Настоящее изобретение относится к IL-17A-связывающим пептидам, ингибиторам связывания IL-17A с рецептором IL-17AR, и, тем самым, образования комплекса IL-17A/IL-17RA/IL17RC. Задачей изобретения также являются биоконъюгаты и димеры, содержащие указанные выше пептиды, фармацевтические композиции и медицинское применение указанных выше пептидов, биоконъюгатов и димеров.
Уровень техники, к которому относится изобретение
Ассоциированный с цитотоксическими T-лимфоцитами антиген 8 (CTLA8), также называемый интерлейкином 17 (IL-17), впервые был клонирован в 1993 году. Первой биологической активностью, описанной для IL-17 человека, была индукция продуцирования интерлейкина 6 (IL-6) и интерлейкина 8 (IL-8) из синовиоцитов при ревматоидном артрите. Эти данные позволили сделать предположение о роли IL-17 в воспалении через IL-6, и в привлечении нейтрофилов через IL-8, Xu S et al, Cell Mol Immunol. 2010, 164-74; Murcia RY et al., PLOS ONE 2016, 11(5): e0154755doi:10.1371/journal.pone.0154755).
Позднее было обнаружено, что эта молекула, позднее названная IL-17A, является частью более крупного семейства, которое включает пять дополнительных представителей, а именно, с IL-17B по F. IL-17F обладает наибольшей гомологией (приблизительно 56%) с первым открытым IL-17, IL-17A, в то время как IL-17E проявляет наиболее низкую консервативность последовательности (приблизительно 16%). Представители семейства IL-17 проявляют их функции, связываясь с их рецепторами в форме гомодимеров, за исключением IL-17A и IL-17F, которые также могут выступать в качестве гетеродимеров (Goepfert A et al, Sci Rep. 2017, 7 (1), 8906).
Наиболее широко исследованный цитокин этого семейства, IL-17A, играет важную роль в защите хозяина против микробных инфекций, и он считается основным пусковым фактором для ряда воспалительных и аутоиммунных состояний. Патологическое продуцирование IL-17A приводит к избыточному воспалению и заметному повреждению тканей.
В частности, посредством индукции различных молекул, включающих цитокины, хемокины, белки острой фазы, противомикробные пептиды, муцины и матриксные металлопротеиназы, IL-17A может усиливать каскады событий, которые приводят к привлечению нейтрофилов, воспалению и защите хозяина. Хотя IL-17A является характерным цитокином, продуцируемым хелперными T-клетками 17 (Th17), IL-17A, как и другие цитокины семейства IL-17, имеет множество источников в диапазоне от иммунных клеток до неиммунных клеток.
IL-17B, IL-17C и IL-17D также считаются провоспалительными цитокинами, однако их роль до конца не известна. IL-17E, также известный как IL-25, имеет наименьшую гомологию и вовлечен в ответы Th2-клеток против паразитов и аллергии. CCL20 запускает привлечение Th17 и дендритных клеток в область воспаления. В свою очередь Th17 клетки активируются, таким образом, продуцируя медиаторы воспаления и вызывая хроническое воспаление.
Цитокины, принадлежащие семейству IL-17, передают сигнал через их родственные рецепторы и активируют нижеследующие каскады, которые включают NFκB, MAPK и C/EBP, индуцируя экспрессию противомикробных пептидов, цитокинов и хемокинов. Проксимальный адаптер Act1 является общим медиатором передачи сигнала всех цитокинов IL-17, и он вовлечен в IL-17-опосредуемую защиту хозяина и в запускаемые IL-17 аутоиммунные состояния.
Семейство рецепторов IL-17 состоит из пяти представителей, IL-17RA, RB, RC, RD и RE, все из которых, подобно их лигандам, обладают гомологией последовательностей.
IL-17RA широко экспрессируется в большом диапазоне тканей и типов клеток и связывается с IL-17-A, C, E и F. Передача сигнала рецептора происходит через гетеродимерные рецепторы, образованные из общей субъединицы IL17-RA и второй субъединицы, которая зависит от лиганда и регулирует специфичность передачи сигнала. При стимуляции лигандом IL-17RA образует гетеродимерный рецепторный комплекс с IL-17RB (для IL-17E), IL-17RC (для IL-17A и IL-17F) или IL-17RE (для IL-17C). Предполагается, что связывание лиганда с первой субъединицей рецептора IL-17RA способствует второму событию связывания, таким образом, индуцируя образование гетеродимерного рецепторного комплекса. В частности, сигнал IL-17A и IL-17F опосредуется комплексом между IL-17RA и IL-17RC. IL-17F связывается с IL-17RA с приблизительно в 100-1000 раз меньшей аффинностью, чем IL-17A, в то время как аффинность связывания в отношении IL-17RC является сравнимой между двумя цитокинами (Onishi RM et al, Immunology 2010, 129(3), 311-321; Gu C et al, Cytokine 2013, 64(2), 477-485).
IL-17A у человека играет ключевую роль в различных аутоиммунных и воспалительных состояниях, таких как ревматоидный артрит, рассеянный склероз, псориаз, болезнь Крона, системная красная волчанка, астма, болезнь Бехчета и гипер-IgE-синдром (Fujino S et al Gut, 2003, 52, 65-70; Shabgah AG et al, Postepy Dermatol Allergol 2014, 31 (4), 256-261). Более того, блокада IL-17A продемонстрировала доклиническую и клиническую эффективность при анкилозирующем спондилите и ревматоидном артрите (Liu S et al, Nature-Scientific Reports 2016, doi:10,1038/srep26071; Lubberts E et al, Arthritis Rheum 2004, 50, 650-659).
Также было показано, что IL-17A вовлечен в заболевания глаз и, в частности, в патогенез заболеваний поверхности глаза и роговицы (DED), вирусный и бактериальный кератит. Антитела, разработанные для нейтрализации IL-17A, показали перспективные результаты в отношении снижения тяжести этих заболеваний (Garbutcheon-Singh KB et al, Curr Eye Res, 2018, DOI:10.1080/02713683.2018.1519834).
DED представляет собой воспалительное и аутоиммунное офтальмологическое заболевание слезной системы и поверхности глаза, которое приводит к дискомфорту, нарушению зрения и нарушению слезной пленки. DED является одной из наиболее частых причин приобретенного нарушения зрения во взрослой популяции (Stern ME et al, Mucosal Immunol 2010, 3(5), 425-442; Stevenson W et al, Arch Ophthalmol 2012, 130(1), 90-100, Parul Singh, Parul Singh 2012, Hah, Chung et al. 2017). Заболевание вовлекает иммунный и воспалительный процесс, который поражает поверхность глаза и в тяжелых случаях может приводить к слепоте. Лечение DED на текущий момент в основном является симптоматическим, состоящим в глазных смазывающих средствах и неспецифических противовоспалительных средствах, таких как кортикостероиды, циклоспорин A и такролимус.
IL-17A ассоциирован с нарушением барьерной функции эпителия роговицы, которое является наиболее угрожающим зрению осложнением DED. Описано увеличение количества Th17-клеток для тканей глаза пациентов с DED, которое индуцирует повышение концентраций IL-6, TGF-β, IL-23 и IL-17A на глазной поверхности, а также повышение концентрации IL-17 в слезах, и количество Th17-клеток на поверхности глаза в экспериментальных моделях DED. Кроме того, также было продемонстрировано, что нейтрализация IL-17A in vivo приводит к значительно ослабленной индукции и тяжести заболевания (De Paiva CS et al, Mucosal Immunol. 2009; 2(3): 243-253, Chauhan et al, Mucosal Immunol, 2009, 2(3), 243-253; Chauhan SK et al, Mucosal Immunol 2009, 2(4), 375-376; Chauhan, El Annan et al. 2009, De Paiva, Chotikavanich et al. 2009, de Paiva, Huang et al. 2014, Subbarayal, Chauhan et al. 2016).
IL-17A также вовлечен в псориаз. Псориаз представляет собой хроническое воспалительное заболевание. Он проявляется как сухие, возвышающиеся красные очаги на коже (бляшки), покрытые серебристыми чешуйками. Существуют многочисленные клинические фенотипы (т.е. бляшковидный, каплевидный, пустулезный, инверсный), причем тяжесть заболевания находится диапазоне от нескольких рассеянных бляшек до вовлечения значительной поверхности тела. Индивидуумы с псориазом имеют увеличенный риск развития других хронических и серьезных заболеваний, таких как псориатический артрит, метаболический синдром, сердечно-сосудистые заболевания и депрессия. Была продемонстрирована центральная роль IL-17A в патофизиологии псориаза (Zeichner JA et al J Clin Aesthet Dermatol 2016, 9 (6 Suppl 1), S3-S6). При псориазе экспрессия мРНК IL-17 является более высокой в пораженной коже по сравнению с непораженной кожей. Кроме того, уровни IL-17A значительно коррелируют с тяжестью заболевания (Arican Oet al. Mediators Inflamm 2005, 2005, 273-279, Takahashi K et al. Clin Exp Dermatol 2010, 35, 645-649). Было показано, что блокада IL-17A снижает гиперпролиферацию кератиноцитов, инфильтрацию T-клеток в дерму и экспрессию мРНК ключевых усиливающих заболевание генов (Krueger JG et al, J Allergy Clin Immunol 2012, 130, 145-154).
Антитела против IL-17A секукинумаб (AIN457, Consentyx™), полностью человеческое mAb IgG1k против IL-17A, и иксекизумаб (LY2439821), гуманизированное IgG4-антитело, были протестированы в клинических испытаниях при псориазе и одобрены для лечения псориаза от умеренного до тяжелого. В ходе клинических испытаний оба антитела были способны снижать тяжесть заболевания по меньшей мере на 75% (PASI75) в 80% популяции пациентов. Более того, современные передовые клинические испытания показали перспективные результаты также для лечения анкилозирующего спондилита и псориатического артрита. Другие антитела в настоящее время проходят клинические испытания по тем же показаниям.
Описанные выше данные подтверждают разработку молекул, способных нацеливаться на IL-17A и ингибировать его передачу сигнала для лечения указанных выше патологий. До недавнего времени нацеливание на IL17-A и его рецепторы оставалось уделом антител. Причиной предпочтительности антител является то, что цитокины, такие как IL-17A, являются мишенью с белок-белковым взаимодействием (PPI), на которую трудно осуществить нацеливание, до настоящего время считавшуюся не поддающейся медикаментозному нацеливанию. Действительно, поверхности контакта PPI, как правило, являются плоскими и лишены глубоких подкарманов и бороздок, которые обычно необходимы для связывания низкомолекулярных (SMW) молекул. Несмотря на это, в последние годы в новом исследовании было идентифицировано несколько SMW-антагонистов IL-17A, способных связывать IL-17A и препятствовать его взаимодействию с рецептором IL-17RA, и, таким образом, ингибировать активацию каскада IL-17A (Espada A et al. J Med Chem 2016, 59(5), 2255-2260; Ting JP et al PLoS One 2018, 13(1), e0190850; Liu S et al, Scientific reports, 6:26071, doi: 10.1038/srep26071). Среди них был описан пептид с последовательностью IHVTIPADLWDWINK, который был назван высокоаффинным пептидом (HAP) (Liu S et al, Scientific reports, 6:26071, doi: 10.1038/srep26071).
Однако все существует необходимость в идентификации и охарактеризации новых молекул, способных связываться с высокой аффинностью с IL-17A и препятствовать активации каскада IL-17A, для разработки новых терапевтических возможностей для лечения нарушений, вовлекающих ось IL-17A/IL-17RA.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения неожиданно обнаружили пептиды, способные связываться с конкретным участком на N-концевой части IL-17A и, тем самым, ингибировать взаимодействие с IL-17RA и образование комплекса IL-17A7IL-17RA/IL17RC.
Ингибирование указанного взаимодействия блокирует каскад передачи сигнала IL-17A. Таким образом, пептиды по изобретению способны снижать связанное с TH17 воспаление и последующее повреждение, наблюдаемое при воспалительных и аутоиммунных заболеваниях, где эти клетки и IL-17A играют ключевую роль.
Как будет показано в экспериментальном разделе, пептиды по изобретению обладают физико-химическими свойствами, которые делают их особенно пригодными для местного лечения офтальмологических и дерматологических заболеваний.
Такие пептиды являются пригодными для конструирования инновационного и специфического местного лечения заболеваний, зависимых от чрезмерного продуцирования или активности IL-17A.
Задачей настоящего изобретения являются пептиды, способные ингибировать связывание IL-17A с IL-17RA, димерами и биоконъюгатами, фармацевтические композиции, содержащие указанные пептиды, димеры или биоконъюгаты, и применение вышеуказанных для предупреждения и/или лечения аутоиммунных и воспалительных заболеваний, зависимых от IL-17A.
Краткое описание чертежей
На фиг. 1 представлены некоторые кривые доза-ответ, полученные в анализе димеризации IL-17RA-IL17RC, описанном в примере 5, с репрезентативными связывающими IL-17A пептидами по изобретению. Указан процент ингибирования димеризации IL-17RA/IL17RC, индуцированной IL-17A, при обработке различными пептидами.
На фиг. 2 представлена активность связывающего IL-17A пептида SEQ ID N.18 в отношении секреции IL6, IL-17A и IL23 дифференцированными и активированными клетками TH17 человека, измеренная, как описано в примере 6. Указана концентрация различных цитокинов в супернатанте клеток TH17 с или без (контрольного) пептида SEQ ID N.18.
На фиг. 3 представлена активность связывающего IL-17A пептида SEQ ID N.18 в отношении секреции металлопротеиназы 3 дифференцированными и активированными клетками TH17 человека, измеренная, как описано в примере 6. Указана концентрация металлопротеиназы 3 в супернатанте клеток TH17 с или без (контрольного) пептида SEQ ID N.18.
На фиг. 4 представлена жизнеспособность клеток TH17 человека через 24 ч в присутствии и в отсутствии (контрольного) пептида SEQ ID N 18, как описано в примере 6.
На фиг. 5 представлен участок связывания на IL-17A для пептидов IL-17A по изобретению.
На фиг. 6 представлены сенсограммы, полученные в эксперименте примера 9 с пептидом SEQ ID N 2 (верхняя панель) или SEQ ID N 1 (нижняя панель).
На фиг. 7 представлена величина константы аффинности при различных концентрациях пептида SEQ ID N 2 (верхняя панель) или SEQ ID N 1 (нижняя панель), полученная в эксперименте согласно примеру 9.
На фиг. 8 представлен уровень экспрессии IL-8, измеренный посредством ПЦР в реальном времени, как описано в примере 10, в клетках HCEC, не обработанных (носитель) или обработанных пептидом SEQ ID N. 2 (панель A) или пептидом HAP (SEQ ID N. 1) (панель B). Планки погрешности соответствуют стандартному отклонению. Для статистического анализа использовали T-критерий. Значение P *<0,05, **<0,005, ***<0,0005.
На фиг.9 представлено количество IL-6, измеренное посредством ПЦР в реальном времени, как описано в примере 10, в клетках HCEC, не обработанных (носитель) или обработанных пептидом SEQ ID N. 2 (панель A) или пептидом HAP (SEQ ID N. 1) (панель B). Планки погрешности соответствуют стандартному отклонению. Для статистического анализа использовали T-критерий. Значение P *<0,05, **<0,005, ***<0,0005.
На фиг.10 представлено количество TNF-α, измеренное посредством ПЦР в реальном времени, как описано в примере 10, в клетках HCEC, не обработанных (носитель) или обработанных пептидом SEQ ID N. 2 (панель A) или пептидом HAP (SEQ ID N. 1) (панель B). Планки погрешности соответствуют стандартному отклонению. Для статистического анализа использовали T-критерий. Значение P *<0,05, **<0,005, ***<0,0005.
Определения
Если не определено иначе, подразумевается, что все термины в данной области, обозначения и другая научная терминология, используемые в настоящем описании, имеют значения, часто подразумеваемые специалистами в области, к которой относится изобретение. В некоторых случаях термины с часто подразумеваемыми значениями определены в настоящем описании для ясности и/или для простоты отсылки; таким образом, включение таких определений в настоящее описание не следует истолковывать как отражение значительного отличия от того, что обычно подразумевается в данной области.
Термин "фармацевтически приемлемый эксципиент" относится к веществам, отличным от активного фармацевтического ингредиента (API), которые были надлежащим образом оценены в отношении безопасности и намеренно включены в систему доставки лекарственного средства. Фармацевтически приемлемые эксципиенты хорошо известны в данной области и описаны, например, в Handbook of Pharmaceutical Excipients, seventh edition 2013, которая включена в настоящее описание в качестве ссылки (Rowe, Sheskey et al. 2012).
Эксципиенты обычно классифицируют в зависимости от функции, которую они выполняют в конечной фармацевтической композиции. Предпочтительно, подходящие эксципиенты в соответствии с настоящим изобретением представляют собой, например, разбавитель, адсорбент, вещество, способствующее скольжению, связующее вещество, смазывающее вещество, поверхностно-активное вещество, разрыхлитель, консерванты, антиоксидант или их смеси.
Термины "примерно" и "приблизительно" в настоящем описании относятся к экспериментальной погрешности, которая может возникать в ходе эксперимента.
Термины "содержащий", "имеющий", "включающий" и "охватывающий" следует истолковывать как открытые термины (т.е. означающие "включая, но не ограничиваясь ими"), и их следует считать дающими основание также для терминов "по существу состоит из", "по существу состоящий из", "состоит из" или "состоящий из".
Термины "по существу состоит из", "по существу состоящий из" следует истолковывать как полузакрытые термины, что означает, что не включены никакие другие ингредиенты, которые значительно влияют на основные и новые характеристики изобретения (таким образом, могут быть включены необязательные эксципиенты).
Термины "состоит из", "состоящий из" следует истолковывать как закрытые термины.
Термин "биоконъюгат", как используют в рамках изобретения, относится к конъюгату, образованному посредством стабильного ковалентного связывания между различными молекулами, необязательно связанными посредством спейсера, по меньшей мере одна из которых представляет собой биомолекулу.
Термин "биомолекула", как используют в рамках изобретения, относится к молекулам биологического происхождения. Термин включает макромолекулы, такие как углеводы, липиды и белки, или небольшие природные продукты. Для целей настоящего изобретения "биомолекулы" предпочтительно выбраны из аскорбиновой кислоты, каприновой кислоты, капроновой кислоты, N-ацетилглюкозамина (также обозначаемого как NAG), N-ацетилмурамовой кислоты (также обозначаемой как NAM), гиалуроновой кислоты, альгиновой кислоты, хитина, (GalNAc)2, Gal-альфа1,3-GalNAc или тригалактуроновой кислоты.
Определение "консервативная замена" в настоящем описании относится к консервативной перестановке (также называемой консервативной мутацией или консервативной заменой) и представляет собой аминокислотную замену, которая изменяет данную аминокислоту на другую аминокислоту со сходными биохимическими свойствами (Simon French 1983).
Описание изобретения
Авторы настоящего изобретения идентифицировали ряд пептидов, которые способны связываться с высокой аффинностью с IL-17A и ингибировать его взаимодействие с рецептором ILRA. Эти соединения действуют в качестве ингибиторов передачи сигнала IL-17A.
Таким образом, первой задачей настоящего изобретения является пептид, способный ингибировать связывание IL-17A с ILRA, имеющий аминокислотную последовательность либо:
формулы (I):
(I) X1- X2-X3-X4- X5- X6-X7-X8-X9-X10-X11-X12-X13-X14-X15
где, независимо друг от друга:
X1 представляет собой I, V, D, K, W, A, G, L или P;
X2 представляет собой H, M, K, N, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, H, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q или он отсутствует;
X11 представляет собой D, E или N или он отсутствует;
X12 представляет собой W, F, V, H или Y или он отсутствует;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y или он отсутствует;
X14 представляет собой N, R, E, F, Q или D или он отсутствует;
X15 представляет собой K, R, E, F, V, W, H или D или он отсутствует;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1) или ее укороченной на C-конце последовательностью длиной 12-14 аминокислот IHVTIPADLWDWIN, IHVTIPADLWDWI или IHVTIPADLWDW;
либо формулы (II):
(II) an-DLSAVCWAFPWDPECH-bn’
где, независимо друг от друга:
a, b выбраны из A, R, N, D, C, E, Q, G, H, I, L, K, M, F, P, S, T, W, Y или V;
n=целое число от 0 до 3;
n’ = целое число от 0 до 3.
Предпочтительно, в формуле (II) каждая из аминокислот, отличных от a или b, может быть заменена консервативной заменой.
Предпочтительным пептидом формулы (II) является DLSAVCWAFPWDPECH (SEQ ID N. 41).
Особенно предпочтительными являются пептиды формулы (I).
Согласно предпочтительному варианту осуществления в указанном пептиде формулы (I):
X1 представляет собой I, V, D, K или W;
X2 представляет собой H, M, K или N;
X3 представляет собой V или F;
X4 представляет собой T, Q, H, V или Y;
X5 представляет собой I или F;
X6 представляет собой P или G;
X7 представляет собой A или Q;
X8 представляет собой D, E, N;
X9 представляет собой L, V, F или W;
X10 представляет собой W, Y, F или он отсутствует;
X11 представляет собой D, E, N или он отсутствует;
X12 представляет собой W, F, V или он отсутствует;
X13 представляет собой I, V, F или он отсутствует;
X14 представляет собой N, R, E, F или он отсутствует;
X15 представляет собой K, R, E, F, V, W или он отсутствует;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1) или ее укороченной на C-конце последовательностью длиной 12-14 аминокислот IHVTIPADLWDWIN, IHVTIPADLWDWI или IHVTIPADLWDW.
В соответствии с альтернативным предпочтительным вариантом осуществления в указанном пептиде формулы (I):
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q или он отсутствует;
X11 представляет собой D, E или N или он отсутствует;
X12 представляет собой W, F, H или Y или он отсутствует;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y или он отсутствует;
X14 представляет собой N, R, E, Q или D или он отсутствует;
X15 представляет собой K, R, E, V, W, H или D или он отсутствует;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1) или ее укороченной на C-конце последовательностью длиной 12-14 аминокислот IHVTIPADLWDWIN, IHVTIPADLWDWI или IHVTIPADLWDW.
Предпочтительно, в указанном выше пептиде формулы (I) все аминокислоты X1-X15 присутствуют и описанные выше пептиды формулы (I) имеют последовательность из 15 аминокислот. Предпочтительно, в пептидах (I), имеющих последовательность менее 15 аминокислот, когда отсутствует одна аминокислота, также отсутствуют все аминокислоты со стороны N-конца от указанной аминокислоты. Например, если аминокислота X10 отсутствует, также отсутствуют аминокислоты X11-X15.
Особенно предпочтительными пептидами формулы (I) являются пептиды, где X1 представляет собой V.
Конкретным предпочтительным пептидом, имеющим в качестве X1 V, является пептид SEQ ID N 2.
Как продемонстрировано в экспериментальном разделе, авторы настоящего изобретения обнаружили, что пептид SEQ ID N 2, где первая аминокислота изолейцин в пептиде HAP SEQ ID N.1 заменена валином, неожиданно демонстрирует поразительно отличающиеся функциональные и физико-химические свойства, а также улучшенную переносимость по сравнению с HAP.
В частности, как продемонстрировано в примерах, пептид демонстрирует улучшенные физико-химические свойства, которые являются особенно преимущественными для местного и офтальмологического применения, лучшую проницаемость и увеличенную аффинность в отношении IL-17A по сравнению с соответствующим пептидом SEQ ID N 1. Также авторы изобретения обнаружили, что пептид HAP, независимо от его активности в отношении передачи сигнала IL-17A, демонстрирует прямой эффект токсичности в отношении клеток роговицы путем индукции экспрессии воспалительных цитокинов. Неожиданно пептид SEQ ID N. 2 не демонстрирует этого эффекта и, таким образом, характеризуется улучшенной переносимостью по сравнению с HAP.
Предпочтительно, в описанном выше пептиде формулы (I):
X1 представляет собой I, V или L;
X2 представляет собой H, M, R, K или E;
X3 представляет собой V, F или I;
X4 представляет собой T, Q, S, Y или N;
X5 представляет собой I, F или V;
X6 представляет собой P;
X7 представляет собой A, Q или L;
X8 представляет собой D, E или Q;
X9 представляет собой L, W, F, V или I;
X10 представляет собой W, Y или F;
X11 представляет собой D, E или N;
X12 представляет собой W или F;
X13 представляет собой I, V, F или L;
X14 представляет собой N, R, Q или E;
X15 представляет собой K, R, H или E;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1);
или
X1 представляет собой I или V;
X2 представляет собой H, M или R,
X3 представляет собой V или F;
X4 представляет собой T или Q;
X5 представляет собой I, F или V;
X6 представляет собой P или G;
X7 представляет собой A или Q;
X8 представляет собой D или E;
X9 представляет собой L;
X10 представляет собой W или Y;
X11 представляет собой D или E;
X12 представляет собой W;
X13 представляет собой I или V;
X14 представляет собой N, R или E;
X15 представляет собой K, R или E;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1);
или
X1 представляет собой I или V;
X2 представляет собой H или M,
X3 представляет собой V;
X4 представляет собой T;
X5 представляет собой I;
X6 представляет собой P;
X7 представляет собой A;
X8 представляет собой D;
X9 представляет собой L, W, F, V или I;
X10 представляет собой W или Y;
X11 представляет собой D или E;
X12 представляет собой W;
X13 представляет собой I или V;
X14 представляет собой N, R или E;
X15 представляет собой K, R или E;
при условии, что указанная последовательность не является IHVTIPADLWDWINK (SEQ ID N. 1);
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой M, K, N, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, N, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой M, K, N, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
Или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой Y, F или Q;
X11 представляет собой E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой V, F, E, K, A, G, L, P или Y;
X14 представляет собой N, R, E, Q или D;
X15 представляет собой K, R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой R, E, Q или D;
X15 представляет собой R, E, V, W, H или D;
или
X1 представляет собой V, A, G, L или P;
X2 представляет собой H, M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой R, E, Q или D;
X15 представляет собой R, E, V, W, H или D;
или
X1 представляет собой I, V, A, G, L или P;
X2 представляет собой M, K, R, E, Q, W или Y;
X3 представляет собой V, F, A, G, L, P, I, Y или W;
X4 представляет собой T, Q, S, N или Y;
X5 представляет собой I, F, A, G, L, P, V или Y;
X6 представляет собой P, G, A, L, V, I или N;
X7 представляет собой A, Q, G, L, P, V, I, N или E;
X8 представляет собой D, E, N, Q или Y;
X9 представляет собой L, V, F, W, A, G, P, I или H;
X10 представляет собой W, Y, F или Q;
X11 представляет собой D, E или N;
X12 представляет собой W, F, H или Y;
X13 представляет собой I, V, F, E, K, A, G, L, P или Y;
X14 представляет собой R, E, Q или D;
X15 представляет собой R, E, V, W, H или D.
Особенно предпочтительными пептидами формулы (I) среди пептидов, описанных выше, являются пептиды, где X1 представляет собой V.
Как продемонстрировано в экспериментальном разделе, когда первая аминокислота в пептиде формулы (I) представляет собой валин, пептид неожиданно демонстрирует физико-химические свойства, особенно преимущественные для местного и офтальмологического применения, и увеличенную аффинность в отношении IL-17A.
Предпочтительно, в описанном выше пептиде формулы (I) присутствуют все аминокислоты с X1 по X15.
Предпочтительные индивидуальные пептиды формулы (I) в соответствии с настоящим изобретением приведены в таблице 1 ниже.
Таблица 1
Другими пептидными ингибиторами IL-17A согласно изобретению являются пептиды, приведенные в таблице 1A
Таблица 1A:
Другие аминокислотные последовательности могут быть добавлены к N- или C-концу пептидов согласно первой задаче настоящего изобретения.
Согласно предпочтительному варианту осуществления, описанные выше пептиды формулы (I) или (II) связаны на их N- и/или C-конце с дополнительной аминокислотной последовательностью, причем последовательность A имеет аминокислотную последовательность формулы (III)
(III) Y1-Y2-Y3- Y4-Y5-Y6- Y7-Y8-Y9- Y10-Y11-Y12- Y13-Y14-Y15-Y16
где:
Y1 представляет собой A, T, V, K, R, I, L, X или G;
Y2 представляет собой R, W, P, E, Q или A;
Y3 представляет собой K, W, G, T, I, R или P;
Y4 представляет собой K, T, E, W, A, R, D, G, X или F;
Y5 представляет собой A, E, T, G, W, I, R, P или V;
Y6 представляет собой A, W, E, R, G, P или отсутствует;
Y7 представляет собой K, S, W, T, F, R, V, G или отсутствует;
Y8 представляет собой A, Q, W, R, G или отсутствует;
Y9 представляет собой N, G или отсутствует;
Y10 представляет собой R или отсутствует;
Y11 представляет собой R или отсутствует;
Y12 представляет собой M или отсутствует;
Y13 представляет собой K или отсутствует;
Y14 представляет собой W или отсутствует;
Y15 представляет собой K или отсутствует;
Y16 представляет собой K или отсутствует.
Предпочтительно, в последовательности формулы (III), когда аминокислота отсутствует, также отсутствуют аминокислоты на N-конце указанных аминокислотных последовательностей. Например, если аминокислота Y6 отсутствует, также отсутствуют аминокислоты Y7-Y16.
Предпочтительные последовательности формулы (III) в соответствии с настоящим изобретением приведены в таблице 2 ниже.
Таблица 2
Таким образом, второй задачей изобретения является пептид, имеющий аминокислотную последовательность, содержащую, предпочтительно состоящую из, последовательности пептида формулы (I) или (II) согласно первой задаче изобретения и на C- и/или N-конце указанной последовательности A, как описано выше.
Аминокислотная последовательность A может быть связана как с N-, так и с C-концом пептида формулы (I) или (II). В таком случае две аминокислотных последовательности A, связанных с N- и C-концом указанного пептида, могут быть одинаковыми или могут различаться.
Альтернативно аминокислотная последовательность A может быть связана либо с N-, либо с C-концом пептида.
Предпочтительные индивидуальные пептиды согласно этой задаче изобретения приведены в таблице 3 ниже.
Таблица 3
В одном варианте осуществления, альтернативном тому, что описан выше, указанные пептиды формулы (I) или (II) согласно первой задаче изобретения могут быть связаны на их N- и/или C-конце с аминокислотной последовательностью (A)m, где A представляет собой аминокислоту, выбранную из R, K, G, E, Q или A, предпочтительно R, K, G, E или A, и m представляет собой целое число от 1 до 10.
Аминокислотная последовательность (A)m может быть связана как с N-, так и с C-концом пептида формулы (I) или (II). В таком случае две аминокислотных последовательности (A)m, связанных с N- и C-концом указанного пептида, могут быть одинаковыми или могут различаться.
Альтернативно аминокислотная последовательность (A)m может быть связана либо с N-, либо с C-концом указанного пептида.
Таким образом, первой задачей изобретения является пептид, имеющий аминокислотную последовательность, которая содержит, предпочтительно состоит из, последовательности пептида согласно первой задаче изобретения и на C- и/или N-конце указанной последовательности (A)m, как определено выше.
Предпочтительные индивидуальные пептиды согласно этому варианту осуществления изобретения приведены в таблице 4 ниже.
Таблица 4
В конкретном варианте осуществления указанные пептиды формулы (I) или (II) связаны как с последовательностью A, так и с последовательностью (A)m. В этом случае указанные пептиды связаны на их N- и/или C-конце с аминокислотной последовательностью (A)m, как определено выше, которая связана на ее N- и/или C-конце с аминокислотной последовательностью A, как описано выше.
Таким образом, четвертой задачей изобретения является пептид, имеющий аминокислотную последовательность, которая содержит, предпочтительно состоит из, последовательности пептида согласно первой задаче изобретения, на C- и/или N-конце указанной последовательности (A)m, как описано выше, или на C- и/или N-конце указанной последовательности A, как описано выше.
Согласно предпочтительному варианту осуществления пептида в соответствии с любой из описанных выше задач изобретения, указанный пептид связан на C- и/или N-конце с защитной кэп-группой, способной препятствовать деградации.
Можно использовать любую защитную кэп-группу, известную в данной области.
Предпочтительно, защитная группа, связанная с C-концом, выбрана из амидов, предпочтительно N-алкиламидов, альдегидов, сложных эфиров, предпочтительно метиловых и этиловых сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина.
Предпочтительно, защитная кэп-группа, связанная с N-концом, выбрана из ацетила, формила, пироглутамила, жирных кислот, мочевины, карбамата, сульфонамида, алкиламина.
Предпочтительно, описанные выше пептиды связаны с защитной кэп-группой только на N-конце. Более предпочтительно, указанные пептиды связаны с защитной кэп-группой только на N-конце, и защитная кэп-группа представляет собой ацетил.
Является особенно преимущественным получение димеров описанных выше пептидов согласно любой из различных задач по изобретению, предпочтительно пептидов формулы (I) или (II) согласно первой задаче изобретения, для повышения аффинности связывания и ингибиторной активности.
В соответствии с вышеуказанным, предпочтительно пептиды согласно различным задачам изобретения, описанные выше, предпочтительно пептиды формулы (I) или (II) согласно первой задаче изобретения, более предпочтительно пептиды формулы (I) согласно первой задаче изобретения, присутствуют в форме димеров.
Таким образом, пятой задачей настоящего изобретения является димер, образованный двумя пептидами согласно любой из указанных выше задач изобретения. Предпочтительно указанные пептиды представляют собой пептиды формулы (I) или (II) согласно первой задаче изобретения, более предпочтительно они представляют собой пептиды формулы (I) согласно первой задаче изобретения.
Предпочтительно, указанный димер представляет собой гомодимер, где два пептида, образующих димер, имеют идентичную последовательность. Предпочтительно, в указанном димере пептиды связаны посредством спейсерной молекулы, предпочтительно спейсера на основе полиэтиленгликоля.
Предпочтительно, согласно этому варианту осуществления, полиэтиленгликоль связан с N-концом пептидов согласно изобретению или с аминокислотами X4, X7 или X14 двух пептидов формулы (I).
Согласно следующему варианту осуществления, в отсутствии защитной кэп-группы пептиды согласно различным задачам изобретения связаны, предпочтительно на их C- и/или N-конце, с биомолекулой с образованием биоконъюгата. Предпочтительно, указанная биомолекула выбрана из каприновой кислоты, капроновой кислоты, аскорбиновой кислоты, NAG-NAM, NAG, NAM, гиалуроновой кислоты, альгиновой кислоты, хитина, (GalNAc)2, Gal-альфа1,3-GalNAc и тригалактуроновой кислоты.
Биолмолекулу связывают с пептидом либо для добавления пептиду конкретной функции, либо для модулирования физико-химических свойств пептида. Например, аскорбиновая кислота сообщает пептиду антиоксидантную активность, капроновая кислота способствует заякориванию пептида на клеточной мембране.
Таким образом, задачей изобретения является биоконъюгат, содержащий, предпочтительно состоящий из пептида согласно различным задачам изобретению, предпочтительно пептида формулы (I) или (II) согласно первой задаче изобретения, более предпочтительно пептида формулы (I) согласно первой задаче изобретения, и по меньшей мере биомолекулы, как описано выше.
Предпочтительно, в указанном биоконъюгате указанная биомолекула связана с N- и/или C-концом указанного пептида.
Предпочтительные биоконъюгаты согласно этому варианту осуществления приведены в таблице 5 ниже.
Таблица 5
Согласно одному варианту осуществления, в описанном выше биоконъюгате описанная выше биомолекула или защитная кэп-группа связана непосредственно с N- и/или C-концом пептида.
Согласно одному варианту осуществления, указанная биомолекула связана как с N-, так и с C-концом пептида. В таком случае две биомолекулы, связанные с N- и C-концом указанного пептида могут быть одинаковыми или могут различаться.
Согласно альтернативному варианту осуществления указанная биомолекула связана либо с N-, либо с C-концом пептида.
Согласно альтернативному варианту осуществления, описанная выше биомолекула или защитная кэп-группа связана с N- и/или C-концом пептида посредством линкера. Предпочтительно, указанный линкер выбран из 4-аминомасляной кислоты, бета-аланина, 2-аминоэтоксиуксусной кислоты, 5-аминовалериановой кислоты, 6-аминокапроновой кислоты, 8-амино-3,6-диоксаоктановой кислоты, 12-амино-4,7,10-триоксадодекановой кислоты, 15-амино-4,7,10,13-тетраоксапентадекановой кислоты и триоксатридекансукцинамовой кислоты. Предпочтительно, когда линкер связан с одной аминокислотой лизином, указанный линкер выбран из NHS-сложного эфира, изоцианатов, бензоилфторидов или карбаматов.
В соответствии другими вариантами осуществления, описанными выше, настоящее изобретение относится к соединению, содержащему или состоящему из связывающего IL-17A пептида формулы (I) или (II), описанного выше, и имеющему следующую формулу (IV):
(IV) [биомолекула или CAP]a - [линкер]b - (последовательность A)n-Am-[пептид] - A’m’-(последовательность A)n’-[линкер]b’-[биомолекула или CAP]a’
где независимо друг от друга:
a=0 или 1;
b=0 или 1;
a’ = 0 или 1;
b’ = 0 или 1;
m=0-10;
m’ = 0-10;
n= 0 или 5;
n’= 0 или 5,
и
пептид представляет собой полипептид формулы (I) или формулы (II);
A или A’ представляет собой одну аминокислоту, выбранную из R, K, G, E или A, повторенную m или m’ раз;
и последовательность A содержит последовательность формулы (III), описанную выше;
биомолекула представляет собой, в каждом случае независимо, капроновую кислоту, аскорбиновую кислоту, NAG-NAM, NAG, NAM, гиалуроновую кислоту, альгиновую кислоту, хитин, (GalNAc)2, Gal-альфа1,3-GalNAc или тригалактуроновую кислоту;
CAP представляет собой, в каждом случае независимо, амид, альдегид, сложный эфир, п-нитроанилид, 7-амино-4-метилкумарин, ацетил, формил, пироглутамил или жирную кислоту;
Линкер представляет собой, в каждом случае независимо, 4-аминомасляную кислоту, бета-аланин, 2-аминоэтоксиуксусную кислоту, 5-аминовалериановую кислоту, 6-аминокапроновую кислоту, 8-амино-3,6-диоксаоктановую кислоту, 12-амино-4,7,10-триоксадодекановую кислоту, 15-амино-4,7,10,13-тетраоксапентадекановую кислоту или триоксатридекансукцинамовую кислоту.
Согласно предпочтительному варианту осуществления, указанная последовательность A связана с N-концом или C-концом пептида формулы (I) или (II).
Некоторые примеры возможных соединений в соответствии с настоящим изобретением описаны в настоящем описании ниже, но могут присутствовать все другие возможные комбинации, соответствующие формуле (IV):
[биомолекула или кэп] - [линкер] - (последовательность A) -A-[пептид]-;
[биомолекула или кэп] - (последовательность A) -A-[пептид]-;
[биомолекула или кэп] -A-[пептид]-;
[биомолекула или кэп] -[пептид]-;
-[пептид] - Am’- (последовательность A)n’- [биомолекула или кэп]a’;
-[пептид] - Am’- [биомолекула или кэп]a’;
-[пептид]- [биомолекула или кэп]a’.
Согласно предпочтительному варианту осуществления, биомолекула в формуле (IV) по настоящему изобретению выбрана из аскорбиновой кислоты, капроновой кислоты, NAG или NAM.
Согласно следующему предпочтительному варианту осуществления кэп в соответствии с настоящим изобретением представляет собой C-концевую модификацию, выбранную из амидов, предпочтительно N-алкиламидов, альдегидов, сложных эфиров, предпочтительно метиловых и этиловых сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина, или N-концевую модификацию, выбранную из ацетила, формила, пироглутамила, жирных кислот, предпочтительно капроновой кислоты, мочевины, карбамата, сульфонамида или алкиламина, предпочтительно указанный кэп выбран из амидов, жирных кислот, таких как капроновая кислота, и ацетила.
В соответствии с настоящим изобретением, указанные линкер, биомолекула или кэп в формуле (IV) могут быть одинаковыми или могут различаться.
В следующем предпочтительном варианте осуществления настоящего изобретения линкер формулы (IV) выбран из 4-аминомасляной кислоты, бета-аланина, 2-аминоэтоксиуксусной кислоты, 5-аминовалериановой кислоты или триоксатридекансукцинамовой кислоты.
Предпочтительно, когда линкер связан с одной аминокислотой лизином, указанный линкер выбран из NHS-сложного эфира, изоцианатов, бензоилфторидов или карбаматов.
Седьмой задачей настоящего изобретения является фармацевтическая композиция, содержащая указанный пептид, димер или биоконъюгат согласно задачам изобретения, как описано выше, и по меньшей мере один фармацевтически приемлемый эксципиент. Седьмой задачей настоящего изобретения является фармацевтическая композиция, содержащая указанный пептид, димер или биоконъюгат согласно задачам изобретения, как описано выше, и по меньшей мере один фармацевтически приемлемый эксципиент.
Фармацевтическая композиция по настоящему изобретению может быть составлена в форме, пригодной для местного или офтальмологического введения.
Предпочтительно, когда введение фармацевтической композиции по изобретению проводят местным путем, фармацевтическую форму выбирают из крема, мази, геля, бальзама, раствора, жидкости для полоскания, суспензии, капель, буфера (буферного раствора), суспензии, глазных капель, капель, спрея, влажной салфетки или порошка, предпочтительно она выбрана из крема, геля, спрея или мази.
Для офтальмологического введения фармацевтическая форма предпочтительно выбрана из глазных капель, офтальмологических гелей, мазей, жидкостей для полоскания, влажных салфеток, спреев или кремов.
Согласно конкретному варианту осуществления пептиды или биоконъюгаты по настоящему изобретению вводят местным путем с использованием микрочастиц или наночастиц.
Согласно изобретению фармацевтическую композицию по настоящему изобретению можно вводить животным и людям, определяемым как взрослые и как "педиатрическая популяция", где термин "педиатрическая популяция" указывает на часть популяции от рождения до восемнадцати лет.
Восьмой задачей настоящего изобретения является описанный выше пептид, димер или биоконъюгат согласно изобретению для применения для лечения и/или предупреждения воспалительного и аутоиммунного заболевания.
Девятой задачей изобретения является способ лечения и/или предупреждения воспалительного и аутоиммунного заболевания, включающий введение пациенту, нуждающемуся в этом, терапевтически эффективного количества описанного выше пептида, димера или биоконъюгата согласно изобретению.
Предпочтительно, указанное воспалительное и аутоиммунное заболевание выбрано из ревматоидного артрита, рассеянного склероза, болезни Крона, системной красной волчанки, астмы, болезни Бехчета, гипер-IgE-синдрома, анкилозирующего спондилита, псориаза, псориатического артрита, ревматоидного артрита, сухого кератоконъюнктивита, весеннего кератоконъюнктивита, стромального герпетического кератита, отторжения аллотрансплантата роговицы, инфекций роговицы, предпочтительно обусловленного вирусом герпеса и Pseudomonas aeruginosa кератита, и болезни сухого глаза. Более предпочтительно указанное заболевание представляет собой аутоиммунное офтальмологическое или дерматологическое заболевание, еще более предпочтительно оно представляет собой болезнь сухого глаза или псориаз.
Общим для всех из указанных выше заболеваний и медицинских состояний является то, что их происхождение и/или симптомы связаны с IL-17A и/или Th-17.
Более конкретно для последнего, болезни сухого глаза (DED), в высокой степени распространенного состояния, которое включает широкий спектр нарушений поверхности глаза, воспаление слизистой оболочки глаза является отличительной чертой, потенциально вызывающей снижение остроты зрения, если его не контролировать, вследствие индуцированной воспалением изъязвления и рубцевания роговицы. При прогрессировании DED, патогенные иммунные клетки, преимущественно Th17-клетки, непрерывно мигрируют на поверхность слизистой оболочки глаза и секретируют провоспалительные медиаторы, включая IL17, вызывая воспаление и эпителиопатию поверхности глаза.
Пептиды, димеры или биоконъюгаты по изобретению удобно и предпочтительно вводить местным путем в качестве глазных капель и офтальмологических гелей и мазей.
В следующем варианте осуществления пептид, димер или биоконъюгат для применения в соответствии с настоящим изобретением вводят в качестве единственного активного вещества или в комбинации с другими активными веществами, и/или в комбинации с медицинскими устройствами для симптоматического лечения офтальмологических состояний, включая, но не ограничиваясь ими, DED, например, глазные смазывающие вещества или "искусственные слезы", местные вызывающие реэпителизацию средства, терапевтические контактные линзы и обтураторы слезных точек.
Предпочтительно, указанное дополнительное активное вещество представляет собой адъювант, иммуносупрессивное средство, иммуномодулирующее средство или противовоспалительное средство.
Например, связывающий IL-17A пептид по настоящему изобретению можно использовать в комбинации с DMSO.
Согласно предпочтительному варианту осуществления, связывающий IL-17A пептид по настоящему изобретению можно использовать в комбинации с иммуносупрессивными моноклональными антителами, такими как моноклональные антитела, обладающие аффинностью к рецепторам лейкоцитов, выбранным из MHC, CD2, CD3, CD4, CD7, CD8, CD25, CD28, CD40, CD45, CD58, CD80, CD86, или их лигандам; другие иммуномодулирующие соединения, предпочтительно рекомбинантная связывающая молекула, имеющая по меньшей мере часть внеклеточного домена CTLA4 или ее мутант, по меньшей мере внеклеточной части CTLA4 или ее мутанта, связанных с белковой последовательностью не из CTLA4, например (например, CTLA4lg, обозначаемая как ATCC 68629) или ее мутантом, например LEA29Y; ингибиторы молекулы адгезии, антагонисты LFA-I, антагонисты ICAM-I или -3, антагонисты VCAM-4 или антагонисты VLA-4.
В следующем предпочтительном варианте осуществления связывающий IL-17A пептид по настоящему изобретению используют в комбинации с DMARD (модифицирующее заболевание противоревматическое лекарственное средство), предпочтительно солями золота, сульфасалазином, противомалярийными средствами, метотрексатом, D-пеницилламином, азатиоприном, микофеноловой кислотой, циклоспорином A, такролимусом, сиролимусом, миноциклином, лефлуномидом, глюкокортикоидами; ингибитором кальциневрина, предпочтительно циклоспорином A или FK 506; модулятором рециркуляции лимфоцитов, предпочтительно FTY720 и аналогами FTY720; ингибитором mTOR, предпочтительно рапамицином, 40-O-(2-гидроксиэтил)рапамицином, CCI779, ABT578, AP23573 или TAFA-93; аскомицином, имеющим иммуносупрессивные свойства, предпочтительно ABT-281, ASM981; кортикостероидами; циклофосфамидом; азатиопреном; метотрексатом; лефлуномидом; мизорибином; микофеноловой кислотой; микофенолатом мофетилом; 15-дезоксиспергуалином или их иммуносупрессивным гомологом, аналогом или производным; или химиотерапевтическим средством, предпочтительно паклитакселом, гемцитабином, цисплатином, доксорубицином или 5-фторурацилом; средствами против TNF, предпочтительно моноклональными антителами к TNF, предпочтительно инфликсимабом, адалимумабом, CDP870 или рецепторными конструкциями к TNF-RI или TNF-RII, предпочтительно этанерцептом, PEG-TNF-RI; блокаторами провоспалительных цитокинов, блокаторами IL-1, предпочтительно анакинрой или ловушкой IL-1, AAL160, ACZ 885, блокаторами IL-6; ингибиторами или активаторами протеаз, предпочтительно металлопротеиназ, антителами против IL-15, антителами против IL-6, антителами против IL-23, антителами против IL-22, антителами против IL-21, антителами против IL-12, антителами против IFN-гамма, антителами против IFN-альфа, антителами против CD20, противовоспалительными средствами, предпочтительно аспирином или противоинфекционным средством.
Естественно, этот перечень средств для совместного введения не является ни ограничивающим, ни полным.
Следующей задачей настоящего изобретения является одни из пептидов, приведенных в таблице 6, для применения для лечения и/или предупреждения воспалительного и аутоиммунного заболевания, описанного выше.
Таблица 6.
Количество и путь введения пептидов в соответствии с настоящим изобретением варьируется в зависимости от конкретного пептидного ингибитора по изобретению, индивидуальной группы пациентов или пациента, присутствия других медицинских активных соединений и природы и тяжести подвергаемого лечению состояния.
Согласно предпочтительному варианту осуществления дозировки для профилактического и/или терапевтического применения составляют приблизительно 5-50 мкг/мл, предпочтительно приблизительно 10-25 мкг/мл.
Предпочтительно, частота введения для профилактического и/или терапевтического применения находится в диапазоне применений от одного до двух раз в сутки, предпочтительно один раз в сутки.
Кроме того, изобретение описывается посредством иллюстрации следующих примеров, ни один из которых не следует интерпретировать как ограничивающий объем изобретения, указанный в прилагаемой формуле изобретения.
Примеры
Пример 1 Синтез и очистка пептидов:
Авторы изобретения разработали модель гомологии IL-17A с использованием доступных кристаллографических данных (PDB 5h13, PDB 5vb9 и PDB 4hsa) (Liu et al. Nat Commun 2013, 4, 1888, Liu et al. Sci Rep 2016, 6: 30859, Liu et al. Sci Rep 2016, 6, 26071, Ting, Tung et al. Plos One 2018, 13(1): e01908502018). В моделях авторов изобретения они также воссоздали петли, нерешенные в кристаллах и оптимизированных моделях посредством имитации молекулярной динамики, проведенной с использованием программного обеспечения Desmond, осуществленного в пакете программ Schrodinger Maestro macromodel suite. Все молекулярные имитации проводили в течение 1 микросекунды для гарантии стабильности системы.
На основе структуры и свойств α-кармана IL-17A, были сконструированы различные пептидные последовательности, чтобы получить i) стабильную вторичную и третичную конформацию, которая вовлекает IL-17A с оптимизированной занятостью участка связывания белка, и ii) физико-химические свойства, пригодные для местного применения при офтальмологической и дерматологической патологии (Liu et al. Nat Commun 2013, 4, 1888, Espada et al J Med Chem 2016, 59(5), 2255-2260).
Кроме того, авторы изобретения оптимизировали взаимодействия лиганд-IL-17A путем внесения водородных связей и оптимизации гидрофобных взаимодействий.
a. Твердофазный синтез пептидов на основе Fmoc
Все химические реагенты приобретали и использовали без дальнейшей очистки.
Пептиды SEQ ID N. 1-43 и 111-257, приведенные в таблице 1, получали путем ручного или автоматического твердофазного синтеза пептидов на амидной MBHA смоле Ринка в качестве твердой подложки, как описано ниже, и с N-концевым амином, кэппированным ацетильной группой.
Группу Fmoc (9-флуоренилметоксикарбонил), использованную для защиты Nα, расщепляли посредством обработки в течение 8 мин 20% пиперидином в диметилформамиде (DMF), за которой следовала дальнейшая обработка тем же реагентом в течение 10 мин. После отщепления Fmoc комплекс пептид-смола промывали DMF (×6). Затем включали следующий остаток с использованием протокола присоединения с использованием гексафторфосфата 3-оксида 1-[бис(диметиламино)метилен]-1H-1,2,3-триазоло[4,5-b]пиридиния (HATU) / N, N-диизопропиламина (DIPEA) [Fmoc-аминокислота (3 экв.), HATU (3 экв.) и DIPEA (6 экв.)]. После осторожного перемешивания (1 ч) и промывания DMF (×6), часть пептидной смолы анализировали посредством теста Кайзера. После завершения сборки пептидную смолу промывали DMF (×3), дихлорметаном (DCM) (×4), а затем сушили в вакууме.
Расщепление и удаление защитной группы
Пептид отщепляли от смолы и при необходимости защитную группу удаляли с использованием в качестве раствора для расщепления смеси трифторуксусная кислота/ 1,2-этандитиол/ тиоанизол/ фенол/ H2O/ триизопропилсилан в соотношении 68,5/ 10/ 10/ 5/ 3,5/ 1 об./об. Использовали 3 мл раствора для расщепления на 100 мг смолы. Полного удаления защитной группы достигали при перемешивании в течение 4 часов при 30°C. После реакции расщепления пептид в смеси преципитировали в холодном диэтиловом эфире и сушили в вакууме при 50°C.
b. Образование дисульфидной связи в пептиде 41
Образование дисульфидной связи в пептиде SEQ ID N. 41, приведенном в таблице 1, проводили путем инкубации пептида в смеси DMSO/H2O (20/80, об./об.), мониторинг которой проводили посредством ОФ-ВЭЖХ, MS и детекции свободных сульфгидрилов (способ с DTNB).
c. Очистка посредством препаративной ОФ-ВЭЖХ и оценка чистоты посредством аналитической ВЭЖХ
Неочищенные образцы пептидов очищали посредством препаративной ОФ-ВЭЖХ (Agilent) с использованием колонки C18 (10 мкм, 100 Å, 50×250 мм). Для элюирования использовали систему растворителей, состоящую из растворителя A (0,1% TFA, 2% CH3CN в воде) и растворителя B (90% CH3CN/H2O) при скорости потока 25 мл/мин, и проводили детекцию поглощения при 220 нм. Растворитель удаляли посредством лиофилизации. Конечные продукты охарактеризовывали посредством MALDI-TOF-MS, и чистоту материала оценивали посредством аналитической ОФ-ВЭЖХ (C18-250 мм × 4,6 мм в.д., скорость потока 1 мл/мин), и детекцию поглощения проводили при 220 нм.
Пример 2: Получение биоконъюгированных с NAM и/или NAG пептидов BIO-1, BIO-2 и BIO-3
Биоконъюгированные пептиды BIO-1, BIO-2 и BIO-3 получали посредством реакции конъюгации пептида 42 или 43 по следующим методикам.
1. Получение донора и акцептора
Все реакции проводили в атмосфере N2. Спектры ЯМР получали на устройстве Brucker 400 МГц. Анализ посредство ВЭЖХ-УФ проводили на системе Agilent 1260 Infinity System, оборудованной насосом для четырехкомпонентных смесей G1311B Agilent 1260, автоматическим пробоотборником G1329B Agilent 1260, детектором на диодной матрице G1315C Agilent 1260, и термостатными модулями колонки G1316A Agilent 1260. Использовали колонку Phenomenex GEMINI C18 150 × 4,6 мм2 (5 мкм). Подвижная фаза A представляла собой воду MilliQ с 0,05% TFA, и B представляла собой ацетонитрил категории ВЭЖХ с 0,05% TFA при скорости потока 1,0 мл/мин. Система ВЭЖХ была сопряжена с масс-спектрометром LC/MS Agilent Quadrupole 6120, работающим в режиме положительных ионов. Ионы получали с использованием источника ионов с электрораспылительной ионизацией. Полученные данные обрабатывали с использованием программного обеспечения Agilent Chemstation Software.
a. Синтез 1-(2,2,2-трихлорацетимин)2-дезокси-3-O-ацетил-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (донор)
Трихлорэтоксикарбонилхлорид (2,1 мл, 15,3 ммоль) капельно добавляли при комнатной температуре к энергично перемешиваемому раствору гидрохлорида D-глюкозамина (3 г, 13,9 ммоль) и NaHCO3, (3,5 г, 41,7 ммоль) в воде (30 мл). Смесь перемешивали в течение 2 ч, затем нейтрализовывали 1 M HCl, концентрировали и сушили в вакууме. Получали 2-дезокси-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозид (промежуточное соединение A).
Полученный неочищенный продукт прямо использовали для следующей стадии реакции без дальнейшей очистки.
В деталях, хлорид цинка (1,9 г, 13,9 ммоль) добавляли к раствору промежуточного соединения A (4,9 г, 13,9 ммоль) в бензальдегиде (24 мл) и молекулярных сит 4 Å (4 Å MS, 600 мг). После перемешивания в течение ночи при комнатной температуре добавляли насыщенный водный NaHCO3 (30 мл) и диэтиловый эфир (90 мл) и реакционную смесь перемешивали в течение 15 мин. Образовавшийся преципитат фильтровали, промывали водой, диэтиловым эфиром и сушили. Остаток растворяли в пиридине (13 мл), охлаждали до 0°C, обрабатывали уксусным ангидридом (6,4 мл, 68 ммоль) и перемешивали в течение ночи при комнатной температуре, затем раствор концентрировали с толуолом, охлаждали до 0°C и экстрагировали DCM и насыщенным водным NaHCO3 (3x), а затем промывали насыщенным раствором NaCl. Объединенные органические слои сушили, концентрировали и неочищенный материал очищали посредством Isolera (EtPet/EtOAc) с получением 1,3-ди-O-ацетил 2-дезокси-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение B), в виде белого твердого вещества (3,05 г, выход 41,7%).
В деталях, хлорид цинка (1,9 г, 13,9 ммоль) добавляли к раствору промежуточного соединения A (4,9 г, 13,9 ммоль) в бензальдегиде (24 мл) и молекулярных сит 4 Å (4 Å MS, 600 мг). После перемешивания в течение ночи при комнатной температуре добавляли насыщенный водный NaHCO3 (30 мл) и диэтиловый эфир (90 мл) и реакционную смесь перемешивали в течение 15 мин. Образовавшийся преципитат фильтровали, промывали водой, диэтиловым эфиром и сушили. Остаток растворяли в пиридине (13 мл), охлаждали до 0°C, обрабатывали уксусным ангидридом (6,4 мл, 68 ммоль) и перемешивали в течение ночи при комнатной температуре, затем раствор концентрировали с толуолом, охлаждали до 0°C и экстрагировали DCM и насыщенным водным NaHCO3 (3x), а затем промывали насыщенным раствором NaCl. Объединенные органические слои сушили, концентрировали и неочищенный материал очищали посредством Isolera (EtPet/EtOAc) с получением 1,3-ди-O-ацетил-2-дезокси-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение B) в виде белого твердого вещества (3,05 г, выход 41,7%).
Затем к раствору промежуточного соединения B (3,05 г, 5,8 ммоль) в сухом этилацетате (EtOAc) (12,2 мл) добавляли морфолин (1,2 мл, 13,9 ммоль). После перемешивания в течение ночи при комнатной температуре реакционную смесь гасили 3 Н раствором HCl (3,5 мл), а затем перемешивали в течение 20 мин. Экстрагировали EtOAc и промывали водой, насыщенным водным NaHCO3 и насыщенным раствором NaCl, сушили и концентрировали. Неочищенный материал очищали посредством Isolera (EtPet/EtOAc) с получением 2-дезокси-3-O-ацетил-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение C) в виде белого твердого вещества (2,2 г, 78,4%). К раствору промежуточного соединения C (2,2 г, 4,54 ммоль) в сухом DCM (44 мл), добавляли Cs2CO3 (680 мг, 2,1 ммоль) и CCl3CN (4,4 мл, 43,8 ммоль). Реакционную смесь перемешивали в течение 2 ч при комнатной температуре, затем фильтровали через целит и концентрировали с получением 1-(2,2,2-трихлорацетимин)2-дезокси-3-O-ацетил-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (донор) в виде светло-желтого твердого вещества (2,2 г, 77%).
b. Синтез 1,6-ди-O-бензил 2-дезокси-3-O-((R)-10-этоксикарбонилэтил)-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (акцептор)
Аллилхлорформиат (6,6 мл, 62,2 ммоль) капельно добавляли при комнатной температуре к энергично перемешиваемому раствору D-глюкозамина гидрохлорида (12,2 г, 56,6 ммоль) и NaHCO3, (14,3 г, 169,7 ммоль) в воде (61 мл). Смесь перемешивали в течение 2 ч, затем нейтрализовывали 1 M HCl, концентрировали и сушили в вакууме. Получали неочищенный 2-дезокси-2-(аллилоксикарбониламино)-a-D-глюкопиранозид (промежуточное соединение D). Продукт использовали для следующей стадии реакции без дальнейшей очистки. Промежуточное соединение D (14,9 г, 56,6 ммоль) растворяли в бензиловом спирте (89 мл) и капельно добавляли ацетилхлорид (15,4 мл, 215,2 ммоль) при 0°C. После перемешивания в течение 3 ч при 80°C реакционную смесь гасили холодным насыщенным водным NaHCO3 (20 мл) и перемешивали в течение дополнительных 30 мин. Добавляли холодную воду и диэтиловый эфир и реакционную смесь перемешивали в течение 30 мин. Две фазы разделяли и водную фазу концентрировали и сушили в вакууме. Добавляли 30 мл воды и 200 мл этилового эфира до образования преципитата, который фильтровали и промывали несколько раз холодным диэтиловым эфиром (до тех пор, пока больше не обнаруживалось следов бензилового спирта). Бензил 2-дезокси-2-(аллилоксикарбониламино)-a-D-глюкопиранозид (промежуточное соединение E) получали в виде белого твердого вещества (11 г, 55%) и использовали непосредственно для следующей стадии реакции без дальнейшей очистки. К раствору промежуточного соединения E (11 г, 31,1 ммоль) в бензальдегиде (55 мл) и 4 Å MS (4,9 г) добавляли хлорид цинка (4,2 г, 31,1 ммоль). Через 2 ч добавляли дополнительный хлорид цинка (4,2 г, 31,1 ммоль). После перемешивания в течение ночи при комнатной температуре реакционную смесь обрабатывали насыщенным водным NaHCO3 (70 мл), петролейным эфиром (420 мл) и перемешивали в течение 10 мин. Преципитат фильтровали, промывали петролейным эфиром и растворяли в DCM. Органический раствор экстрагировали насыщенным водным NaHCO3, водой и насыщенным раствором NaCl, сушили и концентрировали. Неочищенный материал очищали посредством Isolera (DCM/EtOAc) с получением бензил 2-дезокси-4,6-O-бензилиден-2-(аллилоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение F) в виде белого твердого вещества (2,75 г, 20%).
К раствору промежуточного соединения F (2,75 г, 6,2 ммоль) в сухом DCM (40,6 мл), охлажденному до 0°C, добавляли гидрид натрия (NaH, 348,8 мг, 8,7 ммоль, 60% дисперсия в масле), и смесь перемешивали в течение 30 мин при комнатной температуре. Затем смесь капельно обрабатывали чистым (-)-этил (S)-2-трифторметансульфонилпропионатом и перемешивали в течение 2 ч при комнатной температуре. Реакционную смесь гасили добавлением льда и экстрагировали DCM. Органический раствор промывали насыщенным водным NaHCO3, насыщенным раствором NaCl, сушили и концентрировали. Остаток очищали посредством Isolera (DCM/EtOAc) с получением бензил 2-дезокси-3-O-((R)-10-этоксикарбонилэтил)-4,6-O-бензилиден-2-(аллилоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение G) в виде белого твердого вещества (2,35 г, 69,7%).
К раствору промежуточного соединения G (2,35 г, 4,34 ммоль) в сухом DCM (42,7 мл) добавляли тетракис(трифенилфосфин)палладий (1,5 г, 1,3 ммоль) и уксусную кислоту (AcOH, 0,385 мл, 6,725 ммоль). Реакционную смесь перемешивали при комнатной температуре в течение 15 мин, а затем капельно добавляли 2,2,2-трихлорэтилхлорформиат (TrocCl, 1,224 мл, 8,895 ммоль), и полученный раствор перемешивали в течение 1 ч при комнатной температуре. Реакционную смесь гасили насыщенным водным NaHCO3, экстрагировали DCM, промывали H2O и насыщенным раствором NaCl. После концентрирования в вакууме остаток растворяли в диэтиловом эфире (100 мл), и нерастворимые материалы отфильтровывали. Органическую фазу сушили, концентрировали и очищали посредством Isolera (циклогексан/AcOEt) с получением бензил 2-дезокси-3-O-((R)-10-этоксикарбонилэтил)-4,6-O-бензилиден-2-(2,2,2-три-хлорэтоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение H) в виде белого твердого вещества (2,3 г, 83,7%).
К раствору промежуточного соединения H (2,4g, 3,8ммоль) в сухом CH3CN (38 мл) при 0°C капельно добавляли раствор Me3N-BH3 (332 мг, 4,55 ммоль) в CH3CN (2,2 мл), а затем раствор BF3-OEt2 (2,89 мл, 23,4ммоль) в CH3CN (8 мл). После перемешивания в течение 3 ч при 0°C смесь гасили холодным насыщенным водным NaHCO3 (30 мл), разбавляли EtOAc (350 мл) и промывали насыщенным водным NaHCO3 (100 мл), 5% лимонной кислотой (4×50 мл), насыщенным водным NaHCO3 (50 мл) и насыщенным раствором NaCl (40 мл). Органический слой сушили, концентрировали и неочищенный материал очищали посредством Isolera (циклогексан/AcOEt) с получением 1,6-ди-O-бензил 2-дезокси-3-O-((R)-10-этоксикарбонилэтил)-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (акцептор) в виде бесцветной пены (1,2 г, 49,8%).
2. Синтез конъюгированного с NAG-NAM пептида BIO-3
Донор (1,6 г, 2,6 ммоль) и акцептор (1,1 г, 1,73 ммоль), суспендированные в сухом DCM (55 мл) с 4 Å молекулярными ситами (200 мг), обрабатывали триметилсилилтрифторметансульфонатом (TMSOTf, 188 мкл, 1,04 ммоль) при -15°C. После перемешивания в течение 20 мин добавляли дополнительные 0,75 экв. донора и 0,3 экв. TMSOTf. После перемешивания в течение 20 мин добавляли дополнительные 0,75 экв. донора и 0,3 экв. TMSOTf. Затем смесь гасили холодным насыщенным водным раствором NaHCO3 (15 мл) и экстрагировали DCM (60 мл). Органический слой промывали насыщенным водным NaHCO3 и насыщенным раствором NaCl, сушили и концентрировали. Остаток очищали посредством обращенной фазы Isolera (нейтральные фазы H2O/ACN) с получением бензил 6-O-бензил-4-O-[2-дезокси-3-O-ацетил-4,6-O-бензилиден-2-(2,2,2-трихлорэтоксикарбониламино)-b-D-глюкопиранозил]-2-дезокси-3-O-[(R)-1’-этоксикарбонилэтил]-2-(2,2,2-трихлорэтоксикарбониламино)-a-D-глюкопиранозида (промежуточное соединение L) в виде белой пены (1,2 г, 62,9%).
Раствор промежуточного соединения L (1,2 г, 1,09 ммоль) и пары цинк-медь (3,12 г, 24,2 ммоль) в смеси AcOH/Ac2O/THF 1:1:1 (12 мл) перемешивали в течение 4 ч при комнатной температуре. Реакционную смесь фильтровали через целит, промывали EtOAc и концентрировали. Неочищенный материал концентрировали и очищали колоночной хроматографией (циклогексан/AcOEt) с получением бензил 6-O-бензил-4-O-[2-дезокси-3-O-ацетил-4,6-O-бензилиден-2-ацетиламино-b-D-глюкопиранозил]-2-дезокси-3-O-[(R)-10-этоксикарбонилэтил]-2-ацетиламино-a-D-глюкопиранозида (промежуточное соединение M) в виде белого твердого вещества (560 мг, 0,67 ммоль, 61,6%). ВЭЖХ-MS (ESI+) m/z: C44H54N2O14+Na: 857,3472. Найдено: 857,6124.
К раствору промежуточного соединения M (91 мг, 0,109 ммоль) в THF/1,4-диоксане/H2O 4:2:1 (2,8 мл) добавляли LiOH.H2O (56 мг, 1,34 ммоль). После перемешивания в течение 2 ч при комнатной температуре реакционную смесь фильтровали через Dowex H+ (свежеактивированный посредством 1 Н HCl). Остаток очищали посредством хроматографии на колонке diaion HP-20 (2×7 см), предварительно промытой водой, MeOH и водой. Колонку элюировали сначала посредством H2O (50 мл), а затем посредством MeOH (30 мл). Спиртовые фракции концентрировали с получением бензил 6-O-бензил-4-O-[2-дезокси-4,6-O-бензилиден-2-ацетиламино-b-D-глюкопиранозил]-2-дезокси-3-O-[(R)-10-этоксикарбонилэтил]-2-ацетиламино-a-D-глюкопиранозида (промежуточное соединение N), в виде беловатого твердого вещества (57,4 мг, 69%). Mp 102-105 °C, 1H-ЯМР (600 МГц, CD3OD) δH 7,51-7,27 (м, 15H, ArH), 5,59 (с, 1H, CHPh), 5,36 (д, J 3,1 Гц, H-1), 4,85 (м) H2O, H-10, ½ CH2Ph), 4,66-4,57 (м, 3H, J 12,1 Гц, CHCH3, CH2-Ph), 4,46 (д, 1H, J 12,2 Гц, ½ CH2Ph), 4,29 (дд, 1H, J 10,3 Гц, J 5,0 Гц, H-60b), 4,07 (т, 1H, J 9,6 Гц, H-30), 3,95 (т, 1H, J 9,1 Гц, H-4), 3,82-3,77 (м, 2H, H-6b, H-3), 3,72-3,62 (м, 3H, H-60a, H-5,H-6a), 3,55-3,40 (м, 3H, H-20 , H-40,H-2), 3,30-3,27 (м, 1H, H-50),1,98 (с, 3H, COCH3), 1,96 (c, 3H, COCH3), 1,37 (д, J 6,9 Гц, 3H, CHCH3). ВЭЖХ-MS (ESI+) m/z: C40H48N2O13+Na: 787,3054, найдено: 787,7830.
Пептид SEQ ID N. 42 присоединяли к соединению N в условиях, использованных для присоединения аминокислот, и полученный биоконъюгат отщепляли от твердой подложки посредством обработки трифторуксусной кислотой (1% раствор в DCM). Полученный биоконъюгат (0,026 ммоль) растворяли в уксусной кислоте (8 мл) и добавляли Pd(OH)2/C (58 мг). Полученную смесь перемешивали при комнатной температуре в течение 6 ч в атмосфере водорода. Смесь фильтровали через целит и концентрировали с получением биоконъюгата BIO-3 с количественным выходом.
Биоконъюгат отщепляли от смолы и из последнего остатка удаляли защитную группу с использованием в качестве раствора для расщепления смеси трифторуксусная кислота/ 1,2-этандитиол/ тиоанизол/ фенол/ H2O/ триизопропилсилан в соотношении 68,5/10/10/5/3,5/1 об./об. Использовали 3 мл раствора для расщепления на 100 мг смолы. Полного удаления защитной группы достигали после 4 часов при 30°C. После реакции расщепления неочищенный пептид преципитировали добавлением холодного диэтилового эфира и сушили в вакууме при 50°C.
Неочищенный биоконъюгат очищали посредством препаративной ОФ-ВЭЖХ (Agilent) с использованием колонки C18 (10 мкм, 100 Å, 50°250 мм). Для элюирования использовали смесь растворителей, состоящую из растворителя A (0,1% TFA, 2% CH3CN в H2O) и растворителя B (90% CH3CN/H2O) при скорости потока 25 мл/мин, и проводили детекцию поглощения при 220 нм. Растворитель удаляли посредством лиофилизации и конечные продукты охарактеризовывали посредством MALDI-TOF-MS. Чистоту очищенного материала оценивали посредством аналитической ОФ-ВЭЖХ (C18-250 мм × 4,6 мм в.д., скорость потока 1 мл/мин), и проводили детекцию поглощения при 220 нм.
3. Синтез биоконъюгатов с NAG и NAM BIO-1 и BIO-2
Синтез NAM-пептида и пептид-NAG проводили, начиная со смолы с заякоренным пептидом (пример 1), где пептид представлял собой пептид SEQ ID N. 42 (для BIO-1) или пептид SEQ ID N. 43 (для BIO-2), соответственно, и донор и акцептор представляли собой молекулы, описанные выше, в соответствии с методикой, описанной в Swaminathan et al., Proc Natl Acad Sci USA. 2006, 17;103 (3):684-9.
Пример 3: Синтез биоконъюгата с каприновой кислотой BIO-4
Пептиды, кэппированные каприновой кислотой, синтезировали в соответствии с методикой, описанной в примере 1, с использованием N-концевого амина, кэппированного каприновой кислотой.
Пример 4: Синтез биоконъюгата с аскорбиновой кислотой BIO-5
5,6-O-изопропилиден-L-аскорбиновую кислоту (iASA) синтезировали способом Jung (Jung, M. E.; Shaw, T. J. J. Am. Chem. Soc. 1980, 102, 6304). Для активации к смоле с заякоренным пептидом добавляли CDI (10 экв.) в течение 2 ч. Активацию N-концевого амина пептида после отщепления смолы подтверждали посредством ОФ-ВЭЖХ с использованием следующих условий: A-B (A: 0,1% TFA в H2O, B: 0,1% TFA в CH3CN; от 0% до 60% B в течение 30 мин, при скорости потока 1,0 мл/мин); детекция, УФ 230 нм. Далее в активированную смолу с заякоренным пептидом вносили iASA. Продукт отделяли от смолы посредством обработки 50% TFA/DCM (об./об.) в течение 1 ч. Смолу фильтровали, фильтрат концентрировали в высоком вакууме и преципитировали холодным диэтиловым эфиром. Структуру полученного биоконъюгата BIO-5 подтверждали посредством LC/MS с использованием 0,1% муравьиной кислоты/метанола в качестве элюента при скорости потока 250 мкл/мин в течение 30 мин, и проводили мониторинг при 230 нм.
Пример 5: Анализ димеризации IL-17RA/IL17RC в ответ на IL-17A
Пептиды, приведенные в таблице 7, тестировали в отношении их способности ингибировать связывание IL-17A с его рецептором и последующее взаимодействие IL-17RA и IL17RC.
Для измерения взаимодействия цепей рецепторов IL-17RA и IL17RC при активации IL-17A в отсутствии или в присутствии пептидов, приведенных в таблице 7, использовали анализ димеризации рецепторов интерлейкина RA и RC (DiscoverX).
В анализе рецепторов цитокинов PathHunter® одну цепь рецептора цитокина метят небольшим пептидным эпитопом ProLink (PK), а другую цепь метят ферментным акцептором (EA). Связывание лиганда индуцирует димеризацию двух рецепторов, способствуя комплементации фрагментов PK и EA. Это взаимодействие приводит к образованию активной единицы b-галактозидазы, которое определяют с использованием хемилюминесцентного субстрата. Клеточные линии PathHunter увеличивали в количестве из замороженных исходных культур в соответствии со стандартными методиками. Клетки высевали в общем объеме 20 мкл в 384-луночные микропланшеты с белыми стенками и инкубировали в течение надлежащего периода времени перед тестированием. Для ингибирования активности агониста клетки инкубировали с образцом для индукции ответа. Промежуточное разведение исходных культур образца проводили для получения 5X образца в буфере для анализа. 5 мкл 5X образца добавляли к клеткам и инкубировали при 37°C или комнатной температуре в течение 60 минут. Концентрация носителя составляла 1%. К клеткам добавляли 5 мкл 6X агониста EC80 (IL-17A) в буфере для анализа и инкубировали при 37°C в течение 6-16 часов в зависимости от анализа. 1. Сигнал в анализе генерировали посредством однократного добавления 12,5 или 15 мкл (50% об./об.) коктейля реагентов для детекции PathHunter для анализа агонистов (IL-17A) и антагонистов (пептиды, приведенные в таблице 7), соответственно, а затем проводили инкубацию в течение одного часа при комнатной температуре. Для некоторых анализов детекцию активности проводили с использованием высокочувствительного реагента для детекции (набор PathHunter Flash Kit) для повышения эффективности анализа. В этих анализах в лунки добавляли равный объем реагента для детекции (25 или 30 мкл), а затем проводили инкубацию в течение одного часа при комнатной температуре. Считывание микропланшетов проводили после генерирования сигнала с использованием устройства PerkinElmer Envision™ для детекции хемилюминесцентного сигнала. Активность соединения анализировали с использованием пакета программ для анализа данных CBIS (ChemInnovation, CA). % ингибирование =100% × (1 - (среднее количество RLU для тестируемого образца - среднее количество RLU для контроля в виде носителя) / (среднее количество RLU для контроля в виде EC80 - среднее количество RLU для контроля в виде носителя)).
Как описано в таблице 7, ниже, было обнаружено, что все протестированные молекулы являются активными в отношении ингибирования димеризации IL-17RA/IL17RC, таким образом, ингибируя активацию каскада IL-17A с величиной IC50 в среднем наномолярном диапазоне.
Таблица 7
На фиг.1 представлен профиль доза-ответ для иллюстративных пептидов.
Пример 6: Фенотипический анализ в дифференцированных TH17-клетках человека и анализ жизнеспособности
Для оценки эффекта пептидов по изобретению на клетки-мишени, ответственные за воспаление при офтальмологических или дерматологических патологиях, авторы изобретения протестировали способность пептида SEQ ID N. 18 ингибировать in vitro секрецию провоспалительных цитокинов и металлопротеиназ клетками Th17 человека.
Первичные CD4 T-клетки человека дифференцировали в Th17-клетки in vitro посредством культивирования клеток в среде для дифференцировки Th17 в течение 10 суток. Подробно, очищенные CD4 T-клетки периферической крови человека культивировали в среде для дифференцировки Th-17 (CellXVivo Human Th17, Tocris) в соответствии с инструкциями изготовителя. В кратком изложении: 96-луночные планшеты для культивирования тканей покрывали антителами-антагонистами CD3 и CD28. Клетки подсчитывали по исключению трипанового синего и суспендировали в концентрации 1E5 клеток/мл в среде для дифференцировки Th-17. 0,2 мл этой суспензии высевали в индивидуальные лунки 96-луночного планшета. Клетки инкубировали при 37°C с 5% CO2 в течение 10 суток с заменой среды по необходимости. После дифференцировки клеток в течение 10 суток пептиды, синтезированные согласно примеру 1, такие как пептид Seq.ID No 18, разбавляли до надлежащей концентрации в CGM (среда для роста клеток, XVivo-15, Lonza). Планшет с клетками центрифугировали при 300×g в течение 5 мин и среду для дифференцировки Th17 удаляли.
После дифференцировки клетки стимулировали как активирующим коктейлем, содержащим форбол 12-миристат 13-ацетат, так и лономицин, в присутствии варьирующих концентраций тестируемых изделий.
Через 24 ч после стимуляции культуральные супернатанты анализировали в отношении экспрессии IL-17, IL-6, IL-23, MMP3 и исследовали в отношении жизнеспособности с использованием флуоресцентного красителя для определения жизнеспособности, аламарового синего. CD4 клетки секретировали высокие уровни IL-17A в ответ на стимуляцию активирующим коктейлем, что указывает на успешную дифференцировку в Th17-клетки. Кроме того, стимулированные клетки секретировали IL-6, IL23 и MMP3.
Супернатант удаляли и заменяли 200 мкл разбавленного пептида или носителя (CGM+DMSO, разбавленный в соответствии с концентрацией TI) в соответствующих группах. Клетки инкубировали с тестируемыми пептидами в течение 1 часа перед стимуляцией. Получали следующие растворы для стимуляции: 10x коктейль для активации клеток: разбавленный от 500x до 10x рабочего раствора в CGM. В лунки добавляли 22 мкл соответствующих стимуляторов через 1 ч после добавления TI. Наконец, в каждую лунку добавляли 25 мкл реагента аламарового синего (10x) через 20 часов после стимулятора (4 ч перед сбором сыворотки через 24 ч) для определения жизнеспособности клеток. Через 24 ч после добавления стимулятора определяли флуоресценцию аламарового синего в соответствии с инструкциями изготовителя. Остальные супернатанты собирали для последующего мультиплексного анализа белка. Уровни экспрессии IL-6, IL-17 и IL-23, а также MMP3 в супернатантах стимулированной в течение 24 часов культуры оценивали с использованием мультиплексной платформы Luminex на устройстве MAGPIX.
На фиг.2 и 3 показан эффект пептида SEQ ID N. 18. Как можно видеть, этот пептид способен значительно уменьшать секрецию IL-17A, IL-6 и IL-23, а также секрецию MMP3. Как показано на фиг.4 ингибирование секреции цитокинов и металлопротеиназ не снижает жизнеспособность клеток, поскольку обработка клеток пептидом SEQ.ID N. 18 в течение одиннадцати суток не снижает жизнеспособность клеток.
Пример 7: Физико-химический анализ пептидов
Физико-химические свойства пептида HAP уровня техники и некоторых пептидов по изобретению (см. таблицу 8) тестировали, как описано ниже.
i. Способы
a. Изоэлектрическая точка IP log D и logP
Основные физико-химические свойства (изоэлектрическая точка (IP), log D (7,4) и logP) пептидов, приведенных в таблице 8 ниже, определяли с использованием устройства SiriusT3 (Sirius Analytical Instruments Ltd., East Sussex, Великобритания), оборудованного двухконтактным эталонным pH-электродом Ag/AgCl, спектрометра Sirius D-PAS и устройства для определения мутности. pH-электрод калибровали титрометрически в диапазоне pH 1,8-12,2. Использовали верхнеприводную мешалку, и температурный зонд (контроллер Пельтье) осуществлял мониторинг температуры в ходе титрования. Эксперименты по титрованию проводили в 0,15 M растворе KCl со скорректированной ионной силой (вода ISA) в атмосфере азота при температуре 25±1°C. Все тесты проводили с использованием стандартизованного 0,5 M KOH и 0,5 M HCl в качестве реагентов для титрования, для тестирования коэффициентов распределения использовали насыщенный октанол в воде ISA (5% воды ISA) в качестве распределяющего растворителя. pKas определяли потенциометрическим способом посредством pH-метрического титрования. Порошок (around 0,5 мг) растворяли в 1,5 мл воды ISA и титрование проводили в трех экземплярах в диапазоне pH 2,0-12,0. Каждое значение Log P получали в трех экземплярах путем растворения порошка (приблизительно 1 мг) в 1 мл воды ISA с последующим pH-метрическим титрованием при трех различных процентах октанола (как правило, 50%, 60%, 70%).
b. Растворимость в воде и стабильность
Общая методика получения исходных растворов
Каждый лиофилизированный пептид отвешивали во флакон янтарного цвета. Добавляли вычисленный объем фосфатно-солевого буфера при pH 7,8 для прямого получения 10 мМ исходных растворов. Извлеченные суспензии перемешивали в течение 30 посредством инкубатора с орбитальным встряхивателем при 37°C. Последующие разведения проводили для получения более разбавленных исходных растворов для менее растворимых соединений.
Для оценки стабильности и растворимости все исходные растворы хранили при 25°C. Указанное в заголовке соединение оценивали посредством анализа UHPLC-MS по сравнению со стандартными растворами в 3 момента времени: T0, 3 суток, 1 неделя.
Хроматографический профиль оценивали для количественного анализа, в то время как для испытаний стабильности проводили детекцию MS-спектров.
Общая методика получения стандартного раствора
Каждый пептид взвешивали и прямо разбавляли до 100 мкМ фосфатным буфером при pH 7,8. Растворы перемешивали в течение 5 минут с использованием встряхивателя и нагревали при 37°C в течение 20 минут. Полученный стандартный раствор хранили при 25°C в темноте с контролируемой температурой.
Общая методика подготовки образца
Часть исходных растворов собирали во флакон янтарного цвета и разбавляли до 100 мкМ фосфатным буфером при pH 7,8 (фактор разведения 1:100).
Приборный способ
Обращенно-фазовую UHPLC проводили при скорости потока 0,3 мл/мин с использованием колонки Luna Omega Polar C18 column (размер пор 1,6 мкм, 2,1×100 мм). Растворитель A представлял собой 0,5% муравьиную кислоту в воде и растворитель B представлял собой 0,5% муравьиную кислоту в ацетонитриле. Сначала образцы загружали в колонку и промывали изократически при 10% B в течение 3 мин. Затем прогоняли градиент 10-95% B в течение 22 мин, удерживали при 95% в течение 2 мин, а затем снижали до 10% в течение 1 мин и удерживали для стабилизации давления в течение 5 мин. Мониторинг УФ-поглощения проводили посредством DAD (220-400 нм). Для мониторинга TIC использовали ионную ловушку ESI-MS.
ii. Результаты
Все протестированные пептиды продемонстрировали стабильность при 25°C вплоть до 1 недели, поскольку не было обнаружено значительного варьирования хроматографического профиля и спектра масс.
Физико-химические свойства протестированных пептидов представлены в таблице 8 ниже
Таблица 8
Как можно видеть из вышеуказанного, большинство пептидов по изобретению имеют оптимальные физико-химические свойства, демонстрируя хорошее равновесие между растворимостью и липофильностью. Среднее значение LogD для пептидов составляло -2,64±2,06, в то время как для большинства пептидов растворимость, наблюдаемая в воде, была очень хорошей (> 10 мМ). Наблюдаемые свойства делают их особенно пригодными для применения в офтальмологических и дерматологических применениях, поскольку как гидрофобные, так и гидрофильные компоненты присутствуют в дерме и на поверхности глаза. Также они могут объяснять хорошую проницаемость, наблюдаемую для пептидов (см. пример 8).
Пример 8: Тест проницаемости
Протестированные пептиды представляли собой пептиды SEQ ID N. 1, SEQ ID N. 2, SEQ ID N. 7, SEQ ID N. 11, SEQ ID N. 14, SEQ ID N. 38, SEQ ID N. 111, SEQ ID N. 113, SEQ ID N. 115 и SEQ ID N. 121. Их растворяли воде для культивирования клеток (Coming, Manassas, VA, США) до получения 10 мМ исходного раствора.
i. Способы
Модель in vitro
I-HCEC, линия иммотрализованных эпителиальных клеток роговицы человека, происходила из первичных эпителиальных клеток роговицы человека (чистота >99%). Эти клетки являются пригодными для исследований роговицы человека в здоровом состоянии и при заболевании. Клетки культивировали в соответствии с протоколом и с использованием среды, предлагаемой Innoprot, Bizkaia, Испания (IM-Corneal Epithelial Cell Medium, ссылочный номер P60131).
Иммунофлуоресценция
Были разработаны качественные и количественные способы, сначала для подтверждения, а затем и для количественного определения целостности барьера перед продолжения тестирования лекарственного средства. Сначала авторы изобретения проводили иммунофлуоресценцию для охарактеризации белков плотных контактов (Z0-1) (Sugrue SP et al., (1997) Exp Eye Res. Jan;64 (1): 11-20). Клетки высевали на покровные стекла, покрытые коллагеном I 50 мкг/мл (исходный раствор 3 мг/мл от Gibco, NY, США) с плотностью 1*104 клеток/см2 и культивировали в течение 14 DIV и после смыкания монослоя среду обновляли каждые сутки. Клетки фиксировали в метаноле при -20°C в течение 10 мин, ополаскивали фосфатно-солевым буфером (PBS), инкубировали в течение 10 мин при комнатной температуре (к.т.) в PBS, содержавшем 4% бычий сывороточный альбумин (BSA; Sigma-Aldrich), а затем последовательно инкубировали с первичными и вторичными антителами в течение 60 мин в каждом случае при к.т. Первичное антитело представляло собой антитело козы против Z0-1 человека (1:100, Thermo Fisher) в течение ночи при 4°C а затем с антителом против антител козы с Alexafluor 488 (1:10000). Наконец, клетки контрастно окрашивали посредством DAPI (0,5 мг/мл) (Sigma-Aldrich) в течение 5 мин при комнатной температуре, заливали средой Vectashield Mounting Medium (Vector Laboratories Inc, Burlingame, CA) и наблюдали под конфокальным лазерным сканирующим микроскопом (Leica TCS SP5, Wetzlar, Германия).
Анализ MTS
Жизнеспособность клеток определяли через 24 ч с использованием анализа пролиферации клеток Cell Titer One Solution Cell Proliferation Assay (Promega Corporation Madison, WI, CIF), колориметрического способа на основе 3-(4,5-диметилтиазол-2-ил)-5-(3-карбоксиметоксифенил)-2-(4-сульфофенил)-2H-тетразолия (MTS). Количество образовавшегося формазана в качестве функции жизнеспособности измеряли при 492 нм с использованием устройства для считывания планшетов ELISA, Infinite F200 (Tecan, Mannedorf, Швейцария). Анализ проводили при 50 и 100 мкМ (N=9). Результаты выражали в качестве поглощения при 492 нм.
Анализ TEER
Целостность барьера культур эпителиальных роговицы подтверждали с использованием измерения TEER (трансэпителиальное/трансэндотелиальное электрическое сопротивление)) и проницаемости параклеточных проникающих веществ (например, маннит, 6-карбоксифлуоресцеин) (Ronkko S et al., (2016) Drug Deliv Transl Res 6: 660-675). TEER является широко признанным количественным способом измерения динамики целостности плотных контактов в моделях клеточных культур эндотелиальнных и эпителиальных монослоев. Величины TEER являются индикаторами целостности клеточных барьеров до их оценки в отношении транспорта лекарственных веществ или химических веществ. Измерение TEER можно проводить в реальном времени без повреждения клеток, и оно, как правило, основано на измерении осмотического сопротивления или измерении импеданса в широком спектре частот (Snirivasan et al., 2015 J Lab Autom. 20(2): 107-126). Для проведения этого анализа использовали Millicell ERS-2, системы для определения электрического сопротивления от EMD Millipore (Burlington, MA, США) в соответствии с инструкциями изготовителя. Анализ проводили при 50 и 100 мкМ (N=9). Результаты выражали в качестве сопротивления (кл/см2).
Испытание проницаемости
Задачей этого исследования была оценка величины кажущегося коэффициента проницаемости (Papp) для пептидов через пласт эпителиальных клеток роговицы человека. Все пептиды тестировали в концентрации 50 мкМ в двух экземплярах.
В частности, эпителиальные клетки роговицы человека (hCEPIC) наслаивали на вкладыши для культивирования клеток Transwell (Coming, NY, США). Эксперименты по проницаемости проводили, отбирая образцы из приемных камер в моменты времени 5, 30, 60 минут и из донорных камер в те же моменты времени. Каждую полученную аликвоту анализировали с использованием устройства ВЭЖХ (Knauer), оборудованного колонкой C18 Vertex Plus Column (Knauer), термостатически контролируемой при 40°C. Скорость (кажущийся коэффициент проницаемости, Papp) исследуемых соединений вычисляли и сравнивали с величинами Papp для маркеров с низкой и высокой проницаемостью 6-карбоксифлуоресцеина и родамина B, соответственно.
Кажущийся коэффициент проницаемости (Papp) вычисляли по следующему уравнению:
Papp=(l/A *C0)(dM/dt)
где dM/dt представляет собой поток (наномоли в секунду) через клеточные слои;
A (квадратные сантиметры) представляет собой площадь подвергнутой воздействию поверхности мембраны вкладыша;
C0 представляет собой первоначальную концентрацию лекарственного средства (микромолярная) в донорном отделении при t=0 (Xiang et al., Drug Metab Dispos. 2009 May;37(5):992-8.
Статистика
Все группы (исследованное соединение и родамин) сравнивали с контролем с низкой проницаемостью. Все другие экспериментальные результаты выражали в качестве среднего значения ± S.E. и значимость вычисляли посредством t-критерия Стьюдента против контрольных клеток. Отличие считали статистически значимым (p < 0,05) при определении посредством t-критерия Стьюдента.
ii. Результаты
Анализы MTS и TEER
В экспериментальных условиях авторов изобретения пептиды считались не влияющими на значимом уровне на жизнеспособность клеток или TEER при тестируемой концентрации 50 мкМ и 100 мкМ.
Тест проницаемости
Результаты, полученные для каждого пептида, представлены в таблице 9 ниже.
Таблица 9
(10-6 см/с)
Для подтверждения того, что клеточная культура HCE является пригодной для скрининга пептидов, определяли проницаемость липофильного транклеточного маркера родамина B. Как и ожидалось, родамин B демонстрировал высокую проницаемость клеточной культуры HCE (51,26±5,44 10-6 см/с). Затем оценивали проницаемость 6-карбоксифлуоресцеина в соответствии с литературой (0,61±0,33 10-6 см/с). Этот маркер имеет очень низкую липофильность и он проходит через биологические мембраны только через внутриклеточное пространство.
Протестированные пептиды продемонстрировали очень хорошую проницаемость через иммортализованный эпителия роговицы человека по сравнению со стандартом с высокой проницаемостью родамином B. Очевидно, что модель проницаемости HCE не включает строму или эндотелий, однако эти слои не являются ключевыми барьерами для всасывания лекарственного средства через роговицу. Авторы изобретения обнаружили пептиды с высокой проницаемостью (SEQ ID N. 2) и пептиды с более высокой проницаемостью по сравнению с родамином B (SEQ ID N. 115, SEQ ID N. 113 и SEQ ID N. 111).
Пример 9: Испытание аффинности пептид-IL-17A
Пептид уровня техники HAP (SEQ ID N.1) и пептид SEQ ID N.2 тестировали в отношении аффинности к mIL-17A, как описано ниже.
a. Иммобилизация mIL-17A
Для иммобилизации белка IL-17A мыши использовали химию присоединения аминов. На поверхности сенсорного чипа декстрановую матрицу сначала активировали посредством смеси 1-этил-3-карбодиимида (EDC) и N-гидроксисукцинимида (NHS) для получения реакционноспособных сложных сукцинимидоэфиров. 10 мкг/мл IL17 разбавляли в 10 мМ ацетатном буфере при pH=5,0, а затем лиганд пропускали над поверхностью, чтобы позволить сложным эфирам реагировать спонтанно с незаряженными аминогруппами, таким образом, лиганд ковалентно связывается с декстраном. Ковалентная иммобилизация mIL17 приводит к стабильному связыванию 3000 RU лиганда с поверхностью.
b. Охарактеризация взаимодействия пептиды/IL17
Пептиды SEQ ID N. 1 (HAP) и SEQ ID N 2 анализировали в отношении их способности связывать иммобилизованный mIL-17A. Молекулы разбавляли в различных концентрациях (от 62,5 до 1000 нМ) в буфере HBS-EP и инжектировали на сенсорный чип в течение 4 минут, и позволяли диссоциировать в течение 5 минут. После каждого прогона сенсорный чип регенерировали посредством инжектирования 2 M NaCl. На основе сенсограмм (фиг.1, панель A) и с использованием программного обеспечения BIAevaluation, анализ Скэтчарда для величин RU в равновесии позволил определить константу аффинности в качестве функции концентрации анализируемого соединения в растворе (фиг. 6, панель B). Полученные результаты представлены в таблице 10 ниже.
Таблица 10
Как показано в таблице 8, пептид SEQ ID N. 2 связывается с IL-17A с величиной аффинности, равной 130 нМ, в то время как пептид SEQ ID N. 1 связывается с IL17-A с очень низкой величиной аффинности (3700 нМ).
Ввиду результатов, полученных в примере 8 и 9, модификация изолейцина в положении 1 пептида HAP уровня техники на валин, как в пептиде SEQ ID N. 2, приводит к поразительным различиям, касающимся физико-химических свойств, проницаемости и аффинности в отношении лиганда.
Ниже обобщены свойства двух пептидов:
Таблица 11
(HAP)
В таблице 11 показаны данные сравнения пептидов SEQ ID N. 1 и SEQ ID N. 2.
Авторы изобретения обнаружили значительные отличия с точки зрения logP и logD (отличие 6-log и 3-log, соответственно): это объяснило более высокую проницаемость, наблюдаемую для SEQ ID N. 2, по сравнению с SEQ ID N. 1 (28,8 против 19,3). Kd (нМ), измеренная в анализе с использованием поверхностного плазмонного резонанса (SPR), проведенном на IL-17A мыши, показала, что SEQ ID N. 2 был в 28 раз более аффинным, чем SEQ ID N. 1.
Более того, также проводили исследования имитации молекулярной динамики, которые показали, что валин (SEQ ID N. 2) вместо изолейцина (HAP) в значительной степени стабилизирует свернутую структуру: было спрогнозировано, что SEQ ID N. 2 был на 80% более стабильным.
Пример 10: Оценка in vitro экспрессии воспалительных цитокинов
Пептид HAP (SEQ ID N.1) и пептид SEQ ID N.2 тестировали in vitro для оценки их прямого влияния на экспрессию воспалительных цитокинов в роговице.
Тесты проводили на иммортализованной линии клеток эпителия роговицы человека (I-HCEC) (Innoprot, Bizkaia, Испания, ссылочный номер P10871). Она происходит из первичных эпителиальных клеток роговицы человека (чистота >99%) и является пригодной для исследований роговицы человека в здоровом состоянии и при заболевании. Клетки культивировали в соответствии с протоколом и с использованием среды, предложенной Innoprot (IM-Corneal Epithelial Cell Medium, ссылочный номер P60131). Клеточную среду дополняли 5% эмбриональной телячьей сывороткой (FBS).
Клетки высевали в количестве 320000 клеток/лунка в 6-ячеечный планшет и 15000 клеток/лунка в 96-луночный планшет. Когда клетки достигали 80% смыкания монослоя, их обрабатывали соединениями SEQ ID N. 1 и SEQ ID N. 2 в соответствии с кривой дозы (0,01-500 мкМ) в течение 24 часов.
Тотальную РНК выделяли из планшета с 6 ячейками с использованием набора Quick-Start RNeasy Mini Kit (Qiagen, каталожный номер N 74104) в соответствии с инструкциями изготовителя. Обратную транскрипцию проводили с использованием SuperScript Vilo (ссылочный номер 11755050, Life technologies) и ПЦР в реальном времени с использованием протокола TaqMan (ссылочный номер 4444557, Applied Biosystem), адаптированного для системы для исследования в реальном времени CFX96. Эксперименты проводили три раза, каждый раз в двух экземплярах. Использованные зонды от Thermo Fisher Scientific представляют собой: CXCL8 человека (Hs00174103_m1), IL6 человека (Hs00174131_m1), TNFα человека (Hs99999043_m1). В качестве гена домашнего хозяйства использовали GAPDH (зонд Hs02758991_g1).
Экспрессию IL-8, IL-6 и TNF-α в I-HCEC оценивали после обработки в течение 24 часов различными концентрациями пептида SEQ ID N. 1 или SEQ ID N. 2 (0,01, 0,1, 1, 10, 100, 500 мкМ). В качестве отрицательного контроля (носитель) использовали необработанные I-HCEC.
Результаты показывают, что соединение SEQ ID N. 2 не изменяет экспрессию никакого из измеренных воспалительных цитокинов (фиг.8A, 9A и 10A), в то время как обработка посредством 500 мкМ соединения SEQ ID N. 1 в значительной степени индуцирует экспрессию двух воспалительных цитокинов, IL-6 и TNFα (фиг.9B и 10B).
Полученные данные демонстрируют, что пептид SEQ ID N. 1 имеет прямую провоспалительную активность и индуцирует в значительной степени экспрессию воспалительных цитокинов IL-6 и TNF-α посредством HCEC с повышением в 10 и 8 раз, соответственно. Напротив, пептид SEQ ID N. 2 не демонстрирует никакого эффекта на экспрессию цитокинов.
Ввиду вышесказанного, можно сделать заключение, что пептид SEQ ID N. 2 неожиданно демонстрирует значительно лучшую переносимость и отсутствие токсичности по сравнению с пептидом HAP SEQ ID N. 1.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ:
<110> DOMPE
<120> IL-17A-СВЯЗЫВАЮЩИЕ ПЕПТИДЫ И ИХ ПРИМЕНЕНИЕ В МЕДИЦИНЕ
<130> 19LG52E
<140> PCT/EP2019/070265
<141> 2019-07-26
<150> EP18186029.7
<151> 2018-07-27
<160> 257
<170> BiSSAP 1.3.6
<210> 1
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 2
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Val His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 3
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Ile Met Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 4
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Val Met Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 5
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 5
Ile His Val Thr Ile Pro Ala Glu Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 6
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 6
Ile His Val Thr Ile Pro Ala Asp Leu Tyr Asp Trp Ile Asn Lys
1 5 10 15
<210> 7
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Ile His Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Asn Lys
1 5 10 15
<210> 8
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 8
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Val Asn Lys
1 5 10 15
<210> 9
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 9
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Arg Lys
1 5 10 15
<210> 10
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 10
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Arg
1 5 10 15
<210> 11
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Glu Lys
1 5 10 15
<210> 12
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 12
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Glu
1 5 10 15
<210> 13
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 13
Ile His Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Asn Lys
1 5 10 15
<210> 14
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 14
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Val Arg Arg
1 5 10 15
<210> 15
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 15
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Val Glu Glu
1 5 10 15
<210> 16
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 16
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Asn Lys
1 5 10 15
<210> 17
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 17
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Arg Arg
1 5 10 15
<210> 18
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 18
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu
1 5 10 15
<210> 19
<211> 21
<212> БЕЛОК
<213> Homo sapiens
<400> 19
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Arg
1 5 10 15
Arg Arg Arg Arg Arg
20
<210> 20
<211> 18
<212> БЕЛОК
<213> Homo sapiens
<400> 20
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Arg
1 5 10 15
Arg Arg
<210> 21
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 21
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Ala
1 5 10 15
Arg Lys Lys Ala Ala Lys Ala
20
<210> 22
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 22
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Gly
1 5 10 15
Gly Thr Trp Trp Thr Glu Trp Ser Gln
20 25
<210> 23
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 23
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Thr
1 5 10 15
Trp Trp Glu Thr Trp Trp
20
<210> 24
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 24
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Val
1 5 10 15
Pro Gly Trp Gly
20
<210> 25
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 25
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Gly
1 5 10 15
Gly Lys Glu Thr Trp Trp Glu Thr Trp
20 25
<210> 26
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 26
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Val
1 5 10 15
Pro Gly Lys Gly
20
<210> 27
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 27
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Val
1 5 10 15
Pro Gly Ala Gly
20
<210> 28
<211> 31
<212> БЕЛОК
<213> Homo sapiens
<400> 28
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Arg
1 5 10 15
Gln Ile Lys Ile Trp Phe Gln Asn Arg Arg Met Lys Trp Lys Lys
20 25 30
<210> 29
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 29
Arg Arg Arg Arg Arg Arg Arg Arg Val Met Val Thr Ile Pro Ala Asp
1 5 10 15
Leu Tyr Glu Trp Ile Glu Glu
20
<210> 30
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 30
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Arg
1 5 10 15
Arg Arg Arg Arg Arg Arg Arg
20
<210> 31
<211> 23
<212> БЕЛОК
<213> Homo sapiens
<400> 31
Ala Arg Lys Lys Ala Ala Lys Ala Val Met Val Thr Ile Pro Ala Asp
1 5 10 15
Leu Tyr Glu Trp Ile Glu Glu
20
<210> 32
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 32
Gly Gly Thr Trp Trp Thr Glu Trp Ser Gln Val Met Val Thr Ile Pro
1 5 10 15
Ala Asp Leu Tyr Glu Trp Ile Glu Glu
20 25
<210> 33
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 33
Gly Gly Lys Glu Thr Trp Trp Glu Thr Trp Val Met Val Thr Ile Pro
1 5 10 15
Ala Asp Leu Tyr Glu Trp Ile Glu Glu
20 25
<210> 34
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 34
Val Pro Gly Trp Gly Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu
1 5 10 15
Trp Ile Glu Glu
20
<210> 35
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 35
Val Pro Gly Ala Gly Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu
1 5 10 15
Trp Ile Glu Glu
20
<210> 36
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 36
Val Pro Gly Lys Gly Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu
1 5 10 15
Trp Ile Glu Glu
20
<210> 37
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 37
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Val
1 5 10 15
Pro Gly Ala Gly Val Pro Gly Ala Gly
20 25
<210> 38
<211> 25
<212> БЕЛОК
<213> Homo sapiens
<400> 38
Val Pro Gly Ala Gly Val Pro Gly Ala Gly Val Met Val Thr Ile Pro
1 5 10 15
Ala Asp Leu Tyr Glu Trp Ile Glu Glu
20 25
<210> 39
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 39
Val Pro Gly Asp Gly Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu
1 5 10 15
Trp Ile Glu Glu
20
<210> 40
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 40
Val Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Val
1 5 10 15
Pro Gly Asp Gly
20
<210> 41
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 41
Asp Leu Ser Ala Val Cys Trp Ala Phe Pro Trp Asp Pro Glu Cys His
1 5 10 15
<210> 42
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 42
Ile Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu
1 5 10 15
<210> 43
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 43
Ile Met Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Ile Glu Glu Gln
1 5 10 15
<210> 44
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 44
Ala Arg Lys Lys Ala Ala Lys Ala
1 5
<210> 45
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 45
Thr Trp Trp Thr Glu Trp Ser Gln
1 5
<210> 46
<211> 7
<212> БЕЛОК
<213> Homo sapiens
<400> 46
Thr Trp Trp Glu Thr Trp Trp
1 5
<210> 47
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 47
Val Pro Gly Trp Gly
1 5
<210> 48
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 48
Lys Glu Thr Trp Trp Glu Thr Trp
1 5
<210> 49
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 49
Val Pro Gly Lys Gly
1 5
<210> 50
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 50
Val Pro Gly Ala Gly
1 5
<210> 51
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 51
Arg Gln Ile Lys Ile Trp Phe Gln Asn Arg Arg Met Lys Trp Lys Lys
1 5 10 15
<210> 52
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 52
Arg Arg Arg Arg Arg Arg Arg Arg
1 5
<210> 53
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 53
Val Pro Gly Asp Gly
1 5
<210> 54
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полученный пептид
<220>
<221> ВАРИАНТ
<222> 4
<223> Xaa = Любая аминокислота
<400> 54
Val Pro Gly Xaa Gly
1 5
<210> 55
<211> 4
<212> БЕЛОК
<213> Homo sapiens
<400> 55
Ile Pro Gly Gly
1
<210> 56
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 56
Ala Val Gly Val Pro
1 5
<210> 57
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетически полученный пептид
<220>
<221> ВАРИАНТ
<222> 4
<223> Xaa = Любая аминокислота
<400> 57
Ile Pro Gly Xaa Gly
1 5
<210> 58
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 58
Ile Pro Gly Val Gly
1 5
<210> 59
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полученный пептид
<220>
<221> ВАРИАНТ
<222> 4
<223> Xaa = Любая аминокислота
<400> 59
Leu Pro Gly Xaa Gly
1 5
<210> 60
<211> 5
<212> БЕЛОК
<213> Homo sapiens
<400> 60
Leu Pro Gly Val Gly
1 5
<210> 61
<211> 6
<212> БЕЛОК
<213> Homo sapiens
<400> 61
Val Ala Pro Gly Val Gly
1 5
<210> 62
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полученный пептид
<220>
<221> ВАРИАНТ
<222> 1
<223> Xaa = Любая аминокислота
<400> 62
Xaa Pro Gly Val Gly
1 5
<210> 63
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 63
Gly Val Gly Val Pro Gly Val Gly
1 5
<210> 64
<211> 9
<212> БЕЛОК
<213> Homo sapiens
<400> 64
Val Pro Gly Phe Gly Val Gly Ala Gly
1 5
<210> 65
<211> 8
<212> БЕЛОК
<213> Homo sapiens
<400> 65
Val Pro Gly Gly Val Pro Gly Gly
1 5
<210> 66
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 66
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Lys Ala
1 5 10 15
<210> 67
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 67
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 68
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 68
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Lys
1 5 10 15
<210> 69
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 69
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Ala Ala
1 5 10 15
<210> 70
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 70
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ala Arg Ala
1 5 10 15
<210> 71
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 71
Ile Val Val Thr Met Pro Ala Asp Leu Trp Ala Trp Ile Arg Ala
1 5 10 15
<210> 72
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 72
Ile Val Val Thr Met Pro Ala Asp Leu Ala Asp Trp Ile Arg Ala
1 5 10 15
<210> 73
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 73
Ile Val Val Thr Met Pro Ala Asp Ala Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 74
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 74
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 75
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 75
Ile Val Val Thr Ala Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 76
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 76
Ile Val Val Ala Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 77
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 77
Ile Ala Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 78
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 78
Ala Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 79
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 79
Ile His Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 80
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 80
Ile Gln Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 81
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 81
Ile Arg Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 82
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 82
Ile Thr Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 83
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 83
Ile Trp Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 84
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 84
Ile Tyr Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 85
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 85
Ile Val Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 86
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 86
Ile Val Val Thr Leu Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 87
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 87
Ile Val Val Thr Val Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 88
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 88
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Met Ala
1 5 10 15
<210> 89
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 89
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Asn Ala
1 5 10 15
<210> 90
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 90
Ile Val Val Thr Met Pro Ala Asp Leu Trp Asp Trp Ile Gln Ala
1 5 10 15
<210> 91
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 91
Ile Val Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 92
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 92
Ile Val Val Thr Leu Pro Ala Asp Leu Trp Asp Trp Ile Arg Ala
1 5 10 15
<210> 93
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 93
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 94
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 94
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn
1 5 10
<210> 95
<211> 13
<212> БЕЛОК
<213> Homo sapiens
<400> 95
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile
1 5 10
<210> 96
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 96
Asp Ser Ser Ala Val Cys Trp Ala Phe Pro His His Pro Leu Cys His
1 5 10 15
Met Lys Ala Thr
20
<210> 97
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 97
Ala Asp Ala Asp Met Cys Trp Phe Phe Pro Thr Ser Pro Trp Cys His
1 5 10 15
<210> 98
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 98
Asp Leu Ser Ala Val Cys Trp Ala Phe Pro Trp Asp Pro Glu Cys His
1 5 10 15
Met
<210> 99
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 99
Asp Ser Ser Ala Val Cys Trp Ala Phe Pro Tyr Leu Pro Glu Cys His
1 5 10 15
<210> 100
<211> 16
<212> БЕЛОК
<213> Homo sapiens
<400> 100
Asp Ile Ser Ala Val Cys Trp Ala Phe Pro Phe Asp Pro Glu Cys His
1 5 10 15
<210> 101
<211> 20
<212> БЕЛОК
<213> Homo sapiens
<400> 101
Ala Tyr Glu Cys Pro Arg Leu Glu Tyr Asp Met Phe Gly Ala Leu His
1 5 10 15
Cys Leu Pro Ser
20
<210> 102
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 102
Cys Pro Arg Leu Glu Tyr Asp Met Phe Gly Ala Leu His Cys Leu
1 5 10 15
<210> 103
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 103
Cys Leu Asp Leu Gln Tyr Asp Pro Trp Gly Ala Leu His Cys Ile
1 5 10 15
<210> 104
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 104
Cys Phe Asp Leu Gln Tyr Asp Pro Trp Gly Ala Leu His Cys Ile
1 5 10 15
<210> 105
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 105
Cys Leu Asp Leu Gln Tyr Asp Met Phe Gly Ala Leu His Cys Val
1 5 10 15
<210> 106
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 106
Cys Leu Asp Leu Val Tyr Asp Pro Trp Gly Ala Leu His Cys Ile
1 5 10 15
<210> 107
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 107
Cys Trp Val Leu Glu Tyr Asp Met Phe Gly Ala Leu His Cys Arg
1 5 10 15
<210> 108
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 108
Cys Trp Ala Leu Glu Tyr Asp Met Phe Gly Tyr Leu His Cys Arg
1 5 10 15
<210> 109
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 109
Cys Trp Val Leu Glu Tyr Asp Met Phe Gly Phe Leu His Cys Arg
1 5 10 15
<210> 110
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 110
Cys Trp Val Leu Glu Tyr Asp Met Phe Gly Tyr Leu His Cys Arg
1 5 10 15
<210> 111
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 111
Val His Val Thr Ile Pro Ala Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 112
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 112
Val His Phe Thr Ile Pro Ala Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 113
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 113
Val His Val Gln Ile Pro Ala Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 114
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 114
Val His Val Thr Phe Pro Ala Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 115
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 115
Val His Val Thr Ile Pro Gln Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 116
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 116
Val His Val Thr Ile Pro Ala Asn Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 117
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 117
Val His Val Thr Ile Pro Ala Asp Phe Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 118
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 118
Val His Val Thr Ile Pro Ala Asp Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 119
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 119
Val His Val Thr Ile Pro Ala Asp Leu Trp Asn Trp Val Arg Arg
1 5 10 15
<210> 120
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 120
Val His Val Thr Ile Pro Ala Asp Leu Trp Glu Phe Val Arg Arg
1 5 10 15
<210> 121
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 121
Val His Val Thr Ile Pro Ala Asp Leu Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 122
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 122
Val His Val Tyr Ile Pro Ala Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 123
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 123
Val His Val Thr Ile Pro Ala Glu Trp Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 124
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 124
Val His Phe Thr Phe Pro Gln Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 125
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 125
Val His Phe Thr Phe Pro Gln Asp Phe Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 126
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 126
Val His Phe Thr Ile Pro Gln Asp Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 127
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 127
Val His Phe Thr Phe Pro Gln Asp Leu Trp Asn Trp Val Arg Arg
1 5 10 15
<210> 128
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 128
Val His Phe Thr Phe Pro Gln Asp Leu Trp Glu Phe Val Arg Arg
1 5 10 15
<210> 129
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 129
Val His Phe Thr Phe Pro Gln Asp Leu Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 130
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 130
Val His Phe Gln Phe Pro Ala Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 131
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 131
Val His Phe Gln Phe Pro Ala Asp Phe Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 132
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 132
Val His Phe Gln Phe Pro Ala Asp Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 133
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 133
Val His Phe Gln Phe Pro Ala Asp Leu Trp Asn Trp Val Arg Arg
1 5 10 15
<210> 134
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 134
Val His Phe Gln Phe Pro Ala Asp Leu Trp Glu Phe Val Arg Arg
1 5 10 15
<210> 135
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 135
Val His Phe Gln Phe Pro Ala Asp Leu Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 136
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 136
Val His Phe Gln Phe Pro Gln Asp Trp Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 137
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 137
Val His Phe Gln Ile Pro Gln Asp Trp Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 138
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 138
Val His Phe Gln Ile Phe Gln Asp Trp Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 139
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 139
Val His Phe Gln Phe Pro Gln Asp Trp Trp Asn Trp Val Arg Arg
1 5 10 15
<210> 140
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 140
Val His Phe Gln Phe Pro Gln Asp Leu Trp Glu Phe Val Arg Arg
1 5 10 15
<210> 141
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 141
Val His Phe Gln Phe Pro Gln Asp Trp Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 142
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 142
Val His Phe Thr Ile Pro Ala Asp Phe Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 143
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 143
Val His Val Gln Ile Pro Ala Asp Phe Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 144
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 144
Val His Val Thr Phe Pro Ala Asp Leu Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 145
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 145
Val His Val Thr Ile Pro Gln Asp Phe Trp Glu Trp Phe Arg Arg
1 5 10 15
<210> 146
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 146
Val His Phe Thr Ile Pro Gln Asp Trp Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 147
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 147
Val His Phe Thr Phe Pro Gln Asp Leu Tyr Asn Trp Val Arg Arg
1 5 10 15
<210> 148
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 148
Val His Phe Thr Phe Pro Gln Asp Leu Tyr Asn Phe Val Arg Arg
1 5 10 15
<210> 149
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 149
Val His Val Thr Ile Pro Ala Asp Leu Tyr Asn Phe Phe Arg Arg
1 5 10 15
<210> 150
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 150
Val His Phe Gln Phe Pro Gln Asp Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 151
<211> 14
<212> БЕЛОК
<213> Homo sapiens
<400> 151
Val His Phe Thr Ile Pro Gln Asp Leu Tyr Asn Trp Arg Arg
1 5 10
<210> 152
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 152
Val His Phe Thr Ile Pro Ala Asp Leu Tyr Asn Phe Val Arg Arg
1 5 10 15
<210> 153
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 153
Val His Phe Gln Ile Pro Gln Asp Leu Tyr Asn Phe Phe Arg Arg
1 5 10 15
<210> 154
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 154
Val His Phe Gln Phe Pro Gln Glu Trp Tyr Asn Trp Phe Arg Arg
1 5 10 15
<210> 155
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 155
Val Arg Phe Gln Phe Gly Gln Glu Trp Tyr Asn Phe Phe Arg Arg
1 5 10 15
<210> 156
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 156
Val Pro Gly Ala Gly Val Pro Gly Ala Gly Ile His Val Thr Ile
1 5 10 15
<210> 157
<211> 17
<212> БЕЛОК
<213> Homo sapiens
<400> 157
Val Pro Gly Ala Gly Val Pro Gly Ala Gly Ile His Val Thr Ile Pro
1 5 10 15
Ala
<210> 158
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 158
Ala His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 159
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 159
Gly His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 160
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 160
Leu His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 161
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 161
Pro His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 162
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 162
Ile Arg Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 163
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 163
Ile Lys Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 164
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 164
Ile Glu Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 165
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 165
Ile Gln Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 166
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 166
Ile Tyr Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 167
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 167
Ile His Ala Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 168
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 168
Ile His Gly Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 169
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 169
Ile His Leu Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 170
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 170
Ile His Pro Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 171
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 171
Ile His Ile Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 172
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 172
Ile His Tyr Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 173
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 173
Ile His Trp Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 174
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 174
Ile His Phe Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 175
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 175
Ile His Val Ser Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 176
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 176
Ile His Val Tyr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 177
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 177
Ile His Val Asn Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 178
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 178
Ile His Val Gln Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 179
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 179
Ile His Val Thr Ala Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 180
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 180
Ile His Val Thr Gly Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 181
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 181
Ile His Val Thr Leu Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 182
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 182
Ile His Val Thr Pro Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 183
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 183
Ile His Val Thr Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 184
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 184
Ile His Val Thr Phe Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 185
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 185
Ile His Val Thr Tyr Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 186
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 186
Ile His Val Thr Ile Ala Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 187
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 187
Ile His Val Thr Ile Gly Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 188
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 188
Ile His Val Thr Ile Leu Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 189
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 189
Ile His Val Thr Ile Val Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 190
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 190
Ile His Val Thr Ile Ile Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 191
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 191
Ile His Val Thr Ile Asn Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 192
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 192
Ile His Val Thr Ile Pro Gly Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 193
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 193
Ile His Val Thr Ile Pro Leu Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 194
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 194
Ile His Val Thr Ile Pro Pro Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 195
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 195
Ile His Val Thr Ile Pro Val Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 196
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 196
Ile His Val Thr Ile Pro Ile Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 197
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 197
Ile His Val Thr Ile Pro Asn Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 198
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 198
Ile His Val Thr Ile Pro Gln Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 199
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 199
Ile His Val Thr Ile Pro Ala Gln Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 200
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 200
Ile His Val Thr Ile Pro Ala Tyr Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 201
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 201
Ile His Val Thr Ile Pro Ala Asp Ala Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 202
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 202
Ile His Val Thr Ile Pro Ala Asp Gly Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 203
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 203
Ile His Val Thr Ile Pro Ala Asp Pro Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 204
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 204
Ile His Val Thr Ile Pro Ala Asp Val Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 205
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 205
Ile His Val Thr Ile Pro Ala Asp Ile Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 206
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 206
Ile His Val Thr Ile Pro Ala Asp Leu His Asp Trp Ile Asn Lys
1 5 10 15
<210> 207
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 207
Ile His Val Thr Ile Pro Ala Asp Leu Phe Asp Trp Ile Asn Lys
1 5 10 15
<210> 208
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 208
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asn Trp Ile Asn Lys
1 5 10 15
<210> 209
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 209
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp His Ile Asn Lys
1 5 10 15
<210> 210
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 210
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Phe Ile Asn Lys
1 5 10 15
<210> 211
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 211
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Tyr Ile Asn Lys
1 5 10 15
<210> 212
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 212
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Glu Asn Lys
1 5 10 15
<210> 213
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 213
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Lys Asn Lys
1 5 10 15
<210> 214
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 214
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ala Asn Lys
1 5 10 15
<210> 215
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 215
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Gly Asn Lys
1 5 10 15
<210> 216
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 216
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Leu Asn Lys
1 5 10 15
<210> 217
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 217
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Pro Asn Lys
1 5 10 15
<210> 218
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 218
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Phe Asn Lys
1 5 10 15
<210> 219
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 219
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Tyr Asn Lys
1 5 10 15
<210> 220
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 220
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Gln Lys
1 5 10 15
<210> 221
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 221
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asp Lys
1 5 10 15
<210> 222
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 222
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn His
1 5 10 15
<210> 223
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 223
Ile His Val Thr Ile Pro Ala Asp Leu Trp Asp Trp Ile Asn Asp
1 5 10 15
<210> 224
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 224
Val His Val Thr Val Pro Gln Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 225
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 225
Val His Val Thr Val Pro Gln Glu Leu Phe Glu Trp Val Arg Arg
1 5 10 15
<210> 226
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 226
Val His Val Thr Val Pro Gln Glu Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 227
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 227
Val His Val Thr Val Pro Gln Glu Leu Trp Glu Trp Val Glu Glu
1 5 10 15
<210> 228
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 228
Val His Val Thr Val Pro Gln Glu Leu Phe Glu Trp Val Glu Glu
1 5 10 15
<210> 229
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 229
Val His Val Thr Val Pro Gln Glu Leu Tyr Glu Trp Val Glu Glu
1 5 10 15
<210> 230
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 230
Val His Val Ser Ile Pro Gln Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 231
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 231
Val His Val Ser Val Pro Gln Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 232
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 232
Val His Val Ser Val Pro Gln Glu Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 233
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 233
Val Arg Val Thr Ile Pro Gln Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 234
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 234
Val Arg Val Thr Val Pro Gln Glu Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 235
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 235
Val Arg Val Thr Val Pro Gln Glu Leu Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 236
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 236
Val His Val Thr Val Pro Gln Glu Ile Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 237
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 237
Val His Val Thr Ile Pro Gln Glu Ile Trp Glu Trp Val Arg Arg
1 5 10 15
<210> 238
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 238
Val His Phe Thr Val Pro Gln Glu Leu Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 239
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 239
Val Lys Ile Ser Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 240
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 240
Leu Arg Ile Ser Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 241
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 241
Leu Arg Ile Tyr Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 242
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 242
Val Arg Gly Tyr Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 243
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 243
Val Arg Ala Tyr Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 244
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 244
Val Arg Leu Tyr Val Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 245
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 245
Val Arg Ile Tyr Leu Pro Ala Asp Leu Trp Asp Trp Ile Asn Lys
1 5 10 15
<210> 246
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 246
Ile His Val Thr Ile Pro Leu Glu Ile Phe Glu Trp Leu Gln His
1 5 10 15
<210> 247
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 247
Ile His Val Thr Ile Pro Leu Glu Ile Phe Glu Trp Ala Gln His
1 5 10 15
<210> 248
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 248
Ile His Val Thr Ile Pro Leu Glu Ile Phe Glu Trp Leu Gln Arg
1 5 10 15
<210> 249
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 249
Ile His Val Thr Ile Pro Leu Glu Val Phe Glu Trp Leu Gln His
1 5 10 15
<210> 250
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 250
Ile His Val Thr Ile Pro Gly Glu Ile Phe Glu Trp Leu Gln His
1 5 10 15
<210> 251
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 251
Val Arg Phe Ser Val Pro Gln Glu Ile Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 252
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 252
Leu Arg Ile Ser Val Pro Leu Glu Ile Phe Glu Trp Leu Gln His
1 5 10 15
<210> 253
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 253
Leu Arg Gly Ser Val Pro Leu Glu Ile Phe Glu Trp Leu Gln His
1 5 10 15
<210> 254
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 254
Val Lys Ile Ser Val Pro Leu Glu Ile Phe Glu Trp Leu Gln His
1 5 10 15
<210> 255
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 255
Val Glu Phe Asn Phe Pro Gln Gln Val Tyr Glu Trp Phe Asp Asp
1 5 10 15
<210> 256
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 256
Val Glu Phe Asn Phe Pro Gln Gln Val Tyr Glu Trp Val Arg Arg
1 5 10 15
<210> 257
<211> 15
<212> БЕЛОК
<213> Homo sapiens
<400> 257
Thr Trp Tyr Val Phe Asn Glu Gln His Gln Glu Tyr Val Arg Lys
1 5 10 15
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| НЕОАНТИГЕНЫ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2813924C2 |
| ИММУНОДОМИНАНТНЫЕ БЕЛКИ И ФРАГМЕНТЫ ПРИ РАССЕЯННОМ СКЛЕРОЗЕ | 2019 |
|
RU2828590C2 |
| КОМПОЗИЦИИ И СПОСОБЫ ПОЛУЧЕНИЯ КОМПОЗИЦИЙ T-КЛЕТОК И ИХ ПРИМЕНЕНИЕ | 2020 |
|
RU2835034C2 |
| ИСКУССТВЕННЫЕ АНТИГЕНПРЕЗЕНТИРУЮЩИЕ КЛЕТКИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2763798C1 |
| ИНДУКТОР ИММУНИТЕТА | 2016 |
|
RU2835947C2 |
| ИНДУКТОР ИММУНИТЕТА | 2016 |
|
RU2758112C2 |
| T-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ, РАСПОЗНАЮЩИЕ МУТАЦИЮ R175H ИЛИ Y220C В P53 | 2020 |
|
RU2830061C2 |
| ПЕПТИДНЫЕ ИММУНОГЕНЫ, НАЦЕЛЕННЫЕ НА ИНТЕРЛЕЙКИН 6 (IL-6), И ИХ СОСТАВЫ ДЛЯ ИММУНОТЕРАПИИ ЗАБОЛЕВАНИЙ, НА КОТОРЫЕ ВЛИЯЕТ НАРУШЕНИЕ РЕГУЛЯЦИИ IL-6 | 2019 |
|
RU2780161C1 |
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| Т-КЛЕТОЧНЫЕ РЕЦЕПТОРЫ | 2018 |
|
RU2762255C2 |
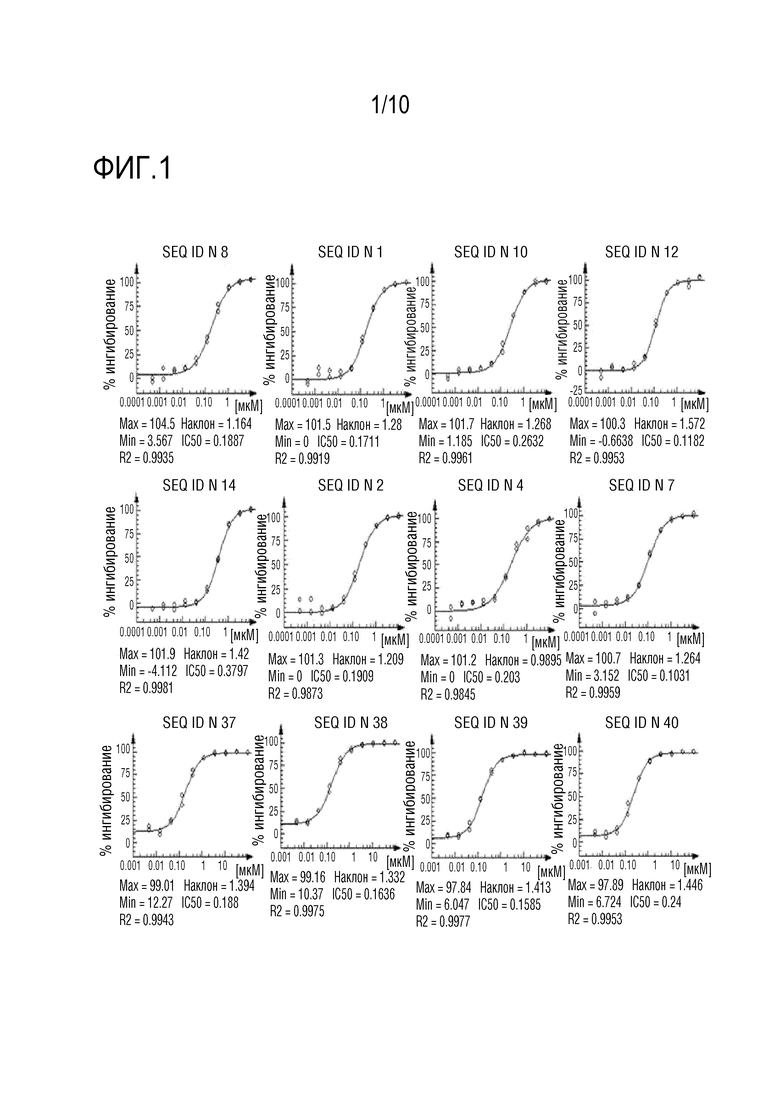
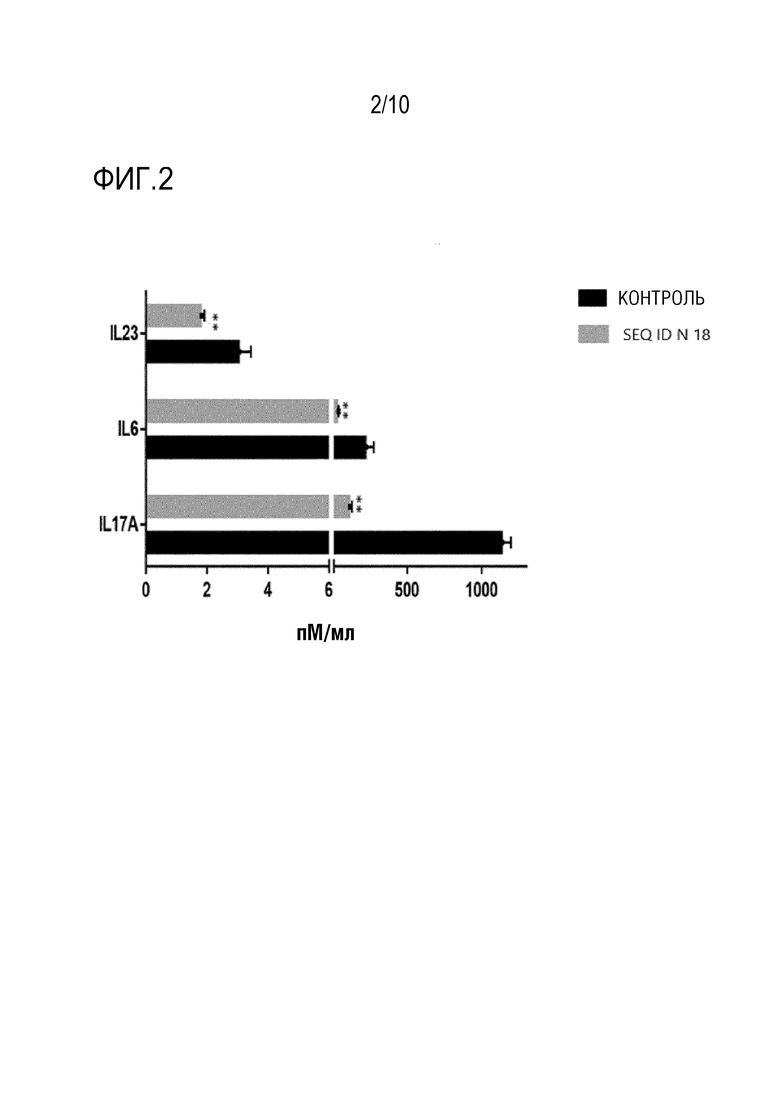
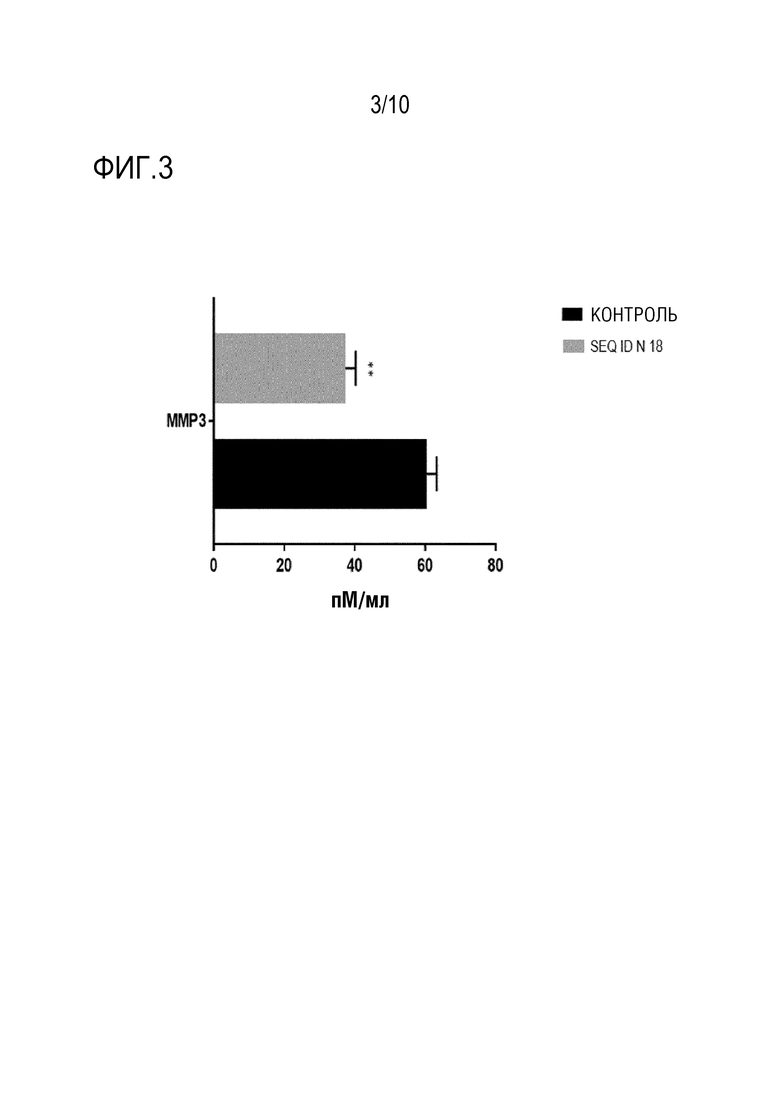
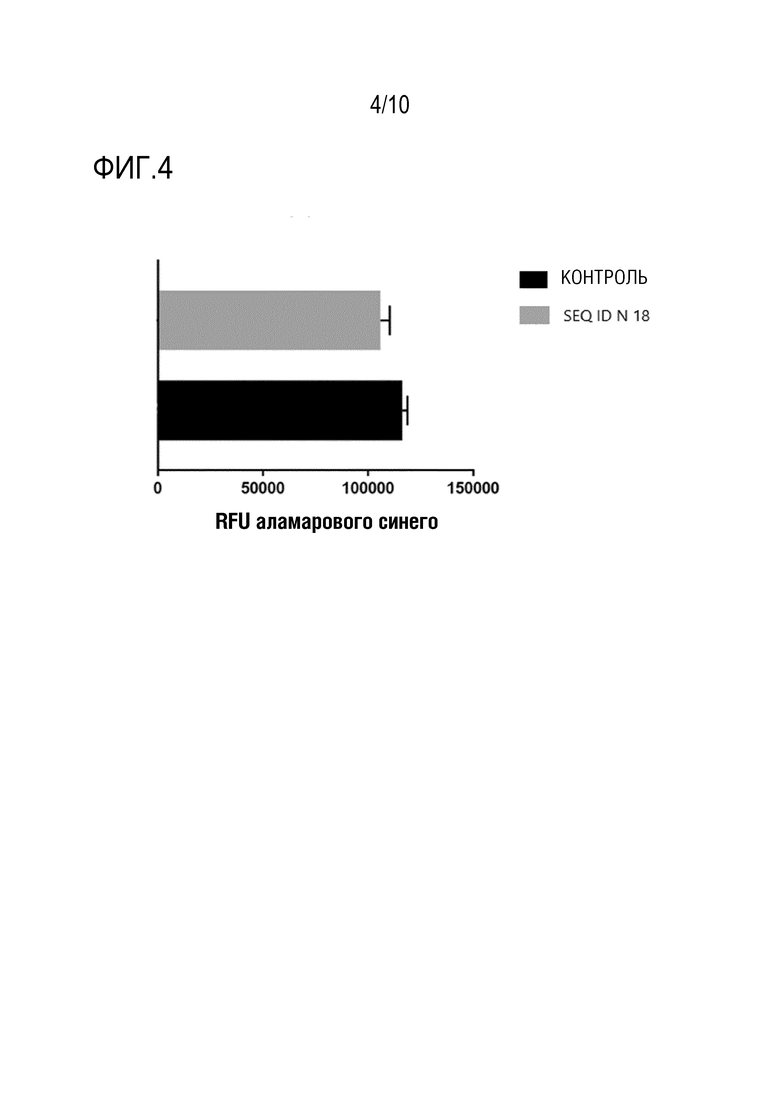

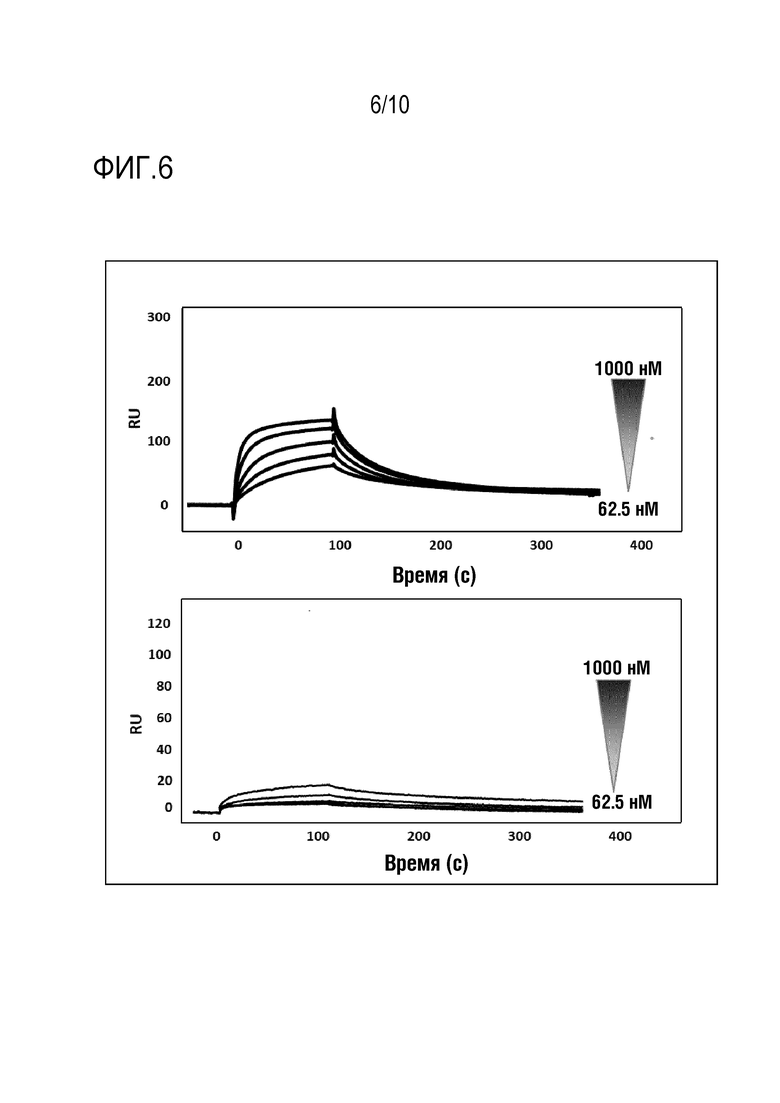
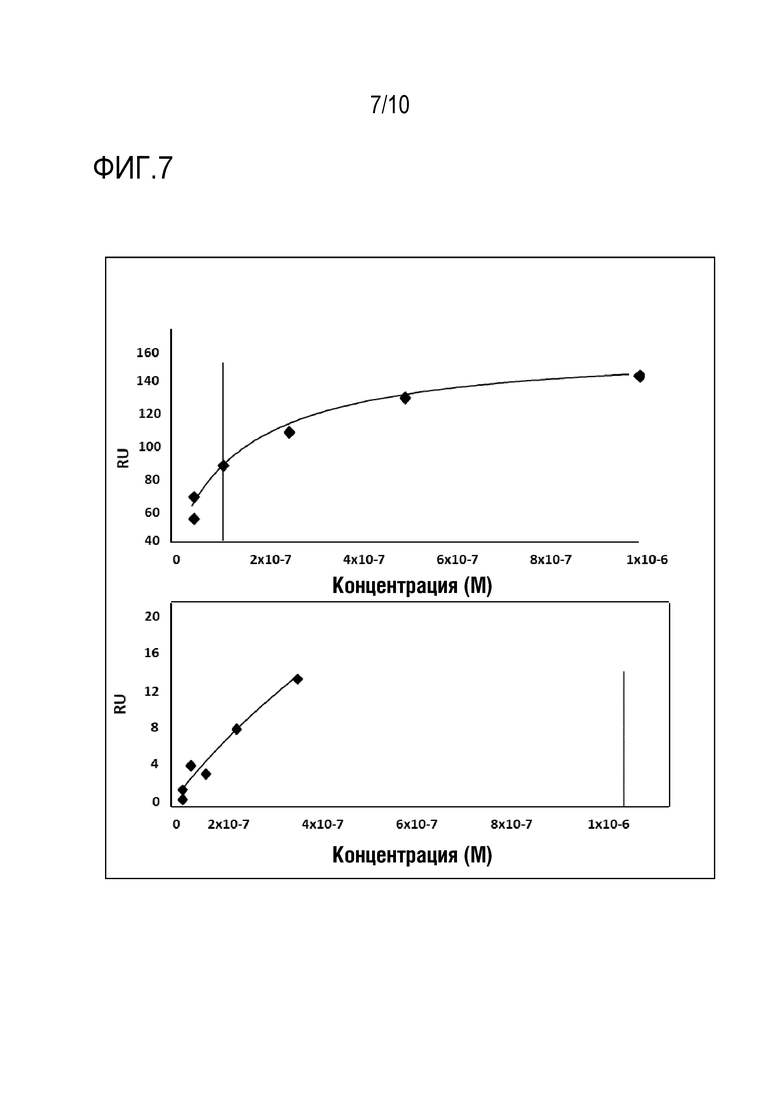
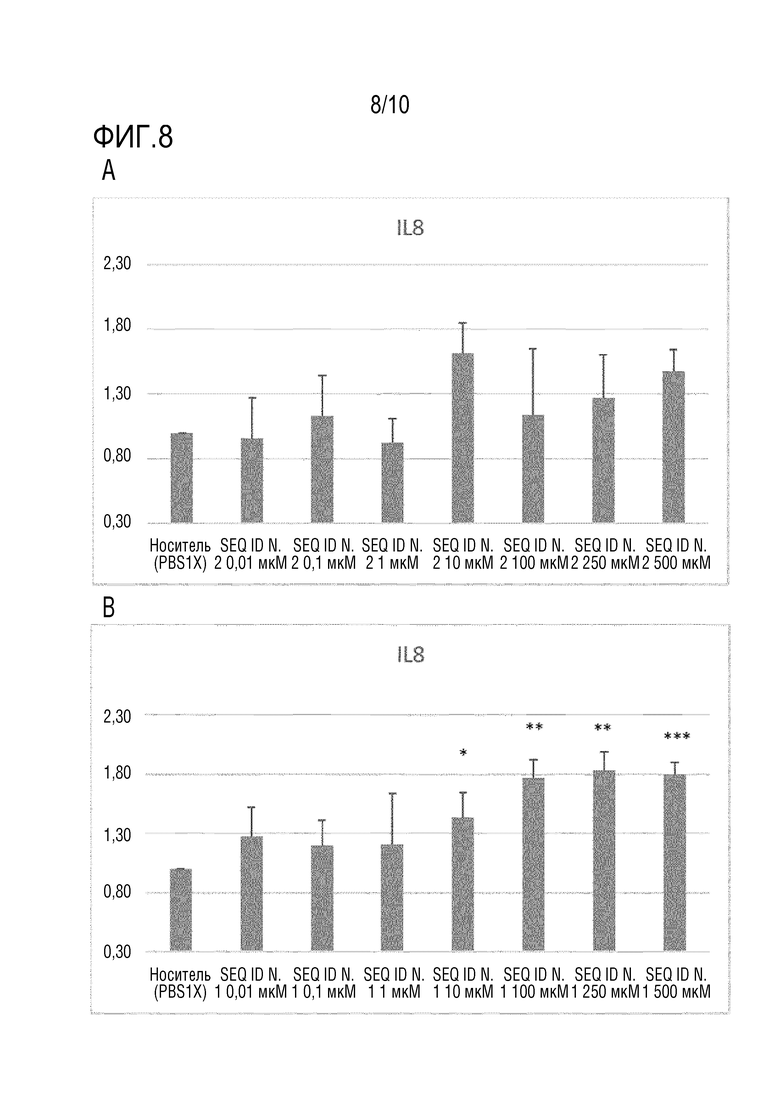
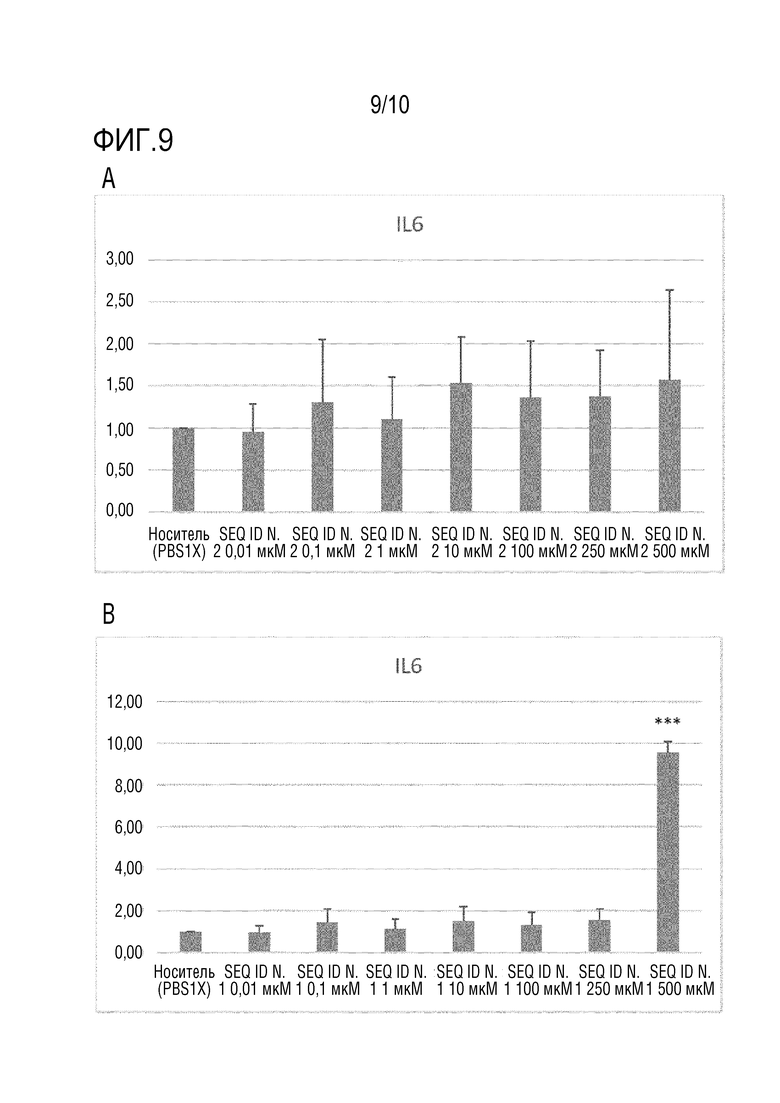
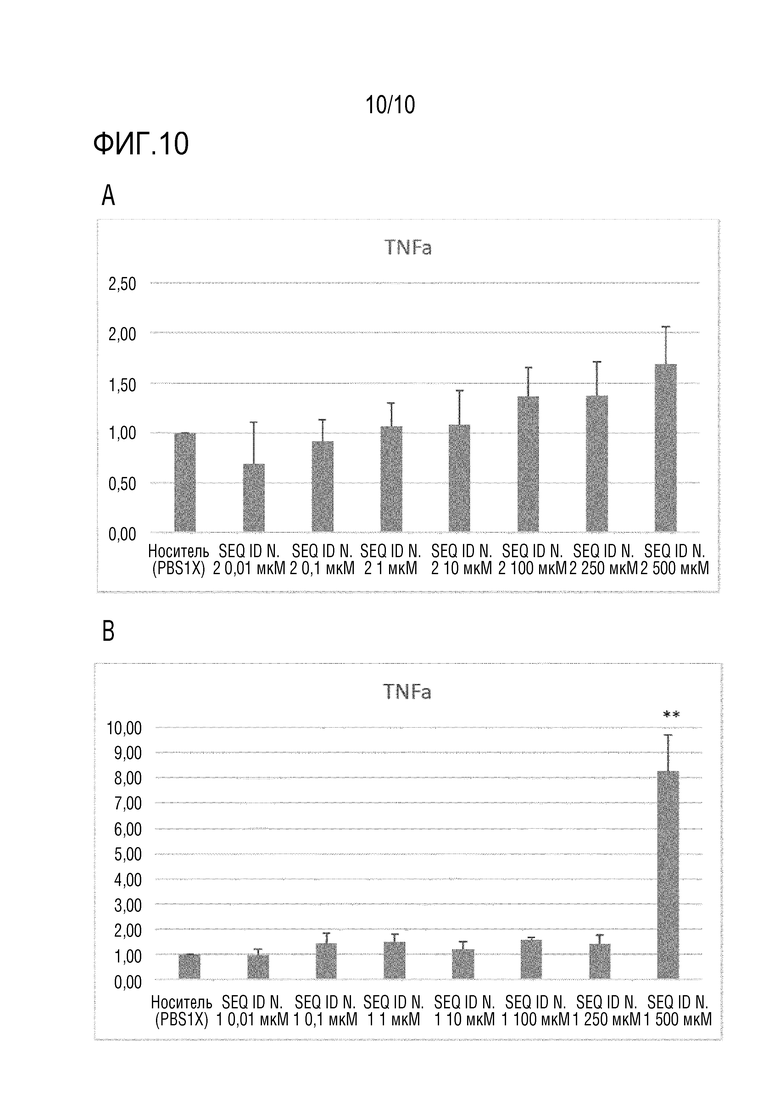
Изобретение относится к связывающим IL-17A пептидам, ингибиторам взаимодействия IL-17A с рецептором IL-17RA, и к биоконъюгатам и фармацевтическим композициям, которые могут быть использованы, например, для лечения и/или предупреждения заболевания такого, как псориаз или болезнь сухого глаза. 5 н. и 2 з.п. ф-лы, 10 ил., 11 табл., 10 пр.
1. Пептид, способный ингибировать связывание IL-17A с ILRA, имеющий аминокислотную последовательность, выбранную из:
SEQ ID N. 2, SEQ ID N. 3, SEQ ID N. 4, SEQ ID N. 5, SEQ ID N. 6, SEQ ID N. 7, SEQ ID N. 8, SEQ ID N. 9, SEQ ID N. 10, SEQ ID N. 11, SEQ ID N. 12, SEQ ID N. 13, SEQ ID N. 14, SEQ ID N. 15, SEQ ID N. 16, SEQ ID N. 17, SEQ ID N. 18, SEQ ID N. 42, SEQ ID N. 111, SEQ ID N. 112, SEQ ID N. 113, SEQ ID N. 114, SEQ ID N. 115, SEQ ID N. 116, SEQ ID N. 117, SEQ ID N. 118, SEQ ID N. 119, SEQ ID N. 120, SEQ ID N. 121, SEQ ID N. 122, SEQ ID N. 123, SEQ ID N. 124, SEQ ID N. 125, SEQ ID N. 126, SEQ ID N. 127, SEQ ID N. 128, SEQ ID N. 129, SEQ ID N. 130, SEQ ID N. 131, SEQ ID N. 132, SEQ ID N. 133, SEQ ID N. 134, SEQ ID N. 135, SEQ ID N. 136, SEQ ID N. 137, SEQ ID N. 138, SEQ ID N. 139, SEQ ID N. 140, SEQ ID N. 141, SEQ ID N. 142, SEQ ID N. 143, SEQ ID N. 144, SEQ ID N. 145, SEQ ID N. 146, SEQ ID N. 147, SEQ ID N. 148, SEQ ID N. 149, SEQ ID N. 150, SEQ ID N. 151, SEQ ID N. 152, SEQ ID N. 153, SEQ ID N. 154, SEQ ID N. 155, SEQ ID N. 158, SEQ ID N. 159, SEQ ID N. 160, SEQ ID N. 161, SEQ ID N. 162, SEQ ID N. 163, SEQ ID N. 164, SEQ ID N. 165, SEQ ID N. 166, SEQ ID N. 167, SEQ ID N. 168, SEQ ID N. 169, SEQ ID N. 170, SEQ ID N. 171, SEQ ID N. 172, SEQ ID N. 173, SEQ ID N. 174, SEQ ID N. 175, SEQ ID N. 176, SEQ ID N. 177, SEQ ID N. 178, SEQ ID N. 179, SEQ ID N. 180, SEQ ID N. 181, SEQ ID N. 182, SEQ ID N. 183, SEQ ID N. 184, SEQ ID N. 185, SEQ ID N. 186, SEQ ID N. 187, SEQ ID N. 188, SEQ ID N. 189, SEQ ID N. 190, SEQ ID N. 191, SEQ ID N. 192, SEQ ID N. 193, SEQ ID N. 194, SEQ ID N. 195, SEQ ID N. 196, SEQ ID N. 197, SEQ ID N. 198, SEQ ID N. 200, SEQ ID N. 201, SEQ ID N. 202, SEQ ID N. 203, SEQ ID N. 204, SEQ ID N. 205, SEQ ID N. 206, SEQ ID N. 207, SEQ ID N. 208, SEQ ID N. 209, SEQ ID N. 210, SEQ ID N. 211, SEQ ID N. 212, SEQ ID N. 213, SEQ ID N. 214, SEQ ID N. 215, SEQ ID N. 216, SEQ ID N. 217, SEQ ID N. 218, SEQ ID N. 219, SEQ ID N. 220, SEQ ID N. 221, SEQ ID N. 222, SEQ ID N. 223, SEQ ID N. 224, SEQ ID N. 225, SEQ ID N. 226, SEQ ID N. 227, SEQ ID N. 228, SEQ ID N. 229, SEQ ID N. 230, SEQ ID N. 231, SEQ ID N. 232, SEQ ID N. 233, SEQ ID N. 234, SEQ ID N. 235, SEQ ID N. 236, SEQ ID N. 237, SEQ ID N. 238, SEQ ID N. 239, SEQ ID N. 240, SEQ ID N. 241, SEQ ID N. 242, SEQ ID N. 243, SEQ ID N. 244, SEQ ID N. 245, SEQ ID N. 246, SEQ ID N. 247, SEQ ID N. 248, SEQ ID N. 249, SEQ ID N. 250, SEQ ID N. 251, SEQ ID N. 252, SEQ ID N. 253, SEQ ID N. 254, SEQ ID N. 255, SEQ ID N. 256, or SEQ ID N. 257;
где пептид может быть связанным на C- и/или N-конце с защитной кэп-группой, причем предпочтительно защитная кэп-группа, связанная с C-концом, выбрана из амидов, альдегидов, сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина, и защитная кэп-группа, связанная с N-концом, выбрана из ацетила, формила, пироглутамила, жирных кислот, мочевины, карбамата, сульфонамида, алкиламина.
2. Пептид по п.1, имеющий последовательность из 15 аминокислот.
3. Пептид, состоящий из аминокислотной последовательности SEQ ID N. 21, SEQ ID N. 22, SEQ ID N. 23, SEQ ID N. 24, SEQ ID N. 25, SEQ ID N. 26, SEQ ID N. 27, SEQ ID N. 28, SEQ ID N. 31, SEQ ID N. 32, SEQ ID N. 33 SEQ ID N. 34, SEQ ID N. 35, SEQ ID N. 36, SEQ ID N. 37, SEQ ID N. 38, SEQ ID N. 39 или SEQ ID N. 40;
где пептид может быть связанным на C- и/или N-конце с защитной кэп-группой, причем предпочтительно защитная кэп-группа, связанная с C-концом, выбрана из амидов, альдегидов, сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина, и защитная кэп-группа, связанная с N-концом, выбрана из ацетила, формила, пироглутамила, жирных кислот, мочевины, карбамата, сульфонамида, алкиламина.
4. Пептид, состоящий из аминокислотной последовательности SEQ ID N. 19, SEQ ID N. 20, SEQ ID N. 29, SEQ ID N. 30 или SEQ ID N. 43;
где пептид может быть связанным на C- и/или N-конце с защитной кэп-группой, причем предпочтительно защитная кэп-группа, связанная с C-концом, выбрана из амидов, альдегидов, сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина, и защитная кэп-группа, связанная с N-концом, выбрана из ацетила, формила, пироглутамила, жирных кислот, мочевины, карбамата, сульфонамида, алкиламина.
5. Пептид по любому из пп.1, 3 или 4, связанный на C- и/или N-конце с защитной кэп-группой, причем предпочтительно защитная кэп-группа, связанная с C-концом, выбрана из амидов, альдегидов, сложных эфиров, п-нитроанилида, 7-амино-4-метилкумарина, и защитная кэп-группа, связанная с N-концом, выбрана из ацетила, формила, пироглутамила, жирных кислот, мочевины, карбамата, сульфонамида, алкиламина.
6. Биоконъюгат, содержащий пептид по любому из пп.1-5 и биомолекулу, причем предпочтительно указанная молекула связана с N- и/или C-концом указанного пептида, где, если указанная молекула находится на N-конце, она представляет собой одну из ацетила, формила, жирных кислот, мочевины, карбамата, сульфонамида или алкиламина; и где, если указанная молекула находится на С-конце, она представляет собой один из амидов, альдегидов, сложных эфиров, п-нитроанилида или 7-амино-4-метилкумарина.
7. Фармацевтическая композиция для лечения и/или предупреждения заболевания, выбранного из ревматоидного артрита, рассеянного склероза, болезни Крона, системной красной волчанки, астмы, болезни Бехчета, гипер-IgE-синдрома, анкилозирующего спондилита, псориаза, псориатического артрита, ревматоидного артрита, сухого кератоконъюнктивита, весеннего кератоконъюнктивита, стромального герпетического кератита, отторжения аллотрансплантата роговицы, инфекций роговицы, обусловленного вирусом герпеса или Pseudomonas aeruginosa кератита и болезни сухого глаза, предпочтительно псориаза или болезни сухого глаза, содержащая в эффективном количестве пептид по любому из пп.1-5 или биоконъюгат по п.6 и по меньшей мере один фармацевтически приемлемый эксципиент.
| JOEY P | |||
| TING ET AL, "Utilization of peptide phage display to investigate hotspots on IL-17A and what it means for drug discovery", PLOS ONE, 2018.01.12, vol | |||
| Насос | 1917 |
|
SU13A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Shenping Liu et al., "Inhibiting complex IL-17A and IL-17RA interactions with a linear peptide", SCIENTIFIC REPORTS, 2016, no | |||
| Приспособление для точного наложения листов бумаги при снятии оттисков | 1922 |
|
SU6A1 |
