Изобретение относится к области биохимии и молекулярной биологии и может быть использовано при создании тест-систем различного назначения, предполагающих использование ПЦР-амплификации ДНК из клеток дрожжей, грибов и грамположительных бактерий с прочной клеточной стенкой.
Из уровня техники известен способ выделения ДНК, включающий использование лизоцима, додецилсульфата натрия, ЭДТА в качестве индукторов лизиса клеточной стенки с последующей депротеинизацией лизата хлороформом или фенолом и осаждением ДНК изопропанолом (Marmur J.A. J.Mol. Biol. 1961.3:208-218). Из уровня техники известно использование литических ферментов (зимолиазы, литиказы и др.) для разрушения клеточных стенок дрожжей при получении образцов высокомолекулярной геномной ДНК (Muller P.M., Werner K.E., Kasai M., Francesconi A., Chanock S.J., Walsh T.J. J. Clin. Microbiol. 1998. 36(6): 1625-1629). Также известен способ выделения ДНК, путем разрушения клеток грамположительных микроорганизмов и дрожжей с помощью 3% SDS (Polaina J., Adam A.C. Nucleic Acid Res. 1991. 19(19): 5443). Все вышеперечисленные способы выделения ДНК достаточно трудоемки, дороги и занимают много времени.
Из уровня техники известен способ выделения ДНК, основанный на использовании буферных растворов, содержащих высокие концентрации солей-хаотропов типа гуанидинийтиоцианата в сочетании с детергентами, способных лизировать оболочки многих грамотрицательных микроорганизмов (Патент США №5234809, МКИ C12N 01/68, опубл. 1993). Недостатками известного способа являются невозможность использования клеток микроорганизмов, обладающих прочной клеточной стенкой, устойчивой к действию хаотропных смесей, длительность и трудоемкость процесса.
В последние годы были разработаны методы лизиса дрожжевых клеток и экстракции ДНК с помощью химических агентов, таких, например, как lithiumacetate (LiOAc) в сочетании с SDS (Looke M., Kristjuhan K., Kristjuhan A. Biotechniques 2011. V.50.P.325–328). К недостатками описанного выше способа является то, что встречаются виды микроорганизмов, в том числе грибов, клеточные стенки которых устойчивы к воздействию указанных реагентов.
Известны способы выделения ДНК, основанные на использовании щелочных реагентов:
- метод экстракции геномной ДНК, эффективный для грамположительных бактерий и дрожжей, включающий обработку клеток смесью 0.1 М КОН и полиэтиленгликоля 200 (60%) и нагрев суспензии (10 мин при 70-90оС), что приводило к хорошим результатам, но не исключало необходимость дополнительной очистки от примесей (Chomczynski P., M. Rymaszewski. Biotechniques. 2006. V. 40. P. 454–458).
- метод, эффективный для бактерий и дрожжей, основан на лизисе клеток в горячей смеси 0.1 M NaOH и 61% этанола (10 мин, 70оС), что позволяет после центрифугирования получать сырые экстракты геномной ДНК, подходящие для ПЦР, или подвергать эти экстракты дальнейшей очистке на колонке с диоксидом кремния (Vingataramin L., Frost E.H. Biotechniques. 2015. V. 58.P.120-125).
Недостатками вышеуказанных способов выделения ДНК: встречаются штаммы микроорганизмов, для которых обязательно необходимо проводить очистку щелочных лизатов, что сильно удлиняет и удорожает процедуру.
Известен способ выделения образцов геномной ДНК из клеток микроорганизмов, состоящий в том, что обработку микроорганизмов, имеющих прочные клеточные стенки, проводят буферным раствором, содержащим соль-хаотроп (производные гуанидина) в концентрации 4-8 М в сочетании с детергентом (саркозилом) при нагревании реакционной смеси при 100оС в течение 3-7 мин (Патент РФ № 2177035, МКИ С12N 15/10, опубл. 2001; Данилевич В.Н., Гришин Е.В. Биоорг. химия. 2002. Т. 28. № 2. С. 156–167). Установлено, что в присутствии высокой концентрации солей-хаотропов и при высокой температуре происходит пермеабилизация клеточных оболочек, солюбилизация мембранных структур и экстракция из клеток значительного количества РНК, белков, липидов, и других компонентов. Образующиеся при этом клеточные оболочки (КО) содержат свыше 97% ДНК и после промывки водой могут быть использованы как источник матричной ДНК в ПЦР. Несколько позже электронная микроскопия ультратонких клеточных срезов показала, что в клетках, обработанных хаотропными смесями (ХС) при 100оС в течение 5 мин, полностью разрушается мембранный аппарат, происходит деградация рибосом, разрушается большинство липидных гранул, резко изменяется строение протоплазмы. Однако клеточная стенка в основном сохраняет свою целостность, ультраструктуру и ригидность, что обуславливает поддержание исходной морфологии клетки. Внутри клеток остается много белков, присутствует РНК и ДНК и некоторая часть липидов. Показано, что одноцепочечная геномная ДНК локализуется в одном из новообразованных компартментов клетки - эктоплазме - периферической гидрофильной зоне (Данилевич В.Н., Дуда В.И., Сузина Н.Е., Гришин Е.В. Микробиология. 2007.T. 76. № 1.C. 72–82.)
Изученные структуры после воздействия температуры и хаотропных смесей лишь условно можно называть КО. Тем не менее, такие структуры не ингибируют ДНК-полимеразу и во многих случаях весьма эффективно отдают свою ДНК в раствор при термоциклировании в ходе ПЦР. Вышерассмотренный способ эффективен в отношении многих дрожжей, мицелиальных грибов и некоторых грамположительных микроорганизмов. Однако его недостатком является то, что он малоэффективен в отношении многих грамположительных бактерий, в том числе микобактерий, патогенных стрептококков, стафиллококков и т.д., почвенных изолятов Bacillus subtilis, Arthrobacter globiformis и Micrococcus luteus, а также многих природных изолятов дрожжей и мицеллиальных грибов, обладающих уникальной очень прочной и плотной клеточной стенкой.
Известен наиболее близкий аналог к заявленному способ выделения образцов геномной ДНК из клеток микроорганизмов (Патент РФ № 2177035 опубл. 2001г). В опубликованном способе (Патент РФ №2317995, МКИ С2, опуб. 01.02.2006) поставленная задача решается за счет того, что клеточные оболочки с ДНК, полученные при обработке клеток хаотропными смесями при нагреве (100оС, 7 мин), дополнительно обрабатывают протеиназой К в течение 1 часа при 37-40оС. В процессе обработки клеток хаотропными смесями и протеиназой К происходит солюбилизация остаточных липидов, протеолитическая деградация внутриклеточных белков и белков клеточной стенки и за счет этого достигается полная депротеинизация хромосомных ДНК и резко увеличивается проницаемость клеточной оболочки по отношению к макромолекулам (ДНК). В то время как белки, пептиды, липиды и другие компоненты уходят из клетки, хромосомная ДНК из-за своей большой длины остается внутри клеточной оболочки. Потери ДНК при последующей промывке клеток водой от компонентов буфера и протеиназы К, как было показано, практически не происходит. Получаемые таким способом клеточные оболочки с ДНК были успешно использованы в ПЦР. К недостаткам этого способа следует отнести длительность и многостадийность процедуры, необходимость использования дорогих реагентов.
Настоящее изобретение раскрывает способ получения образцов геномной ДНК из клеток дрожжей и грамположительных бактерий, ассоциированной с клеточными оболочками и пригодной для непосредственного использования в ПЦР-амплификации. Способ по изобретению предусматривает обработку клеток новыми, неописанными в литературе ХС, содержащими хаотропный агент (мочевину), ионный детергент (додецилсульфат натрия (SDS)), восстановитель дисульфидных связей – (2-меркаптоэтанол), хелатирующий агент (цитрат натрия или ЭДТА) и щелочь (аммиак) в соответствующих концентрациях при соотношении объема ХС к объему клеток, превышающем 10:1. Суспензию клеток в ХС нагревают при температуре кипения воды в течение 10 мин, охлаждают, щелочь нейтрализуют кислым ацетатным буфером, затем смесь центрифугируют. Супернатант отбрасывают, а осадок, содержащий геномную ДНК в составе (внутри) клеточных оболочек микроорганизмов, дважды промывают водой и суспендируют в дистиллированной воде или в буфере ТЕ. Полученную суспензию клеточных оболочек используют непосредственно в ПЦР. Осуществление изобретения позволяет повысить эффективность получения геномной ДНК, добиться большей чистоты получаемых образцов ДНК, снизить содержание в них РНК, белков и иных примесей, другими словами, сделать процедуру выделения ДНК более эффективной и более универсальной.
Заявленное изобретение решает задачу повышения эффективности процесса получения геномной ДНК, ассоциированной с прочными клеточными оболочками дрожжей и грамположительных бактерий.
В изобретении, вместо термальной обработки клеток буферным раствором с нейтральным pH, содержащим хаотропную смесь на основе солей гуанидина и детергента саркозила, используют термальную обработку биомассы (90-100оС, 10 мин) щелочными хаотропными смесями на основе 7-8 М мочевины и 1% SDS с добавкой 100-200 мМ аммиака с последующей нейтрализацией смеси кислым ацетатным буфером и двухкратной промывкой целевого продукта - клеточных оболочек (КО) с ДНК, деионизованной водой.
Использование щелочных хаотропных смесей на основе 7М мочевины и 1% SDS позволяет повысить уровень экстракции из дрожжевых клеток РНК, белков и прочих низкомолекулярных компонентов практически до 100%, что невозможно сделать при использовании нейтральных ХС на основе солей гуанидина и саркозила. Таким образом, за один прием получаются высокоочищенные КО с геномной ДНК. За счет этого повышается чувствительность метода ПЦР.
В другом варианте заявленного изобретения, поставленная задача решается за счет того, что используют обработку биомассы нейтральной хаотропной смесью, содержащей 7М мочевину, 1% SDS с добавкой 25 мМ цитрата натрия. При тех же условиях инкубации (90-100оС, 10 мин) в этом случае достигается близкая к 97% эффективность экстракции РНК. Этот уровень экстракции РНК (а, следовательно, и белков) невозможно получить при использовании хаотропных смесей, содержащих соли гуанидина в сочетании с саркозилом и 25мМ цитратом натрия. Роль цитрата натрия заключается в том, что этот хелатор способен необратимо связывать катионы двухвалентных катионов, в частности Са2+ и Мg2+. Вполне вероятно, что двухвалентные катионы стабилизируют клеточную стенку дрожжей. Полное удаление катионов Me2+ в таком случае может приводить к разрыхлению (пермеабилизации) клеточной стенки и более эффективной экстракции РНК и белков при нагреве с ХС.
Нужно также отметить, что в случае использования нейтральной ХС на основе 7М мочевины и 1% SDS возможно получение клеточных оболочек как с денатурированной ssДНК, так и КО с двухцепочечной dsДНК. Последнее возможно, если температуру нагрева биомассы с нейтральной ХС проводить ниже 70оС, т.е. ниже температуры плавления геномной ДНК дрожжей.
Изобретение решает также вторую задачу – позволяет расширить спектр (круг) микроорганизмов с прочной и плотной клеточной стенкой, которые не поддаются воздействию ХС на основе солей гуанидина в сочетании с саркозилом. Для пока не исследованных микроорганизмов с прочной клеточной оболочкой могут потребоваться некоторые вариации условий пробоподгодотовки (концентрации реагентов, режим нагрева и т.д.). Разработанный способ является гибким и допускает возможность модификаций, в зависимости от сложности объекта.
Состав хаотропных смесей, использованных в заявке, представлен в таблице 1.
Таблица 1.
Состав хаотропных смесей и значение pH раствора до и после добавления аммиака как среднее по данным трех независимых измерений.
§ Саркозил - N-lauroylsarcosine, Na-salt; Меркаптоэтанол - MЭ – 2-mercaptoethanol; GuHCl – гуанидин гидрохлорид; Цитрат Na – Na-citrate.
* Индекс после точки в номере раствора означает количество молей аммиака (0, 0,1, 0,2 соответственно).
В процессе обработки клеток щелочной (или нейтральной) хаотропной смесью в заявленном способе происходит солюбилизация (разрушение) всех мембранных структур, полная денатурация внутриклеточных белков и белков клеточной стенки и за счет этого достигается полная депротеинизация хромосомных ДНК и резко увеличивается проницаемость клеточной оболочки по отношению к макромолекулам (РНК, белкам). Инкубирование клеток при 100оС в щелочных условиях сопровождается щелочным гидролизом всех фракций РНК и резким уменьшением их размеров. По результатам оценки авторов, размер молекул РНК после обработки ХС колеблется от нескольких десятков до 1500 нуклеотидов. Низкомолекулярная РНК вместе с белками и другими низкомолекулярными компонентами легко проникает через поры клеточной стенки и выходит наружу в раствор практически со 100% эффективностью. Что касается геномной ДНК, то в щелочных растворах при нагреве она более стабильна, чем РНК. Тем не менее, в заявленных условиях происходит полная денатурация геномной ДНК и последующее ее расщепление (за счет гидродинамических разрывов) до фрагментов достаточно большой молекулярной массы. По приблизительным оценкам, размер одноцепочечных фрагментов ДНК колеблется в пределах 16-30 т.о. По причине большой длины фрагменты ssДНК остаются внутри КО.
Механизм сохранения ДНК внутри КО, предположительно, состоит в следующем: после высокотемпературной обработки клеток в присутствии ХС и при последующем быстром охлаждении суспензии и нейтрализации, денатурированная ДНК за счет комплементарных взаимодействий с РНК образует трехмерную пространственную сеть, нерастворимую при комнатной температуре - она практически не вымывается из клеточных оболочек в ходе двух промывок дистиллированной водой. В ПЦР-смеси при термоциклировании геномная ssДНК диссоциирует из комплекса с РНК и легко диффундирует через поры пермеабилизованной клеточной оболочки в раствор, где служит матрицей для ДНК-полимеразы.
Таким образом, денатурированная геномная ДНК, получаемая заявленным способом, прочно ассоциирована с КО и представляет собой матричную ДНК, обладающую такой же активностью в ПЦР, как и высокоочищенная одноцепочечная геномная ДНК, не связанная с клеточными стенками. При этом, заявленный способ получения матричной ДНК гораздо дешевле и эффективней.
В работе использовали клинические изоляты дрожжей - из коллекции Института вакцин и сывороток им. И.И. Мечникова (Москва, Россия); а также лабораторные штаммы дрожжей – из коллекции ИБХ РАН. Все они приведены в таблице 2. В работе использовали также штаммы грамположительных бактерий Micrococcus luteus (штамм NCIMB 13267) и Bacillus cereus из коллекции института микробиологии (ИНМИ) им. Виноградского, РАН, Москва).
Таблица 2
Штаммы дрожжей и грибов, использованные в работе.
Клинический изолят № 220
Клинический изолят № 18
Клинический изолят № 3465
Клинический изолят № 132
Лабораторный штамм(ИБХ РАН)
Трансформант штамма GS115
Лабораторный штамм
Изобретение иллюстрируют фигуры.
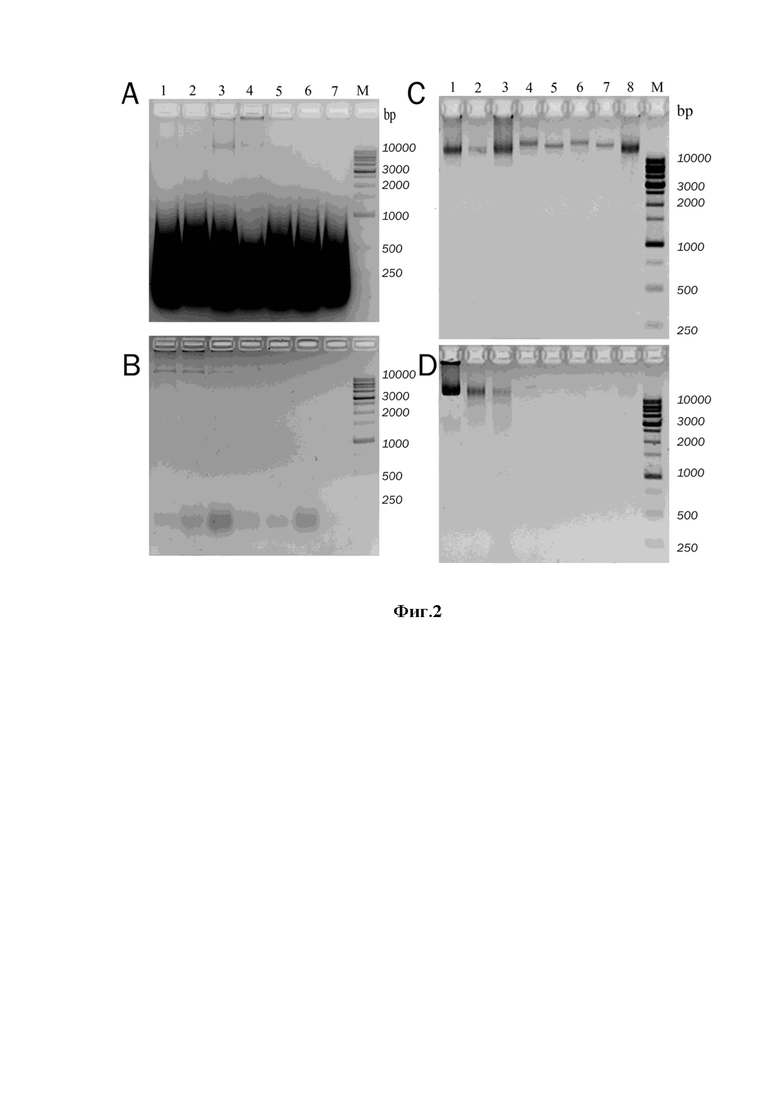
Фиг. 1. Проверка эффективности экстракции НК электрофорезом в 1,5% агарозе. (A) Выход РНК при экстракции из двух штаммов дрожжей P. pastoris (R+) дорожки 1, 2, 5, 6, 8, 9, 10 и P. pastoris (R-) дорожки 3, 4, 7, 12, 14. (B) Выход геномной ДНК из КО двух штаммов дрожжей P. pastoris (R+) дорожки 1, 2, 5, 6, 7, 8 и P. pastoris (R-) дорожки 3, 4, 9, 10, 11, 12. М – стандарты молекулярных масс фрагментов ДНК. Время термической обработки с ХС составляет 10 минут (малый прямоугольник снизу) и 30 минут (большой прямоугольник снизу), цифры обозначают номер ХС.
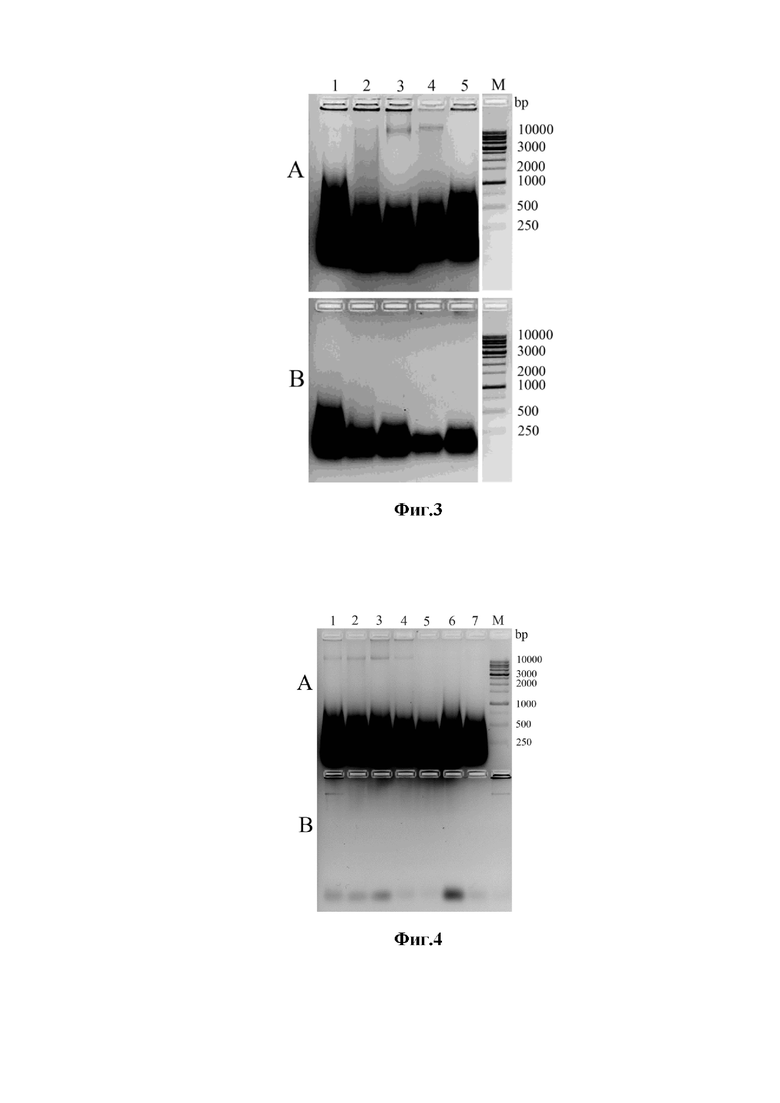
Фиг. 2. Проверка эффективности экстракции РНК (A) и (B), а также последующей диффузии ДНК из клеточных оболочек (C) и (D) электророфорезом в 1,5% агарозе. Данные для смеси 2.1 показаны на панели (A) и (С), смеси 6.2 на панели (B) и (D). Образцы дрожжей: дорожка 1- S. cerevisiae 375; дорожка 2 – P. pastoris; дорожка 3 – C. albicans 927; дорожка 4 - T. cutaneum 18; дорожка 5 – Cr. neoformans 3465; дорожка 6 – Rh. mucilaginosa 132; дорожка 7 –Cr. albidus 220; дорожка 8 - S. cerevisiae Y190. М – стандарты молекулярных масс фрагментов ДНК.
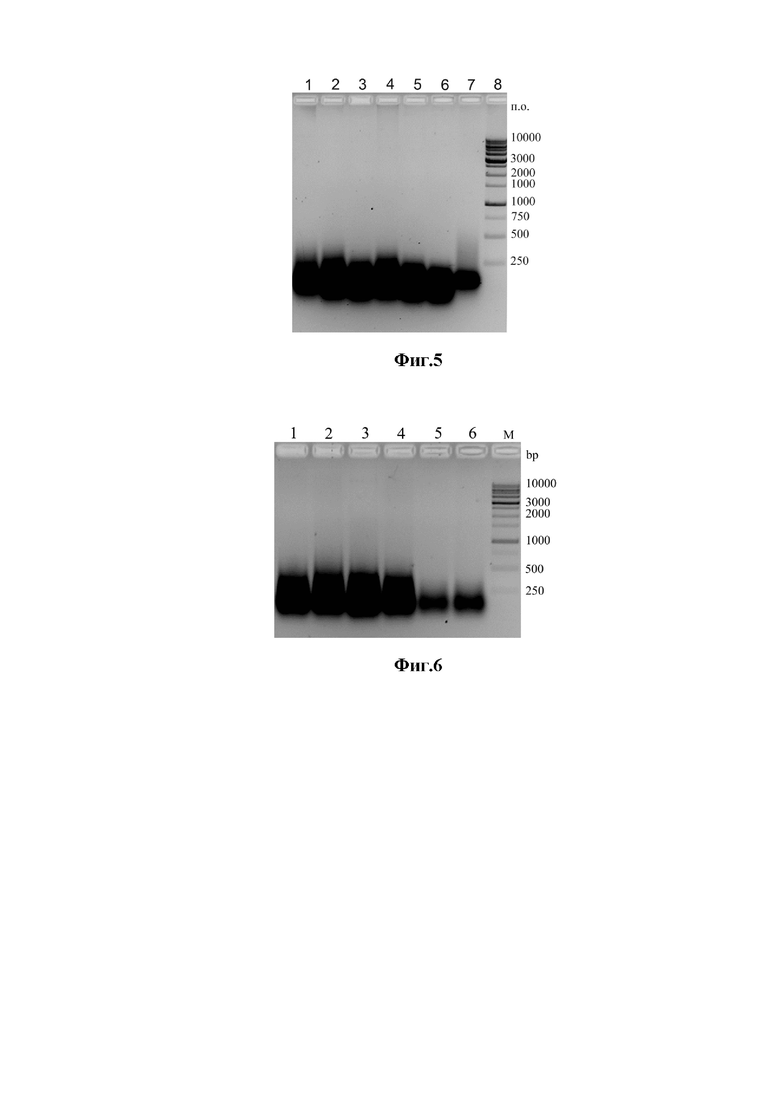
Фиг. 3. Проверка эффективности повышенной концентрации цитрата Na в составе нейтральных ХС на экстракцию РНК из клеток дрожжей электрофорезом в 1,5% агарозе. Панель (А), дорожки 1- 5 - РНК в экстрактах, полученных с помощью ХС1.0, Панель (B), дорожки 1 -5 – РНК в экстрактах, полученных с помощью ХС5.0. Исследовали РНК из дрожжей: дорожка 1- S. cerevisiae 375; дорожка 2 – P. pastoris; дорожка 3 – C. albicans 927; дорожка 4 - T. cutaneum 18; дорожка 5 – Cr. neoformans 3465. М – стандарты молекулярных масс фрагментов ДНК.
Фиг. 4. Проверка эффективности экстракции РНК из клинических штаммов дрожжей с помощью щелочных ХС различного состава, содержащих 25 мМ цитрат натрия электрофорезом в 1,5% агарозе. Панель (А), дорожки 1- 5 - РНК в экстрактах, полученных с помощью ХС1.1, Панель (B), дорожки 1 -5 – РНК в экстрактах, полученных с помощью ХС5.1. Исследовали РНК из дрожжей: дорожка 1- S. cerevisiae 375; дорожка 2 – P. pastoris; дорожка 3 – C. albicans 927; дорожка 4 - T. cutaneum 18; дорожка 5 – Cr. neoformans 3465. М – стандарты молекулярных масс фрагментов ДНК.
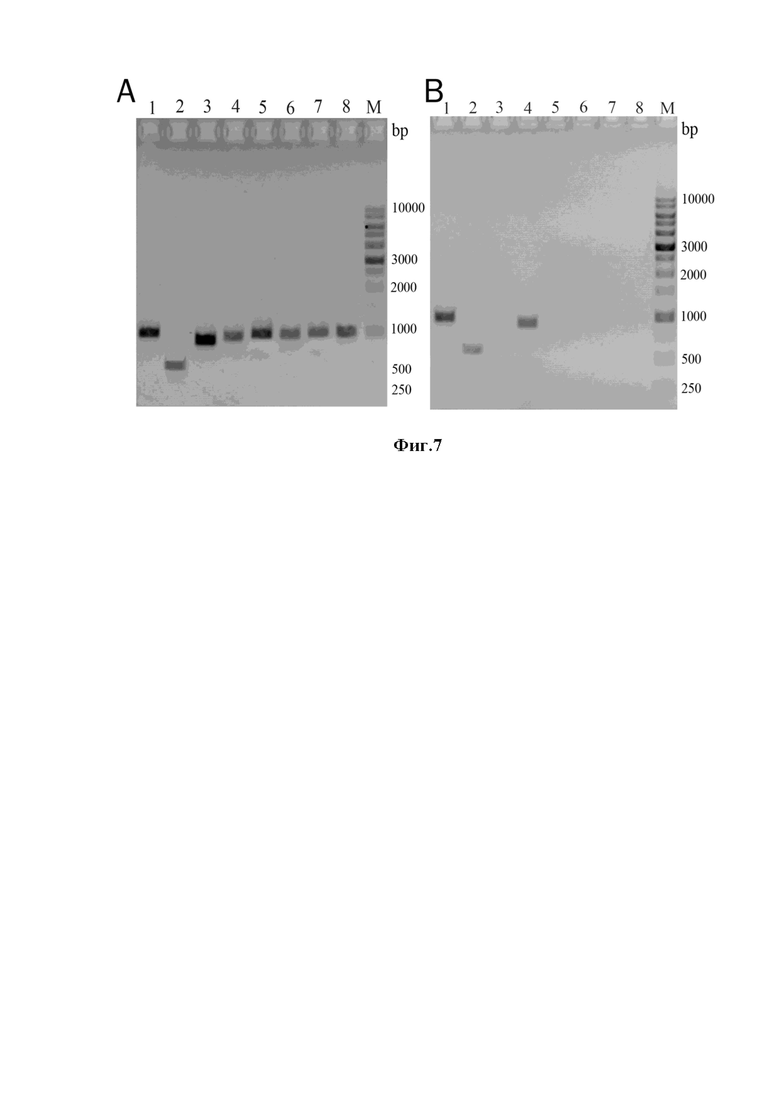
Фиг. 5. Проверка эффективности экстракции РНК из клеток M. luteus (ure+) (дорожки 1-3) и M. luteus (ure-) (дорожки 4-7) электрофорезом в 1,5% агарозном геле. Используют ХС 2.1 с добавками аммиака, 0,1М (дорожка 1 и 4), ХС 2.2 с 0,2М аммиака (дорожка 2 и 5), и ХС 2.0 с добавлением 0,3М аммиака дорожка 3 и 6) и ХС 2.0 дорожка 7. М – стандарты молекулярных масс фрагментов ДНК.
Фиг. 6. Проверка эффективности экстракции РНК из мицеллия гриба A. fumigatus электрофорезом в 1,5% агарозном геле. Используют ХС 1.0 (дорожки 1-2), ХС 1.1 (дорожки 3-4), ХС 5.0 (дорожки 5-6). М – стандарты молекулярных масс фрагментов ДНК.
Фиг. 7. Электрофоретический анализ (0,8% агароза) продуктов ПЦР-амплификации (33 цикла) фрагментов рибосомной ДНК у различных видов дрожжей, где в качестве матрицы используют КО полученные ХС 1.1 (панель A) или 6.0 (панель В). Изученные виды: S. cerevisiae 375 (дорожка 1); P. pastoris GS115 (дорожка 2); C. albicans 927 (дорожка 3); T. cutaneum 18 (дорожка 4); Cr. neoformans 3465 (дорожка 5); Rh. mucilaginosa 132 (дорожка 6); Cr. albidus 220 (дорожка 7); S. cerevisiae Y190 (дорожка 8). М – стандарты молекулярных масс фрагментов ДНК.
Осуществление способа
Клетки дрожжей S. cerevisiae 375, C. albicans 927, Cr. albidus 220, T. cutaneum 18 , Cr. neoformans 3465, Rh. mucilaginosa 132, P. pastoris GS115, S. cerevisiaeY190 (таблица 2) выращивают в жидкой среде YPD (1.0% дрожжевой экстракт, 2.0% бактопептон, 2.0% глюкоза, pH 5.3) или на чашках с агаризованной средой YPD (1.5% агар) при 30оС, или на средах иного состава. Клетки грамположительных бактерий Micrococcus luteus и Bacillus cereus выращивают на чашках с L-агаром при 30оС или 37оС или на жидких и твердых средах другого состава. Биомассу снимают с поверхности чашек стерильной петлей и переносят в пробирку с 0,7 мл буфера TE (10 мМ Tris-HCl, pH 8,0 + 5 мМ EDTA) и затем суспендируют (вортекс) до гомогенного состояния. Аликвоты (100-200 мкл) полученной суспензии далее разливают в 1.5 мл пробирки и центрифугируют при 12000 об /мин в течение 3 мин при комнатной температуре. Надосадочную жидкость аккуратно удаляют, а клеточные осадки обрабатывают хаотропными смесями, как описано ниже.
Экстракция РНК из дрожжей и получение клеточных оболочек с ДНК
Клетки дрожжей S. cerevisiae 375, C. albicans 927, Cr. albidus 220, T. cutaneum 18 , Cr. neoformans 3465, Rh. mucilaginosa 132, P. pastoris GS115, S. cerevisiaeY190 (таблица 2) и бактерий (~20 мкл осадка после центрифуги) суспендируют в 200 мкл соответствующей ХС, содержащей 7 М мочевину, 1% SDS, и другие компоненты, а также различные концентрации аммиака (0,1, 0,2 М). В контрольном образце (смесь Х.0) аммиак отсутствует (образец с нейтральным pH); для любой хаотропной смеси, содержащей 0,1 М аммиака (смесь Х.1), на 1 мл раствора добавляют 17,5 µl 20% водного раствора аммиака; аналогично для любой смеси, содержащей 0,2 М аммиака на 1 мл раствора (смесь Х.2), добавляют 35 µl 20% водного раствора аммиака. Экстракцию РНК (и белков) проводят на кипящей водяной бане или в суховоздушном термостате при 95 - 97оС с минимальным встряхиванием (перемешиванием) в течение 10, 20 или 30 мин. После тепловой обработки пробирки охлаждают на тающем льду (0оC), а щелочные смеси, если планируется анализ РНК, дополнительно нейтрализуют добавлением кислого 3 М Na-ацетатного буфера (pH 5.45), и тотчас центрифугируют. Объем ацетатного буфера для нейтрализации щелочи в образцах определяют предварительно методом титрования до конечного значения pH =7.4, он составляет ~90 мкл 3М Na-ацетатного буфера на 1 мл ХС X.1, или ~ 180 мкл на 1 мл ХС X.2. После центрифугирования (3 мин 12000 об/мин) осадки, содержащие клеточные чехлы с ДНК, дважды промывают 1 мл деионизованной воды и на конечной стадии суспендируют в 200 мкл стерильной деионизированной воды или ТЕ буфера (5 мМ Tris- HCl, pH 8.0, 1 мМ EDTA) и хранят 2 недели при +4оС или при -20оС (для длительного хранения).
Анализ РНК
Ранее и в настоящей работе нами показано, что экстракция РНК из клеток с помощью ХС сопряжена (идет параллельно) с экстракцией белков. Другими словами, чем больше экстрагируется РНК, тем меньше в клетках остается белков и тем эффективнее геномная ДНК в ходе ПЦР (при термоциклировании) будет выходить из КО. Таким образом, анализируя с помощью простого электрофореза количество РНК в дрожжевых экстрактах после обработки хаотропными смесями, можно предсказать результативность ПЦР на ДНК-содержащих клеточных оболочках неизученных ранее штаммов микроорганизмов.
После нейтрализации и центрифугирования дрожжевых суспензий супернатанты переносят в чистые пробирки и к ним добавляют 2 V холодного 96% этанола. Смесь перемешивают и выдерживают 1 час при +4оС для осаждения РНК и ДНК. НК осаждают центрифугированием в течение 3 мин в стандартном режиме, осадки растворяют в 100 мкл стерильной деионизованной воды или ТЕ-буфера. РНК и ДНК в полученных растворах анализируют электрофорезом в 1,0 -1,5% агарозном геле. В лунки геля наносят по 10 µl раствора РНК (1/10 часть).
Получение одноцепочечной (ss) геномной ДНК из клеточных оболочек
Как было установлено авторами, после двукратной промывки клеточных оболочек водой ДНК не теряется. В то же время при промывке могут уходить остатки РНК и белков. Для визуализации ДНК обычно используют 50 мкл аликвоты суспензий клеточных оболочек. Соответствующие пробирки с образцами суспензий инкубируют при 90оС в течение 10 мин в суховоздушном термостате при перемешивании (500 об/мин). В этих условиях трехмерная сеть из ДНК и РНК разрушается и ssДНК диффундирует из КО в раствор. После охлаждения пробирок на воздухе (3-5 мин) и центрифугирования в течение 3 мин в стандартном режиме аликвоты супернатантов (10 мкл), содержащие ssДНК, анализируют электрофорезом в 1.0 -1.5% агарозном геле.
ПЦР-амплификация ДНК
Полученные суспензии КО используют непосредственно в экспериментах по ПЦР-амплификации. В полученных суспензиях титр клеток составляет 5·108 - 1·109. В опыт берут 1 мкл неразведенной суспензии или 1 мкл из 10-1 - 10-2 разведений на 50 мкл реакционной смеси для ПЦР.
ПЦР-амплификацию фрагментов рибосомной ДНК дрожжей проводят с помощью универсальных праймеров 5,8S-R: (5´)TCGATGAAGAACGCAAGC и LR3: (5´)GGTCCGTGTTTCAAGAC (VilgalisR., HesterM.// J. Bacteriol. 1990. V 172. P. 4238-4246).
Условия ПЦР: денатурация – 93оС, 50 с, отжиг – 55оС, 30 с, синтез – 72оС, 40 с; число циклов - 30-40.
ПЦР-амплификацию фрагментов рибосомной ДНК эубактерий осуществляли с помощью универсальных праймеров 27f: (5')AGAGTTTGATCCTGGCTCAG и 1522r: (5') AAGGAGGTGATCCARCCGCA (Johnson J.L. Similarity analysis of RNAs. In P.Herhardt, R.G.E. Murray, W.A. Wood, and N.R. Krieg (ed) Methods for general and molecular bacteriology. American Society for Microbiology, Washington, D.C. 1994. P. 683-700).
Условия ПЦР: денатурация – 93оС, 30 с, отжиг – 60оС, 30с, синтез – 72оС, 60 с; число циклов - 30-40.
Изобретение иллюстрируется примерами.
Пример 1. Отработка условий получения образцов геномной ДНК для ПЦР из дрожжей Pichia pastoris
Клетки двух штаммов дрожжей P. pastoris (R+) и (R-) выращивают на чашках с агаризованной средой YPD (1% дрожжевого экстракта, 2,0% бактопептона , 2% глюкозы, pH 5,3, 1.5% агара) при 30оС.
25 мкл сырой биомассы клеток дрожжей после центрифуги суспендируют в 200 мкл хаотропных смесей различного состава, содержащих:
7 М1) мочевину, 1% SDS, 0,1 М МЭ с добавками 0,1 или 0,2 М аммиака, или
7 М2) мочевину, 1% SDS, 0,1 М МЭ, или
6 M3) GuHCl, 0,5% саркозил, 0,1 М МЭ (контрольная смесь).
Суспензии инкубируют на кипящей водяной бане в течение 10 или 30 мин. Пробирки охлаждают на тающем льду, щелочные смеси нейтрализуют ацетатным буфером, затем клетки осаждают центрифугированием. Супернатанты переносят в чистые пробирки для анализа РНК. Клеточные осадки дважды промывают деионизованной водой. После второй промывки осадки, представляющие собой КО с геномной ДНК, суспендируют в 200 мкл стерильной деионизованной воды и хранят в заморозке при -20оС.
Уровень экстракции РНК из клеток P. pastoris нейтральными и щелочными ХС определяют по результатам электрофореза Фиг. 1(А). Способность ДНК к диффузии в раствор у полученных в эксперименте КО при их нагреве (90оС, 10 мин) определяют по результатам электрофореза Фиг.1(В).
Как видно из Фиг.1, ХС 6.0 с 6М гуанидин гидрохлоридом и 0.5% саркозилом и нейтральным значением рН малоэффективны для экстракции РНК при термальной обработке (10 мин кипячения на водяной бане) (фиг.1A, дорожки 1, 2). Замена основных компонентов контрольной смеси на 7М мочевину и 1% SDS (ХС 4.0) также не обеспечивает заметного повышение выхода РНК из клеток двух штаммов P. pastoris (фиг.1A, дорожки 3, 4, 8), а добавки к ХС 4.0 раствора аммиака до концентрации 0,1 и 0,2 М (до рН 11,32 и 11,50 повышают в 1.5-2 раза эффективность экстракции РНК (Фиг.1A). Пролонгированное кипячение дрожжей (30 мин вместо 10 мин) в ХС 4.1 и 4.2 (в присутствии аммиака) приводит к усилению гидролиза РНК, судя по уменьшению размера пятен на электрофореграммах (Фиг. 1A, дорожки 6, 10, 12, 14).
Как видно из Фиг.1А, при термальной обработке биомассы дрожжей ХС ДНК в супернатантах не детектируется вовсе, присутствует только РНК. Очевидно, вся геномная ДНК находится внутри клеточных оболочек. В осадках, полученных после термальной обработки клеток ХС выявляются КО с ДНК. Однако степень высвобождения ДНК после 10 мин прогрева при 90оС зависит от состава примененной смеси. Так, ДНК не диффундирует в раствор из КО, полученных с использованием ХС 6.0 и ХС4.0 с нейтральным значением рН (Фиг. 1B, дорожки 1-4). Напротив, наблюдают эффективное высвобождение ДНК для ХС 4.1 и 4.2, содержащих 0,1 и 0,2 М аммиака (Фиг. 1B, дорожки 5-12).
Таким образом, добавки аммиака к хаотропным смесям на основе 7М мочевины и 1% SDS в концентрации 0,1 и 0,2 М не только повышают в ~2 раза уровень экстракции РНК из клеток P. pastoris, но также способствуют последующему выходу геномной ДНК из КО в раствор.
Пример 2. Экстракция РНК из клеток клинических штаммов дрожжей щелочными хаотропными смесями
Для того чтобы решить вопрос универсальности щелочных хаотропных смесей 3.1 и 3.2 исследуют широкий круг микроорганизмов (восемь штаммов дрожжей, в том числе, шесть клинических изолятов: S. cerevisiae 375, C. albicans 927, Cr. albidus 220, T. cutaneum 18 , Cr. neoformans 3465, Rh. mucilaginosa 132 и два лабораторных штамма P. pastoris GS115, S. cerevisiaeY190.
Клетки дрожжей выращивают на чашках с агаризованной средой YPD при 30оС. Биомассу снимают с поверхности чашек стерильной петлей и переносят в пробирки с буфером TE (10 мМ Tris-HCl, pH 8,0 + 5 мМ EDTA) и суспендируют до гомогенного состояния. Полученные суспензии разливают в 1.5 мл пробирки и центрифугируют при 12000 об /мин в течение 3 мин. Полученные клеточные осадки 25 мкл суспендируют в 200 мкл щелочной ХС 2.1 и ХС 6.1 с добавлением 5 мМ цитрата натрия. Термальную обработку клеток с хаотропами проводят на кипящей водяной бане в течение 10 мин. После этого клетки охлаждают на тающем льду, щелочь в суспензиях нейтрализуют кислым ацетатным буфером и центрифугируют. Супернатанты отбирают и переносят в чистые пробирки для анализа РНК. Осадки дважды промывают 1 мл деионизованной воды и затем суспендируют в 200 мкл стерильной H2O и хранят при -20оС.
Уровни экстракции РНК использованными хаотропными смесями определяют электрофорезом Фиг. 2, А и В. Изучают также эффективность выхода ДНК из клеточных оболочек дрожжей при нагреве соответствующих суспензий в течение 10 мин при 90оС (Фиг.2, С и D) электророфорезом в 1,5% агарозе.
Устанавливают, что щелочные ХС на основе 7 М мочевины с 5 мМ цитрата (смесь 2.1), эффективно экстрагирует РНК из клеток всех изученных штаммов дрожжей (Фиг. 2А). Напротив, щелочные ХС на основе 6 М GuHCl с 5 мМ цитрата (смесь 6.1) экстрагирует РНК из тех же штаммов на весьма низком уровне (Фиг. 2B).
Геномная ssДНК внутри КО, полученных с помощью щелочных ХС на основе 7М мочевины и 1% SDS, имеет высокое качество. ДНК на электрофорезе детектируют в виде узкой полосы (без шлейфов) в случае ХС 2.1 (Фиг. 2C) для всех штаммов, и частично (Фиг. 2D lines 1-3) для ХС6.1 с 6 M GuHCl, 0,5% саркозилом, 5мМ Na-цитрата и 0,2М NH4OH.
Пример 3. Экстракция РНК из клеток клинических штаммов дрожжей нейтральными хаотропными смесями в присутствии 25 мМ цитрата натрия
Исследовали следующие штаммы дрожжей: S. cerevisiae 375, P. pastoris, C. albicans 927, T. cutaneum 18, Cr. neoformans 3465.
Клетки дрожжей выращивают на чашках с агаризованной средой YPD (1.5% агар) при 30оС. Биомассу дрожжей (везде по ~25 мкл) суспендируют в 200 мкл нейтральной смеси ХС 1.0, содержащей 7 М мочевину, 1% SDS, 25 мМ Na-цитрат и 0,1М МЭ и в 200 мкл ХС 5.0, содержащей 6 М GuHCl, 0,5% саркозил, 25 мМ цитрат, 0,1М МЭ. Термальную обработку клеток с хаотропами проводят на кипящей водяной бане в течение 10 мин. После этого клетки охлаждают на тающем льду, и центрифугируют. Супернатанты отбирают и переносят в чистые пробирки для анализа РНК. Осадки дважды промывают 1 мл деионизованной воды и затем суспендируют в 200 мкл стерильной H2O и хранят при -20оС. Уровни экстракции РНК использованными хаотропными смесями определяют электрофорезом Фиг. 3 (панели А и В).
Как видно на Фиг.3, присутствие 25 мМ цитрата натрия в нейтральной ХС на основе 7М мочевины и 1% SDS резко повышает ее способность экстрагировать РНК при нагреве (100оС). Щелочная ХС на основе 7М мочевины и 1% SDS и 25 мМ цитрата натрия все же имеет определенные преимущества, по сравнению с нейтральной ХС того же состава, - она позволяет практически полностью убрать из дрожжевых клеток все РНК, в том числе длинные матричные РНК.
Пример 4. Экстракция РНК из клеток клинических штаммов дрожжей нейтральными хаотропными смесями, содержащими 8М мочевину, 1% SDS и 25 мМ цитрата натрия.
Использовали те же штаммы дрожжей, что и в примере 3.
Клетки дрожжей выращивают на чашках с агаризованной средой YPD (1.5% агар) при 30оС. Биомассу дрожжей (везде по 25 мкл) суспендируют в 200 мкл нейтральной смеси (близкой по составу с ХС 1.0), содержащей 8 М мочевину, 1% SDS, 25 мМ Na-цитрат и 0,1М МЭ и в 200 мкл ХС 5.0, содержащей 6 М GuHCl, 0,5% саркозил, 25 мМ цитрат, 0,1М МЭ. Термальную обработку клеток с хаотропами проводят на кипящей водяной бане в течение 10 мин. После этого клетки охлаждают на тающем льду, и центрифугируют. Супернатанты отбирают и переносят в чистые пробирки для анализа РНК. Осадки дважды промывают 1 мл деионизованной воды и затем суспендируют в 200 мкл стерильной H2O и хранят при -20оС Уровни экстракции РНК хаотропными смесями определяют электрофорезом. Как было найдено, присутствие 25 мМ цитрата натрия в нейтральной ХС на основе 8М мочевины и 1% SDS резко повышает ее способность экстрагировать РНК при нагреве (100оС).
Пример 5. Экстракция РНК из клеток клинических и лабораторных штаммов дрожжей щелочными хаотропными смесями в присутствии 25 мМ цитрата натрия
Клетки дрожжей (все дрожжевые штаммы из таблицы 2) выращивают на агаризованной среде в тех же условиях, что и в примере 3. По 25 мкл биомассы дрожжей суспендируют в 200 мкл щелочной смеси ХС 1.1, содержащей 7 М мочевину, 1% SDS, 25 мМ Na-цитрат и 0,1М МЭ + 0.1М аммиака, и в 200 мкл ХС 5.0, содержащей 6 М GuHCl, 0,5% саркозил, 25 мМ цитрат, 0,1М МЭ + 0,1 М аммиака. Термальную обработку клеток с ХС проводят на кипящей водяной бане в течение 10 мин. После этого клетки охлаждают на тающем льду, нейтрализуют щелочь и центрифугируют. Супернатанты отбирают и переносят в чистые пробирки для анализа РНК. Осадки дважды промывают 1 мл деионизованной воды и затем суспендируют в 200 мкл стерильной H2O и хранят при -20оС. Уровни экстракции РНК определяют электрофорезом Фиг. 4 (панели А и В).
Как видно из рисунка, уровни экстракции РНК щелочной ХС с 25 мМ цитратом очень высоки у всех изученных образцов, однако не сильно превышают уровни экстракции РНК нейтральной ХС с 25 мМ цитратом. Присутствие аммиака в ХС, содержащей 6М GuHCl, 25 мМ цитрат и другие компоненты, напротив, приводят к резкому снижению уровня экстракции РНК у всех без исключения образцов дрожжей (Фиг.4 В).
Видно, что щелочная ХС 1.1 на основе 7 М мочевины весьма эффективно экстрагирует РНК из всех штаммов. Напротив, защелачивание хаотропной смеси ХС 5.0 на основе 6 M GuHCl уменьшает экстракцию РНК.
Пример 6. Получение образцов геномной ДНК Micrococcus luteus с прочной клеточной стенкой
Клетки грамположительных бактерий M. luteus выращивают на чашках с L-агаром при 37оС. Один из штаммов характеризуется высоким уровнем синтеза уреазы (ure+) – фермента, разлагающего мочевину до углекислого газа и аммиака. Другой штамм неспособен к синтезу уреазы (ure-). Осадки клеток равного объема (20 - 30 мкл) в 1,5 мл пробирках суспендируют в хаотропных смесях объемом 200 мкл, содержащих 7 М мочевину, 1% SDS, 5 мМ цитрат натрия и различные концентрации аммиака (0, 0,1, 0,2 и 0,3 М). Пробирки с суспензией клеток кипятят на водяной бане в течение 10 мин затем охлаждают во льду (0оC), щелочь в смеси нейтрализуют с помощью 3М ацетатного буфера и центрифугируют. Супернатанты отбирают, переносят в чистые пробирки и в них определяют РНК (см. протокол). РНК анализируют электрофорезом в 1,5% агарозном геле Фиг.5.
Осадки – клеточные оболочки с ДНК, дважды промывают деионизованной водой и суспендируют в 200 мкл воды или буфера ТЕ. Затем 1 мкл суспензии клеточных оболочек с ДНК добавляют к 50 мкл смеси для ПЦР, содержащей соответствующий буфер, смеси 4-х dNTPs, праймерные олигонуклеотиды 27f + 1522r и термостабильную ДНК-полимеразу (Taq-полимеразу). Проводят ПЦР с учетом размера амплифицируемого фрагмента и структуры праймеров. Наблюдают синтез линейного фрагмента ДНК размером 1.5 т.п.о, что совпадает с данными литературы (Similarity analysis of RNAs. In P.Herhardt, R.G.E. Murray, W.A. Wood, and N.R. Krieg (ed) Methods for general and molecular bacteriology. American Society for Microbiology, Washington, D.C. 1994. P. 683-700).
Как видно из Фиг. 5, эффективность экстракции РНК из клеток микрококков хаотропной смесью с 0,1М аммиаком, по сравнению с контролем (смесь без аммиака), возрастает, по крайней мере, в 2-3 раза. Дальнейшее увеличение концентрации аммиака (до 0,2 и 0,3М) не приводит к заметному увеличению уровня экстракции РНК. Таким образом, полученные результаты убедительно показывают, что небольшое защелачивание хаотропной смеси с 7М мочевиной и 1% SDS значительно усиливает (ускоряет) экстракцию РНК из клеток обоих штаммов микрококков. Наличие гена ure+ или его отсутствие не влияет на экстракцию РНК. Результат. Клеточные оболочки, полученные с применением щелочных ХС на основе 7М мочевиной и 1% SDS весьма эффективны как матрицы в ПЦР.
Пример 7. Получение образцов геномной ДНК бактерий Bacillus cereus с прочной клеточной стенкой.
Клетки грамположительных бактерий Bacillus cereus выращивают на чашках с L-агаром при 37оС. 25 мкл сырой биомассы клеток суспендируют в 200 мкл щелочной ХС, содержащей 7М мочевину, 1% SDS, 5 мМ цитрат натрия и аммиак (0,1М), и в таком же объеме нейтральной ХС, без аммиака (контроль) и инкубируют на кипящей водяной бане 10 мин. Пробирки далее охлаждают на тающем льду, щелочные смеси нейтрализуют ацетатным буфером, затем клетки осаждают центрифугированием при 12000 об/мин в течение 3 мин. Супернатанты выбрасывают. Клеточные осадки дважды промывают водой и после второй промывки суспендируют в 200 мкл деионизованной воды и хранят в заморозке при -20оС.
По 1 мкл суспензии добавляют к 50 мкл смеси для ПЦР, содержащей соответствующий буфер, смеси 4-х dNTPs, праймерные олигонуклеотиды 27f + 1522r и термостабильную ДНК-полимеразу (Taq-полимеразу). Проводят ПЦР с учетом размера амплифицируемого фрагмента и структуры праймеров. Результат ПЦР – Наблюдают наработку фрагмента ДНК размером ~1,5 т.п.о.
Когда в реакционную смесь для ПЦР вносят по 1 мкл суспензии КО, число необходимых циклов 30. Когда КО разводят в 100, 1000 раз, то число циклов увеличивают до 33. Клеточные оболочки B. cereus, полученные с применением щелочных ХС с 7М мочевиной и 1% SDS весьма эффективны как матрицы в ПЦР.
Пример 8. Получение образцов геномной ДНК для ПЦР-амплификации из мицелия грибов Aspergilus fumigatus.
Используют штамм A. fumigatus RC032. Конидии выращивают 72 ч в жидкой среде Чапека при 37°C при непрерывном встряхивании. Биомассу (мицелий) собирают центрифугированием в 1,5 мл пробирках, используют более высокие обороты - (5мин при 15000 об/мин). Около 25 мкл биомассы суспендируют 1) в нейтральной хаотропной смеси (7М мочевина, 1% SDS, 25 мМ цитрат натрия, 0,1М МЭ), 2) в смеси того же состава, но с добавкой 0,1М аммиака, и 3) в смеси 5.1, содержащей 6 M GuHCl, 0,5% саркозила, 25 мМ цитрата натрия, 0,1 M ME. Полученные суспензии выдерживают 10 мин на кипятящей водяной бане. После этого пробирки охлаждают на тающем льду, щелочные смеси нейтрализуют кислым ацетатным буфером и все пробирки вместе центрифугируют при 15000 об/мин. Супернатанты переносят в чистые пробирки и анализируют содержание в них РНК (Фиг. 6).
Осадки мицеллия дважды промывают водой, суспендируют в 200 мкл стерильной деионизированной воды и затем осадки измельчают с помощью специального пестика (Eppendorf) до гомогенного состояния. Пробирки хранят в заморозке при -20оС. По 1 мкл гомогената добавляют к 50 мкл смеси для ПЦР, содержащей соответствующий буфер, смеси 4-х dNTPs, праймерные олигонуклеотиды 5,8S-R и LR3 и термостабильную ДНК-полимеразу (Taq-полимеразу). Проводят ПЦР с учетом размера амплифицируемого фрагмента и структуры праймеров.
Пример 9. Использование КО дрожжей для ПЦР
КО, полученные из клеток дрожжей S. cerevisiae 375, P. pastoris GS115, C. albicans 927, T. cutaneum 18, Cr. neoformans 3465, Rh. mucilaginosa 132, Cr. albidus 220 с применением щелочных смесей 1.1 и 1.2 с 0.1 М или 0.2 М аммиаком, сравнивают по способности быть источниками ДНК-матриц в ПЦР, (Фиг. 7А).
Синтез ПЦР-продуктов с использованием в качестве матриц КО, полученных из этих же микроорганизмов с использованием нейтральной ХC 6.0, которая характеризуется невысоким уровнем экстракции РНК (см. выше), происходит не во всех случаях (Фиг. 7B). Наработка ПЦР продукта происходит также при разведении суспензии клеточных оболочек в 1000 раз при небольшом увеличении числа термальных циклов до 35. Фиксируют, что ДНК-содержащие клеточные оболочки дрожжей, полученные при использовании щелочных хаотропных смесей на основе 7М мочевины и 1% SDS весьма эффективны как матрицы в ПЦР.
Таким образом, в ходе проведенных исследований выявляют несколько щелочных хаотропных смесей, обладающих способностью к высокоэффективной экстракции РНК, белков и низкомолекулярных компонентов из клеток микроорганизмов с прочной клеточной стенкой. Обязательными компонентами этих смесей являются 7М мочевина, 1% SDS и 0,1М аммиак. Дополнительными компонентами могут быть 0,1 М меркаптоэтанол – восстановитель дисульфидных связей, и 25 мМ цитрат Na в качестве хелатора. Высокую экстракционную эффективность демонстрирует также нейтральная хаотропная смесь следующего состава: 7М мочевина, 1% SDS, 25 мМ цитрат натрия и 0,1 М меркаптоэтанол.
Все эти смеси могут быть использованы для получения ДНК-содержащих клеточных оболочек из бактерий и дрожжей, пригодных для прямого использования в ПЦР.
В итоге предлагаем краткую процедуру получения ДНК-содержащих клеточных оболочек (КО) дрожжей и других микроорганизмов для непосредственного использования в ПЦР.
1. Берут микробиологической петлей одну дрожжевую колонию или небольшую часть колонии с чашки (или клетки, содержащиеся в 10-200 мкл жидкой дрожжевой культуры (OD600 = 0,4)).
2. Суспендируют клетки в 200 мкл хаотропной смеси, содержащей 7-8 М мочевины, 1% SDS, 25 мМ цитрата Na, 0,1М ME и 0,1M NH4OH. Инкубируют в течение 10 мин при 100oC.
3. Добавляют к смеси 18 мкл 3М ацетата натрия (рН 5,45), затем перемешивают для нейтрализации щелочи.
4. Суспензию центрифугируют и удаляют супернатант с РНК и белками при 15000 об/мин в течение 3 мин.
5. ДНК-содержащие клеточные оболочки дважды промывают 500 мкл стерильной деионизованной воды.
6. Осадок ДНК-содержащих клеточных оболочек суспендируют в 100 мкл стерильной деионизированной воды или TE.
7. Используют 1 мкл (или 1/10 мкл) суспензии клеточных оболочек для ПЦР.

Изобретение относится к области биотехнологии. Описана хаотропная смесь для обработки клеток дрожжей или грибов, а также грамположительных бактерий с прочной клеточной стенкой с целью получения образцов геномной ДНК и способ получения образцов геномной ДНК из клеток грамположительных бактерий, дрожжей или грибов с прочной клеточной стенкой для ПЦР-амплификации. Хаотропная смесь в одном варианте реализации содержит 7 М мочевины, 1% додецилсульфата натрия, 0,1-0,2 М аммиака. Изобретение расширяет арсенал средств для получения геномной ДНК, ассоциированной с прочными клеточными оболочками дрожжей или грибов и грамположительных бактерий. 2 н. и 1 з.п. ф-лы, 7 ил., 2 табл., 9 пр.
1. Хаотропная смесь для обработки клеток дрожжей или грибов, а также грамположительных бактерий с прочной клеточной стенкой с целью получения образцов геномной ДНК, содержащая 7 М мочевины, 1% додецилсульфата натрия, 0,1-0,2 М аммиака.
2. Хаотропная смесь по п.1, дополнительно содержащая 25 мМ цитрата натрия и 0,1 М меркаптоэтанола.
3. Способ получения образцов геномной ДНК из клеток грамположительных бактерий, дрожжей или грибов с прочной клеточной стенкой для ПЦР-амплификации, предусматривающий термальную обработку при температуре 95-100°С в течение 10 мин клеток хаотропной смесью по п.1 или 2, в результате которой денатурированная ДНК клеток упомянутых микроорганизмов остается внутри пермеабилизованных клеточных оболочек, а компоненты, затрудняющие проведение ПЦР, удаляются в ходе пробоподготовки.
| Nanasato Y | |||
| et al | |||
| Nuclear isolation and purification using SDS/urea (NIPSU) method for efficient and rapid extraction of high-purity genomic DNAs from Jatropha curcas L: A comparative analysis of DNA isolation protocols // African Journal of Biotechnology, 2018, 17(32), p | |||
| Устройство для избирательного управления с одного конца однопроводной линии несколькими реле | 1918 |
|
SU981A1 |
| Vingataramin L., Frost E.H | |||
| A single protocol for extraction of | |||
