Область техники
Настоящее изобретение относится к области биотехнологии, в частности к аминоацил-тРНК-синтетазе, которая эффективно вводит производное лизина.
Уровень техники
Интегрированный белок (цельный белок) не встречающейся в природе аминокислоты, который получают путем замены аминокислотного остатка в желаемом положении в белке аминокислотами, отличающимися от 20 типов аминокислот, используемых в нормальном синтезе белка (т.е. не встречающейся в природе аминокислот), может использоваться как эффективное средство для анализа структуры и функции белка. Используя пары аминоацил-тРНК-синтетазы (aaRS)/TPHK, полученных из различных биологических видов, было синтезировано более 30 типов цельных белков. Система с самой длинной историей и полезная для введения многих полезных не встречающихся в природе аминокислот представляет собой мутантную тирозил-тРНК-синтетазу (TyrRS) и супрессорную пару тРНКTyr гена, подавляющая амбер-мутацию. Что касается этого способа, ключевым является его ортогональное соотношение, т.е. aaRS в двух группах эубактерий, архей и эукариот может аминоацилировать тРНК в своей соответствующей группе, но не может аминоацилировать тРНК в другой группе.
С другой стороны, пирролизил-тРНК-синтетаза (PylRS) и супрессорный тРНКРу1 ген, подавляющий амбер-мутацию, полученный из Methanosarcina mazei, могут функционировать в качестве ортогональной пары aaRS/тРНК в клетках Escherichia coli. Пирролизин представляет собой производное лизина с объемным метилпирролиновым фрагментом в боковой цепи. PylRS дикого типа может связывать Nε-Boc-L-лизин с тРНКРу1 в Escherichia coli. Однако, поскольку LysRS обладает строгим распознаванием лизина, производные лизина, имеющие функциональные группы различного размера и формы, сложно вводить сайт-специфично в белки.
Следовательно, в данной области желательно модифицировать лизил-тРНК-синтетазу дикого типа и разработать аминоацил-тРНК-синтетазу, которая эффективно вводит производные лизина в белки.
Сущность изобретения
Цель настоящего изобретения - предоставить аминоацил-тРНК-синтетазу и способ эффективного введения производных лизина в белки.
В первом аспекте настоящего изобретения предоставляется мутантный белок аминоацил-тРНК-синтетазы, который включает в себя делеционную мутацию аминокислотного остатка в одном или нескольких положениях, выбранных из группы, состоящей из: положения 102, положений 128-140 и положений 159-179 в аминокислотной последовательности, соответствующей аминоацил-тРНК-синтетазе дикого типа.
В другом предпочтительном варианте осуществления большее количество позиций подразумевает:: 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33 и 34 позиции.
В другом предпочтительном варианте осуществления позиции 128-140 включают в себя позиции 128, 129, 130, 131, 132, 133, 134, 135, 136, 137, 138, 139 и 140.
В другом предпочтительном варианте осуществления позиции 159-179 включают в себя позиции 159, 160, 161, 162, 163, 164, 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178, 179 и 180.
В другом предпочтительном варианте осуществления по меньшей мере один из следующих аминокислотных остатков в аминокислотной последовательности аминоацил-тРНК-синтетазы дикого типа усечен в мутантном белке:
(a) аминокислотный остаток в положении 102;
(b) аминокислотные остатки в положениях от X до Y, где X представляет собой целое положительное число 128-132, a Y представляет собой целое положительное число 133-140; и/или
(c) аминокислотные остатки в положениях от А до В, где А представляет собой положительное целое число 159-164, а В представляет собой положительное целое число 165-179.
В другом предпочтительном варианте осуществления X представляет собой положительное целое число от 128 до 132, например 128, 129, 130, 131 или 132.
В другом предпочтительном варианте осуществления X равен 128.
В другом предпочтительном варианте осуществления Y представляет собой положительное целое число 133-140, например 133, 134, 135, 136, 137, 138, 139 или 140.
В другом предпочтительном варианте осуществления Y равен 140.
В другом предпочтительном варианте осуществления А представляет собой положительное целое число от 159 до 164, например 159, 160, 161, 162, 163 или 164.
В другом предпочтительном варианте осуществления А равно 159.
В другом предпочтительном варианте осуществления В представляет собой положительное целое число 165-179, например 165, 166, 167, 168, 169, 170, 171, 172, 173, 174, 175, 176, 177, 178 или 164.
В другом предпочтительном варианте осуществления В равно 179.
В другом предпочтительном варианте осуществления аминоацил-тРНК-синтетаза дикого типа получена из Methanosarcina mazei, Methanosarcina barkeri или Methanosarcina acetivorans от Methanogenic archaea.
В другом предпочтительном варианте осуществления аминокислотная последовательность аминоацил-тРНК синтетазы дикого типа представлена в SEQ ID №: 1 или 2.
В другом предпочтительном варианте осуществления нумерация удаленного аминокислотного остатка основана на SEQ ID №: 1 или 2.
В другом предпочтительном варианте осуществления мутантный белок представляет собой усеченную аминоацил-тРНК-синтетазу.
В другом предпочтительном варианте осуществления длина аминокислотной последовательности мутантного белка составляет, по меньшей мере, 90%, предпочтительно 92-99% длины последовательности, представленной в SEQ ID №: 1 или 2.
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка (или усеченной формы) имеет, по меньшей мере, 90%, 95%, 96%, 97%, 98% или 99% идентичность последовательности пептидному фрагменту SEQ ID №: 1 или 2 с соответствующей длиной.
В другом предпочтительном варианте осуществления в мутантном белке, по меньшей мере, аминокислотные остатки в положениях 128-140 и положениях 159-179 аминокислотной последовательности аминоацил-тРНК-синтетазы дикого типа усечены.
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка представлена в SEQ ID №: 3.
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка представлена в SEQ ID №: 4.
В другом предпочтительном варианте осуществления по сравнению с последовательностью мутантной аминоацил-тРНК-синтетазы, представленной в SEQ ID №: 4, мутантный белок дополнительно содержит в себе аминокислотную мутацию, выбранную из группы, состоящей из: гистидина (Н) в положении 29, аспарагиновой кислоты (D) в положении 76, серина (S) в положении 89, аспарагина (N) в положении 91, аргинина (R) в положении 96, серина (S) в положении 121, аспарагина (N) в положении 129, серина (S) в положении 145, аланина (А) в положении 148, лейцина (L) в положении 274, цистеина (С) в положении 313, фенилаланина (F) в положении 349 и их комбинаций.
В другом предпочтительном варианте осуществления нумерация удаленных аминокислотных остатков основана на SEQ ID №: 4.
В другом предпочтительном варианте осуществления мутантный белок представляет собой мутантную аминоацил-тРНК-синтазу, имеющую последовательность, представленную в SEQ ID №: 4, и на основе последовательности, предоставленной в SEQ ID №: 4, он дополнительно включает в себя аминокислотную мутацию, выбранную из группы, состоящей из: гистидина (Н) в положении 29, аспарагиновой кислоты (D) в положении 76, серина (S) в положении 89, аспарагина (N) в положении 91, аргинина (R) в положении 96, серина (S) в положении 121, аспарагина (N) в положении 129, серина (S) в положении 145, аланина (А) в положении 148, лейцина (L) в положении 274, цистеина (С) в положении 313, фенилаланина (F) в положении 349 и их комбинаций.
В другом предпочтительном варианте осуществления гистидин (Н) в положении 29 мутировал в тирозин (Y); и/или
аспарагиновая кислота (D) в положении 76 мутировала в глицин (G); и/или
серии (S) в положении 89 мутировал в глицин (G); и/или
аспарагин (N) в положении 91 мутировал на треонин (Т); и/или
аргинин (R) в положении 96 мутировал в лизин (K); и/или
серии (S) в положении 121 мутировал в пролин (Р); и/или
аспарагин (N) в положении 129 мутировал в аспарагиновую кислоту (D); и/или
серии (S) в положении 145 мутировал в пролин (Р); и/или
аланин (А) в положении 148 мутировал в треонин (Т); и/или
лейцин (L) в положении 274 мутировал в аланин (А); и/или
цистеин (С) в положении 313 мутировал в серии (S); и/или
фенилаланин (F) в положении 349 мутировал в тирозин (Y).
В другом предпочтительном варианте осуществления по сравнению с последовательностью мутантной аминоацил-тРНК-синтетазы, представленной в SEQ ID №: 4, мутантный белок дополнительно содержит в себе аминокислотную мутацию, выбранную из группы, состоящей из: H29Y, D76G, S89G, N91T, R96K, S121P, N129D, S145P, А148Т, L274A, C313S, F349Y и их комбинаций.
В другом предпочтительном варианте осуществления по сравнению с последовательностью мутантной аминоацил-тРНК-синтетазы, представленной в SEQ ID №: 4, мутантный белок дополнительно содержит мутацию, выбранную из группы, состоящей из: H29Y, D76G, S89G, N91T, R96K, S121P, N129D, S145P и А148Т.
В другом предпочтительном варианте осуществления по сравнению с последовательностью мутантной аминоацил-тРНК-синтетазы, представленной в SEQ ID №: 4, мутантный белок дополнительно содержит мутацию, выбранную из группы, состоящей из: L274A, C313S и F349Y.
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка представлена в SEQ ID №: 5 или 6.
В другом предпочтительном варианте осуществления за исключением делеции и мутации, другие аминокислоты мутантного белка являются такими же или, по существу, такими же, что и последовательность, представленная в SEQ ID №: 1 или SEQ ID №: 2.
В другом предпочтительном варианте осуществления термин «по существу, такие же» означает, что не более 50 (предпочтительно 1-20, более предпочтительно 1-10) аминокислот отличаются, при этом, отличие включает в себя замену, делецию или добавление аминокислоты, а мутантный белок все еще обладает активностью аминоацил-тРНК-синтетазы.
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка имеет, по меньшей мере, 70%, предпочтительно, по меньшей мере, 75%, 80%, 85%, 90% и более предпочтительно, по меньшей мере, 95%, 96%, 97%, 98%, 99% идентичность последовательности SEQ ID №: 1 или SEQ ID №: 2.
В другом предпочтительном варианте осуществления мутантный белок образован мутацией аминоацил-тРНК-синтетазы дикого типа, представленной в SEQ ID №: 1 или SEQ ID №: 2.
В другом предпочтительном варианте осуществления мутантный белок выбирается из группы, состоящей из:
(1) полипептида, имеющего аминокислотную последовательность, предоставленную в любой из SEQ ID №: 3-6; или
(2) полипептида, полученного из полипептида, имеющего аминокислотную последовательность, представленную в любом из SEQ ID NO: 3-6, который образован заменой, делецией или добавлением одного или нескольких (предпочтительно 1-20, более предпочтительно 1-15, более предпочтительно 1-10, более предпочтительно 1-8, более предпочтительно 1-3 и наиболее предпочтительно 1) аминокислотных остатков в аминокислотной последовательности, представленной в любом из SEQ ID №: 3-6, и выполняет функцию полипептида, как описано в (1).
В другом предпочтительном варианте осуществления аминокислотная последовательность мутантного белка представлена в любом из SEQ ID №: 3-6.
В другом предпочтительном варианте осуществления мутантный белок представляет собой белок искусственного происхождения.
В другом предпочтительном варианте осуществления мутантный белок используется для введения предварительно заданной модифицированной аминокислоты в целевой белок.
В другом предпочтительном варианте осуществления мутантный белок имеет одну или несколько следующих характеристик:
(a) по сравнению с аминоацил-тРНК-синтетазой дикого типа, предварительно заданная модифицированная аминокислота с большими функциональными группами может быть введена в белок;
(b) целевой белок имеет высокий уровень экспрессии; и
(c) мутантный белок имеет более короткую последовательность, что обеспечивает простоту его экспрессии, и обладает высоким уровнем экспрессии.
Во втором аспекте настоящего изобретения предлагается изолированный полинуклеотид, кодирующий мутантный белок согласно первому аспекту настоящего изобретения.
В другом предпочтительном варианте осуществления полинуклеотид кодирует полипептид, предоставленный в любом из SEQ ID №: 3-6.
В другом предпочтительном примере полинуклеотид кодирует полипептид, предоставленный в SEQ ID №: 4, а его нуклеотидная последовательность представлена в SEQ ID №: 9.
В другом предпочтительном варианте осуществления полинуклеотид содержит последовательность ДНК, последовательность РНК и их комбинации.
В другом предпочтительном варианте осуществления полинуклеотид дополнительно содержит вспомогательный элемент на фланкиреумой ORF мутантной аминоацил-тРНК-синтетазы, выбранной из группы, состоящей из: сигнального пептида, секреторного пептида, маркерной последовательности (например 6His) и их комбинаций.
В третьем аспекте настоящего изобретения предлагается вектор, содержащий полинуклеотид по второму аспекту настоящего изобретения.
В другом предпочтительном варианте осуществления вектор включает в себя вектор экспрессии, вектор-челнок и вектор интеграции.
В другом предпочтительном варианте осуществления вектор выбирается из группы, состоящей из рЕТ, pCW, pUC, pPIC9k, рМА5 и их комбинации.
В другом предпочтительном варианте осуществления вектор представляет собой вектор pEvol и/или вектор pBAD.
В другом предпочтительном варианте осуществления вектор используется для экспрессии мутантного белка по первому аспекту настоящего изобретения.
В четвертом аспекте настоящего изобретения предлагается клетка-хозяин, содержащая вектор по третьему аспекту настоящего изобретения, или имеющая полинуклеотид по второму аспекту настоящего изобретения, интегрированный в ее геном, или экспрессирующая мутантный белок по первому аспекту настоящего изобретения.
В другом предпочтительном варианте осуществления клетка-хозяин выбрана из группы, состоящей из прокариотической клетки, эукариотической клетки и их комбинации.
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой эукариотическую клетку, например дрожжевую клетку, растительную клетку или клетку животного (например, клетка млекопитающего).
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой прокариотическую клетку, такую как Escherichia coli
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой Escherichia coli Top 10 или BL21.
В другом предпочтительном варианте осуществления клетка-хозяин содержит:
(a) мутантный белок по первому аспекту настоящего изобретения; и
(b) искусственную тРНК, способную связываться с предварительно заданной модифицированной аминокислотой в присутствии мутантного белка; и, необязательно
(c) первую последовательность нуклеиновой кислоты, кодирующей целевой белок, отличающуюся тем, что первая последовательность нуклеиновой кислоты содержит кодон, распознаваемый искусственной тРНК.
В другом предпочтительном варианте осуществления клетка-хозяин дополнительно содержит:
(d) предварительно заданную модифицированную аминокислоту.
В другом предпочтительном варианте осуществления первая последовательность нуклеиновой кислоты содержит кодон, распознаваемый искусственной тРНК, в положении для введения предварительно заданной модифицированной аминокислоты.
В другом предпочтительном варианте осуществления предварительно заданная модифицированная аминокислота является субстратом мутантного белка.
В другом предпочтительном варианте осуществления предварительно заданная модифицированная аминокислота является лизином с модифицированной группой.
В другом предпочтительном варианте осуществления модифицированная аминокислота выбрана из группы, состоящей из: производного алкинилоксикарбониллизина, производного трет-бутилоксикарборил-L-лизина (ВОС-лизина), ацилированного лизина жирного ряда и их комбинаций.
В другом предпочтительном варианте осуществления структура алкинилоксикарбониллизина представлена в формуле I:

где n равно 0-8.
В другом предпочтительном варианте осуществления искусственная тРНК представляет собой супрессорную тРНК и предпочтительно супрессорную тРНК, подавляющую амбер-мутацию.
В другом предпочтительном варианте осуществления кодирующая последовательность нуклеиновой кислоты искусственной тРНК представлена в SEQ ID №: 7.
GGAAACCTGATCATGTAGATCGAATGGACTCTAAATCCGTTCAGCCGGGTTAGATTCCCGGGGTTTCCGCCA (SEQ ID №: 7)
В другом предпочтительном варианте осуществления кодон, распознаваемый искусственной тРНК, представляет собой UAG (амбер), UAA (охра) или UGA (опал) и предпочтительно амбер-кодон.
В другом предпочтительном варианте осуществления мутантный белок специфически катализирует искусственную тРНК с образованием комплекса «искусственная тРНК-Ха», где Ха представляет собой предварительно заданную модифицированную аминокислоту в аминоацильной форме.
В другом предпочтительном варианте осуществления клетка-хозяин представляет собой прокариотическую клетку или эукариотическую клетку, и предпочтительно Escherichia coli.
В другом предпочтительном варианте осуществления целевой белок выбран из группы, состоящей из: инсулина, белка-предшественника человеческого инсулина, белка-предшественника инсулина лизпро, белка-предшественника инсулина гларгина, паратироидного гормона, кортикорелина, кальцитонина, бивалирудина, глюкагоноподобного пептида и его производных, экзенатида. и лираглутида, сомаглутида, зиконотида, серморелина, соматорелина, секретина, тедуглутида, гирудина, гормона роста, фактора роста, фактора, высвобождающего гормон роста, адренокортикотропного гормона, высвобождающего фактора, дезлорелина, десмопрессина, элкатонина, глюкалинагона, лейпрорелина, фактора, высвобождающего лютеинизирующий гормон, соматостатина, тиреотропин-высвобождающего гормона, трипторелина, вазоактивного интестинального пептида, интерферона, паратироидного гормона, пептида ВН3, амилоидного пептида или фрагмента вышеуказанного пептида, и любой их комбинации.
В пятом аспекте настоящего изобретения предлагается система экспрессии, которая включает в себя:
(a) мутантный белок по первому аспекту настоящего изобретения; и
(b) искусственную тРНК, способную связываться с предварительно заданной модифицированной аминокислотой в присутствии мутантного белка; и, необязательно
(c) первую последовательность нуклеиновой кислоты, кодирующей целевой белок, отличающуюся тем, что первая последовательность нуклеиновой кислоты содержит кодон, распознаваемый искусственной тРНК.
В другом предпочтительном варианте осуществления первая последовательность нуклеиновой кислоты содержит кодон, распознаваемый искусственной тРНК, в положении для введения предварительно заданной модифицированной аминокислоты.
В другом предпочтительном варианте осуществления система обработки дополнительно содержит:
(d) предварительно заданную модифицированную аминокислоту.
В другом предпочтительном варианте осуществления предварительно заданная модифицированная аминокислота является лизином с модифицированной группой.
В другом предпочтительном варианте осуществления модифицированная аминокислота выбрана из группы, состоящей из: производного алкинилоксикарбониллизина, производного трет-бутилоксикарборил-L-лизина (ВОС-лизина), ацилированного лизина жирного ряда и их комбинаций.
В другом предпочтительном варианте осуществления структура алкинилоксикарбониллизина представлена в формуле I:
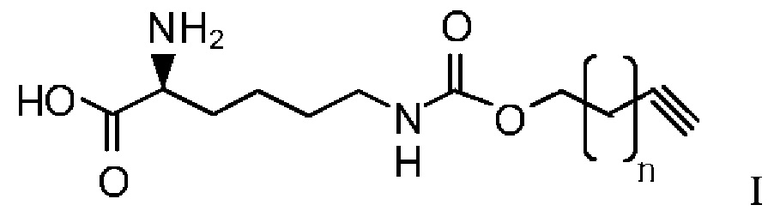
где n равно 0-8.
В другом предпочтительном варианте осуществления искусственная тРНК представляет собой супрессорную тРНК и предпочтительно супрессорную тРНК, подавляющую амбер-мутацию.
В другом предпочтительном варианте осуществления кодирующая последовательность нуклеиновой кислоты искусственной тРНК представлена в SEQ ID №: 7.
В другом предпочтительном варианте осуществления кодон, распознаваемый искусственной тРНК, представляет собой UAG (амбер), UAA (охра) или UGA (опал) и предпочтительно амбер-кодон.
В другом предпочтительном варианте осуществления система экспрессии представляет собой клетку или клеточный экстракт.
В другом предпочтительном варианте осуществления система экспрессии используется для введения предварительно заданной модифицированной аминокислоты в целевой белок или получения целевого белка, содержащего не встречающуюся в природе аминокислоту.
В шестом аспекте настоящего изобретения предлагается плазмидная система, которая включает в себя:
(1) первую кассету экспрессии, содержащую первую кодирующую последовательность для кодирования целевого белка, отличающуюся тем, что первая кодирующая последовательность содержит кодон для введения предварительно заданной модифицированной аминокислоты, а кодон представляет собой UAG (амбер), UAA (охра) или UGA (опал); и
(2) вторую кассету экспрессии, содержащую вторую последовательность нуклеиновой кислоты для кодирования аминоацил-тРНК-синтетазы, отличающуюся тем, что аминоацил-тРНК-синтетаза представляет собой мутантный белок по первому аспекту настоящего изобретения;
и система дополнительно содержит третью кассету экспрессии, которая содержит третью кодирующую последовательность для кодирования искусственной тРНК, отличающуюся тем, что искусственная тРНК содержит антикодон, соответствующий кодону;
причем, аминоацил-тРНК-синтетаза специфически катализирует искусственную тРНК с образованием комплекса «искусственная тРНК-Ха», где Ха представляет собой предварительно заданную модифицированную аминокислоту в аминоацильной форме.
В другом предпочтительном варианте осуществления плазмидная система представляет собой одноплазмидную систему или многоплазмидную систему.
В другом предпочтительном варианте осуществления многоплазмидная система включает в себя двухплазмидную систему, трехплазмидную систему и четырехплазмидную систему.
В другом предпочтительном варианте осуществления кодон представляет собой UAG (амбер) или UGA (опал).
В другом предпочтительном варианте осуществления кодон содержит трехосновную нуклеотидную последовательность, соответствующую аминокислоте в мРНК или ДНК.
В другом предпочтительном варианте осуществления предварительно заданная модифицированная аминокислота является лизином с модифицированной группой.
В другом предпочтительном варианте осуществления модифицированная аминокислота выбрана из группы, состоящей из: производного алкинилоксикарбониллизина, производного трет-бутилоксикарборил-L-лизина (ВОС-лизина), ацилированного лизина жирного ряда и их комбинаций.
В другом предпочтительном варианте осуществления первая кассета экспрессии, вторая кассета экспрессии и третья кассета экспрессии находятся в разных плазмидах, соответственно.
В другом предпочтительном варианте осуществления любые две или три из первой кассеты экспрессии, второй кассеты экспрессии и третьей кассеты экспрессии находятся в одной плазмиде.
В другом предпочтительном варианте осуществления плазмида представляет собой вектор экспрессии, выбранный из группы, состоящей из: pBAD/gIII ABC, pBAD/His ABC, pET28a, pETDuet-1 или вектора pEvol-pBpF.
В другом предпочтительном варианте осуществления плазмида дополнительно содержит в себе ген устойчивости, маркерную последовательность, ген-репрессор (araC), ген-промотор (araBAD) и их комбинации.
В другом предпочтительном варианте осуществления ген устойчивости выбран из группы, состоящей из гена устойчивости к ампициллину (AmpR), гена устойчивости к хлорамфениколу (CmR), гена устойчивости к канамицину (KanaR), гена устойчивости к тетрациклину (TetR) и их комбинаций.
В другом предпочтительном варианте осуществления плазмидная система представляет собой двухплазмидную систему.
В другом предпочтительном варианте осуществления первая кассета экспрессии находится в первой плазмиде, вторая кассета экспрессии находится во второй плазмиде, а третья кассета экспрессии находится в первой плазмиде или во второй плазмиде.
В другом предпочтительном варианте осуществления третья кассета экспрессии находится во второй плазмиде.
В другом предпочтительном варианте осуществления первая плазмида представляет собой вектор экспрессии, выбранный из группы, состоящей из pBAD-His ABC, pBAD/His ABC, pET28a и pETDuet-1.
В другом предпочтительном варианте осуществления первая плазмида, дополнительно, содержит ген устойчивости, маркерную последовательность, ген-репрессор (araC), ген-промотор (araBAD) и их комбинации.
В другом предпочтительном варианте осуществления вторая плазмида представляет собой вектор pEvol-pBpF.
В другом предпочтительном варианте осуществления вторая плазмида, дополнительно, содержит в себе ген устойчивости, маркерную последовательность, ген-репрессор (araC), ген-промотор (araBAD) и их комбинации.
В другом предпочтительном варианте осуществления положения первой кассеты экспрессии, второй кассеты экспрессии и третьей кассеты экспрессии в плазмиде каким бы то ни было образом не ограничиваются.
В другом предпочтительном варианте осуществления любые две или три из первой кассеты экспрессии, второй кассеты экспрессии и третьей кассеты экспрессии могут быть объединены в одну.
В другом предпочтительном варианте осуществления любые две из первой кассеты экспрессии, второй кассеты экспрессии и третьей кассеты экспрессии могут быть связаны посредством линкерной последовательности (например, IRES, Р2А, Т2А и т.д.).
В другом предпочтительном варианте осуществления первая кассета экспрессии, вторая кассета экспрессии и/или третья кассета экспрессии могут содержать или не содержать в себе промотор.
В другом предпочтительном варианте осуществления первая кассета экспрессии, вторая кассета экспрессии и/или третья кассета экспрессии дополнительно содержат один или несколько промоторов, причем промоторы функционально связаны с первой кодирующей последовательностью, второй кодирующей последовательностью, третьей кодирующей последовательностью, энхансерами, сигналами терминации транскрипции, последовательностями полиаденилирования, точками начала репликации, селектируемыми маркерами, сайтами рестрикции нуклеиновых кислот и/или сайтами гомологичной рекомбинации.
В другом предпочтительном варианте осуществления первая кассета экспрессии дополнительно содержит первый промотор, который, предпочтительно, является индуцируемым промотором.
В другом предпочтительном варианте осуществления первый промотор выбран из группы, состоящей из: промотора арабинозы (araBAD), промотора лактозы (Plac), промотора pLacUV5, промотора рТас и их комбинаций.
В другом предпочтительном варианте осуществления первая кассета экспрессии содержит промотор (например araBAD), сайт связывания рибосомы RBS, первую кодирующую последовательность, терминатор или маркерную последовательность в направлении от 5' до 3'.
В другом предпочтительном варианте осуществления вторая кассета экспрессии дополнительно содержит второй промотор, который, предпочтительно, является индуцибельным промотором.
В другом предпочтительном варианте осуществления второй промотор выбран из группы, состоящей из промотора арабинозы (araBAD), промотора glnS, промотора proK и их комбинаций.
В другом предпочтительном варианте осуществления вторая кассета экспрессии содержит промотор (например araBAD или glnS), сайт связывания рибосомы RBS, вторую кодирующую последовательность, терминатор (rrnB или glnS Т) в направлении от 5' до 3'.
В другом предпочтительном варианте осуществления третья кассета экспрессии дополнительно содержит третий промотор, который, предпочтительно, является конститутивным промотором.
В другом предпочтительном варианте осуществления третий промотор представляет собой промотор обратной транскрипции proK.
В другом предпочтительном варианте осуществления третья кассета экспрессии содержит промотор (например, proK), сайт связывания рибосомы RBS, третью кодирующую последовательность, терминатор или маркерную последовательность в направлении от 5' до 3'.
В седьмом аспекте настоящего изобретения предлагается клетка-хозяин или клеточный экстракт, содержащий систему экспрессии согласно пятому аспекту настоящего изобретения или плазмидную систему согласно шестому аспекту настоящего изобретения.
В другом предпочтительном варианте осуществления клетка-хозяин выбирается из группы, состоящей из: Escherichia coli, Bacillus subtilis, дрожжевой клетки, клетки насекомого, клетки млекопитающего и их комбинации.
В другом предпочтительном варианте осуществления клеточный экстракт извлекается из клетки, выбранной из группы, состоящей из: Escherichia coli, Bacillus subtilis, дрожжевой клетки, клетки насекомого, клетки млекопитающего и их комбинации.
В восьмом аспекте настоящего изобретения предлагается способ введения не встречающейся в природе аминокислоты в целевой белок или получения целевого белка, содержащего не встречающуюся в природе аминокислоту, который включает следующие этапы:
(1) предоставление клетки-хозяина согласно четвертому аспекту настоящего изобретения или системы экспрессии согласно пятому аспекту настоящего изобретения, или плазмидной системы согласно шестому аспекту настоящего изобретения; и
(2) в присутствии не встречающейся в природе аминокислоты экспрессия целевого белка с использованием клетки-хозяина, системы экспрессии или плазмидной системы.
В другом предпочтительном варианте осуществления целевой белок выбран из группы, состоящей из: белка-предшественника человеческого инсулина, белка-предшественника инсулина лизпро, белка-предшественника инсулина гларгина, паратироидного гормона, кортикорелина, кальцитонина, бивалирудина, глюкагоноподобного пептида и его производных, эксенатида и лираглутида, зиконотида, серморелина, соматорелина, секретина, тедуглутида, гирудина, гормона роста, фактора роста, фактора, высвобождающего гормон роста, адренокортикотропного гормона, высвобождающего фактора, дезлорелина, десмопрессина, элкатонина, глюкагона, лейпрорелина, фактора, высвобождающего лютеинизирующий гормон, соматостатина, тиреотропин-высвобождающего гормона, трипторелина, вазоактивного интестинального пептида, интерферона, паратироидного гормона, пептида ВНЗ, амилоидного пептида или фрагмента вышеуказанного пептида, и любой их комбинации.
В другом предпочтительном варианте осуществления не встречающаяся в природе аминокислота представляет собой лизин с модифицированной группой.
В другом предпочтительном варианте осуществления не встречающаяся в природе аминокислота выбрана из группы, состоящей из производного алкинилоксикарбониллизина, производного трет-бутилоксикарборил-L-лизина (ВОС-лизина), ацилированного лизина жирного ряда и их комбинаций.
В другом предпочтительном варианте осуществления метод включает в себя этапы:
(1) предоставление клетки-хозяина или клеточного экстракта согласно седьмому аспекту настоящего изобретения; и
(2) культивирование клетки или клеточного экстракта в присутствии не встречающейся в природе аминокислоты, что обеспечивает, таким образом, введение производного лизина в целевой белок посредством мутантного белка согласно первому аспекту настоящего изобретения и искусственной пары тРНК.
В девятом аспекте настоящего изобретения предлагается набор, содержащий (а) контейнер, и (b) мутантный белок согласно первому аспекту настоящего изобретения или полинуклеотид согласно второму аспекту настоящего изобретения, или вектор согласно третьему аспекту изобретения, или систему экспрессии согласно пятому аспекту изобретения, или плазмидную систему согласно шестому аспекту настоящего изобретения, которая расположена в контейнере.
В другом предпочтительном варианте осуществления набор дополнительно содержит клеточный экстракт.
В другом предпочтительном варианте осуществления плазмидная система представляет собой многоплазмидную систему, в которой соответствующие плазмиды расположены в одном или разных контейнерах.
В десятом аспекте настоящего изобретения предлагается система трансляции, которая включает в себя:
(a) мутантный белок согласно первому аспекту настоящего изобретения;
(b) искусственную тРНК, способную связываться с предварительно заданной модифицированной аминокислотой в присутствии мутантного белка; и
(c) заданную модифицированную аминокислоту.
В одиннадцатом аспекте настоящего изобретения предлагается использование мутантного белка в соответствии с первым аспектом настоящего изобретения или клетки-хозяина в соответствии с четвертым аспектом настоящего изобретения или плазмидной системы в соответствии с шестым аспектом настоящего изобретения, или набора согласно девятому аспекту настоящего изобретения для включения не встречающейся в природе аминокислоты в целевой белок или для получения целевого белка, содержащего не встречающуюся в природе аминокислоту.
Двенадцатый объект настоящего изобретения касается способа продуцирования мутантного белка согласно первому объекту настоящего изобретения, содержащего этапы: (i) культивирование клетки-хозяина согласно четвертому аспекту настоящего изобретения, что обеспечивает, таким образом, экспрессию мутантного белка.
В тринадцатом аспекте настоящего изобретения предлагается ферментный препарат, содержащий мутантный белок в соответствии с первым аспектом настоящего изобретения.
В другом предпочтительном варианте осуществления лекарственная форма фармацевтического препарата включает в себя: лиофилизированный препарат, жидкий препарат и их комбинации.
Следует понимать, что в пределах объема настоящего изобретения вышеупомянутые технические признаки настоящего изобретения и технические признаки, конкретно описанные ниже (например, в вариантах реализации), могут быть объединены друг с другом для образования нового или предпочтительного технического решения, которые не приведены поочередно повторно из-за ограничений его объема.
Описание чертежей
На Фигуре 1 показана карта плазмиды pEvol-suppylRs-pylT.
На Фигуре 2 представлена карта плазмиды pEvol-IPYEpylRs-pylT.
На Фигуре 3 показана карта плазмиды pEvol-optpylRs-pylT.
На Фигуре 4 показана карта плазмиды pEvol-IPYEpylRs (L274A, C313S, F349Y)-
pylT.
На Фигуре 5 показана карта плазмиды pBAD-araBAD [Al-u4-u5-TEV-R-MiniINS]-glnS [IPYEpylRs]-proK [pylT].
Подробное описание
После обширных и интенсивных исследований и массового скрининга, изобретатели неожиданно получили мутантную лизил-тРНК-синтетазу. По сравнению с лизил-тРНК-синтетазой дикого типа, мутантная лизил-тРНК-синтетаза по настоящему изобретению обладает высокой активностью, высоким уровнем экспрессии и хорошей растворимостью, а также может значительно увеличивать количество вставок не встречающихся в природе аминокислот и количеств экспрессии целевого белка, содержащего не встречающиеся в природе аминокислоты. Кроме того, мутантная лизил-тРНК-синтетаза по настоящему изобретению также может улучшать стабильность целевого белка, так что он не уязвим для расщепления. Исходя из этого, изобретатели подготовили полное настоящее изобретение.
Термины
Для упрощения понимания настоящего изобретения, сначала предлагаются формулировки определенных терминов. В случае из использования в данной заявке, если здесь явно не указано иное, каждый из следующих терминов имеет указанное ниже значение. Другие определения приводятся в заявке.
Термин «примерно» может относиться к значению или составу в пределах допустимого диапазона погрешностей конкретного значения или состава, определенного специалистом средней квалификации в данной области техники, что будет частично зависеть от того, как значение или состав измеряется или определяется. Например, в контексте данного документа, выражение «около 100» включает в себя все значения от 99 до 101 (такие как, 99,1; 99,2; 99,3; 99,4 и т.д.).
Используемые здесь термины «содержать» или «содержать в себе (или включать в себя)» могут быть раскрытыми, полураскрытыми и закрытыми. Другими словами, термины также включают в себя «в основном состоящий из» или «состоящий из».
Идентичность (или гомология) последовательностей определяется путем сравнения двух выровненных последовательностей в заданном окне сравнения (например, 50%, 60%, 70%, 80%, 90%, 95% или 100% длины эталонной нуклеотидной последовательности или белка) и путем определения количества положений, в которых появляется один и тот же остаток. Как правило, это выражается в процентах. Измерение идентичности нуклеотидных последовательностей является способом, хорошо известным специалистам в данной области техники.
В этой статье термины «аминоацил-тРНК-синтетаза» и «лизил-тРНК-синтетаза» используются как взаимозаменяемые.
Лизил-тРНК-синтетаза дикого типа
Используемый здесь термин «лизил-тРНК-синтетаза дикого типа» относится к естественно присутствующей аминоацил-тРНК-синтетазе, которая не была искусственно модифицирована, а ее нуклеотиды могут быть получены с помощью методов генной инженерии, таких как секвенирование генома, полимеразная цепная реакция (ПЦР) и т.д., а аминокислотная последовательность может быть выведена из нуклеотидной последовательности. Источник лизил-тРНК-синтетазы дикого типа особым образом не ограничивается. Предпочтительный источник включает в себя, но не ограничивается, Methanosarcina mazei, Methanosarcina barkeri и Methanosarcina acetivorans из Methanogenic archaea.
В предпочтительном варианте осуществления настоящего изобретения аминокислотная последовательность лизил-тРНК-синтетазы дикого типа представлена в SEQ ID №: 1.
MDKKPLNTLISATGLWMSRTGTIHKIKHHEVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSESYYNGISTNL (SEQ ID №: 1)
В предпочтительном варианте осуществления настоящего изобретения аминокислотная последовательность лизил-тРНК-синтетазы дикого типа представлена в SEQ ID №: 2.
MDKKPLNTLISATGLWMSRTGTIHKIKHHEVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPTRTKKAMPKSVARAPKPLENTEAAQAQPSGSKFSPAIPVSTQESVSVPASVSTSISSISTGATASALVKGNTNPITSMSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAARSGSYYNGISTNL (SEQ ID №: 2)
Мутантная лизил-тРНК-синтетаза
В контексте данного документа, термины «мутантный белок», «мутантный белок по настоящему изобретению», «мутантная аминоацил-тРНК-синтетаза по настоящему изобретению», «мутантная лизил-тРНК-синтетаза», «мутантный фермент», «мутантная аминоацил-тРНК-синтаза» могут использоваться взаимозаменяемо, и каждая из них относится к не встречающейся в природе мутантной аминоацил-тРНК-синтетазе, а мутантная аминоацил-тРНК-синтетаза представляет собой белок, искусственно модифицированный полипептидом, представленным в SEQ ID №: 1 или SEQ ID №: 2. В частности, мутантная аминоацил-тРНК-синтетаза описана в первом аспекте настоящего изобретения.
Следует понимать, что нумерация аминокислот в мутантной лизил-тРНК-синтетазе по настоящему изобретению основана на лизил-тРНК-синтетазе дикого типа (предпочтительно, SEQ ID №: 1 или SEQ ID №: 2). Когда конкретный мутантный белок имеет 80% или выше гомологию с последовательностью, предоставленной в SEQ ID №: 1 или SEQ ID №: 2, номер аминокислоты мутантного белка может быть неправильным относительно нумерации аминокислот SEQ ID №: 1 или SEQ ID №: 2, например, несовпадение положений 1-5 с N-концом или С-концом аминокислоты. Используя традиционные методы выравнивания последовательностей в данной области техники, специалисты в данной области техники могут, в целом, понять, что такое несовпадение находится в разумных пределах и мутантные белки с такой же или подобной ферментативной активностью с 80% или выше (например, 90%, 95%, 98%) гомологией не следует исключать из объема мутантного белка по настоящему изобретению из-за неправильной нумерации аминокислот.
Мутантный белок по настоящему изобретению представляет собой синтетический белок или рекомбинантный белок, т.е. он может быть химически синтезированным продуктом или получен из прокариотического или эукариотического хозяина (например, бактерий, дрожжей и растений) с использованием рекомбинантной технологии. В зависимости от хозяина, используемого в протоколе рекомбинантного продуцирования, мутантный белок по настоящему изобретению может быть гликозилированным или негликозилированным. Мутантный белок по настоящему изобретению может также включать в себя или не включать в себя исходный остаток метионина.
Настоящее изобретение также включает в себя фрагменты, производные и аналоги мутантного белка. Используемые здесь термины «фрагмент», «производное» и «аналог» относятся к белку, который, по существу, сохраняет ту же биологическую функцию или активность, что и мутантный белок.
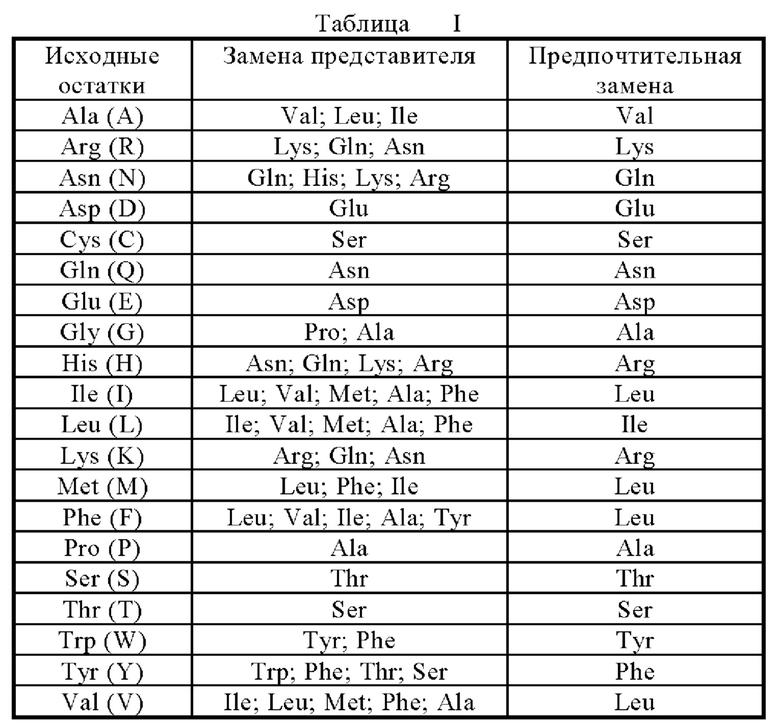
Фрагмент мутантного белка, производное или аналог по настоящему изобретению может быть (i) мутантным белком, в котором заменены один или несколько консервативных или неконсервативных аминокислотных остатков (предпочтительно, консервативных аминокислотных остатков), а такие замененные аминокислотные остатки могут или не могут кодироваться генетическим кодоном, или (ii) мутантным белком с группой замены в одном или нескольких аминокислотных остатках, или (iii) мутантным белком, образованным слиянием зрелого мутантного белка с другим соединением (например, соединение, продлевающее время полужизни мутантного белка, например, полиэтиленгликоль), или (iv) мутантным белком, образованным путем слияния дополнительной аминокислотной последовательности с этой мутантной белковой последовательностью (например, лидерная последовательность или секреторная последовательность или используемая секреторная последовательность для очистки этого мутантного белка или пропротеиновой последовательности, или слитого белка, образованного с фрагментом антигена IgG). Согласно изложенному здесь принципу, эти фрагменты, производные и аналоги находятся в пределах компетенции специалистов в данной области техники. В настоящем изобретении, консервативно замещенные аминокислоты, предпочтительно, получают путем аминокислотных замен согласно таблице I.
Узнавание аминокислотного субстрата PylRS связано со стереохимической структурой каталитически активного функционального домена. Размер производных лизина, которые могут быть активированы PylRS дикого типа, ограничен, а производные лизина с большими функциональными группами не могут быть введены в белки. Следовательно, эффект улучшается путем мутации сайта PylRS, чтобы избежать стерических несоответствий связывания субстрата или взаимодействия мутантной аминокислоты с аминокислотой субстрата или частью основной цепи.
Предпочтительно, мутантный белок представлен в любом из SEQ ID №: 3-6.
MDKKPLNTLISATGLWMSRTGTIHKIKHHEVSRSKIYIEMACGDHLVVNNSRSSRTARALRHHKYRKTCKRCRVSDEDLNKFLTKANEDQTSVKVKVVSAPRTKKAMPKSVARAPKPLENTEAAQASTQESVSVPASVSTSISSSAPVQASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAA RSESYYNGISTNL (SEQ ID №: 3)
MDKKPLDVLISATGLWMSRTGTLHKIKHHEISRSKIYIEMACGDHLVVNNSRSCRPARAFRYHKYRKTCKRCRVSDEDINNFLTRSTESKNSVKVRVVSEPKVKKAMPKSVSRAPKPLENSVSAKASTNTSRSVPSPAKSTPNSSVPASASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVFGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAAR SESYYNGISTNL (SEQ ID №: 4)
MDKKPLDVLISATGLWMSRTGTLHKIKHYEISRSKIYIEMACGDHLVVNNSRSCRPARAFRYHKYRKTCKRCRVSGEDINNFLTRSTEGKTSVKVKVVSEPKVKKAMPKSVSRAPKPLENPVSAKASTDTSRSVPSPAKSTPNSPVPTSASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYLRKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFCQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVFGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAAR SESYYNGISTNL (SEQ ID №: 5)
MDKKPLDVLISATGLWMSRTGTLHKIKHHEISRSKIYIEMACGDHLVVNNSRSCRPARAFRYHKYRKTCKRCRVSDEDINNFLTRSTESKNSVKVRVVSEPKVKKAMPKSVSRAPKPLENSVSAKASTNTSRSVPSPAKSTPNSSVPASASAPALTKSQTDRLEVLLNPKDEISLNSGKPFRELESELLSRRKKDLQQIYAEERENYLGKLEREITRFFVDRGFLEIKSPILIPLEYIERMGIDNDTELSKQIFRVDKNFCLRPMLAPNLYNYARKLDRALPDPIKIFEIGPCYRKESDGKEHLEEFTMLNFSQMGSGCTRENLESIITDFLNHLGIDFKIVGDSCMVYGDTLDVMHGDLELSSAVVGPIPLDREWGIDKPWIGAGFGLERLLKVKHDFKNIKRAAR SESYYNGISTNL (SEQ ID №: 6)
Следует понимать, что по сравнению с последовательностью, предоставленной в SEQ ID №: 1 или SEQ ID №: 2, мутантный белок по настоящему изобретению, как правило, имеет высокую гомологию (или идентичность). Предпочтительно, гомология между мутантным белком и пептидным фрагментом соответствующей длины, присутствующим в SEQ ID №. 1 или 2 составляет, не менее, 90%, 95%, 96%, 97%, 98% или 99%.
Кроме того, мутантный белок по настоящему изобретению также можно модифицировать. Модификация (как правило, не изменяющая первичную структуру) включает в себя: химически полученные формы мутантного белка in vivo или in vitro, например ацетилирование или карбоксилирование. Модификации также включают в себя гликозилирование, например, производимое модификацией гликозилирования во время синтеза и процессинга мутантного белка или во время дальнейших этапов процессинга. Эта модификация может быть достигнута путем воздействия на мутантный белок фермента, выполняющего гликозилирование (например, гликозилазы или дегликозилазы млекопитающих). Модификация также включает в себя последовательности, содержащие фосфорилированные аминокислотные остатки (например фосфотирозин, фосфосерин, фосфотреонин). Он также включает в себя мутантные белки, модифицированные для повышения их устойчивости к протеолизу или для оптимизации их растворимости.
Термин «полинуклеотид, кодирующий мутантный белок по настоящему изобретению» может включать в себя полинуклеотид, кодирующий мутантную лизил-тРНК-синтетазу по настоящему изобретению, или также может быть полинуклеотидом, который также включает в себя дополнительные кодирующие и/или некодирующие последовательности.
Настоящее изобретение также относится к вариантам вышеуказанных полинуклеотидов, кодируютщих фрагменты, аналоги и производные полипептидов или мутантных белков, имеющих такую же аминокислотную последовательность, что и в настоящем изобретении. Эти варианты нуклеотидов включают в себя варианты замены, варианты делеции и варианты вставки. Как известно в данной области техники, аллельный вариант представляет собой альтернативную форму полинуклеотида. Это может быть замена, делеция или вставка одного или нескольких нуклеотидов, но это не приведет к существенному изменению функции кодируемого им мутантного белка.
Настоящее изобретение также относится к полинуклеотидам, гибридизующихся с вышеупомянутыми последовательностями и имеющих, по меньшей мере, 50%, предпочтительно, по меньшей мере, 70% и, более предпочтительно, по меньшей мере, 80% идентичности между двумя последовательностями. Настоящее изобретение, в частности, относится к полинуклеотидам, которые могут гибридизоваться с полинуклеотидом по настоящему изобретению в жестких условиях (или строгих условиях). В настоящем изобретении «жесткое условие» относится к: (1) гибридизации и элюции при более низкой ионной силе и более высокой температуре, например, 0,2×SSC, 0,1% SDS, 60°С; или (2) добавлению денатуранта во время гибридизации, например 50% (об./об.) формамида, 0,1% сыворотки теленка/0,1% фиколла, 42°С и т.д.; или (3) гибридизация происходит только если идентичность между двумя последовательностями составляет, по меньшей мере, 90% или более, а более предпочтительно, 95% или более.
Мутантный белок и полинуклеотид по настоящему изобретению, предпочтительно, представлены в изолированной форме и, более предпочтительно, очищены до гомогенности.
Полноразмерную последовательность полинуклеотида по настоящему изобретению, как правило, можно получить способом ПЦР-амплификации, способом рекомбинации или способом искусственного синтеза. Для способа ПЦР-амплификации, праймеры могут быть сконструированы в соответствии с релевантной нуклеотидной последовательностью, раскрытой в настоящем изобретении, в частности, с последовательностью открытой рамки считывания и с использованием коммерчески доступной библиотеки кДНК или библиотеки кДНК, приготовленной обычным методом, известным специалистам, в данной области техники в качестве шаблона усиления соответствующей последовательности. Если последовательность длинная, часто необходимо выполнить две или более амплификации ПЦР, а затем, полученные каждый раз амплифицированные фрагменты связываются вместе в правильном порядке.
Непосредственно после получения соответствующей последовательности, способ рекомбинации можно использовать для получения соответствующей последовательности в больших количествах. Как правило, это осуществляется путем клонирования в вектор, последующего переноса в клетку и выделения соответствующей последовательности из пролиферированной клетки-хозяина общепринятыми способами.
Кроме того, способы искусственного синтеза также можно использовать для синтеза родственных последовательностей, в частности если длина фрагмента мала. Как правило, фрагменты с очень длинными последовательностями можно получить путем синтеза нескольких небольших фрагментов с последующим их лигированием.
В настоящее время, последовательность ДНК, кодирующая белок (или его фрагмент или производное) по настоящему изобретению, может быть полностью получена посредством химического синтеза. Затем, последовательность ДНК может быть введена в различные существующие молекулы ДНК (или, например, векторы) и клетки, известные в данной области техники. Кроме того, мутации также могут быть введены в последовательность белка по настоящему изобретению посредством химического синтеза.
Способ использования технологии ПЦР для амплификации ДНК/РНК предпочтительно используется для получения полинуклеотида по настоящему изобретению. В частности, в случае трудности получения полноразмерной кДНК из библиотеки, предпочтительно может использоваться способ RACE (способ быстрой амплификации RACE-кДНК конца) а праймеры, используемые для ПЦР, могут быть надлежащим образом выбраны в соответствии с информацией о последовательностях по настоящему изобретению, указанному в этом документе, и которые могут быть синтезированы обычными методами. Амплифицированные фрагменты ДНК/РНК можно разделить и очистить обычными методами, например гель-электрофорезом.
Вектор экспрессии
Используемый здесь термин «конструкция» или «вектор», как правило, относится к нуклеиновой кислоте, способной транспортировать кодирующую последовательность целевого белка, к которому она присоединена. Одним из типов вектора является «плазмида», которая относится к кольцевой двухцепочечной петле ДНК, способной соединять дополнительные сегменты ДНК.
Вектор можно трансформировать или трансфицировать в подходящую клетку-хозяин для обеспечения экспрессии белка. Этот процесс может включать в себя культивирование клетки-хозяина, трансформированной вектором экспрессии, в условиях, обеспечивающих экспрессию вектора, кодирующего последовательность целевой нуклеиновой кислоты белка, и, необязательно, выделение экспрессированного белка.
Вектор может быть, например, плазмидным или вирусным вектором, обеспечивающим начало репликации, необязательно, промотором, используемым для экспрессии последовательности нуклеиновой кислоты-мишени и, необязательно, регулятором промотора. В случае с бактериальными плазмидами вектор может включать в себя один или несколько селективных маркерных генов, например, гены устойчивости к канамицину.
Настоящее изобретение также относится к вектору, содержащему полинуклеотид по настоящему изобретению, к клетке-хозяину, полученной с помощью генной инженерии с использованием вектора по настоящему изобретению или кодирующей последовательности мутантного белка по настоящему изобретению, и к способу получения полипептида по настоящему изобретению с помощью рекомбинантной технологии.
При использовании общепринятой технологии рекомбинантной ДНК полинуклеотидная последовательность по настоящему изобретению может быть использована для экспрессии или получения рекомбинантного мутантного белка. Как правило, к ним относятся следующие этапы:
(1) использование полинуклеотида (или варианта) по настоящему изобретению, кодирующего мутантный белок по настоящему изобретению, или использование рекомбинантного вектора экспрессии, содержащего полинуклеотид, для трансформации или трансфекции подходящей клетки-хозяина;
(2) культивирование клетки-хозяина в подходящей среде;
(3) отделение и очистка белка от культурной среды или клеток.
В настоящем изобретении, полинуклеотидная последовательность, кодирующая мутантный белок, может быть вставлена в рекомбинантный вектор экспрессии. Термин «рекомбинантный вектор экспрессии» относится к бактериальным плазмидам, бактериофагам, дрожжевым плазмидам, вирусам клеток растений, вирусам клеток млекопитающих, например аденовирус, ретровирус или другим векторам, хорошо известным в данной области техники. Можно использовать любую плазмиду и вектор, если они могут реплицироваться и стабилизироваться в хозяине. Важной особенностью вектора экспрессии является то, что он, как правило, включает в себя точку начала репликации, промотор, маркерный ген и элементы управления трансляцией.
Способы, хорошо известные специалистам в данной области техники, можно использовать для конструирования вектора экспрессии, содержащего последовательность ДНК, кодирующую мутантный белок по настоящему изобретению, и соответствующие сигналы контроля транскрипции/трансляции. Эти методы включают в себя технологию рекомбинантной ДНК in vitro, технологию синтеза ДНК и технологию рекомбинации in vivo. Последовательность ДНК может быть эффективно связана с подходящим промотором в векторе экспрессии для управления синтезом мРНК. Характерные примеры этих промоторов включают в себя: Промотор lac или trp Escherichia coli; промотор PL фага лямбда; эукариотические промоторы, включая немедленно-ранний промотор CMV, промотор тимидинкиназы HSV, ранний и поздний промоторы SV40, ретровирусные LTR и некоторые другие известные промоторы, которые могут контролировать экспрессию генов в прокариотических или эукариотических клетках или вирусах. Вектор экспрессии также включает в себя сайт связывания рибосомы для инициации трансляции, а также терминатор транскрипции.
Кроме того, вектор экспрессии, предпочтительно, содержит в себе один или несколько селективных маркерных генов для обеспечения фенотипических признаков для отбора трансформированных клеток-хозяев, например дигидрофолатредуктазу, устойчивость к неомицину и зеленый флуоресцентный белок (GFP) для культуры эукариотических клеток, или устойчивость к тетрациклину или ампициллину для Escherichia coli.
Вектор, содержащий вышеупомянутую подходящую последовательность ДНК и соответствующий промотор или контрольную последовательность, можно использовать для трансформации подходящей клетки-хозяина так, чтобы она могла экспрессировать белок.
Клетка-хозяин может быть прокариотической клеткой, например, бактериальной клеткой; или низшей эукариотической клеткой, например, дрожжевой клеткой; или высшей эукариотической клеткой, например клеткой млекопитающего. Характерные примеры включают в себя: Escherichia coli, Streptomyces; бактериальные клетки Salmonella typhimurium; грибковые клетки, например, дрожжевые и растительные клетки (например, клетки женьшеня).
Когда полинуклеотид по настоящему изобретению экспрессируется в клетках высших эукариот, в случае если в вектор вставлена энхансерная последовательность, транскрипция будет усилена. Энхансеры - это действующие в цис-положении факторы ДНК, как правило, от 10 до 300 пар оснований, действующие на промоторы для усиления транскрипции генов. Примеры включают в себя от 100 до 270 пар оснований энхансера SV40 на участке поздней точки начала репликации, энхансер полиомы на участке поздней точки начала репликации и энхансеры аденовируса и т.п.
Специалисты в данной области техники знают, как выбрать подходящие векторы, промоторы, энхансеры и клетки-хозяева.
Трансформация клеток-хозяев рекомбинантной ДНК может осуществляться обычными способами, хорошо известными специалистам в данной области техники. В случае если хозяином является прокариот, например Escherichia coli, компетентные клетки, способные поглощать ДНК, могут быть собраны после фазы экспоненциального роста и обработаны способом CaCl2. Используемые этапы хорошо известны в данной области техники. Другой способ - использование MgCl2. В случае необходимости трансформация также может быть выполнена электропорацией. В случае если хозяином является эукариот, можно выбрать следующие способы трансфекции ДНК: способ соосаждения фосфата кальция, обычные механические способы, например микроинъекция, электропорация, упаковка липосом и т.д.
Полученные трансформанты можно культивировать обычными способами для экспрессии полипептида, кодируемого геном по настоящему изобретению. В зависимости от используемой клетки-хозяина среда, используемая в культуре, может быть выбрана из различных обычных сред. Культивирование проводят в условиях, подходящих для роста клетки-хозяина. После того, как клетки-хозяева вырастут до подходящей плотности клеток, выбранный промотор индуцируют подходящим способом (например, преобразование температуры или химическая индукция), а клетки культивируют в течение определенного периода времени.
Рекомбинантный полипептид в указанном выше способе может экспрессироваться в клетке или на клеточной мембране или секретироваться из клетки. В случае необходимости, используя физические, химические и другие характеристики, можно отделить и очистить рекомбинантный белок с помощью различных способов разделения. Этим способы хорошо известны специалистам в этой области техники. Примеры этих методов включают в себя, но не ограничиваются: обычная обработка ренатурацией, обработка агентом осаждения белка (способ высаливания), центрифугирование, уничтожение бактерий посредством осмоса, ультраобработка, ультрацентрифугирование, хроматография на молекулярных ситах (гель-фильтрация), адсорбционная хроматография, ионообменная хроматография, высокоэффективная жидкостная хроматография (ВЭЖХ) и различные другие методы жидкостной хроматографии и их комбинации.
Термин «функционально связанный» относится к связыванию транскрибируемого и экспрессируемого целевого гена с его регуляторной последовательностью обычными способами в данной области техники для экспрессии целевого гена.
Плазмидная система
Настоящее изобретение предоставляет плазмидную систему, как это описано в шестом аспекте настоящего изобретения. В частности, плазмидная система включает в себя:
(1) первую кассету экспрессии, содержащую первую кодирующую последовательность для кодирования целевого белка, отличающуюся тем, что первая кодирующая последовательность содержит кодон для введения предварительно заданной модифицированной аминокислоты, а кодон представляет собой UAG (амбер), UAA (охра) или UGA (опал); и
(2) вторую кассету экспрессии, содержащую вторую последовательность нуклеиновой кислоты для кодирования аминоацил-тРНК-синтетазы, отличающуюся тем, что аминоацил-тРНК-синтетаза представляет собой мутантный белок по первому аспекту настоящего изобретения;
и система дополнительно содержит третью кассету экспрессии, которая содержит третью кодирующую последовательность для кодирования искусственной тРНК, отличающуюся тем, что искусственная тРНК содержит антикодон, соответствующий кодону;
причем, аминоацил-тРНК-синтетаза специфически катализирует искусственную тРНК с образованием комплекса «искусственная тРНК-Ха», где Ха представляет собой предварительно заданную модифицированную аминокислоту в аминоацильной форме.
В другом предпочтительном варианте осуществления плазмидная система представляет собой одноплазмидную систему или многоплазмидную систему.
В другом предпочтительном варианте осуществления многоплазмидная система включает в себя двухплазмидную систему, трехплазмидную систему и четырехплазмидную систему.
В другом предпочтительном варианте осуществления первая кассета экспрессии, вторая кассета экспрессии и третья кассета экспрессии находятся в разных плазмидах, соответственно.
В другом предпочтительном варианте осуществления любые две или три из первой кассеты экспрессии, второй кассеты экспрессии и третьей кассеты экспрессии находятся в одной плазмиде.
В другом предпочтительном варианте осуществления плазмида представляет собой вектор экспрессии, выбранный из группы, состоящей из: pBAD/gIII ABC, pBAD/His ABC, pET28a, pETDuet-1 или вектора pEvol-pBpF.
В другом предпочтительном варианте осуществления плазмида дополнительно содержит в себе ген устойчивости, маркерную последовательность, ген-репрессор (araC), ген-промотор (araBAD) и их комбинации.
В другом предпочтительном варианте осуществления ген устойчивости выбран из группы, состоящей из гена устойчивости к ампициллину (AmpR), гена устойчивости к хлорамфениколу (CmR), гена устойчивости к канамицину (KanaR), гена устойчивости к тетрациклину (TetR) и их комбинаций.
В другом предпочтительном варианте осуществления последовательность ДНК промотора glnS соответствует представленной в SEQ ID №: 10.
TTTTAAAAAACTAACAGTTGTCAGCCTGTCCCGCTTTAATATCATACGCCGTTATACGTTGTTTACGCTTTG (SEQ ID №: 10)
В другом предпочтительном варианте осуществления последовательность ДНК терминатора glnS Т соответствует представленной в SEQ ID №: 11.
CAAACAATCCAAAACGCCGCGTTCAGCGGCGTTTTTTCTGCTTTT (SEQ ID №:
И)
В другом предпочтительном варианте осуществления последовательность ДНК промотора proK соответствует представленной в SEQ ID №: 12.
TGTGCTTCTCAAATGCCTGAGGCCAGTTTGCTCAGGCTCTCCCCGTGGAGGTAATAATTGACGATATGATCAGTGCACGGCTAACTAAGCGGCCTGCTGACTTTCTCGCCGATCAAAAGGCATTTTGCTATTAAGGGATTGACGAGGGCGT (SEQ ID №: 12)
В другом предпочтительном варианте осуществления последовательность ДНК терминатора proK Т представлена в SEQ ID №: 13.
AATTCGAAAAGCCTGCTCAACGAGCAGGCTTTTTTGCATG (SEQ ID №: 13)
По сравнению с предшествующим уровнем техники, настоящее изобретение, в основном, имеет следующие преимущества:
После большого количества скринингов и модификаций, в настоящем изобретении впервые было обнаружено, что усеченная мутантная аминоацил-тРНК синтетаза (например, SEQ ID №: 3) полученная делецией аминокислотных остатков в положении 102, положениях 128-140 и положениях 159-179, соответствующих аминоацил-тРНК-синтетазы дикого типа (например, SEQ ID №: 1 или 2) может не только вводить производные лизина с большими функциональными группами в белки, но также значительно увеличивать активность фермента и обладает высоким уровнем экспрессии и хорошей растворимостью. Это может значительно увеличить количество вставок не встречающихся в природе аминокислот и количество экспрессии целевого белка, содержащего не встречающиеся в природе аминокислоты.
Кроме того, мутантная лизил-тРНК-синтетаза по настоящему изобретению также может улучшать стабильность целевого белка, так что его трудно разрушить.
В настоящем изобретении также обнаружено, что мутантный белок (например, SEQ ID №: 4-6), полученный путем мутации в других положениях усеченной мутантной аминоацил-тРНК-синтетазы, может дополнительно увеличить количество вставки не встречающихся в природе аминокислот, количество целевого белка, содержащего не встречающиеся в природе аминокислоты, и/или стабильность целевого белка.
Настоящее изобретение будет дополнительно проиллюстрировано ниже со ссылкой на конкретные примеры. Следует понимать, что эти примеры предназначены исключительно для иллюстрации настоящего изобретения, но не для ограничения объема настоящего изобретения. Экспериментальные методы без конкретных условий, описанные в следующих примерах, обычно выполняются в обычных условиях (например, условиях, описанных Самбрук и соавт., Molecular Cloning: Лабораторное руководство (Нью-Йорк: Cold Spring Harbor Laboratory Press, 1989), или в соответствии с инструкциями производителя. Если не указано иное, все процентные доли и части рассчитываются по массе.
Если не указано иное, экспериментальные материалы и реагенты, используемые в следующих примерах, доступны на рынке.
Пример 1 Конструирование мутантов
Согласно аминокислотной последовательности 1 (SEQ ID №: 1) лизил-тРНК-синтетаза pylR дикого типа и неслучайное использование кодонов Escherichia coli, последовательность pylR ДНК (SEQ ID №: 8) была синтезирована и клонирована в сайт Spel-Sall ниже промотора araBAD плазмиды вектора экспрессии pEvol-pBpF (приобретена у компании NTCC, устойчивость к хлорамфениколу). Сайт рестрикции Spel был увеличен с помощью ПЦР, а сайт Sail был характерен вектору. Исходный глутаминовый промотор glnS плазмиды вектора экспрессии pEvol-pBpF был сохранен. За промотором proK плазмиды вектора экспрессии pEvol-pBpF последовательность ДНК (SEQ ID №: 7) не встречающейся в природе тРНК (pylT) вставляли с помощью ПЦР.
GGAAACCTGATCATGTAGATCGAATGGACTCTAAATCCGTTCAGCCGGGTTAGATTCCCGGGGTTTCCGCC A (SEQ ID №: 7)
Последовательность pylR ДНК (SEQ ID №: 8), 1365bp
ATGGATAAGAAACCGCTGAATACTCTGATTTCTGCAACTGGTCTGTGGATGAGCCGTACCGGCACCATCCACAAGATCAAACACCACGAGGTTTCCCGTAGCAAAATCTACATCGAAATGGCGTGCGGTGACCACCTGGTGGTAAACAACTCCCGTTCTTCTCGTACTGCACGTGCTCTGCGCCACCACAAGTACCGTAAGACCTGCAAGCGCTGTCGCGTGTCTGATGAAGACCTGAACAAATTCCTGACTAAAGCGAACGAAGATCAGACTTCTGTGAAGGTGAAAGTTGTTTCTGCCCCAACCCGCACCAAGAAAGCGATGCCGAAGTCCGTTGCACGCGCTCCGAAACCGCTGGAGAACACCGAAGCCGCACAGGCCCAGCCGTCTGGTTCTAAGTTTTCTCCGGCAATCCCGGTTTCTACTCAGGAGTCTGTGTCTGTGCCAGCTTCTGTTAGCACTTCTATTTCCTCTATCAGCACTGGTGCGACTGCGTCCGCTCTGGTAAAAGGTAACACTAACCCGATCACCAGCATGTCTGCTCCGGTTCAGGCTTCTGCACCGGCACTGACTAAAAGCCAGACTGACCGTCTGGAGGTTCTGCTGAACCCGAAAGATGAAATCAGCCTGAACTCTGGCAAACCGTTCCGTGAACTGGAATCCGAACTGCTGTCTCGTCGTAAGAAAGACCTGCAACAAATCTATGCTGAAGAGCGTGAAAACTACCTGGGTAAACTGGAACGTGAAATCACCCGTTTCTTTGTGGACCGTGGTTTCCTGGAAATCAAGTCTCCGATCCTGATCCCGCTGGAATACATCGAGCGCATGGGTATTGATAACGACACCGAACTGTCCAAGCAGATTTTCCGTGTGGACAAGAACTTCTGCCTGCGTCCGATGCTGGCACCGAACCTGTACAATTACCTGCGTAAACTGGATCGTGCACTGCCGGACCCGATCAAAATCTTTGAAATCGGTCCATGCTATCGTAAGGAGAGCGACGGTAAAGAACACCTGGAAGAGTTCACTATGCTGAACTTTTGTCAGATGGGTTCTGGCTGCACCCGTGAAAATCTGGAATCTATCATCACCGACTTCCTGAACCACCTGGGCATTGACTTCAAAATCGTTGGTGATTCCTGCATGGTTTACGGTGACACTCTGGACGTTATGCATGGTGATCTGGAACTGAGCAGCGCTGTTGTGGGTCCGATTCCGCTGGATCGTGAATGGGGTATCGATAAACCGTGGATTGGTGCTGGCTTCGGTCTGGAACGTCTGCTGAAAGTTAAGCACGACTTTAAGAACATCAAACGTGCTGCGCGTTCCGAGTCCTATTACAACGGCATTAGCACTAACCTGTAA (SEQ ID №: 8)
Последовательность вырезали из вектора клонирования с помощью рестрикционныз ферментов Spel и Sail, а плазмиду pEvol-pylRs-pylT вырезали с помощью Spel и Sail (целевой ДНК-фрагмент представлял собой большой фрагмент размером 4,3 т.п.н.). Затем, продукты разделяли электрофорезом нуклеиновых кислот, экстрагировали набором для восстановления ДНК из агарозного геля, лигировали ДНК-лигазой Т4 и трансформировали в большие компетентные клетки E.coli Тор 10 химическим способом (способ CaCl2). Трансформированные клетки культивировали на среде агара Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl, 1,5% агар), содержащей хлорамфеникол, при 37°С в течение ночи. Отбирали единственную живую колонию и культивировали в жидкой среде Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl), содержащей хлорамфеникол, при 37°С и 220 об/мин в течение ночи. Плазмиду экстрагировали с помощью набора Plasmid Mini Extraction Kit, а полученную плазмиду назвали pEvol-pylRs-pylT.
Согласно аминокислотной последовательности (SEQ ID №: 1) лизил-тРНК-синтетазы pylR дикого типа, 3 исходные последовательности (т.е. 102Т, 128Q-140V и 159I-179M; всего 35 аминокислот) были нокаутированы для получения аминокислотной последовательности (SEQ ID №: 3) синтетазы suppylRs. Плазмида была названа pEvol-suppylRs-pylT, карта плазмиды показана на фигуре 1.
Согласно аминокислотной последовательности (SEQ ID №: 1) лизил-тРНК-синтетазы pylR дикого типа, весь сегмент аминокислот в положениях 1-184 был заменен другим сегментом из 149 аминокислот, при этом 3 аминокислотные последовательности (т.е. 102Т, 128Q-140V и 1591 -179М; всего 35 аминокислот), соответствующие лизил-тРНК-синтетазе pylR дикого типа, также были нокаутированы, как и в плазмиде pEvol-suppylRs-pylT. Была получена аминокислотная последовательность (SEQ ID №: 4) синтетазы IPYEpylRs, а последовательность ДНК (SEQ ID №: 9) IPYEpylR была синтезирована в соответствии с неслучайным использованием кодонов Escherichia coli. Плазмида была названа pEvol-IPYEpylRs-pylT, а карта плазмиды показана на фигуре 2.
Последовательность ДНК IPYEpylR (SEQ ID №: 9), 1260bp
ATGGATAAGAAGCCACTGGATGTTCTGATTTCCGCTACTGGTCTGTGGATGTCTCGCACTGGTACTCTGCACAAGATCAAACACCACGAAATCTCTCGCTCCAAGATCTACATTGAAATGGCTTGTGGTGATCACCTGGTTGTTAACAACTCCCGCTCTTGTCGCCCGGCTCGTGCGTTTCGCTATCACAAATATCGTAAAACCTGCAAACGCTGCCGCGTGAGCGATGAGGATATTAACAACTTCCTGACTCGCTCCACCGAGAGCAAGAACTCTGTGAAAGTTCGCGTAGTTTCTGAACCGAAAGTTAAGAAAGCTATGCCGAAGTCTGTTAGCCGTGCTCCGAAACCGCTGGAGAACTCTGTGTCCGCGAAAGCGAGCACCAACACCAGCCGTTCTGTTCCGTCTCCAGCGAAATCTACTCCGAACTCTAGCGTGCCAGCTTCCGCTTCTGCACCGGCACTGACTAAAAGCCAGACTGACCGTCTGGAGGTTCTGCTGAACCCGAAAGATGAAATCAGCCTGAACTCTGGCAAACCGTTCCGTGAACTGGAATCCGAACTGCTGTCTCGTCGTAAGAAAGACCTGCAACAAATCTATGCTGAAGAGCGTGAAAACTACCTGGGTAAACTGGAACGTGAAATCACCCGTTTCTTTGTGGACCGTGGTTTCCTGGAAATCAAGTCTCCGATCCTGATCCCGCTGGAATACATCGAGCGCATGGGTATTGATAACGACACCGAACTGTCCAAGCAGATTTTCCGTGTGGACAAGAACTTCTGCCTGCGTCCGATGCTGGCACCGAACCTGTACAATTACCTGCGTAAACTGGATCGTGCACTGCCGGACCCGATCAAAATCTTTGAAATCGGTCCATGCTATCGTAAGGAGAGCGACGGTAAAGAACACCTGGAAGAGTTCACTATGCTGAACTTTTGTCAGATGGGTTCTGGCTGCACCCGTGAAAATCTGGAATCTATCATCACCGACTTCCTGAACCACCTGGGCATTGACTTCAAAATCGTTGGTGATTCCTGCATGGTTTTCGGTGACACTCTGGACGTTATGCATGGTGATCTGGAACTGAGCAGCGCTGTTGTGGGTCCGATTCCGCTGGATCGTGAATGGGGTATCGATAAACCGTGGATTGGTGCTGGCTTCGGTCTGGAACGTCTGCTGAAAGTTAAGCACGACTTTAAGAACATCAAACGTGCTGCGCGTTCCGAGTCCTATTACAACGGCATTAGCACTAACCTGTAA (SEQ ID №: 9)
Согласно аминокислотной последовательности (SEQ ID №: 4) лизил-тРНК-синтетазы дикого типа, были введены мутации IPYEpylRs, H29Y, D76G, D89G, N9IT, R96K, D121P, N129D, S145P и А148Т для получения мутантной лизил-тРНК-синтетазы optpylRs с аминокислотной последовательностью, предоставленной в SEQ ID №: 5. Плазмида была названа pEvol-optpylRs-pylT, а карта плазмиды показана на фигуре 3.
Согласно аминокислотной последовательности (SEQ ID №: 4) лизил-тРНК-синтетазы дикого типа, были введены мутации IPYEpylRs, L274A, C313S и F349Y, в результате чего были получена мутантная лизил-тРНК-синтетаза IPYEpylRs (L274A, C313S, F349Y) с аминокислотной последовательностью, предоставленной в SEQ ID №: 6. Плазмида была названа pEvol-IPYEpylRs (L274A, C313S, F349Y)-pylT, а карта плазмиды показана на фигуре 4.
Пример 2 Конструирование штамма для экспрессии одноплазмидного мутанта
Согласно аминокислотной последовательности (SEQ ID №: 4) лизил-тРНК-синтетазы IPYEpylRs дикого типа, в соответствии с последовательностью ДНК (SEQ ID №: 7) не встречающейся в природе тРНК (pylT), в соответствии с последовательностями ДНК (SEQ ID №: 10-13) промотора глутамина glnS и промотора proK, полученных из вектора pEvol-pBpF и их соответствующего терминатора и, в соответствии с неслучайным использованием кодонов Escherichia coli, последовательность ДНК (SEQ ID №: 14) of glnS [pylRs]-proK [pylT] была синтезирована и клонирована в сайт AvrII-XbaI после терминатора rrnB плазмиды pBAD-Al-u4-u5-TEV-R-MiniINS (созданная нашей компанией плазмида устойчивости к канамицину), в которой сайты рестрикции AvrII и XbaI были введены с помощью ПЦР.
Последовательность ДНК glnS [pylRs]-proK [pylT]: (SEQ ID №: 14)
TTTTAAAAAACTAACAGTTGTCAGCCTGTCCCGCTTTAATATCATACGCCGTTATACGTTGTTTACGCTTTGAGGAATCCCATATGGATAAGAAACCGCTGAATACTCTGATTTCTGCAACTGGTCTGTGGATGAGCCGTACCGGCACCATCCACAAGATCAAACACCACGAGGTTTCCCGTAGCAAAATCTACATCGAAATGGCGTGCGGTGACCACCTGGTGGTAAACAACTCCCGTTCTTCTCGTACTGCACGTGCTCTGCGCCACCACAAGTACCGTAAGACCTGCAAGCGCTGTCGCGTGTCTGATGAAGACCTGAACAAATTCCTGACTAAAGCGAACGAAGATCAGACTTCTGTGAAGGTGAAAGTTGTTTCTGCCCCAACCCGCACCAAGAAAGCGATGCCGAAGTCCGTTGCACGCGCTCCGAAACCGCTGGAGAACACCGAAGCCGCACAGGCCCAGCCGTCTGGTTCTAAGTTTTCTCCGGCAATCCCGGTTTCTACTCAGGAGTCTGTGTCTGTGCCAGCTTCTGTTAGCACTTCTATTTCCTCTATCAGCACTGGTGCGACTGCGTCCGCTCTGGTAAAAGGTAACACTAACCCGATCACCAGCATGTCTGCTCCGGTTCAGGCTTCTGCACCGGCACTGACTAAAAGCCAGACTGACCGTCTGGAGGTTCTGCTGAACCCGAAAGATGAAATCAGCCTGAACTCTGGCAAACCGTTCCGTGAACTGGAATCCGAACTGCTGTCTCGTCGTAAGAAAGACCTGCAACAAATCTATGCTGAAGAGCGTGAAAACTACCTGGGTAAACTGGAACGTGAAATCACCCGTTTCTTTGTGGACCGTGGTTTCCTGGAAATCAAGTCTCCGATCCTGATCCCGCTGGAATACATCGAGCGCATGGGTATTGATAACGACACCGAACTGTCCAAGCAGATTTTCCGTGTGGACAAGAACTTCTGCCTGCGTCCGATGCTGGCACCGAACCTGTACAATTACCTGCGTAAACTGGATCGTGCACTGCCGGACCCGATCAAAATCTTTGAAATCGGTCCATGCTATCGTAAGGAGAGCGACGGTAAAGAACACCTGGAAGAGTTCACTATGCTGAACTTTTGTCAGATGGGTTCTGGCTGCACCCGTGAAAATCTGGAATCTATCATCACCGACTTCCTGAACCACCTGGGCATTGACTTCAAAATCGTTGGTGATTCCTGCATGGTTTACGGTGACACTCTGGACGTTATGCATGGTGATCTGGAACTGAGCAGCGCTGTTGTGGGTCCGATTCCGCTGGATCGTGAATGGGGTATCGATAAACCGTGGATTGGTGCTGGCTTCGGTCTGGAACGTCTGCTGAAAGTTAAGCACGACTTTAAGAACATCAAACGTGCTGCGCGTTCCGAGTCCTATTACAACGGCATTAGCACTAACCTGTAAGTCGACCAAACAATCCAAAACGCCGCGTTCAGCGGCGTTTTTTCTGCTTTTGCGGCCGCTGTGCTTCTCAAATGCCTGAGGCCAGTTTGCTCAGGCTCTCCCCGTGGAGGTAATAATTGACGATATGATCAGTGCACGGCTAACTAAGCGGCCTGCTGACTTTCTCGCCGATCAAAAGGCATTTTGCTATTAAGGGATTGACGAGGGCGTATCTGCGCAGTAAGATGCGCCCCGCATTGGAAACCTGATCATGTAGATCGAATGGACTCTAAATCCGTTCAGCCGGGTTAGATTCCCGGGGTTTCGCCAAATTCGAAAAGCCTGCTCAACGAGCAGGCTTTTTTGCATG (SEQ ID №: 14)
Последовательность, представленную в SEQ ID №: 14 вырезали из вектора клонирования pUC57-glnS [pylRs]-proK [pylT] рестрикционными ферментами AvrII и XbaI, а плазмиду pBAD-Al-u4-u5-TEV-R-MiniINS разрезали с помощью AvrII и XbaI. Затем, продукты разделяли электрофорезом нуклеиновых кислот, экстрагировали набором для восстановления ДНК из агарозного геля, лигировали ДНК-лигазой Т4 и трансформировали в большие компетентные клетки Е. coli Тор 10 химическим способом (способ CaCl2). Трансформированные клетки культивировали на среде агара Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl, 1,5% агар), содержащей хлорамфеникол, при 37°С в течение ночи. Отбирали единственную живую колонию и культивировали в жидкой среде Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl), содержащей хлорамфеникол, при 37°С и 220 об/мин в течение ночи. Для сохранения штамма добавляли глицерин в конечной концентрации 20%. Плазмиду экстрагировали с помощью набора для экстракции плазмид Plasmid Mini Extraction Kit, а полученная плазмида была названа pBAD-araBAD[Al-u4-u5-TEV-R-MinilNS]-glnS [IPYEpylRs]-proK[pylT]. Карта плазмиды представлена на фигуре 5.
Пример 3. Конструирование штамма для экспрессии двухплазмидной экспрессии с высокой плотностью модифицированого t-бутилоксикарборилового (Вое) белка слияния.
Плазмида pEvol-pylRs-pylT, плазмида pEvol-suppylRs-pylT, плазмида pEvol-IPYEpylRs-pylT, плазмида pEvol-optpylRs-pylT и плазмида pEvol-IPYEpylRs (L274A) были, соответственно, сотрансформированы с применением вектора экспресии белка силияния инсулина pBAD-INS (плазмида была сконструирована нашей компанией; устойчивость к канамицину) в компетентные клетки Е. coli Тор 10 (компетентные клетки были приобретены у Thermo) химическим способом (способ CaCl2).
Трансформированные клетки культивировали на среде агара Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl, 1,5% агар), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С в течение ночи. Отбирали единственную живую колонию и культивировали в жидкой среде Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С и 220 об/мин в течение ночи. Для сохранения штамма добавляли глицерин до конечной концентрации 20%.
Каждый штамм инокулировали в жидкой среде Луриа-Бертани, культивировали при 37°С и 220 об/мин в течение ночи, затем инокулировали в 1% (об./об.) в ферментере со средой для ферментации (12 г/л пептона жрожжей, 24 г/л порошка экстракта дрожжей, 4 мл/л глицерина, 12,8 г/л гидрофосфата натрия, 3 г/л дигидрофосфата калия, 0,3 мкл пеногасителя) и культивировали при 35 (±3)°С, 200-1000 об/мин, поток воздуха 2-6 л/мин. После культивирования в течение 3-10 часов, питательную среду, содержащую глицерин и пептон дрожжей, подавали постепенно, до завершения ферментации. После культивирования до достижения 25-80 OD600, для индукции добавляли L-ara в конечной концентрации 0,25% и Boc-Lys в конечной концентрации 5 мМ. Культивирование продолжали до достижения 180-220 OD600, затем ферментационный бульон извлекали из ферментера. Различные штаммы собирали центрифугированием (5000 об/мин, 30 мин, 25°С). Электрофорез SDS-полиакриламида использовали для обнаружения экспрессии белков слияния, содержащих Вос-модифицированные лизины в цельных клетках различных штаммов.
Белки слияния экспрессировали в форме нерастворимых «тел включения». Для высвобождения тел включения, клетки Escherichia coli разрушали гомогенизатором высокого давления. Нуклеиновые кислоты, клеточный дебрис и растворимые белки удаляли центрифугированием при 10000 g. Тела включения, содержащие белки слияния, промывали чистой водой, а полученный осадок тел включения использовали в качестве сырья для складывания.
Объем экспрессии белков слияния различных мутантных ферментов показан в следующей таблице.


Для повторной укладки белков слияния, тела включения растворяли в 7,5 М раствора мочевины с рН 10,5, содержащем 2-10 мМ меркаптоэтанола, при этом общая концентрация белка после растворения достигла 10-25 мг/мл. Образец разбавляли 5-10 раз и проводили обычное складывание в течение 16-30 часов при 4-8°С и рН 10,5-11,7. При 18-25°С, значение рН поддерживали в пределах 8,0-9,5, а белки слияния подвергали ферментативному гидролизу с применением трипсина и карбоксипептидазы В в течение 10-20 часов, а затем добавляли 0,45 М сульфата аммония для прекращения ферментолиза. Результаты анализа методом обращенно-фазовой ВЭЖХ показали, что результат процедуры энзимолиза превышал 90%. Аналоги инсулина, полученные после переваривания с применением трипсина и карбоксипептидазы В, были названы ВОС-лизиновыми инсулинами. ВОС-лизиновые инсулины не подвергались ферментативному гидролизу в вышеуказанных условиях. Образцы осветляли мембранной фильтрацией и первоначально очищали гидрофобной хроматографией с применением 0,45 мМ сульфата аммония в качестве буферного раствора, а чистота образца электрофореза геля SDS-полиакриламида достигала 90%. Кроме того, полученные ВОС-человеческие инсулины были проанализированы с помощью масс-спектрометрии MALDI-TOF, и было обнаружено, что их молекулярная масса соответствует теоретической молекулярной массе 5907,7 Да. Образцы смывали и собирали с помощью гидрофобной хроматографии, добавляли соляную кислоту для проведения реакции удаления защитных групп Вос-человеческих инсулинов. Раствор гидроксида натрия добавляли для регулирования значения рН между 2,8-3,2 и прекращения реакции. После двух этапов обращено-фазовой хроматографии высокого давления, результат рекомбинантных человеческих инсулинов превышал 85%.
Объем экспрессии рекомбинантных человеческих инсулинов различных мутантных ферментов показан в следующей таблице:
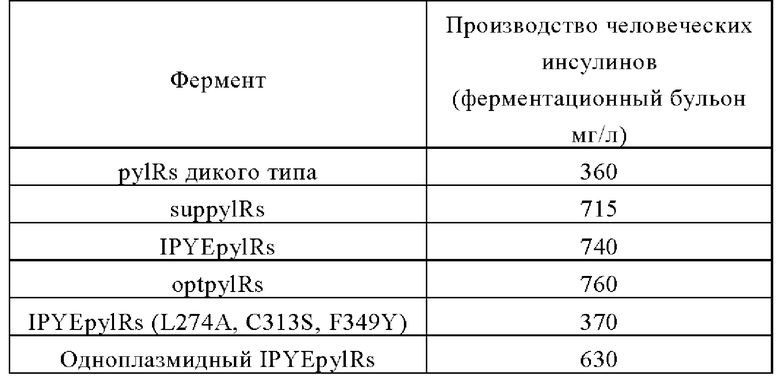
Результаты показали, что использование мутантных ферментов по настоящему изобретению для получения белков-мишеней, содержащих Вос-лизин, может значительно увеличивать количество вставок не встречающихся в природе аминокислот и количество белков-мишеней, содержащих не встречающиеся в природе аминокислоты.
Пример 4 Конструирование штамма для экспрессии двухплазмидной экспрессии с высокой плотностью модифицированного
бутинилоксикарбонилового белка слияния
Плазмида pEvol-pylRs-pylT, плазмида pEvol-suppylRs-pylT, плазмида pEvol-IPYEpylRs-pylT, плазмида pEvol-optpylRs-pylT и плазмида pEvol-IPYEpylRs (L274A, C313S, F349Y) были, соответственно, сотрансформированы с применением вектора экспрессии белка слияния инсулина pBAD-INS (плазмида была сконструирована нашей компанией; устойчивость к канамицину) в компетентные клетки E.coli Тор 10 (компетентные клетки были приобретены у Thermo) химическим способом (способ CaCl2). Трансформированные клетки культивировали на среде агара Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl, 1,5% агар), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С в течение ночи. Отбирали единственную живую колонию и культивировали в жидкой среде Луриа-Бертани (10 г/л пептона дрожжей, 5 г/л порошка экстракта дрожжей, 10 г/л NaCl), содержащей 25 мкг/мл канамицина и 17 мкг/мл хлорамфеникола, при 37°С и 220 об/мин в течение ночи. Для сохранения штамма добавляли глицерин до конечной концентрации 20%.
Каждый штамм инокулировали в жидкой среде Луриа-Бертани, культивировали при 37°С и 220 об/мин в течение ночи, затем инокулировали в 1% (об./об.) в ферментере со средой для ферментации (12 г/л пептона жрожжей, 24 г/л порошка экстракта дрожжей, 4 мл/л глицерина, 12,8 г/л гидрофосфата натрия, 3 г/л дигидрофосфата калия, 0,3 мкл пеногасителя) и культивировали при 35 (±3)°С, 200-1000 об/мин, поток воздуха 2-6 л/мин. После культивирования в течение 3-10 часов, питательную среду, содержащую глицерин и пептон дрожжей, подавали постепенно, до завершения ферментации. После культивирования до достижения 25-80 OD600 для индукции добавляли L-ara в конечной концентрации 0,25% и бутинилоксикарбонил-Lys в конечной концентрации 5 мМ. Культивирование продолжали до достижения 180-220 OD600, затем ферментационный бульон извлекали из ферментера. Различные штаммы собирали центрифугированием (5000 об/мин, 30 мин, 25°С). Электрофорез SDS-полиакриламида использовали для обнаружения экспрессии белков слияния, содержащих бутинилоксикарбонил-модифицированные лизины в цельных клетках различных штаммов.
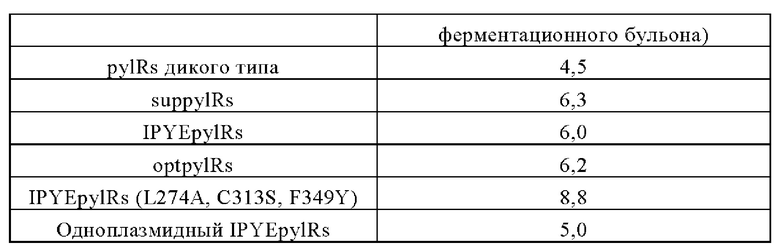
Белки слияния экспрессировали в форме нерастворимых «тел включения». Для высвобождения тел включения, клетки Е. coli разрушали гомогенизатором высокого давления. Нуклеиновые кислоты, клеточный дебрис и растворимые белки удаляли центрифугированием при 10000 g. Тела включения, содержащие белки слияния, промывали чистой водой, а полученный осадок тел включения использовали в качестве сырья для складывания. Для повторной укладки белков слияния, тела включения растворяли в 7,5 М раствора мочевины с рН 10,5, содержащем 2-10 мМ меркаптоэтанола, при этом общая концентрация белка после растворения достигла 10-25 мг/мл. Образец разбавляли 5-10 раз и проводили обычное складывание в течение 16-30 часов при 4-8°С и рН 10,5-11,7. При 18-25°С, значение рН поддерживали в пределах 8,0-9,5, а белки слияния подвергали ферментативному гидролизу с применением трипсина и карбоксипептидазы В в течение 10-20 часов, а затем добавляли 0,45 М сульфата аммония для прекращения ферментолиза. Результаты анализа методом обращенно-фазовой ВЭЖХ показали, что результат процедуры энзимолиза превышал 90%. Аналоги инсулина, полученные после переваривания с применением трипсина и карбоксипептидазы В, были названы бутинилоксикарбонил-лизиновыми инсулинами. Бутинилоксикарбонил-лизиновые инсулины не подвергались ферментативному гидролизу в вышеуказанных условиях. Образцы осветляли мембранной фильтрацией и первоначально очищали гидрофобной хроматографией с применением 0,45 мМ сульфата аммония в качестве буферного раствора, а чистота образца электрофореза геля SDS-полиакриламида достигала 90%. Кроме того, полученные бутинилоксикарбонил-человеческие инсулины были проанализированы с помощью масс-спектрометрии MALDI-TOF, и было обнаружено, что их молекулярная масса соответствует теоретической молекулярной массе 5907,7 Да.


Результаты показали, что использование мутантных ферментов по настоящему изобретению для получения модифицированных бутинилоксикарбонилом целевых белков может значительно увеличить количество вставок не встречающихся в природе аминокислот и количество целевых белков, содержащих не встречающиеся в природе аминокислоты.
Все документы, упомянутые в настоящем изобретении, включены в настоящий документ в виде ссылок, как если бы каждый из документов был отдельно включен в виде ссылки. Кроме того, следует понимать, что после прочтения вышеизложенной идеи изобретения, в настоящее изобретение могут быть внесены различные изменения или модификации специалистами в данной области техники, при этом, данные аналоги также входят в объем формулы изобретения по данной заявке.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> СУЧЖОУ КУНЬПЭН БИОТЕКХ КО., ЛТД.
<120> АМИНОАЦИЛ-тРНК-СИНТЕТАЗА, ЭФФЕКТИВНОЕ ВВЕДЕНИЕ ПРОИЗВОДНЫХ ЛИЗИНА
<130> P2020-0182
<150> CN201910364788.7
<151> 2019-04-30
<160> 14
<170> PatentIn version 3.5
<210> 1
<211> 454
<212> ПРТ
<213> Methanosarcina mazei
<400> 1
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 2
<211> 454
<212> ПРТ
<213> Methanosarcina barkeri
<400> 2
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Thr Arg Thr Lys Lys Ala Met Pro Lys Ser Val
100 105 110
Ala Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Gln
115 120 125
Pro Ser Gly Ser Lys Phe Ser Pro Ala Ile Pro Val Ser Thr Gln Glu
130 135 140
Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser Ile Ser
145 150 155 160
Thr Gly Ala Thr Ala Ser Ala Leu Val Lys Gly Asn Thr Asn Pro Ile
165 170 175
Thr Ser Met Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys
180 185 190
Ser Gln Thr Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile
195 200 205
Ser Leu Asn Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu
210 215 220
Ser Arg Arg Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu
225 230 235 240
Asn Tyr Leu Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp
245 250 255
Arg Gly Phe Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr
260 265 270
Ile Glu Arg Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile
275 280 285
Phe Arg Val Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn
290 295 300
Leu Tyr Asn Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile
305 310 315 320
Lys Ile Phe Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys
325 330 335
Glu His Leu Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser
340 345 350
Gly Cys Thr Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn
355 360 365
His Leu Gly Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr
370 375 380
Gly Asp Thr Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala
385 390 395 400
Val Val Gly Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro
405 410 415
Trp Ile Gly Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His
420 425 430
Asp Phe Lys Asn Ile Lys Arg Ala Ala Arg Ser Gly Ser Tyr Tyr Asn
435 440 445
Gly Ile Ser Thr Asn Leu
450
<210> 3
<211> 419
<212> ПРТ
<213> Искусственная Последовательность
<220>
<223> Мутантная Лизил-тРНК Синтетаза
<400> 3
Met Asp Lys Lys Pro Leu Asn Thr Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Ile His Lys Ile Lys His His Glu Val Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Ser Arg Thr Ala Arg Ala Leu Arg His His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Leu Asn
65 70 75 80
Lys Phe Leu Thr Lys Ala Asn Glu Asp Gln Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Ala Pro Arg Thr Lys Lys Ala Met Pro Lys Ser Val Ala
100 105 110
Arg Ala Pro Lys Pro Leu Glu Asn Thr Glu Ala Ala Gln Ala Ser Thr
115 120 125
Gln Glu Ser Val Ser Val Pro Ala Ser Val Ser Thr Ser Ile Ser Ser
130 135 140
Ser Ala Pro Val Gln Ala Ser Ala Pro Ala Leu Thr Lys Ser Gln Thr
145 150 155 160
Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile Ser Leu Asn
165 170 175
Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu Ser Arg Arg
180 185 190
Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu Asn Tyr Leu
195 200 205
Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp Arg Gly Phe
210 215 220
Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr Ile Glu Arg
225 230 235 240
Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile Phe Arg Val
245 250 255
Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn Leu Tyr Asn
260 265 270
Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile Lys Ile Phe
275 280 285
Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys Glu His Leu
290 295 300
Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser Gly Cys Thr
305 310 315 320
Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn His Leu Gly
325 330 335
Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr Gly Asp Thr
340 345 350
Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala Val Val Gly
355 360 365
Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro Trp Ile Gly
370 375 380
Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His Asp Phe Lys
385 390 395 400
Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn Gly Ile Ser
405 410 415
Thr Asn Leu
<210> 4
<211> 419
<212> ПРТ
<213> Искусственная Последовательность
<220>
<223> Мутантная Лизил-тРНК Синтетаза
<400> 4
Met Asp Lys Lys Pro Leu Asp Val Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Leu His Lys Ile Lys His His Glu Ile Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Cys Arg Pro Ala Arg Ala Phe Arg Tyr His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Ile Asn
65 70 75 80
Asn Phe Leu Thr Arg Ser Thr Glu Ser Lys Asn Ser Val Lys Val Arg
85 90 95
Val Val Ser Glu Pro Lys Val Lys Lys Ala Met Pro Lys Ser Val Ser
100 105 110
Arg Ala Pro Lys Pro Leu Glu Asn Ser Val Ser Ala Lys Ala Ser Thr
115 120 125
Asn Thr Ser Arg Ser Val Pro Ser Pro Ala Lys Ser Thr Pro Asn Ser
130 135 140
Ser Val Pro Ala Ser Ala Ser Ala Pro Ala Leu Thr Lys Ser Gln Thr
145 150 155 160
Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile Ser Leu Asn
165 170 175
Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu Ser Arg Arg
180 185 190
Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu Asn Tyr Leu
195 200 205
Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp Arg Gly Phe
210 215 220
Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr Ile Glu Arg
225 230 235 240
Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile Phe Arg Val
245 250 255
Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn Leu Tyr Asn
260 265 270
Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile Lys Ile Phe
275 280 285
Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys Glu His Leu
290 295 300
Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser Gly Cys Thr
305 310 315 320
Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn His Leu Gly
325 330 335
Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Phe Gly Asp Thr
340 345 350
Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala Val Val Gly
355 360 365
Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro Trp Ile Gly
370 375 380
Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His Asp Phe Lys
385 390 395 400
Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn Gly Ile Ser
405 410 415
Thr Asn Leu
<210> 5
<211> 419
<212> ПРТ
<213> Искусственная Последовательность
<220>
<223> Мутантная Лизил-тРНК Синтетаза
<400> 5
Met Asp Lys Lys Pro Leu Asp Val Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Leu His Lys Ile Lys His Tyr Glu Ile Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Cys Arg Pro Ala Arg Ala Phe Arg Tyr His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Gly Glu Asp Ile Asn
65 70 75 80
Asn Phe Leu Thr Arg Ser Thr Glu Gly Lys Thr Ser Val Lys Val Lys
85 90 95
Val Val Ser Glu Pro Lys Val Lys Lys Ala Met Pro Lys Ser Val Ser
100 105 110
Arg Ala Pro Lys Pro Leu Glu Asn Pro Val Ser Ala Lys Ala Ser Thr
115 120 125
Asp Thr Ser Arg Ser Val Pro Ser Pro Ala Lys Ser Thr Pro Asn Ser
130 135 140
Pro Val Pro Thr Ser Ala Ser Ala Pro Ala Leu Thr Lys Ser Gln Thr
145 150 155 160
Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile Ser Leu Asn
165 170 175
Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu Ser Arg Arg
180 185 190
Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu Asn Tyr Leu
195 200 205
Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp Arg Gly Phe
210 215 220
Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr Ile Glu Arg
225 230 235 240
Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile Phe Arg Val
245 250 255
Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn Leu Tyr Asn
260 265 270
Tyr Leu Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile Lys Ile Phe
275 280 285
Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys Glu His Leu
290 295 300
Glu Glu Phe Thr Met Leu Asn Phe Cys Gln Met Gly Ser Gly Cys Thr
305 310 315 320
Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn His Leu Gly
325 330 335
Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Phe Gly Asp Thr
340 345 350
Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala Val Val Gly
355 360 365
Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro Trp Ile Gly
370 375 380
Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His Asp Phe Lys
385 390 395 400
Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn Gly Ile Ser
405 410 415
Thr Asn Leu
<210> 6
<211> 419
<212> ПРТ
<213> Искусственная Последовательность
<220>
<223> Мутантная Лизил-тРНК Синтетаза
<400> 6
Met Asp Lys Lys Pro Leu Asp Val Leu Ile Ser Ala Thr Gly Leu Trp
1 5 10 15
Met Ser Arg Thr Gly Thr Leu His Lys Ile Lys His His Glu Ile Ser
20 25 30
Arg Ser Lys Ile Tyr Ile Glu Met Ala Cys Gly Asp His Leu Val Val
35 40 45
Asn Asn Ser Arg Ser Cys Arg Pro Ala Arg Ala Phe Arg Tyr His Lys
50 55 60
Tyr Arg Lys Thr Cys Lys Arg Cys Arg Val Ser Asp Glu Asp Ile Asn
65 70 75 80
Asn Phe Leu Thr Arg Ser Thr Glu Ser Lys Asn Ser Val Lys Val Arg
85 90 95
Val Val Ser Glu Pro Lys Val Lys Lys Ala Met Pro Lys Ser Val Ser
100 105 110
Arg Ala Pro Lys Pro Leu Glu Asn Ser Val Ser Ala Lys Ala Ser Thr
115 120 125
Asn Thr Ser Arg Ser Val Pro Ser Pro Ala Lys Ser Thr Pro Asn Ser
130 135 140
Ser Val Pro Ala Ser Ala Ser Ala Pro Ala Leu Thr Lys Ser Gln Thr
145 150 155 160
Asp Arg Leu Glu Val Leu Leu Asn Pro Lys Asp Glu Ile Ser Leu Asn
165 170 175
Ser Gly Lys Pro Phe Arg Glu Leu Glu Ser Glu Leu Leu Ser Arg Arg
180 185 190
Lys Lys Asp Leu Gln Gln Ile Tyr Ala Glu Glu Arg Glu Asn Tyr Leu
195 200 205
Gly Lys Leu Glu Arg Glu Ile Thr Arg Phe Phe Val Asp Arg Gly Phe
210 215 220
Leu Glu Ile Lys Ser Pro Ile Leu Ile Pro Leu Glu Tyr Ile Glu Arg
225 230 235 240
Met Gly Ile Asp Asn Asp Thr Glu Leu Ser Lys Gln Ile Phe Arg Val
245 250 255
Asp Lys Asn Phe Cys Leu Arg Pro Met Leu Ala Pro Asn Leu Tyr Asn
260 265 270
Tyr Ala Arg Lys Leu Asp Arg Ala Leu Pro Asp Pro Ile Lys Ile Phe
275 280 285
Glu Ile Gly Pro Cys Tyr Arg Lys Glu Ser Asp Gly Lys Glu His Leu
290 295 300
Glu Glu Phe Thr Met Leu Asn Phe Ser Gln Met Gly Ser Gly Cys Thr
305 310 315 320
Arg Glu Asn Leu Glu Ser Ile Ile Thr Asp Phe Leu Asn His Leu Gly
325 330 335
Ile Asp Phe Lys Ile Val Gly Asp Ser Cys Met Val Tyr Gly Asp Thr
340 345 350
Leu Asp Val Met His Gly Asp Leu Glu Leu Ser Ser Ala Val Val Gly
355 360 365
Pro Ile Pro Leu Asp Arg Glu Trp Gly Ile Asp Lys Pro Trp Ile Gly
370 375 380
Ala Gly Phe Gly Leu Glu Arg Leu Leu Lys Val Lys His Asp Phe Lys
385 390 395 400
Asn Ile Lys Arg Ala Ala Arg Ser Glu Ser Tyr Tyr Asn Gly Ile Ser
405 410 415
Thr Asn Leu
<210> 7
<211> 72
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК не встречающейся в природе тРНК pylT
<400> 7
ggaaacctga tcatgtagat cgaatggact ctaaatccgt tcagccgggt tagattcccg 60
gggtttccgc ca 72
<210> 8
<211> 1365
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК pylRs
<400> 8
atggataaga aaccgctgaa tactctgatt tctgcaactg gtctgtggat gagccgtacc 60
ggcaccatcc acaagatcaa acaccacgag gtttcccgta gcaaaatcta catcgaaatg 120
gcgtgcggtg accacctggt ggtaaacaac tcccgttctt ctcgtactgc acgtgctctg 180
cgccaccaca agtaccgtaa gacctgcaag cgctgtcgcg tgtctgatga agacctgaac 240
aaattcctga ctaaagcgaa cgaagatcag acttctgtga aggtgaaagt tgtttctgcc 300
ccaacccgca ccaagaaagc gatgccgaag tccgttgcac gcgctccgaa accgctggag 360
aacaccgaag ccgcacaggc ccagccgtct ggttctaagt tttctccggc aatcccggtt 420
tctactcagg agtctgtgtc tgtgccagct tctgttagca cttctatttc ctctatcagc 480
actggtgcga ctgcgtccgc tctggtaaaa ggtaacacta acccgatcac cagcatgtct 540
gctccggttc aggcttctgc accggcactg actaaaagcc agactgaccg tctggaggtt 600
ctgctgaacc cgaaagatga aatcagcctg aactctggca aaccgttccg tgaactggaa 660
tccgaactgc tgtctcgtcg taagaaagac ctgcaacaaa tctatgctga agagcgtgaa 720
aactacctgg gtaaactgga acgtgaaatc acccgtttct ttgtggaccg tggtttcctg 780
gaaatcaagt ctccgatcct gatcccgctg gaatacatcg agcgcatggg tattgataac 840
gacaccgaac tgtccaagca gattttccgt gtggacaaga acttctgcct gcgtccgatg 900
ctggcaccga acctgtacaa ttacctgcgt aaactggatc gtgcactgcc ggacccgatc 960
aaaatctttg aaatcggtcc atgctatcgt aaggagagcg acggtaaaga acacctggaa 1020
gagttcacta tgctgaactt ttgtcagatg ggttctggct gcacccgtga aaatctggaa 1080
tctatcatca ccgacttcct gaaccacctg ggcattgact tcaaaatcgt tggtgattcc 1140
tgcatggttt acggtgacac tctggacgtt atgcatggtg atctggaact gagcagcgct 1200
gttgtgggtc cgattccgct ggatcgtgaa tggggtatcg ataaaccgtg gattggtgct 1260
ggcttcggtc tggaacgtct gctgaaagtt aagcacgact ttaagaacat caaacgtgct 1320
gcgcgttccg agtcctatta caacggcatt agcactaacc tgtaa 1365
<210> 9
<211> 1260
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК IPYEpylRs
<400> 9
atggataaga agccactgga tgttctgatt tccgctactg gtctgtggat gtctcgcact 60
ggtactctgc acaagatcaa acaccacgaa atctctcgct ccaagatcta cattgaaatg 120
gcttgtggtg atcacctggt tgttaacaac tcccgctctt gtcgcccggc tcgtgcgttt 180
cgctatcaca aatatcgtaa aacctgcaaa cgctgccgcg tgagcgatga ggatattaac 240
aacttcctga ctcgctccac cgagagcaag aactctgtga aagttcgcgt agtttctgaa 300
ccgaaagtta agaaagctat gccgaagtct gttagccgtg ctccgaaacc gctggagaac 360
tctgtgtccg cgaaagcgag caccaacacc agccgttctg ttccgtctcc agcgaaatct 420
actccgaact ctagcgtgcc agcttccgct tctgcaccgg cactgactaa aagccagact 480
gaccgtctgg aggttctgct gaacccgaaa gatgaaatca gcctgaactc tggcaaaccg 540
ttccgtgaac tggaatccga actgctgtct cgtcgtaaga aagacctgca acaaatctat 600
gctgaagagc gtgaaaacta cctgggtaaa ctggaacgtg aaatcacccg tttctttgtg 660
gaccgtggtt tcctggaaat caagtctccg atcctgatcc cgctggaata catcgagcgc 720
atgggtattg ataacgacac cgaactgtcc aagcagattt tccgtgtgga caagaacttc 780
tgcctgcgtc cgatgctggc accgaacctg tacaattacc tgcgtaaact ggatcgtgca 840
ctgccggacc cgatcaaaat ctttgaaatc ggtccatgct atcgtaagga gagcgacggt 900
aaagaacacc tggaagagtt cactatgctg aacttttgtc agatgggttc tggctgcacc 960
cgtgaaaatc tggaatctat catcaccgac ttcctgaacc acctgggcat tgacttcaaa 1020
atcgttggtg attcctgcat ggttttcggt gacactctgg acgttatgca tggtgatctg 1080
gaactgagca gcgctgttgt gggtccgatt ccgctggatc gtgaatgggg tatcgataaa 1140
ccgtggattg gtgctggctt cggtctggaa cgtctgctga aagttaagca cgactttaag 1200
aacatcaaac gtgctgcgcg ttccgagtcc tattacaacg gcattagcac taacctgtaa 1260
<210> 10
<211> 72
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК промотора глутамина glnS
<400> 10
ttttaaaaaa ctaacagttg tcagcctgtc ccgctttaat atcatacgcc gttatacgtt 60
gtttacgctt tg 72
<210> 11
<211> 45
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК промотора proK
<400> 11
caaacaatcc aaaacgccgc gttcagcggc gttttttctg ctttt 45
<210> 12
<211> 151
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК терминатора промотора glnS
<400> 12
tgtgcttctc aaatgcctga ggccagtttg ctcaggctct ccccgtggag gtaataattg 60
acgatatgat cagtgcacgg ctaactaagc ggcctgctga ctttctcgcc gatcaaaagg 120
cattttgcta ttaagggatt gacgagggcg t 151
<210> 13
<211> 40
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК терминатора промотора proK
<400> 13
aattcgaaaa gcctgctcaa cgagcaggct tttttgcatg 40
<210> 14
<211> 1798
<212> ДНК
<213> Искусственная Последовательность
<220>
<223> Последовательность ДНК glnS [pylRs]-proK [pylT]
<400> 14
ttttaaaaaa ctaacagttg tcagcctgtc ccgctttaat atcatacgcc gttatacgtt 60
gtttacgctt tgaggaatcc catatggata agaaaccgct gaatactctg atttctgcaa 120
ctggtctgtg gatgagccgt accggcacca tccacaagat caaacaccac gaggtttccc 180
gtagcaaaat ctacatcgaa atggcgtgcg gtgaccacct ggtggtaaac aactcccgtt 240
cttctcgtac tgcacgtgct ctgcgccacc acaagtaccg taagacctgc aagcgctgtc 300
gcgtgtctga tgaagacctg aacaaattcc tgactaaagc gaacgaagat cagacttctg 360
tgaaggtgaa agttgtttct gccccaaccc gcaccaagaa agcgatgccg aagtccgttg 420
cacgcgctcc gaaaccgctg gagaacaccg aagccgcaca ggcccagccg tctggttcta 480
agttttctcc ggcaatcccg gtttctactc aggagtctgt gtctgtgcca gcttctgtta 540
gcacttctat ttcctctatc agcactggtg cgactgcgtc cgctctggta aaaggtaaca 600
ctaacccgat caccagcatg tctgctccgg ttcaggcttc tgcaccggca ctgactaaaa 660
gccagactga ccgtctggag gttctgctga acccgaaaga tgaaatcagc ctgaactctg 720
gcaaaccgtt ccgtgaactg gaatccgaac tgctgtctcg tcgtaagaaa gacctgcaac 780
aaatctatgc tgaagagcgt gaaaactacc tgggtaaact ggaacgtgaa atcacccgtt 840
tctttgtgga ccgtggtttc ctggaaatca agtctccgat cctgatcccg ctggaataca 900
tcgagcgcat gggtattgat aacgacaccg aactgtccaa gcagattttc cgtgtggaca 960
agaacttctg cctgcgtccg atgctggcac cgaacctgta caattacctg cgtaaactgg 1020
atcgtgcact gccggacccg atcaaaatct ttgaaatcgg tccatgctat cgtaaggaga 1080
gcgacggtaa agaacacctg gaagagttca ctatgctgaa cttttgtcag atgggttctg 1140
gctgcacccg tgaaaatctg gaatctatca tcaccgactt cctgaaccac ctgggcattg 1200
acttcaaaat cgttggtgat tcctgcatgg tttacggtga cactctggac gttatgcatg 1260
gtgatctgga actgagcagc gctgttgtgg gtccgattcc gctggatcgt gaatggggta 1320
tcgataaacc gtggattggt gctggcttcg gtctggaacg tctgctgaaa gttaagcacg 1380
actttaagaa catcaaacgt gctgcgcgtt ccgagtccta ttacaacggc attagcacta 1440
acctgtaagt cgaccaaaca atccaaaacg ccgcgttcag cggcgttttt tctgcttttg 1500
cggccgctgt gcttctcaaa tgcctgaggc cagtttgctc aggctctccc cgtggaggta 1560
ataattgacg atatgatcag tgcacggcta actaagcggc ctgctgactt tctcgccgat 1620
caaaaggcat tttgctatta agggattgac gagggcgtat ctgcgcagta agatgcgccc 1680
cgcattggaa acctgatcat gtagatcgaa tggactctaa atccgttcag ccgggttaga 1740
ttcccggggt ttccgccaaa ttcgaaaagc ctgctcaacg agcaggcttt tttgcatg 1798
<---

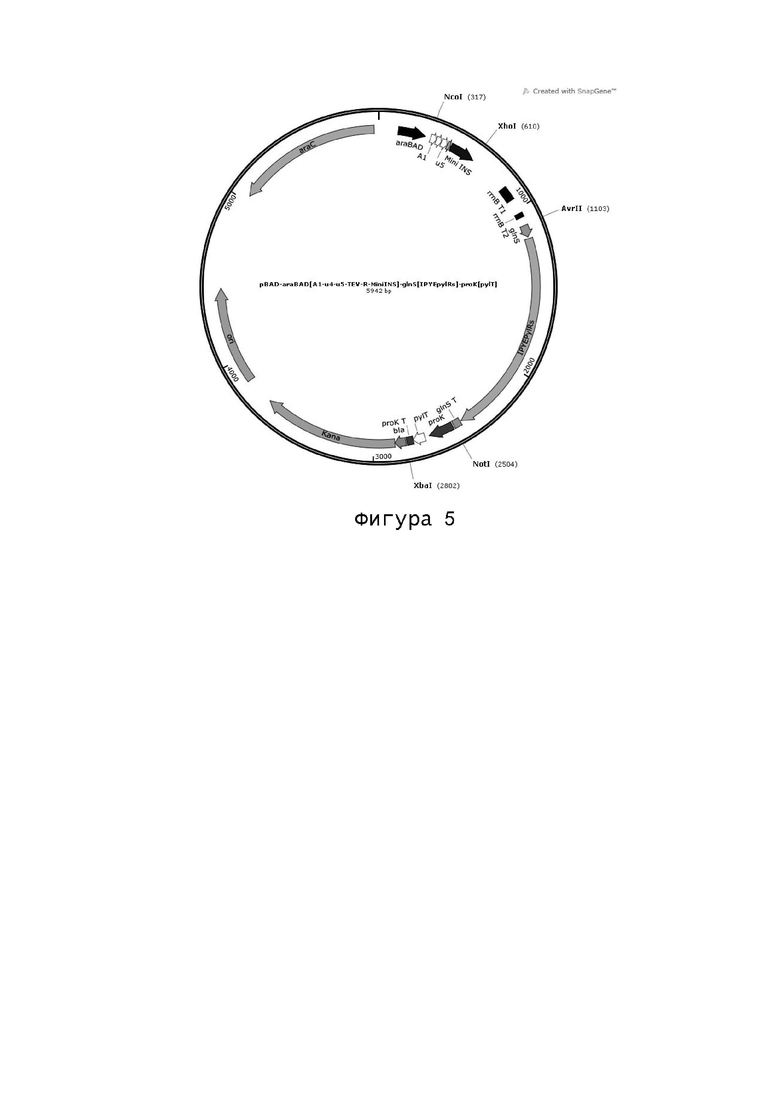
Настоящее изобретение предоставляет мутантную аминоацил-тРНК-синтетазу, имеющую делеционную мутацию в соответствующей аминокислотной последовательности аминоацил-тРНК-синтетазы дикого типа: 102-й, 128-140-й и 159-179-й аминокислотные остатки. По сравнению с лизил-тРНК-синтетазой дикого типа мутантная лизил-тРНК-синтетаза по настоящему изобретению обладает высокой активностью, высоким уровнем экспрессии и хорошей растворимостью, а также может значительно увеличивать количество вставок не встречающейся в природе аминокислоты и уровень экспрессии целевого белка, содержащего не встречающуюся в природе аминокислоту. 7 н. и 4 з.п. ф-лы, 5 ил., 3 табл., 4 пр.
1. Мутантный белок аминоацил-тРНК-синтетазы, который включает
(1) делеционную мутацию аминокислотного остатка в положении 102, положениях 128-140 и положениях 159-179 в аминокислотной последовательности, соответствующей аминоацил-тРНК-синтетазе дикого типа, или
(2) делеционную мутацию аминокислотных остатков в положении 102, положениях 128-140 и положениях 159-179 в аминокислотной последовательности, соответствующей аминоацил-тРНК-синтетазе дикого типа, путем замены положений 1-184 аминоацил-тРНК-синтетазы дикого типа другим сегментом аминокислот в положениях 1-149 SEQ ID №: 4 и аминокислотной мутацией Y384F, соответствующей аминоацил-тРНК-синтетазе дикого типа;
при этом аминокислотная последовательность аминоацил-тРНК-синтетазы дикого типа представлена в SEQ ID №: 1 или 2;
или мутантный белок аминоацил-тРНК-синтетазы дополнительно включает мутацию, выбранную из следующей группы, соответствующей аминокислотной последовательности SEQ ID №: 4:
(3) H29Y, D76G, S89G, N91T, R96K, S121P, N129D, S145P и A148T или
(4) L274A, C313S и F349Y.
2. Мутантный белок по п. 1, в котором аминоацил-тРНК-синтетаза дикого типа представлена в SEQ ID №: 1.
3. Мутантный белок по п. 1, в котором аминокислотная последовательность мутантного белка представлена в SEQ ID №: 3.
4. Мутантный белок по п. 1, в котором аминокислотная последовательность мутантного белка представлена любой из SEQ ID №: 4-6.
5. Изолированный полинуклеотид, кодирующий мутантный белок по п. 1.
6. Вектор для экспрессии мутантного белка по п. 1, который включает полинуклеотид по п. 5.
7. Клетка-хозяин для экспрессии мутантного белка по п. 1, которая включает вектор по п. 6 или имеющая полинуклеотид по п. 5, интегрированный в ее геном.
8. Клетка-хозяин по п. 7, которая содержит:
(а) мутантный белок по п.1; а также
(b) искусственную тРНК, способную связываться с предварительно заданной модифицированной аминокислотой в присутствии мутантного белка; и необязательно
(c) первую последовательность нуклеиновой кислоты, кодирующей целевой белок, где первая последовательность нуклеиновой кислоты содержит кодон, распознаваемый искусственной тРНК.
9. Плазмидная система для экспрессии мутантного белка по п. 1, которая включает:
(1) первую кассету экспрессии, содержащую первую кодирующую последовательность для кодирования целевого белка, при этом первая кодирующая последовательность содержит кодон для введения предварительно заданной модифицированной аминокислоты, а кодон представляет собой UAG (амбер), UAA (охра) или UGA (опал); и
(2) вторую кассету экспрессии, содержащую вторую последовательность нуклеиновой кислоты для кодирования аминоацил-тРНК-синтетазы, при этом аминоацил-тРНК-синтетаза представляет собой мутантный белок по п.1;
при этом система дополнительно содержит третью кассету экспрессии, которая содержит третью кодирующую последовательность для кодирования искусственной тРНК, которая содержит антикодон, соответствующий кодону;
и аминоацил-тРНК-синтетаза специфически катализирует искусственную тРНК с образованием комплекса «искусственная тРНК-Ха», где Ха представляет собой предварительно заданную модифицированную аминокислоту в аминоацильной форме.
10. Набор для повышения экспрессии целевого белка, содержащий не встречающиеся в природе аминокислоты, который содержит (а) контейнер и (b) мутантный белок по п. 1, или полинуклеотид по п. 5, или вектор по п. 6, или плазмидную систему по п. 9, расположенную в контейнере.
11. Применение мутантного белка по п. 1, или клетки-хозяина по п. 7, или плазмидной системы по п. 9, или набора по п. 10 для получения целевого белка, содержащего не встречающуюся в природе аминокислоту.
| WO 2010139948 A2, 09.12.2010, SEQ ID NO: 4,10,12 | |||
| CN 104718285 A, 17.06.2015 | |||
| Сталь | 1986 |
|
SU1315512A1 |
| Прибор для шпаклевки | 1932 |
|
SU30418A1 |

