Клаудины представляют собой интегральные мембранные белки, локализованные в плотных контактах эпителия и эндотелия. Предполагается, что клаудины содержат четыре трансмембранных сегмента с двумя внеклеточными петлями, и N- и C-концы, расположенные в цитоплазме. Клаудин (CLDN), семейство трансмембранных белков, играет важную роль в поддержании эпителиальных и эндотелиальных плотных контактов, а также может участвовать в поддержании цитоскелета и в клеточной сигнализации.
Молекула клаудина 18 (CLDN18) представляет собой интегральный трансмембранный белок (тетраспанин), имеющий в своем составе четыре трансмембранные гидрофобные области и две внеклеточные петли (петлю 1, расположенную между гидрофобной областью 1 и гидрофобной областью 2; петлю 2, расположенную между гидрофобными областями 3 и 4). CLDN18 существует в двух различных сплайсированных вариантах, которые описаны у мышей и у человека (Niimi, Mol. Cell. Biol. 21:7380-90, 2001). Сплайсированные варианты (номер доступа в Genbank: сплайсированный вариант 1 (CLDN18.1): NP_057453, NM_016369, и сплайсированный вариант 2 (CLDN18.2): NM_001002026, NP_001002026) имеют молекулярную массу приблизительно 27,9 / 27,72 кДа. Сплайсированные варианты CLDN18.1 и CLDN18.2 отличаются в N-концевой части, которая содержит первую трансмембранную (TM) область и петлю 1, тогда как первичная белковая последовательность на C-конце является идентичной.
В нормальных тканях отсутствует детектируемая экспрессия CLDN18.2, за исключением желудка, где CLDN18.2 экспрессируется исключительно на короткоживущих дифференцированных клетках эпителия желудка. Экспрессия CLDN18.2 сохраняется в процессе злокачественной трансформации и, таким образом, часто проявляется на поверхности человеческих клеток рака желудка. Более того, этот пан-опухолевый антиген в значительной степени эктопически активируется при аденокарцинаме пищевода, поджелудочной железы и легких. Белок CLDN18.2 также локализован в метастазах в лимфоузлы аденокарциномы желудка и отдаленных метастазах, в особенности, в яичниках (так называемые опухоли Крукенберга).
CLDN6 экспрессируется в ряде различных человеческих раковых клеток, при этом экспрессия в нормальных тканях ограничивается плацентой.
Дифференциальная экспрессия клаудинов, таких как CLDN18.2 и CLDN6, в раковых и нормальных клетках, их локализация в мембране и отсутствие у подавляющего большинства из них токсичности в отношении релевантных нормальных тканей делает эти молекулы привлекательными мишенями для иммунотерапии рака, и использование лекарственных средств на основе антител для направленного воздействия на клаудины в терапии рака обещает высокий уровень терапевтической специфичности.
Подходы, в которых используется потенциал Т-клеток для лечения рака, включают вакцинацию белками, выделенными из опухоли; РНК или пептидными антигенами; инфузию выделенных из опухоли, обогащенных ex-vivo T-клеток (называемую адоптивным переносом); перенос гена T-клеточного рецептора или прямой захват T-клеток би- или триспецифическими антителами. Подобным образом, множество стимуляторов Т-клеточных ответов прошли клиническое тестирование в комбинации или в виде монотерапии, такие как лиганды для Toll-подобных рецепторов; антитела, блокирующие CTLA-4 на Т-клетках; цитокины, стимулирующие иммунный ответ; или антитела, нейтрализующие молекулы, вовлеченные в «ускользание» раковых клеток от иммунного ответа, такие как TGF-бета или B7-H1. Интенсивная разработка терапий на основе Т-клеток продиктована тем наблюдением, что пациенты, по-видимому, живут значительно дольше, если их опухоли инфильтрованы Т-клетками. Более того, многочисленные мышиные модели показали, что захват Т-клеток различными способами может устранить даже крупные опухоли, и ряд Т-клеточных терапий в последнее время добились значительного прогресса в лечении различных симптомов рака.
Целью изобретения является обеспечение новых агентов и способов для терапии раковых заболеваний.
Решение проблемы, лежащее в основе изобретения, основано на концепте создания связывающего агента, содержащего связывающий домен, который является специфическим в отношении опухоль-ассоциированной молекулы клаудина, то есть раковых клеток. Другой связывающий домен является специфическим в отношении CD3, обеспечивая связывание с Т-клетками и вхождение в комплекс, делая, таким образом, возможным направление цитотоксического действия Т-клеток на раковые клетки. Образование этого комплекса может индуцировать передачу сигнала в цитотоксических Т-клетках, либо самостоятельно или в сочетании с А-клетками, что приводит к высвобождению цитотоксических медиаторов.
Авторы впервые сообщают, что связывающие агенты, направленно воздействующие на клаудин и CD3, могут индуцировать сильный опосредованный Т-клетками лизис и являются эффективными в лечении опухолевых заболеваний.
Краткое описание изобретения
В одном аспекте изобретение относится к связывающему агенту, содержащему, по меньшей мере, два связывающих домена, при этом первый связывающий домен связывается с клаудином и второй связывающий домен связывается с CD3. Связывающий агент по изобретению может связываться с цитотоксической клеткой (вовлекая CD3-рецептор) и раковой клеткой, экспрессирующей CLDN, которая является мишенью, подлежащей уничтожению.
В одном варианте осуществления связывающий агент представляет собой биспецифическую молекулу, такую как биспецифическое антитело, в частности, биспецифическое одноцепочечное антитело. В одном варианте осуществления указанный клаудин экспрессируется в раковой клетке. В одном варианте осуществления указанный клаудин экспрессируется на поверхности раковой клетки. В одном варианте осуществления указанный клаудин выбран из группы, состоящей из клаудина 18.2 и клаудина 6. В одном варианте осуществления указанный первый связывающий домен связывается с внеклеточным доменом указанного клаудина. В одном варианте осуществления указанный первый связывающий агент связывается с нативными эпитопами CLDN, присутствующими на поверхности живых клеток. В одном варианте осуществления указанный первый связывающий домен связывается с первой внеклеточной петлей CLDN. В одном варианте осуществления указанный второй связывающий домен связывается с эпсилон-цепью CD3. В одном варианте осуществления указанный CD3 экспрессируется на поверхности Т-клетки. В одном варианте осуществления связывание указанного связывающего агента с CD3 на T-клетках приводит к пролиферации и/или активации указанных Т-клеток, при этом указанные активированные Т-клетки предпочтительно высвобождают цитотоксические факторы, например, перфорины и гранзимы, и инициируют цитолизис и апоптоз раковых клеток. В одном варианте осуществления указанное связывание с клаудином и/или указанное связывание с CD3 представляет собой специфическое связывание.
В одном варианте осуществления связывающий агент находится в формате полноразмерного антитела или фрагмента антитела. В одном варианте осуществления связывающий агент содержит четыре вариабельных домена антитела, по меньшей мере, с двумя связывающими доменами, при этом, по меньшей мере, один связывающий домен связывается с клаудином и, по меньшей мере, один связывающий домен связывается с CD3. В одном варианте осуществления связывающий агент содержит вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью в отношении антигена клаудина (VH(CLDN)), вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью в отношении антигена клаудина (VL(CLDN)), вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью в отношении CD3 (VH(CD3)), и вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью в отношении CD3 (VL(CD3)).
В одном варианте осуществления связывающий агент находится в формате диатела, которое содержит вариабельный домен тяжелой цепи, соединенный с вариабельным доменом легкой цепи на той же самой полипептидной цепи таким образом, что два домена не образуют пару. В одном варианте осуществления диатело содержит две полипептидные цепи, при этом один полипептид содержит VH(CLDN) и VL(CD3) и другая полипептидная цепь содержит VH(CD3) и VL(CLDN).
В одном варианте осуществления связывающий агент находится в формате биспецифического одноцепочечного антитела, которое состоит из двух молекул scFv, соединенных посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) предпочтительно расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-VH(CD3)-VL(CD3), VH(CD3)-VL(CD3)-VH(CLDN)-VL(CLDN) или VH(CD3)-VL(CD3)-VL(CLDN)-VH(CLDN). В одном варианте осуществления указанные вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) соединены посредством длинного пептидного линкера, предпочтительно, пептидного линкера, содержащего аминокислотные последовательности (GGGGS)3 или VE(GGGGS)2GGVD. В одном варианте осуществления указанные две единицы VH-VL или VL-VH scFv соединены посредством короткого пептидного линкера, предпочтительно, пептидного линкера, содержащего аминокислотную последовательность SGGGGS или GGGGS.
В одном варианте осуществления указанный CLDN представляет собой CLDN18.2, и указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 8, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 15, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанный CLDN представляет собой CLDN18.2, и указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 6, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 11, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанный CLDN представляет собой CLDN6, и указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 22, или ее вариант, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 23, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанный CLDN представляет собой CLDN6, и указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 22, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 97, 98, 99 или 100, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанная VH(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 36, 94 или 95, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 37 или 96, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанная VH(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 36, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 37, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанная VH(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 95, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 96 или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном аспекте связывающий агент по изобретению представлен в формате биспецифического одноцепочечного антитела, которое содержит две молекулы scFv, соединенные посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-VH(CD3)-VL(CD3). В одном варианте осуществления указанные VH(CD3) и VL(CD3) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)4. В одном варианте осуществления указанные VH(CLDN) и VL(CLDN) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)4. В одном варианте осуществления указанные две единицы VH-VL scFv соединены посредством линкерного пептида, содержащего аминокислотную последовательность SGGGGS. Одна или обе из указанных единиц VH-VL scFv могут содержать один или более связующих дисульфидных мостиков.
В одном аспекте связывающий агент по изобретению находится в формате биспецифического одноцепочечного антитела, которое содержит две молекулы scFv, соединенные посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VL(CLDN)-VH(CLDN)-VH(CD3)-VL(CD3). В одном варианте осуществления указанные VH(CD3) и VL(CD3) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)4. В одном варианте осуществления указанные VL(CLDN) и VH(CLDN) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)5. В одном варианте осуществления указанные единицы VL-VH и VH-VL scFv соединены посредством линкерного пептида, содержащего аминокислотную последовательность SGGGGS. Одна или обе из указанных двух единиц VL-VH или VH-VL scFv могут содержать один или более связующих дисульфидных мостиков.
В одном аспекте связывающий агент по изобретению находится в формате биспецифического одноцепочечного антитела, которое содержит две молекулы scFv, соединенные посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-VL(CD3)-VH(CD3). Предпочтительно, указанная единица VL(CD3)-VH(CD3) scFv содержит один или более связующих дисульфидных мостиков. В одном варианте осуществления указанные VL(CD3) и VH(CD3) соединены посредством пептидного линкера, состоящего из 20-25, предпочтительно 20 или 25 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)5. В одном варианте осуществления указанные VH(CLDN) и VL(CLDN) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)4. В одном варианте осуществления указанные единицы VH-VL и VL-VH scFv соединены посредством линкерного пептида, содержащего аминокислотную последовательность SGGGGS. Указанная единица VH(CLDN)-VL(CLDN) scFv может содержать один или более связующих дисульфидных мостиков.
В одном аспекте связывающий агент по изобретению находится в формате биспецифического одноцепочечного антитела, которое содержит две молекулы scFv, соединенные посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VL(CLDN)-VH(CLDN)-VL(CD3)-VH(CD3). Предпочтительно, указанная единица VL(CD3)-VH(CD3) scFv содержит один или более связующих дисульфидных мостиков. В одном варианте осуществления указанные VL(CD3) и VH(CD3) соединены посредством пептидного линкера, состоящего из 20-25, предпочтительно 20 или 25 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)5. В одном варианте осуществления указанные VL(CLDN) и VH(CLDN) соединены посредством пептидного линкера, состоящего из 20-25, предпочтительно 20 или 25 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)5. В одном варианте осуществления указанные две единицы VL-VH scFv соединены посредством линкерного пептида, содержащего аминокислотную последовательность SGGGGS. Указанное звено VL(CLDN)-VH(CLDN) scFv может содержать один или более связующих дисульфидных мостиков.
В одном варианте осуществления любого из указанных выше аспектов указанный CLDN представляет собой CLDN18.2. Предпочтительно, указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 8, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 15, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент. Альтернативно, указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 6, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 11, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент. В одном варианте осуществления указанная VH(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 95, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 96, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном аспекте связывающий агент по изобретению находится в формате биспецифического одноцепочечного антитела, которое содержит две молекулы scFv, соединенных посредством линкерного пептида, при этом вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-VH(CD3)-VL(CD3); или в порядке VH(CD3)-VL(CD3)-VH(CLDN)-VL(CLDN). В одном варианте осуществления указанные VH(CLDN) и VL(CLDN) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность (GGGGS)3. В одном варианте осуществления указанные VH(CD3) и VL(CD3) соединены посредством пептидного линкера, состоящего из 15-20, предпочтительно 15 или 20 аминокислот, предпочтительно глицина и/или серина, и предпочтительно соединены посредством пептидного линкера, содержащего аминокислотную последовательность GGGGS(GGS)3GGGS. В одном варианте осуществления указанные две единицы VH-VL scFv соединены посредством линкерного пептида, содержащего аминокислотную последовательность SGGGGS.
В одном варианте осуществления указанного выше аспекта указанный CLDN представляет собой CLDN6. Предпочтительно, указанная VH(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 22, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент. Предпочтительно, указанная VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 98, 99 или 100, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент. Наиболее предпочтительно, указанная VL(CLDN) содержит аминокислотную последовательность, представленную SEQ ID NO: 99, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент. В одном варианте осуществления указанная VH(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 95, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент, и VL(CD3) содержит аминокислотную последовательность, представленную SEQ ID NO: 96, или ее фрагмент, или вариант указанной аминокислотной последовательности или фрагмент.
В одном варианте осуществления указанный CLDN представляет собой CLDN18.2, и указанный связывающий агент по изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 38, 39, 40 и 41, или их фрагментов или вариантов.
В одном варианте осуществления указанный CLDN представляет собой CLDN18.2, и указанный связывающий агент по изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 103, 66, 67, 68, 69, 70, 71 , 72, 73, 74, 75, 76, 77, 78, 79, 80, 81 , 82, 83, 84, 85, 86, 87, 88, 89, 90, 91 , 92 и 93, или ее фрагмент или вариант. В одном варианте осуществления указанный CLDN представляет собой CLDN18.2, и указанный связывающий агент содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 103, 66, 67, 68, 69, 70, 71 , 72, 73, 74, 75, 76, 77, 78, 79, 80, 81 , 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92 и 93, или ее фрагмент или вариант, при этом указанная аминокислотная последовательность лишена сигналов секреции, таких как N-концевые сигналы секреции, в особенности последовательность в соответствии с SEQ ID NO: 51, и/или лишена His-меток, таких как C-концевые His-метки, в особенности последовательность Gly-Gly-Ser-(His)6 или (His)6, в случае присутствия.
В одном варианте осуществления указанный CLDN представляет собой CLDN6, и указанный связывающий агент по изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 42, 43, 44 и 45, или ее фрагмент или вариант.
В одном варианте осуществления указанный CLDN представляет собой CLDN6, и указанный связывающий агент по изобретению содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 101, 102, 60, 61, 62, 63, 64 и 65, или ее фрагмент или вариант. В одном варианте осуществления указанный CLDN представляет собой CLDN6, и указанный связывающий агент содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 101, 102, 60, 61, 62, 63, 64 и 65, или ее фрагмент или вариант, при этом указанная аминокислотная последовательность лишена сигналов секреции, таких как N-концевые сигналы секреции, в особенности последовательность в соответствии с SEQ ID NO: 51, и/или лишена His-меток, таких как C-концевые His-метки, в особенности последовательность Gly-Gly-Ser-(His)6 или (His)6, в случае присутствия.
В одном варианте осуществления указанные раковые клетки, экспрессирующие CLDN18.2, представляют собой раковые клетки рака, выбранного из группы, состоящей из рака желудка, рака пищевода, рака поджелудочной железы, рака легкого, такого как немелкоклеточный рак легкого (NSCLC), рака молочной железы, рак яичников, рака толстой кишки, рака печени, рака головы и шеи, рака желчного пузыря, и их метастазов, опухоли Крукенберга, перитонеальных метастазов и/или метастазов в лимфатические узлы.
В одном варианте осуществления указанные раковые клетки, экспрессирующие CLDN6, представляют собой раковые клетки рака, выбранного из группы, состоящей из рака мочевого пузыря; рака яичника, в частности аденокарциномы яичника и теракарциномы яичника; рака легкого, включая мелкоклеточный рак легкого (SCLC) и немелкоклеточный рак легкого (NSCLC), в частности плоскоклеточный рак легкого и аденокарциному; рака желудка; рака молочной железы; рака печени; рака поджелудочной железы; рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы; злокачественной меланомы; рака головы и шеи, в частности, злокачественной плеоморфной аденомы; саркомы, в частности синовиальной саркомы и карциносаркомы; рака желчного протока; рака мочевого пузыря, в частности переходно-клеточной карциномы и папиллярного рака; рака почки, в частности плоскоклеточного рака, включая светлоклеточный рак почки и папиллярную карциному почки; рака толстой кишки; рака тонкой кишки, включая рак подвздошной кишки, в частности аденокарциномы тонкой кишки и аденокарциномы подвздошной кишки; эмбриональной карциномы яичка; плацентарной хориокарциномы; рака шейки матки; рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка; рака матки; герминомы, такой как тератокарцинома или эмбриональная карцинома, в частности эмбрионально-клеточной опухоли яичка и их метастатических форм.
В одном варианте осуществления связывающий агент содержит N-концевой сигнал секреции и/или C-концевую гистидиновую эпитопную метку, предпочтительно, эпитопную метку из шести гистидинов.
В одном аспекте изобретение относится к рекомбинантной нуклеиновой кислоте, которая кодирует связывающий агент по изобретению. В одном варианте осуществления рекомбинантная нуклеиновая кислота находится в форме вектора. В одном варианте осуществления рекомбинантная нуклеиновая кислота представляет собой РНК.
В одном аспекте изобретение относится к клетке-хозяину, содержащей рекомбинантную нуклеиновую кислоту по изобретению.
В одном аспекте изобретение относится к связывающему агенту по изобретению, рекомбинантной нуклеиновой кислоте по изобретению или клетке-хозяину по изобретению для применения в терапии, в частности, для применения в лечении и предупреждении рака.
В одном аспекте изобретение относится к фармацевтической композиции, содержащей связывающий агент по изобретению, рекомбинантную нуклеиновую кислоту по изобретению или клетку-хозяина по изобретению.
В одном аспекте изобретение относится к способу лечения или предупреждения ракового заболевания, включающему введение пациенту фармацевтической композиции по изобретению.
В одном варианте осуществления клетки указанного рака экспрессируют клаудин, с которым указанный связывающий агент способен связываться.
В одном варианте осуществления указанный клаудин представляет собой CLDN18.2, и указанный рак выбран из группы, состоящей из рака желудка, рака пищевода, рака поджелудочной железы, рака легкого, такого как немелкоклеточный рак легкого (NSCLC), рака молочной железы, рака яичника, рака толстой кишки, рака печени, рака головы и шеи, рака желчного пузыря и их метастаз, опухоли Крукенберга, перитонеальных метастазов и/или метастазов в лимфоузлы.
В одном варианте осуществления указанный клаудин представляет собой CLDN6, и указанный рак выбран из группы, состоящей из рака мочевого пузыря; рака яичника, в частности, аденокарциномы яичника и теракарциномы яичника; рака легкого, включая мелкоклеточный рак легкого (SCLC) и немелкоклеточный рак легкого (NSCLC), в частности, плоскоклеточный рак легкого и аденокарциному; рака желудка; рака молочной железы; рака печени; рака поджелудочной железы; рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы; злокачественной меланомы; рака головы и шеи, в частности злокачественной плеоморфной аденомы; саркомы, в частности, синовиальной саркомы и карциносаркомы; рака желчного протока; рака мочевого пузыря, в частности переходно-клеточной карциномы и папиллярного рака; рака почки, в частности плоскоклеточного рака, включая светлоклеточный рак почки и папиллярную карциному почки; рака толстой кишки; рака тонкой кишки, включая рак подвздошной кишки, в частности аденокарциномы тонкой кишки и аденокарциномы подвздошной кишки; эмбриональной карциномы яичка; плацентарной хориокарциномы; рака шейки матки; рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка; рака матки; герминомы, такой как тератокарцинома или эмбриональная карцинома, в частности эмбрионально-клеточной опухоли яичка и их метастатических форм.
В одном аспекте изобретение обеспечивает связывающий агент или нуклеиновую кислоту, кодирующую связывающий агент, или клетку-хозяина, описанную здесь, для применения в способах лечения, описанных здесь. В одном варианте осуществления изобретение обеспечивает фармацевтическую композицию, описанную здесь, для применения в способах лечения, описанных здесь.
Согласно изобретению, CLDN18.2 предпочтительно содержит аминокислотную последовательность в соответствии с SEQ ID NO: 1, и CLDN6 предпочтительно содержит аминокислотную последовательность в соответствии с SEQ ID NO: 2 или 3.
Другие отличительные признаки и преимущества настоящего изобретения будут очевидны из следующего подробного описания и пунктов формулы изобретения.
Краткое описание чертежей
ФИГУРА 1. Блочная схема, иллюстрирующая дизайн рекомбинантных белков bi-scFv, направленно воздействующих на CLDN18.2 TAA.
Дизайн bi-scFvs в (A) N-концевом и (B) C-концевом положении в отношении вариабельных областей анти-TAA. Области VH и VL анти-CLDN18.2 созданы на основе последовательности моноклонального антитела CLDN18.2 (mCLDN18.2ab). Анти-CD3 является общим для областей VH и VL, созданных из последовательностей следующих моноклональных антител к CD3: UCHT1-HU (гуманизированное mAB), UCHT1, CLB-T3, TR66, 145-2C11. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; His, гистидиновая метка, содержащая 6 остатков гистидина; HU, гуманизированное; LL, длинный линкер (15-18 аминокислот); Sec, сигнал секреции; SL, короткий линкер (5-6 аминокислот); TAA, опухоль-ассоциированный антиген; V, вариабельная область тяжелой (H) и легкой (L) цепи антитела.
ФИГУРА 2. Эффект ориентации доменов и выбора анти-CD3-scFv на специфический лизис клеток-мишеней: 5'-mCLDN18.2ab VH-VL_ TR66 VH-VL -3' 1BiMAB bi-scFvs и no.15 являются наиболее сильными вариантами.
Несколько вариантов bi-scFv, направленных против CLDN18.2 и CD3, были временно экспрессированы в клетках HEK293T и очищены в малом масштабе на колонках Ni-NTA для сравнения их силы в анализе на цитотоксичность. В качестве клеток-мишеней использовали клетки NugC4, эндогенно экспрессирующие CLDN18.2 и стабильно экспрессирующие люциферазу. Человеческие T-клетки и клетки-мишени инкубировали в соотношении E:T, равном 5:1, с 5 нг/мл каждого белка bi-scFv в 96-луночном формате. В качестве отрицательных контролей использовали no.35, направленно воздействующий на неэкспрессированный TAA, no.l1 и no.16 - оба, направленно воздействующие на мышиные, но не человеческие Т-клетки. Каждый тестируемый образец высевали в шестикратной повторности, контрольный образец для Lmin высевали в девятикратной повторности. Время совместной инкубации перед началом анализа составило 8 ч, 16 ч и 24 ч. После добавления раствора люциферина в заданные моменты времени люминесценцию измеряли на ридере Infinite M200 TECAN. Специфический лизис клеток-мишеней рассчитывали путем нормализации к образцам с контрольным no.35 (Lmin) bi-scFv. Наиболее сильные белки bi-scFv - 1BiMAB и no.l5 - имели одинаковую ориентацию доменов и происхождение анти-CD3 mAB TR66, но отличались своей оптимизацией кодонов (HS и CHO, соответственно) и длинными последовательностями линкера. CHO означает клетки яичника китайского хомячка; mAB, моноклональное антитело; HU, гуманизированное; TAA, опухоль-ассоциированный антиген.
ФИГУРА 3. Окрашивание кумасси и вестерн-блот анализ белка 1BiMAB bi-scFv.
Супернатант без FCS моноклональных клеток HEK293, стабильно экспрессирующих 1BiMAB, очищали аффинной хроматографией (IMAC) на колонке Ni-NTA. Аликвоты различных стадий очистки вносили на 4-12% Bis-Tris гели. (A) Окрашивание Кумасси клеточного супернатанта, пропускание и восемь фракций элюата. Фракции первого пика элюирования удаляли, фракции второго пика элюирования объединяли для последующих исследований, диализировали против PBS и затем против 200 мМ аргининового буфера. (Дорожка 1: ΗΕΚ293/1ΒiΜΑΒ SN; дорожка 2: Проточная фракция IMAC; дорожки 3-4: Фракции пика элюирования 1 (удаленные); дорожки 5-10: Фракции пика элюирования 2 (объединенные)); (B) Вестерн-блот анализ 0,5 г 1BiMAB из трех независимых очисток (дорожка 1, 2, 3). Детекцию выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. IMAC означает аффинную хроматографию с использование иммобилизованных металлов; PBS, фосфатно-буферный солевой раствор; SN, супернатант; WB, вестерн-блоттинг.
ФИГУРА 4. Белок 1BiMAB bi-scFv эффективно и специфически связывается с CLDN18.2-экспрессирующими клетками-мишенями и человеческими Т-клетками.
(A) 2,5×105 клеток NugC4, эндогенно экспрессирующих CLDN18.2, инкубировали с 50 мкг/мл 1BiMAB или 10 мкг/мл mCLDN18.2ab в качестве положительного контроля и соответствующими APC-конъюгированными вторичными антителами. Контрольные окрашивания включали только вторичные APC-конъюгированные антитела (g-a-h, g-a-m), анти-His и g-a-m APC, или 1BiMAB и g-a-m APC. Анализ выполняли посредством проточной цитометрии. MFI APC-сигнала рассчитывали с помощью программного обеспечения FlowJo. (B) l×l05 клеток NugC4, эндогенно экспрессирующих CLDN18.2, окрашивали нарастающими концентрациями 1BiMAB (20 пг/мл - 20 мкг/мл), анти-His и g-a-m APC. В качестве отрицательного контроля клетки инкубировали с анти-His и g-a-m APC. В качестве положительного контроля использовали mCLDN18.2ab и g-a-h APC. MFI APC-сигнала рассчитывали с помощью программного обеспечения FlowJo. (C) l×l06 человеческих T-клеток инкубировали с нарастающими концентрациями 1BiMAB (2 нг/мл - 2 мкг/мл), анти-His и g-a-m APC. В качестве отрицательного контроля клетки инкубировали с анти-His и g-a-m APC или только g-a-m APC. MFI APC-сигнала рассчитывали с помощью программного обеспечения FlowJo. (D) l×l05 CLDN18.2-отрицательных клеток PA-1 инкубировали с нарастающими концентрациями 1BiMAB (10 нг/мл - 10 мкг/мл), анти-His и g-a-m APC. В качестве отрицательного контроля клетки окрашивали анти-His и g-a-m APC или только g-a-h APC. 10 мкг/мл mCLDN18.2ab и g-a-h APC использовали для подтверждения негативности клеток в отношении CLDN18.2. G-a-h означает козье-анти-человеческое; g-a-m, козье-анти-мышиное; MFI, средняя интенсивность флуоресценции; TL, T-лимфоцит.
ФИГУРА 5. Белок 1BiMAB bi-scFv вызывает образование кластеров T-клеток на CLDN18.2-положительных клетках-мишенях.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, инкубировали в течение 24 ч с 1 нг/мл и 1 мкг/мл 1BiMAB и человеческими T-клетками в соотношении эффектора к мишени 5:1 в 6-луночных планшетах. В качестве контрольных образцов выбирали только Т-клетки (TL), только клетки-мишени (NugC4) и человеческие Т-клетки с клетками-мишенями (-Контр). Через 24 ч делали снимки образцов с помощью микроскопа Nikon Eclipse Ti при увеличении 200×. Белые стрелки указывают на кластеры Т-клеток на клетках-мишенях. TL означает T лимфоцит.
ФИГУРА 6. 1BiMAB опосредует T-клеточную активацию дозо-зависимым образом.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, инкубировали в течение 24 ч и 48 ч с нарастающими концентрациями белка 1BiMAB bi-scFv (0,001 - 1000 нг/мл) и человеческими Т-клетками в соотношении эффектора к мишени 5:1 в двух повторностях в 24-луночном формате. В качестве контроля человеческие Т-клетки инкубировали с 1-1000 нг/мл 1BiMAB без клеток-мишеней NugC4 для подтверждения зависимой от мишени активации Т-клеток, опосредованной 1BiMAB. Через 24 ч (A) и 48 ч (B) T-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC, и анализировали с помощью проточной цитометрии. TL означает T-лимфоцит.
ФИГУРА 7. 1BiMAB опосредует активацию Т-клеток, строго зависимую от мишени, даже после длительной инкубации с клеточными линиями, высоко-, низко- и не экспрессирующими CLDN18.2.
(A) Показаны данные RT-PCR, полученные из общей РНК шести опухолевых клеточных линий. Величины Ct экспрессии CLDN18.2, нормализованные по отношению к гену домашнего хозяйства HPRT, рассчитывали на основании двух независимых экспериментов. Клеточная линия рака молочной железы MCF7 (серый столбец) не экспрессирующая CLDN18.2 была выбран в качестве контроля. (B) Раковые клеточные линии (A) инкубировали в течение 144 ч с 5 нг/мл белка 1BiMAB bi-scFv с человеческими Т-клетками или без них в соотношении эффектора к мишени 5:1 в двух повторностях в 6-луночном формате. T-клетки метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC для анализа общей популяции Т-клеток (CD3), ранней активации (CD69) и поздней активации (CD25) Т-клеток посредством проточной цитометрии. TL означает Т-лимфоцит.
ФИГУРА 8. 1BiMAB индуцирует T-клеточную пролиферацию и повышающую регуляцию гранзима B только в присутствии CLDN18.2-положительных клеток-мишеней.
(A) Человеческие Т-клетки человека окрашивали CFSE и выращивали отдельно (TL) или в присутствии 1 нг/мл 1BiMAB (TL + 1 нг/мл 1BiMAB), клеток NugC4 (TL + NugC4), или клеток NugC4 и 1 нг/мл 1BiMAB (TL + 1 нг/мл 1BiMAB + NugC4) в течение 120 ч. Было выбрано соотношение эффектора к мишени, равное 5:1. Уменьшение сигнала CFSE, указывающее на Т-клеточную пролиферацию, анализировали посредством проточной цитометрии. (B) Человеческие Т-клетки инкубировали с клетками-мишенями NugC4 или без них, и с 5 нг/мл белка 1BiMAB bi-scFv или без него. Соотношение эффектора к мишени составило 5:1 в 6-луночном формате. Через 96 ч совместной инкубации Т-клетки собирали и внутриклеточно окрашивали анти-GrB-PE и анализировали посредством проточной цитометрии. MFI-сигнал анти-GrB-PE рассчитывали с помощью программного обеспечения FlowJo. Сигнал неокрашенного образца TL + NugC4 + 5 нг/мл 1BiMAB вычитали из всех образцов. CFSE означает сложный эфир сукцинимидила и карбоксифлуоресцеина; GrB, гранзим B; MFI, средняя интенсивность флуоресценции; PE, фикоэритрин; TL, Т-лимфоциты.
ФИГУРА 9. Величина EC50 1BiMAB для специфического лизиса клеток-мишеней через 48 ч составила приблизительно 10 пг/мл.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, которые стабильно экспрессируют люциферазу, инкубировали в течение 24 ч и 48 ч с белком 1BiMAB bi-scFv в нарастающих концентрациях (0,001-1000 нг/мл) с человеческими Т-клетками в соотношении эффектора к мишени 5:1 в трех повторностях в 96-луночном формате. В качестве минимального лизиса (Lmin) контрольные эффекторные клетки и клетки-мишени высевали без 1BiMAB bi-scFv. Максимальный лизис (Lmax) для нормализации по отношению к спонтанному числу импульсов люминесценции был достигнут путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и клетки-мишени в отсутствии bi-scFv непосредственно перед добавлением люциферина. После добавления раствора люциферина люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan через 24 ч и 48 ч. Специфический лизис клеток-мишеней рассчитывали по формуле: % специфического лизиса = [1 - (люминесценциятестируемый образец - Lmax) / (Lmin - Lmax)] × 100. Величины наносили в зависимости от log 10 концентрации 1BiMAB. Величина EC50 означает половину максимальной эффективной концентрации; L, лизис.
ФИГУРА 10. 1BiMAB показывает терапевтическую эффективность in vivo в модели опухоли SC на поздней стадии.
Мышам NOD.Cg-Prkdscid IL2rgtmlWjl/Sz.T (NSG) инъецировали подкожно (SC) клетки HEK293 в количестве l×l07, стабильно экспрессирующие CLDN18.2. Через пять дней клетки PBMC человека в качестве эффекторных клеток в количестве 2×l07 инъецировали интраперитонеально (IP) группам G3 и G4, контрольные группы (Gl и G2) получали только PBS. На следующий день начинали ежедневное IP введение 5 мкг белка 1BiMAB bi-scFv каждому животному или носителя в качестве контроля. Терапию проводили в течение 22 дней, объем опухоли измеряли с использованием калипера и рассчитывали по формуле мм3 = длина мм × ширина мм × (ширина мм/2). (A) Объем опухоли одной мыши и среднее значение на группу показано для лечебных дней 0 и 15 (верхний ряд), и дней 3 и 13 после окончания лечения (нижний ряд). (B) Показан средний объем опухоли двух лечебных групп, привитых человеческими эффекторными клетками. Тире указывает умерщвленных животных. (C) Кривая выживаемости Каплана-Мейера, демонстрирующая все группы от дня инокуляции опухоли до дня 41. Животных умерщвляли после достижения объема опухоли 500 мм3. После дня 41 оставшихся животных умерщвляли для анализа приживления человеческих эффекторных клеток в селезенках мышей. (D) Спленоциты всех мышей изолировали и окрашивали анти-CD45-APC и анти-CD3-FITC для детекции человеческих Т-клеток с помощью проточной цитометрии. Медиана приживления отображена на коробчатой диаграмме. G означает группу; IP, интраперитонеально; PBMC, мононуклеарные клетки периферической крови; PBS, фосфатно-буферный солевой раствор; SC, подкожно.
ФИГУРА 11. Блочная схема, иллюстрирующая дизайн рекомбинантных белков bi-scFv, направленно воздействующих на CLDN6 TAA.
Дизайн bi-scFvs в (A) N-концевом (B) C-концевом положении в отношении вариабельных областей анти-TAA. Области VH и VL анти-CLDN6 созданы на основе последовательности моноклонального антитела CLDN6 (mCLDN6ab). Области VH и VL анти-CD3 созданы на основе последовательности моноклонального антитела CD3 TR66. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; His, гистидиновая метка, содержащая шесть остатков гистидина; LL, длинный линкер (15-18 аминокислот); Sec, сигнал секреции; SL, короткий линкер (5 аминокислот); TAA, опухоль-ассоциированный антиген; V, вариабельная область тяжелой (H) и легкой (L) цепи антитела.
ФИГУРА 12. Белки 6PHU5 и 6PHU3 bi-scFv вызывают образование кластеров Т-клеток на CLDN6-положительных клетках-мишенях.
Клетки PA-1, эндогенно экспрессирующие CLDN6, инкубировали в течение 24 ч с 50 нг/мл 6PHU5 или 6PHU3 и человеческими Т-клетками человека в соотношении эффектора к мишени 5:1 в 6-луночных планшетах. В качестве контрольных образцов были выбраны только Т-клетки (TL), только клетки-мишени (PA-1) и человеческие Т-клетки с клетками-мишенями (-контр). Через 24 ч делали снимки образцов на микроскопе Nikon Eclipse Tj при увеличении 200×. Белые стрелки указывают на Т-клеточные кластеры на клетках-мишенях. TL означает Т-лимфоциты.
ФИГУРА 13. Эффект ориентации доменов на эффективность: белок 6PHU3 bi-scFv является немного более эффективным в отношении индукции активации Т-клеток по сравнению с 6PHU5.
Клетки PA-1, эндогенно экспрессирующие CLDN6, инкубировали в течение 44 ч с нарастающими концентрациями (5 - 200 нг/мл) 6PHU5 или 6PHU3 и человеческими Т-клетками в соотношении эффектора к мишени 5:1 в двух повторностях в 6-луночном формате. В качестве контроля человеческие Т-клетки человека инкубировали с 100 и 200 нг/мл 6PHU5 или 6PHU3 без клеток-мишеней. Через 44 ч Т-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC. Дозозависимую Т-клеточную активацию анализировали с помощью проточной цитометрии. Hu означает человеческие; TL, Т-лимфоциты.
ФИГУРА 14. Окрашивание кумасси и вестерн-блот анализ белка 6PHU3.
Супернатант без FCS поликлональных клеток HEK293, стабильно экспрессирующих 6PHU3, очищали на Ni-NTA аффинной хроматографии (IMAC). Аликвоты различных стадий очистки наносили на 4 - 12% Bis-Tris гели. (A) Окрашивание кумасси клеточного супернатанта, пропускание и девять фракций элюата. Фракции первого пика элюирования удаляли, фракции второго и третьего пиков элюирования объединяли для дальнейших исследований, диализировали против PBS и затем против 200 мМ аргининового буфера. (Дорожка 1: HEK293/6PHU3 SN; дорожка 2: IMAC проточные фракции; дорожки 3-5: Фракции пика элюирования 1 (удаленные); дорожки 6-11: Фракции пика элюирования 2 и 3 (объединенные)) (B) Вестерн-блот анализ 0,5 мкг 6PHU3 из двух независимых очисток. Детекцию выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. IMAC означает аффинную хроматографию с использованием иммобилизованных металлов; PBS; фосфатно-буферный солевой раствор; SN, супернатант; WB, вестерн-блоттинг.
ФИГУРА 15. Белок 6PHU3 bi-scFv эффективно и специфически связывается с CLDN6-экспрессирующими клетками-мишенями и человеческими Т-клетками.
(A) 1×l05 клеток PA-1, эндогенно экспрессирующих CLDN6, и клетки OV-90 инкубировали с нарастающими концентрациями 6PHU3 или контрольным 1BiMAB bi-scFv (10 нг/мл - 10 мкг/мл) и 10 мкг/мл mCLDN6ab или контрольным mAB mCLDN18.2ab с соответствующими APC-конъюгированными вторичными антителами. Контрольные окрашивания представляли собой только вторичные APC-конъюгированные антитела (g-a-h, g-a-m). Анализ выполняли с помощью проточной цитометрии. MFI APC-сигнала рассчитывали с помощью программного обеспечения FlowJo. (B) 5×l05 человеческих Т-клеток инкубировали с нарастающими концентрациями 6PHU3 (100 нг/мл - 10 мкг/мл), анти-His и g-a-m PE. В качестве отрицательного контроля клетки инкубировали с анти-His и g-a-m PE или только g-a-m PE. MFI PE-сигнала рассчитывали с помощью программного обеспечения FlowJo. (C) l×l05 CLDN6-отрицательных клеток NugC4 инкубировали с нарастающими концентрациями 6PHU3 и 1BiMAB (10 нг/мл - 10 мкг/мл), анти-His и g-a-m APC. В качестве отрицательного контроля клетки инкубировали только с g-a-m APC. 10 мкг/мл mCLDN6ab и g-a-h APC использовали для подтверждения негативности CLDN6 клеток. В качестве положительного контроля использовали mCLDN18.2ab и g-a-h APC. MFI APC-сигнала рассчитывали с помощью программного обеспечения FlowJo. APC означает аллофикоцианин; g-a-h, козьи-анти-человеческие; g-a-m, козьи-анти-мышиные; mAB, моноклональное антитело; MFI, средняя интенсивность флуоресценции; PE, фикоэритрин; TL, Т-лимфоцит.
ФИГУРА 16. 6PHU3 опосредует активацию Т-клеток дозо-зависимым образом
Клетки PA-1, эндогенно экспрессирующие CLDN6, инкубировали в течение 24 ч и 48 ч с нарастающими концентрациями белка 6PHU3 bi-scFv (0.001 - 1000 нг/мл) и человеческими Т-клетками в соотношении эффектора к мишени 5:1 в двух повторностях в 24-луночном формате. В качестве контроля человеческие Т-клетки инкубировали с 1-1000 нг/мл 6PHU3 без клеток-мишеней PA-1 для подтверждения зависимой от мишени активации Т-клеток, опосредованной 6PHU3. Через 24 ч (A) и 48 ч (B) Т-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC, и анализировали с помощью проточной цитометрии. TL означает Т-лимфоциты.
ФИГУРА 17. Величина EC50 6PHU3 для специфического лизиса клеток-мишеней через 48 ч составила приблизительно 10 пг/мл.
Клетки PA-1, эндогенно экспрессирующие CLDN6, которые стабильно экспрессируют люциферазу, инкубировали в течение 24 ч и 48 ч с белком 6PHU3 в нарастающих концентрациях (0,001-1000 пг/мл) с человеческими Т-клетками в соотношении эффектора к мишени 5:1 в трех повторностях в 96-луночном формате. В качестве минимального контроля лизиса (Lmin) эффекторные клетки и клетки-мишени высевали без 6PHU3 bi-scFv. Максимальный лизис (Lmax) для нормализации в отношении числа импульсов спонтанной люминесценции был достигнут путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и клетки-мишени при отсутствии bi-scFv непосредственно перед добавлением лициферина. После добавления раствора люциферина люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan через 24 ч и 48 ч. Специфический лизис клеток-мишеней рассчитывали по формуле: % специфического лизиса = [1 - (люминесценция тестируемого образца - Lmax) / (Lmin - Lmax)] × 100. Значения наносили в зависимости от log10 концентрации 6PHU3. Величина EC50 означает половину максимальной эффективной концентрации; L, лизис.
ФИГУРА 18. 6PHU3 показывает терапевтическую in vivo эффективность в модели опухоли SC на поздней стадии.
Мышам NOD.Cg-Prkdscid IL2rgtm1Wj1/SzJ (NSG) инъецировали подкожно (SC) клетки PA-1 в количестве l×l07, эндогенно экспрессирующие CLDN6. Через 15 дней группам G3 и G4 интраперитонеально (IP) инъецировали клетки PBMC человека в количестве 2×l07, контрольные группы (Gl и G2) получали только PBS. Ежедневное IP введение 5 мкг 6PHU3 каждому животному или контрольного 1BiMAB bi-scFv, или только носителя в качестве контроля начинали через пять дней после инъекции PBMC. Терапию проводили в течение 25 дней, объем опухоли измеряли с помощью калипера и рассчитывали по формуле мм3 = длина, мм × ширина, мм × (ширина, мм/2). (A) Объем опухоли одной мыши и среднее значение на группу показано для дней 0 и 14 терапии (верхний ряд), и 21 и 25 (нижний ряд). (B) Показан средний объем опухоли всех лечебных групп. Пунктирные линии означают умерщвленных животных. (C) Показана кривая выживаемости Каплана-Мейера, для всех групп от дня инокуляции опухоли до дня 45. Животных умерщвляли при объеме опухоли >1500 мм3. Через 45 дней всех оставшихся животных умерщвляли для анализа приживления человеческих эффекторных клеток в селезенках мышей. (D) Спленоциты всех мышей выделяли и окрашивали анти-CD45-APC и анти-CD3-FITC для детекции человеческих Т-клеток с помощью проточной цитометрии. Медиана приживления отражена на коробчатой диаграмме. IP означает интраперитонеально; PBMC, мононуклеарные клетки периферической крови; PBS, фосфатно-буферный солевой раствор; SC, подкожно.
ФИГУРА 19. Повышенная инфильтрация Т-клеток в SC PA-1 опухоли в ответ на терапию 6PHU3.
Мышам NSG инъецировали подкожно (SC) клетки PA-1 в количестве 1×107, эндогенно экспрессирующие CLDN6. Через 15 дней группам G3 и G4 интраперитонеально (IP) инъецировали клетки PBMC человека в количестве 2×107 , контрольные группы (Gl и G2) получали только PBS. Ежедневное IP введение 5 мкг 6PHU3 каждому животному или контрольного 1BiMAB bi-scFv, или только носителя в качестве контроля начинали через пять дней после инъекции PBMC. Опухоли вырезали при размере 1500 мм3 или в конце эксперимента, и консервировали в 4% буферном растворе формальдегида для заключения в парафин.
Заключенные в парафин опухолевые ткани SC PA-1 опухолей подвергали иммуногистохимическим окрашиваниям. Последовательные срезы окрашивали поликлональным первичным антителом анти-Клаудин 6 или анти-человеческими CD3. Первичные антитела детектировали с использованием вторичных анти-кроличьих антител, конъюгированных с пероксидазой HRP. Верхние ряды A-E показывают окрашивание CLDN6, нижние ряды окрашивание CD3. Изображения получали с помощью сканера Mirax. (A) и (B) показывают PBS-контрольные группы Gl и G2, которые не получали человеческих эффекторных клеток и получали носитель или 6PHU3 bi-scFv, соответственно, (C) показывает контрольную группу G3, которая получала человеческие эффекторные клетки и носитель в качестве терапии, (D) показывает группу G4, которая получала человеческие эффекторные клетки и 6PHU3 bi-scFv в качестве терапии, и (E) показывает контрольную группу G5, которая получала человеческие эффекторные клетки и контрольные 1BiMAB bi-scFv. Положительные сигналы появляются в виде красного окрашивания. Черные стрелки указывают на примеры сигналов CD3. IP означает интраперитонеально; PBMC, мононуклеарные клетки периферической крови; PBS, фосфатно-буферный солевой раствор; SC, подкожно.
ФИГУРА 20. Схематическое изображение молекул IVT-РНК, кодирующих антитела bi-scFv, направленно воздействующие на CLDN18.2 TAA.
Схема in vitro транскрибированных последовательностей РНК, кодирующих анти-CLDN18.2 антитела bi-scFv. (A) IVT-мРНК в 5'- и 3'-положении относительно вариабельных областей анти-TAA. (B) IVT альфавирусный репликон в 5'-положении относительно вариабельных областей анти-TAA. Области VH и VL анти-CLDN18.2 создавали на основе последовательности моноклонального антитела CLDN18.2 (mCLDN18.2ab). «Кэп» одинаково используется для ARCA, бета-S-ARCA (Dl) или бета-S-ARCA (D2). В (A) «анти-CD3» означает области VH и VL, созданные на основе последовательностей следующих моноклональных CD3 антител: UCHT1-HU (гуманизированное mAB), UCHT1, CLB-T3, TR66, 145-2C11 , в (B) «анти-CD3» описывает только VH и VL из TR66. A означает аденин; bi-scFv, биспецифический одноцепочечный вариабельный фрагмент; hAg, человеческий альфа глобин 5'-UTR; hBg, человеческий бета глобин 3'-UTR; His, гистидиновая метка, содержащая шесть остатков гистидина; IVT, in vitro транскрибированный; LL, длинный линкер (15-18 аминокислот); nsPl-4, неструктурные белки 1-4; Sec, сигнал секреции; sgP, субгеномный промотор; SL, короткий линкер (5-6 аминокислот); TAA, опухоль-ассоциированный антиген; UTR, нетранслируемая область; V, вариабельная область тяжелой (H) цепи и легкой (L) цепи антитела.
ФИГУРА 21. Эффект ориентации доменов и выбора анти-CD3-scFv на зависимую от мишени активацию Т-клеток и специфический лизис клеток-мишеней.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, временно трансфицировали несколькими вариантами bi-scFv, направленными против CLDN18.2 и CD3 для сравнения их силы в анализе на цитотоксичность. Клетки NugC4 в количестве 5×106 на каждый вариант электропорировали с 20 мкг/мл IVT-мРНК. Трансфициованные клетки-мишени подсчитывали, 1×105 клеток засевали на каждый 6-луночный планшет и инкубировали с человеческими цитотоксическими Т-клетками (выбранные T клетки CD8+) в соотношении E:T, равном 5:1. В качестве отрицательных контролей использовали IVT-мРНК bi-scFv, направленно воздействующий на неэкспрессированный TAA (контр), и родительское IgG mAB chCLDN18.2ab (контр IgG), направленно воздействующее на CLDN18.2, но не Т-клетки. Белок 1BiMAB в концентрации 5 нг/мл служил в качестве положительного контроля. В качестве референсных фоновых погибших клеток электропорированные клетки засевали без Т-клеток и, в качестве референсной фоновой активации Т-клетки засевали без клеток-мишеней. Каждый образец высевали в двух повторностях. Через 48 ч Т-клетки и клетки-мишени собирали и метили анти-CD3-FITC, анти-CD25-PE, анти-CD69-APC и 7-AAD для окрашивания живых и погибших клеток и анализировали с помощью проточной цитометрии. (A) TAA-зависимую, опосредованную bi-scFv активацию T-клеток наблюдали для всех вариантов анти-CLDN18.2 bi-scFv. (B) Специфический лизис клеток-мишеней определяли путем вычитания референсной популяции 7-AAD из популяции клеток-мишеней образцов 7-AAD. Антитела bi-scFv, вызывающие немного более высокий лизис клеток-мишеней - 1BiMAB и no.5 - имеют одинаковую ориентацию доменов и анти-CD3-происхождение mAB TR66, но отличаются оптимизацией своих кодонов (HS и CHO, соответственно) и последовательностями длинного линкера. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; контр, контроль; IgG, иммуноглобулин G; IVT, in vitro транскрибированный; мРНК, мессенджер РНК; TL, T-лимфоцит.
ФИГУРА 22. Совместная инкубация клеток-мишеней, трансфицированных IVT-мРНК 1BiMAB, и человеческих Т-клеток приводит к образованию кластеров Т-клеток.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, временно трансфицировали путем электропорации 80 мкг/мл IVT-мРНК 1BiMAB и инкубировали совместно с человеческими цитотоксическими Т-клетками (выбраны T-клетки CD8+ ) в соотношении эффектора к мишени 5:1 в 96-луночных планшетах. В качестве отрицательного контрольного образца использовали клетки-мишени NugC4, трансфицированные IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (-контр), совместно инкубированные с человеческими цитотоксическими Т-клетками (верхний ряд, слева). Нижний ряд показывает клетки NugC4, трансфицированные контрольным bi-scFv (слева) или IVT-мРНК 1BiMAB (справа) без человеческих Т-клеток. Через 24 ч совместной инкубации делали снимки образцов на микроскопе Nikon Eclipse Ti при увеличении 200×. Белые стрелки указывают на Т-клеточные кластеры на клетках-мишенях. CTL означает цитотоксические Т-лимфоциты; контр, контроль; hu, человеческие.
ФИГУРА 23. 1BiMAB, секретируемый клетками-мишенями после трансфекции IVT-мРНК, опосредует активацию Т-клеток зависимым от концентрации образом.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, временно трансфицировали путем электропорации IVT-мРНК в общем количестве 40 мкг/мл, содержащими 0.4-40 мкг/мл IVT-мРНК 1BiMAB плюс соответствующие количества IVT-мРНК люциферазы. Трансфицированные клетки-мишени совместно инкубировали с человеческими цитотоксическими Т-клетками (выбраны T клетки CD8+) в соотношении эффектора и мишени 5:1 в 6-луночных планшетах в двух повторностях. В качестве активации Т-клеток референсные человеческие Т-клетки совместно инкубировали с клетками-мишенями NugC4, трасфицированными 40 мкг/мл IVT-мРНК люциферазы (0.0 мкг/мл IVT-мРНК 1BiMAB). Через 24 ч (A) и 48 ч (B) T-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC, и анализировали с помощью проточной цитометрии. На диаграммах показано процентное содержание положительного окрашенных цитотоксических Т-клеток человека, определенное с помощью программного обеспечения FlowJo. IVT означает in vitro транскрибированные; мРНК, мессенджер РНК; TL, Т-лимфоцит.
ФИГУРА 24. 1BiMAB, секретируемый клетками-мишенями после трансфекции IVT-мРНК, приводит к зависимому от концентрации лизису клеток-мишеней.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, временно трансфицировали путем электропорации IVT-мРНК в общем количестве 40 мкг/мл, содержащем 0,4-40 IVT-мРНК 1BiMAB плюс соответствующие количества IVT-мРНК лициферазы или 40 мкг/мл только IVT-мРНК люциферазы в качестве референсного образца. Трансфицированные клетки-мишени засевали с человеческими цитотоксическими Т-клетками (выбраны T клетки CD8+) в соотношении эффектора и мишени 5:1 или без эффекторных клеток для определения процентного содержания фоновых погибших клеток-мишеней путем индивидуальной электропорации. Все образцы выращивали в 6-луночных планшетах в двух повторностях. Через 24 ч (A) и 48 ч (B) T-клетки собирали, метили иодидом пропидия (PI) для окрашивания живых/погибших клеток и анализировали с помощью проточной цитометрии. Процентное содержание погибших (PI+) клеток-мишеней определяли с помощью программного обеспечения FlowJo. Значения затем нормализовали в отношении каждого индивидуального фонового образца и референсного образца. IVT означает in vitro транскрибированные; мРНК, мессенджер РНК; TL, T-лимфоцит.
ФИГУРА 25. T-клеточная пролиферация специфически индуцирована в ответ на секрецию 1BiMAB клетками-мишенями в присутствие CLDN18.2
Человеческие Т-клетки окрашивали CFSE для анализа. Т-клетки выращивали без клеток-мишеней (T-клетки) в комбинации с 5 мкг/мл OKT3 и 2 мкг/мл αCD28 в качестве положительного контроля активации (+контр), с 5 нг/мл нетаргетного контрольного bi-scFv (белок -контр) или с 5 нг/мл белка 1BiMAB (белок 1BiMAB). Т-клетки и клетки-мишени NugC4, сверхэкспрессирующие CLDN18.2, инкубировали вместе (T-клетки+ CLDN18.2-положительные клетки-мишени) без чего-либо (имитирующий контроль) или с 5 нг/мл белка 1BiMAB (белок 1BiMAB). Для тестирования IVT-мРНК клетки NugC4 трансфицировали 20 мкг/мл IVT-мРНК 1BiMAB (мРНК 1BiMAB) или IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (мРНК -контр) и инкубировали с Т-клетками. Кроме того, клетки NugC4, трансфицированные IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA, объединяли с 5 нг/мл белка 1BiMAB (мРНК -контр + белок 1BiMAB). В качестве дополнительного контроля специфичности включали образцы, содержащие неэкспрессирующую CLDN18.2 линию клеток-мишеней MDA-MB-231 вместе с Т-клетками (T-клетки+ CLDN18.2-отрицательные клетки-мишени). MDA-MB-231 использовали необработанными и инкубировали без чего-либо (имитирующий контроль), с 5 нг/мл контрольного белка bi-scFv (белок -контр) или 5 нг/мл белка 1BiMAB (белок 1BiMAB), или MDA-MB-231 трансфицировали 20 г/мл IVT-мРНК 1BiMAB (мРНК 1BiMAB) или IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (мРНК -контр). Анализ выполняли при соотношении эффектора к мишени 5:1 в 96-луночном планшете, каждый образец готовили в трех повторностях, и время инкубации составило 72 ч. Уменьшение сигнала CFSE, указывающее на пролиферацию Т-клеток, анализировали с помощью проточной цитометрии, рассчитывали с помощью программного обеспечения FlowJo и наносили на график в виде % пролиферирующих Т-клеток. CFSE означает сложный эфир сукцинимидила и карбоксифлуоресцеина; IVT, in vitro транскрибированный; мРНК, мессенджер РНК.
ФИГУРА 26. Активация Т-клеток и опосредованный Т-клетками лизис клеток-мишеней в ответ на секрецию 1BiMAB начинается при соотношении эффектора и мишени 0,3:1.
Клетки NugC4, эндогенно экспрессирующие CLDN18.2, временно трансфицировали путем электропорации 40 мкг/мл IVT-мРНК 1BiMAB. Трансфицированные клетки-мишени совместно инкубировали с человеческими цитотоксическими T-клетками (выбраны T-клетки CD8+) в указанных соотношениях эффектора и мишени в диапазоне от 0,3:1 до 10:1 в 6-луночных планшетах в двух повторностях. В качестве референсных образцов человеческие Т-клетки выращивали в отсутствие клеток-мишеней ((A) 1:0) и клетки-мишени, трансфицированные контрольным IVT-мРНК, выращивали в отсутствие эффекторных клеток ((B) 0:1). В качестве отрицательного контроля человеческие Т-клетки совместно инкубировали с клетками-мишенями NugC4, трансфицированными 40 мкг/мл IVT-мРНК люциферазы (контр IVT-мРНК) в соотношении E:T, равном 10:1, ((A) и (B) контр IVT-мРНК 10:1). Через 48 ч клетки собирали и метили анти-CD3-FITC, анти-CD25-PE, анти-CD69-APC и иодидом пропидия (PI) для окрашивания живых/погибших клеток и анализировали с помощью проточной цитометрии. (A) показывает процентное содержание положительно окрашенных цитотоксических человеческих Т-клеток. (B) демонстрирует процентное содержание погибших (PI+) клеток-мишеней. Все величины определяли с помощью программного обеспечения FlowJo. E:T означает соотношение эффектора и мишени; IVT, in vitro транскрибированный; мРНК, мессенджер РНК; TL, T-лимфоцит.
ФИГУРА 27. Человеческие цитотоксические Т-клетки могут служить в качестве реципиента IVT-мРНК bi-scFv клеток-продуцентов
Человеческие цитотоксические Т-клетки выделяли из свежих PBMC путем CD8-положительного отбора и затем временно трансфицировали путем электропорации 80 или 240 мкг/мл IVT-мРНК 1BiMAB. Трансфицированные эффекторные клетки совместно инкубировали с клетками-мишенями NugC4, эндогенно экспрессирующими CLDN18.2, в соотношении эффектора и мишени 5:1 в 6-луночных планшетах в двух повторностях. В качестве референсных образцов необработанные человеческие Т-клетки выращивали с клетками-мишенями. В качестве отрицательного контроля человеческие Т-клетки, трансфицированные 80 или 240 мкг/мл IVT-мРНК контроля eGFP, совместно инкубировали с клетками-мишенями NugC4. Через 48 ч клетки собирали и метили анти-CD3-FITC, анти-CD25-PE, анти-CD69-APC и иодидом пропидия (PI) для окрашивания живых и погибших клеток, и анализировали с помощью проточной цитометрии. (A) показывает процентное содержание положительно окрашенных цитотоксических человеческих Т-клеток. В (B) показано процентное содержание погибших (PI+) клеток-мишеней, нормализованных к референсному образцу. Все величины определяли с помощью программного обеспечения FlowJo. контр означает контрольный; IVT; in vitro транскрибированные; мРНК, мессенджер РНК; TL, T лимфоцит.
ФИГУРА 28. CLDN18.2-отрицательные клетки-мишени, трансфицированные IVT-мРНК 1BiMAB, не являются лизированными T клетками
CLDN18.2-отрицательная линия клеток PA-1, стабильно экспрессирующая люциферазу, служила в качестве линии клеток-мишеней. 5×106 клеток PA-l/luc трансфицировали путем электропорации IVT-мРНК в общем количестве 40 мкг/мл. В качестве положительного контроля трансфицировали 4 и 40 мкг/мл IVT-мРНК 1BiMAB или 6RHU3, направленно воздействующего на эндогенно экспрессирующийся CLDN6. В качестве bi-scFv-отрицательного контроля трансфицировали 40 мкг/мл IVT-мРНК bi-scFv, направленно воздействующего на неэкспрессированный TAA (- контр). Этот IVT-мРНК служил также в качестве наполнения РНК в 4 мкг/мл образцов IVT-мРНК (4 мкг/мл IVT-мРНК 1BiMAB, 4 мкг/мл IVT-мРНК 6RHU3). Включены контрольные образцы белков, содержащие 1BiMAB и 6PHU3 в комбинации с bi-scFv-отрицательным контролем, трансфицированные клетками PA- l /luc и эффекторными клетками.
Трансфицированные клетки-мишени засевали с человеческими цитотоксическими Т-клетками (пан-Т-клетки) в соотношении эффектора и мишени 5:1. Все образцы засевали в трех повторностях в 96-луночном формате и совместно инкубировали в течение 72 ч. В качестве контроля минимального лизиса (Lmin) каждый индивидуальный образец трансфицированных клеток-мишеней засевали без эффекторных клеток. Максимальный лизис (Lmax) для нормализации по отношению к числу импульсов спонтанной люминесценции достигался путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и необработанные клетки-мишени (Lmax1) или только необработанные клетки-мишени (Lmax2) перед добавлением люциферина. Через 30 мин после добавления раствора люциферина люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan. Специфический лизис клеток-мишеней рассчитывали по формуле: % специфический лизис = [1 - (люминесценция тестируемый образец - Lmax 1 / (Lmin_тестируемый образец - Lmax2)] × 100. контр означает контроль; IVT; in vitro транскрибированный; мРНК, мессенджер РНК.
ФИГУРА 29. Подтверждение продукции 1BiMAB клетками млекопитающих, трансфицированными IVT-мРНК bi-scFv или - репликоном РНК
(A) 5×106 клеток BH 21 временно трансфицировали путем электропорации 40 мкг/мл IVT-мРНК 1BiMAB или - репликоном РНК. В качестве имитирующего контроля клетки электропорировали без РНК. Через 18 ч после трансфекции супернатант и клетки собирали. Клетки лизировали и супернатанты подвергали ~50-кратному концентрированию. Необработанные и концентрированные супернатанты анализировали с помощью ELISA с использованием колонок Ni-NTA, анти-chCLDN18.2ab идиотипического mAB и вторичного AP-конъюгированного антитела. Очищенный белок 1BiMAB в ряде разведений от 2,3 до 37,5 нг/мл с 2 шагами использовали в качестве стандарта. (B) Концентрированный супернатант, клеточные лизаты (A) и 0,1 мкг очищенного белка 1BiMAB в качестве положительного контроля разделяли с помощью SDS-PAGE. Вестерн-блот анализ выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. (C) 5×106 клеток BHK21 временно трансфицировали путем электропорации 40 мкг/мл 1BiMAB или IVT-мРНК no.25. В качестве имитирующего контроля клетки подвергали электропорации без РНК. Через 48 ч после трансфекции супернатант собирали и подвергали 40-кратному концентрированию. SN и 0,1 мкг очищенного белка 1BiMAB в качестве положительного контроля разделяли с помощью SDS-PAGE. Вестерн-блот анализ выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. Контр означает контроль; mAB, моноклональное антитело; SN, супернатант; WB, Вестерн-блоттинг.
ФИГУРА 30. Инъекция IVT-мРНК 1BiMAB bi-scFv или - репликона РНК приводит к in vivo продукции и обнаруживаемым молекулам 1BiMAB bi-scFv у мышей
10 мкг IVT-мРНК 1BiMAB с IVT-мРНК EBK или без него, или 10 мкг IVT-репликона 1BiMAB инъецировали внутримышечно (IM) мышам NSG. Сыворотку из крови, собранной на 2, 4 и 7 дни после инъекции, применяли в анализе на цитотоксичность in vitro. CLDN18.2 и стабильно экспрессирующие люциферазу NugC4-LVT-CLDN18.2/luc клетки-мишени совместно инкубировали с человеческими Т-клетками в соотношении E:T, равном 30:1, с 20 мкл образца сыворотки в течение 48 ч. Стандартный контрольный белок 1BiMAB, Lmin и Lmax содержали 20 мкл сыворотки NSG. EBK означает смесь белков вируса осповакцины E3, B-l 8R, K3; IM, внутримышечно.
ФИГУРА 31. Схематическая иллюстрация молекул IVT-РНК, кодирующих антитела bi-scFv, направленно воздействующие на CLDN6 TAA.
Схема in vitro транскибированных последовательностей РНК, кодирующих анти-CLDN6 антител bi-scFv. (A) IVT мРНК в 5'- и 3'- положении относительно вариабельных областей анти-TAA. (B) альфавирусный IVT-репликон в 3'-положении относительно вариабельных областей анти-TAA. Области VH и VL анти-CLDN6 создавали на основе последовательности моноклонального антитела CLDN6 (mCLDN6ab). "Кэп" одинаково используется для ARCA, бета-S-ARCA (Dl) или бета-S-ARCA (D2). Области VH и VL анти-CD3 создавали на основе последовательности моноклонального CD3 антитела TR66. A означает аденин; bi-scFv, биспецифический одноцепочечный вариабельный фрагмент; hAg, человеческий альфа-глобин 5'-UTR; hBg, человеческий бета-глобин 3'-UTR; His, гистидиновая метка, содержащая шесть остатков гистидина; IVT, in vitro транскрибированный; LL, длинный линкер (15-18 аминокислот); nsPl-4, неструктурные белки 1-4; Sec, сигнал секреции; sgP, субгеномный промотор; SL, короткий линкер (5-6 аминокислот); TAA, опухоль-ассоциированный антиген; UTR, нетранслируемая область; V, вариабельная область тяжелой (H) и легкой (L) цепи антитела.
ФИГУРА 32. Совместная инкубация клеток-мишеней, трансфицированных анти-CLDN6 IVT-мРНК bi-scFv, с человеческими T-клетками приводит образованию кластеров Т-клеток.
Клетки PA-1, эндогенно экспрессирующие CLDN6, временно трансфицировали путем электропорации 20 мкг/мл IVT-мРНК 6RHU5 или 6RHU3 и совместно инкубировали с человеческими Т-клетками (пан-Т-клетки) в соотношении эффектора к мишени 5:1 в 6-луночных планшетах. В качестве отрицательного контрольного образца использовали клетки-мишени PA-1, трансфицированные IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (-контр), совместно инкубированные с человеческими Т-клетками (верхний ряд, фото слева). В среднем ряду показаны необработанные клетки PA-1 и человеческие Т-клетки без белка в качестве отрицательного контроля (имитирующий контроль, фото слева), или с 50 мкг/мл очищенных анти-CLDN6 белков 6PHU5 (в середине) или 6PHU3 (справа) bi-scFv в качестве положительных контролей. В нижнем ряду показаны необработанные клетки PA-1 (слева) и только человеческие Т-клетки (справа). Через 24 ч совместной инкубации делали снимки образцов с помощью микроскопа Nikon Eclipse Ti при увеличении 200×. Белые стрелки указывают на Т-клеточные кластеры на клетках-мишенях. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; контр, контрольный; hu, человеческий; IVT, in vitro транскрибированный; мРНК, мессенджер РНК; TL, T-лимфоцит.
ФИГУРА 33. Эффект ориентации доменов на эффективность: трансфекция клеток-мишеней анти-CLDN6 6RHU3 bi-scFv приводит к более высокому процентному содержанию активированных Т-клеток по сравнению с 6RHU5.
Клетки PA-1, эндогенно экспрессирующие CLDN6, временно трансфицировали двумя вариантами bi-scFv 6RHU5 и 6RHU3, направленными против CLDN6 и CD3 для сравнения их активности в анализе активации Т-клеток. Для каждого варианта 5×106 клеток PA-1 подвергали электропорации 20 мкг/мл IVT-мРНК. Трансфицированные клетки-мишени повторно подсчитывали, 1×105 клеток засевали в каждый 6-луночный планшет и инкубировали с человеческими цитотоксическими Т-клетками (выбраны Т-клетки CD8+) в соотношении E:T, равном 5:1. В качестве отрицательных контролей выбирали необработанные клетки-мишени (hu TL + необработанные PA-1) и клетки-мишени, трасфицированные IVT-мРНК bi-scFv, направленным на неэкспрессированный TAA (hu TL + PA-1 - контр). Белок 6PHU5 служил в качестве положительного контроля в концентрации 50 нг/мл (hu TL + PA-1 белок контр). Кроме того, Т-клетки засевали без клеток-мишеней с белком 6PHU5 и без него в качестве референсной фоновой активации. Каждый образец засевали в двух повторностях. Анализ выполняли через 24 ч и 48 ч: Т-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE, анти-CD69-APC и 7-AAD для окрашивания живых и погибших клеток и анализировали с помощью проточной цитометрии. TAA-зависимую bi-scFv-опосредованную активацию Т-клеток наблюдали в обоих вариантах анти-CLDN6 bi-scFv через 24 ч (A) и 48 ч (B) совместной инкубации. Трансфекция 6RHU3 bi-scFv вызвала приблизительно на 20% более высокую активацию Т-клеток в оба момента времени. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; Контр, контрольный; hu, человеческий; IVT, in vitro транскрибированный; мРНК, мессенджер РНК; TL, T лимфоцит.
ФИГУРА 34. Секреция 6RHU3 опосредует Т-клеточную активацию зависимым от концентрации образом.
Клетки PA-1, эндогенно экспрессирующие CLDN6, временно трансфицировали путем электропорации IVT-мРНК в общем количестве 20 мкг/мл, включающем 0,2-20 мкг/мл IVT-мРНК 6RHU3 плюс соответствующие количества IVT-мРНК bi-scFv, направленно воздействующего на неэкспрессированный TAA. Трансфекция 20 мкг/мл IVT-мРНК bi-scFv, направленно воздействующего на неэкспрессированный TAA (0.0 мкг/мл IVT-мРНК 6RHU3), служила в качестве контроля специфичности. Трасфицированные клетки-мишени совместно инкубировали с человеческими цитотоксическими Т-клетками (пан-Т-клетками) в соотношении эффектора к мишени 5:1 в 6-луночных планшетах в двух повторностях. В качестве референсной активации Т-клеток только человеческие Т-клетки выращивали без белка 6PHU5 (hu TL -) или с белком 6PHU5 (hu TL белковый контр). В качестве отрицательного контроля Т-клетки совместно инкубировали с необработанными клетками-мишенями PA-1 (hu TL + PA-1 - Контр). Белок 6PHU5 служил в качестве положительного контроля в концентрации 50 нг/мл (hu TL + PA-1 белковый контр). Через 48 ч Т-клетки собирали и метили анти-CD3-FITC, анти-CD25-PE и анти-CD69-APC и анализировали с помощью проточной цитометрии. На графиках показано процентное содержание положительно окрашенных человеческих Т-клеток, определенных с помощью программного обеспечения FlowJo. Bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; контр, контрольный; hu, человеческий; IVT, in vitro транскрибированный; мРНК, мессенджер РНК; TL, T лимфоцит.
ФИГУРА 35. Величина EC50 6RHU3 для специфического лизиса клеток-мишеней через 48 ч составила приблизительно 200 нг/мл.
Клетки PA-1, эндогенно экспрессирующие CLDN6, которые стабильно экспрессируют люциферазу, временно трансфицировали путем электропорации IVT-мРНК bi-scFv в общей концентрации 13,3 мкг/мл, включающей 0,004 - 13,3 мкг/мл 6RHU3 и соответствующее количество IVT-мРНК bi-scFv, направленно воздействующего на неэкспрессированный TAA. Трансфицированные клетки-мишени засевали с человеческими Т-клетками в соотношении эффектора к мишени 5:1 в трех повторностях в 96-луночном формате. В качестве контроля минимального лизиса (Lmin) каждый индивидуальный образец трансфицированных клеток-мишеней засевали без эффекторных клеток. Максимальный лизис (Lmax) для нормализации по отношению к числу импульсов спонтанной люминесценции был достигнут путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и необработанные клетки-мишени, непосредственно перед добавлением люциферина. Через 30 мин после добавления раствора люциферина люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan через 24 ч и 48 ч. Специфический лизис клеток-мишеней рассчитывали по формуле: % специфического лизиса = [1 - (люминесценция тестируемого образца - Lmax) (Lmin_тестируемого образца - Lmax)] × 100. Значения наносили на график в зависимости от logl0 концентрации 6RHU3. Величины EC50 означают половину максимальной эффективной концентрации; L, лизис.
ФИГУРА 36. Т-клеточная пролиферация специфически индуцирована в ответ на секрецию 6RHU3 клетками-мишенями в присутствии CLDN6.
Человеческие Т-клетки окрашивали CFSE для анализа. Т-клетки выращивали без клеток-мишеней (Т-клетки) в комбинации с 5 мкг/мл OKT3 и 2 мкг/мл αCD28 в качестве положительного контроля активации (+контр), с 5 нг/мл нетаргетного контрольного bi-scFv (-контр белок) или с 5 нг/мл белка 6PHU3 (белок 6PHU3). Т-клетки и клетки-мишени PA-1, эндогенно экспрессирующие CLDN6, вместе инкубировали (T-клетки + CLDN6-положительные клетки-мишени) без чего-либо (имитирующие) или с 5 нг/мл белка 6PHU3 (белок 6PHU3). Для тестирования IVT-мРНК клетки PA-1 трансфицировали 20 мкг/мл IVT-мРНК 6RHU3 (мРНК 6RHU3) или IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (мРНК -контр), и инкубировали с Т-клетками. Кроме того, клетки PA-1, трансфицированные IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA, объединяли с 5 нг/мл белка 6PHU3 (мРНК -контр + белок 6PHU3). В качестве дополнительного контроля специфичности были включены образцы с неэкспрессирующей CLDN6 линией клеток-мишеней MDA-MB-231 вместе с Т-клетками (T-клетки + CLDN6-отрицательные клетки-мишени). MDA-MB-231 использовали необработанными и инкубировали без чего-либо (имитирующий контроль), с 5 нг/мл контрольного белка bi-scFv (-контр белок) или 5 нг/мл белка 6PHU3 (белок 6PHU3), или MDA-MB-231 трансфицировали 20 мкг/мл IVT-мРНК 6RHU3 (мРНК 6RHU3) или IVT-мРНК bi-scFv, направленно воздействующим на неэкспрессированный TAA (-контр мРНК). Анализ выполняли в соотношении эффектора к мишени 5:1 в 96-луночном планшете, с каждым образцом в трех повторностях и временем инкубации 72 ч. Уменьшение сигнала CFSE, указывающее на Т-клеточную пролиферацию, анализировали с помощью проточной цитометрии, рассчитывали с помощью программного обеспечения FlowJo и наносили на график в виде % пролиферирующих Т-клеток. CFSE означает сложный эфир сукцинимидила и карбоксифлуоресцеина; IVT, in vitro транскрибированный; мРНК, мессенджер РНК.
ФИГУРА 37. Подтверждение 6RHU3 трансляции клетками млекопитающих, трансфицированными IVT-мРНК bi-scFv или - репликоном РНК
(A) 5 x l06 клеток BHK21 временно трансфицировали путем электропорации 40 мкг/мл IVT-мРНК 6RHU3 или -репликона РНК. Трансфекция 40 мкг/мл IVT-мРНК no.25 была включена в качестве дополнительного образца. В качестве имитирующего контроля клетки подвергали электропорации без РНК. Через 18 ч после трансфекции супернатант и клетки собирали. Клетки лизировали и супернатанты подвергали 50-кратному концентрированию. Необработанные и концентрированные супернатанты анализировали с помощью ELISA с использованием колонок Ni-NTA, анти-mCLDN6ab идиотипического mAB и вторичного AP-конъюгированного антитела. Очищенный белок 6PHU3 в разведении от 2,3 до 150 нг/мл за 2 шага использовали в качестве стандарта. (B) Концентрированный супернатант и клеточные лизаты (A) и 0,1 мкг очищенного белка 6PHU3 в качестве положительного контроля разделяли с помощью SDS-PAGE. Вестерн-блот анализ выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. (C) 5×106 клеток BHK21 временно трансфицировали путем электропорации 40 мкг/мл 6RHU3 или IVT-мРНК no.25. В качестве имитирующего контроля клетки подвергали электропорации без РНК. Через 48 ч после трансфекции супернатант собирали и подвергали 40-кратному концентрированию. SN и 0,1 мкг очищенного белка 6PHU3 в качестве положительного контроля разделяли посредством SDS-PAGE. Вестерн-блот анализ выполняли с первичным моноклональным анти-His и вторичным анти-мышиным антителом, конъюгированным с пероксидазой. Контр означает контрольные; mAB, моноклональное антитело; SN, супернатант; WB, Вестерн-блоттинг.
ФИГУРА 38. Инъекция IVT-мРНК 6RHU3 bi-scFv или - репликона РНК приводит к in vivo трансляции и детектируемым молекулам bi-scFv у мышей
10 мкг IVT-мРНК 6RHU3 с IVT-мРНК EBK или без него или 10 мкг IVT-репликона 6RHU3 инъецировали IM мышам NSG. Сыворотку крови, собранной через 7 дней после инъекции, применяли в анализе на цитотоксичность in vitro. Клетки-мишени PA-l/luc, эндогенно экспрессирующие CLDN6 и стабильно экспрессирующие люциферазу, совместно инкубировали с человеческими Т-клетками в соотношении Е:Т, равном 30:1, с 20 мкл образца сыворотки в течение 48 ч. Стандартный контрольный белок 6PHU3, Lmin и Lmax содержащий 20 мкл имитирующей сыворотки NSG. EBK означает смесь белков вируса осповакцины E3, B-18R, K3; IM, внутримышечно.
ФИГУРА 39. Результаты анализа на цитотоксичность анти-CLDN18.2 белков bi-scFv, содержащих анти-CD3 связывающий домен scFv в С-концевой части белка.
Варианты bi-scFv, направленные против CLDN18.2 и CD3, временно экспрессировали в клетках CHO и очищали полимером Protein-L для сравнения их силы в анализе на цитотоксичность. Клетки NugC4, эндогенно экспрессирующие CLDN18.2, которые стабильно экспрессируют люциферазу, были использованы в качестве клеток-мишеней. Человеческие Т-клетки и клетки-мишени инкубировали в соотношении Е:Т, равном 5:1, с 5000, 1000, 200 и 40 нг/мл каждого из белков bi-scFv в 96-луночном формате. Каждый тестируемый образец высевали в трехкратном повторении, контрольный образец для Lmin высевали в трехкратном повторении. Продолжительность совместной инкубации перед анализом составила 24 ч и 48 ч. После добавления раствора люциферина в заданные моменты времени люминесценцию измеряли на ридере Infinite M200 TECAN. Специфический лизис клеток-мишеней рассчитывали для каждой концентрации и записывали.
a. Вариабельные домены анти-CD3 расположены в порядке доменов VH - VL и разделены пептидным линкером LL4.
b. Вариабельные домены анти-CD3 расположены в порядке доменов VH - VL и разделены пептидным линкером LL4. Анти-CD3 scFv содержит связующий дисульфидный мостик между доменами VH и VL.
c. Вариабельные домены анти-CD3 расположены в порядке VL - VH и разделены пептидным линкером LL5.
d. Вариабельные домены анти-CD3 расположены в порядке VL - VH и разделены пептидным линкером LL5. Анти-CD3 scFv содержит связующий дисульфидный мостик между доменами VL и VH.
ФИГУРА 40. Внутрианалитическое сравнение величин EC50, полученных в анализе на цитотоксичность на основе люциферазы с использованием анти-CLDN6 белков bi-scFv.
Анализы на цитотоксичность на основе люциферазы выполняли с помощью трех различных доноров для Т-клеточного препарата. Величины EC50, рассчитанные с использованием 6 тестированных анти-CLDN6 белков bi-scFv через 24 ч и 48 ч инкубации представлены для каждого независимого анализа (A, B и C). Клетки PA-1, эндогенно экспрессирующие CLDN6, инкубировали в течение 24 ч и 48 ч с нарастающими концентрациями (0,025-50000 нг/мл для A и B, 0,0025-5000 нг/мл для C) анти-CLDN6 белков bi-scFv и человеческими Т-клетками в соотношении эффектора и мишени 5:1 в трех повторностях в 96-луночном формате. В качестве контроля минимального лизиса (Lmin) эффекторные клетки и клетки-мишени высевали без белков bi-scFv. Максимальный лизис (Lmax) для нормализации в отношении числа импульсов спонтанной люминесценции достигался путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и клетки-мишени в отсутствие bi-scFv непосредственно перед добавлением люциферина. Через 30 мин после добавления раствора люциферина люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan через 24 ч и 48 ч инкубации клеток-мишеней и эффекторных клеток. Специфический лизист клеток-мишеней рассчитывали по формуле: % специфический лизис = [1 - (люминесценция тестируемый образец - Lmax) / (Lmin - Lmax)] × 100. EC50 означает половину максимальной эффективной концентрации; L, лизис; NA, нет данных.
Подробное описание изобретения
Хотя настоящее изобретение описано подробно далее, следует понимать, что данное изобретение не ограничивается конкретными методиками, протоколами и реагентами, описанными здесь, так как указанные методики, протоколы и реагенты могут изменяться. Также, следует понимать, что терминология, используемая здесь, предназначена только для описания конкретных вариантов осуществления и не предназначена для ограничения объема настоящего изобретения, которое может быть ограничено только прилагаемыми пунктами формулы изобретения. Если не указано иное, все технические и научные термины, используемые здесь, имеют такие же значения, как обычно понимается специалистами в данной области.
Далее будут описаны элементы настоящего изобретения. Эти элементы перечислены вместе с определенными вариантами осуществления, однако, следует понимать, что они могут быть объединены любым способом и в любом количестве для создания дополнительных вариантов осуществления. Различные описанные примеры и предпочтительные варианты осуществления не должны рассматриваться как ограничивающие объем настоящего изобретения только конкретно описанными вариантами осуществления. Следует понимать, что данное описание служит поддержкой и охватывает варианты осуществления, которые объединяют четко описанные варианты осуществления с любым числом раскрытых и/или предпочтительных элементов. Кроме того, любые перестановки и комбинации всех описанных элементов в данной заявке будут считаться раскрытыми с помощью описания по настоящей заявке, если контекст не указывает на иное.
Предпочтительно, термины, используемые здесь, определены, как описано в «A multilingual glossary of biotechnological terms: (IUPAC Recommendations)», H.G.W. Leuenberger, B. Nagel, и H. Kolbl, Eds., Helvetica Chimica Acta, CH-4010 Basel, Switzerland, (1995).
Если не указано иное, осуществление настоящего изобретения будет предусматривать использование стандартных методов химии, биохимии, клеточной биологии, иммунологии и методов рекомбинантных ДНК, описанных в литературе в данной области (смотри, например, Molecular Cloning: A Laboratory Manual, 2nd Edition, J. Sambrook et al. eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor 1989).
Во всем данном описании и прилагаемой формуле изобретения, если контекст не требует иного, слово «содержать» и его вариации, такие как «содержит» и «содержащий», будут пониматься как включение указанного элемента, целого числа или стадии, или групп элементов, целых чисел или стадий, но не как исключение любого другого элемента, целого числа или стадии, или групп элементов, целых чисел или стадий, хотя в некоторых вариантах осуществления такой другой элемент, целое число или стадия, или группы элементов, целых чисел или стадий могут быть исключены, то есть объект изобретения состоит во включении указанного элемента, целого числа или стадии, или групп элементов, целых чисел или стадий. Использование единственного числа в контексте описания изобретения (особенно в контексте прилагаемой формулы изобретения) должно истолковываться так, чтобы охватывать и единственное и множественное число, если иное не указано в данном документе или явно противоречит контексту. Перечисление диапазонов значений в настоящем документе предназначено только для того, чтобы оно служило способом сокращения для указания каждого отдельного значения, попадающего в диапазон. Если не указано иное, каждое отдельное значение включается в техническое описание так, как если бы оно было отдельно перечислено в настоящем документе. Все способы, описанные в настоящем документе, могут быть выполнены в любом надлежащем порядке, если иное не указано в настоящем документе или явно опровергается контекстом. Использование любого или всех примеров, или примерный язык (например, «такой как»), представленный в настоящем документе, предназначен только для лучшего разъяснения изобретения, и не налагает никаких ограничений на область применения изобретения, если не заявлено иное. Никакой язык в данном описании не должен истолковываться как указывающий какой-либо незаявленный в формуле изобретения элемент в качестве важнейшего для осуществления изобретения.
В данном описании цитируются некоторые документы. Каждый из документов, цитируемых здесь (включая все патенты, патентные заявки, научные публикации, спецификации производителя, инструкции и т.д.), выше или ниже, введены здесь ссылками во всей полноте. Ничто здесь не должно быть истолковано как признание того, что изобретатель не имеет права датировать изобретение более ранним числом, вследствие более ранней даты создания изобретения (antedate).
Клаудины представляют собой семейство белков, которые являются наиболее важными компонентами плотных контактов, где они устанавливают параклеточный барьер, который контролирует потоки молекул в межклеточном пространстве между клетками эпителия. Клаудины представляют собой трансмембранные белки, четырежды пересекающие клеточную мембрану, при этом оба конца, N-конец и С-конец, расположены в цитоплазме. Первая внеклеточная петля, названная ECl или ECL1, состоит в среднем из 53 аминокислот, и вторая внеклеточная петля, названная EC2 или ECL2, состоит примерно из 24 аминокислот. Белки клеточной поверхности семейства клаудина, такие как CLDN6 и CLDN18.2, экспрессируются в опухолях различного происхождения, и являются особенно подходящими в качестве структур-мишеней в опосредованной антителами иммунотерапии рака благодаря их селективной экспрессии (экспрессия отсутствует в нормальных тканях, значимых с точки зрения токсичности) и локализации в плазменной мембране.
В контексте настоящего изобретения предпочтительные клаудины представляют собой CLDN6 и CLDN18.2. CLDN6 и CLDN18.2 были идентифицированы как дифференциально экспрессирующиеся в опухолевых тканях, при этом единственными нормальными тканями, экспрессирующими CLDN18.2, является желудок, и единственной нормальной тканью, экспрессирующей CLDN6, является плацента.
CLDN18.2 селективно экспрессируется в нормальных тканях в дифференцированных клетках эпителия слизистой оболочки желудка. CLDN18.2 экспрессируется в раке различного происхождения, таком как карцинома поджелудочной железы, карцинома пищевода, карцинома желудка, бронхиальная карцинома, карцинома молочной железы и опухоли ЛОР-органов. CLDN18.2 является ценной мишенью для предупреждения и/или лечения первичных опухолей, таких как рак желудка, рак пищевода, рак поджелудочной железы, рак легкого, такой как немелкоклеточный рак легкого (NSCLC), рак яичника, рак толстой кишки, рак печени, рак головы и шеи, и рак желчного пузыря, и их метастазов, в частности, метастазов рака желудка, таких как опухоли Крукенберга, перитонеальных метастазов и метастазов в лимфоузлы.
Было обнаружено, что CLDN6 экспрессируется, например, в раке яичника, раке легкого, раке желудка, раке молочной железы, раке печени, раке поджелудочной железы, раке кожи, мелономе, раке головы и шеи, саркоме, раке желчных протоков, почечно-клеточном раке и раке мочевого пузыря. CLDN6 является особенно предпочтительной мишенью для предупреждения и/или лечения рака яичника, в частности, аденокарциномы яичника и тератокарциномы яичника; рака легкого, включая мелкоклеточный рак легкого (SCLC) и немелкоклеточный рак легкого (NSCLC), в частности, плоскоклеточный рак легкого и аденокарциному; рака желудка; рака молочной железы; рака печени; рака поджелудочной железы; рака кожи, в частности, базальноклеточной карциномы и плоскоклеточной карциномы; злокачественной меланомы; рака головы и шеи, в частности, злокачественной плеоморфной аденомы; саркомы, в частности, синовиальной саркомы и карциносаркомы; рака желчного протока; рака мочевого пузыря, в частности переходно-клеточной карциномы и папиллярного рака; рака почки, в частности, плоскоклеточного рака, включая светлоклеточный рак почки и папиллярную карциному почки; рака толстой кишки; рака тонкой кишки, включая рак подвздошной кишки, в частности аденокарциномы тонкой кишки и аденокарциномы подвздошной кишки; эмбриональной карциномы яичка; плацентарной хориокарциномы; рака шейки матки; рака яичка, в частности, семиномы яичка, тератомы яичка и эмбрионального рака яичка; рака матки; герминомы, такой как тератокарцинома или эмбриональная карцинома, в частности, эмбрионально-клеточной опухоли яичка и их метастатических форм. В одном варианте осуществления раковые заболевания, ассоциированные с экспрессией CLDN6, выбраны из группы, состоящей из рака яичников, рака легкого, метастатического рака яичников и метастатического рака легкого. Предпочтительно, рак яичника представляет собой карциному или аденокарциному. Предпочтительно, рак легкого представляет собой карциному или аденокарциному, и предпочтительно представляет собой бронхиолярный рак, такой как бронхиолярная карцинома или бронхиолярная аденокарцинома.
Термин «CLDN», используемый здесь, означает клаудин и включает CLDN18.2 и CLDN6. Предпочтительно, клаудин представляет собой клаудин человека.
Термин «CLDN18» относится к клаудину 18 и включает любые варианты, включая сплайсированный вариант 1 клаудина 18 (клаудин 18.1 (CLDN18.1)) и сплайсированный вариант 2 клаудина 18 (клаудин 18.2 (CLDN 18.2)).
Термин «CLDN18.2» предпочтительно относится к CLDN18.2 человека и, в частности, к белку, содержащему, предпочтительно, состоящему из аминокислотной последовательности согласно SEQ ID NO: 1 перечня последовательностей или варианта указанной аминокислотной последовательности. Первая внеклеточная петля CLDN18.2 предпочтительно содержит аминокислоты с 27 по 81, более предпочтительно аминокислоты с 29 по 78 аминокислотной последовательности, представленной в SEQ ID NO: 1. Вторая внеклеточная петля CLDN18.2 предпочтительно содержит аминокислоты с 140 по 180 аминокислотной последовательности, представленной в SEQ ID NO: 1. Указанные первая и вторая внеклеточные петли предпочтительно образуют внеклеточную часть CLDN18.2.
Термин «CLDN6» предпочтительно относится к CLDN6 человека и, в частности, к белку, содержащему, предпочтительно, состоящему из аминокислотной последовательности SEQ ID NO: 2 или SEQ ID NO: 3 перечня последовательностей или варианта указанной аминокислотной последовательности. Первая внеклеточная петля CLDN6 предпочтительно содержит аминокислоты с 28 по 80, более предпочтительно, аминокислоты с 28 по 76 аминокислотной последовательности, представленной в SEQ ID NO: 2, или аминокислотной последовательности, представленной в SEQ ID NO: 3. Вторая внеклеточная петля CLDN6 предпочтительно содержит аминокислоты с 138 по 160, предпочтительно, аминокислоты с 141 по 159, более предпочтительно, аминокислоты с 145 по 157 аминокислотной последовательности, представленной в SEQ ID NO: 2, или аминокислотной последовательности, представленной в SEQ ID NO: 3. Указанные первая и вторая внеклеточные петли, предпочтительно, образуют внеклеточную часть CLDN6.
Термин «вариант» в соответствии с изобретением относится, в частности, к мутантам, сплайсированным вариантам, конформациям, изоформам, аллельным вариантам, вариантам разновидностей и гомологам разновидностей, в частности, таким, которые присутствуют естественным путем. Аллельный вариант относится к альтерации нормальной последовательности гена, значение которой часто бывает неясным. Полное секвенирование гена часто позволяет идентифицировать многочисленные аллельные варианты заданного гена. Гомолог разновидностей представляет собой нуклеиновую кислоту или аминокислотную последовательность с разновидностями, имеющими происхождение, отличающееся от происхождения заданной нуклеиновой кислоты или аминокислотной последовательности. Термин «вариант» охватывает любые посттрансляционно модифицированные варианты и конформационные варианты.
Второй молекулой-мишенью для связывающих агентов, описанный здесь, является кластер клеточной дифференцировки CD3 (Cluster of Differentiation 3). Комплекс CD3 обозначает антиген, который экспрессируется на зрелых человеческих Т-клетках, тимоцитах и субпопуляции естественных клеток-киллеров, как часть мультимолекулярного комплекса Т-клеточного рецептора (TCR). T-клеточный корецептор представляет собой белковый комплекс и состоит из четырех различных цепей. У млекопитающих комплекс содержит цепь CD3γ, цепь CD35δ и две цепи CD3ε. Эти цепи связаны с молекулой, известной как Т-клеточный рецептор (TCR) и ζ-цепью для генерации сигнала активации в Т-лимфоцитах. Вместе, TCR, ζ-цепь и молекулы CD3 образуют TCR комплекс.
Аминокислотная последовательность CD3-эпсилон человека описана в GenBank под регистрационным номером NM_000733 и содержит SEQ ID NO: 4. Аминокислотная последовательность CD3-гамма человека описана в GenBank под регистрационным номером NM 000073. Аминокислотная последовательность CD3-дельта человека описана в GenBank под регистрационным номером NM_000732. CD3 отвечает за трансдукцию сигнала TCR. Как описано в Lin и Weiss, Journal of Cell Science 114, 243-244 (2001), активация TCR комплекса путем связывания MHC-презентированных специфических эпитопов антигенов приводит к фосфорилированию иммунорецепторных тирозиновых активирующих мотивов (ITAM) киназами Src-семейства, запуская рекрутинг других киназ, что приводит к Т-клеточной активации, включая высвобождение Ca2+. Образование кластеров CD3 на Т-клетках, например, путем иммобилизации анти-CD3-антител, приводит к Т-клеточной активации, аналогичной активации Т-клеточного рецептора, но которая не зависит от типичной для этого клона специфичности.
Используемый здесь термин «CD3» включает CD3 человека и обозначает антиген, который экспрессируется на человеческих Т-клетках как часть мультимолекулярного комплекса Т-клеточного рецептора.
В отношении CD3, связывающий агент по изобретению предпочтительно распознает эпсилон-цепь CD3, в частности, он распознает эпитоп, который соответствует первым 27 N-концевым аминокислотам CD3-эпсилон или функциональным фрагментам этого удлинения из 27 аминокислот.
В соответствии с изобретением, термин «клаудин-позитивный рак», или аналогичные термины, означает рак, в который вовлечены раковые клетки, экспрессирующие клаудин, предпочтительно на поверхности указанных раковых клеток.
«Клеточная поверхность» используется в соответствии с ее обычным значением в данной области и, таким образом, включает наружную часть клетки, которая является доступной для связывания белками и другими молекулами.
Клаудин экспрессируется на клеточной поверхности, если он расположен на поверхности указанных клеток и является доступным для связывания клаудин-специфическими антителами, добавленными в клетки.
Термин «внеклеточная часть» в контексте настоящего изобретения относится к части молекулы, такой как белок, которая обращена к внеклеточному пространству клетки и, предпочтительно, является доступной снаружи указанной клетки, например, для антигенсвязывающих молекул, таких как антитела, локализованные снаружи клетки. Предпочтительно, термин относится к одной или более внеклеточным петлям или доменам, или их фрагментам.
Термин «часть» или «фрагмент» используются здесь взаимозаменяемо и относятся к непрерывному элементу. Например, часть структуры, такой как аминокислотная последовательность или белок, относится к непрерывному элементу указанной структуры. Участок, часть или фрагмент структуры, предпочтительно, обладает одним или несколькими функциональными свойствами указанной структуры. Например, участок, часть или фрагмент эпитопа или пептида является, предпочтительно, иммунологически эквивалентным эпитопу или пептиду, из которого он происходит. Часть или фрагмент белковой последовательности, предпочтительно, содержит последовательность, состоящую по меньшей мере из 6, в частности, по меньшей мере из 8, по меньшей мере из 12, по меньшей мере из 15, по меньшей мере из 20, по меньшей мере из 30, по меньшей мере из 50 или по меньшей мере из 100 последовательных аминокислот белковой последовательности.
В соответствии с изобретением CLDN18.2 значительно не экспрессируется в клетке, если уровень экспрессии ниже по сравнению с экспрессией в клетках желудка или ткани желудка. Предпочтительно, уровень экспрессии составляет менее 10%, предпочтительно, менее 5%, 3%, 2%, 1%, 0,5%, 0,1 % или 0,05%, или даже ниже, от экспрессии в клетках желудка или ткани желудка. Предпочтительно, CLDN18.2 в основном не экспрессируется в клетке, если уровень экспрессии превышает уровень экспрессии в нераковой ткани, отличной от желудка, не более чем в 2 раза, предпочтительно, в 1,5 раза и, предпочтительно, не превышает уровень экспрессии в указанной нераковой ткани. Предпочтительно, CLDN18.2 в основном не экспрессируется в клетке, если уровень экспрессии ниже предела обнаружения и/или если уровень экспрессии является слишком низким для того, чтобы обеспечить связывание CLDN18.2-специфическими антителами, добавленными в клетки.
В соответствии с изобретением, CLDN18.2 экспрессируется в клетке, если уровень экспрессии превышает уровень экспрессии в нераковой ткани, отличной от желудка, предпочтительно, более чем в 2 раза, предпочтительно, в 10 раз, 100 раз, 1000 раз. Предпочтительно, CLDN18.2 экспрессируется в клетке, если уровень экспрессии выше предела обнаружения и/или если уровень экспрессии является достаточно высоким для обеспечения связывания CLDN18.2-специфическими антителами, добавленным в клетки. Предпочтительно, CLDN18.2, экспрессирующийся в клетке, экспрессируется или экспонируется на поверхности указанной клетки.
В соответствии с изобретением, CLDN6 в основном не экспрессируется в клетке, если уровень экспрессии ниже по сравнению с экспрессией в клетках плаценты или ткани плаценты. Предпочтительно, уровень экспрессии составляет менее 10%, предпочтительно, менее 5%, 3%, 2%, 1%, 0,5%, 0,1% или 0,05%, или даже ниже, от экспрессии в клетках плаценты или ткани плаценты. Предпочтительно, CLDN6 в основном не экспрессируется в клетке, если уровень экспрессии превышает уровень экспрессии в нераковой ткани, отличной от плаценты, не более чем в 2 раза, предпочтительно, в 1,5 раза, и предпочтительно, не превышает уровень экспрессии в указанной нераковой ткани. Предпочтительно, CLDN6 в основном не экспрессируется в клетке, если уровень экспрессии ниже предела обнаружения и/или уровень экспрессии является слишком низким для обеспечения связывания CLDN6-специфическими антителами, добавленными в клетки.
В соответствии с изобретением CLDN6 экспрессируется в клетке, если уровень экспрессии превышает уровень экспрессии в нераковой ткани, отличной от плаценты, предпочтительно, более чем 2 раза, предпочтительно, в 10 раз, 100 раз, 1000 раз или 10000 раз. Предпочтительно, CLDN6 экспрессируется в клетке, если уровень экспрессии выше предела обнаружения и/или если уровень экспрессии является достаточно высоким для обеспечения связывания CLDN6-специфическими антителами, добавленными в клетки. Предпочтительно, CLDN6, экспрессирующийся в клетке, экспрессируется или экспонируется на поверхности указанной клетки.
В соответствии с изобретением термин «заболевание» относится к любому патологическому состоянию, включая рак, в частности, таким формам рака, которые описаны в настоящем документе. Любая ссылка в настоящем документе на рак или конкретные формы рака также включает метастазы указанного рака. В предпочтительном варианте осуществления заболевание, подлежащее лечению в соответствии с настоящим изобретением, включает клетки, экспрессирующие клаудин (CLDN), такие как CLDN18.2 и/или CLDN6.
«Заболевание, ассоциированное с клетками, экспрессирующими CLDN» или сходные экспрессии означает в соответствии с изобретением, что CLDN экспрессируется в клетках больной ткани или органа. В одном варианте осуществления экспрессия CLDN в клетках больной ткани или органа является увеличенной по сравнению с состоянием в здоровой ткани или органе. Увеличение относится к увеличению, по меньшей мере, на 10%, в частности, по меньшей мере на 20%, по меньшей мере на 50%, по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 500%, по меньшей мере на 1000%, по меньшей мере на 10000%, или даже больше. В одном варианте осуществления экспрессия обнаруживается только в больной ткани, тогда как экспрессия в здоровой ткани является подавленной. В соответствии с изобретением, заболевания, ассоциированные с клетками, экспрессирующими CLDN, включают раковые заболевания. Кроме того, в соответствии с изобретением раковые заболевания предпочтительно представляют собой такие заболевания, в которых раковые клетки экспрессируют CLDN.
Используемый здесь термин «раковое заболевание» или «рак» включает заболевание, которое характеризуется аберрантно регулированным клеточным ростом, пролиферацией, дифференциацией, адгезией и/или миграцией. Под термином «раковая клетка» понимается аномальная клетка, которая растет путем быстрой неконтролируемой клеточной пролиферации и продолжает расти после стимула, который инициировал новую остановку роста. Предпочтительно, «раковое заболевание» характеризуется клетками, экспрессирующими CLDN, и раковыми клетками, экспрессирующими CLDN. Клетка, экспрессирующая CLDN, предпочтительно представляет собой раковую клетку, предпочтительно типов рака, описанных здесь.
Термин «рак» в соответствии с изобретением включает лейкоз, семиному, меланому, тератому, лимфому, нейробластому, глиому, рак прямой кишки, рак эндометрия, рак почки, рак надпочечной железы, рак щитовидной железы, рак крови, рак кожи, рак головного мозга, рак шейки матки, рак желудочно-кишечного тракта, рак печени, рак толстой кишки, рак желудка, рак кишечника, рак головы и шеи, рак ЖКТ, рак лимфоузлов, рак пищевода, колоректальный рак, рак поджелудочной железы, рак ушей, носа и гортани (ENT), рак молочной железы, рак предстательной железы, рак матки, рак яичника и рак легкого, и их метастазы. Примерами указанного рака является карцинома легкого, карцинома молочной железы, карцинома предстательной железы, карцинома толстой кишки, почечно-клеточная карцинома, карцинома шейки матки или метастазы рака или опухолей, описанных выше. Термин «рак» в соответствии с изобретением также включает метастазы рака.
В соответствии с изобретением «карцинома» представляет собой злокачественную опухоль, происходящую из эпителиальных клеток. Эта группа представляет наиболее распространенные виды рака, включая распространенные формы рака молочной железы, предстательной железы, легкого и толстой кишки.
«Аденокарцинома» представляет собой рак, который происходит из железистой ткани. Эта ткань является также частью более крупной категории ткани, известной как эпителиальная ткань. Эпителиальная ткань включает кожу, железы и целый ряд других тканей, которые выстилают полости и органы организма. Эпителиальная ткань происходит из зародышевых листков, эктодермы, эндодермы и мезодермы. Для классификации рака как аденокарциномы клетки необязательно должны являться частью железы, поскольку они обладают секреторными свойствами. Эта форма карциномы может возникать у некоторых высших млекопитающих, включая человека. Хорошо дифференцированные аденокарциномы имеют сходство с железистой тканью, из которой они происходят, тогда как умеренно дифференцированные аденокарциномы таким свойством не обладают. Путем окрашивания клеток, полученных из биопсии, патолог определяет, представляет ли опухоль аденокарциному или какой-либо другой тип рака. Аденокарцинома может возникать во многих тканях организма за счет убиквитарной природы желез в организме. Несмотря на то, что каждая железа может не секретировать одинаковое вещество, поскольку существует экзокринная функция клетки, рак считается железистым и его злокачественная форма, таким образом, названа аденокарциномой. Злокачественная аденокарцинома инвазирует другие ткани и часто распространяет метастазы, при условии, что ей дали достаточное количество времени. Аденокарцинома яичника является наиболее распространенным типом карциномы яичника. Она включает серозные и муцинозные аденокарциномы, светло-клеточную аденокарциному и эндометриоидную аденокарциному.
Термин «метастазирование» означает распространение раковых клеток из их участка происхождения в другую часть организма. Формирование метастазов представляет собой очень сложный процесс и зависит от открепления злокачественных клеток от первичной опухоли, инвазии внеклеточного матрикса, проникновения сквозь базальные мембраны эндотелия для вхождения в полость организма и сосуды, и затем, после переноса кровью, инфильтрации целевых органов. В заключение, рост новой опухоли в целевом участке зависит от ангиогенеза. Метастазирование опухоли часто возникает даже после удаления первичной опухоли, так как опухолевые клетки или компоненты могут оставаться и развивать метастатический потенциал. В одном варианте осуществления термин «метастазирование» в соответствии с изобретением относится к «отдаленным метастазам», которые относятся к метастазам, удаленным от первичной опухоли и региональной системы лимфатических узлов. В одном варианте осуществления термин «метастазы» в соответствии с изобретением относится к метастазам в лимфатические узлы. Одной конкретной формой метастазов, которая поддается лечению с использованием терапии по изобретению, являют метастазы, происходящее из рака желудка в качестве первичного участка. В предпочтительном варианте осуществления такие метастазы рака желудка представляют собой опухоль Крукенберга, перитонеальные метастазы и/или метастазы в лимфатические узлы.
Опухоль Крукенберга представляет собой редко встречающуюся метастатическую опухоль яичника, на которую приходится от 1% до 2% от всех опухолей яичника. Прогноз при опухоли Крукенберга до сих пор является очень плохим и не существует утвержденного лечения опухолей Крукенберга. Опухоль Крукенберга представляет собой метастатическую перстневидноклеточную аденокарциному яичника. Желудок является первичным участком для большинства случаев опухоли Крукенберга (70%). Карциномы толстой кишки, аппендицита и молочной железы (в основном инвазивная лобулярная карцинома) являются следующими наиболее распространенными первичными участками. Сообщалось о редких случаях опухоли Крукенберга, происходящей из карциномы желчного пузыря, желчного протока, поджелудочной железы, тонкого кишечника, фатерова соска, шейки матки и мочевого пузыря/мочевого протока.
Термин «лечить» означает введение соединения или композиции, или комбинации соединений или композиций субъекту для предупреждения или устранения заболевания, включая уменьшение размера опухоли или количества опухолей у субъекта; купирование или замедление развития заболевания у субъекта; ингибирование или замедление развития нового заболевания у субъекта; уменьшение частоты или тяжести симптомов и/или повторного возникновения у субъекта, который на текущий момент имеет или который ранее имел заболевание, и/или продление, то есть увеличение продолжительности жизни субъекта.
В частности, термин «лечение заболевания» включает излечение, сокращение продолжительности, ослабление, предупреждение, замедление или ингибирование прогрессирования или ухудшения, или предупреждение или задержку начала возникновения заболевания или его симптомов.
В контексте настоящего изобретения термины, такие как «защищать», «предупреждать», «превентивные» или «защитные» относятся к предупреждению или лечению, или и тому и другому, возникновения и/или распространения заболевания у субъекта и, в частности, сведению к минимуму вероятности того, что у субъекта будет развиваться заболевание, или задержке развития заболевания. Например, лицо, имеющее риск развития рака, будет являться кандидатом на лечение или предупреждение рака.
«Имеющий риск» означает субъекта, который идентифицирован как имеющий более высокую, чем нормальная, вероятность развития заболевания, в частности рака, по сравнению с общей популяцией. Кроме того, субъект, который имел или который на текущий момент имеет заболевание, в частности рак, представляет собой субъекта, который имеет повышенный риск развития заболевания, например, у субъекта может продолжать развиваться заболевание. Субъекты, которые на текущий момент имеют, или которые имели рак, также имеют повышенный риск метастазирования рака.
Термин «пациент» в соответствии с изобретением означает субъекта, подлежащего лечению, в частности больного субъекта, включая человека, приматов нечеловеческого происхождения или других животных, в частности млекопитающих, таких как коровы, лошади, свиньи, козы, собаки, кошки или грызуны, такие как мыши и крысы. В особенно предпочтительном варианте осуществления пациентом является человек.
«Клетка-мишень» означает любую нежелательную клетку, такую как раковая клетка. В предпочтительных вариантах осуществления клетка-мишень экспрессирует CLDN.
Термин «антиген» относится к агенту, такому как белок или пептид, содержащему эпитоп, против которого направлен и/или должен быть направлен иммунный ответ. В предпочтительном варианте осуществления антиген представляет собой опухоль-ассоциированный антиген, такой как CLDN18.2 или CLDN6, то есть компонент раковых клеток, который может происходить из цитоплазмы, клеточной поверхности и клеточных ядер, в частности, такие антигены, которые продуцируются, предпочтительно в большом количестве, внутриклеточно или как поверхностные антигены на раковых клетках.
В контексте настоящего изобретения термин «опухоль-ассоциированный антиген» предпочтительно относится к белкам, которые в нормальных условиях специфически экспрессируются в ограниченном числе тканей и/или органов, или на специфических стадиях в процессе развития, и экспрессируются или аберрантно экспрессируются в одной или более опухолей или раковых тканей. В контексте настоящего изобретения опухоль-ассоциированный антиген предпочтительно связан с клеточной поверхностью раковой клетки и предпочтительно не экспрессируется или только редко экспрессируется в нормальных тканях.
Термин «эпитоп» относится к антигенной детерминанте в молекуле, то есть части молекулы, которая распознается иммунной системой, например, которая распознается антителом. Например, эпитопы представляют собой дискретные, трехмерные участки на антигене, которые распознаются иммунной системой. Эпитопы обычно состоят из химически активных групп, расположенных на поверхности молекул, таких как боковые цепи аминокислот или сахаров, и обычно имеют специфические характеристики трехмерной структуры, а также специфические характеристики зарядов. Конформационные и неконформационные эпитопы отличаются тем, что связывание с первым, в отличие от связывания с последним, утрачивается в присутствии денатурирующих растворителей. Эпитоп белка предпочтительно содержит непрерывную или прерывистую часть указанного белка и, предпочтительно, составляет от 5 до 100, предпочтительно от 5 до 50, более предпочтительно от 8 до 30, наиболее предпочтительно от 10 до 25 аминокислот в длину, например, эпитоп может содержать предпочтительно 8, 9, 10, 11 , 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 , 22, 23, 24 или 25 аминокислот в длину.
Термин «антитело» относится к гликопротеину, содержащему, по меньшей мере, две тяжелые (Н) цепи и две легкие (L) цепи, связанные между собой дисульфидными связями. Термин «антитело» включает моноклональные антитела, рекомбинантные антитела, человеческие антитела, гуманизированные антитела и химерные антитела. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначаемой здесь как VH) и константной области тяжелой цепи. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначаемой здесь как VL) и константной области легкой цепи. Области VH и VL могут далее подразделяться на участки гипервариабельности, называемые участками, определяющими комплементарность (CDR), которые перемежаются с участками, являющимися более консервативными, называемыми каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, организованных в направлении от аминоконца к карбоксиконцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включающими различные клетки иммунной системы (например, эффекторные клетки), и первым компонентом (C1q) классической системы комплемента.
Термин «моноклональное антитело», используемый здесь, относится к препарату молекул антитела одной молекулярной композиции. Моноклональное антитело проявляет одну специфичность связывания и аффинность. В одном варианте осуществления моноклональные антитела продуцируются гибридомой, которая включает В-клетку, полученную от животного, отличного от человека, например, мыши, слитую с иммортализованной клеткой.
Термин «рекомбинантное антитело», используемый здесь, предусматривает включение всех антител, которые получены, экспрессированы, созданы или выделены рекомбинантными способами, таких как (а) антитела, выделенные у животного (например, мыши), которое является трансгенным или трансхромосомным по генам иммуноглобулина, или из гибридомы, полученной из него, (b) антитела, выделенные из клетки-хозяина, трансформированной таким образом, чтобы она экспрессировала антитело, например, из трансфектомы, (c) антитела, выделенные из библиотеки рекомбинантных комбинаторных антител, и (d) антитела, полученные, экспрессированные, созданные или выделенные любыми другими способами, которые включают сплайсинг последовательностей гена иммуноглобулина с другими последовательностями ДНК.
Термин «человеческое антитело», используемое здесь, предусматривает включение антител, имеющих вариабельные и константные области, образованные из последовательностей иммуноглобулинов зародышевой линии человека. Человеческие антитела могут включать аминокислотные остатки, не кодируемые последовательностями иммуноглобулинов зародышевой линии человека (например, мутации, внесенные случайным или сайт-специфическим мутагенезом in vitro, или посредством соматической мутации in vivo).
Термин «гуманизированное антитело» относится к молекуле, имеющей антигенсвязывающий сайт, который, по существу, происходит из иммуноглобулина от разновидностей нечеловеческого происхождения, и остальную структуру иммуноглобулина этой молекулы, основанную на структуре и/или последовательности иммуноглобулина человека. Этот антигенсвязывающий сайт может содержать либо полные вариабельные домены, слитые с константными доменами, либо только определяющие комплементарность области (CDR), пересаженные на подходящие каркасные области в вариабельных доменах. Антигенсвязывающие сайты могут быть сайтами дикого типа или модифицированными одной или несколькими аминокислотными заменами, например модифицированными для того, чтобы они имели более близкое сходство с иммуноглобулином человека. Некоторые формы гуманизированных антител сохраняют все CDR-последовательности (например, гуманизированное мышиное антитело, которое содержит все шесть CDR из мышиного антитела). Другие формы имеют одну или более CDR, которые изменены относительно исходного антитела.
Термин «химерное антитело» относится к таким антителам, в которых одна часть каждой из аминокислотных последовательностей тяжелых и легких цепей гомологична соответствующим последовательностям антител, полученных от определенных разновидностей или принадлежащих конкретному классу, тогда как остальной сегмент цепи гомологичен соответствующим последовательностям от других видов. Обычно вариабельная область и легкой, и тяжелой цепей имитирует вариабельные области антител, полученных от одного вида млекопитающих, тогда как константные области гомологичны последовательностям в антителах, полученных от других видов. Одно из явных преимуществ таких химерных форм состоит в том, что вариабельная область может быть легко получена из известных в настоящее время источников с использованием легко доступных В-клеток или гибридом организмов-хозяинов, не являющихся человеком, в комбинации с константными областями, полученными, например, из препаратов клеток человека. В то время как преимущество вариабельной области заключается в простоте получения и на специфичность не влияет ее источник, константная область, если она является человеческой, менее вероятно вызовет иммунный ответ у субъекта-человека при инъекции антител, чем в том случае, когда константная область получена от источника, отличного от человека. Однако определение не ограничено таким конкретным примером.
Антитела могут быть получены от различных видов, включая, но без ограничения, мышей, крыс, кроликов, морских свинок и человека.
Антитела, описанные здесь, включают антитела IgA, такие как IgAl или IgA2, IgGl, IgG2, IgG3, IgG4, IgE, IgM и IgD. В различных вариантах осуществления антитело представляет собой антитело IgGl, в частности, изотип IgGl каппа или лямбда IgGl (то есть. IgGl , κ, λ), антитело IgG2a (например, IgG2a, K, λ), антитело IgG2b (например, IgG2b, κ, λ), антитело IgG3 (например, IgG3, κ, λ) или антитело IgG4 (например, IgG4, κ, λ).
Используемый здесь термин «гетерологичное антитело» определяют относительно трансгенного организма, продуцирующего данное антитело. Этот термин относится к антителу, имеющему последовательность аминокислот или кодирующую последовательность нуклеиновой кислоты, соответствующую последовательностям, обнаруженным в организме, который не является трансгенным животным, и обычно выделенному у разновидностей, отличных от трансгенного животного.
Используемый здесь термин «гетерогибридное антитело» относится к антителу, имеющему легкую и тяжелую цепи организмов различного происхождения. Например, антитело, имеющее человеческую тяжелую цепь, ассоциированную с мышиной легкой цепью, представляет собой гетерогибридное антитело.
Антитела, описанные здесь, предпочтительно, являются выделенными. Термин «выделенное антитело», используемое здесь, относится к антителу, которое практически не содержит другие антитела, имеющих отличные антигенные специфичности (например, выделенное антитело, которое специфически связывается с CLDN18.2, практически не содержит антитела, которые специфически связываются с антигенами, отличными от CLDN18.2). Выделенное антитело, которое специфически связывается с эпитопом, изоформой или вариантом человеческого CLDN18.2, может, однако, обладать перекрестной реактивностью с другими родственными антигенами, например, от других видов (например, гомологи разновидностей CLDN18.2). Кроме того, выделенное антитело может практически не содержать другой клеточный материал и/или химические вещества. В одном варианте осуществления комбинация «выделенных» моноклональных антител относится к антителам, обладающим различными специфичностями, и объединенным в хорошо охарактеризованную композицию или смесь.
Термины «антигенсвязывающая область» антитела (или просто «связывающая область») или «антигенсвязывающий фрагмент» антитела (или просто «связывающий фрагмент»), или аналогичные термины относятся к одному или нескольким фрагментам антитела, которые сохраняют способность к специфическому связыванию с антигеном. Было показано, что антигенсвязывающая функция антитела может осуществляться фрагментами всей последовательности. К примерам связывающих фрагментов, включенных в термин «антигенсвязывающая область» антитела, относятся (i) фрагменты Fab, моновалентные фрагменты, состоящие из доменов VL, VH, CL и CH; (ii) фрагменты F(ab')2 , бивалентные фрагменты, состоящие из двух фрагментов Fab, связанных дисульфидным мостиком в шарнирной области; (iii) фрагменты Fd, состоящие из доменов VH и CH; (iv) фрагменты Fv, состоящие из доменов VL и VH отдельной области антитела; (v) фрагменты dAb (Ward et al., (1989) Nature 341: 544-546), которые состоят из домена VH; (vi) изолированные области, определяющие комплементарность (CDR), и (vii) комбинации двух или более изолированных CDR, которые могут быть необязательно соединены синтетическим линкером. Более того, несмотря на то, что два домена во фрагменте Fv, VL и VH, кодируются различными генами, их можно объединить, используя рекомбинантные технологии, посредством синтетического линкера, который позволяет построить из них единую белковую цепочку, в которой области VL и VH связываются с образованием моновалентных молекул (известных как одиночная цепь Fv (scFv); смотри, например, Bird et al. (1988) Science 242: 423-426; и Huston et al. (1988) Proc. Natl. Acad. Sci. USA 85: 5879-5883). Такие одноцепочечные антитела также предполагаются быть охваченными термином «антигенсвязывающий фрагмент» антитела. Дополнительным примером являются слитые белки связывающий домен-иммуноглобулин, содержащие (i) полипептид связывающего домена, который слит с полипептидом шарнирной области иммуноглобулина, (ii) константную область CH2 тяжелой цепи иммуноглобулина, слитую с шарнирной областью, и (iii) константную область CH3 тяжелой цепи иммуноглобулина, слитую с константной областью CH2. Полипептид связывающего домена может представлять собой вариабельную область тяжелой цепи или вариабельную область легкой цепи. Слитые белки связывающий домен-иммуноглобулин дополнительно раскрыты в US 2003/01 18592 и US 2003/0133939. Такие фрагменты антител получают с использованием общепринятых способов, известных специалистам в данной области, и эти фрагменты подвергают скринингу таким же образом, как и интактные антитела.
Термин «связывающий домен» характеризуется в контексте настоящего изобретения структурой, например, антитела, которая связывается/взаимодействует с заданной целевой структурой/антигеном/эпитопом. Таким образом, связывающий домен в соответствии с изобретением, означает «участок взаимодействия антигена».
Все антитела и производные антител, такие как фрагменты антител, описанные здесь, в целях изобретения обозначены термином «антитело». Термин «производные антитела» относится к любой модифицированной форме антитела, например, конъюгату антитела и другому агенту или антителу, или фрагменту антитела. Кроме того, антитела и производные антител, как описано здесь, являются пригодными для получения связывающих агентов по изобретению, таких как фрагменты антитела.
Антитела природного происхождения, как правило, являются моноспецифическими, то есть они связываются с одним антигеном. Настоящее изобретение обеспечивает связывающие агенты, связывающиеся с цитотоксическими клетками (путем связывания с CD3 рецептором) и раковыми клетками (путем связывания с CLDN). Связывающие агенты по настоящему изобретению являются, по меньшей мере, биспецифическими или мультиспецифическими, такими как триспецифические, тетраспецифические и т.д.
Связывающий агент по изобретению может быть в формате молекулы антитела или молекулы, подобной антителу, или белкового скелета со свойствами, подобными антителу, или циклического пептида, по меньшей мере, с двумя специфичностями связывания. Таким образом, связывающий агент может содержать одно или более антител, описанных здесь, или их фрагменты.
В соответствии с изобретением биспецифическая молекула, в частности, биспецифический белок, такой как биспецифическое антитело, представляет собой молекулу, которая содержит две различные специфичности связывания и, таким образом, может связываться с двумя различными типами антигена, такими как CLDN и CD3. В частности, термин «биспецифическое антитело», используемый здесь, относится к антителу, содержащему два антигенсвязывающих участка, первый участок связывания, обладающий аффинностью в отношении первого антигена или эпитопа, и второй участок связывания, обладающий аффинностью связывания в отношении второго антигена или эпитопа, отличного от первого. В частности, биспецифическое антитело представляет собой искусственный белок, который состоит из фрагментов двух различных антител (указанные фрагменты двух различных антител образуют два связывающих домена) и, таким образом, связывается с двумя различными типами антигена. Биспецифическое антитело в соответствии с изобретением сконструировано для одновременного связывания с клеткой иммунной системы, такой как иммунная эффекторная клетка, в частности, Т-клетка, такая как цитотоксическая клетка (путем связывания с CD3), и клеткой-мишенью, такой как раковая клетка (путем связывания с опухоль-ассоциированным антигеном CLDN), подлежащая уничтожению.
Термин «биспецифическое антитело» также включает диатела. Диатела являются бивалентными, биспецифическими антителами, в которых домены VH и VL экспрессируются на единственной полипептидной цепи, но использование линкера, который является слишком коротким для создания возможности спаривания между двумя доменами на одной и той же цепи, заставляет эти домены спариваться с комплементарными доменами другой цепи и создает два антигенсвязывающих сайта (смотри, например, Holliger, P., et al. (1993) Proc. Natl. Acad. Sci. USA 90: 6444-6448; Poljak, R. J., et al. (1994) Structure 2: 1121- 1123).
«Мультиспецифические связывающие агенты» представляют собой молекулы, которые содержат более двух различных специфичностей связывания.
Особенно предпочтительными в соответствии с изобретением являются биспецифические антитела, включая фрагменты биспецифических антител, в частности, биспецифические одноцепочечные антитела, включая фрагменты биспецифических одноцепочечных антител. Термин «биспецифическое одноцепочечное антитело» означает одну полипептидную цепь, содержащую два связывающих домена. В частности, термин «биспецифическое одноцепочечное антитело» или «одноцепочечное биспецифическое антитело», или родственные термины в соответствии с настоящим изобретением, предпочтительно, означает конструкции антитела, полученные в результате соединения, по меньшей мере, двух вариабельных областей антитела в одной полипептидной цепи, лишенной константной и/или Fc части(ей), присутствующей в полных иммуноглобулинах.
Например, биспецифическое одноцепочечное антитело может представлять собой конструкцию, содержащую всего две вариабельные области антитела, например, две VH-области, при этом каждая область способна к специфическому связыванию с отдельным антигеном и соединены друг с другом посредством короткого полипептидного спейсера, таким образом, что две вариабельные области антитела с их включенным спейсером существуют в виде одной непрерывной полипептидной цепи. Другим примером биспецифического одноцепочечного антитела может являться одна полипептидная цепь с тремя вариабельными областями антитела. В этом случае, две вариабельные области антитела, например, одна VH и одна VL, могут составлять scFv, при этом две вариабельные области антитела соединены друг с другом посредством синтетического полипептидного линкера, причем указанный линкер генетически сконструирован таким образом, чтобы быть минимально иммуногенным, при этом оставаясь максимально устойчивым к протеолизу. Такой scFv способен к специфическому связыванию с конкретным антигеном и соединен с вариабельной областью другого антитела, например, VH-областью, способной к связыванию с антигеном, отличным от антигена, связанного scFv. Еще другим примером биспецифического одноцепочечного антитела может являться единственная полипептидная цепь с четырьмя вариабельными областями антитела. В этом случае, первые две вариабельные области антитела, например, VH-область и VL-область, могут формировать один scFv, способный к связыванию с одним антигеном, тогда как вторая VH-область и VL-область могут формировать второй scFv, способный к связыванию с другим антигеном. В пределах одной непрерывной полипептидной цепи индивидуальные вариабельные области антитела одной специфичности могут быть успешно разделены синтетическим полипептидным линкером, тогда как соответствующие scFv могут быть успешно разделены коротким полипептидным спейсером, как описано выше.
В соответствии с одним вариантом осуществления изобретения первый связывающий домен биспецифического антитела содержит один вариабельный домен антитела, предпочтительно домен VHH. В соответствии с одним вариантом осуществления изобретения первый связывающий домен биспецифического антитела содержит два вариабельных домена антитела, предпочтительно scFv, то есть VH-VL или VL-VH. В соответствии с одним вариантом осуществления изобретения второй связывающий домен биспецифического антитела содержит один вариабельный домен антитела, предпочтительно домен VHH. В соответствии с одним вариантом осуществления изобретения второй связывающий домен биспецифического антитела содержит два вариабельных домена антитела, предпочтительно scFv, то есть VH-VL или VL-VH. В своей минимальной форме общее число вариабельных областей антитела в специфическом антителе в соответствии с изобретением равно, таким образом, только двум. Например, такое антитело может содержать два домена VH или два домена VHH.
В соответствии с одним вариантом осуществления изобретения первый связывающий домен и второй связывающий домен биспецифического антитела, каждый, содержит один вариабельный домен антитела, предпочтительно, домен VHH. В соответствии с одним вариантом осуществления изобретения первый связывающий домен и второй связывающий домен биспецифического антитела, каждый, содержат два вариабельных домена антитела, предпочтительно scFv, то есть VH-VL или VL-VH. В этом варианте осуществления связывающий агент по изобретению предпочтительно содержит (i) вариабельный домен тяжелой цепи (VH) антитела против CLDN, (ii) вариабельный домен легкой цепи (VL) антитела против CLDN, (iii) вариабельный домен тяжелой цепи (VH) антитела против CD3, и (iv) вариабельный домен легкой цепи (VL) антитела против CD3.
Биспецифические антитела полной длины могут быть получены путем ковалентного связывания двух моноклональных антител или с помощью стандартных методов на основе межвидовых гибридом. Ковалентное связывание двух моноклональных антител описано в Anderson, Blood 80 (1992), 2826-34. В контексте данного изобретения одно из антител является специфическим в отношении CLDN и другое в отношении CD3.
В одном варианте осуществления биспецифический связывающий агент находится в формате подобной антителу молекулы, при этом тяжелая цепь содержит два последовательных N-концевых вариабельных домена с различными специфичностями, и легкая цепь содержит два последовательных вариабельных домена с различными специфичностями, приводя к образованию четырех связывающих доменов с двумя различными специфичностями (Wu et al., Nat. Biotechnology, 2007, 25(11)), при этом одной специфичностью является CD3 и другой специфичностью является CLDN.
В предпочтительном варианте осуществления биспецифический связывающий агент по изобретению находится в формате фрагмента антитела.
В одном варианте осуществления биспецифические молекулы в соответствии с изобретением содержат две Fab области, одну, направленную против CLDN, и другую, направленную против CD3. В одном варианте осуществления молекула по изобретению представляет собой комплекс антигенсвязывающих фрагментов (Fab)2. Комплекс Fab2 состоит из двух Fab-фрагментов, при этом один Fab-фрагмент содержит домен Fv, то есть домены VH и VL, специфические в отношении антигена CD3, и другой Fab-фрагмент содержит домен Fv, специфический в отношении CLDN. Каждый из Fab-фрагментов может состоять из двух одиночных цепей, модуля VL-CL и модуля VH-CH. Альтернативно, каждый из индивидуальных Fab-фрагментов может быть составлен в одиночную цепь, предпочтительно VL-CL-CH-VH, и индивидуальные вариабельные и константные домены могут быть соединены с помощью пептидного линкера. В целом, индивидуальные одиночные цепи Fab-фрагментов могут быть соединены посредством дисульфидных связей, адгезивных доменов, связаны химически и/или посредством пептидного линкера. Биспецифическая молекула может также содержать более двух Fab-фрагментов, в частности, молекула может представлять собой Fab3, Fab4 или мультимерный комплекс Fab со специфичностью в отношении 2, 3, 4 или более различных антигенов. Изобретение также включает химически связанные Fab.
В одном варианте осуществления связывающий агент в соответствии с изобретением включает различные типы бивалентных и тривалентных одноцепочечных вариабельных фрагментов (scFv), слитые белки, имитирующие вариабельные домены двух антител. Одноцепочечный вариабельный фрагмент (scFv) представляет собой слитый белок, состоящий из вариабельных областей тяжелой (VH) и легкой цепей (VL) иммуноглобулинов, соединенных коротким линкерным пептидом, состоящим из примерно 10-25 аминокислот. Линкер обычно богат глицином для гибкости, а также серином или треонином для растворимости, и может соединять либо N-конец VH с C-концом VL, или наоборот. Дивалентные (бивалентные) одноцепочечные вариабельные фрагменты (di-scFv, bi-scFv) могут быть сконструированы путем связывания двух scFv. Это может быть сделано путем получения одиночной пептидной цепи с двумя областями VH и двумя областями VL с получением тандемных scFv. Изобретение также включает мультиспецифические молекулы, содержащие более двух scFv-связывающих доменов. Это обеспечивает то, что молекула обладает множеством специфичностей к антигену и является триспецифической, тетраспецифической или мультиспецифической молекулой, или молекула представляет собой биспецифическую молекулу, содержащую более одного связывающего домена scFv, со специфичностью в отношении одного и того же антигена. В частности, молекула по изобретению может представлять собой мультиспецифическую одноцепочечную молекулу Fv.
Другой возможностью является создание scFv с линкерными пептидами, которые являются лишком короткими для соединения двух вариабельных областей (примерно пять аминокислот), инициируя димеризацию scFv. Этот тип известен как диатела. Еще более короткие линкеры (одна или две аминокислоты) приводят к формированию тримеров, так называемых триател или трител. Тетратела также были получены. Они проявляют даже более высокую аффинность по отношению к своим мишеням, чем диатела.
Особенно предпочтительным примером фрагмента биспецифического антитела является диатело (Kipriyanov, Int. J. Cancer 77 (1998), 763-772), которое представляет собой небольшой фрагмент бивалентного и биспецифического антитела. Диатела содержат вариабельный домен тяжелой цепи (VH), соединенный с вариабельным доменом легкой цепи (VL) на одной и той же полипептидной цепи (VH-VL) за счет пептидного линкера, который имеет слишком малую длину, чтобы обеспечивать возможность спаривания между двумя доменами на одной и той же цепи. Это вызывает спаривание с комплементарными доменами другой цепи и способствует сборке димерной молекулы с двумя функциональными антигенсвязывающими участками. Для конструирования биспецифических диател по изобретению V-домены анти-CD3 антитела и анти-CLDN антитела могут быть слиты для создания двух цепей VH(CD3)-VL(CLDN), VH(CLDN)-VL(CD3). Каждая цепь сама по себе неспособна к связыванию с соответствующим антигеном, но воссоздает функциональные антигенсвязывающие участки анти-CD3 антитела и анти-CLDN антитела при спаривании с другой цепью. В этом случае используется пептидный линкер, который является слишком коротким для того, чтобы обеспечить возможность спаривания между двумя доменами на одной и той же цепи. Две молекулы scFv с линкером между вариабельным доменом тяжелой цепи и вариабельным доменом легкой цепи, который является слишком коротким для внутримолекулярной димеризации, соэкспрессируются и самособираются с формированием биспецифических молекул с двумя сайтами связывания на противоположных концах.
В одном варианте осуществления мультиспецифическая молекула в соответствии с изобретением содержит вариабельные (VH, VL) и константные (C) домены иммуноглобулинов. В одном варианте осуществления биспецифическая молекула представляет собой минитело, предпочтительно, минитело, содержащее две одиночные цепи VH-VL-C, которые соединены друг с другом посредством константных доменов (C) каждой цепи. Согласно этому аспекту соответствующие вариабельные области тяжелой цепи (VH), соответствующие вариабельные области легкой цепи (VL) и константные домены (C) расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-(C) и VH(CD3)-VL(CD3)-C, где C предпочтительно представляет собой домен CH3. Спаривание константных доменов приводит к образованию минитела.
В соответствии с другим, особенно предпочтительным аспектом, биспецифический связывающий агент по изобретению находится в формате конструкции биспецифического одноцепочечного антитела, при этом указанная конструкция содержит или состоит, по меньшей мере, из двух связывающих доменов, при этом один из указанных доменов связывается с CLDN и второй домен связывается с CD3. Такие молекулы, также называемые «bispecific Т cell engagers» (BiTE; термин BiTE относится только к биспецифическим молекулам, одно плечо которых является специфическим в отношении CD3), состоят из двух молекул scFv, соединенных посредством линкерного пептида.
Используемый здесь термин «биспецифическое одноцепочечное антитело» означает одиночную полипептидную цепь, содержащую два связывающих домена. Каждый связывающий домен содержит одну вариабельную область из тяжелой цепи антитела («область VH»), при этом область VH первого связывающего домена специфически связывается с CLDN, и область VH второго связывающего домена специфически связывается с CD3. Два связывающих домена необязательно связаны друг с другом коротким полипептидным спейсером. Неограничивающим примером полипептидного спейсера является Gly-Gly-Gly-Gly-Ser (G-G-G-G-S) и ее повторы. Каждый связывающий домен может дополнительно содержать одну вариабельную область из легкой цепи антитела («область VL»), при этом область VH и область VL в пределах каждого из первого и второго связывающих доменов связаны друг с другом посредством полипептидного линкера, имеющего достаточную длину для того, чтобы обеспечить возможность спаривания области VH и области VL первого связывающего домена с областью VH и областью VL второго связывающего домена.
В соответствии с этим аспектом соответствующие вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца к C-концу в порядке VH(CLDN)-VL(CLDN)-VH(CD3)-VL(CD3), VH(CD3)-VL(CD3)-VH(CLDN)-VL(CLDN) или VH(CD3)-VL(CD3)-VL(CLDN)-VH(CLDN). Однако предполагается, что биспецифические одноцепочечные антитела по изобретению содержат другие расположения доменов, такие как VL(CLDN)-VH(CLDN)-VH(CD3)-VL(CD3), VL(CLDN)-VH(CLDN)-VL(CD3)-VH(CD3), VH(CLDN)-VL(CLDN)-VL(CD3)-VH(CD3), VL(CD3)-VH(CD3)-VH(CLDN)-VL(CLDN), VL(CD3)-VH(CD3)-VL(CLDN)-VH(CLDN).
Длинный линкер обычно соединяет соответствующие вариабельные области тяжелой цепи (VL) и соответствующие вариабельные области легкой цепи (VL) с образованием связывающего scFv-домена, тогда как короткий линкер обычно соединяет два связывающих scFv-домена. Линкер обычно разработан для обеспечения гибкости и устойчивости к протеазе и, предпочтительно, линкер содержит аминокислотные остатки глицина и/или серина. Короткие последовательности пептидного линкера могут состоять из 12 или меньше, например, 11, 10, 9, 8, 7, 6, 5, 4, 3 или 2 аминокислот и, предпочтительно, 5 или 6 аминокислот. Короткие пептидные линкеры, предпочтительно, содержат аминокислотные последовательности SGGGGS или GGGGS. Длинные пептидные линкеры могут состоять из 12 или более, например, 15-25 или 15-20, или 15-18 аминокислот. Длинные пептидные линкеры, предпочтительно, содержат аминокислотные последовательности (GGGGS)3 или VE(GGSGGS)2GGVD. Дополнительные длинные пептидные линкеры могут содержать аминокислотные последовательности (GGGGS)4, (GGGGS)5 или GGGGS(GGS)3GGGS.
Связывающие агенты в соответствии с изобретением могут также содержать аминокислотную последовательность для содействия секреции молекулы, например, N-концевого сигнала секреции, и/или одну или более эпитопных меток, содействующих связыванию, очистке или детекции молекулы.
Предпочтительно, сигнал секреции представляет собой сигнальную последовательность (например, выбранную из любой из SEQ ID NO: 51 , 52, 53, 54, 55), которая обеспечивает достаточное прохождение по секреторному пути и/или секрецию связывающего агента во внеклеточное пространство. Предпочтительно, последовательность сигнала секреции является расщепляемой и удаляется из зрелого связывающего агента. Последовательность сигнала секреции, предпочтительно, выбирается в соответствии с клеткой или организмом, в котором связывающий агент продуцируется.
Аминокислотная последовательность эпитопной метки может быть введена в любое положение в пределах аминокислотной последовательности связывающего агента, и может принимать форму петли в пределах кодированной белковой структуры, или может быть слита на N-концах или C-концах со связывающим агентом. Предпочтительно, эпитопная метка слита на С-конце со связывающим агентом. Эпитопная метка может содержать сайт расщепления, который обеспечивает возможность удаление метки из связывающего агента. Указанная эпитопная метка может представлять собой любой вид эпитопной метки, который является функциональным в нативных и/или денатурирующих условиях, предпочтительно, гистидиновую метку, наиболее предпочтительно, метку, содержащую шесть остатков гистидина.
Биспецифический связывающий агент по изобретению может содержать дополнительно к указанному первому и второму связывающим доменам, дополнительный связывающий домен, который служит, например, для повышения селективности в отношении опухолевых клеток. Это может быть достигнуто, например, путем обеспечения связывающих доменов, которые связываются с другими антигенами, экспрессирующимися на опухолевых клетках.
В контексте настоящего изобретения, созданные связывающие агенты, предпочтительно, способны вызывать иммунные эффекторные функции, описанные здесь. Предпочтительно, указанные иммунные эффекторные функции направлены против клеток, несущих опухоль-ассоциированный антиген CLDN на своей поверхности.
Термин «иммунные эффекторные функции» в контексте настоящего изобретения включают любые функции, опосредованные компонентами иммунной системы, которые приводят, например, к ингибированию роста опухоли и/или ингибированию развития опухоли, включая ингибирование распространения опухоли и метастазов. Предпочтительно, иммунные эффекторные функции приводят к гибели опухолевых клеток. Такие функции включают комплемент-зависимую цитотоксичность (CDC), антитело-зависимую клеточную цитотоксичность (ADCC), антитело-зависимый клеточно-опосредованный фагоцитоз (ADCP), индукцию апоптоза в клетках, несущих опухоль-ассоциированный антиген, цитолиз клеток, несущих опухоль-ассоциированный антиген, и/или ингибирование пролиферации клеток, несущих опухоль-ассоциированный антиген. Связывающие агенты могут также проявлять эффект просто путем связывания с опухоль-ассоциированными антигенами на поверхности раковой клетки. Например, антитела могут блокировать функцию опухоль-ассоциированного антигена или индуцировать апоптоз лишь путем связывания с опухоль-ассоциированным антигеном на поверхности раковой клетки.
Связывающие агенты, описанные здесь, могут быть конъюгированы с терапевтическим фрагментом или агентом, таким как цитотоксин, лекарственным средством (например, иммуносупрессантом) или радиоактивным изотопом. Цитотоксин или цитотоксический агент включает любой агент, который является губительным для клеток и, в частности, убивает клетки. Примеры включают таксол, цитохалазин В, грамицидин D, этидиум бромид, эметин, митомицин, этопозид, тенопозид, винкристин, винбластин, колхицин, доксорубицин, даунорубицин, дигидроксиантрациндион, митоксантрон, митрамицин, актиномицин D, 1-дегидротестостерон, глюкокортикоиды, прокаин, татракаин, лидокаин, пропранолол и пуромицин и их аналоги или гомологи. Подходящие терапевтические агенты для формирования конъюгатов включают, но без ограничения, антиметаболиты (например, метотрексат, 6-меркаптопурин, 6-тиогуанин, цитарабин, флударабин, 5-фторурацил, декарбазин), алкилирующие агенты (например, мехлорэтамин, тиоепа хлорамбуцил, мелфалан, кармустин (BSNU) и ломустин (CCNU), циклофосфамид, бусульфан, дибромманитол, стрептозотоцин, митомицин С и цис-дихлордиамин платины (II) (DDP) цисплатин), антрациклины (например, даунорубицин (исходное название дауномицин) и доксорубицин), антибиотики (например, дактиномицин (исходное название актиномицин), блеомицин, митрамицин и антрамицин (AMC), и антимитотические агенты (например, винкристин и винбластин). В предпочтительном варианте осуществления терапевтический агент представляет собой цитотоксический агент или радиотоксический агент. В другом варианте осуществления терапевтический агент представляет собой иммуносуппрессант. В еще другом варианте осуществления терапевтический агент представляет собой GM-CSF. В предпочтительном варианте осуществления терапевтический агент представляет собой доксорубицин, цисплатин, блеомицин, сульфат, кармустин, хлорамбуцил, циклофорамид или рицин А.
Также, связывающие агенты могут быть конъюгированы с радиоактивными изотопами, например, йодом-131, иттрием-90 или индием-111 для создания цитотоксических радиофармацевтических средств.
Методики конъюгации таких терапевтических фрагментов с антителами хорошо известны, смотри, например, Arnon et al., «Monoclonal Antibodies For Immunotargeting Of Drugs In Cancer Therapy» в Monoclonal Antibodies And Cancer Therapy, Reisfeld et al. (eds.), pp. 243-56 (Alan R. Liss, Inc. 1985); Hellstrom et al., «Antibodies For Drug Delivery» в Controlled Drug Delivery (2nd Ed.), Robinson et al. (eds.), pp. 623-53 (Marcel Dekker, Inc. 1987); Thorpe, «Antibody Carriers Of Cytotoxic Agents In Cancer Therapy: A Review» в Monoclonal Antibodies '84: Biological And Clinical Applications, Pincheraet al. (eds. ), pp. 475-506 (1985); «Analysis, Results, And Future Prospective Of The Therapeutic Use Of Radiolabeled Antibody In Cancer Therapy» в Monoclonal Antibodies For Cancer Detection And Therapy, Baldwin et al. (eds.), pp. 303-16 (Academic Press 1985), и Thorpe et al., «The Preparation And Cytotoxic Properties Of Antibody-Toxin Conjugates», Immunol. Rev., 62: 119-58 (1982).
Термин «связывание» в соответствии с изобретением предпочтительно относится к специфическому связыванию.
В соответствии с настоящим изобретением агент, такой как антитело, способен связываться с предварительно определенной мишенью, если он обладает значительной аффинностью в отношении указанной предварительно определенной мишени и связывается с указанной предварительно определенной мишенью в стандартных анализах. «Аффинность» или «аффинность связывания» часто измеряется константой диссоциации (KD). Предпочтительно, термин «значительная аффинность» относится к связыванию с предварительно определенной мишенью с константой диссоциации (KD), равной 10-5 M или ниже, 10-6 M или ниже, 10-7 M или ниже, 10-8 M или ниже, 10-9 M или ниже, 10-10 M или ниже, 10-11 M или ниже, или 10-12 M или ниже.
Агент не способен (значительно) связываться с мишенью, если он не обладает значительной аффинностью в отношении указанной мишени и не связывается в значительной степени, в частности, не связывается детектируемым образом с указанной мишенью в стандартных анализах. Предпочтительно, агент не связывается детектируемым образом с указанной мишенью в случае присутствия при концентрации, составляющей до 2, предпочтительно 10, более предпочтительно 20, в особенности 50 или 100 мг/мл или выше. Предпочтительно, агент не обладает значительной аффинностью в отношении мишени, если он связывается с указанной мишенью с KD, которая, по меньшей мере, в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз или 106 раз выше, чем KD для связывания с предварительно определенной мишенью, с которой агент способен связываться. Например, если KD для связывания агента с мишенью, с которой агент способен связываться, равна 10-7 M, KD для связывания с мишенью, в отношении которой агент не обладает значительной аффинностью, будет составлять, по меньшей мере, 10-6 M, 10-5 M, 10-4 M, 10-3 M, 10-2 M или 10-1 M.
Агент, такой как антитело, является специфическим в отношении предварительно определенной мишени, если он способен связываться с указанной предварительно определенной мишенью, при этом он не способен связываться с другими мишенями, то есть не обладает значительной аффинностью в отношении других мишеней и значительно не связывается с другими мишенями в стандартных анализах. В соответствии с изобретением агент является специфическим в отношении CLDN, если он способен связываться с CLDN, но не способен (значительно) связываться с другими мишенями. Предпочтительно, агент является специфическим в отношении CLDN, если аффинность в отношении таких других мишеней и связывание с такими другими мишенями значительно не превышает аффинность в отношении неродственных CLDN белков или связывание с неродственными CLDN белками, такими как бычий сывороточный альбумин (BSA), казеин, человеческий сывороточный альбумин (HSA), или не относящимися к клаудину трансмембранными белками, такими как молекулы MHC или рецептор трансферрина, или любым другим указанным полипептидом. Предпочтительно, агент является специфическим в отношении предварительно определенной мишени, если он связывается с указанной мишенью с KD, которая, по меньшей мере, в 10 раз, 100 раз, 103 раз, 104 раз, 105 раз, или 106 раз ниже, чем KD для связывания с мишенью, для которой он не является специфическим. Например, если KD для связывания агента с мишенью, в отношении которой он является специфическим, равна 10-7 M, KD для связывания с мишенью, в отношении которой он не является специфическим, будет равна, по меньшей мере, 10-6 M, 10-5 M, 10-4 M, 10-3 M, 10-2 M или 10-1 M.
Связывание агента с мишенью может быть определено экспериментально с использованием любого подходящего способа; смотри, например, Berzofsky et al., «Antibody-Antigen Interactions» в Fundamental Immunology, Paul, W. E., Ed., Raven Press New York, N Y (1984), Kuby, Janis Immunology, W. H. Freeman and Company New York, N Y (1992), и способы, описанные в настоящем документе. Аффинность может быть легко определена стандартными методами, такими как равновесный диализ; анализ с использованием устройства BIAcore 2000 в соответствии с общими процедурами, описанными производителем; радиоиммуноанализ с использованием радиоактивно меченого целевого антигена; или другим методом, известным специалистам в данной области. Данные аффинности могут быть проанализированы, например, методом Scatchard et al., Ann N.Y. Acad. ScL, 51:660 (1949). Измеренная аффинность конкретного антитело-антиген взаимодействия может изменяться при измерении в различных условиях, например, концентрации соли, рН. Таким образом, измерения аффинности и других антигенсвязывающих параметров, например, KD, IC50, предпочтительно проводят в стандартизированных растворах антитела и антигена, и стандартизированном буфере.
Используемый здесь термин «изотип» относится к классу антител (например, IgM или IgGl), кодируемых генами константной области тяжелой цепи.
Используемый здесь термин «переключение изотипа» относится к феномену, вследствие которого класс или изотип антитела переходит из одного класса в один из других классов Ig.
Термин «природный», используемый здесь, применяют к объекту для обозначения того факта, что этот объект может быть обнаружен в естественных условиях. Например, полипептидная или полинуклеотидная последовательность, которая присутствует в организме (включая вирусы), которая может быть выделена из природного источника и которая не была преднамеренно модифицирована человеком в лабораторных условиях, является природной.
Термин «реаранжированный», используемый здесь, относится к конфигурации локуса тяжелой цепи или легкой цепи иммуноглобулина, где положение сегмента V находится в непосредственной близости к сегменту D-J или J в конформации, кодирующей в основном полный домен VH или VL, соответственно. Реаранжированный локус гена иммуноглобулина (антитела) может быть идентифицирован путем сравнения с ДНК зародышевой линии; реаранжированный локус будет содержать, по меньшей мере, один подвергнувшийся рекомбинации гептамерный/нонамерный гомологичный элемент.
Термин «нереаранжированный» или «конфигурация зародышевой линии», используемый здесь в отношении сегмента V, относится к конфигурации, в которой сегмент V не подвергается рекомбинации, при которой он находился бы в непосредственной близости к сегменту D или J.
В одном варианте осуществления связывающий агент по изобретению обладает способностью связываться с CLDN18.2, то есть способностью связываться с эпитопом, присутствующим в CLDN18.2, предпочтительно, эпитопом, расположенным в пределах внеклеточных доменов CLDN18.2, в особенности, первой внеклеточной петле, предпочтительно, в положениях аминокислот с 29 по 78 CLDN18.2. В конкретных вариантах осуществления агент, обладающий способностью связываться с CLDN18.2, связывается с эпитопом на CLDN18.2, который не присутствует на CLDN18.1.
Агент, обладающий способностью связываться с CLDN18.2, предпочтительно связываются с CLDN18.2, но не с CLDN18.1. Предпочтительно, агент, обладающий способностью связываться с CLDN18.2, является специфическим в отношении CLDN18.2. Предпочтительно, агент, обладающий способностью связываться с CLDN18.2, связывается с CLDN1 18.2, экспрессирующимся на клеточной поверхности. В особенно предпочтительных вариантах осуществления агент, обладающий способностью связываться с CLDN18.2, связывается с нативными эпитопами CLDN18.2, присутствующими на поверхности живых клеток.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN18.2, содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 5, 6, 7, 8, 9, 10 и их фрагментов.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN18.2, содержит вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 1 1 , 12, 13, 14, 15, 16, 17, 18, 19 и их фрагментов.
В определенных предпочтительных вариантах осуществления агент, обладающий способностью связываться с CLDN18.2, содержит комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), выбранную из следующих возможных вариантов с (i) по (ix):
(i) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 5, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 12, или ее фрагмент;
(ii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 11, или ее фрагмент;
(iii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 7, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 13, или ее фрагмент;
(iv) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 9, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 16, или ее фрагмент;
(v) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 8, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 15, или ее фрагмент;
(vi) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 10, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 14, или ее фрагмент;
(vii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 10, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 17, или ее фрагмент;
(viii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 10, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 18, или ее фрагмент;
(ix) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 10, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 19, или ее фрагмент.
В особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN18.2, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 8, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 15, или ее фрагмент.
В следующем, особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN18.2, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 6, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 11, или ее фрагмент.
Термин «фрагмент» относится, в частности, к одной или нескольким определяющим комплементарность областям (CDR), предпочтительно, по меньшей мере, вариабельной области CDR3, вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL). В одном варианте осуществления указанная одна или несколько определяющих комплементарность областей (CDR) выбраны из группы определяющих комплементарность областей CDR1, CDR2 и CDR3. В особенно предпочтительном варианте осуществления термин «фрагмент» относится к определяющим комплементарность областям CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL).
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR вместе с расположенными между ними каркасными областями. Предпочтительно, часть будет также содержать, по меньшей мере, примерно 50% одной из двух или обеих первой и четвертой каркасных областей, при этом 50% представляют собой С-концевые 50% первой каркасной области и N-концевые 50% четвертой каркасной области. Конструирование связывающих агентов, осуществляемое способами на основе рекомбинантной ДНК, может приводить к введению N- или C-концевых остатков в вариабельные области, кодированных линкерами, введенными для того, чтобы облегчить клонирование или другие стадии обработки, включая введение линкеров, чтобы соединить вариабельные области согласно изобретению с дополнительными последовательностями белка, включая тяжелые цепи иммуноглобулина, другие вариабельные домены (например, при получении диател) или белковые метки.
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR в каркасной области человеческого антитела.
В одном варианте осуществления связывающий агент по изобретению обладает способностью связываться с CLDN6, то есть способностью связываться с эпитопом, присутствующим в CLDN6, предпочтительно эпитопом, расположенным в пределах внеклеточных доменов CLDN6, в частности, в первой внеклеточной петле, предпочтительно в положениях аминокислот с 28 по 76 CLDN6, или во второй внеклеточной петле, предпочтительно в положениях аминокислот с 141 по 159 CLDN6. В конкретных вариантах осуществления агент, обладающий способностью связываться с CLDN6, связывается с эпитопом на CLDN6, который не присутствует на CLDN9. Предпочтительно, агент, обладающий способностью связываться с CLDN6, связывается с эпитопом на CLDN6, который не присутствует на CLDN4 и/или CLDN3. Наиболее предпочтительно, агент, обладающий способностью связываться с CLDN6, связывается с эпитопом на CLDN6, который не присутствует на белке CLDN, отличном от CLDN6.
Агент, обладающий способностью связываться с CLDN6, предпочтительно связывается с CLDN6, но не с CLDN9, и предпочтительно не связывается с CLDN4 и/или CLDN3. Предпочтительно, агент, обладающий способностью связываться с CLDN6, является специфическим в отношении CLDN6. Предпочтительно, агент, обладающий способностью связываться с CLDN6, связывается с CLDN6, экспрессирующимся на клеточной поверхности. В особенно предпочтительных вариантах осуществления агент, обладающий способностью связываться с CLDN6, связывается с нативными эпитопами CLDN6, присутствующими на поверхности живых клеток.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 20, 22, 24, 26 и их фрагментов.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 21 , 23, 25, 27, 28, 29, 97, 98, 99, 100 и их фрагментов.
В определенных предпочтительных вариантах осуществления агент, обладающий способностью связываться с CLDN6, содержит комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), выбранную из следующих возможных вариантов от (i) до (xi):
(i) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 20, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 21, или ее фрагмент;
(ii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 23, или ее фрагмент;
(iii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 24, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 25, или ее фрагмент;
(iv) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 26, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 27, или ее фрагмент;
(v) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 21, или ее фрагмент;
(vi) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 28, или ее фрагмент;
(vii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 29, или ее фрагмент;
(viii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 97, или ее фрагмент;
(ix) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 98, или ее фрагмент;
(x) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 99, или ее фрагмент;
(xi) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 100, или ее фрагмент.
В особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельную область легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную SEQ ID NO: 22 или ее фрагмент и VL содержит аминокислотную последовательность, представленную SEQ ID NO: 23 или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 97, или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 98, или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 99, или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CLDN6, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 22, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 100, или ее фрагмент.
Термин «фрагмент» относится, в частности, к одной или нескольким определяющим комплементарность областям (CDR), предпочтительно, по меньшей мере, вариабельной области CDR3, вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL). В одном варианте осуществления указанная одна или несколько определяющих комплементарность областей (CDR) выбраны из набора определяющих комплементарность областей CDR1, CDR2 и CDR3. В особенно предпочтительном варианте осуществления термин «фрагмент» относится к определяющим комплементарность областям CDR1, CDR2 и CDR3 вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL).
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR вместе с расположенными между ними каркасными областями. Предпочтительно, часть будет также содержать, по меньшей мере, примерно 50% одной из двух или обеих первой и четвертой каркасных областей, при этом 50% представляют собой С-концевые 50% первой каркасной области и N-концевые 50% четвертой каркасной области. Конструирование связывающих агентов, осуществляемое способами на основе рекомбинантной ДНК, может приводить к введению N- или C-концевых остатков в вариабельные области, кодированных линкерами, введенными для того, чтобы облегчить клонирование или другие стадии обработки, включая введение линкеров, чтобы соединить вариабельные области согласно изобретению с дополнительными последовательностями белка, включая тяжелые цепи иммуноглобулина, другие вариабельные домены (например, при получении диател) или белковые метки..
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR в каркасной области человеческого антитела.
Анти-CD3 антитела, которые являются подходящими для обеспечения связывающих агентов в соответствии с изобретением, включают, но без ограничения, UCHT1-HS (гуманизированное mAB), UCHT1-MM (мышиное mAB), CLB-T3, TR66, 145-2C11.
UCHT1 представляет собой моноклональное IgG1 анти-CD3 моноклональное антитело, которое детектирует CD3 у человека и приматов. CLB-T3 представляет собой мышиное моноклональное анти-CD3 антитело, которое направлено против антигена CD3 и взаимодействует с 80-90% человеческих периферических Т-лимфоцитов и медуллярных тимоцитов. TR66 представляет собой мышиное IgG1 моноклональное анти-CD3 антитело, которое распознает эпсилон-цепь человеческого CD3. 145-2C11 представляет собой моноклональное антитело серого хомячка против мышиного CD3.
Предпочтительно, области VH и VL связывающего домена CD3 получены из молекул антител/антитела и подобных антителу молекул, которые способны специфически распознавать человеческий CD3 в контексте других субъединиц TCR, которые присутствуют на активированных первичных человеческих Т-клетках, экспрессирующих TCR в своей нативной конфигурации. Области VH и VL, полученные из антитела, специфического в отношении CD3-эпсилон цепи, являются наиболее предпочтительными, и указанные (родительские) антитела должны быть способны специфически связывать эпитопы, отражающие нативную или близкую к нативной структуру, или конформационный эпитоп человеческого CD3, представленный в контексте комплекса TCR. В предпочтительном варианте осуществления изобретения области VH и VL связывающего домена CD3 получены из CD3-специфического антитела, выбранного из группы, состоящей из UCHT1-HS, UCHT1-MM, CLB-T3 и TR66, предпочтительно, TR66.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CD3, содержит вариабельную область тяжелой цепи (VH), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 30, 32, 34, 36, 94, 95 и их фрагментов.
В предпочтительном варианте осуществления агент, обладающий способностью связываться с CD3, содержит вариабельную область легкой цепи (VL), содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 31, 33, 35, 37, 96 и их фрагментов.
В определенных предпочтительных вариантах осуществления агент, обладающий способностью связываться с CD3, содержит комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL), выбранную из следующих возможных вариантов с (i) по (ix):
(i) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 30, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 31, или ее фрагмент;
(ii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 32, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 33, или ее фрагмент;
(iii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 34, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 34, или ее фрагмент;
(iv) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 36, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 37, или ее фрагмент;
(v) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 94, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 37, или ее фрагмент;
(vi) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 95, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 37, или ее фрагмент;
(vii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 36, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 96, или ее фрагмент;
(viii) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 94, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 96, или ее фрагмент;
(ix) VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 95, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 96, или ее фрагмент.
В особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CD3, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 36, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 37, или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CD3, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 94, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 37, или ее фрагмент.
В дополнительном особенно предпочтительном варианте осуществления агент, обладающий способностью связываться с CD3, содержит следующую комбинацию вариабельной области тяжелой цепи (VH) и вариабельной области легкой цепи (VL):
VH содержит аминокислотную последовательность, представленную в SEQ ID NO: 95, или ее фрагмент, и VL содержит аминокислотную последовательность, представленную в SEQ ID NO: 96, или ее фрагмент.
Термин «фрагмент» относится, в частности, к одной или нескольким определяющим комплементарность областям (CDR), предпочтительно, по меньшей мере, вариабельной области CDR3, вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL). В одном варианте осуществления указанная одна или несколько из определяющих комплементарность областей (CDR) выбраны из набора определяющих комплементарность областей CDR1, CDR2 и CDR3. В особенно предпочтительном варианте осуществления термин «фрагмент» относится к определяющим комплементарность областям CDR1 , CDR2 и CDR3 вариабельной области тяжелой цепи (VH) и/или вариабельной области легкой цепи (VL).
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR вместе с расположенными между ними каркасными областями. Предпочтительно, часть будет также содержать, по меньшей мере, примерно 50% одной из двух или обеих первой и четвертой каркасных областей, при этом 50% представляют собой С-концевые 50% первой каркасной области и N-концевые 50% четвертой каркасной области. Конструирование связывающих агентов, осуществляемое способами на основе рекомбинантной ДНК, может приводить к введению N- или C-концевых остатков в вариабельные области, кодированных линкерами, введенными для того, чтобы облегчить клонирование или другие стадии обработки, включая введение линкеров, чтобы соединить вариабельные области согласно изобретению с дополнительными последовательностями белка, включая тяжелые цепи иммуноглобулина, другие вариабельные домены (например, при получении диател) или белковые метки.
В одном варианте осуществления связывающий агент, содержащий одну или несколько CDR, набор CDR или комбинацию наборов CDR, как описано здесь, содержит указанные CDR в человеческой каркасной области.
В соответствии с изобретением предпочтительный связывающий агент, направленно воздействующий на CLDN18.2, содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 38, 39, 40 и 41 или их вариантов.
В соответствии с изобретением следующий предпочтительный связывающий агент, направленно воздействующий на CLDN18.2, содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 103, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92 и 93, или их фрагментов или вариантов. В одном варианте осуществления указанная аминокислотная последовательность лишена сигналов секреции, таких как N-концевые сигналы секреции, в частности, последовательности согласно SEQ ID NO: 51, и/или лишена His-меток, таких как C-концевые His-метки, в частности, последовательности Gly-Gly-Ser-(His)6 или (His)6, в случае присутствия.
В соответствии с изобретением предпочтительный связывающий агент, направленно воздействующий на CLDN6, содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 42, 43, 44 и 45 или их вариантов.
В соответствии с изобретением следующий предпочтительный связывающий агент, направленно воздействующий на CLDN6, содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 101, 102, 60, 61, 62, 63, 64 и 65, или их фрагментов или вариантов. В одном варианте осуществления указанная аминокислотная последовательность лишена сигналов секреции, таких как N-концевые сигналы секреции, в частности, последовательности согласно SEQ ID NO: 51, и/или лишена His-меток, таких как C-концевые His-метки, в частности, последовательности Gly-Gly-Ser-(His)6 или (His)6, в случае присутствия.
Следует понимать, что связывающие агенты, описанные здесь, могут быть доставлены пациенту путем введения нуклеиновой кислоты, такой как РНК, кодирующей агент и/или путем введения клетки-хозяина, содержащей нуклеиновую кислоту, такую как РНК, кодирующую агент. Таким образом, нуклеиновая кислота, кодирующая связывающий агент, при введении пациенту, может присутствовать в свободной форме или в подходящем носителе для доставки, например, в форме липосом или вирусных частиц, или в пределах клетки-хозяина. Обеспеченная нуклеиновая кислота может продуцировать агент в течение продолжительных периодов времени пролонгированным образом, подавляя неустойчивость, по меньшей мере частично, наблюдаемую в отношении терапевтических антител, в особенности биспецифических антител. Нуклеиновые кислоты, подлежащие доставке пациенту, могут быть получены с помощью рекомбинантных способов. Если нуклеиновую кислоту вводят пациенту при присутствии в клетке-хозяине, она предпочтительно поглощается клетками пациента для экспрессии связывающего агента, кодированного нуклеиновой кислотой. Если нуклеиновую кислоту вводят пациенту в случае присутствия в клетке-хозяине, она предпочтительно экспрессируется клеткой-хозяином в организме пациента таким образом, чтобы продуцировать связывающий агент, кодированный нуклеиновой кислотой.
Термин «рекомбинантный» в контексте настоящего изобретения означает «изготовленный методом генной инженерии» Предпочтительно, «рекомбинантный объект», такой как рекомбинантная нуклеиновая кислота, в контексте настоящего изобретения не имеет природного происхождения.
Термин «природный», используемый здесь, относится к тому факту, что объект может быть обнаружен в естественных условиях. Например, пептид или нуклеиновая кислота, которая присутствует в организме (включая вирусы), и которая может быть выделена из природного источника, и не была преднамеренно модифицирована человеком в лабораторных условиях, является природной.
Термин «нуклеиновая кислота», используемый здесь, означает включение ДНК и РНК, такую как геномная ДНК, кДНК, мРНК, рекомбинантно полученные и химически синтезированные молекулы. Нуклеиновая кислота может быть однонитевой или двунитевой. РНК включает транскрибированную in vitro РНК (IVT RNA) или синтетическую РНК.
Нуклеиновые кислоты могут быть включены в вектор. Термин «вектор», используемый здесь, включает любые векторы, известные специалисту в данной области, включая плазмидные векторы, козмидные векторы, фаговые векторы, такие как фаг лямбда, вирусные векторы, такие как аденовирусные или бакуловирусные векторы, или векторы на основе искусственных хромосом, таких как бактериальные искусственные хромосомы (BAC), искусственные хромосомы дрожжей (YAC), или искусственные хромосомы на основе PI (PAC). Указанные векторы включают экспрессионные, а также клонирующие векторы. Экспрессионные векторы содержат плазмиды, а также вирусные векторы, и обычно содержат желательную кодирующую последовательность и соответствующие последовательности ДНК, необходимые для экспрессии функционально связанной кодирующей последовательности в конкретном хозяине-организме (например, бактерии, дрожжи, растение, насекомое или млекопитающее) или в экспрессионных системах in vitro. Клонирующие векторы обычно используются для создания и амплификации определенных желательных фрагментов ДНК и могут быть лишены функциональных последовательностей, необходимых для экспрессии желательных фрагментов ДНК.
В контексте настоящего изобретения термин «РНК» относится к молекуле, которая содержит рибонуклеотидные остатки и, предпочтительно, полностью или по существу состоит из рибонуклеоридных остатков. «Рибонуклеотид» относится к нуклеотиду, содержащему гидроксильную группу в 2'-положении β-D-рибофуранозильной группы. Термин включает двухцепочечную РНК, одноцепочечную РНК, выделенную РНК, такую как частично очищенная РНК, по существу чистая РНК, синтетическую РНК, полученную рекомбинантными методами РНК, а также модифицированную РНК, которая отличается от природной РНК за счет внесения в нее добавки, делеции, замещения и/или изменения одного или нескольких нуклеотидов. Такие изменения могут включать добавление материала ненуклеотидной природы, такие как добавки, содержащиеся на одном или обоих концах молекулы РНК или введенные внутрь молекулы, например, локализованные в одном или нескольких нуклеотидах в составе РНК. Нуклеотиды в составе молекул РНК могут также включать нестандартные нуклеотиды, такие как нуклеотиды неприродного происхождения, или химически синтезированные нуклеотиды, или дезоксинуклеотиды. Указанные измененные РНК могут рассматриваться как аналоги или аналоги природных РНК.
В соответствии с настоящим изобретением термин «РНК» включает и предпочтительно относится к «мРНК», что означает «мессенджер РНК» и относится к «транскрипту», который может быть получен с использованием ДНК в качестве матрицы и кодирует пептид или белок. мРНК обычно содержит 5' нетранслируемую область (5'-UTR), область, кодирующую белок или пептид, и 3' нетранслируемую область (3'-UTR). мРНК имеет ограниченное время полужизни в клетках и in vitro. Предпочтительно, мРНК получают путем in vitro транскрипции с использованием матрицы ДНК. В одном варианте осуществления РНК получена путем in vitro транскрипции или химического синтеза. Метод транскрипции in vitro известен для специалистов в данной области. Например, существуют различные коммерчески доступные наборы для vitro транскрипции.
В одном варианте осуществления настоящего изобретения РНК представляет собой самореплицирующуюся РНК, такую как самореплицирующаяся одноцепочечная РНК. В одном варианте осуществления самореплицирующаяся РНК представляет собой одноцепочечную положительно-полярную РНК. В одном варианте осуществления самореплицирующаяся РНК представляет собой вирусную РНК или РНК, полученную из вирусной РНК. В одном варианте осуществления самореплицирующаяся РНК представляет собой альфавирусную геномную РНК или РНК, полученную из геномов альфавирусов. В одном варианте осуществления самореплицирующаяся РНК представляет собой экспрессионный вектор на основе вирусных генов. В одном варианте осуществления вирус представляет собой вирус леса Семлики. В одном варианте осуществления самореплицирующаяся РНК содержит один или несколько трансгенов, при этом, по меньшей мере, один из указанных трансгенов кодирует связывающий агент, описанный здесь. В одном варианте осуществления, если РНК представляет собой вирусную РНК или получена из вирусной РНК, трансгены могут частично или полностью заменять вирусные последовательности, такие как вирусные последовательности, кодирующие структурные белки. В одном варианте осуществления самореплицирующаяся РНК представляет собой in vitro транскрибированную РНК.
Геном альфавирусов представляет собой одноцепочечную положительно-полярную РНК (ssRNA(+)), которая кодирует две открытые рамки считывания (ORF) для крупных полипротеинов. ORF на 5'-конце генома кодирует неструктурные белки с nSP1 по nSP4 (nsPl-4), которые транслируются и процессируются в РНК-зависимую РНК-полимеразу (репликазу); ORF на 3'-конце кодирует структурные белки - капсид и гликопротеины. Обе ORF разделены так называемым субгеномным промотором (SGP), который управляет транскрипцией структурной ORF. При использовании в качестве генных векторов структурные белки, расположенные перед субгенным промотором (SGP) обычно заменены трансгенами. Для упаковки таких векторов в вирусные частицы структурные белки обычно экспрессируются in trans из хелперных конструкций. Репликация альфавирусов происходит в цитоплазме инфицированных клеток, исключительно на уровне РНК. После инфицирования геном ssRNA(+) действует в качестве мРНК для трансляции прекурсора полипротеина nsP1234, который находится на ранних стадиях жизненного цикла вирусов, аутопротеолитически процессированный во фрагменты nsP123 и nsP4. Фрагменты nsP123 и nsP4 формируют (-)нитевый комплекс репликазы, который транскрибирует (-)нитевую РНК из матрицы геномной РНК. На более поздних стадиях полипротеин nsP1234 полностью расщепляется на одиночные белки, которые собираются в (+)нитевый комплекс репликазы, который синтезирует новые (+)нитевые геномы, а также субгеномные транскрипты, которые кодируют структурные белки или трансгены. Субгеномная РНК, а также новая геномная РНК является кэппированной и полиаденилированной и, таким образом, распознается как мРНК после инфицирования клеток-мишеней. Только новая геномная РНК содержит сигнал упаковки, который обеспечивает особую упаковку геномной РНК внутрь почкующихся вирионов. Привлекательность альфавирусных репликонов для векторологии основана на положительной ориентации кэппированного и полиаденилированного генома РНК. Транслируемый репликон РНК может быть легко синтезирован in vitro, при этом кэппирование может быть достигнуто с помощью кэп-аналога, добавленного в реакцию транскрипции in vitro, и поли(А)-хвосты могут быть кодированы как следы политимидиловой кислоты (poly-Т) на плазмидных матрицах. Транскрибированные in vitro (IVT) репликоны трансфицируют традиционными методами трансфекции и даже низкие количества исходной IVT RNA быстро размножались. За несколько часов после переноса трансгены, которые помещают ниже по ходу транскрипции от SGP, транскрибировались в большое число копий, примерно от 40000 до 200000 копий субгеномной РНК на клетку, таким образом, неудивительно, что экспрессия рекомбинантных белков является высокой. В зависимости от определенной цели, IVT репликоны могут быть трансфицированы напрямую в клетки-мишени и упакованы в альфавирусные частицы с хелперными векторами, которые обеспечивают структурные гены in trans. Перенос в кожу или мышцы приводит к высокой и устойчивой локальной экспрессии, наряду с сильной индукцией гуморального и клеточного иммунного ответа.
Для увеличения экспрессии и/или стабильности РНК, используемой в соответствии с настоящим изобретением, РНК может быть модифицирована, предпочтительно без изменения последовательности экспрессированного пептида или белка.
Термин «модификация» в контексте РНК, используемый в соответствии с настоящим изобретением, включает любую модификацию РНК, которая естественным путем не присутствует в указанной РНК.
В одном варианте осуществления изобретения РНК, используемая в соответствии с изобретением, не содержит некэппированных 5'-трифосфатов. Удаление таких некэппированных 5'-трифосфатов может быть достигнуто путем обработки РНК фосфатазой.
РНК в соответствии с изобретением может быть модифицирована рибонуклеотидами природного происхождения или синтетическими для увеличения ее стабильности и/или уменьшения цитотоксичности. Например, в одном варианте осуществления в РНК, используемой в соответствии с изобретением, цитидин заменяли частично или полностью, предпочтительно полностью, на 5-метилцитидин. Альтернативно или дополнительно, в одном варианте осуществления в РНК, используемой в соответствии с изобретением, уридин заменяли частично или полностью, предпочтительно полностью, на псевдоуридин.
В одном варианте осуществления термин «модификация» относится к обеспечению РНК с 5'-кэпом или аналогом 5'-кэпа. Термин «5'-кэп» относится к кэп-структуре, обнаруженной на 5'-конце молекулы мРНК и, как правило, состоит из гуанозинового нуклеотида, соединенного с мРНК посредством необычной 5'-5' трифосфатной связи. В одном варианте осуществления этот гуанозин метилирован в 7-положении. Термин «типичный. 5'-кэп» относится к 5'-кэпу РНК природного происхождения, предпочтительно 7-метилгуанозиновому кэпу (m7G). В контексте настоящего изобретения термин «5'-кэп» включает аналог 5'-кэпа, который имеет сходство с кэп-структурой РНК и является модифицированным для проявления способности стабилизировать РНК в случае присоединения к ней, предпочтительно in vivo и/или в клетке.
Обеспечение РНК 5'-кэпом или аналогом 5'-кэпа может быть достигнуто путем in vitro транскрипции ДНК-матрицы в присутствии указанного 5'-кэпа или аналога 5'-кэпа, при этом указанный 5'-кэп является котранскрипционно встроенным в образованную нить РНК, или РНК может быть образована, например, путем in vitro транскрипции, и 5'-кэп может быть прикреплен к РНК пост-транскрипционно с использованием кэпирующих ферментов, например, кэпирующих ферментов вируса осповакцины.
РНК может содержать дополнительные модификации. Например, дополнительная модификация РНК, используемая в настоящем изобретении, может представлять собой удлинение или укорочение поли(А)-хвоста природного происхождения или изменение 5'- или 3'-нетранслируемых областей (UTR), такое как введение UTR, которая не относится к кодирующей области указанной РНК, например, вставке одной или более, предпочтительно двух копий 3'-UTR, полученных из гена глобина, такого как альфа2-глобин, альфа1-глобин, бета-глобин, предпочтительно бета-глобин, более предпочтительно человеческий бета-глобин.
Таким образом, для повышения стабильности и/или экспрессии РНК, используемой в соответствии с настоящим изобретением, РНК может быть модифицирована таким образом, чтобы присутствовать совместно с последовательностью поли-А, предпочтительно имеющей длину от 10 до 500, более предпочтительно от 30 до 300, даже более предпочтительно от 65 до 200, и особенно предпочтительно от 100 до 150 аденозиновых остатков. В особенно предпочтительном варианте осуществления последовательность поли-А имеет длину приблизительно 120 аденозиновых остатков. Кроме того, встраивание двух или более 3'-нетранслируемых областей (UTR) в 3'-нетранслируемую область молекулы РНК может привести к усилению эффективности трансляции. В одном конкретном варианте осуществления 3'-UTR получена из гена β-глобина человека.
Предпочтительно, РНК при доставке, то есть трансфекции в клетку, в частности клетку, присутствующую in vivo, экспрессирует белок, пептид или антиген, который она кодирует.
Термин “трансфекция» относится к введению нуклеиновых кислот, в частности РНК, в клетку. В целях настоящего изобретения термин «трансфекция» также включает введение нуклеиновой кислоты в клетку или захват нуклеиновой кислоты такой клеткой, при этом клетка может присутствовать у субъекта, например, пациента. Таким образом, в соответствии с настоящим изобретением, клетка, предназначенная для трансфекции нуклеиновой кислоты, описанной здесь, может присутствовать in vitro или in vivo, например, клетка может формировать часть органа, ткани и/или организма пациента. В соответствии с изобретением, трансфекция может быть временной или стабильной. Для некоторых применений трансфекции достаточно, чтобы трансфицированный генетический материал экспрессировался только временно. Поскольку нуклеиновая кислота, введенная в процессе трансфекции, обычно не интегрирована в ядерный геном, чужеродная нуклеиновая кислота будет разбавлена посредством митоза или деградирована. Клетки, обеспечивающие эписомальную амплификацию нуклеиновых кислот, значительно снижают скорость разбавления. Если желательно, чтобы трансфицированная нуклеиновая кислота фактически оставалась в геноме клетки и ее дочерних клетках, должна возникнуть стабильная трансфекция. РНК может быть трансфицирована в клетки для временной экспрессии ее кодированного белка.
Термин «стабильность» РНК относится к «полужизни» РНК. «Полужизнь» относится к периоду времени, который необходим для устранения половины активности, количества или числа молекул. В контексте настоящего изобретения полужизнь РНК является показателем стабильности указанной РНК. Полужизнь РНК может влиять на «продолжительность экспрессии» РНК. Можно предположить, что РНК, имеющая длинный период полужизни, будет экспрессироваться в течение большего периода времени.
В контексте настоящего изобретения термин «транскрипция» относится к процессу, в котором генетический код в последовательности ДНК транскрибирован в РНК. Соответственно, РНК может быть транслирована в белок. В соответствии с настоящим изобретением, термин «транскрипция» включает «in vitro транскрипцию», при этом термин «in vitro транскрипция» относится к процессу, в котором РНК, в частности мРНК, является синтезированной in vitro в бесклеточной системе, предпочтительно с использованием соответствующих клеточных экстрактов. Предпочтительно, клонирующие векторы применяются для образования транскриптов. Эти клонирующие векторы, как правило, разработаны в качестве транскрипционных векторов и в соответствии с настоящим изобретением охвачены термином «вектор».
Термин «трансляция» в соответствии с изобретением относится к процессу в рибосомах клетки, с помощью которого нить мессенджера РНК направляет сборку последовательности аминокислот на получение пептида или белка.
Термин «экспрессия» используется в соответствии с изобретением в наиболее общем значении и включает продукцию РНК и/или пептидов или белков, например, путем транскрипции и/или трансляции. В отношении РНК, термин «экспрессия» или «трансляция» относится, в частности, к продукции пептидов или белков. Указанный термин также включает частичную экспрессию нуклеиновых кислот. Более того, экспрессия может быть временной или стабильной. В соответствии с изобретением, термин «экспрессия» также включает «аберрантную экспрессию» или «аномальную экспрессию».
«Аберрантная экспрессия» или «аномальная экспрессия» означает в соответствии с изобретением, что экспрессия является измененной, предпочтительно увеличенной, по сравнению с референсной экспрессией, например, состоянием субъекта, не имеющего заболевания, связанного с аберрантной или аномальной экспрессией определенного белка, например, опухолевого антигена. Увеличение экспрессии относится к увеличению, по меньшей мере, на 10%, в частности, по меньшей мере на 20%, по меньшей мере на 50% или по меньшей мере на 100%, или более. В одном варианте осуществления экспрессия обнаруживается только в больной ткани, при этом экспрессия в здоровой ткани является подавленной.
Термин «специфически экспрессируется» означает, что белок преимущественно экспрессируется только в специфической ткани или органе. Например, опухолевый антиген, специфически экспрессирующийся в слизистой оболочке желудка, означает, что указанный белок главным образом экспрессируется в слизистой оболочке желудка и не экспрессируется в других тканях, или не экспрессируется в значительной степени в тканях или органах других типов. Таким образом, белок, который экспрессируется исключительно в клетках слизистой оболочки желудка и в значительно меньшей степени в любой другой ткани, такой как семенник, специфически экспрессируется в клетках слизистой оболочки желудка. В некоторых вариантах осуществления опухолевый антиген также может специфически экспрессироваться в нормальных условиях в ткани или органе более чем одного типа, например, в 2 или 3 типах тканей или органах, но предпочтительно не более чем 3 различных типах тканей или органов. В этом случае опухолевый антиген затем специфически экспрессируется в этих органах. Например, если опухолевый антиген экспрессируется в нормальных условиях, предпочтительно, в приблизительно равной степени в легком и желудке, то указанный опухолевый антиген специфически экспрессируется в легком и желудке.
В соответствии с изобретением термин «РНК, кодирующая» означает, что РНК, в случае присутствия в соответствующей среде, предпочтительно в пределах клетки, может экспрессироваться с продуцированием белка или пептида, который она кодирует.
Некоторые аспекты изобретения основаны на адоптивном переносе клеток-хозяев, которые трансфицированы in vitro нуклеиновой кислотой, такой как РНК, кодирующей связывающий агент, описанный здесь, и перенесены реципиентами, таким как пациенты, предпочтительно, после ex vivo экспансии от низкого числа копий прекурсора до клинически значимого количества клеток. Клетки-хозяева, используемые для лечения в соответствии с изобретением, могут быть аутологичными, аллогенными или сингенными в отношении подвергаемого лечению реципиента.
Термин «аутологичный» используется для описания чего-либо, что получено от одного и того же субъекта. Например, «аутологичный трансплантат» относится к ткани или органами, полученным от одного и того же субъекта. Такие процедуры являются предпочтительными, так как они преодолевают иммунологический барьер, что в ином случае приводит к отторжению.
Термин «аллогенный» используется для описания чего-либо, что получено от различных индивидуумов одинаковых разновидностей. Два или более индивидуумов считаются аллогенными друг другу, когда гены в одном или более локусах не являются идентичными.
Термин «сингенный» используется для описания чего-либо, что получено от индивидуумов или тканей, имеющих идентичные генотипы, то есть идентичных близнецов или животных одинакового инбредного штамма, или их тканям.
Термин «гетерологичный» используется для описания чего-либо, состоящего из множества различных элементов. Например, перенос костного мозга одного индивидуума другому индивидууму представляет гетерологичный трансплантат. Гетерологичный ген представляет собой ген, полученный от источника, отличного от субъекта.
Термин «петид» в соответствии с изобретением включает олиго- и полипептиды, и относится к веществам, содержащим две или более, предпочтительно 3 или более, предпочтительно 4 или более, предпочтительно 6 или более, предпочтительно 8 или более, предпочтительно 9 или более, предпочтительно 10 или более, предпочтительно 13 или более, предпочтительно 16 или более, предпочтительно 21 или более и вплоть до предпочтительно 8, 10, 20, 30, 40 или 50, в частности 100 аминокислот, соединенных ковалентно пептидными связями. Термин «белок» относится к крупным пептидам, предпочтительно пептидам, содержащим более 100 аминокислотных остатков, но, как правило, термины «пептиды» и «белки» являются синонимами и используются здесь взаимозаменяемо.
Данные, представленные в настоящем документе в отношении специфических аминокислотных последовательностей, например, приведенных в перечне последовательностей, следует рассматривать как относящиеся также к вариантам указанных специфических последовательностей, приводящих к последовательностям, которые являются функционально эквивалентными указанным специфическим последовательностям, например, аминокислотным последовательностям, проявляющим свойства, идентичные или сходные со свойствами специфических аминокислотных последовательностей. Одним важным свойством является сохранение связывания с мишенью или обеспечение эффекторных функций. Предпочтительно, чтобы последовательность, которая является вариантом по отношению к определенной последовательности, когда она заменяет определенную последовательность в антителе, сохраняла связывание указанного антитела с CLDN и/или CD3, и, предпочтительно, функции указанного антитела, описанные здесь, например, CDC-опосредованный лизис или ADCC-опосредованный лизис.
Например, последовательности, представленные в перечне последовательностей, могут быть модифицированы для удаления одного или нескольких, предпочтительно всех свободных цистеиновых остатков, в частности, путем замены цистеиновых остатков на аминокислоты, отличные от цистеина, предпочтительно серин, аланин, треонин, глицин, тирозин, лейцин или метионин, наиболее предпочтительно аланин или серин. Например, цистеин в положении 103 последовательности, представленной в SEQ ID NO: 36 перечня последовательностей, или соответствующий цистеин в последовательности, содержащей указанную последовательность, может быть, таким образом, модифицирован. Другие цистеины, которые могут быть модифицированы таким способом, представляют собой цистеины в положении 178 в SEQ ID NO: 42, в положении 197 в SEQ ID NO: 43, в положении 427 в SEQ ID NO: 44 или в положении 446 в SEQ ID NO: 45.
Специалистам в данной области следует учесть, что гипервариабельные и вариабельные области последовательности CDR, в частности, могут быть модифицированы без потери способности связывания с CLDN и/или CD3. Например, области CDR будут идентичны или высоко гомологичны областям антител, указанных здесь. Под «высокогомологичными» предполагается, что от 1 до 5, предпочтительно от 1 до 4, например, от 1 до 3, или 1 или 2 замещения может быть сделано в CDR. Кроме того, гипервариабельные и вариабельные области могут быть модифицированы таким образом, что они показывают значительную гомологию с областями антител, специально раскрытых здесь.
В целях настоящего изобретения «варианты» аминокислотной последовательности включают варианты аминокислотных вставок, варианты аминокислотных добавлений, варианты аминокислотных делеций и/или варианты аминокислотных замещений. Варианты аминокислотных делеций, которые содержат делецию на N-конце и/или C-конце белка, также называются укороченными вариантами на N-конце и/или C-конце.
Варианты аминокислотных вставок включают вставки одной или двух, или более аминокислот в конкретную аминокислотную последовательность. В случае вариантов аминокислотной последовательности, имеющих вставку, один или несколько аминокислотных остатков вставлены в конкретный участок в аминокислотной последовательности, хотя случайная вставка при соответствующем скрининге готового продукта также является возможной.
Варианты аминокислотного добавления включают амино- и/или карбокси-концевые слияния одной или более аминокислот, например, 1 , 2, 3, 5, 10, 20, 30, 50, или более аминокислот.
Варианты аминокислотных делеций характеризуются удалением одной или нескольких аминокислот из последовательности, например, путем удаления 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот. Делеции могут быть в любом положении белка.
Варианты аминокислотного замещения характеризуются тем, что, по меньшей мере, один остаток в последовательности удален и на его место вставлен другой остаток. Предпочтение отдается модификациям в положениях в аминокислотной последовательности, которые не являются консервативными между гомологичными белками или пептидами, и/или замене аминокислот другими аминокислотами, обладающими сходными свойствами. Предпочтительно, аминокислотные изменения в белковых вариантах представляют собой консервативные аминокислотные изменения, то есть замещения сходным образом заряженных или незаряженных аминокислот. Консервативное аминокислотное изменение включает замещение одной аминокислоты из семейства аминокислот, которые являются родственными по своим боковым цепям. Аминокислоты природного происхождения, как правило, делятся на четыре семейства: кислотные (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), и незаряженные полярные аминокислоты (глицин, аспарагин, глутамин, цистеин, серин, треонин, тирозин). Фенилаланин, триптофан и тирозин иногда классифицируются как ароматические аминокислоты.
Предпочтительно, степень сходства, предпочтительно, идентичность между заданной аминокислотной последовательностью и аминокислотной последовательностью, которая является вариантом заданной аминокислотной последовательности, будет составлять, по меньшей мере, примерно 60%, 65%, 70%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичность, предпочтительно, представлена для аминокислотной области, которая составляет, по меньшей мере примерно 10%, по меньшей мере примерно 20%, по меньшей мере примерно 30%, по меньшей мере примерно 40%, по меньшей мере примерно 50%, по меньшей мере примерно 60%, по меньшей мере примерно 70%, по меньшей мере примерно 80%, по меньшей мере примерно 90% или примерно 100% от общей длины референсной аминокислотной последовательности. Например, если референсная аминокислотная последовательность состоит из 200 аминокислот, степень сходства или идентичность представлена, предпочтительно, для по меньшей мере 20, по меньшей мере 40, по меньшей мере 60, по меньшей мере 80, по меньшей мере 100, по меньшей мере 120, по меньшей мере 140, по меньшей мере 160, по меньшей мере 180 или примерно 200 аминокислот, предпочтительно непрерывных аминокислот. В предпочтительных вариантах осуществления степень сходства или идентичность представлена для полной длины референсной аминокислотной последовательности. Выравнивание для определения сходства последовательностей, предпочтительно, идентичности последовательностей может быть выполнено с помощью известных в данной области инструментов, предпочтительно, с использованием оптимального выравнивания последовательностей, например, с использованием Align, с использованием стандартных настроек, предпочтительно EMBOSS::needle, Matrix: Blosum62, Gap Open 10.0, Gap Extend 0.5.
«Сходство последовательностей» указывает процентное содержание аминокислот, которые являются идентичным или которые представляют консервативные аминокислотные замещения. «Идентичность последовательностей» между двумя аминокислотным последовательностями указывает процентное содержание аминокислот, которые идентичны между последовательностями.
Термин «процент идентичности» предназначен для обозначения процента идентичных аминокислотных остатков между обеими сравниваемыми последовательностями, полученного после оптимального выравнивания, при этом этот процент является исключительно статистическим и различия между обеими последовательностями распределены случайным образом и по всей их длине. Сравнение двух аминокислотных последовательностей традиционно проводят, сравнивая эти последовательности после выравнивания их оптимальным образом, при этом указанное сравнение может быть проведено посредством сравнения сегментов или с использованием «окна сравнения» для идентификации и сравнения локальных областей сходства последовательности. Оптимальное выравнивание последовательностей для их сравнения может быть осуществлено также и вручную с применением алгоритма локальной гомологии Smith и Waterman, 1981, Ads App. Math. 2, 482, с применением алгоритма локальной гомологии Neddleman и Wunsch, 1970, J. Mol. Biol. 48, 443, с применением аналогичного способа поиска Pearson и Lipman, 1988, Proc. Natl Acad. Sci. USA 85, 2444, или с применением пакетов компьютерных программ, использующих эти алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, Wis.).
Процент идентичности рассчитывают, определяя число идентичных положений, общих для двух сравниваемых последовательностей, и производя деление этого числа на общее число положений в окне сравнения, и производя умножение полученного результата на 100 для получения процента идентичности между обеими этими последовательностями.
Связывающие агенты по изобретению могут быть получены внутиклеточно (например, в цитозоле, периплазме или тельцах включения) и затем выделены из клеток-хозяев и необязательно дополнительно очищены; или они могут быть получены внеклеточно (например, в среде, в которой клетки-хозяева выращиваются) и затем выделены из культуральной среды и необязательно дополнительно очищены. Способы и реагенты, используемые для получения рекомбинантных полипептидов, такие как специфические подходящие экспрессионные векторы, методы трансформации или трансфекции, селективные маркеры, способы индукции экспрессии белка, условия выращивания и тому подобное, являются известными в данной области. Аналогично, технологии выделения и очистки хорошо известны специалистам в данной области.
Термин «клетка» или «клетка-хозяин» предпочтительно относится к интактной клетке, то есть клетке с интактной мембраной, которая не высвобождает свои нормальные внутриклеточные компоненты, такие как фермены, органеллы или генетический материал. Интактная клетка предпочтительно представляет собой жизнеспособную клетку, то есть живую клетку, способную выполнять свои нормальные метаболические функции. Предпочтительно, указанный термин относится в соответствии с изобретением к любой клетке, которая может быть трансфицирована экзогенной нуклеиновой кислотой. Предпочтительно, чтобы клетка при трансфекции экзогенной нуклеиновой кислотой и переносе реципиенту могла экспрессировать нуклеиновую кислоту у реципиента. Термин «клетка» включает бактериальную клетку; другие подходящие клетки представляют собой клетки дрожжей, клетки грибов или клетки млекопитающих. Подходящие бактериальные клетки включают клетки от грамотрицательных штаммов бактерий, таких как штаммы Escherichia coli, Proteus и Pseudomonas, и грамположительные штаммы бактерий, такие как штаммы Bacillus, Streptomyces, Staphylococcus и Lactococcus. Подходящие клетки грибов включают клетки от разновидностей Trichoderma, Neurospora и Aspergillus. Подходящие клетки дрожжей включают клетки от видов Saccharomyces (например, Saccharomyces cerevisiae), Schizosaccharomyces (например, Schizo saccharomyces pombe), Pichia (например, Pichia pastoris и Pichia methanolicd) и Hansenula. Подходящие клетки млекопитающих включают, например, клетки CHO, клетки BHK, клетки HeLa, клетки COS, 293 HEK и тому подобное. Однако клетки земноводных, клетки насекомых, клетки растений и любые другие клетки, используемые в данной области для экспрессии гетерологичных белков, также могут быть использованы. Клетки млекопитающих являются особенно предпочтительными для адоптивного переноса, такие как клетки от человека, мышей, хомяков, свиней, овец и приматов. Клетки могут быть получены из разнообразных типов тканей и включают первичные клетки и клеточные линии, такие как клетки иммунной системы, в частности, антигенпредставляющие клетки, такие как дендритные клетки и Т-клетки, стволовые клетки, такие как гематопоэтические стволовые клетки и мезенхимальные стволовые клетки, и другие типы клеток. Антигенпредставляющая клетка представляют собой клетку, которая представляет антиген посредством его связывания с главным комплексом гистосовместимости на своей поверхности. Т-клетки могут распознавать этот комплекс с использованием своего Т-клеточного рецептора (TCR).
Термин «сокращать», «уменьшать» или «ингибировать», используемый здесь, означает общее уменьшение или способность вызывать общее уменьшение предпочтительно на 5% или более, на 10% или более, на 20% или более, более предпочтительно на 50% или более, и наиболее предпочтительно на 75% или более уровня экспрессии или уровня пролиферации клеток.
Термины, такие как «увеличивает» или «усиливает», предпочтительно, относятся к увеличению или усилению примерно, по меньшей мере на 10%, предпочтительно по меньшей мере на 20%, предпочтительно по меньшей мере на 30%, более предпочтительно по меньшей мере на 40%, более предпочтительно по меньшей мере на 50%, даже более предпочтительно по меньшей мере на 80%, и наиболее предпочтительно по меньшей мере на 100%, по меньшей мере на 200%, по меньшей мере на 500%, по меньшей мере на 1000%, по меньшей мере на 10000%, или даже больше.
Антитело-зависимая клеточно-опосредованная цитотоксичность
ADCC описывает способность эффекторных клеток, в частности лимфоцитов, уничтожать клетки, что предпочтительно требует мечение клетки-мишени антителом.
ADCC предпочтительно возникает, когда антитела связываются с антигенами на опухолевых клетках и домены Fc антитела связываются с Fc-рецепторами (FcR) на поверхности иммунных эффекторных клеток. Идентифицировано несколько семейств Fc-рецепторов, и специфические клеточные популяции характерным образом экспрессируют определенные Fc-рецепторы. ADCC может рассматриваться как механизм прямой индукции изменяющейся степени немедленной деструкции опухоли, что приводит к презентации антигена и индукции опухоль-направленных Т-клеточных ответов. Предпочтительно, индукция ADCC in vivo будет приводить к опухоль-направленным Т-клеточным ответам и гуморальным иммунным ответам организма-хозяина.
Комплементзависимая цитотоксичность
Комплементзависимая цитотоксичность (CDC) представляет собой другой способ уничтожения клеток, который может направляться антителами. IgM является наиболее эффективным изотипом для активации комплемента. Оба, IgG1 и IgG3, также являются эффективными в отношении направления CDC по классическому пути активации системы комплемента. Предпочтительно, в этом каскаде, образование комплексов антиген-антитело приводит к появлению множества участков связывания C1q в непосредственной близости на доменах CH2 принимающих участие молекул антитела, таких как молекулы IgG (C1q является одним из трех субкомпонентов комплемента C1). Предпочтительно, эти связывающие участки C1q переводят ранее низкоаффинные взаимодействия C1q-IgG во взаимодействия с высокой авидностью, которые запускают каскад событий, в которые вовлечены серии других белков комплемента, и приводят к протеолитическому высвобождению хемотаксических/активирующих агентов C3a и C5a. Предпочтительно, каскад комплемента заканчивается образованием мембраноатакующего комплекса, который создает поры в клеточной мембране, способствующие свободному прохождению воды и растворенных веществ внутрь и наружу клетки.
Антитела, описанные здесь, например, для обеспечения областей VL и VH, могут быть получены с использованием множества известных технологий, таких как стандартная технология гибридизации соматических клеток Kohler и Milstein, Nature 256: 495 (1975). Хотя процедуры гибридизации соматических клеток являются предпочтительными, в принципе могут быть использованы другие технологии получения моноклональных антител, например, трансформация В-лимфоцитов вирусами или онкогенами, или технология представления на фагах с использованием библиотек генов антител.
Предпочтительной животной системой получения гибридом, которые продуцируют моноклональные антитела, является мышиная система. Получение гибридомы с использованием мышей хорошо известно в данной области. Протоколы и технологии выделения иммунизированных спленоцитов для слияния известны в данной области. Партнеры слияния (например, клетки мышиной миеломы) и методы слияния также являются известными.
Другими предпочтительными животными системами для получения гибридом, которые продуцируют моноклональные антитела, являются крысиные и кроличьи системы (например, описанные в Spieker-Polet et al., Proc. Natl. Acad. Sci. U.S.A. 92:9348 (1995), смотри также Rossi et al., Am. J. Clin. Pathol. 124: 295 (2005)).
В еще другом предпочтительном варианте осуществления человеческие моноклональные антитела могут быть получены с использованием трансгенных или трансхромосомных мышей, несущих части человеческой иммунной системы, а не мышиной системы. Эти трансгенные и трансхромосомные мыши включают мышей, известных как мыши HuMAb и мыши KM, соответственно, и совместно называются здесь как «трансгенные мыши». Продукция человеческих антител в таких трансгенных мышах может быть выполнена, как описано подробно для CD20 в WO2004 035607.
Еще одной стратегией создания моноклональных антител является непосредственное выделение генов, кодирующих антитела, из лимфоцитов, продуцирующих антитела определенной специфичности, например, смотри Babcock et al., 1996; новая стратегия создания моноклональных антител из одиночных выделенных лимфоцитов, продуцирующих антитела определенных специфичностей. Для более подробной информации относительно конструирования рекомбинантного антитела смотри, также, Welschof и Kraus, Recombinant antibodes for cancer therapy ISBN-0-89603-918-8 и Benny K.C. Lo Antibody Engineering ISBN 1-58829-092-1.
Для генерации антител мышей можно иммунизировать конъюгированными с носителем пептидами, полученными из последовательности антигена, то есть последовательности, против которой должны быть направлены антитела, обогащенным препаратом рекомбинантно экспрессированного антигена или его фрагментов, и/или клетками, экспрессирующими антиген, как описано. Альтернативно, мыши могут быть иммунизированы ДНК, кодирующей антиген или его фрагменты. В случае, когда иммунизации с использованием очищенного или обогащенного препарата антигена не приводят к образованию антител, мышей можно также иммунизировать клетками, экспрессирующими антиген, например, клеточной линией, с целью стимуляции иммунных ответов.
Иммунный ответ может быть подвергнут мониторингу в ходе протокола иммунизации с использованием образцов плазмы и сыворотки, получаемых при отборе крови из хвостовой вены или ретроорбитальной вены. Мыши с достаточными титрами иммуноглобулина могут быть использованы для слияний. Мышам проводят бустерную иммунизацию путем интраперитонеального или внутривенного введения антиген-экспрессирующих клеток за 3 дня до умерщвления и удаления селезенки для увеличения уровня секретирующих специфические антитела гибридом.
Для получения гибридом, продуцирующих моноклональные антитела, спленоциты и клетки лимфатических узлов иммунизированной мыши могут быть выделены и слиты с подходящей иммортализованной клеточной линией, такой клеточная линия мышиной миеломы. Затем проводят скрининг полученных гибридом на продукцию антиген-специфических антител. Затем проводят скрининг отдельных лунок с помощью ELISA на гибридомы, секретирующие антитела. Путем иммунофлуоресценции и анализа FACS с использованием антиген-экспрессирующих клеток, антитела со специфичностью к антигену могут быть идентифицированы. Гибридомы, секретирующие антитела, могут быть повторно высеяны, снова подвергнуты скринингу и, если они остаются положительными в отношении моноклональных антител, их субклонируют путем ограничивающего разведения. Затем стабильные субклоны культивируют in vitro в среде для культур тканей с целью получения антитела для характеризации.
Антитела также можно получить в трансфектоме клетки-хозяина при использовании, например, комбинации технологий рекомбинантной ДНК и способов транфекции гена, как это хорошо известно в данной области (Morrison, S. (1985) Science 229: 1202).
Например, в одном варианте осуществления представляющий интерес ген(ы), например, гены антитела, может быть лигирован в экспрессионный вектор, такой как эукариотическая экспрессионная плазмида, такая как используют в системе экспрессии гена GS, описанной в публикациях WO 87/04462, WO 89/01036 и EP 338 841, или других экспрессионных системах, хорошо известных из уровня техники. Очищенную плазмиду с клонированными генами антитела вводят в эукариотические клетки-хозяева, например клетки CHO, клетки NS/0, клетки HEK 293T или клетки HEK293, или, альтернативно, в другие эукариотические клетки, например клетки, выделенные из растений, грибов или дрожжей. Способ, используемый для введения данных генов, может быть представлен способами, описанными в данной области, такими как электропорация, использование липофектамина, или иными способами. После введения данных генов антител в клетки-хозяева можно идентифицировать и селектировать клетки, экспрессирующие антитело. Данные клетки представляют собой трансфектомы, которые затем могут быть размножены на основе уровня экспрессии и масштабированы для получения антител. Из супернатантов и/или клеток данных культур можно выделить и очистить рекомбинантные антитела.
Альтернативно, данные клонированные гены антител можно экспрессировать в других экспрессионных системах, включая прокариотические клетки, такие как микроорганизмы, например, E. coli. Кроме того, антитела могут быть получены с использованием трансгенных животных, не относящихся к человеку, например, в молоке овец и крыс или в куриных яйцах, или в трансгенных растениях; смотри, например, Verma, R., et al. (1998) J. Immunol. Meth. 216: 165-181; Pollock, et al. (1999) J. Immunol. Meth. 231: 147-157; и Fischer, R., et al. (1999) Biol. Chem. 380: 825-839.
Химеризация
Немеченые мышиные антитела являются высоко иммуногенными у человека при повторяющемся применении, приводя к снижению терапевтического эффекта. Основная иммуногенность опосредована константными областями тяжелой цепи. Иммуногенность мышиных антител у человека может быть снижена или полностью устранена, если соответствующие антитела являются химеризованными или гуманизированными. Химерные антитела представляют собой антитела, различные части которых получены с помощью различных видов животных, например такие, где вариабельная область представляет собой антитело мыши, а константная область - иммуноглобулин человека. Химеризация антител достигается путем соединения вариабельных областей тяжелой и легкой цепи мышиного антитела с константной областью человеческой тяжелой и легкой цепи (например, как описано Kraus et al., в Methods in Molecular Biology series, Recombinant antibodies for cancer therapy ISBN-0-89603-918-8). В предпочтительном варианте осуществления химерные антитела получены путем соединения человеческой константной области каппа-легкой цепи с мышиной вариабельной областью легкой цепи. В также предпочтительном варианте осуществления химерные антитела могут быть получены путем соединения человеческой константной области лямбда-легкой цепи с мышиной вариабельной областью легкой цепи. Предпочтительные константные области тяжелой цепи для создания химерных антител представляют собой IgG1, IgG3 и IgG4. Другие предпочтительные константные области тяжелой цепи для создания химерных антител представляют собой IgG2, IgA, IgD и IgM.
Гуманизация
Антитела взаимодействуют с антигенами-мишенями в основном посредством аминокислотных остатков, которые находятся в шести гипервариабельных участков (CDR) легкой и тяжелой цепей. По этой причине аминокислотные последовательности внутри CDR являются более разнообразными среди отдельных антител, чем последовательности вне CDR. Поскольку последовательности CDR отвечают за большинство взаимодействий антитела и антигена, можно экспрессировать рекомбинантные антитела, которые имитируют свойства специфических природных антител путем конструирования экспрессионных векторов, которые включают последовательности CDR из специфического природного антитела, привитые на каркасные последовательности из другого антитела с другими свойствами (смотри, например, Riechmann, L. et al. (1998) Nature 332: 323-327; Jones, P. et al. (1986) Nature 321: 522-525; и Queen, C. et al. (1989) Proc. Natl. Acad. Sci. U. S. A. 86: 10029-10033). Данные каркасные последовательности могут быть получены из общедоступных баз данных ДНК, которые включают последовательности генов антител зародышевой линии. Данные последовательности зародышевой линии будут отличаться от последовательностей генов зрелого антитела, поскольку они не будут включать полностью собранные вариабельные гены, которые формируются путем связывания V (D) J в процессе созревания В-клеток. Последовательности генов зародышевой линии также будут отличаться от последовательностей высокоаффинного антитела из вторичного репертуара характерным равномерным расположением в вариабельной области.
Способность антител и других связывающих агентов связываться с антигеном может быть определена с использованием стандартных анализов связывания (например, ELISA, Вестерн-блоттинг, иммунофлуоресценция и анализ методом проточной цитометрии).
Для очистки антител отобранные полученные от производителя клеточные линии можно выращивать в двухлитровых роллерных колбах для очистки рекомбинантного антитела. Альтернативно, антитела могут быть получены с помощью диализа в биореакторах. Перед проведением аффинной хроматографии с использованием белка L-сефарозы супернатанты могут быть отфильтрованы и, при необходимости, концентрированы. Элюированный IgG может быть проверен с помощью гель-электрофореза и высокоэффективной жидкостной хроматографии для гарантии чистоты. После замены буферного раствора на PBS концентрацию можно определить по OD280 с использованием соответствующего коэффициента инстинкции. Рекомбинантные антитела могут быть разделены на аликвоты и могут храниться при -80°C.
Для демонстрации связывания моноклональных антител с живыми клетками, экспрессирующими антиген, можно использовать проточную цитометрию. Клеточные линии, экспрессирующие естественным образом или после трансфекции антиген и негативные контроли, лишенные экспрессии антигена (выращенные в условиях стандартного роста), можно смешать с различными концентрациями моноклональных антител в супернатантах гибридомы или в PBS, содержащем 1% FBS, и инкубировать при 4°C в течение 30 мин. После промывания проводят реакцию анти-IgG антитела, меченого APC или Alexa647, с антиген-связанным моноклональным антителом в тех же условиях, что для окрашивания первичного антитела. Образцы можно анализировать методом проточной цитометрии с помощью устройства FACS, с использованием характеристик света и бокового рассеяния дискриминационного окна для отдельных клеток. Для отличия антиген-специфических моноклональных антител от неспецифических связующих в одном измерении, может быть использован способ котрансфекции. Клетки, временно трансфицированные плазмидами, кодирующими антиген, и флуоресцентные маркеры окрашивают, как описано выше. Трансфицированные клетки могут быть детектированы в каналах флуоресценции, отличных от окрашенных антителом клеток. Так как большая часть трансфицированных клеток экспрессирует оба трансгена, антиген-специфические моноклональные антитела связываются преимущественно с клетками, экспрессирующими флуоресцентный маркер, тогда как неспецифические антитела связываются в сопоставимом соотношении с нетрансфицированными клетками. В дополнение или вместо проточного цитометрического анализа может быть использован альтернативный способ анализа с использованием флуоресцентной микроскопии. Клетки окрашивают так, как описано выше, и исследуют с помощью флуоресцентной микроскопии.
Для демонстрации связывания моноклональных антител с живыми клетками, экспрессирующими антиген, может быть использован анализ методом иммунофлуоресцентной микроскопии. Например, клеточные линии, экспрессирующие либо спонтанно или после трансфекции антиген, и отрицательные контроли, лишенные экспрессии антигена, выращивают на предметных стеклах в стандартных условиях роста в среде DMEM/F12, дополненной 10% фетальной бычьей сывороткой (FCS), 2 мМ L-глутамина, 100 ЕД/мл пенициллина и 100 мкг/мл стрептомицина. Клетки затем фиксируют метанолом или параформальдегидом, или оставляют необработанными. Затем проводят реакцию клеток с моноклональными антителами против антигена в течение 30 мин при 25°C. После промывания проводят реакцию клеток с меченым Alexa555 вторичным антимышиным IgG антителом (Molecular Probes) в тех же самых условиях. Затем клетки исследуют с помощью флуоресцентной микроскопии.
Клеточные экстракты клеток, экспрессирующих антиген, и соответствующих отрицательных контролей готовят и подвергают электрофорезу на натрий додецил сульфат-полиакриламидном геле (SDS). После электрофореза разделенные антигены переносят на нитроцеллюлозные мембраны, блокируют и зондируют тестируемыми моноклональными антителами. Связывание IgG можно детектировать с использованием анти-мышиного IgG, конъюгированного с пероксидазой, и проявлять с помощью субстрата ECL.
Антитела могут быть дополнительно тестированы на взаимодействие с антигеном методами иммуногистохимии, хорошо известными специалисту в данной области, например, с использованием фиксированных параформальдегидом или ацетоном криосрезов, или заключенных в парафин срезов тканей, фиксированных параформальдегидом, полученных из образцов нераковой ткани или раковой ткани от пациента во время обычных хирургических процедур, или от мышей, несущих привитые опухоли, инокулированные клеточными линиями, экспрессирующими спонтанно или после трансфекции антиген. Для иммуноокрашивания антитела, реакционноспособные в отношении антигена, могут быть инкубированы с козьим анти-мышиными или козьими анти-кроличьими антителами, конъюгированными с пероксидазой хрена (DAKO) в соответствии с инструкциями изготовителя.
Предклинические исследования
Связывающие агенты, описанные здесь, также могут быть тестированы в модели in vivo, например, на иммунодефицитной мыши, несущей ксенотрансплантированные опухоли, инокулированные клеточными линиями, экспрессирующими CLDN, для определения их эффективности в отношении контроля роста CLDN-экспрессирующих опухолевых клеток.
Исследования in vivo после ксенотрансплантации CLDN-экспрессирующих опухолевых клеток иммунокомпромитированным мышам или другим животным могут быть проведены с использованием связывающих агентов, описанных здесь. Связывающие агенты могут быть введены мышам, не имеющим опухоли, с последующей инъекцией опухолевых клеток для измерения эффектов связывающих агентов в отношении предотвращения формирования опухолей или связанных с опухолью симптомов. Связывающие агенты могут быть введены несущим опухоль мышам для определения терапевтической эффективности соответствующих связывающих агентов в отношении уменьшения роста опухоли, метастазов или связанных с опухолью симптомов. Применение связывающих агентов может быть объединено с применением других веществ, таких как цитостатические лекарственные средства, ингибиторы фактора роста, блокаторы клеточного цикла, ингибиторы антгиогенеза или антитела, для определения синергической эффективности и потенциальной токсичности комбинаций. Для анализа токсических побочных эффектов, опосредованных связывающими агентами, животные могут быть инокулированы связывающими агентами или контрольными реагентами и тщательно исследованы в отношении симптомов, возможно связанных с терапией CLDN-связывающим агентом.
Картирование эпитопов, которые распознали связывающие агенты, может быть выполнено, как описано подробно в «Epitope Mapping Protocols» (Methods in Molecular Biology) by Glenn E. Morris ISBN-089603-375-9 и в «Epitope Mapping: A Practical Approach» Practical Approach Series, 248 by Olwyn M. R. Westwood, Frank C. Hay.
Соединения и агенты, описанные здесь, могут быть введены в форме любой подходящей фармацевтической композиции.
Фармацевтические композиции по изобретению являются предпочтительно стерильными и содержат эффективное количество связывающих агентов, описанных здесь, и необязательно дополнительные агенты, описанные здесь, для достижения желательной реакции или желательного эффекта.
Фармацевтические композиции обычно представлены в виде стандартной лекарственной формы и могут быть приготовлены способами, известными в данной области. Фармацевтическая композиция может, например, находиться в форме раствора или суспензии.
Фармацевтическая композиция может содержать соли, буферные вещества, консерванты, носители, разбавители и/или вспомогательные вещества, все из которых предпочтительно являются фармацевтически приемлемыми. Термин «фармацевтически приемлемый» относится к нетоксичности материала, который не препятствует действию активного компонента фармацевтической композиции.
Соли, не являющиеся фармацевтически приемлемыми, могут быть использованы для приготовления фармацевтически приемлемых солей и включены в изобретение. Фармацевтически приемлемые соли этого вида включают, но без ограничения, соли, полученные из следующих кислот: соляная, бромистоводородная, серная, азотная, фосфорная, малеиновая, уксусная, салициловая, лимонная, муравьиная, малоновая, янтарная кислоты, и т.п. Фармацевтически приемлемые соли могут быть также приготовлены в виде солей щелочных металлов или солей щелочноземельных металлов, таких как соли натрия, соли калия или соли кальция.
Буферные вещества, подходящие для использования в фармацевтической композиции, включают уксусную кислоту в виде соли, лимонную кислоту в виде соли, борную кислоту в виде соли и фосфорную кислоту в виде соли.
Консерванты, подходящие для использования в фармацевтической композиции, включают бензалкония хлорид, хлорбутанол, парабен и тимерозал.
Инъецируемая лекарственная форма может содержать фармацевтически приемлемое вспомогательное вещество, такое как лактат Рингера.
Термин «носитель» относится к органическому или неорганическому компоненту естественного или синтетического происхождения, с которым объединен активный компонент для содействия, усиления или обеспечения возможности применения. В соответствии с изобретением термин «носитель» также включает один или более совместимых твердых или жидких наполнителей, разбавителей или инкапсулирующих веществ, которые являются подходящими для введения пациенту.
Возможные вещества носителя для парентерального введения представляют собой, например, стерильную воду, раствор Рингера, лактат Рингера, стерильный раствор хлорида натрия, полиалкиленгликоли, гидрированные нафталины и, в частности, биосовместимые лактидные полимеры, сополимеры лактида/гликолида или сополимеры полиоксиэтилена/полиоксипропилена.
Термин «вспомогательное вещество», используемый здесь, означает все вещества, которые могут присутствовать в фармацевтической композиции и не являются активными ингредиентами, такие как, например, носители, связующие, лубриканты, загустители, поверхностно-активные вещества, консерванты, эмульгаторы, буферы, вкусовые агенты и красители.
Агенты и композиции, описанные здесь, можно вводить любым обычным способом, таким как парентеральное введение, включая инъекцию или инфузию. Предпочтительно, введение представляет собой парентеральное введение, например, внутривенное, внутриартериальное, подкожное, интрадермальное или внутримышечное.
Композиции, подходящие для парентерального введения, обычно содержат стерильный водный или неводный препарат активного соединения, который предпочтительно является изотоническим по отношению к крови реципиента. Примерами совместимых носителей и растворителей являются раствор Рингера и изотонический раствор хлорида натрия. Кроме того, обычно стерильные нелетучие масла используются в качестве раствора или суспензионной среды.
Агенты и композиции, описанные здесь, вводят в эффективных количествах. «Эффективное количество» относится к количеству, которое достигает желательной реакции или желательного эффекта, отдельно или вместе с дополнительными дозами. При лечении конкретного заболевания или конкретного состояния, желательная реакция предпочтительно относится к ингибированию течения заболевания. Это включает замедление прогрессирования заболевания и, в частности, прерыванию или изменению направления развития заболевания. Желательная реакция при лечении заболевания или состояния может также представлять собой задержку или предупреждение начала развития указанного заболевания или указанного состояния.
Эффективное количество агента или композиции, описанной здесь, будет зависеть от состояния, подлежащего лечению, тяжести заболевания, индивидуальных параметров пациента, включая возраст, физиологическое состояние, размер и вес, продолжительности лечения, типа сопровождающей терапии (в случае присутствия), конкретного пути введения и подобных факторов. Соответствующим образом, дозы вводимых агентов, описанных здесь, могут зависеть от этих различных параметров. В случае, если реакция у пациента недостаточна после введения начальной дозы, могут быть введены более высокие дозы (или, фактически, более высокие дозы достигаются другим, более локализованным способом введения).
Агенты и композиции, описанные здесь, могут быть введены пациентами, например, in vivo, для лечения или предупреждения различных нарушений, таких как описанные здесь нарушения. Предпочтительные пациенты включают пациентов-людей, имеющих нарушения, которые могут быть скорректированы или ослаблены путем введения агентов и композиций, описанных здесь. Это включает нарушения, в которые вовлечены клетки, характеризующиеся измененной картиной экспрессии CLDN, такого как CLDN18.2 и/или CLDN6.
Например, в одном варианте осуществления агенты, описанные здесь, могут быть использованы для лечения пациента с раковым заболеванием, например, раковым заболеванием, описанным здесь, характеризующимся присутствием раковых клеток, экспрессирующих CLDN.
Фармацевтические композиции и способы лечения, описанные в соответствии с изобретением, могут быть также использованы для иммунизации или вакцинации для предупреждения заболевания, описанного здесь.
Фармацевтическая композиция по изобретению может быть введена вместе с дополнительными усиливающими иммунитет веществами, такими как один или несколько адъювантов, и может содержать одно или несколько повышающих иммунитет веществ для дополнительного увеличения их эффективности, предпочтительно, для достижения синергического эффекта иммуностимуляции. Термин «адъювант» относится к соединениям, которые продлевают или усиливают, или ускоряют иммунный ответ. Различные механизмы возможны в этом отношении в зависимости от различных типов адъювантов. Например, соединения, которые обеспечивают возможность созревания DC, например, липополисахариды или лиганд CD40, образуют первый класс подходящих адъювантов. Как правило, любой агент, который оказывает влияние на иммунную систему типа «сигнал опасности» (LPS, GP96, dsRNA и т.д.), или цитокины, такие как GM-CSF, может быть использован в качестве адъюванта, который обеспечивает возможность усиления иммунного ответа и/или влияние на иммунный ответ контролируемым образом. Олигодеоксинуклеотиды могут необязательно использоваться в данном случае, хотя их побочные эффекты, которые возникают при определенных обстоятельствах, как указано выше, подлежат уточнению. Особенно предпочтительные адъюванты представляют собой цитокины, такие как монокины, лимфокины, интерлейкины или хемокины, например, IL-1, IL-2, IL-3, IL-4, IL-5, IL-6, IL-7, IL-8, IL-9, IL-10, IL-12, INFα, INF-γ, GM-CSF, LT-α, или факторы роста, например, hGH. Следующие известные адъюванты представляют собой гидроксид алюминия, адъювант Фрейнда или масло, такое как Montanide®, наиболее предпочтительно Montanide® ISA51. Липопептиды, такие как Pam3Cys, также являются подходящими для использования в качестве адъювантов в фармацевтической композиции по настоящему изобретению.
Агенты и композиции, обеспеченные здесь, могут применяться отдельно или в сочетании с традиционными формами лечения, такими как хирургия, облучение, химиотерапия и/или трансплантация костного мозга (аутологичная, сингенная, аллогенная или неродственная).
Лечение рака представляет область, в которой комбинированные стратегии являются особенно желательными, поскольку часто объединенное действие двух, трех, четырех или даже более противораковых лекарственных средств/терапий создает синергические эффекты, которые являются более сильными по сравнению эффектами при монотерапевтическом подходе. Таким образом, в другом варианте осуществления настоящего изобретения лечение рака, в котором используются основанные на иммунитете или вакцинировании механизмы, такие как способы и фармацевтические композиции по настоящему изобретению, могут быть эффективно объединены с различными другими лекарственными средствами и/или способами, имеющим сходный или другой механизм направленного воздействия. В их число входят, например, комбинации с традиционными терапиями опухолей, стратегии мультиэпитопов, дополнительная иммунотерапия и лечебные подходы, направленно воздействующие на ангиогенез или апоптоз (для обзора смотри, например, Andersen et al., 2008: Cancer treatment: the combination of vaccination with other therapies. Cancer Immunology Immunotherapy, 57(11): 1735-1743.). Последовательное введение различных агентов может ингибировать рост раковых клеток в различные моменты времени, тогда как другие агенты могут, например, ингибировать неоангиогенез, выживаемость злокачественных клеток или метастазов, потенциально переводя рак в хроническое заболевание. Следующий перечень представляет некоторые неограничивающие примеры противораковых лекарственных средств и терапий, которые могут быть использованы в комбинации с настоящим изобретением:
1. Химиотерапия
Химиотерапия представляет собой стандарт лечения множества типов рака. Наиболее распространенные химиотерапевтические агенты действуют путем уничтожения клеток, которые быстро делятся, что является одним из основных особенностей раковых клеток. Таким образом, комбинирование с общепризнанными химиотерапевтическими лекарственными средствами, такими как, например, алкилирующие агенты, антиметаболиты, антрациклины, алкалоиды растений, ингибиторы топоизомеразы и другие противоопухолевые агенты, которые воздействуют на клеточное деление или синтез ДНК, может значительно улучшить терапевтические эффекты по настоящему изобретению путем очистки супрессорных клеток, восстановления иммунной системы путем наделения опухолевых клеток большей восприимчивостью к иммунообусловленному уничтожению, или путем дополнительной активации клеток иммунной системы. Синергическое противораковое действие химиотерапевтических и основанных на вакцинировании иммунотерапевтических лекарственных средств было продемонстрировано во многих исследованиях (смотри, например, Quoix et al. 2011: Therapeutic vaccination with TG4010 and first-line chemotherapy in advanced non-small-cell lung cancer: a controlled phase 2B trial. Lancet Oncol. 12(12): 1125-33.; смотри накже Liseth et al. 2010: Combination of intensive chemotherapy and anticancer vaccines in the treatment of human malignancies: the hematological experience. J Biomed Biotechnol. 2010: 6920979; смотри также Hirooka et al. 2009: A combination therapy of gemcitabine with immunotherapy for patients with inoperable locally advanced pancreatic cancer. Pancreas 38(3): e69-74). Существуют сотни доступных химиотерапевтических средств, которые в основном подходят для комбинированной терапии. Некоторыми (неограничивающие) примерами химиотерапевтических лекарственных средств, которые можно применять в сочетании с настоящим изобретением, являются карбоплатин (параплатин), цисплатин (платинол, платинол-AQ), циклофосфамид (цитоксан, неозар), доцетаксел (таксотер), доксорубицин (адриамицин), эрлотиниб (тарцева), этопозид (вепезид), фторурацил (5-FU), гемцитабин (гемзар), иматиниба мезилат (гливек), иринотекан (камптозар), метотрексат (фолекс, мексат, аметоптерин), паклитаксел (таксол, абраксан), сорафиниб (нексавар), сунитиниб (сутент), топотекан (гикамтин), винкристин (онковин, винказар PFS), и винбластин (велбан).
2. Хирургия
Хирургия рака - операция по удалению опухоли - остается основой для лечения рака. Хирургия может быть комбинирована с другими терапиями рака для удаления любых оставшихся опухолевых клеток. Комбинирование хирургических способов с последующим иммунотерапевтическим лечением является многообещающим подходом, который был неоднократно продемонстрирован.
3. Облучение
Радиационная терапия остается важным компонентом в лечении рака, при этом приблизительно 50% всех раковых пациентов получают радиационную терапию в процессе их заболевания. Основной целью радиационной терапии является лишение раковых клеток их потенциала к размножению (клеточное деление). Типы облучения, используемые для лечения рака, представляют собой облучение фотонами (рентгеновские лучи и гамма лучи) и облучение частицами (электронные, фотонные и нейтронные пучки). Существует два способа доставки излучения в место локализации злокачественной опухоли. Облучение внешним пучком доставляется от источников, расположенных вне организма путем нацеливания высокоэнергетических лучей (фотонов, протонов или облучение частицами) в место локализации опухоли. Внутреннее облучение или брахитерапия доставляется от радиоактивных источников, расположенных вне организма, заключается в герметичные катетеры, или вводится непосредственно в участок опухоли. Методы лучевой терапии, которые можно применять в комбинации с настоящим изобретением, представляют собой фракционирование (сеансы лучевой терапии в режиме фракционирования, например, ежедневные фракции от 1,5 до 3 Гр в течение нескольких недель), 3D-конформную лучевую терапию (3DCRT; доставка излучения в макроскопический объем опухоли), лучевую терапию с модулированной интенсивностью (IMRT; компьютеризированное модулирование интенсивности пучков лучей), лучевую терапию под визуальным контролем (IGRT; метод, заключающийся в создании изображений через короткие промежутки времени в ходе сеанса лучевой терапии, что позволяет делать корректировку), и стереотаксическая радиотерапия (SRBT, доставляет очень высокие дозы излучения лишь за нескольких фракций). Для обзора лучевой терапии смотри Baskar et al. 2012: Cancer and radiation therapy: current advances and future directions. Int. J Med Sci. 9(3): 193-199.
4. Антитела
Антитела (предпочтительно моноклональные антитела) достигают своего терапевтического эффекта против раковых клеток посредством различных механизмов. Антитела могут иметь непосредственное воздействие на апоптоз или программированную гибель клеток. Антитела могут блокировать компоненты путей сигнальной трансдукции, такие как, например, рецепторы факторов роста, эффективно подавляя пролиферацию опухолевых клеток. В клетках, которые экспрессируют моноклональные антитела, они могут способствовать образованию антиидиотипических антител. Непрямые эффекты включают рекрутинг клеток, которые обладают цитотоксичностью, таких как моноциты и макрофаги. Этот тип опосредованной антителом гибели клеток называется антителозависимая клеточно-опосредованная цитотоксичность (ADCC). Антитела также связывают комплемент, что приводит к прямой клеточной токсичности, известной как комплемент-зависимая цитотоксичность (CDC). Комбинирование хирургических методов с иммунотерапевтическими лекарственными средствами и методами является успешным подходом, как продемонстрировано, например, в Gadri et al. 2009: Synergistic effect of dendritic cell vaccination and anti-CD20 antibody treatment in the therapy of murine lymphoma. J Immunother. 32(4): 333-40. Следующий перечень обеспечивает некоторые неограничивающие примеры противораковых антител и потенциальных мишеней антител (в скобках), которые можно применять в комбинации с настоящим изобретением: абаговомаб (CA-125), абциксимаб (CD41), адекатумумаб (EpCAM), афутузумаб (CD20), алацизумаб пегол (VEGFR2), алтумомаб пентетат (CEA), аматуксимаб (MORAb-009), анатумомаб мафенатокс (TAG-72), аполизумаб (HLA-DR), арцитумомаб (CEA), бавитуксимаб (фосфатидилсерин), бектумомаб (CD22), белимумаб (BAFF), бевацизумаб (VEGF-A), биватузумаб мертанзин (CD44 v6), блинатумомаб (CD19), брентуксимаб ведотин (CD30 TNFRSF8), кантузумаб мертанзин (муцин CanAg), кантузумаб равтанзин (MUC1), капромаб пендетид (клетки карциномы предстательной железы), карлумаб (CNT0888), катумаксомаб (EpCAM, CD3), цетуксимаб (EGFR), цитатузумаб богатокс (EpCAM), циксутумумаб (рецептор IGF-1), клаудиксимаб (клаудин), кливатузумаб тетракетан (MUC1), конатумумаб (TRAIL-R2), дацетузумаб (CD40), долатузумаб (рецептор инсулинподобного фактора роста I), деносумаб (RANKL), детумомаб (клетки B-лимфомы), дрозитумаб (DR5), экромексимаб (ганглиозид GD3), эдреколомаб (EpCAM), элотузумаб (SLAMF7), энаватузумаб (PDL192), энситуксимаб (NPC-1 C), эпратузумаб (CD22), эртумаксомаб (HER2/neu, CD3), этарацизумаб (интегрин ανβ3), фарлетузумаб (фолатный рецептор 1), FBTA05 (CD20), фиклатузумаб (SCH 900105), фигитумумаб (IGF-1 рецептор), фланвотумаб (гликопротеин 75), фрезолимумаб (TGF-β), галиксимаб (CD80), ганитумаб (IGF-I), гемтузумаб озогамицин (CD33), гевокизумаб (IL-1β), гирентуксимаб (карбоангидраза 9 (CA-IX)), глембатумумаб ведотин (GPNMB), ибритумомаб тиуксетан (CD20), икрукумаб (VEGFR-1), иговома (CA-125), индатуксимаб равтанзин (SDC1), интетумумаб (CD51), инотузумаб озогамицин (CD22), ипилимумаб (CD 152), иратумумаб (CD30), лабетузумаб (CEA), лексатумумаб (TRAIL-R2), либивирумаб (поверхностный антиген гепатита В), линтузумаб (CD33), лорвотузумаб мертанзин (CD56), лукатумумаб (CD40), лумиликсимаб (CD23), мапатумумаб (TRAIL-R1), матузумаб (EGFR), меполизумаб (IL-5), милатузумаб (CD74), митумомаб (GD3 ганглиозид), могамулизумаб (CCR4), моксетумомаб пасудотокс (CD22), наколомаб тафенатокс (антиген C242), наптумомаб эстафенатокс (5T4), нарнатумаб (RON), нецитумумаб (EGFR), нимотузумаб (EGFR), ниволумаб (IgG4), офатумумаб (CD20), оларатумаб (PDGF-R α), онартузумаб (киназа рецептора фактора роста гепатоцитов человека), опортузумаб монатокс (EpCAM), ореговомаб (CA-125), окселумаб (OX-40), панитумумаб (EGFR), патритумаб (HER3), пемтумома (MUC1), пертузумаб (HER2/neu), пинтумомаб (антиген аденокарциномы), притумумаб (виментин), пакотумомаб (N-гликолилнейраминовая кислота), радретумаб (экстра домен В фибронектина), рафивирумаб (гликопротеин вируса бешенства), рамуципумаб (VEGFR2), рилотумумаб (HGF), ритуксимаб (CD20), робатумумаб (рецептор IGF-1), самализумаб (CD200), сибротузумаб (FAP), силтуксимаб (IL-6), табалумаб (BAFF), такатузумаб тетраксетан (альфа-фетопротеин), таплитумомаб паптокс (CD 19), тенатумомаб (тенасцин C), тепротумумаб (CD221), тицилимумаб (CTLA-4), тигатузумаб (TRAIL-R2), TNX-650 (IL-13), тозитумомаб (CD20), трастузумаб (HER2/neu), TRBS07 (GD2), тремелимумаб (CTLA-4), тукотузумаб целмолейкин (EpCAM), ублитуксимаб (MS4A1), урелумаб (4- IBB), волоциксимаб (интегрин α5β1), вотумумаб (опухолевый антиген CTAA16.88), залутумумаб (EGFR), занолимумаб (CD4).
5. Цитокины, хемокины, костимуляторные молекулы, белки слияния
Комбинированное использование антиген-кодирующих фармацевтических композиций по настоящему изобретению с цитокинами, хемокинами, костимуляторными молекулами и/или их белками слияния для вызывания благоприятного иммунного модулирования или эффектов ингибирования опухоли является другим вариантом осуществления настоящего изобретения. Для увеличения инфильтрации иммунных клеток в опухоли и содействия движению антиген-презентирующих клеток в дренирующие опухоль лимфатические узлы могут быть использованы различные хемокины со структурами C, CC, CXC и CX3C. Некоторыми из самых многообещающих хемокинов являются, например, CCR7 и его лиганды CCL19 и CCL21, кроме того, CCL2, CCL3, CCL5 и CCL16. Другими примерами являются CXCR4, CXCR7 и CXCL12. Кроме того, можно применять костимуляторные или регуляторные молекулы, такие как, например, лиганды B7 (B7.1 и B7.2). Также можно применять другие цитокины, такие как, например, интерлейкины (например, от IL-1 до IL17), интерфероны (например, от IFNalphal до IFNalpha8, IFNalpha10, IFNalpha13, IFNalpha14, IFNalpha16, IFNalpha17, IFNalpha21, IFNbeta1, IFNW, IFNE1 и IFNK), гематопоэтические факторы, TGF (например, TGF-α, TGF-β и другие члены семейства TGF) и, наконец, рецептор семейства фактора некроза опухолей и его лиганды, а также другие стимуляторные молекулы, содержащие, но без ограничения, 4-1BB, 4-1BB-L, CD 137, CD137L, CTLA-4GITR, GITRL, Fas, Fas-L, TNFR1, TRAIL-R1 , TRAIL-R2, p75NGF-R, DR6, LT.beta.R, RANK, EDAR1, XEDAR, Fnl l4, Troy/Trade, TAJ, TNFRII, HVEM, CD27, CD30, CD40, 4- 1BB, OX40, GITR, GITRL, TACI, BAFF-R, BCMA, RELT и CD95 (Fas/APO-1), глюкокортикоид-индуцированные TNFR-связанные белки, TNF рецептор-связанный апоптоз-опосредующий белок (TRAMP) и рецептор клеточной гибели-6 (DR6)., CD40/CD40L и OX40/OX40L являются особенно важными мишенями для комбинированной иммунотерапии благодаря их прямому воздействию на Т-клеточную выживаемость и пролиферацию. Для обзора смотри Lechner et al. 2011: Chemokines, costimulatory molecules and fusion proteins for the immunotherapy of solid tumors. Immunotherapy 3 (11), 1317-1340.
6. Бактериальные терапии
Исследователи использовали анаэробные бактерии, такие как Clostridium novyi, для поглощения внутренней части бедных кислородом опухолей. Бактерии затем погибают при контакте с оксигенированными сторонами опухоли, что означает, что они не будут оказывать вредное воздействие на остальной организм. Другой стратегией является использование анаэробных бактерий, которые были трансформированы ферментом, который может превращать нетоксическое пролекарство в токсическое лекарственное средство. При пролиферации бактерий в некротических и гипоксических областях опухоли фермент экспрессируется только в опухоли. Таким образом, системно применяемое пролекарство метаболизируется в токсическое лекарственное средство только в опухоли. Это было продемонстрировано как эффективное с непатогенными анаэробными спорогенами Clostridium.
7. Ингибиторы киназы
Другая большая группа потенциальных мишеней для комплементарной терапии рака включает ингибиторы киназ, так как рост и выживаемость раковых клеток тесно связана с разрегулированием киназной активности. Для восстановления нормальной киназной активности и, следовательно, уменьшения роста опухоли применяют широкий ряд ингибиторов. Группа целевых киназ включает рецепторные тирозинкиназы, например, BCR-ABL, B-Raf, EGFR, HER-2/ErbB2, IGF-IR, PDGFR-α, PDGFR-β, c-Kit, Flt-4, Flt3, FGFR1, FGFR3, FGFR4, CSF1R, c-Met, RON, c-Ret, ALK, цитоплазматические тирозинкиназы, например, c-SRC, c-YES, Abl, JAK-2, серин/треонинкиназы, например, ATM, Aurora A & B, CDKs, mTOR, PKCi, PLKs, b-Raf, S6K, STK1 1/LKB1 и липидные киназы, например, PI3K, SK1. Низкомолекулярные ингибиторы киназ представляют собой, например, PHA-739358, нилотиниб, дазатиниб и PD166326, NSC 743411, лапатиниб (GW-572016), канертиниб (CI-1033), семаксиниб (SU5416), ваталаниб (PTK787/ZK222584), сутент (SU1 1248), сорафениб (BAY 43-9006) и лефлуномид (SU101). Для дополнительной информации смотри, например, Zhang et al. 2009: Targeting cancer with small molecule kinase inhibitors. Nature Reviews Cancer 9, 28-39.
8. Toll-подобные рецепторы
Члены семейства Toll-подобных рецепторов (TLRs) являются ключевым соединением между врожденным и адаптивным иммунитетом, и эффект многих адъювантов основан на активации TLR. Большое число разработанных вакцин против рака включает лиганды для TLR8, для вызывания ответов на вакцины. Помимо TLR2, TLR3, TLR4, особенно TLR7 и TLR 8 исследовали в отношении терапии рака в пассивных иммунотерапевтических подходах. Близкородственные TLR7 и TLR8 содействуют противоопухолевым ответам путем воздействия на клетки иммунной системы, опухолевые клетки и микроокружение опухоли, и могут быть активированы структурами нуклеозидных аналогов. Все TLR применяются в качестве отдельных иммунотерапевтических средств или адъювантов противораковых вакцин и могут быть синергически комбинированы с лекарственными формами и способами по настоящему изобретению. Для более подробной информации смотри van Duin et al. 2005: Triggering TLR signaling in vaccination. Trends in Immunology, 27(1):49-55.
9. Ингибиторы ангиогенеза
Дополнительно к терапиям, которые направленно воздействуют на иммунные модуляторные рецепторы, находящиеся под воздействием опухоль-опосредованных механизмов ускользания и иммунной супрессии, существуют терапии, которые направленно воздействуют на окружение опухоли. Ингибиторы ангиогенеза предотвращают избыточный рост кровеносных сосудов (ангиогенез), которые требуются опухолям для выживания. Ангиогенез, стимулированный опухолевыми клетками для удовлетворения их растущих потребностей в питательных веществах и кислороде, может быть блокирован путем направленного воздействия на различные молекулы. Неограничивающие примеры ангиогенез-опосредующих молекул или ингибиторов ангиогенеза, которые могут быть комбинированы с настоящим изобретением, включают растворимый VEGF (VEGF, изоформы VEGF121 и VEGF165, рецепторы VEGFR1, VEGFR2 и корецепторы нейропилин-1 и нейропилин-2) 1 и NRP-1, ангиопоэтин 2, TSP-1 и TSP-2, ангиостатин и родственные молекулы, эндостатин, вазостатин, калретикулин, тромбоцитарный фактор-4, TIMP и CDAI, Meth-1 и Meth-2, IFN-α, -β и -γ, CXCL10, IL-4, -12 и -18, протромбин (крингл-домен 2), фрагмент антитромбина III, пролактин, VEGI, SPARC, остеопонтин, маспин, канстатин, пролиферин-связанный белок, рестин и лекарственные средства, такие как, например, бевацизумаб, итраконазол, карбоксиамидотриазол, TNP-470, CM101 , IFN-α тромбоцитарный фактор-4, сурамин, SU5416, тромбоспондин, антагонисты VEGFR, ангиостатические стероиды + гепарин, ингибирующие факторы ангиогенеза в хрящевой ткани, ингибиторы матриксной металлопротеиназы, 2-метоксиэстрадиол, текогалан, тетратиомолибдат, талидомид, тромбоспондин, ингибиторы пролактина ανβ3, линомид, препарат «tasquinimod». Для обзора смотри Schoenfeld и Dranoff 2011: Anti-angiogenesis immunotherapy. Hum Vaccin. (9):976-81.
10. Низкомолекулярные лекарственные средства таргетной терапии
В целом, низкомолекулярные лекарственные средства таргетной терапии представляют собой ингибиторы доменов ферментов на мутировавших, сверхэкспрессирующихся или иным образом важных белках в раковой клетке. Известными и неограничивающими примерами являются ингибиторы тирозинкиназы иматиниб (Gleevec/Glivec) и гефининиб (Iressa). Использование малых молекул, например, сунитиниба малата и/или сорафениба тозилата, направленно воздействующих на некоторые киназы, в сочетании с вакцинами для терапии рака, также описано в более ранней заявке на патент US2009004213.
11. Вакцины на основе вирусов
Существует ряд противораковых вакцин на основе вирусов, доступных или находящихся на стадии разработки, которые можно применять в комбинированном терапевтическом подходе вместе с лекарственными формами по настоящему изобретению. Одним преимуществом использования таких вирусных векторов является их врожденная способность инициировать иммунные ответы, при этом воспалительные реакции, возникающие в результате вирусной инфекции, создают сигнал опасности, необходимый для иммунной активации. Идеальный вирусный вектор должен быть безопасным и не должен приводить к анти-векторному иммунному ответу для того, чтобы обеспечить возможность бустер-иммунизации противоопухолевых специфических ответов. Рекомбинантные вирусы, такие как вирус осповакцины, вирусы простого герпеса, аденовирусы, аденоассоциированные вирусы, ретровирусы и вирусы рода Avipox использовали в животных опухолевых моделях и на основе их обнадеживающих результатов были инициированы клинические исследования на человеке. Особенно важными вакцинами на основе вирусов являются вирусоподобные частицы (VLP), малые частицы, которые содержат определенные белки из внешней оболочки вируса. Вирусоподобные частицы не содержат какого-либо генетического материала вируса и не могут вызвать инфекцию, но они могут быть сконструированы для представления опухолевых антигенов на своей оболочке. VLP могут быть получены из различных вирусов, таких как, например, вирус гепатита В или другие семейства вирусов, включая Parvoviridae (например, аденоассоциированный вирус), Retroviridae (например, ВИЧ) и Flaviviridae (например, вирус гепатита С). Для общего обзора смотри Sorensen и Thompsen 2007: Virus-based immunotherapy of cancer: what do we know and where are we going? APMIS 1 15(11):1 177-93; вирусо-пободные частицы против рака описаны в Buonaguro et al. 2011: Developments in virus-like particle-based vaccines for infectious diseases and cancer. Expert Rev Vaccines 10(1 1): 1569-83; и в Guillen et al. 2010: Virus-like particles as vaccine antigens and adjuvants: application to chronic disease, cancer immunotherapy and infectious disease preventive strategies. Procedia in Vaccinology 2 (2), 128-133.
12. Стратегии на основе мультиэпитопов
Использование мультиэпитопов показало многообещающие результаты в отношении вакцинации. Технологии быстрого секвенирования в сочетании с системами интеллектуального алгоритма позволяют использовать мутаному опухоли и могут обеспечить мультиэпитопы для индивидуальных вакцин, которые могут быть комбинированы с настоящим изобретением. Для более подробной информации смотри 2007: Vaccination of metastatic colorectal cancer patients with matured dendritic cells loaded with multiple major histocompatibility complex class I peptides. J Immunother 30: 762-772; дополнительно, Castle et al. 2012: Exploiting the mutanome for tumor vaccination. Cancer Res 72 (5): 1081 -91.
13. Адоптивный Т-клеточный перенос
Например, комбинация вакцинации опухолевого антигена и Т-клеточного переноса описана в: Rapoport et al. 2011: Combination immunotherapy using adoptive T-cell transfer and tumor antigen vaccination on the basis of hTERT and survivin after ASCT for myeloma. Blood 1 17(3):788-97.
14. Таргетные терапии на основе пептидов
Пептиды могут связываться с рецепторами клеточной поверхности или воздействовать на внеклеточный матрикс, окружающий опухоль. Радионуклиды, которые прикреплены к этим пептидам (например, RGD) в итоге убивают раковые клетки, если нуклид разрушается около клетки. Олиго- или мультимеры этих связывающих мотивов представляют особый интерес, так как это может привести к повышению опухолевой специфичности и авидности. Для обзора неограничивающих примеров смотри Yamada 2011: Peptide-based cancer vaccine therapy for prostate cancer, bladder cancer, and malignant glioma. Nihon Rinsho 69(9): 1657-61.
15. Другие терапии
Существует ряд другие терапий рака, которые могут быть комбинированы с лекарственными формами и способами по настоящему изобретению для создания синергических эффектов. Неограничивающими примерами являются терапии, направленные на апоптоз, гипертермию, гормональную терапию, терапию теломеразой, потенциированную инсулиновую терапию, генную терапию и фотодинамическую терапию.
Настоящее изобретение дополнительно проиллюстрировано следующими примерами, которые не должны рассматриваться как ограничивающие объем изобретения.
Примеры
Пример 1: Создание и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN18.2 и CD3
a. Происхождение последовательности, дизайн конструкций bi-scFv, и клонирование в экспрессионные вектора
Готовили конструкции тандемного биспецифического одноцепочечного антитела (bi-scFv), содержащие связывающие домены, специфические в отношении компонента CD3 человеческого Т-клеточного рецептора и человеческих опухоль-ассоциированных антигенов (TAA). Соответствующие вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) для каждой конструкции специфически расположены в направлении от N- до C-конца в следующем порядке:
N - VHαCLDN18.2 - VLαCLDNl8.2 - VHαCD3 - VLαCD3-C (1BiMAB, 18PHU5, no.11-15)
N - VHαCD3 - VLαCD3 - VHαCLDN18.2 - VLαCLDN18.2-C (18PHU3, no.16-20)
В таблице 1 представлены все конструкции bi-scFv, специфические в отношении TAA CLDN18.2 и PLAC1, которые были созданы в рамках изобретения. Конструкции bi-scFv были созданы путем генного синтеза GeneArt AG (GeneArt/Life Technologies GmbH, Regensburg, Germany) с использованием последовательностей VH и VL соответствующих антител. Оптимизации кодонов, таких как Homo sapiens (HS), Mus musculus (MM) или клетки яичника китайского хомячка (Chinese Hamster Ovary, CHO) были реализованы с помощью программного обеспечения GeneArt's GeneOptimizer®, и перечислены в таблице 1. Информация по специфичности, происхождению последовательности из моноклональных антител (mAB), использованию кодонов, дополнительным отличительным особенностям последовательностей и ссылкам на все применяющиеся домены обобщена в таблице 2. Происхождение последовательностей вариабельного домена соответствующих антител CD3 приведено в таблице 2. Благодаря высокой гомологии человеческих и мышиных TAA, одинаковые последовательности VH и VL анти-TAA могут быть использованы для создания конструкций bi-scFv для тестов на мышах, но в комбинации с последовательностями VH, VL клона 145-2C1 мышиного специфического анти-CD3 антитела.
Клонирование ДНК и конструирование экспрессионного вектора проводили в соответствии со стандартными процедурами (Green/Sambrook, Molecular Cloning, 2012), хорошо известными специалистом в данной области. Вкратце, основные ДНК-последовательности bi-scFv содержали сайты рестрикции 5' HindIII и 3' XhoI (HindIII и XbaI в случае 1BiMAB bi-scFv) для клонирования в экспрессионные плазмиды. Последовательность сигнала секреции вводили на 5'-конце (выше) последовательности bi-scFv для секреции белка из клеточной цитоплазмы в культуральную среду. Последовательность, кодирующую гибкий глицин-сериновый пептидный линкер, состоящий из 15-18 аминокислот, вставляли для соединения доменов VH и VL для композиции одноцепочечных вариабельных фрагментов антитела (scFv), из которых один связывается с CD3 и другой с TAA. Для формирования биспецифического одноцепочечного антитела последовательности двух доменов scFv соединяли последовательностью, кодирующей короткий пептидный линкер (GGGGS). Вместе с этой последовательностью линкера сайт рестрикции BamHI вводили для обменов доменов scFv для клонирования последующих конструкций bi-scFV. В частности, 5'scFv-домены могут быть заменены на сайты рестрикции HindIII и BamHI и 3'scFv-домены на рестрикцию BamHI и XhoI. Схему конструкции смотри также на фигуре 1.
Все используемые конструкции антитела bi-scFv клонировали в стандартный экспрессионный вектор млекопитающего pcDNA™3.1/myc-His (+) (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). C-концевая 6xHis-метка может быть использована для металл-аффинной очистки белка и анализов на обнаружение. Все конструкции подтверждали путем секвенирования через сервис по секвенированию одиночных прочтений (MWG single read sequence service) (Eurofins MWG Operon, Ebersberg, Germany).
Таблица 1. Краткое описание конструкций биспецифических одноцепочечных антител, специфических к TAA и CD3
CHO, клетки яичника китайского хомячка; HS, Homo-sapiens; HU, гуманизированное; MM, Mus.
Таблица 2. Краткая информация по конструкциям bi-scFv
Таблица 2
Продолжение
mAB
ссылки
CHO означает клетки яичника китайского хомячка; HS, Homo-sapiens; mAB, моноклональное антитело; MM, Mus musculus; TAA, опухоль-ассоциированный антиген.
b. Генерация стабильных клеточных линий-продуцентов
Для генерации стабильных клонов клеток-продуцентов CLDN18.2-специфических белков bi-scFv использовали клеточную линию эмбриональных почек человека HEK293 (ATCC CRL-1573) и клеточную линию яичника китайского хомячка CHO-K1 (ATCC CCL-61).
Трансфекция HEK293
1×107 клеток HEK293 засевали за два дня до трансфекции на чашки Петри 14,5 см в 20 мл полной среды DMEM (DMEM/F-12 GlutaMax, дополненной 10% термоинактивированной FBS и 0,5% пенициллина-стрептомицина; все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany). Перед трансфекцией клетки промывали фосфатно-солевым буфером Дульбекко (DPBS), дополненным 2 мМ EDTA, затем добавляли 20 мл чистой среды DMEM без добавления FBS или антибиотиков. 20 мкг линеаризованной ДНК конструкций, описанных в примере 1, разбавляли в 0,5 мл чистой среды DMEM/F-12. 75 мкл раствора линейного PEI при концентрации 1 мг/мл (полиэтиленимин; Polysciences Europe GmbH, Eppelheim, Germany) добавляли в разбавленную ДНК и энергично перемешивали на вортексе. Через 15 мин инкубации при комнатной температуре (RT) в клетки по каплям добавляли комплексы ДНК/PEI, чашки для клеточных культур осторожно перемешивали вращательным движением и затем инкубировали при 37°C, 5% CO2. Через 24 ч после трансфекции среду меняли. Отбор трансфицированных клеток начинали через 48 ч после трансфекции с помощью сульфата G418 (Gibco/Life Technologies GmbH, Darmstadt, Germany) в конечной концентрации 0,8 мг/мл. G418 непрерывно добавляли в культуральную среду для выращивания клеток.
Трансфекция CHO-K1
1×106 клеток CHO-K1 засевали за день до трансфекции в 6-луночные тканевые культуральные планшеты в 2 мл полной среды DMEM (DMEM/F-12 GlutaMax, дополненной 10% термоинактивированной FBS, без антибиотиков; все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany). Перед трансфекцией клетки промывали DPBS, дополненным 2 мМ EDTA, затем добавляли 1,5 мл обычной среды DMEM без добавления FCS или антибиотиков. 4 мкг линеаризованной ДНК конструкций, описанных в примере 1.а, разбавляли в 0,25 мл обычной среды DMEM/F-12 и осторожно перемешивали. Во второй реакционной пробирке 2,5 мкл липофектамина 2000 (Invitrogen/Life Technologies GmbH, Darmstadt, Germany) разбавляли в 0,25 мл обычной среды DMEM/F-12, осторожно перемешивали и инкубировали в течение 5 мин при комнатной температуре (RT). Смесь ДНК и смесь липофектамина объединяли в соотношении 1:1, осторожно перемешивали и инкубировали в течение 20 мин при комнатной температуре (RT). В клетки по каплям добавляли комплексы ДНК/липофектамин 2000, чашки для клеточных культур осторожно перемешивали вращательным движением и затем инкубировали при 37°C, 5% CO2. Через 6 ч после трансфекции среду заменяли полной средой DMEM/F-12. Клетки расщепляли на следующий день в соотношении 1:10. Отбор трансфицированных клеток начинали через 48 ч после трансфекции с помощью сульфата G418 (Gibco/Life Technologies GmbH, Darmstadt, Germany) в конечной концентрации 0,5 мг/мл. G418 непрерывно добавляли в культуральную среду для выращивания клеток.
c. Отбор HEK293 в качестве клеток-продуцентов
Экспрессию белков bi-scFv стабильно трансфицированными клеточными линиями HEK293 и CHO-1, описанными в примере l.b, характеризовали с помощью иммунофлуоресцентного окрашивания для детекции экспрессии bi-scFv в соответствии со стандартными процедурами (Current Protocols in Immunology, 2012). Вкратце, 2×105 клеток выращивали на предметных стеклах в течение 24 ч и затем пермеабилизировали 2% PFA. DPBS, дополненный 5% BSA и 0,2% сапонином, использовали в качестве блокирующего буфера. После промывания с помощью DPBS и блокирования с помощью блокирующего буфера клетки инкубировали с первичным антителом Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в разведении 1:500 в блокирующем буфере в течение 30 мин при RT. После отмывания блокирующим буфером добавляли вторичное конъюгированное с Cy3 козье анти-мышиное IgG (H+L) антитело (Jackson ImmunoResearch Europe, Suffolk, England) в разведении 1:500 в блокирующем буфере и инкубировали в течение 3 ч при RT. После промывания блокирующим буфером и H2O клетки погружали в DAKO-закрепляющую среду (Dako, Carpinteria, CA, USA), дополненную красителем Hoechst 33342 (Pierce/Thermo Fisher Scientific, Rockford, IL, USA). Срезы исследовали и делали снимки с помощью флуоресцентного микроскопа Nikon-Eclipse Ti для выявления присутствия bi-scFv-положительных клеток (данные не показаны). Клетки HEK293 показали лучшую общую экспрессию белков bi-scFv по сравнению с клетками CHO-K1 и, таким образом, были отобраны в качестве клеточной линии-продуцента.
d. Продукция и детекция белка 1BiMAB bi-scFv с помощью клона #28 HEK293
1BiMAB bi-scFv был отобран в качестве первого белка bi-scFv, подлежащего получению, очистке и использованию для проведения различных анализов. Для этой цели клональные клеточные линии всех клеток HEK293, стабильно экспрессирующих 1BiMAB (смотри Пример l.b), получали путем сортинга отдельных клеток с использованием клеточного сортера FACS Aria cell sorter (BD Biosciences, Heidelberg, Germany). После размножения около 40 клональных линий лучший клон-продуцент был отобран с помощью иммунофлуоресценции, как описано в Примере 1.с.
Отобранный клон-продуцент #28 размножали и культивировали в 10-слойной клеточной фабрике (Nunc, Roskilde, Denmark) в среде DMEM/F-12 GlutaMax, дополненной 10% FBS, 0,5% пенициллина-стрептомицина и 0,8 мг/мл G418 (все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany) в соответствии с инструкциями производителя. На конфлюэнтной стадии клетки промывали DPBS и среду заменяли на DMEM/F-12 с антибиотиками, но без FBS. Клеточный супенатант, содержащий белок 1BiMAB bi-scFv, собирали каждые 3-5 дней в течение 4 недель. Супернатант фильтровали с помощью 500 мл Steritop Filter Units (Merck Millipore, Billerica, MA, USA) и хранили при 4°C до FPLC-очистки.
Перед FPLC-очисткой присутствие bi-scFv в супернатанте клеточной культуры тестировали путем электрофореза на полиакриламидном геле с последующим окрашиванием кумасси и вестерн-блот-анализом, выполняемым стандартным способом (Current Protocols in Protein Science, 2012). Супернатант концентрировали 5×-10× с помощью устройств Centricon Centrifugal Filter Devices -10K MWCO (Merck Millipore, Billerica, MA, USA) в соответствии с протоколом производителя. Концентрированные и неконцентрированные супернатанты разделяли на NuPAGE Novex 4-12% Bis-Tris Gels (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). Затем гели окрашивали раствором кумасси брильянтовым синим в соответствии со стандартными процедурами для детекции белка 1BiMAB bi-scFv в диапазоне от 50 до 60 kD и других белков, содержащихся в супернатанте клеточной культуры. Вестерн-блот-анализ выполняли для специфической детекции белка 1BiMAB bi-scFc посредством его 6xHis-метки. Вкратце, после блоттинга белков на PVDF мембране и блокирования с помощью PBST/3% сухого молока мембрану инкубировали в течение 1 ч при 4°C с первичным антителом Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в разведении 1:500 в блокирующем буфере. После промывания блокирующим буфером мембраны инкубировали с Fc-специфическим вторичным козьим-анти-мышиным IgG антитело, конъюгированным с пероксидазой (Sigma Aldrich, Germany), в разведении 1 : 10000 в блокирующем буфере в течение 1 ч при 4°C. После промывания блокирующим буфером сигналы визуализировали с помощью SuperSignal West Femto Chemiluminescent Substrate (Pierce/Thermo Fisher Scientific, Rockford, IL, USA) и записывали при помощи ImageQuant LAS 4000 Imager (GE Healthcare Life Sciences, Munich, Germany). Сигналы bi-scFv детектировали в диапазоне от 50 до 60 kD в сравнении с внутренними стандартами молекулярной массы (смотри фигуру 3 A и B).
e. Очистка и количественное определение белка 1BiMAB bi-scFv
Супернатант клеточной культуры клона #28 HEK293, содержащий белок 1BiMAB bi-scFv (описанный в примере l.d), подвергали аффинной хроматографии с использованием иммобилизованных металлов (IMAC) с использованием стандартных процедур (Current Protocols in Protein Science, 2012). Вкратце, фильтрованный супернатант клеточной культуры наносили на колонку His Trap FF 5 мл, соединенную с системой AКTA Purifier 10 FPLC system (обе фирмы GE Healthcare Life Sciences, Munich, Germany). Промывочный буфер PBS содержал 10 мМ имидазола, элюирующий буфер PBS содержал 500 мМ NaCl, 50 мМ NaH2PO4 и 250 мМ имидазола, pH обоих буферов был доведен до 7,4. Элюирование выполняли с использованием ступенчатого градиента. Элюированный белок 1BiMAB bi-scFv сразу же подвергали диализу против lx PBS с использованием Slide-A-Lyzer G2 Dialysis Cassette 10K MWCO (Pierce/Thermo Fisher Scientific, Rockford, IL, USA). После диализа против l x PBS, bi-scFv подвергали диализу против 200 мМ аргининового буфера на основе H2O (L-аргинин-моногидрохлорид; Roth, Karlsruhe, Germany).
Концентрацию bi-scFv определяли путем измерения при 280 нм с помощью NanoDrop 2000c, учитывая коэффициент экстинкции и молекулярную массу белка 1BiMAB bi-scFv, определенные с помощью инструмента ProtParam tool (http://web.expasy.org/protparam/). Очищенный белок разделяли на аликвоты и хранили при -80°C для длительного хранения или при 4°C для немедленного использования.
Качество и чистоту белка 1BiMAB bi-scFv тестировали путем окрашивания кумасси и вестерн-блот-анализа, как описано в Примере l.d (смотри также фигуры 3А и В). Стандартное разведение BSA было включено в процедуру окрашивания кумасси для предварительного подтверждения концентрации, измеренной с помощью NanoDrop (данные не показаны).
f. Проведение анализа ELISA
Для количественного определения 1BiMAB в супернатанте клеточной культуры клеток HEK 293 проводили специфический анализ ELISA. Для этой цели использовали супернатант из Примера 1.d и очищенный белок IBiMAB bi-scFv, описанный в Примере l.e. Предварительно блокированные BSA колонках Ni-NTA (Thermo Fisher Scientific, Rockford, IL, USA) использовали для иммобилизации белка 1BiMAB bi-scFv посредством его 6xHis-метки. Все стадии промывания выполняли три раза 200 мкл l× PBS/0,05 % Tween (промывочный буфер) на лунку и все стадии выполняли при комнатной температуре. В качестве стандарта использовали очищенный белок 1BiMAB, разбавленный в l× PBS в диапазоне 10-500 нг/мл. Супернатанты разбавляли в соотношении 1:10 в l× PBS. В каждую лунку переносили по 100 мкл разбавленного белка или супернатанта и инкубировали в течение одного часа при встряхивании. После промывания анти-идиотипическое антитело против доменов VH-VL mCLDN18.2ab разбавляли до конечной концентрации 0,5 мкг/мл в l× PBS/3% BSA. В каждую лунку добавляли по 100 мкл раствора анти-mCLDN18.2ab и инкубировали в течение одного часа при встряхивании. После промывания AP-конъюгированное анти-мышиное-Fc антитело (Jackson ImmunoResearch Europe, Suffolk, England) разбавляли до конечной концентрации 300 нг/мл в l× PBS/3% BSA. В каждую лунку добавляли по 100 мкл этого раствора вторичного антитела и инкубировали дополнительно в течение часа при встряхивании. В качестве отрицательных контролей использовали только вторичное антитело, 1BiMAB плюс вторичное антитело, и анти-mCLDN18.2ab плюс вторичное антитело. Кроме того, в анализ был включен супернатант клеток HEK293 без белка bi-scFv. В заключение, после промывания в каждую лунку добавляли по 50 мкл раствора AP-субстрата (1,5 мг pNPP на мл субстратного буфера, AppliChem GmbH, Darmstadt, Germany). Через 5, 15 и 30 мин инкубации в условиях темноты измеряли поглощение при 405 нм с длиной волны испускания 492 нм с помощью микропланшетного ридера Infinite M200 Tecan (Tecan, Mannedorf, Switzerland). Концентрацию белка bi-scFv в супернатанте определяли путем расчета против стандартного ряда (данные не показаны).
g. Временная трансфекция CLDN18.2-специфических белков bi-scFv для сравнительных исследований
Для временной генерации предпочтительно высоких количеств CLDN18.2-специфических белков bi-scFv использовали клеточную линию эмбриональных почек человека HEK293T (ATCC CRL-11268) для трансфекции.
1×107 клеток HEK293T засевали за два дня до трансфекции на чашки Петри 14,5 см в 20 мл полной среды DMEM (DMEM/F-12 GlutaMax, дополненной 10% термоинактивированной FBS и 0,5% пенициллина-стрептомицина; все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany). Перед трансфекцией клетки промывали DPBS, дополненным 2 мМ EDTA, затем добавляли 20 мл обычной среды DMEM без FBS или антибиотиков. 20 мкг кольцевых ДНК-конструкций 1BiMAB, no. 11 - 20 и no.35 (описанных в Примере l.b), разбавляли в 0,5 мл обычной среды DMEM/F-12. В разбавленную ДНК добавляли 75 мкл раствора линейного PEI (полиэтиленимин; Polysciences Europe GmbH, Eppelheim, Germany) при концентрации 1 мг/мл и энергично перемешивали на вортексе. Через 15 мин инкубации при комнатной температуре (RT) в клетки по каплям добавляли комплексы ДНК/PEI, чашки для клеточных культур осторожно перемешивали вращательным движением и затем инкубировали при 37°C, 5% CO2 в течение 24 ч. После замены среды на обычную DMEM/F-12 клетки инкубировали дополнительно в течение 48 ч при 33°C, 5% CO2. Клеточный супернатант собирали после инкубации и стерильно фильтровали с помощью 0,2 мкм шприцевых фильтров Minisart syringe filters (Sigma-Aldrich, Germany). Затем белки bi-scFv очищали в малом масштабе из супернатантов клеточных культур с помощью спин-колонок Ni-NTA в соответствии с протоколом производителя (Qiagen, Hilden, Gemany). Концентрации белка Bi-scFv оценивали с помощью ELISA, как описано в примере 1.f, и подтверждали с помощью вестерн-блот анализа, как описано в примере l.e (данные не показаны). Очищенные белки хранили при 4°C для немедленного использования.
Пример 2: Проведение функциональных анализов для контроля специфической активации Т-клеток и лизиса клеток-мишеней с помощью перенаправленных Т-клеток, опосредованных белками bi-scFv.
FPLC-очищенный белок 1BiMAB bi-scFv использовали для проведения анализов in vitro для контроля способности белков bi-scFv специфически перенаправлять человеческие эффекторные клетки на TAA-положительные клетки-мишени. Целью является визуализация эффектов и количественная оценка активации человеческих Т-клеток и специфического лизиса клеток-мишеней.
a. Микроскопический анализ Т-клеток, перенаправленных на клетки-мишени с помощью белка bi-scFv
Для визуализации функциональности белка bi-scFv проводили анализ для демонстрации перенаправления эффекторных клеток на CLDN18.2-экспрессирующие клетки-мишени белками bi-scFv с помощью микроскопа. Для этой цели в качестве клеточной линии-мишени использовали клеточную линию карциномы желудка NugC4, которая эндогенно экспрессирует сравнительно высокие уровни человеческого CLDN18.2 (Sahin U. et al., Clin Cancer Res. 2008 Dec l;14(23):7624-34).
Человеческие эффекторные клетки свежеизолировали из человеческой крови здоровых доноров в соответствии со стандартными процедурами (Current Protocols in Immunology, 2012): вкратце, кровь разбавляли DPBS, наносили слоями на Ficoll-Paque Plus (GE Healthcare Life Sciences, Munich, Germany) и центрифугировали. Мононуклеарные клетки периферической крови (PBMC) собирали из интерфазы, промывали холодным DPBS, дополненным 2 мМ EDTA и подсчитывали. Человеческие T- клетки затем отделяли путем отделения магнитно-активированных клеток (MACS) от PBMC с помощью набора Pan T Cell Isolation Kit II (Miltenyi Biotec, Teterow, Germany) в соответствии с протоколом производителя.
1×105 клеток NugC4 засевали на лунку в тканевые культуральные 6-луночные планшеты. Человеческие клетки готовили, как описано выше, и добавляли в соотношении эффектора к мишени (E:T) 5:1. Среду RPMI 1640, дополненную 5% термоинактивированной человеческой AB сывороткой, 0,5% пенициллина-стрептомицина, l× NEAA и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany), использовали для всех клеток, и конечный объем каждой лунки доводили до 2 мл на лунку. Контрольные образцы содержали клетки-мишени или Т-клетки с белком bi-scFv или без него. Тканевые культуральные планшеты затем инкубировали при 37°C, 5% CO2. Наблюдение за ходом анализа проводили в непрерывном режиме на инвертационном микроскопе Wilovert S inverted microscope (Hund, Wetzlar, Germany) в интервале от 6 ч до 48 ч совместной инкубации. Значительные эффекты в отношении образования кластеров Т-клеток на клетках-мишенях, формирования иммунологического синапса и уничтожения клеток-мишеней в присутствии белка 1BiMAB bi-scFv наблюдали через 24 ч. Через 48 ч жизнеспособные клетки-мишени с трудом могли быть обнаружены. Снимки делали через 24 ч на инвертационном микроскопе Nikon Eclipse TS 100 (Nikon, Japan). Смотри также фигуру 5.
Этот анализ в дальнейшем использовали в качестве визуального контроля во всех анализах на цитотоксичность в различных форматах лунок.
b. Зависимая от мишени активация Т-клеток белком 1BiMAB bi-scFv
Для детекции специфической активации человеческих Т-клеток белками bi-scFv проводили анализ методом проточной цитометрии. Для детекции активации Т-клеток маркер ранней активации CD69 и маркер поздней активации CD25 были отобраны для окрашивания антителами, конъюгированными с флуоресцентными метками. Для детекции человеческих Т-клеток в смеси клеток-мишеней и Т-клеток, CD3 на Т-клетках окрашивали.
Порядок проведения анализа был выбран из приведенного выше примера (Пример 2.а). Вкратце, клетки-мишени NugC4 засевали с человеческими Т-клетками в соотношении E:T, равном 5:1, в 2 мл полной среды, и белок 1BiMAB bi-scFv добавляли в концентрации с диапазоне 0,001 - 1000 нг/мл. Контрольные образцы содержали клетки-мишени или Т-клетки с белком 1BiMAB bi-scFv или без него. Через 24 ч и/или 48 ч - в зависимости от результата визуального контроля - все клетки собирали путем осторожного соскабливания с помощью Cell Scrapers (Sarstedt AG & Co, Niirmbrecht, Germany) и переносили в пробирки с круглым дном объемом 5 мл (BD Falcon, Heidelberg, Germany). Клетки центрифугировали и промывали DPBS. Для окрашивания клеток использовали мышиные античеловеческие CD3-FITC, мышиные античеловеческие CD69-APC и мышиные античеловеческие CD25-PE антитела (все антитела фирмы BD Biosciences, Heidelberg, Germany). Клеточные осадки ресуспендировали в 50 мкл FACS-буфера (DPBS, дополненный 5% FBS), содержащего антитела, конъюгированные с флуоресцентными метками. После инкубации в течение 20 мин при 4°C в условиях темноты образцы промывали 4 мл DPBS и клеточный осадок ресуспендировали в 200 мкл FACS-буфера, содержащего иодид пропидия (PI) или 7-AAD (обе фирмы Sigma Aldrich, Germany) в конечном разведении 1 : 1000 для детекции погибших клеток. Образцы хранили на льду и в условиях темноты на протяжении проведения измерения. Анализ выполняли с помощью FACSCalibur, последующие измерения выполняли с помощью проточного цитометра FACSCanto II (оба фирмы BD Biosciences, Heidelberg, Germany). Данные анализов оценивали с помощью программного обеспечения FlowJo software (Tree Star, San Carlos, CA, USA).
Как показано на фигурах 6А и В, 1BiMAB-опосредованной активации Т-клеток не обнаруживается в отсутствие клеток-мишеней, подтверждая строгую зависимость функциональности bi-scFv от мишени. Значительная активация Т-клеток в присутствии клеток-мишеней возникала только при концентрации 1BiMAB, составляющей 0,01 нг/мл, через 24 ч. Максимальная эффективность была достигнута с использованием 100 нг/мл 1BiMAB.
Помимо исследования активации Т-клеток, данный анализ обеспечивает также количественные анализы опосредованных bi-scFv эффектов в отношении уничтожения клеток-мишеней, путем гейтирования популяции клеток-мишеней и оценки процентного содержания PI- или 7-AAD-положительных клеток-мишеней (данные отсутствуют). Все анализы выполняли с использованием программного обеспечения FlowJo (Tree Star, San Carlos, CA, USA).
c. Анализ на цитотоксичность с использованием люциферазы
Для определения различий в потенциале белков bi-scFv, направленных против CLDN18.2 и CD3, уничтожать клетки-мишени, должен быть разработан высокочувствительный анализ. Цель состояла в проведении анализа, с помощью которого можно было высокопроизводительным образом количественно контролировать уничтожение клеток-мишеней. Для достижения этой цели был выбран цитотоксический анализ с использованием люциферазы. С помощью данного анализа измерение экспрессии люциферазы жизнеспособными клетками-мишенями обеспечивает возможность непрямого определения лизиса клеток-мишеней, опосредованного цитотоксическими эффекторными клетками в присутствие антитела.
Сначала клетки NugC4 (описанные выше) трансдуцировали лентивирусным вектором, несущим люциферазу светлячка, репортерный ген EGFP и маркер селекции антибиотика. После антибиотического отбора трансдуцированных клеток, клетки, экспрессирующие на высоком уровне EGFP, подвергали сортингу с помощью клеточного сортера FACSAria (BD Biosciences, Heidelberg, Germany), анализировали на высокую экспрессию люциферазы и затем размножали для последующих исследований.
Человеческие эффекторные клетки готовили, как описано в примере 2.а. Анализ проводили при концентрации белка 1BiMAB bi-scFv в диапазоне 1-100 нг/мл, при этом было обнаружено, что концентрация 5 нг/мл обеспечивала высокоэффективные и воспроизводимые результаты, и в дальнейшем была использована в качестве стандартной концентрации. Клетки NugC4, стабильно экспрессирующие люциферазу (описанную выше), использовали в качестве клеток-мишеней. В каждую лунку засевали 1×104 клеток-мишеней в белые плоскодонные 96-луночные планшеты. Человеческие Т-клетки (приготовленные, как описано в Примере 2.а) добавляли в соотношении E:T, равном 5:1. Использовали среду, описанную выше (Пример 2.а), и конечный объем каждой лунки доводили до 100 мкл. Тестируемые образцы и контрольные образцы засевали, по меньшей мере, в трех повторностях. Клеточные культуральные микропланшеты инкубировали в течение 24 ч и 48 ч при 37°C, 5% CO2. Для анализа 50 мкл водного раствора, содержащего 1 мг/мл люциферина (BD Monolight, BD Biosciences, Heidelberg, Germany) и 50 мМ HEPES добавляли в каждую лунку, и планшеты затем инкубировали в течение 30 мин в условиях темноты при 37°C. Люминесценцию, возникающую в результате окисления люциферина люциферазой, экспрессирующей жизнеспособные клетки, измеряли на микропланшетном ридере (Infinite M200, Tecan, Mannedorf, Switzerland). Процент специфического лизиса клеток-мишеней рассчитывали по следующей формуле: % специфического лизиса = [1 - (люминесценциятеституемый образец- Lmax) / (Lmin - Lmax)] × 100, где «L» означает лизис. Lmin относится к минимальному лизису в отсутствие bi-scFv, и Lmax к максимальному лизису (равному числу импульсов спонтанной люминесценции) в отсутствие bi-scFv, достигнутому путем добавления Triton X-100 (конечная концентрация 2%).
Потенциальные прямые эффекты белков bi-scFv на клетки-мишени независимо от эффекторных клеток определяли путем засевания клеток-мишеней без человеческих Т-клеток, включая все контроли, такие как Lmin и Lmax. Данный анализ использовали для последующих исследований для изучения специфического лизиса клеток-мишеней, опосредованного Т-клетками. Модификации осуществляли, например, путем изменения концентраций bi-scFv, белков bi-scFv, отношений E:T или эффекторных клеток (CD8+, CD4+ T-клетки, PBMC).
Пример 3: Отбор лидерного кандидата CLDN18.2-специфического bi-scFv
Анализ на цитотоксичность с использованием люциферазы с различными CLDN18.2-специфическими белками bi-scFv для отбора наиболее сильного варианта bi-scFv
Все 10 кодон-оптимизированных конструкций CHO (no. 11-20), специфических в отношении TAA CLDN18.2, тестировали в сравнении с оптимизированным по кодонам человеческим белком 1BiMAB bi-scFv в анализе на цитотоксичность с использованием люциферазы с клетками-мишенями NugC4, которые эндогенно экспрессируют CLDN18.2 и эктопически экспрессируют люциферазу (смотри также пример 2.с). Характеристики используемых белков bi-scFv представлены в таблице 2. Bi-scFv no.35, специфический в отношении TAA PLAC1, использовали в качестве контроля изотипа, так как PLAC1 не экспрессируется клетками NugC4. Активность связывания с CD3 на человеческих Т-клетках была подтверждена в анализе связывания FACS (данные не показаны). Все белки bi-scFv генерировали, как описано в примере 1.g, и использовали для анализа на цитотоксичность, как описано в примере 2.с.
Все белки bi-scFv использовали в конечной концентрации 5 нг/мл. Для определения Lmin, контрольный белок bi-scFv no.35 засевали с клетками-мишенями и Т-клетками в девяти повторностях, тестируемый образцы засевали в шести повторностях. На каждый момент времени один планшет готовили для анализа.
Специфический лизис в каждый анализируемый момент времени (8 ч, 16 ч, 24 ч) показан в зависимости от используемых белков bi-scFv. Было доказано, что белки 1BiMAB bi-scFv (SEQ ID NO: 39) и no.15 (SEQ ID NO: 41) - которые сконструированы в одинаковой ориентации и содержат одинаковую последовательность анти-CD3 (TR66), и отличаются только по использованию их кодонов на уровне нуклеиновой кислоты и по последовательностям линкера - являются наиболее сильными антителами в отношении опосредования лизиса клеток-мишеней (смотри фигуру 2). Так как 1BiMAB и no.15 равносильны по эффективности, наиболее хорошо изученный на сегодняшний день белок 1BiMAB bi-scFv был отобран для всех последующих анализов. Конструкции 18PHU3 и 18PHU5 (смотри таблицу 1 и 2) сравнивали в более поздний момент времени с 1BiMAB. Эффективность 18PHU5 была эквивалентна 1BiMAB, 18PHU3 был менее сильным (данные не показаны).
Пример 4: Связывающая способность белка 1BiMAB bi-scFv
Проведение анализа на связывание на основе FACS
Анализ методом проточной цитометрии использовали для оценки связывающей способности CLDN18.2 и фрагментов белков bi-scFv, направленно воздействующих на CD3. CLDN18.2, эндогенно экспрессируемый клетками NugC4, использовали для изучения анти-CLDN18.2 сайта и человеческие Т-клетки использовали для изучения анти-CD3 сайта.
Для изучения связывающей способности анти-CLDN18.2, клетки NugC4 трипсинировали, промывали полной средой RPMI 1640 и затем DPBS. Все промывочные стадии выполняли путем центрифугировали при 1200 об/мин в течение 6 мин при 4°C. 2,5 x l05 клеток NugC4 переносили в круглодонные пробирки объемом 5 мл и инкубировали с 50 мкг/мл FPLC-очищенного белка 1BiMAB в FACS-буфере в течение 30 мин при 4°C. Клетки промывали 2 мл FACS-буфера и затем инкубировали с 3,3 мкг/мл моноклонального антитела Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в течение 30 мин при 4°C. После промывания 2 мл FACS-буфера, клеточный осадок инкубировали с APC-конъюгированным козьим-анти-мышиным вторичным антителом (Jackson ImmunoResearch Europe, Suffolk, England) в разведении 1:200 в FACS-буфере в течение 20 мин при 4°C в условиях темноты. Клетки дважды промывали 2 мл FACS-буфера и в заключение ресуспендировали в 150 мкл FACS-буфера, дополненного 1 мкг/мл PI (Sigma Aldrich, Germany) для контрастного окрашивания погибших клеток. Другое окрашивание было включено с использованием такой же процедуры с помощью 50 мкг/мл 1BiMAB и APC-конъюгированного козьего-анти-мышиного вторичного антитела (1:200), но без Anti-HIS Epitope-Tag антитела. Отрицательные контрольные образцы включали вторичное козье-анти-мышиное антитело, конъюгированное с аллофикоцианином (APC), моноклональное антитело Anti-HIS Epitope-Tag плюс вторичное козье-анти-мышиное антитело, конъюгированное с APC. В качестве положительного контроля использовали 10 мкг/мл моноклонального CLDN18.2-специфического антитела mCLDN18.2ab, окрашенного вторичным козьим-анти-человеческим антителом, конъюгированным с APC (Jackson ImmunoResearch Europe, Suffolk, England), и его вторичного антитела.
Образцы измеряли с помощью проточного цитометра FACSCalibur (BD Biosciences, Heidelberg, Germany) и анализировали с помощью программного обеспечения FlowJo Software (Tree Star, San Carlos, CA, USA). Сильные сигналы детектировали путем последовательного окрашивания 1BiMAB, Anti-HIS Epitope-Tag и козьим-анти-мышиным APC. Интенсивность сигнала была сравнима с положительным контролем mCLDN18.2ab с козьим-анти-человеческим APC. Низкий уровень прямого связывания козьего-анти-мышиного APC с 1BiMAB наблюдали в образце, окрашенном 1BiMAB и козьим-анти-мышиным APC без Anti-HIS Epitope-Tag (смотри фигуру 4 A).
Для всех последующих анализов FACS-связывания для исследования связывающей способности белков bi-scFv использовали последовательный протокол, окрашивая bi-scFv, Anti-HIS Epitope-Tag и козьим-анти-мышиным APC (смотри фигуру 4B, C и D). Для исключения неспецифического связывания 1BiMAB, клетки-мишени, которые не экспрессируют CLDN18.2, что подтверждается данными RT-PCR (данные не показаны), подвергали анализу на связывание на основе FACS. Неспецифического связывания 1BiMAB не было обнаружено, как показано на фигуре 4D.
Для изучения связывающей способности анти-CD3 плеча белка 1BiMAB bi-scFv использовали человеческие Т-клетки, 1×106 T-клеток, полученных, как описано в примере 2.а, переносили в круглодонные пробирки объемом 5 мл и инкубировали с FPLC-очищенным белком 1BiMAB при концентрации в диапазоне 0,002 - 2 мкг/мл в FACS-буфере в течение 30 мин при 4°C. Последующая процедура окрашивания описана выше. Контрольные образцы включали только вторичное козье-анти-мышиное APC антитело и моноклональное антитело Anti-HIS Epitope-Tag плюс вторичное козье-анти-мышиное APC антитело. Измерения и анализ проводили, как описано выше. Значительный сигнал был получен при 2 мкг/мл 1BiMAB (смотри фигуру 4C).
Пример 5: Изучение высокоспецифической, зависящей от мишени активации T-клеток белком 1BiMAB bi-scFv
Линии раковых клеток, которые эндогенно экспрессируют высокие или низкие уровни CLDN 18.2, и линии раковых клеток, которые не экспрессируют CLDN18.2, отбирали для подтверждения строгой зависимости от мишени белка 1BiMAB bi-scFv в анализе in vitro цитотоксичности. Отобранные клеточные линии представляли собой два основных типа карциномы, которые экспрессируют CLDN18.2: карциному желудка (NugC4, MKN7, SNU-1) и поджелудочной железы (DanG, KP-4). Клеточную линию рака молочной железы MCF7 использовали в качестве отрицательного контроля.
a. Анализ методом RT-PCR CLDN18.2 клеточных линий злокачественных новообразований
Общую РНК экстрагировали из клеточных линий карциномы, указанных выше, с помощью набора RNEasy Mini Kit согласно процедуре в соответствии с протоколом производителя (Quiagen, Hilden, Germany). 5 мкг РНК использовали для синтеза кДНК с использованием обратной транскриптазы Superscript II (Life Technologies GmbH, Darmstadt, Germany).
Анализы RT-PCR проводили с применением системы ABI Prism 7300 Real Time PCR System (Applied Biosystems/Life Technologies GmbH, Darmstadt, Germany) с использование красителя Sybr Green и следующих праймеров:
CLDN18.2: для TGGCTCTGTGTCGACACTGTG; rev GTGTACATGTTAGCTGTGGAC HPRT: для TGACACTGGCAAAACAATGCA; rev GGTCCTTTTCACCAGCAAGCT
Величину дельта Ct рассчитывали путем вычитания величины Ct гена домашнего хозяйства HPRT из величины Ct CLDN18.2 (результаты показаны на фигуре 7 A).
b. Эксклюзивная активация Т-клеток в присутствие CLDN18.2
Анализ цитотоксичности проводили, как описано в примере 2.а. Клеточные линии карциномы, исследованные в отношении транскриптов CLDN18.2 согласно примеру 5.а посредством количественной RT-PCR, использовали в качестве клеток-мишеней. Концентрация белка 1BiMAB bi-scFv в данном анализе составила 5 нг/мл. Клетки-мишени засевали с человеческими Т-клетками и 1BiMAB в двух повторностях для анализа активации Т-клеток. Для контроля любой потенциальной аллореативности Т-клеток против клеток-мишеней независимо от белка 1BiMAB bi-scF, клетки-мишени и Т-клетки засевали без 1BiMAB в двух повторностях. Клетки непрерывно наблюдали под микроскопом на образование класеров Т-клеток и связывание клеток-мишеней. В случае клеточной линии NugC4, высокоэкспрессирующей CLDN18.2, значительный эффект возникал через 2 ч; через 48 ч жизнеспособные клетки-мишени были еле видимыми. В случае низкоэкспрессирующей CLDN18.2 клеточной линии DanG первые эффекты наблюдались через 96 ч и значительные эффекты через 120 ч. В случае CLDN18.2-отрицательных клеточных линий эффекты, указывающие на какую-либо активацию Т-клеток, отсутствовали даже через 144 ч. Т-клетки всех образцов анализировали через 144 ч совместной инкубации с клетками-мишенями с помощью проточной цитометрии, как описано в примере 2.а для маркера CD69 ранней активации Т-клеток и маркера CD25 поздней активации, контрастно окрашенных CD3 для T-клеточной популяции и PI для погибших клеток. Удивительным образом, до 100% Т-клеток, совместно инкубированных с NugC4 и 1BiMAB, были CD25-положительными, но CD69-отрицательными, что указывало на продолжительную активацию Т-клеток, когда понижающая регуляция CD69 уже возникла. Приблизительно 75% Т-клеток, совместно инкубированных с DanG и 1BiMAB, было активировано, из которых примерно 40% одновременно экспрессировали CD25 и CD69, что указывало на активацию Т-клеток, которая все еще продолжается. Т-клетки, совместно инкубированные с CLDN18.2-отрицательными клеточными линиями, не показали какого-либо признака индукции активации Т-клеток: экспрессия CD69 и CD25 не была значительно повышенной по сравнению с уровнями экспрессии образцов без 1BiMAB (смотри также фигуру 7B).
Пример 6: Исследование индуцированной белком 1BiMAB bi-scFv функции Т-клеток
a. Индукция пролиферации Т-клеток
Пролиферация Т-клеток является признаком активации Т-клеток. Для демонстрации пролиферации Т-клеток в ответ на белок 1BiMAB bi-scFv в присутствии CLDN18.2-положительных клеток-мишеней использовали анализ методом проточной цитометрии. Вкратце, 1×106 человеческих Т-клеток выделяли, как описано в примере 2.а, окрашивали в условиях темноты при 37°C в течение 10 мин с 0,5 мкМ карбоксифлуоресцеина диацетата сукцинимидилового сложного эфира (CellTrace CFSE, Invitrogen/Life Technologies GmbH, Germany), растворенного в DPBS. Окрашивание останавливали путем добавления 5 объемов холодной полной среды RPMI 1640. Клетки хранили на льду в течение 5 мин и промывали 3 раза полной средой RPMI (5% термоинактивированной человеческой сыворотки AB, 0,5% пенициллина-стрептомицина, 1x NEAA и 1 мМ пирувата натрия) и затем ресуспендировали до 1×105 клеток на мл. Анализ на цитотоксичность, описанный в примере 2.b, проводили с CLDN18.2, эндогенно экспрессирующим клетки NugC4, и человеческими Т-клетками в качестве эффекторных клеток. В клетки добавляли 50 ЕД IL-2 на мл среды. Образцы включали только Т-клетки, Т-клетки с 1 нг/мл 1BiMAB, Т-клетки и клетки NugC4, и Т-клетки с 1 нг/мл 1BiMAB и клетками NugC4. Через 120 ч совместной инкубации Т-клетки отбирали, собирали в круглодонные пробирки объемом 5 мл, промывали и окрашивали раствором 7-AAD DPBS в разведении 1:1000 для контрастного окрашивания погибших клеток в течение 15 мин при 4°C. После промывания DPBS клетки ресуспендировали в FACS-буфере и анализировали с помощью FACSCanto II (BD Biosciences, Heidelberg, Germany).
Пролиферацию Т-клеток детектировали по уменьшению CFSE-сигнала только в присутствии клеток-мишеней и белка 1BiMAB bi-scFv (смотри также фигуру 8 A).
b. Индукция серинпротеазы Granzyme B
Для демонстрации повышающего регулирования протеолитических молекул после активации Т-клеток, опосредованной белком 1BiMAB bi-scFv в присутствии CLDN18.2-положительных клеток-мишеней, выбрали детекцию серинпротеазы Granzyme B посредством анализа методом проточной цитометрии. Анализ на цитотоксичность, описанный в примере 2.b, проводили с CLDN18.2, эндогенно экспрессирующим клетки NugC4, и человеческими Т-клетками в качестве эффекторных клеток. Образцы включали только Т-клетки, Т-клетки с 5 нг/мл 1BiMAB, Т-клетки и клетки NugC4, и Т-клетки с 5 нг/мл 1BiMAB и клетками NugC4. Через 96 ч совместной инкубации Т-клетки отбирали, собирали в круглодонные пробирки объемом 5 мл, промывали и окрашивали раствором 7-AAD DPBS в разведении 1 : 1000 для контрастного окрашивания погибших клеток в течение 15 мин при 4°C. После промывания DPBS клетки фиксировали 100 мкл раствора Cytoperm/Cytofix в течение 20 мин при комнатной температуре (RT). Клетки промывали lx Perm/Wash и затем окрашивали PE-конъюгированным мышиным античеловеческим Granzyme B антителом в течение 20 мин при RT. После промывания клетки ресуспендировали в FACS-буфере и анализировали на приборе FACSCanto II (все реагенты и FACS machine BD Biosciences, Heidelberg, Germany).
Повышающую регуляцию Granzyme B в Т-клетках детектировали только в присутствии клеток-мишеней и белка 1BiMAB bi-scFv (смотри также фигуру 8B).
Пример 7: Определение EC50 белка 1BiMAB bi-scFv в анализе in vitro на цитотоксичность
Анализ на цитотоксичность с использованием люциферазы
Для определения половины максимальной эффективной дозы белка 1BiMAB bi-scFv, ряд титрований 1BiMAB тестировали в анализе in vitro на цитотоксичность с использованием люциферазы, главным образом, как описано в примере 2.c.
Стабильно экспрессирующие люциферазу клетки NugC4, описанные в примере 2.c, инкулировали с человеческими Т-клетками и белком 1BiMAB bi-scFv в концентрациях в диапазоне от 1 пг/мл до 1 мкг/мл (10 шагов) и без 1BiMAB для определения величин Lmin. Люминесценцию жизнеспособных клеток измеряли на планшетном ридере Infinite M200 Tecan через 24 ч и 48 ч после начала анализа. Специфический лизис клеток-мишеней рассчитывали по формуле, приведенной в примере 2.c.
Максимальный лизис был достигнут через 48 ч с использованием 1 - 10 нг/мл 1BiMAB. Определенная величина EC50 через 48 ч в этом анализе составила 10 пг/мл (смотри также фигуру 9). Результат этого анализа зависит в значительной степени от активности человеческих Т-клеток, которая различается в зависимости от иммунного статуса донора, как описано в других работах (например, Lutterbuese, R et al., Proc. Natl. Acad. Sci. USA. 2010 Jul 13; 107(28): 12605- 10). Кроме того, используемая линия клеток-мишеней NugC4 показывает изменяющуюся экспрессию CLDN18.2, также влияя на результат. Таким образом, в ходе изобретения наблюдалось отклонение величин EC50 белка 1BiMAB bi-scFv в диапазоне 10 - 300 пг/мл.
Пример 8: Эффективность в мышиной модели ксенотрансплантата
Для исследования терапевтического потенциала белка 1BiMAB bi-scFv in vivo был выбран мышиный штамм NOD.Cg-Prkdscid IL2rgtmlwjl/SzJ или короткий NSG (Jackson laboratory, Bar Harbour, ME, USA). Для описанного исследования прививание человеческих эффекторных клеток и человеческих Т-лимфоцитов у мышей необходимо для исследования эффектов Т-клеточного захвата bi-scFv in vivo. Из-за полного отсутствия B-, T- и NK-клеток мышиный штамм NSG является подходящим для этого вида исследований ксенотрансплантата. Мышиная модель с привитыми, главным образом, человеческими Т-клетками после инъекции PBMC составляет часть изобретения.
a. Лечение с поздним началом опухолей на поздней стадии, высоко экспрессирующих CLDN18.2, у мышей с помощью белка 1BiMAB bi-scFv
В описанном исследовании 40 самкам мышей NSG в возрасте 8 недель подкожно инокулировали 1 x l07 клеток HEK293, стабильно экспрессирующих высокие уровни человеческого CLDN18.2 (HEK293-CLDN18.2). Через 5 дней после инокуляции опухолевых клеток мышей распределяли в соответствии с объемом их опухолей на лечебные группы, при этом мыши, у которых рост опухоли отсутствовал, были исключены. В тот же день мононуклеарные клетки периферической крови (PBMC) выделяли из человеческой крови здоровых доноров методом центрифугирования в градиенте плотности фиколла, как описано в примере 2.а, и использовали в качестве эффекторных клеток in vivo. 2×107 PBMC, разведенных в 300 мкл DPBS, инъецировали интраперитонеально в день выделения экспериментальным лечебным группам, обозначенным «PBMC». Лечебные группы, обозначенные «PBS», получали 300 мкл простого DPBS интраперитонеально и служили в качестве контроля без человеческих эффекторных клеток. С помощью контрольных групп «PBS» можно исследовать потенциальный эффект на рост опухоли самого 1BiMAB или любые потенциальные побочные эффекты, которые вызваны 1BiMAB или носителем, но не человеческими эффекторными клетками, против мышиной ткани (то есть реакция трансплантат против хозяина, вызванная человеческими эффекторными клетками, против мышиной ткани). Группа «PBS/носитель» включала 4 мышей (n=4), «PBS/1BiMAB» 5 мышей (n=5), «PBMC/носитель» 13 мышей (n=13) и «PBMC/1BiMAB» 15 мышей (n=15). Терапию начинали через 1 день после применения DPBS или PBMC: группы «PBS/1BiMAB» и «PBMC/1BiMAB» получали интраперитонеально 5 мкг очищенного белка 1BiMAB bi-scFv, разведенного в 200 мкл DPBS на животное. Группы «PBS/носитель» и «PBMC/носитель» получали интраперитонеально 200 мкл буферного носителя (200 мМ L-аргинина моногидрохлорида, растворенного в H2O, стерильно фильтрованного), разбавленного в DPBS. Лечебные группы кратко описаны в таблице 3. Терапию проводили на ежедневной основе в течение 22 дней. Дважды в неделю измеряли размер опухолей с помощью цифрового калиброванного калипера, и объем опухолей рассчитывали по формуле мм3 = длина × ширина × (ширина/2). На фигурах 10A и B показано ингибирование роста опухоли и уменьшение опухоли в два раза у мышей группы «PBMC/1BiMAB» только с помощью антитела в присутствии человеческих эффекторных клеток. Мышей умерщвляли путем смещения шейных позвонков, когда объем опухоли превышал 500 мм3, или в случае серьезных клинических проявлений (у некоторых мышей наблюдались симптомы реакции трансплантат против хозяина).
Таблица 3. Лечебные группы
(n)
клетки
b. Определение влияния терапии на массу тела
Массу тела каждой мыши измеряли два раза в неделю с использованием лабораторных весов. Ни одна из мышей в любой группе не показала снижение массы тела на протяжении периода лечения (данные не показаны). Некоторые мыши в обеих группах «PBMC» проявили симптомы реакции трансплантат против хозяина через 4 недели после инъекции PBMC и через несколько дней после завершения лечения. Эффекты самого 1BiMAB на массу тела или другие побочные эффекты, относящиеся к состоянию здоровья мышей, не наблюдались.
c. Консервация ткани и выделение спленоцитов
После умерщвления мышей опухоли удаляли, и ткань сразу же фиксировали в 10 мл 4% Roti-Histofix (Carl Roth, Karlsruhe, Germany) для иммуногистохимического анализа. Кроме того, селезенки удаляли для детекции приживления человеческих клеток с помощью проточной цитометрии. Выделение спленоцитов выполняли сразу же после удаления селезенок, продавливая их через клеточный фильтр с размером ячеек 70 мкм, помещенный в реакционную пробирку объемом 50 мл со стерильным поршнем 3-5 мл шприца, и повторной промывки клеточного фильтра теплым DPBS. Выделенные спленоциты центрифугировали, DPBS декантировали и осадок спленоцитов ресуспендировали в 1 мл термоинактивированной фетальной бычьей сыворотки, дополненной 10% DMSO. Образцы немедленно замораживали при -80°C и хранили до получения образцов спленоцитов от всех мышей.
d. Анализ приживления человеческих Т-лимфоцитов в мышиных селезенках
Спленоциты от всех мышей собирали и замораживали, как описано в Примере 8.с. Полную коллекцию образцов спленоцитов оттаивали одновременно, все клетки промывали дважды теплым DPBS, и 1×106 спленоцитов на образец инкубировали с антителами, конъюгированными с флуоресцентными метками, в течение 20 мин при 4°C в условиях темноты для детекции приживления человеческих клеток путем окрашивания анти-CD45 и процентного содержания человеческих клеток путем окрашивания анти-CD3, анти-CD4 и анти-CD8. Анализ методом проточной цитометрии проводили на проточном цитометре FACSCalibur (BD Biosciences, Heidelberg, Germany). Приживление человеческих Т-клеток в обеих группах «PBMC» может подтверждаться высоким процентным содержанием дважды CD45-CD3-положительных спленоцитов, как показано на фигуре 10D.
Пример 9: Генерация и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN6 и CD3
a. Происхождение последовательности, разработка конструкций bi-scFv и клонирование в экспрессионные вектора
Конструкции биспецифического тандемного одноцепочечного антитела (bi-scFv) содержали связывающие домены, специфические в отношении компонента CD3 человеческого Т-клеточного рецептора и человеческих опухоль-ассоциированных антигенов (TAA). Соответствующие вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены для каждой конструкции специфически в направлении от N- до C-конца в следующем порядке:
N-VHαCLDN6 - VLαCLDN6 - VHαCD3 - VLαCD3 - C (6PHU5; SEQ ID NO: 43)
N- VHαCD3 - VLαCD3 -VHαCLDN6- VLαCLDN6 C (6PHU5; SEQ ID NO:45)
В таблице 4 кратко описаны все конструкции bi-scFv, специфические в отношении TAA CLDN6, которые были генерированы в рамках изобретения. CLDN18.2-специфическую конструкцию 1BiMAB bi-scFv использовали в качестве контрольного антитела. Конструкции bi-scFv генерировали путем генного синтеза GeneArt AG (GeneArt/Life Technologies GmbH, Regensburg, Germany) с использованием последовательностей VH и VL соответствующих антител. Оптимизации кодонов, такие как Homo sapiens (HS) или Mus musculus (MM) были реализованы с помощью программного обеспечения GeneArt's GeneOptimizer®, и перечислены в таблице 5. Информация по специфичности, происхождению последовательностей из моноклональных антител (mAB), использованию кодона, дополнительным отличительным признакам последовательностей и ссылкам на все применяющиеся домены кратко описаны в таблице 5. Происхождение последовательностей вариабельного домена соответствующих антител CD3 перечислены в таблице 5. Благодаря высокой гомологии человеческих и мышиных TAA, могут быть использованы последовательности VH и VL одного и того же анти-TAA для генерации конструкций bi-scFv для мышиных анализов, но в комбинации с VH, VL последовательностями мышиного специфического клона 145-2С11 анти-CD3 антитела.
Клонирование ДНК и конструирование экспрессионных векторов проводили в соответствии со стандартными процедурами (Sambrook, 1989), хорошо известными специалисту в данной области. Вкратце, ДНК-последовательности bi-scFv содержали рестрикцию 5' HindIII и 3' BamHI для клонирования в экспрессионные плазмиды. Последовательность сигнала секреции вводили на 5' конце (выше) последовательности bi-scFv для секреции белка из клеточной цитоплазмы в культуральную среду. Последовательность, кодирующую гибкий глицин-сериновый пептидный линкер, состоящий из 15-18 аминокислот, вставляли для соединения доменов VH и VL для композиции одноцепочечных вариабельных фрагментов антитела (scFv), из которых один связывается с CD3 и другой с TAA. Для формирования биспецифического одноцепочечного антитела последовательности двух доменов scFv соединяли последовательностью, кодирующей короткий пептидный линкер (GGGGS). Вместе с этой последовательностью линкера сайт рестрикции BamHI вводили для обменов доменов scFv для клонирования последующих конструкций bi-scFV. В частности, 5'scFv-домены могут быть заменены на сайты рестрикции HindIII и BamHI и 3'scFv-домены на рестрикцию BamHI и XhoI. Схему конструкции смотри также на фигуре 1.
Все используемые конструкции антитела bi-scFv клонировали в стандартный экспрессионный вектор млекопитающего pcDNA™3.1/myc-His (+) (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). C-концевую 6xHis-метку использовали для металл-аффинной очистки белка и анализов на обнаружение. Все конструкции подтверждали путем секвенирования через сервис по секвенированию одиночных прочтений (MWG single read sequence service) (Eurofins MWG Operon, Ebersberg, Germany).
Таблица 4. Краткое описание конструкций биспецифического одноцепочечного антитела, специфического к TAA и CD3
кодонов
HS, Homo-sapiens; ММ, Mus musculus; ТАА, опухоль-ассоциированный антиген.
Таблица 5. Краткая информация по конструкции bi-scFv
Таблица 5
Продолжение
кодонов
mAB ссылка
HS, Homo-sapiens; mAB, моноклональное антитело; ММ, Mus musculus; ТАА, опухоль-ассоциированный антиген.
b. Генерация стабильных клеточных линий-продуцентов
Для генерации стабильных клонов клеток-продуцентов CLDN6-специфических белков bi-scFv использовали клеточную линию эмбриональных почек человека HEK293 (ATCC CRL-1573).
1×107 клеток HEK293 засевали за два дня до трансфекции на чашки Петри 14,5 см в 20 мл полной среды DMEM (DMEM/F-12 GlutaMax, дополненной 10% термоинактивированной FBS и 0,5% пенициллина-стрептомицина; все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany). Перед трансфекцией клетки промывали DPBS, дополненным 2 мМ EDTA, затем добавляли 20 мл обычной среды DMEM без добавления FBS или антибиотиков. 20 мкг линеаризованной ДНК конструкций pcDNA3.1/6PHU5 и pcDNA3.1/6PHU3 (описанных в примере 9.а), разбавляли в 0,5 мл обычной среды DMEM/F-12. 75 мкл раствора линейного PEI при концентрации 1 мг/мл (полиэтиленимин; Polysciences Europe GmbH, Eppelheim, Germany) добавляли в разбавленную ДНК и энергично перемешивали на вортексе. Через 15 мин инкубации при комнатной температуре (RT) в клетки по каплям добавляли комплексы ДНК/PEI, чашки для клеточных культур осторожно перемешивали вращательным движением и затем инкубировали при 37°C, 5% CO2. Через 24 ч после трансфекции среду меняли. Селекцию трансфицированных клеток начинали через 48 ч после трансфекции с помощью сульфата G418 (Gibco/Life Technologies GmbH, Darmstadt, Germany) в конечной концентрации 0,8 мг/мл. G418 непрерывно добавляли в культуральную среду для выращивания клеток.
с. Получение в малом масштабе белков 6PHU5 и 6PHU3 bi-scFv с помощью поликлональных клеток НЕК293
Белки 6PHU5 и 6PHU3 bi-scFv получали в малом масштабе и очищали из супернатантов поликлональных клеток НЕК293 для сравнения in vitro.
Вкратце, в конфлюэнтном состоянии супернатант без FBS собирали из поликлональных клеточных линий, описанных в примере 9.b, и фильтровали через фильтрующую насадку с диаметром пор 0,2 мкм Minisart (Sigma-Aldrich, Germany), используя шприц. Затем белки bi-scFv очищали в малом масштабе из супернатантов клеточных культур на спин-колонках Ni-NTA в соответствии с протоколом производителя (Qiagen, Hilden, Germsny). Концентрации белков bi-scFv определяли путем измерения поглощения при 280 нм при помощи NanoDrop 2000c, учитывая коэффициент экстинкции и молекулярную массу - определенные посредством инструмента ProtParam tool (http://web.expasy.org/protparam/) - белков 6PHU5 и 6PHU3 bi-scFv. Очищенные белки хранили при 4°С для немедленного использования.
Белки bi-scFv тестировали путем подвергания электрофорезу в полиакриламидном геле с последующим окрашиванием кумасси и вестерн-блот-анализом, выполненным с использованием стандартных процедур (Current Protocols in Protein science, 2012). Очищенные в малом масштабе белки разделяли на NuPAGE Novex 4-12% Bis-Tris Gels (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). Затем гели окрашивали раствором кумасси бриллиантовым синим в соответствии со стандартными процедурами (Current Protocols in Protein Science, 2012) для детекции белков 6PHU5 и 6PHU3 bi-scFv, и других белков, содержащихся в супернатанте клеточной культуры. Вестерн-блот анализ выполняли для специфической детекции белков 6PHU5 и 6PHU3 bi-scFv посредством их 6xHis-метки. Вкратце, после блоттинга белков на PVDF мембране и блокирования PBST/3% сухим молоком мембраны инкубировали в течение 1 ч при 4°С с первичным антителом Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany), разведенным в соотношении 1:500 в блокирующем буфере. После промывания блокирующим буфером мембраны инкубировали с Fc-специфическим вторичным козьим-анти-мышиным IgG антителом, конъюгированным с пероксидазой, (Sigma Aldrich, Germany) в разведении 1:10000 в блокирующем буфере в течение 1 ч при 4°С. После повторного промывания блокирующим буфером сигналы визуализировали с помощью SuperSignal West Femto Chemiluminescent Substrate (Pierce/Thermo Fisher Scientific, Rockford, IL, USA) и записывали на ImageQuant LAS 4000 Imager (GE Healthcare life sciences, Munich, Germany). Сигналы белков bi-scFv детектировали в диапазоне от 50 и 60 kD, полученные при сравнении с внутренним стандартом молекулярной массы.
d. Получение в крупном масштабе белков 6PHU3 bi-scFv с помощью поликлональных клеток НЕК293
Поликлональную клеточную линию-продуцент выращивали в 10-слойной клеточной фабрике (Nunc, Roskilde, Denmark) в DMEM/F-12 GlutaMax, дополненной 10% FBS и 0,5% пенициллина-стрептомицина и 0,8 мг/мл G418 (все реагенты от фирмы Gibco/Life Technologies GmbH, Darmstadt, Germany) в соответствии с инструкциями производителя. На конфлюэнтной стадии клетки промывали DPBS и среду заменяли на DMEM/F-12 с антибиотиками, но без FBS. Клеточный супернатант, содержащий белок 6PHU3 bi-scFv, собирали каждые 3-5 дней вплоть до 3 недель. Супернатант фильтровали с помощью 500 мл Steritop Filter Units (Merck Millipore, Billerica, MA, USA) и хранили при 4°С до FPLC-фильтрации.
Перед FPLC-фильтрацией присутствие bi-scFv в супернатанте клеточных культур тестировали при помощи электрофореза в полиакриламидном геле с последующим окрашиванием кумасси и вестерн-блот анализом, выполняемым при помощи стандартных процедур, как кратко описано в примере 9.с.
е. Очистка и количественное определение белка 6PHU3 bi-scFv
Клеточный культуральный супернатант поликлональных клеток НЕК293, содержащий белка 6PHU3 bi-scFv (описан в примере 9.b), подвергали аффинной хроматографии с иммобилизованным металлом (IMAC) с использованием стандартной процедуры (Current Protocol in Protein Science, 2012). Вкратце, супернатант клеточной культуры наносили на колонку His Trap FF 5 мл, соединенную с системой AKTA Purifier 10 FPLC (обе от фирмы GE Healthcare Life Science, Munich, Germany). Промывочный буфер PBS содержал 10 мМ имидазола, PBS-буфер для элюирования содержал 500 мМ NaCl, 50 мМ NaH2PO4 и 250 мМ имидазола, рН обоих буферов доводили до 7,4. Элюирование проводили с использованием ступенчатого градиента. Элюированный белок 6PHU3 bi-scFv немедленно диализировали против 1х PBS с использованием Slide-A-Lyzer G2 Dialysis Cassette 10K MWCO (Pierce/Thermo Fisher Scientific, Rockford, IL, USA). После PBS диализа bi-scFv диализировали против 200 мМ аргининового буфера (L-аргинин-моногидрохлорид; Roth, Karlsruhe, Germany) на основе Н2О.
Концентрацию bi-scFv определяли путем измерения поглощения при 280 нм при помощи NanoDrop 2000c c учетом коэффициента экстинкции и молекулярной массы белка 6PHU3bi-scFv. Очищенный белок разделяли на аликвоты и хранили при -80°С для длительного хранения или при 4°С для немедленного использования.
Качество и чистоту белка 6PHU3 bi-scFv тестировали путем окрашивания кумасси и вестерн-блот-анализа, как описано в примере 9.с. BSA в стандартном разведении включали в процедуру окрашивания кумасси для приблизительного подтверждения концентрации, измеренной при помощи Nano Drop (данные не показаны).
Пример 10: Эффективность кандидатов 6PHU5 и 6PHU3 bi-scFv, направленно воздействующих на CLDN6
а. Микроскопический анализ Т-клеток, перенаправленных на клетки-мишени с помощью белков 6PHU5 и 6PHU3 bi-scFv
Для визуализации перенаправления эффекторных клеток на CLDN6-экспрессирующие клетки-мишени с помощью белков 6PHU5 и 6PHU3 bi-scFv посредством микроскопического анализа, выполняли анализ на цитотоксичность in vitro. Очищенные на колонке NiNTA белки 6PHU3 и 6PHU5 bi-scFv (смотри пример 9.с) использовали для сравнения этих двух вариантов в соответствии с их эффективностью. Использовали линию клеток-мишеней, клеточную линию тератокарциномы яичников РА-1, которая эндогенно экспрессирует высокие уровни человеческого CLDN6.
Человеческие эффекторные клетки выделяли из свежеотобранной человеческой крови здоровых доноров согласно стандартным процедурам (Current Protocols in Protein Science, 2012): вкратце, кровь разводили DPBS, послойно наносили на Ficoll-Paque Plus (GE Healthcare Life Sciences, Munich, Germany) и центрифугировали. Мононуклеарные клетки периферической крови (PBMC) собирали из интерфазы, промывали холодным DPBS, дополненным 2 мМ EDTA, и подсчитывали. Человеческие Т-клетки затем отделяли от PBMC путем магнитно-активированного разделения клеток (MACS) с помощью набора Pan T Cell Isolation Kit II (Miltenyi Biotec, Teterow, Germany) согласно руководствам производителя.
1×105 клеток PA-1 на лунку засевали в тканевые культуральные 6-луночные планшеты. Человеческие клетки готовили, как описано выше, и добавляли в соотношении эффектора к мишени (Е:Т) 5:1. Среду МЕМ, дополненную 10% термоинактивированной FBS, 5% пенициллина-стрептомицина, 1х NEAA, 1 мМ бикарбоната натрия и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany), использовали для всех клеток и конечный объем каждой лунки доводили до 2 мл. Используемая в данном анализе концентрация белка bi-scFv составила 50 нг/мл. Контрольные образцы содержали клетки-мишени или только Т-клетки без белка bi-scFv. Тканевые культуральные планшеты затем инкубировали при 37°С, 5% СО2. В ходе анализа проводили непрерывное наблюдение на инвертационном микроскопе Wilovert S (Hund, Wetzlar, Germany) в интервале от 6 ч до 24 ч совместной инкубации. Значительные эффекты в отношении образования кластеров Т-клеток на клетках-мишенях, формирования иммунологического синапса и гибель клеток-мишеней в присутствии белков 6PHU5 и 6PHU3 bi-scFv наблюдали через 24 ч и делали снимки с помощью инвертационного микроскопа Nikon Eclipse TS100 (Nikon, Japan). Оба белка bi-scFv показали сильную агрегацию Т-клеток и гибель клеток-мишеней, как показано на фигуре 12.
А. Активация Т-клеток, опосредованная белками 6PHU5 и 6PHU3 bi-scFv
Для детекции активации Т-клеток и определения различий в эффективности двух вариантов bi-scFv, специфических в отношении CLDN6, использовали анализ активации Т-клеток на основе FACS. Маркер ранней активации CD69 и маркер поздней активации CD25 были выбраны для окрашивания антител, конъюгированных с флуоресцентными метками. Для детекции человеческих Т-клеток в смеси клеток-мишеней и Т-клеток, проводили окрашивание CD3 на Т-клетках.
В целом, были выбраны условия анализа, указанные выше (Пример 10.а). Вкратце, клетки-мишени РА-1, эндогенно экспрессирующие CLDN6, засевали человеческими Т-клетками в соотношении Е:Т, равном 5:1, в 2 мл полной среды, и белки 6PHU5 и 6PHU3 bi-scFv добавляли при концентрации в диапазоне 5-200 нг/мл. Контрольные образцы содержали только клетки-мишени или Т-клетки вместе с белками bi-scFv или без них. Через 24 ч и 48 ч Т-клетки собирали путем споласкивания и переносили в круглодонные пробирки объемом 5 мл (BD Falcon, Heidelberg, Germany). Клетки центрифугировали и промывали DPBS. Для окрашивания клеток использовали мышиные античеловеческие CD3-FITC, мышиные античеловеческие CD69-APC и мышиные античеловеческие CD25-PE антитела (все антитела фирмы BD Biosciences, Heidelberg, Germany). Клеточный осадок ресуспендировали в 50 мкл FACS-буфера (DPBS, дополненный 5% FBS), содержащего антитела, конъюгированные с флуоресцентными метками, и 2 мкл 7-AAD (BD Biosciences, Heidelberg, Germany). После инкубации в течение 20 мин при 4°С в условиях темноты образцы промывали 4 мл DPBS, и клеточный осадок ресуспендировали в 200 мкл FACS-буфера. Образцы хранили на льду и в условиях темноты на протяжении измерения с помощью проточного цитометра FACSCanto II (оба от фирмы BD Biosciences, Heidelberg, Germany). Результаты анализа оценивали с помощью программного обеспечения FlowJo (Tree Star, San Carlos, CA, USA).
Оба варианта CDLN6-специфических bi-scFv привели к эффективной активации Т-клеток вплоть до 60%. Вариант 6PHU3 (bi-scFv CD3 x CLDN6) был более сильным в диапазоне низких концентраций 5-10 нг/мл (смотри также фигуру 13), и был, следовательно, выбран для дальнейших исследований.
Пример 11: Связывающая способность 6PHU3bi-scFv
Анализ связывания FACS
Для оценки связывающей способности фрагментов белка 6PHU3 bi-scFv, направленно воздействующих на CLDN6 и CD3, использовали анализ методом проточной цитометрии. Клетки РА-1 и OV-90, эндогенно экспрессирующие CLDN6, использовали для изучения сайта анти-CLDN6, и человеческие Т-клетки использовали для изучения сайта анти-CD3. CLDN6-отрицательные клетки NugC4 использовали в качестве контрольных клеток.
Для изучения связывающей способности анти-CLDN6, CLDN6-положительные клетки (РА-1, OV-90) и CLDN6-отрицательные клетки (NugC4) трипсинизировали, промывали полной средой и затем DPBS. Все промывочные стадии выполняли путем центрифугирования при 1200 об/мин в течение 6 мин при 4°С. 1×105 клеток переносили в круглодонные пробирки объемом 5 мл и инкубировали с 0,01-10 мкг/мл FPLC-очищенного белка 6PHU3 или контрольного белка 1BiMAB bi-scFv в FACS-буфере в течение 30 мин при 4°С. Клетки промывали 2 мл FACS-буфера и затем инкубировали с 3,3 мкг/мл моноклонального антитела Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в течение 30 мин при 4°С. После промывания 2 мл FACS-буфера клеточный осадок инкубировали с АРС-конъюгированным козьим-анти-мышиным вторичным антителом (Jackson ImmunoResearch Europe, Suffolk, England) в разведении 1:200 в FACS-буфере в течение 20 мин при 4°С в условиях темноты. Клетки промывали дважды 2 мл FACS-буфера и в заключение ресуспендировали в 150 мкл FACS-буфера, дополненного 1 мкг/мл PI (Sigma Aldrich, Germany) для контрастного окрашивания погибших клеток. Отрицательные контрольные образцы включали только вторичное козье-анти-мышиное АРС антитело. В качестве положительного контроля 10 мкг/мл моноклонального CLDN6-специфического антитела mCLDNab окрашивали вторичным козьим-анти-человеческим АРС антителом (Jackson Immunoresearch Europe, Suffolk, England) и подходящим контрольным вторичным антителом.
Образцы измеряли с помощью проточного цитометра FACSCalibur (BD Biosciences, Heidelberg, Germany) и анализировали с помощью программного обеспечения FlowJo Software (Tree Star, San Carlos, CA, USA). Интенсивность сигнала 10 мкг/мл 6PHU3 была в 4-9 раз ниже, чем положительного контроля mCLDN6ab (смотри фигуру 15А). Неспецифическое связывание 6PHU3 с CLDN6-отрицательной клеточной линией NugC4 не было детектировано (фигура 15С).
Для изучения связывающей способности анти-CD3 плеча белка 6PHU3 bi-scFv использовали человеческие Т-клетки. 5×105 Т-клеток переносили в 5 мл пробирки с круглым дном и инкубировали с FPLC-очищенным белком 6PHU3 при концентрации в диапазоне 100 нг/мл - 10 мкг/мл в FACS-буфере в течение 30 мин при 4°С. Последующая процедура окрашивания была такой же, как описано выше. Контрольные образцы включали только вторичное козье-анти-мышиное АРС антитело и моноклональное антитело Anti-HIS Epitope-Tag плюс вторичное козье-анти-мышиное РЕ антитело. Измерение и анализ выполняли, как описано выше. Значительный сигнал был получен при 100 нг/мл 6PHU3 (смотри также фигуру 15В).
Пример 12: Исследование зависимой от мишени активации Т-клеток с помощью bi-scFv 6PHU3
Анализ на цитотоксичность выполняли, как описано в примере 10.а и b. Вкратце, клетки-мишени РА-1, эндогенно экспрессирующие CLDN6, засевали с человеческими Т-клетками в соотношении Е:Т, равном 5:1, в 2 мл полной среды, и белок 6PHU3 bi-scFv добавляли в концентрации в диапазоне 0,001-1000 нг/мл. Для анализа зависимости от мишени активации Т-клеток, опосредованной bi-scFv, Т-клетки засевали без клеток-мишеней, но инкубировали с такими же концентрациями bi-scFv 6PHU3, что и образцы клеток-мишеней с Т-клетками. Через 24 ч и 48 ч Т-клетки собирали и переносили в 5 мл пробирки с круглым дном (BD Falcon, Heidelberg, Germany). Окрашивание клеток и анализ выполняли, как описано в примере 10.b.
Как показано на фигурах 16А и В, активации Т-клеток, опосредованной 6PHU3, не было детектировано в отсутствие клеток-мишеней, что указывает на прямую строгую зависимость функциональности bi-scFv от мишени. Значительная активация Т-клеток возникала только с 0,1 нг/мл 6PHU3 через 48 ч.
Пример 13: Определение ЕС50 6PHU3 bi-scFv в анализе на цитотоксичность in vitro
Анализ на цитотоксичность с использованием люциферазы
Для определения половины максимальной эффективной дозы белка 6PHU3 bi-scFv ряд титрований 6PHU3 тестировали в анализе на цитотоксичность с использованием люциферазы in vitro.
Стабильно экспрессирующие люциферазу клетки РА-1 и человеческие Т-клетки в соотношении Е:Т, равном 5:1, инкубировали с концентрациями белка 6PHU3 bi-scFv в диапазоне от 1 пг/мл до 1 мкг/мл (10 шагов) или без 6PHU3 для определения величин Lmin.
Клеточные культуральные микропланшеты инкубировали в течение 24 ч и 48 ч при 37°С, 5% СО2. Для анализа 50 мкл водного раствора, содержащего 1 мг/мл люциферина (BD Monolight, BD Biosciences, Heidelberg, Germany) и 50 мМ HEPES добавляли в каждую лунку и планшеты затем инкубировали в течение 30 мин в условиях темноты при 37°С. Люминесценцию, возникающую в результате окисления люциферина люциферазой, экспрессирующей жизнеспособные клетки, измеряли с помощью микропланшетного ридера Inifinite M200 Tecan (Tecan, Mannedorf, Switzerland). Процент специфического лизиса клеток-мишеней рассчитывали по следующей формуле: % специфического лизиса = [1-(люминесценциятеституемый образец - Lmax) / (Lmin - Lmax)] × 100, где «L» означает лизис. Lmin относится к минимальному лизису в отсутствие bi-scFv, и Lmax к максимальному лизису (равному величине спонтанной люминесценции) в отсутствие bi-scFv, достигнутому путем добавления Triton X-100 (в конечной концентрации 2%).
Максимальный лизис был достигнут через 48 ч с помощью 1-10 нг/мл 6PHU3, определенная величина ЕС50 через 48 ч составила приблизительно 10 пг/мл (смотри также фигуру 17). Результат этого анализа в значительной степени зависит от силы человеческих Т-клеток, которая различается в соответствии с иммунным статусом донора, что также было описано в других работах (смотри, например, Lutterbuese, R et al., 2010, Proc. Natl. Acad. Sci USA, 2010 Jul 13;107(28):12605-10). Таким образом, отклонение величины ЕС50 белка 6PHU3 bi-scFv на фактор 3 наблюдалось в ходе изобретения.
Пример 14: Эффективность в мышиной модели ксенотрансплантата
Для исследования терапевтического потенциала белка 6PHU3 bi-scFv in vivo был выбран мышиный штамм NOD.Cg-Prkdscid IL2rgtmlwjl/SzJ или короткий NSG (Jackson laboratory, Bar Harbour, ME, USA). Для описанного исследования прививание человеческих эффекторных клеток и человеческих Т-лимфоцитов у мышей необходимо для исследования эффектов Т-клеточного захвата bi-scFv in vivo. Из-за полного отсутствия B-, T- и NK-клеток мышиный штамм NSG является подходящим для этого вида исследований ксенотрансплантата. Мышиная модель с привитыми, главным образом, человеческими Т-клетками после инъекции PBMC составляет часть изобретения.
а. Лечение с отсроченным началом опухолей на поздней стадии, высоко экспрессирующих CLDN6, у мышей с помощью белка 6PHU3bi-scFv
В описанном исследовании 25 мышиных самок и 25 мышиных самцов линии NSG в возрасте 8-11 подкожно инокулировали 1×107 клеток РА-1, эндогенно экспрессирующих высокие уровни человеческого CLDN6. Через 5 дней после инокуляции опухолевых клеток мышей распределяли в соответствии с объемом их опухолей на лечебные группы, при этом мыши, у которых рост опухоли отсутствовал, были исключены. В тот же день мононуклеарные клетки периферической крови (PBMC) выделяли из человеческой крови здоровых доноров методом центрифугирования в градиенте плотности фиколла и использовали в качестве эффекторных клеток in vivo. 2×107 PBMC, разведенных в 200 мкл DPBS, инъецировали интраперитонеально в день выделения экспериментальным лечебным группам, обозначенным «PBMC». Лечебные группы, обозначенные «PBS», получали 200 мкл обычного DPBS интраперитонеально и служили в качестве контроля без человеческих эффекторных клеток. С помощью контрольных групп «PBS» можно исследовать потенциальный эффект на рост опухоли самого 6PUH3 или любые потенциальные побочные эффекты, которые вызваны 25 мышиным самкам и 25 мышиным самцам линии NSG в возрасте 8-11 или носителем, но не человеческими эффекторными клетками, против мышиной ткани (то есть реакция трансплантат против хозяина, вызванная человеческими эффекторными клетками, против мышиной ткани). Группа «PBS/носитель» включала 8 мышей (n=8), «PBS/6PUH3» 8 мышей (n=8), «PBMC/носитель» 7 мышей (n=7), «PBMC/6PUH3» 7 мышей (n=7) и «PBMC/1BiMAB» 8 мышей (n=8). Терапию начинали через 7 дней после применения DPBS или PBMC: группы «PBS/6PHU3», «PBMC/6PHU3» и «PBMC/1BiMAB» получали интраперитонеально 5 мкг очищенного белка 6PHU3 bi-scFv или 1BiMAB, разведенного в 200 мкл DPBS на животное. Группы «PBS/носитель» и «PBMC/носитель» получали интраперитонеально 200 мкл буферного носителя (200 мМ L-аргинина моногидрохлорида, растворенного в H2O, стерильно фильтрованного), разбавленного в DPBS. Лечебные группы кратко описаны в таблице 3. Терапию проводили на ежедневной основе в течение 26 дней. Дважды в неделю измеряли размер опухолей с помощью цифрового калиброванного калипера, и объем опухолей рассчитывали по формуле мм3 = длина x ширина x (ширина/2). На фигурах 18A и B показано ингибирование роста опухоли у всех мышей группы «PBMC/6PHU3» с помощью антитела в присутствии человеческих эффекторных клеток. Мышей умерщвляли путем смещения шейных позвонков, когда объем опухоли достигал 1500 мм3, или в случае серьезных клинических проявлений (у некоторых мышей наблюдались симптомы реакции трансплантат против хозяина).
Таблица 6. Лечебные группы
b. Определение влияния терапии на массу тела
Массу тела каждой мыши измеряли два раза в неделю с помощью лабораторных весов. Ни одна из мышей к любой группе не показала потерю массы в течение лечения (данные не показаны).
с. Консервация ткани и выделение спленоцитов
После умерщвления мышей опухоли удаляли, и ткань сразу же фиксировали в 10 мл 4% Roti-Histofix (Carl Roth, Karlsruhe, Germany) для иммуногистохимического анализа. Кроме того, селезенки удаляли для детекции приживления человеческих клеток с помощью проточной цитометрии. Выделение спленоцитов выполняли сразу же после удаления селезенок, продавливая их через клеточный фильтр с размером ячеек 70 мкм, помещенный в реакционную пробирку объемом 50 мл со стерильным поршнем 3-5 мл шприца, и повторной промывки клеточного фильтра теплым DPBS. Выделенные спленоциты центрифугировали, DPBS декантировали и осадок спленоцитов ресуспендировали в 1 мл термоинактивированной фетальной бычьей сыворотки, дополненной 10% DMSO. Образцы немедленно замораживали при -80°C и хранили до получения образцов спленоцитов от всех мышей.
d. Анализ приживления человеческих Т-лимфоцитов в мышиных селезенках
Спленоциты от всех мышей собирали и замораживали, как описано в Примере 14.с. Полную коллекцию образцов спленоцитов оттаивали одновременно, все клетки промывали дважды теплым DPBS, и 1×106 спленоцитов на образец инкубировали с антителами, конъюгированными с флуоресцентными метками, в течение 20 мин при 4°C в условиях темноты для детекции приживления человеческих клеток путем окрашивания анти-CD45 и процентного содержания человеческих клеток путем окрашивания анти-CD3, анти-CD4 и анти-CD8. Анализ методом проточной цитометрии проводили на проточном цитометре FACSCalibur (BD Biosciences, Heidelberg, Germany). Приживление человеческих Т-клеток в обеих группах «PBMC» может подтверждаться высоким процентным содержанием дважды CD45-CD3-положительных спленоцитов, как показано на фигуре 18D.
e. Иммуногистохимия для определения экспрессии клеток-мишеней и Т-клеточной инфильтрации
Опухоли фиксировали после удаления с использованием 4% буферного раствора формальдегида (Roti-Histofix, Carl Roth, Karlsruhe, Germany) в течение 48 ч при 4°С. Фиксированные опухоли разделяли на две части и переносили в автоматизированный вакуумный тканевый процессор ASP200 для дегидрирования (Leica Microsystems GmbH, Wetzlar, Germany) с последующим заключением в парафин (Paraplast, Carl Roth, Karlsruhe, Germany) посредством парафиновой диспенсерной станции MPS/C (Slee Medical GmbH, Mainz, Germany). Для иммуногистохимических окрашиваний срезы толщиной 3 мкм фиксированных в формалине и заключенных в парафин тканей получали с использованием роторной микротомы RM2255 (Leica Microsystems GmbH, Wetzlar, Germany). Депарафинизацию и регидрирования выполняли в билинейном устройстве для окрашивания StainMate Max (Thermo Ficher Scientific, Rockford, IL, USA) с последующим извлечением термоиндуцированного эпитопа в 10 мМ лимонном буфере, рН 6, с 0,05% Tween 20 в течение 10 мин при 120°С. Активность эндогенных пероксидаз гасили путем погружения в 0,1% раствор Н2О2 в PBS в течение 15 мин (Carl Roth) с последующей инкубацией с 10% козьей сывороткой в PBS (PAA Laboratories GmbH/GE Healthcare, Pasching, Austria) в течение 30 мин для блокирования неспецифических сайтов связывания антитела. ТАА клаудина 6 детектировали путем инкубации с поликлональным первичным антителом Anti-Mouse Claudin 6 (C) Rabbit (IBL-America, Minneapolis, MN, USA) при 4°С в течение ночи; Т-клетки детектировали на последовательных срезах с использованием поликлонального анти-CD3 AB (Abcam, Cambridge, UK) при 4°С в течение ночи с последующей инкубацией с анти-кроличьим вторичным антителом, конъюгированным с полимером Bright Vision HRP (ImmunoLogic, Duiven, Netherlands). Реакции связывания визуализировали с использованием набора Vector NovaRED kit (Vector Laboratories Ltd., Peterborough, UK) в соответствии с инструкциями производителя с последующим контрастным окрашиванием гематоксилином (Carl Roth), дегидрированием и заключением в среду. Анализы и регистрацию выполняли с использованием установки для визуализации Axio Imager M2 или сканера Mirax scanner (оба от фирмы Carl Zeiss Microscopy GmbH, Goettingen, Germany).
Как показано на фигуре 19, самую высокую инфильтрацию Т-клеток детектировали в опухолях группы «PBMC/6PHU3» путем CD-3 окрашивания, особенно в пограничных областях экспрессии CLDN6. Картина гетерогенной экспрессии ТАА CLDN6 в контрольных группах (фигуры 19А, В, С и D) изменялась в более компактные области экспрессии CLDN6 в опухолях группы «PBMC/6PHU3» в результате терапии (фигура 19D).
Пример 15: Создание и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN18.2 и CD3
а. Исходные последовательности, дизайн конструкций bi-scFv и клонирование матрицу вектора
Готовили конструкции биспецифического тандемного одноцепочечного антитела (bi-scFv), содержащие связывающие домены, специфические в отношении компонента CD3-эпсилон человеческого Т-клеточного рецептора, и опухоль-ассоциированных антигенов (ТАА). Соответствующие вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) для каждой конструкции специфическим образом были расположены в направлении от 5'- до 3'-конца в следующем порядке:
pST1-hAgKozak-VHαCLDN18.2-VLαCLDN18.2-VHαCD3- VLαCD3-His-2hBgUTR-A120
(1BiMAB, 18RHU5, no. 1-5)
pST1-hAgKozak-VHαCD3- VLαCD3- VHαCLDN18.2-VLαCLDN18.2- His-2hBgUTR-A120
(18RHU3, no. 6-10)
В таблице 7 кратко описаны все конструкции bi-scFv, специфические в отношении ТАА CLDN18.2 и PLAC1, которые были генерированы в рамках изобретения. Конструкции bi-scFv генерировали путем генного синтеза GeneArt AG (GeneArt/Life Technologies GmbH, Regensburg, Germany) с использованием последовательностей VH и VL соответствующих антител. Оптимизации кодонов, такие как Homo sapiens (HS) или Mus musculus (MM) или клетки яичника китайского хомячка (СНО), были реализованы с помощью программного обеспечения GeneArt's GeneOptimizer®, и перечислены в таблице 7. Информация по специфичности, происхождению последовательностей из моноклональных антител (mAB), использованию кодона, дополнительным отличительным признакам последовательностей и ссылкам на все применяющиеся домены кратко описаны в таблице 8. Происхождение последовательностей вариабельного домена соответствующих антител CD3 перечислены в таблице 8. Благодаря высокой гомологии человеческих и мышиных TAA, могут быть использованы последовательности VH и VL одного и того же анти-TAA для генерации конструкций bi-scFv для мышиных анализов, но в комбинации с VH, VL последовательностями мышиного специфического клона 145-2С11 анти-CD3 антитела.
Клонирование ДНК и конструирование экспрессионных векторов проводили в соответствии со стандартными процедурами (Sambrook, 1989), хорошо известными специалисту в данной области. Вкратце, основные ДНК-последовательности bi-scFv содержали сайт рестрикции 5'-BsmBI и 3'-XhoI для клонирования в pST1-плазмиды. Последовательность сигнала секреции вводили на 5' конце (выше) последовательности bi-scFv для секреции белка. Последовательность, кодирующую гибкий глицин-сериновый пептидный линкер, состоящий из 15-18 аминокислот, вставляли для соединения доменов VH и VL для композиции одноцепочечных вариабельных фрагментов антитела (scFv), из которых один связывается с CD3 и другой с TAA. Для формирования биспецифического одноцепочечного антитела последовательности двух доменов scFv соединяли последовательностью, кодирующей короткий пептидный линкер (GGGGS). Вместе с этой последовательностью линкера сайт рестрикции BamHI вводили для обменов доменов scFv для клонирования последующих конструкций bi-scFV. Вкратце, 5'scFv-домены заменяли на рестрикцию BsmBI и XhoI и 3'scFv-домены на рестрикцию BamHI и XhoI. C-концевую 6xHis-метку использовали для анализов на обнаружение транслированного белка. Нетранслируемые области человеческого альфа-глобин 5' и человеческого бета-глобин 3' последовательности bi-scFv присутствовали в векторе pST1 (смотри более подробно в WO2007/036366A2: Waggoner, S. et al. (2003) Exp. Biol. Med. (Maywood) 228 (4), pp. 387-395).
Для получения вектора репликона 1BiMAB полную последовательность 1BiMAB, включая сигнал секреции и 6xHis-метку, субклонировали в направлении от 3' от субгеномного промотора вектора репликона вируса леса Семлики (pSFV), любезно предоставленного K. Lundstrom (Lundstrom, K. et al. (2001) Histochem. Cell Biol. 115 (1), pp. 83-91; Ehrengruber, M.U. et al. (1999) Proc. Natl. Acad. Sci. U.S.A. 96 (12), pp. 7041-7046).
Все конструкции подтверждали путем секвенирования через сервис по секвенированию одиночных прочтений (MWG single read sequence service) (Eurofins MWG Operon, Ebersberg, Germany) и использовали конструкции только с правильной последовательностью и поли(А)-хвостом, состоящим из более чем 100 аденинов, для in vitro транскрипции РНК.
Схемы конструкций представлены также на фигуре 20А.
Таблица 7. Краткое описание конструкций мРНК-матриц биспецифического одноцепочечного антитела, специфического в отношении ТАА и CD3
кодонов
bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; СНО, клетки яичника китайского хомячка; HS, Homo-sapiens; HU, гуманизированное; ММ, Mus musculus; ТАА, опухоль-ассоциированный антиген; VH, вариабельный домен тяжелой цепи, VL, вариабельный домен легкой цепи.
Таблица 8. Краткая информация по конструкции мРНК-матрицы bi-scFv
Таблица 8
Продолжение
кодонов
mAB
ссылки
CHO означает клетки яичника китайского хомячка; HS, Homo sapiens; mAB, моноклональное антитело; MM, Mus musculus; TAA, опухоль-ассоциированный антиген.
b. Синтез IVT-РНК
Для создания анти-CLDN18.2-специфических IVT-матриц bi-scFv, плазмиды линеаризовали даунстрим поли(А)-хвоста с использованием эндонуклеазы класса IIs. Линеаризованную матрицу ДНК очищали экстракцией смесью фенол/хлороформ и осаждали ацетатом натрия, как описано в литературных источниках (Holtkamp, S. et al. (2006) Blood 108 (13), pp. 4009-4017).
Линеаризованные матрицы ДНК подвергали in vitro транскрипции с использованием наборов MEGAscript Kits (Ambion/Life Technologies, Darmstadt, Germany) в соответствии с инструкциями производителя: матрицы pST1 транскрибировали с помощью набора MEGAscript Т7 Kit и матрицы pSFV с помощью набора MEGAscript SP6 Kit. Для реакций с кэп-аналогом концентрацию GTP уменьшали до 1,5 мМ и 6 мМ ARCA, бета-S-ARCA(D1) или в реакцию добавляли бета-S-ARCA(D2), синтезированный, как описано в литературе (Kowalska, J. et al. (2008) RNA 14 (6), pp. 1119-1131; Grudzien, E. et al. (2004) RNA 10 (9), pp. 1479-1487; Stepinski, J. et al. (2001) RNA 7 (10), pp. 1486-1495). Очистку IVT-мРНК выполняли с помощью набора MEGAclear Kit (Ambion/Life Technologies, Darmstadt, Germany) в соответствии с руководством. Концентрацию и качество IVT-РНК оценивали с помощью спектрофотометрии и анализировали на биоанализаторе 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA).
Пример 16: Активация Т-клеток в ответ на секрецию bi-scFv клетками-мишенями, трансфицированными различными CLDN18.2-специфическими IVT-мРНК
Для исследования функциональности IVT-мРНК CLDN18.2-специфического bi-scFv клеточную линию карциномы желудка NugC4, которая эндогенно экспрессирует сравнительно высокие уровни человеческого CLDN18.2 (Sahin, U. et al. (2008) Clin. Cancer Res. 14 (23), pp. 7624-7634), использовали в качестве клеточной линии-мишени.
Клетки-мишени NugC4 - которые тестировали обычным способом на экспрессию ТАА с помощью FACS-анализа перед каждым испытанием - дважды промывали ледяной средой X-Vivo 15 (LONZA, Basel, Switzerland) и ресуспендировали до плотности 2×107 клеток/мл. 250 мкл клеточной суспензии переносили в предварительно охлажденные кюветы 0,4 см Gene Pulser/MicroPulser Cuvettes (Bio-Rad, Dreieich, Germany) и добавляли 20 мкг/мл IVT-мРНК. Использовали следующие IVT-мРНК: 1BiMAB, no. 2, no. 3, no. 4, no. 5, no. 7, no. 8, no. 9 и no. 10. Для более подробной информации о вариантах bi-scFv смотри таблицы 7 и 8. После осторожного перемешивания клетки трансфицировали с помощью электропоратора BTX ECM 830 (Harvard Apparatus, Holliston, MA, USA) с использованием следующих условий: 250 В, 2 импульса, длительность импульса 12 мс, пауза между импульсами 40 мс. Сразу же после электропорации кюветы помещали на лед на короткий промежуток времени, и затем клеточные суспензии переносили в нагретую до комнатной температуры аналитическую среду (среда RPMI 1640, дополненная 5% термоинактивированной человеческой сывороткой АВ, 0,5% пенициллина-стрептомицина, 1× NEAA и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany)) в пробирки объемом 15 мл Трансфицированные клетки-мишени подсчитывали и их концентрацию доводили до 1×105 клеток на мл.
Человеческие эффекторные клетки выделяли из свежевзятой крови здоровых доноров-людей в соответствии со стандартными процедурами (Current Protocols in Immunology, 2012): вкратце, кровь разводили DPBS, наносили на Ficoll-Paque Plus (GE Healthcare Life Sciences, Munich, Germany) и центрифугировали. Мононуклеарные клетки периферической крови (PBMC) собирали из интерфазы, отмывали холодным DPBS, дополненным 2 мМ EDTA и подсчитывали. Человеческие цитотоксические Т-клетки выделяли путем магнитно-активированной клеточной сортировки (MACS) из PBMC с помощью набора CD8+ T cell Isolation Kit, human (Miltenyi Biotec, Teterow, Germany) в соответствии с инструкциями производителя. Отделенные эффекторные клетки тестировали обычным способом на успешное выделение Т-клеток посредством FACS-анализа (CD4, CD8 окрашивание). Т-клетки доводили до 5×105 клеток на мл в аналитической среде.
1×105 клеток-мишеней засевали на лунку 6-луночного планшета и человеческие цитотоксические Т-клетки добавляли до соотношения Е:Т, равного 5:1. Конечный объем на лунку составил 2 мл. Контрольные образцы, содержащие эффекторные клетки и клетки-мишени, содержали клетки-мишени, секретирующие no.25, chCLDN18.2ab родительского mAB IgG и белок 1BiMAB в качестве положительного контроля в конечной концентрации 5 нг/мл. Контроли включали только подвергнутые электропорации клетки-мишени или только Т-клетки с белком 1BiMAB и без него. Через 48 ч Т-клетки и клетки-мишени собирали, метили и анализировали методом проточной цитометрии. Вкратце, все клетки собирали путем осторожного соскабливания с помощью клеточного скребка Cell scrapers (Sarstedt AG & Co, Nurmbrecht, Germany) и переносили в 5 мл пробирки с круглым дном (BD Falcon, Heidelberg, Germany). Клетки центрифугировали и отмывали DPBS. Для окрашивания клеток использовали мышиные античеловеческие CD3-FITC, мышиные античеловеческие CD69-APC и мышиные античеловеческие CD25-PE (все антитела фирмы BD Biosciences, Heidelberg, Germany). Клеточные осадки ресуспендировали в 50 мкл FACS-буфера (DPBS, дополненный 5% FBS), содержащего антитела, конъюгированные с флуоресцентными метками. После инкубации в течение 20 мин при 4°С в условиях темноты образцы промывали 4 мл DPBS и клеточные осадки ресуспендировали в 200 мкл FACS-буфера, содержащего йодид пропидия (PI) (Sigma Aldrich, Germany) в конечном разведении 1:1000 для детекции погибших клеток. Образцы хранили на льду и в условиях темноты до измерения. Анализ выполняли с использованием прибора FACSCalibur (BD Biosciences, Heidelberg, Germany). Результаты анализа оценивали с помощью программного обеспечения FlowJo software (Tree Star, San carlos, CA, USA).
Как показано на фигуре 21А, каждый CLDN18.2-специфический IVT-мРНК вызывал секрецию белка bi-scFv, как видно по эффективной активации Т-клеток. Наиболее сильными вариантами лишь с незначительными расхождениями в Т-клеточной активации являлись no.5 > no.8 > no.10 > no.3 > 1BiMAB по убыванию. Все из этих вариантов приводили к общей активации Т-клеток, составляющей более 55%, тогда как варианты no.2, no.4, no.7 и no.9 приводили к общей активации Т-клеток, составляющей от 40 до 50%.
Специфический лизис клеток-мишеней рассчитывали по формуле: % лизис = (% PI+ клеток-мишеней образец - % PI+ клеток-мишеней ЕР сравнения), где «образец» означает совместно инкубированные эффекторные клетки и клетки-мишени, и «ЕР сравнения» только подвергнутые электропорации клетки-мишени каждой индивидуальной электропорации IVT-mRNA. Как показано на фигуре 21В, лизис клеток-мишеней выше 65% был достигнут вариантами 1BiMAB > no.5 > no.8 > no.3 в убывающем порядке. Другие варианты опосредовали лизис клеток-мишеней, составляющий 55-64%. Наиболее сильный CLDN18.2-специфический bi-scFv содержит домены VH и VL TR66 (1BiMAB, no.5, no.10) или UCHT1 (no.3, no.8). В отношении ориентаций доменов значительного различия не наблюдалось в отличие от вариантов белка bi-scFv (смотри пример 3). В соответствии с исследованиями белка, IVT-мРНК, кодирующий 1BiMAB, был выбран для последующих исследований.
Конструкции 18RHU5 и 18RHU3 (смотри таблицы 7 и 8) сравнивали в более поздний момент времени с 1BiMAB. Эффективность 18RHU5 была эквивалентна 1BiMAB, 18RHU3 был менее сильным (данные не показаны).
Пример 17. Микроскопический анализ Т-клеток, перенаправленных на клетки-мишени, секретирующие CLDN18.2-специфический 1BiMAB bi-scFv
Порядок проведения анализа по существу был таким, как описано в примере 2.а.
Человеческие цитотоксические Т-клетки отделяли, используя MACS, от PBMC, выделенных из свежеотобранной крови, с помощью набора CD8+ T Cell Isolation Kit, human (Miltenyi Biotec, Teterow, Germany) в соответствии с инструкциями производителя.
Клетки-мишени NugC4 готовили и трансфицировали, как описано в примере 16, за исключением того, что использовали 80 мкг/мл 1BiMAB IVT-мРНК или 80 мкг/мл no.25 IVT-мРНК. Трансфицированные клетки-мишени подсчитывали и доводили до 2×105 клеток на мл. 1×104 клеток-мишеней засевали в каждую лунку 96-луночного планшета и человеческие цитотоксические Т-клетки добавляли при соотношении Е:Т, равном 5:1. Конечный объем на лунку составил 100 мкл. Контрольные образцы содержали только трансфицированные клетки-мишени для подтверждения лечебных свойств после электропорации и контрольные трансфицированные bi-scFv клетки-мишени с эффекторными клетками. Тканевые культуральные планшеты затем инкубировали при 37°С, 5% СО2. Значительные эффекты в отношении образования кластеров Т-клеток на клетках-мишенях, формирования иммунологического синапса и гибели клеток-мишеней в образцах, содержащих трансфицированные 1BiMAB клетки-мишени, наблюдали через 24 ч и записывали с помощью инвертационного микроскопа Nikon Eclipse TS100 (Nikon, Japan). Образования кластеров Т-клеток или лизиса клеток-мишеней не наблюдалось в контрольном образце с no.25 трансфицированными клетками-мишенями, свидетельствуя о строгой зависимости от экспрессии ТАА для индукции активации Т-клеток. Смотри также фигуру 22.
Пример 18: Анализ методом проточной цитометрии зависимой от концентрации активации Т-клеток CLDN18.2-специфическим 1BiMAB bi-scFv
Для исследования зависимой от концентрации bi-scFv активации Т-клеток в присутствии ТАА-экспрессирующих клеток-мишеней серии трехкратных разведений применяли для процесса трансфекции.
Клетки-мишени NugC4 готовили и трансфицировали, как описано в примере 16, но с конечной концентрацией IVT-мРНК, составляющей 40 мкг/мл. Концентрации 1BiMAB IVT-мРНК изменялись от 0,4 до 40 мкг/мл и были дополнены соответствующими количествами IVT-мРНК люциферазы с целью подвергания всех клеток одинаковому уровню стресса в отношении количеств IVT-мРНК.
1×105 трансфицированных клеток-мишеней и человеческие цитотоксические Т-клетки (выделенные, как описано в примере 16) засевали в соотношении Е:Т, равном 10:1, в 2 мл аналитической среды в 6-луночном формате. Контрольные образцы, содержащие клетки-мишени, трансфицировали 40 мкг/мл IVT-мРНК люциферазы, но не IVT-мРНК 1BiMAB. Через 24 ч и 48 ч совместной инкубации клеток-мишеней и эффекторных клеток, клетки собирали, окрашивали и анализировали, как описано в примере 16.
Как показано на фигурах 23 А и В, значительную активацию Т-клеток наблюдали в образцах, содержащих клетки-мишени, трансфицированные 4 мкг/мл IVT-мРНК 1BiMAB. Максимальная активация Т-клеток в этом анализе была достигнута с помощью 40 мкг/мл IVT-мРНК 1BiMAB. Экспрессия CD69 и CD25 различалась в зависимости от времени инкубации (фигура 23 от А до В) и концентрации IVT-мРНК. Более высокие количества IVT-мРНК 1BiMAB (12 и 40 мкг/мл) приводили к более быстрой инициации механизма активации Т-клеток, наблюдаемой, например, при повышенной экспрессии CD25 по сравнению с экспрессией CD69 на фигуре 23В. Общая активация Т-клеток не превышала ~40%.
Пример 19: Анализ методом проточной цитометрии зависимого от концентрации, опосредованного Т-клетками лизиса клеток-мишеней с помощью CLDN18.2-специфического 1BiMAB bi-scFv
Для исследования зависимого от концентрации bi-scFv опосредованного Т-клетками лизиса клеток-мишеней использовали способ, описанный в примере 18. Помимо образцов совместной инкубированных эффекторных клеток и клеток-мишеней («образец»), также засевали только отдельно индивидуально подвергнутые электропорации клетки-мишени. Последние служили в качестве образцов сравнения («ЕР сравнение») для вычитания клеток-мишеней, погибших в результате процесса электропорации, из клеток-мишеней, лизированных Т-клетками.
Сбор и окрашивание выполняли в соответствии с примером 18. Клетки-мишени в заключение анализировали посредством введения в них пропидия иодида с помощью FACSCalibur (BD Biosciences, Heidelberg, Germany).
Процентный специфический лизис клеток-мишеней определяли путем двухстадийного вычитания исходных погибших клеток:
1. % опосредованного Т-клетками лизиса = (% PI+клеток-мишеней образец - % PI+ клеток- мишеней ЕР сравнения)
2. % специфического лизиса = (% опосредованного Т-клетками лизиса образец - % опосредованного Т-клетками лизиса контроль)
«Опосредованный Т-клетками лизис контроль» вычитается из разницы PI+ клеток-мишеней контрольных образцов (40 мкг/мл только IVT-мРНК люциферазы) с эффекторными клетками и без них.
С помощью этого расчета максимальный специфический лизис, составляющий 55,5% +/- 5,6% достигается с помощью 12 мкг/мл IVT-мРНК 1BiMAB в данном эксперименте. Благодаря чувствительности клеток мишеней NugC4 к стрессу, вызванному электропорацией, и связанной с ним неизбежной гибели исходных клеток-мишеней, которые вычитаются, отмеченный на графике специфический лизис клеток-мишеней может быть ниже в этих экспериментальных условиях, чем фактический процентный лизис.
Пример 20: Пролиферация Т-клеток в ответ на трансфекцию IVT-мРНК 1BiMAB
Пролиферация Т-клеток является признаком активации Т-клеток. Для демонстрации пролиферации Т-клеток bi-scFv, кодирующий 1BiMAB IVT-мРНК в присутствии CLDN18.2-положительных клеток-мишеней, использовали анализ методом проточной цитометрии. Вкратце, 1×107 человеческих Т-клеток, выделенных, как описано в примере 16, окрашивали 1 мкМ карбоксифлуоресцеина диацетата сукцинимидилового сложного эфира (CellTrace CFSE, Invitrogen/Life Technologies GmbH, Germany), растворенного в DPBS, в условиях темноты при комнатной температуре в течение 5 мин.
Клетки промывали дважды DPBS /5%FCS и ресуспендировали в аналитической среде до 2×106 клеток на мл. В качестве клеток-мишеней были выбраны клетки NugC4, лентивирально трандуцированные человеческим CLDN18.2 и - для тестирования специфичности - CLDN18.2-отрицательная клеточная линия рака молочной железы MDA-MB-231. 20 мкг/мл 1BiMAB IVT-мРНК или нетаргетный контроль использовали для электропорации. Электропорацию NugC4 выполняли, как описано в примере 16. Электропорацию MDA-MB-231 выполняли с использованием следующих условий: 400 В, продолжительность импульса 3 мс, 1 импульс, интервал между импульсами 400 мс в 0,4 см кюветах Gene Pulser/MicroPulser Cuvettes (Bio-Rad, dreieich, Germany). Анализ на цитотоксичность, описанный в примере 16, проводили с трансфицированными клетками-мишенями и человеческими CFSE-мечеными Т-клетками в качестве эффекторных клеток. В качестве отрицательных контролей использовали только Т-клетки и нетрансфицированные или контрольные трансфицированные клетки-мишени плюс Т-клетки. В качестве положительного контроля служили только Т-клетки, стимулированные 5 мкг/мл OKT3 (Bio Х Cell, West Lebanon, NH, USA) и 2 мкг/мл анти-CD28 (BioLegend, fell, Germany). Белок 1BiMAB в концентрации 5 нг/мл наносили на нетрансфицированные клетки-мишени плюс Т-клетки для подтверждения валидности анализа. Образцы, объединенные с нетаргетным белком 6PHU3 bi-scFv, включали в качестве контроля специфичности. Все образцы готовили в трех повторностях в общем объеме 0,2 мл аналитической среды в 96 лунках. Через 72 ч совместной инкубации Т-клетки собирали, переносили в 5 мл пробирки с круглым дном, промывали и окрашивали при 4°С в течение 30 мин 2 мкл анти-CD45-APC для дифференциации человеческих Т-клеток от опухолевых клеток, и 0,25 мкл eFluor506 (BD Biosciences, Heidelberg, Germany) для контрастного окрашивания погибших клеток в 200 мкл DPBS. После отмывания DPBS клетки ресуспендировали в FACS-буфере и анализировали с помощью FACSCanto II (BD Biosciences, Heidelberg, Germany).
Пролиферацию Т-клеток детектировали по уменьшению CFSE-сигнала только в присутствии CLDN18.2-положительных клеток-мишеней и 1BiMAB bi-scFv (смотри также фигуру 25). Кроме положительного контроля, пролиферация Т-клеток, составляющую 42-48%, можно было наблюдать в присутствии CLDN18.2-положительных клеток-мишеней в комбинации с белком 1BiMAB. Таргетные положительные клетки, трансфицированные IVT-мРНК 1BiMAB, привели к 22% +/- 3%. Более низкая пролиферация в ответ на IVT-мРНК, чем на белок, возможно обусловлена низкой эффективностью трансфекции, достигаемой клетками-мишенями NugC4. Т-клетки, инкубированные с CLDN18.2-отрицательными клетками-мишенями MDA-MB-231, не показали значительной пролиферации в любых условиях, а также Т-клетки без клеток-мишеней, за исключением положительного контроля.
Пример 21: Титрование соотношений эффектора к мишени для определения эффективного соотношения
Для определения подходящего соотношения Е:Т для проведения анализа цитотоксичности in vitro на основе FACS-анализа, соотношения Е:Т в диапазоне от 0,3:1 до 10:1 выбирали за 3 стадии.
Клетки-мишени NugC4 готовили и трансфицировали, как описано в примере 16, с концентрацией IVT-мРНК, составляющей 40 мкг/мл. Один препарат трансфицированных IVT-мРНК 1BiMAB клеток использовали для всех тестируемых образцов. Трансфекция 40 мкг/мл IVT-мРНК люциферазы была выбрана в качестве негативного контроля.
Человеческие цитотоксические Т-клетки отделяли PBMC, выделенных из свежеотобранной крови, как описано в примере 16, и служили в качестве эффекторных клеток. 1×105 трансфицированных клеток-мишеней совместно инкубировали с цитотоксическими Т-клетками в 6-луночных планшетах в двух повторностях в следующих соотношениях эффектора к мишени: 0,3:1 - 1:1 - 3:1 - 10:1. Кроме того, человеческие цитотоксические Т-клетки выращивали в отсутствие клеток-мишеней для определения исходной активации Т-клеток. Клетки-мишени, трансфицированные контрольным IVT-мРНК или IVT-мРНК 1BiMAB, также выращивали в отсутствие эффекторных клеток для определения исходных погибших клеток путем подвергания стрессу электропорации. Люциферазный отрицательный контроль засевали только в максимальном соотношении Е:Т, равном 10:1. Через 48 ч совместной инкубации клетки отбирали, метили и анализировали, как описано в примере 16.
На фигуре 26А показана специфическая активация цитотоксических Т-клеток в ответ на секрецию 1BiMAB клетками-мишенями NugC4. Независимо от количества Т-клеток в образцах, была обнаружена общая активация 50-60%. Кроме того, распределение экспрессии CD25 и CD69 в образцах является в высокой степени сопоставимым. Это указывает на то, что в популяции цитотоксических Т-клеток имеется фиксированный процент Т-клеток, которые могут быть активирован.
На фигуре 26В зависимость эффективного лизиса клеток-мишеней от количества цитотоксических Т-клеток становится очевидной. Даже если клетки-мишени лизированы в соотношении Е:Т, равном только 0,3:1, активный лизис начинается при соотношении 3:1.
Пример 22: Анализ активации Т-клеток и лизис клеток-мишеней в анализе на основе FACS с использованием трансфицированных IVT-мРНК 1BiMAB эффекторных клеток
В этом эксперименте целью являлось тестирование способности человеческих эффекторных клеток продуцировать и секретировать 1BiMAB после трансфекции IVT-мРНК. Рациональной основой был гипотетический вариант развития событий у пациента, где собственные Т-клетки пациента могут быть трансфицированы bi-scFv с последующим повторным переносом в организм пациента.
Человеческие цитотоксические Т-клетки - изолированные как описано в примере 16 - промывали дважды средой Х-Vivo 15 (LONZA, Basel, Switzerland) и ресуспендировали до плотности 2×107 клеток/мл. 250 мкл клеточной суспензии переносили в предварительно охлажденные 0,4 см кюветы Gene Pulser/MicroPulser Cuvettes (Bio-Rad, dreieich, Germany) и добавляли 80 или 240 мкг/мл IVT-мРНК. Используемыми IVT-мРНК являлись 1BiMAB и eGFP в качестве контроля. После осторожного перемешивания клетки электропорировали с использованием электропоратора BTX ECM 830 (Harvard Apparatus, Holliston, MA, USA) с использованием следующих условий: 500 В, 1 импульс, продолжительность импульса 3 мс, интервал между импульсами 400 мс. Сразу же после электропорации кюветы помещали на лед на короткий промежуток времени и затем клеточные суспензии переносили в аналитическую среду (среда RPMI 1640, дополненная 5% термоинактивированной человеческой сывороткой АВ, 0,5% пенициллина-стрептомицина, 1х NEAA и 1 мМ пирувата натрия (Gibco/Life Nechnologies GmbH, Darmstadt, Germany)), содержащую 10 Ед/мл IL-2. Трансфицированные эффекторные клетки выращивали в течение ночи при 37°С, 5% СО2. На следующий день эффективность трансфекции анализировали с помощью FACS, выявляя эффективность трансфекции выше 70% для обеих концентраций IVT-мРНК eGFP. Каждый образец эффекторных клеток подсчитывали и доводили до 5×105 клеток на мл. Клетки-мишени NugC4 собирали для анализа путем трипсинизации, промывали аналитической средой, подсчитывали и доводили до 1×105 клеток на мл. Эффекторные клетки и клетки-мишени смешивали и засевали в 6-луночных планшетах в двух повторностях в конечном соотношении Е:Т, равном 5:1, и конечном объеме 2 мл. Необработанные Т-клетки засевали без клеток-мишеней для определения исходных сигналов активации. Анализ проводили, как описано в примере 16, через 48 ч совместной инкубации при 37°С, 5% СО2.
Значительная активация цитотоксических Т-клеток, трансфицированных IVT-мРНК 1BiMAB и совместно инкубированных с клетками-мишенями, достигалась, как показано на фигуре 27А. Лизис клеток-мишеней трансфицированными IVT-мРНК 1BiMAB Т-клетками составил более 60%, как показано на фигуре 27В. Эффекты 80 мкг/мл IVT-мРНК 1BiMAB не могли быть увеличены с помощью более высоких количеств РНК.
В результате этого эксперимента можно сделать вывод о том, что эффекторные клетки могут теоретически быть использованы в качестве продуцирующих и секретирующих bi-scFv клеток-реципиентов.
Пример 23: Исследование специфичности в отношении мишени IVT-мРНК 1BiMAB в анализе на цитотоксичность на основе люциферазы с использованием CLDN18.2-отрицательных клеток-мишеней.
Строгая специфичность в отношении мишени является важным вопросом для избежания нежелательных побочных эффектов у пациентов. В этом первичном исследовании CLDN18.2-отрицательную клеточную линию - клеточную линию тератокарциномы РА-1 (ATCC CRL-1572) - использовали для исследования неспецифического цитолитического потенциала 1BiMAB, введенного в виде IVT-мРНК.
Используемая клеточная линия РА-1 была стабильно трансдуцирована лентивирусным вектором на основе люциферазы и, таким образом, могла применяться в анализе на цитотоксичности на основе люциферазы. Клетки-мишени РА-1/luc готовили для электропорации, как описано в примере 16. Всего было трансфицировано 40 мкг/мл IVT-мРНК на образец. Использовали следующие IVT-мРНК: 1BiMAB, no. 25 и 6RHU3. No. 25 направленно воздействует на неэкспрессирующийся PLAC-1 ТАА, и 6RHU3 направленно воздействует на высокоэкспрессирующийся таргетный CLDN6 в клетках РА-1/luc. Для получения более подробной информации, касающейся вариантов bi-scFv, смотри таблицы 7 и 8. No. 25 использовали в качестве дополнения IVT-мРНК в образцах для электропорации 4 мкг/мл IVT-мРНК для обеспечения одинакового уровня стресса, вызванного трансфекцией РНК, для всех образцов клеток-мишеней. После осторожного перемешивания клетки трансфицировали с помощью электропоратора BTX ECM 830 (Harvard Apparatus, Holliston, MA, USA) с использованием следующих условий для 0,4 см кюветты: 200 В, 2 импульса, продолжительность импульса 12 мс, интервал между импульсами 400 мс. Сразу же после электропорации кюветы помещали на лед на короткий промежуток времени, и затем клеточные суспензии переносили в нагретую до комнатной температуры аналитическую среду РА-1 (МЕМ среда, дополненная 10% термоинактивированной FCS, 0,5% пенициллина-стрептомицина, 1х NEAA, 1,5 г/л бикарбоната натрия и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany)) в 15 мл пробирках. Трансфицированные клетки-мишени подсчитывали и доводили до 1 х 105 клеток на мл.
Человеческие эффекторные клетки выделяли, как описано в примере 16. Человеческие цитотоксические Т-клетки выделяли путем магнитно-активированной клеточной сортировки (MACS) из PBMC с помощью набора Pan T Cell Isolation Kit II, human (Miltenyi Biotec, Tererow, Germany) согласно инструкциям производителя. Т-клетки доводили до 5×105 клеток на мл в аналитической среде РА-1.
1×104 клеток-мишеней завевали в каждую лунку 96-луночного планшета и человеческие цитотоксические Т-клетки добавляли до соотношения Е:Т, равного 5:1. Конечный объем каждой лунки составил 100 мкл. Контрольные образцы, содержащие эффекторные клетки и клетки-мишени, содержали в качестве отрицательного контроля клетки-мишени, секретирующие no. 25, в качестве положительного контроля белок 6RHU3 или 6PHU3, и белок 1BiMAB. Концентрации белков доводили до конечной концентрации 100 нг/мл и объединяли с no. 25-трансфицированными клетками-мишенями для обеспечения одинаковых условий для используемых клеток-мишеней. Контроли минимального лизиса (Lmin) включали только каждый образец электропорированных клеток-мишеней. Контроли спонтанного лизиса (Lmax) состояли из необработанных клеток-мишеней и эффекторных клеток (Lmax) для вычитания из Lтестируемого образца или только необработанные клетки-мишени (Lmax2) для вычитания из Lmin. Каждый образец засевали в трех повторностях. Анализ проводили через 72 ч инкубации при 37°С, 5% СО2. Контроли спонтанного лизиса (Lmax) обрабатывали Triton X-100 в конечной концентрации 2%.
Для анализа 50 мкл водного раствора, содержащего 1 мг/мл люциферина (BD Monolight, BD, Biosciences, Heidelberg, Germany) и 50 мМ HEPES, добавляли в каждую лунку, и планшеты затем инкубировали в течение 30 мин в условиях темноты при 37°С. Люминесценцию, возникающую в результате окисления люциферина люциферазой, экспрессирующей жизнеспособные клетки, измеряли на микропланшетном ридере (Infinite M200, Tecan, Mannedorf, Switzerland). Процент специфического лизиса клеток-мишеней рассчитывали по следующей формуле: % специфического лизиса = [1 - (люминесценциятестируемый образец - Lmax) / (Lmin - Lmax2)] × 100.
На фигуре 28 показан процент специфического лизиса клеток-мишеней. CLDN18.2-отрицательные клетки РА-1/luc, трансфицированные отрицательным контролем no. 25 или 1BiMAB, не показывают значительного лизиса даже после инкубации в течение 72 ч. Также, 100 нг/мл белка 1BiMAB не приводит к значительному лизису, тогда как лизис, вызванный bi-scFv-секрецией 6RHU3, направленно воздействующего на ТАА, или 100 нг/мл белка 6RHU3, составил 85-93%. Анализ также проводили через 24 ч и 48 ч, который показал эквивалентные результаты (данные не показаны).
Пример 24: Количественный анализ продукции белка 1BiMAB после трансфекции клеток млекопитающих IVT-мРНК
Для исследования трансляции РНК в белок в клетках млекопитающего выбирали клеточную линию ВНК21 (АТСС CRL-13001) в качестве системы экспрессии. 2×107 клеток ВНК21 на мл трансфицировали путем электропорации, как описано в примере 16, при этом отличие состояло в том, что все стадии выполняли при комнатной температуре. 250 мкл клеточной суспензии переносили в 0,4 см кюветы Gene Pulser/MicroPulser Cuvettes (Bio-rad, Dreiech, Germany) и добавляли 40 мкг/мл IVT-mRNA 1BiMAB или IVT-репликон РНК. Условия электропорации были следующие: 300 В, продолжительность импульса 16 мс, 1 импульс, интервал между импульсами 400 мс.
Электропорированные клетки ресуспендировали в культуральной среде при комнатной температуре (RPMI 1640, 10% FCS) и переносили в 15 см культуральные чашки. Через 5 ч после засевания среду, содержащую FCS, заменяли средой, не содержащей FCS. В примерах 24 а и b супернатант клеточной культуры и клетки собирали для анализа отдельно через 18 ч после электропорации. Клеточные осадки лизировали в 1х LDS буфере для образца (номер по каталогу NP0008; Life technologies, Darmstadt, Germany) и нагревали при 72°С в течение 15 мин. Для анализа ELISA использовали неконцентрированный и приблизительно 50-кратно концентрированный супернатант. Концентрирование проводили с помощью центрифужного фильтра Amicon ultra-15 (Merck Millipore, Billerica, MA, USA) в соответствии с протоколом производителя. В случае примера 24с (только IVT-мРНК) супернатант собирали через 48 ч после трансфекции и подвергали 40-кратному концентрированию, как описано выше.
а. ELISA с использованием супернатанта
Для анализа ELISA планшеты, покрытые никелем (Thermo Fisher Scientific, Bonn, Germany) использовали для захвата аналита через его His-метку. Сначала планшет промывали три раза 200 мкл промывочного буфера (0,01% Tween-20 в 1х PBS) на лунку. В качестве стандарта очищенный белок 1BiMAB разводили в 1,75% Na-Казеин в 1 х PBS (= разбавитель). Ряд разведений изменялся от 2,34 до 37,50 нг/мл с шагом в 2. 100 мкл на стандартное разведение переносили в лунки в трех повторностях каждой концентрации. Соответствующим образом, 100 мкл образцов переносили в трех повторностях. Планшет герметично закрывали адгезивной пленкой и инкубировали при 37°С в течение 30 минут. После этого планшеты промывали три раза 200 мкл промывочного буфера на каждую лунку. Для детекции 1BiMAB использовали анти-идиотипическое моноклональное Ig 8B1F3, которое специфически связывается с VH-VL CLDN18.2ab и, следовательно, также с 1BiMAB. 8B1F3 смешивали с разбавителем до конечной концентрации 1 мкг/мл. 100 мкл раствора анти-идиотипического антитела переносили в каждую лунку с последующей инкубацией в течение 30 мин при 37°С. Затем планшет промывали 3-кратно 200 мкл промывочного буфера на каждую лунку, и 100 мкл АР-конъюгированного анти-мышиного детекторного антитела (Jackson Immuno Research Laboratories, West Grove, PA, USA) - в разведении 1:500 в разбавителе - добавляли в каждую лунку с последующей инкубацией в течение 30 мин при 37°С. После заключительной стадии промывания (3 х 200 мкл промывочного буфера) в каждую лунку добавляли 1,5 мг/мл субстрата pNPP в соответствующем субстратном буфере (1М диэтаноламин, 0,5 мМ MgCl2, 0,01% Na-азид, рН 9,8) с последующей инкубацией в течение 30 мин при комнатной температуре в условиях темноты. Для остановки ферментативной реакции использовали 100 мкл 3М КОН на каждую лунку. Поглощение измеряли с помощью микропланшетного ридера (Infinite M200, Tecan, Mannedorf, Switzerland). Для двухволнового анализа 405 нм устанавливали в качестве длины волны измерения и 492 нм в качестве длины волны сравнения. Величины поглощения рассчитывали путем вычитания длины волны сравнения из длины волны измерения.
На фигуре 29А нанесены средние величины поглощения при 405 нм, включая стандартные отклонения. Концентрированный супернатант из клеток, трансфицированных IVT-мРНК 1BiMAB и IVT-репликоном, вызвал значительный сигнал, подтверждая трансляцию bi-scFv, кодирующего IVT-РНК. Концентрированный супернатант из ложно-трансфицированных клеток не вызвал какого-либо сигнала. Фактические концентрации белка не могут быть предложены из-за различающейся токсичности конструкций IVT-мРНК и IVT-репликона и незначительно различающихся х-кратных концентратов. Определенная приблизительная концентрация белка в неконцентрированном супернатанте находилась в пределах 1,5 нг/мл для образцов IVT-репликона и 2,4 нг/мл для образцов IVT-mRNA.
b. Вестерн-блот анализ супернатанта и клеточного лизата (образцы IVT-мРНК и IVT-репликона)
Для анализа методом Вестерн-блоттинга концентрированные супернатанты и клеточные лизаты разделяли на NuPAGE Novex 4-12% Bis-tris Gels (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). Нагружали супернатант и клеточный лизат клеток ВНК21, трансфицированных IVT-мРНК 1BiMAB, IVT-репликоном РНК, или необработанных клеток и положительного контроля - 0,1 мкг очищенного белка 1BiMAB. Вестерн-блот-анализ выполняли с помощью стандартных процедур (Current Protocols in Protein Science, 2012). Вкратце, после блоттинга белков на PVDF-мембране и блокирования с помощью PBST/3% сухого молока мембрану инкубировали в течение 1 ч при 4°С с первичным антителом Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в разведении 1:500 в блокирующем буфере. После повторного промывания блокирующим буфером мембраны инкубировали с Fc-специфическим вторичным козьим-анти-мышиным IgG антителом, конъюгированным с пероксидазой (Sigma Aldrich, Germany), в разведении 1:10000 в блокирующем буфере в течение 1 ч при 4°С. После повторного промывания блокирующим буфером сигналы визуализировали с помощью SuperSignal West Femto Chemiluminescent Substrate (Pierce/Thermo Fisher Scientific, Rockford, IL, USA) и записывали с помощью ImageQuant LAS 4000 Imager (GE Healthcare Life Sciences, Munich, Germany). Сигналы 1BiMAB детектировали в диапазоне от 50 до 60 кДа в сравнении с молекулярной массой внутреннего стандарта.
Как показано на фигуре 29В слабые сигналы были детектированы в супернатанте клеток, трансфицированных IVT-мРНК (дорожка 2) и IVT-репликоном РНК (дорожка 3), тогда как сигнал отсутствовал в дорожке 4 (супернатант от необработанных клеток). Сильные сигналы могут быть генерированы клеточными лизатами клеток, трансфицированных IVT-мРНК (линия 5) и IVT-репликона РНК (линия 6). Сигнал снова отсутствовал в клеточном лизате необработанных клеток (дорожка 7). Все сигналы возникали на такой же высоте, что и у очищенного контрольного белка 1BiMAB (дорожка 8). Слабый сигнал в супернатанте и слабая секреция 1BiMAB возможно обусловлены относительно коротким временем инкубации после трансфекции. По причине токсического эффекта репликона РНК более длительные периоды инкубации не могут быть тестированы.
Оба анализа являются качественными и не могут быть использованы для определения концентрации белка.
с. Вестерн-блот анализ супернатанта (образцы IVT-мРНК)
Супернатант, собранный через 48 ч после трансфекции, отделяли с помощью SDS-PAGE с последующим вестерн-блот-анализом, как описано в примере 24b. Как показано на фигуре 29С, no.25 и 1BiMAB, транслированный из IVT-мРНК и секретированный в супернатант, детектировали посредством их His-метки. При этом продукция и секреция 1BiMAB, а также no.25, который использовали в качестве контроля специфичности bi-scFv, могут быть подтверждены.
Пример 25: Детекция in vivo транслированного и функционального белка 1BiMAB после внутримышечной инъекции РНК
Мышиных самок и самцов линии NSG в возрасте 8-16 недель отбирали и распределяли на 4 группы по 5 мышей. Каждой мыши в бедренную мышцу инъецировали 40 мкл раствора РНК. В 40 мкл раствора РНК содержалось 1х PBS, 5 мкг D2-кэппированного IVT-мРНК 1BiMAB или репликона, 2 мкг D1-кэпированной люциферазы IVT-мРНК и 0 или 15 мкг D1-кэппированного IVT-мРНК EBK. IVT-мРНК EBK, кодирующий белки E3L, B18R и K3L (EBK) вируса осповакцины, совместно инъецировали для ингибирования ответа IFN и для противодействия активации PKR в целях усиления трансляции РНК (заявка на патент PCT/EP2012/04673). Сигнал люциферазы наблюдали через 24 ч после инъекции Xenogen IVIS 2000 для исключения мышей без сигнала из коллекции образцов.
Кровь собирали через 2, 4 и 7 дней после инъекции. Сыворотку собирали и затем замораживали при -80°С. Мышцы мышей с сильным люминесцентным сигналом вырезали и гистохимически фиксировали для иммуногистохимического анализа (IHC) или замораживали путем шоковой заморозки для вестерн-блот-анализа через 4 дня после инъекции РНК.
Анализ на цитотоксичность
Сыворотку мышей NSG анализировали в анализе на цитотоксичность in vitro. Клетки-мишени NugC4, стабильно трансфектированные люциферазой светлячка и человеческим CLDN18.2 для лучшей экспрессии мишени, засевали человеческими Т-клетками (изолированными как описано в примере 16) в соотношении Е:Т, равном 30:1, для максимальной чувствительности. Аналитическая среда состояла из среды RPMI 1640, дополненной 10% термоинактивированной FCS, 0,5% пенициллина-стрептомицина, 1х NEAA и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany). 20 мкл оттаявшей тестируемой сыворотки добавляли в каждую лунку с тестируемым образцом. Лунки со стандартным контрольным белком 1BiMAB, лунки с Lmin и Lmax наполняли 20 мкл сыворотки необработанных мышей NSG. Конечный объем каждой лунки составил 100 мкл. Lmin засевали двенадцатикратно, Lmax шестикратно и тестируемые образцы в трех повторностях. Через 48 ч инкубации при 37°С и 5% СО2 лунки с Lmax смешивали с 10 мкл 2% раствора Triton X-100 и инкубировали в течение 10 мин. Во все другие лунки добавляли 10 мкл аналитической среды. Добавляли 50 мкл раствора люциферина (смотри пример 23) и планшеты измеряли - через 30 мин стадии инкубации при 37°С в условиях темноты - на микропланшетном ридере Infinite M200 (TECAN, Mannedorf, Switzerland). Расчет специфического лизиса клеток-мишеней выполняли, как описано в примере 23.
На фигуре 30 нанесен процент специфического лизиса. Значительные цитотоксические эффекты детектировали в каждой группе. Цитотоксические эффекты увеличились на фактор 1,7 в лунках, содержащих сыворотку мышей, инъецированных EBK и IVT-mRNA 1BiMAB, и собирали для анализа через 2 дня после инъекции. Значительно более низкие эффекты наблюдались в образцах, собранных на 4 или 7 день после инъекции. В случае образцов 1BiMAB-репликона, сыворотка, собранная в более поздние моменты времени, также генерировали сильные цитолитические эффекты.
Эти данные подтверждают in vivo трансляцию 1BiMAB из IVT-мРНК или -репликона и секрецию в кровоток после внутримышечной инъекции.
Пример 26: Генерация и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN6 и CD3
а. Исходные последовательности, дизайн конструкции bi-scFv и клонирование в матрицу вектора
Готовили конструкции биспецифического тандемного одноцепочечного антитела (bi-scFv), содержащие связывающие домены, специфические в отношении компонента CD3 человеческого Т-клеточного рецептора и человеческих опухоль-ассоциированных антигенов (TAA). Соответствующие вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) для каждой конструкции расположены специфически в направлении от 5'- до 3'-конца в следующем порядке:
pST1-5'AgKozak-VHαCLDN6-VLαCLDN6-VHαCD3- VLαCD3-His-2hBgUTR-A120 (6RHU5)
pST1-5'AgKozak-VHαCD3- VLαCD3- VHαCLDN6-VLαCLDN6- His-2hBgUTR-A120 (6RHU3)
В таблице 9 кратко описаны все конструкции bi-scFv, специфические в отношении TAA CLDN6, которые были генерированы в рамках изобретения. CLDN18.2-специфическую конструкцию 1BiMAB bi-scFv использовали в качестве контрольного антитела. Конструкции bi-scFv генерировали путем генного синтеза GeneArt AG (GeneArt/Life Technologies GmbH, Regensburg, Germany) с использованием последовательностей VH и VL соответствующих антител. Оптимизации кодонов, таких как Homo sapiens (HS) или Mus musculus (MM) были реализованы с помощью программного обеспечения GeneArt's GeneOptimizer®, и перечислены в таблице 9. Информация, касающаяся специфичности, происхождения последовательностей из моноклональных антител (mAB), использования кодонов, дополнительных отличительных признаков последовательностей и ссылок на все применяющиеся домены, обобщена в таблице 10. Исходные последовательности вариабельных доменов соответствующих антител CD3 перечислены в таблице 10. Благодаря высокой гомологии человеческих и мышиных TAA, одинаковые последовательности VH и VL анти-TAA могут быть использованы для создания конструкций bi-scFv для анализов на мышах, но в комбинации с последовательностями VH, VL мышиного специфического клона 145-2С11 анти-CD3 антитела.
Клонирование ДНК и конструирование экспрессионных векторов проводили в соответствии со стандартными процедурами (Green/Sambrook, Molecular Cloning, 2012), которые хорошо известны специалистам в данной области. Вкратце, главные ДНК последовательности bi-scFv содержали сайт рестрикции 5'-BsmBI и 3'-XhoI для клонирования в плазмиды pST1. Последовательность сигнала секреции вводили на 5' конце (выше) последовательности bi-scFv для секреции белка. Последовательность, кодирующую гибкий глицин-сериновый пептидный линкер, состоящий из 15-18 аминокислот, вставляли для соединения доменов VH и VL для композиции одноцепочечных вариабельных фрагментов антитела (scFv), из которых один связывается с CD3 и другой с TAA. Для формирования биспецифического одноцепочечного антитела последовательности двух доменов scFv соединяли последовательностью, кодирующей короткий пептидный линкер (GGGGS). Вместе с этой последовательностью линкера вводили сайт рестрикции BamHI для обмена доменов scFv для клонирования последующих конструкций bi-scFV. Вкратце, домены 5'scFv- заменяли путем рестрикции BsmBI и BamHI и 3'scFv-домены путем рестрикции BamHI и XhoI. C-концевую 6xHis-метку использовали для анализов на обнаружение транслированного белка. Для получения вектора репликона 6RHU3 полную последовательность 6RHU3, включая сигнал секреции и 6xHis-метку, субклонировали в направлении от 3' от субгеномного промотора вектора репликона вируса леса Семлики (pSFV), любезно предоставленного K. Lundstrom (Lundstrom, K. et al. (2001) Histochem. Cell Biol. 115 (1), pp. 83-91; Ehrengruber, M.U. et al. (1999) Proc. Natl. Acad. Sci. U.S.A. 96 (12), pp. 7041-7046).
Все конструкции подтверждали путем секвенирования через сервис по секвенированию одиночных прочтений (MWG single read sequence service) (Eurofins MWG Operon, Ebersberg, Germany) и использовали конструкции только с правильной последовательностью и поли(А)-хвостом, состоящим из более чем 100 аденинов, для in vitro транскрипции РНК.
Схемы конструкций представлены также на фигуре 31А.
Таблица 9. Краткое описание конструкций мРНК-матрицы биспецифического одноцепочечного антитела, специфического в отношении ТАА и CD3
ность
bi-scFv означает биспецифический одноцепочечный вариабельный фрагмент; HS, Homo-sapiens; HU, гуманизированное; ММ, Mus musculus; ТАА, опухоль-ассоциированный антиген; VH, вариабельный домен тяжелой цепи, VL, вариабельный домен легкой цепи.
Таблица 10. Краткое описание конструкции матрицы мРНК- bi-scFv
Таблица 10
mAB
HS, Homo sapience; mAB, моноклональное антитело; ММ, Mus musculus; ТАА, опухоль-ассоциированный антиген.
b. Синтез IVT-РНК
Для создания анти-CLDN6-специфических IVT-матриц bi-scFv, плазмиды линеаризовали ниже поли(А)-хвоста с использованием эндонуклеазы класса IIs. Линеаризованную матрицу ДНК очищали экстракцией смесью фенол/хлороформ и осаждали ацетатом натрия, как описано в литературных источниках (Holtkamp, S. et al. (2006) Blood 108 (13), pp. 4009-4017).
Линеаризованные матрицы ДНК подвергали in vitro транскрипции с использованием наборов MEGAscript Kits (Ambion/Life Technologies, Darmstadt, Germany) в соответствии с инструкциями производителя: матрицы pST1 транскрибировали с помощью набора MEGAscript Т7 Kit и матрицы pSFV с помощью набора MEGAscript SP6 Kit. Для реакций с кэп-аналогом концентрацию GTP уменьшали до 1,5 мМ и 6 мМ ARCA, бета-S-ARCA(D1) или в реакцию добавляли бета-S-ARCA(D2), синтезированный, как описано в литературе (Kowalska, J. et al. (2008) RNA 14 (6), pp. 1119-1131; Grudzien, E. et al. (2004) RNA 10 (9), pp. 1479-1487; Stepinski, J. et al. (2001) RNA 7 (10), pp. 1486-1495). Очистку IVT-мРНК выполняли с помощью набора MEGAclear Kit (Ambion/Life Technologies, Darmstadt, Germany) в соответствии с руководством. Концентрацию и качество IVT-РНК оценивали с помощью спектрофотометрии и анализировали на биоанализаторе 2100 Bioanalyzer (Agilent Technologies, Santa Clara, CA, USA).
Пример 27: Микроскопический анализ Т-клеток, перенаправленных на клетки-мишени, секретирующие CLDN6-специфический bi-scFv 1BiMAB
Порядок проведения анализа по существу был таким, как описано в примере 2.а.
В качестве линии клеток-мишеней использовали субклон клеточной линии тератокарциномы яичников PA-1 (ATCC CRL-1572), которая эндогенно экспрессирует высокие уровни человеческого CLDN6. Человеческие цитотоксические Т-клетки отделяли, используя MACS, от PBMC, выделенных из свежеотобранной крови, с помощью набора Pan T Cell Isolation Kit, human (Miltenyi Biotec, Teterow, Germany) в соответствии с инструкциями производителя.
Клетки-мишени PA-1 - которые тестировали обычным способом в отношении экспрессии ТАА с помощью FACS-анализа перед каждым анализом - дважды промывали ледяной средой X-Vivo 15 (LONZA, Basel, Switzerland) и ресуспендировали до плотности 2×107 клеток/мл. 250 мкл клеточной суспензии переносили в предварительно охлажденные 0,4 см кюветы Gene Pulser/MicroPulser Cuvettes (Bio-Rad, Dreieich, Germany) и добавляли 20 мкг/мл IVT-мРНК. Условия электропоратора BTX ECM 830 (Harvard apparatus, Holliston, MA, USA) были следующие: 200 В, длина импульса 12 мс, 2 импульса, интервал между импульсами 400 мс. Использовали следующие IVT-мРНК: 6RHU5, 6RHU3, no. 25 (для подробной информации смотри таблицы 9 и 10). Трансфицированные клетки-мишени подсчитывали и доводили до 1×105 клеток на мл. 1×105 клеток-мишеней засевали в каждую лунку 6-луночного планшета, и человеческие цитотоксические Т-клетки добавляли в соответствии с соотношением Е:Т, равным 5:1. Конечный объем каждой лунки составил 2 мл. Контрольные образцы содержали только трансфицированные клетки мишени для подтверждения лечебных свойств после электропорации и контрольные трансфицированные bi-scFv клетки-мишени с эффекторными клетками. В качестве положительных контролей соответствующие CLDN6-специфические белки 6RHU5 и 6RHU3 bi-scFv готовили в концентрации 50 мкг/мл. Таким образом, использовали необработанные клетки РА-1 с человеческими Т-клетками. Затем тканевые культуральные планшеты инкубировали при 37°С, 5% СО2. Значительные эффекты с точки зрения образования Т-клеточных кластеров на клетках-мишенях, формирования иммунологического синапса и гибели клеток-мишеней в образцах, содержащих трансфицированные 6RHU5 и 6RHU3 клетки-мишени, наблюдали через 24 ч и записывали с помощью инвертационного микроскопа Nikon Eclipse TS100 (Nikon, Japan). Как показано на фигуре 32, образования кластеров Т-клеток или лизиса клеток-мишеней не наблюдалось в контрольном образце с no. 25 трансфицированными клетками-мишенями, свидетельствуя о строгой зависимости от экспрессии ТАА для индукции активации Т-клеток. Имитирующий контроль без bi-scFv также показывает отсутствие образования кластеров Т-клеток. Контрольные белки наоборот приводят к образованию кластеров Т-клеток и лизису клеток-мишеней.
Пример 28: Активация Т-клеток, индуцированных кандидатами 6RHU5 и 6RHU3 bi-scFv , направленно воздействующими на CLDN6
Для детекции активации Т-клеток и определения различий в эффективности двух вариантов CLDN6-специфических bi-scFv, выполняли анализ активации Т-клеток на основе FACS. Маркер ранней активации CD69 и маркер поздней активации CD25 были выбраны для окрашивания антителами, конъюгированными с флуоресцентной меткой. Для детекции человеческих Т-клеток в смеси клеток-мишеней и Т-клеток их окрашивали CD3, который экспрессируется всем Т-клетками.
Клетки-мишени и эффекторные клетки готовили, как описано выше (пример 27). Вкратце, клетки-мишени РА-1, эндогенно экспрессирующие CLDN6, трансфицировали путем электропорации 20 мкг/мл следующих IVT-мРНК: 6RHU5, 6RHU3 и no. 25. В качестве контроля специфичности использовали no.25, направленно воздействующий на неэкспрессированный ТАА, в качестве имитирующего контроля использовали необработанные клетки-мишени. В качестве положительного контроля использовали 50 нг/мл белка 6RHU5. Кроме того, Т-клетки засевали без клеток-мишеней с белком 6RHU5 или без него в качестве референсных исходных активаций. Каждый образец засевали в двух повторностях в 6-луночные планшеты и инкубировали при 37°С, 5% СО2. Через 24 ч и 48 ч Т-клетки собирали путем соскабливания и переносили в 5 мл пробирки с круглым дном (BD Falcon, Heidelberg, Germany). Клетки центрифугировали и промывали DPBS. Для окрашивания клеток использовали мышиные античеловеческие CD3-FITC, мышиные античеловеческие CD69-APC и мышиные античеловеческие CD25-PE (все антитела BD Biosciences, Heidelberg, Germany). Клеточные осадки ресуспендировали в 50 мкл FACS-буфера (DPBS, дополненный 5% FBS), содержащего антитела, конъюгированные с флуоресцентными метками, и 2 мкл 7-AAD (BD Biosciences, Heidelberg, Germany). После инкубации в течение 20 мин при 4°С в условиях темноты образцы промывали 4 мл DPBS и клеточные осадки ресуспендировали в 200 мкл FACS-буфера. Образцы хранили на льду и условиях темноты на протяжении измерения с помощью проточного цитометра FACSCanto II (оба от фирмы BD Biosciences, Heidelberg, Germany). Результаты оценивали с помощью программного обеспечения FlowJo software (Tree Star, San Carlos, CA, USA).
Как показано на фигуре 33А и В, оба варианта привели к активации Т-клеток, тогда как ни один из отрицательных контролей не показал значительной экспрессии маркеров CD69 и CD25 Т-клеточной активации.
Общая активация Т-клеток в ответ на 6RHU3 была в 1,53 раза выше через 24 ч (А) и в 1,35 раз выше через 48 ч (В) совместной инкубации, по сравнению с ответом на 6RHU5. На основании этих обнаружений все последующие исследования выполняли только с вариантом 6RHU3.
Пример 29: Зависимая от концентрации активация Т-клеток путем совместной инкубации с трансфицированными 6RHU3 клетками-мишенями
Для определения самой низкой концентрации IVT-мРНК 6RHU3, необходимой для индукции зависимой от мишени активации Т-клеток, ряд разведений в диапазоне 0,2-20 мкг/мл IVT-мРНК трансфицировали в клетки-мишени РА-1. Чтобы все образцы подвергнулись одинаковому уровню стресса вызванной электропорацией РНК, трансфицировали IVT-мРНК в общей концентрации 20 мкг/мл. No.25 - направленно воздействующий на неэкспрессированный ТАА - использовали в качестве наполнения IVT-мРНК. Соответственно, 0 мкг/мл 6RHU3 коррелирует с 20 мкг/мл no.25. Электропорацию выполняли, как описано в примере 27. Человеческие Т-клетки использовали в качестве эффекторных клеток. Выделение из PBMC выполняли в соответствии с инструкциями производителя (Pan T Cell Isolation Kit II, Miltenyi, Teterow, Germany). Эффекторные клетки и клетки-мишени смешивали в соотношении 5:1. Необработанные клетки-мишени с Т-клетками служили в качестве имитирующего контроля. Затем Т-клетки засевали без клеток-мишеней с белком 6RHU5 или без него в качестве рефференсных исходных активаций. В качестве положительного контроля необработанные клетки-мишени смешивали с Т-клетками и 50 нг/мл белка 6RHU5. Все образцы готовили в двух повторностях в 6-луночных планшетах.
Клетки-мишени и эффекторные клетки совместно инкубировали в течение 48 ч при 37°С, 5% СО2. Образцы готовили для анализа методом проточной цитометрии, как описано в примере 28.
На фигуре 34 нанесено процентное содержание CD3-положительных Т-клеток, которые экспрессируют маркеры активации. Активации Т-клеток не наблюдалось в контролях. Начало значительной активации Т-клеток детектировали при концентрации 0,7 мкг/мл IVT-мРНК 6RHU3. Эффекты в ответ на 2,0 мкг/мл IVT-мРНК 6RHU3 были сравнимы с эффектами, опосредованными 50 нг/мл контрольного белка 6RHU5.Таким образом, трансляция белка и секреция из трансфицированного IVT-мРНК, по-видимому, является эффективным процессом.
Пример 30: Определение ЕС50 для 6RHU3
Для определения половины максимальной эффективной дозы bi-scfv, кодирующего 6RHU3 IVT-мРНК, ряд титрований 6RHU3 тестировали в in vitro анализе на цитотоксичность на основе люциферазы. Стабильно экспрессирующие люциферазу клетки РА-1, временно трансфицировали путем электропорации, как описано в примере 27. Используемые концентрации IVT-мРНК 6RHU3 находились в диапазоне 0,004-13,3 мкг/мл в 6-ступенчатых разведениях. Общая концентрацию IVT-мРНК была постоянной и составила 13,3 мкг/мл, no. 25 использовали в качестве наполнения IVT-мРНК. В качестве контролей минимального лизиса (Lmin) все образцы трансфицированных клеток-мишеней засевали без эффекторных клеток. С помощью этой процедуры исходно погибшие клетки каждого индивидуального образца электропорации вычитают и получают только эффекты лизиса, опосредованного Т-клетками.
Трансфицированные клетки-мишени засевали с человеческими Т-клетками в соотношении эффектора к мишени, равном 5:1, в трех повторностях в 96-луночном формате и инкубировали при 37°С, 5% СО2. Максимальный лизис (Lmax) для нормализации до показателей спонтанной люминесценции достигался путем добавления Triton X-100 в контрольные лунки, содержащие эффекторные клетки и необработанные клетки-мишени, непосредственно перед добавлением люциферина. После добавления раствора люциферина - в каждую лунку 50 мкл водного раствора, содержащего 1 мг/мл люциферина (BD Monolight, BD Biosciences, Heidelberg, Germany) и 50 мМ HEPES - люминесценцию измеряли на микропланшетном ридере Infinite M200 Tecan через 24 ч и 48 ч. Специфический лизис клеток-мишеней рассчитывали по формуле: % специфического лизиса = [1-(люминесценция тестируемый образец - Lmax) / (Lmin_тестируемый образец - Lmax)] × 100.
На фигуре 35 показана кривая зависимости от концентрации специфического лизиса клеток-мишеней в ответ на 6RHU3. С использованием уравнения GraphPad Prism для расчета величин ЕС50 «log(агонист) против ответа - вариабельный наклон» выявили EC50 (24 ч) = 548,0 нг/мл, и ЕС50 (48 ч) = 194,5 нг/мл. Результат этого анализа зависит в значительной степени от силы человеческих Т-клеток, которая изменяется в соответствии с иммунным статусом донора, как описано также в других источниках (смотри, например, (Lutterbuese, R. et al. (2010) Proc. Natl. Acad. Sci U.S.A. 107 (28), pp. 12605-12610).
Пример 31: Пролиферация Т-клеток в ответ на трансфекцию IVT-мРНК 6RHU3
Пролиферация Т-клеток является признаком активации Т-клеток. Для демонстрации специфической пролиферации Т-клеток в ответ на bi-scFv, кодирующий IVT-мРНК 1BiMAB, в присутствии CLDN6-положительных клеток-мишеней, использовали анализ методом проточной цитометрии. Вкратце, 1×107 человеческих Т-клеток, выделенных, как описано в примере 16, окрашивали 1 мкМ карбоксифлуоресцеина диацетата сукцинимидилового сложного эфира (CellTrace CFSE, Invitrogen/Life Technologies GmbH, Germany), растворенного в DPBS, в условиях темноты при комнатной температуре в течение 5 мин. Клетки промывали дважды DPBS /5%FCS и ресуспендировали в аналитической среде до 2×106 клеток на мл. В качестве клеток-мишеней были выбраны клетки РА-1, эндогенно экспрессирующие CLDN6 и - для тестирования специфичности - CLDN6-отрицательная клеточная линия рака молочной железы MDA-MB-231. 20 мкг/мл IVT-мРНК 6RHU3 или нетаргетный контроль использовали для электропорации. Электропорацию РА-1 выполняли, как описано в примере 27. Электропорацию MDA-MB-231 выполняли с использованием следующих условий: 400 В, продолжительность импульса 3 мс, 1 импульс, интервал между импульсами 400 мс в 0,4 см кюветах Gene Pulser/MicroPulser Cuvettes (Bio-Rad, dreieich, Germany). Анализ на цитотоксичность, описанный в примере 27, проводили с трансфицированными клетками-мишенями и человеческими CFSE-мечеными Т-клетками в качестве эффекторных клеток. В качестве отрицательных контролей использовали только Т-клетки и нетрансфицированные или контрольные трансфицированные клетки-мишени плюс Т-клетки. В качестве положительного контроля служили только Т-клетки, стимулированные 5 мкг/мл OKT3 (Bio Х Cell, West Lebanon, NH, USA) и 2 мкг/мл анти-CD28 (BioLegend, fell, Germany),. Белок 6PHU3 в концентрации 5 нг/мл наносили на нетрансфицированные клетки-мишени плюс Т-клетки для подтверждения валидности анализа. Образцы, объединенные с нетаргетным белком 1BiMAB, включали в качестве контроля специфичности. Все образцы готовили в трех повторностях в общем объеме 0,2 мл аналитической среды в 96-луночных планшетах. Через 72 ч совместной инкубации Т-клетки собирали, переносили в 5 мл пробирки с круглым дном, промывали и окрашивали при 4°С в течение 30 мин 2 мкл анти-CD45-APC для дифференциации человеческих Т-клеток от опухолевых клеток, и 0,25 мкл eFluor506 (BD Biosciences, Heidelberg, Germany) для контрастного окрашивания погибших клеток в 200 мкл DPBS. После отмывания DPBS клетки ресуспендировали в FACS-буфере и анализировали с помощью FACSCanto II (BD Biosciences, Heidelberg, Germany).
Пролиферацию Т-клеток детектировали по уменьшению CFSE-сигнала только в присутствии CLDN6-положительных клеток-мишеней и анти-CLDN6-специфического bi-scFv (смотри также фигуру 36). Кроме положительного контроля, пролиферацию Т-клеток, составляющую 49-61%, можно было наблюдать в присутствии CLDN6-положительных клеток-мишеней в комбинации с белком 6PHU3. Таргетные положительные клетки, трансфицированные 6PHU3 IVT-мРНК, привели к 62% +/- 2%. Т-клетки, инкубированные с CLDN6-отрицательными клетками-мишенями MDA-MB-231, не показали значительной пролиферации в любых условиях, а также Т-клетки без клеток-мишеней, за исключением положительного контроля.
Пример 32: Количественный анализ продукции белка после трансфекции IVT-мРНК 6PHU3 клеток млекопитающего
Для исследования трансляции РНК в белок в клетках млекопитающего в качестве системы экспрессии выбирали клеточную линию ВНК21 (ATCC CRL-13001). 2×107 клеток ВНК на мл трансфицировали путем электропорации, как описано в примере 16, с тем отличием, что все стадии проводили при комнатной температуре. 250 мкл клеточной суспензии переносили в 0,4 см кюветы Gene Pulser/MicroPulser Cuvettes (Bio-Rad, dreieich, Germany) и добавляли 40 мкг/мл no.25 IVT-мРНК, IVT-мРНК 6PHU3 или IVT-репликон РНК 6PHU3. Электропорацию выполняли с использованием следующих условий: 300 В, продолжительность импульса 16 мс, 1 импульс, интервал между импульсами 400 мс.
Электропорированные клетки ресуспендировали в культуральной среде при комнатной температуре (RPMI1640, 10% FCS) и переносили в 15 см культуральные чашки. Через 5 ч после засевания среду, содержащую FCS, заменяли средой, не содержащей FCS. Для примеров 32 а и b через 18 ч после электропорации отдельно собирали супернатант клеточной культуры и клетки. Клеточные осадки лизировали в 1× LDS буфере для образца (номер по каталогу NP0008; Life technologies, Darmstadt, Germany) и нагревали при 72°С в течение 15 мин. Для анализа ELISA использовали неконцентрированный и приблизительно 50-кратно концентрированный супернатант. Концентрирование проводили с помощью центрифужных фильтров Amicon ultra-15 (Merck Millipore, Billerica, MA, USA) в соответствии с протоколом производителя. Для примера 32 с (только IVT-мРНК) супернатант собирали через 48 ч после трансфекции и подвергали 40-кратному концентрированию, как описано выше.
а. ELISA с использованием супернатанта
Для анализа ELISA планшеты, покрытые никелем (Thermo Fisher Scientific, Bonn, Germany) использовали для захвата аналита через его His-метку. Сначала планшет промывали три раза 200 мкл промывочного буфера (0,01% Tween-20 в 1х PBS) на лунку. В качестве стандарта очищенный белок 6PHU3 разводили в 1,75% Na-Казеин в 1× PBS (= разбавитель). Ряд разведений изменялся от 2,34 до 150 нг/мл с шагом в 2. 100 мкл на стандартное разведение переносили в лунки в трех повторностях каждой концентрации. Соответствующим образом, 100 мкл образцов переносили в трех повторностях. Планшет герметично закрывали адгезивной пленкой и инкубировали при 37°С в течение 30 минут. После этого планшеты промывали три раза 200 мкл промывочного буфера на каждую лунку. Для детекции 6PHU3/6RHU3 использовали анти-идиотипическое моноклональное IgG 4F9, которое специфически связывается с VH-VL CLDN6ab и, следовательно, также с 6PHU3/6RHU3. 4F9 смешивали с разбавителем до конечной концентрации 2,5 мкг/мл. 100 мкл раствора анти-идиотипического антитела переносили в каждую лунку с последующей инкубацией в течение 30 мин при 37°С. Затем планшет промывали 3-кратно 200 мкл промывочного буфера на каждую лунку, и 100 мкл АР-конъюгированного анти-мышиного детекторного антитела (Jackson Immuno Research Laboratories, West Grove, PA, USA) - в разведении 1:500 в разбавителе - добавляли в каждую лунку с последующей инкубацией в течение 30 мин при 37°С. После заключительной стадии промывания (3 х 200 мкл промывочного буфера) в каждую лунку добавляли 1,5 мг/мл субстрата pNPP в соответствующем субстратном буфере (1М диэтаноламин, 0,5 мМ MgCl2, 0,01% Na-азид, рН 9,8) с последующей инкубацией в течение 30 мин при комнатной температуре в условиях темноты. Для остановки ферментативной реакции использовали 100 мкл 3М КОН на каждую лунку. Поглощение измеряли с помощью микропланшетного ридера (Infinite M200, Tecan, Mannedorf, Switzerland). Для двухволнового анализа 405 нм устанавливали в качестве длины волны измерения и 492 нм в качестве длины волны сравнения. Величины поглощения рассчитывали путем вычитания длины волны сравнения из длины волны измерения.
На фигуре 37А нанесены средние величины поглощения, включая стандартные отклонения. Концентрированный супернатант из клеток, трансфицированных 6RHU3 IVT-мРНК и IVT-репликоном, вызвал значительный сигнал, подтверждая трансляцию bi-scFv-кодирующей IVT-РНК. Концентрированный супернатант из ложно-трансфицированных клеток и контроля no.25 (-контр) не вызвал какого-либо сигнала, как ожидалось. Фактические концентрации белка не могут быть предположены из-за различающейся токсичности конструкций IVT-мРНК и IVT-репликона и незначительно различающихся х-кратных концентратов. Определенная приблизительная концентрация белка в неконцентрированном супернатанте находилась в пределах 1,4 нг/мл для образцов IVT-репликона и 5,9 нг/мл для образцов IVT-мРНК.
b. Вестерн-блот анализ супернатанта и клеточных лизатов (образцов IVT-mRNA и IVT-репликона)
Для анализа методом Вестерн-блоттинга концентрированные супернатанты и клеточные лизаты разделяли на NuPAGE Novex 4-12% Bis-tris Gels (Invitrogen/Life Technologies GmbH, Darmstadt, Germany). Нагружали супернатант и клеточный лизат клеток ВНК21, трансфицированных IVT-мРНК 6RHU3, IVT-репликоном РНК, IVT-мРНК no.25 или необработанных клеток и положительного контроля - 0,1 мкг очищенного белка 6RHU3. Вестерн-блот-анализ выполняли с помощью стандартных процедур (Current Protocols in Protein Science, 2012). Вкратце, после блоттинга белков на PVDF-мембране и блокирования с помощью PBST/3% сухого молока мембрану инкубировали в течение 1 ч при 4°С с первичным антителом Anti-HIS Epitope-Tag (Dianova GmbH, Hamburg, Germany) в разведении 1:500 в блокирующем буфере. После повторного промывания блокирующим буфером мембраны инкубировали с Fc-специфическим вторичным козьим-анти-мышиным IgG антителом, конъюгированным с пероксидазой (Sigma Aldrich, Germany), в разведении 1:10000 в блокирующем буфере в течение 1 ч при 4°С. После повторного промывания блокирующим буфером сигналы визуализировали с помощью SuperSignal West Femto Chemiluminescent Substrate (Pierce/Thermo Fisher Scientific, Rockford, IL, USA) и записывали с помощью ImageQuant LAS 4000 Imager (GE Healthcare Life Sciences, Munich, Germany). Сигналы 6RHU3 детектировали в диапазоне от 50 до 60 кДа в сравнении с молекулярной массой внутреннего стандарта.
Как показано на фигуре 37В, сигналы были детектированы в супернатантах клеток, трансфицированных no.25 IVT-мРНК (дорожка 1), 6RHU3 IVT-мРНК (дорожка 4) и IVT-репликоном РНК 6RHU3 (дорожка 5), тогда как сигнал отсутствовал в дорожке 6 -супернатант от необработанных клеток. Сильные сигналы могут быть генерированы клеточными лизатами клеток, трансфицированных IVT-мРНК 6RHU3 (дорожка 8) и IVT-репликоном РНК (дорожка 9). Клеточный лизат клеток, трансфицированных no.25, привел к очень слабому сигналу (дорожка 2), необработанные клетки (дорожка 10) показали отсутствие сигнала. Поддавался детекции очищенный контрольный белок 6RHU3 (дорожка 11). Сравнительно слабые сигналы в супернатанте и вместе с тем слабая секреция 6RHU3 возможно обусловлены относительно коротким периодом инкубации после трансфекции. По причине токсического эффекта репликона РНК более длительные периоды инкубации не могут быть тестированы.
Оба анализа являются качественными и не могут быть использованы для определения концентрации белка.
с. Вестерн-блот анализ супернатанта (образцы IVT-мРНК)
Супернатант, собранный через 48 ч после трансфекции, отделяли с помощью SDS-PAGE с последующим вестерн-блот-анализом, как описано в примере 32b. Как показано на фигуре 27С, no.25 и 6RHU3, транслированный из IVT-мРНК и секретированный в супернатант, детектировали посредством их His-метки. При этом может быть подтверждена продукция и секреция 6RHU3, а также no.25, который использовали в качестве контроля специфичности bi-scFv.
Пример 33: Детекция in vivo транслированного и функционального CLDN6-специфического bi-scFv белка после внутримышечной инъекции РНК
Мышиных самок и самцов линии NSG в возрасте 8-16 недель отбирали и распределяли на 4 группы по 5 мышей. Каждой мыши в бедренную мышцу инъецировали 40 мкл раствора РНК. В 40 мкл раствора РНК содержалось 1х PBS, 5 мкг D2-кэппированного IVT-мРНК 6RHU3 или репликона, 2 мкг D1-кэпированного IVT-мРНК люциферазы и 0 или 15 мкг D1-кэппированного IVT-мРНК EBK. IVT-мРНК EBK, кодирующий белки E3L, B18R и K3L (EBK) вируса осповакцины, совместно инъецировали для ингибирования ответа IFN и для противодействия активации PKR в целях усиления трансляции РНК (заявка на патент PCT/EP2012/04673). Сигнал люциферазы наблюдали через 24 ч после инъекции Xenogen IVIS 2000 для исключения мышей без сигнала из коллекции образцов.
Кровь собирали через 2, 4 и 7 дней после инъекции. Сыворотку собирали и затем замораживали при -80°С.
Анализ на цитотоксичность
Сыворотку мышей NSG анализировали в анализе на цитотоксичность in vitro. Клетки-мишени РА-1, стабильно трандуцированные люциферазой светлячка и эндогенно экспрессирующимся CLDN6, засевали человеческими Т-клетками (выделенными, как описано в примере 16) в соотношении Е:Т, равном 30:1, для максимальной чувствительности. Аналитическая среда состояла из среды RPMI 1640, дополненной 10% термоинактивированной FCS, 0,5% пенициллина-стрептомицина, 1х NEAA и 1 мМ пирувата натрия (Gibco/Life Technologies GmbH, Darmstadt, Germany). 20 мкл оттаявшей тестируемой сыворотки добавляли в каждую лунку с тестируемым образцом. Лунки со стандартным контрольным белком 6PHU3, лунки с Lmin и Lmax наполняли 20 мкл сыворотки необработанных мышей NSG. Конечный объем на лунку составил 100 мкл. Lmin засевали двенадцатикратно, Lmax шестикратно и тестируемые образцы в трех повторностях. Через 48 ч инкубации при 37°С и 5% СО2 лунки Lmax смешивали с 10 мкл 2% раствора Triton X-100 и инкубировали в течение 10 мин. Во все другие лунки добавляли 10 мкл аналитической среды. Добавляли 50 мкл раствора люциферина (смотри пример 23) и планшеты измеряли - через 30 мин стадии инкубации при 37°С в условиях темноты - на микропланшетном ридере Infinite M200 (TECAN, Mannedorf, Switzerland). Расчет специфического лизиса клеток-мишеней выполняли, как описано в примере 23.
На фигуре 28 показан процент специфического лизиса. Значительные цитотоксические эффекты детектировали в каждой группе. Концентрация CLDN6-специфического белка bi-scFv была значительно увеличена путем совместной инъекции с EBK в случае IVT-мРНК 6RHU3.
Эти данные подтверждают in vivo трансляцию 6RHU3 из IVT-мРНК или -репликона и секрецию в кровоток после внутримышечной инъекции.
Пример 34: Генерирование и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN18.2 или CLDN6 и CD3
Для дальнейшей разработки анти-CLDN18.2 и анти-CLDN6 специфических фрагментов антитела bi-scFv были рассмотрены другие аспекты для оптимизации оригинального белка. Эти аспекты в основном направлены на получение препаратов с более высокой однородностью в отношении различных разновидностей фолдинга, дисульфидных изомеров и олигомеров, которые могут формироваться при рекомбинантной продукции белка. Эти модификации не должны влиять на функциональную активность белков bi-scFv.
Замещение дополнительного (непарного) цистеинового остатка в анти-CD3 связывающем домене bi-scFv белков.
Для исследования способности непарных цистеиновых остатков, возникающих в пределах первичной последовательности домена Ig, препятствовать правильному формированию внутрицепочечных и/или межцепочечных дисульфидных связей, которые являются важными для надлежащего сворачивания и стабильности полученного фрагмента антитела, было генерировано несколько синтетических конструкций. Такие непарные цистеины могут отрицательно влиять на эффективность, однородность, продуктивность и стабильность конечного белкового продукта и, следовательно, должны избегаться. В дополнение к «стандартному» набору цистеинов, вовлеченных и образование пар дисульфидных связей, в вариабельных доменах могут присутствовать свободные цистеиновые остатки.
Например, в домене VH антитела OKT3 в положении Н92 присутствуют три остатка консервативного цистеина перед началом CDR-H3 и формируют структурную дисульфидную связь с положением Н22. Но в этой молекуле в положении Н100А (CDR-H3) другой цистеин (Cys) может обеспечить неправильное сворачивание, при котором Н100А вместо Н92 является вовлеченным в формирование дисульфидной связи с Н22, генерируя, таким образом, неправильно свернутый, нерастворимый и нефункциональный продукт. Для преодоления этого возможного неправильного спаривания цистеиновых остатков выполняли сайт-направленное замещение свободного цистеина (Kipriyanov, Protein Engineering 10:445-453, 1997). С помощью этого одиночного замещения было достигнуто значительное увеличение продуктивности и стабильности scFv, полученного из OKT3, при сохранении общей связывающей активности.
Домен VH анти-CD3 антитела TR66 (SEQ ID NO: 36), используемый в настоящем исследовании, содержит такой свободный цистеин в положении Н103 первичной последовательности, как показано в SEQ ID NO: 36. Сравнение последовательностей домена VH анти-CD3 антитела TR66 (SEQ ID NO: 36) с доменом VH анти-CD3 антитела ОКТ3 показало гомологию последовательностей, составляющую 96,6%.
Согласно этим результатам, замещение свободного цистеина сериновым остатком в пределах CDR-H3 домена VH анти-CD3 антитела TR66 выполняли для белков bi-scFv, направленно воздействующих на CD3 (SEQ ID NO: 94) и либо для CLDN18.2 или CLDN6. Введение таких заместителей позволило сконструировать белки bi-scFv 1-BiMAB-S (SEQ ID NO: 103) и 6-PHU3-S (SEQ ID NO: 110) (смотри таблицы 11 и 12, соответственно).
Замещение дополнительных (непарных) цистеиновых остатков в анти-CLDN6 белках bi-scFv
Родительское анти-CLDN6 антитело mCLDN6ab, вариабельные домены которого использовали для сборки соответствующих белков bi-scFv 6PHU3 (SEQ ID NO: 45) и 6PHU5 (SEQ ID NO: 43), содержит непарный цистеиновый остаток в пределах фланкированной области CDR-L2 домена L2 в положении 46 соответствующей первичной последовательности. Это соответствует положению 45 в пределах SEQ ID NO: 23, где первая аминокислота VL пропущена. Различные замещения выполняли для замещения этого свободного цистеина:
- сериновым остатком аналогично замещению в домене VH анти-CD3 антитела (TR66 (SEQ ID NO: 100);
- лейциновым остатком, путем сравнения с аминокислотной последовательностью других анти-CLDN6 антител (SEQ ID NO: 97 и 98);
- триптофановым остатком, путем сравнения аминокислотной последовательности с базой данных зародышевой линии (SEQ ID NO: 99).
Оценка длины линкера, порядка V-доменов и искусственных связующих дисульфидных связей в анти-CLDN18.2 bi-scFv белках
В качестве другого примера Arndt et al. (Biochemistry 37:12918-12926, 1998) описывает так называемую замену домена как возможное объяснение появления нековалентно связанных олигомеров фрагментов scFv. Согласно этой модели белок находится в состоянии возможного термодинамического равновесия между мономерной и димерной/олигомерной формой по причине постоянно возникающего внутри- и межмолекулярного обмена пограничных контактов VL/VH. Эти олигомеры могут уже присутствовать в супернатанте клеточной культуры и должны быть удалены во время процесса очистки. Однако эти молекулярные разновидности могут также формироваться во время хранения очищенных мономерных разновидностей.
На предпочтительный энергетический статус сильное влияние оказывает общая конструкция (первичная последовательность, длина линкера, ориентация VL/VH и т.д.).
Worn и Pluckthun (JMB 305:989-1010, 1999) указывают, что формы с более высоким содержанием мономерных разновидностей могут быть получены путем использования линкера, состоящего из 20 или более остатков. В публикации Desplancq et al., (Ptotein Eng. 7:1027-1033, 1994) указано, что ориентация вариабельного домена также может влиять на формирование димеров и высокомолекулярных форм. В этой же публикации Desplancq указано, что линкер, состоящий из 25 или 30 аминокислот (аа) показал лучшее соотношение мономера по сравнению с димером для их конкретного антитела. Расстояние между С-концом VL и N-концом VH составляет около 39-43 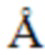 , и расстояние между С-концом VH и N-концом VL составляет 32-34 (Pluckthun et al., From PCR to fermentation. (J. McCafferty, H. R. Hoogenboom, & D. J. Chriswell, Eds.). In: (IRL Press., pp. 203-252, 1996). Для получения аналогичных молекулярных свойств линкер для ориентации VL-VH должен быть длиннее, чем линкер VH-VL. В публикации Pluckthun et al., From PCR to fermentation. (J. McCafferty, H. R. Hoogenboom, & D. J. Chriswell, Eds.). In: (IRL Press., pp. 203-252, 1996) рекомендовано использование линкеров длиной от 15 до 20 аминокислот в ориентации VH/VL и линкеров длиной 20-25 аминокислот в ориентации VL/VH.
, и расстояние между С-концом VH и N-концом VL составляет 32-34 (Pluckthun et al., From PCR to fermentation. (J. McCafferty, H. R. Hoogenboom, & D. J. Chriswell, Eds.). In: (IRL Press., pp. 203-252, 1996). Для получения аналогичных молекулярных свойств линкер для ориентации VL-VH должен быть длиннее, чем линкер VH-VL. В публикации Pluckthun et al., From PCR to fermentation. (J. McCafferty, H. R. Hoogenboom, & D. J. Chriswell, Eds.). In: (IRL Press., pp. 203-252, 1996) рекомендовано использование линкеров длиной от 15 до 20 аминокислот в ориентации VH/VL и линкеров длиной 20-25 аминокислот в ориентации VL/VH.
Другой возможностью направить формирование мономеров и стабилизировать взаимодействие доменов VH/VL является конструирование пограничной дисульфидной связи в пограничную область контакта между двумя доменами. Введение дисульфидного мостика в положении Н44-L100 (нумерация Kabat) наиболее часто использовали в scFv с удовлетворительными результатами (Brinkmann et al., PNAS. 90:7538-7542, 1993; Worn and Pluckthun, Biochemistry 38: 8739-8750, 1999; Weatherill et al., PEDS. 25:321-329, 2012). Эту стратегию успешно использовали для стабилизации IgG-подобных биспецифических антител, объединяя scFv, слитый с IgG полной длины (Michaelson et al., mAbs 1: 128-141, 2009; Schanzre et al., Antimicrob. Agents. Chemother. 55:2369-2378, 2011).
Weatherill et al., (PEDS 25:321-239, 2012) стабилизировал человеческий scFv (VH-(G4S)4-VL и VL-(G4S)4-VH) с дисульфидной связью между положением VH44 и VL-100. Более того, в этой публикации освещена проблема возможной замены домена на scFv, не содержащий пограничной дисульфидной связи, путем выполнения различных экспериментов SE-HPLC при различном объеме нагрузки и концентрации. Анализы показали различные результаты в зависимости от условий нагрузки образца для нестабилизированного scFv, но независимо от условий, в которых использовались дисульфидные стабилизированные молекулы, элюированные аналогично мономеру. В работе Zhao et al., (Int. J. Mol. Sci. 12:1-11, 2011) введена такая же мутация в scFv и наблюдалась более высокая стабильность стабилизированной молекулы после хранения в течение 20 ч при 37°С.
Для биспецифического формата с использованием scFv, слитого с IgG полной длины, Schanzer et al., (Antimicrob. Agents Chemother, 55:2369-2378 2011) сравнивали эффект длины линкера и пограничной дисульфидной связи. Они сливали родительский scFv или scdFv (VH-(G4S)3-VL) на С- или N-концевой части тяжелой цепи или легких цепей. Для различной длины линкера (20, 25 и 30 аминокислот) они сливали родительский scFv с С-концевой частью тяжелых или легких цепей. Результаты, полученные с использованием различной длины линкера, идентифицировали 30 аа пептид в качестве более предпочтительного линкера для получения стабильных мономеров. Уровень агрегатов через 7 дней хранения при 40°С составил 50% для scFv15, 18% для scFv20, 8% для scFv25 и 6% для scFv30, но дисульфид scFv15, стабилизированный пограничной дисульфидной связью, незначительно превосходил scFv30. Такой же подход был использован Michaelson et al., (mAbs. 1:128-141 2009), и они улучшили свое родительское IgG-подобное биспецифическое антитело, содержащее scFv с 15 аа линкером в ориентации VH/VL (генерируя 40% агрегатов), путем увеличения длины линкера до 20 аа scFv и введения пограничной дисульфидной связи между положением VH44 и VL-100. Полученная молекула содержала более 98% мономеров, которые были стабильными через три месяца при 4°С. Авторы решили работать над улучшением молекулы scFv перед переходом к биспецифическому формату.
В случае анти-CLDN18.2-специфических белков bi-scFv неизвестно, может ли возникать формирование димеров и высокомолекулярных форм и каково вовлечение молекул анти-Клаудин и/или анти-CD3 scFv. Для оценки всей оптимальной молекулы для анти-CLDN18.2-специфического белка bi-scFv для каждого отдельного scFv оценивали следующие модификации:
- ориентацию домена;
- длину линкера;
- введение пограничной дисульфиной связи;
- комбинацию трех модификаций.
Для анти-Клаудин 18.2 связывающего домена, дополнительно к вариабельным доменам, полученным из mCLDN18.2ab (VH: SEQ ID NO: 8; VL: SEQ ID NO: 15), последовательности, полученные из mCLDN18.2ab1 (VH: SEQ ID NO: 6; VL: SEQ ID NO: 11), использовали для конструирования анти-CLDN18.2-специфических белков bi-scFv.
В таблице 11 в разделе А.а «Исходные последовательности, дизайн 32 анти-CLDN18.2-специфических конструкций bi-scFv и клонирование в экспрессионные вектора» описаны различные конструкции.
А. Генерация и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN18.2 и CD3
а. Исходные последовательности, дизайн 32 анти-CLDN18.2-специфических конструкций bi-scFv и клонирование в экспрессионные вектора
Конструкции биспецифического тандемного одноцепочечного антитела bi-scFv, представленные здесь, содержат два отличающихся связывающих домена, из которых первый является специфическим в отношении человеческого опухоль-ассоциированного антигена (ТАА), тогда как второй является специфическим в отношении ε-цепи человеческого Т-клеточного рецептора (CD3). Каждый из двух связывающих доменов биспецифической молекулы содержит два вариабельных домена антитела, расположенных как фрагмент scFv в VH-VL или VL-VH ориентации. Вариабельные домены антитела соединены посредством гибкого глицин-серинового пептидного линкера, состоящего из четырех или пяти повторов субъединицы G4S в зависимости от ориентации. Таким образом, фрагменты scFv, расположенные в ориентации VH-VL, соединены линкером из 20 аминокислот (названным «LL4»), VL-VH соединены линкером из 25 аминокислот (названным «LL5»). Два фрагмента scFv, с другой стороны, соединены посредством длинного линкера SG4S, состоящего из шести аминокислот (названного «SL»). Информация по исходным последовательностям, происходящим из моноклональных антител (mAB) и организации доменов обобщена в таблице 11.
Варианты 5504, 5505, 5506, 5507, 5512, 5513, 5514, 5515, 5520, 5521, 5522, 5523, 5528, 5529, 5530, 5531, 5536, 5537, 5538, 5539, 5544, 5545, 5546, 5547, 5552, 5553, 5554, 5555, 5560, 5561, 5562 и 5563 содержат VH анти-CD3 с SEQ ID NO: 95 и VL анти-CD3 с SEQ ID NO: 96.
В конкретном случае 1-BiMAB-S сделано только замещение свободного цистеина на серин в VH анти-CD3 (SEQ ID NO: 94) по сравнению с аминокислотной последовательностью 1-BiMAB последовательности (SEQ ID NO: 39). Следует отметить, что SEQ ID NO: 39 еще содержит аминокислотную последовательность N-концевой сигнальной последовательности, которая опосредует секрецию 1-BiMAB белка bi-scFv в супернатант клеточной культуры при экспрессии у млекопитающего. Эта сигнальная последовательность не является частью секретированного рекомбинантного белка, так как расщепляется сигнальной пептидазой в просвете эндоплазматического ретикулума.
Гены, кодирующие конструкции bi-scFv, были созданы посредством генного синтеза GeneArf® (Life Technologies GmbH, Darmstadt, Germany) с использованием программного обеспечения GeneOptimizer® software для оптимизации использования кодонов для экспрессии в клетках СНО. Помимо общего сигнала секреции, все ДНК-конструкции содержат одинаковую последовательность Kozak и сайт рестрикции HindIII на 5'-конце. На 3'-конце сайты распознавания BsiWI и XhoI добавлены для обеспечения гибкого субклонирования в различные экспрессионные вектора. Субклонирование в предпочтительный экспрессионный вектор pCEP4 (Life Technologies GmbH, Darmstadt, Germany) выполняли с помощью Life Technologies с использованием сайтов рестрикции HindIII и XhoI.
Таблица 11. Краткое описание конструкций анти-CLDN18.2 bi-scFv
варианта
анти-CD3
*) LL4, (G4S)4; LL5; (G4S)5; SL, SG4S.
Следует отметить, что последовательности SEQ ID NO: с 66 по 93 еще содержат аминокислотную последовательность N-концевой сигнальной последовательности, которая опосредует секрецию 1-BiMAB белка bi-scFv в супернатант клеточной культуры при экспрессии у млекопитающего. Эта сигнальная последовательность не является частью секретированного рекомбинантного белка, так как расщепляется сигнальной пептидазой в просвете эндоплазматического ретикулума.
b. Получение и очистка 32 анти-CLDN18.2-специфических белков bi-scFv путем временной трансфекции
Адаптированные к росту в суспензии СНО-клетки субкультивировали в бессывороточной среде в увлажненной атмосфере СО2 на орбитальном встряхивателе. За день до трансфекции клетки засевали в бессывороточную среду в колбы во встряхивателе. В день трансфекции клетки центрифугировали (5 мин при 200×g) и ресуспендировали в свежей среде DMEM (Invitrogen, 41965-039) в колбах стряхивателя. ДНК и реагент для трансфекции добавляли в клетки и осторожно перемешивали путем встряхивания. После инкубации в стандартных условиях в СО2-инкубаторе клетки разбавляли бессывороточной средой для роста и затем выращивали для экспрессии в инкубационном встряхивателе. Клетки подпитывали СНО CD EfficientFeed™ C (Invitrogen, A13275) согласно потребностям в питании. Белки bi-scFv собирали после того, как жизнеспособность клеток начинала снижаться. Конструкции антитела очищали с помощью Capto L сефарозы. Концентрацию белков определяли путем измерения поглощения при 280 нм.
с. Анализ на цитотоксичность на основе люциферазы с использованием 32 анти-CLDN18.2-специфических белков bi-scFv
Для функционального скрининга 32 анти-CLDN18.2-специфических белков bi-scFv четыре титрования (5000, 1000, 200 и 40 нг/мл) тестировали в in vitro анализе на цитотоксичность на основе люциферазы, как описано в примере 2.с.
Стабильно экспрессирующие люциферазу клетки NugC4, описанные в примере 2.с, инкубировали с человеческими Т-клетками и белками bi-scFv или без белка bi-scFv для определения величин Lmin. Люминесценцию жизнеспособных клеток измеряли при помощи планшетного ридера Infinite M200 Tecan через 24 ч и 48 ч после начала анализа. Специфический лизис клеток-мишеней рассчитывали по формуле, приведенной в примере 2.с.
После выполнения количественной оценки результатов анализа на цитотоксичность (фигуры 39 а, b, с и d) на основе scFv анти-CD3-связывающих доменов могут быть сделаны следующие выводы.
Наиболее эффективные анти-CLDN18.2-специфические белки bi-scFv в анализе на цитотоксичность на основе люциферазы содержат фрагмент анти-CD3 в ориентации доменов VH/VL, соединенный линкером «LL4» с пограничным дисульфидным мостиком или без него (5504, 5505, 5506, 5507, 5536, 5537, 5538, 5539, 5512, 5513, 5514, 5515, 5544, 5545, 5546, 5547; фигуры 39 а и b). Более низкая цитотоксическая активность получена для вариантов, содержащих фрагмент анти-CD3 в ориентации доменов VL/VH с пептидным линкером «LL5» и содержащих пограничный дисульфидный мостик (5528, 5529, 5530, 5531, 5560, 5561, 5562, 5563; фигура 39 d). Самая низкая цитотоксическая активность получена для вариантов, содержащих фрагмент анти-CD3 в ориентации доменов VL/VH с пептидным линкером «LL5» без пограничного дисульфидного мостика (5520, 5521, 5522, 5523, 5552, 5553, 5554, 5555; фигура 39 с).
В. Генерация и тестирование биспецифических связывающих агентов, направленно воздействующих на CLDN6 и CD3
а. Происхождение последовательностей, дизайн конструкций bi-scFv и клонирование экспрессионные вектора
Конструкции биспецифического тандемного одноцепочечного антитела bi-scFv, представленные здесь, содержат два отличающихся связывающих домена, один из которых является специфическим в отношении человеческого опухоль-ассоциированного антигена (ТАА), и другой является специфическим в отношении ε-цепи человеческого Т-клеточного рецептора (CD3). Каждый из двух доменов биспецифической молекулы содержит два вариабельных домена антитела, расположенных в виде фрагмента scFv в любой из ориентаций VH-VL или VL-VH. Вариабельные домены антитела соединены посредством гибкого глицин-серинового пептидного линкера. Фрагменты scFv, направленно воздействующие на ТАА, расположены в ориентации VH-VL и соединены линкером из 15 аминокислот (названным «LL3»), состоящим из трех идентичных повторов субъединицы G4S. Фрагменты scFv, направленно воздействующие на CD3, также расположены в ориентации VH-VL, но соединены линкером из 18 аминокислот (названным «LLv1») с последовательностью G4S(G2S)3G3S. Два фрагмента scFv, с другой стороны, соединены посредством длинного линкера SG4S из шести аминокислот (названного «SL»). Информация по происхождению последовательностей из моноклональных антител (mAB) и организации доменов обобщена в таблице 12. Варианты 5454, 5456, 5458, 5460, 5462 и 5464 содержат для анти-CD3 связывающего домена VH анти-CD3 с SEQ ID NO: 95 и VL анти-CD3 с SEQ ID NO: 96. Варианты 5454 и 5458 содержат VL анти-CLDN6 с SEQ ID NO: 98; варианты 5457 и 5460 содержат VL анти-CLDN6 с SEQ ID NO: 99; варианты 5462 и 5464 содержат VL анти-CLDN6 с SEQ ID NO: 100.
В конкретном случае 6PHU3-S (SEQ ID NO: 101) сделано только замещение свободного цистеина на сериновый остаток в VH анти-CD3 (SEQ ID NO: 94) по сравнению с аминокислотной последовательностью 6-PHU3 (SEQ ID NO: 45). Следует отметить, что SEQ ID NO: 45 еще содержит аминокислотную последовательность N-концевой сигнальной последовательности, которая опосредует секрецию 6-PHU-3 белка bi-scFv в супернатант клеточной культуры при экспрессии у млекопитающего. Эта сигнальная последовательность не является частью секретированного рекомбинантного белка, так как расщепляется сигнальной пептидазой в просвете эндоплазматического ретикулума. Более того, для 6PHU3-SL (SEQ ID NO: 102) сделано замещение свободного цистеина на лейциновый остаток в VL анти-CLDN (SEQ ID NO: 97) в положении 46 соответствующей первичной последовательности по сравнению с аминокислотной последовательностью 6PHU3-S. Это соответствует положению 45 в пределах SEQ ID NO: 23, где первая аминокислота VL является пропущенной.
Гены, кодирующие конструкции bi-scFv, были генерированы посредством генного синтеза GeneArf® (Life Technologies GmbH, Darmstadt, Germany) с использованием программного обеспечения GeneOptimizer® software для оптимизации использования кодонов для экспрессии в клетках СНО. Помимо общего сигнала секреции, все ДНК-конструкции содержат одинаковую последовательность Kozak и сайт рестрикции HindIII на 5'-конце. На 3'-конце сайты распознавания BsiWI и XhoI добавлены для обеспечения гибкого субклонирования в различные экспрессионные вектора. Субклонирование в предпочтительный экспрессионный вектор pEE12.4 (LONZA Group Ltd.,Basel, Switzerland) выполняли с помощью стандартных технологий с использованием сайтов рестрикции HindIII и BsiWI.
Таблица 12. Краткое описание конструкций анти-CLDN6 bi-scFv
варианта
анти-CLDN6
*) LL3, (G4S)3; LLv1, G4S(G2S)3G3S; SL; SG4S.
b. Получение и очистка 6 анти-CLDN6-специфических белков bi-scFv путем временной трансфекции
Адаптированные к росту в суспензии СНО-клетки субкультивировали в бессывороточной среде в увлажненной атмосфере СО2 на орбитальном встряхивателе. За день до трансфекции клетки засевали в бессывороточную среду в колбы встряхивателя. В день трансфекции клетки центрифугировали (5 мин при 200×g) и ресуспендировали в свежей среде DMEM (Invitrogen, 41965-039) в колбах встряхивателя. ДНК и реагент для трансфекции добавляли в клетки и осторожно перемешивали путем встряхивания. После инкубации в стандартных условиях в СО2-инкубаторе клетки разбавляли бессывороточной средой для роста и затем выращивали для экспрессии в инкубационном встряхивателе. Клетки подпитывали СНО CD EfficientFeed™ C (Invitrogen, A13275) согласно потребностям в питании. Белки bi-scFv собирали после того, как жизнеспособность клеток начинала снижаться. Конструкции антитела очищали с помощью FPLC с использованием Capto L сефарозы. Концентрацию белков определяли путем измерения поглощения при 280 нм.
с. Определение ЕС50 5 анти-CLDN6-специфических белков bi-scFv
Для определения половины максимальной эффективной дозы анти-CLDN6-специфических белков bi-scFv ряд титрований белков bi-scFv тестировали в анализе на цитотоксичность с использованием люциферазы in vitro, как описано в примере 13.
Стабильно экспрессирующие люциферазу клетки РА-1 и человеческие Т-клетки в соотношении Е:Т, равном 5:1, инкубировали с белком bi-scFv при концентрации в диапазоне от 2,5 пг/мл до 5 мкг/мл (10 шагов) или без анти-CLDN6-специфических белков bi-scFv для определения величин Lmin.
Три независимых анализа препарата человеческих Т-клеток выполняли с различными донорами-людьми. Результаты представлены на фигуре 40.
После выполнения количественного сравнения результатов анализа на цитотоксичность на основе замещения цистеинового остатка в пределах фланкирующей области CDR-L2 серином, лейцином или триптофаном, и на основе положения анти-CLDN6 и анти-CD3 связывающих доменов, могут быть сделаны следующие выводы.
Наиболее эффективные анти-CLDN6-специфические белки bi-scFv в анализе на цитотоксичность на основе люциферазы содержат замещение цистеина на триптофан для анти-CLDN6 связывающего домена в N-концевой части белка bi-scFv (вариант 5456). Незначительно более низкая цитотоксическая активность получена для вариантов, содержащих анти-CLDN6 в С-концевой части с замещением цистеина на триптофан (вариант 5460) или серин (вариант 5464). Для вариантов, содержащих замещение цистеина на лейцин, содержащих анти-CLDN6 в N-концевой части (вариант 5454) или С-концевой части (вариант 5458) белков bi-scFv, более низкая цитотоксическая активность измерена в сравнении с предыдущими вариантами. Неожиданно было обнаружено, что вариант, содержащий замещение цистеина на серин с анти-CLDN6 в N-концевой части (вариант 5462) белка bi-scFv обладал самой низкой цитотоксической активностью.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> BioNTech AG et al.
<120> AGENTS FOR TREATMENT OF CLAUDIN EXPRESSING CANCER DISEASES
<130> 674-86 PCT
<150> PCT/EP2012/004712
<151> 2012-11-13
<150> PCT/EP2013/002270
<151> 2013-07-30
<160> 108
<170> PatentIn version 3.5
<210> 1
<211> 261
<212> PRT
<213> Homo sapiens
<400> 1
Met Ala Val Thr Ala Cys Gln Gly Leu Gly Phe Val Val Ser Leu Ile
1 5 10 15
Gly Ile Ala Gly Ile Ile Ala Ala Thr Cys Met Asp Gln Trp Ser Thr
20 25 30
Gln Asp Leu Tyr Asn Asn Pro Val Thr Ala Val Phe Asn Tyr Gln Gly
35 40 45
Leu Trp Arg Ser Cys Val Arg Glu Ser Ser Gly Phe Thr Glu Cys Arg
50 55 60
Gly Tyr Phe Thr Leu Leu Gly Leu Pro Ala Met Leu Gln Ala Val Arg
65 70 75 80
Ala Leu Met Ile Val Gly Ile Val Leu Gly Ala Ile Gly Leu Leu Val
85 90 95
Ser Ile Phe Ala Leu Lys Cys Ile Arg Ile Gly Ser Met Glu Asp Ser
100 105 110
Ala Lys Ala Asn Met Thr Leu Thr Ser Gly Ile Met Phe Ile Val Ser
115 120 125
Gly Leu Cys Ala Ile Ala Gly Val Ser Val Phe Ala Asn Met Leu Val
130 135 140
Thr Asn Phe Trp Met Ser Thr Ala Asn Met Tyr Thr Gly Met Gly Gly
145 150 155 160
Met Val Gln Thr Val Gln Thr Arg Tyr Thr Phe Gly Ala Ala Leu Phe
165 170 175
Val Gly Trp Val Ala Gly Gly Leu Thr Leu Ile Gly Gly Val Met Met
180 185 190
Cys Ile Ala Cys Arg Gly Leu Ala Pro Glu Glu Thr Asn Tyr Lys Ala
195 200 205
Val Ser Tyr His Ala Ser Gly His Ser Val Ala Tyr Lys Pro Gly Gly
210 215 220
Phe Lys Ala Ser Thr Gly Phe Gly Ser Asn Thr Lys Asn Lys Lys Ile
225 230 235 240
Tyr Asp Gly Gly Ala Arg Thr Glu Asp Glu Val Gln Ser Tyr Pro Ser
245 250 255
Lys His Asp Tyr Val
260
<210> 2
<211> 220
<212> PRT
<213> Homo sapiens
<400> 2
Met Ala Ser Ala Gly Met Gln Ile Leu Gly Val Val Leu Thr Leu Leu
1 5 10 15
Gly Trp Val Asn Gly Leu Val Ser Cys Ala Leu Pro Met Trp Lys Val
20 25 30
Thr Ala Phe Ile Gly Asn Ser Ile Val Val Ala Gln Val Val Trp Glu
35 40 45
Gly Leu Trp Met Ser Cys Val Val Gln Ser Thr Gly Gln Met Gln Cys
50 55 60
Lys Val Tyr Asp Ser Leu Leu Ala Leu Pro Gln Asp Leu Gln Ala Ala
65 70 75 80
Arg Ala Leu Cys Val Ile Ala Leu Leu Val Ala Leu Phe Gly Leu Leu
85 90 95
Val Tyr Leu Ala Gly Ala Lys Cys Thr Thr Cys Val Glu Glu Lys Asp
100 105 110
Ser Lys Ala Arg Leu Val Leu Thr Ser Gly Ile Val Phe Val Ile Ser
115 120 125
Gly Val Leu Thr Leu Ile Pro Val Cys Trp Thr Ala His Ala Ile Ile
130 135 140
Arg Asp Phe Tyr Asn Pro Leu Val Ala Glu Ala Gln Lys Arg Glu Leu
145 150 155 160
Gly Ala Ser Leu Tyr Leu Gly Trp Ala Ala Ser Gly Leu Leu Leu Leu
165 170 175
Gly Gly Gly Leu Leu Cys Cys Thr Cys Pro Ser Gly Gly Ser Gln Gly
180 185 190
Pro Ser His Tyr Met Ala Arg Tyr Ser Thr Ser Ala Pro Ala Ile Ser
195 200 205
Arg Gly Pro Ser Glu Tyr Pro Thr Lys Asn Tyr Val
210 215 220
<210> 3
<211> 220
<212> PRT
<213> Homo sapiens
<400> 3
Met Ala Ser Ala Gly Met Gln Ile Leu Gly Val Val Leu Thr Leu Leu
1 5 10 15
Gly Trp Val Asn Gly Leu Val Ser Cys Ala Leu Pro Met Trp Lys Val
20 25 30
Thr Ala Phe Ile Gly Asn Ser Ile Val Val Ala Gln Val Val Trp Glu
35 40 45
Gly Leu Trp Met Ser Cys Val Val Gln Ser Thr Gly Gln Met Gln Cys
50 55 60
Lys Val Tyr Asp Ser Leu Leu Ala Leu Pro Gln Asp Leu Gln Ala Ala
65 70 75 80
Arg Ala Leu Cys Val Ile Ala Leu Leu Val Ala Leu Phe Gly Leu Leu
85 90 95
Val Tyr Leu Ala Gly Ala Lys Cys Thr Thr Cys Val Glu Glu Lys Asp
100 105 110
Ser Lys Ala Arg Leu Val Leu Thr Ser Gly Ile Val Phe Val Ile Ser
115 120 125
Gly Val Leu Thr Leu Ile Pro Val Cys Trp Thr Ala His Ala Val Ile
130 135 140
Arg Asp Phe Tyr Asn Pro Leu Val Ala Glu Ala Gln Lys Arg Glu Leu
145 150 155 160
Gly Ala Ser Leu Tyr Leu Gly Trp Ala Ala Ser Gly Leu Leu Leu Leu
165 170 175
Gly Gly Gly Leu Leu Cys Cys Thr Cys Pro Ser Gly Gly Ser Gln Gly
180 185 190
Pro Ser His Tyr Met Ala Arg Tyr Ser Thr Ser Ala Pro Ala Ile Ser
195 200 205
Arg Gly Pro Ser Glu Tyr Pro Thr Lys Asn Tyr Val
210 215 220
<210> 4
<211> 207
<212> PRT
<213> Homo sapiens
<400> 4
Met Gln Ser Gly Thr His Trp Arg Val Leu Gly Leu Cys Leu Leu Ser
1 5 10 15
Val Gly Val Trp Gly Gln Asp Gly Asn Glu Glu Met Gly Gly Ile Thr
20 25 30
Gln Thr Pro Tyr Lys Val Ser Ile Ser Gly Thr Thr Val Ile Leu Thr
35 40 45
Cys Pro Gln Tyr Pro Gly Ser Glu Ile Leu Trp Gln His Asn Asp Lys
50 55 60
Asn Ile Gly Gly Asp Glu Asp Asp Lys Asn Ile Gly Ser Asp Glu Asp
65 70 75 80
His Leu Ser Leu Lys Glu Phe Ser Glu Leu Glu Gln Ser Gly Tyr Tyr
85 90 95
Val Cys Tyr Pro Arg Gly Ser Lys Pro Glu Asp Ala Asn Phe Tyr Leu
100 105 110
Tyr Leu Arg Ala Arg Val Cys Glu Asn Cys Met Glu Met Asp Val Met
115 120 125
Ser Val Ala Thr Ile Val Ile Val Asp Ile Cys Ile Thr Gly Gly Leu
130 135 140
Leu Leu Leu Val Tyr Tyr Trp Ser Lys Asn Arg Lys Ala Lys Ala Lys
145 150 155 160
Pro Val Thr Arg Gly Ala Gly Ala Gly Gly Arg Gln Arg Gly Gln Asn
165 170 175
Lys Glu Arg Pro Pro Pro Val Pro Asn Pro Asp Tyr Glu Pro Ile Arg
180 185 190
Lys Gly Gln Arg Asp Leu Tyr Ser Gly Leu Asn Gln Arg Arg Ile
195 200 205
<210> 5
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 5
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Met Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Thr Gly Tyr Thr Phe Ser Ser Tyr
20 25 30
Trp Ile Glu Trp Val Lys Gln Arg Pro Gly His Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Leu Pro Gly Ser Gly Ser Thr Asn Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Phe Thr Ala Asp Thr Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Asp Tyr Pro Trp Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ala
115
<210> 6
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 6
Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu
1 5 10 15
Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr
20 25 30
Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met
35 40 45
Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe
50 55 60
Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr
65 70 75 80
Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys
85 90 95
Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 7
<211> 116
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 7
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Asn Trp Val Lys Gln Arg Thr Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Tyr Pro Gly Ser Gly Asn Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Ser Tyr Gly Ala Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210> 8
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 8
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser
115
<210> 9
<211> 118
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 9
Gln Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Val Ile Ser Trp Val Lys Gln Arg Thr Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Tyr Pro Gly Ser Gly Ser Thr Tyr Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Asn Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Phe Cys
85 90 95
Ala Arg Gly Val Leu Leu Arg Ala Met Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Ser Val Thr Val Ser Ser
115
<210> 10
<211> 120
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 10
Gln Val His Leu Gln Gln Ser Gly Ser Glu Leu Arg Ser Pro Gly Ser
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Asp Phe Asp Ser Glu Val Phe Pro Phe
20 25 30
Ala Tyr Met Ser Trp Ile Arg Gln Lys Pro Gly His Gly Phe Glu Trp
35 40 45
Ile Gly Asp Ile Leu Pro Ser Ile Gly Arg Thr Ile Tyr Gly Glu Lys
50 55 60
Phe Glu Asp Lys Ala Thr Leu Asp Ala Asp Thr Val Ser Asn Thr Ala
65 70 75 80
Tyr Leu Glu Leu Asn Ser Leu Thr Ser Glu Asp Ser Ala Ile Tyr Tyr
85 90 95
Cys Ala Arg Gly Glu Gly Tyr Gly Ala Trp Phe Ala Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ala
115 120
<210> 11
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 11
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 12
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 12
Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ser Tyr Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 13
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 13
Asp Ile Val Met Thr Gln Ser Gln Lys Phe Met Ser Thr Ser Val Gly
1 5 10 15
Asp Arg Val Ser Ile Thr Cys Lys Ala Ser Gln Asn Val Arg Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ser Pro Lys Ala Leu Ile
35 40 45
Tyr Leu Ala Ser Asn Arg His Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Asn Val Gln Ser
65 70 75 80
Glu Asp Leu Ala Asp Tyr Phe Cys Leu Gln His Trp Asn Tyr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 14
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 14
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 15
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn
85 90 95
Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 16
<211> 112
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 16
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Ala Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser
20 25 30
Arg Thr Arg Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Lys Gln
85 90 95
Ser Tyr Asn Leu Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 17
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 17
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Ala Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Asp Tyr His Cys Gly Gln
85 90 95
Gly Tyr Ser Tyr Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile
100 105 110
Lys
<210> 18
<211> 113
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 18
Asp Ile Val Met Ser Gln Ser Pro Ser Ser Leu Ala Val Ser Val Gly
1 5 10 15
Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Tyr Ser
20 25 30
Ser Asn Gln Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Ser Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val
50 55 60
Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Val Lys Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln
85 90 95
Tyr Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu
100 105 110
Lys
<210> 19
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 19
Asn Ile Val Met Thr Gln Ser Pro Lys Ser Met Ser Met Ser Val Gly
1 5 10 15
Glu Arg Val Thr Leu Thr Cys Lys Ala Ser Glu Asn Val Val Thr Tyr
20 25 30
Val Ser Trp Tyr Gln Gln Lys Pro Glu Gln Ser Pro Lys Leu Leu Ile
35 40 45
Tyr Gly Ala Ser Asn Arg Tyr Thr Gly Val Pro Asp Arg Phe Thr Gly
50 55 60
Ser Gly Ser Ala Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Lys Ala
65 70 75 80
Glu Asp Leu Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Ser Tyr Pro Leu
85 90 95
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 20
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 20
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ile Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 21
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 21
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Leu His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Val Tyr Ser
35 40 45
Thr Ser Asn Leu Pro Ser Gly Val Pro Ala Arg Phe Gly Gly Ser Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Ile Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 22
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 22
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 23
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 23
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Cys Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 24
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 24
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Ile Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Phe Tyr Cys
85 90 95
Ala Arg Asp Phe Gly Tyr Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 25
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 25
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Ser Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Thr Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 26
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 26
Glu Val Gln Leu Gln Gln Ser Arg Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Leu Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Ser Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Tyr Val Phe Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 27
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 27
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Met His
20 25 30
Trp Phe Gln Leu Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Met Glu Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Asn Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 28
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 28
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Gly Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 29
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 29
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Ser Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 30
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 30
Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Ser Gly Tyr Thr Lys Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Trp Gln Asp Tyr Asp Val Tyr Phe Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 31
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 31
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 32
<211> 122
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 32
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Met
35 40 45
Gly Leu Ile Asn Pro Tyr Lys Gly Val Ser Thr Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Phe Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Gly Tyr Tyr Gly Asp Ser Asp Trp Tyr Phe Asp Val Trp
100 105 110
Gly Ala Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 33
<211> 107
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 33
Asp Ile Gln Met Thr Gln Thr Thr Ser Ser Leu Ser Ala Ser Leu Gly
1 5 10 15
Asp Arg Val Thr Ile Ser Cys Arg Ala Ser Gln Asp Ile Arg Asn Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Asp Gly Thr Val Lys Leu Leu Ile
35 40 45
Tyr Tyr Thr Ser Arg Leu His Ser Gly Val Pro Ser Lys Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Tyr Ser Leu Thr Ile Ser Asn Leu Glu Gln
65 70 75 80
Glu Asp Ile Ala Thr Tyr Phe Cys Gln Gln Gly Asn Thr Leu Pro Trp
85 90 95
Thr Phe Ala Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 34
<211> 119
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 34
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Phe Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Arg Tyr Ser Arg Tyr Ile Tyr Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Val Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Ser Leu Lys Ser Glu Asp Thr Ala Ile Tyr Tyr Cys
85 90 95
Ala Arg Arg Pro Leu Tyr Gly Ser Ser Pro Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 35
<211> 108
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 35
Asp Ile Glu Asn Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser
1 5 10 15
Pro Gly Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Thr
20 25 30
Tyr Val His Trp Tyr Gln Gln Lys Ser Asn Thr Ser Pro Lys Leu Trp
35 40 45
Ile Tyr Asp Thr Ser Lys Leu Ala Ser Gly Val Pro Gly Arg Val Ser
50 55 60
Gly Ser Gly Ser Gly Asn Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu
65 70 75 80
Ala Glu Asp Val Ala Thr Tyr Tyr Cys Phe Gln Gly Ser Gly Tyr Pro
85 90 95
Leu Thr Phe Gly Ser Gly Thr Lys Leu Glu Met Arg
100 105
<210> 36
<211> 117
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 36
Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val
1 5 10 15
Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met
20 25 30
His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr
35 40 45
Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp
50 55 60
Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln
65 70 75 80
Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg
85 90 95
Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser
115
<210> 37
<211> 106
<212> PRT
<213> Artificial Sequence
<220>
<223> Antibody fragment
<400> 37
Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 38
<211> 495
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 38
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu
130 135 140
Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln
145 150 155 160
Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln
165 170 175
Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr
180 185 190
Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val
210 215 220
Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly
225 230 235 240
Thr Lys Leu Glu Ile Lys Ser Gly Gly Gly Gly Ser Asp Ile Lys Leu
245 250 255
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
260 265 270
Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
275 280 285
Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn
290 295 300
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
305 310 315 320
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
325 330 335
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
340 345 350
Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
355 360 365
Val Ser Ser Val Glu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
370 375 380
Ser Gly Gly Val Asp Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met
385 390 395 400
Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser
405 410 415
Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro
420 425 430
Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr
435 440 445
Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser
450 455 460
Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser
465 470 475 480
Ser Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
485 490 495
<210> 39
<211> 520
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 39
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro
145 150 155 160
Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys
165 170 175
Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr
180 185 190
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp
195 200 205
Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly
210 215 220
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp
225 230 235 240
Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe
245 250 255
Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly Gly Gly Gly Ser Asp
260 265 270
Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser
275 280 285
Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr
290 295 300
Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
305 310 315 320
Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys
325 330 335
Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met
340 345 350
Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala
355 360 365
Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr
370 375 380
Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly Gly Ser Gly Gly
385 390 395 400
Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu Thr Gln Ser Pro
405 410 415
Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg
420 425 430
Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly
435 440 445
Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly
450 455 460
Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu
465 470 475 480
Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln
485 490 495
Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
500 505 510
Leu Lys His His His His His His
515 520
<210> 40
<211> 490
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 40
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
115 120 125
Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu
130 135 140
Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln
145 150 155 160
Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln
165 170 175
Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr
180 185 190
Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr
195 200 205
Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val
210 215 220
Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly
225 230 235 240
Thr Lys Leu Glu Ile Lys Ser Gly Gly Gly Gly Ser Lys Leu Gln Gln
245 250 255
Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys
260 265 270
Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys
275 280 285
Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser
290 295 300
Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu
305 310 315 320
Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu
325 330 335
Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp
340 345 350
His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
355 360 365
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
370 375 380
Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly
385 390 395 400
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met
405 410 415
Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
420 425 430
Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser
435 440 445
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu
450 455 460
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
465 470 475 480
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
485 490
<210> 41
<211> 515
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 41
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro
145 150 155 160
Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys
165 170 175
Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr
180 185 190
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp
195 200 205
Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly
210 215 220
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp
225 230 235 240
Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe
245 250 255
Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly Gly Gly Gly Ser Lys
260 265 270
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
275 280 285
Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
290 295 300
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
305 310 315 320
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
325 330 335
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
340 345 350
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
355 360 365
Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
370 375 380
Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
385 390 395 400
Gly Gly Ser Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala
405 410 415
Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val
420 425 430
Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg
435 440 445
Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe
450 455 460
Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met
465 470 475 480
Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn
485 490 495
Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys His His His
500 505 510
His His His
515
<210> 42
<211> 487
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 42
Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr
20 25 30
Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile
35 40 45
Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr
100 105 110
Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
115 120 125
Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser
130 135 140
Val Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser
145 150 155 160
Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys
165 170 175
Leu Cys Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg
180 185 190
Phe Ser Gly Arg Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg
195 200 205
Val Ala Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn
210 215 220
Tyr Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Ser
225 230 235 240
Gly Gly Gly Gly Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu
245 250 255
Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr
260 265 270
Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln
275 280 285
Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn
290 295 300
Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser
305 310 315 320
Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser
325 330 335
Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp
340 345 350
Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly
355 360 365
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile
370 375 380
Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys
385 390 395 400
Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp
405 410 415
Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr
420 425 430
Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser
435 440 445
Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala
450 455 460
Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly
465 470 475 480
Ala Gly Thr Lys Leu Glu Leu
485
<210> 43
<211> 513
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 43
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe
35 40 45
Thr Gly Tyr Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu
50 55 60
Glu Trp Ile Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Leu Thr Gln Ser Pro Ser
145 150 155 160
Ile Met Ser Val Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala
165 170 175
Ser Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr
180 185 190
Ser Pro Lys Leu Cys Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val
195 200 205
Pro Ala Arg Phe Ser Gly Arg Gly Ser Gly Thr Ser Tyr Ser Leu Thr
210 215 220
Ile Ser Arg Val Ala Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln
225 230 235 240
Arg Ser Asn Tyr Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu
245 250 255
Ile Lys Ser Gly Gly Gly Gly Ser Asp Ile Lys Leu Gln Gln Ser Gly
260 265 270
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr
275 280 285
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
290 295 300
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
305 310 315 320
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
325 330 335
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
340 345 350
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
355 360 365
Cys Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val
370 375 380
Glu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val
385 390 395 400
Asp Asp Ile Gln Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro
405 410 415
Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr
420 425 430
Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile
435 440 445
Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly
450 455 460
Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala
465 470 475 480
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu
485 490 495
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys His His His His His
500 505 510
His
<210> 44
<211> 488
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 44
Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly Gly Ser Gly
115 120 125
Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu Thr Gln Ser
130 135 140
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
145 150 155 160
Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
165 170 175
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser
180 185 190
Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
195 200 205
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
210 215 220
Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu
225 230 235 240
Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu Gln Gln Ser
245 250 255
Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile Ser Cys Lys
260 265 270
Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp Val Lys Gln
275 280 285
Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn Pro Tyr Asn
290 295 300
Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr
305 310 315 320
Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr
325 330 335
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr Gly Phe Val
340 345 350
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
355 360 365
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
370 375 380
Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu Lys Val
385 390 395 400
Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His Trp Phe
405 410 415
Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Cys Ile Tyr Ser Thr Ser
420 425 430
Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly Ser Gly
435 440 445
Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp Ala Ala
450 455 460
Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr Phe Gly
465 470 475 480
Gly Gly Thr Lys Leu Glu Ile Lys
485
<210> 45
<211> 513
<212> PRT
<213> Artificial Sequence
<220>
<223> Bispecific molecule
<400> 45
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Cys Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Asp Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Cys Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys His His His His His
500 505 510
His
<210> 46
<211> 15
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 46
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 47
<211> 16
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 47
Val Glu Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Val Asp
1 5 10 15
<210> 48
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 48
Ser Gly Gly Gly Gly Ser
1 5
<210> 49
<211> 5
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 49
Gly Gly Gly Gly Ser
1 5
<210> 50
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 50
Val Glu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
1 5 10 15
Val Asp
<210> 51
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Secretion signal
<400> 51
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser
<210> 52
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Secretion signal
<400> 52
Met Asn Ser Gly Leu Gln Leu Val Phe Phe Val Leu Thr Leu Lys Gly
1 5 10 15
Ile Gln Gly
<210> 53
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Secretion signal
<400> 53
Met Asn Phe Gly Leu Ser Leu Ile Phe Leu Ala Leu Ile Leu Lys Gly
1 5 10 15
Val Gln Cys
<210> 54
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Secretion signal
<400> 54
Met Glu Trp Ser Trp Ile Phe Leu Phe Leu Leu Ser Val Thr Thr Gly
1 5 10 15
Val His Ser
<210> 55
<211> 19
<212> PRT
<213> Artificial Sequence
<220>
<223> Secretion signal
<400> 55
Met Gly Trp Leu Trp Asn Leu Leu Phe Leu Met Ala Ala Ala Gln Ser
1 5 10 15
Ala Gln Ala
<210> 56
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Oligonucleotide
<400> 56
tggctctgtg tcgacactgt g 21
<210> 57
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Oligonucleotide
<400> 57
gtgtacatgt tagctgtgga c 21
<210> 58
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Oligonucleotide
<400> 58
tgacactggc aaaacaatgc a 21
<210> 59
<211> 21
<212> DNA
<213> Artificial Sequence
<220>
<223> Oligonucleotide
<400> 59
ggtccttttc accagcaagc t 21
<210> 60
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 60
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe
35 40 45
Thr Gly Tyr Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu
50 55 60
Glu Trp Ile Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ser
145 150 155 160
Ile Met Ser Val Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala
165 170 175
Ser Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr
180 185 190
Ser Pro Lys Leu Leu Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val
195 200 205
Pro Ala Arg Phe Ser Gly Arg Gly Ser Gly Thr Ser Tyr Ser Leu Thr
210 215 220
Ile Ser Arg Val Ala Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln
225 230 235 240
Arg Ser Asn Tyr Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu
245 250 255
Ile Lys Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly
260 265 270
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr
275 280 285
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
290 295 300
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
305 310 315 320
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
325 330 335
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
340 345 350
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
355 360 365
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
370 375 380
Gly Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly
385 390 395 400
Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro
405 410 415
Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr
420 425 430
Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile
435 440 445
Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly
450 455 460
Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala
465 470 475 480
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu
485 490 495
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
500 505
<210> 61
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 61
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe
35 40 45
Thr Gly Tyr Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu
50 55 60
Glu Trp Ile Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ser
145 150 155 160
Ile Met Ser Val Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala
165 170 175
Ser Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr
180 185 190
Ser Pro Lys Leu Trp Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val
195 200 205
Pro Ala Arg Phe Ser Gly Arg Gly Ser Gly Thr Ser Tyr Ser Leu Thr
210 215 220
Ile Ser Arg Val Ala Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln
225 230 235 240
Arg Ser Asn Tyr Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu
245 250 255
Ile Lys Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly
260 265 270
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr
275 280 285
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
290 295 300
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
305 310 315 320
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
325 330 335
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
340 345 350
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
355 360 365
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
370 375 380
Gly Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly
385 390 395 400
Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro
405 410 415
Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr
420 425 430
Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile
435 440 445
Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly
450 455 460
Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala
465 470 475 480
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu
485 490 495
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
500 505
<210> 62
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 62
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Glu Val Gln Leu Gln Gln Ser Gly Pro Glu Leu Val Lys
20 25 30
Pro Gly Ala Ser Met Lys Ile Ser Cys Lys Ala Ser Gly Tyr Ser Phe
35 40 45
Thr Gly Tyr Thr Met Asn Trp Val Lys Gln Ser His Gly Lys Asn Leu
50 55 60
Glu Trp Ile Gly Leu Ile Asn Pro Tyr Asn Gly Gly Thr Ile Tyr Asn
65 70 75 80
Gln Lys Phe Lys Gly Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Glu Leu Leu Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Asp Tyr Gly Phe Val Leu Asp Tyr Trp Gly Gln
115 120 125
Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
130 135 140
Gly Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ser
145 150 155 160
Ile Met Ser Val Ser Pro Gly Glu Lys Val Thr Ile Thr Cys Ser Ala
165 170 175
Ser Ser Ser Val Ser Tyr Met His Trp Phe Gln Gln Lys Pro Gly Thr
180 185 190
Ser Pro Lys Leu Ser Ile Tyr Ser Thr Ser Asn Leu Ala Ser Gly Val
195 200 205
Pro Ala Arg Phe Ser Gly Arg Gly Ser Gly Thr Ser Tyr Ser Leu Thr
210 215 220
Ile Ser Arg Val Ala Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln
225 230 235 240
Arg Ser Asn Tyr Pro Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu
245 250 255
Ile Lys Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly
260 265 270
Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr
275 280 285
Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg
290 295 300
Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly
305 310 315 320
Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr
325 330 335
Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser
340 345 350
Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr
355 360 365
Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly
370 375 380
Gly Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly
385 390 395 400
Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro
405 410 415
Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr
420 425 430
Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile
435 440 445
Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly
450 455 460
Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala
465 470 475 480
Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu
485 490 495
Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
500 505
<210> 63
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 63
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly Ser Gln Ile Val Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Gln Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
500 505
<210> 64
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 64
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly Ser Gln Ile Val Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Gln Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
500 505
<210> 65
<211> 507
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 65
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly Ser Gln Ile Val Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Gln Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Ser Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
500 505
<210> 66
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 66
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 67
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 67
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Cys Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 68
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 68
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 69
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 69
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Cys Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 70
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 70
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Cys
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 71
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 71
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Cys Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Cys
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 72
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 72
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Cys
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 73
<211> 530
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 73
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Cys Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala Glu Leu Ala
275 280 285
Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr
290 295 300
Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Cys
305 310 315 320
Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr
325 330 335
Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser
340 345 350
Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala
355 360 365
Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr
370 375 380
Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln
405 410 415
Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu
420 425 430
Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn
435 440 445
Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp
450 455 460
Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly
465 470 475 480
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp
485 490 495
Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe
500 505 510
Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly Ser His His His His
515 520 525
His His
530
<210> 74
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 74
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 75
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 75
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Cys Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 76
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 76
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 77
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 77
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Cys Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 78
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 78
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Gly Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 79
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 79
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Ile Gln Leu Val Gln Ser Gly Pro Glu Leu Lys Lys
20 25 30
Pro Gly Glu Thr Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Asn Tyr Gly Met Asn Trp Val Lys Gln Ala Pro Gly Lys Cys Leu
50 55 60
Lys Trp Met Gly Trp Ile Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala
65 70 75 80
Glu Glu Phe Lys Gly Arg Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser
85 90 95
Thr Ala Tyr Leu Gln Ile Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr
100 105 110
Tyr Phe Cys Ala Arg Leu Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Ser Val Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 80
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 80
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 81
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 81
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Cys Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val
145 150 155 160
Met Thr Gln Ser Pro Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val
165 170 175
Thr Met Ser Cys Lys Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln
180 185 190
Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys
195 200 205
Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg
210 215 220
Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser
225 230 235 240
Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser
245 250 255
Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys Leu Glu Ile Lys Ser Gly
260 265 270
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
275 280 285
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
290 295 300
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
305 310 315 320
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
325 330 335
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
340 345 350
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
355 360 365
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
370 375 380
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser Gly Ala
405 410 415
Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser
420 425 430
Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro
435 440 445
Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr
450 455 460
Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp
465 470 475 480
Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu
485 490 495
Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser
500 505 510
Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 82
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 82
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 83
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 83
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Cys Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 84
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 84
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 85
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 85
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 86
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 86
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 87
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 87
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Cys Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 88
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 88
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 89
<211> 535
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 89
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Val Gln Leu Gln Gln Ser
275 280 285
Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met Ser Cys Lys
290 295 300
Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp Val Lys Gln
305 310 315 320
Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg
325 330 335
Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr
340 345 350
Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr
355 360 365
Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His
370 375 380
Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser Ser
385 390 395 400
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
405 410 415
Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser Pro Ala Ile Met Ser
420 425 430
Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser
435 440 445
Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys
450 455 460
Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly Val Pro Tyr Arg
465 470 475 480
Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Ser
485 490 495
Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser
500 505 510
Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu Glu Leu Lys Gly Gly
515 520 525
Ser His His His His His His
530 535
<210> 90
<211> 540
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 90
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Ala Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Gly Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser
275 280 285
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
290 295 300
Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
305 310 315 320
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser
325 330 335
Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
340 345 350
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
355 360 365
Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu
370 375 380
Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu
405 410 415
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
420 425 430
Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
435 440 445
Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn
450 455 460
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
465 470 475 480
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
485 490 495
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
500 505 510
Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
515 520 525
Val Ser Ser Gly Gly Ser His His His His His His
530 535 540
<210> 91
<211> 540
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 91
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Leu Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Ile Gln
145 150 155 160
Leu Val Gln Ser Gly Pro Glu Leu Lys Lys Pro Gly Glu Thr Val Lys
165 170 175
Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asn Tyr Gly Met Asn
180 185 190
Trp Val Lys Gln Ala Pro Gly Lys Cys Leu Lys Trp Met Gly Trp Ile
195 200 205
Asn Thr Asn Thr Gly Glu Pro Thr Tyr Ala Glu Glu Phe Lys Gly Arg
210 215 220
Phe Ala Phe Ser Leu Glu Thr Ser Ala Ser Thr Ala Tyr Leu Gln Ile
225 230 235 240
Asn Asn Leu Lys Asn Glu Asp Thr Ala Thr Tyr Phe Cys Ala Arg Leu
245 250 255
Gly Phe Gly Asn Ala Met Asp Tyr Trp Gly Gln Gly Thr Ser Val Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser
275 280 285
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
290 295 300
Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
305 310 315 320
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser
325 330 335
Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
340 345 350
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
355 360 365
Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu
370 375 380
Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu
405 410 415
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
420 425 430
Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
435 440 445
Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn
450 455 460
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
465 470 475 480
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
485 490 495
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
500 505 510
Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
515 520 525
Val Ser Ser Gly Gly Ser His His His His His His
530 535 540
<210> 92
<211> 540
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 92
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Ser Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser
275 280 285
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
290 295 300
Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
305 310 315 320
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser
325 330 335
Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
340 345 350
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
355 360 365
Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu
370 375 380
Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu
405 410 415
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
420 425 430
Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
435 440 445
Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn
450 455 460
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
465 470 475 480
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
485 490 495
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
500 505 510
Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
515 520 525
Val Ser Ser Gly Gly Ser His His His His His His
530 535 540
<210> 93
<211> 540
<212> PRT
<213> artificial
<220>
<223> artificial Antibody
<400> 93
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Thr Val
20 25 30
Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys Ser Ser Gln Ser Leu
35 40 45
Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr Trp Tyr Gln Gln Lys
50 55 60
Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Glu
65 70 75 80
Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly Ser Gly Thr Asp Phe
85 90 95
Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp Leu Ala Val Tyr Tyr
100 105 110
Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe Gly Cys Gly Thr Lys
115 120 125
Leu Glu Ile Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln
145 150 155 160
Leu Gln Gln Pro Gly Ala Glu Leu Val Arg Pro Gly Ala Ser Val Lys
165 170 175
Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr Trp Ile Asn
180 185 190
Trp Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Asn Ile
195 200 205
Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
210 215 220
Ala Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
225 230 235 240
Ser Ser Pro Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Thr Arg Ser
245 250 255
Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
260 265 270
Val Ser Ser Ser Gly Gly Gly Gly Ser Gln Ile Val Leu Thr Gln Ser
275 280 285
Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys
290 295 300
Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser
305 310 315 320
Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser
325 330 335
Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser
340 345 350
Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys
355 360 365
Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Cys Gly Thr Lys Leu
370 375 380
Glu Leu Lys Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
385 390 395 400
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gln Val Gln Leu
405 410 415
Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys Met
420 425 430
Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His Trp
435 440 445
Val Lys Gln Arg Pro Gly Gln Cys Leu Glu Trp Ile Gly Tyr Ile Asn
450 455 460
Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys Ala
465 470 475 480
Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu Ser
485 490 495
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr Tyr
500 505 510
Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr
515 520 525
Val Ser Ser Gly Gly Ser His His His His His His
530 535 540
<210> 94
<211> 119
<212> PRT
<213> Mus musculus
<400> 94
Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr
20 25 30
Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe
50 55 60
Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Thr Leu Thr Val Ser Ser
115
<210> 95
<211> 116
<212> PRT
<213> Mus musculus
<400> 95
Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser Val Lys
1 5 10 15
Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr Met His
20 25 30
Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly Tyr Ile
35 40 45
Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys Asp Lys
50 55 60
Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met Gln Leu
65 70 75 80
Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Tyr
85 90 95
Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu
100 105 110
Thr Val Ser Ser
115
<210> 96
<211> 103
<212> PRT
<213> Mus musculus
<400> 96
Leu Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val
1 5 10 15
Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr
20 25 30
Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser
35 40 45
Lys Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly
50 55 60
Thr Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala
65 70 75 80
Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala
85 90 95
Gly Thr Lys Leu Glu Leu Lys
100
<210> 97
<211> 107
<212> PRT
<213> Mus musculus
<400> 97
Asp Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr
35 40 45
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
85 90 95
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 98
<211> 106
<212> PRT
<213> Mus musculus
<400> 98
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 99
<211> 106
<212> PRT
<213> Mus musculus
<400> 99
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Trp Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 100
<211> 106
<212> PRT
<213> Mus musculus
<400> 100
Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly Glu
1 5 10 15
Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met His
20 25 30
Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Ser Ile Tyr Ser
35 40 45
Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg Gly
50 55 60
Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu Asp
65 70 75 80
Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 101
<211> 513
<212> PRT
<213> artificial
<220>
<223> bispecific molecule 107
<400> 101
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Asp Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Cys Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys His His His His His
500 505 510
His
<210> 102
<211> 513
<212> PRT
<213> artificial
<220>
<223> bispecific molecule 123
<400> 102
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Asp Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg
20 25 30
Pro Gly Ala Ser Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe
35 40 45
Thr Arg Tyr Thr Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Ala Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp
115 120 125
Gly Gln Gly Thr Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly
130 135 140
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu
145 150 155 160
Thr Gln Ser Pro Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr
165 170 175
Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln
180 185 190
Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys
195 200 205
Val Ala Ser Gly Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr
210 215 220
Ser Tyr Ser Leu Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr
225 230 235 240
Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly
245 250 255
Thr Lys Leu Glu Leu Lys Ser Gly Gly Gly Gly Ser Glu Val Gln Leu
260 265 270
Gln Gln Ser Gly Pro Glu Leu Val Lys Pro Gly Ala Ser Met Lys Ile
275 280 285
Ser Cys Lys Ala Ser Gly Tyr Ser Phe Thr Gly Tyr Thr Met Asn Trp
290 295 300
Val Lys Gln Ser His Gly Lys Asn Leu Glu Trp Ile Gly Leu Ile Asn
305 310 315 320
Pro Tyr Asn Gly Gly Thr Ile Tyr Asn Gln Lys Phe Lys Gly Lys Ala
325 330 335
Thr Leu Thr Val Asp Lys Ser Ser Ser Thr Ala Tyr Met Glu Leu Leu
340 345 350
Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala Arg Asp Tyr
355 360 365
Gly Phe Val Leu Asp Tyr Trp Gly Gln Gly Thr Thr Leu Thr Val Ser
370 375 380
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
385 390 395 400
Asp Ile Val Leu Thr Gln Ser Pro Ser Ile Met Ser Val Ser Pro Gly
405 410 415
Glu Lys Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Ser Tyr Met
420 425 430
His Trp Phe Gln Gln Lys Pro Gly Thr Ser Pro Lys Leu Leu Ile Tyr
435 440 445
Ser Thr Ser Asn Leu Ala Ser Gly Val Pro Ala Arg Phe Ser Gly Arg
450 455 460
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Ala Ala Glu
465 470 475 480
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Arg Ser Asn Tyr Pro Pro Trp
485 490 495
Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys His His His His His
500 505 510
His
<210> 103
<211> 520
<212> PRT
<213> artificial
<220>
<223> bispecific molecule 124
<400> 103
Met Gly Trp Ser Cys Ile Ile Leu Phe Leu Val Ala Thr Ala Thr Gly
1 5 10 15
Val His Ser Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Arg
20 25 30
Pro Gly Ala Ser Val Lys Leu Ser Cys Lys Ala Ser Gly Tyr Thr Phe
35 40 45
Thr Ser Tyr Trp Ile Asn Trp Val Lys Gln Arg Pro Gly Gln Gly Leu
50 55 60
Glu Trp Ile Gly Asn Ile Tyr Pro Ser Asp Ser Tyr Thr Asn Tyr Asn
65 70 75 80
Gln Lys Phe Lys Asp Lys Ala Thr Leu Thr Val Asp Lys Ser Ser Ser
85 90 95
Thr Ala Tyr Met Gln Leu Ser Ser Pro Thr Ser Glu Asp Ser Ala Val
100 105 110
Tyr Tyr Cys Thr Arg Ser Trp Arg Gly Asn Ser Phe Asp Tyr Trp Gly
115 120 125
Gln Gly Thr Thr Leu Thr Val Ser Ser Gly Gly Gly Gly Ser Gly Gly
130 135 140
Gly Gly Ser Gly Gly Gly Gly Ser Asp Ile Val Met Thr Gln Ser Pro
145 150 155 160
Ser Ser Leu Thr Val Thr Ala Gly Glu Lys Val Thr Met Ser Cys Lys
165 170 175
Ser Ser Gln Ser Leu Leu Asn Ser Gly Asn Gln Lys Asn Tyr Leu Thr
180 185 190
Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro Lys Leu Leu Ile Tyr Trp
195 200 205
Ala Ser Thr Arg Glu Ser Gly Val Pro Asp Arg Phe Thr Gly Ser Gly
210 215 220
Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Val Gln Ala Glu Asp
225 230 235 240
Leu Ala Val Tyr Tyr Cys Gln Asn Asp Tyr Ser Tyr Pro Phe Thr Phe
245 250 255
Gly Ser Gly Thr Lys Leu Glu Ile Lys Ser Gly Gly Gly Gly Ser Asp
260 265 270
Ile Lys Leu Gln Gln Ser Gly Ala Glu Leu Ala Arg Pro Gly Ala Ser
275 280 285
Val Lys Met Ser Cys Lys Thr Ser Gly Tyr Thr Phe Thr Arg Tyr Thr
290 295 300
Met His Trp Val Lys Gln Arg Pro Gly Gln Gly Leu Glu Trp Ile Gly
305 310 315 320
Tyr Ile Asn Pro Ser Arg Gly Tyr Thr Asn Tyr Asn Gln Lys Phe Lys
325 330 335
Asp Lys Ala Thr Leu Thr Thr Asp Lys Ser Ser Ser Thr Ala Tyr Met
340 345 350
Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys Ala
355 360 365
Arg Tyr Tyr Asp Asp His Tyr Ser Leu Asp Tyr Trp Gly Gln Gly Thr
370 375 380
Thr Leu Thr Val Ser Ser Val Glu Gly Gly Ser Gly Gly Ser Gly Gly
385 390 395 400
Ser Gly Gly Ser Gly Gly Val Asp Asp Ile Gln Leu Thr Gln Ser Pro
405 410 415
Ala Ile Met Ser Ala Ser Pro Gly Glu Lys Val Thr Met Thr Cys Arg
420 425 430
Ala Ser Ser Ser Val Ser Tyr Met Asn Trp Tyr Gln Gln Lys Ser Gly
435 440 445
Thr Ser Pro Lys Arg Trp Ile Tyr Asp Thr Ser Lys Val Ala Ser Gly
450 455 460
Val Pro Tyr Arg Phe Ser Gly Ser Gly Ser Gly Thr Ser Tyr Ser Leu
465 470 475 480
Thr Ile Ser Ser Met Glu Ala Glu Asp Ala Ala Thr Tyr Tyr Cys Gln
485 490 495
Gln Trp Ser Ser Asn Pro Leu Thr Phe Gly Ala Gly Thr Lys Leu Glu
500 505 510
Leu Lys His His His His His His
515 520
<210> 104
<211> 20
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 104
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 105
<211> 25
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 105
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser Gly Gly Gly Gly Ser
20 25
<210> 106
<211> 18
<212> PRT
<213> Artificial Sequence
<220>
<223> Linker
<400> 106
Gly Gly Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
1 5 10 15
Gly Ser
<210> 107
<211> 6
<212> PRT
<213> Artificial Sequence
<220>
<223> His Tag
<400> 107
His His His His His His
1 5
<210> 108
<211> 9
<212> PRT
<213> Artificial Sequence
<220>
<223> His Tag
<400> 108
Gly Gly Ser His His His His His His
1 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СПОСОБЫ И КОМПОЗИЦИИ ДЛЯ ПРОГНОЗИРОВАНИЯ ТЕРАПЕВТИЧЕСКОЙ ЭФФЕКТИВНОСТИ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ И ПРОГНОЗА ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ | 2016 |
|
RU2785291C2 |
| КОНЪЮГАТЫ ЛЕКАРСТВЕННЫХ СРЕДСТВ, СОДЕРЖАЩИЕ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2016 |
|
RU2841168C2 |
| КОМБИНИРОВАННАЯ ТЕРАПИЯ С ИСПОЛЬЗОВАНИЕМ АНТИТЕЛ ПРОТИВ КЛАУДИНА 18.2 ДЛЯ ЛЕЧЕНИЯ РАКА | 2014 |
|
RU2792932C2 |
| БИСПЕЦИФИЧЕСКИЕ ТРЕХВАЛЕНТНЫЕ АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С КЛАУДИНОМ 6 ИЛИ КЛАУДИНОМ 18.2, И CD3, ДЛЯ ЛЕЧЕНИЯ ОНКОЛОГИЧЕСКИХ ЗАБОЛЕВАНИЙ С ЭКСПРЕССИЕЙ КЛАУДИНА | 2017 |
|
RU2798988C2 |
| ТЕРАПИЯ ДЛЯ ЛЕЧЕНИЯ РАКА, ВКЛЮЧАЮЩАЯ АНТИТЕЛА ПРОТИВ КЛАУДИНА 18.2 | 2014 |
|
RU2832015C2 |
| МУЛЬТИВАЛЕНТНЫЕ FV-АНТИТЕЛА | 2016 |
|
RU2785766C2 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2714232C2 |
| АНТИТЕЛА К C10ORF54 И ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2819627C1 |
| ИНДУЦИБЕЛЬНЫЕ СВЯЗЫВАЮЩИЕ БЕЛКИ И СПОСОБЫ ИСПОЛЬЗОВАНИЯ | 2017 |
|
RU2800034C2 |
| АНТИТЕЛА ПРОТИВ КЛАУДИНА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2815926C2 |
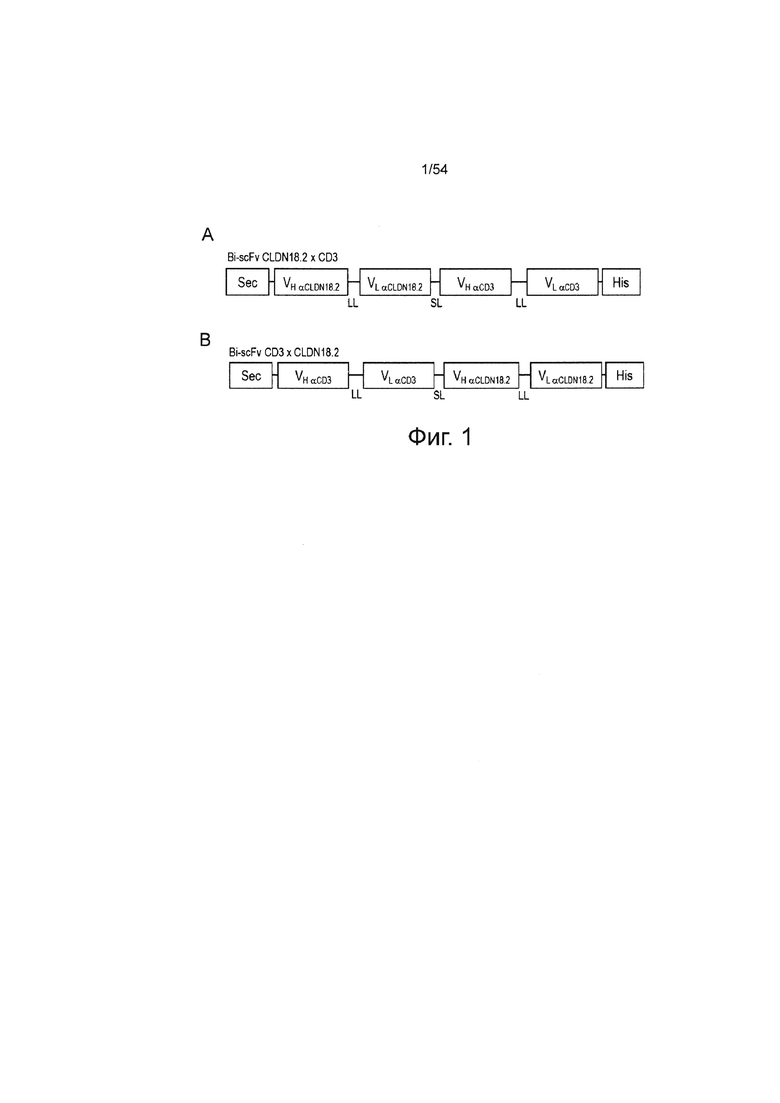
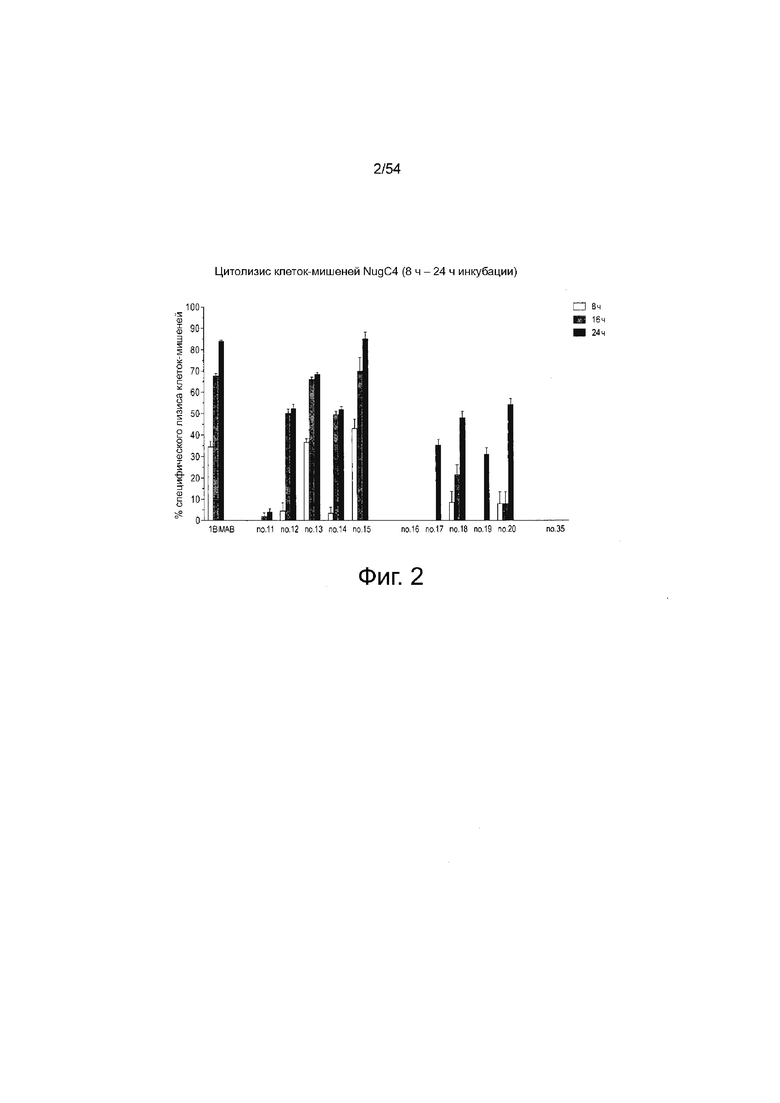
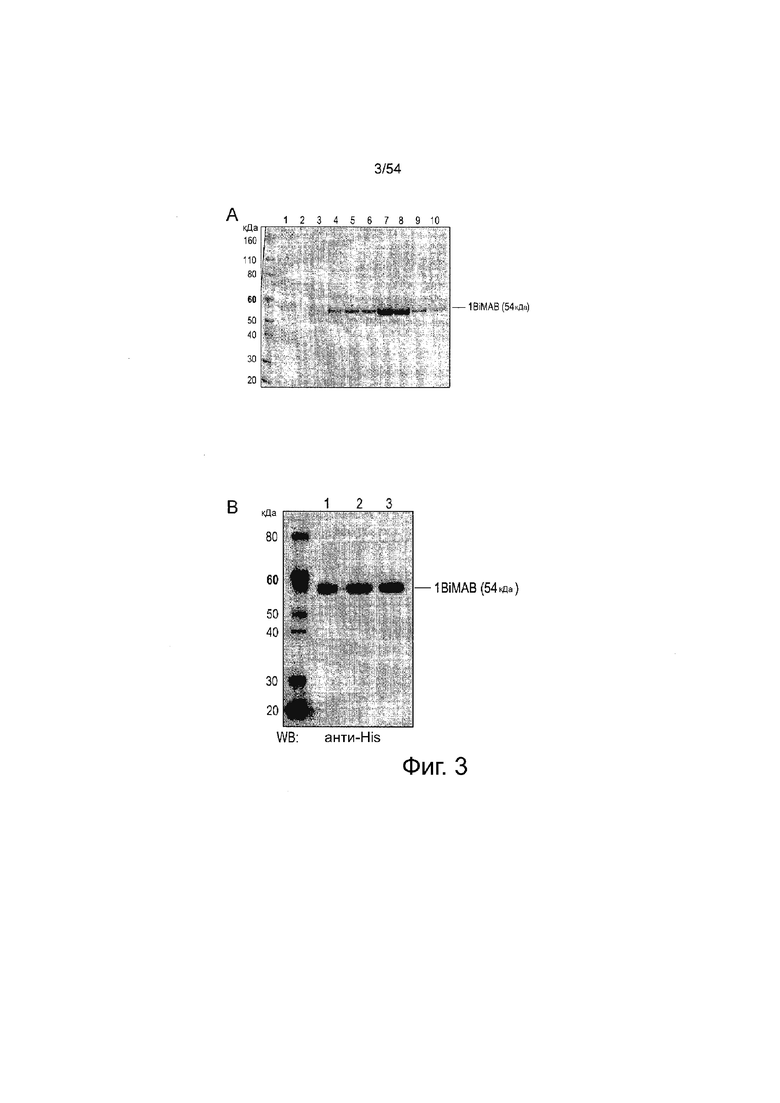
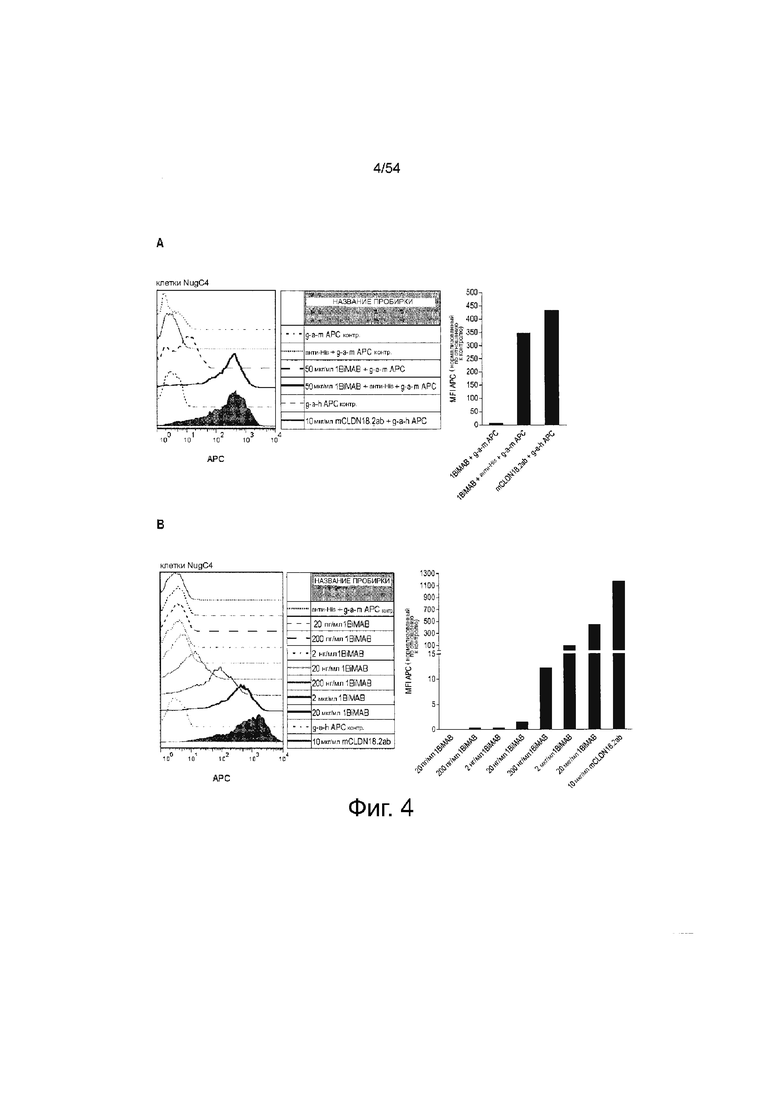


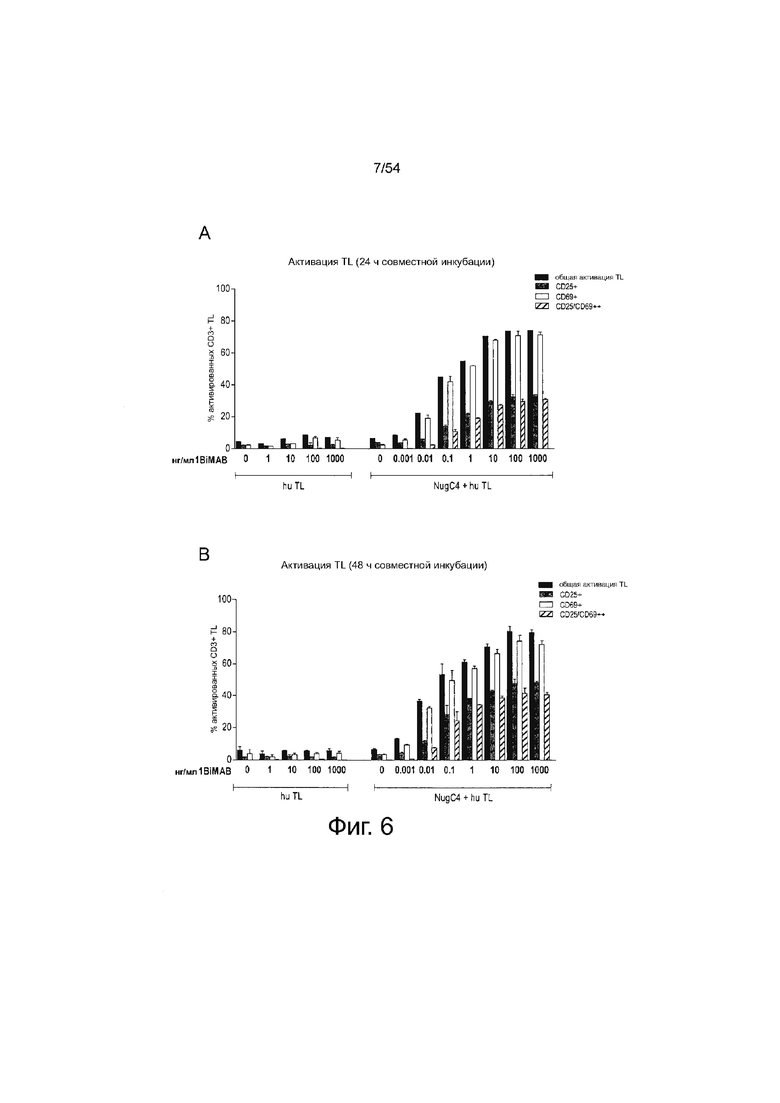
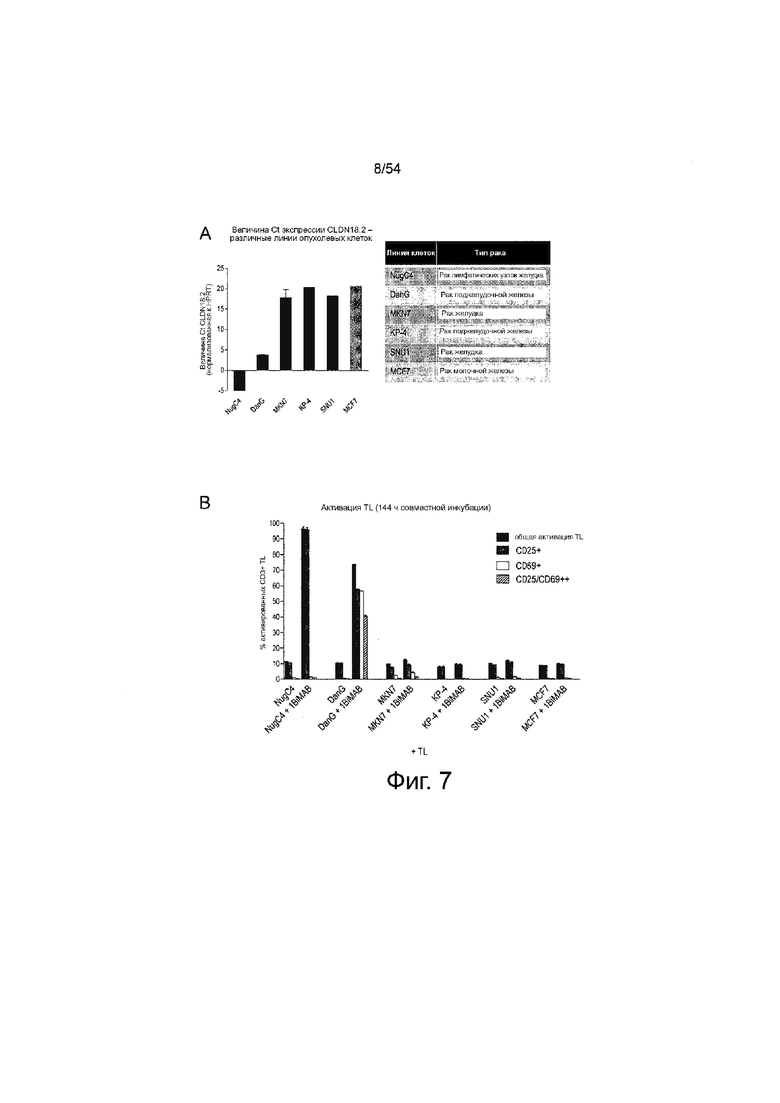
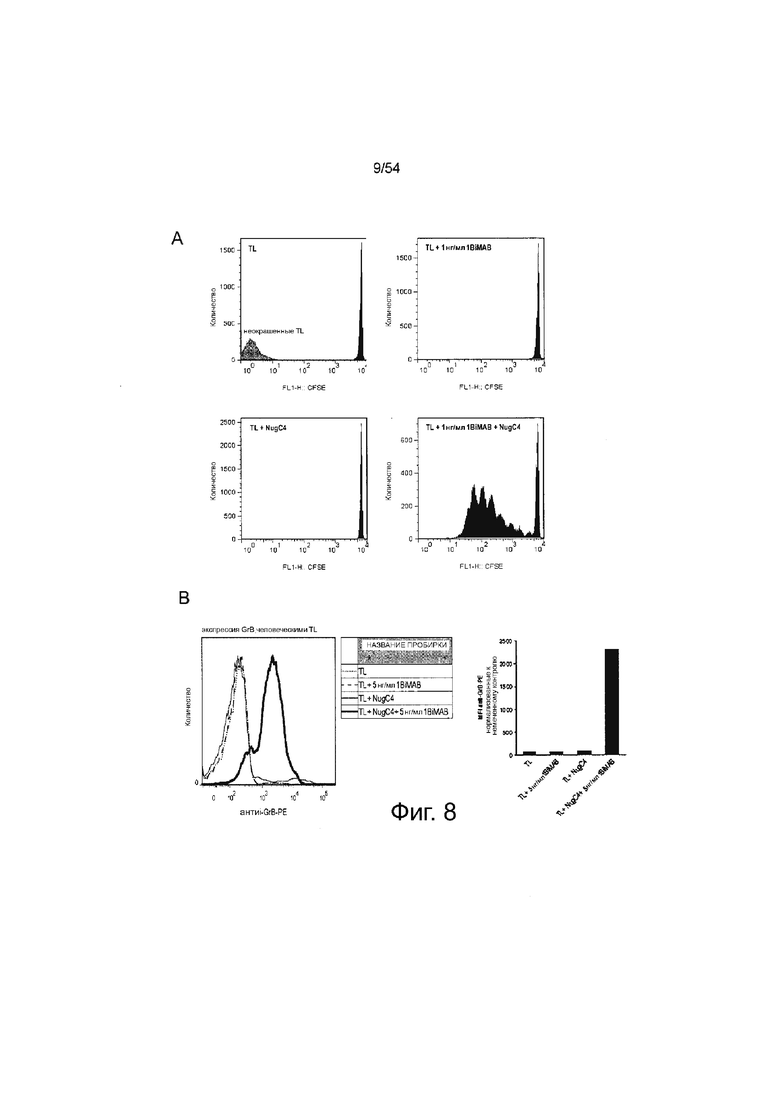
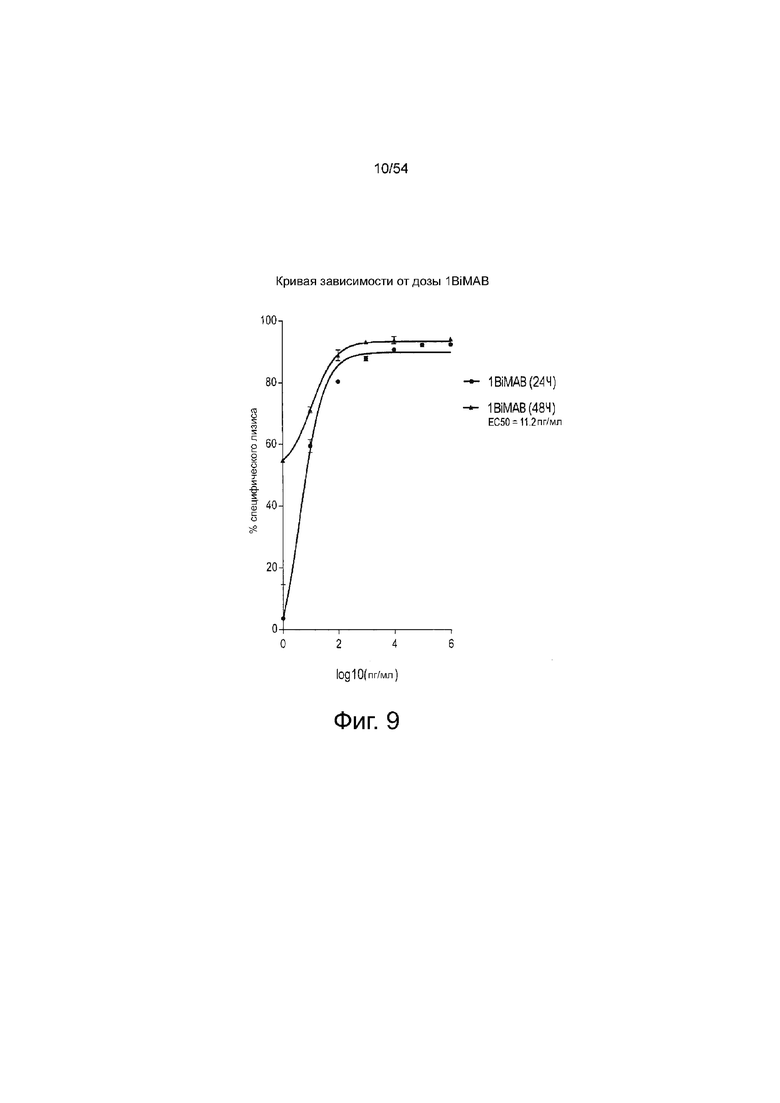
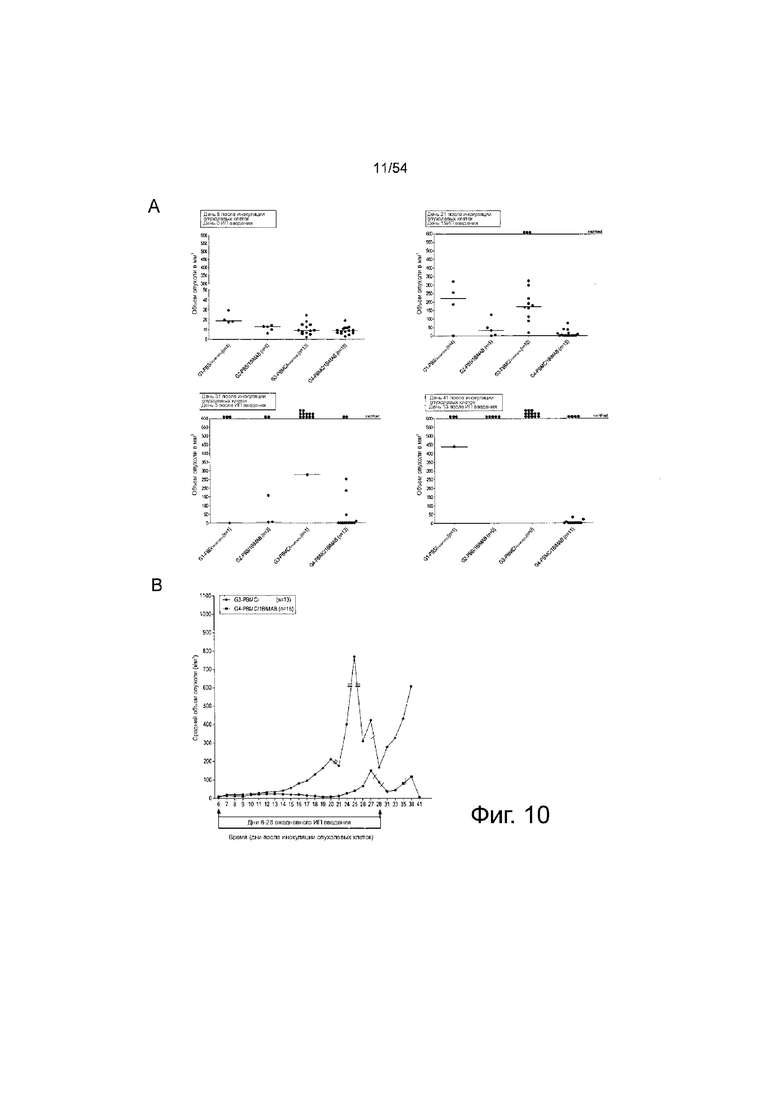
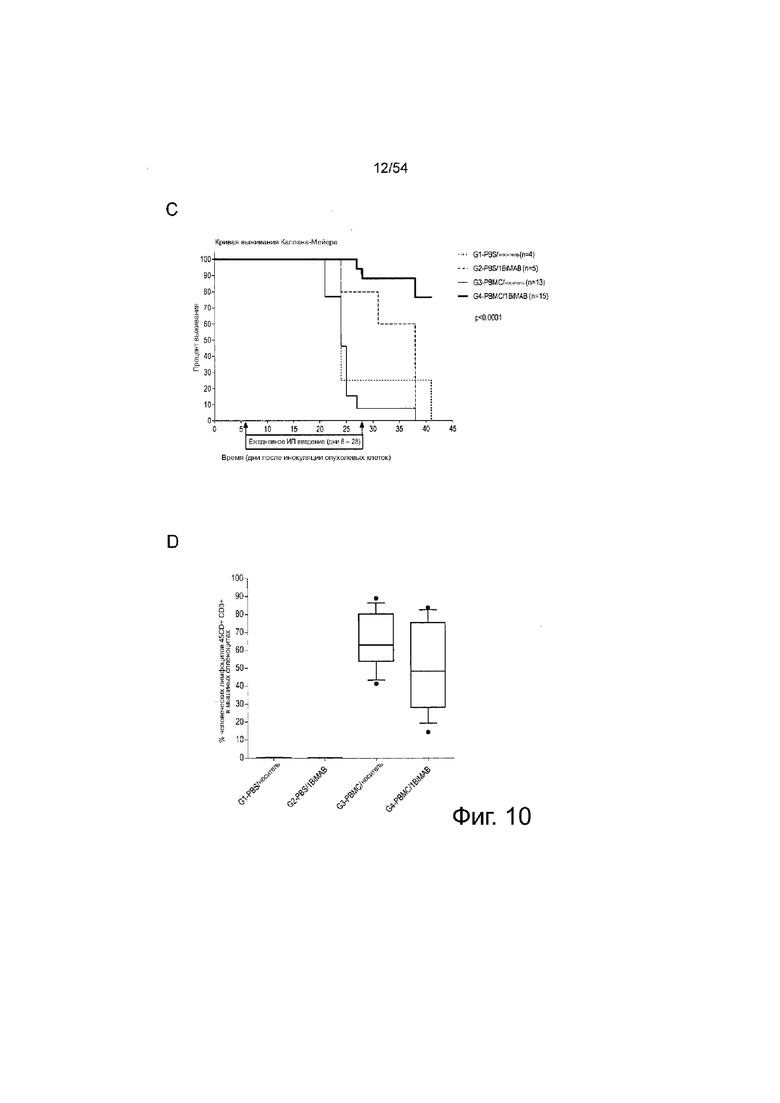


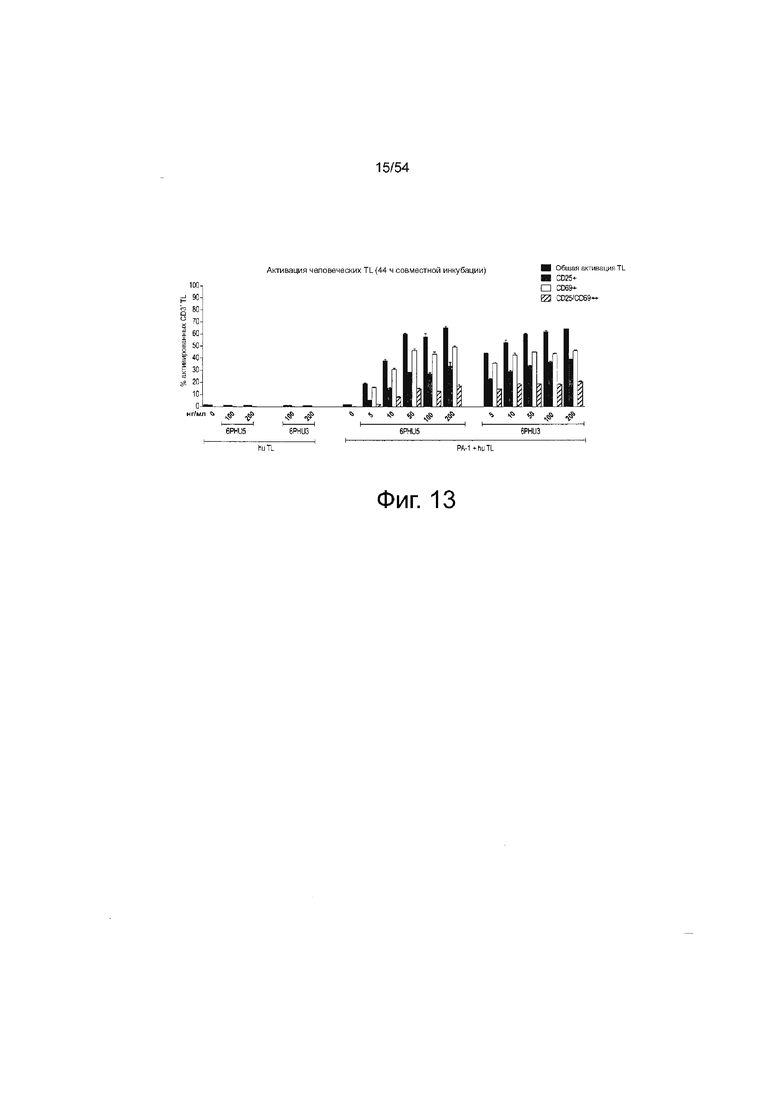

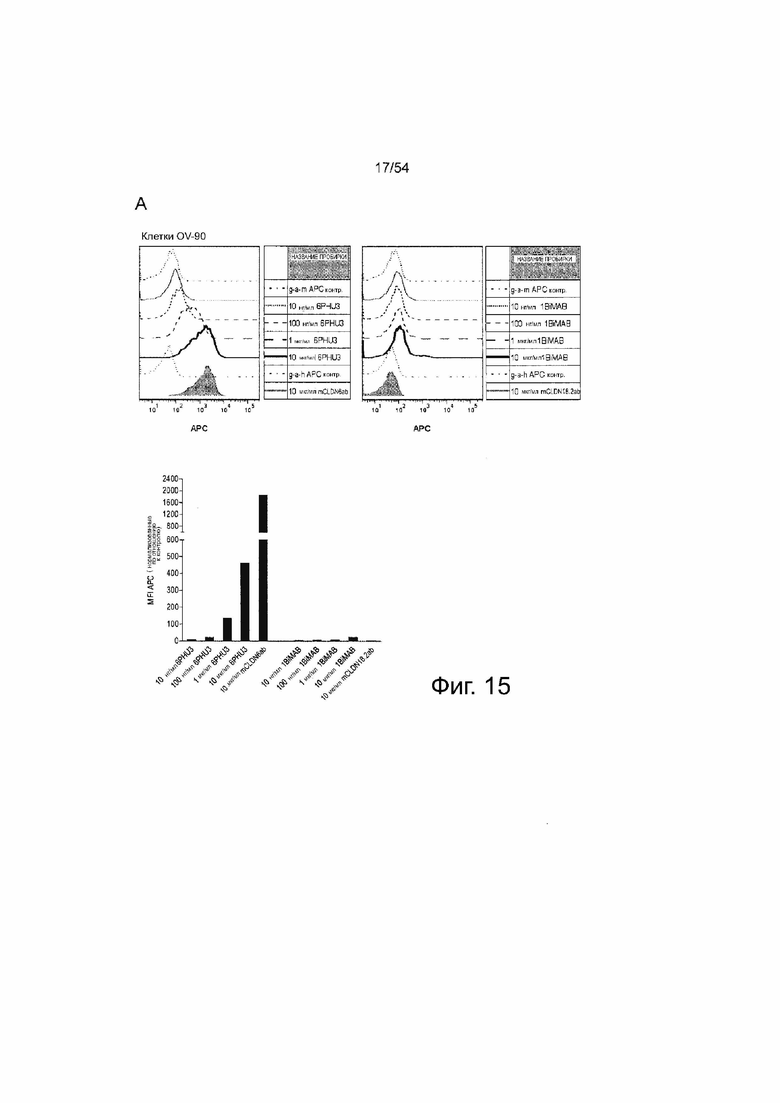
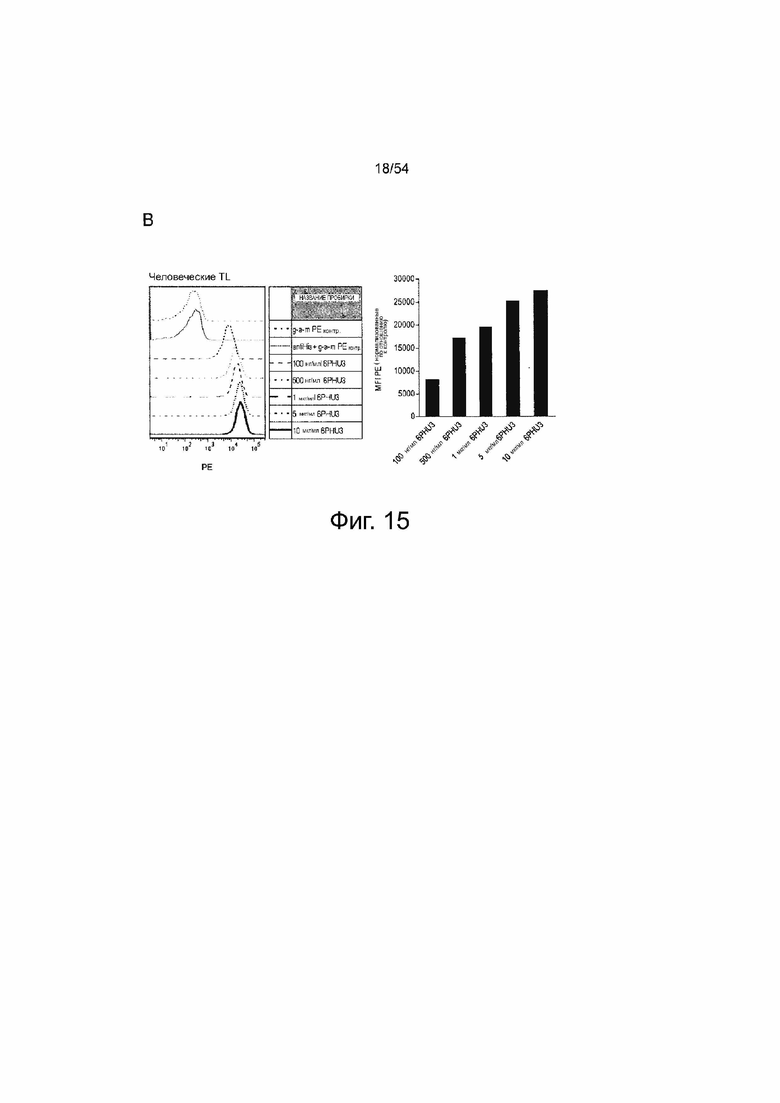
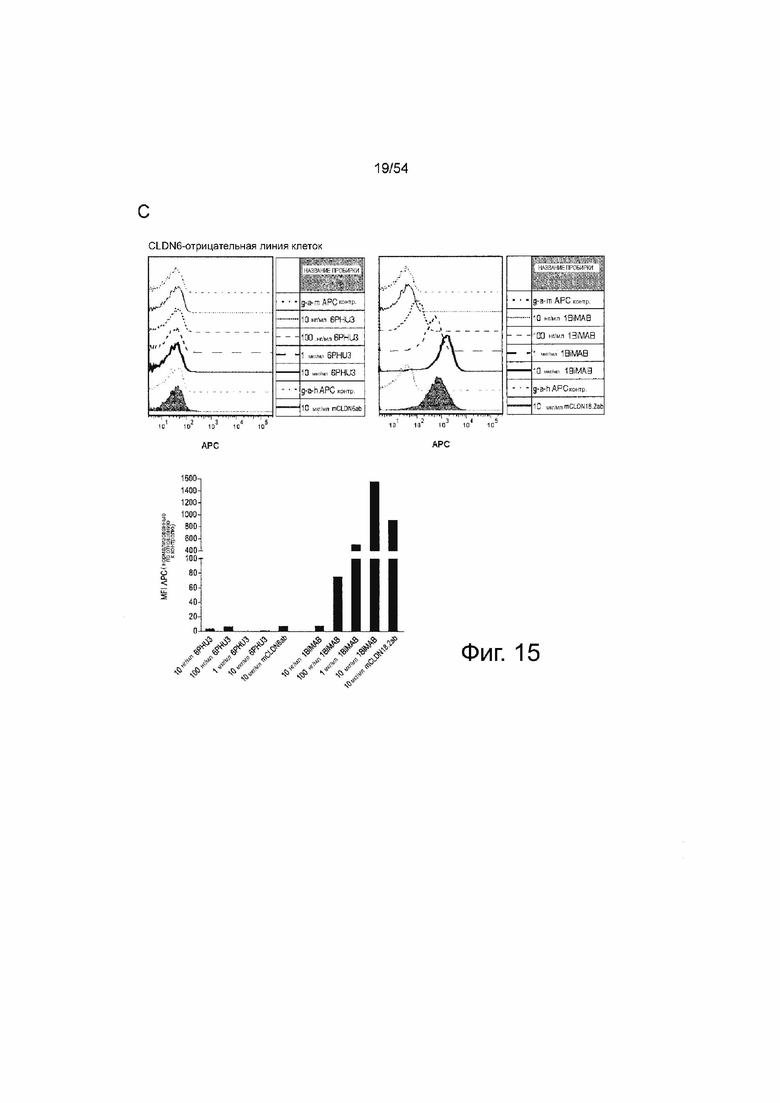
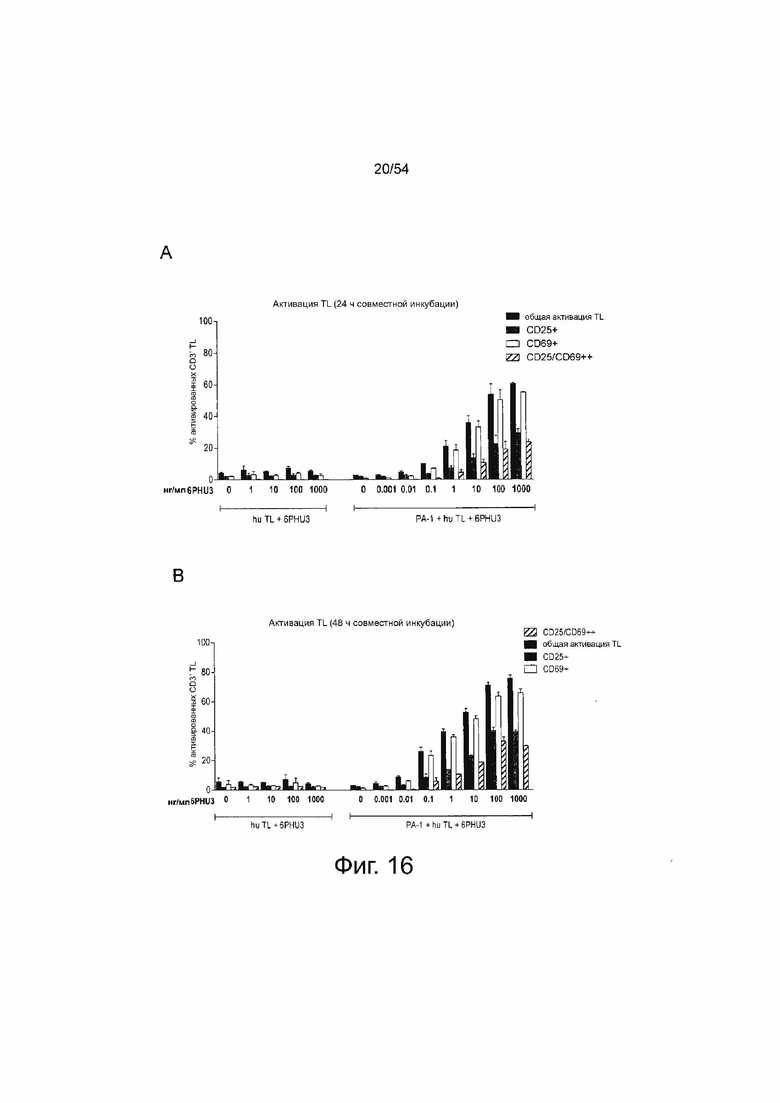
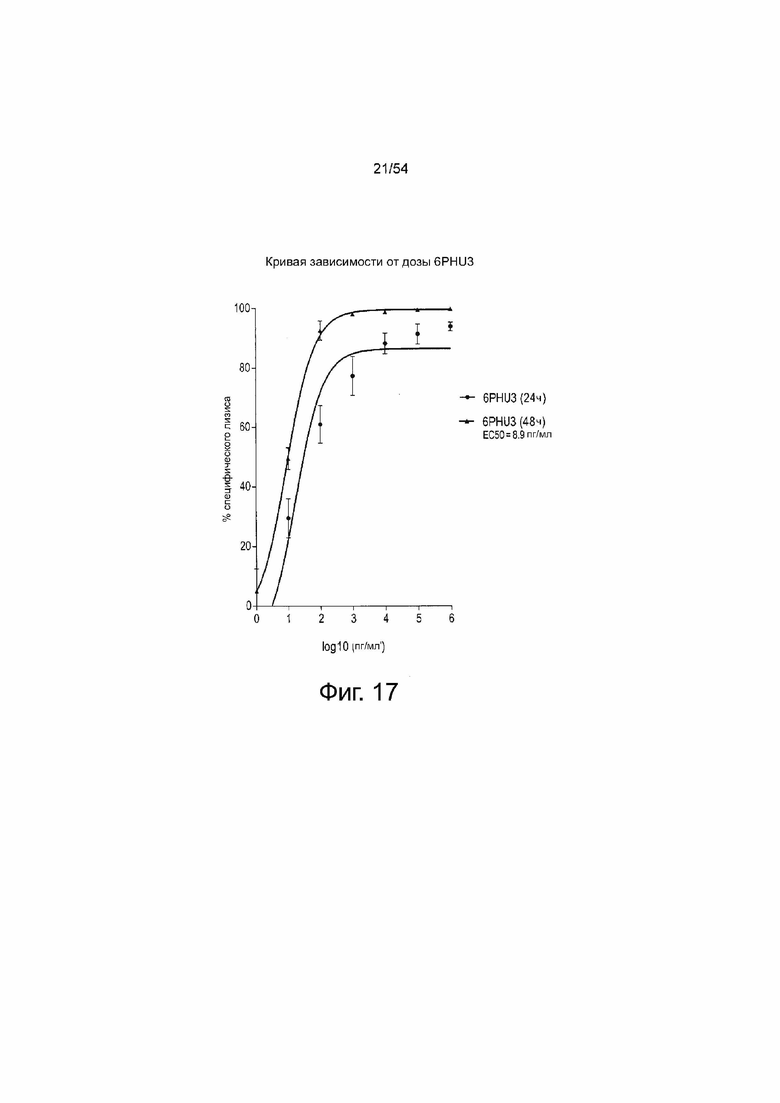
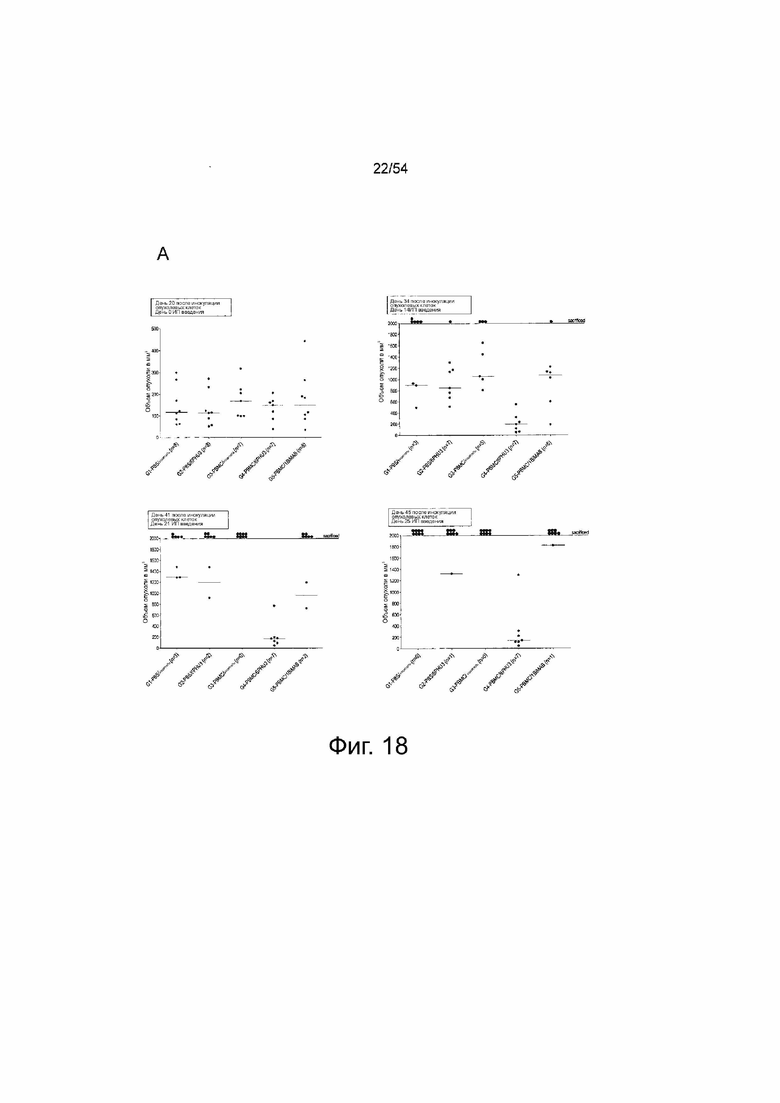
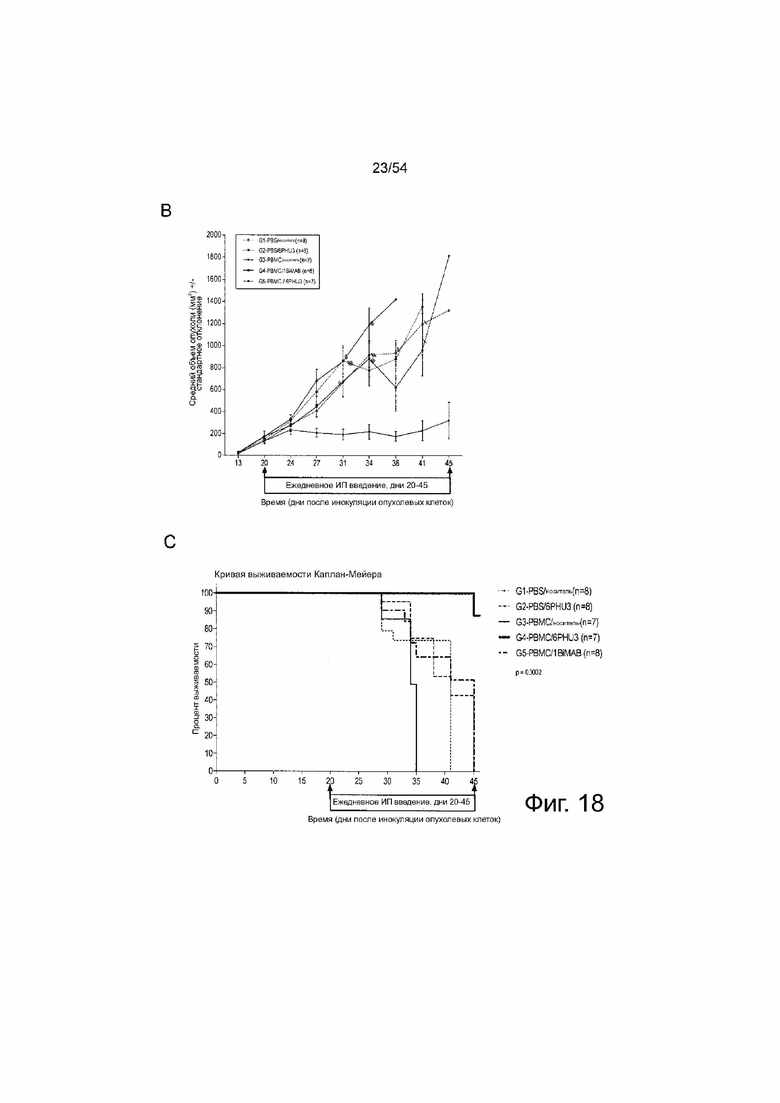
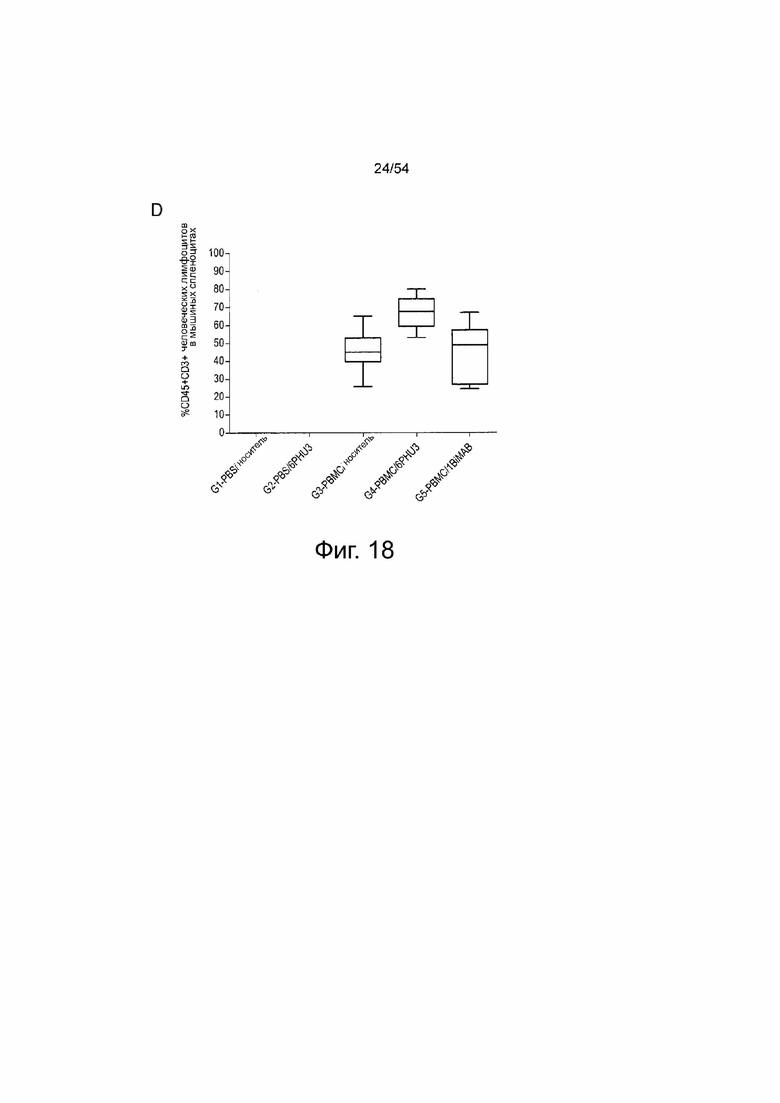

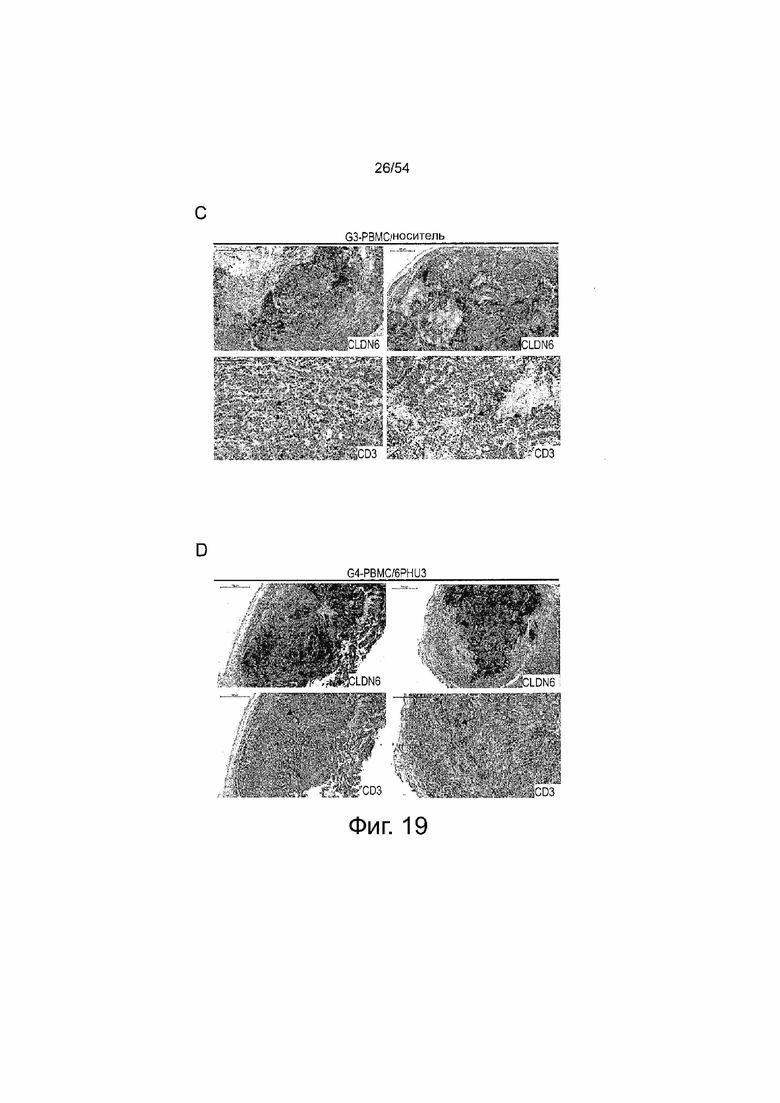
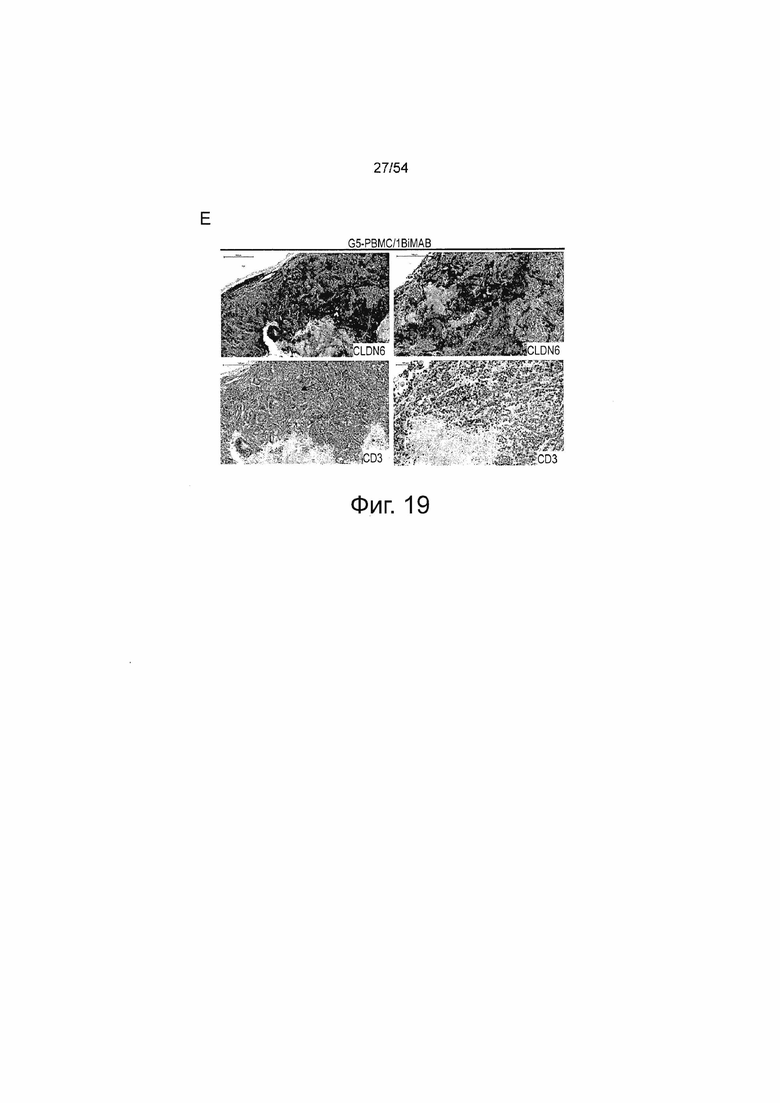

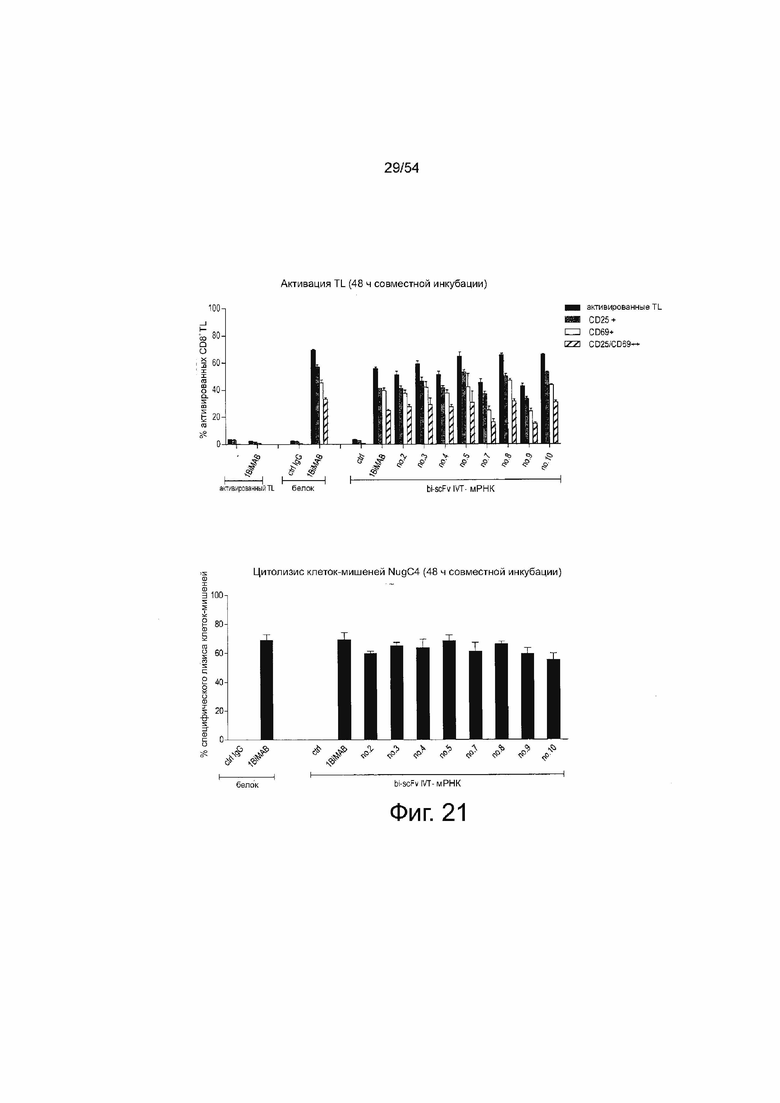

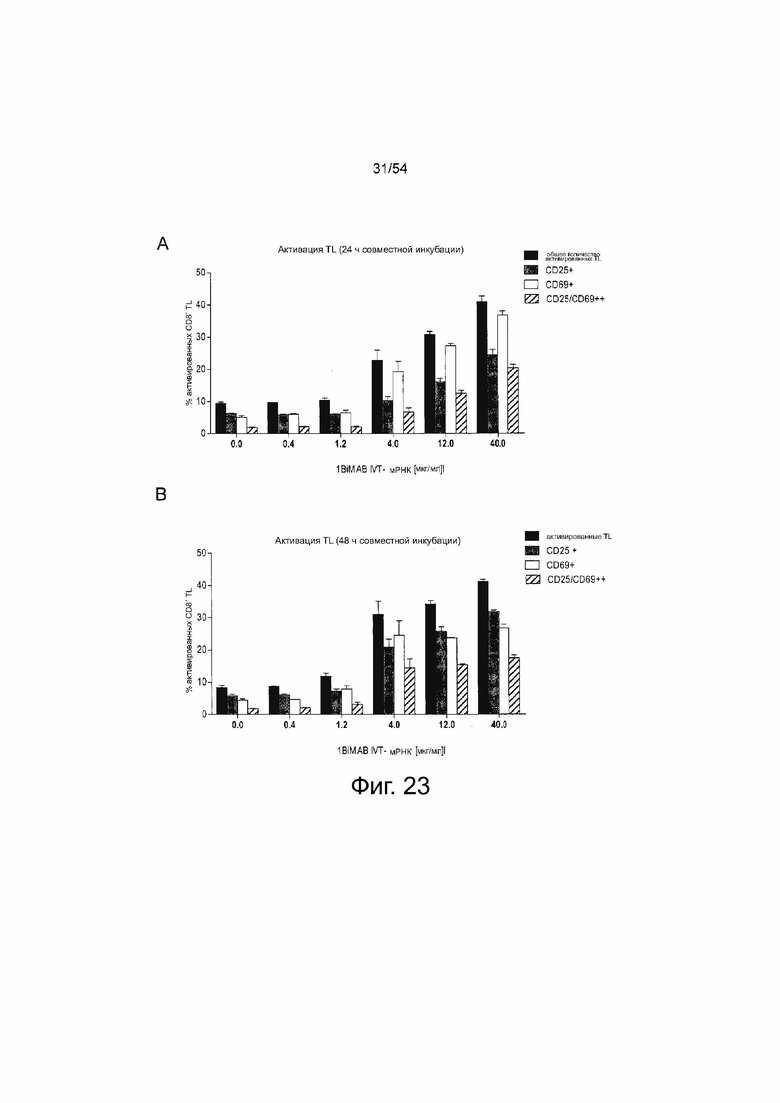
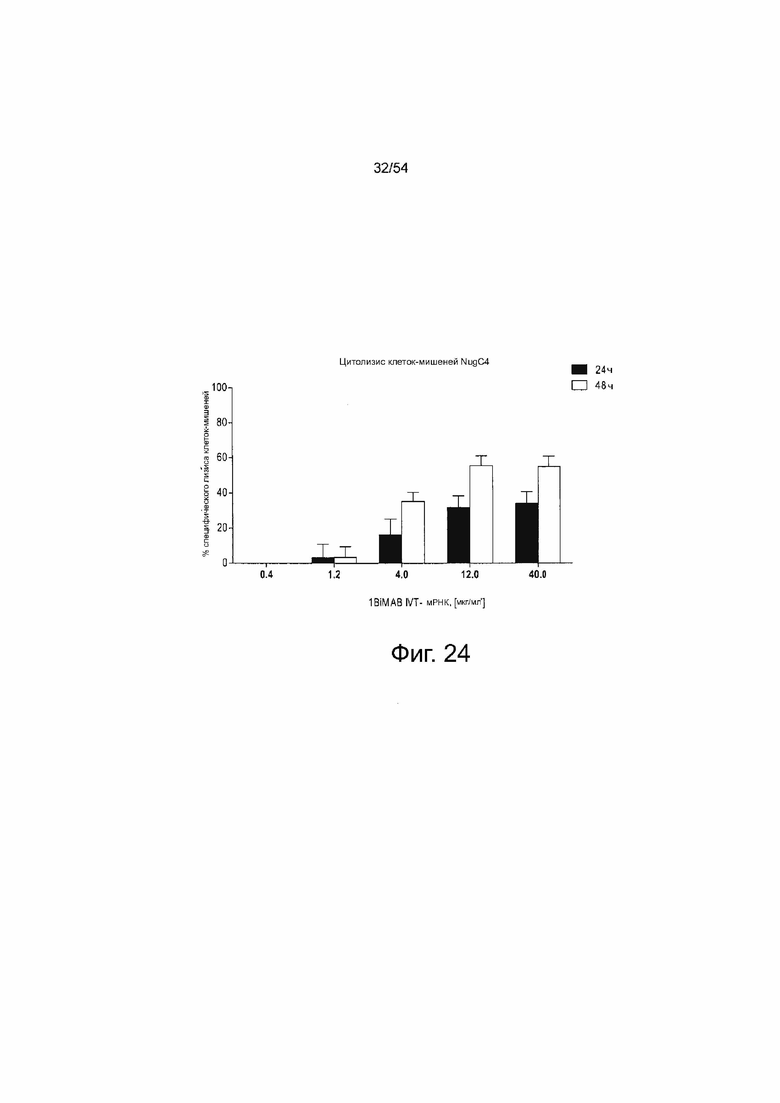
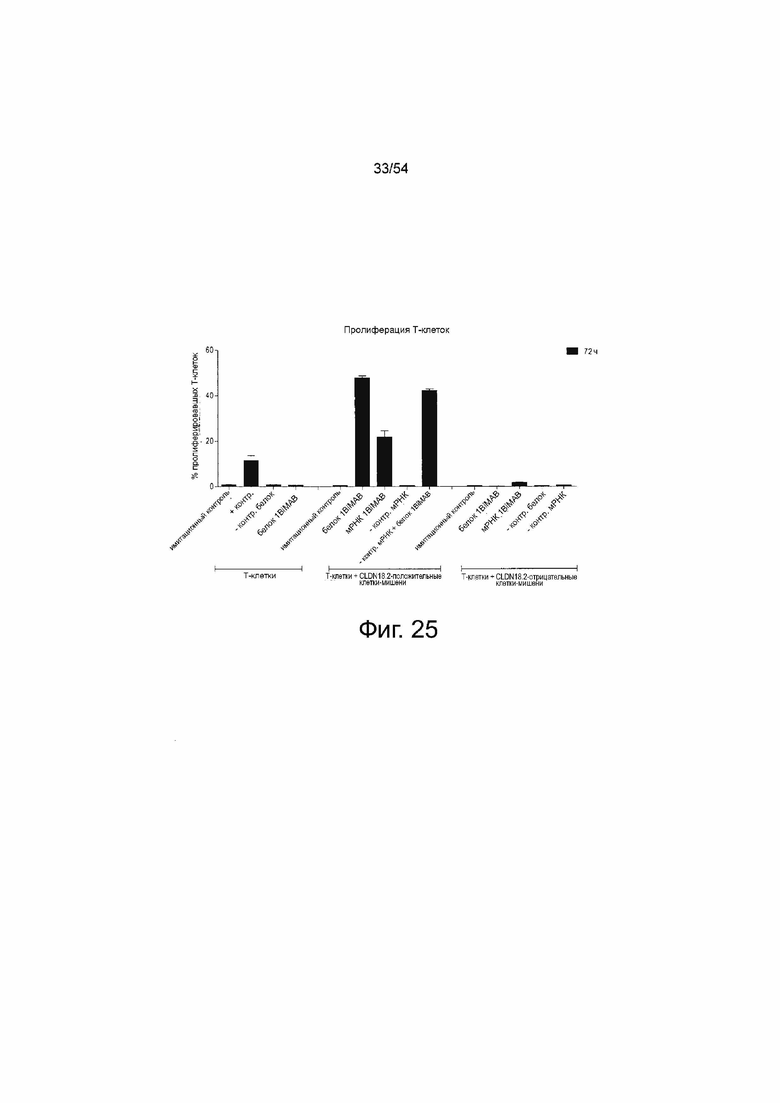
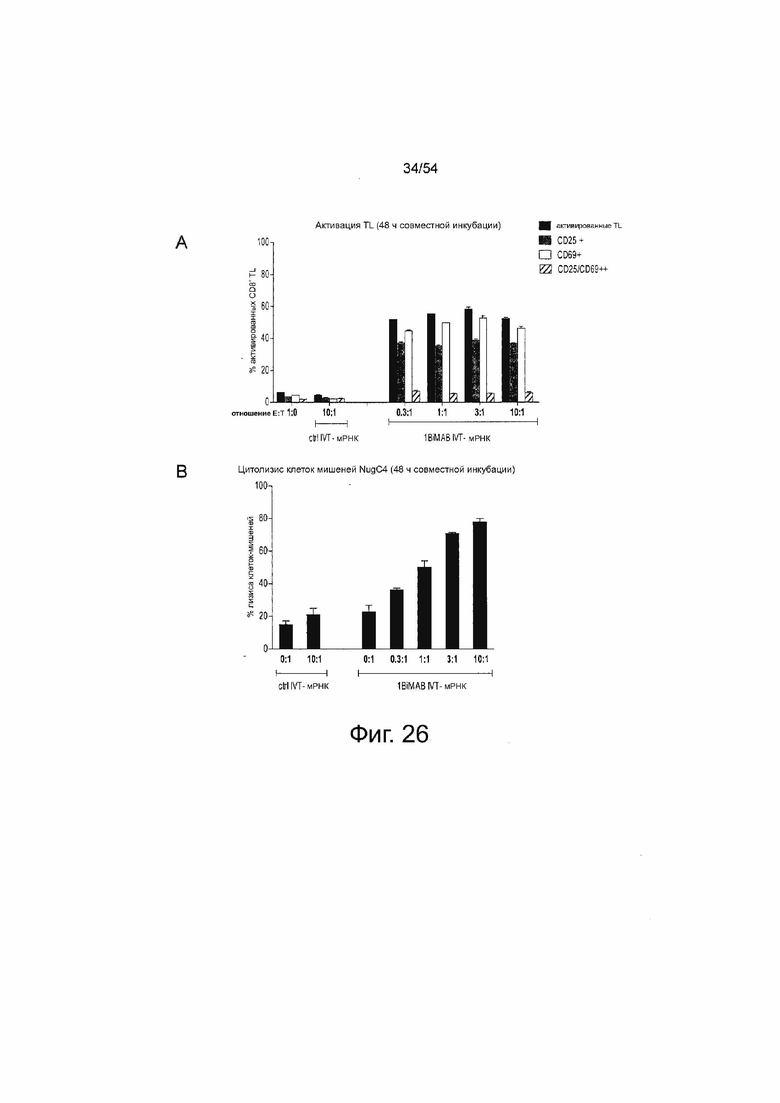
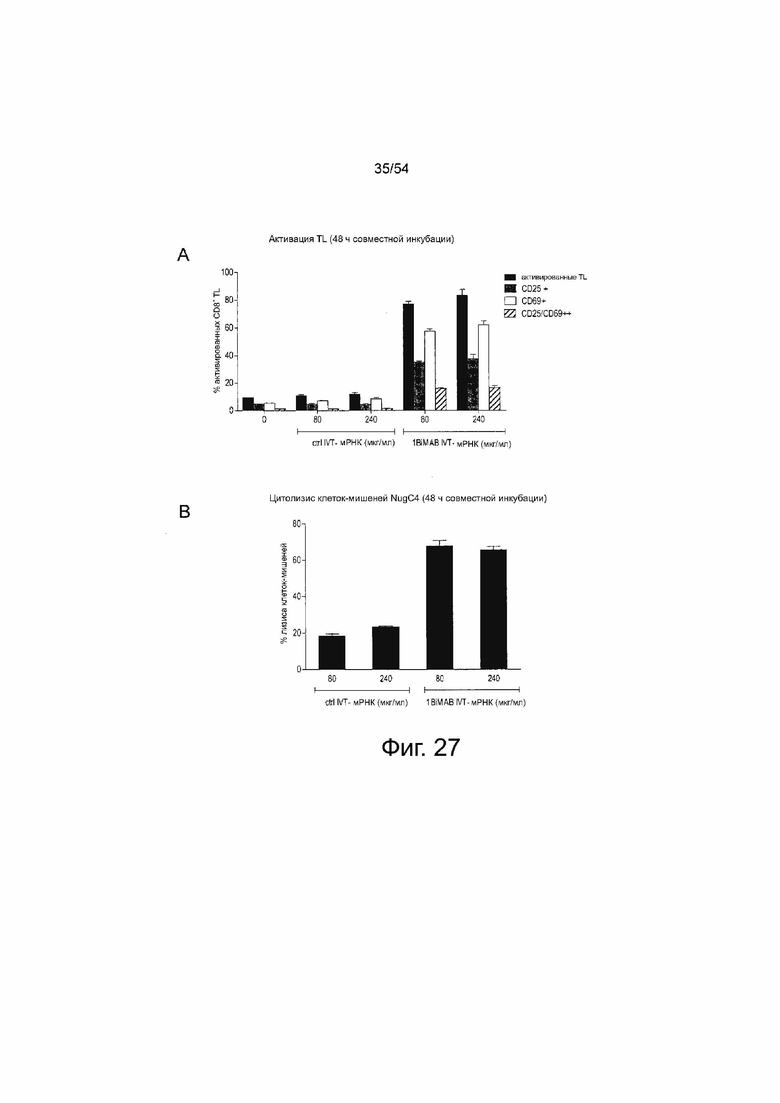

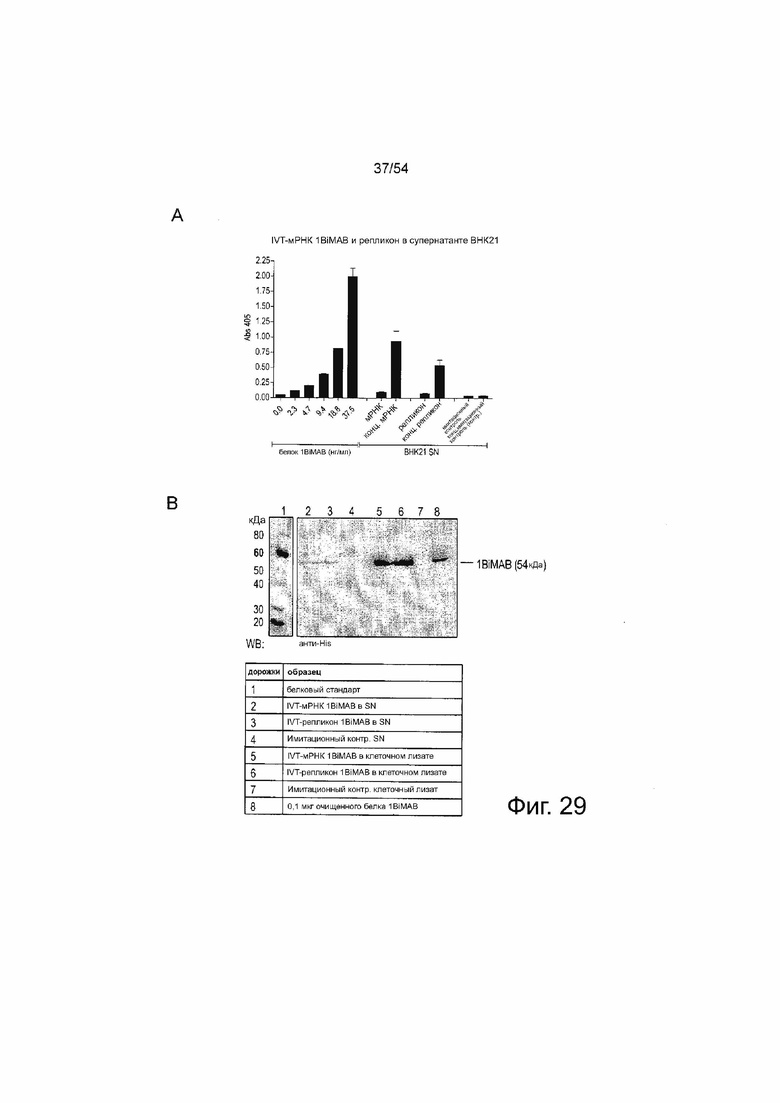

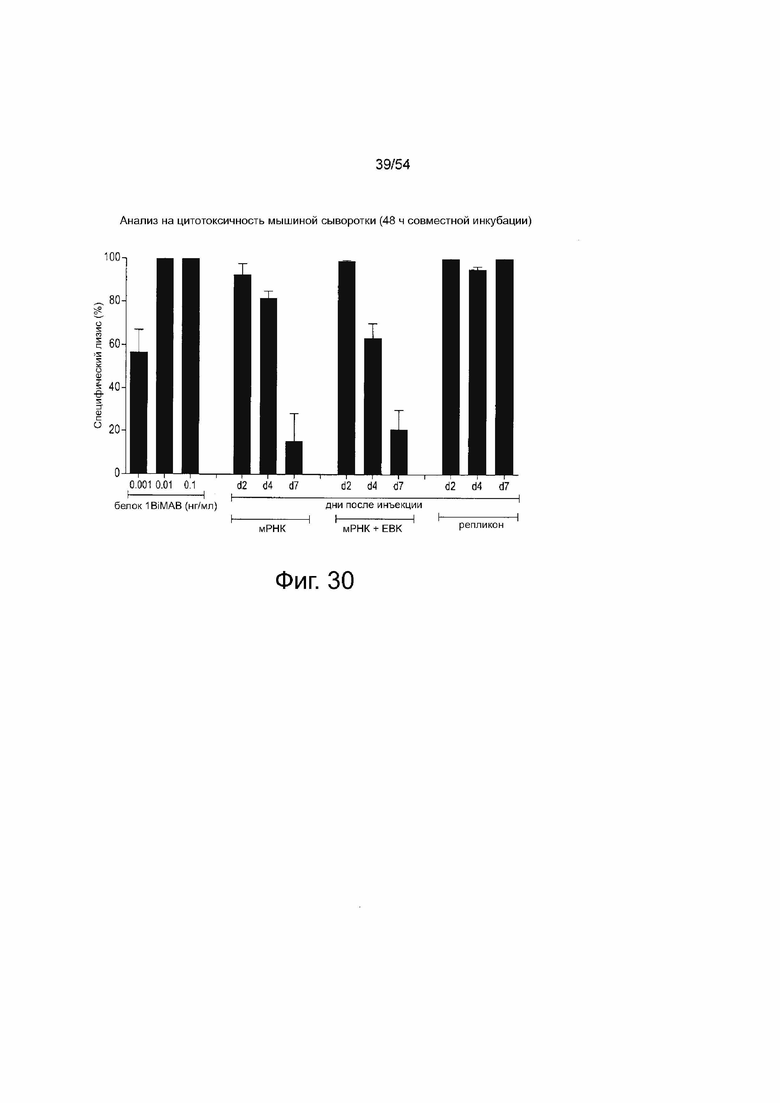
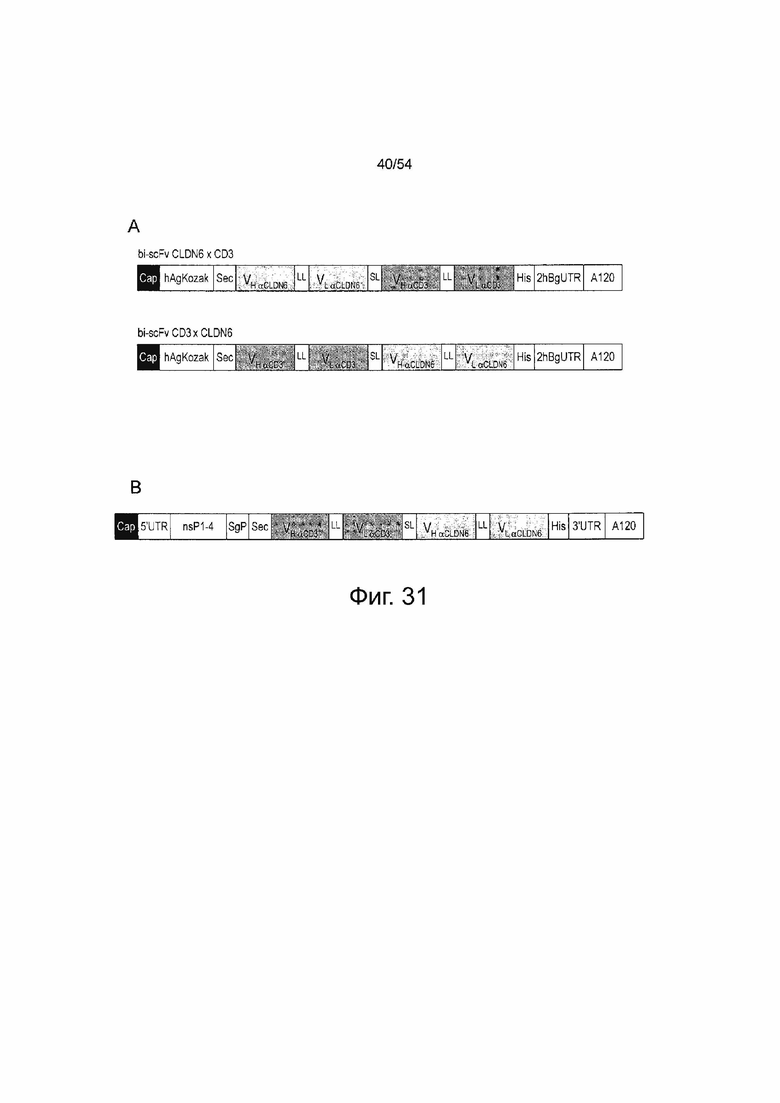

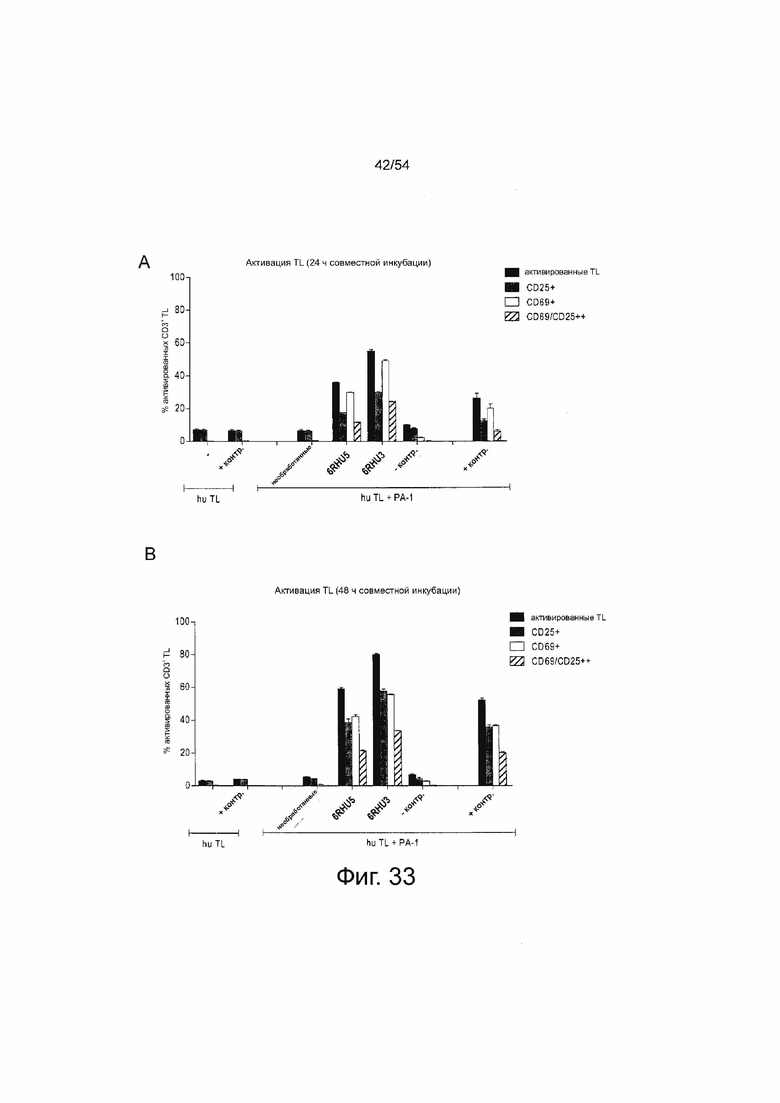
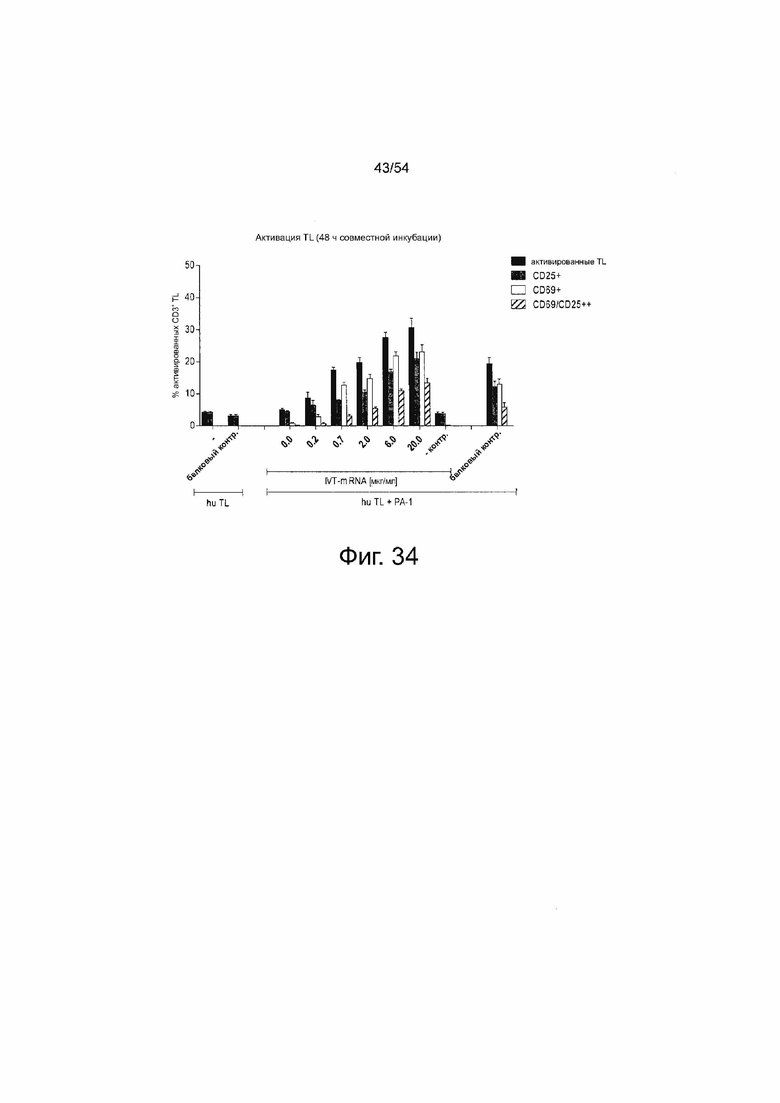
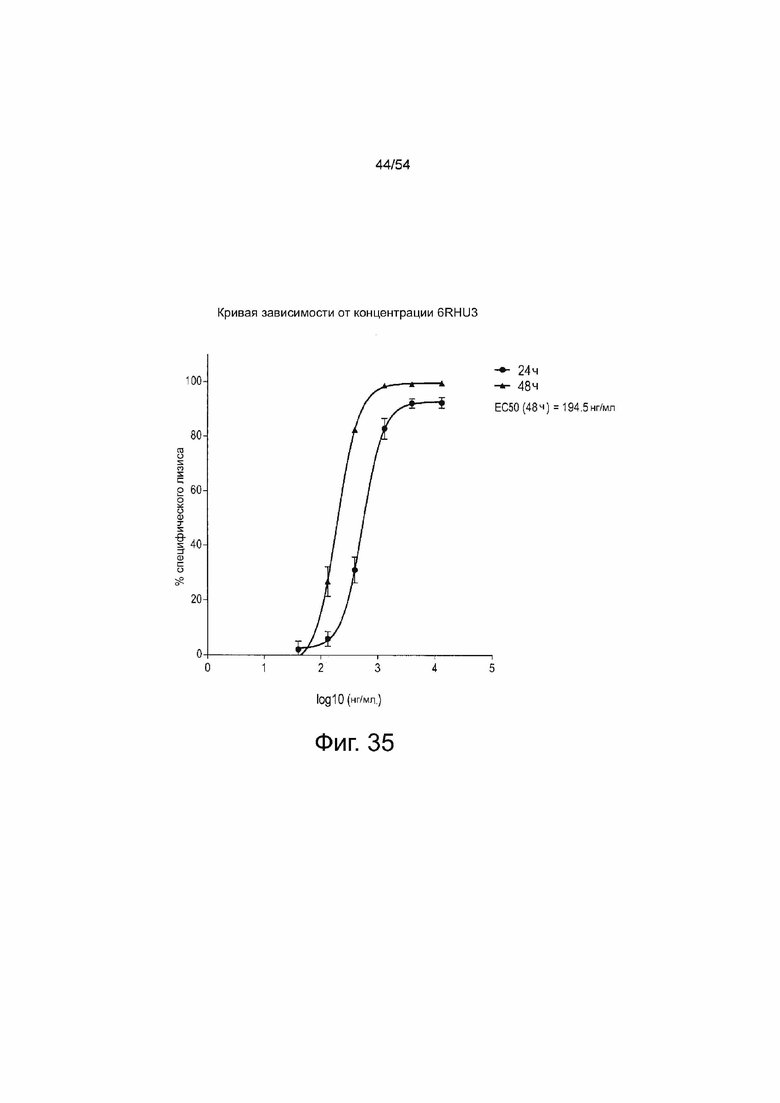
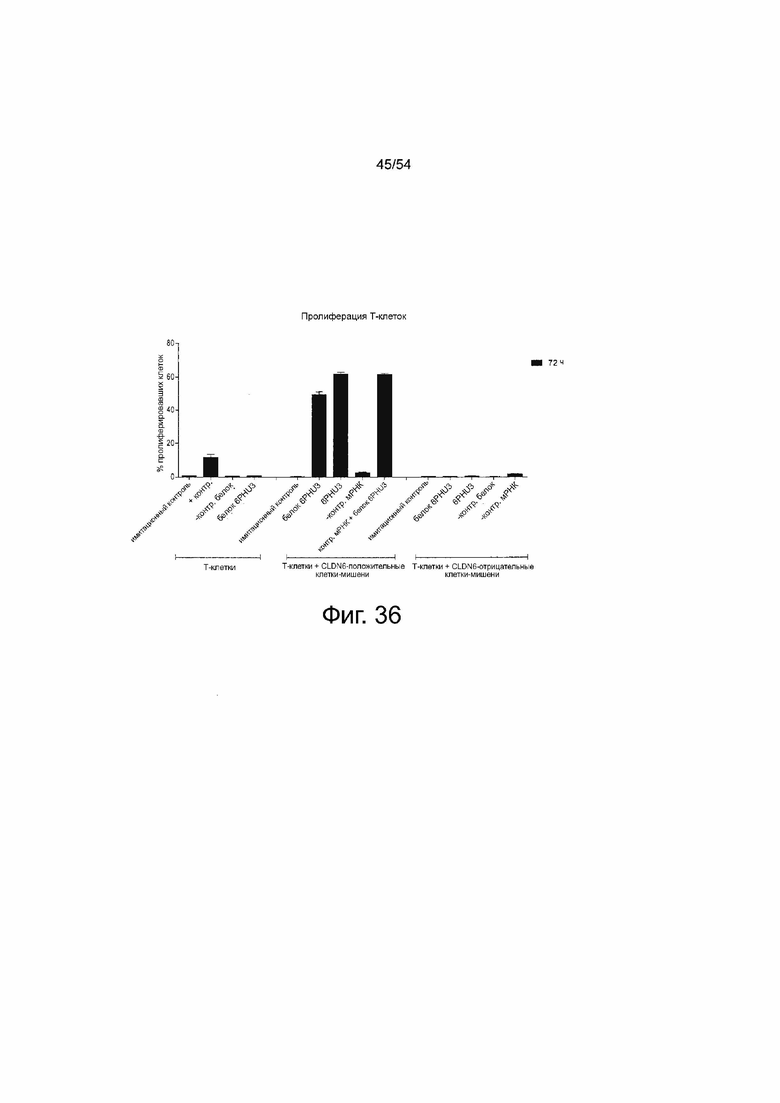
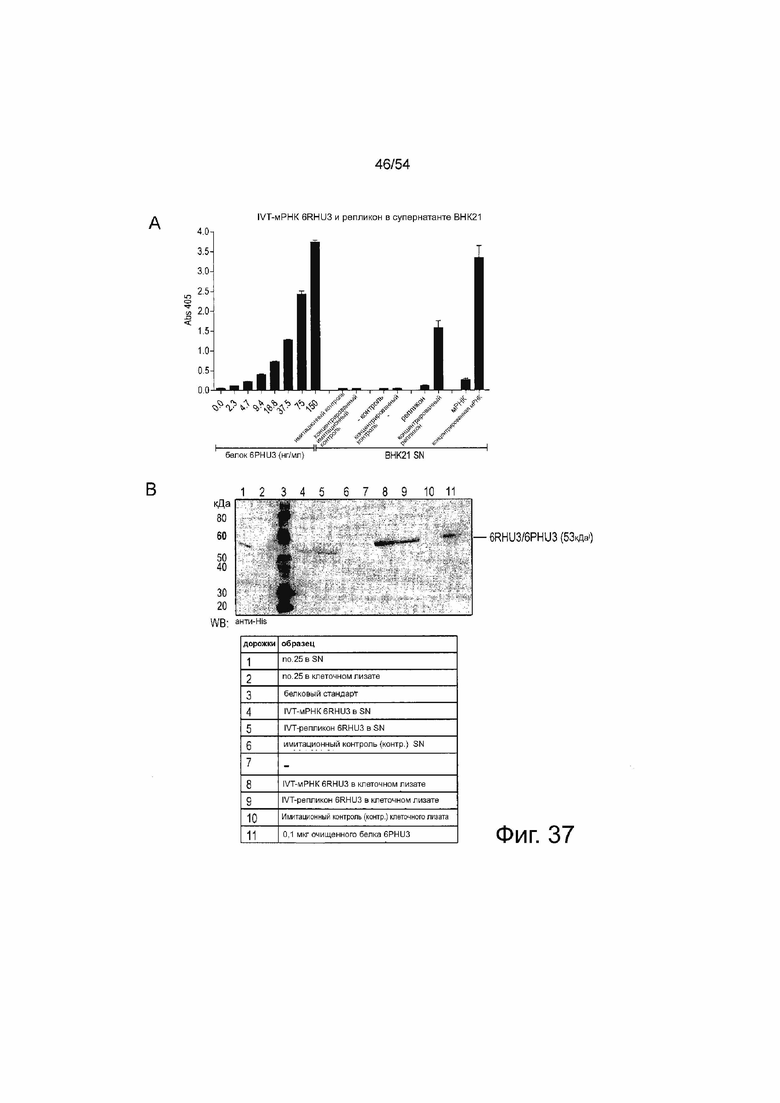

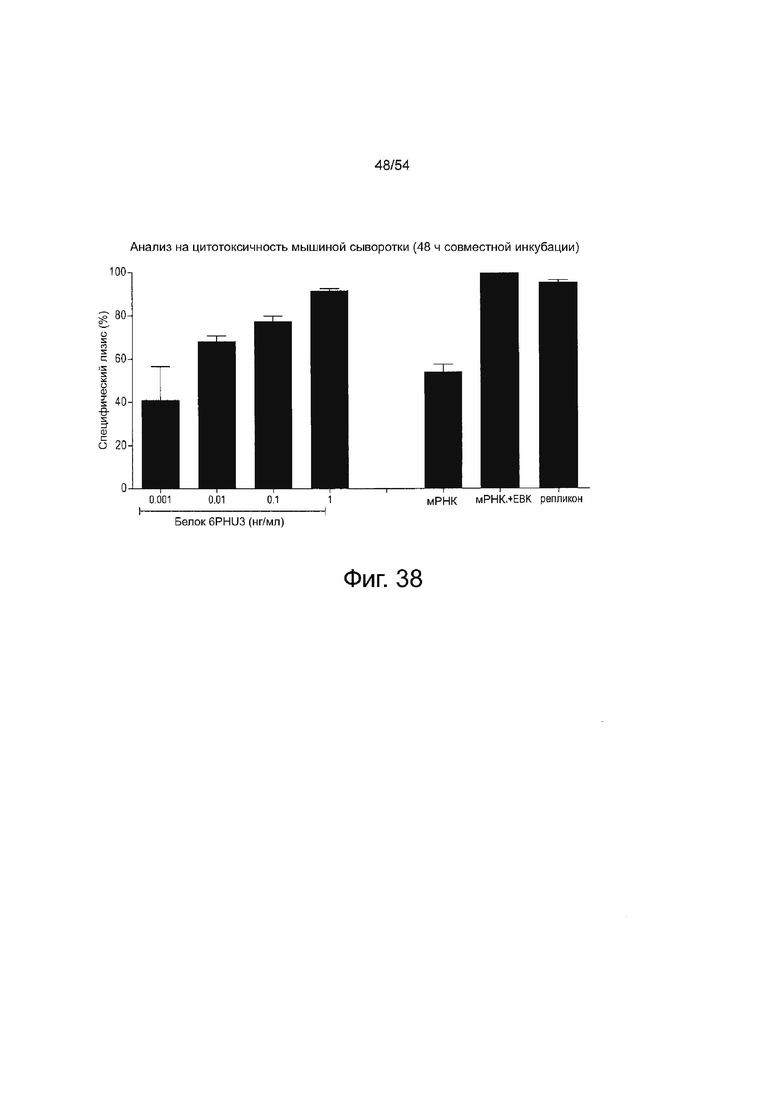


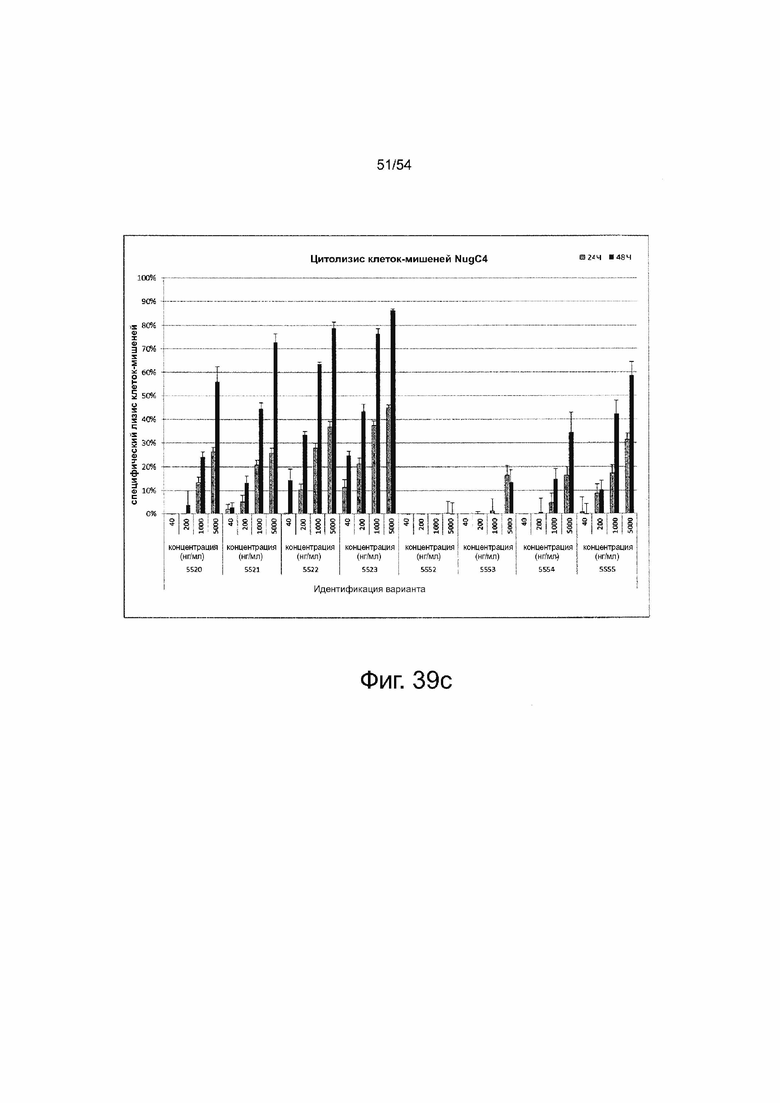

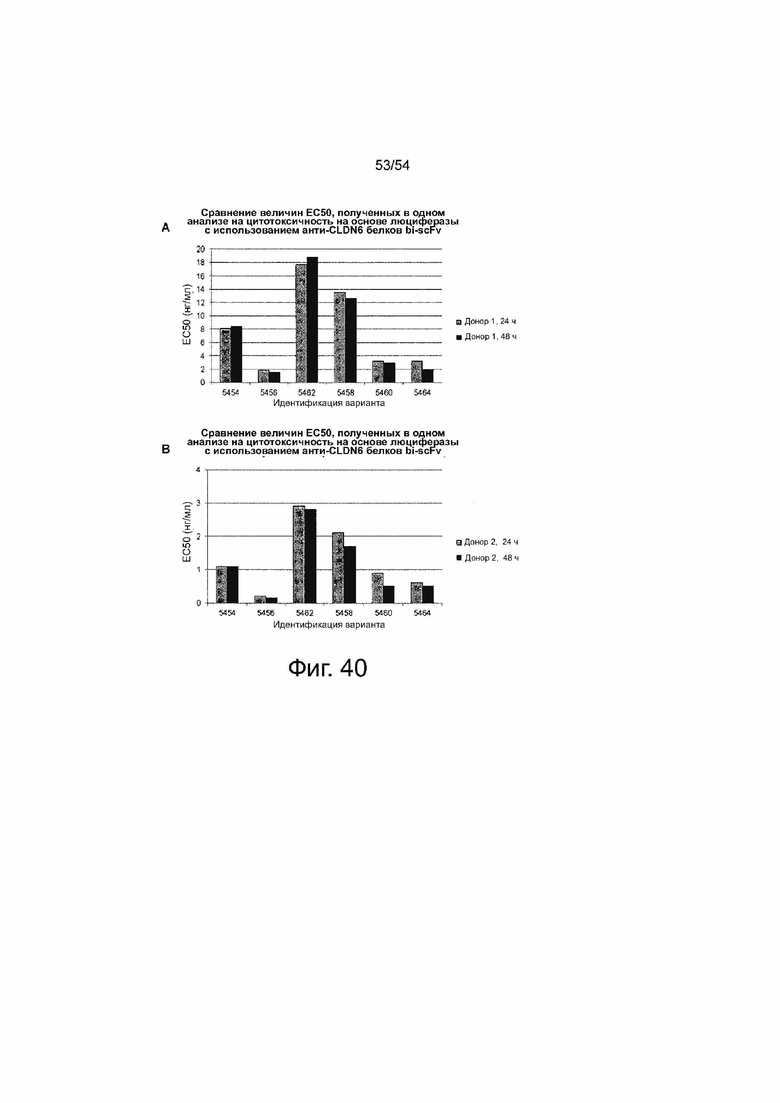
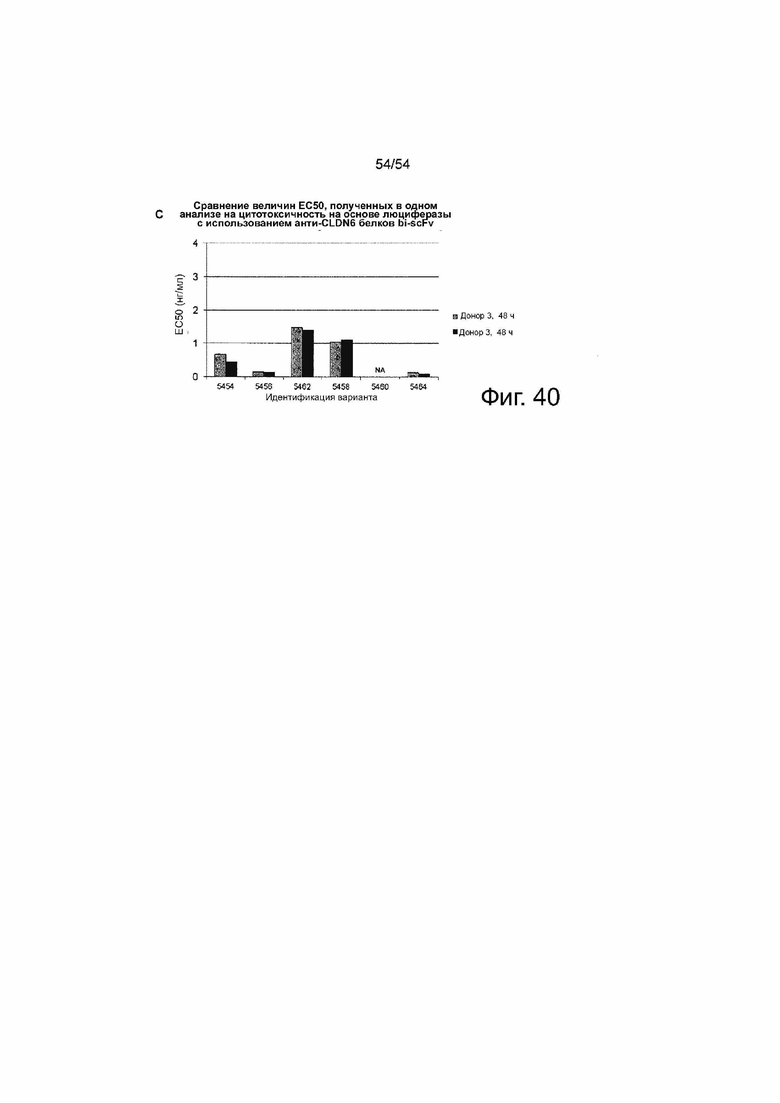
Изобретение относится к области биотехнологии. Описана группа изобретений, включающая биспецифический агент, связывающий клаудин 6 (CLDN6) и CD3, рекомбинантную нуклеиновую кислоту, клетку-хозяин для экспрессии биспецифического агента, применение биспецифического агента, рекомбинантной нуклеиновой кислоты или клетки-хозяина для изготовления фармацевтической композиции для лечения или предупреждения рака, характеризующегося раковыми клетками, экспрессирующими CLDN6, фармацевтическую композицию для лечения или предупреждения рака, характеризующегося раковыми клетками, экспрессирующими CLDN6 и способ лечения или предупреждения ракового заболевания, характеризующегося раковыми клетками, экспрессирующими CLDN6. Изобретение расширяет арсенал средств, связывающихся с клаудином 6 (CLDN6) и CD3. 6 н. и 15 з.п. ф-лы, 54 ил., 12 табл., 34 пр.
1. Биспецифический агент, связывающий клаудин 6 (CLDN6) и CD3, содержащий первый связывающий домен, который связывается с CLDN6, и второй связывающий домен, который связывается с CD3,
где биспецифический агент содержит вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью в отношении антигена клаудина (VH(CLDN6)), вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью в отношении антигена клаудина (VL(CLDN6)), вариабельный домен тяжелой цепи иммуноглобулина (VH) со специфичностью в отношении CD3 (VH(CD3)) и вариабельный домен легкой цепи иммуноглобулина (VL) со специфичностью в отношении CD3 (VL(CD3)), отличающийся тем, что
(i) указанный VH(CLDN6) содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных SEQ ID NO: 20, 22, 24 и 26, или ее фрагмент, или вариант указанной аминокислотной последовательности или указанного фрагмента, где фрагмент содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 указанной VH, а VL(CLDN6) содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных SEQ ID NO: 21, 23, 25 и 27-29 или ее фрагмент, или вариант указанной аминокислотной последовательности или указанного фрагмента, где фрагмент содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 указанной VL, и
(ii) указанный VH(CD3) содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных SEQ ID NO: 30, 32, 34, 36 и 95, или ее фрагмент, или вариант указанной аминокислотной последовательности или указанного фрагмента, где фрагмент содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 указанной VH, и VL(CD3) содержит аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей, представленных SEQ ID NO: 31, 33, 35, 37 и 96, или ее фрагмент, или вариант указанной аминокислотной последовательности или указанного фрагмента, где фрагмент содержит определяющие комплементарность участки CDR1, CDR2 и CDR3 указанной VL,
где вариант включает:
(i) аминокислотную последовательность, которая по меньшей мере на 98% идентична указанной аминокислотной последовательности;
(ii) одну или две консервативные замены аминокислотных остатков.
2. Биспецифический агент по п. 1, где связывающий агент представляет собой биспецифическую молекулу.
3. Биспецифический агент по п. 2, где биспецифическая молекула представляет собой биспецифическое антитело.
4. Биспецифический агент по п. 3, где биспецифическое антитело представляет собой биспецифическое одноцепочечное антитело.
5. Биспецифический агент по любому из пп. 1-4, где связывание указанного биспецифического агента с CD3 на Т-клетках приводит к пролиферации и/или активации указанных Т-клеток, при этом указанные активированные Т-клетки предпочтительно высвобождают цитотоксические факторы, например, перфорины и гранзимы, и инициируют цитолизис и апоптоз раковых клеток.
6. Биспецифический агент по любому из пп. 1-5, где связывающий агент находится в формате полноразмерного антитела, фрагмента антитела, диатела, которое содержит вариабельный домен тяжелой цепи, соединенный с вариабельным доменом легкой цепи на одной и той же полипептидной цепи таким образом, что два домена не образуют пару.
7. Биспецифический агент по п. 6, где диатело содержит две полипептидные цепи, при этом один полипептид содержит VH(CLDN6) и VL(CD3), и другая полипептидная цепь содержит VH(CD3) и VL(CLDN6).
8. Биспецифический агент по любому из пп. 1-6, где биспецифический агент находится в формате биспецифического одноцепочечного антитела, которое состоит из двух молекул scFv, соединенных посредством линкерного пептида.
9. Биспецифический агент по п. 8, где вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) расположены в направлении от N-конца до С-конца в порядке VH(CLDN6)-VL(CLDN6)-VH(CD3)-VL(CD3), VH(CD3)-VL(CD3)-VH(CLDN6)-VL(CLDN6) или VH(CD3)-VL(CD3)-VL(CLDN6)-VH(CLDN6).
10. Биспецифический агент по п. 9, где указанные вариабельные области тяжелой цепи (VH) и соответствующие вариабельные области легкой цепи (VL) соединены посредством длинного пептидного линкера, предпочтительно пептидного линкера, содержащего аминокислотную последовательность, представленную SEQ ID NO: 46, 47 или 104.
11. Биспецифический агент по п. 9 или 10, где указанные две единицы VH-VL или VL-VH scFv соединены посредством короткого пептидного линкера, предпочтительно пептидного линкера, содержащего аминокислотную последовательность, представленную SEQ ID NO: 48 или 49.
12. Биспецифический агент по любому из пп. 1-11, где указанный биспецифический агент содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 42, 43, 44 и 45, или ее фрагмент или вариант.
13. Биспецифический агент по любому из пп. 1-12, где биспецифический агент содержит N-концевой сигнал секреции и/или С-концевую эпитопную гистидиновую метку, предпочтительно эпитопную метку, содержащую шесть гистидиновых остатков.
14. Рекомбинантная нуклеиновая кислота, которая кодирует биспецифический агент по любому из пп. 1-13.
15. Рекомбинантная нуклеиновая кислота по п. 14, которая находится в форме вектора или в форме РНК.
16. Клетка-хозяин для экспрессии биспецифического агента, кодируемого нуклеиновыми кислотами, содержащая рекомбинантную нуклеиновую кислоту по п. 14 или 15.
17. Применение биспецифического агента по любому из пп. 1-13, рекомбинантной нуклеиновой кислоты по п. 14 или 15 или клетки-хозяина по п. 16 для изготовления фармацевтической композиции для лечения или предупреждения рака, характеризующегося раковыми клетками, экспрессирующими CLDN6.
18. Фармацевтическая композиция для лечения или предупреждения рака, характеризующегося раковыми клетками, экспрессирующими CLDN6, содержащая одно из следующего:
i) биспецифический агент по любому из пп. 1-13,
ii) рекомбинантную нуклеиновую кислоту по п. 14 или 15,
iii) рекомбинантные нуклеиновые кислоты, кодирующие биспецифический агент по любому из пп. 1-13,
iv) клетку-хозяина по п. 16 или
v) клетку-хозяина, содержащую рекомбинантные нуклеиновые кислоты, кодирующие биспецифический агент по любому из пп. 1-13.
19. Фармацевтическая композиция по п.18, в которой рекомбинантные нуклеиновые кислоты находятся в форме вектора или в форме РНК.
20. Способ лечения или предупреждения ракового заболевания, характеризующегося раковыми клетками, экспрессирующими CLDN6, включающий введение пациенту фармацевтической композиции по п. 18 или 19.
21. Применение по п. 17, фармацевтическая композиция по п. 18 или 19, или способ по п. 20, в которых указанный рак выбран из группы, состоящей из рака мочевого пузыря; рака яичника, в частности аденокарциномы яичника и теракарциномы яичника; рака легкого, включая мелкоклеточный рак легкого (SCLC) и немелкоклеточный рак легкого (NSCLC), в частности плоскоклеточный рак легкого и аденокарциному; рака желудка; рака молочной железы; рака печени; рака поджелудочной железы; рака кожи, в частности базальноклеточной карциномы и плоскоклеточной карциномы; злокачественной меланомы; рака головы и шеи, в частности злокачественной плеоморфной аденомы; саркомы, в частности синовиальной саркомы и карциносаркомы; рака желчного протока; рака мочевого пузыря, в частности переходно-клеточной карциномы и папиллярного рака; рака почки, в частности почечно-клеточного рака, включая светлоклеточный рак почки и папиллярную карциному почки; рака толстой кишки; рака тонкой кишки, включая рак подвздошной кишки, в частности аденокарциномы тонкой кишки и аденокарциномы подвздошной кишки; эмбриональной карциномы яичка; плацентарной хориокарциномы; рака шейки матки; рака яичка, в частности семиномы яичка, тератомы яичка и эмбрионального рака яичка; рака матки; герминомы, такой как тератокарцинома или эмбриональная карцинома, в частности эмбрионально-клеточной опухоли яичка и их метастатических форм.
| US 8258268 B2 04.09.2012 | |||
| WO 2011057788 A1 19.05.2011 | |||
| МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА К КЛАУДИНУ-18 ДЛЯ ЛЕЧЕНИЯ РАКА | 2006 |
|
RU2445319C2 |
