УРОВЕНЬ ТЕХНИКИ
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[1] Настоящая заявка в целом относится к получению полипептидов, состоящих из модифицированных доменов α1-α2 лигандов NKG2D, которые специфически связываются с неприродным эктодоменом неприродного рецептора NKG2D, и при этом к модифицированным доменам α1-α2 лигандов NKG2D присоединены гетерологичные молекулы.
УРОВЕНЬ ТЕХНИКИ
NKG2D представляет собой активирующий рецептор, экспрессируемый в виде гомодимерного интегрального белка типа II на поверхности естественных киллерных (NK) клеток и некоторых Т-клеток. При связывании с одним из своих восьми природных лигандов, экспрессируемых в основном на поверхности поврежденных клеток, NKG2D активирует NK-клетку на уничтожение подверженной стрессу клетки, или, в случае Т-клеток, NKG2D, связанный с лигандом, костимулирует Т-клетку на выполнение своей эффекторной функции. Для эктодомена человеческого природного NKG2D, некоторых его растворимых природных лигандов и, в некоторых случаях, связанного комплекса растворимого лиганда с эктодоменом рецептора были получены трехмерные структуры. Мономерные домены α1-α2 лигандов NKG2D специфически связываются с двумя эктодоменами природного гомодимера NKG2D.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[2] Настоящее изобретение относится к модифицированным доменам α1-α2 лигандов NKG2D, присоединенным к гетерологичным молекулам, включая полипептиды, в некоторых вариантах осуществления, антитела или фрагменты антител. Изобретение также относится к модифицированным формам рецептора NKG2D, сконструированным для обеспечения комбинации повышенного и пониженного связывания с неприродными и природными версиями лигандов NKG2D, соответственно.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
[3] Фиг. 1a. Выравнивание природного эктодомена NKG2D.wt (SEQ ID NO: 17) с неприродными вариантами NKG2D.YA (SEQ ID NO: 18) и NKG2D.AF (SEQ ID NO: 25). Показаны положения Y152 и Y199, и серым цветом выделены мутированные остатки, присутствующие в неприродных вариантах.
[4] Фиг. 1В. Выравнивание домена α1-α2 ULBP2 природного/дикого типа (SEQ ID NO: 4) и неприродных вариантов ULBP2, включая ULBP2.R80W (SEQ ID NO: 108). Серым цветом выделены остатки, важные для связывания неприродных вариантов ULBP2 с неприродными рецепторами NKG2D.YA или NKG2D.AF. Указаны положения остатка R80, а также область M154-F159, которая была изучена в отношении связывания ортоганальных вариантов с NKG2D.YA (ULBP2.S3, SEQ ID NO: 127) или NKG2D.AF (ULBP2.C, SEQ ID NO: 111; ULBP2.R, SEQ ID NO: 113; ULPB2.AA, SEQ ID NO: 115; и ULBP2.AB, SEQ ID NO: 117).
[5] Фиг. 2. Краткая хараткристика кандидатов, мутаций в вариантах неприродного Fc-eNKG2D и свойств агрегации белков, определенных с помощью эксклюзионной хроматографии (SEC), как показано на фиг.3 и 4.
[6] Фиг. 3. Сравнение результатов эксклюзионной хроматографии неприродных слитых белков Fc-NKG2D, проанализированных на колонке Akta HiLoad 16/600 Superdex 200. Миграция правильно собранного материала проиллюстрирована дискретным симметричным пиком, который элюируется при более высоких объемах, в то время как агрегированный материал элюируется раньше при более низких объемах. Участок и природа модификаций обозначены номерами аминокислот Y152, Y199 или обоими (SEQ ID NO: 48, 43, 42, 58, 41 и 40, начиная с верхней части фигуры).
[7] Фиг. 4. Профили эксклюзионной хроматографии неприродных вариантов Fc-eNKG2D с одной или двумя аминокислотными заменами анализировали на колонке Akta Superdex 200 Increase 10/300 GL. Миграция правильно собранного материала проиллюстрирована дискретным симметричным пиком, элюируемым при более высоких объемах, в то время как агрегированный материал, характеризующийся широким пиком или серией пиков с низкой амплитудой, элюируется при более низких объемах. Буквы в скобках представляют аминокислоты в положениях 152 и 199 (SEQ ID NO: 57, 56, 55, 54, 53, 52, 51, 50, 49, 48, 47, 46, 45, 44 и 40 в порядке сверху вниз), соответственно.
[8] Фиг. 5. Процент насыщения (Rmax) вариантов eNKG2D, нормализованный по связыванию NKG2D дикого типа с MICwed-MicAbody или MIC25-MicAbody. Fc-NKG2D дикого типа и каждый рецептор Fc-eNKG2D иммобилизовали на биосенсорах AHC, а затем подвергали воздействию трастузумаб-специфических MicAbody при 20 нМ. Отслеживали кинетику диссоциации, и ранжировали значения Rmax слитых Fc-eNKG2D. Непротестированные образцы (nt) были результатом либо сильной агрегации, либо недостаточного количества материала, экспрессированного или выделенного после фракционирования SEC.
[9] Фиг. 6. Оценка методом ELISA связывания ULBP2 дикого типа, MICwed- и MIC25-ритуксимаб MicAbody с кандидатами Fc-eNKG2D. Ключ указан в верхней части фигуры, но поскольку многие кривые перекрываются, на каждом графике кривые также помечены индивидуально.
[10] Фиг. 7. Значения EC50 (нМ) для проанализированных методом ELISA Fc-eNKG2D, показанных на фиг.4. nt=не тестировали; nb=связывание отсутствует или связывание очень низкое даже при 300 нМ, поэтому значение EC50 не вычисляли.
[11] Фиг. 8. Связывание вариантов eNKG2D с лигандами дикого типа. Лиганды дикого типа (все в формате Fc-слитых белков) иммобилизовали на биосенсорах Octet AHC, и каждый из природных NKG2D, NKG2D.Y152A или eNKG2D5 (Y152A/Y199F) в виде Fc-слитого белка титровали от 300 нМ до 0,41 нМ. Максимальные реакции связывания оценивали количественно с помощью системы Octet. (Следует обратить внимание на разные ординаты на каждом графике).
[12] Фиг. 9. Подмножество комбинаторных мутаций в ULBP2, которые привели к получению фаговых клонов с селективным связыванием с NKG2D.AF по сравнению с природным NKG2D.wt, оцененное методом точечного ELISA. Мутанты были ранжированы по частоте появления среди выбранных фагов.
[13] Фиг. 10. Титрование ELISA отдельных вариантов фага для подтверждения селективного связывания с Fc-NKG2D.AF и сниженного уровня или отсутствия связывания с Fc-NKG2D.wt. Мутации подробно описаны на фиг. 11.
[14] Фиг. 11. Специфичность связывания NKG2D.AF-селективных вариантов ULBP2 в формате ритуксимаб-MicAbody с NKG2D.AF сохранилась согласно результатам количественного определения с помощью ELISA. Показаны конкретные аминокислотные модификации каждого варианта ULBP2, а также отношение их связывания со слитым белком Fc-NKG2D.wt относительно связывания со слитым белком Fc-NKG2D.AF. Расположение аминокислотных остатков ULBP2 показано на фиг. 1B.
[15] Фиг. 12. Выбранные мутации в указанных аминокислотных положениях ULBP2.R80W (Фиг. 1В; SEQ ID NO: 108), которые привели к образованию Y152A-специфических фаговых клонов.
[16] Фиг. 13. Данные ELISA для четырех неприродных вариантов α1-α2 ULBP2 в формате MicAbody, связывающихся с NKG2D.wt, NKG2D.YA и NKG2D.AF. Варианты Fc-NKG2D использовали в качестве агентов захвата. MicAbody титровали и детектировали с помощью HRP-конъюгированного с антитела к человеческой каппа-цепи.
[17] Фиг. 14: ULBP2.C (SEQ ID NO: 111), ULBP2.R (SEQ ID NO: 113), ULBP2.AA (SEQ ID NO: 115) и ULBP2.AB (SEQ ID NO: 117) изучали на изменение иммуногенности пептид-MHCI относительно ULBP2 дикого типа (SEQ ID NO: 4) с помощью сервера NetMHC4.0 путем запроса на представителей супертипов HLA. Для входной последовательности вводили вариабельную область (остатки 154-159 в соответствии с выравниванием на фиг. 1B) для каждого варианта вместе с девятью остатками, расположенными выше и ниже (всего 24 остатка), и исследовали 9-мерные пептидные окна на предмет предсказанной иммуногенности. Темно-серые прямоугольники соответствуют пептидам, которые, по прогнозам, с большой вероятностью связываются с карманом MHCI (определенные как имеющие % ранга <0,5) и, следовательно, присутствуют с высокой вероятностью. Светло-серые прямоугольники соответствуют слабым, согласно прогнозу, связующим (% ранга <2). Дополнительные сведения см. в тексте описания примера 5.
[18] Фиг. 15: ULBP2.C (SEQ ID NO: 111), ULBP2.R (SEQ ID NO: 113), ULBP2.AA (SEQ ID NO: 115) и ULBP2.AB (SEQ ID NO: 117) изучали на изменение иммуногенности комплекса пептид-MHC класса II по сравнению с ULBP2 дикого типа (SEQ ID NO: 4) с помощью сервера NetMHCII 2.3 путем запроса на HLA-DR, HLA-DQ, HLA-DP. Для входной последовательности вводили вариабельную область (остатки 154-159 согласно выравниванию на фиг. 1B) для каждого варианта вместе с 15 остатками, расположенными выше и ниже (всего 36 остатков), и исследовали 15-мерные пептидные окна на предмет прогнозируемой иммуногенности. Темно-серые прямоугольники соответствуют пептидам, которые, по прогнозам, с большой вероятностью связываются с карманом MHCII и, следовательно, могут быть присутствовать и быть иммуногенными. Светло-серые прямоугольники соответствуют слабым, согласно прогнозу, связующим.
[19] Фиг. 16A и 16B. Измеренное с помощью ELISA связывание ритуксимаб-MicAbody, состоящих из ULBP2.wt (дикого типа), ULBP2.R80W (который имеет повышенную сродство к NKG2D дикого типа), ULPB2.S3 (NKG2D.YA-селективный ортогональный вариант) или ULBP2.R (NKG2D.AF-селективный ортогональный вариант) с природным NKG2D.wt, NKG2D.YA и NKG2D.AF. Фиг. 16А: кривые ELISA. Снижение поглощения при 458 нм в некоторых анализах при более высоких концентрациях является артефактом, который часто наблюдается при взаимодействиях с высоким сродством при более высоких концентрациях из-за осаждения реагента для проявки TMB-Ultra ELISA. Фиг. 16B: Значения EC50 (выраженные в нМ), определенные с помощью GraphPad Prism на основе кривых, приведенных на фиг. 16A. nd=не определено из-за отсутствия связи между повышенной концентрацией и связыванием.
[20] Фиг. 17A, 17B и 17C. Цитолитические in vitro анализы с использованием эффекторных клеток CD8, которые были либо нетрансдуцированы, либо трансдуцированы конструкциями CAR NKG2D.wt, NKG2D.YA или NKG2D.AF, состоящими из шарнирного/трансмембранного домена CD8a и внутриклеточных сигнальных доменов 4-1BB и CD3-дзета. Клетки-мишени предварительно загружали кальцеином и подвергали действию эффекторных клеток при увеличивающемся соотношении эффектор-мишень (E:T). Количество высвобожденного кальцеина определяли через пять часов. Фиг. 17А: лизис клеток HeLa. Фиг. 17B: лизис клеток HeLa, трансфицированных для сверхэкспрессии поверхностного ULBP1. Фиг. 17C: цитолиз клеток HeLa, экспрессирующих на своей поверхности неприродный ULBP2.R. Планки погрешностей соответствуют стандартному отклонению технических повторов в эксперименте.
[21] Фиг. 18A и 18B. MicAbody направлял цитолиз опухолевых линий посредством CD8 NKG2D-CAR-T-клеток. Фиг. 18А: Клетки Ramos, экспрессирующие CD20, мишень ритуксимаба, предварительно загружали кальцеином и подвергали воздействию NKG2D.AF- или NKG2D.YA-CAR клеток при соотношении E:T=20:1 вместе с возрастающими концентрациями либо ULBP2.S2, либо ULBP2.R ритуксимаб-Micabody. Уровень цитолиза количественно определяли через два часа совместной инкубации. Фиг. 18B: линию мышиной опухоли CT26, трансфицированную для экспрессии человеческого Her2, использовали в качестве мишени для цитолиза параллельно с клетками Ramos. CD8 NKG2D.AF-CAR-T-клетки предварительно загружали насыщающей концентрацией (5 нМ) ритуксимаба-ULBP2.R, трастузумаба-ULBP2.R или эквимолярной смесью двух MicAbody. Несвязанные MicAbody удаляли промывкой, и к клеткам-мишеням добавляли CD8 клетки при двух различных соотношениях E:T. Цитолиз измеряли через два часа.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[22] Естественные киллерные (NK) клетки и некоторые (CD8+ αβ и γδ) Т-клетки иммунной системы играют важную роль у людей и других млекопитающих в качестве врожденного (защиты первой линии) иммунитета от неопластических и инфицированных клеток (Cerwenka, A., and L.L. Lanier. 2001. NK cells, viruses and cancer. Nat. Rev. Immunol. 1:41-49). NK-клетки и некоторые T-клетки демонстрируют на своих поверхностях NKG2D, выступающий гомодимерный, поверхностный иммунорецептор, ответственный за распознавание клетки-мишени и активацию врожденной защиты против патологической клетки (Lanier, LL, 1998. NK cell receptors. Ann. Rev. Immunol. 16:359-393; Houchins JP et al. 1991. DNA sequence analysis of NKG2, a family of related cDNA clones encoding type II integral membrane proteins on human NK cells. J. Exp. Med. 173:1017-1020; Bauer, S et al., 1999. Activation of NK cells and T cells by NKG2D, a receptor for stress-inducible MICA. Science 285:727-730). Молекула человеческого NKG2D содержит лектин-подобный внеклеточный (экто-) домен С-типа, который связывается с восемью различными родственными лигандами; наиболее изученные лиганды представляют собой имеющие на 84% идентичные или гомологичные последовательности мономерные MICA и MICB полиморфные аналоги MHC (главный комплекс гистосовместимости)-подобных гликопротеинов (MIC) класса I (Weis et al. 1998. The C-type lectin superfamily of the immune system. Immunol. Rev. 163:19-34; Bahram et al. 1994. A second lineage of mammalian MHC class I genes. PNAS 91:6259-6263; Bahram et al. 1996a. Nucleotide sequence of the human MHC class I MICA gene. Immunogenetics 44: 80-81; Bahram and Spies TA. 1996. Nucleotide sequence of human MHC class I MICB cDNA. Immunogenetics 43:230-233). Непатологическая экспрессия MICA и MICB ограничена кишечным эпителием, кератиноцитами, эндотелиальными клетками и моноцитами, но аберрантная поверхностная экспрессия этих белков MIC происходит в ответ на многие типы клеточного стресса, такие как пролиферация, окисление и тепловой шок, и маркирует клетку как патологическую (Groh et al. 1996. Cell stress-regulated human MHC class I gene expressed in GI epithelium. PNAS 93:12445-12450; Groh et al. 1998. Recognition of stress-induced MHC molecules by intestinal γδ T cells. Science 279:1737-1740; Zwirner et al. 1999. Differential expression of MICA by endothelial cells, fibroblasts, keratinocytes and monocytes. Human Immunol. 60:323-330). Патологическая экспрессия белков MIC также, по-видимому, происходит при некоторых аутоиммунных заболеваниях (Ravetch, JV and Lanier LL. 2000. Immune Inhibitory Receptors. Science 290:84-89; Burgess, SJ. 2008. Immunol. Res. 40:18-34). Дифференциальная регуляция лигандов NKG2D, таких как полиморфные MICA и MICB, важна для обеспечения иммунной системы средствами идентификации и реагирования на широкий спектр аварийных сигналов, защищая при этом здоровые клетки от нежелательной атаки (Stephens HA, (2001) MICA and MICB genes: can the enigma of their polymorphism be resolved? Trends Immunol. 22:378-85; Spies, T. 2008. Regulation of NKG2D ligands: a purposeful but delicate affair. Nature Immunol. 9:1013-1015).
[23] Вирусная инфекция является обычным индуктором экспрессии белка MIC и идентифицирует инфицированную вирусом клетку для NK- или T-клеточной атаки (Groh et al. 1998; Groh et al. 2001. Co-stimulation of CD8+ αβT cells by NKG2D via engagement by MIC induced on virus-infected cells. Nat. Immunol. 2:255-260; Cerwenka, A., and L.L. Lanier. 2001). Фактически, чтобы избежать такой атаки на клетку-хозяина, цитомегаловирус и другие вирусы разработали механизмы, предотвращающие экспрессию белков MIC на поверхности клетки, которую они инфицируют, чтобы избежать нацеливания системы врожденного иммунитета (Lodoen, M., K. Ogasawara, J.A. Hamerman, H. Arase, J.P. Houchins, E.S. Mocarski, and L.L. Lanier. 2003. NKG2D-mediated NK cell protection against cytomegalovirus is impaired by gp40 modulation of RAE-1 molecules. J. Exp. Med. 197:1245-1253; Stern-Ginossar et al., (2007) Host immune system gene targeting by viral miRNA. Science 317:376-381; Stern-Ginossar et al., (2008) Human microRNAs regulate stress-induced immune responses mediated by the receptor NKG2D. Nature Immunology 9:1065-73; Slavuljica, I A Busche, M Babic , M Mitrovic, I Gašparovic, Đ Cekinovic, E Markova Car, EP Pugel, A Cikovic, VJ Lisnic, WJ Britt, U Koszinowski, M Messerle, A Krmpotic and S Jonjic. 2010. Recombinant mouse cytomegalovirus expressing a ligand for the NKG2D receptor is attenuated and has improved vaccine properties. J. Clin. Invest. 120:4532-4545).
[24] Экспрессия белков MIC также индуцируется на многих опухолевых клетках, присутствие которых может сделать их чувствительными к нацеливанию и лизису NK-клетками. Неудивительно, что неспособность экспрессировать белок MIC представляет собой один из способов, с помощью которого многие злокачественные клетки, такие как клетки рака легких и глиобластомы мозга, могут ускользать от врожденной иммунной системы (Busche, A et al. 2006, NK cell mediated rejection of experimental human lung cancer by genetic over expression of MHC class I chain-related gene A. Human Gene Therapy 17:135-146; Doubrovina, ES, MM Doubrovin, E Vider, RB Sisson, RJ O’Reilly, B Dupont, and YM Vyas, 2003. Evasion from NK Cell Immunity by MHC Class I Chain-Related Molecules Expressing Colon Adenocarcinoma (2003) J. Immunology 6891-99; Friese, M. et al. 2003. MICA/NKG2D-mediated immunogene therapy of experimental gliomas. Cancer Research 63: 8996-9006; Fuertes, MB, MV Girart, LL Molinero, CI Domaica, LE Rossi, MM Barrio, J Mordoh, GA Rabinovich and NW Zwirner. (2008) Intracellular Retention of the NKG2D Ligand MHC Class I Chain-Related Gene A in Human Melanomas Confers Immune Privilege and Prevents NK Cell-Mediated Cytotoxicity. J. Immunology, 180:4606-4614).
[25] Определена структура с высоким разрешением человеческого MICA, связанного с NKG2D, которая демонстрирует, что домен α3 MICA не имеет прямого контакта с NKG2D (Li et al. 2001. Complex structure of the activating immunoreceptor NKG2D and its MHC class I-like ligand MICA. Nature Immunol. 2:443-451; Protein Data Bank accession code 1HYR). Домен α3 MICA, как и MICB, связан с доменом α1-α2-платформы посредством короткого гибкого линкерного пептида и сам естественно позиционируется как «спейсер» между платформой и поверхностью клетки, экспрессирующей MIC. Трехмерные структуры доменов α3 человеческих MICA и MICB почти идентичны (среднеквадратичное расстояние <1Å на 94 C-αα) и функционально взаимозаменяемы (Holmes et al. 2001. Structural Studies of Allelic Diversity of the MHC Class I Homolog MICB, a Stress-Inducible Ligand for the Activating Immunoreceptor NKG2D. J Immunol. 169:1395-1400).
[26] Описаны некоторые неприродные домены α1-α2 лигандов NKG2D, модифицированные для связывания человеческих природных рецепторов NKG2D с более высоким сродством, чем природные домены α1-α2 (Candice S. E. Lengyel, Lindsey J. Willis, Patrick Mann, David Baker, Tanja Kortemme, Roland K. Strong and Benjamin J. McFarland. Mutations Designed to Destabilize the Receptor-Bound Conformation Increase MICA-NKG2D Association Rate and Affinity. Journal of Biological Chemistry Vol. 282, no.42, pp.30658-30666, 2007; Samuel H. Henager, Melissa A. Hale, Nicholas J. Maurice, Erin C. Dunnington, Carter J. Swanson, Megan J. Peterson, Joseph J. Ban, David J. Culpepper, Luke D. Davies, Lisa K. Sanders, and Benjamin J. McFarland. Combining different design strategies for rational affinity maturation of the MICA-NKG2D interface. Protein Science 2012 VOL.21:1396-1402). В настоящей заявке описаны неприродные домены α1-α2 лигандов NKG2D, которые модифицированы для связывания неприродных рецепторов NKG2D, которые сам по себе мутированы по двум специфическим сайтам, каждый из которых по отдельности приводит к нарушению или потере связывания со всеми известными в настоящее время природными доменами α1-α2 лигандов человеческого NKG2D (David J. Culpepper, Michael K. Maddox, Andrew B. Caldwell, and Benjamin J. McFarland. Systematic mutation and thermodynamic analysis of central tyrosine pairs in polyspecific NKG2D receptor interactions. (Mol Immunol. 2011 January; 48(4):516-523).
[27] В соответствии с настоящим изобретением созданы биспецифические молекулы, состоящие из (а) специфически модифицированных неприродных доменов α1-α2 и (b) специфически нацеливающих гетерологичных молекул, включая, без ограничения, гетерологичные пептиды или полипептиды. Биспецифические молекулы могут связываться с химерными рецепторами антигена (CAR), причем рецептор CAR состоит из неприродного эктодомена рецептора NKG2D, который связывается с родственными модифицированными доменами α1-α2 с более высоким сродством, чем с природными доменами α1-α2. Вторая специфичность биспецифической молекулы заключается в гетерологичном компоненте, который специфически связывается со своей соответствующей молекулой-мишенью. Генно-модифицированные клетки системы иммунитета, состоящие из таких CAR и родственных биспецифических молекул, могут, таким образом, преодолеть многие недостатки, включая известные серьезную системную токсичность, ускользание антигена, а также ограниченную и неконтролируемую персистентность современных терапевтических средств на основе CAR-T и CAR-NK клеток (Kalos M, Levine, BL, Porter, DL, Katz, S, Grupp, SA, Bagg, A and June, C. T Cells with chimeric antigen receptors have potent antitumor effects and can establish memory in patients with advanced leukemia. Sci Transl Med 2011; 3:95ra73; Morgan RA, Yang JC, Kitano M, Dudley ME, Laurencot CM, Rosenberg SA. Case report of a serious adverse event following the administration of T cells transduced with a chimeric antigen receptor recognizing ERBB2. Mol Ther 2010, 18:843-851).
[28] Т-клетки и NK-клетки могут быть модифицированы с помощью технологий переноса генов для прямой и стабильной экспрессии на поверхности этих клеток трансмембранных сигнальных рецепторов, которые придают новые антигенные специфичности Т-клеткам благодаря связывающим доменам, полученным из антител (Saar Gill & Carl H. June. Going viral: chimeric antigen receptor T cell therapy for hematological malignancies. Immunological Reviews 2015. Vol. 263:68-89; Wolfgang Glienke, Ruth Esser, Christoph Priesner, Julia D. Suerth, Axel Schambach, Winfried S. Wels, Manuel Grez, Stephan Kloess, Lubomir Arseniev and Ulrike Koehl. 2015. Advantages and applications of CAR-expressing natural killer cells. Front. Pharmacol. doi: 10.3389/fphar.2015.00021). Такие Т-клетки, экспрессирующие химерный рецептор антигена (CAR-T-клетки), применяются в подходе, в котором домен распознавания специфического антитела объединен с внутриклеточным доменом цепи CD3-ζ, представляющим собой основной передатчик сигналов от эндогенных T-клеточных рецепторов антигена (TCR), в один химерный белок вместе с последовательностью цитоплазматического полипептида из костимулирующей молекулы, такой как CD27, CD28, ICOS, 4-1BB или OX40. Созданные таким образом CAR могут запускать активацию Т-клеток при связывании антигена-мишени аналогично эндогенному Т-клеточному рецептору, но независимо от главного комплекса гистосовместимости (MHC).
[29] В контексте настоящей заявки термины «растворимый белок MIC», «растворимый белок MICA» и «растворимый белок MICB» относятся к белку MIC, содержащему домены α1, α2 и α3 белка MIC, но без трансмембранных или внутриклеточных доменов. Лиганды NKG2D, ULBP1-6, в природе не содержат домен α3 (Cerwenka A, Lanier LL. 2004. NKG2D ligands: unconventional MHC class I-like molecules exploited by viruses and cancer. Tissue Antigens 61 (5):335-43. doi:10.1034/j.1399-0039.2003.00070.x. PMID 12753652). Термин «домен α1-α2» лиганда NKG2D относится к домену белка лиганда, который связывается с рецептором NKG2D.
[30] В некоторых вариантах осуществления изобретения домены α1-α2 неприродных белков-лигандов NKG2D по изобретению по меньшей мере на 80% идентичны или гомологичны нативному или природному домену α1-α2 лиганда NKG2D (SEQ ID NO: 1-9 для MICA, MICB, ULBP1, ULBP2, ULBP3, ULBP4, ULBP5, ULBP6 и OMCP, соответственно). В других вариантах осуществления модифицированный домен α1-α2 по меньшей мере на 85% идентичен нативному или природному домену α1-α2 лиганда NKG2D. В других вариантах осуществления модифицированный домен α1-α2 по меньшей мере на 90% идентичен нативному или природному домену α1-α2 природного белка-лиганда NKG2D и связывается с неприродным NKG2D.
[31] Предпочтительно модифицированные или неприродные домены α1-α2 неприродных белков MIC по настоящему изобретению по меньшей мере на 80% идентичны или гомологичны нативному или природному домену α1-α2 OMPC (SEQ ID NO: 9) или одному из восьми известных человеческих белков-лигандов NKG2D (SEQ ID NO: 1-8) и связываются с неприродным эктодоменом NKG2D. В некоторых вариантах осуществления неприродный домен α1-α2 по меньшей мере на 85% идентичен нативному или природному домену α1-α2 белка-лиганда NKG2D и связывается с неприродным NKG2D. В других вариантах осуществления неприродный домен α1-α2-платформы по меньшей мере на 90%, 95%, 96%, 97%, 98% или 99% идентичен нативной или природной платформе α1-α2 человеческого природного белка с доменом α1-α2 и связывается с неприродным NKG2D.
[32] В некоторых вариантах осуществления гетерологичная молекулярная метка может быть слита с N-концом или C-концом неприродного домена α1-α2 растворимого белка MIC для облегчения очистки растворимого белка MIC. Последовательности метки включают пептиды, такие как полигистидин, myc-пептид, метку FLAG, стрептавидин-подобную метку или малую молекулу, такую как биотин. Такие метки можно удалить после выделения молекулы MIC методами, известными специалистам в данной области.
[33] В других вариантах осуществления изобретения в доменах α1-α2 лигандов NKG2D могут быть сделаны специфические мутации для создания неприродных доменов α1-α2, которые связываются с неприродными рецепторами NKG2D, которые, в свою очередь, сконструированы так, чтобы они имели пониженное сродство к природным лигандам NKG2D. Это можно сделать, например, с помощью генной инженерии. Модифицированный таким образом неприродный рецептор NKG2D может быть использован для создания на поверхности NK-клеток, Т-клеток или других клеток иммунной системы химерного рецептора антигена (CAR) на основе NKG2D, который предпочтительно может связываться и активироваться молекулами, состоящими из неприродных доменов α1-α2 по изобретению. Эти пары неприродных рецепторов NKG2D и родственных им неприродных лигандов NKG2D по изобретению позволяют обеспечить важные преимущества в вопросах безопасности, эффективности и производства для лечения рака и вирусных инфекций по сравнению с существующими CAR-T-клетками и CAR-NK-клетками, как описано ниже.
[34] Конструирование Т-клеток с помощью CAR стало многообещающим подходом к адоптивной Т-клеточной терапии рака, и в качестве терапевтических средств от злокачественных новообразований на CAR-T-клетках были протестированы CAR, нацеленные на множество различных молекул (Porter DL, Levine BL, Kalos M, Bagg A, June CH., 2011. Chimeric antigen receptor-modified T cells in chronic lymphoid leukemia. N Engl J Med. 365:725-733). Несмотря на хорошую клиническую эффективность, наблюдаемую у нескольких сотен пациентов, получавших адоптивный перенос Т-клеток, экспрессирующих CD19-специфические химерные рецепторы антигена, процессы индивидуальной разработки CAR для нацеливания на конкретный антиген, выделения аутологичных Т-клеток из пациента, генетической инженерии аутологичных Т-клеток для экспрессии персонализированного CAR, размножения модифицированных клеток in vitro и контроля качества их производства являются обременительными и дорогостоящими.
[35] После введения аутологичных CAR-T-клеток обратно пациенту-донору, их размножение in vivo невозможно контролировать, и эффективность созданной таким образом, так называемой «живой терапии», нельзя оценить по показателю доза-ответ (Gill & June, 2015). Кроме того, из-за потери антигена опухолевыми клетками может происходить ускользание опухоли от CAR-T-клеток (Stephan A. Grupp, M.D., Ph.D., Michael Kalos, Ph.D., David Barrett, M.D., Ph.D., Richard Aplenc, M.D., Ph.D., David L. Porter, M.D., Susan R. Rheingold, M.D., David T. Teachey, M.D., Anne Chew, Ph.D., Bernd Hauck, Ph.D., J. Fraser Wright, Ph.D., Michael C. Milone, M.D., Ph.D., Bruce L. Levine, Ph.D., and Carl H. June, M.D. Chimeric Antigen Receptor-Modified T Cells for Acute Lymphoid Leukemia. N Engl J Med 2013; 368:1509-1518). Хотя этот путь ускользания может быть устранен последовательной терапией с использованием CAR-T-клеток, нацеленных на разные мишени, или начальной инфузией продукта Т-клеток, содержащих CAR с двумя или более специфичностями, такие этапы связаны со значительными дополнительными затратами на производственные процессы, контроль качества и увеличением стоимости.
[36] В дополнение к CAR-T-клеткам, нацеленным на опухоли посредством одноцепочечных связывающих доменов антител (scFv), на животных, а недавно и на людях, были изучены CAR-T-клетки, в которых используется лиганд-связывающий домен рецептора NKG2D (Sentman CL, Meehan KR. NKG2D CARs as cell therapy for cancer. Cancer J. 2014 Mar-Apr; 20(2):156-9. doi: 10.1097/PPO.0000000000000029; Manfred Lehner, Gabriel Götz, Julia Proff, Niels Schaft, Jan Dörrie, Florian Full, Armin Ensser, Yves A. Muller, Adelheid Cerwenka, Hinrich Abken, Ornella Parolini, Peter F. Ambros, Heinrich Kovar, Wolfgang Holter. Redirecting T Cells to Ewing's Sarcoma Family of Tumors by a Chimeric NKG2D Receptor Expressed by Lentiviral Transduction or mRNA Transfection. Research Article | published 15 Feb 2012 | PLOS ONE 10.1371/journal.pone.0031210; www.clinicaltrials.gov NCT02203825). Поскольку экспрессия лиганда NKG2D повышается на поверхности подвергшихся стрессу клеток, таких как опухолевые клетки, это семейство природных лигандов NKG2D представляет значительный интерес в качестве мишеней для иммунотерапии рака (Spear P, Wu MR, Sentman ML, Sentman CL. NKG2D ligands as therapeutic targets. Cancer Immun. 2013 May 1; 13:8.; Song DG, Ye Q, Santoro S, Fang C, Best A, Powell DJ Jr., Chimeric NKG2D CAR-expressing T cell-mediated attack of human ovarian cancer is enhanced by histone deacetylase inhibition. Hum Gene Ther. 2013 Mar; 24(3):295-305). Один NKG2D-CAR представлял собой слияние полноразмерного рецептора NKG2D с CD3ζ (NKG2Dζ); другой содержал только эктодомен NKG2D, слитый с каркасом CAR второго поколения, состоящим из трансмембранного и внутриклеточного доменов от CD28 и сигнального домена CD3ζ (NKG2D28ζ). Поскольку активация NKG2D зависит от наличия DAP10, также была сконструирована CAR-T-клетка, в которой DAP10 коэкспрессировался NKG2Dζ (NKG2Dζ10). Т-клетки, экспрессирующие любой из вышеуказанных NKG2D-CAR, продуцировали IFNɣ и TNFα в ответ на стимуляцию лигандом NKG2D и эффективно убивали in vitro опухолевые мишени, экспрессирующие лиганды NKG2D (Heather VanSeggelen, Joanne A. Hammill, Anna Dvorkin-Gheva, Daniela G.M. Tantalo, Jacek M. Kwiecien, Galina F. Denisova, Brian Rabinovich, Yonghong Wan, Jonathan L. Bramson, T cells engineered with chimeric antigen receptors targeting NKG2D ligands display lethal toxicity in mice, Molecular Therapy 2015 Oct; 23(20):1600-10; doi:10.1038/mt.2015.119). Цитотоксический потенциал NK-клеток против широкого спектра подтипов опухолей также может быть заметно усилен за счет экспрессии CAR на основе NKG2D-DAP10-CD3ζ (Yu-Hsiang Chang, John Connolly, Noriko Shimasaki, Kousaku Mimura, Koji Kono, and Dario Campana. Chimeric Receptor with NKG2D Specificity Enhances Natural Killer Cell Activation and Killing of Tumor Cells. Cancer Res; 73(6) March 15, 2013).
[37] Однако после инфузии сингенным мышам-хозяевам наблюдали значительную токсичность этих CAR-T-клеточных конструкций, которые связываются и активируются природными лигандами природного рецептора NKG2D. Признаки токсичности, включая плохое состояние организма, сутулую осанку, затрудненное дыхание и снижение внутренней температуры тела, наблюдали у мышей с опухолью и без опухолей, получавших NKG2D-CAR-T-клетки, по сравнению с необработанными контрольными мышами. Тяжесть токсичности NKG2D-CAR-T-клеток варьировала: NKG2Dζ10 был сильно токсичным, NKG2D28ζ проявлял промежуточную токсичность, а NKG2Dζ был переносимым. Клинические симптомы токсичности и смертности усугублялись, когда мыши получали химиотерапию до адоптивного переноса Т-клеток, экспрессирующих любой из NKG2D-CAR (VanSeggelen et al. 2015). Известно, что химиотерапия и облучение индуцируют лиганды NKG2D на здоровых тканях (Xiulong Xu, Geetha S Rao, Veronika Groh, Thomas Spies, Paolo Gattuso, Howard L Kaufman, Janet Plate and Richard A Prinz. Major histocompatibility complex class I-related chain A/B (MICA/B) expression in tumor tissue and serum of pancreatic cancer: Role of uric acid accumulation in gemcitabine-induced MICA/B expression. BMC Cancer 2011, 11:194 doi:10.1186/1471-2407-11-194; Gannagé M, Buzyn A, Bogiatzi SI, Lambert M, Soumelis V, Dal Cortivo L, Cavazzana-Calvo M, Brousse N, Caillat-Zucman Induction of NKG2D ligands by gamma radiation and tumor necrosis factor-alpha may participate in the tissue damage during acute graft-versus-host disease. Transplantation. 2008 Mar 27; 85(6):911-5. doi: 10.1097/TP.0b013e31816691ef.). Дальнейшая характеристика показала, что токсичность совпадает с системным цитокиновым штормом и летальным уровнем воспаления в легких. Эти данные являются предупреждением того, что следует проявлять крайнюю осторожность при использовании природных лигандов NKG2D в таргетной иммунотерапии, и демонстрируют, что экспрессия CAR Т-клетками, нацеленными на такие лиганды, которые особенно сильно активируют CAR, может оказаться пагубной in vivo (VanSeggelen et al. 2015).
[38] CAR-T-клетки или CAR-NK-клетки, состоящие из эктодоменов неприродных рецепторов NKG2D, которые не связывают природные лиганды NKG2D или демонстрируют слабое связывание, не будут подвергаться вышеуказанной активации и, следовательно, не будут такими токсигенными, как клетки, экспрессирующие CAR на основе природного рецептора NKG2D. Кроме того, эктодомены неприродных рецепторов NKG2D на клетках не будут подавляться природными лигандами NKG2D в растворимом формате или на миелоидных супрессорных клетках (MDSC) (Deng W, Gowen BG, Zhang L, Wang L, Lau S, Iannello A, Xu J, Rovis TL, Xiong N, Raulet DH, 2015. Antitumor immunity. A shed NKG2D ligand that promotes natural killer cell activation and tumor rejection. Science. 2015 Apr 3; 348(6230):136-9. doi: 10.1126/science.1258867. Epub 2015 Mar 5). Однако, при связывании таких CAR клеток, несущих эктодомены неприродных рецепторов NKG2D, с биспецифическими молекулами с родственными неприродными доменами α1-α2 по настоящему изобретению и гетерологичным нацеливающим мотивом, который нашел и связал свою целевую мишень, CAR активируются и проявятся эффекторные функции CAR-клеток.
[39] Поскольку высвобождение цитокинов и цитолитическая активность CAR-T-клеток или CAR-NK-клеток, содержащих эктодомены неприродного рецептора NKG2D, не инициируются, за исключением случаев связывания с биспецифической молекулой, состоящей из родственного неприродного домена α1-α2, причем биспецифическая молекула также связана матрицей своей мишени, их активацию можно контролировать введенными биспецифическими молекулами, которые, в качестве биофармацевтических агентов, будут демонстрировать фармакокинетику и фармакодинамику, хорошо известные в данной области. В случае развития нежелательного явления врач может просто изменить режим дозирования введенной биспецифической молекулы вместо того, чтобы задействовать индуцированный механизм самоубийства для разрушения введенных CAR-клеток, как это делается в настоящее время (Monica Casucci and Attilio Bondanza. Suicide Gene Therapy to Increase the Safety of Chimeric Antigen Receptor-Redirected T Lymphocytes. J Cancer. 2011; 2:378-382). Кроме того, такие биспецифические молекулы с различными специфическими целевыми мотивами можно вводить одновременно или последовательно, что позволяет преодолеть резистентность опухоли и ускользание в результате потери целевого антигена без необходимости создания, размножения и инфузии множества различных аутологичных CAR-клеток (Gill & June, 2015). Поскольку все конструкции CAR могут быть идентичными для всех CAR клеток, а специфичность нацеливания определяется просто нацеливающим мотивом вводимой биспецифической молекулы по настоящему изобретению, это позволит упростить процессы производства CAR-клеток и сделать их менее дорогостоящими.
[40] Примеры родительских или реципиентных белков или полипептидов, которые являются кандидатами для присоединения к неприродным доменам α1-α2 лигандов NKG2D, включают, без ограничения, антитела, белки, состоящие из складок Ig или доменов Ig, включая модифицированные Fc-домены, которые рекрутируют природные молекулы или неспособны рекрутировать или связывать природные молекулы, глобулины, белки, фибронектины и домены фибронектина, интегрины, флуоресцентные белки, ферменты, белки внешней мембраны, рецепторные белки, рецепторы Т-клеток, рецепторы химерных антигенов, вирусные антигены, вирусные капсиды, вирусные лиганды для клеточных рецепторов - гистоны, гормоны, цитокины и модифицированные цитокины, такие как интерлейкины, ноттины, циклические пептиды или полипептиды, белки семейства главного комплекса гистосовместимости (MHC), белки MIC, лектины и лиганды лектинов. К модифицированным доменам α1-α2 лигандов NKG2D также можно присоединить небелковые молекулы, такие как полисахариды, дендримеры, полигликоли, пептидогликаны, антибиотики и поликетиды.
[41] Таким образом, настоящее изобретение позволяет расширить разнообразие и практичность этого замечательного, очень многообещающего иммунологического подхода к лечению рака с помощью CAR-T-клеток, CAR-NK-клеток и CAR-макрофагоподобных клеток, преодолевая при этом многие из признанных в настоящее время трудностей.
[42] В контексте настоящей заявки термины «пептид», «полипептид» и «белок» используются взаимозаменяемо; и «гетерологичная молекула», «гетерологичный пептид», «гетерологичная последовательность» или «гетерологичный атом» представляет собой молекулу, пептид, нуклеиновую кислоту или аминокислотную последовательность или атом, соответственно, которые в природе или обычно не встречаются в физическом соединении с рассматриваемой молекулой. В контексте настоящего описания «неприродный» и «модифицированный» используются взаимозаменяемо. В контексте настоящего описания «естественный», «нативный» и «дикий тип» используются взаимозаменяемо, и термины «NKG2D» и «рецептор NKG2D» используются взаимозаменяемо. Термин «антитело» в настоящем описании используется в самом широком смысле и, в частности, охватывает моноклональные антитела, полиспецифические антитела (например, биспецифические антитела) и фрагменты антител, если они проявляют требуемую биологическую активность. «Фрагменты антитела» включают часть антитела, предпочтительно содержащую его антигенсвязывающую область. Примеры фрагментов антител включают Fab, Fab', F(ab')2, Fv фрагменты и вставляемые Fv-фрагменты; диатела; линейные антитела; молекулы одноцепочечных антител; и полиспецифические антитела, образованные из фрагмента(ов) антитела. «Фрагменты антител» могут также включать Fc-часть антитела.
[43] Термин «содержащий», используемый взаимозаменяемо с терминами «включающий», «состоящий из» или «характеризующийся», является инклюзивным или открытым и не исключает дополнительных, неперечисленных элементов или этапов способа. Фраза «состоящий из» исключает любой элемент, этап или ингредиент, не указанные в пункте формулы изобретения. Фраза «по существу состоящий из» ограничивает объем формулы изобретения указанными материалами или этапами, и тем, что существенно не влияет на основные и новые характеристики заявленного изобретения. Настоящее раскрытие предполагает варианты осуществления композиций и способов по изобретению, соответствующих объему каждой из этих фраз. Таким образом, композиция или способ, содержащие перечисленные элементы или этапы, предполагают конкретные варианты осуществления, в которых композиция или способ, по существу, состоят из этих элементов или этапов или состоят из них.
[44] Все ссылки, цитируемые в настоящей заявке, полностью включены в настоящее описание посредством ссылки, независимо от того, были ли они включены ранее специально или нет. Используемое в настоящем описании единственное число предполагает включение как единственного, так и множественного числа.
[45] После полного описания изобретения специалисты в данной области техники поймут, что то же самое может быть выполнено в широком диапазоне эквивалентных параметров, концентраций и условий без отступления от сущности и объема изобретения и без чрезмерного экспериментирования. Хотя настоящее изобретение описано в связи с конкретными вариантами осуществления, следует понимать, что оно допускает дальнейшие модификации. Настоящая заявка предназначена для охвата любых вариантов, применений или адаптаций изобретения, следуя, в общем, принципам изобретения и включая такие отклонения от настоящего раскрытия, которые входят в известную или обычную практику в области техники, к которой относится изобретение, и могут быть применены к указанным выше существенным признакам.
ПРИМЕРЫ
Модифицированный эктодомен рецептора NKG2D и модифицированные домены α1-α2 лигандов NKG2D
[46] Пример 1. Модификация тирозина 152 на аланин (Y152A) и тирозина 199 на фенилаланин (Y199F) человеческого рецептора NKG2D для создания инертного эктодомена NKG2D
[47] Было показано, что мутации тирозина 152 или тирозина 199 в человеческом NKG2D, эквивалентные положениям 75 и 122 эктодомена NKG2D (фиг. 1A, SEQ ID NO:17), могут существенно снизить связывание с природным лигандом, MICA (David J. Culpepper, Michael K. Maddox, Andrew B. Caldwell, and Benjamin J. McFarland. Systematic mutation and thermodynamic analysis of central tyrosine pairs in polyspecific NKG2D receptor interactions. Mol Immunol. 2011 January; 48(4):516-523). Авторами изобретения было сделано предположение, что, хотя мутация любого остатка тирозина сильно влияет на способность NKG2D связываться со своими природными лигандами, одновременная мутация как тирозина 152 (Y152), так и тирозина 199 (Y199) фактически может привести к исчезновению способности рецептора связываться со всеми нативными лигандами. Поэтому были изучены индивидуальные и комбинаторные замены Y152 и Y199, которые были охарактеризованы с точки зрения биохимического поведения с целью идентификации как одиночных, так и двойных мутантных вариантов, неспособных связываться с какими-либо природными лигандами. Те варианты, которые также хорошо экспрессировались и собирались, представляли особый интерес, как указывающие на инертные лиганды, которые могут быть более легко получены для анализа.
[48] Природный эктодомен NKG2D (дикого типа) (NKG2D.wt, SEQ ID NO: 17) и неприродные варианты-кандидаты эктодомена NKG2D (SEQ ID NO: 18-35), также называемые «сконструированный NKG2D» или «eNKG2D», клонировали в виде слитого белка с C-концом Fc человеческого IgG1 (без Fab-доменов) через короткий линкер Ile-Glu-Gly-Arg, узнаваемый фактором Ха (SEQ ID NO: 38), также (взаимозаменяемо) называемый Fc-NKG2D.wt или NKG2D.wt и Fc-eNKG2D или eNKG2D (SEQ ID NO: 40-58). Фрагменты ДНК gBlocks® (Integrated DNA Technologies, Сан-Диего, Калифорния), соответствующие сигнальной последовательности MHCI (SEQ ID NO: 36 и 37), Fc человеческого IgG1 с линкером (SEQ ID NO: 39) и варианты эктодомена NKG2D (SEQ ID NO: 59-77) синтезировали и вставляли в pD2610-V12 (ATUM, Ньюарк, Калифорния). Конструкции ДНК для изучения замен Y152, Y199 или комбинации Y152/Y199 мутаций (фиг. 2), временно экспрессировали в клетках Expi293TM (ThermoFisher Scientific, Waltham, MA), и секретируемый белок очищали с помощью аффинной хроматографии с белком A (номер по каталогу 20334, Pierce Biotechnology, Рокфорд, Иллинойс). Элюированный материал характеризовали с помощью эксклюзионной хроматографии (SEC) на колонках Akta Pur Superdex, и правильно собранный материал соответствующего размера фракционировали и выделяли из пиков агрегатов перед включением в анализы.
[49] SEC-характеристика очищенного слитого белка NKG2D.Y199A-Fc выявила состав преимущественно агрегированного материала (фиг. 3). Для сравнения, как природный слитый Fc-NKG2D, так и слитый материал Fc-NKG2D.Y152A отличался дискретным, неагрегированным пиком, который можно было легко отличить от более быстро мигрирующего агрегата. Влияние мутации Y199A на агрегацию также было очевидным в варианте слияния Fc-NKG2D с двойной мутацией Y152A/Y199A, свидетельствующее о том, что она имеет преимущественное влияние на неправильную укладку белка (фиг. 3). Таким образом, получение материала, необходимого для последующих структурных изменений становится проблематичным в случае включения Y199A в варианты NKG2D в комбинации с любой мутацией Y152, и вызывает сомнения в вопросах сборки и представления на поверхности клетки. Как следствие, была предпринята попытка изучения других замен Y152 и Y199, которые можно было бы объединить для получения более устойчивой молекулы. Комбинаторные мутанты eNKG2D по Y152 и Y199 исследовали в виде слитых белков с Fc и подробно описали (фиг. 2). Кроме того, с помощью SEC получали профили всех очищенных и экспрессированных слитых бклков-кандидатов Fc-eNKG2D, хроматограммы которых выявили различные уровни образования агрегатов (Фиг. 2-4). Из изученных одиночных аминокислотных замен в остатке 152, аланин, серин, треонин и валин не влияли на сборку молекулы Fc-NKG2D, хотя замена Y152-лейцин (Y152L) давала сильно агрегированный материал. Подобно аланину, ни глутамат, ни аспартат не были толерантными в положении 199, хотя фенилаланин лишь незначительно увеличивал образование агрегатов. Из изученных комбинаций мутации Y152A/Y199F, Y152S/Y199F, Y152T/Y199F и Y152F/Y199F не оказывали отрицательного воздействия на образование требуемого димера, тогда как другие комбинации приводили к усилению агрегации (фиг. 2-4).
[50] Пример 2: Создание биспецифических молекул на основе антител, «MicAbody», с вариантами неприродного лиганда NKG2D.
[51] Для создания неприродных вариантов MicA, слитых с человеческим IgG1, полинуклеотиды ДНК, кодирующие домены α1-α2, например MICwed (SEQ ID NO: 79) и MIC25 (SEQ ID NO: 81), амплифицировали с помощью ПЦР, используя праймеры, которые также вводили полинуклеотид, кодирующий либо линкер APTSSSGGGGS для слияния с C-концевой легкой каппа-цепью (SEQ ID NO: 84), либо линкер GGGS для слияния с C-концевой тяжелой цепью человеческого IgG1 (SEQ ID NO: 82). Кроме того, в домен CH2 тяжелой цепи вводили две мутации - D265A/N297A (нумерация по Кабат), которые уменьшают связывание со всеми рецепторами FcγR, тем самым устраняя функцию антителозависимой клеточной цитотоксичности (ADCC) (Shields et al., 2001 JBC, 276:6591-6604). Полинуклеотид, кодирующий домен α1-α2 ULBP2 дикого типа (ULBP2.wt), не содержащий GPI-связь (SEQ ID NO: 12), клонировали аналогичным образом, и осуществляли слияние с полинуклеотидами ДНК, кодирующими линкеры и тяжелую цепь или легкую цепь IgG1. Эти биспецифические антитела, называемые «MicAbodyTM», являются бивалентными по отношению к слитому домену α1-α2. Примеры антител, используемых для создания MicAbody для изучения конструкций eNKG2D включают, без ограничения, трастузумаб (SEQ ID NO: 94 и 96) и ритиксумаб (SEQ ID NO: 98 и 100), далее упоминаемые как «трастузумаб-MicAbody» (например, SEQ ID NO: 102 и 104) и «ритуксимаб-MicAbody» (например, SEQ ID NO: 106), соответственно. Слитые конструкции вставляли индивидуально в pD2610-V12 (ATUM, Ньюарк, Калифорния), используя метод клонирования по Гибсону (New England Biolabs Inc., Ипсвич, Массачусетс). Для заданного антитела, распознающего специфический антиген, плазмиду, кодирующую тяжелую цепь, и плазмиду, кодирующую легкую цепь, слитую с природным или неприродным лигандом NKG2D, котрансфицировали для временной экспрессии в клетках Expi293TM (ThermoFisher Scientific, Waltham, MA). Альтернативно, котрансфицировали плазмиду, кодирующую тяжелую цепь, слитую с природным или неприродным лигандом NKG2D, и плазмиду, кодирующую легкую цепь. Секретированные биспецифические антитела очищали с помощью аффинной хроматографии с протеином А (номер по каталогу 20334, Pierce Biotechnology, Rockford, IL), элюированный материал характеризовали с помощью эксклюзионной хроматографии (SEC) на колонках Akta Pur Superdex, и по мере необходимости выполняли фракционирование. Кроме того, выполняли SDS-PAGE анализ очищенных образцов для проверки ожидаемых молекулярных масс слитых видов тяжелой цепи и легкой цепи.
[52] Пример 3: Идентификация вариантов модифицированного NK2GD, неспособных связываться ни с природными NKG2D-связывающими лигандами, ни с неприродными лигандами, с усиленным связыванием с NKG2D дикого типа
[53] Сродство связывания α1-α2 вариантов с внеклеточными доменами природных (дикого типа) белков NKG2D и неприродных белков eNKG2D анализировали с помощью метода ELISA. Каждый из SEC-фракционированных слитых белков Fc-природный NKG2D и Fc-неприродный eNKG2D наносили на отдельные лунки 96-луночных планшетов Nunc Maxisorp в течение ночи при 4°C (Thermo Fisher Scientific, Waltham, MA) в концентрации покрытия 1 мкг/мл в фосфатно-солевом буфере (PBS). Планшеты трижды промывали PBS/0,05% Твин-20 (PBS-T) при 20-22°C и блокировали 0,5% бычьим сывороточным альбумином в PBS (PBS-B) в течение 2 часов при 20-22°C. MicAbody титровали против связанных с планшетом слитых белков Fc с природным или неприродным NKG2D в течение 60 минут при 20-22°C в PBS/0,5% бычий сывороточный альбумин (BSA)/0,05% Твин-20 (PBS-BT), промывали PBS-T 3 раза при 20-22°C, и связанные биспецифические белки детектировали с помощью HRP-конъюгированного антитела к человеческой каппа-цепи в PBS-BT (Abcam, Cambridge MA) и проявляли, используя субстратный раствор 1-StepTM Ultra TMB ELISA (Thermo Fisher Scientific, Уолтем, Массачусетс). Связывание с ULBP2.wt ритуксимаб-MicAbody (SEQ ID NO: 98 и 106) различалось между вариантами NKG2D дикого типа и eNKG2D, последний из которых имел более низкий уровень связывания, и варианты лигандов MICwed (SEQ ID NO: 96 и 102) и MIC25 (SEQ ID NO: 96 и 104) оказались более сильными при идентификации вариантов eNKG2D, у которых отсутствовало связывание лиганда. Связывание каждого варианта eNKG2D со всеми тремя биспецифическими лигандами выявило комбинации модификаций NKG2D, которые обеспечили наиболее низкое связывания лигандов дикого типа и вариантных лигандов, что позволило отобрать инертные варианты NKG2D.
[54] Дополнительный биофизический анализ связывания вариантов eNKG2D с лигандами также выполняли с помощью биослойной интерферометрии (BLI) с помощью системы FortéBio Octet (все FortéBio LLC, Fremont, CA). Для этих экспериментов у компании R&D Systems, Inc. (Миннеаполис, Миннесота) приобретали лиганды человеческого NKG2D: MICA-Fc, MICB-Fc, ULBP1-Fc, ULBP2-Fc, ULBP3-Fc и ULBP4-Fc. Лиганды в формате MicAbody связывали с Fc человеческого IgG (AHC) на наконечниках биосенсора. После установления базовых уровней наконечники подвергали серии титрований слитых белков Fc-eNKG2D в диапазоне от 300 нМ до 0,41 нМ, и на всех этапах, выполняемых в PBS-BT, отслеживали кинетику ассоциации/диссоциации. Затем слитые белки Fc-eNKG2D захватывали наконечниками AHC, и MicAbody титровали для характеристики кинетики связывания.
[55] Для определения максимального ответа, определяемого по степени связывания природного NKG2D либо с MICwed, либо с MIC25, слитые белки Fc-природный NKG2D захватывали биосенсорами AHC, и 20 нМ трастузумаб-MICwed или 20 нМ трастузумаб-MIC25 инкубировали в течение двух минут и затем в течение 30 секунд наблюдали кинетику диссоциации. После этого в тех же условиях выполняли анализ связывания со слитыми с Fc рецепторами eNKG2D, в качестве захватывающего агента, и уровень связывания для каждого eNKG2D оценивали в виде процента от максимального ответа связывания, установленного для Fc-NKG2D.wt (фиг. 5). Для MICwed ответы всех вариантов Fc-eNKG2D с одной мутацией, за исключением Y199F, были снижены до 50%. Y199F сохранял 100% ответ связывания. Однако все варианты Fc-eNKG2D с двумя мутациями полностью утратили связывание с MICwed. В случае MIC25, все варианты Fc-eNKG2D с одной мутацией и вариант Y152V/Y199F сохранили 100% ответ связывания относительно связывания Fc-NKG2D дикого типа. Однако связывание было снижено до 50% у нескольких вариантов Fc-eNKG2D с двумя мутациями, включая Y152A/Y199F, Y152S/Y199F и Y152T/Y199F.
[56] Анализ ELISA слитых белков Fc-eNKG2D в качестве захватывающих агентов выполняли для ULBP2.wt, MICwed, MIC25 в формате MicAbody, титрование которых начинали с 300 нМ (фиг. 6). По возможности значения EC50 рассчитывали с помощью GraphPad Prism (фиг. 7). Природный NKG2D связывался с ULBP2, MICwed и MIC25 в формате MicAbody со сродством, вычисленным в виде значения Kd, равном 1,4, 0,007 и 0,005 нМ, соответственно. Хотя у всех кандидатов eNKG2D с одной мутацией сродство к ULBP2 и MICwed в формате MicAbody было снижено, связывание кандидатов eNKG2D с MIC25 сохранялось. Однако у всех кандидатов eNKG2D с двумя мутациями связывание со всеми тремя лигандами, ULBP2, MICwed и MIC25, в форматах Micabody отсутствовало либо было существенно снижено.
[57] Согласно результатам анализов Octet и ELISA, связывание вариантов eNKG2D eNKG2D5 (Y152A/Y199F), eNKG2D7 (Y152S/Y199F), eNKG2D8 (Y152T/Y199F) и eNKG2D9 (Y152V/Y199F) с ULBP, MICwed, MIC25 в формате Micabody было снижено или отсутствовало (фиг. 5 и 7). Кроме того, eNKG2D 5, 7 и 8 имели продемонстрировали наименьшую степень агрегации, что свидетельствует о более надежной сборке белка при экспрессии 293T (фиг. 2). Связывания с eNKG2D5 (SEQ ID NO: 48) с лигандами дикого типа в виде MicAbody, захваченными на наконечниках Octet AHC подвергли более тщательному изучению. Fc-NKG2D.Y152A с одной мутацией (SEQ ID NO: 41) продемонстрировал пониженное связывание со всеми природными лигандами по сравнению с природным NKG2D (SEQ ID NO: 40) (фиг. 8). Кривая ответа на связывание с eNKG2D5 (Y152A/Y199F) была еще более низкой по сравнению с Y152A eNKG2D. eNKG2D5 (Y152A/Y199F, далее обозначаемый как «AF» или «NKG2D.AF») был выбран в качестве ведущего варианта NKG2D, для которого создавали родственные селективные, ортогональные, неприродные лиганды.
[58] Пример 4: Конструирование ортогональных неприродных доменов α1-α2 с селективным связыванием с неприродным эктодоменом NKG2D.AF
[59] Для создания ортогональных неприродных доменов α1-α2, которые проявляют селективное связывание с рецептором Fc-NKG2D AF (SEQ ID NO: 48), использовали фаговый дисплей. В качестве отправной точки был выбран неприродный домен α1-α2 ULBP2.R80W (фиг. 1B; SEQ ID NO: 108) с высоким сродством к природному эктодомену NKG2D (NKG2D.wt) дикого типа в качестве родительского домена для дальнейшего мутагенеза и скрининга библиотеки фагового дисплея. Для домена α1-α2 ULBP2.R80W (SEQ ID NO: 108), который имеет дополнительную мутацию C8S, устраняющую возможность образования дисульфидных связей, создавали библиотеки синтетической ДНК. Кодоны аминокислотных остатков лиганда, которые в связанном состоянии расположены в непосредственной близости к положениям Y152 и Y199 на природном рецепторе NKG2D, заменяли кодонами NNK; библиотеки состояли из кодонов NNK в положениях 154-159 (фиг. 1B; SEQ ID NO: 110). Библиотеки клонировали в виде слитых белков с минорным белком оболочки pIII фага M13, и фаговые частицы, демонстрирующие мутагенизированные варианты домена α1-α2, продуцировали в клетках SS320 E.coli, используя стандартные методы (Andris-Widhopf, J., Steinberger, P., Fuller, R., Rader, C., and Barbas, C. F., 3rd. (2011)). Эти библиотеки α1-α2 фагового дисплея сортировали по силе сродства связывания с неприродным рецептором NKG2D.AF путем селективного захвата фаговых клонов, связанных с биотинилированным белком Fc-NKG2D.AF в присутствии небиотинилированного природного белка-конкурента Fc-NKG2D.wt. Селективные клоны обогащали путем циклического выполнения нескольких раундов конкурентного отбора с увеличивающимися концентрациями небиотинилированного природного Fc-NKG2D.
[60] После четырех раундов отбора фаговые клоны индивидуально распределяли в 96-луночный планшет, выполняли точечный ELISA для проверки предпочтительного дифференциального связывания со связанным с планшетом неприродным NKG2D.AF по сравнению с NKG2D.wt. Связанные фаги обнаруживали с помощью биотинилированного моноклонального антитела E1 к белку оболочки фага M13 (ThermoFisher Scientific, Уолтем, Массачусетс), детектирования с помощью HRP-стрептавидина (R&D Systems, Миннеаполис, Миннесота) и одноэтапной проявки с помощью Ultra TMB ELISA (ThermoFisher Scientific, Уолтем, Массачусетс). Сигнал пятна ELISA для каждого клона выражали в виде отношения связывания фага с NKG2D.AF к связыванию фага с NKG2D.wt. Эти фаги с соотношением больше или равным 14 секвенировали для идентификации специфических мутаций в мутагенизированных областях NNK. На фиг.9 показаны выбранные аминокислотные остатки для каждого варианта фага α1-α2, который селективно связывается с NKG2D.AF. В случаях, когда были идентифицированы несколько клонов, представляющих одну и ту же последовательность, строили график отношения сигналов ELISA, и проверяли совместимость фаговых клонов путем кластеризации точек данных (данные не показаны).
[61] Тридцать вариантов, идентифицированных в ELISA, размножали в отдельных монокультурах для получения микропартий с высоким титром фага. Концентрации очищенных фагов нормализовали до OD268=0,5, затем подвергали серии разведений 1:3 против связанного с планшетом Fc-NKG2D.AF или Fc-NKG2D.wt с детектированием фага и выполнением ELISA, как описано выше. Все тридцать вариантов, проанализированных таким образом, последовательно демонстрировали сопоставимое связывание с NKG2D.AF с незначительным связыванием или отсутствием связывания с NKG2D.wt (фиг. 10) даже при самых высоких концентрациях проанализированного фага. Выбранные фаги также показали сдвиг в концентрации фага на log 2 или более для достижения полумаксимального связывания, между NKG2D.AF и NKG2D.wt.
[62] Для подтвердждения того, что NKG2D.AF-селективные варианты домена α1-α2 сохраняют специфические связывающие свойства в формате слитых антител, клонировали 21 вариант (фиг. 11; например, SEQ ID NO: 111-118) в виде белка с линкером APTSSSGGGGS, слитого на С-конце с легкой цепью антитела ритуксимаба (SEQ ID NO: 119-126). Полученные слитые белки клонировали в вектор экспрессии млекопитающих pD2610-V12 (ATUM, Ньюарк, Калифорния) методом клонирования Гибсона (New England Biolabs Inc., Ипсвич, Массачусетс) и коэкспрессировали с тяжелой цепью родительского антитела (SEQ ID NO: 99) в виде парных полных антител IgG. Временную экспрессию осуществляли в клетках Expi293TM (ThermoFisher Scientific, Уолтем, Массачусетс) в соответствии с протоколом производителя и очищали с помощью стандартной аффинной хроматографии с протеином A (номер по каталогу 20334, Pierce Biotechnology, Rockford, IL). ELISA, измеряющий связывание каждого варианта слитых антител ULBP2 α1-α2 с неприродным Fc-NKG2D.AF и с природным Fc-NKG2D.wt, продемонстрировал значительно более высокое сродство связывания с NKG2D.AF по сравнению с природным NKG2D.wt (фиг. 11). В совокупности эти данные продемонстрировали получение неприродных ортогональных доменов α1-α2 по изобретению, которые обладают высоким сродством связывания с неприродным рецептором NKG2D.AF и существенно сниженным сродством связывания с природным рецептором NKG2D. Кроме того, слитые белки этих ортогональных доменов α1-α2 с полипептидами антител сохраняют свои свойства селективного связывания и могут использоваться, например, в контексте Т-клеток с химерным рецептором антигена (CAR) для перенаправления неприродных рецепторов NKG2D.AF на специфические антигены.
[63] Пример 5: Идентификация неприродных лигандов NKG2D, которые могут различать варианты неприродных рецепторов NKG2D путем селективного связывания одного или другого
[64] Фаговый дисплей для создания ортогональных неприродных доменов α1-α2 с селективным связыванием рецептора NKG2D.Y152A (далее называемым NKG2D.YA) выполняли с неприродным доменом α1-α2 ULBP2.R80W (SEQ ID NO: 108) в качестве отправной точки, как описано выше. Библиотеки фагового дисплея α1-α2 сортировали методом пеннинга по высокому сродству связывания с неприродным рецептором Fc-NKG2D.YA путем селективного захвата фаговых клонов, связанных с биотинилированным Fc-NKG2D.YA (SEQ ID NO: 41) в присутствии небиотинилированного природного белка-конкурента Fc-NKG2D.wt (SEQ ID NO: 40). Дополнительная работа по проверке фаговых клонов привела к идентификации вариантов с преимущественным связыванием с Fc-NKG2D.YA по сравнению с Fc-NKG2D.wt (Фиг. 12). ULBP2.S3 (SEQ ID NO: 127), например, стабильно демонстрировал селективное связывание, согласно результатам ELISA и анализа Octet (как в формате слитого His-меченного моноспецифического, так и биспецифического антитела), с неприродным NKG2D.YA по сравнению с природным NKG2D.wt. Была получена другая форма неприродных ортогональных доменов α1-α2 по изобретению, обладающих высоким сродством связывания с неприродными рецепторами NKG2D (в данном случае NKG2D.YA в отличие от NKG2D.AF, как в Примере 4). Кроме того, слитые конструкции ортогональных доменов α1-α2 с полипептидами антител сохраняли свои свойства селективного связывания и использовались для селективного перенаправления неприродных рецепторов NKG2D на специфические молекулы, определяемые слитыми гетерологичными пептидами, такими как антитела.
[65] Для определения, имеются ли различия между двумя неприродными вариантами рецепторов: неприродным доменом α1-α2 с селективным связыванием с NKG2D.YA (ULBP2.S3, SEQ ID NO: 127) и неприродным доменом α1-α2 с селективным связыванием с NKG2D.AF, выполняли титрование ELISA. Все 21 из выбранных вариантов α1-α2, которые связывались с NKG2D.AF, напрямую сравнивали с NKG2D.AF относительно связывания с NKG2D.YA. Из них четыре продемонстрировали неспособность связывать NKG2D.wt, сильное сродство к NKG2D.AF и существенно сниженное связывание (в 15-20 раз) или отсутствие связывания с NKG2D.YA по сравнению с NKG2D.AF (фиг. 13). Эти четыре неприродных варианта ULBP2 α1-α2: ULBP2.C, ULBP2.R, ULBP2.AA и ULBP2.AB (SEQ ID NO: 111, 113, 115 и 117), также изучали на предмет изменений прогнозируемого профиля иммуногенности относительно последовательности пептида ULBP2 дикого типа (SEQ ID NO: 4) с помощью сервера NetMHC4.0 (с запросом по связыванию пептидов с MHC класса I относительно всех представителей HLA супертипа с помощью 9-мерного пептидного анализа; http://www.cbs.dtu.dk/services/NetMHC/) и NetMHCII 2.3 Server (с запросом по связыванию пептидов с MHC класса II относительно гаплотипов HLA-DR, HLA-DQ, HLA-DP с помощью 15-мерного пептидного анализа; http://www.cbs.dtu.dk/services/NetMHCII/), оба алгоритма разработаны Техническим университетом Дании (http://www.bioinformatics.dtu.dk/; Andreatta M and Nielsen M, Gapped sequence alignment using artificial neural networks: application to the MHC class I system, 2016 Bioinformatics, 32:511, PMID:26515819; Jensen KK, Andreatta M, Marcatili P, Buus S, Greenbaum JA, Yan Z, Sette A, Peters B, and Nielsen M, Improved methods for predicting peptide binding affinity to MHC class I molecules, 2018 Immunology, PMID:29315598). Мутации, включенные в ULBP2.C, ULBP2.R и ULBP2.AB, не увеличивали прогнозируемую иммуногенность, в то время как иммуногенность ULPB2.AA была слегка увеличена для нескольких гаплотипов (фиг. 14 и 15). Вследствие специфичности ULBP2.R к NKG2D.AF и отсутствия предсказуемой иммуногенности для дальнейшего анализа ELISA был выбран ULBP2.R для непосредственного сравнения его поведения связывания с поведением ULBP2.S3 (NKG2D.YA-селективный неприродный ортогональный лиганд), ULBP2.R80W (неприродный лиганд с повышенным сродством к NKG2D дикого типа) и ULBP2 дикого типа (ULBP2.wt). Связывание четырех реагентов ритуксимаб-MicAbody (SEQ ID NO: 98 и 121, 98 и 129, 131 и 100, 98 и 106 как тяжелой цепи, так и легкой цепи для ULBP2.R, ULBP2.S3, ULBP2.R80W и ULBP2.wt, соответственно) тестировали относительно NKG2D дикого типа (NKG2D.wt) и двух инертных неприродных вариантов NKG2D.YA и NKG2D.AF (Фиг. 16A и 16B). Данные продемонстрировали, что NKG2D.YA-селективный вариант ULBP2.S3 в формате MicAbody связывается с высоким сродством с NKG2D.YA, но не взаимодействует с NKG2D.AF или природным NKG2D. Кроме того, NKG2D.AF-селективный вариант ULBP2.R в формате MicAbody связывался с высоким сродством с NKG2D.AF, но не взаимодействовал с NKG2D.YA или природным NKG2D. Эти результаты продемонстрировали высокий потенциал исследования оси NKG2D-MIC лиганда и разработки уникальных пар новых селективных неприродных рецепторов NKG2D и их соответствующих родственных неприродных партнеров по связыванию лигандов MIC.
[66] Пример 6: Таргетирующая и убивающая активность CAR-T-клеток, экспрессирующих неприродный эктодомен NKG2D.AF, контролируется ортогональными доменами α1-α2, слитыми с гетерологичными нацеливающими полипептидами
[67] Средства для селективного контроля терапии CAR-T-клетками очень востребованы для снижения токсичности и повышения эффективности в борьбе с опухолями (Gill and June, op cit). Ранее были предприняты попытки по разработке CAR с использованием эктодомена CD16, который затем можно было бы связать через Fc-домен терапевтических моноклональных антител, что позволило бы контролировать нацеливание CAR-T на основе антител (Chang et al., Op cit). Однако CAR-T-клетки на основе CD16 могут распознавать практически все молекулы эндогенных антител в крови и тканях, и терапевтические антитела, используемые для контроля этих клеток, будут сталкиваться с конкуренцией со стороны эндогенных рецепторов CD16 на NK-клетках, PMN, моноцитах и макрофагах. Обе эти особенности увеличивают проблемы, связанные с токсичностью вне опухоли и плохой фармакокинетикой, соответственно.
[68] Природные лиганды NKG2D присутствуют в некоторых здоровых тканях и многих тканях, подвергшихся стрессу, что создает чрезвычайный риск токсического воздействия при использовании современных подходов к разработке NKG2D-CAR (VanSeggelen et al. 2015). Неприродный рецептор Y152A-NKG2D, специфически связывающийся с неприродным доменом α1-α2 лигандов NKG2D, представляет собой пример средства, с помощью которого можно селективно контролировать активность неприродного NKG2D-CAR с использованием биспецифических белков, состоящих из неприродным доменом α1-α2 лигандов NKG2D.
[69] Конструировали CAR-T-клетки с рецептором, состоящим из Y152A/Y199F-модифицированного эктодомена («AF») NKG2D, который лишен способности связываться со всеми природными лигандами NKG2D, или ранее описанных неприродных доменов α1-α2, ортогональных и родственных Y152A-модифицированному NKG2D (NKG2D.YA). Родственные неприродные α1-α2 домены по изобретению связываются с высоким сродством с неприродным эктодоменом NKG2D.AF и лишены связывания с природными эктодоменами NKG2D и с эктодоменом NKG2D.YA. Таким образом, сконструированные домены α1-α2, которые проявляют высокую селективность в отношении неприродного эктодомена NKG2D.AF по сравнению с природным NKG2D и неприродным NKG2D.YA, представляют собой идеальную систему для селективного контроля неприродных рецепторов NKG2D-CAR или любого рецептора или белка, слитого с неприродными эктодоменами NKG2D, которые могут быть селективно связаны с неприродными доменами α1-α2 по настоящему изобретению. Настоящее изобретение также позволяет экспрессировать в отдельных клетках два разных CAR, один из которых состоит из NKG2D.YA, а другой - из NKG2D.AF, причем каждый из них передает сигналы с помощью четко различающихся внутриклеточных доменов. Эти различающиеся CAR могут обеспечить независимый двойной контроль активности клетки путем внеклеточного воздействия соответствующими родственными ортогональными MicAbody или другими слитыми полипептидами, не являющимися антителами.
[70] Для демонстрации селективного контроля CAR-T-клеток, сконструированных с использованием химерного рецептора, используя неприродный эктодомен NKG2D.AF, создавали CAR либо с природным NKG2D.wt (SEQ ID NO: 135), неприродным NKG2D.YA (SEQ ID NO: 137), либо неприродным эктодоменом NKG2D.AF (SEQ ID NO: 139) на основе предыдущей работы, используя конструкции 4-1BB/CD3-дзета CAR (патент Campana 8,399,645), в которых соответствующие эктодомены NKG2D сливали с CD8 шарнирной областью CAR (SEQ ID NO: 151, 153, 155). Эти конструкции (SEQ ID NO: 152, 154, 156) клонировали в лентивирусный вектор и экспрессировали в первичных человеческих CD8-положительных Т-клетках путем трансдукции лентивируса. Клетки HeLa имеют постоянно повышенные уровни лигандов MIC на своей поверхности, включая MICA, MICB, ULBP3 и ULBP2/5/6 (антитело, используемое для подтверждения этого, не способно различать эти три ULBP; человеческое антитело ULBP-2/5/6, R&D Systems, Миннеаполис, Миннесота). Клетки HeLa трансфицировали также для сверхэкспрессии на своей поверхности либо природного ULBP1, либо NKG2D.AF-селективного варианта ULBP2.R, и эти клетки использовали в качестве мишени для анализа гибели in vitro. Клетки-мишени HeLa предварительно загружали кальцеином и подвергали воздействию NKG2D.wt-CAR, NKG2D.YA-CAR или NKG2D.AF-CAR CD8 клетками при увеличивающемся соотношении эффектор/мишень (E:T) в течение пяти часов, после чего определяли количество кальцеина, высвобожденного в супернатант, и нормализовали по общему количеству кальцеина, высвободившемуся после обработки детергентом (Фиг. 17A-17C). Из-за повышенных уровней лигандов MIC, естественным образом экспрессируемых на поверхности клеток HeLa, клетки CD8, экспрессирующие природный NKG2D (NKG2D.wt) в качестве CAR, взаимодействовали с клетками HeLa через этот сверхэкспрессируемый природный лиганд, вызывая цитолиз. Однако клетки CD8, трансдуцированные как NKG2D.YA-, так и NKG2D.AF-CAR, продемонстрировали очень слабый лизис природных клеток HeLa даже при высоких соотношениях E:T, уровень активности которых находился на одном уровне с нетрансдуцированными CD8 Т-клетками. Если на поверхности клеток HeLa сверхэкспрессировался ULBP1, то только CD8 NKG2D.wt-CAR-T-клетки оказались способны обеспечивать существенный уровень лизиса таких клеток. При высоком соотношении E:T в случае NKG2D.YA-CAR-клеток наблюдали некоторую дополнительную гибель клеток, которая отсутствовала в случае NKG2D.AF-CAR-клеток, демонстрируя, что двойная мутация Y152A/Y199F делает NKG2D даже более инертным, чем единичная мутация Y152A. Клетки NKG2D.wt-CAR направляют лизис (вследствие распознавания эндогенных лигандов MIC) на клетки HeLa, сверхэкспрессирующие NKG2D.AF-селективный неприродный ULBP2.R, в то время как клетки NKG2D.AF-CAR обеспечивают существенный уровень лизиса, сопоставимый со связыванием рецептора и селективного к нему лиганда.
[71] Для демонстрации того, что лизис клеток NKG2D.YA- или NKG2D.AF-CAR может быть направлен только соответствующим родственным нацеливающим MicAbody, в качестве мишени для цитолиза использовали клетки Ramos в комбинации с MicAbody на основе ритуксимаба, связанные либо с неприродными ортогональными лигандами ULBP2.S3, либо с ULBP2.R. Как показано на Фиг. 18A, ритуксимаб-ULBP2.S3 MicAbody может управлять активностью по уничтожению CD8 NKG2D.YA-CAR-клеток, но не NKG2D.AF-CAR-клеток, тогда как ритуксимаб-ULBP2.R MicAbody может управлять активностью NKG2D.AF-CAR-клеток, но не NKG2D.YA-CAR клеток. Это является дополнительным свидетельством селективности двух неприродных вариантов ULBP2 в отношении родственных неприродных вариантов NKG2D, для которых они были сконструированы в качестве предпочтительных партнеров. Для демонстрации специфичности части антитела в MicAbody, оценивали гибель клеток in vitro, используя CD8 NKG2D.AF-CAR-клетки, которые предварительно связывали путем инкубации с ритуксимаб-ULBP2.R, трастузумаб-ULPB2.R (SEQ ID NO: 95 и 133, тяжелая и легкая цепи, соответственно), или их эквимолярной комбинацией при общей насыщающей концентрации MicAbody. После удаления несвязанного MicAbody путем промывки CD8 клетки вносили либо к клеткам Ramos (экспрессирующие CD20, мишень ритуксимаба), либо к клеткам CT26-Her2 (линии мышиных клеток, трансфицированных для экспрессии человеческого Her2), которые были предварительно загружали кальцеином. После двухчасовой инкубации при двух различных соотношениях E:T определяли количество высвободившегося кальцеина. Как показано на фиг. 18B, когда клетки были предварительно связаны с ритуксимаб-MicAbody, наблюдали лизис только клеток Ramos, тогда как трастузумаб-MicAbody направлял цитолитическую активность только против клеток CT26-Her2. Однако, когда CD8 NKG2D.AF-CAR-клетки связывали одновременно как с ритуксимаб-, так и с трастузумаб-ULBP2.R MicAbody, наблюдали лизис обеих линий клеток-мишеней, что свидетельствовало о легкости мультиплексирования этих CAR-клеток, благодаря селективному привилегированному партнерству, которое было создано между рецептором и лигандом, что позволяет направлять их на поражение одновременно различных опухолевых мишеней.
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> XYPHOS BIOSCIENCES INC.
<120> МОДИФИЦИРОВАННЫЕ ДОМЕНЫ АЛЬФА1-АЛЬФА2 НЕПРИРОДНЫХ ЛИГАНДОВ
NKG2D, КОТОРЫЕ СВЯЗЫВАЮТСЯ С НЕПРИРОДНЫМИ РЕЦЕПТОРАМИ NKG2D
<130> F246240
<150> US 62/648,636
<151> 2018-03-27
<160> 156
<170> PatentIn version 3.5
<210> 1
<211> 183
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида MICA
<400> 1
Ala Ala Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val Leu Ser Trp
1 5 10 15
Asp Gly Ser Val Gln Ser Gly Phe Leu Thr Glu Val His Leu Asp Gly
20 25 30
Gln Pro Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala Lys Pro Gln
35 40 45
Gly Gln Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp Asp Arg Glu
50 55 60
Thr Arg Asp Leu Thr Gly Asn Gly Lys Asp Leu Arg Met Thr Leu Ala
65 70 75 80
His Ile Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln Glu Ile Arg
85 90 95
Val Cys Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser Gln His Phe
100 105 110
Tyr Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Lys Glu
115 120 125
Trp Thr Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val
130 135 140
Arg Asn Phe Leu Lys Glu Asp Ala Met Lys Thr Lys Thr His Tyr His
145 150 155 160
Ala Met His Ala Asp Cys Leu Gln Glu Leu Arg Arg Tyr Leu Lys Ser
165 170 175
Gly Val Val Leu Arg Arg Thr
180
<210> 2
<211> 182
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида MICB
<400> 2
Ala Glu Pro His Ser Leu Arg Tyr Asn Leu Met Val Leu Ser Gln Asp
1 5 10 15
Glu Ser Val Gln Ser Gly Phe Leu Ala Glu Gly His Leu Asp Gly Gln
20 25 30
Pro Phe Leu Arg Tyr Asp Arg Gln Lys Arg Arg Ala Lys Pro Gln Gly
35 40 45
Gln Trp Ala Glu Asp Val Leu Gly Ala Lys Thr Trp Asp Thr Glu Thr
50 55 60
Glu Asp Leu Thr Glu Asn Gly Gln Asp Leu Arg Arg Thr Leu Thr His
65 70 75 80
Ile Lys Asp Gln Lys Gly Gly Leu His Ser Leu Gln Glu Ile Arg Val
85 90 95
Cys Glu Ile His Glu Asp Ser Ser Thr Arg Gly Ser Arg His Phe Tyr
100 105 110
Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Gln Glu Ser
115 120 125
Thr Val Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val Thr
130 135 140
Asn Phe Trp Lys Glu Asp Ala Met Lys Thr Lys Thr His Tyr Arg Ala
145 150 155 160
Met Gln Ala Asp Cys Leu Gln Lys Leu Gln Arg Tyr Leu Lys Ser Gly
165 170 175
Val Ala Ile Arg Arg Thr
180
<210> 3
<211> 187
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP1
<400> 3
Ala Ala Glu Pro His Cys Leu Cys Tyr Asp Phe Ile Ile Thr Pro Lys
1 5 10 15
Ser Arg Pro Glu Pro Gln Trp Cys Glu Val Gln Gly Leu Val Asp Glu
20 25 30
Arg Pro Phe Leu His Tyr Asp Cys Val Asn His Lys Ala Lys Ala Phe
35 40 45
Ala Ser Leu Gly Lys Lys Val Asn Val Thr Lys Thr Trp Glu Glu Gln
50 55 60
Thr Glu Thr Leu Arg Asp Val Val Asp Phe Leu Lys Gly Gln Leu Leu
65 70 75 80
Asp Ile Gln Val Glu Asn Leu Ile Pro Ile Glu Pro Leu Thr Leu Gln
85 90 95
Ala Arg Met Ser Cys Glu His Glu Ala His Gly His Gly Arg Gly Ser
100 105 110
Trp Gln Phe Leu Phe Asn Gly Gln Lys Phe Leu Leu Phe Asp Ser Asn
115 120 125
Asn Arg Lys Trp Thr Ala Leu His Pro Gly Ala Lys Lys Met Thr Glu
130 135 140
Lys Trp Glu Lys Asn Arg Asp Val Thr Met Phe Phe Gln Lys Ile Ser
145 150 155 160
Leu Gly Asp Cys Lys Met Trp Leu Glu Glu Phe Leu Met Tyr Trp Glu
165 170 175
Gln Met Leu Asp Pro Thr Lys Pro Pro Met Val
180 185
<210> 4
<211> 189
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP2
<400> 4
Ala Ala Glu Pro His Ser Leu Cys Tyr Asp Ile Thr Val Ile Pro Lys
1 5 10 15
Phe Arg Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu
20 25 30
Lys Thr Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val
35 40 45
Ser Pro Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln
50 55 60
Asn Pro Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Arg
65 70 75 80
Asp Ile Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln
85 90 95
Ala Arg Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser
100 105 110
Trp Gln Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu
115 120 125
Lys Arg Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu
130 135 140
Lys Trp Glu Asn Asp Lys Val Val Ala Met Ser Phe His Tyr Phe Ser
145 150 155 160
Met Gly Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp
165 170 175
Ser Thr Leu Glu Pro Ser Ala Gly Ala Pro Pro Met Val
180 185
<210> 5
<211> 187
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP3
<400> 5
Ala Ala Glu Pro His Ser Leu Trp Tyr Asn Phe Thr Ile Ile His Leu
1 5 10 15
Pro Arg His Gly Gln Gln Trp Cys Glu Val Gln Ser Gln Val Asp Gln
20 25 30
Lys Asn Phe Leu Ser Tyr Asp Cys Gly Ser Asp Lys Val Leu Ser Met
35 40 45
Gly His Leu Glu Glu Gln Leu Tyr Ala Thr Asp Ala Trp Gly Lys Gln
50 55 60
Leu Glu Met Leu Arg Glu Val Gly Gln Arg Leu Arg Leu Glu Leu Ala
65 70 75 80
Asp Thr Glu Leu Glu Asp Phe Thr Pro Ser Gly Pro Leu Thr Leu Gln
85 90 95
Val Arg Met Ser Cys Glu Cys Glu Ala Asp Gly Tyr Ile Arg Gly Ser
100 105 110
Trp Gln Phe Ser Phe Asp Gly Arg Lys Phe Leu Leu Phe Asp Ser Asn
115 120 125
Asn Arg Lys Trp Thr Val Val His Ala Gly Ala Arg Arg Met Lys Glu
130 135 140
Lys Trp Glu Lys Asp Ser Gly Leu Thr Thr Phe Phe Lys Met Val Ser
145 150 155 160
Met Arg Asp Cys Lys Ser Trp Leu Arg Asp Phe Leu Met His Arg Lys
165 170 175
Lys Arg Leu Glu Pro Thr Ala Pro Pro Met Val
180 185
<210> 6
<211> 187
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP4
<400> 6
Ala Ala Glu Pro His Ser Leu Cys Phe Asn Phe Thr Ile Lys Ser Leu
1 5 10 15
Ser Arg Pro Gly Gln Pro Trp Cys Glu Ala Gln Val Phe Leu Asn Lys
20 25 30
Asn Leu Phe Leu Gln Tyr Asn Ser Asp Asn Asn Met Val Lys Pro Leu
35 40 45
Gly Leu Leu Gly Lys Lys Val Tyr Ala Thr Ser Thr Trp Gly Glu Leu
50 55 60
Thr Gln Thr Leu Gly Glu Val Gly Arg Asp Leu Arg Met Leu Leu Cys
65 70 75 80
Asp Ile Lys Pro Gln Ile Lys Thr Ser Asp Pro Ser Thr Leu Gln Val
85 90 95
Glu Met Phe Cys Gln Arg Glu Ala Glu Arg Cys Thr Gly Ala Ser Trp
100 105 110
Gln Phe Ala Thr Asn Gly Glu Lys Ser Leu Leu Phe Asp Ala Met Asn
115 120 125
Met Thr Trp Thr Val Ile Asn His Glu Ala Ser Lys Ile Lys Glu Thr
130 135 140
Trp Lys Lys Asp Arg Gly Leu Glu Lys Tyr Phe Arg Lys Leu Ser Lys
145 150 155 160
Gly Asp Cys Asp His Trp Leu Arg Glu Phe Leu Gly His Trp Glu Ala
165 170 175
Met Pro Glu Pro Thr Val Ser Pro Pro Met Val
180 185
<210> 7
<211> 198
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP5
<400> 7
Gly Leu Ala Asp Pro His Ser Leu Cys Tyr Asp Ile Thr Val Ile Pro
1 5 10 15
Lys Phe Arg Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp
20 25 30
Glu Lys Thr Phe Leu His Tyr Asp Cys Gly Ser Lys Thr Val Thr Pro
35 40 45
Val Ser Pro Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala
50 55 60
Gln Asn Pro Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu
65 70 75 80
Leu Asp Ile Gln Leu Glu Asn Tyr Ile Pro Lys Glu Pro Leu Thr Leu
85 90 95
Gln Ala Arg Met Ser Cys Glu Gln Lys Ala Glu Gly His Gly Ser Gly
100 105 110
Ser Trp Gln Leu Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser
115 120 125
Glu Asn Arg Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys
130 135 140
Glu Lys Trp Glu Asn Asp Lys Asp Met Thr Met Ser Phe His Tyr Ile
145 150 155 160
Ser Met Gly Asp Cys Thr Gly Trp Leu Glu Asp Phe Leu Met Gly Met
165 170 175
Asp Ser Thr Leu Glu Pro Ser Ala Gly Ala Pro Pro Thr Met Ser Ser
180 185 190
Gly Thr Ala Gln Pro Arg
195
<210> 8
<211> 189
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида ULBP6
<400> 8
Ala Ala Glu Pro His Ser Leu Cys Tyr Asp Ile Thr Val Ile Pro Lys
1 5 10 15
Phe Arg Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu
20 25 30
Lys Thr Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val
35 40 45
Ser Pro Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln
50 55 60
Asn Pro Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Leu
65 70 75 80
Asp Ile Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln
85 90 95
Ala Arg Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser
100 105 110
Trp Gln Phe Ser Ile Asp Gly Gln Thr Phe Leu Leu Phe Asp Ser Glu
115 120 125
Lys Arg Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu
130 135 140
Lys Trp Glu Asn Asp Lys Asp Val Ala Met Ser Phe His Tyr Ile Ser
145 150 155 160
Met Gly Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp
165 170 175
Ser Thr Leu Glu Pro Ser Ala Gly Ala Pro Pro Met Val
180 185
<210> 9
<211> 161
<212> Белок
<213> Искусственная последовательность
<220>
<223> Домен альфа1-альфа2 синтетического пептида OMCP
<400> 9
Ala Ala Ala Glu Pro His Lys Leu Ala Phe Asn Phe Asn Leu Glu Ile
1 5 10 15
Asn Gly Ser Asp Thr His Ser Thr Val Asp Val Tyr Leu Asp Asp Ser
20 25 30
Gln Ile Ile Thr Phe Asp Gly Lys Asp Ile Arg Pro Thr Ile Pro Phe
35 40 45
Met Ile Gly Asp Glu Ile Phe Leu Pro Phe Tyr Lys Asn Val Phe Ser
50 55 60
Glu Phe Phe Ser Leu Phe Arg Arg Val Pro Thr Ser Thr Pro Tyr Glu
65 70 75 80
Asp Leu Thr Tyr Phe Tyr Glu Cys Asp Tyr Thr Asp Asn Lys Ser Thr
85 90 95
Phe Asp Gln Phe Tyr Leu Tyr Asn Gly Glu Glu Tyr Thr Val Lys Thr
100 105 110
Gln Glu Ala Thr Asn Lys Asn Met Trp Leu Thr Thr Ser Glu Phe Arg
115 120 125
Leu Lys Lys Trp Phe Asp Gly Glu Asp Cys Ile Met His Leu Arg Ser
130 135 140
Leu Val Arg Lys Met Glu Asp Ser Lys Arg Arg Thr Val Pro Pro Met
145 150 155 160
Val
<210> 10
<211> 543
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен MICA
<400> 10
gagccccaca gtcttcgtta taacctcacg gtgctgtcct gggatggatc tgtgcagtca 60
gggtttctca ctgaggtaca tctggatggt cagcccttcc tgcgctgtga caggcagaaa 120
tgcagggcaa agccccaggg acagtgggca gaagatgtcc tgggaaataa gacatgggac 180
agagagacca gagacttgac agggaacgga aaggacctca ggatgaccct ggctcatatc 240
aaggaccaga aagaaggctt gcattccctc caggagatta gggtctgtga gatccatgaa 300
gacaacagca ccaggagctc ccagcatttc tactacgatg gggagctctt cctctcccaa 360
aacctggaga ctaaggaatg gacaatgccc cagtcctcca gagctcagac cttggccatg 420
aacgtcagga atttcttgaa ggaagatgcc atgaagacca agacacacta tcacgctatg 480
catgcagact gcctgcagga actacggcga tatctaaaat ccggcgtagt cctgaggaga 540
aca 543
<210> 11
<211> 561
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен ULBP1
<400> 11
gctgctgagc cccactgtct ctgctacgac tttattataa ctcctaagtc aagaccagag 60
cctcagtggt gcgaagtaca aggtttggtt gacgaaaggc ctttccttca ctacgattgt 120
gtgaaccata aggcaaaggc tttcgccagc ctgggtaaga aggtaaacgt tactaagacg 180
tgggaggagc agacggaaac cctccgtgat gtggttgact ttcttaaggg tcagctcctc 240
gatatccaag tggagaattt aatccctatc gaaccgctca ctctgcaggc cagaatgtca 300
tgcgaacatg aagcacacgg tcatggaaga ggtagttggc aatttttatt taacggtcaa 360
aaattcctgc tgttcgactc aaacaaccgc aaatggactg cgctgcaccc tggagctaag 420
aagatgactg aaaaatggga gaagaacaga gacgttacca tgttcttcca gaagatttcc 480
ctgggagatt gtaagatgtg gttagaggag ttcttaatgt actgggaaca gatgctggac 540
cccacaaaac cccccatggt g 561
<210> 12
<211> 567
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен ULBP2
<400> 12
gctgctgagc cccatagtct gtgttacgac atcacagtta ttcccaagtt caggcccgga 60
ccgcgctggt gtgccgtgca aggacaagtc gacgaaaaaa cctttcttca ttacgattgc 120
ggaaataaga ctgtaacgcc agtctctcct ttaggtaaga agttaaacgt cactacggcg 180
tggaaggcac aaaaccccgt cctgcgcgag gtcgtcgaca tcctgactga acaattgcgc 240
gacatccagc tcgagaatta cactccaaag gagcctctta ccctgcaggc tagaatgtct 300
tgcgagcaaa aggcagaggg ccactcctcc ggcagctggc agttcagttt cgacggacaa 360
atctttctgt tattcgattc agagaagaga atgtggacta cagttcaccc cggtgcccgt 420
aaaatgaagg agaagtggga aaacgacaaa gtggtggcga tgtcattcca ctatttctcg 480
atgggagact gcatcggttg gctggaagat ttcctcatgg gtatggactc cactttggag 540
ccatcggctg gtgccccccc catggtg 567
<210> 13
<211> 561
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен ULBP3
<400> 13
gctgctgagc cccacagctt gtggtacaac ttcaccatta tccacttgcc gagacatggc 60
cagcagtggt gcgaagtgca atcgcaagtc gaccaaaaaa acttcttatc atacgactgc 120
ggcagcgata aggtcttatc tatgggtcat ttggaggaac agctctacgc gaccgacgcc 180
tggggtaaac agctcgagat gctccgtgag gttggacaga ggctgagact ggaactggct 240
gacactgagc tggaagattt cacacctagt ggtccactca cattgcaagt acgcatgagc 300
tgcgagtgtg aggccgatgg atacattagg ggcagctggc agtttagctt cgacggaagg 360
aaattcctgc tcttcgacag taacaatagg aagtggactg ttgtgcatgc tggtgcgcgc 420
agaatgaagg aaaagtggga gaaagatagc ggcctgacga ccttcttcaa gatggtgtct 480
atgcgtgact gtaagagctg gctcagagat ttcctcatgc atcgcaagaa gaggttagaa 540
cctaccgctc cccccatggt g 561
<210> 14
<211> 561
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен ULBP4
<400> 14
gctgctgagc cccactctct ttgcttcaac ttcaccatta aatccctgag caggcctggt 60
cagccgtggt gtgaggcgca ggtctttctt aacaagaatc tcttcctcca atacaactct 120
gataacaaca tggtaaagcc actgggtctc ctgggtaaaa aagtctatgc tacgagcact 180
tggggagaac tcacccagac tcttggcgag gtaggaagag acctgcgcat gctcctctgc 240
gatataaagc cccaaattaa gaccagtgat ccgtccactt tacaagtcga aatgttctgc 300
caaagggagg ctgaacgctg caccggagcc tcttggcagt tcgcgaccaa tggcgaaaag 360
tccctcttgt tcgatgccat gaatatgacc tggaccgtga tcaatcatga ggcctctaag 420
atcaaggaga cgtggaaaaa ggaccgcggc cttgaaaagt actttaggaa gttgtctaaa 480
ggagactgcg accattggtt acgcgagttc ctcggccatt gggaagcgat gcccgagcca 540
acggttagcc cccccatggt g 561
<210> 15
<211> 567
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен ULBP6
<400> 15
gctgctgagc cccactcctt atgctatgat atcaccgtga ttccaaagtt ccgaccagga 60
ccccgatggt gcgccgtaca gggacaggtc gacgaaaaga cttttttaca ttacgactgc 120
ggtaacaaga cagtcacacc ggtaagtcct ttgggaaaaa agttaaacgt aaccactgct 180
tggaaggccc agaaccccgt ccttcgagaa gtagtggata ttttgactga acagctgctt 240
gacatccagc tggaaaacta cacacccaaa gagcccctga ctcttcaagc gcgtatgtcg 300
tgtgagcaaa aggccgaagg acacagctcc ggatcctggc agttcagtat cgacggtcag 360
accttcctcc tcttcgattc agaaaagcgc atgtggacta ctgtgcaccc cggcgctcgt 420
aagatgaagg aaaagtggga gaatgataag gacgttgcca tgagttttca ttacattagt 480
atgggagatt gcatcggttg gctggaagac ttcctgatgg gtatggatag tacccttgaa 540
cctagtgccg gagctccccc catggtg 567
<210> 16
<211> 483
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий альфа1-альфа2 домен OMCP
<400> 16
gctgctgctg agccccacaa gcttgcgttc aacttcaatc tggaaataaa cggttcagat 60
acccattcaa ccgtggacgt ttatttagac gattcgcaga taatcacctt tgacggcaag 120
gacatccgcc caactatccc gttcatgata ggtgacgaaa tcttccttcc tttttataag 180
aatgtgttct ctgagttctt cagtttgttc cgccgcgtcc ctacctcaac cccctacgaa 240
gacttgactt atttctatga atgcgactac accgacaaca aatctacatt cgatcaattc 300
tacctgtaca acggtgaaga gtacaccgtg aagactcaag aggctactaa caagaacatg 360
tggctgacca cttccgagtt cagactgaag aagtggttcg acggcgagga ctgtatcatg 420
caccttagaa gtttagtgag gaaaatggaa gatagcaaga gaagaacagt gccccccatg 480
gtg 483
<210> 17
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Природный эктодомен NKG2D синтетического пептида
<400> 17
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 18
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152A синтетического пептида
<400> 18
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 19
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y199A синтетического пептида
<400> 19
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Ala Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 20
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152A/Y199A синтетического пептида
<400> 20
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Ala Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 21
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y199F eNKG2D1 синтетического пептида
<400> 21
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 22
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152S eNKG2D2 синтетического пептида
<400> 22
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ser His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 23
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152T eNKG2D3 синтетического пептида
<400> 23
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Thr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 24
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152V eNKG2D4 синтетического пептида
<400> 24
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Val His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 25
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152A/Y199F eNKG2D5 синтетического
пептида
<400> 25
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 26
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152L/Y199F eNKG2D6 синтетического
пептида
<400> 26
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Leu His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 27
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152S/Y199F eNKG2D7 синтетического
пептида
<400> 27
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ser His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 28
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152T/Y199F eNKG2D8 синтетического
пептида
<400> 28
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Thr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 29
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152V/Y199F eNKG2D9 синтетического
пептида
<400> 29
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Val His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 30
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y199D eNKG2D10 синтетического пептида
<400> 30
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Asp Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 31
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y199E eNKG2D11 синтетического пептида
<400> 31
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Glu Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 32
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152D/Y199D eNKG2D12 синтетического
пептида
<400> 32
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Asp His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Asp Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 33
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152E/Y199E eNKG2D13 синтетического
пептида
<400> 33
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Glu His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Glu Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 34
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152L eNKG2D14 синтетического пептида
<400> 34
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Leu His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 35
<211> 139
<212> Белок
<213> Искусственная последовательность
<220>
<223> Неприродный эктодомен NKG2D Y152F/Y199F eNKG2D15 синтетического
пептида
<400> 35
Phe Leu Asn Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu
1 5 10 15
Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn
20 25 30
Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala
35 40 45
Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu
50 55 60
Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Phe His Trp Met Gly Leu
65 70 75 80
Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile
85 90 95
Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys
100 105 110
Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr
115 120 125
Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
130 135
<210> 36
<211> 23
<212> Белок
<213> Искусственная последовательность
<220>
<223> Сигнальная поледовательность синтетического пептида MHCI
<400> 36
Met Gly Leu Gly Pro Val Phe Leu Leu Leu Ala Gly Ile Phe Pro Phe
1 5 10 15
Ala Pro Pro Gly Ala Ala Ala
20
<210> 37
<211> 69
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий сигнальную
поледовательность MHCI
<400> 37
atgggccttg gcccagtgtt tctgctgttg gcaggcattt tcccttttgc tccgcccggc 60
gccgcagcc 69
<210> 38
<211> 237
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид Fc человеческого IgG1 с линкером IEGR
<400> 38
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg
225 230 235
<210> 39
<211> 711
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий Fc человеческого
IgG1 с линкером IEGR
<400> 39
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg c 711
<210> 40
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого
IgG1 с NKG2D
<400> 40
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 41
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1 с
NKG2D Y152A
<400> 41
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 42
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид Fc человеческого IgG1-эктодомен NKG2D Y199A
<400> 42
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Ala Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 43
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетического пептид Fc человеческого IgG1- эктодомен
NKG2D Y152A/Y199A
<400> 43
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Ala Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 44
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y199F eNKG2D1
<400> 44
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 45
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152S eNKG2D2
<400> 45
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 46
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152T eNKG2D3
<400> 46
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Thr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 47
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152V eNKG2D4
<400> 47
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Val His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 48
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152A/Y199F eNKG2D5
<400> 48
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 49
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152L/Y199F eNKG2D6
<400> 49
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Leu His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 50
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152S/Y199F eNKG2D7
<400> 50
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Ser His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 51
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152T/Y199F eNKG2D8
<400> 51
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Thr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 52
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152V/Y199F eNKG2D9
<400> 52
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Val His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 53
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y199D eNKG2D10
<400> 53
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Asp Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 54
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y199E eNKG2D11
<400> 54
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Glu Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 55
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152D/Y199D eNKG2D12
<400> 55
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Asp His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Asp Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 56
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152E/Y199E eNKG2D13
<400> 56
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Glu His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Glu Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 57
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152L eNKG2D14
<400> 57
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Leu His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 58
<211> 376
<212> Белок
<213> Искусственная последовательность
<220>
<223> Слитый белок синтетического пептида Fc человеческого IgG1
с NKG2D Y152F/Y199F eNKG2D15
<400> 58
Met Asp Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1 5 10 15
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
20 25 30
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
35 40 45
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
50 55 60
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
65 70 75 80
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
85 90 95
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
100 105 110
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
115 120 125
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
130 135 140
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
145 150 155 160
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
165 170 175
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
180 185 190
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
195 200 205
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
210 215 220
Lys Ser Leu Ser Leu Ser Pro Gly Lys Ile Glu Gly Arg Phe Leu Asn
225 230 235 240
Ser Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr Glu Ser Tyr Cys
245 250 255
Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn Asn Cys Tyr Gln
260 265 270
Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln Ala Ser Cys Met
275 280 285
Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys Glu Asp Gln Asp
290 295 300
Leu Leu Lys Leu Val Lys Ser Phe His Trp Met Gly Leu Val His Ile
305 310 315 320
Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser Ile Leu Ser Pro
325 330 335
Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp Cys Ala Leu Tyr
340 345 350
Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser Thr Pro Asn Thr
355 360 365
Tyr Ile Cys Met Gln Arg Thr Val
370 375
<210> 59
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D
<400> 59
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcatatcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggctatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 60
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152A
<400> 60
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagctcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggctatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 61
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий
человеческий IgG1 Fc-эктодомен NKG2D Y199A
<400> 61
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcataccatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgctata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 62
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий
человеческтй IgG1 Fc-эктодомен NKG2D Y152A/Y199A
<400> 62
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagctcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgctata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 63
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y199F eNKG2D1
<400> 63
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcataccatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 64
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
Человеческий IgG1 Fc-NKG2D Y152S eNKG2D2
<400> 64
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcataccatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 65
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152T eNKG2D3
<400> 65
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcaactcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggctatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 66
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152V eNKG2D4
<400> 66
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagtgcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggctatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 67
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый D
человеческий IgG1 Fc-NKG2Y152A/Y199F eNKG2D5
<400> 67
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagctcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 68
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152L/Y199F eNKG2D6
<400> 68
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcactgcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 69
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий человеческий IgG1 Fc-NKG2D
Y152S/Y199F eNKG2D7 fusion
<400> 69
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcaagtcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 70
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152T/Y199F eNKG2D8
<400> 70
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcaactcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 71
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152V/Y199F eNKG2D9 fusion
<400> 71
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagtgcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 72
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y199D eNKG2D10
<400> 72
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcatatcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 73
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y199E eNKG2D11
<400> 73
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcataccatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgagata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 74
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152D/Y199D eNKG2D12
<400> 74
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagatcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 75
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152E/Y199E eNKG2D13
<400> 75
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcagagcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcgagata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 76
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152L eNKG2D14
<400> 76
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcactgcatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggctatata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 77
<211> 1128
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий слитый
человеческий IgG1 Fc-NKG2D Y152F/Y199F eNKG2D15
<400> 77
atggacccga aaagctgcga caagactcac acttgtccgc cgtgccccgc ccccgaactg 60
ctgggtggcc cctccgtgtt cctgttcccg cctaagccta aggacaccct tatgatcagc 120
cgcacccctg aagtgacctg tgtcgtcgtg gatgtgtcac acgaggaccc ggaggtcaag 180
ttcaattggt acgtggacgg cgtggaagtg cataacgcaa agaccaagcc tcgggaggaa 240
cagtacaact cgacctaccg cgtggtgtca gtcctgactg tgctgcacca ggactggctg 300
aacgggaagg agtacaagtg caaagtgtcg aacaaggccc tgccggctcc aattgaaaag 360
accatcagca aggccaaggg ccagccaagg gaaccacagg tgtacaccct ccctccttcc 420
cgggacgagc tgaccaaaaa ccaagtgtcc ctgacttgcc ttgtgaaggg gttctaccct 480
tctgacattg ccgtcgaatg ggaatcgaac ggacagcctg aaaacaacta taagactacc 540
ccgcccgtgc tggattccga cggaagcttc ttcctgtact ccaagctgac cgtggacaag 600
tcgagatggc agcagggaaa tgtgttcagc tgctccgtga tgcatgaggc gctgcacaac 660
cactacaccc agaagtcact gagcctctcc cccggaaaga tcgaaggacg cttcttaaac 720
tcattattca accaagaagt tcaaattccc ttgaccgaaa gttactgtgg cccatgtcct 780
aaaaactgga tatgttacaa aaataactgc taccaatttt ttgatgagag taaaaactgg 840
tatgagagcc aggcttcttg tatgtctcaa aatgccagcc ttctgaaagt atacagcaaa 900
gaggaccagg atttacttaa actggtgaag tcattccatt ggatgggact agtacacatt 960
ccaacaaatg gatcttggca gtgggaagat ggctccattc tctcacccaa cctactaaca 1020
ataattgaaa tgcagaaggg agactgtgca ctctatgcct cgagctttaa aggcttcata 1080
gaaaactgtt caactccaaa tacatacatc tgcatgcaaa ggactgtg 1128
<210> 78
<211> 181
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид MICwed альфа1-альфа2
<400> 78
Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val Leu Ser Trp Asp Gly
1 5 10 15
Ser Val Gln Ser Gly Phe Leu Thr Glu Val His Leu Asp Gly Gln Pro
20 25 30
Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala Lys Pro Gln Gly Gln
35 40 45
Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp Asp Arg Glu Thr Arg
50 55 60
Asp Leu Thr Gly Trp Gly Lys Asp Leu Arg Met Thr Leu Ala His Ile
65 70 75 80
Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln Glu Ile Arg Val Cys
85 90 95
Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser Gln His Phe Tyr Tyr
100 105 110
Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Lys Glu Trp Thr
115 120 125
Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val Arg Asn
130 135 140
Phe Leu Lys Glu Asp Ala Met Glu Thr Asp Thr His Tyr His Ala Met
145 150 155 160
His Ala Asp Cys Leu Gln Glu Leu Arg Arg Tyr Leu Lys Ser Gly Val
165 170 175
Val Leu Arg Arg Thr
180
<210> 79
<211> 543
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий MICwed альфа1-альфа2
<400> 79
gagcctcaca gcctccggta taatttgact gtactctctt gggatggctc cgtgcagtcc 60
ggctttctga ctgaagttca tctcgacggt caacctttcc tgcgctgcga ccgacaaaaa 120
tgccgcgcca agccccaagg gcagtgggcc gaagatgtac tgggaaacaa gacctgggac 180
cgggagacac gagacctgac aggctggggc aaggacttgc gcatgacact cgcccatatc 240
aaggaccaga aggaaggatt gcactctttg caagagattc gcgtgtgtga aatccacgag 300
gacaattcaa cgaggagctc ccagcacttc tattacgatg gagaactctt cttgtcacag 360
aacttggaaa ccaaggaatg gactatgcct cagagctctc gggcacagac tctcgctatg 420
aacgttagaa acttccttaa ggaggatgct atggagaccg atactcacta ccacgccatg 480
cacgccgact gcctccagga actgcggaga tatctgaagt ccggcgtggt tttgagaaga 540
acc 543
<210> 80
<211> 181
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид MIC25 альфа1-альфа2
<400> 80
Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val Leu Ser Trp Asp Gly
1 5 10 15
Ser Val Gln Ser Gly Phe Leu Thr Glu Val His Leu Asp Gly Gln Pro
20 25 30
Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala Lys Pro Gln Gly Gln
35 40 45
Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp Asp Arg Glu Thr Arg
50 55 60
Asp Leu Thr Gly Trp Gly Lys Asp Leu Arg Met Thr Leu Ala His Ile
65 70 75 80
Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln Glu Ile Arg Val Cys
85 90 95
Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser Gln His Phe Tyr Tyr
100 105 110
Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu Thr Leu Glu Trp Thr
115 120 125
Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala Met Asn Val Arg Asn
130 135 140
Phe Leu Lys Glu Asp Ala Met Glu Thr Asp Thr His Tyr His Ala Met
145 150 155 160
Arg Ala Asp Cys Leu Ser Glu Leu Arg Arg Tyr Leu Lys Ser Gly Val
165 170 175
Val Leu Arg Arg Thr
180
<210> 81
<211> 543
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий MIC25 альфа1-альфа2
<400> 81
gagcctcaca gcctccggta taatttgact gtactctctt gggatggctc cgtgcagtcc 60
ggctttctga ctgaagttca tctcgacggt caacctttcc tgcgctgcga ccgacaaaaa 120
tgccgcgcca agccccaagg gcagtgggcc gaagatgtac tgggaaacaa gacctgggac 180
cgggagacac gagacctgac aggctggggc aaggacttgc gcatgacact cgcccatatc 240
aaggaccaga aggaaggatt gcactctttg caagagattc gcgtgtgtga aatccacgag 300
gacaattcaa cgaggagctc ccagcacttc tattacgatg gagaactctt cttgtcacag 360
aacttggaaa ccctcgaatg gactatgcct cagagctctc gggcacagac tctcgctatg 420
aacgttagaa acttccttaa ggaggatgct atggagaccg atactcacta ccacgccatg 480
cgcgccgact gcctctctga actgcggaga tatctgaagt ccggcgtggt tttgagaaga 540
acc 543
<210> 82
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид D265A/N297A тяжелой цепи CH1-CH2-CH3
человеческого IgG1
<400> 82
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Ala Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu
225 230 235 240
Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 83
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> D265A/N297A тяжелой цепи CH1-CH2-CH3 человеческого IgG1
<400> 83
gcgtcgacca agggcccgtc agtgttcccg ctggccccgt catccaagtc cacgtctggg 60
ggcacagcag ccctgggatg cttggtcaag gactacttcc ccgagcccgt gactgtgtcc 120
tggaactccg gagcactgac ctccggagtg cacacctttc ccgcggtgct gcagtcctcc 180
ggactgtact ccctgtcgtc ggtcgtgacc gtgccgagct cctcgctcgg aacccagacc 240
tacatctgca acgtgaacca caagccctcg aacaccaaag tggacaagaa ggtcgagccc 300
aaaagctgcg acaagactca cacttgtccg ccgtgccccg cccccgaact gctgggtggc 360
ccctccgtgt tcctgttccc gcctaagcct aaggacaccc ttatgatcag ccgcacccct 420
gaagtgacct gtgtcgtcgt ggcagtgtca cacgaggacc cggaggtcaa gttcaattgg 480
tacgtggacg gcgtggaagt gcataacgca aagaccaagc ctcgggagga acagtacgcc 540
tcgacctacc gcgtggtgtc agtcctgact gtgctgcacc aggactggct gaacgggaag 600
gagtacaagt gcaaagtgtc gaacaaggcc ctgccggctc caattgaaaa gaccatcagc 660
aaggccaagg gccagccaag ggaaccacag gtgtacaccc tccctccttc ccgggacgag 720
ctgaccaaaa accaagtgtc cctgacttgc cttgtgaagg ggttctaccc ttctgacatt 780
gccgtcgaat gggaatcgaa cggacagcct gaaaacaact ataagactac cccgcccgtg 840
ctggattccg acggaagctt cttcctgtac tccaagctga ccgtggacaa gtcgagatgg 900
cagcagggaa atgtgttcag ctgctccgtg atgcatgagg cgctgcacaa ccactacacc 960
cagaagtcac tgagcctctc ccccggaaag 990
<210> 84
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид человеческой легкой каппа-цепи
<400> 84
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 85
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий человеческую
Легкую каппа-цепь
<400> 85
agaaccgtgg ccgccccgag cgtgttcatt ttccctccct ccgacgagca gttgaaatcg 60
ggcaccgcta gcgtggtctg ccttctcaac aatttctatc cacgggaagc caaagtgcag 120
tggaaggtcg acaacgcgct ccaatccggg aactcacagg aatccgtgac tgagcaggat 180
tccaaggact cgacctactc cctgtcatcc acgctgaccc tgagcaaggc agactacgag 240
aagcacaagg tctacgcctg cgaagtgaca caccagggac tgtccagccc cgtgaccaag 300
agcttcaaca gaggagaatg c 321
<210> 86
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид VH трастузумаба
<400> 86
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser
115 120
<210> 87
<211> 360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий VH трастузумаба
<400> 87
gaagtccaat tggtcgaatc aggcggtgga ctcgtgcaac ctggaggttc gttacgctta 60
tcatgtgctg caagtggatt taatattaaa gatacctaca tccactgggt acgtcaagct 120
cccggcaagg gtctcgagtg ggtcgcacgc atttacccca ccaacggata cacgcgctac 180
gccgattcag tgaagggacg tttcacaatc tctgctgata ctagcaaaaa taccgcatac 240
ctccagatga actctcttag ggccgaggac acagctgtgt actactgtag ccgttgggga 300
ggagacggtt tttacgcaat ggattactgg ggccaaggaa ccctggtcac agtttcatcg 360
<210> 88
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид VL трастузумаба
<400> 88
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 89
<211> 321
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий VL трастузумаба
<400> 89
gatatccaaa tgactcaatc accatcttca ctctccgcga gcgtgggtga tcgggtcacc 60
atcacatgta gggcgagcca agatgtgaat accgccgtcg cgtggtatca acaaaagccg 120
ggaaaagcac caaaactgct tatatactct gcatccttcc tgtactctgg ggtgccaagc 180
cggttctccg gtagtagatc tggtactgac tttacactca ctatcagcag tctgcaacct 240
gaggactttg cgacatacta ttgccagcag cactacacaa ccccacctac atttggtcag 300
gggacaaagg tggagatcaa g 321
<210> 90
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид VH ритуксимаба
<400> 90
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Lys Gln Thr Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asn Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ala
115 120
<210> 91
<211> 363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий VH ритуксимаба
<400> 91
caagttcagc ttcagcagcc gggggctgag ttggtgaaac ccggggccag tgtgaagatg 60
agctgtaaag cgagcggcta caccttcact tcttataata tgcattgggt taagcaaacg 120
ccaggaaggg ggctggagtg gatcggcgct atttacccag gtaacggtga cacatcatat 180
aaccaaaagt ttaagggaaa ggcaaccctc acagcggaca agagtagctc aaccgcatac 240
atgcaactgt caagccttac ctccgaagac agcgcagtgt actactgcgc cagaagcacc 300
tactatgggg gtgattggta cttcaacgtc tggggggctg gcaccacagt gactgtaagc 360
gca 363
<210> 92
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид VL ритуксимаба
<400> 92
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105
<210> 93
<211> 318
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий VL ритуксимаба
<400> 93
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaag 318
<210> 94
<211> 450
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид тяжелой цепи трастузумаба
<400> 94
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 95
<211> 1350
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий тяжелую цепь трастузумаба
<400> 95
gaagtccaat tggtcgaatc aggcggtgga ctcgtgcaac ctggaggttc gttacgctta 60
tcatgtgctg caagtggatt taatattaaa gatacctaca tccactgggt acgtcaagct 120
cccggcaagg gtctcgagtg ggtcgcacgc atttacccca ccaacggata cacgcgctac 180
gccgattcag tgaagggacg tttcacaatc tctgctgata ctagcaaaaa taccgcatac 240
ctccagatga actctcttag ggccgaggac acagctgtgt actactgtag ccgttgggga 300
ggagacggtt tttacgcaat ggattactgg ggccaaggaa ccctggtcac agtttcatcg 360
gcgtcgacca agggcccgtc agtgttcccg ctggccccgt catccaagtc cacgtctggg 420
ggcacagcag ccctgggatg cttggtcaag gactacttcc ccgagcccgt gactgtgtcc 480
tggaactccg gagcactgac ctccggagtg cacacctttc ccgcggtgct gcagtcctcc 540
ggactgtact ccctgtcgtc ggtcgtgacc gtgccgagct cctcgctcgg aacccagacc 600
tacatctgca acgtgaacca caagccctcg aacaccaaag tggacaagaa ggtcgagccc 660
aaaagctgcg acaagactca cacttgtccg ccgtgccccg cccccgaact gctgggtggc 720
ccctccgtgt tcctgttccc gcctaagcct aaggacaccc ttatgatcag ccgcacccct 780
gaagtgacct gtgtcgtcgt ggatgtgtca cacgaggacc cggaggtcaa gttcaattgg 840
tacgtggacg gcgtggaagt gcataacgca aagaccaagc ctcgggagga acagtacaac 900
tcgacctacc gcgtggtgtc agtcctgact gtgctgcacc aggactggct gaacgggaag 960
gagtacaagt gcaaagtgtc gaacaaggcc ctgccggctc caattgaaaa gaccatcagc 1020
aaggccaagg gccagccaag ggaaccacag gtgtacaccc tccctccttc ccgggacgag 1080
ctgaccaaaa accaagtgtc cctgacttgc cttgtgaagg ggttctaccc ttctgacatt 1140
gccgtcgaat gggaatcgaa cggacagcct gaaaacaact ataagactac cccgcccgtg 1200
ctggattccg acggaagctt cttcctgtac tccaagctga ccgtggacaa gtcgagatgg 1260
cagcagggaa atgtgttcag ctgctccgtg atgcatgagg cgctgcacaa ccactacacc 1320
cagaagtcac tgagcctctc ccccggaaag 1350
<210> 96
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид легкой цепи трастузумаба
<400> 96
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 97
<211> 642
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий легкую цепь трастузумаба
<400> 97
gatatccaaa tgactcaatc accatcttca ctctccgcga gcgtgggtga tcgggtcacc 60
atcacatgta gggcgagcca agatgtgaat accgccgtcg cgtggtatca acaaaagccg 120
ggaaaagcac caaaactgct tatatactct gcatccttcc tgtactctgg ggtgccaagc 180
cggttctccg gtagtagatc tggtactgac tttacactca ctatcagcag tctgcaacct 240
gaggactttg cgacatacta ttgccagcag cactacacaa ccccacctac atttggtcag 300
gggacaaagg tggagatcaa gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgct ag 642
<210> 98
<211> 451
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид тяжелой цепи ритуксимаба
<400> 98
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Lys Gln Thr Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asn Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Lys
450
<210> 99
<211> 1353
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий тяжелую цепь ритуксимаба
<400> 99
caagttcagc ttcagcagcc gggggctgag ttggtgaaac ccggggccag tgtgaagatg 60
agctgtaaag cgagcggcta caccttcact tcttataata tgcattgggt taagcaaacg 120
ccaggaaggg ggctggagtg gatcggcgct atttacccag gtaacggtga cacatcatat 180
aaccaaaagt ttaagggaaa ggcaaccctc acagcggaca agagtagctc aaccgcatac 240
atgcaactgt caagccttac ctccgaagac agcgcagtgt actactgcgc cagaagcacc 300
tactatgggg gtgattggta cttcaacgtc tggggggctg gcaccacagt gactgtaagc 360
gcagcgtcga ccaagggccc gtcagtgttc ccgctggccc cgtcatccaa gtccacgtct 420
gggggcacag cagccctggg atgcttggtc aaggactact tccccgagcc cgtgactgtg 480
tcctggaact ccggagcact gacctccgga gtgcacacct ttcccgcggt gctgcagtcc 540
tccggactgt actccctgtc gtcggtcgtg accgtgccga gctcctcgct cggaacccag 600
acctacatct gcaacgtgaa ccacaagccc tcgaacacca aagtggacaa gaaggtcgag 660
cccaaaagct gcgacaagac tcacacttgt ccgccgtgcc ccgcccccga actgctgggt 720
ggcccctccg tgttcctgtt cccgcctaag cctaaggaca cccttatgat cagccgcacc 780
cctgaagtga cctgtgtcgt cgtggatgtg tcacacgagg acccggaggt caagttcaat 840
tggtacgtgg acggcgtgga agtgcataac gcaaagacca agcctcggga ggaacagtac 900
aactcgacct accgcgtggt gtcagtcctg actgtgctgc accaggactg gctgaacggg 960
aaggagtaca agtgcaaagt gtcgaacaag gccctgccgg ctccaattga aaagaccatc 1020
agcaaggcca agggccagcc aagggaacca caggtgtaca ccctccctcc ttcccgggac 1080
gagctgacca aaaaccaagt gtccctgact tgccttgtga aggggttcta cccttctgac 1140
attgccgtcg aatgggaatc gaacggacag cctgaaaaca actataagac taccccgccc 1200
gtgctggatt ccgacggaag cttcttcctg tactccaagc tgaccgtgga caagtcgaga 1260
tggcagcagg gaaatgtgtt cagctgctcc gtgatgcatg aggcgctgca caaccactac 1320
acccagaagt cactgagcct ctcccccgga aag 1353
<210> 100
<211> 212
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид легкой цепи ритуксимаба
<400> 100
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Thr Val Ala Ala Pro Ser
100 105 110
Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr Ala
115 120 125
Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys Val
130 135 140
Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu Ser
145 150 155 160
Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser Thr
165 170 175
Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala Cys
180 185 190
Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe Asn
195 200 205
Arg Gly Glu Cys
210
<210> 101
<211> 639
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий легкую цепь ритуксимаба
<400> 101
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagac cgtggccgcc ccgagcgtgt tcattttccc tccctccgac 360
gagcagttga aatcgggcac cgctagcgtg gtctgccttc tcaacaattt ctatccacgg 420
gaagccaaag tgcagtggaa ggtcgacaac gcgctccaat ccgggaactc acaggaatcc 480
gtgactgagc aggattccaa ggactcgacc tactccctgt catccacgct gaccctgagc 540
aaggcagact acgagaagca caaggtctac gcctgcgaag tgacacacca gggactgtcc 600
agccccgtga ccaagagctt caacagagga gaatgctag 639
<210> 102
<211> 634
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид HC_MICwed трастузумаба
<400> 102
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Gly Gly Gly Ser Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val
450 455 460
Leu Ser Trp Asp Gly Ser Val Gln Ser Gly Phe Leu Thr Glu Val His
465 470 475 480
Leu Asp Gly Gln Pro Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala
485 490 495
Lys Pro Gln Gly Gln Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp
500 505 510
Asp Arg Glu Thr Arg Asp Leu Thr Gly Trp Gly Lys Asp Leu Arg Met
515 520 525
Thr Leu Ala His Ile Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln
530 535 540
Glu Ile Arg Val Cys Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser
545 550 555 560
Gln His Phe Tyr Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu
565 570 575
Thr Lys Glu Trp Thr Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala
580 585 590
Met Asn Val Arg Asn Phe Leu Lys Glu Asp Ala Met Glu Thr Asp Thr
595 600 605
His Tyr His Ala Met His Ala Asp Cys Leu Gln Glu Leu Arg Arg Tyr
610 615 620
Leu Lys Ser Gly Val Val Leu Arg Arg Thr
625 630
<210> 103
<211> 1902
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий HC_MICwed трастузумаба
<400> 103
gaagtccaat tggtcgaatc aggcggtgga ctcgtgcaac ctggaggttc gttacgctta 60
tcatgtgctg caagtggatt taatattaaa gatacctaca tccactgggt acgtcaagct 120
cccggcaagg gtctcgagtg ggtcgcacgc atttacccca ccaacggata cacgcgctac 180
gccgattcag tgaagggacg tttcacaatc tctgctgata ctagcaaaaa taccgcatac 240
ctccagatga actctcttag ggccgaggac acagctgtgt actactgtag ccgttgggga 300
ggagacggtt tttacgcaat ggattactgg ggccaaggaa ccctggtcac agtttcatcg 360
gcgtcgacca agggcccgtc agtgttcccg ctggccccgt catccaagtc cacgtctggg 420
ggcacagcag ccctgggatg cttggtcaag gactacttcc ccgagcccgt gactgtgtcc 480
tggaactccg gagcactgac ctccggagtg cacacctttc ccgcggtgct gcagtcctcc 540
ggactgtact ccctgtcgtc ggtcgtgacc gtgccgagct cctcgctcgg aacccagacc 600
tacatctgca acgtgaacca caagccctcg aacaccaaag tggacaagaa ggtcgagccc 660
aaaagctgcg acaagactca cacttgtccg ccgtgccccg cccccgaact gctgggtggc 720
ccctccgtgt tcctgttccc gcctaagcct aaggacaccc ttatgatcag ccgcacccct 780
gaagtgacct gtgtcgtcgt ggcagtgtca cacgaggacc cggaggtcaa gttcaattgg 840
tacgtggacg gcgtggaagt gcataacgca aagaccaagc ctcgggagga acagtacgcc 900
tcgacctacc gcgtggtgtc agtcctgact gtgctgcacc aggactggct gaacgggaag 960
gagtacaagt gcaaagtgtc gaacaaggcc ctgccggctc caattgaaaa gaccatcagc 1020
aaggccaagg gccagccaag ggaaccacag gtgtacaccc tccctccttc ccgggacgag 1080
ctgaccaaaa accaagtgtc cctgacttgc cttgtgaagg ggttctaccc ttctgacatt 1140
gccgtcgaat gggaatcgaa cggacagcct gaaaacaact ataagactac cccgcccgtg 1200
ctggattccg acggaagctt cttcctgtac tccaagctga ccgtggacaa gtcgagatgg 1260
cagcagggaa atgtgttcag ctgctccgtg atgcatgagg cgctgcacaa ccactacacc 1320
cagaagtcac tgagcctctc ccccggagga ggtggcagcg agcctcacag cctccggtat 1380
aatttgactg tactctcttg ggatggctcc gtgcagtccg gctttctgac tgaagttcat 1440
ctcgacggtc aacctttcct gcgctgcgac cgacaaaaat gccgcgccaa gccccaaggg 1500
cagtgggccg aagatgtact gggaaacaag acctgggacc gggagacacg agacctgaca 1560
ggctggggca aggacttgcg catgacactc gcccatatca aggaccagaa ggaaggattg 1620
cactctttgc aagagattcg cgtgtgtgaa atccacgagg acaattcaac gaggagctcc 1680
cagcacttct attacgatgg agaactcttc ttgtcacaga acttggaaac caaggaatgg 1740
actatgcctc agagctctcg ggcacagact ctcgctatga acgttagaaa cttccttaag 1800
gaggatgcta tggagaccga tactcactac cacgccatgc acgccgactg cctccaggaa 1860
ctgcggagat atctgaagtc cggcgtggtt ttgagaagaa cc 1902
<210> 104
<211> 634
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид HC_MIC25 трастузумаба
<400> 104
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Asn Ile Lys Asp Thr
20 25 30
Tyr Ile His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Arg Ile Tyr Pro Thr Asn Gly Tyr Thr Arg Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Ala Asp Thr Ser Lys Asn Thr Ala Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ser Arg Trp Gly Gly Asp Gly Phe Tyr Ala Met Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Gly Gly Gly Ser Glu Pro His Ser Leu Arg Tyr Asn Leu Thr Val
450 455 460
Leu Ser Trp Asp Gly Ser Val Gln Ser Gly Phe Leu Thr Glu Val His
465 470 475 480
Leu Asp Gly Gln Pro Phe Leu Arg Cys Asp Arg Gln Lys Cys Arg Ala
485 490 495
Lys Pro Gln Gly Gln Trp Ala Glu Asp Val Leu Gly Asn Lys Thr Trp
500 505 510
Asp Arg Glu Thr Arg Asp Leu Thr Gly Trp Gly Lys Asp Leu Arg Met
515 520 525
Thr Leu Ala His Ile Lys Asp Gln Lys Glu Gly Leu His Ser Leu Gln
530 535 540
Glu Ile Arg Val Cys Glu Ile His Glu Asp Asn Ser Thr Arg Ser Ser
545 550 555 560
Gln His Phe Tyr Tyr Asp Gly Glu Leu Phe Leu Ser Gln Asn Leu Glu
565 570 575
Thr Leu Glu Trp Thr Met Pro Gln Ser Ser Arg Ala Gln Thr Leu Ala
580 585 590
Met Asn Val Arg Asn Phe Leu Lys Glu Asp Ala Met Glu Thr Asp Thr
595 600 605
His Tyr His Ala Met Arg Ala Asp Cys Leu Ser Glu Leu Arg Arg Tyr
610 615 620
Leu Lys Ser Gly Val Val Leu Arg Arg Thr
625 630
<210> 105
<211> 1902
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий HC_MIC25 трастузумаба
<400> 105
gaagtccaat tggtcgaatc aggcggtgga ctcgtgcaac ctggaggttc gttacgctta 60
tcatgtgctg caagtggatt taatattaaa gatacctaca tccactgggt acgtcaagct 120
cccggcaagg gtctcgagtg ggtcgcacgc atttacccca ccaacggata cacgcgctac 180
gccgattcag tgaagggacg tttcacaatc tctgctgata ctagcaaaaa taccgcatac 240
ctccagatga actctcttag ggccgaggac acagctgtgt actactgtag ccgttgggga 300
ggagacggtt tttacgcaat ggattactgg ggccaaggaa ccctggtcac agtttcatcg 360
gcgtcgacca agggcccgtc agtgttcccg ctggccccgt catccaagtc cacgtctggg 420
ggcacagcag ccctgggatg cttggtcaag gactacttcc ccgagcccgt gactgtgtcc 480
tggaactccg gagcactgac ctccggagtg cacacctttc ccgcggtgct gcagtcctcc 540
ggactgtact ccctgtcgtc ggtcgtgacc gtgccgagct cctcgctcgg aacccagacc 600
tacatctgca acgtgaacca caagccctcg aacaccaaag tggacaagaa ggtcgagccc 660
aaaagctgcg acaagactca cacttgtccg ccgtgccccg cccccgaact gctgggtggc 720
ccctccgtgt tcctgttccc gcctaagcct aaggacaccc ttatgatcag ccgcacccct 780
gaagtgacct gtgtcgtcgt ggcagtgtca cacgaggacc cggaggtcaa gttcaattgg 840
tacgtggacg gcgtggaagt gcataacgca aagaccaagc ctcgggagga acagtacgcc 900
tcgacctacc gcgtggtgtc agtcctgact gtgctgcacc aggactggct gaacgggaag 960
gagtacaagt gcaaagtgtc gaacaaggcc ctgccggctc caattgaaaa gaccatcagc 1020
aaggccaagg gccagccaag ggaaccacag gtgtacaccc tccctccttc ccgggacgag 1080
ctgaccaaaa accaagtgtc cctgacttgc cttgtgaagg ggttctaccc ttctgacatt 1140
gccgtcgaat gggaatcgaa cggacagcct gaaaacaact ataagactac cccgcccgtg 1200
ctggattccg acggaagctt cttcctgtac tccaagctga ccgtggacaa gtcgagatgg 1260
cagcagggaa atgtgttcag ctgctccgtg atgcatgagg cgctgcacaa ccactacacc 1320
cagaagtcac tgagcctctc ccccggagga ggtggcagcg agcctcacag cctccggtat 1380
aatttgactg tactctcttg ggatggctcc gtgcagtccg gctttctgac tgaagttcat 1440
ctcgacggtc aacctttcct gcgctgcgac cgacaaaaat gccgcgccaa gccccaaggg 1500
cagtgggccg aagatgtact gggaaacaag acctgggacc gggagacacg agacctgaca 1560
ggctggggca aggacttgcg catgacactc gcccatatca aggaccagaa ggaaggattg 1620
cactctttgc aagagattcg cgtgtgtgaa atccacgagg acaattcaac gaggagctcc 1680
cagcacttct attacgatgg agaactcttc ttgtcacaga acttggaaac cctcgaatgg 1740
actatgcctc agagctctcg ggcacagact ctcgctatga acgttagaaa cttccttaag 1800
gaggatgcta tggagaccga tactcactac cacgccatgc gcgccgactg cctctctgaa 1860
ctgcggagat atctgaagtc cggcgtggtt ttgagaagaa cc 1902
<210> 106
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.wt ритуксимаба
<400> 106
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Arg Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Met Ser Phe His Tyr Phe Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 107
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.wt ритуксимаба
<400> 107
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgcgcgaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgatgtc attccactat 1140
ttctcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 108
<211> 187
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид варианта R80W ULPB2 альфа1-альфа2
<400> 108
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Met Ser Phe His Tyr Phe Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser Ala Gly Ala Pro Pro Met Val
180 185
<210> 109
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант R80W
ULPB2 альфа1-альфа2
<400> 109
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgatgtcat tccactattt ctcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 110
<211> 513
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий библиотеку NNK варианта
R80W ULPB2 альфа1-альфа2
<220>
<221> misc_feature
<222> (457)..(458)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (460)..(461)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (463)..(464)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (466)..(467)
<223> n представляет собой a, c, g или t
<220>
<221> misc_feature
<222> (469)..(470)
<223> n представляет собой a, c, g или t
<400> 110
gagccccata gcctcagcta tgacattacg gtgattccca aatttcgccc aggaccacgt 60
tggtgcgccg tccagggtca ggtagatgaa aagactttcc tgcattatga ttgcggcaat 120
aaaaccgtga cgccggtatc gccgttaggc aaaaaattga atgtcacgac agcgtggaaa 180
gcacagaacc cggtgttgcg cgaggtagtc gatattttga cggaacaact ctgggacatt 240
cagctcgaga attacacccc aaaagaaccg ctgacgctgc aagcgcgtat gtcgtgcgaa 300
caaaaagcag aaggtcactc tagcgggagt tggcagtttt ccttcgatgg gcagattttt 360
ctgctgtttg attcggagaa acgcatgtgg actacagtcc acccgggtgc ccggaaaatg 420
aaagagaagt gggagaatga taaagtggtg gccactnnkn nknnknnknn ktccatgggc 480
gattgcattg gctggttaga ggattttctc atg 513
<210> 111
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид, вариант ULBP2.C ULBP2 альфа1-альфа2
<400> 111
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Thr Ile Leu Trp Gln Thr Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser
180
<210> 112
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант ULBP2.C
ULBP2 альфа1-альфа2
<400> 112
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgactattc tgtggcagac ttcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 113
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид, вариант ULBP2.R ULBP2 альфа1-альфа2
<400> 113
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Thr Leu Leu Trp Gly Trp Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser
180
<210> 114
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант ULBP2.R
ULBP2 альфа1-альфа2
<400> 114
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgactttgt tgtgggggtg gtcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 115
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид, вариант ULBP2.AA ULBP2 альфа1-альфа2
<400> 115
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Thr Met Phe Trp Ser Trp Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser
180
<210> 116
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант ULBP2.AA
ULBP2 альфа1-альфа2
<400> 116
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgactatgt tttggagttg gtcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 117
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид, вариант ULBP2.AB ULBP2 альфа1-альфа2
<400> 117
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Thr Leu Met Trp Gln Trp Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser
180
<210> 118
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант ULBP2.AB
ULBP2 альфа1-альфа2
<400> 118
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgactctta tgtggcagtg gtcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 119
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.C ритуксимаба
<400> 119
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Thr Ile Leu Trp Gln Thr Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 120
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.C ритуксимаба
<400> 120
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgtgggaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgactat tctgtggcag 1140
acttcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 121
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.R ритуксимаба
<400> 121
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Thr Leu Leu Trp Gly Trp Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 122
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.R ритуксимаба
<400> 122
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgtgggaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgacttt gttgtggggg 1140
tggtcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 123
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.AA ритуксимаба
<400> 123
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Thr Met Phe Trp Ser Trp Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 124
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.AA ритуксимаба
<400> 124
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgtgggaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgactat gttttggagt 1140
tggtcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 125
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.AB ритуксимаба
<400> 125
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Thr Leu Met Trp Gln Trp Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 126
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.AB ритуксимаба
<400> 126
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgtgggaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgactct tatgtggcag 1140
tggtcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 127
<211> 180
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид, вариант ULBP2.S3 ULBP2 альфа1-альфа2
<400> 127
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
1 5 10 15
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
20 25 30
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
35 40 45
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
50 55 60
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
65 70 75 80
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
85 90 95
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
100 105 110
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
115 120 125
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
130 135 140
Glu Asn Asp Lys Val Val Ala Thr Lys Leu Tyr Leu Trp Ser Met Gly
145 150 155 160
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
165 170 175
Leu Glu Pro Ser
180
<210> 128
<211> 540
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий вариант ULBP2.S3
ULBP2 альфа1-альфа2
<400> 128
gagccccata gtctgagcta cgacatcaca gttattccca agttcaggcc cggaccgcgc 60
tggtgtgccg tgcaaggaca agtcgacgaa aaaacctttc ttcattacga ttgcggaaat 120
aagactgtaa cgccagtctc tcctttaggt aagaagttaa acgtcactac ggcgtggaag 180
gcacaaaacc ccgtcctgcg cgaggtcgtc gacatcctga ctgaacaatt gtgggacatc 240
cagctcgaga attacactcc aaaggagcct cttaccctgc aggctagaat gtcttgcgag 300
caaaaggcag agggccactc ctccggcagc tggcagttca gtttcgacgg acaaatcttt 360
ctgttattcg attcagagaa gagaatgtgg actacagttc accccggtgc ccgtaaaatg 420
aaggagaagt gggaaaacga caaagtggtg gcgactaagc tttatctttg gtcgatggga 480
gactgcatcg gttggctgga agatttcctc atgggtatgg actccacttt ggagccatcg 540
<210> 129
<211> 404
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.S3 ритуксимаба
<400> 129
Gln Ile Val Leu Ser Gln Ser Pro Ala Ile Leu Ser Ala Ser Pro Gly
1 5 10 15
Glu Lys Val Thr Met Thr Cys Arg Ala Ser Ser Ser Val Ser Tyr Ile
20 25 30
His Trp Phe Gln Gln Lys Pro Gly Ser Ser Pro Lys Pro Trp Ile Tyr
35 40 45
Ala Thr Ser Asn Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Arg Val Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Thr Ser Asn Pro Pro Thr
85 90 95
Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly Ser
210 215 220
Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe Arg
225 230 235 240
Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys Thr
245 250 255
Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser Pro
260 265 270
Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn Pro
275 280 285
Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp Ile
290 295 300
Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala Arg
305 310 315 320
Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp Gln
325 330 335
Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys Arg
340 345 350
Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys Trp
355 360 365
Glu Asn Asp Lys Val Val Ala Thr Lys Leu Tyr Leu Trp Ser Met Gly
370 375 380
Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser Thr
385 390 395 400
Leu Glu Pro Ser
<210> 130
<211> 1212
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.S3 ритуксимаба
<400> 130
cagatcgtgt tgtcccaatc acccgcaatt ctctctgcga gcccagggga gaaggtgacc 60
atgacttgcc gcgcttcaag ttccgtatcc tacattcact ggttccagca gaagcccgga 120
agttccccta agccctggat ctatgctaca tccaatctgg caagcggtgt tcccgttaga 180
ttttccggaa gcgggtctgg aaccagttac agtctgacta tttccagggt cgaggccgaa 240
gatgcggcta cttattattg ccaacagtgg acctctaacc cacccacatt cggcggcggc 300
actaagttgg aaattaagcg gaccgtggcc gccccgagcg tgttcatttt ccctccctcc 360
gacgagcagt tgaaatcggg caccgctagc gtggtctgcc ttctcaacaa tttctatcca 420
cgggaagcca aagtgcagtg gaaggtcgac aacgcgctcc aatccgggaa ctcacaggaa 480
tccgtgactg agcaggattc caaggactcg acctactccc tgtcatccac gctgaccctg 540
agcaaggcag actacgagaa gcacaaggtc tacgcctgcg aagtgacaca ccagggactg 600
tccagccccg tgaccaagag cttcaacaga ggagaatgcg cacctacctc aagctctgga 660
ggaggtggca gcgagcccca tagtctgagc tacgacatca cagttattcc caagttcagg 720
cccggaccgc gctggtgtgc cgtgcaagga caagtcgacg aaaaaacctt tcttcattac 780
gattgcggaa ataagactgt aacgccagtc tctcctttag gtaagaagtt aaacgtcact 840
acggcgtgga aggcacaaaa ccccgtcctg cgcgaggtcg tcgacatcct gactgaacaa 900
ttgtgggaca tccagctcga gaattacact ccaaaggagc ctcttaccct gcaggctaga 960
atgtcttgcg agcaaaaggc agagggccac tcctccggca gctggcagtt cagtttcgac 1020
ggacaaatct ttctgttatt cgattcagag aagagaatgt ggactacagt tcaccccggt 1080
gcccgtaaaa tgaaggagaa gtgggaaaac gacaaagtgg tggcgactaa gctttatctt 1140
tggtcgatgg gagactgcat cggttggctg gaagatttcc tcatgggtat ggactccact 1200
ttggagccat cg 1212
<210> 131
<211> 634
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид HC_ULBP2.R80W ритуксимаба
<400> 131
Gln Val Gln Leu Gln Gln Pro Gly Ala Glu Leu Val Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Met Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Ser Tyr
20 25 30
Asn Met His Trp Val Lys Gln Thr Pro Gly Arg Gly Leu Glu Trp Ile
35 40 45
Gly Ala Ile Tyr Pro Gly Asn Gly Asp Thr Ser Tyr Asn Gln Lys Phe
50 55 60
Lys Gly Lys Ala Thr Leu Thr Ala Asp Lys Ser Ser Ser Thr Ala Tyr
65 70 75 80
Met Gln Leu Ser Ser Leu Thr Ser Glu Asp Ser Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ser Thr Tyr Tyr Gly Gly Asp Trp Tyr Phe Asn Val Trp Gly
100 105 110
Ala Gly Thr Thr Val Thr Val Ser Ala Ala Ser Thr Lys Gly Pro Ser
115 120 125
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
130 135 140
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
145 150 155 160
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
165 170 175
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
180 185 190
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
195 200 205
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
210 215 220
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
225 230 235 240
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
245 250 255
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
260 265 270
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
275 280 285
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Ala Ser Thr Tyr
290 295 300
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
305 310 315 320
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
325 330 335
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
340 345 350
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
355 360 365
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
370 375 380
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
385 390 395 400
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
405 410 415
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
420 425 430
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
435 440 445
Pro Gly Gly Gly Gly Ser Glu Pro His Ser Leu Ser Tyr Asp Ile Thr
450 455 460
Val Ile Pro Lys Phe Arg Pro Gly Pro Arg Trp Cys Ala Val Gln Gly
465 470 475 480
Gln Val Asp Glu Lys Thr Phe Leu His Tyr Asp Cys Gly Asn Lys Thr
485 490 495
Val Thr Pro Val Ser Pro Leu Gly Lys Lys Leu Asn Val Thr Thr Ala
500 505 510
Trp Lys Ala Gln Asn Pro Val Leu Arg Glu Val Val Asp Ile Leu Thr
515 520 525
Glu Gln Leu Trp Asp Ile Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro
530 535 540
Leu Thr Leu Gln Ala Arg Met Ser Cys Glu Gln Lys Ala Glu Gly His
545 550 555 560
Ser Ser Gly Ser Trp Gln Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu
565 570 575
Phe Asp Ser Glu Lys Arg Met Trp Thr Thr Val His Pro Gly Ala Arg
580 585 590
Lys Met Lys Glu Lys Trp Glu Asn Asp Lys Val Val Ala Met Ser Phe
595 600 605
His Tyr Phe Ser Met Gly Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu
610 615 620
Met Gly Met Asp Ser Thr Leu Glu Pro Ser
625 630
<210> 132
<211> 1902
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий HC_ULBP2.R80W ритуксимаба
<400> 132
caagttcagc ttcagcagcc gggggctgag ttggtgaaac ccggggccag tgtgaagatg 60
agctgtaaag cgagcggcta caccttcact tcttataata tgcattgggt taagcaaacg 120
ccaggaaggg ggctggagtg gatcggcgct atttacccag gtaacggtga cacatcatat 180
aaccaaaagt ttaagggaaa ggcaaccctc acagcggaca agagtagctc aaccgcatac 240
atgcaactgt caagccttac ctccgaagac agcgcagtgt actactgcgc cagaagcacc 300
tactatgggg gtgattggta cttcaacgtc tggggggctg gcaccacagt gactgtaagc 360
gcagcgtcga ccaagggccc gtcagtgttc ccgctggccc cgtcatccaa gtccacgtct 420
gggggcacag cagccctggg atgcttggtc aaggactact tccccgagcc cgtgactgtg 480
tcctggaact ccggagcact gacctccgga gtgcacacct ttcccgcggt gctgcagtcc 540
tccggactgt actccctgtc gtcggtcgtg accgtgccga gctcctcgct cggaacccag 600
acctacatct gcaacgtgaa ccacaagccc tcgaacacca aagtggacaa gaaggtcgag 660
cccaaaagct gcgacaagac tcacacttgt ccgccgtgcc ccgcccccga actgctgggt 720
ggcccctccg tgttcctgtt cccgcctaag cctaaggaca cccttatgat cagccgcacc 780
cctgaagtga cctgtgtcgt cgtggcagtg tcacacgagg acccggaggt caagttcaat 840
tggtacgtgg acggcgtgga agtgcataac gcaaagacca agcctcggga ggaacagtac 900
gcctcgacct accgcgtggt gtcagtcctg actgtgctgc accaggactg gctgaacggg 960
aaggagtaca agtgcaaagt gtcgaacaag gccctgccgg ctccaattga aaagaccatc 1020
agcaaggcca agggccagcc aagggaacca caggtgtaca ccctccctcc ttcccgggac 1080
gagctgacca aaaaccaagt gtccctgact tgccttgtga aggggttcta cccttctgac 1140
attgccgtcg aatgggaatc gaacggacag cctgaaaaca actataagac taccccgccc 1200
gtgctggatt ccgacggaag cttcttcctg tactccaagc tgaccgtgga caagtcgaga 1260
tggcagcagg gaaatgtgtt cagctgctcc gtgatgcatg aggcgctgca caaccactac 1320
acccagaagt cactgagcct ctcccccgga ggaggtggca gcgagcccca tagtctgagc 1380
tacgacatca cagttattcc caagttcagg cccggaccgc gctggtgtgc cgtgcaagga 1440
caagtcgacg aaaaaacctt tcttcattac gattgcggaa ataagactgt aacgccagtc 1500
tctcctttag gtaagaagtt aaacgtcact acggcgtgga aggcacaaaa ccccgtcctg 1560
cgcgaggtcg tcgacatcct gactgaacaa ttgtgggaca tccagctcga gaattacact 1620
ccaaaggagc ctcttaccct gcaggctaga atgtcttgcg agcaaaaggc agagggccac 1680
tcctccggca gctggcagtt cagtttcgac ggacaaatct ttctgttatt cgattcagag 1740
aagagaatgt ggactacagt tcaccccggt gcccgtaaaa tgaaggagaa gtgggaaaac 1800
gacaaagtgg tggcgatgtc attccactat ttctcgatgg gagactgcat cggttggctg 1860
gaagatttcc tcatgggtat ggactccact ttggagccat cg 1902
<210> 133
<211> 405
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид LC_ULBP2.R трастузумаба
<400> 133
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Asp Val Asn Thr Ala
20 25 30
Val Ala Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ser Ala Ser Phe Leu Tyr Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Arg Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln His Tyr Thr Thr Pro Pro
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys Ala Pro Thr Ser Ser Ser Gly Gly Gly Gly
210 215 220
Ser Glu Pro His Ser Leu Ser Tyr Asp Ile Thr Val Ile Pro Lys Phe
225 230 235 240
Arg Pro Gly Pro Arg Trp Cys Ala Val Gln Gly Gln Val Asp Glu Lys
245 250 255
Thr Phe Leu His Tyr Asp Cys Gly Asn Lys Thr Val Thr Pro Val Ser
260 265 270
Pro Leu Gly Lys Lys Leu Asn Val Thr Thr Ala Trp Lys Ala Gln Asn
275 280 285
Pro Val Leu Arg Glu Val Val Asp Ile Leu Thr Glu Gln Leu Trp Asp
290 295 300
Ile Gln Leu Glu Asn Tyr Thr Pro Lys Glu Pro Leu Thr Leu Gln Ala
305 310 315 320
Arg Met Ser Cys Glu Gln Lys Ala Glu Gly His Ser Ser Gly Ser Trp
325 330 335
Gln Phe Ser Phe Asp Gly Gln Ile Phe Leu Leu Phe Asp Ser Glu Lys
340 345 350
Arg Met Trp Thr Thr Val His Pro Gly Ala Arg Lys Met Lys Glu Lys
355 360 365
Trp Glu Asn Asp Lys Val Val Ala Thr Leu Leu Trp Gly Trp Ser Met
370 375 380
Gly Asp Cys Ile Gly Trp Leu Glu Asp Phe Leu Met Gly Met Asp Ser
385 390 395 400
Thr Leu Glu Pro Ser
405
<210> 134
<211> 1215
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий LC_ULBP2.R трастузумаба
<400> 134
gatatccaaa tgactcaatc accatcttca ctctccgcga gcgtgggtga tcgggtcacc 60
atcacatgta gggcgagcca agatgtgaat accgccgtcg cgtggtatca acaaaagccg 120
ggaaaagcac caaaactgct tatatactct gcatccttcc tgtactctgg ggtgccaagc 180
cggttctccg gtagtagatc tggtactgac tttacactca ctatcagcag tctgcaacct 240
gaggactttg cgacatacta ttgccagcag cactacacaa ccccacctac atttggtcag 300
gggacaaagg tggagatcaa gcggaccgtg gccgccccga gcgtgttcat tttccctccc 360
tccgacgagc agttgaaatc gggcaccgct agcgtggtct gccttctcaa caatttctat 420
ccacgggaag ccaaagtgca gtggaaggtc gacaacgcgc tccaatccgg gaactcacag 480
gaatccgtga ctgagcagga ttccaaggac tcgacctact ccctgtcatc cacgctgacc 540
ctgagcaagg cagactacga gaagcacaag gtctacgcct gcgaagtgac acaccaggga 600
ctgtccagcc ccgtgaccaa gagcttcaac agaggagaat gcgcacctac ctcaagctct 660
ggaggaggtg gcagcgagcc ccatagtctg agctacgaca tcacagttat tcccaagttc 720
aggcccggac cgcgctggtg tgccgtgcaa ggacaagtcg acgaaaaaac ctttcttcat 780
tacgattgcg gaaataagac tgtaacgcca gtctctcctt taggtaagaa gttaaacgtc 840
actacggcgt ggaaggcaca aaaccccgtc ctgcgcgagg tcgtcgacat cctgactgaa 900
caattgtggg acatccagct cgagaattac actccaaagg agcctcttac cctgcaggct 960
agaatgtctt gcgagcaaaa ggcagagggc cactcctccg gcagctggca gttcagtttc 1020
gacggacaaa tctttctgtt attcgattca gagaagagaa tgtggactac agttcacccc 1080
ggtgcccgta aaatgaagga gaagtgggaa aacgacaaag tggtggcgac tttgttgtgg 1140
gggtggtcga tgggagactg catcggttgg ctggaagatt tcctcatggg tatggactcc 1200
actttggagc catcg 1215
<210> 135
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид эктодомена NKG2D.wt
<400> 135
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
145 150 155
<210> 136
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий эктодомен NKG2D.wt
<400> 136
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agctaccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggctacatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtg 468
<210> 137
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид эктодомена NKG2D.YA
<400> 137
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
145 150 155
<210> 138
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий эктодомен NKG2D.YA
<400> 138
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agcgcccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggctacatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtg 468
<210> 139
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид эктодомена NKG2D.AF
<400> 139
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val
145 150 155
<210> 140
<211> 468
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий эктодомен NKG2D.AF
<400> 140
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agcgcccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggcttcatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtg 468
<210> 141
<211> 69
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид шарнира CD8альфа и трансмембранного домена
<400> 141
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile
35 40 45
Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val
50 55 60
Ile Thr Leu Tyr Cys
65
<210> 142
<211> 207
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий шарнир CD8альфа и
трансмембранный домен
<400> 142
accaccacac cagctcctag acctccaact cctgctccta caatcgccag ccagcctctg 60
tctctgaggc cagaagcttg tagacctgct gcaggcggag ccgtgcatac aagaggactg 120
gatttcgcct gcgacatcta catctgggcc cctctggctg gaacatgtgg cgtgctgctg 180
ctgagcctgg tcatcaccct gtactgc 207
<210> 143
<211> 42
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид 4-1BB
<400> 143
Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln Pro Phe Met
1 5 10 15
Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser Cys Arg Phe
20 25 30
Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu
35 40
<210> 144
<211> 126
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий 4-1BB
<400> 144
aagcggggca gaaagaagct gctgtacatc tttaagcagc ccttcatgcg gcccgtgcag 60
accacacaag aggaagatgg ctgctcctgc agattccccg aggaagaaga aggcggctgc 120
gagctg 126
<210> 145
<211> 112
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид CD3-дзета
<400> 145
Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly
1 5 10 15
Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr
20 25 30
Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys
35 40 45
Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys
50 55 60
Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg
65 70 75 80
Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala
85 90 95
Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg
100 105 110
<210> 146
<211> 336
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий CD3-дзета
<400> 146
agagtgaagt tcagccgttc tgccgacgct cccgcctata agcagggaca gaaccagctg 60
tacaacgagc tgaacctggg gagaagagaa gagtacgacg tgctggacaa gcggagaggc 120
agagatcctg agatgggcgg caagcccaga cggaagaatc ctcaagaggg cctgtataat 180
gagctgcaga aagacaagat ggccgaggcc tacagcgaga tcggaatgaa gggcgagcgc 240
agaagaggca agggacacga tggactgtac cagggcctga gcaccgccac caaggatacc 300
tatgatgccc tgcacatgca ggccctgcct ccaaga 336
<210> 147
<211> 239
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид EGFP
<400> 147
Met Val Ser Lys Gly Glu Glu Leu Phe Thr Gly Val Val Pro Ile Leu
1 5 10 15
Val Glu Leu Asp Gly Asp Val Asn Gly His Lys Phe Ser Val Ser Gly
20 25 30
Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile
35 40 45
Cys Thr Thr Gly Lys Leu Pro Val Pro Trp Pro Thr Leu Val Thr Thr
50 55 60
Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg Tyr Pro Asp His Met Lys
65 70 75 80
Gln His Asp Phe Phe Lys Ser Ala Met Pro Glu Gly Tyr Val Gln Glu
85 90 95
Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu
100 105 110
Val Lys Phe Glu Gly Asp Thr Leu Val Asn Arg Ile Glu Leu Lys Gly
115 120 125
Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu Gly His Lys Leu Glu Tyr
130 135 140
Asn Tyr Asn Ser His Asn Val Tyr Ile Met Ala Asp Lys Gln Lys Asn
145 150 155 160
Gly Ile Lys Ala Asn Phe Lys Ile Arg His Asn Ile Glu Asp Gly Ser
165 170 175
Val Gln Leu Ala Asp His Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly
180 185 190
Pro Val Leu Leu Pro Asp Asn His Tyr Leu Ser Thr Gln Ser Ala Leu
195 200 205
Ser Lys Asp Pro Asn Glu Lys Arg Asp His Met Val Leu Leu Glu Phe
210 215 220
Val Thr Ala Ala Gly Ile Thr Leu Gly Met Asp Glu Leu Tyr Lys
225 230 235
<210> 148
<211> 717
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий EGFP
<400> 148
atggtgtcta aaggcgagga actgttcacc ggcgtggtgc ccattctggt ggaactggac 60
ggggatgtga acggccacaa gtttagcgtt agcggcgaag gcgaagggga tgccacatac 120
ggaaagctga ccctgaagtt catctgcacc accggcaagc tgcctgtgcc ttggcctaca 180
ctggtcacca cactgacata cggcgtgcag tgctttagca gataccccga ccatatgaag 240
cagcacgact tcttcaagtc cgccatgcct gagggctacg tgcaagagcg gaccatcttc 300
tttaaggacg acggcaacta caagaccagg gccgaagtga agtttgaggg cgacaccctg 360
gtcaaccgga tcgagctgaa gggcatcgac ttcaaagagg atggcaacat cctgggccac 420
aagctcgagt acaactacaa cagccacaac gtgtacatca tggccgacaa gcagaagaac 480
ggcatcaagg ccaacttcaa gatccggcac aacatcgagg acggcagcgt tcagctggcc 540
gatcactacc agcagaacac ccctatcgga gatggccctg tgctgctccc cgacaatcac 600
tacctgagca cacagagcgc cctgagcaag gaccccaacg agaagaggga tcacatggtg 660
ctgctggaat ttgtgaccgc cgcaggcatc accctcggca tggacgaact gtacaaa 717
<210> 149
<211> 473
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 149
Thr Thr Thr Pro Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala
1 5 10 15
Ser Gln Pro Leu Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly
20 25 30
Gly Ala Val His Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile
35 40 45
Trp Ala Pro Leu Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val
50 55 60
Ile Thr Leu Tyr Cys Ser Leu Lys Arg Gly Arg Lys Lys Leu Leu Tyr
65 70 75 80
Ile Phe Lys Gln Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu
85 90 95
Asp Gly Cys Ser Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu
100 105 110
Leu Arg Val Lys Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln
115 120 125
Gly Gln Asn Gln Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu
130 135 140
Tyr Asp Val Leu Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly
145 150 155 160
Lys Pro Arg Arg Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln
165 170 175
Lys Asp Lys Met Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu
180 185 190
Arg Arg Arg Gly Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr
195 200 205
Ala Thr Lys Asp Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro
210 215 220
Arg Ser Gly Ser Gly Ser Gly Ser Gly Ser Met Val Ser Lys Gly Glu
225 230 235 240
Glu Leu Phe Thr Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp
245 250 255
Val Asn Gly His Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala
260 265 270
Thr Tyr Gly Lys Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu
275 280 285
Pro Val Pro Trp Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln
290 295 300
Cys Phe Ser Arg Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys
305 310 315 320
Ser Ala Met Pro Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys
325 330 335
Asp Asp Gly Asn Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp
340 345 350
Thr Leu Val Asn Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp
355 360 365
Gly Asn Ile Leu Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn
370 375 380
Val Tyr Ile Met Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe
385 390 395 400
Lys Ile Arg His Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His
405 410 415
Tyr Gln Gln Asn Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp
420 425 430
Asn His Tyr Leu Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu
435 440 445
Lys Arg Asp His Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile
450 455 460
Thr Leu Gly Met Asp Glu Leu Tyr Lys
465 470
<210> 150
<211> 1419
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 150
accaccacac cagctcctag acctccaact cctgctccta caatcgccag ccagcctctg 60
tctctgaggc cagaagcttg tagacctgct gcaggcggag ccgtgcatac aagaggactg 120
gatttcgcct gcgacatcta catctgggcc cctctggctg gaacatgtgg cgtgctgctg 180
ctgagcctgg tcatcaccct gtactgcagc ctgaagcggg gcagaaagaa gctgctgtac 240
atctttaagc agcccttcat gcggcccgtg cagaccacac aagaggaaga tggctgctcc 300
tgcagattcc ccgaggaaga agaaggcggc tgcgagctga gagtgaagtt cagccgttct 360
gccgacgctc ccgcctataa gcagggacag aaccagctgt acaacgagct gaacctgggg 420
agaagagaag agtacgacgt gctggacaag cggagaggca gagatcctga gatgggcggc 480
aagcccagac ggaagaatcc tcaagagggc ctgtataatg agctgcagaa agacaagatg 540
gccgaggcct acagcgagat cggaatgaag ggcgagcgca gaagaggcaa gggacacgat 600
ggactgtacc agggcctgag caccgccacc aaggatacct atgatgccct gcacatgcag 660
gccctgcctc caagatcagg ctctggttct ggcagcggca gcatggtgtc taaaggcgag 720
gaactgttca ccggcgtggt gcccattctg gtggaactgg acggggatgt gaacggccac 780
aagtttagcg ttagcggcga aggcgaaggg gatgccacat acggaaagct gaccctgaag 840
ttcatctgca ccaccggcaa gctgcctgtg ccttggccta cactggtcac cacactgaca 900
tacggcgtgc agtgctttag cagatacccc gaccatatga agcagcacga cttcttcaag 960
tccgccatgc ctgagggcta cgtgcaagag cggaccatct tctttaagga cgacggcaac 1020
tacaagacca gggccgaagt gaagtttgag ggcgacaccc tggtcaaccg gatcgagctg 1080
aagggcatcg acttcaaaga ggatggcaac atcctgggcc acaagctcga gtacaactac 1140
aacagccaca acgtgtacat catggccgac aagcagaaga acggcatcaa ggccaacttc 1200
aagatccggc acaacatcga ggacggcagc gttcagctgg ccgatcacta ccagcagaac 1260
acccctatcg gagatggccc tgtgctgctc cccgacaatc actacctgag cacacagagc 1320
gccctgagca aggaccccaa cgagaagagg gatcacatgg tgctgctgga atttgtgacc 1380
gccgcaggca tcaccctcgg catggacgaa ctgtacaaa 1419
<210> 151
<211> 629
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид полного химерного рецептора антигена
NKG2D.wt_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 151
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Tyr His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val Thr Thr Thr Pro
145 150 155 160
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
165 170 175
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
180 185 190
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu
195 200 205
Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr
210 215 220
Cys Ser Leu Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
225 230 235 240
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
245 250 255
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
260 265 270
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln
275 280 285
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
290 295 300
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
305 310 315 320
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
325 330 335
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
340 345 350
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
355 360 365
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Ser Gly Ser
370 375 380
Gly Ser Gly Ser Gly Ser Met Val Ser Lys Gly Glu Glu Leu Phe Thr
385 390 395 400
Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn Gly His
405 410 415
Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys
420 425 430
Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp
435 440 445
Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg
450 455 460
Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro
465 470 475 480
Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn
485 490 495
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
500 505 510
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
515 520 525
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Met
530 535 540
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
545 550 555 560
Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn
565 570 575
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
580 585 590
Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
595 600 605
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
610 615 620
Asp Glu Leu Tyr Lys
625
<210> 152
<211> 1887
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий
полный химерный рецептор антигена
NKG2D.wt_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 152
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agctaccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggctacatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtgac caccacacca 480
gctcctagac ctccaactcc tgctcctaca atcgccagcc agcctctgtc tctgaggcca 540
gaagcttgta gacctgctgc aggcggagcc gtgcatacaa gaggactgga tttcgcctgc 600
gacatctaca tctgggcccc tctggctgga acatgtggcg tgctgctgct gagcctggtc 660
atcaccctgt actgcagcct gaagcggggc agaaagaagc tgctgtacat ctttaagcag 720
cccttcatgc ggcccgtgca gaccacacaa gaggaagatg gctgctcctg cagattcccc 780
gaggaagaag aaggcggctg cgagctgaga gtgaagttca gccgttctgc cgacgctccc 840
gcctataagc agggacagaa ccagctgtac aacgagctga acctggggag aagagaagag 900
tacgacgtgc tggacaagcg gagaggcaga gatcctgaga tgggcggcaa gcccagacgg 960
aagaatcctc aagagggcct gtataatgag ctgcagaaag acaagatggc cgaggcctac 1020
agcgagatcg gaatgaaggg cgagcgcaga agaggcaagg gacacgatgg actgtaccag 1080
ggcctgagca ccgccaccaa ggatacctat gatgccctgc acatgcaggc cctgcctcca 1140
agatcaggct ctggttctgg cagcggcagc atggtgtcta aaggcgagga actgttcacc 1200
ggcgtggtgc ccattctggt ggaactggac ggggatgtga acggccacaa gtttagcgtt 1260
agcggcgaag gcgaagggga tgccacatac ggaaagctga ccctgaagtt catctgcacc 1320
accggcaagc tgcctgtgcc ttggcctaca ctggtcacca cactgacata cggcgtgcag 1380
tgctttagca gataccccga ccatatgaag cagcacgact tcttcaagtc cgccatgcct 1440
gagggctacg tgcaagagcg gaccatcttc tttaaggacg acggcaacta caagaccagg 1500
gccgaagtga agtttgaggg cgacaccctg gtcaaccgga tcgagctgaa gggcatcgac 1560
ttcaaagagg atggcaacat cctgggccac aagctcgagt acaactacaa cagccacaac 1620
gtgtacatca tggccgacaa gcagaagaac ggcatcaagg ccaacttcaa gatccggcac 1680
aacatcgagg acggcagcgt tcagctggcc gatcactacc agcagaacac ccctatcgga 1740
gatggccctg tgctgctccc cgacaatcac tacctgagca cacagagcgc cctgagcaag 1800
gaccccaacg agaagaggga tcacatggtg ctgctggaat ttgtgaccgc cgcaggcatc 1860
accctcggca tggacgaact gtacaaa 1887
<210> 153
<211> 629
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид полного химерного рецептора антигена
NKG2D.YA_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 153
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Tyr Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val Thr Thr Thr Pro
145 150 155 160
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
165 170 175
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
180 185 190
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu
195 200 205
Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr
210 215 220
Cys Ser Leu Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
225 230 235 240
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
245 250 255
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
260 265 270
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln
275 280 285
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
290 295 300
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
305 310 315 320
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
325 330 335
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
340 345 350
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
355 360 365
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Ser Gly Ser
370 375 380
Gly Ser Gly Ser Gly Ser Met Val Ser Lys Gly Glu Glu Leu Phe Thr
385 390 395 400
Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn Gly His
405 410 415
Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys
420 425 430
Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp
435 440 445
Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg
450 455 460
Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro
465 470 475 480
Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn
485 490 495
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
500 505 510
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
515 520 525
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Met
530 535 540
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
545 550 555 560
Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn
565 570 575
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
580 585 590
Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
595 600 605
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
610 615 620
Asp Glu Leu Tyr Lys
625
<210> 154
<211> 1887
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий
Полный химерный рецептор антигена
NKG2D.YA_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 154
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agcgcccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggctacatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtgac caccacacca 480
gctcctagac ctccaactcc tgctcctaca atcgccagcc agcctctgtc tctgaggcca 540
gaagcttgta gacctgctgc aggcggagcc gtgcatacaa gaggactgga tttcgcctgc 600
gacatctaca tctgggcccc tctggctgga acatgtggcg tgctgctgct gagcctggtc 660
atcaccctgt actgcagcct gaagcggggc agaaagaagc tgctgtacat ctttaagcag 720
cccttcatgc ggcccgtgca gaccacacaa gaggaagatg gctgctcctg cagattcccc 780
gaggaagaag aaggcggctg cgagctgaga gtgaagttca gccgttctgc cgacgctccc 840
gcctataagc agggacagaa ccagctgtac aacgagctga acctggggag aagagaagag 900
tacgacgtgc tggacaagcg gagaggcaga gatcctgaga tgggcggcaa gcccagacgg 960
aagaatcctc aagagggcct gtataatgag ctgcagaaag acaagatggc cgaggcctac 1020
agcgagatcg gaatgaaggg cgagcgcaga agaggcaagg gacacgatgg actgtaccag 1080
ggcctgagca ccgccaccaa ggatacctat gatgccctgc acatgcaggc cctgcctcca 1140
agatcaggct ctggttctgg cagcggcagc atggtgtcta aaggcgagga actgttcacc 1200
ggcgtggtgc ccattctggt ggaactggac ggggatgtga acggccacaa gtttagcgtt 1260
agcggcgaag gcgaagggga tgccacatac ggaaagctga ccctgaagtt catctgcacc 1320
accggcaagc tgcctgtgcc ttggcctaca ctggtcacca cactgacata cggcgtgcag 1380
tgctttagca gataccccga ccatatgaag cagcacgact tcttcaagtc cgccatgcct 1440
gagggctacg tgcaagagcg gaccatcttc tttaaggacg acggcaacta caagaccagg 1500
gccgaagtga agtttgaggg cgacaccctg gtcaaccgga tcgagctgaa gggcatcgac 1560
ttcaaagagg atggcaacat cctgggccac aagctcgagt acaactacaa cagccacaac 1620
gtgtacatca tggccgacaa gcagaagaac ggcatcaagg ccaacttcaa gatccggcac 1680
aacatcgagg acggcagcgt tcagctggcc gatcactacc agcagaacac ccctatcgga 1740
gatggccctg tgctgctccc cgacaatcac tacctgagca cacagagcgc cctgagcaag 1800
gaccccaacg agaagaggga tcacatggtg ctgctggaat ttgtgaccgc cgcaggcatc 1860
accctcggca tggacgaact gtacaaa 1887
<210> 155
<211> 629
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический пептид полного химерного рецептора антигена
NKG2D.AF_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 155
Met Ala Leu Pro Val Thr Ala Leu Leu Leu Pro Leu Ala Leu Leu Leu
1 5 10 15
His Ala Ala Arg Pro Leu Phe Asn Gln Glu Val Gln Ile Pro Leu Thr
20 25 30
Glu Ser Tyr Cys Gly Pro Cys Pro Lys Asn Trp Ile Cys Tyr Lys Asn
35 40 45
Asn Cys Tyr Gln Phe Phe Asp Glu Ser Lys Asn Trp Tyr Glu Ser Gln
50 55 60
Ala Ser Cys Met Ser Gln Asn Ala Ser Leu Leu Lys Val Tyr Ser Lys
65 70 75 80
Glu Asp Gln Asp Leu Leu Lys Leu Val Lys Ser Ala His Trp Met Gly
85 90 95
Leu Val His Ile Pro Thr Asn Gly Ser Trp Gln Trp Glu Asp Gly Ser
100 105 110
Ile Leu Ser Pro Asn Leu Leu Thr Ile Ile Glu Met Gln Lys Gly Asp
115 120 125
Cys Ala Leu Tyr Ala Ser Ser Phe Lys Gly Phe Ile Glu Asn Cys Ser
130 135 140
Thr Pro Asn Thr Tyr Ile Cys Met Gln Arg Thr Val Thr Thr Thr Pro
145 150 155 160
Ala Pro Arg Pro Pro Thr Pro Ala Pro Thr Ile Ala Ser Gln Pro Leu
165 170 175
Ser Leu Arg Pro Glu Ala Cys Arg Pro Ala Ala Gly Gly Ala Val His
180 185 190
Thr Arg Gly Leu Asp Phe Ala Cys Asp Ile Tyr Ile Trp Ala Pro Leu
195 200 205
Ala Gly Thr Cys Gly Val Leu Leu Leu Ser Leu Val Ile Thr Leu Tyr
210 215 220
Cys Ser Leu Lys Arg Gly Arg Lys Lys Leu Leu Tyr Ile Phe Lys Gln
225 230 235 240
Pro Phe Met Arg Pro Val Gln Thr Thr Gln Glu Glu Asp Gly Cys Ser
245 250 255
Cys Arg Phe Pro Glu Glu Glu Glu Gly Gly Cys Glu Leu Arg Val Lys
260 265 270
Phe Ser Arg Ser Ala Asp Ala Pro Ala Tyr Lys Gln Gly Gln Asn Gln
275 280 285
Leu Tyr Asn Glu Leu Asn Leu Gly Arg Arg Glu Glu Tyr Asp Val Leu
290 295 300
Asp Lys Arg Arg Gly Arg Asp Pro Glu Met Gly Gly Lys Pro Arg Arg
305 310 315 320
Lys Asn Pro Gln Glu Gly Leu Tyr Asn Glu Leu Gln Lys Asp Lys Met
325 330 335
Ala Glu Ala Tyr Ser Glu Ile Gly Met Lys Gly Glu Arg Arg Arg Gly
340 345 350
Lys Gly His Asp Gly Leu Tyr Gln Gly Leu Ser Thr Ala Thr Lys Asp
355 360 365
Thr Tyr Asp Ala Leu His Met Gln Ala Leu Pro Pro Arg Ser Gly Ser
370 375 380
Gly Ser Gly Ser Gly Ser Met Val Ser Lys Gly Glu Glu Leu Phe Thr
385 390 395 400
Gly Val Val Pro Ile Leu Val Glu Leu Asp Gly Asp Val Asn Gly His
405 410 415
Lys Phe Ser Val Ser Gly Glu Gly Glu Gly Asp Ala Thr Tyr Gly Lys
420 425 430
Leu Thr Leu Lys Phe Ile Cys Thr Thr Gly Lys Leu Pro Val Pro Trp
435 440 445
Pro Thr Leu Val Thr Thr Leu Thr Tyr Gly Val Gln Cys Phe Ser Arg
450 455 460
Tyr Pro Asp His Met Lys Gln His Asp Phe Phe Lys Ser Ala Met Pro
465 470 475 480
Glu Gly Tyr Val Gln Glu Arg Thr Ile Phe Phe Lys Asp Asp Gly Asn
485 490 495
Tyr Lys Thr Arg Ala Glu Val Lys Phe Glu Gly Asp Thr Leu Val Asn
500 505 510
Arg Ile Glu Leu Lys Gly Ile Asp Phe Lys Glu Asp Gly Asn Ile Leu
515 520 525
Gly His Lys Leu Glu Tyr Asn Tyr Asn Ser His Asn Val Tyr Ile Met
530 535 540
Ala Asp Lys Gln Lys Asn Gly Ile Lys Ala Asn Phe Lys Ile Arg His
545 550 555 560
Asn Ile Glu Asp Gly Ser Val Gln Leu Ala Asp His Tyr Gln Gln Asn
565 570 575
Thr Pro Ile Gly Asp Gly Pro Val Leu Leu Pro Asp Asn His Tyr Leu
580 585 590
Ser Thr Gln Ser Ala Leu Ser Lys Asp Pro Asn Glu Lys Arg Asp His
595 600 605
Met Val Leu Leu Glu Phe Val Thr Ala Ala Gly Ile Thr Leu Gly Met
610 615 620
Asp Glu Leu Tyr Lys
625
<210> 156
<211> 1887
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический полинуклеотид, кодирующий
полный химерный рецептор антигена
NKG2D.AF_CD8шарнирTM_4-1BB_CD3-дзета_EGFP
<400> 156
atggcattgc ctgttacagc tctgctgctg cccctggctc tgcttctgca tgctgccaga 60
cctctgttca atcaagaggt gcagatccct ctgaccgaga gctactgtgg cccctgtcct 120
aagaactgga tctgctacaa gaacaactgc taccagttct tcgacgagag caagaattgg 180
tacgagagcc aggccagctg catgagccag aatgccagcc tgctgaaggt gtacagcaaa 240
gaggaccagg atctgctgaa gctggtcaag agcgcccact ggatgggact cgtgcacatc 300
cctacaaacg gcagctggca gtgggaggac ggctctatcc tgtctcctaa cctgctgacc 360
atcatcgaga tgcagaaggg cgactgcgcc ctgtacgcca gcagctttaa gggcttcatc 420
gagaactgca gcacccctaa cacctacatc tgtatgcagc ggaccgtgac caccacacca 480
gctcctagac ctccaactcc tgctcctaca atcgccagcc agcctctgtc tctgaggcca 540
gaagcttgta gacctgctgc aggcggagcc gtgcatacaa gaggactgga tttcgcctgc 600
gacatctaca tctgggcccc tctggctgga acatgtggcg tgctgctgct gagcctggtc 660
atcaccctgt actgcagcct gaagcggggc agaaagaagc tgctgtacat ctttaagcag 720
cccttcatgc ggcccgtgca gaccacacaa gaggaagatg gctgctcctg cagattcccc 780
gaggaagaag aaggcggctg cgagctgaga gtgaagttca gccgttctgc cgacgctccc 840
gcctataagc agggacagaa ccagctgtac aacgagctga acctggggag aagagaagag 900
tacgacgtgc tggacaagcg gagaggcaga gatcctgaga tgggcggcaa gcccagacgg 960
aagaatcctc aagagggcct gtataatgag ctgcagaaag acaagatggc cgaggcctac 1020
agcgagatcg gaatgaaggg cgagcgcaga agaggcaagg gacacgatgg actgtaccag 1080
ggcctgagca ccgccaccaa ggatacctat gatgccctgc acatgcaggc cctgcctcca 1140
agatcaggct ctggttctgg cagcggcagc atggtgtcta aaggcgagga actgttcacc 1200
ggcgtggtgc ccattctggt ggaactggac ggggatgtga acggccacaa gtttagcgtt 1260
agcggcgaag gcgaagggga tgccacatac ggaaagctga ccctgaagtt catctgcacc 1320
accggcaagc tgcctgtgcc ttggcctaca ctggtcacca cactgacata cggcgtgcag 1380
tgctttagca gataccccga ccatatgaag cagcacgact tcttcaagtc cgccatgcct 1440
gagggctacg tgcaagagcg gaccatcttc tttaaggacg acggcaacta caagaccagg 1500
gccgaagtga agtttgaggg cgacaccctg gtcaaccgga tcgagctgaa gggcatcgac 1560
ttcaaagagg atggcaacat cctgggccac aagctcgagt acaactacaa cagccacaac 1620
gtgtacatca tggccgacaa gcagaagaac ggcatcaagg ccaacttcaa gatccggcac 1680
aacatcgagg acggcagcgt tcagctggcc gatcactacc agcagaacac ccctatcgga 1740
gatggccctg tgctgctccc cgacaatcac tacctgagca cacagagcgc cctgagcaag 1800
gaccccaacg agaagaggga tcacatggtg ctgctggaat ttgtgaccgc cgcaggcatc 1860
accctcggca tggacgaact gtacaaa 1887
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| МОДИФИЦИРОВАННЫЕ НЕПРИРОДНЫЕ ЛИГАНДЫ NKG2D, КОТОРЫЕ ИЗБИРАТЕЛЬНО ДОСТАВЛЯЮТ ПРИСОЕДИНЕННЫЕ ГЕТЕРОЛОГИЧНЫЕ МОЛЕКУЛЫ К НЕПРИРОДНЫМ РЕЦЕПТОРАМ NKG2D НА CAR-КЛЕТКАХ | 2020 |
|
RU2823728C2 |
| НЕПРИРОДНЫЕ РЕЦЕПТОРЫ NKG2D, КОТОРЫЕ НЕПОСРЕДСТВЕННО НЕ ПЕРЕДАЮТ СИГНАЛ КЛЕТКАМ, С КОТОРЫМИ ОНИ СВЯЗАНЫ | 2019 |
|
RU2832538C2 |
| Белки, связывающие NKG2D, CD16 и опухолеассоциированный антиген | 2018 |
|
RU2816716C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2788531C2 |
| БЕЛОК, СВЯЗЫВАЮЩИЙСЯ С NKG2D, CD16 И С ОПУХОЛЕСПЕЦИФИЧЕСКИМ АНТИГЕНОМ | 2018 |
|
RU2826991C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА NKG2D, CD16 И TROP2 | 2018 |
|
RU2820629C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ ВСМА, NKG2D И CD16 | 2018 |
|
RU2805254C2 |
| БЕЛКИ, СВЯЗЫВАЮЩИЕ CD33, NKG2D И CD16 | 2018 |
|
RU2820603C2 |
| УСЕЧЕННЫЕ ХИМЕРНЫЕ РЕЦЕПТОРЫ NKG2D И ПУТИ ИХ ПРИМЕНЕНИЯ В ИММУНОТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ | 2018 |
|
RU2830556C2 |
| ПОЛИСПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ БЕЛКИ, НАЦЕЛЕННЫЕ НА CAIX, ANO1, МЕЗОТЕЛИН, TROP2, СEA ИЛИ КЛАУДИН-18.2 | 2018 |
|
RU2792671C2 |


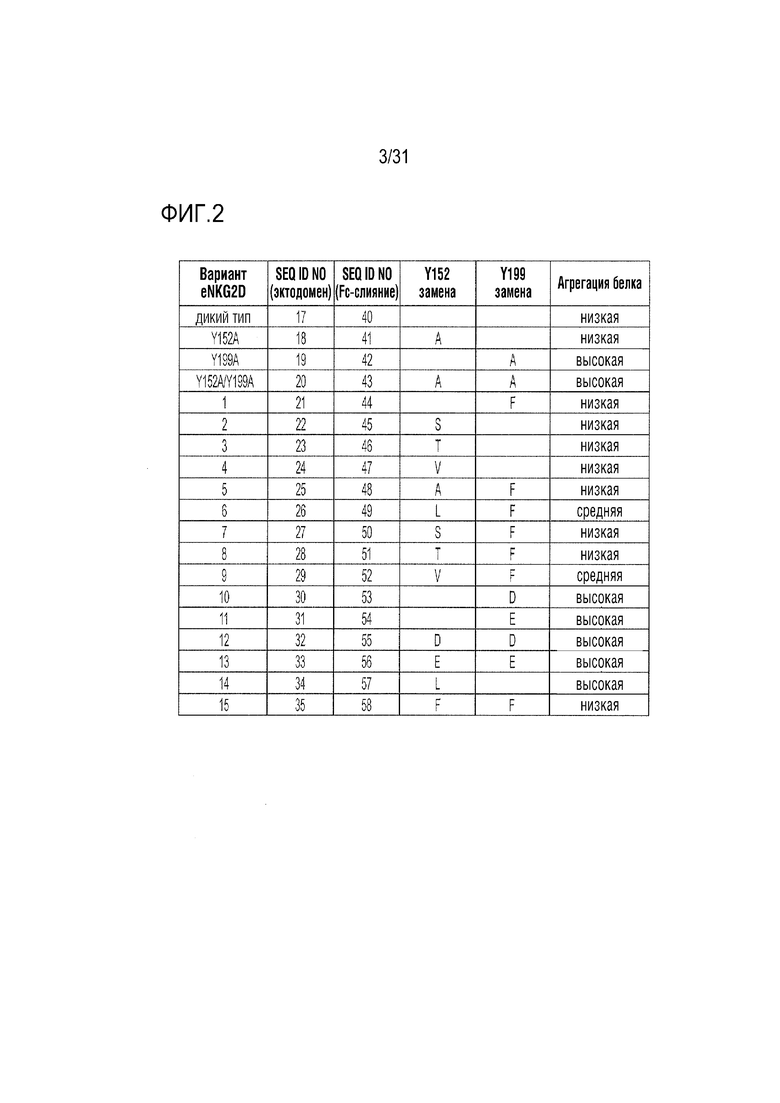
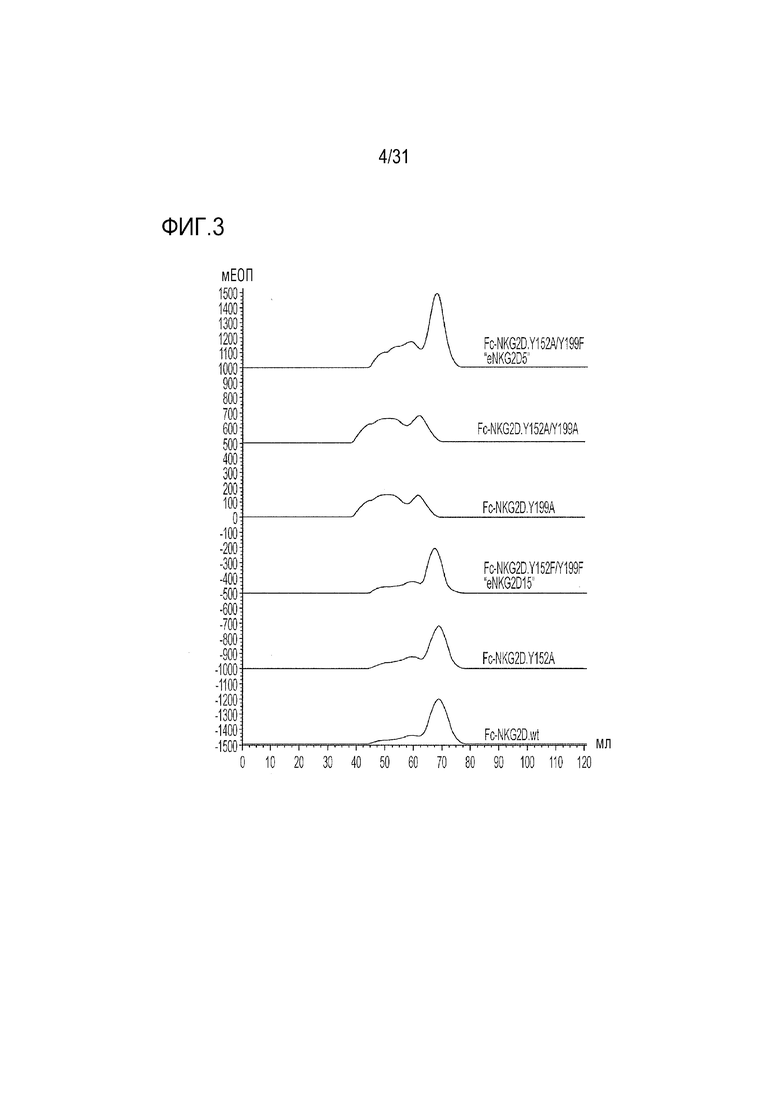
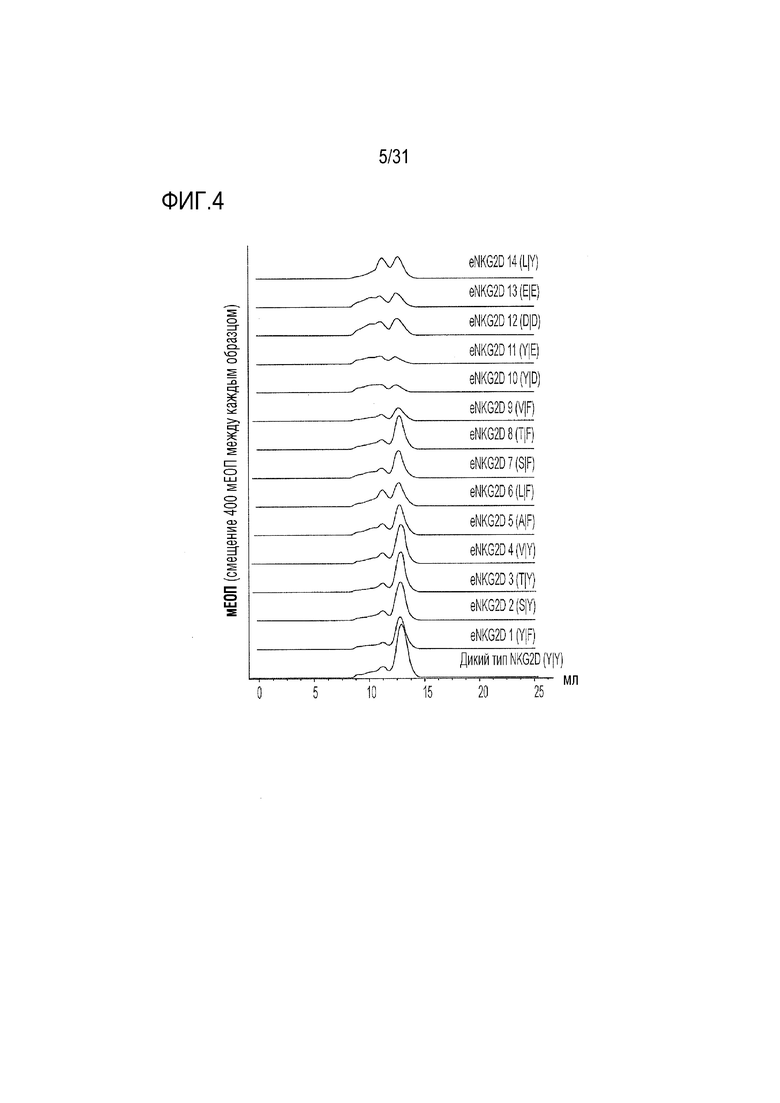

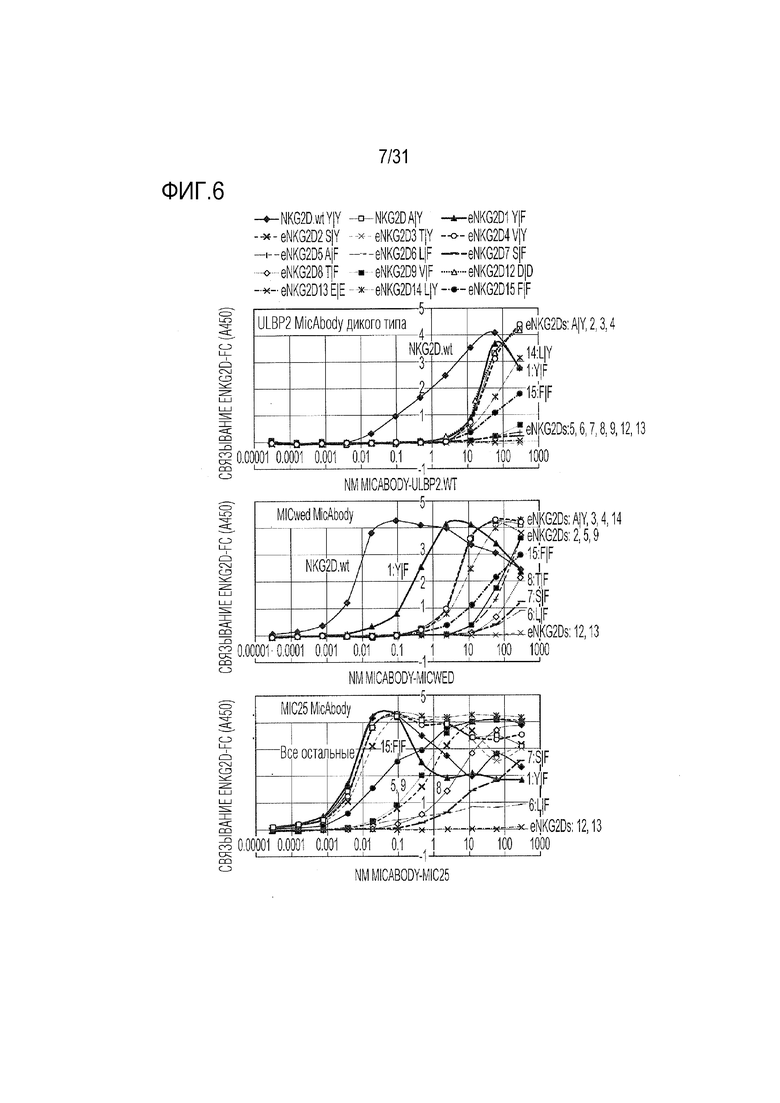
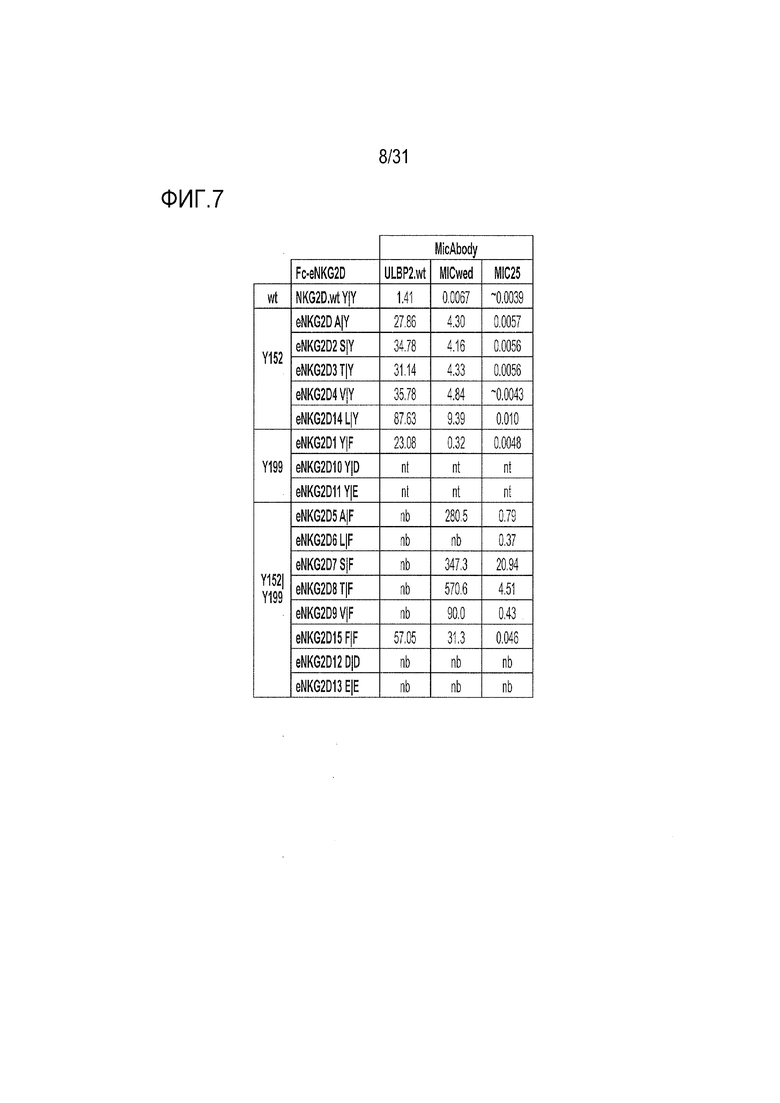
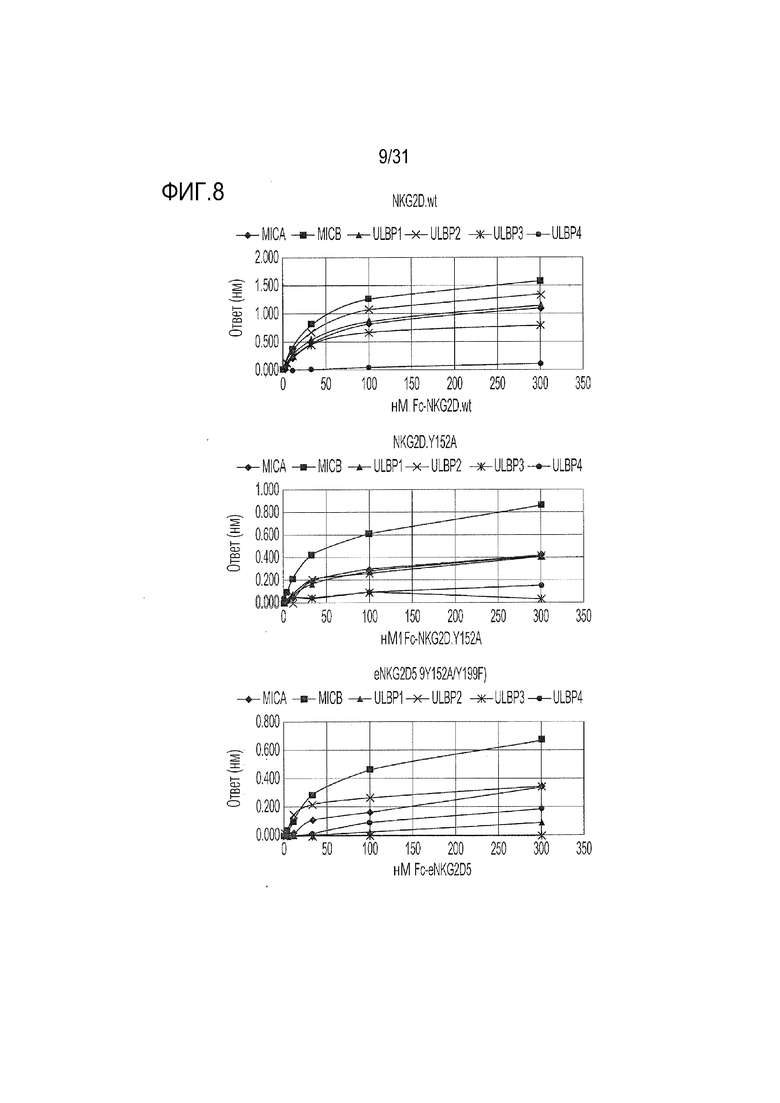
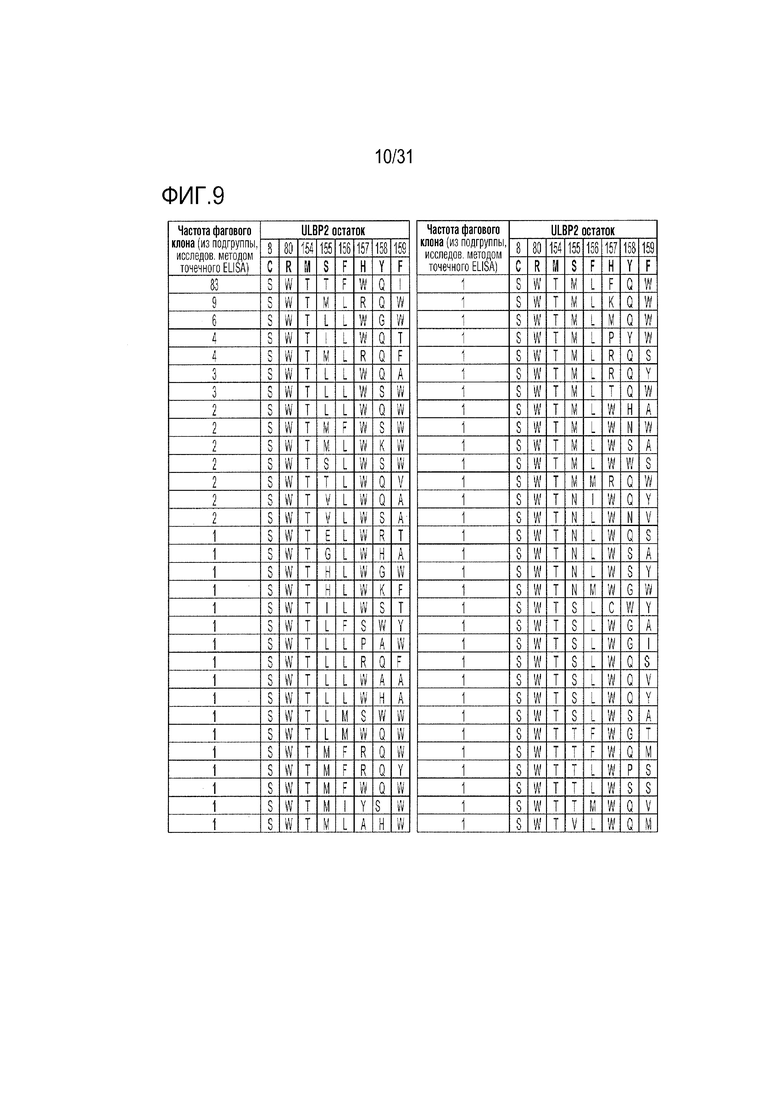
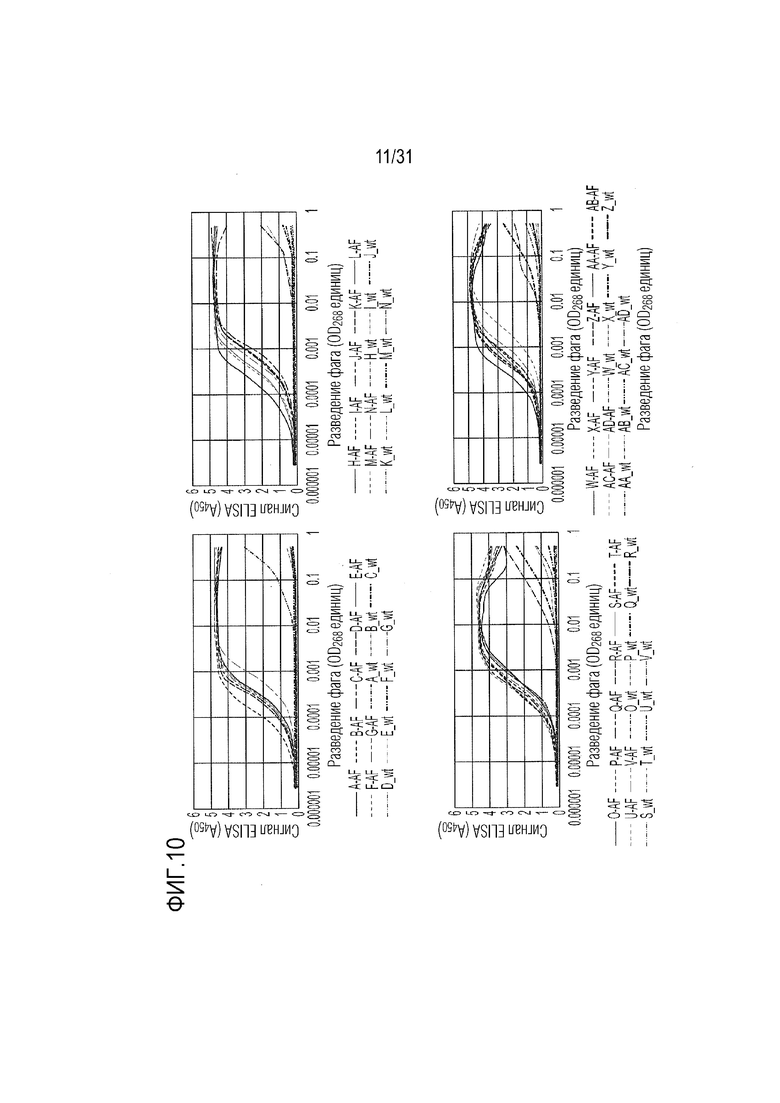
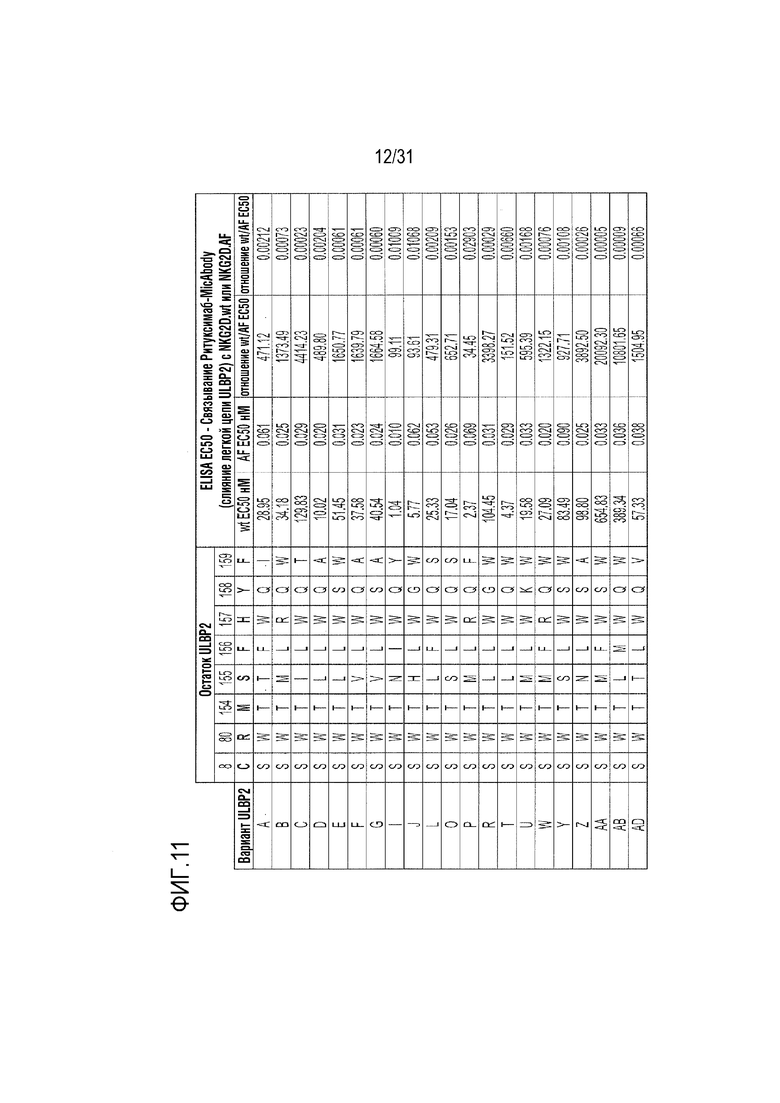

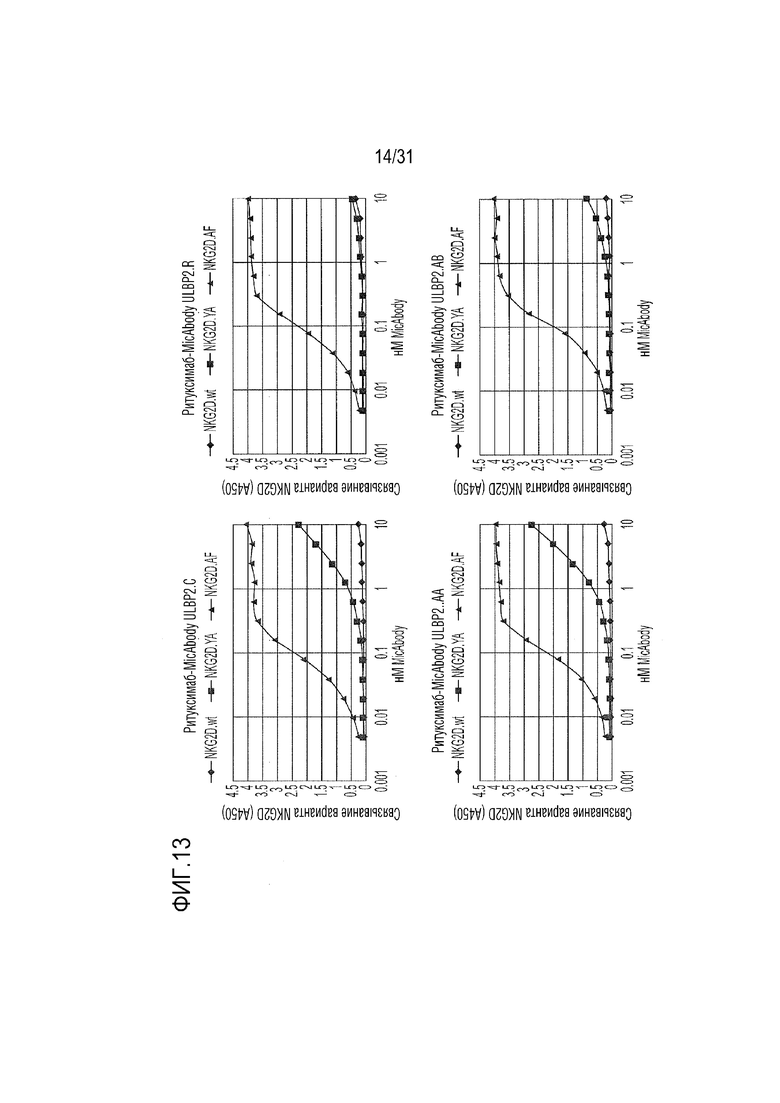



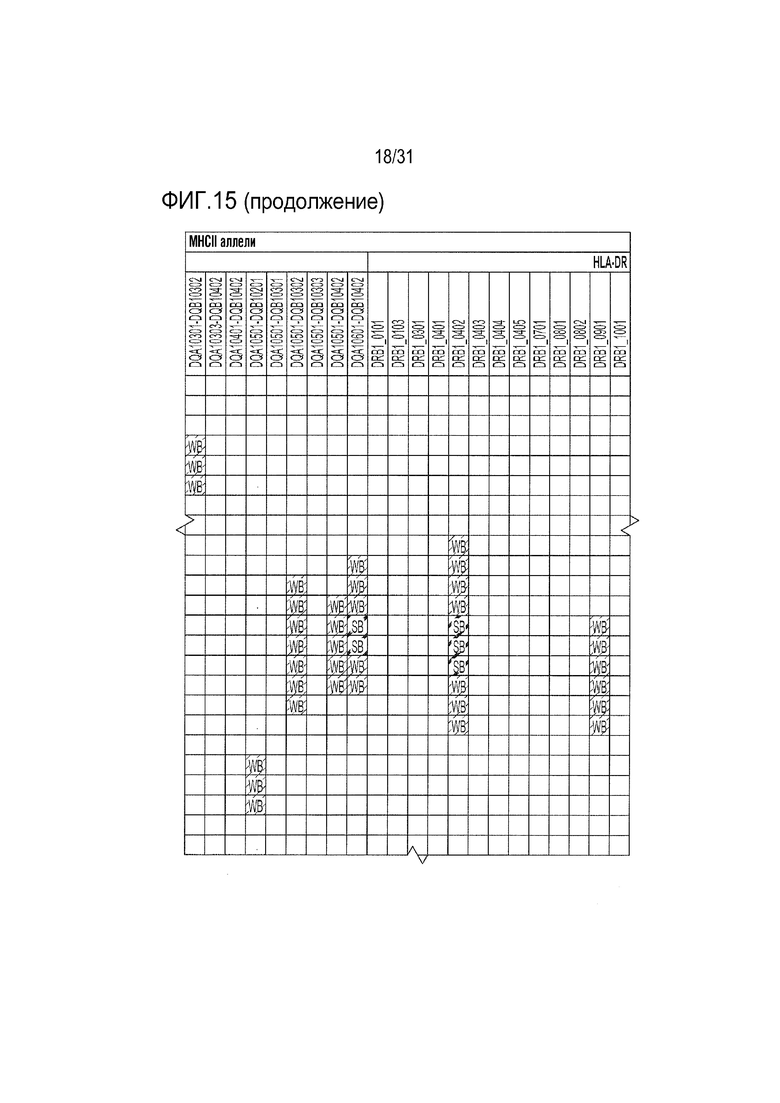
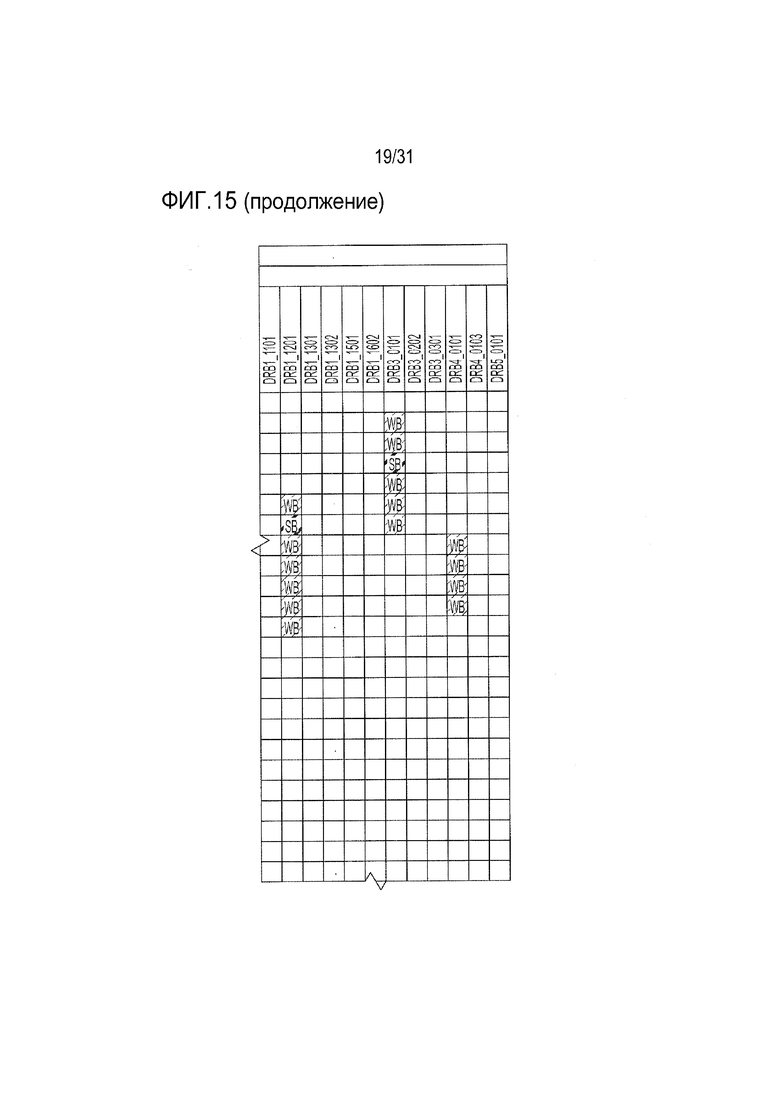
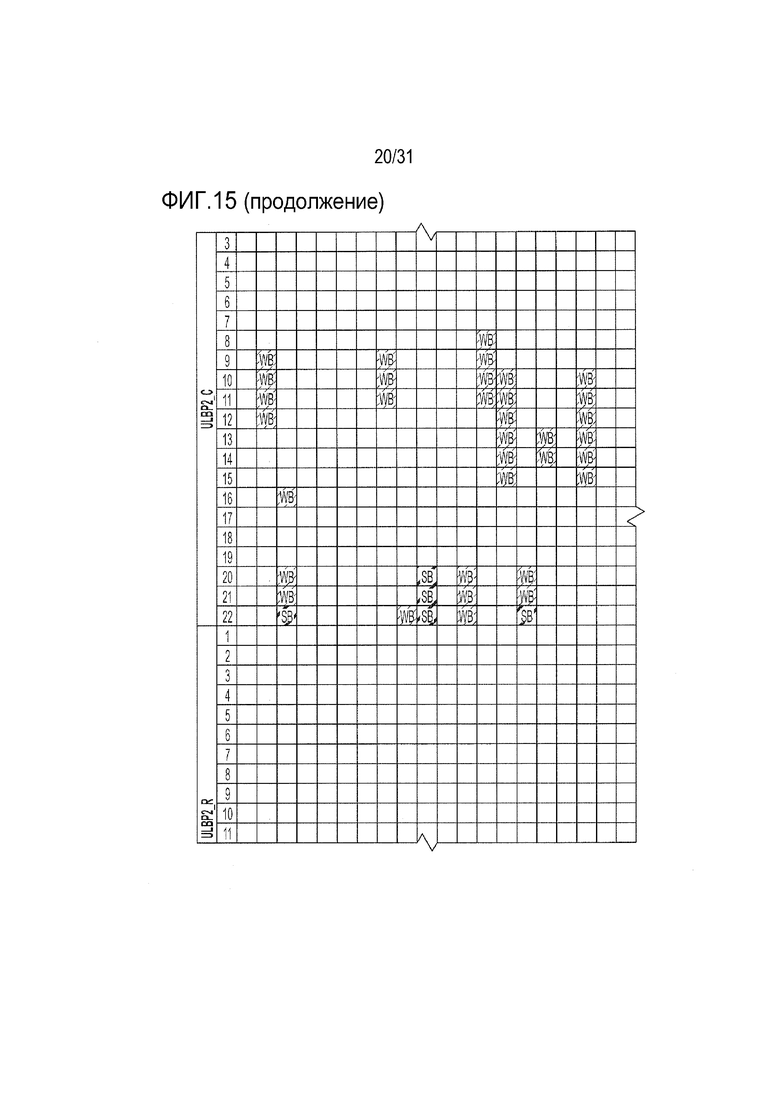
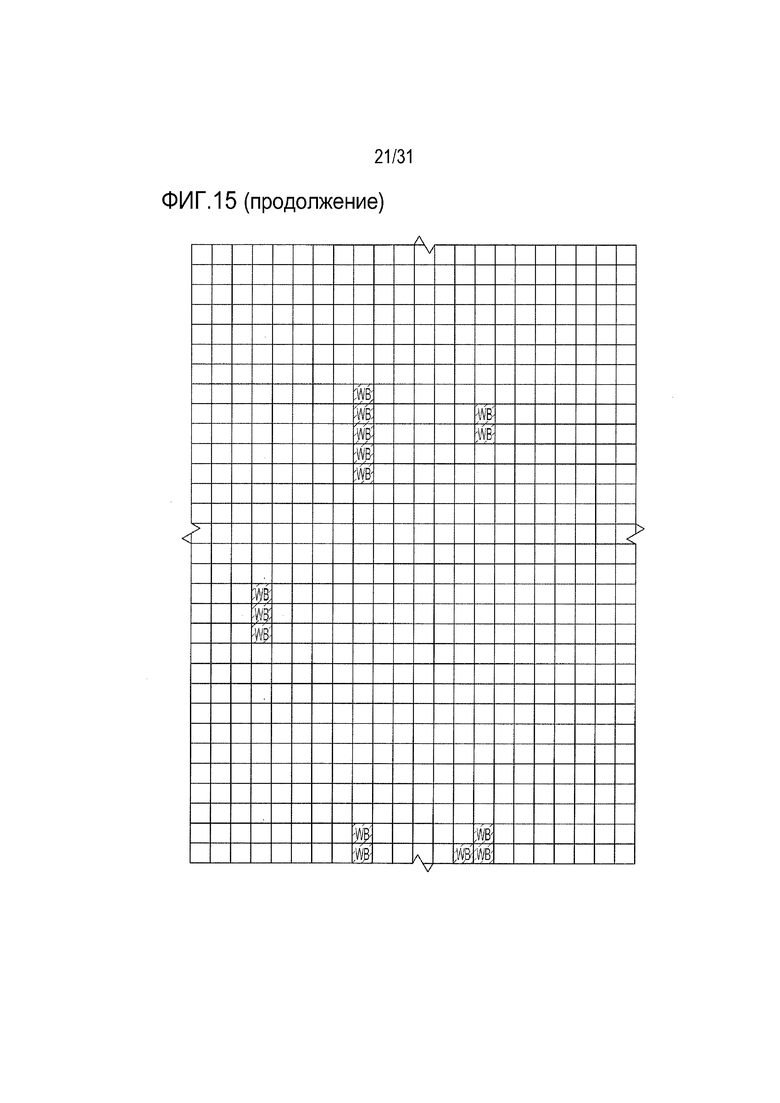
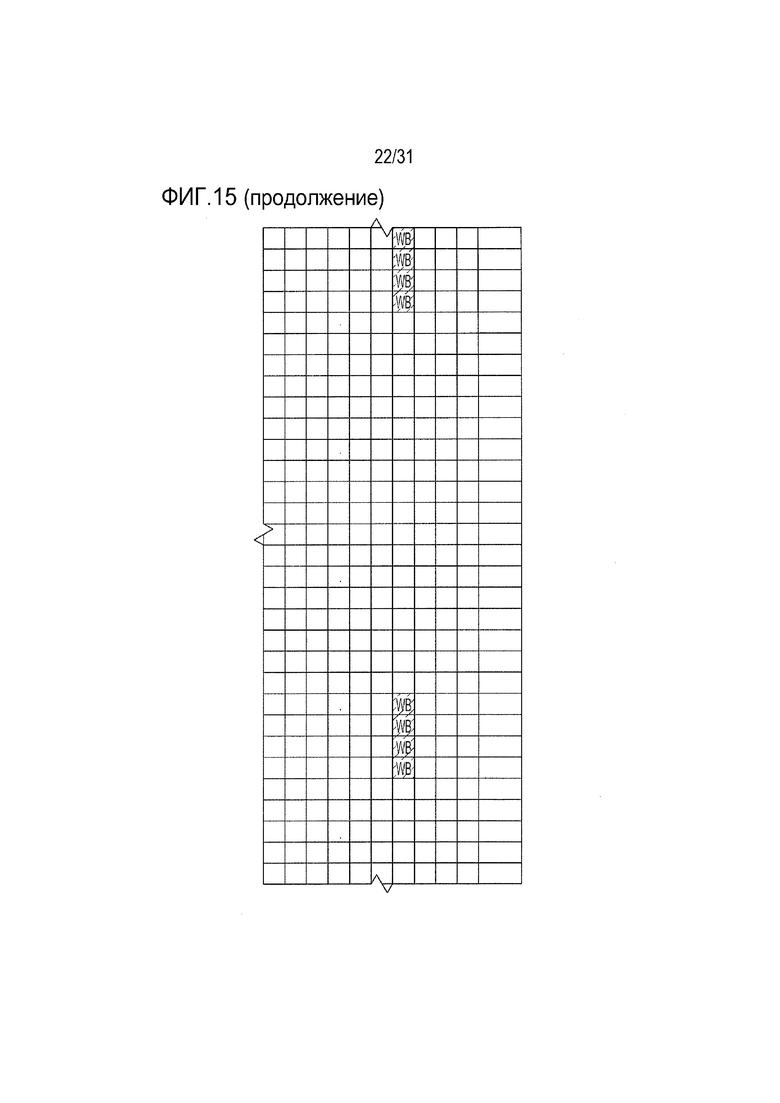
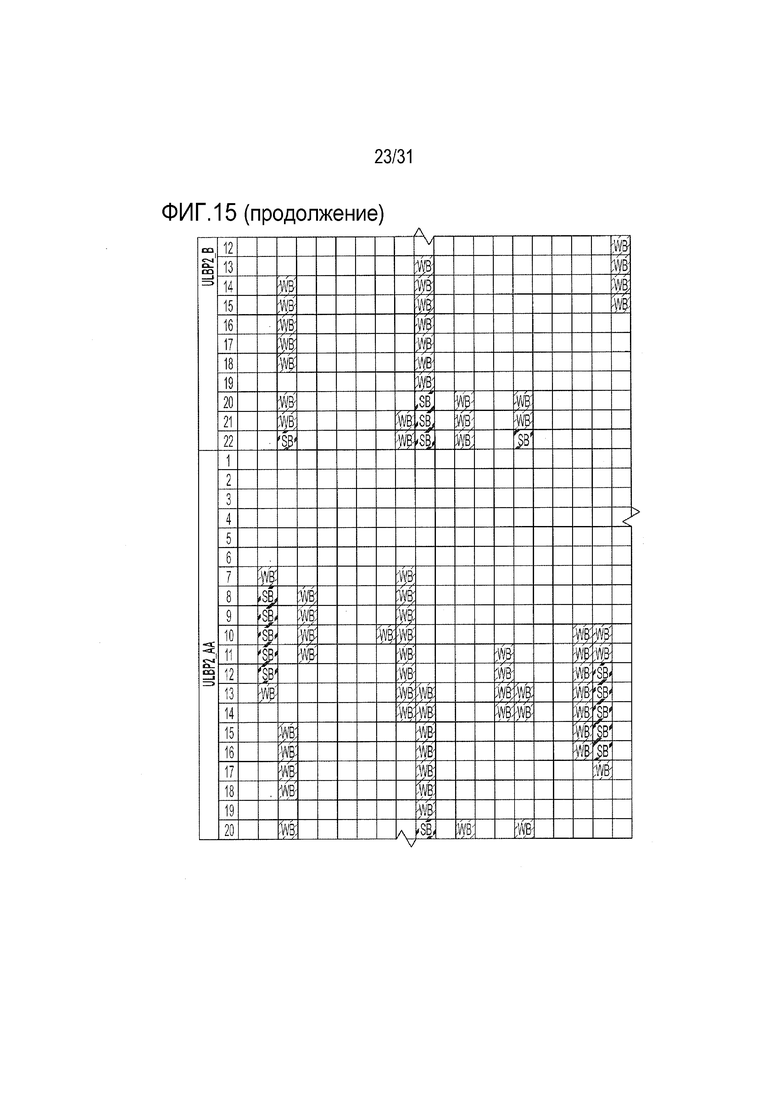


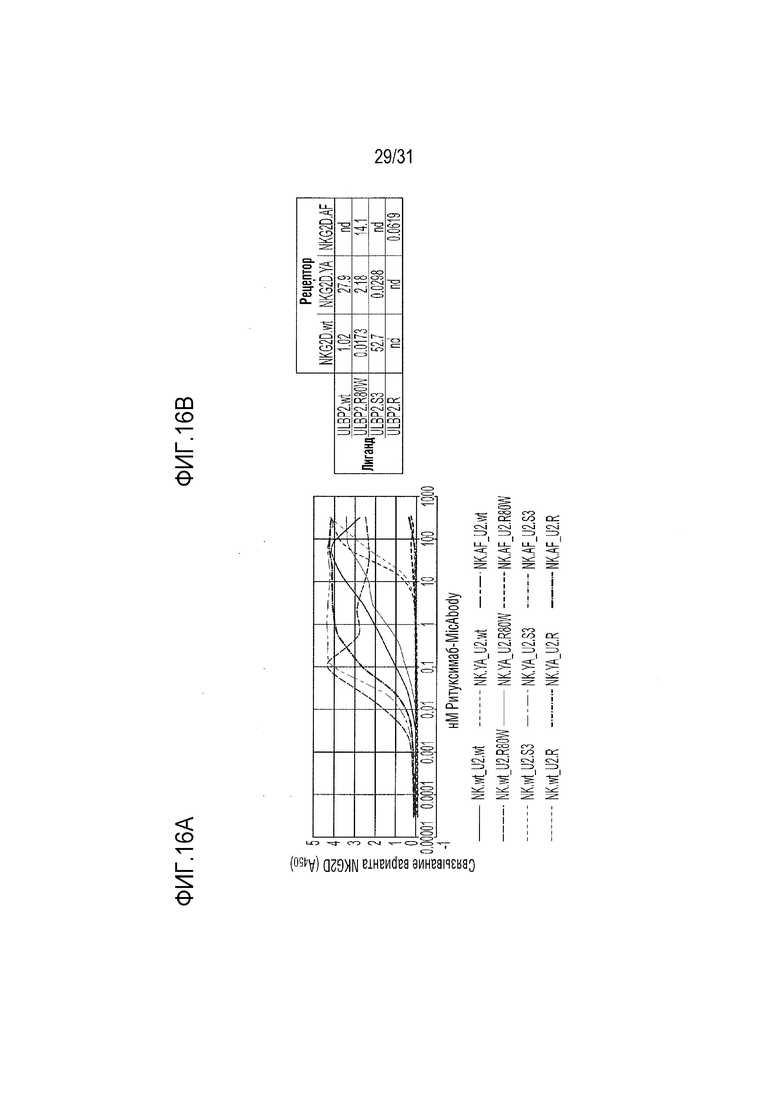
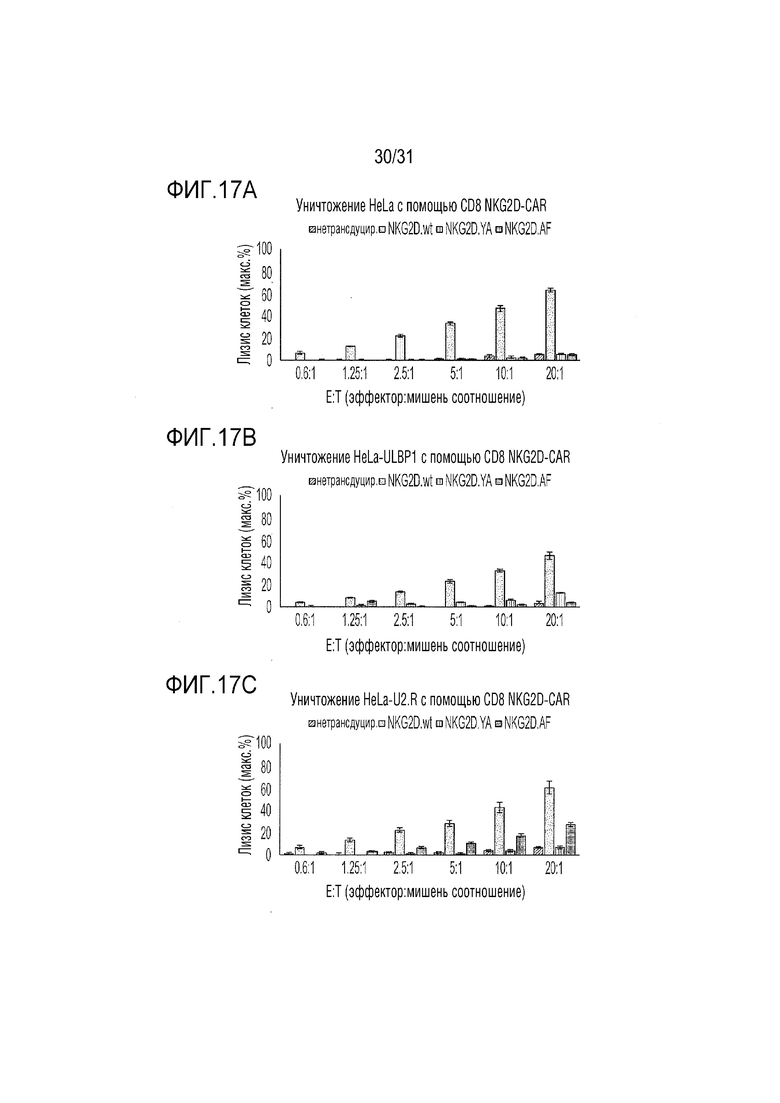
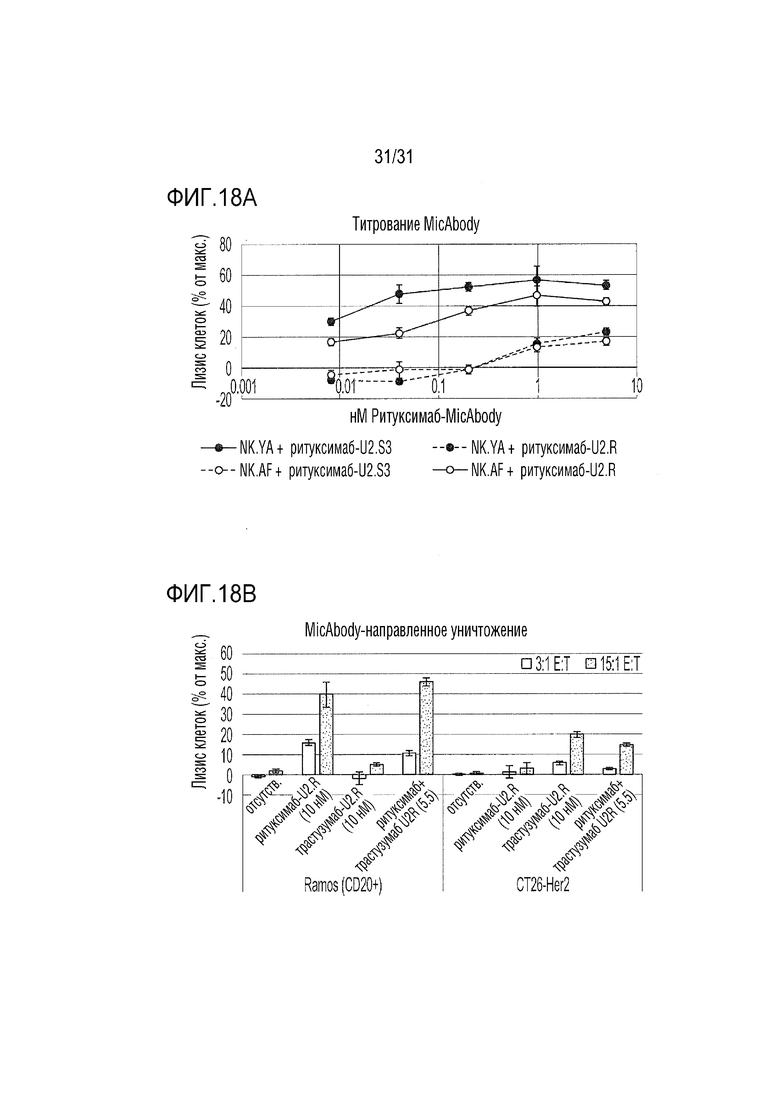
Изобретение относится к биотехнологии, а именно к неприродному модифицированному домену α1-α2 молекулы природного лиганда NKG2D. Указанный модифицированный домен α1-α2 связывает неприродный NKG2D рецептор и содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичность с SEQ ID NO: 4, где в указанной аминокислотной последовательности, имеющей по меньшей мере 90% идентичность с SEQ ID NO: 4, положения, соответствующие положениям 8, 80, 154, 155, 156, 157, 158 и 159 последовательности SEQ ID NO: 4, заменены по сравнению с SEQ ID NO: 4. Настоящее изобретение обеспечивает высокую активность связывания с неприродным эктодоменом рецептора NKG2D, содержащим аминокислотную последовательность SEQ ID NO: 17 с заменами Y75A и Y122F, по сравнению с (а) неприродным эктодоменом рецептора NKG2D, содержащим аминокислотную последовательность SEQ ID NO: 17, имеющую Y75A, и (b) соответствующим природным эктодоменом рецептора NKG2D, содержащим аминокислотную последовательность SEQ ID NO: 17. 10 з.п. ф-лы, 18 ил., 6 пр.
1. Неприродный модифицированный домен α1-α2 молекулы природного лиганда NKG2D, где указанный модифицированный домен α1-α2 связывает неприродный NKG2D рецептор и содержит аминокислотную последовательность, имеющую по меньшей мере 90% идентичность с SEQ ID NO: 4, где в указанной аминокислотной последовательности, имеющей по меньшей мере 90% идентичность с SEQ ID NO: 4, положения, соответствующие положениям 8, 80, 154, 155, 156, 157, 158 и 159 последовательности SEQ ID NO: 4, заменены по сравнению с SEQ ID NO: 4 и где указанная аминокислотная последовательность, имеющая по меньшей мере 90% идентичность с SEQ ID NO: 4, имеет все из следующих признаков:
аминокислота в положении, соответствующем положению 80 последовательности SEQ ID NO: 4, представляет собой W;
аминокислота в положении, соответствующем положению 154 последовательности SEQ ID NO: 4 представляет собой T;
аминокислота в положении, соответствующем положению 155 последовательности SEQ ID NO: 4, представляет собой I, L или M;
аминокислота в положении, соответствующем положению 157 последовательности SEQ ID NO: 4, представляет собой W;
аминокислота в положении, соответствующем положению 158 последовательности SEQ ID NO: 4, представляет собой Q, G или S; и
аминокислота в положении, соответствующем положению 159 последовательности SEQ ID NO: 4, представляет собой T или W.
2. Неприродный модифицированный домен α1-α2 по п.1, который содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 111, 113, 115 и 117.
3. Неприродный модифицированный домен α1-α2 по п.1, где в указанной аминокислотной последовательности, имеющей по меньшей мере 90% идентичность с SEQ ID NO: 4, аминокислота в положении, соответствующем положению 8 последовательности SEQ ID NO: 4, представляет собой S.
4. Неприродный модифицированный домен α1-α2 по п.1, где в указанной аминокислотной последовательности, имеющей по меньшей мере 90% идентичность с SEQ ID NO: 4, аминокислота в положении, соответствующем положению 156 последовательности SEQ ID NO: 4, представляет собой L или M.
5. Неприродный модифицированный домен α1-α2 по п.1, дополнительно содержащий присоединенную гетерологичную молекулу.
6. Неприродный модифицированный домен α1-α2 по п.5, в котором гетерологичная молекула представляет собой пептид или полипептид.
7. Неприродный модифицированный домен α1-α2 по п.6, в котором полипептид представляет собой антитело, фрагмент антитела, цитокин, лимфокин или гормон.
8. Неприродный модифицированный домен α1-α2 по п.5, в котором гетерологичная молекула представляет собой олигосахарид, дендример, ноттин, гормон, нуклеиновую кислоту или липид.
9. Неприродный модифицированный домен α1-α2 по п.1, дополнительно содержащий присоединенную клетку млекопитающего.
10. Неприродный модифицированный домен α1-α2 по п.9, где клетка млекопитающего представляет собой лимфоцит или макрофаг.
11. Неприродный модифицированный домен α1-α2 по п.10, где клетка млекопитающего представляет собой лимфоцит человека или макрофаг человека.
| CANDICE, S.E | |||
| LENGYEL et.al., Mutations designed to destabilize the receptor-bound conformation increase MICA-NKG2D association rate and affinity, J BIOL CHEM., 2007, v.282, n.42, p.30658-30666 | |||
| WITTENBRINK, M | |||
| et.al., Differential NKG2D binding to highly related human NKG2D ligands ULBP2 and RAET1G is determined by a single amino acid in the |
