Ссылка на перечень последовательностей
Настоящая заявка содержит перечень последовательностей в машиночитаемой форме, который включен в данный документ с помощью ссылки.
Область изобретения
Настоящее изобретение относится к способам удаления смолистых веществ из растительного масла и его рафинирования. Настоящее изобретение дополнительно относится к полипептидам, обладающим активностью фосфолипазы A, к полипептидам, обладающим активностью фосфолипазы C, и к полинуклеотидам, кодирующим данные полипептиды. Настоящее изобретение также относится к конструкциям нуклеиновой кислоты, векторам и клеткам-хозяевам, содержащим полинуклеотиды, а также к способам получения и применения данных полипептидов.
Предпосылки изобретения
Независимо от того, предназначено ли растительное масло для употребления человеком или в качестве сырья в получении продуктов переработки масла или биодизеля, его необходимо предварительно обрабатывать с целью удаления примесей, таких как фосфолипиды ("смолистые вещества") и свободные жирные кислоты. Предварительная обработка включает удаление смолистых веществ, рафинирование (также называемые "нейтрализацией", осветлением и дезодорированием).
Целью способа удаления смолистых веществ является удаление гидратируемых и негидратируемых фосфолипидов или смолистых веществ, присутствующих в масле. Традиционно способ удаления смолистых веществ основывался на применении экстрагирования водой ("удалении смолистых веществ с помощью воды"), которое предусматривает обработку масла водой и отделение гидратируемых фосфолипидов или смолистых веществ от триглицеридного масла. В зависимости от источника масла удаление смолистых веществ с помощью воды можно комбинировать с "удалением смолистых веществ с помощью кислоты", при котором масло обрабатывают кислотой и отделяют негидратируемые смолистые вещества от триглицеридного масла.
Удаление смолистых веществ с помощью ферментов проводят для масел, которые подвергали удалению смолистых веществ с помощью воды, а также для неочищенных масел. В способе удаления смолистых веществ с помощью ферментов фосфолипиды гидролизируются в реакции, катализируемой ферментами, обладающими активностью фосфолипазы, и таким образом превращаются в водорастворимые и экстрагируемые водой компоненты.
Существуют два основных типа рафинирования: "химическое рафинирование" (также называемое "рафинированием с помощью щелочи") и "физическое рафинирование". Химическое рафинирование, которое предусматривает обработку масла раствором щелочи или другим раствором для рафинирования, проводят с целью снижения содержания свободных жирных кислот, а также удаления других примесей, таких как фосфолипиды, белковые и вязкие вещества и красящие соединения. Данный способ приводит к значительному снижению содержания свободных жирных кислот посредством их превращения в мыла с высокой удельной плотностью, которые удаляют посредством центрифугирования с некоторой потерей нейтрального масла. Большинство фосфатидов и вязких веществ растворимы в масле только в безводной форме, и их легко отделить после гидратации с помощью раствора каустической соды или другого раствора для рафинирования. После рафинирования с помощью щелочи жир или масло промывают водой с удалением остаточного мыла.
Масла с низким содержанием фосфолипидов (пальмовое и кокосовое) можно рафинировать физическим способом (т.е. десорбировать паром) с удалением свободных жирных кислот. При физическом рафинировании свободные жирные кислоты в неочищенном масле или масле, подвергнутом удалению смолистых веществ с помощью воды, удаляют посредством выпаривания, а не нейтрализуют и удаляют в виде мыла, как в способе щелочного рафинирования.
Хотя удаление смолистых веществ с помощью ферментов недавно стало более распространенным, оно никогда не принималось в промышленности в качестве способа, совмещаемого с химическим рафинированием. Затруднение состояло в том, что получение слишком большого количества FFA может приводить к нежелательному увеличению образования мыла при рафинировании или что технология с применением ферментов не будет отвечать требованиям в отношении условий pH при химическом рафинировании. При химическом рафинировании первая стадия представляет собой этап образования хелатных соединений с кислотой с последующим повышением условий pH путем добавления щелочи. Следовательно, химическое рафинирование, как правило, применяют для неочищенного масла или, как показано на фигуре 2 в данном документе, для масла, которое подвергали удалению смолистых веществ с помощью воды и/или удалению смолистых веществ с помощью кислоты. Специалисту в данной области техники также будет известно, что в традиционных способах химическое рафинирование связано со значительной потерей выхода, если гидратируемые и негидратируемые смолистые вещества не удалить до применения каустической соды или другого(-их) средства(средств) для рафинирования.
Несмотря на достигнутый в последнее время прогресс в отношении удаления смолистых веществ из масла и его рафинирования, существует необходимость в обеспечении новых упрощенных способов удаления смолистых веществ из растительного масла и его рафинирования, в которых потеря масла сведена к минимуму. Настоящее изобретение относится к таким новым способам, к новым полипептидам, обладающим активностью фосфолипазы A, к новым полипептидам, обладающим активностью фосфолипазы C, и к полинуклеотидам, кодирующим данные полипептиды.
Краткое описание изобретения
Вопреки тому, как считалось ранее, авторы настоящего изобретения наблюдали, что при рафинировании растительного масла, содержащего фосфолипиды, обеспечиваются значительные преимущества, когда фосфолипиды подвергают ферментативному гидролизу и затем масло подвергают химическому рафинированию без отделения фазы смолистых веществ между стадиями гидролиза и рафинирования. В частности, авторы настоящего изобретения наблюдали значительное увеличение выхода по сравнению с проведением химического рафинирования неочищенного масла или масла, которое подвергали удалению смолистых веществ с помощью воды.
Соответственно, в первом аспекте настоящее изобретение предусматривает способ рафинирования растительного масла, содержащего фосфолипиды, предусматривающий осуществление ферментативного гидролиза фосфолипидов путем приведения растительного масла в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, и после этого осуществление химического рафинирования растительного масла.
Во втором аспекте настоящее изобретение относится к применению фермента, расщепляющего фосфолипиды, для гидролиза фосфолипидов в растительном масле, где растительное масло приводится в контакт с ферментом, расщепляющим фосфолипиды, и после этого осуществляется химическое рафинирование.
В третьем аспекте настоящее изобретение предусматривает рафинированное растительное масло или соапсток, которые являются получаемыми или получают с помощью способа в соответствии с настоящим изобретением.
В четвертом аспекте настоящее изобретение относится к выделенному или очищенному полипептиду, обладающему активностью фосфолипазы A, выбранному из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 3 и 5,
b. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 4 и 6;
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы A.
В пятом аспекте настоящее изобретение предусматривает выделенный или очищенный полипептид, обладающий активностью фосфолипазы С, выбранный из группы, состоящей из:
i) полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 19, 21, 23,
ii) полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 20, 22, 24, и
iii) фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы С.
В шестом аспекте настоящее изобретение предусматривает композицию, содержащую полипептид в соответствии с настоящим изобретением.
В седьмом аспекте настоящее изобретение предусматривает выделенный или очищенный полинуклеотид, кодирующий полипептид по настоящему изобретению.
В восьмом аспекте настоящее изобретение относится к конструкции нуклеиновой кислоты или вектору экспрессии, содержащим полинуклеотид по настоящему изобретению, где полинуклеотид предпочтительно функционально связан с одной или несколькими регуляторными последовательностями, которые управляют продуцированием полипептида у экспрессирующего хозяина.
В девятом аспекте настоящее изобретение относится к рекомбинантной клетке-хозяину, содержащей полинуклеотид по настоящему изобретению, функционально связанный с одной или несколькими регуляторными последовательностями, которые управляют продуцированием полипептида.
В десятом аспекте настоящее изобретение предусматривает способ получения полипептида по настоящему изобретению, предусматривающий культивирование клетки, которая в своей форме дикого типа продуцирует полипептид, в условиях, способствующих продуцированию полипептида.
В одиннадцатом аспекте настоящее изобретение относится к способу получения полипептида, обладающего активностью фосфолипазы A, или полипептида, обладающего активностью фосфолипазы С, предусматривающему культивирование рекомбинантной клетки-хозяина по настоящему изобретению в условиях, способствующих продуцированию полипептида.
Краткое описание графических материалов
Формулировка "Lipr287" относится к PLC Bacillus macauensis: зрелому полипептиду под SEQ ID NO: 9.
На фиг. 1 показаны места, в которых разные фосфолипазы расщепляют фосфолипид, а также четыре основные функциональные группы фосфолипидов.
На фиг. 2 показаны способы обработки растительного масла, включающие удаление смолистых веществ и рафинирование.
На фиг. 3 показана величина выхода после окончания центрифугирования. Зрелый полипептид под SEQ ID NO: 9 в качестве предварительной обработки для удаления смолистых веществ с помощью щелочи; 70°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, общее содержание NaOH 1141 ppm.
На фиг. 4 показана разница в содержании диглицеридов. Зрелый полипептид под SEQ ID NO: 9 в качестве средства для предварительной обработки для удаления смолистых веществ с помощью щелочи; 0°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, общее содержание NaOH 1141 ppm.
На фиг. 5 показан уровень интактных фосфолипидов. Зрелый полипептид под SEQ ID NO: 9 в качестве предварительной обработки для удаления смолистых веществ с помощью щелочи; 70°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, общее содержание NaOH 1141 ppm.
На фиг. 6 показан уровень гидролизованных фосфолипидов (всех 4). Зрелый полипептид под SEQ ID NO: 9 в качестве предварительной обработки для удаления смолистых веществ с помощью щелочи; 70°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, содержание NaOH 1141 ppm.
На фиг. 7 показан уровень гидролизованных PC+PE. Зрелый полипептид под SEQ ID NO: 9 в качестве предварительной обработки для удаления смолистых веществ с помощью щелочи; 70°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, общее содержание NaOH 1141 ppm.
На фиг. 8 показано общее содержание фосфора после окончания центрифугирования. Зрелый полипептид под SEQ ID NO: 9 в качестве предварительной обработки для удаления смолистых веществ с помощью щелочи; 70°C, ферментативная реакция в течение 1 часа, общее содержание воды 3%, общее содержание NaOH 1141 ppm.
На фиг. 9 показано общее содержание масла после окончания центрифугирования; содержание Ca 650 ppm, общее содержание воды 2,5%, содержание NaOH 3000 ppm для щелочной обработки при 70°C.
На фиг. 10 показано увеличение выхода по сравнению с холостой пробой. Содержание Ca 650 ppm, общее содержание воды 2,5%, содержание NaOH 3000 ppm для щелочной обработки при 70°C для масла с низким содержанием NHP (P 15 ppm).
На фиг. 11 показано содержание FFA до и после щелочной обработки; содержание Ca 650 ppm, общее содержание воды 2,5%, содержание NaOH 3000 ppm для щелочной обработки при 70°C.
На фиг. 12 показана разница в содержании диглицеридов; содержание Ca 650 ppm, общее содержание воды 2,5%, содержание NaOH 3000 ppm для щелочной обработки при 70°C.
Подробное описание изобретения
Определения
Щелочь. В данном контексте термин "щелочь" используется взаимозаменяемо с основанием, которое является растворимым в воде и образует гидроксид-ионы, как, например, NaOH, KOH, карбонат натрия, Ca(OH)2 и Mg(OH)2, и раствором основания в воде.
Осветление. Термин "осветление" относится к способу удаления веществ, обеспечивающих окрашивание, и дополнительной очистки жира или масла. Обычно осветление выполняют после того, как масло было рафинировано.
Химическое рафинирование. В настоящей заявке термин "химическое рафинирование" используется синонимически с терминами "рафинирование с помощью щелочи" и "щелочное рафинирование"; термин также охватывает "рафинирование с помощью каустической соды" и "нейтрализацию с помощью каустической соды".
Неочищенное масло. Термин "неочищенное масло" относится к нерафинированному и необработанному маслу, полученному путем прессования или экстракции из источника растительного происхождения, включая без ограничения масло асаи, миндальное масло, масло из ореха бабассу, масло семян черной смородины, масло семян огуречника, масло канолы, масло из ореха кешью, касторовое масло, кокосовое масло, кориандровое масло, кукурузное масло, хлопковое масло, масло крамбе, масло семян льна, масло виноградных косточек, масло из лесного ореха, конопляное масло, ятрофное масло, масло жожоба, льняное масло, масло из ореха макадамия, масло из косточек манго, масло пенника лугового, горчичное масло, костное масло, оливковое масло, пальмовое масло, пальмоядровое масло, пальмовый олеин, арахисовое масло, масло из ореха пекан, масло из кедрового ореха, фисташковое масло, маковое масло, рапсовое масло, масло из рисовых отрубей, сафлоровое масло, масло сасанквы, кунжутное масло, масло ши, соевое масло, масло семян подсолнечника, талловое масло, масло камелии, масло из грецкого ореха, разновидности "натуральных" масел, характеризующихся измененным составом жирных кислот вследствие генетически модифицированных организмов (GMO) или традиционной "селекции", такие как масла с высоким содержанием олеиновой кислоты, масла с низким содержанием линоленовой кислоты или низконасыщенные масла (масло канолы с высоким содержанием олеиновой кислоты, соевое масло с низким содержанием линоленовой кислоты или виды подсолнечного масла с высоким содержанием стеариновой кислоты). Термин также охватывает смесь нескольких нерафинированных и необработанных масел, полученных из источников, определенных выше, путем прессования или экстракции.
Дезодорирование. "Дезодорирование" представляет собой способ перегонки с паром под вакуумом с целью удаления примесных компонентов, которые являются источником нежелательных привкусов, окрасок и неприятных запахов в жирах и маслах. Обычно данный способ выполняют после рафинирования и осветления.
Фракционирование. Фракционирование представляет собой способ отделения триглицеридов из жиров и масел посредством разности в температурах плавления, растворимости или летучести. Его наиболее часто применяют для отделения жиров, которые находятся в твердом состоянии при комнатной температуре, но его также применяют для отделения триглицеридов, обнаруживаемых в жидких маслах.
Смолистое вещество. В контексте настоящего изобретения "смолистое вещество", "смолистые вещества" или "фракция смолистых веществ" относится к фракции, обогащенной фосфатидами, которую отделяют от основной массы растительного масла в ходе осуществления способа удаления смолистых веществ. "Смолистые вещества" в основном состоят из фосфатидов, но также содержат унесенное масло, азот и сахар, а также тонкоизмельченные частицы.
Гетерологичный. Термин "гетерологичный" в отношении клетки-хозяина означает, что полипептид или нуклеиновая кислота не встречаются в природных условиях у клетки-хозяина. Термин "гетерологичный" в отношении полипептида или нуклеиновой кислоты означает, что регуляторная последовательность, например, промотор или домен полипептида или нуклеиновой кислоты, в природе не ассоциирована с полипептидом или нуклеиновой кислотой, т.е. регуляторная последовательность получена из гена, отличного от гена, кодирующего полипептид под SEQ ID NO: 1.
Клетка-хозяин. Термин "клетка-хозяин" означает любую микробную или растительную клетку, в которую ввели конструкцию нуклеиновой кислоты или вектор экспрессии, содержащие полинуклеотид по настоящему изобретению. Способы введения включают без ограничения слияние протопластов, трансфекцию, трансформацию, электропорацию, конъюгацию и трансдукцию. В некоторых вариантах осуществления клетка-хозяин представляет собой выделенную рекомбинантную клетку-хозяина, которая частично или полностью отделена от по меньшей мере одного другого компонента, включая без ограничения, например, белки, нуклеиновые кислоты, клетки и т.д.
Выделенный. Термин "выделенный" означает полипептид, нуклеиновую кислоту, клетку или другой указанный материал или компонент, которые отделены от по меньшей мере одного другого материала или компонента, с которым они ассоциированы в природных условиях, как обнаружено в природе, включая без ограничения, например, другие белки, нуклеиновые кислоты, клетки и т.д. Выделенный полипептид включает без ограничения культуральный бульон, содержащий секретированный полипептид.
Лизофосфолипаза. "Лизофосфолипаза" (EC 3.1.1.5) представляет собой фермент, который может гидролизовать 2-лизофосфолипиды с высвобождением жирной кислоты.
Активность лизофосфолипазы (LLU) можно измерить с применением набора для анализа NEFA C, используя L-α-лизолецитин из яичного желтка в качестве субстрата. 20 мкл образца смешивают со 100 мкл 20 мM натрий-ацетатного буфера (pH 4,5) и 100 мкл 1% раствора L-α-лизолецитина и инкубируют при 55°C в течение 20 мин. Через 20 мин реакционную смесь переносят в пробирку, содержащую 30 мкл раствора A из набора NEFA, предварительно нагретого при 37°C. После 10 мин инкубирования при 37°C в реакционную смесь добавляют 600 мкл раствора B из набора NEFA и инкубируют при 37°C в течение 10 мин. Активность измеряют при 555 нм на спектрофотометре. Одна единица активности лизофосфолипазы (1 LLU) определяется как количество фермента, при котором величина A550 может увеличиваться на 0,01 в минуту при 55°C.
Зрелый полипептид. Термин "зрелый полипептид" означает полипептид в его конечной форме после трансляции и любых посттрансляционных модификаций, таких как процессинг N-концевой части, усечение C-концевой части, гликозилирование, фосфорилирование и удаление сигнальных пептидов, пропептидов и препропептидов. Из уровня техники известно, что клетка-хозяин может продуцировать смесь двух или более разных зрелых полипептидов (т.е. с отличающейся аминокислотой на C-конце и/или N-конце), экспрессируемых одним и тем же полинуклеотидом. Из уровня техники также известно, что в разных клетках-хозяевах процессинг полипептидов осуществляется разным образом, и поэтому одна клетка-хозяин, экспрессирующая полинуклеотид, может продуцировать другой зрелый полипептид (например, с отличающейся аминокислотой на C-конце и/или N-конце) по сравнению с другой клеткой-хозяином, экспрессирующей тот же полинуклеотид.
Конструкция нуклеиновой кислоты. Термин "конструкция нуклеиновой кислоты" означает одно- либо двухнитевую молекулу нуклеиновой кислоты, которая выделена из встречающегося в природе гена или модифицирована таким образом, что она содержит сегменты нуклеиновых кислот в таком порядке, который в ином случае не существовал бы в природе, или которая является синтетической, которая содержит одну или несколько регуляторных последовательностей.
Функционально связанный. Термин "функционально связанный" означает конфигурацию, при которой регуляторная последовательность размещена в соответствующем положении относительно кодирующей последовательности полинуклеотида таким образом, что регуляторная последовательность управляет экспрессией кодирующей последовательности.
Активность фосфолипазы A. В контексте настоящего изобретения термин "активность фосфолипазы A" предусматривает ферменты, обладающие активностью фосфолипазы A1 и/или фосфолипазы A2 (A1 или A2, EC 3.1.1.32 или EC 3.1.1.4), т.е. гидролитической активностью в отношении одной или обеих сложноэфирных связей с карбоксильной группой в фосфолипидах, таких как лецитин. Фосфолипаза, обладающая активностью как A1, так и A2, также называется фосфолипазой B.
Для целей настоящего изобретения активность фосфолипазы A предпочтительно определяют в соответствии со следующей процедурой.
Активность фосфолипазы A (LEU)
В анализе LEU активность фосфолипазы A определяют исходя из способности гидролизовать лецитин при значении pH 8,0, 40°C. После реакции гидролиза можно осуществлять титрование с помощью NaOH в течение времени реакции, составляющего 2 минуты. Фосфолипаза из Fusarium oxysporum (LIPOPAN F), раскрытая в WO 1998/26057, обладает активностью, составляющей 1540 LEU/мг ферментного белка, и ее можно применять в качестве стандартного образца.
Анализ на планшетах
A) Буфер представляет собой смесь 100 мM HEPES и 100 мM цитратного буфера со значением pH, отрегулированным от pH 3,0 до pH 7,0.
B) 2% агарозу (Litex HSA 1000) получают путем смешивания и подвергания тепловой обработке в буфере (A) в течение 5 минут с последующим охлаждением до примерно 60°C.
C) Субстрат представляет собой 95% L-альфа-фосфатидилхолин из сои (Avanti 441601), диспергированный в воде (MilliQ) при 60°C в течение 1 минуты с помощью Ultra Turrax.
D) Растворы очищенных ферментов от LECITASE ULTRA и зрелой фосфолипазы под SEQ ID NO: 2 разбавляли до 0,4 мг/мл.
В планшеты заливают смесь из 5 мл субстрата (C) и 5 мл агарозы (B), осторожно перемешанную в чашках Петри с диаметром, составляющим 7 см, и охлаждают до комнатной температуры перед проделыванием отверстий диаметром, составляющим примерно 3 мм, под действием вакуума. Десять микролитров разбавленного фермента (D) добавляют в каждую лунку перед герметизацией планшетов с помощью парафильма и помещения в инкубатор при 55°C на 48 часов. Планшеты доставали для фотографирования через равные промежутки времени.
Активность фосфолипазы. В контексте настоящего изобретения термин "активность фосфолипазы" относится к катализу гидролиза глицерофосфолипида или полученного из глицерина фосфолипида.
Условия, способствующие гидролизу фосфолипидов. Выбор условий, которые будут способствовать гидролизу фосфолипидов посредством ферментов, расщепляющих фосфолипиды, находится в компетенции специалиста в данной области техники и включает, например, регулирование значения pH и/или температуры, при которых фермент, расщепляющий фосфолипиды, является активным.
Активность фосфолипазы C. Термин "активность фосфолипазы C" или "активность PLC" относится к ферментативной активности, в результате которой фрагмент сложного эфира фосфорной кислоты удаляется из фосфолипида с получением 1,2-диацилглицерина (см. фигуру 1). Большинство ферментов, представляющих собой PLC, принадлежат к семейству гидролаз и фосфодиэстераз и в общем классифицированы как EC 3.1.4.3, E.C. 3.1.4.11 или EC 4.6.1.13. Активность фосфолипазы C можно определить в соответствии с процедурой, описанной в следующем подразделе "Анализ активности фосфолипазы C".
Анализ активности фосфолипазы C. Реакционные смеси, содержащие 10 мкл 100 мМ раствора п-нитрофенилфосфорилхолина (p-NPPC) в 100 мМ буфере бура-HCl, pH 7,5, и 90 мкл раствора фермента, перемешивают в лунке титрационного микропланшета при температуре окружающей среды. Титрационный микропланшет затем помещают в ридер для титрационных микропланшетов, и количество высвободившегося п-нитрофенола определяют посредством измерения поглощения при 410 нм. Результаты измерений фиксируют в течение 30 мин с интервалами в 1 минуту. Калибровочные кривые в диапазоне 0,01-1 мкл/мл п-нитрофенола получают путем разбавления 10 микромоль/мл исходного раствора п-нитрофенола от Sigma в буфере бура-HCl. Одна единица будет соответствовать высвобождению 1,0 микромоль/минута p-NPPC при температуре окружающей среды.
Специфичность фосфолипазы C. Термин "специфичность фосфолипазы C" относится к полипептиду, обладающему активностью фосфолипазы С, где активность является специфичной в отношении одного или нескольких фосфолипидов, причем четырьмя наиболее важными являются фосфатидилхолин (PC), фосфатидилэтаноламин (PE), фосфатидная кислота (PA) и фосфатидилинозитол (PI) (см. фигуру 1). Специфичность фосфолипазы C можно определить с помощью 31P-ЯМР, как описано выше в отношении термина "активность фосфолипазы".
PC- и PE-специфичная фосфолипаза C. Термины "PC- и PE-специфичная фосфолипаза C", и "фосфолипаза C, обладающая специфичностью в отношении фосфатидилхолина (PC) и фосфатидилэтаноламина (PE)", и "полипептид, обладающий активностью в отношении фосфатидилхолина (PC) и фосфатидилэтаноламина (PE)" используются взаимозаменяемо. Они относятся к полипептиду, обладающему активностью в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE). В дополнение к PC- и PE-специфичности он может также обладать некоторой активностью в отношении фосфатидной кислоты (PA) и фосфатидилинозитола (PI). Предпочтительно PC- и PE-специфичная фосфолипаза C обеспечивает удаление по меньшей мере 30% PC и по меньшей мере 30% PE из масла или жира с по меньшей мере 100 ppm PC и 100 ppm PE при применении анализа посредством P-ЯМР из примера 1 при оптимальном значении pH для фермента и дозе фермента, составляющей 10 мг/кг. Более предпочтительно она обеспечивает удаление 40%, 50%, 60%, 70% или 80%, еще более предпочтительно она обеспечивает удаление 90% и наиболее предпочтительно она обеспечивает удаление от 90% до 100% PC из масла или жира, и 40%, 50%, 60%, 70% или 80%, еще более предпочтительно она обеспечивает удаление 90% и наиболее предпочтительно она обеспечивает удаление от 90% до 100% PE из масла или жира.
PI-специфичная фосфолипаза C. Термины "PI-специфичная фосфолипаза C", "фосфолипаза C, действующая в отношении фосфатидилинозитола" и "полипептид, обладающий активностью в отношении фосфатидилинозитола (PI)" используются взаимозаменяемо. Они относятся к полипептиду, обладающему активностью в отношении фосфатидилинозитола (PI), что означает то, что его активность в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE), фосфатидной кислоты (PA) является низкой по сравнению с активностью в отношении PI. Ферменты, представляющие собой PI-специфичную фосфолипазу C, могут принадлежать к семейству гидролаз и фосфодиэстераз, классифицированных как EC 3.1.4.11, либо к семейству лиаз, классифицированных как EC 4.6.1.13. Активность PI-специфичной фосфолипазы C можно определить в соответствии с процедурой, описанной в примере 5. Предпочтительно PI-специфичная фосфолипаза C обеспечивает удаление по меньшей мере 30% PI из масла или жира с по меньшей мере 50 ppm PI при применении анализа посредством P-ЯМР из примера 1 при оптимальном значении pH для фермента и дозе фермента, составляющей 10 мг/кг. Более предпочтительно она обеспечивает удаление 40%, 50%, 60%, 70% или 80%, еще более предпочтительно она обеспечивает удаление 90% и наиболее предпочтительно она обеспечивает удаление от 90% до 100% PI из масла или жира.
Предпочтительно PI-специфичная фосфолипаза C обеспечивает удаление PI в количестве, на по меньшей мере 20% превышающем количество PC, PE или PA, удаление которого она может обеспечить, более предпочтительно удаление PI в количестве, на по меньшей мере 30%, 40%, еще более предпочтительно на по меньшей мере 50% и наиболее предпочтительно на по меньшей мере 60% превышающем количество PC, PE или PA, удаление которого она может обеспечить.
PC-, PE-, PA- и PI-специфичная фосфолипаза C. Термины "PC-, PE-, PA- и PI-специфичная фосфолипаза C" и "полипептид, обладающий активностью в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE), фосфатидной кислоты (PA) и фосфатидилинозитола (PI)" используются взаимозаменяемо. Они относятся к полипептиду, обладающему активностью в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE), фосфатидной кислоты (PA) и фосфатидилинозитола (PI). Предпочтительно PC-, PE-, PA- и PI-специфичная фосфолипаза C обеспечивает удаление по меньшей мере 30% каждой из четырех молекул фосфолипидов из масла или жира с по меньшей мере 100 ppm PC, 75 ppm PE, 5 ppm PA и 50 ppm PI при применении анализа посредством P-ЯМР из примера 1 при оптимальном значении pH для фермента и дозе фермента, составляющей 10 мг/кг. Более предпочтительно она обеспечивает удаление 40%, 50%, 60%, 70% или 80%, еще более предпочтительно она обеспечивает удаление 90% и наиболее предпочтительно она обеспечивает удаление от 90% до 100% PC из масла или жира, и 40%, 50%, 60%, 70% или 80%, еще более предпочтительно она обеспечивает удаление 90% и наиболее предпочтительно она обеспечивает удаление от 90% до 100% PE из масла или жира.
Очищенный. Термин "очищенный" означает нуклеиновую кислоту или полипептид, которые по сути не содержат других компонентов, с которыми они ассоциированы в природе, как определено с помощью аналитических методик, хорошо известных из уровня техники (например, очищенные полипептид или нуклеиновая кислота могут образовывать дискретную полосу в геле для электрофореза, хроматографическом элюате и/или среде, подвергнутой центрифугированию в градиенте плотности). Чистота очищенных нуклеиновой кислоты или полипептида составляет по меньшей мере приблизительно 50%, обычно чистота составляет по меньшей мере приблизительно 60%, приблизительно 65%, приблизительно 70%, приблизительно 75%, приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99%, приблизительно 99,5%, приблизительно 99,6%, приблизительно 99,7%, приблизительно 99,8% или больше (например, процент по весу в пересчете на моли). В схожем смысле композиция является обогащенной молекулой, если имеет место значительное увеличение концентрации молекулы после применения методики очистки или обогащения. Термин "обогащенный" относится к соединению, полипептиду, клетке, нуклеиновой кислоте, аминокислоте или другому указанному материалу или компоненту, которые присутствуют в композиции в более высокой относительной или абсолютной концентрации, чем в исходной композиции.
Рекомбинантный. Термин "рекомбинантный" при использовании в отношении клетки, нуклеиновой кислоты, белка или вектора, являющихся субъектом, указывает на то, что субъект был получен в результате модификации из его нативного состояния. Таким образом, например, рекомбинантные клетки экспрессируют гены, которые не обнаружены в нативной (нерекомбинантной) форме клетки, или экспрессируют нативные гены на других уровнях или при других условиях, чем те, которые обнаружены в природе. Рекомбинантные нуклеиновые кислоты отличаются от нативной последовательности одним или несколькими нуклеотидами и/или функционально связаны с гетерологичными последовательностями, например, гетерологичным промотором в векторе экспрессии. Рекомбинантные белки могут отличаться от нативной последовательности одной или нескольким аминокислотами и/или быть слитыми с гетерологичными последовательностями. Вектор, содержащий нуклеиновую кислоту, кодирующую полипептид, представляет собой рекомбинантный вектор. Термин "рекомбинантный" является синонимичным с терминами "генетически модифицированный" и "трансгенный".
Скорость реакции. Для цели настоящего изобретения термин "скорость реакции" является синонимичным с термином "скорость протекания реакции" и определен в соответствии с IUPAC, Compendium of Chemical Terminology, 2nd ed. ("Gold Book") (1997): "Скорость протекания реакции".
Идентичность последовательностей. Родство между двумя аминокислотными последовательностями или между двумя нуклеотидными последовательностями описывается параметром "идентичность последовательностей".
Для целей настоящего изобретения идентичность последовательностей для двух аминокислотных последовательностей определяют в виде полученного "наиболее длинного идентичного участка" с применением алгоритма Нидлмана-Вунша (Needleman and Wunsch, 1970, J. Mol. Biol. 48: 443-453), реализованного в программе Needle из пакета программ EMBOSS (EMBOSS: The European Molecular Biology Open Software Suite, Rice et al., 2000, Trends Genet. 16: 276-277), предпочтительно версии 6.6.0 или более поздней. Применяемыми параметрами являются штраф за открытие гэпа, составляющий 10, штраф за продление гэпа, составляющий 0,5, и матрица замен EBLOSUM62 (версия EMBOSS с BLOSUM62). Чтобы программа Needle выдала сообщение о наиболее длинном идентичном участке, опция nobrief должна быть задана в командной строке. Выходные данные Needle, помечающие "наиболее длинный идентичный участок", рассчитывают следующим образом:
(идентичные остатки x 100) / (длина выравниваемого участка - общее число гэпов в выравниваемом участке).
Соапсток. В данном контексте "соапсток" относится к фракции, содержащей мыла, которую отделяют от основной массы растительного масла в ходе осуществления способа химического рафинирования. Мыла образуются посредством реакции химического вещества для рафинирования, такого как щелочь, со свободными жирными кислотами, которые присутствуют в растительном масле. Точный состав соапстоков зависит от источника растительного масла, из которого их получают; например, было обнаружено, что хлопковый соапсток в основном состоит из влаги и растворителя, жирных кислот, органических фосфатов, моноглицеридов, диглицеридов, триглицеридов, стеринов, многоатомных спиртов, углеводов и прочих других компонентов. Большинство таких классов органических соединений обнаружены в соапстоках из других растительных масел.
Стехиометрическое количество. Термин "стехиометрическое количество" в сущности означает измерение количества, требуемого для соблюдения стехиометрии; т. е. оптимального количества, при этом, принимая во внимание, что реакция протекает до завершения, весь реагент расходуется, нет дефицита реагента и нет избытка реагента.
В контексте настоящего изобретения "стехиометрическое количество" относится, в частности, к количеству молей реагента (например, щелочи, такой как NaOH), добавляемого в реакционную смесь, которое равняется количеству молей соединений (например, свободных жирных кислот и/или кислоты, добавляемой в качестве средства, образующего хелатный комплекс с кальцием, такой как лимонная кислота), с которыми реагент реагирует в указанной реакционной смеси.
Удаление смолистых веществ с помощью воды. Термин "удаление смолистых веществ с помощью воды" относится к способу, который предусматривает обработку неочищенного масла некоторым количеством воды, необходимым для гидратации фосфолипидов, присутствующих в масле, и обеспечения возможности их отделения посредством центрифугирования.
В первом аспекте настоящее изобретение предусматривает способ рафинирования растительного масла, содержащего фосфолипиды, предусматривающий осуществление ферментативного гидролиза фосфолипидов путем приведения растительного масла в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, в условиях, способствующих гидролизу фосфолипидов, и после этого осуществление химического рафинирования растительного масла.
В предпочтительных вариантах осуществления настоящего изобретения отделение продуктов реакции ферментативного гидролиза от масла перед указанным химическим рафинированием не осуществляют или осуществляют в небольшом количестве. Следовательно, ферментативный гидролиз фосфолипидов можно осуществлять в первом реакционном сосуде, а химическое рафинирование можно осуществлять во втором реакционном сосуде, при этом два реакционных сосуда сообщены по текучей среде и/или сообщены таким образом, что обеспечивается возможность прохождения жидкости из первого во второй реакционный сосуд.
В дополнительных предпочтительных вариантах осуществления ферментативный гидролиз фосфолипидов и химическое рафинирование осуществляют в одном и том же сосуде. То есть ферментативный гидролиз фосфолипидов проводят в реакционном сосуде и химическое рафинирование проводят после ферментативного гидролиза в том же реакционном сосуде. Такие варианты осуществления позволяют избежать необходимости переносить содержимое первого реакционного сосуда во второй реакционный сосуд, поскольку обе реакции происходят в одном и том же реакционном сосуде.
В дополнительных вариантах осуществления химическое рафинирование проводят одновременно с ферментативным гидролизом или после него, предпочтительно после ферментативного гидролиза.
Способ по настоящему изобретению можно осуществлять в виде периодического способа или в виде непрерывного способа. Таким образом, способ может подходить для существующей схемы способа независимо от того, представляет ли она собой периодический технологический процесс или обычный непрерывный процесс, применяемые в промышленности. Один конкретный вариант осуществления относится к способу в соответствии с настоящим изобретением, где химическое рафинирование осуществляют непосредственно после ферментативного гидролиза; предпочтительно посредством непрерывного технологического процесса.
Кроме того, следует понимать, что ферментативный гидролиз фосфолипидов можно осуществлять в первом реакционном сосуде и химическое рафинирование можно осуществлять во втором реакционном сосуде, где сообщение по текучей среде между реакционными сосудами или прохождение жидкости из первого во второй реакционный сосуд обеспечивается не через разделительное устройство, такое как центрифуга.
В некоторых вариантах осуществления способ в соответствии с настоящим изобретением представляет собой способ, где
- ферментативный гидролиз проводят в реакционной смеси, содержащей тяжелую фазу или водную фазу и легкую фазу или масляную фазу/гидрофобную фаза, и
- до указанного химического рафинирования не происходит снижение или значительное снижение объема тяжелой фазы или не осуществляют отделение смолистых веществ/тяжелой фазы от масла.
При традиционном удалении смолистых веществ две фазы смешивают, например, с применением смесителя с высоким сдвиговым усилием, и создают эмульсию. В эмульсии фермент реагирует с фосфолипидами с получением водорастворимых продуктов реакции. Обеспечивают расслоение эмульсии, например, посредством центрифугирования, отделяя водорастворимые продукты реакции от масла. Способ в соответствии с настоящим изобретением предпочтительно не включает любую стадию для отделения тяжелой фазы/водной фазы или ее части, содержащей водорастворимые продукты реакции, от легкой фазы, масляной фазы или гидрофобной фазы.
Предпочтительно ферментативный гидролиз проводят путем приведения указанного растительного масла в контакт с одним или несколькими ферментами, обладающими активностью фосфолипазы.
Способ в соответствии с настоящим изобретением может предусматривать:
i) получение реакционной смеси, содержащей указанное растительное масло и один или несколько ферментов, обладающих активностью расщепления фосфолипидов, такой как реакционная смесь, содержащая тяжелую фазу или водную фазу и легкую фазу или масляную фазу/гидрофобную фазу;
ii) помещение реакционной смеси в условия, обеспечивающие возможность осуществления ферментативного гидролиза фосфолипидов в масле, с получением прореагировавшей смеси на основе указанного растительного масла и
iii) осуществление химического рафинирования прореагировавшей смеси на основе указанного растительного масла.
Способ в соответствии с настоящим изобретением может дополнительно предусматривать осуществление удаления из растительного масла смолистых веществ с помощью воды перед приведением его в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды.
В дополнительных вариантах осуществления растительное масло выбрано из группы, состоящей из масла асаи, миндального масла, масла из ореха бабассу, масла семян черной смородины, масла семян огуречника, масла канолы, масла из ореха кешью, касторового масла, кокосового масла, кориандрового масла, кукурузного масла, хлопкового масла, масла крамбе, масла семян льна, масла виноградных косточек, масла из лесного ореха, конопляного масла, ятрофного масла, масла жожоба, льняного масла, масла из ореха макадамия, масла из косточек манго, масла пенника лугового, горчичного масла, костного масла, оливкового масла, пальмового масла, пальмоядрового масла, пальмового олеина, арахисового масла, масла из ореха пекан, масла из кедрового ореха, фисташкового масла, макового масла, рапсового масла, масла из рисовых отрубей, сафлорового масла, масла сасанквы, кунжутного масла, масла ши, соевого масла, масла семян подсолнечника, таллового масла, масла камелии и масла из грецкого ореха.
В предпочтительных вариантах осуществления настоящего изобретения растительное масло выбрано из группы, состоящей из рапсового масла, соевого масла, масла семян подсолнечника, пальмового масла, кокосового масла, масла из рисовых отрубей и арахисового масла/масла из земляного ореха. Такие растительные масла считаются важными с коммерческой точки зрения, поскольку они широко распространены и поскольку обрабатывают большие объемы масла для получения пищевого растительного масла очень светлого цвета для соответствия предпочтениям потребителей или применяют в качестве сырья для получения биотоплива.
В некоторых вариантах осуществления настоящего изобретения растительное масло, которое приводят в контакт с указанными одним или несколькими ферментами, расщепляющими фосфолипиды, представляет собой неочищенное растительное масло.
Способ в соответствии с настоящим изобретением может предусматривать приведение растительного масла в контакт с одним или несколькими хелатообразующими средствами, которые могут образовывать комплексы с ионами Ca и/или Mg, до его приведения в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды. Подходящие хелатообразующие средства могут быть выбраны из группы, состоящей из лимонной кислоты, фосфорной кислоты, молочной кислоты и этилендиаминтетрауксусной кислоты (EDTA).
Реакционная смесь может характеризоваться значением pH, которое находится в диапазоне 1,5-7. Как будет понятно специалисту в данной области техники, потребность в регулировании значения pH зависит от требований для применяемого(-ых) фермента(-ов) и от количества любого хелатообразующего средства, которое было добавлено. В частности, значение pH может находиться в пределах диапазона 3-7, такого как 3,5-6,6, в пределах диапазона 3-5, такого как 3,5-4,5, или в пределах диапазона 5-7, такого как 4,5-6,5.
В одном варианте осуществления значение pH регулируют путем добавления основания, например, путем добавления NaOH, KOH, карбоната натрия или их комбинаций. В конкретных вариантах осуществления количество эквивалентов основания, применяемого для нейтрализации кислоты, применяемой для предварительной обработки, находится в диапазоне от 1,2 до 7 эквивалентов, например, от 1,5 до 6, от 1,5 до 5 эквивалентов; или, например, от 2 до 7, от 3 до 7, или, например, от 3 до 7 или от 3 до 5 эквивалентов кислоты; в дополнительных конкретных вариантах осуществления один или несколько ферментов, расщепляющих фосфолипиды, содержат или состоят из SEQ ID NO. 11 и SEQ ID NO: 13.
В способе в соответствии с настоящим изобретением реакционная смесь характеризуется содержанием воды, находящимся в диапазоне 0,5-10% (вес/вес), например, в диапазоне 1-10% (вес/вес), в диапазоне 1-5% (вес/вес), например, в диапазоне 0,5-5% (вес/вес), например, содержанием воды, составляющим 5% (вес/вес) или меньше, например, содержанием воды, составляющим 4% или меньше, или, например, содержанием воды, составляющим 3% или меньше.
Растительное масло приводят в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, при температуре, которая находится в диапазоне 45-90°C, например, в диапазоне 50-90°C, 60-90°C, 60-80°C, 65-75°C, или, например, 65-75°C.
Ферментативный гидролиз фосфолипидов может характеризоваться продолжительностью, составляющей 6 часов или меньше, например, 4 часа или меньше, например, продолжительностью, составляющей 0,5-6 часов или 0,5-4 часа, или, например, продолжительностью, составляющей 5 минут - 4 часа, например, от 5 минут до 2 часов, от 5 минут до 1 часа, или, например, 5-30 минут.
Один или несколько ферментов, обладающих активностью расщепления фосфолипидов, можно вводить в общем количестве, соответствующем 0,1-30 мг ферментного белка.
В способе в соответствии с настоящим изобретением растительное масло предпочтительно приводят в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, в таких условиях, при которых в ходе ферментативного гидролиза обеспечивается снижение количества интактных молекул фосфолипидов на 30-100%, например, на 30-90%, 30-80%, 30-70% или, например, на 30-60%. Процентное содержание интактных молекул фосфолипидов можно определить путем сравнения процентного содержания фосфатидилхолина (PC) + фосфатидилэтаноламина (PE) + фосфатидилинозитола (PI) + фосфатидной кислоты (PA (PC+PE+PI+PA), присутствующих после реакции, с содержанием PC+PE+PI+PA в масле перед реакцией. Содержание фосфолипидов можно определить с помощью анализа посредством 31P-ЯМР или жидкостной хроматографии-масс-спектрометрии (LC-MS). В некоторых вариантах осуществления растительное масло приводят в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, в таких условиях, при которых в результате ферментативной реакции происходит по меньшей мере 10% снижение содержания PC+PE+PI+PA в масле, например, по меньшей мере 25% или по меньшей мере 40% снижение содержания PC+PE+PI+PA в масле. Следует понимать, что для обеспечения преимущества касательно увеличения выхода, обеспечиваемого способом по настоящему изобретению, не будет требоваться полный или почти полный гидролиз фосфолипидов; даже частичный гидролиз фосфолипидов, присутствующих в масле, будет улучшать выход масла и облегчать отделение мыльной фазы после химического рафинирования.
Предпочтительно, чтобы растительное масло после осуществления химического рафинирования в соответствии со способом по настоящему изобретению содержало фосфолипиды в количествах, соответствующих 20 ppm фосфора или меньше, например, 15 ppm или меньше, например, 10 ppm или меньше или, например, 5 ppm или меньше. Предпочтительно количества фосфолипидов определяют в соответствии с AOCS Official Method Ca 20-99 (2009), Analysis for Phosphorous in Oil by Inductively Coupled Plasma Optical Emission Spectroscopy (ICP-OES), Official Methods and Recommended Practices of the AOCS, AOCS Press, Champaign IL. Дополнительное руководство в отношении того, как определять количества фосфора в масле, предоставлено в Z. Benzo et al.: Determination of phosphorus in edible oils by inductively coupled plasma-Atomic emission spectrometry and oil-in-water emulsion of sample introduction, Journal of the American Oil Chemists' Society, September 2000, Volume 77, Issue 9, pp. 997-1000.
В дополнительных предпочтительных вариантах осуществления настоящего изобретения растительное масло и указанные один или несколько ферментов, обладающих активностью расщепления фосфолипидов, инкубируют в течение 0,1-6 часов, как, например, 0,25-6 часов, или, например 0,5-6 часов, при ряде условий, включающем:
a) температуру в диапазоне 45-90°C или, например, в диапазоне 60-80°C;
b) значение pH в диапазоне от приблизительно 1,5 до приблизительно 12,0, например, в диапазоне от 1,5 до 7,0, в диапазоне значений pH 4-7, в диапазоне 3-6, в диапазоне 6-9 или в диапазоне значений pH 7-12;
c) перемешивание или смешивание, например, путем смешивания со сдвиговым усилием, смешивания с высоким сдвиговым усилием, смешивания посредством кавитации или воздействия ультразвука.
В предпочтительных вариантах осуществления настоящего изобретения химическое рафинирование проводят после ферментативного гидролиза. В дополнительных предпочтительных вариантах осуществления химическое рафинирование проводят непосредственно после ферментативного гидролиза без промежуточных стадий разделения. Как указано выше, стадию химического рафинирования можно осуществлять в том же реакционном сосуде, в котором проводили ферментативный гидролиз.
Химическое рафинирование в случае осуществления в соответствии с настоящим изобретением может предусматривать смешивание растительного масла со щелочью, например, смешивание прореагировавшей смеси на основе указанного растительного масла, определенной выше, со щелочью.
Щелочь предпочтительно вводят в количествах, которые превышают стехиометрические количества относительно количеств свободных жирных кислот, присутствующих в масле. Как специалисту в данной области техники будет понятно из контекста настоящего изобретения, количество щелочи, вводимое в способе, предпочтительно превышает количество, которое является достаточным для нейтрализации свободных жирных кислот и любого хелатообразующего средства, такого как лимонная, молочная или фосфорная кислота.
В частности, щелочь может быть выбрана из NaOH, KOH, карбоната натрия и их комбинаций.
Авторы настоящего изобретения неожиданно обнаружили, что взаимозависимость между количеством щелочи, применяемым для нейтрализации любой кислоты для предварительной обработки, и количеством щелочи, вводимом в ходе химического рафинирования, может обладать положительным эффектом в отношении снижения содержания фосфора и FFA-кислоты в конечном образце (см. пример 12). Соответственно, конкретные варианты осуществления настоящего изобретения относятся к способам в соответствии с настоящим изобретением, где количество щелочи, добавляемое в ходе химического рафинирования, составляет по меньшей мере 60% от общего количества щелочи, добавляемого в ходе осуществления способа (т.е. щелочи, добавляемой после предварительной обработки кислотой с целью регулирования значения pH до ферментативного гидролиза, вместе со щелочью, добавляемой для химического рафинирования); предпочтительно указанное количество щелочи, добавляемое на стадии щелочного рафинирования, находится в диапазоне 60-90%, например, 60-85%, 60-80%, от 60 до 78%, или, например, от 62 до 76%.
В качестве альтернативы настоящее изобретение можно описать как такое, где количество щелочи (например, NaOH), добавляемое на стадии регулирования значения pH после предварительной обработки кислотой, составляет не больше 40% от общего количества щелочи (т.е. щелочи, добавляемой после предварительной обработки кислотой с целью регулирования значения pH до ферментативного гидролиза, вместе со щелочью, добавляемой для химического рафинирования); предпочтительно указанное количество основания, добавляемое на стадии регулирования значения pH, находится в диапазоне 10-40%, например, 15-40%, 20-40%, например, 40-22%, или, например, от 24% до 48%.
В способе в соответствии с настоящим изобретением смесь указанного растительного масла и указанной щелочи предпочтительно инкубируют от 1 минуты до 8 часов, например, от 1 минуты до 5 часов, от 1 минуты до 2 часов, от 5 минут до 8 часов, от 5 минут до 5 часов, от 5 минут до 2 часов, от 10 минут до 5 часов, от 10 минут до 2 часов, от 20 минут до 5 часов или от 20 минут до 2 часов.
Авторы настоящего изобретения неожиданно обнаружили, что введение дополнительной стадии подкисления, осуществляемой после ферментативного гидролиза и до химического рафинирования, может обеспечить снижение количества фосфора в конечном образце после удаления смолистых веществ (см. пример 14). Таким образом, некоторые варианты осуществления относятся к способу в соответствии с настоящим изобретением, дополнительно предусматривающему стадию подкисления, осуществляемую после ферментативного гидролиза и до химического рафинирования.
Конкретные варианты осуществления настоящего изобретения относятся к таковым, где количество эквивалентов основания, применяемого для нейтрализации кислоты, применяемой для предварительной обработки, находится в диапазоне от 0,5 до 7 эквивалентов, например, от 0,5 до 6, от 0,5 до 5, или, например, от 1,2 до 7 эквивалентов, например, от 1,5 до 6, от 1,5 до 5 эквивалентов; или, например, от 2 до 7, от 3 до 7, или, например, от 3 до 7 или от 3 до 5 эквивалентов кислоты, и при этом способ предусматривает дополнительную стадию подкисления, как описано.
В конкретных вариантах осуществления настоящего изобретения, предусматривающих дополнительную стадию подкисления, один или несколько ферментов, расщепляющих фосфолипиды, содержат или состоят из SEQ ID NO: 11 и SEQ ID NO: 13.
Химическое рафинирование, осуществляемое в соответствии с настоящим изобретением, предпочтительно предусматривает отделение смолистых веществ и/или соапстока от масла.
Соответственно, способ в соответствии с настоящим изобретением может предусматривать осуществление переноса смеси растительного масла и химического вещества, такой как смесь прореагировавшей смеси на основе указанного растительного масла и химического вещества, в сепаратор, предпочтительно центробежный сепаратор или горизонтальный отстойник.
В промышленности химическое рафинирование обычно осуществляют с применением так называемых способов "длительного перемешивания" или "короткого перемешивания" или их вариантов. В "способе длительного перемешивания" относительно большой избыток каустической соды смешивают с маслом при относительно низкой температуре (например, 20-40°C), при этом применяют время выдержки с перемешиванием, составляющее 3-6 минут, после чего обеспечивают расслоение смеси масло/мыло путем нагревания при 60-80°C. Затем смесь загружают в сепаратор; например, центробежный сепаратор, и поток масла, выходящий из центрифуги, нагревают, промывают водой и высушивают, например, в вакуумной распылительной сушилке. В "способе короткого перемешивания" относительно небольшой избыток щелочи добавляют в масло, после чего смесь почти сразу загружают в сепаратор, например, центробежный сепаратор, промывают водой и высушивают.
В обоих способах масло можно последовательно осветлять с удалением красящих соединений и дезодорировать с удалением летучих соединений, обуславливающих неприятные запахи и вкусы.
Следовательно, в конкретных вариантах осуществления способ в соответствии с настоящим изобретением предусматривает:
i) смешивание растительного масла или прореагировавшей смеси, определенной выше в данном документе, со щелочью при температуре 20-90°C, например, 20-80°C, например, 20-40°C, при этом количества щелочи превышают стехиометрические количества;
ii) инкубирование смеси растительного масла и щелочи или смеси прореагировавшей смеси и щелочи при температуре 20-80°C, например, 20-40°C, в течение 2-15 минут с перемешиванием;
iii) повышение температуры смеси до 55-95°C, например, до 55-85°C; и
iv) после достижения температуры 55-95°C, например, 55-85°C, загрузку смеси в сепаратор с отделением смолистых веществ и/или соапстока от масла.
В альтернативных вариантах осуществления настоящего изобретения способ предусматривает:
i) смешивание растительного масла или прореагировавшей смеси, определенной выше, со щелочью, при этом количества щелочи превышают стехиометрические количества;
ii) загрузку смеси растительного масла, кислоты и щелочи или смеси прореагировавшей смеси, кислоты и щелочи в сепаратор с отделением смолистых веществ и/или соапстока от масла.
Способы рафинирования с помощью каустической соды с "длительным перемешиванием" или "коротким перемешиванием" раскрыты, например, в A.J. Dijkstra: Degumming, Refining, Washing and Drying Fats and Oils; в Proceedings of the World Conference on Oilseed Technology and Utilization (1992), Budapest, Hungary; T. H. Applewhite (ed); pp. 138-151; и в Lipid Handbook, 3rd Edition; F.D. Gunstone, J.L. Harwood, A.J. Dijkstra (Eds.), CRC Press, Taylor & Francis Group, 6000 Broken Sound Parkway NW, Suite 300, Boca Raton, FL 33487-2742, © 2007, Taylor & Francis Group, LLC; см. главу 3: Production and refining of oils and fats, A.J. Dijkstra and J.C. Segers: способ длительного перемешивания описан на стр. 193, способ короткого перемешивания описан на стр. 195.
Рафинирование с помощью каустической соды с применением оборудования, работающего под давлением, с применением способа нанонейтрализации раскрыто на сайте по адресу www.nanoneutralization.com.
Указанные один или несколько ферментов, обладающих активностью расщепления фосфолипидов, могут включать фермент, обладающий активностью фосфолипазы A, фермент, обладающий активностью фосфолипазы С, лизофосфолипазу или их смесь.
Известно несколько типов фосфолипаз, которые отличаются по их специфичности в зависимости от положения связи, на которую они воздействуют в молекуле фосфолипида. Фосфолипаза A1 (PLA1) отщепляет жирную кислоту в положении 1 с образованием свободной жирной кислоты и 1-лизо-2-ацилфосфолипида. Фосфолипаза A2 (PLA2) отщепляет жирную кислоту в положении 2 с образованием свободной жирной кислоты и 1-ацил-2-лизофосфолипида. Термин "фосфолипаза B" (PLB) используется применительно к фосфолипазам, обладающим активностью как A1, так и A2. Фосфолипаза C (PLC) отщепляет фосфатный фрагмент с образованием 1,2-диацилглицерина и сложного эфира фосфорной кислоты. Фосфолипаза D (PLD) обеспечивает образование 1,2-диацилглицерофосфата и основной группы (см. фигуру 1).
Для обзора удаления смолистых веществ с помощью ферментов см. Dijkstra 2010 Eur. J. Lipid Sci. Technol. 112, 1178. Применение фосфолипазы A и/или фосфолипазы C в удалении смолистых веществ, например, описано в Clausen 2001 Eur J Lipid Sci Techno 103 333-340, WO 2003/089620 и WO 2008/094847. Растворы фосфолипазы A обеспечивают получение лизофосфолипида и свободных жирных кислот, в результате чего происходит потеря масла. Фосфолипаза C, напротив, обладает преимуществом в том, что она обеспечивает получение диглицерида (фигура 2), при этом будет оставаться в масле и, следовательно, будет обеспечивать снижение потерь. В растительном масле присутствуют четыре основных фосфолипида: фосфатидилхолин (PC), фосфатидилэтаноламин (PE), фосфатидная кислота (PA) и фосфатидилинозитол (PI). Ферменты, представляющие собой фосфолипазу C, обладают разной специфичностью в отношении данных фосфолипидов. Коммерчески доступной фосфолипазой C является Purifine от Verenium/DSM (Dijkstra, 101-е ежегодное заседание AOCS, 10 мая 2010 года), которая обладает специфичностью в отношении PC и PE. В WO07/059927 описывается термоустойчивая PLC Bacillus, используемая для удаления смолистых веществ. В WO 2012/062817 описывается PLC гриба со специфичностью в отношении всех четырех фосфолипидов.
Для цели настоящего изобретения может быть предпочтительно, чтобы фермент или ферменты, обладающие активностью расщепления фосфолипазы, включали PLC. Вдобавок к увеличению выхода, наблюдаемому авторами настоящего изобретения, также важным является то, что гидролиз фосфолипидов посредством PLC не приводит к образованию свободных жирных кислот. В целом, требуется, чтобы образование свободных жирных кислот было сведено к минимуму в ходе обработки растительного масла.
В рамках настоящего изобретения наблюдали, что увеличение выхода обычно ниже в случае применения PLA для гидролиза фосфолипидов перед химическим рафинированием по сравнению с применением PLC. Однако дополнительные преимущества способа в соответствии с настоящим изобретением, которые включают более низкие уровни фосфолипидов в полученном соапстоке и более низкую вязкость соапстока, также достигаются при применении PLA. В определенных вариантах осуществления настоящего изобретения лизофосфолипаза может быть предпочтительной, поскольку она превращает эмульгирующие лизофосфолипиды в соединения, не обладающие эмульгирующей способностью.
В отношении способа по настоящему изобретению один или несколько ферментов, расщепляющих фосфолипиды, могут обладать одним или несколькими из следующий свойств:
i) температура диссоциации (Td) в диапазоне 50-95°C, например, 60-95°C, 70-95°C, например, в диапазоне 70-90°C;
ii) оптимальное значение pH в диапазоне pH 3-12, например, в диапазоне pH 4-7, или, например, значение pH в диапазоне 3-6, например 3,5-6 или 4-6, или, например, значение pH в диапазоне 5-9, например, 6-9 или 6-8, или, например, в диапазоне pH 7-12, например, 8-12 или 8-10.
Предпочтительно один или несколько ферментов, расщепляющих фосфолипиды, характеризуются скоростью реакции в отношении фосфолипидов в растительном масле, в которое было добавлено одно или несколько хелатообразующих средств, которые могут образовывать комплексы с ионами Ca и/или Mg, при этом указанная скорость реакции составляет по меньшей мере 30%, например, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80% или, например, по меньшей мере 90% от скорости реакции для одного или нескольких ферментов, расщепляющих фосфолипиды, в отношении фосфолипидов в указанном растительном масле, в которое не было добавлено хелатообразующее(-ие) средство(-а). Как указано выше, подходящие хелатообразующие средства могут быть выбраны из группы, состоящей из лимонной кислоты, фосфорной кислоты, молочной кислота и EDTA. В таких вариантах осуществления растительное масло предпочтительно представляет собой неочищенное соевое масло, а хелатообразующее средство предпочтительно представляет собой лимонную кислоту, добавляемую в количествах, соответствующих 500-1000 ppm, например, 650 ppm.
Один или несколько ферментов, расщепляющих фосфолипиды, в частности, могут быть выбраны из группы, состоящей из:
a. фосфолипазы C, обладающей специфичностью в отношении фосфатидилинозитола (PI),
b. фосфолипазы C, обладающей специфичностью в отношении фосфатидилхолина (PC) и фосфатидилэтаноламина (PE), предпочтительно PLC Bacillus macauensis, SEQ ID NO: 9,
c. фосфолипазы C, обладающей специфичностью в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE), фосфатидной кислоты (PA) и фосфатидилинозитола (PI),
d. комбинации фосфолипазы A и фосфолипазы C, такой как фосфолипаза C, определенная в a) или b),
e. комбинации фосфолипазы A и лизофосфолипазы,
f. фосфолипазы A,
g. комбинации a) и b)
или их комбинаций.
В конкретных вариантах осуществления настоящего изобретения фосфолипаза A выбрана из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности, например, по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 1, 4 и 7;
b. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности, например, по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 2, 5 и 8;
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы A.
В дополнительных конкретных вариантах осуществления настоящего изобретения фосфолипаза A выбрана из группы коммерчески доступных PLA, включающей PLA Lecitase® 10L, Lecitase® Novo, Lecitase® Ultra и Quara® LowP, все из которых доступны от Novozymes A/S, и GumZyme™, доступную от DSM, LysoMax® Oil, доступную от DuPont, и ROHALASE® PL-XTRA и ROHALASE® MPL, доступные от AB Enzymes.
Предпочтительно полипептиды по настоящему изобретению обладают по меньшей мере 20%, например, по меньшей мере 40%, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90%, по меньшей мере 95% или по меньшей мере 100% активностью фосфолипазы A по сравнению со зрелым полипептидом под SEQ ID NO: 1 и/или с полипептидом под SEQ ID NO: 3.
В конкретных вариантах осуществления в соответствии с настоящим изобретением один из указанных одного или нескольких ферментов, расщепляющих фосфолипиды, представляет собой вариант зрелого полипептида зрелого полипептида под любым из SEQ ID NO: 1, 4 и 7 или представляет собой вариант полипептида, изложенного под любым из SEQ ID NO: 2, 5 и 8, содержащий замену, делецию и/или вставку в одном или нескольких положениях.
В частности, указанный вариант может содержать замену, делецию и/или вставку в не более чем 20 положениях, например, в не более чем 19, 18, 17, 16, 15, 14, 13, 12, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 положении.
В других вариантах осуществления один из указанных одного или нескольких ферментов, расщепляющих фосфолипиды, содержит последовательность, изложенную под любым из SEQ ID NO: 2, 5 и 8, по сути состоит из нее или состоит из нее.
В способе в соответствии с настоящим изобретением указанная фосфолипаза C может быть выбрана из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности, например, по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 9 (PLC Bacillus macauensis), 11 (PLC Bacillus thuringiensis), 13 (специфичная в отношении PI PLC Pseudomonas sp.), 15 (PLC P. emersonii); 17 (PLC Kionochaeta), 19 (PLC N. mariannaeae), 22 (PLC Rasamsonia), 25 (PLC T. spiralis), 28 (PLC T. harzianum),
b. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности, например, по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NOs: 10, 12, 14, 16, 18, 20, 23, 26, 29; и
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы С.
В частности, один из указанных одного или нескольких ферментов, расщепляющих фосфолипиды, может представлять собой вариант зрелого полипептида зрелого полипептида под любым из SEQ ID NOs: 9, 11, 13, 15, 17, 19, 22, 25 и 28 или представлять собой вариант полипептида, изложенного под любым из SEQ ID NOs: 10, 12, 14, 16, 18, 20, 23, 26 и 29, содержащий замену, делецию и/или вставку в одном или нескольких положениях.
В других вариантах осуществления один из указанных одного или нескольких ферментов, расщепляющих фосфолипиды, содержит последовательность, изложенную под любым из SEQ ID NOs: 10, 12, 14, 16, 18, 20, 23, 26 и 29, по сути состоит из нее или состоит из нее.
В дополнительных конкретных вариантах осуществления настоящего изобретения фосфолипаза C выбрана из группы коммерчески доступных PLC, включающей Purifine®, доступную от DSM, и Quara® Boost, доступную от Novozymes A/S.
Один предпочтительный вариант осуществления относится к таковому, где указанный по меньшей мере один фермент, расщепляющий фосфолипиды, содержит SEQ ID NO: 9 (PLC Bacillus macauensis) или состоит из нее.
Дополнительные предпочтительные варианты осуществления относятся к таким, где указанный по меньшей мере один фермент, расщепляющий фосфолипиды, предусматривает или включает фосфолипазу C, обладающую специфичностью в отношении фосфатидилинозитола (PI), и фосфолипазу C, обладающую специфичностью в отношении фосфатидилхолина (PC) и фосфатидилэтаноламина (PE), предпочтительно PLC Bacillus macauensis, SEQ ID NO. 9.
Лизофосфолипаза может быть выбрана из группы коммерчески доступных лизофосфолипаз, включающей Finizym™, которая доступна от Novozymes.
Способ по настоящему изобретению может дополнительно включать стадии осветления, дезодорирования и фракционирования.
Обычный способ осветления осуществляют путем адсорбции веществ, обеспечивающих окрашивание, на адсорбирующий материал. Активированная кислотой отбеливающая глина или глинистый минерал, иногда называемая бентонитом, представляет собой адсорбирующий материал, который применяется наиболее широко. Данное вещество состоит в основном из гидратированного силиката алюминия. Безводный силикагель и активированный уголь также применяют в качестве осветляющих адсорбирующих веществ в ограниченной степени.
Дезодорирование жиров и масел представляет собой удаление относительно летучих компонентов из жира или масла с применением пара. Это осуществимо благодаря значительным отличиям в летучести у веществ, которые придают нежелательные привкусы, окраски и неприятные запахи жирам и маслам, и триглицеридов. Дезодорирование проводят под вакуумом для облегчения удаления летучих веществ во избежание излишнего гидролиза жира и обеспечения наиболее эффективного применения пара. Что касается растительных масел, достаточное количество токоферолов остается в конечных маслах после дезодорирования для обеспечения стабильности.
Дезодорирование не оказывает какого-либо значительного эффекта в отношении состава жирных кислот большинства жиров или масел. В зависимости от степени ненасыщенности масла, подлежащего дезодорированию, путем изомеризации могут образовываться небольшие количества транс-жирных кислот.
Жиры, которые являются твердыми при комнатной температуре, обычно содержат смесь множества отдельных триглицеридов, все из которых характеризуются разными температурами плавления. Такие компоненты можно отделить друг от друга посредством способа фракционирования.
Результатом фракционирования является получение двух компонентов, называемых фракциями, которые, как правило, в значительной степени отличаются друг от друга по своим физическим свойствам. Фракции можно снова подвергать фракционированию ("двойное" фракционирование) с получением дополнительных фракций, которые будут обладать уникальными физическими свойствами. Способ изначально разработали для фракционирования животных жиров, таких как говяжье сало.
Существует два типа методик фракционирования: сухое и влажное. Сухое фракционирование относится к способу, в котором не применяют растворитель для облегчения разделения компонентов жира. Сначала жир расплавляют, а затем медленно охлаждают с получением больших кристаллов жира с высокой температурой плавления. Взвесь кристаллов, суспендированных в жидком масле, переносят в фильтр-пресс высокого давления, где жидкую (олеинсодержащую) фракцию выжимают, а твердый (стеаринсодержащий) жир остается на фильтре. Данный способ широко применяют для пальмового масла и пальмоядрового масла для получения нескольких уникальных продуктов из одного природного источника без необходимости химической обработки. Фракции, полученные таким образом, можно смешивать вместе или смешивать с жидкими растительными маслами с получением широкого разнообразия функциональных продуктов для многих путей применения в пищевой промышленности.
Во втором аспекте настоящего изобретения предусмотрено применение фермента, расщепляющего фосфолипиды, для гидролиза фосфолипидов в растительном масле, где растительное масло приводится в контакт с ферментом, расщепляющим фосфолипиды, и после этого осуществляется химическое рафинирование.
При применении фермента, расщепляющего фосфолипиды, в соответствии с настоящим изобретением, предпочтительно, чтобы
- ферментативный гидролиз указанных фосфолипидов осуществлялся в реакционной смеси, содержащей тяжелую фазу или водную фазу и легкую фазу или масляную фазу/гидрофобную фазу, и
- до указанного химического рафинирования не происходило снижение или значительное снижение объема тяжелой фазы или не осуществлялось отделение смолистых веществ/тяжелой фазы от масла.
В третьем аспекте настоящее изобретение предусматривает рафинированное растительное масло, отделенную фракцию смолистых веществ или соапсток, которые являются получаемыми или получены с помощью способа, определенного в первом аспекте настоящего изобретения. В конкретных вариантах осуществления масло в соответствии с настоящим изобретением содержит диглицериды в количестве 0,1% (вес/вес), например, 0,2% (вес/вес) или больше, или, например, 0,3% (вес/вес) или больше. Соапсток может характеризоваться более низкой вязкостью и более низким содержанием фосфолипидов, чем соапсток, полученный с помощью традиционного способа химического рафинирования.
В четвертом аспекте настоящее изобретение предусматривает выделенный или очищенный полипептид, обладающий активностью фосфолипазы A, выбранный из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 4 и 7,
b. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, например, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 5 и 8;
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы A.
В определенных вариантах осуществления полипептид представляет собой вариант зрелого полипептида зрелого полипептида под любым из SEQ ID NO: 4 и 7, или он может представлять собой вариант полипептида, изложенного под любым из SEQ ID NO: 5 и 8, содержащий замену, делецию и/или вставку в одном или нескольких положениях. В частности, указанный вариант может содержать замену, делецию и/или вставку в не более чем 20 положениях, например, в не более чем 19, 18, 17, 16, 15, 14, 13, 12, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 положении.
В частности, полипептид может содержать последовательность, изложенную под SEQ ID NO: 5 или SEQ ID NO: 8, по сути состоять из нее или состоять из нее.
В пятом аспекте настоящее изобретение предусматривает выделенный или очищенный полипептид, обладающий активностью фосфолипазы C, выбранный из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 22, 25 и 28,
b. полипептида, характеризующегося по меньшей мере 60% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 23, 26 и 29; и
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы С.
В некоторых вариантах осуществления настоящего изобретения полипептид представляет собой вариант зрелого полипептида зрелого полипептида под любым из SEQ ID NO: 22, 25 и 28 или представляет собой вариант полипептида, изложенного под любым из SEQ ID NO: 23, 26 и 29, содержащий замену, делецию и/или вставку в одном или нескольких положениях. В частности, указанный вариант может содержать замену, делецию и/или вставку в не более чем 20 положениях, например, в не более чем 19, 18, 17, 16, 15, 14, 13, 12, 10, 9, 8, 7, 6, 5, 4, 3, 2 или 1 положении.
Полипептид может содержать последовательность, изложенную под SEQ ID NO: 23, 26 или 29, по сути состоять из нее или состоять из нее.
В шестом аспекте настоящее изобретение предусматривает композицию, содержащую полипептид в соответствии с четвертым или пятым аспектами настоящего изобретения, изложенными выше.
В седьмом аспекте настоящее изобретение предусматривает выделенный или очищенный полинуклеотид, кодирующий полипептид в соответствии с четвертым или пятым аспектами настоящего изобретения, изложенными выше.
Последовательность полинуклеотида, кодирующего полипептид, изложенный под SEQ ID NO: 4 или 5, изложена под SEQ ID NO: 3. Последовательность полинуклеотида, кодирующего полипептид, изложенный под SEQ ID NO: 7 или 8, изложена под SEQ ID NO: 6.
Последовательность полинуклеотида, кодирующего полипептид, изложенный под SEQ ID NO: 22 или 23, изложена под SEQ ID NO: 21. Последовательность полинуклеотида, кодирующего полипептид, изложенный под SEQ ID NO: 25 или 26, изложена под SEQ ID NO: 24. Последовательность полинуклеотида, кодирующего полипептид, изложенный под SEQ ID NO: 28 или 29, изложена под SEQ ID NO: 27
В восьмом аспекте настоящее изобретение предусматривает конструкцию нуклеиновой кислоты или вектор экспрессии, содержащие полинуклеотид, предусмотренный в седьмом аспекте настоящего изобретения, где полинуклеотид предпочтительно функционально связан с одной или несколькими регуляторными последовательностями, которые управляют продуцированием полипептида у экспрессирующего хозяина.
В девятом аспекте настоящее изобретение предусматривает рекомбинантную клетку-хозяина, содержащую полинуклеотид, предусмотренный в седьмом аспекте настоящего изобретения, функционально связанный с одной или несколькими регуляторными последовательностями, которые управляют продуцированием полипептида.
В частности, настоящее изобретение относится к клетке-хозяину, где полипептид является гетерологичным по отношению к рекомбинантной клетке-хозяину.
Рекомбинантная клетка-хозяин может представлять собой такую, где по меньшей мере одна из одной или нескольких регуляторных последовательностей является гетерологичной по отношению к полинуклеотиду, кодирующему полипептид.
В десятом аспекте настоящее изобретение предусматривает способ получения полипептида в соответствии с четвертым или пятым аспектами настоящего изобретения, предусматривающий культивирование клетки, которая в своей форме дикого типа продуцирует полипептид, в условиях, способствующих продуцированию полипептида.
Способ может дополнительно предусматривать стадию извлечения полипептида.
Одиннадцатый аспект настоящего изобретения относится к способу получения полипептида, обладающего активностью фосфолипазы A, или полипептида, обладающего активностью фосфолипазы С, предусматривающему культивирование рекомбинантной клетки-хозяина в соответствии с девятым аспектом настоящего изобретения в условиях, способствующих продуцированию полипептида.
Способ может дополнительно предусматривать стадию извлечения полипептида.
ПРИМЕРЫ
Пример 1. Фермент: PLC Bacillus macauensis; зрелый полипептид под SEQ ID NO: 9
Масло: неочищенное соевое масло. Содержание отдельных компонентов, представляющих собой фосфолипиды, измеряли с помощью количества фосфора (P) из компонентов в виде ppm P.
Анализ с осуществлением ферментативной реакции и щелочного рафинирования
Эффективность действия фермента тестировали в анализе ферментативной реакции, который имитирует условия промышленного масштаба.
Неочищенное соевое масло (75 г) сначала предварительно обрабатывали кислотой путем добавления 650 ppm лимонной кислоты. В тестовых образцах значение pH повышали до ~6 путем добавления 1,5 экв. NaOH; в контрольных образцах (холостая проба; без фермента) значение pH поддерживали при ~4. Обеспечивали смешивание образцов в ультразвуковой ванне (BRANSON 5800) в течение 5 мин и инкубировали их во встряхивающем устройстве в течение 15 мин при 70°C. Ферментативную реакцию проводили в системе с низким содержанием воды (общее содержание воды составляло 2% в пересчете на количество масла) в цилиндрических центрифужных пробирках с коническим дном объемом 100 мл. Образцы подвергали обработке ультразвуком в течение 5 мин с последующим инкубированием во встряхивающем устройстве в камере с подогревом при 70°C с перемешиванием при 20 об/мин в течение 1 часа. После ферментативной реакции в раствор добавляли большое количество щелочи в соответствии со следующей процедурой:
i) в масло добавляли 1141 ppm NaOH;
ii) реакционные пробирки встряхивали вручную в течение 10 с и выдерживали в ультразвуковой ванне в течение 5 мин;
iii) реакционную смесь оставляли на 0,5 ч при 70°C при 20 об/мин;
iv) после окончания реакции образцы по 10 мл центрифугировали в центрифуге Hotspin при 700g при 85°C в течение 5 мин (Koehler Instruments, центрифуга K600X2 для масла).
Образцы и условия
Таблица 1
Таблица 1. Условия для образца; значения pH 4 и 6 при 70°C для ферментативной реакции в течение 1 часа
Величина выхода по объему смолистых веществ
Объем смолистых веществ оценивали визуально с применением шкалы на стеклянных центрифужных пробирках по ASTM D91, и значения объема смолистых веществ затем использовали для расчета выхода масла.
Содержание сухого вещества в смолистых веществах принимали во внимание при расчете выхода масла в целях обеспечения точности. Значения объема смолистых веществ могут быть одинаковыми в обработанном ферментом образце и в образце холостой пробы, но значения содержания сухого вещества в значительной степени отличаются. Таким образом, относительное содержание масла рассчитывали с помощью следующего уравнения:

Масло: исходный объем неочищенного масла перед удалением смолистых веществ (мл)
Смолистые вещества: измеренный объем смолистых веществ (мл)
Сухое вещество: содержание сухого вещества в смолистых веществах (%)
Выход: выход оценивали по объему восстановленного масла (%)
Содержание диглицеридов
Содержание диглицеридов определяли с помощью высокоэффективной жидкостной хроматографии (HPLC) в сочетании с детектором заряженного аэрозоля (Corona Veo) в соответствии с принципами, описанными в официальном способе AOCS Cd 11d-96. Применяли оборудование DIONEX и Kinetex 2,6 мкм HILIC 100A, 150×4,6 мм, колонку Phenomenex.
Количественное определение фосфолипидов
Количество фосфолипидов в масле определяли с помощью 31P ЯМР с применением следующей процедуры. В образец масла добавляли 0,500 мл раствора внутреннего стандарта (IS) с последующим добавлением 0,5 мл CDCl3 и 0,5 мл буфера Cs-EDTA. Образец встряхивали в течение 5 мин и затем центрифугировали (настольная центрифуга, 5 мин, 13400 об/мин) с обеспечением разделения фаз. Нижнюю фазу переносили в пробирку для ЯМР. P-ЯМР проводили со 128 сканированиями и временем задержки 5 с. Все сигналы объединяли. Отнесения (приблизительное значение в ppm): 1,5 (PA), -0,1 (PE), -0,6 (PI), -0,8 (PC). Концентрацию каждого вида молекул рассчитывали в виде "ppm P", т.е. мг элементарного P на кг образца масла. Следовательно, ppm P=I/I(IS)*n(IS)*M(P)/m(масло). Остаточное содержание фосфолипидов рассчитывали в виде соотношения обработанного ферментом образца и холостой пробы. Раствор внутреннего стандарта представляет собой раствор 2 мг/мл трифенилфосфата в метаноле. Буфер Cs-EDTA получали следующим образом. EDTA (17,55 г) диспергировали в воде (примерно 20 мл). Значение pH регулировали до 7,5 с применением 50% вес/вес CsOH. Таким образом получали прозрачный раствор. Воду добавляли до 100 мл с получением концентрации EDTA 0,6 M.
Общее содержание фосфора
Общее содержание фосфора измеряли посредством оптико-эмиссионной спектрометрии с индуктивно-связанной плазмой (ICP-OES) с точностью, составляющей примерно ±1 ppm P.
Результаты
Выход измеряли с помощью уровня содержания смолистых веществ и содержания сухого вещества в смолистых веществах. При применении PLC B. macauensis перед удалением смолистых веществ с помощью щелочи отмечали очевидную и значимую пользу. Выход являлся одинаковым для обеих концентраций фермента. По оценкам, выход масла составил на 1,4% больше, чем у холостой пробы. Результаты показаны на фиг. 3.
Содержание диглицеридов измеряли с течением времени для отслеживания ферментативной активности PLC. В холостой пробе, как ожидалось, не наблюдали увеличения содержания диглицеридов. Образец с PLC B. macauensis характеризовался значительным увеличением содержания диглицеридов по сравнению с холостой пробой и неочищенным маслом, более 1,08-1,33% DG. Две дозы фермента характеризовались более или менее одинаковым увеличением содержания диглицеридов.
Наблюдали небольшое увеличение уровня содержания диглицеридов после этапа с применением щелочи. Дополнительные 0,1-0,2% диглицеридов, которые теоретически образуются в ходе этапа с применением щелочи, не являются настолько высокими, чтобы вызывать обеспокоенность. Результаты показаны на фиг. 4.
Количество интактных фосфолипидов измеряли для отслеживания превращения фосфолипидов. PLC B. macauensis обеспечивала превращение 66% всех четырех фосфолипидов после осуществления реакции в течение 1 часа. PLC B. macauensis является специфичной в отношении PC и PE, и для данных двух молекул фосфолипидов степень гидролиза достигала 92%. Результаты показаны на фигурах 5, 6 и 7.
Выводы
В испытании подтвердили увеличение выхода на высоком уровне (1,4%), обеспечиваемое обработкой PLC B. macauensis. Значимого различия между дозами PLC B. macauensis, составляющими 2 и 4 мг ферментного белка/кг, не отмечали. Доза, составляющая 2 мг EP/кг масла, по-видимому, является достаточной или возможно даже высокой дозой. Ферментативная реакция длительностью всего 1 час обеспечивала высокую степень гидролиза PC и PE (92%). Наблюдали полное удаление смолистых веществ; при этом содержание остаточного фосфора в масле являлось достаточно низким.
Пример 2. Фермент: PLA T. leycettanus; зрелый полипептид под SEQ ID NO: 1
PLC Bacillus macauensis; зрелый полипептид под SEQ ID NO: 9.
Масло: неочищенное соевое масло, описанное в примере 1.
Ферментативная реакция и щелочное рафинирование
Эффективность действия ферментов тестировали в анализе ферментативной реакции, следуя процедуре, изложенной в примере 1. Неочищенное соевое масло (75 г) сначала предварительно обрабатывали кислотой путем добавления 650 ppm лимонной кислоты. В образцах для тестирования PLC Bacillus macauensis значение pH повышали до ~6 путем добавления 1,5 экв. NaOH; в контрольных образцах (холостая проба; без фермента) и образцах для тестирования PLA T. leycettanus значение pH поддерживали при ~4.
После ферментативной реакции добавляли щелочь, следуя процедуре, описанной в примере 1.
Таблица 2. Образцы и условия
Таблица 2. Условия для образца; значения pH 4 и 6 при 70°C для ферментативной реакции в течение 2 часов
Величину выхода по объему смолистых веществ и содержание диглицеридов определяли, как изложено в примере 1.
Содержание свободных жирных кислот определяли в соответствии с официальным способом AOCS Ca 5a-40. Вкратце, известную массу масла растворяли в 2-пропаноле и добавляли фенолфталеин в качестве индикатора. Свободные жирные кислоты затем нейтрализовали путем титрования с помощью раствора NaOH до первого появления стойкого бледно-розового цвета.
Результаты
Выход измеряли с помощью уровня содержания смолистых веществ и содержания сухого вещества в смолистых веществах. При применении 30 ppm PLA T. leycettanus или 2 мг EP/кг масла PLC B. macauensis перед удалением смолистых веществ с помощью щелочи отмечали очевидную и значимую пользу. Выход составляет 0,3% для PLA T. leycettanus и 0,8% для PLC B. macauensis. Результаты показаны на фиг. 9 и 10.
Сама по себе фаза смолистых веществ являлась достаточно неоднородной после центрифугирования и образовывала твердые белые кристаллы. В условиях присутствия гидроксида натрия (NaOH) повторное растворение смолистых веществ с помощью ручного встряхивания и ультразвуковой ванны было почти невозможным. Смешивание надлежащим образом может привести к получению большего выхода для ферментативной обработки.
Содержание FFA измеряли с течением времени для отслеживания ферментативной активности PLA. Результаты показаны на фиг. 11.
Содержание диглицеридов измеряли с течением времени для отслеживания ферментативной активности PLC. Результаты показаны на фиг. 12. Как ожидалось, образец с PLA T. leycettanus и холостая проба не характеризовались увеличением содержания диглицеридов. Образец с PLC B. macauensis характеризовался значительным содержанием диглицеридов по сравнению с холостой пробой. Для образца с PLC B. macauensis наблюдали высокое стандартное отклонение содержания DG. Причина может состоять в том, что пробирки с применяемыми ферментами размораживали и замораживали несколько раз.
Пример 3. PLA Cryphonectria parasitica и PLA Gloeophyllum trabeum; гидролитическая активность и субстратная специфичность, определенные с помощью анализа посредством P-ЯМР
Происхождение
PLA Cryphonectria parasitica клонировали из донора NN008388 Cryphonectria parasitica из Швеции, 1994 год.
PLA Gloeophyllum trabeum клонировали из донора NN050212 Gloeophyllum trabeum из России, 1997 год.
Гидролитическая активность и субстратная специфичность
Методология
Анализ проводили посредством инкубирования фосфолипазы со смесью 10:1 неочищенного растительного масла и водного буфера. Концентрация фермента составляла 10 мг/кг (мг EP на кг масла). Смесь инкубировали с энергичным встряхиванием при 50 C в течение 2 ч. Затем реакционную смесь анализировали посредством 31P-ЯМР и подсчитывали количество остаточного (негидролизованного, интактного) фосфолипида. Результатом является показатель гидролитической активности и субстратной специфичности фермента.
Процедура проведения анализа
Очищенный фермент разбавляли до 0,09 мг/мл в 100 мM цитратном буфере со значением pH 4,0, 5,5 и 7,0. Анализ начинали с добавления 25 мкл разбавленного фермента в 250 мкл неочищенного растительного масла в пробирке Eppendorf объемом 2 мл и инкубирования смеси в термошейкере при 50 C в течение 2 ч. Применяемое масло представляло собой неочищенное соевое масло, содержащее значительное количество каждого из PA, PE, PI и PC (по 100-200 ppm P для каждого).
Анализ посредством ЯМР
В образец масла затем добавляли 0,500 мл раствора внутреннего стандарта (IS) с последующим добавлением 0,5 мл CDCl3 и 0,5 мл буфера Cs-EDTA. Образец встряхивали в течение 5 мин и затем центрифугировали (настольная центрифуга, 5 мин, 13400 об/мин) с обеспечением разделения фаз. Нижнюю фазу переносили в пробирку для ЯМР. 31P-ЯМР проводили со 128 сканированиями и временем задержки 5 с. Все сигналы объединяли. Отнесения (приблизительное значение в ppm): 1,5 (PA), -0,1 (PE), -0,6 (PI), -0,8 (PC). Концентрацию каждого вида молекул рассчитывали в виде "ppm P", т.е. мг элементарного P на кг образца масла. Следовательно, ppm P=I/I(IS)*n(IS)*M(P)/m(масло). Раствор IS представляет собой раствор 2 мг/мл трифенилфосфата в MeOH. Буфер Cs-EDTA получали следующим образом. EDTA (5,85 г) диспергировали в воде (примерно 50 мл). Значение pH регулировали до 7,5 с применением 50% вес/вес CsOH. Таким образом получали прозрачный раствор. Воду добавляли до 100 мл с получением концентрации EDTA 0,2 M.
Результаты
Результаты в таблицах 14 и 15 ниже показывают, что оба фермента активны в отношении всех четырех фосфолипидов и для обоих предпочтительным является значение pH 4, а не значения pH 5,5 и 7,0.
Таблица 14. Содержание фосфолипидов (ppm P) в соевом масле, инкубированном с PLA Gloeophyllum trabeum
Таблица 15. Содержание фосфолипидов (ppm P) в соевом масле, инкубированном с PLA Cryphonectria parasitica
Пример 16. PLC Trichoderma harzianum; клонирование, экспрессия, ферментация и очистка
Геномную ДНК экстрагировали из штамма NN051266 Trichoderma harzianum с применением набора для выделения ДНК из почвы FastDNA под кат. № 6560-200 от MP Biochemicals, следуя протоколу производителя.
Ген D23CR9, P33XXG (SEQ ID NO: 27) амплифицировали с помощью ПЦР из геномной ДНК. Смесь для ПЦР составляли из 1 мкл геномной ДНК штамма; 2,5 мкл прямого праймера для клонирования (SEQ ID NO: 52 и 53) (10 пмоль/мкл), 2,5 мкл обратного праймера для клонирования (SEQ ID NO: 54 и 55) (10 пмоль/мкл), 25 мкл смеси iProof HF Master Mix (BioRad, № по каталогу 172-5310) и 19 мкл воды класса чистоты для ПЦР.
Реакцию амплификации проводили с применением термоциклера с заданной программой: 2 минуты при 98°C с последующими 30 циклами при 98°C в течение 10 секунд и 60°C в течение 10 секунд каждый с последующим одним циклом при 72°C в течение 5 минут.
P8_43-F 5' ACACAACTGGGGATCCACCATGCGTCCCAGCTCGACGC -3'
P8_43-F 5' ACACAACTGGGGATCCACCATGCGTCCCAGCTCGACGC -3'
P8_43-R 5' AGATCTCGAGAAGCTTAAGCCTTGGCTTTCAACTCATTAGCC 3'
P7_30-R 5' AGATCTCGAGAAGCTTAAGCCTTGGCTTTCAACTCATTGGC 3'
Страной происхождения NN051266 Trichoderma harzianum является Китай (2008).
4 мкл продукта ПЦР визуализировали с помощью электрофореза в 1,0% агарозном геле с применением буфера TAE. Оставшийся продукт ПЦР очищали с применением набора для очистки ДНК из полоски геля после проведения ПЦР GFX® (GE Healthcare, Хиллеред, Дания) в соответствии с инструкциями производителя. Очищенный продукт ПЦР, соответствующий гену D23CR9 PLC NN051266 Trichoderma harzianum, клонировали в вектор экспрессии pDAu109 (WO 2005/042735), предварительно линеаризованный с помощью Bam HI и Hind III, с применением набора Dry-Down для ПЦР-клонирования IN-FUSION™ (BD Biosciences, Пало-Альто, Калифорния, США) в соответствии с инструкциями производителя.
Неразбавленную смесь для лигирования объемом 1 мкл применяли для трансформации BD Phusion-Blue (Clontech). Отбирали одну колонию на чашку с агаровой средой LB, содержащей 100 мкг ампициллина на мл, и культивировали в течение ночи в 2 мл среды LB, дополненной 100 мкг ампициллина на мл. Плазмидную ДНК очищали с применением набора Jetquick Plasmid Miniprep Spin Kit (Genomed GmbH, Лоне, Германия) в соответствии с инструкциями производителя. Последовательности гена D23CR9 PLC NN051266 Trichoderma harzianum подтверждали с помощью секвенирования по Сэнгеру перед осуществлением гетерологичной экспрессии. Одну плазмиду, обозначенную как P8_43 (содержащую ген с SEQ ID NO: 27), выбирали для гетерологичной экспрессии генов PLC в клетках-хозяевах Aspergillus oryzae MT3568.
Штамм MT3568 Aspergillus oryzae применяли для гетерологичной экспрессии гена D23CR9, P33XXG. A. oryzae MT3568 является производным Aspergillus oryzae JaL355 (WO 2002/40694) с разрушенным геном amdS (ацетамидазы), у которого ауксотрофия по pyrG была восстановлена путем разрушения гена ацетамидазы (amdS) A. oryzae геном pyrG. Протопласты Aspergillus oryzae MT3568 получали согласно WO 95/002043.
Сто мкл протопластов Aspergillus oryzae MT3568 смешивали с 1-2 мкг вектора экспрессии Aspergillus с клонированным геном D23CR9 и 250 мкл 60% PEG 4000 (Applichem, Дармштадт, Германия) (полиэтиленгликоль, молекулярная масса 4000), 10 мM CaCl2 и 10 мM Tris-HCl pH 7,5 и тщательно перемешивали. После 30 мин инкубирования при 37°C добавляли 4 мл агара для верхнего слоя (темп. 40°C) и протопласты распределяли на планшеты COVE для проведения отбора. После инкубирования в течение 4-7 дней при 37°C споры четырех трансформантов инокулировали в 0,5 мл среды DAP-4C-01 в 96-луночных планшетах с глубокими лунками. Через 4-5 дней культивирования при 30°C культуральные бульоны анализировали с помощью SDS-PAGE для выявления трансформантов, продуцирующих наибольшее количество рекомбинантной фосфолипазы C из NN051266 Trichoderma harzianum.
Споры наилучшего трансформанта с геном D23CR9 PLC NN051266 Trichoderma harzianum распределяли по поверхности планшетов COVE, содержащих 0,01% TRITON® X-100, с целью выделения отдельных колоний. Распределение по поверхности повторяли еще раз перед консервацией клонов.
Ферментация с целью очистки
Трансформант Aspergillus oryzae, сконструированный, как описано выше, подвергали ферментации в 150 мл среды DAP-4C-01 в рифленых встряхиваемых колбах объемом 500 мл, инкубируемых при 30°C в инкубаторе с качающейся платформой, вращающейся со скоростью 150 об/мин в течение 5 дней, и затем применяли в анализах, описанных ниже.
Пример 4. PLC Trichurus spiralis; клонирование, экспрессия и ферментация
Геномную ДНК экстрагировали из штамма NN009739 Trichurus spiralis с применением набора для выделения ДНК из почвы FastDNA под кат. № 6560-200 от MP Biochemicals, следуя протоколу производителя.
Ген D23YRT, P34CUT (SEQ ID NO. 26) амплифицировали с помощью ПЦР из геномной ДНК. Смесь для ПЦР составляли из 1 мкл геномной ДНК штамма; 2,5 мкл прямого праймера для клонирования (P7_37-F) (10 пмоль/мкл), 2,5 мкл обратного праймера для клонирования (P7_37-R) (10 пмоль/мкл), 25 мкл смеси iProof HF Master Mix (BioRad, № по каталогу 172-5310) и 19 мкл воды класса чистоты для ПЦР.
Реакцию амплификации проводили с применением термоциклера с заданной программой: 2 минуты при 98°C с последующими 30 циклами при 98°C в течение 10 секунд и 60°C в течение 10 секунд каждый с последующим одним циклом при 72°C в течение 5 минут.
P7_37-F 5' ACACAACTGGGGATCCACCATGCATCTCACTCGCGTCGC -3'
P7_37-R 5' AGATCTCGAGAAGCTTAGATTAGGAGTCTCTTGTTCTCCTCGACC 3'
Страной происхождения NN009739 Trichurus spiralis является Дания (1996).
4 мкл продукта ПЦР визуализировали с помощью электрофореза в 1,0% агарозном геле с применением буфера TAE. Оставшийся продукт ПЦР очищали с применением набора для очистки ДНК из полоски геля после проведения ПЦР GFX® (GE Healthcare, Хиллеред, Дания) в соответствии с инструкциями производителя. Очищенный продукт ПЦР, соответствующий гену D23YRT PLC NN009739 Trichurus spiralis, клонировали в вектор экспрессии pDAu109 (WO 2005042735), предварительно линеаризованный с помощью Bam HI и Hind III, с применением набора Dry-Down для ПЦР-клонирования IN-FUSION™ (BD Biosciences, Пало-Альто, Калифорния, США) в соответствии с инструкциями производителя.
Неразбавленную смесь для лигирования объемом 1 мкл применяли для трансформации BD Phusion-Blue (Clontech). Отбирали одну колонию на чашку с агаровой средой LB, содержащей 100 мкг ампициллина на мл, и культивировали в течение ночи в 2 мл среды LB, дополненной 100 мкг ампициллина на мл. Плазмидную ДНК очищали с применением набора Jetquick Plasmid Miniprep Spin Kit (Genomed GmbH, Лоне, Германия) в соответствии с инструкциями производителя. Последовательность гена D23YRT PLC NN009739 Trichurus spiralis подтверждали с помощью секвенирования по Сэнгеру перед осуществлением гетерологичной экспрессии. Одну плазмиду, обозначенную как P7_37 (содержащую ген с SEQ ID NO: 14), выбирали для гетерологичной экспрессии гена PLC в клетке-хозяине Aspergillus oryzae MT3568.
Штамм MT3568 Aspergillus oryzae применяли для гетерологичной экспрессии D23YRT, P34CUT. A. oryzae MT3568 является производным Aspergillus oryzae JaL355 (WO 2002/40694) с разрушенным геном amdS (ацетамидазы), у которого ауксотрофия по pyrG была восстановлена путем разрушения гена ацетамидазы (amdS) A. oryzae геном pyrG. Протопласты Aspergillus oryzae MT3568 получали согласно WO 95/002043.
Сто мкл протопластов Aspergillus oryzae MT3568 смешивали с 1-2 мкг вектора экспрессии Aspergillus с клонированным геном D23YRT и 250 мкл 60% PEG 4000 (Applichem, Дармштадт, Германия) (полиэтиленгликоль, молекулярная масса 4000), 10 мM CaCl2 и 10 мM Tris-HCl pH 7,5 и тщательно перемешивали. После 30 мин инкубирования при 37°C добавляли 4 мл агара для верхнего слоя (темп. 40°C) и протопласты распределяли на планшеты COVE для проведения отбора. После инкубирования в течение 4-7 дней при 37°C споры четырех трансформантов инокулировали в 0,5 мл среды DAP-4C-01 в 96-луночных планшетах с глубокими лунками. Через 4-5 дней культивирования при 30°C культуральные бульоны анализировали с помощью SDS-PAGE для выявления трансформантов, продуцирующих наибольшее количество рекомбинантной фосфолипазы C из NN009739 Trichurus spiralis, и культуральные бульоны также анализировали в анализах для подтверждения активности.
Споры наилучшего трансформанта распределяли по поверхности планшетов COVE, содержащих 0,01% TRITON® X-100 с целью выделения отдельных колоний. Распределение по поверхности повторяли еще раз перед консервацией клона.
Ферментация с целью очистки
Трансформант Aspergillus oryzae, сконструированный, как описано выше, подвергали ферментации в 150 мл среды DAP-4C-01 в рифленых встряхиваемых колбах объемом 500 мл, инкубируемых при 30°C в инкубаторе с качающейся платформой, вращающейся со скоростью 150 об/мин в течение 5 дней, и затем применяли в анализах, описанных ниже.
Применяемые среды
Содержимое чашек с LB состояло из 10 г бактотриптона, 5 г дрожжевого экстракта, 10 г хлорида натрия, 15 г бактоагара и деионизированной воды в объеме до 1 литра.
Среда LB состояла из 10 г бактотриптона, 5 г дрожжевого экстракта и 10 г хлорида натрия с добавлением деионизированной воды до 1 литра.
DAP-4C-1
11 г MgSO4⋅7H2O
1 г KH2PO4
2 г C6H8O7⋅H2O
20 г декстрозы
10 г мальтозы
5,2 г K3PO4⋅H2O
0,5 г дрожжевого экстракта
0,5 мл KU6 раств. металлов, содержащихся в следовых количествах (AMG) (MSA-SUB-FS-0042)
Перемешивают до полного растворения
Добавляют 1 мл Dowfax 63N10
Доводят объем до 1000 мл с помощью воды Milli-Q
CaCO3 в табл. по 0,5 г (добавляют 1 табл./200 мл)
Перед инокуляцией в каждую встряхиваемую колбу объемом 150 мл добавляют 3,5 мл 50% диаммония гидрофосфата (NH4)2HPO4 и 5,0 мл 20% молочной кислоты.
KU6 раств. металлов, содержащихся в следовых количествах (AMG) (MSA-SUB-FS-0042)
6,8 г ZnCl2
2,5 г CuSO4⋅5H2O
0,13 г безводного хлорида никеля
13,9 г FeSO4⋅7H2O
8,45 г MnSO4⋅H2O
3 г C6H8O7⋅H2O
Вода, подвергнутая ионообмену, до 1000 мл
Содержимое чашек с сахарозой COVE состояло из 342 г сахарозы, 20 г порошка агара, 20 мл солевого раствора COVE и деионизированной воды в объеме до 1 литра. Среду стерилизовали автоклавированием при давлении 15 фунтов/кв. дюйм в течение 15 минут (Bacteriological Analytical Manual, 8th Edition, Revision A, 1998). Среду охлаждали до 60°C и добавляли 10 мM ацетамида, Triton X-100 (50 мкл/500 мл).
Солевой раствор COVE состоял из 26 г MgSO4⋅7H2O, 26 г KCL, 26 г KH2PO4, 50 мл раствора металлов COVE, содержащихся в следовых количествах, и деионизированной воды в объеме до 1 литра.
Раствор металлов COVE, содержащихся в следовых количествах, состоял из 0,04 г Na2B4O7⋅10H2O, 0,4 г CuSO4⋅5H2O, 1,2 г FeSO4⋅7H2O, 0,7 г MnSO4⋅H2O, 0,8 г Na2MoO4⋅2H2O, 10 г ZnSO4⋅7H2O и деионизированной воды до 1 литра.
Пример 5. PLC Rasamsonia eburnean; клонирование и экспрессия
Ген, кодирующий фосфолипазу, клонировали с помощью традиционных методик из указанного штамма и вставляли в плазмиду pCaHj505 (WO 2013/029496, пример 1: клонирование и экспрессия).
Ген, кодирующий фосфолипазу, клонировали с помощью традиционных методик из указанного штамма и вставляли в плазмиду pCaHj505 (WO 2013/029496). Ген экспрессировался с нативным сигналом секреции, имеющим следующую аминокислотную последовательность MRAFLITALASLATAAGA (аминокислотные остатки 1-18 из SEQ ID NO: 22).
Экспрессия в A. oryzae
Выбирали один клон с правильной последовательностью рекомбинантного гена и обеспечивали интеграцию соответствующей плазмиды в геном клетки-хозяина Aspergillus oryzae MT3568. A. oryzae MT3568 является производным A. oryzae JaL355 (WO 02/40694) с разрушенным геном amdS (ацетамидазы), у которого ауксотрофия по pyrG была восстановлена путем разрушения гена ацетамидазы (amdS) A. oryzae геном pyrG.
Гидролитическую активность фосфолипазы, продуцированной трансформантами Aspergillus, исследовали с применением планшетов с лецитином/агарозой (анализ на планшетах, описанный в аналитическом разделе). Аликвоты культурального бульона объемом 20 мкл от разных трансформантов или буфер (отрицательный контроль) распределяли в проделанных отверстиях диаметром 3 мм и инкубировали в течение 1 часа при 37°C. Затем планшеты проверяли в отношении присутствия или отсутствия темно-фиолетовой зоны вокруг отверстий, соответствующей активности фосфолипазы.
Отбирали рекомбинантный клон Aspergillus oryzae, содержащий интегрированную конструкцию для экспрессии, и культивировали его в 2400 мл среды YPM (10 г дрожжевого экстракта, 20 г бактопептона, 20 г мальтозы и до 1000 мл деионизированной воды) во встряхиваемых колбах в течение 3 дней при температуре 30°C с перемешиванием при 80 об/мин. Культуральный бульон собирали посредством фильтрации с применением фильтрующего устройства с размером пор 0,2 мкм. Профильтрованный ферментационный бульон применяли для определения характеристик фермента. Ген экспрессировался с нативным сигналом секреции, имеющим следующую аминокислотную последовательность MRAFLITALASLATAAGA (аминокислотные остатки 1-18 из SEQ ID NO: 22).
Очистка
Надосадочную жидкость культуры сперва осаждали с помощью (NH4)2SO4, а затем подвергали диализу с использованием 20 мМ Bis-Tris с pH 6,5. Затем образец наносили на хроматографическую колонку Q Sepharose Fast Flow (GE Healthcare), уравновешенную с помощью 20 мМ Bis-Tris с pH 6,5. Применяли увеличение градиента концентрации NaCl от нуля до 0,35 M NaCl с 15 CV (объем колонки), затем до 0,5 M NaCl с 3 CV и, наконец, до 1 M NaCl с 2 CV. Фракции и образцы, проходящие через колонку (проточную фракцию), проверяли посредством SDS-PAGE. На основании картины SDS фракции от № 18 до № 43 собирали и добавляли (NH4)2SO4 до конечной концентрации 1,2 M.
Объединенные фракции загружали в колонку Phenyl Sepharose 6 Fast Flow (GE Healthcare), уравновешенную с помощью 20 мМ Bis-Tris с pH 6,5 с добавлением 1,2 M (NH4)2SO4. Применяли уменьшение градиента концентрации (NH4)2SO4 от 1,2 M до нуля. Элюированные фракции и проточную фракцию собирали и тестировали в отношении активности PLC с применением планшета с лецитином с pH 5,5. Фракции с активностью PLC проверяли посредством SDS-PAGE. Как элюированные фракции от № 1 до № 18, так и проточную фракцию, характеризующиеся надлежащей активностью PLC и чистотой, выбирали в качестве целевых белков.
Процессинг N- и C-концевых частей
N-концевое секвенирование
Анализы с N-концевым секвенированием проводили с применением системы для секвенирования белков Procise® от Applied Biosystems. Образцы очищали на предварительно отлитом полиакриламидном геле Novex® с 4-20% SDS (Life Technologies). Гель прогоняли в соответствии с инструкциями производителя и проводили блоттинг на PVDF-мембране ProBlott® (Applied Biosystems). Для N-концевого сиквенирования аминокислот главную полосу белка вырезали и помещали в картридж для блоттинга системы для секвенирования белков Procise®. N-концевое секвенирование проводили с применением исполняемого файла способа для образцов на PVDF-мембране (Pulsed liquid PVDF) в соответствии с инструкциями производителя. N-концевую аминокислотную последовательность расшифровывали на основании 7 хроматограмм, соответствующих аминокислотным остаткам 1-7, путем сравнения времени удерживания пиков на хроматограммах со значениями времени удерживания PTH-аминокислот на стандартной хроматограмме.
Идентификация белков посредством последовательной масс-спектрометрии (MS/MS)
Идентификацию белков проводили с помощью анализа посредством тандемной масс-спектрометрии (MS/MS) триптических пептидов, полученных путем ферментативного гидролиза в геле. Первый образец восстанавливали с помощью DTT и подвергали алкилированию с помощью йодацетамида. Восстановленный и подвергнутый алкилированию образец затем применяли для электрофореза в геле с SDS.
Гель прогоняли и окрашивали в соответствии с инструкциями производителя (предварительно отлитый полиакриламидный гель Novex® с 4-20% SDS (Life Technologies)). Главную полосу белка вырезали и часть геля подвергали расщеплению трипсином со степенью чистоты для секвенирования (Roche) в течение ночи. После расщепления полученные триптические пептиды экстрагировали и анализировали на масс-спектрометре Orbitrap LTQ XL (Thermo Scientific), где измеряли значения массы пептидов и значения массы фрагментов пептидов. Для идентификации белков экспериментально полученные значения массы сравнивали с теоретическими значениями массы пептидов и значениями массы фрагментов пептидов белков, хранящимися в базах данных, с помощью программы Mascot для поиска массы (Matrix science).
Определение молекулярной массы
Анализы в отношении молекулярной массы интактных молекул проводили с применением масс-спектрометра с электрораспылением MAXIS II (Bruker Daltonik GmbH, Бремен, Германия). Образцы разбавляли до 1 мг/мл в воде MQ. Разбавленные образцы наносили на колонку Aeris Widepore C4 (Phenomenex). Образцы промывали и элюировали из колонки, прогоняя с линейным градиентом ацетонитрила, и вносили в источник электрораспыления со скоростью потока 300 мл/мин с помощью системы Ultimate 3000 LC (Dionex). Анализ данных проводили с помощью DataAnalysis версии 4.2 (Bruker Daltonik GmbH, Бремен, Германия). Молекулярную массу образцов рассчитывали путем свертки необработанных данных в диапазоне от 20000 до 80000 Да.
Субстратная специфичность
Анализ посредством P-ЯМР очищенных ферментов, представляющих собой PLC
Методология
Анализ проводили посредством инкубирования PLC со смесью 10:1 неочищенного растительного масла и водного цитратного буфера с pH 5,5. Концентрация фермента составляла 30 мг/кг (мг EP на кг масла). Смесь инкубировали с энергичным встряхиванием при 50 C в течение 2 ч. Затем реакционную смесь анализировали посредством 31P-ЯМР. Это предусматривало стадию водной экстракции, на которой фосфорсодержащие молекулы, высвобождаемые посредством PLC, удаляли из масляной фазы. Следовательно, выявляют только липофильные P-содержащие молекулы, т.е. непрореагировавшие фосфолипиды.
Ферменты
Зрелый полипептид под SEQ ID NO: 22 (PLC Rasamsonia), SEQ ID NO: 25 (PLC T. spiralis) и SEQ ID NO: 28 (PLC T. harzianum).
Процедура проведения анализа
Очищенный фермент разбавляли до 0,27 мг/мл в 100 мM цитратном буфере с pH 5,5. Анализ начинали с добавления 25 мкл разбавленного фермента в 250 мкл неочищенного растительного масла в пробирке Eppendorf объемом 2 мл и инкубирования смеси в термошейкере при 50 C в течение 2 ч. Применяемое масло представляло собой неочищенное соевое масло, содержащее значительное количество каждого из PA, PE, PI и PC (по 100-200 ppm P для каждого).
Анализ посредством ЯМР
В образец масла затем добавляли 0,500 мл раствора внутреннего стандарта (IS) с последующим добавлением 0,5 мл CDCl3 и 0,5 мл буфера Cs-EDTA. Образец встряхивали в течение 5 мин и затем центрифугировали (настольная центрифуга, 5 мин, 13400 об/мин) с обеспечением разделения фаз. Нижнюю фазу переносили в пробирку для ЯМР. P-ЯМР проводили со 128 сканированиями и временем задержки 5 с. Все сигналы объединяли. Отнесения (приблизительное значение в ppm): 1,5 (PA), -0,1 (PE), -0,6 (PI), -0,8 (PC). Концентрацию каждого вида молекул рассчитывали в виде "ppm P", т.е. мг элементарного P на кг образца масла. Следовательно, ppm P=I/I(IS)*n(IS)*M(P)/m(масло). Остаточное содержание фосфолипидов рассчитывали в виде соотношения обработанного ферментом образца и холостой пробы. Раствор внутреннего стандарта представляет собой раствор 2 мг/мл трифенилфосфата в метаноле. Буфер Cs-EDTA получали следующим образом. EDTA (5,85 г) диспергировали в воде (примерно 50 мл). Значение pH регулировали до 7,5 с применением 50% вес/вес CsOH. Таким образом получали прозрачный раствор. Воду добавляли до 100 мл с получением концентрации EDTA 0,2 M.
Результаты
Ниже в таблице 2 показано остаточное содержание фосфолипидов в процентах (0 обозначает полный гидролиз, 100 обозначает отсутствие гидролиза).
Таблица 2
DCS-температурный профиль
Определение Td посредством дифференциальной сканирующей калориметрии
Термостабильность Harzianum (U49A3) определяли посредством дифференциальной сканирующей калориметрии (DSC) с применением дифференциального сканирующего калориметра с VP-капиллярной ячейкой (MicroCal Inc., Пискатауэй, Нью-Джерси, США). Температуру тепловой денатурации Td (°C) принимали за вершину пика денатурации (главного эндотермического пика) на термограммах (Cp от T), полученных после нагревания растворов фермента (примерно 0,5 мг/мл) в буфере (50 мM ацетатном буфере с pH 5,0) при постоянной заданной в программе скорости нагревания, составляющей 200 K/час.
Растворы образца и эталонные растворы (примерно 0,2 мл) загружали в калориметр (эталон: буфер без фермента) из условий хранения при 10 градусах C и осуществляли предварительное термическое уравновешивание в течение 20 минут при 20°C перед DSC-сканированием при 20°C - 100°C. Значения температуры денатурации определяли с точностью примерно +/-1°C. Td, полученное при таких условиях, для U49A3 составляло 79°C.
Термостабильность Spiralis (U4G2D) определяли посредством дифференциальной сканирующей калориметрии (DSC) с применением дифференциального сканирующего калориметра с VP-капиллярной ячейкой (MicroCal Inc., Пискатауэй, Нью-Джерси, США). Температуру тепловой денатурации Td (°C) принимали за вершину пика денатурации (главного эндотермического пика) на термограммах (Cp от T), полученных после нагревания растворов фермента (примерно 0,5 мг/мл) в буфере (50 мM ацетатном буфере с pH 5,5) при постоянной заданной в программе скорости нагревания, составляющей 200 K/час.
Растворы образца и эталонные растворы (примерно 0,2 мл) загружали в калориметр (эталон: буфер без фермента) из условий хранения при 10 градусах C и осуществляли предварительное термическое уравновешивание в течение 20 минут при 20°C перед DSC-сканированием при 20°C - 100°C. Значения температуры денатурации определяли с точностью примерно +/-1°C. Td, полученное при таких условиях, для U4G2D составляло 67°C.
Термостабильность Rasamsonia (U4BCJ) определяли посредством дифференциальной сканирующей калориметрии (DSC) с применением дифференциального сканирующего калориметра с VP-капиллярной ячейкой (MicroCal Inc., Пискатауэй, Нью-Джерси, США). Температуру тепловой денатурации Td (°C) принимали за вершину пика денатурации (главного эндотермического пика) на термограммах (Cp от T), полученных после нагревания растворов фермента (примерно 0,5 мг/мл) в буфере (50 мM ацетатном буфере с pH 5,5) при постоянной заданной в программе скорости нагревания, составляющей 200 K/час.
Растворы образца и эталонные растворы (примерно 0,2 мл) загружали в калориметр (эталон: буфер без фермента) из условий хранения при 10 градусах C и осуществляли предварительное термическое уравновешивание в течение 20 минут при 20°C перед DSC-сканированием при 20°C - 100°C. Значения температуры денатурации определяли с точностью примерно +/-1°C. Td, полученное при таких условиях, для U4BCJ составляло 82°C.
Эффективность удаления смолистых веществ
Эффективность действия фермента по настоящему изобретению, представляющего собой фосфолипазу C Rasamsonia, Harzianum и Spiralis, тестировали в анализе удаления смолистых веществ, который имитирует условия промышленного масштаба. В анализе измеряли один из следующих параметров или их оба в масляной фазе после процедуры удаления смолистых веществ, описанной ниже в подразделе "Анализ удаления смолистых веществ":
a) Содержание диглицеридов, определяемое с помощью высокоэффективной жидкостной хроматографии (HPLC) в сочетании с детектором заряженного аэрозоля (Corona Veo) в соответствии с принципами, описанными в официальном способе AOCS Cd 11d-96. Применяли оборудование DIONEX и Lichrocart Si-60, 5 мкм, Lichrosphere 250-4 мм, колонку MERCK.
b) Общее содержание фосфора и других металлов, таких как Ca, Mg, Zn, измеряемое посредством оптико-эмиссионной спектрометрии с индуктивно-связанной плазмой (ICP-OES) с точностью примерно ±1 ppm P.
Анализ удаления смолистых веществ
Неочищенное соевое масло (75 г) сначала предварительно обрабатывали кислотой/основанием для облегчения превращения нерастворимых солевых форм фосфолипидов в более гидратируемые формы и обеспечения подходящей для фермента среды. Предварительную обработку кислотой/основанием осуществляли путем добавления кислоты, представляющей собой либо ортофосфорную кислоту (75% раствор), либо лимонную кислоту (50% раствор). Кислоту применяли в количествах, равных либо 0,065%, либо 0,09% (100% чистой ортофосфорной кислоты/100% чистой лимонной кислоты) в пересчете на количество масла, и перемешивали в ультразвуковой ванне (BRANSON 5800) в течение 5 мин и инкубировали во встряхивающем устройстве в течение 15 мин. Затем осуществляли нейтрализацию основанием с применением 1 M NaOH в количествах, эквивалентных (от 0,45 до 5) количествам чистой ортофосфорной кислоты (т.е. кислоты, которую применяли в предварительной обработке), и перемешивали в ультразвуковой ванне в течение 5 мин. Ферментативную реакцию проводили в системе с низким содержанием воды (общее содержание воды составляло 3% в пересчете на количество масла) в цилиндрических центрифужных пробирках с коническим дном объемом 100 мл. Образцы обрабатывали ультразвуком в течение 5 мин с последующим инкубированием в камере с подогревом при выбранной температуре (от 60 до 70°C) с перемешиванием при 20 об/мин в течение выбранного времени инкубирования (от 1 до 24 часов). Для разделения смеси на масляную фазу и тяжелую фазу вода/смолистые вещества образцы центрифугировали при 700g при 85°C в течение 5 мин (Koehler Instruments, центрифуга K600X2 для масла).
Содержание фосфора, кальция, магния и цинка в неочищенном соевом масле, применяемом в экспериментах, указано в таблице 3.
Таблица 3. Содержание металлов в неочищенном масле, измеренное с помощью ICP-OES (мг/кг масло)
Ниже в примерах 6-11 описываются результаты, полученные в анализе удаления смолистых веществ.
Пример 6. Сравнение активности фермента из Harzianum U4AVG с активностью фермента из Mariannaeae U4DB1 при 60°C (идентичность 58,6)
Фермент из Harzianum (U4AVG) (зрелый полипептид под SEQ ID NO: 28) применяли в анализе удаления смолистых веществ при 60°C при сравнении с ферментом из N. mariannaeae (U4DB1) (зрелый полипептид под SEQ ID NO: 19) при дозе фермента, составляющей 10 мг ферментного белка на кг масла, при этом применяли масло 1. Измеряли содержание диглицеридов после удаления смолистых веществ с помощью ферментов в течение 2, 5 и 24 ч (масло предварительно обрабатывали с помощью 0,065% лимонной кислоты/1,5 экв. NaOH), а также посредством ICP измеряли общее содержание фосфора после 2, 5 и 24 часов инкубирования. Результаты (средние значения двукратного определения) и стандартное отклонение (STDEV) представлены в таблицах 4A и 4B.
Таблицы 4A и 4B
Удаление смолистых веществ с помощью фермента из Harzianum (U4AVG) при 60°C приводит к лучшему образованию диглицеридов по сравнению с образованием диглицеридов посредством PLC mariannaeae (U4DB1). Фермент из Harzianum обеспечивает превращение вплоть до 78% фосфолипидов в тестируемых условиях (60°C, 24 часа). Расчет превращения основан на допущении, что общее содержание P, составляющее 743 ppm, измеренное посредством ICP, соответствует 1,86 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему 1,49% (80% молекул фосфолипида).
Пример 7. Исследование влияния величины дозы на эффективность фермента из Harzianum (U4AVG) при 60°C
Фермент из Harzianum (U4AVG) (зрелый полипептид под SEQ ID NO: 28) применяли в анализе удаления смолистых веществ при 60°C при различных дозах фермента, составляющих 1x-2x-5x-10x мг ферментного белка на кг масла, при этом применяли масло 2. Измеряли содержание диглицеридов после удаления смолистых веществ с помощью ферментов в течение 1, 2 и 4 ч (масло предварительно обрабатывали с помощью 0,09% фосфорной кислоты/1,5 экв. NaOH). Результаты представлены в таблице 5.
Таблица 5. Увеличение содержания диглицеридов
Для удаления смолистых веществ с помощью фермента из Harzianum (U4AVG) при 60°C показано увеличение образования диглицеридов при повышенной дозе фермента в тестируемых пределах (1-10X мг EP/кг масла). Обеспечивалось превращение вплоть до 86% фосфолипидов при тестируемых условиях (60°C, 4 часа, 10 мг EP/кг масла). Расчет превращения основан на допущении, что общее содержание P, составляющее 615 ppm, измеренное посредством ICP, соответствует 1,54 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему 1,23% (80% молекул фосфолипида).
Пример 8. Сравнение эффективности действия ферментов из Rasamsonia (U3GPC), P. emersonii (U4DB4) и PLC Kionochaeta (U1A3F) при 60°C с применением фосфорной кислоты для предварительной обработки масла
Фермент из Rasamsonia (U3GPC) (зрелый полипептид под SEQ ID NO: 22) применяли в анализе удаления смолистых веществ при 60°C при сравнении с PLC Kionochaeta (U1A3F) (зрелый полипептид под SEQ ID NO: 17) и ферментом из P. emersonii (U1DW6) (зрелый полипептид под SEQ ID NO: 15) при дозе фермента, составляющей 30 мг ферментного белка на кг масла, при этом применяли масло 3. Измеряли содержание диглицеридов после удаления смолистых веществ с помощью ферментов в течение 2, 4, 6 и 24 ч (масло предварительно обрабатывали с помощью 0,09% фосфорной кислоты (PA) и ±1,5 экв. NaOH), а также посредством ICP измеряли общее содержание фосфора после 2 и 24 часов инкубирования. Результаты представлены в таблице 6.
Таблица 6. Увеличение содержания диглицеридов (% вес/вес)
Для удаления смолистых веществ с помощью фермента из Rasamsonia при 60°C показано более быстрое образование диглицеридов по сравнению с PLC Kion в ходе первых 6 часов (с применением одной и той же предварительной обработки масла) и почти идентичная эффективность действия в сравнении с ферментом из P. emersonii при тестировании без какого-либо добавления каустической соды (NAOH). Фермент из Rasamsonia обеспечивал превращение вплоть до 66% фосфолипидов при тестируемых условиях (60°C, 24 часа, 30 мг EP/кг масла, 0,09% фосфорная кислота+1,5 экв. NaOH). Расчет превращения основан на допущении, что общее содержание P, составляющее 631 ppm, измеренное посредством ICP, соответствует 1,58 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему 1,26% (80% молекул фосфолипида).
Пример 9. Эффективность действия фермента из Rasamsonia (U4BCJ) при 60°C и 70°C с предварительной обработкой масла с применением лимонной кислоты для предварительной обработки масла (70C) и (60C) и без нее
Фермент из Rasamsonia (U4BCJ) (зрелый полипептид под SEQ ID NO: 22) применяли в анализе удаления смолистых веществ при 60°C и 70°C при дозе фермента, составляющей 10 мг ферментного белка на кг масла, при этом применяли масло 4/масло 5, и сравнивали с PLC P. emersonii (зрелый полипептид под SEQ ID NO: 15). Измеряли содержание диглицеридов после удаления смолистых веществ с помощью ферментов в течение 2, 5 и 24 ч. Результаты представлены ниже в таблицах 7A и 7B.
Таблицы 7A и 7B
Для удаления смолистых веществ с помощью фермента из Rasamsonia показано увеличение образования диглицеридов с течением времени и надлежащая эффективность действия при 60°C, а также при 70°C в масле, предварительно обработанном лимонной кислотой и каустической содой, а также в масле без какой-либо предварительной обработки. Полное превращение ~96-100% фосфолипидов получали при 70°C, 24 часа, 10 мг EP/кг масла, 650 ppm CA+0,4 экв. NaOH, а также 60°C, 24 часа, 10 мг EP/кг масла без предварительной обработки масла. Расчет превращения основан на допущении, что общее содержание P, составляющее 465-479 ppm, измеренное посредством ICP, соответствует ~1,2 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему ~0,96% (80% молекул фосфолипида).
Пример 10. Сравнение эффективности действия ферментов из Spiralis (U4G2D) и Marianneaea (U4DB1) при 60°C
Фермент из Spiralis (U4G2D) (зрелый полипептид под SEQ ID NO: 25) применяли в анализе удаления смолистых веществ при 60°C при сравнении с ферментом из Mariannaeae (U4DB1) (зрелый полипептид под SEQ ID NO: 19) при дозе фермента, составляющей 10 мг ферментного белка на кг масла, при этом применяли неочищенное масло 5. Измеряли содержание диглицеридов после удаления смолистых веществ с помощью ферментов в течение 2, 5 и 24 ч (масло предварительно обрабатывали с помощью 0,065 лимонной кислоты и 1,5 экв. NaOH), а также посредством ICP измеряли общее содержание фосфора после 5 и 24 часов инкубирования. Результаты (средние значения двукратного определения) представлены в таблице 8.
Таблица 8
Для фермента из Spiralis показаны хорошая эффективность действия в тестируемых условиях реакции и более быстрое образование диглицеридов после 2 и 5 часов по сравнению с ферментом из Mariannaeae, для которого показано образование DG в наибольшем количестве после 24 ч. Фермент из Spiralis обеспечивал превращение вплоть до 53% фосфолипидов при тестируемых условиях (60°C, 24 часа, 10 мг EP/кг масла, 0,065% фосфорная кислота+1,5 экв. NaOH). Расчет превращения основан на допущении, что общее содержание P, составляющее 622 ppm, измеренное посредством ICP, соответствует 1,56 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему 1,24% (80% молекул фосфолипида).
Пример 11. Сравнение эффективности действия PLC Harzianum, Rasamsonia и Spiralis с PLC Mariannaeae, Kionochaeta sp. и PLC P. emersonii при 60°C
Удаление смолистых веществ
Ферменты из Harzianum, Rasamsonia и Spiralis (зрелые полипептиды под SEQ ID NO: 28, 22 и 25 соответственно) применяли в анализе удаления смолистых веществ при 60°C в сравнении с PLC Kionochaeta sp. (зрелый полипептид под SEQ ID NO: 17), экспрессирующейся в A. niger либо в A. oryzae, PLC Mariannaeae и P. emersonii (зрелые полипептиды под SEQ ID NO: 19 и 15 соответственно) при дозе фермента, составляющей 10 мг ферментного белка на кг масла, при этом применяли неочищенное масло 8. Масло предварительно обрабатывали с помощью 0,065% лимонной кислоты и 0,4 молярных эквивалентов или 1,5 молярных эквивалентов NaOH перед удалением смолистых веществ с помощью PLC P. emersonii и всех остальных ферментов соответственно. Измеряли показатели содержания диглицеридов после удаления смолистых веществ с помощью ферментов в течение 2, 5 и 24 ч посредством HPLC и измеряли общее содержание фосфора после 5 и 24 часов инкубирования посредством ICP. Результаты представлены в таблице 9.
Таблица 9
При указанных условиях реакции (60°C, 10 мг EP/кг масла, 0,065% лимонная кислота+1,5 экв. NaOH) удаление смолистых веществ с помощью PLC Harzianum приводило к более быстрому увеличению содержания диглицеридов и снижению содержания фосфора по сравнению с другими ферментами, представляющими собой PLC. Также наиболее высокого содержания диглицеридов (1,12% вес/вес) после 24 ч достигали с помощью PLC Harzianum, что соответствовало превращению примерно 97% фосфолипидов. Расчет превращения основан на допущении, что общее содержание P, составляющее 574 ppm, измеренное посредством ICP, соответствует 1,44 вес.% фосфолипида (средняя Mw PL ~772 г/моль, Mw P ~31 г/моль), что соответствует максимальному увеличению количества получаемого DG, составляющему 1,15% (80% молекул фосфолипида).
Количественный анализ фосфолипидов посредством LCMS/MS
Жидкостную хроматографию в сочетании с тройным квадрупольным масс-спектрометром (LC/MS/MS) или в сочетании с времяпролетным квадрупольным масс-спектрометром (LC/TOF/MS) применяли для определения количества отдельных молекул фосфолипидов: фосфатидилхолина (PC); фосфатидилинозитола (PI); фосфатидилэтаноламина (PE) и фосфатидной кислоты (фосфатидата) (PA). Чувствительность анализа снижается до менее 1 мг фосфора/кг масла для PC, PE и PI (ppm) и менее 10 мг фосфора/кг для PA. Образец масла растворяли в хлороформе. Затем экстракт анализировали с помощью LC-TOF-MS (или с помощью LC-MS/MS, если требуются более низкие пределы обнаружения) с применением следующих установочных параметров.
Установочные параметры для LC
Элюент A: 50% ацетонитрил, 50% вода, 0,15% муравьиная кислота
Элюент B: 100% изопропионовая кислота, 0,15% муравьиная кислота
Время записи хроматограммы: 26,9 мин
Скорость потока: 0,50 мл/мин
Температура колонки: 50°C
Температура автоматического пробоотборника: 15-25°C
Объем вводимой пробы: 1 мкл
Тип колонки, материал: Гибридный с заряженной поверхностью, длина: 50 мм, размер: 1,7 мкм, ID: 2,1 мм
Установочные параметры для MS
Напряжение на конусе: 28
Напряжение на экстракторе: 2 В
Напряжение на RF-линзах: 0,5 В
Температура источника: 125°C
Температура десольватации: 500°C
Скорость потока газа в конусе: 30 л/час
Скорость потока газа десольватации: 850 л/час
Напряжение на конусе: характерное для компонентов
Напряжение на экстракторе: 2,5 В
Напряжение на RF-линзах:
Температура источника: 150°C
Температура десольватации: 500°C
Скорость потока газа в конусе: 30 л/ч
Скорость потока газа десольватации: 850 л/ч
Данные обрабатывали с применением программного обеспечения MassLynx версии 4.1. В приведенных ниже примерах способ называется просто LCMS.
Результаты Таблица 10
Содержание фосфолипидов в образцах масла после 2, 5 и 24 ч инкубирования показано в таблице 10. Наблюдали, что ферменты, представляющие собой PLC, обеспечивали снижение содержания всех четырех фосфолипидов после инкубирования вплоть до 24 ч. Удаление смолистых веществ с применением PLC Harzianum приводило к наиболее быстрому уменьшению содержания PA, PE и PC.
Пример 12. Влияние нейтрализации с помощью NaOH на выход
Эксперимент проводили, как описано выше (см. подраздел "Удаление смолистых веществ"). В частности, лимонную кислоту вводили в количестве, составляющем 650 ppm, и фермент вводили в количестве, составляющем 200 ppm. Фермент, применяемый во всех образцах, представлял собой комбинацию PLC Bacillus thuringiensis (SEQ ID NO. 11) и специфичной в отношении PI PLC Pseudomonas sp. (SEQ ID NO. 13). Количество эквивалентов NaOH, используемое для нейтрализации CA, применяемой для предварительной обработки, варьировали, см. таблицу 11 ниже. Увеличение количества применяемого NaOH до 3-5 эквивалентов кислоты, применяемой в предварительной обработке, обеспечивает улучшение выхода и снижение потери сухого вещества. Такой эффект наблюдали как для рапсового, так и соевого масла. Таким образом, обеспечение соответствующего значения pH в реакции с PLC увеличивает образование DG.
Образцы были такими, как указано в таблице 11 ниже.
Таблица 11
*потерю сухого вещества и изменение содержания DG измеряли, как описано выше.
Таким образом, в предпочтительных вариантах осуществления настоящее изобретение относится к способу в соответствии с настоящим изобретением, где количество NaOH для обработки составляет по меньшей мере 3,0 экв. кислоты для предварительной обработки, например, от 3,0 до 6,0, например, от 3 до 5,5, от 3 до 5,0, от 3 до 4,5 или от 3 до 4,0 эквивалентов.
Пример 13. Снижение остаточного содержания фосфора и FFA
Как показано в примере 12, увеличение количества NaOH может обеспечивать увеличение выхода, как измерено по изменению содержания DG, и в то же время снижение потери сухого вещества.
Удаление смолистых веществ проводили тем же способом, как описано выше (см. подраздел "Удаление смолистых веществ"), со следующими изменениями.
Масло представляло собой неочищенное рапсовое масло. Предварительную обработку кислотой осуществляли путем добавления 750 или 1500 ppm фосфорной кислоты в течение 15 мин при 70°C. Для нейтрализации кислоты и подготовки для ферментативной обработки добавляли NaOH в количестве, составляющем 1,33, 2,0 или 3,0 эквивалентов кислоты. Ферментативный гидролиз осуществляли с помощью PLC Bacillus thuringiensis (SEQ ID NO. 11) и специфичной в отношении PI PLC Pseudomonas sp. (SEQ ID NO: 13), вводимых в количестве, составляющем 200 ppm, при этом смесь содержала 2% воды. Смесь инкубировали в течение 2 ч при 60°C. По окончании гидролиза осуществляли щелочное рафинирование путем добавления NaOH, 1707 ppm - 2040 ppm с применением 8% NaOH. Общее количество NaOH в каждом образце составляло 2700 ppm, что соответствует 35% избытку по отношению к FFA в неочищенном масле (1,3%).
Образцы получали в соответствии с таблицей 12 ниже. Результаты также приведены в данной таблице.
Таблица 12. Условия для образца и результаты
8% NaOH
25% от всего количества NaOH
37% от всего количества NaOH
33% от всего количества NaOH
49% от всего количества NaOH
8% NaOH
75% от всего количества NaOH
63% от всего количества NaOH
67% от всего количества NaOH
51% от всего количества NaOH
*значения в двойных круглых скобках являются неоптимальными; значения в одинарных круглых скобках являются приемлемыми, хотя не соответствуют целевым значениям; и значения без круглых скобок находятся в пределах целевых значений.
Содержание фосфора. Величину выхода, содержание FFA и изменение содержания удаляемых смолистых веществах определяли, как описано выше.
Как можно увидеть из таблицы 12, для образцов, в которые добавляли одинаковые количества NaOH после обработки кислотой в качестве нейтрализации (регулирование значения pH) и на стадии щелочного рафинирования (каустическая сода, 8% NaOH) (см. ID колб 7 и 8 в таблице 12), содержание FFA в конечном образце являлось неоптимальным. В отличие от этого, для образцов, для которых стадия щелочного рафинирования предусматривала добавление NaOH в количестве, соответствующем более чем 60% от общего количества NaOH, добавляемого в ходе осуществления способа, содержание FFA было приемлемым.
В экспериментах показано, что введение кислоты перед ферментативным гидролизом и после ферментативного гидролиза оказывает влияние на выход.
Пример 14. Удаление смолистых веществ с помощью ферментов в промышленном масштабе с межпроцессным подкислением
Целью данного эксперимента было снижение уровней содержания фосфора в масле, подвергнутом удалению смолистых веществ, до менее чем 30 ppm, предпочтительно ниже 10 ppm, с одновременным достижением приемлемых уровней содержания FFA (т.е. уровней содержания, составляющих 0,1% - 0,2%) с применением одной стадии разделения.
Следующий способ проводили для образца неочищенного масла (180 кг). Масло нагревали до 80°C при слабом перемешивании содержимого резервуара (40% скорости вращения мешалки). Затем добавляли чистую лимонную кислоту (CA) в количестве, соответствующем 650 ppm. CA в виде 30% (вес/вес) раствора. Смесь подвергали смешиванию с большим сдвиговым усилием в течение 15 мин с применением Sylversson HSM (пропускная способность оборудования составляет 1000 кг/ч) и после этого подвергали механическому перемешиванию в течение 15 мин при 80°C и при 70% скорости вращения мешалки, установленной в реакторе. Затем значение pH регулировали путем добавления 6 мол. экв. NaOH. NaOH добавляли в виде 8% (вес/вес) раствора.
Смесь подвергали смешиванию с большим сдвиговым усилием в течение 15 мин с применением Siversson HSM (пропускная способность оборудования составляет 1000 кг/час) и затем охлаждали до 60°C. Добавляли 200 ppm фермента (PLC Bacillus thuringiensis (SEQ ID NO: 11) и специфичной в отношении PI PLC Pseudomonas sp. (SEQ ID NO: 13). Обеспечивали прохождение ферментативной реакции в течение 45 мин. Прекращение активности ферментов в смеси обеспечивали посредством нагревания масла в реакторе до 80°C.
Добавляли фосфорную кислоту (2,5 кг фосфорной кислоты/тонна масла) и смесь подвергали смешиванию с большим сдвиговым усилием в течение 15 мин. Для нейтрализации добавляли NaOH в виде 8% раствора. Нанонейтрализацию проводили (70 бар) в течение приблизительно 7 минут и собирали масло в резервуаре, выполненном с возможностью прямой загрузки в центрифугу GEA. Это добавление кислоты после ферментативного гидролиза, но перед стадией щелочного рафинирования в данном документе называется межпроцессным подкислением.
Вышеизложенная процедура проводила к получению приблизительно 0,099% FFA и 26 ppm фосфора в полученном масле, подвергнутом удалению смолистых веществ. Таким образом, можно сделать вывод, что добавление кислоты после этапа воздействия фермента и до этапа конечной нейтрализации может обеспечивать полное снижение остаточного содержания фосфора и FFA в маслах, где количество щелочи, добавляемое после этапа хелатообразования, обуславливает меньшую эффективность стадии щелочной нейтрализации после воздействия фермента.
Пример 15. Комбинации PLC
Анализ удаления смолистых веществ проводили, как описано выше (см. подраздел "Анализ удаления смолистых веществ"), с изменением, заключающимся в том, что после проведения ферментативного гидролиза добавляли большее количество щелочи.
Как показано в таблице ниже, тестировали различные комбинации ферментов.
Таблица 13. Условия для образца; значения pH 4 и 6 при 70°C для ферментативной реакции в течение 1 часа и результаты
Авторы настоящего изобретения пришли к выводу, что применение удаления смолистых веществ с помощью ферментов в комбинации со щелочным рафинированием приводит к повышению показателя увеличения выхода и снижению потери сухого вещества. Хотя все тестируемые комбинации ферментов приводили к увеличению, комбинация PLC Bacillus macauensis (SEQ ID NO: 9) и специфичной в отношении PI PLC Pseudomonas sp. (SEQ ID NO. 13) приводила к наибольшему увеличению выхода в комбинации с наибольшим снижением потери сухого вещества. Данная комбинация также имела наилучшие показатели в гидролизе фосфолипидов (как измеряли после гидролиза, но перед обработкой с помощью щелочи) и в отношении увеличения содержания диглицеридов.
Таким образом, настоящее изобретение в одном варианте осуществления относится к способу в соответствии с настоящим изобретением, где указанные ферменты, расщепляющие фосфолипиды, включают по меньшей мере специфичную в отношении PI PLC Pseudomonas sp. (SEQ ID NO: 13). Предпочтительные варианты осуществления относятся к таким, где ферменты предусматривают или включают специфичную в отношении PI PLC Pseudomonas sp. (SEQ ID NO: 13) и PLC Bacillus macauensis (зрелый полипептид под SEQ ID NO: 9).
Объем описанного и заявленного в данном документе изобретения не должен ограничиваться конкретными аспектами, раскрытыми в данном документе, поскольку эти аспекты предполагаются как иллюстрации некоторых аспектов настоящего изобретения. Предполагается, что любые эквивалентные аспекты находятся в пределах объема настоящего изобретения. Действительно, различные модификации настоящего изобретения в дополнение к продемонстрированным и описанным в данном документе станут очевидны специалистам в данной области из вышеприведенного описания. Предполагается, что такие модификации также входят в объем прилагаемой формулы изобретения. В случае противоречия настоящее раскрытие, в том числе определения, будет иметь преобладающую силу.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Novozymes A/S
Holm, Hans Christian
Nielsen, Per Munk
Longin, Fanny
Landvik, Sara
Brask, Jesper
Borch, KIm
Olinski, Robert
Noergaard, Allan
Lilbaek, Hanna
Damstrup, Marianne
Li, Ming
Sun, Tianqi
<120> СПОСОБ УДАЛЕНИЯ СМОЛИСТЫХ ВЕЩЕСТВ ИЗ РАСТИТЕЛЬНОГО МАСЛА
И ЕГО РАФИНИРОВАНИЯ
<130> 14265-WO-PCT[2]
<160> 29
<170> PatentIn версия 3.5
<210> 1
<211> 296
<212> БЕЛОК
<213> Talaromyces leycettanus
<400> 1
Met His Arg Pro Leu Gln Leu Trp Ala Leu Ala Ala Leu Thr Ser Leu
1 5 10 15
Val Thr Ala Ala Pro Ala Pro Val Leu Arg Arg Asp Val Ser Ser Ser
20 25 30
Val Leu Ser Glu Leu Asp Leu Phe Ala Gln Tyr Ser Ala Ala Ala Tyr
35 40 45
Cys Ser Ser Asn Ile Gly Ser Pro Gly Thr Lys Leu Thr Cys Ser Val
50 55 60
Gly Asn Cys Pro Arg Val Glu Ala Ala Asp Thr Glu Thr Leu Ile Glu
65 70 75 80
Phe Asn Glu Ser Ser Ser Phe Gly Asp Val Thr Gly Tyr Ile Ala Val
85 90 95
Asp Arg Thr Asn Ser Leu Leu Val Leu Ala Phe Arg Gly Ser Ser Thr
100 105 110
Val Ser Asn Trp Glu Ala Asp Leu Asp Phe Pro Leu Thr Asp Ala Ser
115 120 125
Ser Leu Cys Ser Gly Cys Glu Ile His Ser Gly Phe Trp Ala Ala Trp
130 135 140
Gln Thr Val Gln Ala Ser Ile Thr Ser Thr Leu Glu Ser Ala Ile Ala
145 150 155 160
Ser Tyr Pro Gly Tyr Thr Leu Val Phe Thr Gly His Ser Tyr Gly Ala
165 170 175
Ala Leu Ala Ala Ile Ala Ala Thr Thr Leu Arg Asn Ala Gly Tyr Thr
180 185 190
Ile Gln Leu Tyr Asp Tyr Gly Gln Pro Arg Leu Gly Asn Leu Ala Leu
195 200 205
Ala Gln Tyr Ile Thr Ala Gln Thr Gln Gly Ala Asn Tyr Arg Val Thr
210 215 220
His Thr Asp Asp Ile Val Pro Lys Leu Pro Pro Glu Leu Phe Gly Tyr
225 230 235 240
His His Phe Ser Pro Glu Tyr Trp Ile Thr Ser Gly Asp Asn Val Thr
245 250 255
Val Thr Thr Ser Asp Val Gln Val Val Thr Gly Ile Asp Ser Thr Ala
260 265 270
Gly Asn Asp Gly Thr Leu Leu Asp Ser Thr Ser Ala His Asp Trp Tyr
275 280 285
Ile Val Tyr Ile Asp Gly Cys Asp
290 295
<210> 2
<211> 269
<212> БЕЛОК
<213> Talaromyces leycettanus
<400> 2
Asp Val Ser Ser Ser Val Leu Ser Glu Leu Asp Leu Phe Ala Gln Tyr
1 5 10 15
Ser Ala Ala Ala Tyr Cys Ser Ser Asn Ile Gly Ser Pro Gly Thr Lys
20 25 30
Leu Thr Cys Ser Val Gly Asn Cys Pro Arg Val Glu Ala Ala Asp Thr
35 40 45
Glu Thr Leu Ile Glu Phe Asn Glu Ser Ser Ser Phe Gly Asp Val Thr
50 55 60
Gly Tyr Ile Ala Val Asp Arg Thr Asn Ser Leu Leu Val Leu Ala Phe
65 70 75 80
Arg Gly Ser Ser Thr Val Ser Asn Trp Glu Ala Asp Leu Asp Phe Pro
85 90 95
Leu Thr Asp Ala Ser Ser Leu Cys Ser Gly Cys Glu Ile His Ser Gly
100 105 110
Phe Trp Ala Ala Trp Gln Thr Val Gln Ala Ser Ile Thr Ser Thr Leu
115 120 125
Glu Ser Ala Ile Ala Ser Tyr Pro Gly Tyr Thr Leu Val Phe Thr Gly
130 135 140
His Ser Tyr Gly Ala Ala Leu Ala Ala Ile Ala Ala Thr Thr Leu Arg
145 150 155 160
Asn Ala Gly Tyr Thr Ile Gln Leu Tyr Asp Tyr Gly Gln Pro Arg Leu
165 170 175
Gly Asn Leu Ala Leu Ala Gln Tyr Ile Thr Ala Gln Thr Gln Gly Ala
180 185 190
Asn Tyr Arg Val Thr His Thr Asp Asp Ile Val Pro Lys Leu Pro Pro
195 200 205
Glu Leu Phe Gly Tyr His His Phe Ser Pro Glu Tyr Trp Ile Thr Ser
210 215 220
Gly Asp Asn Val Thr Val Thr Thr Ser Asp Val Gln Val Val Thr Gly
225 230 235 240
Ile Asp Ser Thr Ala Gly Asn Asp Gly Thr Leu Leu Asp Ser Thr Ser
245 250 255
Ala His Asp Trp Tyr Ile Val Tyr Ile Asp Gly Cys Asp
260 265
<210> 3
<211> 1457
<212> ДНК
<213> Cryphonectria parasitica
<400> 3
cagttgagtc cggatgtgtt tggccttcat ccgctccctt tctgcacacg gtctttagcc 60
cagccagcct gccagcctgc tagccagcca gacagacaag atgagactgt cggtcgcctc 120
ctcactttcc ctggcctctg gcctggtggc agcctcgcct cttcaggctc gtggtaagta 180
gagcttggac tcaacctctc cgttgaaggc tatcttgaac ctaattgtga ttatacacag 240
actccgccgt cctggacgag acgagttacg aaaacatgaa gtactacgtg cagtatgctg 300
cgtctgcgta ctgcgacagc gaagatgctg tgggcacact cgtatcctgc ggtgactctg 360
ggtgtcctaa tgtcacggcg aatggtgcca ccatcgtcgg taccatgccg tgagtctttc 420
cttagccccc gaggacttta tgctccttga catcagagag cccagatgct aatatttgcg 480
actccagcac caccaccacc ttcgacctgg aaggctatgt ggccgtcgac cccacccgcg 540
aagaaattgt ggtcgcattc cgtggctcat ctgacctgcg caactggatt gcagacttcg 600
acttcattga ggttgcgtac tcggattgca ctggctgcta tgtccacgat ggattctacg 660
aatcctggaa ggaaattcag acctatgccg tcgggtacgt cgaggcggcg tacgagacgt 720
acccagacta taccctcgtc atcacaggtc actccctggg cgccgccgtc gccaccctcg 780
cgggtgtgca gttcagaatc gacgggtgag taaaaccaac acacaaaagg actcatgtga 840
gacgtccaca tagtaccgtg ctgacggaac tattcaaagc tacccttgtg atatctacac 900
tgtcggctcc ccccgcatcg gcaacctggc atgggccgag tttgtgaccg cccaggatgg 960
cgctgaatac cgggctaccc attacgatga ccccgtgccg cgcctgccac ccatcgtgct 1020
tggctactac cacacgagcc ccgagttctg gctcgccgcc ggtcccgcca ccaacatcga 1080
ctacaccatt gacggaattg atgtctgcgt aggcaacgca aacaccagtt gcaacgccgg 1140
cacgactggc tttgacgccg atgcgcacga atactacttc cagtacatgg gctgtggcga 1200
tgagagcaac ggcattgaca tgcgcaagcg gtcccccaat actctgtttg gtcgtgacgg 1260
cagcagcaac atcaccgatg cggagctggc ccagaagctc accaactaca cgatgcaaga 1320
tatcgccttt tcggagaagc tcgcctccag cagctaagcc tggtaatgct cggatcagag 1380
gcgcggtcgg tatgaggaca attccatgag gtcactggta ggggcgatcg ttcccgtcca 1440
acatcaaaag tggtggg 1457
<210> 4
<211> 333
<212> БЕЛОК
<213> Cryphonectria parasitica
<400> 4
Met Arg Leu Ser Val Ala Ser Ser Leu Ser Leu Ala Ser Gly Leu Val
1 5 10 15
Ala Ala Ser Pro Leu Gln Ala Arg Asp Ser Ala Val Leu Asp Glu Thr
20 25 30
Ser Tyr Glu Asn Met Lys Tyr Tyr Val Gln Tyr Ala Ala Ser Ala Tyr
35 40 45
Cys Asp Ser Glu Asp Ala Val Gly Thr Leu Val Ser Cys Gly Asp Ser
50 55 60
Gly Cys Pro Asn Val Thr Ala Asn Gly Ala Thr Ile Val Gly Thr Met
65 70 75 80
Pro Thr Thr Thr Thr Phe Asp Leu Glu Gly Tyr Val Ala Val Asp Pro
85 90 95
Thr Arg Glu Glu Ile Val Val Ala Phe Arg Gly Ser Ser Asp Leu Arg
100 105 110
Asn Trp Ile Ala Asp Phe Asp Phe Ile Glu Val Ala Tyr Ser Asp Cys
115 120 125
Thr Gly Cys Tyr Val His Asp Gly Phe Tyr Glu Ser Trp Lys Glu Ile
130 135 140
Gln Thr Tyr Ala Val Gly Tyr Val Glu Ala Ala Tyr Glu Thr Tyr Pro
145 150 155 160
Asp Tyr Thr Leu Val Ile Thr Gly His Ser Leu Gly Ala Ala Val Ala
165 170 175
Thr Leu Ala Gly Val Gln Phe Arg Ile Asp Gly Tyr Pro Cys Asp Ile
180 185 190
Tyr Thr Val Gly Ser Pro Arg Ile Gly Asn Leu Ala Trp Ala Glu Phe
195 200 205
Val Thr Ala Gln Asp Gly Ala Glu Tyr Arg Ala Thr His Tyr Asp Asp
210 215 220
Pro Val Pro Arg Leu Pro Pro Ile Val Leu Gly Tyr Tyr His Thr Ser
225 230 235 240
Pro Glu Phe Trp Leu Ala Ala Gly Pro Ala Thr Asn Ile Asp Tyr Thr
245 250 255
Ile Asp Gly Ile Asp Val Cys Val Gly Asn Ala Asn Thr Ser Cys Asn
260 265 270
Ala Gly Thr Thr Gly Phe Asp Ala Asp Ala His Glu Tyr Tyr Phe Gln
275 280 285
Tyr Met Gly Cys Gly Asp Glu Ser Asn Gly Ile Asp Met Arg Lys Arg
290 295 300
Asn Ile Thr Asp Ala Glu Leu Ala Gln Lys Leu Thr Asn Tyr Thr Met
305 310 315 320
Gln Asp Ile Ala Phe Ser Glu Lys Leu Ala Ser Ser Ser
325 330
<210> 5
<211> 315
<212> БЕЛОК
<213> Cryphonectria parasitica
<400> 5
Ser Pro Leu Gln Ala Arg Asp Ser Ala Val Leu Asp Glu Thr Ser Tyr
1 5 10 15
Glu Asn Met Lys Tyr Tyr Val Gln Tyr Ala Ala Ser Ala Tyr Cys Asp
20 25 30
Ser Glu Asp Ala Val Gly Thr Leu Val Ser Cys Gly Asp Ser Gly Cys
35 40 45
Pro Asn Val Thr Ala Asn Gly Ala Thr Ile Val Gly Thr Met Pro Thr
50 55 60
Thr Thr Thr Phe Asp Leu Glu Gly Tyr Val Ala Val Asp Pro Thr Arg
65 70 75 80
Glu Glu Ile Val Val Ala Phe Arg Gly Ser Ser Asp Leu Arg Asn Trp
85 90 95
Ile Ala Asp Phe Asp Phe Ile Glu Val Ala Tyr Ser Asp Cys Thr Gly
100 105 110
Cys Tyr Val His Asp Gly Phe Tyr Glu Ser Trp Lys Glu Ile Gln Thr
115 120 125
Tyr Ala Val Gly Tyr Val Glu Ala Ala Tyr Glu Thr Tyr Pro Asp Tyr
130 135 140
Thr Leu Val Ile Thr Gly His Ser Leu Gly Ala Ala Val Ala Thr Leu
145 150 155 160
Ala Gly Val Gln Phe Arg Ile Asp Gly Tyr Pro Cys Asp Ile Tyr Thr
165 170 175
Val Gly Ser Pro Arg Ile Gly Asn Leu Ala Trp Ala Glu Phe Val Thr
180 185 190
Ala Gln Asp Gly Ala Glu Tyr Arg Ala Thr His Tyr Asp Asp Pro Val
195 200 205
Pro Arg Leu Pro Pro Ile Val Leu Gly Tyr Tyr His Thr Ser Pro Glu
210 215 220
Phe Trp Leu Ala Ala Gly Pro Ala Thr Asn Ile Asp Tyr Thr Ile Asp
225 230 235 240
Gly Ile Asp Val Cys Val Gly Asn Ala Asn Thr Ser Cys Asn Ala Gly
245 250 255
Thr Thr Gly Phe Asp Ala Asp Ala His Glu Tyr Tyr Phe Gln Tyr Met
260 265 270
Gly Cys Gly Asp Glu Ser Asn Gly Ile Asp Met Arg Lys Arg Asn Ile
275 280 285
Thr Asp Ala Glu Leu Ala Gln Lys Leu Thr Asn Tyr Thr Met Gln Asp
290 295 300
Ile Ala Phe Ser Glu Lys Leu Ala Ser Ser Ser
305 310 315
<210> 6
<211> 1824
<212> ДНК
<213> Gloeophyllum trabeum
<400> 6
cattccgtgt cggcatttat accagcaggg tgtatagagg ggccatgcga gccagcccta 60
caacattcag cgcaacccgc tcgaactcca agctctgcag atggcagctt cgttcgcttt 120
agcagttgcg ctcttggttg gtgctgtcgt ggatgctgcc cccaatctac agaagcgtca 180
gtctataacg acgttgtctt cctcgcaagt ctcagctttc acgccttaca catggtatgc 240
gagcgccggg tattgctcac cttccacaac cttgacatgg acctgcggaa gtaagtcggg 300
ttgtgtttcc catcggaaag tagcactgac tgtgtcaact cagccaattg ccaagctaac 360
agtgggttcg aacccgtggc ttctggaggc gatggtgatt ccactcagta ctgtaagctt 420
agtaccgtac accttgcgct tagtcagatt aatgagcgac ttcctaaagg gtttgttggg 480
tatgacccgt ctctggacac cgtgatcgtc tcgcatcaag gcacggacac ttcggagatg 540
taggtgtcgc ctacttaatc gcacctagaa acatacctag ctaaccctgg tgttccctgg 600
tagcctccct ctgctgacag acgccgatat cgagaagacg actcttgatt cgtcactatt 660
cccgggtata agctcctcta ttgaggtgca cgctgggttc gcagatgcgc aagccaacgc 720
cgccacggac gtcttagcgg ctgtccagac agcactgtcg aagtatagca cgaacaccgt 780
tactatggtc ggacactccc ttggtgcgtt gcacatctgc ccacatttct gaattgacgc 840
tcacctgctg tgaaggtgcc gcaatttctc tgttggatct ggtttacctt cctttgcact 900
tgccctccgg gctcacttac aaaatgtacg gttacggaat gcccagggta tgtctacttc 960
ctttgggacc aggggtgaca agttgaccag gacttctagg ttggcaacca ggccttcgct 1020
gactacatcg acgcgaacca tcccatgact cacgtgaaca ataaggaaga ctacatcccc 1080
atcctacccg ggatgttctt aggcttcgtt catccctctg gcgaggtcca catcgaagac 1140
tcgggatcat gggatgcctg cccaggtacg tctgcccagg ggacttatcc atttcattgg 1200
ctcatttcag accgcaggtc aagacaaccc cagcactcta tgcatcgtcg gcgatgtccc 1260
ggagatttgg gatggtgacg aatctgatca tgacggaccc tacgggccag tcactatggg 1320
ttgctaaaat aacctgtact ttcttttgcg gtcggactgg tgccaaatca aaactgtact 1380
atgtcttcac tgttcattgg tacggctgca cgggccgagg tcgtgcgagt ggacttccca 1440
tgttcattgt tggcacgcga acgcgaaggt gctctgacag gaagacatca cccggagggc 1500
gttcaagatg agaaggtagg gaaggtcagg tgcttgtggt cgtttgcgat ctctcagcga 1560
ttgatcgtca tactcttcca catacccata tctcttatca cccatgaaag accgtaggat 1620
tgacgctagc cgcacgcctc cgtagcggtc ctcgaagcag gcccaccaaa tctgaacaac 1680
ccgaaaatca tggtccccgg gcaatttggt gggacattcg ttgaccccaa ggcaagcttc 1740
tccggaatag cctatgtcat ccctgctgag aacaatcctg ttgtagtatg actggtgctt 1800
ctcgacagtc tcgctgaagg attg 1824
<210> 7
<211> 298
<212> БЕЛОК
<213> Gloeophyllum trabeum
<400> 7
Met Ala Ala Ser Phe Ala Leu Ala Val Ala Leu Leu Val Gly Ala Val
1 5 10 15
Val Asp Ala Ala Pro Asn Leu Gln Lys Arg Gln Ser Ile Thr Thr Leu
20 25 30
Ser Ser Ser Gln Val Ser Ala Phe Thr Pro Tyr Thr Trp Tyr Ala Ser
35 40 45
Ala Gly Tyr Cys Ser Pro Ser Thr Thr Leu Thr Trp Thr Cys Gly Thr
50 55 60
Asn Cys Gln Ala Asn Ser Gly Phe Glu Pro Val Ala Ser Gly Gly Asp
65 70 75 80
Gly Asp Ser Thr Gln Tyr Trp Phe Val Gly Tyr Asp Pro Ser Leu Asp
85 90 95
Thr Val Ile Val Ser His Gln Gly Thr Asp Thr Ser Glu Ile Leu Pro
100 105 110
Leu Leu Thr Asp Ala Asp Ile Glu Lys Thr Thr Leu Asp Ser Ser Leu
115 120 125
Phe Pro Gly Ile Ser Ser Ser Ile Glu Val His Ala Gly Phe Ala Asp
130 135 140
Ala Gln Ala Asn Ala Ala Thr Asp Val Leu Ala Ala Val Gln Thr Ala
145 150 155 160
Leu Ser Lys Tyr Ser Thr Asn Thr Val Thr Met Val Gly His Ser Leu
165 170 175
Gly Ala Ala Ile Ser Leu Leu Asp Leu Val Tyr Leu Pro Leu His Leu
180 185 190
Pro Ser Gly Leu Thr Tyr Lys Met Tyr Gly Tyr Gly Met Pro Arg Val
195 200 205
Gly Asn Gln Ala Phe Ala Asp Tyr Ile Asp Ala Asn His Pro Met Thr
210 215 220
His Val Asn Asn Lys Glu Asp Tyr Ile Pro Ile Leu Pro Gly Met Phe
225 230 235 240
Leu Gly Phe Val His Pro Ser Gly Glu Val His Ile Glu Asp Ser Gly
245 250 255
Ser Trp Asp Ala Cys Pro Gly Gln Asp Asn Pro Ser Thr Leu Cys Ile
260 265 270
Val Gly Asp Val Pro Glu Ile Trp Asp Gly Asp Glu Ser Asp His Asp
275 280 285
Gly Pro Tyr Gly Pro Val Thr Met Gly Cys
290 295
<210> 8
<211> 279
<212> БЕЛОК
<213> Gloeophyllum trabeum
<400> 8
Ala Pro Asn Leu Gln Lys Arg Gln Ser Ile Thr Thr Leu Ser Ser Ser
1 5 10 15
Gln Val Ser Ala Phe Thr Pro Tyr Thr Trp Tyr Ala Ser Ala Gly Tyr
20 25 30
Cys Ser Pro Ser Thr Thr Leu Thr Trp Thr Cys Gly Thr Asn Cys Gln
35 40 45
Ala Asn Ser Gly Phe Glu Pro Val Ala Ser Gly Gly Asp Gly Asp Ser
50 55 60
Thr Gln Tyr Trp Phe Val Gly Tyr Asp Pro Ser Leu Asp Thr Val Ile
65 70 75 80
Val Ser His Gln Gly Thr Asp Thr Ser Glu Ile Leu Pro Leu Leu Thr
85 90 95
Asp Ala Asp Ile Glu Lys Thr Thr Leu Asp Ser Ser Leu Phe Pro Gly
100 105 110
Ile Ser Ser Ser Ile Glu Val His Ala Gly Phe Ala Asp Ala Gln Ala
115 120 125
Asn Ala Ala Thr Asp Val Leu Ala Ala Val Gln Thr Ala Leu Ser Lys
130 135 140
Tyr Ser Thr Asn Thr Val Thr Met Val Gly His Ser Leu Gly Ala Ala
145 150 155 160
Ile Ser Leu Leu Asp Leu Val Tyr Leu Pro Leu His Leu Pro Ser Gly
165 170 175
Leu Thr Tyr Lys Met Tyr Gly Tyr Gly Met Pro Arg Val Gly Asn Gln
180 185 190
Ala Phe Ala Asp Tyr Ile Asp Ala Asn His Pro Met Thr His Val Asn
195 200 205
Asn Lys Glu Asp Tyr Ile Pro Ile Leu Pro Gly Met Phe Leu Gly Phe
210 215 220
Val His Pro Ser Gly Glu Val His Ile Glu Asp Ser Gly Ser Trp Asp
225 230 235 240
Ala Cys Pro Gly Gln Asp Asn Pro Ser Thr Leu Cys Ile Val Gly Asp
245 250 255
Val Pro Glu Ile Trp Asp Gly Asp Glu Ser Asp His Asp Gly Pro Tyr
260 265 270
Gly Pro Val Thr Met Gly Cys
275
<210> 9
<211> 280
<212> БЕЛОК
<213> Bacillus macauensis
<400> 9
Met Lys Lys Thr Phe Val Ala Val Ala Thr Ala Ala Leu Leu Val Thr
1 5 10 15
Gly Phe Gln Gly Asn Ala Ser Ala Glu Asp Asp Ala Gln Pro Pro Ile
20 25 30
Thr Ala Lys Trp Ser Ala Glu Asp Pro His His Glu Asp Thr Asn Thr
35 40 45
His Leu Trp Ile Val Arg His Ala Met Glu Ile Met Ala Asn Asn Lys
50 55 60
Asp Val Val Lys Pro Gly Glu Val Glu Gln Leu Lys Gln Trp Gln Ser
65 70 75 80
Asp Leu Glu Gln Gly Ile Tyr Asp Ala Asp His Ala Asn Pro Tyr Tyr
85 90 95
Asp Asn Ala Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Thr Gly Lys
100 105 110
Ser Tyr Ile Pro Leu Ala Ala His Ala Lys Thr Thr Ser Val Lys Tyr
115 120 125
Phe Lys Arg Ala Gly Glu Ala Tyr Gln Lys Gly Asp His Lys Gln Ala
130 135 140
Phe Tyr Asn Leu Gly Leu Ala Leu His Tyr Ile Gly Asp Leu Asn Gln
145 150 155 160
Pro Met His Ala Ala Asn Phe Thr Asn Leu Ser Tyr Pro Gln Gly Phe
165 170 175
His Ser Lys Tyr Glu Asn Tyr Val Asp Ser Phe Lys Glu Asp Tyr Ala
180 185 190
Val Lys Asp Gly Glu Gly Tyr Trp His Trp Lys Gly Thr Asn Pro Glu
195 200 205
Asp Trp Leu His Gly Thr Ala Val Ala Ala Lys Lys Asp Tyr Pro Asp
210 215 220
Ile Val Asn Asp Thr Thr Lys Ala Trp Phe Val Lys Ala Ala Val Ser
225 230 235 240
Asn Ser Tyr Ala Ala Lys Trp Arg Ala Ala Val Val Pro Ala Thr Gly
245 250 255
Lys Arg Leu Thr Glu Ala Gln Arg Ile Leu Ala Gly Tyr Met Gln Leu
260 265 270
Trp Phe Asp Thr Tyr Val Asn Lys
275 280
<210> 10
<211> 245
<212> БЕЛОК
<213> Bacillus macauensis
<400> 10
Trp Ser Ala Glu Asp Pro His His Glu Asp Thr Asn Thr His Leu Trp
1 5 10 15
Ile Val Arg His Ala Met Glu Ile Met Ala Asn Asn Lys Asp Val Val
20 25 30
Lys Pro Gly Glu Val Glu Gln Leu Lys Gln Trp Gln Ser Asp Leu Glu
35 40 45
Gln Gly Ile Tyr Asp Ala Asp His Ala Asn Pro Tyr Tyr Asp Asn Ala
50 55 60
Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Thr Gly Lys Ser Tyr Ile
65 70 75 80
Pro Leu Ala Ala His Ala Lys Thr Thr Ser Val Lys Tyr Phe Lys Arg
85 90 95
Ala Gly Glu Ala Tyr Gln Lys Gly Asp His Lys Gln Ala Phe Tyr Asn
100 105 110
Leu Gly Leu Ala Leu His Tyr Ile Gly Asp Leu Asn Gln Pro Met His
115 120 125
Ala Ala Asn Phe Thr Asn Leu Ser Tyr Pro Gln Gly Phe His Ser Lys
130 135 140
Tyr Glu Asn Tyr Val Asp Ser Phe Lys Glu Asp Tyr Ala Val Lys Asp
145 150 155 160
Gly Glu Gly Tyr Trp His Trp Lys Gly Thr Asn Pro Glu Asp Trp Leu
165 170 175
His Gly Thr Ala Val Ala Ala Lys Lys Asp Tyr Pro Asp Ile Val Asn
180 185 190
Asp Thr Thr Lys Ala Trp Phe Val Lys Ala Ala Val Ser Asn Ser Tyr
195 200 205
Ala Ala Lys Trp Arg Ala Ala Val Val Pro Ala Thr Gly Lys Arg Leu
210 215 220
Thr Glu Ala Gln Arg Ile Leu Ala Gly Tyr Met Gln Leu Trp Phe Asp
225 230 235 240
Thr Tyr Val Asn Lys
245
<210> 11
<211> 283
<212> БЕЛОК
<213> Bacillus thuringiensis
<400> 11
Met Lys Lys Lys Val Leu Ala Leu Ala Ala Ala Ile Thr Leu Val Ala
1 5 10 15
Pro Leu Gln Ser Val Ala Phe Ala His Glu Asn Asp Gly Gly Ser Lys
20 25 30
Ile Lys Ile Val His Arg Trp Ser Ala Glu Asp Lys His Lys Glu Gly
35 40 45
Val Asn Ser His Leu Trp Ile Val Asn Arg Ala Ile Asp Ile Met Ser
50 55 60
Arg Asn Thr Thr Leu Val Lys Gln Asp Arg Val Ala Gln Leu Asn Glu
65 70 75 80
Trp Arg Thr Glu Leu Glu Asn Gly Ile Tyr Ala Ala Asp Tyr Glu Asn
85 90 95
Pro Tyr Tyr Asp Asn Ser Thr Phe Ala Ser His Phe Tyr Asp Pro Asp
100 105 110
Asn Gly Lys Thr Tyr Ile Pro Phe Ala Lys Gln Ala Lys Glu Thr Gly
115 120 125
Ala Lys Tyr Phe Lys Leu Ala Gly Glu Ser Tyr Lys Asn Lys Asp Met
130 135 140
Lys Gln Ala Phe Phe Tyr Leu Gly Leu Ser Leu His Tyr Leu Gly Asp
145 150 155 160
Val Asn Gln Pro Met His Ala Ala Asn Phe Thr Asn Leu Ser Tyr Pro
165 170 175
Gln Gly Phe His Ser Lys Tyr Glu Asn Phe Val Asp Thr Ile Lys Asp
180 185 190
Asn Tyr Lys Val Thr Asp Gly Asn Gly Tyr Trp Asn Trp Lys Gly Thr
195 200 205
Asn Pro Glu Asp Trp Ile His Gly Ala Ala Val Val Ala Lys Gln Asp
210 215 220
Tyr Ser Gly Ile Val Asn Asp Asn Thr Lys Asp Trp Phe Val Lys Ala
225 230 235 240
Ala Val Ser Gln Glu Tyr Ala Asp Lys Trp Arg Ala Glu Val Thr Pro
245 250 255
Met Thr Gly Lys Arg Leu Met Asp Ala Gln Arg Val Thr Ala Gly Tyr
260 265 270
Ile Gln Leu Trp Phe Asp Thr Tyr Gly Asp Arg
275 280
<210> 12
<211> 245
<212> БЕЛОК
<213> Bacillus thuringiensis
<400> 12
Trp Ser Ala Glu Asp Lys His Lys Glu Gly Val Asn Ser His Leu Trp
1 5 10 15
Ile Val Asn Arg Ala Ile Asp Ile Met Ser Arg Asn Thr Thr Leu Val
20 25 30
Lys Gln Asp Arg Val Ala Gln Leu Asn Glu Trp Arg Thr Glu Leu Glu
35 40 45
Asn Gly Ile Tyr Ala Ala Asp Tyr Glu Asn Pro Tyr Tyr Asp Asn Ser
50 55 60
Thr Phe Ala Ser His Phe Tyr Asp Pro Asp Asn Gly Lys Thr Tyr Ile
65 70 75 80
Pro Phe Ala Lys Gln Ala Lys Glu Thr Gly Ala Lys Tyr Phe Lys Leu
85 90 95
Ala Gly Glu Ser Tyr Lys Asn Lys Asp Met Lys Gln Ala Phe Phe Tyr
100 105 110
Leu Gly Leu Ser Leu His Tyr Leu Gly Asp Val Asn Gln Pro Met His
115 120 125
Ala Ala Asn Phe Thr Asn Leu Ser Tyr Pro Gln Gly Phe His Ser Lys
130 135 140
Tyr Glu Asn Phe Val Asp Thr Ile Lys Asp Asn Tyr Lys Val Thr Asp
145 150 155 160
Gly Asn Gly Tyr Trp Asn Trp Lys Gly Thr Asn Pro Glu Asp Trp Ile
165 170 175
His Gly Ala Ala Val Val Ala Lys Gln Asp Tyr Ser Gly Ile Val Asn
180 185 190
Asp Asn Thr Lys Asp Trp Phe Val Lys Ala Ala Val Ser Gln Glu Tyr
195 200 205
Ala Asp Lys Trp Arg Ala Glu Val Thr Pro Met Thr Gly Lys Arg Leu
210 215 220
Met Asp Ala Gln Arg Val Thr Ala Gly Tyr Ile Gln Leu Trp Phe Asp
225 230 235 240
Thr Tyr Gly Asp Arg
245
<210> 13
<211> 320
<212> БЕЛОК
<213> Pseudomonas sp.
<400> 13
Met Thr His Leu Ile Gly Phe Ala Pro Arg Leu Leu Ala Phe Ser Ala
1 5 10 15
Leu Leu Leu Ser Gln Thr Ala Phe Ser Gln Glu Ser Pro Ala Phe Ile
20 25 30
Asp Pro Ala Ser Trp Asn Thr Pro Phe Asn Gly Ile Ala Gln Val Ala
35 40 45
Cys His Asn Cys Tyr Glu Lys Gln Tyr Ala Asn Thr Phe Ser Ser Val
50 55 60
Leu Asp Ser Val Arg Thr Leu Glu Leu Asp Phe Trp Asp Gln Arg Asp
65 70 75 80
Ala Val Ser Gly Gly Ser Pro His His Trp Phe Val Arg His Asn Pro
85 90 95
Gly Thr Leu Phe Gln Ser Gly Asn Asp Asn Asn Cys Thr Gly Asp Gly
100 105 110
Thr Gly Lys Asn Asp Leu Glu Ala Cys Leu Asn Asp Val Lys Asn Trp
115 120 125
Ser Asp Lys His Pro Gly His Phe Pro Ile Thr Leu Ile Leu Asp Lys
130 135 140
Lys Gln Gly Trp Ser Lys Glu Ser Ser Gly Arg Thr Pro Lys Asp Phe
145 150 155 160
Asp Glu Leu Val Ala Arg Val Phe Gln Gly Lys Leu Phe Thr Pro Gln
165 170 175
Asp Leu Ala Thr His Ile Gly Ser Gly Ala Gly Ala Leu Gln Gly Asn
180 185 190
Leu Lys Gly Lys Ser Trp Pro Thr Ala Asn Asp Leu Gln Gly Lys Val
195 200 205
Leu Leu Val Leu Asn His Ser Glu Asn Gln Lys Leu Ser Gln Tyr Ala
210 215 220
Glu Ala Arg Thr Ser Lys Ala Lys Val Phe Ile Ser Pro Val Thr Asn
225 230 235 240
Gly Gln Asn Asp Ile Ser Gly Lys Val Ser Gly Met Ser Ser Gln Ser
245 250 255
Ser Gly Tyr Val Ala Met Asn Asn Met Gly Lys Gly Asp Lys Ser Trp
260 265 270
Ala Lys Gln Ala Phe Ala Tyr Ser His Ile Gly Arg Val Trp Gly Asp
275 280 285
Asp Glu Val Ser Phe Ala Gln His Ile Asn Gln Lys Ile Asn Leu Ser
290 295 300
Ala Tyr Tyr Arg Phe Ala Ala Gln Ser Ala Gly Gly Tyr Arg Ile Arg
305 310 315 320
<210> 14
<211> 297
<212> БЕЛОК
<213> Pseudomonas sp.
<400> 14
Gln Glu Ser Pro Ala Phe Ile Asp Pro Ala Ser Trp Asn Thr Pro Phe
1 5 10 15
Asn Gly Ile Ala Gln Val Ala Cys His Asn Cys Tyr Glu Lys Gln Tyr
20 25 30
Ala Asn Thr Phe Ser Ser Val Leu Asp Ser Val Arg Thr Leu Glu Leu
35 40 45
Asp Phe Trp Asp Gln Arg Asp Ala Val Ser Gly Gly Ser Pro His His
50 55 60
Trp Phe Val Arg His Asn Pro Gly Thr Leu Phe Gln Ser Gly Asn Asp
65 70 75 80
Asn Asn Cys Thr Gly Asp Gly Thr Gly Lys Asn Asp Leu Glu Ala Cys
85 90 95
Leu Asn Asp Val Lys Asn Trp Ser Asp Lys His Pro Gly His Phe Pro
100 105 110
Ile Thr Leu Ile Leu Asp Lys Lys Gln Gly Trp Ser Lys Glu Ser Ser
115 120 125
Gly Arg Thr Pro Lys Asp Phe Asp Glu Leu Val Ala Arg Val Phe Gln
130 135 140
Gly Lys Leu Phe Thr Pro Gln Asp Leu Ala Thr His Ile Gly Ser Gly
145 150 155 160
Ala Gly Ala Leu Gln Gly Asn Leu Lys Gly Lys Ser Trp Pro Thr Ala
165 170 175
Asn Asp Leu Gln Gly Lys Val Leu Leu Val Leu Asn His Ser Glu Asn
180 185 190
Gln Lys Leu Ser Gln Tyr Ala Glu Ala Arg Thr Ser Lys Ala Lys Val
195 200 205
Phe Ile Ser Pro Val Thr Asn Gly Gln Asn Asp Ile Ser Gly Lys Val
210 215 220
Ser Gly Met Ser Ser Gln Ser Ser Gly Tyr Val Ala Met Asn Asn Met
225 230 235 240
Gly Lys Gly Asp Lys Ser Trp Ala Lys Gln Ala Phe Ala Tyr Ser His
245 250 255
Ile Gly Arg Val Trp Gly Asp Asp Glu Val Ser Phe Ala Gln His Ile
260 265 270
Asn Gln Lys Ile Asn Leu Ser Ala Tyr Tyr Arg Phe Ala Ala Gln Ser
275 280 285
Ala Gly Gly Tyr Arg Ile Arg Pro Phe
290 295
<210> 15
<211> 610
<212> БЕЛОК
<213> Penicillium emersonii
<400> 15
Met Arg Val Leu Ala Leu Ile Ala Ala Leu Ala Thr Val Ala Thr Ala
1 5 10 15
Ser Ala Pro Tyr Asp Lys Arg Asp Leu Ala Gln Glu Ile Trp Asp Asp
20 25 30
Ile Lys Asn Ala Val Asp Cys Ala Gly Cys Gln Val Val Leu Thr Ala
35 40 45
Leu Lys Gly Val Ala Asp Leu Gly Thr Thr Ala Leu Val Asp Val Leu
50 55 60
Thr Glu Val Cys Asn Ile Ser Gly Lys Glu Asp Ser Asp Val Cys Ser
65 70 75 80
Gly Ile Ile Ser Arg Glu Gly Pro Val Leu Asp Tyr Val Leu Gln His
85 90 95
Leu Asp Ile Gly Ser His Thr Ser Gln Val Ile Cys Ala Ser Ala Phe
100 105 110
Gly Leu Cys Gln Tyr Pro Glu Val Arg Pro Tyr Asn Leu Thr Phe Pro
115 120 125
Lys Pro Lys Pro Asn Thr Thr Arg Pro Glu Pro Ser Gly Glu Ser Pro
130 135 140
Ile Gln Val Val His Phe Ser Asp Thr His Val Asp Leu Ser Tyr Glu
145 150 155 160
Thr Gly Ser Asn Tyr Asn Cys Thr Lys Pro Ile Cys Cys Arg Pro Tyr
165 170 175
Thr Ala Glu Asp Ala Pro Gly Asn Thr Thr Thr Pro Cys Gly Pro Tyr
180 185 190
Gly Asn Thr Lys Cys Asp Ala Pro Leu Ser Leu Glu Glu Ser Met Phe
195 200 205
Ala Ala Ile Lys Ala Leu Asn Pro Gln Pro Ala Phe Ser Ile Tyr Thr
210 215 220
Gly Asp Val Val Ala His Asp Ile Trp Leu Val Asp Gln Asn Glu Val
225 230 235 240
Ile Glu Asp Leu Asn Ala Thr Tyr Asp Arg Met Ala Gly Leu Gly Leu
245 250 255
Val Tyr Ala Ala Ile Gly Asn His Asp Thr Ala Pro Val Asn Asp Leu
260 265 270
Pro Thr Ser Asn Ile Pro Ser Glu Tyr Ser Ala Asn Trp Thr Tyr Glu
275 280 285
Ala Leu Ser Tyr Asp Phe Thr Met Leu Thr Gln Ser Ala Ser Ala Gln
290 295 300
Thr Ala Ala Asn Tyr Gly Ser Tyr Ser Ala Ile Tyr Pro Gly Ser Tyr
305 310 315 320
Gly Thr Asp Leu Arg Val Ile Ser Tyr Asn Ser Ile Phe Tyr Tyr Val
325 330 335
Asp Asn Phe Trp Ala Tyr Gln Asp Pro Met Glu Phe Asp Pro Asp Gly
340 345 350
Gln Leu Ala Trp Leu Ile Asn Glu Leu Gln Glu Ala Glu Thr Ala Gly
355 360 365
Gln Arg Val Trp Ile Ile Ala His Val Pro Thr Gly Thr Ser Asp His
370 375 380
Phe His Asp Tyr Ser His Tyr Phe Asp Gln Ile Val Gln Arg Tyr Glu
385 390 395 400
Ala Thr Ile Ala Ala Leu Phe Tyr Gly His Thr His Ile Asp Gln Phe
405 410 415
Gln Ile Ser Tyr Ser Asn Tyr Ser Asn Arg Ala Phe Asp Thr Ala Thr
420 425 430
Ala Ile Gly Tyr Ile Met Pro Ser Leu Thr Pro Thr Ser Gly Pro Pro
435 440 445
Thr Phe Arg Val Tyr Asp Val Asp Pro Lys Thr Phe Ala Val Leu Asp
450 455 460
Phe Thr Asn Tyr Ile Ala Asn Ile Ser Asp Pro Ala Phe Gln Ser Gly
465 470 475 480
Pro Ser Trp Gln Lys Tyr Tyr Ser Ala Lys Glu Thr Tyr Gly Ser Leu
485 490 495
Leu Ser Pro Pro Val Thr Asp Pro Thr Ala Glu Leu Thr Pro Ala Phe
500 505 510
Trp His Asn Val Thr Val Ala Phe Glu Gln Asp Asn Ala Thr Phe Gln
515 520 525
Glu Tyr Trp Ala Arg Gln Thr Arg Gly Tyr Asp Val Ser Ser Cys Thr
530 535 540
Gly Ser Cys Ile Thr Gln Ala Ile Cys Gly Leu Arg Ala Gly Asp Ala
545 550 555 560
Gln Tyr Asn Cys Val Thr Pro Thr Pro Gly Phe Asn Phe Ala Lys Arg
565 570 575
Asp Thr Ser Asn Pro Lys Gln Ala Leu Ser His Val Glu Lys Cys Glu
580 585 590
Gly Ser Gly Leu Leu Gly Leu Leu Arg Arg Met Val Ala Asp Ser Lys
595 600 605
Ser Ser
610
<210> 16
<211> 594
<212> БЕЛОК
<213> Penicillium emersonii
<400> 16
Ser Ala Pro Tyr Asp Lys Arg Asp Leu Ala Gln Glu Ile Trp Asp Asp
1 5 10 15
Ile Lys Asn Ala Val Asp Cys Ala Gly Cys Gln Val Val Leu Thr Ala
20 25 30
Leu Lys Gly Val Ala Asp Leu Gly Thr Thr Ala Leu Val Asp Val Leu
35 40 45
Thr Glu Val Cys Asn Ile Ser Gly Lys Glu Asp Ser Asp Val Cys Ser
50 55 60
Gly Ile Ile Ser Arg Glu Gly Pro Val Leu Asp Tyr Val Leu Gln His
65 70 75 80
Leu Asp Ile Gly Ser His Thr Ser Gln Val Ile Cys Ala Ser Ala Phe
85 90 95
Gly Leu Cys Gln Tyr Pro Glu Val Arg Pro Tyr Asn Leu Thr Phe Pro
100 105 110
Lys Pro Lys Pro Asn Thr Thr Arg Pro Glu Pro Ser Gly Glu Ser Pro
115 120 125
Ile Gln Val Val His Phe Ser Asp Thr His Val Asp Leu Ser Tyr Glu
130 135 140
Thr Gly Ser Asn Tyr Asn Cys Thr Lys Pro Ile Cys Cys Arg Pro Tyr
145 150 155 160
Thr Ala Glu Asp Ala Pro Gly Asn Thr Thr Thr Pro Cys Gly Pro Tyr
165 170 175
Gly Asn Thr Lys Cys Asp Ala Pro Leu Ser Leu Glu Glu Ser Met Phe
180 185 190
Ala Ala Ile Lys Ala Leu Asn Pro Gln Pro Ala Phe Ser Ile Tyr Thr
195 200 205
Gly Asp Val Val Ala His Asp Ile Trp Leu Val Asp Gln Asn Glu Val
210 215 220
Ile Glu Asp Leu Asn Ala Thr Tyr Asp Arg Met Ala Gly Leu Gly Leu
225 230 235 240
Val Tyr Ala Ala Ile Gly Asn His Asp Thr Ala Pro Val Asn Asp Leu
245 250 255
Pro Thr Ser Asn Ile Pro Ser Glu Tyr Ser Ala Asn Trp Thr Tyr Glu
260 265 270
Ala Leu Ser Tyr Asp Phe Thr Met Leu Thr Gln Ser Ala Ser Ala Gln
275 280 285
Thr Ala Ala Asn Tyr Gly Ser Tyr Ser Ala Ile Tyr Pro Gly Ser Tyr
290 295 300
Gly Thr Asp Leu Arg Val Ile Ser Tyr Asn Ser Ile Phe Tyr Tyr Val
305 310 315 320
Asp Asn Phe Trp Ala Tyr Gln Asp Pro Met Glu Phe Asp Pro Asp Gly
325 330 335
Gln Leu Ala Trp Leu Ile Asn Glu Leu Gln Glu Ala Glu Thr Ala Gly
340 345 350
Gln Arg Val Trp Ile Ile Ala His Val Pro Thr Gly Thr Ser Asp His
355 360 365
Phe His Asp Tyr Ser His Tyr Phe Asp Gln Ile Val Gln Arg Tyr Glu
370 375 380
Ala Thr Ile Ala Ala Leu Phe Tyr Gly His Thr His Ile Asp Gln Phe
385 390 395 400
Gln Ile Ser Tyr Ser Asn Tyr Ser Asn Arg Ala Phe Asp Thr Ala Thr
405 410 415
Ala Ile Gly Tyr Ile Met Pro Ser Leu Thr Pro Thr Ser Gly Pro Pro
420 425 430
Thr Phe Arg Val Tyr Asp Val Asp Pro Lys Thr Phe Ala Val Leu Asp
435 440 445
Phe Thr Asn Tyr Ile Ala Asn Ile Ser Asp Pro Ala Phe Gln Ser Gly
450 455 460
Pro Ser Trp Gln Lys Tyr Tyr Ser Ala Lys Glu Thr Tyr Gly Ser Leu
465 470 475 480
Leu Ser Pro Pro Val Thr Asp Pro Thr Ala Glu Leu Thr Pro Ala Phe
485 490 495
Trp His Asn Val Thr Val Ala Phe Glu Gln Asp Asn Ala Thr Phe Gln
500 505 510
Glu Tyr Trp Ala Arg Gln Thr Arg Gly Tyr Asp Val Ser Ser Cys Thr
515 520 525
Gly Ser Cys Ile Thr Gln Ala Ile Cys Gly Leu Arg Ala Gly Asp Ala
530 535 540
Gln Tyr Asn Cys Val Thr Pro Thr Pro Gly Phe Asn Phe Ala Lys Arg
545 550 555 560
Asp Thr Ser Asn Pro Lys Gln Ala Leu Ser His Val Glu Lys Cys Glu
565 570 575
Gly Ser Gly Leu Leu Gly Leu Leu Arg Arg Met Val Ala Asp Ser Lys
580 585 590
Ser Ser
<210> 17
<211> 643
<212> БЕЛОК
<213> Kionochaeta sp.
<400> 17
Met Arg Ala Ser Ser Ile Leu Ser Leu Ala Leu Gly Leu Ser Val Ala
1 5 10 15
Gln Ala Ala Val Asn Pro Ala Asp Val Leu Ser Val Val Glu Lys Arg
20 25 30
Val Asp Pro Ala Ser Gly Leu Glu Val Arg Ser Ile Trp Asp Thr Ile
35 40 45
Trp Asn Asp Ile Lys Ser Ala Ala Asp Cys Thr Ala Cys Glu Ala Val
50 55 60
Leu Thr Leu Leu Lys Gly Val Ala Ala Phe Gly Asp Asn Phe Phe Val
65 70 75 80
Glu Val Leu Thr Glu Ile Cys Asp Leu Ser Gly Ala Glu Asp Asp Asp
85 90 95
Val Cys Ser Gly Val Leu Ser Leu Glu Gly Pro Ile Ile Ala Asn Asp
100 105 110
Ile Arg Lys Met Ser Ile Gly Ser Lys Thr Ser Glu Leu Phe Cys Ile
115 120 125
Thr Phe Leu Gly Leu Cys Ser Tyr Pro Ala Val Asp Ala Phe Thr Val
130 135 140
Pro Phe Pro Thr Ala Lys Ser Ala Ala Thr Arg Pro Val Ser Ser Gly
145 150 155 160
Lys Asp Pro Ile Tyr Val Val His Tyr Ser Asp Ile His Ile Asp Pro
165 170 175
Phe Tyr Val Ala Gly Ser Ala Ser Asn Cys Thr Lys Pro Ile Cys Cys
180 185 190
Arg Asp Tyr Thr Ser Ala Ser Ser Pro Gly Asn Asn Asn Ser Pro Ala
195 200 205
Gly Pro Tyr Gly Asp His Asn Cys Asp Val Pro Ile Ser Leu Glu Asp
210 215 220
Ser Met Tyr Ala Ala Ile Lys Lys Leu Val Pro Asp Ala Ala Phe Gly
225 230 235 240
Ile Phe Thr Gly Asp Ile Val Asp His Ala Val Trp Asn Thr Ser Glu
245 250 255
Ser Gln Asn Ile Ile Asp Met Asn Asp Ala Tyr Thr Arg Met Lys Asn
260 265 270
Ser Gly Met Leu Pro Thr Ile Phe Ala Thr Ala Gly Asn His Glu Ala
275 280 285
Ser Pro Val Asn Ser Phe Pro Pro Pro Ala Ile Gly Asn Glu Ser Gln
290 295 300
Trp Val Tyr Asp Thr Leu Ala Ser Asp Trp Ser Gln Trp Ile Gly Thr
305 310 315 320
Ser Gly Ala Ser Ser Val Glu Ser Ile Gly Ala Tyr Ser Val Gln Tyr
325 330 335
Gly Ser Thr Lys Leu Arg Val Ile Ser Leu Asn Thr Asn Met Tyr Tyr
340 345 350
Ile Glu Asn Phe Tyr Leu Tyr Glu Pro Thr Met Glu Gln Asp Pro Ala
355 360 365
Gly Gln Phe Ala Trp Leu Val Ser Glu Leu Ser Ala Ala Glu Ala Ala
370 375 380
Gly Glu Arg Val Trp Ile Ile Gly His Met Pro Leu Gly Leu Ser Asp
385 390 395 400
Ala Phe His Asp Pro Ser Asn Tyr Phe Asp Gln Ile Val Asn Arg Tyr
405 410 415
Glu Ala Thr Ile Ala Ala Met Phe Phe Gly His Thr His Glu Asp His
420 425 430
Phe Gln Ile Ser Tyr Ser Asp Tyr Asn Ala Arg Thr Ala Ala Asn Ala
435 440 445
Arg Ala Val Ser Tyr Ile Met Pro Ser Leu Thr Pro Thr Ser Gly His
450 455 460
Pro Thr Phe Arg Val Tyr Thr Val Asp Pro Glu Thr Phe Gly Val Leu
465 470 475 480
Asp Ala Thr Thr Tyr Tyr Ala Asp Met Ser Gln Pro Thr Tyr Gln Thr
485 490 495
Ala Gly Pro Ala Trp Ser Val Tyr Tyr Ser Ala Lys Ala Ala Tyr Gly
500 505 510
Gly Leu Val Asp Pro Pro Val Ala Ala Asp Asp Ala Ala Ala Glu Leu
515 520 525
Thr Pro Ala Phe Trp His Asn Val Thr Ala Ala Leu Ala Ala Asp Pro
530 535 540
Ala Ser Phe Asp Ala Tyr Tyr Ala Arg Lys Thr Arg Gly Trp Asp Val
545 550 555 560
Ala Ala Cys Ala Gly Ala Cys Ala Ala Ala Glu Val Cys Ala Leu Arg
565 570 575
Ala Ala Arg Ala Gln Asp Asn Cys Val Val Pro Thr Pro Gly Val His
580 585 590
Phe Ser Lys Arg Ala Asp Glu Gly Thr Leu Ala His His Arg Asp Glu
595 600 605
Cys Gly Val Ser Val Ala Arg Asn Ser Leu Ser Ser Leu Val Val Gln
610 615 620
Arg Glu Ala Leu Glu His Leu Glu Gly Arg Leu Ser Glu Lys Arg Arg
625 630 635 640
Met Ala Val
<210> 18
<211> 625
<212> БЕЛОК
<213> Kionochaeta sp.
<400> 18
Ala Val Asn Pro Ala Asp Val Leu Ser Val Val Glu Lys Arg Val Asp
1 5 10 15
Pro Ala Ser Gly Leu Glu Val Arg Ser Ile Trp Asp Thr Ile Trp Asn
20 25 30
Asp Ile Lys Ser Ala Ala Asp Cys Thr Ala Cys Glu Ala Val Leu Thr
35 40 45
Leu Leu Lys Gly Val Ala Ala Phe Gly Asp Asn Phe Phe Val Glu Val
50 55 60
Leu Thr Glu Ile Cys Asp Leu Ser Gly Ala Glu Asp Asp Asp Val Cys
65 70 75 80
Ser Gly Val Leu Ser Leu Glu Gly Pro Ile Ile Ala Asn Asp Ile Arg
85 90 95
Lys Met Ser Ile Gly Ser Lys Thr Ser Glu Leu Phe Cys Ile Thr Phe
100 105 110
Leu Gly Leu Cys Ser Tyr Pro Ala Val Asp Ala Phe Thr Val Pro Phe
115 120 125
Pro Thr Ala Lys Ser Ala Ala Thr Arg Pro Val Ser Ser Gly Lys Asp
130 135 140
Pro Ile Tyr Val Val His Tyr Ser Asp Ile His Ile Asp Pro Phe Tyr
145 150 155 160
Val Ala Gly Ser Ala Ser Asn Cys Thr Lys Pro Ile Cys Cys Arg Asp
165 170 175
Tyr Thr Ser Ala Ser Ser Pro Gly Asn Asn Asn Ser Pro Ala Gly Pro
180 185 190
Tyr Gly Asp His Asn Cys Asp Val Pro Ile Ser Leu Glu Asp Ser Met
195 200 205
Tyr Ala Ala Ile Lys Lys Leu Val Pro Asp Ala Ala Phe Gly Ile Phe
210 215 220
Thr Gly Asp Ile Val Asp His Ala Val Trp Asn Thr Ser Glu Ser Gln
225 230 235 240
Asn Ile Ile Asp Met Asn Asp Ala Tyr Thr Arg Met Lys Asn Ser Gly
245 250 255
Met Leu Pro Thr Ile Phe Ala Thr Ala Gly Asn His Glu Ala Ser Pro
260 265 270
Val Asn Ser Phe Pro Pro Pro Ala Ile Gly Asn Glu Ser Gln Trp Val
275 280 285
Tyr Asp Thr Leu Ala Ser Asp Trp Ser Gln Trp Ile Gly Thr Ser Gly
290 295 300
Ala Ser Ser Val Glu Ser Ile Gly Ala Tyr Ser Val Gln Tyr Gly Ser
305 310 315 320
Thr Lys Leu Arg Val Ile Ser Leu Asn Thr Asn Met Tyr Tyr Ile Glu
325 330 335
Asn Phe Tyr Leu Tyr Glu Pro Thr Met Glu Gln Asp Pro Ala Gly Gln
340 345 350
Phe Ala Trp Leu Val Ser Glu Leu Ser Ala Ala Glu Ala Ala Gly Glu
355 360 365
Arg Val Trp Ile Ile Gly His Met Pro Leu Gly Leu Ser Asp Ala Phe
370 375 380
His Asp Pro Ser Asn Tyr Phe Asp Gln Ile Val Asn Arg Tyr Glu Ala
385 390 395 400
Thr Ile Ala Ala Met Phe Phe Gly His Thr His Glu Asp His Phe Gln
405 410 415
Ile Ser Tyr Ser Asp Tyr Asn Ala Arg Thr Ala Ala Asn Ala Arg Ala
420 425 430
Val Ser Tyr Ile Met Pro Ser Leu Thr Pro Thr Ser Gly His Pro Thr
435 440 445
Phe Arg Val Tyr Thr Val Asp Pro Glu Thr Phe Gly Val Leu Asp Ala
450 455 460
Thr Thr Tyr Tyr Ala Asp Met Ser Gln Pro Thr Tyr Gln Thr Ala Gly
465 470 475 480
Pro Ala Trp Ser Val Tyr Tyr Ser Ala Lys Ala Ala Tyr Gly Gly Leu
485 490 495
Val Asp Pro Pro Val Ala Ala Asp Asp Ala Ala Ala Glu Leu Thr Pro
500 505 510
Ala Phe Trp His Asn Val Thr Ala Ala Leu Ala Ala Asp Pro Ala Ser
515 520 525
Phe Asp Ala Tyr Tyr Ala Arg Lys Thr Arg Gly Trp Asp Val Ala Ala
530 535 540
Cys Ala Gly Ala Cys Ala Ala Ala Glu Val Cys Ala Leu Arg Ala Ala
545 550 555 560
Arg Ala Gln Asp Asn Cys Val Val Pro Thr Pro Gly Val His Phe Ser
565 570 575
Lys Arg Ala Asp Glu Gly Thr Leu Ala His His Arg Asp Glu Cys Gly
580 585 590
Val Ser Val Ala Arg Asn Ser Leu Ser Ser Leu Val Val Gln Arg Glu
595 600 605
Ala Leu Glu His Leu Glu Gly Arg Leu Ser Glu Lys Arg Arg Met Ala
610 615 620
Val
625
<210> 19
<211> 632
<212> БЕЛОК
<213> Nectria mariannaeae
<400> 19
Met Gln Leu Leu Ser Ile Leu Ala Val Gly Leu Gly Leu Ala Gln Asn
1 5 10 15
Ala Phe Cys Gln Glu Val Thr His Asp Leu Ala Gly Ile Lys Arg Ser
20 25 30
Leu Glu Ser Arg Asp Trp Val Glu Asp Leu Trp Asp Lys Phe Glu Ser
35 40 45
Asp Ala Thr Cys Ala Gly Cys Glu Ser Leu Val Leu Val Leu Lys Gly
50 55 60
Leu Ala Ala Ile Ser Asp Gln Ala Phe Ile Asp Val Leu Gln Glu Ile
65 70 75 80
Cys Lys Ile Ser Gly Ala Glu Asp Asp Asp Val Cys Asp Gly Ser Ile
85 90 95
Gln Leu Glu Gly Pro Val Ile Ala Ser Gly Leu Arg Ser Met Ala Ile
100 105 110
Gly Ser Arg Thr Ser Lys Glu Phe Cys Thr Thr Phe Leu Gly Leu Cys
115 120 125
Ala Tyr Pro Ala Val Gln Gln Trp Ser Val Pro Phe Ser Ser Ser Lys
130 135 140
Ser Ser Lys Thr Arg Pro Ser Ser Ser Gly Lys Asp Pro Ile Lys Val
145 150 155 160
Val His Tyr Ser Asp Ile His Ile Asp Pro Leu Tyr Val Gly Gly Ser
165 170 175
Asn Ser Asn Cys Thr Lys Pro Ile Cys Cys Arg Ser Tyr Thr Lys Ala
180 185 190
Asp Gln Pro Gly Asn Asn Lys Tyr Pro Ala Gly Pro Asn Gly Asp His
195 200 205
Asn Cys Asp Ser Pro Val Ser Leu Glu Lys Ser Met Tyr Asn Ala Ile
210 215 220
Lys Glu Ile Val Pro Asp Ala Ala Phe Thr Ile Phe Thr Gly Asp Ile
225 230 235 240
Val Asp His Ala Val Trp Asn Thr Ser Gln Ser Tyr Asn Thr Glu Gln
245 250 255
Ile Thr Asn Ala Tyr Gly Leu Met Ser Asp Asn Leu Gly Thr Ile Tyr
260 265 270
Gly Thr Ala Gly Asn His Glu Ala His Pro Ala Asn Ala Phe Gln Pro
275 280 285
Asn Ser Val Gly Asn Val Ser Gln Trp Val Tyr Asp Leu Leu Ser Gly
290 295 300
Leu Trp Ser Gln Trp Ile Ser Thr Glu Ala Lys Ala Asp Ser Glu Lys
305 310 315 320
Leu Gly Ala Tyr Ser Thr Lys Tyr Pro Gly Gly Asn Leu Arg Ile Ile
325 330 335
Ser Leu Asn Thr Asn Met Tyr Tyr Arg Glu Asn Tyr Trp Leu Tyr Arg
340 345 350
Lys Thr Met Ile Gln Asp Pro Ser Asn Gln Ile Ser Trp Leu Val Asn
355 360 365
Glu Leu Glu Ala Ala Glu Thr Ala Gly Glu Arg Val Tyr Ile Ile Gly
370 375 380
His Met Pro Leu Gly Asp Ser Asn Ser Phe His Asp Gln Ser Asn Tyr
385 390 395 400
Leu Asp Gln Val Ile Asn Arg Tyr Ser Ala Thr Ile Ser Ala Met Phe
405 410 415
Phe Gly His Thr His Asp Asp Gln Phe Gln Ile Ser Tyr Ser Asn Trp
420 425 430
Ser Asn Arg Asn Phe Ser Asn Ala Leu Val Thr Ser Tyr Ile Gly Pro
435 440 445
Ser Leu Thr Pro Thr Ala Gly Met Pro Ala Phe Arg Val Tyr Asp Val
450 455 460
Asp Pro Val Thr Phe Gly Ile Leu Asp Ser Thr Thr Tyr Ile Ala Asp
465 470 475 480
Met Thr Asp Ser Ala Phe Gln Thr Thr Gly Pro Val Trp Lys Lys Tyr
485 490 495
Tyr Ser Ala Lys Glu Val Tyr Gly Ser Leu Leu Ser Pro Ala Val Thr
500 505 510
Asp Ser Ser Ala Glu Leu Thr Ala Ala Phe Trp His Asn Val Thr Thr
515 520 525
Leu Phe Glu Ala Asp Asn Thr Ala Phe Glu Ala Phe Leu Ser Arg Lys
530 535 540
Ser Arg Gly Trp Lys Ser Glu Ser Cys Thr Gly Thr Cys Lys Ala Asn
545 550 555 560
Glu Ile Cys Gln Leu Arg Ala Ala Arg Ser Glu Asn Asn Cys Tyr Thr
565 570 575
Pro Ser Leu Gly Ile Ser Phe Asn Lys Arg Ser Leu Asn Pro Val Glu
580 585 590
Glu Arg Asp Glu Cys Gly Ile Ser Val Thr Arg Ala Thr Val Ser Ala
595 600 605
Met Gly Val Arg Lys Asp Val Leu Arg Leu Leu Lys Lys Arg Phe Ile
610 615 620
Glu Lys Ala Gly Glu Val Arg Gly
625 630
<210> 20
<211> 596
<212> БЕЛОК
<213> Nectria mariannaeae
<400> 20
Asp Trp Val Glu Asp Leu Trp Asp Lys Phe Glu Ser Asp Ala Thr Cys
1 5 10 15
Ala Gly Cys Glu Ser Leu Val Leu Val Leu Lys Gly Leu Ala Ala Ile
20 25 30
Ser Asp Gln Ala Phe Ile Asp Val Leu Gln Glu Ile Cys Lys Ile Ser
35 40 45
Gly Ala Glu Asp Asp Asp Val Cys Asp Gly Ser Ile Gln Leu Glu Gly
50 55 60
Pro Val Ile Ala Ser Gly Leu Arg Ser Met Ala Ile Gly Ser Arg Thr
65 70 75 80
Ser Lys Glu Phe Cys Thr Thr Phe Leu Gly Leu Cys Ala Tyr Pro Ala
85 90 95
Val Gln Gln Trp Ser Val Pro Phe Ser Ser Ser Lys Ser Ser Lys Thr
100 105 110
Arg Pro Ser Ser Ser Gly Lys Asp Pro Ile Lys Val Val His Tyr Ser
115 120 125
Asp Ile His Ile Asp Pro Leu Tyr Val Gly Gly Ser Asn Ser Asn Cys
130 135 140
Thr Lys Pro Ile Cys Cys Arg Ser Tyr Thr Lys Ala Asp Gln Pro Gly
145 150 155 160
Asn Asn Lys Tyr Pro Ala Gly Pro Asn Gly Asp His Asn Cys Asp Ser
165 170 175
Pro Val Ser Leu Glu Lys Ser Met Tyr Asn Ala Ile Lys Glu Ile Val
180 185 190
Pro Asp Ala Ala Phe Thr Ile Phe Thr Gly Asp Ile Val Asp His Ala
195 200 205
Val Trp Asn Thr Ser Gln Ser Tyr Asn Thr Glu Gln Ile Thr Asn Ala
210 215 220
Tyr Gly Leu Met Ser Asp Asn Leu Gly Thr Ile Tyr Gly Thr Ala Gly
225 230 235 240
Asn His Glu Ala His Pro Ala Asn Ala Phe Gln Pro Asn Ser Val Gly
245 250 255
Asn Val Ser Gln Trp Val Tyr Asp Leu Leu Ser Gly Leu Trp Ser Gln
260 265 270
Trp Ile Ser Thr Glu Ala Lys Ala Asp Ser Glu Lys Leu Gly Ala Tyr
275 280 285
Ser Thr Lys Tyr Pro Gly Gly Asn Leu Arg Ile Ile Ser Leu Asn Thr
290 295 300
Asn Met Tyr Tyr Arg Glu Asn Tyr Trp Leu Tyr Arg Lys Thr Met Ile
305 310 315 320
Gln Asp Pro Ser Asn Gln Ile Ser Trp Leu Val Asn Glu Leu Glu Ala
325 330 335
Ala Glu Thr Ala Gly Glu Arg Val Tyr Ile Ile Gly His Met Pro Leu
340 345 350
Gly Asp Ser Asn Ser Phe His Asp Gln Ser Asn Tyr Leu Asp Gln Val
355 360 365
Ile Asn Arg Tyr Ser Ala Thr Ile Ser Ala Met Phe Phe Gly His Thr
370 375 380
His Asp Asp Gln Phe Gln Ile Ser Tyr Ser Asn Trp Ser Asn Arg Asn
385 390 395 400
Phe Ser Asn Ala Leu Val Thr Ser Tyr Ile Gly Pro Ser Leu Thr Pro
405 410 415
Thr Ala Gly Met Pro Ala Phe Arg Val Tyr Asp Val Asp Pro Val Thr
420 425 430
Phe Gly Ile Leu Asp Ser Thr Thr Tyr Ile Ala Asp Met Thr Asp Ser
435 440 445
Ala Phe Gln Thr Thr Gly Pro Val Trp Lys Lys Tyr Tyr Ser Ala Lys
450 455 460
Glu Val Tyr Gly Ser Leu Leu Ser Pro Ala Val Thr Asp Ser Ser Ala
465 470 475 480
Glu Leu Thr Ala Ala Phe Trp His Asn Val Thr Thr Leu Phe Glu Ala
485 490 495
Asp Asn Thr Ala Phe Glu Ala Phe Leu Ser Arg Lys Ser Arg Gly Trp
500 505 510
Lys Ser Glu Ser Cys Thr Gly Thr Cys Lys Ala Asn Glu Ile Cys Gln
515 520 525
Leu Arg Ala Ala Arg Ser Glu Asn Asn Cys Tyr Thr Pro Ser Leu Gly
530 535 540
Ile Ser Phe Asn Lys Arg Ser Leu Asn Pro Val Glu Glu Arg Asp Glu
545 550 555 560
Cys Gly Ile Ser Val Thr Arg Ala Thr Val Ser Ala Met Gly Val Arg
565 570 575
Lys Asp Val Leu Arg Leu Leu Lys Lys Arg Phe Ile Glu Lys Ala Gly
580 585 590
Glu Val Arg Gly
595
<210> 21
<211> 2889
<212> ДНК
<213> Rasamsonia eburnea
<400> 21
cggattagct gggtagacat agcctatcgc gtgtttgccc tggggccacg aggacggctg 60
tccctgacca agattcaatc atcccgcgtc aggaggacac ccgcttcgtg atgtttcatt 120
ccagtcaatc attactgcac tcttgcacag ccaaaagcca cgaccctgac gaatgacgaa 180
cgaacgcgct atctttttcg ttcgttttct tttccacata gtagagcgtt gatttagcgt 240
ccgactaaga agccaagctt ttttgattgc ttcatctacg atcctactct caatgacacg 300
gcgcatcccc cgcccgatca acctcgggca cgtgggagta aggggctccc tgatcaattg 360
atgatgcgtt caggctgtaa taggggcaat atatagacat gggagggcta ggctggcttt 420
gcatccgttc tggaccctgg gtatcccggc gagtccatct ctgacaagtt ctgttcactt 480
caggccagtc aacaagacaa atgagagctt tcctcatcac ggcgctggct tcactagcca 540
ccgccgctgg tgccacctat gacaagcgcg gcctggccca ggatatctgg aacgatatca 600
agaatgcggt ggattgcgct ggctgccagg gcatcctgac tgcattgaag ggcctctcct 660
atctaggcac gactgccttt gtcgacgtac tgaccgaagt ttgcgacatc agtggcgtac 720
gtaggtgaat acccgtgcgc gtggatatac tgactggatt ggctatcagg tggaagattc 780
agatgtctgc tcaggcatca tctccagcga gggtccggcg ctggtctaca tcctaaaaca 840
cctcgatatc ggatcgcaca cctcgcaggt catctgtgct agtgtatttg gtctctgcca 900
gtacccagca gtccgagcct acaacctcac gttcccttcg cccaagcccg acaagacttg 960
tccagagccc agtggagaat cacccgtgca aattgttcac ttcagtgaca cccacgcgga 1020
tctctcgtac gagactggat ccaattacaa ctgtaccaag cccatctgct gtcgctcgta 1080
caccgcggaa gacgctccgg gaaacacgac gactccatgc ggcccatatg gcaaccccaa 1140
gtgcgatgcc cccatgagcc tcgaggagag catgtttgcc gctatcaaag cgctgagccc 1200
gcagccagcc ttttccatct atacgggaga cgtcgtcgca catgacatct ggcttgtgga 1260
ccagaacgaa gttgtcgagg atttgaatgc tacctatgac cgcatggccg gactgggact 1320
ggtctacgcc gcaattggga accacgacac ggcgccggtc aacaatttgc ctaccagcaa 1380
catccctagc cagtacagcg cgaattggac gtacgaagcg cttgagtatc acttctcgct 1440
gctgaccaag tcggcgtccg cccagacagc agagaactac gggtcctatt cctctgtcta 1500
ccggggcaga tacggaacgg atctccgcgt aatttcctac aacagcatct tctattacat 1560
cgccgacttc tgggcctacc aggaccccat gttgtacgac ccagatgggc agctcgcctg 1620
gctgatcaac gagctccagg aggccgaaac ggcgggacaa cgggtctggt tgatagcgca 1680
tgttccgtca ggcacggccg accatttcca cgattactcg cactactttg accagatcgt 1740
gcagcgctac gaggccacca ttgctgcatt gttctatggc cacacgcaca tcgatcagtt 1800
ccagatttcc tactcggact actccaatcg agcgtttgac actgccacag ccatcggcta 1860
catcatgcca tcgatgacgc cgacctcggg acctcctacc tttcgagttt atgatgtcga 1920
tcccaagacc ttcgcagtct tggacttcac taactatatt gccaacatca gtgatccggc 1980
ctaccagtcc ggcccgacgt ggcagaagta ctactcggca aaggaggcgt acggatcatt 2040
gctctctccg cccgtgacgg acgcgacggc ggagctgacg ccagccttct ggcacaacgt 2100
cacggtggct tttgagaacg atgacacggc tttccaggag tactgggcgc ggcagactcg 2160
cggatatgcc gtgtcgagct gcacggggga ctgcatcacg caggcgatct gtggcttgcg 2220
ggcgggagag tcgcagcaca actgcgtgac gccgacacca ggcttcaact tcgccaaacg 2280
agatgtttcg actgatggtc aggctttgcc gcatattgag aaatgtgagg gatcaggatt 2340
gatggctttg ctggccaaga tggtggccag caaccgacaa tcttcctagt tggaattgac 2400
ggatacgaga gtatatataa gtgcagttaa agaattcagt caacaatgta tagtacctag 2460
atttacaaag agtccatagt gtatgatgta cttattaaca ctatggcagt atagcggtgt 2520
tggtgttttc aatagaaacg atgtggaaaa ctagaacatt actatctcag aagatacaga 2580
aataataggg agtttcttct gggagttaga atatttggac ccagaaagct ccacttaatc 2640
agtgctgtat gtgcttggac tcgaatagat caataactaa aggtctattt aatatggcta 2700
agaatcgtga aatccatatt tatataataa tattcacaca tatagttatc tttttaagta 2760
atatatatgt aagaatataa gagcagaggc taatcgatcg gaggtactgt aattctcgtt 2820
gatatttaga ctatgtaaca agctctgccc tatttccttt cttctttttt tttttttaac 2880
aatccctac 2889
<210> 22
<211> 611
<212> БЕЛОК
<213> Rasamsonia eburnea
<400> 22
Met Arg Ala Phe Leu Ile Thr Ala Leu Ala Ser Leu Ala Thr Ala Ala
1 5 10 15
Gly Ala Thr Tyr Asp Lys Arg Gly Leu Ala Gln Asp Ile Trp Asn Asp
20 25 30
Ile Lys Asn Ala Val Asp Cys Ala Gly Cys Gln Gly Ile Leu Thr Ala
35 40 45
Leu Lys Gly Leu Ser Tyr Leu Gly Thr Thr Ala Phe Val Asp Val Leu
50 55 60
Thr Glu Val Cys Asp Ile Ser Gly Val Glu Asp Ser Asp Val Cys Ser
65 70 75 80
Gly Ile Ile Ser Ser Glu Gly Pro Ala Leu Val Tyr Ile Leu Lys His
85 90 95
Leu Asp Ile Gly Ser His Thr Ser Gln Val Ile Cys Ala Ser Val Phe
100 105 110
Gly Leu Cys Gln Tyr Pro Ala Val Arg Ala Tyr Asn Leu Thr Phe Pro
115 120 125
Ser Pro Lys Pro Asp Lys Thr Cys Pro Glu Pro Ser Gly Glu Ser Pro
130 135 140
Val Gln Ile Val His Phe Ser Asp Thr His Ala Asp Leu Ser Tyr Glu
145 150 155 160
Thr Gly Ser Asn Tyr Asn Cys Thr Lys Pro Ile Cys Cys Arg Ser Tyr
165 170 175
Thr Ala Glu Asp Ala Pro Gly Asn Thr Thr Thr Pro Cys Gly Pro Tyr
180 185 190
Gly Asn Pro Lys Cys Asp Ala Pro Met Ser Leu Glu Glu Ser Met Phe
195 200 205
Ala Ala Ile Lys Ala Leu Ser Pro Gln Pro Ala Phe Ser Ile Tyr Thr
210 215 220
Gly Asp Val Val Ala His Asp Ile Trp Leu Val Asp Gln Asn Glu Val
225 230 235 240
Val Glu Asp Leu Asn Ala Thr Tyr Asp Arg Met Ala Gly Leu Gly Leu
245 250 255
Val Tyr Ala Ala Ile Gly Asn His Asp Thr Ala Pro Val Asn Asn Leu
260 265 270
Pro Thr Ser Asn Ile Pro Ser Gln Tyr Ser Ala Asn Trp Thr Tyr Glu
275 280 285
Ala Leu Glu Tyr His Phe Ser Leu Leu Thr Lys Ser Ala Ser Ala Gln
290 295 300
Thr Ala Glu Asn Tyr Gly Ser Tyr Ser Ser Val Tyr Arg Gly Arg Tyr
305 310 315 320
Gly Thr Asp Leu Arg Val Ile Ser Tyr Asn Ser Ile Phe Tyr Tyr Ile
325 330 335
Ala Asp Phe Trp Ala Tyr Gln Asp Pro Met Leu Tyr Asp Pro Asp Gly
340 345 350
Gln Leu Ala Trp Leu Ile Asn Glu Leu Gln Glu Ala Glu Thr Ala Gly
355 360 365
Gln Arg Val Trp Leu Ile Ala His Val Pro Ser Gly Thr Ala Asp His
370 375 380
Phe His Asp Tyr Ser His Tyr Phe Asp Gln Ile Val Gln Arg Tyr Glu
385 390 395 400
Ala Thr Ile Ala Ala Leu Phe Tyr Gly His Thr His Ile Asp Gln Phe
405 410 415
Gln Ile Ser Tyr Ser Asp Tyr Ser Asn Arg Ala Phe Asp Thr Ala Thr
420 425 430
Ala Ile Gly Tyr Ile Met Pro Ser Met Thr Pro Thr Ser Gly Pro Pro
435 440 445
Thr Phe Arg Val Tyr Asp Val Asp Pro Lys Thr Phe Ala Val Leu Asp
450 455 460
Phe Thr Asn Tyr Ile Ala Asn Ile Ser Asp Pro Ala Tyr Gln Ser Gly
465 470 475 480
Pro Thr Trp Gln Lys Tyr Tyr Ser Ala Lys Glu Ala Tyr Gly Ser Leu
485 490 495
Leu Ser Pro Pro Val Thr Asp Ala Thr Ala Glu Leu Thr Pro Ala Phe
500 505 510
Trp His Asn Val Thr Val Ala Phe Glu Asn Asp Asp Thr Ala Phe Gln
515 520 525
Glu Tyr Trp Ala Arg Gln Thr Arg Gly Tyr Ala Val Ser Ser Cys Thr
530 535 540
Gly Asp Cys Ile Thr Gln Ala Ile Cys Gly Leu Arg Ala Gly Glu Ser
545 550 555 560
Gln His Asn Cys Val Thr Pro Thr Pro Gly Phe Asn Phe Ala Lys Arg
565 570 575
Asp Val Ser Thr Asp Gly Gln Ala Leu Pro His Ile Glu Lys Cys Glu
580 585 590
Gly Ser Gly Leu Met Ala Leu Leu Ala Lys Met Val Ala Ser Asn Arg
595 600 605
Gln Ser Ser
610
<210> 23
<211> 552
<212> БЕЛОК
<213> Rasamsonia eburnea
<400> 23
Gly Leu Ala Gln Asp Ile Trp Asn Asp Ile Lys Asn Ala Val Asp Cys
1 5 10 15
Ala Gly Cys Gln Gly Ile Leu Thr Ala Leu Lys Gly Leu Ser Tyr Leu
20 25 30
Gly Thr Thr Ala Phe Val Asp Val Leu Thr Glu Val Cys Asp Ile Ser
35 40 45
Gly Val Glu Asp Ser Asp Val Cys Ser Gly Ile Ile Ser Ser Glu Gly
50 55 60
Pro Ala Leu Val Tyr Ile Leu Lys His Leu Asp Ile Gly Ser His Thr
65 70 75 80
Ser Gln Val Ile Cys Ala Ser Val Phe Gly Leu Cys Gln Tyr Pro Ala
85 90 95
Val Arg Ala Tyr Asn Leu Thr Phe Pro Ser Pro Lys Pro Asp Lys Thr
100 105 110
Cys Pro Glu Pro Ser Gly Glu Ser Pro Val Gln Ile Val His Phe Ser
115 120 125
Asp Thr His Ala Asp Leu Ser Tyr Glu Thr Gly Ser Asn Tyr Asn Cys
130 135 140
Thr Lys Pro Ile Cys Cys Arg Ser Tyr Thr Ala Glu Asp Ala Pro Gly
145 150 155 160
Asn Thr Thr Thr Pro Cys Gly Pro Tyr Gly Asn Pro Lys Cys Asp Ala
165 170 175
Pro Met Ser Leu Glu Glu Ser Met Phe Ala Ala Ile Lys Ala Leu Ser
180 185 190
Pro Gln Pro Ala Phe Ser Ile Tyr Thr Gly Asp Val Val Ala His Asp
195 200 205
Ile Trp Leu Val Asp Gln Asn Glu Val Val Glu Asp Leu Asn Ala Thr
210 215 220
Tyr Asp Arg Met Ala Gly Leu Gly Leu Val Tyr Ala Ala Ile Gly Asn
225 230 235 240
His Asp Thr Ala Pro Val Asn Asn Leu Pro Thr Ser Asn Ile Pro Ser
245 250 255
Gln Tyr Ser Ala Asn Trp Thr Tyr Glu Ala Leu Glu Tyr His Phe Ser
260 265 270
Leu Leu Thr Lys Ser Ala Ser Ala Gln Thr Ala Glu Asn Tyr Gly Ser
275 280 285
Tyr Ser Ser Val Tyr Arg Gly Arg Tyr Gly Thr Asp Leu Arg Val Ile
290 295 300
Ser Tyr Asn Ser Ile Phe Tyr Tyr Ile Ala Asp Phe Trp Ala Tyr Gln
305 310 315 320
Asp Pro Met Leu Tyr Asp Pro Asp Gly Gln Leu Ala Trp Leu Ile Asn
325 330 335
Glu Leu Gln Glu Ala Glu Thr Ala Gly Gln Arg Val Trp Leu Ile Ala
340 345 350
His Val Pro Ser Gly Thr Ala Asp His Phe His Asp Tyr Ser His Tyr
355 360 365
Phe Asp Gln Ile Val Gln Arg Tyr Glu Ala Thr Ile Ala Ala Leu Phe
370 375 380
Tyr Gly His Thr His Ile Asp Gln Phe Gln Ile Ser Tyr Ser Asp Tyr
385 390 395 400
Ser Asn Arg Ala Phe Asp Thr Ala Thr Ala Ile Gly Tyr Ile Met Pro
405 410 415
Ser Met Thr Pro Thr Ser Gly Pro Pro Thr Phe Arg Val Tyr Asp Val
420 425 430
Asp Pro Lys Thr Phe Ala Val Leu Asp Phe Thr Asn Tyr Ile Ala Asn
435 440 445
Ile Ser Asp Pro Ala Tyr Gln Ser Gly Pro Thr Trp Gln Lys Tyr Tyr
450 455 460
Ser Ala Lys Glu Ala Tyr Gly Ser Leu Leu Ser Pro Pro Val Thr Asp
465 470 475 480
Ala Thr Ala Glu Leu Thr Pro Ala Phe Trp His Asn Val Thr Val Ala
485 490 495
Phe Glu Asn Asp Asp Thr Ala Phe Gln Glu Tyr Trp Ala Arg Gln Thr
500 505 510
Arg Gly Tyr Ala Val Ser Ser Cys Thr Gly Asp Cys Ile Thr Gln Ala
515 520 525
Ile Cys Gly Leu Arg Ala Gly Glu Ser Gln His Asn Cys Val Thr Pro
530 535 540
Thr Pro Gly Phe Asn Phe Ala Lys
545 550
<210> 24
<211> 2975
<212> ДНК
<213> Trichurus spiralis
<400> 24
gggggggggg gggggggggg gggaaagcgg ccgaggtcca gtgcaggcgg ccctatacca 60
cccgctaata ttccttatga tgatgacgac cttgcttttt atctccaagc tgagagccct 120
tctcagctct ggtgcttata aggctggcgg gagggccgcg gcttcgcgaa cctgtcgatc 180
ttatttctct tacacgccaa atgctgcttt gtatataacg ggtggtctgt atatatacat 240
atataaatag attctccgtg tgtatataca tcatagcatt tatcccctcc cgcgggtggc 300
gctcttcagg caatgcatct cactcgcgtc gcggccctcg cctttgggct tggcagggcg 360
tttgccacgg ctcccgcgtc tcagctcctc tcgcgcgacg aggccgaact cctcggcagg 420
ggtctcgcgg aagatgtctg ggacgagatc aaggacggtc tctcatgcgc cggctgcgaa 480
gtacgatgcc cctcaccatg gccaactacc caccccctgg acttaagtcc acttacacga 540
gcaacattga ctaacaaccg ccagaccctc gttgtcctgc tcaagggcct cgcgctctta 600
ggcgataacg cctttgtaga tgtcctccaa tcagtctgca agctcacaaa tgtaaacccc 660
ccgtttttga tccctcaccc aaagagcttg gatccgcatc aagactcaca aaaaactgac 720
gttccctcct tgctagagcg ttgacgagga tgtctgccaa ggcgccatag agctcgaggg 780
tcccatcatc gcagacgcca tccggaacat gaaggtcccc agccagacct ccaagctctt 840
ctgcatcaac ttcctcggcg tctgccccta cccgaaggtg acgccctacc aggtcccctt 900
ccccacggag aagaaggcca tcgagggcgg ctcgtcgcgg cccgcggcga gcggcaagga 960
cccgatccgc atcgtccact actccgacgt gcacgtcgac cacctttaca cccccggctc 1020
gaccgcgaac tgcacgaagc ccatatgctg caggccctac accgaggacg acgagccggg 1080
cgtgagcacc tcccccgccg gccccaacgg cgaccacaac tgtgatacgc ccgtgagcct 1140
cgaggagagc atgtacgccg ccattaacga ggtcgccccg gacgcggcct ttgtcatttg 1200
cactggcgac atcgtcgacc acaccgtctg gaacacctcc gtcgagtaca acacattctc 1260
tagtatgtcc accatgccca cctcccccgt ccctctccac cccatctcat gtagcttctc 1320
cgtatcgatc aagcgccaac tactaacacg ccggaacccc cgccgccagt caccgacgcc 1380
tacagccgca tggctgcctc cttcccgctc gtctacggca cggccggcaa ccacgaggcc 1440
cacccgacga acgccttccc gcccacggcc gaggccccga ccagctgggt ctacaacctc 1500
ctctctacga gctggtccca ctggatcggc gacgccgccg cgtcctccac ctccagccgc 1560
ggctcctact ctgtcaaata ccccggcggt aacctgcgcg tcattagcat gaacacgaac 1620
accttttacg tgcagaacta ctacctctac cgccgcgtca tgaccctcga cccggacggc 1680
cagatcgcct ggctcgcgtc cgagctcgac gccgccgaga aggccggcga gagggcgtac 1740
atcatcggcc acatgccccc cggcgacaac gacgccttcc acgaccagag caactacttc 1800
gaccaggtcg tcaaccggta ctccgacacc atcgctgcca tgttcttcgg ccacacccac 1860
ctcgacgagt tccacgtcag ctactcgtcc tacgacgccc agaccgccga caccgctgtc 1920
gccgtgcagt acgtcgcccc gagcctcacg cccacgagcg gccatccctc ctttagggtt 1980
tacgtcgtcg acccggagac ctttgccatc ctggacgccg agacctggat cgccgacatg 2040
gccgacccca acttccagac tacccccacc tggacgaagt tctactccgc cagggaagtc 2100
tacgggcccg accaccctgc cggcgaagag ctctcccccg ccttctggca caacgtcacc 2160
gaggccttcg cctcatcccc cgacctcctg gacgcctacc tctcccggaa gtcgcggggc 2220
tgggaggaca tcgcctgcga cgatgcttgt cgagagcagg agatttgcaa gctacgtggc 2280
gggagggccg aggataactg ctacgagccg aaacctggca tccatttcaa gaagaggagc 2340
ctggagtcta ggggccatgg ggatgactgc ggcgtctcgg tgctgcgcgg tgcggtgggt 2400
tcgctcgctg tcgagaagaa ggccgtggag cttgtgagtg cgagggtcga ggagaacaag 2460
agactcctaa tctagaggca tcattagcgc ttctcggtcc gtaagagaga gatggaggag 2520
ggtggttggt agtgaagagg gatacggatc ttggatgaac agccaatttt tattctggag 2580
ctagaagttg tgctcgggat ataggataac caaaatcgct gtctctcttc ttgaaaagct 2640
gaatcgctgt ccgtctgtct tctcgggtaa ccaaagtcgc tggccgtcac ttccgtggat 2700
aaccaaagcc gttgaccgtc tccgtggata acattgtcta tctccctcct tacaccaccc 2760
gagcttgata gcagtcattc catacgctag caagcccgcc ctatcctcgg tctgcagtac 2820
acaagctcca aaaaaaagaa atgatatcct agcgctcaag gcagcatcac attcctccct 2880
tcgcacctat gaatgctgca atacaaactg attcgggcaa cccactaaca tggccggtaa 2940
aaaaaggctc gtcacaaaac ctgccccgat gataa 2975
<210> 25
<211> 628
<212> БЕЛОК
<213> Trichurus spiralis
<400> 25
Met His Leu Thr Arg Val Ala Ala Leu Ala Phe Gly Leu Gly Arg Ala
1 5 10 15
Phe Ala Thr Ala Pro Ala Ser Gln Leu Leu Ser Arg Asp Glu Ala Glu
20 25 30
Leu Leu Gly Arg Gly Leu Ala Glu Asp Val Trp Asp Glu Ile Lys Asp
35 40 45
Gly Leu Ser Cys Ala Gly Cys Glu Thr Leu Val Val Leu Leu Lys Gly
50 55 60
Leu Ala Leu Leu Gly Asp Asn Ala Phe Val Asp Val Leu Gln Ser Val
65 70 75 80
Cys Lys Leu Thr Asn Ser Val Asp Glu Asp Val Cys Gln Gly Ala Ile
85 90 95
Glu Leu Glu Gly Pro Ile Ile Ala Asp Ala Ile Arg Asn Met Lys Val
100 105 110
Pro Ser Gln Thr Ser Lys Leu Phe Cys Ile Asn Phe Leu Gly Val Cys
115 120 125
Pro Tyr Pro Lys Val Thr Pro Tyr Gln Val Pro Phe Pro Thr Glu Lys
130 135 140
Lys Ala Ile Glu Gly Gly Ser Ser Arg Pro Ala Ala Ser Gly Lys Asp
145 150 155 160
Pro Ile Arg Ile Val His Tyr Ser Asp Val His Val Asp His Leu Tyr
165 170 175
Thr Pro Gly Ser Thr Ala Asn Cys Thr Lys Pro Ile Cys Cys Arg Pro
180 185 190
Tyr Thr Glu Asp Asp Glu Pro Gly Val Ser Thr Ser Pro Ala Gly Pro
195 200 205
Asn Gly Asp His Asn Cys Asp Thr Pro Val Ser Leu Glu Glu Ser Met
210 215 220
Tyr Ala Ala Ile Asn Glu Val Ala Pro Asp Ala Ala Phe Val Ile Cys
225 230 235 240
Thr Gly Asp Ile Val Asp His Thr Val Trp Asn Thr Ser Val Glu Tyr
245 250 255
Asn Thr Phe Ser Ile Thr Asp Ala Tyr Ser Arg Met Ala Ala Ser Phe
260 265 270
Pro Leu Val Tyr Gly Thr Ala Gly Asn His Glu Ala His Pro Thr Asn
275 280 285
Ala Phe Pro Pro Thr Ala Glu Ala Pro Thr Ser Trp Val Tyr Asn Leu
290 295 300
Leu Ser Thr Ser Trp Ser His Trp Ile Gly Asp Ala Ala Ala Ser Ser
305 310 315 320
Thr Ser Ser Arg Gly Ser Tyr Ser Val Lys Tyr Pro Gly Gly Asn Leu
325 330 335
Arg Val Ile Ser Met Asn Thr Asn Thr Phe Tyr Val Gln Asn Tyr Tyr
340 345 350
Leu Tyr Arg Arg Val Met Thr Leu Asp Pro Asp Gly Gln Ile Ala Trp
355 360 365
Leu Ala Ser Glu Leu Asp Ala Ala Glu Lys Ala Gly Glu Arg Ala Tyr
370 375 380
Ile Ile Gly His Met Pro Pro Gly Asp Asn Asp Ala Phe His Asp Gln
385 390 395 400
Ser Asn Tyr Phe Asp Gln Val Val Asn Arg Tyr Ser Asp Thr Ile Ala
405 410 415
Ala Met Phe Phe Gly His Thr His Leu Asp Glu Phe His Val Ser Tyr
420 425 430
Ser Ser Tyr Asp Ala Gln Thr Ala Asp Thr Ala Val Ala Val Gln Tyr
435 440 445
Val Ala Pro Ser Leu Thr Pro Thr Ser Gly His Pro Ser Phe Arg Val
450 455 460
Tyr Val Val Asp Pro Glu Thr Phe Ala Ile Leu Asp Ala Glu Thr Trp
465 470 475 480
Ile Ala Asp Met Ala Asp Pro Asn Phe Gln Thr Thr Pro Thr Trp Thr
485 490 495
Lys Phe Tyr Ser Ala Arg Glu Val Tyr Gly Pro Asp His Pro Ala Gly
500 505 510
Glu Glu Leu Ser Pro Ala Phe Trp His Asn Val Thr Glu Ala Phe Ala
515 520 525
Ser Ser Pro Asp Leu Leu Asp Ala Tyr Leu Ser Arg Lys Ser Arg Gly
530 535 540
Trp Glu Asp Ile Ala Cys Asp Asp Ala Cys Arg Glu Gln Glu Ile Cys
545 550 555 560
Lys Leu Arg Gly Gly Arg Ala Glu Asp Asn Cys Tyr Glu Pro Lys Pro
565 570 575
Gly Ile His Phe Lys Lys Arg Ser Leu Glu Ser Arg Gly His Gly Asp
580 585 590
Asp Cys Gly Val Ser Val Leu Arg Gly Ala Val Gly Ser Leu Ala Val
595 600 605
Glu Lys Lys Ala Val Glu Leu Val Ser Ala Arg Val Glu Glu Asn Lys
610 615 620
Arg Leu Leu Ile
625
<210> 26
<211> 427
<212> БЕЛОК
<213> Trichurus spiralis
<400> 26
Ala Ser Gly Lys Asp Pro Ile Arg Ile Val His Tyr Ser Asp Val His
1 5 10 15
Val Asp His Leu Tyr Thr Pro Gly Ser Thr Ala Asn Cys Thr Lys Pro
20 25 30
Ile Cys Cys Arg Pro Tyr Thr Glu Asp Asp Glu Pro Gly Val Ser Thr
35 40 45
Ser Pro Ala Gly Pro Asn Gly Asp His Asn Cys Asp Thr Pro Val Ser
50 55 60
Leu Glu Glu Ser Met Tyr Ala Ala Ile Asn Glu Val Ala Pro Asp Ala
65 70 75 80
Ala Phe Val Ile Cys Thr Gly Asp Ile Val Asp His Thr Val Trp Asn
85 90 95
Thr Ser Val Glu Tyr Asn Thr Phe Ser Ile Thr Asp Ala Tyr Ser Arg
100 105 110
Met Ala Ala Ser Phe Pro Leu Val Tyr Gly Thr Ala Gly Asn His Glu
115 120 125
Ala His Pro Thr Asn Ala Phe Pro Pro Thr Ala Glu Ala Pro Thr Ser
130 135 140
Trp Val Tyr Asn Leu Leu Ser Thr Ser Trp Ser His Trp Ile Gly Asp
145 150 155 160
Ala Ala Ala Ser Ser Thr Ser Ser Arg Gly Ser Tyr Ser Val Lys Tyr
165 170 175
Pro Gly Gly Asn Leu Arg Val Ile Ser Met Asn Thr Asn Thr Phe Tyr
180 185 190
Val Gln Asn Tyr Tyr Leu Tyr Arg Arg Val Met Thr Leu Asp Pro Asp
195 200 205
Gly Gln Ile Ala Trp Leu Ala Ser Glu Leu Asp Ala Ala Glu Lys Ala
210 215 220
Gly Glu Arg Ala Tyr Ile Ile Gly His Met Pro Pro Gly Asp Asn Asp
225 230 235 240
Ala Phe His Asp Gln Ser Asn Tyr Phe Asp Gln Val Val Asn Arg Tyr
245 250 255
Ser Asp Thr Ile Ala Ala Met Phe Phe Gly His Thr His Leu Asp Glu
260 265 270
Phe His Val Ser Tyr Ser Ser Tyr Asp Ala Gln Thr Ala Asp Thr Ala
275 280 285
Val Ala Val Gln Tyr Val Ala Pro Ser Leu Thr Pro Thr Ser Gly His
290 295 300
Pro Ser Phe Arg Val Tyr Val Val Asp Pro Glu Thr Phe Ala Ile Leu
305 310 315 320
Asp Ala Glu Thr Trp Ile Ala Asp Met Ala Asp Pro Asn Phe Gln Thr
325 330 335
Thr Pro Thr Trp Thr Lys Phe Tyr Ser Ala Arg Glu Val Tyr Gly Pro
340 345 350
Asp His Pro Ala Gly Glu Glu Leu Ser Pro Ala Phe Trp His Asn Val
355 360 365
Thr Glu Ala Phe Ala Ser Ser Pro Asp Leu Leu Asp Ala Tyr Leu Ser
370 375 380
Arg Lys Ser Arg Gly Trp Glu Asp Ile Ala Cys Asp Asp Ala Cys Arg
385 390 395 400
Glu Gln Glu Ile Cys Lys Leu Arg Gly Gly Arg Ala Glu Asp Asn Cys
405 410 415
Tyr Glu Pro Lys Pro Gly Ile His Phe Lys Lys
420 425
<210> 27
<211> 3204
<212> ДНК
<213> Trichoderma harzianum
<400> 27
tcctgatctg ccgagccgca caccatttcc tctatttaac tggtctctgc cgatgcatgt 60
gtgggagcct gcctctgatt ggatctccat ctcggggcct cgctattaga cggccctcag 120
ggaggccgtt gaggggttct tactggacga tgcctcggtc ctgcattcgt tcagcgtggc 180
attggagatg gagcgagaca gagtccggaa ggtcatccgc cctggggtgc gttcttgact 240
tggcacaggg ccggttcctt gtgagctgag tttcccttgt gaagggccca aagttcgtat 300
agttcgagcg gagaggattt gcaacggcct ggagtgcagt tcgtagatgg gtgggcacta 360
acatcatgta ggtaggtgga cgtggctatc gtctatttat atatagctat gatggaagac 420
ggtattcggt tcctggttcc tgatccttgc ggacaaccca ggcatctgaa ggcagctcaa 480
gccagcattt gttaccagct atgcgtccca gctcgacgct ctccctgctg gctctgggct 540
ctgttgccca ggctgcggtt tctcttgaga atagcagtct ccctccccga gatatcgaga 600
acatcgagag ggctattgag gctagaagct tggctgatga tatatggaac gatatcaaga 660
atgctgcgac atgcactgct tgccaggtca gaaaacacca ttcacagtca tccgcatatg 720
tgactctatg ctgacacggc tccagggcat tctcgtcctt ctcaaaggca tcgctgtatt 780
tggagacgga gcattcgtta gcgtagctac ggaactctgc aagctcgcaa aggtacctag 840
atgactacga tggtctggtt ttcatctagc cacggctaac gaaagtcttc aggttgaaga 900
cgatgatgtg tgcgaaggca ccgtaggact agagggtccc atcattgcgg atgctatccg 960
aaacatggat cttgggtctg atacatcaaa actcttctgt ggatcgttcc ttggcctttg 1020
ccctgagccc tccgttcctc agtggaagat tccttttccc tcccccaagc cacagactgg 1080
tcgccctgcg ccgagtggga agacgcctct caaggtggtt caatactcgg atattcatat 1140
tgaccctctt tatgtgtctg gctcaagctc gaattgcacc aagcccatct gctgcaggtg 1200
agtcgattta tacaaacaaa gagtgacgat gaagtcaaaa agtaaagaca ctgacgatac 1260
tgttactata aagaccttat accgccgctg acgagcctgg caagtcaaca tcgcctgctg 1320
ggccaaatgg ggaccacaag tgtgacactc ctgtcggctt agaaattagc atgtatcagg 1380
ccatcaagaa cattgttcca gatgctgccc taacgctatt cacgggcgat attgttgacc 1440
atgcgatctg gaacacctcg cagccttaca accagaagca aagtcagtgc tcgcctttct 1500
tgaatatatt gcgtatgagt attacgtagt tgactaacat actccacttg cagtttccga 1560
cgcatacact tacatgagcc agtatctggg catcgtttac ggtacggctg gtaaccatga 1620
atccaatccc acaaatgctt ttcctcctag gtccatcagc aactcatcac aatgggtata 1680
tgatgcccta tctgaccaat ggacgcgctg ggtgggggcc tcagcagaat ccgagattga 1740
aagtatcggc gcatactcta ccaagtatcc caatggcaac ctgagagtca tctctctcaa 1800
caccaacttt tactaccgca tgaacttttg gctatatcag gagggtattg agcaggaccc 1860
cgatggccaa atccagtggc tcgtaagtga actcgacgct gcggaaaagg caggagagag 1920
agtttacatc attggacaca tgcccctcgg agaaggcgac gctttccacg cagggtcaaa 1980
ctaccttgat caagttgtga accgctactc gtcgaccatt gccgccatgt tcttcggcca 2040
cacacacgtc gatcactttg aggtcagcta ctccgactac aacagccgag atgcttcaca 2100
tgctgtcatg gcctcttaca tctgcccctc gctgacgccc acctcgggca tgccctcgtt 2160
ccgagtatat gatgtcgacc cggagacgtt tgcagtgctc gacactacga cctacattgc 2220
cgacatgacc aacgcggatt tccaaacgac tggccccgtg tggacgaagc tttactctgc 2280
caaggagacg tacggttcca agctgaatcc ccccgtcacc gacgcaagtg tcgagctgac 2340
tcccgcgttt tggcacaacg taacggcctt gttcgactca aactcggacg tgttcaacga 2400
gtacatctcg ctcaagagcc gtggctggaa cgtcgcctcg tgcactgggg attgcaagac 2460
gcaagagctc tgccagcttc gagccggtcg tagcgagaac aactgcgtgg ttccttcaat 2520
tggccttcac cttaacaaga gatcggatga gctgcatgag cacgatcact cgcatcacgg 2580
atcccacgac caccaagaat gcggtatgtc ggcaggtatg aagacgctgg gctctttggc 2640
tgtgaggaag gatttgttgg atgagttgga gactcgggct aatgagttga aagccaaggc 2700
ttgaggcatg agaaggttgt attttattct tgatagagcg taatgaagac gtctttaatc 2760
ccaattacgc gaggtaattg atttccggtc gaggtattat gtcatgctgg aagctatgaa 2820
agcgagatct gtccgaacat cagcgaattc ggaattgtat catgaagtga agccggagtc 2880
tactaatgga ggccgccggc gcaggcaaag tatggagtac tgatgttcgt gctgcctttt 2940
caagcaaact tccaaggtac ctgtacaatt catatgacta gagataacga tgaggatctt 3000
tttcagcttc atttcccgga agtcacaggg atcaacaatg tattgcagta gaaacaaaga 3060
tcttagatga tggagccgat gcgcgtctgc tgtggctttg ttgatattac gttcatgctc 3120
caagtcatgc acatgtacct ctacccaatg ctaccacttg acacaccaac tgtctcgcag 3180
ctgtaaatgt acaggtactg tccg 3204
<210> 28
<211> 645
<212> БЕЛОК
<213> Trichoderma harzianum
<400> 28
Met Arg Pro Ser Ser Thr Leu Ser Leu Leu Ala Leu Gly Ser Ile Ala
1 5 10 15
Gln Ala Ala Val Ser Leu Glu Asn Ser Ser Leu Pro Arg Arg Asp Ile
20 25 30
Glu Asn Phe Glu Arg Ala Ile Glu Ala Arg Ser Leu Ala Asp Asp Ile
35 40 45
Trp Asn Asp Ile Lys Asn Ala Ala Thr Cys Thr Ala Cys Gln Gly Ile
50 55 60
Leu Val Leu Leu Lys Gly Ile Ala Val Phe Gly Asp Gly Ala Phe Val
65 70 75 80
Ser Val Ala Thr Glu Leu Cys Lys Leu Ala Lys Val Glu Asp Asp Asp
85 90 95
Val Cys Glu Gly Thr Val Arg Leu Glu Gly Pro Ile Ile Ala Asp Ala
100 105 110
Ile Arg Asn Met Asp Leu Gly Ser Asp Thr Ser Lys Leu Phe Cys Gly
115 120 125
Ser Phe Leu Gly Leu Cys Pro Glu Pro Ser Val Pro Gln Trp Lys Ile
130 135 140
Pro Phe Pro Ser Pro Lys Pro Gln Thr Gly Arg Pro Ala Pro Ser Gly
145 150 155 160
Lys Thr Pro Leu Lys Val Val Gln Tyr Ser Asp Ile His Ile Asp Pro
165 170 175
Leu Tyr Val Ser Gly Ser Ser Ser Asn Cys Thr Lys Pro Ile Cys Cys
180 185 190
Arg Pro Tyr Thr Ala Ala Asp Glu Pro Gly Lys Ser Thr Ser Pro Ala
195 200 205
Gly Pro Asn Gly Asp His Lys Cys Asp Thr Pro Val Gly Leu Glu Val
210 215 220
Ser Met Tyr Gln Ala Ile Lys Asn Ile Val Pro Asp Ala Ala Leu Thr
225 230 235 240
Leu Phe Thr Gly Asp Ile Val Asp His Ala Ile Trp Asn Thr Ser Gln
245 250 255
Pro Tyr Asn Gln Lys Gln Ile Ser Asp Ala Tyr Thr Tyr Met Ser Gln
260 265 270
Tyr Leu Gly Ile Val Tyr Gly Thr Ala Gly Asn His Glu Ser Asn Pro
275 280 285
Thr Asn Ala Phe Pro Pro Arg Ser Ile Ser Asn Ser Ser Gln Trp Val
290 295 300
Tyr Asp Ala Leu Ser Asp Gln Trp Thr Arg Trp Val Gly Ala Ser Ala
305 310 315 320
Glu Ser Glu Ile Glu Ser Ile Gly Ala Tyr Ser Thr Lys Tyr Pro Asn
325 330 335
Gly Asn Leu Arg Val Ile Ser Leu Asn Thr Asn Phe Tyr Tyr Arg Met
340 345 350
Asn Phe Trp Leu Tyr Gln Glu Gly Ile Glu Gln Asp Pro Asp Gly Gln
355 360 365
Ile Gln Trp Leu Val Ser Glu Leu Asp Ala Ala Glu Lys Ala Gly Glu
370 375 380
Arg Val Tyr Ile Ile Gly His Met Pro Leu Gly Glu Gly Asp Ala Phe
385 390 395 400
His Ala Gly Ser Asn Tyr Leu Asp Gln Val Val Asn Arg Tyr Ser Ser
405 410 415
Thr Ile Ala Ala Met Phe Phe Gly His Thr His Val Asp His Phe Glu
420 425 430
Val Ser Tyr Ser Asp Tyr Asn Ser Arg Asp Ala Ser His Ala Val Met
435 440 445
Ala Ser Tyr Ile Cys Pro Ser Leu Thr Pro Thr Ser Gly Met Pro Ser
450 455 460
Phe Arg Val Tyr Asp Val Asp Pro Glu Thr Phe Ala Val Leu Asp Thr
465 470 475 480
Thr Thr Tyr Ile Ala Asp Met Thr Asn Ala Asp Phe Gln Thr Thr Gly
485 490 495
Pro Val Trp Thr Lys Leu Tyr Ser Ala Lys Glu Thr Tyr Gly Ser Lys
500 505 510
Leu Asn Pro Pro Val Thr Asp Ala Ser Val Glu Leu Thr Ala Ala Phe
515 520 525
Trp His Asn Val Thr Ala Leu Phe Glu Ser Asn Ser Asp Val Phe Asn
530 535 540
Glu Tyr Ile Ser Leu Lys Ser Arg Gly Trp Asn Val Ala Ser Cys Thr
545 550 555 560
Gly Asp Cys Lys Thr Gln Glu Leu Cys Gln Leu Arg Ala Gly Arg Ser
565 570 575
Glu Asn Asn Cys Val Val Pro Ser Ile Gly Leu His Leu Asn Lys Arg
580 585 590
Ser Asp Gly Leu His Glu His Glu His Ser His His Arg Ser His Asp
595 600 605
His Gln Glu Cys Gly Met Ser Ala Gly Met Lys Thr Leu Gly Ser Leu
610 615 620
Ala Val Arg Lys Asp Leu Leu Asp Glu Leu Glu Thr Arg Ala Asn Glu
625 630 635 640
Leu Lys Ala Lys Ala
645
<210> 29
<211> 521
<212> БЕЛОК
<213> Trichoderma harzianum
<400> 29
Ser Leu Ala Asp Asp Ile Trp Asn Asp Ile Lys Asn Ala Ala Thr Cys
1 5 10 15
Thr Ala Cys Gln Gly Ile Leu Val Leu Leu Lys Gly Ile Ala Val Phe
20 25 30
Gly Asp Gly Ala Phe Val Ser Val Ala Thr Glu Leu Cys Lys Leu Ala
35 40 45
Lys Val Glu Asp Asp Asp Val Cys Glu Gly Thr Val Gly Leu Glu Gly
50 55 60
Pro Ile Ile Ala Asp Ala Ile Arg Asn Met Asp Leu Gly Ser Asp Thr
65 70 75 80
Ser Lys Leu Phe Cys Gly Ser Phe Leu Gly Leu Cys Pro Glu Pro Ser
85 90 95
Val Pro Gln Trp Lys Ile Pro Phe Pro Ser Pro Lys Pro Gln Thr Gly
100 105 110
Arg Pro Ala Pro Ser Gly Lys Thr Pro Leu Lys Val Val Gln Tyr Ser
115 120 125
Asp Ile His Ile Asp Pro Leu Tyr Val Ser Gly Ser Ser Ser Asn Cys
130 135 140
Thr Lys Pro Ile Cys Cys Arg Pro Tyr Thr Ala Ala Asp Glu Pro Gly
145 150 155 160
Lys Ser Thr Ser Pro Ala Gly Pro Asn Gly Asp His Lys Cys Asp Thr
165 170 175
Pro Val Gly Leu Glu Ile Ser Met Tyr Gln Ala Ile Lys Asn Ile Val
180 185 190
Pro Asp Ala Ala Leu Thr Leu Phe Thr Gly Asp Ile Val Asp His Ala
195 200 205
Ile Trp Asn Thr Ser Gln Pro Tyr Asn Gln Lys Gln Ile Ser Asp Ala
210 215 220
Tyr Thr Tyr Met Ser Gln Tyr Leu Gly Ile Val Tyr Gly Thr Ala Gly
225 230 235 240
Asn His Glu Ser Asn Pro Thr Asn Ala Phe Pro Pro Arg Ser Ile Ser
245 250 255
Asn Ser Ser Gln Trp Val Tyr Asp Ala Leu Ser Asp Gln Trp Thr Arg
260 265 270
Trp Val Gly Ala Ser Ala Glu Ser Glu Ile Glu Ser Ile Gly Ala Tyr
275 280 285
Ser Thr Lys Tyr Pro Asn Gly Asn Leu Arg Val Ile Ser Leu Asn Thr
290 295 300
Asn Phe Tyr Tyr Arg Met Asn Phe Trp Leu Tyr Gln Glu Gly Ile Glu
305 310 315 320
Gln Asp Pro Asp Gly Gln Ile Gln Trp Leu Val Ser Glu Leu Asp Ala
325 330 335
Ala Glu Lys Ala Gly Glu Arg Val Tyr Ile Ile Gly His Met Pro Leu
340 345 350
Gly Glu Gly Asp Ala Phe His Ala Gly Ser Asn Tyr Leu Asp Gln Val
355 360 365
Val Asn Arg Tyr Ser Ser Thr Ile Ala Ala Met Phe Phe Gly His Thr
370 375 380
His Val Asp His Phe Glu Val Ser Tyr Ser Asp Tyr Asn Ser Arg Asp
385 390 395 400
Ala Ser His Ala Val Met Ala Ser Tyr Ile Cys Pro Ser Leu Thr Pro
405 410 415
Thr Ser Gly Met Pro Ser Phe Arg Val Tyr Asp Val Asp Pro Glu Thr
420 425 430
Phe Ala Val Leu Asp Thr Thr Thr Tyr Ile Ala Asp Met Thr Asn Ala
435 440 445
Asp Phe Gln Thr Thr Gly Pro Val Trp Thr Lys Leu Tyr Ser Ala Lys
450 455 460
Glu Thr Tyr Gly Ser Lys Leu Asn Pro Pro Val Thr Asp Ala Ser Val
465 470 475 480
Glu Leu Thr Pro Ala Phe Trp His Asn Val Thr Ala Leu Phe Asp Ser
485 490 495
Asn Ser Asp Val Phe Asn Glu Tyr Ile Ser Leu Lys Ser Arg Gly Trp
500 505 510
Asn Val Ala Ser Cys Thr Gly Asp Cys
515 520
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВАРИАНТЫ ПОЛИПЕПТИДОВ, РАСЩЕПЛЯЮЩИХ ФУЗАРИОТОКСИНЫ, СОДЕРЖАЩАЯ ИХ ДОБАВКА, ИХ ПРИМЕНЕНИЕ И СПОСОБ РАСЩЕПЛЕНИЯ ФУЗАРИОТОКСИНОВ | 2015 |
|
RU2720517C2 |
| ЭКСПРЕССИЯ ПОЛИПЕПТИДОВ НИТРОГЕНАЗЫ В РАСТИТЕЛЬНЫХ КЛЕТКАХ | 2020 |
|
RU2833880C2 |
| БИСПЕЦИФИЧЕСКИЕ 2+1 КОНТОРСТЕЛА | 2018 |
|
RU2797305C2 |
| ПРОТИВООПУХОЛЕВЫЕ СОЕДИНЕНИЯ | 2018 |
|
RU2773023C2 |
| ДИСПЛЕЙ ИНТЕГРАЛЬНОГО МЕМБРАННОГО БЕЛКА НА ВНЕКЛЕТОЧНЫХ ОБОЛОЧЕЧНЫХ ВИРИОНАХ ПОКСВИРУСА | 2017 |
|
RU2759846C2 |
| РАСТЕНИЯ С МОДИФИЦИРОВАННЫМИ ПРИЗНАКАМИ | 2017 |
|
RU2809117C2 |
| Конструкции для направленного редактирования генов и способы с их применением | 2020 |
|
RU2832109C2 |
| СПОСОБЫ ПОЛУЧЕНИЯ ПРОМЫШЛЕННЫХ ПРОДУКТОВ ИЗ РАСТИТЕЛЬНЫХ ЛИПИДОВ | 2015 |
|
RU2743384C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
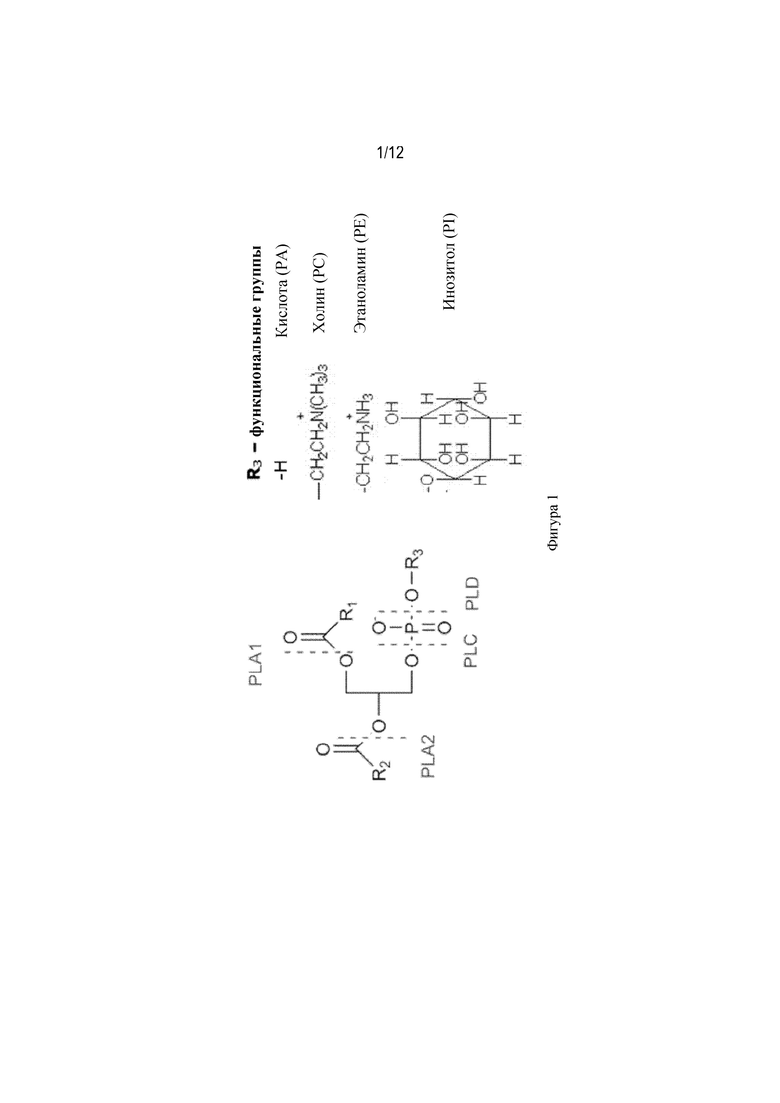
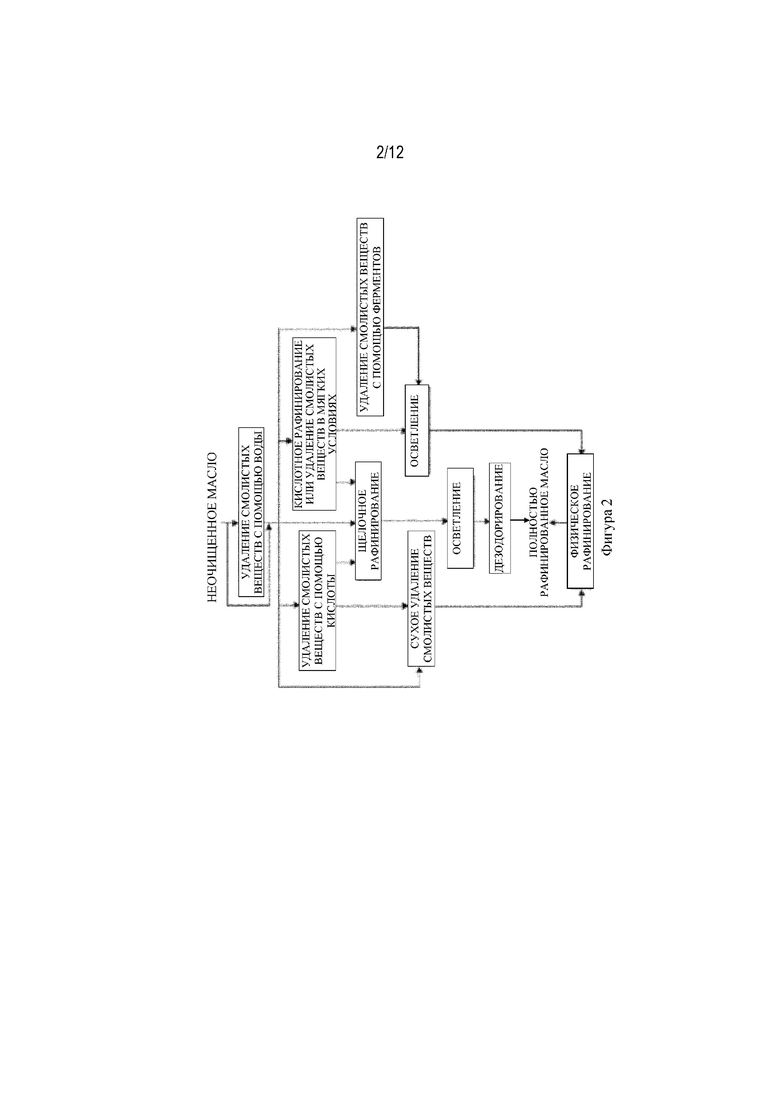


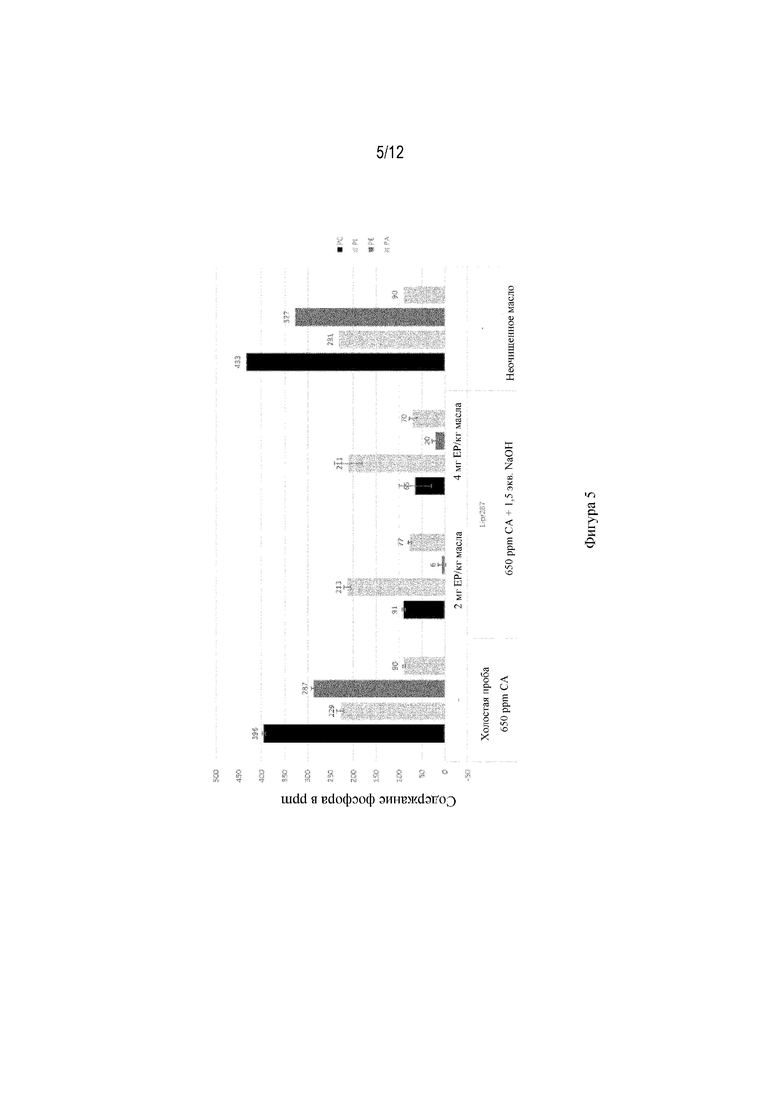






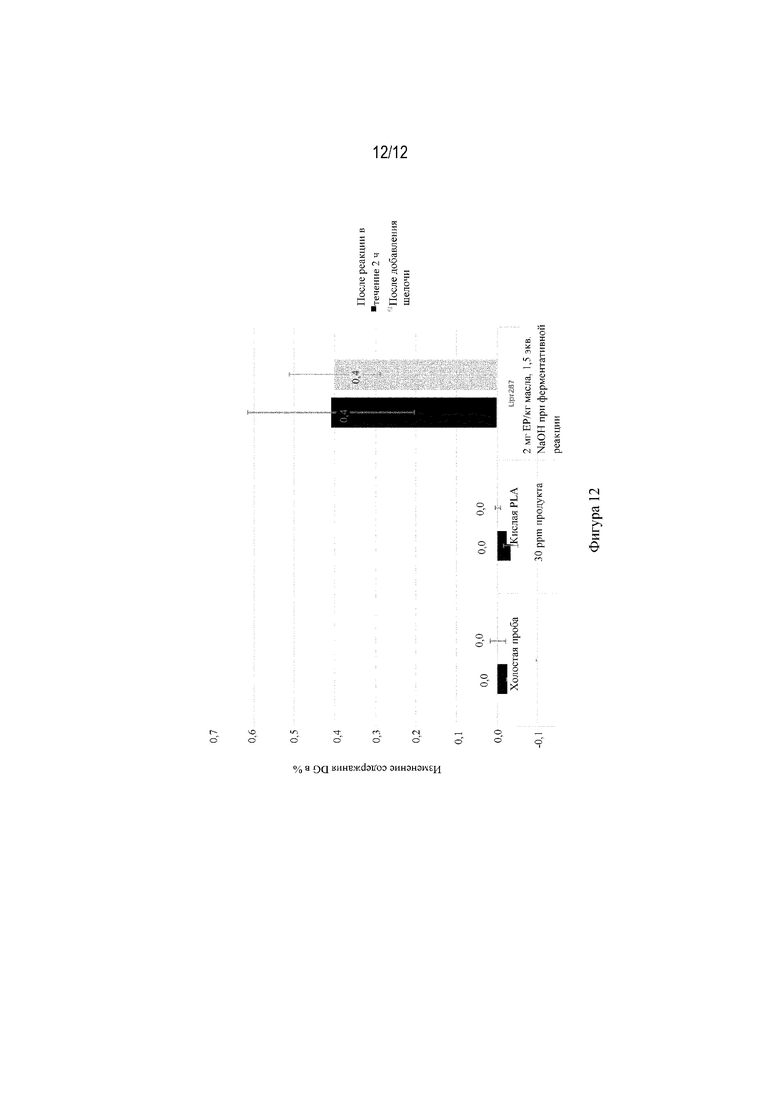
Изобретение относится к масложировой прмышленности. Способ рафинирования растительного масла, содержащего фосфолипиды, предусматривающий: i) получение реакционной смеси, содержащей указанное растительное масло и один или несколько ферментов, обладающих активностью расщепления фосфолипидов; ii) помещение реакционной смеси в условия, обеспечивающие возможность осуществления ферментативного гидролиза фосфолипидов в масле, с получением прореагировавшей смеси на основе указанного растительного масла и iii) осуществление химического рафинирования прореагировавшей смеси на основе указанного растительного масла, где стадию подкисления осуществляют после ферментативного гидролиза и до химического рафинирования. При этом ферментативный гидролиз осуществляют в реакционной смеси, содержащей тяжелую фазу и легкую фазу, и не происходит снижение или по существу не происходит снижение объема тяжелой фазы или не осуществляют отделение смолистых веществ/тяжелой фазы от масла до указанного химического рафинирования. Изобретение позволяет увеличить выход масла по сравнению с проведением химического рафинирования неочищенного масла или масла, которое подвергали удалению смолистых веществ с помощью воды. 19 з.п. ф-лы, 22 табл.,15 пр., 12 ил.
1. Способ рафинирования растительного масла, содержащего фосфолипиды, предусматривающий:
i) получение реакционной смеси, содержащей указанное растительное масло и один или несколько ферментов, обладающих активностью расщепления фосфолипидов;
ii) помещение реакционной смеси в условия, обеспечивающие возможность осуществления ферментативного гидролиза фосфолипидов в масле, с получением прореагировавшей смеси на основе указанного растительного масла и
iii) осуществление химического рафинирования прореагировавшей смеси на основе указанного растительного масла,
где стадию подкисления осуществляют после ферментативного гидролиза и до химического рафинирования, при этом:
ферментативный гидролиз осуществляют в реакционной смеси, содержащей тяжелую фазу и легкую фазу, и
не происходит снижение или по существу не происходит снижение объема тяжелой фазы или не осуществляют отделение смолистых веществ/тяжелой фазы от масла до указанного химического рафинирования.
2. Способ по п. 1, где ферментативный гидролиз фосфолипидов осуществляют в первом реакционном сосуде, а химическое рафинирование осуществляют во втором реакционном сосуде, при этом два реакционных сосуда сообщены по текучей среде и/или сообщены таким образом, что обеспечивается возможность прохождения жидкости из первого во второй реакционный сосуд.
3. Способ по п. 1 или 2, где ферментативный гидролиз фосфолипидов осуществляют в первом реакционном сосуде, а химическое рафинирование осуществляют во втором реакционном сосуде, при этом сообщение по текучей среде между реакционными сосудами или прохождение жидкости из первого во второй реакционный сосуд обеспечивается не через разделительное устройство.
4. Способ по п. 1, где ферментативный гидролиз фосфолипидов и химическое рафинирование осуществляют в одном и том же реакционном сосуде.
5. Способ по п. 3 или 4, где химическое рафинирование осуществляют непосредственно после ферментативного гидролиза.
6. Способ по любому из предыдущих пунктов, где растительное масло выбрано из группы, состоящей из масла асаи, миндального масла, масла из ореха бабассу, масла семян черной смородины, масла семян огуречника, масла канолы, масла из ореха кешью, касторового масла, кокосового масла, кориандрового масла, кукурузного масла, хлопкового масла, масла крамбе, масла семян льна, масла виноградных косточек, масла из лесного ореха, конопляного масла, ятрофного масла, масла жожоба, льняного масла, масла из ореха макадамия, масла из косточек манго, масла пенника лугового, горчичного масла, костного масла, оливкового масла, пальмового масла, пальмоядрового масла, пальмового олеина, арахисового масла/масла из земляного ореха, масла из ореха пекан, масла из кедрового ореха, фисташкового масла, макового масла, рапсового масла, масла из рисовых отрубей, сафлорового масла, масла сасанквы, кунжутного масла, масла ши, соевого масла, масла семян подсолнечника, таллового масла, масла камелии и масла из грецкого ореха.
7. Способ по любому из предыдущих пунктов, предусматривающий приведение растительного масла в контакт с одним или несколькими хелатообразующими средствами, которые могут образовывать комплексы с ионами Ca и/или Mg, до его приведения в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды.
8. Способ по любому из предыдущих пунктов, где из некоторого количества неочищенного масла с помощью указанного способа получают такое количество рафинированного масла, которое превышает на по меньшей мере 0,2% (вес/вес) количество рафинированного масла, получаемое с помощью способа, в котором химическое рафинирование осуществляют с таким же количеством указанного неочищенного масла без предварительного ферментативного гидролиза фосфолипидов, присутствующих в указанном неочищенном масле.
9. Способ по любому из пп. 6-8, где реакционная смесь характеризуется значением pH, которое находится в диапазоне 1,5–12, например, в диапазоне 1,5–7.
10. Способ по любому из пп. 6-9, где реакционная смесь характеризуется содержанием воды, находящимся в диапазоне 0,5–10% (вес/вес), например, в диапазоне 1–10% (вес/вес), в диапазоне 1–5% (вес/вес) или, например, в диапазоне 0,5–5% (вес/вес).
11. Способ по любому из предыдущих пунктов, где растительное масло приводят в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, при температуре, которая находится в диапазоне 45–90ºC.
12. Способ по любому из предыдущих пунктов, где указанный ферментативный гидролиз фосфолипидов характеризуется продолжительностью, составляющей 6 часов или меньше, например, 4 часа или меньше, например, продолжительностью, составляющей 0,5–6 часов или 0,5–4 часа, или, например, продолжительностью, составляющей 5 минут – 4 часа, например, от 5 минут до 2 часов, от 5 минут до 1 часа или, например, 5–30 минут.
13. Способ по любому из предыдущих пунктов, где указанные один или несколько ферментов, обладающих активностью расщепления фосфолипидов, вводят в общем количестве, соответствующем 0,1–30 мг ферментного белка.
14. Способ по любому из предыдущих пунктов, где растительное масло приводят в контакт с одним или несколькими ферментами, расщепляющими фосфолипиды, в таких условиях, при которых в ходе ферментативного гидролиза обеспечивается снижение количества интактных молекул фосфолипидов на 30–100%.
15. Способ по любому из предыдущих пунктов, где растительное масло после осуществления химического рафинирования содержит фосфолипиды в количествах, соответствующих 20 ppm фосфора или меньше.
16. Способ по любому из предыдущих пунктов, где указанные один или несколько ферментов, обладающих активностью расщепления фосфолипидов, предусматривают фермент, обладающий активностью фосфолипазы A, фермент, обладающий активностью фосфолипазы С, лизофосфолипазу или их смесь.
17. Способ по любому из предыдущих пунктов, где указанные один или несколько ферментов, расщепляющих фосфолипиды, выбраны из группы, состоящей из
a. фосфолипазы C, обладающей специфичностью в отношении фосфатидилинозитола (PI),
b. фосфолипазы C, обладающей специфичностью в отношении фосфатидилхолина (PC) и фосфатидилэтаноламина (PE),
c. фосфолипазы C, обладающей специфичностью в отношении фосфатидилхолина (PC), фосфатидилэтаноламина (PE), фосфатидной кислоты (PA) и фосфатидилинозитола (PI),
d. комбинации фосфолипазы A и фосфолипазы C, такой как фосфолипаза C, определенная в a) или b),
e. комбинации фосфолипазы A и лизофосфолипазы,
f. фосфолипазы A,
g. комбинации a) и b)
h. и/или их комбинаций.
18. Способ по п. 17, где указанная фосфолипаза A выбрана из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или по меньшей мере 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NO: 4 и 7;
b. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NO: 5 и 8;
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы A.
19. Способ по любому из предыдущих пунктов, где указанная фосфолипаза C выбрана из группы, состоящей из:
a. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности со зрелым полипептидом под любым из SEQ ID NOs: 9, 11, 13, 15, 17, 19, 22, 25, 28,
b. полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности с полипептидом, изложенным под любым из SEQ ID NOs: 10, 12, 14, 16, 18, 20, 23, 26, 29; и
c. фрагмента полипептида (a) или (b), который обладает активностью фосфолипазы С.
20. Способ по любому из предыдущих пунктов, где один из указанных одного или нескольких ферментов, расщепляющих фосфолипиды, обладает активностью фосфолипазы C и выбран из группы, состоящей из:
i) полипептида, характеризующегося по меньшей мере 75% идентичностью последовательности, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% или 100% идентичностью последовательности со зрелым полипептидом под SEQ ID NO: 9 или с полипептидом, изложенным под SEQ ID NO: 10,
ii) варианта полипептида, определенного в i), содержащего замену, делецию и/или вставку в одном или нескольких положениях,
iii) полипептида, который содержит последовательность, изложенную под SEQ ID NO: 8, по сути состоит из нее или состоит из нее.
| CN 105073985 A, 18.11.2015 | |||
| РИЧАРД ОБРАЙЕН "Жиры и масла | |||
| Производство, состав и свойства, применение", СПб, Профессия, 2007, стр.109-111 | |||
| СПОСОБЫ РАФИНИРОВАНИЯ МАСЛА | 2010 |
|
RU2573916C2 |
| Способ обеззоливания голья | 1928 |
|
SU14985A1 |

