[0001] Настоящая заявка заявляет приоритет предварительной заявки на патент США № 62/652199, поданной 3 апреля 2018 года, которая включена в данный документ посредством ссылки во всей своей полноте.
[0002] Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате с кодировкой ASCII и настоящим включен посредством ссылки во всей своей полноте. Копия указанного файла с кодировкой ASCII, созданная 27 марта 2019 года, имеет название 2019-03-27_01121-0031-00PCT_SL_ST25.txt, а ее размер составляет 187354 байта.
[0003] Даже несмотря на многочисленные успехи в области вакцинологии, необходимы новые открытия для защиты людей от многих опасных для жизни инфекционных заболеваний. Многие лицензированные на данный момент вакцины основаны на технологиях десятилетней давности для получения живых аттенуированных или инактивированных убитых патогенов, которые несут в себе проблемы безопасности и во многих случаях стимулируют лишь кратковременные слабые иммунные ответы, что требует введения нескольких доз. Хотя достижения в генетической и биохимической инженерии сделали возможной разработку терапевтических средств для лечения трудноизлечимых заболеваний, эти пути применения в области вакцинологии не были полностью реализованы. Технологии рекомбинантных белков теперь позволяют разработку оптимальных антигенов. Кроме того, наночастицы все чаще демонстрируют потенциал для оптимального представления антигенов и нацеленной доставки лекарственных средств. Было показано, что наночастицы с множественными прикрепленными антигенами обладают повышенной авидностью связывания, обеспечиваемой поливалентным отображением их молекулярных грузов, и способностью более эффективно преодолевать биологические барьеры благодаря их наноскопическому размеру. Ферритиновые наночастицы Helicobacter pylori (H. pylori), слитые с белком, представляющим собой гемагглютинин (HA) вируса гриппа, позволили улучшить стабильность антигена и повысить иммуногенность на мышиных моделях гриппа (см. Kanekiyo et al., Nature 499:102-106 (2013)). Этот слитый белок самоорганизовался в октаэдрически-симметричную наночастицу с 8 тримерными шиповидными отростками из НА с получением устойчивого иммунного ответа в различных доклинических моделях при применении с адъювантом.
[0004] Респираторно-синцитиальный вирус (RSV) является ведущей причиной тяжелых респираторных заболеваний у младенцев и основной причиной респираторных заболеваний у пожилых людей. Несмотря на десятилетия исследований, потребность в вакцинах остается неудовлетворенной. Хотя потребность в вакцине очевидна, разработка вакцины против RSV остановилась в 1960-х, когда клинические испытания с применением инактивированного формалином вируса RSV у младенцев привели к более тяжелому заболеванию после инфекции, обусловленной RSV. См., Hurwitz (2011) Expert Rev Vaccines 10(10): 1415-1433. Относительно недавно в ходе клинических программ с применением антигена F RSV в его конформации после слияния не смогли добиться достаточной эффективности у взрослых. См., Faloon et al. (2017) JID 216:1362-1370. Однако антигены F RSV, стабилизированные в конформации до слияния, могут вызывать нейтрализующий ответ, превосходящий ответ антигенов после слияния, которые не показали клинических результатов.
[0005] В данном документе представлен ряд новых полипептидов, наночастиц, композиций, способов и вариантов применения, в которых используются полипептиды RSV. Были созданы новые полипептиды F RSV, в том числе полипептиды, в которых эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, заблокирован, например N-гликаном в сайте гликозилирования, добавленном с помощью мутации. Также были созданы антигенные полипептиды и наночастицы, содержащие эти новые полипептиды RSV и ферритин. Также были созданы антигенные полипептиды на основе ферритина, содержащие полипептиды G RSV. Кроме того, были разработаны самоадъювантные антигенные полипептиды, содержащие полипептиды RSV и ферритин, в которых иммуностимулирующие функциональные группы, такие как адъюванты, были непосредственно химически присоединены к антигенному полипептиду. Прямая конъюгация иммуностимулирующей функциональной группы с антигенным полипептидом обеспечивает возможность направленной совместной доставки иммуностимулирующей функциональной группы и полипептида RSV в виде единого макромолекулярного соединения, что может значительно снизить возможность развития системной токсичности, чего опасаются в случае традиционных вакцин, которые содержат антигены и иммуностимулирующие молекулы, такие как адъюванты, в виде отдельных молекул. Совместная доставка иммуностимулирующих функциональных групп вместе с полипептидами RSV в виде макромолекулярного соединения и их поливалентная презентация может также снизить общую дозу, необходимую для обеспечения защиты, снижая производственные затраты и стоимость.
КРАТКОЕ ОПИСАНИЕ
[0006] Целью настоящего раскрытия является обеспечение композиций, наборов, способов и применений, которые способны обеспечить одно или несколько преимуществ, обсуждаемых выше, или по меньшей мере предоставить общественности полезный выбор. Соответственно, в данном документе раскрыты следующие варианты осуществления.
[0007] Вариант осуществления 1 представляет собой антигенный полипептид на основе последовательности RSV, содержащий полипептид F RSV, где эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, является заблокированным.
[0008] Вариант осуществления 2 представляет собой антигенный полипептид на основе последовательности RSV, содержащий полипептид F RSV, где полипептид F RSV содержит аминокислотные остатки 62-69 и 196-209 из SEQ ID NO: 26 и аспарагин, соответствующий положению 328, 348 или 507 из SEQ ID NO: 26.
[0009] Вариант осуществления 3 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 2, где эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, является заблокированным.
[0010] Вариант осуществления 3b представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 1 или 3, где заблокированный эпитоп представляет собой эпитоп антигенного сайта 1 F RSV.
[0011] Вариант осуществления 3c представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 1 или 3-3b, где два или более эпитопов, общих для F RSV до слияния и F RSV после слияния, являются заблокированными.
[0012] Вариант осуществления 3d представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 1 или 3-3c, где два или более эпитопов антигенного сайта 1 F RSV являются заблокированными.
[0013] Вариант осуществления 3e представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 1 или 3-3d, где один, или несколько, или все эпитопы, которые топологически перекрываются с заблокированным эпитопом, также являются заблокированными.
[0014] Вариант осуществления 3f представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 3e, где заблокированный эпитоп представляет собой эпитоп антигенного сайта 1 F RSV.
[0015] Вариант осуществления 4 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, содержащий F RSV до слияния.
[0016] Вариант осуществления 5 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, который распознается антителом, специфичным в отношении F RSV до слияния, выбранным из D25 или AM14.
[0017] Вариант осуществления 6 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 4 или 5, где F RSV до слияния содержит эпитоп не обнаруживаемый на F RSV после слияния.
[0018] Вариант осуществления 7 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 1-3, содержащий F RSV после слияния.
[0019] Вариант осуществления 8 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 1 или 3-6, где эпитоп заблокирован N-гликаном, присоединенным к аспарагину.
[0020] Вариант осуществления 9 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 7, где аспарагин соответствует остатку, отличному от аспарагина, в последовательности F RSV дикого типа (SEQ ID NO: 26), где необязательно остаток, отличный от аспарагина, соответствует положению 328, 348 или 507 из SEQ ID NO: 26.
[0021] Вариант осуществления 10 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, дополнительно содержащий белок ферритин.
[0022] Вариант осуществления 11 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 11, где ферритин содержит мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином.
[0023] Вариант осуществления 12 представляет собой антигенный полипептид на основе последовательности RSV, содержащий полипептид F RSV и белок ферритин, где белок ферритин содержит мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином.
[0024] Вариант осуществления 13 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 11-12, где ферритин содержит одну или несколько из мутаций E12C, S26C, S72C, A75C, K79C, S100C и S111C ферритина из H. pylori или одну или несколько соответствующих мутаций ферритина из организма, отличного от H. pylori, как определяют путем попарного или структурного выравнивания.
[0025] Вариант осуществления 14 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 10-13, содержащий один или несколько иммуностимулирующих функциональных групп, связанных с ферритином посредством экспонированной на поверхности аминокислоты, где необязательно экспонированная на поверхности аминокислота представляет собой цистеин, полученный в результате мутации.
[0026] Вариант осуществления 15 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 10-14, где ферритин содержит мутацию, обеспечивающую замещение экспонированного на поверхности аспарагина отличной от аспарагина аминокислотой, где необязательно аспарагин находится в положении 19 ферритина из H. pylori или аналогичном положении ферритина из организма, отличного от H. pylori, как определяют путем попарного или структурного выравнивания.
[0027] Вариант осуществления 16 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 10-15, где ферритин содержит мутацию, обеспечивающую замещение внутреннего цистеина отличной от цистеина аминокислотой, где необязательно внутренний цистеин находится в положении 31 ферритина из H. pylori или положении, которое соответствует положению 31 ферритина из H. pylori, согласно определению путем попарного или структурного выравнивания.
[0028] Вариант осуществления 17 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 12-16, где полипептид F RSV содержит эпитоп, не обнаруживаемый на F RSV после слияния, который представляет собой эпитоп сайта 0, где необязательно эпитоп сайта 0 содержит аминокислотные остатки 62-69 и 196-209 из SEQ ID NO: 26.
[0029] Вариант осуществления 18 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 328 из SEQ ID NO: 26.
[0030] Вариант осуществления 19 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 348 из SEQ ID NO: 26.
[0031] Вариант осуществления 20 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 507 из SEQ ID NO: 26.
[0032] Вариант осуществления 21 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит лейцин в положении, соответствующем положению лизина 498 из SEQ ID NO: 26.
[0033] Вариант осуществления 22 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит пролин в положении, соответствующем положению изолейцина 217 из SEQ ID NO: 26.
[0034] Вариант осуществления 23 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит аминокислоту, отличную от цистеина, в положении, соответствующем положению 155 из SEQ ID NO: 26, и/или аминокислоту, отличную от цистеина, в положении, соответствующем положению 290 из SEQ ID NO: 26.
[0035] Вариант осуществления 24 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, содержащий серин в положении, соответствующем положению 155 из SEQ ID NO: 26, и/или серин в положении, соответствующем положению 290 из SEQ ID NO: 26.
[0036] Вариант осуществления 25 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV не содержит сайта расщепления фурином, где необязательно вместо сайта расщепления фурином присутствует линкер.
[0037] Вариант осуществления 26 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью аминокислотам 1-478 из SEQ ID NO: 17.
[0038] Вариант осуществления 27 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью последовательности под SEQ ID NO: 17.
[0039] Вариант осуществления 28 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 20 или 21, содержащий аминокислоты 1-478 из SEQ ID NO: 17.
[0040] Вариант осуществления 29 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 1-19, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью аминокислотам 1-478 из SEQ ID NO: 23.
[0041] Вариант осуществления 30 представляет собой антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 1-19 или 23, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью последовательности под SEQ ID NO: 23.
[0042] Вариант осуществления 31 представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 23 или 24, содержащий аминокислоты 1-478 из SEQ ID NO: 23.
[0043] Вариант осуществления 32 представляет собой антигенный полипептид на основе последовательности RSV по любому из предыдущих вариантов осуществления, содержащий последовательность под любой из SEQ ID NO: 3-23.
[0044] Вариант осуществления 32a представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 32, содержащий последовательность из SEQ ID NO: 17.
[0045] Вариант осуществления 32b представляет собой антигенный полипептид на основе последовательности RSV согласно варианту осуществления 32, содержащий последовательность из SEQ ID NO: 23.
[0046] Вариант осуществления 33 представляет собой ферритиновую частицу, содержащую антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 10-32b.
[0047] Вариант осуществления 34 представляет собой композицию, содержащую антигенный полипептид на основе последовательности RSV, или ферритиновую частицу согласно любому из предыдущих вариантов осуществления и полипептид G RSV.
[0048] Вариант осуществления 34b представляет собой композицию согласно варианту осуществления 34, где композиция содержит ферритиновую частицу, и ферритиновая частица содержит полипептид G RSV, где необязательно полипептид G RSV химически конъюгирован с частицей ферритина.
[0049] Вариант осуществления 34c представляет собой композицию согласно варианту осуществления 34 или 34b, где полипептид G RSV не является гликозилированным.
[0050] Вариант осуществления 35 представляет собой композицию, содержащую антигенный полипептид на основе последовательности RSV или ферритиновую частицу согласно любому из вариантов осуществления 1-33, или композицию согласно любому из вариантов осуществления 34-34c, дополнительно содержащую фармацевтически приемлемый носитель.
[0051] Вариант осуществления 36 представляет собой антигенный полипептид на основе последовательности RSV, ферритиновую частицу или композицию согласно любому из вариантов осуществления 1-35 для применения в способе обеспечения формирования иммунного ответа на RSV или в защите субъекта от инфекции, обусловленной RSV.
[0052] Вариант осуществления 37 представляет собой способ обеспечения формирования иммунного ответа на RSV или защиты субъекта от инфекции, обусловленной RSV, включающий введение любого одного или нескольких из антигенного полипептида на основе последовательности RSV, ферритиновой частицы или композиции согласно любому из вариантов осуществления 1-36 субъекту.
[0053] Вариант осуществления 38 представляет собой антигенный полипептид на основе последовательности RSV, ферритиновую частицу, композицию или способ согласно любому из вариантов осуществления 36-37, где субъектом является человек.
[0054] Вариант осуществления 39 представляет собой нуклеиновую кислоту, кодирующую антигенный полипептид на основе последовательности RSV согласно любому из вариантов осуществления 1-32b, где необязательно нуклеиновая кислота представляет собой мРНК.
[0055] Вариант осуществления 39b представляет собой композицию или набор, содержащие нуклеиновую кислоту согласно варианту осуществления 39 и нуклеиновую кислоту, кодирующую полипептид G RSV, где необязательно одна или обе нуклеиновые кислоты представляют собой мРНК.
[0056] Дополнительные цели и преимущества будут частично изложены в последующем описании и частично будут понятны из описания, или их можно будет установить при осуществлении на практике. Цели и преимущества будут реализованы и достигнуты посредством элементов и комбинаций, конкретно указанных в прилагаемой формуле изобретения.
[0057] Следует понимать, что как предшествующее общее описание, так и следующее подробное описание являются лишь иллюстративными и пояснительными и не ограничивают формулу изобретения.
[0058] В прилагаемых графических материалах, которые включены в настоящее описание и составляют его часть, проиллюстрированы несколько вариантов осуществления, и вместе с описанием они служат для объяснения принципов, описанных в данном документе.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
[0059] На фиг. 1A-1D показана структура иллюстративного полипептида Pre-F-NP RSV. (Фиг. 1A) Линейная диаграмма, на которой указаны номера остатков, соответствующие N-концу каждого сегмента. Нумерация соответствует SEQ ID NO: 26. Домены 1-3 обозначены DI, DII и DIII соответственно, и также отмечены область гептадных повторов А (HRA), и область гептадных повторов В (HRB). Отмечен С-концевой ферритин (ферритиновая наночастица). Фрагменты F1 и F2 функциональной группы, представляющей собой F RSV, помечены под рисунком. Область между фрагментами F1 и F2, где фрагмент, представляющий собой пептид 27 (p27), являющийся слитым пептидом (FP), и сайты расщепления фурином (сайты фурина) были удалены и замещены гибким линкером с образованием одноцепочечных конструкций F, изображена линией и отмечена над рисунком. Звездочки над диаграммой указывают приблизительное расположение сконструированных сайтов гликозилирования E328N, S348N и R507N. (Фиг. 1В) Структурная модель функциональной группы, представляющей собой F RSV до слияния, на которой указаны ключевые эпитопы, подлежащие нейтрализации антителами (Nab) D25, AM14, 101F и паливизумабом. Примерная область структурных эпитопов, общих для конформации до слияния и после слияния, обозначена белым треугольником. Отмечены положения иллюстративных сконструированных сайтов гликозилирования E328N, S348N и R507N. Сконструированные сайты гликозилирования соответствуют областям, которые являются структурно общими для конформаций до слияния и после слияния, и находятся вдали от ключевых эпитопов, подлежащих нейтрализации, распознаваемых антителами, такими как D25, AM14, 101F и паливизумаб. По сути, конструкции, содержащие эти сконструированные сайты гликана, сохраняют способность связываться с вышеуказанными нейтрализующими антителами (данные не показаны). (Фиг. 1C) Структурная модель наночастицы белка F RSV до слияния (Pre-F-NP) с областями HRA и HRB, заштрихованными более темным цветом. Полученные в результате свернутые конструкции Pre-F-NP могут образовывать 24-меры, которые экспонируют ключевые эпитопы, перечисленные на фиг. 1B. (Фиг. 1D) Средние значения класса 2D электронных микрофотографий конструкции RF8085 Pre-F-NP RSV (SEQ ID NO: 1), демонстрирующие симметрию расположения тримерных функциональных групп F RSV на 24-мерной ферритиновой наночастице.
[0060] На фиг. 2 показана мелкомасштабная экспрессия нескольких конструкций Pre-F-NP, экспрессированных в кондиционированной среде клеток 293, согласно измерению посредством вестерн-блоттинга с применением антитела D25. RF8090 представляет собой SEQ ID NO: 2, который представляет собой вариант для клонирования, используемый при экспрессии в CHO, имеющий ту же последовательность, что и RF8085, т. е. SEQ ID NO:1. RF8085 и RF8090 представляют собой конструкции Pre-F-NP, несущие дисульфидные мутации и мутации заполнения полости DS-CAV с делециями и одноцепочечным линкером, описанным на фиг. 1A, слитым с N-концом ферритина. RF8100-RF8105 и RF8108-RF8112 имеют последовательности под SEQ ID NO: 3-8 и 11-14 соответственно. scF-pFerr=слитый белок на основе полипептида F RSV и ферритина. Мутации, которые, как оказалось, обеспечивают улучшение экспрессии конструкции по сравнению с эталоном RF8090, указаны под результатами вестерн-блоттинга. Заслуживающие упоминания мутации предусматривают добавление сайтов гликана посредством мутаций E328N, S348N и R507N и мутацию I327P. обеспечивающую кэппирование центральной спирали, которые обеспечивали увеличение уровня экспрессии и секреции наночастиц F RSV в кондиционированной среде, согласно измерению с помощью вестерн-блоттинга.
[0061] На фиг. 3 показана экспрессия RF8085 (SEQ ID NO: 1; контрольная конструкция) и RF8106 (SEQ ID NO: 9; содержащая мутацию I217P, как в RF8108, и не содержащая мутации дисульфидной связи (DS) в DS-CAV1), согласно измерению с помощью анализа методом вестерн-блоттинга в кондиционированной среде при экспрессии в 293. Замещение DS мутацией I217P, обеспечивающей кэппирование центральной спирали, обеспечивало существенное увеличение уровня экспрессии. Замещение DS мутацией, обеспечивающей кэппирование центральной спирали, не влияло на связывание конструкции с антителами D25 и AM14, специфичными в отношении белка в конформации до слияния.
[0062] На фиг. 4 показаны результаты очистки конструкции RF8106 с помощью эксклюзионной хроматографии (SEQ ID NO: 9). Объем удержания наночастицы RF8106, составляющий приблизительно 65 мл на препаративной колонке SEC Superose 6, соответствует свернутой 24-мерной наночастице, что свидетельствует о том, что мутации в RF8106 не препятствуют образованию наночастиц.
[0063] На фиг. 5A-5B показаны результаты анализа методом динамического рассеяния света (DLS) для невосстановленного (5A) и восстановленного (5B) RF8106. Как и анализ методом SEC, DLS демонстрирует, что Pre-F-NP RSV образовала ожидаемую свернутую наночастицу. Данные для восстановленного белка также показывают, что частица не была разрушена восстановлением, которое выполняли перед конъюгацией адъюванта с экспонированным на поверхности цистеином, введенным в ферритин путем мутации (см. фиг. 6).
[0064] На фиг. 6 показаны результаты анализа на геле SDS-PAGE с окрашиванием кумасси для RF8106 с конъюгацией с CpG, являющейся агонистом TLR9, и без таковой. Увеличение сдвига на геле для наночастицы, обработанной CpG, продемонстрировало, что может быть добавлено до приблизительно 40-50% адъюванта CpG к наночастице F RSV. Конъюгация CpG или других иммуностимулирующих функциональных групп, таких как агонист TLR7/8 SM7/8, не ингибировала способность частицы связывать антитела D25 и AM14, специфичные в отношении белка в конформации до слияния.
[0065] На фиг. 7 показаны результаты вестерн-блоттинга наночастиц, содержащих F RSV с дополнительными гликанами (RF8117, SEQ ID NO: 17) и без дополнительных гликанов (RF8085, SEQ ID NO: 1; и RF8113, SEQ ID NO: 16). RF8113 аналогичен RF8106, но экспонированный на поверхности цистеин S111C (с применением нумерации остатков ферритина, т. е. соответствующий положениям в последовательности ферритина SEQ ID NO: 208) из RF8106 был замещен экспонированным на поверхности цистеином K79C (также с применением нумерации остатков ферритина), чтобы разместить сайт конъюгации дальше от функциональной группы, представляющей собой Pre-F. Как и RF8106, RF8113 обеспечивает сохранение улучшенной экспрессии по сравнению с эталонной молекулой RF8085. RF8117 аналогичен RF8113, но дополнительно включает три мутации, обеспечивающие гликозилирование, идентифицированные на фиг. 2, т. е. E328N, S348N и R507N, для дополнительного улучшения экспрессии и блокирования эпитопов, не подлежащих нейтрализации, общих для F в конформации до слияния и F в конформации после слияния, как описано на фиг. 1B.
[0066] На фиг. 8 показана экспрессия конструкций F RSV с разными заменами по потенциальным сайтам расщепления трипсиноподобной протеазой. При экспрессии RF8090 в линии клеток СНО (та же последовательность белка, что и у RF8085, но с другой последовательностью ДНК, адаптированной к вектору экспрессии в СНО) наблюдали, что в полипептиде происходило вырезание участка между функциональными группами, представляющими собой F и ферритин, что приводило к снижению уровня экспрессии. По полученным значениям массы функциональной группы, представляющей собой F, установили, что протеолиз может происходить рядом с HRB, областью линкера лягушки-быка из конструкции Pre-F-NP. Мутации остатков лизина и аргинина в этой области (остатки ~450-550) исследовали для устранения потенциального протеолиза конструкции по трипсиновому типу. Мутации в RF8122 (SEQ ID NO: 18) по сравнению с RF8117 (K498L и K508Q) обеспечивали улучшенную экспрессию в клетках 293 и могут обеспечивать снижение или устранение протеолиза в клетках CHO. Альтернативные мутации ограничивали экспрессию.
[0067] Фиг. 9A-B. Экспрессия RF8090, RF8117 и RF8140 в стабильно трансфицированных клетках CHO. Выход экспрессии RF8090 (SEQ ID NO: 2) наблюдали на низких уровнях. Мутации для замещения дисульфида в DS-CAV1 и мутации в линкере между функциональной группой, представляющей собой F, и функциональной группой, представляющей собой ферритин, для устранения потенциальных сайтов расщепления трипсином вводили, как описано выше, в конструкции RF8117 (SEQ ID NO: 17) и RF8140 (SEQ ID NO: 23), которые клонировали в стабильно экспрессирующие клетки CHO. (Фиг. 9A) Экспрессию RF8117 и RF8140 из трех и четырех пулов клеток СНО соответственно в кондиционированной среде СНО сравнивали с выходами RF8090 в кондиционированной среде СНО с помощью анализа методом вестерн-блоттинга с применением D25. Все три пула CHO для RF8117 и все четыре пула CHO в случае RF8140 обеспечивали экспрессию с более высокими значениями выхода, чем в случае RF8090. (Фиг. 9В) Экспрессия RF8117 в кондиционированной среде СНО, измеренная с применением антитела D25, специфичного в отношении F до слияния, с помощью Octet. На левой панели показан график зависимости ответа для известных концентраций RF8140, очищенного из среды 293, и ответа на связывание с D25 на наконечнике с белком А, обеспечивающий стандартную кривую. Отдельные точки представляют ответы на связывание RF8117 из кондиционированной среды CHO с помощью D25. На правой панели показан рассчитанный выход RF8117 или RF8140 в объединенной кондиционированной среде CHO на основании ответа на связывание D25. В среде экспрессировались как RF8117, так и RF8140, согласно измерению по связыванию D25 и AM14, демонстрируя, что подобно клеткам 293, клетки CHO способны экспрессировать Pre-F-NP в свернутом виде, что позволяет сохранить тримерную структуру F до слияния.
[0068] Фиг. 10A-B. Ответ нейтрализующих антител на Pre-F-NP RF8117. (Фиг. 10A) Сравнение титров нейтрализующих антител к RSV, полученных при иммунизации высокой дозой (1 мкг) и низкой дозой (0,1 мкг) DS-CAV1 (тримера Pre-F, SEQ ID NO: 25), тримера F после слияния (тримера Post-F; SEQ ID NO: 24) или Pre-F-NP с гликозилированием посредством конструирования (Pre-F-NP; RF8117, SEQ ID NO: 17), измеряли с помощью анализа клеток VERO. Все полипептиды RSV вводили с адъювантом AF03, как описано в данном документе. Во всех случаях, если не указано иное, AF03 вводили с полипептидом или наночастицей RSV, но без конъюгации с ними. Полипептиды RSV и дозы отмечены под осью x. Показан статистический анализ ответов при высоких дозах относительно иммунизации Pre-F-NP. (Фиг. 10B) Сравнение титров нейтрализующих антител к RSV, полученных при иммунизации высокой дозой (1 мкг) и низкой дозой (0,1 мкг) DS-CAV1 (тример Pre-F), Pre-F-NP без гликозилирования посредством конструирования (RF8113, SEQ ID NO: 16) или Pre-F-NP с гликозилированием посредством конструирования (RF8117, SEQ ID NO: 17), согласно измерению с помощью анализа на основе клеток VERO. Все полипептиды RSV вводили с адъювантом AF03 (не конъюгированным ни с какими полипептидом или наночастицей), как описано в данном документе. Полипептиды RSV и дозы отмечены под осью x.
[0069] Фиг. 11A-D. Сравнение антитела, связывающего тример F RSV до слияния (DS-CAV1), и нейтрализующих антител к RSV, образование которых вызвано иммунизацией с помощью тримера F после слияния (SEQ ID NO: 24) или Pre-F-NP (RF8140 SEQ ID NO: 23) в модели на мыши или модели на примате, отличном от человека. (Фиг. 11A) Сравнение ответ в виде образования антитела, связывающего тример F до слияния, вызванных у мышей в результате иммунизации с помощью F после слияния и Pre-F-NP (RF8140, SEQ ID NO: 23). (Фиг. 11B) Показаны ответы с образованием нейтрализующих антител, вызванные у мышей в результате иммунизации с помощью F после слияния и Pre-F-NP (RF8140, SEQ ID NO: 23). (Фиг. 11C) Сравнение ответ в виде образования антитела, связывающего тример F до слияния, вызванных у приматов, отличных от человека, с помощью Pre-F-NP с адъювантом или без такового (AF03, указывается в скобках ниже). (Фиг. 11D) Сравнение титров нейтрализующих антител к RSV, полученных при иммунизации с помощью Pre-F-NP (RF8140, SEQ ID NO: 23) с адъювантом AF03 и без такового. У мышей Pre-F-NP вызывает ответ в виде образования антител, связывающих F до слияния, и ответ в виде образования нейтрализующих антител к RSV на более высоком уровне по сравнению с тримером после слияния. У приматов, отличных от человека, Pre-F-NP вызывает мощный нейтрализующий ответ.
[0070] На фиг. 12A-12B показано, что сконструированные сайты гликозилирования блокируют эпитопы, присутствующие в конформации после слияния. (Фиг. 12A) Показан ответ в виде образования антитела к F до слияния (DS-CAV1), вызванный иммунизацией с помощью Pre-F-NP без гликозилирования посредством конструирования (RF8113) или Pre-F-NP с гликозилированием посредством конструирования (частица со сконструированными Gly) при высокой (1 мкг) и низкой (0,1 мкг) дозе, согласно измерению с помощью Octet. (Фиг. 12В) Показан ответ в виде образования антитела к тримеру после слияния, вызванный иммунизацией с помощью Pre-F-NP без гликозилирования посредством конструирования (RF8113) или Pre-F-NP с гликозилированием посредством конструирования (RF8117) при высокой (1 мкг) и низкой (0,1 мкг) дозе, согласно измерению с помощью Octet. Как указано выше, все полипептиды RSV смешивали с AF03 во время иммунизации. Хотя как RF8113, так и RF8117 вызывают устойчивые ответы в виде образования антител против F до слияния, ответ в виде образования антител против F после слияния, вызванный RF8117, значительно снижен. Это обусловлено наличием сконструированных гликанов, которые картируются в эпитопах, общих для конформации до слияния и после слияния (фиг. 2B).
[0071] На фиг. 13A-C показано блокирование эпитопов, не подлежащих нейтрализации, с помощью сконструированных сайтов гликозилирования. (Фиг. 13A) Сравнение титров нейтрализующих антител к RSV, полученных при иммунизации с помощью pre-F-NP с сайтами гликозилирования дикого типа ("частица с гликаном Wt"; RF8113, SEQ ID NO: 16) и pre-F-NP с добавленными сконструированными сайтами гликозилирования ("частица+гликан"; RF8117, SEQ ID NO: 17) при дозе 0,1 мкг в исследованиях на мышах, согласно измерению с помощью анализа на основе клеток VERO. (Фиг. 13B) Сравнение ответов в виде образования антитела, связывающего тример F после слияния RSV, вызванных иммунизацией с помощью частицы с гликаном Wt (RF8113, SEQ ID NO: 16) и частицы с+гликаном (RF8117, SEQ ID NO: 17) при дозе 0,1 мкг в исследованиях на мышах. (Фиг. 13C) Отношение измеренных титров нейтрализующих антител к титрам связывающих антител из панелей A и B, демонстрирующее, что сконструированные гликаны не снижали ответ в виде образования функциональных нейтрализующих антител, но снижали уровень антител, отличных от нейтрализующих, образующихся в ответ на эпитопы, общие для конформации до слияния/после слияния (фиг. 1B), тем самым обеспечивая улучшение в отношении отношения нейтрализующие/связывающие антитела.
[0072] Фиг. 14A-D. Характеристика пептида центрального домена G RSV (Gcc), конъюгированного с ферритиновой наночастицей. (Фиг. 14A) Гель SDS-PAGE, окрашенный кумасси, демонстрирующий клик-конъюгацию центрального домена G RSV (SEQ ID NO: 29) с ферритиновой наночастицей с образованием антигена Gcc-NP. (Фиг. 14B) Структурная модель Gcc-NP. (Фиг. 14C) Сравнение ответов в виде образования антитела, связывающего Gcc, вызванных иммунизацией только пептидом Gcc (пептид Gcc, SEQ ID NO. 29) и пептидом Gcc, конъюгированным с наночастицей (Gcc-NP) в исследованиях на мышах. Репрезентативный ответ в не подвергавшихся воздействию образцах сыворотки крови показан с помощью белого прямоугольника, тогда как ответы в результате второй иммунизации показаны с помощью светло-серых прямоугольников, а ответы в результате процедур третей иммунизации показаны с помощью темно-серых прямоугольников. (Фиг. 14D) Сравнение титров нейтрализующих антител к RSV, полученных при иммунизации пептидом Gcc (SEQ ID NO. 29) и Gcc-NP в исследованиях на мышах после третьей инъекции, согласно измерению с помощью анализа на основе клеток HAE. Образцы сыворотки крови от не подвергавшихся воздействию животных и образцы сыворотки крови от животных, иммунизированных пептидом Gcc, объединяли, и титры показаны в виде столбиков.
[0073] Фиг. 15A-C. Совместное введение Pre-F-NP RSV (RF8140) и Gcc-NP вызывает нейтрализующий ответ. Мышей иммунизировали с помощью Pre-F-NP (RF8140) отдельно, Gcc-NP отдельно или объединенных Pre-F-NP и Gcc-NР при дозе 1 мкг на антиген. Все процедуры иммунизации осуществляли с применением адъюванта AF03, как показано выше. (Фиг. 15A) Иммунизация мышей с помощью RF8140 отдельно (Pre-F-NP) или RF8140 и Gcc-NP (Pre-F-NP+Gcc-NP) вызывала образование антител, которые связывают тример F до слияния. (Фиг. 15В) Иммунизация мышей с помощью Gcc-NP отдельно (Gcc-NP) или RF8140 и Gcc-NP (Pre-F-NO+Gcc-NP) вызывала образование антител, которые связывают пептид Gcc. (FIG 15C) У животных иммунизация с помощью Pre-F-NP отдельно, Gcc-NP отдельно или совместного введения Pre-F-NP и Gcc-NP вызывает нейтрализующий ответ после второй и после третьей иммунизации, согласно измерению с помощью анализа нейтрализации с клетками HAE. Совместное введение Pre-F-NP+Gcc-NP вызывало нейтрализующий ответ, превосходящий таковой, вызванный иммунизацией с помощью только Pre-F-NP.
[0074] Фиг. 16A-B. Совместное введение Pre-F-NP и Gcc-NP не влияет на обеспечение образования антител, которые связывают тример F до слияния или Gcc-наночастицу. Титры нейтрализующих антител, измеренные с помощью анализа на основе клеток VERO, чувствительных к F, показаны слева на фиг. 16A, тогда как титры нейтрализующих антител, измеренные с помощью анализа на основе клеток HAE чувствительных к F- и G, показаны справа на фиг. 16B. Процедуры иммунизации животных были такими же, как на фиг. 15. Полипептиды RSV, используемые при иммунизации, показаны под горизонтальной осью. Черные столбики представляют образцы сыворотки крови, собранные из групп иммунизации, описанных на фиг. 15, и отмечены аналогичным образом. Образцы сыворотки крови от не подвергавшихся воздействию животных также показаны черными столбиками и отмечены для сравнения. Образцы сыворотки крови, истощенные по тримеру F до слияния, показаны белым, непосредственно справа от соответствующего черного столбика. Образцы сыворотки крови, истощенные по эктодомену G, показаны заштрихованными по диагонали столбиками, непосредственно справа от соответствующего черного столбика. Образцы сыворотки крови, истощенные по тримеру F до слияния с последующим истощением по эктодомену G, показаны вертикально заштрихованными столбиками. (Фиг. 16A) Титры нейтрализующих антител наблюдали в анализах на основе клеток VERO для образцов сыворотки крови, полученных при иммунизации с помощью RF8140 и совместном введении RF8140+Gcc-NP, но не для образцов сыворотки крови от не подвергавшихся воздействию животных или образцов сыворотки крови, полученных при иммунизации с помощью Gcc-NP отдельно. Истощение образцов сыворотки крови от групп, получавших RF8140 или RF8140+Gcc-NP, по тримеру F до слияния обеспечивало снижение поддающихся измерению титров нейтрализующих антител. (Фиг. 16B) Титры нейтрализующих антител наблюдали в анализах на основе клеток HAE для образцов сыворотки крови от животных, иммунизированных с помощью RF8140, Gcc-NP или RF8140, совместно вводимых с Gcc-NP. Образцы сыворотки крови от не подвергавшихся воздействию животных не демонстрировали нейтрализующего ответа. Сыворотка крови от животных, иммунизированных с помощью RF8140, истощенная по тримеру F до слияния, характеризуется снижением поддающегося измерению титра нейтрализующих антител. Сыворотка крови от животных, иммунизированных с помощью Gcc-NP, истощенная по эктодомену G, характеризуется снижением поддающегося измерению титра нейтрализующих антител. Сыворотка крови от животных, иммунизированных посредством совместного введения RF8140 и Gcc-NP, не характеризуется снижением поддающегося измерению титра нейтрализующих антител при истощении по тримеру F до слияния отдельно, но характеризуется снижением поддающегося измерению титра нейтрализующих антител при истощении как по тримеру F до слияния, так и по эктодомену G. Вместе эти данные говорят о том, что совместное введение Pre-F-NP и Gcc-NP не влияет на соответствующие способности антигенов обеспечивать образование нейтрализующих антител к F до слияния или G.
[0075] Фиг. 17A-B. Применение RF8117 или RF8140 с адъювантом, представляющим собой AF03, SPA09 или алюминиевые квасцы, вызывает превосходящий нейтрализующий ответ у мышей по сравнению с применением RF8117 без адъюванта. (Фиг. 17А) Показаны титры нейтрализующих антител в сыворотке крови от мышей, иммунизированных с помощью RF8117, либо без применения адъюванта (без адъюванта), либо с применением адъюванта, представляющего собой алюминиевые квасцы, либо с применением адъюванта AF03, согласно измерению с помощью анализа на основе клеток VERO. (Фиг. 17B) Показаны титры нейтрализующих антител в сыворотке крови от мышей, иммунизированных с помощью RF8117 либо без применения адъюванта (без адъюванта), с помощью RF8117 с применением адъюванта SPA09, либо с помощью RF8140 с применением адъюванта AF03, согласно измерению с помощью анализа на основе клеток VERO. Во всех случаях RF8117 или RF8140 у получавших адъювант групп не подвергавшихся воздействию мышей обеспечивали получение более высокого титра нейтрализующих антител, чем у не получавших адъювант групп.
[0076] Фиг. 18A-B. Применение RF8140 с адъювантом AF03 или SPA09 вызывает превосходящий нейтрализующий ответ у отличных от человека приматов (NHP) по сравнению с таковым при процедурах иммунизации с помощью RF8140 без применения адъюванта. (Фиг. 18A) Ответы на тример F до слияния, измеренные в сыворотке крови от NHP после иммунизации с помощью RF8140 без применения адъюванта (без адъюванта), либо с применением адъюванта AF03, либо с применением адъюванта SPA09 (использовали две дозы SPA09, как указано ниже), согласно измерению с помощью ELISA. Во все моменты времени применение адъюванта AF03 или SPA09 вызывает превосходящий нейтрализующий ответ. (Фиг. 18B) Титры нейтрализующих антител в образцах сыворотки крови от NHP, иммунизированных с помощью RF8140 без применения адъюванта (без адъюванта), либо с применением адъюванта AF03 или с применением адъюванта SPA09 (использовали две дозы SPA09, как указано ниже), согласно измерению с помощью анализа на основе клеток VERO. Во всех случаях иммунизация с помощью RF8140 с адъювантом обеспечивает получение более высокого титра нейтрализующих антител, чем у не получавших адъювант групп во все моменты времени.
[0077] Фиг. 19A-B. Конъюгация RF8140 с агонистом TLR7/8 SM7/8 или агонистом TLR9 CpG обеспечивает получение превосходящего титра антител, связывающих F до слияния, по сравнению с применением RF8140 отдельно без адъюванта. (Фиг. 19A) Показан ответ в виде образования антител, связывающих тример F до слияния, измеряемый в сыворотке крови от не подвергавшихся воздействию мышей, мышей, иммунизированных с помощью RF8140 без применения адъюванта, мышей, иммунизированных с помощью RF8140, конъюгированным с адъювантом SM7/8, RF8140, с применением адъюванта в виде 130 нг SM7/8, или RF8140, с применением адъюванта в виде 20 мкг SM7/8. RF8140, конъюгированный с SM7/8, обеспечивает получение более высокого титра антител, связывающих тример F до слияния, чем в случае групп, не получавших адъюванта или получавших адъювант SM7/8. (Фиг. 19В) Показан ответ в виде образования антител, связывающих тример F до слияния, измеренный в образцах сыворотки крови от не подвергавшихся воздействию мышей, мышей, иммунизированных с помощью RF8140 без применения адъюванта, мышей, иммунизированных с помощью RF8140, конъюгированным с адъювантом CpG, RF8140 с применением адъюванта в виде 680 нг CpG или RF8140 с применением адъюванта в виде 20 мкг SM7/8. RF8140, конъюгированный с SM7/8, обеспечивает получение более высокого титра антител, связывающих тример F до слияния, чем группы, не получавшие адъюванта или получавшие адъювант SM7/8.
[0078] Фиг. 20A-G. F-субъединичные вакцины-кандидаты вызывают образование нейтрализующих антител, направленных против pre-F, и Th1 CD4+ T-клеточный ответ в системе MIMIC. (Фиг. 20A) Титры антител против pre-F в системе MIMIC измеряли с помощью AF после праймирования каждым Ag в молярной эквивалентной концентрации F с 10 нг/мл pre-F-NP (n=48-49 доноров на группу). (Фиг. 20B) Титры микронейтрализации были измерены и представлены в международных единицах/мл (МЕ/мл). (Фиг. 20C) Отношение антител к pre-F к антителам к post-F, составляющее > 1, соответствует более высокому уровню антитела, связывающего pre-F, по сравнению с антителом, связывающим post-F, в то время как значение отношения < 1 соответствует большему уровню ответа в виде образования Ab к post-F. (Фиг. 20D) Продуцирование TNFα в активированных CD154+/CD4+ T-клетках, повторно стимулированных целевыми клетками, нагруженными белком F, измеряли с применением проточной цитометрии, n=48. Статистическую значимость определяли посредством множественного сравнения Тьюки-Крамера-HSD. (Фиг. 20E) Титр ранее существовавших антител у субъектов-людей (серостатус) сильно коррелировал с величиной иммунного ответа на RSV в системе MIMIC. График линейной регрессии, показывающий зависимость уровня IgG против pre-F в образцах сыворотки крови от каждого донора и общего ответа в виде образования IgG против pre-F, создавали с помощью программного обеспечения или алгоритма, и p-значение для общего наклона анализировали статистическим методом (n=50). Ось Y представляет уровни IgG к pre-F, полученные после праймирования с помощью RSV. (Фиг. 20 F) Как и на фиг. 20Е, график линейной регрессии, показывающий зависимость уровня IgG к pre-F в сыворотке крови от каждого донора и общего уровня IgG к pre-F после примирования F-субъединичными вакцинами-кандидатами (post-F - квадраты, pre-F-NP - круги и DC-Cav1 - ромбы). Ранее существующие титры циркулирующих IgG к pre-F варьировали от 199800 до 3037600000. Каждая точка представляет значение IgG каждого отдельного донора. (Фиг. 20G) Сравнение ответов в виде образования антител, связывающих Gcc, вызванных обработкой пептидом Gcc отдельно (пептид Gcc), с таковыми в случае пептида Gcc, конъюгированного с наночастицей (Gcc-NP), в B-клетках человека. Группа без лечения показана для сравнения, как указано выше.
[0079] Фиг. 21A-C. Титры нейтрализующих антител, полученных при низкой дозе (0,5 мкг) наночастиц на основе Gcc RSV-ферритина ("Gcc-NP"). Показаны титры нейтрализующих антител к штамму A RSV в HAE, вызванные иммунизацией с помощью Gcc RSV-NP, содержащих последовательность Gcc RSV A2 (составлены с AF03), из образцов сыворотки крови, отобранных через две недели после второй иммунизации (2wp2) (фиг. 21A) или через две недели после третьей иммунизации (2wp3) (фиг. 21B), с образцами не подвергавшейся воздействию и гипериммунной сыворотки крови в качестве отрицательного и положительного контроля. Также показан титр нейтрализующих антител к штамму В RSV в HAE, вызванный иммунизацией с помощью Gcc RSV-NP, содержащих последовательность Gcc RSV A2 (составлены с AF03), из образцов сыворотки крови, взятых через две недели после третьей иммунизации (2wp3) (фиг. 21C).
[0080] Фиг. 22A-B. Ответы в виде образования антитела, связывающего антиген штамма А2 RSV, вызванные с помощью Gcc RSV-NP. (Фиг. 22A) Ответы в виде образования антитела, связывающего Gcc, вызванные к Gcc штамму A2, измеренные через две недели после второй инъекция (светло-серые прямоугольники) и через две недели после третьей инъекции (темно-серые прямоугольники), вызванные высокой дозой (5 мкг) Gcc RSV-NP. Ответ в сыворотке крови от не подвергавшейся воздействию мыши показан в качестве отрицательного контроля. (Фиг. 22В) Ответы в виде образования антитела, связывающего Gcc, вызванные к штамму Gcc A2, измеренные через две недели после второй инъекция (светло-серые прямоугольники) и через две недели после третьей инъекции (темно-серые прямоугольники), вызванные низкой дозой (0,5 мкг) Gcc RSV-NP.
[0081] Фиг. 23A-B. Ответы в виде образования антитела, связывающего антиген штамма B1 RSV, вызванные с помощью Gcc RSV-NP. (Фиг. 23A) Ответы в виде образования антитела, связывающего Gcc, вызванные к штамму Gcc B1, измеренные через две недели после второй инъекция (светло-серые прямоугольники) и через две недели после третьей инъекции (темно-серые прямоугольники), вызванные высокой дозой (5 мкг) Gcc RSV-NP. Ответ в сыворотке крови от не подвергавшейся воздействию мыши показан в качестве отрицательного контроля. (Фиг. 23В) Ответы в виде образования антитела, связывающего Gcc, вызванные к штамму Gcc B1, измеренные через две недели после второй инъекция (светло-серые прямоугольники) и через две недели после третьей инъекции (темно-серые прямоугольники), вызванные низкой дозой (0,5 мкг) Gcc RSV-NP.
ПОДРОБНОЕ ОПИСАНИЕ
[0082] Предусмотрены полипептиды RSV, которые могут быть антигенными при введении отдельно, с адъювантом в виде отдельной молекулы и/или в виде части наночастицы (например, ферритиновой частицы или лумазинсинтазной частицы), которая может быть самоадъювантной. В некоторых вариантах осуществления антигенные полипептиды на основе последовательности RSV содержат полипептид F RSV и ферритин и/или полипептид F RSV, в котором эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, является заблокированным. Полипептиды F RSV, которые управляют продуцированием антител против F RSV в конформации до слияния, индуцировали более сильный ответ in vivo в виде образования антител к F RSV до слияния по сравнению с F RSV после слияния. Также в данном документе описываются полипептиды G RSV, содержащие весь или часть G RSV и которые могут дополнительно содержать ферритин. Белки G RSV и F RSV необходимы для прикрепления RSV к клеткам-хозяевам и слияния с ними.
[0083] F RSV существует в двух конформационных состояниях - в конформациях до слияния и после слияния. В своем нативном состоянии до слияния F RSV представляет собой тример, состоящий из 3 протомеров. Таким образом, иммунизация полипептидами F RSV в конформации до слияния может обладать улучшенными свойствами. В некоторых вариантах осуществления полипептид F RSV разработан таким образом, чтобы индуцировать иммунитет против F RSV в конформации до слияния. G RSV представляет собой белок прикрепления, отвечающий за ассоциацию RSV с эпителиальными клетками дыхательных путей человека.
Определения
[0084] Термин "антигенный сайт 0" или "эпитоп сайта 0", используемый в данном документе, относятся к сайту, расположенному на вершине тримера F RSV до слияния, содержащему аминокислотные остатки 62-69 и 196-209 F RSV дикого типа (SEQ ID NO: 26). Эпитоп сайта 0 представляет собой сайт связывания для антител, которые обладают специфичностью в отношении F RSV до слияния, таких как D25 и AM14, и связывание антител с эпитопом сайта 0 блокирует прикрепление RSV к поверхности клетки (см. McLellan et al., Science 340(6136):1113-1117 (2013)).
[0085] Термин "стабильность антигена", используемый в данном документе, относится к стабильности антигена во времени или в растворе.
[0086] Термин "замены, обеспечивающие заполнение полости", используемый в данном документе, относится к сконструированным гидрофобным заменам для заполнения полостей, присутствующих в тримере F RSV до слияния.
[0087] Термин "белок F" или "белок F RSV" относится к белку RSV, ответственному за слияние вирусной оболочки с мембраной клетки-хозяина во время проникновения вируса.
[0088] Термин "полипептид F RSV" или "полипептид F" относится к полипептиду, содержащему по меньшей мере один эпитоп белка F.
[0089] Термин "добавление гликана", используемый в данном документе, относится к добавлению мутаций, которые вводят сайты гликозилирования, не присутствующие в F RSV дикого типа, которые могут быть сконструированы для увеличения уровня экспрессии конструкции, повышения стабильности конструкции или блокирования эпитопов, общих для конформации до слияния и после слияния. Модифицированный белок, предусматривающий добавления гликана, будет характеризоваться большим уровнем гликозилирования и, следовательно, более высокой молекулярной массой. Добавление гликана может уменьшить степень, в которой полипептид F RSV вызывает образование антител против F RSV в конформации после слияния.
[0090] Термин "белок G" или "белок G RSV", используемый в данном документе, относится к белку прикрепления, ответственному за ассоциацию RSV с эпителиальными клетками дыхательных путей человека. Иллюстративная аминокислотная последовательность G RSV дикого типа представлена в виде SEQ ID NO: 27. Белок G RSV содержит эктодомен (приблизительно аминокислоты 66-297 из G RSV (SEQ ID NO: 27)), который находится вне клетки. В пределах эктодомена G RSV находится центральная консервативная область (Gcc или CCR, приблизительно аминокислоты 151-193 из SEQ ID NO: 27). CCR в G RSV содержит мотив CX3C. Мотив CX3C опосредует связывание белка G с рецептором CX3CR1.
[0091] Термин "кэппирование спирали с помощью PRO" или "кэппирование спирали пролином", используемый в данном документе, относятся к случаю, когда кэп спирали предусматривает пролин, который может стабилизировать образование спирали.
[0092] Термин "замены, обеспечивающие внутрипротомерную стабилизацию", используемый в данном документе, описывает аминокислотные замены в F RSV, которые стабилизируют конформацию до слияния путем стабилизации взаимодействия внутри протомера тримера F RSV.
[0093] Термин "замены, обеспечивающие межпротомерную стабилизацию", используемый в данном документе, описывает аминокислотные замены в F RSV, которые стабилизируют конформацию до слияния путем стабилизации взаимодействия протомеров тримера F RSV друг с другом.
[0094] Термин "расщепление протеазой", используемый в данном документе, относится к протеолизу (иногда в уровне техники называемому "вырезание") чувствительных остатков (например, лизина или аргинина) в полипептидной последовательности.
[0095] Термин "после слияния", используемый в данном документе, в отношении F RSV, относится к стабильной конформации F RSV, которая возникает после слияния вируса и мембран клетки.
[0096] Термин "до слияния", используемый в данном документе в отношении F RSV, относится к конформации F RSV, которая существует до взаимодействия вируса с клеткой.
[0097] Термин "протомер", используемый в данном документе, относится к структурной единице олигомерного белка. В случае F RSV отдельной единицей тримера F RSV является протомер.
[0098] Применяемые в данном документе термины "ферритин" или "белок ферритин" относятся к белку с выявляемой идентичностью последовательности с ферритином из H. pylori (SEQ ID NO: 208 или 209) или другим ферритином, обсуждаемым в данном документе, таким как ферритин P. furiosus, ферритин Trichoplusia ni или человеческий ферритин, который служит для хранения железа, например, внутриклеточно или в тканях, или для переноса железа в кровотоке. Такие иллюстративные ферритины, в том числе те, которые встречаются в виде двух полипептидных цепей, известные как тяжелая и легкая цепи (например, ферритин T. ni и человеческий ферритин), подробно обсуждаются ниже. В некоторых вариантах осуществления ферритин включает последовательность с по меньшей мере 15%, 20%, 30%, 40%, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью с последовательностью ферритина, раскрытой в данном документе, например, в таблице 1 (таблица последовательностей). Ферритин может представлять собой фрагмент полноразмерной встречающейся в природе последовательности.
[0099] Применяемый в данном документе термин "ферритин дикого типа" относится к ферритину, последовательность которого состоит из встречающейся в природе последовательности. Ферритины также включают полноразмерный ферритин или фрагмент ферритина с одним или несколькими отличиями его аминокислотной последовательности от ферритина дикого типа.
[00100] Применяемый в данном документе термин "мономер ферритина" относится к одной молекуле ферритина (или, если применимо, к одной тяжелой или легкой цепи ферритина), которая не была собрана с другими молекулами ферритина. "Ферритиновый мультимер" содержит несколько ассоциированных мономеров ферритина. "Белок ферритин" включает мономерный ферритин и мультимерный ферритин.
[00101] Применяемый в данном документе термин "ферритиновая частица" относится к ферритину, который самоорганизовался в глобулярную форму. Ферритиновые частицы иногда называют "ферритиновыми наночастицами" или просто "наночастицами". В некоторых вариантах осуществления ферритиновая частица содержит 24 мономера ферритина (или, если применимо, 24 полных тяжелых и легких цепей).
[00102] Применяемый в данном документе термин "гибридный ферритин" относится к ферритину, включающему ферритин из H. pylori с аминоконцевым удлинением из ферритина лягушки-быка. Иллюстративная последовательность, применяемая в качестве аминоконцевого удлинения из ферритина лягушки-быка, представлена как SEQ ID NO: 217. В гибридном ферритине аминоконцевое удлинение из ферритина лягушки-быка может быть слито с ферритином из H. pylori таким образом, что сайты присоединения иммуностимулирующих фрагментов равномерно распределяются на поверхности ферритиновой частицы. Применяемый в данном документе термин "линкер лягушки-быка" представляет собой линкер, содержащий последовательность под SEQ ID NO: 217. Гибридный ферритин также иногда называют "bfpFerr" или "bfp-ферритин". Любая из конструкций, включающих последовательность лягушки-быка, может быть предусмотрена без последовательности лягушки-быка, например, без линкера или с альтернативным линкером. Иллюстративные линкерные последовательности лягушки-быка предусмотрены в таблице 1. Если в таблице 1 показан линкер лягушки-быка, та же конструкция может быть получена без линкера или с альтернативным линкером.
[00103] Применяемый в данном документе термин "N-гликан" относится к сахаридной цепи, присоединенной к белку по амидному азоту остатка N (аспарагина) белка. Таким образом, N-гликан образуется в процессе N-гликозилирования. Этот гликан может представлять собой полисахарид.
[00104] Применяемый в данном документе термин "гликозилирование" относится к добавлению сахаридного звена к белку.
[00105] Применяемый в данном документе термин "иммунный ответ" относится к ответу клетки иммунной системы, такой как В-клетка, Т-клетка, дендритная клетка, макрофаг или полиморфноядерный лейкоцит, на стимул, такой как антиген или вакцина. Иммунный ответ может включать любую клетку организма, участвующую в защитном ответе хозяина, включая, например, эпителиальную клетку, которая секретирует интерферон или цитокин. Иммунный ответ включает без ограничения врожденный и/или адаптивный иммунный ответ. Применяемый в данном документе термин "защитный иммунный ответ" относится к иммунному ответу, который защищает субъекта от инфекции (например, предупреждает инфекцию или предупреждает развитие заболевания, связанного с инфекцией). Способы измерения иммунных ответов хорошо известны из уровня техники и включают, например, измерение пролиферации и/или активности лимфоцитов (таких как B- или Т-клетки), секреции цитокинов или хемокинов, воспаления, образования антител и т. п. "Ответ с образованием антител" представляет собой иммунный ответ, при котором образуются антитела.
[00106] Применяемый в данном документе термин "антиген" относится к средству, которое вызывает иммунный ответ, и/или к средству, связываемому Т-клеточным рецептором (например, когда оно представляется с помощью молекулы МНС) или антителом (например, продуцируемым В-клеткой) при экспонировании или введении в организм. В некоторых вариантах осуществления антиген вызывает гуморальный ответ (например, включающий образование антигенспецифических антител) в организме. В качестве альтернативы или дополнительно, в некоторых вариантах осуществления антиген вызывает клеточный ответ (например, с участием T-клеток, рецепторы которых специфически взаимодействуют с антигеном) в организме. Конкретный антиген может вызывать иммунный ответ у одного или нескольких представителей целевого организма (например, мышей, кроликов, приматов, людей), но не у всех представителей вида целевого организма. В некоторых вариантах осуществления антиген вызывает иммунный ответ у по меньшей мере приблизительно 25%, 30%, 35%, 40%, 45%, 50%, 55%, 60%, 65%, 70%, 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% представителей вида целевого организма. В некоторых вариантах осуществления антиген связывается с антителом и/или Т-клеточным рецептором и может индуцировать или не индуцировать конкретный физиологический ответ в организме. В некоторых вариантах осуществления, например, антиген может связываться с антителом и/или с Т-клеточным рецептором in vitro, независимо от того, происходит ли такое взаимодействие in vivo. В некоторых вариантах осуществления антиген реагирует с продуктами специфического гуморального или клеточного иммунитета, в том числе с индуцируемыми гетерологичными иммуногенами. Антигены включают антигенные белки на основе последовательности ферритина, содержащие полипептид ферритина (например, содержащий одну или несколько мутаций), и полипептид белка, отличного от ферритина (например, полипептид RSV), как описано в данном документе.
[00107] Применяемый в данном документе термин "иммуностимулирующий фрагмент" относится к фрагменту, который ковалентно присоединяется к ферритину или антигенному полипептиду на основе последовательности ферритина и который способен активировать компонент иммунной системы (либо отдельно, либо при присоединении к ферритину или антигенному полипептиду на основе последовательности ферритина). Иллюстративные иммуностимулирующие фрагменты включают агонистов toll-подобных рецепторов (TLR), например, TLR 4, 7, 8 или 9. В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой адъювант.
[00108] Применяемый в данном документе термин "адъювант" относится к веществу или среде-носителю, которые неспецифически усиливают иммунный ответ на антиген. Адъюванты могут включать без ограничения суспензию минералов (например, алюминиевые квасцы, гидроксид алюминия или фосфат), на которых адсорбируется антиген; эмульсию типа вода-в-масле или масло-в-воде, в которой раствор антигена эмульгируется в минеральном масле или в воде (например, неполный адъювант Фрейнда). Иногда включают убитые микобактерии (например, полный адъювант Фрейнда) для дополнительного усиления антигенности. В качестве адъювантов также можно применять иммуностимулирующие олигонуклеотиды (например, см. патенты США №№ 6194388, 6207646, 6214806, 6218371, 6239116, 6339068, 6406705 и 6429199). Адъюванты также могут включать биологические молекулы, такие как агонисты Toll-подобного рецептора (TLR) и костимулирующие молекулы. Адъювант можно вводить в виде отдельной молекулы в композиции или ковалентно связанным (конъюгированным) с ферритином или антигенным полипептидом на основе последовательности ферритина.
[00109] Термин "антигенный полипептид на основе последовательности RSV" используют в данном документе в отношении полипептида, содержащего всю аминокислотную последовательность RSV или ее часть достаточной длины, чтобы молекула была антигенной в отношении RSV. Антигенность может быть признаком последовательности RSV в виде части конструкции, дополнительно содержащей гетерологичную последовательность, такую как ферритин и/или иммуностимулирующая функциональная группа. То есть, если последовательность RSV является частью конструкции, дополнительно содержащей гетерологичную последовательность, то достаточно, чтобы конструкция могла служить в качестве антигена, который обеспечивает образование антител против RSV, независимо от того, может ли это делать последовательность RSV без гетерологичной последовательности.
[00110] Термины "антигенный полипептид на основе ферритина" и "антигенный белок на основе ферритина" используются в данном документе взаимозаменяемо для обозначения полипептида, содержащего ферритин и полипептид, отличный от ферритина (например, полипептид RSV) достаточной длины, чтобы молекула была антигенной в отношении полипептида, отличного от ферритина. Антигенный полипептид на основе последовательности ферритина может дополнительно содержать иммуностимулирующий фрагмент. Антигенность может быть признаком последовательности, отличной от последовательности ферритина, как части более крупной конструкции. То есть достаточно, чтобы конструкция могла служить в качестве антигена против полипептида, отличного от ферритина, независимо от того, способен ли на это полипептид, отличный от ферритина, без ферритина (и иммуностимулирующего фрагмента, если применимо). В некоторых вариантах осуществления полипептид, отличный от ферритина, представляет собой полипептид RSV, в этом случае антигенный полипептид на основе последовательности ферритина также является "антигенным полипептидом на основе последовательности RSV". Однако для ясности следует отметить, что антигенный полипептид на основе последовательности RSV не обязательно должен содержать ферритин. Термин "антигенный полипептид" используют в данном документе для обозначения полипептида, который представляет собой любой из антигенного полипептида на основе последовательности ферритина и антигенного полипептида на основе последовательности RSV или оба.
[00111] Применяемый в данном документе термин "самоадъювантный" относится к композиции или полипептиду, включающим ферритин и иммуностимулирующий фрагмент, непосредственно конъюгированный с ферритином таким образом, что ферритин и иммуностимулирующий фрагмент находятся в составе одного молекулярного объекта. Антигенный полипептид на основе последовательности ферритина, включающий отличный от ферритина полипептид, может быть конъюгирован с иммуностимулирующим фрагментом с образованием самоадъювантного полипептида.
[00112] Применяемый в данном документе термин "экспонированная на поверхности" аминокислота относится к аминокислотному остатку в белке (например, ферритине) с боковой цепью, которая способна контактировать с молекулами растворителя, когда белок находится в своей нативной трехмерной конформации после мультимеризации, если применимо. Таким образом, например, в случае ферритина, который образует 24-мер, экспонированный на поверхности аминокислотный остаток представляет собой остаток, боковая цепь которого способна контактировать с растворителем, когда ферритин собирается как 24-мер, например, как ферритиновый мультимер или ферритиновая частица.
[00113] Применяемый в данном документе термин "субъект" относится к любому представителю царства животных. В некоторых вариантах осуществления "субъект" относится к людям. В некоторых вариантах осуществления "субъект" относится к отличным от человека животным. В некоторых вариантах осуществления субъекты включают без ограничения млекопитающих, птиц, рептилий, амфибий, рыб, насекомых и/или червей. В определенных вариантах осуществления отличный от человека субъект представляет собой млекопитающее (например, грызуна, мышь, крысу, кролика, обезьяну, собаку, кошку, овцу, крупный рогатый скот, примата и/или свинью). В некоторых вариантах осуществления субъект может представлять собой трансгенное животное, животное, полученное с помощью способов генетической инженерии, и/или клон. В определенных вариантах осуществления настоящего изобретения субъект представляет собой взрослого, подростка или младенца. В некоторых вариантах осуществления применяются термины "индивидуум" или "пациент", и подразумевается, что они взаимозаменяемы с термином "субъект".
[00114] Применяемые в данном документе термины "вакцинация" или "вакцинировать" относятся к введению композиции, предназначенной для генерации иммунного ответа, например, на фактор, вызывающий заболевание. Вакцинацию можно осуществлять до, во время и/или после контакта с фактором, вызывающим заболевание, и/или развития одного или нескольких симптомов и, в некоторых вариантах осуществления до, во время и/или вскоре после контакта с фактором. В некоторых вариантах осуществления вакцинация включает многократное введение композиции для вакцинации с соответствующими интервалами времени.
[00115] В настоящем изобретении описаны последовательности нуклеиновых кислот и аминокислотные последовательности, характеризующиеся определенной степенью идентичности с данной последовательностью нуклеиновой кислоты или аминокислотной последовательностью, соответственно (эталонная последовательность).
[00116] "Идентичность последовательностей" двух последовательностей нуклеиновых кислот указывает на процентное содержание нуклеотидов, которые являются идентичными в двух последовательностях. "Идентичность последовательностей" двух аминокислотных последовательностей указывает на процентное содержание аминокислот, которые являются идентичными в двух последовательностях.
[00117] Термины "идентичный %", "% идентичности" или подобные термины предназначены для обозначения, в частности, процентного содержания нуклеотидов или аминокислот, которые являются идентичными при оптимальном выравнивании в сравниваемых последовательностях. Указанное процентное содержание является чисто статистическим, и различия между двумя последовательностями могут быть, но не обязательно, случайным образом распределены по всей длине сравниваемых последовательностей. Сравнение двух последовательностей обычно осуществляют посредством сравнения указанных последовательностей после оптимального выравнивания относительно сегмента или "окна сравнения", чтобы идентифицировать локальные участки соответствующих последовательностей. Оптимальное выравнивание для сравнения можно осуществлять вручную или с помощью алгоритма поиска локальной гомологии по Smith and Waterman, 1981, Ads App. Math. 2, 482, с помощью алгоритма поиска локальной гомологии по Neddleman and Wunsch, 1970, J. Mol. Biol. 48, 443, с помощью алгоритма поиска совпадений по Pearson and Lipman, 1988, Proc. Natl Acad. Sci. USA 88, 2444, или с помощью компьютерных программ, в которых применяются указанные алгоритмы (GAP, BESTFIT, FASTA, BLAST P, BLAST N и TFASTA в пакете программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Drive, Мэдисон, Висконсин).
[00118] Процент идентичности получают посредством определения количества идентичных положений, в которых сравниваемые последовательности совпадают, деления этого количества на количество сравниваемых положений (например, количество положений в эталонной последовательности) и умножения этого результата на 100.
[00119] В некоторых вариантах осуществления степень идентичности дана для участка, который составляет по меньшей мере приблизительно 50%, по меньшей мере приблизительно 60%, по меньшей мере приблизительно 70%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 90% или приблизительно 100% от полной длины эталонной последовательности. Например, если эталонная последовательность нуклеиновой кислоты состоит из 200 нуклеотидов, то степень идентичности дана для по меньшей мере приблизительно 100, по меньшей мере приблизительно 120, по меньшей мере приблизительно 140, по меньшей мере приблизительно 160, по меньшей мере приблизительно 180 или приблизительно 200 нуклеотидов, в некоторых вариантах осуществления для непрерывных нуклеотидов. В некоторых вариантах осуществления степень идентичности дана для полной длины эталонной последовательности.
[00120] Последовательности нуклеиновых кислот или аминокислотные последовательности, характеризующиеся определенной степенью идентичности с данной последовательностью нуклеиновой кислоты или аминокислотной последовательностью соответственно, могут иметь по меньшей мере одно функциональное свойство данной указанной последовательности, например, и в некоторых случаях функционально эквивалентны данной указанной последовательности. Одно важное свойство включает способность действовать в качестве цитокина, в частности, при введении субъекту. В некоторых вариантах осуществления последовательность нуклеиновой кислоты или аминокислотная последовательность, характеризующаяся определенной степенью идентичности с данной последовательностью нуклеиновой кислоты или аминокислотной последовательностью, функционально эквивалентна данной указанной последовательности.
[00121] Применяемый в данном документе термин "набор" относится к упакованному набору связанных компонентов, таких как одно или несколько соединений или композиций и один или несколько связанных материалов, таких как растворители, растворы, буферы, инструкции или высушивающие средства.
Антигенные полипептиды на основе последовательности RSV, содержащие полипептид F RSV, содержащий один или несколько остатков аспарагина в определенных положениях
[00122] В данном документе предусмотрены антигенные полипептиды на основе последовательности RSV, содержащие полипептид F RSV. Полипептид F RSV может содержать целую последовательность F RSV или часть F RSV. В некоторых вариантах осуществления эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, является заблокированным. Блокирование эпитопа снижает или устраняет образование антител против эпитопа при введении субъекту антигенного полипептида на основе последовательности RSV. Это может обеспечить увеличение доли антител, нацеленных на эпитоп, специфичный в отношении конкретной конформации F, такой как конформация до слияния. Поскольку F имеет конформацию до слияния в вирусах, которые еще не вошли в клетки, повышенная доля антител, нацеленных на F до слияния, может обеспечить большую степень нейтрализации (например, выраженную как отношение нейтрализации к связыванию, как описано в данном документе). Блокирование может быть достигнуто путем конструирования объемной функциональной группы, такой как N-гликан, вблизи общего эпитопа. Например, может быть добавлен сайт N-гликозилирования, отсутствующий в F дикого типа, например, путем мутации соответствующего остатка с заменой на аспарагин. В некоторых вариантах осуществления заблокированный эпитоп представляет собой эпитоп антигенного сайта 1 F RSV. В некоторых вариантах осуществления два или более эпитопов, общих для F RSV до слиянием и F RSV после слияния, являются заблокированными. В некоторых вариантах осуществления два или более эпитопов антигенного сайта 1 F RSV заблокированы. В некоторых вариантах осуществления один, или несколько, или все эпитопы, которые топологически перекрываются с заблокированным эпитопом, также блокируются, при этом заблокированный эпитоп необязательно представляет собой эпитоп антигенного сайта 1 F RSV.
[00123] В некоторых вариантах осуществления полипептид F RSV содержит аспарагин, соответствующий положению 328, 348 или 507 из SEQ ID NO: 26. В некоторых вариантах осуществления полипептид содержит остатки аспарагина, которые соответствуют по меньшей мере двум из положений 328, 348 или 507 из SEQ ID NO: 26. В некоторых вариантах осуществления полипептид содержит остатки аспарагина, которые соответствуют положениям 328, 348 или 507 из SEQ ID NO: 26. Как описано в примерах, было обнаружено, что такие остатки аспарагина могут функционировать в качестве сайтов гликозилирования. Кроме того, без ограничения какой-либо конкретной теорией, при введении полипептида субъекту наличие гликанов в этих сайтах может ингибировать развитие антител против близлежащих эпитопов, которые включают эпитопы, общие для белка F RSV до и после слияния. В некоторых вариантах осуществления гликозилирование аспарагина, соответствующего положению 328, 348 или 507 из SEQ ID NO: 26, обеспечивает блокирование по меньшей мере одного эпитопа, общего для F RSV до слияния и F RSV после слияния, такого как эпитоп антигенного сайта 1. Ингибирование развития антител против эпитопов, общих для белка F RSV до и после слияния, может быть благоприятным, поскольку это может обеспечить направление в сторону развития антител против эпитопов, специфичных в отношении белка F RSV до слияния, таких как эпитоп сайта 0, которые могу обладать более эффективной нейтрализующей активностью по сравнению с антителами против других эпитопов F RSV. Эпитоп сайта 0 содержит аминокислотные остатки 62-69 и 196-209 из SEQ ID NO: 26. Следовательно, в некоторых вариантах осуществления полипептид F RSV содержит аминокислотные остатки 62-69 и 196-209 из SEQ ID NO: 26.
[00124] Следует отметить, что конструкции, описанные в данном документе, могут иметь делеции или замены разной длины относительно F RSV дикого типа. Например, в конструкции под SEQ ID NO: 23 и других положения 98-144 последовательности дикого типа (SEQ ID NO: 26) замещены GSGNVGL (положения 98-104 из SEQ ID NO: 23; также SEQ ID NO: 31), что приводит к суммарному удалению 40 аминокислот, таким образом, что положения 328, 348 или 507 из SEQ ID NO: 26 соответствуют положениям 288, 308 и 467 из SEQ ID NO: 23. В целом, положения в конструкциях, описанных в данном документе, могут быть картированы в последовательности дикого типа из SEQ ID NO: 26 путем попарного выравнивания, например, с применением алгоритма Нидлмана-Вунша со стандартными параметрами (матрица EBLOSUM62, штраф за гэп 10, штраф за продление гэпа 0,5). См. также обсуждение структурного выравнивания, представленное в данном документе в качестве альтернативного подхода к идентификации соответствующих положений.
[00125] В некоторых вариантах осуществления полипептид F RSV содержит мутации, которые обеспечивают добавление гликанов для блокирования эпитопов на антигене до слияния, которые структурно аналогичны таковым на поверхности F RSV после слияния. В некоторых вариантах осуществления гликаны добавляют для специфичного блокирования эпитопов, которые могут присутствовать в F RSV в конформации после слияния. В некоторых вариантах осуществления добавляют гликаны, которые блокируют эпитопы, которые могут присутствовать в F RSV в конформации после слияния, но не оказывают эффекта в отношении одного или нескольких эпитопов, присутствующих в F RSV в конформации до слияния, таких как эпитоп сайта 0.
[00126] В некоторых вариантах осуществления гликаны, добавленные в один или несколько сайтов гликозилирования, обсуждаемых выше, обеспечивают увеличение уровня секреции в системах экспрессии, таких как клетки млекопитающих, по сравнению с другими конструкциями.
[00127] В некоторых вариантах осуществления полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью аминокислотам 1-478 из SEQ ID NO: 17. В некоторых вариантах осуществления полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью последовательности под SEQ ID NO: 17. В некоторых вариантах осуществления полипептид F RSV содержит аминокислоты 1-478 из SEQ ID NO: 17. В некоторых вариантах осуществления полипептид F RSV содержит последовательность из SEQ ID NO: 17.
[00128] В некоторых вариантах осуществления полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью аминокислотам 1-478 из SEQ ID NO: 23. В некоторых вариантах осуществления полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 85%, 90%, 95%, 97%, 98%, 99% или 99,5% идентичностью последовательности под SEQ ID NO: 23. В некоторых вариантах осуществления полипептид F RSV содержит аминокислоты 1-478 из SEQ ID NO: 23. В некоторых вариантах осуществления полипептид F RSV содержит последовательность из SEQ ID NO: 23.
[00129] В некоторых вариантах осуществления полипептид F RSV содержит последовательность DS-CAV1 (описанную, например, в McLellan, J.S., et al., Science 342(6158):592-598 (2013)) (SEQ ID NO: 25), в которой осуществлены дополнительные модификации, включая по меньшей мере один, два или три из остатков аспарагина, описанных выше.
[00130] В некоторых вариантах осуществления полипептид дополнительно содержит белок ферритин. Белок ферритин может дополнительно обладать любым из признаков, описанных ниже в разделе, касающемся ферритина, или их комбинацией.
[00131] Полипептид F RSV может дополнительно обладать любым из дополнительных признаков, представленных в следующем обсуждении, или любой возможной комбинацией таких признаков.
Одноцепочечные конструкции
[00132] В некоторых вариантах осуществления полипептид RSV представляет собой одноцепочечную конструкцию, например полипептид RSV, который не содержит сайтов расщепления фурином. В некоторых вариантах осуществления F RSV не содержит одного или нескольких сайтов расщепления фурином. Конструкции, которые не содержат сайты расщепления фурином, экспрессируются в виде целых полипептидов, которые не расщепляются на биологические фрагменты F1/F2 нативного белка F.
Аминокислотные замены
[00133] В некоторых вариантах осуществления F RSV содержит одну аминокислотную замену относительно последовательности дикого типа. В некоторых вариантах осуществления F RSV содержит более чем одну аминокислотную замену, например 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 замен, относительно последовательности дикого типа. Иллюстративной последовательностью дикого типа является SEQ ID NO: 26.
[00134] В некоторых вариантах осуществления аминокислотная замена или пара аминокислотных замен представляет собой замену(-ы), обеспечивающую(-ие) межпротомерную стабилизацию. Иллюстративные замены, которые могут обеспечивать межпротомерную стабилизацию, представляют собой V207L; N228F; I217V и E218F; I221L и E222M; или Q224A и Q225L, при применении нумерации положений из SEQ ID NO: 26.
[00135] В некоторых вариантах осуществления аминокислотная замена или пара аминокислотных замен являются заменами, обеспечивающими внутрипротомерную стабилизацию. Иллюстративными заменами, которые могут обеспечивать внутрипротомерную стабилизацию, являются V220I, а также A74L и Q81L, при применении нумерации положений из SEQ ID NO: 26.
[00136] В некоторых вариантах осуществления аминокислотная замена обеспечивает стабилизацию спирали, т.е. предсказано, что она будет обеспечивать стабилизацию спирального домена F RSV. Стабилизация спирального домена может вносить вклад в стабильность эпитопа сайта 0 и в целом F RSV в конформации до слияния. Иллюстративными заменами, которые могут обеспечивать стабилизацию спирали, являются N216P или I217P, при применении нумерации положений из SEQ ID NO: 26.
[00137] В некоторых вариантах осуществления аминокислотная замена представляет собой замену, обеспечивающую кэппирование спирали. В некоторых вариантах осуществления аминокислотная замена представляет собой замену, обеспечивающую кэппирование спирали с помощью PRO. Кэппирование спирали основано на наблюдении с помощью биофизических методов того, что, в то время как наличие мутации в месте остатка пролина в альфа-спирали может нарушить формирование спирали, наличие пролина на N-конце спиральной области может способствовать индуцированию образования спирали за счет стабилизации углов связи PHI/PSI. Иллюстративными заменами, которые могут обеспечивать кэппирование спирали, являются N216P или I217P, при применении нумерации положений из SEQ ID NO: 26.
[00138] В некоторых вариантах осуществления аминокислотная замена обеспечивает замещение дисульфидной мутации DS-CAV1. В некоторых вариантах осуществления сконструированный дисульфид в DS-CAV1 возвращается к дикому типу (мутации C69S и/или C212S в DS-CAV1, при применении нумерации положений из SEQ ID NO: 26). В некоторых вариантах осуществления один или несколько остатков C в DS-CAV1 замещаются остатком S для устранения дисульфидной связи. В некоторых вариантах осуществления замена C69S или C212S, при применении нумерации положений из SEQ ID NO: 26, обеспечивает устранение дисульфидной связи. В некоторых вариантах осуществления полипептид F RSV содержит как C69S, так и C212S, при применении нумерации положений из SEQ ID NO: 26. В некоторых вариантах осуществления замещение таких остатков цистеина с устранением тем самым дисульфидной связи обеспечивает блокирование восстановления (т. е. прием электронов от восстанавливающего средства) полипептида F RSV. В некоторых вариантах осуществления замена I217P, при применении нумерации положений из SEQ ID NO: 26, включена в антиген вместо замены по C69 и/или C212. Положение 217 в SEQ ID NO: 26 соответствует положению 177 в SEQ ID NO: 23.
[00139] В некоторых вариантах осуществления аминокислотная замена обеспечивает предотвращение протеолиза трипсином или трипсиноподобными протеазами. В некоторых вариантах осуществления аминокислотная замена, которая обеспечивает предотвращение такого протеолиза, находится в области гептадных повторов В (HRB) в F RSV. Появление фрагментов, разрешающих протеолиз, в конструкции F RSV-ферритин, которая содержала область HRB дикого типа, свидетельствует о том, что лизин или аргинин в этой области являлся мишенью для протеолиза. Аминокислотная замена, обеспечивающая удаление остатка K или R, может быть названа нокаутом (KO). В некоторых вариантах осуществления K или R заменены на L или Q. В некоторых вариантах осуществления K заменен на L или Q. В некоторых вариантах осуществления полипептид F RSV содержит K498L и/или K508Q, при применении нумерации положений из SEQ ID NO: 26. Соответствующими положениями в SEQ ID NO: 23 являются 458 и 468 соответственно. В некоторых вариантах осуществления полипептид F RSV содержит как K498L, так и K508Q.
[00140] В некоторых вариантах осуществления аминокислотная замена обеспечивает добавление гликанов. В некоторых вариантах осуществления аминокислотная замена обеспечивает увеличение уровня гликозилирования путем добавления гликанов в полипептиды F RSV. Замены, обеспечивающие добавление гликанов, также можно называть гликозилированием посредством конструирования по сравнению с естественным гликозилированием (без дополнительных гликанов).
[00141] В некоторых вариантах осуществления аминокислотная замена для добавления гликанов представляла собой замену на N. В некоторых вариантах осуществления аминокислотная замена на N обеспечивает N-связанное гликозилирование. В некоторых вариантах осуществления замена на N сопровождается заменой на T или S аминокислоты, находящейся во втором положении от N в направлении С-конца, что образует мотив гликозилирования NxT/S. В некоторых вариантах осуществления N экспонирован на поверхности. Как показано в приведенных ниже примерах, мутации, которые обеспечивают увеличение уровня гликозилирования, могут обеспечивать увеличение уровня экспрессии полипептида, содержащего полипептид F RSV.
Изменения свойств полипептида F RSV в зависимости от модификаций
[00142] Модификации в аминокислотной последовательности F RSV могут изменять свойства полипептида F RSV. Свойство полипептида F RSV может включать любую структурную или функциональную характеристику полипептида F RSV.
[00143] В некоторых вариантах осуществления одна модификация аминокислотной последовательности изменяет несколько свойств полипептида F RSV. В некоторых вариантах осуществления полипептид F RSV может содержать несколько модификаций, которые изменяют различные свойства полипептида F RSV. В некоторых вариантах осуществления несколько модификаций вызывают большее изменение свойств полипептида F RSV.
[00144] В некоторых вариантах осуществления несколько модификаций могут оказывать аддитивный эффект в отношении конкретного свойства. Например, две аминокислотные замены для добавления гликанов могут вызвать большее усиление гликозилирования полипептида F RSV по сравнению с любой одной аминокислотной заменой.
[00145] В некоторых вариантах осуществления несколько модификаций оказывают эффект в отношении различных свойств полипептида F RSV. Например, одна или несколько аминокислотных замен для увеличения уровня гликозилирования могут быть осуществлены вместе с одной или несколькими аминокислотными заменами для блокирования восстановления.
[00146] В некоторых вариантах осуществления модификации полипептида СSV F обеспечивают стабилизацию конформации до слияния.
[00147] В некоторых вариантах осуществления модификации обеспечивают стабилизацию эпитопа сайта 0 (также известного как антигенный сайт 0) F RSV до слияния, как описано, например, в McLellan et al., Science 340(6136):1113-1117 (2013). В некоторых вариантах осуществления модификация, которая обеспечивает стабилизацию эпитопа сайта 0, представляет собой модификацию, обеспечивающую межпротомерную стабилизацию. В некоторых вариантах осуществления модификация, которая обеспечивает стабилизацию эпитопа сайта 0, обеспечивает стабилизацию F до слияния, согласно данным измерения связывания сайта 0 и сайта V, согласно данным измерения связывания с антителами D25 или AM14 соответственно.
[00148] В некоторых вариантах осуществления модификации обеспечивают увеличение уровня экспрессии F RSV в системах экспрессии. В некоторых вариантах осуществления модификации обеспечивают увеличение уровня секреции F RSV в системах экспрессии. В некоторых вариантах осуществления модификации обеспечивают увеличение стабильности рекомбинантного F RSV после экспрессии. Это изменение может происходить в системе экспрессии любого типа, такой как система экспрессии на основе бактерии, гриба, насекомого или млекопитающего.
[00149] В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают введение пролина, обеспечивают увеличение уровня экспрессии по сравнению с другими конструкциями. В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают добавление гликанов, обеспечивают увеличение уровня экспрессии по сравнению с другими конструкциями. В некоторых вариантах осуществления аминокислота замены, которые обеспечивают замену K или R на другие аминокислоты, обеспечивают увеличение уровня экспрессии по сравнению с другими конструкциями. Наблюдаемое увеличение уровня экспрессии может обеспечиваться в результате действия любого механизма, который обеспечивает увеличение выхода процесса ферментации или другого процесса получения, включая соответствующее ингибирование расщепления или разложения протеазой и/или повышение стабильности в клетке-хозяине или во внеклеточной среде. В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают замену одного или нескольких остатков K в области HRB F RSV на другие аминокислоты, обеспечивают увеличение уровня экспрессии по сравнению с другими конструкциями.
[00150] В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают замену K на другие аминокислоты, обеспечивают повышение стабильности полипептидов F RSV. В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают замену одного или нескольких остатков К в области HRB F RSV на другие аминокислоты, обеспечивают повышение стабильности полипептидов F RSV. В некоторых вариантах осуществления такая повышенная стабильность обусловлена снижением уровня расщепления протеазой.
[00151] В некоторых вариантах осуществления F RSV содержит мутацию(-и), которая(-ые) обеспечивает(-ют) удаление дисульфида, например для предотвращения конъюгации после восстановления. В некоторых вариантах осуществления замена I217P обеспечивают блокирование восстановления полипептида F RSV. В некоторых вариантах осуществления аминокислотные замены, которые обеспечивают замену K на другие аминокислоты, блокируют восстановление полипептида F RSV в присутствии восстанавливающего средства.
[00152] В некоторых вариантах осуществления одноцепочечные конструкции обеспечивают увеличение уровня экспрессии по сравнению с другими конструкциями.
[00153] В некоторых вариантах осуществления полипептид F RSV содержит последовательность DS-CAV1 (SEQ ID NO: 25) (как описано в McLellan, J.S., et al., Science 342(6158):592-598 (2013)). В некоторых вариантах осуществления полипептид F RSV содержит последовательность DS-CAV1, в которой выполнены дополнительные модификации, например, включая по меньшей мере один, два или три остатка аспарагина, описанных выше.
Полипептиды G RSV
[00154] Как используется в данном документе, полипептид G RSV может содержать целую последовательность G RSV или часть G RSV. Полипептид G RSV может содержать модификации по сравнению с последовательностью дикого типа. В некоторых вариантах осуществления полипептид G RSV представляет собой G RSV, модифицированный по сравнению с G RSV дикого типа (SEQ ID NO: 27).
[00155] В некоторых вариантах осуществления такие модификации представляют собой изменения аминокислоты полипептида G RSV по сравнению с G RSV дикого типа.
[00156] В некоторых вариантах осуществления полипептид G RSV содержит весь эктодомен G RSV (SEQ ID NO: 28 или положения, соответствующие им) или его часть. В некоторых вариантах осуществления полипептид G RSV содержит всю область Gcc (аминокислоты 151-193 в G RSV (SEQ ID NO: 27)) или ее часть. В некоторых вариантах осуществления полипептид G RSV содержит мотив CX3C. В некоторых вариантах осуществления полипептид G RSV связывается с рецептором CX3CR1. Область Gcc является как консервативной, так и иммуногенной, поэтому ее можно использовать для обеспечения образования антител с широкой активностью против штаммов RSV. В некоторых вариантах осуществления представлен Gcc RSV штамма A, как показано в SEQ ID NO: 32. В некоторых вариантах осуществления представлен Gcc RSV штамма В, как показано в SEQ ID NO: 33.
[00157] В некоторых вариантах осуществления полипептид G RSV не является гликозилированным. Например, полипептид G RSV может не содержать сайтов гликозилирования NXS/TX либо из-за усечения, либо из-за мутации остатков N или S/T (например, с заменой на Q или A соответственно), или из-за их комбинации.
[00158] В некоторых вариантах осуществления полипептид G RSV является частью антигенного полипептида на основе последовательности ферритина. Например, полипептид G RSV может быть конъюгирован с ферритином, как описано в данном документе, например посредством экспонированного на поверхности цистеина в ферритине. В некоторых вариантах осуществления такая ферритиновая наночастица представляет собой слитый белок, также содержащий полипептид F RSV, такой как любой из полипептидов, содержащих полипептид F RSV и белок ферритин, описанных выше.
Антигенные полипептиды на основе последовательности RSV, содержащие ферритин
[00159] Также в данном документе представлен антигенный полипептид на основе последовательности RSV, содержащий ферритин и полипептид RSV. Полипептид RSV может представлять собой полипептид F RSV, такой как любой из полипептидов F RSV, описанных в данном документе. Полипептид F RSV может содержать целую последовательность F RSV или часть F RSV. Полипептид F RSV может содержать одну или несколько модификаций (например, аминокислотную замену) по сравнению с последовательностью дикого типа. Полипептид RSV может представлять собой полипептид G RSV, такой как любой из полипептидов G RSV, описанных в данном документе.
[00160] В некоторых вариантах осуществления ферритин в полипептиде представляет собой ферритин дикого типа. В некоторых вариантах осуществления ферритин получен из бактерий, насекомых, грибов, птиц или млекопитающих. В некоторых вариантах осуществления ферритин является человеческим. В некоторых вариантах осуществления ферритин является бактериальным.
[00161] В некоторых вариантах осуществления ферритин представляет собой легкую цепь и/или тяжелую цепь ферритина. В некоторых вариантах осуществления ферритин представляет собой ферритин насекомого, такой как тяжелая цепь ферритина из Trichoplusia ni (SEQ ID NO: 211) или легкая цепь ферритина из Trichoplusia ni (SEQ ID NO: 212). В некоторых вариантах осуществления ферритин представляет собой ферритин человека, такой как тяжелая цепь ферритина человека (SEQ ID NO: 214 или FTH1, GENE ID No: 2495) или легкая цепь ферритина человека (SEQ ID NO: 215 или FTL, GENE ID No: 2512). В некоторых вариантах осуществления ферритиновая наночастица содержит всего 24 субъединицы тяжелой цепи ферритина и легкой цепи ферритина, такие как в ферритиновых наночастицах человека или Trichoplusia ni. Ферритиновые наночастицы из T. ni могут содержать 12 субъединиц тяжелой цепи ферритина и 12 субъединиц легкой цепи ферритина.
[00162] В некоторых вариантах осуществления антигенный полипептид на основе последовательности RSV содержит легкую цепь ферритина и полипептид RSV. В некоторых вариантах осуществления антигенный полипептид на основе последовательности RSV содержит тяжелую цепь ферритина и полипептид RSV. В некоторых вариантах осуществления антигенный полипептид на основе последовательности RSV, содержащий легкую цепь ферритина и полипептид RSV, может собираться с тяжелой цепью ферритина, которая не связана с полипептидом RSV. В некоторых вариантах осуществления антигенный полипептид на основе последовательности RSV, содержащий тяжелую цепь ферритина и полипептид RSV, может собираться с легкой цепью ферритина, которая не связана с полипептидом RSV. Ферритин, не связанный с полипептидом RSV (или, в более широком смысле, полипептидом, отличным от ферритина), может называться "голым ферритином".
[00163] В некоторых вариантах осуществления антигенный полипептид, содержащий тяжелую цепь ферритина и полипептид, может собираться с антигенным полипептидом, содержащим легкую цепь ферритина и полипептид RSV, с обеспечением презентирования двух одинаковых или разных полипептидов, отличных от ферритина, на одной ферритиновой наночастице. В некоторых вариантах осуществления два разных полипептида, отличные от ферритина, представляют собой полипептиды RSV. В некоторых вариантах осуществления два разных полипептида, отличные от ферритина, кодируются RSV и другим возбудителем инфекции. В некоторых вариантах осуществления другой полипептид, отличный от ферритина, из другого возбудителя инфекции происходит из вируса или бактерии.
[00164] В некоторых вариантах осуществления антигенный полипептид, содержащий тяжелую цепь ферритина и полипептид, отличный от ферритина, может собираться с полипептидом, содержащим легкую цепь ферритина и полипептид, отличный от ферритина, с получением двухвалентной композиции, в которой один или оба полипептида, отличные от ферритина, представляют собой полипептиды RSV, такие как полипептиды F или G RSV, например полипептид F или G RSV, описанные в данном документе.
[00165] В некоторых вариантах осуществления ферритин представляет собой ферритин из H. pylori (см. SEQ ID NO: 208 или 209 для иллюстративной последовательности ферритина из H. pylori), необязательно с одной или несколькими мутациями, такими как описанные в данном документе. В некоторых вариантах осуществления более низкая степень гомологии последовательностей между ферритином из H. pylori (или другими вариантами бактериального ферритина) и ферритином человека может снижать возможность развития аутоиммунитета при использовании в качестве платформы для вакцины (см. Kanekiyo et al., Cell 162, 1090-1100 (2015)).
[00166] В некоторых вариантах осуществления ферритин представляет собой ферритин из Pyrococcus furiosus (NCBI seq WP_011011871.1) с одной или несколькими мутациями, описанными в данном документе.
[00167] В некоторых вариантах осуществления ферритин включает последовательность, характеризующуюся более чем 70%, более чем 75%, более чем 80%, более чем 85%, более чем 90%, более чем 95%, более чем 97%, более чем 98% или более чем 99% идентичностью с ферритином дикого типа.
[00168] В некоторых вариантах осуществления представлена наночастица, содержащая антигенный полипептид на основе последовательности RSV, раскрытый в данном документе, содержащий полипептид RSV и ферритин.
[00169] В некоторых вариантах осуществления другой белок, способный формировать наночастицу, заменяет ферритин. В некоторых вариантах осуществления этим белком является лумазинсинтаза (см. Ra et al., Clin Exp Vaccine Res 3:227-234 (2014)). В некоторых вариантах осуществления этот белок представляет собой лумазинсинтазу серотипа 1, 2, 3, 4, 5, 6 или 7. Иллюстративные последовательности лумазинсинтазы предусмотрены как SEQ ID NO: 216 и 219. В некоторых вариантах осуществления лумазинсинтаза включает последовательность с 80%, 85%, 90%, 95%, 98% или 99% идентичностью с SEQ ID NO: 216 или 219.
1. Цистеин для конъюгации
[00170] В некоторых вариантах осуществления ферритин подвергают мутированию для обеспечения химической основы для конъюгации иммуностимулирующей функциональной группы и/или полипептида RSV. Это может быть достигнуто с помощью мутации, обеспечивающей замену экспонированной на поверхности отличной от цистеина аминокислоты на цистеин. Во избежание сомнений такая формулировка как "замена экспонированной на поверхности аминокислоты на цистеин" обязательно подразумевает, что экспонированная на поверхности аминокислота в последовательности дикого типа или премутационной последовательности не представляет собой цистеин. Другой подход для обеспечения химической основы для конъюгации иммуностимулирующей функциональной группы или полипептида RSV заключается во включении сегмента из аминокислот, такого как линкер, с N- или C-конца ферритина, при этом сегмент из аминокислот содержит цистеин. В некоторых вариантах осуществления этот цистеин (независимо от того, заменяет ли он аминокислоту, экспонированную на поверхности или в N- или C-концевом линкере) является непарным, что означает, что он не имеет подходящего цистеина-партнера для образования дисульфидной связи. В некоторых вариантах осуществления этот цистеин не изменяет вторичную структуру ферритина. В некоторых вариантах осуществления этот цистеин не изменяет третичную структуру ферритина.
[00171] В некоторых вариантах осуществления этот цистеин можно применять для конъюгации средств, таких как иммуностимулирующие фрагменты, с ферритином. В некоторых вариантах осуществления этот цистеин обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с этим цистеином на ферритине, экспонированы на поверхности собранной ферритиновой частицы. В некоторых вариантах осуществления этот цистеин способен взаимодействовать с молекулами и клетками субъекта после введения в ходе сборки ферритиновой частицы.
[00172] В некоторых вариантах осуществления присутствие этого цистеина обеспечивает конъюгацию одного или нескольких иммуностимулирующих фрагментов, например, адъювантов. В некоторых вариантах осуществления конъюгация иммуностимулирующего фрагмента не происходила бы в отсутствие этого цистеина.
[00173] В некоторых вариантах осуществления отличная от цистеина аминокислота, которая заменена на цистеин, выбрана из E12, S72, A75, K79, S100 и S111 из ферритина H. pylori. Таким образом, в некоторых вариантах осуществления экспонированная на поверхности аминокислота, которая заменена на цистеин, представляет собой аминокислотный остаток, который соответствует E12, S26, S72, A75, K79, S100 или S111 из ферритина H. pylori. Аналогичные аминокислоты можно найти в ферритине, отличном от ферритина из H. pylori, посредством парного или структурного выравнивания. В некоторых вариантах осуществления отличная от цистеина аминокислота, которая заменена на цистеин, выбрана из аминокислоты, которая соответствует S3, S19, S33, I82, A86, A102 и A120 из легкой цепи человеческого ферритина. В некоторых вариантах осуществления экспонированная на поверхности аминокислота, подлежащая замене на цистеин, выбрана на основании понимания того, что если бы нативная аминокислота была заменена на цистеин, она была бы реакционноспособна в собранном ферритиновом мультимере или ферритиновой частице, и/или что этот цистеин не нарушает стабильность ферритинового мультимера или ферритиновой частицы, и/или что этот цистеин не приводит к снижению уровней экспрессии ферритина.
[00174] В некоторых вариантах осуществления ферритин содержит мутацию E12C. В некоторых вариантах осуществления остаток E12C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток E12C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком E12C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка E12C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00175] В некоторых вариантах осуществления ферритин содержит мутацию S26C. В некоторых вариантах осуществления остаток S26C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток S26C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком S26C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка S26C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00176] В некоторых вариантах осуществления ферритин содержит мутацию S72C. В некоторых вариантах осуществления остаток S72C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток S72C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком S72C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка S72C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00177] В некоторых вариантах осуществления ферритин содержит мутацию A75C. В некоторых вариантах осуществления остаток A75C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток A75C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком A75C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка A75C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00178] В некоторых вариантах осуществления ферритин содержит мутацию K79C. В некоторых вариантах осуществления остаток K79C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток K79C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком K79C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка K79C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00179] В некоторых вариантах осуществления ферритин содержит мутацию S100C. В некоторых вариантах осуществления остаток S100C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток S100C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком S100C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка S100C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
[00180] В некоторых вариантах осуществления ферритин содержит мутацию S111C. В некоторых вариантах осуществления остаток S111C может быть использован для конъюгации средств (например, иммуностимулирующих функциональных групп и/или полипептидов RSV) с ферритином. В некоторых вариантах осуществления остаток S111C обеспечивает свободную тиольную группу, которая является реакционноспособной. В некоторых вариантах осуществления средства, конъюгированные с остатком S111C на мономерах ферритина, экспрессируются на поверхности собранного ферритинового мультимера или ферритиновой частицы. В некоторых вариантах осуществления двадцать четыре остатка S111C (по одному из каждого мономера) присутствуют на поверхности ферритинового мультимера или ферритиновой частицы.
2. Удаление внутреннего цистеина
[00181] В некоторых вариантах осуществления ферритин содержит мутацию с заменой внутреннего цистеина на отличную от цистеина аминокислоту. Удаление нативного внутреннего остатка цистеина может гарантировать наличие только одного неспаренного цистеина в мономере ферритина, что позволяет избежать нежелательных реакций, таких как образование дисульфидных связей, и может привести к более стабильному и эффективному результату (например, представлению адъюванта). В некоторых вариантах осуществления C31 из ферритина H. pylori заменен на отличную от цистеина аминокислоту. В некоторых вариантах осуществления C31 из ферритина H. pylori заменен на серин (C31S), хотя можно применять любой отличный от цистеина остаток, например, аланин, глицин, треонин или аспарагин. Аналогичные аминокислоты можно найти в ферритине, отличном от ферритина H. pylori, посредством парного или структурного выравнивания. Таким образом, в некоторых вариантах осуществления внутренний цистеин, который заменен на отличную от цистеина аминокислоту, представляет собой аминокислотный остаток, который выравнивается с C31 из ферритина H. pylori. Иллюстративные последовательности ферритина, демонстрирующие мутацию C31S, показаны в SEQ ID NO: 201-207. В некоторых вариантах осуществления, когда в ферритине присутствует более одного внутреннего цистеина, два или более (например, каждый) внутренних цистеина заменены на отличную от цистеина аминокислоту, такую как серин, или аминокислоту, выбранную из серина, аланина, глицина, треонина или аспарагина.
3. Гликозилирование
[00182] Гликозилирование, совместимое с человеческим гликозилированием, может способствовать безопасности и эффективности рекомбинантных лекарственных продуктов. Разрешение регулирующих органов может зависеть от демонстрации соответствующего гликозилирования как критического атрибута качества (см. Zhang et al., Drug Discovery Today 21(5):740-765 (2016)). N-гликаны могут образовываться в результате гликозилирования боковых цепей аспарагина и могут различаться по структуре у людей и других организмов, таких как бактерии и дрожжи. Таким образом, может быть желательно уменьшить или устранить гликозилирование, отличное от человеческого гликозилирования, и/или образование N-гликанов в ферритине в соответствии с настоящим изобретением. В некоторых вариантах осуществления контроль гликозилирования ферритина улучшает эффективность и/или безопасность композиции, особенно при применении для вакцинации человека.
[00183] В некоторых вариантах осуществления ферритин мутирован для ингибирования образования N-гликана. В некоторых вариантах осуществления мутированный ферритин характеризуется пониженным гликозилированием по сравнению с соответствующим ему ферритином дикого типа.
[00184] В некоторых вариантах осуществления ферритин содержит мутацию с заменой экспонированного на поверхности аспарагина на отличную от аспарагина аминокислоту. В некоторых вариантах осуществления экспонированный на поверхности аспарагин представляет собой N19 из ферритина H. pylori или находится в положении, которое соответствует положению 31 ферритина H. pylori, как определено посредством парного или структурного выравнивания. В некоторых вариантах осуществления мутирование такого аспарагина, например, N19 из ферритина H. pylori, снижает гликозилирование ферритина. В некоторых вариантах осуществления мутация заменяет аспарагин на глутамин. В некоторых вариантах осуществления ферритин представляет собой ферритин из H. pylori, включающий мутацию N19Q. SEQ ID NO: 201-207 представляют собой иллюстративные последовательности ферритина, включающие мутации N19Q.
[00185] Млекопитающее, подвергшееся воздействию гликозилированного белка, полученного в бактериях или дрожжах, может генерировать иммунный ответ на гликозилированный белок, потому что паттерн гликозилирования данного белка у бактерий или дрожжей может отличаться от паттерна гликозилирования того же белка у млекопитающих. Таким образом, некоторые гликозилированные терапевтические белки могут не подходить для получения в бактериях или дрожжах.
[00186] В некоторых вариантах осуществления снижение гликозилирования ферритина за счет мутации аминокислоты облегчает получение белка в бактериях или дрожжах. В некоторых вариантах осуществления снижение гликозилирования ферритина снижает возможность проявления нежелательных эффектов у млекопитающих при введении мутантного ферритина, который экспрессируется в бактериях или дрожжах. В некоторых вариантах осуществления реактогенность мутантного ферритина, полученного в бактериях или дрожжах, у субъекта-человека ниже, поскольку гликозилирование снижено. В некоторых вариантах осуществления частота ответов с гиперчувствительностью у субъектов-людей ниже после лечения мутантным ферритином с пониженным гликозилированием по сравнению с ферритином дикого типа.
[00187] В некоторых вариантах осуществления у субъекта деградация композиции, содержащей мутантный ферритин с пониженным гликозилированием, происходит медленнее по сравнению с композицией, содержащей ферритин дикого типа, или композицией, содержащей соответствующий ферритин с гликозилированием дикого типа. В некоторых вариантах осуществления композиция, содержащая мутантный ферритин с пониженным гликозилированием, характеризуется пониженным клиренсом у субъекта по сравнению с композицией, содержащей ферритин дикого типа, или композицией, содержащей соответствующий ферритин с гликозилированием дикого типа. В некоторых вариантах осуществления композиция, содержащая мутантный ферритин с пониженным гликозилированием, характеризуется более длительным периодом полужизни в сыворотке крови по сравнению с ферритином дикого типа или композицией, содержащей соответствующий ферритин с гликозилированием дикого типа.
4. Комбинации мутаций
[00188] В некоторых вариантах осуществления ферритин включает более одного типа мутаций, описанных в данном документе. В некоторых вариантах осуществления ферритин содержит одну или несколько мутаций, независимо выбранных из мутации для снижения гликозилирования, мутации для удаления внутреннего цистеина и мутации для образования экспонированного на поверхности цистеина. В некоторых вариантах осуществления ферритин содержит мутацию для снижения гликозилирования, мутацию для удаления внутреннего цистеина и мутацию для образования экспонированного на поверхности цистеина.
[00189] В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию для образования экспонированного на поверхности цистеина. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию E12C. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию S72C. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию A75C. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию K79C. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию S100C. В некоторых вариантах осуществления ферритин содержит мутацию N19Q, мутацию C31S и мутацию S111C. В некоторых вариантах осуществления ферритин включает мутации, соответствующие любой из упомянутых выше групп мутаций, где соответствующие мутации изменяют N на Q, C на S и отличную от цистеина экспонированную на поверхности аминокислоту на цистеин в положениях, определенных посредством парного выравнивания аминокислотной последовательности ферритина с аминокислотной последовательностью ферритина из H. pylori (SEQ ID NO: 208 или 209).
[00190] Иллюстративные ферритины, включающие более одного типа мутаций, представлены под SEQ ID NO: 201-207.
5. Структурное выравнивание
[00191] Как обсуждается в данном документе, положения мутаций, соответствующие положениям мутаций, описанным по отношению к данному полипептиду (например, ферритину из H. pylori), можно идентифицировать посредством попарного или структурного выравнивания. Структурное выравнивание подходит для больших семейств белков, таких как ферритин, где белки имеют сходные структуры, несмотря на значительные вариации последовательности, и многие представители этого семейства были структурно охарактеризованы, и его можно применять для идентификации соответствующих положений в различных версиях других полипептидов, описанных в данном документе, таких как полипептиды RSV (например, F или G RSV). База данных "Protein databank" (PDB) содержит 3D-структуры многих ферритинов, включая перечисленные ниже с их номерами доступа.
[00192] 2jd6, 2jd7 - PfFR - Pyrococcus furiosus. 2jd8 - PfFR+Zn. 3a68 - soFR из гена SferH4 - соя. 3a9q - soFR из гена SferH4 (мутантного). 3egm, 3bvf, 3bvi, 3bvk, 3bvl - HpFR - Heliobacter pylori. 5c6f - HpFR (мутантный) + Fe. 1z4a, 1vlg - FR - Thermotoga maritime. 1s3q, 1sq3, 3kx9 - FR - Archaeoglubus fulgidus. 1krq - FR - Campylobacter jejuni. 1eum - EcFR - Escherichia coli. 4reu - EcFR+Fe. 4xgs - EcFR (мутантный) + Fe2O2. 4ztt - EcFR (мутантный) + Fe2O+Fe2+Fe+O2. 1qgh - LiFR - Listeria innocua. 3qz3 - VcFR - Vibrio cholerae. 3vnx - FR - Ulva pertusa. 4ism, 4isp, 4itt, 4itw, 4iwj, 4iwk, 4ixk, 3e6s - PnmFR - смешанный род Pseudo-nitschia. 4zkh, 4zkw, 4zkx, 4zl5, 4zl6, 4zlw, 4zmc - PnmFR (мутантный) + Fe. 1z6o - FR - Trichoplusia ni. 4cmy - FR+Fe - Chlorobaculum tepidum. Легкая цепь ферритина (FTL). 1lb3, 1h96 - mFTL - мышь. 1rcc, 1rcd, 1rci - bFTL+тартрат+Mg. 1rce, 1rcg - bFTL+тартрат+Mn. 3noz, 3np0, 3np2, 3o7r - hoFTL (мутантный) - лошадь. 3o7s, 3u90 - hoFTL. 4v1w - hoFTL - cryo EM. 3rav, 3rd0 - hoFTL+барбитурат. Легкие цепи ферритина+тяжелые цепи ферритина: 5gn8 - hFTH+Ca.
[00193] Структурное выравнивание включает идентификацию соответствующих остатков в двух (или более) полипептидных последовательностях посредством (i) моделирования структуры первой последовательности с применением известной структуры второй последовательности или (ii) сравнения структур первой и второй последовательностей, где обе известны, и идентификацию остатка в первой последовательности, расположенного наиболее сходно с представляющим интерес остатком во второй последовательности. Соответствующие остатки идентифицируют с помощью некоторых алгоритмов на основе минимизации расстояния между альфа-атомами углерода в перекрывающихся структурах (например, какой набор парных альфа-атомов углерода обеспечивает минимизированное среднеквадратичное отклонение для выравнивания). При идентификации положений в ферритине, отличном от ферритина из H. pylori, соответствующих положениям, описанным по отношению к ферритину из H. pylori, ферритин из H. pylori может представлять собой "вторую" последовательность. Если представляющий интерес ферритин, отличный от ферритина из H. pylori, не имеет доступной известной структуры, но является близкородственным по отношению к другому ферритину, отличному от ферритина из H. pylori, который имеет известную структуру по сравнению с ферритином из H. pylori, то может быть наиболее эффективным моделирование представляющего интерес ферритина, отличного от ферритина из H. pylori, с применением известной структуры близкородственного ферритина, отличного от ферритина из H. pylori, а затем сравнение этой модели со структурой ферритина из H. pylori для идентификации необходимого соответствующего остатка в представляющем интерес ферритине. Имеется обширная литература по структурному моделированию и выравниванию; иллюстративные раскрытия включают US 6859736; US 8738343; и раскрытия, которые цитируются в Aslam et al., Electronic Journal of Biotechnology 20 (2016) 9-13. Для обсуждения моделирования структуры на основании известной родственной структуры или структур см., например, Bordoli et al., Nature Protocols 4 (2009) 1-13, и цитируемые в нем ссылки.
6. Иммуностимулирующие функциональные группы, адъюванты, конъюгированные полипептиды на основе RSV
[00194] В некоторых вариантах осуществления полипептид RSV и/или иммуностимулирующая функциональная группа, такая как адъювант, прикрепляются к экспонированной на поверхности аминокислоте. В некоторых вариантах осуществления экспонированная на поверхности аминокислота представляет собой цистеин, например, полученный в результате мутации, обсуждаемой выше. В некоторых вариантах осуществления экспонированная на поверхности аминокислота представляет собой лизин, аспартат или глутамат. Процедуры конъюгации с применением глутаральдегида (для конъюгации лизина с несущим аминогруппу линкером или функциональной группой) или карбодиимида (например, 1-циклогексил-3-(2-морфолин-4-ил-этил)карбодиимида или 1-этил-3-(3-диметил-аминопропил)карбодиимида (EDC; EDAC) для конъюгирования аспартата или глутамата с несущим аминогруппу линкером или функциональной группой или лизина с несущим карбоксильную группу линкером или функциональной группой) описаны, например, в разделе 4 в Holtzhauer, M., Basic Methods for the Biochemical Lab, Springer 2006, ISBN 978-3-540-32785-1, доступной на www.springer.com.
[00195] В некоторых вариантах осуществления иммуностимулирующий фрагмент, такой как адъювант, присоединен к экспонированной на поверхности аминокислоте в ферритине. В некоторых вариантах осуществления более чем один иммуностимулирующий фрагмент, такой как адъювант, присоединен к экспонированной на поверхности аминокислоте в ферритине. В некоторых вариантах осуществления двадцать четыре иммуностимулирующих фрагмента присоединены к ферритиновому мультимеру или ферритиновой частице (например, по одному фрагменту для каждого мономера в частице с ферритином из H. pylori). В некоторых вариантах осуществления с множеством иммуностимулирующих фрагментов, присоединенных к ферритиновой наночастице, все иммуностимулирующие фрагменты идентичны. В некоторых вариантах осуществления с множеством иммуностимулирующих фрагментов, присоединенных к ферритиновой наночастице, все иммуностимулирующие фрагменты не идентичны.
Типы иммуностимулирующих функциональных групп, адъюванты
[00196] Любая иммуностимулирующая функциональная группа, которая может быть прикреплен к экспонированной на поверхности аминокислоте (например, цистеину), может применяться в вариантах ферритина в соответствии с настоящим изобретением. В некоторых вариантах осуществления иммуностимулирующий фрагмент является B-клеточным агонистом.
[00197] В некоторых вариантах осуществления иммуностимулирующий фрагмент не является гидрофобным. В некоторых вариантах осуществления иммуностимулирующий фрагмент является гидрофильным. В некоторых вариантах осуществления иммуностимулирующих фрагмент является полярным. В некоторых вариантах осуществления иммуностимулирующий фрагмент способен к образованию водородной связи или ионной связи, например, содержит донор водородной связи, акцептор водородной связи, катионный фрагмент или анионный фрагмент. Фрагмент считается катионным или анионным, если он будет ионизироваться в водном растворе при физиологически релевантном pH, таком как pH 6, 7, 7,4 или 8.
[00198] В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой адъювант. В некоторых вариантах осуществления адъювант включает молекулярный паттерн, связанный с патогеном (PAMP). В некоторых вариантах осуществления адъювант является агонистом Toll-подобного рецептора (TLR) или стимулятором агониста генов интерферонов (STING). В некоторых вариантах осуществления адъювант активирует передачу сигнала TLR в B- и/или T-клетках. В некоторых вариантах осуществления адъювант регулирует адаптивный иммунный ответ.
(1) Агонисты TLR2
[00199] В некоторых вариантах осуществления иммуностимулирующий фрагмент является агонистом TLR2. В некоторых вариантах осуществления иммуностимулирующий фрагмент стимулирует передачу сигнала TLR2. В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой синтетический низкомолекулярный лиганд TLR2. В некоторых вариантах осуществления иммуностимулирующий фрагмент является синтетическим низкомолекулярным агонистом передачи сигнала TLR2.
[00200] В некоторых вариантах осуществления агонист TLR2 представляет собой PAM2CSK4, FSL-1 или PAM3CSK4.
(2) Агонисты TLR7/8
[00201] В некоторых вариантах осуществления иммуностимулирующий фрагмент является агонистом TLR7 и/или TLR8 (т. е. агонистом по меньшей мере одного из TLR7 и TLR8). В некоторых вариантах осуществления иммуностимулирующий фрагмент стимулирует передачу сигнала TLR7 и/или TLR8. В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой синтетический низкомолекулярный лиганд TLR7 и/или TLR8. В некоторых вариантах осуществления иммуностимулирующий фрагмент является синтетическим низкомолекулярным агонистом передачи сигнала TLR7 и/или TLR8.
[00202] В некоторых вариантах осуществления агонист TLR7 и/или TLR8 представляет собой однонитевую РНК (ssRNA). В некоторых вариантах осуществления агонист TLR7 и/или TLR8 представляет собой имидазохинолин. В некоторых вариантах осуществления агонист TLR7 и/или TLR8 представляет собой аналог нуклеозида.
[00203] В некоторых вариантах осуществления агонист TLR7 и/или TLR8 является имидазохинолинаминовым агонистом Toll-подобного рецептора (TLR), таким как 3M-012 (3M Pharmaceuticals). Структура свободного 3M-012 представляет собой:
 . Следует понимать, что иммуностимулирующий фрагмент, такой как 3M-012 или любой фрагмент, обсуждаемый в данном документе, может быть конъюгирован с ферритином посредством замещения соответствующего периферического атома фрагмента (например, водорода) связью с ферритином, описанным в данном документе, например, по атому серы экспонированного на поверхности цистеина или по линкеру, присоединенному к такому атому серы. Таким образом, при конъюгации с ферритином структура иммуностимулирующего фрагмента будет немного отличаться от структуры свободной молекулы.
. Следует понимать, что иммуностимулирующий фрагмент, такой как 3M-012 или любой фрагмент, обсуждаемый в данном документе, может быть конъюгирован с ферритином посредством замещения соответствующего периферического атома фрагмента (например, водорода) связью с ферритином, описанным в данном документе, например, по атому серы экспонированного на поверхности цистеина или по линкеру, присоединенному к такому атому серы. Таким образом, при конъюгации с ферритином структура иммуностимулирующего фрагмента будет немного отличаться от структуры свободной молекулы.
[00204] В некоторых вариантах осуществления агонистом TLR7 и/или TLR8 представляет собой SM 7/8a. Структура свободного SM 7/8a является следующей:
 .
.
[00205] См., например, Nat Biotechnol. 2015 Nov;33(11):1201-10. doi: 10.1038/nbt.3371.
(3) Агонисты TLR9
[00206] В некоторых вариантах осуществления иммуностимулирующий фрагмент является агонистом TLR9. В некоторых вариантах осуществления иммуностимулирующий фрагмент стимулирует передачу сигнала TLR9. В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой синтетический низкомолекулярный лиганд TLR9. В некоторых вариантах осуществления иммуностимулирующий фрагмент является синтетическим низкомолекулярным агонистом передачи сигнала TLR9.
[00207] В некоторых вариантах осуществления агонист TLR9 представляет собой олигодезоксинуклеотид CpG (ODN). В некоторых вариантах осуществления агонист TLR9 представляет собой неметилированный ODN CpG. В некоторых вариантах осуществления ODN CpG включает частичный или полный фосфоротиоатный (PS) остов вместо природного фосфодиэфирного (PO) остова, обнаруженного в обычной ДНК.
[00208] В некоторых вариантах осуществления ODN CpG представляет собой ODN класса B, который включает один или несколько 6-мерных мотивов CpG, содержащих 5'-пурин (Pu)-пиримидин (Py)-C-G-Py-Pu-3'; имеет полностью фосфоротиоатный (т. е. PS-модифицированный) остов, и его длина составляет 18-28 нуклеотидов. В некоторых вариантах осуществления ODN CpG включает последовательность под SEQ ID NO: 210, необязательно содержащую фосфоротиоатные связи в остове.
[00209] В некоторых вариантах осуществления агонист TLR9 включает иммуностимулирующую последовательность (ISS). В некоторых вариантах осуществления агонист TLR9 представляет собой ISS-1018 (Dynavax) (SEQ ID NO: 210).
(4) Агонисты STING
[00210] В некоторых вариантах осуществления иммуностимулирующий фрагмент является агонистом STING (стимулятор белка генов интерферонов, также известный как стимулятор IFN эндоплазматического ретикулума). В некоторых вариантах осуществления иммуностимулирующий фрагмент стимулирует передачу сигнала STING. В некоторых вариантах осуществления иммуностимулирующий фрагмент представляет собой синтетический низкомолекулярный лиганд STING. В некоторых вариантах осуществления иммуностимулирующий фрагмент является синтетическим низкомолекулярным агонистом передачи сигнала STING.
[00211] В некоторых вариантах осуществления агонист STING представляет собой циклический динуклеотид (CDN). См., например, Danilchanka et al., Cell 154:962-970 (2013). Иллюстративные CDN включают cdA, cdG, cAMP-cGMP и 2'-5',3'-5'-cGAMP (структуры см. в Danilchanka et al.). Агонисты STING также включают синтетические агонисты, такие как DMXAA
 .
.
Конъюгированные полипептиды на основе RSV
[00212] В некоторых вариантах осуществления полипептид RSV конъюгирован с экспонированной на поверхности аминокислотой ферритина. В некоторых вариантах осуществления полипептид RSV делает белок ферритин антигенным. В некоторых вариантах осуществления полипептид RSV является антигенным сам по себе, тогда как в некоторых вариантах осуществления полипептид RSV является антигенным благодаря его ассоциации с ферритином. В некоторых вариантах осуществления полипептид RSV является любым из полипептидов F или G RSV, описанных в данном документе.
Конъюгация
[00213] В некоторых вариантах осуществления цистеин, экспонированный на поверхности (например, полученный в результате мутации, описанной в данном документе), или цистеин в пептидном линкере, присоединенном к ферритину (например, к N-концу ферритина), используют для конъюгации иммуностимулирующей функциональной группы, такой как адъювант, или полипептида RSV с ферритином. В некоторых вариантах осуществления линкер конъюгирован с таким цистеином, при этом линкер затем может быть конъюгирован с иммуностимулирующей функциональной группой, такой как адъювант, или полипептидом RSV. В некоторых вариантах осуществления такой цистеин создает химическую основу для реакций конъюгации с присоединением адъюванта, линкера или полипептида RSV. В некоторых вариантах осуществления получают биоконъюгаты, где иммуностимулирующая функциональная группа, такая как адъювант, или полипептид RSV связываются с ферритином после восстановления такого цистеина. В некоторых вариантах осуществления цистеин представляет собой непарный экспонированную на поверхности цистеин, т.е. в котором отсутствует цистеин-партнер в подходящем положении для образования дисульфидной связи. В некоторых вариантах осуществления цистеин представляет собой непарный цистеин, который содержит свободную тиольную боковую цепь.
(1) Типы химии конъюгации
[00214] Химические реакции любого типа могут применяться для конъюгации иммуностимулирующей функциональной группы, такой как адъювант, или полипептида RSV с ферритином, например, путем реакции с экспонированной на поверхности аминокислотой, такой как цистеин или другая аминокислота, такая как Lys, Glu или Asp.
[00215] В некоторых вариантах осуществления конъюгацию осуществляют с применением клик-химии. Применяемый в данном документе термин "клик-химия" относится к реакции между парой функциональных групп, которые быстро и селективно реагируют (т. е. "клик") друг с другом. В некоторых вариантах осуществления клик-химию можно осуществлять в мягких водных условиях. В некоторых вариантах осуществления в реакции клик-химии применяется преимущество цистеина на поверхности ферритина, такого как цистеин, возникающий в результате мутации экспонированной на поверхности аминокислоты, для осуществления клик-химии с применением функциональной группы, которая способна реагировать с цистеином.
[00216] Из уровня техники известны различные реакции, удовлетворяющие критериям клик-химии, и специалист в данной области техники может применять любую из ряда опубликованных методик (см., например, Hein et al., Pharm Res 25(10):2216-2230 (2008)). Можно применять широкий спектр коммерчески доступных реагентов для клик-химии, например, от Sigma Aldrich, Jena Bioscience или Lumiprobe. В некоторых вариантах осуществления конъюгацию осуществляют с применением клик-химии, как описано в примерах ниже.
[00217] В некоторых вариантах осуществления реакция клик-химии происходит после восстановления ферритина.
[00218] В некоторых вариантах осуществления клик-химия может представлять собой 1-стадийную клик-реакцию. В некоторых вариантах осуществления клик-химия может представлять собой 2-стадийную клик-реакцию.
[00219] В некоторых вариантах осуществления реакция(-и) включает(-ют) клик-химию без участия металла. В некоторых вариантах осуществления реакция(-и) включает(-ют) тиол-малеимидный и/или дисульфидный обмен.
Клик-химия без участия металла
[00220] Клик-химию без участия металла можно применять для реакций конъюгации во избежание потенциального окисления белков. Клик-химию без участия металла применяли для образования конъюгатов антител (см van Geel et al., Bioconjugate Chem. 2015, 26, 2233-2242).
[00221] В некоторых вариантах осуществления клик-химию без участия металла применяют в реакциях для присоединения адъюванта к ферритину. В некоторых вариантах осуществления конъюгацию без участия меди применяют в реакциях для присоединения адъюванта к ферритину. В некоторых вариантах осуществления в клик-химии без участия металла применяют бицикло[6.1.0]нонин (BCN). В некоторых вариантах осуществления в клик-химии без участия металла применяют дибензоазациклооктин (DBCO). В некоторых вариантах осуществления BCN или DBCO реагируют с азидной группой.
[00222] DBCO характеризуется высокой специфичностью к азидным группам за счет стимулируемой напряжением клик-реакции в отсутствие катализатора, что приводит к высокому выходу стабильного триазола. В некоторых вариантах осуществления DBCO реагирует с азидом в отсутствие медного катализатора.
[00223] В некоторых вариантах осуществления клик-химию без участия металла применяют в 1-стадийной клик-реакции. В некоторых вариантах осуществления клик-химию без участия металла применяют в 2-стадийной клик-реакции.
Тиол-малеимидный и дисульфидный обмен
[00224] Ферритины, описанные в данном документе, могут включать цистеин, включающий тиол, также известный как сульфгидрил, который доступен для реакции с реагирующими с сулфгидрилом химическими группами (или который можно сделать доступным посредством восстановления). Таким образом, цистеин обеспечивает хемоселективную модификацию с добавлением иммуностимулирующего фрагмента, такого как адъювант, к ферритину. В основных условиях цистеин будет депротонирован с образованием тиолатного нуклеофила, который способен реагировать с мягкими электрофилами, такими как малеимиды и йодацетамиды. Реакция цистеина с малеимидом или йодацетамидом приводит к образованию связи углерод-сера.
[00225] В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа реагирует с экспонированным на поверхности цистеином или цистеином в линкере ферритина. В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа представляет собой галогенацетил, малеимид, азиридин, акрилоил, арилирующее средство, винилсульфон, пиридилдисульфид или TNB-тиол.
[00226] В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа конъюгирует с сульфгидрилом цистеина посредством алкилирования (т. е. образования тиоэфирной связи). В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа конъюгирует с сульфгидрилом цистеина посредством дисульфидного обмена (т. е. образования дисульфидной связи).
[00227] В некоторых вариантах осуществления реакция для конъюгации иммуностимулирующего фрагмента, такого как адъювант, с ферритином, представляет собой реакцию тиола с малеимидом.
[00228] В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа представляет собой малеимид. В некоторых вариантах осуществления реакция малеимида с цистеином приводит к образованию стабильной тиоэфирной связи, например, которая является необратимой. В некоторых вариантах осуществления малеимид не реагирует с тирозинами, гистидинами или метионинами в ферритине. В некоторых вариантах осуществления непрореагировавшие малеимиды гасят по окончанию реакции посредством добавления свободного тиола, например, в избытке.
[00229] В некоторых вариантах осуществления реакция для конъюгации иммуностимулирующего фрагмента, такого как адъювант, с ферритином, представляет собой тиол-дисульфидный обмен, также известный как дисульфидный обмен. В некоторых вариантах осуществления реакция включает образование смешанного дисульфида, содержащего часть исходного дисульфида. В некоторых вариантах осуществления исходный дисульфид представляет собой цистеин, встроенный в ферритин посредством мутации экспонированной на поверхности аминокислоты или добавления N-концевого линкера.
[00230] В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа представляет собой пиридилдитиол. В некоторых вариантах осуществления реагирующая с сульфгидрилом химическая группа представляет собой TNB-тиольную группу.
(2) Конъюгированные линкеры
[00231] В некоторых вариантах осуществления иммуностимулирующая функциональная группа, такая как адъювант, или полипептид RSV присоединяется к ферритину через линкер, который ковалентно связывается с экспонированной на поверхности аминокислотой, такой как цистеин. В некоторых вариантах осуществления линкер включает полиэтиленгликоль, например, PEG-линкер. В некоторых вариантах осуществления полиэтиленгликолевый (например, PEG) линкер усиливает растворимость в воде и эффективность лигирования ферритина, связанного с иммуностимулирующим фрагментом, таким как адъювант. Длина PEG-линкера составляет 2-18 PEG, например, PEG4, PEG5, PEG6, PEG7, PEG8, PEG9, PEG10, PEG11, PEG12, PEG13, PEG14, PEG15, PEG16, PEG17 и PEG18.
[00232] В некоторых вариантах осуществления линкер содержит малеимид. В некоторых вариантах осуществления линкер содержит компоненты иммуностимулирующего фрагмента, представляющего собой (ISM)-линкер-малеимид. В некоторых вариантах осуществления ISM-линкер-малеимид конъюгируют с ферритином в 1-стадийной реакции клик-химии посредством осуществления реакции малеимида с цистеином в ферритине. В некоторых вариантах осуществления ISM из адъювант-линкер-малеимида представляет собой SM7/8a. В некоторых вариантах осуществления линкер из ISM-линкер-малеимида представляет собой PEG4. В некоторых вариантах осуществления ISM-линкер-малеимид представляет собой SM7/8a-PEG4-малеимид.
[00233] В некоторых вариантах осуществления протокол 2-стадийной клик-химии применяют с линкером, включающим реагирующую с сульфгидрилом химическую группу на одном конце и реагирующую с амином группу на другом конце. В таком протоколе 2-стадийной клик-химии реагирующая с сульфгидрилом химическая группа реагирует с цистеином в ферритине, тогда как реагирующая с амином группа реагирует с реагентом, присоединенным к ISM. Таким образом, ISM конъюгируют с ферритином с помощью набора из 2 реагентов для клик-химии.
[00234] В некоторых вариантах осуществления протокола 2-стадийной клик-химии реагирующая с сульфгидрилом химическая группа представляет собой малеимид. В некоторых вариантах осуществления протокола 2-стадийной клик-химии малеимид реагирует с цистеином, встроенным в ферритин посредством мутации экспонированной на поверхности аминокислоты или добавления N-концевого линкера.
[00235] В некоторых вариантах осуществления протокола 2-стадийной клик-химии реагирующая с амином группа представляет собой DBCO. В некоторых вариантах осуществления протокола 2-стадийной клик-химии DBCO реагирует с азидной группой, присоединенной к ISM.
[00236] В некоторых вариантах осуществления применяется малеимид-линкер-DBCO. В некоторых вариантах осуществления малеимид-линкер-DBCO конъюгируют с ферритином после восстановления ферритина. В некоторых вариантах осуществления малеимид-линкер-реагент конъюгируют с ферритином посредством реакции малеимида с цистеином в ферритине на первой стадии. В некоторых вариантах осуществления DBCO применяется для связывания с ISM, присоединенным к азиду. В некоторых вариантах осуществления ISM, связанный с азидом, представляет собой ISS-1018. В некоторых вариантах осуществления адъювант, связанный с азидом, представляет собой 3M-012 или CpG.
[00237] В некоторых вариантах осуществления к ISM добавляют линкер с реакционноспособной группой. В некоторых вариантах осуществления линкер представляет собой PEG4-азидный линкер или PEG4-малеимидный линкер.
[00238] В некоторых вариантах осуществления PEG4-азидный линкер конъюгирован с 3M-012. Иллюстративная структура 3M-012, конъюгированного с PEG4-азидным линкером, представляет собой:
[00239] В некоторых вариантах осуществления PEG4-азидный линкер конъюгирован с SM7/8a. Иллюстративная структура SM7/8a, конъюгированного с PEG4-азидным линкером, представляет собой:
 .
.
[00240] В некоторых вариантах осуществления PEG4-малеимидный линкер конъюгирован с SM7/8a. Иллюстративная структура SM7/8a, конъюгированного с PEG4-малеимидным линкером, представляет собой:
[00241] В некоторых вариантах осуществления азидная группа конъюгирована с ISS-1018. Иллюстративная структура ISS-1018, конъюгированного с NHS-эфир-азидным линкером, представляет собой:
Линкеры
[00242] В некоторых вариантах осуществления линкер отделяет аминокислотную последовательность полипептида RSV от аминокислотной последовательности ферритина. Можно применять любой линкер. В некоторых вариантах осуществления линкер представляет собой пептидный линкер, который может способствовать экспрессии антигенного полипептида на основе последовательности ферритина в виде слитого белка (например, из одной открытой рамки считывания). В некоторых вариантах осуществления линкер представляет собой глицин-сериновый линкер. В некоторых вариантах осуществления глицин-сериновый линкер представляет собой GS, GGGS (SEQ ID NO: 226), 2XGGGS (SEQ ID NO: 227) (т. е., GGGSGGGS (SEQ ID NO: 227)) или 5XGGGS (SEQ ID NO: 228). Линкер может быть расположен N- или C-терминально относительно ферритина.
[00243] В некоторых вариантах осуществления длина линкера составляет 2, 3, 4, 5, 6, 7, 8, 9 или 10 аминокислот. В некоторых вариантах осуществления длина линкера составляет приблизительно 2-4, 2-6, 2-8, 2-10, 2-12 или 2-14 аминокислот. В некоторых вариантах осуществления длина линкера составляет по меньшей мере 15 аминокислот. В некоторых вариантах осуществления длина линкера составляет по меньшей мере 25 аминокислот. В некоторых вариантах осуществления длина линкера составляет по меньшей мере 30 аминокислот. В некоторых вариантах осуществления длина линкера составляет по меньшей мере 35 аминокислот. В некоторых вариантах осуществления длина линкера составляет по меньшей мере 40 аминокислот. В некоторых вариантах осуществления длина линкера составляет менее 60 аминокислот или равна 60 аминокислотам. В некоторых вариантах осуществления длина линкера составляет менее 50 аминокислот или равна 50 аминокислотам. В некоторых вариантах осуществления длина линкера составляет приблизительно 16, 28, 40, 46 или 47 аминокислот. В некоторых вариантах осуществления линкер является гибким. В некоторых вариантах осуществления линкер включает цистеин, например, для применения в качестве сайта для конъюгации иммуностимулирующего фрагмента (например, адъюванта); иллюстративный линкер, включающий цистеин, предусмотрен как SEQ ID NO: 225. В некоторых вариантах осуществления линкер включает последовательность с по меньшей мере 75%, 80%, 85%, 90% или 95% идентичностью с SEQ ID NO: 225 и дополнительно включает цистеин, соответствующий цистеину в SEQ ID NO: 225. В некоторых вариантах осуществления линкер включает по меньшей мере 25 аминокислот (например, 25-60 аминокислот), где цистеин расположен в положении, варьирующем от 8-ой аминокислоты с N-конца до 8-ой аминокислоты с С-конца, или в пределах 10 аминокислот от центрального остатка или связи с линкером.
[00244] В некоторых вариантах осуществления линкер включает аминокислоты глицин (G) и/или серин (S). В некоторых вариантах осуществления линкер включает аминокислоты глицин (G), серин (S), аспарагин (N) и/или аланин (A) и необязательно цистеин, или состоит из них, как обсуждалось выше. В некоторых вариантах осуществления линкер включает аминокислотную последовательность с по меньшей мере 80%, 85%, 90%, 95%, 97%, 98%, 99% или 100% идентичностью с SEQ ID NO: 222. В некоторых вариантах осуществления линкер содержит ggggsggggsggggsg (SEQ ID NO: 220), ggsgsgsnssassgassggasggsggsg (SEQ ID NO: 221), ggsgsassgasasgssngsgsgsgsnssassgassggasggsggsg (SEQ ID NO: 222) или GS. В некоторых вариантах осуществления линкер содержит FR1 (SEQ ID NO: 223) или FR2 (SEQ ID NO: 224).
[00245] В некоторых вариантах осуществления ферритин включает ферритин из H. pylori с аминоконцевым удлинением из ферритина лягушки-быка (который будет называться гибридным ферритином). В некоторых вариантах осуществления этот гибридный ферритин образует мультимеры с сайтами присоединения полипептида RSV, равномерно распределенными по поверхности (см. Kanekiyo 2015). В некоторых вариантах осуществления N-концевые слитые белки с гибридным ферритином обеспечивают презентирование полипептида RSV на поверхности ферритиновой наночастицы. В некоторых вариантах осуществления ферритин включает глутамат в положении, соответствующем положению 13 из SEQ ID NO: 208 (гибридный ферритин, который включает этот глутамат), или в положении 6 в SEQ ID NO: 209 (ферритин из H. pylori дикого типа, в котором в положении 6 находится изолейцин). Считается, что в комбинации с линкером лягушки-быка этот глутамат сохраняет консервативный солевой мостик, обнаруженный в ферритинах человека и лягушки-быка (6R и 14E как в легкой цепи человеческого ферритина, так и в ферритинах лягушки-быка с меньшим количеством субъединиц). См. Kanekiyo et al., Cell 162, 1090-1100 (2015)).
[00246] В некоторых вариантах осуществления полипептид RSV связывается с ферритином с помощью цистеин-тромбин-гистидинового линкера. В некоторых вариантах осуществления этот линкер применяют для непосредственного конъюгирования функциональной группы (например, иммуностимулирующей функциональной группы или полипептида RSV) с ферритином посредством клик-химии. Иллюстративная последовательность, включающая цистеин-тромбин-гистидиновый линкер, представляет собой SEQ ID NO: 218. Клик-химия, подходящая для реакций конъюгации с участием цистеин-тромбин-гистидинового линкера, обсуждается выше.
[00247] В некоторых вариантах осуществления линкер, содержащий цистеин в качестве сайта конъюгации для иммуностимулирующего фрагмента, такого как адъювант, применяется в конструкции, содержащей молекулу ферритина, не имеющую непарного экспонированного на поверхности цистеина, или в конструкции, содержащей молекулу ферритина, содержащую неспаренный экспонированный на поверхности цистеин.
[00248] В некоторых вариантах осуществления конструкция не включает линкер. В некоторых вариантах осуществления конструкция включает один линкер. В некоторых вариантах осуществления конструкция включает два линкера или более двух линкеров.
Композиции, применения и способы вакцинация
[00249] В некоторых вариантах осуществления настоящее изобретения предусматривает способы иммунизации субъекта против инфекции, обусловленной RSV. Кроме того, настоящее изобретения предусматривает способы обеспечения формирования иммунного ответа против RSV у субъекта. В некоторых вариантах осуществления способы в соответствии с настоящим изобретением включают введение субъекту эффективного количества фармацевтической композиции, описанной в данном документе. В некоторых вариантах осуществления способы в соответствии с настоящим изобретением предусматривают введение субъекту эффективного количества антигенного полипептида на основе последовательности RSV, антигенного полипептида на основе последовательности ферритина или наночастицы, описанных в данном документе.
[00250] В некоторых вариантах осуществления предусмотрена композиция, содержащая любое одно или несколько из полипептидов, наночастиц или слитых белков, описанных в данном документе, и фармацевтически приемлемые среду-носитель, адъювант или вспомогательное вещество.
[00251] В некоторых вариантах осуществления полипептид, наночастицу или композицию, описанные в данном документе, вводят субъекту, такому как человек, для иммунизации против инфекции, вызываемой RSV. В некоторых вариантах осуществления полипептид или слитый белок, описанные в данном документе, вводят субъекту, такому как человек, для получения защитного иммунного ответа на инфекцию, обусловленную RSV, в будущем. В некоторых вариантах осуществления предусмотрены любое одно или несколько из полипептидов, наночастиц или композиций, описанных в данном документе, для применения в иммунизации против инфекции, вызываемой RSV. В некоторых вариантах осуществления предусмотрены любое одно или несколько из полипептидов, наночастиц или композиций, описанных в данном документе, для применения в получении защитного иммунного ответа на инфекцию, обусловленную RSV, в будущем. В некоторых вариантах осуществления защитный иммунный ответ обеспечивает снижение частоты возникновения инфекции, обусловленной RSV, пневмонии, бронхита или астмы.
[00252] В некоторых вариантах осуществления композиция содержит полипептид F RSV, описанный в данном документе. В некоторых вариантах осуществления композиция содержит полипептид G RSV, описанный в данном документе. В некоторых вариантах осуществления композиция содержит полипептид F RSV, описанный в данном документе, и полипептид G RSV. В некоторых вариантах осуществления композиция содержит полипептид G RSV, описанный в данном документе, и полипептид F RSV. В некоторых вариантах осуществления композиция содержит полипептид F RSV, описанный в данном документе, и полипептид G RSV, описанный в данном документе.
[00253] В некоторых вариантах осуществления композиция, содержащая полипептид F RSV, описанный в данном документе, вызывает нейтрализующий ответ на RSV, являющийся превосходящим по сравнению с таковым при иммунизации полипептидом F RSV после слияния. В некоторых вариантах осуществления иммунизация полипептидом F RSV, описанным в данном документе (например, полипептидом или наночастицей, содержащими полипептид F RSV, описанный в данном документе), обеспечивает получение более высокого титра антител, направленных против F RSV до слияния, по сравнению с таковым при иммунизации с помощью F RSV после слияния. В некоторых вариантах осуществления иммунизация полипептидом F RSV, описанным в данном документе, обеспечивает получение более низкого титра антител, направленных против F RSV после слияния, по сравнению с таковым при иммунизации с помощью F RSV после слияния. В некоторых вариантах осуществления иммунизация полипептидом F RSV, описанным в данном документе, обеспечивает образование большей доли суммарного антитела, направленного против F RSV до слияния, по сравнению с таковой при иммунизации с помощью F RSV после слияния. Иммунизация антигеном RSV, описанным в данном документе, может обеспечивать лучшую защиту против RSV по сравнению с иммунизацией с помощью F RSV после слияния. Эпитопы, присутствующие в F RSV после слияния и общие с F до слияния, могут представлять собой эпитопы, не подлежащие нейтрализации, и, как предположили, в некоторых случаях обеспечивают образование антител, которые усиливают инфекцию, обусловленную RSV. В некоторых вариантах осуществления композиция, содержащая полипептид F RSV, описанный в данном документе, вызывает более сильный нейтрализующий ответ на RSV, при этом обеспечивая уменьшение уровня антител, направленных против F RSV после слияния. Таким образом, в некоторых вариантах осуществления композиция, содержащая полипептид F RSV, описанный в данном документе, обеспечивает получение более высокого титра нейтрализующих антител к RSV, в ответ на связывание F после слияния.
[00254] В некоторых вариантах осуществления иммунизация антигеном RSV, описанным в данном документе, позволяет получить улучшенный профиль безопасности по сравнению с иммунизацией с помощью F RSV после слияния. Этот улучшенный профиль безопасности может быть связан с блокированием эпитопов, не подлежащих нейтрализации, или эпитопов, подлежащих нейтрализации в небольшой степени, присутствующих на белке в конформации после слияния. Сообщалось о том, что антитела, которые связывают белок в конформации после слияния, могут обеспечивать усиление инфекции, обусловленной RSV, из-за опосредованной антителом вирусной инфекции. Таким образом, антитела против белка в конформации после слияния, которые не нейтрализуют вирус RSV в значительной степени, могут усиливать инфекцию, обусловленную RSV, например антитела, которые распознают белки в конформации как до слияния, так и после слияния.
[00255] В некоторых вариантах осуществления композиция, содержащая полипептид G RSV, описанный в данном документе, вызывает нейтрализующий ответ на RSV.
[00256] В некоторых вариантах осуществления композиция, содержащая полипептиды F RSV и G RSV, описанные в данном документе, вызывает нейтрализующий ответ на RSV. В некоторых вариантах осуществления композиция, содержащая полипептиды F RSV и G RSV, описанные в данном документе, обеспечивает улучшенную защиту от RSV, например более высокий титр нейтрализующих антител, чем композиция, которая не содержит оба антигена.
1. Субъекты
[00257] В некоторых вариантах осуществления субъект представляет собой млекопитающее. В некоторых вариантах осуществления субъект является человеком.
[00258] В некоторых вариантах осуществления субъект является взрослым (в возрасте 18 лет или старше). В некоторых вариантах осуществления субъектом является ребенок или подросток (в возрасте младше 18 лет). В некоторых вариантах осуществления субъектом является пожилой человек (в возрасте старше 60 лет). В некоторых вариантах осуществления субъект является взрослым не пожилого возраста (в возрасте от 18 лет или старше до 60 лет или старше).
[00259] В некоторых вариантах осуществления субъекту осуществляют более одного введения композиции. В некоторых вариантах осуществления бустерное введение улучшает иммунный ответ.
[00260] В некоторых вариантах осуществления любые один или несколько из антигенных полипептидов или композиций, описанных в данном документе, предназначены для применения у млекопитающего, такого как примат (например, отличный от человека примат, такой как обезьяна (например, макак, такой как макак-резус или яванский макак) или человекообразная обезьяна), грызун (например, мышь или крыса) или одомашненное млекопитающее (например, собака, кролик, кот, лошадь, овца, корова, коза, верблюд или осел). В некоторых вариантах осуществления любое одно или несколько из антигенных полипептидов или композиций, описанных в данном документе, предназначены для применения для птиц, таких как домашняя птица (например, курица, индейка, утка, гусь, цесарка или лебедь).
2. Адъюванты
[00261] Как описано в данном документе, адъюванты могут быть конъюгированы с ферритином посредством экспонированной на поверхности аминокислоты, например цистеина. Неконъюгированный адъювант также можно вводить субъекту вместе с антигенными полипептидами на основе последовательности ферритина, описанными в данном документе. В некоторых вариантах осуществления введение адъюванта вместе с антигенным полипептидом на основе последовательности ферритина обеспечивает продуцирование более высокого титра антител против полипептида RSV у субъекта по сравнению с введением полипептида RSV отдельно или антигенного полипептида на основе ферритина отдельно без адъюванта. Адъювант может способствовать более раннему, более сильному или более стойкому иммунному ответу на антигенный полипептид.
[00262] В некоторых вариантах осуществления композиция содержит один адъювант. В некоторых вариантах осуществления композиция содержит более одного адъюванта. В некоторых вариантах осуществления композиция не содержит адъювант.
[00263] В некоторых вариантах осуществления адъювант включает алюминий. В некоторых вариантах осуществления адъювант представляет собой фосфат алюминия. В некоторых вариантах осуществления адъювант представляет собой алюминиевые квасцы (Alyhydrogel '85 2%; Brenntag, № по каталогу 21645-51-2).
[00264] В некоторых вариантах осуществления адъювант представляет собой органический адъювант. В некоторых вариантах осуществления адъювант представляет собой адъювант на основе масла. В некоторых вариантах осуществления адъювант включает наноэмульсию типа масло-в-воде.
[00265] В некоторых вариантах осуществления адъювант включает сквален. В некоторых вариантах осуществления адъювант, содержащий сквален, представляет собой Ribi (Sigma adjuvant system, № по каталогу S6322-1vl), Addavax™ MF59, AS03 или AF03 (см. US9703095). В некоторых вариантах осуществления адъювант, включающий сквален, представляет собой наноэмульсию.
[00266] В некоторых вариантах осуществления адъювант включает полимер полиакриловой кислоты (PAA). В некоторых вариантах осуществления адъювант, включающий PAA, представляет собой SPA09 (см. WO 2017218819).
[00267] В некоторых вариантах осуществления адъювант включает неметаболизируемые масла. В некоторых вариантах осуществления адъювант представляет собой неполный адъювант Фрейнда (IFA).
[00268] В некоторых вариантах осуществления адъювант включает неметаболизируемые масла и убитую Mycobacterium tuberculosis. В некоторых вариантах осуществления адъювант представляет собой полный адъювант Фрейнда (CFA).
[00269] В некоторых вариантах осуществления адъювант представляет собой липополисахарид. В некоторых вариантах осуществления адъювант представляет собой монофосфорил A (MPL или MPLA).
3. Фармацевтические композиции
[00270] В различных вариантах осуществления предусмотрена фармацевтическая композиция, содержащая антигенный полипептид на основе последовательности ферритина, описанный в данном документе, и/или родственные объекты. В некоторых вариантах осуществления фармацевтическая композиция представляет собой иммуногенную композицию (например, вакцину), способную вызывать иммунный ответ, такой как защитный иммунный ответ против патогена.
[00271] Например, в некоторых вариантах осуществления фармацевтические композиции могут содержать один или несколько из следующего: (1) антигенный белок на основе последовательности ферритина, содержащий (i) мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином, и (ii) полипептид RSV; (2) антигенный белок на основе последовательности ферритина, содержащий (i) мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином, и иммуностимулирующую функциональную группу, связанную с цистеином; и (ii) полипептид RSV; (3) антигенный белок на основе последовательности ферритина, содержащий (i) экспонированный на поверхности цистеин, (ii) пептидный линкер, N-концевой по отношению к белку ферритина, и (iii) полипептид RSV, N-концевой по отношению к пептидному линкеру; (4) антигенный белок на основе последовательности ферритина, содержащий (i) мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином, и иммуностимулирующую функциональную группу, связанную с цистеином, (ii) мутацию, обеспечивающую замещение внутреннего цистеина в положении 31 ферритина из H. pylori, или мутацию внутреннего цистеина в положении, аналогичном положению 31 ферритина из организма, отличного от H. pylori, согласно определению путем попарного или структурного выравнивания, аминокислотой, отличной от цистеина, (iii) мутацию, обеспечивающую замещение экспонированного на поверхности остатка аспарагина аминокислотой, отличной от аспарагина, и (iv) полипептид RSV; или (5) ферритиновую частицу, содержащую любой из вышеупомянутых белков ферритина. В некоторых вариантах осуществления фармацевтическая композиция содержит антигенный полипептид на основе последовательности RSV, содержащий полипептид F RSV, где эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния, является заблокированным, и/или антигенный полипептид на основе последовательности RSV, содержащий полипептид F RSV, где полипептид F RSV содержит аминокислотные остатки 62-69 и 196-209 из SEQ ID NO: 26 и аспарагин, соответствующий положению 328, 348 или 507 из SEQ ID NO: 26, где необязательно антигенный полипептид на основе последовательности RSV, дополнительно содержит ферритин.
[00272] В некоторых вариантах осуществления в настоящем изобретении предусмотрены фармацевтические композиции, содержащие антитела или другие средства, родственные антигенным полипептидам, описанным в данном документе. В одном варианте осуществления фармацевтическая композиция содержит антитела, которые связываются с и/или конкурируют с антигенным полипептидом, описанным в данном документе. В качестве альтернативы, антитела могут распознавать вирусные частицы, содержащие компонент полипептида RSV антигенного полипептида, описанного в данном документе.
[00273] В некоторых вариантах осуществления фармацевтические композиции, описанные в данном документе, вводят отдельно или в комбинации с одним или несколькими средствами для усиления иммунного ответа, например, с адъювантом, описанным выше. В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит адъювант, описанный выше.
[00274] В некоторых вариантах осуществления фармацевтическая композиция дополнительно содержит фармацевтически приемлемый носитель или вспомогательное вещество. Применяемый в данном документе термин "носитель" относится к разбавителю, адъюванту, вспомогательному веществу или среде-носителю, с которыми вводят фармацевтическую композицию. В иллюстративных вариантах осуществления носители могут включать стерильные жидкости, такие как, например, вода и масла, в том числе масла нефтяного, животного, растительного или синтетического происхождения, такие как, например, арахисовое масло, соевое масло, минеральное масло, кунжутное масло и т. п. В некоторых вариантах осуществления носители представляют собой или включают один или несколько твердых компонентов. Фармацевтически приемлемые носители также могут включать без ограничения солевой раствор, буферный солевой раствор, декстрозу, глицерин, этанол и их комбинации. Применяемый в данном документе термин вспомогательное вещество относится к любому нетерапевтическому средству, которое можно включить в фармацевтическую композицию, например, для обеспечения или содействия достижению необходимой консистенции или стабилизирующего эффекта. Подходящие фармацевтические вспомогательные вещества включают без ограничения крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропилен, гликоль, воду, этанол и т. п. В различных вариантах осуществления фармацевтическая композиция является стерильной.
[00275] В некоторых вариантах осуществления фармацевтическая композиция содержит незначительные количества смачивающих или эмульгирующих средств или буферных средств для доведения pH. В некоторых вариантах осуществления фармацевтические композиции могут содержать любую из множества добавок, таких как стабилизаторы, буферы или консерванты. Кроме того, можно включать вспомогательные, стабилизирующие, загущающие, смазывающие и окрашивающие средства.
[00276] В различных вариантах осуществления фармацевтическая композиция может быть составлена для соответствия любому требуемому способу введения. Например, фармацевтическая композиция может иметь форму растворов, суспензий, эмульсии, капель, таблеток, пилюль, пеллет, капсул, капсул, содержащих жидкости, желатиновых капсул, порошков, составов с замедленным высвобождением, суппозиториев, эмульсий, аэрозолей, спреев, суспензий, лиофилизированного порошка, замороженной суспензии, высушенного порошка или любую другую форму, подходящую для применения. Общие соображения по составлению и изготовлению фармацевтических средств можно найти, например, в Remington's Pharmaceutical Sciences, 19th ed., Mack Publishing Co., Easton, PA, 1995; включенном в данный документ посредством ссылки.
[00277] Фармацевтическую композицию можно вводить любым путем введения. Пути введения включают, например, пероральный, интрадермальный, внутримышечный, интраперитонеальный, внутривенный, подкожный, интраназальный, чресслизистый, эпидуральный, сублингвальный, интраназальный, интрацеребральный, интравагинальный, трансдермальный, ректальный, посредством интратрахеальной инстилляции, бронхиальной инстилляции, ингаляции, или местный. Введение может быть локальным или системным. В некоторых вариантах осуществления введение осуществляют перорально. В другом варианте осуществления введение осуществляют посредством парентеральной инъекции. В некоторых случаях введение приводит к высвобождению антигенного полипептида на основе последовательности ферритина, описанного в данном документе, в кровоток. Способ введения может быть оставлен на усмотрение практикующего специалиста.
[00278] В некоторых вариантах осуществления фармацевтическая композиция подходит для парентерального введения (например, внутривенного, внутримышечного, интраперитонеального и подкожного). Такие композиции могут быть составлены, например, в виде растворов, суспензий, дисперсий, эмульсий и т. п. Они также могут быть изготовлены в форме стерильных твердых композиций (например, лиофилизированной композиции), которые могут быть растворены или суспендированы в стерильной инъекционной среде непосредственно перед применением. Например, парентеральное введение можно осуществлять путем инъекции. В таких вариантах осуществления инъекционные препараты получают в общепринятых формах, т. е. либо в виде жидких растворов или суспензий, твердых форм, подходящих для растворения или суспендирования в жидкости перед инъекцией, либо в виде эмульсий. В некоторых вариантах осуществления инъекционные растворы и суспензии получают из стерильных порошков, лиофилизированных порошков или гранул.
[00279] В дополнительном варианте осуществления фармацевтическую композицию составляют для доставки посредством ингаляции (например, для прямой доставки в легкие и дыхательную систему). Например, композиция может иметь форму назального спрея или любого другого известного аэрозольного состава. В некоторых вариантах осуществления препараты для ингаляционной или аэрозольной доставки содержат множество частиц. В некоторых вариантах осуществления такие препараты могут характеризоваться средним размером частиц, составляющим приблизительно 1, приблизительно 2, приблизительно 3, приблизительно 4, приблизительно 5, приблизительно 6, приблизительно 7, приблизительно 8, приблизительно 9, приблизительно 10, приблизительно 11, приблизительно 12 или приблизительно 13 микрон. В некоторых вариантах осуществления препараты для ингаляционной или аэрозольной доставки составлены в виде сухого порошка. В некоторых вариантах осуществления препараты для ингаляционной или аэрозольной доставки составлены в виде влажного порошка, например, посредством включения смачивающего средства. В некоторых вариантах осуществления смачивающее средство выбрано из группы, состоящей из воды, солевого раствора или другой жидкости с физиологическим значением pH.
[00280] В некоторых вариантах осуществления фармацевтическую композицию в соответствии с настоящим изобретением вводят в виде капель в носовую или ротовую полость. В некоторых вариантах осуществления доза может содержать множество капель (например, 1-100, 1-50, 1-20, 1-10, 1-5 и т. д.)
[00281] Фармацевтическую композицию по настоящему изобретению можно вводить в любой дозе, подходящей для достижения требуемого результата. В некоторых вариантах осуществления требуемым результатом является индуцирование длительного адаптивного иммунного ответа против источника полипептида RSV, присутствующего в антигенном полипептиде на основе последовательности ферритина, присутствующем в композиции. В некоторых вариантах осуществления требуемый результат представляет собой снижение интенсивности, тяжести, частоты и/или задержки проявления одного или нескольких симптомов инфекции. В некоторых вариантах осуществления требуемый результат представляет собой ингибирование или предупреждение инфекции. Необходимая доза будет варьироваться от субъекта к субъекту в зависимости от вида, возраста, веса и общего состояния субъекта, тяжести подлежащей предупреждению или лечению инфекции, конкретной применяемой композиции и способа ее введения.
[00282] В некоторых вариантах осуществления фармацевтические композиции в соответствии с настоящим изобретением вводят в виде одной или нескольких доз. В некоторых вариантах осуществления фармацевтические композиции вводят несколькими дозами, вводимыми в разные дни (например, прайм-буст стратегии вакцинации). В некоторых вариантах осуществления фармацевтическую композицию вводят как часть бустерного режима.
[00283] В различных вариантах осуществления фармацевтическую композицию вводят совместно с одним или несколькими дополнительными терапевтическими средствами. Совместное введение не требует одновременного введения терапевтических средств, если время их введения таково, что фармакологические активности дополнительного терапевтического средства и активного(-ых) ингредиента(-ов) в фармацевтической композиции перекрываются во времени, тем самым оказывая комбинированный терапевтический эффект. Как правило, каждое средство будут вводить в дозе и в соответствии с графиком, установленными для данного средства.
4. Нуклеиновая кислота/мРНК
[00284] Также представлена нуклеиновая кислота, кодирующая антигенный полипептид, описанный в данном документе. В некоторых вариантах осуществления нуклеиновая кислота представляет собой мРНК. Любая нуклеиновая кислота, способная подвергаться трансляции, в результате которой образуется полипептид, считается мРНК для целей настоящего изобретения.
5. Наборы
[00285] В данном документе также представлены наборы, содержащие один или несколько антигенных полипептидов, нуклеиновых кислот, антигенных ферритиновых частиц, антигенных лумазинсинтазных частиц, композиций или фармацевтических композиций, описанных в данном документе. В некоторых вариантах осуществления набор дополнительно содержит один или несколько из растворителя, раствора, буфера, инструкций или высушивающего средства.
Таблица 1 (таблица последовательностей): Описание последовательностей
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTNNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKNGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPTYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGNVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFSDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFINKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
PETVIEFQQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFLQKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFLVKNNRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNLRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
IETVIEFQQKNFRLLEITREFSVNAGVTTPVSTYMLTNSELLSLINDMPITNDQKKLMSNNVQIVRQQSYSIMCIIKEEVLAYVVQLPLYGVIDTPCWKLHTSPLCTTNTKEGSNICLTRTDRGWYCDNAGSVSFFPQAETCKVQSNRVFCDTMNSRTLPSEVNLCNVDIFNPKYDCKIMTSKTDVSSSVITSLGAIVSCYGKTKCTASNKNRGIIKTFSNGCDYVSNKGVDTVSVGNTLYYVNKQEGKSLYVKGEPIINFYDPLVFPSDEFDASISQVNEKINQSLAFIRKSDELLSGSGSESQVRQQFSKDIEKLLNEQVNKEMQSSNLYMSMSSWSYTHSLDGAGLFLFDHAAEEYEHAKKLIIFLNENNVPVQLTSISAPEHKFEGLTQIFQKAYEHEQHISESINNIVDHAIKCKDHATFNFLQWYVAEQHEEEVLFKDILDKIELIGNENHGLYLADQYVKGIAKSRKS
STTTIIQDATNQIKNTTPTYLTQNPQLGISPSNPSDITSLITTILDSTTPGVKSTLQSTTVGTKNTTTTQ
AQPNKPTTKQRQNKPPSKPNNDFHFEVFNFVPCSICSNNPTCWAICKRIPNKKPGKRTTTKPTKKPTPKT
TKKGPKPQTTKSKEAPTTKPTEEPTINTTKTNIITTLLTSNTTRNPELTSQMETFHSTSSEGNPSPSQVS
ITSEYPSQPSSPPNTPR
IFDKKLLGIDV
IYDRNGRVELDEIPKPPKEWESPLKAFEAAYEHEKFISKSIYELAALAEEEKDYSTRAFL
EWFINEQVEEEASVKKILDKLKFAKDSPQILFMLDKELSARAPKLPGLLMQGGE
[00286] Это описание и иллюстративные варианты осуществления не должны рассматриваться как ограничивающие. Для целей настоящего описания и прилагаемой формулы изобретения, если не указано иное, все числа, выражающие количества, процентные доли или пропорциональные доли и другие числовые значения, применяемые в описании и формуле изобретения, следует понимать как модифицируемые во всех случаях термином "приблизительно" в той степени, в которой они еще не были модифицированы. "Приблизительно" указывает на степень вариации, которая не оказывает существенного влияния на свойства описанного заявленного объекта, например, в пределах 10%, 5%, 2% или 1%. Соответственно, если не указано иное, числовые параметры, изложенные в следующем описании и прилагаемой формуле изобретения, являются примерными значениями, которые могут варьироваться в зависимости от требуемых свойств, которые должны быть получены. В крайнем случае, и не как попытка ограничить применение доктрины эквивалентов к объему формулы изобретения, каждый числовой параметр должен по меньшей мере истолковываться с учетом количества сообщаемых значащих цифр и путем применения обычных способов округления.
[00287] Следует отметить, что, как применяется в данном описании и прилагаемой формуле изобретения, форма единственного числа, а также любое применение любого слова в единственном числе включают объекты во множественном числе, если только однозначно и недвусмысленно не ограничены одним объектом. Применяемый в данном документе термин "включать" и его грамматические варианты не предназначены для ограничения, так что перечисление элементов в списке не исключает другие подобные элементы, которые могут быть заменены или добавлены к перечисленным элементам.
ПРИМЕРЫ
[00288] Следующие примеры предусмотрены для иллюстрации определенных раскрытых вариантов осуществления и не должны рассматриваться как ограничивающие объем этого раскрытия каким-либо образом.
1. Дизайн и характеристика модификаций полипептидов F RSV
[00289] Подобно другим белкам F парамиксовируса, F RSV экспрессируется в виде белка-предшественника с N-концевым сигнальным пептидом и С-концевой трансмембранной областью, которая прикрепляет белок к вирусной поверхности. RSV-F подвергается внутриклеточному расщеплению протеазой фурин с высвобождением гидрофобного слитого пептида («FP» на фиг. 1A), роль которого заключается в прикреплении к клетке-мишени во время инфицирования. К слитому пептиду примыкает область гептадных повторов А (HRA), тогда как область гептадных повторов В (HRB) прилегает к трансмембранному домену.
[00290] Кристаллические структуры тримеров эктодомена F RSV в их конформациях до слияния и после слияния демонстрируют, что области HRA и HRB претерпевают значительные перегруппировки для обеспечения управления событием клеточного слияния (фиг. 1B) (см. Swanson, K.A., et al., Proc Natl Acad Sci U S A 108(23): p. 9619-24 (2011); McLellan, J.S., et al., Science 342(6158):592-598 (2013); McLellan, J.S., et al., J Virol 85(15):7788-96 (2011); и McLellan, J.S., et al., Science 342(6158): p. 592-8 (2013)). В конформации до слияния, область гептадных повторов A (HRA) связана с глобулярной концевой группой, а кончик слитого пептида в основном погружен в центр белка. Конформация до слияния содержит ряд спиралей и предусматривает определенные контакты между протомерами с образованием тримера до слияния.
[00291] Разрабатывали ряд аминокислотных замен для межпротомерной стабилизации. Иллюстративные замены включают V207L; N228F; I217V и E218F; I221L и E222M; или Q224A и Q225L. При нумерации всех аминокислотных последовательностей F RSV в примерах используется нумерация SEQ ID NO: 26.
[00292] Разрабатывали аминокислотные замены для стабилизации спирали. Предполагается, что как таковые эти замены стабилизируют спиральный домен F RSV. Иллюстративные замены включают N216P или I217P.
[00293] Разрабатывали аминокислотные замены для внутрипротомерной стабилизации. Иллюстративные замены включают V220I; или A74L и Q81L.
[00294] Разрабатывали аминокислотные замены для кэппирования спирали. Иллюстративные замены включают N216P или I217P.
[00295] Разрабатывали аминокислотные замены для уменьшения агрегации. Иллюстративные замены включают V192E и L61Q.
[00296] Другие аминокислотные замены разрабатывали для заполнения полости путем введения гидрофобных аминокислот, таких как N228F.
[00297] Аминокислотные замещения E328N, S348N и R507N были разработаны для добавления сайтов гликозилирования путем замещения остатков, отличных от остатков аспарагина, аспарагином. Было высказано предположение, что добавление ненативных гликанов можно применять для блокирования эпитопов, которые экспонируются в F RSV после слияния (фиг. 1B), на поверхности белка F до слияния.
[00298] Представляющие интерес конструкции F RSV создавали в виде одноцепочечных (scF) белков, слитых с гибридным ферритином, содержащим N-концевой ферритиновый линкер лягушки-быка и ферритин из H. pylori (pFerr) (фиг. 1A). Ферритин содержал экспонированный на поверхности цистеин, полученный в результате мутации K79C или S111C (нумерация последовательности ферритина соответствует SEQ ID NO: 208).
[00299] Создание различных последовательностей, кодирующих Pre-F-NP RSV и ферритин, выполняли с применением стандартных методов клонирования, известных в данной области техники. В общих чертах, ДНК для конструкций F RSV с описанными заменами синтезировали и клонировали в вектор экспрессии млекопитающих с помощью Genscript. F RSV DS-CAV1 и тримеры F после слияния получали аналогично ранее опубликованным протоколам (см. McLellan, J.S., et al., Science 342(6158):592-598 (2013)). В конструкции DS-CAV1 сохраняется С-концевой домен тримеризации F RSV и он объединяется с гидрофобными заменами, обеспечивающими заполнение полости. F RSV DS-CAV1 содержит дисульфидную мутацию (DS) S155C-S290C и S190F-V207L (CAV1).
[00300] Векторы, кодирующие наночастицы на основе F RSV-ферритина, голый ферритин (т. е. не соединенный с F RSV) и тримеры F RSV, трансфицировали в клетки 293EXPI и собирали продукты экспрессии из кондиционированной среды через 4 дня. Наночастицы F RSV очищали серией очисток на анионной колонке Q (GE Healthcare, № по каталогу 17-1154-01) при pH 7,0 и 8,5 с последующей очисткой с помощью Superose 6 SEC в PBS (GE Healthcare, № по каталогу 90-1000-42) с применением общепринятых способов хроматографии. Тримеры DS-CAV1 до слияния и тримеры после слияния хранили при -80°C, а наночастицы F RSV хранили при 4°C.
[00301] Для определения конформации наночастиц F RSV проводили электронную микроскопию. Препараты наночастиц F RSV (30 мкг/мл в 25 мМ Tris, 50 мМ NaCl) абсорбировали на сетке с углеродным покрытием с размером пор 400 меш (Electron Microscopy Sciences) и окрашивали 0,75% уранилформиатом. Для анализа образцов использовался микроскоп JEOL 1200EX, работающий при 80 кВ. Микрофотографии были получены при увеличении 65000× и средние значения класса 2D были получены с применением общепринятых способов в области техники компанией, специализирующейся на EM, Nanoimaging Services, INC (Сан-Диего, Калифорния) (фиг. 1D).
[00302] Экспрессию и секрецию полипептидов, содержащих эти полипептиды F RSV и ферритин (SEQ ID NO: 1-8 и 11-15) транзиентно трансфицированными клетками 293 EXPI (Invitrogen), оценивали с помощью вестерн-блоттинга с антителами против F RSV. Во всех вестерн-блоттингах с антителами против F RSV использовали антитело D25, специфическое в отношении сайта 0, описанное в McLellan et al., Science 340(6136):1113-1117 (2013) и в патенте США № 8562996. Как показано на фигуре 2, многие конструкции были успешно экспрессированы и секретированы.
[00303] Полипептид RF8085 (SEQ ID NO: 1) представляет собой одноцепочечный мутантный вариант опубликованного DS-CAV1 F RSV (см. McLellan, J.S., et al., Science 342(6158):592-598 (2013)), слитый на N-конце с ферритиновой наночастицей. Эта конструкция содержит мутантный вариант (DS) F RSV с двумя мутациями, S155C-S290C, в котором сохраняется антигенный сайт 0.
[00304] Полипептид RF8106 (SEQ ID NO: 9) содержит замену I217P вместо 2 остатков цистеина, замененных в DS-CAV1. Как показано на фигуре 3, конструкция RF8106 характеризовалась значительно лучшей экспрессией в транзиентно трансфицированных клетках 293 EXPI, согласно оценке в кондиционированной среде через 4 дня с помощью вестерн-блоттинга с антителом против F RSV.
[00305] В результате эксклюзионной хроматографии (SEC) RF8106 показано элюирование основного пика при времени удерживания, соответствующем собранной частице ферритина, слитой с антигеном RSV, соответствующей наночастице слитого белка (Pre-F-NP, фигура 4). Анализ методом динамического рассеяния света (DLS) RF8106 проводили в восстановленном (фигура 5B) и невосстановленном состояниях (фигура 5A). Восстановление предусматривало обработку с помощью 2 мМ TCEP. RF8106 имел радиус приблизительно 15 нм, что соответствует включению в наночастицу (24-мерную) как в восстановленном, так и в невосстановленном состояниях. Стабильность слитого белка в присутствии восстанавливающих средств облегчает конъюгацию адъювантов со слитыми белками с образованием самоадъювантных наночастиц, как описано ниже.
[00306] Затем оценивали конъюгацию адъюванта со слитым белком на основе полипептида F RSV и ферритина (Pre-F-NP). Выяснили, что свободный поверхностный цистеин на ферритине может быть использован для присоединения дополнительной функциональной группы к слитому белку scF-pFerr. На фигуре 6 показана успешная конъюгация олигодезоксинуклеотида CpG (ODN) с последовательностью T*G*A*C*T*G*T*G*A*A*C*G*T*T*C*G*A*G*A*T*G*A (SEQ ID NO: 30; звездочки указывают фосфоротиоатные связи) с RF8106, о чем свидетельствует увеличение молекулярной массы, согласно оценке на геле SDS-PAGE, окрашенном кумасси.
[00307] Эффект добавления сайтов гликозилирования с применением замен E328N, S348N и R507N (RF8117, SEQ ID NO: 17) оценивали в клетках 293EXPI, транзиентно трансфицированных этой конструкцией в виде белка, слитого с ферритином (т. е. в виде конструкции Pre-F-NP). RF8117 также содержит замену I217P, как в RF8113. Как показано на фигуре 7, увеличенный уровень экспрессии отмечали для RF8117 по сравнению как с контрольной конструкцией RF8085, так и с конструкцией RF8113 (SEQ ID NO: 16, которая содержит замену на пролин I217P, но не замены E328N, S348N и R507N). RF8113 аналогичен RF8106, описанному ранее, за исключением того, что сконструированный цистеин ферритина находится на остатке ферритина K79C, а не на S111C. Конструкция RF8117 также показала увеличение молекулярной массы по сравнению с RF8113 и RF8117, что указывает на успешное добавление гликанов.
[00308] На фигуре 8 кратко описываются модификации наночастиц F RSV, которые обеспечивали повышение протеолитической стабильности Pre-F-NP. Исходной конструкцией служил RF8117 (выше). Когда более раннюю конструкцию RF8085 клонировали в вектор для CHO в виде RF8090 и трансфицировали в клетки CHO, наблюдали, что некоторое количество материала было вырезано между F и функциональной группой, представляющей собой ферритин. Предполагалось, что остатки аргинина или лизина в области HRB или линкере между F и функциональной группой, представляющей собой ферритин, разрезаются трипсиноподобными протеазами. Мутации остатков лизина и аргинина в пределах области тестировали на предмет экспрессии в клетках 293. На фигуре 8 мутации K498L и R508Q (в RF8122, SEQ ID NO: 18) идентифицированы как не оказывающие влияния или не обеспечивающие увеличения уровня экспрессии по сравнению с RF8117. Эти мутации с R523Q объединяли с упомянутыми в данном документе мутациями RF8117 с образованием конструкции RF8140 (SEQ ID NO: 23).
[00309] Более значительное улучшение экспрессии (приблизительно в 5 раз) отмечали при комбинации одноцепочечных и пролиновых (I217P) модификаций при экспрессии в клетках 293 (иллюстративные конструкции с этими заменами включают RF8106 (SEQ ID NO: 9) и RF8113 (SEQ ID NO: 16)) с дальнейшим улучшением экспрессии и растворимости в результате модификаций, обеспечивающих добавление сайта гликозилирования, в F RSV (иллюстративные конструкции RF8117 (SEQ ID NO: 17) и RF8140 (SEQ ID NO: 23). Во всех этих конструкциях области слитого пептида и пептида p27 (аминокислоты 98-144 SEQ ID NO: 26) замещены последовательностью GSGNVGL (SEQ ID NO: 31). Однако при экспрессии RF8090 в линиях клеток-продуцентов СНО наблюдали дополнительные полосы F RSV в вестерн-блоттинге, что позволяет предположить, что конструкция была чувствительна к протеолизу, возможно, к расщеплению, подобному расщеплению трипсином, по остатку аргинина или лизина.
[00310] Также исследовали потенциальную роль чувствительности к протеазам. Выполняли замену остатков K (нокаут или KO) в области HRB и в линкере между функциональной группой, представляющей собой F, и функциональной группой, представляющей собой ферритин, поскольку они были предсказаны в качестве возможных сайтов опосредованного К расщепления, первоначально наблюдаемого в линии клеток-продуцентов CHO. Как показано на фиг. 9A и 9B, как RF8117, так и RF8140 экспрессируются на высоких уровнях по сравнению с RF8090 в линии продуцирующих клеток CHO, согласно измерению с помощью вестерн-блоттинга с D25 или анализа Octet с D25 и AM14.
[00311] Эти данные показывают, что одноцепочечные конструкции и модификации аминокислот для кэппирования спирали, увеличения уровня гликозилирования и элиминации остатков лизина или остатков аргинина, чувствительных к расщеплению протеазой, могут обеспечивать улучшение в отношении экспрессии полипептидов F RSV, включая антигены Pre-F-NP RSV.
2. Характеристика слитых белков на основе F RSV и ферритиновых наночастиц
[00312] Перед исследованиями на животных концентрацию наночастиц DS-CAV1 и F RSV анализировали путем связывания с применением Octet. Связывание антигенов до слияния с антителами D25 и AM14, специфическими в отношении конформации до слияния, также измеряли с помощью устройства Octet от FortéBio. Все анализы проводили в PBS при 30°C. Антитела загружали на наконечники сенсора с белком A (ProA) (FortéBio, № 18-5013) на 400 секунд для обеспечения уровня захвата, близкого к состоянию насыщения. Наконечники биосенсора затем уравновешивали в течение 90 секунд в PBS с последующим связыванием антигена при известных концентрациях в PBS в течение 300 секунд с последующей диссоциацией антигена в PBS. Анализ данных и аппроксимация кривой, с предположением взаимодействия 1:1, проводили с помощью программного обеспечения Octet Data Analysis HT10.0 с применением кривой внешнего стандарта для связывания очищенного Pre-F-NP при известной концентрации. Результат иллюстративного анализа для определения концентрации Pre-F-NP в кондиционированной среде СНО показан на фиг. 9В.
3. Характеристика иммунного ответа на полипептиды F RSV in vivo
[00313] Для оценивания in vivo ответа на антигены RSV у мышей, самок мышей BALBc внутримышечно иммунизировали антигенами RSV при определенных дозах на неделе 0, 3 и 6. Если не указано иное, антигены RSV (например, в экспериментах на фигурах 10A-B и 12A-B, среди прочих) применяли с адъювантом AF03 с применением стратегии смешивания перед использованием. То есть 50 мкл раствора соответствующего белка смешивали с 50 мкл адъюванта AF03 от Sanofi (эмульсия на основе сквалена; см. Klucker et al., J Pharm Sci. 2012 Dec;101(12):4490-500) непосредственно перед инъекцией 50 мкл в каждую заднюю лапу. Для групп без применения адъюванта антигены смешивали, как указано выше, но AF03 замещали эквивалентным объемом PBS. Для антигенов, смешанных с SPA09 или алюминиевыми квасцами, описанную выше процедуру выполняли с замещением AF03 эквивалентным объемом SPA09 или алюминиевые квасцы соответственно. Никаких побочных эффектов иммунизации не наблюдали ни для одного состава. Кровь собирали за 1 день до первой иммунизации и по меньшей мере через 2 недели после каждой инъекции (т. е. недели 2, 5 и 8). Если не указано иное, показаны данные для 2 недель после третьей инъекции (неделя 8, также обозначаемая как 2wp3). Обычно анализировали образцы сыворотки крови от животных до иммунизации (обозначенных как не подвергавшиеся воздействию) через две недели после второй инъекции (после 2 или 2wp2) или через две недели после третьей инъекции (после 3-ей или 2wp3).
[00314] Для анализа нейтрализации на клетках Vero сыворотку крови инактивировали нагреванием в течение 30 минут при 56°C. Выполняли серию четырехкратных серийных разведений инактивированной сыворотки крови в среде Игла, модифицированной по Дульбекко (DMEM), дополненной 2% фетальной бычьей сыворотки (FBS), 1% GlutaMAX и 1% антибиотика-антимитотического средства. Исходные растворы вируса RSV объединяли 1:1 с разведениями сыворотки крови и инкубировали в течение 1,5 часа при 37°C. Затем смесь вируса с сывороткой крови добавляли в 24-луночные планшеты, содержащие конфлюентные монослои клеток Vero в количестве 100 мкл на лунку, и инкубировали в течение 1,5 часа при 37°C, 5% CO2. Затем на инокулюм наносили 1 мл на лунку 0,75% метилцеллюлозы в DMEM, дополненной 2% FBS, 2% GlutaMAX и 2% антибиотика-антимитотического средства. После 5 дней инкубации при 37°C, 5% CO2, верхний слой удаляли, а монослои фиксировали ледяным метанолом в течение 20 минут.
[00315] Затем планшеты один раз промывали водой и блокировали 5% обезжиренным сухим молоком в фосфатно-солевом буфере (PBS) в течение 30 минут при комнатной температуре с осторожным встряхиванием. Затем блокирующий раствор замещали из расчета 200 мкл на лунку с помощью 2% сухого молока в PBS, содержащего разведение 1:2000 антитела против RSV, конъюгированного с пероксидазой хрена (Abcam AB20686). После 3 часов инкубации при комнатной температуре планшеты промывали 2 раза водой, проявляли субстратом для HRP от TrueBlue, еще дважды промывали водой и сушили на воздухе.
[00316] Окрашенные бляшки подсчитывали с помощью препаровального микроскопа. Титры нейтрализующих антител определяли в конечной точке снижения на 60% относительно контролей с имитацией нейтрализации вируса по формуле: Титр, при котором количество бляшек снижается на 60% = (C/V x 0,4 - низкое)/(высокое - низкое) x (HSD - LSD) + LSD, где C/V=среднее число бляшек RSV в контрольных лунках с имитацией нейтрализации вируса, «низкое» и «высокое» представляют собой среднее число бляшек RSV в двух разведениях, которые ограничивают значение C/V x 0,4 для образца сыворотки крови, а HSD и LSD представляют собой более высокие и более низкие разведения сыворотки крови.
[00317] Для анализа нейтрализации в HAE, сыворотку крови инактивировали нагреванием в течение 30 минут при 56°C. Выполняли серию четырехкратных серийных разведений инактивированной сыворотки крови в базовой среде PneumaCult™-ALI (Stem Cell Technologies; 05002), дополненной PneumaCult™-ALI 10X Supplement (Stem Cell Technologies; 05003) и 1% антибиотика/противогрибкового средства (далее среда). Исходные растворы вируса RSV объединяли 1:1 с разведениями сыворотки крови и инкубировали в течение 1,5 часа при 37°C. Затем смесь вируса с сывороткой добавляли в 24-луночные планшеты, содержащие полностью дифференцированные клетки HAE при 50 мкл на лунку, и инкубировали в течение 1,5 часа при 37°C, 5% CO2. После инкубации инокулюм удаляли, лунки дважды промывали средой для удаления несвязанного вируса и инкубировали еще 20 часов при 37°C, 5% CO2. События инфицирования в культурах, инфицированных RSV, экспрессирующих репортер mKate (TagFP635), подсчитывали на флуоресцентном микроскопе.
[00318] Для выявления инфекции, обусловленной RSV, при которой не экспрессируется репортер mKate, псевдостратифицированный эпителий тщательно промывали средой для удаления слизи, затем фиксировали 4% параформальдегидом в течение 30 минут при комнатной температуре, пермеабилизировали с помощью 0,25% Triton X-100 в течение 30 минут и блокировали с помощью DMEM, дополненной 2% FBS, в течение 1 часа при 37°C. Блокирующий раствор замещали из расчета 100 мкл на лунку смесью моноклональных Ab мыши против RSV (Millipore; MAB 858-4), разведенной 1:200 в DMEM, дополненной 2% FBS, и планшеты инкубировали при 37°C в течение 2 часов. Затем планшеты промывали 3 раза с помощью PBS с добавлением 0,05% Tween 20. В каждую лунку добавляли по 100 мкл антитела козы против IgG мыши (H+L) (Invitrogen; A11001), разведенного 1:200 в DMEM, дополненной 2% FBS, и планшеты инкубировали в течение ночи при 4°C. На следующее утро планшеты промывали 3 раза с помощью PBS с добавлением 0,05% Tween 20, флуоресцентный сигнал стабилизировали с помощью ProLong Gold AntiFade с DAPI (Thermo Fisher Scientific; P36935) и подсчитывали на флуоресцентном микроскопе. Титры нейтрализующих антител определяли при конечной точке снижения на 60%, как указано выше.
[00319] Для связывания антителами против F, либо F до слияния (DS-CAV1), либо F после слияния связывали с наконечниками, покрытыми антителом против HIS на Octet. Если не указано иное, все связывание антител против F относится к связыванию антител против тримера F до слияния (DS-CAV1). His6-меченный (SEQ ID NO: 230) тример F RSV (DS-CAV1 или F после слияния предварительно загружали на наконечники сенсора Anti-Penta-HIS (HIS1K) (FortéBio, №18-5122) на 400 секунд для обеспечения уровня захвата, близкого к состоянию насыщения. Наконечники биосенсора затем уравновешивали в течение 90 секунд в промывочном буфере Octet, с последующим связыванием с разбавленной сывороткой крови в течение 300 секунд. Окончательные ответы кривой связывания измеряли с применением программного обеспечения Octet Data Analysis HT10.0, и ответ умножали на коэффициент разведения (100 или 300) для получения окончательного регистрируемого ответа.
[00320] Для связывания антитела против Gcc, тримеризованный димер пептида Gcc с C-концевой меткой HIS использовали на наконечнике Octet, как описано выше. His6-меченный (SEQ ID NO: 230) гексамер Gcc (штамма A2) предварительно загружали на наконечники сенсоров Anti-Penta-HIS (HIS1K) (FortéBio, №18-5122) на 400 секунд для обеспечения уровня захвата, близкого к состоянию насыщения. Наконечники биосенсора затем уравновешивали в течение 90 секунд в промывочном буфере Octet, с последующим связыванием с разбавленной сывороткой крови в течение 300 секунд. Окончательные ответы кривой связывания измеряли с применением программного обеспечения Octet Data Analysis HT10.0, и ответ умножали на коэффициент разведения (100 или 300) для получения окончательного регистрируемого ответа.
[00321] В исследованиях на приматах, отличных от человека (NHP), NHP предварительно отбирали на предмет ответа на RSV (исходные уровни оказались ниже пределов выявления для всех анализов). NHP иммунизировали с помощью 50 мкг RF8140 с обозначенным адъювантом по протоколу, аналогичному описанному выше для мышей, но с большим объемом адъюванта (фиг. 11C-D и фиг. 18).
[00322] В исследовании на приматах, отличных от человека, анализы нейтрализации в VERO проводили, как описано выше. Связывание pre-F оценивали с помощью нижеописанного анализа ELISA.
[00323] Образцы сыворотки крови от NHP серийно разводили в 2 раза (начальное разведение 1:100) и инкубировали в планшетах, покрытых блокированным растворимым F RSV (Sinobiological, № 11049-V08B) (1 мкг/мл, 100 мкл/лунка), в течение 1 часа при 37°C. Специфические в отношении F RSV IgG выявляли с применением конъюгированных с пероксидазой хрена антител против IgG обезьяны (BioRad AAI42P, разведение 1:10000) в течение 90 минут при 37°C. Планшеты проявляли с применением 3,3',5,5'-тетраметилбензидина (TMB Tebu-Bio) и останавливали с помощью 1 н. соляной кислоты (Prolabo, № 30024290). Оптическую плотность (OD) измеряли при 450-650 нм с помощью устройства для считывания микропланшетов (SpectraMax). Титры IgG, специфических в отношении sF RSV, рассчитывали с применением программного обеспечения SoftmaxPro для диапазона значений OD от 0,2 до 3,0, исходя из кривой титрования (на каждый планшет наносили стандартную гипериммунную сыворотку мыши).
[00324] Титры IgG этого эталона, выраженные в произвольных единицах ELISA (EU), соответствовали log10 кратности разведения, дающего OD 1,0. Порог выявления антител составлял 20 (1,3 log10) EU. Все окончательные титры выражали в log10 для построения графика. Каждому титру < 1,3 log10 назначали произвольный титр 1,0 log10.
[00325] Для оценки клеточного иммунитета в исследовании на NHP использовали набор IFNγ/IL-2 FluoroSpot (FS-2122-10, Mabtech) в соответствии с инструкциями производителя. Вкратце, мембраны планшетов IPFL предварительно смачивали 35% этанолом и покрывали иммобилизованными антителами (антителами против IFNγ и антителами против IL-2) в течение ночи при 4°C.
[00326] Затем планшеты блокировали в течение 2 часов при 37°C из расчета 200 мкл/лунка средой для инкубации клеток, содержащей 10% фетальной телячьей сыворотки (FCS). Среду удаляли и в лунки добавляли стимулы: полноразмерный антиген F (антиген-специфическая стимуляция), антитело против CD3 (положительный контроль) или среду для культивирования клеток (не подвергнутый стимулированию контроль). Мононуклеарные клетки периферической крови (PBMC) макака размораживали и подсчитывали. Добавляли 400000 клеток на лунку и инкубировали в течение 24 часов при 37°C в увлажненном инкубаторе с 5% CO2.
[00327] Для выявления клетки удаляли и добавляли антитела для выявления (конъюгированные антитела против IFNγ и антитела против IL-2) и инкубировали 2 часа при комнатной температуре. Затем добавляли реагенты, конъюгированные с флуорофором, и инкубировали 1 час при к. т. Планшеты освобождали от содержимого, сушили и хранили в темноте при к. т. до анализа. MAb против CD3 использовали в качестве положительного контроля, и во всех образцах были получены ответы > 500 случаев образования пятен (SFC)/миллион PBMC, что подтверждало приемлемое качество образца. Пятна, выявляемые в лунках без стимуляции (среда для культивирования клеток), вычитали из значений для клеток, стимулированных F-антигеном.
[00328] Для анализа клеток человека (или В-клеток) выполняли эксперименты, аналогичные эксперименту, упомянутому в Dauner, et al. Vaccine 2017 Oct 4;35(41):5487-5494 фиг. 20). Клетки либо не обрабатывали (обрабатывали с помощью PBS), либо обрабатывали полипептидами F RSV или G RSV, как указано, при дозах 100 нг. Ответы в виде связывания F и связывания G анализировали с применением описанного в литературе анализа Luminex с гранулами, покрытыми тримером pre-F (DS-CAV1) или эктодоменом G соответственно.
[00329] RF8117 (SEQ ID NO: 17) содержит сконструированные сайты гликозилирования в положениях E328N, S348N и R507N, которые, как упоминалось выше, не предотвращают связывание с D25 или AM14. Чтобы продемонстрировать, что эта наночастица на основе белка до слияния вызывает иммунный ответ, аналогичный таковому на другие антигены до слияния (DS-CAV1), авторы настоящего изобретения иммунизировали мышей в группах по 5 особей либо тримером pre-F (DS-CAV1), либо F после слияния, либо RF8117, при дозах 1 мкг или 0,1 мкг, при этом их все применяли с адъювантом AF03, три раза с трехнедельным интервалами между инъекциями. Образцы сыворотки крови тестировали на предмет титра нейтрализующих антител через две недели после третьей иммунизации с применением анализа на основе клеток VERO. RF8117 в более высокой дозе обеспечивал получение титра нейтрализующих антител, подобного титру контроля до слияния, и превосходящего титр контроля после слияния. При более низкой дозе RF8117 обеспечивал получение более высокого титра нейтрализующих антител, чем контроль до слияния и контроль после слияния (фигура 10A).
[00330] Pre-F-NP RSV содержит сайты гликозилирования, сконструированные для блокирования эпитопов, общих для конформации до слияния и после слияния. Оценивали ингибирование этими гликанами нейтрализующего ответа. RF8117 со сконструированными гликанами (SEQ ID NO: 17) сравнивали с RF8113 (аналогичным RF8117, но без сконструированных гликанов; SEQ ID NO: 16) и контролем тримера до слияния (DS-CAV1). Мышей в группах по 5 особей иммунизировали дозами 1 мкг или 0,1 мкг, при этом их все применяли с адъювантом AF03, три раза с трехнедельными интервалами между инъекциями. Образцы сыворотки крови тестировали на предмет титра нейтрализующих антител через две недели после третьей иммунизации с применением анализа на основе клеток VERO. Не было существенной разницы в обеих дозах между конструкциями RF8113 и RF8117, о чем судили по титру нейтрализующих антител (фигура 10B).
[00331] Чтобы продемонстрировать, что упомянутые в данном документе нокауты лизина и аргинина в RF8140 (SEQ ID NO: 23) не нарушают способность антигена вызывать нейтрализующий ответ, авторы настоящего изобретения сравнивали иммуногенность RF8140 (SEQ ID NO: 25) с иммуногенностью тримера F после слияния (SEQ ID NO: 24) у мышей (фиг. 11A и B). При низкой дозе (0,1 мкг) RF8140 (SEQ ID NO: 25) обеспечивает получение титра нейтрализующих антител, превосходящего титр для тримера после слияния (SEQ ID NO: 24). Для демонстрации того, что RF8140 (SEQ ID NO: 23) вызывает иммунный ответ у NHP, авторы настоящего изобретения иммунизировали NHP с помощью RF8140 (SEQ ID NO: 25) с адъювантом (AF03) или без такового. На фигуре 11C показан ответ в виде связывания F RSV (титр ELISA), а на фигуре 11D сравниваются титры нейтрализующих антител к RSV, полученные при иммунизации с помощью Pre-F-NP (RF8140, SEQ ID NO: 23). Как без применения адъюванта, так и с применением адъюванта RF8140 (SEQ ID NO: 25) вызывает иммунный ответ у NHP.
[00332] Показывая то, что сконструированные сайты гликозилирования RF8117 (SEQ ID NO: 17) и RF8140 (SEQ ID NO: 23) не препятствуют тому, чтобы эти антигены вызывали нейтрализующий ответ, авторы настоящего изобретения хотели продемонстрировать, что они действительно блокируют эпитопы, не подлежащие нейтрализации или подлежащие нейтрализации в небольшой степени, общие для конформации до слияния и после слияния (фигура 12). Ответ в виде образования антител к F до слияния (DS-CAV1, SEQ ID NO: 25) вызывали иммунизацией с помощью Pre-F-NP без гликозилирования посредством конструирования (RF8113, SEQ ID NO: 16) или с помощью Pre-F-NP с гликозилированием посредством конструирования (частица со сконструированным Gly, RF8117 SEQ ID NO: 17) при высокой (1 мкг) и низкой (0,1 мкг) дозе, согласно измерению с помощью Octet (фиг. 12A). Ответы, вызванные любой из Pre-F-NP, были подобными. Ответ в виде образования антител к тримеру после слияния вызывали иммунизацией с помощью Pre-F-NP без гликозилирования посредством конструирования (RF8113, SEQ ID NO: 16) или с помощью Pre-F-NP с гликозилированием посредством конструирования (RF8117 SEQ ID NO: 17) при высокой (1 мкг) и низкой (0,1 мкг) дозе, согласно измерению с помощью Octet (фиг. 12В). Ответы в виде связывания F после слияния, вызванные RF8117 (SEQ ID NO:17), были значительно ниже вызванных RF8113 (SEQ ID NO: 16). Следовательно, хотя и RF8113, и RF8117 вызывают устойчивые Ответы в виде образования антител против F до слияния, ответ в виде образования антител против F после слияния, вызванный посредством RF8117, значительно подавлен. Это обеспечивается наличием сконструированных гликанов, которые картируются в эпитопах, общих для конформации до слияния и после слияния (фиг. 2B).
[00333] Чтобы дополнительно продемонстрировать, что сконструированные сайты гликозилирования блокируют эпитопы, не подлежащие нейтрализации, но обеспечивают смещение соотношения титра нейтрализующих антител и не нейтрализующих антител, авторы настоящего изобретения анализировали приведенные выше данные другим способом (фиг. 13). Измеренные с помощью анализа на основе клеток VERO титры нейтрализующих антител к RSV, полученные при иммунизации с помощью pre-F-NP с сайтами гликозилирования дикого типа (частица с гликаном Wt; RF8113, SEQ ID NO: 16) по сравнению с Pre-F-NP с добавленными сконструированными сайтами гликозилирования (частица+гликан; RF8117, SEQ ID NO: 17) в исследованиях на мышах, были измерены и не продемонстрировали существенной разницы (фиг. 13A). Сравнение Ответ в виде образования антитела, связывающего тример F после слияния RSV, вызванных иммунизацией с помощью частицы с гликаном Wt (RF8113, SEQ ID NO: 16) и с помощью частицы с+гликаном (RF8117, SEQ ID NO: 17), в исследованиях на мышах показывало подавление ответа в виде связывания F после слияния в случае Pre-F-NP со сконструированными гликанами (фиг. 13B). Чтобы дополнительно продемонстрировать, что сконструированные гликаны не уменьшают ответ в виде образования функциональных нейтрализующих антител, но уменьшают уровень антител, отличных от нейтрализующих, образование которых происходит в ответ на общие эпитопы до слияния/после слияния, что тем самым обеспечивает улучшение в отношении соотношения нейтрализующих и суммарных антител, образующихся под воздействием конструкций со сконструированными гликанами, отношение титра нейтрализующих антител к ответу в виде связывания F наносили на график (фиг. 13C). Следовательно, в исследованиях на мышах Pre-F-NP со сконструированными гликанами обеспечивают получение профиля нейтрализующих антител, превосходящего профиль связывающих антител.
[00334] Чтобы продемонстрировать возможность использования ферритиновых наночастиц для улучшения иммуногенности антигена центрального домена G RSV, авторы настоящего изобретения разрабатывали способ химического конъюгирования пептида Gcc (SEQ ID NO: 29) с ферритиновой наночастицей. Ферритин, несущий мутацию S111C, описанную в данном документе, может быть конъюгирован с пептидом Gcc (SEQ ID NO: 29), синтезированным с малеимидной группой на линкере PEG4, присоединенном к N-концу с помощью группы NHS. Пептид Gcc с N-концевым малеимидом синтезировали и очищали с помощью HPLC в компании Peptides International (Луисвилл, Кентукки, США). При добавлении антигена малеимид-Gcc к частице ферритина с S111C, малеимид конъюгируется со свободным цистеином и образует Gcc-NP, что можно наблюдать с помощью анализа на геле SDS-PAGE, окрашенном кумасси (фиг. 14A). Хотя эффективность конъюгации обычно составляет от 50% до 90%, модель наночастицы на основе пептида Gcc и ферритина (конъюгированной на 100%) показана на фиг. 14B.
[00335] Чтобы определить, вызывает ли Gcc-NP иммунный ответ, превосходящий таковой на пептид Gcc (SEQ ID NO: 29), иммунизировали по 5 мышей на группу либо пептидом Gcc, либо Gcc-NP (доза 1,3 мкг, смесь 1:1 с RIBI для каждой иммунизации). Ответ в виде связывания Gcc (Octet) через две недели после второй и две недели после третьей процедур иммунизации сравнивали с репрезентативной сывороткой крови от группы мышей, не подвергавшихся воздействию (фиг. 14C). Также нейтрализующий ответ, вызванный иммунизацией пептидом Gcc (SEQ ID NO. 29), сравнивали с Gcc-NP в исследованиях на мышах после третьей инъекции в анализах нейтрализации с клетками HAE (фиг. 14D). Gcc-NP вызывает иммунный ответ, превосходящий таковой на пептид Gcc отдельно, о чем судят как по ответу в виде связывания Gcc, так и по нейтрализующему ответу.
[00336] Чтобы продемонстрировать, что совместное введение Pre-F-NP RSV (RF8140) и Gcc-NP не влияет на способность любого из антигенов вызывать иммунный ответ, мышей иммунизировали с помощью либо Pre-F-NP отдельно (RF8140, SEQ ID NO: 23), либо Gcc-NP (ферритина, конъюгированного с пептидом Gcc SEQ ID NO: 29), либо Pre-F-NP (RF8140, SEQ ID NO: 23) в сочетании с Gcc-NP (FIG 15A-C). Все процедуры иммунизации осуществляли с применением адъюванта AF03. У мышей, иммунизированных с помощью RF8140 отдельно (Pre-F-NP) или RF8140 и Gcc-NP (Pre-F-NP+Gcc-NP), вырабатывались антитела, которые связывают тример F до слияния (DS-CAV1, SEQ ID:25), тогда как у мышей, иммунизированных с помощью Gcc-NP, не вырабатывались. У мышей, иммунизированных с помощью Gcc-NP отдельно (Gcc-NP) или RF8140 и Gcc-NP, вырабатывались антитела, которые связывают пептид Gcc, тогда как у мышей, иммунизированных с помощью только RF8140, не вырабатывались. У всех животных, иммунизированных с помощью Pre-F-NP отдельно, Gcc-NP отдельно или совместного введения Pre-F-NP и Gcc-NP, вырабатывался нейтрализующий ответ после второй и после третьей иммунизации, согласно измерению в анализе нейтрализации с клетками HAE.
[00337] Чтобы определить, влияет ли совместное введение Pre-F-NP RSV и Gcc-NP на способность любого антигена вызывать нейтрализующие антитела, нейтрализующие антитела как против F, так и против G изучали в анализе с истощением (фиг. 16A-B). Чтобы продемонстрировать, что добавление Gcc-NP не влияет на способность Pre-F-NP вызывать нейтрализующий ответ, титры нейтрализующих антител измеряли с помощью анализа на основе клеток VERO, чувствительных к F, для упомянутых выше групп (фиг. 16A). Образцы сыворотки крови от не подвергавшихся воздействию животных также тестировали, чтобы судить о качестве истощения по антигенам. В анализе на основе VERO образцы сыворотки крови от мышей, иммунизированных RF8140 (SEQ ID NO: 23) отдельно или RF8140, смешанным с Gcc-NP, вызывали аналогичные нейтрализующие ответы, тогда как Gcc-NP, как оказалось, не вызывала нейтрализующий ответ в анализе на основе клеток VERO, чувствительных к антителу против F. Когда объединенные образцы сыворотки крови животных, иммунизированных с помощью RF8140 (SEQ ID NO:23) отдельно или иммунизированных с помощью RF8140 (SEQ ID: 23) и Gcc-NP, истощали по антителам, связывающим тример до слияния (DS-CAV1, SEQ ID: 25), наблюдали снижение поддающихся измерению титров нейтрализующих антител в анализе на основе клеток VERO. Когда в вышеуказанных группах измеряли титр нейтрализующих антител в анализе на основе клеток HAE, наблюдали, что у всех групп иммунизации вырабатывался нейтрализующий ответ в анализе чувствительности к F и G (фиг. 16B). Объединенные образцы сыворотки крови от животных, иммунизированных с помощью RF8140 (SEQ ID NO: 23) отдельно, вызывали нейтрализующий ответ в анализе на основе клеток HAE, которые могли быть истощены по тримеру F до слияния (DS-CAV1, SEQ ID NO: 25). Объединенные образцы сыворотки крови от животных, иммунизированных с помощью Gcc-NP отдельно, вызывали нейтрализующих ответ в анализе на основе клеток HAE, которые могли быть истощены по эктодомену G (SEQ ID NO: 28). Объединенные образцы сыворотки крови от животных, иммунизированных с помощью как RF8140 (SEQ ID NO: 23), так и Gcc-NP, вызывали нейтрализующих ответ в анализе на основе клеток HAE, которые не были полностью истощены по DS-CAV1 (SEQ ID NO: 25), но были полностью истощены путем последовательных процедур истощения по DS-CAV1, а затем по эктодомену G (SEQ ID NO: 28). Вместе эти данные говорят о том, что совместное введение с Pre-F-NP и Gcc-NP не влияет на способность любого антигена вызывать образование нейтрализующих антител против F до слияния или G соответственно.
[00338] Чтобы продемонстрировать эффект применения с адъювантом для RF8117 (SEQ ID NO: 17) или RF8140 (SEQ ID NO: 23), мышам вводили дозу этих конструкций, смешанных с AF03, SPA09 или алюминиевыми квасцами. На фигуре 17A показано, что мышей иммунизировали с помощью 10 мкг антигена, смешанного с адъювантом, тогда как на фигуре 17В показано, что мышей иммунизировали с помощью 1 мкг антигена, смешанного с адъювантом. На фигуре 17A показаны титры нейтрализующих антител, измеряемые с помощью анализа на основе клеток VERO в момент времени через две недели после третьей иммунизации. Показаны образцы сыворотки крови от мышей, иммунизированных с помощью RF8117 (SEQ ID NO: 17) либо без применения адъюванта (без адъюванта), с применением адъюванта, представляющего собой алюминиевые квасцы, либо с применением адъюванта AF03. На фигуре 17B показано, что титры нейтрализующих антител измеряли с помощью анализа на основе клеток VERO для образцов сыворотки крови от мышей, иммунизированных с помощью RF8117 (SEQ ID NO: 17) с применением адъюванта AF03, RF8117 (SEQ ID NO: 17) с применением адъюванта SPA09, или RF8140 с применением адъюванта AF03. Во всех случаях для RF8117 (SEQ ID NO: 17) или RF8140 (SEQ ID NO: 23) у получавших адъювант групп не подвергавшихся воздействию мышей получали более высокий титр нейтрализующих антител, чем у не получавших адъювант групп. У мышей, иммунизированных с помощью RF8117 (SEQ ID NO: 17) или RF8140 (SEQ ID NO: 23), смешанного с AF03, получали аналогичный нейтрализующий ответ, что позволяет предположить, что мутации, обеспечивающие добавление лизина и аргинина в RF8140 (SEQ ID NO: 23), не влияют на способность Pre-F-NP вызывать нейтрализующий ответ.
[00339] Для дальнейшего изучения эффекта без применения адъюванта AF03 и SPA09, приматов, отличных от человека (NHP), иммунизировали с помощью RF8140 без применения адъюванта, с применением адъюванта AF03 или с применением адъюванта в виде двух доз SPA09 (фиг. 18A). NHP иммунизировали с помощью 50 мкг антигена, смешанного с указанным адъювантом, в дни 0 и 29, и иммунный ответ измеряли с помощью ELISA или нейтрализующего ответа в клетках VERO в указанные моменты времени. Ответы на тример F до слияния с помощью ELISA измеряли в образцах сыворотки крови от NHP после иммунизации с помощью RF8140, без применения адъюванта (без адъюванта), с применением адъюванта AF03 или с применением адъюванта SPA09 (использовали дозы 500 мкг и 2000 мкг SPA09). Во все моменты времени применение адъюванта AF03 или SPA09 вызывает превосходящий нейтрализующий ответ. Также измеряли титры нейтрализующих антител в образцах сыворотки крови для упомянутых выше групп NHP с помощью анализа на основе клеток VERO (фиг. 18B). Во всех случаях иммунизация с помощью RF8140 с адъювантом обеспечивает получение более высокого титра нейтрализующих антител, чем у не получавших адъювант групп во все моменты времени.
[00340] Тестировали эффект прямой конъюгации RF8140 (SEQ ID NO: 23) с агонистом TLR7/8 SM7/8 или агонистом TLR9 CpG. Антиген конъюгировали с малыми молекулами SM7/8 или CpG и вводили мышам в виде дозы 10 мкг. В своей последовательности ферритина RF8140 содержит мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином (K79C), что можно использовать для конъюгации с помощью клик-химии. Для сравнения мышам вводили дозу RF8140 либо без применения адъюванта (без адъюванта), либо с применением адъюванта путем смешивания с малой молекулой при высокой или низкой дозе (без конъюгирования), как показано на фиг. 19. Образцы сыворотки крови от животных после второй и после третьей иммунизации тестировали на предмет связывания с тримером F до слияния.
[00341] На фигуре 19A показан результат измерения ответа в виде связывания тримера F до слияния в образцах сыворотки крови от любых из не подвергавшихся воздействию мышей, мышей, иммунизированных с помощью RF8140 (SEQ ID NO: 23) без применения адъюванта, мышей, иммунизированных с помощью RF8140 (SEQ ID NO: 23), конъюгированного с адъювантом SM7/8, RF8140 (SEQ ID NO: 23) с применением адъюванта в виде 130 нг SM7/8 или RF8140 (SEQ ID NO: 23) с применением адъюванта в виде 20 мкг SM7/8. RF8140 (SEQ ID NO: 23), конъюгированный с SM7/8, обеспечивает получение более высокого титра антител, связывающих тример F до слияния, чем в случае групп, получавших RF8140 без применения адъюванта или с применением адъюванта SM7/8.
[00342] На фигуре 19В показан также результат измерения ответа в виде связывания тримера F до слияния в образцах сыворотки крови от любых из не подвергавшихся воздействию мышей, мышей, иммунизированных с помощью RF8140 (SEQ ID NO: 23) без применения адъюванта, мышей, иммунизированных с помощью RF8140 (SEQ ID NO: 23), конъюгированного с адъювантом CpG, RF8140 (SEQ ID NO: 23) с применением адъюванта в виде 680 нг CpG, или RF8140 (SEQ ID NO: 23) с применением адъюванта в виде 20 мкг CpG. RF8140 (SEQ ID NO: 23), конъюгированный с SM7/8, обеспечивает получение более высокого титра антител, связывающих тример F до слияния, чем в случае групп, получавших RF8140 без применения адъюванта или с применением адъюванта SM7/8.
[00343] Чтобы продемонстрировать способность антигена Pre-F-NP и антигена Gcc-NP вызывать ответ в клетках человека, проводили эксперименты с платформой MIMIC (фиг. 20A-D). Платформа MIMIC состоит исключительно из аутологичных иммунных клеток человека, способных быстро и воспроизводимо вырабатывать антигенспецифические врожденные и адаптивные ответы при заражении. В предыдущей работе продемонстрирована способность системы MIMIC воспроизводить иммунные профили in vivo против таких разнообразных мишеней, как HBV, столбнячный анатоксин, моноклональные антитела, YF-VAX и В-клеточные ответы на вирус гриппа. Сравнивали Ответы в виде образования антитела, связывающего тример F до слияния RSV, вызванные обработкой с помощью Pre-F-NP RF8140 (SEQ ID NO: 23) и с помощью тримера F после слияния (SEQ ID NO: 24), в человеческих B-клетках, и репрезентативный исходный ответ показан для сравнения (без обработки) (фиг. 20A). Соотношения измеренных ответов в виде связывания с тримером F до слияния (DS-CAV1, SEQ ID NO: 25) и с тримером F после слияния (SEQ ID NO: 24), вызванных путем обработки с помощью Pre-F-NP (RF8140, SEQ ID NO: 23) по сравнению с F после слияния (SEQ ID NO: 24) в человеческих B-клетках, показаны на фиг. 20C. Антитела из MIMIC, образование которых вызвано обработкой различными антигенами F, измеряли с применением анализа на основе клеток VERO (фиг. 20B). Титры нейтрализующих антител, полученные при обработке с помощью pre-F-NP (RF8140, SEQ ID NO: 23) и с помощью тримера F после слияния (SEQ ID NO: 24) в человеческих B-клетках, сравнивали с титрами в группе без обработки, в результате чего было показано, что RF8140 (SEQ ID NO: 23) вызывал превосходящий нейтрализующий ответ в клетках человека.
[00344] Величину ответа в виде образования Ab на инфекцию, обусловленную RSV, или на кандидатную вакцину на основе субъединицы F определяли на основании серологического статуса субъектов-людей, изучаемых в исследованиях с применением MIMIC, который оценивали с помощью анализа линейной регрессии. Доноры с более высокими ранее существовавшими титрами циркулирующих IgG против pre-F продуцировали значительно больше IgG против pre-F после обработки с помощью RSV (фиг. 20E, p=0,0041) и после праймирования с помощью post-F (фиг. 20F, p=0,0019). Хотя для показателя корреляция не была достигнута статистическая значимость, для pre-F также показали взаимосвязь между уровнем индуцированного Ab и уровнем ранее существовавшего Ab. Примечательно, что в отличие от других методов лечения, pre-F-NP индуцировал сравнительно высокий уровень IgG против pre-F у доноров с низким ранее существующим IgG против pre-F, как и у доноров с высоким уровнем уже существующих Ab (фиг. 20F). Это указывает на то, что pre-F-NP способен эффективно сохранять (или усиливать) ответ в виде образования Ab даже у доноров с низким уровнем ранее существующего IgG.
[00345] Чтобы продемонстрировать, что Gcc-NP вызывает ответ в виде образования антитела против G, превосходящий ответ в случае пептида Gcc (SEQ ID NO: 29) отдельно, клетки человека обрабатывали пептидом Gcc отдельно (SEQ ID NO: 29) или пептидом Gcc, конъюгированным с наночастицей (Gcc-NP) в B-клетках человека. Gcc-NP вызывал превосходящий ответ в виде образования антитела, связывающего G (фиг. 20G). В совокупности эти данные позволяют предположить, что Pre-F-NP и Gcc-NP будут вызывать иммунные ответы при иммунизации человека.
4. Характеристика иммунного ответа на наночастицы на основе Gcc RSV-ферритина in vivo
[00346] Gcc RSV-NP получали, как описано выше. Для оценивания ответа in vivo на Gcc RSV-NP у мышей, самок мышей BALBc внутримышечно иммунизировали антигенами RSV при определенных дозах на неделе 0, 3 и 6 либо высокой дозой (5 мкг), либо низкой дозой (0,5 мкг) антигена. Если не указано иное, Gcc RSV-NP применяли с адъювантом AF03 с применением стратегии смешивания перед использованием. То есть 50 мкл раствора белка смешивали с 50 мкл адъюванта AF03 Sanofi (эмульсия на основе сквалена; см. Klucker et al., J Pharm Sci. 2012 Dec;101(12):4490-500) непосредственно перед инъекцией 50 мкл в каждую заднюю лапу. Никаких побочных эффектов иммунизации не наблюдали. Кровь собирали за 1 день до первой иммунизации и по меньшей мере через 2 недели после каждой инъекции (т. е. недели 2, 5 и 8). Если не указано иное, показаны данные для 2 недель после третьей инъекции (неделя 8, также обозначаемая как 2wp3). Обычно анализировали образцы сыворотки крови от животных до иммунизации (обозначенных как не подвергавшиеся воздействию), через две недели после второй инъекции (после 2 или 2wp2) или через две недели после третьей инъекции (после 3-ей или 2wp3).
[00347] Для анализа нейтрализации в HAE сыворотку крови инактивировали нагреванием в течение 30 минут при 56°C. Выполняли серию четырехкратных серийных разведений инактивированной сыворотки в базовой среде PneumaCult™-ALI (Stem Cell Technologies; 05002), дополненной PneumaCult™-ALI 10X Supplement (Stem Cell Technologies; 05003) и 1% антибиотика/противогрибкового средства (далее среда). Исходные растворы вируса RSV объединяли 1:1 с разведениями сыворотки крови и инкубировали в течение 1,5 часа при 37°C. Затем смесь вируса с сывороткой добавляли в 24-луночные планшеты, содержащие полностью дифференцированные клетки HAE из расчета 50 мкл на лунку и инкубировали в течение 1 часа при 37°C, 5% CO2. После инкубации инокулюм удаляли, лунки дважды промывали средой для удаления несвязанного вируса и инкубировали еще 20 часов при 37°C, 5% CO2. События инфицирования в культурах, инфицированных RSV, экспрессирующих репортер mKate (TagFP635), подсчитывали на флуоресцентном микроскопе.
[00348] Для выявления инфекции, обусловленной RSV, при которой не экспрессируется репортер mKate (нейтрализация RSV штамма B), образцы псевдостратифицированного эпителия тщательно промывали средой для удаления слизи, затем фиксировали 4% параформальдегидом в течение 30 минут при комнатной температуре, пермеабилизировали с помощью 0,25% Triton X-100 в течение 30 минут и блокировали с помощью DMEM, дополненной 2% FBS, в течение 1 часа при 37° C. Блокирующий раствор замещали из расчета 100 мкл на лунку смесью моноклональных Ab мыши против RSV (Millipore; MAB 858-4), разведенной 1:200 в DMEM, дополненной 2% FBS, и планшеты инкубировали при 37°C в течение 2 часов. Затем планшеты промывали 3 раза с помощью PBS с добавлением 0,05% Tween 20. В каждую лунку добавляли по 100 мкл антитела козы против IgG мыши (H+L) (Invitrogen; A11001), разведенного 1:200 в DMEM, дополненной 2% FBS, и планшеты инкубировали в течение ночи при 4°C. На следующее утро планшеты промывали 3 раза с помощью PBS с добавлением 0,05% Tween 20, флуоресцентный сигнал стабилизировали с помощью ProLong Gold AntiFade с DAPI (Thermo Fisher Scientific; P36935) и подсчитывали на флуоресцентном микроскопе. Титры нейтрализующих антител определяли при конечной точке снижения на 60%.
[00349] Чтобы продемонстрировать, что более высокая степень поливалентности улучшает способность вызывать нейтрализующий ответ антигенами G RSV, мышей иммунизировали антигенами F RSV. Все процедуры иммунизации осуществляли с применением адъюванта AF03. Мышей иммунизировали с помощью Gcc RSV-NP, составленных с AF03, и измеряли титры нейтрализующих антител через 2 недели после второй и через 2 недели после третьей инъекций (фиг. 21A-C). Gcc RSV-NP вызывали нейтрализующий ответ по сравнению с сывороткой крови от не подвергавшейся воздействию мыши. Как через 2 недели после второй (фиг. 21A), так и через 2 недели после третьей (фиг. 21B) иммунизации мыши, иммунизированные с помощью Gcc-NP, содержащих Gcc из штамма A2, демонстрировали нейтрализующие ответы на штамм А RSV. Через 2 недели после третьей инъекции Gcc-NP также вызывали нейтрализующий ответ на штамм В1 RSV (фиг. 21C).
[00350] Для связывания антитела против Gcc тримеризованный димер пептида Gcc с C-концевой меткой HIS использовали на наконечнике Octet. His6-меченный гексамер Gcc (штамма A2) или His6-меченный гексамер Gcc (штамма B1) предварительно загружали на наконечники сенсоров Anti-Penta-HIS (HIS1K) (FortéBio, №18-5122) на 400 секунд для обеспечения уровня захвата, близкого к состоянию насыщения. Наконечники биосенсора затем уравновешивали в течение 90 секунд в промывочном буфере Octet, с последующим связыванием с разбавленной сывороткой крови в течение 300 секунд. Окончательные ответы кривой связывания измеряли с применением программного обеспечения Octet Data Analysis HT10.0, и ответ умножали на коэффициент разведения (100 или 300) для получения окончательного регистрируемого ответа.
[00351] Чтобы определить, вызывает ли Gcc RSV-NP иммунный ответ в виде связывания Gcc, образцы сыворотки крови, полученные в результате процедур иммунизации, описанных выше, тестировали на предмет их способности связывать гексамер Gcc A2 или гексамер Gcc B1. Ответ в виде связывания Gcc при высокой дозе (фиг. 22A и фиг. 23A) и низкой дозе (фиг. 22B и фиг. 23B) тестировали через 2 недели после второй и 2 недели после третьей процедур иммунизации. Как в случае штамма A2 (фиг. 22A-B), так и в случае штамма B1 (фиг. 23A-B) Gcc RSV-NP вызывала ответ в виде связывания в отличие от образцов сыворотки крови от не подвергавшихся воздействию мышей.
5. Ответ на Pre-F-NP и Gcc-NP в клетках человека
[00352] Чтобы продемонстрировать способность Pre-F-NP и Gcc-NP вызывать ответ в клетках человека, проводили эксперименты с платформой MIMIC. Платформа MIMIC состоит исключительно из аутологичных иммунных клеток человека, способных быстро и воспроизводимо вырабатывать антигенспецифические врожденные и адаптивные ответы при заражении. В предыдущей работе продемонстрирована способность системы MIMIC воспроизводить иммунные профили in vivo против таких разнообразных мишеней, как HBV, столбнячный анатоксин, моноклональные антитела, YF-VAX и В-клеточные ответы на вирус гриппа. Сравнивали ответы в виде образования антитела, связывающего тример F до слияния RSV, вызванные обработкой с помощью Pre-F-NP RF8140 (SEQ ID NO: 23) и с помощью тримера F после слияния (SEQ ID NO: 24) в B-клетках человека, а также сравнивали с репрезентативным исходным ответом. Соотношения измеренных ответов в виде связывания с тримером F до слияния (DS-CAV1, SEQ ID NO: 25) и с тримером F после слияния (SEQ ID NO: 24), вызванных путем обработки с помощью Pre-F-NP (RF8140, SEQ ID NO: 23) по сравнению с обработкой с помощью F после слияния (SEQ ID NO: 24), определяли в B-клетках человека. Антитела из MIMIC, образование которых вызвано обработкой различными антигенами F, измеряли с применением анализа на основе клеток VERO. Титры нейтрализующих антител, полученные при обработке с помощью pre-F-NP (RF8140, SEQ ID NO: 23) и с помощью тримера F после слияния (SEQ ID NO: 24) в B-клетках человека, сравнивали с титрами в группе без обработки, в результате чего было показано, что RF8140 (SEQ ID NO: 23) вызывал превосходящий нейтрализующий ответ в клетках человека. Чтобы продемонстрировать, что Gcc-NP вызывает ответ в виде образования антитела против G, превосходящий ответ в случае пептида Gcc (SEQ ID NO: 29) отдельно, клетки человека обрабатывали пептидом Gcc отдельно (SEQ ID NO: 29) или пептидом Gcc, конъюгированным с наночастицей (Gcc-NP) в B-клетках человека. Gcc-NP вызывал превосходящий ответ в виде образования антитела, связывающего G. Таким образом, Pre-F-NP и Gcc-NP будут вызывать иммунные ответы при иммунизации человека.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> SANOFI
<120> АНТИГЕННЫЕ ПОЛИПЕПТИДЫ НА ОСНОВЕ ПОСЛЕДОВАТЕЛЬНОСТИ
РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА
<130> 01121-0031-00PCT
<150> 62/652,199
<151> 2018-04-03
<160> 230
<170> PatentIn версия 3.5
<210> 1
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 1
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 2
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 2
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 3
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 3
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Asn Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 4
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 4
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 5
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 5
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Thr Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 6
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 6
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 7
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 7
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Ser Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 8
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 8
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 9
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 9
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 10
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 10
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Leu Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 11
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 11
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 12
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 12
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Leu Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 13
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 13
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Leu Val Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 14
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 14
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Leu Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 15
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 15
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Cys Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Ile Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Phe Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Cys Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 16
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 16
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Arg Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 17
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 17
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 18
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 18
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Leu Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Gln Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 19
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 19
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Val Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Val Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Lys Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 20
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 20
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Leu Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Gln Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Asn Asp Ile Glu Asn Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 21
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 21
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Asn Ser Leu Tyr Val Asn Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Leu Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Gln Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 22
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 22
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Asn Ser Leu Tyr Val Asn Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Leu Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Gln Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Arg Gln Gln Phe Ser Asn Asp Ile Glu Asn Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 23
<211> 651
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 23
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gly Ser Gly Asn Val Gly Leu Gly Gly Ala Ile Ala Ser Gly Val
100 105 110
Ala Val Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys
115 120 125
Ser Ala Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly
130 135 140
Val Ser Val Leu Thr Phe Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp
145 150 155 160
Lys Gln Leu Leu Pro Ile Leu Asn Lys Gln Ser Cys Ser Ile Ser Asn
165 170 175
Pro Glu Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu
180 185 190
Ile Thr Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser
195 200 205
Thr Tyr Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met
210 215 220
Pro Ile Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile
225 230 235 240
Val Arg Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val
245 250 255
Leu Ala Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro
260 265 270
Cys Trp Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Asn
275 280 285
Gly Ser Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp
290 295 300
Asn Ala Gly Asn Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val
305 310 315 320
Gln Ser Asn Arg Val Phe Cys Asp Thr Met Asn Ser Arg Thr Leu Pro
325 330 335
Ser Glu Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp
340 345 350
Cys Lys Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr
355 360 365
Ser Leu Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala
370 375 380
Ser Asn Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp
385 390 395 400
Tyr Val Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu
405 410 415
Tyr Tyr Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu
420 425 430
Pro Ile Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe
435 440 445
Asp Ala Ser Ile Ser Gln Val Asn Glu Leu Ile Asn Gln Ser Leu Ala
450 455 460
Phe Ile Asn Gln Ser Asp Glu Leu Leu Ser Gly Ser Gly Ser Glu Ser
465 470 475 480
Gln Val Gln Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu Asn Glu
485 490 495
Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser Met Ser
500 505 510
Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe
515 520 525
Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe
530 535 540
Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro
545 550 555 560
Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu
565 570 575
His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala
580 585 590
Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val
595 600 605
Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys
610 615 620
Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln
625 630 635 640
Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
645 650
<210> 24
<211> 530
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 24
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gln Ser Thr Pro Ala Thr Asn Asn Arg Ala Arg Arg Glu Leu Pro
100 105 110
Arg Phe Met Asn Tyr Thr Leu Asn Asn Ala Lys Lys Thr Asn Val Thr
115 120 125
Leu Ser Lys Lys Arg Lys Arg Arg Ala Ile Ala Ser Gly Val Ala Val
130 135 140
Ser Lys Val Leu His Leu Glu Gly Glu Val Asn Lys Ile Lys Ser Ala
145 150 155 160
Leu Leu Ser Thr Asn Lys Ala Val Val Ser Leu Ser Asn Gly Val Ser
165 170 175
Val Leu Thr Ser Lys Val Leu Asp Leu Lys Asn Tyr Ile Asp Lys Gln
180 185 190
Leu Leu Pro Ile Val Asn Lys Gln Ser Cys Ser Ile Ser Asn Ile Glu
195 200 205
Thr Val Ile Glu Phe Gln Gln Lys Asn Asn Arg Leu Leu Glu Ile Thr
210 215 220
Arg Glu Phe Ser Val Asn Ala Gly Val Thr Thr Pro Val Ser Thr Tyr
225 230 235 240
Met Leu Thr Asn Ser Glu Leu Leu Ser Leu Ile Asn Asp Met Pro Ile
245 250 255
Thr Asn Asp Gln Lys Lys Leu Met Ser Asn Asn Val Gln Ile Val Arg
260 265 270
Gln Gln Ser Tyr Ser Ile Met Ser Ile Ile Lys Glu Glu Val Leu Ala
275 280 285
Tyr Val Val Gln Leu Pro Leu Tyr Gly Val Ile Asp Thr Pro Cys Trp
290 295 300
Lys Leu His Thr Ser Pro Leu Cys Thr Thr Asn Thr Lys Glu Gly Ser
305 310 315 320
Asn Ile Cys Leu Thr Arg Thr Asp Arg Gly Trp Tyr Cys Asp Asn Ala
325 330 335
Gly Ser Val Ser Phe Phe Pro Gln Ala Glu Thr Cys Lys Val Gln Ser
340 345 350
Asn Arg Val Phe Cys Asp Thr Met Asn Ser Leu Thr Leu Pro Ser Glu
355 360 365
Val Asn Leu Cys Asn Val Asp Ile Phe Asn Pro Lys Tyr Asp Cys Lys
370 375 380
Ile Met Thr Ser Lys Thr Asp Val Ser Ser Ser Val Ile Thr Ser Leu
385 390 395 400
Gly Ala Ile Val Ser Cys Tyr Gly Lys Thr Lys Cys Thr Ala Ser Asn
405 410 415
Lys Asn Arg Gly Ile Ile Lys Thr Phe Ser Asn Gly Cys Asp Tyr Val
420 425 430
Ser Asn Lys Gly Val Asp Thr Val Ser Val Gly Asn Thr Leu Tyr Tyr
435 440 445
Val Asn Lys Gln Glu Gly Lys Ser Leu Tyr Val Lys Gly Glu Pro Ile
450 455 460
Ile Asn Phe Tyr Asp Pro Leu Val Phe Pro Ser Asp Glu Phe Asp Ala
465 470 475 480
Ser Ile Ser Gln Val Asn Glu Lys Ile Asn Gln Ser Leu Ala Phe Ile
485 490 495
Arg Lys Ser Asp Glu Leu Leu Gly Leu Glu Val Leu Phe Gln Gly Pro
500 505 510
His His His His His His His His Ser Ala Trp Ser His Pro Gln Phe
515 520 525
Glu Lys
530
<210> 25
<211> 689
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 25
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gln Ser Thr Pro Ala Thr Asn Asn Arg Ala Arg Arg Glu Leu Pro
100 105 110
Arg Phe Met Asn Tyr Thr Leu Asn Asn Ala Lys Lys Thr Asn Val Thr
115 120 125
Leu Ser Lys Lys Arg Lys Arg Arg Phe Leu Gly Phe Leu Leu Gly Val
130 135 140
Gly Ser Ala Ile Ala Ser Gly Val Ala Val Cys Lys Val Leu His Leu
145 150 155 160
Glu Gly Glu Val Asn Lys Ile Lys Ser Ala Leu Leu Ser Thr Asn Lys
165 170 175
Ala Val Val Ser Leu Ser Asn Gly Val Ser Val Leu Thr Phe Lys Val
180 185 190
Leu Asp Leu Lys Asn Tyr Ile Asp Lys Gln Leu Leu Pro Ile Leu Asn
195 200 205
Lys Gln Ser Cys Ser Ile Ser Asn Ile Glu Thr Val Ile Glu Phe Gln
210 215 220
Gln Lys Asn Asn Arg Leu Leu Glu Ile Thr Arg Glu Phe Ser Val Asn
225 230 235 240
Ala Gly Val Thr Thr Pro Val Ser Thr Tyr Met Leu Thr Asn Ser Glu
245 250 255
Leu Leu Ser Leu Ile Asn Asp Met Pro Ile Thr Asn Asp Gln Lys Lys
260 265 270
Leu Met Ser Asn Asn Val Gln Ile Val Arg Gln Gln Ser Tyr Ser Ile
275 280 285
Met Cys Ile Ile Lys Glu Glu Val Leu Ala Tyr Val Val Gln Leu Pro
290 295 300
Leu Tyr Gly Val Ile Asp Thr Pro Cys Trp Lys Leu His Thr Ser Pro
305 310 315 320
Leu Cys Thr Thr Asn Thr Lys Glu Gly Ser Asn Ile Cys Leu Thr Arg
325 330 335
Thr Asp Arg Gly Trp Tyr Cys Asp Asn Ala Gly Ser Val Ser Phe Phe
340 345 350
Pro Gln Ala Glu Thr Cys Lys Val Gln Ser Asn Arg Val Phe Cys Asp
355 360 365
Thr Met Asn Ser Leu Thr Leu Pro Ser Glu Val Asn Leu Cys Asn Val
370 375 380
Asp Ile Phe Asn Pro Lys Tyr Asp Cys Lys Ile Met Thr Ser Lys Thr
385 390 395 400
Asp Val Ser Ser Ser Val Ile Thr Ser Leu Gly Ala Ile Val Ser Cys
405 410 415
Tyr Gly Lys Thr Lys Cys Thr Ala Ser Asn Lys Asn Arg Gly Ile Ile
420 425 430
Lys Thr Phe Ser Asn Gly Cys Asp Tyr Val Ser Asn Lys Gly Val Asp
435 440 445
Thr Val Ser Val Gly Asn Thr Leu Tyr Tyr Val Asn Lys Gln Glu Gly
450 455 460
Lys Ser Leu Tyr Val Lys Gly Glu Pro Ile Ile Asn Phe Tyr Asp Pro
465 470 475 480
Leu Val Phe Pro Ser Asp Glu Phe Asp Ala Ser Ile Ser Gln Val Asn
485 490 495
Glu Lys Ile Asn Gln Ser Leu Ala Phe Ile Arg Lys Ser Asp Glu Leu
500 505 510
Leu Ser Gly Gly Ser Ser Gly Ser Ser Gly Gly Ser Asp Ile Ile Lys
515 520 525
Leu Leu Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr
530 535 540
Met Ser Met Ser Ser Trp Cys Tyr Thr His Ser Leu Asp Gly Ala Gly
545 550 555 560
Leu Phe Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys
565 570 575
Leu Ile Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser
580 585 590
Ile Ser Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln
595 600 605
Lys Ala Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile
610 615 620
Val Asp His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu
625 630 635 640
Gln Trp Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp
645 650 655
Ile Leu Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr
660 665 670
Leu Ala Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser Gly
675 680 685
Ser
<210> 26
<211> 574
<212> БЕЛОК
<213> Респираторно-синцитиальный вирус человека
<400> 26
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1. 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gln Ser Thr Pro Pro Thr Asn Asn Arg Ala Arg Arg Glu Leu Pro
100 105 110
Arg Phe Met Asn Tyr Thr Leu Asn Asn Ala Lys Lys Thr Asn Val Thr
115 120 125
Leu Ser Lys Lys Arg Lys Arg Arg Phe Leu Gly Phe Leu Leu Gly Val
130 135 140
Gly Ser Ala Ile Ala Ser Gly Val Ala Val Ser Lys Val Leu His Leu
145 150 155 160
Glu Gly Glu Val Asn Lys Ile Lys Ser Ala Leu Leu Ser Thr Asn Lys
165 170 175
Ala Val Val Ser Leu Ser Asn Gly Val Ser Val Leu Thr Ser Lys Val
180 185 190
Leu Asp Leu Lys Asn Tyr Ile Asp Lys Gln Leu Leu Pro Ile Val Asn
195 200 205
Lys Gln Ser Cys Ser Ile Ser Asn Ile Glu Thr Val Ile Glu Phe Gln
210 215 220
Gln Lys Asn Asn Arg Leu Leu Glu Ile Thr Arg Glu Phe Ser Val Asn
225 230 235 240
Ala Gly Val Thr Thr Pro Val Ser Thr Tyr Met Leu Thr Asn Ser Glu
245 250 255
Leu Leu Ser Leu Ile Asn Asp Met Pro Ile Thr Asn Asp Gln Lys Lys
260 265 270
Leu Met Ser Asn Asn Val Gln Ile Val Arg Gln Gln Ser Tyr Ser Ile
275 280 285
Met Ser Ile Ile Lys Glu Glu Val Leu Ala Tyr Val Val Gln Leu Pro
290 295 300
Leu Tyr Gly Val Ile Asp Thr Pro Cys Trp Lys Leu His Thr Ser Pro
305 310 315 320
Leu Cys Thr Thr Asn Thr Lys Glu Gly Ser Asn Ile Cys Leu Thr Arg
325 330 335
Thr Asp Arg Gly Trp Tyr Cys Asp Asn Ala Gly Ser Val Ser Phe Phe
340 345 350
Pro Gln Ala Glu Thr Cys Lys Val Gln Ser Asn Arg Val Phe Cys Asp
355 360 365
Thr Met Asn Ser Leu Thr Leu Pro Ser Glu Ile Asn Leu Cys Asn Val
370 375 380
Asp Ile Phe Asn Pro Lys Tyr Asp Cys Lys Ile Met Thr Ser Lys Thr
385 390 395 400
Asp Val Ser Ser Ser Val Ile Thr Ser Leu Gly Ala Ile Val Ser Cys
405 410 415
Tyr Gly Lys Thr Lys Cys Thr Ala Ser Asn Lys Asn Arg Gly Ile Ile
420 425 430
Lys Thr Phe Ser Asn Gly Cys Asp Tyr Val Ser Asn Lys Gly Met Asp
435 440 445
Thr Val Ser Val Gly Asn Thr Leu Tyr Tyr Val Asn Lys Gln Glu Gly
450 455 460
Lys Ser Leu Tyr Val Lys Gly Glu Pro Ile Ile Asn Phe Tyr Asp Pro
465 470 475 480
Leu Val Phe Pro Ser Asp Glu Phe Asp Ala Ser Ile Ser Gln Val Asn
485 490 495
Glu Lys Ile Asn Gln Ser Leu Ala Phe Ile Arg Lys Ser Asp Glu Leu
500 505 510
Leu His Asn Val Asn Ala Gly Lys Ser Thr Thr Asn Ile Met Ile Thr
515 520 525
Thr Ile Ile Ile Val Ile Ile Val Ile Leu Leu Ser Leu Ile Ala Val
530 535 540
Gly Leu Leu Leu Tyr Cys Lys Ala Arg Ser Thr Pro Val Thr Leu Ser
545 550 555 560
Lys Asp Gln Leu Ser Gly Ile Asn Asn Ile Ala Phe Ser Asn
565 570
<210> 27
<211> 297
<212> БЕЛОК
<213> Респираторно-синцитиальный вирус человека
<400> 27
Met Ser Lys Asn Lys Asp Gln Arg Thr Ala Lys Thr Leu Glu Arg Thr
1. 5 10 15
Trp Asp Thr Leu Asn His Leu Leu Phe Ile Ser Ser Cys Leu Tyr Lys
20 25 30
Leu Asn Leu Lys Ser Val Ala Gln Ile Thr Leu Ser Ile Leu Ala Met
35 40 45
Ile Ile Ser Thr Ser Leu Ile Ile Val Ala Ile Ile Phe Ile Ala Ser
50 55 60
Ala Asn His Lys Ile Thr Ser Thr Thr Thr Ile Ile Gln Asp Ala Thr
65 70 75 80
Asn Gln Ile Lys Asn Thr Thr Pro Thr Tyr Leu Thr Gln Asn Pro Gln
85 90 95
Leu Gly Ile Ser Pro Ser Asn Pro Ser Asp Ile Thr Ser Leu Ile Thr
100 105 110
Thr Ile Leu Asp Ser Thr Thr Pro Gly Val Lys Ser Thr Leu Gln Ser
115 120 125
Thr Thr Val Gly Thr Lys Asn Thr Thr Thr Thr Gln Ala Gln Pro Asn
130 135 140
Lys Pro Thr Thr Lys Gln Arg Gln Asn Lys Pro Pro Ser Lys Pro Asn
145 150 155 160
Asn Asp Phe His Phe Glu Val Phe Asn Phe Val Pro Cys Ser Ile Cys
165 170 175
Ser Asn Asn Pro Thr Cys Trp Ala Ile Cys Lys Arg Ile Pro Asn Lys
180 185 190
Lys Pro Gly Lys Arg Thr Thr Thr Lys Pro Thr Lys Lys Pro Thr Pro
195 200 205
Lys Thr Thr Lys Lys Gly Pro Lys Pro Gln Thr Thr Lys Ser Lys Glu
210 215 220
Ala Pro Thr Thr Lys Pro Thr Glu Glu Pro Thr Ile Asn Thr Thr Lys
225 230 235 240
Thr Asn Ile Ile Thr Thr Leu Leu Thr Ser Asn Thr Thr Arg Asn Pro
245 250 255
Glu Leu Thr Ser Gln Met Glu Thr Phe His Ser Thr Ser Ser Glu Gly
260 265 270
Asn Pro Ser Pro Ser Gln Val Ser Ile Thr Ser Glu Tyr Pro Ser Gln
275 280 285
Pro Ser Ser Pro Pro Asn Thr Pro Arg
290 295
<210> 28
<211> 233
<212> БЕЛОК
<213> Респираторно-синцитиальный вирус человека
<400> 28
Asn His Lys Val Thr Leu Thr Thr Ala Ile Ile Gln Asp Ala Thr Ser
1. 5 10 15
Gln Ile Lys Asn Thr Thr Pro Thr Tyr Leu Thr Gln Asp Pro Gln Leu
20 25 30
Gly Ile Ser Phe Ser Asn Leu Ser Glu Ile Thr Ser Gln Thr Thr Thr
35 40 45
Ile Leu Ala Ser Thr Thr Pro Gly Val Lys Ser Asn Leu Gln Pro Thr
50 55 60
Thr Val Lys Thr Lys Asn Thr Thr Thr Thr Gln Thr Gln Pro Ser Lys
65 70 75 80
Pro Thr Thr Lys Gln Arg Gln Asn Lys Pro Pro Asn Lys Pro Asn Asn
85 90 95
Asp Phe His Phe Glu Val Phe Asn Phe Val Pro Cys Ser Ile Cys Ser
100 105 110
Asn Asn Pro Thr Cys Trp Ala Ile Cys Lys Arg Ile Pro Asn Lys Lys
115 120 125
Pro Gly Lys Lys Thr Thr Thr Lys Pro Thr Lys Lys Pro Thr Phe Lys
130 135 140
Thr Thr Lys Lys Asp His Lys Pro Gln Thr Thr Lys Pro Lys Glu Val
145 150 155 160
Pro Thr Thr Lys Pro Thr Glu Glu Pro Thr Ile Asn Thr Thr Lys Thr
165 170 175
Asn Ile Ile Thr Thr Leu Leu Thr Asn Asn Thr Thr Gly Asn Pro Lys
180 185 190
Leu Thr Ser Gln Met Glu Thr Phe His Ser Thr Ser Ser Glu Gly Asn
195 200 205
Leu Ser Pro Ser Gln Val Ser Thr Thr Ser Glu His Pro Ser Gln Pro
210 215 220
Ser Ser Pro Pro Asn Thr Thr Arg Gln
225 230
<210> 29
<211> 56
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 29
Ser Gly Gly Ser Ser Gly Ser Ser Glu Glu Glu Gly Gly Ser Arg Gln
1. 5 10 15
Asn Lys Pro Asn Asn Asp Phe His Phe Glu Val Phe Asn Phe Val Pro
20 25 30
Cys Ser Ile Cys Ser Asn Asn Pro Thr Cys Trp Ala Ile Cys Lys Arg
35 40 45
Ile Pro Asn Lys Lys Glu Glu Glu
50 55
<210> 30
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 30
tgactgtgaa cgttcgagat ga 22
<210> 31
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 31
Gly Ser Gly Asn Val Gly Leu
1. 5
<210> 32
<211> 43
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 32
Arg Gln Asn Lys Pro Pro Asn Lys Pro Asn Asn Asp Phe His Phe Glu
1. 5 10 15
Val Phe Asn Phe Val Pro Cys Ser Ile Cys Ser Asn Asn Pro Thr Cys
20 25 30
Trp Ala Ile Cys Lys Arg Ile Pro Asn Lys Lys
35 40
<210> 33
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 33
Arg Lys Asn Pro Pro Lys Lys Pro Lys Asp Asp Tyr His Phe Glu Val
1. 5 10 15
Phe Asn Phe Val Pro Cys Ser Ile Cys Gly Asn Asn Gln Leu Cys Lys
20 25 30
Ser Ile Cys Lys Thr Ile Pro Asn Lys Lys
35 40
<210> 34
<400> 34
000
<210> 35
<400> 35
000
<210> 36
<400> 36
000
<210> 37
<400> 37
000
<210> 38
<400> 38
000
<210> 39
<400> 39
000
<210> 40
<400> 40
000
<210> 41
<400> 41
000
<210> 42
<400> 42
000
<210> 43
<400> 43
000
<210> 44
<400> 44
000
<210> 45
<400> 45
000
<210> 46
<400> 46
000
<210> 47
<400> 47
000
<210> 48
<400> 48
000
<210> 49
<400> 49
000
<210> 50
<400> 50
000
<210> 51
<400> 51
000
<210> 52
<400> 52
000
<210> 53
<400> 53
000
<210> 54
<400> 54
000
<210> 55
<400> 55
000
<210> 56
<400> 56
000
<210> 57
<400> 57
000
<210> 58
<400> 58
000
<210> 59
<400> 59
000
<210> 60
<400> 60
000
<210> 61
<400> 61
000
<210> 62
<400> 62
000
<210> 63
<400> 63
000
<210> 64
<400> 64
000
<210> 65
<400> 65
000
<210> 66
<400> 66
000
<210> 67
<400> 67
000
<210> 68
<400> 68
000
<210> 69
<400> 69
000
<210> 70
<400> 70
000
<210> 71
<400> 71
000
<210> 72
<400> 72
000
<210> 73
<400> 73
000
<210> 74
<400> 74
000
<210> 75
<400> 75
000
<210> 76
<400> 76
000
<210> 77
<400> 77
000
<210> 78
<400> 78
000
<210> 79
<400> 79
000
<210> 80
<400> 80
000
<210> 81
<400> 81
000
<210> 82
<400> 82
000
<210> 83
<400> 83
000
<210> 84
<400> 84
000
<210> 85
<400> 85
000
<210> 86
<400> 86
000
<210> 87
<400> 87
000
<210> 88
<400> 88
000
<210> 89
<400> 89
000
<210> 90
<400> 90
000
<210> 91
<400> 91
000
<210> 92
<400> 92
000
<210> 93
<400> 93
000
<210> 94
<400> 94
000
<210> 95
<400> 95
000
<210> 96
<400> 96
000
<210> 97
<400> 97
000
<210> 98
<400> 98
000
<210> 99
<400> 99
000
<210> 100
<400> 100
000
<210> 101
<400> 101
000
<210> 102
<400> 102
000
<210> 103
<400> 103
000
<210> 104
<400> 104
000
<210> 105
<400> 105
000
<210> 106
<400> 106
000
<210> 107
<400> 107
000
<210> 108
<400> 108
000
<210> 109
<400> 109
000
<210> 110
<400> 110
000
<210> 111
<400> 111
000
<210> 112
<400> 112
000
<210> 113
<400> 113
000
<210> 114
<400> 114
000
<210> 115
<400> 115
000
<210> 116
<400> 116
000
<210> 117
<400> 117
000
<210> 118
<400> 118
000
<210> 119
<400> 119
000
<210> 120
<400> 120
000
<210> 121
<400> 121
000
<210> 122
<400> 122
000
<210> 123
<400> 123
000
<210> 124
<400> 124
000
<210> 125
<400> 125
000
<210> 126
<400> 126
000
<210> 127
<400> 127
000
<210> 128
<400> 128
000
<210> 129
<400> 129
000
<210> 130
<400> 130
000
<210> 131
<400> 131
000
<210> 132
<400> 132
000
<210> 133
<400> 133
000
<210> 134
<400> 134
000
<210> 135
<400> 135
000
<210> 136
<400> 136
000
<210> 137
<400> 137
000
<210> 138
<400> 138
000
<210> 139
<400> 139
000
<210> 140
<400> 140
000
<210> 141
<400> 141
000
<210> 142
<400> 142
000
<210> 143
<400> 143
000
<210> 144
<400> 144
000
<210> 145
<400> 145
000
<210> 146
<400> 146
000
<210> 147
<400> 147
000
<210> 148
<400> 148
000
<210> 149
<400> 149
000
<210> 150
<400> 150
000
<210> 151
<400> 151
000
<210> 152
<400> 152
000
<210> 153
<400> 153
000
<210> 154
<400> 154
000
<210> 155
<400> 155
000
<210> 156
<400> 156
000
<210> 157
<400> 157
000
<210> 158
<400> 158
000
<210> 159
<400> 159
000
<210> 160
<400> 160
000
<210> 161
<400> 161
000
<210> 162
<400> 162
000
<210> 163
<400> 163
000
<210> 164
<400> 164
000
<210> 165
<400> 165
000
<210> 166
<400> 166
000
<210> 167
<400> 167
000
<210> 168
<400> 168
000
<210> 169
<400> 169
000
<210> 170
<400> 170
000
<210> 171
<400> 171
000
<210> 172
<400> 172
000
<210> 173
<400> 173
000
<210> 174
<400> 174
000
<210> 175
<400> 175
000
<210> 176
<400> 176
000
<210> 177
<400> 177
000
<210> 178
<400> 178
000
<210> 179
<400> 179
000
<210> 180
<400> 180
000
<210> 181
<400> 181
000
<210> 182
<400> 182
000
<210> 183
<400> 183
000
<210> 184
<400> 184
000
<210> 185
<400> 185
000
<210> 186
<400> 186
000
<210> 187
<400> 187
000
<210> 188
<400> 188
000
<210> 189
<400> 189
000
<210> 190
<400> 190
000
<210> 191
<400> 191
000
<210> 192
<400> 192
000
<210> 193
<400> 193
000
<210> 194
<400> 194
000
<210> 195
<400> 195
000
<210> 196
<400> 196
000
<210> 197
<400> 197
000
<210> 198
<400> 198
000
<210> 199
<400> 199
000
<210> 200
<400> 200
000
<210> 201
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 201
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Cys
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 202
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 202
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Cys Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 203
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 203
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Cys Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 204
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 204
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Cys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 205
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 205
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Cys Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 206
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 206
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Cys Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 207
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 207
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Cys Gln Val Asn Lys Glu Met Gln Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Ser Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 208
<211> 173
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 208
Glu Ser Gln Val Arg Gln Gln Phe Ser Lys Asp Ile Glu Lys Leu Leu
1. 5 10 15
Asn Glu Gln Val Asn Lys Glu Met Asn Ser Ser Asn Leu Tyr Met Ser
20 25 30
Met Ser Ser Trp Cys Tyr Thr His Ser Leu Asp Gly Ala Gly Leu Phe
35 40 45
Leu Phe Asp His Ala Ala Glu Glu Tyr Glu His Ala Lys Lys Leu Ile
50 55 60
Ile Phe Leu Asn Glu Asn Asn Val Pro Val Gln Leu Thr Ser Ile Ser
65 70 75 80
Ala Pro Glu His Lys Phe Glu Gly Leu Thr Gln Ile Phe Gln Lys Ala
85 90 95
Tyr Glu His Glu Gln His Ile Ser Glu Ser Ile Asn Asn Ile Val Asp
100 105 110
His Ala Ile Lys Ser Lys Asp His Ala Thr Phe Asn Phe Leu Gln Trp
115 120 125
Tyr Val Ala Glu Gln His Glu Glu Glu Val Leu Phe Lys Asp Ile Leu
130 135 140
Asp Lys Ile Glu Leu Ile Gly Asn Glu Asn His Gly Leu Tyr Leu Ala
145 150 155 160
Asp Gln Tyr Val Lys Gly Ile Ala Lys Ser Arg Lys Ser
165 170
<210> 209
<211> 166
<212> БЕЛОК
<213> Helicobacter pylori
<400> 209
Leu Ser Lys Asp Ile Ile Lys Leu Leu Asn Glu Gln Val Asn Lys Glu
1. 5 10 15
Met Asn Ser Ser Asn Leu Tyr Met Ser Met Ser Ser Trp Cys Tyr Thr
20 25 30
His Ser Leu Asp Gly Ala Gly Leu Phe Leu Phe Asp His Ala Ala Glu
35 40 45
Glu Tyr Glu His Ala Lys Lys Leu Ile Ile Phe Leu Asn Glu Asn Asn
50 55 60
Val Pro Val Gln Leu Thr Ser Ile Ser Ala Pro Glu His Lys Phe Glu
65 70 75 80
Gly Leu Thr Gln Ile Phe Gln Lys Ala Tyr Glu His Glu Gln His Ile
85 90 95
Ser Glu Ser Ile Asn Asn Ile Val Asp His Ala Ile Lys Ser Lys Asp
100 105 110
His Ala Thr Phe Asn Phe Leu Gln Trp Tyr Val Ala Glu Gln His Glu
115 120 125
Glu Glu Val Leu Phe Lys Asp Ile Leu Asp Lys Ile Glu Leu Ile Gly
130 135 140
Asn Glu Asn His Gly Leu Tyr Leu Ala Asp Gln Tyr Val Lys Gly Ile
145 150 155 160
Ala Lys Ser Arg Lys Ser
165
<210> 210
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический олигонуклеотид
<400> 210
tgactgtgaa cgttcgagat ga 22
<210> 211
<211> 191
<212> БЕЛОК
<213> Trichoplusia ni
<400> 211
Thr Gln Cys Asn Val Asn Pro Val Gln Ile Pro Lys Asp Trp Ile Thr
1. 5 10 15
Met His Arg Ser Cys Arg Asn Ser Met Arg Gln Gln Ile Gln Met Glu
20 25 30
Val Gly Ala Ser Leu Gln Tyr Leu Ala Met Gly Ala His Phe Ser Lys
35 40 45
Asp Val Val Asn Arg Pro Gly Phe Ala Gln Leu Phe Phe Asp Ala Ala
50 55 60
Ser Glu Glu Arg Glu His Ala Met Lys Leu Ile Glu Tyr Leu Leu Met
65 70 75 80
Arg Gly Glu Leu Thr Asn Asp Val Ser Ser Leu Leu Gln Val Arg Pro
85 90 95
Pro Thr Arg Ser Ser Trp Lys Gly Gly Val Glu Ala Leu Glu His Ala
100 105 110
Leu Ser Met Glu Ser Asp Val Thr Lys Ser Ile Arg Asn Val Ile Lys
115 120 125
Ala Cys Glu Asp Asp Ser Glu Phe Asn Asp Tyr His Leu Val Asp Tyr
130 135 140
Leu Thr Gly Asp Phe Leu Glu Glu Gln Tyr Lys Gly Gln Arg Asp Leu
145 150 155 160
Ala Gly Lys Ala Ser Thr Leu Lys Lys Leu Met Asp Arg His Glu Ala
165 170 175
Leu Gly Glu Phe Ile Phe Asp Lys Lys Leu Leu Gly Ile Asp Val
180 185 190
<210> 212
<211> 212
<212> БЕЛОК
<213> Trichoplusia ni
<400> 212
Ala Asp Thr Cys Tyr Asn Asp Val Ala Leu Asp Cys Gly Ile Thr Ser
1. 5 10 15
Asn Ser Leu Ala Leu Pro Arg Cys Asn Ala Val Tyr Gly Glu Tyr Gly
20 25 30
Ser His Gly Asn Val Ala Thr Glu Leu Gln Ala Tyr Ala Lys Leu His
35 40 45
Leu Glu Arg Ser Tyr Asp Tyr Leu Leu Ser Ala Ala Tyr Phe Asn Asn
50 55 60
Tyr Gln Thr Asn Arg Ala Gly Phe Ser Lys Leu Phe Lys Lys Leu Ser
65 70 75 80
Asp Glu Ala Trp Ser Lys Thr Ile Asp Ile Ile Lys His Val Thr Lys
85 90 95
Arg Gly Asp Lys Met Asn Phe Asp Gln His Ser Thr Met Lys Thr Glu
100 105 110
Arg Lys Asn Tyr Thr Ala Glu Asn His Glu Leu Glu Ala Leu Ala Lys
115 120 125
Ala Leu Asp Thr Gln Lys Glu Leu Ala Glu Arg Ala Phe Tyr Ile His
130 135 140
Arg Glu Ala Thr Arg Asn Ser Gln His Leu His Asp Pro Glu Ile Ala
145 150 155 160
Gln Tyr Leu Glu Glu Glu Phe Ile Glu Asp His Ala Glu Lys Ile Arg
165 170 175
Thr Leu Ala Gly His Thr Ser Asp Leu Lys Lys Phe Ile Thr Ala Asn
180 185 190
Asn Gly His Asp Leu Ser Leu Ala Leu Tyr Val Phe Asp Glu Tyr Leu
195 200 205
Gln Lys Thr Val
210
<210> 213
<211> 174
<212> БЕЛОК
<213> Pyrococcus furiosus
<400> 213
Met Leu Ser Glu Arg Met Leu Lys Ala Leu Asn Asp Gln Leu Asn Arg
1. 5 10 15
Glu Leu Tyr Ser Ala Tyr Leu Tyr Phe Ala Met Ala Ala Tyr Phe Glu
20 25 30
Asp Leu Gly Leu Glu Gly Phe Ala Asn Trp Met Lys Ala Gln Ala Glu
35 40 45
Glu Glu Ile Gly His Ala Leu Arg Phe Tyr Asn Tyr Ile Tyr Asp Arg
50 55 60
Asn Gly Arg Val Glu Leu Asp Glu Ile Pro Lys Pro Pro Lys Glu Trp
65 70 75 80
Glu Ser Pro Leu Lys Ala Phe Glu Ala Ala Tyr Glu His Glu Lys Phe
85 90 95
Ile Ser Lys Ser Ile Tyr Glu Leu Ala Ala Leu Ala Glu Glu Glu Lys
100 105 110
Asp Tyr Ser Thr Arg Ala Phe Leu Glu Trp Phe Ile Asn Glu Gln Val
115 120 125
Glu Glu Glu Ala Ser Val Lys Lys Ile Leu Asp Lys Leu Lys Phe Ala
130 135 140
Lys Asp Ser Pro Gln Ile Leu Phe Met Leu Asp Lys Glu Leu Ser Ala
145 150 155 160
Arg Ala Pro Lys Leu Pro Gly Leu Leu Met Gln Gly Gly Glu
165 170
<210> 214
<211> 183
<212> БЕЛОК
<213> Homo sapiens
<400> 214
Met Thr Thr Ala Ser Thr Ser Gln Val Arg Gln Asn Tyr His Gln Asp
1. 5 10 15
Ser Glu Ala Ala Ile Asn Arg Gln Ile Asn Leu Glu Leu Tyr Ala Ser
20 25 30
Tyr Val Tyr Leu Ser Met Ser Tyr Tyr Phe Asp Arg Asp Asp Val Ala
35 40 45
Leu Lys Asn Phe Ala Lys Tyr Phe Leu His Gln Ser His Glu Glu Arg
50 55 60
Glu His Ala Glu Lys Leu Met Lys Leu Gln Asn Gln Arg Gly Gly Arg
65 70 75 80
Ile Phe Leu Gln Asp Ile Lys Lys Pro Asp Cys Asp Asp Trp Glu Ser
85 90 95
Gly Leu Asn Ala Met Glu Cys Ala Leu His Leu Glu Lys Asn Val Gln
100 105 110
Gln Ser Leu Leu Glu Leu His Lys Leu Ala Thr Asp Lys Asn Asp Pro
115 120 125
His Leu Cys Asp Phe Ile Glu Thr His Tyr Leu Asn Glu Gln Val Lys
130 135 140
Ala Ile Lys Glu Leu Gly Asp His Val Thr Asn Leu Arg Lys Met Gly
145 150 155 160
Ala Pro Glu Ser Gly Leu Ala Glu Tyr Leu Phe Asp Lys His Thr Leu
165 170 175
Gly Asp Ser Asp Gln Glu Ser
180
<210> 215
<211> 204
<212> БЕЛОК
<213> Homo sapiens
<400> 215
Met Asp Ser Lys Gly Ser Ser Gln Lys Gly Ser Arg Leu Leu Leu Leu
1. 5 10 15
Leu Val Val Ser Asn Leu Leu Leu Pro Gln Gly Val Leu Ala Ser Ser
20 25 30
Gln Ile Arg Gln Asn Tyr Ser Thr Asp Val Glu Ala Ala Val Asn Ser
35 40 45
Leu Val Asn Leu Tyr Leu Gln Ala Ser Tyr Thr Tyr Leu Ser Leu Gly
50 55 60
Phe Tyr Phe Asp Arg Asp Asp Val Ala Leu Glu Gly Val Ser His Phe
65 70 75 80
Phe Arg Glu Leu Ala Glu Glu Lys Arg Glu Gly Tyr Glu Arg Leu Leu
85 90 95
Lys Met Gln Asn Gln Arg Gly Gly Arg Ala Leu Phe Gln Asp Ile Lys
100 105 110
Lys Pro Ala Glu Asp Glu Trp Gly Lys Thr Pro Asp Ala Met Lys Ala
115 120 125
Ala Met Ala Leu Glu Lys Lys Leu Asn Gln Ala Leu Leu Asp Leu His
130 135 140
Ala Leu Gly Ser Ala Arg Thr Asp Pro His Leu Cys Asp Phe Leu Glu
145 150 155 160
Thr His Phe Leu Asp Glu Glu Val Lys Leu Ile Lys Lys Met Gly Asp
165 170 175
His Leu Thr Asn Leu His Arg Leu Gly Gly Pro Glu Ala Gly Leu Gly
180 185 190
Glu Tyr Leu Phe Glu Arg Leu Thr Leu Lys His Asp
195 200
<210> 216
<211> 154
<212> БЕЛОК
<213> Aquifex aeolicus
<400> 216
Met Gln Ile Tyr Glu Gly Lys Leu Thr Ala Glu Gly Leu Arg Phe Gly
1. 5 10 15
Ile Val Ala Ser Arg Phe Asn His Ala Leu Val Asp Arg Leu Val Glu
20 25 30
Gly Ala Ile Asp Cys Ile Val Arg His Gly Gly Arg Glu Glu Asp Ile
35 40 45
Thr Leu Val Arg Val Pro Gly Ser Trp Glu Ile Pro Val Ala Ala Gly
50 55 60
Glu Leu Ala Arg Lys Glu Asp Ile Asp Ala Val Ile Ala Ile Gly Val
65 70 75 80
Leu Ile Arg Gly Ala Thr Pro His Phe Asp Tyr Ile Ala Ser Glu Val
85 90 95
Ser Lys Gly Leu Ala Asn Leu Ser Leu Glu Leu Arg Lys Pro Ile Thr
100 105 110
Phe Gly Val Ile Thr Ala Asp Thr Leu Glu Gln Ala Ile Glu Arg Ala
115 120 125
Gly Thr Lys His Gly Asn Lys Gly Trp Glu Ala Ala Leu Ser Ala Ile
130 135 140
Glu Met Ala Asn Leu Phe Lys Ser Leu Arg
145 150
<210> 217
<211> 8
<212> БЕЛОК
<213> Неустановленное
<220>
<223> Последовательность линкера лягушки-быка
<400> 217
Glu Ser Gln Val Arg Gln Gln Phe
1. 5
<210> 218
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 218
Cys Leu Val Pro Arg Gly Ser Leu Glu His His His His His His
1. 5 10 15
<210> 219
<211> 156
<212> БЕЛОК
<213> Escherichia coli
<400> 219
Met Asn Ile Ile Glu Ala Asn Val Ala Thr Pro Asp Ala Arg Val Ala
1. 5 10 15
Ile Thr Ile Ala Arg Phe Asn Asn Phe Ile Asn Asp Ser Leu Leu Glu
20 25 30
Gly Ala Ile Asp Ala Leu Lys Arg Ile Gly Gln Val Lys Asp Glu Asn
35 40 45
Ile Thr Val Val Trp Val Pro Gly Ala Tyr Glu Leu Pro Leu Ala Ala
50 55 60
Gly Ala Leu Ala Lys Thr Gly Lys Tyr Asp Ala Val Ile Ala Leu Gly
65 70 75 80
Thr Val Ile Arg Gly Gly Thr Ala His Phe Glu Tyr Val Ala Gly Gly
85 90 95
Ala Ser Asn Gly Leu Ala His Val Ala Gln Asp Ser Glu Ile Pro Val
100 105 110
Ala Phe Gly Val Leu Thr Thr Glu Ser Ile Glu Gln Ala Ile Glu Arg
115 120 125
Ala Gly Thr Lys Ala Gly Asn Lys Gly Ala Glu Ala Ala Leu Thr Ala
130 135 140
Leu Glu Met Ile Asn Val Leu Lys Ala Ile Lys Ala
145 150 155
<210> 220
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 220
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1. 5 10 15
<210> 221
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 221
Gly Gly Ser Gly Ser Gly Ser Asn Ser Ser Ala Ser Ser Gly Ala Ser
1. 5 10 15
Ser Gly Gly Ala Ser Gly Gly Ser Gly Gly Ser Gly
20 25
<210> 222
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 222
Gly Gly Ser Gly Ser Ala Ser Ser Gly Ala Ser Ala Ser Gly Ser Ser
1. 5 10 15
Asn Gly Ser Gly Ser Gly Ser Gly Ser Asn Ser Ser Ala Ser Ser Gly
20 25 30
Ala Ser Ser Gly Gly Ala Ser Gly Gly Ser Gly Gly Ser Gly
35 40 45
<210> 223
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 223
Gly Gly Ser Gly Ser Ala Ser Ala Glu Ala Ala Ala Lys Glu Ala Ala
1. 5 10 15
Ala Lys Ala Gly Gly Ser Gly Gly Ser Gly
20 25
<210> 224
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 224
Gly Gly Ser Gly Ser Ala Ser Ala Glu Ala Ala Ala Lys Glu Ala Ala
1. 5 10 15
Ala Lys Glu Ala Ala Ala Lys Ala Ser Gly Gly Ser Gly Gly Ser Gly
20 25 30
<210> 225
<211> 47
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический полипептид
<400> 225
Ser Gly Gly Gly Ser Gly Ser Ala Ser Ser Gly Ala Ser Ala Ser Gly
1. 5 10 15
Ser Ser Cys Ser Gly Ser Gly Ser Gly Ser Ser Ser Ala Ser Ser Gly
20 25 30
Ala Ser Ser Gly Gly Ala Ser Gly Gly Gly Ser Gly Gly Ser Gly
35 40 45
<210> 226
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 226
Gly Gly Gly Ser
1
<210> 227
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 227
Gly Gly Gly Ser Gly Gly Gly Ser
1. 5
<210> 228
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 228
Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser Gly Gly Gly Ser
1. 5 10 15
Gly Gly Gly Ser
20
<210> 229
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Синтетический пептид
<400> 229
Ser Gly Ser Gly Ser
1. 5
<210> 230
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 6xHis-метка
<400> 230
His His His His His His
1. 5
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИГЕННЫЕ ПОЛИПЕПТИДЫ НА ОСНОВЕ ПОСЛЕДОВАТЕЛЬНОСТИ OspA | 2019 |
|
RU2816208C2 |
| БЕЛКИ RSV В ПРЕДШЕСТВУЮЩЕЙ СЛИЯНИЮ КОНФОРМАЦИИ И ИХ ПРИМЕНЕНИЕ | 2014 |
|
RU2761631C2 |
| МУТАНТЫ БЕЛКА F RSV | 2016 |
|
RU2788403C2 |
| МУТАНТНЫЙ БЕЛОК F RSV И ЕГО ПРИМЕНЕНИЕ | 2020 |
|
RU2807742C1 |
| МУТАНТЫ БЕЛКА F RSV | 2016 |
|
RU2723039C2 |
| СТАБИЛИЗИРОВАННЫЕ БЕЛКИ F RSV И ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2795459C2 |
| СКОНСТРУИРОВАННЫЕ МУЛЬТИСПЕЦИФИЧЕСКИЕ АНТИТЕЛА И ДРУГИЕ МУЛЬТИМЕРНЫЕ БЕЛКИ С АСИММЕТРИЧНЫМИ МУТАЦИЯМИ В ОБЛАСТИ CH2-CH3 | 2018 |
|
RU2804031C2 |
| МУЛЬТИВАЛЕНТНЫЕ PD-L1-СВЯЗЫВАЮЩИЕ СОЕДИНЕНИЯ ДЛЯ ЛЕЧЕНИЯ ЗЛОКАЧЕСТВЕННЫХ НОВООБРАЗОВАНИЙ | 2020 |
|
RU2816646C2 |
| АНТАГОНИСТЫ NGF ДЛЯ МЕДИЦИНСКОГО ИСПОЛЬЗОВАНИЯ | 2020 |
|
RU2829812C2 |
| ВАРИАНТЫ FC-ОБЛАСТИ С ИЗМЕНЕННЫМ СВЯЗЫВАНИЕМ С НЕОНАТАЛЬНЫМ FC-РЕЦЕПТОРОМ (FCRN) ДЛЯ ПРИМЕНЕНИЯ В ВЕТЕРИНАРИИ | 2019 |
|
RU2830231C2 |
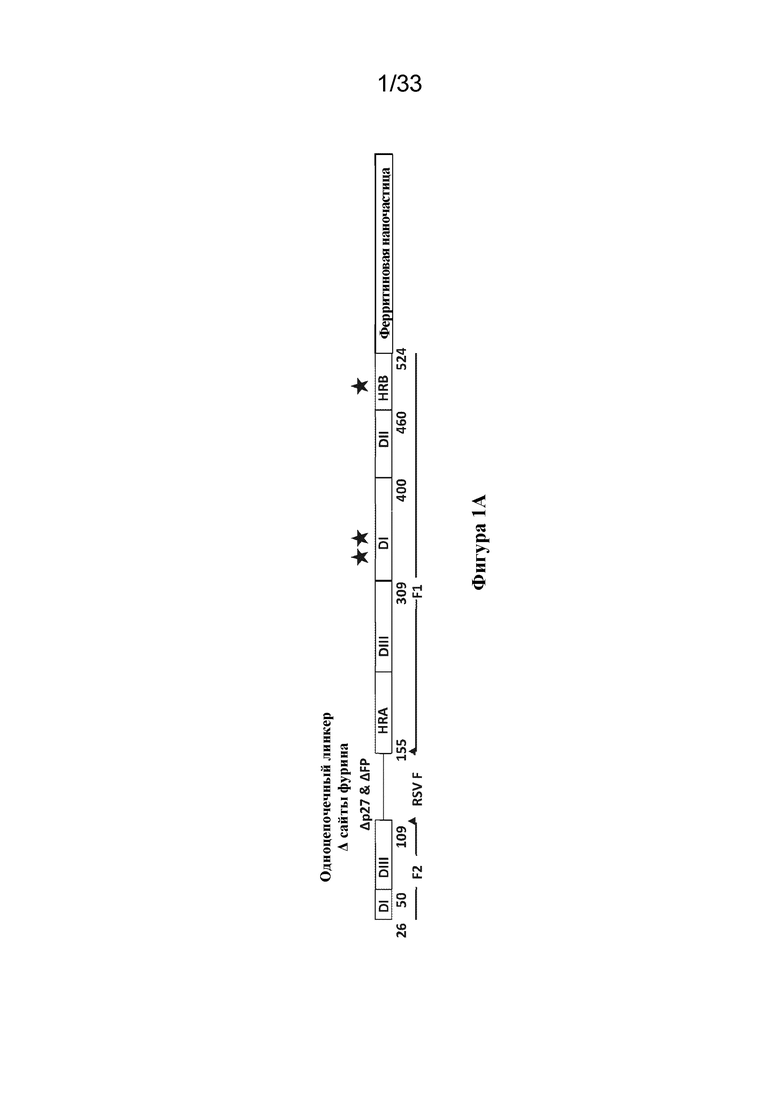
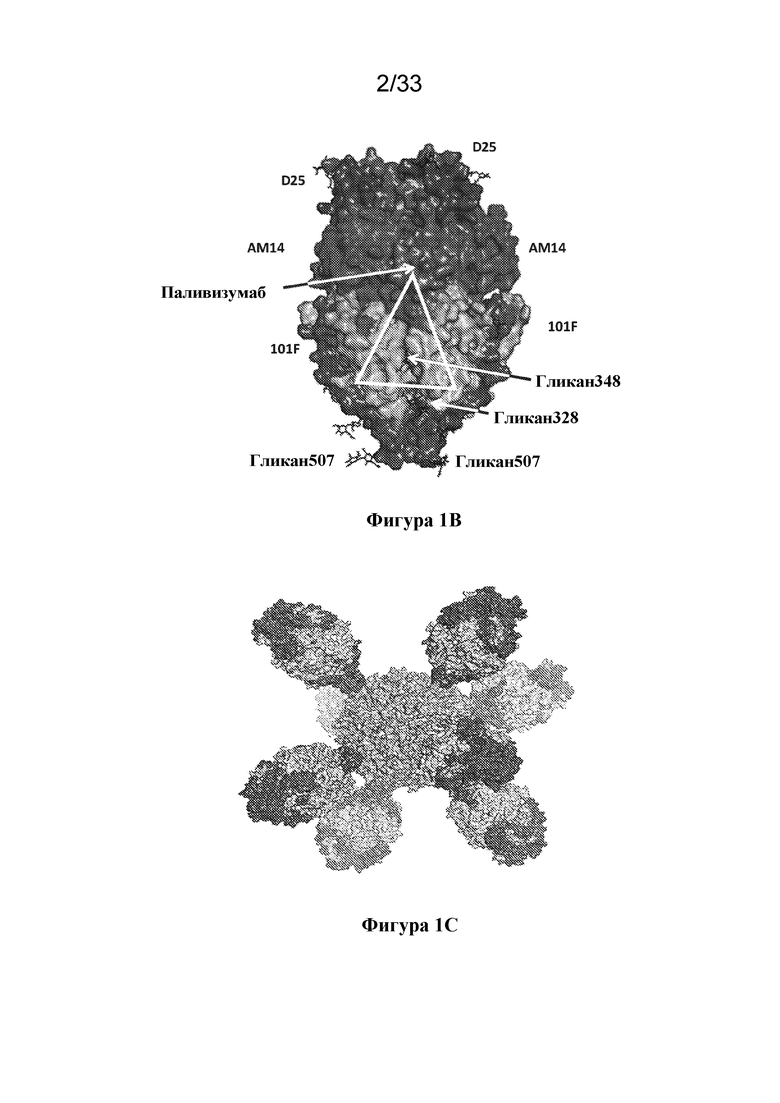

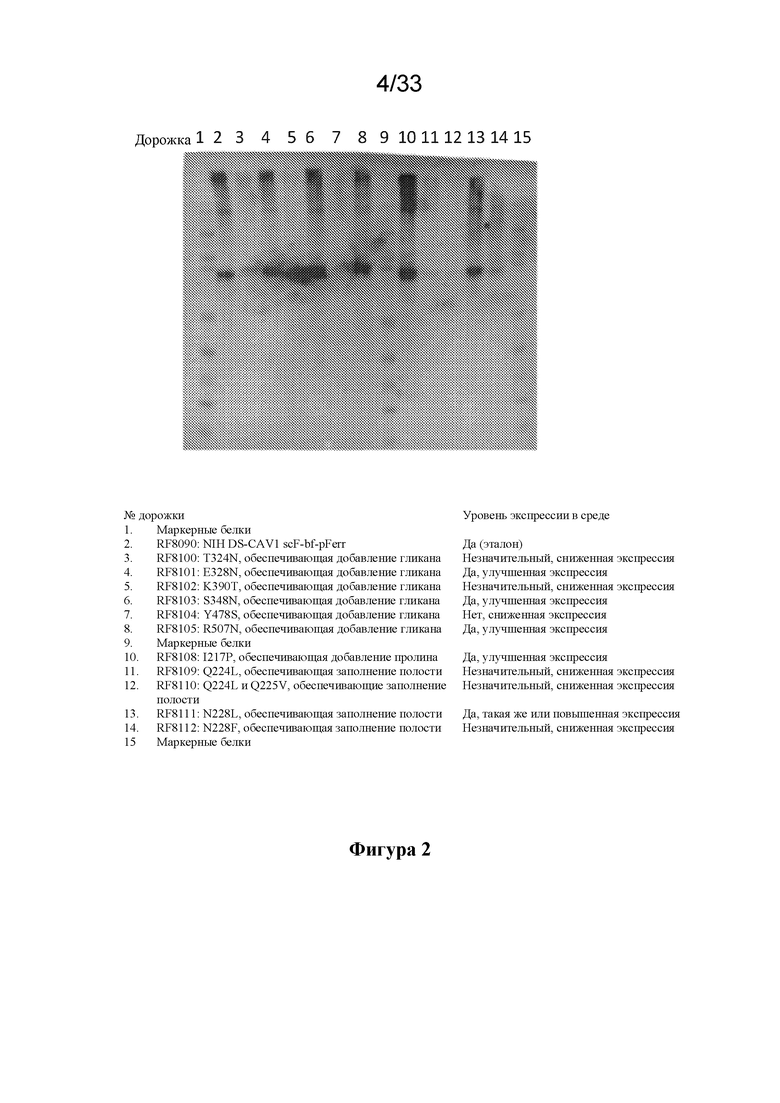
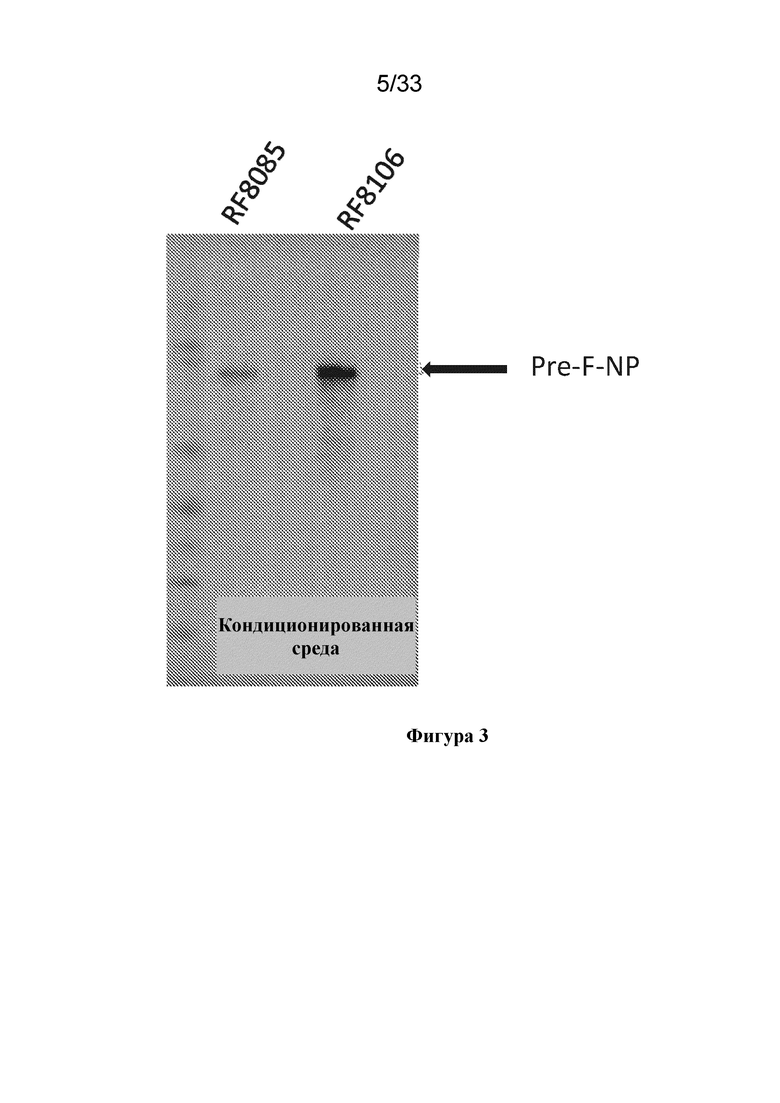
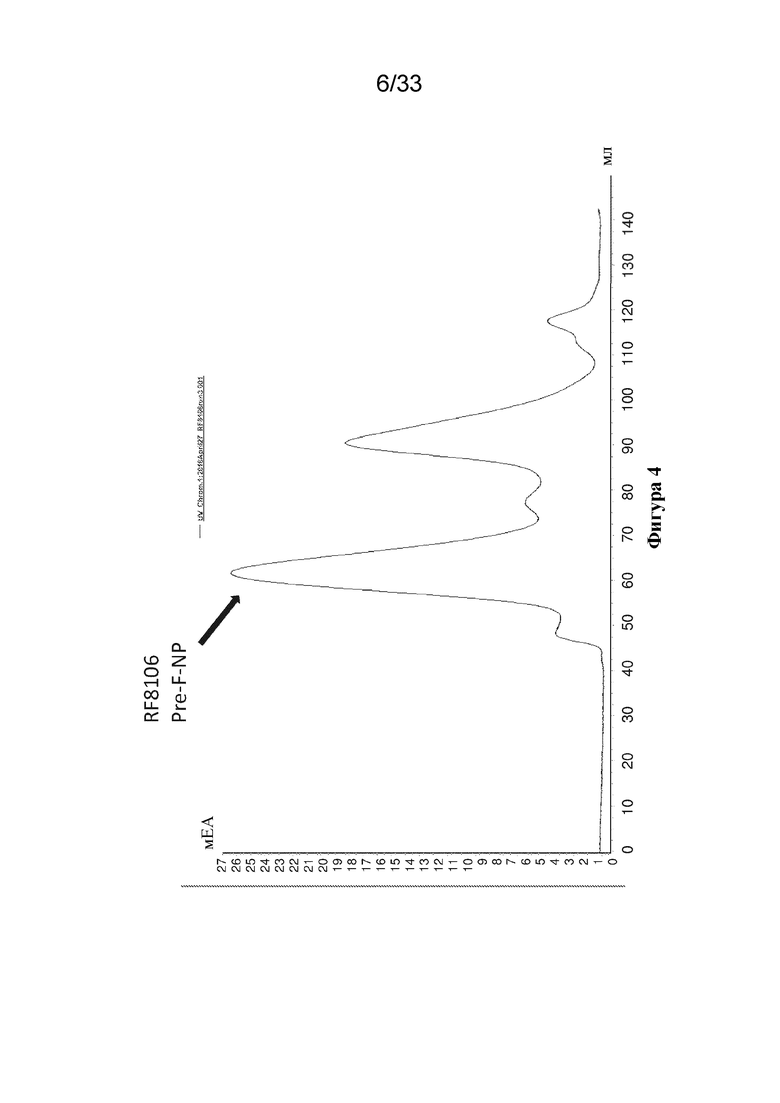
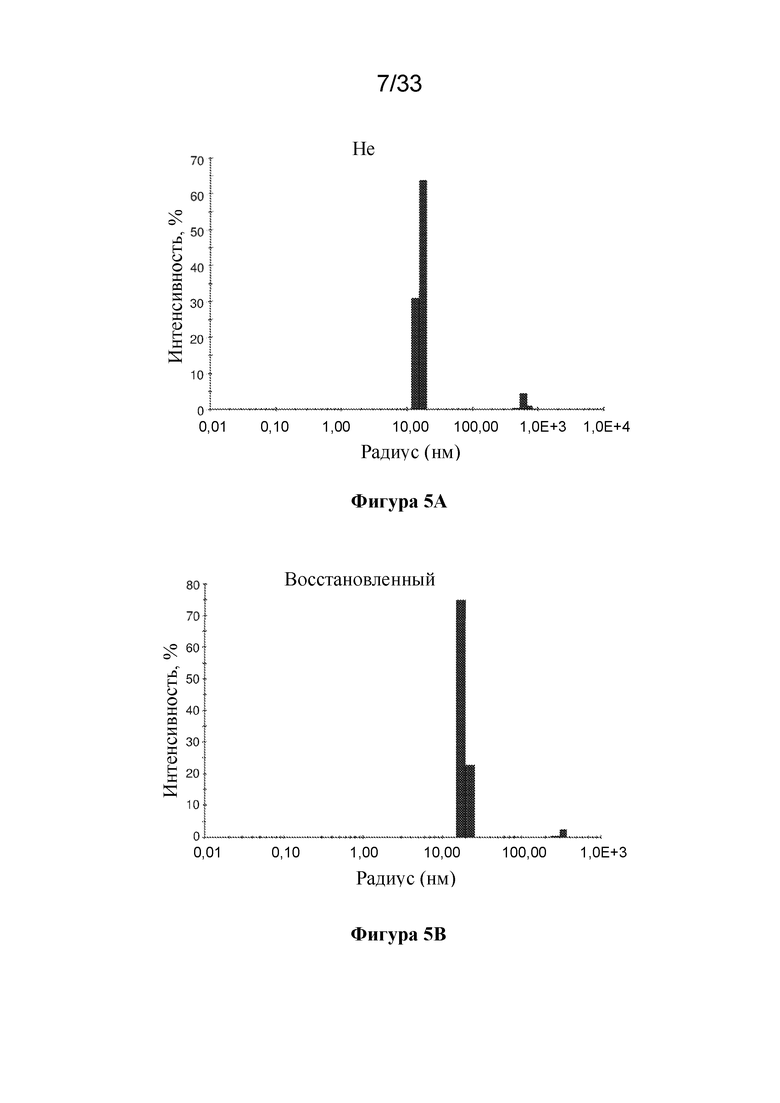
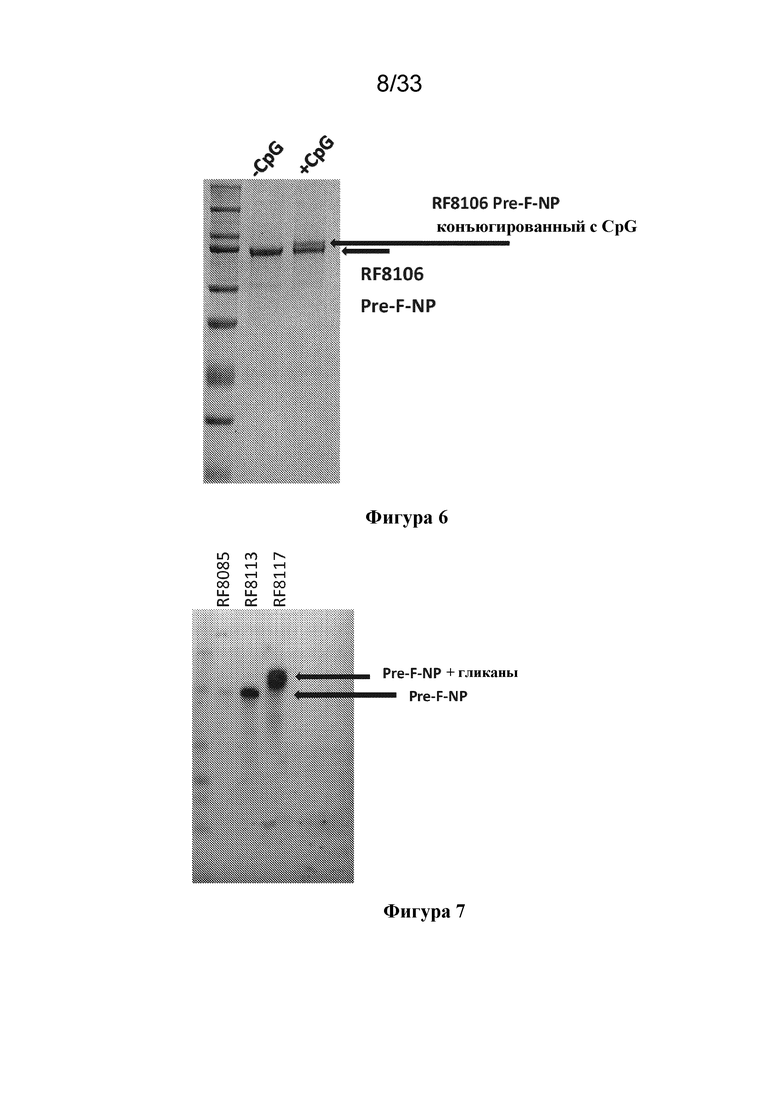
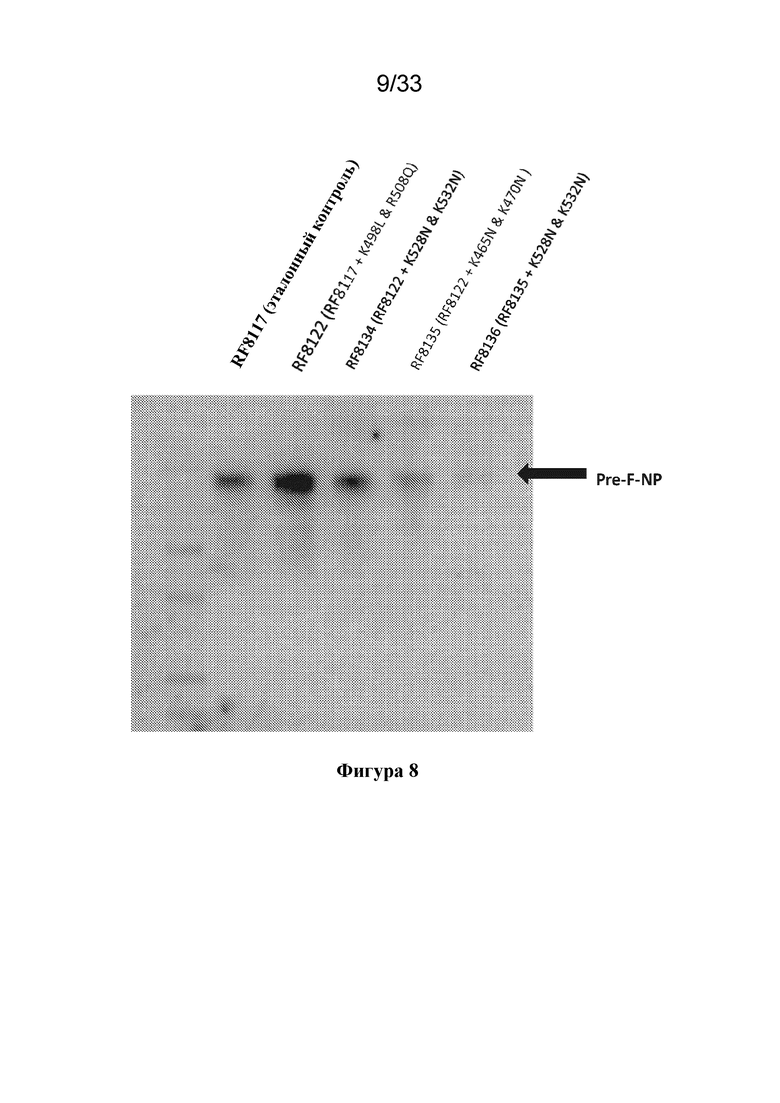
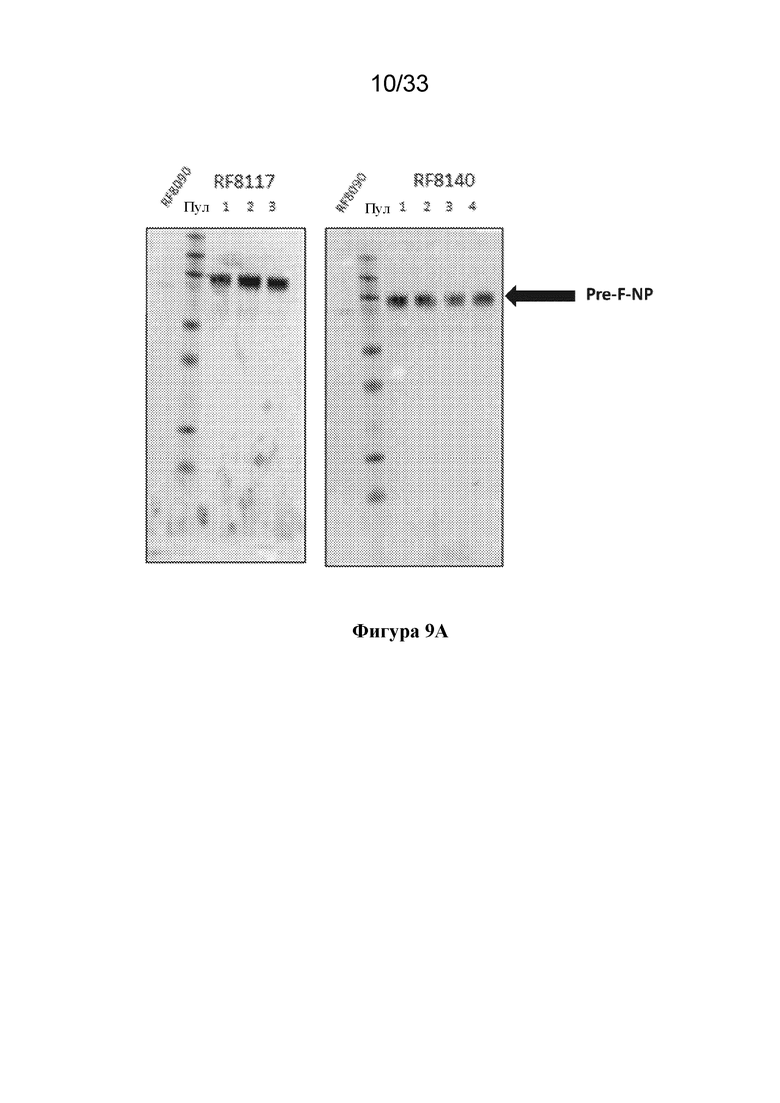
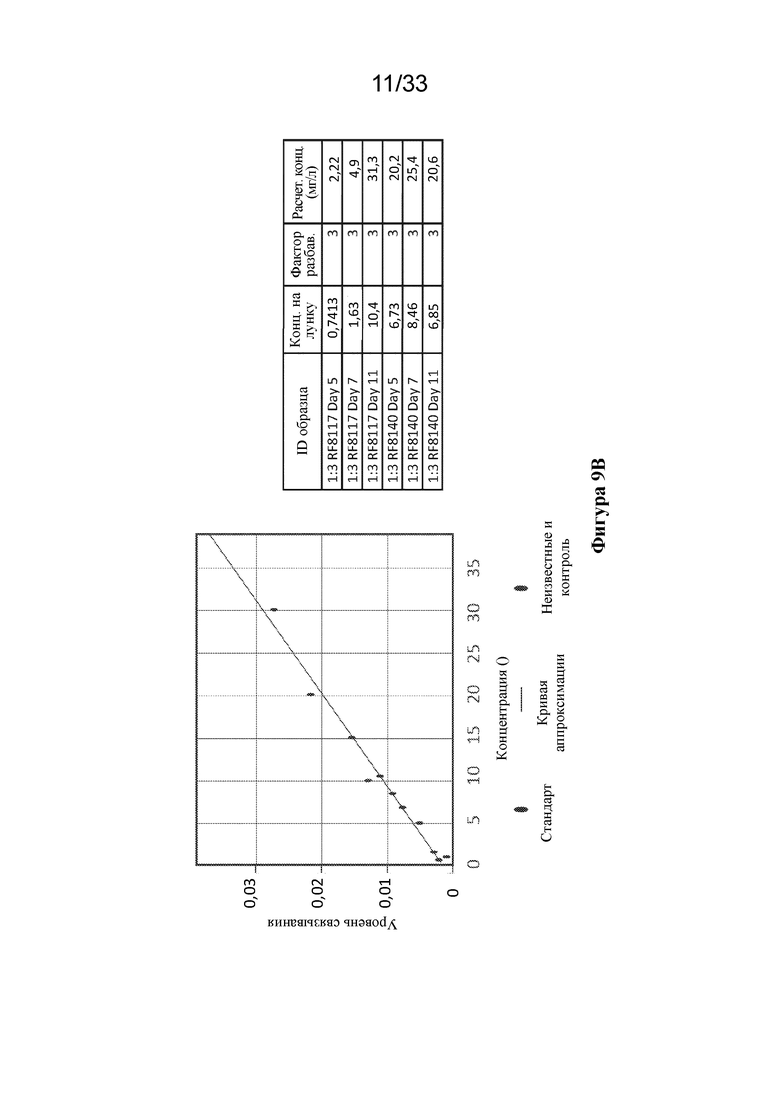
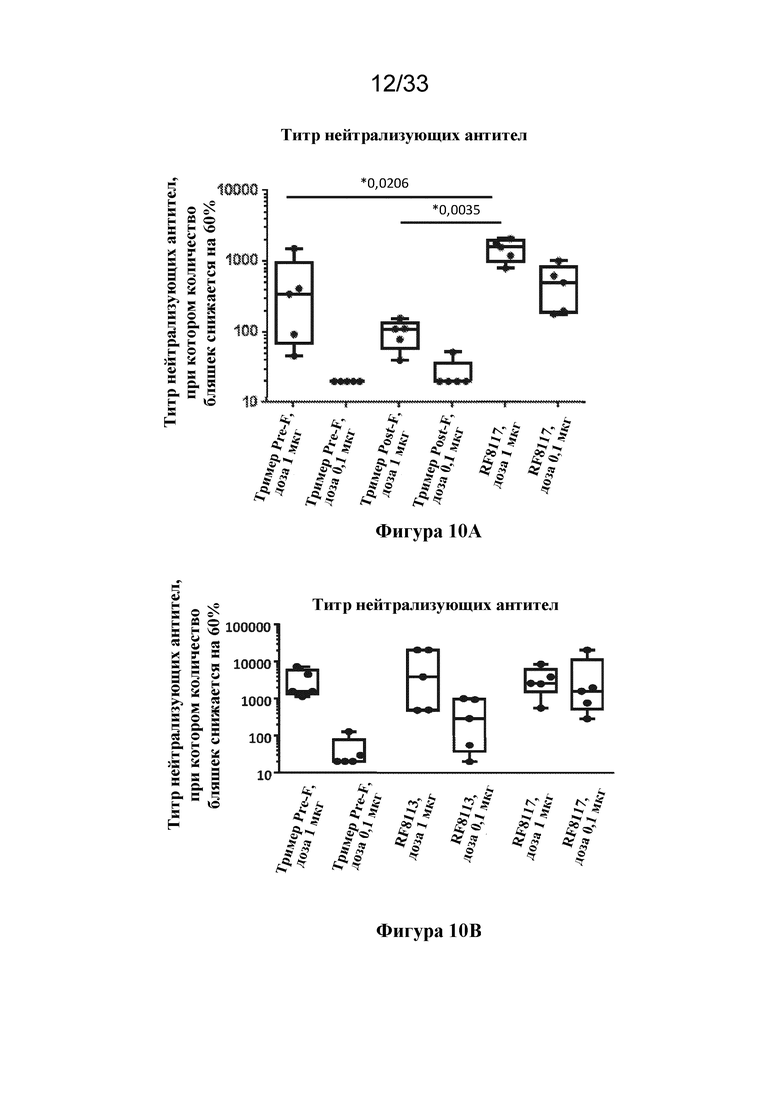
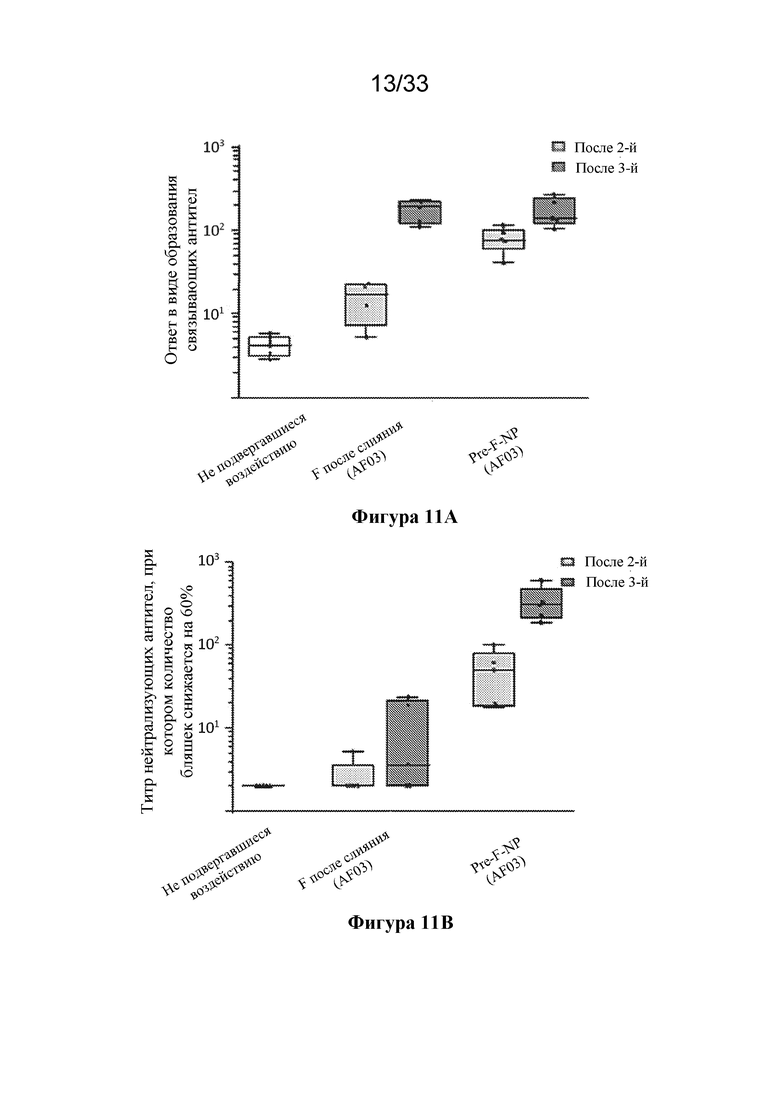
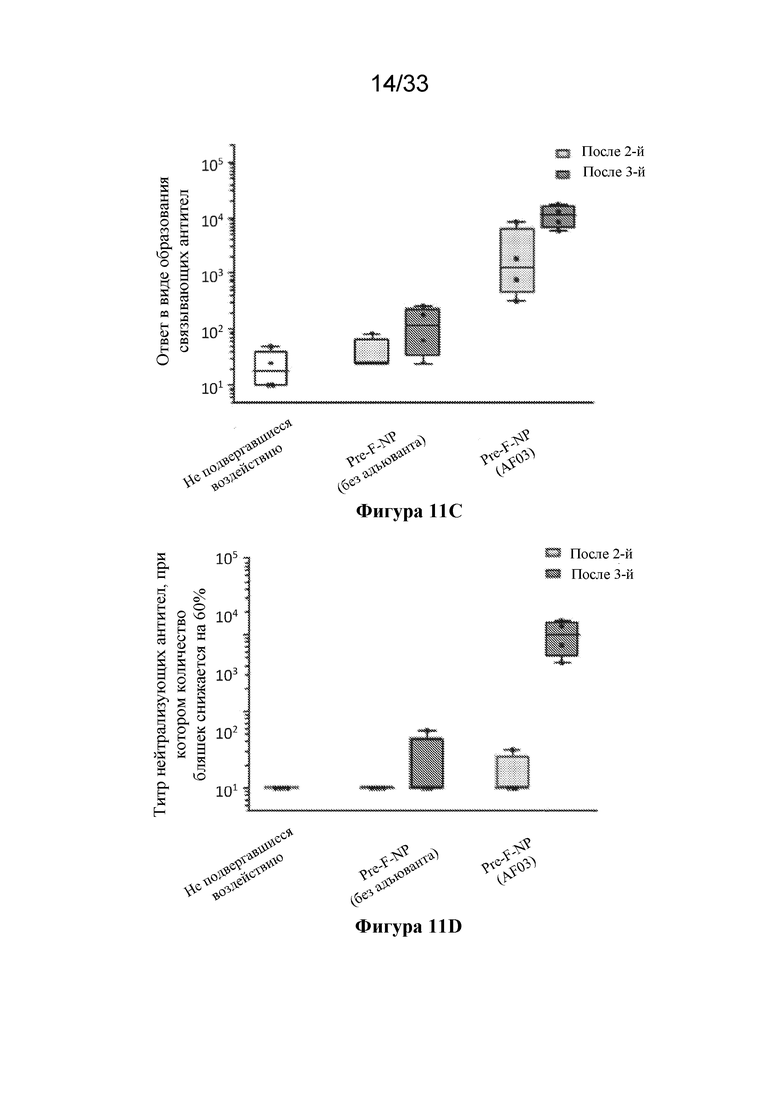
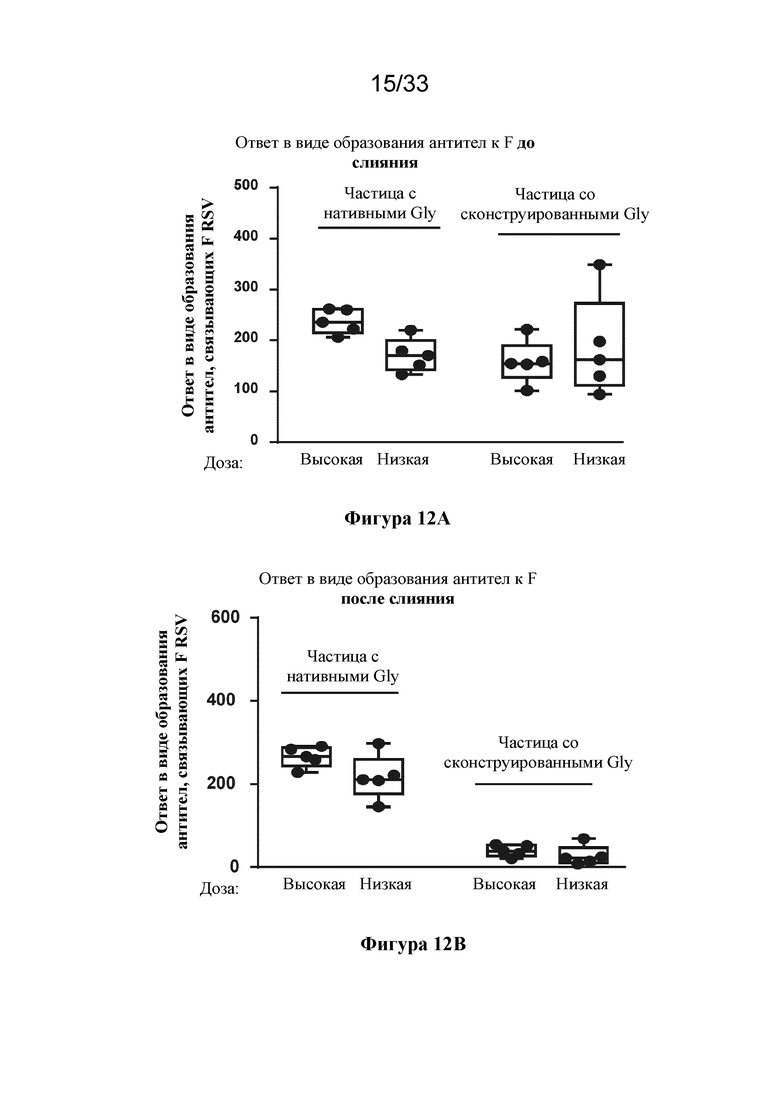
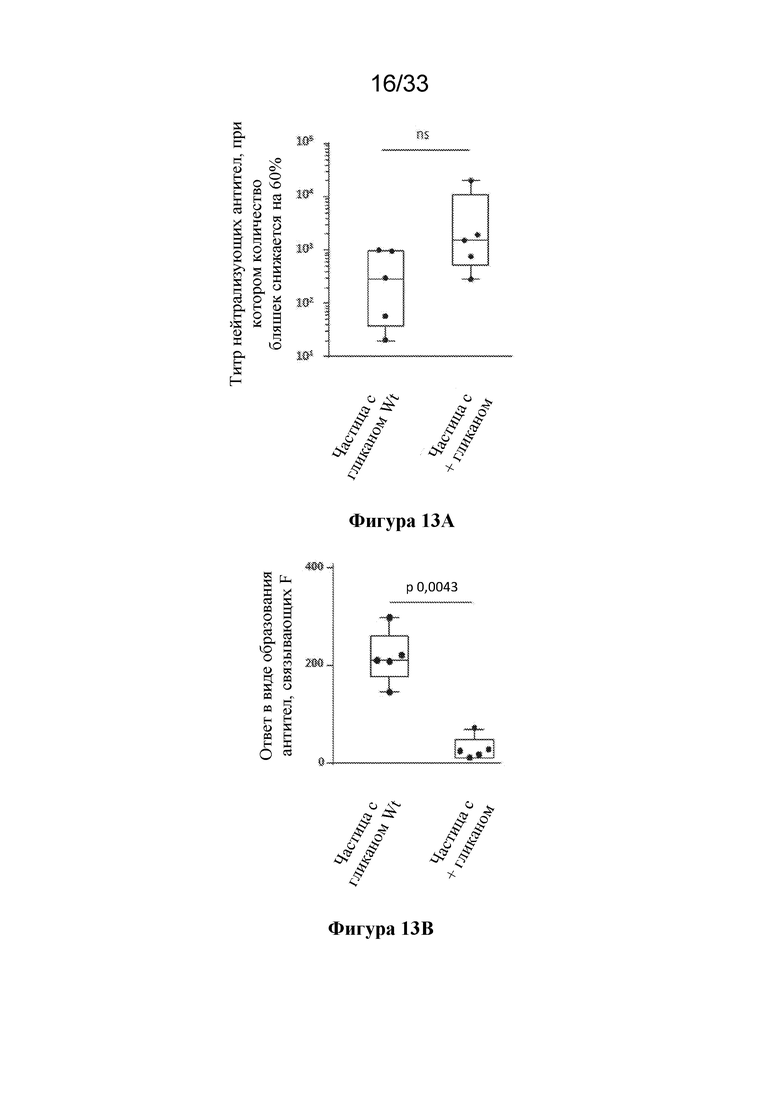
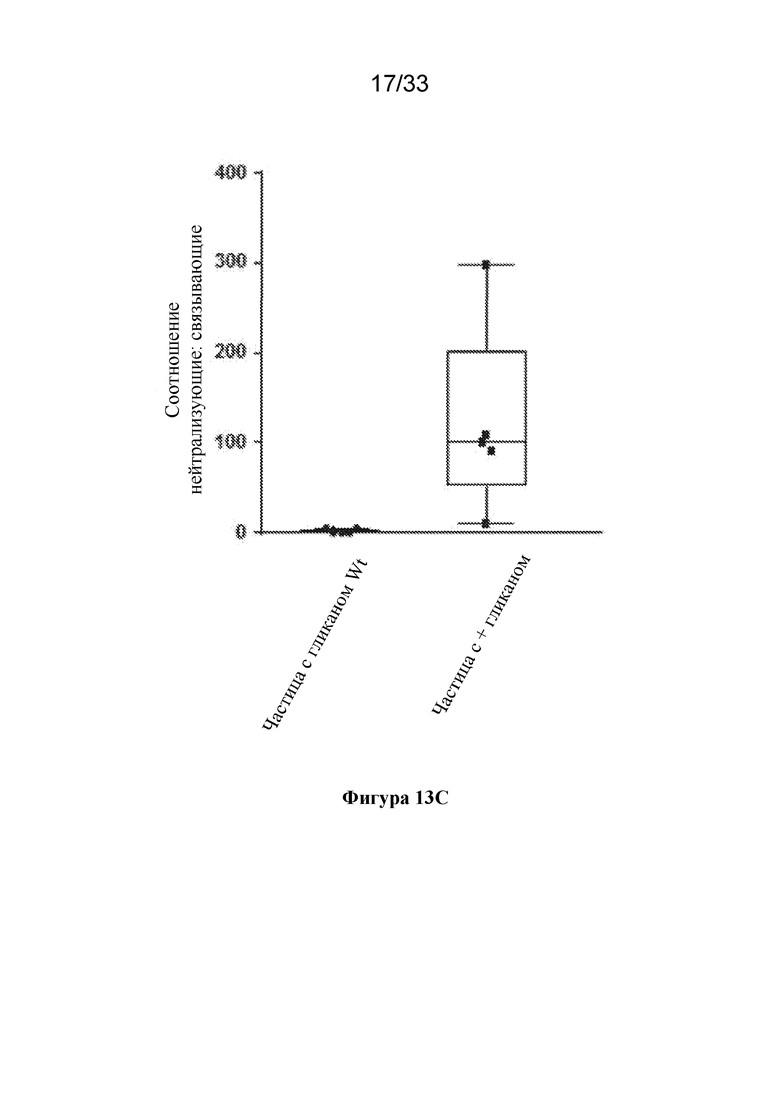
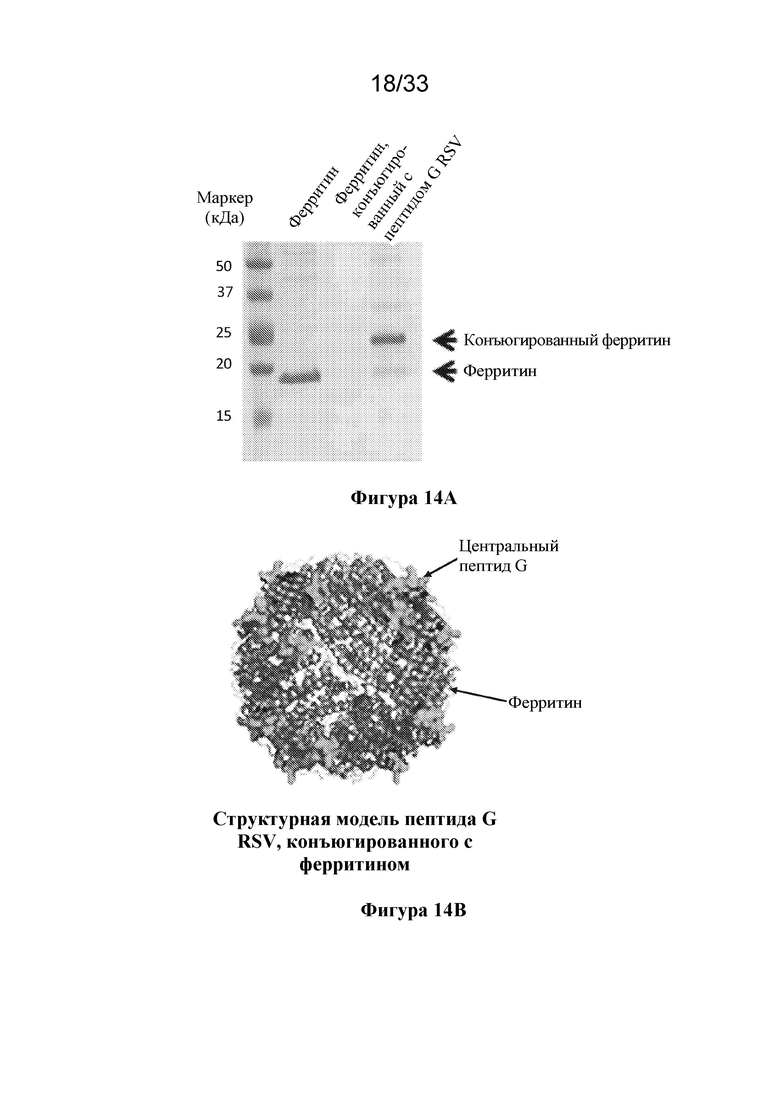
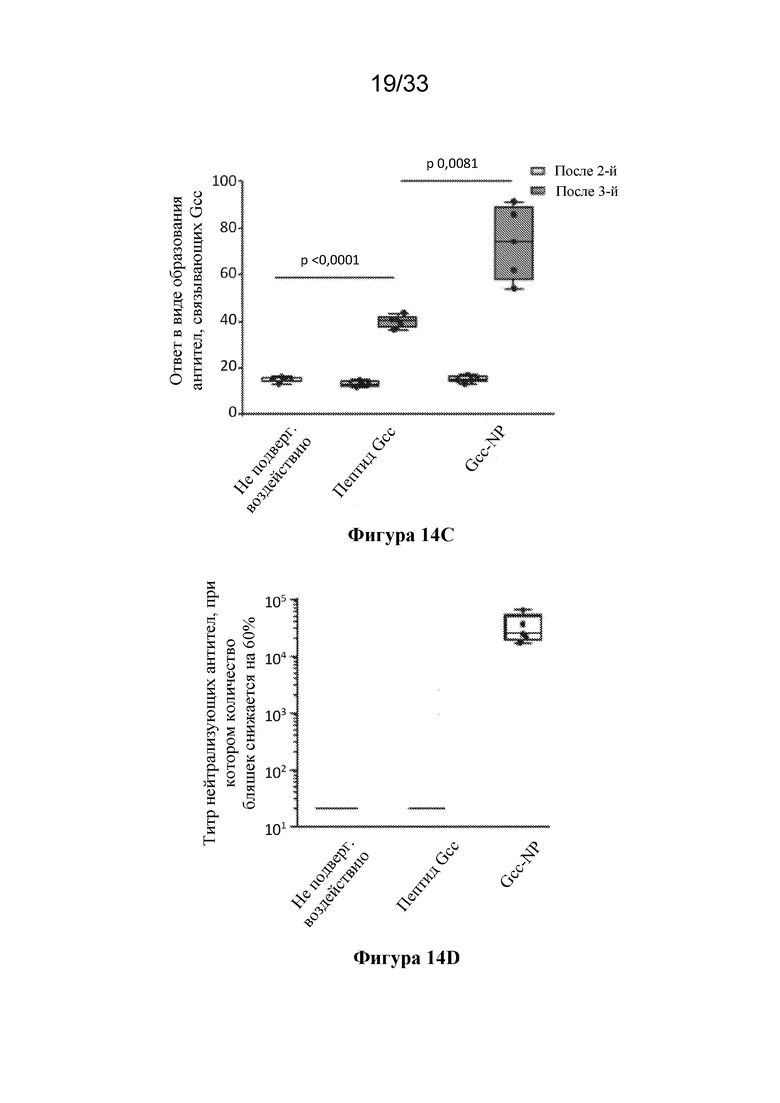
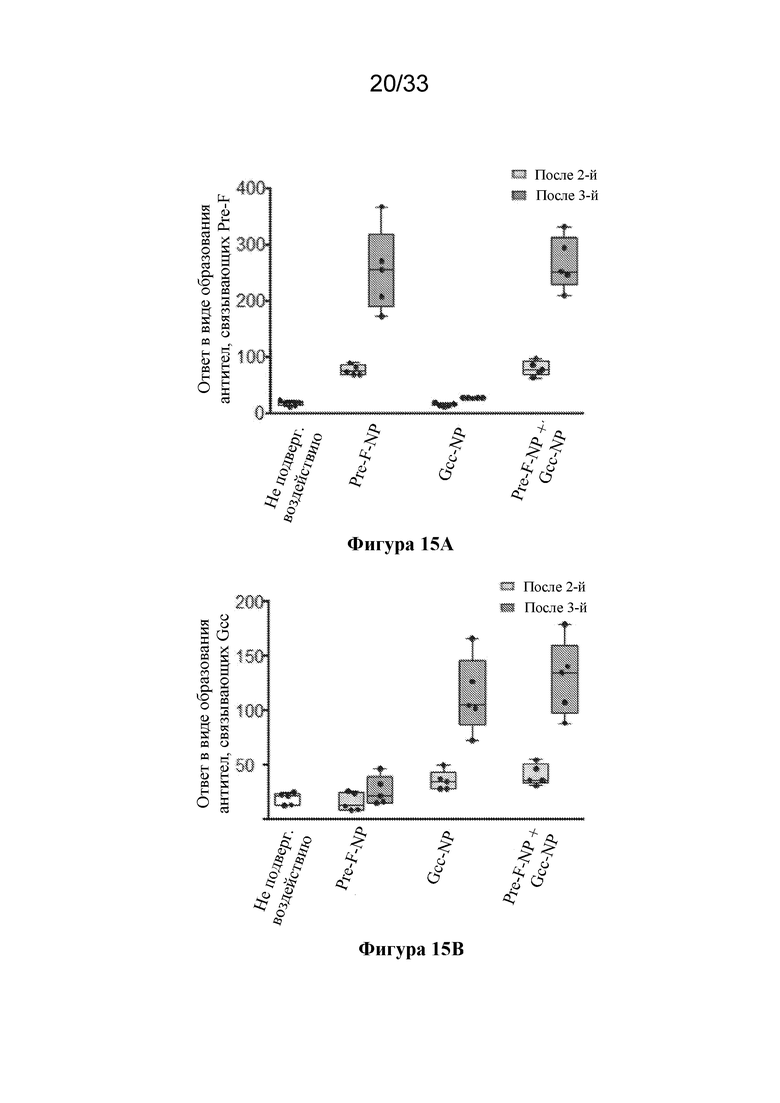
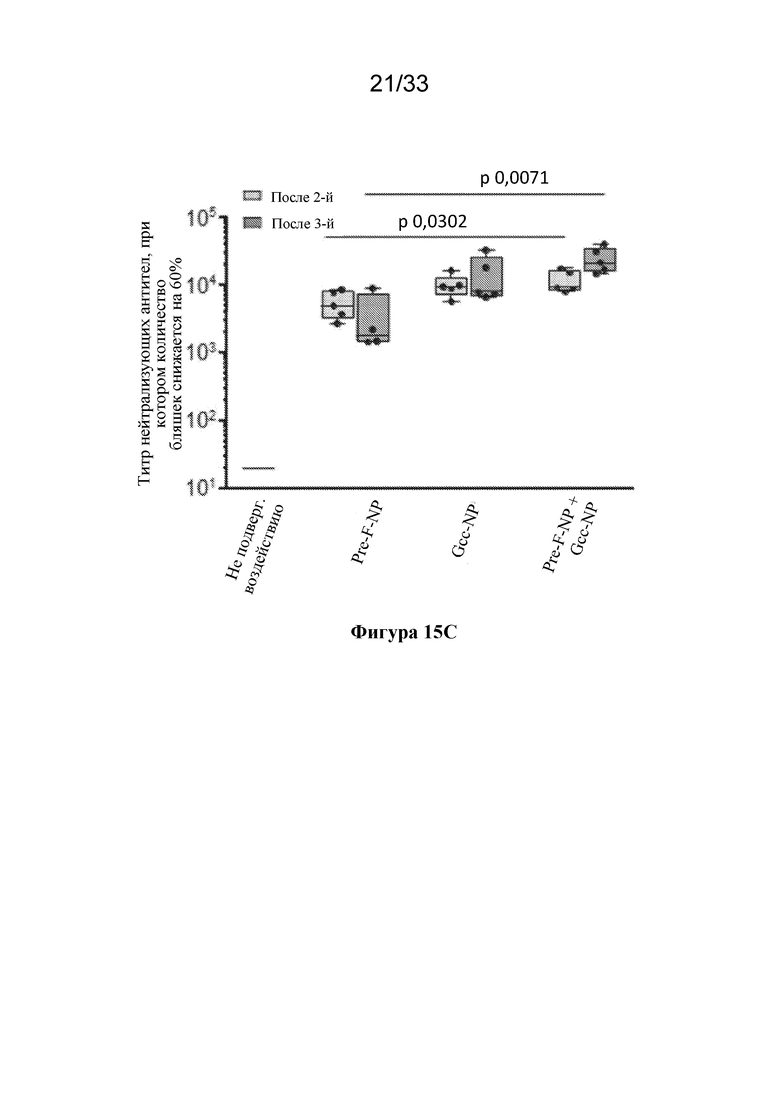
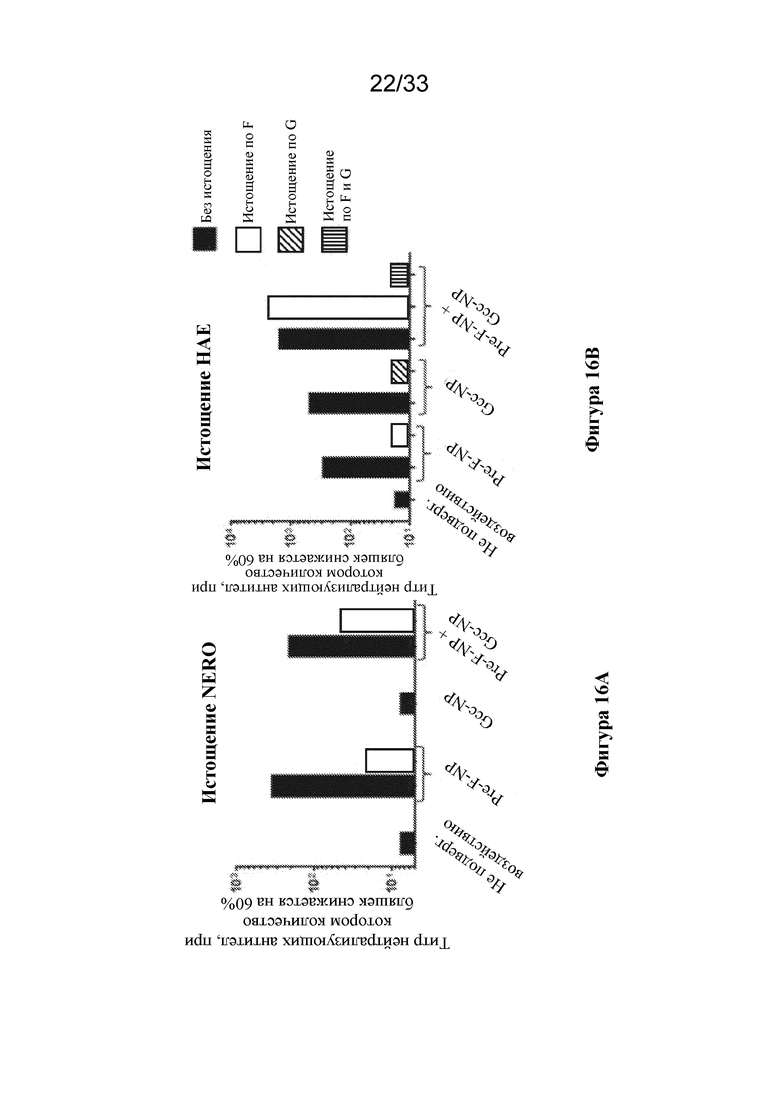
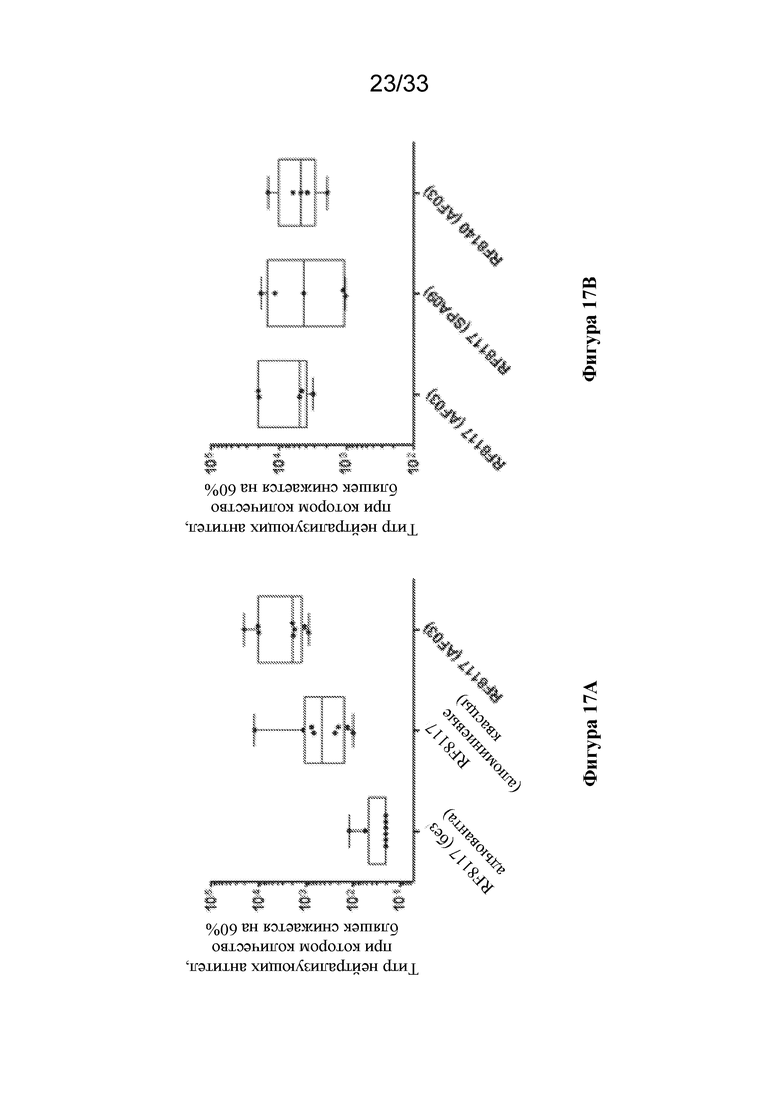
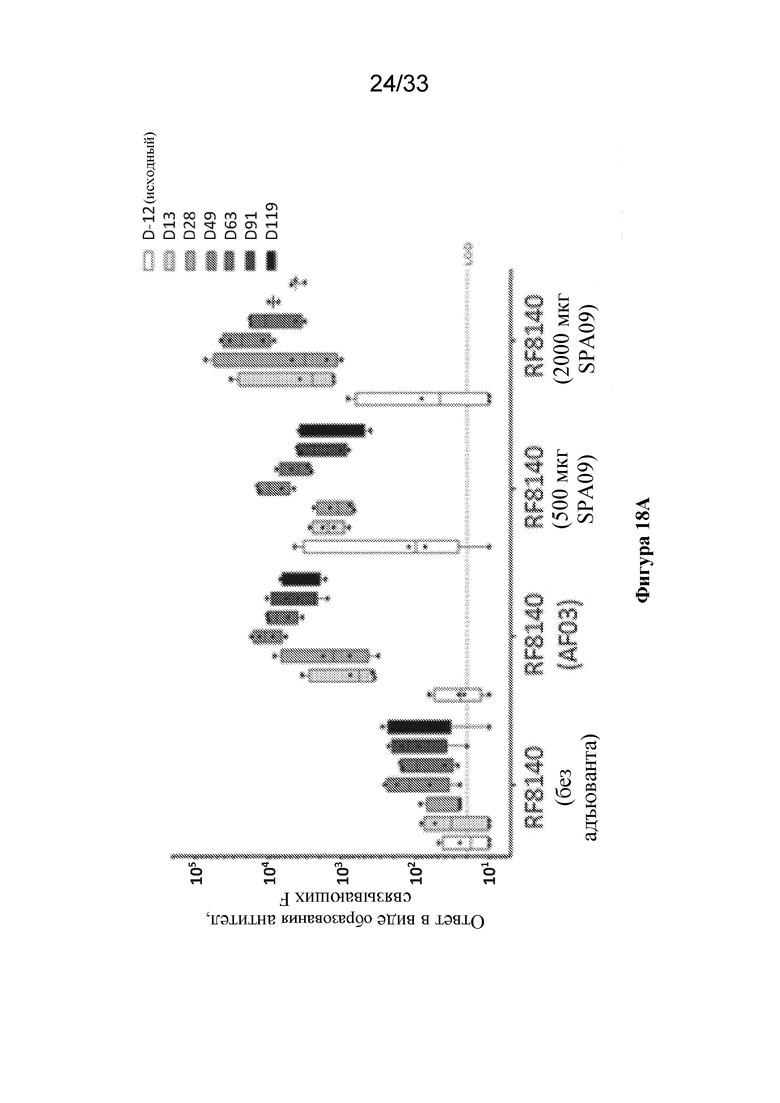
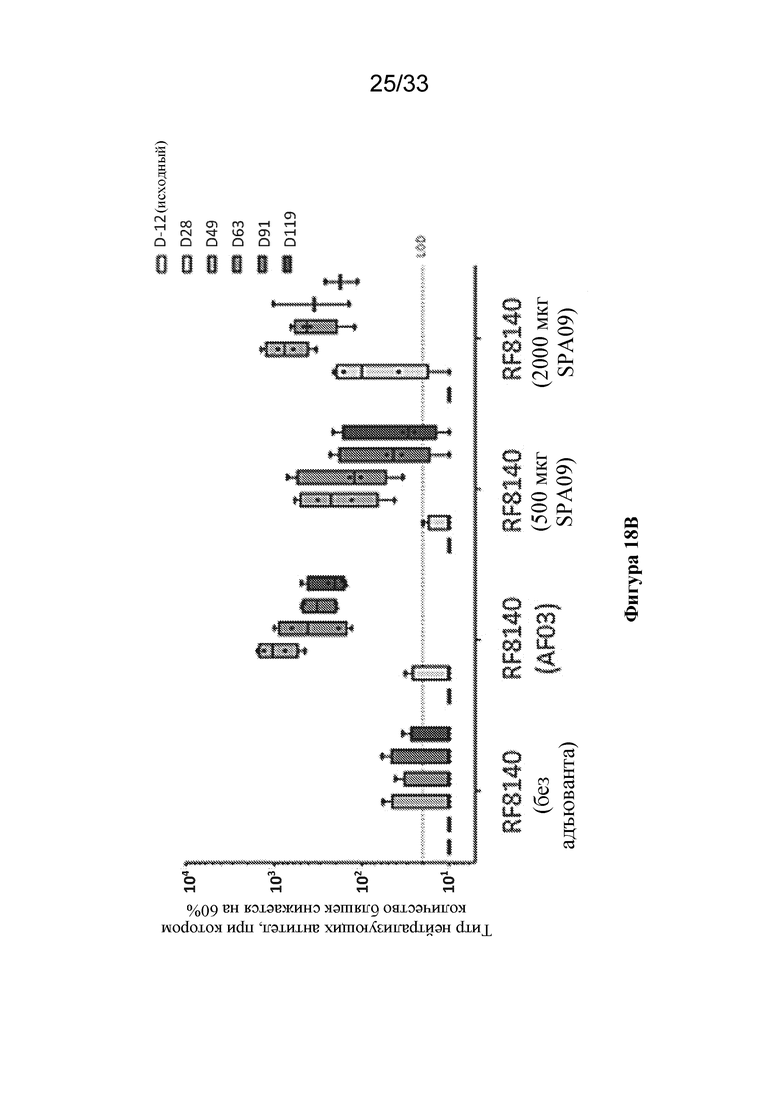
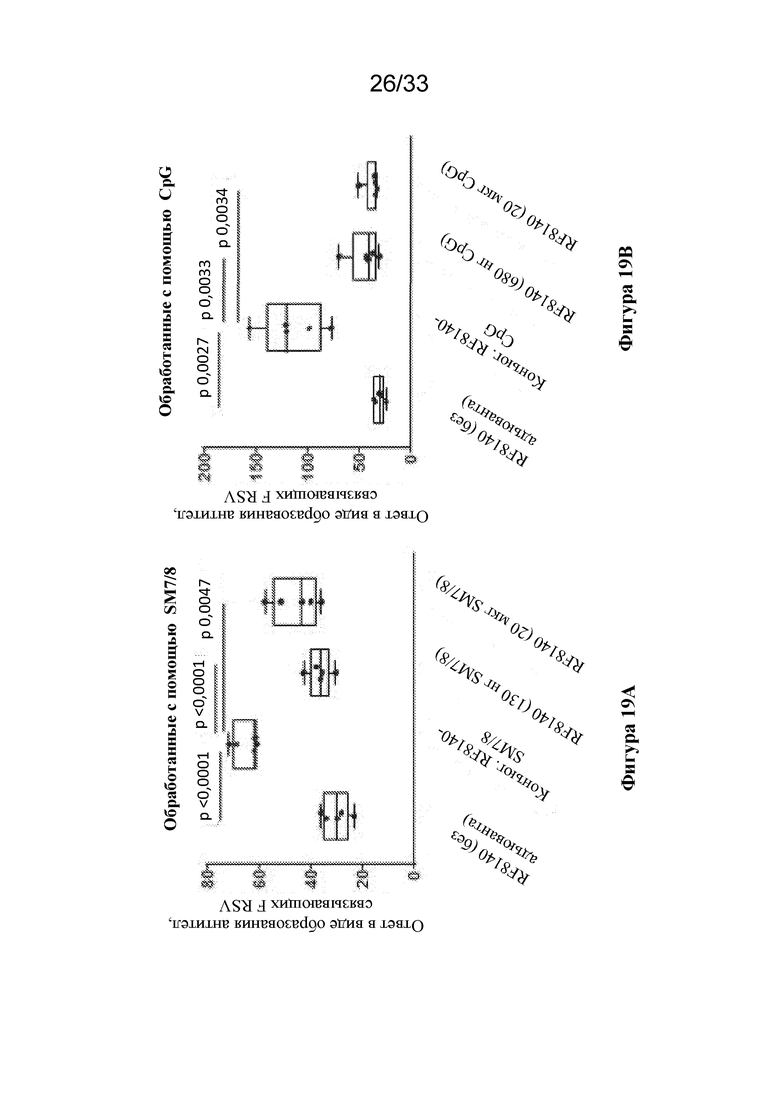
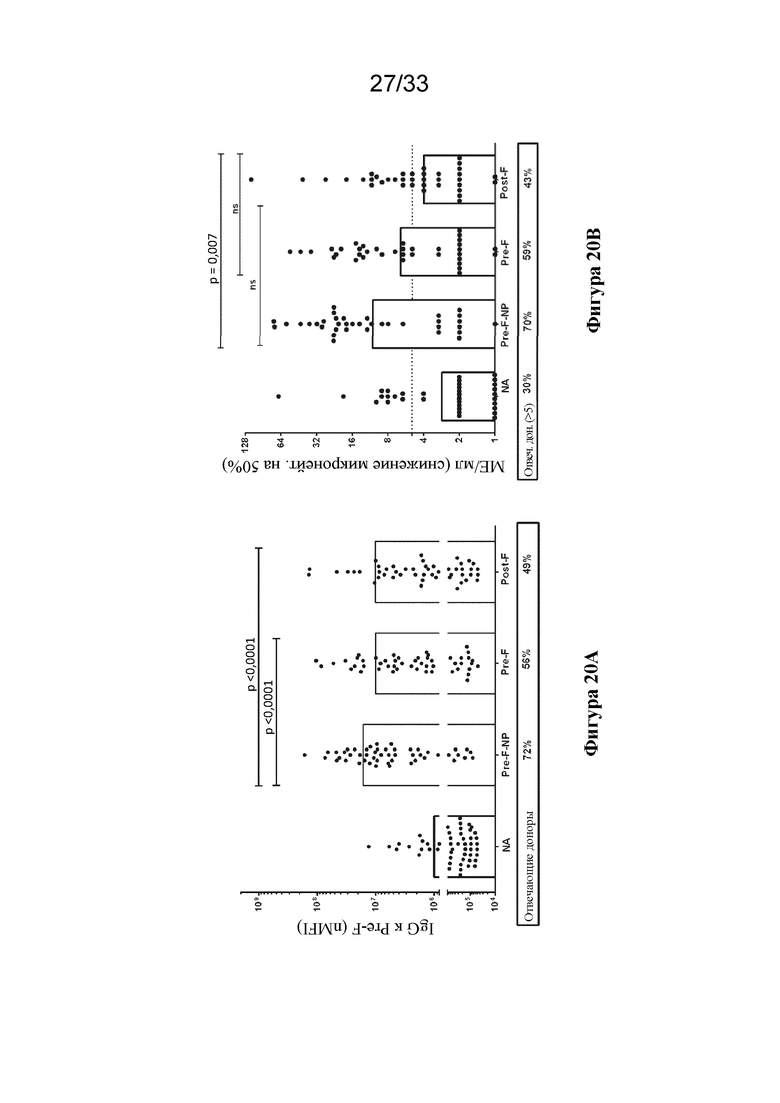
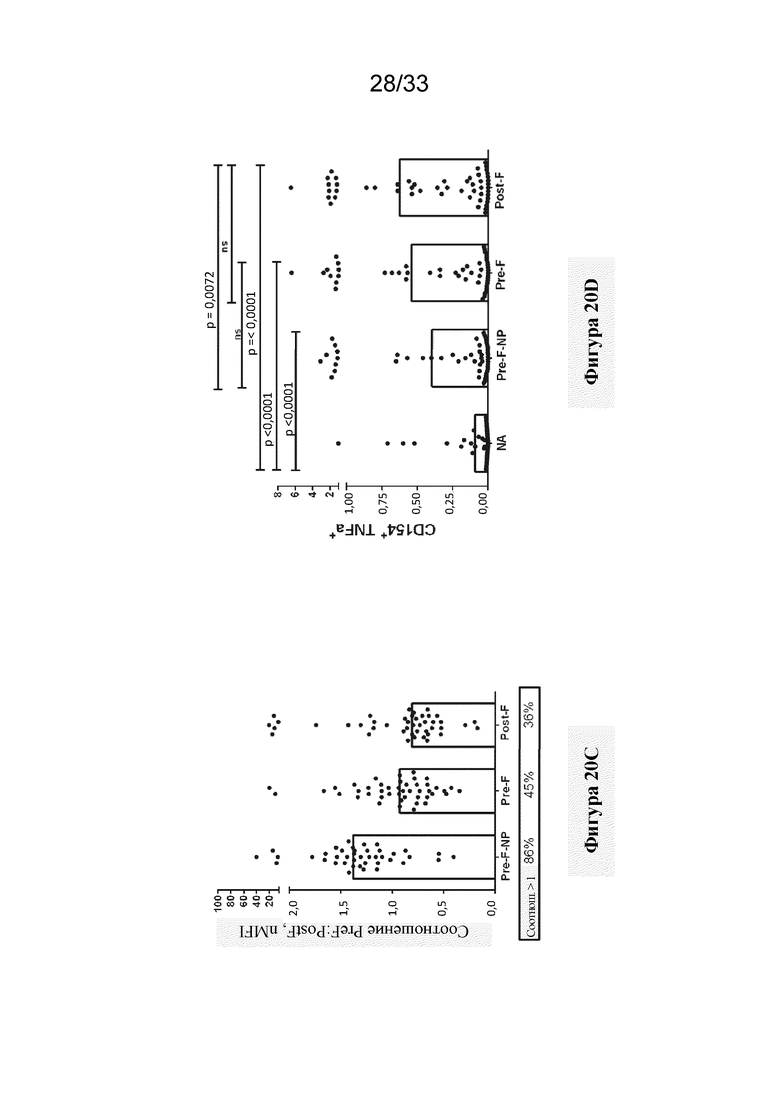
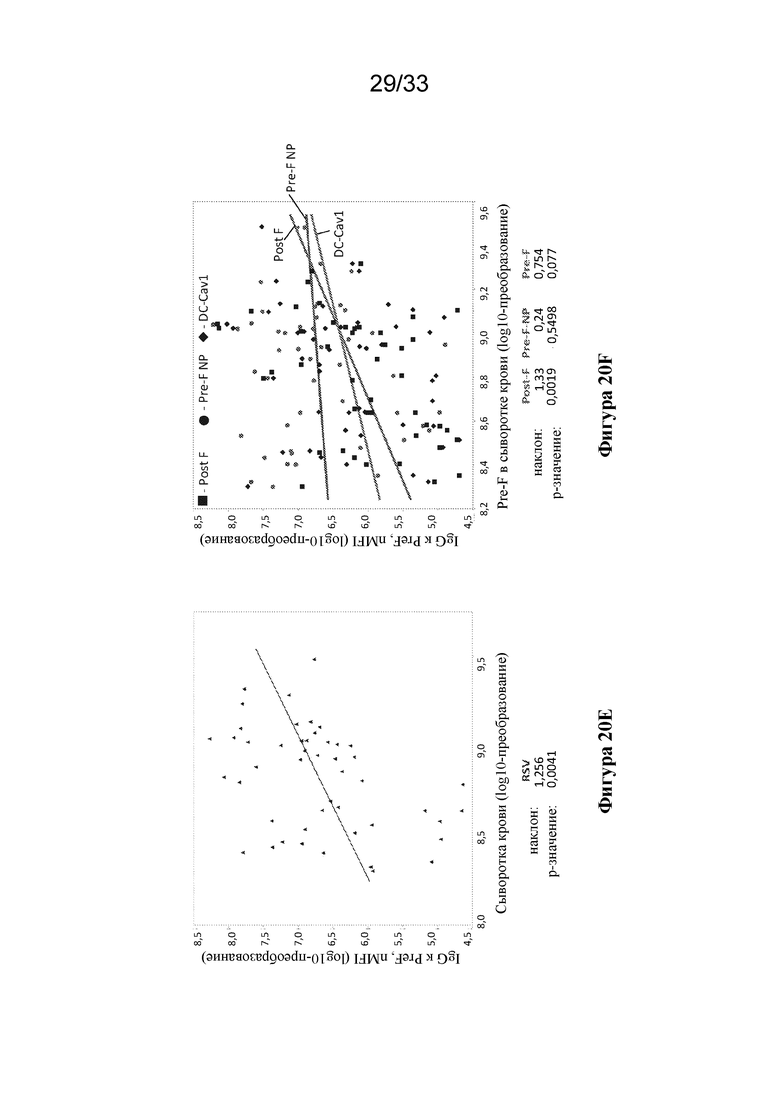
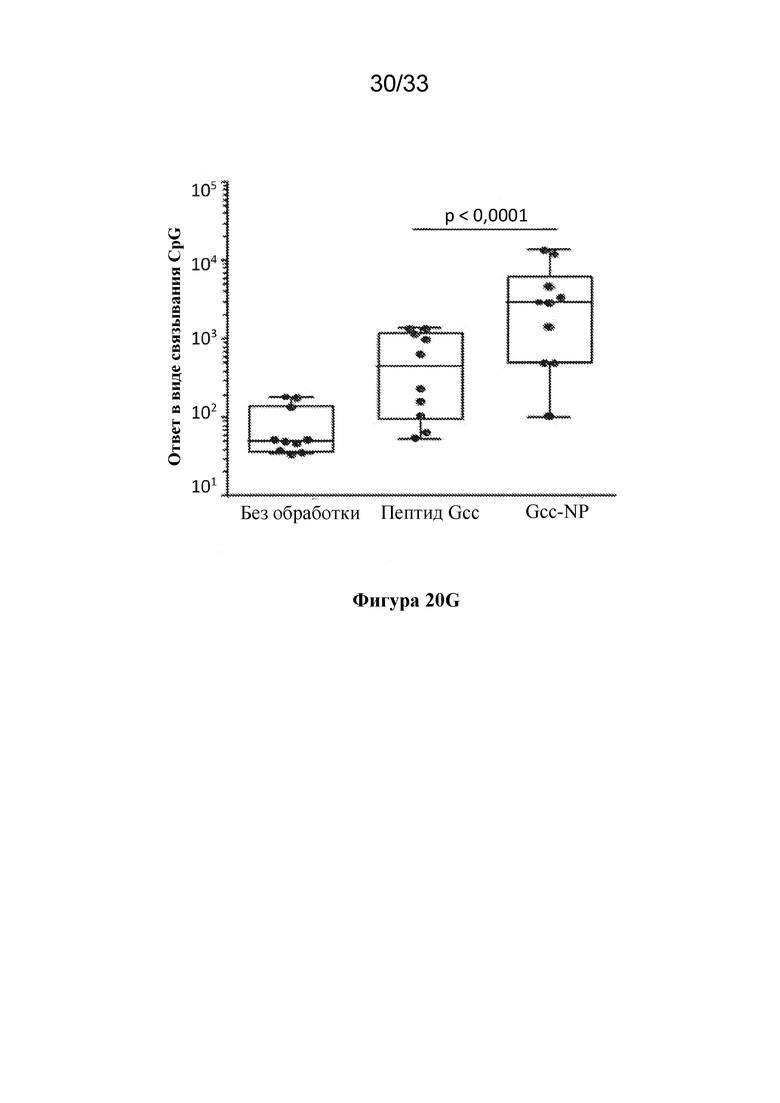
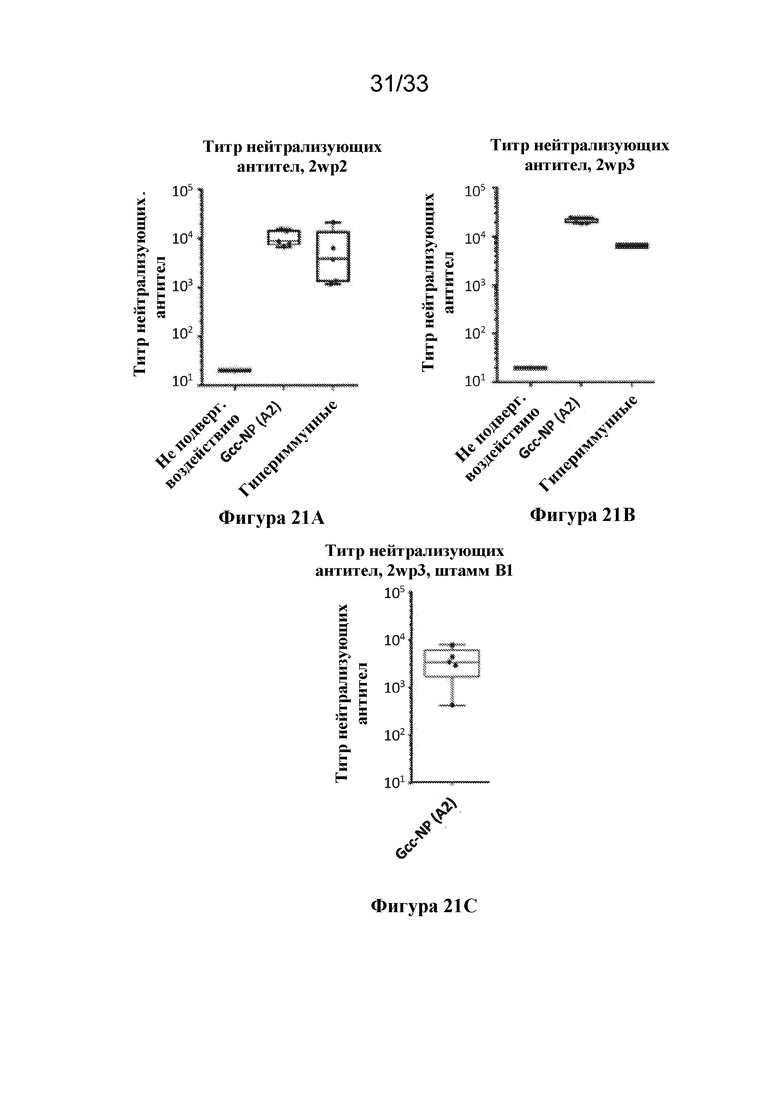
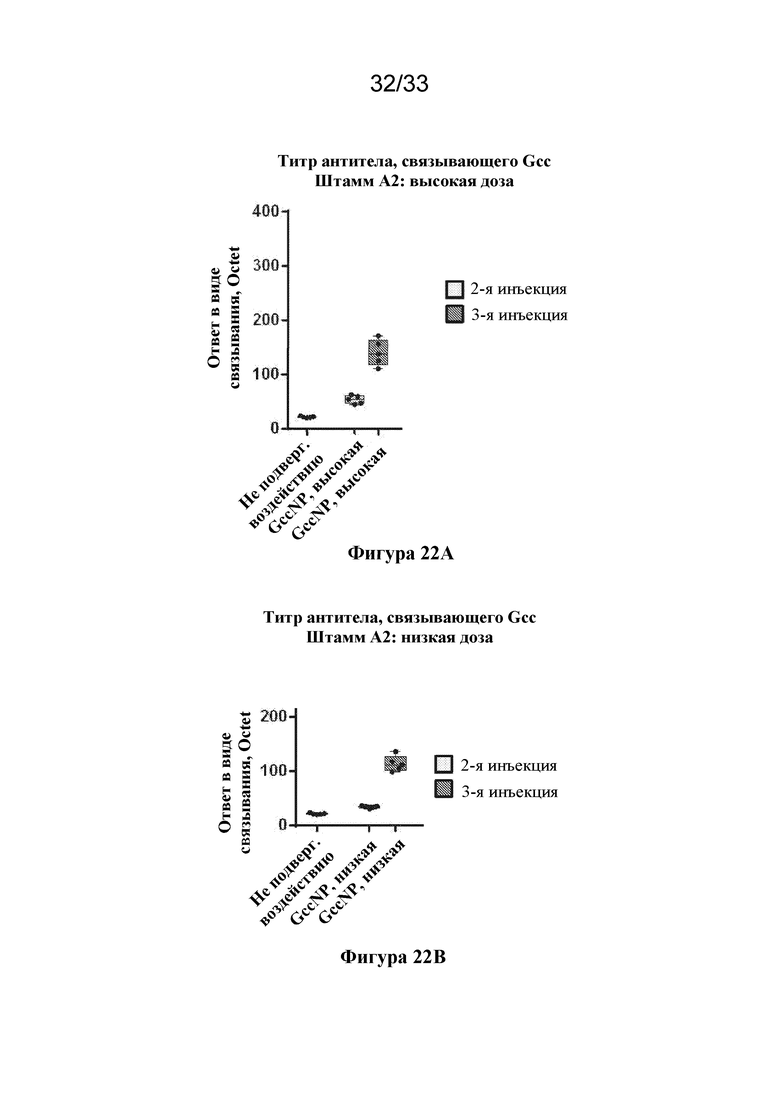
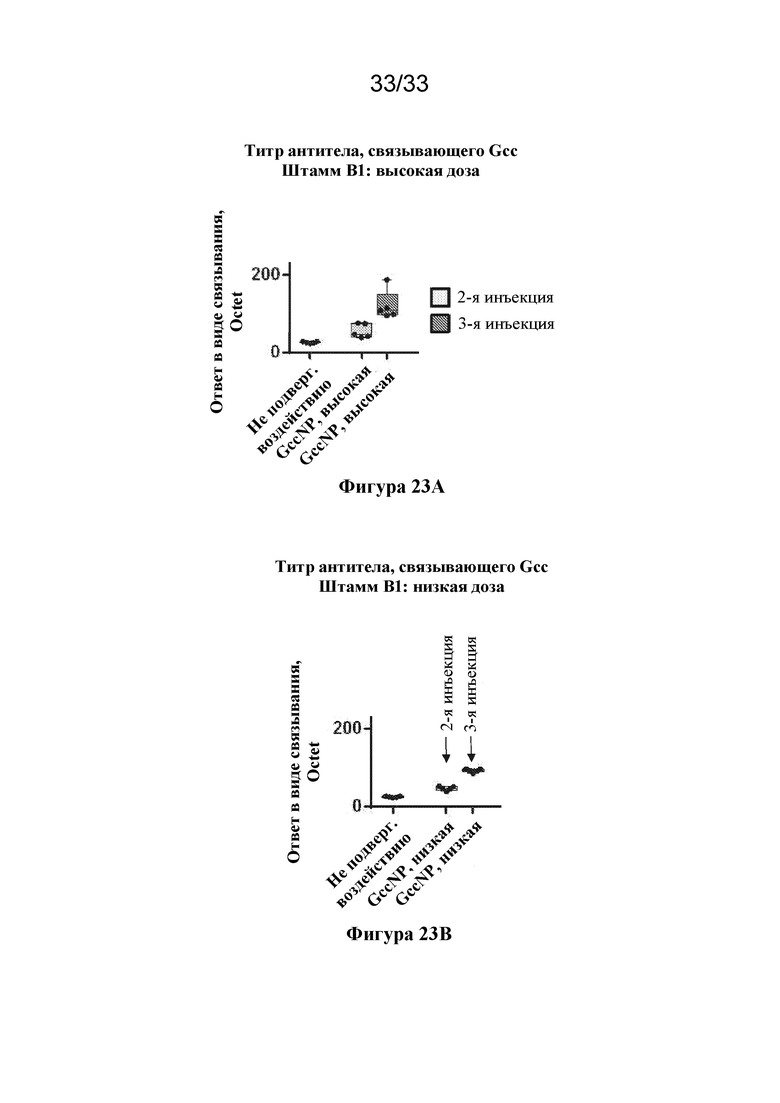
Изобретение относится к области биотехнологии, в частности к антигенному полипептиду респираторно-синцитиального вируса (RSV), а также ферритиновой частице и композиции, его содержащим. Также раскрыта нуклеиновая кислота, кодирующая вышеуказанный полипептид. Изобретение эффективно для индукции иммунного ответа на RSV у субъекта. 7 н. и 25 з.п. ф-лы, 23 ил., 1 табл.
1. Антигенный полипептид респираторно-синцитиального вируса (RSV), содержащий последовательность, имеющую по меньшей мере 90% идентичности аминокислотам 1-478 SEQ ID NO: 23.
2. Антигенный полипептид RSV, содержащий полипептид F RSV, где полипептид F RSV содержит последовательность, имеющую по меньшей мере на 90% идентичности аминокислотам 1-478 SEQ ID NO: 23, и где полипептид F RSV сохраняет аспарагин в по меньшей мере одном из положений 288, 308 или 467 SEQ ID NO: 23.
3. Антигенный полипептид RSV по п. 2, где аспарагин полипептида RSV гликозилирован, тем самым блокируя эпитоп полипептида RSV, который является общим для F RSV до слияния и F RSV после слияния.
4. Антигенный полипептид RSV по любому из пп. 1-3, содержащий F RSV до слияния.
5. Антигенный полипептид RSV по любому из пп. 1-4, который распознается антителом, специфичным в отношении F RSV до слияния, выбранным из D25 или AM14.
6. Антигенный полипептид RSV по п. 4 или 5, где F RSV до слияния содержит эпитоп, не обнаруживаемый на F RSV после слияния.
7. Антигенный полипептид RSV по любому из пп. 1-3, содержащий F RSV после слияния.
8. Антигенный полипептид RSV по любому из пп. 1 или 3-6, где эпитоп заблокирован N-гликаном, присоединенным к аспарагину.
9. Антигенный полипептид RSV по любому из пп. 1-8, дополнительно содержащий ферритин.
10. Антигенный полипептид RSV по п. 9, где ферритин содержит мутацию, обеспечивающую замещение экспонированной на поверхности аминокислоты цистеином.
11. Антигенный полипептид RSV по п. 10, где ферритин содержит одну или более из мутаций E12C, S26C, S72C, A75C, K79C, S100C и S111C ферритина из H. pylori или одну или более соответствующих мутаций ферритина из организма, отличного от H. pylori, согласно определению путем попарного или структурного выравнивания.
12. Антигенный полипептид RSV по любому из пп. 9-11, содержащий одну или более иммуностимулирующих функциональных групп, связанных с ферритином посредством экспонированной на поверхности аминокислоты, где необязательно экспонированная на поверхности аминокислота представляет собой цистеин, полученный в результате мутации.
13. Антигенный полипептид RSV по любому из пп. 9-12, где ферритин содержит мутацию, обеспечивающую замещение экспонированного на поверхности аспарагина аминокислотой, отличной от аспарагина, где необязательно аспарагин находится в положении 19 ферритина из H. pylori или аналогичном положении ферритина из организма, отличного от H. pylori, согласно определению путем попарного или структурного выравнивания.
14. Антигенный полипептид RSV по любому из пп. 9-13, где ферритин содержит мутацию, обеспечивающую замещение внутреннего цистеина аминокислотой, отличной от цистеина, где необязательно внутренний цистеин находится в положении 31 ферритина из H. pylori или в положении, которое соответствует положению 31 ферритина из H. pylori, согласно определению путем попарного или структурного выравнивания.
15. Антигенный полипептид RSV по любому из пп. 9-14, где полипептид F RSV содержит эпитоп, не обнаруживаемый на F RSV после слияния, который представляет собой эпитоп сайта 0, где необязательно эпитоп сайта 0 содержит аминокислотные остатки 62-69 и 156-169 из SEQ ID NO: 23.
16. Антигенный полипептид RSV по любому из пп. 1-15, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 288 из SEQ ID NO: 23.
17. Антигенный полипептид RSV по любому из пп. 1-16, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 308 из SEQ ID NO: 23.
18. Антигенный полипептид RSV по любому из пп. 1-17, где полипептид F RSV содержит аспарагин в положении, соответствующем положению 467 из SEQ ID NO: 23.
19. Антигенный полипептид RSV по любому из пп. 1-18, где полипептид F RSV содержит лейцин в положении, соответствующем положению лизина 458 из SEQ ID NO: 23.
20. Антигенный полипептид RSV по любому из пп. 1-19, где полипептид F RSV содержит пролин в положении, соответствующем положению изолейцина 177 из SEQ ID NO: 23.
21. Антигенный полипептид RSV по любому из пп. 1-20, где полипептид F RSV не содержит сайта расщепления фурином, где необязательно вместо сайта расщепления фурином присутствует линкер.
22. Антигенный полипептид RSV по любому из пп. 1-21, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 95%, 97%, 98%, 99% или 99,5% идентичностью аминокислотам 1-478 из SEQ ID NO: 23.
23. Антигенный полипептид RSV по любому из пп. 1-21, где полипептид F RSV содержит последовательность, характеризующуюся по меньшей мере 95%, 97%, 98%, 99% или 99,5% идентичностью последовательности под SEQ ID NO: 23.
24. Антигенный полипептид RSV по любому из пп. 1-23, содержащий аминокислоты 1-478 из SEQ ID NO: 23.
25. Антигенный полипептид RSV по любому из пп. 1-21 или 23, содержащий последовательность SEQ ID NO: 23.
26. Ферритиновая частица для применения в способе индукции иммунного ответа на RSV у субъекта, содержащая антигенный полипептид RSV по любому из пп. 9-25.
27. Композиция для применения в способе индукции иммунного ответа на RSV у субъекта, содержащая эффективное количество антигенного полипептида RSV по любому из пп. 1-25 и полипептид G RSV.
28. Композиция для применения в способе индукции иммунного ответа на RSV у субъекта, содержащая эффективное количество антигенного полипептида RSV по любому из пп. 1-25 и фармацевтически приемлемый носитель.
29. Антигенный полипептид RSV по любому из пп. 1-25 для применения в способе индукции иммунного ответа на RSV у субъекта.
30. Способ индукции иммунного ответа на RSV у субъекта, включающий введение субъекту любого одного или более из антигенного полипептида RSV по любому из пп. 1-25.
31. Антигенный полипептид RSV по любому из пп. 1-25, где субъектом является человек.
32. Нуклеиновая кислота, кодирующая антигенный полипептид RSV по любому из пп. 1-25, 29 или 31, где полипептид F RSV антигенного полипептида RSV содержит последовательность, имеющую по меньшей мере 90%, 95%, 97%, 98 %, 99% или 99,5% идентичности аминокислотам 1-478 SEQ ID NO: 23, где необязательно нуклеиновая кислота представляет собой мРНК.
| WO2014160463 A1, 02.10.2014 | |||
| WO2015183969 A1, 03.12.2015 | |||
| WO2016138160 A1, 01.09.2016 | |||
| JASON S | |||
| MCLELLAN et al., Structure-Based Design of a Fusion Glycoprotein Vaccine for Respiratory Syncytial Virus, SCIENCE, 2013, 342(6158), 592-598 | |||
| ИНАКТИВИРОВАННЫЕ ВАКЦИНЫ РЕСПИРАТОРНО-СИНЦИТИАЛЬНОГО ВИРУСА | 1994 |
|
RU2142817C1 |
