По данной заявке испрашивается приоритет согласно патентной заявке 202011105063.5, поданной в Национальное управление интеллектуальной собственности Китая 15 октября 2020 года с названием "Рекомбинантный штамм для продуцирования L-аминокислоты, способ его конструирования и его применение"; патентной заявке 202010790887.4, поданной в Национальное управление интеллектуальной собственности Китая 7 августа 2020 года с названием "Рекомбинантный штамм для продуцирования L-аминокислоты, способ его конструирования и его применение"; и патентной заявке 202010514037.1 поданной в Национальное управление интеллектуальной собственности Китая 8 июня 2020 года с названием "Рекомбинантный штамм для модификации гена lysc, способ его конструирования и его применение". Все эти три предшествующие заявки включены в данной описание посредством ссылки во всей их полноте.
Область техники
Данное изобретение относится к области генной инженерии и микробных технологий, в частности относится к рекомбинантному штамму, продуцирующему L-аминокислоту, к способу его конструирования и к его применению.
Предшествующий уровень техники
L-лизин имеет широкий спектр применений, включая медицину, питание, корма и другие аспекты. Среди них L-лизин, используемый в качестве пищевой добавки, составляет более чем 90% от общего количества. В настоящее время Китай является вторым по величине потребительским рынком и крупнейшим производителем L-лизина.
В настоящее время L-лизин главным образом получают посредством прямой ферментации, для чего используют штаммы с полным путем биосинтетза L-лизина и используют отбросную мелассу, гидролизат крахмала и тому подобное, в качестве субстратов для продуцирования L-лизина посредством аэробной ферментации. На этот способ сегодня приходится 2/3 производства L-лизина во всем мире и его процесс является очень отработанным. Этот способ в основном применяется к дрожжам, бактериям и плесени, и широкого представлен в микроорганизмах. В настоящее время, производственные штаммы, используемые для ферментации L-лизина в промышленности, представляют собой главным образом размножаемые посредством мутагенеза мутантные штаммов рода Corynebacterium и Brevibacterium. С разработкой метаболической инженерии и генной инженерии, генные мутации стали контролируемыми. Следовательно, в процессе конструирования исходного штамма с помощью метаболической инженерии можно точно определить ключевые гены ферментов для продуцирования L-лизина в метаболическом процессе, и затем улучшить экспрессию таких ключевых генов ферментов, чтобы достичь увеличения продуцирования L-лизина.
L-глутаминовую кислоту главным образом используют в производстве однозамещенного глутамата натрия и специй, и используется в качестве замены соли, пищевой добавки и биохимического реагента и тому подобного. Сама L-глутаминовая кислота может быть использована в качестве лекарственного средства для участия в метаболизме белка и сахара в головном мозге, чтобы способствовать процессу окисления. Этот продукт объединяется с аммиаком в организме для синтеза нетоксичного глутамина, который может уменьшать количество аммиака в крови и облегчать симптомы печеночной комы. В прошлом получение глутамата натрия в основном осуществляли посредством гидролиза клейковины пшеницы (глутенина), а теперь для крупномасштабного производства используют способ микробной ферментации.
Краткое изложение сущности изобретения
Цель настоящего изобретения состоит в том, чтобы разработать новый штамм со способностью продуцировать L-аминокислоту, тем самым предложить способ эффективного получения L-аминокислоты.
Для достижения вышеуказанной цели, автор изобретения обнаружил посредством исследования, что ген 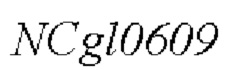 и/или ген
и/или ген 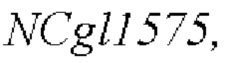 способный продуцировать аминокислоты посредством ферментации, может иметь высокоэффективную способность продуцировать L-аминокислоты при модификации этого гена или улучшения его экспрессии, что было неизвестно в предшествующем уровне техники; кроме того, автор изобретения также обнаружил, что мутация определенной промоторной последовательности также может улучшать способность соответствующих микроорганизмов продуцировать L-аминокислоты. На основании полученных результатов изобретение было завершено.
способный продуцировать аминокислоты посредством ферментации, может иметь высокоэффективную способность продуцировать L-аминокислоты при модификации этого гена или улучшения его экспрессии, что было неизвестно в предшествующем уровне техники; кроме того, автор изобретения также обнаружил, что мутация определенной промоторной последовательности также может улучшать способность соответствующих микроорганизмов продуцировать L-аминокислоты. На основании полученных результатов изобретение было завершено.
В изобретении предлагается бактерия, продуцирующая L-аминокислоты, где экспрессия полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3, улучшена, и/или экспрессия полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 31, улучшена, и/или основания в положениях 45 п. о. и -47п.о. промоторной области, представленные в SEQ ID NO: 57, мутированы. В изобретении также предложен способ продуцирования L-аминокислоты посредством применения такого микроорганизма.
Согласно изобретению, улучшение экспрессии состоит в том, что экспрессия полинуклеотида усилена, или полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3 или SEQ ID NO: 31, имеет точечные мутации, или полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3 или SEQ ID NO: 31, имеет точечные мутации и экспрессия усилена.
В первом аспекте изобретения предлагается бактерия, которая продуцирует L-аминокислоту, в которой экспрессия полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3, улучшена. Предпочтительно, L-аминокислота представляет собой L-лизин или L-глутаминовую кислоту.
Аминокислотная последовательность SEQ ID NO: 3 представляет собой белок, кодируемый геном 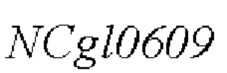 .
.
Бактерия имеет повышенную способность продуцировать L-аминокислоту.
Бактерия со способностью продуцировать L-аминокислоту может быть бактерией, которая может накапливать целевую L-аминокислоту в культуральной среде в количестве, предпочтительно превышающем 0,5 г/л, более предпочтительно превышающем 1,0 г/л.
Полинуклеотиды могут кодировать аминокислотные последовательности с гомологией последовательности примерно 90% или более, примерно 92% или более, примерно 95% или более, примерно 97% или более, примерно 98% или более, примерно 99% или более с аминокислотной последовательностью SEQ ID NO: 3.
В одном конкретном воплощении изобретения полинуклеотид с улучшенной экспрессией содержит нуклеотидную последовательность SEQ ID NO: 1.
В одном воплощении изобретения улучшение экспрессии означает, что полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, имеет точечные мутации, так что аргинин в положении 334 аминокислотной последовательности SEQ ID NO: 3 заменен терминатором.
Согласно изобретению, аминокислотная последовательность, в которой аргинин в положении 334 аминокислотной последовательности, показанной в SEQ ID NO: 3, заменен терминатором, представлена в SEQ ID NO: 4.
В одном воплощении изобретения последовательность полинуклеотида с точечной мутацией образована посредством мутации 1000-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1.
Согласно изобретению, мутация включает мутацию 1000-ого основания полинуклеотидной последовательности, представленной в SEQ ID NO: 1, с цитозина (С) на тимин (Т).
В одном воплощении изобретения полинуклеотидная последовательность с точечной мутацией включает полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
В изобретении также предлагается бактерия, которая продуцирует L-аминокислоты, которые имеют улучшенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 31. Предпочтительно, L-аминокислота представляет собой L-лизин. Предпочтительно, бактерия представляет собой бактерию, которая принадлежит к роду Corynebacterium.
Аминокислотная последовательность SEQ ID NO: 31 представляет собой белок, кодируемый геном
Микроорганизм обладает повышенной способностью продуцировать L-лизин по сравнению с диким типом или родительским штаммом.
Полинуклеотиды могут кодировать аминокислотные последовательности с гомологией последовательности примерно 90% или более, примерно 92% или более, примерно 95% или более, примерно 97% или более, примерно 98% или более, или примерно 99% или более с аминокислотной последовательностью SEQ ID NO: 31.
В одном конкретном воплощении изобретения полинуклеотид может содержать нуклеотидную последовательность SEQ ID NO: 29.
В одном воплощении изобретения полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 31, имеет точечные мутации, так что тирозин в положении 592 аминокислотной последовательности SEQ ID NO: 31 заменен другими аминокислотами.
Согласно изобретению, предпочтительно, чтобы тирозин в положении 592 был заменен фенилаланином.
Согласно изобретению, аминокислотная последовательность, в которой тирозин (Y) в положении 592 аминокислотной последовательности, показанной в SEQ ID NO: 31, заменен фенилаланином (F), показана в SEQ ID NO: 32.
В одном воплощении изобретения полинуклеотидная последовательность с точечной мутацией образуется посредством мутации 1775-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29.
Согласно изобретению, мутация включает мутацию 1775-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29, с аденина (А) на тимин (Т).
В одном воплощении изобретения, полинуклеотидная последовательность с точечной мутацией включает полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
Согласно изобретению, бактерия может быть микроорганизмом, который принадлежит к роду Corynebacterium, такому как Corynebacterium glutamicum, Brevibacterium flavum, Brevibacterium lactofermentum, Corynebacterium ammoniagenes, Corynebacterium pekinense.
В одном воплощении изобретения, микроорганизм, принадлежащий к роду Corynebacterium представляет собой Corynebacterium glutamicum YP97158, с номером депонирования CGMCC No.12856, который депонирован 16 августа 2016 года, депозитарий представляет собой General Microbiology Center of China Microbial Species Conservation and Management Commission (Центр общей микробиологии Китайской комиссии по сохранению и управлению микробными видами), No. 3, Yard. 1 Beichen West Road, Chaoyang District, Beijing, Tel: 010-64807355, зафиксированный в китайской патентной заявке CN106367432A (дата подачи: 1 сентября 2016 г., дата публикации: 1 февраля 2017 г ).
В одном воплощении изобретения, микроорганизм, принадлежащий к роду Corynebacterium, представляет собой Corynebacterium glutamicum АТСС 13869.
Экспрессия полинуклеотидов может быть усилена следующим образом: замена или мутация в последовательностях регуляции экспрессии, введение мутации в полинуклеотидные последовательности, увеличение количества копий полинуклеотидов, введенных посредством хромосомной вставки, или вектора, или их комбинации.
Последовательности регуляции экспрессии полинуклеотидов могут быть модифицированы. Последовательности регуляции экспрессии контролируют экспрессию полинуклеотидов, с которыми они функционально связаны, и могут включать, например, промоторы, терминаторы, энхансеры, сайленсеры и тому подобное. Полинуклеотиды могут иметь изменения в стартовом кодоне. Полинуклеотиды могут быть включены в конкретные сайты хромосом, чтобы увеличить количество копий. Здесь, специфические сайты могут включать, например, сайты транспозонов или межгенные сайты. Кроме того, полинуклеотиды могут быть включены в экспрессионный вектор, и экспрессионный вектор может быть введен в клетки-хозяева для увеличения числа копий.
В одном воплощении изобретения число копий увеличено путем включения полинуклеотидов или полинуклеотидов с точечными мутациями в конкретные сайты микробных хромосом.
В одном воплощении изобретения последовательность нуклеиновой кислоты сверхэкспрессируется путем включения полинуклеотидов с промоторными последовательностями или полинуклеотидов с промоторными последовательностями и точечными мутациями в конкретные сайты микробных хромосом.
В одном воплощении изобретения число копий увеличено в результате включения полинуклеотидов или полинуклеотидов с точечными мутациями в экспрессионные векторы и введения этих экспрессионных векторов в клетки-хозяев а.
В одном воплощении изобретения, аминокислотная последовательность сверхэкспрессируется путем включения полинуклеотидов с промоторными последовательностями или полинуклеотидов с промоторными последовательностями и точечными мутациями в экспрессионные векторы и введения этих экспрессионных векторов в клетки-хозяев а.
В одном конкретном воплощении изобретения, промотор представляет собой промотор полинуклеотида (ген 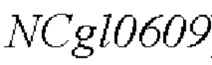 ), кодирующего аминокислотную последовательность SEQ ID NO: 3.
), кодирующего аминокислотную последовательность SEQ ID NO: 3.
В одном конкретном воплощении изобретения, промотор представляет собой промотор полинуклеотида (ген 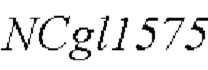 ), кодирующего аминокислотную последовательность SEQ ID NO: 31.
), кодирующего аминокислотную последовательность SEQ ID NO: 31.
В некоторых конкретных воплощениях изобретения, используемые векторы представляют собой плазмиду pK18mobsacB и плазмиду pXMJ19.
Согласно изобретению, бактерия также может иметь другие улучшения, связанные с увеличением продуцирования L-аминокислот.
Во втором аспекте изобретения предлагается полинуклеотидная последовательность, аминокислотная последовательность, кодируемая полинуклеотидной последовательностью, рекомбинантный вектор, включающий полинуклеотидную последовательность, и рекомбинантный штамм, содержащий полинуклеотидную последовательность.
Согласно изобретению, полинуклеотидная последовательность имеет улучшенную экспрессию и улучшение включает точечные мутации полинуклеотида, кодирующего полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 3, где аргинин в положении 334 аминокислотной последовательности заменен терминатором.
Согласно изобретению, аминокислотная последовательность, в которой аргинин в положении 334 аминокислотной последовательности, представленной в SEQ ID NO: 3, заменен на терминатор, представлена в SEQ ID NO: 4.
Согласно изобретению, полинуклеотидная последовательность, кодирующая полипептид, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 3, содержит полинуклеотидную последовательность, представленную в SEQ ID NO: 1.
В одном воплощении изобретения, мутантная полинуклеотидная последовательность, представленная в изобретении, образована посредством мутации 1000-ого основания полинуклеотидной последовательности, представленной в SEQ ID NO: 1.
Согласно изобретению, мутация включает мутацию 1000-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1, с цитозина (С) на тимин (Т).
В одном воплощении изобретения, мутантная полинуклеотидная последовательность включает полинуклеотидную последовательность, представленную в SEQ ID NO: 2.
Согласно изобретению, аминокислотная последовательность с заменой включает аминокислотную последовательность, представленную в SEQ ID NO: 4.
Согласно изобретению, полинуклеотидная последовательность включает полинуклеотид, кодирующий полипептиды, содержащие аминокислотную последовательность, представленную в SEQ ID NO: 31, где тирозин в положении 592 заменен другими аминокислотами.
Согласно изобретению, предпочтительно, тирозин в положении 592 заменен фенилаланином.
Согласно изобретению, аминокислотная последовательность, в которой тирозин (Y) в положении 592 аминокислотной последовательности, показанной в SEQ ID NO: 31, заменен фенилаланином (F), представлена в SEQ ID NO: 32.
Согласно изобретению, предпочтительно, полинуклеотидная
последовательность, кодирующая полипептид, содержащий аминокислотную последовательность, показанную в SEQ ID NO: 31, содержит полинуклеотидную последовательность, показанную в SEQ ID NO: 29.
В одном воплощении изобретения, полинуклеотидная последовательность образована мутацией 1775-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29.
Согласно изобретению, мутация включает мутацию 1775-ого основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29, с аденин (А) на тимин (Т).
В одном воплощении изобретения, полинуклеотидная последовательность включает полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
Согласно изобретению, аминокислотная последовательность включает аминокислотную последовательность, показанную в SEQ ID NO: 32.
Согласно изобретению, мутация относится к изменению основания/нуклеотида сайта. Способ мутации может быть выбран из по меньшей мере одного из мутагенеза, способа ПЦР-сайт-направленной мутации, и/или способа гомологичной рекомбинации. В данном изобретении предпочтительными являются способ ПЦР-сайт-направленной мутации и/или способ гомологичной рекомбинации.
Согласно изобретению, рекомбинантный вектор конструируют путем введения полинуклеотидной последовательности в плазмиду.
В одном воплощении изобретения плазмида представляет собой плазмиду pK18mobsacB.
В другом воплощении изобретения плазмида представляет собой плазмиду pXMJ19.
В частности, полинуклеотидная последовательность и плазмида может быть сконструирована в виде рекомбинантного вектора посредством системы рекомбинации NEBuider.
Согласно изобретению, рекомбинантный штамм содержит полинуклеотидную последовательность.
В качестве одного воплощения изобретения, исходный штамм рекомбинантного штамма представляет собой YP97158.
В качестве одного воплощения изобретения, исходный штамм рекомбинантного штамма представляет собой АТСС 13869.
В третьем аспекте изобретения также предложен способ конструирования рекомбинантного штамма для продуцирования L-аминокислоты.
Согласно изобретению, способ конструирования включает следующие стадии:
Модификацию полинуклеотидной последовательности 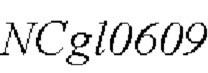 дикого типа, показанной в SEQ ID NO: 1, в штамме-хозяине для мутации его 1000-ого основания, с получением рекомбинантного штамма, содержащего мутированный кодирующий ген
дикого типа, показанной в SEQ ID NO: 1, в штамме-хозяине для мутации его 1000-ого основания, с получением рекомбинантного штамма, содержащего мутированный кодирующий ген 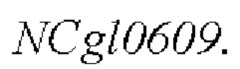
Согласно способу конструирования по изобретению, модификация включает по меньшей мере один из способа мутагенеза, способа ПЦР-сайт-направленной мутации и/или способа гомологичной рекомбинации.
Согласно способу конструирования по изобретению, мутация относится к мутации 1000-ого основания в SEQ ID NO: 1 с цитозина (С) на тимин (Т). В частности, полинуклеотидная последовательность, содержащая мутантный кодирующий ген 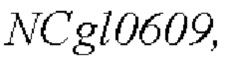 показана в SEQ ID NO: 2. Кроме того, способ конструирования включает следующие стадии:
показана в SEQ ID NO: 2. Кроме того, способ конструирования включает следующие стадии:
(1) Модификацию нуклеотидной последовательности гена 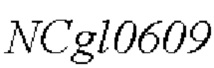 дикого типа, как показано в SEQ ID NO: 1, для мутации его 1000-ого основания, с получением мутантной полинуклеотидной последовательности гена
дикого типа, как показано в SEQ ID NO: 1, для мутации его 1000-ого основания, с получением мутантной полинуклеотидной последовательности гена 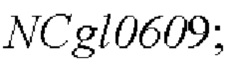
(2) Связывание мутантной полинуклеиновокислотной последовательности с плазмидой для конструирования рекомбинантного вектора; и
(3) Введение рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма, содержащего мутантный кодирующий ген .
Согласно способу конструирования по изобретению, стадия (1) включает: конструирование гена с точечной мутацией: синтез двух пар праймеров Р1 и Р2, РЗ и Р4 для амплификации фрагментов гена на основе последовательности генома немодифицированного штамма, и введение точечной мутации в последовательность SEQ ID NO: 1 гена дикого типа посредством способа ПЦР-сайт-направленной мутации, с получением нуклеотидной последовательности SEQ ID NO: 2 гена с точечной мутацией, который зафиксирован как 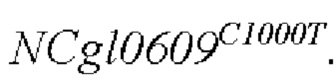
В одном воплощении изобретения геном немодифицированного штамма может происходит из штамма АТСС13032 и последовательность его генома может быть получена с веб-сайта NCBI.
В одном воплощении изобретения на стадии (1), праймеры представляют собой:
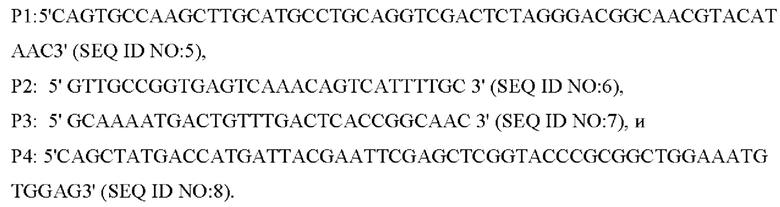
В одном воплощении изобретения ПЦР-амплификацию выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 40 секунд при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
В одном воплощении изобретения ПЦР-амплификацию с перекрывающимися праймерами выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
Согласно способу конструирования по изобретению, стадия (2) включает конструирование рекомбинантной плазмиды, включающее: объединение выделенных и очищенных плазмид 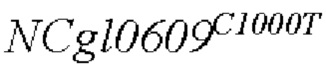 и pK18mobsacB посредством системы рекомбинации NEBuider с получением рекомбинантной плазмиды.
и pK18mobsacB посредством системы рекомбинации NEBuider с получением рекомбинантной плазмиды.
Согласно способу конструирования по изобретению, стадия (3) включает конструирование рекомбинантного штамма: трансформацию рекомбинантной плазмиды в штамм-хозяин с получением рекомбинантного штамма.
В одном воплощении изобретения, трансформация на стадии (3) представляет собой способ электрической трансформации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158. В одном воплощении изобретения рекомбинация достигается посредством гомологичной рекомбинации.
В четвертом аспекте изобретения также предложен способ конструирования рекомбинантного штамма для продуцирования L-аминокислоты.
Согласно изобретению, способ конструирования включает следующие стадии: амплификацию вышерасположенных и нижерасположенных фрагментов гомологичного плеча гена , кодирующей области гена и последовательности его промоторной области, или амплификацию кодирующей области гена или 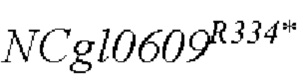 и последовательности его промоторной области, с последующим введением гена или
и последовательности его промоторной области, с последующим введением гена или 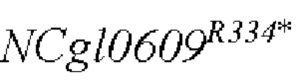 в геном штамма-хозяина посредством гомологичной рекомбинации, чтобы получить сверхэкспрессию гена или
в геном штамма-хозяина посредством гомологичной рекомбинации, чтобы получить сверхэкспрессию гена или 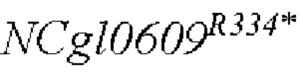 в штамме.
в штамме.
В одном воплощении изобретения, праймеры для амплификации вышерасположенного фрагмента гомологичного плеча представляют собой:
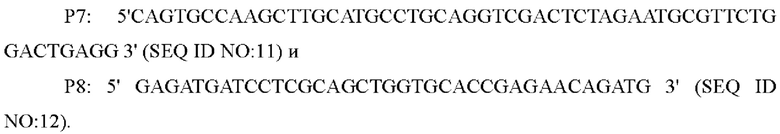
В одном воплощении изобретения, праймеры для амплификации нижерасположенного фрагмента гомологичного плеча представляют собой:
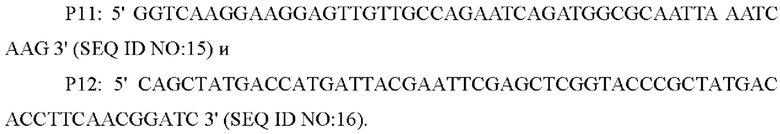
В одном воплощении изобретения, праймеры для амплификации последовательности кодирующей области гена и его промоторной области представляют собой:
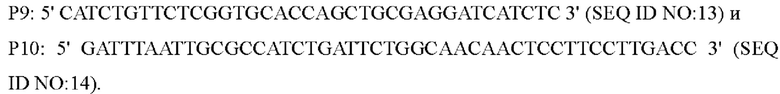
В одном воплощении изобретения, вышеуказанные Р7-Р12 используют в качестве праймеров и вышерасположенный гомологичный фрагмент, нижерасположенный гомологичный фрагмент и фрагмент или с его собственным промотором, полученным посредством амплификации, смешивают в качестве матриц для амплификации, чтобы получить интегрированный фрагмент гомологичного плеча.
В одном воплощении изобретения, используемая система ПЦР представляет собой: 10×Ех Taq буфер 5 мкл, смесь dNTP (дезоксинуклеотиды) (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5 ед/мкл) 0,25 мкл, общий объем: 50 мкл; ПЦР-амплификацию выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С.
В одном воплощении изобретения, шаттл-плазмиду PK18mobsacB объединяют с вышерасположенными и нижерасположенными фрагментами гомологичного плеча, фрагментами кодирующей области гена и промоторной области, посредством использования системы рекомбинации NEBuider, для получения интегрированной плазмиды.
В одном воплощении изобретения, интегрированную плазмиду трансфицируют в штамм-хозяин, и ген или 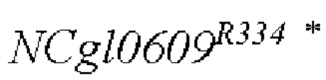 вводят в геном штамма-хозяина посредством гомологичной рекомбинации.
вводят в геном штамма-хозяина посредством гомологичной рекомбинации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения, штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
В пятом аспекте изобретения также предложен способ конструирования рекомбинантного штамма для продуцирования L-аминокислоты.
Согласно изобретению, способ конструирования включает следующие стадии:
Амплификацию кодирующей области гена и последовательности промоторной области, или кодирующей области гена 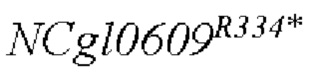 и последовательности промоторной области, конструирование сверхэкспрессирующего плазмидного вектора, и трансфекция вектора в штамм-хозяин для получения сверхэкспрессии или в штамме.
и последовательности промоторной области, конструирование сверхэкспрессирующего плазмидного вектора, и трансфекция вектора в штамм-хозяин для получения сверхэкспрессии или в штамме.
В одном воплощении изобретения праймеры для амплификации последовательности кодирующей области гена и его промоторной области представляют собой:
В одном воплощении изобретения система ПЦР представляет: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5 ед./мкл) 0,25 мкл, общий объем 50 мкл; ПЦР-амплификацию выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С.
В одном воплощении изобретения шаттл-плазмиду рХМJ19 объединяют с фрагментами и 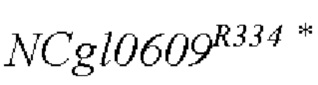 с их собственными промоторами посредством использования системы рекомбинации NEBuider для получения сверхэкспрессирующей плазмиды.
с их собственными промоторами посредством использования системы рекомбинации NEBuider для получения сверхэкспрессирующей плазмиды.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158. В одном воплощении изобретения штамм-хозяин представляет собой АТСС 13869.
В одном воплощении изобретения штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
В изобретении также предложен способ конструирования рекомбинантного штамма коринебактерии.
Согласно изобретению, способ конструирования включает следующие стадии:
модификацию полинуклеотидной последовательности 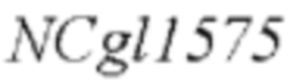 дикого типа в штамме-хозяине, как показано в SEQ ID NO: 29 для мутации его 1775-ого основания, с получением рекомбинантного штамма Corynebacterium, содержащего мутантный кодирующий ген NCgl1575.
дикого типа в штамме-хозяине, как показано в SEQ ID NO: 29 для мутации его 1775-ого основания, с получением рекомбинантного штамма Corynebacterium, содержащего мутантный кодирующий ген NCgl1575.
Согласно способу конструирования по изобретению, модификация включает по меньшей мере один способ из мутагенеза, способа ПЦР-сайт-направленной мутации и/или способа гомологичной рекомбинации.
Согласно способу конструирования по изобретению, мутация относится к мутации 1775-ого основания в SEQ ID NO: 29 с аденина (А) на тимин (Т); в частности, полинуклеотидная последовательность, содержащая мутантный кодирующий ген NCgl1575, показана в SEQ ID NO: 30.
Кроме того, способ конструирования включает следующие стадии:
(1) модификация нуклеотидной последовательности гена NCgl1575 дикого типа, показанной в SEQ ID NO: 29 путем мутации его 1775-ого основания с получением мутантной полинуклеотидной последовательности гена NCgl1575;
(2) связывание мутантной полинуклеиновокислотной последовательности с плазмидой для конструирования рекомбинантного вектора; и
(3) введение рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма Corynebacterium, содержащего мутантный кодирующий ген NCgl1575.
Согласно способу конструирования по изобретению, стадия (1) включает: конструирование гена NCgl1575 с точечной мутацией: синтез двух пар праймеров Р1' и Р2', а также Р3' и Р4' для амплификации фрагментов гена NCgl1575, на основе последовательности генома Corynebacterium glutamicum, и введение точечной мутации в SEQ ID NO: 29 гена NCgl1575 дикого типа при помощи способа ПЦР-сайт-направленной мутации, с получением нуклеотидной последовательности SEQ ID NO: 30 гена NCgl1575 с точечной мутацией, который зарегистрирован, как NCgl1575A1775T.
В одном воплощении изобретения геном Corynebacterium glutamicum может происходить из штамма АТСС13032 и последовательность его генома может быть получена с веб-сайта NCBI.
В одном воплощении изобретения на стадии (1), праймеры представляют собой:
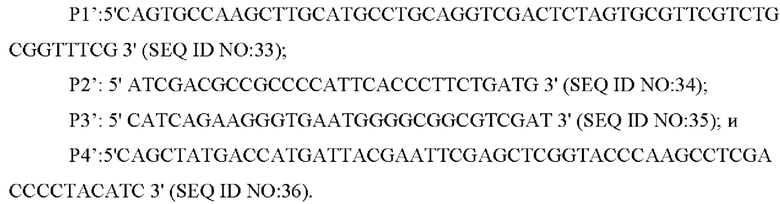
В одном воплощении изобретения ПЦР-амплификацию выполняют следующим образом: денатурацию в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С и элонгацию в течение 40 секунд при 72°С (30 циклов).
В одном воплощении изобретения ПЦР-амплификацию с перекрывающимися праймерами выполняют следующим образом: денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С и элонгация в течение 90 сек при 72°С (30 циклов).
Согласно способу конструирования по изобретению, стадия (2) включает конструирование рекомбинантной плазмиды, включающее: объединение выделенных и очищенных плазмид NCgl1575A1775T и pK18mobsacB при помощи системы рекомбинации NEBuider с получением рекомбинантной плазмиды pK18-NCgl1575A1775T.
Согласно способу конструирования по изобретению, стадия (3) включает конструирование рекомбинантного штамма: трансформацию рекомбинантной плазмиды pK18-NCgl1575A1775T в штамм-хозяин с получением рекомбинантного штамма.
В одном воплощении изобретения, трансформация на стадии (3) представляет метод электрической трансформации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения рекомбинацию осуществляют посредством гомологичной рекомбинации.
В изобретении также предложен способ конструирования рекомбинантного штамма Corynebacterium.
Согласно изобретению, способ конструирования включает следующие стадии:
амплификацию вышерасположенных и нижерасположенных фрагментов гомологичного плеча гена NCgl1575, кодирующей области гена NCgl1575 и последовательности его промоторной области, или кодирующей области гена NCgl1575A1775T и последовательности его промоторной области, и введение гена NCgl1575 или NCgl1575A1775T в геном штамма-хозяина посредством гомологичной рекомбинации, чтобы достичь сверхэкспрессии гена NCgl1575 или NCgl1575A17751 в штамме.
В одном воплощении изобретения праймеры для амплификации вышерасположенного фрагмента гомологичного плеча представляют собой:
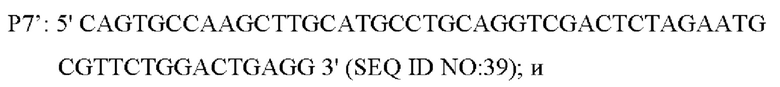
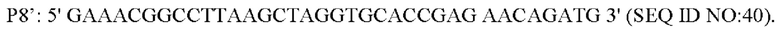
В одном воплощении изобретения, праймеры для амплификации нижерасположенного фрагмента гомологичного плеча представляют собой:

В одном воплощении изобретения, праймеры для амплификации последовательности кодирующей области гена и его промоторной области представляют собой:
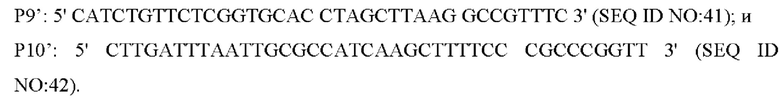
В одном воплощении изобретения, вышеуказанные Р77Р12' используют в качестве праймеров, а вышерасположенный гомологичный фрагмент, нижерасположенный гомологичный фрагмент и фрагмент NCgl1575 или NCgl1575A1775T с его собственным промотором, полученные путем амплификации, смешивают в качестве матриц для амплификации, с получением интегрированного фрагмента гомологичного плеча.
В одном воплощении изобретения, используемая система ПЦР представляет собой: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5 ед/мкл) 0,25 мкл, общий объем 50 мкл; ПЦР-амплификацию выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 180 сек при 72 С (30 циклов) и удлинение в течение 10 минут при 72°С.
В одном воплощении изобретения шаттл-плазмиду PK18mobsacB объединяют с интегрированным фрагментом гомологичного плеча путем использования рекомбинантной системы NEBuider для получения интегрированной плазмиды.
В одном воплощении изобретения интегрированную плазмиду трансфицируют в штамм-хозяин и ген NCgl1575 или NCgl1575A1775T вводят в геном штамма-хозяина посредством гомологичной рекомбинации.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
В изобретении также предложен способ конструирования рекомбинантного штамма Corynebacterium.
Согласно изобретению, способ конструирования включает следующие стадии:
амплификацию кодирующей области гена NCgl1575 и последовательности промоторной области, или кодирующей области гена NCgl1575A1775T и последовательности промоторной области, конструирование сверхэкспрессирующегося плазмидного вектора, и трансфекцию вектора в штамм-хозяин для достижения сверхэкспрессии гена NCgl1575 или NCgl1575A1775T в этом штамме.
В одном воплощении изобретения праймеры для амплификации последовательности кодирующей области гена и промоторной области представляют собой:

В одном воплощении изобретения система ПЦР представляет собой: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5 ед/мкл) 0,25 мкл, общий объем 50 мкл; ПЦР-амплификацию выполняют следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 120 сек при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
В одном воплощении изобретения шаттл-плазмиду рХМJ19 объединяют с фрагментами NCgl1575 или NCgl1575A1775T с их собственными промоторами путем использования рекомбинантной системы NEBuider, чтобы получить сверхэкспрессирующую плазмиду.
В одном воплощении изобретения штамм-хозяин представляет собой YP97158.
В одном воплощении изобретения штамм-хозяин представляет собой штамм, несущий полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
Другой аспект изобретения заключается в предложении нуклеотидной последовательности промотора, которая включает нуклеотидную последовательность, образованную посредством мутации оснований в положениях -45 п. о. и 47 п. о. в промоторной области, показанной в SEQ ID NO: 57.
Согласно изобретению, нуклеотид гуанин (G) в положении -45 п. о. мутирован на аденин (А), а нуклеотид гуанин (G) в положении -47 п. о. мутирован на тимин (Т) в промоторной области, показанной в SEQ ID NO: 57.
Согласно изобретению, нуклеотидная последовательность промотора представляет собой следующее:
(а) нуклеотидную последовательностью, показанную в SEQ ID NO: 58; или
(б) нуклеотидную последовательность, имеющую идентичность последовательности более 90%, предпочтительно более 95%, 98% с нуклеотидной последовательностью, показанной в SEQ ID NO: 58, и сохраняющую повышенную активность промотора (а), при сохранении аденина (А) в положении 45 п. о., и тимина (Т) в положении 47 п. о.
В изобретении также предлагается экспрессионная кассета, содержащая вышеприведенный промотор, включающая промотор и кодирующую последовательность, которая может быть функционально присоединена за промотором. В одном воплощении изобретения, кодирующая последовательность представляет собой кодирующую последовательность гена  .
.
В изобретении также предложен рекомбинантный вектор, содержащий нуклеотидную последовательность промотора по изобретению.
Согласно изобретению, рекомбинантный вектор конструируют посредством соединения нуклеотидной последовательности промотора по изобретению с шаттл-плазмидой; в качестве одного воплощения изобретения шаттл-плазмида представляет собой плазмиду pK18mobsacB.
В изобретении также предложен рекомбинантный штамм, содержащий нуклеотидную последовательность промотора или вышеуказанный рекомбинантный вектор.
Рекомбинантный штамм по изобретению содержит нуклеотидную последовательность, показанную в SEQ ID NO: 58. Нуклеотидная последовательность, показанная в SEQ ID NO: 58, представляет собой промоторную область гена . Кроме того, нуклеотидная последовательность, показанная в SEQ ID NO: 58, соединена с кодирующей последовательностью гена . В частности, рекомбинантный штамм может включать экспрессионную кассету или рекомбинантный вектор, как описано в изобретении выше. В частности, рекомбинантный штамм по изобретению получают посредством трансформации экспрессионной кассеты или рекомбинантного вектора. Рекомбинантный штамм по изобретению образуется посредством введения нуклеотидной последовательности вышеупомянутого мутантного промотора в штамм-хозяин для рекомбинации; штамм-хозяин может быть выбран из штаммов, которые продуцируют L-аминокислоту, особенно L-лизин, как известно в данной области, например по меньшей мере штамм, выбранный из Corynebacterium. Corynebacterium может быть Corynebacterium glutamicum, Corynebacterium flavum, Corynebacterium crenatum и Corynebacterium pekinene; Corynebacterium glutamicum является предпочтительным. В качестве одного воплощения изобретения штамм-хозяин представляет собой YP97158.
В рекомбинантном штамме по изобретению используется плазмида pK18mobsacB в качестве вектора.
Рекомбинантный штамм по изобретению может дополнительно включать другие модификации.
В изобретении также предложен способ конструирования рекомбинантного штамма, продуцирующего L-лизин, который включает следующие стадии:
(1) модификацию промоторной области, показанной в SEQ ID NO:57 посредством мутации положении -45п.о. и -47п.о. с получением нуклеотидной последовательности, содержащей мутированныую промоторную область.
Согласно изобретению, мутация относится к мутации нуклеотида гуанин (г) в положении -45 п. о. на аденин (а) и нуклеотида гуанин (г) в положении -47 п. о. на тимин (Т) в промоторной области, показанного в SEQ ID NO: 57. В частности, нуклеотидная последовательность мутантной промоторной области показана в SEQ ID NO: 58. Кроме того, способ конструирования дополнительно включает следующие стадии:
(2) Связывание нуклеотидной последовательности мутантной промоторной области с плазмидой для конструирования рекомбинантного вектора; и
(3) Введение рекомбинантного вектора в штамм-хозяин с получением рекомбинантного штамма, продуцирующего L-лизин, содержащего мутантную промоторную область.
Согласно изобретению, способ мутации на стадии (1) включает мутагенез, ПЦР-сайт-направленную мутацию или гомологичную рекомбинацию, предпочтительно ПЦР- сайт-направленную мутацию.
Согласно изобретению, стадия (1) включает:
Конструирование двух пар праймеров для амплификации промоторной области гена  с последующим получением нуклеотидной последовательности мутантной промоторной области посредством ПЦР-технологии.
с последующим получением нуклеотидной последовательности мутантной промоторной области посредством ПЦР-технологии.
В одном воплощении настоящего изобретения праймеры на стадии (1) представляют собой:
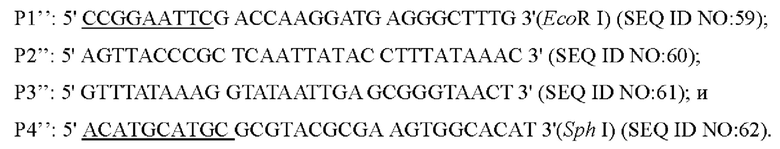
В одном воплощении настоящего изобретения стадия (1) включает: использование Corynebacterium glutamicum АТСС13032 в качестве матрицы и использование праймеров Р1'' и Р2'', Р3'' и Р4'' соответственно для выполнения ПЦР-амплификации с получением двух фрагментов ДНК; использование вышеуказанных двух фрагментов ДНК в качестве матриц и Р1'' и Р4'' в качестве праймеров с получением фрагмента ДНК, содержащего нуклеотидную последовательность промоторной области (SEQ ID NO: 58) по настоящему изобретению посредством ПЦР-амплификации с перекрывающимися праймерами.
Согласно изобретению, на стадии (1), фрагмент ДНК, полученный посредством ПЦР-амплификации с перекрывающимися праймерами, содержит сайты ферментативного переваривания EcoR I и Sph I на обоих концах соответственно.
Согласно изобретению, стадия (2) включает: электрофорез в агарозном геле продукта, амплифицированного посредством реакции ПЦР с перекрывающимися праймерами и разделение и очистка, соединение фрагмента посредством двойного ферментативного переваривания (EcoR I/Sph I) с шаттл-плазмидой посредством такого же переваривания двумя ферментами ((EcoR I/Sph I)) с получением рекомбинантного вектора с аллельной заменой.
Согласно изобретению, шаттл-плазмида представляет собой плазмиду pK18mobsacB; и сконструированный рекомбинантный вектор представляет собой 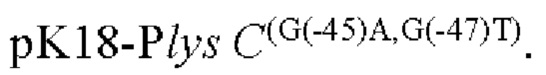
В одном воплощении настоящего изобретения рекомбинантная плазмида имеет маркер резистентности к канамицину.
В одном воплощении настоящего изобретения стадия трансформации (3) представляет собой метод электрической трансформации; например, на стадии (3), рекомбинантный вектор трансформируют в штамм YP97158.
Вышеприведенные различные рекомбинантные штаммы, полученные посредством изобретения, могут быть использованы в ферментации для получения L-аминокислот по отдельности или в комбинации, или могут быть смешаны с другими бактериями, продуцирующими L-аминокислоты, для ферментации, чтобы продуцировать L-аминокислоты.
В другом аспекте изобретения предложен способ продуцирования L-аминокислот, который включает культивирование бактерий; и получение L-аминокислот из культуры.
Бактерии можно культивировать в подходящей среде в условиях культивирования, известных в данной области. Культуральная среда может содержать источник углерода, источник азота, микроэлементы и их комбинации. При культивировании рН культуры может быть скорректирован. Кроме того, при культивировании может быть включено предупреждение образования пузырьков, например посредством использования пеногасителей для предупреждения образования пузырьков. Кроме того, при культивировании может быть включена введение газа в культуру. Газы могут включать любой газ, способный поддерживать аэробные условия в культуре. При культивировании температура культуры может составлять от 20 до 45°. Образовавшиеся L-аминокислоты могут быть извлечены из культуры, то есть культуру обрабатывают серной или соляной кислотой и т.д., с последующим комбинированием методов, таких как анионообменная хроматография, концентрирование, кристаллизация и осаждение в изоэлектрической точке.
В изобретении:
SEQ ID NO:1: последовательность ORF (открытая рамка считывания) дикого типа:
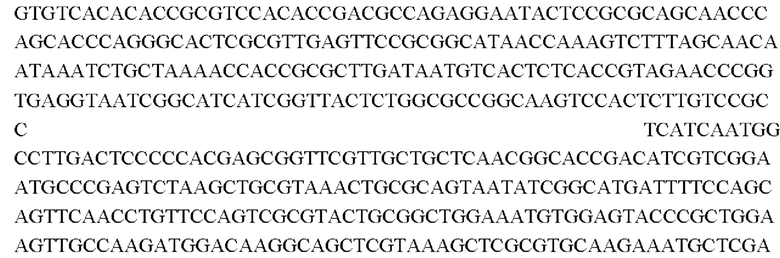

SEQ ID NO:2: последовательность ORE NCgl0609334*:

SEQ ID NO:3: аминокислотная последовательность белка, кодируемая NCg10609 дикого типа:

SEQ ID NO:4: аминокислотная последовательность белка, кодируемая NCg10609R334*:

SEQ ID NO:29: последовательность ORF NCgl1575 дикого типа:


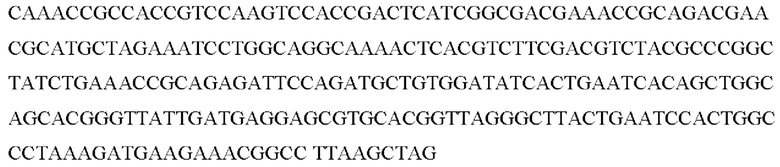
SEQ ID NO:30: последовательность ORF NCgl1575A1775T:


SEQ ID NO:31: последовательность белка, кодируемая NCgl1575 дикого типа:


SEQ ID NO:32: последовательность белка, кодируемая NCgl1575Y592F:
 SEQ ID NO:57: промоторная последовательность дикого типа:
SEQ ID NO:57: промоторная последовательность дикого типа:
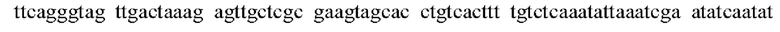
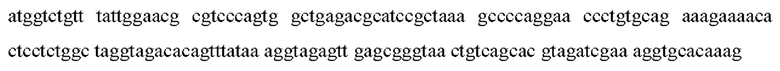
SEQ ID NO:58: мутантная промоторная последовательность:
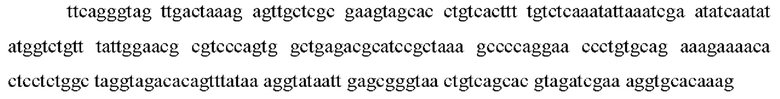
Определение терминов:
В настоящем изобретении термин "бактерия со способностью продуцировать L-аминокислоты" относится к способности продуцировать и накапливать представляющие интерес L-аминокислоты в культуральной среде и/или в клетках бактерий до указанной ниже величины, так что бактерия, продуцирующая L-аминокислоту, может быть собрана, когда бактерия культивируется в культуральной среде. Бактерией со способность продуцировать L-аминокислоту может быть бактерия, которая может накапливать представляющие интерес L-аминокислоты в культуральной среде и/или в клетках бактерий в количестве, которое больше, чем может быть получено при помощи немодифицированного штамма.
Примеры L-аминокислот включают основные аминокислоты, такие как L-лизин, L-орнитин, L-аргинин, L-гистидин и L-цитруллин; алифатические аминокислоты, такие как L-изолейцин, L-аланин, L-валин, L-лейцин и глицин; аминокислоты, такие как гидрокси-моноаминокарбоновые кислоты, такие как L-треонин и L-серин; циклические аминокислоты, такие как L-пролин; ароматические аминокислоты, такие как L-фенилаланин, L-тирозин и L-триптофан; серосодержащие аминокислоты, такие как L-цистеин, L-цистин и L-метионин; кислые аминокислоты, такие как L-глутамат и L-аспартат; и аминокислоты с амидными группами в боковой цепи, такие как L-глутамин и L-аспарагин. Конкретные примеры L-аминокислот включают L-глутаминовую кислоту, L-лизин, L-треонин, L-аргинин, L-гистидин, L-изолейцин, L-валин, L-лейцин, L-фенилаланин, L-тирозин, L-триптофан и L-цистеин. Более конкретные примеры L-аминокислот включают L-глутамат, L-лизин, l-треонин и L-триптофан. Еще более конкретные примеры L-аминокислот включают L-глутамат и L-лизин.
В настоящем изобретении, если не указано иное, термин "аминокислота" относится к L-аминокислоте. В настоящем изобретении, если не указано иное, термин "L-аминокислота" относится к L-аминокислоте в свободной форме, ее соли или их смеси.
Термин "немодифицированный штамм" относится к контрольному штамму, который не был модифицирован так что он приобрел конкретные характеристики. То есть примеры немодифицированных штаммов включают штаммы дикого типа и родительские штаммы.
Термин "гомология" относится к процентной идентичности двух полинуклеотидов или двух полипептидных элементов. Гомологию последовательности одного элемента с другим элементом можно определить с использованием способов, известных в данной области. Например, такую гомологию последовательности можно определить с помощью алгоритма BLAST.
Термин "функциональное связывание" относится к функциональному связыванию регуляторной последовательности с полинуклеотидной последовательностью, при этом регуляторная последовательность контролирует транскрипцию и/или трансляцию полинуклеотидной последовательности. Регуляторная последовательность может быть сильным промотором, который может улучшать уровень экспрессии полинуклеотидов. Регуляторная последовательность может быть промотором, происходящим из микроорганизма, принадлежащего к роду Corynebacterium или может быть промотором, происходящим из других микроорганизмов. Например, промотор может быть промотором trc, промотором gap, промотором tac, промотором Т7, промотором lac, промотором trp, промотором araBAD, или промотором cj7.
Термин "вектор" относится к полинуклеотидной конструкции, которая содержит регуляторную последовательность гена и последовательность гена и конфигурирована для экспрессии целевого гена в подходящей клетке-хозяине. В качестве альтернативы, вектор также может относиться к полинуклеотидной конструкции, содержащей последовательности, которые могут быть использованы для гомологичной рекомбинации, так что, благодаря вектору, введенному в клетку-хозяин а, регуляторная последовательность эндогенного гена в геноме клетки-хозяина может быть изменена, или целевой ген, который может быть экспрессирован, может быть введен в конкретный сайт генома-хозяина. При этом вектор, используемый в настоящем изобретении, может дополнительно содержать селективный маркер для определения введения вектора в клетку-хозяина или встраивания вектора в хромосому клетки-хозяина. Селективные маркеры могут включать маркеры, придающие селективные фенотипы, такие как резистентность к лекарственному средству, ауксотрофный тип, резистентность к цитотоксичным агентам или экспрессию поверхностных белков. В средах, обработанных селективными агентами, трансформированные клетки могут быть отобраны, поскольку клетки, экспрессирующие только селективные маркеры, могут выживать или демонстрировать различные фенотипические признаки.
Используемый здесь термин "трансформация" относится к введению полинуклеотидов в клетки-хозяева, так что полинуклеотиды можно использовать в качестве внегеномных элементов или встраивать в геном клеток-хозяев для репликации. Способ трансформации векторов, используемых в настоящем изобретении, может включать способ введения нуклеиновых кислот в клетки. Кроме того, как описано в соответствующей технологии, способ электрического импульса может быть реализован на основе клеток-хозяев.
Полезные эффекты
В данном изобретении было обнаружено, что продукт, кодируемый геном, оказывает влияние на уровень производства аминокислот в результате ослабления или инактивирования гена или гена NCgl1575. Рекомбинантный штамм получают посредством введения точечных мутаций в кодирующую последовательность или увеличения числа копий или сверхэкспрессии гена. По сравнению со штаммом дикого типа, полученный штамм приводит к продуцированию высоких концентраций аминокислот. Кроме того, рекомбинантный штамм был получен посредством введения точечных мутаций в промоторную область гена lysC. По сравнению с немутантным штаммом, полученный штамм также может значительно улучшать продуципрование L-лизина, кроме того, улучшать эффективность генерации, снижать стоимость генерации и облегчать популяризацию и применение.
Подробные воплощения
Ниже техническое решение настоящего изобретения будет дополнительно подробно описано в сочетании с конкретными примерами. Следует понимать, что следующие примеры являются только иллюстративными и поясняющими данное изобретения и не должны рассматриваться как ограничение объема изобретения. Все технологии, реализованные на основе вышеупомянутого содержания настоящего изобретения, попадают в объем настоящего изобретения. Если не указано иное, все сырье и реагенты, используемые в следующих примерах, являются коммерчески доступными продуктами или могут быть получены известными способами. Все операции известны в данной области или выполняются в соответствии с руководством пользователя имеющихся в продаже продуктов.
В следующих примерах основная среда, используемая для культивирования штаммов, имеет одинаковую композицию, и сахарозу, канамицин или хлорамфеникол и т.д. добавляют к такой композиции основной среды, если это необходимо. Состав основной среды является следующим:
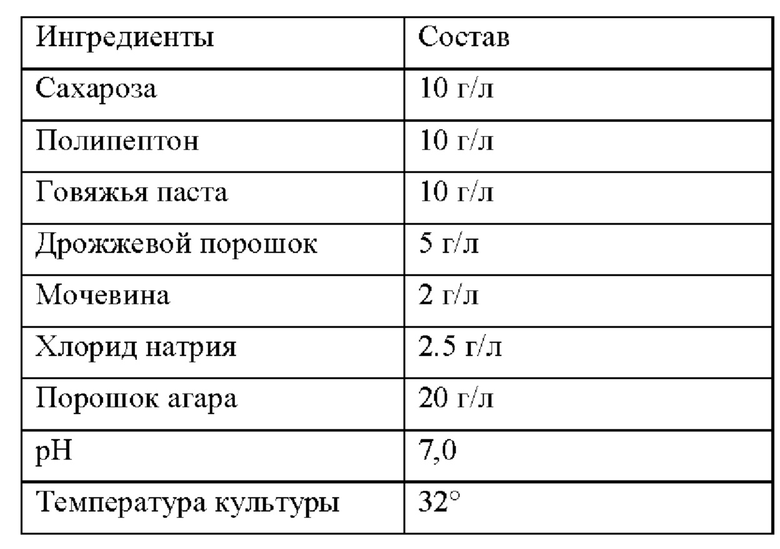
Приготовление и условия электрофореза PAGE (электрофорез в полиакриламидном геле) с SSCP (анализ конформационного полиморфизма однонитевой ДНК) в следующих примерах были следующими:
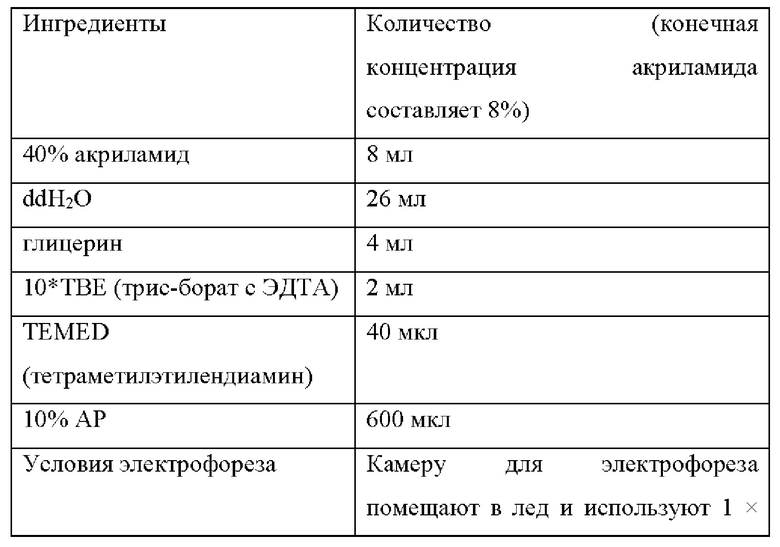

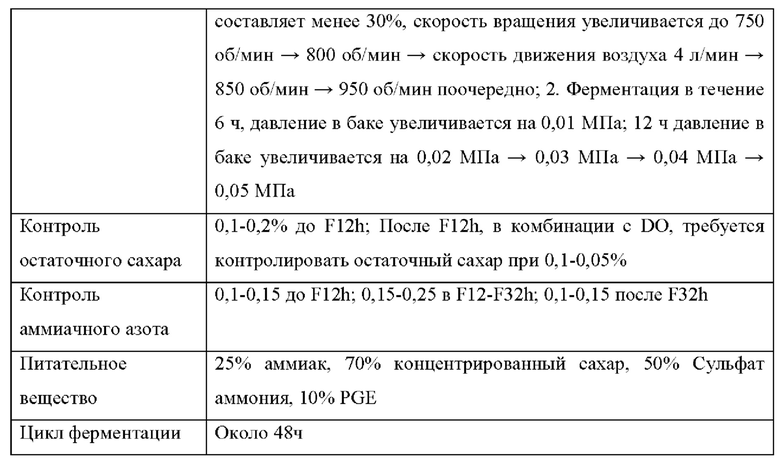
В следующих примерах среда для ферментации и процесс ферментации L-лизина показаны в Таблице 1 и 2 ниже:

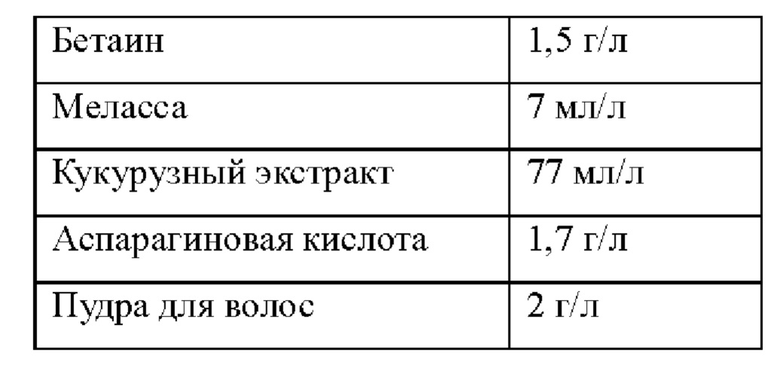
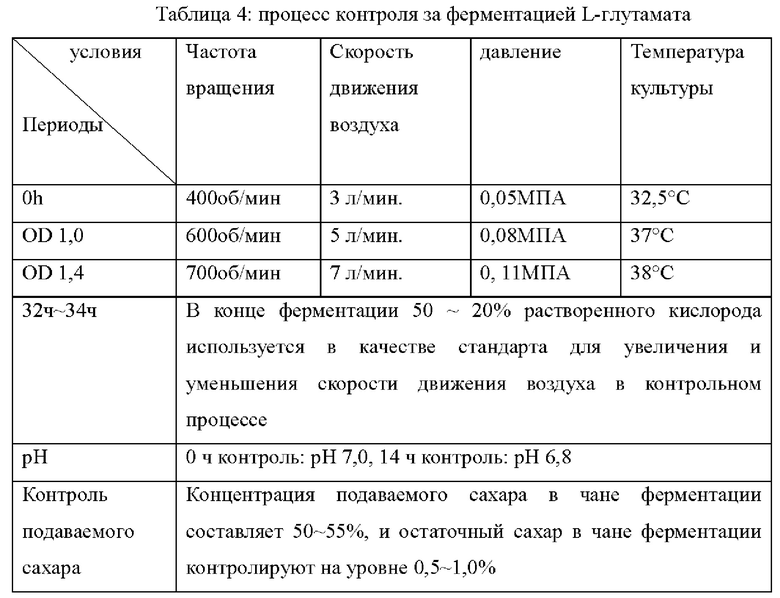
В следующих примерах, ферментационная среда и процесс ферментации L-глутамата показаны в Таблице 3 и 4 ниже:

Пример 1: Конструирование трансформированного вектора 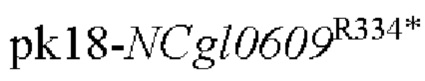 , содержащего кодирующую область гена NCgl0609 с точечной мутацией
, содержащего кодирующую область гена NCgl0609 с точечной мутацией
На основе последовательности генома Corynebacterium glutamicum АТСС13032, опубликованной NCBI, были спроектированы и синтезированы две пары праймеров для амплификации последовательности кодирующей области гена NCgl0609. Точечная мутация была введена в кодирующую область гена (SEQ ID NO: 1, и соответствующая аминокислотная последовательность, кодирующая белки, представляет собой SEQ ID NO: 3) на основе штамма YP97158 [номер депонирования: CGMCC No. 12856, дата депонирования: August 16, 2016, депозиторий: Институт Микробиологии, Китайская академия наук, No. 3, Yard. 1, Beichen West Road, Chaoyang District, Beijing, Tel: 010-64807355, зафиксирован в китайской патентной заявке CN106367432A (дата подачи: 1 сентября 2016 г, и дата публикации: 1 февраля 2017), и посредством секвенирования было подтверждено, что ген 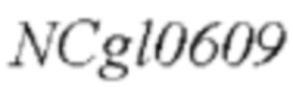 дикого типа сохранялся в хромосоме штамма] посредством аллельной замены, и, таким образом, нуклеотидная последовательность гена в положении 1000 была заменена с С на Т (SEQ ID NO: 2), и соответствующая аминокислотная последовательность, кодирующая белки в положении 334, была замена с аргинина на терминатор (SEQ ID NO: 4:
дикого типа сохранялся в хромосоме штамма] посредством аллельной замены, и, таким образом, нуклеотидная последовательность гена в положении 1000 была заменена с С на Т (SEQ ID NO: 2), и соответствующая аминокислотная последовательность, кодирующая белки в положении 334, была замена с аргинина на терминатор (SEQ ID NO: 4: 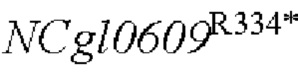 ). Праймеры были сконструированы следующим образом
). Праймеры были сконструированы следующим образом
(синтезированы компанией Shanghai Invitrogen):
Способ конструирования: Corynebacterium glutamicum АТСС13032 был использован в качестве матрицы, и праймеры Р1 и Р2, Р3 и Р4 использовали, соответственно, для ПЦР-амплификации. Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (по 2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) каждый по 2 мкл, Ex Taq(5 ед/мкл) 0,25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, (денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 40 секунд при 72°С, 30 циклов), и удлинение в течение 10 минут при 72°С, и затем были получены два фрагмента ДНК, содержащие кодирующую область гена , с размером 698 п. о. и 648 п. о., соответственно ( выше и ниже). После разделения двух фрагментов ДНК и очистки с помощью электрофореза в агарозном геле, два фрагмента ДНК в качестве матрицы амплифицировали в 1317 п. о. фрагменты посредством ПЦР с перекрыванием праймеров, с использованием Р1 и Р4 в качестве праймеров.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5ед/мкл) 0,25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, (денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С, 30 циклов), и удлинение в течение 10 минут при 72°С. В этом фрагменте ДНК происходила замена цитозина (С) в положении 1000 в кодирующей области гена YP97158 на тимин (Т), и, наконец, в замене 334ой аминокислоты кодируемого белка с аргинина (R) на терминатор. Этот фрагмент ДНК очищали посредством электрофореза в агарозном геле и соединяли с плазмидой pK18mobsacB (приобретенной в компании Addgene, расщепленной двумя ферментами  I/BamH I, соответственно), которую расщепляли двумя ферментами и затем очищали с помощью фермента NEBuilder (приобретенного в компании NEB) при 50°С в течение 30 мин, и положительный вектор
I/BamH I, соответственно), которую расщепляли двумя ферментами и затем очищали с помощью фермента NEBuilder (приобретенного в компании NEB) при 50°С в течение 30 мин, и положительный вектор 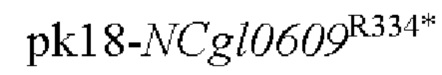 был получен из моноклона, выросшего после трансформации соединенного продукта посредством ПЦР-идентификации, и эта плазмида содержала маркер резистентности к канамицину. Вектор с правильным расщеплением ферментом отправляли в секвенирующую компанию для секвенирования и идентификации, и вектор
был получен из моноклона, выросшего после трансформации соединенного продукта посредством ПЦР-идентификации, и эта плазмида содержала маркер резистентности к канамицину. Вектор с правильным расщеплением ферментом отправляли в секвенирующую компанию для секвенирования и идентификации, и вектор 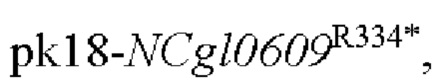 содержащий правильную точечную мутацию (С-Т), хранили для использования.
содержащий правильную точечную мутацию (С-Т), хранили для использования.
Пример 2: Конструирование спроектированных штаммов , содержащих точечную мутацию.
Способ конструирования: плазмиду с аллельной заменой трансформировали в L-лизин-продуцирующий штамм YP97158 посредством электрического шока (см. WO2014121669A1 в отношении способа конструирования; секвенированием подтверждено, что кодирующая область гена дикого типа сохраняется в хромосоме этого штамма). Единичную колонию, полученную посредством культивирования, идентифицировали с помощью праймера Р1 и универсального праймера M13R, и штамм, который мог амплифицировать бэнды размером 1375 п. о., являлся положительным штаммом. Положительный штамм культивировали в среде, содержащей 15% сахарозы, единичную колонию, полученную в результате культивирования, выращивали на среде, содержащий канамицин, и на среде без канамицина, соответственно, и штаммы, которые росли на среде без канамицина, но не росли на среде, содержащей канамицин, затем идентифицировали с помощью ПЦР с использованием следующих праймеров (синтезированных компанией Shanghai mvitrogen):

Вышеописанный продукт ПЦР-амплификации имел 264 п. о., и его денатурировали при 95°С в течение 10 минут и подвергали воздействию ледяной бани в течение 5 минут с последующим sscp электрофорезом (амплифицированный фрагмент плазмиды использовали в качестве положительного контроля, амплифицированный фрагмент YP97158 использовали в качестве отрицательного контроля, а воду использовали в качестве холостого контроля). Благодаря разным структурам фрагментов и положениям на электрофорезе, штаммы с положениями на электрофорезе, отличными от положений фрагментов отрицательного контроля и совпадающими с положениями фрагментов положительного контроля, представляют собой штаммы с успешной аллельной заменой. Фрагмент положительного штамма подвергали ПЦР-амплификации с использованием праймера Р5/Р6, и соединяли с вектором PMD19-T для секвенирования. Согласно выравниванию последовательностей, штамм с мутацией (С-Т) в исходной последовательности представлял собой положительный штамм с удачной аллельной заменой и был назван YPL-4-041.
Пример 3: Конструирование спроектированных штаммов, сверхэкспрессирующих гены и в геноме.
На основе последовательности генома Corynebacterium glutamicum АТСС13032 дикого типа, опубликованного NCBI, были спроектированы и синтезированы три пары праймеров для амплификации вышерасположенного и нижерасположенного фрагментов гомологичного плеча и последовательностей кодирующей области и промоторной области гена и , и ген или был введен в штамм YP97158 посредством гомологичной рекомбинации.
Праймеры были сконструированы следующим образом (синтезированы компанией Shanghai Invitrogen):
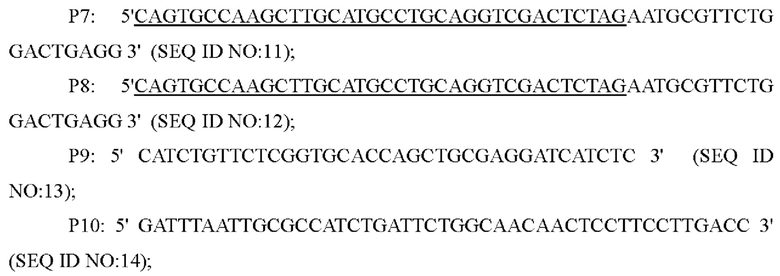
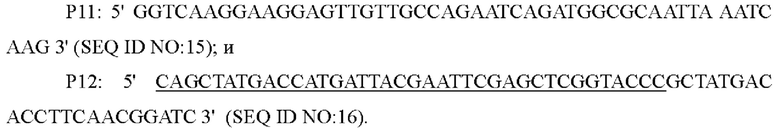
Способ конструирования: Corynebacterium glutamicum АТСС13032 или YPL-4-041 использовали в качестве матрицы, соответственно, для ПЦР-амплификации с праймерами Р7/Р8, Р9/Р10, Р11/Р12 с получением вышерасположенного фрагмента гомологичного плеча размером 768 п. о., гена или и его промоторного фрагмента размером 1626 п. о. и нижерасположенного фрагмента гомологичного плеча размером 623 п. о. После завершения реакции ПЦР три амплифицированных фрагмента были выделены с помощью электрофореза с использованием набор для извлечения ДНК на гелевой колонке, соответственно. Три выделенных фрагмента были соединены с плазмидой pK18mobsacB (приобретенной у Addgene Company, переваренной двумя ферментами  I/BamH I, соответственно), которая была переварена двумя ферментами, и затем очищены с помощью фермента NEBuilder (приобретенного у NEB Company) при 50°С в течение 30 минут, и положительная интегрированная плазмида была получена из моноклона, выросшего после трансформации соединенного продукта посредством ПЦР-идентификацмм. Эта плазмида содержала маркер резистентности к канамицину и рекомбинант с плазмидой, интегрированной в геном, может быть получен посредством отбора на канамицине.
I/BamH I, соответственно), которая была переварена двумя ферментами, и затем очищены с помощью фермента NEBuilder (приобретенного у NEB Company) при 50°С в течение 30 минут, и положительная интегрированная плазмида была получена из моноклона, выросшего после трансформации соединенного продукта посредством ПЦР-идентификацмм. Эта плазмида содержала маркер резистентности к канамицину и рекомбинант с плазмидой, интегрированной в геном, может быть получен посредством отбора на канамицине.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq(5ед/мкл) 0.25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С. Правильно секвенированную интегрированную плазмиду электротрансформировали в L-лизин-продуцирующий штамм YP97158. Единичная колония, полученная посредством культивирования, была идентифицирована посредством ПЦР с помощью праймеров Р13/Р14. Штамм с амплифицированный посредством ПЦР фрагментом размером 1317 п. о. был положительным штаммом, а штамм без фрагмента, при амплификации, был исходным штаммом. Положительный штамм культивировали в среде, содержащей 15% сахарозы и единичную колонию, полученную при культивировании, затем идентифицировали посредством ПЦР с помощью праймеров Р15/Р16. Бактерии, амплифицирующие фрагмент размером 1352 п. о., являются положительными штаммами с геном или , интегрированным в геном YP97158, и они были названы YPL-4-042 (без сайта мутации) и YPL-4-043 (с сайтом мутации).
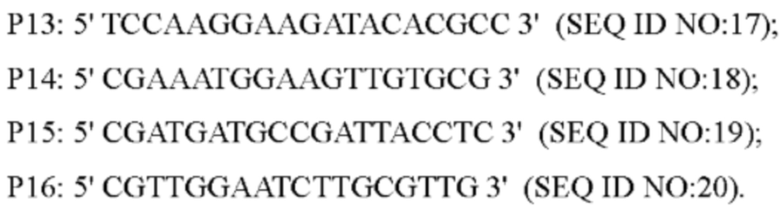
Пример 4: Конструирование спроектированных штаммов, сверхэкспрессирующих гены или плазмиде.
На основе последовательности генома Corynebacterium glutamicum АТСС13032 дикого типа, опубликованной NCBI, была разработана и синтезирована пара праймеров для амплификации последовательностей кодирующей области и промоторной области гена или . Праймеры были разработаны следующим образом (синтезированы компанией Shanghai Invitrogen):
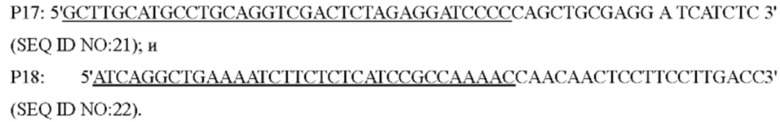
Способ конструирования: Corynebacterium glutamicum АТСС 13032 и YPL-4-041 были использованы в качестве матрицы, соответственно, для ПЦР-амплификации с праймерами Р17/Р18 для получения генов и и их промоторных фрагментов размером 1582 п. о. Амплифицированные продукты были подвергнуты электрофорезу и очищены с использованием колонки набора для извлечения ДНК на гелевой колонке. Выделенный фрагмент ДНК и шаттл-плазмиду рХМJ19 , полученные посредством переваривания ферментом EcoR I, соединяли при 50°С с ферментом NEBuilder (приобретенным у NEB) в течение 30 минут, и положительные сверхэкспрессирующие плазмиды pXMJ19- и pXMJ19- были получены из моноклонов, выросших после трансформации связанных продуктов посредством ПЦР-идентификации с праймером М13, и затем эти плазмиды были отправлены на секвенирование. Поскольку плазмида содержит маркер резистентности к хлорамфениколу, хлорамфеникол может быть использован для определения, была ли плазмида трансформирована в штамм.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq(5ед/мкл) 0,25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 60 секунд при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
Правильно секвенированные плазмиды pXMJ19- и pXMJ19- электротрансформировали в L-лизин-продуцирующий штамм YP97158, соответственно. Единичную колонию, полученную посредством культивирования, идентифицировали с помощью ПЦР с праймерами M13F/P18. Штаммы с амплификацией фрагмента размером 1585 п. о. посредством ПЦР были положительными штаммами, и они были названы YPL-4-044 (без сайта мутации) и YPL-4-045 (с сайтом мутации).
Пример 5: Конструирование спроектированных штаммов с геном , удаленным из генома.
На основании последовательности генома Corynebacterium glutamicum АТСС13032 опубликованного NCBI, были разработаны и синтезированы две пары праймеров для амплификации фрагментов на двух концах кодирующей области гена , в качестве вышерасположенного и нижерасположенного фрагментов гомологичного плеча. Праймеры были спроектированы следующим образом (синтезированы компанией Shanghai Invitrogen):
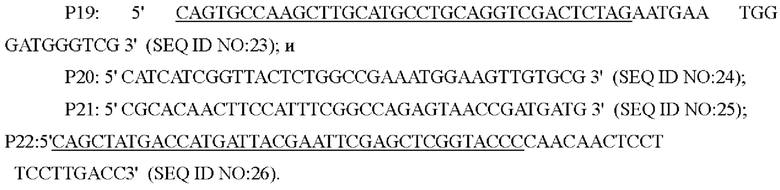
Способ конструирования: Corynebacterium glutamicum АТСС 13032 использовали в качестве матрицы для ПЦР-амплификации с праймерами Р19/Р20 и Р21/Р22, соответственно, для получения вышерасположенного фрагмента гомологичного плеча размером 661 п. о. и нижерасположенного фрагмента гомологичного плеча размером 692 п. о. Затем праймеры Р19/Р22 использовали для ПЦР с перекрывающимися праймерами с получением всего фрагмента гомологичного плеча размером 1334 п. о. Амплифицированные продукты были подвергнуты электрофорезу и очищены с использованием набора для извлечения ДНК на гелевой колонке. Выделенные фрагменты ДНК были соединены с плазмидой pK18mobsacB (приобретенной в Addgene Company, переваренной двумя ферментами  I/BamH I, соответственно), которая была переварена двумя ферментами, и затем очищены с помощью фермента NEBuilder (приобретенного в NEB Company) при 50°С в течение 30 минут. Положительный нокаутный вектор
I/BamH I, соответственно), которая была переварена двумя ферментами, и затем очищены с помощью фермента NEBuilder (приобретенного в NEB Company) при 50°С в течение 30 минут. Положительный нокаутный вектор 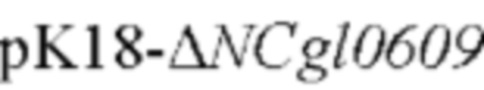 был получен из моноклонов, выросших после трансформации соединенными продуктами посредством ПЦР-идентификации с помощью праймера М13, и зхатем эти плазмиды подвергали секвенированию. Плазмида имела резистентность к канамицину в качестве маркера для отбора.
был получен из моноклонов, выросших после трансформации соединенными продуктами посредством ПЦР-идентификации с помощью праймера М13, и зхатем эти плазмиды подвергали секвенированию. Плазмида имела резистентность к канамицину в качестве маркера для отбора.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5ед/мкл) 0,25 мкл, общий объем 50 мкл.
ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 90 сек при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
Правильно секвенированная нокаутная плазмида была электротрансформирована в лизин-продуцирующий штамм YP97158, и единичную колонию, полученную посредством культивирования, идентифицировали с помощью ПЦР со следующими праймерами (синтезированными компанией Shanghai Invitrogen Company):
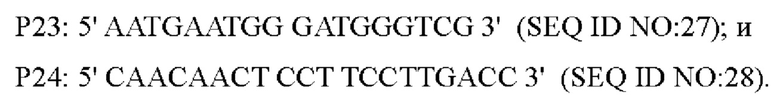
Штаммы с одновременной амплификацией бэндов 1334 п. о. и 1788 п. о. посредством вышеуказанной ПЦР, были положительными штаммами, а штаммы только с амплификацией бэнда размером 1788 п. о. были исходными штаммами. После отбора на среде с 15% сахарозой, положительные штаммы культивировали в среде, содержащей канамицин, и в среде без канамицина, соответственно. Штаммы, которые росли в среде без канамицина, но не росли в среде, не содержащей канамицин, затем идентифицировали с помощью ПЦР с использованием праймеров Р23/Р24. Штаммы с амплификацией бэнда 1334 п. о., были положительными штаммами, у которых кодирующая область гена была нокаутирована. Снова, фрагмент положительного штамма был ПЦР-амплифицирован с праймерами Р23/Р24 и соединен с вектором PMD19-T для секвенирования. Правильно секвенированный штамм был назван YPL-4-046.
Пример 6: эксперимент по ферментации L-лизина
На штаммах, сконструированных в Примерах 2-5, и исходном штамме YP97158, был выполнен эксперимент по ферментации в ферментационном чане BLBIO-5GC-4-H (приобретенном у Shanghai Bailun Biotechnology Co., Ltd.) с культуральной средой, показанной в Таблице 1, и с контрольным процессом, показанным в Таблице 2. Эксперимент с каждым штаммом повторяли три раза и результаты показаны в Таблице 5.
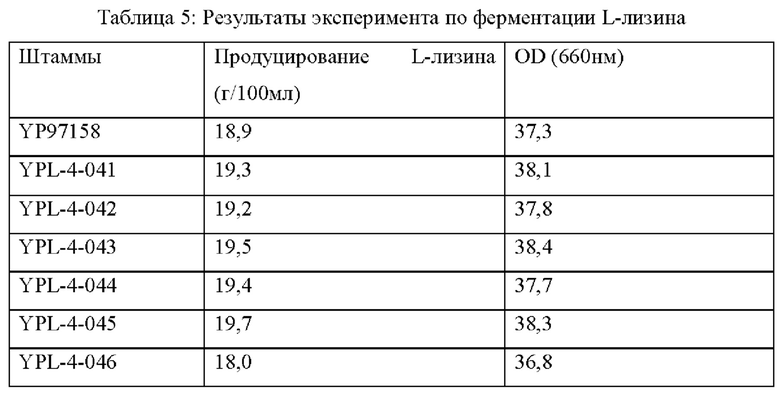
Результаты показаны в Таблице 5. Точечная мутация NCgl0609R334* и сверхэкспрессия кодирующей области гена у Corynebacterium glutamicum способствует увеличению продуцирования L-лизина и темпа роста, в то время как ослабление или нокаутирование гена не способствует накоплению L-лизина и снижает скорость роста штамма.
Пример 7: Введение сверхэкспрессии гена в глутамат-продуцирующий штамм или точечной мутации и сверхэкспрессии в кодирующую область гена и выполнение экспериментов по ферментации.
Согласно способам Примеров 1-5, с использованием тех же праймеров и экспериментальных условий, Corynebacterium АТСС 13 869 использовали в качестве исходной бактерии и бактерию АТСС 13869 использовали в качестве экспрессирующих бактерий для получения глутамат-продуцирующих сконструированных штаммов YPG-013, содержащих 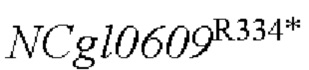 с точечной мутацией, глутамат-продуцирующих конструированных штаммов YPG-014 и YPG-015, сверхэкспрессирующих гены и
с точечной мутацией, глутамат-продуцирующих конструированных штаммов YPG-014 и YPG-015, сверхэкспрессирующих гены и 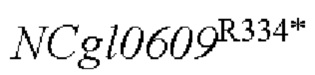 в геноме, глутамат-продуцирующих сконструированных штаммов YPG-016 и YPG-017, сверхэкспрессирующих гены и в плазмиде, и глутамат-продуцирующего сконструированного штамма YPG-018 с отсутствием гена в геноме.
в геноме, глутамат-продуцирующих сконструированных штаммов YPG-016 и YPG-017, сверхэкспрессирующих гены и в плазмиде, и глутамат-продуцирующего сконструированного штамма YPG-018 с отсутствием гена в геноме.
На штаммах, сконструированных в Примерах, и исходном штамме выполняли эксперимент по ферментации (с бактерией АТСС 13869 в качестве экспрессирующей бактерии) в ферментационном чане BLBIO-5GC-4-H (приобретенном у Shanghai Bailun Biotechnology Co., Ltd.) с культуральной средой, показанной в Таблице 3 и с контрольным процессом, показанным в Таблице 4. Эксперимент с каждым штаммом повторяли три раза и результаты показаны в Таблице 6.
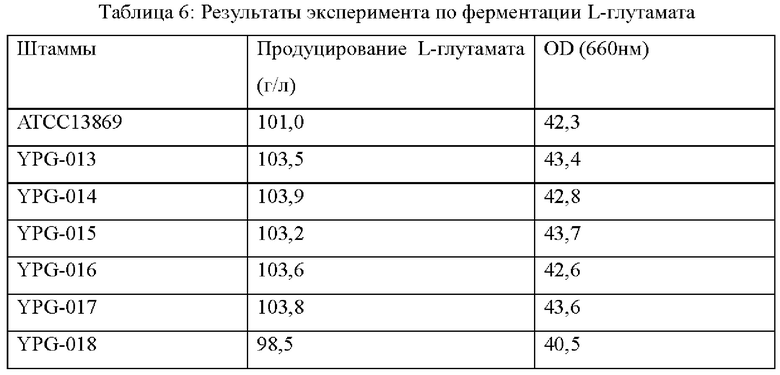
Результаты показаны в Таблице 6. Точечная мутация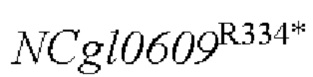 и сверхэкспрессия кодирующей области гена у Corynebacterium glutamicum способствует увеличению продуцированию L-глутамата и скорости роста, в то время как ослабление или инактивация гена не способствует накоплению L- глутаминовой кислоты и снижает скорость роста штамма.
и сверхэкспрессия кодирующей области гена у Corynebacterium glutamicum способствует увеличению продуцированию L-глутамата и скорости роста, в то время как ослабление или инактивация гена не способствует накоплению L- глутаминовой кислоты и снижает скорость роста штамма.
Пример 8: Конструирование трансформационного вектора pK18-NCgl1575A1775T содержащего кодирующую область гена NCgl1575 с точечной мутацией
На основе последовательности генома Corynebacterium glutamicum АТСС 13032 дикого типа, опубликованной NCBI, были сконструированы и синтезированы две пары праймеров для амплификации последовательности кодирующей области гена NCgl1575. Точечную мутацию вводили в кодирующую область гена NCgl1575 (SEQ ID NO:29) на основе штамма YP97158 (было подтвержено посредством секвенирования, что ген дикого типа NCgl1575 сохранялся в хромосоме штамма), посредством аллельной замены. Соответствующая аминокислотная последовательность, кодирующая белки, была SEQ ID NO:31, и в нуклеотидной последовательности гена NCgl1575 в положении 1775 нуклеотид А был заменен на Т (SEQ ID N0:30: NCgl1575A1775T), и в соответствующей аминокислотной последовательности, кодирующей белки, в положении 592 тирозин был заменен на фенилаланин (SEQ ID NO:32: NCgl1575 Y592F).
Праймеры были разработаны следующим образом (синтезированы компанией Shanghai mvitrogen Company):

Способ конструирования: Corynebacterium glutamicum АТСС13032 использовали в качестве матрицы для ПЦР-амплификации с праймерами Р1' и Р2', Р3' и Р4', соответственно.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq(5ед/мкл) 0,25 мкл, общий объем 50 мкл.
ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 40 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С.Были получены два фрагмента ДНК, содержащих кодирующую область гена NCgl1575 размером 766 п. о. и 759 п. о., соответственно (NCgl1575 вышерасположенный и NCgl1575 нижерасположенный).
После выделения и очистки вышеуказанных двух фрагментов ДНК с помощью электрофореза в агарозном геле, вышеуказанные два фрагмента ДНК были использованы в качестве матриц, и Р1' и Р4' использовали в качестве праймеров, для амплификации фрагмента длиной примерно 1495 п. о. посредством ПЦР с перекрывающимися праймерами.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5ед/мкл) 0,25 мкл, общий объем 50 мкл.
ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 90 секунд при 72°С (30 циклов) и удлинение в течение 10 минут при 72°С.
Этот фрагмент ДНК (NCgl1575A1775T) был получен в результате замены аденина (А) в положении 1775 в кодирующей области гена YP97158 NCgl1575 на тимин (Т), и наконец приводило к замене аминокислоты в положении 592 кодирующего белка с тирозина (Y) на фенилаланин (F).
NCgl1575A1775T, выделенный и очищенный посредством электрофореза в агарозном геле и плазмиду pK18mobsacB (приобретенную у Addgene), выделенную посредством переваривания ферментом Xba I, объединили при помощи рекомбинантной системы NEBuider с получением вектора pK18-NCgl1575A1775T и плазмида содержала маркер резистентности к канамицину. Вектор pK18-NCgl1575A1775T отправляли в компанию для секвенирования и идентификации, а вектор рК18-NCgl1575A1775T; содержащий правильную точечную мутацию (А-Т), был сохранен для использования.
Пример 9: Конструирование спроектированных штаммов, содержащих NCgl1575A1775T с точечной мутацией
Способ конструирования: Плазмиду pK18-NCgl1575A1775T с аллельной заменой трансформировали в L-лизин-продуцирующий штамм YP97158 посредством электрического шока. Единичную колонию, полученную посредством культивирования, идентифицировали с помощью праймера Р1' и универсального праймера M13R, соответственно. Штамм, который может амплифицировать бэнд 1502 п. о., является положительным штаммом. Положительные штаммы культивировали в среде, содержащей 15% сахарозу, и единичную колонию, полученную посредством культивирования, выращивали в среде, содержащей канамицин, и в среде без канамицина, соответственно. Штаммы, которые росли в среде без канамицина, но не росли в среде, содержащей канамицин, затем были идентифицированы с помощью ПЦР со следующими праймерами (синтезированными компанией Shanghai mvitrogen Company):
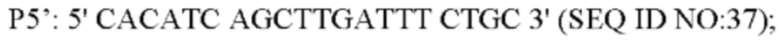 и
и
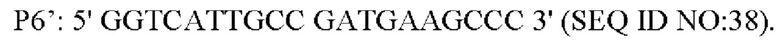
Вышеописанный продукт ПЦР-амплификации имел размер 256 п. о. и был денатурирован при высокой температуре и помещен в ледяную баню, за этим следовал sscp электрофорез (амплифицированный фрагмент плазмиды pK18-NCgl1575A1775T использовали в качестве положительного контроля, амплифицированный фрагмент YP97158 использовали в качестве отрицательного контроля, а воду использовали в качестве холостого контроля). Из-за разных структур фрагментов и положений при электрофорезе, штаммы, положения электрофореза которых отличаются от положений фрагментов отрицательного контроля и совпадают с положениями фрагментов положительного контроля, представляют собой штаммы с успешными аллельными заменами. Представляющий интерес фрагмент штаммов с удачной аллельной заменой был еще раз подвергнут ПЦР-амплификации с использованием праймера Р5' и Р6' и был соединен с вектором PMD19-T для секвенирования. Посредством выравнивания последовательности, для последовательности, в которой мутирована последовательность оснований, подтверждено, что аллельная замена штамма является успешной и он назван YPL-4-023.
Пример 10: Конструирование спроектированных штаммов, сверхэкспрессирующих гены NCgl1575 или NCgl1575A1775T в геноме.
На основе последовательности генома Corynebacterium glutamicum АТСС 13032 дикого типа, опубликованной NCBI, были сконструированы и синтезированы три пары праймеров для амплификации вышерасположенного и нижерасположенного фрагментов гомологичного плеча и последовательностей кодирующей области гена NCgl1575 или NCgl1575A17751 и промоторной области, а ген NCgl1575 или NCgl1575A1775T был введен в штамм YP97158 посредством гомологичной рекомбинации.
Праймер был разработан следующим образом (синтезирован компанией Shanghai mvitrogen Company):

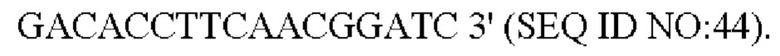
Способ конструирования: Corynebacterium glutamicum АТСС 13032 или YPL-4-023 использовали в качестве матрицы, соответственно, для ПЦР-амплификации с праймерами Р7'/Р8', Р9'/Р10', Р11'/Р12', для получения вышерасположенного фрагмента гомологичного плеча размером 802 п. о., гена NCgl1575 и его промоторного фрагмента размером 2737 п. о., или гена NCgl1575A1775T и его промоторного фрагмента размером 2737 п. о., и нижерасположенного фрагмента гомологичного плеча размером 647п.о. Затем вышеуказанные три амплифицированные фрагмента (вышерасположенный фрагмент гомологичного плеча, ген NCgl1575 и его промоторный фрагмент и нижерасположенный фрагмент гомологичного плеча; или вышерасположенный фрагмент гомологичного плеча, ген NCgl1575A1775T и его промоторный фрагмент, и нижерасположенный фрагмент гомологичного плеча) были смешаны в качестве матрицы для амплификации с праймерами Р7'/Р12' для получения интегрированного фрагмента гомологичного плеча размером 4111 п. о.
После завершения реакции ПЦР, амплифицированный продукт электрофоретически выделяли и требуемый фрагмент ДНК размером 4111 п. о. выделяли с помощью набора для выделения ДНК на гелевой колонке (TIANGEN), и соединяли с шаттл-плазмидой PK18mobsacB, выделенной посредством переваривания ферментом Xba I, с использованием системы рекомбинации NEBBuider с получением интегрированной плазмиды PK18mobsacB-NCgl1575 или PK18mobsacB-NCgl1575A1775T. Плазмида, содержащая маркер резистентности к канамицину, и рекомбинант с плазмидой, интегрированной в геном, могут быть получены посредством отбора с канамицином.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25mM) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq(5ед/мкл) 0.25 мкл, общий объем 50 мкл.
ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 180 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С.
Две объединенные плазмиды электротрансформировали в L-лизин-продуцирующий штамм YP97158, соответственно, и единичную колонию, полученную посредством культивирования, идентифицировали с помощью ПЦР с праймерами Р13'/Р14'. Штамм с ПЦР-амплифицированными фрагментами размером 1778 п. о. был положительным штаммом, а штамм без амплифицированных фрагментов был исходным штаммом. Положительные штаммы отбирали на среде с 15% сахарозой и затем культивировали на среде, содержащей канамицин, и на среде без канамицина, соответственно. Штаммы, которые росли на среде без канамицина, но не росли на среде, содержащей канамицин, затем были идентифицированы с помощью ПЦР с праймерами Р15'/Р16'. Бактерии с амплификацией фрагмента размером 1756 п. о. представляли собой штаммы с геном NCgl1575 или NCgl1575A1775T, интегрированным в геном YP97158, которые были назвааны YPL-4-024 (без сайта мутации) и YPL-4-025 (с сайтом мутации) соответственно.
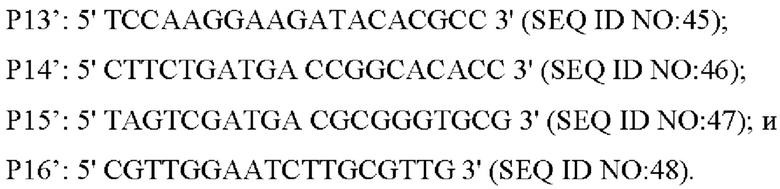
Пример 11: Конструирование спроектированных штаммов, сверхэкспрессирующих ген NCgl1575 или NCgl1575A1775TB плазмиде.
На основе последовательности генома Corynebacterium glutamicum АТСС 13032 дикого типа, опубликованной NCBI, была сконструирована и синтезирована пара праймеров для амплификации последовательностей кодирующей области и промоторной области гена NCgl1575 или NCgl1575A1775T. Праймеры были сконструированы следующим образом (синтезированы компанией Shanghai mvitrogen Company):
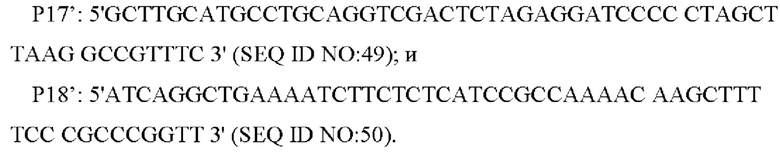
Способ конструирования: АТСС13032 и YPL-4-023 были использованы в качестве матриц, соответственно, для ПЦР-амплификации с праймерами Р17'/Р18', для получения гена NCgl1575 или NCgl1575A1775T и их промоторных фрагментов размером 2749 п. о. Амплифицированные продукты были выделены посредством электрофореза. Необходимые 2749 п. о. фрагменты ДНК были выделены с помощью набора для выделения ДНК на гелевой колонке и были соединены с шаттл-плазмидой рХМJ19 , выделенной посредством переваривания ферментом EcoR I с использованием рекомбинантной системы NEBuider с получением сверхэкспрессирующих плазмид pXMJ19-NCgl1575 и pXMJ19-NCgl1575A1775T. Плазмиды, содержащие маркеры резистентности к хлорамфениколу, могут быть получены посредством резистентности к хлармафениколу и трансформированы в штаммы.
Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq(5ед/мкл) 0,25 мкл, общий объем 50 мкл.
ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 120 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С.
Плазмиды pXMJ19-NCgl1575 и pXMJ19-NCgl1575A1775T электротрансформировали в L-лизин-продуцирующий штамм YP97158 соответственно. Отдельную колонию, полученную посредством культивирования, идентифицировали с помощью ПЦР с праймерами М13 (-48) и Р18'. Отдельную колонию с ПЦР-амплифицированным фрагментом размером 2752п.о. трансформировали в штаммы, которые были названы YPL-4-026 (без сайта мутации) и YPL-4-027 (с сайтом мутации).
Пример 12: Конструирование спроектированных штаммов с геном NCgl1575. удаленным из генома
На основании последовательности генома Corynebacterium glutamicum АТСС13032, опубликованной NCBI, были синтезированы две пары праймеров для амплификации фрагментов на двух концах кодирующей области гена NCgl1575 в качестве вышерасположенного и нижерасположенного фрагментов гомологичного плеча. Праймеры были сконструированы следующим образом (синтезированы компанией Shanghai Invitrogen Company):

Corynebacterium glutamicum АТСС 13032 использовали в качестве матрицы для ПЦР-амплификации с праймерами Р19'/Р20' и Р21'/Р22' соответственно, для получения вышерасположенного фрагмента гомологичного плеча размером 775 п. о. и нижерасположенного фрагмента гомологичного плеча размером 807 п. о. Затем они были подвергнуты ПЦР с перекрывающимися праймерами с праймерами Р19'/Р22' для получения всего фрагмента гомологичного плеча размером 1545 п. о. После завершения ПЦР реакции, амплифицированный продукт был электрофоретически выделен и необходимый 1545 п. о. фрагмент ДНК извлечен с использованием набора для выделения ДНК на гелевой колонке и соединен с шаттл-плазмидой pk18mobsacB, выделенной посредством переваривания энзимом Xba I, посредством рекомбинантной системы NEBuider с получения нокаутной плазмиды. Эта плазмида содержала маркер резистентности к канамицину.
Нокаутную плазмиду электротрансформировали в лизин-продуцирующий штамм YP97158 и единичную колонию, полученную посредством культивирования, идентифицировали с помощью ПЦР со следующими праймерами (синтезированными компанией Shanghai Invitrogen):
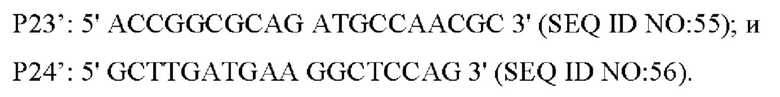
Штаммы с амплифицированными бэндами размером 1471 п. о. и 4150 п. о. при помощи вышеописанной ПЦР, были положительными штаммами, а штаммы с амплификацией только бэнда размером 4150, были исходными бактериями. После отбора в среде с 15% сахарозы, положительные штаммы культивировали в среде, содержащей канамицин, и в среде без канамицина соответственно, и штаммы, которые росли в среде без канамицина, но не росли в среде, содержащей канамицин, затем идентифицировали с помощью ПЦР с использованием праймеров Р237Р24'. Штамм с амплификацией бэнда размером 1471 п. о был спроектированным штаммом с удаленной кодирующей последовательностью гена NCgl1575, который был назван YPL-4-028.
Пример 13: эксперимент по ферментации L-лизина
Сконструированные штаммы из Примеров 9-12 и исходный штамм YP97158 подвергали эксперименту по ферментации в ферментационном чане BLBIO-5GC-4-H (приобретенном у Shanghai Bailun Biotechnology Co., Ltd.) с культуральной средой, показанной в Таблице 1, и контрольным процессом, показанным в Таблице 2. Эксперимент с каждым штаммом повторяли три раза и результаты показаны в Таблице 7.
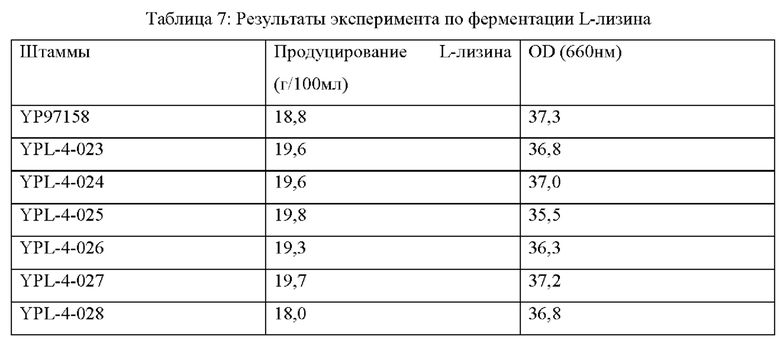
Результаты показаны в Таблице 7. Сверхэкспрессия гена NCgl1575 в Corynebacterium glutamicum, или точечная мутация NCgl1575A1775T и сверхэкспрессия кодирующей области гена NCgl1575 способствуют увеличению продуцирования L-лизина, а ослабление или нокаутирование гена не способствует накоплению L-лизина.
Пример 14: Конструирование трансформирующего вектора 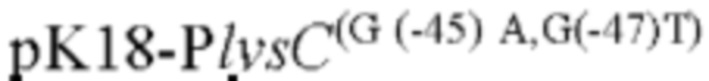 , содержащего промоторную область гена
, содержащего промоторную область гена  gene с точечной мутацией
gene с точечной мутацией
На основании последовательности генома Corynebacterium glutamicum АТСС13032, опубликованной NCBI были сконструированы и синтезированы две пары праймеров для амплификации последовательностей промоторной области гена , и точечная мутация была введена в промоторную область гена lysC (SEQ ID NO: 57) на основе штамма YP97158 посредством аллельной замены. G в положении -45 п. о. нуклеотидной последовательности промоторной области гена lysC заменяли на А, и G в положении -47 п. о. заменяли на Т (SEQ ID NO: 58).
Праймеры были сконстрированы следующим образом (синтезированы компанией Shanghai mvitrogen Company):
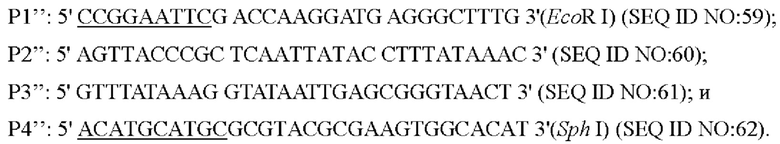
Способ конструирования: Corynebacterium glutamicum АТСС13032 использовали в качестве матрицы для ПЦР-амплификации с праймерами Р1'' и Р2'', Р3'' и Р4'' соответственно. Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ex Taq (5ед/мкл) 0,25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 сек при 94°С, отжиг в течение 30 сек при 52°С, элонгация в течение 40 сек при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С. Были получены два фрагмента ДНК с точечной мутацией размером 729 п. о. и 760 п. о. соответственно (фрагменты промотора в восходящем и нисходящем направлении). После разделения и очистки вышеупомянутых двух фрагментов с помощью электрофореза в агарозном геле, два очищенные фрагмента ДНК были использованы в качестве матриц, и Р1'' и Р4'' были использованы в качестве праймеров для амплификации фрагмента длиной примерно 1459 п. о. (фрагмент в восходящем направлении) посредством ПЦР с перекрывающимися праймерами. Система ПЦР: 10×Ех Taq буфер 5 мкл, смесь dNTP (2,5 мМ каждого) 4 мкл, Mg2+ (25 мМ) 4 мкл, праймеры (10 пМ) 2 мкл каждого, Ех Taq(5ед/мкл) 0,25 мкл, общий объем 50 мкл. ПЦР-амплификацию выполняли следующим образом: предварительная денатурация в течение 5 минут при 94°С, денатурация в течение 30 секунд при 94°С, отжиг в течение 30 секунд при 52°С, элонгация в течение 90 секунд при 72°С (30 циклов), и удлинение в течение 10 минут при 72°С. Вышеописанный фрагмент в восходящем направлении был выделен и очищен с помощью электрофореза в агарозном геле и фрагмент содержал промоторную область гена  и его вышерасположенную и нижерасположенную последовательности и оба конца фрагмента, содержащие сайты переваривания ферментами EcoR I и Sph I соответственно. Этот фрагмент ДНК вызывает замену нуклеотида гуанин (G) в положении -45 п. о. в промоторной области YP97158 гена
и его вышерасположенную и нижерасположенную последовательности и оба конца фрагмента, содержащие сайты переваривания ферментами EcoR I и Sph I соответственно. Этот фрагмент ДНК вызывает замену нуклеотида гуанин (G) в положении -45 п. о. в промоторной области YP97158 гена  на аденин (А), и нуклеотида гуанин (G) в положении -47 п. о. на тимин (Т). Фрагмент был очищен и выделен после переваривания двумя ферментами (EcoR I/Sph I) и был соединен с шаттл-плазмидой pK18mobsacB (приобретенной у Addgene) после двойного переваривания теми же ферментами (EcoR I/Sph I) с получением плазмиды
на аденин (А), и нуклеотида гуанин (G) в положении -47 п. о. на тимин (Т). Фрагмент был очищен и выделен после переваривания двумя ферментами (EcoR I/Sph I) и был соединен с шаттл-плазмидой pK18mobsacB (приобретенной у Addgene) после двойного переваривания теми же ферментами (EcoR I/Sph I) с получением плазмиды 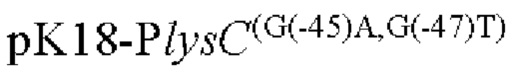 с аллельной заменой, содержащей маркер резистентности к канамицину. Вектор был отправлен в секвенирующую компанию для секвенирования и идентификации, и вектор
с аллельной заменой, содержащей маркер резистентности к канамицину. Вектор был отправлен в секвенирующую компанию для секвенирования и идентификации, и вектор 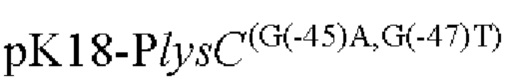 содержащий правильную точечную мутацию был сохранен для использования.
содержащий правильную точечную мутацию был сохранен для использования.
Пример 15: Конструирование спроектированных штаммов, содержащих 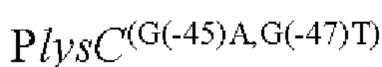 с точечной мутацией
с точечной мутацией
Плазмиду 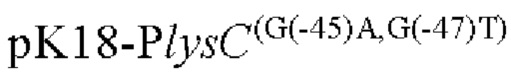 с аллельной заменой трансформировали в L-лизин-продуцирующий штамм YP97158 посредством электрошока. Единичную колонию, полученную посредством культивирования, идентифицировали с помощью праймера Р1'' и универсального праймера M13F соответственно, и штаммы, которые могут амплифицировать бэнд размером 1500 п. о., были положительными штаммами. Положительные штаммы культивировали в среде, содержащей 15% сахарозу, и отдельную колонию, полученную посредством культивирования, выращивали в среде, содержащей канамицин, и в среде без канамицина соответственно; штаммы, которые росли на среде без канамицина, но не росли на среде, содержащей канамицин, затем были идентифицированы посредством ПЦР с использованием следующих праймеров (синтезированные компанией Shanghai Invitrogen Company):
с аллельной заменой трансформировали в L-лизин-продуцирующий штамм YP97158 посредством электрошока. Единичную колонию, полученную посредством культивирования, идентифицировали с помощью праймера Р1'' и универсального праймера M13F соответственно, и штаммы, которые могут амплифицировать бэнд размером 1500 п. о., были положительными штаммами. Положительные штаммы культивировали в среде, содержащей 15% сахарозу, и отдельную колонию, полученную посредством культивирования, выращивали в среде, содержащей канамицин, и в среде без канамицина соответственно; штаммы, которые росли на среде без канамицина, но не росли на среде, содержащей канамицин, затем были идентифицированы посредством ПЦР с использованием следующих праймеров (синтезированные компанией Shanghai Invitrogen Company):
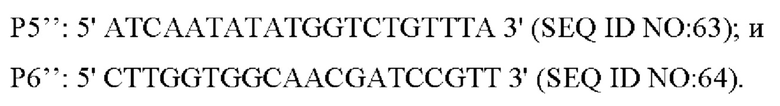
Вышеупомянутый продукт ПЦР-амплификации денатурировали при высокой температуре и подвергали воздействию ледяной бани с последующим sscp электрофорезом (амплифицированный фрагмент плазмиды использовали в качестве положительного контроля, амплифицированный фрагмент YP97158 использовали в качестве отрицательного контроля, а воду использовали в качестве холостого контроля). Из-за разных структур фрагментов и положений при электрофорезе, штаммы, положения электрофореза которых отличались от положений фрагментов отрицательного контроля и совпадали с положениями фрагментов положительного контроля, представляли собой штаммы с успешными аллельными заменами. Целевой фрагмент положительного штамма еще раз ПЦР-амплифицировали и соединяли с вектором PMD19-T для секвенирования. Посредством выравнивания последовательностей, последовательность, в которой последовательность оснований мутирована, подтверждает, что аллельная замена штамма является успешной, и штамм назван YPL-4-009.
Пример 16: эксперимент по ферментации L-лизина
Эксперимент по ферментации штамма YPL-4-009, конструированного в Примере 15, и исходного штамма YP97158 выполняли в ферментационном чане BLBIO-5GC-4-H (приобретенном у Shanghai Bailun Biotechnology Co., Ltd.) с культуральной средой, показанной в Таблице 1, и контрольным процессом, показанным в Таблице 2. Эксперимент с каждым штаммом повторяли три раза и результаты показаны в Таблице 8.
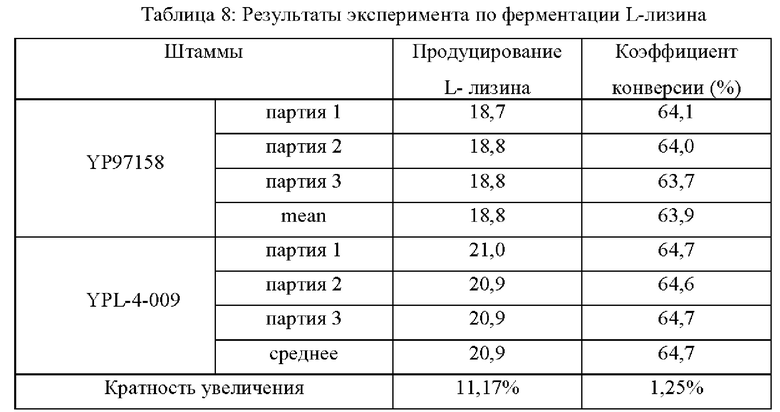
Вышеуказанный коэффициент конверсии = общая масса лизина/общее потребление глюкозы * 100%
Результаты показаны в Таблице 8. Точечная мутация 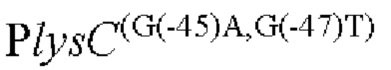 промотора гена в Corynebacterium glutamicum способствует увеличению продуцирования L-лизина.
промотора гена в Corynebacterium glutamicum способствует увеличению продуцирования L-лизина.
Воплощение изобретения описано выше. Однако настоящее изобретение не ограничено вышеприведенными воплощениями. Любая модификация, эквивалентная замена, улучшение и т.д., сделанные в соответствии с сущностью и принципом изобретения, включены в объем правовой защиты данного изобретения.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> INNER MONGOLIA EPPEN BIOTECH CO., LTD.
<120> РЕКОМБИНАНТНЫЙ ШТАММ ДЛЯ ПРОДУЦИРОВАНИЯ
L-АМИНОКИСЛОТЫ, МЕТОД ЕГО КОНСТРУИРОВАНИЯ И ЕГО ПРИМЕНЕНИЕ
<130> CPWO20411699
<160> 64
<170> PatentIn version 3.3
<210> 1
<211> 1083
<212> DNA
<213> Corynebacterium glutamicum
<400> 1
gtgtcacaca ccgcgtccac accgacgcca gaggaatact ccgcgcagca acccagcacc 60
cagggcactc gcgttgagtt ccgcggcata accaaagtct ttagcaacaa taaatctgct 120
aaaaccaccg cgcttgataa tgtcactctc accgtagaac ccggtgaggt aatcggcatc 180
atcggttact ctggcgccgg caagtccact cttgtccgcc tcatcaatgg ccttgactcc 240
cccacgagcg gttcgttgct gctcaacggc accgacatcg tcggaatgcc cgagtctaag 300
ctgcgtaaac tgcgcagtaa tatcggcatg attttccagc agttcaacct gttccagtcg 360
cgtactgcgg ctggaaatgt ggagtacccg ctggaagttg ccaagatgga caaggcagct 420
cgtaaagctc gcgtgcaaga aatgctcgag ttcgtcggcc tgggcgacaa aggcaaaaac 480
taccccgagc agctgtcggg cggccagaag cagcgcgtcg gcattgcccg tgcactggcc 540
accaatccaa cgcttttgct tgccgacgaa gccacctccg ctttggaccc agaaaccacc 600
catgaagttc tggagctgct gcgcaaggta aaccgcgaac tgggcatcac catcgttgtg 660
atcacccacg aaatggaagt tgtgcgttcc atcgcagaca aggttgctgt gatggaatcc 720
ggcaaagttg tggaatacgg cagcgtctac gaggtgttct ccaatccaca aacacaggtt 780
gctcaaaagt tcgtggccac cgcgctgcgt aacaccccag accaagtgga atcggaagat 840
ctgcttagcc atgagggacg tctgttcacc attgatctga ctgaaacgtc cggcttcttt 900
gcagcaaccg ctcgtgctgc cgaacaaggt gcttttgtca acatcgttca cggtggcgtg 960
accaccttgc aacgccaatc atttggcaaa atgactgttc gactcaccgg caacaccgct 1020
gcgattgaag agttctatca aaccttgacc aagaccacga ccatcaagga gatcacccga 1080
tga 1083
<210> 2
<211> 1083
<212> DNA
<213> artifical sequence
<400> 2
gtgtcacaca ccgcgtccac accgacgcca gaggaatact ccgcgcagca acccagcacc 60
cagggcactc gcgttgagtt ccgcggcata accaaagtct ttagcaacaa taaatctgct 120
aaaaccaccg cgcttgataa tgtcactctc accgtagaac ccggtgaggt aatcggcatc 180
atcggttact ctggcgccgg caagtccact cttgtccgcc tcatcaatgg ccttgactcc 240
cccacgagcg gttcgttgct gctcaacggc accgacatcg tcggaatgcc cgagtctaag 300
ctgcgtaaac tgcgcagtaa tatcggcatg attttccagc agttcaacct gttccagtcg 360
cgtactgcgg ctggaaatgt ggagtacccg ctggaagttg ccaagatgga caaggcagct 420
cgtaaagctc gcgtgcaaga aatgctcgag ttcgtcggcc tgggcgacaa aggcaaaaac 480
taccccgagc agctgtcggg cggccagaag cagcgcgtcg gcattgcccg tgcactggcc 540
accaatccaa cgcttttgct tgccgacgaa gccacctccg ctttggaccc agaaaccacc 600
catgaagttc tggagctgct gcgcaaggta aaccgcgaac tgggcatcac catcgttgtg 660
atcacccacg aaatggaagt tgtgcgttcc atcgcagaca aggttgctgt gatggaatcc 720
ggcaaagttg tggaatacgg cagcgtctac gaggtgttct ccaatccaca aacacaggtt 780
gctcaaaagt tcgtggccac cgcgctgcgt aacaccccag accaagtgga atcggaagat 840
ctgcttagcc atgagggacg tctgttcacc attgatctga ctgaaacgtc cggcttcttt 900
gcagcaaccg ctcgtgctgc cgaacaaggt gcttttgtca acatcgttca cggtggcgtg 960
accaccttgc aacgccaatc atttggcaaa atgactgttt gactcaccgg caacaccgct 1020
gcgattgaag agttctatca aaccttgacc aagaccacga ccatcaagga gatcacccga 1080
tga 1083
<210> 3
<211> 360
<212> PRT
<213> Corynebacterium glutamicum
<400> 3
Met Ser His Thr Ala Ser Thr Pro Thr Pro Glu Glu Tyr Ser Ala Gln
1 5 10 15
Gln Pro Ser Thr Gln Gly Thr Arg Val Glu Phe Arg Gly Ile Thr Lys
20 25 30
Val Phe Ser Asn Asn Lys Ser Ala Lys Thr Thr Ala Leu Asp Asn Val
35 40 45
Thr Leu Thr Val Glu Pro Gly Glu Val Ile Gly Ile Ile Gly Tyr Ser
50 55 60
Gly Ala Gly Lys Ser Thr Leu Val Arg Leu Ile Asn Gly Leu Asp Ser
65 70 75 80
Pro Thr Ser Gly Ser Leu Leu Leu Asn Gly Thr Asp Ile Val Gly Met
85 90 95
Pro Glu Ser Lys Leu Arg Lys Leu Arg Ser Asn Ile Gly Met Ile Phe
100 105 110
Gln Gln Phe Asn Leu Phe Gln Ser Arg Thr Ala Ala Gly Asn Val Glu
115 120 125
Tyr Pro Leu Glu Val Ala Lys Met Asp Lys Ala Ala Arg Lys Ala Arg
130 135 140
Val Gln Glu Met Leu Glu Phe Val Gly Leu Gly Asp Lys Gly Lys Asn
145 150 155 160
Tyr Pro Glu Gln Leu Ser Gly Gly Gln Lys Gln Arg Val Gly Ile Ala
165 170 175
Arg Ala Leu Ala Thr Asn Pro Thr Leu Leu Leu Ala Asp Glu Ala Thr
180 185 190
Ser Ala Leu Asp Pro Glu Thr Thr His Glu Val Leu Glu Leu Leu Arg
195 200 205
Lys Val Asn Arg Glu Leu Gly Ile Thr Ile Val Val Ile Thr His Glu
210 215 220
Met Glu Val Val Arg Ser Ile Ala Asp Lys Val Ala Val Met Glu Ser
225 230 235 240
Gly Lys Val Val Glu Tyr Gly Ser Val Tyr Glu Val Phe Ser Asn Pro
245 250 255
Gln Thr Gln Val Ala Gln Lys Phe Val Ala Thr Ala Leu Arg Asn Thr
260 265 270
Pro Asp Gln Val Glu Ser Glu Asp Leu Leu Ser His Glu Gly Arg Leu
275 280 285
Phe Thr Ile Asp Leu Thr Glu Thr Ser Gly Phe Phe Ala Ala Thr Ala
290 295 300
Arg Ala Ala Glu Gln Gly Ala Phe Val Asn Ile Val His Gly Gly Val
305 310 315 320
Thr Thr Leu Gln Arg Gln Ser Phe Gly Lys Met Thr Val Arg Leu Thr
325 330 335
Gly Asn Thr Ala Ala Ile Glu Glu Phe Tyr Gln Thr Leu Thr Lys Thr
340 345 350
Thr Thr Ile Lys Glu Ile Thr Arg
355 360
<210> 4
<211> 333
<212> PRT
<213> artifical sequence
<400> 4
Met Ser His Thr Ala Ser Thr Pro Thr Pro Glu Glu Tyr Ser Ala Gln
1 5 10 15
Gln Pro Ser Thr Gln Gly Thr Arg Val Glu Phe Arg Gly Ile Thr Lys
20 25 30
Val Phe Ser Asn Asn Lys Ser Ala Lys Thr Thr Ala Leu Asp Asn Val
35 40 45
Thr Leu Thr Val Glu Pro Gly Glu Val Ile Gly Ile Ile Gly Tyr Ser
50 55 60
Gly Ala Gly Lys Ser Thr Leu Val Arg Leu Ile Asn Gly Leu Asp Ser
65 70 75 80
Pro Thr Ser Gly Ser Leu Leu Leu Asn Gly Thr Asp Ile Val Gly Met
85 90 95
Pro Glu Ser Lys Leu Arg Lys Leu Arg Ser Asn Ile Gly Met Ile Phe
100 105 110
Gln Gln Phe Asn Leu Phe Gln Ser Arg Thr Ala Ala Gly Asn Val Glu
115 120 125
Tyr Pro Leu Glu Val Ala Lys Met Asp Lys Ala Ala Arg Lys Ala Arg
130 135 140
Val Gln Glu Met Leu Glu Phe Val Gly Leu Gly Asp Lys Gly Lys Asn
145 150 155 160
Tyr Pro Glu Gln Leu Ser Gly Gly Gln Lys Gln Arg Val Gly Ile Ala
165 170 175
Arg Ala Leu Ala Thr Asn Pro Thr Leu Leu Leu Ala Asp Glu Ala Thr
180 185 190
Ser Ala Leu Asp Pro Glu Thr Thr His Glu Val Leu Glu Leu Leu Arg
195 200 205
Lys Val Asn Arg Glu Leu Gly Ile Thr Ile Val Val Ile Thr His Glu
210 215 220
Met Glu Val Val Arg Ser Ile Ala Asp Lys Val Ala Val Met Glu Ser
225 230 235 240
Gly Lys Val Val Glu Tyr Gly Ser Val Tyr Glu Val Phe Ser Asn Pro
245 250 255
Gln Thr Gln Val Ala Gln Lys Phe Val Ala Thr Ala Leu Arg Asn Thr
260 265 270
Pro Asp Gln Val Glu Ser Glu Asp Leu Leu Ser His Glu Gly Arg Leu
275 280 285
Phe Thr Ile Asp Leu Thr Glu Thr Ser Gly Phe Phe Ala Ala Thr Ala
290 295 300
Arg Ala Ala Glu Gln Gly Ala Phe Val Asn Ile Val His Gly Gly Val
305 310 315 320
Thr Thr Leu Gln Arg Gln Ser Phe Gly Lys Met Thr Val
325 330
<210> 5
<211> 55
<212> DNA
<213> artifical sequence
<400> 5
cagtgccaag cttgcatgcc tgcaggtcga ctctagggac ggcaacgtac ataac 55
<210> 6
<211> 29
<212> DNA
<213> artifical sequence
<400> 6
gttgccggtg agtcaaacag tcattttgc 29
<210> 7
<211> 29
<212> DNA
<213> artifical sequence
<400> 7
gcaaaatgac tgtttgactc accggcaac 29
<210> 8
<211> 56
<212> DNA
<213> artifical sequence
<400> 8
cagctatgac catgattacg aattcgagct cggtacccgc ggctggaaat gtggag 56
<210> 9
<211> 18
<212> DNA
<213> artifical sequence
<400> 9
ctagccggtt ccagtcag 18
<210> 10
<211> 19
<212> DNA
<213> artifical sequence
<400> 10
ggacgtctgt tcaccattg 19
<210> 11
<211> 55
<212> DNA
<213> artifical sequence
<400> 11
cagtgccaag cttgcatgcc tgcaggtcga ctctagaatg cgttctggac tgagg 55
<210> 12
<211> 55
<212> DNA
<213> artifical sequence
<400> 12
cagtgccaag cttgcatgcc tgcaggtcga ctctagaatg cgttctggac tgagg 55
<210> 13
<211> 37
<212> DNA
<213> artifical sequence
<400> 13
catctgttct cggtgcacca gctgcgagga tcatctc 37
<210> 14
<211> 46
<212> DNA
<213> artifical sequence
<400> 14
gatttaattg cgccatctga ttctggcaac aactccttcc ttgacc 46
<210> 15
<211> 49
<212> DNA
<213> artifical sequence
<400> 15
ggtcaaggaa ggagttgttg ccagaatcag atggcgcaat taaatcaag 49
<210> 16
<211> 60
<212> DNA
<213> artifical sequence
<400> 16
cagctatgac catgattacg aattcgagct cggtacccgc tatgacacct tcaacggatc 60
<210> 17
<211> 19
<212> DNA
<213> artifical sequence
<400> 17
tccaaggaag atacacgcc 19
<210> 18
<211> 18
<212> DNA
<213> artifical sequence
<400> 18
cgaaatggaa gttgtgcg 18
<210> 19
<211> 19
<212> DNA
<213> artifical sequence
<400> 19
cgatgatgcc gattacctc 19
<210> 20
<211> 18
<212> DNA
<213> artifical sequence
<400> 20
cgttggaatc ttgcgttg 18
<210> 21
<211> 55
<212> DNA
<213> artifical sequence
<400> 21
gcttgcatgc ctgcaggtcg actctagagg atcccccagc tgcgaggatc atctc 55
<210> 22
<211> 54
<212> DNA
<213> artifical sequence
<400> 22
atcaggctga aaatcttctc tcatccgcca aaaccaacaa ctccttcctt gacc 54
<210> 23
<211> 54
<212> DNA
<213> artifical sequence
<400> 23
cagtgccaag cttgcatgcc tgcaggtcga ctctagaatg aatgggatgg gtcg 54
<210> 24
<211> 37
<212> DNA
<213> artifical sequence
<400> 24
catcatcggt tactctggcc gaaatggaag ttgtgcg 37
<210> 25
<211> 37
<212> DNA
<213> artifical sequence
<400> 25
cgcacaactt ccatttcggc cagagtaacc gatgatg 37
<210> 26
<211> 58
<212> DNA
<213> artifical sequence
<400> 26
cagctatgac catgattacg aattcgagct cggtacccca acaactcctt ccttgacc 58
<210> 27
<211> 18
<212> DNA
<213> artifical sequence
<400> 27
aatgaatggg atgggtcg 18
<210> 28
<211> 20
<212> DNA
<213> artifical sequence
<400> 28
caacaactcc ttccttgacc 20
<210> 29
<211> 2679
<212> DNA
<213> Corynebacterium glutamicum
<400> 29
atggcagaat caaacgctat ggaccgggca caaatctctg cactgctaga tagagcacag 60
cacacaatca accttgccga acaagcaaac aacgtgctcc gactgttgaa aacacccgga 120
acggccacag taggggacaa cgggacactc ggcaccgata cctatctgat cccatcccgc 180
aacatcacct ggcctgacaa cctgtatgtc aacgtctttc tagacggcat gaatgcagaa 240
gccaccctta ccgattacgt cgcatcagtc gcttcgatcc cacgcctatg ccagatcatc 300
aacgagggcc aaggcggcat gttccgcaga ctattcaacc ccaccaaggt ccaagccggc 360
gaccaagctg tcttcgacct catggtcaaa ctcgacgaga tttcatctac cacccacgaa 420
gtctcccgca tgctcgaggg cgtccacgct gcccgcaccc gccaacaaca aggcgttgca 480
cttttcccag gtattcatgg agtgggagag cgctacatcg aacgcgcaca acaggtactc 540
gcctcagccc tcggtatcgc tggattcggt gccgaaccct gggacggaca tacccttgcc 600
caagcgcgcc gggtagtcca acgctacgcc caagatccta actccgaata ccggctgaaa 660
agcgaagccg agaaacacct cacatccatc aacgagctcc gcgtacagat actcctcgaa 720
caactccccg ttgatgccct acgcatggct accgaccacc gcctgcgctt tggatccctc 780
gattccatcc acgtcgcaac cgtcgccgac gtcctaaaaa cacacacctc catcctcacc 840
accgtgcaag gtatcggcgc ccaaaccgcg gggcggatga aagccgcagc agaaacactc 900
aaacaagaag cactacgccg ccaaaacacc tccatcggcg acgaacctac ccaacccgcc 960
atgcgtctaa tcaacgtgct ggcccgcttc gaccaaaccg aaaccatcac gcccgaagaa 1020
cgcgcccgcc gcacccgcgt catcgactac gtagaacaca tacccccaag cctcgacccc 1080
tacatcgtca tcaacccagc aacgcctgag ttcaacaact tcaccgacga cctccgctgg 1140
atcgacgcaa accccaacct cttccaccca caaacaatca ccaccccacc cgccgacatc 1200
tgggacgact acatctcccg tcccgctcac taccaaggcc tgctagccac gctgctcggc 1260
cgcgacatcg aaggcgcaga cgaactcctc gacgccacca ccctccaaaa aatcagagac 1320
ctcaccctcg acaaaactca tctcaccgac ctccacctcc gcggatacca atcattcggc 1380
gcccgcttcg ccatcatcca aaagaaaacc ctcctcggcg acgacatggg actcggcaaa 1440
acagtccaag ccctctccgc agctgcacac cttgccgcca ccgaaaaaga cttccgcacc 1500
ctcgtcgtcg tacccgcatc cgtcattgtt aactggaccc gcgaatgcaa acgcttcctc 1560
aacctccccg tattcatcgc ccacggagac aacaaacaag acgccatcaa cgcctggtct 1620
aacaccaacg gaatcgcaat ctgcacctac gacggcgtcc gcaccatgga catccccgcg 1680
ccgggtctgg tcattgccga tgaagcccac ctgatcaaaa acccctccac caaacgcacc 1740
caagcactgc gcaaacttat cgacgccgcc ccatacaccc ttctgatgac cggcacacca 1800
ctagaaaaca aagtggaaga gtttgtaaat ctcgtgcgct acatccaacc ggagctgatc 1860
acccgtggca tgtccaaaat gcaggccgag aatttccgcg agcgcatcgc accagcctat 1920
ctgcgcagaa atcaagctga tgtgcttgac gaactcccag agcgcaccga ctccatcgac 1980
tggatcgacc tcaccccaga agaccgcagc gcctacgacg accaagtccg ccaaggcagc 2040
tggatgggca tgcgccgctc cgccatgctc tcaccaacac cacgcctaac ttccgcaaaa 2100
atgcaacgca tcctagaact cttcgaagaa gcagaagaac acggccgcaa agccctcatc 2160
ttcacctact tcctcgacgt cctcgacgaa ctggaaaagc atctaggcga gcgcgtcatc 2220
ggccgcattt ccggcgacgt gccagccacc aagcgccaat tgcttgtcga cgccctgtcc 2280
cactccaaac ccggatccgc cctcattgcc caaatcaccg ccgggggagt aggcctaaac 2340
atccaatccg cgagcctatg cattatttgt gaacctcaag taaagccaac catcgaacag 2400
caggccgtcg cccgagtcca ccgcatgggc caaaccgcca ccgtccaagt ccaccgactc 2460
atcggcgacg aaaccgcaga cgaacgcatg ctagaaatcc tggcaggcaa aactcacgtc 2520
ttcgacgtct acgcccggct atctgaaacc gcagagattc cagatgctgt ggatatcact 2580
gaatcacagc tggcagcacg ggttattgat gaggagcgtg cacggttagg gcttactgaa 2640
tccactggcc ctaaagatga agaaacggcc ttaagctag 2679
<210> 30
<211> 2679
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 30
atggcagaat caaacgctat ggaccgggca caaatctctg cactgctaga tagagcacag 60
cacacaatca accttgccga acaagcaaac aacgtgctcc gactgttgaa aacacccgga 120
acggccacag taggggacaa cgggacactc ggcaccgata cctatctgat cccatcccgc 180
aacatcacct ggcctgacaa cctgtatgtc aacgtctttc tagacggcat gaatgcagaa 240
gccaccctta ccgattacgt cgcatcagtc gcttcgatcc cacgcctatg ccagatcatc 300
aacgagggcc aaggcggcat gttccgcaga ctattcaacc ccaccaaggt ccaagccggc 360
gaccaagctg tcttcgacct catggtcaaa ctcgacgaga tttcatctac cacccacgaa 420
gtctcccgca tgctcgaggg cgtccacgct gcccgcaccc gccaacaaca aggcgttgca 480
cttttcccag gtattcatgg agtgggagag cgctacatcg aacgcgcaca acaggtactc 540
gcctcagccc tcggtatcgc tggattcggt gccgaaccct gggacggaca tacccttgcc 600
caagcgcgcc gggtagtcca acgctacgcc caagatccta actccgaata ccggctgaaa 660
agcgaagccg agaaacacct cacatccatc aacgagctcc gcgtacagat actcctcgaa 720
caactccccg ttgatgccct acgcatggct accgaccacc gcctgcgctt tggatccctc 780
gattccatcc acgtcgcaac cgtcgccgac gtcctaaaaa cacacacctc catcctcacc 840
accgtgcaag gtatcggcgc ccaaaccgcg gggcggatga aagccgcagc agaaacactc 900
aaacaagaag cactacgccg ccaaaacacc tccatcggcg acgaacctac ccaacccgcc 960
atgcgtctaa tcaacgtgct ggcccgcttc gaccaaaccg aaaccatcac gcccgaagaa 1020
cgcgcccgcc gcacccgcgt catcgactac gtagaacaca tacccccaag cctcgacccc 1080
tacatcgtca tcaacccagc aacgcctgag ttcaacaact tcaccgacga cctccgctgg 1140
atcgacgcaa accccaacct cttccaccca caaacaatca ccaccccacc cgccgacatc 1200
tgggacgact acatctcccg tcccgctcac taccaaggcc tgctagccac gctgctcggc 1260
cgcgacatcg aaggcgcaga cgaactcctc gacgccacca ccctccaaaa aatcagagac 1320
ctcaccctcg acaaaactca tctcaccgac ctccacctcc gcggatacca atcattcggc 1380
gcccgcttcg ccatcatcca aaagaaaacc ctcctcggcg acgacatggg actcggcaaa 1440
acagtccaag ccctctccgc agctgcacac cttgccgcca ccgaaaaaga cttccgcacc 1500
ctcgtcgtcg tacccgcatc cgtcattgtt aactggaccc gcgaatgcaa acgcttcctc 1560
aacctccccg tattcatcgc ccacggagac aacaaacaag acgccatcaa cgcctggtct 1620
aacaccaacg gaatcgcaat ctgcacctac gacggcgtcc gcaccatgga catccccgcg 1680
ccgggtctgg tcattgccga tgaagcccac ctgatcaaaa acccctccac caaacgcacc 1740
caagcactgc gcaaacttat cgacgccgcc ccattcaccc ttctgatgac cggcacacca 1800
ctagaaaaca aagtggaaga gtttgtaaat ctcgtgcgct acatccaacc ggagctgatc 1860
acccgtggca tgtccaaaat gcaggccgag aatttccgcg agcgcatcgc accagcctat 1920
ctgcgcagaa atcaagctga tgtgcttgac gaactcccag agcgcaccga ctccatcgac 1980
tggatcgacc tcaccccaga agaccgcagc gcctacgacg accaagtccg ccaaggcagc 2040
tggatgggca tgcgccgctc cgccatgctc tcaccaacac cacgcctaac ttccgcaaaa 2100
atgcaacgca tcctagaact cttcgaagaa gcagaagaac acggccgcaa agccctcatc 2160
ttcacctact tcctcgacgt cctcgacgaa ctggaaaagc atctaggcga gcgcgtcatc 2220
ggccgcattt ccggcgacgt gccagccacc aagcgccaat tgcttgtcga cgccctgtcc 2280
cactccaaac ccggatccgc cctcattgcc caaatcaccg ccgggggagt aggcctaaac 2340
atccaatccg cgagcctatg cattatttgt gaacctcaag taaagccaac catcgaacag 2400
caggccgtcg cccgagtcca ccgcatgggc caaaccgcca ccgtccaagt ccaccgactc 2460
atcggcgacg aaaccgcaga cgaacgcatg ctagaaatcc tggcaggcaa aactcacgtc 2520
ttcgacgtct acgcccggct atctgaaacc gcagagattc cagatgctgt ggatatcact 2580
gaatcacagc tggcagcacg ggttattgat gaggagcgtg cacggttagg gcttactgaa 2640
tccactggcc ctaaagatga agaaacggcc ttaagctag 2679
<210> 31
<211> 892
<212> PRT
<213> Corynebacterium glutamicum
<400> 31
Met Ala Glu Ser Asn Ala Met Asp Arg Ala Gln Ile Ser Ala Leu Leu
1 5 10 15
Asp Arg Ala Gln His Thr Ile Asn Leu Ala Glu Gln Ala Asn Asn Val
20 25 30
Leu Arg Leu Leu Lys Thr Pro Gly Thr Ala Thr Val Gly Asp Asn Gly
35 40 45
Thr Leu Gly Thr Asp Thr Tyr Leu Ile Pro Ser Arg Asn Ile Thr Trp
50 55 60
Pro Asp Asn Leu Tyr Val Asn Val Phe Leu Asp Gly Met Asn Ala Glu
65 70 75 80
Ala Thr Leu Thr Asp Tyr Val Ala Ser Val Ala Ser Ile Pro Arg Leu
85 90 95
Cys Gln Ile Ile Asn Glu Gly Gln Gly Gly Met Phe Arg Arg Leu Phe
100 105 110
Asn Pro Thr Lys Val Gln Ala Gly Asp Gln Ala Val Phe Asp Leu Met
115 120 125
Val Lys Leu Asp Glu Ile Ser Ser Thr Thr His Glu Val Ser Arg Met
130 135 140
Leu Glu Gly Val His Ala Ala Arg Thr Arg Gln Gln Gln Gly Val Ala
145 150 155 160
Leu Phe Pro Gly Ile His Gly Val Gly Glu Arg Tyr Ile Glu Arg Ala
165 170 175
Gln Gln Val Leu Ala Ser Ala Leu Gly Ile Ala Gly Phe Gly Ala Glu
180 185 190
Pro Trp Asp Gly His Thr Leu Ala Gln Ala Arg Arg Val Val Gln Arg
195 200 205
Tyr Ala Gln Asp Pro Asn Ser Glu Tyr Arg Leu Lys Ser Glu Ala Glu
210 215 220
Lys His Leu Thr Ser Ile Asn Glu Leu Arg Val Gln Ile Leu Leu Glu
225 230 235 240
Gln Leu Pro Val Asp Ala Leu Arg Met Ala Thr Asp His Arg Leu Arg
245 250 255
Phe Gly Ser Leu Asp Ser Ile His Val Ala Thr Val Ala Asp Val Leu
260 265 270
Lys Thr His Thr Ser Ile Leu Thr Thr Val Gln Gly Ile Gly Ala Gln
275 280 285
Thr Ala Gly Arg Met Lys Ala Ala Ala Glu Thr Leu Lys Gln Glu Ala
290 295 300
Leu Arg Arg Gln Asn Thr Ser Ile Gly Asp Glu Pro Thr Gln Pro Ala
305 310 315 320
Met Arg Leu Ile Asn Val Leu Ala Arg Phe Asp Gln Thr Glu Thr Ile
325 330 335
Thr Pro Glu Glu Arg Ala Arg Arg Thr Arg Val Ile Asp Tyr Val Glu
340 345 350
His Ile Pro Pro Ser Leu Asp Pro Tyr Ile Val Ile Asn Pro Ala Thr
355 360 365
Pro Glu Phe Asn Asn Phe Thr Asp Asp Leu Arg Trp Ile Asp Ala Asn
370 375 380
Pro Asn Leu Phe His Pro Gln Thr Ile Thr Thr Pro Pro Ala Asp Ile
385 390 395 400
Trp Asp Asp Tyr Ile Ser Arg Pro Ala His Tyr Gln Gly Leu Leu Ala
405 410 415
Thr Leu Leu Gly Arg Asp Ile Glu Gly Ala Asp Glu Leu Leu Asp Ala
420 425 430
Thr Thr Leu Gln Lys Ile Arg Asp Leu Thr Leu Asp Lys Thr His Leu
435 440 445
Thr Asp Leu His Leu Arg Gly Tyr Gln Ser Phe Gly Ala Arg Phe Ala
450 455 460
Ile Ile Gln Lys Lys Thr Leu Leu Gly Asp Asp Met Gly Leu Gly Lys
465 470 475 480
Thr Val Gln Ala Leu Ser Ala Ala Ala His Leu Ala Ala Thr Glu Lys
485 490 495
Asp Phe Arg Thr Leu Val Val Val Pro Ala Ser Val Ile Val Asn Trp
500 505 510
Thr Arg Glu Cys Lys Arg Phe Leu Asn Leu Pro Val Phe Ile Ala His
515 520 525
Gly Asp Asn Lys Gln Asp Ala Ile Asn Ala Trp Ser Asn Thr Asn Gly
530 535 540
Ile Ala Ile Cys Thr Tyr Asp Gly Val Arg Thr Met Asp Ile Pro Ala
545 550 555 560
Pro Gly Leu Val Ile Ala Asp Glu Ala His Leu Ile Lys Asn Pro Ser
565 570 575
Thr Lys Arg Thr Gln Ala Leu Arg Lys Leu Ile Asp Ala Ala Pro Tyr
580 585 590
Thr Leu Leu Met Thr Gly Thr Pro Leu Glu Asn Lys Val Glu Glu Phe
595 600 605
Val Asn Leu Val Arg Tyr Ile Gln Pro Glu Leu Ile Thr Arg Gly Met
610 615 620
Ser Lys Met Gln Ala Glu Asn Phe Arg Glu Arg Ile Ala Pro Ala Tyr
625 630 635 640
Leu Arg Arg Asn Gln Ala Asp Val Leu Asp Glu Leu Pro Glu Arg Thr
645 650 655
Asp Ser Ile Asp Trp Ile Asp Leu Thr Pro Glu Asp Arg Ser Ala Tyr
660 665 670
Asp Asp Gln Val Arg Gln Gly Ser Trp Met Gly Met Arg Arg Ser Ala
675 680 685
Met Leu Ser Pro Thr Pro Arg Leu Thr Ser Ala Lys Met Gln Arg Ile
690 695 700
Leu Glu Leu Phe Glu Glu Ala Glu Glu His Gly Arg Lys Ala Leu Ile
705 710 715 720
Phe Thr Tyr Phe Leu Asp Val Leu Asp Glu Leu Glu Lys His Leu Gly
725 730 735
Glu Arg Val Ile Gly Arg Ile Ser Gly Asp Val Pro Ala Thr Lys Arg
740 745 750
Gln Leu Leu Val Asp Ala Leu Ser His Ser Lys Pro Gly Ser Ala Leu
755 760 765
Ile Ala Gln Ile Thr Ala Gly Gly Val Gly Leu Asn Ile Gln Ser Ala
770 775 780
Ser Leu Cys Ile Ile Cys Glu Pro Gln Val Lys Pro Thr Ile Glu Gln
785 790 795 800
Gln Ala Val Ala Arg Val His Arg Met Gly Gln Thr Ala Thr Val Gln
805 810 815
Val His Arg Leu Ile Gly Asp Glu Thr Ala Asp Glu Arg Met Leu Glu
820 825 830
Ile Leu Ala Gly Lys Thr His Val Phe Asp Val Tyr Ala Arg Leu Ser
835 840 845
Glu Thr Ala Glu Ile Pro Asp Ala Val Asp Ile Thr Glu Ser Gln Leu
850 855 860
Ala Ala Arg Val Ile Asp Glu Glu Arg Ala Arg Leu Gly Leu Thr Glu
865 870 875 880
Ser Thr Gly Pro Lys Asp Glu Glu Thr Ala Leu Ser
885 890
<210> 32
<211> 892
<212> PRT
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 32
Met Ala Glu Ser Asn Ala Met Asp Arg Ala Gln Ile Ser Ala Leu Leu
1 5 10 15
Asp Arg Ala Gln His Thr Ile Asn Leu Ala Glu Gln Ala Asn Asn Val
20 25 30
Leu Arg Leu Leu Lys Thr Pro Gly Thr Ala Thr Val Gly Asp Asn Gly
35 40 45
Thr Leu Gly Thr Asp Thr Tyr Leu Ile Pro Ser Arg Asn Ile Thr Trp
50 55 60
Pro Asp Asn Leu Tyr Val Asn Val Phe Leu Asp Gly Met Asn Ala Glu
65 70 75 80
Ala Thr Leu Thr Asp Tyr Val Ala Ser Val Ala Ser Ile Pro Arg Leu
85 90 95
Cys Gln Ile Ile Asn Glu Gly Gln Gly Gly Met Phe Arg Arg Leu Phe
100 105 110
Asn Pro Thr Lys Val Gln Ala Gly Asp Gln Ala Val Phe Asp Leu Met
115 120 125
Val Lys Leu Asp Glu Ile Ser Ser Thr Thr His Glu Val Ser Arg Met
130 135 140
Leu Glu Gly Val His Ala Ala Arg Thr Arg Gln Gln Gln Gly Val Ala
145 150 155 160
Leu Phe Pro Gly Ile His Gly Val Gly Glu Arg Tyr Ile Glu Arg Ala
165 170 175
Gln Gln Val Leu Ala Ser Ala Leu Gly Ile Ala Gly Phe Gly Ala Glu
180 185 190
Pro Trp Asp Gly His Thr Leu Ala Gln Ala Arg Arg Val Val Gln Arg
195 200 205
Tyr Ala Gln Asp Pro Asn Ser Glu Tyr Arg Leu Lys Ser Glu Ala Glu
210 215 220
Lys His Leu Thr Ser Ile Asn Glu Leu Arg Val Gln Ile Leu Leu Glu
225 230 235 240
Gln Leu Pro Val Asp Ala Leu Arg Met Ala Thr Asp His Arg Leu Arg
245 250 255
Phe Gly Ser Leu Asp Ser Ile His Val Ala Thr Val Ala Asp Val Leu
260 265 270
Lys Thr His Thr Ser Ile Leu Thr Thr Val Gln Gly Ile Gly Ala Gln
275 280 285
Thr Ala Gly Arg Met Lys Ala Ala Ala Glu Thr Leu Lys Gln Glu Ala
290 295 300
Leu Arg Arg Gln Asn Thr Ser Ile Gly Asp Glu Pro Thr Gln Pro Ala
305 310 315 320
Met Arg Leu Ile Asn Val Leu Ala Arg Phe Asp Gln Thr Glu Thr Ile
325 330 335
Thr Pro Glu Glu Arg Ala Arg Arg Thr Arg Val Ile Asp Tyr Val Glu
340 345 350
His Ile Pro Pro Ser Leu Asp Pro Tyr Ile Val Ile Asn Pro Ala Thr
355 360 365
Pro Glu Phe Asn Asn Phe Thr Asp Asp Leu Arg Trp Ile Asp Ala Asn
370 375 380
Pro Asn Leu Phe His Pro Gln Thr Ile Thr Thr Pro Pro Ala Asp Ile
385 390 395 400
Trp Asp Asp Tyr Ile Ser Arg Pro Ala His Tyr Gln Gly Leu Leu Ala
405 410 415
Thr Leu Leu Gly Arg Asp Ile Glu Gly Ala Asp Glu Leu Leu Asp Ala
420 425 430
Thr Thr Leu Gln Lys Ile Arg Asp Leu Thr Leu Asp Lys Thr His Leu
435 440 445
Thr Asp Leu His Leu Arg Gly Tyr Gln Ser Phe Gly Ala Arg Phe Ala
450 455 460
Ile Ile Gln Lys Lys Thr Leu Leu Gly Asp Asp Met Gly Leu Gly Lys
465 470 475 480
Thr Val Gln Ala Leu Ser Ala Ala Ala His Leu Ala Ala Thr Glu Lys
485 490 495
Asp Phe Arg Thr Leu Val Val Val Pro Ala Ser Val Ile Val Asn Trp
500 505 510
Thr Arg Glu Cys Lys Arg Phe Leu Asn Leu Pro Val Phe Ile Ala His
515 520 525
Gly Asp Asn Lys Gln Asp Ala Ile Asn Ala Trp Ser Asn Thr Asn Gly
530 535 540
Ile Ala Ile Cys Thr Tyr Asp Gly Val Arg Thr Met Asp Ile Pro Ala
545 550 555 560
Pro Gly Leu Val Ile Ala Asp Glu Ala His Leu Ile Lys Asn Pro Ser
565 570 575
Thr Lys Arg Thr Gln Ala Leu Arg Lys Leu Ile Asp Ala Ala Pro Phe
580 585 590
Thr Leu Leu Met Thr Gly Thr Pro Leu Glu Asn Lys Val Glu Glu Phe
595 600 605
Val Asn Leu Val Arg Tyr Ile Gln Pro Glu Leu Ile Thr Arg Gly Met
610 615 620
Ser Lys Met Gln Ala Glu Asn Phe Arg Glu Arg Ile Ala Pro Ala Tyr
625 630 635 640
Leu Arg Arg Asn Gln Ala Asp Val Leu Asp Glu Leu Pro Glu Arg Thr
645 650 655
Asp Ser Ile Asp Trp Ile Asp Leu Thr Pro Glu Asp Arg Ser Ala Tyr
660 665 670
Asp Asp Gln Val Arg Gln Gly Ser Trp Met Gly Met Arg Arg Ser Ala
675 680 685
Met Leu Ser Pro Thr Pro Arg Leu Thr Ser Ala Lys Met Gln Arg Ile
690 695 700
Leu Glu Leu Phe Glu Glu Ala Glu Glu His Gly Arg Lys Ala Leu Ile
705 710 715 720
Phe Thr Tyr Phe Leu Asp Val Leu Asp Glu Leu Glu Lys His Leu Gly
725 730 735
Glu Arg Val Ile Gly Arg Ile Ser Gly Asp Val Pro Ala Thr Lys Arg
740 745 750
Gln Leu Leu Val Asp Ala Leu Ser His Ser Lys Pro Gly Ser Ala Leu
755 760 765
Ile Ala Gln Ile Thr Ala Gly Gly Val Gly Leu Asn Ile Gln Ser Ala
770 775 780
Ser Leu Cys Ile Ile Cys Glu Pro Gln Val Lys Pro Thr Ile Glu Gln
785 790 795 800
Gln Ala Val Ala Arg Val His Arg Met Gly Gln Thr Ala Thr Val Gln
805 810 815
Val His Arg Leu Ile Gly Asp Glu Thr Ala Asp Glu Arg Met Leu Glu
820 825 830
Ile Leu Ala Gly Lys Thr His Val Phe Asp Val Tyr Ala Arg Leu Ser
835 840 845
Glu Thr Ala Glu Ile Pro Asp Ala Val Asp Ile Thr Glu Ser Gln Leu
850 855 860
Ala Ala Arg Val Ile Asp Glu Glu Arg Ala Arg Leu Gly Leu Thr Glu
865 870 875 880
Ser Thr Gly Pro Lys Asp Glu Glu Thr Ala Leu Ser
885 890
<210> 33
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 33
cagtgccaag cttgcatgcc tgcaggtcga ctctagtgcg ttcgtctgcg gtttcg 56
<210> 34
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 34
atcgacgccg ccccattcac ccttctgatg 30
<210> 35
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 35
catcagaagg gtgaatgggg cggcgtcgat 30
<210> 36
<211> 57
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 36
cagctatgac catgattacg aattcgagct cggtacccaa gcctcgaccc ctacatc 57
<210> 37
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 37
cacatcagct tgatttctgc 20
<210> 38
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 38
ggtcattgcc gatgaagccc 20
<210> 39
<211> 55
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 39
cagtgccaag cttgcatgcc tgcaggtcga ctctagaatg cgttctggac tgagg 55
<210> 40
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 40
gaaacggcct taagctaggt gcaccgagaa cagatg 36
<210> 41
<211> 36
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 41
catctgttct cggtgcacct agcttaaggc cgtttc 36
<210> 42
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 42
cttgatttaa ttgcgccatc aagcttttcc cgcccggtt 39
<210> 43
<211> 39
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 43
aaccgggcgg gaaaagcttg atggcgcaat taaatcaag 39
<210> 44
<211> 60
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 44
cagctatgac catgattacg aattcgagct cggtacccgc tatgacacct tcaacggatc 60
<210> 45
<211> 19
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 45
tccaaggaag atacacgcc 19
<210> 46
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 46
cttctgatga ccggcacacc 20
<210> 47
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 47
tagtcgatga cgcgggtgcg 20
<210> 48
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 48
cgttggaatc ttgcgttg 18
<210> 49
<211> 54
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 49
gcttgcatgc ctgcaggtcg actctagagg atccccctag cttaaggccg tttc 54
<210> 50
<211> 53
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 50
atcaggctga aaatcttctc tcatccgcca aaacaagctt ttcccgcccg gtt 53
<210> 51
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 51
cagtgccaag cttgcatgcc tgcaggtcga ctctagaccg gcgcagatgc caacgc 56
<210> 52
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 52
cccagaactg aaggtctaat tgcctaaggc cggaatt 37
<210> 53
<211> 37
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 53
aattccggcc ttaggcaatt agaccttcag ttctggg 37
<210> 54
<211> 56
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 54
cagctatgac catgattacg aattcgagct cggtacccgc ttgatgaagg ctccag 56
<210> 55
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 55
accggcgcag atgccaacgc 20
<210> 56
<211> 18
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 56
gcttgatgaa ggctccag 18
<210> 57
<211> 242
<212> DNA
<213> Corynebacterium glutamicum
<400> 57
ttcagggtag ttgactaaag agttgctcgc gaagtagcac ctgtcacttt tgtctcaaat 60
attaaatcga atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc 120
atccgctaaa gccccaggaa ccctgtgcag aaagaaaaca ctcctctggc taggtagaca 180
cagtttataa aggtagagtt gagcgggtaa ctgtcagcac gtagatcgaa aggtgcacaa 240
ag 242
<210> 58
<211> 242
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 58
ttcagggtag ttgactaaag agttgctcgc gaagtagcac ctgtcacttt tgtctcaaat 60
attaaatcga atatcaatat atggtctgtt tattggaacg cgtcccagtg gctgagacgc 120
atccgctaaa gccccaggaa ccctgtgcag aaagaaaaca ctcctctggc taggtagaca 180
cagtttataa aggtataatt gagcgggtaa ctgtcagcac gtagatcgaa aggtgcacaa 240
ag 242
<210> 59
<211> 29
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 59
ccggaattcg accaaggatg agggctttg 29
<210> 60
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 60
agttacccgc tcaattatac ctttataaac 30
<210> 61
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 61
gtttataaag gtataattga gcgggtaact 30
<210> 62
<211> 30
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 62
acatgcatgc gcgtacgcga agtggcacat 30
<210> 63
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 63
atcaatatat ggtctgttta 20
<210> 64
<211> 20
<212> DNA
<213> Artificial Sequence
<220>
<223> искусственная последовательность
<400> 64
cttggtggca acgatccgtt 20
<---
Изобретение относится к области биотехнологии. Предложена бактерия для продуцирования L-аминокислоты. Бактерия имеет улучшенную экспрессию полинуклеотида для кодирования белка, представленного SEQ ID NO: 3, и улучшенную экспрессию полинуклеотида для кодирования белка, представленного SEQ ID NO: 31, и/или имеет мутации оснований в положениях -45 п.о. и -47 п.о. промоторной области, представленной SEQ ID NO: 57. Изобретение позволяет получать аминокислоты, в том числе лизин, с высокой степенью эффективности. 4 н. и 15 з.п. ф-лы, 8 табл., 16 пр.
1. Бактерия, продуцирующая L-аминокислоту, выбранную из L-лизина и L-глутаминовой кислоты, причем указанная бактерия имеет улучшенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3, в которой аргинин в положении 334 заменен терминатором.
2. Бактерия, продуцирующая L-лизин, причем указанная бактерия имеет улучшенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 3, в которой аргинин в положении 334 заменен терминатором, или улучшенную экспрессию полинуклеотида, кодирующего аминокислотную последовательность SEQ ID NO: 31, в которой тирозин в положении 592 заменен другими аминокислотами, или мутации оснований в положениях -45 п.о. и -47 п.о. в промоторной области, показанной в SEQ ID NO: 57.
3. Бактерия по п. 1 или 2, характеризующаяся тем, что полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 3, содержит нуклеотидную последовательность SEQ ID NO: 1;
или полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 31, содержит нуклеотидную последовательность SEQ ID NO: 29.
4. Бактерия по любому из пп. 1-3, характеризующаяся тем, что полинуклеотидная последовательность с точечными мутациями, кодирующая аминокислотную последовательность SEQ ID NO: 3, образована посредством мутации 1000-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1; или мутация включает мутацию 1000-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1, с цитозина (C) на тимин (T); или полинуклеотидная последовательность с точечными мутациями включает полинуклеотидную последовательность, показанную в SEQ ID NO: 2;
или полинуклеотидная последовательность с точечными мутациями, кодирующая аминокислотную последовательность SEQ ID NO: 31, образована посредством мутации 1775-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29; или мутация включает мутацию 1775-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29, с аденина (A) на тимин (T); или полинуклеотидная последовательность с точечными мутациями включает полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
5. Бактерия по любому из пп. 2-4, характеризующаяся тем, что мутации оснований в положениях -45 п.о. и -47 п.о. в промоторном участке, показанном в SEQ ID NO: 57, выбраны из замены гуанина (G) в положении -45 п.о. на аденин (A) и замены гуанина (G) в положении -47 п.о. на тимин (T) в промоторном участке, показанном в SEQ ID NO: 57;
или нуклеотидная последовательность промотора содержит:
(a) нуклеотидную последовательность, показанную в SEQ ID NO: 58; или
(б) нуклеотидную последовательность, имеющую идентичность последовательности более чем 90%, предпочтительно более чем 95%, 98% с нуклеотидной последовательностью, показанной в SEQ ID NO: 58, и сохраняющую повышенную промоторную активность (a), при сохранении аденина (A) в положении -45 п.о. и тимина (T) в положении -47 п.о.
6. Бактерия по любому из пп. 1-5, отличающаяся тем, что указанная бактерия представляет собой Corynebacterium glutamicum.
7. Бактерия по п. 6, отличающаяся тем, что указанная бактерия представляет собой YP97158 или ATCC 13869.
8. Рекомбинантный штамм бактерии для получения L-лизина, содержащий любое из (I) и (IV), (II) и (IV), (III) и (IV), (V), (VI) или (VII), указанных далее:
(I) полинуклеотидная последовательность, характеризующаяся тем, что указанная полинуклеотидная последовательность содержит полинуклеотид, кодирующий аминокислотную последовательность, показанную в SEQ ID NO: 3, и аргинин в положении 334 заменен на терминатор;
(II) полинуклеотидная последовательность, характеризующаяся тем, что указанная полинуклеотидная последовательность содержит полинуклеотид, кодирующий аминокислотную последовательность, показанную в SEQ ID NO: 31, где тирозин в положении 592 заменен другими аминокислотами;
(III) рекомбинантный вектор, характеризующийся тем, что указанный рекомбинантный вектор содержит полинуклеотидную последовательность, как определено в (I) или (II);
(IV) нуклеотидная последовательность промотора, содержащая нуклеотидную последовательность, образованную посредством мутации оснований в положениях -45 п.о. и -47 п.о. промоторной области, как показано в SEQ ID NO: 57;
(V) экспрессионная кассета, содержащая нуклеотидную последовательность промотора, как определено в (IV), и кодирующую последовательность, функционально присоединенную за нуклеотидной последовательностью промотора;
(VI) рекомбинантный вектор, содержащий нуклеотидную последовательность промотора, как определено в (IV);
и
(VII) нуклеотидная последовательность промотора, как определено в (IV), или рекомбинантный вектор, как определено в (VI).
9. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что
указанная полинуклеотидная последовательность (I) включает полинуклеотид, кодирующий аминокислотную последовательность, показанную в SEQ ID NO: 4;
указанная полинуклеотидная последовательность (I) образована посредством мутации 1000-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1;
указанная полинуклеотидная последовательность (I) включает мутацию 485-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 1, с цитозина (C) на тимин (T); или
указанная полинуклеотидная последовательность (I) включает полинуклеотидную последовательность, показанную в SEQ ID NO: 2.
10. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что
указанная полинуклеотидная последовательность (II) включает полинуклеотид, кодирующий аминокислотную последовательность, показанную в SEQ ID NO: 32;
указанная полинуклеотидная последовательность (II) образована посредством мутации 1775-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29;
указанная полинуклеотидная последовательность (II) включает мутацию 1775-го основания полинуклеотидной последовательности, показанной в SEQ ID NO: 29, с аденина (A) на тимин (T); и
указанная полинуклеотидная последовательность (II) включает полинуклеотидную последовательность, показанную в SEQ ID NO: 30.
11. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что
нуклеотид гуанин (G) в положении -45 п.о. в промоторном участке мутирован на аденин (A) и нуклеотид гуанин (G) в положении -47 п.о. мутирован на тимин (T) в промоторном участке, показанном в SEQ ID NO: 57.
12. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что
указанная нуклеотидная последовательность промотора (IV) дополнительно содержит (а) или (б):
(а) нуклеотидную последовательность, показанную в SEQ ID NO:58; или
(б) нуклеотидную последовательность, имеющую идентичность последовательности более чем 90% нуклеотидной последовательности, показанной в SEQ ID NO: 58, и сохраняющую повышенную промоторную активность (a), при сохранении аденина (A) в положении -45 п.о. и тимина (T) в положении -47 п.о.
13. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что кодирующая последовательность представляет собой кодирующую последовательность гена lysC.
14. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что указанная нуклеотидная последовательность промотора (VI) связана с шаттл-плазмидой для конструирования рекомбинантного вектора; и указанная шаттл-плазмида представляет собой плазмиду pK18mobsacB.
15. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что указанная нуклеотидная последовательность (VII) представляет собой нуклеотидную последовательность, показанную в SEQ ID NO: 58; или указанная нуклеотидная последовательность представляет собой нуклеотидную последовательность, показанную в SEQ ID NO: 58, связанную с кодирующей последовательностью гена lysC.
16. Рекомбинантный штамм бактерии по п. 8, отличающийся тем, что тирозин в положении 592 заменен фенилаланином.
17. Рекомбинантный штамм бактерии по п. 12, отличающийся тем, что указанная нуклеотидная последовательность промотора (IV) дополнительно содержит нуклеотидную последовательность, имеющую идентичность последовательности более чем 95% нуклеотидной последовательности, показанной в SEQ ID NO: 58.
18. Рекомбинантный штамм бактерии по п. 12, отличающийся тем, что указанная нуклеотидная последовательность промотора (IV) дополнительно содержит нуклеотидную последовательность, имеющую идентичность последовательности более чем 98% нуклеотидной последовательности, показанной в SEQ ID NO: 58.
19. Способ продуцирования L-аминокислоты, включающий культивирование бактерии по любому из пп. 1-5 или рекомбинантного штамма бактерии по п. 8 и выделение L-аминокислоты из культуры, где L-аминокислота представляет собой L-лизин.
| CN 110951662 A, 03.04.2020 | |||
| Микроорганизм, обладающий продуктивностью по L-лизину, и способ получения L-лизина с использованием этого микроорганизма | 2016 |
|
RU2683551C1 |
| ФЕРМЕНТАЦИОННЫЙ СПОСОБ ПОЛУЧЕНИЯ L-ЛИЗИНА | 1994 |
|
RU2204605C2 |
| МИКРООРГАНИЗМЫ CORYNEBACTERIUM SP., ОБЛАДАЮЩИЕ СПОСОБНОСТЬЮ ПРОДУЦИРОВАТЬ L-ЛИЗИН, И СПОСОБ ПРОДУЦИРОВАНИЯ L-ЛИЗИНА С ИСПОЛЬЗОВАНИЕМ ЭТИХ МИКРООРГАНИЗМОВ | 2016 |
|
RU2685482C1 |
| Микроорганизм Corynebacterium sp. с повышенной способностью продуцировать L-лизин и способ получения L-лизина с его помощью | 2014 |
|
RU2667425C2 |

