ОБЛАСТЬ ТЕХНИКИ
Настоящее изобретение относится к композиции для культивирования Т-клеток и к способу культивирования регуляторных Т-кпеток с ее применением.
ПРЕДШЕСТВУЮЩИЙ УРОВЕНЬ ТЕХНИКИ
Т-клетки играют центральную роль в клеточно-опосредованном иммунитете. Т-клетки отличаются от других лимфоцитов, включая В-клетки, рецепторами на поверхности Т-клеток, такими как рецепторы Т-кпеток (TCR). Кроме того, Т-клетки включают разные типы Т-клеток, такие как хелперные Т-клетки (ТН-клетки), цитотоксические Т-клетки (ТС-кпетки или CTL), Т-клетки памяти (ТМ-клетки), включающие центральные Т-клетки памяти (ТСМ-клетки) и эффекторные Т-клетки памяти (ТЕМ-клетки), Т-клетки-природные киллеры (NKT-клетки), гамма дельта Т-клетки (γδ Т-клетки) и регуляторные Т-клетки (Treg-клетки).
Среди них регуляторные Т-клетки (Treg) представляют собой Т-клетки, которые действуют для подавления иммунного ответа других клеток. Например, Treg-клетки действуют для подавления опосредованного Т-клетками иммунитета во время иммунного ответа и для подавления аутореактивных Т-клеток, которые избежали отрицательного отбора в тимусе. Treg-клетки, главным образом, могут классифицироваться на природные регуляторные Т-клетки (nTreg) и индуцированные регуляторные Т-клетки (iTreg). Природные регуляторные Т-клетки, известные как CD4+CD25+FoxP3+ регуляторные Т-клетки, развиваются в тимусе. Индуцированные регуляторные Т-клетки имеют целый ряд общих характеристик со встречающимися в природе Treg-клетками, но характеристика превращения из CD4+CD25-FoxP3- Т-клеток в CD4+CD25+FoxP3+ регуляторные Т-клетки известна как репрезентативное отличие.
Активно проводили исследования по важности регуляторных Т-клеток как таковых в связи с заболеванием, вызванным нарушениями в разных аутоиммунных системах. С момента введения и первого предложения Gershon идеи Т-клеток-супрессоров (R K Gershon and K Kondo, immunology, 1970, 18: 723-37) в начале 1970 гг. были проведены исследования во многих областях иммунологии для выяснения биологических свойств и функций регуляторных Т-клеток. В частности, с момента предложения в 1995 г. Sakaguchi того, что CD25 может функционировать в качестве важного фенотипического маркера для встречающихся в природе регуляторных Т-клеток CD4+ (S Sakaguchi et, al., J Immunol, 1995, 155: 1151-1164), проводили исследования, сосредоточенные на роли и важности регуляторных Т-клеток в индуцировании периферческой толерантности каутоантигенам.
Известно, что регуляторные Т-клетки способны секретировать IL-10 (интерлейкин-10), TGF-β (трансформирующий фактор роста-бета), IL-35 или тому подобные, известные в качестве иммунодепрессивных цитокинов (Н Nishikawa et, al., Int. J. Cancer, 2010, 127: 759-767). Провели исследование, которое подтверждает то, что регуляторные Т-клетки, которые секретируют иммунодепрессивные цитокины, могут секретировать IL-10 или тому подобные с индукцией антигенспецифичных Т-клеток, которые вызывают аутоиммунное заболевание в иммунотолерантных антигенпрезентирующих клетках, индуцируя, посредством этого, иммунологическую толерантность.
Применения регуляторных клеток как таковых для лечения аутоиммунного заболевания широко известно, но конкретные способы, которые могут амплифицировать кпетки-природные киллеры для их эффективного применения, до сих пор недостаточны.
Подробное описание изобретения
Техническая проблема
Соответственно, авторы настоящего изобретения исследовали способ эффективного культивирования регуляторных Т-клеток и, в результате, выяснили то, что новый димер слитого белка, содержащий белок IL-2 и белок CD80 в одной молекуле, может эффективно пролиферировать регуляторные Т-клетки, и осуществили настоящее изобретение.
Решение проблемы
Для достижения приведенной выше цели согласно типичному воплощению композиция или среда для культивирования регуляторных Т-клеток содержит, в качестве активного ингредиента, слитый белок-димер, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент.
Согласно другому типичному воплощению предложен способ культивирования регуляторных Т-клеток с использованием димера слитого белка, содержащего белок IL-2 или его вариант и белок CD80 или его фрагмент.
Согласно еще одному другому типичному воплощению фармацевтическая композиция содержит, в качестве активного ингредиента, регуляторные Т-клетки, культивируемые в среде, содержащей димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент.
Эффект изобретения
Культуральная композиция согласно настоящему изобретению может не только эффективно пролиферировать Т-клетки, но, в частности, эффективно пролиферировать регуляторные Т-клетки. В частности, подтвердили то, что жизнеспособность регуляторных Т-клеток значимо возрастала по сравнению с традиционно используемым способом культивирования с IL-2. Также подтвердили то, что у полученных регуляторных Т-кпеток увеличвался уровень экспрессии Foxp3+. Следовательно, данный способ пролиферации можно использовать в области клеточной терапии с использованием регуляторных Т-клеток.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Типичные воплощения могут быть поняты с большими подробностями из следующего описания, взятого в сочетании с сопровождающими графическими материалами, в которых:
ФИГ. 1А представляет собой схематическую диаграмму димера слитого белка, используемого в настоящем изобретении;
На ФИГ. 1В показано изображение геля SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), подтверждающее полученный димер слитого белка (GI-101);
На ФИГ. 1С показано содержание димера слитого белка (GI-101) согласно поглощению;
На ФИГ. 1D показан анализ гель-фильтрацией (SEC) полученного димера слитого белка (GI-101);
На ФИГ. 2А показано изображение геля SDS-PAGE, подтверждающее полученный димер слитого белка hCD80-Fc;
На ФИГ. 2В показан анализ гель-фильтрацией (SEC) полученного димера слитого белка hCD80-Fc;
На ФИГ. 3А показано изображение геля SDS-PAGE, подтверждающее полученный димер слитого белка Fc-IL2v2;
На ФИГ. 3В показан анализ гель-фильтрацией (SEC) полученного димера слитого белка Fc-IL2v2;
На ФИГ. 3С показано изображение геля SDS-PAGE, подтверждающее полученный димер слитого белка Fc-IL2wt;
На ФИГ. 3D показан анализ гель-фильтрацией (SEC) полученного димера слитого белка Fc-IL2wt;
На ФИГ. 4А показано изображение геля SDS-PAGE, подтверждающее полученный димер слитого белка hCD80-Fc-IL2wt;
На ФИГ. 4В показан анализ гель-фильтрацией (SEC) полученного димера слитого белка hCD80-Fc-IL2wt;
На ФИГ. 5 показана схематическая диаграмма способа культивирования Treg-клеток с использованием димера слитого белка;
На ФИГ. 6 показано число регуляторных Т-клеток, культивируемых в композиции, содержащей среду RPMI1640;
На ФИГ. 7 показана жизнеспособность регуляторных Т-кпеток, культивируемых в композиции, содержащей среду RPMI1640;
На ФИГ. 8 показано число регуляторных Т-клеток, культивируемых в композиции, содержащей среду TexMACS;
На ФИГ. 9 показана жизнеспособность регуляторных Т-кпеток, культивируемых в композиции, содержащей среду TexMACS;
На ФИГ. 10 показана способность к секреции IL-10 регуляторных Т-клеток, культивируемых с использованием композиции, содержащей среду RPMI1640;
На ФИГ. 11 показана способность к секреции IL-10 регуляторных Т-клеток, культивируемых с использованием композиции, содержащей среду TexMACS;
На ФИГ. 12А и 12В показано число пролиферировавших регуляторных Т-клеток при культивировании в композиции, содержащей среду RPMI1640, в оптимизированном способе;
На ФИГ. 13А и 13В показана жизнеспособность регуляторных Т-клеток при культивировании в композиции, содержащей среду RPMH640, в оптимизированном способе;
На ФИГ. 14А и 14В показано число регуляторных Т-клеток при культивировании в композиции, содержащей среду TexMACS, в оптимизированном способе;
На ФИГ. 15А и 15В показана жизнеспособность регуляторных Т-клеток при культивировании в композиции, содержащей среду TexMACS, в оптимизированном способе;
На ФИГ. 16А-16С показаны результаты анализа FACS (флуоресцентная сортировка клеток) свойств клеток, культивируемых в композиции, содержащей среду RPMI1640, в оптимизированном способе;
На ФИГ. 17А-17С показано число регуляторных Т-клеток, экспрессирующих Foxp3, при культивировании в композиции, содержащей среду RPMH640, в оптимизированном способе;
На ФИГ. 18А-18С показаны результаты анализа FACS свойств клеток, культивируемых в композиции, содержащей среду TexMACS, в оптимизированном способе;
На ФИГ. 19А-19С показано число регуляторных Т-клеток, экспрессирующих Foxp3, при культивировании в композиции, содержащей среду TexMACS, в оптимизированном способе;
На ФИГ. 20 показана способность к секреции IL-10 регуляторных Т-кпеток при культивировании в композиции, содержащей среду RPMH640, в оптимизированном способе; и
На ФИГ. 21 показана способность к секреции IL-10 регуляторных Т-кпеток при культивировании в композиции, содержащей среду TexMACS.
Наилучший способ осуществления изобретения
Композиция и среда для пролиферации регуляторных Т-клеток
Согласно одному аспекту настоящего изобретения предложена композиция для пролиферации регуляторных Т-клеток, содержащая, в качестве активного ингредиента, димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент.Кроме того, предложена культуральная среда для регуляторных Т-клеток, содержащая димер слитого белка в качестве активного ингредиента.
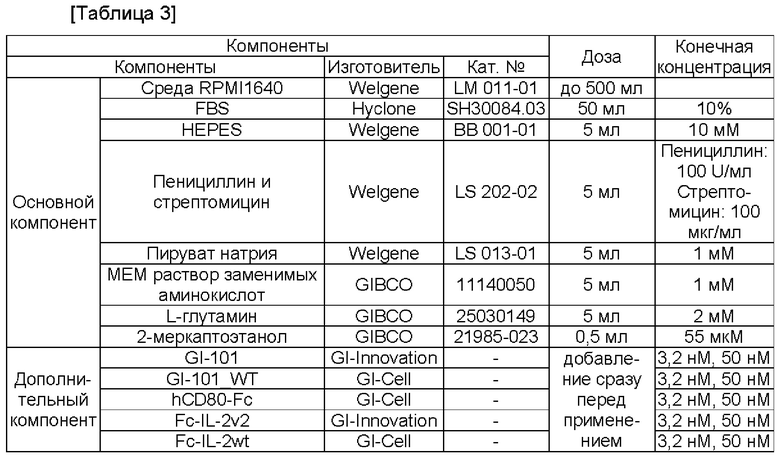
Среда пролиферации регуляторных Т-кпеток может представлять собой среду, в которой димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, добавляют в среду для культивирования Т-клеток. В данном случае культуральная среда Т-клеток может содержать любой компонент, выбранный из группы, состоящей из аминокислот, Сахаров, неорганических солей и витаминов. Предпочтительно культуральная среда Т-кпеток может содержать любую аминокислоту, любой сахар, любую неорганическую соль и любой витамин. Кроме того, данная среда может дополнительно содержать фетальную телячью сыворотку (FBS), гидроксиэтилпиперазинэтансульфоновую кислоту (HEPES), белки, углеводы, меркаптоэтанол и факторы роста. Также культуральная среда регуляторных Т-клеток может дополнительно содержать ретиноевую кислоту. В одном воплощении среда пролиферации регуляторных Т-клеток может может содержать базовые компоненты, описанные в следующих Таблице 3 или Таблице 4.
Термин «культуральная среда клеток» в том виде, как он используется в данном документе, означает среду, используемую для культивирования клеток, в частности, регуляторных Т-клеток, и, более конкретно, означает среду для культивирования CD4+CD25+CD127- клеток. Она содержит компоненты, требующиеся клетками для роста и выживания клеток in vitro, или содержит компоненты, которые помогают росту и выживанию клеток. В частности, данные компоненты могут представлять собой витамины, незаменимые или заменимые аминокислоты и микроэлементы. Данная среда может представлять собой среду, используемую для культивирования клеток, предпочтительно эукариотических клеток, наиболее предпочтительно регуляторных Т-клеток и намного более предпочтительно Т-клеток CD4+CD25+CD127- или Т-клеток CD4+CD25+Foxp3+.
Культуральная среда клеток согласно настоящему изобретению содержит любой аминокислотный компонент, любой витаминный компонент, любой компонент в виде неорганической соли, любой другой компонент и очищенную воду, где:
a) аминокислотный компонент представляет собой по меньшей мере одну аминокислоту, выбранную из группы, состоящей из глицина, L-аланина, L-валина, L-лейцина, L-изолейцина, L-треонина, L-серина, L-цистеина, L-метионина, L-аспарагиновой кислоты, L-глутаминовой кислоты, L-глутамина, L-лизина, L-аргинина, L-гистидина, L-фенилаланина, L-тирозина, L-триптофана, L-пролина, β-аланина, γ-аминомасляной кислоты, орнитина, цитруллина, гомосерина, трийодтиронина, тироксина и диоксифенилаланина или их комбинации, и предпочтительно по меньшей мере одну аминокислоту, выбранную из группы, состоящей из глицина, L-аланина, L-аргинина, L-цистеина, L-глутамина, L-гистидина, L-лизина, L-метионина, L-пролина, L-серина, L-треонина и L-валина или их комбинации;
b) витаминный компонент представляет собой по меньшей мере один витамин, выбранный из группы, состоящей из биотина, кальция D-пантотената, фолиевой кислоты, ниацинамида, пиридоксина гидрохлорида, рибофлавина, тиамина гидрохлорида, витамина В12, холина хлорида, i-инозита и аскорбиновой кислоты или их комбинации, и предпочтительно по меньшей мере один витамин, выбранный из группы, состоящей из i-инозита, тиамина гидрохлорида, ниацинамида и пиридоксина гидрохлорида или их комбинации;
c) компонент в виде неорганической соли представляет собой по меньшей мере одну неорганическую соль, выбранную из группы, состоящей из хлорида кальция (CaCl2) (безводный), сульфата меди пентагидрата (CuSO4-5H2O), сульфата железа (II) гептагидрата (FeSO4-7H2O), хлорида магния (безводный), сульфата магния (MgSO4) (безводный), хлорида калия (KCl), хлорида натрия (NaCl), динатрия гидрофосфата (Na2HPO4), натрия дигидрофосфата моногидрата (NaH2PO4-H2O), сульфата цинка гептагидрата (ZnSO4-7H2O), нитрата железа (III) нонагидрата (Fe(NO3)3⋅9H2O) и гидрокарбоната натрия (NaHCO3) или их комбинации, и предпочтительно по меньшей мере одну неорганическую соль, выбранную из группы, состоящей из хлорида натрия (NaCl), гидрокарбоната натрия (NaHCO3), хлорида калия (KCl), хлорида кальция (CaCl2) (безводный) и натрия дигидрофосфата моногидрата (NaH2PO4-H2O) или их комбинации;
d) другой компонент представляет собой по меньшей мере один другой компонент, выбранный из группы, состоящей из D-глюкозы (декстрозы), пирувата натрия, гипоксантина Na, тимидина, линолевой кислоты, липоевой кислоты, аденозина, цитидина, гуанозина, уридина, 2'-дезоксиаденозина, 2'-дезоксицитидина HCl и 2'-дезоксигуанозина или их комбинации, и он предпочтительно может представлять собой пируват натрия; и
e) очищенная вода используется для растворения аминокислот, витаминов, неорганических солей и других компонентов, и может быть получена посредством одного или более чем одного способа дистилляции, или очищена через фильтр.
Кроме того, культуральная среда клеток согласно настоящему изобретению может дополнительно содержать факторы роста или цитокины. Данный фактор роста может представлять собой IGF (инсулиноподобный фактор роста), bFGF (основной фактор роста фибробластов), TGF (трансформирующий фактор роста), HGF (фактор роста гепатоцитов), EGF (эпидермальный фактор роста), VEGF (фактор роста эндотелия сосудов), PDGF (фактор роста тромбоцитов) или тому подобный, один или по меньшей мере два из них, но не ограничиваясь ими. Цитокин может представлять собой IL-1 (интерлейкин-1), IL-4, IL-6, IFN-γ (интерферон-гамма), IL-10, IL-17 или тому подобные, один или по меньшей мере два из них, но не ограничиваясь ими.
Термин «Т-клетка» в том виде, как он используется в данном документе, относится к одному из лимфоцитов, ответственных за антигенспецифичный адаптивный иммунитет. Т-клетки классифицируются на нативные Т-клетки, которые еще не встретили антиген, и зрелые Т-клетки, и Т-клетки памяти, которые встречали антиген. При этом зрелые эффекторные Т-клетки содержат Т-клетки-хелперы, цитотоксические Т-клетки и Т-клетки-природные киллеры.
Термин «Т-клетка-хелпер или Th клетка» в том виде, как он используется в данном документе, относится к клетке, которая стимулирует гуморальный иммунитет посредством осуществления регуляции дифференциации и активации других лейкоцитов. Она также называется Т-клетка CD4+, так как она имеет белок CD4 на поверхности клетки. Т-клетки-хелперы могут быть дополнительно классифицированы на Th1, Th2, Th17 и Treg согласно их детальным функциям. Клетки Th1 секретируют интерферон-гамма (IFN-γ) и фактор некроза опухолей бета (TNF-β), индуцируя, посредством этого, слияние эндосом и лизосом с образованием эндолизосом внутри макрофагов. Тем временем, клетки Th2 секретируют несколько типов интерлейкина (IL), обеспечивая дифференциацию В-клеток в плазматические клетки. Клетки Th17 секретируют интерлейкин-17 (IL-17) для рекрутирования нейтрофилов.
Термин «регуляторная Т-клетки (Treg)» в том виде, как он используется в данном документе, включает природные регуляторные Т-клетки (nTreg) или индуцированные регуляторные Т-клетки (iTreg). Регуляторные Т-клетки в данном документе включают Т-клетки CD4+CD25+, Т-клетки CD4+CD25+CD127 низкая - или Т-клетки CD4+CD25+Foxp3+. Регуляторные Т-клетки поддерживают иммунный гомеостаз и блокируют аутоиммунный ответ и тому подобное посредством ингибирования иммунного ответа.
Термин «цитотоксическая Т-клетка» в том виде, как он используется в данном документе, относится к клетке, которая умерщвляет инфицированные вирусом клетки или опухолевые клетки или тому подобное посредством осуществления секреции цитотоксичных веществ, таких как гранзим или перфорин. Она также называется Т-клетка CD8, так как она имеет белок CD8 на поверхности клетки. В отличие от хелперных Т-клеток она устраняет вирус и раковые клетки опосредованием клеточного иммунитета.
Термин «Т-кпетка-природный киллер» в том виде, как он используется в данном документе, относится к одной из эффекторных Т-клеток, которая распостранена в малой доле по сравнению с Т-клетками-хелперами и цитотоксическими Т-клетками. Т-клетки-природные киллеры имеют такие же рецепторы Т-клеток (TCR) на поверхности клеток, что и Т-клетки, но также имеют специфичные для клетки-природного киллера молекулы, такие как NK1.1. Т-клетки-природные киллеры секретируют гамма-интерферон, интерлейкин-4 или тому подобные для регуляции иммунного ответа.
Термин «Т-клетка памяти» в том виде, как он используется в данном документе, относится к Т-клетке, которая имеет потенциальную способность функционировать в качестве эффекторной Т-клетки, так как она распознала антиген и выжила в течение длительного времени после процессов дифференциации и отбора, и позднее, при повторном вторжении антигена, быстро активируется. Наивные Т-клетки дифференцируются в активированные клетки посредством распознавания антигена, или эффекторные Т-клетки дифференцируются в долгоживущие Т-клетки памяти посредством влияния интерлейкина-7 и интерлейкина-15.
В данном случае димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент, может содержаться в культуральной среде в количестве от 1 нМ до 2000 нМ. Кроме того, данный димер может содержаться в количестве от 1 нМ до 1000 нМ или от 1 нМ до 500 нМ. Кроме того, данный димер может содержаться в количестве от 2 нМ до 300 нМ, от 5 нМ до 100 нМ, от 10 нМ до 80 нМ, от 20 нМ до 70 нМ или от 40 нМ до 50 нМ. В частности, слитый белок-димер может содержаться в среде в количестве 1 нМ, 3,2 нМ, 10 нМ или 50 нМ.
Димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент
Термин «IL-2» или «интерлейкин-2» в том виде, как он используется в данном документе, если не утверждается иное, относится к любому IL-2 дикого типа, полученному из любого источника-позвоночного, включая млекопитающих, например, приматов (таких как человек) и грызунов (таких как мыши и крысы). IL-2 может быть получен из животных клеток и также включает IL-2, полученный из рекомбинантных клеток, способных к продукции IL-2. Кроме того, IL-2 может представлять собой IL-2 дикого типа или его вариант.
В настоящем описании изобретения IL-2 или его вариант могут быть совместно выражены термином «белок IL-2» или «полипептид IL-2». IL-2, белок IL-2, полипептид IL-2 и вариант IL-2 специфично связываются, например, с рецептором IL-2. Это специфичное связывание может быть идентифицировано способами, известными специалистам в данной области.
Одно воплощение IL-2 может иметь аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36. Здесь IL-2 также может находиться в зрелой форме. В частности, зрелый IL-2 может не содержать сигнальную последовательность и может иметь аминокислотную последовательность SEQ ID NO: 10. Здесь IL-2 может использоваться согласно конфигурации, охватывающей фрагмент IL-2 дикого типа, в котором усечена часть N-конца или С-конца IL-2 дикого типа.
Кроме того, данный фрагмент IL-2 может находиться в форме, в которой 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 смежных аминокислот усечены с N-конца белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36. Кроме того, фрагмент IL-2 может находиться в форме, в которой 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 смежных аминокислот усечены с С-конца белка, имеющего аминокислотную последовательность SEQ ID NO: 35 или SEQ ID NO: 36.
Термин «вариант IL-2» в том виде, как он используется в данном документе, относится к форме, в которой часть аминокислот в полноразмерном IL-2 или вышеописанном фрагменте IL-2 заменена. То есть, вариант IL-2 может иметь отличную аминокислотную последоватеьность от IL-2 дикого типа или его фрагмента. Однако вариант IL-2 может иметь эквивалентную или аналогичную активность относительно IL-2 дикого типа. Здесь «активность IL-2» может, например, относиться к специфичному связыванию с рецептором IL-2, причем данное специфичное связывание можно измерять способами, известными специалистам в данной области.
В частности, вариант IL-2 можно получать заменой части аминокислот в IL-2 дикого типа. Одно воплощение варианта IL-2, полученного аминокислотной заменой, может быть получено заменой по меньшей мере одной из 38-ой, 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен заменой по меньшей мере одной из 38-ой, 42-ой, 45-ой, 61-ой или 72-ой аминокислоты в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой. Кроме того, когда IL-2 находится в форме, в которой часть N-конца аминокислотной последовательности SEQ ID NO: 35 усечена, аминокислота в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10, может быть заменена другой аминокислотой. Например, когда IL-2 имеет аминокислотную последовательность SEQ ID NO: 35, его вариант IL-2 может быть получен заменой по меньшей мере одной из 58-ой, 62-ой, 65-ой, 81-ой или 92-ой аминокислоты в аминокислотной последовательности SEQ ID NO: 35 другой аминокислотой. Данные аминокислотные остатки соответствуют 38-му, 42-му, 45-му, 61-му и 72-му аминокислотному остатку в аминокислотной последовательности SEQ ID NO: 10 соответственно. Согласно одному воплощению могут быть заменены одна, две, три, четыре, пять, шесть, семь, восемь, девять или десять аминокислот, при условии, что такой вариант IL-2 сохраняет активность IL-2. Согласно другому воплощению могут быть заменены от одной до пяти аминокислот.
В одном воплощении вариант IL-2 может находиться в форме, в которой заменены две аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой и 42-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются три аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой, 42-ой и 45-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются четыре аминокислоты. В частности, вариант IL-2 может быть получен заменой 38-ой, 42-ой, 45-ой и 61-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой, 45-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 38-ой, 42-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10. Кроме того, в одном воплощении вариант IL-2 может быть получен заменой 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10.
Кроме того, вариант IL-2 может находится в форме, в которой заменяются пять аминокислот. В частности, вариант IL-2 может быть получен заменой каждой из 38-ой, 42-ой, 45-ой, 61-ой и 72-ой аминокислот в аминокислотной последовательности SEQ ID NO: 10 другой аминокислотой.
Здесь «другая аминокислота», вводимая заменой, может представлять собой любую аминокислоту, выбранную из группы, состоящей из аланина, аргинина, аспарагина, аспарагиновой кислоты, цистеина, глутаминовой кислоты, глутамина, гистидина, изолейцина, лейцина, лизина, метионина, фенилаланина, пролина, серина, треонина, триптофана, тирозина и валина. Однако относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота не может быть заменена аргинином, 42-ая аминокислота не может быть заменена фенилаланином, 45-ая аминокислота не может быть заменена тирозином, 61-ая аминокислота не может быть заменена глутаминовой кислотой, и 72-ая аминокислота не может быть заменена лейцином.
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота - аргинин - может быть заменена аминокислотой, отличной от аргинина. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 38-ая аминокислота - аргинин - может быть заменена аланином (R38A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 42-ая аминокислота - фенилаланин - может быть заменена другой аминокислотой, чем фенилаланин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 42-ая аминокислота - фенилаланин - может быть заменена аланином (F42A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 45-ая аминокислота - тирозин - может быть заменена другой аминокислотой, чем тирозин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 45-ая аминокислота - тирозин - может быть заменена аланином (Y45A).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 61-ая аминокислота - глутаминовая кислота -может быть заменена другой аминокислотой, чем глутаминовая кислота. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 61-ая аминокислота -глутаминовая кислота - может быть заменена аргинином (E61R).
Относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 72-ая аминокислота - лейцин - может быть заменена другой аминокислотой, чем лейцин. Предпочтительно относительно аминокислотной замены для варианта IL-2 в аминокислотной последовательности SEQ ID NO: 10, 72-ая аминокислота - лейцин - может быть заменена глицином (L72G).
В частности, вариант IL-2 может быть получен по меньшей мере одной заменой, выбранной из группы, состоящей из R38A, F42A, Y45A, E61R и L72G в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может быть получен аминокислотными заменами в двух, трех, четырех или пяти положениях среди положений, выбранных в группе, состоящей из R38A, F42A, Y45A, E61R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменены две аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A и F42A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами E61R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменяются три аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A, F42A и Y45A. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами Y45A, E61R и L72G.
Кроме того, вариант IL-2 может находиться в форме, в которой заменяются четыре аминокислоты. В частности, вариант IL-2 может быть получен заменами R38A, F42A, Y45A и E61R. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A, Y45A и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, F42A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами R38A, Y45A, E61R и L72G. Кроме того, в одном воплощении вариант IL-2 может быть получен заменами F42A, Y45A, E61R и L72G.
Кроме того, вариант IL-2 может быть получен заменами R38A, F42A, Y45A, E61R и L72G.
Предпочтительно одно воплощение варианта IL-2 может содержать замену, которая представляет собой любую замену, выбранную из следующих комбинаций замен (a)-(d) в аминокислотной последовательности SEQ ID NO: 10:
(a) R38A/F42A
(b) R38A/F42A/Y45A
(c) R38A/F42A/E61R
(d) R38A/F42A/L72G
Здесь, когда IL-2 имеет аминокислотную замену SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10. Кроме того, даже когда IL-2 представляет собой фрагмент аминокислотной последовательности SEQ ID NO: 35, аминокислотная замена может присутствовать в положении, комплементарно соответствующем положению в аминокислотной последовательности SEQ ID NO: 10.
В частности, вариант IL-2 может иметь аминокислотную последовательность SEQ ID NO: 6, 22, 23 или 24.
Кроме того, вариант IL-2 может отличаться наличием низкой токсичности in vivo. Здесь низкая токсичность in vivo может представлять собой побочный эффект, вызванный связыванием IL-2 с альфа цепью рецептора IL-2 (IL-2Rα). Были разработаны разные варианты IL-2 для уменьшения интенсивности побочного эффекта, вызванного связыванием IL-2 с IL-2Rα, и такие варианты IL-2 могут представлять собой варианты, раскрытые в патенте США №5229109 и корейском патенте №1667096. В частности, варианты IL-2, описанные в настоящей заявке, имеют низкую аффинность связывания в отношении альфа цепи рецептора IL-2 (IL-2Rα) и, таким образом, имеют более низкую токсичность in vivo, чем IL-2 дикого типа.
Термин «CD80» в том виде, в котором он используется в данном документе, также именуемый «В7-1», относится к мембранному белку, присутствующему в дендритных клетках, активированных В-клетках и моноцитах. CD80 предоставляет костимулирующие сигналы, важные для активации и выживания Т-клеток. CD80 известен в качестве лиганда двух разных белков - CD28 и CTLA-4, присутствующих на поверхности Т-клеток. CD80 состоит из 288 аминокислот и может, в частности, иметь аминокислотную последовательность SEQ ID NO: 11. Кроме того, термин «белок CD80» в том виде, в котором он используется в данном документе, относится к полноразмерному CD80 или к фрагменту CD80.
Термин «фрагмент CD80» в том виде, в котором он используется в данном документе, относится к усеченной форме CD80. Кроме того, фрагмент CD80 может представлять собой внеклеточный домен CD80. Одно воплощение фрагмента CD80 может быть получено устранением 1-ой-34-ой аминокислот с N-конца, которые представляют собой сигнальную последовательность CD80. В частности, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 288-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 242-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 232-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 35-ой - 139-ой аминокислот в SEQ ID NO: 11. Кроме того, одно воплощение фрагмента CD80 может представлять собой белок, состоящий из 142-ой - 242-ой аминокислот в SEQ ID NO: 11. В одном воплощении фрагмент CD80 может иметь аминокислотную последовательность SEQ ID NO: 2.
Кроме того, белок IL-2 и белок CD80 могут присоединяться друг к другу через линкер или носитель. В частности, IL-2 или его вариант и CD80 (В7-1) или его фрагмент могут присоединяться друг к другу через линкер или носитель. В настоящем описании термины «линкер» и «носитель» могут использоваться взаимозаменяемо.
Линкер связывает два белка. Одно воплощение линкера может содержать от 1 до 50 аминокислот, альбумин или его фрагмент, домен Fc иммуноглобулина или тому подобное. Здесь домен Fc иммуноглобулина относится к белку, который содержит константную область 2 тяжелой цепи (СН2) и константную область 3 тяжелой цепи (СН3) иммуноглобулина, и не содержит вариабельные области тяжелой и легкой цепи, и константную область 1 легкой цепи (CL1) иммуноглобулина. Данный иммуноглобулин может представлять собой IgG, IgA, IgE, IgD или IgM и предпочтительно может представлять собой IgG4. Здесь домен Fc иммуноглобулина G4 дикого типа может иметь аминокислотную последовательность SEQ ID NO: 4.
Кроме того, домен Fc иммуноглобулина может представлять собой вариант домена Fc, а также домен Fc дикого типа. Кроме того, термин «вариант домена Fc» в том виде, как он используется в данном документе, может относиться к форме, которая отличается от домена Fc дикого типа в показателях картины гликозилирования, имеет высокое гликозилирование по сравнению с доменом Fc дикого типа или имеет низкое гликозилирование по сравнению с доменом Fc дикого типа, или представляет собой дегликозилированную форму. Кроме того, в данный документ включается негликозилированный домен Fc. Домен Fc или его вариант может быть адаптирован для наличия скорректированного числа сиаловых кислот, фукозилирований или гликозилирований посредством культуральных условий или генетических манипуляций с хозяином.
Кроме того, гликозилирование домена Fc иммуноглобулина может быть модифицировано традиционными способами, такими как химические способы, ферментативные способы и способы генной инженерии с использованием микроорганизмов. Кроме того, вариант домена Fc может находиться в смешанной форме соответствующих областей Fc иммуноглобулинов - IgG, IgA, IgE, IgD и IgM. Кроме того, вариант домена Fc может находиться в форме, в которой некоторые аминокислоты домена Fc заменяются другими аминокислотами. Одно воплощение варианта домена Fc может иметь аминокислотную последовательность SEQ ID NO: 12.
Данный слитый белок может иметь структуру, в которой с использованием домена Fc в качестве линкера (или носителя) белок CD80 и белок IL-2 или белок IL-2 и белок CD80 связываются с N-концом и С-концом линкера или носителя соответственно (Фиг. 1А). Связь между N-концом или С-концом домена Fc и CD-80 или IL-2 возможно может достигаться посредством линкерного пептида.
В частности, слитый белок может состоять из следующей структурной формулы (I) или (II):
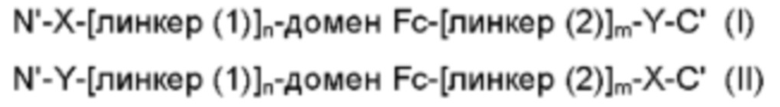
Здесь в структурных формулах (I) и (II)
N' представляет собой N-конец слитого белка,
С представляет собой С-конец слитого белка,
X представляет собой белок CD80,
Y представляет собой белок IL-2,
линкеры (1) и (2) представляют собой пептидные линкеры и
n и m каждый независимо равен 0 или 1.
Предпочтительно данный слитый белок может состоять из структурной формулы (I). Белок IL-2 является таким, как описано выше. Кроме того, белок CD80 является таким, как описано выше. Согласно одному воплощению белок IL-2 может представлять собой вариант IL-2 с одной-пятью аминокислотными заменами по сравнению с IL-2 дикого типа. Белок CD80 может представлять собой фрагмент, полученный усечением вплоть до примерно 34 смежных аминокислотных остатков от N-конца или С-конца CD80 дикого типа. В качестве альтернативы, белок CD может представлять собой внеклеточный иммуноглобулиноподобный домен, имеющий активность связывания с рецепторами поверхности Т-клеток CTLA-4 и CD28.
В частности, данный слитый белок может иметь аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30. Согласно другому воплощению данный слитый белок содержит полипептид, имеющий идентичность последовательности 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% с аминокислотной последовательностью SEQ ID NO: 9, 26, 28 или 30. Здесь идентичность представляет собой, например, процент гомологии и может определяться посредством программы для сравнения гомологии, такой как программа BlastN Национального центра биотехнологической информации (NCBI).
Пептидный линкер (1) может содержаться между белком CD80 и доменом Fc. Пептидный линкер (1) может состоять из 5-80 смежных аминокислот, 20-60 смежных аминокислот, 25-50 смежных аминокислот или 30-40 смежных аминокислот. В одном воплощении пептидный линкер (1) может состоять из 30 аминокислот. Кроме того, пептидный линкер (1) может содержать по меньшей мере один цистеин. В частности, пептидный линкер (1) может содержать один, два или три цистеина. Кроме того, пептидный линкер (1) может происходить из шарнира иммуноглобулина. В одном воплощении пептидный линкер (1) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 3.
Пептидный линкер (2) может состоять из 1-50 смежных аминокислот, 3-30 смежных аминокислот или 5-15 смежных аминокислот. В одном воплощении пептидный линкер (2) может представлять собой (G4S)n (где n представляет собой целое число от 1 до 10). Здесь в (G4S)n n может быть равен 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10. В одном воплощении пептидный линкер (2) может представлять собой пептидный линкер, состоящий из аминокислотной последовательности SEQ ID NO: 5.
В другом аспекте настоящего изобретения предложен димер, полученный связыванием двух слитых белков, каждый из которых содержит белок IL-2 и белок CD80. Слитый белок, содержащий IL-2 или его вариант и CD80 или его фрагмент, является таким, как описано выше.
Здесь связывание между слитыми белками, составляющими данный димер, может достигаться посредством дисульфидной связи, образованной цистеинами, присутствующими в линкере, но не ограничиваясь ей. Слитые белки, составляющие димер, могут быть одинаковыми или отличающимися друг от друга слитыми белками. Предпочтительно данный димер может представлять собой гомодимер. Одним воплощением слитого белка, составляющего димер, может быть белок, имеющий аминокислотную последовательность SEQ ID NO: 9.
Способ культивирования регуляторной Т-клетки
Согласно другому аспекту настоящего изобретения предложен способ культивирования регуляторных Т-клеток, включающий культивирование Т-клеток CD4+CD25+CD127- в среде, содержащей димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент.
В настоящее время Т-клетки CD4+CD25+CD127- можно получать из клеток крови. В настоящее время Т-клетки CD4+CD25+CD127- могут быть выделены из одноядерных клеток периферической крови (РВМС). В качестве альтернативы, Т-клетки CD4+CD25+CD127- можно получать посредством осуществления специфичной пролиферации Т-клеток CD4+CD25+CD127- из клеток крови. В качестве альтернативы, Т-клетки CD4+CD25+CD127- можно получать из клеток CD4+ после удаления РВМС от Т-клеток CD4-. Кроме того, Т-клетки CD4+ могут быть выделены с использованием антител против CD4, и в одном воплощении они были выделены с использованием шариков, с которыми связываются антитела против CD4. Кроме того, Т-клетки CD25+ могут быть выделены с использованием антител против CD25+. В частности, регуляторные Т-клетки могут быть выделены посредством выделения Т-клеток CD4+, CD25+ и CD127-.
Среда может представлять собой традиционно используемую среду. Предпочтительно она может представлять собой среду, оптимизированную для Т-клеток CD4+CD25+CD127-. В конкретном воплощении она может представлять собой среду, в которой FBS, HEPES, L-глутамин и 2-меркаптоэтанол добавлены в среду RPMI1640 или в среду TexMACS, как раскрыто в Таблицах 3 и 4. Кроме того, данная среда может дополнительно содержать ретиноевую кислоту. Кроме того, данная среда может дополнительно содержать пенициллин и/или стерптомицин.
При этом регуляторные Т-клетки CD4+ можно культивировать в среде в течение 1-30 суток или 2-20 суток. Кроме того, они могут культивироваться в течение 3-10 суток или 4-6 суток.
Тем временем, Т-клетки CD4+CD25+CD127- могут быть получены посредством стадии культивирования Т-клеток CD4+ или стадии культивирования Т-клеток CD25+.
В настоящее время Т-клетки CD4+ и Т-клетки CD25+ могут быть получены из клеток крови соответственно. В настоящее время Т-клетки CD4+ и Т-клетки CD25+ могут быть выделены из одноядерных клеток периферической крови (РВМС) или могут быть получены из клеток крови посредством осуществления специфичной пролиферации Т-клеток CD4+ или Т-клеток CD25+ соответственно. В качестве альтернативы, Т-клетки CD4+ или Т-клетки CD25+ могут быть получены после удаления Т-клеток CD4- или Т-клеток CD25- от РВМС. Кроме того, Т-клетки CD4+ могут быть выделены с использованием антител против CD4, и в одном воплощении они могут быть выделены с использованием шариков, с которыми связываются антитела против CD4. Кроме того, Т-клетки CD25+ могут быть выделены с использованием антител против CD25+. В частности, регуляторные Т-клетки могут быть выделены посредством осуществления выделения Т-клеток CD4+, CD25+ и CD127- из Т-клеток CD4+ или Т-клеток CD25+.
Полученные регуляторные Т-клетки и их применение
Согласно другому аспекту настоящего изобретения предложены регуляторные Т-клетки, полученные данным способом культивирования.
Согласно еще одному другому аспекту настоящего изобретения предложена композиция для лечения заболевания, опосредованного регуляторными Т-клетками, содержащая регуляторные Т-клетки, полученные вышеописанным способом, в качестве активного ингредиента.
В настоящее время регуляторные Т-клетки, полученные данным способом культивирования, могут иметь повышенное количество экспрессии Foxp3+. Термин «Foxp3» в том виде, как он используется в данном документе, относится к белку, также именуемому скурфин. Данный белок представляет собой белок, участвующий в пути регуляторного механизма регуляторных Т-клеток и известен как маркер регуляторных Т-клеток. Предпочтительно регуляторные Т-клетки, полученные выше, могут представлять собой Т-клетки CD4+CD25+CD127-Foxp3+.
Термин «заболевание, опосредованное регуляторными Т-клетками» в том виде, как он используется в данном документе, относится к заболеванию, индуцированному ненормальностью или недостаточностью регуляторных Т-клеток, и может конкретно характеризоваться как воспалительное заболевание или аутоиммунное заболевание.
Конкретное воплощение настоящего изобретения может отличаться тем, что данное воспалительное заболевание представляет собой по меньшей мере одно, выбранное из группы, состоящей из волчанки, синдрома Шегрена, ревматоидного артрита, фибромиозита, склеродермии, анкилозирующего спондилита, болезни Бехчета, автозного стоматита, синдрома Гийена-Барре, гнездной алопеции, полимиозита, болезни Крона, колита, узелкового полиартериита, рецидивирующего полихондрита и аутоиммунной тромбоцитопении. Кроме того, конкретное воплощение настоящего изобретения может отличаться тем, что аутоиммунное заболевание представляет собой по меньшей мере одно, выбранное из группы, состоящей из ревматоидного артрита, системного склероза, инсулинозависимого юношеского диабета, вызванного антителами против клеток поджелудочной железы, гнездной алопеции, псориаза, пузырчатки, астмы, молочницы полости рта, хронического тиреоидита, некоторой приобретенной апластической анемии, первичного цирроза, язвенного колита, болезни Бехчета, болезни Крона, силикоза, асбестоза, IgA нефропатии, постстрептококкового гломерулонефрита, синдрома Шегрена, синдрома Гийена-Барре, полимиозита, множественного миозита, рассеянного склероза, аутоиммунной гемолитической анемии, аутоиммунного энцефаломиелита, тяжелой миастении, гипертиреоза Грейвса, узелкового полиартериита, анкилозирующего спондилита, фибромиалгии, височного артериита, болезни Вильсона, синдрома Фанкони, множественной миеломы и системной красной волчанки.
Композиция по настоящему изобретению может содержать фармацевтически приемлемый носитель и/или добавку, или тому подобное. Например, она может содержать стерильную воду, нормальный физиологический раствор, традиционный буфер (например, на основе фосфорной кислоты, лимонной кислоты и другой органической кислоты), стабилизатор, соль, антиоксидант, поверхностно-активное вещество, суспендирующий агент, изотоничный агент или консервант. Кроме того, она может содержать органическое вещество, такое как биополимер, и неорганическое вещество, такое как гидроксиапатит, в частности, коллагеновую матрицу, полимер полимолочной кислоты или ее сополимер, полимер полиэтиленгликоля или его сополимер, его химическое производное и их смесь, но не ограничиваясь ими. Примеры стабилизатора могут включать декстран 40, метил целлюлозу, желатин, сульфит натрия, метасульфат натрия или тому подобные. Примеры антиоксиданта могут включать хелатор, такой как эриторбовая кислота, дибутилгидрокситолуол, бутилгидроксианизол, α-токоферол, токоферилацеат, L-аскорбиновой кислоты пальмитат, L-аскорбиновой кислоты стеарат, гидросульфит натрия, сульфит натрия, триамил галловой кислоты, пропил галловой кислоты или натрий этилендиаминтетрауксусную кислоту (EDTA), пирофосфат натрия, метафосфат натрия и тому подобные. Примеры суспендирующего агента могут включать метилцеллюлозу, полисорбат 80, гидроксиэтилцеллюлозу, аравийскую камедь, трагакантовую камедь, натрий карбоксиметилцеллюлозу, полиоксиэтилена сорбитана монолаурат и тому подобные. Примеры изотоничного агента могут включать D-маннит и сорбит. Примеры консерванта могут включать метилпараоксибензоат, этилпараоксибензоат, сорбиновую кислоту, фенол, крезол, хлоркрезол или тому подобные.
Способ лечения с использованием полученных регуляторных Т-клеток
Согласно другому аспекту настоящего изобретения предложен способ лечения заболевания, опосредованного регуляторными Т-клетками, включающий введение регуляторных Т-клеток индивидууму, имеющему заболевание, опосредованное регуляторными Т-клетками. В настоящее время регуляторные Т-клетки и заболевание, опосредованное регуляторными Т-клетками, являются такими, как описано выше.
Согласно еще одному другому аспекту настоящего изобретения предложено применение регуляторных Т-клеток для лечения заболевния, опосредованного регуляторными Т-клетками.
Способ осуществления данного изобретения
Ниже настоящее изобретение будет более подробно описано посредством следующих примеров. Однако следующие примеры предназначены только для осуществления иллюстрации настоящего изобретения, и объем настоящего изобретения не ограничивается ими.
Пример получения 1. Получение варианта hCD80-Fc-IL-2 (2М): GI-101
Для того, чтобы получать слитый белок, содержащий фрагмент человеческого CD80, домен Fc и вариант IL-2, синтезировали полинукпеотид, содержащий нуклеотидную последовательность (SEQ ID NO: 8), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, связанный с линкером (SEQ ID NO: 3), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2M), в котором заменены две аминокислоты (R38A, F42A) (SEQ ID NO: 6), в данном порядке от N-конца посредством службы синтеза генов GeneArt Invitrogen Thermo Fisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, данный вектор вводили в клетки СНО (яйцеклетки китайского хомяка) (EXPI-СНО™) для экспрессии слитого белка SEQ ID NO: 9. После введения вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин и 8% СО2 в течение 7 суток и затем отбирали для очистки слитого белка. Очищенный димер слитого белка называли «GI-101)).
Очистку проводили с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался при условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения в сборную пробирку 20% 1 М Tris-HCl с рН 9 отбирали слитый белок. Отобранный слитый белок диализировали в буфер PBS (фосфатно-солевой буферный раствор) в течение 16 часов для замены буфера.
Затем измеряли поглощение при длине волны 280 нм с течением времени посредством применения гель-фильтрации с колонкой TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом выделенный и очищенный слитый белок подвергали SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия) при восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим для подтверждения его чистоты (ФИГ. 1В). Подтвердили то, что слитый белок содержался в концентрации 2,78 мг/мл при выявлении с использованием NanoDrop (ФИГ. 1С). Также результат, проанализированный с использованием гель-фильтрации, является таким, как показано на ФИГ. 1D.
Пример получения 2. Получение димера варианта Fc-IL-2 (2М): Fc-IL-2v2
Для того, чтобы получить слитый белок, содержащий домен Fc и вариант IL-2, синтезировали полинуклеотид, содержащий нуклеотидную последовательность (SEQ ID NO: 45), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), шарнир Ig (SEQ ID NO: 38), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и вариант IL-2 (2М), в котором заменяются две аминокислоты (R38A, F42A) (SEQ ID NO: 6), в данном порядке от N-конца посредством службы синтеза генов GeneArt Invitrogen ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, данный вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 44. После введения данного вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин и 8% СО2 в течение 7 суток и затем отбирали для очистки димера слитого белка. Очищенный димер слитого белка называли «Fc-IL2v2».
Очистку и отбор данного слитого белка проводили таким же способом, как и в Примере получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE при восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим с подтверждением его чистоты (ФИГ. 3А). В результате, подтвердили то, что слитый белок образует димер. Также результат, проанализированный с использованием гель-фильтрации, является таким, как показано на ФИГ. 3В.
Пример получения 3. Получение димера Fc-IL-2: Fc-IL-2wt
Для того, чтобы получить слитый белок, содержащий домен Fc и IL-2 дикого типа, синтезировали полинуклеотид, содержащий нуклеотидную последовательность (SEQ ID NO: 43), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), шарнир Ig (SEQ ID NO: 38), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и IL-2 дикого типа (SEQ ID NO: 10), в данном порядке от N-конца посредством службы синтеза генов GeneArt Invitrogen Thermo Fisher Scientific inc. и клонировали в вектор pcDNA3_4. Кроме того, данный вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 42. После введения вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин и 8% СО2 в течение 7 суток и затем отбирали для очистки димера слитого белка. Очищенный димер слитого белка называли «Fc-IL2wt».
Очистку и отбор данного слитого белка проводили таким же способом, как и в Примере получения 1. Выделенный и очищенный слитый белок подвергали SDS-PAGE при восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим с подтверждением его чистоты (ФИГ. 3С). В результате, подтвердили то, что слитый белок образует димер. Также результат, проанализированный с использованием гель-фильтрации, является таким, как показано на ФИГ. 3D.
Пример получения 4. Получение димера hCD80-Fc-IL-2 дикого типа: hCD80-Fc-IL-2wt
Для того, чтобы получать слитый белок, содержащий фрагмент человеческого CD80, домен Fc и белок IL-2 дикого типа, синтезировали полинуклеотид, содержащий нукпеотидную последовательность (SEQ ID NO: 41), кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, связанный с линкером (SEQ ID NO: 3), домен Fc (SEQ ID NO: 4), линкер (SEQ ID NO: 5) и IL-2 дикого типа (SEQ ID NO: 10) в данном порядке от N-конца, посредством службы синтеза генов GeneArt Invitrogen ThermoFisher Scientific Inc. и клонировали в вектор pcDNA3_4. Кроме того, данный вектор вводили в клетки СНО (EXPI-CHO™) для экспрессии слитого белка SEQ ID NO: 46. После введения вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин и 8% СО2 в течение 7 суток и затем отбирали для очистки димера слитого белка. Очищенный димер слитого белка называли «hCD80-Fc-IL2wt».
Очистку проводили с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался при условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения в сборную пробирку 20% 1 М Tris-HCl с рН 9 отбирали слитый белок. Отобранный слитый белок диализировали в буфер PBS в течение 16 часов для замены буфера.
Затем измеряли поглощение при длине волны 280 нм с течением времени посредством применения гель-фильтрации с колонкой TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом выделенный и очищенный слитый белок подвергали SDS-PAGE при восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим с подтверждением его чистоты (ФИГ. 4А). В результате подтвердили то, что данный слитый белок образует димер. Также результат, проанализированный с использованием гель-фильтрации, является таким, как показано на ФИГ. 4В.
Пример получения 5. Получение димера hCD80-Fc: hCD80-Fc
Для того, чтобы получить слитый белок, содержащий фрагмент человеческого CD80 и домен Fc, синтезировали полинуклеотид (SEQ ID NO: 39), содержащий нуклеотидную последовательность, кодирующую слитый белок, содержащий сигнальный пептид (SEQ ID NO: 1), фрагмент CD80 (SEQ ID NO: 2), шарнир Ig, связанный с линкером (SEQ ID NO: 3), и домен Fc (SEQ ID NO: 4) в данном порядке от N-конца, посредством службы синтеза генов GeneArt invitrogen ThermoFisher Scientific inc. и клонировали в вектор pcDNA3_4. Кроме того, данный вектор вводили в клетки СНО (EXPi-CHO™) для экспрессии слитого белка SEQ ID NO: 40. После введения вектора культуральный раствор культивировали в окружающей среде с 37°С, 125 об./мин и 8% СО2 в течение 7 суток и затем отбирали для очистки димера слитого белка. Очищенный димер слитого белка называли «hCD80-Fc».
Очистку проводили с использованием хроматографии, включающей смолу с белком A MabSelect SuRe. Слитый белок связывался при условиях 25 мМ Tris, 25 мМ NaCl и рН 7,4. Затем его элюировали 100 мМ NaCl и 100 мМ уксусной кислотой при рН 3. После помещения в сборную пробирку 20% 1 М Tris-HCl с рН 9 отбирали слитый белок. Отобранный слитый белок диализировали в буфер PBS в течение 16 часов для замены буфера.
Затем измеряли поглощение при длине волны 280 нм с течением времени посредством применения гель-фильтрации с колонкой TSKgel G3000SWXL (TOSOH Bioscience) с получением высокой концентрации слитого белка. При этом выделенный и очищенный слитый белок подвергали SDS-PAGE при восстанавливающих (R) или невосстанавливающих (NR) условиях и окрашивали кумасси синим с подтверждением его чистоты (ФИГ. 2А). В результате подтвердили то, что данный слитый белок образует димер. Также результат, проанализированный с использованием гель-фильтрации, является таким, как показано на ФИГ. 2В.
Пример получения 1. Культуральная композиция для культивирования регуляторных Т-клеток
Пример получения 1.1. Культуральная композиция клеток CD4+
Культуральную среду клеток CD4+ получали в виде следующей композиции. При этом получали среду из основных компонентов, и затем перед применением добавляли дополнительные компоненты Gl-101, Gl-101WT, hCD80-Fc, Fc-IL-2v2 или Fc-IL-2wt.
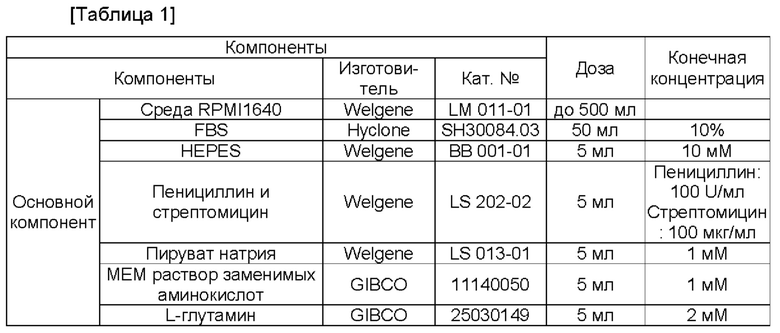
Пример получения 1.2. Культуральная композиция клеток CD4+CD25+CD127-
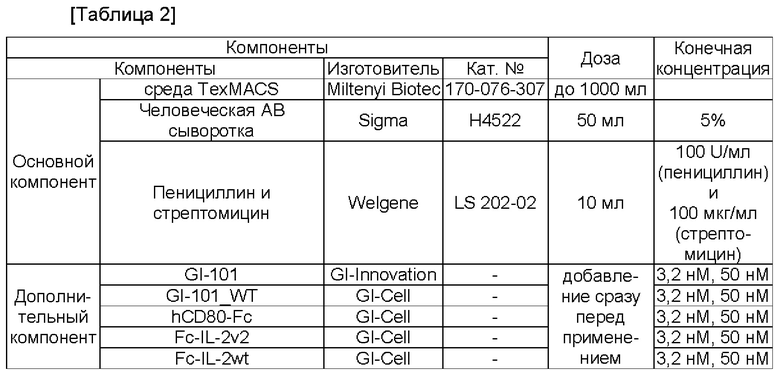
Культуральную композицию клеток CD4+CD25+CD127- получали следующим образом.
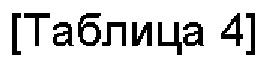
Пример 1. Определение уровня пролиферации регуляторных Т-клеток посредством димера слитого белка, содержащего белок IL-2 и белок CD80
Пример 1.1. Получение шариков для стимулирования пролиферации регуляторных Т-клеток
Для того, чтобы стимулировать пролиферацию регуляторных Т-клеток в выделенных клетках CD4+, получали шарики для стимулирования пролиферации регуляторных Т-клеток с использованием набора MACS GMP ExpAct Treg (кат. №:170-076-119) (Miltenyi Biotec, Bergisch Giadbach, Германия). В частности, реактив в наборе MACS GMP ExpAct Treg, который содержит шарики для стимулирования пролиферации регуляторных Т-клеток, переносили в новую пробирку, давали данной пробирке находиться на магните в течение 1 минуты, и затем супернатант удаляли для отделения шариков. При этом использовали 1 мкл реактива в наборе MACS GMP ExpAct Treg на 2×105 клеток. После выделения шариков добавляли от 0,5 мл до 1 мл культуральной композиции клеток CD4+ в Таблице 1 или Таблице 2, которая не содержит какой-либо дополнительный компонент (содержащей только основные компоненты), для ресуспендирования шариков.
Пример 1.2. Выделение Т-клеток CD4+
Измеряли число приобретенных РВМС (Кат. №: SER-PBMC-200-F) (Zen-Bio. Inc, NC 27709, США). Затем их центрифугировали при 300 g в течение 10 минут. Затем, после удаления буферного раствора супернатанта, добавляли 80 мкл буфера MAC на 1×107 клеток для ресуспендирования клеточных осадков. Затем дозировали 20 мкл микрошариков CD4 (кат. №130-045-101) (Miltenyi Biotec, Bergisch Gladbach, Германия) на 1×107 клеток, постукивали и достаточно перемешивали. Затем проводили взаимодействие при 4°С-8°С в течение 15 минут.
Для промывки добавляли 10 мл буфера MAC и затем центрифугировали при 300 g в течение 10 минут. Затем после удаления супернатанта добавляли 500 мкл буфера MAC на 1×108 клеток для ресуспендирования клеточных осадков. Затем готовили колонку LS, и затем пропускали 3 мл буфера MAC. Полученную выше суспензию клеток пропускали через колонку LS (кат. №130-042-401) (Miltenyi Biotec, Bergisch Gladbach, Германия). 3 мл буфера MAC пропускали 3 раза таким образом, что клетки, присоединенные к колонке LS, могли быть в достаточной степени промыты. Затем после снятие колонки LS с магнитного столика добавляли 3 мл буфера MACS и прилагали давление поршнем для выделения клеток CD4+. Затем осуществляли центрифугирование при 300 g в течение 5 минут. Затем супернатант удаляли, и затем измеряли число клеток.
Кроме того, культуральный раствор регуляторных Т-клеток, содержащий шарики, полученные в Примере 1.1, инокулировали на Т-клетки CD4+, выделенные выше.
Пример 1.3. Культура Т-клеток CD4+
В 6-луночном планшете клетки CD4+, полученные в Примере 1.2, высевали в концентрации 1×107 клеток/мл, и клетки CD4+ культивировали при условиях культуральной композиции клеток в Таблице 1 или Таблице 2, соответственно, содержащей GI-101 (50 нМ), GI-101_WT (50 нМ), димер CD80-Fc (50 нМ) + димер Fc-IL-2-2v2 (50 нМ) или димер CD80-Fc (50 нМ) + димер Fc-IL-2wt (50 нМ) в качестве добавок. При демонстрировании клетками в 6-луночном планшете больше, чем 80%-ной конфлюентности их субкультивировали во флаконе 25Т. При демонстрировании клетками во флаконе 25Т больше, чем 80%-ной конфлюентности их субкультивировали во флаконе 75Т. Наконец, при демонстрировании клетками во флаконе 75Т больше, чем 80%-ной конфлюентности их отбирали.
Пример 1.4. Выделение клеток CD4+CD25+CD127-
Клетки CD4+, выделенные в Примере 1.3, центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Кроме того, удаляли супернатант. Добавляли 1 мл блокирующего Fc реактива (biolegend, кат. №422302), разведенного в буфере FACS до 1:200, и оставляли на льду на 10 минут, и затем добавляли 50 мкл CD4-Pacific Blue (BioLegend, кат. №317429), 50 мкл CD25-PE/Cy7 (BioLegend, кат. №356108) и 50 мкл CD127-PE (BD, кат. №557938) и оставляли на льду на 20 минут. Затем дополнительно добавляли 4 мл буфера FACS и центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Затем, после удаления супернатанта, добавляли 3 мл буфера FACS и центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Затем удаляли супернатант, и добавляли 3 мл буфера FACS для ресуспендирования клеток. Наконец, клетки CD4+CD25+CD127- выделяли с использованием BD FACS Aria.
Пример 1.5. Культивирование клеток CD4+CD25+CD127-
Клетки CD4+CD25+CD127-, выделенные в Примере 1.4, высевали в концентрации 3×105 клеток/мл в 48-луночный планшет, и в то же самое время выделяли шарики из 1 мкл набора MACS GMP ExpAct Treg (кат. №:170-076-119) (Miltenyi Biotec, Bergisch Gladbach, Германия) на 2×106 клеток для стимулирования пролиферации регуляторных Т-клеток, как в Примере 1.1. Выделенные шарики ресуспендировали в культуральной композиции клеток (от 0,5 мл до 1 мл) в Таблице 3 или Таблице 4 (содержащей только базовые компоненты), которая не содержит какой-либо дополнительный компонент, и затем добавляли в луночный планшет, в который высевали клетки CD4+CD25+CD127-, полученные в Примере 1.4. Затем клетки CD4+CD25+CD127- культивировали при условиях культуральной композиции клеток в Таблице 3 или Таблице 4, соответственно, содержащей GI-101 (50 нМ), GI-101_WT (50 нМ), CD80-Fc (50 нМ) + Fc-IL-2-2v2 (50 нМ) или CD80-Fc (50 нМ) + Fc-IL-2wt (50 нМ) в качестве добавки.
В культуре при демонстрировании клетками больше, чем 80%-ной конфлюентности в 48-луночном планшете их субкультивировали в 24-луночном планшете. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 24-луночном планшете их субкультивировали в 12-луночном планшете. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 12-луночном планшете их субкультивировали в 6-луночном планшете. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 6-луночном планшете их субкультивировали в 25Т флаконе. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 25Т флаконе их субкультивировали в 75Т флаконе. Наконец, клетки отбирали, когда они демонстрировали больше, чем 80%-ную конфлюентность после субкультивирования в 75Т флаконе.
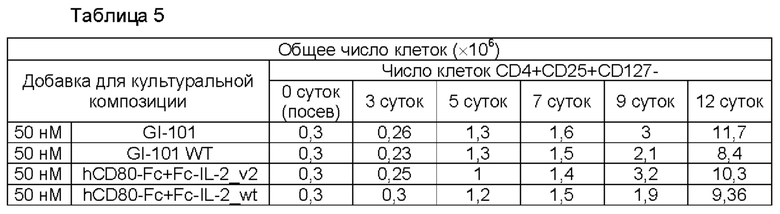
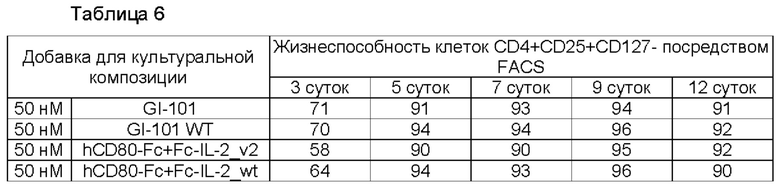
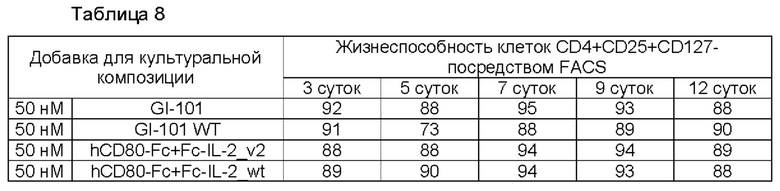
В результате, результаты пролиферации регуляторных Т-клеток в композиции культуральной среды для регуляторных Т-клеток, содержащей среду RPMI1640, являются такими, как показано в Таблице 5 и на ФИГ. 6, и жизнеспособности клеток являются такими, как показано в Таблице 6 и на ФИГ. 7. Кроме того, результаты пролиферации регуляторных Т-клеток в культуральной композиции, содержащей среду TexMACS, показаны в Таблице 7 и на ФИГ. 8, и жизнеспособности клеток показаны в Таблице 8 и на ФИГ. 9.

Пример 1.6. Определение секреторных способностей иммунодепрессивных цитокинов: интерлейкин-10
Для того, чтобы оценить секреторные способности IL-10 регуляторных Т-клеток, полученных в Примере 1,5, данные клетки культивировали таким образом, что число клеток корректировали до 1×106 клеток/мл, и затем получали супернатант культуры для анализа посредством ELISA (твердофазный иммуноферментный анализ).
Во-первых, 100 мкл инкубационного буфера набора ELISA для человеческого IL-10 (Invitrogen, кат. № KAC1321) дозировали в каждый микропланшет, и затем добавляли для дозирования 100 мкл калибровки, контроля или образца супернатанта культуры регуляторных Т-клеток соответственно. Затем данные микропланшеты покрывали клейкой пленкой и осуществляли взаимодействие со встряхиванием при 700 об./мин при комнатной температуре (от 18°С до 25°С) в течение 2 часов на шейкере. Затем ряды микролунок 3 раза промывали примерно 150 мкл промывочного буфера на лунку.
После промывки добавляли 100 мкл разбавителя образцов. Затем добавляли 50 мкл антитела против IL-10, конъюгированного с HRP (пероксидаза хрена), и затем осуществляли взаимодействие со встряхиванием при 700 об./мин при комнатной температуре в течение 2 часов, с последующей промывкой 3 раза с использованием примерно 150 мкл промывочного буфера на лунку. После промывки добавляли 100 мкл ТМВ (тетраметилбензидин) и затем осуществляли взаимодействие при комнатной температуре в течение 15 минут, с последующим завершением реакции посредством добавления 100 мкл останавливающего раствора. Затем значения флуоресценции каждой микролунки измеряли при 450 нм.
В результате, секреторные способности интерлейкина-10 клеток, культивируемых в композиции, содержащей среду RPMH640, являются такими, как показано на ФИГ. 10, и секреторные способности интерлейкина-10 клеток, культивируемых в композиции, содержащей среду TexMACS, являются такими, как показано на ФИГ. 11. На ФИГ. 10 и 11 подтверждено, что интерлейкин-10 высоко экспрессировался в регуляторных Т-клетках, культивируемых в культуральной композиции, содержащей GI-101.
Пример 2. Определение пролиферации регуляторных Т-клеток посредством GH01 в оптимизированном способе культивирования
Пример 2.1. Выделение Т-клеток CD4+
Измеряли число приобретенных РВМС (Кат. №: SER-PBMC-200-F) (Zen-Bio Inc, NC 27709, США). Затем их центрифугировали при 300 g в течение 10 минут. Затем, после удаления буферного раствора супернатанта, добавляли 80 мкл буфера MAC на 1×107 клеток для ресуспендирования клеточных осадков. Затем дозировали 20 мкл микрошариков CD4 (кат. №130-045-101) (Miltenyi Biotec, Bergisch Gladbach, Германия) на 1×107 клеток, постукивали и достаточно перемешивали. Затем проводили взаимодействие при 4°С-8°С в течение 15 минут.
Для промывки добавляли 10 мл буфера MAC и затем центрифугировали при 300 g в течение 10 минут. Затем после удаления супернатанта добавляли 500 мкл буфера MAC на 1×108 клеток для ресуспендирования клеточных осадков. Затем готовили колонку LS, и затем пропускали 3 мл буфера MAC. Полученную выше суспензию клеток пропускали через колонку LS (кат. №130-042-401) (Miltenyi Biotec, Bergisch Gladbach, Германия). 3 мл буфера MAC пропускали 3 раза таким образом, что клетки, присоединенные к колонке LS, могли быть в достаточной степени промыты. Затем после снятие колонки LS с магнитного столика добавляли 3 мл буфера MAC и прилагали давление поршнем для выделения клеток CD4+. Затем осуществляли центрифугирование при 300 g в течение 5 минут. После центрифугирования супернатант удаляли, и измеряли число клеток.
Для того, чтобы стимулировать пролиферацию регуляторных Т-клеток в выделенных клетках CD4+ добавляли от 0,5 мл до 1 мл культуральных композиций клеток CD4+ в Таблице 1 или Таблице 2 (содержащих только базовые компоненты), которые не содержат какого-либо дополнительного компонента, для ресуспендирования шариков, которые выделяли и готовили как в Примере 1.1, и это инокулировали в клетки CD4+, выделенные из РВМС.
Пример 2.2. Культивирование Т-клеток CD4+
В 6-луночном планшете клетки CD4+, полученные в Примере 2.1, высевали в концентрации 1×107 клеток/мл и культивировали при условиях композиции культуры клеток в Таблице 1 или Таблице 2, соответственно, содержащей GI-101 (3,2 нМ/50 нМ), GI-101_WT (3,2 нМ/50 нМ), димер CD80-Fc (3,2 нМ/50 нМ) + димер Fc-IL-2v2 (3,2 нМ/50 нМ) или димер CD80-Fc (3,2 нМ/50 нМ) + димер Fc-IL-2wt (3,2 нМ/50 нМ) в качестве добавки. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 6-луночном планшете их субкультивировали во флаконе 25Т. При демонстрировании клетками во флаконе 25Т больше, чем 80%-ной конфлюентности их субкультивировали во флаконе 75Т. Наконец, при демонстрировании клетками во флаконе 75Т больше, чем 80%-ной конфлюентности их отбирали.
Пример 2.3. Выделение Т-клеток CD4+CD25+CD127-
Клетки CD4+, выделенные в Примере 2.2, центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Кроме того, удаляли супернатант и добавляли 1 мл блокирующего Fc реактива (biolegend, кат. №422302), разведенного в буфере FACS до 1:200, с последующим оставлением на льду на 10 минут. Добавляли в образец 2 50 мкл CD4-Pacific Blue (BioLegend, кат. №317429), 50 мкл CD25-PE/Cy7 (BioLegend, кат. №356108) и 50 мкл CD127-PE (BD, кат. №557938) и оставляли на льду на 20 минут.
Затем дополнительно добавляли 4 мл буфера FACS и центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Затем удаляли супернатант и добавляли 3 мл буфера FACS, с последующим центрифугированием при 1300 об./мин и 4°С в течение 5 минут. Затем удаляли супернатант, и добавляли 3 мл буфера FACS для ресуспендирования клеток. Наконец, клетки CD4+CD25+CD127- выделяли с использованием BD FACS Aria.
Пример 2.4. Культивирование Т-клеток CD4+CD25+CD127-
Все клетки CD4+CD25+CD127-, выделенные в Примере 2.3, высевали в 24-луночный планшет и, в то же самое время, для стимуляции пролиферации регуляторных Т-клеток, выделяли шарики из 1 мкл набора MACS GMP ExpAct Treg (кат. №:170-076-119) (Miltenyi Biotec, Bergisch Gladbach, Германия) на 2×105 клеток, как и в Примере 2.1, и данные шарики ресуспендировали в культуральной композиции клеток (от 0,5 мл до 1 мл) в Таблице 3 или Таблице 4 (содержащей только базовые компоненты), которая не содержит какого-либо дополнительного компонента. Затем их добавляли в луночный планшет, в который были посеяны клетки CD4+CD25+CD127-.
Затем клетки CD4+CD25+CD127- культивировали при условиях культуральной композиции клеток в Таблице 3 или Таблице 4, соответственно, содержащей GI-101 (3,2 нМ/50 нМ), GI-101_WT (3,2 нМ/50 нМ), димер CD80-Fc (3,2 нМ/50 нМ) + димер Fc-IL-2v2 (3,2 нМ/50 нМ) или димер CD80-Fc (3,2 нМ/50 нМ) + Fc-IL-2wt (3,2 нМ/50 нМ) в качестве добавки. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 24-луночном планшете их субкультивировали в 12-луночном планшете. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 12-луночном планшете их субкультивировали в 6-луночном планшете. При демонстрировании клетками больше, чем 80%-ной конфлюентности в 6-луночном планшете их субкультивировали во флаконе 25Т. При демонстрировании клетками во флаконе 25Т больше, чем 80%-ной конфлюентности их субкультивировали во флаконе 75Т.
Для группы, обработанной 3,2 нМ добавок в Таблице 3 или Таблице 4, клетки наконец отбирали при демонстрировании ими больше, чем 80%-ной конфлюентности во флаконе 75Т. Для группы, обработанной 50 нМ добавок в Таблице 3 или Таблице 4, клетки наконец отбирали при демонстрировании ими больше, чем 80%-ной конфлюентности после субкультивирования во флаконе 175Т.
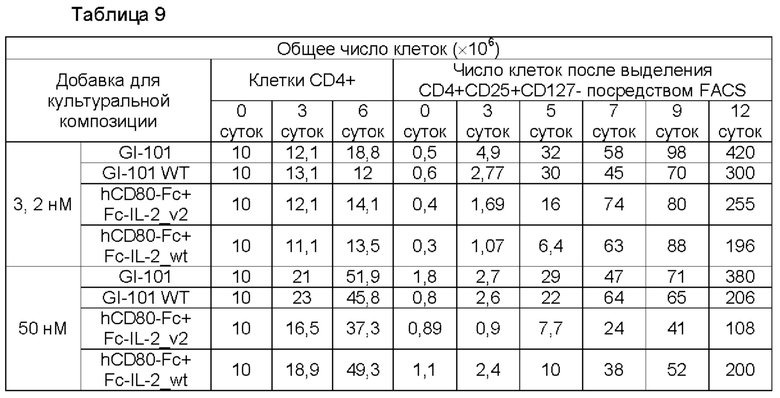
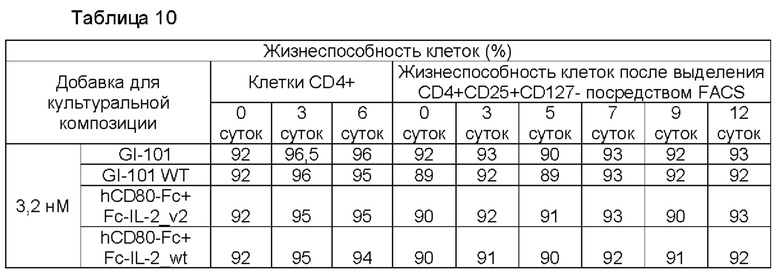
В результате, в культуральной композиции, содержащей среду RPMH640, результаты пролиферации клеток являются такими, как показано в Таблице 9, на ФИГ. 12А и ФИГ. 12В, и жизнеспособности клеток являются такими, как показано в Таблице 10, на ФИГ. 13А и ФИГ. 13В. Кроме того, в культуральной композиции, содержащей среду TexMACS, результаты пролиферации клеток показаны в Таблице 11, на ФИГ. 14А и ФИГ. 14В, и жизнеспособности клеток показаны в Таблице 12, на ФИГ. 15А и ФИГ. 15В.
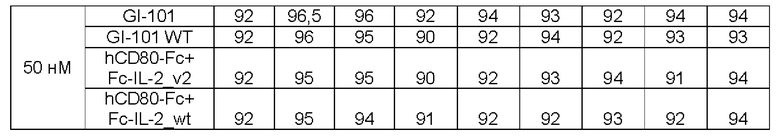
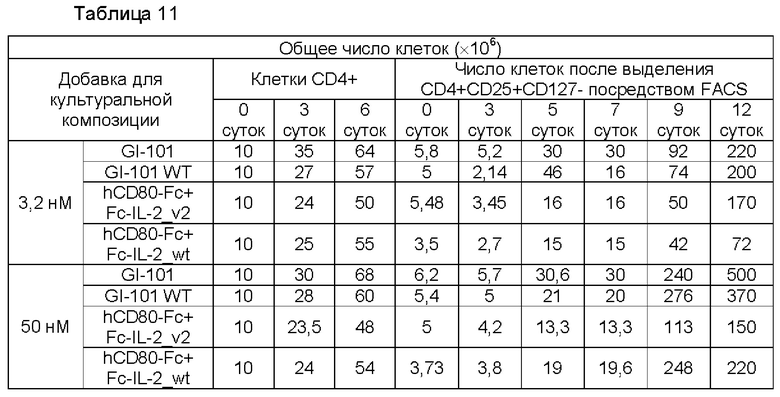
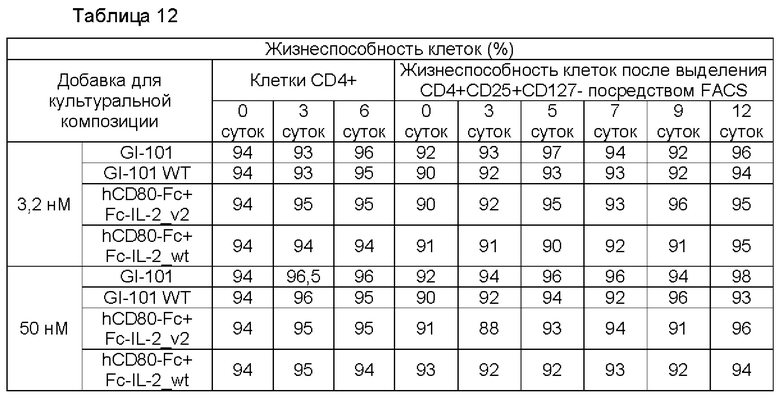
Пример 2.5. Характеристика клеток
Клетки CD4+CD25+CD127-, выделенные в Примере 2.4, центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Кроме того, супернатант удаляли. Затем добавляли 100 мкл блокирующего Fc реактива (biolegend, кат. №422302), разведенного в буфере FACS до 1:200, оставляли на льду на 10 минут и дополнительно добавляли 2,5 мкл CD4-PerCP-Cy5.5 (eBioscience, кат. №45-0048-42), 2,5 мкл CD25-APC/Cy7 (BD, кат. №557753) и 2,5 мкл CD127-бриллиантовый фиолетовый 785 (Biolegend, кат. №351330) и оставляли на льду на 20 минут.
Затем дополнительно добавляли 1 мл буфера FACS и центрифугировали при 1300 об./мин и 4°С в течение 5 минут. Получали раствор смеси 1:3 концентрата фиксатора/пермеабилизатора и разбавителя фиксатора/пермеабилизатора, и 100 мкл добавляли после удаления супернатанта от клеток, полученных центрифугированием, и оставляли на льду на 30 минут. Затем дополнительно добавляли 1 мл буфера FACS, и дважды повторяли центрифугирование при 1300 об./мин и 4°С в течение 5 минут. Затем удаляли супернатант, добавляли 100 мкл буфера FACS и 2,5 мкл Foxp3-Pacific Blue (Biolegend, кат. №320116) и оставляли на льду на 20 минут. Затем дополнительно добавляли 1 мл буфера FACS и центрифугирование при 1300 об./мин и 4°С в течение 5 минут. Затем удаляли супернатант и добавляли 200 мкл буфера FACS для ресуспендирования клеток. Наконец, клетки характеризовали с использованием оборудования Cytek Aurora.
В результате результаты анализа FACS свойств клеток, культивируемых в композиции, содержащей среду RPIVH1640, являются такими, как показано на ФИГ. 16А-16С, и число Т-клеток, экспрессирующих CD4+CD25+Foxp3+, подтвержденное в приведенных выше культуральных условиях, является таким, как показано в Таблице 13 и на ФИГ. 17А-17С. Кроме того, результаты анализа FACS свойств клеток, культивируемых в композиции, содержащей среду TexMACS, являются такими, как показано на ФИГ. 18А-18С, и число Т-клеток, экспрессирующих CD4+CD25+Foxp3+, подтвержденное в приведенных выше культуральных условиях, является таким, как показано в Таблице 14 и на ФИГ. 19А-19С.
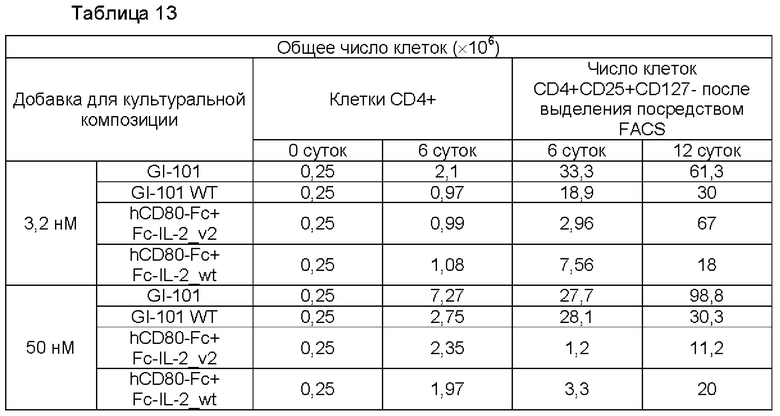
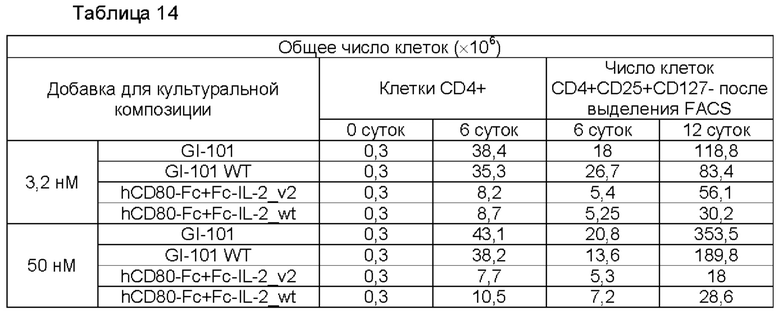
Пример 2.6. Определение секреторных способностей иммунодепрессивных цитокинов: интерлейкин-10
Для того, чтобы оценить секреторные способности IL-10 регуляторных Т-клеток, полученных в Примере 2.4, данные клетки культивировали таким образом, что число клеток корректировали до 1×106 клеток/мл, и затем получали супернатант культуры для анализа посредством ELISA (твердофазный иммуноферментный анализ).
Во-первых, 100 мкл инкубационного буфера набора ELiSA для человеческого IL-10 (Invitrogen, кат. № KAC1321) дозировали в каждый микропланшет, и затем добавляли для дозирования 100 мкл калибровки, контроля или образца культурального супернатанта регуляторных Т-клеток соответственно. Затем данные микропланшеты покрывали клейкой пленкой и осуществляли взаимодействие со встряхиванием при 700 об./мин при комнатной температуре (от 18°С до 25°С) в течение 2 часов на шейкере. Затем ряды микролунок 3 раза промывали примерно 150 мкл промывочного буфера на лунку.
После промывки добавляли 100 мкл разбавителя образцов. Затем добавляли 50 мкл антитела против IL-10, конъюгированного с HRP, и затем осуществляли взаимодействие со встряхиванием при 700 об./мин при комнатной температуре в течение 2 часов, с последующей промывкой 3 раза с использованием примерно 150 мкл промывочного буфера на лунку. После промывки добавляли 100 мкл ТМВ и затем осуществляли взаимодействие прикомнатной температуре в течение 15 минут, с последующим завершением реакции посредством добавления 100 мкл останавливающего раствора. Затем значения флуоресценции каждой микролунки измеряли при 450 нм.
В результате, секреторные способности интерлейкина-10 клеток, культивируемых в композиции, содержащей среду RPMI1640, являются такими, как показано на ФИГ. 20, и секреторные способности интерлейкина-10 клеток, культивируемых в композиции, содержащей среду TexMACS, являются такими, как показано на ФИГ. 21. На ФИГ. 20 и 21 подтверждено, что интерлейкин-10 высоко экспрессировался в регуляторных Т-клетках, культивируемых в культуральной композиции, содержащей GI-101.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> GI Cell, Inc.
<120> КОМПОЗИЦИЯ ДЛЯ КУЛЬТИВИРОВАНИЯ РЕГУЛЯТОРНЫХ Т-КЛЕТОК И ЕЕ ПРИМЕНЕНИЕ
<130> SPO20-038-PCT-GIC
<150> KR 10-2019-0149779
<151> 2019-11-20
<150> KR 10-2020-0033229
<151> 2020-03-18
<160> 46
<170> KopatentIn 3.0
<210> 1
<211> 25
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> сигнальный пептид (TPA)
<400> 1
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala
20 25
<210> 2
<211> 208
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hB7-1:35-242
<400> 2
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
<210> 3
<211> 30
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> шарнир с линкером
<400> 3
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1 5 10 15
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
20 25 30
<210> 4
<211> 216
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> fc иммуноглобулина
<400> 4
Ala Pro Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Phe Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Leu Gly
210 215
<210> 5
<211> 5
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> linker
<400> 5
Gly Gly Gly Gly Ser
1 5
<210> 6
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2M
<400> 6
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 7
<211> 617
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок, содержащий варианты IL-2 и фрагменты CD80
<400> 7
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Ile His Val Thr Lys Glu
20 25 30
Val Lys Glu Val Ala Thr Leu Ser Cys Gly His Asn Val Ser Val Glu
35 40 45
Glu Leu Ala Gln Thr Arg Ile Tyr Trp Gln Lys Glu Lys Lys Met Val
50 55 60
Leu Thr Met Met Ser Gly Asp Met Asn Ile Trp Pro Glu Tyr Lys Asn
65 70 75 80
Arg Thr Ile Phe Asp Ile Thr Asn Asn Leu Ser Ile Val Ile Leu Ala
85 90 95
Leu Arg Pro Ser Asp Glu Gly Thr Tyr Glu Cys Val Val Leu Lys Tyr
100 105 110
Glu Lys Asp Ala Phe Lys Arg Glu His Leu Ala Glu Val Thr Leu Ser
115 120 125
Val Lys Ala Asp Phe Pro Thr Pro Ser Ile Ser Asp Phe Glu Ile Pro
130 135 140
Thr Ser Asn Ile Arg Arg Ile Ile Cys Ser Thr Ser Gly Gly Phe Pro
145 150 155 160
Glu Pro His Leu Ser Trp Leu Glu Asn Gly Glu Glu Leu Asn Ala Ile
165 170 175
Asn Thr Thr Val Ser Gln Asp Pro Glu Thr Glu Leu Tyr Ala Val Ser
180 185 190
Ser Lys Leu Asp Phe Asn Met Thr Thr Asn His Ser Phe Met Cys Leu
195 200 205
Ile Lys Tyr Gly His Leu Arg Val Asn Gln Thr Phe Asn Trp Asn Thr
210 215 220
Thr Lys Gln Glu His Phe Pro Asp Asn Gly Ser Gly Gly Gly Gly Ser
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly
245 250 255
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
290 295 300
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly
465 470 475 480
Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
485 490 495
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
500 505 510
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe
515 520 525
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
530 535 540
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
545 550 555 560
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
565 570 575
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
580 585 590
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
595 600 605
Cys Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 8
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101)
<400> 8
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga
1857
<210> 9
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101)
<400> 9
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 10
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2
<400> 10
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 11
<211> 288
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80
<400> 11
Met Gly His Thr Arg Arg Gln Gly Thr Ser Pro Ser Lys Cys Pro Tyr
1 5 10 15
Leu Asn Phe Phe Gln Leu Leu Val Leu Ala Gly Leu Ser His Phe Cys
20 25 30
Ser Gly Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu
35 40 45
Ser Cys Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile
50 55 60
Tyr Trp Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp
65 70 75 80
Met Asn Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr
85 90 95
Asn Asn Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly
100 105 110
Thr Tyr Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg
115 120 125
Glu His Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr
130 135 140
Pro Ser Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile
145 150 155 160
Ile Cys Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu
165 170 175
Glu Asn Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp
180 185 190
Pro Glu Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met
195 200 205
Thr Thr Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg
210 215 220
Val Asn Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro
225 230 235 240
Asp Asn Leu Leu Pro Ser Trp Ala Ile Thr Leu Ile Ser Val Asn Gly
245 250 255
Ile Phe Val Ile Cys Cys Leu Thr Tyr Cys Phe Ala Pro Arg Cys Arg
260 265 270
Glu Arg Arg Arg Asn Glu Arg Leu Arg Arg Glu Ser Val Arg Pro Val
275 280 285
<210> 12
<211> 215
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> модифицированный Fc
<400> 12
Ser His Thr Gln Pro Leu Gly Val Phe Leu Phe Pro Pro Lys Pro Lys
1 5 10 15
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
20 25 30
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
35 40 45
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
50 55 60
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
65 70 75 80
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
85 90 95
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
100 105 110
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
115 120 125
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
130 135 140
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
145 150 155 160
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
165 170 175
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
180 185 190
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
195 200 205
Leu Ser Leu Ser Leu Gly Lys
210 215
<210> 13
<211> 306
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> mCD80
<400> 13
Met Ala Cys Asn Cys Gln Leu Met Gln Asp Thr Pro Leu Leu Lys Phe
1 5 10 15
Pro Cys Pro Arg Leu Ile Leu Leu Phe Val Leu Leu Ile Arg Leu Ser
20 25 30
Gln Val Ser Ser Asp Val Asp Glu Gln Leu Ser Lys Ser Val Lys Asp
35 40 45
Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His Glu Asp Glu Ser
50 55 60
Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val Val Leu Ser Val
65 70 75 80
Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys Asn Arg Thr Leu
85 90 95
Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly Leu Val Leu Ser
100 105 110
Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys Glu Arg Gly Thr
115 120 125
Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser Ile Lys Ala Asp
130 135 140
Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro Ser Ala Asp Thr
145 150 155 160
Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro Lys Pro Arg Phe
165 170 175
Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile Asn Thr Thr Ile
180 185 190
Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser Ser Gln Leu Asp
195 200 205
Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu Ile Lys Tyr Gly
210 215 220
Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys Pro Pro Glu Asp
225 230 235 240
Pro Pro Asp Ser Lys Asn Thr Leu Val Leu Phe Gly Ala Gly Phe Gly
245 250 255
Ala Val Ile Thr Val Val Val Ile Val Val Ile Ile Lys Cys Phe Cys
260 265 270
Lys His Arg Ser Cys Phe Arg Arg Asn Glu Ala Ser Arg Glu Thr Asn
275 280 285
Asn Ser Leu Thr Phe Gly Pro Glu Glu Ala Leu Ala Glu Gln Thr Val
290 295 300
Phe Leu
305
<210> 14
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101)
<400> 14
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg
120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa
180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag
240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc
300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag
360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag
420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct
480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt
540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc
600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt
660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc
720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca
780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag
840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa
900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag
960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg
1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg
1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt
1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg
1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag
1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct
1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg
1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt
1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg
1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc
1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag
1620
tgcctggaag aggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac
1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa
1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt
1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc
1848
<210> 15
<211> 616
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101)
<400> 15
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 16
<211> 1437
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C1)
<400> 16
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctaggtgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc cctgggc
1437
<210> 17
<211> 454
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C1)
<400> 17
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly
450
<210> 18
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101C2)
<400> 18
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccatctc acgccgctga gtctaagtac ggccctcctt gtcctccatg tcctgctcca
120
gaagctgctg gcggaccctc tgtgttcctg tttcctccaa agcctaagga ccagctcatg
180
atctctcgga cccctgaagt gacctgcgtg gtggtggatg tgtctcaaga ggaccctgag
240
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga
300
gaggaacagt tcaactccac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat
360
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca agggcctgcc ttccagcatc
420
gaaaagacca tctccaaggc taagggccag cctagggaac cccaggttta caccctgcct
480
ccaagccaag aggaaatgac caagaaccag gtgtccctga cctgcctggt caagggcttc
540
tacccttccg acattgccgt ggaatgggag tccaatggcc agcctgagaa caactacaag
600
accacacctc ctgtgctgga ctccgacggc tccttctttc tgtactctcg cctgaccgtg
660
gacaagtcta ggtggcaaga gggcaacgtg ttctcctgct ctgtgctgca cgaggccctg
720
cacaatcact acacccagaa gtccctgtct ctgtctcttg gcggaggcgg aggatctgct
780
cctacctcca gctccaccaa gaaaacccag ctccagttgg agcatctgct gctggacctc
840
cagatgatcc tgaatggcat caacaattac aagaacccca agctgaccgc catgctgacc
900
gctaagttct acatgcccaa gaaggccacc gagctgaagc acctccagtg cctggaagag
960
gaactgaagc ccctggaaga agtgctgaat ctggcccagt ccaagaactt ccacctgagg
1020
cctagggacc tgatctccaa catcaacgtg atcgtgctgg aactgaaagg ctccgagaca
1080
accttcatgt gcgagtacgc cgacgagaca gccaccatcg tggaatttct gaaccggtgg
1140
atcaccttct gccagtccat catctccaca ctgacc
1176
<210> 19
<211> 367
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101C2)
<400> 19
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
1 5 10 15
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
20 25 30
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
35 40 45
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
50 55 60
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
85 90 95
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
100 105 110
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
115 120 125
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
130 135 140
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
145 150 155 160
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
165 170 175
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
180 185 190
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
195 200 205
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
210 215 220
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
225 230 235 240
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
245 250 255
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Ala
260 265 270
Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
275 280 285
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
290 295 300
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
305 310 315 320
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
325 330 335
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
340 345 350
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
355 360 365
<210> 20
<211> 1434
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI101C1)
<400> 20
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg
120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa
180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag
240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc
300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag
360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag
420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct
480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt
540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc
600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt
660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc
720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca
780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag
840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa
900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag
960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg
1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg
1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt
1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg
1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag
1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct
1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg
1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtccct gggc
1434
<210> 21
<211> 478
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI101C1)
<400> 21
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
465 470 475
<210> 22
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M45)
<400> 22
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 23
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M61)
<400> 23
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 24
<211> 133
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> варианты IL-2 (3M, M72)
<400> 24
Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His
1 5 10 15
Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys
20 25 30
Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys
35 40 45
Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys
50 55 60
Pro Leu Glu Glu Val Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu
65 70 75 80
Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu
85 90 95
Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala
100 105 110
Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile
115 120 125
Ile Ser Thr Leu Thr
130
<210> 25
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M45)
<400> 25
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
accgccatgc tgaccgctaa gttcgccatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c
1851
<210> 26
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M45)
<400> 26
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Ala Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 27
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M61)
<400> 27
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aaagggaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c
1851
<210> 28
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M61)
<400> 28
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Arg Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 29
<211> 1857
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI102-M72)
<400> 29
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
accgccatgc tgaccgctaa gttctacatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatggggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac ctgatga
1857
<210> 30
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI102-M72)
<400> 30
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Ala Met Leu Thr Ala Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Gly Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 31
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (GI101w)
<400> 31
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
acccgcatgc tgacctttaa gttctacatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c
1851
<210> 32
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (GI101w)
<400> 32
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<210> 33
<211> 1848
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (mGI102-M61)
<400> 33
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgga cgagcagctc tccaagtccg tgaaggataa ggtcctgctg
120
ccttgccggt acaactctcc tcacgaggac gagtctgagg accggatcta ctggcagaaa
180
cacgacaagg tggtgctgtc cgtgatcgcc ggaaagctga aagtgtggcc tgagtacaag
240
aacaggaccc tgtacgacaa caccacctac agcctgatca tcctgggcct cgtgctgagc
300
gatagaggca cctattcttg cgtggtgcag aagaaagagc ggggcaccta cgaagtgaag
360
cacctggctc tggtcaagct gtccatcaag gccgacttca gcacccctaa catcaccgag
420
tctggcaacc cttccgccga caccaagaga atcacctgtt tcgcctctgg cggcttccct
480
aagcctcggt tctcttggct ggaaaacggc agagagctgc ccggcatcaa taccaccatt
540
tctcaggacc cagagtccga gctgtacacc atctccagcc agctcgactt taacaccacc
600
agaaaccaca ccatcaagtg cctgattaag tacggcgacg cccacgtgtc cgaggacttt
660
acttgggaga aacctcctga ggaccctcct gactctggat ctggcggcgg aggttctggc
720
ggaggtggaa gcggaggcgg aggatctgct gagtctaagt atggccctcc ttgtcctcca
780
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag
840
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa
900
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag
960
accaagccta gagaggaaca gttcaactcc acctatagag tggtgtccgt gctgaccgtg
1020
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg
1080
ccttccagca tcgaaaagac catcagcaag gctaagggcc agcctaggga accccaggtt
1140
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg
1200
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag
1260
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct
1320
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg
1380
cacgaggctc tgcacaacca ctacacccag aagtccctgt ctctgtctct tggaggtggt
1440
ggcggttctg cccctacctc cagctctacc aagaaaaccc agctccagtt ggagcatctg
1500
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc
1560
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacttgcag
1620
tgcctggaaa gggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac
1680
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa
1740
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt
1800
ctgaaccggt ggatcacctt ctgccagagc atcatctcca cactgacc
1848
<210> 34
<211> 616
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> слитый белок (mGI102-M61)
<400> 34
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Asp Glu Gln Leu Ser Lys
20 25 30
Ser Val Lys Asp Lys Val Leu Leu Pro Cys Arg Tyr Asn Ser Pro His
35 40 45
Glu Asp Glu Ser Glu Asp Arg Ile Tyr Trp Gln Lys His Asp Lys Val
50 55 60
Val Leu Ser Val Ile Ala Gly Lys Leu Lys Val Trp Pro Glu Tyr Lys
65 70 75 80
Asn Arg Thr Leu Tyr Asp Asn Thr Thr Tyr Ser Leu Ile Ile Leu Gly
85 90 95
Leu Val Leu Ser Asp Arg Gly Thr Tyr Ser Cys Val Val Gln Lys Lys
100 105 110
Glu Arg Gly Thr Tyr Glu Val Lys His Leu Ala Leu Val Lys Leu Ser
115 120 125
Ile Lys Ala Asp Phe Ser Thr Pro Asn Ile Thr Glu Ser Gly Asn Pro
130 135 140
Ser Ala Asp Thr Lys Arg Ile Thr Cys Phe Ala Ser Gly Gly Phe Pro
145 150 155 160
Lys Pro Arg Phe Ser Trp Leu Glu Asn Gly Arg Glu Leu Pro Gly Ile
165 170 175
Asn Thr Thr Ile Ser Gln Asp Pro Glu Ser Glu Leu Tyr Thr Ile Ser
180 185 190
Ser Gln Leu Asp Phe Asn Thr Thr Arg Asn His Thr Ile Lys Cys Leu
195 200 205
Ile Lys Tyr Gly Asp Ala His Val Ser Glu Asp Phe Thr Trp Glu Lys
210 215 220
Pro Pro Glu Asp Pro Pro Asp Ser Gly Ser Gly Gly Gly Gly Ser Gly
225 230 235 240
Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly Pro
245 250 255
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
260 265 270
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
275 280 285
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
290 295 300
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
305 310 315 320
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
325 330 335
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
340 345 350
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
355 360 365
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
370 375 380
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
385 390 395 400
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
405 410 415
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
420 425 430
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
435 440 445
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
450 455 460
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
465 470 475 480
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
485 490 495
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
500 505 510
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
515 520 525
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Arg
530 535 540
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
545 550 555 560
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
565 570 575
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
580 585 590
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
595 600 605
Gln Ser Ile Ile Ser Thr Leu Thr
610 615
<210> 35
<211> 153
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> hIL-2 дикого типа
<400> 35
Met Tyr Arg Met Gln Leu Leu Ser Cys Ile Ala Leu Ser Leu Ala Leu
1 5 10 15
Val Thr Asn Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu
20 25 30
Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile
35 40 45
Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe
50 55 60
Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu
65 70 75 80
Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys
85 90 95
Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile
100 105 110
Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala
115 120 125
Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe
130 135 140
Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150
<210> 36
<211> 158
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> IL-2 с сигнальной последовательностью
<400> 36
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Ala Pro Thr Ser Ser Ser Thr
20 25 30
Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln Met
35 40 45
Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg Met
50 55 60
Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys His
65 70 75 80
Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu Asn
85 90 95
Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile Ser
100 105 110
Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe
115 120 125
Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu Asn
130 135 140
Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
145 150 155
<210> 37
<211> 474
<212> ДНК
<213> Искусственная последовательность
<220>
<223> последовательность нуклеотидов, кодирующая IL-2 с сигнальной
последовательностью
<400> 37
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgcccc taccagctcc tctaccaaga aaacccagct ccagttggag
120
catctgctgc tggacctcca gatgattctg aacgggatca acaactataa gaaccccaag
180
ctgacccgca tgctgacctt taagttctac atgcccaaga aggccaccga gctgaagcac
240
ctccagtgcc tggaagaaga actgaagccc ctggaagagg tgctgaatct ggcccagtcc
300
aagaacttcc acctgaggcc acgggacctg atcagcaaca tcaacgtgat cgtgctggaa
360
ctgaagggct ccgagacaac ctttatgtgc gagtacgccg acgagacagc caccatcgtg
420
gaatttctga accggtggat caccttctgc cagagcatca tctccacact gacc
474
<210> 38
<211> 13
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> шарнир
<400> 38
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro
1 5 10
<210> 39
<211> 1461
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие CD80-Fc белок
<400> 39
ggatccgcca ccatggatgc tatgctgaga ggcctgtgtt gcgtgctgct gctgtgtggc
60
gctgtgttcg tgtctccttc tcacgctgtg atccacgtga ccaaagaagt gaaagaggtc
120
gccacactgt cctgcggcca caacgtttca gtggaagaac tggcccagac caggatctac
180
tggcagaaag aaaagaaaat ggtgctgacc atgatgtccg gcgacatgaa catctggcct
240
gagtacaaga accggaccat cttcgacatc accaacaacc tgtccatcgt gattctggcc
300
ctgaggcctt ctgatgaggg cacctatgag tgcgtggtgc tgaagtacga gaaggacgcc
360
ttcaagcgcg agcacctggc tgaagtgaca ctgtccgtga aggccgactt tcccacacct
420
tccatctccg acttcgagat ccctacctcc aacatccggc ggatcatctg ttctacctct
480
ggcggctttc ctgagcctca cctgtcttgg ctggaaaacg gcgaggaact gaacgccatc
540
aacaccaccg tgtctcagga ccccgaaacc gagctgtacg ctgtgtcctc caagctggac
600
ttcaacatga ccaccaacca cagcttcatg tgcctgatta agtacggcca cctgagagtg
660
aaccagacct tcaactggaa caccaccaag caagagcact tccctgacaa tggatctggc
720
ggcggaggtt ctggcggagg tggaagcgga ggcggaggat ctgctgagtc taagtatggc
780
cctccttgtc ctccatgtcc tgctccagaa gctgctggcg gaccctctgt gttcctgttt
840
cctccaaagc ctaaggacca gctcatgatc tctcggacac ccgaagtgac ctgcgtggtg
900
gtggatgtgt ctcaagagga ccctgaggtg cagttcaatt ggtacgtgga cggcgtggaa
960
gtgcacaacg ccaagaccaa gcctagagag gaacagttca actccaccta cagagtggtg
1020
tccgtgctga ccgtgctgca ccaggattgg ctgaacggca aagagtacaa gtgcaaggtg
1080
tccaacaagg gcctgccttc cagcatcgaa aagaccatct ccaaggctaa gggccagcct
1140
agggaacccc aggtttacac cctgcctcca agccaagagg aaatgaccaa gaaccaggtg
1200
tccctgacct gcctggtcaa gggcttctac ccttccgaca ttgccgtgga atgggagtcc
1260
aatggccagc ctgagaacaa ctacaagacc acacctcctg tgctggactc cgacggctcc
1320
ttctttctgt actctcgcct gaccgtggac aagtctaggt ggcaagaggg caacgtgttc
1380
tcctgctctg tgctgcacga ggccctgcac aatcactaca cccagaagtc cctgtctctg
1440
tccctgggct gatgactcga g
1461
<210> 40
<211> 479
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80-Fc protein
<400> 40
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Val Ile His Val Thr Lys Glu
20 25 30
Val Lys Glu Val Ala Thr Leu Ser Cys Gly His Asn Val Ser Val Glu
35 40 45
Glu Leu Ala Gln Thr Arg Ile Tyr Trp Gln Lys Glu Lys Lys Met Val
50 55 60
Leu Thr Met Met Ser Gly Asp Met Asn Ile Trp Pro Glu Tyr Lys Asn
65 70 75 80
Arg Thr Ile Phe Asp Ile Thr Asn Asn Leu Ser Ile Val Ile Leu Ala
85 90 95
Leu Arg Pro Ser Asp Glu Gly Thr Tyr Glu Cys Val Val Leu Lys Tyr
100 105 110
Glu Lys Asp Ala Phe Lys Arg Glu His Leu Ala Glu Val Thr Leu Ser
115 120 125
Val Lys Ala Asp Phe Pro Thr Pro Ser Ile Ser Asp Phe Glu Ile Pro
130 135 140
Thr Ser Asn Ile Arg Arg Ile Ile Cys Ser Thr Ser Gly Gly Phe Pro
145 150 155 160
Glu Pro His Leu Ser Trp Leu Glu Asn Gly Glu Glu Leu Asn Ala Ile
165 170 175
Asn Thr Thr Val Ser Gln Asp Pro Glu Thr Glu Leu Tyr Ala Val Ser
180 185 190
Ser Lys Leu Asp Phe Asn Met Thr Thr Asn His Ser Phe Met Cys Leu
195 200 205
Ile Lys Tyr Gly His Leu Arg Val Asn Gln Thr Phe Asn Trp Asn Thr
210 215 220
Thr Lys Gln Glu His Phe Pro Asp Asn Gly Ser Gly Gly Gly Gly Ser
225 230 235 240
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Ala Glu Ser Lys Tyr Gly
245 250 255
Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser
260 265 270
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg
275 280 285
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro
290 295 300
Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
305 310 315 320
Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val
325 330 335
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
340 345 350
Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr
355 360 365
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
370 375 380
Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys
385 390 395 400
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
405 410 415
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp
420 425 430
Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser
435 440 445
Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala
450 455 460
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly
465 470 475
<210> 41
<211> 1851
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие слитый белок (hCD80-Fc-IL2wt)
<400> 41
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgtgat ccacgtgacc aaagaagtga aagaggtcgc cacactgtcc
120
tgcggccaca acgtttcagt ggaagaactg gcccagacca ggatctactg gcagaaagaa
180
aagaaaatgg tgctgaccat gatgtccggc gacatgaaca tctggcctga gtacaagaac
240
cggaccatct tcgacatcac caacaacctg tccatcgtga ttctggccct gaggccttct
300
gatgagggca cctatgagtg cgtggtgctg aagtacgaga aggacgcctt caagcgcgag
360
cacctggctg aagtgacact gtccgtgaag gccgactttc ccacaccttc catctccgac
420
ttcgagatcc ctacctccaa catccggcgg atcatctgtt ctacctctgg cggctttcct
480
gagcctcacc tgtcttggct ggaaaacggc gaggaactga acgccatcaa caccaccgtg
540
tctcaggacc ccgaaaccga gctgtacgct gtgtcctcca agctggactt caacatgacc
600
accaaccaca gcttcatgtg cctgattaag tacggccacc tgagagtgaa ccagaccttc
660
aactggaaca ccaccaagca agagcacttc cctgacaatg gatctggcgg cggaggttct
720
ggcggaggtg gaagcggagg cggaggatct gctgagtcta agtatggccc tccttgtcct
780
ccatgtcctg ctccagaagc tgctggcgga ccctctgtgt tcctgtttcc tccaaagcct
840
aaggaccagc tcatgatctc tcggacaccc gaagtgacct gcgtggtggt ggatgtgtct
900
caagaggacc ctgaggtgca gttcaattgg tacgtggacg gcgtggaagt gcacaacgcc
960
aagaccaagc ctagagagga acagttcaac tccacctaca gagtggtgtc cgtgctgacc
1020
gtgctgcacc aggattggct gaacggcaaa gagtacaagt gcaaggtgtc caacaagggc
1080
ctgccttcca gcatcgaaaa gaccatctcc aaggctaagg gccagcctag ggaaccccag
1140
gtttacaccc tgcctccaag ccaagaggaa atgaccaaga accaggtgtc cctgacctgc
1200
ctggtcaagg gcttctaccc ttccgacatt gccgtggaat gggagtccaa tggccagcct
1260
gagaacaact acaagaccac acctcctgtg ctggactccg acggctcctt ctttctgtac
1320
tctcgcctga ccgtggacaa gtctagatgg caagagggca acgtgttctc ctgctctgtg
1380
ctgcacgagg ccctgcacaa tcactacacc cagaagtccc tgtctctgtc tcttggaggt
1440
ggtggcggtt ctgcccctac cagctcctct accaagaaaa cccagctcca gttggagcat
1500
ctgctgctgg acctccagat gattctgaac gggatcaaca actataagaa ccccaagctg
1560
acccgcatgc tgacctttaa gttctacatg cccaagaagg ccaccgagct gaagcacctc
1620
cagtgcctgg aagaagaact gaagcccctg gaagaggtgc tgaatctggc ccagtccaag
1680
aacttccacc tgaggccacg ggacctgatc agcaacatca acgtgatcgt gctggaactg
1740
aagggctccg agacaacctt tatgtgcgag tacgccgacg agacagccac catcgtggaa
1800
tttctgaacc ggtggatcac cttctgccag agcatcatct ccacactgac c
1851
<210> 42
<211> 367
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Fc-IL2wt
<400> 42
Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu
1 5 10 15
Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp
20 25 30
Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp
35 40 45
Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly
50 55 60
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn
65 70 75 80
Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
85 90 95
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro
100 105 110
Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
115 120 125
Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn
130 135 140
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
145 150 155 160
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
165 170 175
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg
180 185 190
Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys
195 200 205
Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
210 215 220
Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser Ser
225 230 235 240
Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu Gln
245 250 255
Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr Arg
260 265 270
Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu Lys
275 280 285
His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val Leu
290 295 300
Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu Ile
305 310 315 320
Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr Thr
325 330 335
Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe Leu
340 345 350
Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
355 360 365
<210> 43
<211> 1176
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Fc-IL2wt
<400> 43
atggatgcta tgctgagagg cctgtgttgc gtgctgctgc tgtgtggcgc tgtgttcgtg
60
tctccttctc acgctgctga gtctaagtat ggccctcctt gtcctccatg tcctgctcca
120
gaagctgctg gcggaccctc tgtgttcctg tttcctccaa agcctaagga ccagctcatg
180
atctctcgga cacccgaagt gacctgcgtg gtggtggatg tgtctcaaga ggaccctgag
240
gtgcagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga
300
gaggaacagt tcaactccac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat
360
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca agggcctgcc ttccagcatc
420
gaaaagacca tctccaaggc taagggccag cctagggaac cccaggttta caccctgcct
480
ccaagccaag aggaaatgac caagaaccag gtgtccctga cctgcctggt caagggcttc
540
tacccttccg acattgccgt ggaatgggag tccaatggcc agcctgagaa caactacaag
600
accacacctc ctgtgctgga ctccgacggc tccttctttc tgtactctcg cctgaccgtg
660
gacaagtcta gatggcaaga gggcaacgtg ttctcctgct ctgtgctgca cgaggccctg
720
cacaatcact acacccagaa gtccctgtct ctgtctcttg gaggtggtgg cggttctgcc
780
cctaccagct cctctaccaa gaaaacccag ctccagttgg agcatctgct gctggacctc
840
cagatgattc tgaacgggat caacaactat aagaacccca agctgacccg catgctgacc
900
tttaagttct acatgcccaa gaaggccacc gagctgaagc acctccagtg cctggaagaa
960
gaactgaagc ccctggaaga ggtgctgaat ctggcccagt ccaagaactt ccacctgagg
1020
ccacgggacc tgatcagcaa catcaacgtg atcgtgctgg aactgaaggg ctccgagaca
1080
acctttatgt gcgagtacgc cgacgagaca gccaccatcg tggaatttct gaaccggtgg
1140
atcaccttct gccagagcat catctccaca ctgacc
1176
<210> 44
<211> 392
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> Fc-IL2v2
<400> 44
Met Asp Ala Met Leu Arg Gly Leu Cys Cys Val Leu Leu Leu Cys Gly
1 5 10 15
Ala Val Phe Val Ser Pro Ser His Ala Ala Glu Ser Lys Tyr Gly Pro
20 25 30
Pro Cys Pro Pro Cys Pro Ala Pro Glu Ala Ala Gly Gly Pro Ser Val
35 40 45
Phe Leu Phe Pro Pro Lys Pro Lys Asp Gln Leu Met Ile Ser Arg Thr
50 55 60
Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu Asp Pro Glu
65 70 75 80
Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
85 90 95
Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg Val Val Ser
100 105 110
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
115 120 125
Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu Lys Thr Ile
130 135 140
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
145 150 155 160
Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
165 170 175
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
180 185 190
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
195 200 205
Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp Lys Ser Arg
210 215 220
Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Leu His Glu Ala Leu
225 230 235 240
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu Gly Gly Gly
245 250 255
Gly Gly Ser Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln
260 265 270
Leu Glu His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn
275 280 285
Asn Tyr Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr
290 295 300
Met Pro Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu
305 310 315 320
Glu Leu Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn
325 330 335
Phe His Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val
340 345 350
Leu Glu Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp
355 360 365
Glu Thr Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys
370 375 380
Gln Ser Ile Ile Ser Thr Leu Thr
385 390
<210> 45
<211> 1200
<212> ДНК
<213> Искусственная последовательность
<220>
<223> нуклеотиды, кодирующие Fc-IL2v2
<400> 45
ggatccgcca ccatggatgc tatgctgaga ggcctgtgtt gcgtgctgct gctgtgtggc
60
gctgtgttcg tgtctccatc tcacgccgct gagtctaagt acggccctcc ttgtcctcca
120
tgtcctgctc cagaagctgc tggcggaccc tctgtgttcc tgtttcctcc aaagcctaag
180
gaccagctca tgatctctcg gacccctgaa gtgacctgcg tggtggtgga tgtgtctcaa
240
gaggaccctg aggtgcagtt caattggtac gtggacggcg tggaagtgca caacgccaag
300
accaagccta gagaggaaca gttcaactcc acctacagag tggtgtccgt gctgaccgtg
360
ctgcaccagg attggctgaa cggcaaagag tacaagtgca aggtgtccaa caagggcctg
420
ccttccagca tcgaaaagac catctccaag gctaagggcc agcctaggga accccaggtt
480
tacaccctgc ctccaagcca agaggaaatg accaagaacc aggtgtccct gacctgcctg
540
gtcaagggct tctacccttc cgacattgcc gtggaatggg agtccaatgg ccagcctgag
600
aacaactaca agaccacacc tcctgtgctg gactccgacg gctccttctt tctgtactct
660
cgcctgaccg tggacaagtc taggtggcaa gagggcaacg tgttctcctg ctctgtgctg
720
cacgaggccc tgcacaatca ctacacccag aagtccctgt ctctgtctct tggcggaggc
780
ggaggatctg ctcctacctc cagctccacc aagaaaaccc agctccagtt ggagcatctg
840
ctgctggacc tccagatgat cctgaatggc atcaacaatt acaagaaccc caagctgacc
900
gccatgctga ccgctaagtt ctacatgccc aagaaggcca ccgagctgaa gcacctccag
960
tgcctggaag aggaactgaa gcccctggaa gaagtgctga atctggccca gtccaagaac
1020
ttccacctga ggcctaggga cctgatctcc aacatcaacg tgatcgtgct ggaactgaaa
1080
ggctccgaga caaccttcat gtgcgagtac gccgacgaga cagccaccat cgtggaattt
1140
ctgaaccggt ggatcacctt ctgccagtcc atcatctcca cactgacctg atgactcgag
1200
1200
<210> 46
<211> 592
<212> ПРТ
<213> Искусственная последовательность
<220>
<223> CD80-Fc-IL2wt
<400> 46
Val Ile His Val Thr Lys Glu Val Lys Glu Val Ala Thr Leu Ser Cys
1 5 10 15
Gly His Asn Val Ser Val Glu Glu Leu Ala Gln Thr Arg Ile Tyr Trp
20 25 30
Gln Lys Glu Lys Lys Met Val Leu Thr Met Met Ser Gly Asp Met Asn
35 40 45
Ile Trp Pro Glu Tyr Lys Asn Arg Thr Ile Phe Asp Ile Thr Asn Asn
50 55 60
Leu Ser Ile Val Ile Leu Ala Leu Arg Pro Ser Asp Glu Gly Thr Tyr
65 70 75 80
Glu Cys Val Val Leu Lys Tyr Glu Lys Asp Ala Phe Lys Arg Glu His
85 90 95
Leu Ala Glu Val Thr Leu Ser Val Lys Ala Asp Phe Pro Thr Pro Ser
100 105 110
Ile Ser Asp Phe Glu Ile Pro Thr Ser Asn Ile Arg Arg Ile Ile Cys
115 120 125
Ser Thr Ser Gly Gly Phe Pro Glu Pro His Leu Ser Trp Leu Glu Asn
130 135 140
Gly Glu Glu Leu Asn Ala Ile Asn Thr Thr Val Ser Gln Asp Pro Glu
145 150 155 160
Thr Glu Leu Tyr Ala Val Ser Ser Lys Leu Asp Phe Asn Met Thr Thr
165 170 175
Asn His Ser Phe Met Cys Leu Ile Lys Tyr Gly His Leu Arg Val Asn
180 185 190
Gln Thr Phe Asn Trp Asn Thr Thr Lys Gln Glu His Phe Pro Asp Asn
195 200 205
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
210 215 220
Ser Ala Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
225 230 235 240
Glu Ala Ala Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
245 250 255
Asp Gln Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
260 265 270
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
275 280 285
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
290 295 300
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
305 310 315 320
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
325 330 335
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
340 345 350
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
355 360 365
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
370 375 380
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
385 390 395 400
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
405 410 415
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
420 425 430
Cys Ser Val Leu His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
435 440 445
Leu Ser Leu Ser Leu Gly Gly Gly Gly Gly Ser Ala Pro Thr Ser Ser
450 455 460
Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu His Leu Leu Leu Asp Leu
465 470 475 480
Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr Lys Asn Pro Lys Leu Thr
485 490 495
Arg Met Leu Thr Phe Lys Phe Tyr Met Pro Lys Lys Ala Thr Glu Leu
500 505 510
Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu Glu Val
515 520 525
Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg Asp Leu
530 535 540
Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser Glu Thr
545 550 555 560
Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val Glu Phe
565 570 575
Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr Leu Thr
580 585 590
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| ПОЛИВАЛЕНТНЫЕ МОДУЛЯТОРЫ РЕГУЛЯТОРНЫХ T-КЛЕТОК | 2017 |
|
RU2769871C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ ДЛЯ ПРЕДОТВРАЩЕНИЯ ИЛИ ЛЕЧЕНИЯ ГЕПАТИТА, ФИБРОЗА ПЕЧЕНИ И ЦИРРОЗА ПЕЧЕНИ, ВКЛЮЧАЮЩАЯ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2795548C2 |
| МУЛЬТИФУНКЦИОНАЛЬНЫЙ МУЛЬТИСПЕЦИФИЧЕСКИЙ МУЛЬТИМЕРНЫЙ БИОМОЛЕКУЛЯРНЫЙ ПОЛИМЕР, ИМЕЮЩИЙ ПРОЛОНГИРОВАННУЮ ЭФФЕКТИВНОСТЬ IN VIVO | 2020 |
|
RU2805307C1 |
| БЕЛКИ С ДВОЙНОЙ ФУНКЦИЕЙ И СОДЕРЖАЩАЯ ИХ ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ | 2016 |
|
RU2741345C2 |
| КОМПОЗИЦИЯ ДЛЯ ОБЛЕГЧЕНИЯ ИЛИ ЛЕЧЕНИЯ БОЛИ | 2017 |
|
RU2725830C1 |
| МОДИФИЦИРОВАННЫЕ ДОМЕНЫ α1-α2 НЕПРИРОДНЫХ ЛИГАНДОВ NKG2D, КОТОРЫЕ СВЯЗЫВАЮТСЯ С НЕПРИРОДНЫМИ РЕЦЕПТОРАМИ NKG2D | 2019 |
|
RU2815278C2 |
| КОМПОЗИЦИЯ ДЛЯ ПРОТИВОРАКОВОГО ЛЕЧЕНИЯ, СОДЕРЖАЩАЯ NK-КЛЕТКИ И СЛИТЫЙ БЕЛОК, КОТОРЫЙ СОДЕРЖИТ БЕЛОК IL-2 И БЕЛОК CD80 | 2020 |
|
RU2832136C1 |
| МОДИФИЦИРОВАННЫЕ МИТОХОНДРИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2811467C2 |
| КОМПОЗИЦИЯ ДЛЯ КУЛЬТИВИРОВАНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ И СПОСОБ ПОЛУЧЕНИЯ ЕСТЕСТВЕННЫХ КЛЕТОК-КИЛЛЕРОВ С ЕЕ ИСПОЛЬЗОВАНИЕМ | 2020 |
|
RU2824216C1 |
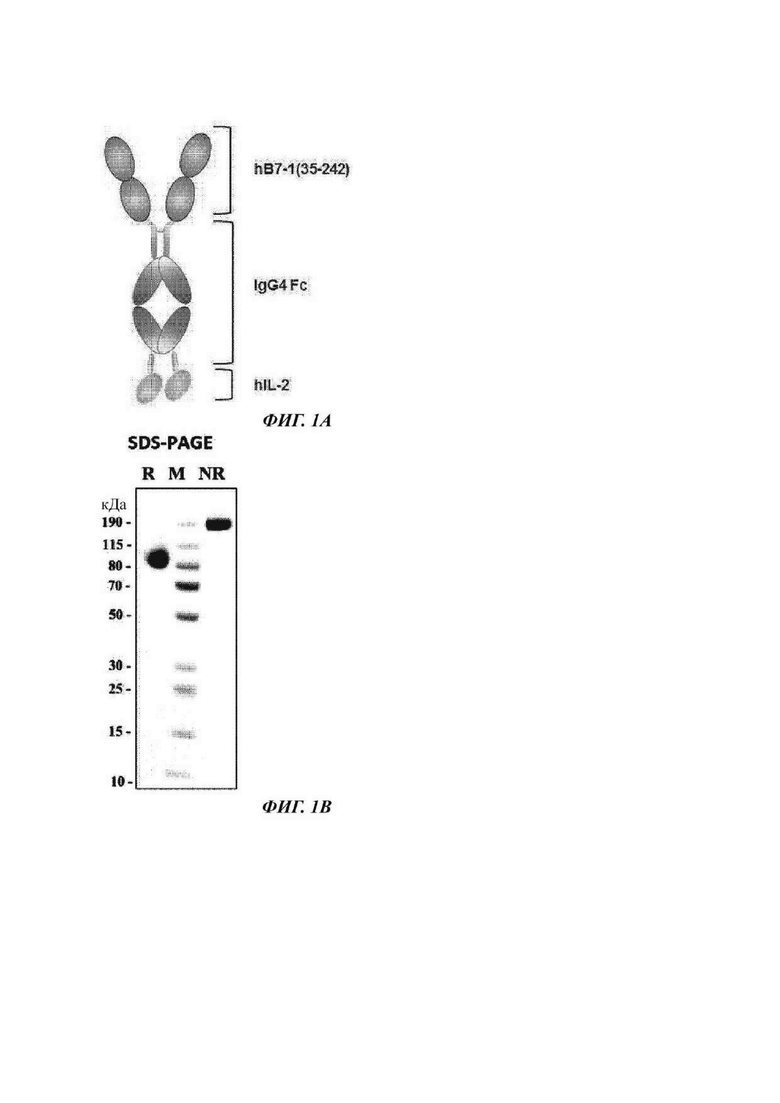
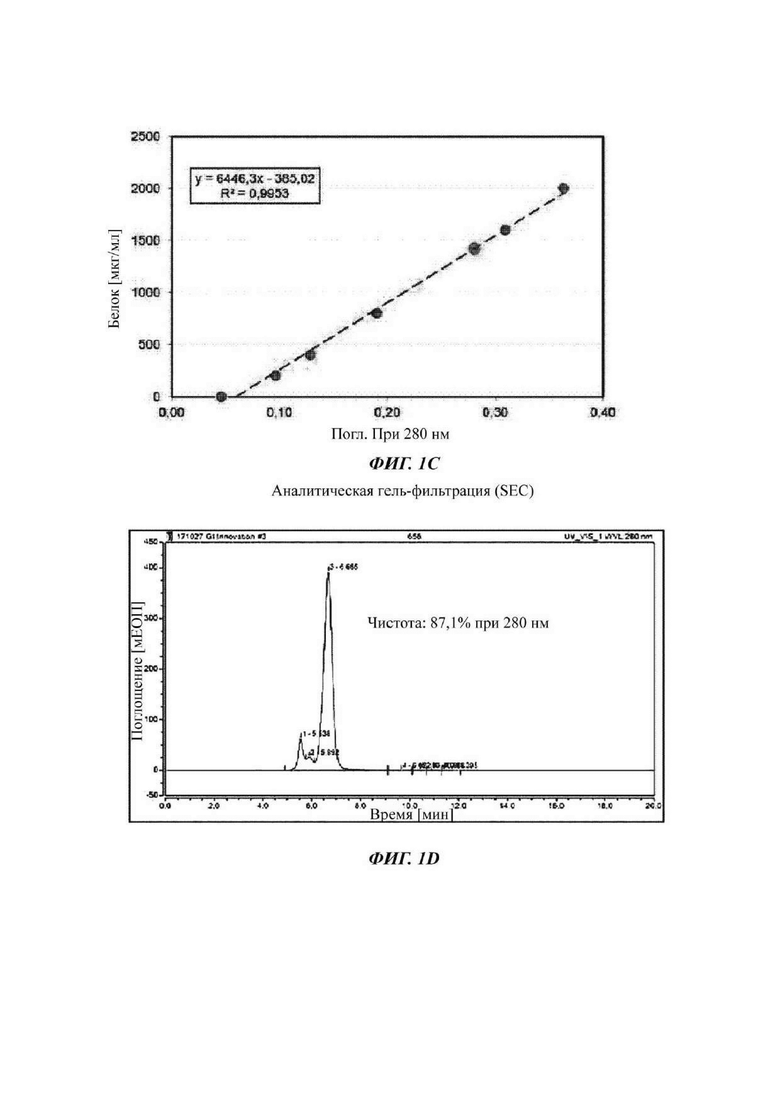
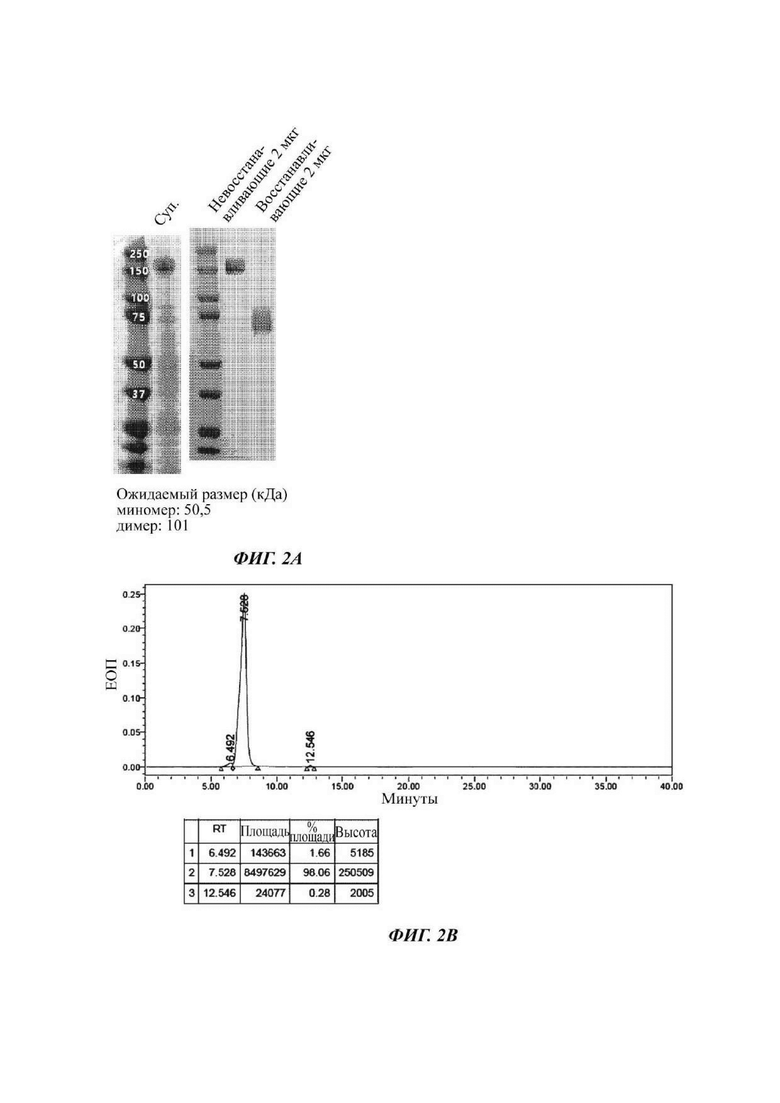
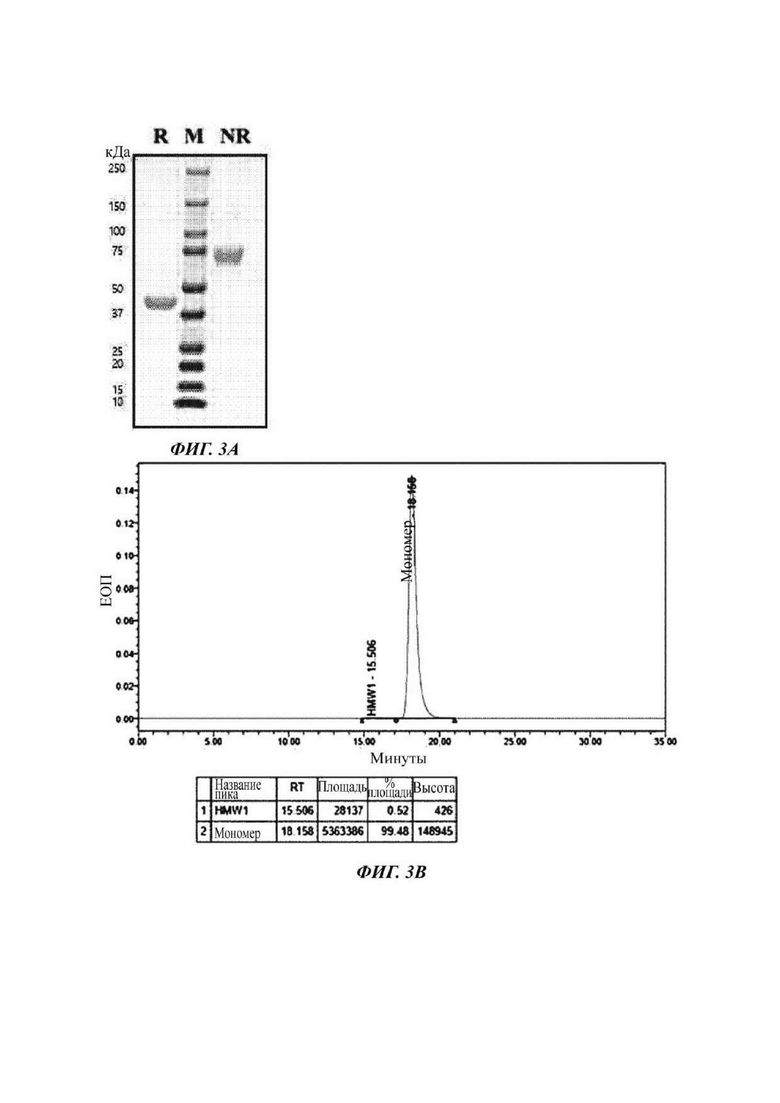

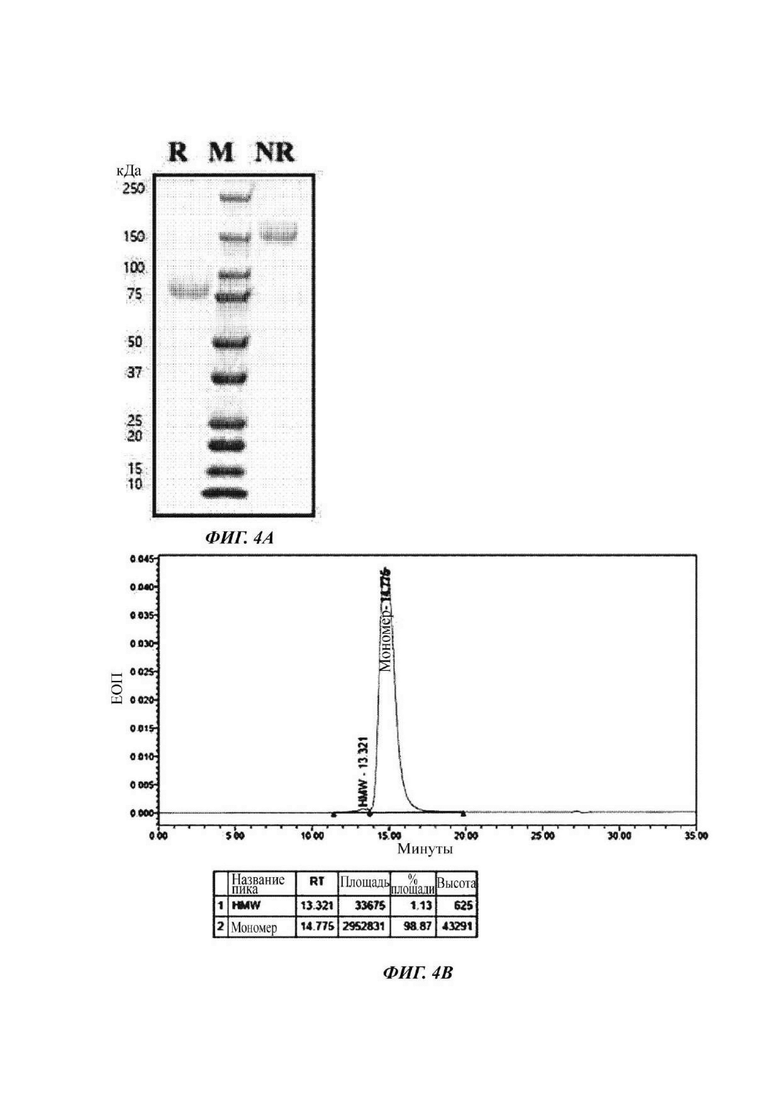
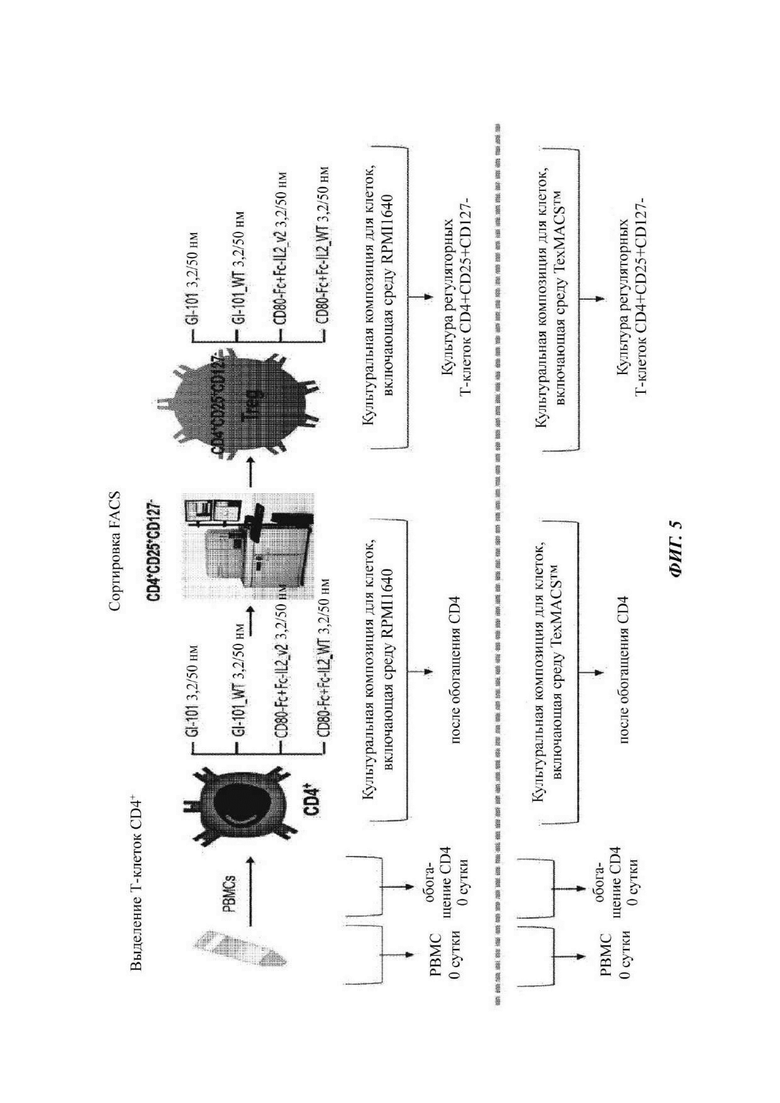
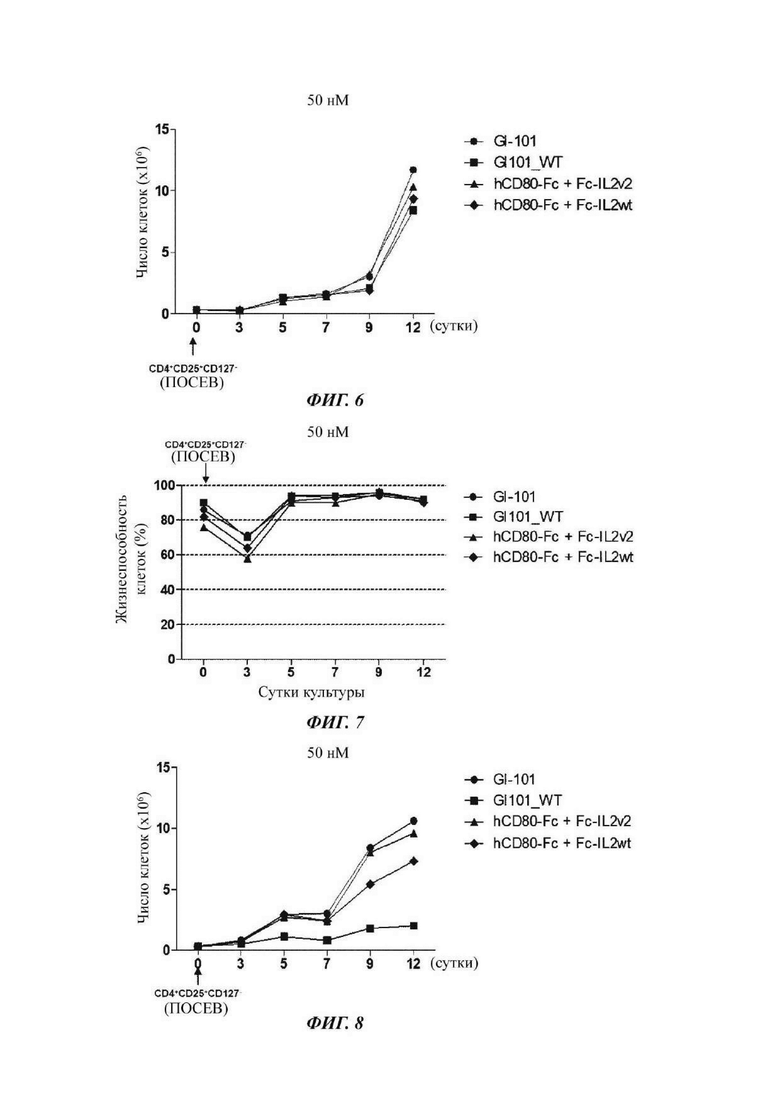
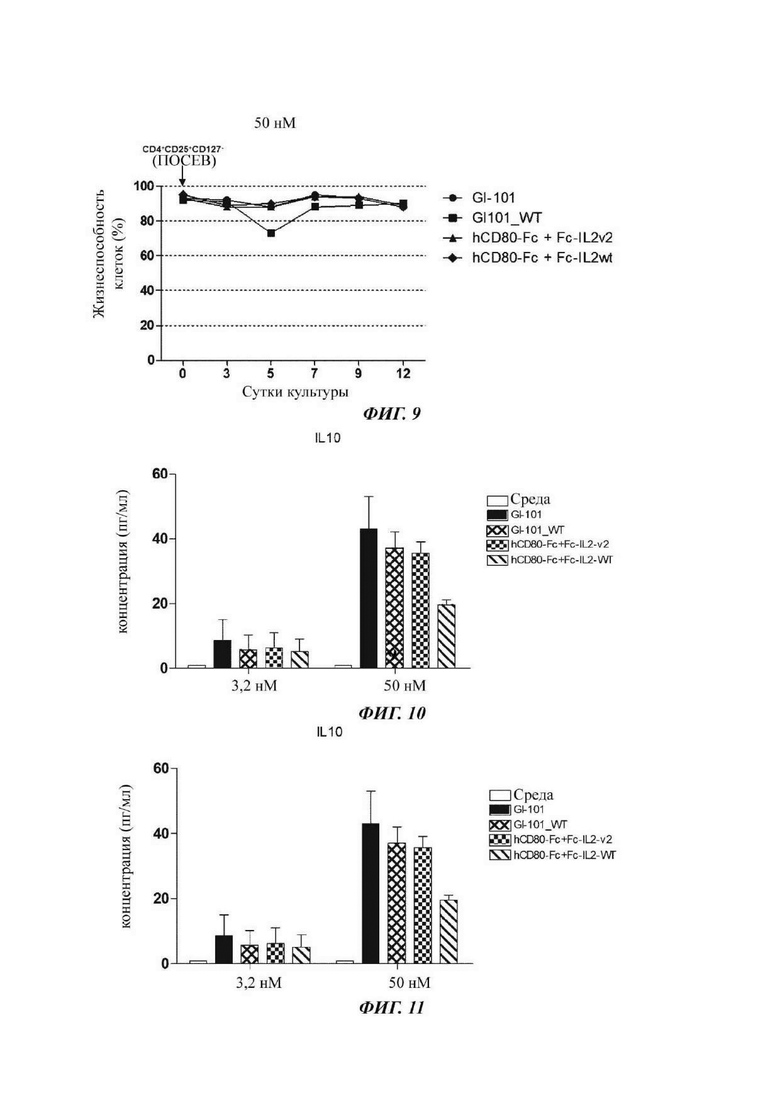
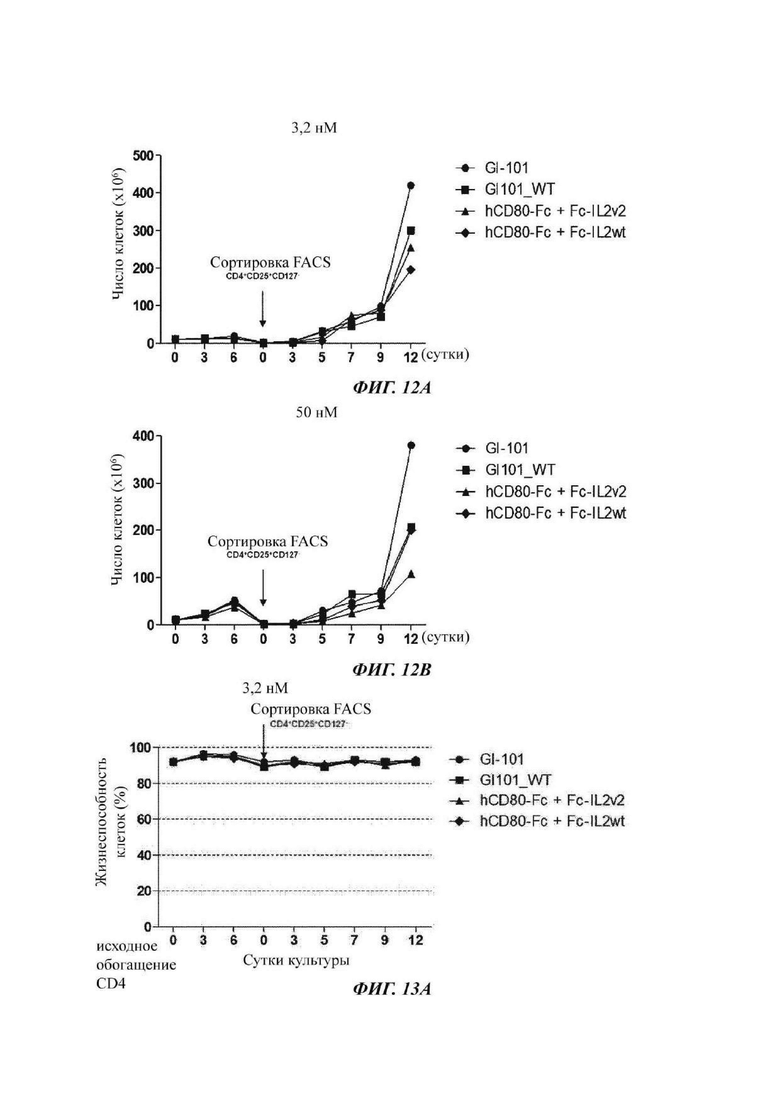
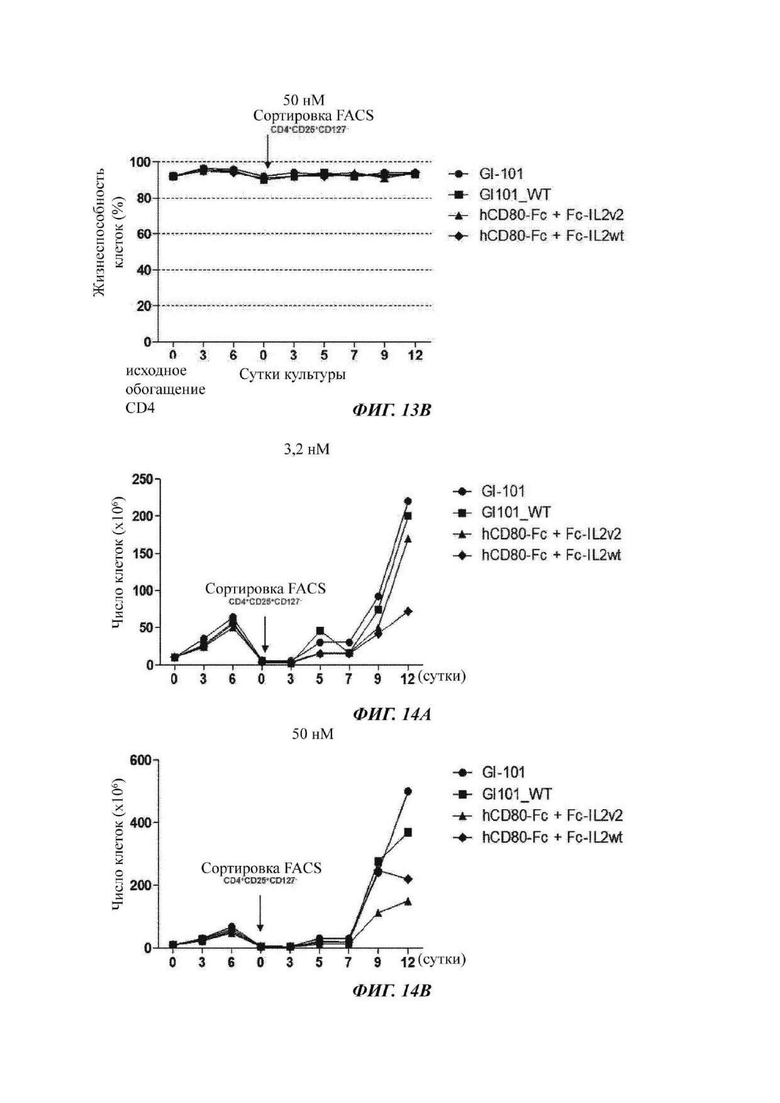
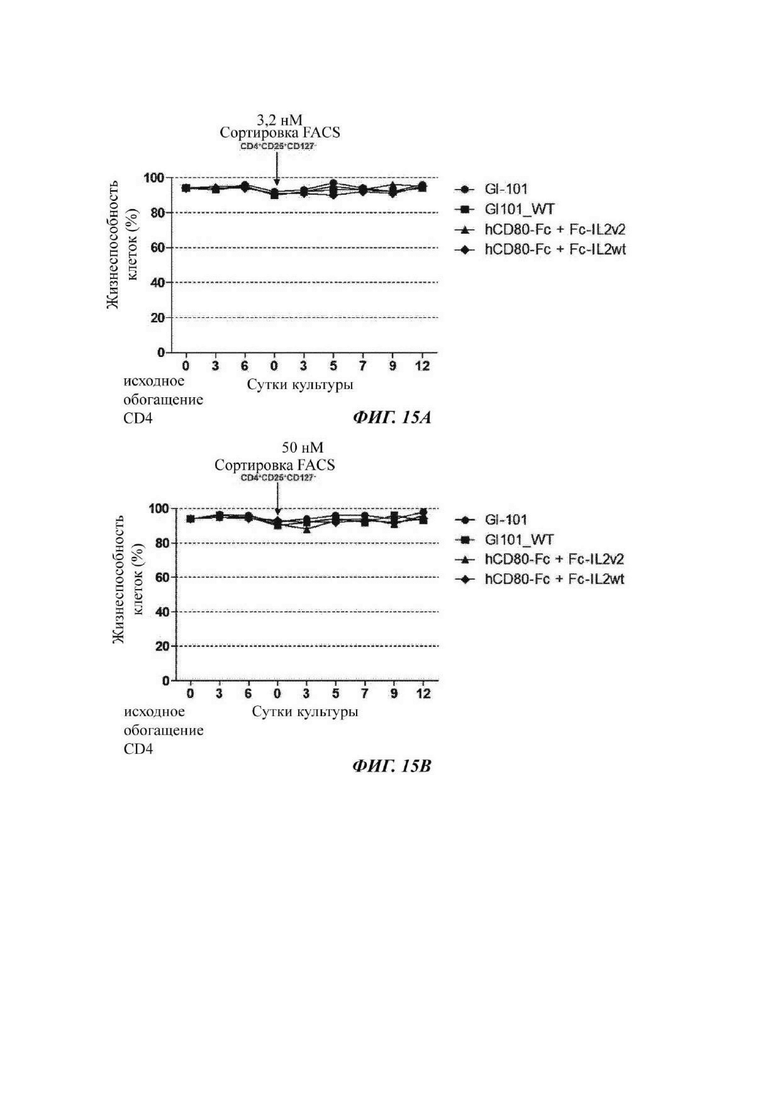

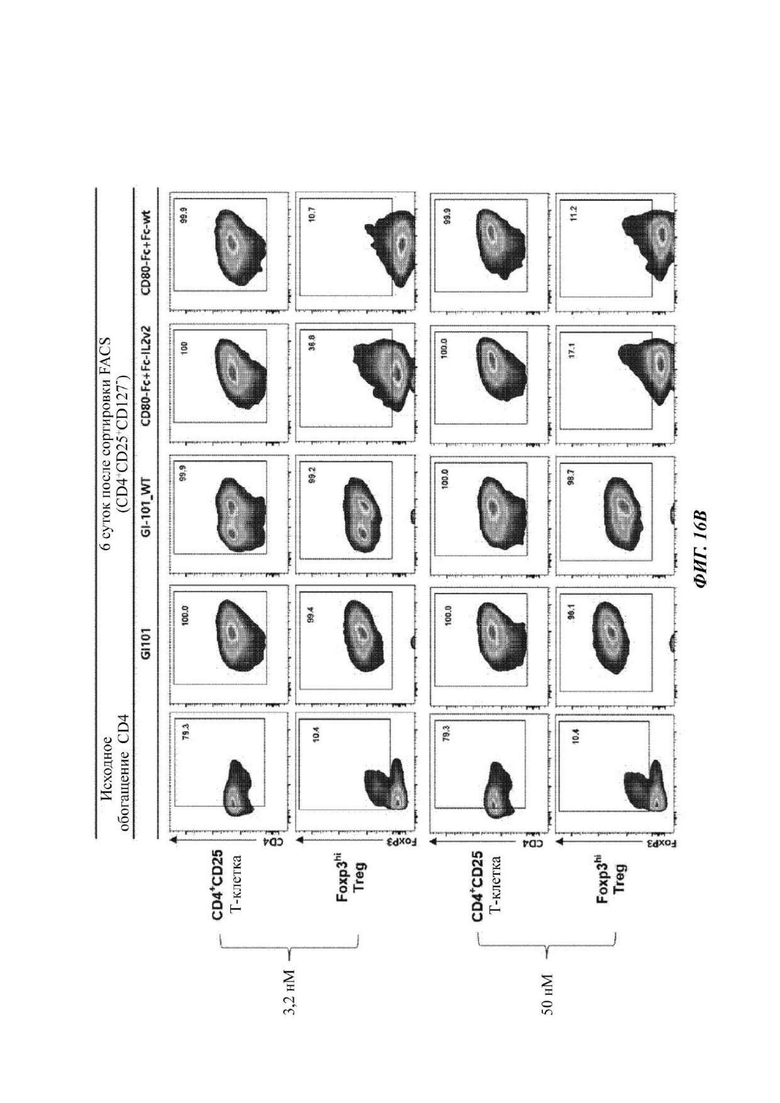

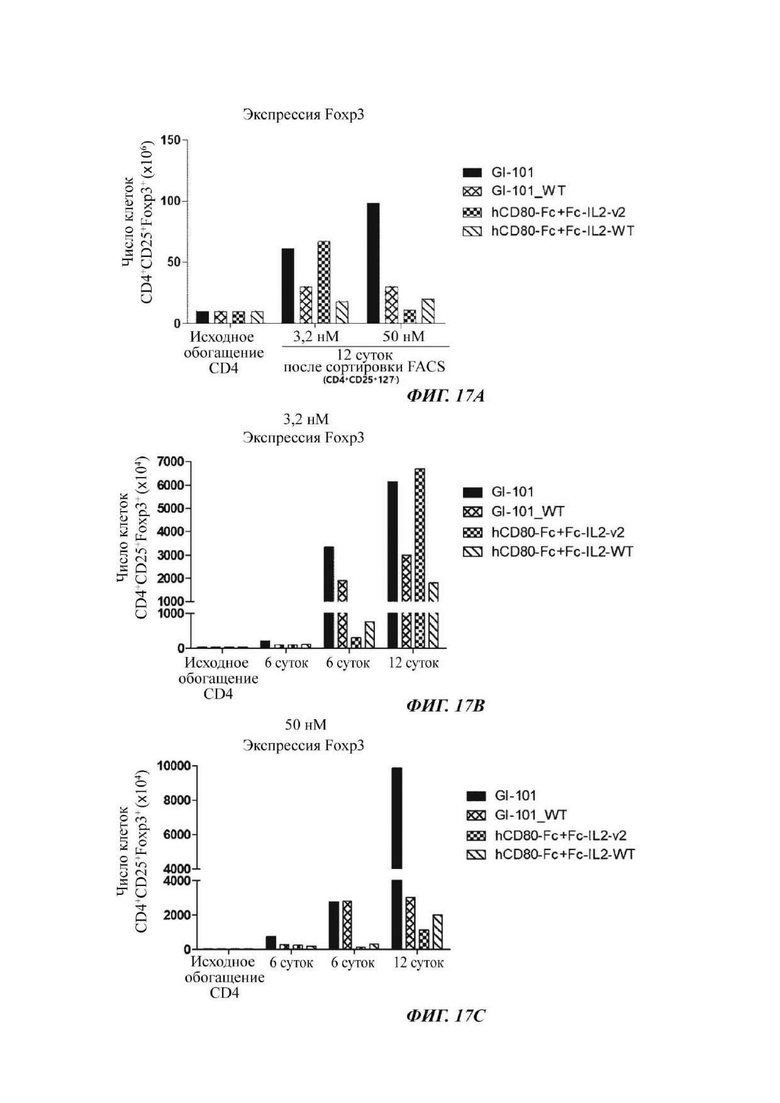

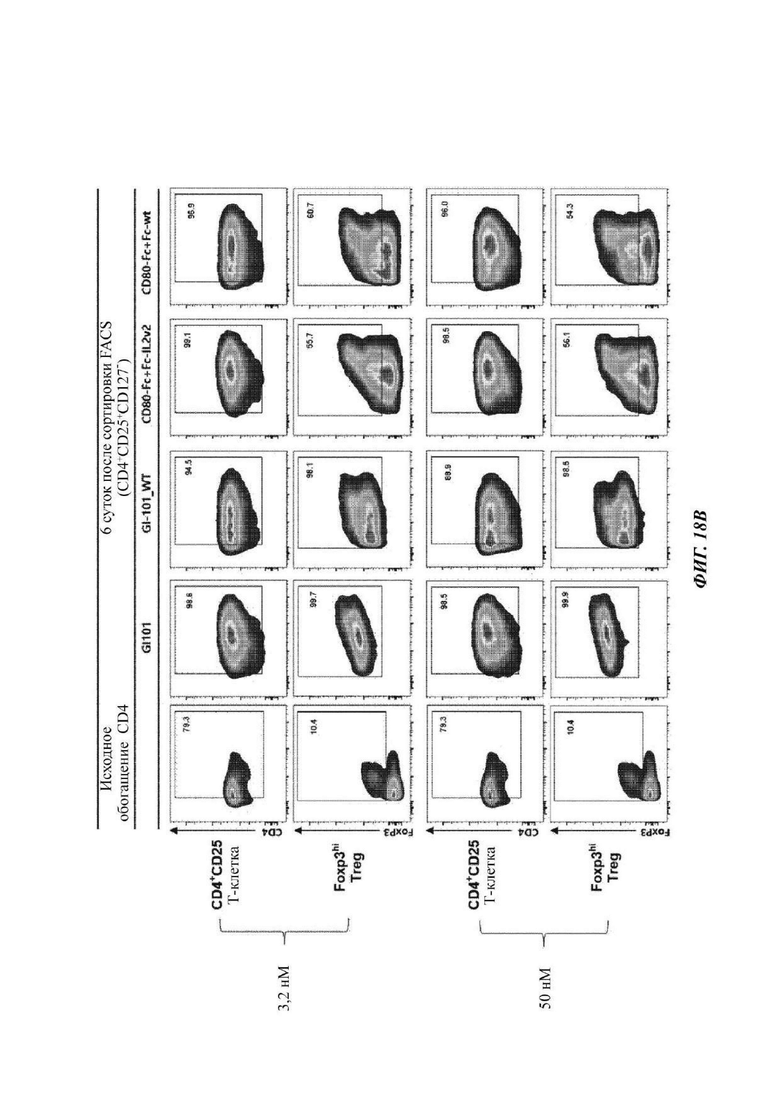

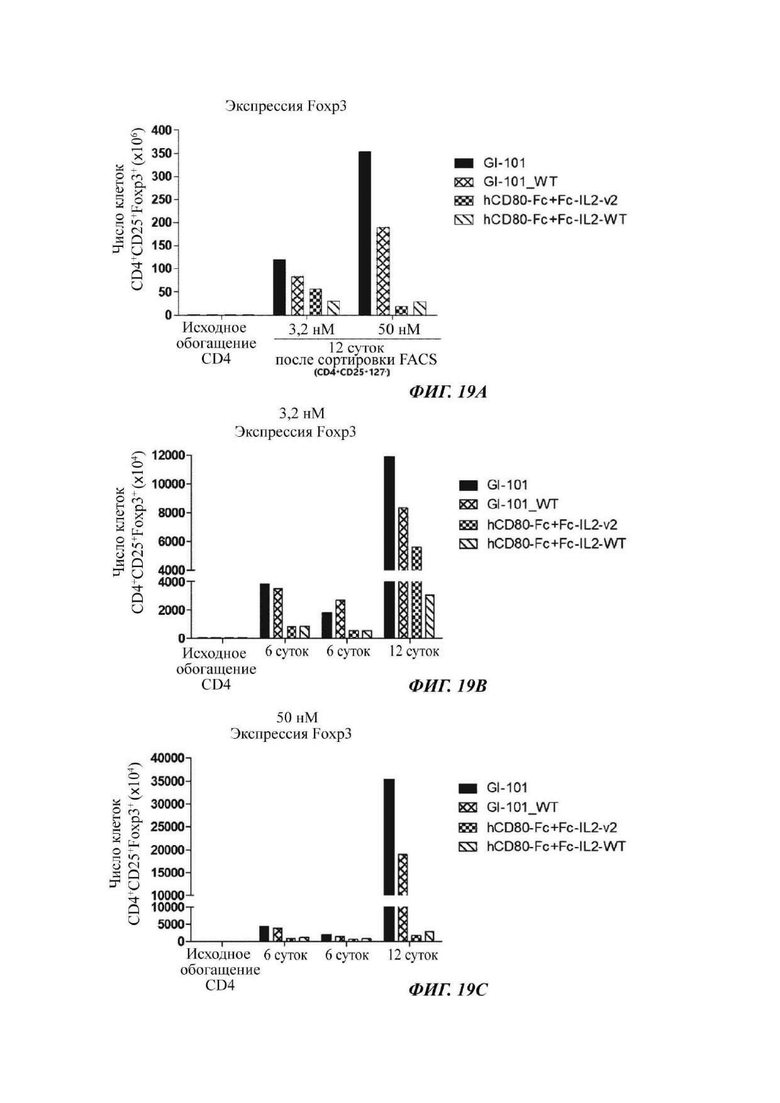
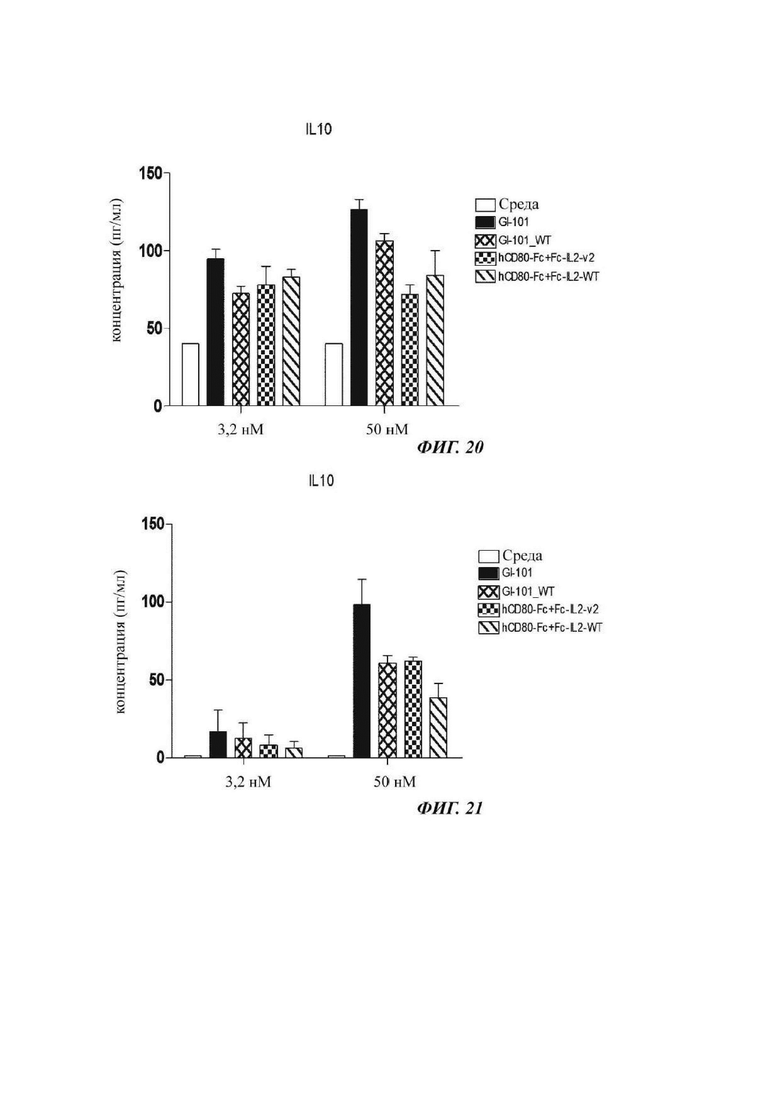
Изобретение относится к области биотехнологии. Предложен способ культивирования регуляторных Т-клеток, включающий культивирование выделенных регуляторных Т-клеток в среде, содержащей димер слитого белка. Указанный димер слитого белка содержит белок IL-2 или его вариант и белок CD80 или его фрагмент. Изобретение обеспечивает увеличение эффективности пролиферации и жизнеспособности полученных регуляторных Т-клеток Foxp3+. 8 з.п. ф-лы, 41 ил., 14 табл., 2 пр.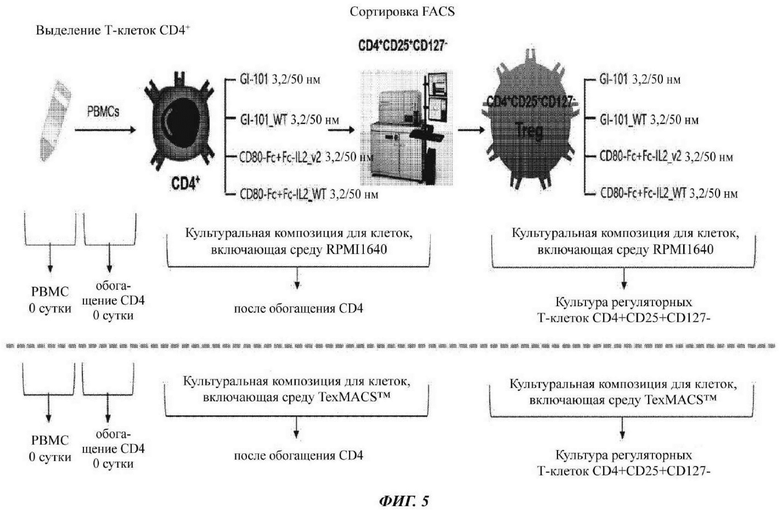
1. Способ культивирования регуляторных Т-клеток, включающий:
культивирование выделенных регуляторных Т-клеток в среде, содержащей димер слитого белка, содержащий белок IL-2 или его вариант и белок CD80 или его фрагмент,
где слитый белок характеризуется следующими структурными формулами (I) или (II):
N'-X-[линкер (1)]n-домен Fc-[линкер (2)]m-Y-C' – формула (I),
N'-Y-[линкер (1)]n-домен Fc-[линкер (2)]m-X-C' – формула (II),
где N' представляет собой N-конец слитого белка,
C' представляет собой С-конец слитого белка,
X представляет собой белок CD80 или его фрагмент,
Y представляет собой белок IL-2 или его вариант,
линкеры (1) и (2) представляют собой пептидные линкеры и
n и m каждый независимо равен 0 или 1,
при этом вариант белка IL-2 содержит замену 38-й и 42-й аминокислот в аминокислотной последовательности SEQ ID NO: 10, при этом фрагмент белка CD80 содержит внеклеточный домен CD80.
2. Способ культивирования регуляторных Т-клеток по п. 1, в котором регуляторные Т-клетки представляют собой Т-клетки CD4+CD25+CD127-.
3. Способ культивирования регуляторных Т-клеток по п. 1, в котором культивирование осуществляют в течение 1-20 суток.
4. Способ культивирования регуляторных Т-клеток по п. 2, в котором Т-клетки CD4+CD25+CD127- получают культивированием Т-клеток CD4+ или культивированием Т-клеток CD25+.
5. Способ культивирования регуляторных Т-клеток по п. 1, в котором белок IL-2 имеет аминокислотную последовательность SEQ ID NO: 10.
6. Способ культивирования регуляторных Т-клеток по п. 1, в котором вариант белка IL-2 имеет аминокислотную последовательность SEQ ID NO: 6.
7. Способ культивирования регуляторных Т-клеток по п. 1, в котором CD80 имеет аминокислотную последовательность SEQ ID NO: 11.
8. Способ культивирования регуляторных Т-клеток по п. 1, в котором вариант белка IL-2 содержит любую замену, выбранную из следующих комбинаций замен (а)-(d) в аминокислотной последовательности SEQ ID NO: 10:
(a) R38A/F42A,
(b) R38A/F42A/Y45A,
(c) R38A/F42A/E61R,
(d) R38A/F42A/L72G.
9. Способ культивирования регуляторных Т-клеток по п. 1, в котором слитый белок имеет аминокислотную последовательность SEQ ID NO: 9, 26, 28 или 30.
| WEIHONG LIU et al | |||
| Способ получения морфия из опия | 1922 |
|
SU127A1 |
| MARCO ROMANO et al | |||
| Treg therapy in transplantation: a general overview, Transplant International, 2017, vol.30, pp.745-753 | |||
| CHAN L | |||
| et al., IL-2/B7.1 (CD80) Fusagene Transduction of | |||

