Область техники
Изобретение относится к биотехнологии и генной инженерии. Предложенный вектор может применяться для разработки вакцин против инфекционных заболеваний и противоопухолевых вакцин, лекарственных средств для иммунотерапии опухолей. Вектор на основе мРНК можно использовать для создания терапевтических лекарственных средств, активным действующим веществом которых являются антитела, синтезируемые in vivo. Также существует вариант использования изобретения для синтеза других терапевтических белков in vivo.
Уровень техники
Как известно, в 1961 году S.Brenner и соавторы обнаружили матричную рибонуклеиновую кислоту (мРНК), которая является ключевой промежуточной молекулой, необходимой для экспрессии генов в виде белков. В 1990 г. J.Wolff и соавторы впервые показали, что целевой белок может быть эффективно синтезирован in vivo путем внутримышечного введения мышам чистой РНК [He Q, Gao H, Tan D, Zhang H, Wang JZ. mRNA cancer vaccines: Advances, trends and challenges. Acta Pharm Sin B. 2022 Jul;12(7):2969-2989. doi: 10.1016/j.apsb.2022.03.011. Epub 2022 Mar 23. PMID: 35345451; PMCID: PMC8942458.]. Эти события послужили стимулом для развития технологических платформ на основе мРНК, которые в настоящее время активно используются в различных областях медицины для разработки профилактических и терапевтических средств.
Технология на основе мРНК имеет ряд преимуществ, которые выделяют ее среди других технологических платформ. Средства на основе рибонуклеиновой кислоты могут быть разработаны и протестированы в короткие сроки. Технология позволяет синтезировать белки непосредственно в организме, устраняя необходимость очистки и долговременной стабилизации белка. Природные свойства РНК позволяют избежать противовекторного иммунитета, таким образом обеспечивая возможность многократного введения препарата. Вакцины, полученные на основе мРНК, способны индуцировать как клеточный, так и гуморальный иммунный ответ. Возможность получения мРНК в бесклеточных системах позволяет организовать масштабируемое и экономически эффективное производство.
В настоящее время технология на основе мРНК используется для разработки различных лекарственных средств. В 2020 году в ответ на пандемию, вызванную распространением вируса SARS-CoV-2, были разработаны две вакцины на основе мРНК: BNT162b2 производитель Pfizer/BioNTech (США/Германия) и mRNA-1273 производитель Moderna (США). Обе вакцины показали высокую эффективность и были одобрены для применения во многих странах мира. Кроме того, на различных стадиях клинических исследований находятся вакцины против гриппа, СПИД, бешенства, лихорадки Зика, против цитомегаловирусной инфекции, против метапневмовирусной инфекции и других инфекционных заболеваний. Технология на основе мРНК также применяется для создания противоопухолевых лекарственных средств (против острого миелоидного лейкоза, колоректального рака, меланомы, глиобластомы, злокачественной мезотелиомы для лечения метастазов в печени и мозге и др.), которые в настоящее время проходят клинические испытания. Проводятся исследования терапевтических препаратов на основе мРНК для лечения генетических заболеваний (муковисцидоз, пропионовая ацидемия, метилмалоновая ацидемия, фенилкетонурия, гемофилия), аутоиммунных заболеваний, метаболических нарушений (диабет второго типа), сердечно-сосудистых заболеваний (гиперхолестеринемия, ишемия миокарда), фиброза (гипертрофические рубцы, фиброз печени, фиброз легких, первичный склерозирующий холангит, анемия) и др. [Xu S, Yang K, Li R, Zhang L. mRNA Vaccine Era-Mechanisms, Drug Platform and Clinical Prospection. Int J Mol Sci. 2020 Sep 9;21(18):6582. doi: 10.3390/ijms21186582. PMID: 32916818; PMCID: PMC7554980; Qin S, Tang X, Chen Y, Chen K, Fan N, Xiao W, Zheng Q, Li G, Teng Y, Wu M, Song X. mRNA-based therapeutics: powerful and versatile tools to combat diseases. Signal Transduct Target Ther. 2022 May 21;7(1):166. doi: 10.1038/s41392-022-01007-w. PMID: 35597779; PMCID: PMC9123296].
Таким образом, мРНК является одной из наиболее перспективных и востребованных технологических платформ. Однако ее огромный потенциал может быть ограничен из-за низкой стабильности и недостаточно эффективной продукции целевого белка.
В течение нескольких десятилетий были предприняты различные попытки получения стабильного вектора на основе мРНК, обеспечивающего высокую эффективность продукции целевого белка.
В патенте CN105849269B описан РНК-вектор для эффективного синтеза целевого белка, содержащий трансляционный энхансер (translational Enhancer), транскрибируемую последовательность и стабилизирующий элемент, в роли которого выступает: сигнал удержания мРНК в ядре (nuclear retention element). При этом сигнал удержания мРНК в ядре предпочтительно является «экспрессионным и удерживающим в ядре элементом» (expression and nuclear retention element, ENE) вируса герпеса, ассоциированного с саркомой Капоши. Транскрибируемая последовательность выбирается из списка, содержащего рибонуклеиновую кислоту, кодирующую CD40L, CD70, caTLR4 или рибонуклеиновую кислоту, специфичную для антигена/заболевания. Описано применение вектора для терапевтических целей.
В патенте RU2650795C2 описана нуклеиновая кислота, содержащая: 1) область, кодирующую по меньшей мере один пептид или белок; по меньшей мере одну шпильку из мРНК гистона и поли(А)-последовательность или сигнал полиаденилирования; или 2) область, кодирующую по меньшей мере один пептид или белок; поли(А)-последовательность и, по меньшей мере, одну шпильку из мРНК гистона. При этом вышеупомянутый пептид или белок содержит специфичный для опухоли антиген или его фрагмент.
В патенте EP3424524A2 описана синтетическая нуклеиновая кислота, кодирующая по меньшей мере один антигенный пептид или белок и по меньшей мере одну дополнительную последовательность, нацеливающую антигенные пептиды или белки на представляющие интерес клеточные компартменты. Кроме того, изобретение включает фармацевтические композиции или вакцины, а также наборы, содержащие указанные молекулы нуклеиновой кислоты. Данное изобретение может быть использовано в терапии различных заболеваний: опухолевых, инфекционных, аутоиммунных, реакции отторжения трансплантата.
Из патента US8785611B2 известна изолированная полинуклеотидная последовательность, содержащая по меньшей мере два элемента усилителя трансляции мРНК (TEE), в которой по меньшей мере один TEE состоит из полноразмерной последовательности, выбранной из группы, состоящей из 5'-CGCGGCTGA-3', 5'-AGCCGCCGCA-3' и 5'-ACGCCGCCGA-3'. Кроме того известен вектор для продукции рекомбинантного полипептида в эукариотической клетке, содержащий эукариотический промотор, функционально связанный с полинуклеотидной последовательностью, содержащей по меньшей мере один элемент усилителя трансляции, который выбран из группы, состоящей из 5'-CGCGGCTGA-3', 5'-AGCCGCCGCA-3' и 5'-ACGCCGCCGA-3'.
В патенте JP6759196B2 описана нуклеиновая кислота, обладающая повышенной стабильностью. Указанный эффект достигается за счет фрагментации 3' поли(А)-содержащей кассеты (поли(dA:dT) области) случайной последовательностью из 10 нуклеотидов. Более того, ни последовательность, ни положение линкера в последовательности 3' поли(A) не приводили к снижению эффективности трансляции и стабильности транскрибируемой in vitro РНК
В патенте US10335486B2 описан способ лечения рака у субъекта путем индукции или усиления противоопухолевого иммунного ответа, в котором субъекту на первом этапе вводят мРНК, кодирующую полипептид IL-23 и полипептид IL-12p40, функционально связанный (линкером или без него) с полипептидом IL-23p19, на втором этапе вводят мРНК, кодирующую полипептид IL-18, а на третьем этапе вводят мРНК, кодирующую полипептид OX40L.
Известно изобретение JP6704850B2, в котором описана молекула синтетической нуклеиновой кислоты, содержащая открытую рамку считывания, элемент 3'-нетранслируемой области (3'-НТО), представляющий собой последовательность 3'-НТО гена рибосомного белка и, необязательно, поли(А)-последовательность и/или сигнал полиаденилирования. Изобретение дополнительно относится к вектору, содержащему данную синтетическую нуклеиновую кислоту, а также к клетке, содержащей данный вектор или данную синтетическую нуклеиновую кислоту. Изобретение может быть использовано для генной терапии и/или вакцинации.
Из патента CA2621444C известно техническое решение, в котором молекула нуклеиновой кислоты содержит в направлении 5'→3':
промотор; транскрибируемую последовательность нуклеиновой кислоты, одну или две последовательности 3'-НТО гена глобина.
Известен вектор на основе мРНК, который содержит 5´-нетраслируемую область (5´-НТО) мРНК альфа-глобина человека с оптимизированной последовательностью Козак и 3´-НТО, содержащую две последовательности, полученные из мРНК гена AES и митохондриально-кодируемой рибосомной 12S РНК (https://web.archive.org/web/20210105162941/ https://mednet-communities.net/inn/db/media/docs/11889.doc). Данное решение, как наиболее близкое к заявляемому, выбрано за прототип.
На основе прототипа была получена вакцина против COVID-19 BNT-162b2, Pfizer/BioNTech, которая показала высокую эффективность против различных вариантов SARS-CoV-2 (от 65,5% до 95%) [Andrews N et al. Covid-19 Vaccine Effectiveness against the Omicron (B.1.1.529) Variant. N Engl J Med. 2022 Apr 21;386(16):1532-1546. doi: 10.1056/NEJMoa2119451. Epub 2022 Mar 2. PMID: 35249272; PMCID: PMC8908811]. Однако существует мнение, что вектор на основе мРНК был сконструирован неоптимальным образом, что могло негативно повлиять на наработку целевого белка [Xia X. Detailed Dissection and Critical Evaluation of the Pfizer/BioNTech and Moderna mRNA Vaccines. Vaccines (Basel). 2021 Jul 3;9(7):734. doi: 10.3390/vaccines9070734. PMID: 34358150; PMCID: PMC8310186].
Таким образом, в настоящее время существует острая потребность в разработке вектора на основе мРНК, который позволил бы преодолеть существующие технические проблемы, связанные с недостаточной эффективностью продукции целевого белка.
Раскрытие изобретения
Целью настоящей разработки является создание вектора на основе матричной рибонуклеиновой кислоты, обеспечивающего высокую эффективность продукции целевого белка.
Технический результат заключается в создании вектора на основе мРНК, обладающего стабильностью и обеспечивающего повышенные уровни продукции целевого белка.
Указанный технический результат достигается тем, что создан вектор на основе мРНК для увеличенной продукции целевого белка, содержащий 5'-нетраслируемую область, выбранную из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, SEQ ID NO:4, область, кодирующую целевой белок, и 3'-нетраслируемую область, выбранную из SEQ ID NO:5.
Также технический результат достигается тем, что создан вектор на основе мРНК для увеличенной продукции целевого белка, содержащий 5'-нетраслируемую область, выбранную из SEQ ID NO:1, SEQ ID NO:2, SEQ ID NO:3, область, кодирующую целевой белок, и 3'-нетраслируемую область, выбранную из SEQ ID NO:6.
При этом в разработанных вариантах вектора на основе мРНК область, кодирующая целевой белок, может представлять собой последовательность, кодирующую вакцинный антиген. Также область, кодирующая целевой белок, может представлять собой последовательность, кодирующую антитело. Кроме того, существует вариант изобретения, в котором область, кодирующая целевой белок, представляет собой последовательность, кодирующую терапевтический белок. Также область, кодирующая целевой белок, может представлять собой последовательность, кодирующую фермент.
Указанный технический результат также достигается тем, что разработан способ использования вариантов вектора на основе мРНК для синтеза целевого белка в клетках млекопитающих. Кроме того, предложен способ использования вектора на основе мРНК для синтеза целевого белка in vivo.
Осуществление изобретения
Краткое описание чертежей.
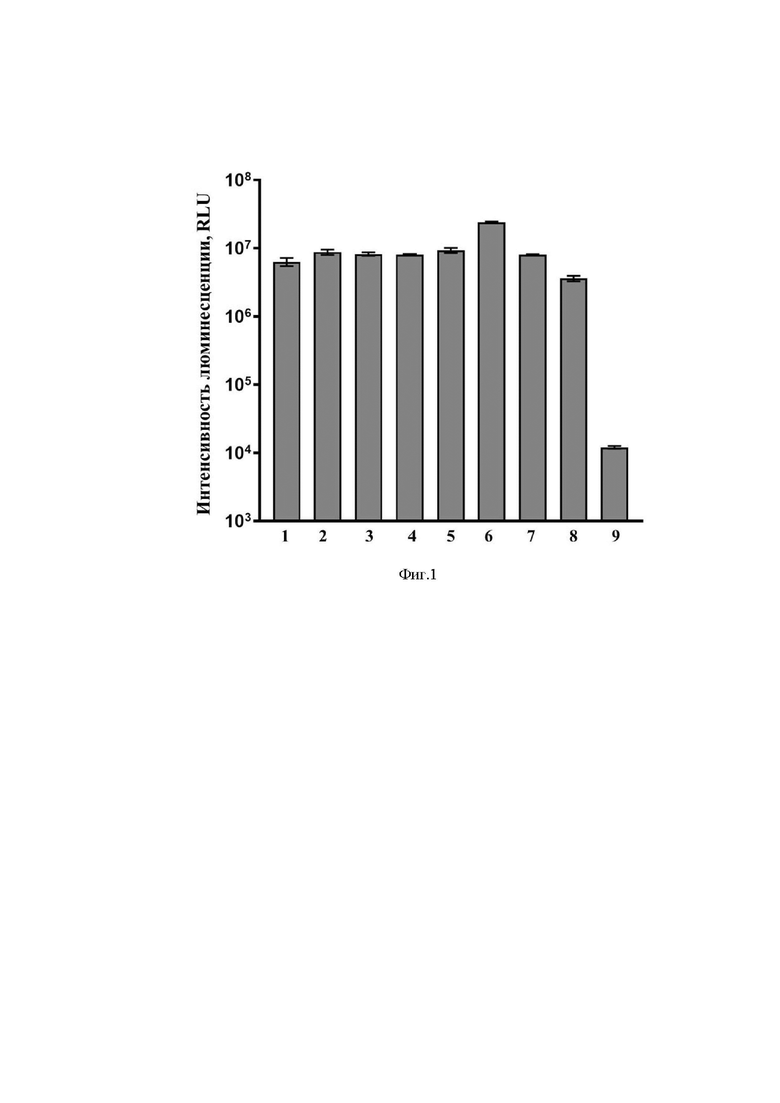
На Фиг. 1 представлены результаты оценки активности люциферазы в клетках HeLa после добавления к ним вариантов разработанного вектора на основе мРНК или прототипа методом липофекции.
На оси ординат - интенсивность люминесценции, отн. ед.,
На оси абсцисс - названия экспериментальных групп:
1) mRNA-cas-luc-glo
2) mRNA-hsp-luc-glo
3) mRNA-tpl-luc-glo
4) mRNA-his-luc-glo
5) mRNA-cas-luc-cyt
6) mRNA-hsp-luc-cyt
7) mRNA-tpl-luc-cyt
8) Контрольная мРНК (содержащая последовательность, кодирующую люциферазу)
9) Отрицательный контроль (буферный раствор).
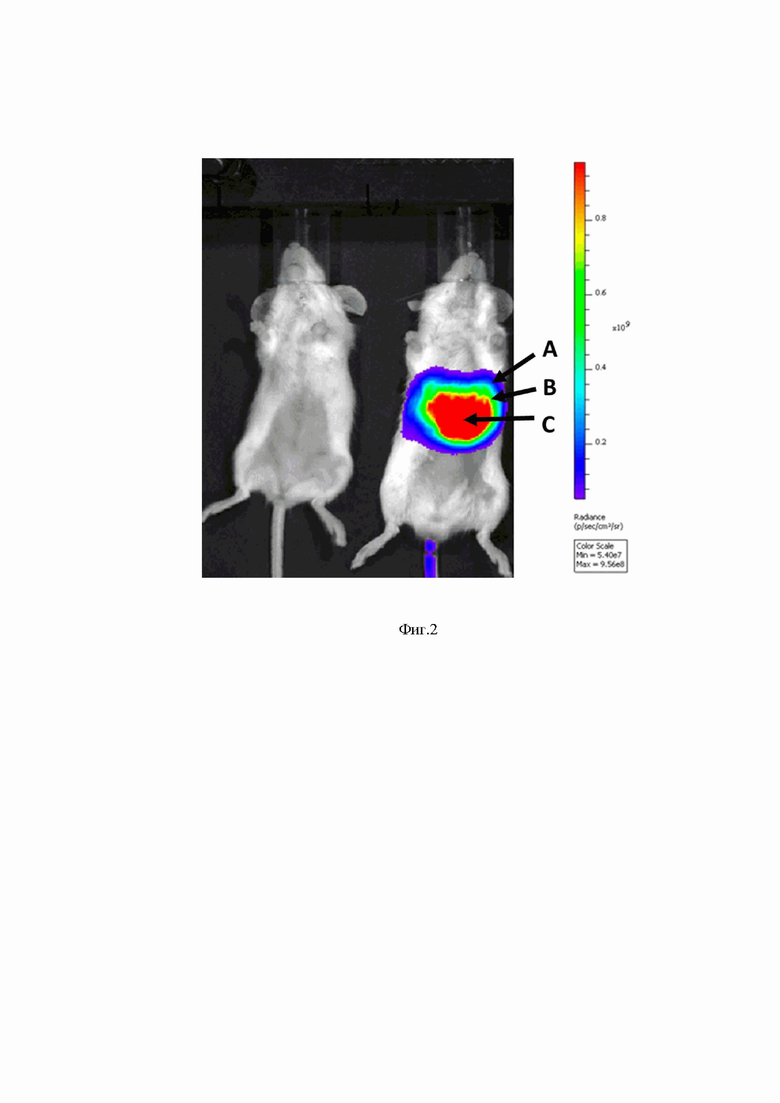
На Фиг. 2 представлены результаты оценки активности люциферазы у мышей через 3 часа после внутривенного введения разработанного вектора на основе мРНК (mRNA-cas-luc-glo), упакованного в липидные частицы.
Слева расположено контрольное животное, которому вводили буферный раствор.
Справа расположено животное, которому внутривенно вводили вектор mRNA-cas-luc-glo, упакованный в липидные частицы.
А - интенсивность люминесценции  от 0,1*109 отн. ед. до 0,3*109 отн. ед.;
от 0,1*109 отн. ед. до 0,3*109 отн. ед.;
В - интенсивность люминесценции от 0,4*109 отн. ед. до 0,7*109 отн. ед.;
С - интенсивность люминесценции от 0,8*109 отн. ед. до 0,9*109 отн. ед.;
На шкале отображается интенсивность люминесценции (отн. ед. люминесценции).
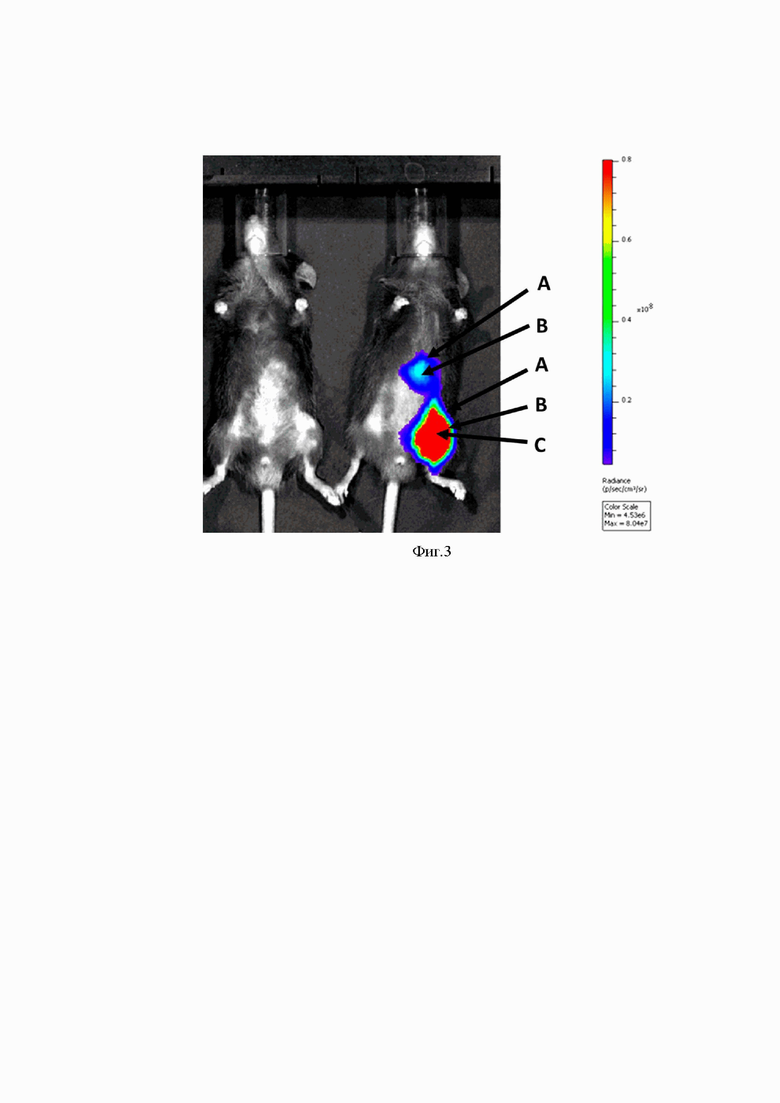
На Фиг. 3 представлены результаты оценки активности люциферазы у мышей через 3 часа после внутримышечного введения разработанного вектора на основе мРНК (mRNA-cas-luc-glo), упакованного в липидные частицы.
Слева расположено контрольное животное, которому вводили буферный раствор.
Справа расположено животное, которому внутримышечно вводили вектор mRNA-cas-luc-glo, упакованный в липидные частицы.
А - интенсивность люминесценции от 0,1*108 отн. ед. до 0,2*108 отн. ед.;
В - интенсивность люминесценции от 0,3*108 отн. ед. до 0,6*108 отн. ед.;
С - интенсивность люминесценции от 0,7*108 отн. ед. до 0,9*108 отн. ед.;
На шкале отображается интенсивность люминесценции (отн. ед. люминесценции).
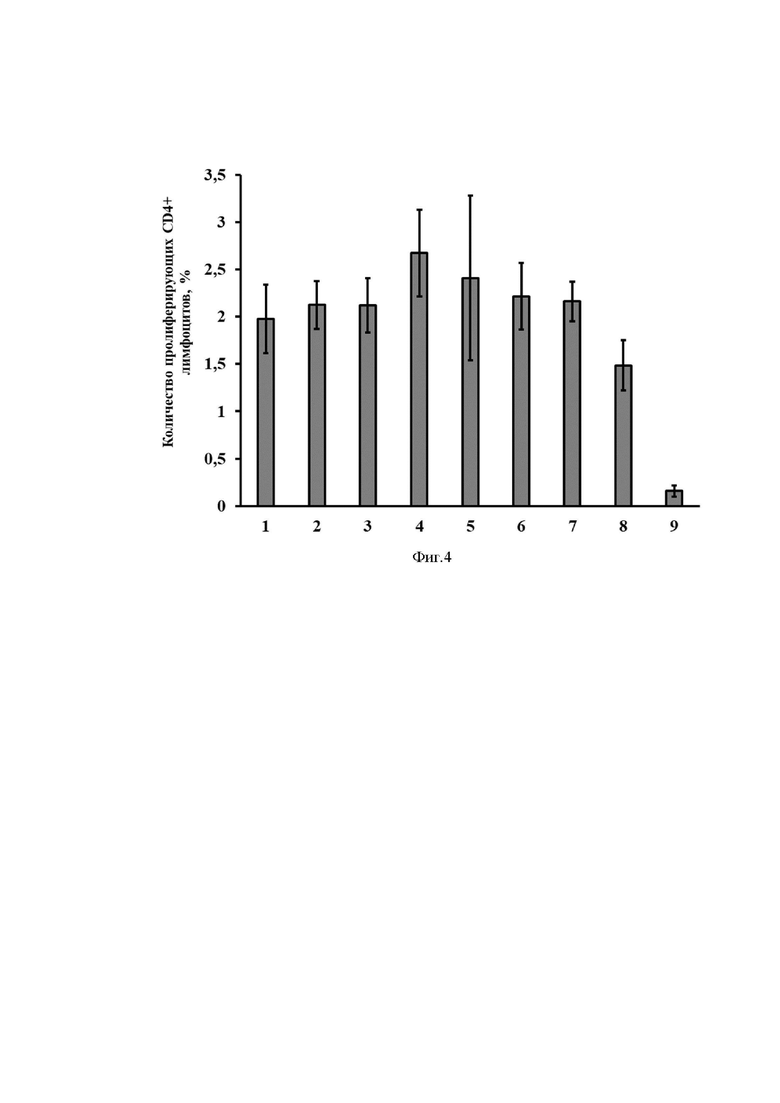
На Фиг. 4 представлены результаты оценки эффективности иммунизации разработанным вектором на основе мРНК по оценке доли пролиферирующих CD4+ лимфоцитов в ответ на стимуляцию антигеном (S белок вируса SARS-CoV-2).
Ось ординат - количество пролиферирующих клеток, %.
Ось абсцисс - экспериментальные группы.
1) mRNA-cas-S-glo, 3 мкг/мышь
2) mRNA-hsp-S-glo, 3 мкг/мышь
3) mRNA-tpl-S-glo, 3мкг/мышь
4) mRNA-his-S-glo, 3 мкг/мышь
5) mRNA-cas-S-cyt, 3 мкг/мышь
6) mRNA-hsp-S-cyt, 3 мкг/мышь
7) mRNA-tpl-S-cyt, 3 мкг/мышь
8) Контрольная мРНК (содержащая последовательность S белка SARS-CoV-2)
9) Буферный раствор
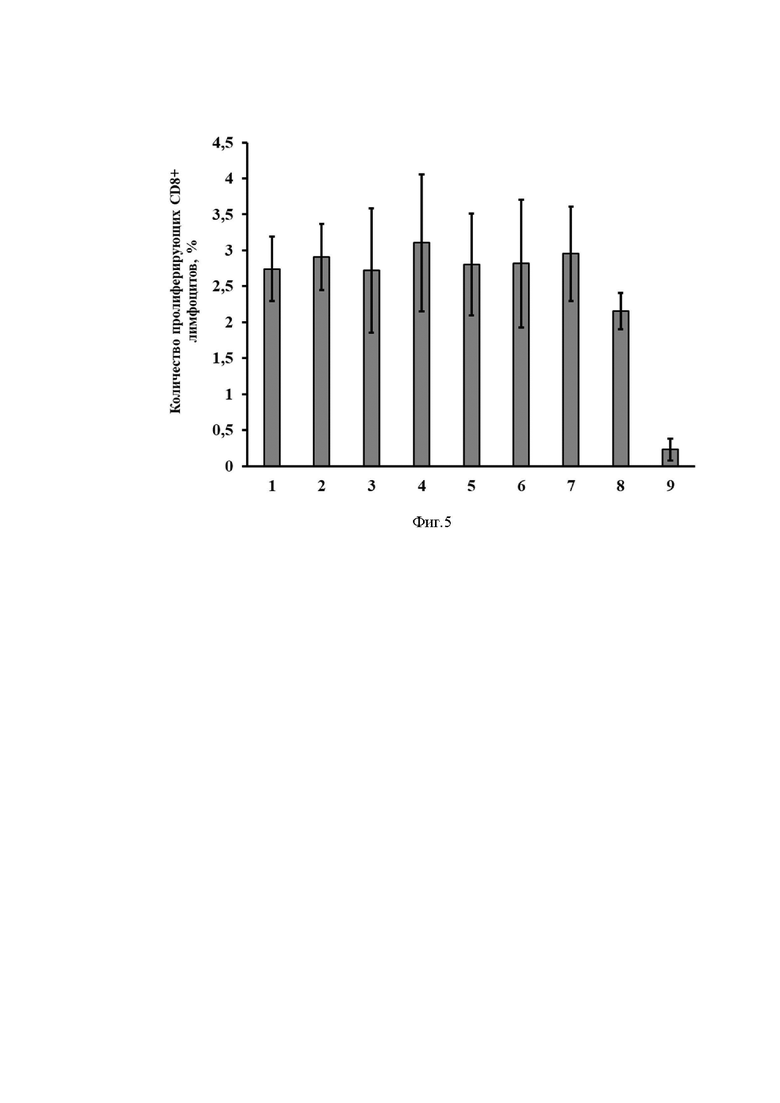
На Фиг. 5 представлены результаты оценки эффективности иммунизации разработанным вектором на основе мРНК по оценке доли пролиферирующих CD8+ лимфоцитов в ответ на стимуляцию антигеном (S белок вируса SARS-CoV-2).
Ось ординат - количество пролиферирующих клеток, %.
Ось абсцисс - экспериментальные группы.
1) mRNA-cas-S-glo, 3 мкг/мышь
2) mRNA-hsp-S-glo, 3 мкг/мышь
3) mRNA-tpl-S-glo, 3мкг/мышь
4) mRNA-his-S-glo, 3 мкг/мышь
5) mRNA-cas-S-cyt, 3 мкг/мышь
6) mRNA-hsp-S-cyt, 3 мкг/мышь
7) mRNA-tpl-S-cyt, 3 мкг/мышь
8) Котнрольная мРНК (содержащая последовательность S белка SARS-CoV-2)
9) Буферный раствор
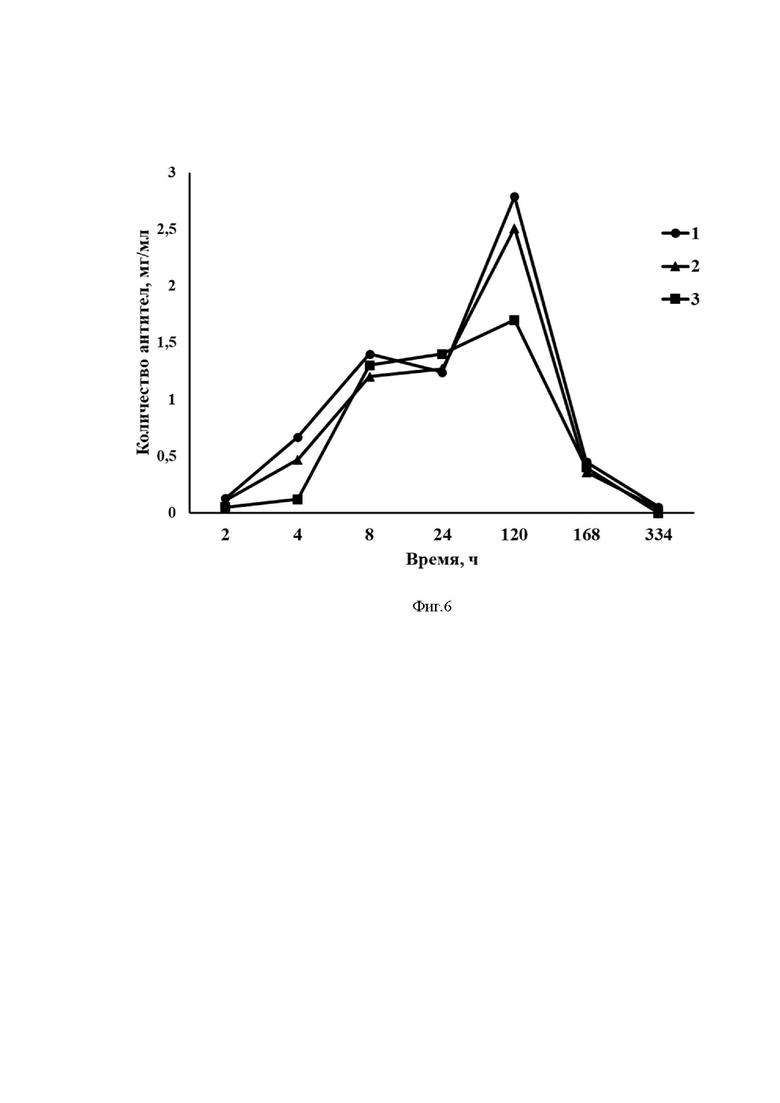
На Фиг. 6 представлены результаты оценки продукции однодоменного антитела против RBD SARS-CoV-2 у мышей после введения разработанного вектора на основе мРНК.
Ось ординат - количество антител, мкг/мл
Ось абсцисс - экспериментальные группы.
1) mRNA-cas-C5-glo, 10 мкг/мышь
2) mRNA-hsp-C5-glo, 10 мкг/мышь
3) mRNA-tpl-C5-glo, 10 мкг/мышь
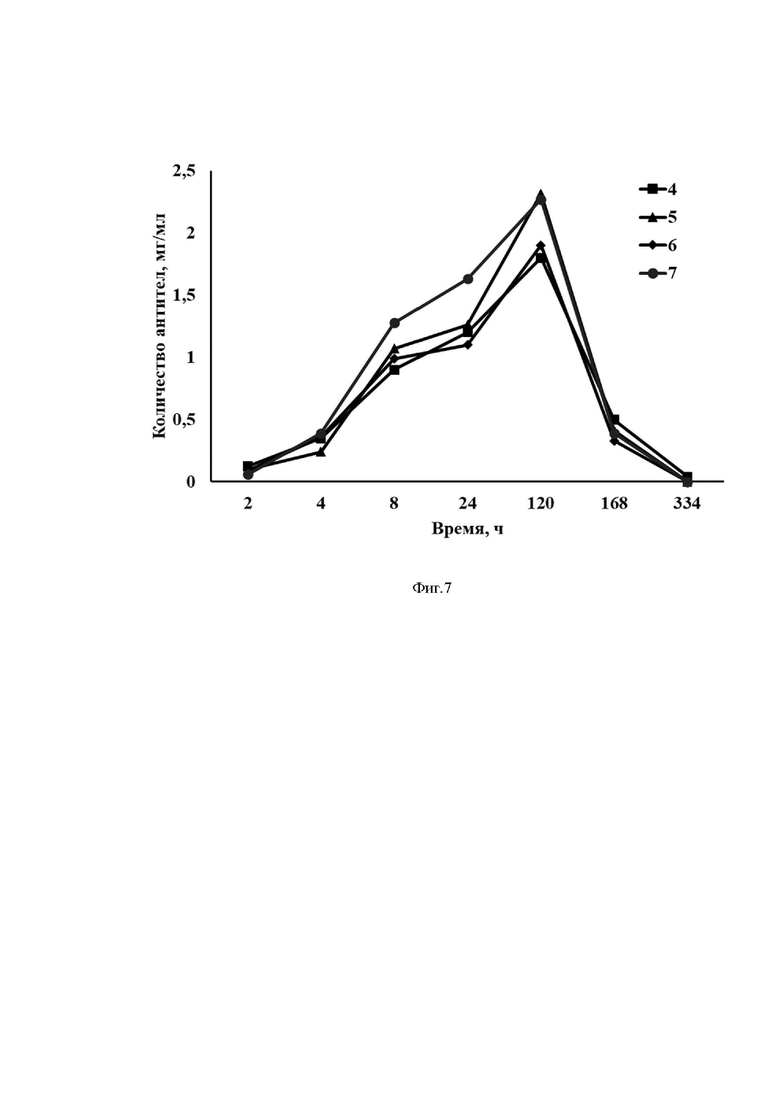
На Фиг. 7 представлены результаты оценки продукции однодоменного антитела против RBD SARS-CoV-2 у мышей после введения разработанного вектора на основе мРНК.
Ось ординат - количество антител, мкг/мл
Ось абсцисс - экспериментальные группы.
4) mRNA-his-C5-glo, 10 мкг/мышь
5) mRNA-cas-C5-cyt, 10 мкг/мышь
6) mRNA-hsp-C5-cyt, 10 мкг/мышь
7) mRNA-tpl-C5-cyt. 10 мкг/мышь
Перечень последовательностей, поясняющих сущность изобретения
SEQ ID NO:1 последовательность рибонулеиновой кислоты, представляющая собой 5'-НТО из мРНК каспазы 8 человека;
SEQ ID NO:2 последовательность рибонулеиновой кислоты, представляющая собой 5'-НТО из мРНК белка теплового шока HSP70;
SEQ ID NO:3 последовательность рибонулеиновой кислоты, представляющая собой 5'-НТО из поздних аденовирусных РНК (TPL);
SEQ ID NO:4 последовательность рибонулеиновой кислоты, представляющая собой 5'-НТО на основе мРНК гистона h1.2 человека;
SEQ ID NO:5 последовательность рибонулеиновой кислоты, представляющая собой 3'-НТО из мРНК альфа-субъединицы гемоглобина;
SEQ ID NO:6 последовательность рибонулеиновой кислоты, представляющая собой мРНК альфа-цепи цитохрома b-245.
Матричная РНК (мРНК, информационная РНК) является промежуточным звеном в потоке генетической информации от гена к белку. Каждая встречающаяся в природе мРНК имеет свой индивидуальный период полураспада. Различные скорости деградации природных мРНК являются одним из инструментов регуляции экспрессии генов. Нестабильные мРНК обеспечивают временную экспрессию гена в различные моменты времени, тогда как долгоживущие мРНК могут быть связаны с непрерывной экспрессией генов или накоплением отдельных белков.
Для синтетических мРНК, которые используются в медицинских целях, стабильность и эффективность трансляции имеют первостепенное значение, поскольку данные характеристики существенно влияют на фармакокинетические и фармакодинамические свойства лекарственных средств. Синтетическая мРНК должна сохранять свою структурную целостность и оставаться функциональной в процессе производства, хранения и использования лекарственных средств. Кроме того, она должна обеспечивать высокую эффективность продукции целевого белка in vivo.
При конструировании синтетических мРНК используют природные механизмы регуляции генов. Было обнаружено, что встречающиеся в природе молекулы мРНК эукариот содержат характерные регуляторные элементы. Например, только при секвенировании мРНК дрожжей было обнаружено около 560 стабилизирующих и 851 дестабилизирующих элементов [Geisberg JV, Moqtaderi Z, Fan X, Ozsolak F, Struhl K. Global analysis of mRNA isoform half-lives reveals stabilizing and destabilizing elements in yeast. Cell. 2014 Feb 13;156(4):812-24. doi: 10.1016/j.cell.2013.12.026. PMID: 24529382; PMCID: PMC3939777.].
Для получения вектора на основе мРНК для увеличенной продукции целевого белка была разработана синтетическая мРНК-конструкция. Под увеличением продукции целевого белка авторы подразумевают его наработку в количествах, превышающих те, которые способен обеспечить аналог. Синтетическая мРНК сконструирована по принципу природной мРНК и содержит следующие основные элементы:
1) 5'-кэп. У подавляющего большинства эукариот 5'-конец транскриптов, синтезируемых РНК-полимеразой II, во время транскрипции модифицируется путем присоединения 7-метилгуанозина, который называется 5'кэп. Данная структура обеспечивает эффективную трансляцию мРНК, направляет сплайсинг пре-мРНК и экспорт мРНК из ядра, ограничивает деградацию мРНК клеточными 5'→3' экзонуклеазами и позволяет распознавать чужеродные РНК. Минимальной кэпирующей структурой является m7GpppNp (7-метилгуанозин, соединенный 5',5'-трифосфатным мостиком с первым нуклеотидом РНК), который также называют кэп 0. Данная структура характерна для мРНК растений и грибов. Для мРНК животных более характерен кэп 1 (отличается от кэпа 0 метилированием первого нуклеотида транскрипта по 2'-O-положению рибозы) и кэп 2 (отличается от кэпа 0 метилированием первого и второго нуклеотидов транскрипта по 2'-O-положению рибозы).
Альтернативой 5'-кэпу для привлечения трансляционного аппарата могут служить кэп-независимые вирусные последовательности, такие как, например, 5'-нетранслируемая область вируса табачной мозаики (омега-последовательность) или внутренние сайты посадки рибосомы (IRES).
В данном изобретении авторы использовали кэп 1.
2) 5'-нетранслируемая область (5'-НТО, синонимы: 5'-untranslated region, 5'-UTR). Известно, что НТО играют важную роль в посттранскрипционной регуляции экспрессии генов, включая модуляцию транспорта мРНК из ядра, эффективность трансляции, субклеточную локализацию и стабильность мРНК.
Посттранскрипционная регуляция экспрессии генов связана в том числе с первым этапом инициации трансляции - образованием на мРНК преинициаторного комплекса 48S (48S PIC). У подавляющего большинства эукариотических мРНК эта стадия связана с механизмом кэп-зависимого сканирования. Считается, что оно состоит из трех этапов:
- присоединения к кэпированному 5'-концу мРНК преинициаторного комплекса 43S, состоящего из малой субчастицы рибосомы, инициаторной Мет-тРНК и факторов инициации трансляции eIF1, eIF1A, eIF2, eIF3 и eIF5;
- сканирования мРНК в направлении 5'→3' в поисках стартового кодона при участии факторов группы eIF4;
- узнавания стартового кодона, остановки сканирования и диссоциации факторов.
Стартовым кодоном часто является триплет AUG, ближайший к 5'-концу. В процессе узнавания стартового кодона важную роль играет его окружение. Первый AUG может быть пропущен, если он окружен неблагоприятной последовательностью - процесс, называемый “пропускающее сканирование”. Консенсусная последовательность Козак, играющая важную роль в инициации трансляции у эукариот, включает 3-6 нуклеотидов, предшествующих старт-кодону, и один-два нуклеотида непосредственно после старт-кодона. Считается, что у млекопитающих оптимальный контекст стартового кодона имеет вид GCCRCCAUGG.
Вторичная структура 5'-НТО также может влиять на эффективность инициации. Существует мнение, что необходимым условием для эффективной трансляции является отсутствие стабильных элементов вторичной структуры внутри 5'-НТО. Однако было показано, что фактор eIF4F совместно с фактором eIF4B обеспечивает эффективное сканирование лидеров, имеющих вторичную структуру умеренной стабильности.
Важным фактором, определяющим эффективность трансляции, также является длина 5'-НТО. Обнаружено, что удлинение 5'-НТО не приводит к снижению эффективности трансляции, однако чрезмерно короткий 5'-НТО (<20 нт) может провоцировать пропускающее сканирование. Поэтому мРНК с короткими 5'-НТО, как правило, показывают эффективность трансляции ниже среднего.
В 2008 году был открыт регуляторный элемент - Инициатор трансляции на коротких 5'-НТО, который присутствует в значительном количестве генов с основными клеточными функциями и очень короткой 5'-НТО. Данный регуляторный элемент обеспечивает стабильную эффективную кэп-зависимую инициацию, когда внутриклеточные уровни eIF1 и eIF4A подвергаются колебаниям.
В данном изобретении для увеличения эффективности продукции целевого белка авторы использовали 5'-НТО, взятые из мРНК каспазы 8 человека (SEQ ID NO:1), или мРНК белка теплового шока (HSP70) (SEQ ID NO:2), или поздних аденовирусных РНК (TPL) (SEQ ID NO:3), или мРНК полученную на основе мРНК гистона h1.2 человека (SEQ ID NO:4).
3) Последовательность, кодирующую целевой белок. Вектор на основе мРНК, являющийся предметом настоящего изобретения, может содержать любую открытую рамку считывания (ORF). В этом качестве может быть использована последовательность, кодирующая антиген патогена человека. Например, последовательность, кодирующая S-белок вируса SARS-CoV-2. Также в качестве последовательности, кодирующей целевой белок, может быть использована последовательность, кодирующая антитело. Например, последовательность, кодирующая однодоменное антитело против RBD S-белка вируса SARS-CoV-2. Кроме того, в качестве последовательности, кодирующей целевой белок, может быть использована последовательность, кодирующая терапевтический белок. Например, последовательность, кодирующая однодоменное антитело против ботулинического нейротоксина. Также в качестве последовательности, кодирующей целевой белок, может быть использована последовательность, кодирующая фермент. Например, последовательность, кодирующая люциферазу.
4) 3'-нетранслируемая область (3'-НТО, 3'-untranslated region, 3'-UTR). Известно, что 3'-НТО играют решающую роль в посттранскрипционной регуляции экспрессии генов. В мРНК млекопитающих данная область зачастую содержит множество регуляторных цис-действующих элементов, которые специфично распознаются РНК-связывающими белками или микроРНК. Их взаимодействие имеет решающее значение для посттранскрипционной регуляции экспрессии генов.
Наиболее распространенными регуляторными последовательностями в 3'-НТО мРНК являются AU-богатые регуляторные элементы (AREs). Эти элементы имеют длину от 50 до 150 нуклеотидов и обычно содержат многочисленные копии последовательности AUUUA. Показано, что они влияют на стабильность мРНК, альтернативный процессинг пре-мРНК и эффективность трансляции. Клеточные факторы, которые связываются с данными элементами, называются ARE-связывающие белки (ARE-BP). Большинство известных ARE-BP (например, TTP, BRF1, KSRP) стимулируют деаденилирование и тем самым способствуют дестабилизации и деградации мРНК. Однако, некоторые ARE-BP, например, HuR, ELAV, могут, наоборот, стабилизировать мРНК. Конечный эффект на мРНК сильно зависит от ARE-BP и от их конкуренции в пределах одного и того же транскрипта, а также от состояния клетки, что может привести либо к стабилизации мРНК и усилению трансляции, либо, наоборот, к дестабилизации мРНК и репрессии трансляции.
Около 5% мРНК человека содержат в 3'-НТО GU-богатые регуляторные элементы (GRE). Они напоминают ARE по характеру мотива базовой последовательности, однако минимальная функциональная длина элемента составляет 11 нуклеотидов (UGUUUGUUUGU). Наблюдается некоторое функциональное сходство GRE с ARE. Данные элементы способствуют регуляции посттранскрипционных событий, таких как деаденилирование, распад мРНК и сплайсинг пре-мРНК.
Другими регуляторными элементами, встречающимися в 3'-НТО, являются CU-богатые элементы (CURE). Одним из широко охарактеризованных факторов, взаимодействующих с CURE, является белок, связывающий полипиримидиновый тракт (PTB). PTB может связываться с удаленными участками в одной и той же молекуле РНК и, таким образом, может модулировать ее вторичную структуру. Например, в случае вирусных IRES-элементов PTB участвует в стабилизации правильной ориентации петель РНК-доменов, критически важных для привлечения трансляционных факторов и рибосом, а в случае некоторых клеточных пре-мРНК он обеспечивает взаимодействие с факторами, необходимыми для альтернативного сплайсинга.
CA-богатые элементы (CAREs) оказывают стабилизирующее действие на мРНК. Основным партнером, взаимодействующим с CARE является белок hnRNP L, который изменяет восприимчивость мРНК к деградации эндо- или экзо-нуклеазами. В 3'-НТО отдельных мРНК могут встречаться особые, характерные для них регуляторные элементы например, элементы, чувствительные к концентрации ионов железа; элемент, необходимый для вставки селеноцистеина и др.
Кроме того, 3'-НТО содержит последовательности, комплементарные микроРНК. Это особые регуляторные молекулы РНК, взаимодействие с которыми подавляет трансляцию мРНК. Зрелая микроРНК является частью активного РНК-индуцируемого комплекса выключения гена (RISC). Центральную роль в функционировании RISC играют белки семейства Argonaute. Некоторые из них могут непосредственно разрезать транскрипт-мишень, другие привлекают дополнительные белки для осуществления репрессии трансляции.
Вторичная структура 3'-НТО также имеет большое значение для трансляции. Модуляция вторичной структуры 3'-НТО при помощи белков или иных средств может изменять ее специфичность к связыванию с различными транс-факторами, тем самым регулируя экспрессию генов на посттранскрипционном уровне.
Результат коллективного эффекта всех факторов, связанных с 3'-НТО одной и той же мРНК, зависит от многих обстоятельств, таких как их сродство к сайтам связывания, представленность и локализация в клетке, которые могут варьировать в зависимости от физиологического состояния.
В данном изобретении для увеличения эффективности продукции целевого белка авторы использовали 3'-НТО, взятый из мРНК альфа-субъединицы гемоглобина (SEQ ID NO:5) или мРНК альфа-цепи цитохрома b-245 (SEQ ID NO:6).
5) Поли(А)-хвост (poly(A)-tail). Это регуляторная последовательность, состоящая в основном из аденозиновых нуклеотидов. Длина данной последовательности влияет на время полураспада молекулы мРНК и может регулироваться рядом ферментов. Поли(А)-хвост имеет важное значение для транспорта мРНК из ядра, ее трансляции и стабильности.
Поли(А)-хвост часто находится в комплексе с поли(А)-связывающим белком (РАВР), молекулы которого выполняют сразу несколько функций. РАВР содержит четыре РНК-связывающих домена, и его комплекс с поли(А)-хвостом не только предохраняет мРНК от деградации, но и выступает в качестве энхансера инициации трансляции и терминации. РАВР может взаимодействовать с 5'-концом мРНК через факторы инициации трансляции, такие как eIF4G и eIF4E, что, в свою очередь, регулирует ее стабильность и эффективность трансляции. Однако PABP также может связываться с комплексами аденилирования или участвовать в процессе ингибирования трансляции, опосредованном микроРНК. Противоречивая функция PABP указывает на то, что различная длина последовательности поли(A) может по-разному влиять на эффективность трансляции мРНК.
В данном изобретении авторы использовали поли(А)-хвост, состоящий из 30 остатков аденозина, 10-тинуклеотидной линкерной последовательности и еще 70 остатков аденозина.
Пример 1. Получение молекул мРНК.
На первом этапе работы было необходимо разработать дизайн мРНК. Синтетическая мРНК содержит следующие элементы:
1) Котранскрипционно встраиваемый кэп-аналог (cap analog). В данном изобретении авторы использовали кэп 1.
2) 5'-нетранслируемая область (5'-НТО, 5'-untranslated region, 5'-UTR). Было показано, что данная область имеет ключевое значение для регуляции экспрессии генов на посттранскрипционном уровне. В ходе работы было получено несколько вариантов вектора на основе мРНК, в которых использовался один из следующих элементов:
- 5'-НТО из мРНК каспазы 8 человека (SEQ ID NO:1);
- 5'-НТО из мРНК белка теплового шока HSP70 (SEQ ID NO:2);
- 5'-НТО из поздних аденовирусных РНК (TPL) (SEQ ID NO:3);
- 5'-НТО на основе мРНК гистона h1.2 человека (SEQ ID NO:4).
3) Область, кодирующая целевой белок. Разработанный вектор на основе мРНК может использоваться для разработки средств профилактики заболеваний, вызываемых инфекционными агентами, а также терапевтических средств. К инфекционным агентам могут относиться в том числе патогены, являющиеся триггерами развития опухолевых заболеваний. В частном случае область, кодирующая целевой белок, может представлять собой последовательность, кодирующую вакцинный антиген, антитело, терапевтический белок или фермент. Для примера авторами были получены векторы на основе мРНК, содержащие:
- последовательность, кодирующую S-белок вируса SARS-CoV-2 (вакцинный антиген),
- последовательность, кодирующую однодоменное антитело против RBD S-белка вируса SARS-CoV-2 (антитело),
- последовательность, кодирующую однодоменное антитело против ботулинического нейротоксина (для терапии ботулизма),
- последовательность, кодирующую люциферазу (фермент).
4) 3'-нетранслируемая область (3'-НТО, 3'-untranslated region, 3'-UTR). Данный элемент играет важную роль в стабилизации мРНК и регуляции трансляции. Для получения вариантов вектора на основе мРНК для увеличенной продукции целевого белка использовали один из следующих элементов:
- 3'-НТО из мРНК альфа-субъединицы гемоглобина (SEQ ID NO:5);
- 3'-НТО из мРНК альфа-цепи цитохрома b-245 (SEQ ID NO:6).
5) Поли(А)-хвост (poly(A)-tail). Авторы использовали поли(А)-хвост, состоящий из 30 остатков аденозина, 10-тинуклеотидной линкерной последовательности и еще 70 остатков аденозина.
Таким образом, был осуществлен дизайн 28 векторов:
1. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-S-glo, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
2. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-S-glo, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
3. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-S-glo, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
4. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-his-S-glo, содержащий 5'-нетраслируемую область SEQ ID NO:4, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
5. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-S- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO: 6.
6. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-S- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO: 6.
7. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-S- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую S белок SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:6.
8. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
9. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
10. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
11. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-his-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:4, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:5.
12. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO: 1.
13. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO: 6.
14. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело C5 против RBD SARS-CoV-2 и 3'-нетраслируемую область SEQ ID NO:6.
15. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-B11-glo, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:5.
16. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-B11-glo, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:5.
17. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-B11-glo, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:5.
18. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-his-B11-glo, содержащий 5'-нетраслируемую область SEQ ID NO:4, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:5.
19. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-B11-cyt, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:6.
20. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-B11-cyt, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:6.
21. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-B11- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело B11 против ботулинического нейротоксина и 3'-нетраслируемую область SEQ ID NO:6.
22. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-luc-glo, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:5.
23. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-luc-glo, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:5.
24. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-luc-glo, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:5.
25. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-his-luc-glo, содержащий 5'-нетраслируемую область SEQ ID NO:4, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:5.
26. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-luc-cyt, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:6.
27. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-luc-cyt, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:6.
28. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-luc- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую люциферазу и 3'-нетраслируемую область SEQ ID NO:6.
На основании дизайна мРНК была разработана последовательность матричной ДНК, которая затем была получена с помощью методов генной инженерии. Далее полученная матричная ДНК, которая представляла собой плазмиду, была линеаризована с использованием рестриктазы BsmBI-v2 (New England Biolabs) при температуре 55°C в течение 3 часов. После этого матричную ДНК очищали из реакционной смеси осаждением смесью фенола, хлороформа и изоамилового спирта (25:24:1), насыщенной 20 мМ Трис-HCl рН 7.9 (ФХ).
Далее проводили реакцию транскрипции in vitro. Для этого в пробирку добавляли: ДНК-матрицу, буфер для T7-транскрипции, ДТТ, смесь рНТФ, T7 РНК-полимеразу, ингибитор РНКаз, стерильную воду. Далее проводили инкубацию в течение 1,5 часов при температуре 37°С. Для удаления ДНК-матрицы в реакционную смесь добавляли ДНКазу (из расчета 1 е.а. на 1 мкг ДНК-матрицы) и инкубировали 10 минут при температуре 37°С. Очистку мРНК из реакционной смеси проводили путем ВЭЖХ. Кэпирование проводили ко-транскрипционно.
Таким образом, в результате проведенной работы было получено 28 вариантов вектора на основе мРНК для увеличенной продукции целевого белка. Кроме того, аналогичным способом были получены 4 контрольные молекулы мРНК, сконструированные по аналогии с прототипом и содержащие последовательности, кодирующие целевые белки (люциферазу, или S-белок вируса SARS-CoV-2, или однодоменное антитело против RBD S-белка вируса SARS-CoV-2, или однодоменное антитело против ботулинического нейротоксина).
Пример 2. Упаковка молекул мРНК в липидные частицы.
Препараты мРНК, получение которых описано в примере 1 инкапсулировали в липидные наночастицы (ЛНЧ) с использованием процесса микрофлюидного смешивания раствора мРНК (pH 3,0) с раствором липидов, растворенных в спирте. Для этого липиды растворяли в 96 % этаноле при молярных соотношениях 46,3:9:42,7:1,6 (ионизируемый липид (ALC-0315): дистеароилфосфатидилхолин: холестерин: пегилированный липид (ПЭГ-липид)). Раствор липидов объединяли с 10 мМ цитратного буфера (pH 3,0), содержащим мРНК (0,2 мг/мл) в объемном соотношении 3:1 (водный раствор:этанол), используя микрожидкостное смешивание с помощью Nanoassmblr Benchtop (Precision NanoSystems). Отношение ионизируемых атомов азота в ионизируемом липиде к количеству фосфатных групп в мРНК (соотношение N:P) равно 6 для каждой композиции. Препараты подвергали диализу против раствора PBS (фосфатно-солевой буферный раствор), pH 7,2, в диализных кассетах Slide-A-Lyzer (Thermo Fisher Scientific) в течение не менее 24 часов. Полученные формуляции затем стерилизовали через фильтр 0,22 мкм и хранили при 4°C (PBS) до использования. Размер и дзета-потенциал частиц измеряли с помощью динамического светорассеяния с использованием прибора Zetasizer Nano ZS (Malvern Panalytical). Инкапсуляция мРНК составляла около 90% (измерено с помощью анализа RiboGreen - Quant-iT™ RiboGreen™ RNA Reagent, Thermo Fisher Scientific).
В результате проведенной работы было получено 32 препарата липидных частиц, содержащих мРНК (28 экспериментальных образцов и 4 контрольных образца). Полученные липидные частицы использовались для доставки мРНК в клетки млекопитающих.
Пример 3. Оценка стабильности липидных частиц, содержащих разработанный вектор на основе мРНК.
Целью данного эксперимента являлась оценка стабильности полученных липидных частиц, содержащих разработанный вектор на основе мРНК, при хранении в холодильнике в диапазоне температур от +4°С до +8°С.
Для эксперимента было выбрано по две аликвоты от каждого из 28 экспериментальных образцов полученных липидных частиц, содержащих разработанный вектор на основе мРНК. Одна аликвота использовалась для оценки уровня продукции целевого белка, которую определяли in vitro на клетках HeLa. Другую аликвоту поместили в холодильник, который поддерживает температуру в диапазоне от +4°С до +8°С. Через 3 месяца данные аликвоты также использовали для оценки уровня продукции целевого белка in vitro на клетках HeLa в аналогичных условиях.
Результаты эксперимента показали, что хранение аликвот липидных частиц, содержащих разработанный вектор на основе мРНК, в течение 3 месяцев в диапазоне температур +4°С до +8°С не приводило к достоверному снижению уровня продукции целевого белка.
Пример 4. Сравнительная оценка эффективности продукции целевого белка в клетках млекопитающих после добавления вариантов разработанного вектора на основе мРНК и контрольного препарата.
Клетки карциномы шейки матки человека HeLa выращивали в культуральной среде DMEM (Gibco), 10% FBS (Sigma-Aldrich) с добавлением 2 мМ L-глутамина (Gibco) и смеси антибиотиков (пенициллина/стрептомицина) 1% (Gibco) в инкубаторе в условиях 37°C, 5% CO2.
На следующий день клеточную культуру при конфлюентности ~70% трансфицировали вариантами разработанного вектора на основе мРНК. За 24 ч до трансфекции клетки высевали в 96-луночный планшет из расчета 3*104 клеток/лунку. В день эксперимента меняли среду в планшете на среду для липофекции OptiMEM (Gibco). Далее 30 нг мРНК (один из перечисленных ниже вариантов) в 20 мкл Opti-MEM (Gibco) на лунку смешивали с раствором 0,06 мкл GenJectorTM-U (Molecta) в 3 мкл Opti-MEM (на лунку), инкубировали 15 мин, затем добавляли 0,4 мкл 100 мМ D-люциферина (Promega) на лунку, а затем 23 мкл полученной смеси добавляли к клеткам. В эксперименте были использованы следующие варианты мРНК:
1) mRNA-cas-luc-glo
2) mRNA-hsp-luc-glo
3) mRNA-tpl-luc-glo
4) mRNA-his-luc-glo
5) mRNA-cas-luc-cyt
6) mRNA-hsp-luc-cyt
7) mRNA-tpl-luc-cyt
8) Контрольная мРНК (с последовательностью люциферазы)
9) Отрицательный контроль (буферный раствор).
Измерения люминесценции в режиме реального времени проводились в течение ночи в планшет-ридере CLARIOstar (BMG Labtech), оснащенном блоком управления атмосферой для поддержания 5% CO2, при 37°C (время интегрирования сигнала - 5 с). Все эксперименты повторяли не менее трех раз (в том числе с разными пассажами клеток) и затем рассчитывали средние значения.
В данном примере описан способ использования разработанного вектора для синтеза целевого белка в клетках млекопитающих. Синтез белка происходит внутри клеток млекопитающих, в условиях, обеспечивающих жизнеспособность клеток (например, температура 37оС и 5% СО2) и реализуется благодаря конструктивным особенностям вектора на основе мРНК. Вектор обеспечивает доставку в клетки млекопитающих последовательности РНК, кодирующей белок и обеспечивает ее успешную трансляцию.
Результаты эксперимента представлены на Фиг. 1. Как видно из полученных результатов, уровень экспрессии люциферазы в клетках, к которым добавляли варианты разработанного вектора достоверно выше, чем в контрольных клетках, к которым добавляли прототип.
Таким образом, в результате проведенной работы был разработан вектор на основе мРНК, обеспечивающий увеличенную продукцию целевого белка.
Пример 5. Оценка способности разработанного вектора на основе мРНК, кодирующего фермент люциферазу, обеспечивать синтез целевого белка in vivo.
В данном эксперименте были использованы мыши линии BALB/c весом около 18 г. Животные были разделены на несколько экспериментальных групп, которым вводили:
1) mRNA-cas-luc-glo, упакованная в липидные частицы (пример 2), внутривенно 3,3 мкг/100 мкл;
2) Фосфатный буфер, внутривенно, 100 мкл;
3) mRNA-cas-luc-glo, упакованная в липидные частицы (пример 2), внутримышечно, 3,3 мкг/100 мкл;
4) Фосфатный буфер, внутримышечно, 100 мкл.
Через 3 часа после введения экспериментальных средств мышам внутрибрюшинно вводили 100 мкл раствора D-люциферина в PBS (25 мг/мл). Через 5 минут после инъекции животных анестезировали 1-2% изофлураном и помещали в систему визуализации IVIS Lumina III (Perkin Elmer). Фотографии мышей были обработаны с помощью программного обеспечения Living Image (Perkin Elmer).
В данном примере описан способ использования разработанного вектора для синтеза целевого белка в клетках млекопитающих, отличающийся тем, что синтез целевого белка происходит in vivo. Синтез белка происходит внутри клеток млекопитающих и обеспечивается благодаря конструктивным особенностям вектора на основе мРНК. Вектор обеспечивает доставку в клетки млекопитающих последовательности РНК, кодирующей белок и обеспечивает ее успешную трансляцию in vivo (в условиях живого организма).
В рамках данной работы были исследованы дозы вектора на основе мРНК от 2 мкг/дозу до 5 мкг/дозу, однако специалисту среднего уровня очевидно, что дозы могут отличаться в зависимости от конкретного вида млекопитающих. Нижняя граница диапазона доз определяется уровнем, при котором возможно детектировать экспрессию целевого белка, а верхняя граница диапазона будет определяться экономической целесообразностью и концентрацией, при которой достигается токсичность.
Результаты эксперимента представлены на Фиг. 2 (группы 1,2) и Фиг. 3 (группы 3,4). Как видно из полученных данных, после внутривенного введения вектора на основе мРНК максимальные уровни экспрессии люциферазы визуализируются в верхней части брюшной полости животного, предположительно в районе печени. Тогда как после внутримышечного введения вектора на основе мРНК максимальные уровни экспрессии люциферазы визуализируются в основном в районе места введения и в меньшей степени в верхней части брюшной полости животного.
Таким образом, результаты эксперимента показывают, что разработанный вектор на основе мРНК, обеспечивает эффективную продукцию фермента люциферазы in vivo. Кроме того, можно сделать вывод, что фармакокинетика средства, содержащего разработанный вектор на основе мРНК, может отличаться в зависимости от способа введения.
Пример 6. Оценка способности различных вариантов разработанного вектора на основе мРНК, кодирующего S-белок вируса SARS-CoV-2, индуцировать гуморальный иммунный ответ против вируса SARS-CoV-2 у животных.
В эксперименте использовали мышей линии BALB/c весом 18-20 г. Животные были рандомно разделены на группы по 5 мышей, которым двукратно внутримышечно вводили различные варианты вектора, которые были предварительно упакованы в липидные частицы (пример 2):
1) mRNA-cas-S-glo, 2 мкг/мышь
2) mRNA-hsp-S-glo, 2 мкг/мышь
3) mRNA-tpl-S-glo, 2 мкг/мышь
4) mRNA-his-S-glo, 2 мкг/мышь
5) mRNA-cas-S-cyt, 2 мкг/мышь
6) mRNA-hsp-S-cyt, 2 мкг/мышь
7) mRNA-tpl-S-cyt, 2 мкг/мышь
8) Контрольная мРНК (с последовательность S белка SARS-CoV-2), 2 мкг/мышь
9) mRNA-cas-S-glo, 5 мкг/мышь
10) mRNA-hsp-S-glo, 5 мкг/мышь
11) mRNA-tpl-S-glo, 5 мкг/мышь
12) mRNA-his-S-glo, 5 мкг/мышь
13) mRNA-cas-S-cyt, 5 мкг/мышь
14) mRNA-hsp-S-cyt, 5 мкг/мышь
15) mRNA-tpl-S-cyt, 5 мкг/мышь
16) Контрольная мРНК (с последовательность S белка SARS-CoV-2), 2 мкг/мышь
17) Отрицательный контроль (буферный раствор).
Через три недели у животных отбирали кровь из хвостовой вены и выделяли сыворотку крови. Титр антител определяли методом иммуноферментного анализа (ИФА) по следующему протоколу:
1) Антиген - S-белок вируса SARS-CoV-2 - адсорбировали на лунках 96-луночного планшета для ИФА в течение 16 часов при температуре +4°С.
2) Далее для избавления от неспецифического связывания в планшет добавляли 5%-молоко, растворенное в буфере для блокирования неспецифического сигнала, в объеме 100 мкл на лунку. Инкубировали на шейкере при температуре 37°С на протяжении часа.
3) Образцы сывороток иммунизированных мышей разводили вначале в 100 раз, а затем делали серию двукратных разведений. Всего было приготовлено 12 разведений каждого образца.
4) Добавляли по 50 мкл каждого разведенного образца сыворотки в лунки планшета.
5) Далее проводили инкубацию в течение 1 часа при 37°С.
6) После инкубации проводилась трехкратная промывка лунок фосфатным буфером.
7) Затем добавляли вторичные антитела против IgG мыши, конъюгированные с пероксидазой хрена.
8) Далее проводили инкубацию в течение 1 часа при 37°С.
9) После инкубации проводилась трехкратная промывка лунок фосфатным буфером.
10) Затем добавили раствор тетраметилбензидина (ТМВ), который является субстратом пероксидазы хрена и в результате реакции превращается в окрашенное соединение.
11) Реакцию останавливали через 15 минут добавлением серной кислоты. Далее с помощью спектрофотометра измеряли оптическую плотность раствора (OD) в каждой лунке при длине волны 450 нм.
12) Титр антител определяли как последнее разведение, в котором оптическая плотность раствора была достоверно выше, чем в группе отрицательного контроля. Полученные результаты (среднее геометрическое значение) представлены в таблице 1.
Как видно из результатов эксперимента, двукратное внутримышечное введение всех вариантов разработанного вектора на основе мРНК, кодирующего S-белок вируса SARS-CoV-2, в концентрациях от 2 мкг/мышь до 5 мкг/мышь индуцирует развитие гуморального иммунного ответа.
Пример 7. Определение эффективности иммунизации мышей различными вариантами вектора на основе мРНК, кодирующего S-белок вируса SARS-CoV-2, по оценке доли пролиферирующих лимфоцитов.
Пролиферативный анализ позволяет оценивать способность лимфоцитов усиленно делиться после встречи с антигеном. Для того чтобы оценить пролиферацию, авторы использовали окраску лимфоцитов флуоресцентным красителем CFSE (carboxyfluorescein diacetate succinimidyl ester). Данный краситель связывается с клеточными белками, сохраняется долгое время и при этом никогда не передается соседним клеткам в популяции. Однако флуоресцентная метка передается дочерним клеткам. Концентрация метки, а следовательно, и интенсивность флуоресценции, снижаются ровно в два раза. Поэтому делящиеся клетки легко отслеживать по уменьшению их флуоресценции.
В эксперименте использовались мыши линии C57BL/6. Мыши были рандомно разделены на группы по 3 животных, которым двукратно внутримышечно вводили различные варианты вектора, которые были предварительно упакованы в липидные частицы (пример 2):
1) mRNA-cas-S-glo, 3 мкг/мышь
2) mRNA-hsp-S-glo, 3 мкг/мышь
3) mRNA-tpl-S-glo, 3 мкг/мышь
4) mRNA-his-S-glo, 3 мкг/мышь
5) mRNA-cas-S-cyt, 3 мкг/мышь
6) mRNA-hsp-S-cyt, 3 мкг/мышь
7) mRNA-tpl-S-cyt, 3 мкг/мышь
8) Контрольная мРНК (с последовательность S белка SARS-CoV-2), 3 мкг/мышь
9) Буферный раствор (отрицательный контроль)
Через две недели животных усыпляли. Из селезенки выделяли лимфоциты методом центрифугирования в градиенте фиколла-урографина. Затем выделенные клетки окрашивали CFSE по методике [Quah BJ, Warren HS, Parish CR. Monitoring lymphocyte proliferation in vitro and in vivo with the intracellular fluorescent dye carboxyfluorescein diacetate succinimidyl ester. Nat Protoc. 2007;2(9):2049-56. doi: 10.1038/nprot.2007.296. PMID: 17853860.]и культивировали в присутствии антигена. Далее клетки анализировали методом проточной цитофлуориметрии. На Фиг. 4 представлены результаты оценки эффективности иммунизации разработанным вектором на основе мРНК по оценке доли пролиферирующих CD4+ лимфоцитов в ответ на стимуляцию антигеном. На Фиг. 5 представлены результаты оценки эффективности иммунизации разработанным вектором на основе мРНК по оценке доли пролиферирующих CD8+ лимфоцитов в ответ на стимуляцию антигеном. Как видно из результатов эксперимента, все варианты разработанного вектора на основе мРНК, кодирующего S-белок вируса SARS-CoV-2, в данной дозе эффективно стимулируют пролиферацию лимфоцитов.
Таким образом, можно заключить, что полученные конструкции на основе мРНК индуцируют формирование антиген-специфичного иммунного ответа (как CD4+, так и CD8+).
Пример 8. Оценка экспрессии однодоменных антител против RBD S-белка вируса SARS-CoV-2 у животных после введения вариантов вектора на основе мРНК.
Одним из перспективных подходов к лечению вирусных инфекций является терапия на основе антител. Введение антител одобрено для иммунопрофилактики и лечения вирусных инфекций, вызванных респираторно-синцитиальным вирусом и вирусом Эбола. На сегодняшний день несколько моноклональных антител уже используются для лечения COVID-19 (бамланивимаб плюс этесевимаб, казиривимаб плюс имдевимаб и сотровимаб). Было показано, что введение этих антител негоспитализированным пациентам с симптомами COVID-19 и факторами риска прогрессирования заболевания снижало риск госпитализации и смерти.
Перспективным направлением в данной области является использование однодоменных антител. Небольшой размер и природа данных антител позволяют им связывать эпитопы, недоступные для обычных антител, в частности, вогнутые эпитопы, такие как бороздки и щели. Однодоменные антитела демонстрируют сильное сродство к связыванию, а также очень стабильны в широком диапазоне температур и pH.
Авторами патента были получены варианты вектора на основе мРНК, содержащие область, кодирующую однодоменное антитело против RBD S-белка вируса SARS-CoV-2 (Пример 1):
1. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO:5.
2. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO:5.
3. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO:5.
4. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-his-C5-glo, содержащий 5'-нетраслируемую область SEQ ID NO:4, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO:5.
5. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-cas-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:1, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO: 1.
6. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-hsp-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:2, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO: 6.
7. Вектор на основе мРНК для увеличенной продукции целевого белка mRNA-tpl-C5- cyt, содержащий 5'-нетраслируемую область SEQ ID NO:3, область, кодирующую однодоменное антитело C5 против RBD S-белка вируса SARS-CoV-2, и 3'-нетраслируемую область SEQ ID NO:6.
Для оценки экспрессии однодоменного антитела против RBD S-белка вируса SARS-CoV-2 проводили эксперимент с использованием мышей линии BALB/c, самки 16-18 г. Животные были разделены на 8 групп, которым внутримышечно вводили один из 7 вариантов разработанного вектора на основе мРНК, кодирующего однодоменное антитело (10 мкг/мышь), упакованного в липидные частицы (пример 2) или буферный раствор (отрицательный контроль). Через 2 часа, 4 часа, 8 часов, 24 часа, 5 дней (120 часов), 7 дней (168 часов), 14 дней (334 часов) осуществляли отбор периферической крови из хвостовой вены. Кровь инкубировали 30 мин при 37°С, центрифугировали 10 мин при 1100 об/мин для отделения сыворотки.
Полученные сыворотки анализировали с помощью ИФА с использованием тест-системы SARS-CoV-2 FRT (производства Медгамал).
Сыворотки титровали в буфере для разведения, параллельно титровали антитело p2c5-Fc для построения калибровочной кривой в тех же условиях. Раститрованные сыворотки и антитело вносили в планшеты и инкубировали 1 час 37°С при 350 об/мин на планшетном термошейкере, далее промывали 4 раза промывочным буфером (ФСБТ с 0,05% твин-20), вносили конъюгат "polyclonal goat anti-human IgG HRP-conjugated antibodies" (MilliporeSigma, Burlington, MA, USA) в разведении 1:20000 в буфере для разведения и инкубировали 1 час на 37°С при 350 об/мин, промывали 4 раза промывочным буфером и вносили по 100 мкл однокомпонентный ТМБ "НВО Иммунотех", инкубировали 15 мин. Реакцию останавливали 1М раствором серной кислоты и измеряли оптическую плотность на планшетном спектрофотометре при длине волны 450 нм. Далее по калибровочной кривой определяли концентрацию однодоменного антитела в крови животных в экспериментальных группах.
Результаты эксперимента представлены на Фиг. 6, 7.
Как видно из представленных результатов, после внутримышечного введения всех десяти вариантов разработанного вектора на основе мРНК у животных выработались однодоменные антитела против RBD SARS-CoV-2.
Пример 9. Оценка терапевтических свойств различных вариантов разработанного вектора на основе мРНК, кодирующего однодоменное антитело против ботулинического нейротоксина.
Ботулинический нейротоксин вырабатывается анаэробной грамположительной бактерией Clostridium botulinum. Это сильнейший из известных науке органических токсинов и одно из самых ядовитых веществ в мире. Основной механизм действия ботулинического нейротоксина реализуется на пресинаптическом уровне через торможение высвобождения ацетилхолина в терминалях холинэргических нейронов, что приводит к периферическому нейропараличу скелетной и автономной нервной систем.
Вакцинация хотя и является эффективным методом борьбы с ботулизмом, остается нежелательной стратегией из-за широкого терапевтического использования ботулинического нейротоксина при различных патологических состояниях. Поэтому актуальным направлением является разработка различных терапевтических средств против ботулизма.
Целью данного эксперимента является оценка терапевтических свойств различных вариантов разработанного вектора на основе мРНК, кодирующего однодоменное антитело против ботулинического нейротоксина.
В эксперименте использовали мышей линии BALB/c весом 18-20 г. Животным вводили ботулинический нейротоксин (BoNT/A) от Clostridium botulinum штамма A98 в дозе 5LD50.
Далее животные были разделены на несколько экспериментальных групп (по 3 мыши в группе), которым однократно внутримышечно вводили:
1) mRNA-cas-B11-glo
2) mRNA-hsp-B11-glo
3) mRNA-tpl-B11-glo
4) mRNA-his-B11-glo
5) mRNA-cas-B11-cyt
6) mRNA-hsp-B11-cyt
7) mRNA-tpl-B11-cyt
8) Буферный раствор
Мышей контролировали на наличие клинических симптомов с интервалами - 0, 4, 8, 26 и 48 часов после заражения. Оценку клинических симптомов проводили по шкале: 0 баллов (отсутствие симптомов), 1 балл (легкие симптомы), 2 балла (умеренные симптомы), 3 балла (тяжелые симптомы = гуманная конечная точка). Время смерти определяли как время, когда мышь была найдена мертвой или была подвергнута эвтаназии в конечной точке. Результаты эксперимента приведены в Таблице 2.
Как видно из результатов эксперимента, все варианты разработанного вектора на основе мРНК, кодирующего однодоменное антитело В11, оказывают терапевтический эффект и предотвращают смерть животных от ботулинического нейротоксина.
Промышленная применимость
Все приведенные примеры подтверждают эффективность всех вариантов векторов на основе мРНК и возможность их применения для создания профилактических и терапевтических средств, а также их промышленную применимость.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3" fileName="вектор на основе
мРНК.xml" softwareName="WIPO Sequence" softwareVersion="2.1.2"
productionDate="2022-10-21">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2022-10-21</ApplicationNumberText>
<FilingDate>2022-10-21</FilingDate>
</ApplicationIdentification>
<ApplicantFileReference>2022-10-21</ApplicantFileReference>
<ApplicantName languageCode="ru">федеральное государственное
бюджетное учреждение "Национальный исследовательский центр
эпидемиологии и микробиологии имени почетного академика
Н.Ф.Гамалеи" Министерства здравоохранения Российской
Федерации</ApplicantName>
<ApplicantNameLatin>The National Research Center for Epidemiology
and Microbiology named after Honorary Academician N.F. Gamaleya of
the Ministry of Health of the Russian Federation</ApplicantNameLatin>
<InventionTitle languageCode="ru">Вектор на основе мРНК для
увеличенной продукции целевого белка в клетках млекопитающих
(варианты)</InventionTitle>
<SequenceTotalQuantity>6</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>208</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..208</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q7">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>tgctctgagtttttggtttctgtttcaccttgtgtctgagctggtctga
aggctggttgttcagactgagcttcctgcctgcctgtaccccgccaacagcttcagaagaaggagcagcc
cctgggtgcgtccactttctgggcacgtgaggttgggccttggccgcctgagcccttgagttggtcactt
gaaccttgggaatattgag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>214</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..214</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q8">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>aacggctagcctgaggagctgctgcgacagtccactacctttttcgaga
gtgactcccgttgtcccaaggcttcccagagcgaacctgtgcggctgcaggcaccggcgcgtcgagtttc
cggcgtccggaaggaccgagctcttctcgcggatccagtgttccgtttccagcccccaatctcagagccg
agccgacagagagcagggaaccggc</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="3">
<INSDSeq>
<INSDSeq_length>202</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..202</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q9">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Human adenovirus</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>actctcttcgcatcgctgtctgcgagggccagctgttgggctcgcggtt
gaggacaaactcttcgcggtctttccagtactcttggatcggaaacccgtcggcctccgaacggtactcc
gccaccgagggacctgagcgagtccgcatcgaccggatcggaaaacctctcgagaaaggcgtctaaccag
tcacagtcgcaag</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="4">
<INSDSeq>
<INSDSeq_length>51</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..51</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other RNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q10">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atcggcgctttgccacttgtacccgagtttttgattctcaagatggcgg
ca</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="5">
<INSDSeq>
<INSDSeq_length>111</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..111</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q11">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>gctggagcctcggtggcctagcttcttgccccttgggcctccccccagc
ccctcctccccttcctgcacccgtacccccgtggtctttgaataaagtctgagtgggcggca</INSDSe
q_sequence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="6">
<INSDSeq>
<INSDSeq_length>64</INSDSeq_length>
<INSDSeq_moltype>RNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..64</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>mRNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q6">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>cctcgccccggacctgccctcccgccaggtgcacccacctgcaataaat
gcagcgaagccggga</INSDSeq_sequence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Иммунобиологическое средство для индукции комплексного иммунного ответа против вируса SARS-CoV-2 и вируса гриппа и его применение | 2024 |
|
RU2836687C1 |
| Плазмидная ДНК-кассета pVAX-C3-PolyA, плазмидная ДНК-матрица pVAX-C3-RBD-PolyA для синтеза мРНК и полинуклеотидная вакцина против SARS-CoV-2, представляющая собой молекулы мРНК-C3-RBD-PolyA, индуцирующие SARS-CoV-2-специфические антитела | 2024 |
|
RU2839841C1 |
| Плазмидная ДНК-матрица и молекула мРНК, используемая для создания вакцины против коронавируса, полученная с использованием ДНК-матрицы и обеспечивающая экспрессию гена искусственного полиэпитопного белка-иммуногена, содержащего консервативные эпитопы антигенов вируса SARS-Cov-2 и индуцирующего SARS-Cov-2-специфический иммунитет | 2024 |
|
RU2839372C1 |
| Катионный липид для доставки нуклеиновых кислот в клетки млекопитающих, способ его получения и его применение. | 2024 |
|
RU2836309C1 |
| Экспрессионный вектор на основе аденоассоциированного вируса, несущий гены рекомбинантных антител, и его применение для профилактики заболеваний, вызываемых вирусом гриппа А и вирусом SARS-CoV-2 | 2023 |
|
RU2817792C1 |
| Тяжелоцепочечные моноклональные антитела, специфически связывающиеся с S белком вируса SARS-CoV-2, и способ их применения для терапии заболеваний, вызываемых различными вариантами вируса SARS-CoV-2 | 2024 |
|
RU2836313C1 |
| Экспрессионный вектор на основе аденоассоциированного вируса и способ его применения для экстренной профилактики и профилактики заболеваний, вызываемых вирусом SARS-CoV-2 (варианты) | 2022 |
|
RU2777404C1 |
| Нуклеиновая кислота, содержащая регуляторные элементы TPL, AES и mtRNR1 | 2023 |
|
RU2831163C1 |
| Средство и способ терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 на основе рекомбинантного антитела и гуманизированного моноклонального антитела | 2021 |
|
RU2769223C1 |
| Однодоменное антитело и его модификации, специфически связывающиеся с RBD S белка вируса SARS-CoV-2, и способ их применения для терапии и экстренной профилактики заболеваний, вызываемых вирусом SARS-CoV-2 | 2021 |
|
RU2763001C1 |
Изобретение относится к области биотехнологии, иммунологии и микробиологии, в частности к векторам на основе мРНК, содержащим определенные 5’- и 3’-нетраслируемые области, для увеличенной продукции целевого белка, а также к способу использования упомянутых векторов для синтеза целевого белка в клетках млекопитающих. Изобретение обеспечивает создание вектора на основе мРНК, обладающего стабильностью и обеспечивающего повышенные уровни продукции целевого белка, например вакцинного антигена, антитела, терапевтического белка или фермента. 3 н. и 5 з.п. ф-лы, 7 ил., 2 табл., 9 пр.
1. Вектор на основе мРНК для увеличенной продукции целевого белка, содержащий 5’-нетраслируемую область, выбранную из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 4, область, кодирующую целевой белок, и 3’-нетраслируемую область, выбранную из SEQ ID NO: 5.
2. Вектор на основе мРНК для увеличенной продукции целевого белка, содержащий 5’-нетраслируемую область, выбранную из SEQ ID NO: 1, SEQ ID NO: 2, SEQ ID NO: 3, область, кодирующую целевой белок, и 3’-нетраслируемую область, выбранную из SEQ ID NO: 6.
3. Вектор по п.1 или 2, в котором область, кодирующая целевой белок, представляет собой последовательность, кодирующую вакцинный антиген.
4. Вектор по п.1 или 2, в котором область, кодирующая целевой белок, представляет собой последовательность, кодирующую антитело.
5. Вектор по п.1 или 2, в котором область, кодирующая целевой белок, представляет собой последовательность, кодирующую терапевтический белок.
6. Вектор по п.1 или 2, в котором область, кодирующая целевой белок, представляет собой последовательность, кодирующую фермент.
7. Способ использования вектора по п.1 или 2 для синтеза целевого белка в клетках млекопитающих.
8. Способ использования вектора по п.7, отличающийся тем, что синтез целевого белка происходит in vivo.
| Xia X | |||
| Detailed dissection and critical evaluation of the Pfizer/BioNTech and Moderna mRNA vaccines | |||
| Vaccines (Basel) | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Vol | |||
| Разборный с внутренней печью кипятильник | 1922 |
|
SU9A1 |
| No | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Art | |||
| ИСКУССТВЕННАЯ АКТИВНАЯ ВЕРХНЯЯ КОНЕЧНОСТЬ | 1922 |
|
SU734A1 |
| WO 2021213945 A1, 28.10.2021 | |||
| MATEUS J | |||
| et al | |||
| Способ расщепления слюды | 1924 |
|
SU1273A1 |
| Science | |||
| Способ регенерирования сульфо-кислот, употребленных при гидролизе жиров | 1924 |
|
SU2021A1 |
| Vol | |||
| Устройство для телефонирования по проводам токами высокой частоты | 1921 |
|
SU374A1 |
| No | |||
| МНОГОХОДОВОЙ РАСПРЕДЕЛИТЕЛЬНЫЙ КРАН | 1927 |
|
SU6566A1 |
| Art | |||
| Выключательное приспособление к мотальным машинам | 1927 |
|
SU9853A1 |
| RU | |||

