Область техники, к которой относится настоящее изобретение
Настоящее изобретение относится к человеческим антителам, связывающимся с ROR2, предпочтительно, с человеческим ROR2 (hROR2), и к их фрагментам и конъюгатам, а также к их применениям.
Предшествующий уровень техники настоящего изобретения
Рак является одной из основных причин смерти. Он представляет собой класс заболеваний, вызванных злокачественной трансформацией здоровых клеток, являющейся результатом генетических изменений, подобных хромосомным транслокациям, мутациям в генах-супрессорах опухолевого роста, воздействий транскрипционных факторов или рецепторов ростовых факторов, приводящих к иммортализации клеток. Если иммортализация сочетается с чрезмерной пролиферацией, иммортализированные клетки образуют опухоли с метастазированием или без него (в случае солидных опухолей) или лейкозы и лимфомы (злокачественные новообразования из клеток крови). Дефектный апоптоз, или программируемая гибель клеток, может вносить дополнительный вклад в злокачественную трансформацию клеток, приводящую к раку.
Было описано, что семейство мембраноассоциированных рецепторных тирозинкиназ, состоящее из подобных рецепторной тирозинкиназе орфанных рецепторов 1 и 2 (ROR1 и ROR2), специфически ассоциировано с конкретными видами рака (Rebagay et al. (2012) Front Oncol. 2(34): 1-8; doi 10.3389/onc.2012.00034), при этом их экспрессия преимущественно отсутствует в здоровой ткани за редкими исключениями, например, в случае ROR1 (Balakrishnan et al. (2016) Clin Cancer Res. doi: 10.1158/1078-0432). Остается неясным, является ли экспрессия ROR функционально связанной с образованием опухолей. Тем не менее, вследствие очень селективной экспрессии членов семейства ROR в опухолях они являются значимыми мишенями для противораковых терапевтических средств. Важно, что экспрессия человеческого ROR2 (hROR2) была описана на опухолевых клетках в нейробластоме, саркоме (и, в особенности, остеосаркоме), почечно-клеточной карциноме, при раке молочной железы, раке яичка, раке яичника, раке поджелудочной железы, раке почки, раке желудка, раке предстательной железы, раке головы и шеи, меланоме, плоскоклеточной карциноме, множественной миеломе и других видах рака.
Подобные рецепторной тирозинкиназе орфанные рецепторы 1 и 2, ROR1 и ROR2, представляют собой единственные два члена, определяющие новое семейство рецепторных тирозинкиназ на основании общего плана структуры и некоторых функциональных сходств. Оба из белков ROR1 и ROR2 представляют собой однократно пронизывающие мембрану трансмембранные рецепторы I типа с внеклеточным доменом (ECD), состоящим из иммуноглобулинового домена, богатого цистеином frizzled-домена и kringle-домена. За этими тремя внеклеточными доменами следует трансмембранный домен, соединяющий ECD с внутриклеточной частью белка, содержащей киназные домены (Rebagay et al. (2012) Frontiers Oncol. 2: 1-8).
Человеческие белки ROR1 и ROR2 являются на 58% гомологичными друг другу, но каждый из белков ROR является очень консервативным между видами. Это представляет сложность для разработки специфичных в отношении человеческого ROR2 моноклональных антител, и известно очень немного антител.
В нормальных физиологических условиях hROR2 является ответственным за аспекты роста костей и хрящей во время эмбрионального развития. После рождения экспрессия hROR2 снижается, и hROR2 в норме не выявляется или экспрессируется на очень низких уровнях во взрослых тканях. Сообщалось только о слабой экспрессии hROR2 в желудке и ткани щитовидной железы (Morioka et al. (2009) Cancer Sci. 100: 1227-1233). hROR2 ранее рассматривали как мишень для разработки специфичных в отношении hROR2 антител (международная заявка WO 2013/103637 А1, международная заявка WO 2016/142768 А1).
Существует потребность в высококачественных антителах к ROR2, которые можно применять в качестве основы для разработки целенаправленно воздействующих терапевтических средств на основе антител для экспрессирующих ROR2 видов рака. В частности, также существует потребность в антителах к ROR2, характеризующихся отличными параметрами удобства разработки, в том числе низкой склонностью к агрегации, и демонстрирующих высокую термостабильность, которые, как правило, получают с помощью В-клеток от иммунизированных животных. Кроме того, существует потребность в антителах, которые дополнительно к связыванию с человеческим ROR2 также связываются с ROR2 у стандартных видов животных, используемых для исследования токсикологических параметров, таких как мыши, крысы, кролики и/или яванские макаки, и, в особенности, связываются с ROR2 как яванского макака, так и с мышиным ROR2, обеспечивая возможность токсикологической характеристики целенаправленно воздействующих терапевтических средств на основе антител в преддверии клинических испытаний на людях. Кроме того, в случае терапии у человека существует потребность в антителах, которые представляют собой, по существу, полностью человеческие антитела с наименьшим риском иммуногенности при системном введении субъектам-людям. Также существует потребность в дополнительных диагностических инструментах для выявления экспрессии ROR2 при связанных с ROR2 болезненных состояниях. Настоящее изобретение направлено на удовлетворение этих и других потребностей.
Краткое раскрытие настоящего изобретения
Настоящее изобретение относится к полностью человеческим антителам, которые специфично связываются с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2). Настоящее изобретение и общие преимущества его признаков будут подробно обсуждаться ниже.
Краткое описание чертежей
Фиг. 1. Схематическое изображение процесса, применяемого для разработки новых человеческих антител, целенаправленно воздействующих на hROR2. (А) На первой стадии трансгенных по человеческому антителу мышей (H2L2 мышей) (описанных в международной заявке WO 2010/070263 А1) иммунизировали исключительно внеклеточным доменом, несущим на С-конце метку Twin-Strep (hROR2-TwinStrep). (В) Затем количество В-клеток селезенки, связывающихся с hROR2-TwinStrep, увеличивали посредством обогащения клеток от иммунизированных H2L2 мышей с помощью магнитно-активированного клеточного сортинга (MACS). (С) Из выделенных В-клеток селезенки создавали библиотеки транспозируемых экспрессионных векторов для человеческого IgG1 с помощью ПЦР-клонирования вариабельных доменов тяжелой и легкой цепи из выделенных связывающихся с hROR2 В-клеток селезенки. (D) За этим следовало опосредуемое транспозицией создание клеточных библиотек, обеспечивающих дисплей библиотек полноразмерных человеческих IgG1, устойчиво экспрессирующих ПЦР-клонированные библиотеки антител в иммортализированных пре-В-клетках. (Е) Скрининг на основе метода FACS представленных на поверхности клетки антител IgG1 в отношении связывания с hROR2 и выделение клонов отдельных клеток с помощью FACS. (F) Функциональный скрининг антител IgG1 из выбранных клональных супернатантов. (G) ПЦР-амплификация антител из выбранных клонов.
Фиг. 2. Выявление титров антитела к hROR2-ECD после иммунизации H2L2 мышей, называемых 1357, 1368 и 1359, с ECD-hROR2-TwinStrep.(А) перед иммунизацией и на 7 сутки (В), на 28 сутки (С) и на 49 сутки (D) после первоначальной иммунизации.
Фиг. 3. Антиген-специфичный сортинг методом FACS в расчете на мышь на основе библиотек IgG/IgK и IgM/IgK: (A) H2L2 мышь 1357, (В) H2L2 мышь 1363 и (С) H2L2 мышь 1359.
Фиг. 4. Определение аффинностей связывания с hROR2-ECD для отдельных моноклональных антител, клонированных и экспрессированных с помощью методик генетической рекомбинации, которые указаны, измеренных с помощью метода поверхностного плазмонного резонанса (SPR).
Фиг. 5. Определение межвидовой перекрестной реактивности рекомбинантных моноклональных антител к hROR2 с мышиным ROR2 и ROR2 яванского макака, которую оценивают с помощью метода ELISA.
Фиг. 6. (А) Химическая структура модифицированного пентаглицином производного PNU («G5-PNU»), (В) модифицированного триглицином производного PNU («G3-PNU»).
Фиг. 7. Анализ методом FACS экспрессии hROR2 в линии клеток злокачественной опухоли молочной железы ЕМТ-6, подвергнутых воздействию методов генной инженерии для того, чтобы устойчиво сверхэкспрессировать hROR2.
Фиг. 8. Оценка in vitro цитолитической активности конъюгатов антитело-лекарственное средство, содержащих токсин PNU (G5-PNU), сайт-специфически конъюгированный с IgH и IgL цепями выбранных антител к hROR2, которые указаны, с применением клеток ЕМТ-6, подвергнутых воздействию методов генной инженерии для того, чтобы устойчиво экспрессировать hROR2, в соответствии с примером 8.
Фиг. 9. Оценка in vitro цитолитической активности конъюгатов антитело-лекарственное средство, содержащих клон МК-ЗВ12 нового моноклонального антитела к hROR2, конъюгированного с одним из токсинов PNU G3-PNU или G5-PNU, с применением клеток ЕМТ-6, подвергнутых воздействию методов генной инженерии для того, чтобы стабильно экспрессировать hROR2, в соответствии с примером 9. Соответствующий по изотипу контрольный ADC, содержащий клон специфичного в отношении CD30 антитела - брентуксимаб (клон Ас10) и сайт-специфически конъюгированный G5-PNU, а также экспрессирующие CD30 клетки Karpas-299 применяли для контрольных экспериментов, как указано.
Фиг. 10. Оценка in vitro цитолитической активности конъюгатов антитело-лекарственное средство, содержащих токсин PNU (G5-PNU), сайт-специфически конъюгированный с IgH и IgL цепями выбранных антител к hROR2, которые указаны, с применением клеток ЕМТ-6, подвергнутых воздействию методов генной инженерии для того, чтобы устойчиво экспрессировать hROR2, в соответствии с примером 10.
Фиг. 11. Определение межвидовой перекрестной реактивности рекомбинантных моноклональных антител к hROR2 с мышиным ROR2 и ROR2 яванского макака, которую оценивают с помощью метода ELISA.
Подробное раскрытие настоящего изобретения
Несмотря на то что настоящее изобретение было подробно проиллюстрировано и описано в описании чертежей и вышеизложенном описании, такая иллюстрация и описание считаются иллюстративными или примерными, а не ограничительными; настоящее изобретение не ограничивается раскрытыми вариантами осуществления. Другие варианты раскрытых вариантов осуществления могут быть понятны специалистам в данной области техники и осуществлены ими при практической реализации заявленного изобретения в результате изучения чертежей, раскрытия и прилагаемой формулы изобретения. В пунктах формулы изобретения слово «содержащий» не исключает другие элементы или стадии, и формы единственного числа не исключают множественного числа. Сам по себе тот факт, что определенные измеряемые величины перечислены в отличающихся друг от друга зависимых пунктах формулы изобретения, не указывает на то, что комбинация этих измеряемых величин не может быть применена для получения преимущества. Любые ссылочные позиции в пунктах формулы изобретения не следует толковать как ограничивающие объем.
Все аминокислотные последовательности, раскрытые в данном документе, приведены в ориентации от N-конца к С-концу; все последовательности нуклеиновой кислоты, раскрытые в данном документе, приведены в ориентации от 5' к 3'.
В соответствии с одним аспектом настоящее изобретение относится к новым высокоаффинным связывающим доменам полностью человеческих моноклональных антител, которые специфично связываются с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2). В соответствии с одним вариантом осуществления ROR2 представляет собой человеческий ROR2 (hROR2).
Такие моноклональные антитела были отобраны из разнообразных библиотек антител, созданных посредством стимуляции трансгенных по вариабельному домену легкой и тяжелой цепи человеческого антитела мышей (международная заявка WO 2010/070263 А1, H2L2 мышей) рекомбинантным внеклеточным доменом антигена hROR2 с последующим созданием и скринингом клеточных библиотек, представляющих IgG1, от стимулируемых hROR2 H2L2 мышей с применением ДНК-транспозируемых векторов для трансфекции в 63-12 мышиных пре-В-клеток в соответствии с международной заявкой WO 2014/013026 А1. Клетки млекопитающих, представляющие антитела согласно международной заявке WO 2014/013026 А1, подвергали скринингу с применением антиген-специфического клеточного сортинга (FACS), в том числе сортинга отдельных клеток. В конечном итоге антитела, секретируемые в супернатант отсортированными клонами отдельных клеток, подвергали оценке связывания с антигеном с помощью метода ELISA и анализам цитолиза раковых клеток вторичными антителами (т.е. в которых вторичное антитело, конъюгированное с токсином, связывается с антигеном, связанным с антителом к hROR2 на клеточной поверхности, и интернализируется для осуществления цитолиза). Кодирующие последовательности вариабельных доменов тяжелой и легкой цепи были идентифицированы из выбранных клонов клеток, проявляющих hROR2-специфичное связывание, и на основании функциональной характеристики, включающей цитолиз экспрессирующих hROR2 клеток-мишеней с помощью конъюгированного с токсином вторичного реактива на основе Fab. С помощью этой стратегии были идентифицированы новые полностью человеческие моноклональные антитела к hROR2 высокого качества и с подходящими функциональными свойствами.
В соответствии со вторым аспектом настоящего изобретения предусмотрены конъюгаты антитело-лекарственное средство (ADC) на основе указанных антител к ROR2, связывающих белков на основе антител или их антигенсвязывающих фрагментов с одним или несколькими токсинами и, в частности, с сильнодействующим антрациклиновым токсином. В частности, такие ADC получают посредством сайт-специфического конъюгирования, обеспечиваемого посредством ферментативного конъюгирования с применением фермента-сортазы, как раскрыто в международной заявке WO 2014/140317 А1, которая включена в данный документ посредством ссылки. Один конкретный сильнодействующий антрациклиновый токсин с высокой активностью был раскрыт в международной заявке WO 2016/102679 А1, которая включена в данный документ посредством ссылки.
В соответствии с третьим аспектом настоящего изобретения предусмотрены конъюгаты антитело-эффекторная молекула (АЕС) на основе указанных антител к ROR2, связывающих белков на основе антител или их антигенсвязывающих фрагментов, конъюгированных с одной или несколькими метками.
Кроме того, настоящее изобретение относится к химерным антигенным рецепторам (CAR) и Т-клеткам, подвергнутым воздействию методов генной инженерии с целью экспрессии этих CAR, т.е. Т-клеткам с CAR, использующим указанные антитела к ROR2, производные, модифицированные форматы или фрагменты.
Кроме того, настоящее изобретение относится к би- или мультиспецифичным антителам, содержащим связывающие домены раскрытых антител к ROR2, к би- или мультиспецифичным форматам антител, содержащим по меньшей мере один связывающий домен, специфичный в отношении другой мишени, например, без ограничения, в отношении мишеней, которые осуществляют рекрутмент и/или активацию клеток иммунной системы, подобных Т-клеткам или NK-клеткам. Такие другие связывающие домены могут быть специфичными в отношении CD3, CD16, CD32, CD56, CD64 или других маркеров, специфических для Т- и NK-клеток.
В соответствии с еще одним аспектом настоящее изобретение относится к выделенным или практически очищенным полинуклеотидам, кодирующим вариабельный участок тяжелой цепи иммуноглобулина или легкой цепи иммуноглобулина в полностью человеческом антителе к ROR2, связывающем белке на основе антитела или его антигенсвязывающем фрагменте, и к экспрессионным векторам, несущим такие полинуклеотиды, а также к клеткам-хозяевам, трансформированным или трансфицированным этими полинуклеотидами, и к способам получения антител к ROR2, связывающих белков на основе антител и антигенсвязывающих фрагментов.
В соответствии с еще одним аспектом настоящее изобретение относится к полностью человеческому антителу к ROR2, связывающему белку на основе антитела или его антигенсвязывающему фрагменту, или к ADC, би- или мультиспецифичному антителу, или к химерному антигенному рецептору (CAR), которые описаны в данном документе, для применения в лечении субъекта, который страдает от неопластического заболевания, имеет риск развития неопластического заболевания, и/или у которого диагностировано неопластическое заболевание.
В соответствии с еще одним аспектом настоящее изобретение относится к способу лечения субъекта, страдающего от неопластического заболевания, имеющего риск развития неопластического заболевания, и/или у которого диагностировано неопластическое заболевание, с помощью человеческого антитела к hROR2, связывающего белка на основе антитела или его антигенсвязывающего фрагмента, или ADC, или би- или мультиспецифичного антитела, или клетки, в которой с помощью методов генной инженерии обеспечена экспрессия CAR, как описано в данном документе.
В соответствии с еще одним аспектом настоящее изобретение относится к способу выявления неопластического заболевания или иммунопатологического заболевания или нарушения, для лечения которого подходит антитело к ROR2, связывающий белок на основе антитела или его антигенсвязывающий фрагмент, или АЕС.
Определения
Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понятно квалифицированным специалистам в области техники, к которой относится настоящее изобретение. Кроме того, следующие определения представлены для помощи читателю в практическом осуществлении настоящего изобретения.
Термин «человеческое антитело» относится к антителу, связывающему белку на основе антитела, модифицированному формату антитела, сохраняющему способность к связыванию с мишенью, или к производному или фрагменту антитела, сохраняющим способность к связыванию с мишенью, которые содержат последовательности, полученные из человеческих иммуноглобулинов, вследствие чего практически все из участков CDR1 и CDR2 тяжелой и легкой цепи имеют человеческое происхождение, и практически все из участков 1, 2, 3 и 4 FR тяжелой и легкой цепи соответствуют таковым в последовательности человеческого иммуноглобулина с ограниченным количеством соматических мутаций или без соматических мутаций, которые можно вводить в отдельные последовательности вариабельного домена CDR1 и CDR2 и FR1, 2, 3 и 4 тяжелой и легкой цепи.
Термины «антитело», «связывающий белок на основе антитела», «модифицированный формат антитела, сохраняющий способность к связыванию с мишенью», «производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью» относятся к полипептидной(полипептидным) цепи(цепям), которая(которые) проявляет(проявляют) сильное моновалентное, бивалентное или поливалентное связывание с заданным антигеном, эпитопом или эпитопами. Антитела, связывающие белки на основе антител и антигенсвязывающие фрагменты, применяемые в настоящем изобретении, могут быть получены с применением любой подходящей технологии, например, технологии гибридом, рибосомного дисплея, фагового дисплея, библиотек для шаффлинга генов, полусинтетических или полностью синтетических библиотек или их комбинаций. Антитела, связывающие белки на основе антител и антигенсвязывающие фрагменты согласно настоящему изобретению включают в себя интактные антитела и фрагменты антител или антигенсвязывающие фрагменты, которые содержат антигенсвязывающие участки интактного антитела и сохраняют способность к связыванию с когнатным антигеном. Если в данном документе не определено иное, все пептидные последовательности, в том числе все последовательности антитела и антигенсвязывающего фрагмента, представлены в порядке N->С.
Интактное антитело, как правило, содержит по меньшей мере две тяжелых (Н) цепи (приблизительно 50-70 кДа) и две легких (L) цепи (приблизительно 25 кДа), соединенные между собой дисульфидными связями. Известные гены иммуноглобулинов, кодирующие цепи антител, включают в себя гены константных участков каппа, лямбда, альфа, гамма, дельта, эпсилон и мю цепей, а также гены несметного числа вариабельных участков иммуноглобулина. Легкие цепи относятся либо к типу каппа, либо к типу лямбда. Тяжелые цепи относятся к типу гамма, мю, альфа, дельта или эпсилон, которые, в свою очередь, определяют классы иммуноглобулинов, IgG, IgM, IgA, IgD и IgE, соответственно. Каждая тяжелая цепь антитела состоит из вариабельного участка тяжелой цепи (VH) и константного участка тяжелой цепи. В случае IgG константный участок тяжелой цепи состоит из трех доменов, CH1, СН2 и СН3. Каждая легкая цепь состоит из вариабельного участка легкой цепи (VL) и константного участка легкой цепи. Константный участок легкой цепи состоит из одного домена, CL. Вариабельные участки тяжелой и легкой цепей содержат связывающий домен, который взаимодействует с антигеном. Константные участки антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, в том числе с различными клетками иммунной системы и первым компонентом (Clq) классической системы комплемента. Моноклональные антитела (mAb) состоят из идентичных молекул антител.
Участки VH и VL антитела можно дополнительно подразделять на гипервариабельные участки, также называемые определяющими комплементарность участками (CDR), в промежутках между которыми находятся более консервативные каркасные участки (FR). Каждый VH и VL состоит из трех CDR и четырех FR, расположенных от амино-кольца к карбокси-кольцу в следующем порядке: FR1, CDR1, FR2, CDR2, FR3, CDR3, FR4. Были определены положения участков CDR и FR и система нумерации, например, согласно системе IMGT (Lefranc MP et al., 2015) или схеме нумерации Kabat.
Антитела, связывающие белки на основе антител и антигенсвязывающие фрагменты согласно настоящему изобретению также охватывают одноцепочечные антитела. Термин «одноцепочечное антитело» относится к полипептиду, который содержит домен VH и домен VL в полипептидном мостике, обычно связанные посредством спейсерного пептида, и который может содержать дополнительные домены или аминокислотные последовательности на амино- и/или карбокси-концах. Например, одноцепочечное антитело может содержать ограничивающий сегмент для связывания с кодирующим полинуклеотидом. В качестве примера одноцепочечный фрагмент вариабельного участка (scFv) представляет собой одноцепочечное антитело. По сравнению с доменами VL и VH Fv фрагмента, которые кодируются отдельными генами, scFv имеет два домена, соединенных (например, с помощью методов на основе генетической рекомбинации) синтетическим линкером. Это позволяет получать их в виде одной белковой цепи, в которой участки VL и VH образуют пару с образованием моновалентных молекул.
Примерами связывающих белков на основе антител являются полипептиды, в которых связывающие домены антитела объединены с другими полипептидами или полипептидными доменами, например, альтернативными молекулярными остовами, Fc-участками, другими функциональными или связывающими доменами других полипептидов или антител, что дает в результате молекулы с дополнительными свойствами связывания, например, би- или мультиспецифичные белки или антитела. Такие полипептиды могут создавать характер расположения связывающих или функциональных доменов, который обычно не обнаруживается во встречающихся в естественных условиях антителах или фрагментах антител.
Примеры антигенсвязывающих фрагментов включают в себя (i) Fab фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CHI; (ii) F(ab')2 фрагмент, бивалентный фрагмент, содержащий два Fab фрагмента, связанных дисульфидным мостиком в шарнирном участке; (iii) Fd фрагмент, состоящий из доменов VH и CH1; (iv) Fv фрагмент, состоящий из доменов VL и VH одного плеча интактного антитела; (v) стабилизированные дисульфидными связями Fv (dsFv), которые имеют дисульфидную связь между цепями, встроенную с помощью методов генной инженерии между структурно консервативными каркасными участками; (vi) однодоменное антитело (dAb), которое состоит из домена VH или VL (см., например, Ward et al., Nature 341:544-546, 1989); и (vii) выделенный определяющий комплементарность участок (CDR) в виде линейного или циклического пептида.
Антитела к ROR2, связывающие белки на основе антител и антигенсвязывающие фрагменты, описанные в данном документе, можно получать посредством ферментативной или химической модификации интактных антител или синтезировать de novo с применением методов на основе использования рекомбинантной ДНК. Способы получения этих антител, связывающих белков на основе антител и антигенсвязывающих молекул являются хорошо известными в уровне техники. В частности, scFv антитела можно получать с применением способов, описанных, например, в Bird et al., Science 242:423-426, 1988; и Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883, 1988. Fv фрагменты антител можно получать, как описано в Skerra and 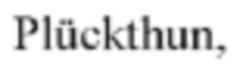 Science 240:1038-41, 1988. Стабилизированные дисульфидными связями Fv фрагменты (dsFv) можно получать с применением способов, описанных, например, в Reiter et al., Int. J. Cancer 67:113-23, 1996. Аналогично, однодоменные антитела (dAb) можно получать с помощью ряда способов, описанных, например, в Ward et al., Nature 341:544-546, 1989; и Cai and Garen, Proc. Natl. Acad. Sci. USA 93:6280-85, 1996. Однодоменные антитела верблюдовых можно получать с применением способов, хорошо известных в уровне техники, например, Dumoulin et al., Nat. Struct. Biol. 11:500-515, 2002; Ghahroudi et al., FEBS Letters 414:521-526, 1997; и Bond et al., J. Mol. Biol. 332:643-55, 2003. Другие типы антигенсвязывающих фрагментов (например, Fab, F(ab')2 или Fd фрагменты) также можно легко получать с помощью обычно используемых в практике иммунологических методов. См., например, Harlow & Lane, Using Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1998.
Science 240:1038-41, 1988. Стабилизированные дисульфидными связями Fv фрагменты (dsFv) можно получать с применением способов, описанных, например, в Reiter et al., Int. J. Cancer 67:113-23, 1996. Аналогично, однодоменные антитела (dAb) можно получать с помощью ряда способов, описанных, например, в Ward et al., Nature 341:544-546, 1989; и Cai and Garen, Proc. Natl. Acad. Sci. USA 93:6280-85, 1996. Однодоменные антитела верблюдовых можно получать с применением способов, хорошо известных в уровне техники, например, Dumoulin et al., Nat. Struct. Biol. 11:500-515, 2002; Ghahroudi et al., FEBS Letters 414:521-526, 1997; и Bond et al., J. Mol. Biol. 332:643-55, 2003. Другие типы антигенсвязывающих фрагментов (например, Fab, F(ab')2 или Fd фрагменты) также можно легко получать с помощью обычно используемых в практике иммунологических методов. См., например, Harlow & Lane, Using Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1998.
Антитела к ROR2, связывающие белки на основе антител или антигенсвязывающие фрагменты согласно настоящему изобретению можно получать с помощью любой подходящей методики, например, с применением любой подходящей эукариотической экспрессионной системы или экспрессионной системы, отличной от эукариотической. В соответствии с определенными вариантами осуществления антитело, связывающий белок на основе антитела или антигенсвязывающий фрагмент получают с применением экспрессионной системы на основе млекопитающего. Некоторые конкретные методики для получения антител, связывающих белков на основе антител или антигенсвязывающих фрагментов или антигенсвязывающих фрагментов согласно настоящему изобретению проиллюстрированы примером в данном документе. В соответствии с некоторыми вариантами осуществления антитела, связывающие белки на основе антител или антигенсвязывающие фрагменты согласно настоящему изобретению можно получать с применением подходящей экспрессионной системы, отличной от эукариотической, такой как бактериальная экспрессионная система. Бактериальные экспрессионные системы можно применять для получения фрагментов, таких как F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2, и диател. Методики изменения кодирующих последовательностей ДНК для получения таких фрагментов, являются известными в уровне техники.
Термин «консервативно модифицированный вариант» относится как к аминокислотным последовательностям, так и к последовательностям нуклеиновой кислоты. Применительно к конкретным последовательностям нуклеиновой кислоты термин «консервативно модифицированные варианты» относится к тем нуклеиновым кислотам, которые кодируют идентичные или, по существу, идентичные аминокислотные последовательности, или в случаях, когда нуклеиновая кислота не кодирует аминокислотную последовательность, к, по существу, идентичным последовательностям. Вследствие вырожденности генетического кода большое число функционально идентичных нуклеиновых кислот кодируют любой заданный белок. Например, все из кодонов GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором аланин определен кодоном, кодон можно изменять на любые соответствующие описанные кодоны без изменения кодируемого полипептида. Такие вариации нуклеиновой кислоты представляют собой «молчащие вариации», которые являются одной разновидностью консервативно модифицированных вариаций. Каждая последовательность нуклеиновой кислоты в данном документе, которая кодирует полипептид, также описывает каждую молчащую вариацию нуклеиновой кислоты. Квалифицированному специалисту будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) можно модифицировать с получением на выходе функционально идентичной молекулы. Соответственно, в каждой описанной последовательности предполагается каждая молчащая вариация нуклеиновой кислоты, которая кодирует полипептид.
В случае полипептидных последовательностей «консервативно модифицированные варианты» относятся к варианту, который имеет консервативные аминокислотные замены, при этом аминокислотные остатки заменяют на другой аминокислотный остаток, имеющий боковую цепь с аналогичным зарядом. В уровне техники были определены семейства аминокислотных остатков, имеющих боковые цепи с аналогичными зарядами. Эти семейства включают в себя аминокислоты с основными боковыми цепями (например, лизин, аргинин, гистидин), кислотными боковыми цепями (например, аспарагиновая кислота, глутаминовая кислота), незаряженными полярными боковыми цепями (например, глицин, аспарагин, глутамин, серии, треонин, тирозин, цистеин), неполярными боковыми цепями (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленными боковыми цепями (например, треонин, валин, изолейцин) и ароматическими боковыми цепями (например, тирозин, фенилаланин, триптофан, гистидин).
Термины «идентичный» или процентная «идентичность» в контексте двух или более нуклеиновых кислот или полипептидных последовательностей относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми. Две последовательности являются «практически идентичными», если две последовательности имеют определенный процент аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т.е. 60% идентичность, необязательно, 65%, 70%, 75%, 80%, 85%, 90%, 95% или 99% идентичность в определенном участке, или в случае, когда он не определен, во всей последовательности) при сравнении и выравнивании для максимального соответствия в окне сравнения или обозначенном участке при измерении с применением одного из следующих алгоритмов для сравнения последовательностей или ручного выравнивания и визуального просмотра. Необязательно, идентичность существует в участке, который имеет длину по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот), или, более предпочтительно, в участке, который имеет длину 100-500 или 1000 или больше нуклеотидов (или 20, 50, 200 или больше аминокислот).
Способы выравнивания последовательностей для сравнения являются хорошо известными в уровне техники. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии из Smith and Waterman, Adv. Appl. Math. 2:482c, 1970; с помощью алгоритма выравнивания гомологичных участков из Needleman and Wunsch, J. Mol. Biol. 48:443, 1970; с помощью способа поиска сходства из Pearson and Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444, 1988; с помощью реализаций этих алгоритмов в компьютерных программах (GAP, BESTFIT, FASTA и TFASTA в составе пакета программного обеспечения Wisconsin Genetics Software Package, Genetics Computer Group, Мадисон, Висконсин, США); или с помощью ручного выравнивания и визуального просмотра (см., например, Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (Ringbou ed., 2003)). Двумя примерами алгоритмов, которые являются подходящими для определения процентной идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977; и Altschul et al., J. Mol. Biol. 215:403-410, 1990, соответственно.
Процентная (%) идентичность пептидных последовательностей можно рассчитать, например, как 100 × [(идентичные положения)/min(TGA, TGB)], где TGA и TGB представляют собой сумму числа остатков и положений внутренних гэпов в пептидных последовательностях А и В при выравнивании, которое сводит к минимуму TGA и TGB. См., например, Russell et al, J. Mol. Biol, 244: 332-350 (1994).
Искусственные Т-клеточные рецепторы (также известные как химерные Т-клеточные рецепторы, химерные иммунорецепторы, химерные антигенные рецепторы (CAR) или Т-клеточные тельца (T-bodies)) представляют собой сконструированные рецепторы, которые придают произвольную специфичность эффекторной иммунной клетке. Как правило, эти рецепторы применяют для придания специфичности моноклонального антитела Т-клетке; причем перенос их кодирующей последовательности облегчается ретровирусными или лентивирусными векторами или транспозонами. Т-клетки, подвергнутые воздействию методов генной инженерии для экспрессии CAR, представляют собой подвергнутые воздействию методов генной инженерии Т-клетки, снаряженные химерными рецепторами, чья внеклеточная распознающая часть состоит из происходящего из антитела распознающего домена, и чей внутриклеточный участок происходит из лимфоцит-стимулирующего фрагмента(фрагментов). Структура прототипных CAR является модульной, она предназначена для обеспечения размещения различных функциональных доменов и, благодаря этому, создания возможности выбора специфичности и контролируемой активации Т-клеток. Предпочтительная происходящая из антитела распознающая часть представляет собой одноцепочечный вариабельный фрагмент (scFv), который объединяет обеспечивающие специфичность и связывание остатки в вариабельных участках как тяжелой, так и легкой цепи моноклонального антитела, а также другие происходящие из антитела форматы, подобные Fab фрагментам, V-доменам и т.д., можно использовать для придания желаемой специфичности CAR Т-клеткам, подвергнутым воздействию методов генной инженерии для экспрессии CAR. Наиболее распространенные лимфоцит-активирующие фрагменты включают в себя Т-клеточный костимулирующий домен (например, CD28) в тандеме с запускающим Т-клетки (например, CD3zeta) фрагменты, а также другие сигнальные домены, подобные 4-1 ВВ, можно использовать во внутриклеточной части CAR. При снаряжении эффекторных лимфоцитов (таких как Т-клетки и клетки-натуральные киллеры) такими химерными рецепторами подвергнутая воздействию методов генной инженерии клетка пере нацеливается с предварительно определенной специфичностью на любой желаемый антиген-мишень без ограничения по HLA. Конструкции CAR вводят ex vivo в Т-клетки из периферических лимфоцитов заданного пациента с применением ретровирусных или лентивирусных векторов или транспозонов. После инфузии полученных в результате Т-клеток, подвергнутых воздействию методов генной инженерии для экспрессии CAR, обратно пациенту они мигрируют, достигают своего сайта-мишени и при взаимодействии со своей клеткой- или тканью-мишенью они подвергаются активации и выполняют свою предварительно определенную эффекторную функцию. Терапевтические мишени для CAR-подхода включают в себя злокачественную опухоли и ВИЧ-инфицированные клетки или аутоиммунные эффекторные клетки.
Термины «лечить», «осуществление лечения», «лечение» и «терапевтически эффективный», используемые в контексте данного документа, не обязательно предполагают 100% или полное лечение. Точнее, существуют переменные степени лечения, известные квалифицированному специалисту в данной области техники как оказывающие потенциальное благоприятное воздействие или терапевтический эффект. В связи с этим способ согласно настоящему изобретению может обеспечивать любую величину любого уровня лечения. Более того, лечение, обеспечиваемое с помощью способа согласно настоящему изобретению, может включать в себя лечение одного или нескольких состояний или симптомов заболевания, подлежащего лечению.
«Вектор» представляет собой репликон, такой как плазмида, фаг или космида, к которому может быть прикреплен другой полинуклеотидный сегмент для того, чтобы осуществлять репликацию прикрепленного сегмента. Векторы, способные к управлению экспрессией генов, кодирующих один или несколько полипептидов, называются «экспрессионными векторами».
Подробное раскрытие вариантов осуществления
Аспекты настоящего изобретения, относящиеся к связыванию с человеческим ROR2
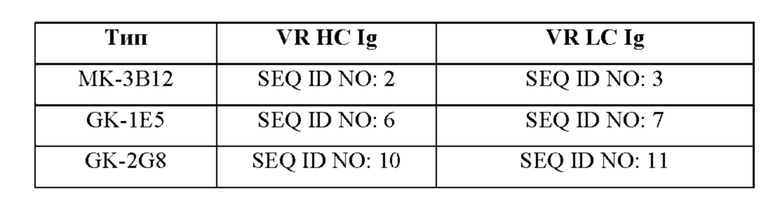
В соответствии с одним аспектом настоящее изобретение относится к человеческим антителам к ROR2, связывающим белкам на основе антител (в том числе би- или мультиспецифичным антителам), их антигенсвязывающим фрагментам, АЕС, ADC или CAR, имеющим одинаковую специфичность связывания в отношении ROR2 (и, в особенности, в отношении hROR2, содержащего или состоящего из SEQ ID NO. 1), т.е. связывающимся с таким же эпитопом ROR2 и/или конкурирующим за связывание с ROR2 со специфичными в отношении ROR2 антителами, содержащими пару из последовательности вариабельного участка тяжелой цепи иммуноглобулина и последовательности вариабельного участка легкой цепи иммуноглобулина согласно таблице 2. В частности, настоящее изобретение относится к человеческим антителам к ROR2, связывающим белкам на основе антител (в том числе би- или мультиспецифичным антителам), их антигенсвязывающим фрагментам, АЕС, ADC или CAR, имеющим одинаковую специфичность связывания в отношении ROR2 (и, в особенности, в отношении hROR2, содержащего или состоящего из SEQ ID NO. 1).
В соответствии с одним аспектом предусмотрено полностью человеческое антитело или его производное, модифицированный формат или фрагмент, которые специфично связываются с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2).
В соответствии с одним вариантом осуществления антитело, производное, модифицированный формат или фрагмент представляют собой антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью.
В соответствии с одним вариантом осуществления антитело, производное, модифицированный формат или фрагмент (i) связываются с тем же эпитопом ROR2 и/или (ii) конкурируют за связывание с ROR2 с антителом, содержащим последовательность вариабельного участка тяжелой цепи Ig и последовательность вариабельного участка легкой цепи Ig, соответственно, представленные в:
(i) SEQ ID NO:2 и SEQ ID NO:3;
(ii) SEQ ID NO:4 и SEQ ID NO: 5;
(iii) SEQ ID NO:6 и SEQ ID NO:7;
(iv) SEQ ID NO:8 и SEQ ID NO:9;
(v) SEQ ID NO: 10 и SEQ ID NO: 11;
(vi) SEQ ID NO: 12 и SEQ ID NO: 13;
(vii) SEQ ID NO:14 и SEQ ID NO: 15;
(viii) SEQ ID NO:16 и SEQ ID NO:17;
(ix) SEQ ID NO: 18 и SEQ ID NO: 19;
(x) SEQ ID N0:20 и SEQ ID NO:21;
(xi) SEQ ID NO:22 и SEQ ID NO:23 или
(xii) SEQ ID NO:24 и SEQ ID NO:25.
В соответствии с одним другим вариантом осуществления антитело, производное, модифицированный формат или фрагмент содержат последовательность вариабельного участка тяжелой цепи и последовательность вариабельного участка легкой цепи, соответственно, одна или или обе из которых являются по меньшей мере на 90%, предпочтительно, по меньшей мере на 95%, более предпочтительно, по меньшей мере на 98%, еще более предпочтительно, по меньшей мере на 99% и, наиболее предпочтительно, на 100% идентичными парам последовательности вариабельного участка тяжелой цепи/последовательности вариабельного участка легкой цепи, представленным выше.








Анализы для оценки конкуренции за связывание включают в себя, без ограничения, иммунологические анализы с использованием метки из радиоактивного материала (RIA), твердофазные иммуноферментные анализы (ELISA), сэндвич-анализы ELISA, анализы методом флуоресцентно-активированного клеточного сортинга (FACS) и анализы методом Biacore (SPR). При проведении конкурентного анализа антител между контрольным антителом и исследуемым антителом можно вначале пометить эталонный образец выявляемой меткой, такой как флуорофор, биотин или ферментативная (или даже радиоактивная) метка, для обеспечения возможности последующей идентификации. Если исследуемое антитело конкурирует с меченым контрольным антителом, интенсивность будет снижаться относительно контрольной реакции, которую проводят без исследуемого антитела.
Способы определения связывающего эпитопа включают в себя, без ограничения, оценку связывания с массивом олигопептидов, имеющих (перекрывающиеся) аминокислотные последовательности из последовательности ROR2, расшифровку кристаллической или ЯМР структуры антитела, связывающего белка на основе антитела или его антигенсвязывающего фрагмента с ROR2, с помощью оценки потери связывания антител, связывающих белков на основе антител, их антигенсвязывающих фрагментов с ROR2, содержащим одну или несколько мутаций аминокислот («мутагенез с высокой пропускной способностью»), и с помощью водородно-дейтериевого обмена для оценки доступной для растворителя поверхности у комплекса ROR2 с антителом, связывающим белком на основе антитела или антигенсвязывающим фрагментом (Abbott М. et al, 2014).
В соответствии с предпочтительным вариантом осуществления полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, АЕС, ADC или CAR, имеющие одинаковую специфичность связывания в отношении ROR2 (и, в особенности, в отношении hROR2, содержащего или состоящего из SEQ ID NO. 1), т.е. связывающиеся с таким же эпитопом ROR2 и/или конкурирующие за связывание с ROR2 со специфичными в отношении ROR2 антителами, содержащими пару из последовательности вариабельного участка тяжелой цепи иммуноглобулина (Ig HCVR) и последовательности вариабельного участка легкой цепи иммуноглобулина (Ig LCVR), соответственно, представленными в: (i) SEQ ID NO:2 и SEQ ID NO:3; (ii) SEQ ID NO:6 и SEQ ID NO:7; (iii) SEQ ID NO:10 и SEQ ID NO:ll; (iv) SEQ ID NO:12 и SEQ ID NO:13; (v) SEQ ID NO:14 и SEQ ID NO:15; (vi) SEQ ID NO:16 и SEQ ID NO:17; (vii) SEQ ID NO:4 и SEQ ID NO:5; (viii) SEQ ID NO:8 и SEQ ID NO:9; (ix) SEQ ID NO:18 и SEQ ID NO:19; (x) SEQ ID NO:24 и SEQ ID NO:25; (xi) SEQ ID NO:20 и SEQ ID NO:21; и (xii) SEQ ID NO:22 и SEQ ID NO:23, и, более предпочтительно, которые представлены в (i) SEQ ID NO:2 и SEQ ID NO:3; (ii) SEQ ID NO:8 и SEQ ID NO:9 и (iii) SEQ ID N0:20 и SEQ IDNO:21.
В соответствии с различными вариантами осуществления антитело, связывающий белок на основе антитела, его антигенсвязывающий фрагмент, АЕС, ADC или CAR считают конкурирующими с контрольным антителом, если они снижают связывание контрольного антитела по меньшей мере приблизительно на 20% или больше, например, по меньшей мере приблизительно на 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90%, 95% или еще больше, или на процентное значение, находящееся в диапазоне между любым из вышеизложенных значений, при концентрации контрольного антитела, которая составляет 80% от максимального связывания в условиях конкретного применяемого анализа, и концентрации исследуемого антитела или антигенсвязывающего фрагмента, которая является 10-кратно большей, чем концентрация контрольного антитела.
Предпочтительные варианты осуществления, относящиеся к вышеизложенным аспектам настоящего изобретения
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческое антитело к ROR2, связывающий белок на основе антитела, его антигенсвязывающий фрагмент, АЕС, ADC или CAR предпочтительно связываются с ROR2, содержащим или состоящим из SEQ ID NO. 1, с Kd, составляющим менее 100 нМ, более предпочтительно, менее 75 нМ, еще более предпочтительно, менее 50 нМ и, наиболее предпочтительно, менее 25 нМ.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческое антитело к ROR2, связывающий белок на основе антитела, его антигенсвязывающий фрагмент, АЕС, ADC или CAR предпочтительно связываются с ROR2, содержащим или состоящим из SEQ ID NO. 1, с kon, составляющей более 1×104 М-1 с-1, более предпочтительно, более 1×105 М-1 с-1, еще более предпочтительно, более l×106 M-1c-1.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческое антитело к ROR2, связывающий белок на основе антитела, его антигенсвязывающий фрагмент, АЕС, ADC или CAR предпочтительно связываются с ROR2, содержащим или состоящим из SEQ ID NO. 1, с k0ff, составляющим менее 1×10-2 с-1, более предпочтительно, менее 1×10-3 с-1 и, еще более предпочтительно, менее 1×10-4 с-1.
Аффинность связывания обычно выражают через константы равновесия ассоциации или диссоциации (Ka или Kd, соответственно), которые, в свою очередь, представляют собой обратные величины констант скорости диссоциации и ассоциации (k0ff и kon, соответственно). Таким образом, эквивалентные аффинности могут соответствовать различным константам скорости, поскольку соотношение констант скорости остается одинаковым. Аффинности связывания и/или константы скорости можно определять с применением методик, хорошо известных в уровне техники или описанных в данном документе, таких как, например, ELISA, изотермическая титрационная калориметрия (ITC), Biacore (SPR), биослойная интерферометрия или поляризация флуоресценции.
Настоящее изобретение также относится к полностью человеческим антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающий фрагментам, ADC, АЕС или CAR, содержащим цепь VH, содержащую три CDR, причем:
CDR №1 VH содержит или состоит из пептидной последовательности GYSISSGYY (SEQ ID NO. 26) или GX1X2FX3X4X5X6 (SEQ ID NO. 67), в которой X1=F или G, Х2=Т или S, X3=R или S, X4=S, Т, R или G, X5=H, Y или Q, X6=G, Y или R; и/или
CDR №2 VH содержит или состоит из пептидной последовательности IYQSGST (SEQ ID NO. 27), INHSRTT (SEQ ID NO. 59), INHSGIT (SEQ ID NO. 93) или IX7X8DGX9X10K (SEQ ID NO. 68), в которой X7=W или K, X8=Y, N, F или Q, X9=S или T, Х10=K, N или E; и/или
VH CDR №3 содержит или состоит из пептидной последовательности, выбранной из: CAREDRAGWYPFDCW (SEQ ID NO. 28), CARVGAGLYLDYW (SEQ ID NO. 33), CAREGSGWYDYYYGMDVW (SEQ ID NO. 40), CQHYNTYSRTF (SEQ ID NO. 62), CARPGIAMTGLDYW (SEQ ID NO. 37), CARGGDQWLVPFDNW (SEQ ID NO. 95), CARVAAALHFHYW (SEQ ID NO. 47), CARGGEQWLVPFDYW (SEQ ID NO. 60), CARPGVAMTGLDLW (SEQ ID NO. 43), CIRVKFGDLYFQHW (SEQ ID NO. 49), CVRVRFGELYFQHW (SEQ ID NO. 44), CARDGYRNGWHIPEDYW (SEQ ID NO. 65), CARMGAINRGGGGFDYW (SEQ ID NO. 52) и CARDKGEWFGELRYYYYGMDVW (SEQ ID NO. 55).
Настоящее изобретение также относится к полностью человеческим антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающим фрагментам, ADC, AEC или CAR, содержащим цепь VH, содержащую три CDR, причем:
CDR №1 VH содержит или состоит из пептидной последовательности GYSISSGYY (SEQ ID NO. 26) или GX1X2FX3X4X5X6 (SEQ ID NO. 67), в которой X1=F или G, Х2=Т или S, X3=R или S, X4=S, Т, R или G, Х5=Н, Y или Q, X6=G, Y или R; и
CDR №2 VH содержит или состоит из пептидной последовательности IYQSGST (SEQ ID NO. 27), INHSRTT (SEQ ID NO. 59) или IX7X8DGX9X10K (SEQ ID NO. 68), в которой X7=W или K, X8=Y, N, F или Q, X9=S или T, X10=K, N или E; и
VH CDR №3 содержит или состоит из пептидной последовательности, выбранной из: CAREDRAGWYPFDCW (SEQ ID NO. 28), CARVGAGLYLDYW (SEQ ID NO. 33), CAREGSGWYDYYYGMDVW (SEQ ID NO. 40), CQHYNTYSRTF (SEQ ID NO. 62), CARPGIAMTGLDYW (SEQ ID NO. 37), CARGGDQWLVPFDNW (SEQ ID NO. 95), CARVAAALHFHYW (SEQ ID NO. 47), CARGGEQWLVPFDYW (SEQ ID NO. 60), CARPGVAMTGLDLW (SEQ ID NO. 43), CIRVKFGDLYFQHW (SEQ ID NO. 49), CVRVRFGELYFQHW (SEQ ID NO. 44), CARDGYRNGWHIPEDYW (SEQ ID NO. 65), CARMGAINRGGGGFDYW (SEQ ID NO. 52) и CARDKGEWFGELRYYYYGMDVW (SEQ ID NO. 55).
Настоящее изобретение также относится к полностью человеческим антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающим фрагментам, ADC, АЕС или CAR, содержащим цепь VH, содержащую три CDR, причем:
CDR №1 VH содержит или состоит из пептидной последовательности GYSISSGYY (SEQ ID NO. 26), GFTFRSHG (SEQ ID NO. 31), GGSFSGYY (SEQ ID NO. 58), GFTFSSQR (SEQ ID NO. 63) или GFTFX3X4YG (SEQ ID NO. 71), в которой X3=R или S, и X4=S, T или R, и, предпочтительно, содержит или состоит из пептидной последовательности GYSISSGYY (SEQ ID NO. 26), GFTFRSYG (SEQ ID NO 36), GFTFRTYG (SEQ ID NO 94), GFTFSRYG (SEQ ID NO 45), GFTFRSHG (SEQ ID NO. 31), GFTFSSYG (SEQ ID NO. 38), GGSFSGYY (SEQ ID NO. 58) или GFTFSSQR (SEQ ID NO. 63), и, более предпочтительно, содержит или состоит из пептидная последовательность GYSISSGYY (SEQ ID NO. 26) или GFTFSSYG (SEQ ID NO. 38); и
CDR №2 VH содержит или состоит из пептидной последовательности IYQSGST (SEQ ID NO. 27), IWYDGSKK (SEQ ID NO 32), IWYDGSNK (SEQ ID NO 46), IWNDGSNK (SEQ ID NO. 39), IWFDGTNK (SEQ ID NO. 54), INHSRTT (SEQ ID NO. 59) или IKQDGSEK (SEQ ID NO. 64) и, предпочтительно, содержит или состоит из пептидной последовательности IYQSGST (SEQ ID NO. 27), IWNDGSNK (SEQ ID NO. 39) или IWFDGTNK (SEQ ID NO. 54); и
VH CDR №3 содержит или состоит из пептидной последовательности, выбранной из: CAREDRAGWYPFDCW (SEQ ID NO. 28), CARPGIAMTGLDYW, CARPGVAMTGLDLW (DEQ ID NO. 43), CVRVRFGELYFQHW (SEQ ID NO. 44), CARVAAALHFHYW (SEQ ID NO. 47), CIRVKFGDLYFQHW (SEQ ID NO. 49), CARVGAGLYLDYW (SEQ ID NO. 33), CARPGIAMTGLDYW (SEQ ID NO. 37), CAREGSGWYDYYYGMDVW (SEQ ID NO. 40), CARDKGEWFGELRYYYYGMDVW (SEQ ID NO. 55), CARGGEQWLVPFDYW (SEQ ID NO. 60) или CARDGYRNGWHIPEDYW (SEQ ID NO. 65) и, предпочтительно, содержит или состоит из пептидной последовательности, выбранной из: CAREDRAGWYPFDCW (SEQ ID NO. 28), CAREGSGWYDYYYGMDVW (SEQ ID NO. 40) или CARDKGEWFGELRYYYYGMDVW (SEQ ID NO. 55).
В соответствии с любым из вышеуказанных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VH, содержащую три CDR, причем:
CDR №1 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR1 НС, перечисленных в таблице 3; и/или
CDR №2 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR2 НС, перечисленных в таблице 3; и/или
CDR №3 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR3 НС, перечисленных в таблице 3.
В соответствии с любым из вышеуказанных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VH, содержащую три CDR, причем:
CDR №1 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR1 НС, перечисленных в таблице 3; и
CDR №2 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR2 НС, перечисленных в таблице 3; и
CDR №3 VH содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR3 НС, перечисленных в таблице 3.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VH, содержат три CDR, причем CDR №1 VH, CDR №2 VH и CDR №3 VH содержит или состоит из пептидной последовательности, соответствующей заданному набору из CDR1, CDR2 и CDR3 НС, представленному в таблице 3.
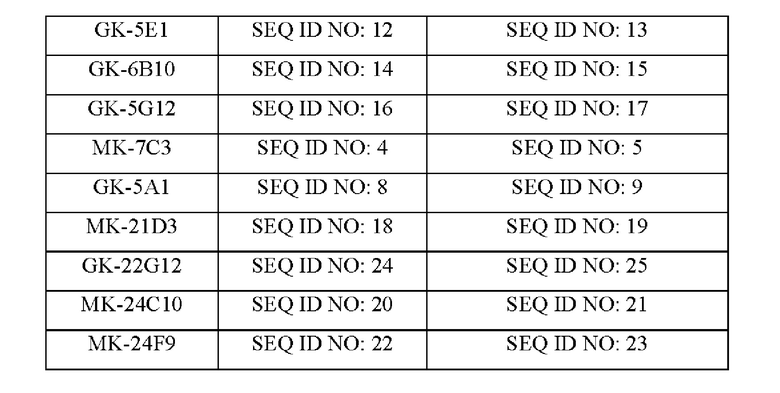
В соответствии с любым из вышеизложенных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VH, содержат три CDR, причем CDR №1 VH, CDR №2 VH и CDR №3 VH содержат или состоят из пептидной последовательности, соответствующей заданному набору из CDR1, CDR2 и CDR3 НС из MK-3 В12, GK-5A1, GK-21D3, MK-24С10, MK-24F9 или GK-22G12 и, более предпочтительно, из MK-3 В12, GK-5A1 или MK-24С10, представленных в таблице 3.
Настоящее изобретение также относится к антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающих фрагментов, ADC, АЕС или CAR, содержащим цепь VL, содержащую три CDR, причем:
CDR №1 VL содержит или состоит из пептидной последовательности QSLLHSNGYNY (SEQ ID NO. 56), QSIDNW (SEQ ID NO. 48) или QX11ISX12X13 (SEQ ID NO. 69), в которой X11=S, T или G, X12=S, N или H и X13=W или Y; и/или
CDR №2 VL содержит или состоит из пептидной последовательности KAS, AAS или LGS; и/или
CDR №3 VL содержит или состоит из пептидной последовательности CX14X15X16X17X18X19X20X21X22F (SEQ ID NO. 70), в которой X14=Q или М, X15=K, Н или Q, X16=H, Y или A, X17=N или L, X18=R, Т, Q, S или N; X19=A, Y или Т; Х20=Р, S или W, X21=W, R, Y или отсутствует, X22=S или Т.
Настоящее изобретение также относится к антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающих фрагментов, ADC, АЕС или CAR, содержащих цепь VL, содержащих три CDR, причем:
CDR №1 VL содержит или состоит из пептидной последовательности QSLLHSNGYNY (SEQ ID NO. 56), QSIDNW (SEQ ID NO. 48) или QX11ISX12X13 (SEQ ID NO. 69), в которой X11=S, T или G, X12=S, N или H и X13=W или Y; и
CDR №2 VL содержит или состоит из пептидной последовательности KAS, AAS или LGS; и
CDR №3 VL содержит или состоит из пептидной последовательности CX14X15X16X17X18X19X20X21X22F (SEQ ID NO. 70), в которой X14=Q или М, X15=K, Н или Q, X16=H, Y или A, X17=N или L, X18=R, T, Q, S или N; X19=A, Y или T; X20=P, S или W, X21=W, R, Y или отсутствует, X22=S или Т.
Настоящее изобретение также относится к антителам к ROR2, связывающим белкам на основе антител, их антигенсвязывающих фрагментов, ADC, АЕС или CAR, содержащих цепь VL, содержащих три CDR, причем:
CDR №1 VL содержит или состоит из пептидной последовательности QSISSW (SEQ ID NO. 29), QTISNW (SEQ ID NO. 34), QSIDNW (SEQ ID NO. 48), QGISNY (SEQ ID NO. 50), QSLLHSNGYNY (SEQ ID NO. 56) или QSISHW (SEQ ID NO. 61) и, предпочтительно, содержит или состоит из пептидной последовательности QSISSW (SEQ ID NO. 29) или QSLLHSNGYNY (SEQ ID NO. 56); и
CDR №2 VL содержит или состоит из пептидной последовательности KAS, LGS или AAS и, предпочтительно, содержит или состоит из пептидной последовательности KAS или LGS; и
CDR №3 VL содержит или состоит из пептидной последовательности CQQYNNYWTF (SEQ ID NO. 30), CQQYNSYWTF (SEQ ID NO. 41), CQQYNSYSYSF (SEQ ID NO. 42), CQQYNSYSYSF (SEQ ID NO. 44), CQQYNSYSYTF (SEQ ID NO. 35), CQKYNSAPYTF (SEQ ID NO. 51), CARMGAINRGGGGFDYW (SEQ ID NO. 52), CQKYNSAPWTF (SEQ ID NO. 53), CMQALQTPYTF (SEQ ID NO. 57), CQHYNTYSRTF (SEQ ID NO. 62) или CQKHNRAPWTF (SEQ ID NO. 66) и предпочтительно, содержит или состоит из, пептидной последовательности CQQYNNYWTF (SEQ ID NO. 30), CQQYNSYWTF (SEQ ID NO. 41) или CMQALQTPYTF (SEQ ID NO. 57).
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VL, содержащую три CDR, причем:
CDR №1 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR1 LC, перечисленных в таблице 3; и/или
CDR №2 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR2 LC, перечисленных в таблице 3; и/или
CDR №3 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR3 LC, перечисленных в таблице 3.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VL, содержащую три CDR, причем:
CDR №1 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR1 LC, перечисленных в таблице 3; и
CDR №2 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR2 LC, перечисленных в таблице 3; и
CDR №3 VL содержит или состоит из пептидной последовательности, выбранной из последовательностей CDR3 LC, перечисленных в таблице 3.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающих фрагментов, ADC, АЕС или CAR, содержащих цепь VL, содержат три CDR, причем CDR №1 VL, CDR №2 VL и CDR №3 VL содержат или состоят из пептидной последовательности, соответствующей заданному набору из CDR1, CDR2 и CDR3 LC, представленному в таблице 3.
В соответствии с любым из вышеизложенных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержащие цепь VL, содержат три CDR, причем CDR №1 VL, CDR №2 VL и CDR №3 VL содержат или состоят из пептидной последовательности, соответствующей набору из CDR1, CDR2 и CDR3 LC из MK-3 В12, GK-5A1, GK-21D3, MK-24С10, MK-24F9 или GK-22G12, и, более предпочтительно, из MK-3 В12, GK-5A1 или MK-24С10, представленных в таблице 3.
В соответствии с любым из вышеуказанных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержат цепь VL и VH, каждая из которых содержит три CDR, содержащих или состоящих, в соответствии с таблицей 3, из пептидной последовательности соответствующих CDR из: MK-3 В12, MK-7С3, GK-1E5, GK-5E1, GK-2G8, GK-5A1, GK-6B10, GK-5G12, GK-21D3, MK-24С10, MK-24F9 или GK-22G12 и, предпочтительно, из: MK-3 В12, GK-5A1, GK-21D3, MK-24С10, MK-24F9 или GK-22G12, и, более предпочтительно, из: Mk-3 В12, GK-5A1 или МК-24С10.
В соответствии с любым из вышеуказанных аспектов настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR, содержат цепь VL и VH, каждая из которых содержит три CDR, содержащих или состоящих, в соответствии с таблицей 3, из пептидной последовательности соответствующих CDR из: MK-3 В12, GK-1E5, GK-2G8, GK-5E1, GK-6 В10 или GK-5G12, или из: GK-5A1, GK-21D3, MK-24С10, MK-24F9, MK-7С3 или GK-22G12, или, более предпочтительно, из: MK-3 В12, GK-5A1, GK-21D3, MK-24С10, MK-24F9 или GK-22G12, или, еще более предпочтительно, из: MK-3 В12, GK-5A1 или MK-24С10.
В соответствии с одним вариантом осуществления антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью, обладают перекрестной реактивностью в отношении (i) человеческого ROR2 (hROR2) и (ii) по меньшей мере одного из ROR2 яванского макака (cROR2) и мышиного RoR2 (mROR2).
При использовании в контексте данного документа термин «перекрестная реактивность» означает, что антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью, являются способными к связыванию (i) с человеческим ROR2 (hROR2) и (ii) по меньшей мере с одним из ROR2 яванского макака (cROR2) и мышиного RoR2 (mROR2) с достаточной аффинностью.
Указанная перекрестная-реактивность имеет значительные преимущества в применениях в доклинических исследованиях, поскольку то же самое антитело, которое применяли для исследований на мышах и яванских макаках, можно в дальнейшем применять для клинических испытаний. Кроме того, указанная перекрестная реактивность имеет значительные преимущества в диагностике, а также в научных применениях.
В соответствии с одним вариантом осуществления антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью, содержат один из следующих наборов CDR:
a) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 38, 39 и 40, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 29 и 41, с CDR 2 легкой цепи, имеющим последовательность KAS;
b) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 26-28, и CDR 1-3 легкой цепи, которые изложены в SEQ ID NO 29, 30 и 73;
c) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 94, 32 и 44, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 29 и 42, с CDR 2 легкой цепи, имеющим последовательность KAS;
d) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 38, 46 и 52, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 50 и 53, с CDR 2 легкой цепи, имеющим последовательность AAS;
e) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 38, 54 и 55, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 56 и 57, с CDR 2 легкой цепи, имеющим последовательность LGS;
f) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 58-60, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 61 и 62, с CDR 2 легкой цепи, имеющим последовательность KAS;
g) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO 63-65, с CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO 50 и 66, с CDR 2 легкой цепи, имеющим последовательность AAS.
CDR содержатся в подходящем белковом каркасе для того, чтобы они были способны к связыванию с hROR2, а также по меньшей мере с одним из ROR2 яванского макака (cROR2) и мышиного RoR2 (mROR2) с достаточной аффинностью.
В соответствии с одним вариантом осуществления антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью, содержат одну из следующих пар последовательностей:
a) последовательность вариабельного участка тяжелой цепи антитела GK-5A1, представленную в SEQ ID NO. 8, и последовательность вариабельного участка легкой цепи антитела GK-5A1, представленную в SEQ ID NO. 9,
b) последовательность вариабельного участка тяжелой цепи антитела MK-3 В12, представленную в SEQ ID NO. 2, и последовательность вариабельного участка легкой цепи антитела МК-ЗВ12, представленную в SEQ ID NO. 3,
c) последовательность вариабельного участка тяжелой цепи антитела GK-5E1, представленную в SEQ ID NO. 12, и последовательность вариабельного участка легкой цепи антитела GK-5E1, представленную в SEQ ID NO. 13,
d) последовательность вариабельного участка тяжелой цепи антитела GK-21D3, представленную в SEQ ID NO. 18, и последовательность вариабельного участка легкой цепи антитела GK-5E1, представленную в SEQ ID NO. 19,
e) последовательность вариабельного участка тяжелой цепи антитела МК-24С10, представленную в SEQ ID NO. 20, и последовательность вариабельного участка легкой цепи антитела MK-24С10, представленную в SEQ ID NO. 21,
f) последовательность вариабельного участка тяжелой цепи антитела MK-24F9, представленную в SEQ ID NO. 22, и последовательность вариабельного участка легкой цепи антитела MK-24F9, представленную в SEQ ID NO. 23,
g) последовательность вариабельного участка тяжелой цепи антитела GK-22G12, представленную в SEQ ID NO. 24, и последовательность вариабельного участка легкой цепи антитела GK-22G12, представленную в SEQ ID NO. 25.
В соответствии с одним вариантом осуществления антитело, связывающий белок на основе антитела, модифицированный формат антитела, сохраняющий способность к связыванию с мишенью, производное или фрагмент антитела, сохраняющие способность к связыванию с мишенью, би- или мультиспецифичные молекулы содержат:
• первую часть, которая связывается с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2), и
• по меньшей мере вторую часть, которая связывается с эффекторным антигеном, выбранным из группы, состоящей из CD3, CD16, NKG2D, NKp46, CD2, CD28 и/или CD25.
Некоторые последовательности представлены ниже:





В соответствии со всеми вышеизложенными аспектами настоящего изобретения антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR предпочтительно содержат пару вариабельных участков тяжелой цепи/легкой цепи, последовательности которых являются практически идентичными вариабельному участку из любой пары из таблицы 2 и, в особенности, последовательностям, выбранным из следующих пар (таблица 2): (i) SEQ ID NO:2 и SEQ ID NO:3; (ii) SEQ ID NO:4 и SEQ ID NO:5; (iii) SEQ ID NO:6 и SEQ ID NO:7; (iv) SEQ ID NO:8 и SEQ ID NO:9; (v) SEQ ID N0:10 и SEQ ID NO:11; (vi) SEQ ID NO:12 и SEQ ID NO:13; (vii) SEQ ID NO:14 и SEQ ID NO:15; (vii) SEQ ID NO:16 и SEQ ID NO:17; (ix) SEQ ID NO:18 и SEQ ID NO:19; (x) SEQ ID N0:20 и SEQ ID NO:21; (xi) SEQ ID NO:22 и SEQ ID NO:23 или (xii) SEQ ID NO:24 и SEQ ID NO:25. В соответствии с предпочтительным вариантом осуществления полностью человеческое антитело к ROR2 или антигенсвязывающий фрагмент содержит пару вариабельных участков тяжелой цепи/легкой цепи, последовательности которых являются практически идентичными последовательностям, выбранным из следующих пар (таблица 2): (i) SEQ ID NO:2 и SEQ ID NO:3; (ii) SEQ ID NO:8 и SEQ ID NO:9; (iii) SEQ ID NO:18 и SEQ ID NO:19; (iv) SEQ ID NO:20 и SEQ ID NO:21; (v) SEQ ID NO:22 и SEQ ID NO:23 или (vi) SEQ ID NO:24 и SEQ ID NO:25. В соответствии с предпочтительным вариантом осуществления полностью человеческое антитело к ROR2 или антигенсвязывающий фрагмент содержит пару вариабельных участков тяжелой цепи/легкой цепи, последовательности которых являются практически идентичными последовательностям, выбранным из следующих пар (таблица 2): (i) SEQ ID NO:2; SEQ ID NO:3; (ii) SEQ ID NO:8 и SEQ ID NO:9 и (iii) SEQ ID NO:20 и SEQ ID NO:21.
В соответствии с предпочтительным вариантом осуществления «практически идентичный» означает по меньшей мере на 90%, предпочтительно, на 95%, более предпочтительно, на 100% идентичный соответствующей аминокислотной последовательности вариабельного участка, идентифицированной в соответствующей SEQ ID NO.
В соответствии с некоторыми вариантами осуществления настоящим изобретением предусмотрены полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, которые представляют собой консервативно модифицированные варианты человеческих антител к ROR2, проиллюстрированных в данном документе. Как правило, вариабельные участки этих вариантов имеют аминокислотную последовательность, которая является идентичной одной из этих проиллюстрированных последовательностей за исключением консервативных замен по одному или нескольким аминокислотным остаткам.
В соответствии со всеми вышеизложенными аспектами настоящего изобретения человеческие антитела, связывающие белки на основе антител, их антигенсвязывающие фрагменты предпочтительно представляют собой IgA1, IgA2, IgD, IgE, IgG1, IgG2, IgG3, IgG4, IgM, F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, scFv-Fc, (scFv)2, необедняющий IgG, диатело или бивалентное антитело. В соответствии с более предпочтительным вариантом осуществления человеческие антитела, связывающие белки на основе антител, их антигенсвязывающие фрагменты представляют собой IgG1, IgG2, IgG3, IgG4, IgM.
В случае, когда человеческие антитела, связывающие белки на основе антител или их антигенсвязывающие фрагменты (или ADC, или CAR, полученные с ними) являются бивалентными, предпочтительно, чтобы они связывались как с человеческим ROR2, так и с человеческим CD3.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с человеческим ROR2 и с ROR2 яванского макака и конкурируют за связывание с антителом, содержащим вариабельные домены GK-5A1, MK-3 В12, GK-5E1, MK-21D3, MK-24С10, MK-24F9 или MK-22G12 и, предпочтительно, с антителом, содержащим вариабельные домены GK-5A1, MK-3 В12, МК-24С10, MK-24F9 или MK-22G12.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающий белок на основе антитела, их антигенсвязывающие фрагменты, ADC, AEC или CAR согласно настоящему изобретению связываются с человеческим ROR2 и с ROR2 яванского макака. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR связываются с ROR2 яванского макака и с человеческим ROR2 и содержат CDR из GK-5A1, MK-3В12, GK-5E1, MK-21D3, MK-24С10, MK-24F9 или MK-22G12 и, предпочтительно, из GK-5A1, MK-3В12, MK-24С10, MK-24F9 или MK-22G12. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с ROR2 яванского макака и с человеческим ROR2 и содержат вариабельные домены из GK-5A1, MK-3В12, GK-5E1, MK-21D3, MK-24С10, MK-24F9 или MK-22G12 и, предпочтительно, из GK-5A1, MK-3 В12, MK-24С10, MK-24F9 или MK-22G12.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с человеческим ROR2 и с мышиным ROR2 и конкурируют за связывание с антителом, содержащим вариабельные домены из GK-5A1, MK-24С10 или MK-24F9, и, предпочтительно, с антителом, содержащим вариабельные домены из GK-5A1.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с мышиным ROR2 и с человеческим ROR2. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающим белкам на основе антитела, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с мышиным ROR2 и с человеческим ROR2 и содержат CDR из GK-5A1, MK-24С10 или MK-24F9 и, предпочтительно, из GK-5A1. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с мышиным ROR2 и с человеческим ROR2 и содержат вариабельные домены из GK-5A1, MK-24С10 или MK-24F9 и, предпочтительно, из GK-5А1.
Перекрестная-реактивность антител согласно настоящему изобретению с мышиным ROR2 является особенно неожиданной с учетом того, что эти антитела происходят из мыши.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с человеческим ROR2, с ROR2 яванского макака и с мышиным ROR2 и конкурируют за связывание с антителом, содержащим вариабельные домены из GK-5A1, MK-24С10 или MK-24F9, и, предпочтительно, с антителом, содержащим вариабельные домены из GK-5A1.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с ROR2 яванского макака, с мышиным ROR2 и с человеческим ROR2. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с ROR2 яванского макака, с мышиным ROR2 и с человеческим ROR2 и содержат CDR из GK-5A1, MK-24С10 или MK-24F9 и, предпочтительно, из GK-5А1. В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению связываются с ROR2 яванского макака, с мышиным ROR2 и с человеческим ROR2 и содержат вариабельные домены из GK-5A1, MK-24С10 или MK-24F9 и, предпочтительно, из GK-5A1.
В соответствии с предпочтительным вариантом осуществления антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению происходят из мыши. Антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, ADC, АЕС или CAR согласно настоящему изобретению могут проявлять благоприятную более низкую склонность к агрегации, улучшенную технологичность производства и/или более высокие уровни экспрессии по сравнению с полностью человеческими антителами к ROR2, связывающими белками на основе антител, их антигенсвязывающими фрагментами, ADC, АЕС или CAR, полученными с помощью способов, включающих фаговый дисплей.
Также предусмотрены полинуклеотиды кодирующие полностью человеческие антитела к ROR2, связывающие белки на основе антител, их антигенсвязывающие фрагменты, описанные в данном документе, клетки-хозяева, трансформированные или трансфицированные этими полинуклеотидами, и способы получения различных полностью человеческих антител к ROR2, связывающих белков на основе антител, их антигенсвязывающих фрагментов, описанных в данном документе. Настоящим изобретением также предусмотрены практически очищенные полинуклеотиды (ДНК или РНК), которые являются идентичными или комплементарными последовательностям, кодирующим полипептиды, содержащие сегменты или домены цепей антител или антигенсвязывающих фрагментов, описанных в данном документе. В случае экспрессии с соответствующих экспрессионных векторов полипептиды, кодируемые этими полинуклеотидами, способны проявлять антигенсвязывающую способность. В соответствии с настоящим изобретением также предусмотрены полинуклеотиды, которые кодируют по меньшей мере один участок CDR и обычно все три участка CDR из тяжелой или легкой цепи в полностью человеческих антителах к ROR2, связывающих белках на основе антител, их антигенсвязывающих фрагментах, описанных в данном документе. Некоторые другие полинуклеотиды кодируют всю или практически всю из последовательности вариабельного участка тяжелой цепи и/или легкой цепи проиллюстрированных антителах. Вследствие вырожденности кода различные последовательности нуклеиновой кислоты будут кодировать каждую из аминокислотных последовательностей иммуноглобулина. Полинуклеотиды согласно настоящему изобретению могут кодировать только последовательность вариабельного участка проиллюстрированного антитела. Они также могут кодировать как вариабельный участок, так и константный участок антитела. Некоторые из полинуклеотидных последовательностей нуклеиновых кислот согласно настоящему изобретению кодируют зрелую последовательность вариабельного участка тяжелой цепи антитела, который является практически идентичным (например, по меньшей мере на 80%, 90% или 99%) представленной зрелой последовательности вариабельного участка тяжелой цепи антитела. Некоторые другие полинуклеотидные последовательности кодируют зрелую последовательность вариабельного участка легкой цепи, которая является практически идентичной представленной зрелой последовательности вариабельного участка легкой цепи. Некоторые из полинуклеотидных последовательностей кодируют полипептид, который содержит вариабельные участки обеих из тяжелой цепи и легкой цепи одного из проиллюстрированных антител. Некоторые другие полинуклеотиды кодируют два полипептидных сегмента, которые, соответственно, являются практически идентичными вариабельным участкам тяжелой цепи и легкой цепи одного из проиллюстрированных антител.
Полинуклеотидные последовательности можно получать посредством твердофазного синтеза ДНК de novo или посредством мутагенеза с помощью ПЦР существующей последовательности, кодирующей проиллюстрированное функциональное антитело. Прямой химический синтез нуклеиновых кислот можно осуществлять с помощью методов, известных в уровне техники, таких как фосфотриэфирный метод из Narang et al., Meth. Enzymol. 68:90, 1979; фосфодиэфирный метод из Brown et al., Meth. Enzymol. 68:109, 1979; диэтилфосфорамидитный метод из Beaucage et al., Tetra. Lett., 22:1859, 1981; и способ с применением твердофазной подложки из патента США №4458066. Введение мутаций в полинуклеотидную последовательность с помощью ПЦР можно осуществлять, как описано, например, в PCR Technology: Principles and Applications for DNA Amplification, H.A. Erlich (Ed.), Freeman Press, NY, NY, 1992; PCR Protocols: A Guide to Methods and Applications, mnis etal. (Ed.), Academic Press, San Diego, CA, 1990; Manila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
В соответствии с настоящим изобретением также предусмотрены векторы экспрессии и клетки-хозяева для получения полностью человеческих антител к ROR2, связывающих белков на основе антител, их антигенсвязывающих фрагментов, описанных в данном документе. Различные экспрессионные векторы можно использовать для экспрессии полинуклеотидов, кодирующих функциональные цепи антител или связывающих фрагментов. Для получения антител в клетке-хозяине млекопитающего можно применять как векторы на основе вирусов, так и невирусные экспрессионные векторы. Невирусные векторы и системы включают в себя плазмиды, эписомные векторы, как правило, с экспрессионной кассетой для экспрессии белка или РНК, и человеческие искусственные хромосомы (см., например, Harrington et al., Nat. Genet. 15:345, 1997). Например, невирусные векторы, пригодные для экспрессии полинуклеотидов и полипептидов антитела в клетках млекопитающего (например, человека), включают в себя рСЕР4, pREP4, pThioHis А, В & С, pcDNA3.1/His, pEBVHis А, В & С (Invitrogen, Сан-Диего, Калифорния, США), MPSV векторы и многочисленные другие векторы, известные в области экспрессии других белков. Другие пригодные невирусные векторы включают в себя векторы, содержащие любую из Sleeping Beauty, PiggyBac и других транспозонных систем или последовательностей, обеспечивающих возможность транспозиции компонентов вектора с помощью указанных Sleeping Beauty, PiggyBac и других транспозонных систем. Пригодные вирусные векторы включают в себя векторы на основе лентивирусов или других ретровирусов, аденовирусов, аденоассоциированных вирусов, герпесвирусов, векторы на основе SV40, вируса папилломы, НВР вируса Эпштейна-Барр, векторы на основе вируса коровьей оспы и вируса леса Семлики (SFV). See, Brent et al., выше; Smith, Annu. Rev. Microbiol. 49:807, 1995; и Rosenfeld et al., Cell 68:143, 1992.
Выбор экспрессионного вектора зависит от предполагаемых клеток-хозяев, в которых должен экспрессироваться вектор. Как правило, экспрессионные векторы содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально соединены с полинуклеотидами, кодирующими функциональную цепь или антигенсвязывающий фрагмент антитела. В соответствии с некоторыми вариантами осуществления индуцируемый промотор используют для предотвращения экспрессии введенных последовательностей в условиях, отличных от условий, обеспечивающих индукцию. Индуцируемые промоторы включают в себя, например, индуцируемые тетрациклином, арабинозой, lacZ, металлотионеиновые промоторы или промотор белков теплового шока. Культуры трансформированных или трансфицированных клеток-хозяев можно разращивать в не индуцирующих условиях без изменения популяции кодирующих последовательностей, чьи продукты экспрессии лучше переносятся клетками-хозяевами. Для эффективной экспрессии функциональной цепи или фрагмента антитела также могут требоваться или быть желательными другие регуляторные элементы помимо промоторов. Эти элементы, как правило, включают в себя инициирующий кодон ATG и смежный сайт связывания рибосомы или другие последовательности. Кроме того, эффективность экспрессии можно повысить посредством включения энхансеров, подходящих для применяемой клеточной системы (см., например, Scharf et al., Results Probl. Cell Differ. 20:125, 1994; и Bittner et al., Meth. Enzymol, 153:516, 1987). Например, энхансер SV40 или энхансер CMV можно применять для повышения экспрессии в клетках-хозяевах млекопитающих.
Экспрессионные векторы также могут обеспечивать положение для сигнальной последовательности для секреции с образованием гибридного белка с полипептидами, кодируемыми вставленными функциональными последовательностями антител. Чаще, вставленные функциональные последовательности антител являются связанными с сигнальными последовательностями до включения в вектор. Векторы, которые нужно применять для получения последовательностей, кодирующих функциональные вариабельные домены легкой и тяжелой цепи антител, иногда также кодируют константные участки или их части. Такие векторы обеспечивают экспрессию вариабельных участков в виде гибридных белков с константными участками, тем самым приводя к выработке интактных антител или их фрагментов. Как правило, такие константные участки являются человеческими.
Клетки-хозяева для сохранения и экспрессии функциональных цепей антитела могут быть либо прокариотическими, либо эукариотическими. В соответствии с некоторыми предпочтительными вариантами осуществления клетки-хозяева млекопитающих применяют для экспрессии и получения полипептидов антител согласно настоящему изобретению. Например, они могут представлять собой либо клеточную линию гибридомы, экспрессирующую эндогенные гены иммуноглобулинов, либо линию клеток млекопитающего, несущую экзогенный экспрессионный вектор. Они включают в себя любую нормальную смертную или нормальную или ненормальную иммортализированную клетку животного или человека. Помимо клеток, проиллюстрированных в данном документе, в уровне техники также известен ряд других подходящих линий клеток-хозяев, способных к секреции интактных иммуноглобулинов. Они включают в себя, например, клеточные линии СНО, различные клеточные линии НЕК 293, различные клеточные линии Cos, клетки HeLa, линии клеток миеломы, трансформированные клетки В-линии и гибридомы. Применение культуры клеток ткани млекопитающего для экспрессии полипептидов, в целом, обсуждается, например, в Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987. Экспрессионные векторы для клеток-хозяев млекопитающих могут включать в себя последовательности для управления экспрессией, такие как точка начала репликации, промотор и энхансер, а также необходимые сайты для процессинга информационной РНК, такие как сайты связывания рибосом, сайты сплайсинга РНК, сайты полиаденилирования и последовательности терминатора транскрипции. Эти экспрессионные векторы обычно содержат промоторы, полученные из генов млекопитающего или из вирусов млекопитающего. Подходящие промоторы могут являться конститутивными, специфическими для типа клетки, специфическими для стадии развития и/или модулируемыми или регулируемыми. Пригодные промоторы включают в себя, без ограничения, промоторы EF1α и человеческого UbC, проиллюстрированные в данном документе, металлотионеиновый промотор, конститутивный большой поздний промотор аденовируса, индуцируемый дексаметазоном промотор MMTV, промотор SV40, промотор MRP polIII, конститутивный промотор MPSV, индуцируемый тетрациклином промотор CMV (такой как немедленный ранний промотор CMV человека), конститутивный промотор CMV и комбинации промотор-энхансер, известные в уровне техники.
Способы введения экспрессионных векторов, содержащих полинуклеотидные последовательности, представляющие интерес, варьируют в зависимости от типа клеточного хозяина. Например, трансформацию с использованием хлорида кальция или электропорацию зачастую используют для прокариотических клеток, в то время как обработку фосфатом кальция или электропорацию можно применять в отношении других клеточных хозяев (в целом, см., Sambrook et al., выше). Другие способы включают в себя, например, электропорацию, обработку фосфатом кальция, опосредуемую липосомами трансфекцию, инъекцию и микроинъекцию, баллистические методы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, депротеинизированную ДНК, искусственные вирионы, гибридные белки со структурным белком VP22 вируса герпеса(Elliot and O'Hare, Cell 88:223, 1997), усиленное средствами поглощение ДНК и трансдукцию ex vivo. В случае продолжительной выработки рекомбинантных белков с высоким выходом часто будет желательной устойчивая экспрессия. Например, клеточные линии, которые устойчиво экспрессируют цепи или связывающие фрагменты антител, можно получать с применением экспрессионных векторов согласно настоящему изобретению, которые содержат вирусные точки начала репликации или эндогенные экспрессионные элементы и ген селектируемого маркера. После введения вектора клеткам могут позволять расти в течение 1-2 суток в обогащенной среде до того, как их переводили на селективную среду. Цель селектируемого маркера заключается в придании устойчивости к отбору, и его присутствие обеспечивает возможность роста клеток, которые успешно экспрессируют введенные последовательности в селективной среде. Пролиферацию устойчивых клеток, подвергнутых устойчивой трансфекции, можно обеспечивать с применением методик ведения тканевой культуры, подходящих для типа клеток.
Аспекты настоящего изобретения, относящиеся к новым ADC и АЕС
Настоящее изобретение также относится к конъюгатам антитело-эффекторная молекула (АЕС) или конъюгатам антитело-лекарственное средство (ADC).
Термин «конъюгаты антитело-эффекторная молекула» (АЕС) используют в контексте данного документа для описания липосомы или метящих средств, связанных с полностью человеческим антителом к ROR2, связывающим белком на основе антитела или его антигенсвязывающим фрагментом, которые описаны в данном документе. Метящее средство обеспечивает возможность прямого или косвенного выявления антитела к ROR2 или антигенсвязывающего фрагмента, к которому оно прикреплено. Липосома, как описано в Bendas, BioDrugs, 15: 215-224, 2001, может обеспечивать возможность контролируемой доставки средства в пораженные заболеванием клетки. При получении липосомного конъюгата, например, иммунолипосомы, средство, такое как химиотерапевтическое или другое лекарственное средство, может быть заключено в липосому для доставки в клетку-мишень.
Термин «конъюгаты антитело-лекарственное средство» (ADC) используют в контексте данного документа для описания цитотоксических и/или цитостатических средств («токсин»), связанных с полностью человеческим антителом к ROR2, связывающим белком на основе антитела или антигенсвязывающим фрагментом, которые описаны в данном документе. Цитотоксические и/или цитостатические средства представляют собой любое средство, о котором известно, что оно ингибирует рост и/или ингибирует деление клеток и/или уничтожает клетки, в особенности, злокачественные клетки. Ковалентные конъюгаты низкомолекулярных цитотоксических и/или цитостатических средств (с молекулярной массой предпочтительно <2500 Дальтон) с антителами или антигенсвязывающими фрагментами, специфичными в отношении опухолевых клеток, являются эффективными инструментами для специфичного целенаправленного воздействия на раковые клетки с целью их уничтожения.
В ADC полезная нагрузка в виде токсина может быть конъюгирована с антителом, связывающим белком на основе антитела или фрагментом антитела не сайт-специфично через боковые цепи аминокислоты лизина или цистеина с использованием классических химических линкеров с малеимидной функциональной группой или других химических продуктов, известных в уровне техники, которые могут опосредовать конъюгацию боковых цепей аминокислоты лизина или цистеина. В ADC согласно настоящему изобретению низкомолекулярную полезную нагрузку также можно конъюгировать сайт-специфично с помощью химического, химико-ферментативного или ферментативного механизма конъюгации, известного в уровне техники, как например, с использованием бифункциональных линкеров, линкеров, обеспечивающих возможность протекания химической реакции Пикте-Шпенглер а на формил-глицине с образованием модифицированных ферментом антител, с помощью антител с ремоделированным гликановым компонентом, или с помощью бактериальных ферментов трансглутаминазы или сортазы.
В соответствии с одним вариантом осуществления конъюгат антитело-лекарственное средство (ADC) имеет общую формулу A-(L)n-(T)m, в которой
 А представляет собой антитело, производное, модифицированный формат или фрагмент, которые описаны в данном документе,
А представляет собой антитело, производное, модифицированный формат или фрагмент, которые описаны в данном документе,
 L представляет собой линкер,
L представляет собой линкер,
 Т представляет собой полезную нагрузку в виде цитотоксического или цитостатического средства,
Т представляет собой полезную нагрузку в виде цитотоксического или цитостатического средства,
и в которой n и m представляют собой целые числа от >1 до <10.
В соответствии с одним вариантом осуществления конъюгат антитело-эффекторная молекула (АЕС) обычно имеет общую формулу A-(L)n-(T)m, в которой
 А представляет собой антитело, производное, модифицированный формат или фрагмент по любому из п. 1,
А представляет собой антитело, производное, модифицированный формат или фрагмент по любому из п. 1,
 L представляет собой линкер,
L представляет собой линкер,
 Т представляет собой метку, и в которой n и m представляют собой целые числа от >1 до <10.
Т представляет собой метку, и в которой n и m представляют собой целые числа от >1 до <10.
В соответствии с предпочтительным вариантом осуществления линкер содержит или состоит из по меньшей мере одного, выбранного из группы, состоящей из: олигопептидного линкера (в том числе расщепляемых и нерасщепляемых олигопептидных линкеров), гидразинового линкера, тиомочевинного линкера, саморасщепляющегося линкера, сукцинимидил-транс-4-(малеимидилметил)циклогексан-1-карбоксилатного (SMCC) линкера, дисульфидного линкера, тиоэфирного линкера и/или малеимидного линкера.
Малеимидный линкер, необязательно, содержит расщепляемые спейсеры, которые могут расщепляться при изменениях рН, окислительно-восстановительного потенциала и или конкретными внутриклеточными ферментами.
Специалисту в данной области техники понятно, что дополнительные линкеры могут быть подходящими. Такие линкеры могут быть нерасщепляемыми или могут расщепляться под действием изменений рН, окислительно-восстановительного потенциала или конкретных внутриклеточных ферментов. Расщепляемые олигопептидные линкеры включают в себя расщепляемые протеазой линкеры. Понятно, что линкер может содержать комбинации вышеизложенного. Например, линкер может представлять собой валино-цитрулиновый РАВ линкер.
В соответствии с одним вариантом осуществления линкер имеет по меньшей мере одну из следующих аминокислотных последовательностей: - LPXTGn-, -LPXAGn-, -LPXSGn-, -LAXTGn-, -LPXTGn-, -LPXTAn- или -NPQTGn-, причем Gn представляет собой олиго- или полиглицин, при этом n представляет собой целое число от ≥1 до ≤21, An представляет собой олиго- или полиаланин, при этом n представляет собой целое число от ≥1 до ≤21, и X представляет собой любую возможную аминокислотную последовательность.
В соответствии с предпочтительным вариантом осуществления линкер конъюгирован с С-концом по меньшей мере одного субдомена антитела, антитела, производного, модифицированного формата или фрагмента.
В соответствии с одним вариантом осуществления до конъюгирования
 антитело, производное, модифицированный формат или фрагмент несет мотив для распознавания сортазой, слитый или конъюгированный с С-концом по меньшей мере одного его субдомена, и
антитело, производное, модифицированный формат или фрагмент несет мотив для распознавания сортазой, слитый или конъюгированный с С-концом по меньшей мере одного его субдомена, и
 токсин или метка содержит глициновый отрезок длиной от ≥1 до ≤21 глициновых остатков, предпочтительно, длиной от ≥2 до ≤5 глициновых остатков.
токсин или метка содержит глициновый отрезок длиной от ≥1 до ≤21 глициновых остатков, предпочтительно, длиной от ≥2 до ≤5 глициновых остатков.
Цитостатическое средство в ADC может представлять собой растительную, грибную или бактериальную молекулу. В соответствии с некоторыми вариантами осуществления цитотоксическое средство для конъюгирования с антителом или антигенсвязывающим фрагментом согласно настоящему изобретению представляет собой клеточный токсин-малую молекулу, пептидный токсин или белковый токсин. Многие конкретные примеры этих токсинов являются хорошо известными в уровне техники. См., например, Dyba et al., Curr. Pharm. Des. 10:2311-34, 2004; Kuyucak et al., Future Med. Chem. 6:1645-58, 2014; Beraud et al., Inflamm. Allergy Drug Targets. 10:322-42, 2011; и Middlebrook et al., Microbiol. Rev. 48:199-221, 1984. В соответствии с некоторыми вариантами осуществления терапевтическое средство конъюгировано с антителом. Например, терапевтическое средство может представлять собой майтансиноид (например, майтансинол или майтансиноид DM1), таксан, калихеамицин, цематодин, монометилауристатин (например, монометилауристатин Е или монометилауристатин F), пирролобензодиазепин (PBD), псевдодимер индилино-бензодиазепина или антрациклин. Терапевтические средства также включают в себя винкристин и преднизон. В соответствии с различными вариантами осуществления терапевтическое средство, которое можно использовать в настоящем изобретении, может представлять собой антиметаболит (например, антифолат, такой как метотрексат, фторпиримидин, такой как 5-фторурацил, цитозин-арабинозид, или аналог пурина или аденозина); интеркалирующее средство (например, антрациклин, такой как доксорубицин, PNU-159682, дауномицин, эпирабицин, идарубицин, митомицин-С, дактиномицин или митрамицин, или другие интеркалирующие средства, такие как пирролобензодиазепин); реагирующее с ДНК средство, такое как калихеамицины, тианцимицины и другие ендиины; производное платины (например, цисплатин или карбоплатин); акилирующее средство (например, азотистый иприт, мелфалан, хлорамбуцил, бусульфан, циклофосфамид, ифосфамид, нитрозомочевины или тиотепа); ингибитор РНК-полимеразы, такой как а-аманитин; антимитотическое средство (например, алкалоид барвинка, такой как винкристин, или анатоксин, такой как паклитаксел или доцетаксел); ингибитор топоизомеразы (например, этопозид, тенипозид, амсакрин, топотекан); ингибитор клеточного цикла (например, флавопиридол); или средство, нарушающее образование микротрубочек (например, эпотилон, тубулизин, претубулизин, аналог дискодермолида или аналог элеутеробина). Терапевтическое средство может представлять собой ингибитор протеосомы или ингибитор топоизомеразы, такой как бортезомиб, амсакрин, этопозид, фосфат этопозида, тенипозид или доксорубицин. Терапевтические радиоактивные изотопы включают в себя йод (131I), иттрий (90Y), лютеций (177Lu), актиний (225Ас), празеодим, астат (At), рений (Re), висмут (Bi или Bi) и родий (Rh). Антиангиогенные средства включают в себя линомид, бевацизумаб, ангиостатин и разоксан.
В соответствии с предпочтительным вариантом осуществления ADC цитотоксическая или цитостатическая полезная нагрузка представляет собой по меньшей мере одну, выбранную из группы, состоящей из:
 майтансиноидов,
майтансиноидов,
 ауристатинов,
ауристатинов,
 антрациклинов,
антрациклинов,
 калихеамицинов,
калихеамицинов,
 тубулизинов,
тубулизинов,
 дуокармицинов,
дуокармицинов,
 радиоизотопов,
радиоизотопов,
 липосом, содержащих полезную нагрузку в виде анатоксина,
липосом, содержащих полезную нагрузку в виде анатоксина,
 белковых токсинов,
белковых токсинов,
 таксанов,
таксанов,
 псевдодимеров индилино-бензодиазепина и/или
псевдодимеров индилино-бензодиазепина и/или
 пирролобензодиазепинов, или представляет собой их производное.
пирролобензодиазепинов, или представляет собой их производное.
В соответствии с предпочтительным вариантом осуществления ADC токсин является выбранным из PNU-159682, который описан в Quintieri et al. (2005), и его производных, майтансина, монометил-ауристатин ММАЕ и монометил-ауристатин MMAF. В соответствии с предпочтительным вариантом осуществления ADC токсин, соединенный с линкером, что показано волнистой линией, имеет формулу (i), как описано в международной заявке WO 2016/102679:
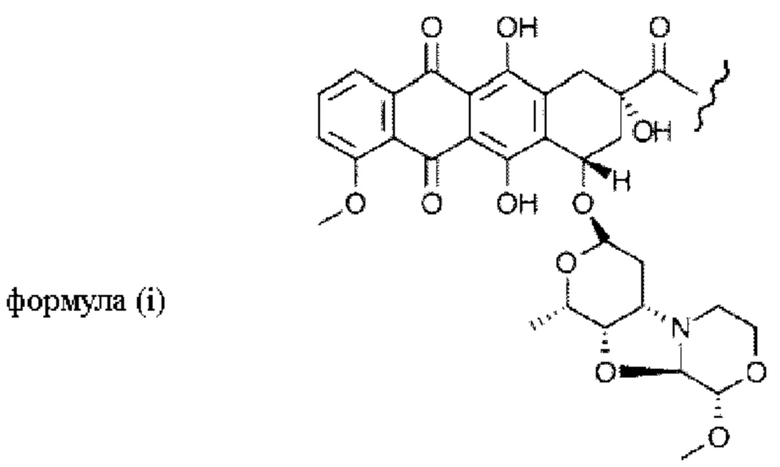
В соответствии с вариантом осуществления, в котором токсин имеет формулу (i), предпочтительно, чтобы линкер содержал алкилдиамино-группу формы NH2-(CH2)m-NH2, где m ≥1 и ≤11, предпочтительно, m=2, в результате чего одна аминогруппа непосредственно связана, как показано волнистой линией в формуле (i), с образованием амидной связи. Кроме того, предпочтительно, чтобы вторая аминогруппа была связана с олигопептидным линкером, который, более предпочтительно, представляет собой олигоглицин.
В соответствии с некоторыми вариантами осуществления молекула для коньюгирования с антителом представляет собой белок (например, антитело) или аптамер РНК или ДНК.
В соответствии с предпочтительным вариантом осуществления АЕС метка представляет собой по меньшей мере одну, выбранную из группы, состоящей из: флуоресцентной метки (в том числе флуоресцентного красителя или флуоресцентного белка), хромофорной метки, радиоизотопной метки, содержащей йод (например, 125I), галлий (67Ga), индий (111I), технеций (99mTc), фосфор (32Р), углерод (14С), тритий (3Н), другой радиоактивный изотоп (например, радиоактивный ион), и/или белковой метки, такой как авидин или стрептавидин.
В соответствии с предпочтительным вариантом осуществления ADC или АЕС ADC или АЕС характеризуются стехиометрическим соотношением между антителом и полезной нагрузкой, составляющей любое целое число от ≥1 до ≤10, предпочтительно, от 2 до 4. В случае ADC это соотношение также может называться соотношением лекарственного средства и антитела («DAR»). В случае смеси или набора ADC DAR может представлять собой рациональное число, представляющее собой среднее значение отдельных DAR b виде целых чисел.
В соответствии с одним вариантом осуществления ADC или АЕС только одна или несколько тяжелых цепей антитела или соответствующего антигенсвязывающего фрагмента конъюгированы с полезной нагрузкой.
В соответствии с одним вариантом осуществления ADC или АЕС только одна или несколько легких цепей антитела или соответствующего антигенсвязывающего фрагмента конъюгированы с полезной нагрузкой.
Понятно, что данный ADC или АЕС может одновременно содержать различные связанные токсины или метки; также возможно, чтобы антитело к ROR2 было связано с комбинацией как одной или нескольких меток, так и одного или нескольких токсинов.
Антитела или антигенсвязывающие фрагменты согласно настоящему изобретению можно конъюгировать с синтетической молекулой с применением любого типа подходящего конъюгирования. Методы генной инженерии на основе рекомбинации и включение селеноцистеина (например, как описано в патенте США №8916159) можно применять для конъюгирования синтетической молекулы. Другие способы конъюгирования могут включать в себя ковалентное связывание с нативными или сконструированными аминами боковой цепи лизина или тиолами боковой цепи цистеина. См., например, Wu et al., Nat. Biotechnol, 23: 1 137-1 146 (2005). Синтетическая молекула может представлять собой любую молекулу, такую как молекула, оказывающая целенаправленное воздействие на опухоль.
В соответствии с предпочтительным вариантом осуществления ADC или АЕС получают с помощью опосредуемого ферментом-сортазой сайт-специфического конъюгирования антитела (SMAC-технология), которое всесторонне раскрыто в международной заявке WO 2014140317, права по которой принадлежат тому же заявителю. Содержание этой публикации включено в данный документ посредством ссылки. Сортазы (также называемые транспептидазами-сортазами) из группы прокариотических ферментов, которые модифицируют поверхностные белки посредством распознавания и расщепления специфического пептидного мотива, называемого «мотивом для распознавания сортазой» или «тэгом для сортазы». Обычно, данный фермент-сортаза распознает один или несколько мотивов для распознавания сортазой. Ферменты-сортазы могут встречаться в естественных условиях, или они могли быть подвергнуты воздействию методов генной инженерии (Dorr et al., 2014), и их применяют в технологии SMAC для конъюгирования двух белков, один из которых несет такой мотив для распознавания, тогда как другой несет олиголициновый пептид (Gly)n. Важно понимать, что в соответствии с одним конкретным вариантом осуществления (в котором применяют сортазу A Streptococcus pyogenes, см. ниже) олигоглицин (Gly)n необязательно может быть заменен олигоаланином (Ala)n.
В соответствии с предпочтительным вариантом осуществления ADC или АЕС получают с помощью опосредуемого ферментом-сортазой сайт-специфического конъюгирования:
a) антитела или антигенсвязывающго фрагмента, которые описаны в данном документе и несут один или несколько мотивов для распознавания сортазой, и
b) одной или нескольких молекул-полезной нагрузки, несущих олигоглициновый тэг;
или
a) антитела или антигеневязывающего фрагмента, которые описаны в данном документе и несут один или несколько олигоглициновых тэгов, и
b) одной или нескольких молекул-полезной нагрузки, несущих мотив для распознавания сортазой.
Важно понимать, что в соответствии с одним конкретным вариантом осуществления (в котором применяют сортазу A Streptococcus pyogenes, см. ниже) олигоглицин (Gly)n необязательно может быть заменен олигоаланином (Ala)n.
Предпочтительно, мотив для распознавания сортазой слит или конъюгирован с С-концом по меньшей мере одного субдомена антитела.
Предпочтительно, олигоглициновый тэг имеет длину от ≥1 до ≤21 глициновых остатков, предпочтительно, с длиной от ≥2 до ≤5 аминокислот.
Настоящее изобретение также относится к способу получения АЕС или ADC, причем способ предусматривает следующие стадии:
а) обеспечения антитела, производного, модифицированного формата или фрагмента, которые описаны в данном документе, причем антитело или антигенсвязывающий фрагмент несут один или несколько мотивов для распознавания сортазой,
b) обеспечения одной или нескольких молекул-полезной нагрузки, несущих олигоглициновый тэг, и
c) конъгирования антитела или антигенсвязывающего фрагмента и одной или нескольких молекул-полезной нагрузки с помощью опосредуемого сортазой конъюгирования.
Термин «олигоглициновый тэг» (также называемый Gn или G(n)) относится к олигоглицину с длиной, составляющей n, которое может составлять от ≥1 до ≤21, предпочтительно, от ≥1 до ≤5.
Настоящее изобретение также относится к способу получения АЕС или ADC, причем способ предусматривает следующие стадии:
a) обеспечения антитела к ROR2 или антигенсвязывающего фрагмента, которые описаны в данном документе, причем антитело или антигенсвязывающий фрагмент несут один или несколько олигоглициновых тэгов,
b) обеспечения одной или нескольких молекул-полезной нагрузки, несущих мотив для распознавания сортазой, и
c) конъгирования антитела или антигенсвязывающего фрагмента и одной или нескольких молекул-полезной нагрузки с помощью опосредуемого сортазой конъюгирования.
В соответствии с еще одним вариантом осуществления указанный мотив для распознавания ферментом-сортазой содержит по меньшей мере одну из следующих аминокислотных последовательностей: LPXTG, LPXAG, LPXSG, LAXTG, LPXTA или NPQTN, причем X представляет собой любую возможную аминокислотную последовательность.
В соответствии с еще одним вариантом осуществления полученный в результате линкер имеет по меньшей мере одну из следующих аминокислотных последовательностей: -LPXTGn-, -LPXAGn-, -LPXSGn-, -LAXTGn-, -LPXTGn-, -LPXTAn- или -NPQTGn-, причем Gn представляет собой олиго- или полиглицин, при этом n представляет собой целое число от ≥1 до ≤21, An представляет собой олиго- или полиаланин, при этом n представляет собой целое число от ≥1 до ≤21, и X представляет собой любую возможную аминокислотную последовательность.
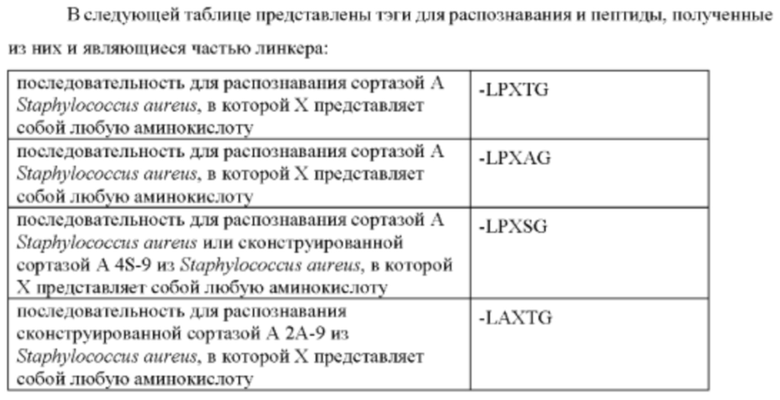
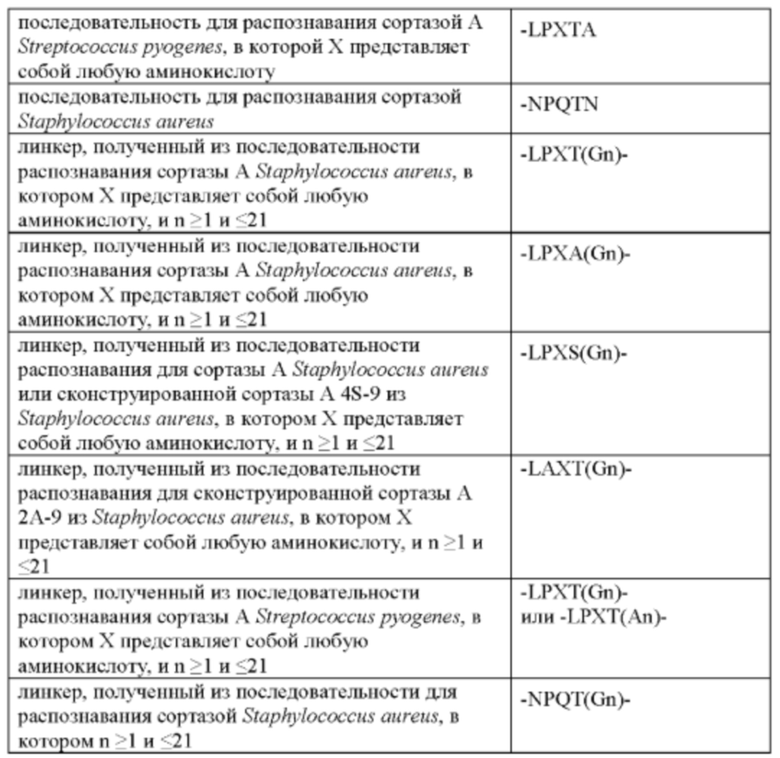
Таблица 4. Мотивы для распознавания сортазой и полученные в результате линкеры
Подвергнутые воздействию методов генной инженерии сортазы, в том числе, без ограничения, мутант 2А-9 сортазы А и мутант 4S-9 сортазы А из Staphylococcus aureus, описаны в Dorr et al. (2014), а также мутанты, описанные в Chen et al. (2011).
В качестве справочных сведений и для иллюстрирования общей идеи транспептидирования под действием сортазы сортаза А использует олигоглициновый отрезок в качестве нуклеофила для катализа транспептидирования, при котором концевая аминогруппа олигоглицина производит нуклеофильную атаку на пептидную связь, соединяющую последние два С-концевых остатка в тэге для сортазы. Это приводит в результате к разрыву пептидной связи и образованию новой пептидной связи между вторым с С-конца остатком в тэге для сортазы и N-концевым глицином в олигоглициновом пептиде, т.е. приводит в результате к транспептидированию.
Важно понимать, что в соответствии с одним конкретным вариантом осуществления (в котором применяют сортазу A Streptococcus pyogenes, см. выше) олигоглицин (Gly)n, необязательно, может быть заменен олигоаланином (А1а) n.
Перед конъюгированием с использованием сортазы распознаваемый сортазой мотив может дополнительно нести на своем С-конце другие тэги, подобные His-тэгам, Мус-тэгам или Strep-тэгам (см. фиг. 4а в международной заявке WO 2014/140317, содержание которой включено в данный документ посредством ссылки). Тем не менее, ввиду того что пептидная связь между 4-ой и 5-ой аминокислотами в тэге для сортазы расщепляется при опосредованной сортазой А конъюгации, эти дополнительные тэги не появляются в конъюгированном продукте.
Тэг для сортазы, например, может быть слит с С-концом связывающего белка или с его доменом или субъединицей посредством генетического слияния и коэкспрессироваться с ним. В соответствии с еще одним предпочтительным вариантом осуществления тэг для сортазы может быть напрямую присоединен к последней встречающейся в естественных условиях С-концевой аминокислоте в легких цепях или тяжелых цепях иммуноглобулинов, которая в случае легкой каппа-цепи человеческого иммуноглобулина представляет собой С-концевой цистеиновый остаток, и которая в случае тяжелой цепи человеческого иммуноглобулина IgG1 может представлять собой С-концевой лизиновый остаток, кодируемый кДНК человеческого Fcγ1. Тем не менее другой предпочтительный вариант осуществления также заключается в непосредственном присоединении тэга для сортазы ко второму от С-конца глициновому остатку, кодируемому кДНК человеческого Fcγ1, поскольку обычно концевые лизиновые остатки в тяжелых цепях антитела вырезаются при посттрансляционной модификации в клетках млекопитающих. Таким образом, в более чем 90% случаев у встречающегося в естественных условиях человеческого IgG1 отсутствуют С-концевые лизиновые остатки в тяжелых цепях IgG1.
Следовательно, один предпочтительный вариант осуществления настоящего изобретения заключается в отсутствии включения С-концевых лизиновых аминокислотных остатков в константных участках тяжелой цепи человеческого IgG1 в экспрессионных конструкциях для тяжелых цепей Igγ1, меченных мотивом для распознавания сортазой. Другой предпочтительный вариант осуществления заключается во включении С-концевых лизиновых аминокислотных остатков в константных участках тяжелой цепи человеческого IgG1 в экспрессионных конструкциях для тяжелых цепей Igγ1, меченных мотивом для распознавания сортазой.
В соответствии с еще одним предпочтительным вариантом осуществления тэг для сортазы или олигоглициновый тэг могут быть присоединены к С-концу тяжелой цепи человеческого иммуноглобулина IgG1, в котором С-концевой лизиновый остаток, кодируемый кДНК человеческого Fcγ1, заменен на аминокислотный остаток, отличный от лизина, для предотвращения непродуктивных реакций сортазы с ε-аминогруппой указанного С-концевого лизинового остатка, приводящих к сшиванию тяжелых цепей между собой.
Авторами настоящего изобретения ранее было описано, что в некоторых случаях (например, на С-конце легких каппа-цепей Ig, см.: Beerli et al. (2015) PloS One 10, e 131177) является выгодным добавление дополнительных аминокислот между С-концом связывающего белка и тэгом для сортазы. Было показано, что это улучшает эффективность конъюгирования ферментом сортазой «полезной нагрузки» со связывающим белком. В случае легких каппа-цепей Ig наблюдали, что при добавлении 5 аминокислот между последней С-концевой цистеиновой аминокислотой в легкой каппа-цепи Ig и пентапептидным мотивом для сортазы улучшались кинетические характеристики конъюгации, в результате чего С-концы легких каппа-цепей Ig и тяжелых цепей Ig могут конъюгировать с аналогичными характеристиками (см.: Beerli et al. (2015) PloS One 10, e131177). Таким образом, еще один предпочтительный вариант осуществления заключается в том, что необязательно ≥1 и ≤11 аминокислот добавляют между последней С-концевой аминокислотой в связывающем белке или субъединице антитела и тэгом для сортазы.
В соответствии с предпочтительным вариантом осуществления пептид GnS (причем n составляет от ≥1 до ≤21, предпочтительно, от ≥1 до ≤5) добавляют между последней С-концевой аминокислотой в связывающем белке или субъединице антитела и тэгом для сортазы.
Наконец, в соответствии с еще одним предпочтительным вариантом осуществления могут быть благоприятно включены дополнительные аминокислоты между С-концом связывающего белка и тэгом для сортазы или олигоглициновым тэгом, которые содержат последовательность и/или линкер, которые могут расщепляться посредством гидролиза, посредством изменения рН или посредством изменения окислительно-восстановительного потенциала, или которые могут расщепляться под действием отличного от сортазы фермента, например, под действием протеаз.
В соответствии с еще одним аспектом настоящего изобретения предусмотрен химерный антигенный рецептор (CAR) ROR2, использующий антитело, производное, модифицированный формат или фрагмент в соответствии с вышеизложенным описанием, слитый или конъюгированный по меньшей мере с одним трансмембранным участком и по меньшей мере одним внутриклеточным доменом.
Кроме того, предусмотрена клетка, содержащая такой химерный антигенный рецептор, причем клетка предпочтительно представляет собой подвергнутую воздействию методов генной инженерии Т-клетку.
Кроме того, предусмотрено применение антитела, производного, модифицированного формата или фрагмента, конъюгата антитело-лекарственное средство или CAR или клетки в соответствии с вышеизложенным описанием для лечения пациента, который
 страдает от неопластического заболевания,
страдает от неопластического заболевания,
 имеет риск развития неопластического заболевания, и/или
имеет риск развития неопластического заболевания, и/или
 у которого диагностировано неопластическое заболевание.
у которого диагностировано неопластическое заболевание.
В качестве альтернативы, предусмотрен способ лечения пациента, страдающего от неопластического заболевания, имеющего риск развития неопластического заболевания, и/или у которого диагностировано неопластическое заболевание, причем способ предусматривает введение одной или нескольких терапевтически активных доз антитела, конъюгата антитело-лекарственное средство, или CAR, или клетки в соответствии с вышеизложенным описанием.
В соответствии с одним вариантом осуществления неопластическое заболевание представляет собой по меньшей мере одно, выбранное из группы, состоящей из
 почечно-клеточной карциномы,
почечно-клеточной карциномы,
 остеосаркомы,
остеосаркомы,
 рака почки.
рака почки.
В соответствии с одним аспектом предусмотрена фармацевтическая композиция, содержащая антитело, производное, модифицированный формат или фрагмент, конъюгат антитело-лекарственное средство, или CAR, или клетку в соответствии с вышеизложенным описанием вместе с одним или несколькими фармацевтически приемлемыми ингредиентами.
В соответствии с еще одним аспектом настоящим изобретением предусмотрены способы уничтожения или ингибирования роста клетки, экспрессирующей ROR2 in vitro или у пациента, причем способ предусматривает введение в клетку фармацевтически эффективного количества или дозы антитела, производного, модифицированного формата или фрагмента, конъюгата антитело-лекарственное средство, или CAR, или клетки в соответствии с вышеизложенным описанием или фармацевтической композиции в соответствии с вышеизложенным описанием.
Предпочтительно, клетка, экспрессирующая ROR2, представляет собой раковую клетку.
Способы предполагают введение терапевтически эффективное количество антитела к ROR2, связывающего белка на основе антитела, фрагмента этого антитела, ADC или CAR, раскрытых в данном документе, субъекту, нуждающемуся в лечении. Это обеспечивает возможность уничтожения или ингибирования роста клетки, экспрессирующей ROR2 у субъекта. В соответствии с различными вариантами осуществления клетка, экспрессирующая ROR2, представляет собой опухолевую клетку. В соответствии со связанным аспектом настоящим изобретением предусмотрены способы лечения заболевания или состояния, ассоциированного с повышенной экспрессией ROR2 у субъекта. Эти способы включают введение терапевтически эффективного количества антитела к ROR2, связывающего белка на основе антитела, фрагмента этого антитела, ADC или CAR согласно настоящему изобретению субъекту, который поражен заболеванием или состоянием, ассоциированным с повышенной экспрессией ROR2. Это обеспечивает возможность лечения заболевания или состояния у субъекта. Некоторые из этих способов направлены на лечение рака у субъекта. Виды рака, которые восприимчивы к лечению с помощью способов согласно настоящему изобретению, включают в себя, например, нейробластому, саркому (и, в особенности, остеосаркому), почечно-клеточную карциному, рак молочной железы, рак яичка, рак яичника, рак поджелудочной железы, рак почки, рак желудка, рак предстательной железы, рак головы и шеи, меланому, плоскоклеточную карциному, множественную миелому и другие виды рака.
В соответствии с различными вариантами осуществления вводимые антитело к ROR2, связывающий белок на основе антитела, фрагмент этого антитела или ADC или CAR на их основе представляют собой F(ab)2, Fv, scFv, IgGACH2, F(ab')2, scFv2CH3, Fab, VL, VH, scFv4, scFv3, scFv2, dsFv, Fv, (scFv)2 или синтетический IgG. В соответствии с некоторыми вариантами осуществления вводимые антитело к ROR2, связывающий белок на основе антитела или фрагмент этого антитела конъюгированы с синтетической молекулой. В соответствии с некоторыми из этих вариантов осуществления антитело к ROR2, связывающий белок на основе антитела, фрагмент этого антитела конъюгированы с трансмембранным участком и внутриклеточным сигнальным доменом Т-клеточного рецептора (TCR) с образованием химерного антигенного рецептора (CAR). В соответствии с некоторыми вариантами осуществления химерный антигенный рецептор (CAR) присутствует на Т-клетке, которую нужно вводить субъекту. В соответствии с некоторыми другими вариантами осуществления антитело может быть конъюгировано с цитотоксическим средством, радиоактивным изотопом или липосомой.
Термины «осуществление лечения» или «лечение», используемые в контексте данного документа, не обязательно предполагают 100% или полное лечение. Точнее, существуют переменные степени лечения, известные квалифицированному специалисту в данной области техники как оказывающие потенциальное благоприятное воздействие или терапевтический эффект. В связи с этим способ согласно настоящему изобретению может обеспечивать любую величину любого уровня лечения. Более того, лечение, обеспечиваемое с помощью способа согласно настоящему изобретению, может включать в себя лечение одного или нескольких состояний или симптомов заболевания, подлежащего лечению. В частности, лечение можно осуществлять в виде внутривенной инфузии.
В соответствии с одним вариантом осуществления полностью человеческое антитело к ROR2, связывающий белок на основе антитела или фрагмент этого антитела, би- или мультиспецифичное антитело, ADC или CAR вводят в виде монотерапии. В соответствии с альтернативным вариантом осуществления полностью человеческое антитело к ROR2, связывающий белок на основе антитела или фрагмент этого антитела, би- или мультиспецифичное антитело, ADC или CAR вводят совместно, или параллельно, или последовательно с дополнительными терапевтическими средствами.
В частности, полностью человеческое антитело к ROR2, связывающий белок на основе антитела или фрагмент этого антитела, би- или мультиспецифичное антитело, ADC или CAR можно вводить в дозе, составляющей приблизительно 0,1-20 мг/кг.
Термин «субъект» относится к человеку и отличным от человека животным (в особенности, отличным от человека млекопитающим) и, предпочтительно, к людям. Помимо связывания с ROR2 антитела, связывающего белка на основе антитела или фрагмента этого антитела, би- или мультиспецифичного антитела, ADC или CAR, они также могут связываться с ROR2 из других видов, что делает их эффективными для лечения этих видов.
Настоящее изобретение также относится к способу лечения субъекта, страдающего от, имеющего риск развития, и/или у которого диагностировано неопластическое заболевание, предусматривающему введение полностью человеческого антитела к ROR2, связывающего белка на основе антитела или фрагмента этого антитела, ADC или CAR, которые описаны в данном документе.
В частности, указанное неопластическое заболевание является выбранным из группы, состоящей из: нейробластомы, саркомы (и, в особенности, остеосаркомы), почечно-клеточной карциномы, рака молочной железы, рака яичка, рака яичника, рака поджелудочной железы, рака почки, рака желудка, рака предстательной железы, рака головы и шеи, меланомы, плоскоклеточной карциномы, множественной миеломы и других видов рака.
В соответствии с некоторыми связанными аспектами настоящим изобретением предусмотрены фармацевтические композиции или наборы, которые содержат терапевтически эффективное количество полностью человеческого антитела к ROR2 или антитела, связывающего белка на основе антитела или фрагмента этого антитела, би- или мультиспецифического антитела или ADC, описанных в данном документе, и фармацевтически приемлемый носитель. Некоторые наборы согласно настоящему изобретению могут дополнительно содержать один или несколько буферов для иммунологического анализа.
В соответствии с некоторыми вариантами осуществления композиции согласно настоящему изобретению содержат носитель, желательно, фармацевтически приемлемый носитель. Фармацевтически приемлемый носитель может представлять собой один или несколько совместимых твердых или жидких наполнителей, разбавителей, других вспомогательных веществ или инкапсулирующих веществ, которые являются подходящими для введения человеку или субъекту, получающему ветеринарную помощь, (например, физиологически приемлемый носитель или фармакологически приемлемый носитель). Термин «носитель» определяет органический или неорганический ингредиент, натуральный или синтетический, с которым активный ингредиент объединяют для облегчения применения активного ингредиента, например, введения активного ингредиента субъекту. Фармацевтически приемлемый носитель может быть смешан с одним или несколькими из активных компонентов, например, с гибридной молекулой, и друг с другом, если более чем один фармацевтически приемлемый носитель присутствует в композиции, таким образом, чтобы он не ухудшал существенно желаемую фармацевтическую эффективность. Фармацевтически приемлемые материалы, как правило, можно вводить субъекту, например, пациенту, без получения значительных нежелательных физиологических эффектов, таких как тошнота, головокружение, высыпание или расстройство желудка. Например, желательно, чтобы композиция, содержащая фармацевтически приемлемый носитель, не являлась иммуногенной при введении пациенту-человеку для терапевтических целей.
Фармацевтические композиции согласно настоящему изобретению могут дополнительно содержать подходящие буферные средства, в том числе, например, уксусную кислоту в соли, лимонную кислоту в соли, борную кислоту в соли и фосфорную кислоту в соли. Композиции также могут необязательно содержать подходящие консерванты, такие как хлорид бензалкония, хлорбутанол, парабены и тимеросал. Фармацевтические композиции согласно настоящему изобретению могут присутствовать в единичной лекарственной форме, и их можно получать с помощью любого подходящего способа, многие из которых являются хорошо известными в области фармацевтики. Такие способы включают в себя стадию объединения антитела или антигенсвязывающего фрагмента согласно настоящему изобретению с носителем, который составляют один или несколько дополнительных ингредиентов. Обычно композицию получают посредством равномерного и тщательного объединения активного средства с жидким носителем, тонкоизмельченным твердым носителем или обоими из них, а затем, если необходимо, придания продукту определенной формы.
Композиция, подходящая для парентерального введения, в подходящем случае содержит стерильный водный препарат композиции согласно настоящему изобретению, который, предпочтительно, является изотоничным с кровью реципиента. Этот водный препарат можно составлять согласно известным способам с применением подходящих диспергирующих или смачивающих средств и суспендирующих средств. Стерильный инъекционный препарат также может представлять собой стерильный инъекционный раствор или суспензию в нетоксичном приемлемом для парентерального введения разбавителе или растворителе, например, в виде раствора в 1,3-бутандиоле. В числе приемлемых сред и растворителей, которые можно использовать, находятся вода, раствор Рингера и изотоничный раствор хлорида натрия. Кроме того, стерильные жирные масла обычно используют в качестве растворителя или суспендирующей среды. Для этой цели можно можно использовать любое легкое жирное масло, такое как синтетические моно- или диглицериды. Кроме того, жирные кислоты, такие как олеиновая кислота, можно применять в получении инъекционных растворов. Составы носителей, подходящие для перорального, подкожного, внутривенного, внутримышечного и других введений, можно найти в Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, PA.
Получение фармацевтических композиций согласно настоящему изобретению, а также их введение различными путями можно осуществлять в соответствии со способами, хорошо известными в уровне техники. См., например, Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000; и Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978. Системы для доставки, пригодные в контексте настоящего изобретения включают в себя системы доставки с медленным высвобождением, замедленным высвобождением и пролонгированным высвобождением, благодаря которым доставка композиции согласно настоящему изобретению происходит до сенсибилизации места, подлежащего лечению, и в течение достаточного периода времени для того, чтобы вызвать сенсибилизацию. Композицию согласно настоящему изобретению можно применять совместно с другими терапевтическими средствами или методами терапии. Такие системы могут избегать повторных введений композиции согласно настоящему изобретению, тем самым повышая удобство для субъекта и врача, и могут являться особенно подходящими для определенных композиций согласно настоящему изобретению.
Многие типы систем для доставки с высвобождением являются доступными и известны квалифицированным специалистам в данной области техники. Подходящие системы для доставки с высвобождением включают в себя системы на основе полимеров, таких как поли(лактид-гликолид), сополимеры оксалатов, поликапролактоны, полиэфирамиды, полиортоэфиры, полигидроксимасляная кислота и полиангидриды. Микрокапсулы из вышеизложенных полимеров, содержащие лекарственные средства, описаны, например, в патенте США №5075109. Системы для доставки также включают в себя неполимерные системы, которые представляют собой липиды, в том числе стерины, такие как холестерин, сложные эфиры холестерина, и жирные кислоты или нейтральные жиры, такие как моно-, ди- и триглицериды; системы для высвобождения на основе гидрогеля; системы на основе материала sylastic; системы на основе пептидов; восковые покрытия; прессованные таблетки с применением традиционных связующих и вспомогательных веществ; частично слитые имплантаты и т.п. Конкретные примеры включают в себя, без ограничения: (а) эрозионные системы, в которых активная композиция содержится в форме внутри матрицы, такой как описанные в патентах США №№4452775, 4667014, 4748034 и 5239660; и (b) диффузионные системы, в которых активный компонент проникает с контролируемой скоростью из полимера, такого как описанный в патентах США №3832253 и №3854480. Кроме того, можно применять аппаратные системы для доставки на основе насоса, некоторые из которых адаптированы для имплантации.
Настоящим изобретением также предусмотрены наборы, подходящие для осуществления способов согласно настоящему изобретению. Как правило, наборы содержат два или более компонентов, требующихся для осуществления терапевтических или диагностических способов согласно настоящему изобретению. Компоненты набора включают в себя, без ограничения, одно или несколько антител или антигенсвязывающих фрагментов согласно настоящему изобретению, соответствующие реактивы и/или оборудование. В соответствии с некоторыми вариантами осуществления наборы могут содержать антитело или антигенсвязывающий фрагмент согласно настоящему изобретению и буфер для иммунологического анализа, подходящий для выявления ROR2 (например, с помощью метода ELISA, проточной цитометрии, магнитного сортинга или FACS). Набор также может содержать один или несколько микротитровальных планшетов, стандартов, разбавителей для анализа, промывочных буферов, клеящихся крышек для планшета, магнитных гранул, магнитов и/или инструкций по осуществлению способа согласно настоящему изобретению с применением набора. Набор может включать в себя антитело или антигенсвязывающий фрагмент согласно настоящему изобретению, связанные с подложкой (например, многолуночный планшет или чип), которые подходящим образом упакованы и пригодны для выявления ROR2. В соответствии с некоторыми вариантами осуществления набор включает в себя антитело или антигенсвязывающий фрагмент согласно настоящему изобретению, которые конъюгированы с меткой, такой как флуоресцентная метка, биологически активная ферментная метка, люминесцентная метка или хромофорная метка. Наборы могут дополнительно включать в себя реактивы для визуализации конъюгированного антитела, например, субстрат для фермента. В соответствии с некоторыми вариантами осуществления наборы включают в себя антитело или антигенсвязывающий фрагмент согласно настоящему изобретению, которые конъюгированы с контрастным средством, и, необязательно, один или несколько реактивов или узлов оборудования, пригодных для визуализации антитела у субъекта.
Обычно антитело или антигенсвязывающий фрагмент согласно настоящему изобретению в наборе подходящим образом упакованы, например, в пузырек, мешок, ампулу и/или любой контейнер, подходящий для терапевтического способа или способа выявления. Компоненты набора могут быть обеспечены в виде концентратов (в том числе в виде лиофилизированных композиций), которые можно дополнительно разбавить перед применением, или они могут быть обеспечены в концентрации для применения. В случае применения антитела, производного, модифицированного формата или фрагмента согласно настоящему изобретению in vivo однократные дозы могут быть обеспечены в стерилизованных контейнерах, содержащих желаемое количество и концентрацию компонентов.
Аспекты настоящего изобретения, относящиеся к способам выявления
В соответствии с еще одним аспектом настоящего изобретения предусмотрен способ определения того,
 страдает ли предполагаемый пациент от неопластического заболевания или иммунного заболевания,
страдает ли предполагаемый пациент от неопластического заболевания или иммунного заболевания,
 имеет ли предполагаемый пациент риск развития неопластического заболевания или иммунного заболевания, и/или
имеет ли предполагаемый пациент риск развития неопластического заболевания или иммунного заболевания, и/или
 диагностировано ли у предполагаемого пациента неопластическое заболевание или иммунное заболевание,
диагностировано ли у предполагаемого пациента неопластическое заболевание или иммунное заболевание,
причем указанный способ предусматривает обработку образца, взятого у этого субъекта, конъюгатом антитело-эффекторная молекула в соответствии с вышеизложенным описанием.
В соответствии с одним вариантом осуществления неопластическое заболевание или иммунное заболевание является подходящим для лечения антителом к ROR2, конъюгатом антитело к ROR2-лекарственное средство или Т-клеткой с CAR, содержащей антитело к ROR2.
В соответствии с некоторыми другими вариантами осуществления настоящее изобретение относится к способу выявления в биологическом образце измененного уровня ROR2 (например, ROR2 клеточной поверхности), например, по сравнению с контролем с помощью любого из FACS, иммуногистохимического анализа (IHC) или Вестерн-блоттинга. Обычно способ включает в себя обеспечение контакта биологического образца с антителом, связывающим белком на основе антитела, фрагментом этого антитела согласно настоящему изобретению и определение количества антитела, которое селективно связывается с материалом (например, клетками) в образце, чтобы таким образом определить уровень ROR2 в биологическом образце. Биологический образец может быть получен из клеточной культуры или от исследуемого субъекта, например, плазма крови или образец ткани от субъекта, который имеет заболевание или состояние, ассоциированное с повышенным уровнем ROR2 у субъекта, предположительно имеет заболевание или состояние, ассоциированное с повышенным уровнем ROR2 у субъекта, или имеет риск развития заболевания или состояния, ассоциированного с повышенным уровнем ROR2 у субъекта. Контрольный уровень желательно соответствует уровню ROR2, выявляемому с применением того же антитела в соответствующем(соответствующих) образце(образцах) из одной или нескольких контрольных культур или от субъектов, не страдающих заболеванием. Способы применения антитела согласно настоящему изобретению для определения уровней ROR2 могут включать в себя любой иммунологический анализ, такой как иммуно- (вестерн) блоттинг, твердофазный иммуноферментный анализ (ELISA), иммуногистохимический анализ (IHC) и проточную цитометрию, например, анализ методом флуоресцентно-активированного клеточного сортинга (FACS).
Способы выявления можно применять для скрининга в отношении присутствия нарушения, ассоциированного с повышенным уровнем ROR2. Способы включают в себя получение образца от исследуемого субъекта, нуждающегося в скрининге, например, от субъекта, который имеет нарушение, ассоциированное с повышенным уровнем ROR2, предположительно имеет нарушение, ассоциированное с повышенным уровнем ROR2, или имеет риск развития нарушения, ассоциированного с повышенным уровнем ROR2. Уровень ROR2 (например, количество или концентрация) в образце измеряют с применением антитела, связывающего белка на основе антитела, фрагмента этого антитела согласно настоящему изобретению, и уровень в образце сравнивают с контрольным уровнем ROR2. Контрольный уровень представляет собой, например, средний уровень (например, количество или концентрацию) в образце (образцах) из одной или, предпочтительно, нескольких субъектов в контрольной группе, которые не имеют нарушение, ассоциированное с повышенным уровнем ROR2. В качестве альтернативы, контрольный уровень может соответствовать уровню или среднему уровню ROR2 в одном или нескольких образцах, взятых у исследуемого субъекта один или несколько раз до исследования, как например, когда исследуемый субъект не имел состояние, ассоциированное с повышенным уровнем ROR2, или у него не проявлялось состояние, ассоциированное с повышенным уровнем ROR2. Значительно более высокий уровень ROR2 в биологическом образце по сравнению с контрольным уровнем, указывает на нарушение, ассоциированное с повышенным уровнем ROR2 у субъекта. У субъектов, таких как люди, в случаях, когда экспрессия ROR2 на клеточной поверхности преимущественно ограничивается эмбриональным развитием, контрольный уровень ROR2 может составлять ноль или отсутствовать. Таким образом, в соответствии с некоторыми вариантами осуществления способа выявления, предусмотренного настоящим изобретением, любое значимое и выявляемое количество ROR2 в биологическом образце может указывать на нарушение, ассоциированное с повышенным уровнем ROR2 у субъекта.
Кроме того, способы выявления можно применять для мониторинга развития нарушения, ассоциированного с повышенным уровнем ROR2. Способ включает в себя получение образца от субъекта, нуждающегося в скрининге, например, от субъекта, у которого было диагностировано, нарушение, ассоциированное с повышенным уровнем ROR2, или который предположительно имеет нарушение, ассоциированное с повышенным уровнем ROR2. Уровень ROR2 в образце измеряют с применением антитела, связывающего белка на основе антитела, фрагмента этого антитела согласно настоящему изобретению, и уровень в образце сравнивают с контрольным уровнем, соответствующим уровню или среднему уровню ROR2 в одном или нескольких образцах, взятых у исследуемого субъекта один или несколько раз до исследования. Уровни ROR2, которые являются значительно повышенными или пониженными по сравнению с контрольным, указывают на то, что нарушение у субъекта усугубляется или улучшается, соответственно. Вышеизложенные способы выявления можно применять для скрининга в отношении присутствия или для мониторинга развития нарушений, в том числе, например, CLL, ALL, лимфомы из клеток мантийной зоны, нейробластомы, саркомы, почечно-клеточной карциномы, рака молочной железы, рака легкого, рака толстой кишки, рака головы и шеи, меланомы и других видов рака.
В соответствии с некоторыми вариантами осуществления настоящим изобретением предусмотрены способы скрининга субъекта в отношении измененного уровня ROR2. Обычно способы предполагают введение субъекту антитела, связывающего белка на основе антитела, фрагмента этого антитела согласно настоящему изобретению, которые конъюгированы с меткой (например, контрастным средством), визуализацию субъекта способом, подходящим для выявления метки, и определения того, имеет ли область у субъекта измененную плотность или концентрацию метки по сравнению с фоновым уровнем метки в ближайшей ткани. В качестве альтернативы, способы включают в себя определение того, присутствует ли измененная плотность или концентрация метки в области по сравнению с плотностью или концентрацией метки, выявленной ранее в той же области субъекта. Способы визуализации субъекта могут включать в себя рентгенографическую визуализацию, визуализацию с помощью рентгеновской компьютерной томографии (КТ) (например, визуализацию с помощью КТ-ангиографии (СТА)), магнитно-резонансную (MR) визуализацию, магнитно-резонансную ангиографию (MRA), визуализацию с помощью методов медицинской радиологии, ультразвуковую (US) визуализацию, оптическую визуализацию, эластографию, визуализацию в инфракрасных лучах, СВЧ-визуализацию и т.п., которые являются подходящими для выявления метки, конъюгированной с антителом. В соответствии с предпочтительным вариантом осуществления субъект имеет, предположительно имеет или имеет риск развития экспрессирующей ROR2 опухоли, такой как CLL, ALL, лимфома из клеток мантийной зоны, нейробластома, саркома, почечно-клеточная карцинома, рак молочной железы, рак легкого, рак толстой кишки, рак головы и шеи, меланома и другие виды рака, и способ применяют для скрининга в отношении присутствия опухоли или выявления присутствия опухоли. В соответствии с еще одним вариантом осуществления способ можно применять для мониторинга размера или плотности экспрессирующей ROR2 опухоли со временем, например, в течение курса лечения.
Таким образом, настоящее изобретение относится к способам выявления измененного уровня ROR2 у субъекта. Способы включают (а) получение биологического образца от субъекта, (b) обеспечение контакта образца с антителом к ROR2, связывающим белком на основе антитела, фрагментом этого антитела, которые раскрыты в данном документе, (с) определение уровня ROR2 в биологическом образце и (d) сравнение уровня ROR2 в биологическом образце с контрольным уровнем ROR2. Это обеспечивает возможность определения того, является ли уровень ROR2 в биологическом образце измененным по сравнению с контрольным уровнем ROR2. В этих способах повышенный уровень ROR2 у субъекта по сравнению с контрольным уровнем указывает на заболевание или состояние, ассоциированное с повышенной экспрессией ROR2 у субъекта. Например, выявление повышенной экспрессии ROR2 может указывать на присутствие нейробластомы, остеосаркомы, почечно-клеточной карциномы, рака молочной железы, рака желудка, рака предстательной железы, меланомы, плоскоклеточной карциномы или множественной миеломы у субъекта.
В соответствии с еще одним вариантом осуществления настоящее изобретение относится к способам выявления экспрессирующей ROR2 опухоли у субъекта. Эти способы предполагают (а) введение антитела к ROR2, связывающего белка на основе антитела, фрагмента этого антитела согласно настоящему изобретению субъекту, который имеет, предположительно имеет или имеет риск развития экспрессирующей ROR2 опухоли, и (b) визуализацию субъекта для выявления области с измененной плотностью или концентрацией конъюгированной метки, причем плотность или концентрация является относительной по сравнению (i) с фоном в ближайшей ткани или с (ii) с плотностью или концентрацией, выявленными ранее в той же области у субъекта. Существование области с измененной плотностью или концентрацией конъюгированной метки указывает на присутствие экспрессирующей ROR2 опухоли у субъекта.
Аспекты настоящего изобретения, относящиеся к клеткам и Т-клеткам
В соответствии с некоторыми вариантами осуществления настоящим изобретением предусмотрены способы лечения субъекта, который имеет нарушение, ассоциированное с повышенными уровнями ROR2, предположительно имеет нарушение, ассоциированное с повышенными уровнями ROR2, или имеет риск развития нарушения, ассоциированного с повышенными уровнями ROR2, с помощью адоптивного переноса подвергнутых воздействию методов генной инженерии Т-клеток, описанных в данном документе, которые экспрессируют антитело или антигенсвязывающий фрагмент согласно настоящему изобретению в виде химерного антигенного рецептора (CAR), который селективно связывает ROR2. Технологию рекомбинантной ДНК можно применять для введения кодирующего CAR генетического материала в любые подходящие Т-клетки, например, центральные Т-клетки памяти от субъекта, подлежащего лечению. Т-клетки, несущие генетический материал, можно разращивать (например, в присутствии цитокинов). Подвергнутые воздействию методов генной инженерии Т-клетки переносят пациенту, как правило, посредством инфузии. Перенесенные Т-клетки согласно настоящему изобретению могут затем вызывать иммунный ответ в отношении экспрессирующих ROR2 клеток у субъекта. Способ адоптивного переноса можно применять, например, для лечения субъектов, которые имеют или предположительно имеют любой из видов рака, ассоциированных с ROR2, в том числе CLL, ALL, лимфомы из клеток мантийной зоны, нейробластомы, саркомы, почечно-клеточной карциномы, рака молочной железы, рака легкого, рака толстой кишки, рака головы и шеи, меланомы и других видов рака. В соответствии с некоторыми вариантами осуществления вышеизложенные способы лечения могут дополнительно включать в себя совместное введение второго терапевтического средства для лечения нарушения, ассоциированного с повышенным уровнем ROR2. Например, если нарушение, подлежащее лечению, включает экспрессирующую ROR2 злокачественную опухоль, способ может дополнительно включать в себя совместное введение цитотоксического, цитостатического, или антиангиогенного, или иммуностимулирующего средства (например, антител-ингибиторов иммунных контрольных точек, например, без ограничения, антител, связывающихся с PD1, PDL1, CTLA4, ОХ40, TIM3, GITR, LAG3 и т.п.), подходящего для лечения рака. Если рак представляет собой В-клеточную злокачественную опухоль, способ может дополнительно включать в себя, например, совместное введение ритуксимаба, алемтузумаба, офатумумаба, окрелизумаба или химиотерапевтическую схему CHOP.
Таким образом, настоящим изобретением дополнительно предусмотрены эукариотические клетки или клетки, отличные от эукариотических (например, Т-лимфоциты), которые были подвергнуты воздействию генной инженерии с помощью технологии рекомбинантной ДНК для выработки полностью человеческих антител к ROR2 или антигенсвязывающих фрагментов согласно настоящему изобретению. Эукариотические клетки или клетки, отличные от эукариотических, можно применять в качестве экспрессионных систем для получения антитела согласно настоящему изобретению. В соответствии с некоторыми вариантами осуществления настоящим изобретением предусмотрены нацеленные на ROR2 иммунные клетки, которые подвергнуты воздействию методов генной инженерии для рекомбинантной экспрессии специфичного в отношении ROR2 отличного от человеческого или гуманизированного антитела или антигенсвязывающего фрагмента согласно настоящему изобретению. Например, настоящим изобретением предусмотрена Т-клетка, подвергнутая воздействию методов генной инженерии для экспрессии человеческого антитела к ROR2 или антигенсвязывающего фрагмента согласно настоящему изобретению (например, scFv, scFv-Fc или (scFv)2), которые связаны с синтетической молекулой, содержащей один или несколько из следующих доменов: спейсерный или шарнирный участок (например, последовательность CD28 или последовательность шарнирного участка-Fc IgG4), трансмембранный участок (например, канонический трансмембранный домен) и внутриклеточный сигнальный домен Т-клеточного рецептора (TCR), таким образом образуя химерный антигенный рецептор (CAR) или Т-клеточное тельце. Внутриклеточные сигнальные домены TCR, которые могут быть включены в CAR (или Т-клеточное тельце) включают в себя, без ограничения, сигнальные домены CD3ξ, FcR-γ и Syk-PT, а также совместно участвующие в передаче сигнала домены CD28, 4-1 ВВ и CD134. Способы конструирования Т-клеток, экспрессирующих CAR (или Т-клеточных телец), являются известными в уровне техники. См., например, Marcu-Malina et al., Expert Opinion on Biological Therapy, Vol.9, No. 5 (размещенный в Интернете 16 апреля 2009 года).
ПРИМЕРЫ
Несмотря на то что настоящее изобретение было подробно проиллюстрировано и описано в описании чертежей и вышеизложенном описании, такая иллюстрация и описание считаются иллюстративными или примерными, а не ограничительными; настоящее изобретение не ограничивается раскрытыми вариантами осуществления. Другие варианты раскрытых вариантов осуществления могут быть понятны специалистам в данной области техники и осуществлены ими при практической реализации заявленного изобретения в результате изучения чертежей, раскрытия и прилагаемой формулы изобретения. В пунктах формулы изобретения слово «содержащий» не исключает другие элементы или стадии, и формы единственного числа не исключают множественного числа. Сам по себе тот факт, что определенные измеряемые величины перечислены в отличающихся друг от друга зависимых пунктах формулы изобретения, не указывает на то, что комбинация этих измеряемых величин не может быть применена для получения преимущества. Любые ссылочные позиции в пунктах формулы изобретения не следует толковать как ограничивающие объем.
Все аминокислотные последовательности, раскрытые в данном документе, приведены в ориентации от N-конца к С-концу; все последовательности нуклеиновой кислоты, раскрытые в данном документе, приведены в ориентации от 5' к 3'.
Пример 1. Создание и скрининг библиотеки антител (фиг. 1)
Экспрессия антигена hROR2-TwinStrep. EBNA экспрессионный вектор pCB14b-hROR2-Thr-ECD-TwinStrep (где Thr представляет собой треонин), управляющий экспрессией внеклеточного домена hROR2 (ECD), меченного на С-конце меткой TwinStrep, трансфицировали в НЕК293Т с применением Lipofectamine® LTX с реактивом PLUS® (Thermo Fisher Scientific, 15388100). После 1 суток инкубирования (37°С, 5% СО2, ростовая среда: среда Игла в модификации Дульбекко (DMEM) с высоким содержанием глюкозы (4,5 г/л), с L-глутамином с 10% (объем/объем) фетальной телячьей сыворотки (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином (все от Bioconcept)) клетки разращивали в условиях отбора (2 мкг/мл пуромицина (Sigma-Aldrich, Р8833-25 мг маточный раствор в концентрации 2 мг/мл)). Клетки разделяли и дополнительно разращивали (37°С, 5% СО2); по достижению конфлюентности флаконы hyperflasks покрывали 20 мкг/мл поли-L-лизина (Sigma-Aldrich, P1524) в течение 2 часов при 37°С и промывали дважды PBS. Затем клетки трипсинизировали, промывали PBS и высевали на покрытые поли-L-лизином флаконы hyperflasks. После повторного достижения конфлюентности клетки промывали PBS с последующей заменой среды с применением среды для продуцирования (DMEM/F-12, Gibco/Thermo Fisher Scientific, 31330-038), дополненной 1 мг/мл пуромицина (Sigma-Aldrich, Р8833), 100 МЕ/мл пенициллина-стрептомицина-фунгизона (Bioconcept, 4-02F00-H), 161 мкг/мл N-ацетил-L-цистеина (Sigma-Aldrich, А8199) и 10 мкг/мл L-глутатиона в восстановленном состоянии (Sigma-Aldrich, G6529). Супернатант, который собирали раз в две недели и фильтровали (0,22 мкм) для удаления клеток, хранили при 4°С до очистки. Удаленный супернатант заменяли свежей средой для продуцирования. Экспресс-анализ всех собранных супернатантов методом электрофореза в полиакриламидном геле подтверждал присутствие полос, соответствующих hROR2.
Для очистки фильтрованный супернатант загружали на колонку, подходящую для связывания strep-меток; очистку и элюирование осуществляли в соответствии с протоколами производителя на системе АЕКТА pure (GE Healthcare). Фракции анализировали в отношении чистоты и целостности белка с помощью электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Содержащие белок фракции смешивали и подвергали замене буфера с применением фильтровальных установок Amicon (Millipore, Шаффхаузен, Швейцария, UFC901008) до достижения степени разведения ≥1:100 в PBS, а затем стерилизовали фильтрованием с применением фильтра с низким удержанием (0,20 мкм, Carl Roth, Карлсруэ, Германия, РА49.1).
Иммунизация H2L2 мышей hROR2-TwinStrep.Каждую из пяти трансгенных гуманизированных H2L2 мышей (полученных от Harbour Biomed; H2L2 мыши представляли собой результат скрещивания следующих линий мышей: F129, fvb/n и C57BL6, и при иммунизации они вырабатывают антитела с человеческим вариабельным доменом и крысиным константным доменом, раскрытые в международной заявке WO 2010/070263 А1) возрастом 6-10 недель иммунизировали посредством двух внутрибрюшинных инъекций (IP) с последующей одной внутривенной инъекцией 100 мкл hROR2-TwinStrep (из примера 1), составленной согласно таблице 5.

Таблица 5. Режим иммунизации H2L2 мышей
Образцы крови забирали у каждой мыши из хвостовой вены в Дни: -7, 7 и 28 относительно первой инъекции и посредством прокола сердца в День 49. Все процедуры, описанные в данном документе, в которых были задействованы животные, согласовались со швейцарскими директивами по уходу и содержанию животных.
Определение титров антител. Собранные образцы крови инкубировали при комнатной температуре в течение 15-60 минут, чтобы позволить крови свернуться, а затем центрифугировали для получения сыворотки, в которой оценивали титры антитела. Для этого планшеты для ELISA покрывали 100 мкл 2 мкг/мл hROR2-TwinStrep (из примера 1) в буфере на основе бикарбоната натрия для нанесения покрытия (0,1 М Na2CO3, 0,1 М NaHCO3, рН 9,6). После инкубирования в течение 1 суток при 4°С планшеты промывали PBS, дополненным 0,05% (объем/объем) Tween 20, и блокировали 150 мкл PBS, дополненным 0,05% (объем/объем) Tween 20 и 3% бычьего сывороточного альбумина (BSA). Образцы сыворотки разводили 100-кратно и из них готовили серии 2,5-кратных серий разведений, добавляя 50 мкл каждого разведенного образца в соответствующие планшеты для ELISA. После 1 часа инкубирования при температуре 37°С планшеты промывали PBS, дополненным 0,05% (объем/объем) Tween 20, перед добавлением конъюгированного с HRP антитела к крысиному FC-гамма фрагменту (Jackson Immunoresearch, 112-036-071). После дополнительного промывания планшета планшеты промывали 50 мкл Sigmafast OPD Tablet set (Sigma, P9187) и реакцию останавливали посредством добавления 2 М серной кислоты. Планшеты считывали на планшет-ридере ELISA (OD 490), результаты представлены на фиг. 2.
Выделение с помощью MACS специфичных к антигену hROR2 В-клеток из H2L2 мышей. Мышей умерщвляли через 49 суток после первоначальной инъекции антигена и селезенки мышей собирали. Селезенки переносили в пробирку gentleMACS C-tube (№в каталоге 130-093-237), содержащий 2,4 мл RPMI-10%FCS, и гомогенизировали с применением гомогенизатора gentleMACS Octo Dissociator (Miltenyi Biotec) перед фильтрованием через клеточное сито (пробирки для FACS, BD Flacon, 734-0001) с получением суспензий отдельных клеток.
Специфичные в отношении hROR2 В-клетки из мышиных спленоцитов затем подвергали селекции с помощью MACS сортинга: вначале В-клетки выделяли с использованием набора для выделения мышиных В-клеток Pan В Cell Isolation Kit, human (Miltenyi Biotec, 130-095-813) посредством отрицательной селекции. Для этого клетки спленоцитов промывали и суспендировали в буфере для MACS (2-4×106 клеток в 600 мкл) перед добавлением 2 мкл мышиного IgG (ChromPure, Jackson Immunoresearch, №в каталоге 015-000-003 в концентрации 20 мкг/мл), икубируя на льду в течение 15 минут. Клетки промывали и ресуспендировали в 40 мкл холодного буфера для MACS. Клетки, отличные от В-клеток, метили магнитной меткой посредством добавления 10 мкл коктейля меченых биотином пан-В-клеточных антител при 4°С с последующим добавлением 30 мкл холодного буфера для MACS и 20 мкл микрогранул с антителами к биотину. Конечный объем доводили до 500 мкл холодным буфером для MACS.
Клетки, отличные от В-клеток, удаляли, пропуская суспензию клеток через LD-колонку (Miltenyi Biotec, 130-042-901) в магнитном поле с применением сепаратора QuadroMACS™ (Miltenyi Biotec, 130-091-051) и собирая элюат.
Параллельно, hROR2-TwinStrep (12 мкг, из примера 1) инкубировали с магнитными наногранулами Strep-Tactin® (IBA 6-5500-005, 50 мкл в концентрации 7,044 мг/мл) и буфером для MACS при температуре 4°С и загружали на LS колонку (Miltenyi Biotec, 130-042-401) в магнитном поле для вымывания несвязанного антигена. Нагруженные антигеном гранулы затем ресуспендировали с элюатом из LD-колонки (содержащим В-клетки) и инкубировали 45 минут на льду. После центрифугирования и промывания буфером для MACS смесь помещали в LS-колонку, промывая в магнитном поле. Клетки, которые впоследствии вымывались с гранулами за пределами воздействия магнитного поля, представляют собой антиген-положительные В-клетки.
Создание библиотеки кДНК антитела. Антиген-положительные В-клетки ресуспендировали в 500 мкл реактива TRI; 100 мкл хлороформа добавляли после 5 минутного инкубирования. После применения вихревой мешалки и центрифугирования прозрачную верхнюю фазу переносили в пробирку Eppendorf, в которую добавляли 1 мкл гликогена и 250 мкл изопропанола. После смешивания, инкубирования, промывания и центрифугирования супернатант удаляли посредством декантирования и 500 мкл ледяного этанола (75%) добавляли к осадку РНК с перемешиванием посредством переворачивания. Осадок РНК затем центрифугировали и супернатант декантировали перед высушиванием на воздухе.
РНК подвергали обратной транскрипции в кДНК с применением стандартных методик. Вариабельные домены амплифицировали на ПЦР-циклере с использованием человеческих праймеров в две стадии: первый набор праймеров в соответствии с таблицей 6, (а) достроенный 5'-конец вариабельного домена с включением вышележащей нуклеотидной последовательности, соответствующей части лидерного пептида и (b) достроенный 5'-конец крысиного константного домена. Второй набор праймеров включал в себя: (а) для достройки 5'-конца нуклеотидные последовательности, обеспечивающие возможность получения полного лидерного пептида, а также сайт рестрикции NotI; (b) для амплификации полностью человеческого вариабельного домена относительно 3'-конца с применением нуклеотидной последовательности, связывающейся с крайним 3'-концом J-домена, а также сайты рестрикции Nhel или BsiWI для IgG и IgM или IgK, соответственно.

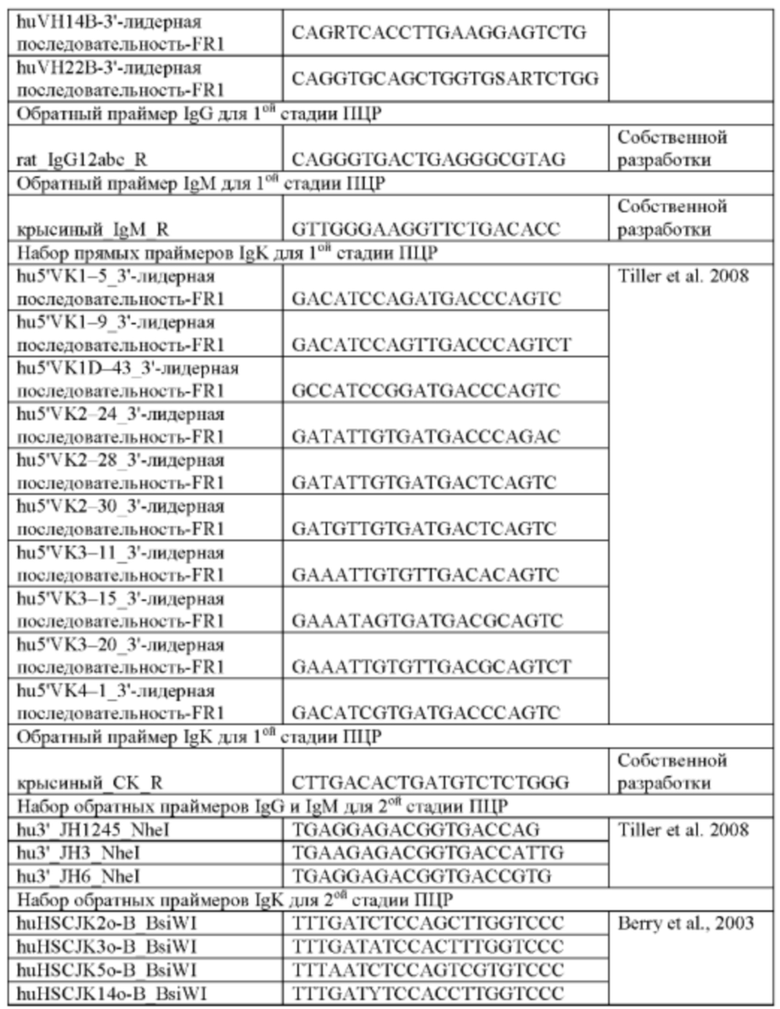
Таблица 6. Праймеры, применяемые для создания библиотеки кДНК.
Опосредуемое-транспозицией создание клеточных библиотек и характеристика библиотек. Кодирующие вариабельный домен антитела кДНК затем клонировали в векторы с помощью кодируемых сайтов рестрикции рядом с нуклеотидами, кодирующими человеческие константные домены, и между нуклеотидами, кодирующими последовательности, функциональные с транспозазой PiggyBac, а также устойчивость к ампициллину. Библиотеку характеризовали с помощью Miniprep с последующим секвенированием. Библиотека состояла из от 6×106 до 5×107 членов.
Их совместно трансфицировали в 63-12 мышиные трансформированные A-MuLV пре-В-клетки со вторым вектором, кодирующим транспозазу PiggyBac, для опосредуемого транспозицией дисплея на поверхности В-клеток и секреции антител («Transpo-mAb Display»), как описано в Waldmeier et al., 2016.
Функциональный скрининг супернатантов от клонов и отбор клонов. В-клетки представляли собой отдельные клетки, отсортированные методом FACS с использованием FACSAria, и их отбирали на основании положительного двойного окрашивания антителом к Strep-метке (IBA, 2-1555-050) и поликлональным конъюгированным с РЕ антителом к IgG (ebioscience, 12-4998-82); до 288 клонов на библиотеку IgG/IgK и IgM/IgK на мышь отбирали и выращивали в среде SF-IMDM с 2% (объем/объем) фетальной телячьей сыворотки (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона, 50 мкМ бета-меркаптоэтанолом и 2 мМ L-глутамином (все от Bioconcept, Альшвиль, Швейцария)) при температуре 37°С и 7,5% СО2 (фиг. 3). Антитела в супернатантах от отобранных клонов В-клеток затем оценивали с помощью двух средств: (1) в отношении связывания с hROR2 с помощью метода ELISA и (2) в отношении опосредования цитолиза сверхэкспрессирующих hROR2 клеток ЕМТ-6 после связывания с hROR2 с применением вторичного ADC, связывающегося с антителами к ROR2.
Связывание hROR2 с помощью ELISA одного пятна Половину лунок в 96-луночных планшетах покрывали 50 мкл 2 мкг/мл антитела к человеческому Fc (Jackson Immunoresearch, 109-006-008), а вторую половину покрывали 50 мкл 2 мкг/мл hROR2-Twin-Strep (из примера 1), оба в промывочном буфере (0,1 М Na2CO3, 0,1 М NaHCO3, рН 9,6), и хранили в течение ночи при температуре 4°С. После блокирования каждой лунки с добавлением 150 мкл PBS, дополненного 0,05% (объем/объем) Tween 20 и 3% (масса/объем) бычьего сывороточного альбумина (BS А), в течение 1 часа при температуре 37°С в каждую лунку добавляли 50 мкл разведенных супернатантов (разведенного 5-кратно в PBS, дополненном 1% (масса/объем) бычьего сывороточного альбумина (BSA) и 0,05% (объем/объем) Tween 20). Для сравнения на планшет добавляли серийно разведенные 1:20 образцы очищенного антитела к ROR2, а также антитела изотипического контроля, начиная с 0,5 мкг/мл в PBS, дополненном 1% (масса/объем) бычьего сывороточного альбумина (BSA) и 0,05% (объем/объем) Tween 20. После 1 часа инкубирования при температуре 37°С планшеты промывали PBS, дополненным 0,05% (объем/объем) Tween 20, перед добавлением конъюгированного с HRP антитела к человеческому IgG (Jackson Immunoresearch, 109-036-008). После дополнительного промывания планшета планшеты промывали 50 мкл Sigmafast OPD Tablet set (Sigma, P9187) и реакцию останавливали посредством добавления 2 М серной кислоты. Планшеты считывали на планшет-ридере ELISA (OD 490).
Анализ цитолиза клеток вторичными антителами с одного пятна: Клетки ЕМТ6, подвергнутые воздействию методов генной инженерии для сверхэкспрессии hROR2 из примера 7, высевали в количестве 1000 клеток в 100 мкл на лунку в полной среде DMEM (среда Игла в модификации Дульбекко (DMEM) с высоким содержанием глюкозы (4,5 г/л), с L-глутамином с 10% (объем/объем) фетальной телячьей сыворотки (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином (все от Bioconcept, Альшвиль, Швейцария)) и инкубировали при температуре 37°С и 5% СО2. На следующие сутки 15 мкл неразбавленного супернатанта или 15 мкл серийно разведенного 2-кратно образца очищенного антитела к ROR2, начиная с концентрации 10 мкг/мл, добавляли к клеткам. После инкубирования в течение 30 минут 35 мкл антитела anti-hu-IgG-CL-PNU (Moradec, AH-102PN-50) добавляли в концентрации 0,32 мкг/мл. После дополнительных трех суток планшеты удаляли из инкубатора и уравновешивали при комнатной температуре. Спустя примерно 30 минут 50 мкл удаляли из каждой лунки, а затем 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243) добавляли в каждую лунку. После встряхивания планшетов при 750 об./мин. в течение 5 минут с последующим инкубированием в течение 20 минут без встряхивания люминесценцию измеряли на планшет-ридере Spark 10М с продолжительностью интегрирования, составляющей 1 секунду на лунку.
Антитела, демонстрирующие высокий уровень связывания с ROR2, а также низкую жизнеспособность клеток, что, таким образом, указывало на них как на хороших кандидатов для ADC, подвергали дополнительному анализу методом ELISA с титрованием и анализам жизнеспособности клеток. В анализе ELISA с титрованием использовали те же условия, которые описаны выше в анализе ELISA одного пятна, за исключением того, что 5-кратно разведенный супернатант серийно разводили 3-кратно на общее количество 8 лунок. В анализе цитолиза вторичным антителом с титрованием использовали условия, подобные описанным выше. В данном случае концентрации IgG во всех супернатантах корректировали к концентрации IgG у клона с самым низким уровнем экспрессии (измеренным методом ELISA с титрованием при покрытии антителом к IgG). С этой концентрации все супернатанты серийно разводили 2-кратно, при этом общее количество этапов разведения составляло 6, и 15 мкл супернатантов добавляли в планшет. Анализ осуществляли с применением программного обеспечения Graphpad Prism.
Пример 2. Экспрессия и очистка антител к ROR2
Экспрессионные векторы: Последовательности антител, которые, как определено выше, связываются с hROR2, синтезировали в виде ДНК с помощью GenScript (Пискатауэй, США) и включали в экспрессионный вектор, содержащий подходящие сайты рестрикции и соответствующий константный домен.
Экспрессия и очистка антител kROR2: Экспрессионные векторы трансфицировали в клетки HEK293T с применением реактива Lipofectamine® LTX с реактивом PLUS™ (Thermo Fisher Scientific, Райнах, Швейцария, 15388100); после инкубирования в течение 1 суток (37°С, 5% СО2, ростовая среда: среда Игла в модификации Дульбекко (DMEM) с высоким содержанием глюкозы (4,5 г/л), с L-глутамином с 10% (объем/объем) фетальной телячьей сыворотки (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином (все от Bioconcept, Альшвиль, Швейцария)) клетки разращивали в условиях отбора (2 мкг/мл пуромицина (Sigma-Aldrich, Букс, Санкт-Галлен, Швейцария, Р8833-25 мг маточный раствор в концентрации 2 мг/мл)). Клетки разделяли и дополнительно разращивали (37°С, 5% СО2); по достижению конфлюентности чашки для тканевых культур покрывали 20 мкг/мл поли-L-лизина (Sigma-Aldrich, P1524) в течение 2 часов при 37°С и промывали дважды PBS. Затем клетки трипсинизировали и рассаживали в соотношении 1:3 на покрытые поли-L-лизином планшеты. После повторного достижения конфлюентности клетки промывали PBS с последующей заменой среды на среду для продуцирования (DMEM/F-12, Gibco/Thermo Fisher Scientific, 31330-03), дополненной 1 мкг/мл пуромицина (Sigma, Р8833), 100 МЕ/мл пенициллина-стрептомицина-фунгизона (Bioconcept), 161 мкг/мл N-ацетил-L-цистеина (Sigma-Aldrich, А8199) и 10 мкг/мл L-глутатиона в восстановленном состоянии (Sigma-Aldrich, G6529). Супернатант, который собирали раз в две недели и фильтровали (0,22 мкм) для удаления клеток, хранили при 4°С до очистки.
Для очистки профильтрованный супернатант загружали на уравновешенную PBS колонку HiTrap с белком A (GE Healthcare, Франкфурт-на-Майне, Германия, 17-0405-01) или колонку JSR Amsphere™ с белком A (JSR Life Sciences, Левен, Бельгия, JWT203CE) и промывали PBS; элюирование осуществляли с использованием 0,1 М глицина (рН 2,5) на системе АЕКТА pure (GE Healthcare). Фракции немедленно нейтрализовали 1 М буфером Tris-HCl (pH 8,0) и анализировали в отношении чистоты и целостности белка методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия. Содержащие белок фракции смешивали и подвергали замене буфера с применением фильтровальных установок Amicon (Millipore, Шаффхаузен, Швейцария, UFC 901008) до достижения степени разведения 1:100, а затем стерилизовали фильтрованием с применением фильтра с низким удержанием (0,20 мкм, Carl Roth, Карлсруэ, Германия, РА49.1).
Чистоту и целостность рекомбинантных антител анализировали методом электрофореза в полиакриламидном геле в присутствии додецилсульфата натрия.


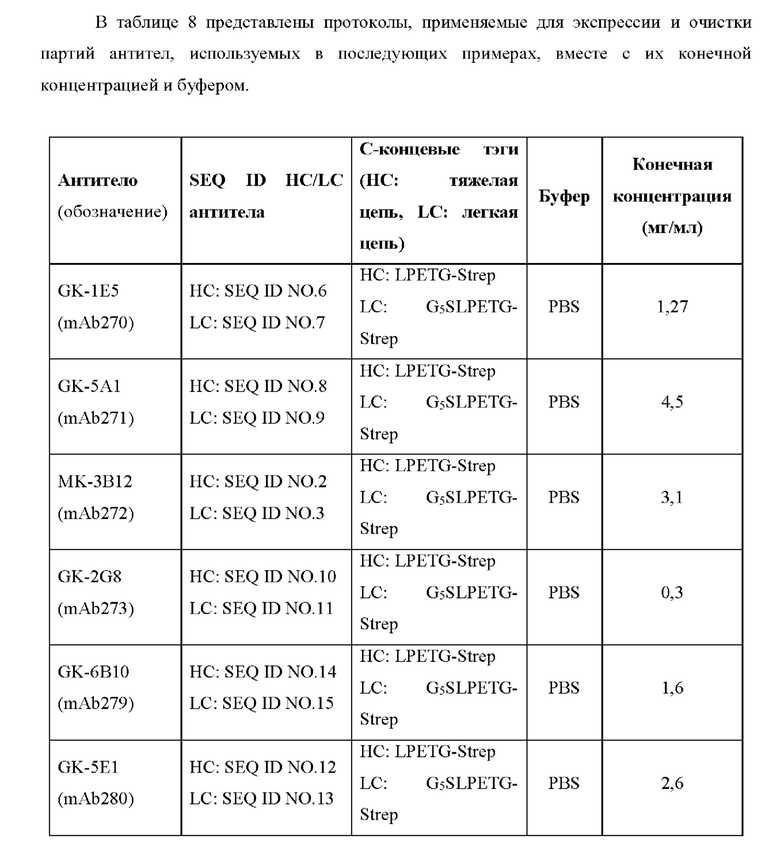
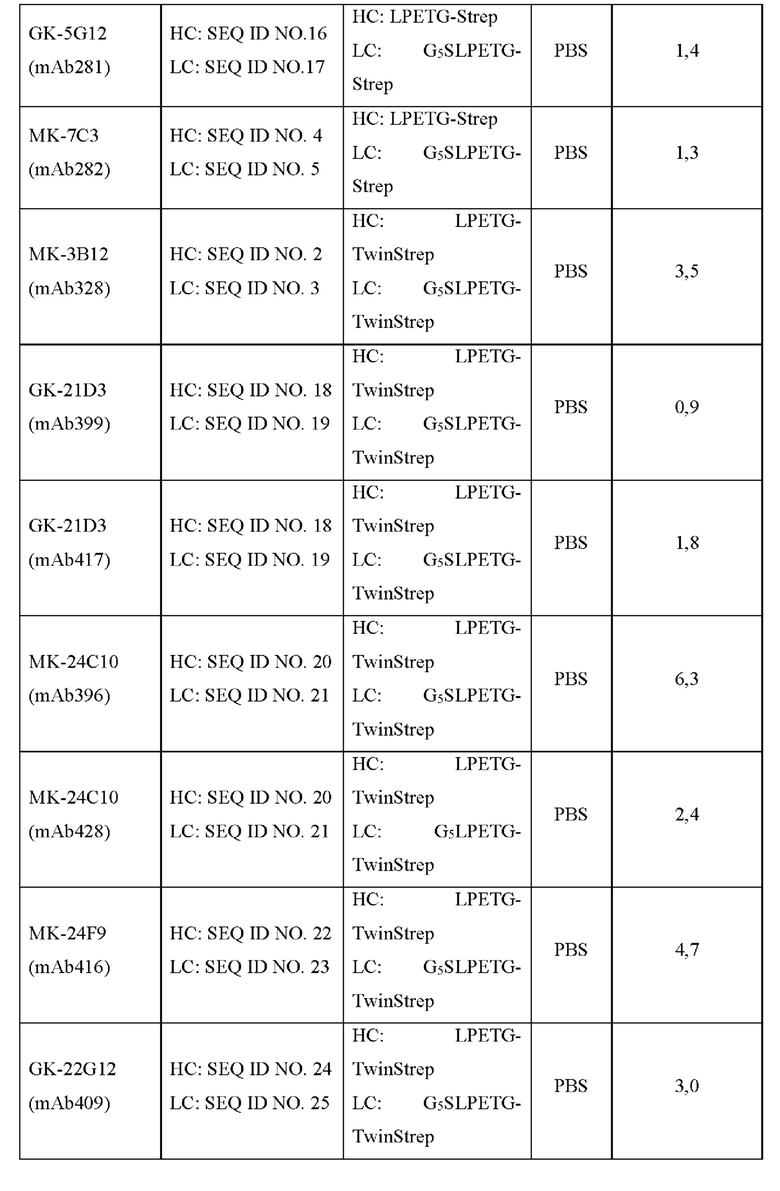
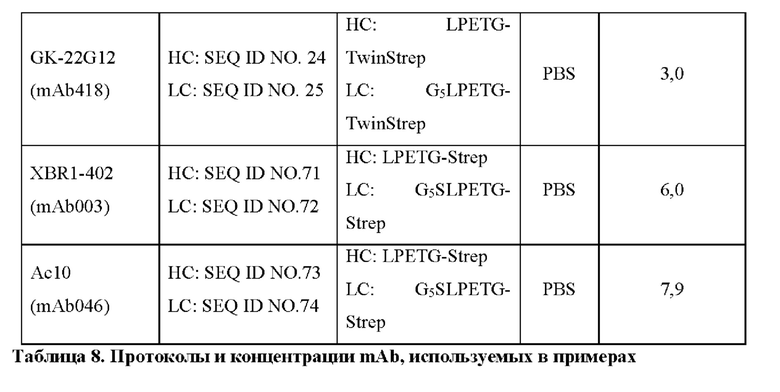
Пример 3. Связывание антитела с hROR2 с помощью SPR
Поверхностный плазмонный резонанс для измерения аффинностей антител к hROR2 в отношении hROR2 осуществляли на инструменте Biacore Т200 (GE Healthcare). Антитела захватывали с использованием сенсорного чипа СМ5 с белком A (GE Healthcare, 29127556) или белок G иммобилизировали на сенсорном чипе СМ5. Для измерений аффинности применяли либо очищенные антитела к hROR2, либо супернатанты 293Т, содержащие антитела к hROR2. Во всех случаях антитела к hROR2 разводили до концентрации 1-3 мкг/мл в 1х подвижном буфере HBS-EP+ (10 мМ HEPES, 150 мМ NaCl, 3 мМ EDTA (рН 7,4) и 0,05% (объем/объем) Tween 20) и подвергали захвату в течение 30 секунд со скоростью потока 30 мкл/мин. hROR2-TwinStrep (из примера 1) разводили в подвижном буфере с применением 2-кратных серийных разведений в диапазоне от 40 нМ до 2,5 нМ.
Ассоциацию и диссоциацию измеряли при скорости потока 30 мкл/мин. в течение 120 секунд и 200 секунд, соответственно. Расчет констант скорости ассоциации (kon) и диссоциации (koff) основывался на ленгмюровской модели связывания 1:1. Равновесную константу диссоциации (Kd) рассчитывали на основании koff/kon. SPR сенсограммы для антител из таблицы 9 представлены на фиг. 4, причем значения Kd, kon и koff представлены в таблице 9.
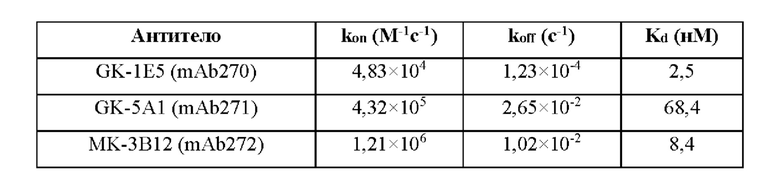

Пример 4. Экспрессия мышиного ROR2 и ROR2 яванского макака
Экспрессию каждого из мышиного ROR2 (mROR2) и ROR2 яванского макака (cROR2) осуществляли в соответствии с протоколом, аналогичным обеспеченному для экспрессии hROR2 в примере 1.
Пример 5. Перекрестная реактивность mAb с мышиным ROR2 и RQR2 яванского макака
Связывание антител с hROR2, мышиным ROR2 (mROR2) и ROR2 яванского макака (cROR2) оценивали в анализе на основе ELIS А. Для этого планшеты для ELIS А покрывали 50 мкл 2 мкг/мл следующих молекул в буфере на основе бикарбоната натрия для нанесения покрытия (0,1 М Na2CO3, 0,1 М NaHCO3, рН 9,6): (а) антитело к Fc человека (Jackson Immunoresearch, 109-006-098), (b) hROR2-TwinStrep, (с) ROR2 яванского макака-TwinStrep или (d) мышиный ROR2-TwinStrep. После инкубирования в течение 1-2 суток при температуре 4°С планшеты промывали PBS, дополненным 0,05% (объем/объем) Tween 20, и блокировали 150 мкл PBS, дополненным 0,05% (объем/объем) Tween 20 и 3% бычьего сывороточного альбумина (BSA). После этого добавляли 3-кратные или 4-кратные разведения очищенных антител, начиная с концентраций 0,5 или 2 мкг/мл, соответственно, в PBS, дополненном 1% (масса/объем) бычьего сывороточного альбумина (BSA) и 0,05% (объем/объем) Tween 20. После 1 часа инкубирования при температуре 37°С планшеты промывали PBS, дополненным 0,05% (объем/объем) Tween 20, перед добавлением конъюгированного с HRP антитела к человеческому IgG (Jackson Immunoresearch, 109-036-008). После дополнительного промывания планшета планшеты промывали 50 мкл Sigmafast OPD Tablet set (Sigma, P9187) и реакцию останавливали посредством добавления 2 М серной кислоты. Планшеты считывали на планшет-ридере ELIS A (OD 490).
На фиг. 5 и фиг. 11 представлены профили ELISA для mAb из таблицы 10. В этой таблице также обобщен статус связывания для каждого mAb с ROR2 из каждого из оцениваемых видов.
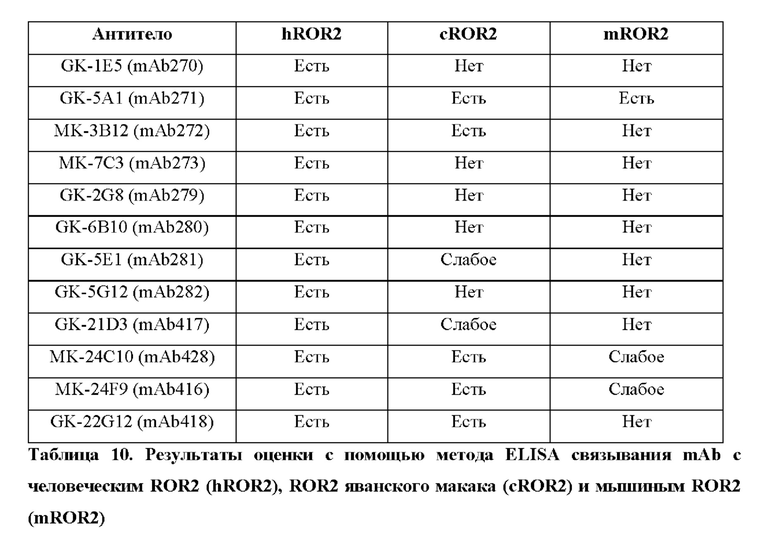
Пример 6. Конъюгирование mAb с модифицированными глицином токсинами с образованием ADC с применением технологии SMAC™
Сортаза А. Рекомбинантный и очищенный с помощью аффинной очистки фермент сортазу А из Staphylococcus aureus продуцировали в Е. coli, как раскрыто в международной заявке WO 2014140317 A1.
Создание модифицированных глицином токсинов. Модифицированное пентаглицином EDA-антрациклиновое производное G5-PNU и модифицированное триглицином EDA-антрациклиновое производное G3-PNU (фиг.6 (А) и (В), соответственно) производилось Concords, Сан-Диего, США. Идентичность и чистоту модифицированных глицином токсинов подтверждали с помощью масс-спектрометрии и ВЭЖХ. Каждый из модифицированных глицином токсинов демонстрировал чистоту >95%, определенную с помощью хроматографии методом ВЭЖХ.
Опосредуемая сортазой конъюгация антител. Вышеупомянутые токсины конъюгировали с антителами к ROR2 и сравнительными антителами, как указано в таблице 7, посредством инкубирования меченых LPETG mAb [5-10 мкМ] с модифицированным глицином токсином [100-200 мкМ] и 2,5-3 мкМ сортазой А в представленном буфере для конъюгирования в течение 3,5 часов при температуре 25°С. Реакцию останавливали посредством пропускания через колонку BioRad GraviTrap с белком А, как указано в таблице 11. Связавшийся конъюгат элюировали 5 объемами колонки буфера для элюирования (0,1 М глицин, рН 2,5, 50 нМ NaCl), причем фракции в количестве 1 объема колонки собирали в пробирки, содержащие 25% объем/объем 1 М Tris-основания для нейтрализации кислоты. Содержащие белок фракции объединяли и получали состав с ними в буфере для получения состава из таблицы 11 с использованием ZebaSpin колонки для обессоливания.
Анализы ADC. Значения DAR оценивали с помощью обращенно-фазовой хроматографии, осуществляемой на колонке Polymer Labs PLRP 2,1 мм × 5 см, 5 мкм со скоростью потока 1 мл/мин./80°С с 25-минутным линейным градиентом от 0,05 к 0,1% TFA/H2O и от 0,04 до 0,1% TFA/CH3CN. Образцы вначале восстанавливали посредством инкубирования с DTT при рН 8,0 и при температуре 37°С в течение 15 минут. Значения DAR, определенные с помощью обращенно-фазовой хроматографии, обобщены в таблице 7 ниже.

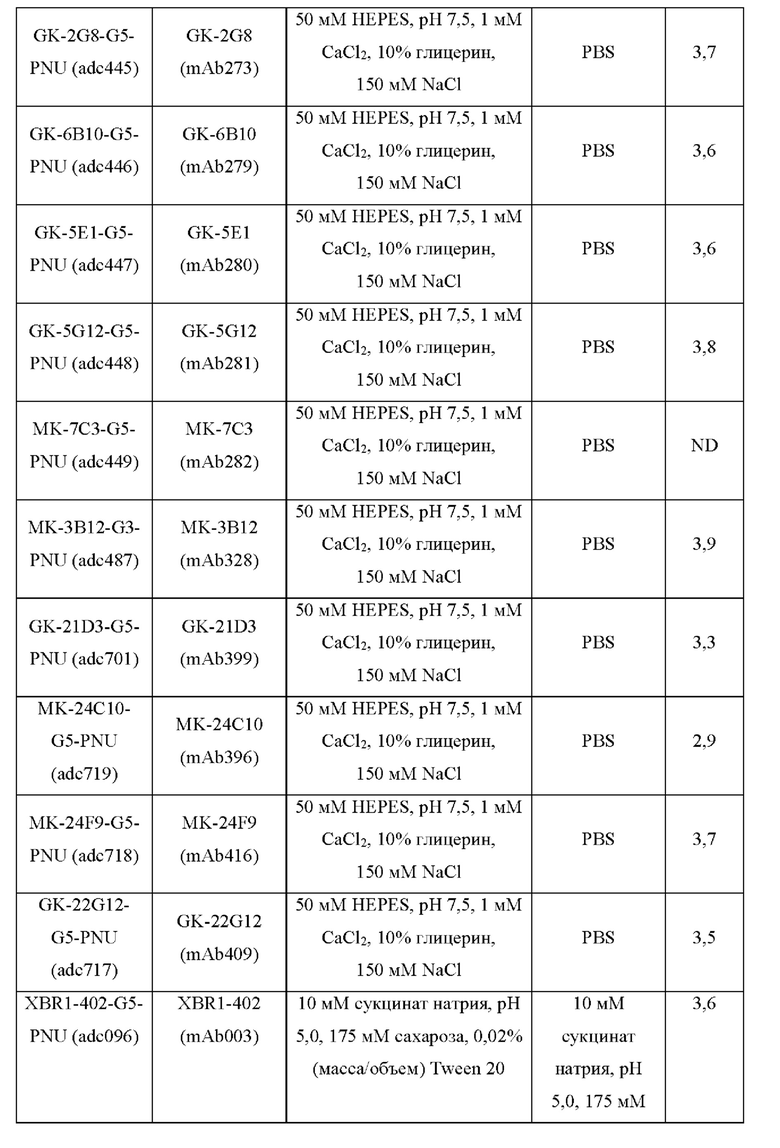

Из этих анализов можно сделать вывод, что конъюгирование по технологии SMAC™ протекало с высокой эффективностью, обеспечивая в результате общие средние значения DAR в диапазоне примерно от 3,5 до 4,0 для каждой из комбинаций антитело к hROR2 -токсин.
Пример 7. Воздействие на клетки ЕМТ-6 с помощью методов генной инженерии для устойчивой экспрессии hROR2
Клетки ЕМТ-6, культивируемые в полной среде DMEM (среда Игла в модификации Дульбекко (DMEM) с высоким содержанием глюкозы (4,5 г/л), с L-глутамином с 10% (объем/объем) фетальной телячьей сыворотки (FCS), 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином (все от Bioconcept, Альшвиль, Швейцария)) при температуре 37°С и 5% СО2, центрифугировали (6 минут, 1200 об./мин., 4°С) и суспендировали в среде RPMI-1640 (5×106 клеток/мл). 400 мкл этой клеточной суспензии затем добавляли к 400 мкл RPMI, содержащей 10,2 мкг транспозируемого вектора pPB-PGK-Puro-hROR2-Thr, управляющего коэкспрессией полноразмерного ROR2 и гена устойчивости к пуромицину, и 3,6 мкг содержащего транспозазу вектора pCDNA3.1_hy_mPB. Смесь клеток DNA/EMT-6 трансфицировали в кюветах для электропорации (просвет 0,4 см, 165-2088, BioRad, Кресье, Швейцария) и подвергали электропорации с применением электропоратора Biorad Gene Pulser II с модулем дополнительной емкости на 300 В и 950 мкФ. Затем клетки инкубировали в течение 5-10 минут при комнатной температуре. После инкубирования клетки центрифугировали при 1200 об./мин. в течение 6 минут, промывали один раз и затем ресуспендировали в полной среде DMEM перед инкубированием при температуре 37°С в увлажняемом инкубаторе в атмосфере с 5% СО2. Через сутки после электропорации пулы клеток, устойчиво экспрессирующих hROR2, подвергали отбору по устойчивости к пуромицину с применением 3 мкг/мл пуромицина (Sigma-Aldrich, Р8833-25 мг маточный раствор в концентрации 2 мг/мл).
Экспрессию ROR2 на отобранных клетках ЕМТ-6 ROR2 подтверждали с помощью проточной цитометрии. Кратко, после трипсинизации 10б клеток центрифугировали в пробирках для FACS; полученные осадки ресуспендировали в буфере (PBS с 2% (объем/объем) FCS). Клетки затем инкубировали с mAb Orb38364 к ROR2 (Biorbyt; 30 минут, 4°С, конечная концентрация 2 мкг/мл) с последующим центрифугированием и промыванием. Клетки затем ресуспендировали, как описано выше, и инкубировали с конъюгированным с РЕ мышиным антителом к кроличьему IgG (Abcam, ab99704) в разведении 1:250 в темноте (30 минут, 4°С) перед промыванием.
При использовании FACS Aria II клетки сортировали в виде отдельных клеток в 96-луночный плоскодонный планшет, содержащий 200 мкл полной среды DMEM на лунку. Этот планшет инкубировали при 37°С и разращивали в 6 лунках перед скринингом. Клетки анализировали с применением инструмента FACSCalibur (BD Biosciences) и аналитического программного обеспечения FlowJo (Tree Star, Ашленд, Орегон, США). На фиг. 7 представлены данные FACS анализа клона 14, экспрессирующего ROR2 на высоком уровне.
Пример 8. In vitro анализы цитотоксичности ADC на основе PNU в отношении экспрессирующих hROR2 раковых клеток ЕМТ-6
Цитотоксичность ADC к ROR2 из таблицы 12 изучали с применением подвергнутых воздействию методов генной инженерии клеток линии ЕМТ-6 из примера 7.
Для этого 1000 подвергнутых воздействию методов генной инженерии клеток ЕМТ6 (клон 14) на лунку высевали на 96-луночные планшеты (за исключением крайних лунок, которые содержали воду) в 75 мкл среды DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином, с плотностью 1,3 хЮ5 клеток на лунку и выращивали при температуре 37°С в увлажняемом инкубаторе в атмосфере 5% СО2. После 1 суток инкубирования каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в ростовой среде (начиная с концентрации ADC 80 мкг/мл, с получением конечных концентраций ADC в диапазоне от около 20 мкг/мл до 0,3 нг/мл). Каждое разведение выполняли в двух параллелях. После 4 дополнительных суток планшеты удаляли из инкубатора и уравновешивали при комнатной температуре. Спустя примерно 30 минут 50 мкл удаляли из каждой лунки, а затем 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243) добавляли в каждую лунку. После встряхивания планшетов при 750 об/мин в течение 5 минут с последующим инкубированием в течение 20 минут без встряхивания люминесценцию измеряли на планшет-ридере Spark 10 М с продолжительностью интегрирования, составляющей 1 секунду на лунку. Кривые зависимости люминесценции от концентрации ADC (нг/мл) аппроксимировали с использованием программного обеспечения Graphpad Prism. Измерение повторяли дважды. Значения IC50, определенные с использованием встроенной функции определения IC50 «log(inhibitor) vs. response -- Variable slope (four parameters))) в программном обеспечении Prism Software, представлены в таблице 12.

На фиг. 8 представлена кривая зависимости доза-эффект в in vitro анализах цитолиза на клетках ЕМТ-6 злокачественной опухоли молочной железы, устойчиво экспрессирующих hROR2, с использованием ADC на основе PNU из таблицы 12. Согласно таблице 12 и фиг. 8 ADC на основе новых антител к ROR2 согласно настоящему изобретению уничтожают экспрессирующие ROR2 клетки.
Пример 9. In vitro анализы цитотоксичности ADC на основе PNU в отношении экспрессирующих hROR2 клеток ЕМТ-6 злокачественной опухоли молочной железы и в отношении ROR2-отрицатель.ных раковых клеток
Цитотоксичность ADC к ROR2 из таблицы 13 изучали с применением подвергнутых воздействию методов генной инженерии клеток линии ЕМТ-6 из примера 7. Экспрессирующую ROR2 на низком уровне линию клеток человека Karpas-299 применяли в качестве контроля. ADC Ac10-G5-PNU включали в качестве изотипического контроля.
Для этого 1×103 подвергнутых воздействию методов генной инженерии клеток EMT6-ROR2 (клон 14) и 2,5×103 клеток Karpas-299 на лунку высевали на 96-луночные планшеты (за исключением крайних лунок, которые содержали воду) в 75 мкл среды DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином, с плотностью 1,3×105 клеток на лунку и выращивали при 37°С в увлажняемом инкубаторе в атмосфере 5% СО2. После 1 суток инкубирования каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в ростовой среде (начиная с концентрации ADC 80 мкг/мл, с получением конечных концентраций ADC в диапазоне от 20 мкг/мл до 0,89 нг/мл). Каждое разведение выполняли в двух параллелях. После 4 дополнительных суток планшеты удаляли из инкубатора и уравновешивали при комнатной температуре. Спустя примерно 30 минут 50 мкл удаляли из каждой лунки, а затем 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243) добавляли в каждую лунку. После встряхивания планшетов при 750 об./мин. в течение 5 минут с последующим инкубированием в течение 20 минут без встряхивания люминесценцию измеряли на планшет-ридере Spark 10 М с продолжительностью интегрирования, составляющей 1 секунду на лунку. Кривые зависимости люминесценции от концентрации ADC (нг/мл) аппроксимировали с использованием программного обеспечения Graphpad Prism. Значения IC50, определенные с использованием встроенной функции определения IC50 «log(inhibitor) vs. response -- Variable slope (four parameters))) в программном обеспечении Prism Software, представлены в таблице 13.
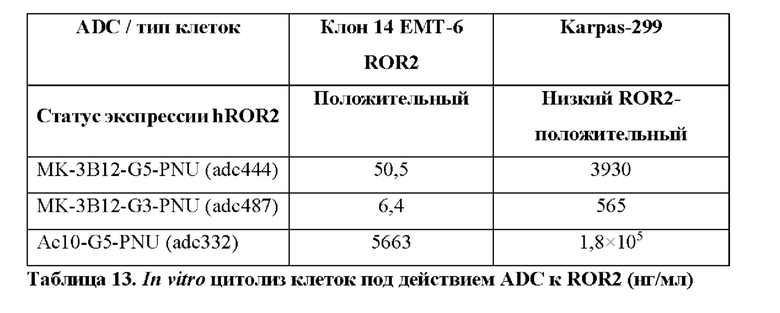
Пример 10. In vitro анализы цитотоксичности ADC на основе PNU в отношении экспрессирующих hRQR2 раковых клеток ЕМТ-6
Цитотоксичность ADC к ROR2 из таблицы 14 изучали с применением подвергнутых воздействию методов генной инженерии клеток линии ЕМТ-6 из примера 7.
Для этого 1×103 подвергнутых воздействию методов генной инженерии клеток EMT6-ROR2 (клон 14) на лунку высевали на 96-луночные планшеты (за исключением крайних лунок, которые содержали воду) в 75 мкл среды DMEM, дополненной 10% по объему FCS, 100 МЕ/мл пенициллина-стрептомицина-фунгизона и 2 мМ L-глутамином, с плотностью 1,3×105 клеток на лунку и выращивали при температуре 37°С в увлажняемом инкубаторе в атмосфере 5% СО2. После 1 суток инкубирования каждый ADC добавляли в соответствующие лунки в количестве 25 мкл 3,5-кратных серийных разведений в ростовой среде (начиная с концентрации ADC 80 мкг/мл, с получением конечных концентраций ADC в диапазоне от 20 мкг/мл до 0,89 нг/мл). Каждое разведение выполняли в двух параллелях. После 4 дополнительных суток планшеты удаляли из инкубатора и уравновешивали при комнатной температуре. Спустя примерно 30 минут 50 мкл люминесцентного раствора CellTiter-Glo® 2.0 (Promega, G9243) добавляли в каждую лунку. После встряхивания планшетов при 750 об/мин. В течение 5 минут с последующим инкубированием в течение 20 минут без встряхивания люминесценцию измеряли на планшет-ридере Spark 10 М с продолжительностью интегрирования, составляющей 1 секунду на лунку. Кривые зависимости люминесценции от концентрации ADC (нг/мл) аппроксимировали с использованием программного обеспечения Graphpad Prism. Значения IC50, определенные с использованием встроенной функции определения IC50 «log(inhibitor) vs. response -- Variable slope (four parameters))) в программном обеспечении Prism Software, представлены в таблице 14.

Несмотря на то что вышеизложенное изобретение было описано довольно подробно с целью иллюстрации и подтверждения примерами с целью ясности понимания, квалифицированному специалисту в данной области техники с учетом идей настоящего изобретения будет очевидно, что в нем можно производить определенные изменения и модификации без отступления от идеи или объема прилагаемой формулы изобретения.
Все публикации, базы данных, последовательности в GenBank, патенты и патентные заявки, цитируемые в данном описании, включены в данный документ посредством ссылки, как если бы было указано, что каждый из них был специально и отдельно включен посредством ссылки.
ПОСЛЕДОВАТЕЛЬНОСТИ
С этой заявкой также представлен перечень последовательностей в электронной форме согласно ST 25 WIPO. Во избежание сомнений, если существуют несоответствия между последовательностями в следующей таблице и перечнем последовательностей в электронной форме, последовательности в данной таблице будут считаться правильными.










СПИСОК ИСПОЛЬЗОВАННЫХ ИСТОЧНИКОВ
Непатентная литература
Abbott М. et al., "Current approaches to fine mapping of antigen-antibody interactions";
Immunology, 2014; 142(4); 526-35.
Altschul et al., J. Mol. Biol. 215:403-410, 1990.
Altschul et al., Nuc. Acids Res. 25:3389-3402, 1977.
Balakrishnan et al. (2016) Clin Cancer Res. doi: 10.1158/1078-0432.
Beaucage et al., Tetra. Lett, 22:1859, 1981.
Beerli et al. (2015) PloS One 10, e131177.
Bendas, BioDrugs, 15: 215-224, 200.1
Beraud et al., Inflamm. Allergy Drag Targets. 10:322-42, 2011.
Berry et al., 2003 Hybridoma and Hybridomics 329(1-2): 112-124.
Bird et al., Science 242:423-426, 1988.
Bittner et al., Meth. Enzymol., 153:516, 1987.
Bond et al., J. Mol. Biol. 332:643-55, 2003.
Brent et al., Current Protocols in Molecular Biology, John Wiley & Sons, Inc. (Ringbou ed., 2003).
Brown et al., Meth. Enzymol. 68:109, 1979.
Cai and Garen, Proc. Natl. Acad. Sci. USA 93:6280-85, 1996.
Chen et al., "A general strategy for the evolution of bond-forming enzymes using yeast display".
PNAS 2011; 108(28); 11399-11404.
Dorr BM et al., "Reprogramming the specificity of sortase enzymes"; PNAS 2014; 111, 13343-8.
Dumoulin et al., Nat. Struct. Biol. 11:500-515, 2002.
Dyba et al., Curr. Pharm. Des. 10:2311-34, 2004.
Elliot and O'Hare, Cell 88:223, 1997.
Ghahroudi et al., FEBS Letters 414:521 526, 1997.
Harlow & Lane, Using Antibodies, A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1998.
Lefranc MP et al. "IMGT®, the International ImMunoGeneTics information system® 25 years on"; Nucleic Acids Res 2015; 43; D413-22.
Harrington et al., Nat. Genet. 15:345, 1997.
Huston et al., Proc. Natl. Acad. Sci. USA 85:5879-5883, 1988.
Mattila et al., Nucleic Acids Res. 19:967, 1991; и Eckert et al., PCR Methods and Applications 1:17, 1991.
Kuyucak et al., Future Med. Chem. 6:1645-58, 2014.
Marcu-Malina et al., Expert Opinion on Biological Therapy, Vol. 9, No. 5.
Middlebrook et al., Microbiol. Rev. 48:199-221, 1984.
Morioka et al., Cancer Sci. 100: 1227-1233, 2009.
Narang et al., Meth. Enzymol. 68:90, 1979.
Needleman and Wunsch, J. Mol. Biol. 48:443, 1970.
Pearson and Lipman, Proc. Nat'l. Acad. Sci. USA 85:2444, 1988.
Quintieri et al., Clin. Cancer Res 11:1608-1617 (2005).
Rebagay et al. (2012) Front Oncol. 2(34).
Reiter et al., Int. J. Cancer 67:113-23, 1996.
Remington: The Science and Practice of Pharmacy, Mack Publishing Co., 20th ed., 2000; and Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
Remington's Pharmaceutical Sciences, Mack Publishing Co., Easton, PA.
Rosenfeld et al., Cell 68:143, 1992.
Russell et al, J. Mol. Biol., 244: 332-350 (1994).
Sblattero and Bradbury 1998 hnmunotechnology3, 271-278.
Scharf et al., Results Probl. Cell Differ. 20:125, 1994.
Skerra and Pluckthun, Science 240:1038-41, 1988.
Smith and Waterman, Adv. Appl. Math. 2:482c, 1970.
Smith, Annu. Rev. Microbiol. 49:807, 1995.
Tiller et al., J Immunol Methods. 2008 May 20; 334(1-2): 142.
Waldmeier L et al. "Transpo-mAb display: Transposition-mediated В cell display and functional screening of full-length IgG antibody libraries"; MAbs 2016; 8(4), 726-40.
Ward et al., Nature 341:544-546, 1989.
Ward et al., Nature 341:544-546, 1989.
Winnacker, From Genes to Clones, VCH Publishers, N.Y., N.Y., 1987.
Wu et al., Nat. Biotechnol, 23: 1 137-1 146 (2005).
Патентная литература
Патент США №5075109.
Патент США №4452775.
Патент США №4667014.
Патент США №4748034.
Патент США №5239660.
Патент США №3832253.
Патент США №3854480.
Патент США №4458066.
Патент США №8916159.
Международная заявка WO 2010/070263 А1.
Международная заявка WO 2014/013026 А1.
Международная заявка WO 2014/140317 А1.
Международная заявка WO 2016/102679 А1.
Международная заявка WO 2013/103637 А1.
Международная заявка WO 2016/142768 А1.
Международная заявка WO 2014/140317 А1.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> НБЕ-ТЕРАПОЙТИКС АГ
<120> Человеческие антитела, связывающиеся с ROR2
<130> ND 40646
<160> 142
<170> BiSSAP 1.3.6
<210> 1
<211> 370
<212> Белок
<213> Искусственная последовательность
<220>
<223> внеклеточный домен человеческого ROR2
<400> 1
Glu Val Glu Val Leu Asp Pro Asn Asp Pro Leu Gly Pro Leu Asp Gly
1 5 10 15
Gln Asp Gly Pro Ile Pro Thr Leu Lys Gly Tyr Phe Leu Asn Phe Leu
20 25 30
Glu Pro Val Asn Asn Ile Thr Ile Val Gln Gly Gln Thr Ala Ile Leu
35 40 45
His Cys Lys Val Ala Gly Asn Pro Pro Pro Asn Val Arg Trp Leu Lys
50 55 60
Asn Asp Ala Pro Val Val Gln Glu Pro Arg Arg Ile Ile Ile Arg Lys
65 70 75 80
Thr Glu Tyr Gly Ser Arg Leu Arg Ile Gln Asp Leu Asp Thr Thr Asp
85 90 95
Thr Gly Tyr Tyr Gln Cys Val Ala Thr Asn Gly Met Lys Thr Ile Thr
100 105 110
Ala Thr Gly Val Leu Phe Val Arg Leu Gly Pro Thr His Ser Pro Asn
115 120 125
His Asn Phe Gln Asp Asp Tyr His Glu Asp Gly Phe Cys Gln Pro Tyr
130 135 140
Arg Gly Ile Ala Cys Ala Arg Phe Ile Gly Asn Arg Thr Ile Tyr Val
145 150 155 160
Asp Ser Leu Gln Met Gln Gly Glu Ile Glu Asn Arg Ile Thr Ala Ala
165 170 175
Phe Thr Met Ile Gly Thr Ser Thr His Leu Ser Asp Gln Cys Ser Gln
180 185 190
Phe Ala Ile Pro Ser Phe Cys His Phe Val Phe Pro Leu Cys Asp Ala
195 200 205
Arg Ser Arg Thr Pro Lys Pro Arg Glu Leu Cys Arg Asp Glu Cys Glu
210 215 220
Val Leu Glu Ser Asp Leu Cys Arg Gln Glu Tyr Thr Ile Ala Arg Ser
225 230 235 240
Asn Pro Leu Ile Leu Met Arg Leu Gln Leu Pro Lys Cys Glu Ala Leu
245 250 255
Pro Met Pro Glu Ser Pro Asp Ala Ala Asn Cys Met Arg Ile Gly Ile
260 265 270
Pro Ala Glu Arg Leu Gly Arg Tyr His Gln Cys Tyr Asn Gly Ser Gly
275 280 285
Met Asp Tyr Arg Gly Thr Ala Ser Thr Thr Lys Ser Gly His Gln Cys
290 295 300
Gln Pro Trp Ala Leu Gln His Pro His Ser His His Leu Ser Ser Thr
305 310 315 320
Asp Phe Pro Glu Leu Gly Gly Gly His Ala Tyr Cys Arg Asn Pro Gly
325 330 335
Gly Gln Met Glu Gly Pro Trp Cys Phe Thr Gln Asn Lys Asn Val Arg
340 345 350
Met Glu Leu Cys Asp Val Pro Ser Cys Ser Pro Arg Asp Ser Ser Lys
355 360 365
Met Gly
370
<210> 2
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 HC
<400> 2
Glu Val Gln Leu Val Glu Ser Gly Pro Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Ser Gly Tyr Ser Ile Ser Ser Gly
20 25 30
Tyr Tyr Trp Gly Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp
35 40 45
Ile Gly Ser Ile Tyr Gln Ser Gly Ser Thr His Tyr Asn Pro Ser Leu
50 55 60
Lys Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser
65 70 75 80
Leu Lys Leu Thr Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Asp Arg Ala Gly Trp Tyr Pro Phe Asp Cys Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Arg
450
<210> 3
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 LC
<400> 3
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 4
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 HC
<400> 4
Glu Val Gln Leu Leu Glu Thr Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Val Ala Ser Gly Phe Thr Phe Arg Ser His
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Gly Ala Gly Leu Tyr Leu Asp Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 5
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 LC
<400> 5
Ala Ile Arg Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Thr Ile Ser Asn Trp
20 25 30
Leu Ala Trp Phe Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Ser Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Arg Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 6
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1E5 HC
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Thr Asp Ser Val
50 55 60
Gln Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Ile Ala Met Thr Gly Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 7
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1E5 LC
<400> 7
Asp Ile Gln Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 8
<211> 453
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 HC
<400> 8
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met Tyr Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Asn Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Glu Gly Ser Gly Trp Tyr Asp Tyr Tyr Tyr Gly Met Asp Val
100 105 110
Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala Ser Thr Lys Gly
115 120 125
Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly
130 135 140
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
145 150 155 160
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
165 170 175
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
180 185 190
Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val
195 200 205
Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys
210 215 220
Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu
225 230 235 240
Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr
245 250 255
Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
260 265 270
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val
275 280 285
Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
290 295 300
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
305 310 315 320
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
325 330 335
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
340 345 350
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
355 360 365
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
370 375 380
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
385 390 395 400
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
405 410 415
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
420 425 430
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
435 440 445
Leu Ser Pro Gly Lys
450
<210> 9
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 LC
<400> 9
Glu Ile Val Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 10
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-2G8 HC
<400> 10
Gln Val Gln Leu Gln Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Val Ala Met Thr Gly Leu Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 11
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-2G8 LC
<400> 11
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 12
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5E1 HC
<400> 12
Gln Val Thr Leu Lys Glu Ser Gly Gly Asp Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Pro Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Val Arg Val Arg Phe Gly Glu Leu Tyr Phe Gln His Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 13
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5E1 LC
<400> 13
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr
85 90 95
Ser Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 14
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-6B10 HC
<400> 14
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Arg Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Val Ala Ala Ala Leu His Phe His Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro
115 120 125
Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly
130 135 140
Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn
145 150 155 160
Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln
165 170 175
Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser
180 185 190
Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser
195 200 205
Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr
210 215 220
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
225 230 235 240
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
245 250 255
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
260 265 270
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
275 280 285
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
290 295 300
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
305 310 315 320
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
325 330 335
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
340 345 350
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
355 360 365
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
370 375 380
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
385 390 395 400
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser
405 410 415
Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
420 425 430
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 15
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-6B10 LC
<400> 15
Asp Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Asp Asn Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Val Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 16
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5G12 HC
<400> 16
Gln Ile Thr Leu Lys Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ile Arg Val Lys Phe Gly Asp Leu Tyr Phe Gln His Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 17
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5G12 LC
<400> 17
Glu Ile Val Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ala Thr Tyr Tyr Cys Gln Lys Tyr Asn Ser Ala Pro Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 18
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21D3 HC
<400> 18
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Gly Ala Ile Asn Arg Gly Gly Gly Gly Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 19
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21D3 LC
<400> 19
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Ile Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys Tyr Asn Ser Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 20
<211> 457
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 HC
<400> 20
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Thr Asn Lys His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Glu Trp Phe Gly Glu Leu Arg Tyr Tyr Tyr Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala
115 120 125
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
130 135 140
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
145 150 155 160
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
165 170 175
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
210 215 220
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 21
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 LC
<400> 21
Glu Ile Val Leu Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Thr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 22
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 HC
<400> 22
Glu Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Asp Ile Asn His Ser Arg Thr Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Val Thr Ile Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Glu Gln Trp Leu Val Pro Phe Asp Tyr Trp Asp Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 23
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 LC
<400> 23
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser His Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Lys Ser Gly Val Pro Ser Arg Phe Asn Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Asn Thr Tyr Ser Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 24
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 HC
<400> 24
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Gln
20 25 30
Arg Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Asn Tyr Val Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Ile Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Arg Asn Gly Trp His Ile Pro Glu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 25
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 LC
<400> 25
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys His Asn Arg Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 26
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 HC CDR1
<400> 26
Gly Tyr Ser Ile Ser Ser Gly Tyr Tyr
1 5
<210> 27
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 HC CDR2
<400> 27
Ile Tyr Gln Ser Gly Ser Thr
1 5
<210> 28
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 HC CDR3
<400> 28
Cys Ala Arg Glu Asp Arg Ala Gly Trp Tyr Pro Phe Asp Cys Trp
1 5 10 15
<210> 29
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 LC CDR1; GK-1E5 LC CDR1; GK-5A1 LC CDR1; GK-2G8 LC CDR1;
GK-5E1 LC CDR1; GK-1H2 LC CDR1; GK-2A9 LC CDR1; GK-5A6 LC CDR1
<400> 29
Gln Ser Ile Ser Ser Trp
1 5
<210> 30
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 LC CDR3; GK-1E5 LC CDR3; GK-2G8 LC CDR3; GK-1H2 LC CDR3;
GK-2A9 LC CDR3
<400> 30
Cys Gln Gln Tyr Asn Asn Tyr Trp Thr Phe
1 5 10
<210> 31
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 HC CDR1
<400> 31
Gly Phe Thr Phe Arg Ser His Gly
1 5
<210> 32
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 HC CDR2; GK-1E5 HC CDR2; GK-2G8 HC CDR2; GK-5E1 HC CDR2;
GK-1H2 HC CDR2; GK-2A9 HC CDR2
<400> 32
Ile Trp Tyr Asp Gly Ser Lys Lys
1 5
<210> 33
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 HC CDR3
<400> 33
Cys Ala Arg Val Gly Ala Gly Leu Tyr Leu Asp Tyr Trp
1 5 10
<210> 34
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 LC CDR1
<400> 34
Gln Thr Ile Ser Asn Trp
1 5
<210> 35
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-7C3 LC CDR3; GK-6B10 LC CDR3; GK-5A6 LC CDR3
<400> 35
Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr Thr Phe
1 5 10
<210> 36
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1E5 HC CDR1; GK-2G8 HC CDR1; GK-1H2 HC CDR1; GK-2A9 HC CDR1
<400> 36
Gly Phe Thr Phe Arg Ser Tyr Gly
1 5
<210> 37
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1E5 HC CDR3; GK-1H2 HC CDR3
<400> 37
Cys Ala Arg Pro Gly Ile Ala Met Thr Gly Leu Asp Tyr Trp
1 5 10
<210> 38
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 HC CDR1; GK-21D3 HC CDR1; MK-24C10 HC CDR1; GK-21F1 HC
CDR1; MK-24C12 HC CDR1
<400> 38
Gly Phe Thr Phe Ser Ser Tyr Gly
1 5
<210> 39
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 HC CDR2
<400> 39
Ile Trp Asn Asp Gly Ser Asn Lys
1 5
<210> 40
<211> 18
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 HC CDR3
<400> 40
Cys Ala Arg Glu Gly Ser Gly Trp Tyr Asp Tyr Tyr Tyr Gly Met Asp
1 5 10 15
Val Trp
<210> 41
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A1 LC CDR3
<400> 41
Cys Gln Gln Tyr Asn Ser Tyr Trp Thr Phe
1 5 10
<210> 42
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5E1 LC CDR3
<400> 42
Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr Ser Phe
1 5 10
<210> 43
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-2G8 HC CDR3; GK-5E1 HC CDR1; GK-5G12 HC CDR1; GK-2A9 HC CDR3;
GK-5A6 HC CDR1
<400> 43
Cys Ala Arg Pro Gly Val Ala Met Thr Gly Leu Asp Leu Trp
1 5 10
<210> 44
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5E1 HC CDR3
<400> 44
Cys Val Arg Val Arg Phe Gly Glu Leu Tyr Phe Gln His Trp
1 5 10
<210> 45
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-6B10 HC CDR1
<400> 45
Gly Phe Thr Phe Ser Arg Tyr Gly
1 5
<210> 46
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-6B10 HC CDR2; GK-5G12 HC CDR2; GK-21D3 HC CDR2; GK-5A6 HC
CDR2; GK-21F1 HC CDR2
<400> 46
Ile Trp Tyr Asp Gly Ser Asn Lys
1 5
<210> 47
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-6B10 HC CDR3
<400> 47
Cys Ala Arg Val Ala Ala Ala Leu His Phe His Tyr Trp
1 5 10
<210> 48
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> >GK-6B10 LC CDR1
<400> 48
Gln Ser Ile Asp Asn Trp
1 5
<210> 49
<211> 14
<212> Белок
<213> Искусственная последовательность
<220>
<223> >GK-5G12 HC CDR3; GK-5A6 HC CDR3
<400> 49
Cys Ile Arg Val Lys Phe Gly Asp Leu Tyr Phe Gln His Trp
1 5 10
<210> 50
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5G12 LC CDR1; GK-21D3 LC CDR1; GK-22G12 LC CDR1; GK-21F1 LC
CDR1; GK-21G5 LC CDR1; GK-21E6 LC CDR1; GK-22E12 LC CDR1
<400> 50
Gln Gly Ile Ser Asn Tyr
1 5
<210> 51
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5G12 LC CDR3
<400> 51
Cys Gln Lys Tyr Asn Ser Ala Pro Tyr Thr Phe
1 5 10
<210> 52
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21D3 HC CDR3; GK-21F1 HC CDR3
<400> 52
Cys Ala Arg Met Gly Ala Ile Asn Arg Gly Gly Gly Gly Phe Asp Tyr
1 5 10 15
Trp
<210> 53
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21D3 LC CDR3; GK-21F1 LC CDR3
<400> 53
Cys Gln Lys Tyr Asn Ser Ala Pro Trp Thr Phe
1 5 10
<210> 54
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 HC CDR2; MK-24C12 HC CDR2
<400> 54
Ile Trp Phe Asp Gly Thr Asn Lys
1 5
<210> 55
<211> 22
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 HC CDR3; MK-24C12 HC CDR3
<400> 55
Cys Ala Arg Asp Lys Gly Glu Trp Phe Gly Glu Leu Arg Tyr Tyr Tyr
1 5 10 15
Tyr Gly Met Asp Val Trp
20
<210> 56
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 LC CDR1; MK-24C12 LC CDR1
<400> 56
Gln Ser Leu Leu His Ser Asn Gly Tyr Asn Tyr
1 5 10
<210> 57
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 LC CDR3; MK-24C12 LC CDR3
<400> 57
Cys Met Gln Ala Leu Gln Thr Pro Tyr Thr Phe
1 5 10
<210> 58
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 HC CDR1; GK-21G5 HC CDR1; GK-23A8 HC CDR1
<400> 58
Gly Gly Ser Phe Ser Gly Tyr Tyr
1 5
<210> 59
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> >MK-24F9 HC CDR2
<400> 59
Ile Asn His Ser Arg Thr Thr
1 5
<210> 60
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 HC CDR3
<400> 60
Cys Ala Arg Gly Gly Glu Gln Trp Leu Val Pro Phe Asp Tyr Trp
1 5 10 15
<210> 61
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 LC CDR1; GK-23A8 LC CDR1
<400> 61
Gln Ser Ile Ser His Trp
1 5
<210> 62
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24F9 LC CDR3; GK-23A8 LC CDR3
<400> 62
Cys Gln His Tyr Asn Thr Tyr Ser Arg Thr Phe
1 5 10
<210> 63
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 HC CDR1; GK-21E6 HC CDR1; GK-22E12 HC CDR1
<400> 63
Gly Phe Thr Phe Ser Ser Gln Arg
1 5
<210> 64
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 HC CDR2; GK-21E6 HC CDR2; GK-22E12 HC CDR2
<400> 64
Ile Lys Gln Asp Gly Ser Glu Lys
1 5
<210> 65
<211> 17
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 HC CDR3; GK-21E6 HC CDR3; GK-22E12 HC CDR3
<400> 65
Cys Ala Arg Asp Gly Tyr Arg Asn Gly Trp His Ile Pro Glu Asp Tyr
1 5 10 15
Trp
<210> 66
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22G12 LC CDR3; GK-21G5 LC CDR3; GK-21E6 LC CDR3; GK-22E12 LC
CDR3
<400> 66
Cys Gln Lys His Asn Arg Ala Pro Trp Thr Phe
1 5 10
<210> 67
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1; X1=F или G, X2=T или S, X3=R или S, X4=S, T, R или G, X5=H, Y
или Q, X6=G, Y или R
<400> 67
Gly Xaa Xaa Phe Xaa Xaa Xaa Xaa
1 5
<210> 68
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR2; X7=W или K, X8=Y, N, F или Q, X9=S или T, X10=K, N или E
<400> 68
Ile Xaa Xaa Asp Gly Xaa Xaa Lys
1 5
<210> 69
<211> 6
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1; X11=S, T или G, X12=S, N или H and X13=W или Y
<400> 69
Gln Xaa Ile Ser Xaa Xaa
1 5
<210> 70
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR3; X14=Q или M, X15=K, H или Q, X16=H, Y или A, X17=N или L,
X18=R, T, Q, S или N; X19=A, Y или T; X20=P, S или W, X21=W, R, Y или
отсутствует, X22=S или T.
<400> 70
Cys Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Xaa Phe
1 5 10
<210> 71
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> CDR1; X3=R или S, и X4=S, T
<400> 71
Gly Phe Thr Phe Xaa Xaa Tyr Gly
1 5
<210> 72
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5G12 LC CDR2; GK-21D3 LC CDR2; GK-22G12 LC CDR2; GK-21F1 LC
CDR2; GK-21G5 LC CDR2; GK-21E6 LC CDR2; GK-22E12 LC CDR2
(последовательность имеет только 3 AA (X отсуствует)
<400> 72
Ala Ala Ser Xaa
1
<210> 73
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-3B12 LC CDR2; MK-7C3 LC CDR2; GK-1E5 LC CDR2; GK-5A1 LC CDR2;
GK-2G8 LC CDR2; GK-5E1 LC CDR2; GK-6B10 LC CDR2; MK-24F9 LC CDR2;
GK-1H2 LC CDR2; GK-2A9 LC CDR2; GK-5A6 LC CDR2; GK-23A8 LC CDR2
(последовательность имеет только 3 AA (X отсуствует)
<400> 73
Lys Ala Ser Xaa
1
<210> 74
<211> 4
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C10 LC CDR2, MK-24C12 LC CDR2 (последовательность имеет только
3 AA (X отсуствует)
<400> 74
Leu Gly Ser Xaa
1
<210> 75
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1H2 HC
<400> 75
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Ile Ala Met Thr Gly Leu Asp Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 76
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-1H2 LC
<400> 76
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 77
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-2A9 HC
<400> 77
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Lys Lys Tyr Tyr Thr Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Pro Gly Val Ala Met Thr Gly Leu Asp Leu Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 78
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-2A9 LC
<400> 78
Glu Ile Val Leu Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Asn Tyr Trp Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 79
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A6 HC
<400> 79
Glu Val Gln Leu Gln Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Thr Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Leu Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ile Arg Val Lys Phe Gly Asp Leu Tyr Phe Gln His Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 80
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5A6 LC
<400> 80
Asp Val Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Glu Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Tyr Asn Ser Tyr Ser Tyr
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 81
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21F1 HC
<400> 81
Gln Val Gln Leu Val Gln Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Ile Ile Trp Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Asp Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Met Gly Ala Ile Asn Arg Gly Gly Gly Gly Phe Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 82
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21F1 LC
<400> 82
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Ile Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys Tyr Asn Ser Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 83
<211> 457
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C12 HC
<400> 83
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Phe Asp Gly Thr Asn Lys His Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Lys Gly Glu Trp Phe Gly Glu Leu Arg Tyr Tyr Tyr Tyr
100 105 110
Gly Met Asp Val Trp Gly Gln Gly Thr Thr Val Thr Val Ser Ser Ala
115 120 125
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
130 135 140
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
145 150 155 160
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
165 170 175
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
180 185 190
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
195 200 205
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
210 215 220
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
225 230 235 240
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
245 250 255
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
260 265 270
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
275 280 285
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
290 295 300
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
305 310 315 320
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
325 330 335
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
340 345 350
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
355 360 365
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
370 375 380
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
385 390 395 400
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
405 410 415
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
420 425 430
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
435 440 445
Lys Ser Leu Ser Leu Ser Pro Gly Lys
450 455
<210> 84
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> MK-24C12 LC
<400> 84
Glu Ile Val Leu Thr Gln Ser Pro Leu Ser Leu Pro Val Thr Pro Gly
1 5 10 15
Glu Pro Ala Ser Ile Ser Cys Arg Ser Ser Gln Ser Leu Leu His Ser
20 25 30
Asn Gly Tyr Asn Tyr Leu Asp Trp Tyr Leu Gln Lys Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Leu Gly Ser Asn Arg Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Lys Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln Ala
85 90 95
Leu Gln Thr Pro Tyr Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 85
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21G5 HC
<400> 85
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Leu Thr Val Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Asp Gln Trp Leu Val Pro Phe Asp Asn Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 86
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21G5 LC
<400> 86
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys His Asn Arg Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 87
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-23A8 HC
<400> 87
Gln Val Gln Leu Gln Gln Trp Gly Ala Gly Leu Leu Lys Pro Ser Glu
1 5 10 15
Thr Leu Ser Leu Thr Cys Ala Val Tyr Gly Gly Ser Phe Ser Gly Tyr
20 25 30
Tyr Trp Ser Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Ile
35 40 45
Gly Glu Ile Asn His Ser Gly Ile Thr Asn Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Leu Thr Val Ser Val Asp Thr Ser Lys Asn Gln Phe Ser Leu
65 70 75 80
Lys Leu Ser Ser Val Thr Ala Ala Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Gly Asp Gln Trp Leu Val Pro Phe Asp Asn Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
Lys
<210> 88
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-23A8 LC
<400> 88
Glu Ile Val Met Thr Gln Ser Pro Ser Thr Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser His Trp
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Lys Ala Ser Ser Leu Lys Ser Gly Val Pro Ser Arg Phe Asn Gly
50 55 60
Ser Gly Ser Gly Thr Glu Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Asp Asp Phe Ala Thr Tyr Tyr Cys Gln His Tyr Asn Thr Tyr Ser Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Asp Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 89
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21E6 HC
<400> 89
Arg Val Gln Leu Val Gln Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Gln
20 25 30
Arg Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Asn Tyr Val Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Ile Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Arg Asn Gly Trp His Ile Pro Glu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 90
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21E6 LC
<400> 90
Asp Val Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys His Asn Arg Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 91
<211> 452
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22E12 HC
<400> 91
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Ser Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Gln
20 25 30
Arg Leu Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Asn Tyr Val Asp Ser Val
50 55 60
Arg Gly Arg Phe Thr Ile Ser Arg Asp Ile Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Asp Gly Tyr Arg Asn Gly Trp His Ile Pro Glu Asp Tyr Trp
100 105 110
Gly Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro
115 120 125
Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr
130 135 140
Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr
145 150 155 160
Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro
165 170 175
Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr
180 185 190
Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn
195 200 205
His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser
210 215 220
Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
225 230 235 240
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
245 250 255
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
260 265 270
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
275 280 285
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
290 295 300
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
305 310 315 320
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
325 330 335
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
340 345 350
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
355 360 365
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
370 375 380
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
385 390 395 400
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
405 410 415
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
420 425 430
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
435 440 445
Ser Pro Gly Lys
450
<210> 92
<211> 214
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-22E12 LC
<400> 92
Ala Ile Arg Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Ser Asn Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Lys Val Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Val Ser Thr Tyr Tyr Cys Gln Lys His Asn Arg Ala Pro Trp
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 93
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21G5 HC CDR2, GK-23A8 HC CDR2
<400> 93
Ile Asn His Ser Gly Ile Thr
1 5
<210> 94
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-5E1 HC CDR1; GK-5G12 HC CDR1; GK-5A6 HC CDR1
<400> 94
Gly Phe Thr Phe Arg Thr Tyr Gly
1 5
<210> 95
<211> 15
<212> Белок
<213> Искусственная последовательность
<220>
<223> GK-21G5 HC CDR3, GK-23A8 HC CDR3
<400> 95
Cys Ala Arg Gly Gly Asp Gln Trp Leu Val Pro Phe Asp Asn Trp
1 5 10 15
<210> 96
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Последовательность для распознавания сортазой A Staphylococcus aureus,
в которой X представляет собой любую аминокислоту
<400> 96
Leu Pro Xaa Ser Gly
1 5
<210> 97
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сортазой A Staphylococcus aureus,
в которой X представляет собой любую аминокислоту
<400> 97
Leu Pro Xaa Ala Gly
1 5
<210> 98
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сортазой A Staphylococcus aureus
или сконструированной сортазой A 4S-9 из Staphylococcus aureus, в которой X
представляет собой любую аминокислоту
<400> 98
Leu Pro Xaa Thr Gly
1 5
<210> 99
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сконструированной сортазой A 2A-9
из Staphylococcus aureus, в которой X представляет собой любую аминокислоту
<400> 99
Leu Ala Xaa Thr Gly
1 5
<210> 100
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сортазой A Staphylococcus aureus
или сконструированной сортазой A 4S-9 из Staphylococcus aureus
<400> 100
Leu Ala Glu Thr Gly
1 5
<210> 101
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сортазой A Streptococcus pyogenes,
в которой X представляет собой любую аминокислоту
<400> 101
Leu Pro Xaa Thr Ala
1 5
<210> 102
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> последовательность для распознавания сортазой Staphylococcus aureus
<400> 102
Asn Pro Gln Thr Asn
1 5
<210> 103
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> >huVH4B-3'Leader-FR1
<400> 103
caggtgcagc tgcaggagtc sg 22
<210> 104
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH5B-3'-лидерная последовательность-FR1
<400> 104
caggtacagc tgcagcagtc a 21
<210> 105
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH6B-3'-лидерная последовательность-FR1
<400> 105
caggtgcagc tacagcagtg gg 22
<210> 106
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH10Ba-3'-лидерная последовательность-FR1
<400> 106
gaggtgcagc tgktggagwc t 21
<210> 107
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH10Bb-3'-лидерная последовательность-FR1
<400> 107
gaggtgcagc tgktggagwc c 21
<210> 108
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH12B-3'-лидерная последовательность-FR1
<400> 108
caggtccagc tkgtrcagtc tgg 23
<210> 109
<211> 22
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH14B-3'-лидерная последовательность-FR1
<400> 109
cagrtcacct tgaaggagtc tg 22
<210> 110
<211> 23
<212> DNA
<213> Искусственная последовательность
<220>
<223> huVH22B-3'-лидерная последовательность-FR1
<400> 110
caggtgcagc tggtgsartc tgg 23
<210> 111
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> rat_IgG12abc_R
<400> 111
cagggtgact gagggcgtag 20
<210> 112
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> rat_IgM_R
<400> 112
gttgggaagg ttctgacacc 20
<210> 113
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK1-5_3'-лидерная последовательность-FR1
<400> 113
gacatccaga tgacccagtc 20
<210> 114
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK1-9_3'-лидерная последовательность-FR1
<400> 114
gacatccagt tgacccagtc t 21
<210> 115
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK1D-43_3'-лидерная последовательность-FR1
<400> 115
gccatccgga tgacccagtc 20
<210> 116
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK2-24_3'-лидерная последовательность-FR1
<400> 116
gatattgtga tgacccagac 20
<210> 117
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK2-28_3'-лидерная последовательность-FR1
<400> 117
gatattgtga tgactcagtc 20
<210> 118
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK2-30_3'-лидерная последовательность-FR1
<400> 118
gatgttgtga tgactcagtc 20
<210> 119
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK3-11_3'-лидерная последовательность-FR1
<400> 119
gaaattgtgt tgacacagtc 20
<210> 120
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK3-15_3'-лидерная последовательность-FR1
<400> 120
gaaatagtga tgacgcagtc 20
<210> 121
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK3-20_3'-лидерная последовательность-FR1
<400> 121
gaaattgtgt tgacgcagtc t 21
<210> 122
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu5'VK4-1_3'-лидерная последовательность-FR1
<400> 122
gacatcgtga tgacccagtc 20
<210> 123
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> rat_CK_R
<400> 123
cttgacactg atgtctctgg g 21
<210> 124
<211> 18
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu3'_JH1245_NheI
<400> 124
tgaggagacg gtgaccag 18
<210> 125
<211> 20
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu3'_JH3_NheI
<400> 125
tgaagagacg gtgaccattg 20
<210> 126
<211> 19
<212> DNA
<213> Искусственная последовательность
<220>
<223> hu3'_JH6_NheI
<400> 126
tgaggagacg gtgaccgtg 19
<210> 127
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huHSCJK2o-B_BsiWI
<400> 127
tttgatctcc agcttggtcc c 21
<210> 128
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huHSCJK3o-B_BsiWI
<400> 128
tttgatatcc actttggtcc c 21
<210> 129
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huHSCJK5o-B_BsiWI
<400> 129
tttaatctcc agtcgtgtcc c 21
<210> 130
<211> 21
<212> DNA
<213> Искусственная последовательность
<220>
<223> huHSCJK14o-B_BsiWI
<400> 130
tttgatytcc accttggtcc c 21
<210> 131
<211> 212
<212> Белок
<213> Искусственная последовательность
<220>
<223> XBR1-402 LC
<400> 131
Ser Tyr Glu Leu Thr Gln Leu Pro Ser Val Ser Val Ser Leu Gly Gln
1 5 10 15
Thr Ala Arg Ile Thr Cys Glu Gly Asn Asn Ile Gly Ser Lys Ala Val
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Leu Ala Pro Gly Leu Leu Ile Tyr
35 40 45
Asp Asp Asp Glu Arg Pro Ser Gly Val Pro Asp Arg Phe Ser Gly Ser
50 55 60
Asn Ser Gly Asp Thr Ala Thr Leu Thr Ile Ser Gly Ala Gln Ala Gly
65 70 75 80
Asp Glu Ala Asp Tyr Tyr Cys Gln Val Trp Asp Ser Ser Ala Tyr Val
85 90 95
Phe Gly Gly Gly Thr Gln Leu Thr Val Thr Gly Gln Pro Lys Ala Ala
100 105 110
Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu Leu Gln Ala Asn
115 120 125
Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr Pro Gly Ala Val
130 135 140
Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys Ala Gly Val Glu
145 150 155 160
Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr Ala Ala Ser Ser
165 170 175
Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His Lys Ser Tyr Ser
180 185 190
Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys Thr Val Ala Pro
195 200 205
Thr Glu Cys Ser
210
<210> 132
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> XBR1-402 HC
<400> 132
Gln Glu Gln Gln Lys Glu Ser Gly Gly Gly Leu Phe Lys Pro Thr Asp
1 5 10 15
Thr Leu Thr Leu Thr Cys Thr Ala Ser Gly Phe Asp Ile Ser Ser Tyr
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Asn Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Gly Ile Ser Gly Asn Ala Tyr Tyr Ala Ser Trp Ala Lys
50 55 60
Ser Arg Ser Thr Ile Thr Arg Asn Thr Asn Leu Asn Thr Val Thr Leu
65 70 75 80
Lys Met Thr Ser Leu Thr Ala Ala Asp Thr Ala Thr Tyr Phe Cys Ala
85 90 95
Arg Asp His Pro Thr Tyr Gly Met Asp Leu Trp Gly Pro Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 133
<211> 447
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ac10 LC
<400> 133
Gln Ile Gln Leu Gln Gln Ser Gly Pro Glu Val Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Ile Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Asp Tyr
20 25 30
Tyr Ile Thr Trp Val Lys Gln Lys Pro Gly Gln Gly Leu Glu Trp Ile
35 40 45
Gly Trp Ile Tyr Pro Gly Ser Gly Asn Thr Lys Tyr Asn Glu Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Val Asp Thr Ser Ser Ser Thr Ala Phe
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Thr Ala Val Tyr Phe Cys
85 90 95
Ala Asn Tyr Gly Asn Tyr Trp Phe Ala Tyr Trp Gly Gln Gly Thr Gln
100 105 110
Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
435 440 445
<210> 134
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Ac10 LC
<400> 134
Asp Ile Val Leu Thr Gln Ser Pro Ala Ser Leu Ala Val Ser Leu Gly
1 5 10 15
Gln Arg Ala Thr Ile Ser Cys Lys Ala Ser Gln Ser Val Asp Phe Asp
20 25 30
Gly Asp Ser Tyr Met Asn Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Val Leu Ile Tyr Ala Ala Ser Asn Leu Glu Ser Gly Ile Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Glu Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Ser Asn
85 90 95
Glu Asp Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 135
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания сортазой A
Staphylococcus aureus, в котором X представляет собой любую аминокислоту, и
n≥1 и ≤21
<400> 135
Leu Pro Xaa Thr Gly
1 5
<210> 136
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания сортазой A
Staphylococcus aureus, в котором X представляет собой любую аминокислоту, и
n≥1 и ≤21
<400> 136
Leu Pro Xaa Ala Gly
1 5
<210> 137
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания сортазой А
Staphylococcus aureus или сконструированной сортазы A 4S-9 из Staphylococcus
aureus, в котором X представляет собой любую аминокислоту, и n≥1 и ≤21
<400> 137
Leu Pro Xaa Ser Gly
1 5
<210> 138
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания
сконструированной сортазой A 2A-9 из Staphylococcus aureus, в котором X
представляет собой любую аминокислоту, и n≥1 и ≤21
<400> 138
Leu Ala Xaa Thr Gly
1 5
<210> 139
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания сортазой А
Streptococcus pyogenes, в котором X представляет собой любую аминокислоту, и
n≥1 и ≤21
<400> 139
Leu Pro Xaa Thr Gly
1 5
<210> 140
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> линкер, полученный из последовательности для распознавания сортазой
Staphylococcus aureus, в котором n≥1 и ≤21
<400> 140
Asn Pro Gln Thr Gly
1 5
<210> 141
<211> 370
<212> Белок
<213> Искусственная последовательность
<220>
<223> внеклеточный домен ROR2 яванского макака (cROR2)
<400> 141
Glu Val Glu Val Pro Asp Pro Asn Asp Pro Leu Gly Pro Leu Asp Gly
1 5 10 15
Gln Asp Gly Pro Ile Pro Thr Leu Lys Gly Tyr Phe Leu Asn Phe Leu
20 25 30
Glu Pro Val Asn Asn Ile Thr Ile Val Gln Gly Gln Thr Ala Ile Leu
35 40 45
His Cys Lys Val Ala Gly Asn Pro Pro Pro Asn Val Arg Trp Leu Lys
50 55 60
Asn Asp Ala Pro Val Val Gln Glu Pro Arg Arg Ile Ile Ile Arg Lys
65 70 75 80
Thr Glu Tyr Gly Ser Arg Leu Arg Ile Gln Asp Leu Asp Thr Thr Asp
85 90 95
Thr Gly Tyr Tyr Gln Cys Val Ala Thr Asn Gly Met Lys Thr Ile Thr
100 105 110
Ala Thr Gly Val Leu Phe Val Arg Leu Gly Pro Thr His Ser Pro Asn
115 120 125
His Asn Phe Gln Asp Asp Tyr His Glu Asp Gly Phe Cys Gln Pro Tyr
130 135 140
Arg Gly Ile Ala Cys Ala Arg Phe Ile Gly Asn Arg Thr Ile Tyr Val
145 150 155 160
Asp Ser Leu Gln Met Gln Gly Glu Ile Glu Asn Arg Ile Thr Ala Ala
165 170 175
Phe Thr Met Ile Gly Thr Ser Thr His Leu Ser Asp Gln Cys Ser Gln
180 185 190
Phe Ala Ile Pro Ser Phe Cys His Phe Val Phe Pro Leu Cys Asp Ala
195 200 205
Arg Ser Arg Ala Pro Lys Pro Arg Glu Leu Cys Arg Asp Glu Cys Glu
210 215 220
Val Leu Glu Ser Asp Leu Cys Arg Gln Glu Tyr Thr Ile Ala Arg Ser
225 230 235 240
Asn Pro Leu Ile Leu Met Arg Leu Gln Leu Pro Lys Cys Glu Ala Leu
245 250 255
Pro Met Pro Glu Ser Pro Asp Ala Ala Asn Cys Met Arg Ile Gly Ile
260 265 270
Pro Ala Glu Arg Leu Gly Arg Tyr His Gln Cys Tyr Asn Gly Ser Gly
275 280 285
Thr Asp Tyr Arg Gly Thr Ala Ser Thr Thr Lys Ser Gly His Gln Cys
290 295 300
Gln Pro Trp Ala Leu Gln His Pro His Ser His His Leu Ser Ser Thr
305 310 315 320
Asp Phe Pro Glu Leu Gly Gly Gly His Ala Tyr Cys Arg Asn Pro Gly
325 330 335
Gly Gln Met Glu Gly Pro Trp Cys Phe Thr Gln Asn Lys Asn Val Arg
340 345 350
Met Glu Leu Cys Asp Val Pro Ser Cys Ser Pro Arg Asp Ser Ser Lys
355 360 365
Met Gly
370
<210> 142
<211> 370
<212> Белок
<213> Искусственная последовательность
<220>
<223> внеклеточный домен мышиного ROR2 (mROR2)
<400> 142
Glu Val Glu Asp Ser Glu Ala Ile Asp Thr Leu Gly Gln Pro Asp Gly
1 5 10 15
Pro Asp Ser Pro Leu Pro Thr Leu Lys Gly Tyr Phe Leu Asn Phe Leu
20 25 30
Glu Pro Val Asn Asn Ile Thr Ile Val Gln Gly Gln Thr Ala Ile Leu
35 40 45
His Cys Lys Val Ala Gly Asn Pro Pro Pro Asn Val Arg Trp Leu Lys
50 55 60
Asn Asp Ala Pro Val Val Gln Glu Pro Arg Arg Val Ile Ile Arg Lys
65 70 75 80
Thr Glu Tyr Gly Ser Arg Leu Arg Ile Gln Asp Leu Asp Thr Thr Asp
85 90 95
Thr Gly Tyr Tyr Gln Cys Val Ala Thr Asn Gly Leu Lys Thr Ile Thr
100 105 110
Ala Thr Gly Val Leu Tyr Val Arg Leu Gly Pro Thr His Ser Pro Asn
115 120 125
His Asn Phe Gln Asp Asp Asp Gln Glu Asp Gly Phe Cys Gln Pro Tyr
130 135 140
Arg Gly Ile Ala Cys Ala Arg Phe Ile Gly Asn Arg Thr Ile Tyr Val
145 150 155 160
Asp Ser Leu Gln Met Gln Gly Glu Ile Glu Asn Arg Ile Thr Ala Ala
165 170 175
Phe Thr Met Ile Gly Thr Ser Thr Gln Leu Ser Asp Gln Cys Ser Gln
180 185 190
Phe Ala Ile Pro Ser Phe Cys His Phe Val Phe Pro Leu Cys Asp Ala
195 200 205
Arg Ser Arg Ala Pro Lys Pro Arg Glu Leu Cys Arg Asp Glu Cys Glu
210 215 220
Val Leu Glu Asn Asp Leu Cys Arg Gln Glu Tyr Thr Ile Ala Arg Ser
225 230 235 240
Asn Pro Leu Ile Leu Met Arg Leu Gln Leu Pro Lys Cys Glu Ala Leu
245 250 255
Pro Met Pro Glu Ser Pro Asp Ala Ala Asn Cys Met Arg Ile Gly Ile
260 265 270
Pro Ala Glu Arg Leu Gly Arg Tyr His Gln Cys Tyr Asn Gly Ser Gly
275 280 285
Ala Asp Tyr Arg Gly Met Ala Ser Thr Thr Lys Ser Gly His Gln Cys
290 295 300
Gln Pro Trp Ala Leu Gln His Pro His Ser His Arg Leu Ser Ser Thr
305 310 315 320
Glu Phe Pro Glu Leu Gly Gly Gly His Ala Tyr Cys Arg Asn Pro Gly
325 330 335
Gly Gln Met Glu Gly Pro Trp Cys Phe Thr Gln Asn Lys Asn Val Arg
340 345 350
Val Glu Leu Cys Asp Val Pro Pro Cys Ser Pro Arg Asp Gly Ser Lys
355 360 365
Met Gly
370
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ АНТИТЕЛ К ROR1 И СООТВЕТСТВУЮЩИЕ СПОСОБЫ | 2017 |
|
RU2766190C2 |
| АНТИТЕЛА К РЕЦЕПТОРУ IL4 ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2837141C2 |
| АНТИ-IL31 АНТИТЕЛА ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2020 |
|
RU2835543C1 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| АНТИТЕЛО ПРОТИВ ФАКТОРА РОСТА СОЕДИНИТЕЛЬНОЙ ТКАНИ И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2819228C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ ВИРУСА БЕШЕНСТВА И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2764740C1 |
| ИММУНОЦИТОКИНЫ И ИХ ПРИМЕНЕНИЕ | 2021 |
|
RU2818371C1 |
| АНТИТЕЛО, СПОСОБНОЕ СВЯЗЫВАТЬСЯ С ТИМИЧЕСКИМ СТРОМАЛЬНЫМ ЛИМФОПОЭТИНОМ, И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2825304C2 |
| Биспецифическое антитело анти-HER2/анти-4-1BB и его применение | 2020 |
|
RU2831838C2 |
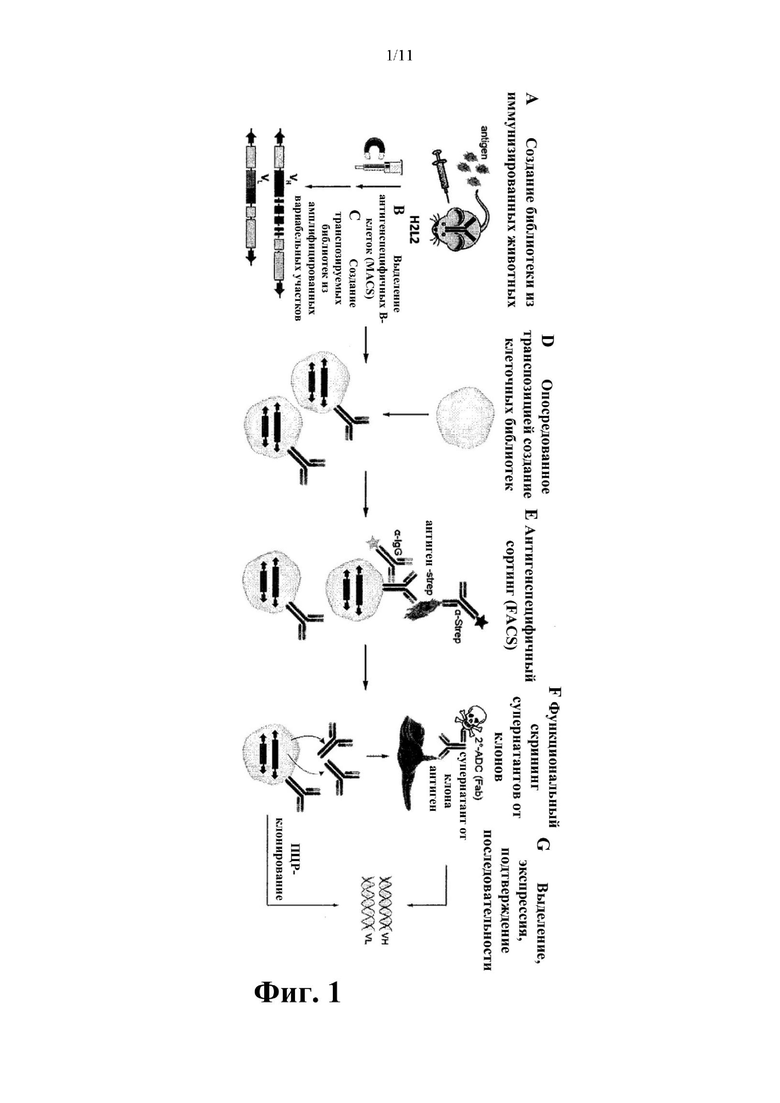

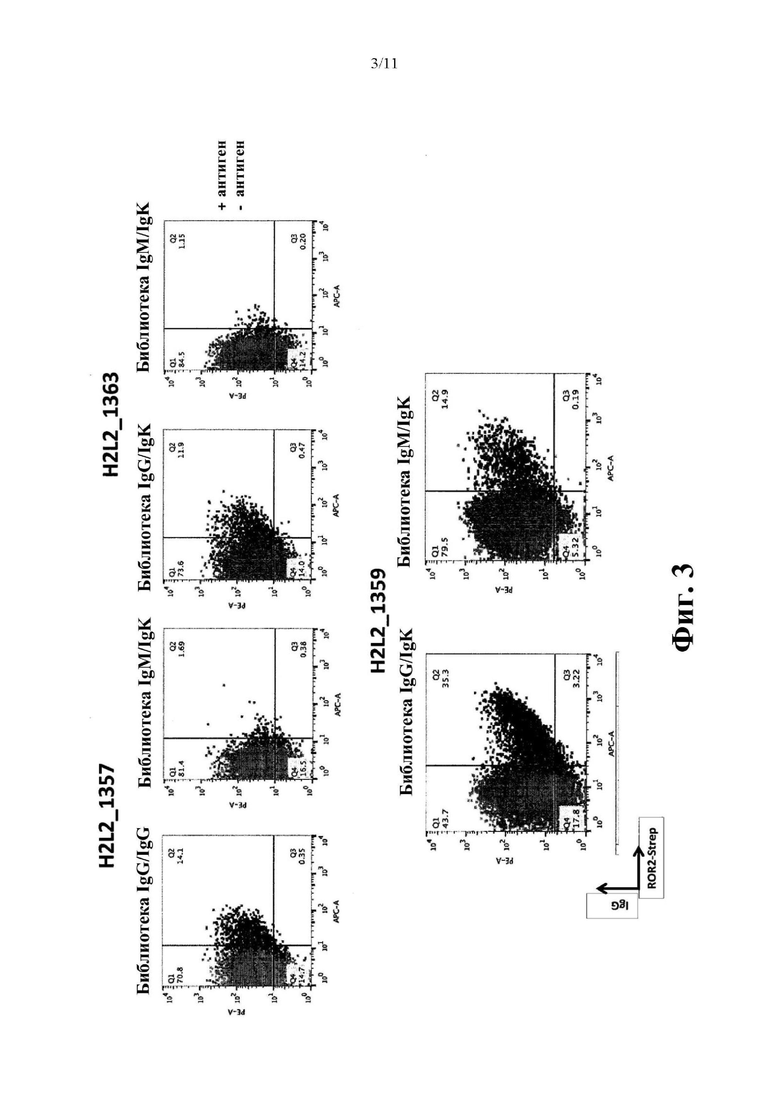
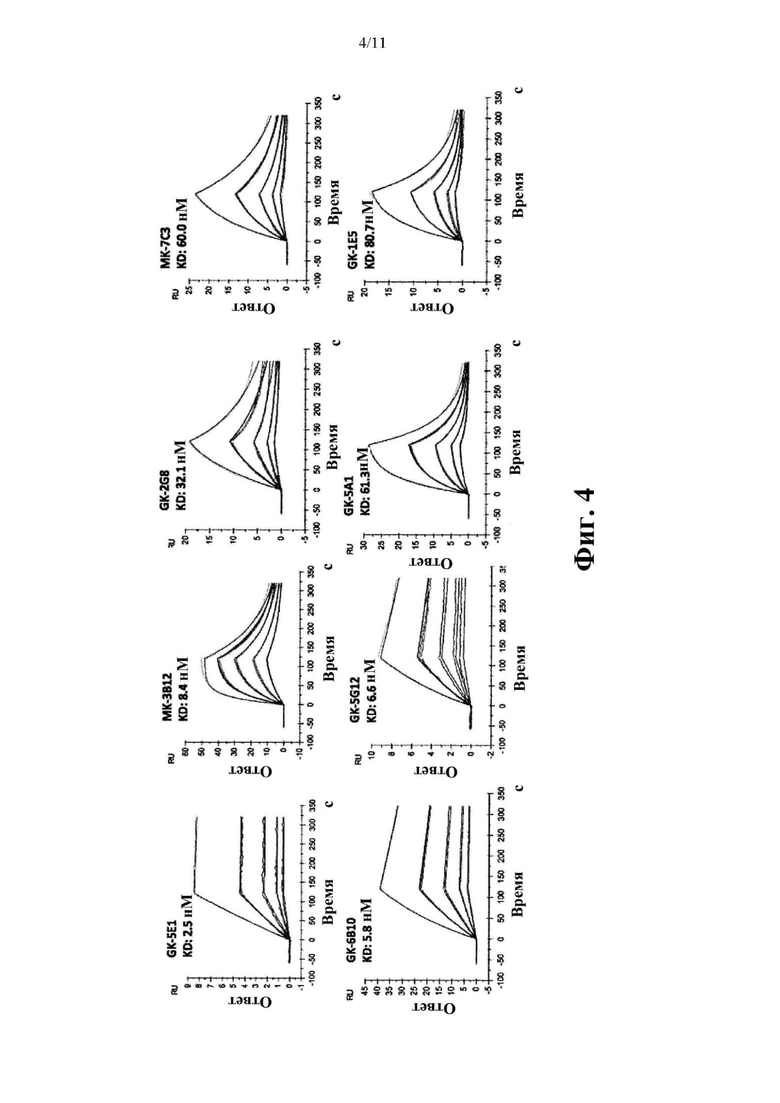

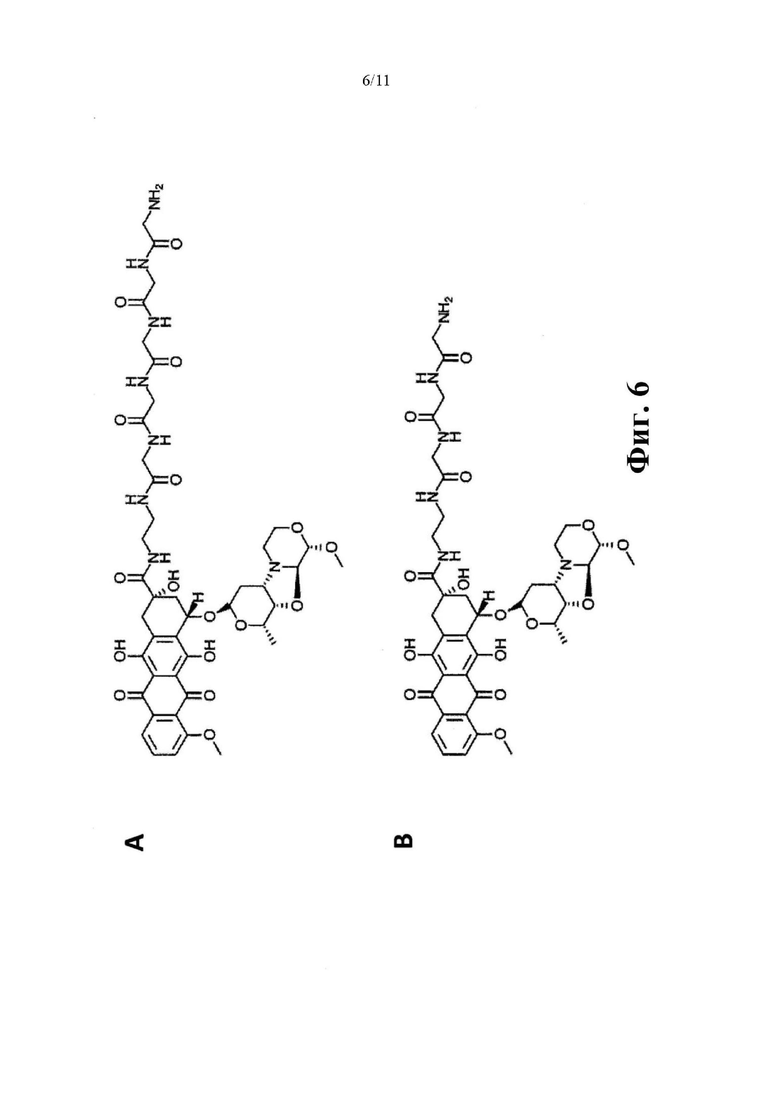
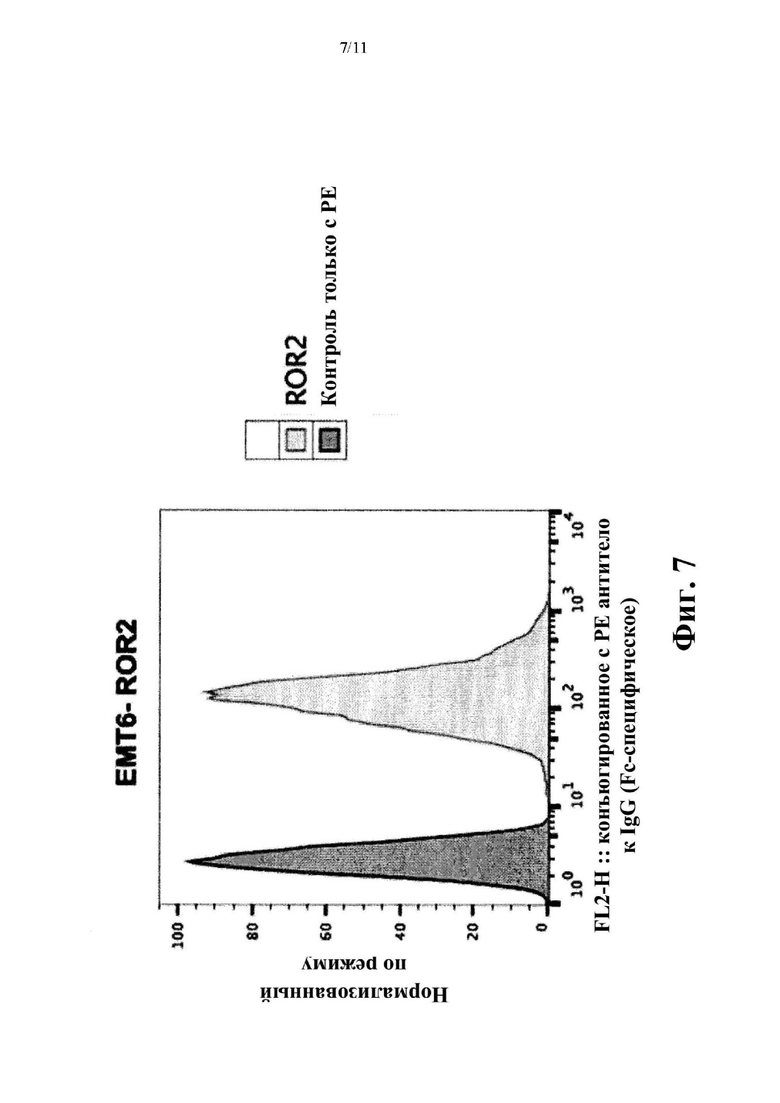
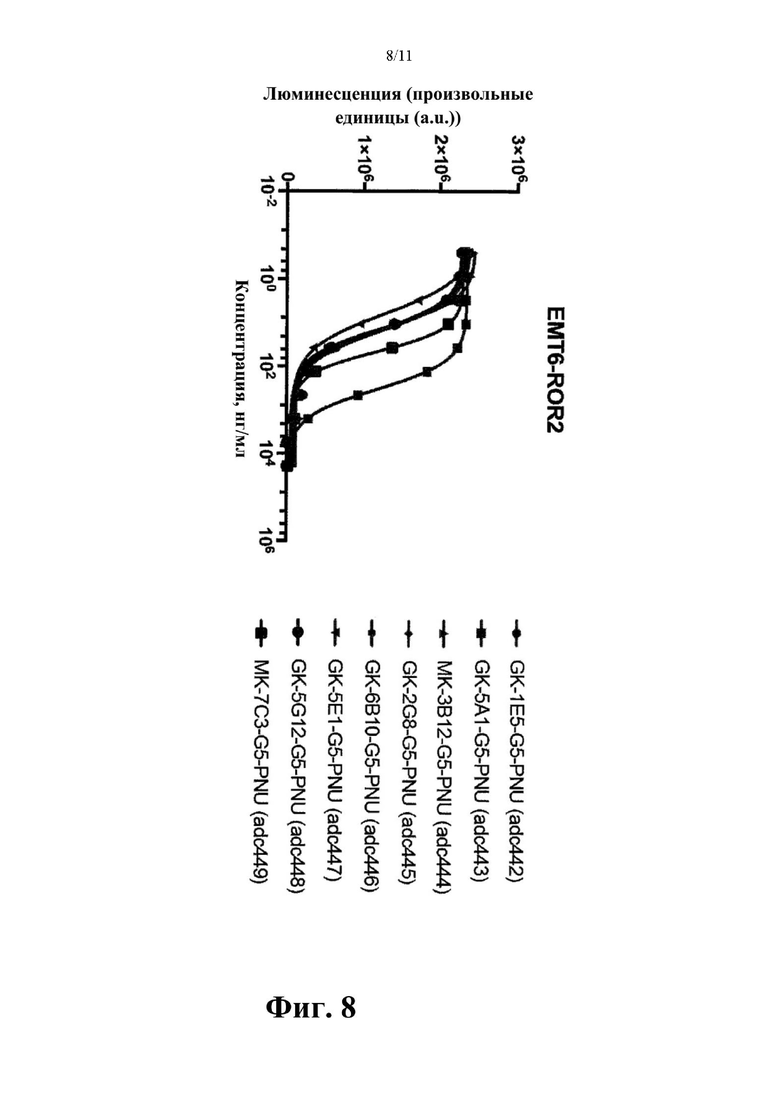
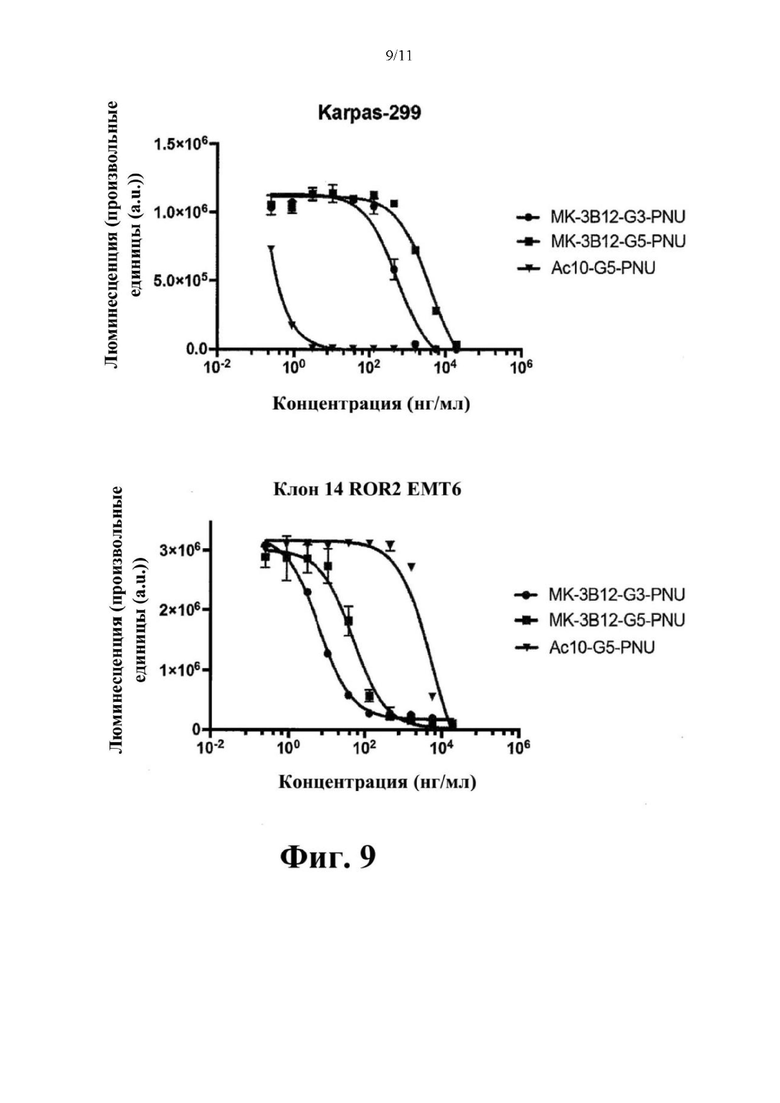
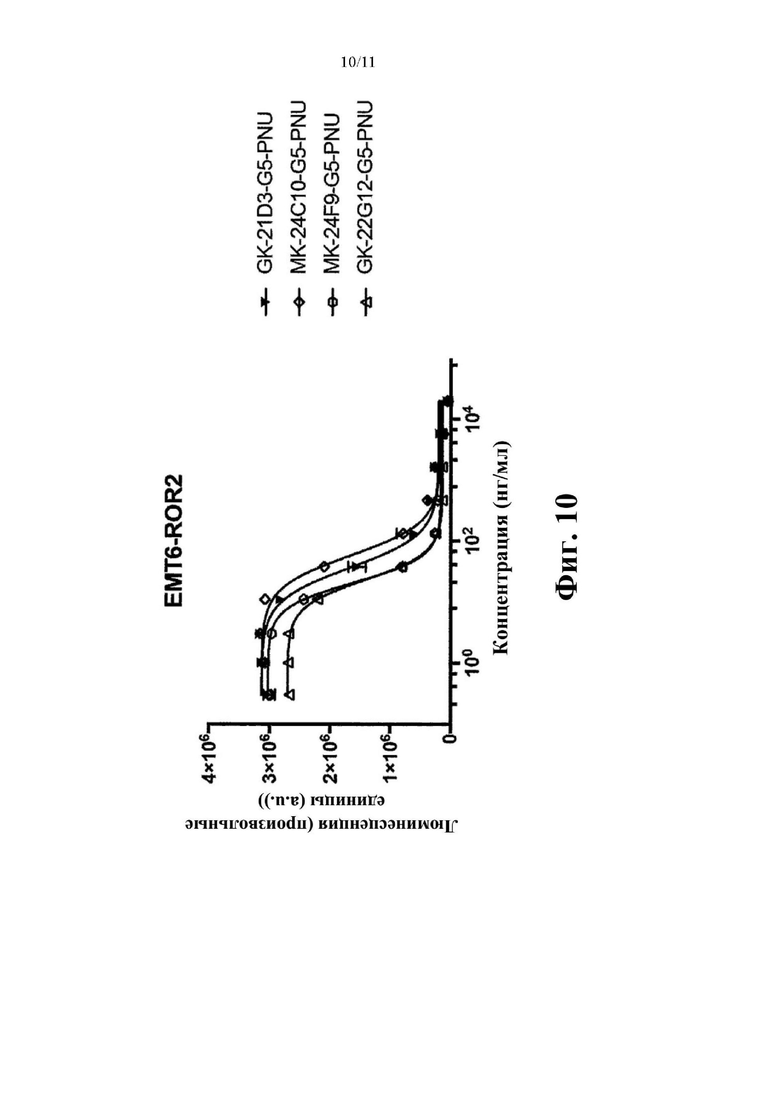
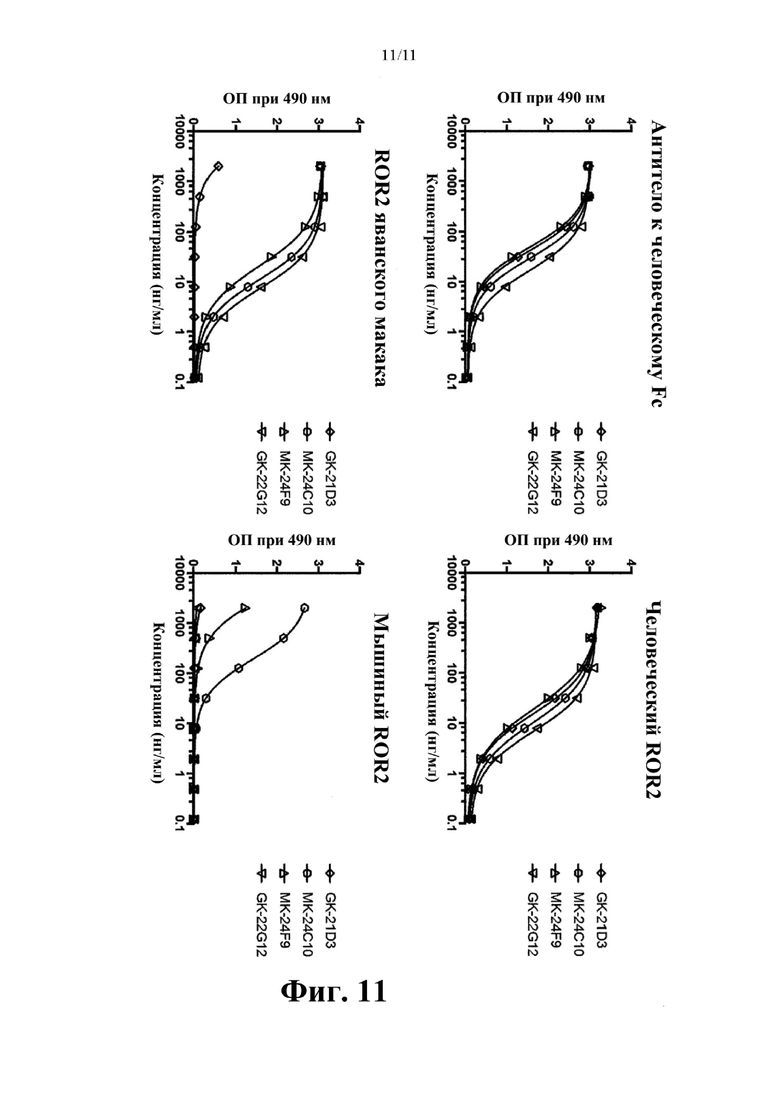
Изобретение относится к области биотехнологии. Предложено полностью человеческое антитело или его фрагмент, которые специфично связываются с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2). Также предложены конъюгаты антитело-лекарственное средство, включающие антитела по изобретению. Кроме того, раскрыто применение указанных конъюгатов в способе лечения неопластического заболевания, а также в способе определения неопластического заболевания или иммунного заболевания у пациента или риска его развития. Изобретение может быть использовано в терапии рака. 8 н. и 9 з.п. ф-лы, 11 ил., 14 табл., 10 пр.
1. Полностью человеческое антитело или его фрагмент, которые специфично связываются с внеклеточным доменом подобного рецепторной тирозинкиназе орфанного рецептора 2 (ROR2),
которые характеризуются перекрестной реактивностью с (i) человеческим ROR2 (hROR2) и (ii) с ROR2 яванского макака (cROR2), которые содержат один из следующих наборов CDR:
a) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 38, 39 и 40, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 29 и 41, с CDR 2 легкой цепи, имеющим последовательность KAS,
b) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 26-28, и CDR 1-3 легкой цепи, которые изложены в SEQ ID NO: 29, 73 и 30,
c) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 94, 32 и 44, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 29 и 42, с CDR 2 легкой цепи, имеющим последовательность KAS,
d) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 38, 46 и 52, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 50 и 53, с CDR 2 легкой цепи, имеющим последовательность AAS,
e) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 38, 54 и 55, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 56 и 57, с CDR 2 легкой цепи, имеющим последовательность LGS,
f) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 58-60, и CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 61 и 62, с CDR 2 легкой цепи, имеющим последовательность KAS,
g) CDR 1-3 тяжелой цепи, которые изложены в SEQ ID NO: 63-65, с CDR 1 и 3 легкой цепи, которые изложены в SEQ ID NO: 50 и 66, с CDR 2 легкой цепи, имеющим последовательность AAS,
и при этом CDR содержатся в подходящем белковом каркасе для того, чтобы они были способны к связыванию с hROR2, а также с ROR2 яванского макака с достаточной аффинностью.
2. Антитело или его фрагмент по п. 1, которые содержат одну из следующих пар последовательностей:
a) последовательность вариабельного участка тяжелой цепи антитела GK-5A1, представленную в SEQ ID NO: 8, и последовательность вариабельного участка легкой цепи антитела GK-5A1, представленную в SEQ ID NO: 9,
b) последовательность вариабельного участка тяжелой цепи антитела MK-3B12, представленную в SEQ ID NO: 2, и последовательность вариабельного участка легкой цепи антитела MK-3B12, представленную в SEQ ID NO: 3,
c) последовательность вариабельного участка тяжелой цепи антитела GK-5E1, представленную в SEQ ID NO: 12, и последовательность вариабельного участка легкой цепи антитела GK-5E1, представленную в SEQ ID NO: 13,
d) последовательность вариабельного участка тяжелой цепи антитела GK-21D3, представленную в SEQ ID NO: 18, и последовательность вариабельного участка легкой цепи антитела GK-5E1, представленную в SEQ ID NO: 19,
e) последовательность вариабельного участка тяжелой цепи антитела MK-24C10, представленную в SEQ ID NO: 20, и последовательность вариабельного участка легкой цепи антитела MK-24C10, представленную в SEQ ID NO: 21,
f) последовательность вариабельного участка тяжелой цепи антитела MK-24F9, представленную в SEQ ID NO: 22, и последовательность вариабельного участка легкой цепи антитела MK-24F9, представленную в SEQ ID NO: 23,
g) последовательность вариабельного участка тяжелой цепи антитела GK-22G12, представленную в SEQ ID NO: 24, и последовательность вариабельного участка легкой цепи антитела GK-22G12, представленную в SEQ ID NO: 25.
3. Конъюгат антитело-лекарственное средство (ADC) для лечения неопластического заболевания, имеющий общую формулу A-(L)n-(T)m, в которой
A представляет собой антитело или его фрагмент по любому из пп. 1, 2,
L представляет собой линкер,
T представляет собой полезную нагрузку в виде цитотоксического или цитостатического средства
и в которой n и m представляют собой целые числа от >1 до <10.
4. Конъюгат антитело-эффекторная молекула (AEC) для лечения неопластического заболевания, имеющий общую формулу A-(L)n-(T)m, в которой
A представляет собой антитело или его фрагмент по любому из пп. 1, 2,
L представляет собой линкер,
T представляет собой метку
и в которой n и m представляют собой целые числа от >1 до <10.
5. Конъюгат по любому из пп. 3 и 4, причем линкер содержит или состоит по меньшей мере из одного, выбранного из группы, состоящей из
олигопептидного линкера,
малеимидного линкера, необязательно содержащего расщепляемые спейсеры, которые могут расщепляться при изменениях pH, окислительно-восстановительного потенциала и/или конкретными внутриклеточными ферментами.
6. Конъюгат по любому из пп. 3-5, причем линкер имеет по меньшей мере одну из следующих аминокислотных последовательностей: -LPXTGn-, -LPXAGn-, -LPXSGn-, -LAXTGn-, -LPXTGn-, -LPXTAn- или -NPQTGn-, причем Gn представляет собой олиго- или полиглицин, при этом n представляет собой целое число от ≥1 до ≤21, An представляет собой олиго- или полиаланин, при этом n представляет собой целое число от ≥1 до ≤21, и X представляет собой любую возможную аминокислотную последовательность.
7. Конъюгат по любому из пп. 3-6, причем линкер конъюгирован с C-концом по меньшей мере одного субдомена антитела или его фрагмента.
8. Конъюгат по любому из пп. 3-7, причем перед конъюгированием
антитело или его фрагмент несет мотив для распознавания сортазой, слитый или конъюгированный с C-концом по меньшей мере одного его субдомена, и
токсин или метка содержит глициновый отрезок длиной от ≥1 до ≤21 глициновых остатков, предпочтительно длиной от ≥2 до ≤5 глициновых остатков.
9. Конъюгат антитело-лекарственное средство по любому из пп. 3-8, причем токсин представляет собой по меньшей мере один, выбранный из группы, состоящей из:
майтансиноидов,
ауристатинов,
антрациклинов, предпочтительно, антрациклинов, полученных из PNU-159682,
калихеамицинов,
тубулизинов,
дуокармицинов,
радиоактивных изотопов,
липосом, содержащих полезную нагрузку в виде анатоксина,
белковых токсинов,
таксанов,
псевдодимеров индилино-бензодиазепина и/или
пирролобензодиазепинов,
или представляет собой их производное.
10. Конъюгат по любому из пп. 3-9, который получен посредством опосредованного сортазой конъюгирования (i) антитела, несущего один или несколько мотивов для распознавания сортазой, и (ii) одного или нескольких токсинов или меток, несущих олигоглициновый тэг.
11. Способ получения конъюгата по любому из пп. 3-10, причем способ предусматривает следующие стадии:
a) обеспечение антитела или его фрагмента по любому из пп. 1, 2, причем антитело несет один или несколько мотивов для распознавания сортазой,
b) обеспечение одного или нескольких токсинов или меток, несущих олигоглициновый тэг, и
c) конъюгирование антитела и токсина или метки с помощью опосредованного сортазой конъюгирования.
12. Применение конъюгата антитело-лекарственное средство по любому из пп. 3 или 5-10 для изготовления лекарственного средства для лечения неопластического заболевания.
13. Применение по п. 12, причем неопластическое заболевание представляет собой по меньшей мере одно, выбранное из группы, состоящей из
почечно-клеточной карциномы,
остеосаркомы,
рака почки.
14. Фармацевтическая композиция для лечения неопластического заболевания, содержащая конъюгат антитело-лекарственное средство по любому из пп. 3 или 5-10 вместе с одним или несколькими фармацевтически приемлемыми ингредиентами.
15. Способ уничтожения или ингибирования роста клетки, экспрессирующей ROR2 у пациента, причем способ предусматривает введение в клетку фармацевтически эффективного количества или дозы (i) конъюгата антитело-лекарственное средство по любому из пп. 3 или 5-10 или (ii) фармацевтической композиции по п. 14, причем клетка, экспрессирующая ROR2, представляет собой раковую клетку.
16. Способ определения того,
страдает ли предполагаемый пациент от неопластического заболевания или иммунного заболевания и/или
имеет ли предполагаемый пациент риск развития неопластического заболевания или иммунного заболевания, причем указанный способ предусматривает обработку образца, взятого у субъекта, конъюгатом антитело-эффекторная молекула по любому из пп. 4-8 или 10.
17. Способ по п. 16, причем неопластическое заболевание или иммунное заболевание является подходящим для лечения антителом к ROR2 или его фрагментом или конъюгатом антитело к ROR2-лекарственное средство.
| WO 2016142768 A1, 15.09.2016 | |||
| WO 2013103637 A1, 11.07.2013 | |||
| WO 2014140317 A2, 18.09.2014 | |||
| АНТИТЕЛА И ИММУНОКОНЪЮГАТЫ И ИХ ПРИМЕНЕНИЕ | 2007 |
|
RU2483080C2 |
| DEBEBE Z., RATHMELL W | |||
| K., Ror2 as a Therapeutic Target in Cancer, Pharmacology & Therapeutics, 2015, Volume 150, pp.143-148. | |||

