ОБЛАСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к способам получения молочного продукта и к способам снижения содержания лактозы в молочном продукте с использованием новых пептидов или димерных пептидов, проявляющих активность бета-галактозидазы, с улучшенной активностью при низких температурах.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Для роста молочнокислых бактерий на молоке оптимальным путем получения ими глюкозы и галактозы в качестве источника углерода является гидролиз лактозы. Лактаза (бета-галактозидаза; ЕС 3.2.1.23) представляет собой фермент, который осуществляет стадию гидролиза молочного сахара лактозы до моносахаридов. В коммерческих целях лактазу используют для расщепления лактозы в молочных продуктах. Лица с непереносимостью лактозы испытывают затруднения при переваривании молочных продуктов с высоким содержанием лактозы. По расчетам примерно 70% населения мира обладает ограниченной способностью к перевариванию лактозы. Соответственно, растет потребность в молочных пищевых продуктах, не содержащих или содержащих лишь низкие концентрации лактозы.
Лактазы выделены из широкого ряда организмов, включая такие микроорганизмы, как Kluyveromyces и Bacillus. Распространенным источником грибных лактаз являются дрожжи Kluyveromyces, в частности K. fragilis и K. lactis, и другие грибы, например родов Candida, Torula и Torulopsis, при этом хорошо известными источниками бактериальных лактаз являются В. coagulans и В. circulans. В продаже имеется несколько полученных из этих организмов коммерческих препаратов, таких как Lactozym® (производства компании Novozymes, Дания), HA-Lactase (производства компании Chr. Hansen, Дания) и Maxilact® (производства компании DSM, Нидерланды), все из K. lactis. Все эти лактазы представляют собой так называемые нейтральные лактазы, имеющие оптимум рН от рН 6 до рН 8, а также температурный оптимум примерно 37°С. При применении таких лактаз в производстве, например, йогурта с низким содержанием лактозы обработку ферментом либо необходимо проводить на отдельной стадии, предшествующей ферментации, либо чаще всего приходится использовать высокие дозы ферментов, поскольку их активность будет падать по мере снижения рН в процессе ферментации.
Характерный способ получения пастеризованного молока с пониженным содержанием лактозы включает добавление к молоку фермента лактазы с последующей длительной инкубацией (10-48 ч, часто 24 ч) при температурах примерно 6°С. Поскольку активность Ha-Lactase и NOLA® Fit находится в диапазоне 45-70 мкмоль в минуту на мг фермента, для достижения желаемого содержания остаточной лактозы требуются дозы фермента 55-70 мг/л и 45-60 мг/л соответственно. Ферменты Ha-Lactase и NOLA® Fit имеют оптимум температуры примерно 37°С. Более длительная инкубация молока при 37°С может приводить в результате к росту микроорганизмов
Эти лактазы также не подходят для гидролиза лактозы в молоке, проводимого при высокой температуре, что в некоторых случаях было бы полезно для поддержания микроорганизмов в малом количестве и, следовательно, гарантии высокого качества молока. Кроме того, известные лактазы не подойдут для применения, если желателен процесс производства ультрапастеризованного молока (UHT), в котором ферменты добавляют до обработки UHT.
WO 92/13068 относится к композициям, обладающим активностью лактазы, полученным в результате обработки ультразвуком клеток микроорганизмов - бактерий или дрожжей. WO 2010092057 и WO 0104276 относятся к бета-галактозидазам, активным при низкой температуре. WO 07110619 относится к бета-галактозидазе с высокой активностью транс-галактозилирования, где WO 2009071539 относится к бета-галактозидазе со сниженной активностью транс-галактозилирования.
ЗАДАЧА ИЗОБРЕТЕНИЯ
Задача воплощений изобретения состоит в разработке способов применения бета-галактозидаз, обеспечивающих получение усовершенствованных безлактозных или низколактозных продуктов при низких температурах.
Дополнительная задача воплощений изобретения состоит в разработке способов применения бета-галактозидаз со свойствами, улучшающими снижение содержания лактозы в продукте, таком как безлактозные или низколактозные продукты.
КРАТКОЕ ИЗЛОЖЕНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Автор(-ы) настоящего изобретения идентифицировал(-и) бета-галактозидазы, обладающие прежде не описанными свойствами, обеспечивающими получение усовершенствованных безлактозных или низколактозных продуктов, а также обеспечивающими усовершенствованные способы получения таких безлактозных или низколактозных продуктов. В частности, показано, что эти бета-галактозидазы обладают высокой стабильностью при относительно высокой активности в очень широких диапазонах значений температуры, а также рН. Их также можно применять при определенных температурах, например, при высоких температурах, и значениях рН, обычно не рассматриваемых для этих ферментов. Прежде всего, это обеспечивает возможность использования бета-галактозидаз при определенных значениях рН и температуры, которая прежде не была известна. Это также обеспечивает применение одного и того же конкретного фермента в нескольких различных областях применения с высокой потребностью в промышленности.
В первом аспекте настоящего изобретения предложены способы получения молочного продукта, включающие:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида или димерного пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л, например, от 20 до 55 мг/л;
(б) инкубацию смеси при температуре от 1°С до 10°С в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 0,2 г/л.
В родственном воплощении настоящего изобретения предложены способы снижения содержания лактозы в субстрате на молочной основе, включающие:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида или димерного пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л, например, от 20 до 55 мг/л;
(б) инкубацию смеси при температуре от 1°С до 10°С в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 0,2 г/л.
Описанные выше способы можно осуществлять с помощью пептида или димерного пептида, проявляющего активность бета-галактозидазы, который может быть дополнительно охарактеризован как:
(1) пептид, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 7, 9, 11, 30 и 1, или пептид, имеющий идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(2) пептид, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 13, 19, 7, 9, 11, 26 и 27, 30 и 1, или пептид, имеющий идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(3) пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, или его обладающие ферментативной активностью фрагменты, или аминокислотная последовательность любого из них, имеющая не более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 замен, добавлений или делеций аминокислот.
Преимущество способов по настоящему изобретению состоит в том, что для них требуется лишь низкая концентрация пептида или димерного пептида, проявляющего активность бета-галактозидазы, и при этом они значительно снижают концентрацию лактозы. В предпочтительной альтернативе пептид или димерный пептид, проявляющий активность бета-галактозидазы, добавляют в концентрации от 35 до 52 мг/л, в концентрации от 40 до 52 мг/л или в концентрации от 45 до 52 мг/л.
Субстрат на молочной основе может представлять собой любой субстрат, содержащий молоко. В одном аспекте описанных выше способов используют субстрат на молочной основе, представляющий собой:
(1) коровье молоко, овечье молоко, козье молоко, буйволиное молоко, верблюжье молоко или указанное молоко в пастеризованной и/или фильтрованной форме; или
(2) кисломолочный продукт, полученный из (1) путем ферментации.
В особенно предпочтительном воплощении изобретения в описанных выше способах в качестве субстрата на молочной основе используют коровье молоко, содержащее лактозу в концентрации от примерно 37 до 50 г/л, или это молоко в термически обработанной, пастеризованной, сырой и/или фильтрованной форме.
Описанные выше способы обеспечивают значительное снижение концентрации лактозы за короткий период времени. В некоторых воплощениях изобретения концентрация лактозы снижается до значения менее 0,2 г/л после инкубации в течение по меньшей мере 4 часов, по меньшей мере 8 часов, по меньшей мере 12 часов или по меньшей мере 24 часов.
Одно из преимуществ способов по настоящему изобретению заключается в снижении концентрации лактозы при низких температурах. Например, температура инкубации на стадии (б) описанных выше способов может находиться в диапазоне от 2°С до 7°С или в диапазоне от 3°С до 6°С.
Эти способы обеспечивают значительное снижение концентрации лактозы и предпочтительно при инкубации на стадии (б) концентрация лактозы в смеси снижается до менее 0,05 г/л, до менее 0,02 г/л или до менее 0,01 г/л.
Конкретный пептид или димерный пептид, проявляющий активность бета-галактозидазы, для применения в способах по изобретению обладает высокой активностью не только при низких температурах, но и при высоких температурах. Таким образом, в одном аспекте в изобретении предложен способ, как описано выше, в котором до или после инкубации при температуре от 1°С до 10°С смесь, содержащую субстрат на молочной основе и пептид или димерный пептид, проявляющий активность бета-галактозидазы, нагревают до температуры по меньшей мере 60°С в течение по меньшей мере четырех секунд. В частности, способ может включать в себя стадию нагревания, включающую нагревание до температуры 72°С в течение примерно 15 секунд до или после инкубации смеси при низких температурах на стадии (б) или нагревание до температуры 140°С в течение примерно четырех секунд до или после инкубации смеси при температуре от 1°С до 10°С.
В одной альтернативе способы по настоящему изобретению используют для получения молочного продукта. Эти способы могут дополнительно включать стадию ферментации субстрата на молочной основе молочнокислыми бактериями. Стадию ферментации выполняют до или после инкубации с пептидом или димерным пептидом, проявляющим активность бета-галактозидазы,
Эти способы особенно подходят для получения молочных продуктов, таких как кисломолочный продукт, сыр, йогурт, сливочное масло, молочный спред, пахта, напиток из сквашенного молока, сметана, напиток на основе молочной сыворотки, мороженое, сгущенное молоко, карамелизированное сгущенное молоко или ароматизированный молочный напиток.
В особенно предпочтительном воплощении настоящего изобретения предложены способы получения молока или молочного продукта, включающие:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 35 до 52 мг/л, который имеет аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 7, 9, 11, 30 и 1, или идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(б) инкубацию смеси при температуре от 1°С до 10°С в течение 12 часов в условиях, достаточных для снижения концентрации лактозы в смеси до менее 0,2 г/л.
В дополнительном предпочтительном воплощении настоящего изобретения предложены способы получения молока или молочного продукта, включающие:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 35 до 52 мг/л, который имеет аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 7, 9, 11, 30 и 1, или идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(б) инкубацию смеси при температуре от 1°С до 10°С в течение 12 часов в условиях, достаточных для снижения концентрации лактозы в смеси до менее 0,2 г/л,
где до или после инкубации при температуре от 1°С до 10°С смесь, содержащую субстрат на молочной основе и пептид, проявляющий активность бета-галактозидазы, нагревают до температуры по меньшей мере 60°С в течение по меньшей мере четырех секунд.
В дополнительном воплощении настоящее изобретение относится к применению пептида или димерного пептида, проявляющего активность бета-галактозидазы, для получения молочного продукта с пониженным содержанием лактозы при температуре от 1°С до 10°С в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 0,2 г/л, где пептид или димерный пептид, проявляющий активность бета-галактозидазы, представляет собой:
(1) пептид, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 7, 9, 11, 30 и 1, или пептид, имеющий идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(2) пептид, имеющий аминокислотную последовательность, выбранную из SEQ ID NO: 22, 33, 14, 13, 19, 7, 9, 11, 26 и 27, 30 и 1, или пептид, имеющий идентичность аминокислотной последовательности более 85% с любой из этих последовательностей;
(3) пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33 или его обладающие ферментативной активностью фрагменты, или аминокислотная последовательность любого из них, имеющую не более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 замен, добавлений или делеций аминокислот.
ПОДПИСИ К ФИГУРАМ
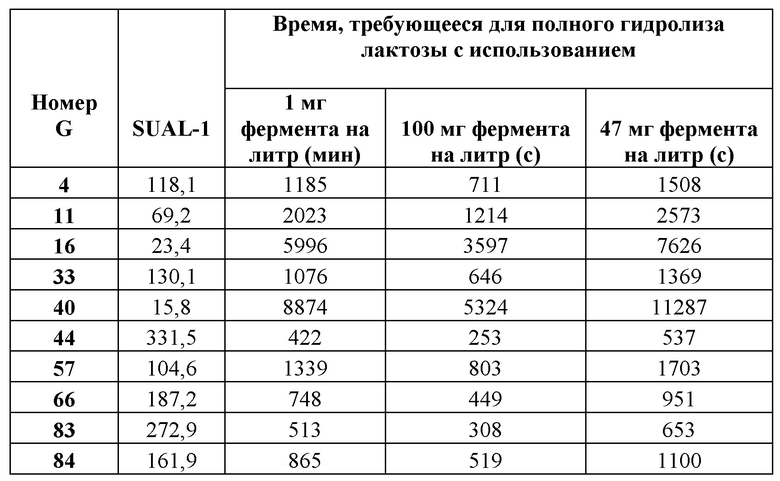
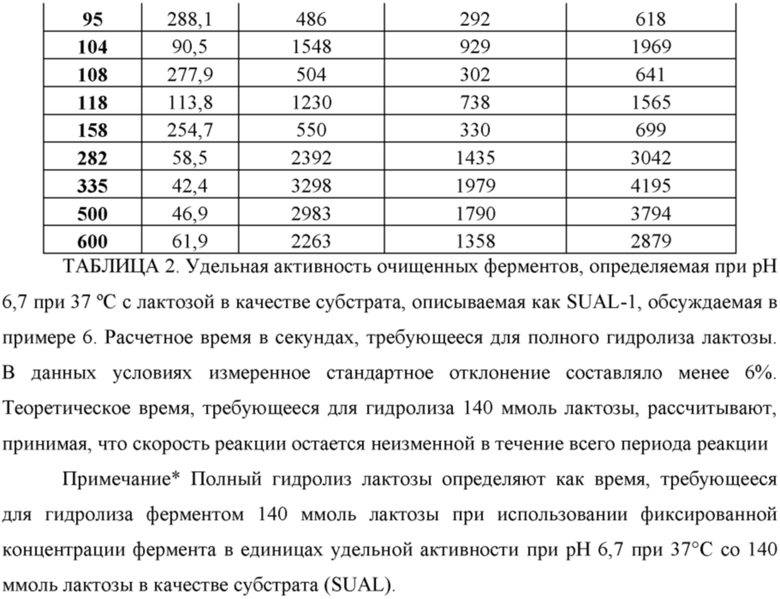
Фиг. 1. Удельная активность очищенных ферментов, определяемая при рН 6,7 при 37°С с лактозой в качестве субстрата, описываемая как SUAL-1, обсуждаемая в примере 6. В данных условиях измеренное стандартное отклонение составляло менее 6%.
Фиг. 2. Удельная активность очищенных ферментов, определяемая при рН 6,7 при 37°С в присутствии галактозы, описываемая как SUAG, обсуждаемая в примере 7. В данных условиях измеренное стандартное отклонение составляло менее 15%.
Фиг. 3. Удельная активность очищенных ферментов, определяемая при рН 6,7 при 4°С с лактозой в качестве субстрата, описываемая как SUAL-2, обсуждаемая в примере 8. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 4. Удельная активность очищенных ферментов, определяемая при рН 6,7 при 43°С с лактозой в качестве субстрата, описываемая как SUAL-3, обсуждаемая в примере 9. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 5. Удельная активность очищенных ферментов, определяемая при рН 5,5 при 4°С с лактозой в качестве субстрата, описываемая как SUAL-4, обсуждаемая в примере 10. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 6. Удельная активность очищенных ферментов, определяемая при рН 5,5 при 37°С с лактозой в качестве субстрата, описываемая как SUAL-5, обсуждаемая в примере 11. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 7. Удельная активность очищенных ферментов, определяемая при рН 5,5 при 43°С с лактозой в качестве субстрата, описываемая как SUAL-6, обсуждаемая в примере 12. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 8. Удельная активность очищенных ферментов, определяемая при рН 4,5 при 4°С с лактозой в качестве субстрата, описываемая как SUAL-7, обсуждаемая в примере 13. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 9. Удельная активность очищенных ферментов, определяемая при рН 4,5 при 37°С с лактозой в качестве субстрата, описываемая как SUAL-8, обсуждаемая в примере 14. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 10. Удельная активность очищенных ферментов, определяемая при рН 4,5 при 43°С с лактозой в качестве субстрата, описываемая как SUAL-9, обсуждаемая в примере 15. В данных условиях измеренное стандартное отклонение составляло менее 5%.
Фиг. 11. Процентное содержание остаточной лактозы в пастеризованном молоке после обработки фиксированным количеством фермента, определенное через 24 ч при 5°С с помощью высокоэффективной жидкостной хроматографии (ВЭЖХ).
Фиг. 12. Процентное содержание остаточной лактозы в ультрапастеризованном (UHT) молоке после обработки фиксированным количеством фермента, определенное через 24 ч при 25°С с помощью ВЭЖХ.
Фиг. 13. Процент остаточной активности очищенных ферментов при повышенных температурах, определенной с использованием лактозы в качестве субстрата. Активность при рН 6,7 при 37°С считали 100%-ной.
Фиг. 14. Процентное содержание остаточной лактозы, присутствующей в пастеризованном молоке после инкубации с ферментами лактазами при различной температуре, 37°С, 55°С или 60°С. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 15. Процентное содержание остаточной лактозы, присутствующей в пастеризованном молоке после инкубации с ферментами лактазами в концентрации 0,047 мг/мл. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг.16. Процентное содержание остаточной лактозы, присутствующей в пастеризованном молоке после инкубации с ферментами лактазами в течение различного времени реакции, в частности 15 или 30 минут. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 17. Процентное содержание остаточной лактозы, присутствующей в пастеризованном молоке после инкубации с ферментами лактазами при различных дозах фермента, в частности 0,047 мг/мл или 0,024 мг/мл. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 18. Процентное содержание остаточной лактозы, присутствующей в пастеризованном молоке после инкубации с ферментами лактазами при использовании различных доз и различного времени реакции. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 19. Процентное содержание остаточной лактозы, присутствующей в фильтрованном молоке после инкубации с ферментами лактазами при 55°С. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 20. Процентное содержание остаточной лактозы, присутствующей в фильтрованном молоке после инкубации с ферментами лактазами при 55°С и при различных дозах фермента. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 21. Процентное содержание остаточной лактозы, присутствующей в фильтрованном молоке после инкубации с ферментами лактазами при 55°С и в течение различного времени реакции. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 22. Кинетика гидролиза лактозы в пастеризованном молоке при 4°С с Ha-Lactase и NOLA® Fit в дозе 50 мг/л. Фермент смешивали с молоком и хранили при 4°С в течение различных интервалов времени. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 23. Процентное содержание остаточной лактозы, измеренное через 12 и 24 ч после добавления ферментов. Фермент смешивали с молоком и хранили при 4°С в течение различных интервалов времени. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 24. Кинетика гидролиза лактозы в пастеризованном молоке при 4°С с новыми лактазами в дозе 0,050 мг/мл. Фермент смешивали с молоком и хранили при 4°С в течение различных интервалов времени. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). NOLA® Fit и Ha-Lactase использовали в качестве контрольных образцов. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 25. Кинетика гидролиза лактозы в пастеризованном молоке при 4°С с отдельными новыми лактазами в дозе 0,050 мг/л. Измеренные значения содержания остаточной лактозы показаны на графике. Фермент смешивали с молоком и хранили при 4°С в течение различных интервалов времени. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). NOLA® Fit и Ha-Lactase использовали в качестве контрольных образцов. Измеренные значения содержания остаточной лактозы показаны на графике. Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 26. Влияние дозы фермента на гидролиз лактозы. Молоко инкубировали с различными дозами ферментов, смешивали и хранили при 4°С в течение 24 ч. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет либо от 0,01% до 0,2%, либо от 0,02% до 1,0% лактозы.
Фиг. 27. Сравнение характеристик ферментов в различных типах молока. Молоко инкубировали с 0,052 мг/л фермента в пастеризованном и фильтрованном молоке, смешивали и хранили при 4°С в течение 24 ч. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Предел обнаружения используемого в анализе набора реактивов LactoSens® составляет от 0,01% до 0,2% лактозы.
Фиг. 28. Измеренная удельная активность очищенных ферментов, определенная при рН 6,7 при различных температурах. Значения удельной активности определяли как количество микромоль (мкмоль) образующейся за минуту глюкозы на один миллиграмм фермента в данных условиях. В данных условиях измеренное стандартное отклонение составляло 5-20%.
Фиг. 29. Измеренная удельная активность очищенных ферментов, определенная при рН 5,5 при различных температурах. Значения удельной активности определяли как количество микромоль (мкмоль) образующейся за минуту глюкозы на один миллиграмм фермента в данных условиях. В данных условиях измеренное стандартное отклонение составляло примерно 5%.
Фиг. 30. Измеренная удельная активность очищенных ферментов, определенная при рН 4,5 при различных температурах. Значения удельной активности определяли как количество микромоль (мкмоль) образующейся за минуту глюкозы на один миллиграмм фермента в данных условиях. В данных условиях измеренное стандартное отклонение составляло примерно 5%.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Авторы настоящего изобретения обнаружили, что некоторые пептиды и димерные пептиды, проявляющие ферментативную активность бета-галактозидазы, неожиданно оказались стабильными при различных физических условиях и обладают относительно высокой активностью за пределами диапазонов, которые обычно рассматривают как оптимальные для этого класса ферментов.
Соответственно, авторами настоящего изобретения были идентифицированы ферменты, обладающие относительно высокой активностью при температуре примерно 4°С или 5°С, и поэтому их можно применять для гидролиза лактозы в производстве, например, свежего молока. Таким образом, эти новые ферменты особенно подходят для снижения содержания лактозы в продуктах на основе молока, таких как молочные продукты, при низких температурах.
Дополнительное преимущество этих новых улучшенных пептидов, проявляющих ферментативную активность бета-галактозидазы, состоит в том, что они характеризуются относительно низкой степенью ингибирования галактозой. Пониженное ингибирование галактозой этих новых ферментов в высокой степени актуально для областей применения, где желательны очень низкие концентрации лактозы.
Что касается применимости для кисломолочных продуктов, значительное преимущество состоит в том, что описанные в настоящем документе ферменты обладают высокой ферментативной активностью бета-галактозидазы в относительно широком температурном диапазоне от 4°С до 43°С, например, примерно 37°С, где ферментация в норме должна быть оптимальной, но эта ферментативная активность бета-галактозидазы также присутствует при низком рН, например, пониженном до 4,5, или пониженном до 4,0, или пониженном до 3,5, или даже пониженном до рН 3.
В итоге, авторами настоящего изобретения обнаружено, что некоторые пептиды, проявляющие ферментативную активность бета-галактозидазы, активны в широком диапазоне температуры, активны в широком диапазоне рН, обладают общей высокой гидролитической активностью без побочных активностей, что ингибирование этих пептидов галактозой отсутствует или происходит незначительно, например, менее чем на 60%, и что они стабильны в течение длительного срока хранения.
Активность бета-галактозидазы можно определить путем измерения количества высвобождаемой глюкозы после инкубации с лактозой в заданных условиях. Высвобождаемую глюкозу можно обнаружить с помощью цветной реакции.
Определения
Используемый в настоящем документе и в контексте настоящего изобретения термин «молоко» следует понимать как выделяемое молоко, полученное в результате доения любого млекопитающего, такого как корова, овца, коза, буйвол или верблюд.
Используемый в настоящем документе термин «содержащая лактозу композиция» относится к любой композиции, такой как любая жидкость, содержащая лактозу в значительной измеримой степени, например, с содержанием лактозы выше 0,002% (0,002 г/100 мл). Этот термин охватывает молоко и субстраты на молочной основе.
В контексте настоящего изобретения термин «субстрат на молочной основе» может означать любой материал из сырого и/или обработанного молока. Полезные субстраты на молочной основе включают, но не ограничены ими, растворы/суспензии любого молока или подобных молоку продуктов, содержащих лактозу, такие как цельное молоко или молоко низкой жирности, обезжиренное молоко, пахта, низколактозное молоко, восстановленное сухое молоко, сгущенное молоко, растворы сухого молока, молоко UHT, молочная сыворотка, пермеат молочной сыворотки, кислая молочная сыворотка, сливки, кисломолочные продукты, такие как йогурт, сыр, диетическая добавка и пробиотические диетические продукты. Как правило, термин «субстрат на молочной основе» относится к материалу из сырого и/или обработанного молока, который обрабатывают дополнительно в целях получения молочного продукта.
Используемый в настоящем документе термин «пастеризация» относится к процессу снижения или устранения в субстрате на молочной основе присутствия живых организмов, таких как микроорганизмы. Предпочтительно пастеризация достигается путем поддержания определенной температуры в течение определенного периода времени. Определенная температура обычно достигается путем нагревания. Температура и продолжительность времени может быть выбрана так, чтобы уничтожить или инактивировать некоторые бактерии, такие как вредные бактерии, и/или инактивировать ферменты в молоке. За этим может следовать стадия быстрого охлаждения.
Используемый в настоящем документе термин «молочный продукт» может означать любой пищевой продукт, один из основных компонентов которого основан на молоке. Обычно основной компонент основан на молоке, и в некоторых воплощениях изобретения основной компонент представляет собой субстрат на молочной основе, обработанный в соответствии со способом по настоящему изобретению ферментом, обладающим активностью бета-галактозидазы.
Молочный продукт в соответствии с изобретением может представлять собой, например, обезжиренное молоко, молоко низкой жирности, цельное молоко, сливки, молоко UHT, молоко с длительным сроком хранения, кисломолочный продукт, сыр, йогурт, сливочное масло, молочный спред, пахту, кисломолочный напиток, сметану, напиток на основе молочной сыворотки, мороженое, сгущенное молоко, карамелизованное сгущенное молоко или ароматизированный молочный напиток.
Молочный продукт может дополнительно содержать отличные от молока компоненты, например растительные компоненты, такие как, например, растительное масло, растительный белок и/или растительные углеводы. Молочные продукты могут также содержать дополнительные добавки, такие как, например, ферменты, вкусоароматические добавки, культуры микроорганизмов, такие как пробиотические культуры, соли, подсластители, сахара, кислоты, фрукты, приготовленные фрукты, фруктовые соки или любой другой известный в данной области техники компонент в качестве компонента или добавки молочного продукта.
Используемые в настоящем документе термины «кисломолочный продукт» или «продукт из сквашенного молока» следует понимать как любой молочный продукт, в состав технологического процесса которого входит ферментация любого типа. Примерами кисломолочных продуктов являются такие продукты, как йогурт, пахта, крем-фреш, мягкий творог и свежеприготовленный сыр. Кисломолочный продукт может быть получен любым известным в данной области техники способом или способом, включающим его стадии.
Используемый в настоящем документе термин «ферментация» относится к конверсии углеводов в спирты или кислоты под действием микроорганизмов. В некоторых воплощениях изобретения ферментация в соответствии с настоящим изобретением включает конверсию лактозы в молочную кислоту. В контексте настоящего изобретения «микроорганизм» может включать любую бактерию или гриб, способные ферментировать молочный субстрат.
Используемый в настоящем документе термин «повышенная ферментативная активность бета-галактозидазы» относится к относительно повышенной активности фермента бета-галактозидазы по сравнению с эталонной последовательностью.
Используемый в настоящем документе термин «пептид, проявляющий повышенную ферментативную активность бета-галактозидазы» относится к любому пептиду, который обладает ферментативной активностью катализа гидролиза дисахарида лактозы до его моносахаридных компонентов глюкозы и галактозы. Этот пептид может также называться лактазой или просто бета-галактозидазой (ЕС: 3.2.1.23).
В предпочтительном воплощении изобретения активность бета-галактозидазы определяют путем инкубации 13 мкл раствора, содержащего известное количество очищенного фермента лактазы, с раствором, содержащим 140 мМ лактозы, при рН 6,7 и 37°С в течение 10 мин, завершения реакции с лактазой путем повышения температуры до 95°С в течение 10 мин. Количество образовавшейся глюкозы определяли путем инкубации продукта реакции при 30°С в течение 40 мин с 80 мкл раствора глюкозооксидазы (0,6 г/л), диаммониевой соли 2,2'-азино-бис(3-этилбензолтиазолин-6-сульфокислоты) (1,0 г/л ABTS) и пероксидазы хрена (0,02 г/л) и определения оптической плотности при 610 нм с помощью фотометра FLUO.
Оптическая плотность коррелирует с концентрацией образующейся за минуту глюкозы, и ее определенное максимальное значение (в мкмоль образовавшейся глюкозы/мин) определяют как единицу активности лактазы 1 (также обозначена в настоящем документе как UAL-1; Unit of Lactase Activity). Удельную активность лактазы (также обозначена в настоящем документе как SUAL-1; Specific Activity of Lactase) при рН 6,7 при 37°С определяют в мкмоль образовавшейся глюкозы/мин/мг фермента и рассчитывают путем деления UAL-1 на концентрацию белка лактазы в мг. Полное подробное описание предпочтительной альтернативы выполнения этого анализа проиллюстрировано в примере 6.
Хотя стандартным подходом к определению активности является характеристика активности бета-галактозидазы по соотнесению со значениями в единицах, представляющих собой мкмоль образовавшейся глюкозы/мин/мг фермента, для характеристики активности ферментов лактаз с использованием описанного выше теста можно в равной степени использовать другие единицы. Соответственно, в некоторых из примеров активность фермента лактазы характеризуют по соотнесению с мкмоль образующейся за секунду глюкозы на мкмоль фермента.
В альтернативных воплощениях изобретения этот анализ можно выполнять с использованием различных значений температуры или различных значений рН для инкубации с лактазой.
Используемые в контексте настоящей заявки термины «пептид» и «олигопептид», как общепризнанно, считают синонимами, и каждый термин можно использовать взаимозаменяемо в зависимости от требований контекста как указывающий на цепь из по меньшей мере двух аминокислот, связанных пептидильными связями. Слово «полипептид» используют в настоящем документе для цепей, содержащих более десяти аминокислотных остатков. Все приведенные в настоящем документе формулы или последовательности пептида и полипептида написаны слева направо и в направлении от амино-конца к карбокси-концу. Используемый в настоящем документе термин «белки» относится к пептидным последовательностям, продуцируемым некоторым организмом-хозяином, и могут включать посттрансляционные модификации, такие как присоединенные гликаны.
Используемые в настоящем документе термины «аминокислота» или «аминокислотная последовательность» относятся к последовательности олигопептида, пептида, полипептида или белка или к любому ее фрагменту и к природным или синтетическим молекулам. В данном контексте «фрагмент» относится к фрагментам пептида, проявляющим ферментативную активность бета-галактозидазы, которые сохраняют некоторую ферментативную активность. Если указанная в настоящем документе «аминокислотная последовательность» относится к аминокислотной последовательности встречающейся в природе молекулы белка, «аминокислотная последовательность» и другие подобные термины не подразумевают как ограничивающие аминокислотную последовательность полноразмерной нативной аминокислотной последовательностью, связанной с молекулой указанного пептида.
Примеры пептидов по изобретению также включают фрагменты длиной по меньшей мере примерно 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800 или более остатков, либо превышающей длину полноразмерного фермента. Соответственно «пептидный фрагмент» или «ферментативно-активный фрагмент» по изобретению представляют собой фрагменты, сохраняющие по меньшей мере некоторую функциональную ферментативную активность. Как правило, пептидный фрагмент по изобретению будет по-прежнему содержать функциональный каталитический домен или другие существенные активные центры пептида, проявляющего ферментативную активность бета-галактозидазы. Другие домены могут быть делетированы.
Как правило, удельная ферментативная активность бета-галактозидазы будет измерена и указана в мкмоль образовавшейся глюкозы/мин/мг используемого фермента. Однако это конкретное значение будет изменяться в зависимости от применяемых условий, таких как температура и рН. Соответственно, значения ферментативной активности бета-галактозидазы могут быть также указаны как относительные по отношению к известному эталонному ферменту, такому как фермент бета-галактозидаза, определенный SEQ ID NO: 34 или SEQ ID NO: 35.
Если не указано иное, используемый в настоящем документе термин «идентичность последовательности» для аминокислот относится к идентичности последовательности, рассчитываемой как (nref - ndif)⋅100/nref, где ndif представляет собой общее число неидентичных остатков в двух последовательностях при выравнивании, и где nref представляет собой число остатков в одной из последовательностей.
В некоторых воплощениях изобретения идентичность последовательности определяют традиционными способами, например, Smith and Waterman, 1981, Adv. Appl. Math. 2:482, путем поиска с применением метода подобия Pearson & Lipman, 1988, Proc. Natl. Acad. Sci. USA 85:2444, применения алгоритма CLUSTAL W Thompson et al., 1994, Nucleic Acids Res 22:467380, с помощью компьютерных приложений этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в пакете программ Wisconsin Genetics Software Package, Genetics Computer Group). Можно также применять алгоритм BLAST (Altschul et al., 1990, Mol. Biol. Biol. 215: 403-10), для которого можно получить программное обеспечение через Национальный центр биотехнологической информации (National Center for Biotechnology Information, www.ncbi.nlm.nih.gov/). При использовании любого из упомянутых выше алгоритмов для «окна» длины, штрафа на гэп и т.д. используют параметры по умолчанию.
Пептид с определенной аминокислотной последовательностью, как описано в настоящем документе, может отличаться от последовательности эталонного пептида заменой, добавлением/вставкой или делецией любой из аминокислот.
Некоторые воплощения в соответствии с настоящим изобретением относятся к применению пептида с аминокислотной последовательностью, представленной SEQ ID NO: 1-33, или последовательностью по меньшей мере с 80%-ной идентичностью любой из указанных последовательностей. В некоторых воплощениях изобретения эта идентичность последовательности может составлять по меньшей мере приблизительно 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, например, пептид, имеющий не более 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21 или 22 замен, добавлений или делеций по сравнению с любой одной эталонной аминокислотной последовательностью, представленной SEQ ID NO: 1-33. Признаками изобретения также являются биологически активные фрагменты пептидов в соответствии с изобретением. Биологически активные фрагменты пептида по изобретению включают пептиды, содержащие аминокислотные последовательности, по существу идентичные аминокислотной последовательности пептида по изобретению или полученные из нее, которые включают в себя меньше аминокислот, чем полноразмерный белок, но проявляют существенную часть биологической активности соответствующего полноразмерного пептида. Как правило, биологически активные фрагменты содержат домен или мотив по меньшей мере с одной активностью варианта белка по изобретению. Биологически активный фрагмент пептида по изобретению может представлять собой пептид, длина которого составляет, например, 100, 200, 300, 400, 500, 600, 700, 800, 900, 1000 или более аминокислот.
Используемый в настоящем документе термин «клетка-хозяин» включает клетку любого типа, подверженную трансформации, трансфекции, трансдукции и т.п. нуклеиново-кислотной конструкцией или экспрессионным вектором, которые содержат полинуклеотид, кодирующий пептиды по настоящему изобретению. Клетка-хозяин может представлять собой клетку любого типа, из которой получают конкретный фермент, или альтернативно клетку такого типа, который склонен к продуцированию конкретного фермента. Этот термин включает и штаммы дикого типа, и аттенуированные штаммы.
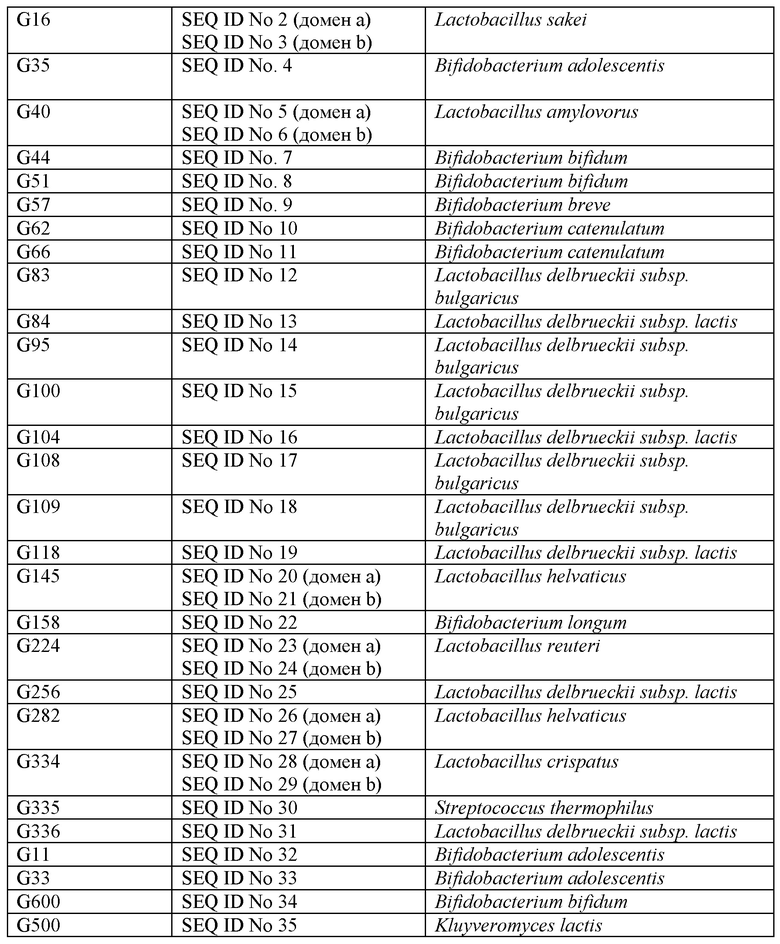
Подходящей клеткой-хозяином может быть любая бактерия, включающая молочнокислые бактерии в пределах порядка Lactobacillales, который включает Lactococcus spp., Streptococcus spp., Lactobacillus spp., Leuconostoc spp., Pseudoleuconostoc spp., Pediococcus spp., Brevibacterium spp., Enterococcus spp. и Propionibacterium spp. Он также включает продуцирующие молочную кислоту бактерии, относящиеся к группе анаэробных бактерий, бифидобактерий, т.е. Bifidobacterium spp., часто используемых в качестве пищевых культур отдельно или в комбинации с молочнокислыми бактериями. В это определение также включают Lactococcus lactis, Lactococcus lactis subsp. cremoris, Leuconostoc mesenteroides subsp. cremoris, Pseudoleuconostoc mesenteroides subsp. cremoris, Pediococcus pentosaceus, Lactococcus lactis subsp. lactis biovar. diacetylactis, Lactobacillus casei subsp. casei и Lactobacillus paracasei subsp. Paracasei и виды термофильных молочнокислых бактерий, включающие в качестве примеров Streptococcus thermophilus, Enterococcus faecium, Lactobacillus delbrueckii subsp. lactis, Lactobacillus helveticus, Lactobacillus delbrueckii subsp. bulgaricus и Lactobacillus acidophilus. Другие конкретные бактерии в пределах этого определения включают бактерии семейства Bifidobacteriaceae, например из рода Bifidobacterium, например штаммы Bifidobacterium animalis или Bifidobacterium longum, Bifidobacterium adolescentis, Bifidobacterium bifodum, Bifidobacterium breve, Bifidobacterium catenulatum, Bifidobacterium infantus, или из рода Lactobacillus, например L. sakei, L. amylovorus, L. delbrueckii subsp. Lactis, и L. helveticus.
В это определение клеток-хозяев также включают штамм Agaricus, например А. bisporus; Ascovaginospora; Aspergillus, например A. niger, A. awamori, A. foetidus, А. japonicus, A. oryzae; Candida; Chaetomium; Chaetotomastia; Dictyostelium, например D. discoideum; Kluveromyces, например K. fragilis, K. lactis; Mucor, например M. javanicus, M. mucedo, M. subtilissimus; Neurospora, например N. crassa; Rhizomucor, например R. pusillus; Rhizopus, например R arrhizus, R. japonicus, R. stolonifer; Sclerotinia, например S. libertiana; Torula; Torulopsis; Trichophyton, например Т. rubrum; Whetzelinia, например W. sclerotiorum; Bacillus, например В. coagulans, В. circulans, В. megaterium, В. novalis, В. subtilis, В. pumilus, В. stearothermophilus, В. thuringiensis; Bifidobacterium, например В. Iongum, В. bifidum, В. animalis; Chryseobacterium; Citrobacter, например С. freundii; Clostridium, например С perfringens; Diplodia, например D. gossypina; Enterobacter, например E. aerogenes, E. cloacae Edwardsiella, E. tarda; Erwinia, например E. herbicola; Escherichia, например E. coli; Klebsiella, например K. pneumoniae; Miriococcum; Myrothesium; Mucor; Neurospora, например N. crassa; Proteus, например P. vulgaris; Providencia, например P. stuartii; Pycnoporus, например Pycnoporus cinnabarinus, Pycnoporus sanguineus; Ruminococcus, например R torques; Salmonella, например S. typhimurium; Serratia, например S. liquefasciens, S. marcescens; Shigella, например S. flexneri; Streptomyces, например S. antibioticus, S. castaneoglobisporus, S. violeceoruber; Trametes; Trichoderma, например Т. reesei, Т. viride; Yersinia, например Y. enterocolitica.
Для получения безлактозного пастеризованного молока (содержание остаточной лактозы менее 0,01%) при низких температурах (4-5°С) за 24 ч рекомендуемая доза Ha-Lactase и NOLA® составляет 55-70 мг/л (10000 NLU/л) и 45-60 мг/л (10000 BLU/л) соответственно. Ферменты по настоящему изобретению обеспечивают очень низкие остаточные концентрации лактозы при низких температурах (от менее 0,01% до 0,2%). Измерения удельной активности показывают, что новые ферменты обладают в 2-5 раз более высокой активностью по сравнение с Ha-Lactase и NOLA® Fit, поэтому для получения безлактозного молока требуется меньше времени.
В приведенных ниже примерах показано, что новые лактазы действуют быстрее, чем Ha-Lactase и NOLA® Fit, и в результате безлактозное пастеризованное молоко получают за значительно более короткое время. Эти новые ферменты могут уменьшить общее время технологического процесса. Дополнительно с помощью новых ферментов можно дополнительно уменьшить дозу фермента для получения безлактозного/низколактозного пастеризованного молока до 25-50%.

ПРИМЕРЫ
Общие материалы и методы
Молекулярное клонирование и генетические методики
Методики расщепления ферментами рестрикции, лигирования, трансформации и другие стандартные молекулярно-биологические манипуляции были основаны на методах, описанных в литературе (Maniatis et al. "Molecular cloning: a laboratory manual, 2nd edition" Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY, 1989; Sambrook and Russell "Molecular Cloning: A Laboratory Manual, 3rd edition" Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY 2001; Miller "Experiment in molecular genetics" Cold Spring Harbor Laboratory Press, 1972); или предложенных производителем. ПЦР проводили в термоциклере ДНК производства компании Bio-Rad, США. Секвенирование ДНК было выполнено компанией LGC, г. Берлин, Германия. Белки анализировали с помощью электрофореза в полиакриламидном геле (ПААГ-электрофореза) в денатурирующих условиях, используя додецилсульфат натрия, в гелях, содержащих 10% ДСН (гель Mini-PROTEAN® TGX stain-free™, Biorad, США). Концентрации белка определяли, используя метод с бицинхониновой кислотой (ВСА), следуя протоколу, поставляемому с набором реактивов.
Штаммы бактерий, плазмиды и условия выращивания
Для клонирования и выделения плазмид использовали штамм Escherichia coli ТОР10 (Invitrogen). Для получения рекомбинантного белка использовали штамм Е. coli BW25113 с дефицитом бета-галактозидазы (Δ(araD-araB)567, ΔlacZ4787(::rrnB-3), λ-, rph-1, Δ(rhaD-rhaB)568, hsdR514) (Datsenko KA, Wanner BL; 2000, Proc Natl Acad Sci U.S.A. 97: 6640-6645) в комбинации с вектором pBAD/His, полученным от компании Invitrogen™ Life Technologies Corporation Europe BV.
Питательная среда для выращивания для экспрессии белка
Для продукции рекомбинантного белка использовали питательную среду 2xPY, содержащую 16 г/л пептона BD BBL™ Phyton ТМ, 10 г/л дрожжевого экстракта, 5 г/л NaCl. Среда для роста содержала добавку ампициллина (100 мкг/мл) для поддержания плазмиды. Продукцию белка инициировали добавлением в культуральную среду 0,05% арабинозы.
Пример 1. Конструирование экспрессионного вектора для продукции лактаз
Геномную ДНК молочнокислых бактерий или бифидобактерий выделяли с использованием коммерческого набора реактивов для выделения геномной ДНК, следуя протоколу поставщика (DNeasy, Qiagen, Германия). Ген лактазы амплифицировали с помощью полимеразной цепной реакции (ПЦР) с использованием двух синтетических праймеров, очищенной геномной ДНК в качестве источника биомассы и реактивы, поставляемые в наборе с ДНК-полимеразой «горячего старта» Phusion U Hot start (Thermo Scientific, США). Ген лактазы клонировали в стартовом кодоне экспрессионного вектора pBAD/His с использованием метода клонирования USER (Nour-Eldin НН, Geu-Flores F, Halkier BA, Plant Secondary Metabolism Engineering, Methods in Molecular Biology, 643; 2010) с получением в результате экспрессионной конструкции. При использовании метода USER и в продукте ПЦР, и в векторе доставки образуются длинные комплементарные выступающие концы. Эти выступающие концы можно подвергать отжигу друг с другом с образованием стабильного продукта гибридизации, который используют для трансформации Е. coli без лигирования. Для создания выступающих концов в продукте ПЦР в расположенный выше участок каждого праймера для амплификации целевой ДНК включали единственный остаток дезоксиуридина. Ген лактазы амплифицировали с использованием прямого праймера (5'-ATTAACCAUGCGACGCAACTTCGAATGGCC-3') и обратного праймера (ATCTTCTCUTTACCGCCTTACCACGAGCACG), содержащих уридин в 9-ом положении (показано полужирным шрифтом), за которым следовала последовательность гена лактазы. Параллельно векторную ДНК амплифицировали с помощью ПЦР, используя пару прямого (5'-AGAGAAGAUTTTCAGCCTGATACAGATTAAATC-3') и обратного праймера (5'-ATGGTTAAUTCCTCCTGTTAGCCCAAAAAACGG-3'), содержащих единственный остаток дезоксиурацила в 9-ом положении (выделено жирным шрифтом), за которым следовала последовательность векторной ДНК. Продукты ПЦР очищали с помощью коммерческого набора для очистки продуктов ПЦР (Qiagen, Дания). Очищенные продукты ПЦР (ген лактазы и векторную ДНК) смешивали в эквимолярном количестве и инкубировали с коммерческой смесью ферментов USER (New England Biolabs, США), следуя протоколу поставщика. Эти ферменты удаляют остаток урацила, а также расположенный выше уридина короткий фрагмент, в результате чего в продуктах ПЦР образуются комплементарные выступающие концы. Эти комплементарные выступающие концы претерпевают отжиг друг с другом с получением в результате экспрессионного вектора лактазы pBAD. Аликвоты этой лигированной смеси трансформировали в полученные химическим способом компетентные клетки Е. coli ТОР 10. Трансформанты отбирали при 37°С на чашках Петри со средой LB-Amp (LB; среда Лурия - Бертани, Amp; 100 мкг/мл ампициллина). На следующий день проводили ПЦР отдельных колоний, используя небольшую биомассу выросших за ночь трансформантов, используя праймеры к вектору (праймер 1; 5'-CGGCGTCACACTTTGCTATGCC-3' и праймер 2; 5'-CCGCGCTACTGCCGCCAGGC-3'). Положительные клоны в ПЦР отдельных колоний культивировали в 5 мл среды LB-Amp и выделяли из клеток плазмидную ДНК. Клонированный ген лактазы секвенировали, чтобы подтвердить, что в процессе амплификации гена не было внесено дополнительных мутаций. Плазмидной ДНК трансформировали экспрессионный штамм-хозяин BW25113.
Пример 2. Экспрессия лактаз в экспрессионном штамме-хозяине Е. coli
Фермент лактазу продуцировали в Е. coli BW25113 с использованием экспрессионной системы pBAD. Свежетрансформированные клетки Е. coli BW25113, несущие плазмидную ДНК, собирали с чашки LB-Amp с помощью стерильной петли и использовали для инокуляции 5 мл среды LB-Amp. Выращенную ночную культуру (200 мкл) использовали для инокуляции 50 мл среды 2х PY, содержащей 100 мкг/мл ампициллина, в колбу емкостью 250 мл и помещали в устройство для встряхивания (Innova® 42). Культуру выращивали при 37°С при 220 об/мин до достижения оптической плотности при 600 нм (OD600) 0,6-0,8. Экспрессию лактазы инициировали добавлением в культуральную среду 0,05% арабинозы, и клетки культивировали дополнительно в течение 16-20 часов при 18°С при 180 об/мин. Клетки собирали центрифугированием (5000 об/мин, 10 мин при 4°С) и хранили при -20°С до дальнейшего использования.
Пример 3. Очистка белка с помощью аффинной хроматографии на иммобилизованных металлах
Клетки из культуры объемом 50 мл оттаивали на льду и лизировали, используя 10 мл смеси буфера для лизиса BugBuster® (Novagen), содержащей 2 мг/мл лизоцима (Sigma Aldrich), 1 единицу бензоназы (Sigma Aldrich) и 1-кратную полную смесь ингибиторов протеаз (без ЭДТА, Roche), путем инкубации клеток при комнатной температуре в течение 30 мин. Через 30 мин клеточный дебрис удаляли центрифугированием при 16000 об/мин в течение 20 мин при 4°С. Полученную надосадочную жидкость фильтровали через фильтр с диаметром пор 0,45 мкм. Колонку с гравитационным течением с Ni-сефарозой (GE Healthcare) готовили с 1 мл суспензии путем промывания этанолом и водой. Затем колонку уравновешивали отмывочным буфером 50 ммоль/л NaH2PO4, рН 8,0, содержащим 300 ммоль/л NaCl и 20 ммоль/л имидазола. Бесклеточный экстракт наносили на колонку, и несвязанные белки элюировали из колонки. Колонку промывали 20 мл отмывочного буфера, и удержавшиеся на колонке белки элюировали 3,5 мл буфера для элюции 50 ммоль/л NaH2PO4, рН 8,0, содержащего 300 ммоль/л NaCl и 250 ммоль/л имидазола. Собранные фракции анализировали методом электрофореза в ДСН-ПААГ в гелях, содержащих 10% акриламид, и фракции, содержащие очищенные ферменты лактазы, объединяли вместе. Проводили обмен буфера на буфер для хранения (буферный раствор 50 ммоль/л KH2PO4, рН 7,0, содержащий 10 ммоль/л NaCl, 1 ммоль/л MgCl2), используя предварительно упакованную колонку PD-10 для обессоливающей гель-фильтрации G-25 (GE Healthcare). Очищенные ферменты хранили при 4°С до дальнейшего использования.
Пример 4. Очистка белка с использованием гель-фильтрационной хроматографии
Клетки из культуры объемом 50 мл оттаивали на льду и лизировали, используя 10 мл смеси буфера для лизиса BugBuster® (Novagen), содержащей 2 мг/мл лизоцима (Sigma Aldrich), 1 единицу бензоназы (Sigma Aldrich) и 1-кратную полную смесь ингибиторов протеаз (без ЭДТА, Roche), путем инкубации клеток при комнатной температуре в течение 30 мин. Через 30 мин клеточный дебрис удаляли центрифугированием при 16000 об/мин в течение 20 мин при 4°С. Полученную надосадочную жидкость фильтровали через фильтр с диаметром пор 0,45 мкм. Осветленный бесклеточный экстракт концентрировали фильтрованием через фильтр с порогом отсечения 30000 Дальтон (Vivaspin 20, GE Healthcare), следуя протоколу поставщика. Колонку Sephadex G50 superfine с гравитационным потоком (Pharmacia Chemicals, Швеция) готовили с 1 г материала колонки, подготовленного путем кипячения в 100 мл воды в течение 1 часа, охлажденного до комнатной температуры. Колонку готовили путем нанесения 20 мл охлажденной суспензии на фильтрационную колонку объемом 30 мл. Колонку промывали водой MilliQ и уравновешивали отмывочным буфером В (буферный раствор 50 ммоль/л NaH2PO4, рН 7,0). 500 мкл концентрированной надосадочной жидкости наносили на колонку и оставляли для вхождения в неподвижную фазу. Отмывочный буфер (буферный раствор 50 ммоль/л NaH2PO4, рН 7,0) наносили на колонку сверху и собирали фракции элюента по отдельности. Собранные фракции анализировали в геле с ДСН-ПААГ, содержащем 10% акриламид. Фракции белка объединяли вместе и проводили обмен буфера на буфер для хранения (буферный раствор 50 ммоль/л KH2PO4, рН 7,0, содержащий 10 ммоль/л NaCl, 1 ммоль/л MgCl2) с помощью обессоливающей колонки, как описано в предыдущем разделе. Очищенные ферменты хранили при 4°С до дальнейшего использования.
Пример 5. Измерение концентрации белка методом с бицинхониновой кислотой (ВСА)
Концентрацию очищенных лактаз определяли с помощью набора реактивов для анализа белков Pierce™ ВСА (Thermo Fisher Scientific, Германия), следуя протоколу, поставляемому с набором.
Пример 6. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 6,7 и температуре 37°С
Для измерения активности бета-галактозидазы очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере В (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, охлаждение при 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Для определения количества глюкозы, образовавшейся в ходе реакции, 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета (Thermo Fischer Scientific, Дания), содержащую 80 мкл буфера С (буферный раствор 100 ммоль/л NaH2PO4, рН 7,0, содержащий глюкозооксидазу, 0,6 г/л (Sigma Aldrich); диаммониевую соль 2,2'-азино-бис(3-этилбензотиазолин-6-сульфокислоты), ABTS, 1,0 г/л (Sigma Aldrich); пероксидазу хрена, 0,02 г/л (Sigma Adrich)) и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega (BMG Labtech, Германия). Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 10-кратной концентрации буфером А. Для каждого очищенного фермента реакции проводили в трех повторностях и использовали для расчетов среднее значение трех повторных измерений. Для нормализации использовали очистку белка, проведенную на клетках Е. coli, трансформированных вектором pBAD/His без вставки. Строили стандартную кривую, используя известную концентрацию глюкозы (0-2,5 ммоль/л), и наклон кривой использовали для расчета количества глюкозы, образовавшейся в ходе реакции. Для определения количества мкмоль образующейся за минуту глюкозы (например, путем приведения значения оптической плотности в соответствие с концентрацией образовавшейся глюкозы с помощью стандартной или калибровочной кривой) и также обозначенного как единица активности лактазы 1 (или UAL-1) при рН 6,7 при 37°С, использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность (обозначенную как SUAL-1) при рН 6,7 при 37°С определяют как количество мкмоль образующейся за минуту глюкозы на один мг фермента (мкмоль глюкозы/мин/мг фермента) и определяют путем деления UAL-1 на концентрацию белка в мг. Удельная активность SEQ ID NO: 34 и SEQ ID NO: 35 была определена по существу в таких же условиях. Высокая удельная активность при рН 6,7 в высокой степени желательна для устойчивости фермента в областях применения для свежего и ферментированного молока. Подробные результаты определения удельной активности ферментов при рН 6,7 при 37°С описаны на Фиг. 28. Дополнительно активность была описана в виде количества мкмоль образующейся за секунду глюкозы на один мкмоль добавленного фермента. Результаты представлены на Фиг. 1.
Удельную активность ферментов определяли при рН 6,7 и при 37°С и использовали для расчета приблизительного времени, требующегося для гидролиза лактозы с использованием фиксированной дозы фермента в единицах активности при рН 6,7 при 37°С и 140 ммоль/л лактозы в качестве субстрата (SUAL-1). Результаты определения расчетного времени для гидролиза лактозы представлены в таблице 2.
Пример 7. Определение активности с использованием очищенных ферментов в присутствии галактозы при рН 6,7 и температуре 37°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельных реакциях разведенные ферменты инкубировали с буфером D (раствор лактозы 140 ммоль/л и галактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционная смесь состояла из 13 мкл разведенного фермента и 37 мкл буфера D в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере при следующих параметрах инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, охлаждение 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Для определения количества глюкозы, образовавшейся в ходе реакции, 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета (Thermo Fischer Scientific, Дания), содержащую 80 мкл буфера С (буферный раствор 100 ммоль/л NaH2PO4, рН 7,0, содержащий глюкозооксидазу, 0,6 г/л (Sigma Aldrich); диаммониевую соль 2,2'-азино-бис(3-этилбензотиазолин-6-сульфокислоты), ABTS, 1,0 г/л (Sigma Aldrich); пероксидазу хрена, 0,02 г/л (Sigma Adrich)) и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega (BMG Labtech, Германия). Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 10-кратной концентрации буфером А. Для каждого очищенного фермента реакции проводили в трех повторностях и использовали для расчетов среднее значение трех повторных измерений. Для нормализации использовали очистку белка, проведенную на клетках Е. coli, трансформированных вектором pBAD/His без вставки. Строили стандартную кривую, используя известную концентрацию глюкозы (0-2,5 ммоль/л), и наклон кривой использовали для расчета оптической плотности, соответствующей 1 мкмоль глюкозы, образовавшейся в ходе реакции. Для определения количества мкмоль образующейся за секунду глюкозы описываемого как 1 единица активности с галактозой при рН 6,7 при 37°С (UAG), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 6,7 при 37°С в присутствии галактозы, описываемую как SUAG, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAG на концентрацию белка в мкмоль.
Процент ингибирования ферментов галактозой рассчитывают по формуле
% ингибирования = 100*(А-В)/А,
где А представляет собой удельную активность ферментов с лактозой при рН 6,7 при 37°С (SUAL), как описано в примере 6, а В означает удельную активность ферментов в присутствии галактозы при рН 6,7 при 37°С (SUAG), как описано в примере 7. Подробные результаты % ингибирования галактозой описаны на Фиг. 2 и Фиг. 28. Пониженное ингибирование галактозой в высокой степени актуально для областей применения, где желательны очень низкие концентрации лактозы.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 28.
Примечание: относительно высокие стандартные отклонения измерения ингибирования галактозой связаны со следовыми количествами примесей глюкозы в приобретенной галактозе.
Пример 8. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 6,7 и температуре 4°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере В (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4. Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 60 мин при 4°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили в морозильной камере при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 6. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 6,7 при 4°С (UAL-2), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 6,7 при 4°С, описываемую как SUAL-2, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAG на концентрацию белка в мкмоль. Высокая удельная активность при рН 6,7 при 4°С весьма желательна для гидролиза лактозы в областях применения для свежего/пастеризованного молока. Подробные результаты определения удельной активности ферментов при рН 6,7 при 4°С описаны на Фиг. 3.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 28.
Пример 9. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 6,7 и температуре 43°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере В (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 43°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили в морозильной камере при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 6. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 6,7 при 43°С (UAL-3), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 6,7 при 43°С, описываемую как SUAL-3, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-3 на концентрацию белка в мкмоль. Высокая удельная активность при рН 6,7 при 43°С весьма желательна для гидролиза лактозы в областях применения для свежего/пастеризованного молока. Подробные результаты определения удельной активности ферментов при рН 6,7 при 43°С описаны на Фиг. 4.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 28.
Пример 10. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 5,5 и температуре 4°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере Е (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 5,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 5,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 5,5 до 5,7.
Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 60 мин при 4°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили в морозильной камере при -20°С до дальнейшего использования. Для определения количества глюкозы, образовавшейся в ходе реакции, 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета, содержащую 80 мкл буфера С, и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega (BMG Labtech, Германия). Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 10-кратной концентрации буфером А. Для каждого очищенного фермента реакции проводили в трех повторностях и использовали для расчетов среднее значение трех повторных измерений. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 5,5 при 4°С (UAL-4), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 5,5 при 4°С, описываемую как SUAL-4, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-4 на концентрацию белка в мкмоль. Высокая удельная активность при рН 5,5 при 4°С весьма желательна для гидролиза лактозы в областях применения для ферментированного молока. Подробные результаты определения удельной активности ферментов при рН 5,5 при 4°С описаны на Фиг. 5.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 29.
Пример 11. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 5,5 и температуре 37°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере Е (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 5,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 5,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 5,5 до 5,7.
Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 10. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 5,5 при 37°С (UAL-5), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 5,5 при 37°С, описываемую как SUAL-5, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-5 на концентрацию белка в мкмоль. Высокая удельная активность при рН 5,5 при 37°С актуальна для гидролиза лактозы в областях применения для ферментированного молока и гидролиза лактозы в сладкой молочной сыворотке. Подробные результаты определения удельной активности ферментов при рН 5,5 при 37°С описаны на Фиг. 6.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 29.
Пример 12. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 5,5 и температуре 43°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере Е (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 5,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 5,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 5,5 до 5,7.
Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 43°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 10. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 5,5 при 43°С (UAL-6), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 5,5 при 43°С, описываемую как SUAL-6, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-6 на концентрацию белка в мкмоль. Высокая удельная активность при рН 5,5 при 43°С актуальна для гидролиза лактозы в областях применения для ферментированного молока и гидролиза лактозы в сладкой молочной сыворотке. Подробные результаты определения удельной активности ферментов при рН 5,5 при 43°С описаны на Фиг. 7.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 29.
Пример 13. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 4,5 и температуре 4°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере F (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 4,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 4,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 4,5 до 4,7.
Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 60 мин при 4°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Для определения количества глюкозы, образовавшейся в ходе реакции, 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета, содержащую 80 мкл буфера С (как описано в примере 6), и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega (BMG Labtech, Германия). Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 5-кратной концентрации буфером А. Для каждого очищенного фермента реакции проводили в трех повторностях и использовали для расчетов среднее значение трех повторных измерений. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 4,5 при 4°С (UAL-7), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 4,5 при 4°С, описываемую как SUAL-7, определяют как количество мкмоль глюкозы, образующейся за секунду, на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-7 на концентрацию белка в мкмоль. Высокая удельная активность при рН 4,5 при 4°С актуальна для гидролиза лактозы в областях применения для ферментированного молока. Подробные результаты определения удельной активности ферментов при рН 4,5 при 4°С описаны на Фиг. 8.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 30.
Пример 14. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 4,5 и температуре 37°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере F (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 4,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 4,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 4,5 до 4,7.
Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 13. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 4,5 при 37°С (UAL-8), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 4,5 при 37°С, описываемую как SUAL-8, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-8 на концентрацию белка в мкмоль. Высокая удельная активность при рН 4,5 при 37°С актуальна для гидролиза лактозы в областях применения для ферментированного молока и гидролиза лактозы в кислой молочной сыворотке. Подробные результаты определения удельной активности ферментов при рН 4,5 при 37°С описаны на Фиг. 9. Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 30.
Пример 15. Определение активности с использованием очищенных ферментов на лактозе в качестве субстрата при рН 4,5 и температуре 43°С
Очищенные лактазы разводили до 40-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере F (раствор лактозы 140 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 4,5, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Раствор субстрата готовили в буфере рН 4,5, в раствор фермента имел рН 6,7. Для инициации реакции добавляли 13 мкл фермента к 37 мкл раствора субстрата. Таким образом, при смешивании этих двух буферов конечный рН реакции повышался от 4,5 до 4,7. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 43°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Очищенные ферменты хранили при -20°С до дальнейшего использования. Количество глюкозы, образовавшейся в ходе реакции, определяли, следуя протоколу, описанному в примере 13. Для определения количества мкмоль образующейся за секунду глюкозы, описываемого как 1 единица активности с лактозой при рН 4,5 при 43°С (UAL-9), использовали максимальное значение оптической плотности для каждой лактазы. Удельную активность при рН 4,5 при 43°С, описываемую как SUAL-9, определяют как количество мкмоль образующейся за секунду глюкозы на один мкмоль фермента (мкмоль глюкозы/с/мкмоль фермента) и рассчитывают путем деления UAL-9 на концентрацию белка в мкмоль. Высокая удельная активность при рН 4,5 при 43°С актуальна для гидролиза лактозы в областях применения для ферментированного молока и гидролиза лактозы в кислой молочной сыворотке. Подробные результаты определения удельной активности ферментов при рН 4,5 при 43°С описаны на Фиг. 10.
Дополнительно активность была описана в виде количества мкмоль образующейся за минуту глюкозы на один миллиграмм добавленного фермента. Результаты представлены на Фиг. 30.
Пример 16. Определение активности в единицах BLU
Готовили разведения имеющегося в продаже фермента NOLA® Fit (Chr-Hansen, Дания) в диапазоне концентрации от 0,5 BLU/мл до 2,5 BLU/мл в буфере G (буферный раствор 50 ммоль/л NaH2PO4, рН 7,0, содержащий 100 мкмоль/л MgSO4, 0,045% красителя Бридж, Sigma Aldrich). Разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере Н (раствор лактозы 105 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. Конверсию глюкозы определяли количественно путем переноса 10 мкл реакционной смеси в одну лунку стандартного микротитрационного планшета, содержащую 80 мкл буфера С, и инкубации при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega (BMG Labtech, Германия). Стандартную кривую строили с использованием измеренных значений оптической плотности против BLU/мл. Максимальный наклон кривой использовали для определения активности новых ферментов в BLU/мл.
Пример 17. Определение активности новых лактаз в BLU/мл с использованием лактозы в качестве субстрата
Очищенные лактазы разводили до 50-кратной концентрации в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции разведенный фермент инкубировали с раствором лактозы, приготовленном в буфере Н (раствор лактозы 105 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл разведенного очищенного фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. После проведения реакции 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета, содержащую 80 мкл буфера С (как описано в примере 6), и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega. Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 5-кратной концентрации буфером А. Для расчета активности фермента в BLU/мл с помощью стандартной кривой, описанной в примере 16, использовали максимальные значения оптической плотности.
Пример 18. Измерение процентного содержания остаточной лактозы в свежем молоке при температуре охлаждения
2 мл имеющегося в продаже пастеризованного молока (пастеризованное молоко 1,5%-ной жирности, Arla Food) смешивали в стеклянной пробирке емкостью 10 мл с 10-125 мкл фермента (эквивалент 10 BLU/мл), как определено в примере 17. Образцы инкубировали в постоянных условиях в течение 24 часов при 4°С. После инкубации реакцию останавливали путем инактивации нагреванием при 95°С в течение 7 мин с последующим хранением при -20°С до дальнейшего использования. Количество остаточной лактозы в молоке анализировали, используя метод высокоэффективной жидкостной хроматографии (ВЭЖХ). Образцы для анализа обрабатывали 1,8 мл раствора для осаждения белка (0,083 моль/л РСА и 2 ммоль/л Na-ЭДТА) и 2 мл воды Milli-Q (MQW), после чего центрифугировали при 2800 об/мин в течение 30 мин при 4°С. Аликвоту надосадочной жидкости разводили в общей сложности в 200 раз, используя автоматическую систему разведения JANUS (PerkinElmer, Уолтем, штат Массачусетс, США). Разведенные образцы анализировали в системе Dionex ICS-5000 (Thermo Fischer Scientific, Уолтем, штат Массачусетс, США), используя аналитическую колонку 4 × 250 мм CarboPac SA20 (Thermo Fischer Scientific, Уолтем, штат Массачусетс, США) и импульсный амперометрический детектор. Детектор был настроен на определение простой трехступенчатой потенциальной формы колебательного сигнала для избирательного обнаружения углеводов. Элюент был настроен на 1 ммоль/л KOH, и его постоянно регенерировали пропусканием через предколонку (CR-TC, Thermo Fischer Scientific, Уолтем, штат Массачусетс, США). Расход элюента составлял 1,2 мл/мин, а время анализа на одно введение пробы составляло 10 мин. Для количественного определения лактозы в каждом образце использовали внешнюю калибровочную кривую, построенную по трем точкам путем добавления известных количеств моногидрата лактозы (Sigma-Aldrich, Сент-Луис, штат Миссури, США) к MQW. Концентрации рассчитывали по высоте хроматографических пиков. Измеренное процентное содержание остаточной лактозы в свежем молоке представлено на Фиг. 11.
Пример 19. Определение активности в UHT молоке при комнатной температуре 2 мл имеющегося в продаже UHT молока (UHT молоко 1,5%-ной жирности, Arla Food) смешивали в стеклянной пробирке емкостью 2 мл с 2-25 мкл фермента (эквивалент 10 BLU/мл), как определено в примере 17. Образцы инкубировали в постоянных условиях в течение 24 часов при 25°С. После инкубации реакцию останавливали путем инактивации нагреванием при 95°С в течение 7 мин с последующим хранением при -20°С до дальнейшего использования. Количество остаточной лактозы в UHT молоке анализировали методом ВЭЖХ, следуя протоколу, описанному в примере 18. Измеренное процентное содержание остаточной лактозы в свежем молоке после гидролиза представлено на Фиг. 12.
Пример 20. Характеристики фермента при высокой температуре в буфере
Очищенный фермент разводили до 5 BLU/мл в буфере А (буферный раствор 50 ммоль/л NaH2PO4 рН 6,7, содержащий 100 мкмоль/л MgSO4). В отдельной реакции 13 мкл разведенного фермента инкубировали в термоциклере для ДНК с раствором лактозы (раствор лактозы 105 ммоль/л, приготовленный в 100 мМ натрийцитратном буфере рН 6,7, содержащем 100 мкмоль/л MgSO4). Реакционную смесь готовили путем смешивания 13 мкл фермента и 37 мкл раствора лактозы в пробирке для ПЦР. Реакционную смесь инкубировали в термоциклере ДНК с использованием следующих параметров инкубации: время реакции 10 мин при 37°С, инактивация фермента 10 мин при 95°С, хранение при 4°С. После проведения реакции 10 мкл реакционной смеси переносили в одну лунку стандартного микротитрационного планшета, содержащую 80 мкл буфера С (как описано в примере 6), и инкубировали при 30°С в течение 40 мин. Через 40 мин определяли оптическую плотность при 610 нм, используя сканирующий УФ-спектрофотометр для прочтения планшетов FLUOStar Omega. Для расчетов использовали значения оптической плотности от 0,1 до 1,5; если значение А610 нм превышало 1,5, реакционную смесь разводили до 5-кратной концентрации буфером А. Измеренную оптическую плотность обозначили как Abs37°C и считали эталонным значением при расчетах.
Для измерения влияния термической обработки на активность фермента в отдельной реакции инкубировали 13 мкл разведенного фермента (5 BLU/мл) в термоциклере для ДНК, используя следующие параметры инкубации: при 72°С в течение 15 с, либо при 74°С в течение 15 с, либо при 76°С в течение 6 с, либо 78°С в течение 6 с, либо при 80°С в течение 4 с, либо при 85°С в течение 5 с, либо при 90°С в течение 5 с, либо при 95°С в течение 5 с, с последующим хранением при 4°С. Активность обработанного нагреванием фермента определяли путем инкубации с раствором лактозы (раствор 105 ммоль/л лактозы, приготовленный в натрийцитратном буфере 100 ммоль/л при рН 6,7, содержащем 100 мкмоль/л MgSO4), как описано выше. Оптическую плотность, измеренную при различной температуре (например, при 72°С, 74°С, 76°С, 78°С, 80°С, 85°С, 90°С или 95°С), обозначали как Abs72°C, Abs74°C, Abs76°C, Abs78°C, Abs80°C, Abs85°C, Abs90°C, Abs95°C.
Процент остаточной активности при высокой температуре определяли по формуле
% остаточной активности = (Abs72°C/Abs37°C)*100
Проценты остаточной активности различных ферментов при различной температуре описаны на Фиг. 13.
Пример 21. Процентное содержание остаточной лактозы после высокотемпературной обработки
Влияние термической обработки на характеристики фермента в пастеризованном молоке определяли путем инкубации фиксированного количества фермента в молоке с последующей термической обработкой. В отдельных реакциях 50 мкл пастеризованного молока смешивали с 10 BLU/мл очищенного фермента (как определено в примере 17) в пробирке для ПЦР. Образец молока инкубировали при 72°С в течение 15 или при 76°С в течение 10 с, либо при 85°С в течение 5 с и при 90°С в течение 5 с, с последующей инкубацией при 5°С в течение 24 ч. После 24 ч при 5°С реакцию останавливали нагреванием при 95°С в течение 7 мин с последующим хранением при -20°С. Количество остаточной лактозы измеряли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания), следуя протоколу поставщика. Измеренное количество остаточной лактозы, определенное при различной температуре, выражали в г/л. Предел обнаружения для набора реактивов LactoSens® составляет от 0,2 г/л до 10 г/л. Результаты описаны в таблице 3.
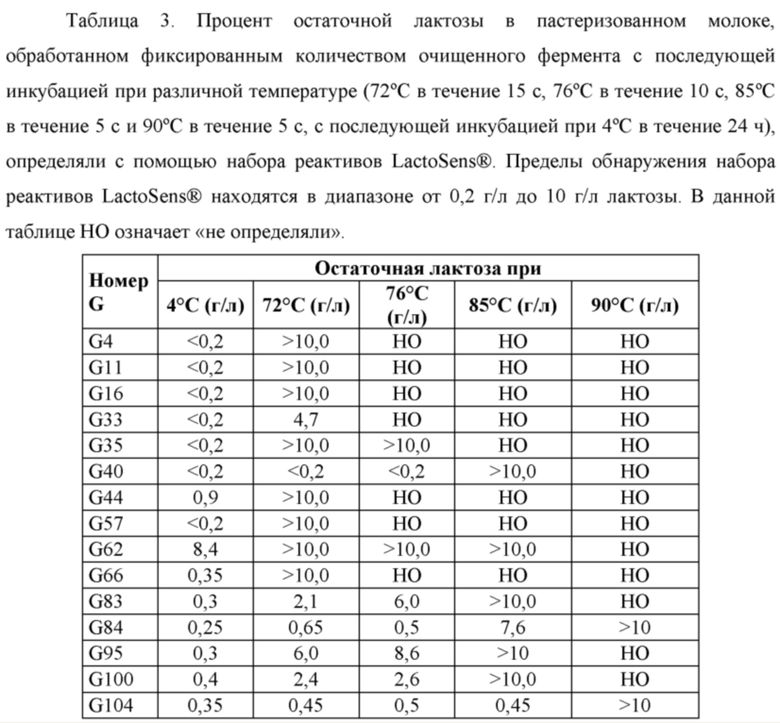
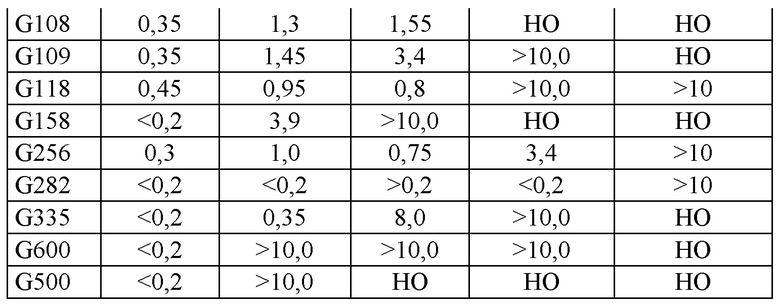
Пример 22. Процентное содержание остаточной лактозы в пастеризованном молоке после инкубации при различных температурах
1 мл имеющегося в продаже пастеризованного молока (молоко 1,5% жирности, содержащее 4,7% лактозы, Arla Foods, Дания) смешивали с 0,047 мг/мл фермента в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком мягким вращением или перемешиванием пипеткой. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 37°С в течение 30 мин, или 55°С в течение 30 мин, или 60°С в течение 30 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы молока либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Для анализа остаточной лактозы в молоке использовали набор реактивов LactoSens® (Chr. Hansen, Дания), следуя протоколу, поставляемому с набором. Измеренное процентное содержание остаточной лактозы в пастеризованном молоке представлено на Фиг. 14.
Для испытания этих новых лактаз на потенциальную способность к гидролизу лактозы авторы изобретения инкубировали 0,047 мг фермента на один миллилитр пастеризованного молока и проводили инкубацию при 37°С, 55°С и 60°С в течение 30 мин. После 30-минутной инкубации ферменты инактивировали нагреванием при 95°С. Количество остаточной лактозы определяли с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Для обоих ферментов, Ha-Lactase и NOLA® fit, при их оптимальной температуре (37°С) показано высокое содержание остаточной лактозы (более 1% остаточной лактозы), поэтому можно предположить, что эти ферменты обладают пониженной активностью, и в данной временной рамке получить безлактозное пастеризованное молоко не удается. Кроме того, аналогичное содержание остаточной лактозы было измерено при 55°С и 60°С. Напротив, для ферментов G33, G44, G95 и G158 было показано содержание остаточной лактозы менее 0,1% при 37°С, Фиг. 15. Новые ферменты благодаря их высокой активности при повышенной температуре (55°С или 60°С) показали содержание остаточной лактозы менее 0,01% после 30 мин инкубации. Таким образом, показано, что при использовании действующей дозы фермента можно получить по существу безлактозное пастеризованное и фильтрованное молоко менее чем за 30 мин. Фильтрованное молоко больше напоминает сырое молоко, чем пастеризованное молоко. Гидролиз лактозы при повышенной температуре (55°С - 60°С) за короткое время уменьшает вероятность роста микроорганизмов, не влияя на качество молока.
Пример 23. Процентное содержание остаточной лактозы в пастеризованном молоке после инкубации в течение различных периодов времени
1 мл имеющегося в продаже пастеризованного молока (молоко 1,5% жирности, содержащее 4,7% лактозы, Arla Foods, Дания) смешивали с 0,047 мг/мл фермента в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком мягким вращением или с помощью пипетки. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 15 мин или 55°С в течение 30 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры и времени на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы молока либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Для анализа остаточной лактозы в молоке использовали набор реактивов LactoSens® (Chr. Hansen, Дания), как описано в примере 22. Измеренное процентное содержание остаточной лактозы в пастеризованном молоке представлено на Фиг. 16.
Пример 24. Процентное содержание остаточной лактозы в пастеризованном молоке после инкубации с различными дозами фермента
1 мл имеющегося в продаже пастеризованного молока (молоко 1,5% жирности, содержащее 4,7% лактозы, Arla Foods, Дания) смешивали с различными дозами фермента (0,024 мг/мл или 0,047 мг/мл) в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком аккуратным вращением или с помощью пипетки. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 30 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. После проведения реакции образцы либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Количество остаточной лактозы в молоке анализировали, следуя тому же протоколу, который описан в примере 22. Измеренное процентное содержание остаточной лактозы в пастеризованном молоке представлено на Фиг. 17.
Пример 25. Процентное содержание остаточной лактозы в пастеризованном молоке после инкубации с различными дозами фермента и в течение различных периодов времени
1 мл имеющегося в продаже пастеризованного молока (молоко 1,5% жирности, содержащее 4,7% лактозы, Arla Foods, Дания) смешивали с различными дозами фермента (0,024 мг/мл или 0,047 мг/мл) в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком аккуратным вращением или перемешиванием пипеткой. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 15 мин или 55°С в течение 30 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры и времени на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Количество остаточной лактозы определяли с использованием протокола, описанного в примере 22. Измеренное процентное содержание остаточной лактозы в пастеризованном молоке представлено на Фиг. 18.
Пример 26. Процентное содержание остаточной лактозы в фильтрованном молоке
1 мл имеющегося в продаже микрофильтрованного, частично обезжиренного молока (молоко 1,5% жирности, содержащее 4,8% лактозы, молоко «Маргарита», Франция) смешивали с 0,047 мг/мл фермента в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком аккуратным вращением или перемешиванием пипеткой. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 30 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры и времени на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Для количественного анализа остаточной лактозы в молоке использовали набор реактивов LactoSens® (Chr. Hansen, Дания), следуя протоколу, поставляемому с набором. Измеренное процентное содержание остаточной лактозы в фильтрованном молоке представлено на Фиг. 19.
Таким образом, показано, что при использовании действующей дозы фермента можно получить безлактозное фильтрованное молоко (фильтрованное молоко больше напоминает сырое молоко, чем пастеризованное) менее чем за 30 мин. Гидролиз лактозы при повышенной температуре (55°С - 60°С) за короткое время уменьшает вероятность роста микроорганизмов, не влияя на качество молока.
Пример 27. Процентное содержание остаточной лактозы в фильтрованном молоке после инкубации с различными дозами фермента
1 мл имеющегося в продаже микрофильтрованного, частично обезжиренного молока (молоко 1,5% жирности, содержащее 4,8% лактозы, молоко «Маргарита», Франция) смешивали с различными дозами фермента (0,055 мг/мл, 0,55 мкмоль/л, или 0,11 мг/мл, 0,11 мкмоль/л) в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком аккуратным вращением или перемешиванием пипеткой. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 5 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры и времени на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Количество остаточной лактозы в молоке анализировали, следуя тому же протоколу, который описан в примере 22. Измеренное процентное содержание остаточной лактозы в фильтрованном молоке представлено на Фиг. 20.
Пример 28. Процентное содержание остаточной лактозы в фильтрованном молоке после инкубации в течение различных периодов времени
1 мл имеющегося в продаже микрофильтрованного, частично обезжиренного молока (молоко 1,5% жирности, содержащее 4,8% лактозы, молоко «Маргарита», Франция) смешивали с 0,11 мг/мл (1 мкмоль/л) фермента в пробирке Eppendorf емкостью 1,5 мл. Фермент смешивали с молоком мягким вращением или перемешиванием пипеткой. 50 мкл содержащего фермент молока переносили в пробирку для ПЦР. Для каждого фермента реакцию проводили в объеме реакционной смеси 2 по 50 мкл. Реакционную смесь инкубировали в термоциклере ДНК при следующих параметрах инкубации: температура и время реакции 55°С в течение 5 мин, или 55°С в течение 6 мин, или 55°С в течение 7 мин; температура и время инактивации фермента 95°С в течение 10 мин; температура хранения 4°С. Чтобы свести к минимуму влияние температуры и времени на производительность фермента, в процессе добавления, переноса пипеткой и смешивания с молоком образцы выдерживали в смеси льда с водой. После проведения реакции образцы либо сразу использовали для измерения остаточной лактозы, либо хранили при -20°С до дальнейшего использования. Для количественного анализа остаточной лактозы в молоке использовали набор реактивов LactoSens® (Chr. Hansen, Дания), следуя протоколу, поставляемому с набором. Измеренное процентное содержание остаточной лактозы в фильтрованном молоке представлено на Фиг. 21. Таким образом, показано, что при использовании действующей дозы фермента можно получить безлактозное пастеризованное и фильтрованное молоко (фильтрованное молоко больше напоминает сырое молоко, чем пастеризованное) менее чем за 5-30 мин. Гидролиз лактозы при повышенной температуре (55°С - 60°С) за короткое время уменьшает вероятность роста микроорганизмов, не влияя на качество молока.
Пример 29. Активность фермента при 4-5°С
Для анализа кинетики гидролиза лактозы новыми ферментами в пастеризованном молоке добавляли 0,05 мг фермента на один миллилитр имеющегося в продаже пастеризованного молока (молоко 1,5% жирности, содержащее 4,7% лактозы, Arla Foods, Дания). Фермент хорошо перемешивали аккуратным вращением и переносили в пробирку для ПЦР, 10 по 100 мкл каждая. Реакционные смеси инкубировали при 4°С и отбирали образцы после фиксированного интервала времени. Реакцию останавливали нагреванием при 95°С в течение 10 мин в устройстве для ПЦР. Образцы охлаждали до комнатной температуры и измеряли содержание остаточной лактозы с помощью набора реактивов LactoSens® (Chr. Hansen, Дания). Измеренное значение содержания остаточной лактозы наносили на график против времени реакции.
Как показано на Фиг. 22 и 23 при 4-5°С для снижения концентрации остаточной лактозы в коровьем молоке до менее 0,1% и до менее 0,01% с помощью известных коммерческих продуктов NOLA® Fit (G600) и Ha-Lactase™ (G500) требуется от 8-12 ч до 18-24 ч соответственно.
Лактазы по настоящему изобретению в этих условиях значительно более активны. Например, при использовании самого активного фермента G95 концентрация остаточной лактозы достигает уровня менее 0,1% (4 ч). G158 и G33 способны снизить концентрацию остаточной лактозы до уровня менее 0,1% за 5-6 ч и до уровня менее 0,01% лактозы за 8-12 ч. Как показано на Фиг. 24 и 25, для нескольких лактаз после 12 ч инкубации показана сниженная концентрация остаточной лактозы по сравнению с контрольными ферментами. Эти результаты показывают, что новые лактазы действуют быстрее, чем Ha-Lactase™ и NOLA® Fit, и в результате безлактозное пастеризованное молоко получают за значительно более короткое время. Эти новые ферменты могут уменьшить общее время технологического процесса на 50%. Кроме того, как показано на Фиг. 26, с помощью новых ферментов можно дополнительно уменьшить дозу фермента для получения безлактозного/низколактозного пастеризованного молока на 25-50%.
Таким образом, эти результаты показывают, что при использовании новых лактаз можно получить безлактозное пастеризованное молоко за значительно более короткое время (8-12 ч) при дозе фермента 50 мг/л. Кроме того, в зависимости от требуемой концентрации лактозы дозу фермента можно уменьшить на 25-50%.
Пример 30. Ферментативная активность в различных типах молока при 4-5°С
Для сравнения ферментативной активности в различных типах молока пастеризованное и фильтрованное молоко инкубировали, используя фермент лактазу в концентрации 0,052 мг/л. Образцы смешивали и хранили при 4°С в течение 24 ч.
Содержание остаточной лактозы было определено с помощью набора реактивов LactoSens® (Chr. Hansen, Denmark) и представлено на Фиг. 27, где показано, что многие из новых ферментов лактаз обладают высокой активностью расщепления лактозы в пастеризованном и фильтрованном молоке при 4°С.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Chr. Hansen A/S
<120> ФЕРМЕНТЫ ЛАКТАЗЫ С УЛУЧШЕННЫМИ СВОЙСТВАМИ ПРИ
ПОНИЖЕННЫХ ТЕМПЕРАТУРАХ
<130> P 102838
<160> 41
<170> PatentIn version 3.5
<210> 1
<211> 1049
<212> PRT
<213> Bifidobacterium adolescentis
<400> 1
Met Ala Asp Thr Ala Glu Leu Ala Ile Val His Ala Thr Thr Ala Ser
1 5 10 15
Ala Ser Trp Leu Thr Asp Pro Thr Val Phe Ala Ala Asn Arg Lys Pro
20 25 30
Ala His Ser Ser His Arg Tyr Val Ile Gly Glu Thr Ser Glu Pro Lys
35 40 45
Gln Ser Leu Asp Gly Glu Trp Lys Val Arg Ile Glu Gln Ala Arg Asn
50 55 60
Val Asp Val Glu Ser Ala Pro Phe Ala Ala Val Asp Phe Glu Asp Gly
65 70 75 80
Asp Phe Gly Ala Ile Glu Val Pro Gly His Leu Gln Met Ala Gly Tyr
85 90 95
Leu Lys Asn Lys Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asp Pro Gln Ala Pro Asn Ile Pro Glu Asn Asn His Val Ala Ile Tyr
115 120 125
Arg Arg Arg Phe Ala Leu Asp Ala Gln Leu Ala Arg Thr Leu Glu Asn
130 135 140
Asp Gly Thr Val Ser Leu Thr Phe His Gly Ala Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Leu Asp Gly Thr Phe Val Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Ser Glu Phe Asp Val Thr Glu Ala Leu Arg Asn Gly Asn Gly Asn
180 185 190
Ala Ala Asp Ser Pro Glu Ala Glu His Thr Leu Thr Val Ala Cys Tyr
195 200 205
Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu
210 215 220
His Gly Leu Phe Arg Thr Val Glu Leu Ala Ala Gln Pro His Thr His
225 230 235 240
Val Glu Thr Val Gln Leu Glu Ala Asp Tyr Thr Ala Ala Asp Thr Ala
245 250 255
Gly Thr Ala Asp Thr Ala Glu Leu Asn Ala Ala Leu Thr Leu Arg Asn
260 265 270
Ser Ala Asp Ala Met Thr Ile Glu Ser Thr Leu Arg Asp Gly Asp Gly
275 280 285
Asn Val Val Trp Glu Ser Thr Gln Ala Cys Asn Gly Glu Ile Ala Leu
290 295 300
Asn Ser Gly Lys Met Thr Asn Ile Ala Pro Trp Ser Ala Glu Ser Pro
305 310 315 320
Thr Leu Tyr Thr Leu Thr Val Arg Val Val Gly His Asp Gly Ala Ile
325 330 335
Ile Glu Thr Val Thr Gln Lys Ile Gly Phe Arg Thr Phe Arg Ile Glu
340 345 350
Asn Gly Ile Met Thr Leu Asn Gly Lys Arg Ile Val Phe Lys Gly Ala
355 360 365
Asp Arg His Glu Phe Asp Ala Lys Arg Gly Arg Ala Ile Thr Arg Glu
370 375 380
Asp Met Leu Ser Asp Val Val Phe Cys Lys Arg His Asn Ile Asn Ala
385 390 395 400
Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Tyr Trp Tyr Asp Leu Cys
405 410 415
Asp Glu Tyr Gly Leu Tyr Leu Ile Asp Glu Thr Asn Met Glu Thr His
420 425 430
Gly Thr Trp Val Ala Asn Asn Val Glu Arg Pro Glu Asp Gly Ile Pro
435 440 445
Gly Ser Arg Pro Glu Trp Glu Gly Ala Cys Val Asp Arg Ile Asn Ser
450 455 460
Met Met Arg Arg Asp Tyr Asn His Pro Ser Val Leu Ile Trp Ser Leu
465 470 475 480
Gly Asn Glu Ser Ser Ala Gly Glu Val Phe Arg Ala Met Tyr Arg His
485 490 495
Ala His Thr Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly Ser Val
500 505 510
His Met Arg Glu Phe Glu Asp Val Thr Asp Ile Glu Ser Arg Met Tyr
515 520 525
Ala His Ala Asp Glu Ile Glu Arg Tyr Leu Asn Asp Gly Ser Pro Ala
530 535 540
His Thr Asp Gly Pro Lys Lys Pro Tyr Ile Ser Cys Glu Tyr Met His
545 550 555 560
Ala Met Gly Asn Ser Cys Gly Asn Met Asp Glu Tyr Thr Ala Leu Glu
565 570 575
Arg Tyr Pro Met Tyr Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln
580 585 590
Ala Ile Glu Thr Lys Leu Pro Asp Gly Thr Thr Arg Met Cys Tyr Gly
595 600 605
Gly Asp Phe Gly Asp Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asp Gly
610 615 620
Leu Leu Phe Ala Asp Arg Thr Pro Ser Pro Lys Ala Gln Glu Val Lys
625 630 635 640
Gln Leu Tyr Ala Asn Val Lys Ile Val Val Ser Val Asp Glu Ala Arg
645 650 655
Ile Thr Asn Asp Asn Leu Phe Val Ser Thr Gly Asp Tyr Arg Phe Val
660 665 670
Leu Arg Ile Leu Ala Asp Gly Lys Pro Val Trp Ser Thr Thr Arg Arg
675 680 685
Phe Asp Val Ala Ala Gly Glu Ser Ala Ser Phe Glu Val Asp Trp Pro
690 695 700
Val Asp Asp Tyr Arg Ser Asn Ala Glu Glu Leu Val Leu Glu Val Ser
705 710 715 720
Gln Gln Leu Gly Asn Ala Cys Asp Trp Ala Pro Ala Gly Tyr Glu Leu
725 730 735
Ala Phe Gly Gln Cys Val Val Ala Gly Ala Lys Thr Thr Ala Asp Ala
740 745 750
Val Asp Ala Ala Gly Ala Pro Ala Asp Gly Thr Val Thr Leu Gly Arg
755 760 765
Trp Asn Ala Gly Val Arg Gly Gln Gly Arg Glu Ala Leu Phe Ser Arg
770 775 780
Thr Gln Gly Gly Met Val Ser Tyr Thr Phe Gly Glu Arg Glu Phe Val
785 790 795 800
Leu Arg Arg Pro Ser Ile Thr Thr Phe Arg Pro Leu Thr Asp Asn Asp
805 810 815
Arg Gly Ala Gly His Ala Phe Glu Arg Ala Ala Trp Ala Val Ala Gly
820 825 830
Lys Tyr Ala Arg Cys Val Asp Cys Ala Ile Ala Asn Arg Gly Glu Asn
835 840 845
Ala Val Glu Ala Thr Tyr Thr Tyr Glu Leu Ala Ile Pro Gln Arg Thr
850 855 860
Lys Val Thr Val Arg Tyr Val Ala Asp Thr Ala Gly Leu Val Ser Leu
865 870 875 880
Asp Val Glu Tyr Pro Gly Glu Lys Asn Gly Asp Leu Pro Thr Ile Pro
885 890 895
Ala Phe Gly Ile Glu Trp Ala Leu Pro Val Glu Tyr Ala Asn Leu Arg
900 905 910
Phe Tyr Gly Ala Gly Pro Glu Glu Thr Tyr Ala Asp Arg Arg His Ala
915 920 925
Lys Leu Gly Val Trp Ser Thr Thr Ala Gly Asp Asp Cys Ala Pro Tyr
930 935 940
Leu Leu Pro Gln Glu Thr Gly Asn His Glu Asp Val Arg Trp Ala Glu
945 950 955 960
Ile Thr Asp Asp Ser Gly His Gly Val Arg Val Lys Arg Gly Ala Gly
965 970 975
Ala Lys Pro Phe Ala Met Ser Leu Leu Pro Tyr Ser Ser Thr Met Leu
980 985 990
Glu Glu Ala Leu His Gln Asp Glu Leu Pro Lys Pro Arg His Met Phe
995 1000 1005
Leu Arg Leu Leu Ala Ala Gln Met Gly Val Gly Gly Asp Asp Ser
1010 1015 1020
Trp Met Ser Pro Val His Glu Gln Tyr Gln Leu Pro Ala Asp Gln
1025 1030 1035
Pro Leu Ser Leu Asn Val Gln Leu Lys Leu Phe
1040 1045
<210> 2
<211> 625
<212> PRT
<213> Lactobacillus sakei
<400> 2
Met Gln Pro Asn Ile Gln Trp Leu Asp Thr Pro Ala Val Phe Arg Val
1 5 10 15
Gly Gln Leu Pro Ala His Ser Asp His Arg Tyr Tyr Ala Thr Leu Ala
20 25 30
Glu Met Ala Gln Gln Gln Ser Ser Phe Glu Gln Ser Leu Asn Gly Thr
35 40 45
Trp Gln Phe His Tyr Ser Val Asn Ala Ala Ser Arg Pro Lys Ser Phe
50 55 60
Tyr Glu Leu Ala Phe Asp Ala Gln Asp Phe Glu Pro Ile Thr Val Pro
65 70 75 80
Gln His Ile Glu Leu Ala Gly Tyr Glu Gln Leu His Tyr Ile Asn Thr
85 90 95
Met Tyr Pro Trp Glu Gly His Tyr Tyr Arg Arg Pro Ala Phe Ser Thr
100 105 110
Ser Asp Asp Lys Gln His Leu Gly Met Phe Ser Glu Ala Asp Tyr Asn
115 120 125
Pro Val Gly Ser Tyr Leu His His Phe Asp Leu Thr Pro Ala Leu Arg
130 135 140
Asn Gln Arg Val Ile Ile Arg Phe Glu Gly Val Glu Gln Ala Met Tyr
145 150 155 160
Val Trp Leu Asn Gly Gln Phe Ile Gly Tyr Ala Glu Asp Ser Phe Thr
165 170 175
Pro Ser Glu Phe Asp Leu Thr Pro Tyr Leu Lys Glu Thr Asp Asn Cys
180 185 190
Leu Ala Val Glu Val His Lys Arg Ser Ser Ala Ala Phe Ile Glu Asp
195 200 205
Gln Asp Phe Phe Arg Phe Phe Gly Ile Phe Arg Asp Val Lys Leu Leu
210 215 220
Ala Lys Pro Arg Thr His Leu Glu Asp Leu Trp Val Ile Pro Glu Tyr
225 230 235 240
Asp Val Val Gln Gln Thr Gly Gln Val Lys Leu Arg Leu Gln Phe Ser
245 250 255
Gly Asp Glu Asn Arg Val His Leu Arg Ile Arg Asp Gln His Gln Ile
260 265 270
Ile Leu Thr Ala Asp Leu Thr Ser Ala Ala Gln Val Asn Gly Leu Tyr
275 280 285
Lys Met Pro Glu Leu Val Gln Ala Trp Ser Asn Gln Thr Pro Asn Leu
290 295 300
Tyr Thr Leu Glu Leu Glu Val Val Asp Gln Ala Gly Glu Thr Ile Glu
305 310 315 320
Ile Ser Gln Gln Pro Phe Gly Phe Arg Lys Ile Glu Ile Lys Asp Lys
325 330 335
Val Met Leu Leu Asn Gly Lys Arg Leu Val Ile Asn Gly Val Asn Arg
340 345 350
His Glu Trp His Pro Glu Thr Gly Arg Thr Ile Thr Ala Glu Asp Glu
355 360 365
Ala Trp Asp Ile Ala Cys Met Gln Arg Asn His Ile Asn Ala Val Arg
370 375 380
Thr Ser His Tyr Pro Asp Arg Leu Ser Phe Tyr Asn Gly Cys Asp Gln
385 390 395 400
Ala Gly Ile Tyr Met Met Ala Glu Thr Asn Leu Glu Ser His Gly Ser
405 410 415
Trp Gln Lys Met Gly Ala Val Glu Pro Ser Trp Asn Val Pro Gly Ser
420 425 430
Tyr Asp Glu Trp Glu Ala Ala Thr Leu Asp Arg Ala Arg Thr Asn Phe
435 440 445
Glu Thr Phe Lys Asn His Val Ser Ile Leu Phe Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Ser Val Leu Glu Lys Met Asn Ala Tyr Tyr Lys
465 470 475 480
Gln Gln Asp Pro Thr Arg Leu Val His Tyr Glu Gly Val Phe Arg Ala
485 490 495
Pro Glu Tyr Lys Ala Thr Ile Ser Asp Val Glu Ser Arg Met Tyr Ala
500 505 510
Thr Pro Ala Glu Ile Lys Ala Tyr Leu Asp Asn Ala Pro Gln Lys Pro
515 520 525
Phe Ile Leu Cys Glu Tyr Met His Asp Met Gly Asn Ser Leu Gly Gly
530 535 540
Met Gln Ser Tyr Ile Asp Leu Leu Ser Gln Tyr Asp Met Tyr Gln Gly
545 550 555 560
Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val Thr Asp Pro
565 570 575
Val Thr Gly Gln Arg Glu Leu Arg Tyr Gly Gly Asp Phe Asp Asp Arg
580 585 590
Pro Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Val Phe Ala Thr Arg
595 600 605
Asp Glu Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr Gly Glu His
610 615 620
Lys
625
<210> 3
<211> 330
<212> PRT
<213> Lactobacillus sakei
<400> 3
Met Lys Asn Gln Gln Cys Arg Arg Leu Asp Thr Ile Met Ala Asn Thr
1 5 10 15
Asn Lys Arg Leu Ala Val Ile Phe Gly Asp Val Thr Leu Gly Leu Lys
20 25 30
Gly Pro Asp Phe His Tyr Leu Phe Ser Tyr Gln Thr Gly Gly Pro Glu
35 40 45
Ser Leu Arg Ile Gln Gly Lys Glu Trp Leu Tyr Arg Ser Pro Lys Pro
50 55 60
Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly Asn Gln Phe Pro
65 70 75 80
Leu Lys Ser Gly Met Trp Leu Ala Ala Asp Gln Phe Ile Ala Cys Gln
85 90 95
Ser Ile Thr Val Ala Ile Asp Gly Gln Thr Ile Pro Leu Pro Ile Ala
100 105 110
Pro Glu Asn Asn Arg Tyr Ser Gly Gln Glu Thr Ala Gln Glu Val Thr
115 120 125
Val Thr Tyr Thr Tyr Gln Thr Ile Thr Thr Pro Gln Thr Thr Val Glu
130 135 140
Val Ser Tyr Thr Ile Gln Ala Ser Gly Lys Ile Arg Val Ala Val Thr
145 150 155 160
Tyr His Gly Gln Ala Gly Leu Pro Ser Leu Pro Val Phe Gly Leu Arg
165 170 175
Phe Val Met Pro Thr Pro Ala Thr Arg Phe Ile Tyr Gln Gly Leu Ser
180 185 190
Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Gly Met Ala Gly Glu Tyr
195 200 205
Glu Val Thr Gly Leu Pro Val Thr Pro Tyr Leu Val Pro Gln Asp Cys
210 215 220
Gly Val His Met Ala Thr Asp Trp Val Thr Ile Tyr Arg Gln Ala Val
225 230 235 240
Leu Asp Asn Arg Leu Arg Glu Pro Val Glu Thr Gly Leu Lys Phe Lys
245 250 255
Met Val Asp Gln Pro Phe Ala Phe Ser Cys Leu Pro Tyr Thr Ala Glu
260 265 270
Glu Leu Glu Asn Ala Thr His His Ser Glu Leu Pro Ala Pro His Arg
275 280 285
Thr Val Leu Ser Leu Leu Gly Ala Val Arg Gly Val Gly Gly Ile Asp
290 295 300
Ser Trp Gly Ser Asp Val Glu Ala Ala Tyr Gln Ile Asp Ala Thr Gln
305 310 315 320
Asp His His Leu Glu Phe Glu Ile Ser Phe
325 330
<210> 4
<211> 1049
<212> PRT
<213> Bifidobacterium adolescentis
<400> 4
Met Ala Asp Thr Ala Glu Leu Ala Ile Val His Ala Thr Thr Ala Ser
1 5 10 15
Ala Ser Trp Leu Thr Asp Pro Thr Val Phe Ala Ala Asn Arg Lys Pro
20 25 30
Ala His Ser Ser His Arg Tyr Val Ile Gly Glu Thr Ser Glu Pro Lys
35 40 45
Gln Ser Leu Asp Gly Glu Trp Lys Val Arg Ile Glu Gln Ala Arg Asn
50 55 60
Val Asp Val Glu Ser Ala Pro Phe Ala Ala Val Asp Phe Glu Asp Gly
65 70 75 80
Asp Phe Gly Ala Ile Glu Val Pro Gly His Leu Gln Met Ala Gly Tyr
85 90 95
Leu Lys Asn Lys Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asp Pro Gln Ala Pro Asn Ile Pro Glu Asn Asn His Val Ala Ile Tyr
115 120 125
Arg Arg Arg Phe Ala Leu Asp Ala Gln Leu Ala Arg Thr Leu Glu Asn
130 135 140
Asp Gly Thr Val Ser Leu Thr Phe His Gly Ala Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Leu Asp Gly Thr Phe Val Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Ser Glu Phe Asp Val Thr Glu Ala Leu Arg Asn Gly Asn Gly Asn
180 185 190
Ala Ala Asp Ser Pro Glu Ala Glu His Thr Leu Thr Val Ala Cys Tyr
195 200 205
Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu
210 215 220
His Gly Leu Phe Arg Thr Val Glu Leu Ala Ala Gln Pro His Thr His
225 230 235 240
Val Glu Thr Val Gln Leu Glu Ala Asp Tyr Thr Ala Ala Asp Thr Ala
245 250 255
Gly Thr Ala Asp Thr Ala Glu Leu Asn Ala Ala Leu Thr Leu Arg Asn
260 265 270
Pro Ala Asp Ala Met Thr Ile Glu Ser Thr Leu Arg Asp Gly Asp Gly
275 280 285
Asn Val Val Trp Glu Ser Thr Gln Ala Cys Asn Gly Glu Ile Ala Leu
290 295 300
Asn Ser Gly Lys Met Thr Asn Ile Ala Pro Trp Ser Ala Glu Ser Pro
305 310 315 320
Thr Leu Tyr Thr Leu Thr Val Arg Val Val Gly His Asp Gly Ala Ile
325 330 335
Ile Glu Thr Val Thr Gln Lys Ile Gly Phe Arg Thr Phe Arg Ile Glu
340 345 350
Asn Gly Ile Met Thr Leu Asn Gly Lys Arg Ile Val Phe Lys Gly Ala
355 360 365
Asp Arg His Glu Phe Asp Ala Lys Arg Gly Arg Ala Ile Thr Arg Glu
370 375 380
Asp Met Leu Ser Asp Val Val Phe Cys Lys Arg His Asn Ile Asn Ala
385 390 395 400
Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Tyr Trp Tyr Asp Leu Cys
405 410 415
Asp Glu Tyr Gly Leu Tyr Leu Ile Asp Glu Thr Asn Met Glu Thr His
420 425 430
Gly Thr Trp Val Ala Asn Asn Val Glu Arg Pro Glu Asp Gly Ile Pro
435 440 445
Gly Ser Arg Pro Glu Trp Glu Gly Ala Cys Val Asp Arg Ile Asn Ser
450 455 460
Met Met Arg Arg Asp Tyr Asn His Pro Ser Val Leu Ile Trp Ser Leu
465 470 475 480
Gly Asn Glu Ser Ser Ala Gly Glu Val Phe Arg Ala Met Tyr Arg His
485 490 495
Ala His Thr Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly Ser Val
500 505 510
His Met Arg Glu Phe Glu Asp Val Thr Asp Ile Glu Ser Arg Met Tyr
515 520 525
Ala His Ala Asp Glu Ile Glu Arg Tyr Leu Asn Asp Gly Ser Pro Ala
530 535 540
His Thr Asp Gly Pro Lys Lys Pro Tyr Ile Ser Cys Glu Tyr Met His
545 550 555 560
Ala Met Gly Asn Ser Cys Gly Asn Met Asp Glu Tyr Thr Ala Leu Glu
565 570 575
Arg Tyr Pro Met Tyr Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln
580 585 590
Ala Ile Glu Thr Lys Leu Pro Asp Gly Thr Thr Arg Met Cys Tyr Gly
595 600 605
Gly Asp Phe Gly Asp Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asp Gly
610 615 620
Leu Leu Phe Ala Asp Arg Thr Pro Ser Pro Lys Ala Gln Glu Val Lys
625 630 635 640
Gln Leu Tyr Ala Asn Val Lys Ile Ala Val Ser Val Asp Glu Ala Arg
645 650 655
Ile Thr Asn Asp Asn Leu Phe Val Ser Thr Gly Asp Tyr Arg Phe Val
660 665 670
Leu Arg Ile Leu Ala Asp Gly Lys Pro Val Trp Ser Thr Thr Arg Arg
675 680 685
Phe Asp Val Ala Ala Gly Glu Ser Ala Ser Phe Glu Val Asp Trp Pro
690 695 700
Val Asp Asp Tyr Arg Ser Asn Ala Glu Glu Leu Val Leu Glu Val Ser
705 710 715 720
Gln Gln Leu Gly Asn Ala Cys Asp Trp Ala Pro Ala Gly Tyr Glu Leu
725 730 735
Ala Phe Gly Gln Cys Val Val Ala Gly Ala Lys Thr Thr Ala Asp Ala
740 745 750
Val Asp Ala Ala Gly Ala Pro Ala Asp Gly Thr Val Thr Leu Gly Arg
755 760 765
Trp Asn Ala Gly Val Arg Gly Gln Gly Arg Glu Ala Leu Phe Ser Arg
770 775 780
Thr Gln Gly Gly Met Val Ser Tyr Thr Phe Gly Glu Arg Glu Phe Val
785 790 795 800
Leu Arg Arg Pro Ser Ile Thr Thr Phe Arg Pro Leu Thr Asp Asn Asp
805 810 815
Arg Gly Ala Gly His Ala Phe Glu Arg Ala Ala Trp Ala Val Ala Gly
820 825 830
Lys Tyr Ala Arg Cys Val Asp Cys Ala Ile Ala Asn Arg Gly Glu Asn
835 840 845
Ala Val Glu Ala Thr Tyr Thr Tyr Glu Leu Ala Ile Pro Gln Arg Thr
850 855 860
Lys Val Thr Val Arg Tyr Val Ala Asp Thr Ala Gly Leu Val Ser Leu
865 870 875 880
Asp Val Glu Tyr Pro Gly Glu Lys Asn Gly Asp Leu Pro Thr Ile Pro
885 890 895
Ala Phe Gly Ile Glu Trp Ala Leu Pro Val Glu Tyr Ala Asn Leu Arg
900 905 910
Phe Tyr Gly Ala Gly Pro Glu Glu Thr Tyr Ala Asp Arg Arg His Ala
915 920 925
Lys Leu Gly Val Trp Ser Thr Thr Ala Gly Asp Asp Cys Ala Pro Tyr
930 935 940
Leu Leu Pro Gln Glu Thr Gly Asn His Glu Asp Val Arg Trp Ala Glu
945 950 955 960
Ile Thr Asp Asp Ser Gly His Gly Val Arg Val Lys Arg Gly Ala Gly
965 970 975
Ala Lys Pro Phe Ala Met Ser Leu Leu Pro Tyr Ser Ser Thr Met Leu
980 985 990
Glu Glu Ala Leu His Gln Asp Glu Leu Pro Lys Pro Arg His Met Phe
995 1000 1005
Leu Arg Leu Leu Ala Ala Gln Met Gly Val Gly Gly Asp Asp Ser
1010 1015 1020
Trp Met Ser Pro Val His Glu Gln Tyr Gln Leu Pro Ala Asp Gln
1025 1030 1035
Pro Leu Ser Leu Asn Val Gln Leu Lys Leu Phe
1040 1045
<210> 5
<211> 626
<212> PRT
<213> Lactobacillus amylovorus
<400> 5
Met Lys Ala Asn Ile Lys Trp Leu Asp Asp Pro Glu Val Phe Arg Ile
1 5 10 15
Asn Gln Leu Pro Ala His Ser Asp His Pro Phe Tyr Lys Asp Tyr Arg
20 25 30
Glu Trp Gln Asn His Ser Ser Ser Phe Lys Gln Ser Leu Asn Gly Ala
35 40 45
Trp Gln Phe His Phe Ser Lys Asp Pro Gln Ser Arg Pro Ile Asp Phe
50 55 60
Tyr Lys Arg Ser Phe Asp Ser Ser Ser Phe Asp Thr Ile Pro Val Pro
65 70 75 80
Ser Glu Ile Glu Leu Asn Gly Tyr Ala Gln Asn Gln Tyr Thr Asn Ile
85 90 95
Leu Tyr Pro Trp Glu Ser Lys Ile Tyr Arg Lys Pro Ala Tyr Thr Leu
100 105 110
Gly Arg Gly Ile Lys Asp Gly Asp Phe Ser Gln Gly Lys Asp Asn Thr
115 120 125
Val Gly Ser Tyr Leu Lys His Phe Asp Leu Asn Pro Ala Leu Ala Gly
130 135 140
His Asp Ile His Ile Gln Phe Glu Gly Val Glu Arg Ala Met Tyr Val
145 150 155 160
Tyr Leu Asn Gly His Phe Ile Gly Tyr Ala Glu Asp Ser Phe Thr Pro
165 170 175
Ser Glu Phe Asp Leu Thr Pro Tyr Ile Gln Ala Lys Asp Asn Ile Leu
180 185 190
Ala Val Glu Val Phe Lys His Ser Thr Ala Ser Trp Leu Glu Asp Gln
195 200 205
Asp Met Phe Arg Phe Ser Gly Ile Phe Arg Ser Val Glu Leu Leu Ala
210 215 220
Leu Pro Arg Thr His Leu Met Asp Leu Asp Ile Lys Pro Thr Val Val
225 230 235 240
Asn Asp Tyr His Asp Gly Val Phe Asn Ala Lys Leu His Phe Met Gly
245 250 255
Lys Thr Ser Gly Asn Val His Val Leu Ile Glu Asp Ile Asp Gly Lys
260 265 270
Thr Leu Leu Asn Lys Lys Leu Pro Leu Lys Ser Thr Val Glu Ile Glu
275 280 285
Asn Glu Thr Phe Ala Asn Val His Leu Trp Asp Asn His Asp Pro Tyr
290 295 300
Leu Tyr Gln Leu Ile Ile Glu Val His Asp Gln Asp Gly Lys Leu Val
305 310 315 320
Glu Leu Ile Pro Tyr Gln Phe Gly Phe Arg Lys Ile Glu Ile Thr Lys
325 330 335
Asp His Val Val Leu Leu Asn Gly Lys Arg Leu Ile Ile Asn Gly Val
340 345 350
Asn Arg His Glu Trp Asp Ala Lys Arg Gly Arg Ser Ile Thr Leu Ala
355 360 365
Asp Met Lys Gln Asp Ile Ala Thr Phe Lys His Asn Asn Ile Asn Ala
370 375 380
Val Arg Thr Cys His Tyr Pro Asn Gln Ile Pro Trp Tyr Tyr Leu Cys
385 390 395 400
Asp Gln Asn Gly Ile Tyr Met Met Ala Glu Asn Asn Leu Glu Ser His
405 410 415
Gly Thr Trp Gln Lys Leu Gly Gln Val Glu Ala Thr Ser Asn Val Pro
420 425 430
Gly Ser Ile Pro Glu Trp Arg Glu Val Val Val Asp Arg Ala Arg Ser
435 440 445
Asn Tyr Glu Thr Phe Lys Asn His Thr Ala Ile Leu Phe Trp Ser Leu
450 455 460
Gly Asn Glu Ser Tyr Ala Gly Ser Asn Ile Ala Ala Met Asn Lys Leu
465 470 475 480
Tyr Lys Asp His Asp Ser Ser Arg Leu Thr His Tyr Glu Gly Val Phe
485 490 495
His Ala Pro Glu Phe Lys Lys Glu Ile Ser Asp Leu Glu Ser Cys Met
500 505 510
Tyr Leu Pro Pro Lys Glu Ala Glu Glu Tyr Leu Gln Asn Pro Lys Lys
515 520 525
Pro Leu Val Glu Cys Glu Tyr Met His Asp Met Gly Thr Pro Asp Gly
530 535 540
Gly Met Gly Ser Tyr Ile Lys Leu Ile Asp Lys Tyr Pro Gln Tyr Met
545 550 555 560
Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val His Asp
565 570 575
Pro Val Ser Gly Gln Asp Val Leu Arg Tyr Gly Gly Asp Phe Asp Asp
580 585 590
Arg His Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Met Phe Ala Asp
595 600 605
Arg Thr Pro Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr Gly Leu
610 615 620
His Lys
625
<210> 6
<211> 316
<212> PRT
<213> Lactobacillus amylovorus
<400> 6
Met Ala Tyr Thr Asn Asn Leu His Val Val Tyr Gly Glu Ala Ser Leu
1 5 10 15
Gly Val Asn Gly Gln Asp Phe Ala Tyr Leu Phe Ser Tyr Glu Arg Gly
20 25 30
Gly Leu Glu Ser Leu Lys Ile Lys Asp Lys Glu Trp Leu Tyr Arg Thr
35 40 45
Pro Thr Pro Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly Ser
50 55 60
Gly Phe Asn Gln Lys Ala Ala Gln Trp Leu Gly Ala Asp Met Phe Thr
65 70 75 80
Lys Cys Val Gly Ile His Val Gln Val Asp Asp His Arg Phe Asp Glu
85 90 95
Leu Pro Val Ala Pro Ile Asn Asn Gln Phe Ser Asn Gln Glu Phe Ala
100 105 110
His Glu Val Lys Val Ala Phe Asp Tyr Glu Thr Leu Thr Thr Pro Ala
115 120 125
Thr Lys Val Lys Ile Ile Tyr Asn Ile Asn Asp Phe Gly His Met Thr
130 135 140
Ile Thr Met His Tyr Phe Gly Lys Lys Gly Leu Pro Pro Leu Pro Val
145 150 155 160
Ile Gly Met Arg Phe Ile Met Pro Thr Lys Ala Lys Ser Phe Asp Tyr
165 170 175
Thr Gly Leu Ser Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Ala Glu
180 185 190
Arg Gly Thr Phe His Ile Asp Gly Leu Pro Val Thr Lys Tyr Leu Val
195 200 205
Pro Gln Glu Asn Gly Met His Met Gln Thr Asn Glu Leu Val Ile Thr
210 215 220
Arg Asn Ser Thr Gln Asn Asn Ala Asp Lys Asp Gly Asp Phe Ser Leu
225 230 235 240
Lys Ile Thr Gln Thr Lys Gln Pro Phe Asn Phe Ser Leu Leu Pro Tyr
245 250 255
Thr Ala Glu Glu Leu Glu Asn Ala Thr His Ile Glu Glu Leu Pro Leu
260 265 270
Ala Arg Arg Ser Val Leu Val Ile Ala Gly Ala Val Arg Gly Val Gly
275 280 285
Gly Ile Asp Ser Trp Gly Ser Asp Val Glu Glu Gln Tyr His Ile Asp
290 295 300
Pro Glu Gln Asp His Glu Phe Ser Phe Thr Leu Asn
305 310 315
<210> 7
<211> 1052
<212> PRT
<213> Bifidobacterium bifidum
<400> 7
Met Asn Thr Thr Asp Asp Gln Arg Lys Asn Gly Asp Pro Ile Val Ser
1 5 10 15
Pro Ser Ile Pro Thr Thr Ala Trp Leu Ala Asp Pro Arg Val Tyr Ala
20 25 30
Val His Arg Leu Asp Ala His Ser Asp His Ala Cys Trp Ser Arg Ser
35 40 45
Pro Val Asp Gly Glu Ser Thr Asp Leu Arg Gln Ser Leu Asp Gly Glu
50 55 60
Trp Arg Val Arg Val Glu Thr Ala Pro Thr Gly Arg Phe Pro Asp Gly
65 70 75 80
Thr Ser Asp Gly Pro Asp Trp Ile Ser Asp Val Ser Pro Leu Phe Ala
85 90 95
Ala Pro Gly Phe Asp Asp Ser Ser Phe Ser Arg Val Gln Val Pro Ser
100 105 110
His Leu Glu Thr Ala Gly Leu Leu Ala Pro Gln Tyr Val Asn Val Gln
115 120 125
Tyr Pro Trp Asp Gly His Glu Asp Pro Lys Ala Pro Ala Ile Pro Glu
130 135 140
His Gly His Val Ala Val Tyr Arg Arg Glu Phe Asp Ala Asp Gly Glu
145 150 155 160
Val Ala Gln Ala Val Arg Glu Gly Arg Pro Val Thr Leu Thr Phe Gln
165 170 175
Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu Asn Gly Ser Phe Ile Gly
180 185 190
Tyr Ala Glu Asp Ser Phe Thr Pro Ser Glu Phe Asp Val Thr Asp Ala
195 200 205
Ile Lys Val Asp Gly Asn Val Leu Ala Val Ala Cys Tyr Glu Tyr Ser
210 215 220
Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu His Gly Leu
225 230 235 240
Phe Arg Ser Val Glu Leu Asn Ala Arg Pro Ala Ala His Val Ala Asp
245 250 255
Leu His Ala Asp Ala Asp Trp Asp Leu Ala Thr Ser Arg Gly Ser Leu
260 265 270
Ser Leu Asp Val Leu Ile Asp Gly Ala Ala Asn Ala Ala Thr Ala Asp
275 280 285
Phe Ala Leu Arg Asp Lys Asn Gly Thr Ile Val Trp Arg Thr Ala Thr
290 295 300
Lys Ala Asp Gly Thr Leu His Ala Glu Ala Glu Ile Asp Asp Ala Ala
305 310 315 320
Pro Trp Ser Ala Glu Arg Pro Asp Leu Tyr Glu Leu Ser Val Thr Leu
325 330 335
Leu Asp Ala Asp Gly Lys Val Leu Glu Thr Ala Arg Thr Arg Ile Gly
340 345 350
Phe Arg His Val Ala Ile Glu Asp Gly Ile Leu Lys Leu Asn Gly Lys
355 360 365
Arg Leu Val Phe Arg Gly Val Asn Arg His Glu Phe Asp Cys Arg Arg
370 375 380
Gly Arg Ala Ile Thr Glu Glu Asp Met Leu Trp Asp Ile Arg Phe Met
385 390 395 400
Lys Arg His Asn Ile Asn Ala Val Arg Thr Ser His Tyr Pro Asn Gln
405 410 415
Ser Arg Trp Tyr Glu Leu Cys Asp Glu Tyr Gly Ile Tyr Leu Ile Asp
420 425 430
Glu Thr Asn Leu Glu Thr His Gly Ser Trp Asn Ser Pro Gly Asp Ile
435 440 445
Pro Val Gly Thr Ser Val Pro Gly Asp Asp Glu Ala Trp Leu Gly Ala
450 455 460
Cys Ile Asp Arg Leu Asp Ser Met Ile Leu Arg Asp Arg Asn His Pro
465 470 475 480
Ser Val Leu Val Trp Ser Leu Gly Asn Glu Ser Tyr Ala Gly Glu Val
485 490 495
Leu Lys Ala Met Ser Ala His Ala His Gln Leu Asp Pro Gly Arg Pro
500 505 510
Val His Tyr Glu Gly Val Asn Trp Asn His Ala Tyr Asp Gly Ile Ser
515 520 525
Asp Phe Glu Ser Arg Met Tyr Ala Lys Pro Ala Glu Ile Gln Asp Trp
530 535 540
Leu Glu His Gly Asp Glu Arg Gly Glu Ala Ser Lys Pro Phe Val Ser
545 550 555 560
Cys Glu Tyr Met His Ala Met Gly Asn Ser Cys Gly Gly Leu Ser Glu
565 570 575
Phe Ile Asp Leu Glu Arg Tyr Glu Arg Tyr Ser Gly Gly Phe Ile Trp
580 585 590
Asp Tyr Ile Asp Gln Gly Leu Val Gln Arg Leu Pro Asp Gly Ser Glu
595 600 605
Arg Leu Ser Val Gly Gly Glu Trp Gly Asp Arg Pro Thr Asp Tyr Glu
610 615 620
Phe Val Gly Asn Gly Ile Val Phe Ala Asp Arg Thr Pro Ser Pro Lys
625 630 635 640
Ala Gln Glu Val Lys Gln Leu Tyr Ser Pro Val Lys Leu Ala Pro Asp
645 650 655
Gly His Gly Val Thr Ile Glu Asn Arg Asn Leu Phe Ala Gly Thr Asp
660 665 670
Gly Tyr Val Phe Ala Ala Arg Leu Leu Glu Asp Gly His Glu Ile Trp
675 680 685
His Ala Asp Tyr Arg Phe Asp Val Ala Ala Gly Asp Thr Gln His His
690 695 700
Asp Ile Ala Phe Pro Asp Ile Asp Ala Asp Gly Asp Thr Arg Glu Val
705 710 715 720
Thr Tyr Glu Val Asp Leu Leu Leu Ala Glu Ala Thr Ala Trp Ala Pro
725 730 735
Ala Gly Tyr Glu Leu Ala Phe Gly Gln Leu Thr Gly Thr Leu Asn Pro
740 745 750
Glu Gln Asp Ile Thr Glu Thr Ser His Asp Asp Asp Gly Arg Ala Thr
755 760 765
Arg Thr Leu Ser Arg Trp Asn Ala Gly Ile Arg Arg Asp Asp Glu Glu
770 775 780
Ile Leu Leu Ser Arg Thr Gln Gly Gly Ile Val Ser Trp Lys Arg Asp
785 790 795 800
Asp Arg Glu Met Val Ile Arg Arg Pro Glu Leu Val Thr Phe Arg Pro
805 810 815
Leu Thr Asp Asn Asp Arg Gly Asn His Ser Gly Phe Asp Arg Ala Ala
820 825 830
Trp Phe Ala Ala Gly Arg Tyr Ala Ile Val Thr Glu Thr Lys Ile His
835 840 845
Glu Ser Asp Asp Gly Leu Val Ala Glu Tyr Gln Tyr Glu Leu Ala Asp
850 855 860
Pro Asn His Thr Pro Val Ser Val Thr Tyr His Val Asn Ser Asp Met
865 870 875 880
Arg Met Gln Leu Thr Val Glu Tyr Pro Gly Asn Ala Thr Asp Met Ala
885 890 895
Ser Leu Pro Ala Phe Gly Ile Glu Trp Glu Leu Pro Gly Glu Tyr Asp
900 905 910
Arg Leu Arg Tyr Tyr Gly Pro Gly Pro Glu Glu Thr Tyr Arg Asp Arg
915 920 925
Lys Gln Gly Gly Lys Leu Gly Ile Trp Asp Ala Thr Ala Lys Ala Ser
930 935 940
Met Ala Pro Tyr Leu Met Val Gln Glu Thr Gly Ser His Glu Asp Val
945 950 955 960
Arg Trp Leu Glu Ala Thr Asp Ile Gln Gly His Gly Leu Arg Val Thr
965 970 975
Gln Arg Gly Asp Arg His Phe Thr Ala Ser Leu Leu Pro Trp Asn Thr
980 985 990
Tyr Thr Ile Glu Ala Ala Arg Arg His Glu Asp Leu Pro Lys Pro Arg
995 1000 1005
His Asn Tyr Leu Arg Leu Leu Ala Ala Gln Met Gly Val Gly Gly
1010 1015 1020
Asp Asp Ser Trp Gly Ala Pro Val His Thr Ala Tyr Gln Leu Pro
1025 1030 1035
Ala Gly Arg Pro Leu Thr Leu Asp Val Asn Leu Glu Leu Ile
1040 1045 1050
<210> 8
<211> 1052
<212> PRT
<213> Bifidobacterium bifidum
<400> 8
Met Asn Thr Thr Asp Asp Gln Arg Lys Asn Gly Asp Pro Ile Val Ser
1 5 10 15
Pro Ser Ile Pro Thr Thr Ala Trp Leu Ala Asp Pro Arg Val Tyr Ala
20 25 30
Val His Arg Leu Asp Ala His Ser Asp His Ala Cys Trp Ser Arg Ser
35 40 45
Pro Val Asp Gly Glu Ser Thr Asp Leu Arg Gln Ser Leu Asp Gly Glu
50 55 60
Trp Arg Val Arg Val Glu Thr Ala Pro Thr Gly Arg Phe Pro Asp Gly
65 70 75 80
Thr Ser Asp Gly Pro Asp Trp Ile Ser Asp Val Ser Pro Leu Phe Ala
85 90 95
Ala Pro Gly Phe Asp Asp Ser Ser Phe Ser Arg Val Gln Val Pro Ser
100 105 110
His Leu Glu Thr Ala Gly Leu Leu Ala Pro Gln Tyr Val Asn Val Gln
115 120 125
Tyr Pro Trp Asp Gly His Glu Asp Pro Lys Ala Pro Ala Ile Pro Glu
130 135 140
His Gly His Val Ala Val Tyr Arg Arg Glu Phe Asp Ala Asp Gly Glu
145 150 155 160
Val Ala Gln Ala Val Arg Glu Gly Arg Pro Val Thr Leu Thr Phe Gln
165 170 175
Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu Asn Gly Ser Phe Ile Gly
180 185 190
Tyr Ala Glu Asp Ser Phe Thr Pro Ser Glu Phe Asp Val Thr Asp Ala
195 200 205
Ile Lys Val Asp Gly Asn Val Leu Ala Val Ala Cys Tyr Glu Tyr Ser
210 215 220
Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu His Gly Leu
225 230 235 240
Phe Arg Ser Val Glu Leu Asn Ala Arg Pro Ala Ala His Val Ala Asp
245 250 255
Leu His Ala Asp Ala Asp Trp Asp Leu Ala Thr Ser Arg Gly Ser Leu
260 265 270
Ser Leu Asp Val Leu Ile Asp Gly Ala Ala Asn Ala Ala Thr Ala Asp
275 280 285
Phe Ala Leu Trp Asp Lys Asn Gly Thr Ile Val Trp His Ile Val Thr
290 295 300
Lys Ala Asp Gly Thr Leu His Ala Glu Ala Glu Ile Asp Asp Ala Ala
305 310 315 320
Pro Trp Ser Ala Glu Arg Pro Asp Leu Tyr Glu Leu Ser Val Thr Leu
325 330 335
Leu Asp Ala Asp Gly Lys Val Leu Glu Thr Ala Arg Thr Arg Ile Gly
340 345 350
Phe Arg His Val Ala Ile Glu Asp Gly Ile Leu Lys Leu Asn Gly Lys
355 360 365
Arg Leu Val Phe Arg Gly Val Asn Arg His Glu Phe Asp Cys Arg Arg
370 375 380
Gly Arg Ala Ile Thr Glu Glu Asp Met Leu Trp Asp Ile Arg Phe Met
385 390 395 400
Lys Arg His Asn Ile Asn Ala Val Arg Thr Ser His Tyr Pro Asn Gln
405 410 415
Ser Arg Trp Tyr Glu Leu Cys Asp Glu Tyr Gly Ile Tyr Leu Ile Asp
420 425 430
Glu Thr Asn Leu Glu Thr His Gly Ser Trp Asn Ser Pro Gly Asp Ile
435 440 445
Pro Val Gly Thr Ser Val Pro Gly Asp Asp Glu Ala Trp Leu Gly Ala
450 455 460
Cys Ile Asp Arg Leu Asp Ser Met Ile Leu Arg Asp Arg Asn His Pro
465 470 475 480
Ser Val Leu Val Trp Ser Leu Gly Asn Glu Ser Tyr Ala Gly Glu Val
485 490 495
Leu Lys Ala Met Ser Ala His Ala His Arg Leu Asp Pro Gly Arg Pro
500 505 510
Val His Tyr Glu Gly Val Asn Trp Asn His Ala Tyr Asp Gly Ile Ser
515 520 525
Asp Phe Glu Ser Arg Met Tyr Ala Lys Pro Ala Glu Ile Gln Asp Trp
530 535 540
Leu Glu His Gly Asp Glu Arg Gly Glu Ala Ser Lys Pro Phe Val Ser
545 550 555 560
Cys Glu Tyr Met His Ala Met Gly Asn Ser Cys Gly Gly Leu Ser Glu
565 570 575
Phe Ile Asp Leu Glu Arg Tyr Glu Arg Tyr Ser Gly Gly Phe Ile Trp
580 585 590
Asp Tyr Ile Asp Gln Gly Leu Val Gln Arg Leu Pro Asp Gly Ser Glu
595 600 605
Arg Leu Ser Val Gly Gly Glu Trp Gly Asp Arg Pro Thr Asp Tyr Glu
610 615 620
Phe Val Gly Asn Gly Ile Val Phe Ala Asp Arg Thr Pro Ser Pro Lys
625 630 635 640
Ala Gln Glu Val Lys Gln Leu Tyr Ser Pro Val Lys Leu Ala Pro Asp
645 650 655
Gly His Gly Val Thr Ile Glu Asn Arg Asn Leu Phe Ala Gly Thr Asp
660 665 670
Gly Tyr Val Phe Ala Ala Arg Leu Leu Glu Asp Gly His Glu Ile Trp
675 680 685
His Ala Asp Tyr Arg Phe Asp Val Ala Ala Gly Asp Thr Gln His His
690 695 700
Asp Ile Ala Phe Pro Asp Ile Asp Ala Asp Gly Asp Thr Arg Glu Val
705 710 715 720
Thr Tyr Glu Val Asp Leu Leu Leu Ala Glu Ala Thr Ala Trp Ala Pro
725 730 735
Ala Gly Tyr Glu Leu Ala Phe Gly Gln Leu Thr Gly Thr Leu Asn Pro
740 745 750
Glu Gln Asp Ile Thr Glu Thr Ser His Asp Asp Asp Gly Arg Ala Thr
755 760 765
Arg Thr Leu Ser Arg Trp Asn Ala Gly Ile Arg Arg Asp Asp Lys Glu
770 775 780
Ile Leu Leu Ser Arg Thr Gln Gly Gly Ile Val Ser Trp Lys Arg Asp
785 790 795 800
Asp Arg Glu Met Val Ile Arg Arg Pro Glu Leu Val Thr Phe Arg Pro
805 810 815
Leu Thr Asp Asn Asp Arg Gly Asn His Ser Gly Phe Asp Arg Ala Ala
820 825 830
Trp Phe Ala Ala Gly Arg Tyr Ala Ile Val Thr Glu Thr Lys Ile His
835 840 845
Glu Ser Asp Asp Gly Leu Val Ala Glu Tyr Gln Tyr Glu Leu Ala Asp
850 855 860
Pro Asn His Thr Pro Val Ser Val Thr Tyr His Val Asn Ser Asp Met
865 870 875 880
Arg Met Gln Leu Thr Val Glu Tyr Pro Gly Asn Ala Thr Asp Met Ala
885 890 895
Ser Leu Pro Ala Phe Gly Ile Glu Trp Glu Leu Pro Gly Glu Tyr Asp
900 905 910
Arg Leu Arg Tyr Tyr Gly Pro Gly Pro Glu Glu Thr Tyr Arg Asp Arg
915 920 925
Lys Gln Gly Gly Lys Leu Gly Ile Trp Asp Ala Thr Ala Lys Ala Ser
930 935 940
Met Ala Pro Tyr Leu Met Val Gln Glu Thr Gly Ser His Glu Asp Val
945 950 955 960
Arg Trp Leu Glu Ala Thr Asp Ile Gln Gly His Gly Leu Arg Val Thr
965 970 975
Gln Arg Gly Asp Arg His Phe Thr Ala Ser Leu Leu Pro Trp Asn Thr
980 985 990
Tyr Met Ile Glu Ala Ala Arg Arg His Glu Asp Leu Pro Glu Pro Arg
995 1000 1005
His Asn Tyr Leu Arg Leu Leu Ala Ala Gln Met Gly Val Gly Gly
1010 1015 1020
Asp Asp Ser Trp Gly Ala Pro Val His Thr Ala Tyr Gln Leu Pro
1025 1030 1035
Ala Gly Arg Pro Leu Thr Leu Asp Val Asn Leu Glu Leu Ile
1040 1045 1050
<210> 9
<211> 1055
<212> PRT
<213> Bifidobacterium breve
<400> 9
Met Thr Asn Ser Met Gln Gly Lys Ala Lys Thr Ile Met Thr Asn Leu
1 5 10 15
Gln Ser Ala Gln Gln Phe Ser Gln Ala Trp Leu Thr Asp Pro Arg Val
20 25 30
Phe Ala Val Asn Arg Leu Ala Ala His Ser Ser His Lys Phe Tyr Asp
35 40 45
His Ser Pro Gln Cys Gly Glu Ala Met Asp Leu Lys Gln Ser Leu Asp
50 55 60
Gly Gln Trp Arg Val Gln Met Leu Asp Leu Ala Asp Leu Ala Asp Asn
65 70 75 80
Glu Leu Ala Glu Ala Ala Phe Ala Gln Pro Gly Tyr Asp Ala Ala Gly
85 90 95
Phe Ser Pro Ile Glu Val Pro Ser Ala Leu Glu Thr Lys Gly Phe Leu
100 105 110
Asn His Gln Tyr Val Asn Gln Gln Tyr Pro Trp Ser Gly His Glu Ser
115 120 125
Pro Val Ala Pro Asp Val Pro Lys His Asn His Val Ala Leu Tyr Arg
130 135 140
His Glu Phe Ser Leu Glu Pro Lys Ala Ala Ala Val Leu Glu Ala Asn
145 150 155 160
Lys Thr Ala Ala Asp Asp Ala Ala Lys Arg Arg Val Thr Leu Thr Phe
165 170 175
Gln Gly Ala Ala Thr Ala Ile Val Val Trp Leu Asn Gly Ala Phe Ile
180 185 190
Gly Tyr Ala Glu Asp Ser Phe Thr Pro Ser Glu Phe Asp Val Thr Asp
195 200 205
Val Leu Arg Asp Gly Val Asn Thr Leu Ala Val Ala Cys Phe Glu Phe
210 215 220
Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu His Gly
225 230 235 240
Ile Phe Arg Ser Val Glu Leu Glu Ala Gln Pro Leu Val His Val Asn
245 250 255
Asp Leu Arg Val Leu Ala Asp Tyr Asp His Thr Thr Gly Glu Gly Ser
260 265 270
Leu Asp Val Val Ala Leu Leu Arg Asn Ala Gly Thr Ala Ala Ala Val
275 280 285
Ala Ala Thr Val Leu Asp Ala Ala Gly Asn Thr Val Trp His Ser Lys
290 295 300
Leu Thr Ala Gly Ala Asp Ala Glu Thr Leu Thr Val Lys Ala Asn Val
305 310 315 320
Gly Lys Val Asn Pro Trp Ser Ala Glu Glu Pro Thr Leu Tyr Thr Leu
325 330 335
Gln Val Val Ala Thr Asp Ala Ala Gly Gln Val Ile Glu Ala Ala Leu
340 345 350
Gln Arg Ile Gly Phe Arg His Phe Ala Ile Glu Asp Gly Leu Met Lys
355 360 365
Leu Asn Gly Lys Arg Ile Val Phe Lys Gly Val Asp Arg His Glu Phe
370 375 380
Asp Ala Arg Thr Gly Arg Thr Ile Ala Glu Ala Asp Met Ile Glu Asp
385 390 395 400
Ile His Ser Phe Lys Arg Leu Asn Ile Asn Ala Val Arg Thr Ser His
405 410 415
Tyr Pro Asn Glu Thr Arg Trp Tyr Glu Leu Cys Asp Glu Tyr Gly Ile
420 425 430
Tyr Val Leu Asp Glu Thr Asn Leu Glu Thr His Gly Ser Trp Thr Asp
435 440 445
Pro Gly Asp Val Phe Gln Pro Ala Arg Ala Ile Pro Gly Ser Lys Asp
450 455 460
Glu Trp Arg Ala Ala Cys Val Asp Arg Thr Ala Ser Met Val Arg Arg
465 470 475 480
Asp Tyr Asn His Pro Ser Val Val Ile Trp Ser Leu Gly Asn Glu Ala
485 490 495
Phe Gly Gly Asp Val Phe Tyr Ser Met Arg Asp Phe Val His Glu Asn
500 505 510
Asp Pro Phe Arg Pro Val His Tyr Glu Gly Thr Phe Asn Asp Pro Glu
515 520 525
Phe Ser Ala Ala Thr Asp Ile Met Ser Arg Met Tyr Ala Lys Pro Asp
530 535 540
Glu Ile Val Lys Leu Tyr Leu Gly Glu Asp Gly Lys Lys Pro Tyr Ile
545 550 555 560
Ser Cys Glu Tyr Ser His Ser Met Gly Asn Ser Thr Gly Gly Leu His
565 570 575
Leu Tyr Thr Glu Leu Glu Arg Tyr Pro Leu Tyr Gln Gly Gly Phe Ile
580 585 590
Trp Asp Tyr Val Asp Gln Ala Leu Trp Gln Asp Cys Gly Asn Gly Thr
595 600 605
Glu Arg Leu Ala Tyr Gly Gly Asp Phe Glu Asp Arg Pro Asn Asp Tyr
610 615 620
Glu Phe Ser Gly Asp Gly Val Met Phe Ala Asp Arg Thr Pro Ser Pro
625 630 635 640
Lys Ala Gln Glu Val Lys Gln Leu Tyr Ala Asn Val Lys Leu Val Pro
645 650 655
Asp Glu Ser Gly Val Thr Ile Thr Asn Asp Asn Leu Phe Ile Ser Thr
660 665 670
Ala Ser Ser Leu Phe Thr Ala Arg Val Leu Val Asp Gly Ala Glu Arg
675 680 685
Trp His Ala Asn Tyr Arg Phe Asp Val Pro Ala Gly Glu Thr Val Arg
690 695 700
Glu Pro Ile Ala Phe Pro Lys Val Thr Asp Leu Val Ala Leu Ser Gly
705 710 715 720
Ser Ala Glu Val Thr Tyr Glu Val Asp Gln Arg Leu Ala Glu Ala Thr
725 730 735
Asp Trp Ala Pro Ala Gly Tyr Glu Leu Thr Phe Gly Gln Tyr Val Ala
740 745 750
Ala Val Ser Phe Asp Asp Gly Ala Ala Asp Ala Val Val Ala Gly Asp
755 760 765
Ala Glu Val Ala Ala Asp Gly Phe Asn Ala Gly Ile His Thr Asp Phe
770 775 780
Gly Glu Val Leu Leu Ser Lys Thr Gln Gly Gly Met Val Ser Phe Lys
785 790 795 800
Arg Asp Gly Arg Glu Met Val Ile Arg Arg Pro Asn Leu Thr Thr Phe
805 810 815
Arg Ala Leu Thr Asp Asn Asp Arg Gly Asn Gly Ser Gly Phe Glu Arg
820 825 830
Ala Gln Trp Met Ala Ala Gly Arg Tyr Ala Arg Val Thr Gly Thr Ser
835 840 845
Val Glu Glu Thr Ala Asp Gly Lys Gly Leu Lys Ala Thr Tyr Ser Tyr
850 855 860
Glu Leu Ala Asp Ala Lys His Thr Pro Val Thr Val His Tyr Glu Val
865 870 875 880
Asp Ala Ala Leu Arg Val His Leu Thr Val Glu Tyr Pro Gly Glu Ala
885 890 895
Asp Ala Ala Thr Leu Pro Ala Phe Gly Leu Glu Trp Ile Leu Pro Lys
900 905 910
Gln Tyr Asp Arg Leu Arg Phe Tyr Gly Leu Gly Pro Glu Glu Thr Tyr
915 920 925
Ala Asp Arg Leu His Gly Ala Lys Leu Gly Val Phe Ser Arg Thr Ala
930 935 940
Ala Glu Asp Cys Ala Pro Tyr Leu Leu Pro Gln Glu Thr Gly Asn His
945 950 955 960
Glu Gln Val Arg Trp Ala Glu Ile Thr Asp Glu Tyr Gly His Gly Met
965 970 975
Arg Val Thr Ala Ala Gly Gly Thr Arg Phe Ala Thr Ser Leu Leu Pro
980 985 990
Tyr Ser Ser Leu Met Phe Glu Asp Ala Leu His Gln Asn Glu Leu Pro
995 1000 1005
Lys Pro Arg His Thr Phe Leu Arg Leu Leu Ala Ala Gln Met Gly
1010 1015 1020
Val Gly Gly Asp Asp Thr Trp Gly Ala Pro Val His Asp Glu Phe
1025 1030 1035
Gln Val Pro Ala Asp Gln Pro Leu Lys Leu Asp Val Thr Leu Glu
1040 1045 1050
Leu Ile
1055
<210> 10
<211> 689
<212> PRT
<213> Bifidobacterium catenulatum
<400> 10
Met Thr Gln Arg Arg Ser Tyr Arg Trp Pro Gln Pro Leu Ala Gly Gln
1 5 10 15
Gln Ala Arg Ile Trp Tyr Gly Gly Asp Tyr Asn Pro Asp Gln Trp Pro
20 25 30
Glu Glu Val Trp Asp Asp Asp Val Arg Leu Met Lys Lys Ala Gly Val
35 40 45
Asn Leu Val Ser Val Gly Ile Phe Ser Trp Ala Lys Ile Glu Thr Ser
50 55 60
Glu Gly Val Tyr Asp Phe Asp Trp Leu Asp Arg Ile Ile Asp Lys Leu
65 70 75 80
Gly Glu Ala Gly Ile Ala Val Asp Leu Ala Ser Ala Thr Ala Ser Pro
85 90 95
Pro Met Trp Leu Thr Gln Ala His Pro Glu Val Leu Trp Lys Asp Tyr
100 105 110
Arg Gly Asp Val Cys Gln Pro Gly Ala Arg Gln His Trp Arg Pro Thr
115 120 125
Ser Pro Val Phe Arg Glu Tyr Ala Leu Lys Leu Cys Arg Ala Met Ala
130 135 140
Glu His Tyr Lys Gly Asn Pro Tyr Val Val Ala Trp His Val Ser Asn
145 150 155 160
Glu Tyr Gly Cys His Asn Arg Phe Asp Tyr Ser Glu Asp Ala Glu Arg
165 170 175
Ala Phe Arg Lys Trp Cys Glu Glu Arg Tyr Gly Thr Ile Asp Ala Val
180 185 190
Asn Asp Ala Trp Gly Thr Ala Phe Trp Ala Gln Arg Met Asn Asp Phe
195 200 205
Thr Glu Ile Val Pro Pro Arg Phe Ile Gly Asp Gly Asn Phe Met Asn
210 215 220
Pro Gly Lys Leu Leu Asp Phe Lys Arg Phe Ser Ser Asp Ala Leu Lys
225 230 235 240
Ala Phe Tyr Val Ala Glu Arg Asp Ala Leu Ala Glu Ile Thr Pro Asp
245 250 255
Leu Pro Leu Thr Thr Asn Phe Met Val Ser Ala Ala Gly Ser Val Leu
260 265 270
Asp Tyr Asp Asp Trp Gly Arg Glu Val Asp Phe Val Ser Asn Asp His
275 280 285
Tyr Phe Ile Pro Gly Glu Ala His Leu Asp Glu Leu Ala Phe Ser Ala
290 295 300
Ser Leu Val Asp Gly Ile Ala Arg Lys Asp Pro Trp Phe Leu Met Glu
305 310 315 320
His Ser Thr Ser Ala Val Asn Trp Arg Pro Val Asn Tyr Arg Lys Glu
325 330 335
Pro Gly Gln Leu Val Arg Asp Ser Leu Ala His Val Ala Met Gly Ala
340 345 350
Asp Ala Val Cys Tyr Phe Gln Trp Arg Gln Ser Lys Ala Gly Ala Glu
355 360 365
Lys Phe His Ser Ala Met Val Pro His Thr Gly Glu Asp Ser Ala Val
370 375 380
Phe Arg Asp Val Cys Glu Leu Gly Ala Asp Leu Asn Thr Leu Ala Asp
385 390 395 400
Asn Gly Leu Leu Gly Thr Lys Leu Ala Lys Ser Lys Val Ala Val Val
405 410 415
Phe Asp Tyr Glu Ser Glu Trp Ala Thr Glu His Thr Ala Thr Pro Thr
420 425 430
Gln Lys Val His His Val Asp Glu Pro Leu Gln Trp Phe Arg Ala Leu
435 440 445
Ala Asp His Gly Val Thr Ala Asp Val Val Pro Val Ser Ser Asn Trp
450 455 460
Asp Glu Tyr Glu Val Val Val Leu Pro Ser Val Tyr Ile Leu Ser Glu
465 470 475 480
Glu Thr Thr Arg Arg Val Arg Asp Tyr Val Val Asn Gly Gly Arg Leu
485 490 495
Ile Val Thr Tyr Tyr Thr Gly Leu Ser Asp Glu Lys Asp His Val Trp
500 505 510
Leu Gly Gly Tyr Pro Gly Ser Ile Arg Asp Val Val Gly Val Arg Val
515 520 525
Glu Glu Phe Met Pro Met Gly Asp Asp Phe Pro Gly Val Pro Asp Cys
530 535 540
Leu Gly Leu Ser Asn Gly Ala Val Ala His Asp Ile Ala Asp Val Ile
545 550 555 560
Gly Ser Val Asp Gly Thr Ala Thr Val Leu Glu Thr Phe Arg Asp Asp
565 570 575
Pro Trp Thr Gly Met Asp Gly Ala Pro Ala Ile Val Ala Asn Thr Phe
580 585 590
Gly Glu Gly Arg Ser Val Tyr Val Gly Ala Arg Leu Gly Arg Asp Gly
595 600 605
Ile Ala Lys Ser Leu Pro Glu Ile Phe Glu Ser Leu Gly Met Ala Glu
610 615 620
Thr Gly Glu Asn Asp Ser Arg Val Leu Arg Val Glu Arg Glu Gly Ser
625 630 635 640
Asp Gly Ser Arg Phe Val Phe Ser Phe Asn Arg Thr His Glu Ala Val
645 650 655
Gln Ile Pro Phe Glu Gly Lys Ile Val Val Ser Ser Phe Ala Glu Val
660 665 670
Ser Gly Glu Asn Val Ser Ile Lys Pro Asn Gly Val Ile Val Thr Lys
675 680 685
Gln
<210> 11
<211> 1023
<212> PRT
<213> Bifidobacterium catenulatum
<400> 11
Met Ala Asn Ser Asn Arg Val Glu His Ala Ser Glu Thr Trp Leu Thr
1 5 10 15
Asp Ala Thr Val Phe Glu Val Asn Arg Thr Pro Ala His Ser Asn His
20 25 30
Lys Cys Phe Thr His Asp Pro Gln Ser Gly Glu His Ser Asp Leu Thr
35 40 45
Gln Ser Leu Asp Gly Glu Trp Arg Val Glu Ile Val Gln Ala Ser Asp
50 55 60
Ile Asp Phe Asn Glu Glu Pro Phe Val Ala Glu Asn Phe Asp Asp Ser
65 70 75 80
Ser Phe Cys Arg Ala Gln Val Pro Gly His Leu Gln Met Ala Gly Leu
85 90 95
Leu Lys Asn Lys Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asn Pro Leu Glu Pro Asn Val Pro Glu Asn Asn His Val Ala Leu Tyr
115 120 125
Arg Arg Lys Phe Val Val Ser Lys Arg Leu Ala Asp Thr Lys Glu Ser
130 135 140
Glu Gly Ser Val Ser Ile Val Phe His Gly Met Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Val Asn Gly Leu Phe Ala Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Asn Glu Phe Asp Ile Thr Asp Leu Leu His Asp Gly Glu Asn Val
180 185 190
Val Ala Val Ala Cys Tyr Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp
195 200 205
Gln Asp Phe Trp Arg Leu His Gly Leu Phe Arg Ser Val Glu Leu Thr
210 215 220
Ala Gln Pro His Val His Val Glu Asn Met Gln Leu Glu Ala Asp Trp
225 230 235 240
Asp Ala Glu Ser Gly Thr Ala Ser Leu Asp Ala Ala Leu Ser Val Arg
245 250 255
Asn Ala Ser Asp Ala Ala Thr Ile Ser Ala Thr Leu Lys Asp Ser Glu
260 265 270
Gly Asn Val Val Trp Glu Ala Ser Thr Asn Ala Asp Ala Asn Thr Thr
275 280 285
Phe Ala Ser Gly Ser Leu Gln Gly Leu Glu Pro Trp Ser Ala Glu Ser
290 295 300
Pro Ser Leu Tyr Glu Leu Glu Val Asn Val Ile Asp Gln Ala Gly Asn
305 310 315 320
Ile Val Glu Ala Ala Val Gln Lys Val Gly Phe Arg Arg Phe Arg Ile
325 330 335
Glu Asn Gly Ile Met Thr Leu Asn Gly Lys Arg Ile Val Phe Lys Gly
340 345 350
Ala Asp Arg His Glu Phe Asp Ala Lys Arg Gly Arg Ser Ile Thr Glu
355 360 365
Gln Asp Met Ile Asp Asp Val Ile Phe Cys Lys Arg His Asn Ile Asn
370 375 380
Ala Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Arg Trp Tyr Asp Leu
385 390 395 400
Cys Asp Glu Tyr Gly Ile Tyr Leu Ile Asp Glu Thr Asn Leu Glu Thr
405 410 415
His Gly Ser Trp Cys Leu Pro Gly Asp Val Val Thr Ala Glu Thr Ala
420 425 430
Val Pro Gly Ser Lys Ala His Trp Glu Gly Ala Cys Val Asp Arg Val
435 440 445
Asn Ser Met Val Arg Arg Asp Tyr Asn His Pro Ser Val Val Ile Trp
450 455 460
Ser Leu Gly Asn Glu Ser Tyr Thr Gly Asp Val Phe Arg Ala Met Tyr
465 470 475 480
Lys His Val His Asp Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly
485 490 495
Val Thr Lys Asn Arg Asp Tyr Asp Asp Val Thr Asp Ile Glu Thr Arg
500 505 510
Met Tyr Glu His Ala Asp Val Val Glu Glu Tyr Leu Lys Asn Asp Pro
515 520 525
Gln Lys Pro Tyr Ile Ser Cys Glu Tyr Met His Ala Met Gly Asn Ser
530 535 540
Val Gly Asn Leu Asp Glu Tyr Thr Ala Leu Glu Arg Tyr Pro His Tyr
545 550 555 560
Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Ile Tyr Ala Thr
565 570 575
Gln Pro Asp Gly Ser Thr Arg Leu Cys Tyr Gly Gly Asp Phe Gly Asp
580 585 590
Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asn Gly Leu Val Phe Ala Asp
595 600 605
Arg Thr Pro Thr Pro Lys Ala Gln Glu Val Lys Gln Leu Tyr Ser Asn
610 615 620
Val His Ile Asp Val Thr Asp Arg Ser Val Ser Ile Lys Asn Asp Asn
625 630 635 640
Leu Phe Ile Ser Thr Gly Gly Tyr Gln Phe Val Leu Arg Ile Leu Ala
645 650 655
Asp Gly Glu Pro Val Trp Gln Ser Glu Arg Arg Phe Asp Val Pro Ala
660 665 670
Asp Ser Ala Cys Thr Phe Asp Val Glu Trp Pro Val Asp Leu Tyr Arg
675 680 685
Ala Asn Ala Asp Glu Leu Val Leu Glu Val Ser Gln Arg Leu Ala Glu
690 695 700
Ala Thr Asp Trp Ala Pro Ala Gly Tyr Glu Leu Ala Phe Gly Gln Thr
705 710 715 720
Ile Val Ala Gly Thr Lys Ala Ala Glu Asp Ala Ala Leu Pro Ala Asp
725 730 735
Gly Ile Val Thr Val Gly Arg Trp Asn Ala Gly Val Gln Gly Ser Gly
740 745 750
Arg Glu Ile Leu Leu Ser Arg Thr Gln Gly Gly Leu Val Ser Tyr Thr
755 760 765
Phe Asp Gly His Glu Phe Val Leu Arg Arg Pro Ala Ile Thr Thr Phe
770 775 780
Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly His Gly Phe Glu Arg
785 790 795 800
Ala Gln Trp Met Val Ala Gly Arg Tyr Ala Arg Cys Val Asp Asn Val
805 810 815
Ile Glu Gln Val Asp Glu Asp Thr Leu Lys Ala Val Tyr Thr Tyr Glu
820 825 830
Leu Ala Thr Pro Gln Cys Thr Lys Val Thr Val Gly Tyr Thr Ala Asp
835 840 845
Thr Thr Gly Arg Leu Asn Leu His Val Glu Tyr Pro Gly Glu Ser Gly
850 855 860
Glu Leu Pro Thr Ile Pro Ala Phe Gly Ile Glu Trp Thr Leu Pro Val
865 870 875 880
Gln Tyr Ser Asn Leu Arg Phe Phe Gly Ala Gly Pro Glu Glu Thr Tyr
885 890 895
Gln Asp Arg Lys His Ala Lys Leu Gly Val Trp Ser Thr Asp Ala Phe
900 905 910
Lys Asp His Ala Pro Tyr Leu Met Pro Gln Glu Thr Gly Asn His Glu
915 920 925
Glu Val Arg Trp Ala Glu Ile Thr Asp Glu Asn Gly His Gly Leu Arg
930 935 940
Val Ser Arg Ala Asn Gly Ala Ala Pro Phe Ala Val Ser Leu Gln Pro
945 950 955 960
Tyr Ser Ser Phe Met Ile Glu Glu Ala Gln His Gln Asp Glu Leu Pro
965 970 975
Ala Pro Lys His Met Phe Leu Arg Val Leu Ala Ala Gln Met Gly Val
980 985 990
Gly Gly Asp Asp Ser Trp Met Ser Pro Val His Ser Gln Tyr His Ile
995 1000 1005
Thr Ala Asp Gln Pro Ile Ser Leu Asp Val Asn Leu Glu Leu Ile
1010 1015 1020
<210> 12
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. bulgaricus
<400> 12
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Ile Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Asp Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Arg Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Arg Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Ala Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Lys Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Val Ile Glu Glu Tyr Leu Thr Asn Lys Pro Ala Lys Pro Phe
515 520 525
Ile Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Glu Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ala Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ser Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Arg
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Ala Asp Glu Lys Gly Glu Val Val Tyr Arg Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Val Ala Gln Lys Leu Pro Glu Phe Lys Pro
705 710 715 720
Glu Gly Arg Pro Asp Leu Val Asp Ser Asp Tyr Asn Leu Gly Leu Lys
725 730 735
Gly Asn Asn Phe Gln Ile Leu Phe Ser Lys Val Lys Gly Trp Pro Val
740 745 750
Ser Leu Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Arg Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Lys
785 790 795 800
Asp Ile Ser Cys Glu Val Lys Glu Asp Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Val Thr Tyr Glu
820 825 830
Val Asp Gly Arg Gly Lys Ile Ala Val Thr Ala Asp Phe Pro Gly Ala
835 840 845
Glu Glu Ala Gly Leu Leu Pro Ala Phe Gly Leu Asn Leu Ala Leu Pro
850 855 860
Lys Glu Leu Thr Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Pro Asp Arg Leu Glu Gly Asn Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Lys Lys Asn Phe Ser Pro Tyr Leu Arg Pro Gln Glu Thr Gly Asn
900 905 910
Arg Ser Lys Val Arg Trp Tyr Gln Leu Phe Asp Glu Lys Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Asp Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ser Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Lys Ala Arg Gln Leu Arg Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 13
<211> 1007
<212> PRT
<213> Lactobacillus delbrueckii subsp. lactis
<400> 13
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asn Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Ile Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Val Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Leu Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Ala Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Gly Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Leu Ser Gly Leu Phe Arg Ser Val Thr Leu Glu Ala Lys Pro
210 215 220
Leu Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Val Gly Pro Ile Arg Ser Glu Lys Leu Gly Phe Ser
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Ala Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Gln Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp Asp
420 425 430
Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met Ala
435 440 445
Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn Glu
450 455 460
Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg Lys
465 470 475 480
Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn Arg
485 490 495
Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro Ala
500 505 510
Lys Glu Ile Glu Glu Tyr Leu Thr Lys Lys Pro Ala Lys Pro Phe Ile
515 520 525
Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu Ala
530 535 540
Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe Ile
545 550 555 560
Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu Tyr
565 570 575
Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly Asp
580 585 590
Gly Leu Val Phe Ala Asp Arg Thr Thr Ser Pro Lys Leu Ala Asn Val
595 600 605
Lys Ala Leu Tyr Ser Asn Leu Lys Leu Glu Val Lys Asp Gly Gln Leu
610 615 620
Phe Ile Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ala Tyr Tyr Phe
625 630 635 640
Leu Ala Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Gln Pro
645 650 655
Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Val Leu Pro
660 665 670
Trp Pro Glu Val Glu Asp Glu Lys Gly Glu Ile Val Tyr Gln Val Thr
675 680 685
Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr Val
690 695 700
Ala Glu Ala Glu Glu Ala Val Thr Lys Leu Pro Glu Phe Tyr Pro Ala
705 710 715 720
Gly Arg Pro Glu Leu Val Asp Ser Asp Phe Asn Leu Gly Leu Lys Gly
725 730 735
Asn Gly Phe Arg Ile Leu Phe Ser Lys Ala Lys Gly Trp Pro Val Ser
740 745 750
Ile Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe Thr
755 760 765
Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly Tyr
770 775 780
Asp Leu Ala Lys Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Gln Asp
785 790 795 800
Ile Ser Tyr Glu Ile Lys Glu Asn Ser Ala Leu Val Lys Thr Thr Phe
805 810 815
Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Ile Thr Tyr Glu Val
820 825 830
Asp Ser Leu Gly Lys Ile Ala Val Thr Ala Asn Phe Pro Gly Ala Val
835 840 845
Glu Asn Gly Leu Leu Pro Ala Phe Gly Leu Asn Phe Ala Leu Pro Lys
850 855 860
Glu Leu Ser Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser Tyr
865 870 875 880
Ala Asp Arg Leu Glu Gly Ser Tyr Leu Gly Ile Tyr Gln Gly Ala Val
885 890 895
Glu Lys Asn Phe Thr Pro Tyr Leu Arg Pro Gln Glu Ala Gly Asn Arg
900 905 910
Ser Lys Val Arg Tyr Tyr Gln Leu Phe Asp Glu Glu Gly Gly Leu Glu
915 920 925
Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr Ser
930 935 940
Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn Asn
945 950 955 960
Tyr Thr Trp Val Arg Ala Leu Ala Ala Gln Met Gly Val Gly Gly Asp
965 970 975
Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala Gln
980 985 990
Glu Ala Arg Gln Leu Lys Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 14
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. bulgaricus
<400> 14
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Phe Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Ile Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Asp Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Arg Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Arg Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Ala Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Lys Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Val Ile Glu Glu Tyr Leu Thr Asn Lys Pro Ala Lys Pro Phe
515 520 525
Ile Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Glu Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ala Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ser Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Arg
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Ala Asp Glu Lys Gly Glu Val Val Tyr Arg Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Val Ala Gln Lys Leu Pro Glu Phe Lys Pro
705 710 715 720
Glu Gly Arg Pro Asp Leu Val Asp Ser Asp Tyr Asn Leu Gly Leu Lys
725 730 735
Gly Asn Asn Phe Gln Ile Leu Phe Ser Lys Val Lys Gly Trp Pro Val
740 745 750
Ser Leu Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Arg Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Lys
785 790 795 800
Asp Ile Ser Cys Glu Val Lys Glu Asp Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Val Thr Tyr Glu
820 825 830
Val Asp Gly Arg Gly Lys Ile Ala Val Thr Ala Asp Phe Pro Gly Ala
835 840 845
Glu Glu Ala Gly Leu Leu Pro Ala Phe Gly Leu Asn Leu Ala Leu Pro
850 855 860
Lys Glu Leu Thr Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Pro Asp Arg Leu Glu Gly Asn Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Lys Lys Asn Phe Ser Pro Tyr Leu Arg Pro Gln Glu Thr Gly Asn
900 905 910
Arg Ser Lys Val Arg Trp Tyr Gln Leu Phe Asp Glu Lys Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ser Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Lys Ala Arg Gln Leu Arg Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 15
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. bulgaricus
<400> 15
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Ile Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Asp Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Arg Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Arg Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Ala Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Lys Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Val Ile Glu Glu Tyr Leu Thr Asn Lys Pro Ala Lys Pro Phe
515 520 525
Ile Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Glu Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ala Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ser Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Arg
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Ala Asp Glu Lys Gly Glu Val Val Tyr Arg Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Val Ala Gln Lys Leu Pro Glu Phe Lys Pro
705 710 715 720
Glu Gly Arg Pro Asp Leu Val Asp Ser Asp Tyr Asn Leu Gly Leu Lys
725 730 735
Gly Asn Asn Phe Gln Ile Leu Phe Ser Lys Val Lys Gly Trp Pro Val
740 745 750
Ser Leu Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Arg Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Lys
785 790 795 800
Asp Ile Ser Cys Glu Val Lys Glu Asp Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Val Thr Tyr Glu
820 825 830
Val Asp Gly Arg Gly Lys Ile Ala Val Thr Ala Asp Phe Pro Gly Ala
835 840 845
Glu Glu Ala Gly Leu Leu Pro Ala Phe Gly Leu Asn Leu Ala Leu Pro
850 855 860
Lys Glu Leu Thr Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Pro Asp Arg Leu Glu Gly Asn Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Lys Lys Asn Phe Ser Pro Tyr Leu Arg Pro Gln Glu Thr Gly Asn
900 905 910
Arg Ser Lys Val Arg Trp Tyr Gln Leu Phe Asp Glu Lys Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ser Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Lys Ala Arg Gln Leu Arg Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 16
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. lactis
<400> 16
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Val Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Ala Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Leu Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Leu Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Arg Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp Asp
420 425 430
Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met Ala
435 440 445
Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn Glu
450 455 460
Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg Lys
465 470 475 480
Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn Arg
485 490 495
Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro Ala
500 505 510
Lys Glu Ile Glu Glu Tyr Leu Thr Lys Lys Pro Ala Lys Pro Phe Ile
515 520 525
Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu Ala
530 535 540
Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe Ile
545 550 555 560
Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu Tyr
565 570 575
Gly Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Thr Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ser Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ala Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Gln
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Val Leu
660 665 670
Pro Trp Pro Glu Val Glu Asp Glu Lys Gly Glu Ile Val Tyr Gln Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Ala Val Thr Lys Leu Pro Glu Phe Tyr Pro
705 710 715 720
Ala Gly Arg Pro Glu Leu Val Asp Ser Asp Phe Asn Leu Gly Leu Lys
725 730 735
Gly Asn Gly Phe Arg Ile Leu Phe Ser Lys Ala Lys Gly Trp Pro Val
740 745 750
Ser Ile Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Lys Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Gln
785 790 795 800
Asp Ile Ser Tyr Glu Ile Lys Glu Asn Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Ile Thr Tyr Glu
820 825 830
Val Asp Ser Leu Gly Lys Ile Ala Val Thr Ala Asn Phe Pro Gly Ala
835 840 845
Val Glu Asn Gly Leu Leu Pro Ala Phe Gly Leu Asn Phe Ala Leu Pro
850 855 860
Lys Glu Leu Ser Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Ala Asp Arg Leu Glu Gly Ser Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Glu Lys Asn Phe Thr Pro Tyr Leu Arg Pro Gln Glu Ala Gly Asn
900 905 910
Arg Ser Lys Val Arg Tyr Tyr Gln Leu Phe Asp Glu Glu Ser Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Ser Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ala Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Glu Ala Arg Gln Leu Lys Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 17
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. bulgaricus
<400> 17
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Ile Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Asp Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Arg Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Arg Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Ile Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Lys Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Val Ile Glu Glu Tyr Leu Thr Asn Lys Pro Ala Lys Pro Phe
515 520 525
Ile Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Glu Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ala Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ser Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Arg
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Ala Asp Glu Lys Gly Glu Val Val Tyr Arg Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Val Ala Gln Lys Leu Pro Glu Phe Lys Pro
705 710 715 720
Glu Gly Arg Pro Asp Leu Val Asp Ser Asp Tyr Asn Leu Gly Leu Lys
725 730 735
Gly Asn Asn Phe Gln Ile Leu Phe Ser Lys Val Lys Gly Trp Pro Val
740 745 750
Ser Leu Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Arg Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Lys
785 790 795 800
Asp Ile Ser Cys Glu Val Lys Glu Asp Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Val Thr Tyr Glu
820 825 830
Val Asp Gly Arg Gly Lys Ile Ala Val Thr Ala Asp Phe Pro Gly Ala
835 840 845
Glu Glu Ala Gly Leu Leu Pro Ala Phe Gly Leu Asn Leu Ala Leu Pro
850 855 860
Lys Glu Leu Thr Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Pro Asp Arg Leu Glu Gly Asn Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Lys Lys Asn Phe Ser Pro Tyr Leu Arg Pro Gln Glu Thr Gly Asn
900 905 910
Arg Ser Lys Val Arg Trp Tyr Gln Leu Phe Asp Glu Lys Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Asp Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ser Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Lys Ala Arg Gln Leu Arg Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 18
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. bulgaricus
<400> 18
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Val Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Ile Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Asp Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Asn Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Gln Ala Lys Pro
210 215 220
Arg Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Leu Gly Pro Ile Gly Ser Glu Gln Leu Glu Phe Thr
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Glu Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Ile Lys Arg Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Lys Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Val Ile Glu Glu Tyr Leu Thr Asn Lys Pro Ala Lys Pro Phe
515 520 525
Ile Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly
580 585 590
Asn Gly Leu Val Phe Ala Asp Arg Thr Glu Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ala Asn Leu Lys Leu Glu Val Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ser Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Arg
645 650 655
Pro Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Ala Asp Glu Lys Gly Glu Val Val Tyr Arg Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Val Ala Gln Lys Leu Pro Glu Phe Lys Pro
705 710 715 720
Glu Gly Arg Pro Asp Leu Val Asp Ser Asp Tyr Asn Leu Gly Leu Lys
725 730 735
Gly Asn Asn Phe Gln Ile Leu Phe Ser Lys Val Lys Gly Trp Pro Val
740 745 750
Ser Leu Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Arg Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Lys
785 790 795 800
Asp Ile Ser Cys Glu Val Lys Glu Asp Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Val Thr Tyr Glu
820 825 830
Val Asp Gly Arg Gly Lys Ile Ala Val Thr Ala Asp Phe Pro Gly Ala
835 840 845
Glu Glu Ala Gly Leu Leu Pro Ala Phe Gly Leu Asn Leu Ala Leu Pro
850 855 860
Lys Glu Leu Thr Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Pro Asp Arg Leu Glu Gly Asn Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Lys Lys Asn Phe Ser Pro Tyr Leu Arg Pro Gln Glu Thr Gly Asn
900 905 910
Arg Ser Lys Val Arg Trp Tyr Gln Leu Phe Asp Glu Lys Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ser Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Lys Ala Arg Gln Leu Arg Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 19
<211> 1007
<212> PRT
<213> Lactobacillus delbrueckii subsp. lactis
<400> 19
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asn Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Ile Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Val Pro Ser Lys Asn Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Leu Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Ala Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Gly Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Leu Ser Gly Leu Phe Arg Ser Val Thr Leu Glu Ala Lys Pro
210 215 220
Leu Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Val Gly Pro Ile Arg Ser Glu Lys Leu Asp Phe Ser
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Ala Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Gln Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp Asp
420 425 430
Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met Ala
435 440 445
Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn Glu
450 455 460
Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg Lys
465 470 475 480
Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn Arg
485 490 495
Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro Ala
500 505 510
Lys Glu Ile Glu Glu Tyr Leu Thr Lys Lys Pro Ala Lys Pro Phe Ile
515 520 525
Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu Ala
530 535 540
Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe Ile
545 550 555 560
Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu Tyr
565 570 575
Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly Asp
580 585 590
Gly Leu Val Phe Ala Asp Arg Thr Thr Ser Pro Lys Leu Ala Asn Val
595 600 605
Lys Ala Leu Tyr Ser Asn Leu Lys Leu Glu Val Lys Asp Gly Gln Leu
610 615 620
Phe Ile Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ala Tyr Tyr Phe
625 630 635 640
Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Gln Pro
645 650 655
Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu Pro
660 665 670
Trp Pro Glu Val Glu Asp Glu Lys Gly Glu Ile Val Tyr Gln Val Thr
675 680 685
Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr Val
690 695 700
Ala Glu Ala Glu Glu Ala Val Thr Lys Leu Pro Glu Phe Tyr Pro Ala
705 710 715 720
Gly Arg Pro Glu Leu Val Asp Ser Asp Phe Asn Leu Gly Leu Lys Gly
725 730 735
Asn Gly Phe Arg Ile Leu Phe Ser Lys Ala Lys Gly Trp Pro Val Ser
740 745 750
Ile Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe Thr
755 760 765
Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly Tyr
770 775 780
Asp Leu Ala Lys Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Gln Asp
785 790 795 800
Ile Ser Tyr Glu Ile Lys Glu Asn Ser Ala Leu Val Lys Thr Ala Phe
805 810 815
Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Ile Thr Tyr Glu Val
820 825 830
Asp Ser Leu Gly Lys Ile Ala Val Thr Ala Asn Phe Pro Gly Ala Val
835 840 845
Glu Asn Gly Leu Leu Pro Ala Phe Gly Leu Asn Phe Ala Leu Pro Lys
850 855 860
Glu Leu Ser Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser Tyr
865 870 875 880
Ala Asp Arg Leu Glu Gly Ser Tyr Leu Gly Ile Tyr Gln Gly Met Val
885 890 895
Glu Lys Asn Phe Thr Pro Tyr Leu Arg Pro Gln Glu Ala Gly Asn Arg
900 905 910
Ser Lys Val Arg Tyr Tyr Gln Leu Phe Asp Glu Glu Gly Gly Leu Glu
915 920 925
Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr Ser
930 935 940
Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn Asn
945 950 955 960
Tyr Thr Trp Val Arg Ala Leu Ala Ala Gln Met Gly Val Gly Gly Asp
965 970 975
Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala Gln
980 985 990
Glu Ala Arg Gln Leu Lys Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 20
<211> 628
<212> PRT
<213> Lactobacillus helvaticus
<400> 20
Met Gln Ala Asn Ile Asn Trp Leu Asp Asn Pro Glu Val Phe Arg Val
1 5 10 15
Asn Gln Leu Pro Ala His Ser Asp His Pro Phe Phe Arg Asp Tyr Arg
20 25 30
Glu Trp Gln Lys Gln His Ser Ser Tyr Gln Gln Ser Leu Asn Gly Lys
35 40 45
Trp Lys Phe His Phe Ser Ala Asn Pro Met Asp Arg Pro Gln Asp Phe
50 55 60
Tyr Gln Arg Asp Phe Asp Ser Ser Asn Phe Asp Ser Ile Pro Val Pro
65 70 75 80
Ser Glu Ile Glu Leu Ser Asn Tyr Thr Gln Asn Gln Tyr Ile Asn Val
85 90 95
Leu Phe Pro Trp Glu Gly Lys Ile Phe Arg Arg Pro Ala Tyr Ala Leu
100 105 110
Asp Pro Asn Asp His Glu Glu Gly Ser Phe Ser Lys Gly Ala Asp Asn
115 120 125
Thr Val Gly Ser Tyr Leu Lys Arg Phe Asp Leu Ser Ser Ala Leu Ile
130 135 140
Gly Lys Asp Val His Ile Lys Phe Glu Gly Val Glu Gln Ala Met Tyr
145 150 155 160
Val Trp Leu Asn Gly His Phe Val Gly Tyr Ala Glu Asp Ser Phe Thr
165 170 175
Pro Ser Glu Phe Asp Leu Thr Pro Tyr Ile Gln Asp Lys Asp Asn Leu
180 185 190
Leu Ala Val Glu Val Phe Lys His Ser Thr Ala Ser Trp Leu Glu Asp
195 200 205
Gln Asp Met Phe Arg Phe Ser Gly Ile Phe Arg Ser Val Glu Leu Leu
210 215 220
Gly Ile Pro Ala Thr His Leu Met Asp Met Asp Leu Lys Pro Arg Val
225 230 235 240
Ala Asp Asn Tyr Gln Asp Gly Ile Phe Asn Leu Lys Leu His Phe Ile
245 250 255
Gly Lys Lys Ala Gly Ser Phe His Leu Leu Val Lys Asp Ile Lys Gly
260 265 270
His Thr Leu Leu Glu Lys Asn Glu Asp Ile Lys Glu Asn Val Gln Ile
275 280 285
Asn Asn Glu Lys Phe Glu Asn Val His Leu Trp Asn Asn His Asp Pro
290 295 300
Tyr Leu Tyr Gln Leu Leu Ile Glu Val Tyr Asp Glu Gln Gln Asn Leu
305 310 315 320
Leu Glu Leu Ile Pro Phe Gln Phe Gly Phe Arg Arg Ile Glu Ile Ser
325 330 335
Pro Glu Lys Val Val Leu Leu Asn Gly Lys Arg Leu Ile Ile Asn Gly
340 345 350
Val Asn Arg His Glu Trp Asp Ala Lys Arg Gly Arg Ser Ile Thr Met
355 360 365
Ser Asp Met Thr Thr Asp Ile Asn Thr Phe Lys Glu Asn Asn Ile Asn
370 375 380
Ala Val Arg Thr Cys His Tyr Pro Asn Gln Ile Pro Trp Tyr Tyr Leu
385 390 395 400
Cys Asp Gln Asn Gly Ile Tyr Val Met Ala Glu Asn Asn Leu Glu Ser
405 410 415
His Gly Thr Trp Gln Lys Met Gly Glu Ile Glu Pro Ser Asp Asn Val
420 425 430
Pro Gly Ser Ile Pro Gln Trp Lys Glu Ala Val Ile Asp Arg Ala Arg
435 440 445
Asn Asn Tyr Glu Thr Phe Lys Asn His Thr Ser Ile Leu Phe Trp Ser
450 455 460
Leu Gly Asn Glu Ser Tyr Ala Gly Asp Asn Ile Ile Ala Met Asn Glu
465 470 475 480
Phe Tyr Lys Ser His Asp Asp Thr Arg Leu Val His Tyr Glu Gly Val
485 490 495
Val His Arg Pro Glu Leu Lys Asp Lys Ile Ser Asp Val Glu Ser Cys
500 505 510
Met Tyr Leu Pro Pro Lys Lys Val Glu Glu Tyr Leu Gln Asn Asp Pro
515 520 525
Pro Lys Pro Phe Met Glu Cys Glu Tyr Met His Asp Met Gly Asn Ser
530 535 540
Asp Gly Gly Met Gly Ser Tyr Ile Lys Leu Leu Asp Lys Tyr Pro Gln
545 550 555 560
Tyr Phe Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val
565 570 575
His Asp Glu Ile Ser Gly His Asp Val Leu Arg Tyr Gly Gly Asp Phe
580 585 590
Asp Asp Arg His Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Met Phe
595 600 605
Ala Asp Arg Thr Pro Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr
610 615 620
Gly Leu His Lys
625
<210> 21
<211> 318
<212> PRT
<213> Lactobacillus helvaticus
<400> 21
Met Asp Tyr Thr Asn Asn Gln Leu His Ile Ile Tyr Gly Asp Ala Thr
1 5 10 15
Phe Gly Val Asn Gly Lys Asp Phe Gln Tyr Ile Phe Ser Tyr Glu Arg
20 25 30
Gly Gly Leu Glu Ser Leu Lys Val His Gly Lys Glu Trp Leu Tyr Arg
35 40 45
Val Pro Thr Pro Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly
50 55 60
Ser Gly Phe Asn Leu Lys Ala Ala Gln Trp Leu Gly Ala Asp Met Phe
65 70 75 80
Thr Lys Cys Thr Asp Ile His Leu Lys Val Asp Arg His Asp Phe Ala
85 90 95
Glu Leu Pro Ile Ala Pro Phe Asn Asn Lys Phe Ser Asn His Glu Tyr
100 105 110
Ala Lys Ser Ala Glu Ile Ser Phe Thr Tyr Gln Thr Leu Thr Thr Pro
115 120 125
Ala Thr Asn Ala Lys Ile Ile Tyr Asn Ile Asp Asp Val Gly His Ile
130 135 140
Lys Val Thr Met Arg Tyr Tyr Gly Lys Lys Gly Leu Pro Pro Leu Pro
145 150 155 160
Val Ile Gly Ile Arg Leu Ile Met Pro Thr Ala Ala Thr Gly Phe Asp
165 170 175
Tyr Glu Gly Leu Ser Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Ala
180 185 190
Lys Glu Gly Lys Phe His Ile Asp Gly Leu Pro Val Thr Glu Tyr Leu
195 200 205
Val Pro Gln Glu Asn Gly Met His Met Gln Thr Lys Lys Leu Thr Ile
210 215 220
Asn Arg Glu Thr Thr Gln Asn Asn Val Asp Arg Thr Asn Glu Lys Phe
225 230 235 240
Ser Leu Ser Ile Gln Gln Ala Glu Lys Pro Phe Asn Phe Ser Cys Leu
245 250 255
Pro Tyr Thr Ala Glu Glu Leu Glu Asn Ala Thr His Ile Glu Glu Leu
260 265 270
Pro Leu Val Arg Arg Thr Val Leu Val Ile Ala Gly Ala Val Arg Gly
275 280 285
Val Gly Gly Ile Asp Ser Trp Gly Thr Asp Val Glu Ser Ala Tyr His
290 295 300
Ile Asn Pro Glu Leu Asp His Glu Phe Ser Phe Ile Leu Asn
305 310 315
<210> 22
<211> 1023
<212> PRT
<213> Bifidobacterium longum
<400> 22
Met Thr Asp Val Thr His Val Asp Arg Ala Ser Gln Ala Trp Leu Thr
1 5 10 15
Asp Pro Thr Val Phe Glu Val Asn Arg Thr Pro Ala His Ser Ser His
20 25 30
Lys Trp Tyr Ala Arg Asp Pro Gln Ser Gly Gln Trp Ser Asp Leu Lys
35 40 45
Gln Ser Leu Asp Gly Glu Trp Arg Val Glu Val Val Gln Ala Ala Asp
50 55 60
Ile Asn Leu Glu Glu Glu Pro Ala Thr Ala Glu Ser Phe Asp Asp Ser
65 70 75 80
Ser Phe Glu Arg Ile Gln Val Pro Gly His Leu Gln Thr Ala Gly Leu
85 90 95
Met Asn His Lys Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asn Pro Leu Glu Pro Asn Ile Pro Glu Asn Asn His Val Ala Leu Tyr
115 120 125
Arg Arg Lys Phe Thr Val Ser Ala Pro Val Ala Asn Ala Lys Gln Ala
130 135 140
Gly Gly Ser Val Ser Ile Val Phe His Gly Met Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Val Asn Gly Ala Phe Val Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Asn Glu Phe Asp Ile Thr Glu Leu Leu His Asp Gly Glu Asn Val
180 185 190
Val Ala Val Ala Cys Tyr Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp
195 200 205
Gln Asp Phe Trp Arg Leu His Gly Leu Phe Arg Ser Val Glu Leu Ala
210 215 220
Ala Arg Pro His Val His Ile Glu Asn Thr Gln Ile Glu Ala Asp Trp
225 230 235 240
Asp Pro Glu Ala Gly Thr Ala Ser Leu Asp Ala Ala Leu Thr Val Leu
245 250 255
Asn Ala Ala Asp Ala Ala Thr Val Arg Ala Thr Leu Lys Asp Ala Asp
260 265 270
Gly Asn Thr Val Trp Gln Thr Thr Gly Asp Ala Glu Ala Gln Thr Ala
275 280 285
Ile Ser Ser Gly Pro Leu Gln Gly Ile Ala Pro Trp Ser Ala Glu Ser
290 295 300
Pro Thr Leu Tyr Glu Leu Asp Val Asp Val Ile Asp Gln Ala Gly Asp
305 310 315 320
Val Ile Glu Cys Thr Ser Gln Lys Val Gly Phe Arg Arg Phe Arg Ile
325 330 335
Glu Asp Gly Ile Leu Thr Ile Asn Gly Lys Arg Ile Val Phe Lys Gly
340 345 350
Ala Asp Arg His Glu Phe Asp Ala Glu Gln Gly Arg Ala Ile Thr Glu
355 360 365
Gln Asp Met Ile Asp Asp Val Val Phe Cys Lys Arg His Asn Ile Asn
370 375 380
Ser Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Arg Trp Tyr Glu Leu
385 390 395 400
Cys Asp Glu Tyr Gly Ile Tyr Leu Ile Asp Glu Ala Asn Leu Glu Ala
405 410 415
His Gly Ser Trp Ser Leu Pro Gly Asp Val Leu Thr Glu Asp Thr Ile
420 425 430
Val Pro Gly Ser Lys Arg Glu Trp Glu Gly Ala Cys Val Asp Arg Val
435 440 445
Asn Ser Met Met Arg Arg Asp Tyr Asn His Pro Ser Val Leu Ile Trp
450 455 460
Ser Leu Gly Asn Glu Ser Tyr Val Gly Asp Val Phe Arg Ala Met Tyr
465 470 475 480
Lys His Val His Asp Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly
485 490 495
Val Thr His Asn Arg Asp Tyr Asp Asp Val Thr Asp Ile Glu Thr Arg
500 505 510
Met Tyr Ser His Ala Asp Glu Ile Glu Lys Tyr Leu Lys Asp Asp Pro
515 520 525
Lys Lys Pro Tyr Leu Ser Cys Glu Tyr Met His Ala Met Gly Asn Ser
530 535 540
Val Gly Asn Met Asp Glu Tyr Thr Ala Leu Glu Arg Tyr Pro Lys Tyr
545 550 555 560
Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Ile Tyr Ala Thr
565 570 575
Gln Pro Asp Gly Thr Arg Ser Leu Arg Tyr Gly Gly Asp Phe Gly Asp
580 585 590
Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Leu Phe Ala Asn
595 600 605
Arg Lys Pro Ser Pro Lys Ala Gln Glu Val Lys Gln Leu Tyr Ser Asn
610 615 620
Val His Ile Asp Val Thr Lys Asp Ser Val Ser Val Lys Asn Asp Asn
625 630 635 640
Leu Phe Thr Ala Thr Gly Asp Tyr Val Phe Val Leu Ser Val Leu Ala
645 650 655
Asp Gly Lys Pro Val Trp Gln Ser Thr Arg Arg Phe Asp Val Pro Ala
660 665 670
Gly Glu Thr Arg Thr Phe Asp Val Ala Trp Pro Val Ala Ala Tyr Arg
675 680 685
Ala Asp Ala Arg Glu Leu Val Leu Gln Val Ser Gln Arg Leu Ala Lys
690 695 700
Ala Thr Asp Trp Ala Glu Ser Gly Tyr Glu Leu Ala Phe Gly Gln Thr
705 710 715 720
Val Val Pro Ala Asp Ala Thr Ala Thr Pro Asp Thr Lys Pro Ala Asp
725 730 735
Gly Thr Ile Thr Val Gly Arg Trp Asn Ala Gly Val Arg Gly Ala Gly
740 745 750
Arg Glu Val Leu Leu Ser Arg Thr Gln Gly Gly Met Val Ser Tyr Thr
755 760 765
Phe Ala Gly Asn Glu Phe Val Leu Arg Arg Pro Ala Ile Thr Thr Phe
770 775 780
Arg Pro Leu Thr Asp Asn Asp Arg Gly Ala Gly His Gly Phe Glu Arg
785 790 795 800
Val Gln Trp Leu Gly Ala Gly Arg Tyr Ala Arg Cys Val Asp Asn Val
805 810 815
Leu Glu Gln Ile Asp Asp Ser Thr Leu Lys Gly Thr Tyr Thr Tyr Glu
820 825 830
Leu Ala Thr Ala Gln Arg Thr Lys Val Thr Val Ser Tyr Thr Ala His
835 840 845
Thr Asp Gly Arg Val Asn Leu His Val Glu Tyr Pro Gly Glu Gln Gly
850 855 860
Asp Leu Pro Thr Ile Pro Ala Phe Gly Ile Glu Trp Thr Leu Pro Val
865 870 875 880
Gln Tyr Thr Asn Leu Arg Phe Phe Gly Thr Gly Pro Ala Glu Thr Tyr
885 890 895
Leu Asp Arg Lys His Ala Lys Leu Gly Val Trp Ser Thr Asn Ala Phe
900 905 910
Ala Asp His Ala Pro Tyr Leu Met Pro Gln Glu Thr Gly Asn His Glu
915 920 925
Asp Val Arg Trp Ala Glu Ile Thr Asp Asp His Gly His Gly Met Arg
930 935 940
Val Ser Arg Ala Asp Gly Ala Ala Pro Phe Ala Val Ser Leu Leu Pro
945 950 955 960
Tyr Ser Ser Phe Met Leu Glu Glu Ala Gln His Gln Asp Glu Leu Pro
965 970 975
Lys Pro Lys His Met Phe Leu Arg Val Leu Ala Ala Gln Met Gly Val
980 985 990
Gly Gly Asp Asp Ser Trp Met Ser Pro Val His Pro Gln Tyr His Ile
995 1000 1005
Pro Ala Asp Lys Pro Ile Ser Leu Asp Val Asp Leu Glu Leu Ile
1010 1015 1020
<210> 23
<211> 628
<212> PRT
<213> Lactobacillus reuteri
<400> 23
Met Asp Ala Asp Ile Lys Trp Leu Asp Glu Pro Glu Thr Phe Arg Val
1 5 10 15
Asn Gln Leu Pro Ala His Ser Asp His Tyr Tyr Tyr Gly Asn Tyr Asp
20 25 30
Glu Trp Arg His Asn Asn Ser Arg Phe Ala Gln Asn Leu Asp Gly Gln
35 40 45
Trp Gln Phe Asn Phe Ala Glu Asn Leu Arg Glu Arg Glu Asn Asp Phe
50 55 60
Tyr Lys Met Asp Tyr Asp Ser Ser Ser Phe Gly Thr Ile Glu Val Pro
65 70 75 80
Ser Glu Ile Glu Leu Asn Asn Tyr Ala Gln Asn Asn Tyr Ile Asn Thr
85 90 95
Leu Ile Pro Trp Glu Gly Lys Ile Tyr Arg Arg Pro Ala Tyr Thr Leu
100 105 110
Ser Pro Asp Asp Ala Gln Glu Gly Ser Phe Ser Asp Gly Asp Asp Asn
115 120 125
Thr Ile Gly Glu Tyr Leu Lys His Phe Asp Leu Asp Pro Ser Leu Arg
130 135 140
Gly Lys Gln Val Arg Ile Arg Phe Asp Gly Val Glu Arg Ala Met Tyr
145 150 155 160
Val Trp Leu Asn Gly His Phe Ile Gly Tyr Ala Glu Asp Ser Phe Thr
165 170 175
Pro Ser Glu Phe Asp Leu Thr Pro Tyr Ile Gln Asp Glu Gly Asn Val
180 185 190
Leu Ala Val Glu Val Phe Lys His Ser Thr Ala Ser Trp Ile Glu Asp
195 200 205
Gln Asp Met Phe Arg Phe Ser Gly Ile Phe Arg Ser Val Asn Leu Leu
210 215 220
Ala Gln Pro Leu Val His Val Glu Asp Leu Asn Ile Arg Pro Ile Val
225 230 235 240
Thr Asp Asn Tyr Gln Asp Gly Ile Phe Asn Val Glu Leu Gln Leu His
245 250 255
Gly Glu Lys Thr Gly Asn Val Asn Val Arg Val Ile Asp Asn Asp Gly
260 265 270
Asn Thr Leu Val Asn Glu Thr His Pro Val Asp Ser Thr Val Lys Val
275 280 285
Gln Asp Gln Phe Leu Glu Asn Val His Leu Trp Asp Asn His Asp Pro
290 295 300
Tyr Leu Tyr Gln Leu Leu Ile Glu Ile Arg Asp Asp Glu Gly Asn Leu
305 310 315 320
Val Glu Leu Val Pro Tyr Arg Phe Gly Phe Arg Arg Ile Glu Ile Asn
325 330 335
Lys Asp His Val Val Leu Leu Asn Gly Gln Arg Leu Ile Ile Asn Gly
340 345 350
Val Asn Arg His Glu Trp Asp Ala Arg Arg Gly Arg Ala Ile Thr Met
355 360 365
Asp Asp Met Thr Ser Asp Ile His Thr Phe Lys Glu Asn Asn Ile Asn
370 375 380
Ala Val Arg Thr Cys His Tyr Pro Asp Gln Ile Pro Trp Tyr Tyr Leu
385 390 395 400
Cys Asp Asp Asn Gly Ile Tyr Met Met Ala Glu Asn Asn Leu Glu Ser
405 410 415
His Ala Thr Trp Gln Lys Met Gly Ala Ile Glu Pro Ser Tyr Asn Val
420 425 430
Pro Gly Ser Val Pro Gln Trp Arg Asp Val Val Val Asp Arg Ala Arg
435 440 445
Thr Asn Tyr Glu Thr Phe Lys Asn His Pro Ser Ile Leu Phe Trp Ser
450 455 460
Leu Gly Asn Glu Ser Tyr Ala Gly Asp Asn Ile Val Lys Met Asn Glu
465 470 475 480
Phe Tyr Lys Lys His Asp Asp Ser Arg Leu Val His Tyr Glu Gly Val
485 490 495
Cys His Thr Pro Glu Tyr Arg Asp Arg Ile Ser Asp Val Glu Ser Trp
500 505 510
Met Tyr Leu Pro Pro Lys Glu Val Glu Glu Tyr Leu Lys Asn Asn Pro
515 520 525
Asp Lys Pro Phe Met Glu Cys Glu Tyr Met His Asp Met Gly Asn Ser
530 535 540
Asp Gly Gly Met Gly Ser Tyr Ile Ser Leu Leu Asp Lys Tyr Pro Gln
545 550 555 560
Tyr Phe Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val
565 570 575
Lys Asp Pro Val Ser Gly Gln Glu Val Met Arg Tyr Gly Gly Asp Phe
580 585 590
Asp Asp Arg His Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Met Phe
595 600 605
Ala Asp Arg Thr Pro Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr
610 615 620
Gly Leu His Lys
625
<210> 24
<211> 319
<212> PRT
<213> Lactobacillus reuteri
<400> 24
Met Ala Tyr Thr Asn Lys Leu Arg Val Ile Tyr Gly Asp Ala Thr Leu
1 5 10 15
Gly Leu Ser Gly Asp Gly Phe His Tyr Ile Phe Ser Tyr Glu Arg Gly
20 25 30
Gly Leu Glu Ser Leu Lys Leu Asn Gly Lys Glu Trp Leu Tyr Arg Glu
35 40 45
Pro Met Pro Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly Ser
50 55 60
Gly Phe Asn Ile Arg Ser Ala Gln Trp Leu Ala Ala Asp Thr Phe His
65 70 75 80
Lys Cys Val Gly Ile Asp Leu Thr Val Asp Asn Gln His Phe Ala Glu
85 90 95
Leu Pro Ile Ala Pro Ile Thr Asn Glu Phe Ser Asp Pro Val Ser Ala
100 105 110
Glu Ser Val Lys Ile Lys Tyr Thr Phe Ala Thr Leu Thr Val Pro Ala
115 120 125
Thr Gln Val Thr Val Ile Tyr Glu Val Asn Gly Gln Gly Glu Ile Lys
130 135 140
Val Thr Met His Tyr Tyr Gly His Glu Asp Leu Pro Gly Leu Pro Val
145 150 155 160
Val Gly Met Arg Phe Ile Met Pro Thr Val Ala Thr Gly Phe Asp Tyr
165 170 175
Gln Gly Leu Ser Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Ala Thr
180 185 190
Glu Gly Thr Phe His Val Asp Gly Leu Pro Val Thr Lys Tyr Leu Val
195 200 205
Pro Gln Glu Asn Gly Met His Met Ala Thr His Ala Leu Thr Ile Thr
210 215 220
Arg Asp Ser Thr Gln Asn Asn Ala Asp His Ser Arg Glu Pro Phe Ser
225 230 235 240
Leu Thr Val Lys Gln Asp Ala Gln Pro Phe Ala Phe Ser Cys Leu Pro
245 250 255
Tyr Thr Ala Glu Glu Leu Glu Asn Ala Thr His Ile Glu Glu Leu Pro
260 265 270
Leu Ala Arg Arg Thr Val Leu Val Val Ala Gly Ala Val Arg Gly Val
275 280 285
Gly Gly Ile Asp Ser Trp Gly Ala Asp Val Glu Glu Gln Tyr His Ile
290 295 300
Pro Ala Asp Arg Asp Val Glu Phe Ser Phe Val Leu Asn Ala Lys
305 310 315
<210> 25
<211> 1007
<212> PRT
<213> Lactobacillus delbrueckii subsp. lactis
<400> 25
Met Ser Asn Lys Leu Val Lys Glu Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Val Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Gln Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asn Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Gln Gly Pro Ile Asn Phe Tyr Ala Glu Asp Phe Asp
65 70 75 80
Asp Ser Asn Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Glu Glu Ile Phe Pro Pro Gln Val Pro Ser Lys Ile Pro Leu Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Asp Glu Ala Leu Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Ala Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Gly Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Leu Ser Gly Leu Phe Arg Ser Val Thr Leu Glu Ala Lys Pro
210 215 220
Leu Leu His Leu Glu Asp Leu Lys Leu Thr Ala Ser Leu Thr Asp Asn
225 230 235 240
Tyr Gln Lys Gly Lys Leu Glu Val Glu Ala Asn Ile Ala Tyr Arg Leu
245 250 255
Pro Asn Ala Ser Phe Lys Leu Glu Val Arg Asp Ser Glu Gly Asp Leu
260 265 270
Val Ala Glu Lys Val Gly Pro Ile Arg Ser Glu Lys Leu Asp Phe Ser
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Lys Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Tyr Leu Tyr Gln Ala Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Gln Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Ala Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Gln Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Leu Phe Tyr Arg Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp Asp
420 425 430
Gln His Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met Ala
435 440 445
Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn Glu
450 455 460
Ser Tyr Ala Gly Thr Val Phe Ala Gln Met Ala Asp Tyr Val Arg Lys
465 470 475 480
Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn Arg
485 490 495
Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro Ala
500 505 510
Lys Glu Ile Glu Glu Tyr Leu Thr Lys Lys Pro Ala Lys Pro Phe Ile
515 520 525
Ser Val Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu Ala
530 535 540
Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe Ile
545 550 555 560
Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Asp Gly His Leu Leu Tyr
565 570 575
Gly Gly Asp Phe Asp Asp Arg Pro Thr Asp Tyr Glu Phe Cys Gly Asp
580 585 590
Gly Leu Val Phe Ala Asp Arg Thr Thr Ser Pro Lys Leu Ala Asn Val
595 600 605
Lys Ala Leu Tyr Ser Asn Leu Lys Leu Glu Val Lys Asp Gly Gln Leu
610 615 620
Phe Ile Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ala Tyr Tyr Phe
625 630 635 640
Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Gln Pro
645 650 655
Leu Thr Phe Gly Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu Pro
660 665 670
Trp Pro Glu Val Glu Asp Glu Lys Gly Glu Ile Val Tyr Gln Val Thr
675 680 685
Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr Val
690 695 700
Ala Glu Ala Glu Glu Ala Val Thr Lys Leu Pro Glu Phe Tyr Pro Ala
705 710 715 720
Gly Arg Pro Glu Leu Val Asp Ser Asp Phe Asn Leu Gly Leu Lys Gly
725 730 735
Asn Gly Phe Arg Ile Leu Phe Ser Lys Ala Lys Gly Trp Pro Val Ser
740 745 750
Ile Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe Thr
755 760 765
Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly Tyr
770 775 780
Asp Leu Ala Lys Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Gln Asp
785 790 795 800
Ile Ser Tyr Glu Ile Lys Glu Asn Ser Ala Leu Val Lys Thr Thr Phe
805 810 815
Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Ile Thr Tyr Glu Val
820 825 830
Asp Ser Leu Gly Lys Ile Ala Val Thr Ala Asn Phe Pro Gly Ala Val
835 840 845
Glu Asn Gly Leu Leu Pro Ala Phe Gly Leu Asn Phe Ala Leu Pro Lys
850 855 860
Glu Leu Ser Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser Tyr
865 870 875 880
Ala Asp Arg Leu Glu Gly Ser Tyr Leu Gly Ile Tyr Gln Gly Met Val
885 890 895
Glu Lys Asn Phe Thr Pro Tyr Leu Arg Pro Gln Glu Ala Gly Asn Arg
900 905 910
Ser Lys Val Arg Tyr Tyr Gln Leu Phe Asp Glu Glu Gly Gly Leu Glu
915 920 925
Phe Thr Ala Asn Gly Ala Asp Leu Asn Leu Ser Ala Leu Pro Tyr Ser
930 935 940
Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn Asn
945 950 955 960
Tyr Thr Trp Val Arg Ala Leu Ala Ala Gln Met Gly Val Gly Gly Asp
965 970 975
Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala Gln
980 985 990
Glu Ala Arg Gln Leu Lys Leu Val Ile Gln Pro Leu Leu Leu Lys
995 1000 1005
<210> 26
<211> 628
<212> PRT
<213> Lactobacillus helvaticus
<400> 26
Met Gln Ala Asn Ile Asn Trp Leu Asp Asn Pro Glu Val Phe Arg Val
1 5 10 15
Asn Gln Leu Pro Ala His Ser Asp His Pro Phe Phe Arg Asp Tyr Arg
20 25 30
Glu Trp Gln Lys Gln His Ser Ser Tyr Gln Gln Ser Leu Asn Gly Lys
35 40 45
Trp Lys Phe His Phe Ser Ala Asn Pro Met Asp Arg Pro Gln Asp Phe
50 55 60
Tyr Gln Arg Asp Phe Asp Ser Ser Asn Phe Asp Ser Ile Pro Val Pro
65 70 75 80
Ser Glu Ile Glu Leu Ser Asn Tyr Thr Gln Asn Gln Tyr Ile Asn Val
85 90 95
Leu Phe Pro Trp Glu Gly Lys Ile Phe Arg Arg Pro Ala Tyr Ala Leu
100 105 110
Asp Pro Asn Asp His Glu Glu Gly Ser Phe Ser Lys Gly Ala Asp Asn
115 120 125
Thr Val Gly Ser Tyr Leu Lys Arg Phe Asp Leu Ser Ser Ala Leu Ile
130 135 140
Gly Lys Asp Val His Ile Lys Phe Glu Gly Val Glu Gln Ala Met Tyr
145 150 155 160
Val Trp Leu Asn Gly His Phe Val Gly Tyr Ala Glu Asp Ser Phe Thr
165 170 175
Pro Ser Glu Phe Asp Leu Thr Pro Tyr Ile Gln Glu Lys Asp Asn Leu
180 185 190
Leu Ala Val Glu Val Phe Lys His Ser Thr Ala Ser Trp Leu Glu Asp
195 200 205
Gln Asp Met Phe Arg Phe Ser Gly Ile Phe Arg Ser Val Glu Leu Leu
210 215 220
Gly Ile Pro Ala Thr His Leu Met Asp Met Asp Leu Lys Pro Arg Val
225 230 235 240
Ala Asp Asn Tyr Gln Asp Gly Ile Phe Asn Leu Lys Leu His Phe Ile
245 250 255
Gly Lys Lys Ala Gly Ser Phe His Leu Leu Val Lys Asp Ile Lys Gly
260 265 270
His Thr Leu Leu Glu Lys Asn Glu Asp Ile Lys Glu Asn Val Gln Ile
275 280 285
Asn Asn Glu Lys Phe Glu Asn Val His Leu Trp Asn Asn His Asp Pro
290 295 300
Tyr Leu Tyr Gln Leu Leu Ile Glu Val Tyr Asp Glu Gln Gln Asn Leu
305 310 315 320
Leu Glu Leu Ile Pro Phe Gln Phe Gly Phe Arg Arg Ile Glu Ile Ser
325 330 335
Pro Glu Lys Val Val Leu Leu Asn Gly Lys Arg Leu Ile Ile Asn Gly
340 345 350
Val Asn Arg His Glu Trp Asp Ala Lys Arg Gly Arg Ser Ile Thr Met
355 360 365
Ser Asp Met Thr Thr Asp Ile Asn Thr Phe Lys Glu Asn Asn Ile Asn
370 375 380
Ala Val Arg Thr Cys His Tyr Pro Asn Gln Ile Pro Trp Tyr Tyr Leu
385 390 395 400
Cys Asp Gln Asn Gly Ile Tyr Val Met Ala Glu Asn Asn Leu Glu Ser
405 410 415
His Gly Thr Trp Gln Lys Met Gly Glu Ile Glu Pro Ser Asp Asn Val
420 425 430
Pro Gly Ser Ile Pro Gln Trp Lys Glu Ala Val Ile Asp Arg Ala Arg
435 440 445
Asn Asn Tyr Glu Thr Phe Lys Asn His Thr Ser Ile Leu Phe Trp Ser
450 455 460
Leu Gly Asn Glu Ser Tyr Ala Gly Asp Asn Ile Ile Ala Met Asn Glu
465 470 475 480
Phe Tyr Lys Ser His Asp Asp Thr Arg Leu Val His Tyr Glu Gly Val
485 490 495
Val His Arg Pro Glu Leu Lys Asp Lys Ile Ser Asp Val Glu Ser Cys
500 505 510
Met Tyr Leu Pro Pro Lys Lys Val Glu Glu Tyr Leu Gln Asn Asp Pro
515 520 525
Pro Lys Pro Phe Met Glu Cys Glu Tyr Met His Asp Met Gly Asn Ser
530 535 540
Asn Gly Gly Met Asp Ser Tyr Ile Lys Leu Leu Asp Lys Tyr Pro Gln
545 550 555 560
Tyr Phe Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val
565 570 575
His Asp Glu Ile Ser Gly His Asp Val Leu Arg Tyr Gly Gly Asp Phe
580 585 590
Asp Asp Arg His Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Met Phe
595 600 605
Ala Asp Arg Lys Pro Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr
610 615 620
Gly Leu His Lys
625
<210> 27
<211> 318
<212> PRT
<213> Lactobacillus helvaticus
<400> 27
Met Asp Tyr Thr Asn Asn Gln Leu His Ile Ile Tyr Gly Asp Ala Thr
1 5 10 15
Phe Gly Val Asn Gly Lys Asp Phe Gln Tyr Ile Phe Ser Tyr Glu Arg
20 25 30
Gly Gly Leu Glu Ser Leu Lys Val His Gly Lys Glu Trp Leu Tyr Arg
35 40 45
Val Pro Thr Pro Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly
50 55 60
Ser Gly Phe Asn Leu Lys Ala Ala Gln Trp Leu Gly Ala Asp Met Phe
65 70 75 80
Thr Lys Cys Thr Asp Ile His Leu Lys Val Asp Arg His Asp Phe Ala
85 90 95
Glu Leu Pro Ile Ala Pro Phe Asn Asn Lys Phe Ser Asn His Glu Tyr
100 105 110
Ala Lys Ser Ala Glu Ile Ser Phe Thr Tyr Gln Thr Leu Thr Thr Pro
115 120 125
Ala Thr Asn Ala Lys Ile Ile Tyr Asn Ile Asp Asp Gly Gly His Ile
130 135 140
Lys Val Thr Met Arg Tyr Tyr Gly Lys Lys Gly Leu Pro Pro Leu Pro
145 150 155 160
Val Ile Gly Ile Arg Leu Ile Met Pro Thr Ala Ala Thr Gly Phe Asp
165 170 175
Tyr Glu Gly Leu Ser Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Ala
180 185 190
Lys Glu Gly Lys Phe His Ile Asp Gly Leu Pro Val Thr Glu Tyr Leu
195 200 205
Val Pro Gln Glu Asn Gly Met His Met Gln Thr Lys Lys Leu Thr Ile
210 215 220
Asn Arg Glu Thr Thr Gln Asn Asn Val Asp Arg Thr Asn Glu Lys Phe
225 230 235 240
Ser Leu Ser Ile Gln Gln Ala Glu Lys Pro Phe Asn Phe Ser Cys Leu
245 250 255
Pro Tyr Thr Ala Glu Glu Leu Glu Asn Ala Thr His Ile Glu Glu Leu
260 265 270
Pro Leu Val Arg Arg Thr Val Leu Val Ile Ala Gly Ala Val Arg Gly
275 280 285
Val Gly Gly Ile Asp Ser Trp Gly Thr Asp Val Glu Ser Ala Tyr His
290 295 300
Ile Asn Pro Asp Leu Asp His Glu Phe Ser Phe Ile Leu Asn
305 310 315
<210> 28
<211> 626
<212> PRT
<213> Lactobacillus crispatus
<400> 28
Met Lys Ala Asn Ile Lys Trp Leu Asp Asp Pro Glu Val Phe Arg Ile
1 5 10 15
Asn Gln Leu Pro Ala His Ser Asp His Pro Phe Tyr Lys Asp Tyr Arg
20 25 30
Glu Trp Gln Lys His Ser Ser Ser Phe Lys Gln Ser Leu Asn Gly Ala
35 40 45
Trp Gln Phe His Phe Ser Lys Asp Pro Gln Ser Arg Pro Ile Asp Phe
50 55 60
Tyr Lys Leu Ser Phe Asp Ser Ser Ser Phe Asp Thr Ile Pro Val Pro
65 70 75 80
Ser Glu Ile Glu Leu Asn Gly Tyr Ala Gln Asn Gln Tyr Thr Asn Ile
85 90 95
Leu Tyr Pro Trp Glu Ser Lys Ile Tyr Arg Lys Pro Ala Tyr Thr Leu
100 105 110
Gly Arg Gly Ile Lys Asp Gly Asp Phe Ser Gln Gly Lys Asp Asn Thr
115 120 125
Val Gly Ser Tyr Leu Lys His Phe Asp Leu Asn Pro Ala Leu Ala Gly
130 135 140
His Asp Ile His Ile Gln Phe Glu Gly Val Glu Arg Ala Met Tyr Val
145 150 155 160
Tyr Leu Asn Gly His Phe Ile Gly Tyr Ala Glu Asp Ser Phe Thr Pro
165 170 175
Ser Glu Phe Asp Leu Thr Pro Tyr Ile Gln Ala Lys Asp Asn Ile Leu
180 185 190
Ala Val Glu Val Phe Lys His Ser Thr Ala Ser Trp Leu Glu Asp Gln
195 200 205
Asp Met Phe Arg Phe Ser Gly Ile Phe Arg Ser Val Glu Leu Leu Ala
210 215 220
Leu Pro Arg Thr His Leu Met Asp Leu Asp Ile Lys Pro Thr Val Val
225 230 235 240
Asn Asp Tyr His Asp Gly Val Phe Asn Ala Lys Leu His Phe Met Gly
245 250 255
Lys Thr Ser Gly Asn Val His Val Leu Ile Glu Asp Ile Asp Gly Lys
260 265 270
Thr Leu Leu Asn Lys Lys Leu Pro Leu Lys Ser Thr Val Glu Ile Glu
275 280 285
Asn Glu Thr Phe Ala Asn Val His Leu Trp Asp Asn His Asp Pro Tyr
290 295 300
Leu Tyr Gln Leu Ile Ile Glu Val His Asp Gln Asp Gly Lys Leu Val
305 310 315 320
Glu Leu Ile Pro Tyr Gln Phe Gly Phe Arg Lys Ile Glu Ile Thr Lys
325 330 335
Asp His Val Val Leu Leu Asn Gly Lys Arg Leu Ile Ile Asn Gly Val
340 345 350
Asn Arg His Glu Trp Asp Ala Lys Arg Gly Arg Ser Ile Thr Leu Ala
355 360 365
Asp Met Lys Gln Asp Ile Ala Thr Phe Lys His Asn Asn Ile Asn Ala
370 375 380
Val Arg Thr Cys His Tyr Pro Asn Gln Ile Pro Trp Tyr Tyr Leu Cys
385 390 395 400
Asp Gln Asn Gly Ile Tyr Met Met Ala Glu Asn Asn Leu Glu Ser His
405 410 415
Gly Thr Trp Gln Lys Leu Gly Gln Val Glu Ala Thr Ser Asn Val Pro
420 425 430
Gly Ser Ile Pro Glu Trp Arg Glu Val Val Val Asp Arg Ala Arg Ser
435 440 445
Asn Tyr Glu Thr Phe Lys Asn His Thr Ala Ile Leu Phe Trp Ser Leu
450 455 460
Gly Asn Glu Ser Tyr Ala Gly Ser Asn Ile Ala Ala Met Asn Lys Leu
465 470 475 480
Tyr Lys Asp His Asp Ser Ser Arg Leu Thr His Tyr Glu Gly Val Phe
485 490 495
His Ala Pro Glu Phe Lys Lys Glu Ile Ser Asp Leu Glu Ser Cys Met
500 505 510
Tyr Leu Pro Pro Lys Glu Ala Glu Glu Tyr Leu Gln Asn Pro Lys Lys
515 520 525
Pro Leu Val Glu Cys Glu Tyr Met His Asp Met Gly Asn Ser Asp Gly
530 535 540
Gly Ile Gly Ser Tyr Ile Lys Leu Ile Asp Lys Tyr Pro Gln Tyr Met
545 550 555 560
Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Leu Leu Val His Asp
565 570 575
Pro Val Ser Gly Gln Asp Val Leu Arg Tyr Gly Gly Asp Phe Asp Asp
580 585 590
Arg His Ser Asp Tyr Glu Phe Ser Gly Asp Gly Leu Met Phe Ala Asp
595 600 605
Arg Thr Pro Lys Pro Ala Met Gln Glu Val Arg Tyr Tyr Tyr Gly Leu
610 615 620
His Lys
625
<210> 29
<211> 316
<212> PRT
<213> Lactobacillus crispatus
<400> 29
Met Ala Tyr Thr Asn Asn Leu His Val Val Tyr Gly Glu Ala Ser Leu
1 5 10 15
Gly Val Asn Gly Gln Asp Phe Ala Tyr Leu Phe Ser Tyr Glu Arg Gly
20 25 30
Val Leu Glu Ser Leu Lys Ile Lys Asp Lys Glu Trp Leu Tyr Arg Thr
35 40 45
Pro Thr Pro Thr Phe Trp Arg Ala Thr Thr Asp Asn Asp Arg Gly Ser
50 55 60
Gly Phe Asn Gln Lys Ala Ala Gln Trp Leu Gly Ala Asp Met Phe Thr
65 70 75 80
Lys Cys Val Gly Ile His Val Gln Val Asp Asp His Gln Phe Asp Glu
85 90 95
Leu Pro Ile Ala Pro Ile Asn Asn Gln Phe Ser Asn Gln Glu Phe Ala
100 105 110
His Glu Val Lys Val Ala Phe Asp Tyr Glu Thr Leu Thr Thr Pro Ala
115 120 125
Thr Lys Val Lys Ile Ile Tyr Asn Ile Asn Asp Ala Gly His Met Thr
130 135 140
Ile Thr Met His Tyr Phe Gly Lys Lys Gly Leu Pro Pro Leu Pro Val
145 150 155 160
Ile Gly Met Arg Phe Ile Met Pro Thr Lys Ala Lys Ser Phe Asp Tyr
165 170 175
Thr Gly Leu Ser Gly Glu Thr Tyr Pro Asp Arg Met Ala Gly Ala Glu
180 185 190
Arg Gly Thr Phe His Ile Asp Gly Leu Pro Val Thr Lys Tyr Leu Val
195 200 205
Pro Gln Glu Asn Gly Met His Met Gln Thr Asn Glu Leu Val Ile Thr
210 215 220
Arg Asn Ser Thr Gln Asn Asn Ala Asp Lys Asp Gly Asp Phe Ser Leu
225 230 235 240
Lys Ile Thr Gln Thr Lys Gln Pro Phe Asn Phe Ser Leu Leu Pro Tyr
245 250 255
Thr Ala Glu Glu Leu Glu Asn Ala Thr His Ile Glu Glu Leu Pro Leu
260 265 270
Ala Arg Arg Ser Val Leu Val Ile Ala Gly Ala Val Arg Gly Val Gly
275 280 285
Gly Ile Asp Ser Trp Gly Ser Asp Val Glu Glu Gln Tyr His Ile Asp
290 295 300
Pro Glu Gln Asp His Glu Phe Ser Phe Thr Leu Asn
305 310 315
<210> 30
<211> 1025
<212> PRT
<213> Streptococcus thermophilus
<400> 30
Met Asn Met Thr Lys Ile Gln Thr Tyr Leu Asn Asp Pro Lys Ile Val
1 5 10 15
Ser Val Asn Thr Val Asp Ala His Ser Asp His Lys Tyr Phe Glu Ser
20 25 30
Leu Glu Glu Phe Ser Glu Gly Glu Met Lys Leu Arg Gln Ser Leu Asn
35 40 45
Gly Lys Trp Lys Ile His Tyr Ala Gln Asn Thr Asn Gln Val Leu Lys
50 55 60
Asp Phe Tyr Lys Thr Glu Phe Asp Glu Thr Asp Leu Asn Phe Ile Asn
65 70 75 80
Val Pro Gly His Leu Glu Leu Gln Gly Phe Gly Ser Pro Gln Tyr Val
85 90 95
Asn Thr Gln Tyr Pro Trp Asp Gly Lys Glu Phe Leu Arg Pro Pro Gln
100 105 110
Val Pro Gln Glu Ser Asn Ala Val Ala Ser Tyr Val Lys His Phe Thr
115 120 125
Leu Asn Asp Ala Leu Lys Asp Lys Lys Val Phe Ile Ser Phe Gln Gly
130 135 140
Val Ala Thr Ser Ile Phe Val Trp Val Asn Gly Asn Phe Val Gly Tyr
145 150 155 160
Ser Glu Asp Ser Phe Thr Pro Ser Glu Phe Glu Ile Ser Asp Tyr Leu
165 170 175
Val Glu Gly Asp Asn Lys Leu Ala Val Ala Val Tyr Arg Tyr Ser Thr
180 185 190
Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu Tyr Gly Ile Phe
195 200 205
Arg Asp Val Tyr Leu Tyr Ala Ile Pro Lys Val His Val Gln Asp Leu
210 215 220
Phe Val Lys Gly Asp Tyr Asp Tyr Gln Thr Lys Ala Gly Gln Leu Asp
225 230 235 240
Ile Asp Leu Lys Thr Val Gly Asp Tyr Glu Asp Lys Lys Ile Lys Tyr
245 250 255
Val Leu Ser Asp Tyr Glu Gly Ile Val Thr Glu Gly Asp Ala Ser Val
260 265 270
Asn Gly Asp Gly Glu Leu Ser Val Ser Leu Glu Asn Leu Lys Ile Lys
275 280 285
Pro Trp Ser Ala Glu Ser Pro Lys Leu Tyr Asp Leu Ile Leu His Val
290 295 300
Leu Asp Asp Asp Gln Val Val Glu Val Val Pro Val Lys Val Gly Phe
305 310 315 320
Arg Arg Phe Glu Ile Lys Asp Lys Leu Met Leu Leu Asn Gly Lys Arg
325 330 335
Ile Val Phe Lys Gly Val Asn Arg His Glu Phe Asn Ala Arg Thr Gly
340 345 350
Arg Cys Ile Thr Glu Glu Asp Met Leu Trp Asp Ile Lys Val Met Lys
355 360 365
Gln His Asn Ile Asn Ala Val Arg Thr Ser His Tyr Pro Asn Gln Thr
370 375 380
Arg Trp Tyr Glu Leu Cys Asp Glu Tyr Gly Leu Tyr Val Ile Asp Glu
385 390 395 400
Ala Asn Leu Glu Thr His Gly Thr Trp Gln Lys Leu Gly Leu Cys Glu
405 410 415
Pro Ser Trp Asn Ile Pro Ala Ser Glu Pro Glu Trp Leu Pro Ala Cys
420 425 430
Leu Asp Arg Ala Asn Asn Met Phe Gln Arg Asp Lys Asn His Ala Ser
435 440 445
Val Ile Ile Trp Ser Cys Gly Asn Glu Ser Tyr Ala Gly Lys Asp Ile
450 455 460
Ala Asp Met Ala Asp Tyr Phe Arg Ser Val Asp Asn Thr Arg Pro Val
465 470 475 480
His Tyr Glu Gly Val Ala Trp Cys Arg Glu Phe Asp Tyr Ile Thr Asp
485 490 495
Ile Glu Ser Arg Met Tyr Ala Lys Pro Ala Asp Ile Glu Glu Tyr Leu
500 505 510
Thr Thr Gly Lys Leu Val Asp Leu Ser Ser Val Ser Asp Lys His Phe
515 520 525
Ala Ser Gly Asn Leu Thr Asn Lys Pro Gln Lys Pro Tyr Ile Ser Cys
530 535 540
Glu Tyr Met His Thr Met Gly Asn Ser Gly Gly Gly Leu Gln Leu Tyr
545 550 555 560
Thr Asp Leu Glu Lys Tyr Pro Glu Tyr Gln Gly Gly Phe Ile Trp Asp
565 570 575
Phe Ile Asp Gln Ala Ile Tyr Lys Thr Leu Pro Asn Gly Ser Glu Phe
580 585 590
Leu Ser Tyr Gly Gly Asp Trp His Asp Arg Pro Ser Asp Tyr Glu Phe
595 600 605
Cys Gly Asn Gly Ile Val Phe Ala Asp Arg Thr Leu Thr Pro Lys Leu
610 615 620
Gln Thr Val Lys His Leu Tyr Ser Asn Ile Lys Ile Ala Val Asp Glu
625 630 635 640
Lys Ser Val Thr Ile Lys Asn Asp Asn Leu Phe Glu Asp Leu Ser Ala
645 650 655
Tyr Thr Phe Leu Ala Arg Val Tyr Glu Asp Gly Arg Lys Val Ser Glu
660 665 670
Ser Glu Tyr His Phe Asp Val Lys Pro Gly Glu Glu Ala Thr Phe Pro
675 680 685
Val Asn Phe Val Val Glu Ala Ser Asn Ser Glu Gln Ile Tyr Glu Val
690 695 700
Ala Cys Val Leu Arg Glu Ala Thr Lys Trp Ala Pro Lys Gly His Glu
705 710 715 720
Ile Val Arg Gly Gln Tyr Val Val Glu Lys Ile Ser Thr Glu Thr Pro
725 730 735
Val Lys Ala Pro Leu Asn Val Val Glu Gly Asp Phe Asn Ile Gly Ile
740 745 750
Gln Gly Gln Asn Phe Ser Ile Leu Leu Ser Arg Ala Gln Asn Thr Leu
755 760 765
Val Ser Ala Lys Tyr Asn Gly Val Glu Phe Ile Glu Lys Gly Pro Lys
770 775 780
Leu Ser Phe Thr Arg Ala Tyr Thr Asp Asn Asp Arg Gly Ala Gly Tyr
785 790 795 800
Pro Phe Glu Met Ala Gly Trp Lys Val Ala Gly Asn Tyr Ser Lys Val
805 810 815
Thr Asp Thr Gln Ile Gln Ile Glu Asp Asp Ser Val Lys Val Thr Tyr
820 825 830
Val His Glu Leu Pro Gly Leu Ser Asp Val Glu Val Lys Val Thr Tyr
835 840 845
Gln Val Asp Tyr Lys Gly Arg Ile Phe Val Thr Ala Asn Tyr Asp Gly
850 855 860
Lys Ala Gly Leu Pro Asn Phe Pro Glu Phe Gly Leu Glu Phe Ala Ile
865 870 875 880
Gly Ser Gln Phe Thr Asn Leu Ser Tyr Tyr Gly Tyr Gly Ala Glu Glu
885 890 895
Ser Tyr Arg Asp Lys Leu Pro Gly Ala Tyr Leu Gly Arg Tyr Glu Thr
900 905 910
Ser Val Glu Lys Thr Phe Ala Pro Tyr Leu Met Pro Gln Glu Ser Gly
915 920 925
Asn His Tyr Gly Thr Arg Glu Phe Thr Val Ser Asp Asp Asn His Asn
930 935 940
Gly Leu Lys Phe Thr Ala Leu Asn Lys Ala Phe Glu Phe Ser Ala Leu
945 950 955 960
Arg Asn Ser Thr Glu Gln Ile Glu Asn Ala Arg His Gln Tyr Glu Leu
965 970 975
Gln Glu Ser Asp Ala Thr Trp Ile Lys Val Leu Ala Ala Gln Met Gly
980 985 990
Val Gly Gly Asp Asp Ser Trp Gly Ala Pro Val His Asp Glu Phe Leu
995 1000 1005
Leu Ser Ser Ala Asp Ser Tyr Gln Leu Ser Phe Met Ile Glu Pro
1010 1015 1020
Leu Asn
1025
<210> 31
<211> 1008
<212> PRT
<213> Lactobacillus delbrueckii subsp. indicus
<400> 31
Met Asn Asn Lys Leu Ala Gln Val Lys Arg Val Asp Gln Ala Asp Leu
1 5 10 15
Ala Trp Leu Thr Asp Pro Glu Ile Tyr Glu Val Asn Thr Ile Pro Pro
20 25 30
His Ser Asp His Glu Ser Phe Gln Ser Leu Glu Glu Leu Glu Glu Gly
35 40 45
Lys Ser Ser Leu Val Gln Ser Leu Asp Gly Asp Trp Leu Ile Asp Tyr
50 55 60
Ala Glu Asn Gly Glu Gly Pro Ala Asn Phe Tyr Glu Glu Asn Phe Asp
65 70 75 80
Asp Ser Ser Phe Lys Ser Val Lys Val Pro Gly Asn Leu Glu Leu Gln
85 90 95
Gly Phe Gly Gln Pro Gln Tyr Val Asn Val Gln Tyr Pro Trp Asp Gly
100 105 110
Ser Asp Glu Ile Phe Pro Pro Met Ile Pro Ser Lys Asn Pro Val Ala
115 120 125
Ser Tyr Val Arg Tyr Phe Asp Leu Glu Glu Ala Phe Trp Asp Lys Glu
130 135 140
Val Ser Leu Lys Phe Ala Gly Ala Ala Thr Ala Ile Tyr Val Trp Leu
145 150 155 160
Asn Gly His Phe Val Gly Tyr Gly Glu Asp Ser Phe Thr Pro Ser Glu
165 170 175
Phe Met Val Thr Lys Phe Leu Lys Lys Glu Gly Asn Arg Leu Ala Val
180 185 190
Ala Leu Tyr Lys Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe
195 200 205
Trp Arg Met Ser Gly Leu Phe Arg Ser Val Thr Leu Glu Ala Lys Pro
210 215 220
Leu Leu His Leu Gln Asp Leu Lys Leu Thr Ala Ser Leu Thr Asn Asp
225 230 235 240
Tyr Gln Lys Gly Ser Leu Gln Val Glu Ala Asp Ile Asp Tyr Arg Leu
245 250 255
Pro Asn Ser Ser Phe Lys Leu Glu Leu Arg Asp Ser Ala Gly Glu Leu
260 265 270
Val Ala Glu Lys Val Gly Pro Ile Arg Ser Glu Lys Leu Asp Phe Ser
275 280 285
Leu Ala Asp Leu Pro Val Ala Ala Trp Ser Ala Glu Glu Pro Asn Leu
290 295 300
Tyr Gln Val Arg Leu Ser Leu Tyr Gln Gln Gly Ser Leu Leu Glu Val
305 310 315 320
Ser Arg Gln Glu Val Gly Phe Arg Asn Phe Glu Leu Lys Asp Gly Ile
325 330 335
Met Tyr Leu Asn Gly Lys Arg Ile Val Phe Lys Gly Val Asn Arg His
340 345 350
Glu Phe Asp Ser Lys Leu Gly Arg Ala Ile Thr Glu Ala Asp Met Ile
355 360 365
Trp Asp Ile Lys Thr Met Lys Gln Ser Asn Ile Asn Ala Val Arg Cys
370 375 380
Ser His Tyr Pro Asn Gln Ser Ile Phe Tyr His Leu Cys Asp Lys Tyr
385 390 395 400
Gly Leu Tyr Val Ile Asp Glu Ala Asn Leu Glu Ser His Gly Thr Trp
405 410 415
Glu Lys Val Gly Gly His Glu Asp Pro Ser Phe Asn Val Pro Gly Asp
420 425 430
Asp Gln Arg Trp Leu Gly Ala Ser Leu Ser Arg Val Lys Asn Met Met
435 440 445
Ala Arg Asp Lys Asn His Ala Ser Ile Leu Ile Trp Ser Leu Gly Asn
450 455 460
Glu Ser Tyr Ala Gly Lys Val Phe Ala Gln Met Ala Asp Tyr Val Arg
465 470 475 480
Gln Ala Asp Pro Thr Arg Val Gln His Tyr Glu Gly Val Thr His Asn
485 490 495
Arg Lys Phe Asp Asp Ala Thr Gln Ile Glu Ser Arg Met Tyr Ala Pro
500 505 510
Ala Lys Glu Ile Glu Glu Tyr Leu Thr Lys Lys Pro Ala Lys Pro Phe
515 520 525
Val Ser Cys Glu Tyr Ala His Ala Met Gly Asn Ser Val Gly Asp Leu
530 535 540
Ala Ala Tyr Thr Ala Leu Glu Lys Tyr Pro His Tyr Gln Gly Gly Phe
545 550 555 560
Ile Trp Asp Trp Ile Asp Gln Gly Leu Glu Lys Glu Gly His Leu Leu
565 570 575
Tyr Gly Gly Asp Phe Asp Asp Arg Pro Ser Asp Tyr Glu Phe Cys Gly
580 585 590
Asp Gly Leu Val Phe Ala Asp Arg Thr Thr Ser Pro Lys Leu Ala Asn
595 600 605
Val Lys Ala Leu Tyr Ser Asn Leu Lys Leu Glu Leu Lys Asp Gly Gln
610 615 620
Leu Phe Leu Lys Asn Asp Asn Leu Phe Thr Asn Ser Ser Ala Tyr Tyr
625 630 635 640
Phe Leu Thr Ser Leu Leu Val Asp Gly Lys Leu Thr Tyr Gln Ser Gln
645 650 655
Pro Leu Thr Phe Ala Leu Glu Pro Gly Glu Ser Gly Thr Phe Ala Leu
660 665 670
Pro Trp Pro Glu Val Glu Asp Glu Lys Gly Glu Ile Val Tyr Gln Val
675 680 685
Thr Ala His Leu Lys Glu Asp Leu Pro Trp Ala Asp Glu Gly Phe Thr
690 695 700
Val Ala Glu Ala Glu Glu Ala Val Thr Lys Leu Pro Glu Phe Tyr Pro
705 710 715 720
Ala Gly Arg Pro Glu Leu Val Asp Ser Asp Tyr Asn Leu Gly Ile Lys
725 730 735
Gly Asn Gly Phe Arg Ile Leu Phe Ser Lys Ala Lys Gly Trp Pro Val
740 745 750
Ser Ile Lys Tyr Ala Gly Arg Glu Tyr Leu Lys Arg Leu Pro Glu Phe
755 760 765
Thr Phe Trp Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly Tyr Gly
770 775 780
Tyr Asp Leu Ala Lys Trp Glu Asn Ala Gly Lys Tyr Ala Arg Leu Gln
785 790 795 800
Asp Ile Ser Tyr Glu Ile Lys Glu Asn Ser Val Leu Val Lys Thr Ala
805 810 815
Phe Thr Leu Pro Val Ala Leu Lys Gly Asp Leu Thr Ile Thr Tyr Glu
820 825 830
Val Asp Ser Leu Gly Lys Ile Ala Val Thr Ala Asn Phe Pro Gly Ala
835 840 845
Val Glu Asn Gly Leu Leu Pro Ala Phe Gly Leu Asn Phe Ala Leu Pro
850 855 860
Lys Glu Leu Ser Asp Tyr Arg Tyr Tyr Gly Leu Gly Pro Asn Glu Ser
865 870 875 880
Tyr Ala Asp Arg Leu Glu Gly Ser Tyr Leu Gly Ile Tyr Gln Gly Ala
885 890 895
Val Glu Lys Asn Phe Thr Pro Tyr Leu Arg Pro Gln Glu Val Gly Asn
900 905 910
Arg Ser Lys Val Arg Tyr Tyr Gln Leu Phe Asp Glu Glu Gly Gly Leu
915 920 925
Glu Phe Thr Ala Asn Gly Ala Asn Leu Asn Leu Ser Ala Leu Pro Tyr
930 935 940
Ser Ala Ala Gln Ile Glu Ala Ala Asp His Ala Phe Glu Leu Thr Asn
945 950 955 960
Asn Tyr Thr Trp Val Arg Ala Leu Ala Ala Gln Met Gly Val Gly Gly
965 970 975
Asp Asp Ser Trp Gly Gln Lys Val His Pro Glu Phe Cys Leu Asp Ala
980 985 990
Gln Glu Ala Arg Gln Leu Lys Leu Val Ile Gln Pro Leu Phe Thr Glu
995 1000 1005
<210> 32
<211> 1049
<212> PRT
<213> Bifidobacterium adolescentis
<400> 32
Met Ala Asp Thr Ala Glu Leu Ala Ile Val His Ala Thr Thr Ala Ser
1 5 10 15
Ala Ser Trp Leu Thr Asp Pro Thr Val Phe Ala Ala Asn Arg Lys Pro
20 25 30
Ala His Ser Ser His Arg Tyr Val Ile Gly Glu Thr Ser Glu Pro Lys
35 40 45
Gln Ser Leu Asp Gly Glu Trp Lys Val Arg Ile Glu Gln Ala Arg Asn
50 55 60
Val Asp Val Glu Ser Ala Pro Phe Ala Ala Val Asp Phe Glu Asp Gly
65 70 75 80
Asp Phe Gly Ala Ile Glu Val Pro Gly His Leu Gln Met Ala Gly Tyr
85 90 95
Leu Lys Asn Lys Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asp Pro Gln Ala Pro Asn Ile Pro Glu Asn Asn His Val Ala Ile Tyr
115 120 125
Arg Arg Arg Phe Ala Leu Asp Ala Gln Leu Ala Arg Thr Leu Glu Asn
130 135 140
Asp Gly Thr Val Ser Leu Thr Phe His Gly Ala Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Leu Asp Gly Thr Phe Val Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Ser Glu Phe Asp Val Thr Glu Ala Leu Arg Asn Gly Asn Gly Asn
180 185 190
Ala Ala Asp Ser Pro Glu Ala Glu His Thr Leu Thr Val Ala Cys Tyr
195 200 205
Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp Gln Asp Phe Trp Arg Leu
210 215 220
His Gly Leu Phe Arg Thr Val Glu Leu Ala Ala Gln Pro His Thr His
225 230 235 240
Val Glu Thr Val Gln Leu Glu Ala Asp Tyr Thr Ala Ala Asp Thr Ala
245 250 255
Gly Thr Ala Asp Thr Ala Glu Leu Asn Ala Ala Leu Thr Leu Arg Asn
260 265 270
Pro Ala Asp Ala Met Thr Ile Glu Ser Thr Leu Arg Asp Gly Asp Gly
275 280 285
Asn Val Val Trp Glu Ser Thr Gln Ala Cys Asn Gly Glu Ile Ala Leu
290 295 300
Asn Ser Gly Lys Met Thr Asn Ile Ala Pro Trp Ser Ala Glu Ser Pro
305 310 315 320
Thr Leu Tyr Thr Leu Thr Val Arg Val Val Gly His Asp Gly Ala Ile
325 330 335
Ile Glu Thr Val Thr Gln Lys Ile Gly Phe Arg Thr Phe Arg Ile Glu
340 345 350
Asn Gly Ile Met Thr Leu Asn Gly Lys Arg Ile Val Phe Lys Gly Ala
355 360 365
Asp Arg His Glu Phe Asp Ala Lys Arg Gly Arg Ala Ile Thr Arg Glu
370 375 380
Asp Met Leu Ser Asp Val Val Phe Cys Lys Arg His Asn Ile Asn Ala
385 390 395 400
Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Tyr Trp Tyr Asp Leu Cys
405 410 415
Asp Glu Tyr Gly Leu Tyr Leu Ile Asp Glu Thr Asn Met Glu Thr His
420 425 430
Gly Thr Trp Val Ala Asn Asn Val Glu Arg Pro Glu Asp Gly Ile Pro
435 440 445
Gly Ser Arg Pro Glu Trp Glu Asp Ala Cys Val Asp Arg Ile Asn Ser
450 455 460
Met Met Arg Arg Asp Tyr Asn His Pro Ser Val Leu Ile Trp Ser Leu
465 470 475 480
Gly Asn Glu Ser Ser Ala Gly Glu Val Phe Arg Ala Met Tyr Arg His
485 490 495
Ala His Thr Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly Ser Val
500 505 510
His Met Arg Glu Phe Glu Asp Val Thr Asp Ile Glu Ser Arg Met Tyr
515 520 525
Ala His Ala Asp Glu Ile Glu Arg Tyr Leu Asn Asp Gly Ser Pro Ala
530 535 540
His Thr Asp Gly Pro Lys Lys Pro Tyr Ile Ser Cys Glu Tyr Met His
545 550 555 560
Ala Met Gly Asn Ser Cys Gly Asn Met Asp Glu Tyr Thr Ala Leu Glu
565 570 575
Arg Tyr Pro Met Tyr Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln
580 585 590
Ala Ile Glu Thr Lys Leu Pro Asp Gly Thr Thr Arg Met Cys Tyr Gly
595 600 605
Gly Asp Phe Gly Asp Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asp Gly
610 615 620
Leu Leu Phe Ala Asp Arg Thr Pro Ser Pro Lys Ala Gln Glu Val Lys
625 630 635 640
Gln Leu Tyr Ala Asn Val Lys Ile Ala Val Ser Val Asp Glu Ala Arg
645 650 655
Ile Thr Asn Asp Asn Leu Phe Val Ser Thr Gly Asp Tyr Arg Phe Val
660 665 670
Leu Arg Ile Leu Ala Asp Gly Lys Pro Val Trp Ser Thr Thr Arg Arg
675 680 685
Phe Asp Val Ala Ala Gly Glu Ser Ala Ser Phe Glu Val Asp Trp Pro
690 695 700
Val Asp Asp Tyr Arg Ser Asn Ala Glu Glu Leu Val Leu Glu Val Ser
705 710 715 720
Gln Gln Leu Gly Asn Ala Cys Asp Trp Ala Pro Ala Gly Tyr Glu Leu
725 730 735
Ala Phe Gly Gln Cys Val Val Ala Gly Ala Lys Thr Thr Ala Asp Ala
740 745 750
Val Asp Ala Ala Gly Ala Pro Ala Asp Gly Thr Val Thr Leu Gly Arg
755 760 765
Trp Asn Ala Gly Val Arg Gly Gln Gly Arg Glu Ala Leu Phe Ser Arg
770 775 780
Thr Gln Gly Gly Met Val Ser Tyr Thr Phe Gly Glu Arg Glu Phe Val
785 790 795 800
Leu Arg Arg Pro Ser Ile Thr Thr Phe Arg Pro Leu Thr Asp Asn Asp
805 810 815
Arg Gly Ala Gly His Ala Phe Glu Arg Ala Ala Trp Ala Val Ala Gly
820 825 830
Lys Tyr Ala Arg Cys Val Asp Cys Ala Ile Ala Asn Arg Gly Glu Asn
835 840 845
Ala Val Glu Ala Thr Tyr Thr Tyr Glu Leu Ala Ile Pro Gln Arg Thr
850 855 860
Lys Val Thr Val Arg Tyr Val Ala Asp Thr Ala Gly Leu Val Ser Leu
865 870 875 880
Asp Val Glu Tyr Pro Gly Glu Lys Asn Gly Asp Leu Pro Thr Ile Pro
885 890 895
Ala Phe Gly Ile Glu Trp Ala Leu Pro Val Glu Tyr Ala Asn Leu Arg
900 905 910
Phe Tyr Gly Ala Gly Pro Glu Glu Thr Tyr Ala Asp Arg Arg His Ala
915 920 925
Lys Leu Gly Val Trp Ser Thr Thr Ala Gly Asp Asp Cys Ala Pro Tyr
930 935 940
Leu Leu Pro Gln Glu Thr Gly Asn His Glu Asp Val Arg Trp Ala Glu
945 950 955 960
Ile Thr Asp Asp Ser Gly His Gly Val Arg Val Lys Arg Gly Ala Gly
965 970 975
Ala Lys Pro Phe Ala Met Ser Leu Leu Pro Tyr Ser Ser Thr Met Leu
980 985 990
Glu Glu Ala Leu His Gln Asp Glu Leu Pro Lys Pro Arg His Met Phe
995 1000 1005
Leu Arg Leu Leu Ala Ala Gln Met Gly Val Gly Gly Asp Asp Ser
1010 1015 1020
Trp Met Ser Pro Val His Glu Gln Tyr Gln Leu Pro Ala Asp Gln
1025 1030 1035
Pro Leu Ser Leu Asn Val Gln Leu Lys Leu Phe
1040 1045
<210> 33
<211> 1023
<212> PRT
<213> Bifidobacterium adolescentis
<400> 33
Met Ala Asn Glu Thr Arg Ile Glu His Ala Ser Glu Thr Trp Leu Ala
1 5 10 15
Asp Ser Thr Val Phe Glu Val Asn Arg Val Pro Ala His Ser Asp His
20 25 30
Lys Cys Tyr Ala His Asp Ser Gln Thr Asn Glu Trp Ser Asp Leu Arg
35 40 45
Gln Ser Leu Asp Gly Glu Trp Arg Val Glu Val Val Gln Ala Ser Asp
50 55 60
Ile Glu Phe Asn Glu Glu Pro Phe Val Arg Glu Asn Phe Asp Asp Ser
65 70 75 80
Ala Phe Glu Arg Ile Gln Val Pro Gly His Leu Gln Met Ala Gly Leu
85 90 95
Met Asn Asn Lys Tyr Val Asn Ile Gln Tyr Pro Trp Asp Gly His Glu
100 105 110
Asn Pro Ala Glu Pro Asn Ile Pro Glu Asn Asn His Val Ala Leu Tyr
115 120 125
Arg Lys Thr Phe Thr Met Ala Asn Arg Leu Ala Asp Thr Lys Asn Ala
130 135 140
Gly Gly Thr Val Ser Ile Val Phe His Gly Met Ala Thr Ala Ile Tyr
145 150 155 160
Val Trp Val Asn Gly Met Phe Val Gly Tyr Gly Glu Asp Gly Phe Thr
165 170 175
Pro Asn Glu Phe Asp Ile Thr Glu Met Leu His Asp Gly Glu Asn Val
180 185 190
Val Ala Val Ala Cys Tyr Glu Tyr Ser Ser Ala Ser Trp Leu Glu Asp
195 200 205
Gln Asp Phe Trp Arg Leu His Gly Leu Phe Arg Ser Val Glu Leu Ala
210 215 220
Ala Gln Pro His Val His Ile Glu Asn Met Gln Ile Glu Ser Asp Trp
225 230 235 240
Asp Pro Glu Ser Gly Ser Ala Ser Leu Asp Ala Ala Leu Thr Val Arg
245 250 255
Asn Ala Ala Asp Ala Ala Thr Ile Ser Ala Thr Leu Lys Asp Ser Asp
260 265 270
Gly Asn Val Val Trp Glu Thr Ala Asn Cys Ala Asp Pro Asp Thr Ser
275 280 285
Ile Ser Thr Gly Ser Leu Asn Gly Ile Arg Pro Trp Ser Ala Glu Asp
290 295 300
Pro Val Leu Tyr Glu Phe Glu Val Thr Val Ile Asp His Ala Gly Asn
305 310 315 320
Ile Ala Glu Val Ala Val Gln Lys Val Gly Phe Arg Arg Phe Arg Ile
325 330 335
Glu Asp Gly Ile Met Thr Ile Asn Gly Lys Arg Ile Val Phe Lys Gly
340 345 350
Ala Asp Arg His Glu Phe Asp Pro Lys Arg Gly Arg Ala Ile Thr Glu
355 360 365
Gln Asp Met Ile Asp Asp Val Val Phe Cys Lys Arg His Asn Leu Asn
370 375 380
Ala Ile Arg Thr Ser His Tyr Pro Asn Gln Glu Arg Trp Tyr Glu Leu
385 390 395 400
Cys Asp Glu Tyr Gly Ile Tyr Leu Ile Asp Glu Thr Asn Leu Glu Thr
405 410 415
His Gly Ser Trp Cys Leu Pro Gly Asp Val Leu Thr Glu Glu Thr Ala
420 425 430
Val Pro Gly Ser Lys Ala His Trp Glu Gly Ala Cys Val Asp Arg Val
435 440 445
Asn Ser Met Val Arg Arg Asp Tyr Asn His Pro Ser Val Leu Ile Trp
450 455 460
Ser Leu Gly Asn Glu Ser Tyr Thr Gly Asp Val Phe Arg Ala Met Tyr
465 470 475 480
Lys Arg Val His Asp Ile Asp Pro Asn Arg Pro Val His Tyr Glu Gly
485 490 495
Val Thr His Asn Arg Asp Tyr Asn Asp Val Thr Asp Ile Glu Thr Arg
500 505 510
Met Tyr Ala His Ala Asp Ala Ile Glu Glu Tyr Leu Lys Asn Asp Pro
515 520 525
Gln Lys Pro Tyr Ile Ser Cys Glu Tyr Met His Ala Met Gly Asn Ser
530 535 540
Cys Gly Asn Met Asp Glu Tyr Thr Ala Leu Glu Arg Tyr Pro Lys Tyr
545 550 555 560
Gln Gly Gly Phe Ile Trp Asp Phe Ile Asp Gln Ala Ile Tyr Ala Thr
565 570 575
Gln Pro Asp Gly Thr Thr Ser Leu Arg Tyr Gly Gly Asp Phe Gly Asp
580 585 590
Arg Pro Ser Asp Tyr Glu Phe Ser Gly Asn Gly Leu Val Phe Ala Asp
595 600 605
Arg Lys Pro Thr Pro Lys Ala Gln Glu Val Lys Gln Leu Tyr Ser Asn
610 615 620
Val His Ile Asp Val Ala Glu Asp Ser Val Thr Ile Lys Asn Asp Asn
625 630 635 640
Leu Phe Thr Ser Thr Gly Glu Tyr Thr Phe Val Leu Arg Val Leu Ala
645 650 655
Asp Gly Glu Pro Val Trp Gln Ser Glu Arg Arg Phe Asp Val Pro Ala
660 665 670
Gly Ser Thr Glu Lys Leu Asp Val Asp Trp Pro Leu Asp Leu Tyr Arg
675 680 685
Asp Gly Ala Ser Glu Leu Val Leu Glu Val Ser Gln Arg Leu Ala Lys
690 695 700
Ala Thr Asn Trp Ala Val Ala Gly Tyr Glu Leu Ala Phe Gly Gln Thr
705 710 715 720
Val Val Ala Gly Ser Lys Lys Ala Ser Ala Pro Val Lys Pro Val Asp
725 730 735
Gly Ile Val Thr Val Gly Arg Trp Asn Val Gly Val Gln Gly Ser Gly
740 745 750
Arg Glu Val Leu Leu Ser Arg Thr Gln Gly Gly Leu Val Ser Tyr Thr
755 760 765
Phe Asn Asn Arg Glu Phe Val Leu Arg Arg Pro Ala Val Thr Thr Phe
770 775 780
Arg Ala Leu Thr Asp Asn Asp Arg Gly Ala Gly His Gly Phe Glu Arg
785 790 795 800
Ala Gln Trp Leu Gly Ala Gly Arg Tyr Ala Arg Cys Ile Gly Asn Glu
805 810 815
Ile Glu Gln Ile Asp Glu Asn Thr Val Lys Ala Ser Tyr Thr Tyr Glu
820 825 830
Leu Ala Thr Pro Gln Arg Thr Lys Val Thr Val Ser Tyr Thr Ala Asp
835 840 845
Thr Thr Gly Arg Val Asn Leu His Val Glu Tyr Pro Gly Glu Pro Gly
850 855 860
Asp Leu Pro Thr Ile Pro Ala Phe Gly Ile Glu Trp Thr Leu Pro Val
865 870 875 880
Gln Tyr Ser Asn Leu Arg Phe Phe Gly Ala Gly Pro Glu Glu Thr Tyr
885 890 895
Gln Asp Arg Lys His Ala Lys Leu Gly Val Trp Ser Thr Asp Ala Phe
900 905 910
Lys Asp His Ala Pro Tyr Leu Met Pro Gln Glu Thr Gly Asn His Glu
915 920 925
Asp Val Arg Trp Ala Glu Ile Thr Asp Glu Lys Gly His Gly Leu Arg
930 935 940
Ile Ser Arg Ala Glu Gly Ala Glu Pro Phe Ala Met Ser Leu Gln Pro
945 950 955 960
Tyr Ser Ser Phe Met Leu Glu Glu Ala Gln His Gln Asp Glu Leu Pro
965 970 975
Ala Pro Lys His Met Phe Leu Arg Val Leu Ala Glu Gln Met Gly Val
980 985 990
Gly Gly Asp Asp Ser Trp Met Ser Pro Val His Pro Gln Tyr His Ile
995 1000 1005
Pro Ala Asp Gln Pro Ile Ser Leu Asp Val Asp Leu Asp Leu Ile
1010 1015 1020
<210> 34
<211> 1305
<212> PRT
<213> Bifidobacterium bifidum
<400> 34
Met Val Glu Asp Ala Thr Arg Ser Asp Ser Thr Thr Gln Met Ser Ser
1 5 10 15
Thr Pro Glu Val Val Tyr Ser Ser Ala Val Asp Ser Lys Gln Asn Arg
20 25 30
Thr Ser Asp Phe Asp Ala Asn Trp Lys Phe Met Leu Ser Asp Ser Val
35 40 45
Gln Ala Gln Asp Pro Ala Phe Asp Asp Ser Ala Trp Gln Gln Val Asp
50 55 60
Leu Pro His Asp Tyr Ser Ile Thr Gln Lys Tyr Ser Gln Ser Asn Glu
65 70 75 80
Ala Glu Ser Ala Tyr Leu Pro Gly Gly Thr Gly Trp Tyr Arg Lys Ser
85 90 95
Phe Thr Ile Asp Arg Asp Leu Ala Gly Lys Arg Ile Ala Ile Asn Phe
100 105 110
Asp Gly Val Tyr Met Asn Ala Thr Val Trp Phe Asn Gly Val Lys Leu
115 120 125
Gly Thr His Pro Tyr Gly Tyr Ser Pro Phe Ser Phe Asp Leu Thr Gly
130 135 140
Asn Ala Lys Phe Gly Gly Glu Asn Thr Ile Val Val Lys Val Glu Asn
145 150 155 160
Arg Leu Pro Ser Ser Arg Trp Tyr Ser Gly Ser Gly Ile Tyr Arg Asp
165 170 175
Val Thr Leu Thr Val Thr Asp Gly Val His Val Gly Asn Asn Gly Val
180 185 190
Ala Ile Lys Thr Pro Ser Leu Ala Thr Gln Asn Gly Gly Asn Val Thr
195 200 205
Met Asn Leu Thr Thr Lys Val Ala Asn Asp Thr Glu Ala Ala Ala Asn
210 215 220
Ile Thr Leu Lys Gln Thr Val Phe Pro Lys Gly Gly Lys Thr Asp Ala
225 230 235 240
Ala Ile Gly Thr Val Thr Thr Ala Ser Lys Ser Ile Ala Ala Gly Ala
245 250 255
Ser Ala Asp Val Thr Ser Thr Ile Thr Ala Ala Ser Pro Lys Leu Trp
260 265 270
Ser Ile Lys Asn Pro Asn Leu Tyr Thr Val Arg Thr Glu Val Leu Asn
275 280 285
Gly Asp Thr Val Leu Asp Thr Tyr Asp Thr Glu Tyr Gly Phe Arg Trp
290 295 300
Thr Gly Phe Asp Ala Thr Ser Gly Phe Ser Leu Asn Gly Glu Lys Val
305 310 315 320
Lys Leu Lys Gly Val Ser Met His His Asp Gln Gly Ser Leu Gly Ala
325 330 335
Val Ala Asn Arg Arg Ala Ile Glu Arg Gln Val Glu Ile Leu Gln Lys
340 345 350
Met Gly Val Asn Ser Ile Arg Thr Thr His Asn Pro Ala Ala Lys Ala
355 360 365
Leu Ile Asp Val Cys Asn Glu Lys Gly Val Leu Val Val Glu Glu Val
370 375 380
Phe Asp Met Trp Asn Arg Ser Lys Asn Gly Asn Thr Glu Asp Tyr Gly
385 390 395 400
Lys Trp Phe Gly Gln Thr Ile Ala Gly Asp Asn Ala Val Leu Gly Gly
405 410 415
Asp Lys Asp Glu Thr Trp Ala Lys Phe Asp Leu Thr Ser Thr Ile Asn
420 425 430
Arg Asp Arg Asn Ala Pro Ser Val Ile Met Trp Ser Leu Gly Asn Glu
435 440 445
Met Met Glu Gly Ile Ser Gly Ser Val Ser Asp Phe Pro Ala Thr Ser
450 455 460
Ala Lys Leu Val Ala Trp Thr Lys Ala Ala Asp Ser Thr Arg Pro Met
465 470 475 480
Thr Tyr Gly Asp Asn Lys Ile Lys Ala Asn Trp Asn Glu Ser Asn Thr
485 490 495
Met Gly Asp Asn Leu Thr Ala Asn Gly Gly Val Val Gly Thr Asn Tyr
500 505 510
Ser Asp Gly Ala Asn Tyr Asp Lys Ile Arg Thr Thr His Pro Ser Trp
515 520 525
Ala Ile Tyr Gly Ser Glu Thr Ala Ser Ala Ile Asn Ser Arg Gly Ile
530 535 540
Tyr Asn Arg Thr Thr Gly Gly Ala Gln Ser Ser Asp Lys Gln Leu Thr
545 550 555 560
Ser Tyr Asp Asn Ser Ala Val Gly Trp Gly Ala Val Ala Ser Ser Ala
565 570 575
Trp Tyr Asp Val Val Gln Arg Asp Phe Val Ala Gly Thr Tyr Val Trp
580 585 590
Thr Gly Phe Asp Tyr Leu Gly Glu Pro Thr Pro Trp Asn Gly Thr Gly
595 600 605
Ser Gly Ala Val Gly Ser Trp Pro Ser Pro Lys Asn Ser Tyr Phe Gly
610 615 620
Ile Val Asp Thr Ala Gly Phe Pro Lys Asp Thr Tyr Tyr Phe Tyr Gln
625 630 635 640
Ser Gln Trp Asn Asp Asp Val His Thr Leu His Ile Leu Pro Ala Trp
645 650 655
Asn Glu Asn Val Val Ala Lys Gly Ser Gly Asn Lys Val Pro Val Val
660 665 670
Val Tyr Thr Asp Ala Ala Lys Val Lys Leu Tyr Phe Thr Pro Lys Gly
675 680 685
Ser Thr Glu Lys Arg Leu Ile Gly Glu Lys Ser Phe Thr Lys Lys Thr
690 695 700
Thr Ala Ala Gly Tyr Thr Tyr Gln Val Tyr Glu Gly Thr Asp Lys Asp
705 710 715 720
Ser Thr Ala His Lys Asn Met Tyr Leu Thr Trp Asn Val Pro Trp Ala
725 730 735
Glu Gly Thr Ile Ser Ala Glu Ala Tyr Asp Glu Asn Asn Arg Leu Ile
740 745 750
Pro Glu Gly Ser Thr Glu Gly Asn Ala Ser Val Thr Thr Thr Gly Lys
755 760 765
Ala Ala Lys Leu Lys Ala Asp Ala Asp Arg Lys Thr Ile Thr Ala Asp
770 775 780
Gly Lys Asp Leu Ser Tyr Ile Glu Val Asp Val Thr Asp Ala Asn Gly
785 790 795 800
His Ile Val Pro Asp Ala Ala Asn Arg Val Thr Phe Asp Val Lys Gly
805 810 815
Ala Gly Lys Leu Val Gly Val Asp Asn Gly Ser Ser Pro Asp His Asp
820 825 830
Ser Tyr Gln Ala Asp Asn Arg Lys Ala Phe Ser Gly Lys Val Leu Ala
835 840 845
Ile Val Gln Ser Thr Lys Glu Ala Gly Glu Ile Thr Val Thr Ala Lys
850 855 860
Ala Asp Gly Leu Gln Ser Ser Thr Val Lys Ile Ala Thr Thr Ala Val
865 870 875 880
Pro Gly Thr Ser Thr Glu Lys Thr Val Arg Ser Phe Tyr Tyr Ser Arg
885 890 895
Asn Tyr Tyr Val Lys Thr Gly Asn Lys Pro Ile Leu Pro Ser Asp Val
900 905 910
Glu Val Arg Tyr Ser Asp Gly Thr Ser Asp Arg Gln Asn Val Thr Trp
915 920 925
Asp Ala Val Ser Asp Asp Gln Ile Ala Lys Ala Gly Ser Phe Ser Val
930 935 940
Ala Gly Thr Val Ala Gly Gln Lys Ile Ser Val Arg Val Thr Met Ile
945 950 955 960
Asp Glu Ile Gly Ala Leu Leu Asn Tyr Ser Ala Ser Thr Pro Val Gly
965 970 975
Thr Pro Ala Val Leu Pro Gly Ser Arg Pro Ala Val Leu Pro Asp Gly
980 985 990
Thr Val Thr Ser Ala Asn Phe Ala Val His Trp Thr Lys Pro Ala Asp
995 1000 1005
Thr Val Tyr Asn Thr Ala Gly Thr Val Lys Val Pro Gly Thr Ala
1010 1015 1020
Thr Val Phe Gly Lys Glu Phe Lys Val Thr Ala Thr Ile Arg Val
1025 1030 1035
Gln Arg Ser Gln Val Thr Ile Gly Ser Ser Val Ser Gly Asn Ala
1040 1045 1050
Leu Arg Leu Thr Gln Asn Ile Pro Ala Asp Lys Gln Ser Asp Thr
1055 1060 1065
Leu Asp Ala Ile Lys Asp Gly Ser Thr Thr Val Asp Ala Asn Thr
1070 1075 1080
Gly Gly Gly Ala Asn Pro Ser Ala Trp Thr Asn Trp Ala Tyr Ser
1085 1090 1095
Lys Ala Gly His Asn Thr Ala Glu Ile Thr Phe Glu Tyr Ala Thr
1100 1105 1110
Glu Gln Gln Leu Gly Gln Ile Val Met Tyr Phe Phe Arg Asp Ser
1115 1120 1125
Asn Ala Val Arg Phe Pro Asp Ala Gly Lys Thr Lys Ile Gln Ile
1130 1135 1140
Ser Ala Asp Gly Lys Asn Trp Thr Asp Leu Ala Ala Thr Glu Thr
1145 1150 1155
Ile Ala Ala Gln Glu Ser Ser Asp Arg Val Lys Pro Tyr Thr Tyr
1160 1165 1170
Asp Phe Ala Pro Val Gly Ala Thr Phe Val Lys Val Thr Val Thr
1175 1180 1185
Asn Ala Asp Thr Thr Thr Pro Ser Gly Val Val Cys Ala Gly Leu
1190 1195 1200
Thr Glu Ile Glu Leu Lys Thr Ala Thr Ser Lys Phe Val Thr Asn
1205 1210 1215
Thr Ser Ala Ala Leu Ser Ser Leu Thr Val Asn Gly Thr Lys Val
1220 1225 1230
Ser Asp Ser Val Leu Ala Ala Gly Ser Tyr Asn Thr Pro Ala Ile
1235 1240 1245
Ile Ala Asp Val Lys Ala Glu Gly Glu Gly Asn Ala Ser Val Thr
1250 1255 1260
Val Leu Pro Ala His Asp Asn Val Ile Arg Val Ile Thr Glu Ser
1265 1270 1275
Glu Asp His Val Thr Arg Lys Thr Phe Thr Ile Asn Leu Gly Thr
1280 1285 1290
Glu Gln Glu Phe Pro Ala Asp Ser Asp Glu Arg Asp
1295 1300 1305
<210> 35
<211> 1025
<212> PRT
<213> Kluyveromyces lactis
<400> 35
Met Ser Cys Leu Ile Pro Glu Asn Leu Arg Asn Pro Lys Lys Val His
1 5 10 15
Glu Asn Arg Leu Pro Thr Arg Ala Tyr Tyr Tyr Asp Gln Asp Ile Phe
20 25 30
Glu Ser Leu Asn Gly Pro Trp Ala Phe Ala Leu Phe Asp Ala Pro Leu
35 40 45
Asp Ala Pro Asp Ala Lys Asn Leu Asp Trp Glu Thr Ala Lys Lys Trp
50 55 60
Ser Thr Ile Ser Val Pro Ser His Trp Glu Leu Gln Glu Asp Trp Lys
65 70 75 80
Tyr Gly Lys Pro Ile Tyr Thr Asn Val Gln Tyr Pro Ile Pro Ile Asp
85 90 95
Ile Pro Asn Pro Pro Thr Val Asn Pro Thr Gly Val Tyr Ala Arg Thr
100 105 110
Phe Glu Leu Asp Ser Lys Ser Ile Glu Ser Phe Glu His Arg Leu Arg
115 120 125
Phe Glu Gly Val Asp Asn Cys Tyr Glu Leu Tyr Val Asn Gly Gln Tyr
130 135 140
Val Gly Phe Asn Lys Gly Ser Arg Asn Gly Ala Glu Phe Asp Ile Gln
145 150 155 160
Lys Tyr Val Ser Glu Gly Glu Asn Leu Val Val Val Lys Val Phe Lys
165 170 175
Trp Ser Asp Ser Thr Tyr Ile Glu Asp Gln Asp Gln Trp Trp Leu Ser
180 185 190
Gly Ile Tyr Arg Asp Val Ser Leu Leu Lys Leu Pro Lys Lys Ala His
195 200 205
Ile Glu Asp Val Arg Val Thr Thr Thr Phe Val Asp Ser Gln Tyr Gln
210 215 220
Asp Ala Glu Leu Ser Val Lys Val Asp Val Gln Gly Ser Ser Tyr Asp
225 230 235 240
His Ile Asn Phe Thr Leu Tyr Glu Pro Glu Asp Gly Ser Lys Val Tyr
245 250 255
Asp Ala Ser Ser Leu Leu Asn Glu Glu Asn Gly Asn Thr Thr Phe Ser
260 265 270
Thr Lys Glu Phe Ile Ser Phe Ser Thr Lys Lys Asn Glu Glu Thr Ala
275 280 285
Phe Lys Ile Asn Val Lys Ala Pro Glu His Trp Thr Ala Glu Asn Pro
290 295 300
Thr Leu Tyr Lys Tyr Gln Leu Asp Leu Ile Gly Ser Asp Gly Ser Val
305 310 315 320
Ile Gln Ser Ile Lys His His Val Gly Phe Arg Gln Val Glu Leu Lys
325 330 335
Asp Gly Asn Ile Thr Val Asn Gly Lys Asp Ile Leu Phe Arg Gly Val
340 345 350
Asn Arg His Asp His His Pro Arg Phe Gly Arg Ala Val Pro Leu Asp
355 360 365
Phe Val Val Arg Asp Leu Ile Leu Met Lys Lys Phe Asn Ile Asn Ala
370 375 380
Val Arg Asn Ser His Tyr Pro Asn His Pro Lys Val Tyr Asp Leu Phe
385 390 395 400
Asp Lys Leu Gly Phe Trp Val Ile Asp Glu Ala Asp Leu Glu Thr His
405 410 415
Gly Val Gln Glu Pro Phe Asn Arg His Thr Asn Leu Glu Ala Glu Tyr
420 425 430
Pro Asp Thr Lys Asn Lys Leu Tyr Asp Val Asn Ala His Tyr Leu Ser
435 440 445
Asp Asn Pro Glu Tyr Glu Val Ala Tyr Leu Asp Arg Ala Ser Gln Leu
450 455 460
Val Leu Arg Asp Val Asn His Pro Ser Ile Ile Ile Trp Ser Leu Gly
465 470 475 480
Asn Glu Ala Cys Tyr Gly Arg Asn His Lys Ala Met Tyr Lys Leu Ile
485 490 495
Lys Gln Leu Asp Pro Thr Arg Leu Val His Tyr Glu Gly Asp Leu Asn
500 505 510
Ala Leu Ser Ala Asp Ile Phe Ser Phe Met Tyr Pro Thr Phe Glu Ile
515 520 525
Met Glu Arg Trp Arg Lys Asn His Thr Asp Glu Asn Gly Lys Phe Glu
530 535 540
Lys Pro Leu Ile Leu Cys Glu Tyr Gly His Ala Met Gly Asn Gly Pro
545 550 555 560
Gly Ser Leu Lys Glu Tyr Gln Glu Leu Phe Tyr Lys Glu Lys Phe Tyr
565 570 575
Gln Gly Gly Phe Ile Trp Glu Trp Ala Asn His Gly Ile Glu Phe Glu
580 585 590
Asp Val Ser Thr Ala Asp Gly Lys Leu His Lys Ala Tyr Ala Tyr Gly
595 600 605
Gly Asp Phe Lys Glu Glu Val His Asp Gly Val Phe Ile Met Asp Gly
610 615 620
Leu Cys Asn Ser Glu His Asn Pro Thr Pro Gly Leu Val Glu Tyr Lys
625 630 635 640
Lys Val Ile Glu Pro Val His Ile Lys Ile Ala His Gly Ser Val Thr
645 650 655
Ile Thr Asn Lys His Asp Phe Ile Thr Thr Asp His Leu Leu Phe Ile
660 665 670
Asp Lys Asp Thr Gly Lys Thr Ile Asp Val Pro Ser Leu Lys Pro Glu
675 680 685
Glu Ser Val Thr Ile Pro Ser Asp Thr Thr Tyr Val Val Ala Val Leu
690 695 700
Lys Asp Asp Ala Gly Val Leu Lys Ala Gly His Glu Ile Ala Trp Gly
705 710 715 720
Gln Ala Glu Leu Pro Leu Lys Val Pro Asp Phe Val Thr Glu Thr Ala
725 730 735
Glu Lys Ala Ala Lys Ile Asn Asp Gly Lys Arg Tyr Val Ser Val Glu
740 745 750
Ser Ser Gly Leu His Phe Ile Leu Asp Lys Leu Leu Gly Lys Ile Glu
755 760 765
Ser Leu Lys Val Lys Gly Lys Glu Ile Ser Ser Lys Phe Glu Gly Ser
770 775 780
Ser Ile Thr Phe Trp Arg Pro Pro Thr Asn Asn Asp Glu Pro Arg Asp
785 790 795 800
Phe Lys Asn Trp Lys Lys Tyr Asn Ile Asp Leu Met Lys Gln Asn Ile
805 810 815
His Gly Val Ser Val Glu Lys Gly Ser Asn Gly Ser Leu Ala Val Val
820 825 830
Thr Val Asn Ser Arg Ile Ser Pro Val Val Phe Tyr Tyr Gly Phe Glu
835 840 845
Thr Val Gln Lys Tyr Thr Ile Phe Ala Asn Lys Ile Asn Leu Asn Thr
850 855 860
Ser Met Lys Leu Thr Gly Glu Tyr Gln Pro Pro Asp Phe Pro Arg Val
865 870 875 880
Gly Tyr Glu Phe Trp Leu Gly Asp Ser Tyr Glu Ser Phe Glu Trp Leu
885 890 895
Gly Arg Gly Pro Gly Glu Ser Tyr Pro Asp Lys Lys Glu Ser Gln Arg
900 905 910
Phe Gly Leu Tyr Asp Ser Lys Asp Val Glu Glu Phe Val Tyr Asp Tyr
915 920 925
Pro Gln Glu Asn Gly Asn His Thr Asp Thr His Phe Leu Asn Ile Lys
930 935 940
Phe Glu Gly Ala Gly Lys Leu Ser Ile Phe Gln Lys Glu Lys Pro Phe
945 950 955 960
Asn Phe Lys Ile Ser Asp Glu Tyr Gly Val Asp Glu Ala Ala His Ala
965 970 975
Cys Asp Val Lys Arg Tyr Gly Arg His Tyr Leu Arg Leu Asp His Ala
980 985 990
Ile His Gly Val Gly Ser Glu Ala Cys Gly Pro Ala Val Leu Asp Gln
995 1000 1005
Tyr Arg Leu Lys Ala Gln Asp Phe Asn Phe Glu Phe Asp Leu Ala
1010 1015 1020
Phe Glu
1025
<210> 36
<211> 30
<212> ДНК
<213> Праймер
<220>
<221> прочий_признак
<222> (9)..(9)
<223> n представляет собой остаток дезоксиурацила в 9-х положениях
<400> 36
attaaccang cgacgcaact tcgaatggcc 30
<210> 37
<211> 31
<212> ДНК
<213> Праймер
<220>
<221> прочий_признак
<222> (9)..(9)
<223> n представляет собой остаток дезоксиурацила в 9-х положениях
<400> 37
atcttctcnt taccgcctta ccacgagcac g 31
<210> 38
<211> 33
<212> ДНК
<213> Праймер
<220>
<221> прочий_признак
<222> (9)..(9)
<223> n представляет собой остаток дезоксиурацила в 9-х положениях
<400> 38
agagaagant ttcagcctga tacagattaa atc 33
<210> 39
<211> 33
<212> ДНК
<213> Праймер
<220>
<221> прочий_признак
<222> (9)..(9)
<223> n представляет собой остаток дезоксиурацила в 9-х положениях
<400> 39
atggttaant cctcctgtta gcccaaaaaa cgg 33
<210> 40
<211> 22
<212> ДНК
<213> Праймер
<400> 40
cggcgtcaca ctttgctatg cc 22
<210> 41
<211> 20
<212> ДНК
<213> Праймер
<400> 41
ccgcgctact gccgccaggc 20
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ФЕРМЕНТЫ ЛАКТАЗЫ С УЛУЧШЕННЫМИ СВОЙСТВАМИ | 2018 |
|
RU2788608C2 |
| Ферменты лактазы с улучшенными свойствами при кислом pH | 2019 |
|
RU2814542C2 |
| Ферменты лактазы с улучшенными свойствами | 2018 |
|
RU2806709C2 |
| УЛУЧШЕННЫЕ ПОЛИПЕПТИДЫ, СПОСОБНЫЕ ПРЕВРАЩАТЬ СУБСТРАТ 3-КЕТОДЕЗОКСИНИВАЛЕНОЛ В 3-ЭПИДЕЗОКСИНИВАЛЕНОЛ | 2020 |
|
RU2836544C1 |
| ФЕРМЕНТАТИВНОЕ ПОЛУЧЕНИЕ ТАГАТОЗЫ | 2019 |
|
RU2820606C2 |
| ФЕРМЕНТАТИВНЫЙ СИНТЕЗ N-АЦЕТИЛНЕЙРАМИНОВОЙ КИСЛОТЫ | 2018 |
|
RU2809787C2 |
| АBC-ТРАНСПОРТЕРЫ ДЛЯ ВЫСОКОЭФФЕКТИВНОГО ПРОИЗВОДСТВА РЕБАУДИОЗИДОВ | 2020 |
|
RU2795855C2 |
| СПОСОБЫ ПРИГОТОВЛЕНИЯ ИНТЕНСИВНЫХ ПОДСЛАСТИТЕЛЕЙ | 2018 |
|
RU2767792C2 |
| СИАЛИЛТРАНСФЕРАЗЫ И ИХ ПРИМЕНЕНИЕ В ПОЛУЧЕНИИ СИАЛИРОВАННЫХ ОЛИГОСАХАРИДОВ | 2018 |
|
RU2822039C2 |
| ВАРИАНТЫ ПОЛИПЕПТИДОВ, РАСЩЕПЛЯЮЩИХ ФУЗАРИОТОКСИНЫ, СОДЕРЖАЩАЯ ИХ ДОБАВКА, ИХ ПРИМЕНЕНИЕ И СПОСОБ РАСЩЕПЛЕНИЯ ФУЗАРИОТОКСИНОВ | 2015 |
|
RU2720517C2 |

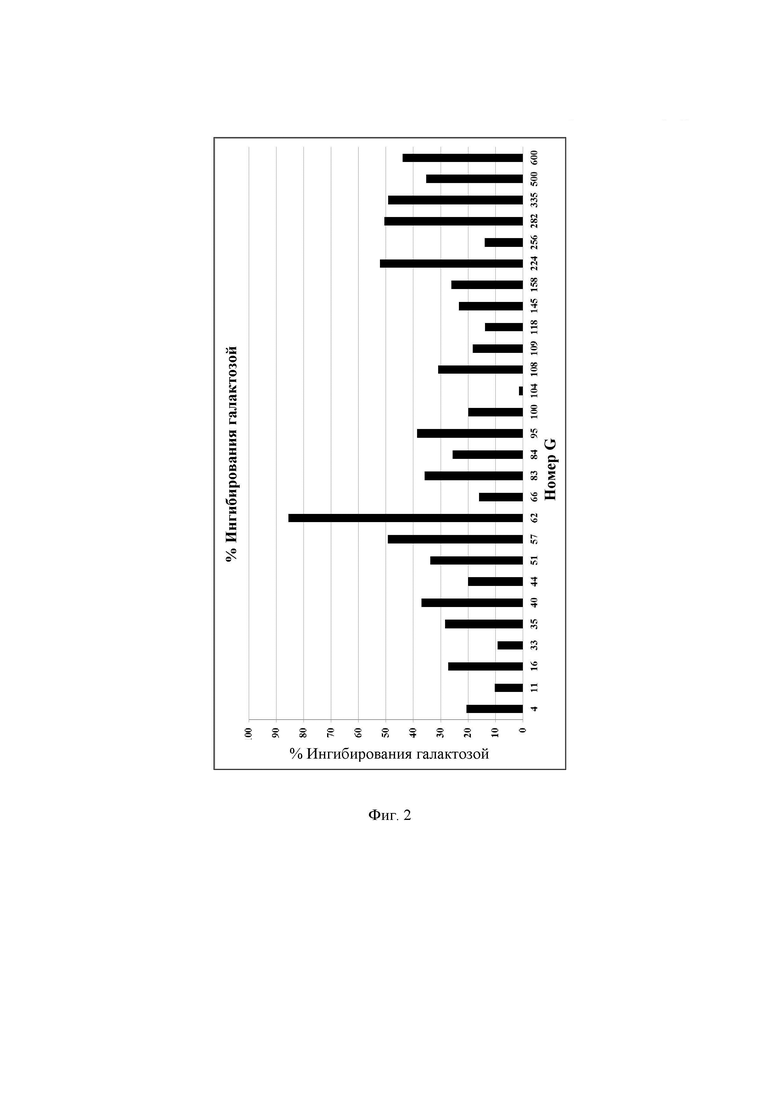
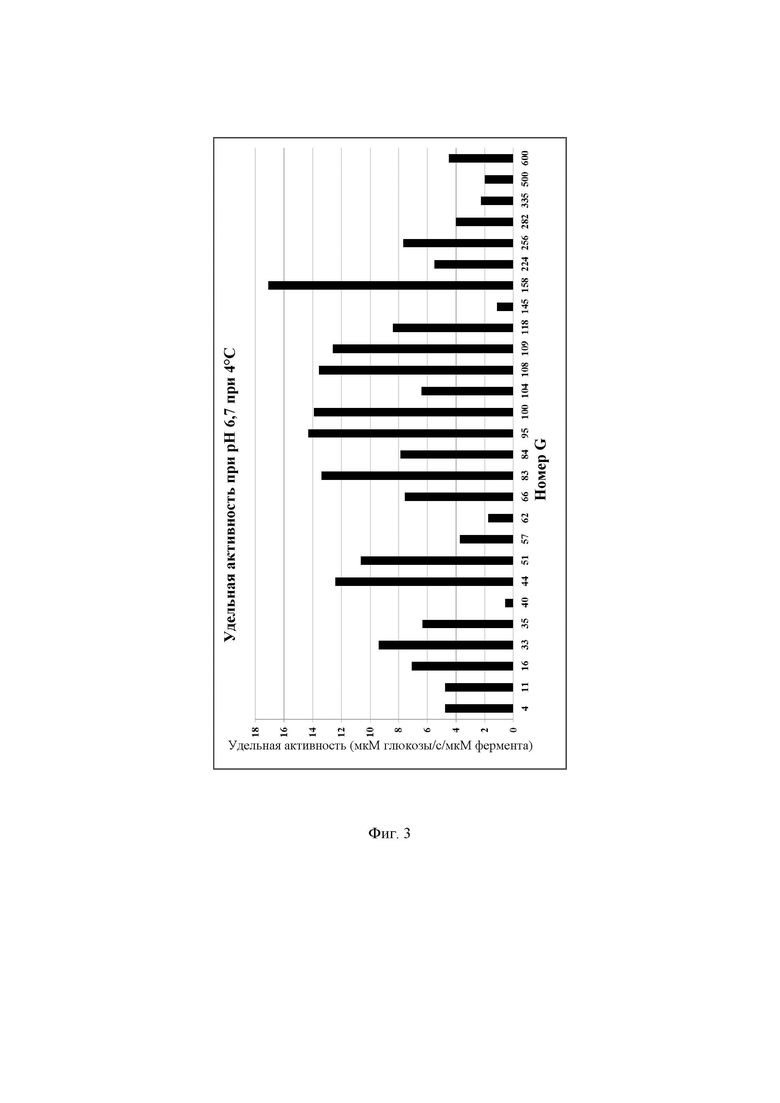
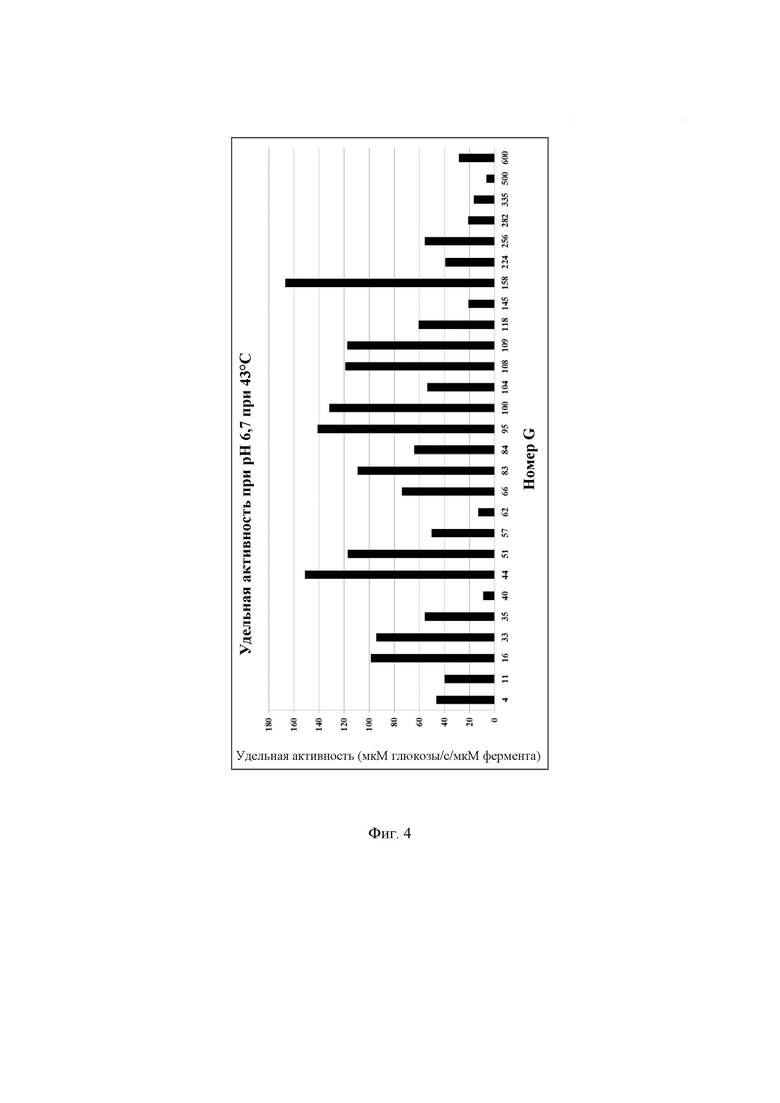
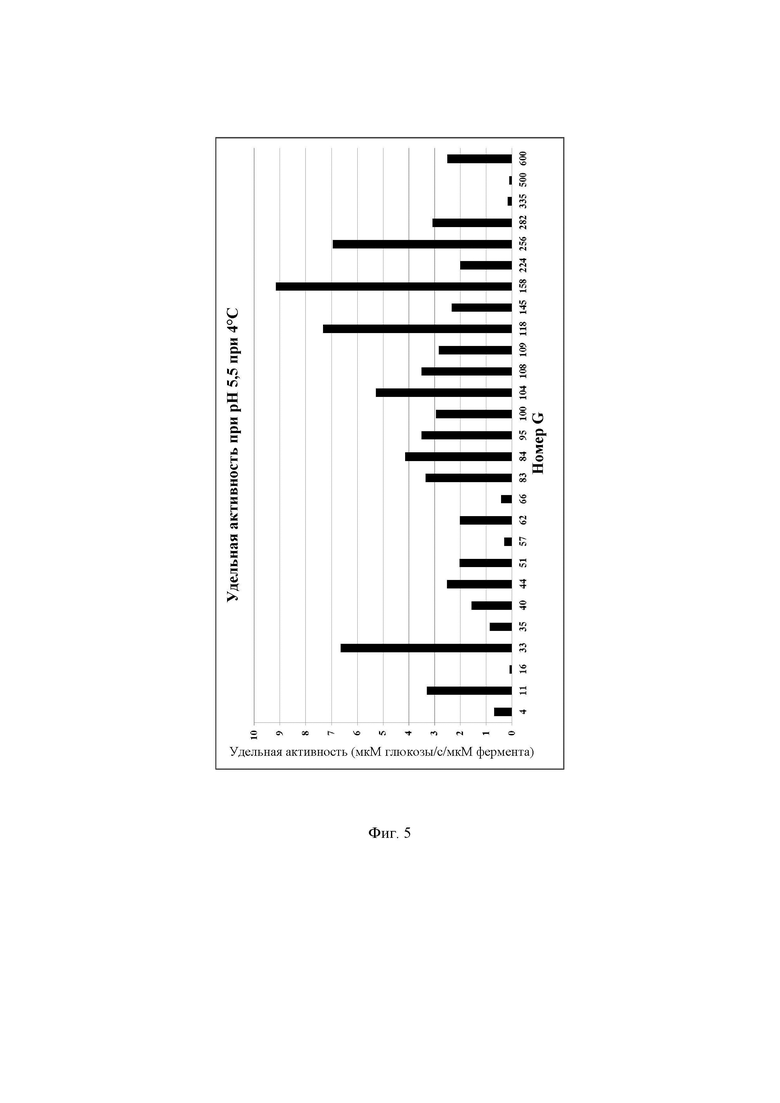

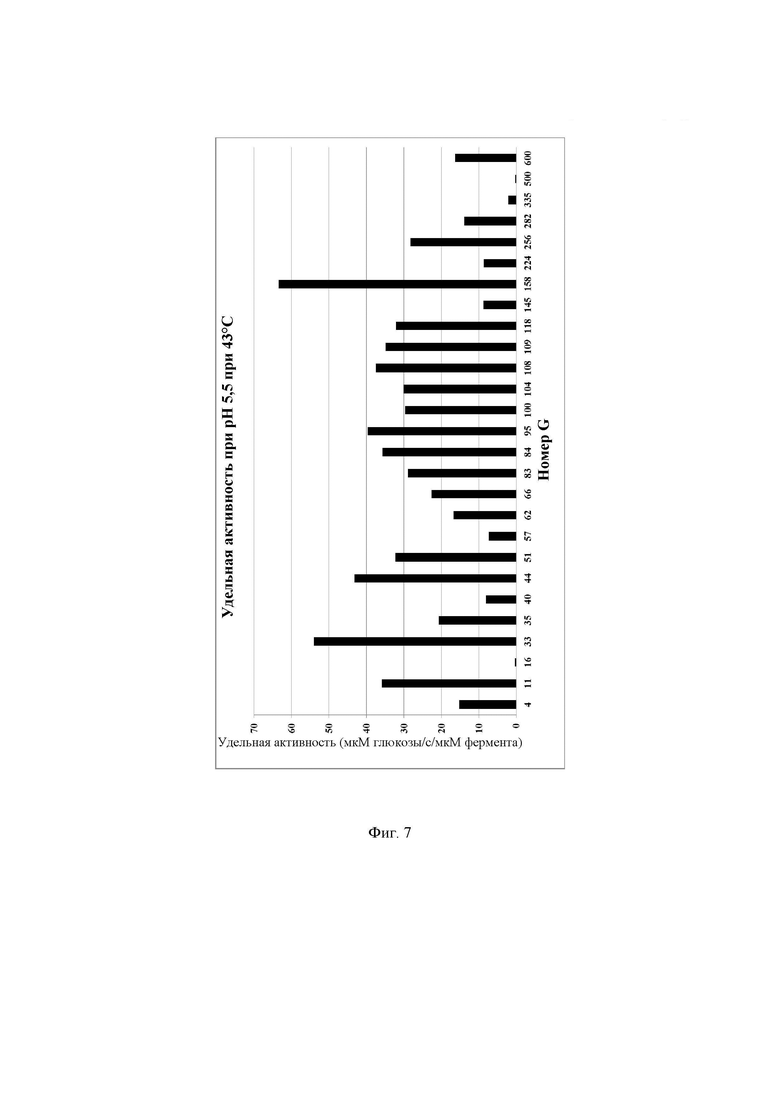


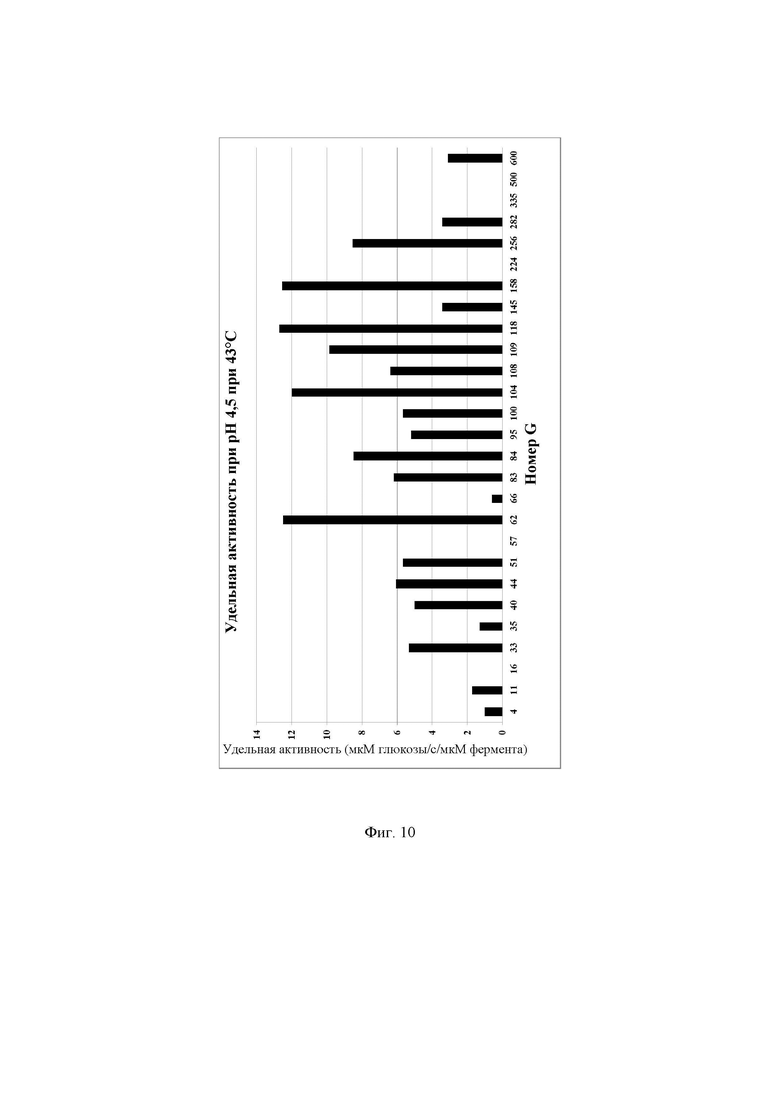


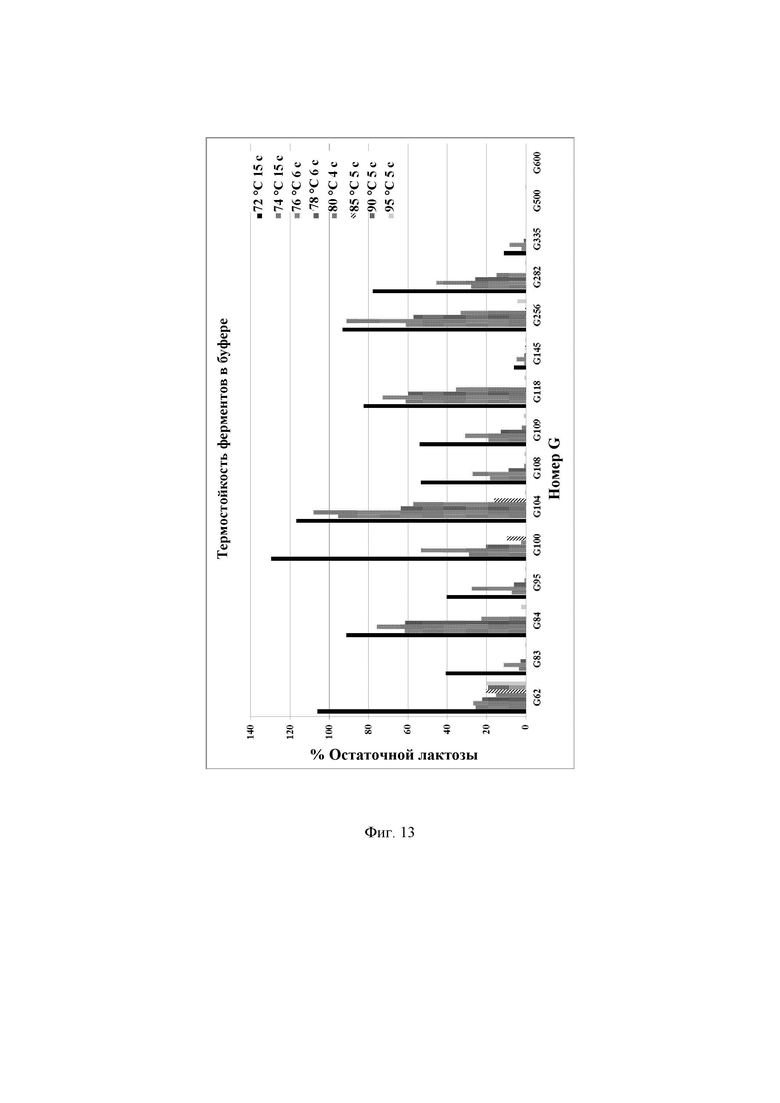

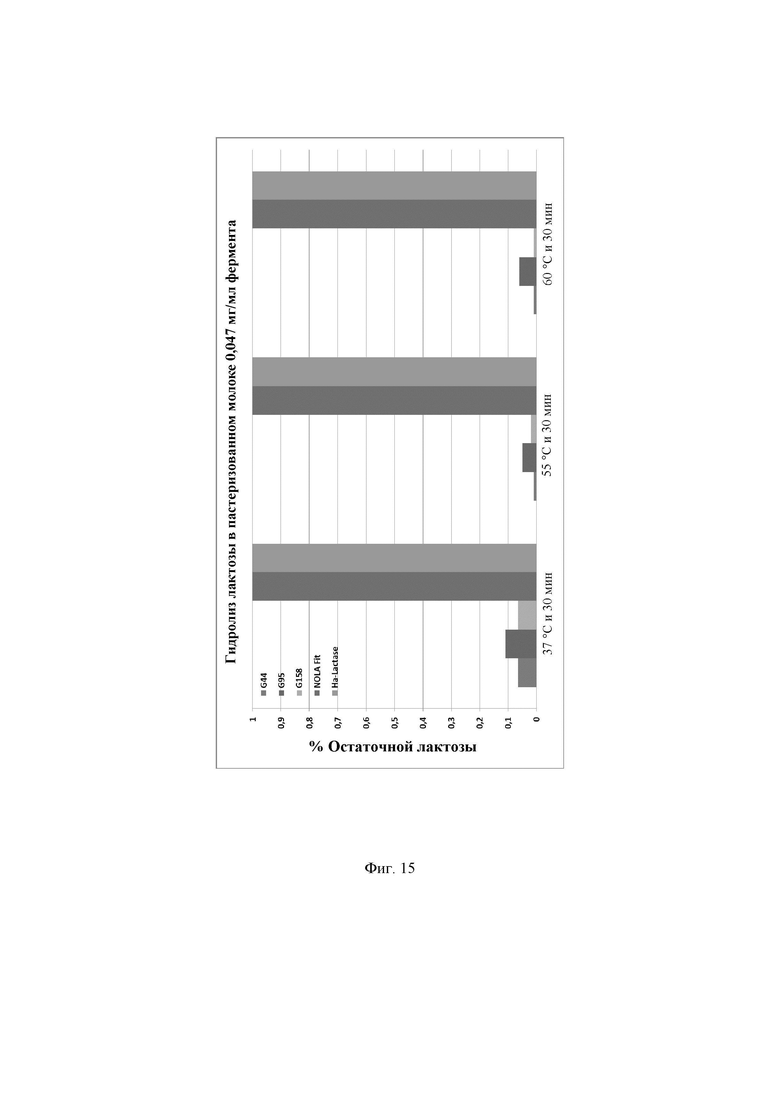
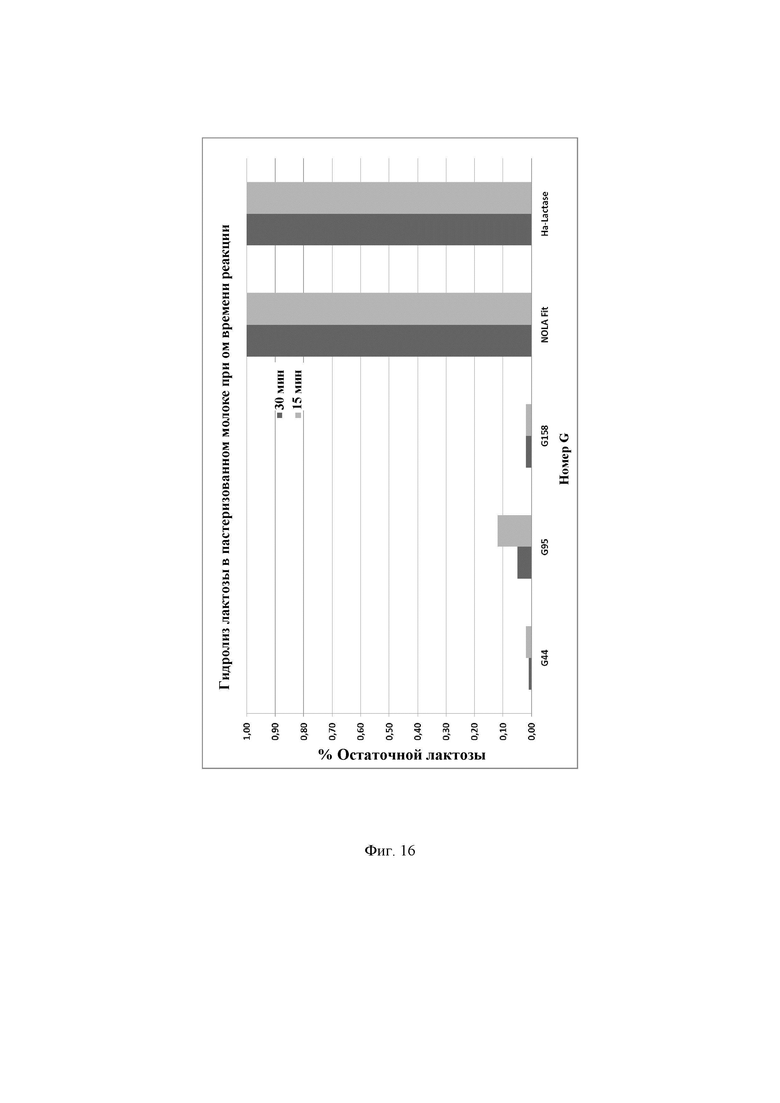
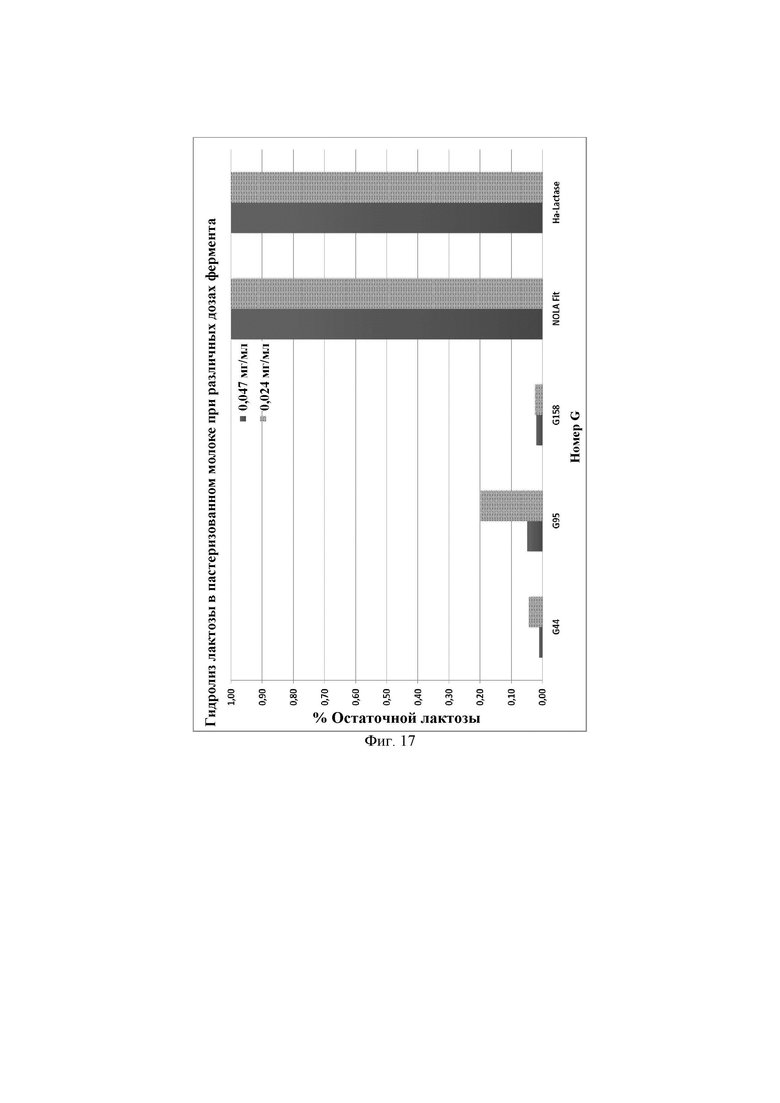

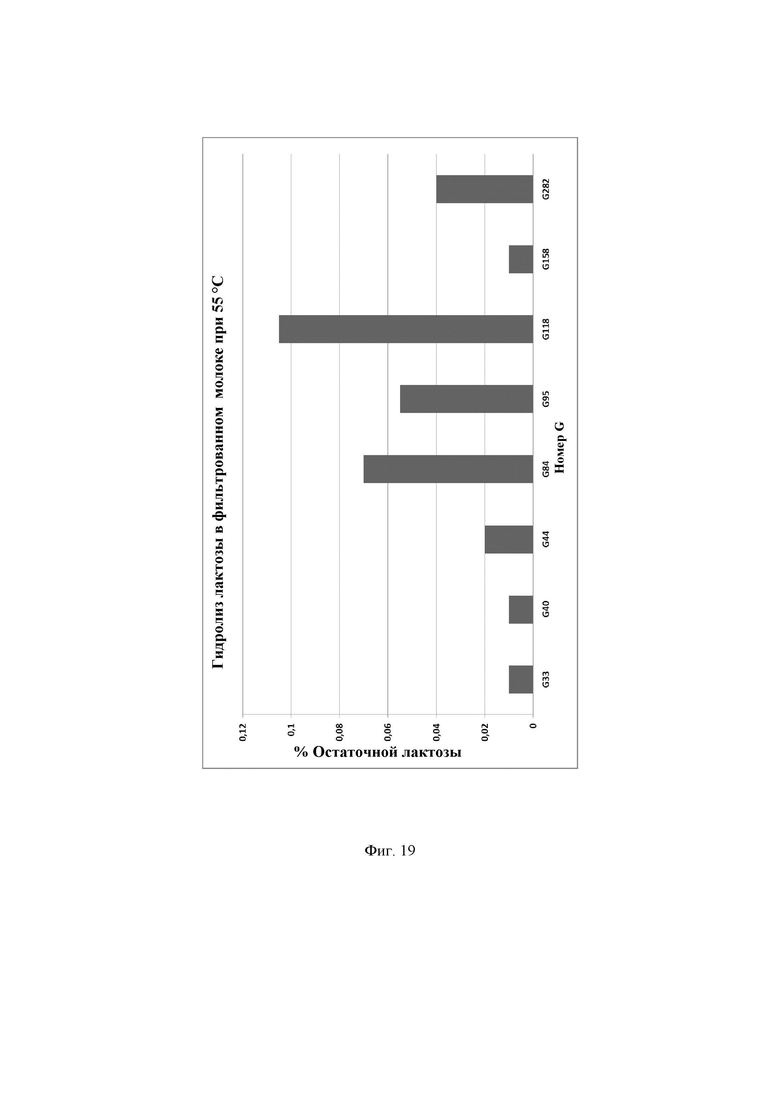
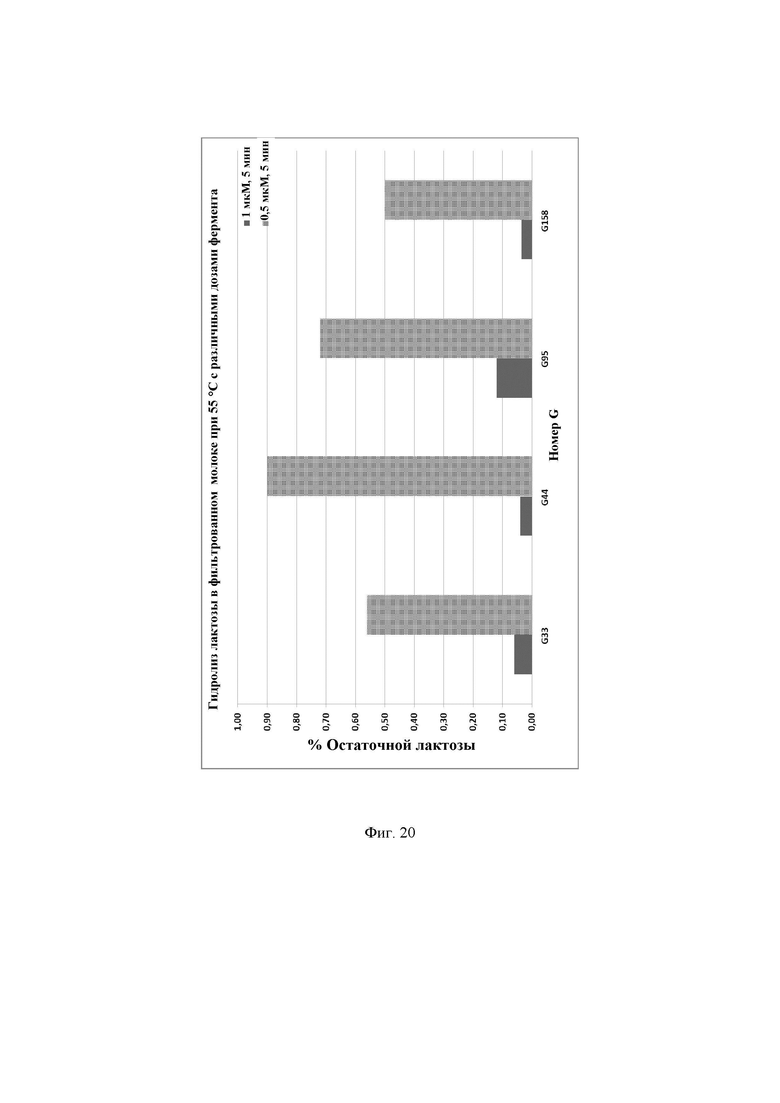

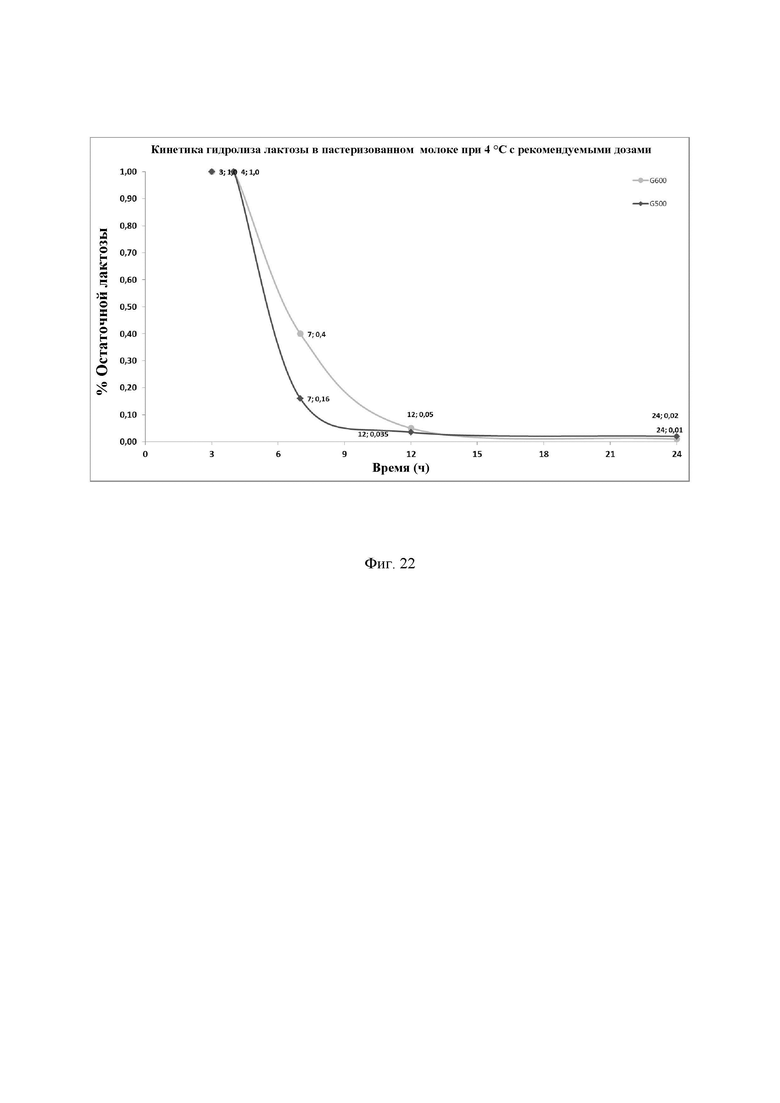

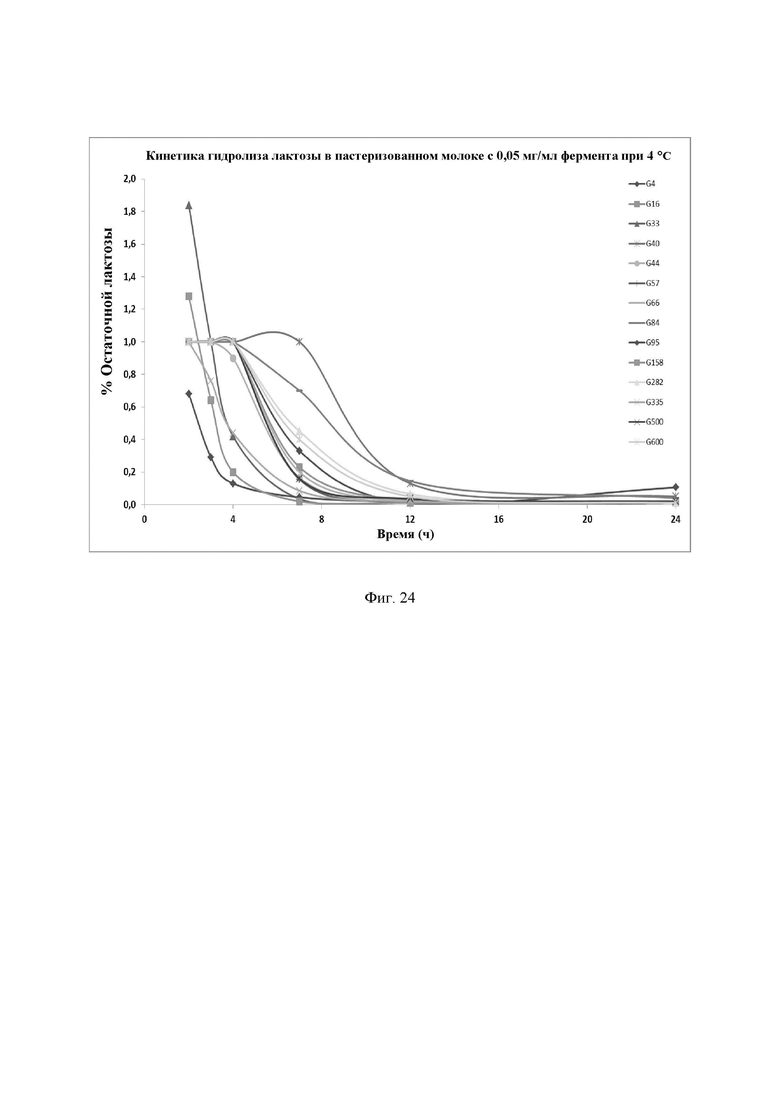
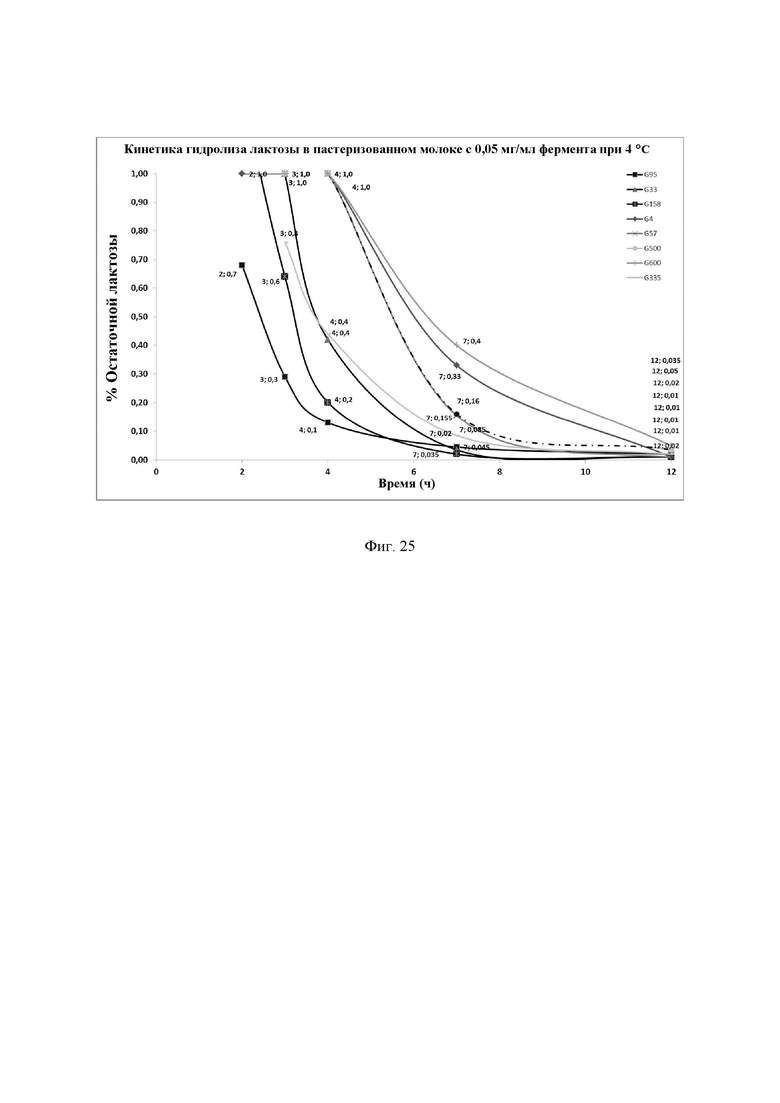
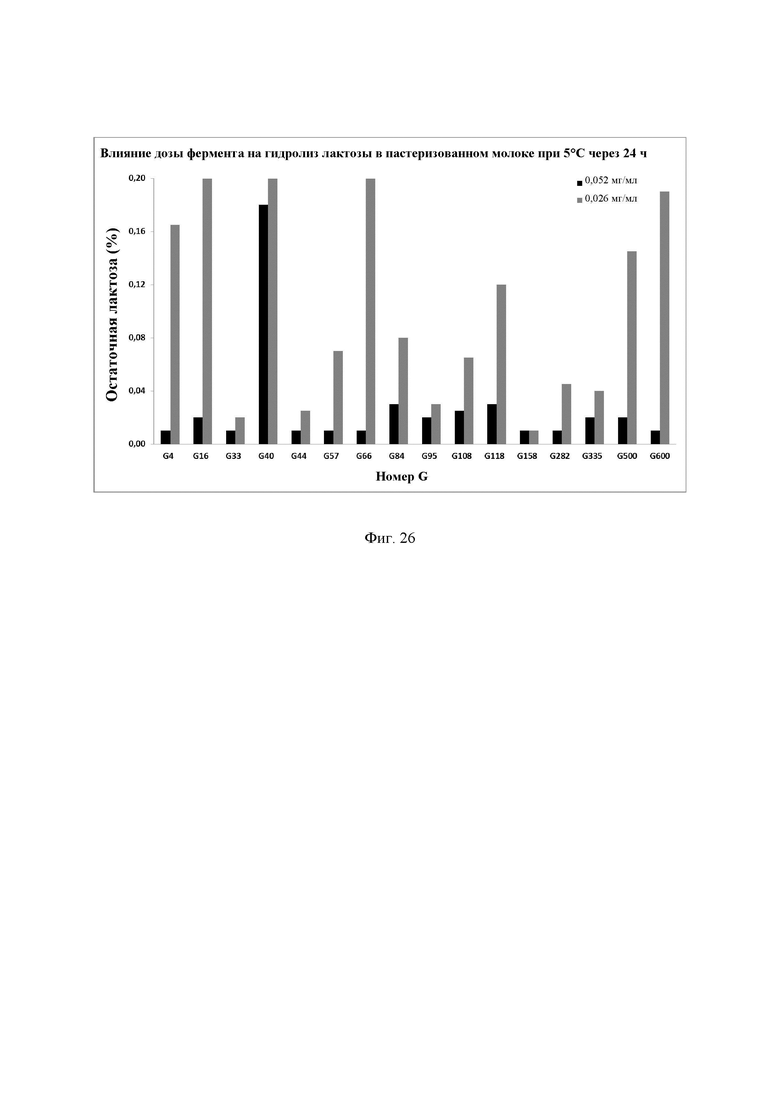
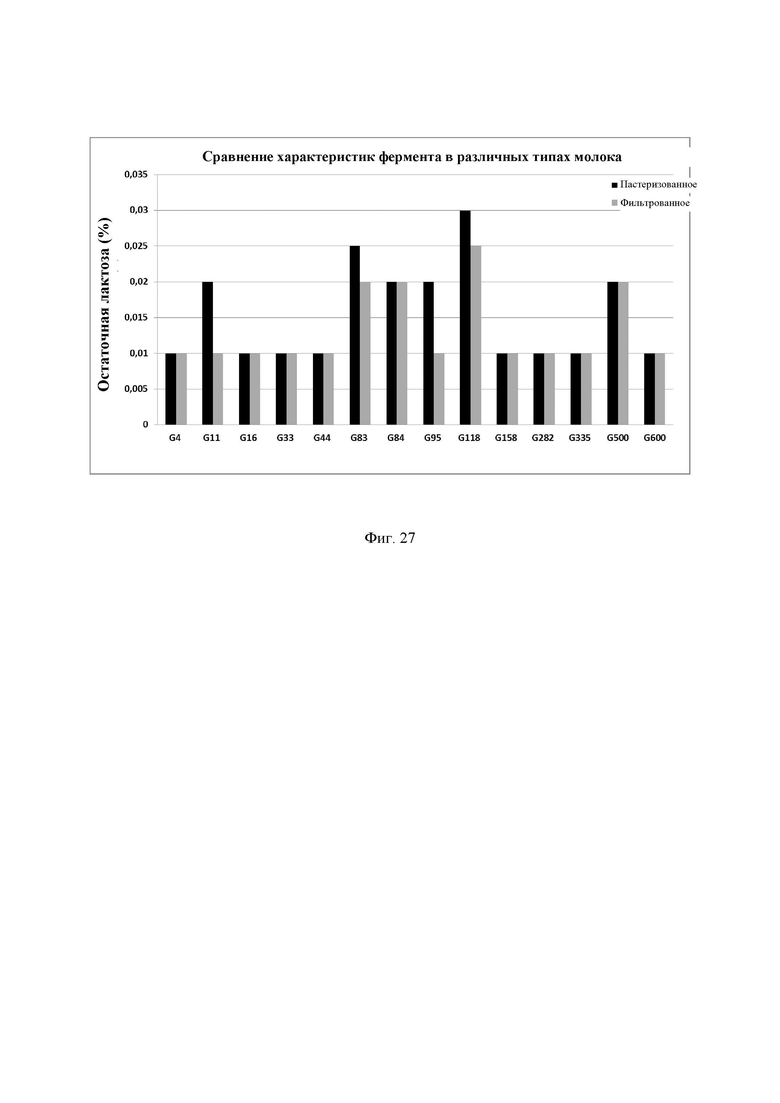
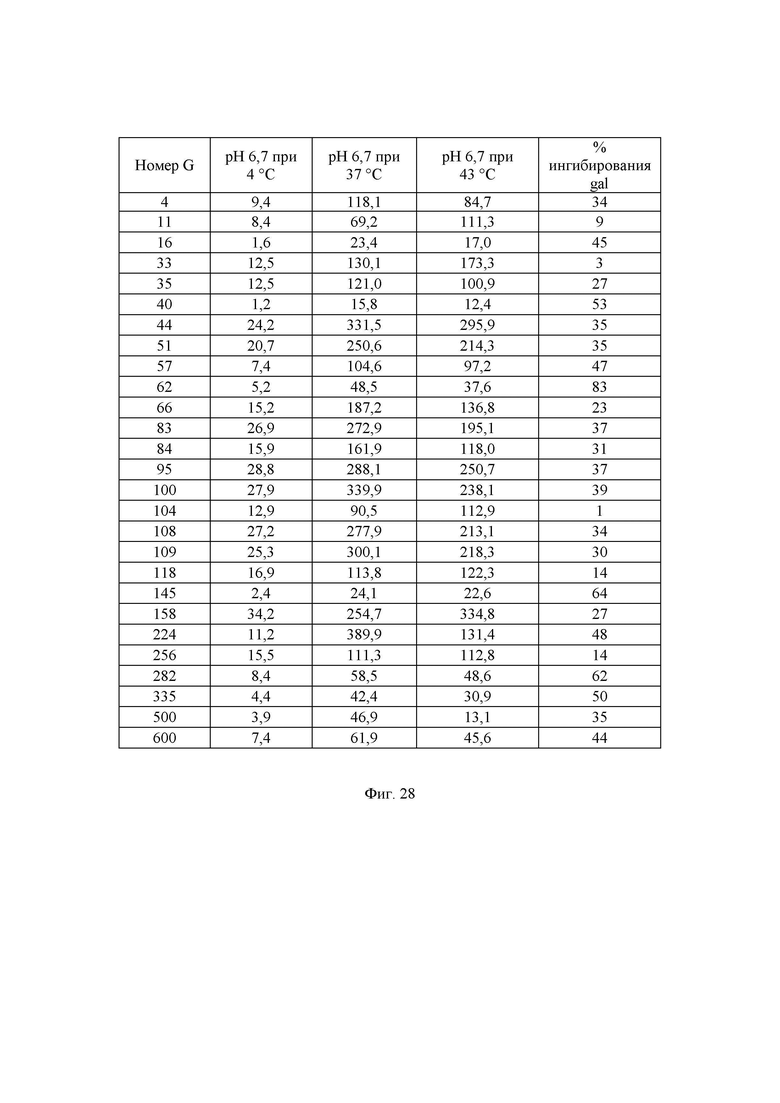
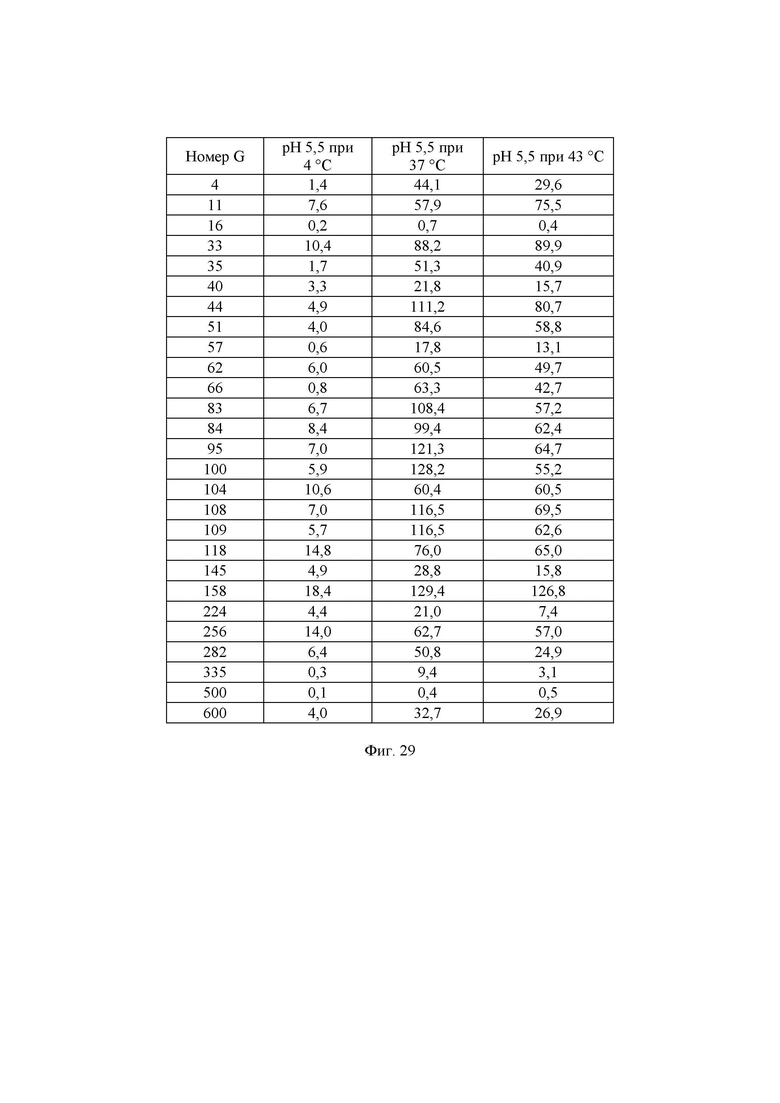
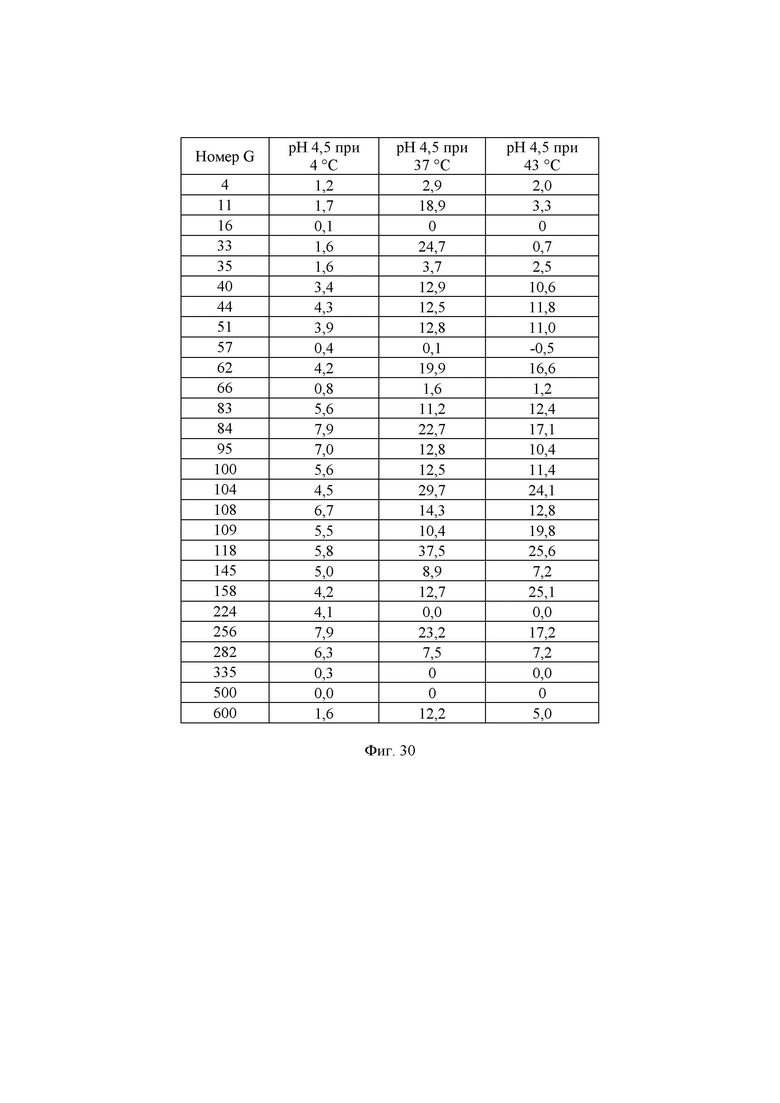
Группа изобретений относится к молочной промышленности и биотехнологии. Способ получения молочного продукта включает: (а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л; (б) инкубацию смеси при температуре от 1°C до 10°C в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 2 г/л, где pH на стадии (б) составляет от 4 до 7, и где пептид, проявляющий активность бета-галактозидазы, представляет собой: пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25, 31, или его обладающие ферментативной активностью фрагменты, или аминокислотную последовательность любого из них, имеющую идентичность последовательности по меньшей мере 90% с SEQ ID NO: 12, 13,14, 15, 16, 17, 18, 19, 25 или 31. Также описан способ содержания лактозы в субстрате на молочной основе с использованием указанного пептида, проявляющего активность бета-галактозидазы, и его применение в способе получения молочного продукта. Группа изобретений позволяет эффективно использовать ферменты, обладающие бета-галактозидазной активностью для снижения содержания лактозы в молочных продуктах. 3 н. и 12 з.п. ф-лы, 30 ил., 3 табл., 30 пр.
1. Способ получения молочного продукта, включающий:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л;
(б) инкубацию смеси при температуре от 1°C до 10°C в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 2 г/л,
где pH на стадии (б) составляет от 4 до 7, и
где пептид, проявляющий активность бета-галактозидазы, представляет собой: пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25, 31, или его обладающие ферментативной активностью фрагменты, или аминокислотную последовательность любого из них, имеющую идентичность последовательности по меньшей мере 90% с SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25 или 31.
2. Способ снижения содержания лактозы в субстрате на молочной основе, включающий:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л;
(б) инкубацию смеси при температуре от 1°C до 10°C в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 2 г/л,
где pH на стадии (б) составляет от 4 до 7, и
где пептид, проявляющий активность бета-галактозидазы, представляет собой: пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25, 31, или его обладающие ферментативной активностью фрагменты, или аминокислотную последовательность любого из них, имеющую идентичность последовательности по меньшей мере 90% с SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25 или 31.
3. Способ по любому из пп. 1, 2, где пептид, проявляющий активность бета-галактозидазы, добавляют в концентрации от 35 до 52 мг/л, в концентрации от 40 до 52 мг/л или в концентрации от 45 до 52 мг/л.
4. Способ по любому из пп. 1-3, где субстрат на молочной основе, содержащий лактозу, представляет собой:
(1) коровье молоко, овечье молоко, козье молоко, буйволиное молоко, верблюжье молоко или указанное молоко в пастеризованной, сырой и/или фильтрованной форме; или
(2) кисломолочный продукт, полученный из молока (1) путем ферментации.
5. Способ по п. 4, где субстрат на молочной основе, содержащий лактозу, представляет собой коровье молоко, содержащее лактозу в концентрации от примерно 37 до 50 г/л, или указанное молоко в термически обработанной, пастеризованной и/или фильтрованной форме.
6. Способ по любому из пп. 1-5, где стадия инкубации (б) снижает концентрацию лактозы в смеси до от менее 0,1 г/л до 2 г/л или до от 0,1 г/л до 2 г/л.
7. Способ по любому из пп. 1-4, где концентрации лактозы менее 0,2 г/л достигают после инкубации в течение по меньшей мере 4 часов, по меньшей мере 8 часов, по меньшей мере 12 часов или по меньшей мере 24 часов.
8. Способ по любому из пп. 1-7, где температура инкубации на стадии (б) находится в диапазоне от 2°C до 7°C или в диапазоне от 3°C до 6°C.
9. Способ по любому из пп. 1-8, где при инкубации на стадии (б) концентрация лактозы в смеси снижается до менее 0,05 г/л, до менее 0,02 г/л или до менее 0,01 г/л.
10. Способ по любому из пп. 1-9, где смесь, содержащую субстрат на молочной основе и пептид, проявляющий активность бета-галактозидазы, нагревают до температуры по меньшей мере 60°C в течение по меньшей мере четырех секунд до или после инкубации смеси при температуре от 1°C до 10°C.
11. Способ по п. 10, при котором смесь, содержащую субстрат на молочной основе и пептид, проявляющий активность бета-галактозидазы, нагревают до температуры 72°C в течение примерно 15 секунд до или после инкубации смеси при низких температурах на стадии (б) или нагревают до температуры 140°C в течение примерно четырех секунд до или после инкубации смеси при температуре от 1°C до 10°C.
12. Способ получения молочного продукта по любому из пп. 1-9, включающий стадию ферментации субстрата на молочной основе с молочнокислыми бактериями.
13. Способ по п. 12, где стадию ферментации выполняют до или после инкубации с пептидом, проявляющим активность бета-галактозидазы.
14. Способ по любому из пп. 1-9, где молочный продукт представляет собой кисломолочный продукт, сыр, йогурт, сливочное масло, молочный спред, пахту, напиток из сквашенного молока, сметану, напиток на основе молочной сыворотки, мороженое, сгущенное молоко, карамелизированное сгущенное молоко или ароматизированный молочный напиток.
15. Применение пептида, проявляющего активность бета-галактозидазы, в способе получения молочного продукта, включающем:
(а) смешивание субстрата на молочной основе, содержащего лактозу в концентрации по меньшей мере 10 г/л, и пептида, проявляющего активность бета-галактозидазы, в концентрации от 10 до 55 мг/л;
(б) инкубацию смеси при температуре от 1°C до 10°C в течение периода времени, достаточного для снижения концентрации лактозы в смеси до менее 2 г/л,
где pH на стадии (б) составляет от 4 до 7, и
где пептид, проявляющий активность бета-галактозидазы, представляет собой: пептид, имеющий аминокислотную последовательность, представленную SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25, 31, или его обладающие ферментативной активностью фрагменты, или аминокислотную последовательность любого из них, имеющую идентичность последовательности по меньшей мере 90% с SEQ ID NO: 12, 13, 14, 15, 16, 17, 18, 19, 25 или 31.
| СПОСОБ ПОЛУЧЕНИЯ ГРАНУЛИРОВАННОГО МИНЕРАЛЬНОГО УДОБРЕНИЯ | 2012 |
|
RU2530148C2 |
| RHIMI M | |||
| et al | |||
| Exploring the acidotolerance of b-galactosidase from Lactobacillus delbrueckii subsp | |||
| bulgaricus: an attractive enzyme for lactose bioconversion, Research in Microbiology, 2009, Vol.160, N 10, pp.775-784 | |||
| Оконечный каскад лампового усилителя | 1951 |
|
SU98003A1 |
| HORNER T.W | |||
| et al | |||
