вариантах осуществления по меньшей мере приблизительно 85%, 90%, 95%, 98% или 99% его биологической активности у человека. В одном варианте осуществления индивидуумы представляют собой детей.
[00237] В некоторых аспектах состав, содержащий рекомбинантные вирионы AAV, дополнительно содержит одно или более буферных средств. Например, в различных вариантах осуществления состав, описанный в настоящем описании, содержит двухосновный фосфат натрия в концентрации от приблизительно 0,1 мг/мл до приблизительно 3 мг/мл, от приблизительно 0,5 мг/мл до приблизительно 2,5 мг/мл, от приблизительно 1 мг/мл до приблизительно 2 мг/мл, или от приблизительно 1,4 мг/мл до приблизительно 1,6 мг/мл. В одном варианте осуществления состав AAV, описанный в настоящем описании, содержит приблизительно 1,42 мг/мл двухосновного фосфата натрия (высушенного). Другое буферное средство, которое может быть использовано в составах рекомбинантного AAV, предусматриваемых в рамках настоящего изобретения, представляет собой моногидрат одноосновного фосфата натрия, который в некоторых вариантах осуществления может использоваться в концентрации от приблизительно 0,1 мг/мл до приблизительно 3 мг/мл, от приблизительно 0,5 мг/мл до приблизительно 2,5 мг/мл, от приблизительно 1 мг/мл до приблизительно 2 мг/мл, или от приблизительно 1,3 мг/мл до приблизительно 1,5 мг/мл. В одном варианте осуществления состав AAV в соответствии с настоящим вариантом осуществления содержит приблизительно 1,38 мг/мл моногидрата одноосновного фосфата натрия. В другом варианте осуществления рекомбинантный состав AAV, описанный в настоящем описании, содержит приблизительно 1,42 мг/мл двухосновного фосфата натрия и приблизительно 1,38 мг/мл моногидрата одноосновного фосфата натрия.
[00238] В другом варианте осуществления состав рекомбинантного AAV, предусматриваемый в рамках настоящего изобретения, может содержать одно или более обеспечивающих изотоничность средств, таких как хлорид натрия, в одном варианте осуществления в концентрации от приблизительно 1 мг/мл до приблизительно 20 мг/мл, например, от приблизительно 1 мг/мл до приблизительно 10 мг/мл, от приблизительно 5 мг/мл до приблизительно 15 мг/мл, или от приблизительно 8 мг/мл до приблизительно 20 мг/мл. В другом варианте осуществления состав, описанный в настоящем описании, содержит приблизительно 8,18 мг/мл хлорида натрия. Другие буферные средства и обеспечивающие изотоничность средства, известные в данной области, являются пригодными и могут быть стандартным образом использованы для применения в составах, предусматриваемых в рамках настоящего изобретения.
[00239] В другом варианте осуществления составы рекомбинантного AAV, предусматриваемые в рамках настоящего изобретения, могут содержать один или более наполнителей. Иллюстративные наполнители включают, но не ограничиваются ими, маннит, сахарозу, декстран, лактозу, трегалозу и повидон (PVP K24). В некоторых вариантах осуществления составы, предусматриваемые в рамках настоящего изобретения, содержат маннит, который может присутствовать в количестве от приблизительно 5 мг/мл до приблизительно 40 мг/мл, или от приблизительно 10 мг/мл до приблизительно 30 мг/мл, или от приблизительно 15 мг/мл до приблизительно 25 мг/мл. В другом варианте осуществления маннит присутствует в концентрации приблизительно 20 мг/мл.
[00240] В другом варианте осуществления составы рекомбинантного AAV по изобретению могут содержать одно или более поверхностно-активных веществ, которые могут представлять собой неионные поверхностно-активные вещества. Иллюстративные поверхностно-активные вещества включают ионные поверхностно-активные вещества, неионные поверхностно-активные вещества и их комбинации. Например, поверхностно-активное вещество может представлять собой, но не ограничиваясь ими, TWEEN 80 (также известный как полисорбат 80, или его химическое наименование полиоксиэтилена сорбитана моноолеат), додецилсульфат натрия, стеарат натрия, лаурилсульфат аммония, TRITON AG 98 (Rhone-Poulenc), полоксамер 407, полоксамер 188 и т.п., и их комбинации. В одном варианте осуществления состав согласно настоящему варианту осуществления содержит полоксамер 188, который может присутствовать в концентрации от приблизительно 0,1 мг/мл до приблизительно 4 мг/мл, или от приблизительно 0,5 мг/мл до приблизительно 3 мг/мл, от приблизительно 1 мг/мл до приблизительно 3 мг/мл, от приблизительно 1,5 мг/мл до приблизительно 2,5 мг/мл, или от приблизительно 1,8 мг/мл до приблизительно 2,2 мг/мл. В другом варианте осуществления полоксамер 188 присутствует в концентрации приблизительно 2,0 мг/мл.
[00241] Составы, содержащие рекомбинантный экспрессирующий терапевтический белок вирус AAV, описанные в настоящем описании, являются стабильными и могут храниться в течение длительных периодов времени без неприемлемого изменения качества, эффективности или чистоты. В одном аспекте состав является стабильным при температуре приблизительно 5°C (например, от 2°C до 8°C) в течение по меньшей мере 1 месяца, например, по меньшей мере 1 месяца, по меньшей мере 3 месяцев, по меньшей мере 6 месяцев, по меньшей мере 12 месяцев, по меньшей мере 18 месяцев, по меньшей мере 24 месяцев, или более. В другом варианте осуществления состав является стабильным при температуре, меньшей или равной приблизительно -20°C, в течение по меньшей мере 6 месяцев, например, по меньшей мере 6 месяцев, по меньшей мере 12 месяцев, по меньшей мере 18 месяцев, по меньшей мере 24 месяцев, по меньшей мере 36 месяцев или более. В другом варианте осуществления состав является стабильным при температуре, меньшей или равной приблизительно -40°C, в течение по меньшей мере 6 месяцев, например, по меньшей мере 6 месяцев, по меньшей мере 12 месяцев, по меньшей мере 18 месяцев, по меньшей мере 24 месяцев, по меньшей мере 36 месяцев, или более. В другом варианте осуществления состав является стабильным при температуре, меньшей или равной приблизительно -60°C, в течение по меньшей мере 6 месяцев, например, по меньшей мере 6 месяцев, по меньшей мере 12 месяцев, по меньшей мере 18 месяцев, по меньшей мере 24 месяца, по меньшей мере 36 месяцев, или более.
[00242] Фармацевтические композиции, как правило, являются стерильными и стабильными в условиях изготовления и хранения. Фармацевтические композиции могут быть составлены в качестве раствора, микроэмульсии, липосомы или другой упорядоченной структуры, пригодной для вмещения высокой концентрации лекарственного средства. Носитель может представлять собой растворитель или дисперсионную среду, включающие, например, воду, этанол, многоатомный спирт (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль, и т.п.), и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, с использованием покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае дисперсии и с использованием поверхностно-активных веществ. В некоторых вариантах осуществления в композицию включены обеспечивающие изотоничность средства, например, сахара, многоатомные спирты, такие как маннит, сорбит или хлорид натрия. Длительного всасывания инъекционных композиций можно достигать путем включения в композицию средства, которое замедляет всасывание, например, моностеаратов и желатина. В некоторых вариантах осуществления нуклеиновую кислоту или вектор, предусматриваемые в рамках настоящего изобретения, можно вводить в составе с замедленным или контролируемым высвобождением, например, в композиции, которая включает полимер для замедленного высвобождения или другие носители, которые защищают соединение от быстрого высвобождения, включая имплантаты и микроинкапсулированные системы доставки. Можно использовать, например, биодеградируемые биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры, полимолочная кислота и сополимеры молочной и гликолевой кислот (PLG).
[00243] В некоторых вариантах осуществления фармацевтическая композиция, содержащая вектор на основе AAV, предусматриваемый в рамках настоящего изобретения, может использоваться для переноса генетического материала в клетку. Такой перенос может происходить in vitro, ex vivo или in vivo. Таким образом, один вариант осуществления относится к способу доставки нуклеотидной последовательности в клетку, который включает приведение с ней в контакт нуклеиновой кислоты, вектора или фармацевтической композиции, как описано в настоящем описании, в условиях, в которых нуклеиновая кислота или вектор, описанные в настоящем описании, входят в клетку. Клетка может представлять собой клетку in vitro, ex vivo или in vivo.
5.5 СПОСОБЫ ЛЕЧЕНИЯ
[00244] В некоторых вариантах осуществления изобретение относится к способам лечения индивидуума, страдающего генетическим нарушением, включающим введение индивидууму терапевтически эффективного количества вектора на основе AAV, экспрессирующего терапевтический белок, или фармацевтической композиции, содержащей его. В этом случае "терапевтически эффективное количество" представляет собой количество вектора на основе AAV, которое после введения приводит к экспрессии терапевтического белка на уровне, достаточным для по меньшей мере частичного и предпочтительно полного смягчения симптомов генетического нарушения.
[00245] В одном варианте осуществления изобретение относится к способу лечения ФКУ, включающему введение терапевтически эффективного количества нуклеиновой кислоты, белка, вектора или клеток или фармацевтической композиции по изобретению пациенту, страдающему ФКУ. В одном варианте осуществления пациентом является человек. В одном варианте осуществления рассматриваемая популяция пациентов представляет собой пациентов с гиперфенилаланинемией от умеренной до тяжелой, включая пациентов с ФКУ, вариантом ФКУ или гиперфенилаланинемией, не являющейся ФКУ. В одном варианте осуществления целью введения вектора на основе AAV является переход у пациентов с тяжелой ФКУ либо к умеренной, либо к мягкой ФКУ, таким образом, снижая нагрузку, ассоциированную с диетой со строго ограниченным содержанием фенилаланина.
[00246] В одном варианте осуществления изобретение относится к способам повышения уровней циркулирующего белка PAH у индивидуума, нуждающегося в этом, включающим введение индивидууму любого из векторов на основе AAV, описанных в настоящем описании, или вирусных частиц, описанных в настоящем описании, или вирусных частиц, полученных способом, описанным в настоящем описании, которые экспрессируют белок PAH. В одном варианте осуществления доза вектора доставляет PAH, что приводит к снижению уровней фенилаланина в плазме до менее чем 160 мкМ.
[00247] В другом варианте осуществления изобретение относится к применению эффективного количества рекомбинантного вируса AAV PAH для получения лекарственного средства для лечения индивидуума, страдающего ФКУ. В одном варианте осуществления индивидуумом, страдающим ФКУ, является человек. В одном варианте осуществления лекарственное средство вводят посредством внутривенного (в/в) введения. В другом варианте осуществления введение лекарственного средства приводит к экспрессии белка PAH в кровотоке индивидуума, достаточной для изменения уровней метаболита нейротрансмиттера или нейротрансмиттера у индивидуума. В некоторых вариантах осуществления лекарственное средство также включает профилактический и/или терапевтический кортикостероид для предупреждения и/или лечения какой-либо гепатотоксичности, ассоциированной с введением вируса AAV PAH. Лекарственное средство, содержащее профилактический или терапевтический стероид, может включать по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60 или более мг кортикостероида/сутки. В некоторых вариантах осуществления лекарственное средство, включающее профилактический или терапевтический кортикостероид, можно вводить на протяжении непрерывного периода времени в течение по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10 недель или более. В другом варианте осуществления терапия ФКУ, описанная в настоящем описании, необязательно включает дополнение тирозином.
[00248] "Терапевтически эффективное количество" вектора на основе AAV, или вируса, или фармацевтической композиции, содержащей их, для лечения, как описано в настоящем описании, может быть определено эмпирически или стандартным образом. Однако в некоторых вариантах осуществления "терапевтически эффективное количество" рекомбинантного вируса AAV находится в диапазоне от приблизительно 1E12 вг/кг массы тела до приблизительно 1E14 вг/кг массы тела, в одном варианте осуществления от приблизительно 6E12 вг/кг массы тела до приблизительно 6E13 вг/кг массы тела. В другом варианте осуществления a терапевтически эффективное количество рекомбинантного вируса AAV составляет приблизительно 2E13 вг/кг массы тела. В другом варианте осуществления a терапевтически эффективное количество рекомбинантного вируса AAV составляет приблизительно 6E13 вг/кг массы тела.
[00249] В одном варианте осуществления рекомбинантные векторы/вирусы AAV, описанные в настоящем описании, можно вводить индивидууму, в одном варианте осуществления млекопитающему или человеку, различными известными способами введения. В другом варианте осуществления рекомбинантный вирус AAV для генной терапии вводят посредством внутривенной инъекции либо в качестве однократного болюсного введения, либо на протяжении длительного периода времени, который может составлять по меньшей мере приблизительно 1, 5, 10, 15, 30, 45, 60, 75, 90, 120, 150, 180, 210 или 240 минут, или более. В других вариантах осуществления вводимый рекомбинантный вирус AAV экспрессирует PAH.
[00250] В некоторых вариантах осуществления, в которых используют вектор на основе AAV, экспрессирующий PAH, для лечения ФКУ у индивидуумов, можно проводить мониторинг эффективности вектора на основе AAV путем измерения уровней фенилаланина в крови подвергаемого лечению индивидуума. Конкретные способы количественного анализа для определения уровней фенилаланина в кровотоке хорошо известны в данной области и включают флуориметрические способы анализа (см., McCaman, M.W. and Robins, E., (1962) J. Lab. Clin. Med., vol. 59, pp. 885-890); способы анализа на основе тонкослойной хроматографии (см., Tsukerman, G. L. (1985) Laboratornoe delo, vol. 6, pp. 326-327); ферментные способы анализа (см., La Du, B. N., et al. (1963) Pediatrics, vol. 31, pp. 39-46; и Peterson, K., et al. (1988) Biochem. Med. Metab. Biol., vol. 39, pp. 98-104); способы с использованием высокоэффективной жидкостной хроматографии (ВЭЖХ) (см., Rudy, J. L., et al. (1987) Clin. Chem., vol. 33, pp. 1152-1154); и высокопроизводительные автоматизированные способы (см., Hill, J. B., et al. (1985) Clin. Chem., vol. 5, pp. 541-546).
[00251] Введение вируса AAV по настоящему изобретению в некоторых случаях может приводить к поддающейся выявлению степени гепатотоксичности. Гепатотоксичность можно измерять различными хорошо известными и часто используемыми способами, например, путем измерения концентраций определенного ассоциированного с печенью фермента(ов) (например, аланинтрансаминаза, ALT) в кровотоке индивидуума как до введения AAV (т.е. исходный уровень), так и после введения AAV. Поддающееся выявлению повышение концентрации ALT после введения AAV (по сравнению с концентрацией до введения) указывает на индуцируемую лекарственным средством гепатотоксичность. В некоторых вариантах осуществления в дополнение к введению терапевтически эффективного количества вируса AAV индивидуума можно лечить либо профилактически, либо терапевтически, либо как профилактически, так и терапевтически, с использованием кортикостероида для предупреждения и/или лечения какой-либо гепатотоксичности, ассоциированной с введением вируса AAV.
[00252] "Профилактическое" лечение кортикостероидом относится к введению кортикостероида для предупреждения гепатотоксичности и/или для предупреждения повышения измеренных уровней ALT у индивидуума. "Терапевтическое" лечение кортикостероидом относится к введению кортикостероида для снижения гепатотоксичности, вызванной введением вируса AVV, и/или для снижения повышенной концентрации ALT в кровотоке индивидуума, вызванной введением вируса AAV. В некоторых вариантах осуществления профилактическое или терапевтическое лечение кортикостероидом может включать введение индивидууму по меньшей мере 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60 или более мг кортикостероида/сутки. В некоторых вариантах осуществления профилактическое или терапевтическое лечение индивидуума кортикостероидом может происходить на протяжении непрерывного периода времени, составляющего по меньшей мере приблизительно 3, 4, 5, 6, 7, 8, 9, 10 недель, или более. Кортикостероиды, пригодные в способах, описанных в настоящем описании, включают любые известные или обычно используемые кортикостероиды, включая, например, дексаметазон, преднизон, флудрокортизон, гидрокортизон и т.п.
5.6 ДЕТЕКЦИЯ АНТИТЕЛ К AAV
[00253] Для максимального повышения вероятности успешной трансдукции печени посредством системного AAV-опосредуемого переноса терапевтических генов перед введением вектора на основе AAV в терапевтическом режиме пациенту-человеку, как описано выше, предполагаемого пациента можно оценивать в отношении присутствия антител к капсиду AAV, которые способны блокировать трансдукцию клеток или иным образом снижать общую эффективность терапевтического режима. Такие антитела могут присутствовать в сыворотке предполагаемого пациента и могут быть направлены против капсида AAV любого серотипа. В одном варианте осуществления серотип, против которого направлены предсуществующие антитела, представляет собой AAV5.
[00254] Способы выявления предсуществующего иммунитета против AAV хорошо известны и часто используются в данной области, и включают клеточные способы анализа ингибирования трансдукции in vitro (TI), способы анализа TI in vivo (например, у мышей), и детекцию на основе ELISA тотальных антител против капсида (TAb) (см., например, Masat et al., Discov. Med., vol. 15, pp. 379-389 и Boutin et al., (2010) Hum. Gene Ther., vol. 21, pp. 704-712). В способах анализа TI могут использоваться клетки-хозяева, в которые предварительно введен индуцируемый AAV репортерный вектор. Репортерный вектор может включать индуцибельный репортерный ген, такой как GFP и т.д., экспрессия которого индуцируется при трансдукции клетки-хозяина вирусом AAV. Антитела против капсида AAV, присутствующие в сыворотке человека, которые способны предупреждать/снижать трансдукцию клетки-хозяина, таким образом, будут снижать общую экспрессию репортерного гена в системе. Таким образом, такие способы анализа можно использовать для выявления присутствия антител против капсида AAV в сыворотке человека, которые способны предупреждать/снижать трансдукцию клеток посредством терапевтического вируса AAV PAH.
[00255] В способах анализа с TAb для детекции антител к капсиду AAV может использоваться связанный с твердой фазой капсид AAV в качестве "улавливающего агента", над которой пропускают сыворотку человека, тем самым позволяя антителам против капсида, присутствующим в сыворотке, связываться со связанным с твердой подложкой капсидным "улавливающим агентом". После промывания для удаления неспецифического связывания можно использовать "агент детекции" для выявления присутствия антикапсидных антител, связанных с улавливающим агентом. Агент детекции может представлять собой антитело, капсид AAV и т.п., и он может быть меченным поддающейся детекции меткой для облегчения детекции и количественного определения связанного антитела против капсида. В одном варианте осуществления агент детекции является меченным рутением или комплексом рутения, детекцию которого можно проводить с использованием электрохемилюминесцентных способов и оборудования.
[00256] Ту же описанную выше методологию можно использовать для оценки и детекции формирования иммунного ответа против капсида AAV у пациента, которому ранее вводили представляющий интерес терапевтический вирус AAV. По существу эти способы можно использовать не только для оценки присутствия антител против капсида AAV перед лечением терапевтическим вирусом AAV, но также их можно использовать для оценки и количественного определения индукции иммунного ответа против введенного терапевтического вируса AAV после введения. По существу, в рамках настоящего изобретения предусматриваются способы, которые сочетают в себе способы детекции антител против капсида AAV в сыворотке человека и введение терапевтического вируса AAV для лечения ФКУ, где способы выявления антител против капсида AAV в сыворотке человека можно проводить либо до, либо после введения терапевтического вируса AAV.
[00257] Другие аспекты и преимущества настоящего изобретения станут понятными при изучении следующих иллюстративных примеров.
6. ПРИМЕРЫ
6.1 ПРИМЕР 1: МЕТАБОЛИЧЕСКИЙ КАСКАД фЕНИЛАЛАНИНА И МЕТОДОЛОГИЯ АНАЛИЗА
[00258] PAH-опосредуемое конвертирование фенилаланина в тирозин служит в качестве реакции-предшественника для продуцирования ряда нейротрансмиттеров, как показано на схеме 1.
Схема 1:
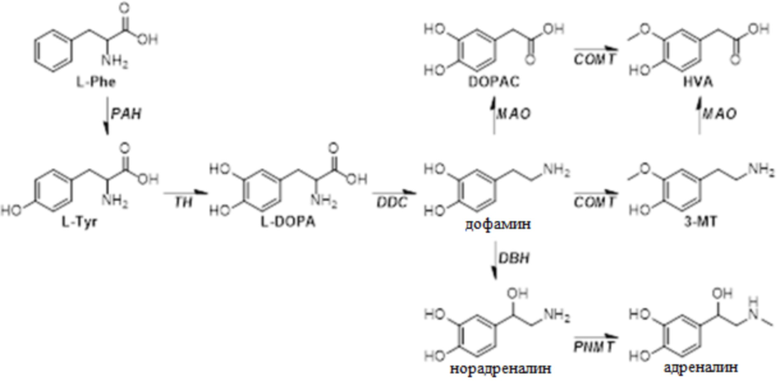
Схема 1 (продолжение):
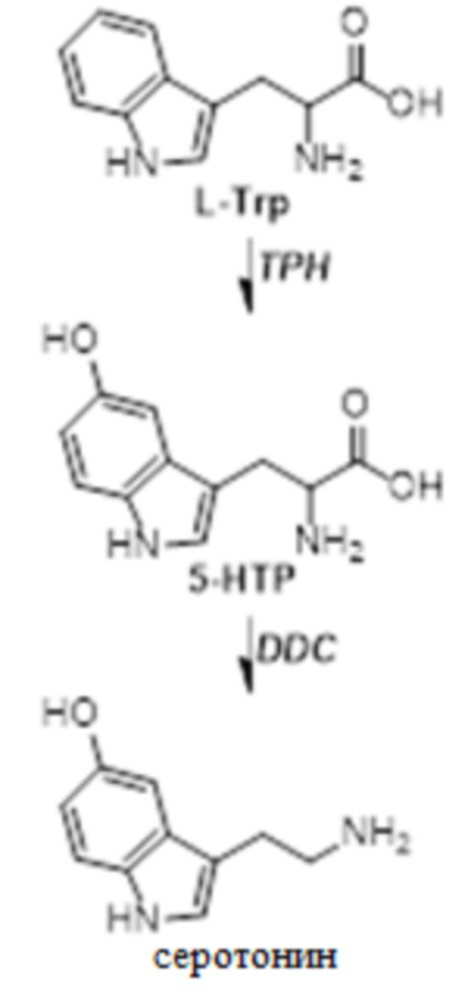
[00259] Потеря активности PAH у пациентов с ФКУ приводит к накоплению фенилаланина, как и отмечалось, посредством повышения уровней фенилаланиина в плазме. Однако нейрокогнитивные эффекты ФКУ по большей части вызываются утратой предшествующей метаболической стадии, которая приводит к продуцированию ряда нейротрансмиттеров и метаболитов.
[00260] Чувствительный анализ на основе жидкостной хроматографии, сопряженной с масс-спектрометрией (LC/MS), использовали для измерения уровней различных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров. В кратком изложении, когда аминокислоты, метаболиты нейротрансмиттеров и нейротрансмиттеры подвергают реакции с этилхлорформиатом и пиридином или бензоилхлоридом и карбонатом натрия, они продуцируют продукты реакции, которые имеют определенное увеличение молекулярной массы, которое позволяет их идентификацию и количественное определение с использованием LC/MS. Этот процесс реакции подробно проиллюстрирован на схеме 2 для нейротрансмиттера дофамина, где дофамин массой 153,08 Да конвертируется в дофамин-ECF массой 369,14 Да.
Схема 2:
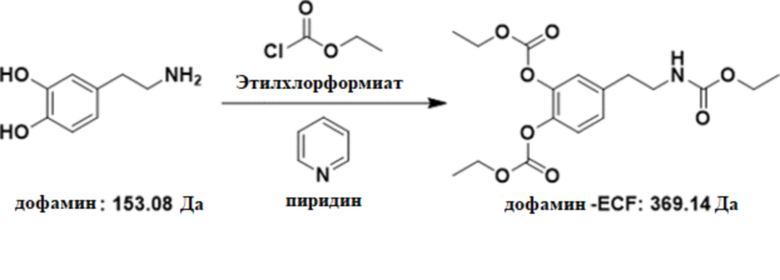
[00261] Продукты реакции этилхлорформиата и пиридина и их точные массы представлены для аминокислот, нейротрансмиттеров, метаболитов нейротрансмиттеров, которые продуцируются после реакции фенилаланина с участием фермента PAH, представлены на схеме 3.
Схема 3:
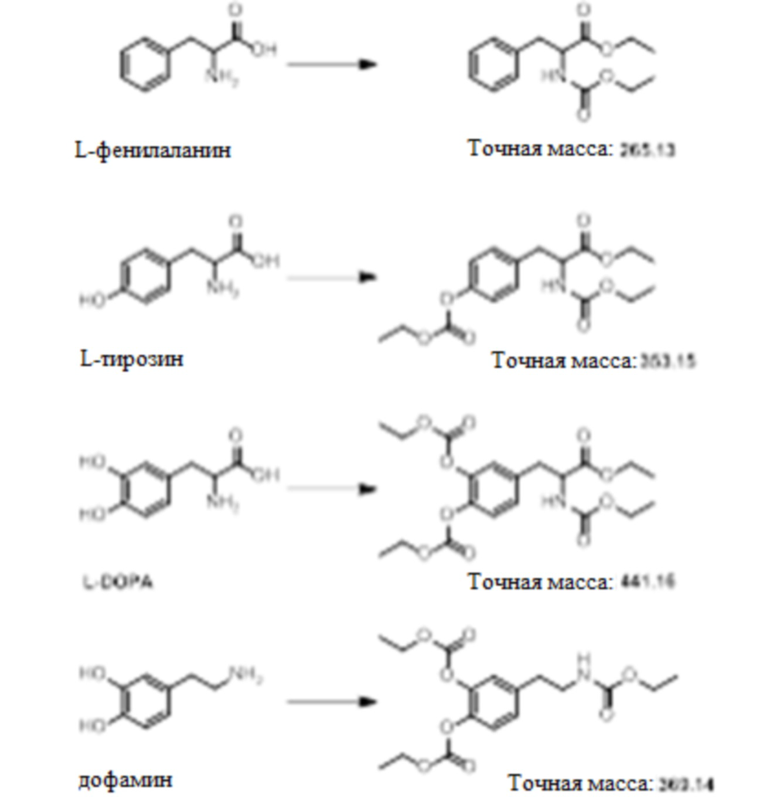
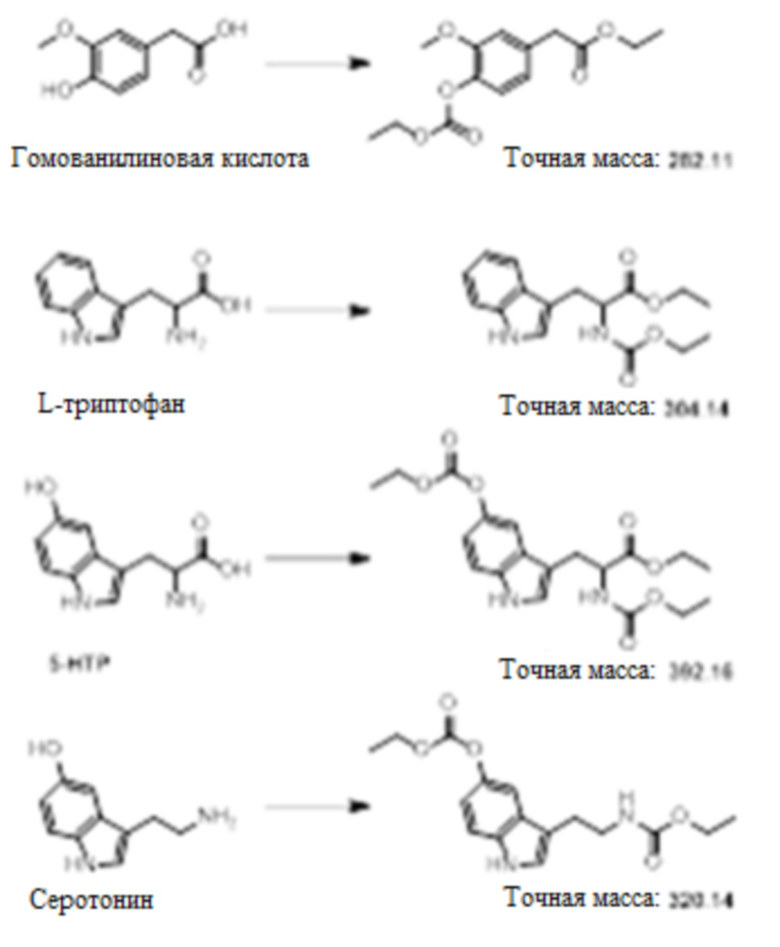
[00262] На фиг.1 представлен иллюстративный пример хроматограммы жидкостной хроматографии - масс-спектрометрии (LC/MS) для происходящих из фенилаланина аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров после реакции с этилхлорформиатом и пиридином или бензоилхлоридом и карбонатом натрия. Как видно на фиг.1, каждый из продуктов реакции может быть отчетливо установлен и количественно определен.
6.2 ПРИМЕР 2: УРОВНИ НЕЙРОТРАНСМИТТЕРОВ НЕИЗМЕННО НИЖЕ В ОБРАЗЦАХ МЫШЕЙ ENU2 ПО СРАВНЕНИЮ С ОБРАЗЦАМИ МЫШЕЙ ДИКОГО ТИПА
[00263] Анализ, описанный выше в примере 1, использовали для количественного определения уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в биологических образцах, полученных от мышей Enu2 и мышей дикого типа, чтобы определить эффект утраты PAH на продуцирование нейротрансмиттеров. Как показано на фиг.2, мыши Enu2 неизменно демонстрировали более низкие уровни нейротрансмиттеров и метаболитов нейротрансмиттеров по сравнению с мышами дикого типа в образцах головного мозга. Мыши Enu2 имели сниженные уровни тирозина, дофамина, 3-метокситирамина (3-MT), DOPAC, гомованилиновой кислоты, норадреналина, триптофана и серотонина в их головном мозге по сравнению с уровнями в головном мозге мышей дикого типа. Эти данные демонстрируют, что утрата активности PAH приводит к снижению уровня ряда ключевых нейротрансмиттеров.
6.3 ПРИМЕР 3: УРОВНИ НЕЙРОТРАНСМИТТЕРОВ СЛУЖАТ В КАЧЕСТВЕ СУРРОГАТНЫХ МАРКЕРОВ ДЛЯ ИДЕНТИФИКАЦИИ ЭФФЕКТИВНОЙ ДОЗЫ ТЕРАПЕВТИЧЕСКОГО СРЕДСТВА ПРОТИВ ФКУ.
[00264] Мышам Enu2 вводили две дозы (2E13 вг/кг и 2E14 вг/кг) аденоассоциированного вирусного вектора, экспрессирующего PAH человека (AAV-PAH). AAV конструировали с ITR серотипа AAV2 и генами капсида серотипа AAV5. Мышам Enu2 и дикого типа также вводили носитель в качестве контроля. Через двенадцать недель после введения вируса проводили измерение уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в образцах головного мозга, полученных от каждой группы животных. Как показано на фиг.3, мыши Enu2, которым вводили носитель, имели повышенные уровни фенилаланина и сниженные уровни тирозина, дофамина, DOPAC, гомованилиновой кислоты, норадреналина, триптофана и серотонина относительно уровней, измеренных у мышей дикого типа, которым вводили носитель. Введение более высокой дозы AAV-PAH (2E14 вг/кг) мышам Enu2, но не более низкой дозы (2E13 вг/кг), восстанавливало уровни фенилаланина, тирозина, дофамина, DOPAC, гомованилиновой кислоты, норадреналина, триптофана и серотонина до уровней, наблюдаемых в случае контролей дикого типа, которым вводили носитель. Эти данные демонстрируют, что уровни определенных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в головном мозге можно использовать для определения эффективной дозы терапевтического средства против ФКУ, требуемого для восстановления путей метаболизма нейротрансмиттеров до состояния, наблюдаемого у контролей дикого типа.
6.4 ПРИМЕР 4: УРОВНИ ФЕНИЛАЛАНИНА В ПЛАЗМЕ КОРРЕЛИРУЮТ С УРОВНЯМИ, ОБНАРУЖЕННЫМИ В ГОЛОВНОМ МОЗГЕ
[00265] Уровни фенилаланина в моче, крови, сыворотке и плазме являются известным биомаркером для диагностики ФКУ и мониторинга хода лечения заболевания. Уровни фенилаланина в плазме и головном мозге измеряют и сравнивают с мышами дикого типа и Enu2, которым вводили носитель, и мышами Enu2, которым вводили 2E13 вг/кг и 2E14 вг/кг AAV-PAH. Как показано на фиг.4, существует строгая линейная корреляция между уровнями фенилаланина в плазме и головном мозге у индивидуальных животных. Более того, корреляция сохраняется, независимо от статуса PAH и дозы введения AAV-PAH. Эти данные обеспечивают дополнительное подтверждение того, что величина уровня фенилаланина в плазме является суррогатным биомаркером нейрокогнитивных симптомов ФКУ, поскольку уровни фенилаланина в головном мозге могут определять уровни образующихся нейротрансмиттеров.
6.5 ПРИМЕР 5: УРОВНИ НЕЙРОТРАНСМИТТЕРОВ И МЕТАБОЛИТОВ НЕЙРОТРАНСМИТТЕРОВ В ПЛАЗМЕ МЫШЕЙ ENU2 И МЫШЕЙ ДИКОГО ТИПА
[00266] Уровни метаболитов нейротрансмиттеров DOPAC и гомованилиновой кислоты измеряли у мышей дикого типа и мышей Enu2. Как показано на фиг.5, мыши Enu2 имеют более низкие уровни метаболитов нейротрансмиттера в их плазме по сравнению с мышами дикого типа. Более того, уровни этих метаболитов в плазме коррелируют с уровнями, обнаруженными в головном мозге (фиг.6 и 8). Эти данные демонстрируют, что уровни DOPAC и гомованилиновой кислоты в плазме могут выступать в качестве суррогатных для понимания уровней метаболитов в головном мозге и могут служить в качестве определяемых биомаркеров нейрокогнитивных симптомов.
[00267] Проводили измерение уровней фенилаланина, тирозина, дофамина, DOPAC, 3MT, гомованилиновой кислоты, норадреналина, триптофана и серотонина в плазме и головном мозге мышей дикого типа и мышей Enu2, которым вводили носитель, и у мышей Enu2, которым вводили 2E13 вг/кг и 2E14 вг/кг AAV-PAH. Как показано на фиг.7, доза 2E14 вг/кг AAV-PAH увеличивала уровни в плазме измеряемых аминокислот (за исключением уровней фенилаланина, которые снижались до уровней дикого типа), нейротрансмиттеров и метаболитов нейротрансмиттераов до уровней, наблюдаемых у мышей дикого типа. Напротив, доза 2E13 вг/кг AAV-PAH не восстанавливала в полной мере уровни измеренных аминокислот, метаболитов нейротрансмиттеров и нейротрансмиттеров. Кроме того, существует обратная корреляция между уровнями фенилаланина и уровнями метаболитов нейротрансмиттеров DOPAC и гомованилиновой кислоты (фиг.9). Это согласуется с мнением, что утрата активности PAH приводит к выраженному снижению уровней метаболитов, которое является основой для нейрокогнитивных симптомов ФКУ. Таким образом, уровни в плазме или головном мозге этих нейротрансмиттеров или их конечных метаболитов могут служить в качестве сопутствующих диагностических средств для определения эффективного количества терапевтического средства на основе ФКУ, требуемого для лечения или смягчения нейрокогнитивных симптомов ФКУ. Это показано в качестве неограничивающих иллюстративных примеров на фиг.10 и 11. В этом случае, уровни в головном мозге дофамина, норадреналина и DOPAC измеряли у мышей дикого типа и мышей Enu2, которым вводили носитель, и у мышей Enu2, которым вводили 2E13 вг/кг и 2E14 вг/кг AAV-PAH. Как показано на фиг.10 и 11, более низкая доза 2E13 вг/кг была достаточной для восстановления уровней дофамина, однако была недостаточной для восстановления уровней DOPAC и норадреналина. Напротив, более высокая доза 2E14 вг/кг полностью восстанавливала уровни дофамина, DOPAC и норадреналина у животных Enu2 до уровней, наблюдаемых у контролей дикого типа. Этот набор суррогатных маркеров был достаточным для определения того, что доза 2E14 вг/кг была эффективной для восстановления этих нейрокогнитивных каскадов, в то время как более низкая доза не была эффективной.
[00268] В качестве дополнительного примера, использование уровней в плазме метаболитов нейротрансмиттеров в качестве суррогатного маркера эффективной дозы показано на фиг.12 и 13. В данном случае уровни в плазме DOPAC и гомованилиновой кислоты измеряли у мышей дикого типа и Enu2, которым вводили носитель, и у мышей Enu2, которым вводили 2E13 вг/кг и 2E14 вг/кг AAV-PAH. Как показано на фиг.12, высокая доза 2E14 вг/кг была способна восстановить уровни в плазме DOPAC и гомованилиновой кислоты до уровней, наблюдаемых в случае дикого типа, в то время как низкая доза 2E13 вг/кг не была. Более того, как показано на фиг.13, уровни этих маркеров в плазме и головном мозге имели обратную корреляцию с уровнями в плазме и головном мозге их аминокислотного предшественника фенилаланина.
6.6 ПРИМЕР 6: ЭФФЕКТИВНОЕ КОЛИЧЕСТВО PALYNZIQ® (ПЕГВАЛИАЗА -PQPZ) ВОССТАНАВЛИВАЕТ УРОВНИ НЕЙРОТРАНСМИТТЕРОВ
[00269] Мышам Enu2 вводили терапевтическое средство против ФКУ, PALYNZIQ® (пегвалиаза-pqpz), или носитель. После дозирования в течение трех суток PALYNZIQ® (пегвалиаза-pqpz) мышей умерщвляли, и проводили взятие крови и головного мозга и исследование в отношении присутствия отдельных нейротрансмиттеров и метаболитов нейротрансмиттеров. Как показано на фиг.14, 15 и 16, мыши Enu2, которым вводили PALYNZIQ® (пегвалиаза-pqpz), демонстрировали увеличенные уровни нейротрансмиттеров и метаболитов нейротрансмиттеров как в плазме, так и в головном мозге, по сравнению с мышами, которым вводили носитель. Введение PALYNZIQ® (пегвалиаза-pqpz) в течение трех суток восстанавливало уровни фенилаланина, тирозина и триптофана в головном мозге до уровней, наблюдаемых у дикого типа, и уровни в плазме фенилаланина, тирозина и триптофана служили в качестве приблизительных заместителей соответствующих уровней в головном мозге (фиг.14). Кроме того, PALYNZIQ® (пегвалиаза-pqpz) в течение трех суток также была способна нормализовать уровни в головном мозге дофамина, норадреналина и серотонина (фиг.15). Введение PALYNZIQ® (пегвалиаза-pqpz) также восстанавливало уровни DOPAC, гомованилиновой кислоты и 5-гидроксилиндолуксусной кислоты в головном мозге до уровней, наблюдаемых в головном мозге нормальной мыши, уровни в плазме всех трех метаболитов служили в качестве суррогатных для уровней, наблюдаемых в головном мозге. Эти данные далее демонстрируют ценность использования уровней нейротрансмиттеров и метаболитов в плазме в качестве суррогатных маркеров для идентификации эффективной дозы терапевтического средства против ФКУ.
[00270] Различные уровни дозы PALYNZIQ® (пегвалиаза-pqpz) (10-80 мг/кг) или носителя (забуференный tris солевой раствор/2 мМ транс-циннамат; 4 мл/кг)) вводили самцам мышей C57BL6-Pahenu2 (n=7) посредством подкожной (п/к) инъекции на 0 сутки. После кратковременного нагревания мыши под нагревательной лампой для индукции расширения сосудов проводили взятие образцов крови от каждого животного до и через 24 часа после введения путем проведения поперечного разреза на коже хвоста длиной приблизительно 0,1 см с использованием стерильного лезвия скальпеля. Выходящую кровь собирали в одну капиллярную пробирку с литий-гепарином, которую сразу центрифугировали с получением плазмы. Плазму сливали в чистые обозначенные пробирки и хранили при -80°C. Через семьдесят два часа после введения PALYNZIQ® (пегвалиаза-pqpz), каждое животное подвергали глубокой анестезии с использованием изофлурана, доставляемого через назальный конус (4% для индукции, 1,5% для поддержания). Взятие последнего образца крови проводили посредством пункции сердца, и его помещали в пробирку, содержавшую литий-гепарин. Затем животных умерщвляли посредством кровопускания, проведения трахеотомии перед полной перфузией организма PBS через сердце в течение ~10 минут, а затем смещения шейных позвонков. Затем головной мозг каждой мыши извлекали, разрезали пополам в сагиттальном направлении, быстро замораживали и хранили в обозначенных пробирках при -80°C. Образцы крови центрифугировали с получением плазмы, которую сливали в чистые обозначенные пробирки и хранили при -80°C.
[00271] Способы анализа, описанные в примерах 12 и 13, использовали для измерения уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров. Как показано на фиг.17-18, через 72 часа после однократной дозы PALYNZIQ® (пегвалиаза-pqpz) 40 мг/кг или 80 мг/кг уровни фенилаланина как в головном мозге, так и в плазме, и уровни триптофана в головном мозге нормализовывались до уровней, наблюдаемых у мышей дикого типа. На фиг.19 показано, что однократная доза PALYNZIQ® (пегвалиаза-pqpz) 40 мг/кг или 80 мг/кг нормализовывала уровни фенэтиламина, дофамина и серотонина и увеличивала уровни норадреналина через 72 часа после введения. Уровни метаболитов фенэтиламина и серотонина в плазме также нормализовывались до уровней дикого типа, как видно на фиг.20. При введении PALYNZIQ® уровни всех метаболитов нейротрансмиттеров в головном мозге возрастали относительно мышей Enu2, которым вводили носитель (пегвалиаза-pqpz) (фиг.21A-C).
6.7 ПРИМЕР 7: УРОВНИ МАРКЕРОВ КОРРЕЛИРУЮТ У МЫШЕЙ ENU
6.7.1 Уровни аминокислот в головном мозге коррелируют со снижением уровня Phe в плазме у мышей Enu
[00272] Уровни аминокислот количественно определяли в плазме и гомогенатах головного мозга мышей PAHEnu2 и PAHWT через 72 часа после введения носителя или PALYNZIQ® (пегвалиаза-pqpz) (фиг.17 и 18A). PALYNZIQ® (пегвалиаза-pqpz) выше 10 мг/кг снижала уровни Phe в плазме мышей PAHEnu2 с 1999,0±85,0 мкМ (среднее значение±SEM) до < 1,0 мкМ (PAHWT=60,4±4,4 мкМ) (фиг.17, слева). В противоположность плазме, уровни Phe в головном мозге нормализовывались относительно PAHWT при введении PALYNZIQ® (пегвалиаза-pqpz) и снижались с 743,7±37,4 до 107,7+6,6 мкМ (PAHWT=108,8±6,7 мкМ) (фиг.18A, слева).
[00273] Хотя уровни Phe в головном мозге не снижались ниже PAHWT, как наблюдалось для Phe в плазме, была выявлена строгая корреляция между этими двумя компартментами с величиной r 0,89 (95% доверительный интервал (CI) 0,81-0,94, p < 0,0001) (фиг.18B, слева).
[00274] В то время как уровень Tyr в плазме оставался низким во всех группах PAHEnu2 относительно PAHWT (фиг.17, середина), Tyr в головном мозге у мышей PAHEnu2, которым вводили PALYNZIQ® (пегвалиаза-pqpz), возрастал от 68,8±2,9 до 85,1±6,1 мкМ (PAHWT=99,0±2,6 мкМ) (фиг.18A, середина). Tyr в головном мозге коррелировал со снижением Phe в головном мозге ниже 400 мкМ (фиг.18B, середина).
[00275] Уровни Trp в плазме оставались нормальными во всех группах введения (фиг.17, справа). Интересно, что Trp в головном мозге у мышей PAHEnu2, которым вводили PALYNZIQ® (пегвалиаза-pqpz) нормализовался относительно PAHWT, и возрастал от 20,4±1,1 мкМ до 30,5±0,8 мкМ (PAHWT=29,2±0,7 мкМ) (фиг.18A, справа). Уровни Trp в головном мозге нормализовывались, когда уровень Phe в головном мозге снижался ниже 400 мкМ (фиг.18B, справа).
6.7.2 Уровни нейротрансмиттеров коррелируют с коррекцией аминокислот в головном мозге мышей
[00276] Уровни всех нейротрансмиттеров нормализовывались относительно PAHWT при введении PALYNZIQ® (пегвалиаза-pqpz); уровень PEA в плазме снижался от 53,6±1,8 до 2,7±0,8 нМ (PAHWT=3,7±1,0 нМ), уровень дофамина в головном мозге возрастал от 1115±78,6 до 1506±96,5 нМ (PAHWT=1492±138,4 нМ), уровень норадреналина в головном мозге возрастал от 1335±54,0 до 1855±78,0 нМ (PAHWT=2247±54,9 нМ), и уровень серотонина в головном мозге возрастал от 166±12,5 до 583,8±45,6 (PAHWT=502±47,6 нМ) (фиг.19A).
[00277] Наблюдалась строгая положительная корреляция между уровнями PEA в плазме и Phe в головном мозге с величиной r 0,95 (95% CI 0,89-0,97, p < 0,0001). Уровень дофамина, норадреналина и серотонина в головном мозге обратно коррелировал с уровнем Phe в головном мозге с величинами r -0,61 (95% CI от -0,79 до -0,34, p=0,0002), -0,63 (95% CI от -0,80 до -0,35, p=0,0001) и -0,92 (95% CI от -0,96 до -0,84, p < 0,0001), соответственно (фиг.19B).
6.7.3 Уровни метаболитов нейротрансмиттера коррелируют с уровнями нейротрансмиттеров у мышей
[00278] Уровни всех метаболитов нейротрансмиттеров в головном мозге возрастали относительно PAHEnu2, которым вводили носитель, при введении PALYNZIQ® (пегвалиаза-pqpz); уровень HVA в головном мозге возрастал от 869±41,01 до 1073±67,5 нМ, уровень MOPEG в головном мозге возрастал от 640,5±55,22 до 746±40,7 нМ, и уровень 5HIAA в головном мозге возрастал от 280,2±20,2 до 1318,0±37,4 нМ (фиг.21).
[00279] Уровни PAGly в плазме нормализовывались относительно PAHWT при введении PALYNZIQ® (пегвалиаза-pqpz) и снижались от 205,7±24,8 нМ до 14,69±2,2 нМ (PAHWT=11,9±0,7 нМ) (фиг.20A, слева). Уровни HVA и MOPEG в плазме были сходными с их соответствующими уровнями в головном мозге; мало мышей PAHEnu2 достигали уровней PAHWT в плазме (фиг.20A, середина). Уровни 5HIAA в плазме были сходными с уровнями 5HIAA до достижения плато при 400 нМ как в группе высокой дозы, так и в группе PAHWT (фиг.20A, справа).
[00280] Уровень PAGly в плазме прямо коррелировал с уровнем PEA в плазме с величиной r 0,93 (95% CI 0,86-0,96, p < 0,0001), указывая на прямую взаимосвязь с метаболитом (фиг.20B, слева). HVA, MOPEG и 5HIAA в плазме коррелировали с соответствующими уровнями метаболитов нейротрансмиттеров в головном мозге с величинами r 0,62 (95% CI 0,37-0,78, p < 0,0001), 0,556 (95% CI 0,29-0,74, p=0,0003) и 0,66 (95% CI 0,40-0,79, p < 0,0001), соответственно (фиг.20B, в середине и справа).
6.8 ПРИМЕР 8: ДОПОЛНЕНИЕ ТИРОЗИНОМ В КОМБИНАЦИИ С ВВЕДЕНИЕМ PALYNZIQ® (ПЕГВАЛИАЗА-PQPZ) ПОВЫШАЕТ УРОВНИ НОРАДРЕНАЛИНА В ГОЛОВНОМ МОЗГЕ У МЫШЕЙ PAHEnu2
[00281] В отдельном испытании самцом мышей C57BL6-PAHEnu2 проводили дополнение тирозином, суспендированном в фосфатно-солевом буфере (PBS), 60 мг/кг, три раза в сутки через желудочный зонд, или контролем в виде носителя, начиная за одни сутки до п/к введения PALYNZIQ® (пегвалиаза-pqpz), 40 мг/кг, или носителя, и продолжали каждые сутки в течение 4 суток после введения дозы PALYNZIQ® (пегвалиаза-pqpz). Взятие образцов крови проводили до начала дозирования Tyr, и через 24 и 96 часов после введения PALYNZIQ® (пегвалиаза-pqpz). Головной мозг извлекали через 96 часов после введения дозы PALYNZIQ® (пегвалиаза-pqpz) и через 1 час после последней дозы Tyr.
[00282] Нарезанный головной мозг взвешивали и гомогенизировали в PBS (4:1 (об./масс.), содержавшем 1% NP-40, 0,5% дезоксихолат натрия и 0,1% додецилсульфат натрия) с использованием стандартных протоколов на устройстве FastPrep-24 5G (MP Biomedicals 116005500). Супернатанты получали путем центрифугирования гомогенатов головного мозга при 21000 g в течение 15 минут при 4°C, а затем хранили аликвотами при -80°C.
[00283] Дополнение тирозином (180 мг/кг/сутки) увеличивало уровни в плазме Tyr у мышей PAHEnu2, которым вводили носитель и PALYNZIQ® (пегвалиаза-pqpz) (фиг.22C). Дополнение Tyr несколько увеличивало уровни Tyr в головном мозге у мышей PAHEnu2, которым вводили носитель, но не имело эффекта у мышей PAHEnu2, которым вводили PALYNZIQ® (пегвалиаза-pqpz) (фиг.22D). Уровень дофамина в головном мозге достигал уровней PAHWT у мышей PAHEnu2, которым вводили PALYNZIQ® (пегвалиаза-pqpz), независимо от дополнения Tyr (фиг.22E). Наиболее неожиданно, дополнение Tyr у мышей PAHEnu2, которым вводили, увеличивало уровни норадреналина в головном мозге относительно дополнения PBS (p=0,0002) от 1481±34,8 до 1818±62,7 нМ (фиг.22F).
6.9 ПРИМЕР 9: УРОВНИ МЕТАБОЛИТОВ НЕЙРОТРАНСМИТТЕРОВ ВОЗРАСТАЛИ У ИНДИВИДУУМОВ С ФКУ СО СНИЖЕННЫМ УРОВНЕМ PHE В ПЛАЗМЕ
[00284] Данные у мышей PAHEnu2, которым проводили введение PALYNZIQ® (пегвалиаза-pqpz), указывают на то, что снижение Phe в плазме восстанавливает нейротрансмиттеры до уровней PAHWT в головном мозге, причем метаболиты нейротрансмиттеров в плазме демонстрируют сходные тенденции. Для определения того, можно ли эти данные транслировать на людей с ФКУ, проводили оценку уровней метаболитов нейротрансмиттеров в образцах плазмы от подгруппы индивидуумов в клинических испытаниях фазы 3 PALYNZIQ® (пегвалиаза-pqpz) с уровнями Phe в крови до лечения >900 мкМ. Оценивали уровни в плазме от 23 индивидуумов, которые участвовали в клинических испытаниях PALYNZIQ® (пегвалиаза-pqpz) фазы 3 (PRISM-1 NCT01819727 или PRISM-2, NCT01889862). Все индивидуумы предоставляли письменное информированное согласие. Контрольные образцы плазмы взрослых были получены от Discovery Life Sciences (Los Osos, CA) и не отличались на статистически значимом уровне по возрасту и полу. BMI не был доступен для контролей. Контрольные образцы представляли собой деидентифицированные остатки образцов от диагностической процедуры, не связанной с ФКУ. Способы оценки уровней Phe и ADHD-RS IV в плазме в клинических испытаниях PALYNZIQ® (пегвалиаза-pqpz) были описаны ранее (Thomas et al).
[00285] Индивидуумов разделяли на две группы: индивидуумы, которые имели уровни Phe в плазме >900 мкМ, или индивидуумы, которые имели уровни Phe в плазме <360 мкМ после лечения в течение 12 месяцев. Уровень Phe в плазме <360 мкМ является верхней границей целевого диапазона Phe, рекомендованного в руководстве по лечению ФКУ ACMG (Vockley et al., 2014), в то время как уровень Phe в плазме >900 мкМ представляет собой уровень Phe, существенно превышающий уровни в руководствах по лучению ФКУ ACMG и EU (Vockley et al., 2014; van Spronsen et al 2018). Исходные характеристики соответствовали для двух групп 2 по возрасту, полу, ADHD-RS IV и уровням Phe в плазме, описанным в таблице 1 ниже:
Таблица 1. Исходные характеристики индивидуумов:
(Phe <360 мкМ)
(Phe >900 мкМ)
[00286] Уровни Phe в плазме пациентов, которых лечили PALYNZIQ® (пегвалиаза-pqpz), сравнивали с образцами, полученными от контрольной группы без ФКУ. Способы анализа, описанные в примерах 10 и 11, использовали для оценки уровней метаболитов нейротрансмиттеров во всех образцах плазмы после лечения в течение 0, 6 и 12 месяцев.
[00287] После лечения PALYNZIQ® (пегвалиаза-pqpz) в течение 12 месяцев, уровень фенилацетилглутамина (PAG) в плазме группы >900 мкМ Phe оставался неизмененным от 4177±446,3 до 3565±481,9 нМ; уровни PAG в группе <360 мКМ Phe нормализовывались относительно контролей и снижались от 3133±452,2 до 1133±317,3 нМ (контроль=1167±150,0 нМ) (фиг.23A). Уровни HVA оставались неизменными (от 27,6±3,2 до 24,35±1,72 нМ) и ниже контролей (42,1±2,45 нМ) в группе >900 мкМ; уровни HVA в плазме нормализовывались относительно контролей в группе <360 мкМ, возрастая от 30,6±3,9 до 44,8±5,6 нМ (фиг.23B). Уровень MOPEG в плазме возрастал от 38,2±2,9 до 50,9±5,3 нМ в группе >900 мкМ; уровень MOPEG в плазме нормализовывался относительно контролей в группе <360 мкМ, увеличиваясь от 49±6,4 до 70±5,4 нМ (контроль=61,0±4,1 нМ) (фиг.23C). Наконец, уровень 5HIAA в плазме оставался неизменным (от 13,4±0,8 до 15,9±1,22 нМ) и ниже уровня у контролей (43,1±2,5 нМ) в группе >900 мкМ; в то время как уровень 5HIAA в плазме приближался к контрольным уровням в группе <360 мкМ, возрастая от 18,1±3,6 до 34,9±2,8 нМ (p=0,0017) (фиг.23D).
6.10 ПРИМЕР 10: УРОВНИ НЕЙРОТРАНСМИТТЕРОВ КОРРЕЛИРУЮТ С УРОВНЯМИ PHE В ПЛАЗМЕ У ИНДИВИДУУМОВ С ФКУ
[00288] Уровень PAG в плазме коррелировал с Phe в плазме в группах >900 мкМ (r=0,68; 95% CI 0,48-0,72; p < 0,0001) и <360 мкМ (r=0,72; 95% CI 0,52-0,84; p < 0,0001; фиг.23E), указывая на то, что снижение уровня Phe в плазме при всех концентрациях приводило к снижению уровня Phe в головном мозге, а затем PAG. Аналогично мышам PAHEnu2, у индивидуумов с уровнями Phe в плазме <30 мкМ поддерживались уровни PAG в том же диапазоне, что и у контролей.
[00289] Уровни HVA и MOPEG в плазме коррелировали с Phe в плазме только в группе <360 мкМ (HVA: r = -0,42; 95% CI -0,64 - -0,12; p=0,0075; MOPEG: r = -0,35; 95% CI -0,60 - -0,049; p=0,0246), указывая на то, что более низкие уровни Phe в плазме необходимы для существенного влияния на уровни метаболитов дофамина и норадреналина. Уровень 5-HIAA в плазме коррелировал со снижением уровня Phe в плазме как в группе >900 мкМ (r = -0,38; 95% CI от -0,61 до -0,088; p=0,0126), так и в группе <360 мкМ (r = -0,72; 95% CI от -0,84 до -0,53; p < 0,0001) (фиг.23E-L). Уровень 5-HIAA в плазме более строго коррелировал с уровнем Phe в плазме в группе <360 мкМ, где уровни 5-HIAA достигали диапазона контрольных образцов.
[00290] Траектории уровней у индивидуумов каждого из метаболита нейротрансмиттера и Phe анализировали для образцов от исходного уровня до 12 месяцев. В группе >900 мкМ траектории уровней у индивидуумов для PAG и 5-HIAA относительно Phe были умеренными по величине и наблюдались у меньшинства индивидуумов, и не наблюдалось устойчивых тенденций для траекторий для HVA или MOPEG в этой группе (фиг.23M-P). Наиболее сильные и наиболее устойчивые изменения уровней метаболитов нейротрансмиттеров относительно изменений Phe наблюдались в группе <360 мкМ. Более неожиданно, в группе <360 мкМ индивидуальные тенденции траекторий у индивидуумов были более высокими по величине и у всех кроме одного индивидуума происходило снижение уровня PAG при снижении уровня Phe в плазме (фиг.23Q). Для HVA наблюдали положительную тенденцию траекторий; у всех кроме трех индивидуумов происходило повышение уровня HVA со снижением уровня Phe (фиг.23R). Аналогично, для MOPEG, у всех кроме трех индивидуумов происходило повышение уровня MOPEG со снижением уровня Phe в плазме (фиг.23S). Для 5HIAA, у всех кроме одного индивидуума происходило повышение уровня 5HIAA со снижением уровня Phe в плазме через 12 месяцев (фиг.23T).
[00291] В некоторых вариантах осуществления можно проводить измерение комбинации двух или более аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров от индивидуума для диагностики и/или управления течением ФКУ. В качестве неограничивающего примера, можно проводить измерение уровней Phe и PAG для диагностики и/или управления течением ФКУ.
6.11 ПРИМЕР 11: УЛУЧШЕНИЕ ПОКАЗАТЕЛЕЙ ОСЛАБЛЕНИЯ ВНИМАНИЯ КОРРЕЛИРУЮТ С УРОВНЯМИ MOPEG В ПЛАЗМЕ У ИНДИВИДУУМОВ С ФКУ
[00292] Домен подшкалы ослабления внимания Оценочной шкалы IV синдрома дефицита внимания и гиперактивности (ADHD RS-IV IA) использовали для оценки нейропсихологических симптомов ослабления внимания в клинических испытаниях у индивидуумов с ФКУ, которым вводили PALYNZIQ® (пегвалиаза-pqpz). Улучшение средних показателей подшкалы ослабления внимания было ассоциировано со снижением средних уровней Phe в плазме у субъектов, которые продолжали длительное лечение PALYNZIQ® (пегвалиаза-pqpz). Ассоциация, в частности, была очевидной у индивидуумов, у которых были описаны симптомы ослабления внимания на исходном уровне, указанные как оценка ≥ 9 по ADHD RS-IV IA (Thomas et al 2018).
[00293] Снижение уровней норадреналина и дофамина ассоциировано с симптомами ADHD, и данные указывают на то, что снижение уровня норадреналина строго ассоциировано с симптомами ослабления внимания. Таким образом, способы анализа, описанные в примерах 10 и 11, использовали для изучения ассоциаций между изменениями показателей IA ADHD RS-IV и уровнями метаболитов нейротрансмиттеров дофамина и норадреналина, HVA и MOPEG, соответственно (фиг.24). Не было обнаружено корреляции между изменением уровней HVA и ADHD RS-IV IA у индивидуумов с исходными показателями ADHD RS-IV IA ≧9 или <9 (фиг.24A-B). Была обнаружена корреляция между изменением уровня MOPEG и изменением ADHD RS-IV IA у индивидуумов с исходным показателем ADHD RS-IV IA ≥9 (r = -0,5434, p=0,0242). Поскольку уровни MOPEG возрастали (как для группы >900 мкМ, так и для группы <360 мкМ), изменение показателей ADHD RS-IV IA было отрицательными, что указывает на улучшение показателей. Не было обнаружено корреляции между HVA или MOPEG и изменением показателей ADHD RS-IV IA у индивидуумов с исходным показателем ADHD RS-IV IA <9 (фиг.24C-D).
[00294] В некоторых вариантах осуществления можно определять комбинацию двух или более аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуумов для диагностики и/или управления течением ADHD. В качестве неограничивающего примера, можно измерять уровни дофамина/HVA, норадреналина/MOPEG, и/или серотонина/5HIAA для диагностики и/или управления течением ADHD.
6.12 ПРИМЕР 12: ПРЕОБРАЗОВАНИЕ ГОМОГЕНАТОВ ПЛАЗМЫ ИЛИ ГОЛОВНОГО МОЗГА
[00295] В 96-луночном планшете, 20 мкл гомогената головного мозга или плазмы преципитировали с 80 мкл ледяного ацетонитрила, содержащего внутренние стандарты. После центрифугирования 20 мкл супернатанта преобразовывали в новом планшете посредством 10 мкл 100 мМ карбоната натрия и 10 мкл BzCl (6% (об./об.) в ацетонитриле). Реакции гасили 60 мкл муравьиной кислоты (0,16% (об./об.) в воде).
6.13 ПРИМЕР 13: АНАЛИЗ НЕЙРОТРАНСМИТТЕРОВ ПОСРЕДСТВОМ LC/MS
[00296] Образцы, преобразованные, как описано в примере 12, анализировали посредством LC/MS на Acquity UPLC (Waters H-Class), сопряженном с масс-спектрометром Sciex 6500 Q-Trap с использованием позитивного программного режима MRM. 10 мкл инжектировали в колонку C18 Acquity HSS (2,1 мм × 150 мм, 1,8 мкМ, размер пор 100 A, Waters 186003534) и элюировали с использованием градиента из 10 мМ формиата аммония с 0,15% муравьиной кислотой (MPA) и ацетонитрилом (MPB) при скорости потока 0,5 мл/мин в течение 8 минут.
[00297] Параметры для каждого анализируемого образца представлены в таблице 2. Интеграцию пиков проводили с использованием программного обеспечения MultiQuant. Фильтры масс 1 (Q1) и Q3 квадрупольного масс-анализатора, время элюирования, потенциал декластеризации (DP), потенциал на входе (EP), энергию столкновений (CE) и выходной потенциал столкновения клеток (CXP) оптимизировали для каждого анализируемого соединения. 5HIAA, 5-гидроксииндолуксусная кислота; DOPEG, 3,4-дигидроксифенилэтиленгликоль; HVA, гомованилиновая кислота; MOPEG, 3-метокси-4-гидроксифенилгликоль; PAGlu, фенилацетилглутамат; PAGly, фенилацетилглицин; PEA-IS, фенилэтиламин-изотоп; PHE, фенилаланин; SRO, серотонин; TYR-IS, тирозингидроксилаза; TRP-IS, триптофангидроксилаза.
Таблица 2. Параметры LC/MS для каждого анализируемого соединения.
[00298] Способы анализа, описанные в примерах 12 и 13, использовали для количественного определения уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в биологических образцах, описанных в примерах 6-11.
6.14 ПРИМЕР 14: ОЦЕНКА ЭКСПРЕССИРУЮЩЕЙ КАССЕТЫ PAH, ВСТРОЕННОЙ В ВЕКТОР rAAV
6.14.1 Получение векторов, экспрессирующих PAH человека дикого типа или мыши
[00299] Первоначальные исследования проводили с использованием рекомбинантного вектора на основе AAV для генной терапии, содержащего кДНК PAH дикого типа (вектор 1; ApoE-HCR-hAAT.GI.hPAH.bGH; SEQ ID NO: 15). В вектор была встроена последовательность PAH либо мыши (muPAH), либо человека (hPAH). Геном вектора был фланкирован происходящими из серотипа 2 AAV (AAV2) инвертированными концевыми повторами (ITR) и экспрессия PAH контролировалась гибридным энхансером аполипопротеина E (ApoE)/HCR человека/ промотором альфа анти-трипсина (AAT) человека. Векторный геном также включал последовательность интрона бета-глоблина (GI) и сигнал полиаденилирования бычьего гормона роста (bGH). (фиг.26). Последовательность векторного генома с кДНК PAH человека имеет длину 2800 п.о., как показано в SEQ ID NO: 15. Вектор получали с использованием общепринятых способов клонирования, как описано, например, Gibson et al. (2009). "Enzymatic assembly of DNA molecules up to several hundred kilobases". Nature Methods. 6 (5): 343-345, и Gibson DG. (2011). "Enzymatic assembly of overlapping DNA fragments". Methods in Enzymology. 498: 349-361, которые включены в настоящее описание в качестве ссылок.
6.14.2 Способы анализа для тестирования экспрессии и активности векторов AAV-PAH
[00300] Способы анализа для тестирования рекомбинантных векторов AAV PAH, описанных в настоящем описании, включают, например, (1) временную трансфекцию двухцепочечными ДНК-плазмидами, содержащими нуклеиновые кислоты вектора на основе AAV, клеток HepG2 - клеточной линии, происходящей из печени человека - для проверки специфической для печени экспрессии и сплайсинга мРНК и продуцирования и секреции белка PAH in vitro; (2) продуцирование вирионов AAV, содержащих векторы AAV PAH, в клетках 293 и инфицированных бакуловирусом клетках насекомых; (3) оценку нуклеиновых кислот вектора на основе AAV посредством анализа в щелочном геле и анализа репликации; и (4) оценку экспрессии PAH, активности PAH и уровней Phe у мышей ENU2.
6.14.3 Анализ временной трансфекции
[00301] Предварительный анализ in vitro проводили для сравнения экспрессии и активности PAH из векторов AAV PAH согласно настоящему варианту осуществления. В одном варианте осуществления плазмидами векторов AAV PAH временно трансфицируют клеточную линию печени человека HepG2. После трансфекции, например, через 24, или 48, или 72 часов, измеряют внутриклеточную экспрессию PAH. С использованием этого анализа было продемонстрировано, что вектор ApoE-HCR-hAAT.GI.hPAH.bGH способен экспрессировать белок PAH во временно трансифицированных клетках HepG2 (см., например; фиг.47), и его количественное определение представлено на фиг.51 (вектор, идентифицированный в качестве WT-hPAH)).
6.14.4 Продуцирование вирионов AAV PAH в клетках 293 и инфицированных бакуловирусом клетках насекомых
[00302] Для демонстрации, того, что рекомбинантные векторы AAV PAH согласно настоящему варианту осуществления действительно упаковывают нуклеиновые кислоты, кодирующие PAH, двухцепочечные формы векторов AAV PAH, полученные, как описано выше (например, SEQ ID NO: 15-23), вводят в клетки, способны продуцировать вирионы AAV. В первой системе продуцирования на основе вируса AAV, плазмидами, содержащими нуклеиновые кислоты вектора AAV PAH в двухцепочечной форме, совместно трансфицируют клетки 293 вместе с плазмидой, которая экспрессирует белки Cap и Rep AAV, и плазмидой, которая экспрессирует функции помощника аденовируса, необходимые для продуцирования вирионов AAV. Во второй системе продуцирования вируса AAV получают бакуловирусные конструкции, экспрессирующие нуклеиновые кислоты вектора AAV PAH и белки Cap и Rep AAV, а затем совместно трансфицируют в клетки насекомых Sf9. Полученные вирионы AAV, продуцированные во временно трансфицированных клетках 293 или инфицированных бакуловирусом клетках Sf9, очищают и анализируют стандартными способами, известными в данной области.
6.14.5 Оценка с использованием анализа в щелочном геле и анализа репликации
[00303] Анализ с использованием щелочного гель-электрофореза используют для определения размера упакованной нуклеиновой кислоты. Анализ репликационного центра используют для определения того, какие векторы AAV PAH упаковываются в интактной форме посредством обоих способов упаковывания. Анализ удлинения праймеров используют для количественного определения нуклеиновых кислот вектора AAV PAH, которые имеют завершенные концы, т.е. терминируются на 5′-конце шпилечной петли 5′-ITR (смысловая цепь) или 3′-ITR (антисмысловая цепь) AAV2. Альтернативно анализ с использованием ПЦР используют для определения того, имеют ли нуклеиновые кислоты векторов AAV PAH завершенные концы, т.е. терминируются ли они на 5′-конце шпилечной петли в 5′-ITR (смысловая цепь) или 3′ ITR (антисмысловая цепь) AAV2.
[00304] Как показано на фиг.27 и 28 наблюдали множество полос, когда эту охарактеризацию проводили для первоначального вектора AAV-PAH (ApoE-HCR-hAAT.GI.hPAH.bGH) посредством щелочного гель-электрофореза. Вектор ApoE-HCR-hAAT.GI.hPAH.bGH обозначают как либо hPAH-sc-AAV-DNA (фиг.27), либо AAV-hPAHv1 (фиг.28). Анализ удлинения праймеров и анализ ПЦР проводили для подтверждения того, что две популяции были самокомплементарными и одноцепочечными. Было обнаружено, что области бакуловируса, соседние с hPAH, имеют в ~100-300 раз более низкий титр VG hPAH. Таким образом, дополнительные последовательности в полосе размером ~4,2 т.п.н. не происходят из соседней бакуловирусной ДНК.
6.14.6 Эффект AAV-PAH у мышей с ФКУ
[00305] Мышь C57BL6-ENU2 или BTBRenu2 имеет гомозиготную мутацию локуса гена фенилаланингидроксилазы (PAH), которая приводит к тяжелой гиперфенилаланинемии у животного. Высокие уровни фенилаланина (Phe) в плазме делают это животное подходящей моделью для оценки способности AAV-PAH снижать уровень Phe в плазме, а также другие параметры, такие как Tyr в плазме, скорость роста, аминокислоты и нейротрансмиттеры в головном мозге, и биомаркеры безопасности, такие как ALT в плазме и гистопатология печени.
[00306] Различные уровни доз вектора (2e12-6e14 вг/кг) или носителя (фосфатно-солевой буфер; 4 мл/кг) вводили самцам или самкам мышей C57BL6-Pahenu2 или контрольным мышам C57Bl6 (n=7-10/группа) посредством внутривенной инъекции через хвостовую вену на 0 сутки. Массу каждого животного измеряли и регистрировали до и каждые две недели после введения дозы. В те же моменты времени проводили взятие образцов крови каждого животного. В конечный момент времени (5 или 12 недель после ведения дозы) проводили взятие образцов крови и тканей.
6.14.7 Уровни Phe и Tyr в плазме
[00307] Мышам Enu2 вводили две дозы (2E13 вг/кг и 2E14 вг/кг) вектора на основе аденоассоциированного вируса (AAV), экспрессирующего PAH либо мыши (muPAH), либо человека (hPAH). AAV-PAH сконструирован с ITR серотипа AAV2 и генами капсида серотипа AAV5, как описано выше для вектора ApoE-HCR-hAAT.GI.mu/hPAH.bGH. Мышам Enu2 и дикого типа также вводили носитель в качестве контроля. В различные моменты времени после введения вируса проводили измерение плазменных уровней фенилаланина (Phe) и тирозина (Tyr) в образцах крови, полученных от каждой группы животных, с использованием способа LC-MS/MS с использованием масс-спектрометра QTRAP 4500. Как показано на фиг.29A, мыши Enu2, которым вводили носитель, имели увеличенные уровни фенилаланина относительно уровней, измеренных у мышей дикого типа, которым вводили носитель.
[00308] Введение более высокой дозы AAV-PAH (2E14 вг/кг) мышам Enu2, но не более низкой дозы (2E13 вг/кг), восстанавливало уровни фенилаланина до уровней, наблюдаемых у контролей дикого типа, которым вводили носитель. Эти данные демонстрируют, что уровни Phe в плазме можно использовать для определения эффективной дозы терапевтического средства против ФКУ, необходимой для восстановления метаболических каскадов нейротрансмиттеров до состояния, наблюдаемого у контролей дикого типа. Дополнительные исследования уровней дозы показали, что доза AAV5-PAH ≥ 6e13 вг/кг также приводила к нормализации уровней Phe в плазме (фиг.29B). По существу, 2e13 вг/кг и 6e13 вг/кг установлены в качестве ориентиров для повышения эффективности при конструировании векторов.
6.14.8 Скорость роста
[00309] Скорость роста также измеряли у мышей Enu2 без введения и мышей дикого типа, которым вводили носитель, в качестве контролей и сравнивали с мышами Enu2, которым вводили AAV-PAH, на основании измерений массы тела до получения образца плазмы (фиг. 30). Введение AAV-PAH в обеих протестированных дозах значительно повышало скорость повышения массы тела.
6.14.9 Цвет шерсти
[00310] Гиперфенилаланинемия у мышей Enu2 ассоциирована с относительной гипопигментацией окраски шерсти. Этот эффект, вероятно, является следствием конкурентного ингибирования фермента тирозиназы в меланоцитах посредством фенилаланина (Miyamoto M, Fitzpatrick T. Competitive inhibition of mammalian tyrosinase by phenylalanine and its relationship to hair pigmentation in phenylketonuria. Nature. 1957;179:199-200). После введения AAV-PAH и нормализации сывороточных уровней Phe цвет шерсти постепенно темнел у мышей, которым проводили введение в обеих тестируемых дозах (2E13 вг/кг и 2E14 вг/кг) (фиг.31). Хотя появление этого эффекта было заметно вскоре после инъекции, оно завершилось через 8 недель.
6.14.10 Аминокислоты и нейротрансмиттеры в головном мозге
[00311] Опосредуемое PAH конвертирование фенилаланина в тирозин служит в качестве реакции-предшественника для продуцирования ряда нейротрансмиттеров, как показано на фиг.32. Потеря активности PAH у пациентов с ФКУ приводит к накоплению фенилаланина, как и отмечалось, посредством повышения уровней фенилаланиина в плазме. Однако нейрокогнитивные эффекты ФКУ по большей части вызываются утратой предшествующей метаболической стадии, которая приводит к продуцированию ряда нейротрансмиттеров и метаболитов.
[00312] Чувствительный анализ на основе жидкостной хроматографии, сопряженной с масс-спектрометрией (LC/MS), использовали для измерения уровней различных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров. В кратком изложении, когда аминокислоты, метаболиты нейротрансмиттеров и нейротрансмиттеры подвергают реакции с этилхлорформиатом и пиридином или бензоилхлоридом и карбонатом натрия, они продуцируют продукты реакции, которые имеют определенное увеличение молекулярной массы, которое позволяет их идентификацию и количественное определение с использованием LC/MS. Этот анализ использовали для измерения уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в биологических образцах от мышей Enu2, которым не проводили введение, контрольных мышей дикого типа и мышей Enu2, которым вводили две дозы (2E13 вг/кг и 2E14 вг/кг) вектора на основе аденоассоциированного вируса, экспрессирующего PAH человека (AAV-PAH), для определения эффекта утраты PAH на продуцирование нейротрансмиттеров. AAV конструируют с вектором ApoE-HCR-hAAT.GI.mu/hPAH.bGH и генами капсида серотипа AAV5.
[00313] Как показано на фиг.32, мыши Enu2, которым не проводили введение, неизменно демонстрировали более низкие уровни нейротрансмиттеров и метаболитов нейротрансмиттеров в образцах головного мозга по сравнению с мышами дикого типа. Мыши Enu2 имели сниженные уровни тирозина, дофамина и норадреналина в их головном мозге по сравнению с уровнями, встречающимися в головном мозге мышей дикого типа. Эти данные подтверждают, что утрата активности PAH приводит к снижению уровня ключевых нейротрансмиттеров. Введение более высокой дозы AAV-PAH (2E14 вг/кг) мышам Enu2, но не более низкой дозы (2E13 вг/кг), восстанавливало уровни фенилаланина, тирозина, дофамина и норадреналина до уровней, наблюдаемых в случае контролей дикого типа, которым вводили носитель. Эти данные демонстрируют, что уровни определенных аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров в головном мозге можно использовать для определения эффективной дозы терапевтического средства против ФКУ, требуемого для восстановления путей метаболизма нейротрансмиттеров до состояния, наблюдаемого у контролей дикого типа.
6.14.11 Длительность эффективности
[00314] Для определения длительности эффективности после введения AAV5-muPAH (вектор ApoE-HCR-hAAT.GI.muPAH.bGH) мышам ENU2, проводили мониторинг уровней Phe в плазме каждые 2 недели после дозирования вектора на основе AAV. Через 60 недель (исследование продолжается) наблюдали снижение эффективности в случае субоптимальной дозы 2e13 вг/кг, однако длительная эффективность сохранялась в случае дозы 2e14 вг/кг (фиг.33).
6.14.12 Предварительные данные безопасности
[00315] Предварительные исследования безопасности, проведенные на животных дикого типа, которым вводили AAV5-hPAH в дозе 2E13 или 2E14 вг/кг, продемонстрировали отсутствие повышения уровня фермента печени аланинаминотрансферазы (ALT) (данные не представлены), минимальные изменения уровней Phe по сравнению с WT (фиг.34) и отсутствие гистологических признаков повреждения или гибели гепатоцитов (например, отсутствует окрашивание TUNEL) (фиг.36), несмотря на увеличенные уровни белка PAH у мышей, которым проводили введение, при измерении посредством вестерн-блоттинга (фиг.35).
6.15 ПРИМЕР 15: ПОЛУЧЕНИЕ КОНСТРУКЦИЙ AAV-PAH, КОТОРЫЕ ВКЛЮЧАЮТ ИНТРОННЫЕ ПОСЛЕДОВАТЕЛЬНОСТИ
[00316] Для получения дополнительного рекомбинантных векторов на основе AAV, сконструированных близко к упаковывающей способности AAV, которые как лишены гетерогенного процессинга бакуловирусной системы продуцирования, так и имеют повышенную экспрессию функциональной PAH, получали конструкции с чередующимися интронными последовательностями. В некоторых вариантах осуществления конструкции содержали укороченные версии 2-го интрона hPAH. В альтернативных вариантах осуществления конструкции содержали составной интрон глобина/AIAT (как указано в SEQ ID NO: 14) или его укороченные версии. Эти конструкции получали с использованием стандартных способов клонирования ДНК, как упоминалось в примере 1, и их репрезентативные последовательности представлены в SEQ ID NO: 19 и 17, соответственно.
[00317] В первоначальных попытках увеличить экспрессию последовательности hPAH в векторе ApoE-HCR-hAAT.GI.hPAH.bGH, укороченный 2-й интрон hPAH размером 2116 пар оснований (п.о.) встраивали между экзонами 2 и 3 в кодирующей последовательности hPAH (ApoE-HCR-hAAT.hPAH-tI2.bGH) (фиг.37). Известно, что встраивание интрона, возможно, может приводить к увеличенному уровню экспрессии мРНК в генах, которые в остальных случаях не имеют интронов, например, таких как гены интерферонов. Однако в этом случае интрон 1 не использовался, поскольку известно, что он содержит промотор, который активен в печени. Также средство поиска ESE не выявило на укороченном 2-ом интроне hPAH экзонных энхансеров сплайсинга в точке присоединения укорочения. Этот вектор также был сконструирован близко к упаковочной способности AAV.
6.15.1 Измерение уровней Phe в плазме у мышей ENU2, которым вводили вектор ApoE-HCR-hAAT.hPAH-tI2.bGH
[00318] Мышам инъецировали различные концентрации вектора ApoE-HCR-hAAT.hPAH-tI2.bGH и проводили взятие образцов крови еженедельно для оценки концентрации фенилаланина в плазме (фиг.38). Более высокий уровень фенилаланина был обнаружен у мышей PAH KO/Enu2 до инъекции по сравнению с однопометными контролями, и через 2 суток после инъекции вектора уровень фенилаланина в плазме снижался у мышей, которым вводили дозу 2E14 вг/кг, и в меньшей степени в группах 6E13 и 2E13 вг/кг, в то время как контроли оставались стабильными. Этот результат продемонстрировал, что однократная инъекция вектора ApoE-HCR-hAAT.hPAH-tI2.bGH в дозе 2E14 вг/кг может восстанавливать дефицитную PAH и снижать патологическое накопление фенилаланина в крови. Однако вектор ApoE-HCR-hAAT.hPAH-tI2.bGH не был настолько же эффективным, как и вектор ApoE-HCR-hAAT.GI.hPAH.bGH, в отношении снижения уровней Phe в плазме, который достигал снижения Phe до уровней дикого типа через 2 суток в дозе 6E13 вг/кг (см. фиг. 29B).
6.15.2 Уровни аминокислот и нейротрансмиттеров в головном мозге вектора ApoE-HCR-hAAT.hPAH-tI2.bGH мышей, которым вводили ENU2
[00319] С использованием анализа нейротрансмиттеров, описанного выше в примере 14, уровни аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров также измеряли у мышей, которым вводили вектор ApoE-HCR-hAAT.hPAH-tI2.bGH. Как показано на фиг.39, у мышей Enu2, которым вводили две наиболее высоких дозы (2E14 вг/кг и 6E13 вг/кг) вектора ApoE-HCR-hAAT.hPAH-tI2.bGH, но не наиболее низкую дозу (≤2E13 вг/кг), восстанавливались уровни аминокислот фенилаланина, тирозина и триптофана (фиг.39A), и уровни нейротрансмиттера норадреналина (фиг.39B), как видно у контролей дикого типа, которым вводили носитель.
6.15.3 Вектор 1 с Large Intron (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH)
[00320] Для наблюдения составной интрон глобина/AIAT из 1815 п.о. (SEQ ID NO: 14) встраивали перед кодирующей последовательностью hPAH (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH) (фиг.40). Этот интрон также был сконструирован для доведения общего размера вектора до размера, близкого к упаковывающей способности AAV. Нуклеотидная последовательность включает промотор ApoE-hAAT с составным интроном глобина/AIAT, кодирующую последовательность дикого типа для PAH человека и последовательность поли(A) bGH, как указано в SEQ ID NO: 17.
[00321] Оценка этого вектора in vitro и in vivo включена в пример 17 со сравнениями с кодон-оптимизированными векторами.
6.16 ПРИМЕР 16 - ПОЛУЧЕНИЕ КОДОН-ОПТИМИЗИРОВАННЫХ КОНСТРУКЦИЙ AAV-PAH
[00322] Далее пытаясь увеличить экспрессию hPAH в векторе ApoE-HCR-hAAT.GI.hPAH.bGH, конструировали дополнительные рекомбинантные векторы на основе AAV с кодон-оптимизированными кодирующими последовательностями PAH (см. SEQ ID NO: 7-13). Как описано выше, эти конструкции получали с использованием стандартных способов клонирования ДНК, и репрезентативная последовательность вектора, содержащая кодон-оптимизированную последовательность SEQ ID NO: 7, представлена в SEQ ID NO: 16.
6.16.1 Получение кодон-оптимизированной PAH
[00323] Авторы изобретения сконструировали альтернативные конструкции hPAH (codop-PAH) для тестирования гипотезы, что замена нечасто используемых кодонов в кДНК кодонами, более часто встречающимися в генах млекопитающих ("оптимизация кодонов") приведет к увеличенной экспрессии hPAH после переноса генов. Подобное действие для факторов свертывания VII, VIII и IX повышало экспрессию вплоть до 10 раз по сравнению с аналогами дикого типа (см., например, US 9393323). Стратегия для конструирования codop-hPAH вовлекала обратную трансляцию аминокислотной последовательности hPAH с набором кодонов, которые наиболее часто встречаются в генах млекопитающих с высокой экспрессией. Затем эту модифицированную последовательность тщательно сканировали и кодоны далее модифицировали для повышения стабильности мРНК и устранения нежелательных последовательностей, таких как избыточные CpG-динуклеотиды, и криптические участки сплайсинга.
[00324] Авторы изобретения использовали 6 различных способов оптимизации кодонов, как описано выше, которые включают:
[00325] GENEius 1/2 (Operon 1) - Программное обеспечение для оптимизации кодонов Operon/Eurofins Genomincs с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00326] Nathwani (NW2-Cop) - оптимизацию кодонов проводили с использованием таблицы использования кодонов, которая использовалась Amit Nathwani для оптимизации кодонов фактора VIII.
[00327] Nathwani-RCG (NW-RCG) - оптимизацию кодонов проводили с использованием таблицы использования кодонов, которая использовалась Amit Nathwani для оптимизации кодонов фактора VIII, совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00328] ATUM (DNA2.0-op/D20-P) - проводили оптимизацию кодонов с использованием алгоритма оптимизации кодонов DNA2.0
[00329] ATUM-RCG (DNA2.0-Cop1) - проводили оптимизацию кодонов с использованием алгоритма оптимизации кодонов DNA2.0 совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00330] JCAT (JCAT) - проводили оптимизацию кодонов с использованием Java Codon Adaptation Tool (www.jcat.de) совместно с уменьшением вручную содержания динуклеотидов CpG и удалением каких-либо ORF в смысловом и антисмысловом направлении.
[00331] Выравнивание последовательностей codop-hPAH приведено на фиг.41, где сравниваются процентная идентичность последовательностей, содержание GC и взаимосвязи согласно кладрограмме, приведенной на фиг.42.
6.16.2 Конструирование кодон-оптимизированного вектора hPAH
[00332] Конечные сконструированные последовательности codop-hPAH содержат единичные изменения в диапазоне 46-316 относительно последовательности hPAH дикого типа, и, как правило, содержание G+C 47-57%, относительно содержания G+C 47% в последовательности дикого типа. Авторы изобретения клонировали варианты codop PAH в их стандартный вектор rAAV (Nathwani A. et al. Blood. 2007 February 15; 109(4): 1414-1421) с промотором ApoE/hAAT в различных конфигурациях (см. фиг.43). В предпочтительном варианте осуществления codop-PAH клонирована в вектор, который включает промотор ApoE-hAAT с составным интроном глобина/AIAT, кодон-оптимизированной последовательностью для PAH человека, и последовательностью поли(A) bGH. Предпочтительный составной интрон глобина/AIAT имеет последовательность SEQ ID NO: 14.
6.16.3 Трансдукция HepG2 векторами с кодон-оптимизированной hPAH на основе AAV
[00333] Оценка этих векторов с кодон-оптимизированной hPAH на основе AAV в линии клеток печени HepG2 (фиг.44) продемонстрировала, что GENEius1, GENEius2 и JCAT продемонстрировали значительное усиление экспрессии относительно WT-hPAH. В совокупности, таким образом, эти данные указывают на то, что молекула codop-PAH авторов изобретения является более эффективной, чем WT-hPAH. Примечательно, что векторные плазмиды GENEius1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH) и GENEius2 (ApoE-HCR-hAAT.GI.hPAHco2.bGH) неизменно обеспечивают на 50-100% более высокий выход вектора, чем rAAV-WT-hPAH.
[00334] Клетки HepG2 высевали на 0 сутки в количестве 5E5 клеток/ячейка 12-ячеечного планшета. Клетки трансдуцировали на 1 сутки с MOI в диапазоне от 1,25E5 до 1E6 с 20 мкМ этопозидом. Среду заменяли на 2 сутки. Клетки собирали на 4 сутки путем удаления среды, открепления с использованием TrypLE, последующей нейтрализации содержащей сыворотку средой, промывания клеток DPBS и замораживания клеточного осадка. Анализ экспрессии проводили посредством лизиса клеток TPER+4 мМ BMe, удаления нерастворимого клеточного дебриса, измерения концентрации белка по A280, SDS-PAGE с использованием загрузки белка 20 мкг и WB с использованием первичного Ab мыши против PAH и конъюгированного с HRP вторичного Ab кролика против антител мыши.
6.16.4 Трансдукция AML12 векторами AAV-codop hPAH
[00335] Клетки AML12 представляют собой клетки, происходящие из гепатоцитов мыши. Клетки высевали на 0 сутки в количестве 5E5 клеток/ячейка в 12-ячеечном планшете. Клетки трансдуцировали на 1 сутки с MOI 1E6 с 20 мкМ этопозидом. Среду заменяли на 2 сутки. Клетки собирали на 4 сутки путем удаления среды, открепления с использованием TrypLE, последующей нейтрализации содержащей сыворотку средой, промывания клеток DPBS и замораживания клеточного осадка. Анализ экспрессии проводили посредством лизиса клеток TPER+4 мМ BMe, удаления нерастворимого клеточного дебриса, измерения концентрации белка по A280, SDS-PAGE с использованием загрузки белка 80 мкг и WB с использованием первичного Ab мыши против PAH и конъюгированного с HRP вторичного Ab кролика против антител мыши. Результаты, представленные на фиг.45 сравнивают использование содержащей сыворотку среды и не содержащей сыворотку среды на сутки 1-4, причем для последней из них ранее было показано, что она индуцирует подобную гепатоцитам регуляцию транскрипции. При сравнении паттерна экспрессии кодон-оптимизированной hPAH, клетки, которые поддерживали в SFM, соответствовали результатам авторов изобретения на клетках HepG2, а также в экспериментах на мышах in vivo.
6.16.5 Измерение уровней Phe в плазме у мышей ENU2, которым вводили кодон-оптимизированные векторы AAV-PAH
[00336] Мышам Enu2 инъецировали 2E13 вг/кг каждого кодон-оптимизированного вектора AAV-PAH, носителя или вектора на основе AAV с последовательностью PAH дикого типа. Взятие образцов крови проводили каждую неделю для оценки концентрации фенилаланина в плазме (фиг.46A-B). Более высокий уровень фенилаланина был обнаружен у мышей ENU2 до инъекции по сравнению с однопометными контролями. Через шесть недель после инъекции вектора уровни фенилаланина в плазме мышей ENU2 снижались, в то время как у контролей они оставались стабильными. Этот результат продемонстрировал, что однократная инъекция кодон-оптимизированного AAV-PAH мышам PAH KO/ENU2 может восстанавливать дефицитную PAH и снижать патологическое накопление фенилаланина в крови.
[00337] Эффекты введения конструкций AAV-codop PAH далее оценивали в качестве массы тела относительно снижения Phe между различными конструкциями (фиг.47). Конструкции GENEius1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH), GENEius2 (ApoE-HCR-hAAT.GI.hPAHco2.bGH) и JCAT (ApoE-HCR-hAAT.GI.hPAHco3.bGH) оказались наиболее эффективными в отношении снижения уровней Phe, которые также коррелировали с более значительным повышением % массы тела.
6.17 ПРИМЕР 17 - СРАВНЕНИЕ КОНСТРУКЦИЙ AAV-PAH С ОПТИМИЗАЦИЕЙ КОДОНОВ ПРОТИВ ИНСЕРЦИИ ИНТРОНА
[00338] Проводили серию экспериментов для дальнейшего сравнения функциональности и эффективности лучшего кодон-оптимизированного вектора (ApoE-HCR-hAAT.GI.hPAHco1.bGH) и лучшего вектора с встроенным интроном (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH).
6.17.1 Временная трансфекция HepG2
[00339] Клетки HepG2 высевали на 0 сутки в количестве 5E5 клеток/ячейка 12-ячеечного планшета. Клетки трансдуцировали на 1 сутки с использованием 1 мкг ДНК/ячейка с соотношением FuGene HD:ДНК 4.5:1. Среду заменяли на 2 сутки. Клетки собирали на 4 сутки путем удаления среды, открепления с использованием TrypLE, последующей нейтрализации содержащей сыворотку средой, промывания клеток DPBS и замораживания клеточного осадка. Анализ экспрессии проводили посредством лизиса клеток TPER+4 мМ BMe, удаления нерастворимого клеточного дебриса, измерения концентрации белка по A280, SDS-PAGE с использованием загрузки белка 20 мкг и WB с использованием первичного Ab мыши против PAH и конъюгированного с HRP вторичного Ab кролика против антител мыши. Данные на фиг.48A-B относятся к сравнению V1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH), V1+ интрон LG (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH), Geneius1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH) и вектора 2 (ApoE-HCR-hAAT.cG-AIATI.hPAHco1.bGH). Результаты показывают пользу включения интрона LG в V1, и вектор Geneius1+интрон LG является по меньшей мере настолько же эффективным, как и Geneius1 отдельно.
6.17.2 Ответ на дозу для векторов с улучшенной эффективностью
[00340] Возрастающие дозы ApoE-HCR-hAAT.GI.hPAHco1.bGH или ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH доставляли мышам ENU2 для определения эффективности снижения уровней Phe. Векторы AAV5-PAH модифицировали включением кодон-оптимизированного гена PAH, Large Intron, и эффект этих модификаций тестировали в модели на мышах ENU2/ФКУ.
[00341] Кривая доза-эффект для AAV5-PAH, модифицированного посредством Geneius-1 или Large Intron, представлена на фиг.49A. Варианты AAV5-PAH вводили мышам ENU2 в дозах в вг/кг, указанных на графике, и определяли Phe в плазме. На фиг.49B представлен ответ на дозу через 2 недели после введения AAV (результаты мышей ENU2 не нанесены на график), и на фиг.49C представлен ответ на дозу через 4 недели после введения AAV с сужением оси y для акцентирования на изменениях ответа на дозу (результаты мышей ENU2 не нанесены на график).
6.17.3 Сравнение биораспределения PAH между конструкциями AAV-PAH с GENEius-1 и Large Intron
[00342] На фиг.50 охарактеризованы изменения биораспределения PAH в ткани печени при введении AAV-PAH. Векторы AAV5-PAH модифицировали включением кодон-оптимизированного гена PAH, Large Intron и экспрессию белка PAH в гепатоцитах печени определяли посредством IHC с использованием стандартных способов. Белок PAH окрашивали с использованием стандартных способов IHC, и % положительных клеток определяли и наносили на график против доз AAV5-PAH в вг/кг (фиг.50A). Возрастающий процент гепатоцитов, экспрессирующих PAH, наблюдали при повышении дозы конструкций либо Large Intron, либо GENEius-1. Однако не было значимых отличий в % положительных по PAH гепатоцитов между конструкциями GENEius-1 и Large Intron для каждой дозы. Изображения IHC печени от мышей ENU2, которым дозировали эти векторы AAV5-PAH в дозе 2e13 вг/кг (фиг.50B) и 1e14 вг/кг (фиг.50C), продемонстрировали, что в большинстве случаев основная часть PAH локализовывалась в области вокруг центральной вены, и в периферической области наблюдали минимальное распределение. Таким образом, оказалось, что экспрессия белка PAH наблюдается в гепатоцитах, окружающих центральные вены, при всех уровнях доз.
6.18 ПРИМЕР 18 - ПОЛУЧЕНИЕ КОНСТРУКЦИЙ AAV-PAH, СОЧЕТАЮЩИХ В СЕБЕ ОПТИМИЗАЦИЮ КОДОНОВ И ИНСЕРЦИЮ ИНТРОНА
6.18.1 Ответ на дозу для вектора 2 у мышей ENU2
[00343] Для оценки кинетики ответа на дозу использовали Phe в плазме в качестве суррогатного маркера для активности PAH в печени. После введения вариантов AAV5-PAH в модели на мышах ENU2/ФКУ наблюдали снижение Phe в плазме в течение двух недель после дозирования AAV. Ответ оставался стабильным в течение вплоть до 5 недель и оставался стабильным на протяжении более длительного периода, исходя из других исследований (фиг.51A). Ответ на дозу вариантов AAV5-PAH, которые вводили мышам ENU2, также наносили на график в момент времени 2 недели, чтобы более отчетливо идентифицировать различия в эффективности различных доз (фиг.51B). Gene1 (вариант Geneius-1) и LgIntron (вариант Large Intron) использовали в качестве эталонных контролей в дозе 4e13 вг/кг. Вектор 2 вводили в различных дозах самцам и самкам мышей ENU2, и эффективность сравнивали с другими вариантами AAV5-PAH.
[00344] Корреляция дозы вектора 2 с эффективностью у самцов и самок мышей ENU2 далее проиллюстрирована на фиг.52, причем только незначительные отличия наблюдались в эффективности вектора 2 между самцами и самками мышей ENU2.
[00345] Сравнение ответа на дозу между вариантами AAV5-PAH (GENEius-1 и Large Intron), и вектором 2 представлено на фиг.53. На график нанесен ответ векторов в отношении снижения Phe в плазме у мышей ENU2 через 2 недели при введении в дозе 2e13 вг/кг. На графике указано, что вектор 2 имеет увеличенную эффективность в отношении снижения уровня Phe в плазме в дозе 2e13 вг/кг по сравнению с другими вариантами AAV5-PAH.
6.19 ПРИМЕР 19 - АНАЛИЗ БЕЗОПАСНОСТИ КОНСТУРКЦИЙ AAV-PAH
6.19.1 Уровни ALT в плазме
[00346] Активность аланинаминотрансферазы (ALT) в плазме можно использовать в качестве индикатора здоровья гепатоцитов, и более высокие уровни ALT указывают на токсичность для гепатоцитов. Образцы плазмы отбирали от мышей до введения AAV5-PAH (фиг.54A) и через 2 недели после введения (фиг.54B), и измеряли уровень ALT в плазме с использованием коммерческого набора (Sigma). На графиках указано, что введение AAV5-PAH в различных дозах не приводило к заметному изменению уровней ALT.
6.19.2 Окрашивание TUNEL, использованное для измерения токсичности
[00347] Терминальное разрезание и мечение дезоксинуклеотидилтрансферазой dUTP (TUNEL) представляет собой ферментный анализ, используемый для детекции фрагментации ДНК в результате гибели клеток вследствие апоптотических сигнальных каскадов. Положительные по TUNEL клетки оценивали в срезах печени мышей, которым вводили варианты AAV5-PAH, и минимальное изменение положительных по TUNEL клеток наблюдали после введения AAV5-PAH в различных дозах (фиг.55).
6.19.3 Окрашивание IBA и H&E, использованное для определения уровней воспаления
[00348] В качестве следующего показателя здоровья печени окрашивание IBA и H&E использовали для детекции инфильтрации макрофагов у животных, которым дозировали AAV-hPAH. На активированных макрофагах присутствуют воспалительный фактор 1 аллотрансплантата (AIF-1), также известный как связывающая ионизированный кальций адаптерная молекула 1 (IBA1). Активированные макрофаги присутствуют в тканях в ходе воспаления (фиг.56A). Положительные по IBA1 клетки/очаги оценивали в срезах печени мышей, которым вводили варианты AAV5-PAH для определения степени воспаления и инфильтрации макрофагов после введения AAV5-PAH (фиг.56B). Минимальное изменение клеток положительных по IBA1 очагов наблюдали после введения AAV5-PAH в различных дозах.
[00349] Окрашивание H&E (гематоксилин и эозин) ткани печени проводили у мышей, которым вводили варианты AAV5-PAH, и сравнивали с паттерном окрашивания у мышей дикого типа. Паттерн окрашивания показал минимальные различия в гистологии тканей между мышами дикого типа и мышами, которым вводили варианты AAV5-PAH (фиг.57A-C).
[00350] Ввиду вышеуказанного, использованные векторы AAV5-PAH и дозы, вероятно, являются непатогенными у мышей ENU2, исходя из отсутствия выраженной гистопатологии печени.
6.20 ПРИМЕР 20: ПОЛУЧЕНИЕ КОНСТУРКЦИЙ С УЛУЧШЕННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ ПРОМОТОРА/ЭНХАНСЕРА
[00351] Для получения дополнительных рекомбинантных векторов на основе AAV с сильными промоторами, которые повышают экспрессию функциональной PAH, получали конструкции с модифицированными последовательностями энхансеров и/или промоторов. В частности, конструкции конструировали с использованием стандартных способов клонирования ДНК с альтернативными специфическими для печени промоторами, такими как промотор тироксин-связывающего глобулина (TBG) или промотор транстиретина (TTR) (см., например, SEQ ID NO: 22 и 23). Другие пригодные промоторы включают промотор альбумина человека (Miyatake et al., J. Virol, 71:5124 32 (1997)) humAlb; специфический для печени промотор (LSP), и основной промотор вируса гепатита B (Sandig et al, Gene Ther., 3: 1002 9 (1996). См., например, The Liver Specific Gene Promoter Database, Cold Spring Harbor, rulai.schl.edifLSPD). Хотя и менее желательно, в векторах, описанных в настоящем описании, можно использовать другие промоторы, такие как вирусные промоторы, синтетические промоторы, конститутивные промоторы, регулируемые промоторы (см., например, WO 2011/126808 и WO 2013/04943) или промотор, отвечающий на физиологические сигналы.
6.21 ПРИМЕР 21: ГЕННАЯ ТЕРАПИЯ ВОССТАНАВЛИВАЕТ АКТИВНОСТЬ PAH У МЫШЕЙ ENU2
6.21.1 Конструирование вектора AAV5-PAH
[00352] Для этих исследований использовали опосредуемую аденоассоциированным вирусом серотипа 5 (AAV5) доставку гена PAH человека (hPAH) или PAH мыши (muPAH) (под специфическим для печени промотором).
6.21.2 Исследования in vivo
[00353] Все исследования проводили в Translational Biology Department BioMarin в виварии, находящемся в Buck Institute, Novato CA. Разведение животных и протоколы эксперимента были одобрены институциональным комитетом по уходу за животными и их использованию (IACUC). Самцов мышей ENU2 (C57BL/6-Pahenu2) и контрольных мышей дикого типа (WT) (C57BL/6J) получали от Jackson Laboratory.
[00354] Мышам ENU2 вводили в возрасте 8 недель посредством инъекции через хвостовую вену AAV5-muPAH или контроля в виде носителя. Мышам дикого типа вводили носитель. Взятие образцов крови проводили каждые две недели после дозирования посредством надреза в хвостовой вене. В заключение исследования проводили взятие крови посредством пункции сердца и перфузию тканей (головной мозг и печень) фосфатно-солевым буфером перед извлечением.
[00355] Уровни Phe и Tyr в плазме мышей измеряли посредством LC-MS/MS, и уровень аланинаминотрансферазы (ALT) в плазме измеряли посредством иммуноанализа (Sigma Aldrich) в соответствии с инструкциями изготовителя. Как видно на фиг.58A, уровни Phe в плазме у мышей ENU2, которым вводили AAV5-muPAH, восстанавливались до уровней WT к моменту времени 2 недели после дозирования. Этот эффект сохранялся на протяжении исследования (80 недель). Отсутствовали статистические отличия уровней ALT между мышами WT, ENU2, которым вводили носитель, и ENU2, которым вводили AAV5-muPAH, на протяжении исследования в течение 80 недель (фиг.58B). Мыши ENU2, которым дозировали носитель, сохраняли светло-коричневый окрас, в то время как у мышей ENU2, которым вводили AAV5-muPAH, происходило изменение окраски шерсти на темно-коричневый аналогично окраске шерсти, которую демонстрировали контроли WT (фиг.58C). Эти данные демонстрируют, что посредством генной терапии в однократной дозе достигалось длительное снижение Phe в плазме до уровней, близких к уровням WT.
[00356] Для исследования эффекта PAH человека на уровень аминокислот и метаболитов нейротрансмиттеров в головном мозге мышам ENU2 вводили AAV5-hPAH. Через 12 недель замороженные полушария головного мозга гомогенизировали и полученные лизаты обрабатывали для измерения посредством LC-MS/MS уровней Phe, Tyr, триптофана, 5-гидроксииндолуксусной кислоты (5-HIAA), гомованилиновой кислоты (HVA) и 3-метокси-4-гидроксифенилгликоля (MOPEG). Как показано на фиг.59, введение AAV5-hPAH нормализовывало уровни аминокислот (Phe, Tyr и триптофан) в головном мозге (фиг.59A), в дополнение к уровням в головном мозге метаболитов нейротрансмиттеров, 5-гидроксииндолуксусной кислоты (5-HIAA), гомованилиновой кислоты (HVA) и 3-метокси-4-гидроксифенилгликоля (MOPEG), происходящих из серотонина, дофамина и норадреналина, соответственно (фиг.59B).
[00357] Для исследования эффекта PAH человека на экспрессию белка PAH в печени, мышей WT, которым вводили AAV5-hPAH, исследовали в отношении признаков гипофенилаланина в течение 12 недель. На фиг.60A показано, что мыши WT, которым вводили AAV5-hPAH, имели нормальные уровни Phe на протяжении исследования. В заключение исследования в течение 12 недель, замороженные срезы печени гомогенизировали, и полученные лизаты восстанавливали, алкилировали и расщепляли трипсином (Promega). Уровни PAH измеряли посредством LC-MS/MS с использованием пептидов, общих для PAH как человека, так и мыши. У мышей дикого типа, которым вводили AAV5-hPAH, происходило 2-кратное повышение уровней белка PAH, как показано на фиг.60B.
[00358] Эти данные показывают, что длительного снижения уровней Phe в плазме практически до уровней у WT можно достигать посредством генной терапии в однократной дозе. Кроме того, уровни аминокислот и метаболитов нейротрансмиттеров в головном мозге при введении также нормализовывались до уровней WT, и уровень Phe не снижался ниже эталонного диапазона у животных WT, которым вводили AAV5-hPAH. В совокупности, эти эксперименты обеспечивают доказательство, что восстановление на основе генной терапии активности PAH является перспективным терапевтическим подходом для лечения ФКУ.
6.22 ПРИМЕР 22: РАСПРЕДЕЛЕНИЕ ФЕНИЛАЛАНИНГИДРОКСИЛАЗЫ (PAH) В ПЕЧЕНИ И ОХАРАКТЕРИЗАЦИЯ ПОСЛЕ ГЕННОЙ ТЕРАПИИ AAV5-hPAH У МЫШЕЙ PAHenu2
6.22.1 Биораспределение трансгена PAH человека
[00359] Мышам ENU2 вводили однократную дозу AAV5-hPAH посредством инъекции через хвостовую вену. Животных умерщвляли через 12 недель после введения вектора, подвергали перфузии посредством солевого раствора, а затем извлекали печень. Все образцы средней доли печени фиксировали в 4% параформальдегиде в течение 48 часов и обрабатывали стандартным образом для заливки парафином в течение одного месяца после получения.
[00360] Печень мыши нарезали на срезы и проводили гибридизацию in situ в соответствии с инструкцией изготовителя (ACD) с использованием Roche Ventana Autostainer. Получали десять изображений 40x на животное и гепатоциты оценивали либо как отрицательные, либо как положительные по сигналу DAB. На фиг. 61 представлено биораспределение векторных геномов AAV5-hPAH и происходящего из трансгена белка PAH в ткани печени в модели ФКУ на мышах. Дозозависимое увеличение количества гепатоцитов с положительным окрашиванием как на ДНК, так и на белок hPAH, было выявлено в печени мышей ENU2 (фиг. 61A-C). Содержание как ДНК, так белка, было более высоким в околоцентральных областях доли печени по сравнению с перипортальной областью.
6.22.2 Оценка воспаления и апоптоза после введения AAV5-hPAH
[00361] Как описано выше, мышам ENU2 вводили однократную дозу AAV5-hPAH и печень извлекали через 12 недель после введения вектора. Для оценки эффекта генной терапии AAV5-PAH и происходящего из трансгена белка PAH на безопасность печени, печень обрабатывали для FFPE и иммуногистохимии для детекции провоспалительных маркеров. На фиг. 62A-C показано, что происходило небольшое увеличение количества макрофагов IBA1 (тотальные) и CD68 (M1/M2 активированные) при введении AAV5-hPAH и не было обнаружено значительного окрашивания на iNOS (M1 активированные), маркер провоспалительных макрофагов. Кроме того, не наблюдали таких иммуногистологических признаков, как гепатоцеллюлярный некроз, гиперплазия купферовских клеток, и воспаление или фиброз портальной триады, что указывает на то, что здоровье печени сохранялось после введения AAV5-hPAH. На фиг. 62D представлено количественное определение окрашенных срезов.
[00362] Окрашивание посредством терминального разрезания и мечения дезоксинуклеотидилтрансферазой dUTP (TUNEL) печени FFPE продемонстрировало отсутствие значимого повышения апоптотической гибели клеток в любой из групп, которым проводили генную терапию, по сравнению с мышами WT, которым вводили носитель, или мышами ENU2, которым вводили носитель. На фиг. 63 представлены визуализированные окрашенные TUNEL срезы печени, и анализ изображений проводили с использованием ImageJ. Было определено приблизительно 14000-17000 ядер на животное. Эти данные демонстрируют, что не наблюдали отсутствия гистопатологических признаков увеличенной гибели клеток после введения вектора AAV5-hPAH, и также не существовало выраженных признаков воспаления (фиг. 62-63). Эти данные подтверждают, что опосредуемая AAV5 доставка гена hPAH может быть перспективной возможностью в качестве потенциального терапевтического подхода для пациентов с ФКУ.
6.23 ПРИМЕР 23: ЭФФЕКТ ВЕКТОРА 2
[00363] Вектор 2, описанный в примере 17 (фиг. 48) и примере 18 (фиг. 51-53), был отобран для дальнейшего исследования.
6.23.1 Биораспределение ДНК вектора 2 у яванских макаков
[00364] Вектор 2 доставляли на двух уровнях дозы (высокая и низкая) посредством в/в инъекции. Самцов и самок яванских макаков умерщвляли через 4 недели после введения. Анализ с использованием количественной полимеразной цепной реакции (кПЦР) использовали для определения количества векторной ДНК вектора 2 в геномной ДНК (гДНК), экстрагированной из крови и тканей яванских макаков. На фиг. 64 показано, что биораспределение ДНК вектора 2 после в/в введения вектора 2 согласовывалось с опубликованной литературой по капсидам AAV5 с печенью в качестве основного органа-мишени. Количества ДНК, обнаруженной в других тканях, представлено в качестве процента ДНК, обнаруженной в печени. Селезенка имеет наиболее высокое количество ДНК вектора после печени (закрашенные квадраты), причем семенники и яичники демонстрировали наиболее низкое количество относительно печени (закрашенные круги).
6.23.2 Эффект вектора 2 на фенилаланин и тирозин у яванских макаков
[00365] Вектор 2 доставляли на двух уровнях доз (высокая и низкая) посредством в/в инъекции, как описано выше. Взятие плазмы от самцов и самок яванских макаков проводили с регулярными интервалами в течение 4 недель после введения. Преципитацию с ацетонитрилом ВЭЖХ/MS/MS (колонка ВЭЖХ C18, после которой следовал тройной квадрупольный масс-спектрометр) использовали для определения концентрации фенилаланина и тирозина в плазме яванского макака с литий-гепарином.
[00366] Уровень фенилаланина и тирозина в плазме оставался неизменным на протяжении исследования (4 недели) (фиг. 65). Известно, что эндогенная PAH аллостерически регулируется, и эти данные указывают на то, что PAH, доставляемая посредством генной терапии, также подвергается аллостерической регуляции. Эта регуляция минимизирует риск гипофенилаланинемии.
[00367] Самкам и самцам мышей ENU2 вводили различные дозы вектора 2. Через 12 недель мышей умерщвляли и гомогенаты головного мозга анализировали для определения уровней конкретных аминокислот. Как показано на фиг. 66, как самцы, так и самки продемонстрировали очевидное зависимое от дозы снижение уровней Phe до уровней WT при введении вектора 2. Полную коррекцию Phe в головном мозге наблюдали, когда вектор 2 вводили в дозе 6E13 вг/кг у самцов мышей и 2E14 вг/кг у самок мышей (фиг. 66A). Вектор 2 в дозе 2E13 вг/кг у самцов ENU2 и 6E13 вг/кг у самок ENU2 повышал уровни Tyr в головном мозге до уровней WT (фиг. 66B). Вектор 2 в дозе 2E13 вг/кг у самцов ENU2 и 2E14 вг/кг у самок ENU2 повышал уровень Trp в головном мозге практически до уровней WT (фиг. 66C).
6.23.4 Введение мышам ENU2 вектора 2 приводило к коррекции уровней нейротрансмиттеров в гомогенатах головного мозга
[00368] Как описано выше, самкам и самцам мышей ENU2 вводили различные дозы вектора 2, и через 12 недель мышей умерщвляли и гомогенаты головного мозга анализировали для определения уровней нейротрансмиттеров. Уровни в головном мозге дофамина и норадреналина, образованных из Tyr, и серотонина, образованного из Trp, возрастали до уровней WT при дозах 2E13 вг/кг у самцов ENU2 и 6E13 вг/кг у самок ENU2 (фиг. 67A-C). Кроме того, уровни PEA, образованного из Phe, снижались до уровней, наблюдаемых у мышей WT, когда мышам ENU2 вводили 6E13 вг/кг (самцы) или 2E14 вг/кг (самки) вектора 2 (фиг.67D).
6.23.5 Коррекция метаболитов нейротрансмиттеров, наблюдаемая при высокой дозе вектора 2
[00369] Уровни метаболитов нейротрансмиттеров измеряли в гомогенатах головного мозга мышей ENU2 после введения вектора 2, как описано выше. Все из метаболитов HVA, MOPEG и 5-HIAA корректировались до уровней WT, когда вектор 2 вводили в дозе 2E13 вг/кг самцам мышей ENU2 и 6E13 или 2E14 самкам мышей ENU2 (фиг. 68A-C).
[00370] Варианты осуществления, описанные в настоящем описании, являются только иллюстративными, и специалистам в данной области будут понятны, или они будут установить с использованием не более чем стандартного экспериментирования, многочисленные эквиваленты конкретных соединений, материалов и методик. Все такие эквиваленты считаются входящими в объем настоящего изобретения.
[00371] Все из патентов, патентных заявок и публикаций, упоминаемых в настоящем описании, включены в настоящее описание в качестве ссылок в полном объеме. Цитирование или идентификация какой-либо ссылки в настоящей заявке не является допущением, что такая ссылка доступна в качестве документа уровня техники для настоящей заявки. Полный объем изобретения станет более понятным с помощью прилагаемой формулы изобретения.
7. ПОСЛЕДОВАТЕЛЬНОСТИ
SEQ ID NO: 1 - последовательность нуклеиновой кислоты PAH человека дикого типа
ATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAA
SEQ ID NO: 2 - аминокислотная последовательность PAH человека дикого типа
MSTAVLENPG LGRKLSDFGQ ETSYIEDNCN QNGAISLIFS LKEEVGALAK VLRLFEENDV NLTHIESRPS RLKKDEYEFF THLDKRSLPA LTNIIKILRH DIGATVHELS RDKKKDTVPW FPRTIQELDR FANQILSYGA ELDADHPGFK DPVYRARRKQ FADIAYNYRH GQPIPRVEYM EEEKKTWGTV FKTLKSLYKT HACYEYNHIF PLLEKYCGFH EDNIPQLEDV SQFLQTCTGF RLRPVAGLLS SRDFLGGLAF RVFHCTQYIR HGSKPMYTPE PDICHELLGH VPLFSDRSFA QFSQEIGLAS LGAPDEYIEK LATIYWFTVE FGLCKQGDSI KAYGAGLLSS FGELQYCLSE KPKLLPLELE KTAIQNYTVT EFQPLYYVAE SFNDAKEKVR NFAATIPRPF SVRYDPYTQR IEVLDNTQQL KILADSINSE IGILCSALQK IK
SEQ ID NO: 3 - последовательность промотора hAAT
GATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagta
SEQ ID NO: 4 - последовательность энхансера HCR или энхансера ApoE
AGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGG
SEQ ID NO: 6 - последовательность синтетического промотора (промотор hAAT, энхансер HCR/энхансер ApoE)
AGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGgtcgacGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtatgcctttcactgcgaggggttctggagaggcttctgagctccccatggcccaggcaggcagcaggtctggggcaggaggggggttgtggagtgggtatccgcctgctgaggtgcagggcagatggagaggctgcagctgagctcctattttcataataacagcagccatgagggttgtgtcctgtttcccagtcctgcccggtcccccctcggtacctcctggtggatacactggttcctgtaagcagaagtggatgagggtgtctaggtctgcagtcctggcaccccaggatgggggacaccagccaagatacagcaacagcaacaaagcgcagccatttctttctgtttgcacagctcctctgtctgtcgggggctcctgtctgttgtctcctataagcctcaccacctctcctactgcttgggcatgcatctttctccccttctatagatgaggaggttaaggtccagagaggggtggggaggaacgccggctcacattctccatcccctccagatatgaccaggaacagacctgtgccaggcctcagccttacatcaaaatgggcctccccatgcaccgtggacctctgggccctcctgtcccagtggaggacaggaagctatgaggggcactgtcacccagggctcaagctggcattcctgaataatcgctctgcaccaggccacggctaagctcagtgcgtgattaagcctcataaccctccaaggcagttactagtgtgattcccattttacagatgaggaagatggggacagagaggtgaataactggccccaaatcacacaccatccataattcgggctcaggcacctggctccagtccccaaactcttgaacctggccctagtgtcactgtttctcttgggtctcaggcgctggatggggaacaggaaacctgggctggacttgaggcctctctgatgctcggtgacttcagacagttgctcaacctctctgttctcttgggcaaaacatgataacctttgacttctgtcccctcccctcaccccacccgaccttgatctctgaagtgttggaaggatttaatttttcctgcactgagttttggagacaggtcaaaaagatgaccaaggccaaggtggccagtttcctatagaacgcctctaaaagacctgcagcaatagcagcaagaactggtattctcgagaacttgctgcgcagcaggcacttcttggcattttatgtgtatttaatttcacaatagctctatgacaaagtccacctttctcatctccaggaaactgaggttcagagaggttaagtaacttgtccaaggtcacacagctaatagcaagttgacgtggagcaatctggcctcagagcctttaattttagccacagactgatgctcccctcttcatttagccaggctgcctctgaagttttctgattcaagacttctggcttcagctttgtacacagagatgattcaatgtcaggttttggagtgaaatctgtttaatcccagacaaaacatttaggattacatctcagttttgtaagcaagtagctctgtgatttttagtgagttatttaatgctctttggggctcaatttttctatctataaaatagggctaataatttgcaccttatagggtaagctttgaggacagattagatgatacggtgcctgtaaaacaccaggtgttagtaagtgtggcaatgatggtgacgctgaggctgatgtttgcttagcatagggttaggcagctggcaggcagtaaacagttggataatttaatggaaaatttgccaaactcagatgcTAGCAGCTACaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagctcctgggcaacgtgctggtctgtgtgctggcccatcactttggcaaagaattgcgatCGCCACC
SEQ ID NO: 7 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (GENEius 1/Operon1)
ATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGAGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGAGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCTGCCCTGACCAACATCATCAAGATCCTGAGACACGATATTGGAGCCACTGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACTGTGCCCTGGTTCCCCAGAACTATCCAGGAGCTGGACAGATTTGCCAACCAGATCCTGAGCTATGGAGCTGAGCTGGATGCTGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGAAGAAAGCAGTTTGCTGACATTGCCTACAACTACAGACACGGCCAGCCCATCCCCAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACTGGCTTCAGACTGAGACCTGTGGCTGGCCTGCTGAGCAGCAGAGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTACATCAGACACGGCAGCAAGCCCATGTACACCCCTGAGCCTGATATCTGCCACGAGCTGCTGGGCCACGTGCCCCTGTTCTCTGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGAGCCCCTGATGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACTGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGAGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCTGAGAAGCCCAAGCTGCTGCCCCTGGAGCTGGAGAAGACTGCCATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCTGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGACCCTTCTCTGTGAGATATGACCCCTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCTGACAGCATCAACTCTGAGATTGGCATCCTGTGCTCTGCCCTGCAGAAGATCAAGTAA
SEQ ID NO: 8 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (GENEius 2/Operon2)
ATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGGGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGGGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCTGCCCTGACCAACATCATCAAGATCCTGAGACACGATATTGGGGCCACTGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACTGTGCCCTGGTTCCCCAGAACTATCCAGGAGCTGGACAGATTTGCCAACCAGATCCTGAGCTATGGGGCTGAGCTGGATGCTGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGAAGAAAGCAGTTTGCTGACATTGCCTACAACTACAGACACGGCCAGCCCATCCCCAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACTGGCTTCAGACTGAGACCTGTGGCTGGCCTGCTGAGCAGCAGAGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTACATCAGACACGGCAGCAAGCCCATGTACACCCCTGAGCCTGATATCTGCCACGAGCTGCTGGGCCACGTGCCCCTGTTCTCTGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGGGCCCCTGATGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACTGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGGGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCTGAGAAGCCCAAGCTGCTGCCCCTGGAGCTGGAGAAGACTGCCATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCTGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGACCCTTCTCTGTGAGATACGACCCCTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCTGACAGCATCAACTCTGAGATTGGCATCCTGTGCTCTGCCCTGCAGAAGATCAAGTGA
SEQ ID NO: 9 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (Nathwani/NW2-Cop)
ATGTCCACTGCTGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTACTGAGGTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTAGGTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAAAGGAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCACGACATTGGTGCCACTGTCCACGAGCTTTCAAGGGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACTATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCTGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACAGGGCAAGAAGGAAGCAGTTTGCTGACATTGCCTACAACTACAGGCATGGGCAGCCCATCCCTAGGGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCACGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCACGAAGATAACATTCCCCAGCTGGAAGATGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCAGGCTCAGGCCTGTAGCTGGCCTGCTTTCCTCTAGGGATTTCTTGGGTGGCCTGGCCTTCAGGGTCTTCCACTGCACACAGTACATCAGACACGGCTCCAAGCCCATGTATACCCCTGAACCTGACATCTGCCACGAGCTGTTGGGACACGTGCCCTTGTTTTCAGATAGGAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATATATTGAAAAGCTGGCCACAATCTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACTGAGTTCCAGCCCCTGTATTATGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTAGGCCCTTCTCAGTTAGGTATGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATCAAGTAA
SEQ ID NO: 10 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (Nathwani-RCG/NW-RCG)
ATGTCCACTGCTGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTACTGAGGTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTAGGTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAAAGGAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCACGACATTGGTGCCACTGTCCACGAGCTTTCAAGGGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACTATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCTGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACAGGGCAAGAAGGAAGCAGTTTGCTGACATTGCCTACAACTACAGGCATGGGCAGCCCATCCCTAGGGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCACGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCACGAAGATAACATTCCCCAGCTGGAAGATGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCAGGCTCAGGCCTGTAGCTGGCCTGCTTTCCTCTAGGGATTTCTTGGGTGGCCTGGCCTTCAGGGTCTTCCACTGCACACAGTACATCAGACACGGCTCCAAGCCCATGTATACCCCTGAACCTGACATCTGCCACGAGCTGTTGGGACACGTGCCCTTGTTTTCAGATAGGAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATATATTGAAAAGCTGGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACTGAGTTCCAGCCCCTGTATTATGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTAGGCCCTTCTCAGTTAGGTATGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAA
SEQ ID NO: 11 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (ATUM (DNA2.0-op/D20-P))
ATGTCCACTGCGGTGTTGGAAAACCCCGGACTGGGCAGAAAGTTGTCCGACTTCGGCCAGGAAACCTCTTACATTGAGGACAACTGCAACCAGAACGGGGCCATCTCACTGATCTTTTCGCTGAAAGAAGAAGTCGGAGCTCTGGCCAAAGTGCTGCGCCTGTTCGAGGAAAACGACGTCAACCTGACCCACATCGAGTCAAGACCGAGCAGGCTGAAGAAGGATGAGTACGAGTTTTTCACCCATCTCGACAAGAGATCCCTGCCTGCCCTGACCAATATTATCAAGATTTTGCGGCACGACATTGGCGCAACCGTGCATGAACTCTCCCGGGACAAGAAGAAGGACACCGTGCCGTGGTTCCCCCGAACCATCCAGGAACTCGACCGCTTCGCTAACCAGATCCTGTCCTACGGCGCCGAACTGGATGCCGATCACCCTGGATTCAAGGACCCAGTGTACAGAGCCCGGCGCAAGCAGTTCGCCGATATCGCCTACAATTATCGGCACGGACAGCCAATCCCGAGGGTGGAGTACATGGAGGAGGAAAAGAAAACCTGGGGAACTGTGTTCAAGACCTTGAAGTCCCTGTACAAGACTCACGCCTGCTACGAGTACAACCACATCTTCCCCCTGCTCGAAAAGTACTGCGGGTTCCATGAGGACAACATCCCCCAACTGGAAGATGTGTCGCAGTTCCTGCAAACCTGTACCGGATTCCGGCTGAGGCCTGTCGCGGGACTTCTGTCCTCCCGGGATTTTCTTGGCGGTCTGGCCTTCCGGGTGTTCCACTGTACTCAGTACATTAGACACGGGAGCAAGCCTATGTACACTCCTGAACCCGACATTTGCCACGAACTCCTGGGTCATGTGCCCCTCTTCTCGGATCGGAGCTTCGCCCAGTTCAGCCAAGAGATCGGTCTGGCTAGCTTGGGAGCACCCGACGAGTACATCGAGAAGCTGGCCACTATCTACTGGTTTACCGTGGAATTCGGACTGTGCAAGCAGGGGGACTCAATCAAGGCCTATGGCGCGGGACTCCTGAGCTCCTTCGGGGAGCTGCAGTACTGCCTGTCCGAAAAGCCAAAGCTGCTCCCTCTTGAACTGGAGAAAACGGCCATCCAGAACTACACCGTGACCGAATTCCAGCCGCTCTACTACGTCGCGGAGTCCTTCAACGATGCCAAGGAGAAGGTCCGCAACTTCGCCGCAACTATCCCGCGGCCGTTTTCCGTGCGCTATGACCCGTACACACAACGCATCGAAGTGCTGGACAACACCCAGCAACTTAAGATTCTGGCCGACTCGATCAACTCCGAGATTGGCATTCTGTGCTCGGCGCTGCAGAAGATCAAGTAA
SEQ ID NO: 12 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (ATUM-RCG (DNA2.0-Cop1))
ATGTCCACTGCGGTCCTGGAAAACCCTGGTCTGGGCCGCAAGCTTTCTGACTTTGGACAGGAAACCTCATACATTGAGGACAACTGTAACCAAAATGGTGCAATCAGCCTGATCTTCAGCCTCAAGGAAGAAGTGGGAGCCCTGGCCAAGGTCCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGTCCAGGCCCTCCAGACTGAAGAAGGATGAATACGAATTCTTCACCCACCTGGACAAGCGCTCCCTCCCTGCCCTCACCAACATCATTAAGATCCTGCGGCACGACATTGGAGCCACTGTGCATGAGTTGAGCCGGGACAAGAAGAAGGATACTGTGCCCTGGTTCCCGAGGACCATCCAGGAACTGGACCGGTTTGCCAACCAAATTCTGTCCTATGGAGCTGAATTGGATGCAGACCACCCTGGCTTCAAGGACCCAGTGTACAGAGCAAGGAGAAAGCAGTTTGCAGACATAGCCTACAACTACAGACATGGACAGCCCATCCCGAGGGTGGAGTACATGGAAGAGGAGAAGAAAACCTGGGGCACTGTGTTTAAGACCCTCAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCACTCCTGGAGAAATACTGTGGCTTCCATGAGGACAACATCCCACAGCTGGAGGATGTGTCCCAGTTCCTGCAGACTTGCACTGGCTTCCGGTTGAGGCCTGTGGCTGGCCTGCTCTCCTCAAGGGACTTCTTGGGGGGACTGGCATTCAGGGTGTTCCACTGCACCCAATACATCAGACACGGCAGCAAGCCAATGTACACCCCTGAACCGGACATCTGCCACGAACTCCTGGGCCACGTCCCTCTGTTCTCGGACAGAAGCTTTGCCCAGTTCTCCCAAGAGATTGGCCTTGCCTCCCTGGGGGCCCCTGATGAATACATTGAAAAGCTGGCCACAATCTACTGGTTCACTGTGGAATTTGGACTTTGCAAGCAGGGAGATAGCATCAAGGCCTACGGGGCTGGACTTCTGTCCTCCTTCGGTGAACTGCAGTACTGTCTGTCAGAGAAGCCCAAGCTGCTGCCCCTGGAACTGGAGAAAACTGCCATCCAAAACTACACTGTGACTGAGTTCCAGCCCCTCTACTATGTGGCTGAGTCCTTCAATGATGCCAAAGAAAAGGTCAGAAATTTTGCGGCCACCATTCCTAGGCCTTTCTCAGTCCGCTATGACCCTTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTCAAGATCCTGGCAGACTCCATCAACTCAGAAATTGGGATCTTGTGCTCGGCCCTCCAAAAGATCAAGTAA
SEQ ID NO: 13 - кодон-оптимизированная последовательность нуклеиновой кислоты PAH человека (JCAT (JCAT))
ATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGGGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGGGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCAGCCCTGACCAATATTATCAAGATCCTGAGGCACGATATTGGGGCCACAGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACAGTGCCCTGGTTCCCCAGGACTATCCAGGAGCTGGACAGGTTTGCCAACCAGATCCTGAGCTACGGAGCTGAGCTGGATGCAGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGGAGAAAGCAGTTTGCAGATATAGCCTACAACTACAGGCACGGCCAGCCAATCCCTAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTACGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACAGGCTTCAGGCTGAGACCAGTGGCAGGCCTGCTGAGCAGCAGGGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTATATCAGGCACGGCAGCAAGCCAATGTACACCCCTGAGCCAGATATCTGCCACGAGCTGCTGGGCCACGTTCCCCTGTTCTCAGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGGGCCCCTGACGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACAGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGGGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCAGAGAAGCCAAAGCTGCTGCCCCTGGAGCTGGAGAAGACAGCTATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCAGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGGCCCTTCTCTGTGAGATATGACCCCTACACCCAGAGGATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCAGACAGCATCAACAGTGAGATTGGTATCCTGTGCTCTGCCCTGCAGAAGATCAAGTAA
SEQ ID NO: 14 - последовательность нуклеиновой кислоты составного интрона глобина/AIAT
TgcctttcactgcgaggggttctggagaggcttctgagctccccatggcccaggcaggcagcaggtctggggcaggaggggggttgtggagtgggtatccgcctgctgaggtgcagggcagatggagaggctgcagctgagctcctattttcataataacagcagccatgagggttgtgtcctgtttcccagtcctgcccggtcccccctcggtacctcctggtggatacactggttcctgtaagcagaagtggatgagggtgtctaggtctgcagtcctggcaccccaggatgggggacaccagccaagatacagcaacagcaacaaagcgcagccatttctttctgtttgcacagctcctctgtctgtcgggggctcctgtctgttgtctcctataagcctcaccacctctcctactgcttgggcatgcatctttctccccttctatagatgaggaggttaaggtccagagaggggtggggaggaacgccggctcacattctccatcccctccagatatgaccaggaacagacctgtgccaggcctcagccttacatcaaaatgggcctccccatgcaccgtggacctctgggccctcctgtcccagtggaggacaggaagctatgaggggcactgtcacccagggctcaagctggcattcctgaataatcgctctgcaccaggccacggctaagctcagtgcgtgattaagcctcataaccctccaaggcagttactagtgtgattcccattttacagatgaggaagatggggacagagaggtgaataactggccccaaatcacacaccatccataattcgggctcaggcacctggctccagtccccaaactcttgaacctggccctagtgtcactgtttctcttgggtctcaggcgctggatggggaacaggaaacctgggctggacttgaggcctctctgatgctcggtgacttcagacagttgctcaacctctctgttctcttgggcaaaacatgataacctttgacttctgtcccctcccctcaccccacccgaccttgatctctgaagtgttggaaggatttaatttttcctgcactgagttttggagacaggtcaaaaagatgaccaaggccaaggtggccagtttcctatagaacgcctctaaaagacctgcagcaatagcagcaagaactggtattctcgagaacttgctgcgcagcaggcacttcttggcattttatgtgtatttaatttcacaatagctctatgacaaagtccacctttctcatctccaggaaactgaggttcagagaggttaagtaacttgtccaaggtcacacagctaatagcaagttgacgtggagcaatctggcctcagagcctttaattttagccacagactgatgctcccctcttcatttagccaggctgcctctgaagttttctgattcaagacttctggcttcagctttgtacacagagatgattcaatgtcaggttttggagtgaaatctgtttaatcccagacaaaacatttaggattacatctcagttttgtaagcaagtagctctgtgatttttagtgagttatttaatgctctttggggctcaatttttctatctataaaatagggctaataatttgcaccttatagggtaagctttgaggacagattagatgatacggtgcctgtaaaacaccaggtgttagtaagtgtggcaatgatggtgacgctgaggctgatgtttgcttagcatagggttaggcagctggcaggcagtaaacagttggataatttaatggaaaatttgccaaactcagatgcTAGCAGCTACaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagctcctgggcaacgtgctggtctgtgtgctggcccatcactttggcaaagaattgcgatCGCCACC
SEQ ID NO: 15 - Последовательность нуклеиновой кислоты вектора 1 (ApoE-HCR-hAAT.GI.hPAH.bGH)
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGGTCGACGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtactagcagctacaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagCTCCTGGGCAACGTGCTGGTCTGTGTGCTGGCCCATCACTTTGGCAAAGAATTgcgatCGCCACCATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGCggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg
SEQ ID NO: 16 - последовательность нуклеиновой кислоты GENEius1 (ApoE-HCR-hAAT.GI.hPAHco1.bGH)
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGGTCGACGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtactagcagctacaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagCTCCTGGGCAACGTGCTGGTCTGTGTGCTGGCCCATCACTTTGGCAAAGAATTgcgatCGCCACCATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGAGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGAGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCTGCCCTGACCAACATCATCAAGATCCTGAGACACGATATTGGAGCCACTGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACTGTGCCCTGGTTCCCCAGAACTATCCAGGAGCTGGACAGATTTGCCAACCAGATCCTGAGCTATGGAGCTGAGCTGGATGCTGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGAAGAAAGCAGTTTGCTGACATTGCCTACAACTACAGACACGGCCAGCCCATCCCCAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACTGGCTTCAGACTGAGACCTGTGGCTGGCCTGCTGAGCAGCAGAGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTACATCAGACACGGCAGCAAGCCCATGTACACCCCTGAGCCTGATATCTGCCACGAGCTGCTGGGCCACGTGCCCCTGTTCTCTGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGAGCCCCTGATGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACTGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGAGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCTGAGAAGCCCAAGCTGCTGCCCCTGGAGCTGGAGAAGACTGCCATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCTGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGACCCTTCTCTGTGAGATATGACCCCTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCTGACAGCATCAACTCTGAGATTGGCATCCTGTGCTCTGCCCTGCAGAAGATCAAGTAACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGCggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg
SEQ ID NO: 17 - Последовательность нуклеиновой кислоты Large Intron-вектор 1 (ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH) cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGgtcgacGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtatgcctttcactgcgaggggttctggagaggcttctgagctccccatggcccaggcaggcagcaggtctggggcaggaggggggttgtggagtgggtatccgcctgctgaggtgcagggcagatggagaggctgcagctgagctcctattttcataataacagcagccatgagggttgtgtcctgtttcccagtcctgcccggtcccccctcggtacctcctggtGgatacactggttcctgtaagcagaagtggatgagggtgtctaggtctgcagtcctggcaccccaggatgggggacaccagccaagatacagcaacagcaacaaagcgcagccatttctttctgtttgcacagctcctctgtctgtcgggggctcctgtctgttgtctcctataagcctcaccacctctcctactgcttgggcatgcatctttctccccttctatagatgaggaggttaaggtccagagaggggtggggaggaacgccggctcacattctccatcccctccagatatgaccaggaacagacctgtgccaggcctcagccttacatCaaaatgggcctccccatgcaccgtggacctctgggccctcctgtcccagtggaggacaggaagctatgaggggcactgtcacccagggctcaagctggcattcctgaataatcgctctgcaccaggccacggctaagctcagtgcgtgattaagcctcataaccctccaaggcagttactagtgtgattcccattttacagatgaggaagatggggacagagaggtgaataactggccccaaatcacacaccatccataattcgggctcaggcacctggctccagtccccaaactcttgaacctggccctagtgtcactgtttctcttgggtCtcaggcgctggatggggaacaggaaacctgggctggacttgaggcctctctgatgctcggtgacttcagacagttgctcaacctctctgttctcttgggcaaaacatgataacctttgacttctgtcccctcccctcaccccacccgaccttgatctctgaagtgttggaaggatttaatttttcctgcactgagttttggagacaggtcaaaaagatgaccaaggccaaggtggccagtttcctatagaacgcctctaaaagacctgcagcaatagcagcaagaactggtattctcgagaacttgctgcgcagcaggcacttcttggcattttAtgtgtatttaatttcacaatagctctatgacaaagtccacctttctcatctccaggaaactgaggttcagagaggttaagtaacttgtccaaggtcacacagctaatagcaagttgacgtggagcaatctggcctcagagcctttaattttagccacagactgatgctcccctcttcatttagccaggctgcctctgaagttttctgattcaagacttctggcttcagctttgtacacagagatgattcaatgtcaggttttggagtgaaatctgtttaatcccagacaaaacatttaggattacatctcagttttgtaagcaagtagctctgtgatttttagtgagttatttaatgctctttggggctcaatttttctatctataaaatagggctaataatttgcaccttatagggtaagctttgaggacagattagatgatacggtgcctgtaaaacaccaggtgttagtaagtgtggcaatgatggtgacgctgaggctgatgtttgcttagcatagggttaggcagctggcaggcagtaaacagttggataatttaatggaaaatttgccaaactcagatgcTAGCAGCTACaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagctcctgggcaacgtgctggtctgtgtgctggcccatcactttggcaaagaattgcgatcgccaccATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAAcctcgagctgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctAATAAAatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggaccggtgcggccgcAGGAACCCCTAGTGATGGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCTTTGCCCGGGCGGCCTCAGTGAGCGAGCGAGCGCGCAGCTGCCTGCAGG
SEQ ID NO: 18- Последовательность нуклеиновой кислоты Large Intron-GENEius 1 (ApoE-HCR-hAAT.cG-AIATI.hPAHco1.bGH)
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGgtcgacGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtatgcctttcactgcgaggggttctggagaggcttctgagctccccatggcccaggcaggcagcaggtctggggcaggaggggggttgtggagtgggtatccgcctgctgaggtgcagggcagatggagaggctgcagctgagctcctattttcataataacagcagccatgagggttgtgtcctgtttcccagtcctgcccggtcccccctcggtacctcctggtggatacactggttCctgtaagcagaagtggatgagggtgtctaggtctgcagtcctggcaccccaggatgggggacaccagccaagatacagcaacagcaacaaagcgcagccatttctttctgtttgcacagctcctctgtctgtcgggggctcctgtctgttgtctcctataagcctcaccacctctcctactgcttgggcatgcatctttctccccttctatagatgaggaggttaaggtccagagaggggtggggaggaacgccggctcacattctccatcccctccagatatgaccaggaacagacctgtgccaggcctcagccttacatcaaaatgggcctccccAtgcaccgtggacctctgggccctcctgtcccagtggaggacaggaagctatgaggggcactgtcacccagggctcaagctggcattcctgaataatcgctctgcaccaggccacggctaagctcagtgcgtgattaagcctcataaccctccaaggcagttactagtgtgattcccattttacagatgaggaagatggggacagagaggtgaataactggccccaaatcacacaccatccataattcgggctcaggcacctggctccagtccccaaactcttgaacctggccctagtgtcactgtttctcttgggtctcaggcgctggatggggaAcaggaaacctgggctggacttgaggcctctctgatgctcggtgacttcagacagttgctcaacctctctgttctcttgggcaaaacatgataacctttgacttctgtcccctcccctcaccccacccgaccttgatctctgaagtgttggaaggatttaatttttcctgcactgagttttggagacaggtcaaaaagatgaccaaggccaaggtggccagtttcctatagaacgcctctaaaagacctgcagcaatagcagcaagaactggtattctcgagaacttgctgcgcagcaggcacttcttggcattttatgtgtatttaatttcacaatagctCtatgacaaagtccacctttctcatctccaggaaactgaggttcagagaggttaagtaacttgtccaaggtcacacagctaatagcaagttgacgtggagcaatctggcctcagagcctttaattttagccacagactgatgctcccctcttcatttagccaggctgcctctgaagttttctgattcaagacttctggcttcagctttgtacacagagatgattcaatgtcaggttttggagtgaaatctgtttaatcccagacaaaacatttaggattacatctcagttttgtaagcaagtagctctgtgatttttagtgagttatttaatgctctttggggctcaatttttctatctataaaatagggctaataatttgcaccttatagggtaagctttgaggacagattagatgatacggtgcctgtaaaacaccaggtgttagtaagtgtggcaatgatggtgacgctgaggctgatgtttgcttagcatagggttaggcagctggcaggcagtaaacagttggataatttaatggaaaatttgccaaactcagatgcTAGCAGCTACaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagctcctgggcaacgtgctggtctgtgtgctggcccatcactttggcaaagaattgcgatCGCCACCATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGAGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGAGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCTGCCCTGACCAACATCATCAAGATCCTGAGACACGATATTGGAGCCACTGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACTGTGCCCTGGTTCCCCAGAACTATCCAGGAGCTGGACAGATTTGCCAACCAGATCCTGAGCTATGGAGCTGAGCTGGATGCTGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGAAGAAAGCAGTTTGCTGACATTGCCTACAACTACAGACACGGCCAGCCCATCCCCAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACTGGCTTCAGACTGAGACCTGTGGCTGGCCTGCTGAGCAGCAGAGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTACATCAGACACGGCAGCAAGCCCATGTACACCCCTGAGCCTGATATCTGCCACGAGCTGCTGGGCCACGTGCCCCTGTTCTCTGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGAGCCCCTGATGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACTGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGAGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCTGAGAAGCCCAAGCTGCTGCCCCTGGAGCTGGAGAAGACTGCCATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCTGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGACCCTTCTCTGTGAGATATGACCCCTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCTGACAGCATCAACTCTGAGATTGGCATCCTGTGCTCTGCCCTGCAGAAGATCAAGTAACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGCggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg
SEQ ID NO: 19 - последовательность нуклеиновой кислоты hPAH+укороченный 2-й интрон (ApoE-HCR-hAAT.hPAH-tI2.bGH)
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcggccgcacgcgtaggctcagaggcacacaggagtttctgggctcaccctgcccccttccaacccctcagttcccatcctccagcagctgtttgtgtgctgcctctgaagtccacactgaacaaacttcagcctactcatgtccctAaaatgggcaaacattgcaagcagcaaacagcaaacacacagccctccctgcctgctgaccttggagctggggcagaggtcagagacctctctgggcccatgccacctccaacatccactcgaccccttggaatttcggtggagaggagcagaggttgtcctggcgtggtttaggtagtgtgagaggggtcgacgatcttgctaccagtggaacagccactaaggattctgcagtgagagcagagggccagctaagtggtactctcccagagactgtctgactcacgccaccccctccaccttggacacaggacgctgtggtttctgagCcaggtacaatgactcctttcggtaagtgcagtggaagctgtacactgcccaggcaaagcgtccgggcagcgtaggcgggcgactcagatcccagccagtggacttagcccctgtttgctcctccgataactggggtgaccttggttaatattcaccagcagcctcccccgttgcccctctggatccactgcttaaatacggacgaggacagggccctgtctcctcagcttcaggcaccaccactgacctgggacAGTGAATCagccagagacctcactcccggggagccagcATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGgtcagtgctacaatcatgtttgtcttggataatgtcgtagcaaacttccatgttcttttctagttagatgcaatgaaaagaacacaggatctggaacaggcagatttgaatttgagctcaagtttttacattaaccaactgtgtgaccttaGttaagtcattcaatctctctgagcttcagtttttccattcataatatagtgctgataatatgtgccttgtcagtttcaacaggaactttgtgatgaaataatgtgttttgtaaaatgcctaatcatatgtgtgattaggcatgattaactataagtcatatgtgacacgtgattacatttcacataacatgtaatcacatgtgtcacatgtgattacagttaaagagtaaacaagaaataaatattaattcttttattcactaaattaatattgattgtcttctatgtataagtgtaaaataatatgcaagacaccgtccctttcttcaagtagcttAacccaaaactatgctttagaaaatagctaatgttcttcaagacattggtaaatgtcttatgattaaagtggttccataattaataaacttgggaaattcgggcatattattttacctgatttctttattataggacttttagagtctataaaactaattatactaagttgtttacagagagaaatgtatgtattattttccaaacttacttgactgtggatcctttttattttaagaacatgtattcacatctgggaacagagtttaggaaacgttgatttgggtgtttgctgggcagccaaacttcacagaactccaaataggtttcctcagccagAttccttccgagtactatgctaatatttttggtgggattttgtcccacctgaaaatacattgctttatgctaagattcctgtgacctctctagctgattgggaggcaggggtagcacattggtcagggctgtgcacatacttagtgctcagtgtgtttgggcatgcaatcaacaaatccttttggggcatgactggatacgattagatatcttgtgaaaacctatgatgttctcagcactgtgggtggggaggaagatgggggacacatagaaattgtaaggaagagaaactgcctcctttaggatctaacaggggaagcaaatattctgagcaGctggggagggaaggcagagcatgagaagttatctcagtaaaaggtagctttatagacatgatcttattcagtctaatgaaaataacatgaggtaggagctatttctgcccccatttttcagagatagaaactgatgcttagaggggttaggtaaatgtgcccaaggtcaaacagctagtaaatacggaagaaatcatttaaacccaggcattctgagtctagaacctacatacactcttaactgctattcagtactgccccagacagaacgggcttcatagtttgcagggcgcagtgcaaaatgaaaacgtggggtctcttcctcaataAcaggaaataatgctgttaaaagtactaaaatatataactttatcctttcttctgtggtctctctcatgtcctgttatggtgtttttgatttgccatttagttgtgctctcctcccctgggcaccttggccctctaactgctgggttccctgtccagccagggctgggctgacagccaagcccctaccaaggatggggaaatccatcttccatttccgtggacctgctgcaccaacccacagtgatgagctatcccccaagagattgtaacctctggaatgagaatggaaaggaagcTTGCCTGAACTTGGTTTTAaagaagaaaaagtaaacaacttttgcaaatttaaggacttttttatactcttataaaaataattgtattcttgaactctccattttgttgcgttaggttttcctgttctggttctgcatctttggcctgcgttagttccagtgactgtctcctcAccctccccattctctcttctagGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTaGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTgTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAAcctcgagctgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctAATAAAatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggaccggtgcggccgcAGGAACCCCTAGTGATGGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCTTTGCCCGGGCGGCCTCAGTGAGCGAGCGAGCGCGCAGCTGCCTGCAGG
SEQ ID NO: 20 - последовательность вектора pFB-ApoE-hAAT-hPAH-Genius2
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGGTCGACGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtactagcagctacaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagCTCCTGGGCAACGTGCTGGTCTGTGTGCTGGCCCATCACTTTGGCAAAGAATTgcgatCGCCACCATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGGGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGGGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCTGCCCTGACCAACATCATCAAGATCCTGAGACACGATATTGGGGCCACTGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACTGTGCCCTGGTTCCCCAGAACTATCCAGGAGCTGGACAGATTTGCCAACCAGATCCTGAGCTATGGGGCTGAGCTGGATGCTGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGAAGAAAGCAGTTTGCTGACATTGCCTACAACTACAGACACGGCCAGCCCATCCCCAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTATGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACTGGCTTCAGACTGAGACCTGTGGCTGGCCTGCTGAGCAGCAGAGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTACATCAGACACGGCAGCAAGCCCATGTACACCCCTGAGCCTGATATCTGCCACGAGCTGCTGGGCCACGTGCCCCTGTTCTCTGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGGGCCCCTGATGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACTGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGGGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCTGAGAAGCCCAAGCTGCTGCCCCTGGAGCTGGAGAAGACTGCCATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCTGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGACCCTTCTCTGTGAGATACGACCCCTACACCCAGAGAATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCTGACAGCATCAACTCTGAGATTGGCATCCTGTGCTCTGCCCTGCAGAAGATCAAGTGACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGCggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg
SEQ ID NO: 21 - последовательность вектора pFB-ApoE-hAAT-hPAH-JCAT
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcGGCCGCACGCGTAGGCTCAGAGGCACACAGGAGTTTCTGGGCTCACCCTGCCCCCTTCCAACCCCTCAGTTCCCATCCTCCAGCAGCTGTTTGTGTGCTGCCTCTGAAGTCCACACTGAACAAACTTCAGCCTACTCATGTCCCTAAAATGGGCAAACATTGCAAGCAGCAAACAGCAAACACACAGCCCTCCCTGCCTGCTGACCTTGGAGCTGGGGCAGAGGTCAGAGACCTCTCTGGGCCCATGCCACCTCCAACATCCACTCGACCCCTTGGAATTTCGGTGGAGAGGAGCAGAGGTTGTCCTGGCGTGGTTTAGGTAGTGTGAGAGGGGTCGACGATCTTGCTACCAGTGGAACAGCCACTAAGGATTCTGCAGTGAGAGCAGAGGGCCAGCTAAGTGGTACTCTCCCAGAGACTGTCTGACTCACGCCACCCCCTCCACCTTGGACACAGGACGCTGTGGTTTCTGAGCCAGGTACAATGACTCCTTTCGGTAAGTGCAGTGGAAGCTGTACACTGCCCAGGCAAAGCGTCCGGGCAGCGTAGGCGGGCGACTCAGATCCCAGCCAGTGGACTTAGCCCCTGTTTGCTCCTCCGATAACTGGGGTGACCTTGGTTAATATTCACCAGCAGCCTCCCCCGTTGCCCCTCTGGATCCACTGCTTAAATACGGACGAGGACAGGGCCCTGTCTCCTCAGCTTCAGGCACCACCACTGACCTGGGACAGTGAATCgtaagtactagcagctacaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagCTCCTGGGCAACGTGCTGGTCTGTGTGCTGGCCCATCACTTTGGCAAAGAATTgcgatCGCCACCATGAGCACTGCTGTGCTGGAGAACCCTGGCCTGGGCAGAAAGCTGTCTGACTTTGGCCAGGAGACCAGCTACATTGAGGACAACTGCAACCAGAATGGGGCCATCAGCCTGATCTTCAGCCTGAAGGAGGAGGTGGGGGCCCTGGCCAAGGTGCTGAGACTGTTTGAGGAGAATGATGTGAACCTGACCCACATTGAGAGCAGACCCAGCAGACTGAAGAAGGATGAGTATGAGTTCTTCACCCACCTGGACAAGAGAAGCCTGCCAGCCCTGACCAATATTATCAAGATCCTGAGGCACGATATTGGGGCCACAGTGCACGAGCTGAGCAGAGACAAGAAGAAGGACACAGTGCCCTGGTTCCCCAGGACTATCCAGGAGCTGGACAGGTTTGCCAACCAGATCCTGAGCTACGGAGCTGAGCTGGATGCAGACCACCCTGGCTTCAAGGACCCTGTGTACAGAGCCAGGAGAAAGCAGTTTGCAGATATAGCCTACAACTACAGGCACGGCCAGCCAATCCCTAGAGTGGAGTACATGGAGGAGGAGAAGAAGACCTGGGGCACTGTGTTCAAGACCCTGAAGAGCCTGTACAAGACCCACGCCTGCTACGAGTACAACCACATCTTCCCCCTGCTGGAGAAGTACTGTGGCTTCCACGAGGACAACATCCCCCAGCTGGAGGATGTGAGCCAGTTCCTGCAGACCTGCACAGGCTTCAGGCTGAGACCAGTGGCAGGCCTGCTGAGCAGCAGGGACTTCCTGGGGGGCCTGGCCTTCAGAGTGTTCCACTGCACCCAGTATATCAGGCACGGCAGCAAGCCAATGTACACCCCTGAGCCAGATATCTGCCACGAGCTGCTGGGCCACGTTCCCCTGTTCTCAGACAGAAGCTTTGCCCAGTTCAGCCAGGAGATTGGCCTGGCCAGCCTGGGGGCCCCTGACGAGTATATTGAGAAGCTGGCCACTATCTACTGGTTCACAGTGGAGTTTGGCCTGTGCAAGCAGGGGGACAGCATCAAGGCCTATGGGGCTGGCCTGCTGAGCAGCTTTGGGGAGCTGCAGTACTGCCTGTCAGAGAAGCCAAAGCTGCTGCCCCTGGAGCTGGAGAAGACAGCTATCCAGAACTACACTGTGACTGAGTTCCAGCCCCTGTACTATGTGGCAGAGAGCTTCAATGATGCCAAGGAGAAGGTGAGAAACTTTGCTGCCACCATCCCCAGGCCCTTCTCTGTGAGATATGACCCCTACACCCAGAGGATTGAGGTGCTGGACAACACCCAGCAGCTGAAGATCCTGGCAGACAGCATCAACAGTGAGATTGGTATCCTGTGCTCTGCCCTGCAGAAGATCAAGTAACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGCggccgcaggaacccctagtgatggagttggccactccctctctgcgcgctcgctcgctcactgaggccgggcgaccaaaggtcgcccgacgcccgggctttgcccgggcggcctcagtgagcgagcgagcgcgcagctgcctgcagg
SEQ ID NO: 22- последовательность вектора pFB-hPAHV1-TBG2uGlob
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcggccgcaCGCGTCAGGTTAATTTTTAAAAAGCAGTCAAAAGTCCAAGTGGCCCTTGGCAGCATTTACTCTCTCTGTTTGCTCTGGTTAATAATCTCAGGAGCACAAACATTCCAGATCCAGGTTAATTTTTAAAAAGCAGTCAAAAGTCCAAGTGGCCCTTGGCAGCATTTACTCTCTCTGTTTGCTCTGGTTAATAATCTCAGGAGCACAAACATTCCAGATCCGGCGCGCCAGGGCTGGAAGCTACCTTTGACATCATTTCCTCTGCGAATGCATGTATAATTTCTACAGAACCTATTAGAAAGGATCACCCAGCCTCTGCTTTTGTACAACTTTCCCTTAAAAAACTGCCAATTCCACTGCTGTTTGGCCCAATAGTGAGAACTTTTTCCTGCTGCCTCTTGGTGCTTTTGCCTATGGCCCCTATTCTGCCTGCTGAAGACACTCTTGCCAGCATGGACTTAAACCCCTCCAGCTCTGACAATCCTCTTTCTCTTTTGTTTTACATGAAGGGTCTGGCAGCCAAAGCAATCACTCAAAGTTCAAACCTTATCATTTTTTGCTTTGTTCCTCTTGGCCTTGGTTTTGTACATCAGCTTTGAAAATACCATCCCAGGGTTAATGCTGGGGTTAATTTATAACTAAGAGTGCTCTAGTTTTGCAATACAGGACATGCTATAAAAATGGAAAGATGTTGCTTTCTGAGAGATGCAGGTGGATTCTTGGGCATTTGCTGTAAGTACTAGCAGCTACAATCCAGCTACCATTCTGCTTTTATTTTATGGTTGGGATAAGGCTGGATTATTCTGAGTCCAAGCTAGGCCCTTTTGCTAATCATGTTCATACCTCTTATCTTCCTCCCACAGCTCCTGGGCAACGTGCTGGTCTGTGTGCTGGCCCATCACTTTGGCAAAGAATTGCGATCGCCACCATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAACCTCGAGCTGTGCCTTCTAGTTGCCAGCCATCTGTTGTTTGCCCCTCCCCCGTGCCTTCCTTGACCCTGGAAGGTGCCACTCCCACTGTCCTTTCCTAATAAAATGAGGAAATTGCATCGCATTGTCTGAGTAGGTGTCATTCTATTCTGGGGGGTGGGGTGGGGCAGGACAGCAAGGGGGAGGATTGGGAAGACAATAGCAGGCATGCTGGGGATGCGGTGGGCTCTATGGACCGGTGC
SEQ ID NO: 23- последовательность вектора pFB-hPAHV1-TTR
cctgcaggcagctgcgcgctcgctcgctcactgaggccgcccgggcaaagcccgggcgtcgggcgacctttggtcgcccggcctcagtgagcgagcgagcgcgcagagagggagtggccaactccatcactaggggttcctgcggccgcGTCTGTCTGCACATTTCGTAGAGCGAGTGTTCCGATACTCTAATCTCCCTAGGCAAGGTTCATATTgactTAGGTTACTTATTCTCCTTTTGTTGACTAAGTCAATAATCAGAATCAGCAGGTTTGGAGTCAGCTTGGCAGGGATCAGCAGCCTGGGTTGGAAGGAGGGGGTATAAAAGCCCCTTCACCAGGAGAAGCCGTCACACAGATCCACAAGCTCCTGACAGGagGCTTCCCTTCGACTCTTCCTCCTTTGCCTCGCTGGACTGGTATTTGTGTCTGAAGCTGGCCCCGCGgtaagtactagcagctacaatccagctaccattctgcttttattttatggttgggataaggctggattattctgagtccaagctaggcccttttgctaatcatgttcatacctcttatcttcctcccacagctcctgggcaacgtgctggtctgtgtgctggcccatcactttggcaaagaattgcgatcgccaccATGTCCACTGCGGTCCTGGAAAACCCAGGCTTGGGCAGGAAACTCTCTGACTTTGGACAGGAAACAAGCTATATTGAAGACAACTGCAATCAAAATGGTGCCATATCACTGATCTTCTCACTCAAAGAAGAAGTTGGTGCATTGGCCAAAGTATTGCGCTTATTTGAGGAGAATGATGTAAACCTGACCCACATTGAATCTAGACCTTCTCGTTTAAAGAAAGATGAGTATGAATTTTTCACCCATTTGGATAAACGTAGCCTGCCTGCTCTGACAAACATCATCAAGATCTTGAGGCATGACATTGGTGCCACTGTCCATGAGCTTTCACGAGATAAGAAGAAAGACACAGTGCCCTGGTTCCCAAGAACCATTCAAGAGCTGGACAGATTTGCCAATCAGATTCTCAGCTATGGAGCGGAACTGGATGCTGACCACCCTGGTTTTAAAGATCCTGTGTACCGTGCAAGACGGAAGCAGTTTGCTGACATTGCCTACAACTACCGCCATGGGCAGCCCATCCCTCGAGTGGAATACATGGAGGAAGAAAAGAAAACATGGGGCACAGTGTTCAAGACTCTGAAGTCCTTGTATAAAACCCATGCTTGCTATGAGTACAATCACATTTTTCCACTTCTTGAAAAGTACTGTGGCTTCCATGAAGATAACATTCCCCAGCTGGAAGACGTTTCTCAGTTCCTGCAGACTTGCACTGGTTTCCGCCTCCGACCTGTAGCTGGCCTGCTTTCCTCTCGGGATTTCTTGGGTGGCCTGGCCTTCCGAGTCTTCCACTGCACACAGTACATCAGACATGGATCCAAGCCCATGTATACCCCCGAACCTGACATCTGCCATGAGCTGTTGGGACATGTGCCCTTGTTTTCAGATCGCAGCTTTGCCCAGTTTTCCCAGGAAATTGGCCTTGCCTCTCTGGGTGCACCTGATGAATACATTGAAAAGCTCGCCACAATTTACTGGTTTACTGTGGAGTTTGGGCTCTGCAAACAAGGAGACTCCATAAAGGCATATGGTGCTGGGCTCCTGTCATCCTTTGGTGAATTACAGTACTGCTTATCAGAGAAGCCAAAGCTTCTCCCCCTGGAGCTGGAGAAGACAGCCATCCAAAATTACACTGTCACGGAGTTCCAGCCCCTGTATTACGTGGCAGAGAGTTTTAATGATGCCAAGGAGAAAGTAAGGAACTTTGCTGCCACAATACCTCGGCCCTTCTCAGTTCGCTACGACCCATACACCCAAAGGATTGAGGTCTTGGACAATACCCAGCAGCTTAAGATTTTGGCTGATTCCATTAACAGTGAAATTGGAATCCTTTGCAGTGCCCTCCAGAAAATAAAGTAAcctcgagctgtgccttctagttgccagccatctgttgtttgcccctcccccgtgccttccttgaccctggaaggtgccactcccactgtcctttcctAATAAAatgaggaaattgcatcgcattgtctgagtaggtgtcattctattctggggggtggggtggggcaggacagcaagggggaggattgggaagacaatagcaggcatgctggggatgcggtgggctctatggaccggtgcggccgcAGGAACCCCTAGTGATGGAGTTGGCCACTCCCTCTCTGCGCGCTCGCTCGCTCACTGAGGCCGGGCGACCAAAGGTCGCCCGACGCCCGGGCTTTGCCCGGGCGGCCTCAGTGAGCGAGCGAGCGCGCAGCTGCCTGCAGG
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
<110> BioMarin Pharmaceutical Inc.
<120> СПОСОБЫ ЛЕЧЕНИЯ ФЕНИЛКЕТОНУРИИ
<130> 11808-407-228
<140> PCT/US2019/031252
<141> 2019-05-08
<150> 62/819,414
<151> 2019-03-15
<150> 62/802,608
<151> 2019-02-07
<150> 62/755,207
<151> 2018-11-02
<150> 62/669,292
<151> 2018-05-09
<160> 23
<170> PatentIn version 3.5
<210> 1
<211> 1359
<212> ДНК
<213> Homo sapiens
<220>
<223> PAH человека дикого типа
<400> 1
atgtccactg cggtcctgga aaacccaggc ttgggcagga aactctctga ctttggacag 60
gaaacaagct atattgaaga caactgcaat caaaatggtg ccatatcact gatcttctca 120
ctcaaagaag aagttggtgc attggccaaa gtattgcgct tatttgagga gaatgatgta 180
aacctgaccc acattgaatc tagaccttct cgtttaaaga aagatgagta tgaatttttc 240
acccatttgg ataaacgtag cctgcctgct ctgacaaaca tcatcaagat cttgaggcat 300
gacattggtg ccactgtcca tgagctttca cgagataaga agaaagacac agtgccctgg 360
ttcccaagaa ccattcaaga gctggacaga tttgccaatc agattctcag ctatggagcg 420
gaactggatg ctgaccaccc tggttttaaa gatcctgtgt accgtgcaag acggaagcag 480
tttgctgaca ttgcctacaa ctaccgccat gggcagccca tccctcgagt ggaatacatg 540
gaggaagaaa agaaaacatg gggcacagtg ttcaagactc tgaagtcctt gtataaaacc 600
catgcttgct atgagtacaa tcacattttt ccacttcttg aaaagtactg tggcttccat 660
gaagataaca ttccccagct ggaagacgtt tctcagttcc tgcagacttg cactggtttc 720
cgcctccgac ctgtagctgg cctgctttcc tctcgggatt tcttgggtgg cctggccttc 780
cgagtcttcc actgcacaca gtacatcaga catggatcca agcccatgta tacccccgaa 840
cctgacatct gccatgagct gttgggacat gtgcccttgt tttcagatcg cagctttgcc 900
cagttttccc aggaaattgg ccttgcctct ctgggtgcac ctgatgaata cattgaaaag 960
ctcgccacaa tttactggtt tactgtggag tttgggctct gcaaacaagg agactccata 1020
aaggcatatg gtgctgggct cctgtcatcc tttggtgaat tacagtactg cttatcagag 1080
aagccaaagc ttctccccct ggagctggag aagacagcca tccaaaatta cactgtcacg 1140
gagttccagc ccctgtatta cgtggcagag agttttaatg atgccaagga gaaagtaagg 1200
aactttgctg ccacaatacc tcggcccttc tcagttcgct acgacccata cacccaaagg 1260
attgaggtct tggacaatac ccagcagctt aagattttgg ctgattccat taacagtgaa 1320
attggaatcc tttgcagtgc cctccagaaa ataaagtaa 1359
<210> 2
<211> 452
<212> БЕЛОК
<213> Homo sapiens
<220>
<223> PAH человека дикого типа
<400> 2
Met Ser Thr Ala Val Leu Glu Asn Pro Gly Leu Gly Arg Lys Leu Ser
1 5 10 15
Asp Phe Gly Gln Glu Thr Ser Tyr Ile Glu Asp Asn Cys Asn Gln Asn
20 25 30
Gly Ala Ile Ser Leu Ile Phe Ser Leu Lys Glu Glu Val Gly Ala Leu
35 40 45
Ala Lys Val Leu Arg Leu Phe Glu Glu Asn Asp Val Asn Leu Thr His
50 55 60
Ile Glu Ser Arg Pro Ser Arg Leu Lys Lys Asp Glu Tyr Glu Phe Phe
65 70 75 80
Thr His Leu Asp Lys Arg Ser Leu Pro Ala Leu Thr Asn Ile Ile Lys
85 90 95
Ile Leu Arg His Asp Ile Gly Ala Thr Val His Glu Leu Ser Arg Asp
100 105 110
Lys Lys Lys Asp Thr Val Pro Trp Phe Pro Arg Thr Ile Gln Glu Leu
115 120 125
Asp Arg Phe Ala Asn Gln Ile Leu Ser Tyr Gly Ala Glu Leu Asp Ala
130 135 140
Asp His Pro Gly Phe Lys Asp Pro Val Tyr Arg Ala Arg Arg Lys Gln
145 150 155 160
Phe Ala Asp Ile Ala Tyr Asn Tyr Arg His Gly Gln Pro Ile Pro Arg
165 170 175
Val Glu Tyr Met Glu Glu Glu Lys Lys Thr Trp Gly Thr Val Phe Lys
180 185 190
Thr Leu Lys Ser Leu Tyr Lys Thr His Ala Cys Tyr Glu Tyr Asn His
195 200 205
Ile Phe Pro Leu Leu Glu Lys Tyr Cys Gly Phe His Glu Asp Asn Ile
210 215 220
Pro Gln Leu Glu Asp Val Ser Gln Phe Leu Gln Thr Cys Thr Gly Phe
225 230 235 240
Arg Leu Arg Pro Val Ala Gly Leu Leu Ser Ser Arg Asp Phe Leu Gly
245 250 255
Gly Leu Ala Phe Arg Val Phe His Cys Thr Gln Tyr Ile Arg His Gly
260 265 270
Ser Lys Pro Met Tyr Thr Pro Glu Pro Asp Ile Cys His Glu Leu Leu
275 280 285
Gly His Val Pro Leu Phe Ser Asp Arg Ser Phe Ala Gln Phe Ser Gln
290 295 300
Glu Ile Gly Leu Ala Ser Leu Gly Ala Pro Asp Glu Tyr Ile Glu Lys
305 310 315 320
Leu Ala Thr Ile Tyr Trp Phe Thr Val Glu Phe Gly Leu Cys Lys Gln
325 330 335
Gly Asp Ser Ile Lys Ala Tyr Gly Ala Gly Leu Leu Ser Ser Phe Gly
340 345 350
Glu Leu Gln Tyr Cys Leu Ser Glu Lys Pro Lys Leu Leu Pro Leu Glu
355 360 365
Leu Glu Lys Thr Ala Ile Gln Asn Tyr Thr Val Thr Glu Phe Gln Pro
370 375 380
Leu Tyr Tyr Val Ala Glu Ser Phe Asn Asp Ala Lys Glu Lys Val Arg
385 390 395 400
Asn Phe Ala Ala Thr Ile Pro Arg Pro Phe Ser Val Arg Tyr Asp Pro
405 410 415
Tyr Thr Gln Arg Ile Glu Val Leu Asp Asn Thr Gln Gln Leu Lys Ile
420 425 430
Leu Ala Asp Ser Ile Asn Ser Glu Ile Gly Ile Leu Cys Ser Ala Leu
435 440 445
Gln Lys Ile Lys
450
<210> 3
<211> 405
<212> ДНК
<213> Homo sapiens
<220>
<223> последовательность промотора hAAT
<400> 3
gatcttgcta ccagtggaac agccactaag gattctgcag tgagagcaga gggccagcta 60
agtggtactc tcccagagac tgtctgactc acgccacccc ctccaccttg gacacaggac 120
gctgtggttt ctgagccagg tacaatgact cctttcggta agtgcagtgg aagctgtaca 180
ctgcccaggc aaagcgtccg ggcagcgtag gcgggcgact cagatcccag ccagtggact 240
tagcccctgt ttgctcctcc gataactggg gtgaccttgg ttaatattca ccagcagcct 300
cccccgttgc ccctctggat ccactgctta aatacggacg aggacagggc cctgtctcct 360
cagcttcagg caccaccact gacctgggac agtgaatcgt aagta 405
<210> 4
<211> 321
<212> ДНК
<213> Homo sapiens
<220>
<223> последовательность энхансера HCR или энхансера ApoE
<400> 4
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg g 321
<210> 5
<400> 5
000
<210> 6
<211> 2734
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность синтетического промотора (промотор hAAT,
энхансер HCR /ApoE энхансер)
<400> 6
aggctcagag gcacacagga gtttctgggc tcaccctgcc cccttccaac ccctcagttc 60
ccatcctcca gcagctgttt gtgtgctgcc tctgaagtcc acactgaaca aacttcagcc 120
tactcatgtc cctaaaatgg gcaaacattg caagcagcaa acagcaaaca cacagccctc 180
cctgcctgct gaccttggag ctggggcaga ggtcagagac ctctctgggc ccatgccacc 240
tccaacatcc actcgacccc ttggaatttc ggtggagagg agcagaggtt gtcctggcgt 300
ggtttaggta gtgtgagagg ggtcgacgat cttgctacca gtggaacagc cactaaggat 360
tctgcagtga gagcagaggg ccagctaagt ggtactctcc cagagactgt ctgactcacg 420
ccaccccctc caccttggac acaggacgct gtggtttctg agccaggtac aatgactcct 480
ttcggtaagt gcagtggaag ctgtacactg cccaggcaaa gcgtccgggc agcgtaggcg 540
ggcgactcag atcccagcca gtggacttag cccctgtttg ctcctccgat aactggggtg 600
accttggtta atattcacca gcagcctccc ccgttgcccc tctggatcca ctgcttaaat 660
acggacgagg acagggccct gtctcctcag cttcaggcac caccactgac ctgggacagt 720
gaatcgtaag tatgcctttc actgcgaggg gttctggaga ggcttctgag ctccccatgg 780
cccaggcagg cagcaggtct ggggcaggag gggggttgtg gagtgggtat ccgcctgctg 840
aggtgcaggg cagatggaga ggctgcagct gagctcctat tttcataata acagcagcca 900
tgagggttgt gtcctgtttc ccagtcctgc ccggtccccc ctcggtacct cctggtggat 960
acactggttc ctgtaagcag aagtggatga gggtgtctag gtctgcagtc ctggcacccc 1020
aggatggggg acaccagcca agatacagca acagcaacaa agcgcagcca tttctttctg 1080
tttgcacagc tcctctgtct gtcgggggct cctgtctgtt gtctcctata agcctcacca 1140
cctctcctac tgcttgggca tgcatctttc tccccttcta tagatgagga ggttaaggtc 1200
cagagagggg tggggaggaa cgccggctca cattctccat cccctccaga tatgaccagg 1260
aacagacctg tgccaggcct cagccttaca tcaaaatggg cctccccatg caccgtggac 1320
ctctgggccc tcctgtccca gtggaggaca ggaagctatg aggggcactg tcacccaggg 1380
ctcaagctgg cattcctgaa taatcgctct gcaccaggcc acggctaagc tcagtgcgtg 1440
attaagcctc ataaccctcc aaggcagtta ctagtgtgat tcccatttta cagatgagga 1500
agatggggac agagaggtga ataactggcc ccaaatcaca caccatccat aattcgggct 1560
caggcacctg gctccagtcc ccaaactctt gaacctggcc ctagtgtcac tgtttctctt 1620
gggtctcagg cgctggatgg ggaacaggaa acctgggctg gacttgaggc ctctctgatg 1680
ctcggtgact tcagacagtt gctcaacctc tctgttctct tgggcaaaac atgataacct 1740
ttgacttctg tcccctcccc tcaccccacc cgaccttgat ctctgaagtg ttggaaggat 1800
ttaatttttc ctgcactgag ttttggagac aggtcaaaaa gatgaccaag gccaaggtgg 1860
ccagtttcct atagaacgcc tctaaaagac ctgcagcaat agcagcaaga actggtattc 1920
tcgagaactt gctgcgcagc aggcacttct tggcatttta tgtgtattta atttcacaat 1980
agctctatga caaagtccac ctttctcatc tccaggaaac tgaggttcag agaggttaag 2040
taacttgtcc aaggtcacac agctaatagc aagttgacgt ggagcaatct ggcctcagag 2100
cctttaattt tagccacaga ctgatgctcc cctcttcatt tagccaggct gcctctgaag 2160
ttttctgatt caagacttct ggcttcagct ttgtacacag agatgattca atgtcaggtt 2220
ttggagtgaa atctgtttaa tcccagacaa aacatttagg attacatctc agttttgtaa 2280
gcaagtagct ctgtgatttt tagtgagtta tttaatgctc tttggggctc aatttttcta 2340
tctataaaat agggctaata atttgcacct tatagggtaa gctttgagga cagattagat 2400
gatacggtgc ctgtaaaaca ccaggtgtta gtaagtgtgg caatgatggt gacgctgagg 2460
ctgatgtttg cttagcatag ggttaggcag ctggcaggca gtaaacagtt ggataattta 2520
atggaaaatt tgccaaactc agatgctagc agctacaatc cagctaccat tctgctttta 2580
ttttatggtt gggataaggc tggattattc tgagtccaag ctaggccctt ttgctaatca 2640
tgttcatacc tcttatcttc ctcccacagc tcctgggcaa cgtgctggtc tgtgtgctgg 2700
cccatcactt tggcaaagaa ttgcgatcgc cacc 2734
<210> 7
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(GENEius 1/Operon1)
<400> 7
atgagcactg ctgtgctgga gaaccctggc ctgggcagaa agctgtctga ctttggccag 60
gagaccagct acattgagga caactgcaac cagaatggag ccatcagcct gatcttcagc 120
ctgaaggagg aggtgggagc cctggccaag gtgctgagac tgtttgagga gaatgatgtg 180
aacctgaccc acattgagag cagacccagc agactgaaga aggatgagta tgagttcttc 240
acccacctgg acaagagaag cctgcctgcc ctgaccaaca tcatcaagat cctgagacac 300
gatattggag ccactgtgca cgagctgagc agagacaaga agaaggacac tgtgccctgg 360
ttccccagaa ctatccagga gctggacaga tttgccaacc agatcctgag ctatggagct 420
gagctggatg ctgaccaccc tggcttcaag gaccctgtgt acagagccag aagaaagcag 480
tttgctgaca ttgcctacaa ctacagacac ggccagccca tccccagagt ggagtacatg 540
gaggaggaga agaagacctg gggcactgtg ttcaagaccc tgaagagcct gtacaagacc 600
cacgcctgct atgagtacaa ccacatcttc cccctgctgg agaagtactg tggcttccac 660
gaggacaaca tcccccagct ggaggatgtg agccagttcc tgcagacctg cactggcttc 720
agactgagac ctgtggctgg cctgctgagc agcagagact tcctgggggg cctggccttc 780
agagtgttcc actgcaccca gtacatcaga cacggcagca agcccatgta cacccctgag 840
cctgatatct gccacgagct gctgggccac gtgcccctgt tctctgacag aagctttgcc 900
cagttcagcc aggagattgg cctggccagc ctgggagccc ctgatgagta tattgagaag 960
ctggccacta tctactggtt cactgtggag tttggcctgt gcaagcaggg ggacagcatc 1020
aaggcctatg gagctggcct gctgagcagc tttggggagc tgcagtactg cctgtctgag 1080
aagcccaagc tgctgcccct ggagctggag aagactgcca tccagaacta cactgtgact 1140
gagttccagc ccctgtacta tgtggctgag agcttcaatg atgccaagga gaaggtgaga 1200
aactttgctg ccaccatccc cagacccttc tctgtgagat atgaccccta cacccagaga 1260
attgaggtgc tggacaacac ccagcagctg aagatcctgg ctgacagcat caactctgag 1320
attggcatcc tgtgctctgc cctgcagaag atcaagtaa 1359
<210> 8
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(GENEius 2/Operon2)
<400> 8
atgagcactg ctgtgctgga gaaccctggc ctgggcagaa agctgtctga ctttggccag 60
gagaccagct acattgagga caactgcaac cagaatgggg ccatcagcct gatcttcagc 120
ctgaaggagg aggtgggggc cctggccaag gtgctgagac tgtttgagga gaatgatgtg 180
aacctgaccc acattgagag cagacccagc agactgaaga aggatgagta tgagttcttc 240
acccacctgg acaagagaag cctgcctgcc ctgaccaaca tcatcaagat cctgagacac 300
gatattgggg ccactgtgca cgagctgagc agagacaaga agaaggacac tgtgccctgg 360
ttccccagaa ctatccagga gctggacaga tttgccaacc agatcctgag ctatggggct 420
gagctggatg ctgaccaccc tggcttcaag gaccctgtgt acagagccag aagaaagcag 480
tttgctgaca ttgcctacaa ctacagacac ggccagccca tccccagagt ggagtacatg 540
gaggaggaga agaagacctg gggcactgtg ttcaagaccc tgaagagcct gtacaagacc 600
cacgcctgct atgagtacaa ccacatcttc cccctgctgg agaagtactg tggcttccac 660
gaggacaaca tcccccagct ggaggatgtg agccagttcc tgcagacctg cactggcttc 720
agactgagac ctgtggctgg cctgctgagc agcagagact tcctgggggg cctggccttc 780
agagtgttcc actgcaccca gtacatcaga cacggcagca agcccatgta cacccctgag 840
cctgatatct gccacgagct gctgggccac gtgcccctgt tctctgacag aagctttgcc 900
cagttcagcc aggagattgg cctggccagc ctgggggccc ctgatgagta tattgagaag 960
ctggccacta tctactggtt cactgtggag tttggcctgt gcaagcaggg ggacagcatc 1020
aaggcctatg gggctggcct gctgagcagc tttggggagc tgcagtactg cctgtctgag 1080
aagcccaagc tgctgcccct ggagctggag aagactgcca tccagaacta cactgtgact 1140
gagttccagc ccctgtacta tgtggctgag agcttcaatg atgccaagga gaaggtgaga 1200
aactttgctg ccaccatccc cagacccttc tctgtgagat acgaccccta cacccagaga 1260
attgaggtgc tggacaacac ccagcagctg aagatcctgg ctgacagcat caactctgag 1320
attggcatcc tgtgctctgc cctgcagaag atcaagtga 1359
<210> 9
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(Nathwani / NW2-Cop)
<400> 9
atgtccactg ctgtcctgga aaacccaggc ttgggcagga aactctctga ctttggacag 60
gaaacaagct atattgaaga caactgcaat caaaatggtg ccatatcact gatcttctca 120
ctcaaagaag aagttggtgc attggccaaa gtactgaggt tatttgagga gaatgatgta 180
aacctgaccc acattgaatc tagaccttct aggttaaaga aagatgagta tgaatttttc 240
acccatttgg ataaaaggag cctgcctgct ctgacaaaca tcatcaagat cttgaggcac 300
gacattggtg ccactgtcca cgagctttca agggataaga agaaagacac agtgccctgg 360
ttcccaagaa ctattcaaga gctggacaga tttgccaatc agattctcag ctatggagct 420
gaactggatg ctgaccaccc tggttttaaa gatcctgtgt acagggcaag aaggaagcag 480
tttgctgaca ttgcctacaa ctacaggcat gggcagccca tccctagggt ggaatacatg 540
gaggaagaaa agaaaacatg gggcacagtg ttcaagactc tgaagtcctt gtataaaacc 600
cacgcttgct atgagtacaa tcacattttt ccacttcttg aaaagtactg tggcttccac 660
gaagataaca ttccccagct ggaagatgtt tctcagttcc tgcagacttg cactggtttc 720
aggctcaggc ctgtagctgg cctgctttcc tctagggatt tcttgggtgg cctggccttc 780
agggtcttcc actgcacaca gtacatcaga cacggctcca agcccatgta tacccctgaa 840
cctgacatct gccacgagct gttgggacac gtgcccttgt tttcagatag gagctttgcc 900
cagttttccc aggaaattgg ccttgcctct ctgggtgcac ctgatgaata tattgaaaag 960
ctggccacaa tctactggtt tactgtggag tttgggctct gcaaacaagg agactccata 1020
aaggcatatg gtgctgggct cctgtcatcc tttggtgaat tacagtactg cttatcagag 1080
aagccaaagc ttctccccct ggagctggag aagacagcca tccaaaatta cactgtcact 1140
gagttccagc ccctgtatta tgtggcagag agttttaatg atgccaagga gaaagtaagg 1200
aactttgctg ccacaatacc taggcccttc tcagttaggt atgacccata cacccaaagg 1260
attgaggtct tggacaatac ccagcagctt aagattttgg ctgattccat taacagtgaa 1320
attggaatcc tttgcagtgc cctccagaaa atcaagtaa 1359
<210> 10
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(Nathwani-RCG / NW-RCG)
<400> 10
atgtccactg ctgtcctgga aaacccaggc ttgggcagga aactctctga ctttggacag 60
gaaacaagct atattgaaga caactgcaat caaaatggtg ccatatcact gatcttctca 120
ctcaaagaag aagttggtgc attggccaaa gtactgaggt tatttgagga gaatgatgta 180
aacctgaccc acattgaatc tagaccttct aggttaaaga aagatgagta tgaatttttc 240
acccatttgg ataaaaggag cctgcctgct ctgacaaaca tcatcaagat cttgaggcac 300
gacattggtg ccactgtcca cgagctttca agggataaga agaaagacac agtgccctgg 360
ttcccaagaa ctattcaaga gctggacaga tttgccaatc agattctcag ctatggagct 420
gaactggatg ctgaccaccc tggttttaaa gatcctgtgt acagggcaag aaggaagcag 480
tttgctgaca ttgcctacaa ctacaggcat gggcagccca tccctagggt ggaatacatg 540
gaggaagaaa agaaaacatg gggcacagtg ttcaagactc tgaagtcctt gtataaaacc 600
cacgcttgct atgagtacaa tcacattttt ccacttcttg aaaagtactg tggcttccac 660
gaagataaca ttccccagct ggaagatgtt tctcagttcc tgcagacttg cactggtttc 720
aggctcaggc ctgtagctgg cctgctttcc tctagggatt tcttgggtgg cctggccttc 780
agggtcttcc actgcacaca gtacatcaga cacggctcca agcccatgta tacccctgaa 840
cctgacatct gccacgagct gttgggacac gtgcccttgt tttcagatag gagctttgcc 900
cagttttccc aggaaattgg ccttgcctct ctgggtgcac ctgatgaata tattgaaaag 960
ctggccacaa tttactggtt tactgtggag tttgggctct gcaaacaagg agactccata 1020
aaggcatatg gtgctgggct cctgtcatcc tttggtgaat tacagtactg cttatcagag 1080
aagccaaagc ttctccccct ggagctggag aagacagcca tccaaaatta cactgtcact 1140
gagttccagc ccctgtatta tgtggcagag agttttaatg atgccaagga gaaagtaagg 1200
aactttgctg ccacaatacc taggcccttc tcagttaggt atgacccata cacccaaagg 1260
attgaggtct tggacaatac ccagcagctt aagattttgg ctgattccat taacagtgaa 1320
attggaatcc tttgcagtgc cctccagaaa ataaagtaa 1359
<210> 11
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(ATUM (DNA2.0-op/D20-P))
<400> 11
atgtccactg cggtgttgga aaaccccgga ctgggcagaa agttgtccga cttcggccag 60
gaaacctctt acattgagga caactgcaac cagaacgggg ccatctcact gatcttttcg 120
ctgaaagaag aagtcggagc tctggccaaa gtgctgcgcc tgttcgagga aaacgacgtc 180
aacctgaccc acatcgagtc aagaccgagc aggctgaaga aggatgagta cgagtttttc 240
acccatctcg acaagagatc cctgcctgcc ctgaccaata ttatcaagat tttgcggcac 300
gacattggcg caaccgtgca tgaactctcc cgggacaaga agaaggacac cgtgccgtgg 360
ttcccccgaa ccatccagga actcgaccgc ttcgctaacc agatcctgtc ctacggcgcc 420
gaactggatg ccgatcaccc tggattcaag gacccagtgt acagagcccg gcgcaagcag 480
ttcgccgata tcgcctacaa ttatcggcac ggacagccaa tcccgagggt ggagtacatg 540
gaggaggaaa agaaaacctg gggaactgtg ttcaagacct tgaagtccct gtacaagact 600
cacgcctgct acgagtacaa ccacatcttc cccctgctcg aaaagtactg cgggttccat 660
gaggacaaca tcccccaact ggaagatgtg tcgcagttcc tgcaaacctg taccggattc 720
cggctgaggc ctgtcgcggg acttctgtcc tcccgggatt ttcttggcgg tctggccttc 780
cgggtgttcc actgtactca gtacattaga cacgggagca agcctatgta cactcctgaa 840
cccgacattt gccacgaact cctgggtcat gtgcccctct tctcggatcg gagcttcgcc 900
cagttcagcc aagagatcgg tctggctagc ttgggagcac ccgacgagta catcgagaag 960
ctggccacta tctactggtt taccgtggaa ttcggactgt gcaagcaggg ggactcaatc 1020
aaggcctatg gcgcgggact cctgagctcc ttcggggagc tgcagtactg cctgtccgaa 1080
aagccaaagc tgctccctct tgaactggag aaaacggcca tccagaacta caccgtgacc 1140
gaattccagc cgctctacta cgtcgcggag tccttcaacg atgccaagga gaaggtccgc 1200
aacttcgccg caactatccc gcggccgttt tccgtgcgct atgacccgta cacacaacgc 1260
atcgaagtgc tggacaacac ccagcaactt aagattctgg ccgactcgat caactccgag 1320
attggcattc tgtgctcggc gctgcagaag atcaagtaa 1359
<210> 12
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(ATUM-RCG (DNA2.0-Cop1))
<400> 12
atgtccactg cggtcctgga aaaccctggt ctgggccgca agctttctga ctttggacag 60
gaaacctcat acattgagga caactgtaac caaaatggtg caatcagcct gatcttcagc 120
ctcaaggaag aagtgggagc cctggccaag gtcctgagac tgtttgagga gaatgatgtg 180
aacctgaccc acattgagtc caggccctcc agactgaaga aggatgaata cgaattcttc 240
acccacctgg acaagcgctc cctccctgcc ctcaccaaca tcattaagat cctgcggcac 300
gacattggag ccactgtgca tgagttgagc cgggacaaga agaaggatac tgtgccctgg 360
ttcccgagga ccatccagga actggaccgg tttgccaacc aaattctgtc ctatggagct 420
gaattggatg cagaccaccc tggcttcaag gacccagtgt acagagcaag gagaaagcag 480
tttgcagaca tagcctacaa ctacagacat ggacagccca tcccgagggt ggagtacatg 540
gaagaggaga agaaaacctg gggcactgtg tttaagaccc tcaagagcct gtacaagacc 600
cacgcctgct atgagtacaa ccacatcttc ccactcctgg agaaatactg tggcttccat 660
gaggacaaca tcccacagct ggaggatgtg tcccagttcc tgcagacttg cactggcttc 720
cggttgaggc ctgtggctgg cctgctctcc tcaagggact tcttgggggg actggcattc 780
agggtgttcc actgcaccca atacatcaga cacggcagca agccaatgta cacccctgaa 840
ccggacatct gccacgaact cctgggccac gtccctctgt tctcggacag aagctttgcc 900
cagttctccc aagagattgg ccttgcctcc ctgggggccc ctgatgaata cattgaaaag 960
ctggccacaa tctactggtt cactgtggaa tttggacttt gcaagcaggg agatagcatc 1020
aaggcctacg gggctggact tctgtcctcc ttcggtgaac tgcagtactg tctgtcagag 1080
aagcccaagc tgctgcccct ggaactggag aaaactgcca tccaaaacta cactgtgact 1140
gagttccagc ccctctacta tgtggctgag tccttcaatg atgccaaaga aaaggtcaga 1200
aattttgcgg ccaccattcc taggcctttc tcagtccgct atgaccctta cacccagaga 1260
attgaggtgc tggacaacac ccagcagctc aagatcctgg cagactccat caactcagaa 1320
attgggatct tgtgctcggc cctccaaaag atcaagtaa 1359
<210> 13
<211> 1359
<212> ДНК
<213> Искусственная последовательность
<220>
<223> кодон-оптимизированная последовательность PAH человека
(JCAT (JCAT))
<400> 13
atgagcactg ctgtgctgga gaaccctggc ctgggcagaa agctgtctga ctttggccag 60
gagaccagct acattgagga caactgcaac cagaatgggg ccatcagcct gatcttcagc 120
ctgaaggagg aggtgggggc cctggccaag gtgctgagac tgtttgagga gaatgatgtg 180
aacctgaccc acattgagag cagacccagc agactgaaga aggatgagta tgagttcttc 240
acccacctgg acaagagaag cctgccagcc ctgaccaata ttatcaagat cctgaggcac 300
gatattgggg ccacagtgca cgagctgagc agagacaaga agaaggacac agtgccctgg 360
ttccccagga ctatccagga gctggacagg tttgccaacc agatcctgag ctacggagct 420
gagctggatg cagaccaccc tggcttcaag gaccctgtgt acagagccag gagaaagcag 480
tttgcagata tagcctacaa ctacaggcac ggccagccaa tccctagagt ggagtacatg 540
gaggaggaga agaagacctg gggcactgtg ttcaagaccc tgaagagcct gtacaagacc 600
cacgcctgct acgagtacaa ccacatcttc cccctgctgg agaagtactg tggcttccac 660
gaggacaaca tcccccagct ggaggatgtg agccagttcc tgcagacctg cacaggcttc 720
aggctgagac cagtggcagg cctgctgagc agcagggact tcctgggggg cctggccttc 780
agagtgttcc actgcaccca gtatatcagg cacggcagca agccaatgta cacccctgag 840
ccagatatct gccacgagct gctgggccac gttcccctgt tctcagacag aagctttgcc 900
cagttcagcc aggagattgg cctggccagc ctgggggccc ctgacgagta tattgagaag 960
ctggccacta tctactggtt cacagtggag tttggcctgt gcaagcaggg ggacagcatc 1020
aaggcctatg gggctggcct gctgagcagc tttggggagc tgcagtactg cctgtcagag 1080
aagccaaagc tgctgcccct ggagctggag aagacagcta tccagaacta cactgtgact 1140
gagttccagc ccctgtacta tgtggcagag agcttcaatg atgccaagga gaaggtgaga 1200
aactttgctg ccaccatccc caggcccttc tctgtgagat atgaccccta cacccagagg 1260
attgaggtgc tggacaacac ccagcagctg aagatcctgg cagacagcat caacagtgag 1320
attggtatcc tgtgctctgc cctgcagaag atcaagtaa 1359
<210> 14
<211> 2002
<212> ДНК
<213> Искусственная последовательность
<220>
<223> составная последовательность нуклеиновой кислоты интрона
глобина/AIAT
<400> 14
tgcctttcac tgcgaggggt tctggagagg cttctgagct ccccatggcc caggcaggca 60
gcaggtctgg ggcaggaggg gggttgtgga gtgggtatcc gcctgctgag gtgcagggca 120
gatggagagg ctgcagctga gctcctattt tcataataac agcagccatg agggttgtgt 180
cctgtttccc agtcctgccc ggtcccccct cggtacctcc tggtggatac actggttcct 240
gtaagcagaa gtggatgagg gtgtctaggt ctgcagtcct ggcaccccag gatgggggac 300
accagccaag atacagcaac agcaacaaag cgcagccatt tctttctgtt tgcacagctc 360
ctctgtctgt cgggggctcc tgtctgttgt ctcctataag cctcaccacc tctcctactg 420
cttgggcatg catctttctc cccttctata gatgaggagg ttaaggtcca gagaggggtg 480
gggaggaacg ccggctcaca ttctccatcc cctccagata tgaccaggaa cagacctgtg 540
ccaggcctca gccttacatc aaaatgggcc tccccatgca ccgtggacct ctgggccctc 600
ctgtcccagt ggaggacagg aagctatgag gggcactgtc acccagggct caagctggca 660
ttcctgaata atcgctctgc accaggccac ggctaagctc agtgcgtgat taagcctcat 720
aaccctccaa ggcagttact agtgtgattc ccattttaca gatgaggaag atggggacag 780
agaggtgaat aactggcccc aaatcacaca ccatccataa ttcgggctca ggcacctggc 840
tccagtcccc aaactcttga acctggccct agtgtcactg tttctcttgg gtctcaggcg 900
ctggatgggg aacaggaaac ctgggctgga cttgaggcct ctctgatgct cggtgacttc 960
agacagttgc tcaacctctc tgttctcttg ggcaaaacat gataaccttt gacttctgtc 1020
ccctcccctc accccacccg accttgatct ctgaagtgtt ggaaggattt aatttttcct 1080
gcactgagtt ttggagacag gtcaaaaaga tgaccaaggc caaggtggcc agtttcctat 1140
agaacgcctc taaaagacct gcagcaatag cagcaagaac tggtattctc gagaacttgc 1200
tgcgcagcag gcacttcttg gcattttatg tgtatttaat ttcacaatag ctctatgaca 1260
aagtccacct ttctcatctc caggaaactg aggttcagag aggttaagta acttgtccaa 1320
ggtcacacag ctaatagcaa gttgacgtgg agcaatctgg cctcagagcc tttaatttta 1380
gccacagact gatgctcccc tcttcattta gccaggctgc ctctgaagtt ttctgattca 1440
agacttctgg cttcagcttt gtacacagag atgattcaat gtcaggtttt ggagtgaaat 1500
ctgtttaatc ccagacaaaa catttaggat tacatctcag ttttgtaagc aagtagctct 1560
gtgattttta gtgagttatt taatgctctt tggggctcaa tttttctatc tataaaatag 1620
ggctaataat ttgcacctta tagggtaagc tttgaggaca gattagatga tacggtgcct 1680
gtaaaacacc aggtgttagt aagtgtggca atgatggtga cgctgaggct gatgtttgct 1740
tagcataggg ttaggcagct ggcaggcagt aaacagttgg ataatttaat ggaaaatttg 1800
ccaaactcag atgctagcag ctacaatcca gctaccattc tgcttttatt ttatggttgg 1860
gataaggctg gattattctg agtccaagct aggccctttt gctaatcatg ttcatacctc 1920
ttatcttcct cccacagctc ctgggcaacg tgctggtctg tgtgctggcc catcactttg 1980
gcaaagaatt gcgatcgcca cc 2002
<210> 15
<211> 2822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты вектора 1
(ApoE-HCR-hAAT.GI.hPAH.bGH)
<400> 15
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtacta gcagctacaa 900
tccagctacc attctgcttt tattttatgg ttgggataag gctggattat tctgagtcca 960
agctaggccc ttttgctaat catgttcata cctcttatct tcctcccaca gctcctgggc 1020
aacgtgctgg tctgtgtgct ggcccatcac tttggcaaag aattgcgatc gccaccatgt 1080
ccactgcggt cctggaaaac ccaggcttgg gcaggaaact ctctgacttt ggacaggaaa 1140
caagctatat tgaagacaac tgcaatcaaa atggtgccat atcactgatc ttctcactca 1200
aagaagaagt tggtgcattg gccaaagtat tgcgcttatt tgaggagaat gatgtaaacc 1260
tgacccacat tgaatctaga ccttctcgtt taaagaaaga tgagtatgaa tttttcaccc 1320
atttggataa acgtagcctg cctgctctga caaacatcat caagatcttg aggcatgaca 1380
ttggtgccac tgtccatgag ctttcacgag ataagaagaa agacacagtg ccctggttcc 1440
caagaaccat tcaagagctg gacagatttg ccaatcagat tctcagctat ggagcggaac 1500
tggatgctga ccaccctggt tttaaagatc ctgtgtaccg tgcaagacgg aagcagtttg 1560
ctgacattgc ctacaactac cgccatgggc agcccatccc tcgagtggaa tacatggagg 1620
aagaaaagaa aacatggggc acagtgttca agactctgaa gtccttgtat aaaacccatg 1680
cttgctatga gtacaatcac atttttccac ttcttgaaaa gtactgtggc ttccatgaag 1740
ataacattcc ccagctggaa gacgtttctc agttcctgca gacttgcact ggtttccgcc 1800
tccgacctgt agctggcctg ctttcctctc gggatttctt gggtggcctg gccttccgag 1860
tcttccactg cacacagtac atcagacatg gatccaagcc catgtatacc cccgaacctg 1920
acatctgcca tgagctgttg ggacatgtgc ccttgttttc agatcgcagc tttgcccagt 1980
tttcccagga aattggcctt gcctctctgg gtgcacctga tgaatacatt gaaaagctcg 2040
ccacaattta ctggtttact gtggagtttg ggctctgcaa acaaggagac tccataaagg 2100
catatggtgc tgggctcctg tcatcctttg gtgaattaca gtactgctta tcagagaagc 2160
caaagcttct ccccctggag ctggagaaga cagccatcca aaattacact gtcacggagt 2220
tccagcccct gtattacgtg gcagagagtt ttaatgatgc caaggagaaa gtaaggaact 2280
ttgctgccac aatacctcgg cccttctcag ttcgctacga cccatacacc caaaggattg 2340
aggtcttgga caatacccag cagcttaaga ttttggctga ttccattaac agtgaaattg 2400
gaatcctttg cagtgccctc cagaaaataa agtaacctcg agctgtgcct tctagttgcc 2460
agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca 2520
ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta 2580
ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc 2640
atgctgggga tgcggtgggc tctatggacc ggtgcggccg caggaacccc tagtgatgga 2700
gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac caaaggtcgc 2760
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 2820
gg 2822
<210> 16
<211> 2822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеиновой кислоты GENEius 1
(ApoE-HCR-hAAT.GI.hPAHco1.bGH)
<400> 16
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtacta gcagctacaa 900
tccagctacc attctgcttt tattttatgg ttgggataag gctggattat tctgagtcca 960
agctaggccc ttttgctaat catgttcata cctcttatct tcctcccaca gctcctgggc 1020
aacgtgctgg tctgtgtgct ggcccatcac tttggcaaag aattgcgatc gccaccatga 1080
gcactgctgt gctggagaac cctggcctgg gcagaaagct gtctgacttt ggccaggaga 1140
ccagctacat tgaggacaac tgcaaccaga atggagccat cagcctgatc ttcagcctga 1200
aggaggaggt gggagccctg gccaaggtgc tgagactgtt tgaggagaat gatgtgaacc 1260
tgacccacat tgagagcaga cccagcagac tgaagaagga tgagtatgag ttcttcaccc 1320
acctggacaa gagaagcctg cctgccctga ccaacatcat caagatcctg agacacgata 1380
ttggagccac tgtgcacgag ctgagcagag acaagaagaa ggacactgtg ccctggttcc 1440
ccagaactat ccaggagctg gacagatttg ccaaccagat cctgagctat ggagctgagc 1500
tggatgctga ccaccctggc ttcaaggacc ctgtgtacag agccagaaga aagcagtttg 1560
ctgacattgc ctacaactac agacacggcc agcccatccc cagagtggag tacatggagg 1620
aggagaagaa gacctggggc actgtgttca agaccctgaa gagcctgtac aagacccacg 1680
cctgctatga gtacaaccac atcttccccc tgctggagaa gtactgtggc ttccacgagg 1740
acaacatccc ccagctggag gatgtgagcc agttcctgca gacctgcact ggcttcagac 1800
tgagacctgt ggctggcctg ctgagcagca gagacttcct ggggggcctg gccttcagag 1860
tgttccactg cacccagtac atcagacacg gcagcaagcc catgtacacc cctgagcctg 1920
atatctgcca cgagctgctg ggccacgtgc ccctgttctc tgacagaagc tttgcccagt 1980
tcagccagga gattggcctg gccagcctgg gagcccctga tgagtatatt gagaagctgg 2040
ccactatcta ctggttcact gtggagtttg gcctgtgcaa gcagggggac agcatcaagg 2100
cctatggagc tggcctgctg agcagctttg gggagctgca gtactgcctg tctgagaagc 2160
ccaagctgct gcccctggag ctggagaaga ctgccatcca gaactacact gtgactgagt 2220
tccagcccct gtactatgtg gctgagagct tcaatgatgc caaggagaag gtgagaaact 2280
ttgctgccac catccccaga cccttctctg tgagatatga cccctacacc cagagaattg 2340
aggtgctgga caacacccag cagctgaaga tcctggctga cagcatcaac tctgagattg 2400
gcatcctgtg ctctgccctg cagaagatca agtaacctcg agctgtgcct tctagttgcc 2460
agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca 2520
ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta 2580
ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc 2640
atgctgggga tgcggtgggc tctatggacc ggtgcggccg caggaacccc tagtgatgga 2700
gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac caaaggtcgc 2760
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 2820
gg 2822
<210> 17
<211> 4635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность Large Intron-вектор 1
(ApoE-HCR-hAAT.cG-AIATI.hPAH.bGH)
<400> 17
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtatgc ctttcactgc 900
gaggggttct ggagaggctt ctgagctccc catggcccag gcaggcagca ggtctggggc 960
aggagggggg ttgtggagtg ggtatccgcc tgctgaggtg cagggcagat ggagaggctg 1020
cagctgagct cctattttca taataacagc agccatgagg gttgtgtcct gtttcccagt 1080
cctgcccggt cccccctcgg tacctcctgg tggatacact ggttcctgta agcagaagtg 1140
gatgagggtg tctaggtctg cagtcctggc accccaggat gggggacacc agccaagata 1200
cagcaacagc aacaaagcgc agccatttct ttctgtttgc acagctcctc tgtctgtcgg 1260
gggctcctgt ctgttgtctc ctataagcct caccacctct cctactgctt gggcatgcat 1320
ctttctcccc ttctatagat gaggaggtta aggtccagag aggggtgggg aggaacgccg 1380
gctcacattc tccatcccct ccagatatga ccaggaacag acctgtgcca ggcctcagcc 1440
ttacatcaaa atgggcctcc ccatgcaccg tggacctctg ggccctcctg tcccagtgga 1500
ggacaggaag ctatgagggg cactgtcacc cagggctcaa gctggcattc ctgaataatc 1560
gctctgcacc aggccacggc taagctcagt gcgtgattaa gcctcataac cctccaaggc 1620
agttactagt gtgattccca ttttacagat gaggaagatg gggacagaga ggtgaataac 1680
tggccccaaa tcacacacca tccataattc gggctcaggc acctggctcc agtccccaaa 1740
ctcttgaacc tggccctagt gtcactgttt ctcttgggtc tcaggcgctg gatggggaac 1800
aggaaacctg ggctggactt gaggcctctc tgatgctcgg tgacttcaga cagttgctca 1860
acctctctgt tctcttgggc aaaacatgat aacctttgac ttctgtcccc tcccctcacc 1920
ccacccgacc ttgatctctg aagtgttgga aggatttaat ttttcctgca ctgagttttg 1980
gagacaggtc aaaaagatga ccaaggccaa ggtggccagt ttcctataga acgcctctaa 2040
aagacctgca gcaatagcag caagaactgg tattctcgag aacttgctgc gcagcaggca 2100
cttcttggca ttttatgtgt atttaatttc acaatagctc tatgacaaag tccacctttc 2160
tcatctccag gaaactgagg ttcagagagg ttaagtaact tgtccaaggt cacacagcta 2220
atagcaagtt gacgtggagc aatctggcct cagagccttt aattttagcc acagactgat 2280
gctcccctct tcatttagcc aggctgcctc tgaagttttc tgattcaaga cttctggctt 2340
cagctttgta cacagagatg attcaatgtc aggttttgga gtgaaatctg tttaatccca 2400
gacaaaacat ttaggattac atctcagttt tgtaagcaag tagctctgtg atttttagtg 2460
agttatttaa tgctctttgg ggctcaattt ttctatctat aaaatagggc taataatttg 2520
caccttatag ggtaagcttt gaggacagat tagatgatac ggtgcctgta aaacaccagg 2580
tgttagtaag tgtggcaatg atggtgacgc tgaggctgat gtttgcttag catagggtta 2640
ggcagctggc aggcagtaaa cagttggata atttaatgga aaatttgcca aactcagatg 2700
ctagcagcta caatccagct accattctgc ttttatttta tggttgggat aaggctggat 2760
tattctgagt ccaagctagg cccttttgct aatcatgttc atacctctta tcttcctccc 2820
acagctcctg ggcaacgtgc tggtctgtgt gctggcccat cactttggca aagaattgcg 2880
atcgccacca tgtccactgc ggtcctggaa aacccaggct tgggcaggaa actctctgac 2940
tttggacagg aaacaagcta tattgaagac aactgcaatc aaaatggtgc catatcactg 3000
atcttctcac tcaaagaaga agttggtgca ttggccaaag tattgcgctt atttgaggag 3060
aatgatgtaa acctgaccca cattgaatct agaccttctc gtttaaagaa agatgagtat 3120
gaatttttca cccatttgga taaacgtagc ctgcctgctc tgacaaacat catcaagatc 3180
ttgaggcatg acattggtgc cactgtccat gagctttcac gagataagaa gaaagacaca 3240
gtgccctggt tcccaagaac cattcaagag ctggacagat ttgccaatca gattctcagc 3300
tatggagcgg aactggatgc tgaccaccct ggttttaaag atcctgtgta ccgtgcaaga 3360
cggaagcagt ttgctgacat tgcctacaac taccgccatg ggcagcccat ccctcgagtg 3420
gaatacatgg aggaagaaaa gaaaacatgg ggcacagtgt tcaagactct gaagtccttg 3480
tataaaaccc atgcttgcta tgagtacaat cacatttttc cacttcttga aaagtactgt 3540
ggcttccatg aagataacat tccccagctg gaagacgttt ctcagttcct gcagacttgc 3600
actggtttcc gcctccgacc tgtagctggc ctgctttcct ctcgggattt cttgggtggc 3660
ctggccttcc gagtcttcca ctgcacacag tacatcagac atggatccaa gcccatgtat 3720
acccccgaac ctgacatctg ccatgagctg ttgggacatg tgcccttgtt ttcagatcgc 3780
agctttgccc agttttccca ggaaattggc cttgcctctc tgggtgcacc tgatgaatac 3840
attgaaaagc tcgccacaat ttactggttt actgtggagt ttgggctctg caaacaagga 3900
gactccataa aggcatatgg tgctgggctc ctgtcatcct ttggtgaatt acagtactgc 3960
ttatcagaga agccaaagct tctccccctg gagctggaga agacagccat ccaaaattac 4020
actgtcacgg agttccagcc cctgtattac gtggcagaga gttttaatga tgccaaggag 4080
aaagtaagga actttgctgc cacaatacct cggcccttct cagttcgcta cgacccatac 4140
acccaaagga ttgaggtctt ggacaatacc cagcagctta agattttggc tgattccatt 4200
aacagtgaaa ttggaatcct ttgcagtgcc ctccagaaaa taaagtaacc tcgagctgtg 4260
ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa 4320
ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt 4380
aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa 4440
gacaatagca ggcatgctgg ggatgcggtg ggctctatgg accggtgcgg ccgcaggaac 4500
ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact gaggccgggc 4560
gaccaaaggt cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 4620
gcagctgcct gcagg 4635
<210> 18
<211> 4635
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность Large Intron-GENEius 1
(ApoE-HCR-hAAT.cG-AIATI.hPAHco1.bGH)
<400> 18
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtatgc ctttcactgc 900
gaggggttct ggagaggctt ctgagctccc catggcccag gcaggcagca ggtctggggc 960
aggagggggg ttgtggagtg ggtatccgcc tgctgaggtg cagggcagat ggagaggctg 1020
cagctgagct cctattttca taataacagc agccatgagg gttgtgtcct gtttcccagt 1080
cctgcccggt cccccctcgg tacctcctgg tggatacact ggttcctgta agcagaagtg 1140
gatgagggtg tctaggtctg cagtcctggc accccaggat gggggacacc agccaagata 1200
cagcaacagc aacaaagcgc agccatttct ttctgtttgc acagctcctc tgtctgtcgg 1260
gggctcctgt ctgttgtctc ctataagcct caccacctct cctactgctt gggcatgcat 1320
ctttctcccc ttctatagat gaggaggtta aggtccagag aggggtgggg aggaacgccg 1380
gctcacattc tccatcccct ccagatatga ccaggaacag acctgtgcca ggcctcagcc 1440
ttacatcaaa atgggcctcc ccatgcaccg tggacctctg ggccctcctg tcccagtgga 1500
ggacaggaag ctatgagggg cactgtcacc cagggctcaa gctggcattc ctgaataatc 1560
gctctgcacc aggccacggc taagctcagt gcgtgattaa gcctcataac cctccaaggc 1620
agttactagt gtgattccca ttttacagat gaggaagatg gggacagaga ggtgaataac 1680
tggccccaaa tcacacacca tccataattc gggctcaggc acctggctcc agtccccaaa 1740
ctcttgaacc tggccctagt gtcactgttt ctcttgggtc tcaggcgctg gatggggaac 1800
aggaaacctg ggctggactt gaggcctctc tgatgctcgg tgacttcaga cagttgctca 1860
acctctctgt tctcttgggc aaaacatgat aacctttgac ttctgtcccc tcccctcacc 1920
ccacccgacc ttgatctctg aagtgttgga aggatttaat ttttcctgca ctgagttttg 1980
gagacaggtc aaaaagatga ccaaggccaa ggtggccagt ttcctataga acgcctctaa 2040
aagacctgca gcaatagcag caagaactgg tattctcgag aacttgctgc gcagcaggca 2100
cttcttggca ttttatgtgt atttaatttc acaatagctc tatgacaaag tccacctttc 2160
tcatctccag gaaactgagg ttcagagagg ttaagtaact tgtccaaggt cacacagcta 2220
atagcaagtt gacgtggagc aatctggcct cagagccttt aattttagcc acagactgat 2280
gctcccctct tcatttagcc aggctgcctc tgaagttttc tgattcaaga cttctggctt 2340
cagctttgta cacagagatg attcaatgtc aggttttgga gtgaaatctg tttaatccca 2400
gacaaaacat ttaggattac atctcagttt tgtaagcaag tagctctgtg atttttagtg 2460
agttatttaa tgctctttgg ggctcaattt ttctatctat aaaatagggc taataatttg 2520
caccttatag ggtaagcttt gaggacagat tagatgatac ggtgcctgta aaacaccagg 2580
tgttagtaag tgtggcaatg atggtgacgc tgaggctgat gtttgcttag catagggtta 2640
ggcagctggc aggcagtaaa cagttggata atttaatgga aaatttgcca aactcagatg 2700
ctagcagcta caatccagct accattctgc ttttatttta tggttgggat aaggctggat 2760
tattctgagt ccaagctagg cccttttgct aatcatgttc atacctctta tcttcctccc 2820
acagctcctg ggcaacgtgc tggtctgtgt gctggcccat cactttggca aagaattgcg 2880
atcgccacca tgagcactgc tgtgctggag aaccctggcc tgggcagaaa gctgtctgac 2940
tttggccagg agaccagcta cattgaggac aactgcaacc agaatggagc catcagcctg 3000
atcttcagcc tgaaggagga ggtgggagcc ctggccaagg tgctgagact gtttgaggag 3060
aatgatgtga acctgaccca cattgagagc agacccagca gactgaagaa ggatgagtat 3120
gagttcttca cccacctgga caagagaagc ctgcctgccc tgaccaacat catcaagatc 3180
ctgagacacg atattggagc cactgtgcac gagctgagca gagacaagaa gaaggacact 3240
gtgccctggt tccccagaac tatccaggag ctggacagat ttgccaacca gatcctgagc 3300
tatggagctg agctggatgc tgaccaccct ggcttcaagg accctgtgta cagagccaga 3360
agaaagcagt ttgctgacat tgcctacaac tacagacacg gccagcccat ccccagagtg 3420
gagtacatgg aggaggagaa gaagacctgg ggcactgtgt tcaagaccct gaagagcctg 3480
tacaagaccc acgcctgcta tgagtacaac cacatcttcc ccctgctgga gaagtactgt 3540
ggcttccacg aggacaacat cccccagctg gaggatgtga gccagttcct gcagacctgc 3600
actggcttca gactgagacc tgtggctggc ctgctgagca gcagagactt cctggggggc 3660
ctggccttca gagtgttcca ctgcacccag tacatcagac acggcagcaa gcccatgtac 3720
acccctgagc ctgatatctg ccacgagctg ctgggccacg tgcccctgtt ctctgacaga 3780
agctttgccc agttcagcca ggagattggc ctggccagcc tgggagcccc tgatgagtat 3840
attgagaagc tggccactat ctactggttc actgtggagt ttggcctgtg caagcagggg 3900
gacagcatca aggcctatgg agctggcctg ctgagcagct ttggggagct gcagtactgc 3960
ctgtctgaga agcccaagct gctgcccctg gagctggaga agactgccat ccagaactac 4020
actgtgactg agttccagcc cctgtactat gtggctgaga gcttcaatga tgccaaggag 4080
aaggtgagaa actttgctgc caccatcccc agacccttct ctgtgagata tgacccctac 4140
acccagagaa ttgaggtgct ggacaacacc cagcagctga agatcctggc tgacagcatc 4200
aactctgaga ttggcatcct gtgctctgcc ctgcagaaga tcaagtaacc tcgagctgtg 4260
ccttctagtt gccagccatc tgttgtttgc ccctcccccg tgccttcctt gaccctggaa 4320
ggtgccactc ccactgtcct ttcctaataa aatgaggaaa ttgcatcgca ttgtctgagt 4380
aggtgtcatt ctattctggg gggtggggtg gggcaggaca gcaaggggga ggattgggaa 4440
gacaatagca ggcatgctgg ggatgcggtg ggctctatgg accggtgcgg ccgcaggaac 4500
ccctagtgat ggagttggcc actccctctc tgcgcgctcg ctcgctcact gaggccgggc 4560
gaccaaaggt cgcccgacgc ccgggctttg cccgggcggc ctcagtgagc gagcgagcgc 4620
gcagctgcct gcagg 4635
<210> 19
<211> 4640
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Последовательность hPAH+укороченный 2-й интрон
(ApoE-HCR-hAAT.hPAH-tI2.bGH)
<400> 19
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc agccagagac ctcactcccg 900
gggagccagc atgtccactg cggtcctgga aaacccaggc ttgggcagga aactctctga 960
ctttggacag gaaacaagct atattgaaga caactgcaat caaaatggtg ccatatcact 1020
gatcttctca ctcaaagaag aagttggtgc attggccaaa gtattgcgct tatttgaggt 1080
cagtgctaca atcatgtttg tcttggataa tgtcgtagca aacttccatg ttcttttcta 1140
gttagatgca atgaaaagaa cacaggatct ggaacaggca gatttgaatt tgagctcaag 1200
tttttacatt aaccaactgt gtgaccttag ttaagtcatt caatctctct gagcttcagt 1260
ttttccattc ataatatagt gctgataata tgtgccttgt cagtttcaac aggaactttg 1320
tgatgaaata atgtgttttg taaaatgcct aatcatatgt gtgattaggc atgattaact 1380
ataagtcata tgtgacacgt gattacattt cacataacat gtaatcacat gtgtcacatg 1440
tgattacagt taaagagtaa acaagaaata aatattaatt cttttattca ctaaattaat 1500
attgattgtc ttctatgtat aagtgtaaaa taatatgcaa gacaccgtcc ctttcttcaa 1560
gtagcttaac ccaaaactat gctttagaaa atagctaatg ttcttcaaga cattggtaaa 1620
tgtcttatga ttaaagtggt tccataatta ataaacttgg gaaattcggg catattattt 1680
tacctgattt ctttattata ggacttttag agtctataaa actaattata ctaagttgtt 1740
tacagagaga aatgtatgta ttattttcca aacttacttg actgtggatc ctttttattt 1800
taagaacatg tattcacatc tgggaacaga gtttaggaaa cgttgatttg ggtgtttgct 1860
gggcagccaa acttcacaga actccaaata ggtttcctca gccagattcc ttccgagtac 1920
tatgctaata tttttggtgg gattttgtcc cacctgaaaa tacattgctt tatgctaaga 1980
ttcctgtgac ctctctagct gattgggagg caggggtagc acattggtca gggctgtgca 2040
catacttagt gctcagtgtg tttgggcatg caatcaacaa atccttttgg ggcatgactg 2100
gatacgatta gatatcttgt gaaaacctat gatgttctca gcactgtggg tggggaggaa 2160
gatgggggac acatagaaat tgtaaggaag agaaactgcc tcctttagga tctaacaggg 2220
gaagcaaata ttctgagcag ctggggaggg aaggcagagc atgagaagtt atctcagtaa 2280
aaggtagctt tatagacatg atcttattca gtctaatgaa aataacatga ggtaggagct 2340
atttctgccc ccatttttca gagatagaaa ctgatgctta gaggggttag gtaaatgtgc 2400
ccaaggtcaa acagctagta aatacggaag aaatcattta aacccaggca ttctgagtct 2460
agaacctaca tacactctta actgctattc agtactgccc cagacagaac gggcttcata 2520
gtttgcaggg cgcagtgcaa aatgaaaacg tggggtctct tcctcaataa caggaaataa 2580
tgctgttaaa agtactaaaa tatataactt tatcctttct tctgtggtct ctctcatgtc 2640
ctgttatggt gtttttgatt tgccatttag ttgtgctctc ctcccctggg caccttggcc 2700
ctctaactgc tgggttccct gtccagccag ggctgggctg acagccaagc ccctaccaag 2760
gatggggaaa tccatcttcc atttccgtgg acctgctgca ccaacccaca gtgatgagct 2820
atcccccaag agattgtaac ctctggaatg agaatggaaa ggaagcttgc ctgaacttgg 2880
ttttaaagaa gaaaaagtaa acaacttttg caaatttaag gactttttta tactcttata 2940
aaaataattg tattcttgaa ctctccattt tgttgcgtta ggttttcctg ttctggttct 3000
gcatctttgg cctgcgttag ttccagtgac tgtctcctca ccctccccat tctctcttct 3060
aggagaatga tgtaaacctg acccacattg aatctagacc ttctcgttta aagaaagatg 3120
agtatgaatt tttcacccat ttggataaac gtagcctgcc tgctctgaca aacatcatca 3180
agatcttgag gcatgacatt ggtgccactg tccatgagct ttcacgagat aagaagaaag 3240
acacagtgcc ctggttccca agaaccattc aagagctgga cagatttgcc aatcagattc 3300
tcagctatgg agcggaactg gatgctgacc accctggttt taaagatcct gtgtaccgtg 3360
caagacggaa gcagtttgct gacattgcct acaactaccg ccatgggcag cccatccctc 3420
gagtggaata catggaggaa gaaaagaaaa catggggcac agtgttcaag actctgaagt 3480
ccttgtataa aacccatgct tgctatgagt acaatcacat ttttccactt cttgaaaagt 3540
actgtggctt ccatgaagat aacattcccc agctggaaga cgtttctcag ttcctgcaga 3600
cttgcactgg tttccgcctc cgacctgtag ctggcctgct ttcctctcgg gatttcttgg 3660
gtggcctggc cttccgagtc ttccactgca cacagtacat cagacatgga tccaagccca 3720
tgtatacccc cgaacctgac atctgccatg agctgttggg acatgtgccc ttgttttcag 3780
atcgcagctt tgcccagttt tcccaggaaa ttggccttgc ctctctgggt gcacctgatg 3840
aatacattga aaagctcgcc acaatttact ggtttactgt ggagtttggg ctctgcaaac 3900
aaggagactc cataaaggca tatggtgctg ggctcctgtc atcctttggt gaattacagt 3960
actgcttatc agagaagcca aagcttctcc ccctggagct ggagaagaca gccatccaaa 4020
attacactgt cacggagttc cagcccctgt attacgtggc agagagtttt aatgatgcca 4080
aggagaaagt aaggaacttt gctgccacaa tacctcggcc cttctcagtt cgctacgacc 4140
catacaccca aaggattgag gtcttggaca atacccagca gcttaagatt ttggctgatt 4200
ccattaacag tgaaattgga atcctttgca gtgccctcca gaaaataaag taacctcgag 4260
ctgtgccttc tagttgccag ccatctgttg tttgcccctc ccccgtgcct tccttgaccc 4320
tggaaggtgc cactcccact gtcctttcct aataaaatga ggaaattgca tcgcattgtc 4380
tgagtaggtg tcattctatt ctggggggtg gggtggggca ggacagcaag ggggaggatt 4440
gggaagacaa tagcaggcat gctggggatg cggtgggctc tatggaccgg tgcggccgca 4500
ggaaccccta gtgatggagt tggccactcc ctctctgcgc gctcgctcgc tcactgaggc 4560
cgggcgacca aaggtcgccc gacgcccggg ctttgcccgg gcggcctcag tgagcgagcg 4620
agcgcgcagc tgcctgcagg 4640
<210> 20
<211> 2822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pFB-ApoE-hAAT-hPAH-Genius 2 (последовательность вектора)
<400> 20
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtacta gcagctacaa 900
tccagctacc attctgcttt tattttatgg ttgggataag gctggattat tctgagtcca 960
agctaggccc ttttgctaat catgttcata cctcttatct tcctcccaca gctcctgggc 1020
aacgtgctgg tctgtgtgct ggcccatcac tttggcaaag aattgcgatc gccaccatga 1080
gcactgctgt gctggagaac cctggcctgg gcagaaagct gtctgacttt ggccaggaga 1140
ccagctacat tgaggacaac tgcaaccaga atggggccat cagcctgatc ttcagcctga 1200
aggaggaggt gggggccctg gccaaggtgc tgagactgtt tgaggagaat gatgtgaacc 1260
tgacccacat tgagagcaga cccagcagac tgaagaagga tgagtatgag ttcttcaccc 1320
acctggacaa gagaagcctg cctgccctga ccaacatcat caagatcctg agacacgata 1380
ttggggccac tgtgcacgag ctgagcagag acaagaagaa ggacactgtg ccctggttcc 1440
ccagaactat ccaggagctg gacagatttg ccaaccagat cctgagctat ggggctgagc 1500
tggatgctga ccaccctggc ttcaaggacc ctgtgtacag agccagaaga aagcagtttg 1560
ctgacattgc ctacaactac agacacggcc agcccatccc cagagtggag tacatggagg 1620
aggagaagaa gacctggggc actgtgttca agaccctgaa gagcctgtac aagacccacg 1680
cctgctatga gtacaaccac atcttccccc tgctggagaa gtactgtggc ttccacgagg 1740
acaacatccc ccagctggag gatgtgagcc agttcctgca gacctgcact ggcttcagac 1800
tgagacctgt ggctggcctg ctgagcagca gagacttcct ggggggcctg gccttcagag 1860
tgttccactg cacccagtac atcagacacg gcagcaagcc catgtacacc cctgagcctg 1920
atatctgcca cgagctgctg ggccacgtgc ccctgttctc tgacagaagc tttgcccagt 1980
tcagccagga gattggcctg gccagcctgg gggcccctga tgagtatatt gagaagctgg 2040
ccactatcta ctggttcact gtggagtttg gcctgtgcaa gcagggggac agcatcaagg 2100
cctatggggc tggcctgctg agcagctttg gggagctgca gtactgcctg tctgagaagc 2160
ccaagctgct gcccctggag ctggagaaga ctgccatcca gaactacact gtgactgagt 2220
tccagcccct gtactatgtg gctgagagct tcaatgatgc caaggagaag gtgagaaact 2280
ttgctgccac catccccaga cccttctctg tgagatacga cccctacacc cagagaattg 2340
aggtgctgga caacacccag cagctgaaga tcctggctga cagcatcaac tctgagattg 2400
gcatcctgtg ctctgccctg cagaagatca agtgacctcg agctgtgcct tctagttgcc 2460
agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca 2520
ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta 2580
ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc 2640
atgctgggga tgcggtgggc tctatggacc ggtgcggccg caggaacccc tagtgatgga 2700
gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac caaaggtcgc 2760
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 2820
gg 2822
<210> 21
<211> 2822
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pFB-ApoE-hAAT-hPAH-JCAT (последовательность вектора)
<400> 21
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtaggct cagaggcaca caggagtttc 180
tgggctcacc ctgccccctt ccaacccctc agttcccatc ctccagcagc tgtttgtgtg 240
ctgcctctga agtccacact gaacaaactt cagcctactc atgtccctaa aatgggcaaa 300
cattgcaagc agcaaacagc aaacacacag ccctccctgc ctgctgacct tggagctggg 360
gcagaggtca gagacctctc tgggcccatg ccacctccaa catccactcg accccttgga 420
atttcggtgg agaggagcag aggttgtcct ggcgtggttt aggtagtgtg agaggggtcg 480
acgatcttgc taccagtgga acagccacta aggattctgc agtgagagca gagggccagc 540
taagtggtac tctcccagag actgtctgac tcacgccacc ccctccacct tggacacagg 600
acgctgtggt ttctgagcca ggtacaatga ctcctttcgg taagtgcagt ggaagctgta 660
cactgcccag gcaaagcgtc cgggcagcgt aggcgggcga ctcagatccc agccagtgga 720
cttagcccct gtttgctcct ccgataactg gggtgacctt ggttaatatt caccagcagc 780
ctcccccgtt gcccctctgg atccactgct taaatacgga cgaggacagg gccctgtctc 840
ctcagcttca ggcaccacca ctgacctggg acagtgaatc gtaagtacta gcagctacaa 900
tccagctacc attctgcttt tattttatgg ttgggataag gctggattat tctgagtcca 960
agctaggccc ttttgctaat catgttcata cctcttatct tcctcccaca gctcctgggc 1020
aacgtgctgg tctgtgtgct ggcccatcac tttggcaaag aattgcgatc gccaccatga 1080
gcactgctgt gctggagaac cctggcctgg gcagaaagct gtctgacttt ggccaggaga 1140
ccagctacat tgaggacaac tgcaaccaga atggggccat cagcctgatc ttcagcctga 1200
aggaggaggt gggggccctg gccaaggtgc tgagactgtt tgaggagaat gatgtgaacc 1260
tgacccacat tgagagcaga cccagcagac tgaagaagga tgagtatgag ttcttcaccc 1320
acctggacaa gagaagcctg ccagccctga ccaatattat caagatcctg aggcacgata 1380
ttggggccac agtgcacgag ctgagcagag acaagaagaa ggacacagtg ccctggttcc 1440
ccaggactat ccaggagctg gacaggtttg ccaaccagat cctgagctac ggagctgagc 1500
tggatgcaga ccaccctggc ttcaaggacc ctgtgtacag agccaggaga aagcagtttg 1560
cagatatagc ctacaactac aggcacggcc agccaatccc tagagtggag tacatggagg 1620
aggagaagaa gacctggggc actgtgttca agaccctgaa gagcctgtac aagacccacg 1680
cctgctacga gtacaaccac atcttccccc tgctggagaa gtactgtggc ttccacgagg 1740
acaacatccc ccagctggag gatgtgagcc agttcctgca gacctgcaca ggcttcaggc 1800
tgagaccagt ggcaggcctg ctgagcagca gggacttcct ggggggcctg gccttcagag 1860
tgttccactg cacccagtat atcaggcacg gcagcaagcc aatgtacacc cctgagccag 1920
atatctgcca cgagctgctg ggccacgttc ccctgttctc agacagaagc tttgcccagt 1980
tcagccagga gattggcctg gccagcctgg gggcccctga cgagtatatt gagaagctgg 2040
ccactatcta ctggttcaca gtggagtttg gcctgtgcaa gcagggggac agcatcaagg 2100
cctatggggc tggcctgctg agcagctttg gggagctgca gtactgcctg tcagagaagc 2160
caaagctgct gcccctggag ctggagaaga cagctatcca gaactacact gtgactgagt 2220
tccagcccct gtactatgtg gcagagagct tcaatgatgc caaggagaag gtgagaaact 2280
ttgctgccac catccccagg cccttctctg tgagatatga cccctacacc cagaggattg 2340
aggtgctgga caacacccag cagctgaaga tcctggcaga cagcatcaac agtgagattg 2400
gtatcctgtg ctctgccctg cagaagatca agtaacctcg agctgtgcct tctagttgcc 2460
agccatctgt tgtttgcccc tcccccgtgc cttccttgac cctggaaggt gccactccca 2520
ctgtcctttc ctaataaaat gaggaaattg catcgcattg tctgagtagg tgtcattcta 2580
ttctgggggg tggggtgggg caggacagca agggggagga ttgggaagac aatagcaggc 2640
atgctgggga tgcggtgggc tctatggacc ggtgcggccg caggaacccc tagtgatgga 2700
gttggccact ccctctctgc gcgctcgctc gctcactgag gccgggcgac caaaggtcgc 2760
ccgacgcccg ggctttgccc gggcggcctc agtgagcgag cgagcgcgca gctgcctgca 2820
gg 2822
<210> 22
<211> 2674
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pFB-hPAHV1-TBG2uGlob (последовательность вектора)
<400> 22
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgca cgcgtcaggt taatttttaa aaagcagtca 180
aaagtccaag tggcccttgg cagcatttac tctctctgtt tgctctggtt aataatctca 240
ggagcacaaa cattccagat ccaggttaat ttttaaaaag cagtcaaaag tccaagtggc 300
ccttggcagc atttactctc tctgtttgct ctggttaata atctcaggag cacaaacatt 360
ccagatccgg cgcgccaggg ctggaagcta cctttgacat catttcctct gcgaatgcat 420
gtataatttc tacagaacct attagaaagg atcacccagc ctctgctttt gtacaacttt 480
cccttaaaaa actgccaatt ccactgctgt ttggcccaat agtgagaact ttttcctgct 540
gcctcttggt gcttttgcct atggccccta ttctgcctgc tgaagacact cttgccagca 600
tggacttaaa cccctccagc tctgacaatc ctctttctct tttgttttac atgaagggtc 660
tggcagccaa agcaatcact caaagttcaa accttatcat tttttgcttt gttcctcttg 720
gccttggttt tgtacatcag ctttgaaaat accatcccag ggttaatgct ggggttaatt 780
tataactaag agtgctctag ttttgcaata caggacatgc tataaaaatg gaaagatgtt 840
gctttctgag agatgcaggt ggattcttgg gcatttgctg taagtactag cagctacaat 900
ccagctacca ttctgctttt attttatggt tgggataagg ctggattatt ctgagtccaa 960
gctaggccct tttgctaatc atgttcatac ctcttatctt cctcccacag ctcctgggca 1020
acgtgctggt ctgtgtgctg gcccatcact ttggcaaaga attgcgatcg ccaccatgtc 1080
cactgcggtc ctggaaaacc caggcttggg caggaaactc tctgactttg gacaggaaac 1140
aagctatatt gaagacaact gcaatcaaaa tggtgccata tcactgatct tctcactcaa 1200
agaagaagtt ggtgcattgg ccaaagtatt gcgcttattt gaggagaatg atgtaaacct 1260
gacccacatt gaatctagac cttctcgttt aaagaaagat gagtatgaat ttttcaccca 1320
tttggataaa cgtagcctgc ctgctctgac aaacatcatc aagatcttga ggcatgacat 1380
tggtgccact gtccatgagc tttcacgaga taagaagaaa gacacagtgc cctggttccc 1440
aagaaccatt caagagctgg acagatttgc caatcagatt ctcagctatg gagcggaact 1500
ggatgctgac caccctggtt ttaaagatcc tgtgtaccgt gcaagacgga agcagtttgc 1560
tgacattgcc tacaactacc gccatgggca gcccatccct cgagtggaat acatggagga 1620
agaaaagaaa acatggggca cagtgttcaa gactctgaag tccttgtata aaacccatgc 1680
ttgctatgag tacaatcaca tttttccact tcttgaaaag tactgtggct tccatgaaga 1740
taacattccc cagctggaag acgtttctca gttcctgcag acttgcactg gtttccgcct 1800
ccgacctgta gctggcctgc tttcctctcg ggatttcttg ggtggcctgg ccttccgagt 1860
cttccactgc acacagtaca tcagacatgg atccaagccc atgtataccc ccgaacctga 1920
catctgccat gagctgttgg gacatgtgcc cttgttttca gatcgcagct ttgcccagtt 1980
ttcccaggaa attggccttg cctctctggg tgcacctgat gaatacattg aaaagctcgc 2040
cacaatttac tggtttactg tggagtttgg gctctgcaaa caaggagact ccataaaggc 2100
atatggtgct gggctcctgt catcctttgg tgaattacag tactgcttat cagagaagcc 2160
aaagcttctc cccctggagc tggagaagac agccatccaa aattacactg tcacggagtt 2220
ccagcccctg tattacgtgg cagagagttt taatgatgcc aaggagaaag taaggaactt 2280
tgctgccaca atacctcggc ccttctcagt tcgctacgac ccatacaccc aaaggattga 2340
ggtcttggac aatacccagc agcttaagat tttggctgat tccattaaca gtgaaattgg 2400
aatcctttgc agtgccctcc agaaaataaa gtaacctcga gctgtgcctt ctagttgcca 2460
gccatctgtt gtttgcccct cccccgtgcc ttccttgacc ctggaaggtg ccactcccac 2520
tgtcctttcc taataaaatg aggaaattgc atcgcattgt ctgagtaggt gtcattctat 2580
tctggggggt ggggtggggc aggacagcaa gggggaggat tgggaagaca atagcaggca 2640
tgctggggat gcggtgggct ctatggaccg gtgc 2674
<210> 23
<211> 2387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pFB-hPAHV1-TTR (последовательность вектора)
<400> 23
cctgcaggca gctgcgcgct cgctcgctca ctgaggccgc ccgggcaaag cccgggcgtc 60
gggcgacctt tggtcgcccg gcctcagtga gcgagcgagc gcgcagagag ggagtggcca 120
actccatcac taggggttcc tgcggccgcg tctgtctgca catttcgtag agcgagtgtt 180
ccgatactct aatctcccta ggcaaggttc atattgactt aggttactta ttctcctttt 240
gttgactaag tcaataatca gaatcagcag gtttggagtc agcttggcag ggatcagcag 300
cctgggttgg aaggaggggg tataaaagcc ccttcaccag gagaagccgt cacacagatc 360
cacaagctcc tgacaggagg cttcccttcg actcttcctc ctttgcctcg ctggactggt 420
atttgtgtct gaagctggcc ccgcggtaag tactagcagc tacaatccag ctaccattct 480
gcttttattt tatggttggg ataaggctgg attattctga gtccaagcta ggcccttttg 540
ctaatcatgt tcatacctct tatcttcctc ccacagctcc tgggcaacgt gctggtctgt 600
gtgctggccc atcactttgg caaagaattg cgatcgccac catgtccact gcggtcctgg 660
aaaacccagg cttgggcagg aaactctctg actttggaca ggaaacaagc tatattgaag 720
acaactgcaa tcaaaatggt gccatatcac tgatcttctc actcaaagaa gaagttggtg 780
cattggccaa agtattgcgc ttatttgagg agaatgatgt aaacctgacc cacattgaat 840
ctagaccttc tcgtttaaag aaagatgagt atgaattttt cacccatttg gataaacgta 900
gcctgcctgc tctgacaaac atcatcaaga tcttgaggca tgacattggt gccactgtcc 960
atgagctttc acgagataag aagaaagaca cagtgccctg gttcccaaga accattcaag 1020
agctggacag atttgccaat cagattctca gctatggagc ggaactggat gctgaccacc 1080
ctggttttaa agatcctgtg taccgtgcaa gacggaagca gtttgctgac attgcctaca 1140
actaccgcca tgggcagccc atccctcgag tggaatacat ggaggaagaa aagaaaacat 1200
ggggcacagt gttcaagact ctgaagtcct tgtataaaac ccatgcttgc tatgagtaca 1260
atcacatttt tccacttctt gaaaagtact gtggcttcca tgaagataac attccccagc 1320
tggaagacgt ttctcagttc ctgcagactt gcactggttt ccgcctccga cctgtagctg 1380
gcctgctttc ctctcgggat ttcttgggtg gcctggcctt ccgagtcttc cactgcacac 1440
agtacatcag acatggatcc aagcccatgt atacccccga acctgacatc tgccatgagc 1500
tgttgggaca tgtgcccttg ttttcagatc gcagctttgc ccagttttcc caggaaattg 1560
gccttgcctc tctgggtgca cctgatgaat acattgaaaa gctcgccaca atttactggt 1620
ttactgtgga gtttgggctc tgcaaacaag gagactccat aaaggcatat ggtgctgggc 1680
tcctgtcatc ctttggtgaa ttacagtact gcttatcaga gaagccaaag cttctccccc 1740
tggagctgga gaagacagcc atccaaaatt acactgtcac ggagttccag cccctgtatt 1800
acgtggcaga gagttttaat gatgccaagg agaaagtaag gaactttgct gccacaatac 1860
ctcggccctt ctcagttcgc tacgacccat acacccaaag gattgaggtc ttggacaata 1920
cccagcagct taagattttg gctgattcca ttaacagtga aattggaatc ctttgcagtg 1980
ccctccagaa aataaagtaa cctcgagctg tgccttctag ttgccagcca tctgttgttt 2040
gcccctcccc cgtgccttcc ttgaccctgg aaggtgccac tcccactgtc ctttcctaat 2100
aaaatgagga aattgcatcg cattgtctga gtaggtgtca ttctattctg gggggtgggg 2160
tggggcagga cagcaagggg gaggattggg aagacaatag caggcatgct ggggatgcgg 2220
tgggctctat ggaccggtgc ggccgcagga acccctagtg atggagttgg ccactccctc 2280
tctgcgcgct cgctcgctca ctgaggccgg gcgaccaaag gtcgcccgac gcccgggctt 2340
tgcccgggcg gcctcagtga gcgagcgagc gcgcagctgc ctgcagg 2387
<---
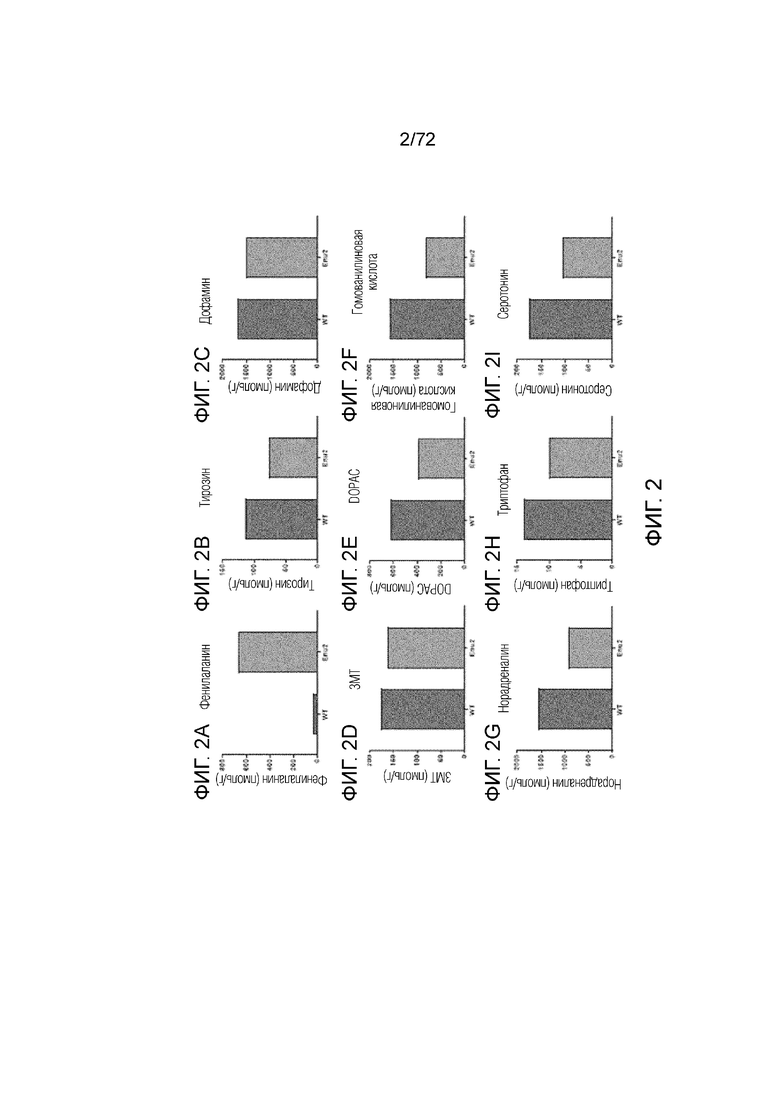
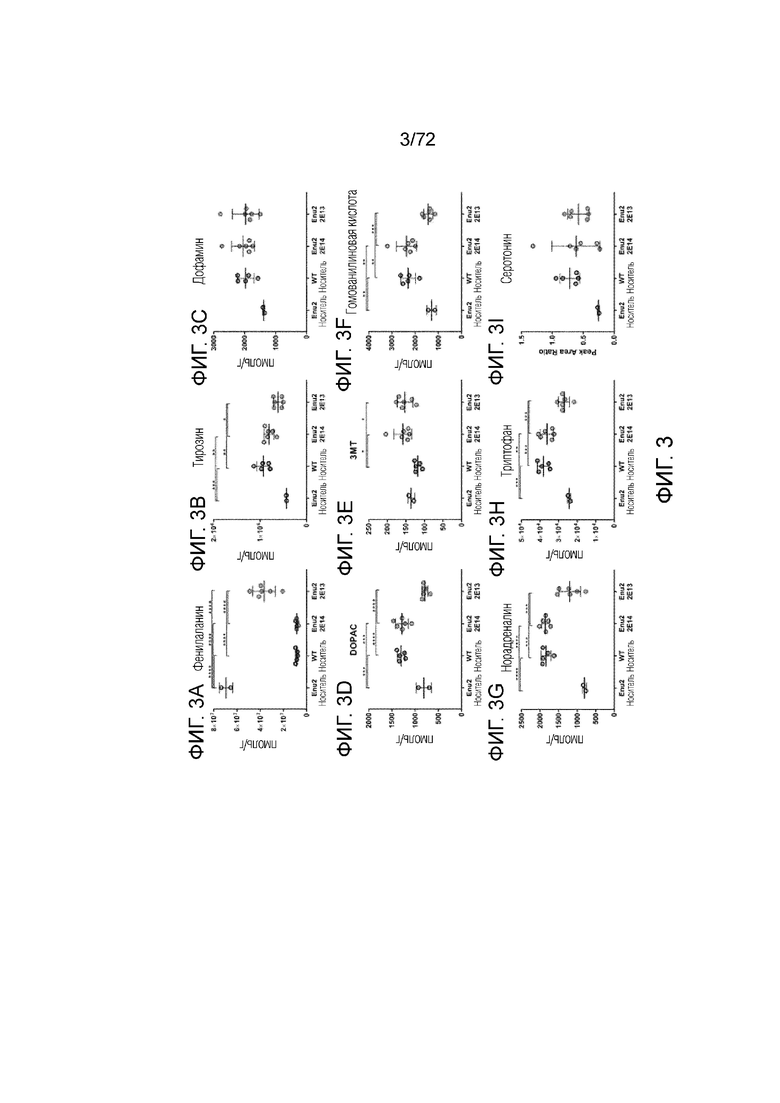
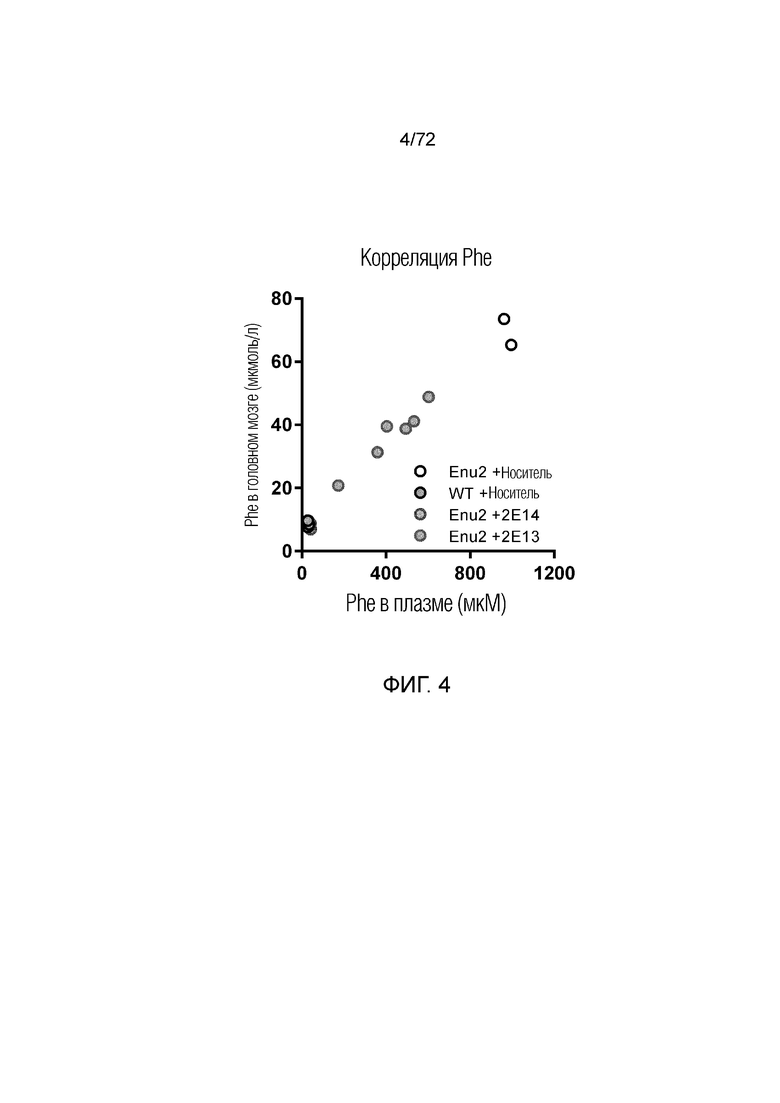
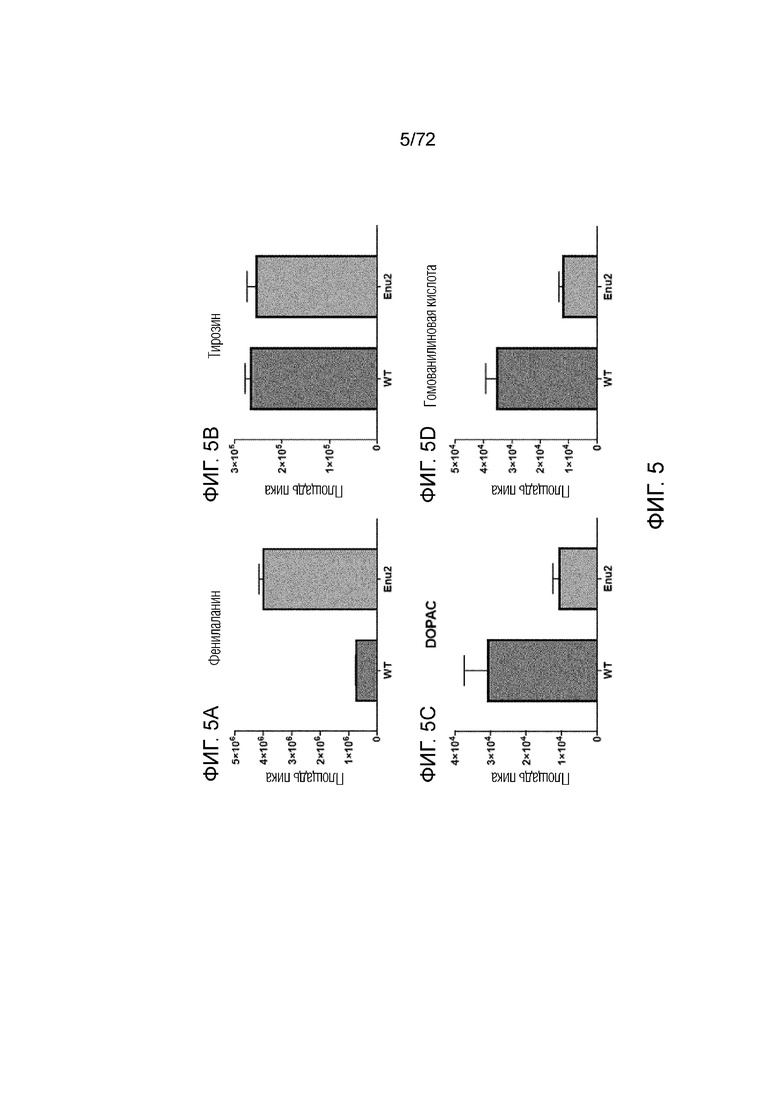

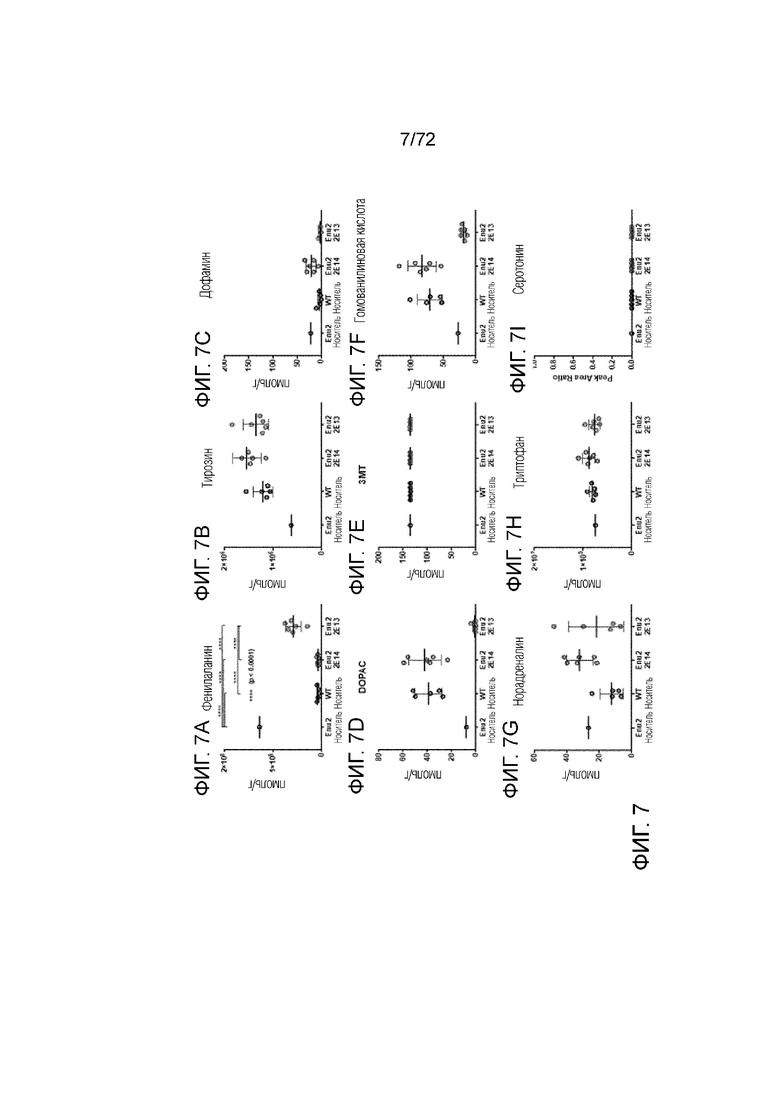
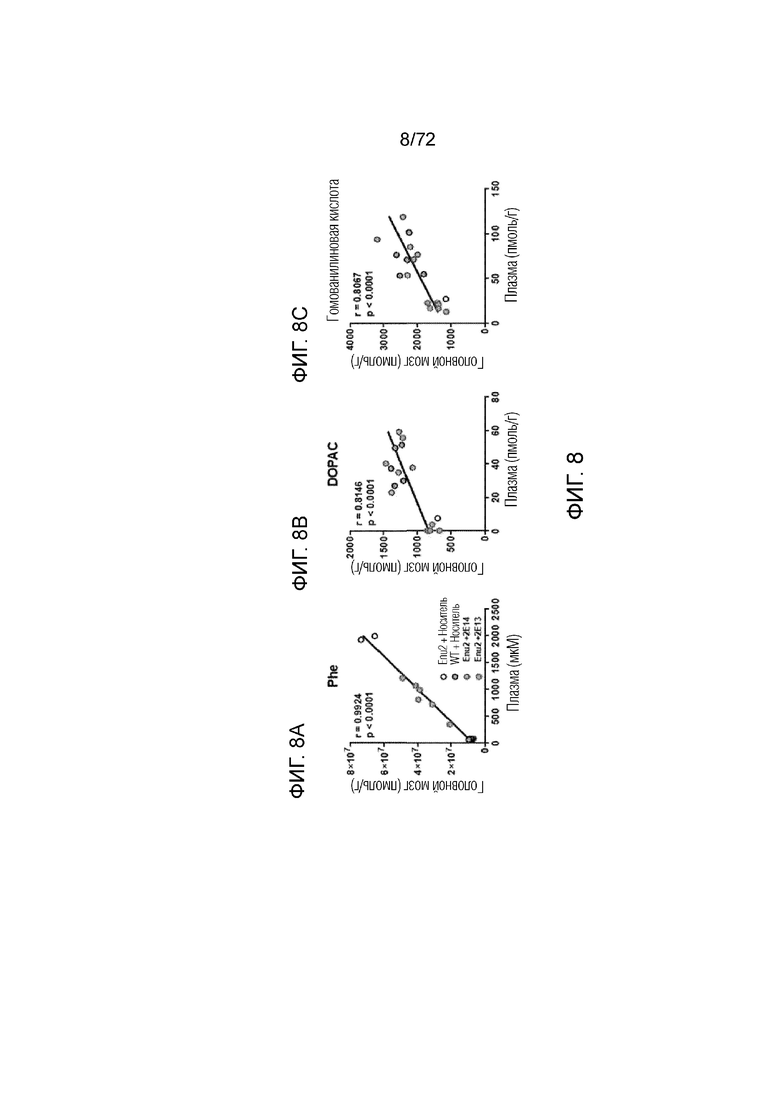
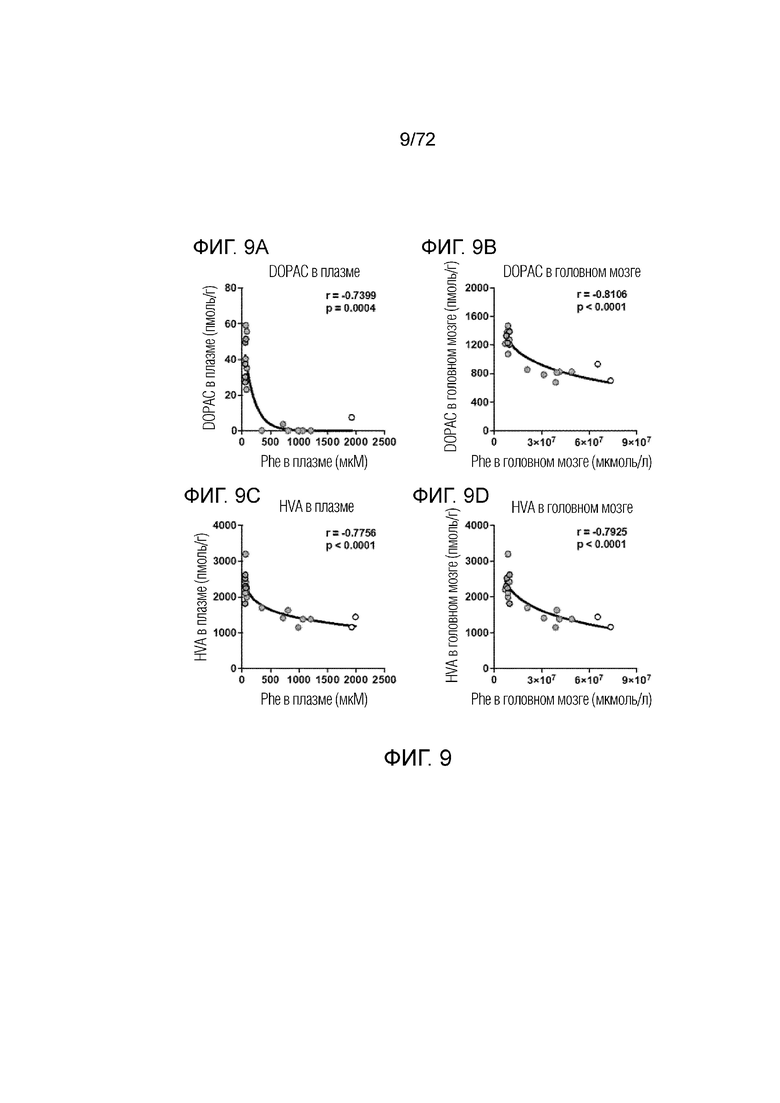
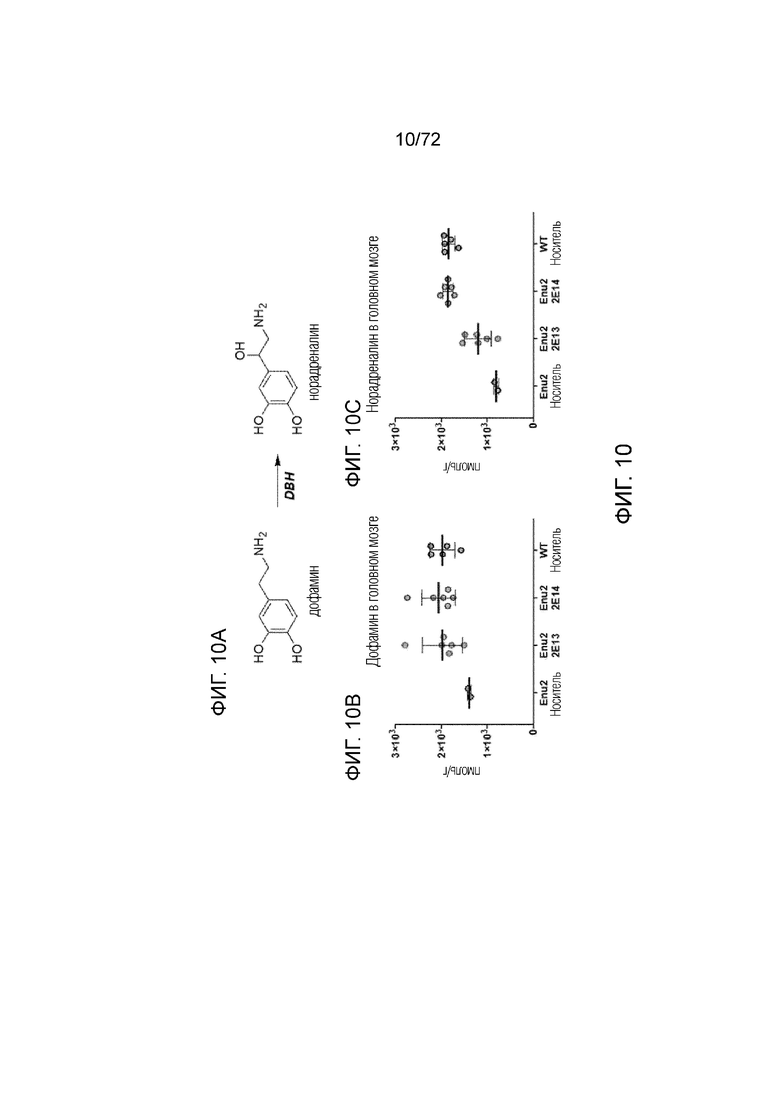
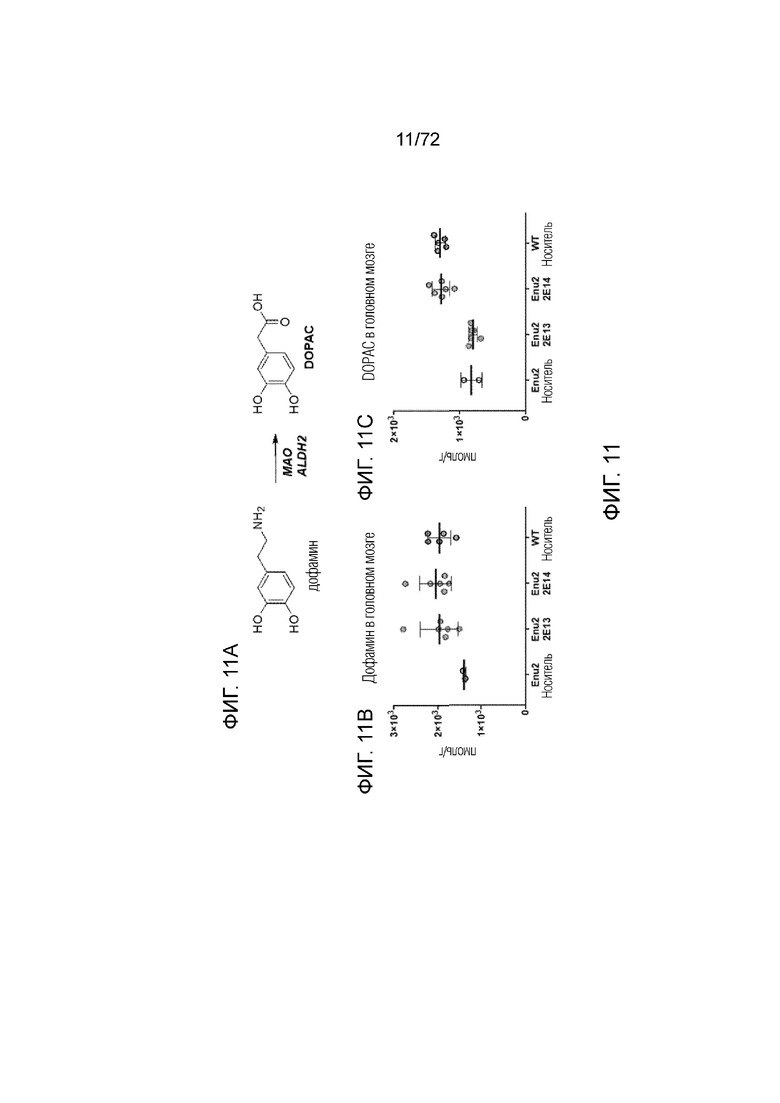

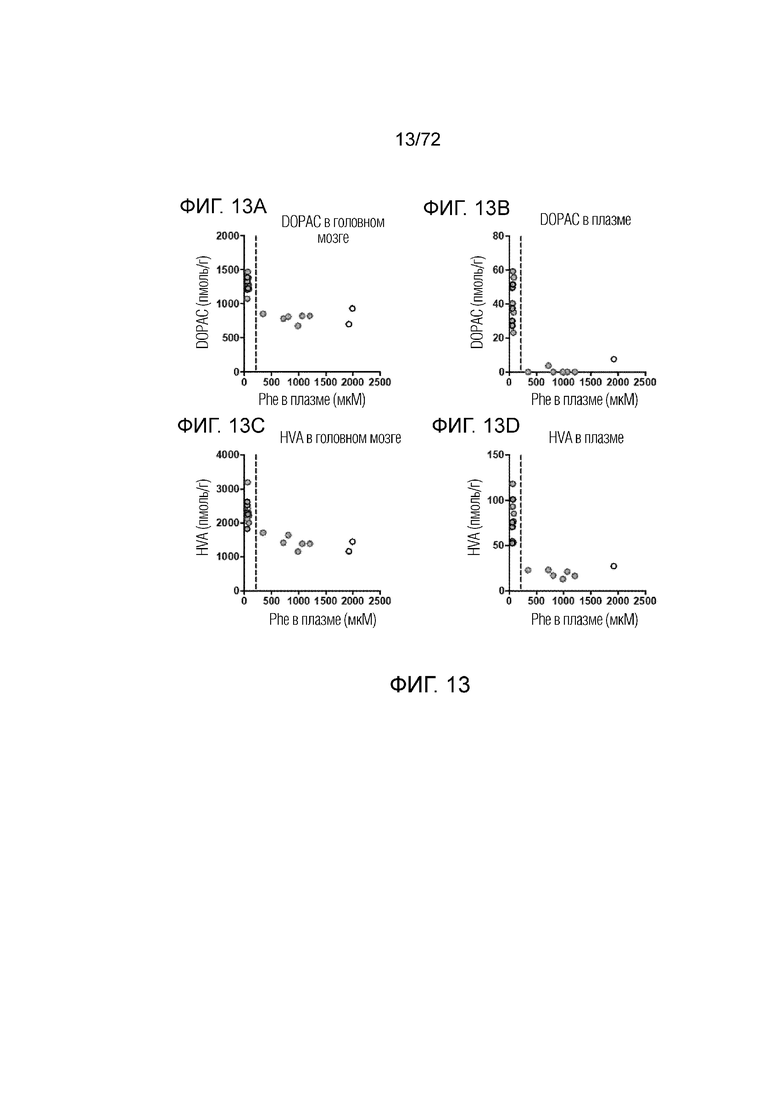
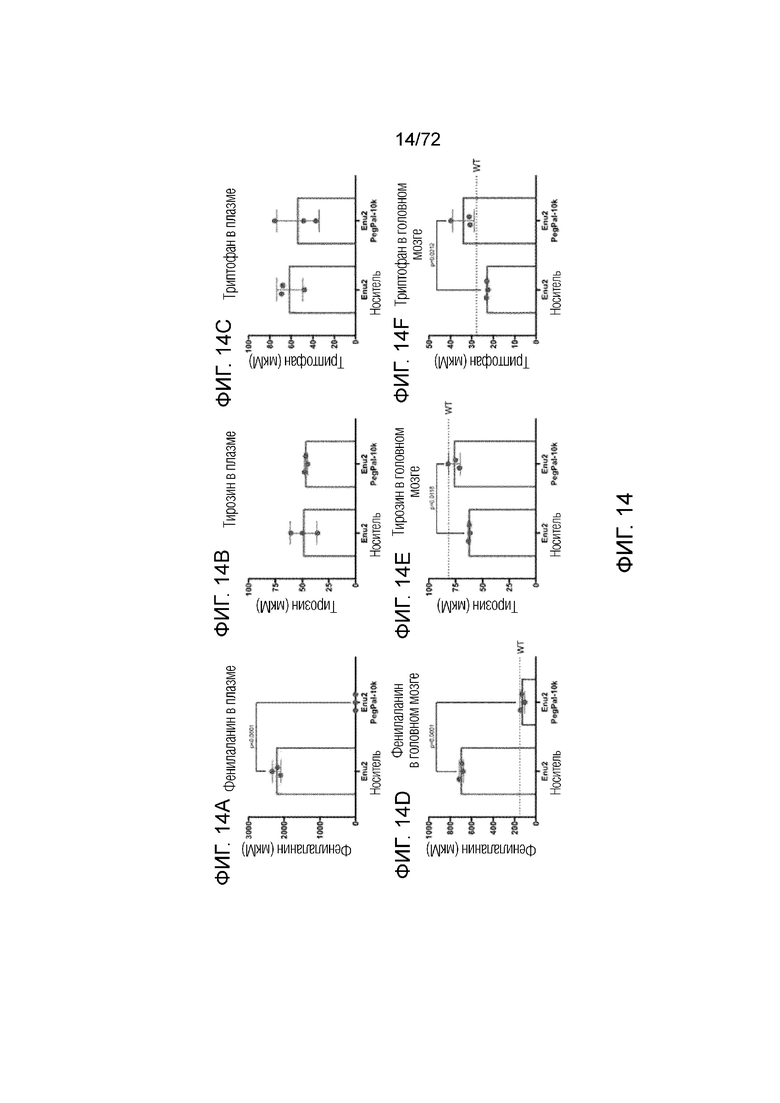
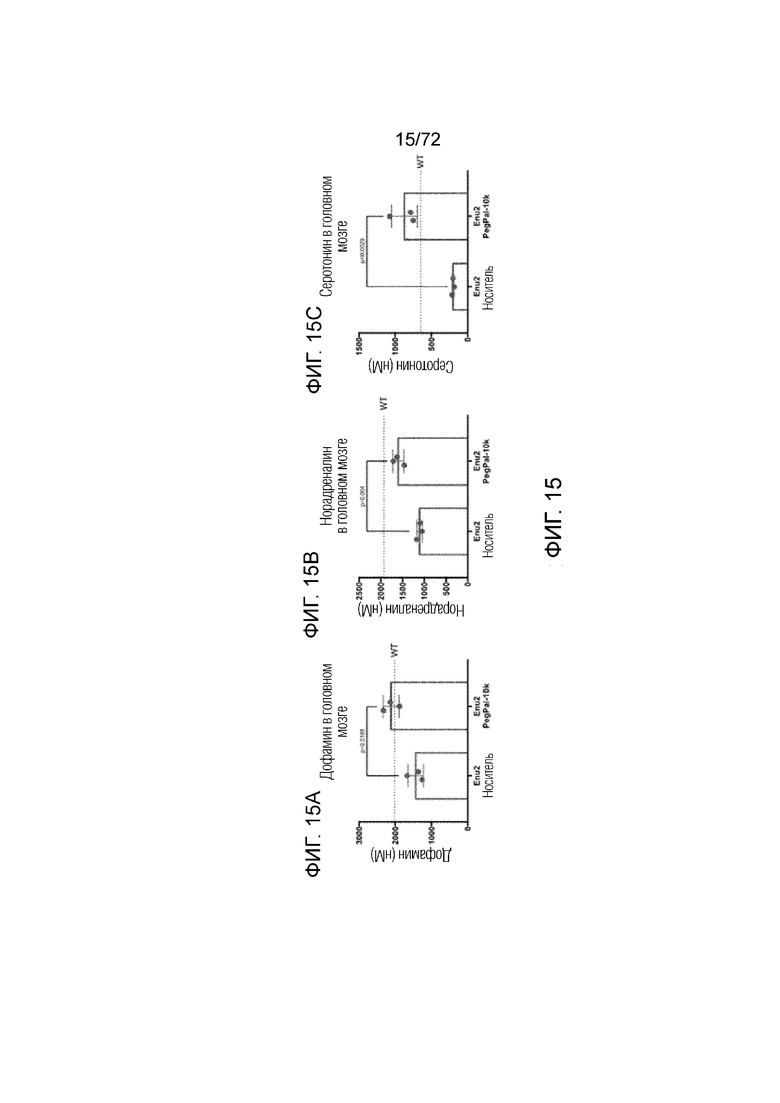
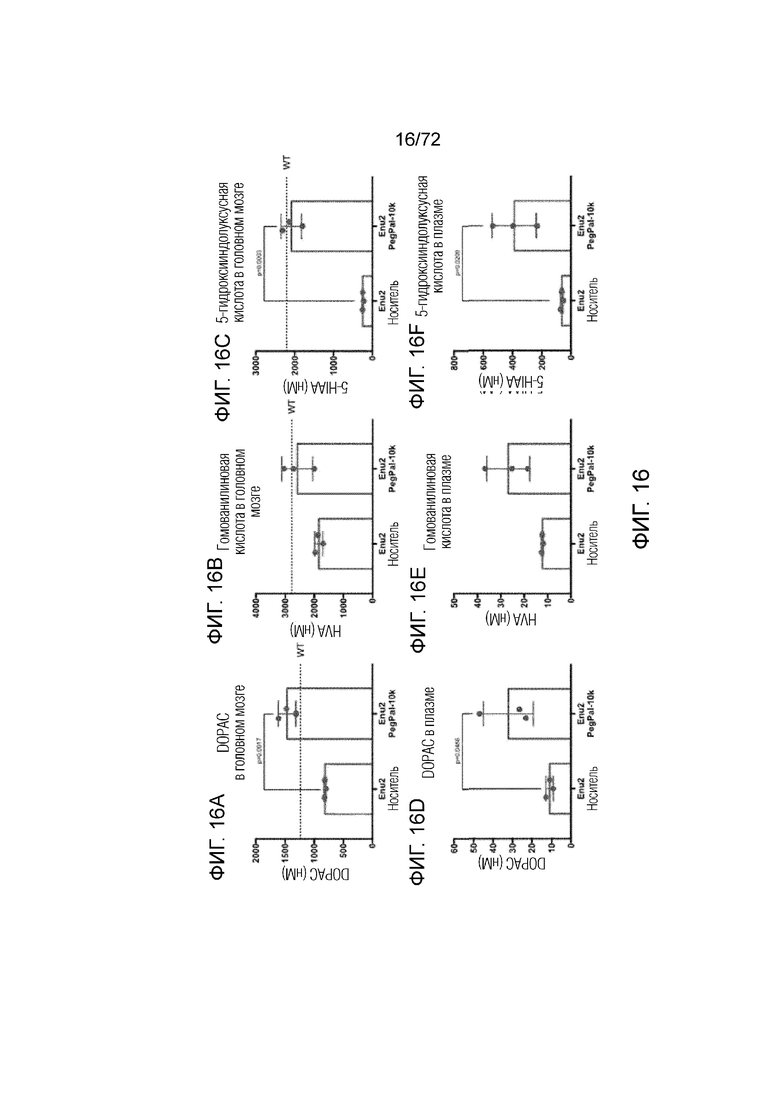
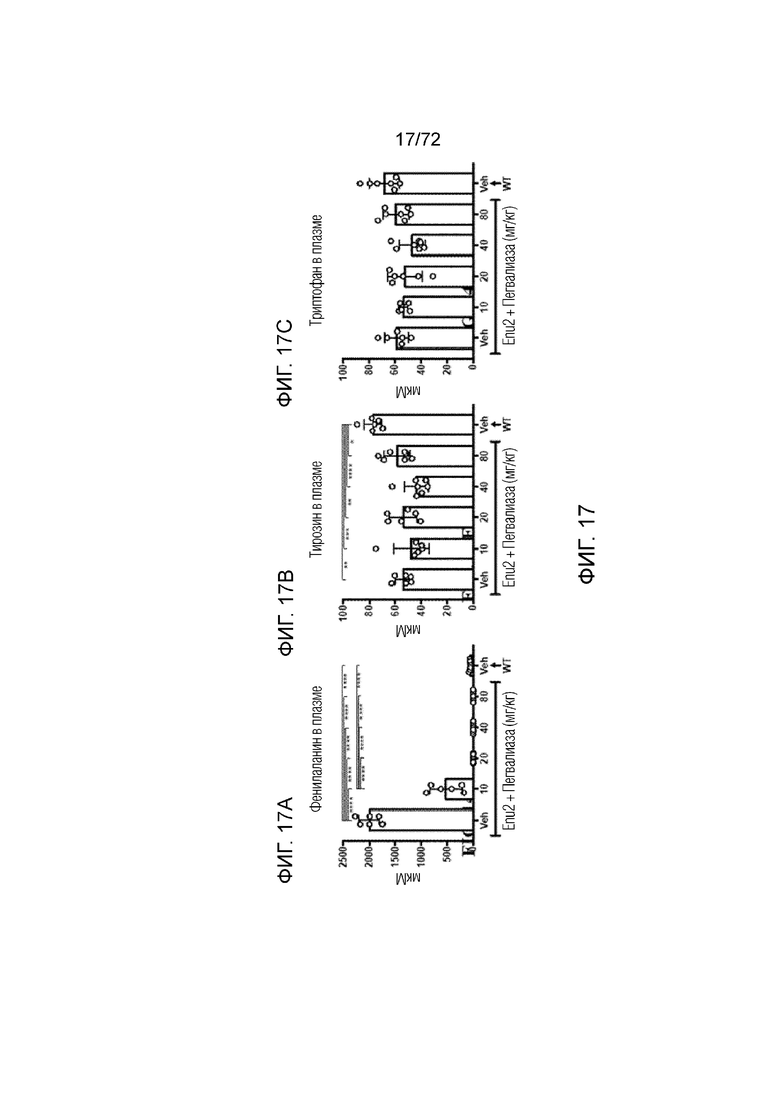
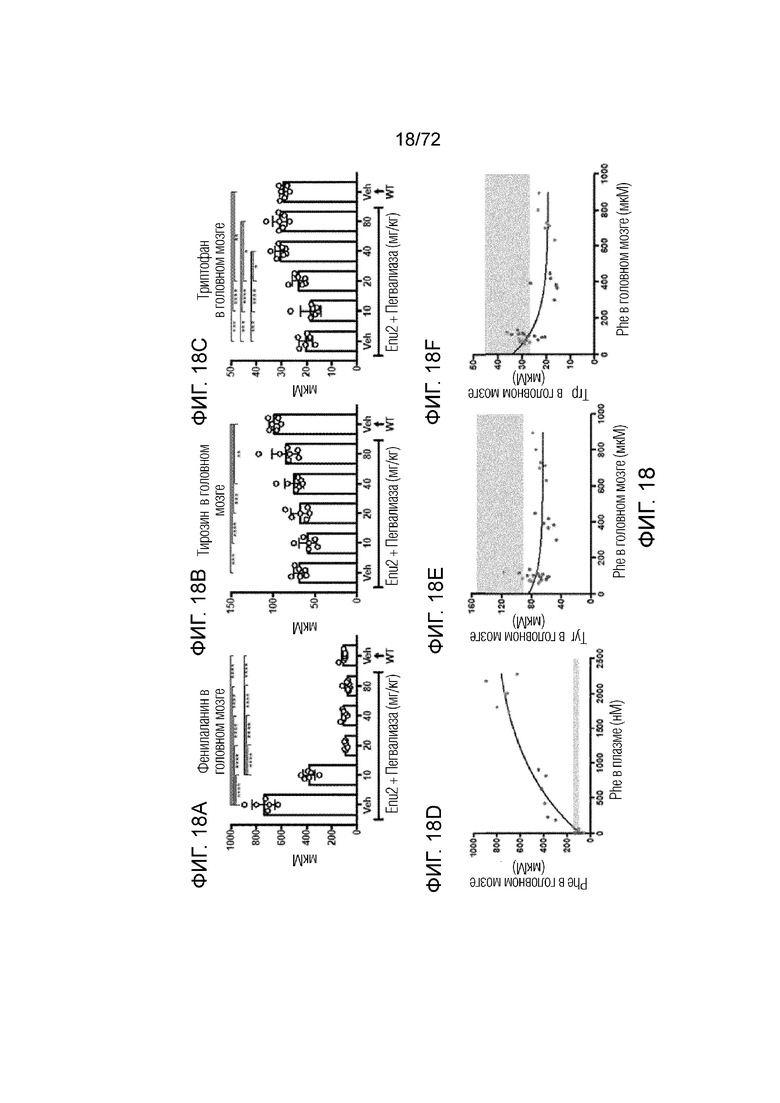
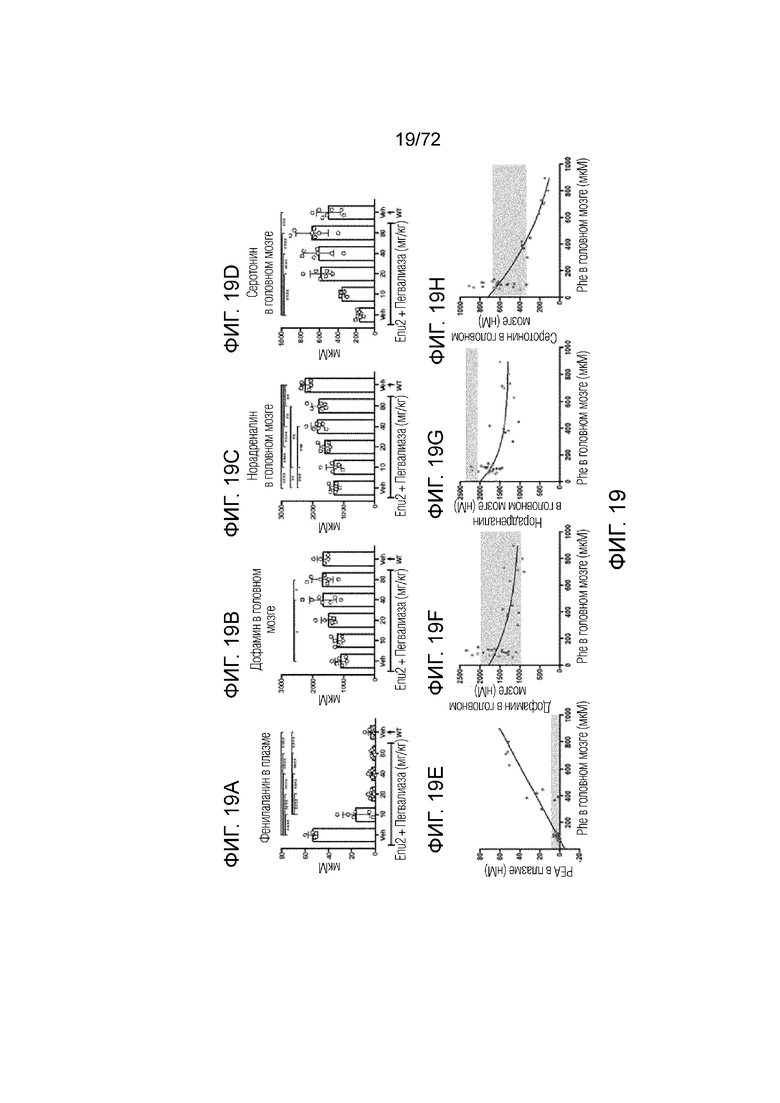
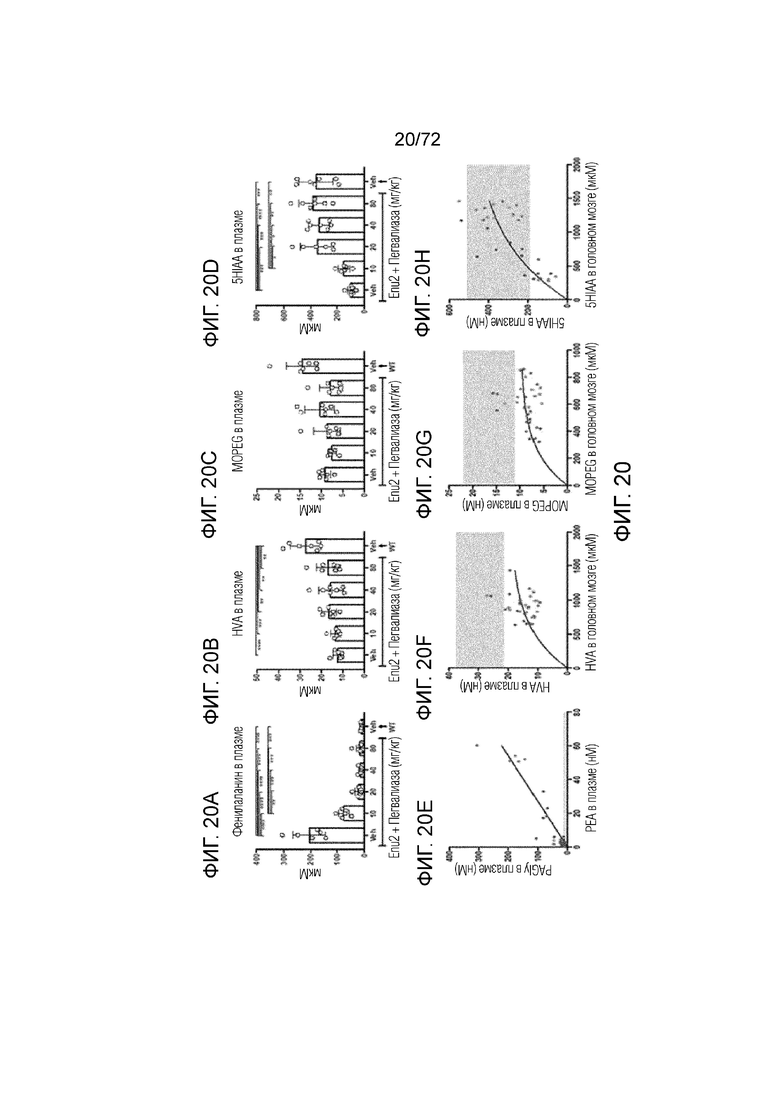
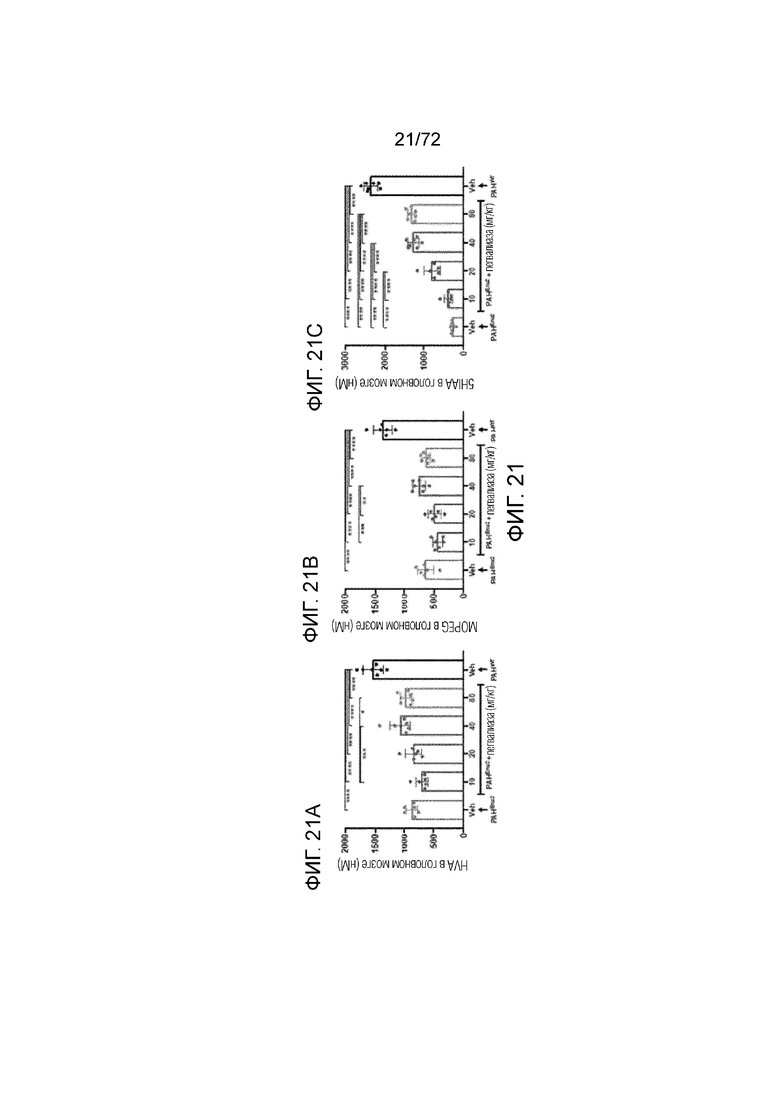
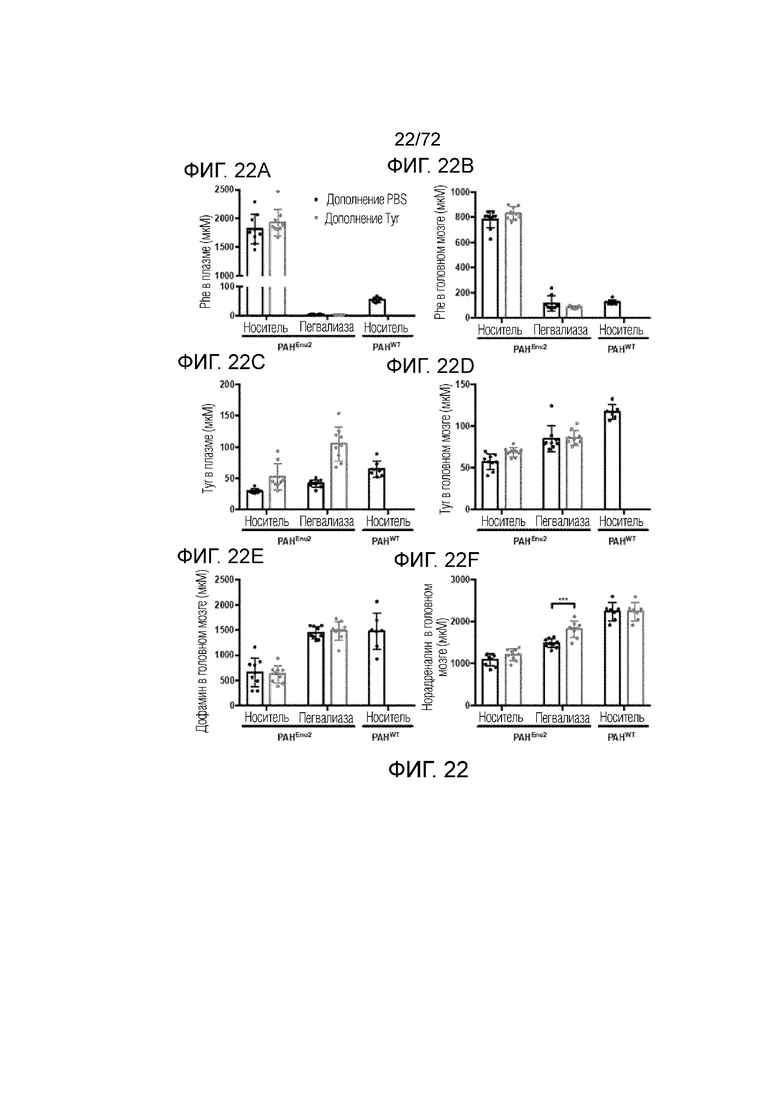
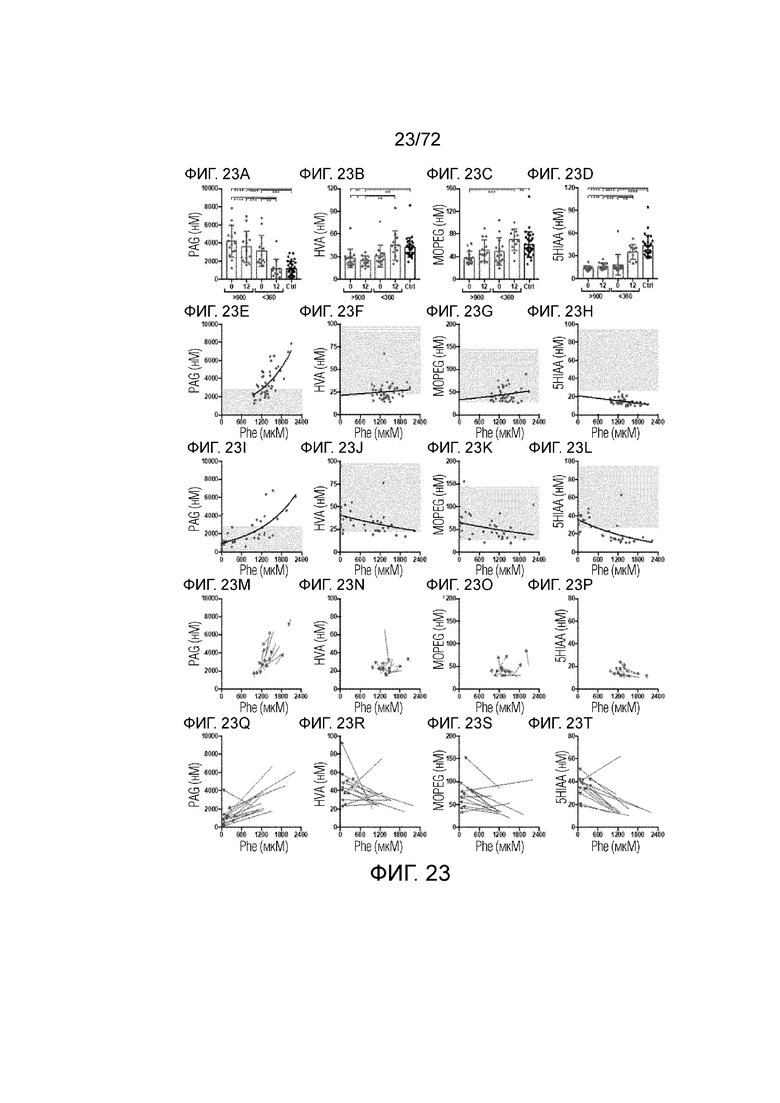
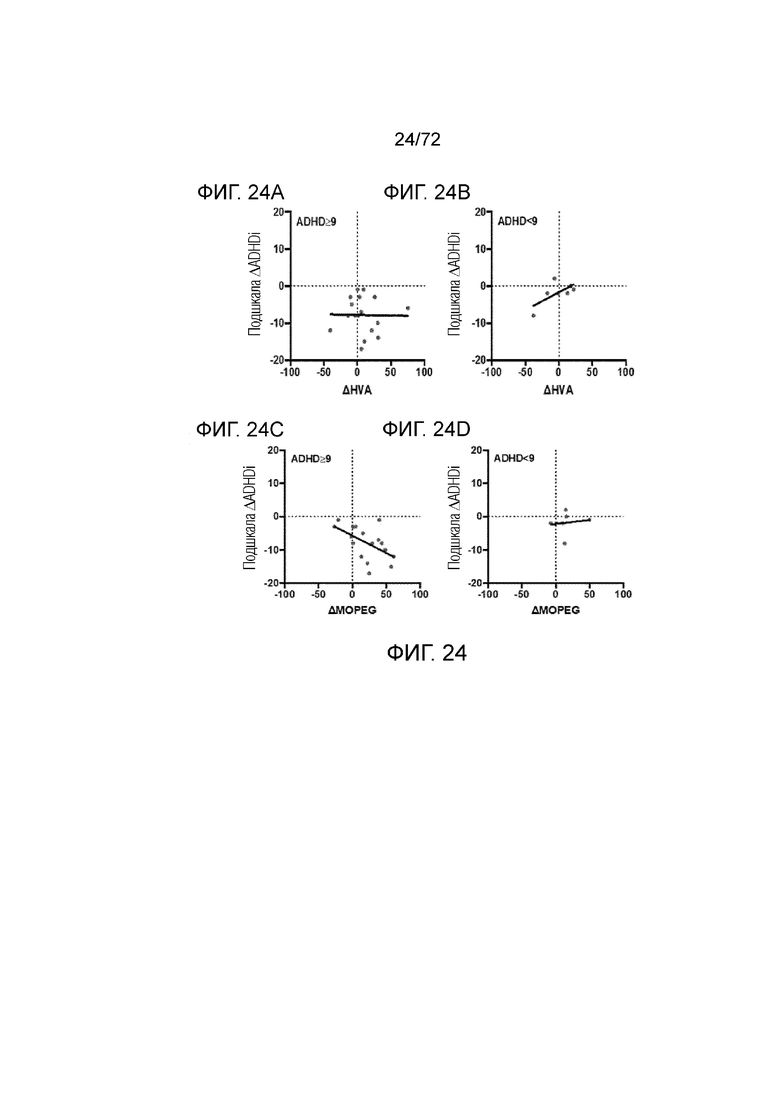

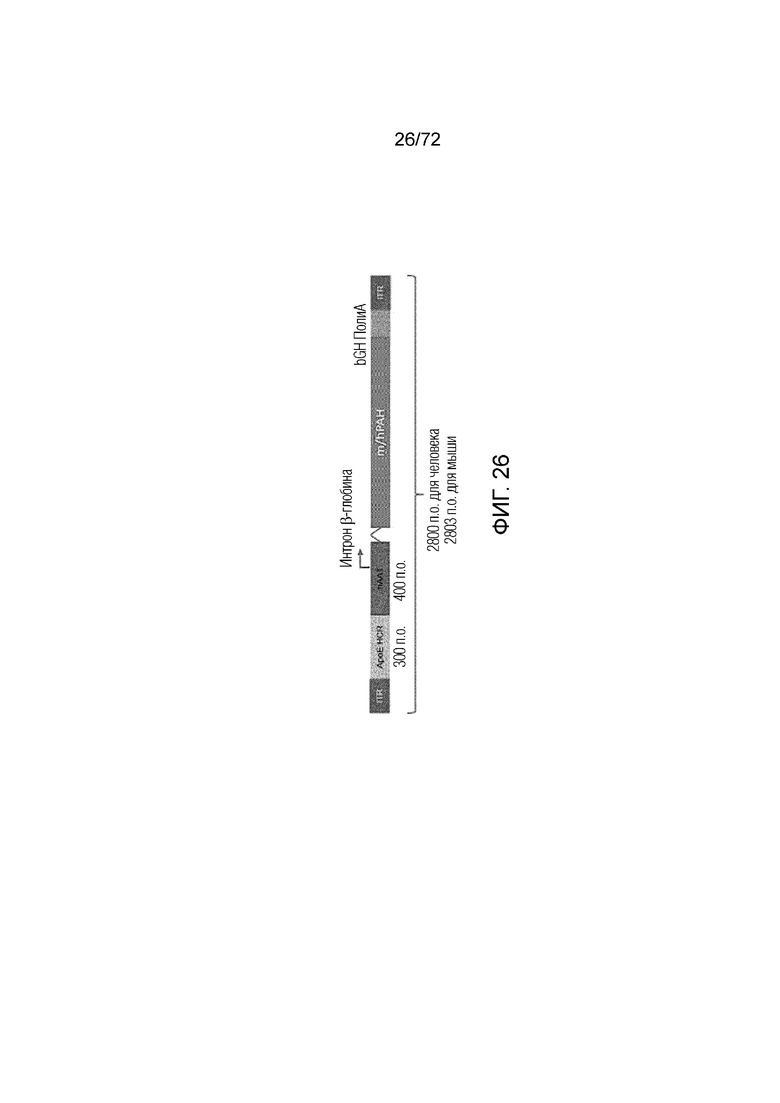
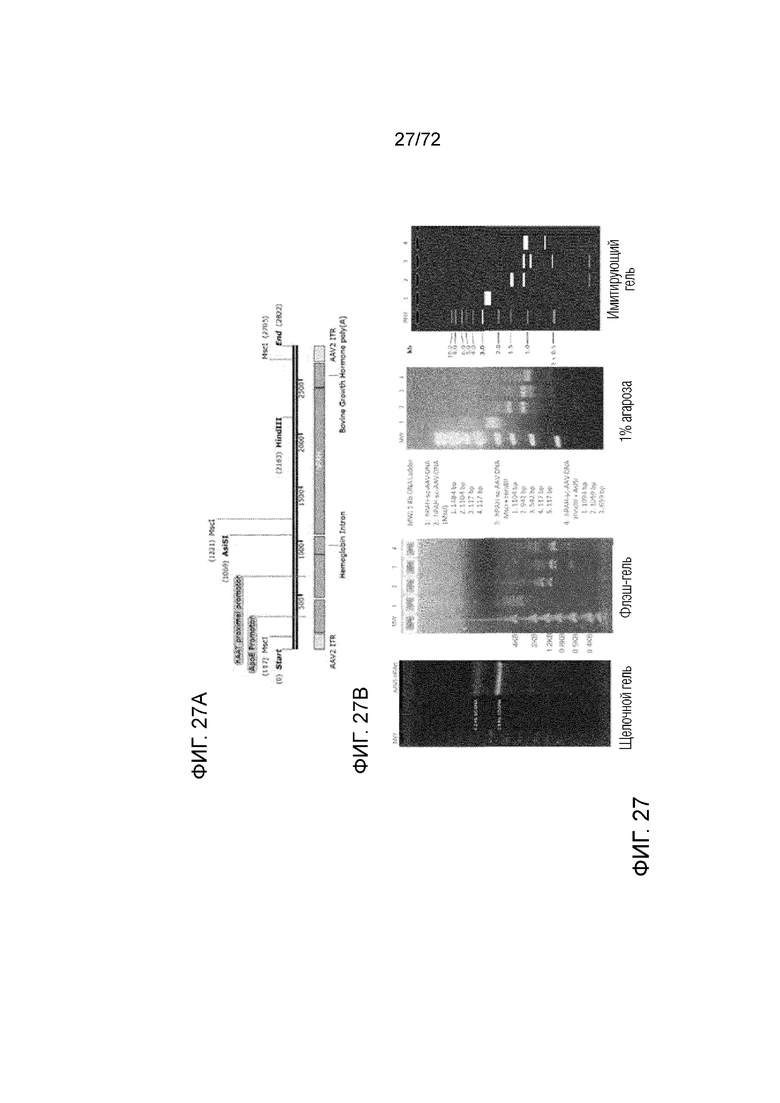
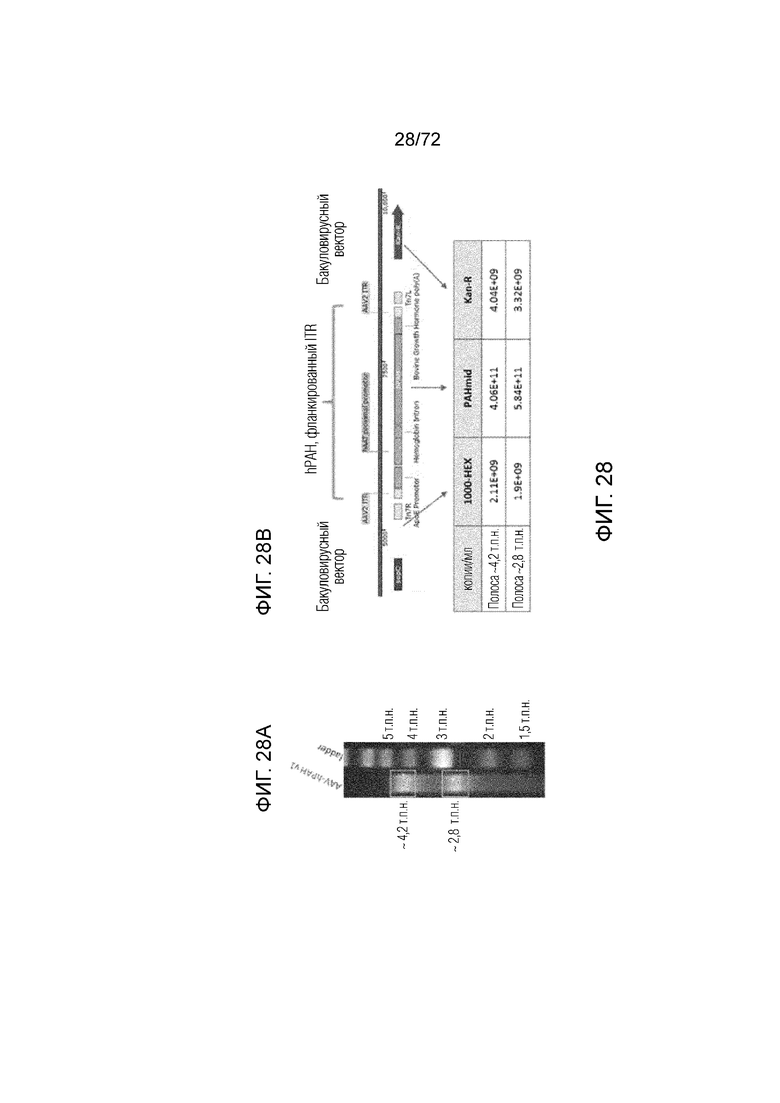
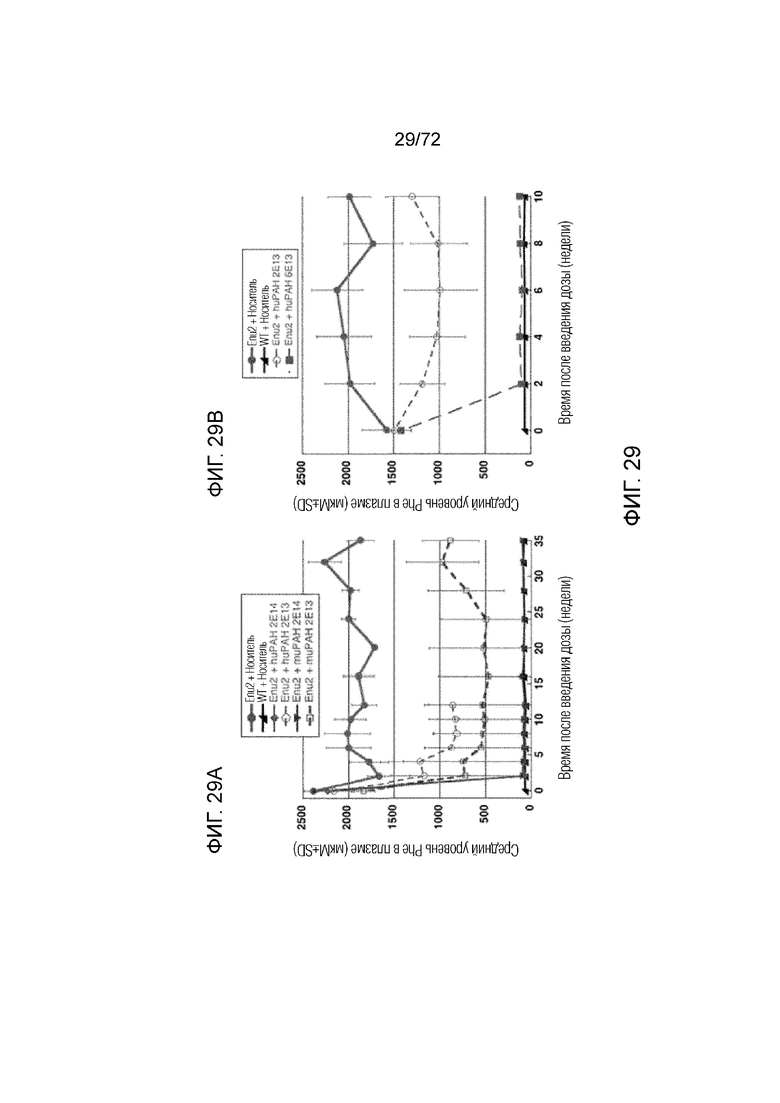
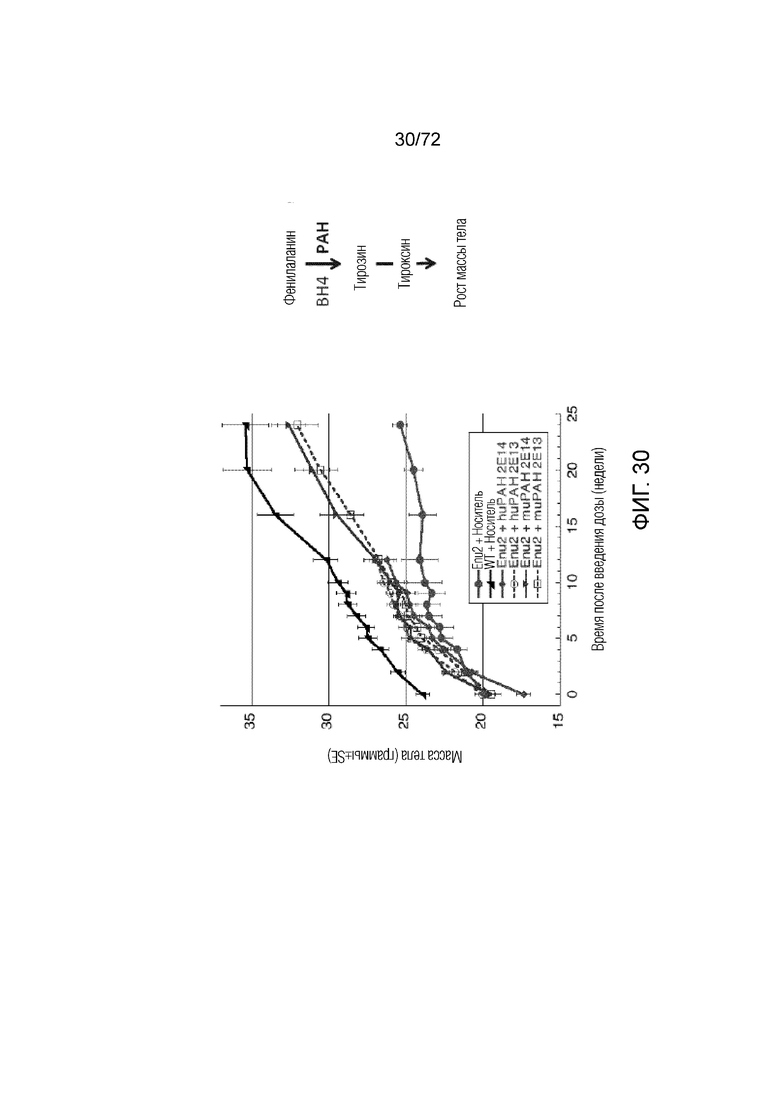

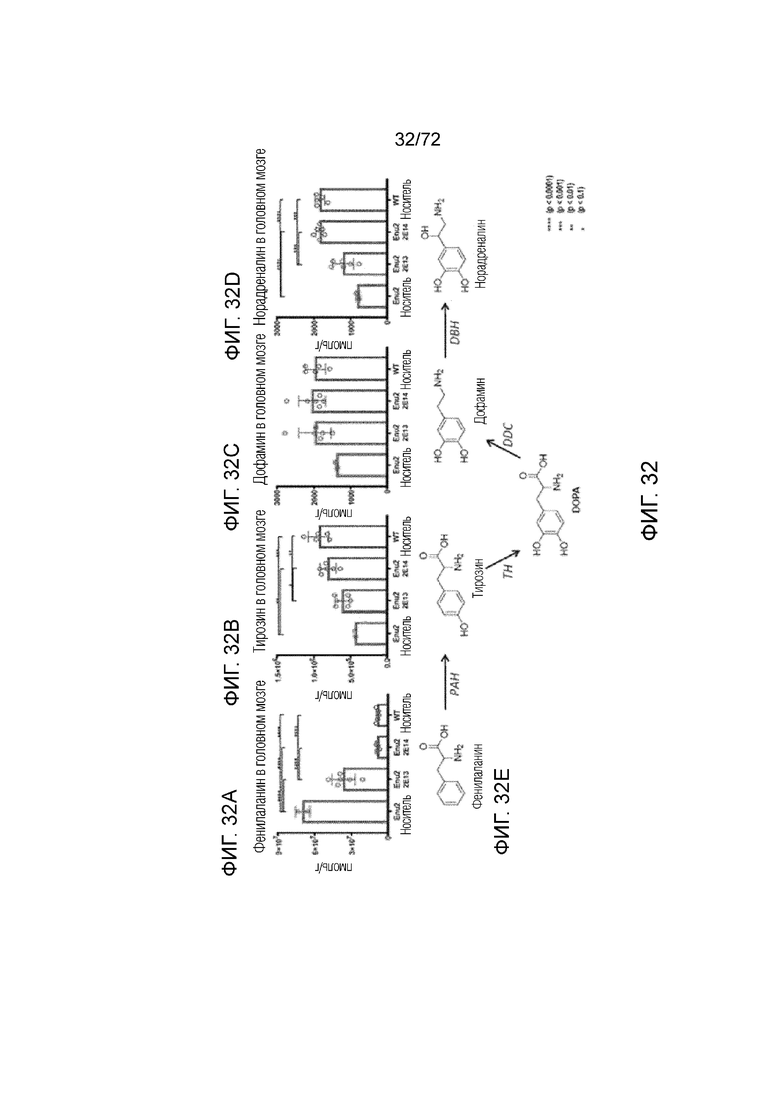
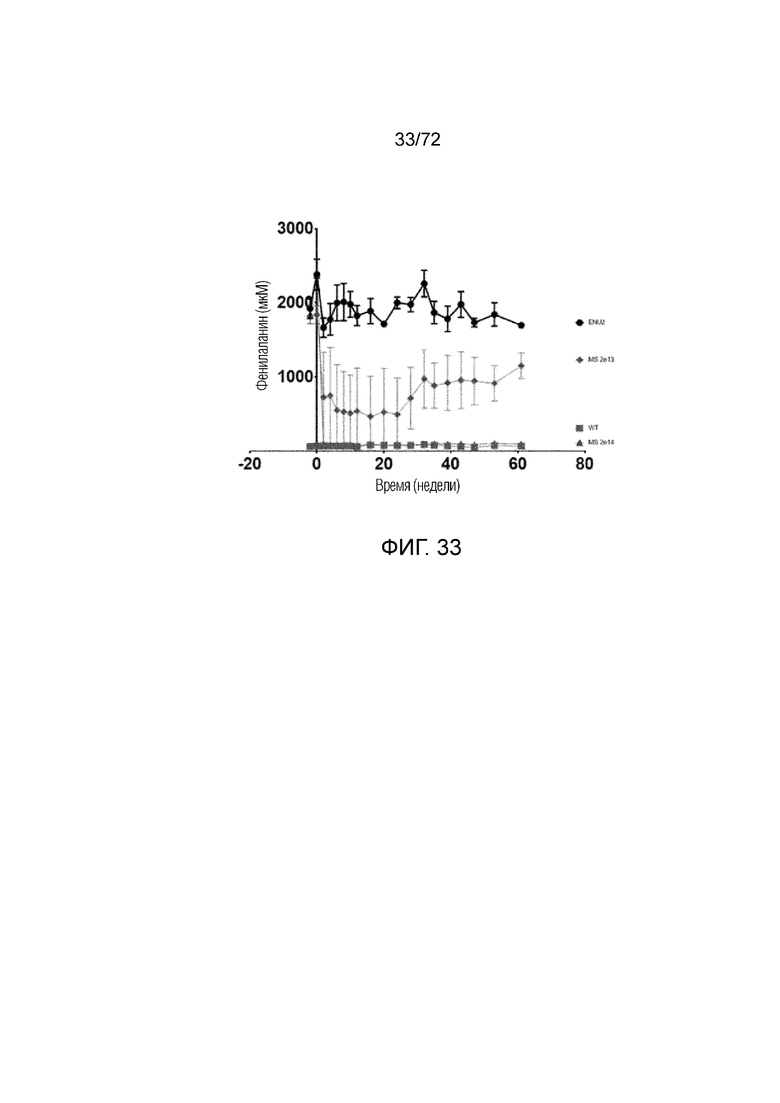
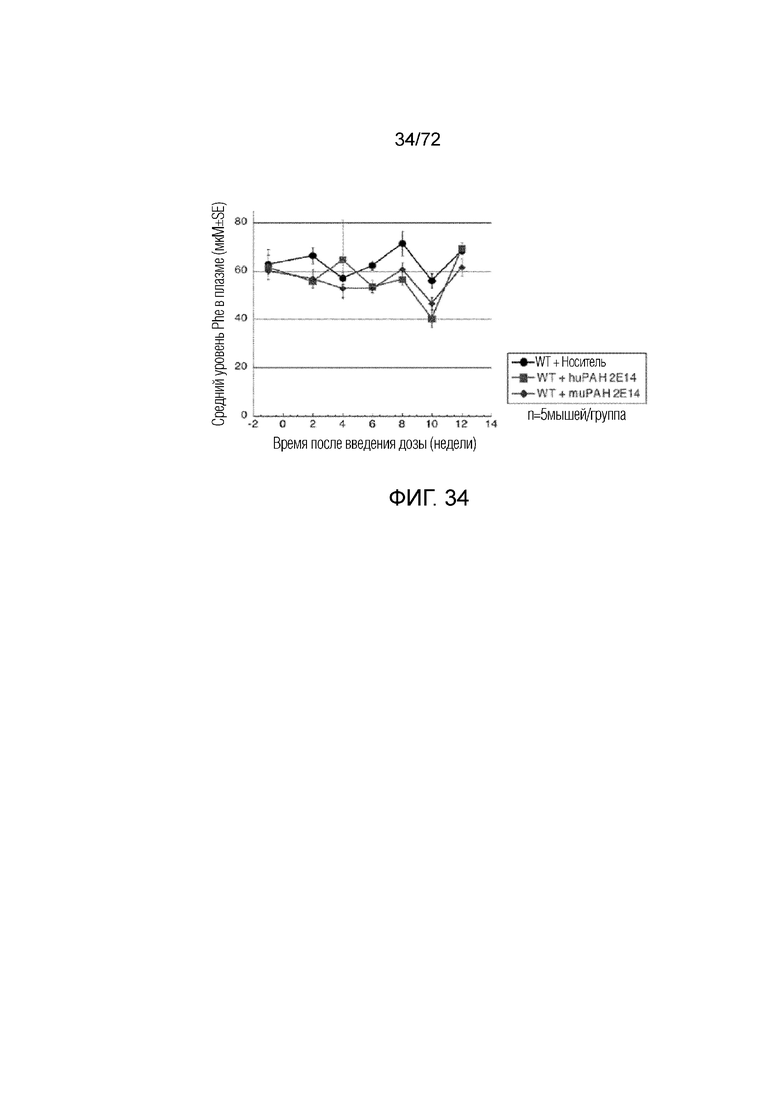

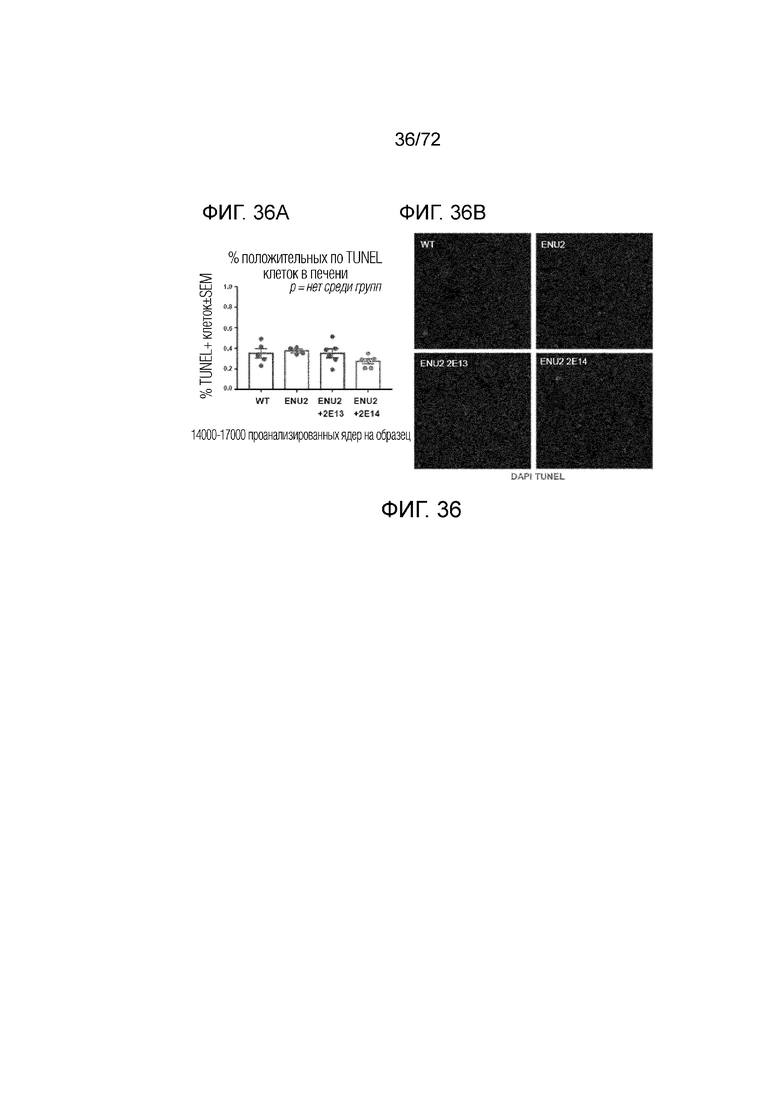
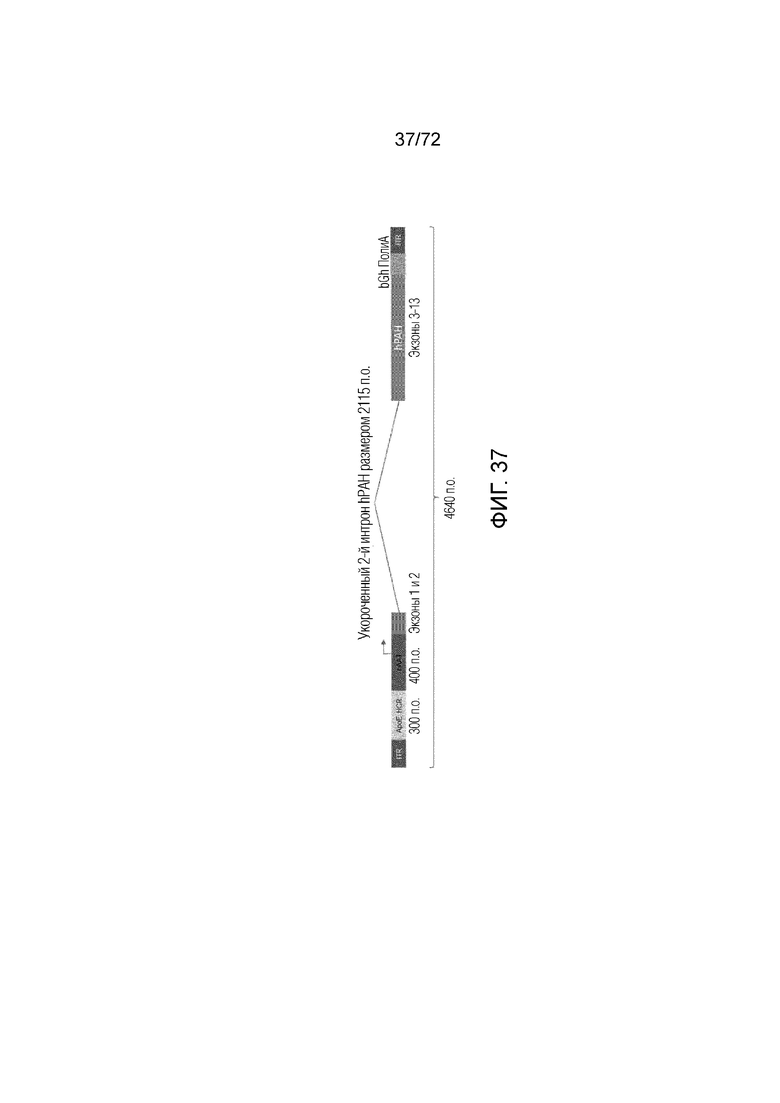
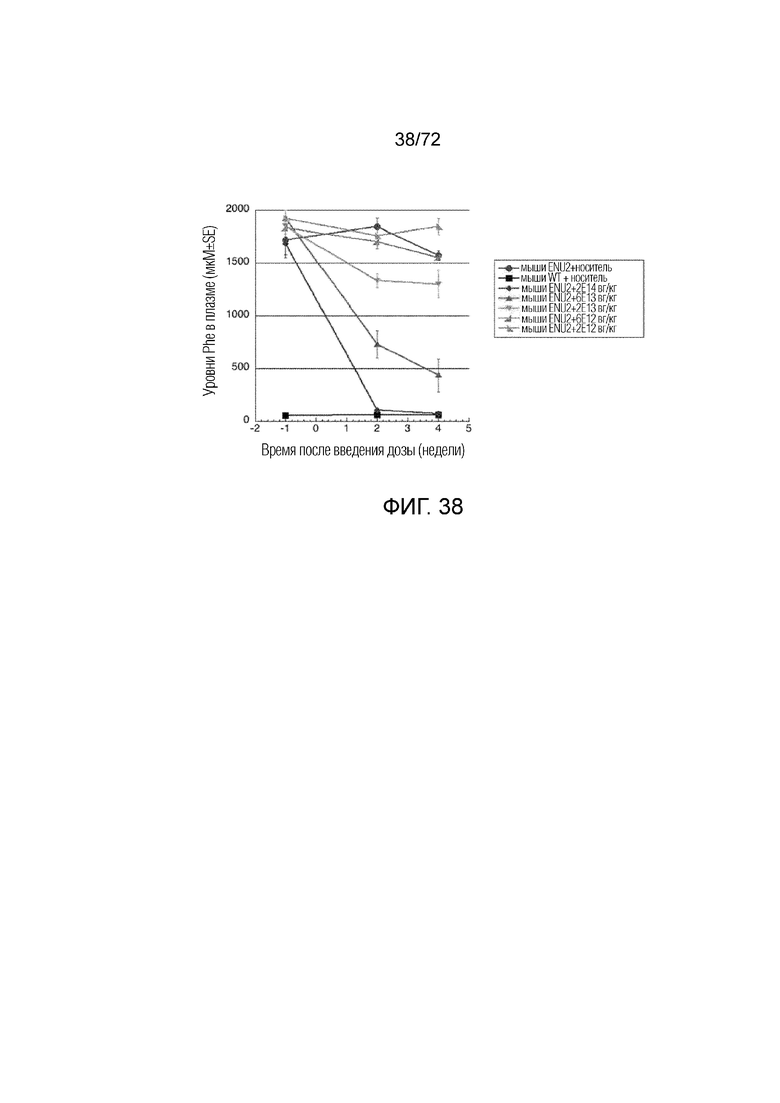
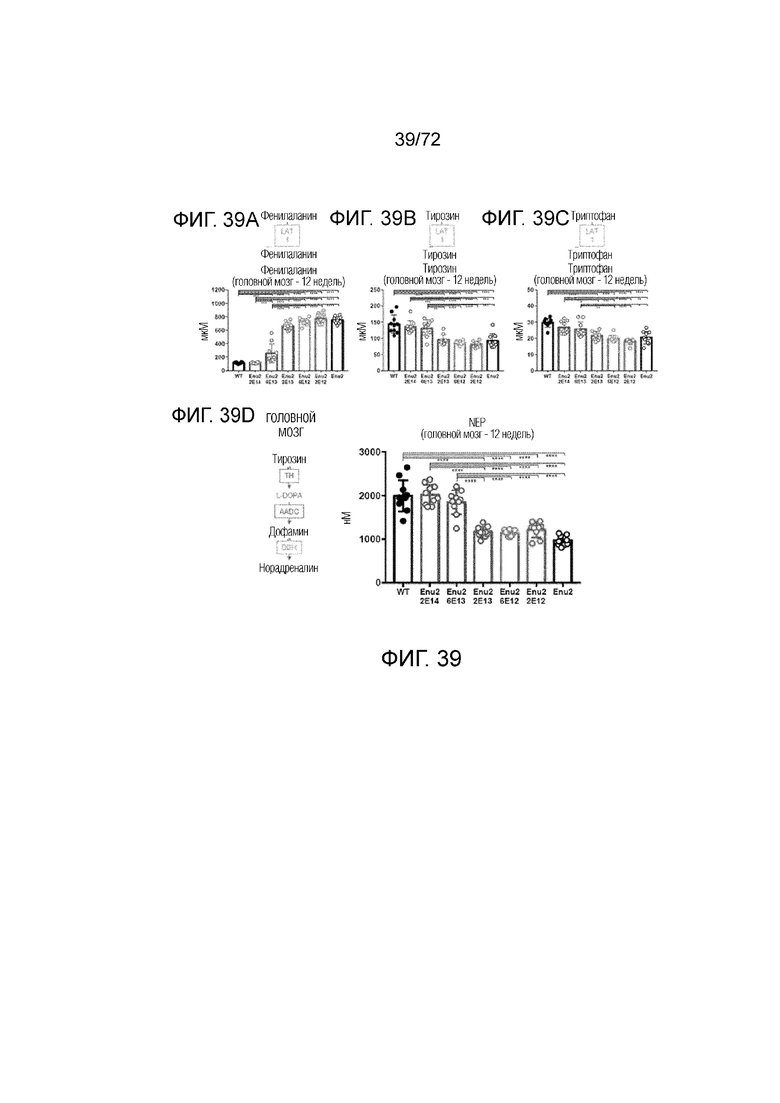
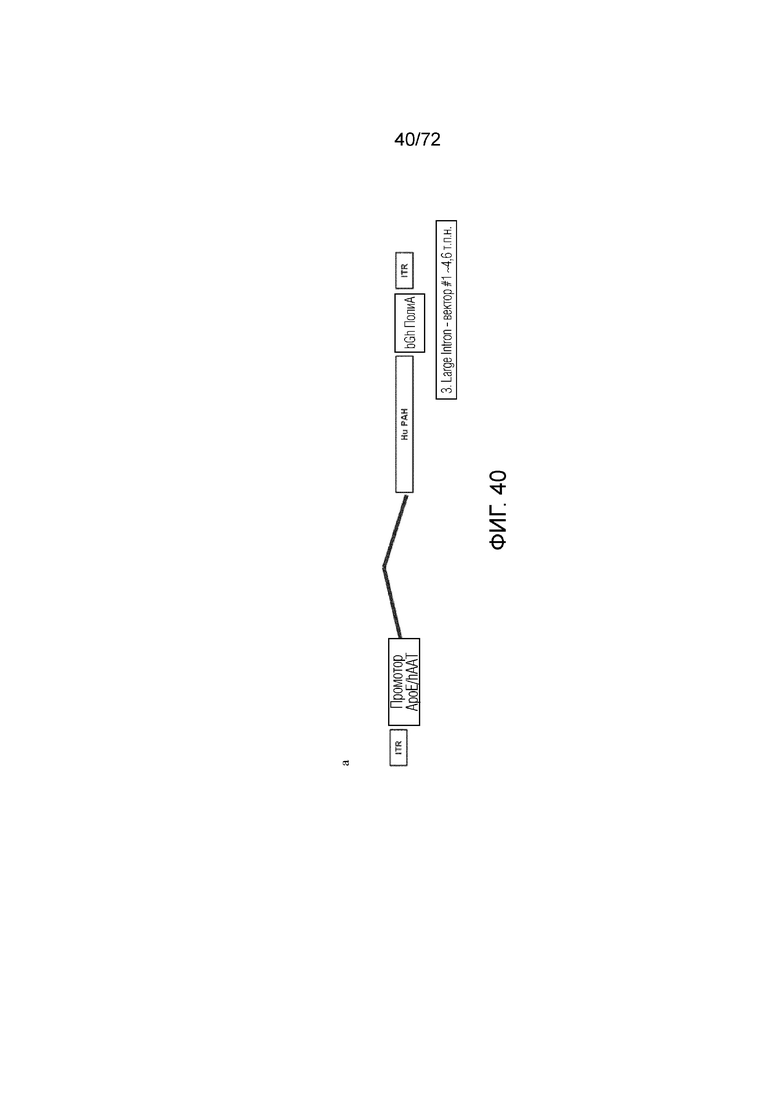





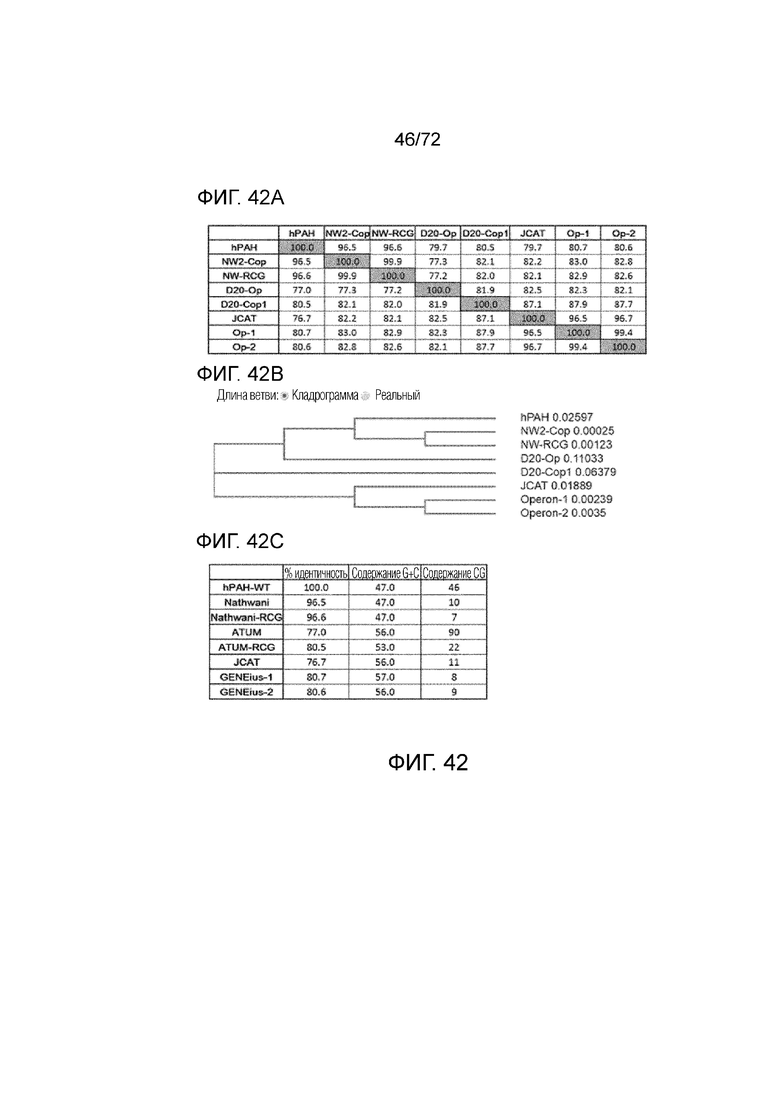
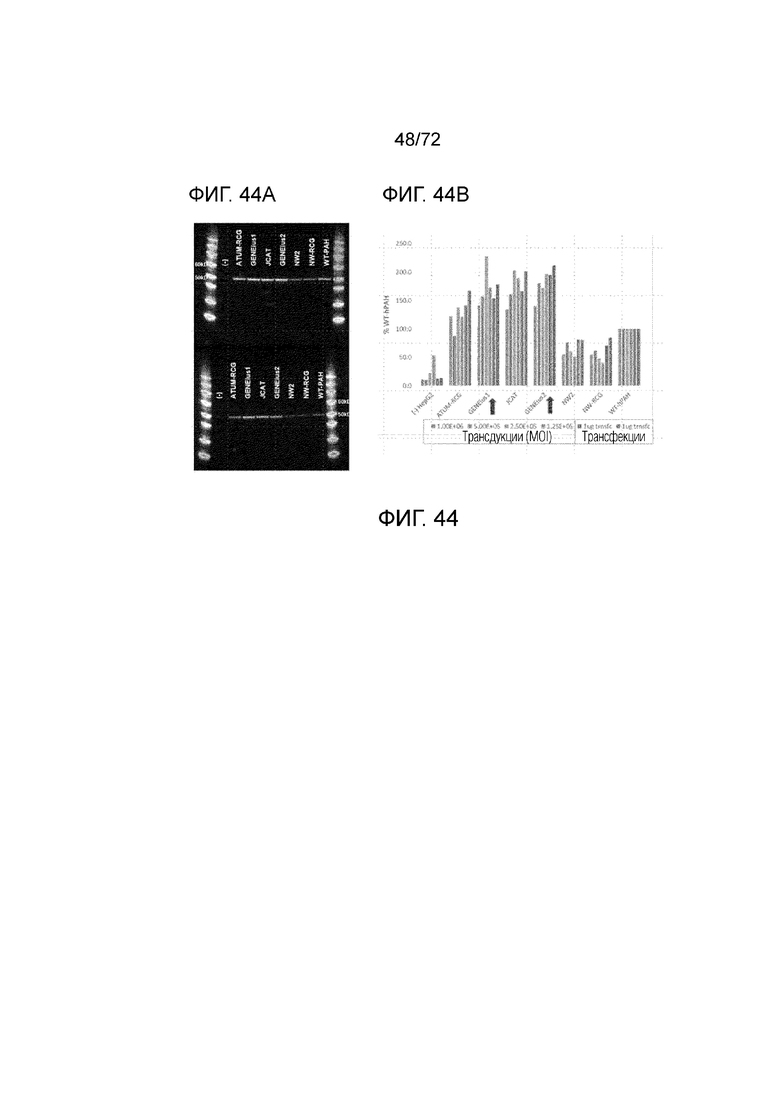
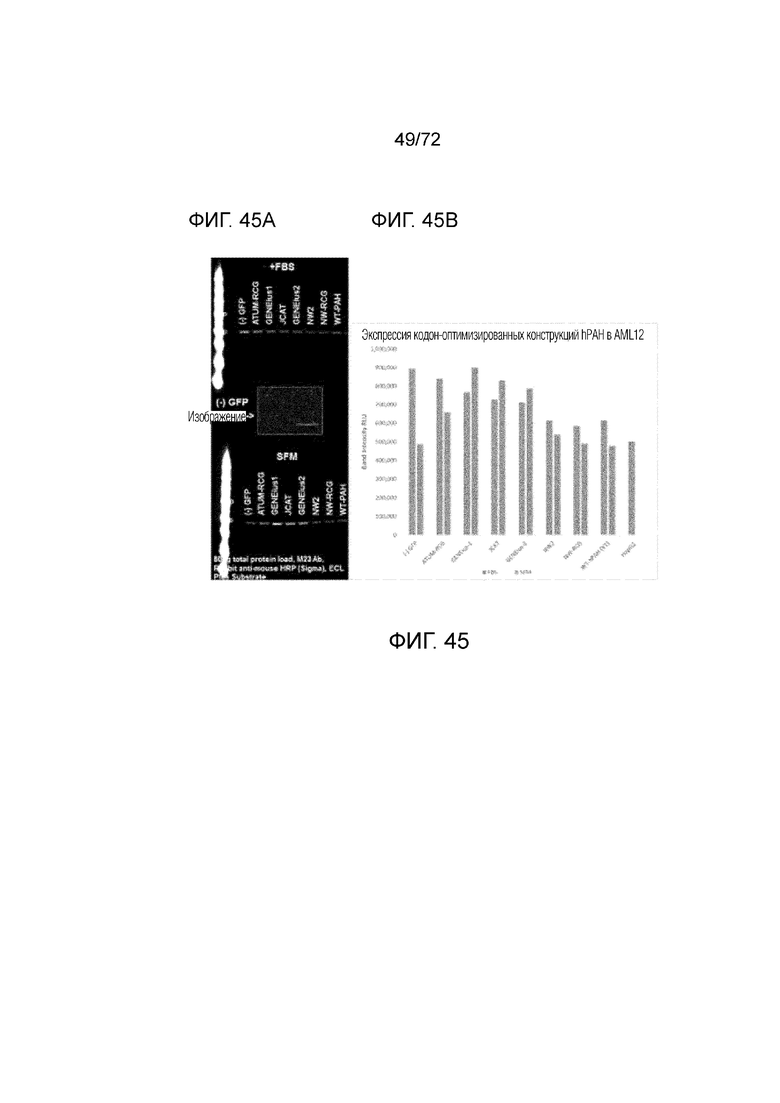
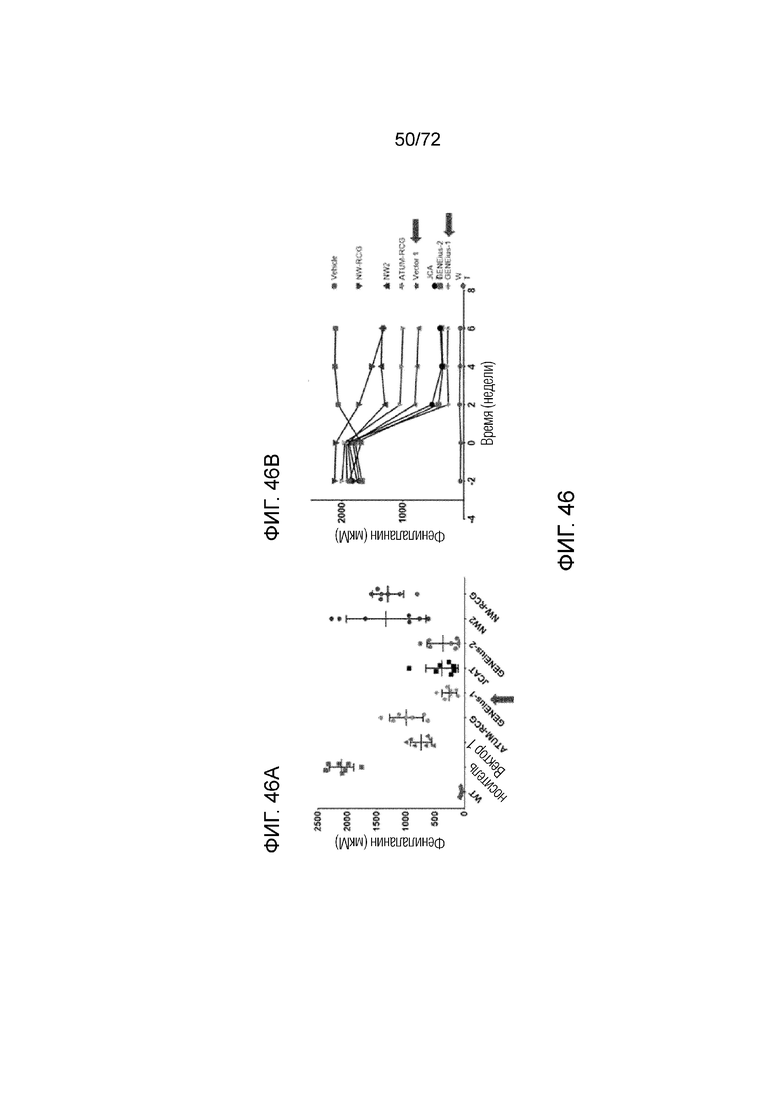
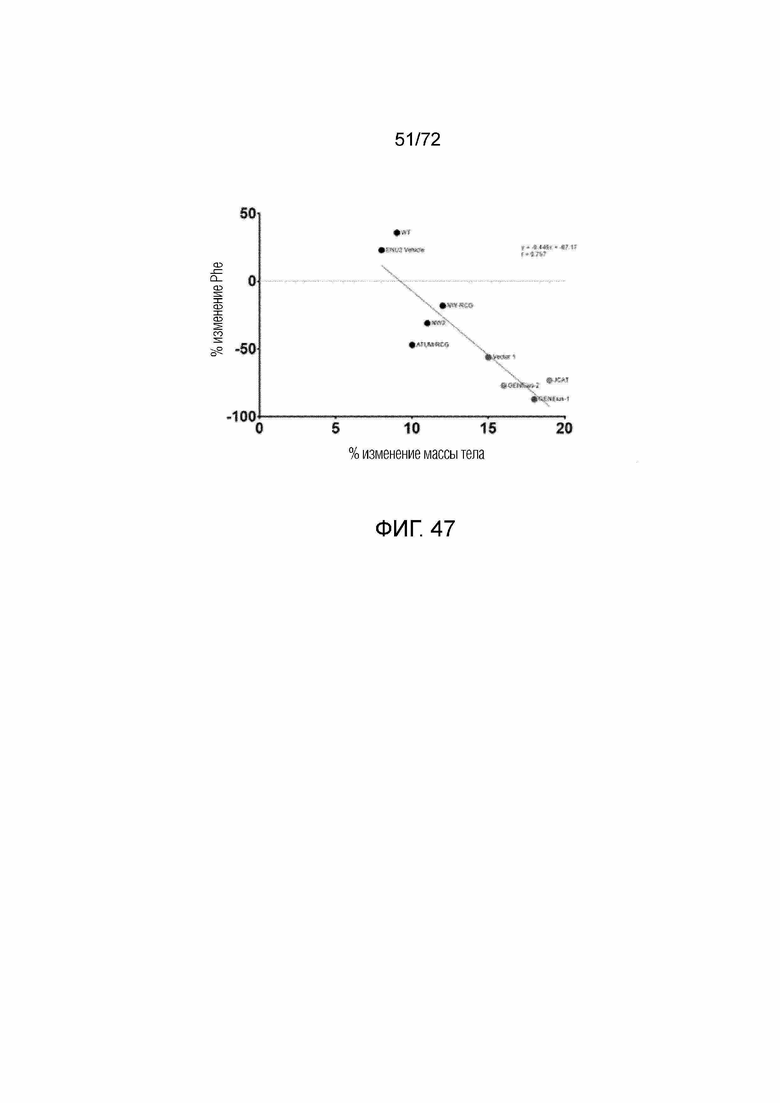

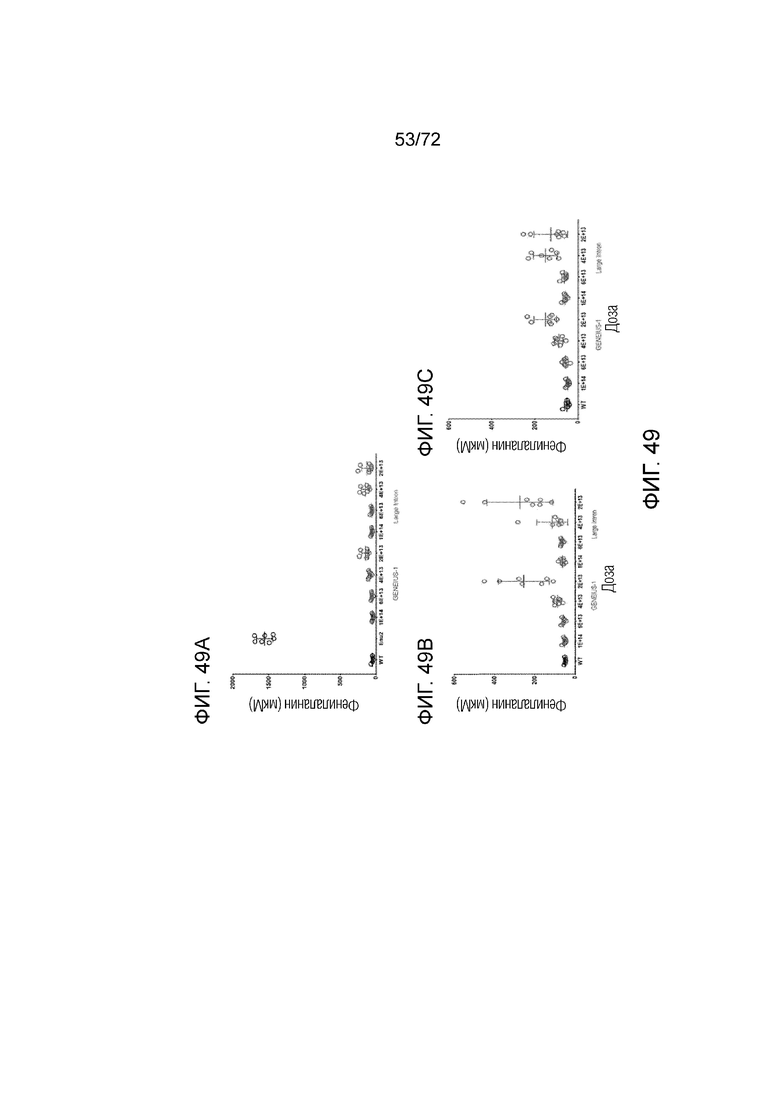
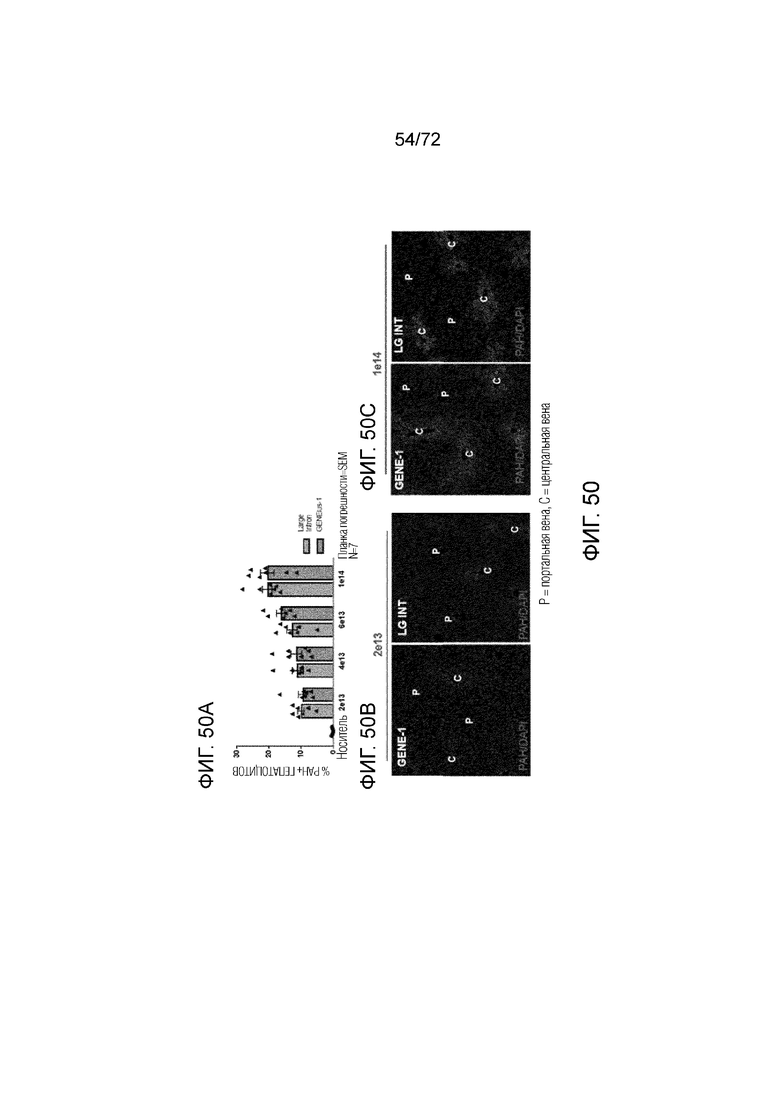
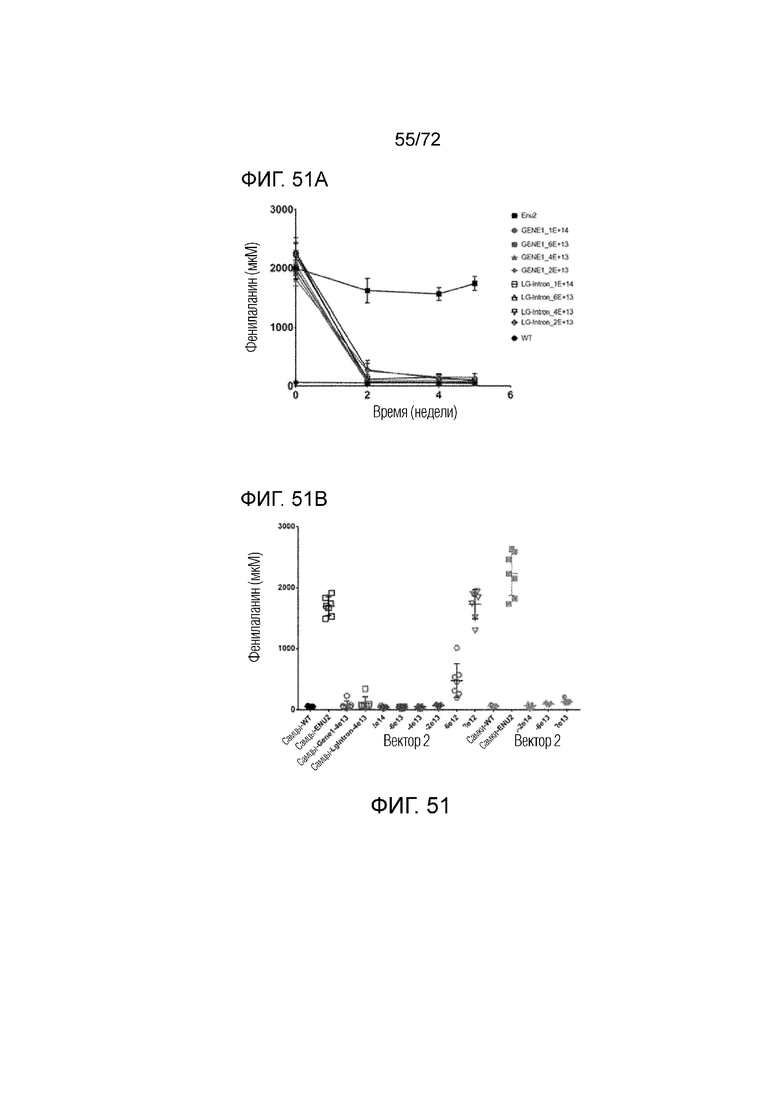
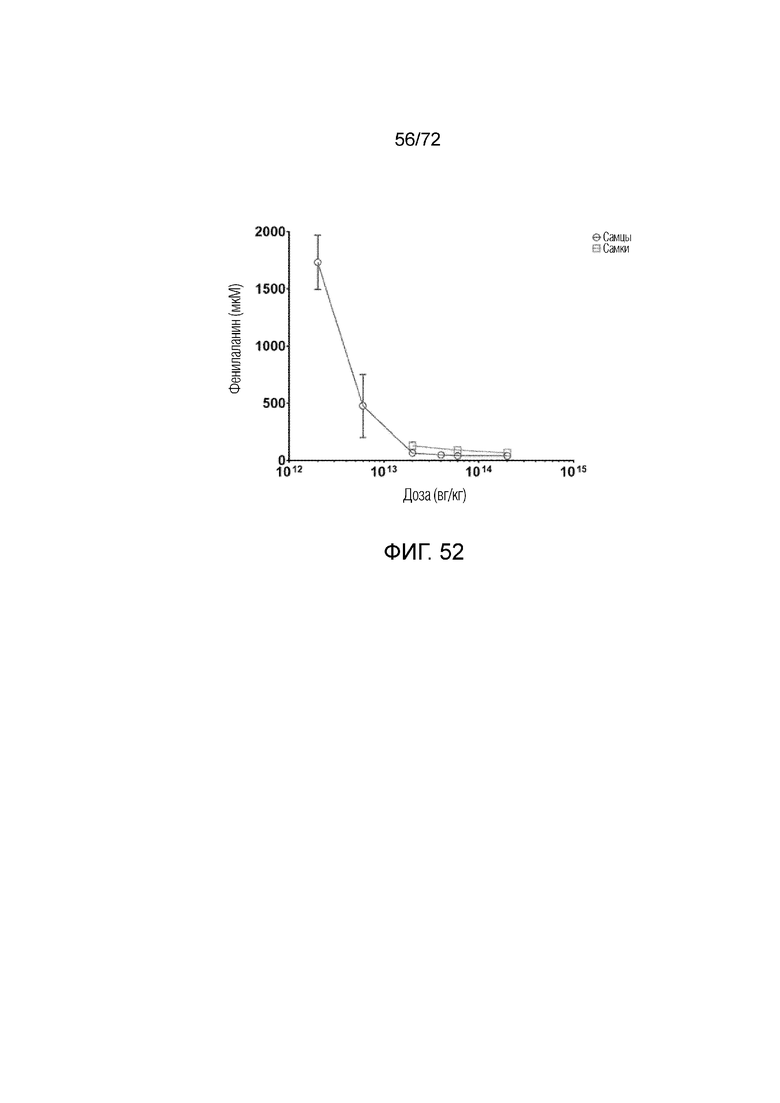
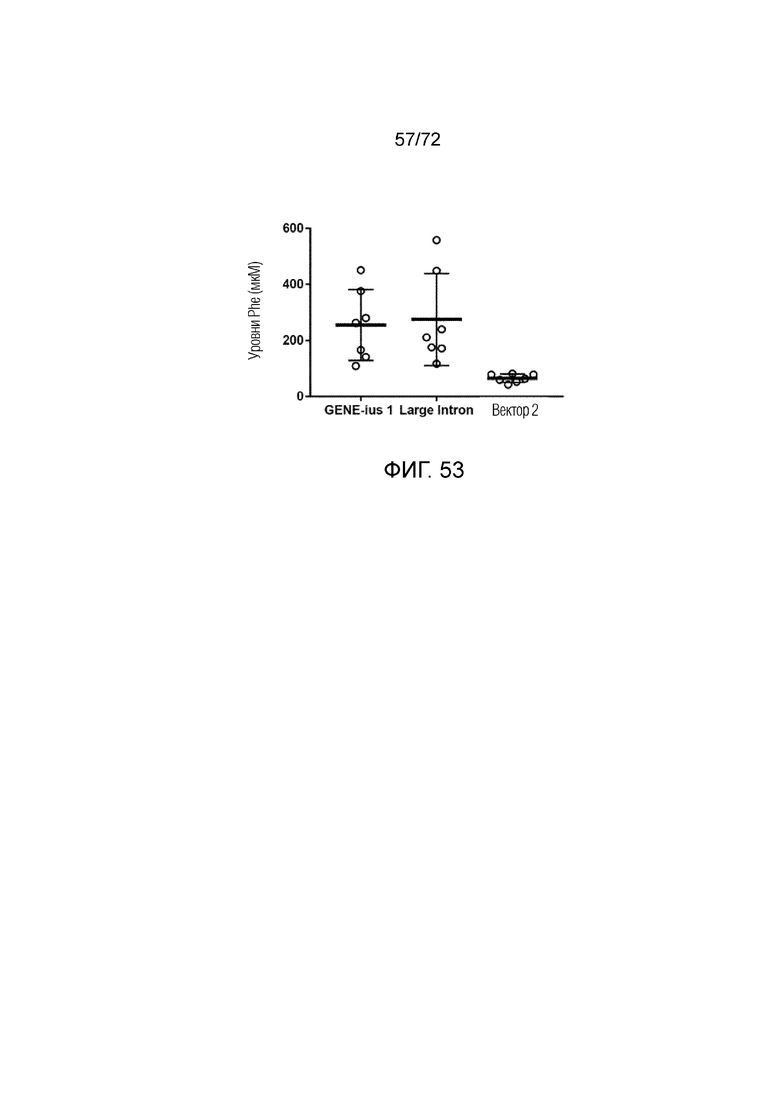
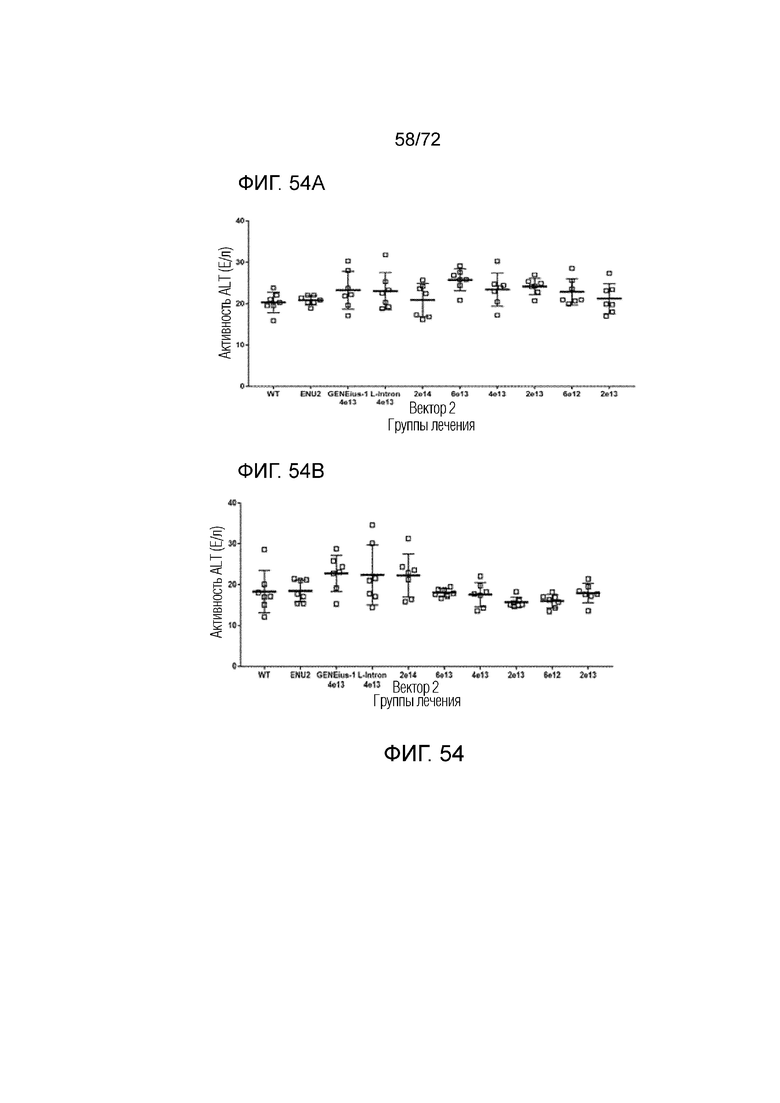
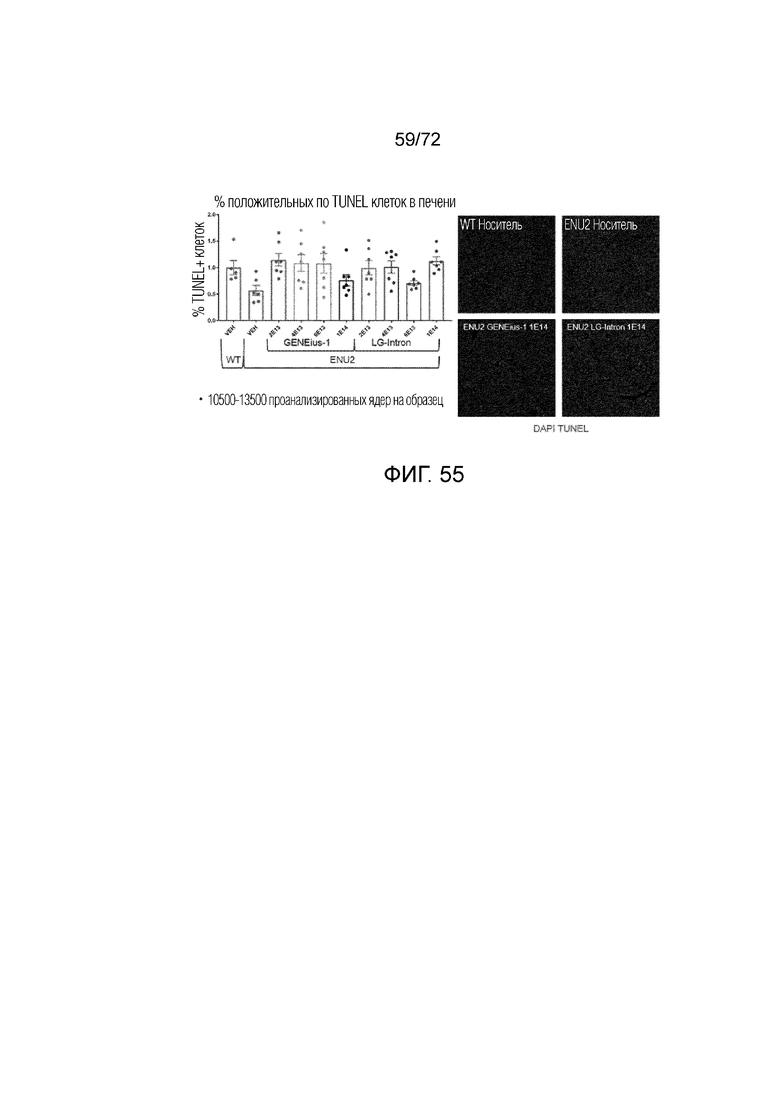
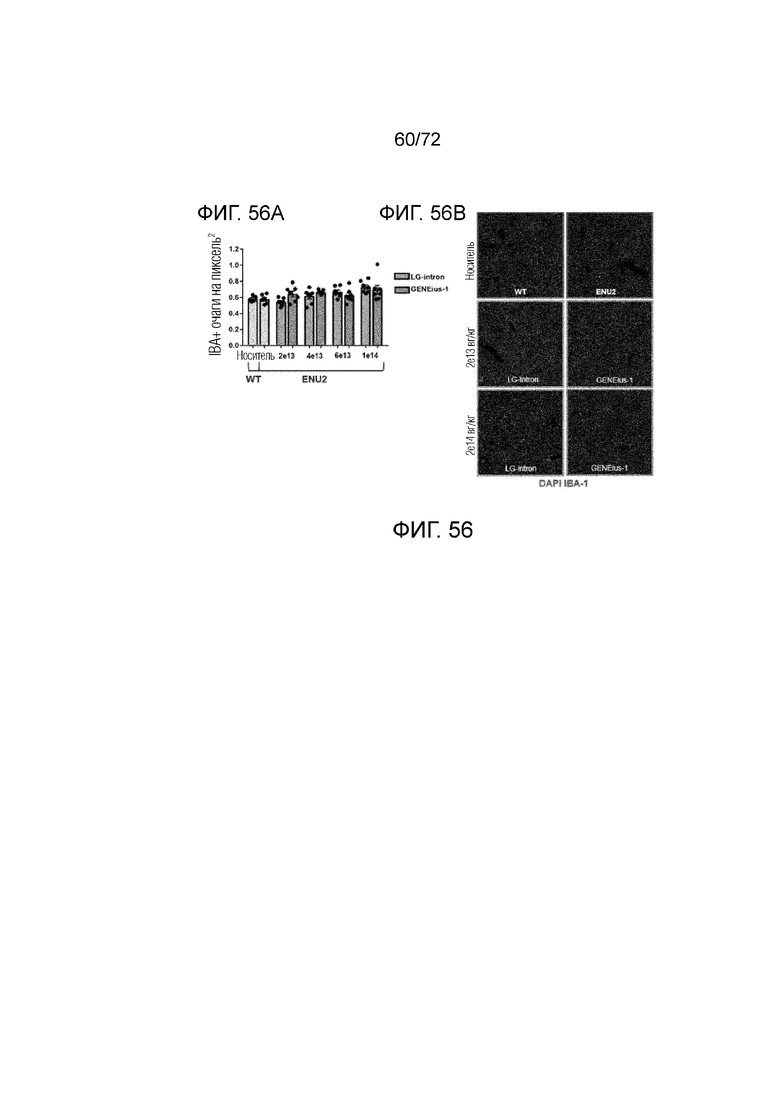
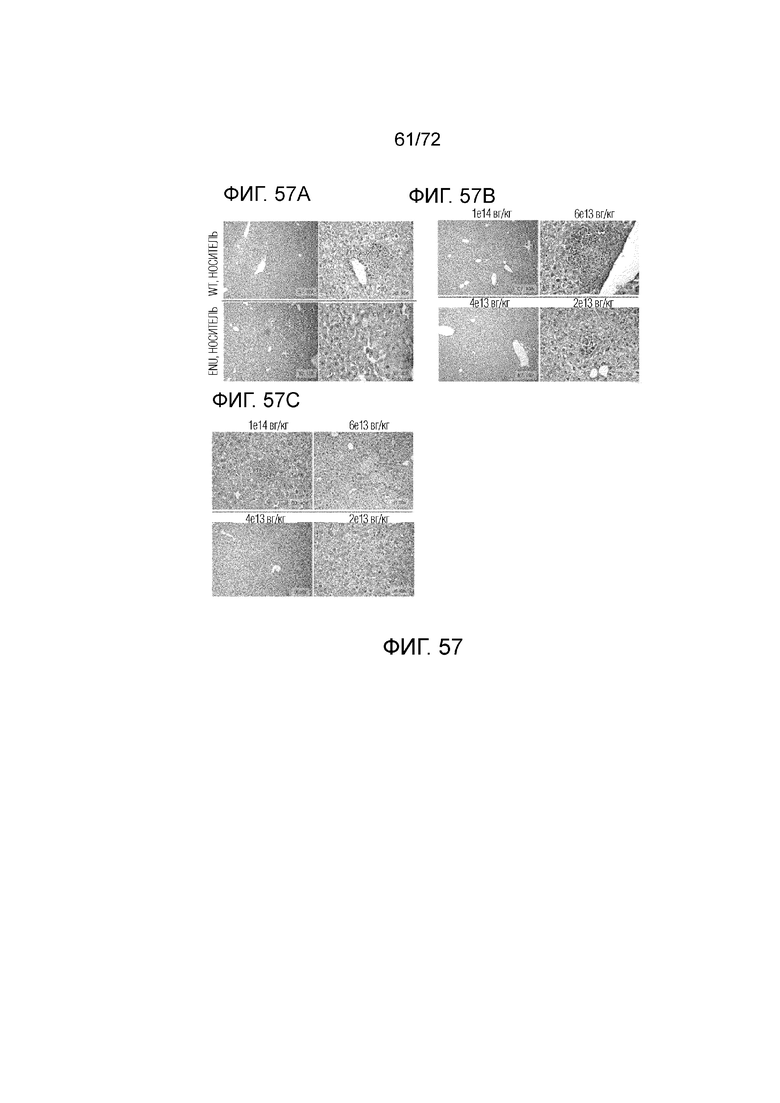
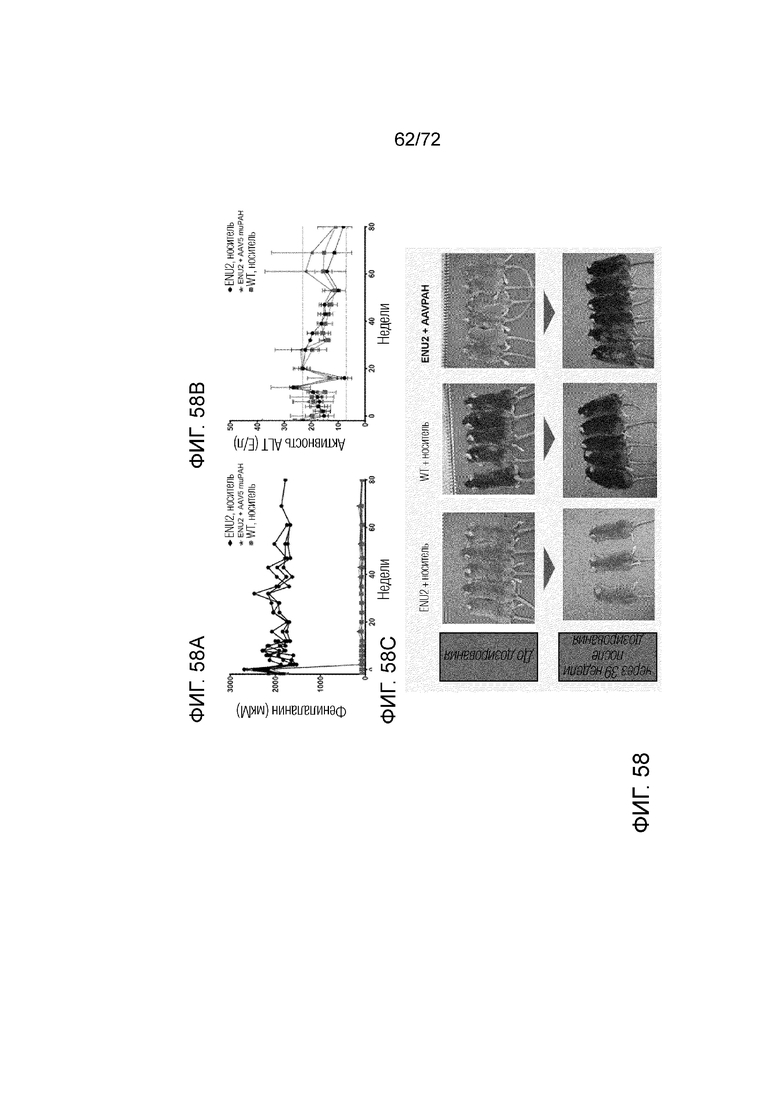
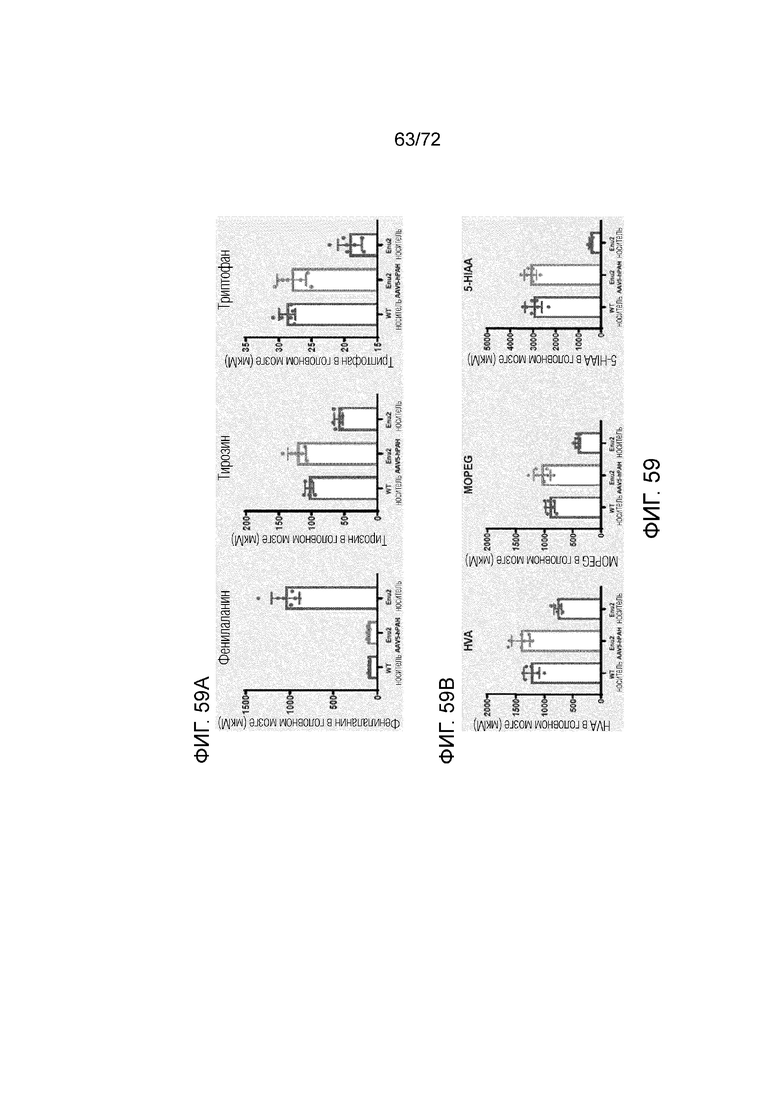
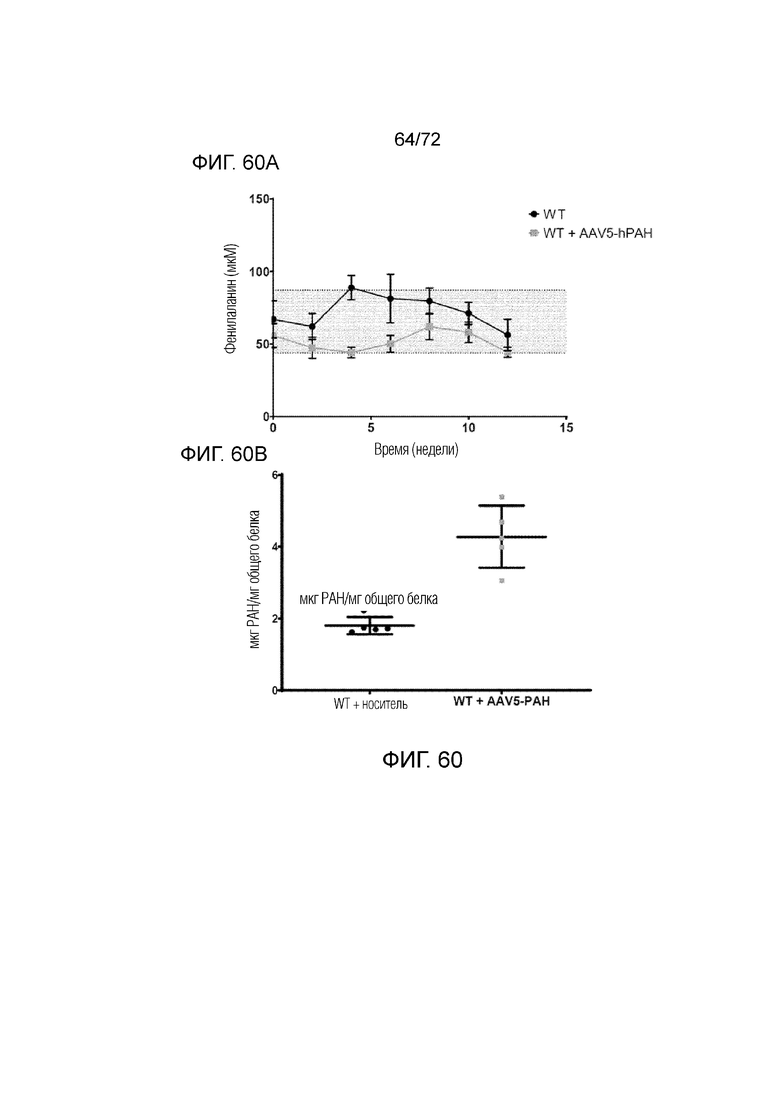
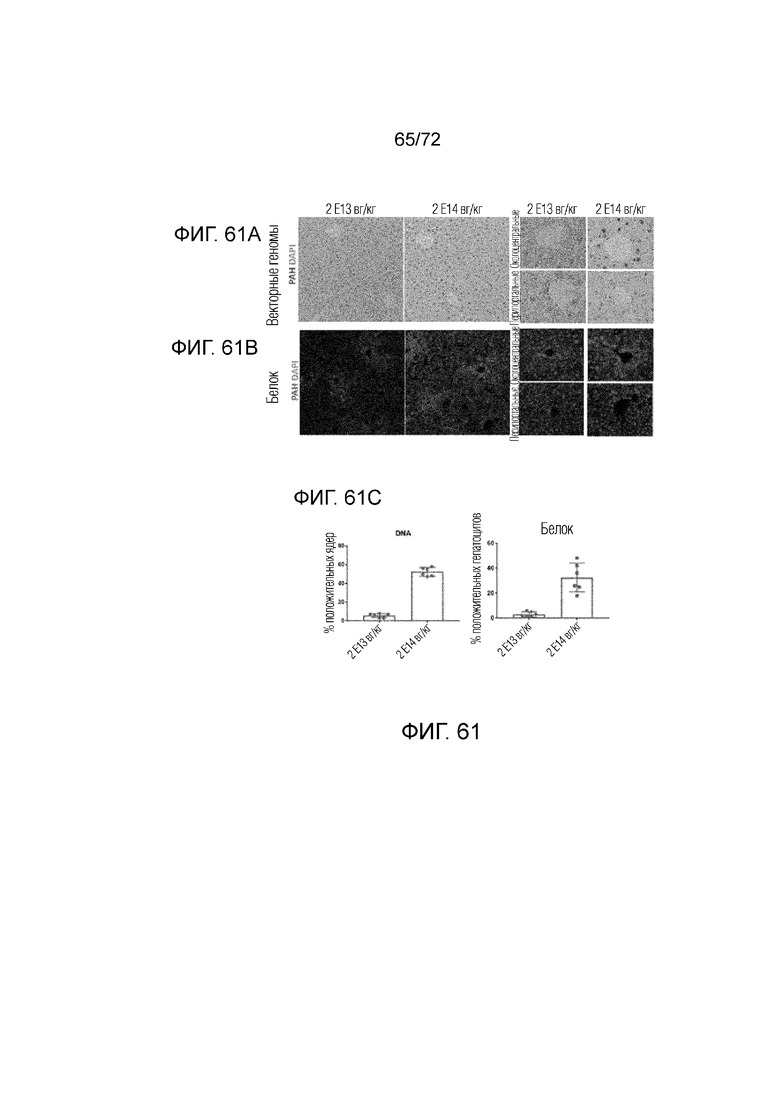
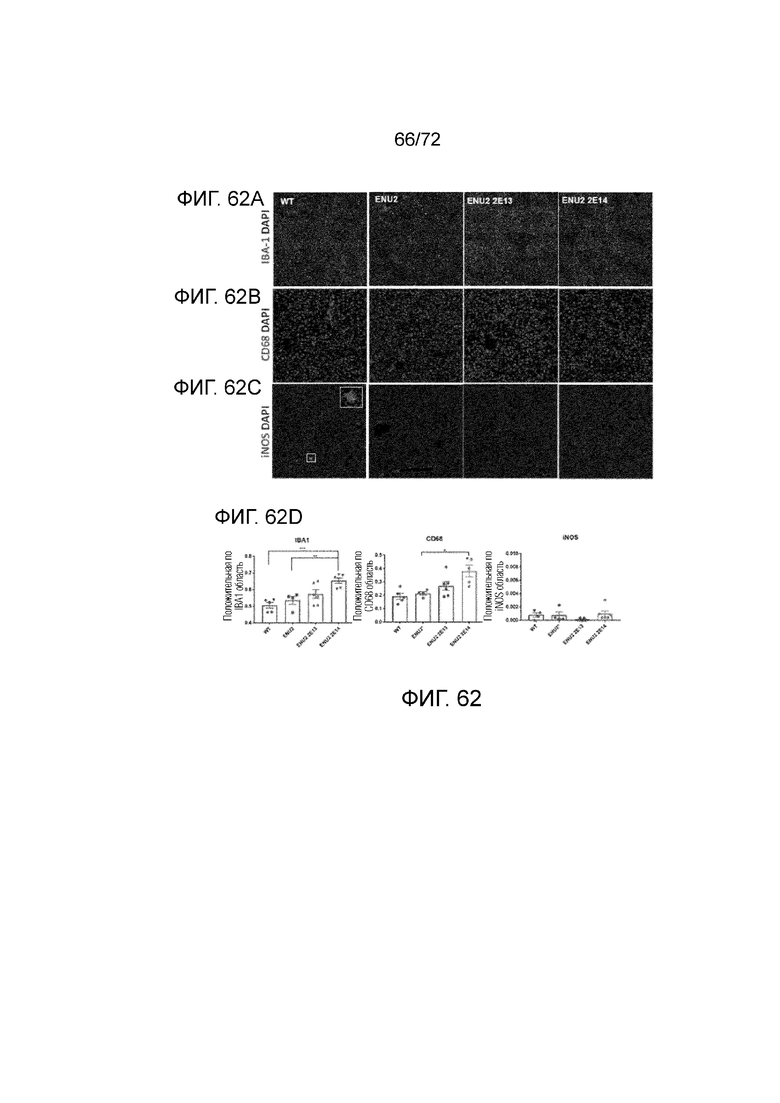

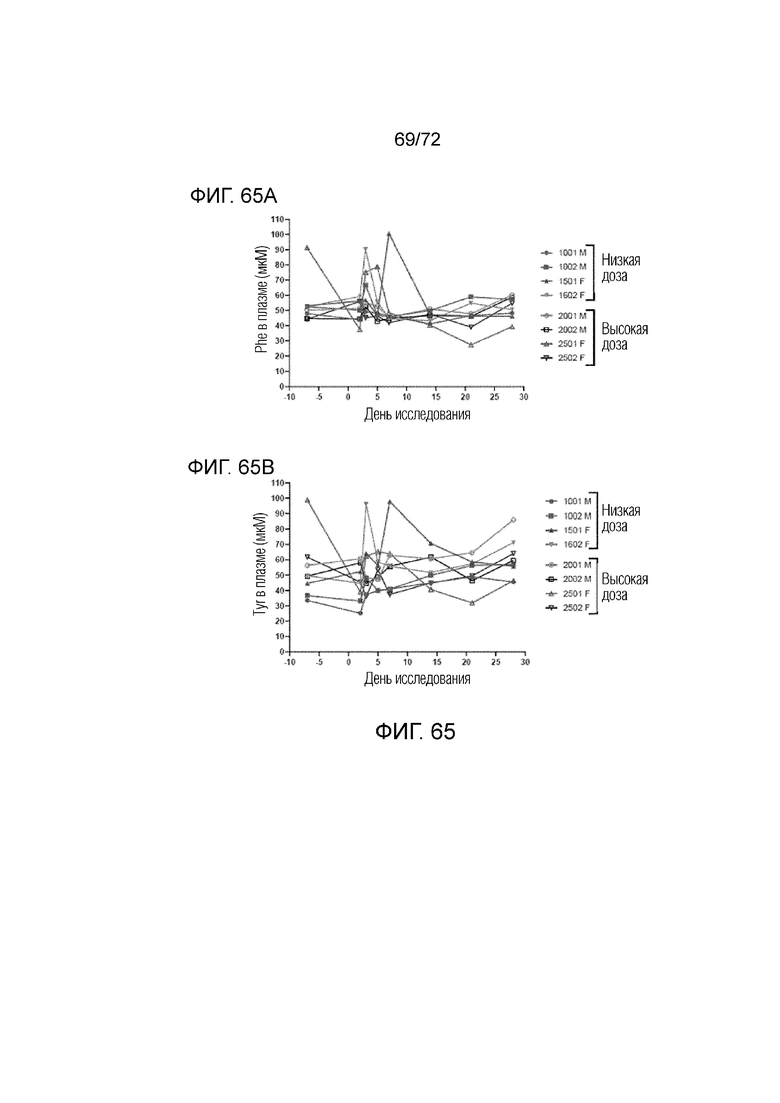
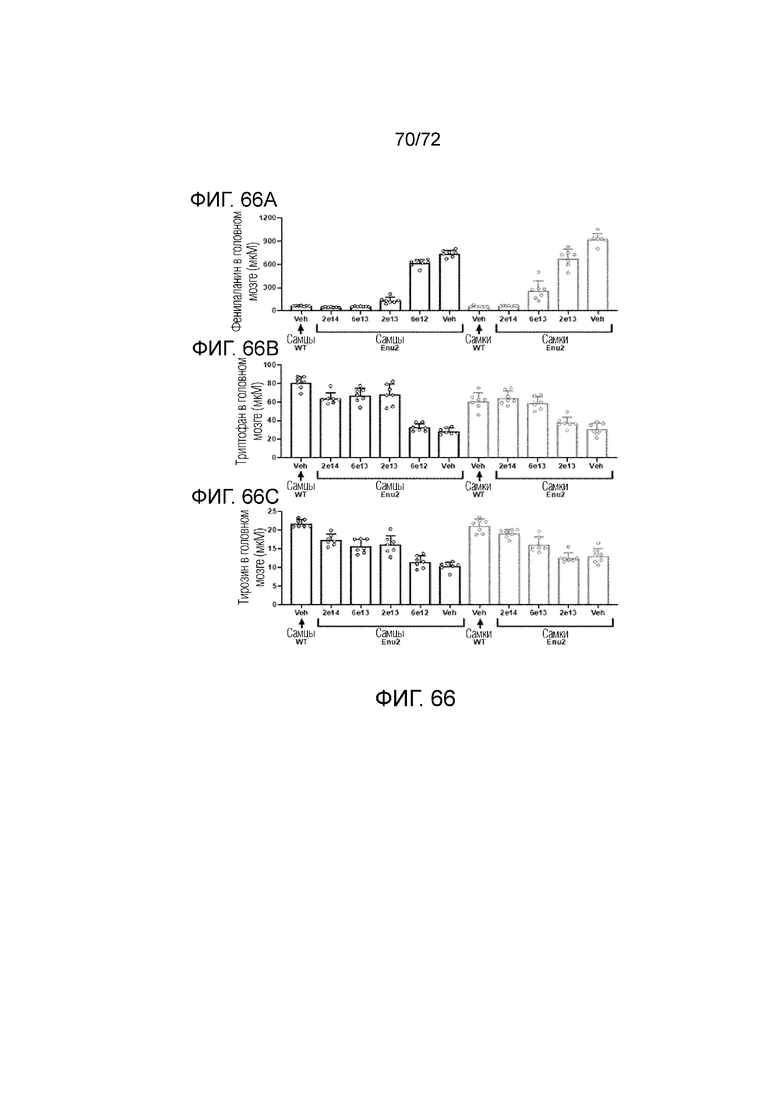
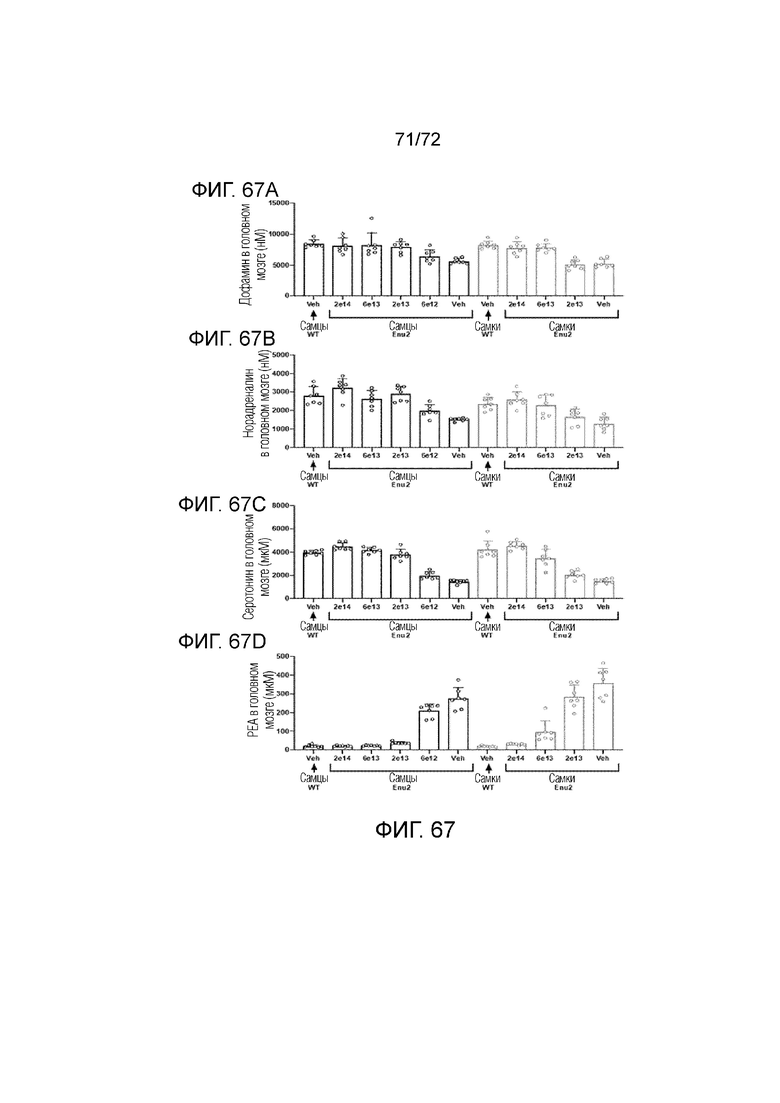
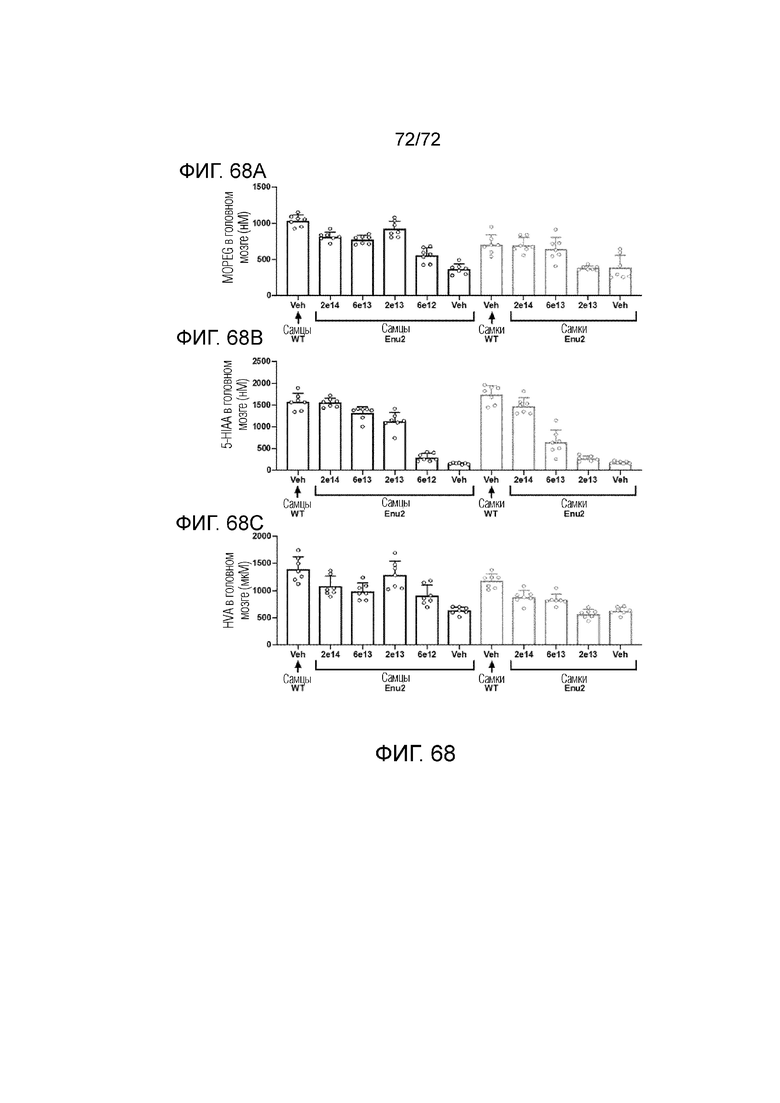
Изобретение относится к биотехнологии. Описан векторный геном на основе рекомбинантного аденоассоциированного вируса (rAAV) для лечения фенилкетонурии (ФКУ), содержащий: промотор; и кодон-оптимизированную последовательность, кодирующую фенилаланингидроксилазу человека (hPAH); причем кодон-оптимизированная последовательность содержит SEQ ID NO: 7; или ее вариант, имеющий по меньшей мере 95% идентичности последовательности с SEQ ID NO: 7. Представлена частица рекомбинантного аденоассоциированного вируса (rAAV) для лечения ФКУ, содержащая капсид AAV и описанный векторный геном. Также представлена фармацевтическая композиция для лечения ФКУ, содержащая векторный геном. Описан способ экспрессии белка у индивидуума, включающий введение индивидууму композиции, содержащей представленную частицу rAAV. Также описан способ лечения ФКУ у индивидуума, где указанный способ включает введение индивидууму описанного векторного генома rAAV, или представленной частицы, или соответствующей фармацевтической композиции. Также представлено применение векторного генома rAAV или частицы, указанной выше, для получении лекарственного средства для лечения ФКУ у индивидуума. Изобретение расширяет арсенал средств лечения фенилкетонурии посредством нормализации уровней аминокислот, нейротрансмиттеров и метаболитов нейротрансмиттеров у индивидуумов, имеющих фенилкетонурию. 12 н. и 37 з.п. ф-лы, 2 табл., 68 ил., 23 пр.
1. Векторный геном на основе рекомбинантного аденоассоциированного вируса (rAAV) для лечения фенилкетонурии (ФКУ), содержащий:
(a) промотор; и
(b) кодон-оптимизированную последовательность, кодирующую фенилаланингидроксилазу человека (hPAH);
причем кодон-оптимизированная последовательность содержит SEQ ID NO: 7; или ее вариант, имеющий по меньшей мере 95% идентичности последовательности с SEQ ID NO: 7.
2. Вектор rAAV по п.1, где кодон-оптимизированная последовательность содержит вариант, имеющий по меньшей мере 98% идентичности последовательности с SEQ ID NO: 7.
3. Вектор rAAV по п.1, где кодон-оптимизированная последовательность содержит SEQ ID NO: 7.
4. Вектор rAAV по п.1, где кодон-оптимизированная последовательность содержит SEQ ID NO: 8.
5. Вектор rAAV по п.1, где кодон-оптимизированная последовательность содержит SEQ ID NO: 13.
6. Вектор rAAV по любому из пп. 1-5, где промотор представляет собой специфичный для печени промотор.
7. Вектор rAAV по любому из пп. 1-5, где промотор содержит синтетический промотор, конститутивный промотор, регулируемый промотор или промотор, реагирующий на физиологические сигналы.
8. Вектор rAAV по п.6, где промотор представляет собой промотор тироксинсвязывающего глобулина (TBG), промотор транстиретина (TTR), промотор альбумина человека (humAlb), промотор, специфичный для печени (LSP), или основной промотор вируса гепатита В.
9. Вектор rAAV по любому из пп. 1-5, где промотор представляет собой последовательность синтетического промотора, содержащую части промотора hAAT, энхансер печеночной области контроля (HCR) и энхансер ApoE.
10. Вектор rAAV по любому из пп. 1-5, где промотор содержит SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6 или их вариант, имеющий по меньшей мере 80% идентичности последовательности с SEQ ID NO: NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6.
11. Вектор rAAV по п.10, где промотор содержит его вариант, последовательность которого по меньшей мере на 85% идентична SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6.
12. Вектор rAAV по п.10, где промотор содержит его вариант, последовательность которого по меньшей мере на 90% идентична SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6.
13. Вектор rAAV по п.10, где промотор содержит его вариант, последовательность которого по меньшей мере на 95% идентична SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6.
14. Вектор rAAV по п.10, где промотор содержит SEQ ID NO: 3, SEQ ID NO: 4 или SEQ ID NO: 6.
15. Вектор rAAV по любому из пп. 1-14, где геном вектора дополнительно содержит последовательность 5'-инвертированного концевого повтора (ITR) AAV или 3'-ITR AAV.
16. Вектор rAAV по любому из пп. 1-14, где геном вектора дополнительно содержит последовательность 5'-инвертированного концевого повтора (ITR) AAV и 3'-ITR AAV.
17. Вектор rAAV по п.15, где 5'-ITR AAV и/или 3'-ITR AAV происходят из AAV2.
18. Вектор rAAV по п.16, где 5'-ITR и 3'-ITR AAV происходят из AAV2.
19. Вектор rAAV по любому из пп. 1-18, где векторный геном дополнительно содержит сигнальную последовательность полиаденилирования.
20. Вектор rAAV по п.19, где сигнал полиаденилирования представляет собой последовательности сигнала полиаденилирования бычьего гормона роста (bGH) поли(А), позднего поли(А) SV40, поли(А) бета-глобина кролика (rBG), поли(А) тимидинкиназы (ТК) или любые их варианты.
21. Вектор rAAV по п.19, где сигнал полиаденилирования представляет собой сигнал полиаденилирования бычьего гормона роста (bGH).
22. Вектор rAAV по любому из пп. 1-21, где геном вектора дополнительно содержит интрон.
23. Вектор rAAV по п.22, где интрон представляет собой составную последовательность интрона глобина/A1AT.
24. Вектор rAAV по п.22, где интрон содержит SEQ ID NO: 14 или ее вариант, имеющий по меньшей мере 80% идентичности последовательности с SEQ ID NO: 14.
25. Вектор rAAV по п.24, где интрон содержит его вариант, последовательность которого по меньшей мере на 85% идентична SEQ ID NO: 14.
26. Вектор rAAV по п.24, где интрон содержит его вариант, имеющий по меньшей мере 90% идентичности последовательности с SEQ ID NO: 14.
27. Вектор rAAV по п.24, где интрон содержит его вариант, последовательность которого по меньшей мере на 95% идентична SEQ ID NO: 14.
28. Вектор rAAV по п.24, где последовательность интрона содержит SEQ ID NO: 14.
29. Вектор rAAV по любому из пп. 1-28, где векторный геном имеет размер от приблизительно 2 т.п.н. до приблизительно 3 т.п.н., от приблизительно 3 т.п.н. до приблизительно 4 т.п.н., или от приблизительно 4 т.п.н. до приблизительно 5 т.п.н.
30. Вектор rAAV по п.1, где последовательность векторного генома содержит SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21 или его вариант, имеющий по меньшей мере 80% идентичности последовательности с SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
31. Вектор rAAV по п.1, где последовательность генома вектора содержит SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
32. Вектор rAAV по п.1, где последовательность генома вектора содержит вариант, имеющий по меньшей мере 80% идентичности последовательности с SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
33. Вектор rAAV по п.1, где последовательность генома вектора содержит вариант, имеющий по меньшей мере 85% идентичности последовательности с SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
34. Вектор rAAV по п.1, где последовательность генома вектора содержит вариант, по меньшей мере на 90% идентичный SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
35. Вектор rAAV по п.1, где последовательность генома вектора содержит вариант, имеющий по меньшей мере 95% идентичности последовательности с SEQ ID NO: 16, SEQ ID NO: 18, SEQ ID NO: 20 или SEQ ID NO: 21.
36. Частица рекомбинантного аденоассоциированного вируса (rAAV) для лечения ФКУ, содержащая капсид AAV и векторный геном по любому из пп. 1-35.
37. Частица rAAV по п.36, где капсид AAV представляет собой капсид AAV5.
38. Фармацевтическая композиция для лечения ФКУ, содержащая векторный геном по любому из пп. 1-35.
39. Фармацевтическая композиция для лечения ФКУ, содержащая частицу rAAV по п.36 или 37 и фармацевтически приемлемый носитель.
40. Способ экспрессии белка у индивидуума, включающий введение индивидууму композиции, содержащей частицу rAAV по п.36 или 37, с обеспечением, тем самым, экспрессии кодируемого белка PAH в печени индивидуума.
41. Способ лечения ФКУ у индивидуума, где указанный способ включает введение индивидууму векторного генома rAAV по любому из пп. 1-35.
42. Способ лечения ФКУ у индивидуума, включающий введение индивидууму частицы rAAV по п.36 или 37.
43. Способ по п.42, где частицу rAAV доставляют в дозе от приблизительно 1×1012 до приблизительно 1×1015 г/кг в водной суспензии.
44. Способ лечения ФКУ у индивидуума, включающий введение индивидууму фармацевтической композиции по п.38 или 39.
45. Применение векторного генома rAAV по любому из пп. 1-35 в получении лекарственного средства для лечения ФКУ у индивидуума.
46. Применение частицы rAAV по п.36 или 37 в получении лекарственного средства для лечения ФКУ у индивидуума.
47. Применение частицы rAAV по п.36 или 37 в получении лекарственного средства для экспрессии hPAH у индивидуума.
48. Применение фармацевтической композиции по п.39 в получении лекарственного средства для экспрессии hPAH у индивидуума.
49. Способ по любому из пп. 40-44 или применение по любому из пп. 45-48, где индивидуумом является человек.
| WO 2003087383 A1, 23.10.2003 | |||
| 0 |
|
SU323618A1 | |
| 0 |
|
SU323618A1 | |
| Припой для пайки магниевых сплавов | 1957 |
|
SU111928A1 |
| Припой для пайки магниевых сплавов | 1957 |
|
SU111928A1 |
| 0 |
|
SU323618A1 | |
| 0 |
|
SU323618A1 | |
| RU 2015144234 A, 24.04.2017. | |||

