ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] В данной заявке испрашивается приоритет заявки на патент Китая №201910591280.0, поданной в Национальное управление интеллектуальной собственности Китая 02 июля 2019 г, содержание которой включено во всей полноте путем ссылки.
ОБЛАСТЬ ТЕХНИКИ
[0002] Данное изобретение относится к биологически активному слитому белку с увеличенным временем полужизни in vivo и его конъюгату с полимером, и в частности, относится к человеческому фактору свертывания IX с увеличенным временем полужизни in vivo, содержащей их фармацевтической композиции и их применению в лечении геморрагических заболеваний (таких как гемофилия В).
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
[0003] Гемофилия, тяжелая коагулопатия, вызванная дефицитом определенных факторов свертывания в крови, представляет собой группу наследственных геморрагических заболеваний, включающих гемофилию А (дефицит фактора VIII, антигемофильного глобулина) и гемофилию В (дефицит фактора IX, плазменного компонента тромбопластина).
[0004] Гемофилия В представляет собой тяжелую коагулопатию, вызванную дефицитом человеческого фактора свертывания IX (hFIX). Ген, кодирующий hFIX (hFIX), расположен в области Xq27.1 на дальнем конце длинного плеча X хромосомы человека, его полная длина составляет 34 тпн, включая 8 экзонов и 7 интронов. Полная длина кДНК hFIX составляет 2802 пн, из которых 1383 пн приходится на кодирующую область (регистрационный номер в Genbank: NM_000133). У пациентов с тяжелой форой заболевания активность фактора IX обычно составляет менее 1% от нормы, они часто страдают от спонтанных кровотечений, которые становятся причиной гематом в мышцах или деформации суставов.
[0005] Рекомбинантные композиции для предупреждения и лечения гемофилии получают все большее признание в медицине и среди пациентов благодаря своей безопасности. В случае гемофилии В введение препаратов фактора IX (который в настоящее время обычно представляет себя рекомбинантный белок фактора IX, экспрессируемый in vitro) является единственным эффективным способом лечения для поддержания необходимого уровня фактора свертывания IX в организме пациента. Однако, поскольку пептидные лекарственные средства обычно характеризуются коротким временем полужизни, плохой физической и химической стабильностью и легко разрушаются различными протеазами в организме, эти лекарственные средства требуют многократных инъекций в течения дня, в результате частое введение медикаментов становится большим физическим, психологическим и финансовым бременем для пациентов, ограничивая их приверженность лечению. Таким образом, в данной области существует неудовлетворенная потребность в новых лекарственных средствах для увеличения времени полужизни лекарственного средства в плазме и увеличения его системного воздействия.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
[0006] После многолетних исследований авторы изобретения создали слитый белок фактора свертывания IX и Fc, который для образования конъюгата может быть дополнительно модифицирован группой, включая ПЭГ, и обладает значительно большим временем полужизни в кровотоке по сравнению с полипептидами FIX.
[0007] В частности, в первом аспекте изобретения предложен слитый белок фактора свертывания IX (FIX), содержащий активную группу фактора свертывания IX и партнер по слиянию (FP), способный увеличивать время полужизни слитого белка, где активная группа фактора свертывания IX и партнер по слиянию связаны непосредственно или через первый линкер L1; предпочтительно, первый линкер L1 представляет собой пептид, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот; более предпочтительно, первый линкер L1 содержит гибкий пептид и/или жесткий элемент, и гибкий пептид представляет собой пептид, содержащий глицин (Gly, G), серии (Ser, S), аланин (Ala, А) и/или треонин (Thr, Т), например, гибкий пептид представляет собой (GS)m(GGS)n(GGGS)o(GGGGS)p, где m, n, о и р независимо выбраны из целых чисел от 0 до 50, таких как 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, предпочтительно, не все из m, n, о и р равны 0.
[0008] В одном воплощении данного изобретения активная группа фактора свертывания IX происходит из человеческого фактора свертывания IX, такого как полноразмерный или укороченный фактор свертывания IX; полноразмерный или укороченный человеческий фактор свертывания IX может содержать одну или более аминокислотных мутаций, при условии что он по-прежнему сохраняет активность фактора IX, например, активная группа фактора свертывания IX содержит аминокислотную последовательность, приведенную в SEQ ID NO: 1, или идентичную по меньшей мере на 80%, 85%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или более аминокислотной последовательности, приведенной в SEQ ID NO: 1.
[0009] В другом воплощении жесткий элемент представляет собой пептид, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более пролинов (Pro, Р), предпочтительно, жесткий элемент содержит аминокислотную последовательность, выбранную из группы, состоящей из:
VAPPPALPAPVRLPGPA (SEQ ID NO: 3),
VAPPPALPAVAPPPALPA (SEQ ID NO: 4),
VAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPA (SEQ ID NO: 5),
VAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPA (SEQ ID NO: 6) и
VAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPA (SEQ ID NO: 7).
[0010] Предпочтительно, первый линкер L1 выбран из группы, состоящей из:
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPA (SEQ ID NO: 8),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPA (SEQ ID NO: 9),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPA (SEQ ID NO 10),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPA (SEQ ID NO: 11) и
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPA (SEQ ID NO 12).
[0011] В еще одном воплощении партнер по слиянию представляет собой полноразмерную форму, укороченную форму или вариант Fc-фрагмента иммуноглобулина, альбумина, трансферрина или XTEN; например, партнер по слиянию происходит из Fc-фрагмента человеческого иммуноглобулина; предпочтительно, Fc-фрагмент состоит из одного - четырех доменов, выбранных из группы, состоящей из домена СН1, домена СН2, домена СН3 и домена СН4; предпочтительно, Fc-фрагмент происходит из Fc-фрагмента IgG IgA, IgD, IgE или IgM, более предпочтительно, из Fc-фрагмента IgG; более предпочтительно, Fc-фрагмент происходит из Fc-фрагмента IgG1, IgG2, IgG3 или IgG4, и более предпочтительно, Fc-фрагмент IgG опосредует менее выраженную ADCC (антителозависимую клеточную цитотоксичность) и/или CDC (комплементзависимую цитотоксичность) и/или обладает большей аффинностью связывания с FcRn.
[0012] В еще одном воплощении, когда партнер по слиянию также связан непосредственно или через второй линкер L2 с пептидом Р, содержащим сайт распознавания сортазой, а сортаза представляет собой, например, сортазу А или сортазу В; предпочтительно, Р содержит основной сайт распознавания сортазой A LPXTG, где X представляет собой любую аминокислоту, и последовательность Р представляет собой, например, LPETG, LPETGG или LPETGGG; более предпочтительно, аминокислотная последовательность Р дополнительно содержит аффинную метку, присоединенную к сайту распознавания сортазой, и аминокислотная последовательность Р, например, представляет собой LPETGGHHHHHH или LPETGGWSHPQFEK.
[0013] В другом воплощении второй линкер L2 представляет собой пептид, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот; предпочтительно, второй линкер L2 представляет собой гибкий пептидный фрагмент, содержащий глицин (Gly G), серии (Ser, S), аланин (Ala, А) и/или треонин (Thr, Т), такой как (GS)w(GGS)x(GGGS)y(GGGGS)z, где w, х, у и z независимо выбраны из целых чисел от 0 до 50, таких как 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, предпочтительно, не все из w, х, у и z равны 0; предпочтительно, второй линкер L2 выбран из группы, состоящей из GGGGS (SEQ ID NO: 14), GGGGSGGGGS (SEQ ID NO: 15) и GSGGSGGGGS (SEQ ID NO: 16).
[0014] В другом воплощении слитый белок имеет структуру FLX-L1-FP-L2-P, где FIX, L1, L2, FP и Р имеют такие же определения, как в указанных выше воплощениях, и где один или оба из L1 и L2 могут отсутствовать.
[0015] В еще одном воплощении слитый белок содержит последовательность, приведенную в SEQ ID NO: 18, 20, 22, 24, 26 или 28, или кодируется нуклеиновокислотной последовательностью, приведенной в SEQ ID NO: 19, 21, 23, 25, 27 или 29.
[0016] Кроме того, в данном изобретении также предложена молекула нуклеиновой кислоты, кодирующая слитый белок по любому из указанных выше воплощений, и экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты.
[0017] Во втором аспекте данного изобретения предложен конъюгат, образованный присоединением гидрофильного полимера к концу слитого белка по любому из воплощений первого аспекта. В составе конъюгата один, два или более гидрофильных полимеров присоединены к концу слитого белка по любому из воплощений первого аспекта, предпочтительно, гидрофильный полимер присоединяют к сайту распознавания сортазы посредством реакции транспептидации; в частности, слитый белок может быть в форме мономера или димера.
[0018] В одном воплощении один, два или более гидрофильных полимеров независимо выбраны из полисахарида и полиалкиленгликоля, такого как полипропиленгликоль и полиэтиленгликоль; полиалкиленгликоль может быть кэпирован на конце, например, кэпирован алкоксигруппой, такой как метоксигруппа; и/или полиалкиленгликоль является линейным или разветвленным, например, полиалкиленгликоль является разветвленным, таким как разветвленный полиэтиленгликоль, в частности, разветвленный полиэтиленгликоль, копированный метокси группой; молекулярная масса полиалкиленгликоля может быть 1 или более, 10 или более, 20 или более, 30 или более, 40 или более, 50 или более, 60 или более, 70 или более, 80 или более, 90 или более, 100 или более, 110 или более, 120 или более, 130 или более, 140 или более, 150 или более или 160 или более кДа, например, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 кДа, или иметь значение, находящееся между любыми двумя значениями; и предпочтительно, молекулярная масса гидрофильного полимера составляет 20 кДа или 40 кДа.
[0019] В третьем аспекте данного изобретения предложен способ получения конъюгата по второму аспекту, включающий приведение слитого белка по первому аспекту данного изобретения в контакт с сортазой (такой как сортаза А или сортаза В) и гидрофильным полимером, где один конец гидрофильного полимера имеет аминогруппу, которая может быть амидирована сортазой.
[0020] В одном воплощении N-конец гидрофильного полимера содержит поли-Gly такой как GGGAA.
[0021] В четвертом аспекте данного изобретения предложена фармацевтическая композиция, содержащая эффективное количество слитого белка, описанного в первом аспекте, и/или конъюгата, описанного во втором аспекте, и возможно фармацевтически приемлемый носитель.
[0022] В одном воплощении фармацевтическую композицию применяют для предупреждения и/или лечения геморрагического заболевания, где геморрагическое заболевание предпочтительно выбрано из геморрагических заболеваний у пациентов с врожденным или приобретенным дефицитом FIX и спонтанного или послеоперационного кровотечения у пациентов с гемофилией В.
[0023] В еще одном воплощении фармацевтическая композиция находится в форме жидкого препарата или лиофилизированного препарата.
[0024] В пятом аспекте данного изобретения предложено применение слитого белка, конъюгата или фармацевтической композиции по аспектам, указанным выше, для изготовления лекарственного средства для предупреждения и/или лечения геморрагического заболевания, где геморрагическое заболевание предпочтительно выбрано из геморрагических заболеваний у пациентов с врожденным или приобретенным дефицитом FIX и спонтанного или послеоперационного кровотечения у пациентов с гемофилией В.
[0025] В шестом аспекте данного изобретения предложен способ предупреждения и/или лечения геморрагического заболевания, включающий введение слитого белка, конъюгата или фармацевтической композиции по аспектам, указанным выше, субъекту, которому это необходимо. Геморрагическое заболевание предпочтительно выбрано из геморрагических заболеваний у пациентов с врожденным или приобретенным дефицитом FIX и спонтанного или послеоперационного кровотечения у пациентов с гемофилией В.
[0026] В седьмом аспекте данного изобретения предложен набор, включающий слитый белок, конъюгат или фармацевтическую композицию по аспектам, указанным выше, и возможно инструкции по применению.
[0027] В восьмом и девятом аспектах данного изобретения, соответственно, предложена молекула нуклеиновой кислоты, кодирующая слитый белок по первому аспекту, и экспрессирующий вектор, содержащий молекулу нуклеиновой кислоты.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0028] На Фиг. 1А показана структура бицистронного экспрессирующего вектора FIX/PACE (фурин).
[0029] На Фиг. 1В показана карта плазмиды pcDNA3.1-Hyg-VKGC.
[0030] На Фиг. 2 приведены результаты очистки слитого белка FIX-C1-Fc-L2-P, полученные при помощи электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия (SDS-PAGE) (А) и эксклюзионной высокоэффективной жидкостной хроматографии (SEC-HPLC) (В).
[0031] На Фиг. 3 показаны результаты определения активности слитого белка FIX-L1-FC-L2-P и конъюгата FIX-C1-Fc-L2-P in vitro.
[0032] На Фиг. 4 показаны результаты анализа конъюгата массой 40 кДа методом SEC-HPLC после реакции конъюгации с ПЭГ.
[0033] На Фиг. 5 показаны результаты анализа конъюгата методом SEC-HPLC после очистки и выделения.
[0034] На Фиг. 6 показаны результаты исследования фармакокинетики конъюгата FIX-C1-Fc-L2-PEG на модели гемофилии у мышей, а в таблице ниже показаны Т1/2 (период полувыведения в конечной фазе), Cmax, AUC0-inf (площадь под кривой), MRT (среднее время удерживания) и CL (скорость выведения).
СВЕДЕНИЯ, ПОДТВЕРЖДАЮЩИЕ ВОЗМОЖНОСТЬ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
[0035] Для более ясного понимания поставленных задач, технических решений и преимуществ данного изобретения ниже будет приведено более подробное описание данного изобретения со ссылкой на графические материалы и воплощения. Очевидно, что описанные воплощения являются лишь частью всех возможных воплощений данного изобретения. Все другие воплощения, к которым специалист в области техники придет на основании воплощений данного изобретения без изобретательской активности, будут входить в объем правовой защиты данного изобретения.
Определения
[0036] Фактор свертывания IX, также обозначаемый фактором IX, FIX, представляет собой витамин K-зависимый фактор свертывания, обладающий структурным сходством с фактором VII, протромбином, фактором X и протеином С.
[0037] В данном изобретении термин «активная группа фактора свертывания IX» относится к группе слитого белка, которая проявляет свертывающую активность фактора IX. Термин «фактор свертывания IX» может относиться к природному белку дикого типа (аминокислотная последовательность дикого типа приведена в SEQ ID NO: 1), а референтная кодирующая нуклеиновокислотная последовательность может быть найдена в базе данных Genbank под регистрационным номером: NM_000133, и в частности, приведена в SEQ ID NO: 2), и также охватывает формы его вариантов, например, вариант белка, образованный в результате одной или более (например, 2, 3, 4 или 5) аминокислотных замен, делеций или вставок, при сохранении активности фактора свертывания IX.
[0038] MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAAINKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLT (SEQ ID NO: 1).
[0039] ATGCAGCGCGTGAACATGATCATGGCAGAATCACCAGGCCTCATCACCATCTGCCTTTTAGGATATCTACTCAGTGCTGAATGTACAGTTTTTCTTGATCATGAAAACGCCAACAAAATTCTGAATCGGCCAAAGAGGTATAATTCAGGTAAATTGGAAGAGTTTGTTCAAGGGAACCTTGAGAGAGAATGTATGGAAGAAAAGTGTAGTTTTGAAGAAGCACGAGAAGTTTTTGAAAACACTGAAAGAACAACTGAATTTTGGAAGCAGTATGTTGATGGAGATCAGTGTGAGTCCAATCCATGTTTAAATGGCGGCAGTTGCAAGGATGACATTAATTCCTATGAATGTTGGTGTCCCTTTGGATTTGAAGGAAAGAACTGTGAATTAGATGTAACATGTAACATTAAGAATGGCAGATGCGAGCAGTTTTGTAAAAATAGTGCTGATAACAAGGTGGTTTGCTCCTGTACTGAGGGATATCGACTTGCAGAAAACCAGAAGTCCTGTGAACCAGCAGTGCCATTTCCATGTGGAAGAGTTTCTGTTTCACAAACTTCTAAGCTCACCCGTGCTGAGACTGTTTTTCCTGATGTGGACTATGTAAATTCTACTGAAGCTGAAACCATTTTGGATAACATCACTCAAAGCACCCAATCATTTAATGACTTCACTCGGGTTGTTGGTGGAGAAGATGCCAAACCAGGTCAATTCCCTTGGCAGGTTGTTTTGAATGGTAAAGTTGATGCATTCTGTGGAGGCTCTATCGTTAATGAAAAATGGATTGTAACTGCTGCCCACTGTGTTGAAACTGGTGTTAAAATTACAGTTGTCGCAGGTGAACATAATATTGAGGAGACAGAACATACAGAGCAAAAGCGAAATGTGATTCGAATTATTCCTCACCACAACTACAATGCAGCTATTAATAAGTACAACCATGACATTGCCCTTCTGGAACTGGACGAACCCTTAGTGCTAAACAGCTACGTTACACCTATTTGCATTGCTGACAAGGAATACACGAACATCTTCCTCAAATTTGGATCTGGCTATGTAAGTGGCTGGGGAAGAGTCTTCCACAAAGGGAGATCAGCTTTAGTTCTTCAGTACCTTAGAGTTCCACTTGTTGACCGAGCCACATGTCTTCGATCTACAAAGTTCACCATCTATAACAACATGTTCTGTGCTGGCTTCCATGAAGGAGGTAGAGATTCATGTCAAGGAGATAGTGGGGGACCCCATGTTACTGAAGTGGAAGGGACCAGTTTCTTAACTGGAATTATTAGCTGGGGTGAAGAGTGTGCAATGAAAGGCAAATATGGAATATATACCAAGGTATCCCGGTATGTCAACTGGATTAAGGAAAAAACAAAGCTCACTTAA (SEQ ID NO: 2).
[0040] В одном воплощении вариант белка по меньшей мере на 90% идентичен последовательности SEQ ID NO: 1. В другом воплощении вариант белка по меньшей мере на 95%, например, на 96%, 97%, 98% или 99% идентичен последовательности SEQ ID NO: 1. В данном описании, если не указано иное, указание любого конкретного положения относится к соответствующему положению в SEQ ID NO: 1.
[0041] Данное изобретение не ограничивается описанными в нем последовательностями. Вариантами FIX, например, являются изложенные в US5521070 (где тирозин в первом положении замещен на аланин) и WO2007/135182 (где один или более природных аминокислотных остатков в FIX замещены остатками цистеина). Процитированные документы включены в данное описание во всей полноте путем ссылки. Таким образом, варианты FIX хорошо известны в области техники, и данное изобретение охватывает формы вариантов, которые известны или которые будут созданы или открыты в будущем.
Слитый белок
[0042] Термин «партнер по слиянию», FP, относится к полипептиду, который слит с целевым полипептидом (полипептидом, у которого желательно увеличить время полужизни в кровотоке). Партнер по слиянию может влиять на функциональные свойства слитого белка посредством некоторых различных механизмов, таких как увеличение времени полужизни целевого полипептида in vivo.
[0043] В одном воплощении партнер по слиянию замедляет клиренс FIX in vivo путем взаимодействия с неонатальным Fc рецептором (FcRn). В другом воплощении партнер по слиянию представляет собой Fc-фрагмент иммуноглобулина, альбумин, трансферрин, XTEN или их часть.
[0044] Fc-фрагмент иммуноглобулина безопасен для применения в качестве фармацевтического носителя, поскольку он является биодеградируемым полипептидом, который может метаболизироваться в организме. Кроме того, по сравнению с молекулой целого иммуноглобулина, Fc область иммуноглобулина имеет относительно низкую молекулярную массу, что благоприятно для получения, очистки и продуцирования конъюгата. Поскольку Fc-фрагмент не содержит Fab-фрагмента (его аминокислотная последовательность варьирует в зависимости от подкласса антитела и поэтому является очень гетерогенной), ожидается, что Fc-область иммуноглобулина может значительно повысить гомогенность вещества и обладать низкой антигенностью.
[0045] Термин «Fc-область/фрагмент иммуноглобулина» относится к белку, включающему 2-ую константную область тяжелой цепи (СН2) и 3-ью константную область тяжелой цепи (СН3) иммуноглобулина, но не включающему вариабельные области тяжелой цепи и легкой цепи иммуноглобулина. Также, Fc-фрагмент, используемый в данном изобретении, может дополнительно содержать шарнирную область константной области тяжелой цепи и/или всю или часть 1-й константной области тяжелой цепи (СН1) и/или 1-й константной области легкой цепи (CL1), не включая вариабельные области тяжелой и легкой цепи, при условии, что он обладает физиологической функцией, по существу схожей или превосходящей таковую естественной формы.
[0046] В некоторых воплощениях Fc-фрагмент иммуноглобулина по данному изобретению может содержать 1) СН1 домен, СН2 домен, СН3 домен и СН4 домен; 2) СН1 домен и СН2 домен; 3) СН1 домен и СН3 домен; 4) СН2 домен и СН3 домен; 5) комбинацию одного или более доменов с (всей или частью) шарнирной областью иммуноглобулина; или 6) димер любых доменов константной области тяжелой цепи и константной области легкой цепи.
[0047] Также Fc-фрагмент иммуноглобулина может представлять собой Fc-область, происходящую из IgG, IgA, IgD, IgE и IgM, или быть получен путем их комбинации или быть их гибридом. Предпочтительно, он происходит из IgG или IgM (поскольку они являются двумя наиболее распространенными белками в крови человека). В одном воплощении с учетом того, что антитела IgG имеют большее время полужизни, предпочтителен Fc домен IgG.
[0048] Fc-фрагмент также может быть модифицирован для улучшения других функций, например, для изменения посредством мутаций его способности индуцировать иммунные ответы, такой как способность связываться с комплементом и/или некоторыми Fc-рецепторами, поскольку это не обязательно при применении Fc для увеличения времени полужизни. Известно, что мутации в положениях 234, 235 и 237 Fc-области IgG обычно приводят к уменьшению связывания с рецепторами FcγRI, а также могут приводить к уменьшению связывания с рецепторами FcγRIIa и FcγRIII, но эти мутации не изменяют его связывания с рецепторами FcRn и поэтому могут по-прежнему способствовать длительному времени полужизни в кровотоке благодаря рециркуляции путем эндоцитоза. Предпочтительно, модифицированный Fc-фрагмент IgG слитого белка по данному изобретению содержит одну или более следующих мутаций: мутации L234A, L235E и G237A приводят к уменьшению аффинности к некоторым Fc-рецепторам, a A330S и P331S приводят к уменьшению Clq-опосредованного связывания комплемента.
[0049] В одном воплощении слитый белок фактора свертывания IX дополнительно связан с фрагментом, содержащим сайт распознавания сортазы, т.е. «пептид Р», описанный в данном изобретении.
[0050] Впервые обнаружили, что «сортаза», включающая, в частности, сортазу А и сортазу В, обладает функцией прикрепления бактериальных поверхностных белков к клеточной стенке. Обнаружили, что «сигнал сортировки» поверхностных белков в большинстве случаев имеет три ключевых составляющих: последовательности LPXTG гидрофобную последовательность и хвост из положительно заряженных остатков. Последовательность LPXTG является очень консервативной и может распознаваться сортазой, после чего цистеин (Cys) сортазы действует как нуклеофильная группа, атакуя пептидную связь между треонином и глицином С-концевого мотива LPXTG субстрата (такого как поверхностный белок), вызывая расщепление пептидной связи (ацилирование), что в свою очередь приводит к образованию ацильного промежуточного соединения, и наконец, пептид ковалентно присоединяется к клеточной стенке или субъединице пилей для выполнения функций (деацилирование).
[0051] В одном воплощении данного изобретения сортазу А применяют для сайт-специфического присоединения ПЭГ. В конкретном воплощении используют распознаваемые сортазой А сайты LPETG LPETGG и LPETGGG. В другом конкретном воплощении к сайту распознавания можно добавлять различные аффинные метки, такие как LPETGGHHHHHH, LPETGGWSHPQFEK и т.д.
[0052] В другом воплощении слитый белок фактора свертывания IX по данному изобретению продуцируют рекомбинантными способами, включая, например, экспрессию белка в подходящих прокариотических или эукариотических клетках-хозяевах, и выделяют слитый белок фактора свертывания IX по данному изобретению стандартными способами. Например, нуклеотидная последовательность, кодирующая пептид, может быть вначале синтезирована методами химического синтеза, а затем последовательность может быть клонирована в подходящий экспрессирующий вектор для экспрессии под контролем подходящего промотора. В альтернативном варианте, для получения нуклеотидной последовательности, кодирующей фактор свертывания IX, из фактора свертывания IX дикого типа могут применяться методы мутагенеза, а затем последовательность других элементов для конструирования слитого белка может быть клонирована в подходящий экспрессирующий вектор для экспрессии под контролем подходящего промотора. Эти технологии находятся в рамках компетенции специалиста в области техники, и в области техники существует множество руководств.
[0053] Подходящие эукариотические клетки включают клетки млекопитающих, такие как клетки СНО, COS, НЕK 293, ВНK, SK-Hep и HepG2. Предпочтительно, клетки выращивают в условиях, подходящих для экспрессии слитого белка фактора свертывания IX по данному изобретению. Что касается реагентов и композиций, использованных для получения или выделения слитого белка фактора свертывания IX по данному изобретению, не существует каких-либо определенных ограничений, и можно использовать любую систему, известную в области техники или доступную для приобретения. В предпочтительном воплощении слитый белок фактора свертывания IX получают способами, описанными в области техники.
[0054] Существуют разнообразные экспрессирующие векторы, которые можно использовать для получения слитого белка фактора свертывания IX, которые могут быть выбраны из векторов для экспрессии в эукариотических и прокариотических клетках. Векторы для экспрессии в прокариотических клетках включают, например, плазмиды, такие как pRSET, рЕТ и pBAD, в которых могут применяться промоторы, включающие, например, lac, trc, trp, rесА или araBAD. Векторы для экспрессии в эукариотических клетках включают: (1) векторы, применяющиеся для экспрессии в дрожжах, такие как pAO, pPIC, pYES и рМЕТ, в которых могут применяться такие промоторы, как АОХ1, GAP, GAL1, AUG1 и так далее; (2) векторы, применяющиеся для экспрессии в клетках насекомых, такие как рМТ, pAc[delta], plB, pMIB, рВАС и так далее, в которых могут применяться такие промоторы, как РН, р10, МТ, Ас5, Ор1Е2, gp64, polh и так далее и (3) векторы, применяющиеся для экспрессии в клетках млекопитающих, такие как pSVL, pCMV, pRc/RSV, pcDNA3, pBPV и так далее, и векторы, являющиеся производными вирусных систем, таких как вирус осповакцины, адено-ассоциированный вирус, вирус герпеса, ретровирус и тому подобные, в которых могут применяться такие промоторы, как промоторы CMV, SV40, EF-1, UbC, RSV, ADV. BPV и β-актина. В предпочтительном воплощении слитый белок фактора свертывания IX экспрессируют в прокариотических или эукариотических клеточных системах и используют кодон-оптимизированные последовательности.
[0055] В предпочтительном воплощении последовательность для экспрессии слитого белка фактора свертывания IX содержит пропептид и/или сигнальный пептид для улучшения секреции слитого белка фактора свертывания IX из клетки за ее пределы для выделения и очистки; для улучшения эффективности отщепления пропептида, выхода FIX и должного процессинга производят оптимизацию экспрессирующего вектора; в конкретном воплощении конструируют экспрессирующий вектор FIX/PACE (фурин).
[0056] В другом предпочтительном воплощении последовательность, экспрессирующая слитый белок фактора свертывания IX, не содержит пропептид и/или сигнальный пептид, и вместо секреции за пределы клетки для выделения и очистки слитый белок фактора свертывания IX получают посредством лизиса клетки.
[0057] В одном воплощении в слитом белке по данному изобретению белок фактора свертывания IX и Fc-фрагмент связаны непосредственно. В другом воплощении данного изобретения белок фактора свертывания IX и Fc-фрагмент связаны через первый линкер L1. В другом воплощении Fc-фрагмент и фрагмент, содержащий сайт распознавания сортазой, связаны непосредственно. В другом воплощении Fc-фрагмент и фрагмент, содержащий сайт распознавания сортазой, связаны через второй линкер L2.
[0058] В предпочтительном воплощении первый линкер L1 представляет собой пептид, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот; более предпочтительно, первый линкер L1 содержит гибкий пептид и/или жесткий элемент, и гибкий пептид представляет собой пептид, содержащий глицин (Gly, G), серии (Ser, S), аланин (Ala, А) и/или треонин (Thr, Т), например, гибкий пептид представляет собой (GS)m(GGS)n(GGGS)o(GGGGS)p, где m, n, о и р независимо выбраны из целых чисел от 0 до 50, таких как 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, предпочтительно, не все из m, n, о и р равны 0.
[0059] Жесткий элемент представляет собой пептид, содержащий один или более пролинов (Pro, Р). Пролин имеет особую циклическую структуру, которая накладывает больше ограничений на структуру пептидной цепи. Авторы изобретения неожиданно обнаружили, что включение жесткой единицы в состав линкера может значительно улучшить активность слитого белка. Предпочтительно, жесткий элемент содержит аминокислотную последовательность, выбранную из группы, состоящей из:
VAPPPALPAPVRLPGPA (SEQ ID NO: 3),
VAPPPALPAVAPPPALPA (SEQ ID NO: 4),
VAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPA (SEQ ID NO: 5),
VAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPA (SEQ ID NO: 6) и
VAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPA (SEQ ID NO: 7).
[0060] В другом предпочтительном воплощении последовательность первого линкера
L1 выбрана из группы, состоящей из:
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPA (SEQ ID NO: 8),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPA (SEQ ID NO: 9),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPA (SEQ ID NO 10),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPA (SEQ ID NO: 11) и
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPA (SEQ ID NO 12).
[0061] Второй линкер L2 представляет собой пептид, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 или более аминокислот, и в основном применяется для разделения Fc и сайта распознавания сортазой для облегчения распознавания и действия сортазы; более предпочтительно, второй линкер L2 представляет собой гибкий пептид, содержащий глицин (Gly, G), серии (Ser, S), аланин (Ala, А) и/или треонин (Thr, Т), такой как (GS)w(GGS)x(GGGS)y(GGGGS)z, где w, х, у и z независимо выбраны из целых чисел от 0 до 50, таких как 0, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19 или 20, предпочтительно, не все из w, х, у и z равны 0.
[0062] В предпочтительном воплощении последовательность второго линкера L2 выбрана из группы, состоящей из GGGGS (SEQ ID NO: 14), GGGGSGGGGS (SEQ ID NO: 15) и GSGGSGGGGS (SEQ ID NO: 16). Конъюгат слитого белка фактора свертывания IX
[0063] Термин «конъюгат» относится к продукту, образованному полипептидом или вариантом полипептида, ковалентно или нековалентно связанными с модифицирующей группой гидрофильного полимера, описанной в данном документе, причем гидрофильный полимер и полипептид могут быть связаны в любом положении, например, соответствующем положении на N-конце, С-конце или в средней части полипептида.
[0064] Как правило, термин «полимер», используемый в данном описании, имеет обычное значение, известное специалистам в области техники, и относится как к самому полимеру, так и к его производным с модифицированным концом, если явным образом не указано иное.
[0065] Кроме того, в случае таких полимеров, как полиэтиленгликоль, существует множество способов определения их молекулярной массы. Поскольку полимеры состоят из молекул с различной степенью полимеризации в пределах определенного диапазона распределения, молекулярные массы полимеров обычно представляют их средней молекулярной массой, в частности, среднечисловой молекулярной массой или среднемассовой молекулярной массой. Среднечисловая молекулярная масса относится к сумме произведений числовой доли каждой молекулы с отличной молекулярной массой и ее соответствующей молекулярной массы; среднемассовая молекулярная масса относится к сумме произведений массовой доли каждой молекулы с отличной молекулярной массой и ее соответствующей молекулярной массы. Хотя среднечисловая молекулярная масса и среднемассовая молекулярная масса могут несколько расходиться, когда степень полимеризации полимеров существенно различается, для полимеров с узким диапазоном распределения оба показателя обычно стремятся к одному значению. В случае таких полимеров, как упомянутый здесь полиэтиленгликоль, его молекулярная масса может быть указана либо как среднечисловая молекулярная масса, либо как среднемассовая молекулярная масса.
[0066] Слитый белок фактора свертывания IX по данному изобретению может быть конъюгирован с одним или более гидрофильными полимерами с образованием конъюгата слитого белка фактора свертывания IX. Предпочтительно, полимер является физиологически приемлемым, что включает растворимые в водном растворе или суспензии и не оказывающие на млекопитающих негативного влияния, такого как побочные эффекты после введения конъюгата слитого белка фактора свертывания IX в фармацевтически приемлемом количестве. Полимеры, которые могут применяться в данном изобретении, не ограничены какими-либо конкретными. Обычно предпочтительны полимеры, имеющие от 2 до приблизительно 3000 повторяющихся единиц. Группа полимеров может быть выбрана из природных или синтетических полимеров, примеры которых включают, без ограничения, полисахариды, полиалкиленгликоли, такие как полиэтиленгликоль (ПЭГ), полипропиленгликоль (ППГ), полиэтиленоксид (ПЭО), сополимер этиленгликоля и пропиленгликоля, поливиниловый спирт и любую их комбинацию. В предпочтительном воплощении конъюгат слитого белка фактора свертывания IX по данному изобретению конъюгирован с одной или более группами ПЭГ для модификации.
[0067] В данном изобретении полимер не ограничен конкретной структурой, он может быть линейным (таким как алкоксиПЭГ или бифункциональный ПЭГ), разветвленным или многолучевым (таким как раздвоенный ПЭГ или ПЭГ, связанный с полиольным ядром), древовидным или может иметь деградируемые связи. Кроме того, внутренняя структура полимеров может быть организована в любое число различных паттернов, которые могут быть выбраны из гомополимеров, чередующихся сополимеров, случайных сополимеров, сополимеров из трех чередующихся мономеров, случайных полимеров из трех мономеров, блок-полимеров из трех мономеров и тому подобного. Полимер может также включать полимеры поли(алкиленоксида), полималеиновой кислоты, поли(D, L-аланина) и тому подобного.
[0068] В некоторых воплощениях полимер представляет собой полиэтиленгликоль (ПЭГ) или его производные, такие как метоксиполиэтиленгликоль (мПЭГ). В данном документе, если не указано иное, полиэтиленгликоли (ПЭГ) включают имеющие в качестве концевой группы либо гидроксильные группы, либо другие группы. Другие группы включают алкокси, циклоалкокси, циклоалкилалкокси, алкенил, арилалкокси или аралкилокси, без ограничения. Эти молекулярные виды ПЭГ известны в уровне техники и используются рутинным образом в модификации полипептидов. Боковая цепь ПЭГ может быть линейной, разветвленной, раздвоенной или состоять из нескольких плечей. Различные полиэтиленгликоли могут иметь различную длину полимерных цепей и различные полимерные структуры.
[0069] В данном изобретении молекулярная масса ПЭГ не ограничена конкретными значениями, и его молекулярная масса может варьировать от 0,1 до 200 кДа, например, от 1 до 150 кДа, от 2 до 100 кДа, от 3 до 80 кДа, от 4 до 50 кДа или от 5 до 40 кДа. Другие полезные ПЭГ включают, например, описанные в WO 03/040211, US 6566506, US 6864350 и US 6455639. В частности, ПЭГ имеет общую формулу НО-СH2СH2О-(СH2СH2O)n-СH2СH2-ОН, где n находится в диапазоне от приблизительно 5 до приблизительно 4000. Как упоминалось выше, ПЭГ по данному изобретению включает ПЭГ с другими концевыми группами, такой как метоксиПЭГ, разветвленный ПЭГ, раздвоенный ПЭГ и тому подобное. Подходящие разветвленные ПЭГи могут быть получены, как описано в патенте US 5932462, содержание которого включено в данное описание во всей полноте путем ссылки. Раздвоенный ПЭГ относится к ПЭГ, который имеет ответвление вблизи одного конца полимерной цепи, и основная цепь раздвоенного ПЭГ может быть линейной или разветвленной.
[0070] Специалистам в области техники известно, что по мере увеличения молекулярной массы полимерной группы в биологически активной молекуле, конъюгированной с полимерной группой, биологическая активность конъюгата обычно постепенно снижается. Специалистам в области техники также известно, что по мере увеличения молекулярной массы полимерной группы время биологической полужизни конъюгата и/или время полужизни конъюгата в плазме, соответственно увеличивается. [0071] Для обеспечения стабильного терапевтического эффекта в течение длительного периода времени и для снижения частоты введения для улучшения комплаентности пациента, желательно как можно больше увеличить время биологической полужизни конъюгатов слитого белка фактора свертывания IX при сохранении существенной активности агониста рецептора фактора свертывания IX. Таким образом, в данном изобретении предложен конъюгат слитого белка фактора свертывания IX с увеличенным периодом биологической полужизни и существенной активностью агониста рецептора фактора свертывания IX.
[0072] В конкретном воплощении молекулярная масса одной или более полимерных групп (таких как ПЭГ) в конъюгате слитого белка фактора свертывания ГХ составляет 1 или более, 10 или более, 20 или более, 30 или более, 40 или более, 50 или более, 60 или более, 70 или более, 80 или более, 90 или более, 100 или более, 110 или более, 120 или более, 130 или более, 140 или более, 150 или более или 160 или более кДа, например, 5, 10, 20, 30, 40, 50, 60, 70, 80, 90 или 100 кДа, или имеет значение, находящееся между любыми двумя значениями. Следует отметить, что если в конъюгате имеется множество конъюгированных полимерных групп, при описании молекулярной массы полимерной группы в конъюгате слитого белка фактора свертывания IX подсчитывают сумму молекулярных масс всех конъюгированных полимерных групп в конъюгате, если не указано иное.
[0073] Полимер, используемый в данном изобретении, известен в уровне техники и может быть получен различными способами, включая, например, приобретение коммерческим путем, или он может быть получен собственными силами согласно способам, известным в области техники. Данное изобретение не ограничивается полимерами, полученными каким-либо определенным способом.
[0074] После реакции конъюгации конъюгат может быть выделен подходящим способом, включая, например, ультрафильтрацию, диализ или хроматографию и так далее, все из которых находятся в рамках компетенции специалиста в области техники. Фармацевтическая композиция
[0075] Слитый белок фактора свертывания IX или конъюгат слитого белка фактора свертывания IX по данному изобретению может иметь различные применения, включая, например, применение в профилактике и/или лечении геморрагических заболеваний. Таким образом, в данном изобретении также предложена фармацевтическая композиция для предупреждения и/или лечения геморрагических заболеваний, содержащая терапевтически эффективное количество слитого белка или конъюгата по данному изобретению и возможно фармацевтически приемлемый носитель. Предпочтительно, фармацевтическая композиция может применяться для предупреждения и/или лечения геморрагических заболеваний, более предпочтительно, для предупреждения и/или лечения геморрагических заболеваний у пациентов с врожденным или приобретенным дефицитом FIX и спонтанного или послеоперационного кровотечения у пациентов с гемофилией В.
[0076] Терапевтически эффективное количество слитого белка или конъюгата по данному изобретению зависит от пути введения, категории субъектов и физических характеристик конкретного млекопитающего, о котором идет речь. Эти факторы и их связь с определением количества хорошо известны специалистам в области медицины. Количество и способ введения можно корректировать для достижения оптимальной эффективности для доставки пептида субъекту, в зависимости от факторов, хорошо известных специалистам в области медицины, таких как масса тела, диета, сопутствующие лекарственные средства, а также других факторов.
[0077] Фармацевтическую композицию по данному изобретению можно вводить в составе комбинированной терапии, то есть в комбинации с одним или более других веществ, при этом фармацевтическую композицию и другие вещества вводят вместе или последовательно. В других воплощениях другие вещества можно вводить до, во время или после введения одного или более из: слитого белка, конъюгата или их фармацевтической композиции по данному изобретению.
[0078] В данном документе «фармацевтически приемлемый носитель» или «физиологически приемлемый носитель» могут использоваться взаимозаменяемо, включая одно или более из всевозможных физиологически совместимых солей, растворителей, диспергирующих сред, покрытий, антибактериальных и противогрибковых агентов, изотонических и замедляющих абсорбцию веществ и т.д. В некоторых воплощениях носитель подходит для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, посредством инъекции или инфузии). В зависимости от способа введения терапевтический агент может быть покрыт некоторыми материалами для защиты терапевтического агента от воздействия кислот и других естественных условий, которые могут привести к инактивации терапевтического агента.
[0079] При введении фармацевтического препарата по данному изобретению его вводят в фармацевтически приемлемом количестве в составе фармацевтически приемлемой композиции. Термин «фармацевтически приемлемый» означает нетоксичное вещество, которое не влияет на биологически активную эффективность активных компонентов. Такие препараты обычно содержат соли, буферы, консерванты, совместимые носители и возможно другие терапевтические агенты, такие как дополнительные иммуномодуляторы, включая адъюванты, хемокины и цитокины. При применении в составе лекарственного средства соль должна быть фармацевтически приемлемой. Однако соли, не являющиеся фармацевтически приемлемыми, могут успешно применяться для получения фармацевтически приемлемых солей, поэтому они не исключены из объема данного изобретения.
[0080] При необходимости слитый белок фактора свертывания IX или конъюгат слитого белка фактора свертывания IX по данному изобретению могут быть скомбинированы с фармацевтически приемлемыми носителями. Термин «фармацевтически приемлемые носители», используемый в данном документе, относится к одному или более совместимым твердым или жидким наполнителям, разбавителям или инкапсулирующим веществам, которые подходят для введения млекопитающим, таким как человек. Термин «носитель» означает органические или неорганические, естественные или синтетические компоненты, которые комбинируют с активными компонентами для выполнения вспомогательной функции. Компоненты фармацевтической композиции также могут быть смешаны в форме, при которой отсутствуют взаимодействия, которые могут существенно повредить терапевтическому действию желаемого лекарства.
[0081] Предпочтительно, фармацевтическая композиция по данному изобретению может содержать буферную систему и, предпочтительно, буферная система представляет собой раствор ацетатного буфера с рН от приблизительно 3,0 до приблизительно 6,0 или раствор фосфатного буфера с рН от приблизительно 5,0 до приблизительно 9,0. В некоторых конкретных воплощениях подходящие буферы включают ацетатный, цитратный, боратный и фосфатный.
[0082] Возможно, фармацевтическая композиция может также содержать подходящие консерванты, такие как хлорид бензалкония, хлор-трет-бутанол, парабены и мертиолят.
[0100] Фармацевтическая композиция может быть для удобства представлена в виде дозированной лекарственной формы и может быть получена любым известным в области фармакологии способом. Способ включает стадии объединения активного агента с носителем, который содержит один или более вспомогательных ингредиентов. В целом, композицию получат путем тщательного смешивания активного соединения с жидким носителем или тонко измельченным твердым носителем или тем и другим, после чего, при необходимости, придают продукту необходимую форму.
[0083] Фармацевтические композиции, подходящие для парентерального введения, могут быть стерильными водными или неводными составами, содержащими один или более слитых белков или конъюгатов. В некоторых воплощениях состав является изотоническим по отношению к крови субъекта. Препарат может быть составлен в соответствии с известными способами с применением подходящего диспергирующих или смачивающих веществ и суспендирующих веществ. Стерильный инъекционный препарат может также представлять собой стерильный инъекционный раствор или суспензию в нетоксичном приемлемом для парентерального применения разбавителе или растворителе, например, раствор в 1,3-бутандиоле. Приемлемые носители и растворители, которые могут применяться, включают воду, раствор Рингера и изотонический раствор хлорида натрия. Кроме того, в качестве растворителей или суспендирующих сред обычно применяют стерильные нелетучие масла. По этой причине можно использовать любое инертное нелетучее масло, включая синтетические моно- или диглицериды. Также, в качестве инъекционных составов можно использовать жирные кислоты, такие как олеиновая кислота. Составы носителей, подходящих для перорального, подкожного, внутривенного, внутримышечного и тому подобного введения, могут быть получены согласно руководствам в области техники.
[0084] Слитый белок или конъюгат по данному изобретению может быть приготовлен с носителями, которые защищают его от быстрого высвобождения, такими как состав с контролируемым высвобождением, включая имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Можно использовать биоразлагаемые и биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, полиортоэфиры и полимолочная кислота. В области техники известно множество способов для получения таких составов.
[0085] Фармацевтическую композицию по данному изобретению можно вводить любым стандартным путем, включая инъекцию или медленную инфузию в течение определенного времени. Например, введение может быть пероральным, внутривенным, внутрибрюшинным, внутримышечным, внутриполостным, внутриопухолевым или чрескожным.
[0086] Фармацевтическую композицию по данному изобретению вводят в фармацевтически эффективном количестве. «Эффективное количество» представляет собой количество любого слитого белка или конъюгата, предложенного в данном изобретении, обеспечивающее желаемый ответ в отдельности или с дополнительными дозами и/или другими терапевтическими агентами.
[0087] Несомненно, такое количество будет зависеть от конкретного заболевания, лечение которого осуществляют, тяжести заболевания, индивидуальных характеристик пациента (включая возраст, физическое состояние, рост и массу тела), продолжительность лечения, природу сопутствующего лечения (если такое проводится), конкретного пути введения и аналогичных факторов, известных медицинским специалистам и работникам здравоохранения. Эти факторы хорошо известны специалистам в области техники и могут быть установлены в ходе рутинных экспериментов. Обычно предпочтительно использовать максимальную дозу каждого компонента или их комбинации, которую на основании врачебного суждения можно назвать максимальной безопасной дозой. Однако, специалисты в области техники поймут, что по медицинским, психологическим или практически любым другим причинам пациентам могут потребоваться более низкие дозы или допустимые дозы.
[0088] Фармацевтическая композиция, применяемая в описанном выше способе, предпочтительно является стерильной и содержит эффективное количество слитого белка или конъюгата в отдельности или в комбинации с другим препаратом в единицах массы или единицах объема, подходящих для введения пациентам для получения желаемого ответа, такого как снижение глюкозы крови.
[0089] Дозировка слитого белка или конъюгата, вводимого субъекту, может быть выбрана на основании различных параметров, в частности, в зависимости от способа введения и состояния субъекта. Другие факторы включают необходимую продолжительность лечения. Если ответ у субъекта на первоначально введенную дозу недостаточен, можно использовать более высокую дозу (или более высокую эффективную дозу, достигаемую другим, более локальным способом введения), переносимую пациентом.
[0090] В некоторых воплощениях диапазон дозировок слитого белка фактора свертывания IX или конъюгата слитого белка фактора свертывания IX может составлять от 30 мг/кг массы тела в сутки до 0,00001 мг/кг массы тела в сутки, или от 3 мг/кг в сутки до 0,0001 мг/кг в сутки или от 0,3 мг/кг в сутки до 0,01 мг/кг в сутки.
[0091] В некоторых воплощениях фармацевтическая композиция по данному изобретению содержит 0,20-5 мг/мл слитого белка фактора свертывания IX и/или 4-40 мг/мл конъюгата слитого белка фактора свертывания IX, предпочтительно 0,20-5 мг/мл слитого белка фактора свертывания IX и/или 4-40 мг/мл конъюгата слитого белка фактора свертывания IX, более предпочтительно, 0,5-2 мг/мл слитого белка фактора свертывания IX и/или 10-20 мг/мл конъюгата слитого белка фактора свертывания IX. Как правило, дозировка слитого белка фактора свертывания IX или конъюгата слитого белка фактора свертывания IX по данному изобретению может варьировать от приблизительно 10 мкг/кг массы тела пациента до приблизительно 100000 мкг/кг массы тела пациента. В некоторых воплощениях дозировка может варьировать от приблизительно 0,1 мг/кг до приблизительно 20 мг/кг. В некоторых воплощениях дозировка может варьировать от приблизительно 0,1 мг/кг до 5 мг/кг, от 0,1 мг/кг до 10 мг/кг, или от 0,1 мг/кг до 15 мг/кг.В некоторых воплощениях дозировка может варьировать от приблизительно 1 мг/кг до 5 мг/кг, от 5 мг/кг до 10 мг/кг, от 10 мг/кг до 15 мг/кг или от 15 мг/кг до 20 мг/кг. В некоторых воплощениях дозировка составляет приблизительно 0,1 мг/кг, 0,5 мг/кг, 1 мг/кг, 2 мг/кг, 3 мг/кг, 5 мг/кг, 7 мг/кг, 10 мг/кг, 12 мг/кг, 15 мг/кг, 17 мг/кг, 20 мг/кг, 25 мг/кг или 30 мг/кг. В некоторых воплощениях дозировка составляет приблизительно 1 мг/кг, 3 мг/кг, 5 мг/кг или 6 мг/кг. В зависимости от свойств композиции дозирование может осуществляться непрерывно (например, при помощи непрерывного введения) или в определенные периоды времени. В некоторых воплощениях при внутривенном введении дозировка слитого белка фактора свертывания IX или конъюгата слитого белка фактора свертывания IX по данному изобретению может составлять от 0,1 до 20 мг/кг или равняться любому значению в указанном интервале. Идеальный временной промежуток для многократных введений конкретной композиции может установить специалист в области техники без излишних экспериментов. Специалистам в области техники известны другие схемы введения предложенной композиции, где дозировка, схема введения, место введения, способ введения и так далее могут различаться в зависимости от следующего. В одном воплощении введение осуществляется внутривенно. В другом воплощении схема введения представляет собой однократное внутривенное введение.
[0092] Набор, содержащий слитый белок фактора свертывания IX или конъюгат слитого белка фактора свертывания IX (например, в фармацевтической композиции) и инструкции по применению, также входит в объем данного изобретения. Набор может дополнительно содержать по меньшей мере один из других агентов, таких как один или более других агентов, обеспечивающих профилактику и/или лечение геморрагических заболеваний. В другом воплощении набор может включать носитель, который имеет отсеки для прочного удержания одного или более контейнеров или ряда контейнеров (таких как пробирки для анализов, пробирки, колбы, флаконы, шприцы и так далее). Компоненты набора могут быть упакованы в водной среде или находиться в лиофилизированной форме.
[0093] Композиция может находиться в лиофилизированной форме или в водной среде.
[0094] Предпочтительно, субъект является позвоночным, более предпочтительно, млекопитающим, наиболее предпочтительно, человеком, однако субъект может также быть другим животным, таким как домашние животные (например, собаки, кошки и так далее), домашним скотом (например, крупным рогатым скотом, овцы и козы, свиньи, лошади и так далее) или лабораторными животными (например, обезьянами, крысами, мышами, кроликами, морскими свинками и так далее).
[0095] Термин «улучшенное время полужизни в кровотоке» или «увеличенное время полужизни в кровотоке» означает, что молекула по данному изобретению имеет измененное время полужизни в кровотоке, предпочтительно, увеличенное время полужизни в кровотоке по сравнению с молекулой фактора IX дикого типа. Время полужизни в кровотоке предпочтительно увеличивается по меньшей мере на 10%, предпочтительно по меньшей мере на 15%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 25%, предпочтительно по меньшей мере на 30%, предпочтительно по меньшей мере на 35%, предпочтительно по меньшей мере на 40%, предпочтительно по меньшей мере на 45%, предпочтительно по меньшей мере на 50%, предпочтительно по меньшей мере на 55%, предпочтительно по меньшей мере на 60%, предпочтительно по меньшей мере на 65%, предпочтительно по меньшей мере на 70%, предпочтительно по меньшей мере на 75%, предпочтительно по меньшей мере на 80%, предпочтительно по меньшей мере на 85%, предпочтительно по меньшей мере на 90%, предпочтительно по меньшей мере на 95%, предпочтительно по меньшей мере на 100%, более предпочтительно по меньшей мере на 125%, более предпочтительно по меньшей мере на 150%, более предпочтительно по меньшей мере на 175%, более предпочтительно по меньшей мере на 200% и более предпочтительно по меньшей мере на 250% или 300%. Еще более предпочтительно, молекула имеет время полужизни в кровотоке, увеличенное по меньшей мере на 400%, 500%, 600% или даже 700%.
[0096] Данное изобретение дополнительно проиллюстрировано следующими примерами, которые не следует рассматривать как ограничивающие объем изобретения каким-либо образом. Все содержание документов, процитированных в данной заявке (включая публикации в журналах, выданные патенты, опубликованные заявки на патенты и находящиеся на рассмотрении заявки на патенты) явным образом включено в данное описание путем ссылки. В следующих ниже примерах, если не указано иное, использованные реагенты и материалы представляют собой доступные для приобретения продукты по меньшей мере аналитической степени чистоты или эквивалентной степени.
ОПИСАНИЕ ПРИМЕРОВ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Пример 1. Получение слитого белка FIX-L1-Fc-L2-P
1. Конструирование экспрессирующего вектора
[0097] Слитые белки FIX, сконструированные в данном примере, имеют общую молекулярную структуру FIX-L1-Fc-L2-P, где FIX обозначает естественный фактор свертывания IX человеческого происхождения, последовательность которого приведена в SEQ ID NO: 1.
[0098] L1, обозначающий первый линкер между FLX и Fc, представляет собой гибкий пептид или линкерный пептид, состоящий из гибкого пептида и жесткой структуры на основании множества пролинов (Pro, Р). В данном примере использовали в общей сложности 6 последовательностей L1, обозначенных как C1, С2, С3, С4, С5 и GS, соответственно, и их конкретные последовательности выглядят следующим образом:
C1: GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPA (SEQ ID NO: 8);
C2: GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPA (SEQ ID NO: 9);
C3: GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPA (SEQ ID NO 10),
C4: GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPA (SEQ ID NO 11),
C5: GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPA (SEQ ID NO: 12) и
GS: GGGGSGGGGSGGGGSGGGGSGGGGS (SEQ ID NO 13).
[0099] Fc происходит из человеческого IgGl, имеющего точечную мутацию N297A, опосредующего пониженную ADCC и CDC активность (см. Lund J. et al. Multiple binding sites on the CH2 domain of IgG for mouse Fc gamma R11. Mol Immunol. 1992 Jan; 29(1):53-9.).
[0100] L2 представляет собой второй линкер, который в основном применяется для разделения Fc и сайта распознавания сортазы. В частности, последовательность L2 в данном примере представляет собой GSGGSGGGS (SEQ ID NO: 16).
[0101] Р представляет собой аминокислотный фрагмент, содержащий сайт распознавания сортазой.
[0102] В Таблице 1 представлена сводная информация по всем слитым белкам FIX-L1-Fc-L2-P, сконструированным в данном Примере.

[0103] Большая часть рекомбинантного FIX, экспрессируемого в клетках СНО, существует в виде белка-предшественника, и только приблизительно 30% пропептида отщепляется от белка-предшественника с образованием зрелого FIX. Процессинг зрелого FIX требует участия фермента РАСЕ/фурин, который представляет собой подобную субтилизину кальций-зависимую сериновую пропептидазу располагающуюся в транс-зоне комплекса Гольджи, и может отщеплять С-конец R46 в белке-предшественнике FIX, чтобы катализировать превращение белка-предшественника FIX в зрелый FIX. Для улучшения выхода и обеспечения надлежащего процессинга FIX в данном Примере дополнительно сконструировали бицистронный экспрессирующий вектор FIX/PACE (фурин), структура которого приведена на Фиг. 1 А.
Конструирование бицистронного экспрессирующего вектора FIX/PACE (фурин)
а. Конструирование экспрессирующего вектора pFRL-rPS, экспрессирующего РАСЕ
[0104] Последовательность РАСЕ/фурин имеет регистрационный номер NP 002560.1 в базе GenBank. Для экспрессии в хозяине выполняли оптимизацию кодонов последовательности, кодирующей аминокислоты 1-715 РАСЕ/фурин (SEQ ID NO: 17). После синтеза полного гена последовательность клонировали в вектор pcDNA3.1 (+) и полученную плазмиду обозначали pcDNA3.1-P-SOL (Shanghai Generay Biotech Co., Ltd.). Вся кассета, экспрессирующая ген, включая промотор SV40, ген РАСЕ/фурин и поли-А, была получена при помощи ПЦР-амплификации с праймерами F1 и R1 (полные последовательности приведены ниже) с использованием плазмиды pcDNA3.1-P-SOL в качестве матрицы. После расщепления по сайтам рестрикции BglII на обоих концах фрагмент встраивали в вектор pFRL-DHFR, расщепленный по одному сайту рестрикции BamHI. Отбирали положительные клоны, верифицировали при помощи секвенирования, и обозначали вектор pFRL-rPS.
F1: 5'-TCAGGAAGATCTCGCGAATTAATTCTGTGGAATGTGT-3' (SEQ ID NO: 30);
R1: 5'-TATCCTAGATCTGCTGGCACGACAGGTTTCCCGACT-3' (SEQ ID NO: 31).
Последовательность аминокислот 1-715 РАСЕ/фурин приведена ниже:
MELRPWLLWVVAATGTLVLLAADAQGQKVFTNTWAVRIPGGPAVANSVARKHGFLNLGQIFGDYYHFWHRGVTKRSLSPHRPRHSRLQREPQVQWLEQQVAKRRTKRDVYQEPTDPKFPQQWYLSGVTQRDLNVKAAWAQGYTGHGIVVSILDDGIEKNHPDLAGNYDPGASFDVNDQDPDPQPRYTQMNDNRHGTRCAGEVAAVANNGVCGVGVAYNARIGGVRMLDGEVTDAVEARSLGLNPNHIHIYSASWGPEDDGKTVDGPARLAEEAFFRGVSQGRGGLGSIFVWASGNGGREHDSCNCDGYTNSIYTLSISSATQFGNVPWYSEACSSTLATTYSSGNQNEKQIVTTDLRQKCTESHTGTSASAPLAAGIIALTLEANKNLTWRDMQHLVVQTSKPAHLNANDWATNGVGRKVSHSYGYGLLDAGAMVALAQNWTTVAPQRKCIIDILTEPKDIGKRLEVRKTVTACLGEPNHITRLEHAQARLTLSYNRRGDLAIHLVSPMGTRSTLLAARPHDYSADGFNDWAFMTTHSWDEDPSGEWVLEIENTSEANNYGTLTKFTLVLYGTAPEGLPVPPESSGCKTLTSSQACVVCEEGFSLHQKSCVQHCPPGFAPQVLDTHYSTENDVETIRASVCAPCHASCATCQGPALTDCLSCPSHASLDPVEQTCSRQSQSSRESPPQQQPPRLPPEVEAGQRLRAGLLPSHLPE (SEQ ID NO 17).
б. Конструирование бицистронного экспрессирующего вектора для экспрессии FIX-L1-Fc-L2-P
b1) pFRL-rPS-FLX-C1-Fc-L2-P
[0105] Выполняли оптимизацию кодонов нуклеотидной последовательности, кодирующей FIX-C1-Fc-L2-P (то есть последовательность L1 в FIX-L1-Fc-L2-P представляет собой С1 и так далее) (аминокислотная последовательность приведена в SEQ ID NO: 18, а кодон-оптимизированная кодирующая нуклеотидная последовательность приведена в SEQ ID NO: 19). После синтеза полного гена в компании Shanghai Generay Biotech Co., Ltd., фрагмент встраивали в pFRL-rPS путем расщепления по двум сайтам рестрикции HindIII/EcoRI для конструирования бицистронного экспрессирующего вектора pFRL-rPS-FIX-C1-Fc-L2-P, совместно экспрессирующего FIX-C1-Fc-L2-P/фypин.
Аминокислотная последовательность FIX-C1-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAAINKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGSGGSGGGGSLPETGG(SEQ ID NO 18).
Нуклеотидная последовательность FIX-C1-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCCGTCGCTCCACCACCTGCACTCCCCGCCCCTGTCAGACTGCCTGGACCCGCTGATAAGACCCACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCCAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACCAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGAATCAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCCAACGGACAGCCAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACААТСACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCAAAGGCAGCGGCGGАТСTGGAGGCGGCGGATCCCTCCCAGAGACCGGCGGATGA (SEQ ID NO 19).
b2) PFRL-rPS-FIX-C2-Fc-L2-P
[0106] Используя кодирующую последовательность FIX-C1-Fc-L2-P в качестве матрицы, добавляли последовательность С2 к 5' концу праймера и амплифицировали последовательность Fc-L2-P при помощи ПЦР с получением последовательности C2-Fc-L2-P. Последовательности праймеров F2 и R2, использованных для амплификации, приведены ниже: Прямой праймер F2:
5'-TTCGCGGATCCGTCGCTCCACCACCTGCACTCCCCGCCGTGGCTCCTCCACCT
GCTTTGCCCGCTGATAAGАСССACACATGTCCAC- 3' (SEQ ID NO: 32);
Обратный праймер R2:
5'-ТАACCGGAATTCATTATCCGCCGGTCTCTGGGAGAGATCCGCCGCCTCCAGATC
CGCCGCTGCCGCCAGGGCTGAGGGACAGGGACT-3' (SEQ ID NO: 33).
[0107] Затем после расщепления по двум рестрикционным сайтам BamHI/EcoRI последовательность C1-Fc-L2-P в векторе pFRL-rPS-FIX-C1-Fc-L2-P заменяли амплифицированным продуктом C2-Fc-L2-P, таким образом получая бицистронный экспрессирующий вектор pFRL-rPS-FIX-C2-Fc-L2-P, совместно экспрессирующий FIX-C2-Fc-L2-P/фурин.
Аминокислотная последовательность FIX-C2-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAAFNKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGGSGGSGGGGSLPETGG (SEQ ID NO: 20).
Нуклеотидная последовательность FIX-C2-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCCGTCGCTCCACCACCTGCACTCCCCGCCGTGGCTCCТССACCTGCTTTGCCCGCTGATAAGACCCACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCСAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACСAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGAATCAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCCAACGGACAGCCAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACААТСACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCGGCAGCGGCGGАТСTGGAGGCGGCGGATCTCTCCCAGAGACCGGCGGATAA (SEQ ID NO: 21).
63) Конструирование pFRL-rPS-FIX-C3-Fc-L2-P
[0108] Способ конструирования вектора был таким же, как и для pFRL-rPS-FIX-C2-Fc-L2-P. Праймеры для ПЦР для амплификации кодирующей последовательности C3-Fc-L2-P включали прямые праймеры F3: 5'-TGGCTCCTCCACCTGCTTTGCCCGCTGTCGCTCCACCACCTGCACTCC-3' (SEQ ID NO: 34) и F4: 5'-TAGATCTGTCGCTCCACCACCTGCACTCCCCGCCGTGGCTCCTCCACCTGCTTTGCC-3' (SEQ ID NO: 35) и обратный праймер R3: 5'-TGAATTCTTATCATCCGCCGGTCTCTGGGAG-3' (SEQ ID NO: 36). Последовательность C3-Fc-L2-P получали в двух раундах ПЦР-амплификации.
Аминокислотная последовательность FIX-C3-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAArNKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAVAPPPALPAPVRLPGPADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGKGSGGSGGGGSLPETGG (SEQ ID NO: 22).
Нуклеотидная последовательность FIX-C3-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCTGTCGCTCCACСACCTGCACTCCCCGCCGTGGCTCCTCСACCTGCTTTGCCCGCTGTCGCTCCACCACCTGCACTCCCCGCCCCTGTCAGACTGCCTGGACCCGCTGATAAGACCCACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCСAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACCAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGAATCAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCCAACGGACAGCСAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACAATCACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCAAAGGCAGCGGCGGATCTGGAGGCGGCGGATCCCTCC С AG AG AC CGGCGG ATG A (SEQ ID NO: 23). 64) Конструирование pFRL-rPS-FIX-C4-Fc-L2-P
[0109] Способ конструирования был таким же, как и для pFRL-rPS-FIX-C2-Fc-P. Праймеры для ПЦР для амплификации гена C4-Fc-L2-P включали прямые праймеры F5: 5'-CCCTGT CAGACTGCCTGGACCCGCTGTCGCTCCACCACCTGCACTCC-3' (SEQ ID NO: 37) и F6:
5-TAGATCTGTCGCTCCACCACCTGCACTCCCCGCCCCTGTCAGACTGCCTGGACC-3' (SEQ ID NO: 38) и обратный праймер R3. Последовательность C4-Fc-L2-P получали в двух раундах ПЦР-амплификации.
Аминокислотная последовательность FIX-C4-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAAINKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPAVAPPPALPAVAPPPALPADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGGSGGS GGGGS LPETGG (SEQ ID NO: 24).
Нуклеотидная последовательность FIX-C4-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCTGTCGCTCCACCACCTGCACTCCCCGCCCCTGTCAGACTGCCTGGACCCGCTGTCGCTCCACCACCTGCACTCCCCGCCGTGGCTCCTCCACCTGCTTTGCCCGCTGATAAGACCCACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCCAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACCAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGААТСAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCCAACGGACAGCCAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACAATCACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCGGCAGCGGCGGATCTGGAGGCGGCGGATCTCTCCCAGAGACCGGCGGATGA (SEQ ID NO: 25).
65) Конструирование pFRL-rPS-FIX-C5-Fc-L2-P
[0110] С использованием FIX-C2-Fc-L2-P в качестве матрицы, проводили ПЦР-амплификацию фрагмента FIX-C2 (прямой праймер F7: 5'-ATATAAGCTTCCGCCACCATGCAGCGC-3' (SEQ ID NO: 39) и обратный праймер R4: 5'-ATATGGATCCAGCGGGCAAAGCAGGTGG AGGAGC-3' (SEQ ID NO: 40)). Полученный фрагмент клонировали в вектор pFRL-rPS-FIX-C2-Fc-L2-P с использованием двух сайтов рестрикции HindIII/BamHI с получением бицистронного экспрессирующего вектора pFRL-rPS-FIX-C5-Fc-L2-P, совместно экспрессирующего FIX-C5-Fc-L2-P/фypин (С5 на самом деле представляет собой тандем двух С2).
Аминокислотная последовательность FIX-C5-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEHTEQKRNVIRIIPHHNYNAAFNKYNHDIALLELDEPLVLNSYVTPICIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAGSVAPPPALPAVAPPPALPADKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGGSGGSGGGGSLPETGG (SEQ ID NO: 26).
Нуклеотидная последовательность FIX-C5-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCTGTCGCTCCACСACCTGCACTCCCCGCCGTGGCTCCTCСACCTGCTTTGCCCGCTGGATCCGTCGCTCCACCACCTGCACTCCCCGCCGTGGCTCCTCCACCTGCTTTGCCCGCTGATAAGACCCACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCCAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACCAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGААТСAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCСAACGGACAGCCAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACAATCACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCGGCAGCGGCGGATCTGGAGGCGGCGGATCTCTCCCAGAGACCGGCGGATAA (SEQ ID NO: 27).
66) Конструирование pFRL-rPS-FLX-GS-Fc-L2-P
[0111] Способ конструирования бицистронного экспрессирующего вектора pFRL-rPS-FIX-GS-Fc-L2-P, совместно экспрессирующего FIX-GS-Fc-L2-P/фypин, был таким же, как и для pFRL-rPS-FIX-C2-Fc-L2-P. Праймеры для ПЦР-амплификации гена Fc-L2-P включали прямой праймер F8:
5'-GATTTGGATCCGATAAGACCCACACATGTCCACCTTG-3' (SEQ ID NO: 41) и обратный праймер R5:
5'-ТААССGGAATTCATTATCCGCCGGTCTCTGGGAGAGATCCGCCGCCTCCAGAT
CCGCCGCTGCCGCCAGGGCTGAGGGACAGGGАСТ-3' (SEQ ID NO: 42).
Аминокислотная последовательность FIX-GS-Fc-L2-P: MQRVNMIMAESPGLITICLLGYLLSAECTVFLDHENANKILNRPKRYNSGKLEEFVQGNLERECMEEKCSFEEAREVFENTERTTEFWKQYVDGDQCESNPCLNGGSCKDDINSYECWCPFGFEGKNCELDVTCNIKNGRCEQFCKNSADNKVVCSCTEGYRLAENQKSCEPAVPFPCGRVSVSQTSKLTRAETVFPDVDYVNSTEAETILDNITQSTQSFNDFTRVVGGEDAKPGQFPWQVVLNGKVDAFCGGSIVNEKWIVTAAHCVETGVKITVVAGEHNIEETEНТЕQKRNVIRIIPHHNYNAAINКYNHDIALLELDEPLVLNSYVTPIСIADKEYTNIFLKFGSGYVSGWGRVFHKGRSALVLQYLRVPLVDRATCLRSTKFTIYNNMFCAGFHEGGRDSCQGDSGGPHVTEVEGTSFLTGIISWGEECAMKGKYGIYTKVSRYVNWIKEKTKLTGGGGSGGGGSGGGGSGGGGSGGGGSDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYASTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSRDELTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGGSGGSGGGGSLPETGG (SEQ ID NO: 28).
Нуклеотидная последовательность FIX-GS-Fc-L2-P: ATGCAGCGCGTCAATATGATTATGGCCGAGTCTCCAGGCCTCATTACCATTTGTCTGCTGGGCTACCTGCTGTCCGCAGAGTGTACCGTCTTCCTCGACCACGAGAACGCAAATAAGATTCTCAATAGGCCTAAGCGCTACAATTCCGGAAAGCTCGAAGAGTTTGTCCAGGGAAATCTGGAGCGCGAGTGTATGGAGGAAAAGTGTAGCTTTGAAGAAGCTAGAGAGGTGTTTGAGAATACTGAGCGCACTACTGAGTTTTGGAAGCAGTATGTGGATGGCGATCAGTGCGAGTCCAATCCATGCCTGAATGGCGGATCTTGCAAGGATGATATTAATTCTTACGAGTGTTGGTGCCCATTCGGCTTTGAGGGAAAGAATTGCGAACTGGATGTCACATGTAACATCAAGAACGGCAGATGCGAGCAGTTTTGTAAGAACAGCGCCGATAATAAGGTGGTGTGTAGTTGTACTGAGGGATACCGCCTCGCAGAGAATCAGAAGTCCTGTGAGCCCGCTGTGCCATTTCCTTGTGGCCGGGTCAGCGTCAGCCAGACTTCTAAGCTGACACGCGCCGAGACCGTCTTTCCAGATGTGGATTATGTCAACTCTACTGAGGCCGAGACCATTCTCGATAATATTACTCAGTCTACACAGTCCTTTAACGACTTCACTCGGGTGGTGGGCGGAGAGGACGCAAAGCCTGGCCAGTTTCCTTGGCAGGTCGTGCTCAACGGAAAGGTGGACGCATTTTGTGGCGGATCTATCGTGAATGAGAAGTGGATTGTGACCGCCGCTCACTGCGTCGAGACAGGCGTCAAGATTACCGTGGTCGCTGGAGAGCACAACATTGAGGAAACAGAGCACACCGAGCAGAAGCGCAATGTGATTCGCATCATCCCACACCACAACTACAACGCCGCTATTAATAAGTACAACCACGACATCGCCCTGTTAGAACTAGATGAGCCTCTGGTCCTGAATAGTTACGTCACTCCTATTTGTATCGCCGATAAAGAATACACTAACАТСTTTCTCAAGTTCGGCTCTGGCTATGTATCTGGATGGGGACGGGTGTTTCACAAGGGACGGTCCGCACTCGTGCTACAATACCTCCGGGTGCCACTGGTGGATCGGGCAACATGCCTCCGGTCTACCAAGTTTACTATATACAATAACATGTTTTGCGCCGGCTTTCACGAGGGCGGACGGGATAGTTGCCAGGGCGATAGTGGAGGCCCTCATGTCACAGAGGTGGAGGGAACTAGCTTTCTGACAGGCATTATCTCTTGGGGCGAAGAGTGCGCTATGAAGGGAAAGTACGGCATCTATACTAAGGTCTCGCGTTACGTCAATTGGATCAAGGAAAAGACAAAGCTGACAGGCGGCGGAGGGAGCGGCGGCGGAGGCTCTGGCGGCGGCGGATCTGGCGGAGGCGGCTCTGGCGGAGGCGGATCCGATAAGACCСACACATGTCCACCTTGCCCAGCACCTGAGCTGCTGGGCGGACCTAGCGTGTTTCTGTTCCCACCAAAGCCAAAGGACACCCTCATGATTAGTAGAACCCCAGAGGTGACTTGCGTCGTGGTCGATGTCAGTCACGAGGACCCCGAGGTGAAGTTTAATTGGTATGTAGATGGCGTCGAGGTGCACAACGCTAAGACCAAGCCACGCGAAGAGCAGTACGCATCTACTTACCGGGTCGTCTCTGTGCTGACTGTGCTGCACCAGGATTGGCTCAATGGCAAAGAGTACAAGTGTAAGGTCAGTAATAAGGCCCTCCCCGCACCTATTGAAAAGACTATTAGCAAGGCAAAGGGCCAGCCTAGGGAGCCCCAGGTCTACACCCTGCCCCCTTCCCGCGATGAGCTGACTAAGAATCAGGTCTCTCTCACTTGCCTCGTGAAGGGCTTTTACCCATCCGACATTGCCGTGGAGTGGGAGTCСAACGGACAGCCAGAGAATAACTACAAGACAACTCCTCCCGTGCTCGATAGCGACGGTTCATTCTTCCTGTACTCCAAGCTCACCGTTGATAAGTCTCGGTGGCAGCAGGGAAATGTGTTCTCTTGTTCCGTCATGCACGAGGCACTGCACAATCACTACACCCAGAAGTCCCTGTCCCTCAGCCCTGGCGGCAGCGGCGGATCTGGAGGCGGCGGATCTCTCCCAGAGACCGGCGGATAA (SEQ ID NO: 29).
[0112] Структурная диаграмма вышеуказанного бицистронного экспрессирующего вектора FIX-Ll-Fc-L2-P/фузин приведена на Фиг. 1 А.
Конструирование вектора, совместно экспрессирующего GGCX и VKORC1 [0113] FIX представляет собой витамин К (VK)-зависимый гликопротеин, и с помощью VK, остаток глутамата в его N-концевом домене G1a карбоксилируется с образованием γ-карбоксиглутамата, обеспечивающего свертывающую активность FIX. В ходе модификации Gla двумя ключевыми ферментами являются γ-глутамилкарбоксилаза (GGCX) и 1 субъединица комплекса витамин К-эпоксидредуктазы (VKORC l). Таким образом, в данном изобретении сконструировали плазмиду pcDNA3.1-Hyg-VKGC для увеличения уровня экспрессии функционального FLX путем совместной трансфекции плазмидой с FIX. Структура плазмиды pcDNA3.1-Hyg-VKGC показана на Фиг. 1 В, а способ конструирования был следующим: гамма-глутамилкарбоксилазу (GGCX), 1 субъединицу комплекса витамин К-эпоксидредуктазы (VKORCl) и внутренний участок посадки рибосомы (IRES) ДНК синтезировали путем синтеза полного гена; IRES клонировали в вектор pcDNA3.1(+)/hygro путем расщепления по двум сайтам рестрикции NheI/XhoI с конструированием плазмиды p3.1hyg-IRES; затем путем рестрикции по двум другим сайтам клонировали ген VKORCl (сайты Nhel/BamHI) и ген GGCX (сайты NotI/XhoI) в р3.1hyg-IRES с конструированием вектора pcDNA3.1-Hyg-VKGC, совместно экспрессирующего GGCX и VKORCl.
2. Конструирование стабильных клеточных линий
[0114] Каждым типом плазмиды, экспрессирующей слитый белок FIX, сконструированной, как описано выше, трансфицировали клетки-хозяева СНО DG44 путем электропорации. Для электропорации 1×107 клеток, 40 мкг плазмиды, совместно экспрессирующей FIX-L1-Fc-L2-P/фурин, и 20 мкг экспрессирующей плазмиды pcDNA3.1-Hyg-VKGC осторожно смешивали и добавляли в кюветы для электропорации (Bio-Rad, 4 мм). Использовали устройство для электропорации Bio-Rad (GENE PULSER XCELL) и устанавливали следующие параметры для электропорации:
[0115] После электропорации клетки переносили в среду для восстановления. Через 24 часа культивирования клетки высаживали в пятнадцать 96-луночных планшетов и заменяли среду на среду для скрининга, содержавшую Opti-MEM I+50 нМ МТХ. После культивирования клеток в течение 4-5 дней, 2-3 раза добавляли среду, содержавшую 50 нМ МТХ. При достижении клетками конфлюентности 50% или более проводили скрининг клонов с высокой экспрессией целевого белка методом дот-блот, а в качестве антитела использовали мышиное антитело к Fc-фрагменту IgG человека. Отобранные клоны с относительно высокими уровнями экспрессии последовательно переносили для размножения в 24-луночные планшеты, 6-луночные планшеты, чашки для культивирования клеток и встряхиваемые колбы для культивирования клеток.
[0116] Для повышения выхода слитого белка клетки культивировали под давлением увеличивающихся концентраций МТХ. Таким образом, за счет ингибирования гена DHFR при помощи МТХ, добивались совместной амплификации гена DHFR и гена слитого белка.
3. Продуцирование слитого белка 3.1 Размораживание и пассаж клеток
[0117] Клетки размораживали и культивировали в культуральных флаконах, а после увеличения плотности клетки переносили во встряхиваемые флаконы, содержавшие среду CD DG44+5 мг/л VK для культивирования и пассажей. Пассаж проводили, когда клетки достигали плотности в диапазоне 2,5-3,5x106 клеток/мл, и высаживали с плотностью 0,8-1,0×106 клеток/мл. Когда культура клеток достигала 1,8-2 л, а клеточная плотность достигала приблизительно 3,5x106 клеток/мл, клетки культивировали в резервуарах.
3.2 Ферментация
[0118] Для ферментации использовали биореактор объемом 15 л (Applikon, Biobundle 15L), объем инокулума составлял 8 л, а клеточная плотность составляла 1,0×106 клеток/мл. Устанавливали следующие параметры культивирования клеток: 1) рН 6,8-7,4; 2) растворенный кислород (DO) 40%; 3) режим подачи кислорода: вентилирование крупными пузырьками; 4) методика охлаждения: культивирование при 37°С на ранней стадии и охлаждение до 33°С на 5й день; 5) скорость перемешивания: 80-140 об/мин; 6) методика подачи питательной среды: подача питательной среды на четвертый день в количестве 2%, 3%, 4%, 3%, 2%, 2%, 2%, 2% и 2%, общий объем подаваемой питательной среды составлял 24%, клетки подсчитывали при помощи счетчика клеток ежедневно; 7) сбор: после культивирования в течение примерно 13 дней собирали надосадочную жидкость путем центрифугирования при 3000 g в течение 10 мин.
3.3 Анализ образцов и очистка
[0119] Кондиционированную надосадочную жидкость очищали в три стадии: аффинной хроматографией, хроматографией со смешанным режимом и анионо-обменной хроматографией. Очищенный слитый белок анализировали при помощи SDS-PAGE и SEC-HPLC. Как показано на Фиг. 2А, результаты SDS-PAGE демонстрируют, что размер белков, представляющих интерес, в основном совпадал с расчетной молекулярной массой.
[0120] SEC-HPLC проводили на колонке Agilent Advancebio SEC с размером пор 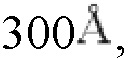 размером частиц 2,7 мкм, внутренним диаметром 7,8 мм и длиной 300 мм, элюировали градиентом 50 мМ Tris+150 мМ NaCl, рН 7,2, при этом температура колонки составляла 30°С, скорость потока составляла 0,5 мл/мин, температура устройства для ввода образца составляла 4,0°С, а объем вводимого образца составлял 30 мкг.На Фиг. 2 В показаны результаты очистки на примере белка FIX-C1-Fc-L2-P, у которого время удерживания составляло 15,1 мин, а чистота по данным SEC после трехстадийной очистки составляла 95%.
размером частиц 2,7 мкм, внутренним диаметром 7,8 мм и длиной 300 мм, элюировали градиентом 50 мМ Tris+150 мМ NaCl, рН 7,2, при этом температура колонки составляла 30°С, скорость потока составляла 0,5 мл/мин, температура устройства для ввода образца составляла 4,0°С, а объем вводимого образца составлял 30 мкг.На Фиг. 2 В показаны результаты очистки на примере белка FIX-C1-Fc-L2-P, у которого время удерживания составляло 15,1 мин, а чистота по данным SEC после трехстадийной очистки составляла 95%.
Пример 2. Определение активности FIX-Ll-Fc-L2-P in vitro и сравнение
[0121] В данном Примере исследовали активность in vitro FIX-C1-Fc-L2-P, FIX-C2-Fc-L2-P, FIX-C3-Fc-L2-P, FIX-C4-Fc-L2-P и FIX-C5-Fc-L2-P, полученных в Примере 1, и сравнивали с активностью in vitro FIX-GS-Fc-L2-P, не имеющего жесткого элемента в первом линкере L1.
[0122] Активность человеческого фактора свертывания IX (hFIX) измеряли одностадийным способом на основе активированного парциального тромбопластинового времени (АПТВ), в котором в качестве активатора использовали эллаговую кислоту и для определения АПТВ использовали 8й международный стандарт 2009 концентрата FIX (стандарт ВОЗ для активности FIX). Для построения линейной стандартной кривой откладывали логарифмы активности известного стандарта и измеренного времени АПТВ. Исследуемый образец разводили, чтобы попасть в диапазон стандартной кривой, и смешивали с дефицитной по FIX плазмой для измерения значения АПТВ. Биологическую активность исследуемого образца рассчитывали по стандартной кривой.
[0123] Аликвоты активного стандарта FIX или образца доводили до комнатной температуры, затем отбирали пипеткой по 10 мкл каждого из них и разводили приблизительно до 1 МЕ/мл при помощи дефицитной плазмы с концентрацией FIX 5%, а затем разводили в 10 раз и 20 раз (и дополнительно разводили в 40 раз и 80 раз для активного стандарта) при помощи дефицитной плазмы с концентрацией FIX 5% и тщательно смешивали при помощи вортекса. Разведенный стандарт или образец помещали в штатив для анализа образцов автоматического коагулометра и в соответствующие позиции для реагентов помещали используемые реагенты. Затем запускали соответствующую программу измерений на автоматическом анализаторе коагуляции для определения времени свертывания и повторяли измерения дважды для каждого разведения. Для построения стандартной кривой по оси абсцисс откладывали логарифм стандартной активности, а по оси ординат откладывали логарифм времени свертывания. При помощи стандартной кривой рассчитывали активность, соответствующую измеренному времени свертывания разведенного исследуемого продукта, которую затем умножали на фактор разведения для получения соответствующей активности.
[0124] Неожиданно обнаружили, что, как показано на Фиг. 3, специфическая активность молекул FIX-Cl-Fc-L2-P, FIX-C2-Fc-L2-P, FIX-C3-Fc-L2-P, FIX-C4-Fc-L2-P и FIX-C5-Fc-L2-P, в которых первый линкер L1 содержал жесткий элемент, была значительно выше, чем у FIX-GS-Fc-L2-P, в которой первый линкер L1 не имел жесткого элемента, что указывает на то, что первый линкер L1, содержащий жесткий элемент, может существенно увеличивать специфическую активность молекулы.
[0125] Кроме того, неожиданно оказалось, что среди всех слитых белков, содержащих жесткий элемент в L1, FIX-C1-Fc-L2-P обладал существенно более высокой активностью, которая почти вдвое или более превышала специфическую активность других слитых белков (Фиг. 3). Таким образом, для более углубленного изучения в следующих Примерах отбирали FIX-C1-Fc-L2-P. Пример 3. Получение конъюгата слитого белка FIX-Cl-Fc-L2-PEG
[0126] Белок FIX-C1-Fc-L2-P содержит LPETGG на С-конце, что может специфически распознаваться сортазой А. Разрезали амидную связь между Т и G путем воздействия сортазы А, и Т взаимодействовал с сульфгидрильной группой С в положении 184 сортазы с образованием промежуточной тиоэфирной связи, которую затем атаковал GGGAA-PEG с поли-Gly на N-конце. В итоге, ПЭГ определенной молекулярной массы мог связаться с С-концом белка.
[0127] Реакционная смесь для ПЭГилирования: 40KD-GGGAA-PEG растворяли в буфере и доводили рН до 7,5. Затем белок и 40KD-GGGAA-PEG добавляли к буферной системе 50 мМ Tris 150 мМ NaCl рН 7,5 в соотношении 1:20, и затем добавляли сортазу А и 10 мМ СаСl2. Через 30 минут добавляли ЕDТА для остановки реакции.
[0128] Степень конъюгации конъюгата (FIX-C1-Fc-L2-PEG) проверяли при помощи SEC-HPLC, которую проводили на колонке TOSOH TSKgel UItraSW Aggregate, с размером частиц 3 мкм, внутренним диаметром 7,8 мм и длиной 300 мм, элюировали градиентом элюирующего буфера 50 мМ Tris+150 мМ NaCl, рН 7,2, при этом температура колонки составляла 25°С, скорость потока составляла 0,5 мл/мин, температура устройства для ввода образца составляла 4,0°С, объем вводимого образца составлял 50 мкг, время удерживания конъюгированного белка составляло 12-14,5 мин, а время удерживания субстрата составляло 15 мин. На Фиг. 14 показаны результаты анализа FIX-C1-Fc-L2-PEG где доля конъюгированного с ПЭГ продукта достигала 70% через 30 мин.
Очистка и анализ конъюгированного с ПЭГ продукта
[0129] Проводили анионообменную хроматографию конъюгированных с ПЭГ продуктов для удаления непрореагировавших GGGAA-PEG сортазы А и неконъюгированного субстрата (FIX-C1-Fc-L2-P), и остаточное количество неконъюгированного субстрата составляло менее 3%.
[0130] Конъюгированные образцы разделяли при помощи анионообменной хроматографии, и собирали пиковые фракции на различных стадиях. Образцы, отобранные на каждом пике, анализировали при помощи SDS-HPLC и SEC, а затем образцы с долей неконъюгированного субстрата менее 5% объединяли для определения чистоты при помощи SEC. Результаты анализа FIX-C1-Fc-L2-PEG при помощи SEC показаны на Фиг. 5, где чистота ПЭГилированного продукта по результатам SEC составляла 97,8%. Среди них на продукт двойной конъюгации, время удерживания которого составляло 13,20 мин, приходилось 15,7%, на продукт одинарной конъюгации, время удерживания которого составляло 13,70 мин, приходилось 82,2%, а на непрореагировавший субстрат FIX-C1-Fc-L2-P, время удерживания которого составляло 15,94 мин, приходилось 1,24%.
Пример 4. Сохранение превосходной активности конъюгата FIX-Cl-Fc-L2-PEG in vitro
[0131] В данном Примере дополнительно исследовали активность конъюгата FIX-Cl-Fc-L2-PEG in vitro. Активность человеческого фактора свертывания IX (hFIX) измеряли одностадийным способом на основе активированного парциального тромбопластинового времени (АПТВ), в котором в качестве активатора использовали эллаговую кислоту, а для определения АПТВ использовали 8-й международный стандарт 2009 концентрата FIX (стандарт ВОЗ для активности FIX). Для построения линейной стандартной кривой использовали логарифмы активности известного стандарта и измеренного времени АПТВ. Исследуемый образец разводили, чтобы попасть в диапазон стандартной кривой и смешивали с дефицитной по FIX плазмой для измерения значения АПТВ. Биологическую активность исследуемого образца рассчитывали по стандартной кривой.
[0132] Аликвоты активного стандарта FIX или образца доводили до комнатной температуры, затем отбирали пипеткой по 10 мкл каждого из них и разводили приблизительно до 1 МЕ/мл при помощи дефицитной плазмы с концентрацией FIX 5%, а затем разводили в 10 раз и 20 раз (и дополнительно разводили в 40 раз и 80 раз для активного стандарта) при помощи дефицитной плазмы с концентрацией FIX 5% и тщательно смешивали при помощи вортекса. Разведенный стандарт или образец помещали в штатив для анализа образцов автоматического коагулометра и в соответствующие позиции для реагентов помещали используемые реагенты. Затем запускали соответствующую программу измерений на автоматическом анализаторе коагуляции для определения времени свертывания и повторяли измерения дважды для каждого разведения. Для построения стандартной кривой по оси абсцисс откладывали логарифм стандартной активности, а по оси ординат откладывали логарифм времени свертывания. При помощи стандартной кривой рассчитывали активность, соответствующую измеренному времени свертывания разведенного исследуемого продукта, которую затем умножали на фактор разведения для получения соответствующей активности.
[0133] Как показано на Фиг. 3, специфическая активность конъюгата FIX-Cl-Fc-L2-PEG была ниже, чем у FIX-C1-Fc-L2-P, что отражает, что конъюгирование с PEG до некоторой степени уменьшает специфическую активность молекулы in vitro, и приведенные выше результаты согласовывались с общими представлениями в данной области. Однако, следует отметить, что активность FIX-C1-Fc-L2-PEG in vitro по-прежнему сохранялась на относительно высоком уровне, не только гораздо более высоком, чем у FIX-GS-Fc-L2-P, не конъюгированного с ПЭГ, но также была сопоставима с активностью других слитых белков FIX-L1-Fc-L2-P, не конъюгированных с ПЭГ, за исключением FIX-C1-Fc-L2-P
Пример 5. FIX-C1-Fc-L2-PEG демонстрирует значительное увеличение времени полужизни in vivo
[0134] В данном Примере дополнительно исследовали активность конъюгата FIX-C1-Fc-L2-PEG у мышей. В частности, FIX-C1-Fc-L2-PEG вводили мышам НВ и в различные моменты времени брали образцы крови из ретроорбитального пространства. Затем отделяли плазму для определения активности hFIX в плазме одностадийным способом и рассчитывали фармакокинетические параметры с применением программного обеспечения WinNonlin 5.2.1.
[0135] Животные, на которых проводили эксперименты в данном Примере, охарактеризованы в Таблице, приведенной ниже.

[0136] В данном Примере использовали следующие образцы:

[0137] В эксперименте принимали участие 2 группы. Препарат сравнения BeneFIX и образец конъюгата FIX-C1-Fc-L2-PEG вводили через хвостовую вену по 300 МЕ/кг. Кровь забирали в следующие моменты времени: 1) BeneFIX: 0,083, 0,5, 1, 4, 8, 24, 48 и 72 ч; 2) FIX-C1-Fc-L2-PEG: 0,083, 0,5, 1, 4, 8, 24, 48, 72, 96, 120, 144, 168, 192, 216 и 240 ч. В каждый момент взятия крови в каждой группе у 5 животных НВ (2 самцов, 3 самок) брали кровь из ретроорбитального пространства и определяли активность hFIX.
[0138] Как показано на Фиг. 6, BeneFIX поддерживал уровень hFIX в плазме выше 5% в течение 2 дней после введения, но через 3 дня после введения уровень hFIX в плазме составлял всего 1%-2%; при этом FIX-C1-Fc-L2-PEG по-прежнему поддерживал уровень hFIX в плазме выше 5% в течение 3 дней после введения, и уровень hFIX в плазме составлял выше 2% через 6 дней после введения, в частности, через 10 дней после введения hFIX в плазме все еще поддерживался на уровне 1%-2%. Результаты эксперимента показывают, что период полувыведения в конечной фазе (Т1/2) FIX-C1-Fc-L2-PEG достигает 70 ч, что значительно превышает Т1/2 препарата сравнения BeneFIX (14 ч; что согласуется со значениями периода полувыведения, описанными в литературе) и T1/2 Alprolix (46 ч), описанными в литературе (Peters et al. Prolonged activity of factor IX as a monomeric Fc fusion protein. Blood 2010; 115(10): 2057-64.). Максимальная концентрация в крови (Cmax) составляла 119 МЕ/дл, площадь под кривой зависимости концентрации лекарственного средства от времени (AUClast) составляла 1475 ч* МЕ/дл, скорость выведения (CL) составляла 0,2033 дд/ч/кг, а среднее время удерживания в организме (MRT) составляло 72 ч, обеспечивая превосходный фармакодинамический эффект.Можно заметить, что сайт-специфическое конъюгирование hFIX, слитого с Fc, и ПЭГ существенно увеличивает время полужизни молекулы hFIX и обеспечивает значительно более продолжительное действие in vivo по сравнению с существующими препаратами, что открывает хорошие перспективы для практического применения.
--->
Перечень последовательностей
SEQUENCE LISTING
<110> JIANGSU GENSCIENCES INC.
<120> IMPROVED FIX FUSION PROTEIN AND CONJUGATE AND USE THEREOF
<130> PCT/CN2020/097911
<150> CN 201910591280.0
<151> 2019-07-02
<150> PCT/CN2020/097911
<151> 2020-06-24
<160> 42
<170> PatentIn version 3.2
<210> 1
<211> 461
<212> PRT
<213> Homo sapiens
<400> 1
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr
450 455 460
<210> 2
<211> 1386
<212> DNA
<213> Homo sapiens
<400> 2
atgcagcgcg tgaacatgat catggcagaa tcaccaggcc tcatcaccat ctgcctttta 60
ggatatctac tcagtgctga atgtacagtt tttcttgatc atgaaaacgc caacaaaatt 120
ctgaatcggc caaagaggta taattcaggt aaattggaag agtttgttca agggaacctt 180
gagagagaat gtatggaaga aaagtgtagt tttgaagaag cacgagaagt ttttgaaaac 240
actgaaagaa caactgaatt ttggaagcag tatgttgatg gagatcagtg tgagtccaat 300
ccatgtttaa atggcggcag ttgcaaggat gacattaatt cctatgaatg ttggtgtccc 360
tttggatttg aaggaaagaa ctgtgaatta gatgtaacat gtaacattaa gaatggcaga 420
tgcgagcagt tttgtaaaaa tagtgctgat aacaaggtgg tttgctcctg tactgaggga 480
tatcgacttg cagaaaacca gaagtcctgt gaaccagcag tgccatttcc atgtggaaga 540
gtttctgttt cacaaacttc taagctcacc cgtgctgaga ctgtttttcc tgatgtggac 600
tatgtaaatt ctactgaagc tgaaaccatt ttggataaca tcactcaaag cacccaatca 660
tttaatgact tcactcgggt tgttggtgga gaagatgcca aaccaggtca attcccttgg 720
caggttgttt tgaatggtaa agttgatgca ttctgtggag gctctatcgt taatgaaaaa 780
tggattgtaa ctgctgccca ctgtgttgaa actggtgtta aaattacagt tgtcgcaggt 840
gaacataata ttgaggagac agaacataca gagcaaaagc gaaatgtgat tcgaattatt 900
cctcaccaca actacaatgc agctattaat aagtacaacc atgacattgc ccttctggaa 960
ctggacgaac ccttagtgct aaacagctac gttacaccta tttgcattgc tgacaaggaa 1020
tacacgaaca tcttcctcaa atttggatct ggctatgtaa gtggctgggg aagagtcttc 1080
cacaaaggga gatcagcttt agttcttcag taccttagag ttccacttgt tgaccgagcc 1140
acatgtcttc gatctacaaa gttcaccatc tataacaaca tgttctgtgc tggcttccat 1200
gaaggaggta gagattcatg tcaaggagat agtgggggac cccatgttac tgaagtggaa 1260
gggaccagtt tcttaactgg aattattagc tggggtgaag agtgtgcaat gaaaggcaaa 1320
tatggaatat ataccaaggt atcccggtat gtcaactgga ttaaggaaaa aacaaagctc 1380
acttaa 1386
<210> 3
<211> 17
<212> PRT
<213> Artificial sequence
<220>
<223> Жесткий элемент
<400> 3
Val Ala Pro Pro Pro Ala Leu Pro Ala Pro Val Arg Leu Pro Gly Pro
1 5 10 15
Ala
<210> 4
<211> 18
<212> PRT
<213> Artificial sequence
<220>
<223> Жесткий элемент
<400> 4
Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala Leu
1 5 10 15
Pro Ala
<210> 5
<211> 35
<212> PRT
<213> Artificial sequence
<220>
<223> Жесткий элемент
<400> 5
Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala Leu
1 5 10 15
Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala Pro Val Arg Leu Pro
20 25 30
Gly Pro Ala
35
<210> 6
<211> 35
<212> PRT
<213> Artificial sequence
<220>
<223> Жесткий элемент
<400> 6
Val Ala Pro Pro Pro Ala Leu Pro Ala Pro Val Arg Leu Pro Gly Pro
1 5 10 15
Ala Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala
20 25 30
Leu Pro Ala
35
<210> 7
<211> 38
<212> PRT
<213> Artificial sequence
<220>
<223> Жесткий элемент
<400> 7
Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala Leu
1 5 10 15
Pro Ala Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro
20 25 30
Pro Pro Ala Leu Pro Ala
35
<210> 8
<211> 42
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: C1
<400> 8
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu
20 25 30
Pro Ala Pro Val Arg Leu Pro Gly Pro Ala
35 40
<210> 9
<211> 43
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: C2
<400> 9
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu
20 25 30
Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala
35 40
<210> 10
<211> 60
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: C3
<400> 10
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu
20 25 30
Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro
35 40 45
Ala Leu Pro Ala Pro Val Arg Leu Pro Gly Pro Ala
50 55 60
<210> 11
<211> 60
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: C4
<400> 11
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu
20 25 30
Pro Ala Pro Val Arg Leu Pro Gly Pro Ala Val Ala Pro Pro Pro Ala
35 40 45
Leu Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala
50 55 60
<210> 12
<211> 63
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: C5
<400> 12
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu
20 25 30
Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala Gly Ser Val Ala Pro
35 40 45
Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala
50 55 60
<210> 13
<211> 25
<212> PRT
<213> Artificial sequence
<220>
<223> Первый линкер L1: GS
<400> 13
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 14
<211> 5
<212> PRT
<213> Artificial sequence
<220>
<223> Второй линкер L2
<400> 14
Gly Gly Gly Gly Ser
1 5
<210> 15
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Второй линкер L2
<400> 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 16
<211> 10
<212> PRT
<213> Artificial sequence
<220>
<223> Второй линкер L2
<400> 16
Gly Ser Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 17
<211> 715
<212> PRT
<213> Artificial sequence
<220>
<223> PACE/Фурин 1-715
<400> 17
Met Glu Leu Arg Pro Trp Leu Leu Trp Val Val Ala Ala Thr Gly Thr
1 5 10 15
Leu Val Leu Leu Ala Ala Asp Ala Gln Gly Gln Lys Val Phe Thr Asn
20 25 30
Thr Trp Ala Val Arg Ile Pro Gly Gly Pro Ala Val Ala Asn Ser Val
35 40 45
Ala Arg Lys His Gly Phe Leu Asn Leu Gly Gln Ile Phe Gly Asp Tyr
50 55 60
Tyr His Phe Trp His Arg Gly Val Thr Lys Arg Ser Leu Ser Pro His
65 70 75 80
Arg Pro Arg His Ser Arg Leu Gln Arg Glu Pro Gln Val Gln Trp Leu
85 90 95
Glu Gln Gln Val Ala Lys Arg Arg Thr Lys Arg Asp Val Tyr Gln Glu
100 105 110
Pro Thr Asp Pro Lys Phe Pro Gln Gln Trp Tyr Leu Ser Gly Val Thr
115 120 125
Gln Arg Asp Leu Asn Val Lys Ala Ala Trp Ala Gln Gly Tyr Thr Gly
130 135 140
His Gly Ile Val Val Ser Ile Leu Asp Asp Gly Ile Glu Lys Asn His
145 150 155 160
Pro Asp Leu Ala Gly Asn Tyr Asp Pro Gly Ala Ser Phe Asp Val Asn
165 170 175
Asp Gln Asp Pro Asp Pro Gln Pro Arg Tyr Thr Gln Met Asn Asp Asn
180 185 190
Arg His Gly Thr Arg Cys Ala Gly Glu Val Ala Ala Val Ala Asn Asn
195 200 205
Gly Val Cys Gly Val Gly Val Ala Tyr Asn Ala Arg Ile Gly Gly Val
210 215 220
Arg Met Leu Asp Gly Glu Val Thr Asp Ala Val Glu Ala Arg Ser Leu
225 230 235 240
Gly Leu Asn Pro Asn His Ile His Ile Tyr Ser Ala Ser Trp Gly Pro
245 250 255
Glu Asp Asp Gly Lys Thr Val Asp Gly Pro Ala Arg Leu Ala Glu Glu
260 265 270
Ala Phe Phe Arg Gly Val Ser Gln Gly Arg Gly Gly Leu Gly Ser Ile
275 280 285
Phe Val Trp Ala Ser Gly Asn Gly Gly Arg Glu His Asp Ser Cys Asn
290 295 300
Cys Asp Gly Tyr Thr Asn Ser Ile Tyr Thr Leu Ser Ile Ser Ser Ala
305 310 315 320
Thr Gln Phe Gly Asn Val Pro Trp Tyr Ser Glu Ala Cys Ser Ser Thr
325 330 335
Leu Ala Thr Thr Tyr Ser Ser Gly Asn Gln Asn Glu Lys Gln Ile Val
340 345 350
Thr Thr Asp Leu Arg Gln Lys Cys Thr Glu Ser His Thr Gly Thr Ser
355 360 365
Ala Ser Ala Pro Leu Ala Ala Gly Ile Ile Ala Leu Thr Leu Glu Ala
370 375 380
Asn Lys Asn Leu Thr Trp Arg Asp Met Gln His Leu Val Val Gln Thr
385 390 395 400
Ser Lys Pro Ala His Leu Asn Ala Asn Asp Trp Ala Thr Asn Gly Val
405 410 415
Gly Arg Lys Val Ser His Ser Tyr Gly Tyr Gly Leu Leu Asp Ala Gly
420 425 430
Ala Met Val Ala Leu Ala Gln Asn Trp Thr Thr Val Ala Pro Gln Arg
435 440 445
Lys Cys Ile Ile Asp Ile Leu Thr Glu Pro Lys Asp Ile Gly Lys Arg
450 455 460
Leu Glu Val Arg Lys Thr Val Thr Ala Cys Leu Gly Glu Pro Asn His
465 470 475 480
Ile Thr Arg Leu Glu His Ala Gln Ala Arg Leu Thr Leu Ser Tyr Asn
485 490 495
Arg Arg Gly Asp Leu Ala Ile His Leu Val Ser Pro Met Gly Thr Arg
500 505 510
Ser Thr Leu Leu Ala Ala Arg Pro His Asp Tyr Ser Ala Asp Gly Phe
515 520 525
Asn Asp Trp Ala Phe Met Thr Thr His Ser Trp Asp Glu Asp Pro Ser
530 535 540
Gly Glu Trp Val Leu Glu Ile Glu Asn Thr Ser Glu Ala Asn Asn Tyr
545 550 555 560
Gly Thr Leu Thr Lys Phe Thr Leu Val Leu Tyr Gly Thr Ala Pro Glu
565 570 575
Gly Leu Pro Val Pro Pro Glu Ser Ser Gly Cys Lys Thr Leu Thr Ser
580 585 590
Ser Gln Ala Cys Val Val Cys Glu Glu Gly Phe Ser Leu His Gln Lys
595 600 605
Ser Cys Val Gln His Cys Pro Pro Gly Phe Ala Pro Gln Val Leu Asp
610 615 620
Thr His Tyr Ser Thr Glu Asn Asp Val Glu Thr Ile Arg Ala Ser Val
625 630 635 640
Cys Ala Pro Cys His Ala Ser Cys Ala Thr Cys Gln Gly Pro Ala Leu
645 650 655
Thr Asp Cys Leu Ser Cys Pro Ser His Ala Ser Leu Asp Pro Val Glu
660 665 670
Gln Thr Cys Ser Arg Gln Ser Gln Ser Ser Arg Glu Ser Pro Pro Gln
675 680 685
Gln Gln Pro Pro Arg Leu Pro Pro Glu Val Glu Ala Gly Gln Arg Leu
690 695 700
Arg Ala Gly Leu Leu Pro Ser His Leu Pro Glu
705 710 715
<210> 18
<211> 746
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-C1-Fc-L2-P
<400> 18
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Pro
485 490 495
Val Arg Leu Pro Gly Pro Ala Asp Lys Thr His Thr Cys Pro Pro Cys
500 505 510
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
515 520 525
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
530 535 540
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
545 550 555 560
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
565 570 575
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
580 585 590
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
595 600 605
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
610 615 620
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
625 630 635 640
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
645 650 655
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
660 665 670
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
675 680 685
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
690 695 700
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
705 710 715 720
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Ser Gly Gly Ser Gly
725 730 735
Gly Gly Gly Ser Leu Pro Glu Thr Gly Gly
740 745
<210> 19
<211> 2241
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-C1-Fc-L2-P
<400> 19
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatccgt cgctccacca cctgcactcc ccgcccctgt cagactgcct 1500
ggacccgctg ataagaccca cacatgtcca ccttgcccag cacctgagct gctgggcgga 1560
cctagcgtgt ttctgttccc accaaagcca aaggacaccc tcatgattag tagaacccca 1620
gaggtgactt gcgtcgtggt cgatgtcagt cacgaggacc ccgaggtgaa gtttaattgg 1680
tatgtagatg gcgtcgaggt gcacaacgct aagaccaagc cacgcgaaga gcagtacgca 1740
tctacttacc gggtcgtctc tgtgctgact gtgctgcacc aggattggct caatggcaaa 1800
gagtacaagt gtaaggtcag taataaggcc ctccccgcac ctattgaaaa gactattagc 1860
aaggcaaagg gccagcctag ggagccccag gtctacaccc tgcccccttc ccgcgatgag 1920
ctgactaaga atcaggtctc tctcacttgc ctcgtgaagg gcttttaccc atccgacatt 1980
gccgtggagt gggagtccaa cggacagcca gagaataact acaagacaac tcctcccgtg 2040
ctcgatagcg acggttcatt cttcctgtac tccaagctca ccgttgataa gtctcggtgg 2100
cagcagggaa atgtgttctc ttgttccgtc atgcacgagg cactgcacaa tcactacacc 2160
cagaagtccc tgtccctcag ccctggcaaa ggcagcggcg gatctggagg cggcggatcc 2220
ctcccagaga ccggcggatg a 2241
<210> 20
<211> 746
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-C2-Fc-L2-P
<400> 20
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Val
485 490 495
Ala Pro Pro Pro Ala Leu Pro Ala Asp Lys Thr His Thr Cys Pro Pro
500 505 510
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
515 520 525
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
530 535 540
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
545 550 555 560
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
565 570 575
Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
580 585 590
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser
595 600 605
Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys
610 615 620
Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
625 630 635 640
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe
645 650 655
Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
660 665 670
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
675 680 685
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
690 695 700
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
705 710 715 720
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Ser Gly Gly Ser Gly
725 730 735
Gly Gly Gly Ser Leu Pro Glu Thr Gly Gly
740 745
<210> 21
<211> 2241
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-C2-Fc-L2-P
<400> 21
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatccgt cgctccacca cctgcactcc ccgccgtggc tcctccacct 1500
gctttgcccg ctgataagac ccacacatgt ccaccttgcc cagcacctga gctgctgggc 1560
ggacctagcg tgtttctgtt cccaccaaag ccaaaggaca ccctcatgat tagtagaacc 1620
ccagaggtga cttgcgtcgt ggtcgatgtc agtcacgagg accccgaggt gaagtttaat 1680
tggtatgtag atggcgtcga ggtgcacaac gctaagacca agccacgcga agagcagtac 1740
gcatctactt accgggtcgt ctctgtgctg actgtgctgc accaggattg gctcaatggc 1800
aaagagtaca agtgtaaggt cagtaataag gccctccccg cacctattga aaagactatt 1860
agcaaggcaa agggccagcc tagggagccc caggtctaca ccctgccccc ttcccgcgat 1920
gagctgacta agaatcaggt ctctctcact tgcctcgtga agggctttta cccatccgac 1980
attgccgtgg agtgggagtc caacggacag ccagagaata actacaagac aactcctccc 2040
gtgctcgata gcgacggttc attcttcctg tactccaagc tcaccgttga taagtctcgg 2100
tggcagcagg gaaatgtgtt ctcttgttcc gtcatgcacg aggcactgca caatcactac 2160
acccagaagt ccctgtccct cagccctggc ggcagcggcg gatctggagg cggcggatct 2220
ctcccagaga ccggcggata a 2241
<210> 22
<211> 764
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-C3-Fc-L2-P
<400> 22
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Val
485 490 495
Ala Pro Pro Pro Ala Leu Pro Ala Val Ala Pro Pro Pro Ala Leu Pro
500 505 510
Ala Pro Val Arg Leu Pro Gly Pro Ala Asp Lys Thr His Thr Cys Pro
515 520 525
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
530 535 540
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
545 550 555 560
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
565 570 575
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
580 585 590
Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr
595 600 605
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
610 615 620
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
625 630 635 640
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
645 650 655
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
660 665 670
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
675 680 685
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
690 695 700
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
705 710 715 720
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
725 730 735
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys Gly Ser Gly Gly
740 745 750
Ser Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly Gly
755 760
<210> 23
<211> 2295
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-C3-Fc-L2-P
<400> 23
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatctgt cgctccacca cctgcactcc ccgccgtggc tcctccacct 1500
gctttgcccg ctgtcgctcc accacctgca ctccccgccc ctgtcagact gcctggaccc 1560
gctgataaga cccacacatg tccaccttgc ccagcacctg agctgctggg cggacctagc 1620
gtgtttctgt tcccaccaaa gccaaaggac accctcatga ttagtagaac cccagaggtg 1680
acttgcgtcg tggtcgatgt cagtcacgag gaccccgagg tgaagtttaa ttggtatgta 1740
gatggcgtcg aggtgcacaa cgctaagacc aagccacgcg aagagcagta cgcatctact 1800
taccgggtcg tctctgtgct gactgtgctg caccaggatt ggctcaatgg caaagagtac 1860
aagtgtaagg tcagtaataa ggccctcccc gcacctattg aaaagactat tagcaaggca 1920
aagggccagc ctagggagcc ccaggtctac accctgcccc cttcccgcga tgagctgact 1980
aagaatcagg tctctctcac ttgcctcgtg aagggctttt acccatccga cattgccgtg 2040
gagtgggagt ccaacggaca gccagagaat aactacaaga caactcctcc cgtgctcgat 2100
agcgacggtt cattcttcct gtactccaag ctcaccgttg ataagtctcg gtggcagcag 2160
ggaaatgtgt tctcttgttc cgtcatgcac gaggcactgc acaatcacta cacccagaag 2220
tccctgtccc tcagccctgg caaaggcagc ggcggatctg gaggcggcgg atccctccca 2280
gagaccggcg gatga 2295
<210> 24
<211> 763
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-C4-Fc-L2-P
<400> 24
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Pro
485 490 495
Val Arg Leu Pro Gly Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala
500 505 510
Val Ala Pro Pro Pro Ala Leu Pro Ala Asp Lys Thr His Thr Cys Pro
515 520 525
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
530 535 540
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
545 550 555 560
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
565 570 575
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
580 585 590
Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr
595 600 605
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
610 615 620
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala
625 630 635 640
Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg
645 650 655
Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
660 665 670
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro
675 680 685
Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
690 695 700
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
705 710 715 720
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
725 730 735
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Ser Gly Gly Ser
740 745 750
Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly Gly
755 760
<210> 25
<211> 2292
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-C4-Fc-L2-P
<400> 25
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatctgt cgctccacca cctgcactcc ccgcccctgt cagactgcct 1500
ggacccgctg tcgctccacc acctgcactc cccgccgtgg ctcctccacc tgctttgccc 1560
gctgataaga cccacacatg tccaccttgc ccagcacctg agctgctggg cggacctagc 1620
gtgtttctgt tcccaccaaa gccaaaggac accctcatga ttagtagaac cccagaggtg 1680
acttgcgtcg tggtcgatgt cagtcacgag gaccccgagg tgaagtttaa ttggtatgta 1740
gatggcgtcg aggtgcacaa cgctaagacc aagccacgcg aagagcagta cgcatctact 1800
taccgggtcg tctctgtgct gactgtgctg caccaggatt ggctcaatgg caaagagtac 1860
aagtgtaagg tcagtaataa ggccctcccc gcacctattg aaaagactat tagcaaggca 1920
aagggccagc ctagggagcc ccaggtctac accctgcccc cttcccgcga tgagctgact 1980
aagaatcagg tctctctcac ttgcctcgtg aagggctttt acccatccga cattgccgtg 2040
gagtgggagt ccaacggaca gccagagaat aactacaaga caactcctcc cgtgctcgat 2100
agcgacggtt cattcttcct gtactccaag ctcaccgttg ataagtctcg gtggcagcag 2160
ggaaatgtgt tctcttgttc cgtcatgcac gaggcactgc acaatcacta cacccagaag 2220
tccctgtccc tcagccctgg cggcagcggc ggatctggag gcggcggatc tctcccagag 2280
accggcggat ga 2292
<210> 26
<211> 766
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-C5-Fc-L2-P
<400> 26
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Val Ala Pro Pro Pro Ala Leu Pro Ala Val
485 490 495
Ala Pro Pro Pro Ala Leu Pro Ala Gly Ser Val Ala Pro Pro Pro Ala
500 505 510
Leu Pro Ala Val Ala Pro Pro Pro Ala Leu Pro Ala Asp Lys Thr His
515 520 525
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
530 535 540
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
545 550 555 560
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
565 570 575
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
580 585 590
Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser
595 600 605
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
610 615 620
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
625 630 635 640
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
645 650 655
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
660 665 670
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
675 680 685
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
690 695 700
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
705 710 715 720
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
725 730 735
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Gly Ser
740 745 750
Gly Gly Ser Gly Gly Gly Gly Ser Leu Pro Glu Thr Gly Gly
755 760 765
<210> 27
<211> 2301
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-C5-Fc-L2-P
<400> 27
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatctgt cgctccacca cctgcactcc ccgccgtggc tcctccacct 1500
gctttgcccg ctggatccgt cgctccacca cctgcactcc ccgccgtggc tcctccacct 1560
gctttgcccg ctgataagac ccacacatgt ccaccttgcc cagcacctga gctgctgggc 1620
ggacctagcg tgtttctgtt cccaccaaag ccaaaggaca ccctcatgat tagtagaacc 1680
ccagaggtga cttgcgtcgt ggtcgatgtc agtcacgagg accccgaggt gaagtttaat 1740
tggtatgtag atggcgtcga ggtgcacaac gctaagacca agccacgcga agagcagtac 1800
gcatctactt accgggtcgt ctctgtgctg actgtgctgc accaggattg gctcaatggc 1860
aaagagtaca agtgtaaggt cagtaataag gccctccccg cacctattga aaagactatt 1920
agcaaggcaa agggccagcc tagggagccc caggtctaca ccctgccccc ttcccgcgat 1980
gagctgacta agaatcaggt ctctctcact tgcctcgtga agggctttta cccatccgac 2040
attgccgtgg agtgggagtc caacggacag ccagagaata actacaagac aactcctccc 2100
gtgctcgata gcgacggttc attcttcctg tactccaagc tcaccgttga taagtctcgg 2160
tggcagcagg gaaatgtgtt ctcttgttcc gtcatgcacg aggcactgca caatcactac 2220
acccagaagt ccctgtccct cagccctggc ggcagcggcg gatctggagg cggcggatct 2280
ctcccagaga ccggcggata a 2301
<210> 28
<211> 728
<212> PRT
<213> Artificial sequence
<220>
<223> FIX-GS-Fc-L2-P
<400> 28
Met Gln Arg Val Asn Met Ile Met Ala Glu Ser Pro Gly Leu Ile Thr
1 5 10 15
Ile Cys Leu Leu Gly Tyr Leu Leu Ser Ala Glu Cys Thr Val Phe Leu
20 25 30
Asp His Glu Asn Ala Asn Lys Ile Leu Asn Arg Pro Lys Arg Tyr Asn
35 40 45
Ser Gly Lys Leu Glu Glu Phe Val Gln Gly Asn Leu Glu Arg Glu Cys
50 55 60
Met Glu Glu Lys Cys Ser Phe Glu Glu Ala Arg Glu Val Phe Glu Asn
65 70 75 80
Thr Glu Arg Thr Thr Glu Phe Trp Lys Gln Tyr Val Asp Gly Asp Gln
85 90 95
Cys Glu Ser Asn Pro Cys Leu Asn Gly Gly Ser Cys Lys Asp Asp Ile
100 105 110
Asn Ser Tyr Glu Cys Trp Cys Pro Phe Gly Phe Glu Gly Lys Asn Cys
115 120 125
Glu Leu Asp Val Thr Cys Asn Ile Lys Asn Gly Arg Cys Glu Gln Phe
130 135 140
Cys Lys Asn Ser Ala Asp Asn Lys Val Val Cys Ser Cys Thr Glu Gly
145 150 155 160
Tyr Arg Leu Ala Glu Asn Gln Lys Ser Cys Glu Pro Ala Val Pro Phe
165 170 175
Pro Cys Gly Arg Val Ser Val Ser Gln Thr Ser Lys Leu Thr Arg Ala
180 185 190
Glu Thr Val Phe Pro Asp Val Asp Tyr Val Asn Ser Thr Glu Ala Glu
195 200 205
Thr Ile Leu Asp Asn Ile Thr Gln Ser Thr Gln Ser Phe Asn Asp Phe
210 215 220
Thr Arg Val Val Gly Gly Glu Asp Ala Lys Pro Gly Gln Phe Pro Trp
225 230 235 240
Gln Val Val Leu Asn Gly Lys Val Asp Ala Phe Cys Gly Gly Ser Ile
245 250 255
Val Asn Glu Lys Trp Ile Val Thr Ala Ala His Cys Val Glu Thr Gly
260 265 270
Val Lys Ile Thr Val Val Ala Gly Glu His Asn Ile Glu Glu Thr Glu
275 280 285
His Thr Glu Gln Lys Arg Asn Val Ile Arg Ile Ile Pro His His Asn
290 295 300
Tyr Asn Ala Ala Ile Asn Lys Tyr Asn His Asp Ile Ala Leu Leu Glu
305 310 315 320
Leu Asp Glu Pro Leu Val Leu Asn Ser Tyr Val Thr Pro Ile Cys Ile
325 330 335
Ala Asp Lys Glu Tyr Thr Asn Ile Phe Leu Lys Phe Gly Ser Gly Tyr
340 345 350
Val Ser Gly Trp Gly Arg Val Phe His Lys Gly Arg Ser Ala Leu Val
355 360 365
Leu Gln Tyr Leu Arg Val Pro Leu Val Asp Arg Ala Thr Cys Leu Arg
370 375 380
Ser Thr Lys Phe Thr Ile Tyr Asn Asn Met Phe Cys Ala Gly Phe His
385 390 395 400
Glu Gly Gly Arg Asp Ser Cys Gln Gly Asp Ser Gly Gly Pro His Val
405 410 415
Thr Glu Val Glu Gly Thr Ser Phe Leu Thr Gly Ile Ile Ser Trp Gly
420 425 430
Glu Glu Cys Ala Met Lys Gly Lys Tyr Gly Ile Tyr Thr Lys Val Ser
435 440 445
Arg Tyr Val Asn Trp Ile Lys Glu Lys Thr Lys Leu Thr Gly Gly Gly
450 455 460
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
465 470 475 480
Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro
485 490 495
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
500 505 510
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
515 520 525
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
530 535 540
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
545 550 555 560
Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
565 570 575
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
580 585 590
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
595 600 605
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
610 615 620
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
625 630 635 640
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
645 650 655
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
660 665 670
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
675 680 685
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
690 695 700
Lys Ser Leu Ser Leu Ser Pro Gly Gly Ser Gly Gly Ser Gly Gly Gly
705 710 715 720
Gly Ser Leu Pro Glu Thr Gly Gly
725
<210> 29
<211> 2187
<212> DNA
<213> Artificial sequence
<220>
<223> FIX-GS-Fc-L2-P
<400> 29
atgcagcgcg tcaatatgat tatggccgag tctccaggcc tcattaccat ttgtctgctg 60
ggctacctgc tgtccgcaga gtgtaccgtc ttcctcgacc acgagaacgc aaataagatt 120
ctcaataggc ctaagcgcta caattccgga aagctcgaag agtttgtcca gggaaatctg 180
gagcgcgagt gtatggagga aaagtgtagc tttgaagaag ctagagaggt gtttgagaat 240
actgagcgca ctactgagtt ttggaagcag tatgtggatg gcgatcagtg cgagtccaat 300
ccatgcctga atggcggatc ttgcaaggat gatattaatt cttacgagtg ttggtgccca 360
ttcggctttg agggaaagaa ttgcgaactg gatgtcacat gtaacatcaa gaacggcaga 420
tgcgagcagt tttgtaagaa cagcgccgat aataaggtgg tgtgtagttg tactgaggga 480
taccgcctcg cagagaatca gaagtcctgt gagcccgctg tgccatttcc ttgtggccgg 540
gtcagcgtca gccagacttc taagctgaca cgcgccgaga ccgtctttcc agatgtggat 600
tatgtcaact ctactgaggc cgagaccatt ctcgataata ttactcagtc tacacagtcc 660
tttaacgact tcactcgggt ggtgggcgga gaggacgcaa agcctggcca gtttccttgg 720
caggtcgtgc tcaacggaaa ggtggacgca ttttgtggcg gatctatcgt gaatgagaag 780
tggattgtga ccgccgctca ctgcgtcgag acaggcgtca agattaccgt ggtcgctgga 840
gagcacaaca ttgaggaaac agagcacacc gagcagaagc gcaatgtgat tcgcatcatc 900
ccacaccaca actacaacgc cgctattaat aagtacaacc acgacatcgc cctgttagaa 960
ctagatgagc ctctggtcct gaatagttac gtcactccta tttgtatcgc cgataaagaa 1020
tacactaaca tctttctcaa gttcggctct ggctatgtat ctggatgggg acgggtgttt 1080
cacaagggac ggtccgcact cgtgctacaa tacctccggg tgccactggt ggatcgggca 1140
acatgcctcc ggtctaccaa gtttactata tacaataaca tgttttgcgc cggctttcac 1200
gagggcggac gggatagttg ccagggcgat agtggaggcc ctcatgtcac agaggtggag 1260
ggaactagct ttctgacagg cattatctct tggggcgaag agtgcgctat gaagggaaag 1320
tacggcatct atactaaggt ctcgcgttac gtcaattgga tcaaggaaaa gacaaagctg 1380
acaggcggcg gagggagcgg cggcggaggc tctggcggcg gcggatctgg cggaggcggc 1440
tctggcggag gcggatccga taagacccac acatgtccac cttgcccagc acctgagctg 1500
ctgggcggac ctagcgtgtt tctgttccca ccaaagccaa aggacaccct catgattagt 1560
agaaccccag aggtgacttg cgtcgtggtc gatgtcagtc acgaggaccc cgaggtgaag 1620
tttaattggt atgtagatgg cgtcgaggtg cacaacgcta agaccaagcc acgcgaagag 1680
cagtacgcat ctacttaccg ggtcgtctct gtgctgactg tgctgcacca ggattggctc 1740
aatggcaaag agtacaagtg taaggtcagt aataaggccc tccccgcacc tattgaaaag 1800
actattagca aggcaaaggg ccagcctagg gagccccagg tctacaccct gcccccttcc 1860
cgcgatgagc tgactaagaa tcaggtctct ctcacttgcc tcgtgaaggg cttttaccca 1920
tccgacattg ccgtggagtg ggagtccaac ggacagccag agaataacta caagacaact 1980
cctcccgtgc tcgatagcga cggttcattc ttcctgtact ccaagctcac cgttgataag 2040
tctcggtggc agcagggaaa tgtgttctct tgttccgtca tgcacgaggc actgcacaat 2100
cactacaccc agaagtccct gtccctcagc cctggcggca gcggcggatc tggaggcggc 2160
ggatctctcc cagagaccgg cggataa 2187
<210> 30
<211> 37
<212> DNA
<213> Artificial sequence
<220>
<223> F1
<400> 30
tcaggaagat ctcgcgaatt aattctgtgg aatgtgt 37
<210> 31
<211> 36
<212> DNA
<213> Artificial sequence
<220>
<223> R1
<400> 31
tatcctagat ctgctggcac gacaggtttc ccgact 36
<210> 32
<211> 87
<212> DNA
<213> Artificial sequence
<220>
<223> F2
<400> 32
ttcgcggatc cgtcgctcca ccacctgcac tccccgccgt ggctcctcca cctgctttgc 60
ccgctgataa gacccacaca tgtccac 87
<210> 33
<211> 87
<212> DNA
<213> Artificial sequence
<220>
<223> R2
<400> 33
taaccggaat tcattatccg ccggtctctg ggagagatcc gccgcctcca gatccgccgc 60
tgccgccagg gctgagggac agggact 87
<210> 34
<211> 48
<212> DNA
<213> Artificial sequence
<220>
<223> F3
<400> 34
tggctcctcc acctgctttg cccgctgtcg ctccaccacc tgcactcc 48
<210> 35
<211> 57
<212> DNA
<213> Artificial sequence
<220>
<223> F4
<400> 35
tagatctgtc gctccaccac ctgcactccc cgccgtggct cctccacctg ctttgcc 57
<210> 36
<211> 31
<212> DNA
<213> Artificial sequence
<220>
<223> R3
<400> 36
tgaattctta tcatccgccg gtctctggga g 31
<210> 37
<211> 47
<212> DNA
<213> Artificial sequence
<220>
<223> F5
<400> 37
ccctgtcaga ctgcctggac ccgctgtcgc tccaccacct gcactcc 47
<210> 38
<211> 54
<212> DNA
<213> Artificial sequence
<220>
<223> F6
<400> 38
tagatctgtc gctccaccac ctgcactccc cgcccctgtc agactgcctg gacc 54
<210> 39
<211> 27
<212> DNA
<213> Artificial sequence
<220>
<223> F7
<400> 39
atataagctt ccgccaccat gcagcgc 27
<210> 40
<211> 34
<212> DNA
<213> Artificial sequence
<220>
<223> R4
<400> 40
atatggatcc agcgggcaaa gcaggtggag gagc 34
<210> 41
<211> 37
<212> DNA
<213> Artificial sequence
<220>
<223> F8
<400> 41
gatttggatc cgataagacc cacacatgtc caccttg 37
<210> 42
<211> 87
<212> DNA
<213> Artificial sequence
<220>
<223> R5
<400> 42
taaccggaat tcattatccg ccggtctctg ggagagatcc gccgcctcca gatccgccgc 60
tgccgccagg gctgagggac agggact 87
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Слитый белок человеческого фактора свертывания IХ (FIX), способ его получения и применения | 2017 |
|
RU2736339C1 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| ПРОКОАГУЛЯНТНЫЕ АНТИТЕЛА | 2018 |
|
RU2810094C2 |
| УЛУЧШЕННЫЙ БЕЛОК СЛИЯНИЯ FVIII И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2789085C2 |
| ГЛИКОЗИЛИРОВАННЫЕ СЛИТЫЕ БЕЛКИ VWF С УЛУЧШЕННОЙ ФАРМАКОКИНЕТИКОЙ | 2017 |
|
RU2782212C2 |
| Антитела против ингибитора пути тканевого фактора и их применения | 2016 |
|
RU2815681C1 |
| СВЯЗЫВАНИЕ СЛИТОГО БЕЛКА С БЕЛКОМ CD47 И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2787521C2 |
| IL-12 ГЕТЕРОДИМЕРНЫЕ СЛИТЫЕ БЕЛКИ FC | 2019 |
|
RU2819097C2 |
| МУЛЬТИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА, ОБЛАДАЮЩАЯ ЗАМЕЩАЮЩЕЙ ФУНКЦИОНАЛЬНОЙ АКТИВНОСТЬЮ КОФАКТОРА КОАГУЛИРУЮЩЕГО ФАКТОРА КРОВИ VIII, И ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, СОДЕРЖАЩАЯ УКАЗАННУЮ МОЛЕКУЛУ В КАЧЕСТВЕ АКТИВНОГО ИНГРЕДИЕНТА | 2018 |
|
RU2812909C2 |
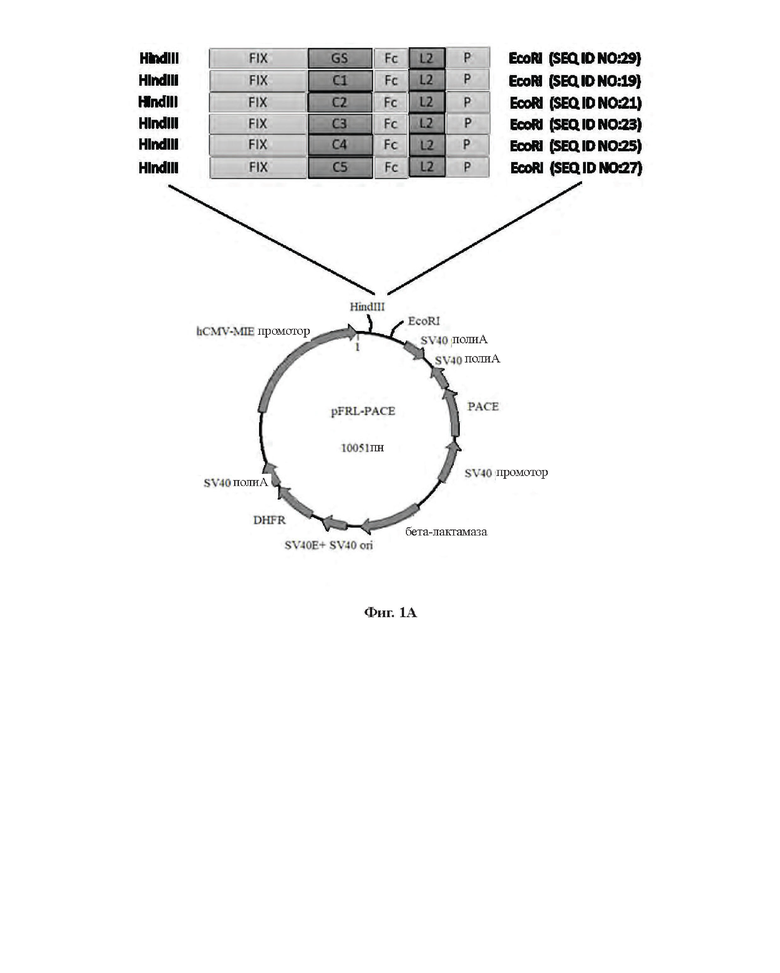
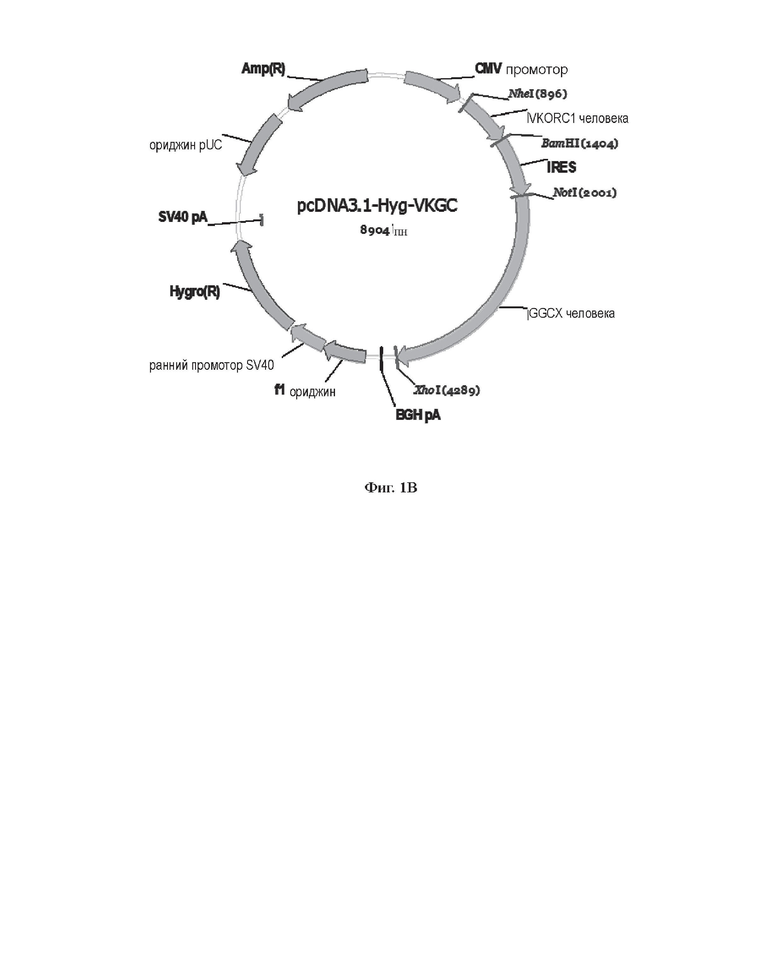
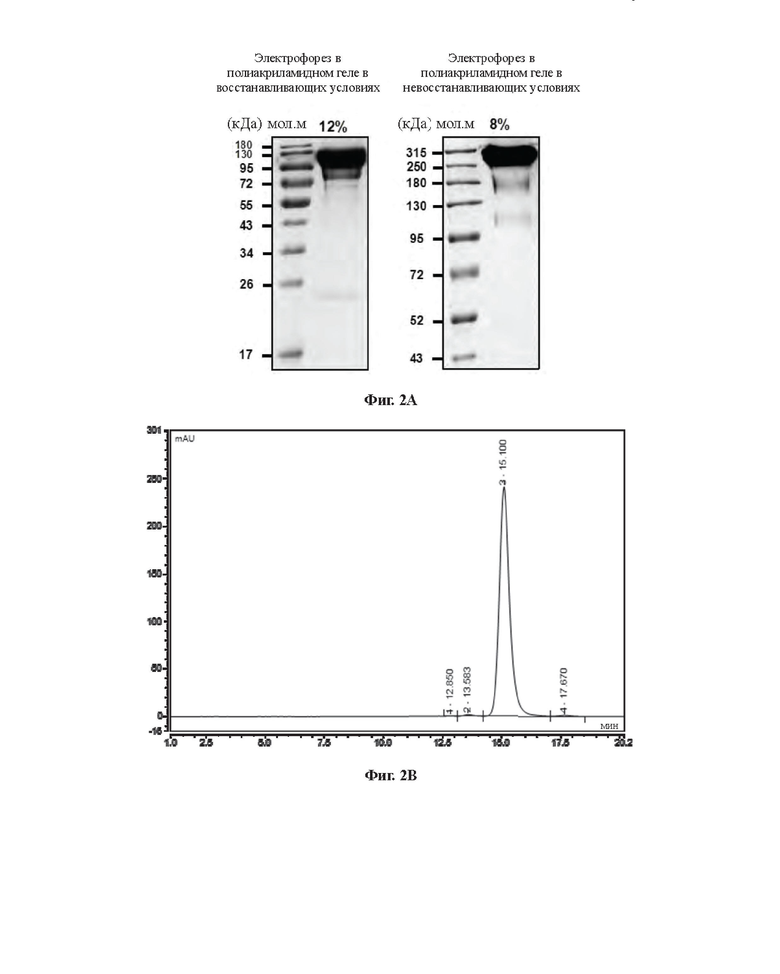
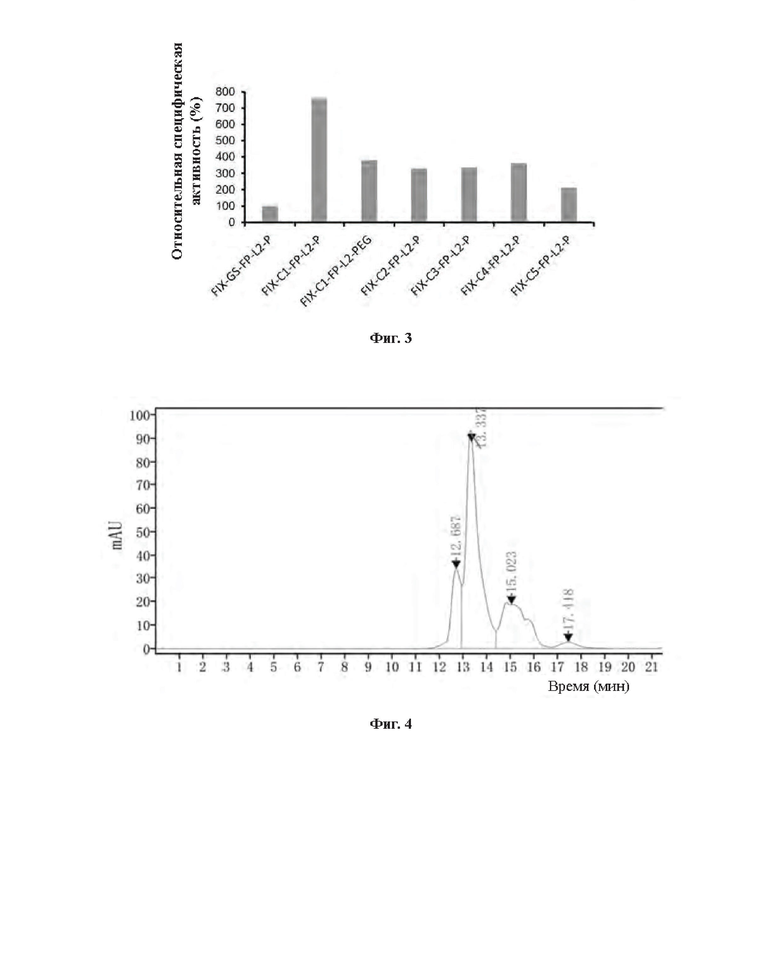
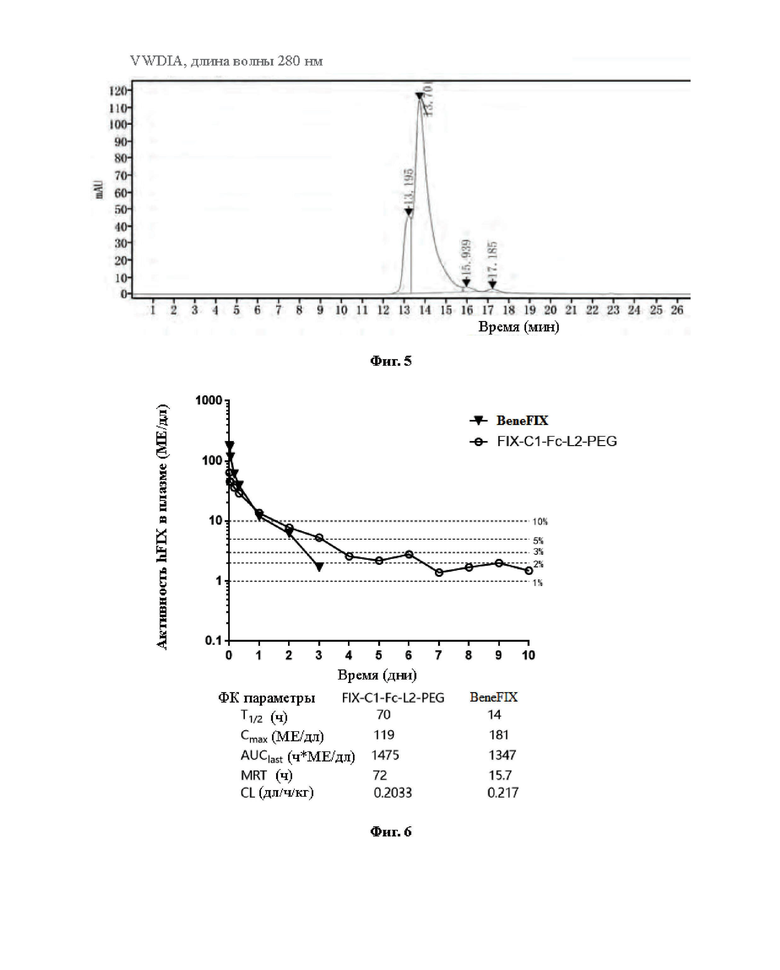
Изобретение относится к области биотехнологии, конкретно к конъюгатам фактора свертывания IX (FIX), и может быть использовано в медицине для предупреждения или лечения заболеваний, ассоциированных с врожденным или приобретенным дефицитом FIX. Предложен конъюгат, содержащий слитый белок FIX и полиэтиленгликоль с молекулярной массой 40 кДа, присоединенный к С-концу слитого белка. При этом слитый белок содержит активную группу FIX и партнер по слиянию (FP) и представлен структурой FIX-L1-Fс-L2-P, где FIX представляет собой полноразмерный человеческий фактор свертывания IX, партнер по слиянию представляет собой Fc-фрагмент иммуноглобулина из человеческого IgG1 c точечной мутацией N297A, L1 и L2 - пептидные линкеры, а P - аминокислотный фрагмент, содержащий сайт распознавания сортазой LPETGG. Изобретение обеспечивает получение конъюгата на основе FIX с увеличенным временем полужизни в кровотоке. 5 н. и 4 з.п. ф-лы, 6 ил., 2 табл., 5 пр. 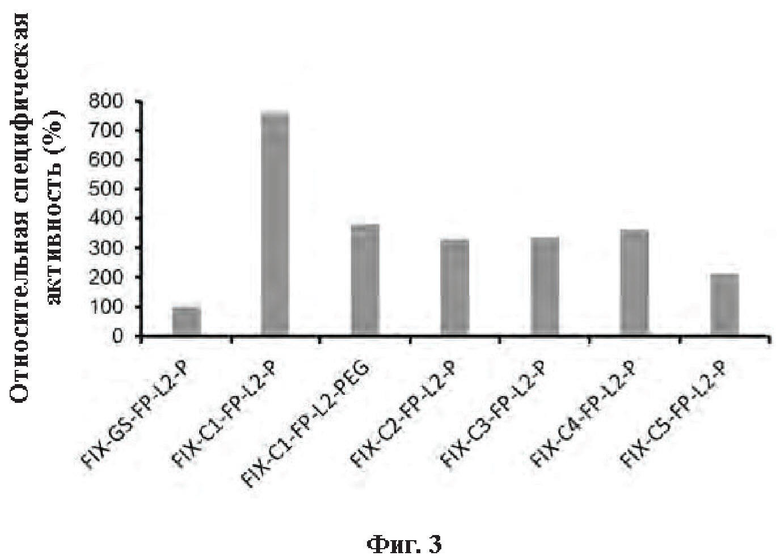
1. Конъюгат для предупреждения или лечения заболеваний, ассоциированных с врожденным или приобретенным дефицитом белка фактора свертывания IX (FIX), содержащий слитый белок фактора свертывания IX (FIX) и гидрофильный полимер, где гидрофильный полимер представляет собой полиэтиленгликоль, присоединенный к С-концу слитого белка, и указанный полиэтиленгликоль имеет молекулярную массу 40 кДа; и слитый белок содержит активную группу фактора свертывания IX и партнер по слиянию (FP), представленный структурой FIX-L1-Fс-L2-P, где
активная группа фактора свертывания IX представляет собой полноразмерный человеческий фактор свертывания IX, содержащий последовательность, приведенную в SEQ ID NO: 1; и
партнер по слиянию представляет собой Fc-фрагмент иммуноглобулина из человеческого IgG1 c точечной мутацией N297A,
L1 представляет собой первый линкер между FIX и Fс, L1 выбран из группы, состоящей из:
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPA (SEQ ID NO: 8),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPA (SEQ ID NO: 9),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAVAPPPALPAPV RLPGPA (SEQ ID NO: 10),
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPAVAPPPALPAVAP PPALPA (SEQ ID NO: 11) и
GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAVAPPPALPAGSVAPPPALPA VAPPPALPA (SEQ ID NO: 12);
L2 представляет собой второй линкер между Fс и сайтом распознавания сортазой Р, L2 содержит последовательность GSGGSGGGGS (SEQ ID NO: 16); и
P представляет собой аминокислотный фрагмент, содержащий сайт распознавания сортазой LPETGG.
2. Конъюгат по п. 1, в котором первый линкер L1 представляет собой GGGGSGGGGSGGGGSGGGGSGGGGSVAPPPALPAPVRLPGPA (SEQ ID NO: 8).
3. Конъюгат по п. 1, в котором слитый белок находится в форме мономера или димера.
4. Способ получения конъюгата по любому из пп. 1-3, включающий
экспрессию слитого белка фактора свертывания IX (FIX) в клетке-хозяине;
очистку слитого белка фактора свертывания IX (FIX);
приведение слитого белка фактора свертывания IX (FIX) в контакт с сортазой, предпочтительно сортазой А или сортазой B, и гидрофильным полимером, где один конец гидрофильного полимера имеет аминогруппу, которая может быть амидирована сортазой, где гидрофильный полимер представляет собой полиэтиленгликоль (PEG) с молекулярной массой 40 кДа.
5. Способ по п. 4, где N-конец гидрофильного полимера содержит поли-Gly, предпочтительно GGGAA.
6. Фармацевтическая композиция для предупреждения или лечения заболеваний, ассоциированных с врожденным или приобретенным дефицитом FIX, содержащая эффективное количество конъюгата по любому из пп. 1-3 и фармацевтически приемлемый носитель.
7. Фармацевтическая композиция по п. 6, которая находится в форме жидкого препарата или лиофилизированного препарата.
8. Применение конъюгата по любому из пп. 1-3 для изготовления лекарственного средства для предупреждения или лечения геморрагического заболевания, где геморрагическое заболевание выбрано из геморрагических заболеваний у пациентов с врожденным или приобретенным дефицитом FIX и спонтанного или послеоперационного кровотечения у пациентов с гемофилией B.
9. Набор для предупреждения или лечения заболеваний, ассоциированных с врожденным или приобретенным дефицитом FIX, содержащий конъюгат по любому из пп. 1-3 и инструкции по применению.
| CN 108137708 A, 08.06.2018, RU 2232192 C2, 10.07.2004, EA 201791134 A1, 28.02.2018, ORLANDO M., Modification of proteins and low molecular weight substances with hydroxyethyl starch (HES), Inauguraldissertation, Giesen, 2003, p.166, с.15 ACCHIONE M | |||
| et al., Impact of linker and conjugation chemistry on antigen binding, Fc receptor binding and |

