Изобретение относится к биотехнологии, биологии и медицине и предназначено для выявления ядерного белка, являющегося транскрипционным фактором, в неопластических и нормальных клетках миелоидного и лимфоидного происхождения иммуноферментным, иммуноцитохимическим, иммуногистохимическим и иммунофлуоресцентным методами, а также методом иммуноблоттинга для научно-исследовательских и клинико-лабораторных целей.
На сегодняшний день антитела являются мощным инструментом для изучения фундаментальных вопросов, направленных на выяснение функций белков человека в норме и при патологиях.
Как известно, иммуноцитохимические методы анализа с использованием антител к известным белкам-маркерам биоптата опухоли позволяют классифицировать тип новообразований и оценить степень злокачественности (М.А. Пальцев, Н.М. Аничков. Патологическая анатомия. - М.: Медицина, 2001). Иммуногистохимическая окраска моноклональными антителами клеточных белков в сочетании с иммунопероксидазным методом позволяет не только оценивать опухоли низкой степени дифференцировки, опухоли неизвестного происхождения, более точно различать опухоли различного тканевого происхождения (Клиническая онкология: учебное пособие / под ред. П.Г. Брюсова, П.Н. Зубаревой. - СПб.: СпецЛит, 2012. - 455 с.: ил.), но и позволяет выбрать соответствующие методы лечения. Антитела к белкам, служащим специфическими онкомаркерами клеток, повсеместно применяются для клинической оценки прогноза опухолевой прогрессии (Lea P., Ling M. "New molecular assays for cancer diagnosis and targeted therapy". Curr. Opin. Mol. Ther. 2008,10: 251-259).
PU.1 относится к семейству транскрипционных факторов E26, необходимых для развития иммунной системы и кодируется геном Spi1. PU.1 участвует в гемопоэзе. Его экспрессия определяет развитие лимфоидных и миелоидных клеток, в особенности макофагов, гранулоцитов, дендритных клеток, а также играет важную роль в дифференцировке иммунных клеток по лимфоидному типу. Высокий уровень экспрессии PU.1 характерен для миелоидных клеток (Heinz S, Benner C, Spann N, Bertolino E, Lin YC, Laslo P, et al. Simple combinations of lineage-determining transcription factors prime cis-regulatory elements required for macrophage and B cell identities. Mol Cell (2010) 38(4):576-89), в то время как B-клетки экспрессируют его в меньшем количестве (Pang SHM, de Graaf CA, Hilton DJ, Huntington ND, Carotta S, Wu L, Nutt SL. PU.1 Is Required for the Developmental Progression of Multipotent Progenitors to Common Lymphoid Progenitors. Front Immunol. 2018 Jun 11;9:1264). Несколько более низкий уровень его экспрессии наблюдается в гранулоцитах, мегакариоцитах, тучных клетках и незрелых клетках эритроидного ряда. Зрелые Т-лимфоциты PU.1 не экспрессируют.
В онкологии PU.1 используется в основном для дифференциальной диагностики классической лимфомы Ходжкина (отсутствие экспрессии PU.1) от узловой лимфомы Ходжкина с лимфоидным преобладанием (присутствие экспрессии PU.1). Также высокий уровень экспресии PU.1 наблюдаются в фолликулярной лимфоме, лимфоме мантийной зоны, B-клеточной лимфоме маргинальной зоны, в малой лимфоцитарной лимфоме, гистиоцитарной / дендритно-клеточной саркоме (Shao H, Xi L, Raffeld M, Feldman AL, Ketterling RP, Knudson R, Rodriguez-Canales J, Hanson J, Pittaluga S, Jaffe ES. Clonally related histiocytic/dendritic cell sarcoma and chronic lymphocytic leukemia/small lymphocytic lymphoma: a study of seven cases. Mod Pathol. 2011 Nov;24(11):1421-32).
Для выявления белка PU.1 у человека на сегодняшний день существует несколько доступных коммерческих как поликлональных, так и моноклональных антител. Известные мышиные гибридомы, продуцирующие моноклональные антитела к ядерному антигену PU.1, были получены при иммунизации мышей полноразмерным рекомбинантным белком PU.1 человека (коммерчески доступные клоны 9G7, E8I8L, MA5-15064); различными рекомбинантными полипептидными участками белка PU.1 человека (коммерчески доступный клон E388.3).
К недостаткам описанных клонов относятся дороговизна моноклональных антител, продуцируемых данными гибридомами, ввиду их зарубежного происхождения, а также отсутствие для некоторых из них данных о тестировании в реакциях иммуногистохимии, иммуноцитохимии, иммунофлуоресценции, использующихся в ежедневной клинико-диагностической практике, а также иммуноблоттинге и иммуноферментном анализе.
Задачей изобретения является расширение ассортимента моноклональных антител, обладающих специфичностью к белку PU.1 человека.
Поставленная задача решается за счет мышиной гибридомы PU.1, клон 4G6, с регистрационным номером ВКПМ: H-187 от 22 сентября 2021 года, - продуцента моноклонального антитела, выявляющего белок PU.1 методами иммуноферментного анализа, иммуноцитохимии, иммуногистохимии, иммунофлуоресценции и иммуноблоттинга, полученной путем иммунизации мышей линии Balb/c рекомбинантым белком, и слиянием сенсибилизированных лимфоцитов и спленоцитов иммунизированных мышей с клетками мышиной миеломы линии sp2/0 с помощью полиэтиленгликоля с молекулярной массой 1450.
Продуцируемые гибридомой 4G6 моноклональные антитела к белку PU.1 человека обладают селективной способностью связывать белок PU.1 человека на иммуноблотах (Фиг.5), на клеточном (Фиг.2, Фиг.3, Фиг. 4) и тканевом уровнях (на парафиновых срезах) (Фиг.1) и расширяют существующий доступный ассортимент моноклональных антител к белку PU.1. Также следует отметить, что каждая вновь полученная гибридома уникальна. Моноклональные антитела, продуцируемые разными гибридомами, различаются по своей первичной структуре, по специфичности к различным антигенным детерминантам, аффинности и другим свойствам. Поэтому получение как можно большего количества моноклональных антител к белку PU.1 человека важно с научной и практической точки зрения.
Технический результат изобретения заключается в получении линии гибридных культивируемых клеток мыши 4G6, продуцирующей доступные для отечественных клинико-диагностических лабораторий моноклональные антитела к ядерному антигену PU.1 человека для иммуноферментного анализа, иммуноцитохимии, иммуногистохимии, иммуноблоттинга и иммунофлуоресценции, позволяющие использоваться для дифференциальной диагностики лимфом.
Мышиную гибридому PU.1, клон 4G6 получали следующим образом:
Для получения бактериального штамма, экспрессирующего рекомбинантный белок PU.1, последовательность, кодирующая 264 аминокислоты (CDS 7…270) изоформы 2 белка PU.1 человека, была клонирована в вектор pET45b(+). Ген SPI1 человека (нуклеотидная последовательность мРНК SPI1 человека доступна в базе данных GeneBank по адресу http://www.ncbi.nlm.nih.gov/genbank/ с порядковым номером NM_003120.3) амплифицируют с использованием в качестве матрицы плазмиду pINDUCER-21-SPI1 (http://www.addgene.org/97039/) в присутствии ген-специфических праймеров (SPI1_F3147_SalI: AAGGGTCGACATGGAAGGGTTTCCCCT, SPI1_R3130_HindIII: AGAGAAGCTTTTAGTGGGGCGGGTGGCGCCGCT; встроенные сайты эндонуклеаз рестрикции SalI и HindIII). Амплифицированные фрагменты ДНК очищали и клонировали в вектор pET45b(+) по сайтам, созданным эндонуклеазами рестрикции. В результате была получена плазмида, несущая последовательность белка PU.1 человека. Плазмиду тестировали на наличие нуклеотидных замен методом автоматического секвенирования ДНК. Компетентные клетки E. coli штамм BL21 DE3 химически трансформировали полученной плазмидой pET45b(+) SPI1, высевали на чашки с LB-агаром, содержащим 50 мкг/мл ампициллина, и инкубировали при 37°С в течение 16 ч. Далее в 20 мл среды LB переносили единичную колонию и наращивали при 37°С в течение 16 ч при постоянном перемешивании. Затем ночную культуру добавляли к 200 мл среды Terrific, содержащей лактозу, и инкубировали в течение 8 часов при 37°C. Стимуляция экспрессии белка PU.1 в бактериях E.coli BL21 DE3 происходила за счёт присутствия в среде галактозы. По истечении времени стимуляции клетки бактерий осаждали, выдерживали при -20°С в течение 16 ч, лизировали, сонифицировали с помощью ультразвука и подвергали процедуре очистки.
Для лизиса бактериальных клеток был использован буфер следующего состава (pH 8,0): мочевина - 8 М, NaH2PO4 - 100 мМ, трис - 10 мМ, имидазол - 20 мМ, бета-меркаптоэтанол - 20 мМ, глицин - 50 мМ, Tween 20 - 0,5%. Лизис осуществляли в течение 1 часа при комнатной температуре. Раствор центрифугировали при 14000 × g в течение 30 минут при +20°С. Дальнейшую очистку белков производили метал-хелатной хроматографией (IMAC) на колонке Ni-NTA Fast Start (Qiagen, 30600). Белок наносили на колонку Ni-NTA Fast Start, уравновешенную буфером, приведенного выше состава. Колонку промывали ступенчатым градиентом pH. Для первой промывки был использован буфер следующего состава (pH 7,0): мочевина - 8 М, NaH2PO4 - 100 мМ, трис - 10 мМ, имидазол - 20 мМ, глицин - 50 мМ, Tween 20 - 0,5%. Для второй промывки был использован буфер следующего состава (pH 6,8): мочевина - 8 М, NaH2PO4 - 100 мМ, трис - 10 мМ, глицин - 50 мМ. Эффективная элюция белка PU.1 с минимальными примесями происходит буфером, приведенного выше состава, при pH 4,8. Полученный в результате элюции белок PU.1 был дополнительно очищен от примесных бактериальных белков на колонке Superdex 200. Собранные фракции проверены методом электрофореза на 10% полиакриламидном геле (ПААГ). Наиболее чистая белковая фракция PU.1 была использована в качестве антигена для дальнейшей иммунизации мышей.
Первичную иммунизацию проводили мышам Balb/c (самкам, 20 гр), вводя около 30 мкг антигена (фракционированного белка PU.1) в виде эмульсии с полным адъювантом Фрейнда (1:1) в подушечки задних лап и в холку. Бустирование антигеном с неполным адъювантом Фрейнда (1:1) проводили через две недели после первичной иммунизации. Гибридизацию (слияние) иммунных спленоцитов и лимфоцитов, полученных из селезенки и лимфоузлов мыши, соответственно, с клеточной линией Sp 2/0 делали на третий день после бустирования в присутствии 50% полиэтиленгликоля (PEG 1450). В среду для посадки гибридом (RPMI-1640 с 15% FetalClone I) добавляли HAT (гипоксантин, аминоптерин, тимидин) для удаления неслившихся миеломных клеток, в соответствии с рекомендациями производителя. Полученные гибридомы рассадили на четыре 96-луночных планшета по 150 мкл суспензии в лунку и на 10-й день кондиционированные среды протестировали методом иммуноферментного анализа (ИФА) на фракционированном антигене PU.1. В результате 25 первичных гибридомных клонов, продуцирующих антитела, связывающие антиген, отсадили в лунки 12-луночных планшетов в среду RPMI-1640, 15% FetalClone I, с добавлением HT (гипоксантина, тимидина). По результатам ИФА на неродственном антигене, содержащим His-tag элиминировали 6 клонов. Кондиционированную среду с оставшихся 19 первичных клонов тестировали методом иммуногистохимии (ИГХ) на срезах миндалины. Четыре первичных клона, продуцирующих антитела, специфично связывающихся с тканью миндалин в ИГХ клонировали на мышином фидере из селезенки: 2A10, 1C10, 4E6, 4G6. Субклоны клонов 4G6 и 1С10 выбрали как клоны с наиболее специфично работающими антителами. Полученные антитела фракционировали на Superose 6. Собранные фракции для каждого клона гибридом тестировали в ИФА, в серийных разведениях начиная от 1:10 на исходном бактериальном антигене и после объединяли. Полученные антитела к PU.1 обоих клонов 4G6 и 1С10 тестировали методами иммуноферментного анализа, в результате которого для дальнейшей работы оставлен клон 4G6. Антитела клона 4G6 проверены методами иммуногистохимии на миндалине и на клеточных блоках лимфобластоподобных клеток Raji, иммуноцитохимии и иммунофлуоресценции на моноцитарной клеточной линии THP1, а также методом иммуноблоттинга. Антитела клона 4G6 показали специфичное связывание с антигеном во всех этих методах.
В иммуноферментном анализе (Таб. 1) антитела 4G6 были проверены как на самом антитене (PU.1) так и на неродственном рекомбинантном белке, содержащим His-tag. Перекрестной реакции на His-tag не обнаружено.
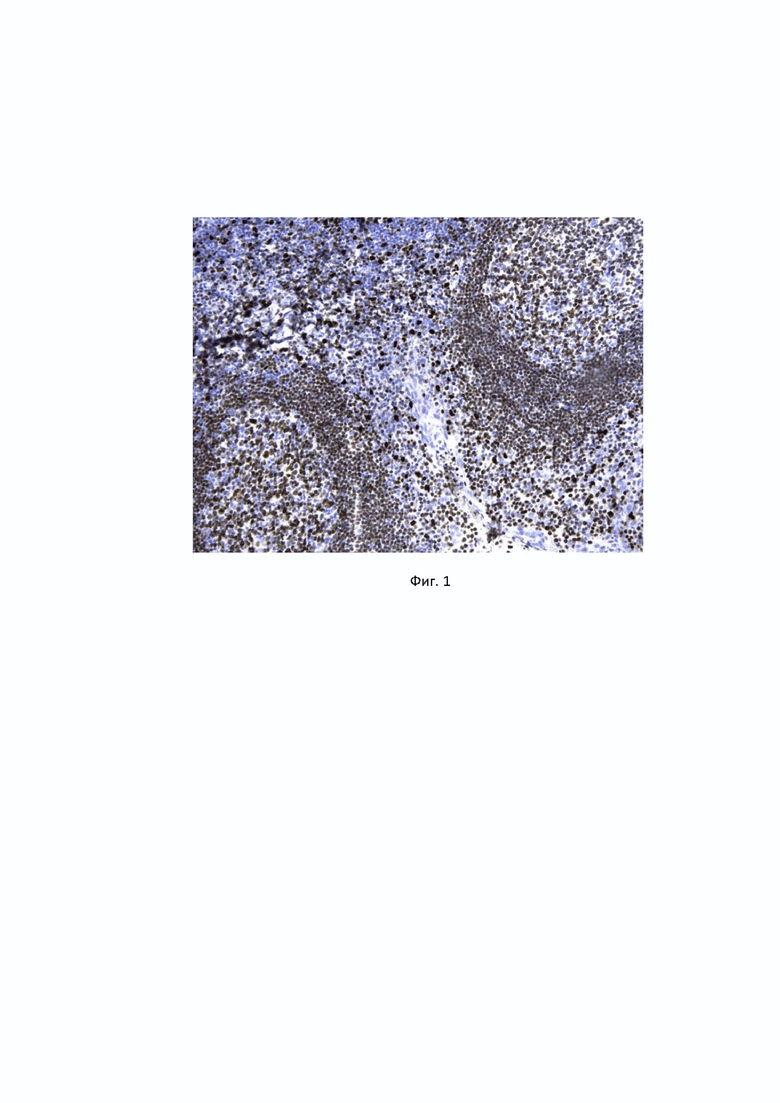
В иммуногистохимическом анализе антител к PU.1, клона 4G6 на срезах миндалин наблюдалась четкая ядерная окраска B-лимфоцитов герминативного центра, B-клеток мантийной зоны, макрофагов, плазматические клетки при этом оставались неокрашенными (Фиг. 1)
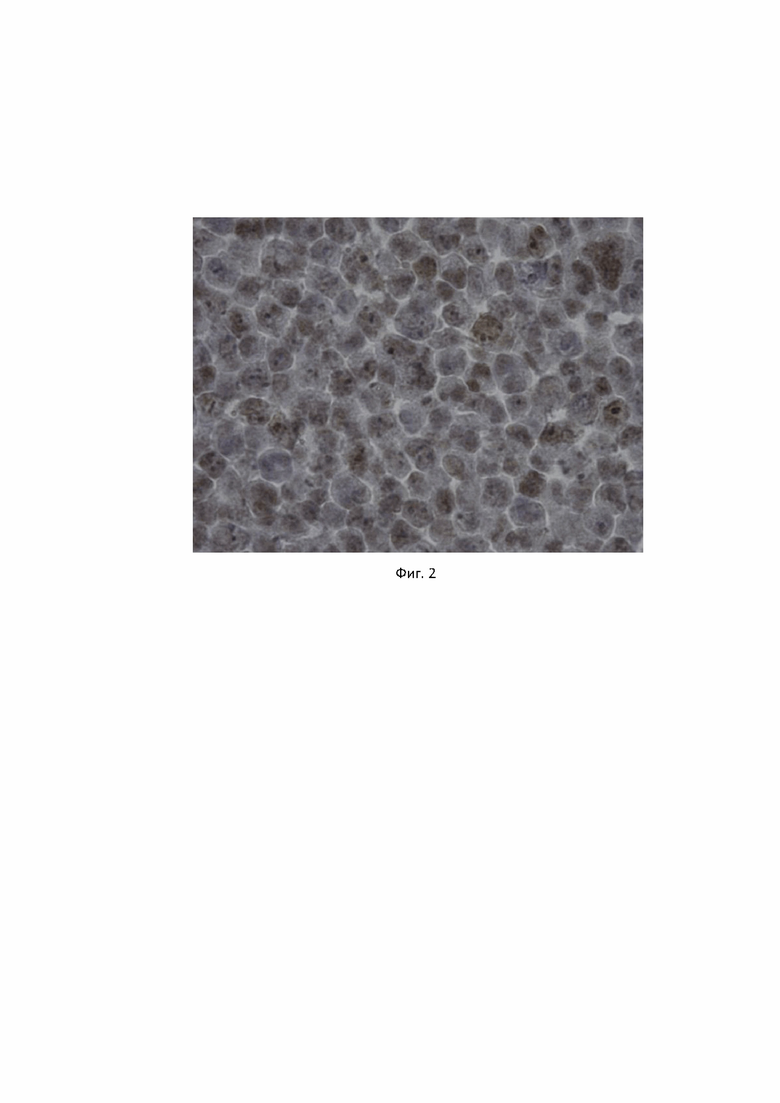
Антитела клона 4G6 были протестированы так же иммуногистохимическим методом на клеточных блоках, подготовленных из клеточной линии Raji. В результате у клеток наблюдалась четкая ядерная окраска (Фиг. 2)
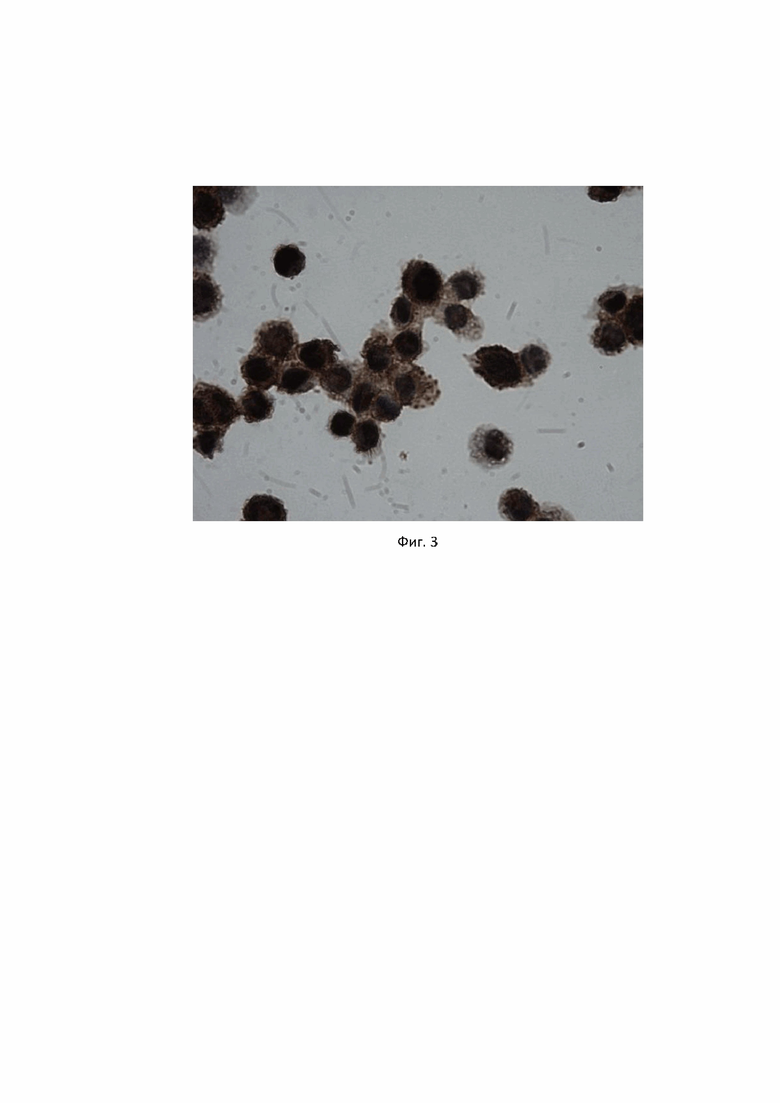
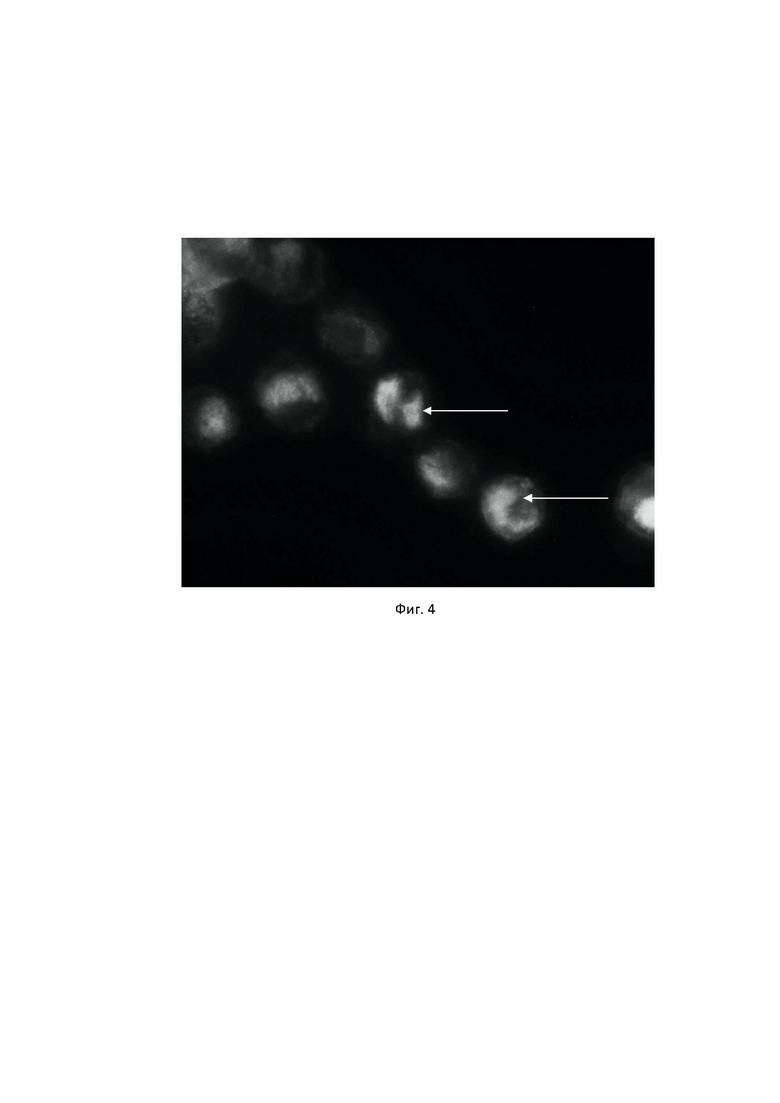
Антитела клона 4G6 тестировали методами иммуноцитохимии и иммунофлуоресценции на моноцитарной клеточной линии THP1. Антитела показали четкое связывание с ядерным антигеном PU.1 (Фиг. 3, Фиг. 4).
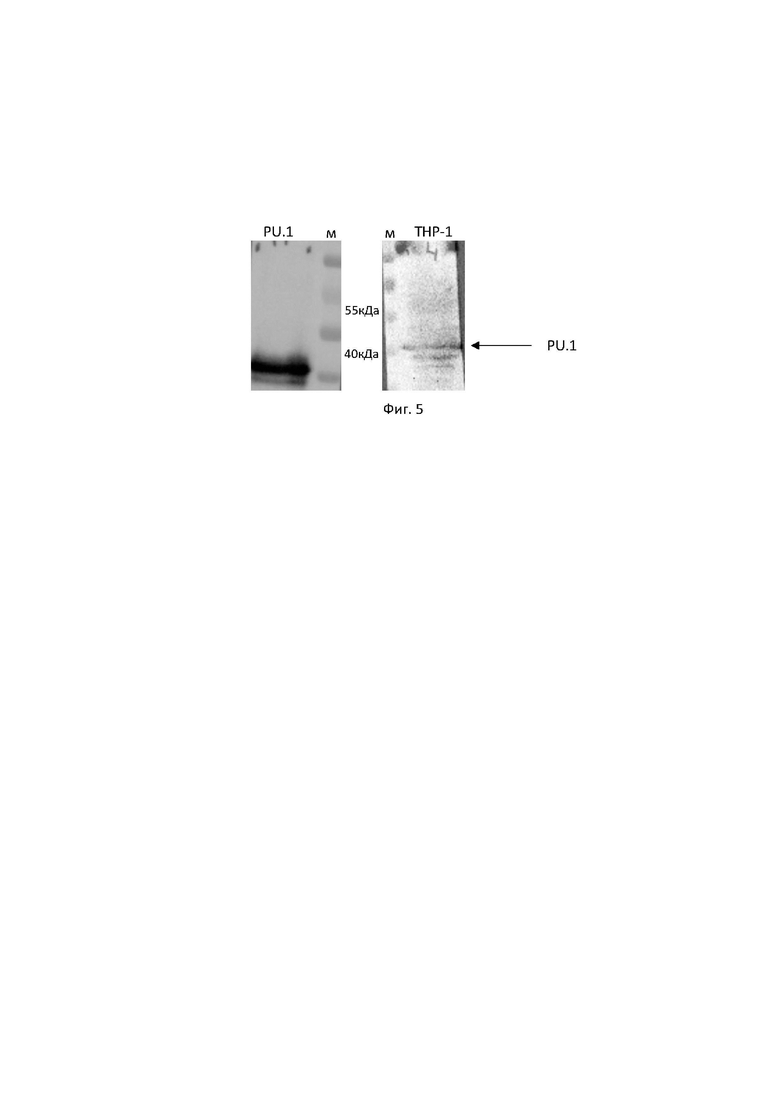
Дополнительно был проведен иммуноблоттинг (Фиг. 5) для проверки антител на тотальных клеточных лизатах моноцитарной клеточной линии THP1 и рекомбинантном белке PU.1. Было показано, что антитела к 4G6 связываются с нативным белком PU.1 (42 кДа).
Таким образом антитела, продуцируемые гибридомой PU.1, клон 4G6 специфически работают в иммуноферментном анализе, иммуногистохимии (на тканях и на клеточных блоках), иммуноцитохимии, иммунофлуоресценции и в иммуноблоттинге.
Мышиная гибридома PU.1, клон 4G6 - продуцент моноклонального антитела к PU.1, имеет следующие характеристики:
- морфологические признаки: культура имеет вид суспензии;
- условия рутинного культивирования клеток гибридомы: среда для культивирования - RPMI-1640 («Gibco», США), 7,5% эмбриональной телячьей сыворотки FetalClone 1 («HyClone», США), 4 мМ L-глутамина, 50 мкг/мл гентамицина, 37°C, абсолютная влажность и 5% СО2 в атмосфере;
- частота пассирования: каждые 3-4 суток, кратность рассева 1:5-1:10;
- способ криоконсервации клеток гибридомы: в суспензию клеток в кондиционированной среде добавляют 7-10% ДМСО («ПанЭко», Россия), криопробирки с суспензией клеток перемешивают и помещают на сутки в холодильник на -80°С, затем переносят в жидкий азот для длительного хранения. Жизнеспособность клеток после размораживания 80%. После размораживания клетки культивируют при плотности 0.2-0.3⋅106 кл/мл.
Бактерии, грибы, дрожжи и микоплазма в культуре не обнаружены.
Изотип моноклональных антител, производимых гибридомой 4G6: IgG2a.
Специфичность моноклональных антител, продуцируемых гибридомой PU.1, клон 4G6: в иммуноферментном анализе связывается с рекомбинантым белком PU.1, в иммуногистохимическом анализе связывается с PU.1 в тканях и клетках, экспрессирующих данный транскрипционный фактор, например с B-клетками миндалин и с лимфобластоподобными клетками Raji, в иммуноцитохимическом и иммунофлуоресцентном анализе - выявляет антиген PU.1 в моноцитарной клеточной линии человека THP1, выявляет нативный белок PU.1 в тотальных клеточных лизатах THP1 на иммуноблотах.
Оптимальные разведения антител к PU.1, клон 4G6, определяемые в супернатанте методом иммуногистохимии - 1:5.
Продуцируемое гибридомой 4G6 моноклональное антитело рекомендуется для специфического выявления антигена PU.1 в тотальных клеточных лизатах методом иммуноферментного анализа, в тканях и клетках, содержащих данный антиген - методом иммуногистохимии, а также в клетках человека иммуноцитохимии, иммунофлуоресценции, иммуноблоттинга.
Изобретение иллюстрируют следующие фотографии и таблица:
Табл. 1. График, представляющий результаты иммуноферментного анализа (ИФА), описанного в Примере 1, показывающий связывание антител к PU.1, продуцируемых гибридомой 4G6 с рекомбинантым белком PU.1 (5 мкг/мл). В качестве отрицательного контроля (К-) взят рекомбинантный белок PMS2-His (5 мкг/мл). Перекрестной реакции на His-tag не обнаружено.
Фиг. 1. Специфичность антител к PU.1, клон 4G6, продуцируемых гибридомой 4G6, к транскрипционному фактору PU.1 в B-лимфоцитах герминативного центра, B-клетках мантийной зоны, макрофагах, выявляемая методом иммуногистохимии. Для детекции связавшихся с антигеном антител, была использована система детекции PrimeVision Ms/Rb HRP DAB (ПраймБиоМед, Россия) на основе вторичных антител к иммуноглобулинам мыши/кролика, конъюгированных с полимерной пероксидазой хрена и субстрата ДАБ, как описано в Примере 2. Наблюдается специфическая ядерная окраска B-клеток герминативного центра и мантийной зоны и макрофагов.
Фиг. 2. Специфичность антител, продуцируемого гибридомой PU.1, клон 4G6, к ядерному белку PU.1 в клетках человека лимфобластоподобной линии Raji, выявляемая методом иммуногистохимии на срезах клеточных блоков. Для детекции связавшихся с антигеном антител, была использована система детекции PrimeVision Ms/Rb HRP DAB (ПраймБиоМед, Россия) на основе вторичных антител к иммуноглобулинам мыши/кролика, конъюгированных с полимерной пероксидазой хрена и субстрата ДАБ, как описано в Примере 2. Наблюдается специфическая ядерная окраска клеток.
Фиг. 3. Специфичность антител, продуцируемых гибридомой PU.1, клон 4G6, к транскрипционному фактору PU.1 в моноцитарной клеточной линии человека THP1, выявляемая методом иммуноцитохимии, описанном в Примере 3. Для детекции связавшихся с антигеном антител, была использована система детекции PrimeVision Ms/Rb HRP DAB (ПраймБиоМед, Россия) на основе вторичных антител к иммуноглобулинам мыши/кролика, конъюгированных с полимерной пероксидазой хрена и субстрата ДАБ. Темные ядра представляют собой окрашенный антителами ядерный белок - транскрипционный фактор PU.1.
Фиг. 4. Специфичность антител, продуцируемых гибридомой PU.1, клон 4G6 к транскрипционному фактору PU.1 в моноцитарной клеточной линии человека THP1, выявляемая методом иммунофлуоресценции. Для выявления связавшихся с антигеном антител использовались вторичные антитела к иммуноглобулинам мыши, конъюгированные с флуорохромом Alexa Fluor 488 (JacksonImmunoresearch, США), как описано в Примере 3. Стрелками отмечены окрашенные ядра клеток.
Фиг. 5. Специфичность антител, продуцируемых гибридомой PU.1, клон 4G6, к транскрипционному фактору PU.1 на тотальных клеточных лизатах моноцитарной клеточной линии человека THP1, и рекомбинантном белке PU.1, выявляемая на иммуноблотах, как описано в Примере 4. Электрофоретическая подвижность белка соответствует расчетной - 42 кДа.
Изобретение иллюстрируют следующие примеры:
Пример 1. Процедура иммуноферментного анализа (ИФА) с использованием мышиной гибридомы PU.1, клон 4G6.
Для ИФА на 96-луночные планшеты (MaxiSorb, SPL, США) сорбируют антиген в концентрации 5 мкг/мл в карбонат-бикарбонатном буфере (67мМ NaHCO3 и 28мМ Na2CO3), pH 9,5 по 100 мкл на лунку в течение 1 часа при комнатной температуре на шейкере. Блокировка свободной поверхности пластика осуществляется добавлением по 100 мкл в лунку 1% раствора бычьего сывороточного альбумина (BSA) на 30 мин. Промывают 4 раза фосфатно-солевым буфером с добавлением 0,05% Твин 20. Антитела добавляют по 100 мкл в лунки в концентрации менее 10 мкг/мл в 0,5% BSA в PBS-T и инкубируют 1 час при комнатной температуре на шейкере. Промывают 4 раза фосфатно-солевым буфером с добавлением 0,05% Твин 20.
Вторичные антитела, козьи анти-мышиные, конъюгированные с пероксидазой хрена (HRP) растворяют в PBS-T 1 к 30000 (JacksonImmunoResearch, США), добавляют в лунки по 100 мкл и инкубируют 40 минут - 1 час при комнатной температуре на шейкере. Промывают 6 раз фосфатно-солевым буфером с добавлением 0,05% Твин 20. Реакцию проявляют с помощью субстрата TMB (Иммунотех, Россия) по 100 мкл в лунку 5 минут, остановка реакции происходит при добавлении 25 мкл 10% серной кислоты. Оптическую плотность снимают при длине волны 450 нм, дифференциальный фильтр 630 нм. Результаты представлены в Таб. 1.
Пример 2. Процедура иммуногистохимического выявления белка PU.1 на срезах миндалины и клеточных блоков Raji.
Иммуногистохимическое исследование проводят на операционном материале и клеточном осадке, фиксированном 10%-ным нейтральным формалином, забуференным фосфатным буфером, в течение 24 часов. После гистологической проводки материал заливают в парафин и затем готовят срезы толщиной 4 мкм. Срезы монтируют на специальные высокоадгезивные стекла (SuperFrost Plus, ApexLab) и высушивают в течение 15 минут при температуре 70°С.
Непосредственно перед иммуногистохимическим исследованием парафин удаляют со срезов в двух сменах ксилола, по 10 мин в каждой смене. Срезы проводят через спирты с объемной долей изопропанола 100% (I), 100% (II), 70%, 50% по 5 мин в каждом и промывают в дистиллированной воде. Температурную демаскировку проводят 10 минут при температуре 110°С в цитратном или трис-ЭДТА буфере. Блокировку эндогенной пероксидазы проводят в 3% перекиси водорода 5-10 минут. Срезы инкубируют с антителами PU.1, клон 4G6 в течение 30 минут, а затем с антителами к иммуноглобулинам мыши, конъюгированными с полимерной пероксидазой хрена (PrimeVision, ПраймБиоМед, Россия). Коричневое окрашивание проявляется при добавлении субстрата и ДАБ хромогена на 5-10 минут (ПраймБиоМед, Россия). Результат представлен на Фиг. 2 и Фиг. 3.
Пример 3. Процедура выявления белка PU.1 в реакции иммуноцитохимии и иммунофлуоресценции.
Моноцитарную культуру THP1 выращивают на стеклах в чашки Петри с добавлением 67 нмоль PMA для лучшей клеточной адгезии. Клетки фиксируют 3,7% параформальдегидом 15 мин, с последующей промывкой в PBS (ПанЭко, Россия) 3 раза по 2 мин. После фиксации клетки инкубируют с антителом к PU.1, клон 4G6 30 минут при комнатной температуре, а затем с козьими антителами к иммуноглобулинам мыши, конъюгированными с флуорохромом Alexa488 (JacksonImmunoresearch, США), или полимерной пероксидазой хрена (ПраймБиоМед, Россия) 40 или 30 мин, соответственно, при комнатной температуре. Для визуализации окрашивания в нефлуоресцентном методе добавляют субстрат ДАБ хромоген на 5-10 минут (ПраймБиоМед, Россия). Препараты заключают в среду для заключения на водной основе и изучают в микроскоп Olympus BX53 (Cheminst, Германия), сопряженный с 2 мп камерой Infinity 2 (Lumenera, США), используя объективы U PlanFL N ×40/ЧА 0.75 и U PlanFL N ×100/ЧА 1.30. Для иммунофлуоресценции используются фильтры, U-FUNA и U-FGNA (Olympus, Япония). Результаты представлены на Фиг. 3 и Фиг. 4.
Пример 4. Способ получения тотальных клеточных лизатов и выявления нативного белка PU.1 методом иммуноблоттинга.
Клетки THP1 в количестве около 4*106 клеток промывают от среды фосфатно-солевым буфером (PBS), и лизируют при +4°C в буфере, содержащем 50 mM Трис-(гидроксиметил)-аминометана (рН 7.5), 150 mM NaCl, 10% глицерина, 0.5% Тритона Х-100 и коктейль протеазных ингибиторов (Roche, США). Суммарную концентрацию белков в лизатах определяют по методу Бредфорд с помощью Protein Assay Kit (Bio-Rad, США), следуя рекомендациям производителя.
Перед электрофоретическим разделением к приготовленным лизатам добавляют 5-ный буфер Лэммли, содержащий 250 мМ Трис-(гидроксиметил)-аминометана (рН 6.8), 50% глицерина, 10% додецилсульфата натрия (ДСН), 500 мМ бета-меркаптоэтанола, 0.5% бромфенолового синего. Образцы подвергают термической обработке при 95°С 5 мин. В каждую лунку 10% полиакриламидного геля с ДСН (ПААГ-ДСН) наносят лизаты, содержащие не менее 30 мкг суммарного белка. Электрофоретическое разделение белков в ПААГ-ДСН проводят 30 мин при 60 В и 60 мин при 120 В для дальнейшего разделения в электрофорезной камере. Перенос белков с геля на нитроцеллюлозную мембрану (0.22 мкм, Millipore, США) проводят при постоянной силе тока 250 мА 60 мин в буфере для проведения блоттинга, содержащем 50 мМ Трис-(гидроксиметил)-аминометана, 38 мМ глицина и 20% этанола. Мембрану блокируют 5% бычьим сывороточным альбумином в PBS-T (ПанЭко, Россия) 1 час, инкубируют с антителами к PU.1, клон 4G6 в том же растворе (ночь при +4°С), а затем с антителами к иммуноглобулинам мыши, конъюгированными с пероксидазой хрена (разведение 1:50000, JacksonImmunoResearch, США) -1 час. Мембрану проявляют методом хемилюминесценции, используя набор Novex ECL (Invitrogen, США) согласно инструкции производителя. Хемилюминесцентную реакцию регистрируют на приборе ChemiDoc MP (Bio-Rad, UK) с последующей обработкой с помощью программы ChemiDoc XRS+ imaging systems. Результаты представлены на Фиг. 5.
Табл. 1
| название | год | авторы | номер документа |
|---|---|---|---|
| Мышиная гибридома SI-CLP, клон 3D4 - продуцент моноклонального антитела, обладающего специфичностью к белку SI-CLP | 2019 |
|
RU2714685C1 |
| Мышиная гибридома YKL-40, клон 2G8 C10 - продуцент моноклонального антитела, обладающего специфичностью к цитоплазмотическим антигенам YKL-39, YKL-40 и SI-CLP человека | 2019 |
|
RU2728228C1 |
| МЫШИНАЯ ГИБРИДОМА YKL-39, КЛОН 1B2 G4 - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА, ОБЛАДАЮЩЕГО СПЕЦИФИЧНОСТЬЮ К ЦИТОПЛАЗМАТИЧЕСКОМУ АНТИГЕНУ YKL-39 ЧЕЛОВЕКА | 2017 |
|
RU2667423C1 |
| Мышиная гибридома AMACR, клон G8 - продуцент моноклонального антитела, обладающего специфичностью к альфа-метилацил-коэнзим A рацемазе (AMACR) человека | 2016 |
|
RU2636042C1 |
| МЫШИНАЯ ГИБРИДОМА Р56 - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА, ОБЛАДАЮЩЕГО СПЕЦИФИЧНОСТЬЮ К ЯДЕРНОМУ АНТИГЕНУ ПРОЛИФЕРИРУЮЩИХ КЛЕТОК PCNA ЧЕЛОВЕКА | 2010 |
|
RU2435850C1 |
| МЫШИНАЯ ГИБРИДОМА S79 - ПРОДУЦЕНТ МОНОКЛОНАЛЬНОГО АНТИТЕЛА, ОБЛАДАЮЩЕГО СПЕЦИФИЧНОСТЬЮ К БЕЛКУ ЯДРЫШКА SURF-6 ЧЕЛОВЕКА | 2010 |
|
RU2422516C1 |
| Штамм гибридных клеток животных Mus musculus 1F1 - продуцент моноклонального антитела к нуклеокапсидному белку N вируса SARS-CoV-2 | 2021 |
|
RU2769817C1 |
| Штамм культивируемых гибридных клеток животных Mus. musculus - продуцент моноклональных антител к мембранному белку, общему для типичных и атипичных холерных вибрионов 01 серогруппы | 2022 |
|
RU2785463C1 |
| ШТАММ ГИБРИДНЫХ КУЛЬТИВИРУЕМЫХ КЛЕТОК ЖИВОТНЫХ Mus musculus L. - ПРОДУЦЕНТ МОНОКЛОНАЛЬНЫХ АНТИТЕЛ ПРОТИВ РАЗНЫХ ИЗОФОРМ АЛЬФА2-МИКРОГЛОБУЛИНА ФЕРТИЛЬНОСТИ (АМГФ)/ГЛИКОДЕЛИНА | 2007 |
|
RU2355762C2 |
| Штамм гибридных клеток животных Mus musculus 2E1B5 - продуцент моноклонального антитела к рецептор-связывающему домену белка S вируса SARS-CoV-2 | 2021 |
|
RU2771288C1 |
Изобретение относится к области биотехнологии, биологии и медицине, в частности к мышиной гибридоме PU.1, клону 4G6, которая является продуцентом моноклонального антитела, выявляющего ядерный белок человека PU.1 методами иммуноферментного анализа, иммуноцитохимии, иммуногистохимии, иммуноблоттинга и иммунофлуоресценции. Гибридоме присвоен регистрационный номер ВКПМ: H-187 от 22 сентября 2021 года. Продуцируемые гибридомой антитела могут быть использованы для дифференциальной диагностики лимфом. 5 ил., 1 табл., 4 пр.
Мышиная гибридома PU.1, клон 4G6, регистрационный номер ВКПМ: H-187, – продуцент моноклональных антител, выявляющих белок PU.1 человека в различных типах клеток нормальных и опухолевых тканей, содержащих данный антиген, методами иммуноферментного анализа, иммуноцитохимии, иммуногистохимии, иммуноблоттинга и иммунофлуоресценции, полученная путем иммунизации мышей линии Balb/c рекомбинантным белком PU.1 человека и слиянием сенсибилизированных спленоцитов и лимфоцитов иммунизированных мышей с клетками мышиной миеломы линии Sp2/0 с помощью 50%-ного раствора полиэтиленгликоля с молекулярной массой 1450 и отличающаяся уникальной эпитопной специфичностью.
| WILL B | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Nat Med | |||
| Устройство для закрепления лыж на раме мотоциклов и велосипедов взамен переднего колеса | 1924 |
|
SU2015A1 |
| Vol | |||
| Выбрасывающий ячеистый аппарат для рядовых сеялок | 1922 |
|
SU21A1 |
| No | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Pp | |||
| Автоматический огнетушитель со спринклером | 1924 |
|
SU1172A1 |
| Мышиная гибридома SI-CLP, клон 3D4 - продуцент моноклонального антитела, обладающего специфичностью к белку SI-CLP | 2019 |
|
RU2714685C1 |
| LEA P., LING M | |||
| New molecular assays for cancer diagnosis and targeted therapy | |||
| Curr | |||
| Opin | |||
| Mol | |||

