Область техники
Настоящее изобретение относится к области аминокислотной ферментации, в частности, к способу получения L-лизина ферментацией, к способам и применениям по названному способу, а также к бактериям, используемым в данных способах и применениях.
Предшествующий уровень техники
Получение L-лизина путем ферментации из продуцирующих L-лизин бактерий (например, E.Coli из Escherichia и бактерий Corynebacterium) уже широко используется в промышленности. Такие бактерии могут быть выделены из природной среды и/или получены путем мутагенеза или генно-инженерной модификации. Предыдущий уровень техники для производства L-лизина использовал генно-инженерную модификацию генов, таких как pnt, dap, ррс и т.п., игнорируя регуляторный элемент кодирующего аконитазу гена (например, аконитаза А из Escherichia coli, аконитаза из Corynebacterium glutamicum или Corynebacterium pekinense).
Аконитаза является ферментом цикла трикарбоновых кислот, катализирующим двухступенчатые химические реакции: превращение лимонной кислоты в аконитовую кислоту и превращение аконитовой кислоты в изолимонную кислоту. В известных бактериях Escherichia, ген acnA, нуклеотидная последовательность которого показана на SEQ ID No: 1, кодирует аконитазу А. Тем не менее, возможно, из-за его метаболита, далекого от конечного продукта L-лизина, а также ввиду многих и сложных промежуточных ветвей метаболизма, его роль в ферментации L-лизина не была выяснена. Отсутствуют сообщения о влиянии аконитазы из других продуцирующих L-лизин бактерий, таких как Corynebacterium glutamicum и т.п., на ферментацию L-лизина. Аконитазы, описанные в таких документах, как CN 1289368А и CN 101631871 А, относятся к ферментации L-глутаминовой кислоты, и не затрагивают ферментацию L-лизина.
После долгих исследований и практики, в силу определенной удачи, изобретатели случайно обнаружили, что модификация гена аконитазы и его регуляторного элемента способствует увеличению выхода L-лизина.
Кроме того, ранее уровень экспрессии и/или активность фермента повышали за счет увеличения числа копий и введения сайт-направленной мутации полезного ферментного гена, или же активность и/или уровень экспрессии фермента элиминировали нокаутом вредного ферментного гена. Однако изобретатели обнаружили, что ген аконитазы и его регуляторный элемент отличаются от таковых, описанных в предшествующем уровне техники, и не могут быть просто увеличены числом копий или геннонокаутированы. В частности, элиминирование экспрессии гена acnA путем генного нокаута приводит к медленному росту бактерий и трудностям в практическом использовании, поэтому был изобретен новый способ модификации гена аконитазы и/или его регуляторного элемента для повышения выхода L-лизина.
Кроме того, настоящий способ не влияет на модифицируемые сайты в хромосомах большого числа продуцирующих L-лизин бактерий, модифицированных в соответствии с известным уровнем техники, что дополнительно усиливает этот эффект и является полезным для практического производства L-лизина с помощью большого разнообразия бактерий.
Раскрытие изобретения
Проблема, решаемая с помощью настоящего изобретения, - это создание нового способа получения L-лизина ферментацией и соответствующими методами, включая способ повышения ферментационного выхода L-лизина по сравнению с немодифицированной бактерией, использование модифицированной бактерии для получения L-лизина с помощью ферментации, вариант использования модифицированной бактерии для увеличения ферментационного выхода L-лизина по сравнению с немодифицированной бактерией, и/или способ модификации бактерии. Кроме того, настоящее изобретение описывает полинуклеотид, вектор и/или бактерию, используемые в указанных выше способах.
В частности, в первом аспекте настоящее изобретение обеспечивает способ получения L-лизина ферментацией, который включает следующие стадии:
(1) модификация гена аконитазы и/или его регуляторного элемента в хромосоме бактерии таким образом, что активность и/или уровень экспрессии аконитазы этой бактерии уменьшаются, но не элиминируются; и
(2) продуцирование L-лизина ферментацией бактерией, полученной с помощью модификации на стадии (1).
Термин «модификация» в данном описании означает изменение объекта, нуждающегося в модификации, в результате чего достигается определенный эффект. Способы модификации гена или его регуляторного элемента в хромосоме включают (но не ограничиваются) мутагенез, сайт-направленную мутацию и/или гомологичную рекомбинацию, предпочтительно - два последних. Модифицирование гена или регуляторного элемента в хромосоме - это вставка, делеция или замена одного или нескольких нуклеотидов в нуклеотидной последовательности этого гена или его регуляторного элемента. Эти способы широко описаны в документах молекулярной биологии и микробиологии, и многие из них применяются промышленно. В вариантах осуществления настоящего изобретения модификация может быть осуществлена в соответствии с принципом гомологичной рекомбинации с использованием плазмиды pKOV, коммерчески доступной из Addgene, или с использованием плазмиды pK18mobsacB, так что немодифицированный ген аконитазы и/или его регуляторный элемент в бактериальной хромосоме модифицируются, становясь новым геном аконитазы и/или его регуляторным элементом, и уровень экспрессии и/или ферментная активность этой аконитазы в модифицированной бактерии уменьшаются, но не элиминируются. Поэтому предпочтительно, чтобы используемая здесь модификация достигалась гомологичной рекомбинацией.
После долгих исследований изобретатели обнаружили, что в Escherichia coli, бактерии из семейства Corynebacterium и т.п., ликвидации/элиминирование экспрессии аконитазы тем или иным способом, например, генным нокаутом, приводит к замедлению роста бактерий, и даже не вырабатываются аминокислоты. Таким образом, модификация по настоящему изобретению действительно уменьшает и не элиминирует уровень экспрессии аконитазы бактерии, полученной с помощью этой модификации, по сравнению с немодифицированной бактерией. Предпочтительно, уровень экспрессии аконитазы А в бактерии, полученной с помощью этой модификации, снижается на 20%~95%, более предпочтительно на 50%~90%, например, на 65%, 70% или 80%.
Настоящее изобретение также обеспечивает другие применения или способы. Например, во втором аспекте настоящее изобретение дает способ повышения ферментационного выхода L-лизина, который включает следующие стадии:
(1) модификация гена аконитазы и/или его регуляторного элемента в хромосоме бактерии таким образом, чтобы активность и/или уровень экспрессии аконитазы бактерии уменьшались, но не элиминировались; и
(2) продуцирование L-лизина путем ферментации в бактерии, полученной с помощью модификации на стадии (1).
L-лизин является важным бактериальным метаболитом, и большинство бактерий вырабатывает большее или меньшее количество L-лизина. Хотя бактерия, продуцирующая L-лизин с низким выходом, не подходит для экономически эффективного производства L-лизина, способы по настоящему изобретению могут повысить выход L-лизина и могут быть использованы в области, где экономичность неважна. Несомненно, предпочтительной бактерией, используемой здесь, является бактерия, продуцирующая L-лизин с высоким выходом, и этот выход может быть дополнительно повышен с помощью способов по настоящему изобретению. Кроме того, в способах или использованиях настоящего изобретения не могут быть осуществлены никакие другие модификации, кроме модификации гена аконитазы и/или его регуляторного элемента в хромосоме бактерии. Например, ген аконитазы в бактериальной хромосоме не может быть модифицирован, в то время как регуляторный элемент гена аконитазы модифицируется, и наоборот. Например, только что-то одно может быть модифицировано: ген аконитазы или его регуляторный элемент в хромосоме бактерии, особенно в бактерии, продуцирующей L-лизин с высоким выходом.
Например, в третьем аспекте настоящее изобретение обеспечивает применение бактерии, полученной путем модификации, для получения L-лизина путем ферментации, где модификацией является модификация гена аконитазы и/или его регуляторного элемента в хромосоме бактерии, и активность фермента и/или уровень экспрессии аконитазы этой бактерии, полученной модификацией, снижается, но не элиминируется.
Полученная модификацией бактерия может быть использована отдельно, с другими продуцирующими L-лизин бактериями, или с другими средствами ферментационного продуцирования L-лизина. Если не указано иное (например, без определения «полученное путем настоящей модификации»), используемый здесь термин «бактерия» означает немодифицированную бактерию, или бактерию без этой модификации, имеющую ген аконитазы и регуляторный элемент, расположенный вокруг локуса этого гена в хромосоме, которой принадлежат ген аконитазы и его регуляторный элемент, без снижения активности фермента и/или уровня экспрессии аконитазы, например, ген аконитазы и регуляторный элемент бактерии дикого типа.
Например, в четвертом аспекте настоящее изобретение обеспечивает применение бактерии, полученной путем модификации для повышения ферментационного выхода L-лизина, где модификация - модификация гена аконитазы и/или его регуляторного элемента в хромосоме бактерии и активность фермента и/или уровень экспрессии аконитазы бактерии, полученной модификацией, снижаются, но не элиминируются.
Здесь бактерия может быть продуцирующей L-лизин бактерией, например бактерией Escherichia или Corynebacterium. В одном предпочтительном аспекте бактерия может быть бактерией Escherichia, более предпочтительно - Escherichia coli, например, филиальные штаммы Escherichia coli К-12, в том числе штаммы, полученные из штамма W3110; В другом предпочтительном аспекте бактерией может быть бактерия Corynebacterium, более предпочтительно - Corynebacterium glutamicum или Corynebacterium pekinense. Поскольку ранее имелось мало свидетельств о роли гена аконитазы и/или его регуляторного элемента в бактерии в производстве/ферментации L-лизина и они сосредоточивались на локусах, таких как pnt, dap, ррс и т.п., чтобы модифицировать эти гены в хромосомах, отсутствуют сообщения о модификации гена аконитазы и/или регуляторного элемента вокруг него в производящих L-лизин бактериях (особенно бактериях Escherichia или Corynebacterium, например, Е.coli, Corynebacterium glutamicum или Corynebacterium pekinense), таким образом, продуцирующие L-лизин бактерии, что существенно, имеют гены аконитазы и регуляторные элементы вокруг них дикого типа. Поэтому они могут быть модифицированы с помощью способов по настоящему изобретению для повышения ферментационного выхода L-лизина. В вариантах осуществления настоящего изобретения каждая из бактерий, продуцирующих L-лизин с низким или высоким выходом, может быть модифицирована с помощью способов по настоящему изобретению для повышения ферментационного выхода L-лизина.
Существенно, что в пятом аспекте настоящее изобретение обеспечивает способ модификации бактерии, который включает стадию модификации гена аконитазы и/или его регуляторного элемента в хромосоме бактерии таким образом, что активность и/или уровень экспрессии аконитазы этой бактерии, полученной модификацией, снижаются, но не элиминируются.
Бактерия, полученная по способу пятого аспекта изобретения, может быть использована для продуцирования или получения L-лизина путем ферментации. Поэтому в шестом аспекте настоящее изобретение описывает бактерию, полученную способом по пятому аспекту настоящего изобретения.
В соответствии с экспериментами изобретателей, многие из нуклеотидных последовательностей генов аконитазы (acnA) в бактериях Escherichia (например, E.coli), приведны на SEQ ID No: 1, в то время как многие нуклеотидные последовательности генов аконитазы в бактериях Corynebacterium (например, Corynebacterium glutamicum или Corynebacterium pekinense) приведены на SEQ ID No: 2. Модифицирование этих генов аконитазы таким образом, чтобы снизить, но не элиминировать ферментную активность и/или экспрессию аконитаз в модифицированных бактериях, может повысить ферментационный выход L-лизина. Поэтому в вариантах осуществления настоящего изобретения нуклеотидная последовательность гена аконитазы приведены на SEQ ID No: 1 или 2. Для других бактерий нуклеотидные последовательности генов аконитазы могут быть получены с помощью таких методик, как секвенирование и сравнение идентичностей последовательностей для модификации способами по настоящему изобретению.
В настоящем изобретении предпочтительная стадия модификации гена аконитазы в хромосоме бактерии - вставка, делеция или замена одного или нескольких нуклеотидов в нуклеотидной последовательности гена аконитазы, при условии, что ферментная активность и/или уровень экспрессии аконитазы в модифицированной бактерии уменьшаются, но не элиминируются.
Согласно опыту авторов изобретения, хотя, как правило, повысить активность и/или уровень экспрессии фермента бактерии трудно, поскольку эта бактерия развивались в течение длительного времени и модифицируемые сайты для такого увеличения должны быть исследованы в частностях, тем не менее, исследования по снижению активности и/или уровня экспрессии фермента проводятся гораздо проще. Например, нуклеотидная последовательность внедоменов фермента может быть мутирована. В настоящем изобретении предпочтительная стадия модификации гена аконитазы в хромосоме бактерии может быть заменой одного или нескольких нуклеотидов в нуклеотидной последовательности гена аконитазы. Например, замена включает в себя замену инициирующего кодона гена аконитазы, предпочтительно замену GTG. Это может быть осуществлено для Escherichia coli, Corynebacterium glutamicum, Corynebacterium pekinense и т.п. Кроме того, в настоящем изобретении предпочтительной стадией модификации гена аконитазы в хромосоме бактерии может быть делеция одного или нескольких нуклеотидов в нуклеотидной последовательности гена аконитазы. Например, делеция включает делецию в нуклеотидной последовательности гена аконитазы, предпочтительно делецию 1-120 нуклеотидов, более предпочтительно делецию 1-90 нуклеотидов, наиболее предпочтительно делецию 90 нуклеотидов, например, делеция 90 нуклеотидов до терминального кодона в нуклеотидной последовательности гена аконитазы. Это может быть осуществлено для Escherichia coli, Corynebacterium glutamicum, Corynebacterium pekinense и т.п.
Здесь, регуляторный элемент означает какой-либо полинуклеотид, расположенный выше или ниже по порядку от гена (например, гена аконитазы) и регулирующий транскрипцию и/или экспрессию этого гена, и поэтому влияющий на уровень экспрессии этого гена. Он может включать в себя некодирующие и кодирующие последовательности. Регуляторным элементом может быть промотор, энхансер, репрессор или другой полинуклеотид, связанный с транскрипцией и/или контролем экспрессии. Предпочтительным регуляторным элементом может быть промотор. В одном из вариантов осуществления настоящего изобретения нуклеотидная последовательность промотора приведена на SEQ ID No: 4 или 6. Предпочтительным регуляторным элементом может также быть репрессор, например, транскрипционный репрессор. В одном из вариантов осуществления настоящего изобретения нуклеотидная последовательность репрессора транскрипции приведена на SEQ ID No: 7. Путем модифицирования промотора и/или репрессора ферментативная активность и/или уровень экспрессии аконитазы в модифицированной бактерии уменьшаются, но не элиминируются.
В настоящем изобретении предпочтительная стадия модификации регуляторного элемента гена аконитазы в хромосоме бактерии - вставка, делеция или замена одного или нескольких нуклеотидов в нуклеотидной последовательности регуляторного элемента гена аконитазы до тех пор, пока ферментная активность и/или уровень экспрессии аконитазы в модифицированной бактерии уменьшаются, но не элиминируются.
Согласно опыту авторов изобретения, хотя, как правило, повысить транскрипционную активность промотора или энхансера бактерии трудно, поскольку эта бактерия развивалась в течение длительного времени и сайты, подлежание модификации для такого увеличения, должны быть тщательно исследована, исследование для снижения транскрипционной активности промотора или энхансера проводится намного легче. Например, могут быть добавлены одна или несколько нуклеотидных последовательностей для увеличения расстояния между промотором и инициирующим кодоном гена, или промотор со слабой транскрипционной активностью заменяется оригинальным промотором. В настоящем изобретении предпочтительной стадией модификации регуляторного элемента гена аконитазы в хромосоме бактерии может быть замена одного или нескольких нуклеотидов в промоторе гена аконитазы, например, замена промотора со слабой транскрипционной активностью, более предпочтительно замена нуклеотидной последовательности, данной на SEQ ID No: 3 или 5.
Кроме того, в настоящем изобретении предпочтительной стадией модификации регуляторного элемента гена аконитазы в хромосоме бактерии может быть вставка нуклеотидной последовательности транскрипционного репрессора для гена аконитазы, например, добавление нуклеотидной последовательности нового репрессора транскрипции или увеличение числа копий нуклеотидной последовательности оригинального транскрипционного репрессора. Репрессор транскрипции может быть добавлен за промотором (особенно за промотором с сильной транскрипционной активностью) для того, чтобы более эффективно повысить его уровень экспрессии и ингибировать транскрипцию гена аконитазы. В одном варианте осуществления настоящего изобретения добавляется нуклеотидная последовательность, приведенная на SEQ ID No: 8 и 7 в тандеме.
Кроме того, настоящее изобретение обеспечивает промежуточные продукты, такие как полинуклеотид и/или вектор, используемые в указанных выше методах, а также использования продуктов. Например, в седьмом аспекте настоящее изобретение предоставляет полинуклеотид, нуклеотидная последовательность которого выбирается из
(a) нуклеотидной последовательности, полученной путем замены (предпочтительно GTG) для инициирующего кодона нуклеотидной последовательности, данной на SEQ ID No: 1;
(b) нуклеотидной последовательности, полученной путем делеции (предпочтительно 1-120 нуклеотидов, более предпочтительно - 1-90 нуклеотидов, наиболее предпочтительно - 90 нуклеотидов) в нуклеотидной последовательности, приведенной на SEQ ID No: 1 или 2, например, делеция 90 нуклеотидов до терминального кодона в нуклеотидной последовательности, данной на SEQ ID No: 1 или 2; и
(с) нуклеотидной последовательности, приведенной на SEQ ID No: 8 или 7 в тандеме.
В восьмом аспекте настоящее изобретение обеспечивает вектор, который включает полинуклеотид по седьмому аспекту изобретения.
В девятом аспекте настоящее изобретение описывает применение полинуклеотида по седьмому аспекту настоящего изобретения и/или вектора по восьмому аспекту настоящего изобретения в способе или использовании по первому, второму, третьему и/или четвертому аспекту изобретения. То есть, в способах или использованиях по первому, второму, третьему и/или четвертому аспекту настоящего изобретения используется полинуклеотид по седьмому аспекту настоящего изобретения и/или вектор по восьмому аспекту настоящего изобретения.
В десятом аспекте настоящее изобретение описывает применение полинуклеотида по седьмому аспекту настоящего изобретения и/или вектора по восьмому аспекту настоящего изобретения для получения бактерии согласно пятому аспекту настоящего изобретения. То есть, в процессе получения бактерии по пятому аспекту изобретения используются полинуклеотид по седьмому аспекту изобретения и/или вектор по восьмому аспекту настоящего изобретения.
Положительный эффект настоящего изобретения состоит в том, что изобретены и подтверждены новые способы увеличения ферментационного выхода L-лизина при использовании бактерий, продуцирующих L-лизин с высоким и низким выходом; и они не интерферируют в модификационных сайтах в хромосомах большого числа бактерий, продуцирующих L-лизин с высоким выходом, модифицированных согласно известным техническим решениям, так, чтобы дополнительно увеличить выход, быть полезными для практического производства L-лизина в результате бактериальной ферментации и быть легкими во внедрении.
Для лучшего понимания настоящего изобретения оно теперь будет описано более подробно со ссылкой на конкретные примеры. Следует отметить, что эти примеры только иллюстрируют изобретение и не должны истолковываться как ограничивающие его объем.
В соответствии с описанием изобретения, возможны различные модификации и изменения/варианты изобретения, очевидные для специалиста в данной области.
Кроме того, публикации, цитируемые в настоящем изобретении, используются для иллюстрации этого изобретения, и их содержание включается здесь посредством ссылки, как если бы они были записаны в данном документе.
Примеры
Варианты осуществления настоящего изобретения дополнительно иллюстрируется следующими примерами. Если не указано иное, методы, используемые в этих примерах, хорошо известны специалистам в данной области; как и коммерчески доступные устройства и реагенты (см. Molecular Cloning: Alaboratory Manual (3rd Edition), Science Press, Microbiology Experiments (4th Edition), Higher Education Press, и инструкции изготовителя устройств и реагентов).
Пример 1. Замена GTG для инициирующего кодона ATG аконитазы
Геномную хромосому штамма Escherichia coli дикого типа, E.coli К12 W3110 (коммерчески доступен в Biological Resource Center, National Institute of Technology and Evaluation (NITE Biological Resource Center, NBRC)) экстрагировали в качестве шаблона для ПЦР-амплификации с помощью праймеров Р1/Р2 и Р3/Р4 соответственно. Были получены два фрагмента ДНК 510 bp и 620 bp и названы фрагментами Up1 и Down1 соответственно. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 30 с (секунд) при 72°С, и повтор 30 раз циклически. Последовательности праймеров были следующие:
Р1: 5'-CGCGGATCCGGAGTCGTCACCATTATGCC-3'
Р2: 5'-TCTCGTAGGGTTGACGACACAGCTCCTCCTTAATGACAGG-3'
Р3: 5'-CCTGTCATTAAGGAGGAGCTGTGTCGTCAACCCTACGAGA-3'
Р4: 5'-ATTGCGGCCGCTCCATTCACCGTCCTGCAAT-3'
Эти два фрагмента ДНК очищали электрофорезом в агарозном геле и смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с использованием праймеров Р1/Р4. Был получен фрагмент 1200bp (приблизительно) и назван фрагментом Up-Down1. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 60 с (секунд) при 72°С, и повтор 30 раз циклически.
Фрагмент Up-Down1, очищенный электрофорезом в агарозном геле, и плазмиду pKOV (коммерчески доступна из Addgene) расщепляли (подвергали ферментативному гидролизу) посредством BamHI/NotI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и лигировали в вектор pKOV-Up-Down1 для дальнейшего преобразования. Вектор pKOV-Up-Down1 был изучен секвенированием в секвенирующей компании, он имел правильную точечную мутацию (A-G) во фрагменте гена acnA, и его хранили для дальнейшего использования.
Полученная плазмида pKOV-Up-Downl была трансформирована электропорацией в штамм, продуцирующий L-лизин с низким выходом, Е.Coli NRRLB-12185 (коммерчески доступен в Agricultural Research Service Culture Collection (NRRL), см. также патент US 4346170 A для способа его построения), и штамм, продуцирующий L-лизин с высоким выходом, Е.Col iK12 W3110 Δ3 (коммерчески доступен в Институте микробиологии Академии наук Китая; штамм является L-лизин-продуцирующим инженерным штаммом, мутагенизированным и мутировавшим из Е.Coli K12 W3110) соответственно. Эти два штамма были определены с помощью секвенирования, что они имеют ген дикого типа acnA (т.е. сайты от 1333855 до 1336530 в GenBank, инвентарный номер U00096.2) и элементы вверх и вниз от него в их хромосомах. В соответствии с инструкцией источника плазмиды pKOV из Addgene, отбирали гомологичные рекомбинационно-положительные клоны после восстановления культуры в среде LB при условиях 30°С и 100 об/мин в течение 2 ч, и с помощью секвенирования было установлено, что они имеют мутацию инициирующего кодона ATGвGTG в гене дикого типа acnA в их хромосомах. Наконец, были получены два штамма E.coli, продуцирующие L-лизин с низким и высоким выходом, имеющие мутацию инициирующего кодона acnA, и названы YP-13633 и YP-13664 соответственно. Уровень экспрессии аконитаз в этих двух модифицированных штаммах был измерен и оказался снижен примерно на 75-85% (разные значения в различных средах).
Пример 2. Мутация последовательности гена аконитазы для снижения активности аконитазы
(1) Получение штаммов E.coli
Удаляли (делецировали) 90 нуклеотидов до окончания кодона в гене acnA E.coli для снижения активности аконитазы. В частности, экстрагировали геномную хромосому штамма Escherichia coli дикого типа, E.coli К12 W3110, в качестве шаблона для ПЦР-амплификации, используя праймеры Р5/Р6 и Р7/Р8 соответственно. Были получены два фрагмента ДНК 752bp и 657bp и названы фрагментами Up2 и Down2 соответственно. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 30 с (секунд) при 72°С, и повтор 30 раз циклически. Последовательности праймеров были следующими:
Р5: 5'-CGCGGATCCCGTCACACGATCCGATACCT-3'
Р6: 5'-CGGCAAGCAAATAGTTGTTATACGACTTCCTGGCTACCAT-3'
Р7: 5'-ATGGTAGCCAGGAAGTCGTATAACAACTATTTGCTTGCCG-3'
Р8: 5'-ATTGCGGCCGCCATGGGGCGATTTCCTGATG-3'
Эти два фрагмента ДНК очищали электрофорезом в агарозном геле и смешивали в качестве шаблона для перекрывающейся ПЦР-амплификации с помощью праймеров Р5/Р8. Был получен фрагмент 1400 bp (приблизительно) и назван фрагментом Up-Down2. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 60 с (секунд) при 72°С, и повтор 30 раз циклически.
Up-Down2, очищенный электрофорезом в агарозном геле, и плазмиду pKOV (коммерчески доступна из Addgene) расщепляли (подвергали ферментативному гидролизу) посредством BamHI/NotI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и лигировали в вектор pKOV-Up-Down2 для дальнейшего преобразования. В секвенирующей компании секвенированием было установлено, что вектор pKOV-Up-Down2 имеет делецию оснований 90bp до терминального кодона во фрагменте гене acnA, и его хранили для дальнейшего использования.
Согласно инструкции производителя плазмиды pKOV из Addgene, построенная плазмида pKOV-Up-Down2 была преобразована электропорацией в штамм, продуцирующий L-лизин с низким выходом, Е. Coli NRRL В-12185 (см. также патент US4346170A для способа его получения), и штамм, продуцирующий L-лизин с высоким выходом, Е. Coli K2 W3110 Δ3 соответственно. С помощью секвенирования было определено, что эти два штамма имеют ген acnA дикого типа и элементы вверх и вниз от него в их хромосомах. Были выбраны гомологичные рекомбинационно-положительные клоны и путем секвенирования определено, что они имеют делецию оснований 90bp до терминального кодона в гене acnA в их хромосомах. Наконец, были получены два штамма E.coli, продуцирующих L-лизин с низким и высоким выходом, в которых были понижены ферментные активности acnA, и названы YP-13675 и YP-13699 соответственно. Активности аконитазы в этих двух модифицированных штаммах были измерены и оказались уменьшены примерно на 60-80% (разные значения в различных средах).
(2) Получение штаммов Corynebacterium
Удаляли 90 нуклеотидов до терминального кодона в гене acnCorynebacterium для снижения активности аконитазы. Процесс был по существу таким же, что и на стадии (1), описанной выше, за исключением того, что использовали геном Corynebacterium pekinense AS1.299 (который похож на Corynebacterium glutamicum и иногда ошибочно рассматривается как Corynebacterium glutamicum, коммерчески доступен в China General Microbiological Culture Collection Center (CGMCC)) в качестве шаблона для амплификации с помощью праймеров Р9-Р12 (соответствующих выше упомянутым праймерам Р4-Р8 соответственно), и был получен фрагмент 542bp (Up2), фрагмент 527bp (Down2) и фрагмент 1069bp (примерно) (Up-Down). Последовательности праймеров были следующие:
Р9: 5'-CGGGATCCTGCAGCTCAGTACTTGGAT-3'
Р10: 5'-AAAGTCTTCTAATTACTTACTGCGTCGAACTCGACG-3'
Р11: 5'-GTTCGACGCAGTAAGTAATTAGAAGACTTTTGAT-3'
Р12: 5'-TCCCCCGGGGAATACCGGGTCGGTGCG-3'
Затем фрагмент Up-Down2, очищенный электрофорезом в агарозном геле, и плазмиду pK18mobsacB (коммерчески доступна в American Type Culture Collection (АТСС)) расщепляли посредством BamHI/SmaI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и лигировали до рекомбинантного вектора pK18mobsacB-Up-P-Down, имеющего делецию оснований 90bp до терминального кодона в гене acn. Полученную плазмиду pK18mobsacB-Up-Down2 подтверждали секвенированием и трансформировали электропорацией в штамм, продуцирующий L-лизин с низким выходом, Corynebacterium pekinense AS1.299, в штамм, продуцирующий L-лизин с низким выходом, Corynebacterium glutamicum АТСС 13032 (коммерчески доступен в АТСС) и штамм, продуцирующий L-лизин с высоким выходом, Corynebacterium pekinense AS1.563 (коммерчески доступен в China General Microbiological Culture Collection Center). Для последних двух штаммов секвенирование показало, что они имеют в своих хромосомах ген сап из Corynebacterium pekinense AS1.299, нуклеотидная последовательность которого дана в SEQ ID No: 2. Отбирали гомологичные рекомбинационно-положительные клоны после восстановления культуры в среде BHIS при условиях 30°С и 120 об/мин в течение 2 ч, и подтверждали секвенированием. Наконец, были получены три штамма-продуцента L-лизина Corynebacterium с низким, низким и высоким выходом, имеющие мутацию гена acn, и названы YP-14808, YP-14852 и YP-14837.
Пример 3. Замена промотора со слабой транскрипционной активностью на промотор гена аконитазы
(1) Получение штаммов E.coli
Анализируя выше расположенную последовательность гена acnA в E.coli К12 W3110, мы выявили промотор со слабой транскрипционной активностью (последовательность на SEQ ID No: 3), чтобы заменить область (домен) промотора дикого типа (последовательность которого дана на SEQ ID No: 4), выше ORF гена acnA для уменьшения экспрессии гена acnA дикого типа.
В частности, геномную хромосому штамма Escherichia coli дикого типа, E.coli К12 W3110, экстрагировали в качестве шаблона для ПЦР-амплификации, используя праймеры Р13/Р14 и Р15/Р16 соответственно. Было получены два фрагмента ДНК, 580bp и 618bp, и названы фрагментами Up3 и Down3 соответственно. Плазмиду, содержащую промотор со слабой транскрипционной активностью, упомянутую выше, использовали в качестве шаблона для ПЦР-амплификации с использованием Р17/Р18, и получили фрагмент промотора 161bp со слабой транскрипционной активностью, назвав фрагментом Р. Процесс ПЦР-амплификации был следующим: денатурирование в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 30 с (секунд) при 72°С, и повтор 30 раз циклически.
Три выше указанных фрагмента ДНК очищали электрофорезом в агарозном геле. Затем фрагменты Up3 и Р смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с помощью праймеров Р13/Р18. Получили фрагмент 716bp (приблизительно) и назвали фрагментом Up-P. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 60 с (секунд) при 72°С, и повтор 30 раз циклически.
Фрагменты Up-P и Down3, очищенные электрофорезом в агарозном геле, смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с помощью праймеров Р13/Р16. Получили фрагмент 1334bp (приблизительно)и назвали фрагментом Up-P-down. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 60 с (секунд) при 72°С, и повтор 30 раз циклически.
Последовательности выше упомянутых праймеров были следующие:
Р13: 5'-CGCGGATCCGAAGAAATTGAGGTCATGTT-3'
Р14: 5'-GGTTTCTTAGACGTCGGATTCGTTTCGTTTCTGTTTCATT-3'
Р15: 5'-ATCAGCAGGACGCACTGACCCATTAAGGAGGAGCTATGTCG-3'
Р16: 5'-ATTGCGGCCGCTCCATTCACCGTCCTGCAATT-3'
Р17: 5'-AATGAAACAGAAACGAAACGCAATCCGACGTCTAAGAAACC-3'
Р18: 5'-CGACATAGCTCCTCCTTAATGGGTCAGTGCGTCCTGCTGAT-3'
Фрагмент Up-Р-down, очищенный электрофорезом в агарозном геле, и плазмиду pKOV (коммерчески доступна в Addgene) расщепляли посредством BamHI/NotI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и лигировали в вектор pKOV-Up-P-Down для дальнейшего преобразования. В секвенирующей компании определили секвенированием, что вектор pKOV-Up-P-Down имеет правильную последовательность промотора со слабой транскрипционной активностью, и его сохраняли для дальнейшего использования.
Полученную плазмиду pKOV-Up-P-Down преобразовали электропорацией в штамм, продуцирующий L-лизин с низким выходом, E.coli NRRL В-12185, и штамм, продуцирующий L-лизин с высоким выходом, E.coli K12 W3110 Δ3, соответственно. Секвенированием определили, что эти два штамма имеют ген acnA дикого типа и его элементы вверх и вниз по их хромосомам, где выше лежащий промотор содержит сайты с инвентарными номерами от 2102518 до 2102713 в Gen Bank СР004009.1. Согласно инструкции производителя плазмиды pKOV из Addgene, отбирали гомологичные рекомбинационно-положительные клоны после восстановления культуры в LB среде при условиях 30°С и 100 об/мин в течение 2 ч, и секвенированием определили, что в них промотор со слабой транскрипционной активностью замещен на промотор дикого типа выше по гену acnA в их хромосомах. Наконец, были получены два штамма E.coli, продуцирующих L-лизин с низким и высоким выходом, имеющие мутацию промотора acnA, и названы YP-13627 и YP-13682 соответственно. Уровень экспрессии аконитаз в этих двух модифицированных штаммах был измерен и оказался снижен на 65-80% в различных средах.
(2) Получение штаммов Corynebacterium
Анализом вышележащей последовательности can в Corynebacterium мы выявилили промотор со слабой транскрипционной активностью (последовательность которого представлена в SEQ ID No: 5), чтобы заменить область промотора 166bp дикого типа (последовательность которого дана в SEQ ID No: 6) выше ORF гена can из Corynebacterium. Процесс был по существу таким же, что и на стадии (1), описанной выше, за исключением того, что:
Использовали геном Corynebacterium pekinense AS1.299 (коммерчески доступен в China General Microbiological Culture Collection Center) в качестве шаблона для амплификации с помощью праймеров Р19-Р24 (соответствующих выше упомянутым праймерам Р13-Р18 соответственно), и был получен фрагмент 573bp (Up3), фрагмент 581bp (Down3), фрагмент 130bp (Р) и фрагмент 1284bp (примерно) (Up-P-down). Последовательности праймеров были следующие:
Р19: 5'-CGGGATCCGCCAAAGCAACCAACCCC-3'
Р20: 5'-CTTTTTAGTTTTCAACGGTCGGATTTGCTCGAAAT-3'
Р21: 5'-GCCGAAACAAAGTAGCCGAAGCAGACGCCGTCG-3'
Р22: 5'-CGGAATTCTGACCTGGTGGACGATAC-3'
Р23: 5'-CGAGCAAATCCGACCGTTGAAAACTAAAAAGCTGG-3'
Р24: 5'-GCGTCTGCTTCGGCTACTTTGTTTCGGCCACCC-3'
Затем фрагмент Up-P-down, очищенный электрофорезом в агарозном геле, и плазмиду pK18mobsac5 расщепляли посредством BamHI/EcoRI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и дотировали до рекомбинантного вектора pK18mobsacB-Up-P-Down для дальнейшего замещения промотора. Полученный plasmid pK18mobsacB-Up-P-Down подтверждали секвенированием и трансформировали электропорацией в штамм, продуцирующий L-лизин с низким выходом, Corynebacterium pekinense AS1.299, штамм, продуцирующий L-лизин с низким выходом, Corynebacterium glutamicum АТСС 13032, а также штамм, продуцирующий L-лизин с высоким выходом, Corynebacterium pekinense AS1.563 соответственно. Секвенированием было определено, что последние два штамма имеют в своих хромосомах ген acn и его вышележащие и нижележащие элементы из Corynebacterium pekinense AS1.299. Выбрали Гомологичные рекомбинационно-положительные клоны после восстановления культуры в среде BHIS при условиях 30°С и 120 об/мин в течение 2 ч, и подтверждены секвенированием. Наконец, были получены три штамма Corynebacterium, продуцирующие L-лизин с низким, низким и высоким выходом, имеющие мутацию промотора acn, и названы YP-14755, YP-14732 и YP-14780.
Пример 4. Увеличение числа копий гена acnR, кодирующего транскрипционный репрессор гена аконитазы
Число копий гена acnR, кодирующего транскрипционный репрессор acn, увеличивали для снижения уровня транскрипции гена асn. В частности, использовали геном Corynebacterium pekinense AS1.299 в качестве шаблона для ПЦР-амплификации с использованием праймеров Р25/Р26 и Р27/Р28 соответственно. Были получены два фрагмента ДНК, 715 bp и 797 bp, и названы фрагментами Up4 nDown4. Этот шаблон использовали для ПЦР-амплификации с использованием праймеров Р29/Р30. Получили фрагмент 567bpacnR и назвали фрагментом R, нуклеотидная последовательность которого показана на SEQ ID No: 7. Кроме того, использовали экспрессионнную плазмиду pXMJ19 (коммерчески доступна в Biovector Science Lab, Inc) в качестве шаблона для ПЦР-амплификации с использованием праймеров Р31/Р32. Получили промотор Ptac(164bp) с сильной транскрипционной активностью и назвали фрагментом Ptac, нуклеотидная последовательность которого показана на SEQ ID No: 8. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 30 с (секунд) при 72°С, и повтор 30 раз циклически.
Последовательности вышеупомянутых праймеров были следующими:
Р25: 5'-CGGGATCCTTCGCAACCGATAGAGCA-3'
Р26: 5'-CACGAATTATGCAGAATAAGCCTTTAAGTAACAA-3'
Р27: 5'-TAAACGCGACTAAGCGTGACCATTAAAAGGCT-3'
P28: 5'-CGGAATTCAAAAGCCTATTAAGTGTC-3'
P29: 5'-TTCACACAGGAAAGTGTCCGTAGCGGCAGGCGA-3'
P30: 5'-TTTAATGGTCACGCTTAGTCGCGTTTACGGACAG-3'
P31: 5'-TTAAAGGCTTATTCTGCATAATTCGTGTCGCTC-3'
P32: 5'-GCCGCTACGGACACTTTCCTGTGTGAAATTGTTA-3'
Фрагменты RandPtac очищали электрофорезом в агарозном геле и смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с помощью праймеров Р31/Р30. Получили фрагмент 731bp (приблизительно) и назвали фрагментом Ptac-R. Фрагменты Up4 and Ptac-R очищали электрофорезом в агарозном геле и смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с использованием праймеров Р25/Р30. Получили фрагмент 1446bp (приблизительно) и назвали фрагментом Up4-Ptac-R. Фрагменты Up4-Ptac-R и Down4 очищали электрофорезом в агарозном геле и смешивали в качестве шаблона для перекрывающей ПЦР-амплификации с использованием праймеров Р25/Р28. Получили фрагмент 2243bp (приблизительно) и назвали фрагментом Up-Ptac-R-Down. Процесс ПЦР-амплификации был следующим: денатурация в течение 30 с (секунд) при 94°С, отжиг в течение 30 с (секунд) при 52°С, элонгация в течение 60 с (секунд) при 72°С и повтор 30 раз циклически.
Затем фрагмент Up-Ptac-R-Down, очищенный электрофорезом в агарозном геле, и плазмиду pK18mobsacB расщепляли посредством BamHI/EcoRI соответственно. Продукты ферментативного гидролиза очищали электрофорезом в агарозном геле и лигировали до рекомбинантного вектора pK18mobsacB-Up-Ptac-R-Down для дальнейшего введения дополнительных копий acnR в некодирующую область хромосомы. В секвенирующей компании определили секвенированием, что вектор pK18mobsacB-Up-Ptac-R-Down имеет фрагменты гена Ptac-acnR, и его сохраняли для дальнейшего использования.
Полученную плазмиду pK18mobsacB-Up-Ptac-R-Down преобразовали электропорацией в штамм, продуцирующий L-лизин с низким выходом, Corynebacterium pekinense AS1.299, в штамм, продуцирующий L-лизин с низким выходом, Corynebacterium glutamicum АТСС 13032, и штамм-продуцент L-лизинас высоким выходом, Corynebacterium pekinense AS1.563 соответственно. Отбирали гомологичные рекомбинационно-положительные клоны после восстановления культуры в среде BHIS при условиях 30°С и 100 об/мин в течение 2 ч, и подтверждали секвенированием. Наконец, получили три штамма Corynebacterium, продуцирующих L-лизин с низким, низким и высоким выходом, в некодирующих регионах хромосом которых были вставлены дополнительные копии гена acnR, и названы YP-14857, YP-14896 и YP-14860.
Пример 5. Ферментационные эксперименты cL-лизином
Для ферментации E.coli 25 мл посевной среды, приведенной в табл. 1, инокулировали каждым из штаммов E.coli К12 W3110 Δ 3, E.coli NRRLB-12185 и штаммами E.coli, полученными в примерах 1-3 соответственно, и культивировали при условиях 37°С и 220 об/мин в течение 9 ч. Затем 25 мл ферментивной среды, показанной в табл. 1, инокулировали посредством 1 мл культурального продукта посевной среды и культивировали при условиях 37°С и 220 об/мин в течение 48 часов. Выход L-лизина измеряли с помощью ВЭЖХ после процесса культивирования.
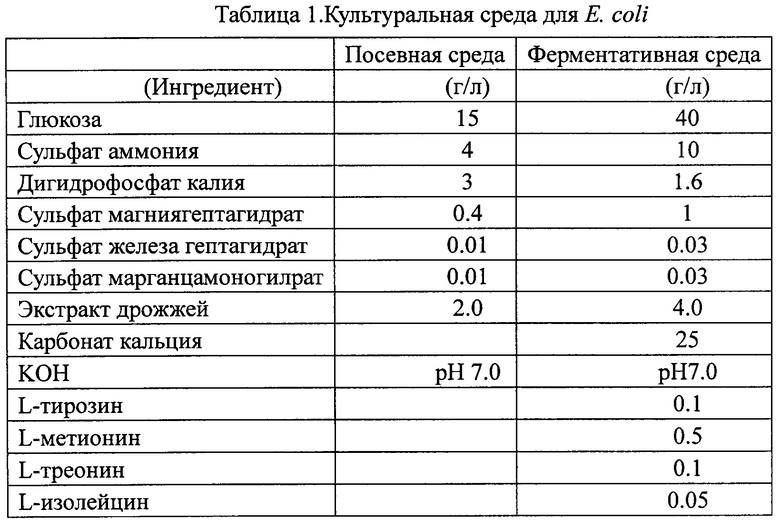
Для ферментации Corynebacterium 30 мл посевной среды, приведенной в табл. 2, инокулировали каждым из штаммов Corynebacterium AS1.299, 13032, AS1.563 и штаммами Corynebacterium, полученными в примерах 2-4 соответственно, и культивировали при условиях 30°С и 220 об/мин в течение 8 часов. Затем 30 мл ферментативной среды, показанной в табл. 2, инокулировали 1 мл культурального продукта посевной среды и культивировали при условиях 30°С и 220 об/мин в течение 48 часов. Выход L-лизина измеряли с помощью ВЭЖХ после процесса культивирования.
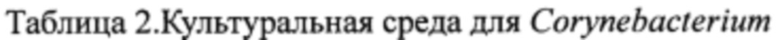
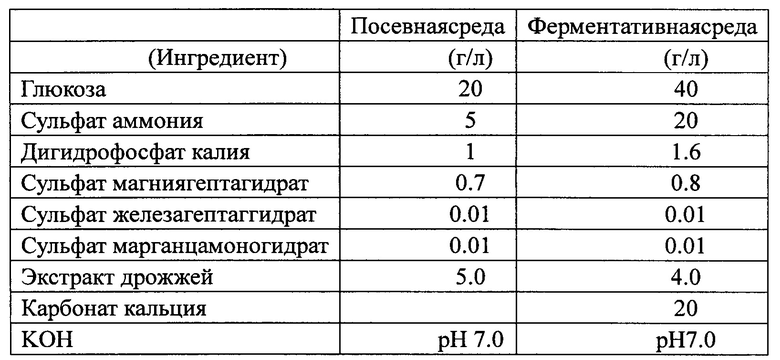
Результаты показаны в табл. 3. Мутирование (например, делеция или замена) структуры аконитазы штаммов Escherichia coli и Corynebacterium для уменьшения активности фермента, или модификация (например, замена и вставка) регуляторных элементов генов аконитазы штаммов Escherichia coli и Corynebacterium для уменьшения уровня экспрессии фермента, способствуют повышению выхода L-лизина.
--->
Перечень последовательностей
<110> Ningxia EPPEN Biotech Co., Ltd
<120> Method for Producing L-Lysine by Modifying Aconitase Gene and/or
Regulatory Elements thereof
<130> YP-PCT-1
<150> CN 201310050196.0
<151> 2013-02-08
<150> CN 201310050144.3
<151> 2013-02-08
<160> 8
<170> PatentIn version 3.5
<210> 1
<211> 2676
<212> DNA
<213> Escherichia coli
<400> 1
atgtcgtcaa ccctacgaga agccagtaag gacacgttgc aggccaaaga taaaacttac 60
cactactaca gcctgccgct tgctgctaaa tcactgggcg atatcacccg tctacccaag 120
tcactcaaag ttttgctcga aaacctgctg cgctggcagg atggtaactc ggttaccgaa 180
gaggatatcc acgcgctggc aggatggctg aaaaatgccc atgctgaccg tgaaattgcc 240
taccgcccgg caagggtgct gatgcaggac tttaccggcg tacctgccgt tgttgatctg 300
gcggcaatgc gcgaagcggt taaacgcctc ggcggcgata ctgcaaaggt taacccgctc 360
tcaccggtcg acctggtcat tgaccactcg gtgaccgtcg atcgttttgg tgatgatgag 420
gcatttgaag aaaacgtacg cctggaaatg gagcgcaacc acgaacgtta tgtgttcctg 480
aaatggggaa agcaagcgtt cagtcggttt agcgtcgtgc cgccaggcac aggcatttgc 540
catcaggtta acctcgaata tctcggcaaa gcagtgtgga gtgaattgca ggacggtgaa 600
tggattgctt atccggatac actcgttggt actgactcgc acaccaccat gatcaacggc 660
cttggcgtgc tggggtgggg cgttggtggg atcgaagcag aagccgcaat gttaggccag 720
ccggtttcca tgcttatccc ggatgtagtg ggcttcaaac ttaccggaaa attacgtgaa 780
ggtattaccg ccacagacct ggttctcact gttacccaaa tgctgcgcaa acatggcgtg 840
gtggggaaat tcgtcgaatt ttatggtgat ggtctggatt cactaccgtt ggcggatcgc 900
gccaccattg ccaatatgtc gccagaatat ggtgccacct gtggcttctt cccaatcgat 960
gctgtaaccc tcgattacat gcgtttaagc gggcgcagcg aagatcaggt cgagttggtc 1020
gaaaaatatg ccaaagcgca gggcatgtgg cgtaacccgg gcgatgaacc aatttttacc 1080
agtacgttag aactggatat gaatgacgtt gaagcgagcc tggcagggcc taaacgccca 1140
caggatcgcg ttgcactgcc cgatgtacca aaagcatttg ccgccagtaa cgaactggaa 1200
gtgaatgcca cgcataaaga tcgccagccg gtcgattatg ttatgaacgg acatcagtat 1260
cagttacctg atggcgctgt ggtcattgct gcgataacct cgtgcaccaa cacctctaac 1320
ccaagtgtgc tgatggccgc aggcttgctg gcgaaaaaag ccgtaactct gggcctcaag 1380
cggcaaccat gggtcaaagc gtcgctggca ccgggttcga aagtcgtttc tgattatctg 1440
gcaaaagcga aactgacacc gtatctcgac gaactggggt ttaaccttgt gggatacggt 1500
tgtaccacct gtattggtaa ctctgggccg ctgcccgatc ctatcgaaac ggcaatcaaa 1560
aaaagcgatt taaccgtcgg tgcggtgctg tccggcaacc gtaactttga aggccgtatc 1620
catccgctgg ttaaaactaa ctggctggcc tcgccgccgc tggtggttgc ctatgcgctg 1680
gcgggaaata tgaatatcaa cctggcttct gagcctatcg gccatgatcg caaaggcgat 1740
ccggtttatc tgaaagatat ctggccatcg gcacaagaaa ttgcccgtgc ggtagaacaa 1800
gtctccacag aaatgttccg caaagagtac gcagaagttt ttgaaggcac agcagagtgg 1860
aagggaatta acgtcacacg atccgatacc tacggttggc aggaggactc aacctatatt 1920
cgcttatcgc ctttctttga tgaaatgcag gcaacaccag caccagtgga agatattcac 1980
ggtgcgcgga tcctcgcaat gctgggggat tcagtcacca ctgaccatat ctctccggcg 2040
ggcagtatta agcccgacag cccagcgggt cgatatctac aaggtcgggg tgttgagcga 2100
aaagacttta actcctacgg ttcgcggcgt ggtaaccatg aagtgatgat gcgcggcacc 2160
ttcgccaata ttcgcatccg taatgaaatg gtgcctggcg ttgaaggggg gatgacgcgg 2220
catttacctg acagcgacgt agtctctatt tatgatgctg cgatgcgcta taagcaggag 2280
caaacgccgc tggcggtgat tgccgggaaa gagtatggat caggctccag tcgtgactgg 2340
gcggcaaaag gtccgcgtct gcttggtatt cgtgtggtga ttgccgaatc gtttgaacga 2400
attcaccgtt cgaatttaat tggcatgggc atcctgccgc tggaatttcc gcaaggcgta 2460
acgcgtaaaa cgttagggct aaccggggaa gagaagattg atattggcga tctgcaaaac 2520
ctacaacccg gcgcgacggt tccggtgacg cttacgcgcg cggatggtag ccaggaagtc 2580
gtaccctgcc gttgtcgtat cgacaccgcg acggagttga cctactacca gaacgacggc 2640
attttgcatt atgtcattcg taatatgttg aagtaa 2676
<210> 2
<211> 2832
<212> DNA
<213> Corynebacterium glutamicum
<400> 2
ttggagctca ctgtgactga aagcaagaac tccttcaatg ctaagagcac ccttgaagtt 60
ggcgacaagt cctatgacta cttcgccctc tctgcagtgc ctggcatgga gaagctgccg 120
tactccctca aggttctcgg agagaacctt cttcgtaccg aagacggcgc aaacatcacc 180
aacgagcaca ttgaggctat cgccaactgg gatgcatctt ccgatccaag catcgaaatc 240
cagttcaccc cagcccgtgt tctcatgcag gacttcaccg gtgtcccttg tgtagttgac 300
ctcgcaacca tgcgtgaggc agttgctgca ctcggtggcg accctaacga cgtcaaccca 360
ctgaacccag ccgagatggt cattgaccac tccgtcatcg tggaggcttt cggccgccca 420
gatgcactgg ctaagaacgt tgagatcgag tacgagcgca acgaggagcg ttaccagttc 480
ctgcgttggg gttccgagtc cttctccaac ttccgcgttg ttcctccagg aaccggtatc 540
gtccaccagg tcaacattga gtacttggct cgcgtcgtct tcgacaacga gggccttgca 600
tacccagata cctgcatcgg taccgactcc cacaccacca tggaaaacgg cctgggcatc 660
ctgggctggg gcgttggtgg cattgaggct gaagcagcaa tgctcggcca gccagtgtcc 720
atgctgatcc ctcgcgttgt tggcttcaag ttgaccggcg agatcccagt aggcgttacc 780
gcaactgacg ttgtgctgac catcaccgaa atgctgcgcg accacggcgt cgtccagaag 840
ttcgttgagt tctacggctc cggtgttaag gctgttccac tggctaaccg tgcaaccatc 900
ggcaacatgt ccccagagtt cggctccacc tgtgcgatgt tcccaatcga cgaggagacc 960
accaagtacc tgcgcctcac cggccgccca gaagagcagg ttgcactggt cgaggcttac 1020
gccaaggcgc agggcatgtg gctcgacgag gacaccgttg aagctgagta ctccgagtac 1080
ctcgagctgg acctgtccac cgttgttcct tccatcgctg gccctaagcg cccacaggac 1140
cgcatccttc tctccgaggc aaaggagcag ttccgtaagg atctgccaac ctacaccgac 1200
gacgctgttt ccgtagacac ctccatccct gcaacccgca tggttaacga aggtggcgga 1260
cagcctgaag gcggcgtcga agctgacaac tacaacgctt cctgggctgg ctccggcgag 1320
tccttggcta ctggcgcaga aggacgtcct tccaagccag tcaccgttgc atccccacag 1380
ggtggcgagt acaccatcga ccacggcatg gttgcaattg catccatcac ctcttgcacc 1440
aacacctcta acccatccgt gatgatcggc gctggcctga tcgcacgtaa ggcagcagaa 1500
aagggcctca agtccaagcc ttgggttaag accatctgtg caccaggttc ccaggttgtc 1560
gacggctact accagcgcgc agacctctgg aaggaccttg aggccatggg cttctacctc 1620
tccggcttcg gctgcaccac ctgtattggt aactccggcc cactgccaga ggaaatctcc 1680
gctgcgatca acgagcacga cctgaccgca accgcagttt tgtccggtaa ccgtaacttc 1740
gagggacgta tctcccctga cgttaagatg aactacctgg catccccaat catggtcatt 1800
gcttacgcaa tcgctggcac catggacttc gacttcgaga acgaagctct tggacaggac 1860
caggacggca acgacgtctt cctgaaggac atctggcctt ccaccgagga aatcgaagac 1920
accatccagc aggcaatctc ccgtgagctt tacgaagctg actacgcaga tgtcttcaag 1980
ggtgacaagc agtggcagga actcgatgtt cctaccggtg acaccttcga gtgggacgag 2040
aactccacct acatccgcaa ggcaccttac ttcgacggca tgcctgtcga gccagtggca 2100
gtcaccgaca tccagggcgc acgcgttctg gctaagctcg gcgactctgt caccaccgac 2160
cacatctccc ctgcttcctc cattaagcca ggtacccctg cagctcagta cttggatgag 2220
cacggtgtgg aacgccacga ctacaactcc ctgggttcca ggcgtggtaa ccacgaggtc 2280
atgatgcgcg gcaccttcgc caacatccgc ctccagaacc agctggttga catcgcaggt 2340
ggctacaccc gcgacttcac ccaggagggt gctccacagg cgttcatcta cgacgcttcc 2400
gtcaactaca aggctgctgg cattccgctg gtcgtcttgg gcggcaagga gtacggcacc 2460
ggttcttccc gtgactgggc agctaagggc actaacctgc tcggaattcg cgcagttatc 2520
accgagtcct tcgagcgtat tcaccgctcc aacctcatcg gtatgggcgt tgtcccactg 2580
cagttccctg caggcgaatc ccacgagtcc ctgggccttg acggcaccga gaccttcgac 2640
atcaccggac tgaccgcact caacgagggc gagactccta agactgtcaa ggtcaccgca 2700
accaaggaga acggcgacgt cgtcgagttc gacgcagttg tccgcatcga caccccaggt 2760
gaggctgact actaccgcca cggcggcatc ctgcagtacg tgctgcgtca gatggctgct 2820
tcttctaagt aa 2832
<210> 3
<211> 161
<212> DNA
<213> Escherichia coli
<400> 3
caatccgacg tctaagaaac cattattatc atgacattaa cctataaaaa taggcgtatc 60
acgaggccct ttcgtcttca cctcgagtcc ctatcagtga tagagatgga catccctatc 120
agtgatagag atactgagca catcagcagg acgcactgac c 161
<210> 4
<211> 454
<212> DNA
<213> Escherichia coli
<400> 4
ggaatgttcc gtcgttattc cagacgactg gcaactaaca tcgcagcagc aagcctttat 60
agaactgttt gctgaagatg atcagccgaa acaataatta tcatcattct tattacccat 120
ttttaatgaa ttaaagggct tttaatacac cgcagcaata acagcttgag ttatctcaac 180
acaaaataat aaccgttaag ggtgtagcct atgatcaaca caaatatgaa atattggtcc 240
tggatgggcg cgttttctct gtcgatgctc ttctgggccg aactcctctg gatcattact 300
cactgatcct tgaccccgct gcggcggggt tgtcatttgc tttgccacaa ggtttctcct 360
cttttatcaa tttgggttgt tatcaaatcg ttacgcgatg tttgtgttat ctttaatatt 420
caccctgaag agaatcaggg cttcgcaacc ctgt 454
<210> 5
<211> 130
<212> DNA
<213> Corynebacterium glutamicum
<400> 5
ccgttgaaaa ctaaaaagct gggaaggtga atcgaatttc ggggctttaa agcaaaaatg 60
aacagcttgg tctatagtgg ctaggtaccc tttttgtttt ggacacatgt agggtggccg 120
aaacaaagta 130
<210> 6
<211> 166
<212> DNA
<213> Corynebacterium glutamicum
<400> 6
acgcgccaag aaccccaact ttcccgccag aacgcttgta ctgttaggat aatgaagacg 60
tagggtcctt ttccacagtt ctgtggaatg agaatccgat gtttttctca cgccggctca 120
gccgaagcag acgccgtcgc gaaatctcac cctaaaaaag ttagaa 166
<210> 7
<211> 567
<212> DNA
<213> Corynebacterium glutamicum
<400> 7
gtgtccgtag cggcaggcga caaaccaaca aatagccgtc aagaaatcct cgaaggtgcc 60
cgacggtgct tcgctgagca cggctatgaa ggcgcaaccg tacgccgact ggaagaagca 120
acaggtaaat cacgcggagc gatctttcat cacttcggtg acaaagaaaa cctgttccta 180
gccctcgcgc gggaagatgc agcccgcatg gcggaggtgg tgtctgaaaa tggcctcgtt 240
gaagtgatgc gaggaatgct ggaagatcct gaacgatatg actggatgtc agtacgcctg 300
gagatctcca agcagctgcg caccgacccg gtattccgcg caaaatggat tgatcaccaa 360
agtgttctag acgaagctgt ccgcgtgcgt ttgtcccgca acgtggataa gggacaaatg 420
cgcactgacg tcccgatcga agtgctgcac accttcttag agactgttct cgacggtttc 480
atctcccgtc ttgctaccgg cgcatccaca gaaggactgt ccgaagtatt ggatctggtc 540
gagggaactg tccgtaaacg cgactaa 567
<210> 8
<211> 164
<212> DNA
<213> Corynebacterium glutamicum
<400> 8
ctgcataatt cgtgtcgctc aaggcgcact cccgttctgg ataatgtttt ttgcgccgac 60
atcataacgg ttctggcaaa tattctgaaa tgagctgttg acaattaatc atcggctcgt 120
ataatgtgtg gaattgtgag cggataacaa tttcacacag gaaa 164
<---
Изобретение относится к способу получения L-лизина ферментацией. Способ предусматривает модификацию гена аконитазы и/или его регуляторного элемента в хромосоме бактерии таким образом, что активность и/или уровень экспрессии аконитазы бактерии уменьшаются, но не элиминируются, и продуцирование L-лизина путём ферментации в бактерии, полученной с помощью указанной модификации. Изобретение обеспечивает повышение выхода L-лизина. 3 табл., 5 пр.
Способ получения L-лизина ферментацией, включающий следующие стадии:
(1) модификацию гена аконитазы и/или его регуляторного элемента в хромосоме бактерии таким образом, что активность и/или уровень экспрессии аконитазы бактерии уменьшаются, но не элиминируются; и
(2) продуцирование L-лизина путём ферментации в бактерии, полученной с помощью модификации на стадии (1), где модификация гена аконитазы и/или его регуляторного элемента включает: замену GTG на инициирующий кодон ATG последовательности нуклеиновой кислоты SEQ ID NO: 1 гена аконитазы в хромосоме Escherichia coli, продуцирующей L-лизин, замену нуклеотидной последовательности SEQ ID NO: 3 на последовательность нуклеиновой кислоты SEQ ID NO: 4 промотора гена аконитазы в хромосоме Escherichia coli, продуцирующей L-лизин, замену нуклеотидной последовательности SEQ ID NO: 5 на последовательность нуклеиновой кислоты SEQ ID NO: 6 промотора гена аконитазы в хромосоме, продуцирующей L-лизин Corynebacterium glutamicum или Corynebacterium pekinense, удаление кодонов 2584-2673 из последовательности нуклеиновой кислоты SEQ ID NO: 1 гена аконитазы в хромосоме Escherichia coli, продуцирующей L-лизин, или удаление кодонов 2740-2829 из последовательности нуклеиновой кислоты SEQ ID NO: 2 гена аконитазы в хромосоме, продуцирующей L-лизин Corynebacterium glutamicum или Corynebacterium pekinense.
| CN 101631871 A, 20.01.2010 | |||
| CN 102191289 A, 21.09.2011 | |||
| СПОСОБ КОНСТРУИРОВАНИЯ БАКТЕРИАЛЬНЫХ ШТАММОВ, ПРОДУЦИРУЮЩИХ АМИНОКИСЛОТЫ (ВАРИАНТЫ), И СПОСОБ ПОЛУЧЕНИЯ АМИНОКИСЛОТЫ ПУТЕМ ФЕРМЕНТАЦИИ С ИСПОЛЬЗОВАНИЕМ СКОНСТРУИРОВАННЫХ БАКТЕРИАЛЬНЫХ ШТАММОВ, ПРОДУЦИРУЮЩИХ АМИНОКИСЛОТЫ | 1999 |
|
RU2214456C2 |
| NCBI, GenBank Accession No | |||
| Способ обкладки резиной сердечниковтранспортерных лент | 1952 |
|
SU99535A1 |
| Найдено онлайн: www.ncbi.nlm.nih.gov/protein/446021680?sat=17&satkey=19951853 Дата обращения 10.01.2023 | |||
| MITSUHASHI S | |||
| ET AL | |||
| Disruption of malate:quinone oxidoreductase | |||

