ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННУЮ ЗАЯВКУ
Эта заявка заявляет приоритет по патентной заявке Китая №201611180971,4, которая называется "Анализ связывания", поданной в патентный офис Китая 19 декабря 2016 года, и содержание которой включено в данное описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ
Данное изобретение относится к способам определения активности связывания с ГКГС класса II средств, содержащий белок гена активации лимфоцитов-3 (LAG-3) или его фрагментов, производных или аналогов, а также к сенсорам и наборам для применения в указанных способах.
УРОВЕНЬ ТЕХНИКИ
Белок LAG-3 представляет собой мембранный белок типа I гомолог CD4 с четырьмя внеклеточными доменами суперсемейства иммуноглобулинов. Подобно CD4, LAG-3 олигомеризуется на поверхностях Т-клеток и связывается с молекулами ГКГС класса II на антиген-презентирующих клетках (АПК), но со значительно более высокой аффинностью, чем CD4. LAG-3 экспрессируется на активированных CD4+ и CD8+ Т-лимфоцитах, где он связывается с рецепторным комплексом CD3/Т-клеток на поверхности клетки и отрицательно регулирует передачу сигнала. Как следствие, он отрицательно регулирует пролиферацию, функцию и гомеостаз Т-клеток. LAG-3 экспрессируется в значительных количествах на истощенных Т-клетках по сравнению с эффекторными или Т-клетками памяти. LAG-3 также экспрессируется в значительных количествах на инфильтрирующих в опухоль лимфоцитах (ОИЛ), и блокада LAG-3 с использованием анти-LAG-3 антитела может усиливать противоопухолевые Т-клеточные ответы.
IMP321 является рекомбинантным растворимым слитым белком LAG-3Ig, который связывается с ГКГС класса II с высокой авидностью. Он является первым в своем классе иммунопотенциатором, нацеленным на ГКГС класса II-положительные антигенпрезентирующие клетки (АПК) (Fougeray et al.: A soluble LAG-3 protein as an immu no potentiator for therapeutic vaccines: Preclinical evaluation of IMP321. Vaccine 2006, 24:5426-5433; Brignone et at. IMP321 (sLAG-3) safety and T cell response potentiation using an influenza vaccine as a model antigen: A single-blind phase I study. Vaccine 2007, 25:4641-4650; Brignone et al.: IMP321 (sLAG-3), an immunopotentiator for T cell responses against a HBsAg antigen in healthy adults: a single blind randomised controlled phase I study. J Immune Based Ther Vaccines 2007, 5:5; Brignone etai: A soluble form of lymphocyte activation gene-3 (IMP321) induces activation of a large range of human effector cytotoxic cells. J Immunol 2007, 179:4202-4211). IMP321 был протестирован на ранее лечившихся пациентах с прогрессирующей почечно-клеточной карциномой, у которых известен факт ослабления иммунитета и показано увеличение процента циркулирующих активированных CD8 Т-клеток и длительно существующих CD8 Т-клеток эффекторной памяти у всех пациентов, которых лечили повторными инъекциями в течение 3 месяцев без какой-либо обнаруживаемой токсичности (Brignone et al.: A phase I pharmacokinetic and biological correlative study of IMP321, a novel MHC class II agonist in patients with advanced renal cell carcinoma. Clin Cancer Res 2009, 15:6225-6231). Было показано, что концентрация только в несколько нг/мл IMP321 активна in vitro на АПК, демонстрируя высокую эффективность IMP321 как агониста иммунной системы (Brignone, et al., 2009, выше).
В исследовании у пациентов с метастатическим раком молочной железы (мРМЖ) Brignone et al. (First-line chemoimmunotherapy in metastatic breast carcinoma: combination of paclitaxel and IMP321 (LAG-3Ig) enhances immune responses and antitumor activity. Journal of Translational Medicine 2010, 8:71) продемонстрировано, что IMP321 увеличивал и активировал в течение нескольких месяцев как первичные клетки-мишени (ГКГС класса II-позитивные моноциты/дендритные клетки), с которыми связывается IMP321, так и вторичные клетки-мишени (HKK/CD8+эффекторные Т-клетки памяти), которые активируются впоследствии. Объединяя результаты всех 30 пациентов и сравнивая регресс опухоли с соответствующей ретроспективной контрольной группой, они увидели удвоение частоты объективного ответа, что свидетельствует о том, что IMP321 является мощным агонистом эффективных противораковых клеточных иммунных ответов в данной клинической ситуации.
В WO 99/04810 описано использование белка LAG-3 или его фрагментов или производных в качестве вспомогательных средств для вакцинации и
для лечения рака. Использование белка LAG-3 или его фрагментов или производных для лечения рака и инфекционных заболеваний описано в WO 2009/044273.
Принимая во внимание медицинское применения LAG-3 и его фрагментов или производных, существует необходимость в создании препаратов таких соединений, в соответствии с правилами надлежащей производственной практики (GMP). Такая практика требуется для того, чтобы соответствовать стандартам, рекомендованным агентствами, которые контролируют утверждение и лицензирование производства и продажи активных лекарственных средств. Эти стандарты содержат минимальные требования, которым должен соответствовать производитель лекарственных препаратов, чтобы гарантировать, что продукция высокого качества и не представляет никакого риска для потребителя или общественности. В рамках процедуры контроля качества при производстве белков GMP-стандарта необходимо определить, сохраняют ли препараты таких соединений высокий уровень биологической активности.
Однако мы обнаружили, что некоторые общепринятые способы определения межбелковых взаимодействий не подходят для определения специфического связывания производного LAG-3 IMP321 с молекулами ГКГС класса II, экспрессируемыми на поверхности иммунных клеток. В частности, способ анализа сортировки клеток с активированной флуоресценцией (FACS), не подходил для установления различий между препаратами IMP321 с различными способностями связывания с клетками, экспрессирующими ГКГС класса II. Верхнее плато не наблюдали при повышении концентрации IMP321 для кривых связывания, полученных с использованием FACS. Это не позволяет рассчитать относительную активность разных препаратов, что требует конвергентных плато (параллелизм).
Мы также обнаружили, что IMP321 неспецифично связывается на планшетах, используемых для электрохемилюминесцентных (ECL) анализов MesoScale Discovery (MSD) и энзим-связанных иммуносорбентных анализов (ELISA). Хотя неспецифическое связывание IMP321 на планшетах, используемых для анализов ELISA и MSD, было значительно снижено при использовании казеина в качестве блокирующего реагента, это снизило абсолютный сигнал в анализе MSD. Не наблюдалось никаких верхних плато на кривых связывания, полученных с использованием анализов, в которых клетки, экспрессирующие молекулы ГКГС класса II, не были иммобилизованы на планшетах MSD. Был также протестирован другой метод ELISA, в котором клетки, экспрессирующие молекулы ГКГС класса II, переносили в другой планшет после связывания IMP321, чтобы минимизировать эффект неспецифического связывания IMP321 в планшетах. Однако изменение сигнала от лунки к лунке оказалось нежелательным. Учитывая это, был сделан вывод о том, что ни анализы MSD ECL, ни анализы ELISA не могли быть использованы для определения специфического связывания IMP321 с иммобилизованными клетками в анализе контроля качества для тестирования продукта по GMP-стандарту.
Следовательно, существует необходимость в разработке способа определения активности связывания с ГКГС класса II средств, содержащих белок LAG-3 или его фрагментов, производных или аналогов, который был бы пригоден для использования в анализе контроля качества таких соединений по GMP-стандарту.
КРАТКОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Согласно изобретению, предложен способ определения активности связывания с ГКГС класса II средства, содержащего белок гена активации лимфоцитов 3 (LAG-3) или его фрагмент, производное или аналог, при этом способ включает определение связывания белка LAG-3, его фрагмента, производного или аналога молекул ГКГС класса II с использованием биослойной интерферометрии (BLI).
Термин «биослойная интерферометрия (BLI)» используется в данном документе для обозначения волоконно-оптического анализа, основанного на фазовой интерферометрии, например, как описано в патенте США №5804453 (Chen). Разработки в технике BLI, включая разработки, направленные на повышение чувствительности и точности обнаружения аналита, описаны в WO 2005/047854 и WO 2006/138294 ForteBio, Inc.
Патент США 5804453 описывает зонды, способы, и системы для детектирования связывания аналита с волоконно-оптической торцевой поверхностью. Детектирование аналита основано на изменении толщины на торцевой поверхности оптического волокна в результате связывания молекул аналита с поверхностью, причем большее количество аналита вызывает в интерференционном сигнале изменение, связанное с толщиной. Изменение интерференционного сигнала происходит из-за фазового сдвига между светом, отраженным от конца волокна и от связующего слоя, переносимого на конце волокна, как показано, в частности, на Фиг. 7а и 7b патента США 5804453.
Зонд, описанный в патенте США №5804453, включает волоконно-оптическую часть, имеющую проксимальный конец и дистальный конец, и слой реагента, расположенный на дистальном конце. Слой реагента взаимодействует (или связывается) с обнаруживаемым веществом (аналитом). Волоконно-оптическая часть имеет первый показатель преломления, а слой реагента имеет второй показатель преломления. Когда какое-либо вещество связывается со слоем реагента, образуется результирующий слой, включающий слой реагента и вещество. Полученный слой можно анализировать как имеющий однородный показатель преломления.
Способ позволяет определять концентрацию вещества в растворе образца с помощью волоконнооптического зонда. Способ включает этапы (i) погружения дистального конца волоконно-оптического зонда в раствор образца, (ii) оптического соединения источника света с проксимальным концом волоконно-оптического зонда, (iii) детектирования по меньшей мере первого светового пучка отраженного от границы раздела между поверхностью дистального конца волоконно-оптической части и слоем реагента, и второй луч света, отраженный от границы раздела между слоем реагента и раствором образца, отраженный от дистального конца волоконно-оптического зонда, (iv) детектирования интерференционной картины, сформированной первым и вторым световыми лучами в первый раз, (v) детектирования интерференционной картины, сформированной первым и вторым световыми лучами во второй раз, и (vi) определение, присутствует ли вещество в образце раствора, основанное на том, происходит ли сдвиг в интерференционных картинах. Концентрация вещества может быть определена на основе сдвига в интерференционных картинах и на основе разности между первой и второй повторностью.
Система для детектирования концентрации вещества в растворе образца имеет источник света для подачи светового луча, волоконно-оптический зонд, детектор, волоконно-оптический соединитель, волоконно-оптический соединитель и процессор. Волоконно-оптический соединитель включает в себя первую волоконно-оптическую часть, имеющую проксимальный конец для приема падающего светового пучка, вторую волоконно-оптическую часть, имеющую проксимальный конец для доставки отраженного интерференционного светового пучка в детектор, и третью волоконно-оптическую часть, имеющую дистальный конец для подключения к волоконно-оптическому зонду. Волоконно-оптический зонд включает проксимальный конец для соединения с волоконно-оптическим соединителем и наконечник дистального конца с расположенным на нем слоем реагента. Волоконно-оптический зонд формирует по меньшей мере первый отраженный луч и второй отраженный луч из падающего светового луча. Детектор обнаруживает интерференционную картину, сформированную первым и вторым отраженными лучами. Соединяющее устройство оптически соединяет источник света с волоконно-оптическим зондом и оптически соединяет волоконно-оптический зонд с детектором. Процессор определяет фазу, связанную с интерференционной картиной, обнаруженной детектором в первый раз, определяет фазу, связанную с интерференционной картиной, обнаруженной детектором во второй раз, и определяет концентрацию вещества на основе сдвига фаз связанного с интерференционными картинами, обнаруженными детектором в первый и второй раз.
Мы установили, что метод BLI можно использовать для определения активности связывания ГКГС класса II препаратов белка LAG-3 или его фрагментов, производных или аналогов, и что такие способы особенно полезны как контроль качества при производстве таких веществ по GMP-стандарту.
В конкретных вариантах реализации, способы согласно изобретению включают определение связывания белка, фрагмента, производного или аналога LAG-3 с молекулами ГКГС класса II, присутствующими в клетках, экспрессирующих ГКГС класса II. В таких вариантах реализации, белок, фрагмент, производное или аналог LAG-3 могут быть иммобилизованы в слое реагента зонда BLI, и клетки, экспрессирующие ГКГС класса II, находятся в растворе.
Зонд, способ и система, описанные в патенте США 5804453, могут быть использованы в соответствии с данным изобретением для определения активности связывания ГКГС класса II препарата белка LAG-3 или его фрагмента, производного или аналога, как проиллюстрировано ниже путем связывания рекомбинантного производного белка LAG-3 IMP321 с клетками Raji, экспрессирующими ГКГС класса II.
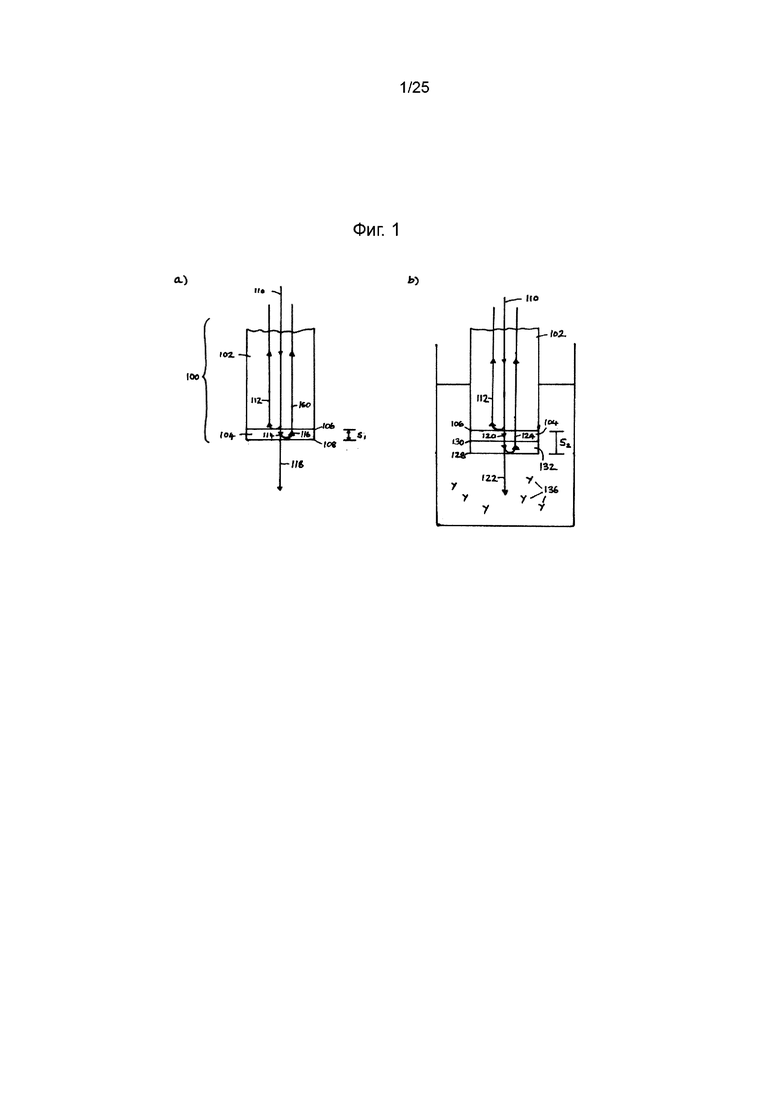
Ссылаясь на Фиг. 1а ниже, биосенсорный зонд 100 включает оптическое волокно 102 и слой реагентов 104, содержащий блокирующий реагент (например, БСА) и IMP321, на дистальном конце оптического волокна 102. Блокирующий реагент и IMP321 могут быть связаны с концом оптического волокна 102 путем смачивания конца в растворе, имеющем заранее определенную концентрацию IMP321, или блокирующего реагента, в течение предварительно определенного периода.
Падающий световой луч 110 направляется через оптическое волокно 102 к его дистальному концу. На границе раздела 106, определенной между оптическим волокном 102, которое имеет первый показатель преломления, и слоем 104 реагента, который имеет второй показатель преломления, первая часть 112 падающего светового пучка 110 будет отражаться, а вторая часть 114 падающего светового пучка 110 будет проходить через слой реагента 104. Как правило, блокирующий реагент и IMP321 будут малы относительно длины волны падающего светового пучка 110 с оптической точки зрения, поэтому блокирующий реагент и IMP321 можно рассматривать как формирующий один слой реагента 104. На границе 108, определенной на открытой поверхности слоя реагента 104 второй части 114 падающего луча 110, будет отражена первая часть 116, а вторая часть 118 пройдет в соседнюю среду. Из первой части 116 второй части 114 падающего луча 110 первая часть 160 будет передаваться обратно через оптическое волокно 102, а вторая часть (не показана) будет отражаться на границе 106 обратно в слой реагента 104.
На проксимальном конце оптического волокна 102 отраженные лучи 112 и 160 детектируются и анализируются. В любой заданной точке вдоль оптического волокна 102, включая его проксимальный конец, отраженные лучи 112 и 160 будут иметь разность фаз. На основании этой разности фаз можно определить толщину Si слоя реагента 104.
Ссылаясь на Фиг. 1b ниже, зонд 100 погружают в раствор 134, содержащий клетки 136 Raji для определения связывание клеток с иммобилизованным IMP321. Клетки 136 будут связываться с иммобилизованным IMP321 в слое реагента 104, тем самым формируя слой клеток 132 в течение определенного периода времени. Толщина S2 слоя будет зависеть от времени погружения зонда 100 в жидкость образца 134, а также от концентрации клеток 136 в жидкости образца 134. Другие молекулы 138 (не показаны) в растворе образца не будут связываться со слоем реагента 104.
Общая толщина S2 этого объединенного слоя будет больше, чем толщина S1 только слоя реагента 104. Таким образом, аналогично зонду 100 на Фиг. 1а, когда падающий луч 110 направлен к дистальному концу оптического волокна 102, на границе 106 между оптическим волокном 102 и объединенным слоем - первая часть 112 падающего луча 110 отражается, в то время как вторая часть 120 падающего луча 110 проходит через объединенный слой. Когда вторая часть 120 достигает клеток клеточного слоя 132, первая его часть (не показана) будет отражаться, когда она встречается с клеточной мембраной и цитоскелетными структурами клеток.
На второй границе 128 между объединенным слоем и образцом раствора 134 отражается вторая часть 124 второй части 120 падающего луча 110, а третья часть 122 второй части 120 падающего луча 110 проходит через образец раствора 134. Из второй части 124 второй части 120 падающего луча 110 первая часть 126 продолжается обратно через оптическое волокно 102, тогда как вторая часть (не показана) отражается обратно в объединенный слой на границе 106.
На проксимальном конце оптического волокна 102 отраженные лучи 112 и 126 детектируются и анализируются. В любой заданной точке вдоль оптического волокна 102, включая его проксимальный конец, отраженные лучи 112 и 126 будут иметь разность фаз. На основании этой разности фаз можно определить толщину S2 объединенного слоя.
Путем определения разницы между толщиной S2 объединенного слоя и толщиной S1 слоя реагента 104 можно определить толщину слоя клеток 132. Толщина S2 объединенного слоя определяется (или «отбирается») в дискретные моменты времени. Таким образом, может быть определена скорость увеличения разности между толщиной S2 объединенного слоя и толщиной S1 слоя реагента 104 (то есть скорость увеличения толщины слоя клеток 132). На основании этой скорости можно определить скорость связывания иммобилизованного IMP321 с молекулами ГКГС класса II на клетках Raji в течение очень короткого периода инкубации.
Диаметр клеток Raji составляет примерно 5-7 мкм, что в 1000 раз больше длины волны света, поэтому можно ожидать, что это повлияет на полученные результаты. Тем не менее, считывание сигнала составляет около 1-2 нМ, что указывает на то, что свет отражается вблизи поверхности клеток. Мы обнаружили, что изменение сигнала повторяется, коррелирует со связыванием клеток и что изменение скорости связывания находится в пределах диапазона измерений, поэтому его можно использовать для определения связывания клеток Raji с IMP321, иммобилизованными на конце оптического волокна.
Активность связывания ГКГС класса II препарата может быть определена как скорость связывания белка LAG-3, его фрагмента, производного или аналога с молекулами ГКГС класса II.
Мы обнаружили, что скорость связывания, полученная с использованием анализа BLI, зависит от плотности клеток, экспрессирующих ГКГС класса II, в растворе, тогда как скорость связывания является низкой и относительно прямой, когда плотность клеток, не экспрессирующих ГКГС класса II, увеличивается. Более высокая скорость, а также более высокое верхнее плато кривой связывания получают, если клетки, экспрессирующие ГКГС класса II, присутствуют при плотности, по меньшей мере, 4Е6/мл, предпочтительно, по меньшей мере, 6Е6/мл или 8Е6/мл.
Мы обнаружили, что специфичность анализа BLI улучшается, когда слой реагента зонда BLI предварительно обрабатывают блокирующим реагентом, чтобы минимизировать неспецифическое связывание клеток, экспрессирующих ГКГС класса II, со слоем реагента. Может быть использован любой подходящий блокирующий реагент, например блокирующие реагенты, содержащие инертный белок, такой как альбумин, например бычий сывороточный альбумин (БСА).
Клетки, экспрессирующие ГКГС класса II, могут быть иммунными клетками, экспрессирующими молекулы ГКГС класса II. Подходящие примеры включают антигенпрезентирующие клетки или клетки клеточных линий, полученных из иммунных клеток. В конкретных вариантах реализации, клетки, экспрессирующие ГКГС класса II, представляют собой В-клетки или клетки В-клеточной линии, например клетки Raji.
Мы обнаружили, что клетки, экспрессирующие ГКГС класса II, используемые для способов согласно изобретению, могут быть размороженными, готовыми к использованию клетками, полученными из замороженного исходного раствора. Использование таких клеток устраняет необходимость культивирования клеток непосредственно перед осуществлением способа согласно изобретению, может помочь обеспечить надежность и воспроизводимость результатов, полученных способами согласно изобретению, а также позволяет сравнивать результаты, полученные в разное время.
Способы согласно изобретению могут включать определение скорости связывания белка, фрагмента, производного или аналога LAG-3 с молекулами ГКГС класса II для множества различных концентраций белка, фрагмента, производного или аналога LAG-3 и создание кривой доза-ответ для скоростей связывания, например, как описано в Примере 6 ниже.
Способы согласно изобретению могут дополнительно включать определение активности связывания ГКГС класса II контрольного образца белка LAG-3 или его фрагмента, производного или аналога путем определения связывания белка, фрагмента, производного или аналога эталонного образца LAG-3 с молекулами ГКГС класса II с использованием BLI в тех же условиях, которые используются для определения связывания белка LAG-3, его фрагмента, производного или аналога препарата, и сравнение активности связывания ГКГС класса II, определенной для эталонного образца, с активностью связывания ГКГС класса II, определенной для препарата.
Активность связывания ГКГС класса II в контрольном образце при заданной концентрации может быть установлена равной 100% и разбавлена до различных желаемых концентраций, например, для обеспечения определения или подтверждения измерений активности связывания ГКГС класса II препарата, содержащего белок LAG-3, или его фрагмент, производное или аналог, полученный с использованием способа согласно изобретению.
В некоторых вариантах реализации, эталонный образец содержит белок LAG-3 или его фрагмент, производное или аналог, который обрабатывался для снижения своей активности связывания ГКГС класса II. Подходящие обработки включают, например, дегликозилирование (например, обработка ПНГазой), хранение при 37°C в течение по меньшей мере 12 дней, окисление (например, обработка 1% или 0,1% перекисью водорода), обработку кислотой или щелочью или выдержку на свету не менее 5 дней.
В приведенном ниже Примере 6 подробно описан анализ BLI для определения активности связывания ГКГС класса II иммобилизованного IMP321 с клетками Raji в растворе.
В соответствии с изобретением также предлагается зонд BLI для определения активности связывания ГКГС класса II белка LAG-3 или его фрагмента, производного или аналога, который содержит слой реагента, к которому относится белок или фрагмент LAG-3, производное или его аналог, являющийся иммобилизованным.
Дополнительно предлагается набор для определения активности связывания ГКГС класса II белка LAG-3 или его фрагмента, производного или аналога, который включает зонд BLI, имеющий слой реагента, к которому относится белок или фрагмент производного белка LAG-3 или его аналог, являющийся иммобилизованным, и клетки, экспрессирующие ГКГС класса II.
В некоторых вариантах реализации, слой реагента зонда BLI предварительно обрабатывали блокирующим реагентом, чтобы минимизировать неспецифическое связывание клеток, экспрессирующих ГКГС класса II, со слоем реагента. Может быть использован любой подходящий блокирующий реагент, например блокирующие реагенты, содержащие инертный белок, такой как альбумин, например бычий сывороточный альбумин (БСА).
В некоторых вариантах реализации, клетки, экспрессирующие ГКГС класса II, представляют собой замороженные клетки.
В некоторых вариантах реализации, клетки, экспрессирующие ГКГС класса II, представляют собой клетки Raji.
Клетки, экспрессирующие ГКГС класса II, могут присутствовать с плотностью, по меньшей мере, 1Е6/мл, предпочтительно, по меньшей мере, 4Е6/мл или 8Е6/мл.
Набор согласно изобретению может дополнительно включать контрольный образец, например, как описано выше, содержащий белок LAG-3 или его фрагмент, производное или аналог. Предпочтительно, активность связывания ГКГС класса II эталонного образца известна (например, как определено с помощью анализа высвобождения CCL4, описанного ниже).
Зонды и наборы согласно изобретению могут быть использованы в способах изобретения.
Белок LAG-3 может быть изолированным природным или рекомбинантным белком LAG-3. Белок LAG-3 может содержать аминокислотную последовательность белка LAG-3 из любых подходящих видов, таких как белок LAG-3 приматов или мыши, но предпочтительно белок LAG-3 человека. Аминокислотная последовательность белка LAG-3 человека и мыши представлена на Фиг. 1 в работе Huard et al. (Proc. Natl. Acad. Sci. USA, 11: 5744-5749, 1997). Последовательность белка LAG-3 человека повторяется на Фиг. 25 ниже (SEQ ID NO: 1). Аминокислотные последовательности четырех внеклеточных доменов суперсемейства Ig (D1, D2, D3 и D4) человеческого LAG-3 также показаны на Фиг. 1 в работе Huard et al., в аминокислотных остатках: 1-149 (D1); 150-239 (D2); 240-330 (D3); и 331-412 (D4).
Производные белка LAG-3 включают растворимые фрагменты, варианты или мутанты белка LAG-3, которые способны связывать молекулы ГКГС класса II. Известно несколько производных белка LAG-3, способных связываться с молекулами ГКГС класса II. Многие примеры таких производных описаны в работе Huard et al. (Proc. Natl. Acad. Sci. USA, 11: 5744-5749, 1997). Этот документ описывает характерные признаки сайта связывания ГКГС класса II на белке LAG-3. Описаны способы получения мутантов LAG-3, а также количественный анализ клеточной адгезии для определения способности мутантов LAG-3 связывать позитивные клетки Daudi класса II. Определено связывание нескольких различных мутантов LAG-3 с молекулами ГКГС класса II. Некоторые мутации были способны уменьшать связывание класса II, тогда как другие мутации увеличивали сродство LAG-3 к молекулам класса II. Многие из остатков, необходимых для связывания белков ГКГС класса II, кластеризованы у основания сверхпетлевой структуры из 30 аминокислот в домене LAG-3 D1. Аминокислотная последовательность сверхпетлевой структуры домена D1 белка LAG-3 человека представляет собой GPPAAAPGHPLAPGPHPAAPSSWGPRPRRY (SEQ ID NO: 2), подчеркнутая последовательность на Фигуре 25.
Производное белка LAG-3 может содержать 30-аминокислотную последовательность сверхпетли домена D1 человеческого LAG-3 или вариант такой последовательности с одной или несколькими консервативными аминокислотными заменами. Вариант может содержать аминокислотную последовательность, которая по меньшей мере на 70, 80, 90 или 95% идентична последовательности из 30-аминокислотной последовательности домена D1 человеческого LAG-3.
Производное белка LAG-3 может содержать аминокислотную последовательность домена D1 и, необязательно, домена D2 белка LAG-3, предпочтительно человеческого белка LAG-3.
Производное белка LAG-3 может содержать аминокислотную последовательность, которая имеет идентичность по аминокислотам по меньшей мере на 70, 80, 90 или 95% с доменом D1 или с доменом D1 и D2 белка LAG-3, предпочтительно белка LAG-3 человека.
Производное белка LAG-3 может содержать аминокислотную последовательность доменов D1, D2, D3 и, необязательно, D4 белка LAG-3, предпочтительно человеческого белка LAG-3.
Производное белка LAG-3 может содержать аминокислотную последовательность, которая имеет идентичность аминокислот по меньшей мере на 70, 80, 90 или 95% с доменом D1, D2 и D3 или с доменом D1, D2, D3, и D4 белка LAG-3, предпочтительно человеческого LAG-3.
Идентичность последовательностей между аминокислотными последовательностями можно определить путем сравнения выравнивания последовательностей. Когда эквивалентная позиция в сравниваемых последовательностях занята одной и той же аминокислотой, то молекулы в этой позиции идентичны. Оценка выравнивания как процента идентичности является функцией числа идентичных аминокислот в положениях, общих для сравниваемых последовательностей. При сравнении последовательностей для оптимального выравнивания может потребоваться ввести пробелы в одну или несколько последовательностей, чтобы учесть возможные вставки и удаления в последовательностях. Способы сравнения последовательностей могут использовать штрафы за пропуск последовательности, так что при одинаковом количестве идентичных молекул в сравниваемых последовательностях выравнивание последовательностей с как можно меньшим количеством пропусков, отражающее более высокую взаимосвязь между двумя сравниваемыми последовательностями, приведет к более высокой оценке, чем у одной со многими пропусками. Расчет максимального процента идентичности включает создание оптимального выравнивания с учетом штрафов за пропуск последовательности.
Подходящие компьютерные программы для сравнения последовательностей широко доступны в коммерческом и общественном секторах. Примеры включают MatGat (Campanella et al., 2003, BMC Bioinformatics 4: 29; программа доступна по ссылке http://bitincka.com/ledion/matgat), Gap (Needleman & Wunsch, 1970, J. Mol. Biol. 48: 443-453), FASTA (Altschul et al., 1990, J. Mol. Biol. 215 403-410; программа доступна по ссылке http://www.ebi.ac.uk/fasta), Clustal W 2.0 and X 2.0 (Larkin et al., 2007, Bioinformatics 23: 2947-2948; программа доступна по ссылке http://www.ebi.ac.uk/tools/clustalw2) and EMBOSS Pairwise Alignment Algorithms (Needleman & Wunsch, 1970, supra; Kruskal, 1983, In: Time warps, string edits and macromolecules: the theory and practice of sequence comparison, Sankoff & Kruskal (eds), pp 1-44, Addison Wesley; программа доступна по ссылке http://www.ebi.ac.uk/tools/emboss/align). Все программы могут быть запущены с использованием параметров по умолчанию.
Например, сравнение последовательностей может быть выполнено с использованием «needle» способа в алгоритме парного выравнивания EMBOSS, который определяет оптимальное выравнивание (включая промежутки) двух последовательностей при рассмотрении по всей их длине и обеспечивает процент идентичности. Параметрами по умолчанию для сравнения аминокислотных последовательностей (опция «Протеиновая молекула») может быть штраф за продолжение гэпа: 0,5, штраф за открытие гэпа: 10,0, матрица: Blosum 62.
Сравнение последовательностей может быть выполнено по всей длине эталонной последовательности.
Производное белка LAG-3 может быть присоединено к аминокислотной последовательности Fc-иммуноглобулина, предпочтительно аминокислотной последовательности Fc человеческого lgG1, необязательно, с помощью аминокислотной последовательности линкера.
Способность производного белка LAG-3 связываться с молекулами ГКГС класса II может быть определена с использованием количественного анализа клеточной адгезии, как описано в работе Huard et al (выше). Аффинность производного белка LAG-3 к молекулам ГКГС класса II может составлять, по меньшей мере, 20%, 30%, 40%, 50%, 60%, 70%, 80%, 90% или 100% от сродства белка LAG-3 человека для молекул класса II. Предпочтительно аффинность производного белка LAG-3 к молекулам ГКГС класса II составляет, по меньшей мере, 50% сродства человеческого белка LAG-3 к молекулам класса II.
Примеры подходящих производных белка LAG-3, которые способны связывать молекулы ГКГС класса II, включают производные, включающие:
аминокислотные остатки с 23 по 448 в последовательности LAG-3 человека;
аминокислотную последовательность доменов D1 и D2 LAG-3;
аминокислотную последовательность доменов D1 и D2 LAG-3 с аминокислотной заменой в одном или нескольких из следующих положений: положение 73, где ARG замещен на GLU; позиция 75, где ARG замещен на ALA или GLU; позиция 76, где ARG замещен на GLU; положение 30, где ASP замещен на ALA; положение 56, где HIS замещен на ALA; положение 77, где TYR замещен на РНЕ; положение 88, где ARG замещен на ALA; положение 103, где ARG замещен на ALA; положение 109, где ASP замещен на GLU; положение 115, где ARG замещен на ALA;
аминокислотную последовательность домена D1 LAG-3 с делецией аминокислотных остатков с 54 по 66;
рекомбинантный растворимый слитый белок LAG-3Ig человека (IMP321) - димер 200 кДа, продуцируемый в клетках яичника китайского хомяка, трансфицированных плазмидой, кодирующей внеклеточный домен hLAG-3, слитый с Fc человеческого IgG1. Последовательность IMP321 приведена в SEQ ID NO: 17 в документе US 2011/0008331.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Варианты реализации изобретения описаны ниже только в качестве примера со ссылкой на следующие фигуры, в которых:
На Фиг. 1 изображено действие зонда, используемого для определения активности связывания ГКГС класса II белка LAG-3 или его фрагментов, производных или аналогов в соответствии с вариантом реализации изобретения (фигура взята из патента США №5804453);
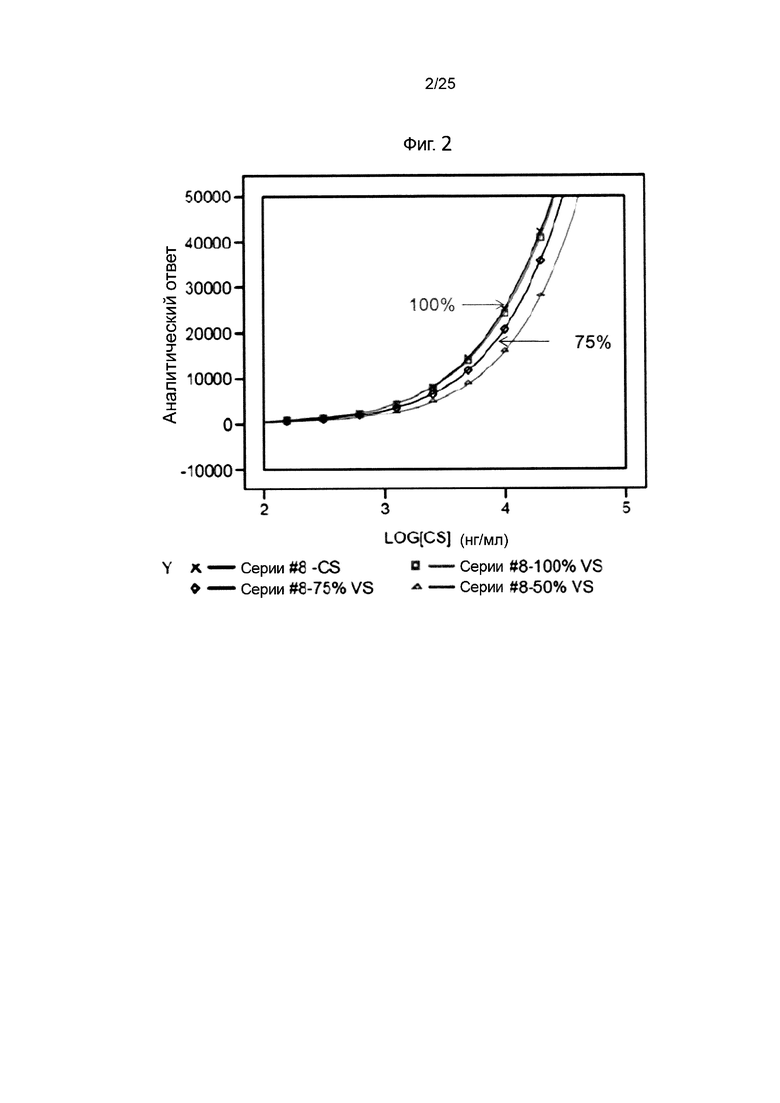
На Фиг. 2 изображены результаты анализа FACS для определения связывания IMP321 с клетками Raji;
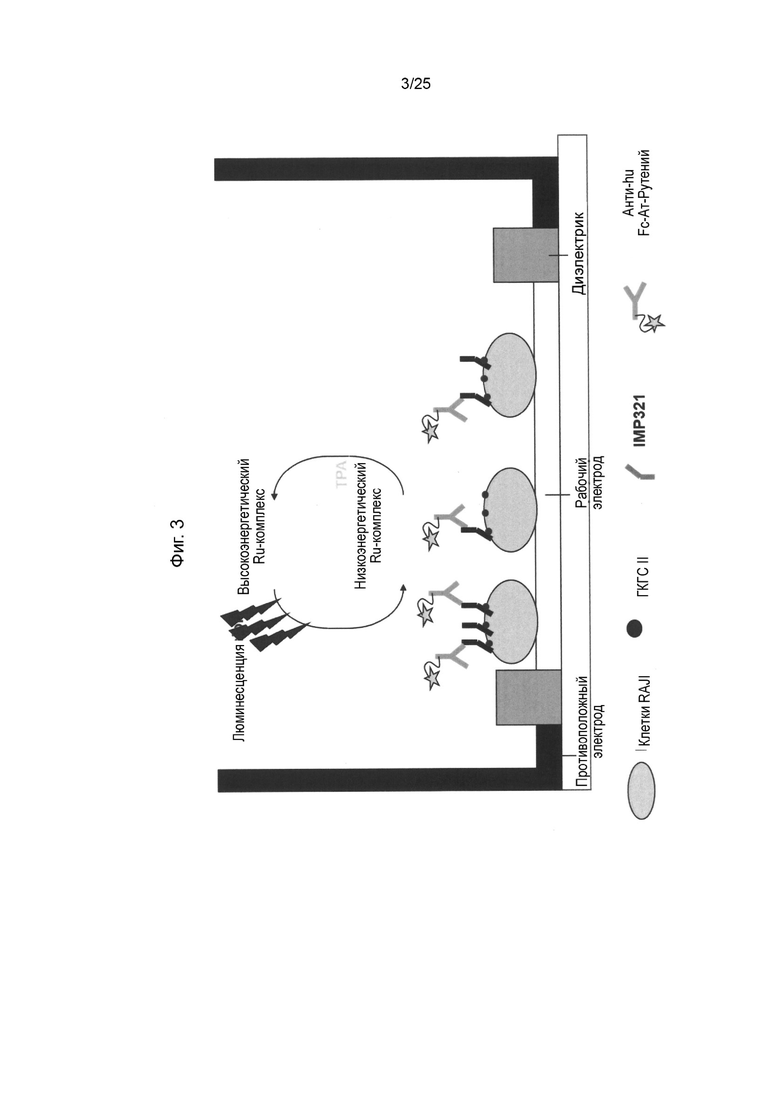
На Фиг. 3 схематически изображен электрохемилюминесцентный (ECL) анализ MesoScale Discovery (MSD) для определения связывания IMP321 с клетками Raji;
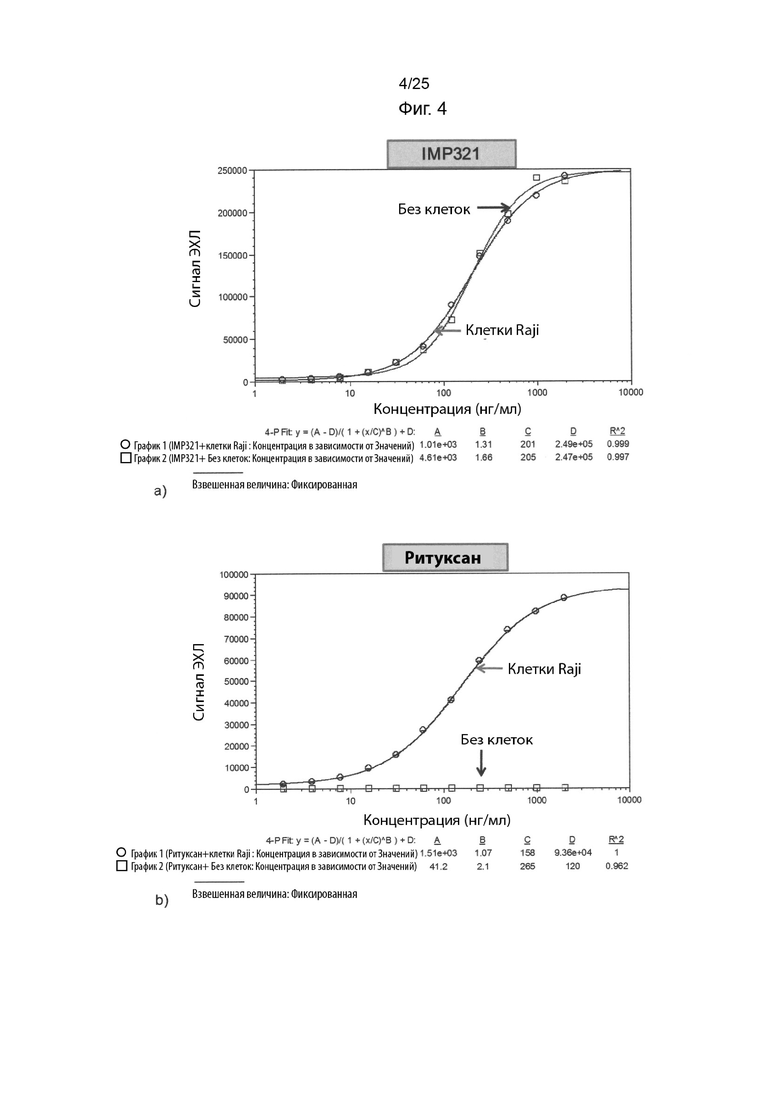
На Фиг. 4(a) изображен график ECL сигнала, полученного в случае анализа MSD при различных концентрациях IMP321 в присутствии и в отсутствие клеток Raji; На Фиг. 4(b) изображен график ECL сигнала, полученного для анализа MSD при различных концентрациях ритуксана в присутствии и в отсутствие клеток Raji;
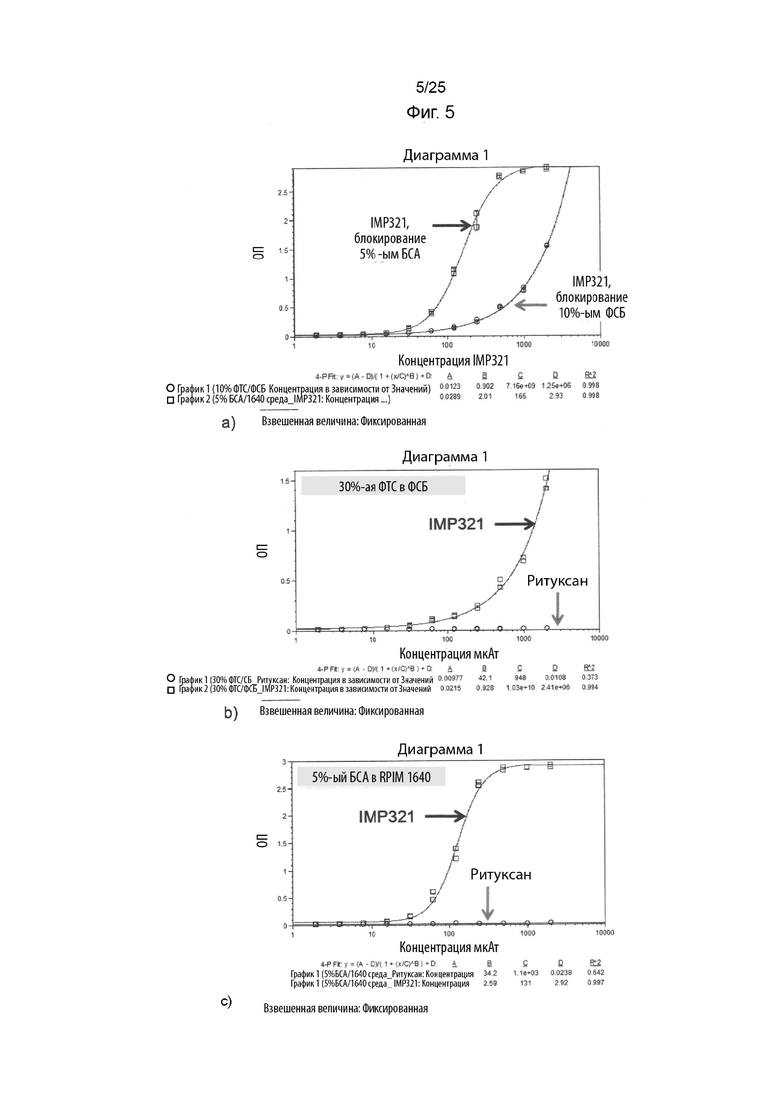
На Фиг. 5(a) изображен график сигнала ОП, полученного в ELISA при различных концентрациях IMP321 после блокирования планшета ELISA с 5% БСА или 10% ФТС; На Фиг. 5(b) изображен график сигнала ОП, полученного для ELISA при различных концентрациях IMP321 или ритуксана после блокирования планшета ELISA с 30% ФТС в ФСБ; На Фиг. 5(c) изображен график сигнала ОП, полученного для ELISA при различных концентрациях IMP321 или ритуксана после блокирования планшета ELISA с 5% БСА в RPIM1640;
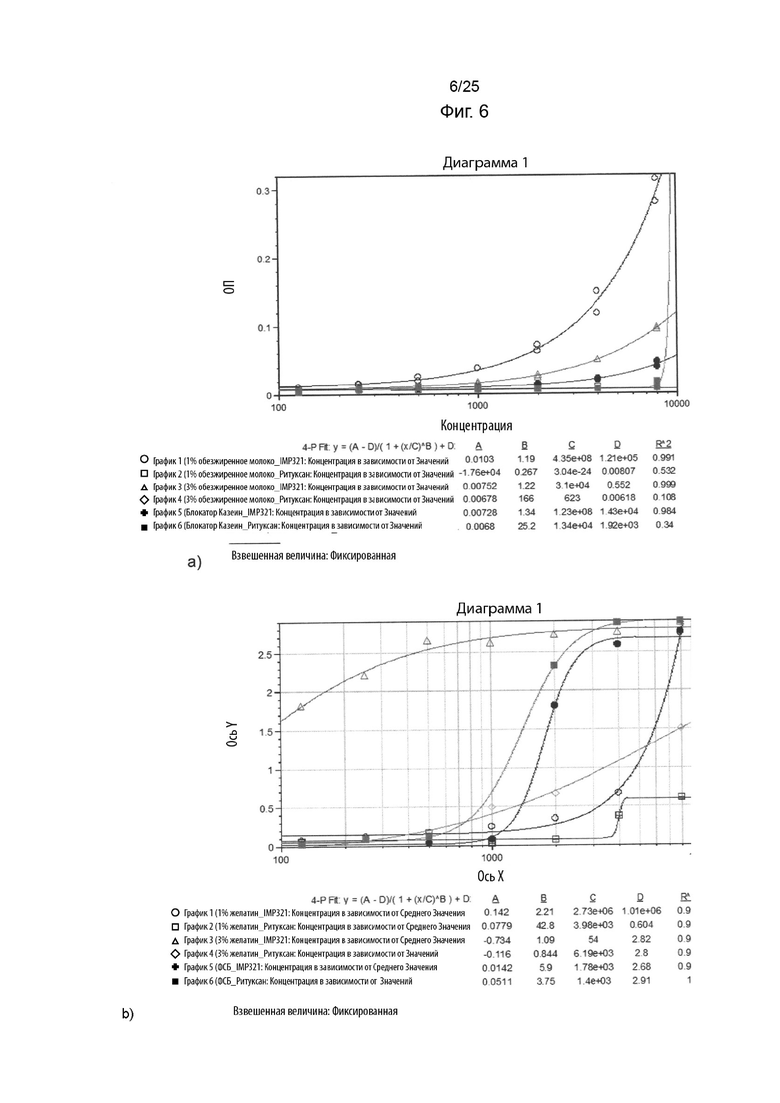
На Фиг. 6(a) изображен график сигнала ОП, полученного в ELISA при различных концентрациях IMP321 или ритуксана после блокирования планшета ELISA различными блокирующими реагентами (1%-ым обезжиренным молоком, 3%-ым обезжиренным молоком, казеином); На Фиг. 6(b) изображен график сигнала ОП, полученного в ELISA при различных концентрациях IMP321 или ритуксана после блокирования планшета ELISA различными блокирующими реагентами (1%-ым желатином, 3%-ым желатином или ФСБ);
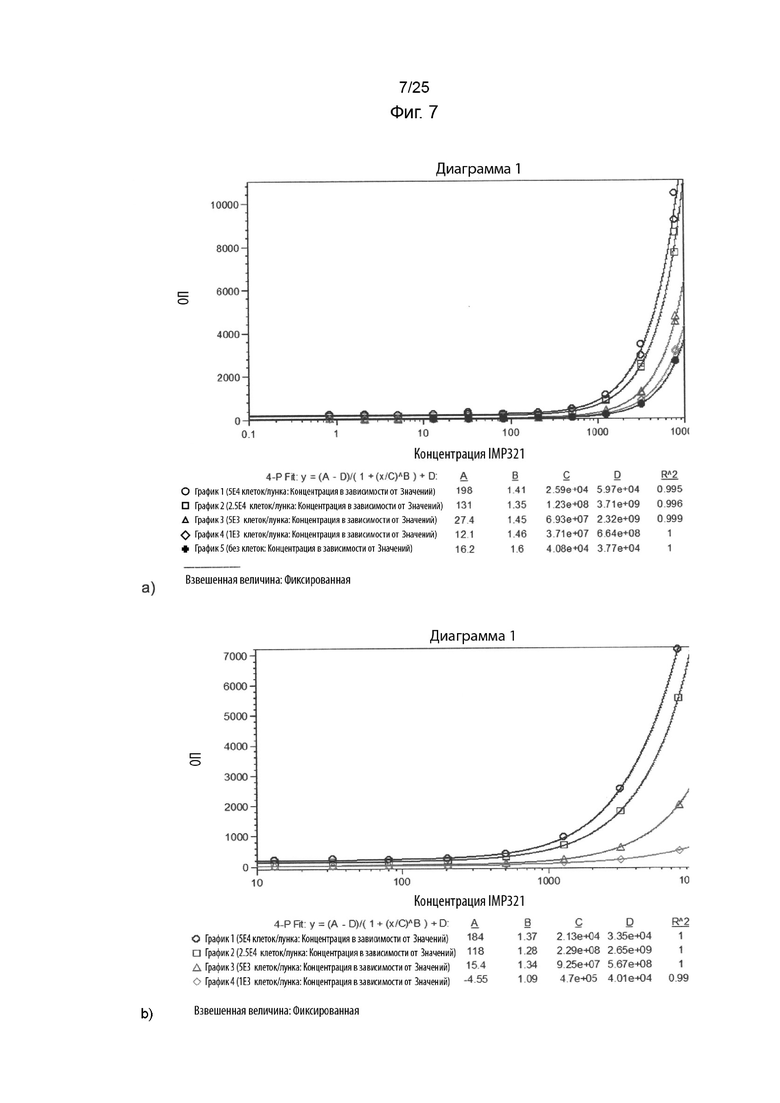
На Фиг. 7(a) изображен график исходного ECL сигнала, полученного в случае анализа MSD при различных концентрациях IMP321 для посевов клеток Raji с разной плотностью; На Фиг. 7(b) изображен график специфического сигнала ECL, полученного для анализа MSD при различных концентрациях IMP321 для посевов клеток Raji с разной плотностью;
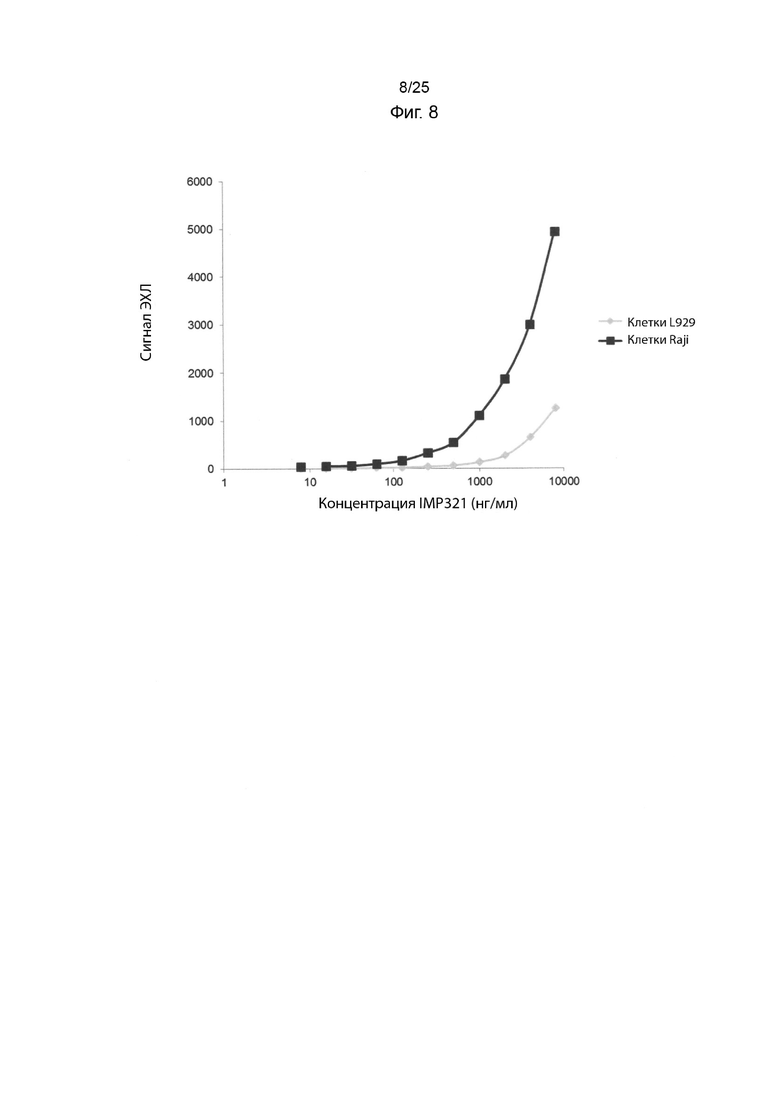
На Фиг. 8 изображен график ECL сигнала, полученного в случае анализа MSD для связывания различных концентраций IMP321 с клетками Raji или клетками HLA-DRdim L929 после блокировки планшета MSD казеином;
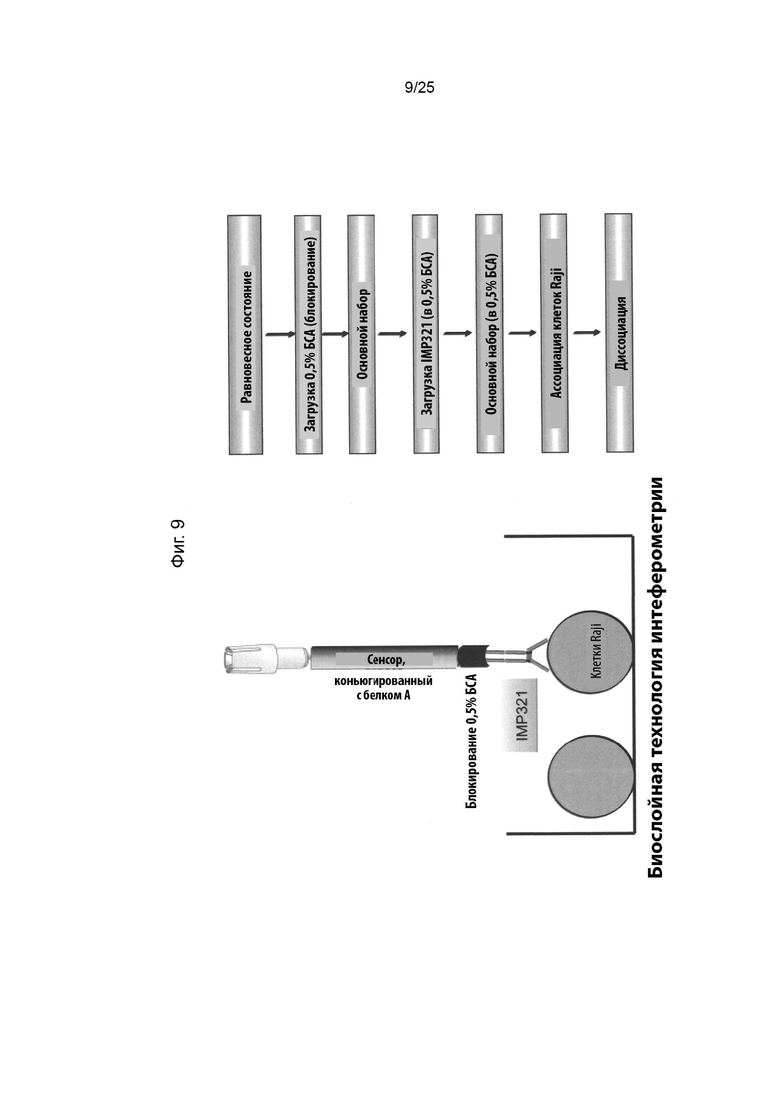
На Фиг. 9 слева схематически изображен зонд BLI с сенсором, конъюгированным с белком А, и IMP321, иммобилизованный на дистальном конце оптического волокна сенсора, с концом сенсора, погруженным в раствор образца, содержащий клетки Raji. Основные этапы способа приведены в правой части фигуры;
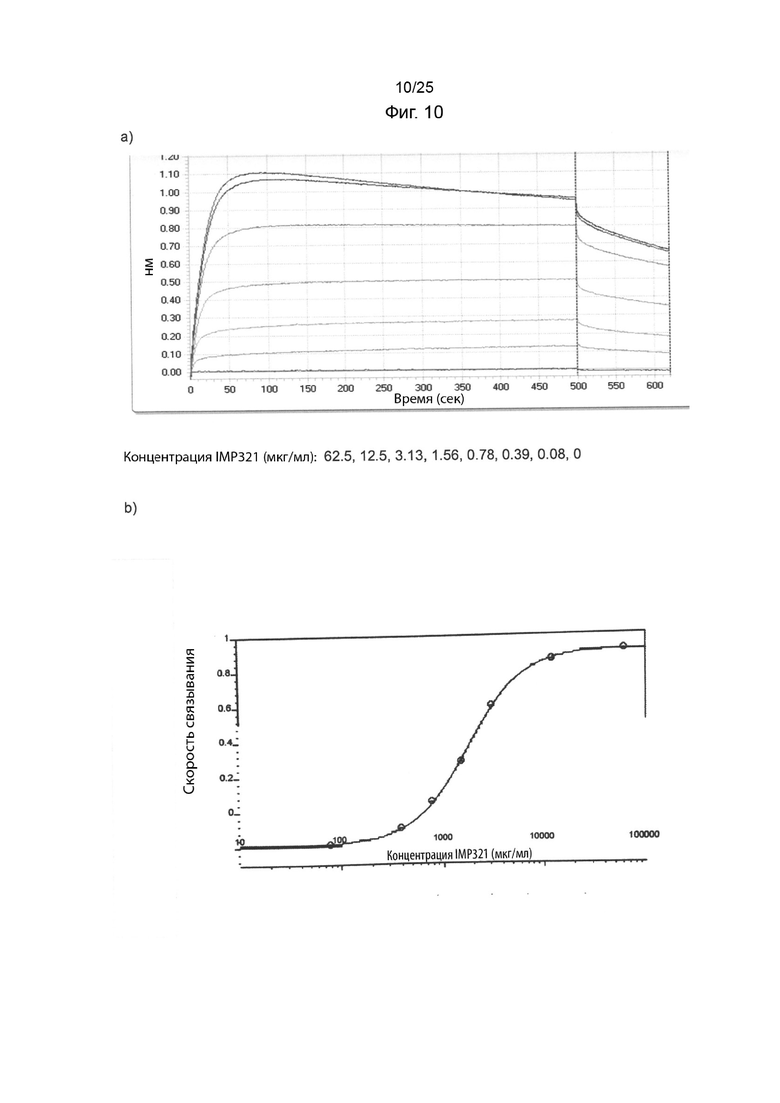
На Фиг. 10(a) изображен график сигнала связывания, полученного в анализе BLI для дозозависимого связывания иммобилизованного IMP321 с клетками Raji в растворе на этапе ассоциации; На Фиг. 10(b) изображена стандартная кривая дозозависимого связывания IMP321 с клетками Raji в анализе BLI;
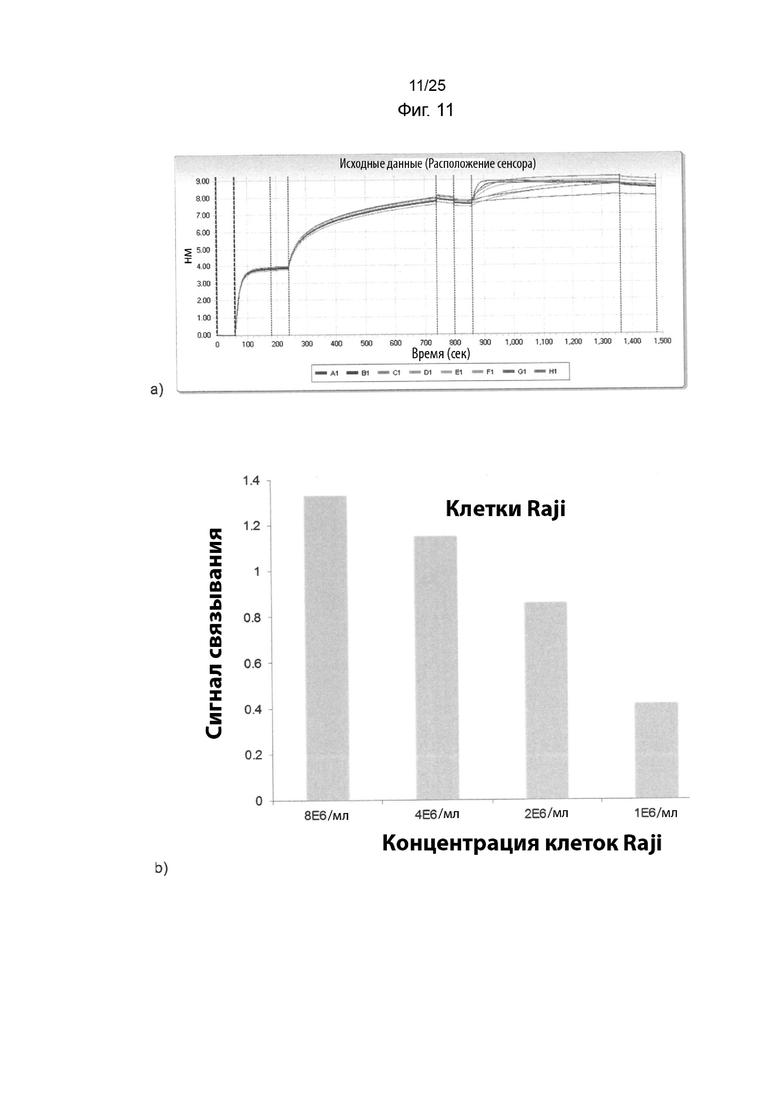
На Фиг. 11(a) изображены кривые ассоциации и диссоциации при связывании иммобилизованного IMP321 с различными концентрациями клеток Raji (которые экспрессируют ГКГС класса II) или клеток Jurkat (которые не экспрессируют ГКГС класса II) в растворе в анализе BLI; На Фиг. 11 (b) изображен график сигнала связывания, полученного для различных концентраций клеток Raji;
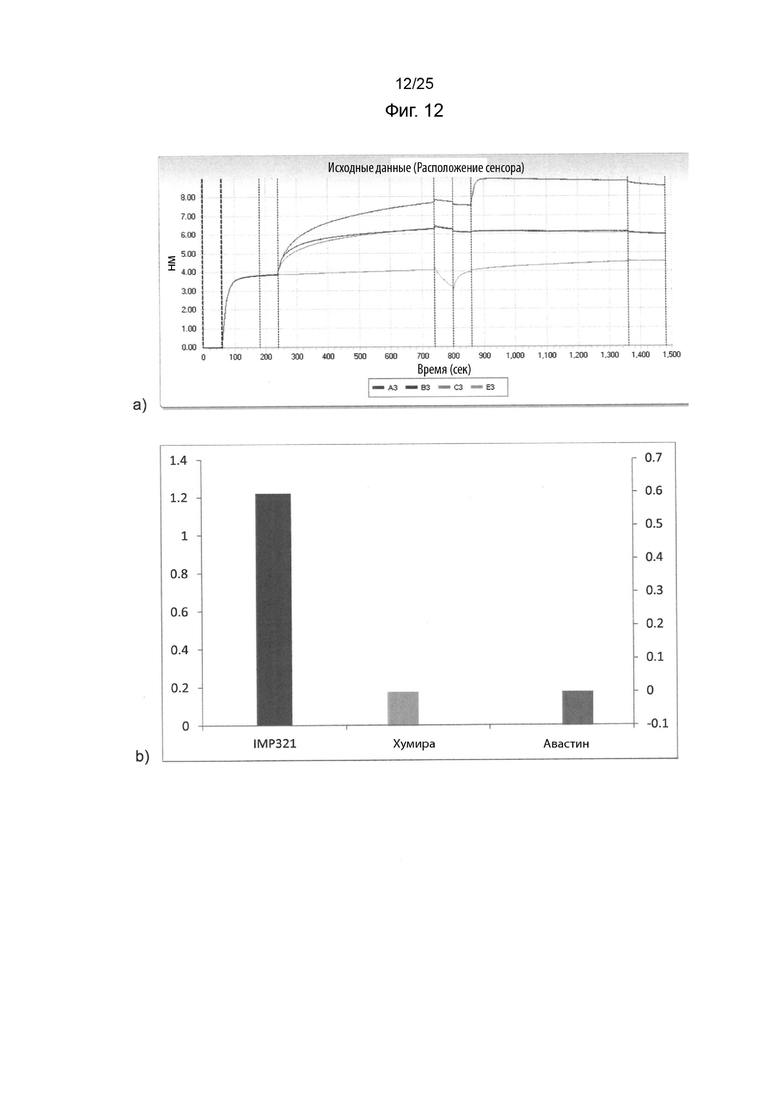
На Фиг. 12(a) изображены кривые ассоциации и диссоциации при связывании иммобилизованного IMP321, Хумира или Авастина с клетками Raji в растворе в анализе BLI; На Фиг. 12(b) изображен график сигнала связывания, полученного для различных иммобилизованных белков;
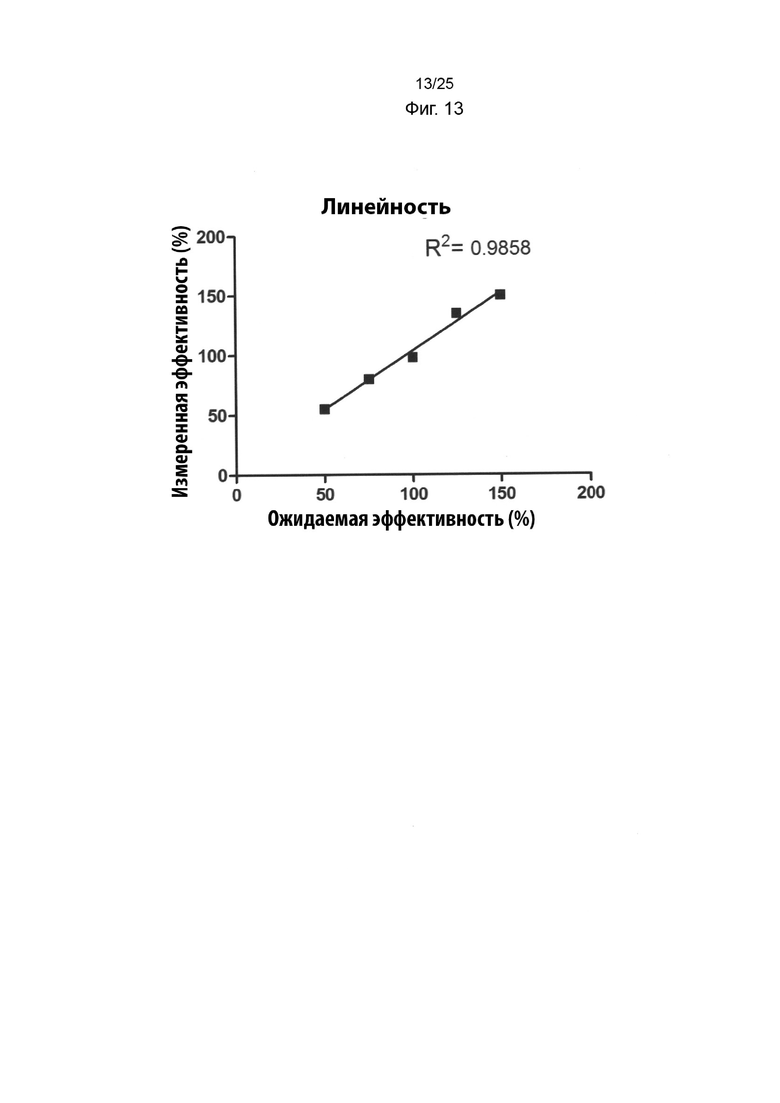
На Фиг. 13 изображен график с процентами активности связывания, измеренной с помощью анализа BLI, для связывания различных иммобилизованных препаратов IMP321 с клетками Raji в растворе в зависимости от их ожидаемой активности;
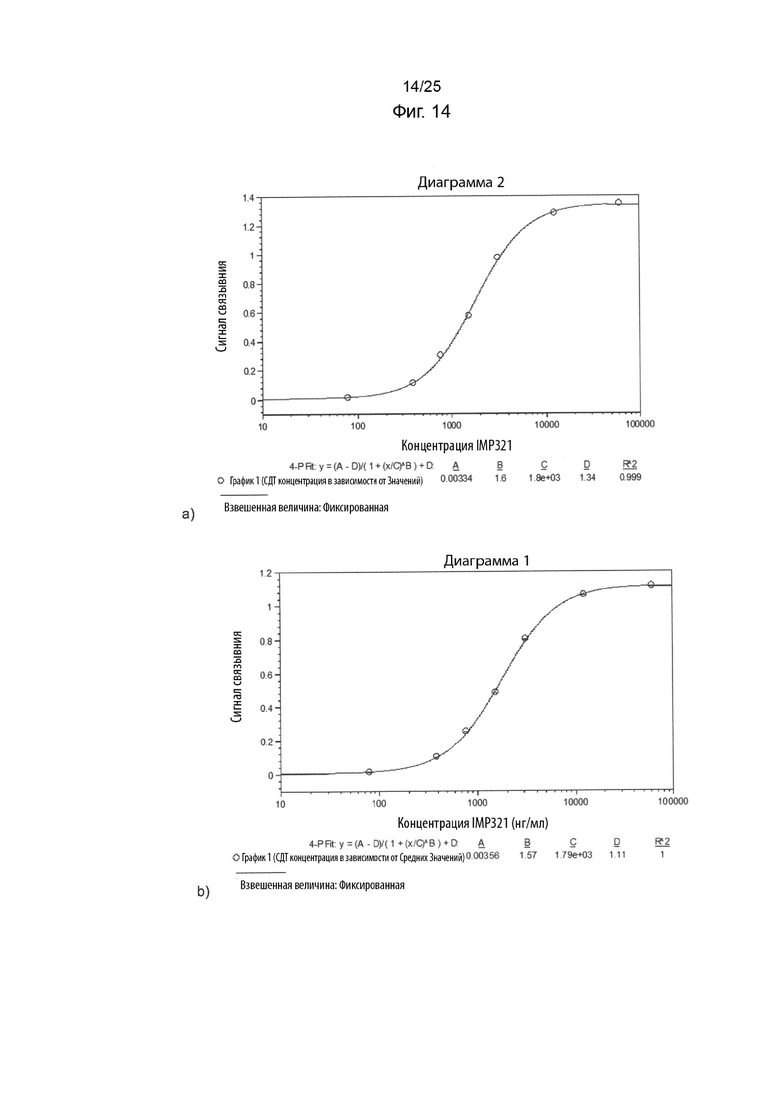
На Фиг. 14(a) изображен график сигнала связывания, полученного с помощью анализа BLI при связывании различных концентраций иммобилизованного IMP321 с предварительно культивированными клетками Raji в растворе;
На Фиг. 14(b) изображен график сигнала связывания, полученного с помощью анализа BLI при связывании различных концентраций иммобилизованного IMP321 с ранее замороженными клетками Raji в растворе;
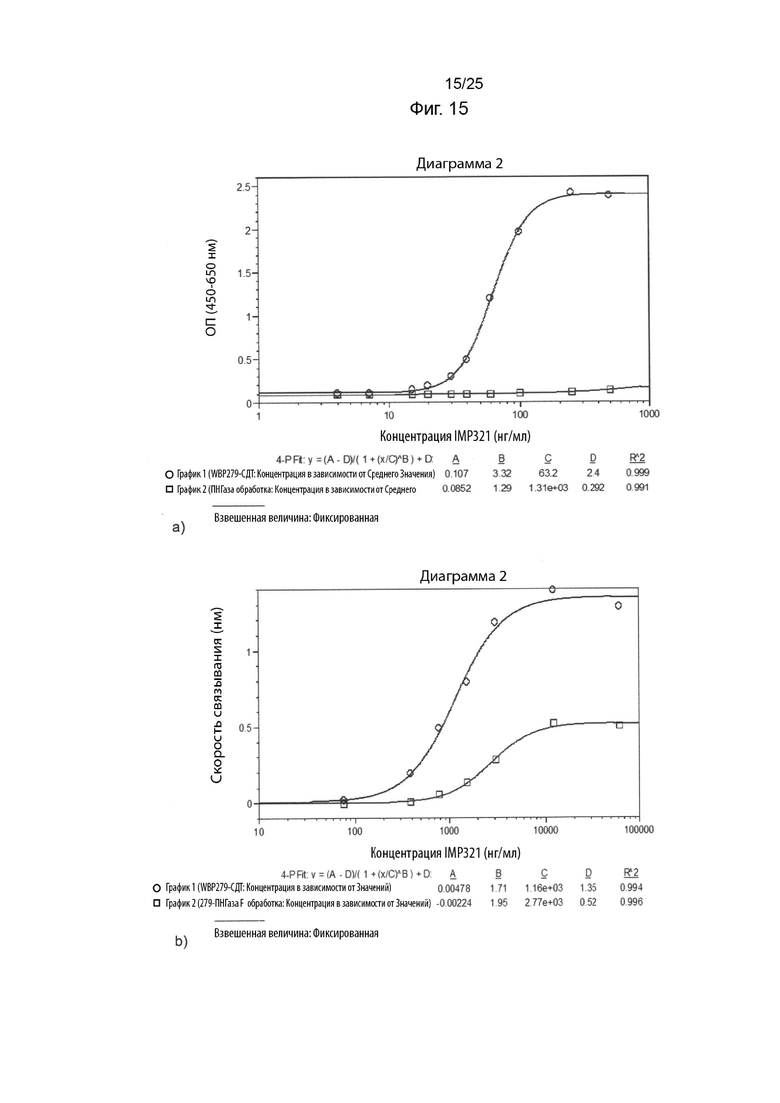
На Фиг. 15(a) изображен график снижения высвобождения CCL4, полученного с помощью клеточного анализа при связывании различных концентраций иммобилизованного IMP321 или дегликозилированного IMP321 с клетками Raji;
На Фиг. 15(b) изображен график сигнала связывания, полученный с помощью анализа BLI при связывании различных концентраций иммобилизованного IMP321 или дегликозилированного IMP321 с клетками Raji;
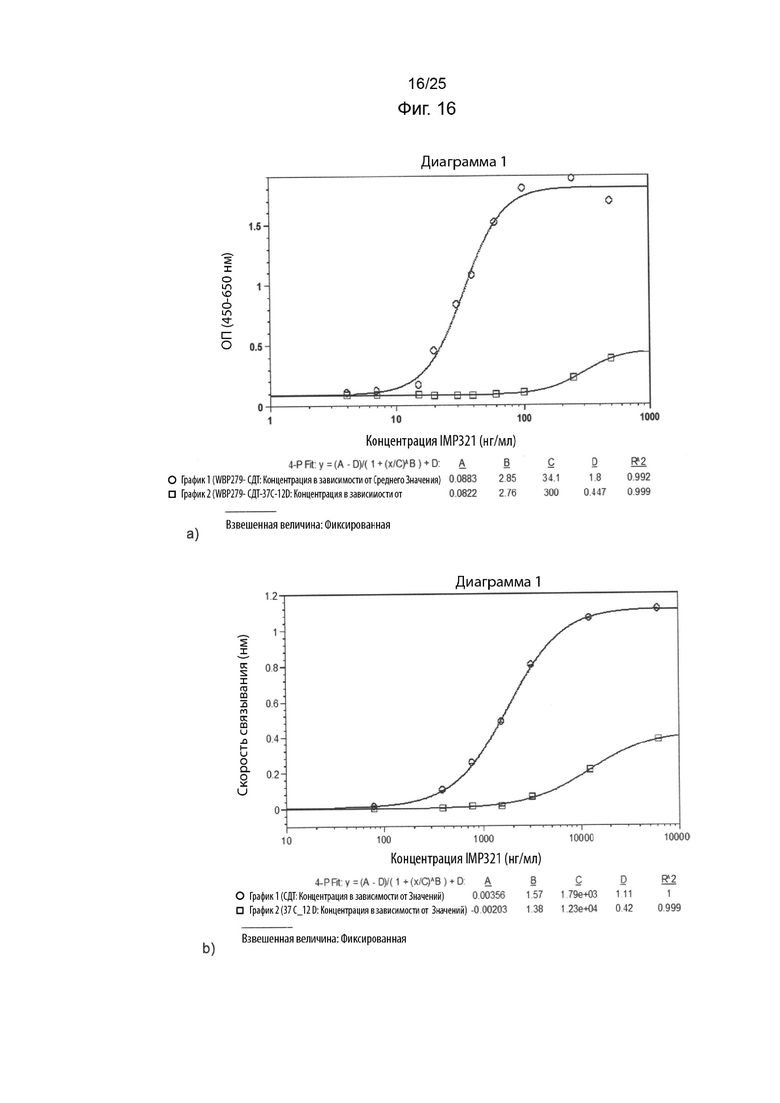
На Фиг. 16 изображены графики сигнала связывания иммобилизованных IMP321 или IMP321 различных концентраций, которые хранятся ненадлежащим образом (при 37°C в течение 12 дней), с клетками Raji. Результаты, изображенные на Фиг. 16(a), были получены с помощью клеточного анализа, измеряющего высвобождение CCL4, а результаты, изображенные на Фиг. 16(b), были получены с помощью анализа BLI;
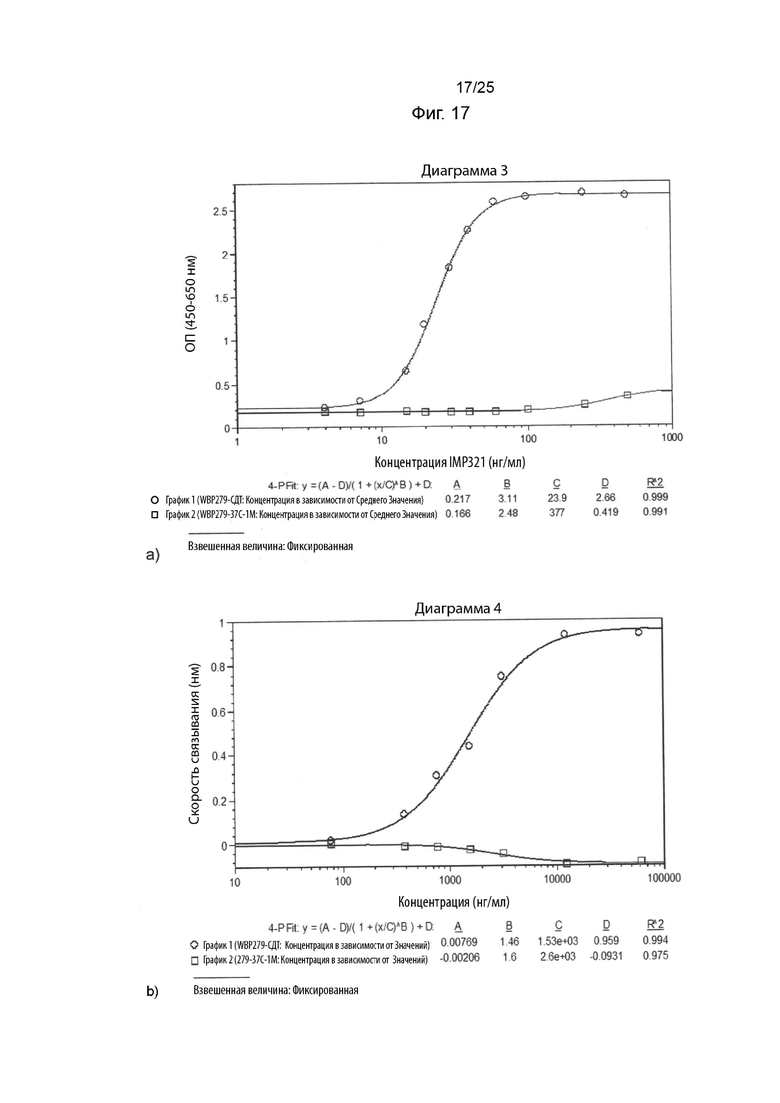
На Фиг. 17 изображены графики сигнала связывания различных концентраций иммобилизованного IMP321 или IMP321, хранящихся ненадлежащим образом (при 37°C в течение 1 месяца), с клетками Raji. Результаты, изображенные на Фиг. 17(a), были получены с помощью клеточного анализа, измеряющего высвобождение CCL4, а результаты, изображенные на Фиг. 17(b), были получены с помощью анализа BLI;
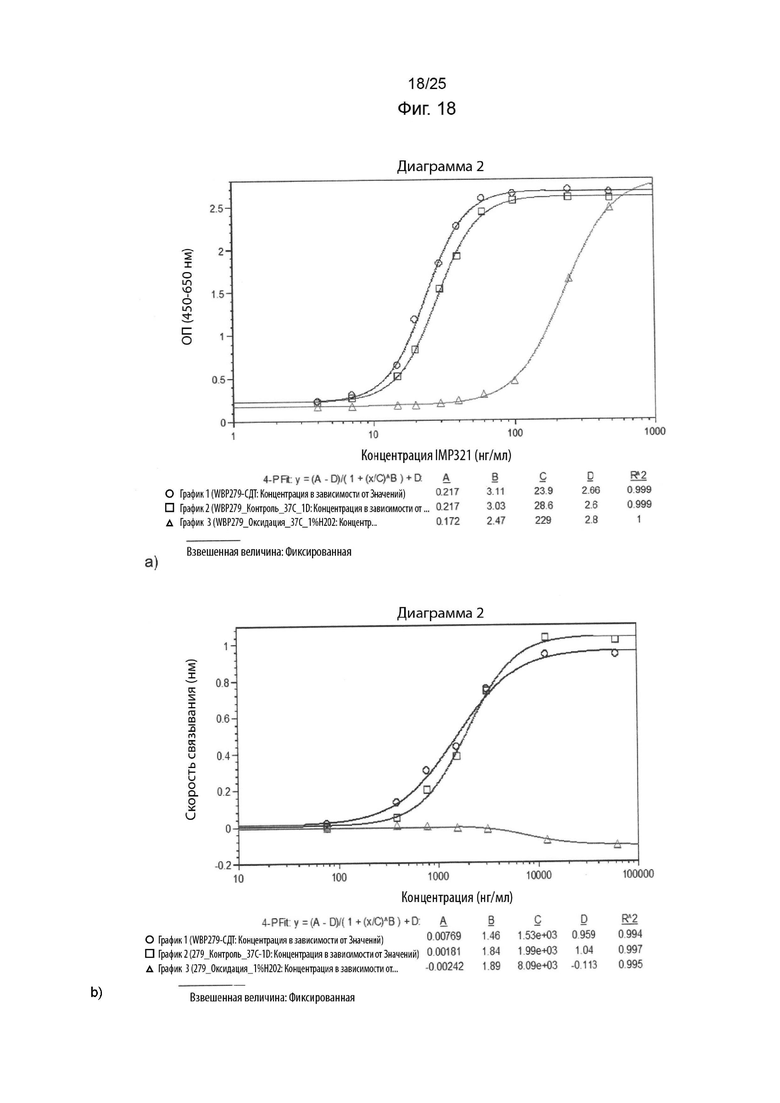
На Фиг. 18 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 18а), или с помощью анализа BLI (Фиг. 18b), при связывании различных концентраций иммобилизованного необработанного IMP321 или окисленного IMP321 (с 1%-ой перекисью водорода) с клетками Raji;
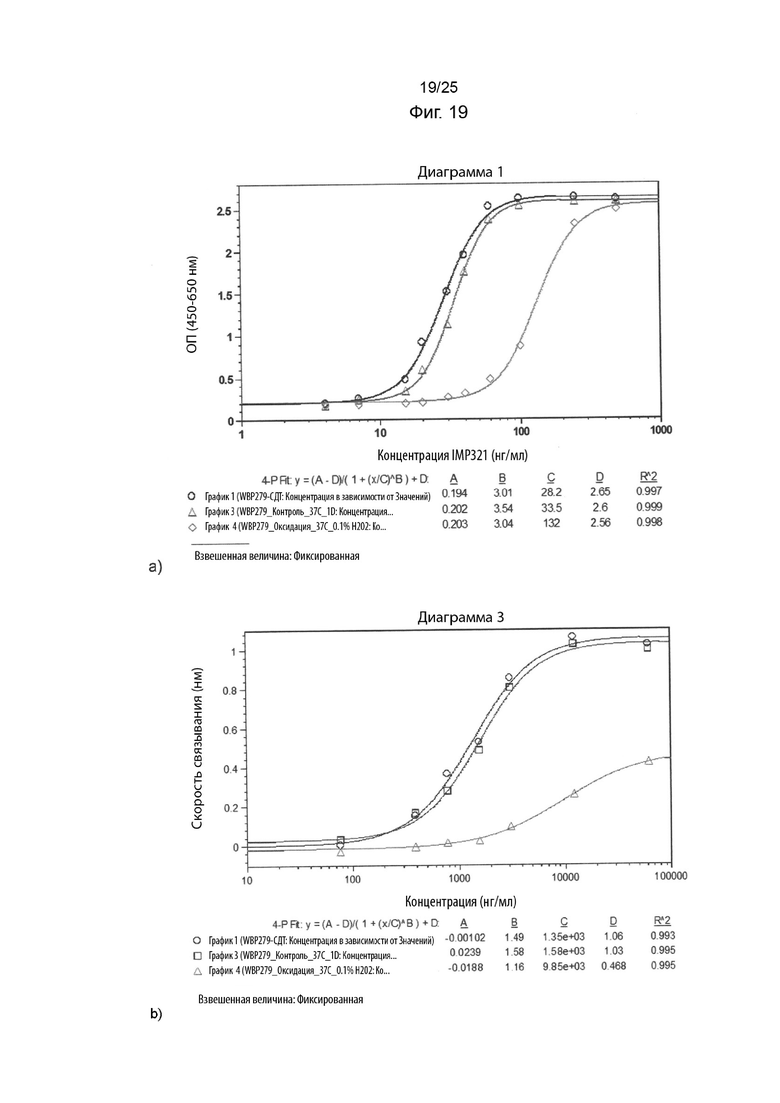
На Фиг. 19 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 19а), или с помощью анализа BLI (Фиг. 19b), при связывании различных концентраций иммобилизованного необработанного IMP321 или окисленного IMP321 (с 0,1%-ой перекисью водорода) с клетками Raji;
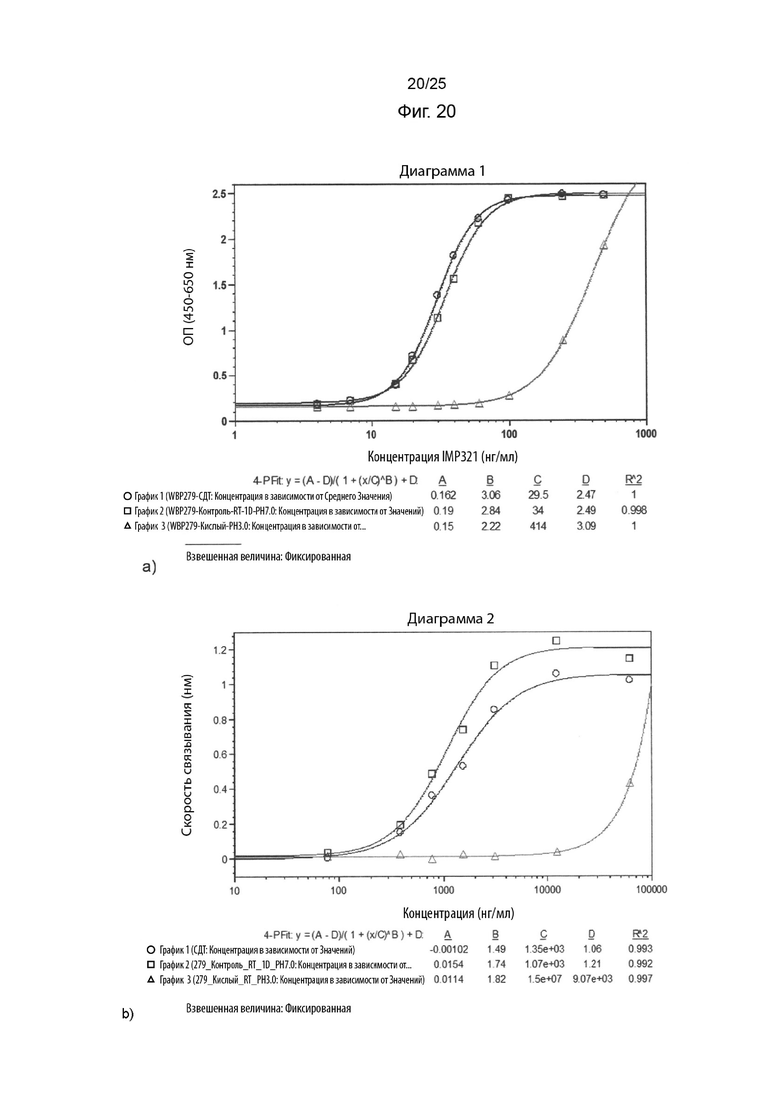
На Фиг. 20 изображен графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 20а), или с помощью анализа BLI (фигура 20b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного кислотой (при рН 3,0), с клетками Raji;
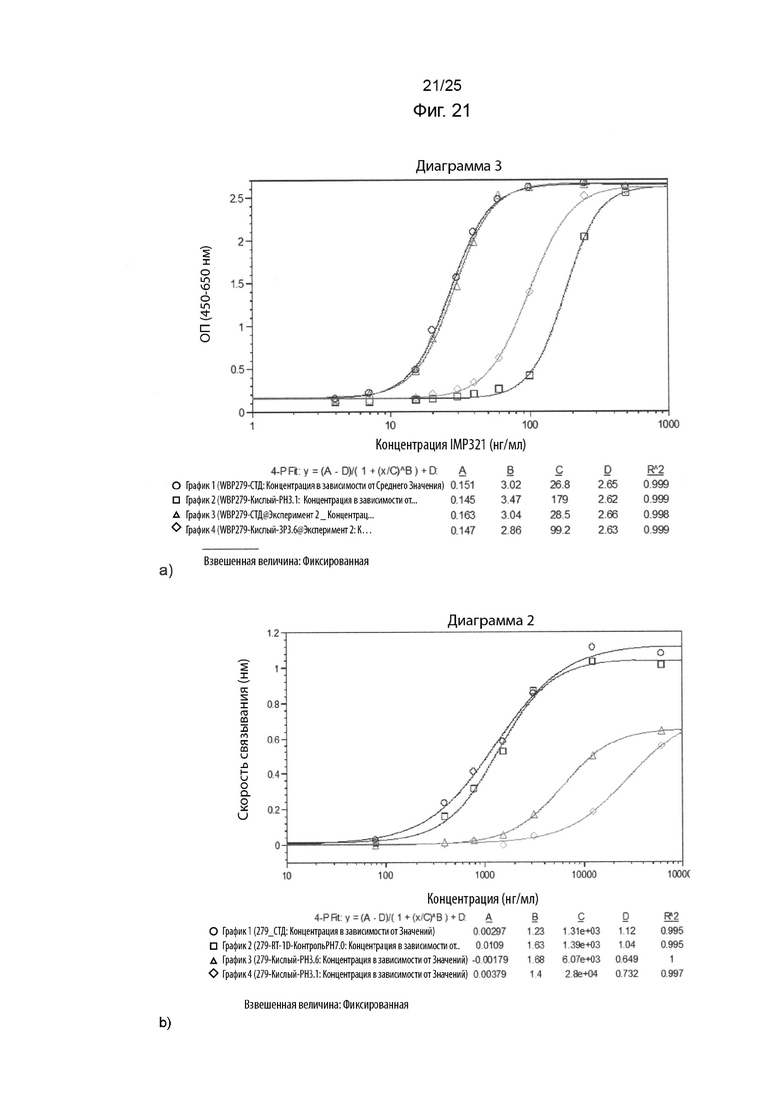
На Фиг. 21 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 21а), или с помощью анализа BLI (Фиг. 21b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного кислотой (при рН 3,1, или рН 3,6), с клетками Raji;
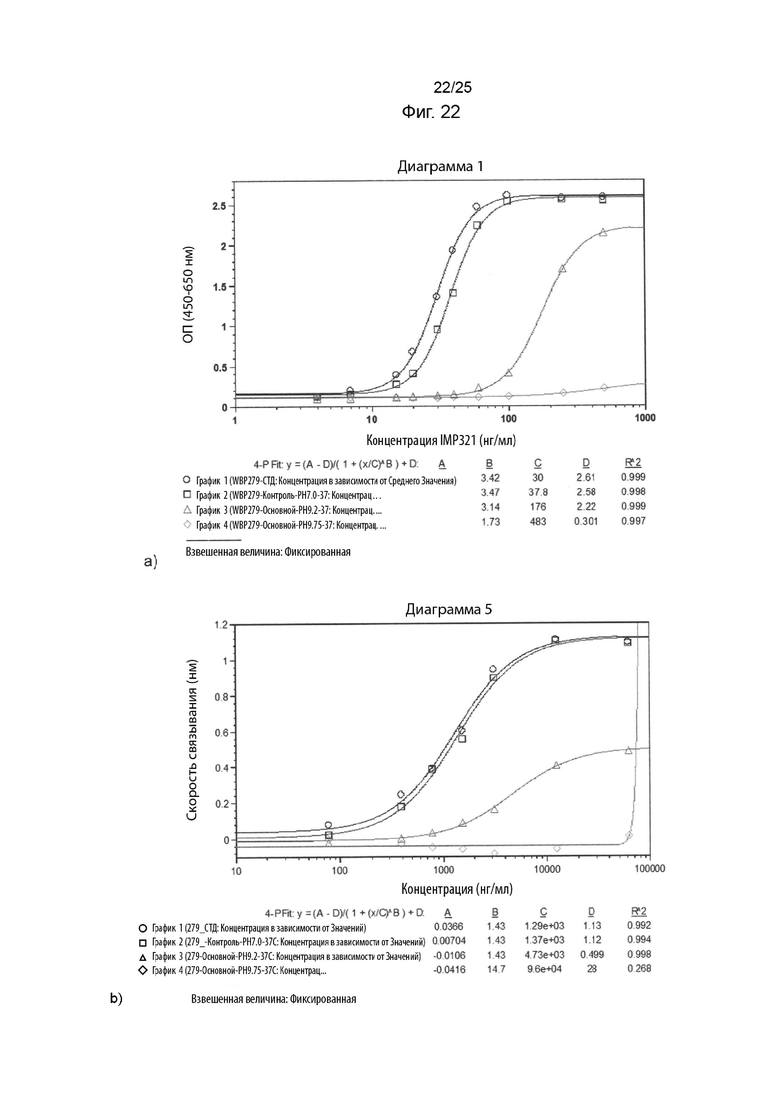
На Фиг. 22 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 22а) или с помощью анализа BLI (Фиг. 22b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного щелочами (при рН 9,2 или рН). 9,75), с клетками Raji;
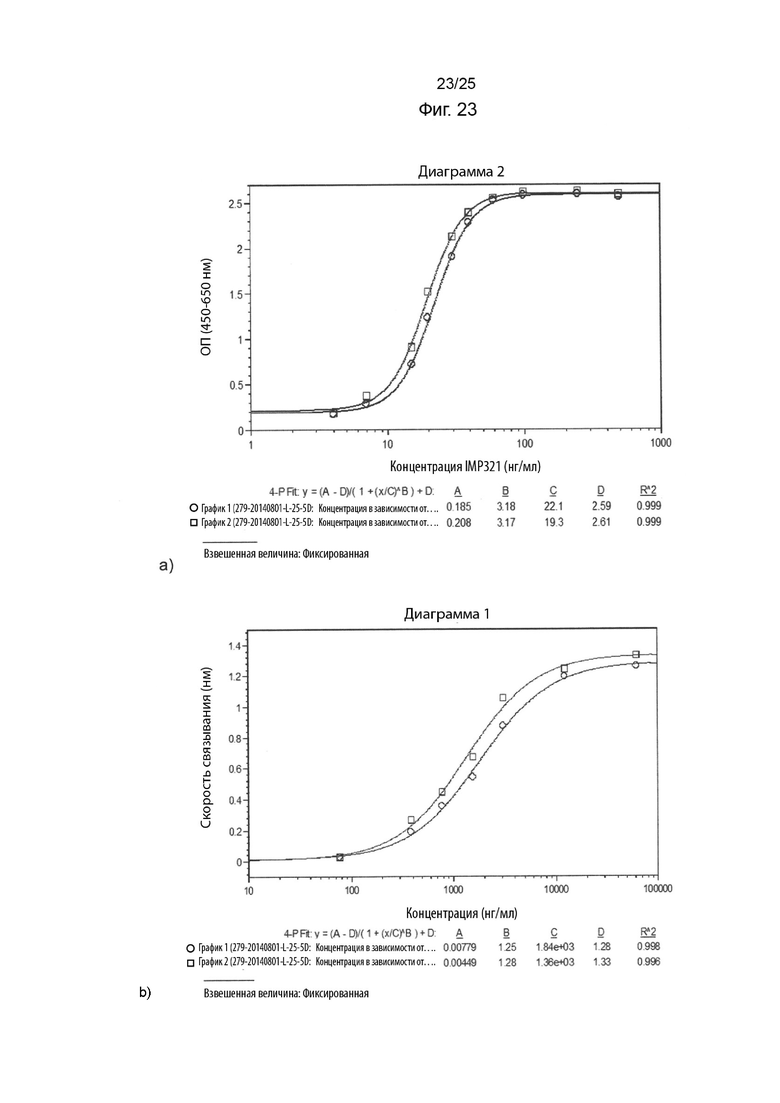
На Фиг. 23 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 23а), или с помощью анализа BLI (Фиг. 23b), при связывании различных концентраций иммобилизованного IMP321, необработанного или находившегося на свету (при 25°C в течение 5 дней).) с клетками Raji;
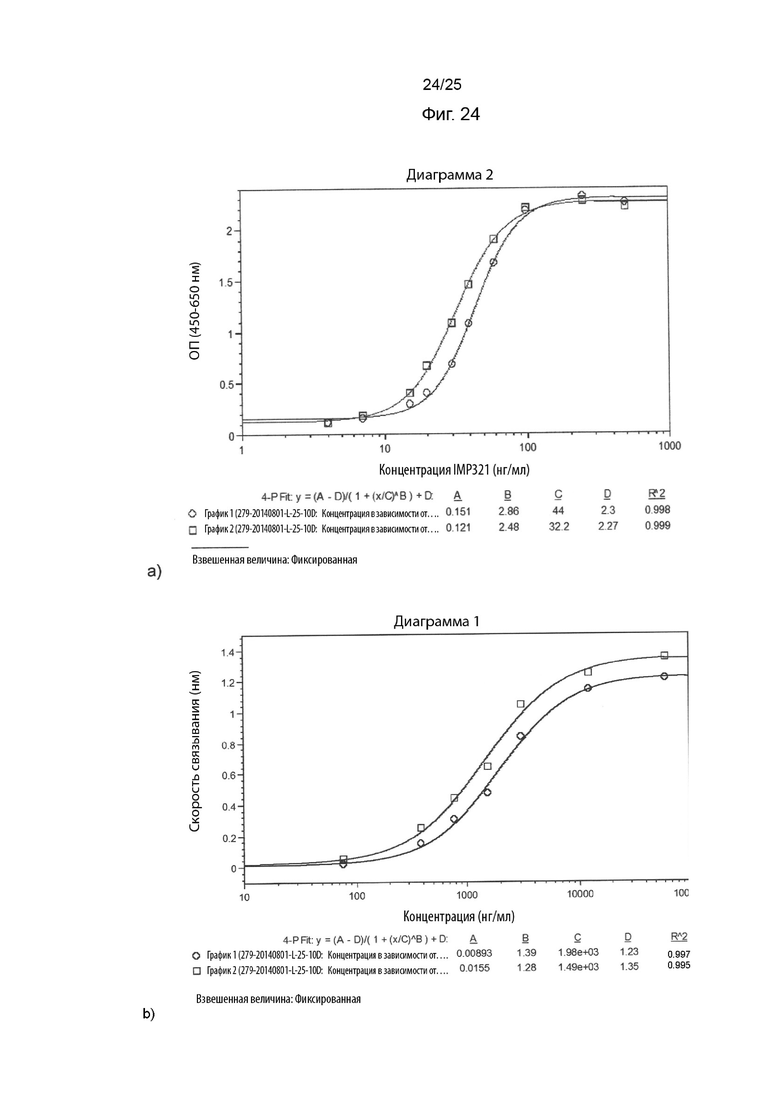
На Фиг. 24 изображены графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 24а), или с помощью анализа BLI (Фиг. 24b), при связывании различных концентраций иммобилизованного IMP321, необработанного или находившегося на свету (при 25°C в течение 10 дней); и
На Фиг. 25 изображена аминокислотная последовательность зрелого человеческого белка LAG-3. Четыре внеклеточных домена суперсемейства Ig находятся в аминокислотных остатках: 1-149 (D1); 150-239 (D2); 240-330 (D3); и 331-412 (D4). Аминокислотная последовательность сверхпетлевой структуры домена D1 белка LAG-3 человека изображена подчеркнутой жирным шрифтом.
ПОДРОБНОЕ ОПИСАНИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
В Примерах 1-5, приведенных ниже, описано исследование различных отличающихся друг от друга анализов связывания, для определения того, подходят ли они для использования в качестве анализов контроля качества согласно GMP-стандарта для производства производного IMP321 от рекомбинантного белка LAG-3. Ни один из анализов не был признан подходящим. Примеры 6-11 описывают клеточные способы анализа BLI и демонстрируют их пригодность для определения активности связывания ГКГС класса II препаратов IMP321.
Пример 1
Оценка использования анализа сортировки клеток с активированной флуоресценцией (FACS) для определения связывания IMP321 с клетками Raji.
Для определения связывания IMP321 с клетками Raji был проведен анализ FACS. Были протестированы образцы IMP321 с активностью связывания ГКГС класса II 100%, 75% и 50%. Образец со 100%-ой активностью представлял собой контрольный образец с известной связывающей активностью ГКГС класса II в заранее определенной концентрации. Образцы с активностью 75% и 50% готовили разведением контрольного образца.
Полученные кривые связывания показаны на Фиг. 2. Они демонстрируют, что верхние плато не были достигнуты, поэтому не было параллелизма между кривой связывания эталонного образца со 100% активностью и других образцов. Это препятствовало вычислению относительной активности различных образцов.
Пример 2
Оценка использования анализа Meso Scale Discovery (MSD) для определения связывания IMP321 с клетками Raji.
В этом примере описана оценка анализа Meso Scale Discovery (MSD) для определения связывания IMP321 с клетками Raji.
Платформа Meso Scale Discovery (MSD-ECL) использует электрохемилюминесцентные метки, которые конъюгированы с антителами для обнаружения. Эти метки инициируют свечение при стимуляции электричеством в соответствующей химической среде, которую затем можно использовать для измерения ключевых белков и молекул.
Электричество подается на пластинчатые электроды платформой Meso Scale Discovery (MSD-ECL), что приводит к излучению света метками. Затем измеряют интенсивность свечения для количественного определения аналитов в образце.
Процесс детектирования инициируется на электродах, расположенных в нижней части микропланшетов Meso Scale Discovery (MSD-ECL), и только метки рядом с электродом возбуждаются и детектируются. В системе используются буферы с высокими концентрациями трипропиламина в качестве катализатора для реакции двойного восстановления с рутением, испускающего свечение при 620 нм.
Используемый анализ MSD схематически представлен на Фиг. 3. Вкратце, приблизительно 2×104 клеток Raji на лунку в ФСБ высевали в 96-луночный MSD-планшет Single-SPOT (Meso Scale Discovery, Gaithersburg, MD) в объеме 25 мкл/лунку. Планшет инкубировали при комнатной температуре в течение 1-1,5 часов, а затем блокировали с помощью блокирующего буфера (25 мкл/лунку). Затем серийные разведения эталонного стандарта IMP321 или образцов загружали в двойные лунки в объеме 50 мкл/лунку. После примерно 1 часа инкубации при комнатной температуре связанный IMP321 детектировали с использованием конъюгированного с рутением античеловеческого Fc в объеме 50 мкл/лунку. Электрохемилюминесцентный сигнал получали с использованием MSD-считывающего буфера без поверхностно-активного вещества. Число ECL сигналов должно быть пропорциональным связыванию IMP321 с поверхностью клетки в пределах диапазона анализа.
Углеродные электроды с высокой степенью связывания в нижней части микропланшетов позволяют легко прикреплять клетки Raji. В анализе используются электрохемилюминесцентные метки, которые конъюгированы с анти-1МР321 антителами. Электричество подается на пластинчатые электроды с помощью прибора MSD, что приводит к излучению свечения метками. Затем измеряют интенсивность свечения для количественного определения присутствия IMP321, связанного с молекулами класса ГКГС, на поверхности иммобилизованных клеток Raji.
Результаты, полученные при использовании образцов, содержащих IMP321 с и без клеток Raji, представлены на Фиг. 4(a), а для образцов, содержащих ритуксан с и без клеток Raji, представлены на Фиг. 4(b).
Результаты показывают, что неспецифическое связывание IMP321 с планшетами MSD наблюдалось в отсутствие клеток Raji. Для сравнения наблюдали специфическое связывание ритуксана с клетками Raji.
Клетки Raji представляют собой клетки клеточной линии, полученной в 1963 году из В-лимфоцита 11-летнего уроженца Нигерии мужского пола больного лимфомой Беркитта. Ритуксан (Ритуксимаб) представляет собой химерное моноклональное антитело против белка CD20, которое преимущественно находится на поверхности В-клеток.
Пример 3
Оценка неспецифического связывания IMP321 в планшетах для ELISA
В данном примере описывается оценка неспецифического связывания IMP321 и ритуксана на планшетах, используемых для энзим-связанных иммуносорбентных анализов (ELISA) с использованием различных блокирующих реагентов.
Вкратце, микропланшеты блокировали реагентом для блокирования при 25°C в течение 2 часов. Образцы и контроль ритуксана разбавляли буфером для разведения до 2 мкг/мл, затем дополнительно разбавляли двукратным серийным разведением. Микропланшеты промывали и хорошо просушивали до и после добавления разбавленных образцов и инкубации. После инкубации со вторичным антителом сигнал измеряли с помощью спектрометрического анализа с использованием SpectraMax М2 (450-650 нм).

Результаты показаны на Фиг. 5. На Фиг. 5(a) показаны результаты ELISA с использованием возрастающих концентраций IMP321 и блокированием 5%-ым БСА или 10%-ым ФСБ планшетов для ELISA. На Фиг. 5(b) показаны результаты ELISA с использованием возрастающих концентраций IMP321 или ритуксана и блокированием 30%-ым ФТС в ФСБ планшетов для ELISA. На Фиг. 5(c) показаны результаты ELISA с использованием возрастающих концентраций IMP321 или ритуксана и блокированием 5%-ым БСА в RPIM 1640 планшетов для ELISA.
Результаты показывают, что имело место сильное неспецифическое связывание IMP321, но не ритуксана, с планшетами для ELISA при использовании БСА или ФТС в качестве блокирующих реагентов.
Различные отличающиеся друг от друга типы блокирующих агентов были затем протестированы с IMP321 или ритуксаном, чтобы определить, можно ли устранить неспецифическое связывание IMP321 с планшетами для ELISA.
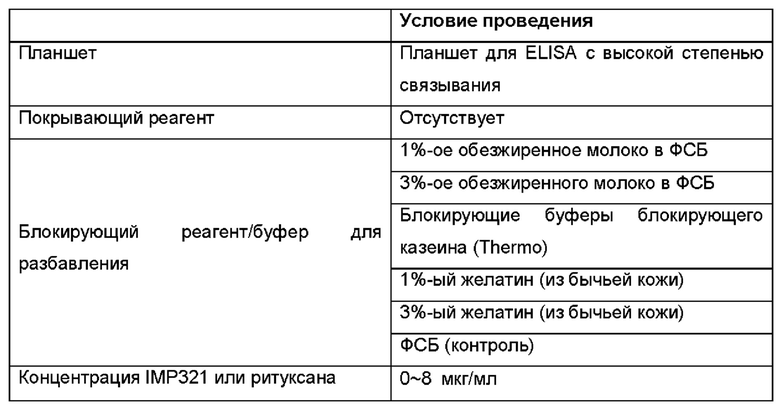
Результаты показаны на Фиг. 6. На Фиг. 6(a) показаны результаты для IMP321 или Ритуксана с использованием 1%-го обезжиренного молока, 3%-го обезжиренного молока или блокирующих буферов блокирующего казеина (Thermo) в качестве блокирующего реагента. На Фиг. 6(b) показаны результаты для IMP321 или ритуксана с использованием 1%-го желатина, 3%-го желатина или фСБ в качестве блокирующего реагента.
Результаты показывают, что казеин был лучшим блокирующим реагентом в отношении снижения неспецифического связывания IMP321 с планшетами для ELISA.
Пример 4
Оценка использования анализа Meso Scale Discovery (MSD) с блокирующим казеиновым буфером, для определения связывания IMP321 с клетками Raji
В данном примере описана оценка анализа MSD для определения связывания IMP321 с клетками Raji при различной плотности посева с использованием блокирующего казеинового буфера.
Был проведен анализ MSD, аналогичный описанному в Примере 2, чтобы оценить, можно ли свести к минимуму неспецифическое связывание IMP321 с планшетом MSD, наблюдаемое в данном примере, используя блокирующий казеиновый буфер.
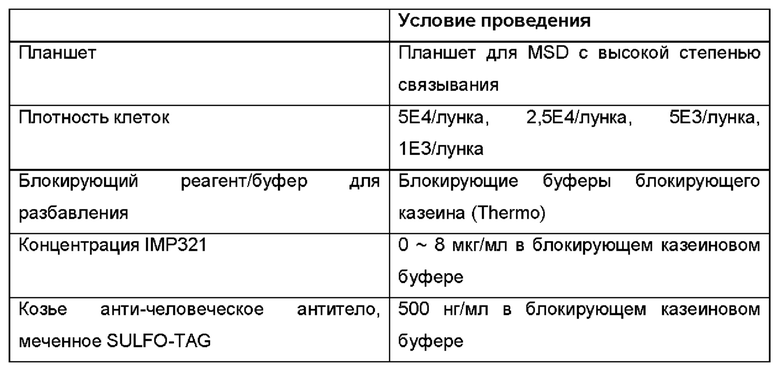
Результаты показаны на Фиг. 7. На Фиг. 7(a) показаны результаты связывания IMP321 с клетками Raji с различной плотностью посева (0-5×104 клеток/лунка) при различных концентрациях IMP321. Результаты показывают увеличение максимальной степени связывания IMP321 в зависимости от плотности клеток. На Фиг. 7(b) показаны результаты специфического связывания IMP321 с клетками Raji при различной плотности посева (1×103-5×104 клеток/лунка). Результаты показывают зависимое от плотности клеток увеличение специфического связывания IMP321.
Связывание IMP321 с клетками Raji сравнивали со связыванием IMP321 с клетками HLA-DRdim L929 (эти клетки не экспрессируют ГКГС класса II) при разных концентрациях IMP321 с использованием анализа MSD с блокирующим казеиновым буфером. L929 представляет собой фибробластоподобную клеточную линию, клонированную из штамма L. Результаты показаны на Фиг. 8. Результаты показывают, что неспецифическое связывание IMP321 с планшетами MSD было значительно снижено в присутствии блокирующего казеина. Однако сигнал специфического связывания был низким, и не наблюдалось никакого верхнего плато кривой в зависимости от дозы IMP321.
Был сделан вывод, что анализ MSD с использованием блокирующего казеинового буфера не может быть использован для демонстрации специфического связывания IMP321 с иммобилизованными на планшетах клетками Raji.
Пример 5
Оценка использования анализов ELISA для определения связывания IMP321 с клетками Raji.
В данном примере описана оценка способности прямого клеточного ELISA и ELISA с перенесением клеток определять связывание IMP321 с клетками Raji.
Прямой ELISA (аналогично анализу, описанному в Примере 3) проводили в присутствии различных блокирующих реагентов (5%-го БСА, 10%-го ФТС, 0,5%-го казеина или 3%-го желатина) с различными количествами иммобилизованных на планшетах клеток Raji (10000, 5000 или 2500 клеток), а также различных концентраций IMP321 или IMP321, обработанных пептид-N-гликозидазой F (ПНГаза F, амидаза, которая расщепляет между внутренним GIcNAc и остатками аспарагина с высоким содержанием маннозы, гибридными и сложными олигосахаридами из числа N-связанных гликопротеинов). Условия, используемые для прямого анализа ELISA, приведены в таблицах ниже:

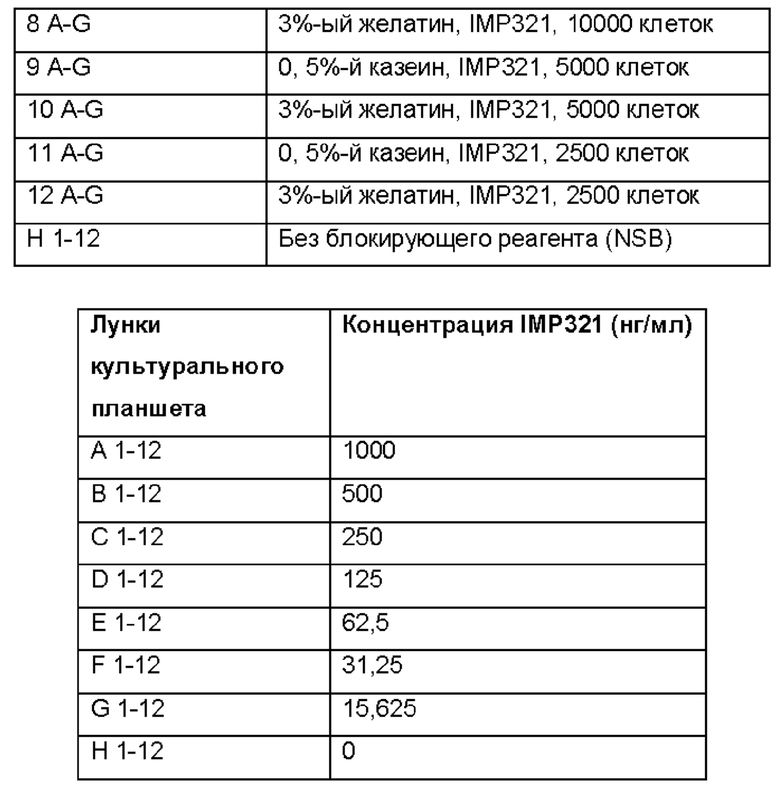
Результаты показаны в таблице ниже.
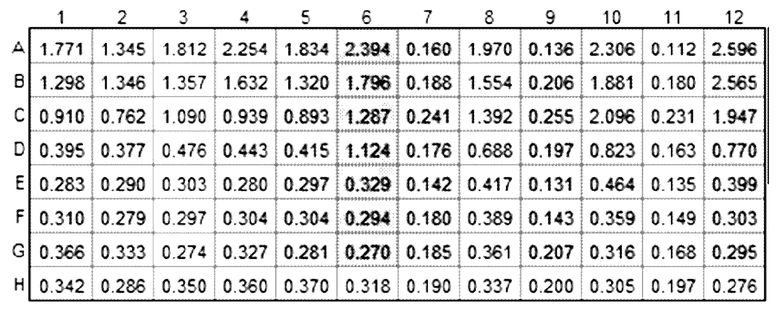
Результаты показывают зависимое от дозы связывание IMP321 с клетками Raji, иммобилизованными на планшетах.
Чтобы проверить, связывается ли IMP321 неспецифично с планшетами для ELISA, проводили прямой ELISA в отсутствие клеток Raji в условиях, приведенных в таблице ниже:
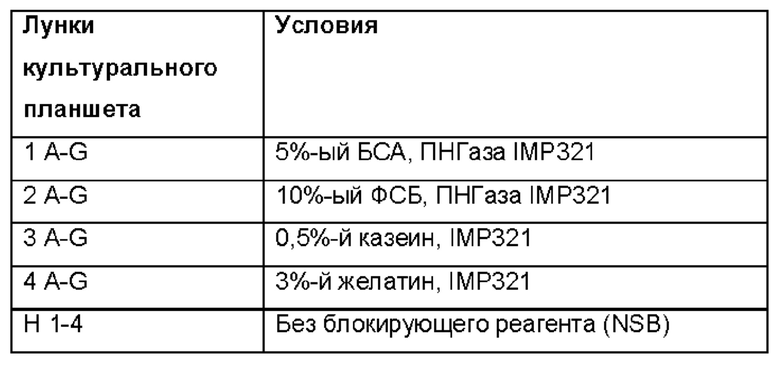
Результаты показаны в таблице ниже:
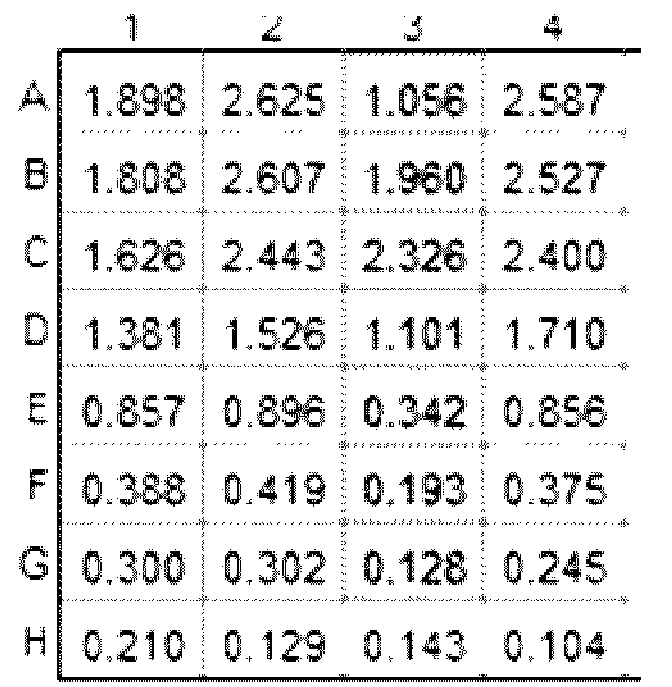
Результаты показывают сильное неспецифическое связывание IMP321 с планшетом для ELISA в отсутствие иммобилизованных на планшете клеток Raji. Ни казеиновые, ни желатиновые блокирующие реагенты, ни обработка ПНГазой IMP321 не устраняли неспецифического связывания.
Был сделан вывод, что прямой клеточный ELISA нельзя использовать для демонстрации специфического связывания IMP321 с клетками Raji, иммобилизованными на планшетах.
Для определения связывания различных концентраций IMP321 или IMP321, обработанных ПНГазой, с иммобилизованными клетками Raji, проводили ELISA с перенесением клеток. Клетки Raji переносили в другой планшет после связывания с IMP321 или обрабатывали IMP321. Условия, использованные для анализа, приведены в таблицах ниже.
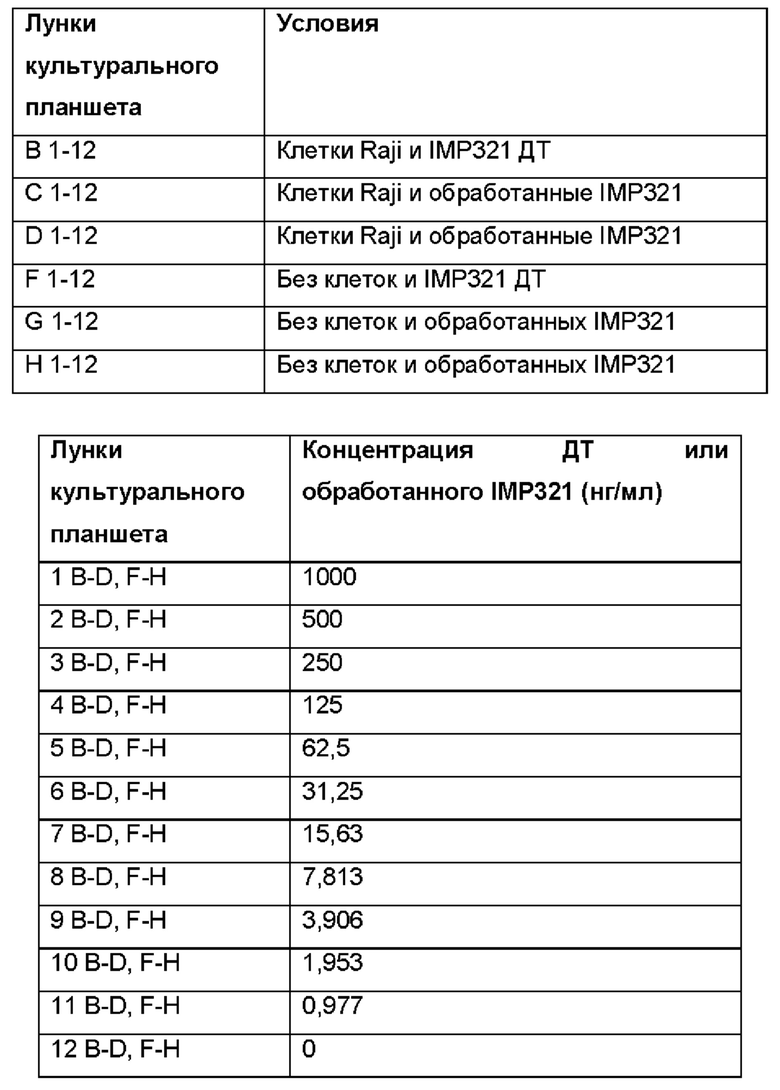
Результаты показаны в таблице ниже:
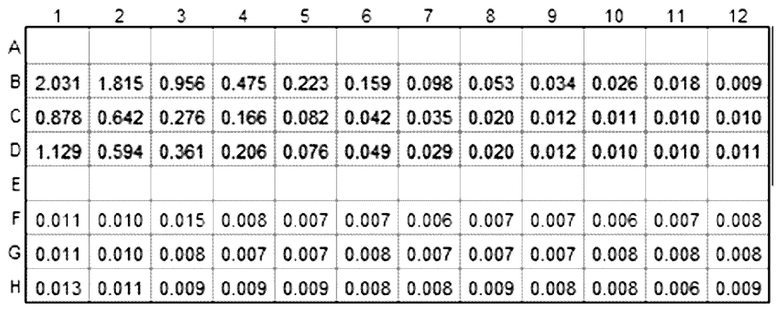
Результаты показывают, что изменение сигнала от лунки к лунке неприемлемо для способа контроля качества. Метод также трудоемкий. Был сделан вывод, что перенос клеток в ELISA нельзя использовать для демонстрации специфического связывания IMP321 с клетками Raji, иммобилизованными на планшетах.
Пример 6
Клеточный анализ для измерения активности связывания препарата производного белка LAG-3 IMP321 с использованием биослойной интерферометрии (BLI)
IMP321 представляет собой растворимое рекомбинантное производное белка LAG-3 с высокой аффинностью к молекулам ГКГС класса II. В данном примере описан клеточнм анализ для измерения активности связывания IMP321 с клетками Raji, экспрессирующими ГКГС класса II, с использованием BLI. Анализ является простым и быстрым и позволяет сравнивать эталонные стандарты и образцы.
На Фиг. 9 слева схематически показан зонд BLI с сенсором, конъюгированным с белком А, и IMP321, иммобилизованный на дистальном конце оптического волокна сенсора, с концом сенсора, погруженным в раствор образца, содержащий клетки Raji. Основные этапы способа изложены в правой части фигуры. Анализ описан более подробно ниже.
Материалы:
1) клетки Raji: АТСС / CCL-86
2) RPMI 1640: Invitrogen / 22400-089
3) ИН-ФТС: Invitrogen /10100147
4) ДФСБ: Hyclone / SH30028.01В
5) БСА: Sigma / А3032
6) Аналитический стандарт IMP321
7) Среда для роста клеток Raji: RPMI 1640, 10%-ая ИН-ФТС
8) Разбавитель для анализа связывания: ДФСБ, 0,5%-ый БСА
9) Планшет с белком A (ForteBio-18-5010)
10) черные 96-луночные планшеты с плоским дном (Greiner-655209)
11) Одно- и многоканальные пипетки: Sartorius и Eppendorf / различные
12) Счетчик клеток: Roche / Cedex HiRes и Beckman/ViCell
13) Биослойный интерферометр: Fortebio / Octet Red с програмным обеспечением версии 7.0 или более поздней
Способы:
1. Приготовление готовых к использованию клеток Raji.
1) Извлеките N флакон клеток Raji из морозильника с жидким азотом и быстро разморозьте на водяной бане при 37°C.
2) В асептических условиях переносите содержимое флакона в стерильную центрифужную пробирку, содержащую приблизительно N × 9 мл среды для выращивания клеток Raji. Хорошо перемешайте, аккуратно пипеткой.
3) Центрифугируйте клетки 5 мин при 300х g. Ресуспендируйте клетки в разбавителе для анализа связывания и подсчитайте их с помощью счетчика клеток или гемоцитометра.
4) Добавьте объем исходной суспензии клеток к достаточному объему разбавителя для анализа связывания, чтобы довести плотность клеток до 4,0Е6-8,0Е6 клеток на мл, и оставьте на льду для использования.
2. Подготовка эталонного образца IMP321, контроля и образцов
ПРИМЕЧАНИЕ: 1) Используйте способ обратного пипитерования для обеспечения точности.
2) Аккуратно перемешайте, чтобы избежать образования пены и пузырьков. 1) Подготовка аналитического стандарта:
1.1) При необходимости разморозьте флакон с аналитическим стандартом IMP321. Хранить при 2-8°C. Срок годности 7 дней от даты оттаивания.
1.2) Разведите аналитический стандарт IMP321 примерно до 1,0 мг/мл в буфере для состава. Готовьте и используйте свежие реактивы. Определите концентрацию белка спектрофотометрически с использованием буфера для составав качестве холостого раствора.
1.3) Исходя из измеренной концентрации белка, разбавьте АС, чтобы подготовить стандартную кривую до соответствующих концентраций, как описано ниже. Перемешайте разведения на вортексе.
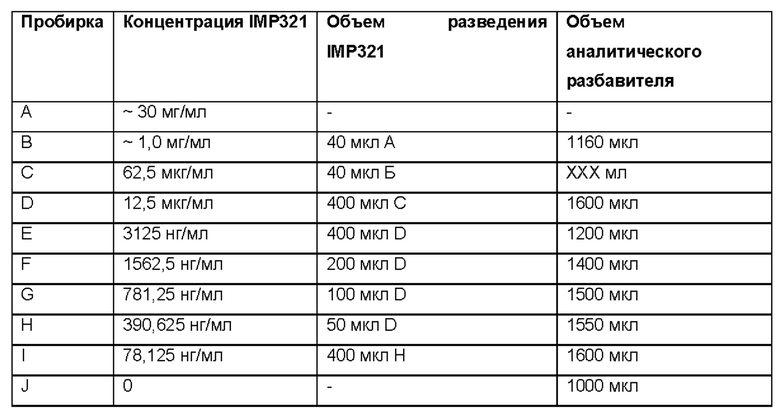
1.4) Используйте разведения C-J для стандартной кривой. При необходимости можно использовать дополнительные концентрации, чтобы включить линейную часть кривой и верхнее и нижнее плато.
2) Подготовка контроля
2.1) Контролем является независимое разбавление аналитического стандарта из пробирки С, приготовленного на шаге 1.3 выше. Далее разбавьте, как описано в таблице выше. Перемешайте разведения на вортексе.
2.2) Используйте разведения C-J для контроля.
3) Подготовка образцов.
3.1) Исходя из концентрации белка, разведите образцы IMP321 до приблизительно 1,0 мг/мл в аналитическом разбавителе. Готовьте и используйте свежие реактивы.
3.2) Для приготовления стандартной кривой, далее разведите до соответствующих концентраций, как описано в таблице выше. Перемешайте разведения на вортексе.
3.3) Используйте разведения C-J для образцов. При необходимости можно использовать дополнительные концентрации, чтобы включить линейную часть кривой и верхнее и нижнее плато.
3. Этапы детектирования в системе Octet
1) Гидратируйте биосенсоры в ФСБ по меньшей мере 10 мин
2) Подготовьте аналитический планшет. В черный полипропиленовый микропланшет перенесите 200 мкл на лунку ФСБ, аналитического разбавителя, титры IMP321 в АР или клетки Raji, соответственно, в соответствующие лунки согласно карте образцов планшета ниже:
Карта образцов планшета

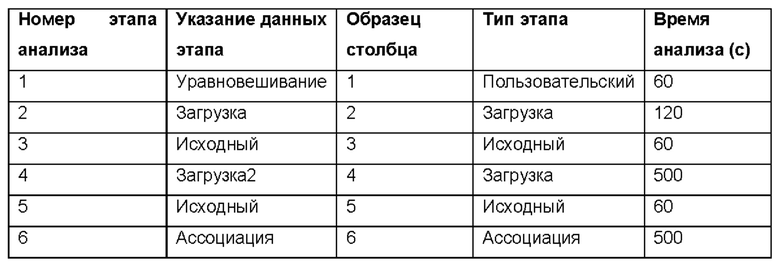
3) Настройте кинетический анализ путем ввода параметров, описанных ниже.
4) Введите расположение и имя файла для сохранения данных.
5) Нажмите Пуск, чтобы начать анализ.

4. Анализ данных
1) В программе анализа данных Octet загрузите папку данных для анализа.
2) На вкладке Обработка выберите этап Ассоциация. Затем нажмите «quantitate the Selected Step».
3) Введите информацию о соответствующей концентрации.
4) На вкладке Results выберите R equilibrium (Req) в качестве уравнения скорости связывания. Это уравнение будет соответствовать кривой связывания, сгенерированной в ходе эксперимента, и рассчитает ответ в отношении равновесия в качестве выходного сигнала.
5) Нажмите на Calculate Binding Rate. Результаты будут автоматически отображаться в таблице.
6) Нажмите кнопку Save Report, чтобы создать файл отчета MS Excel.
7) Используйте SoftMax Pro, программу подбора логистической кривой с 4 параметрами, чтобы сгенерировать стандартную кривую или кривую образца по скорости связывания (нм) в зависимости от концентрации IMP321, выраженной в мкг/мл. Пример показан на Фиг. 10.
8) Рассчитайте относительную силу связывания образца с использованием соотношения ЕК50 для аналитического стандарта и образца.
5. Пригодность системы и критерии соответствия анализа.
Анализ эффективен, если он соответствует ВСЕМ следующим критериям:
1) Готовность использовать клетки Raji с жизнеспособностью >= 60%
2) Относительная активность контроля находится в пределах 80%-120%
3) Соотношение сигнал/ фон в контроле (параметр D/параметр А)>=2.
4) Параллелизм (сопоставимость): угловой коэффициент в стандарте составляет от 0,8 до 1,4.
5) Если результат контроля анализа не соответствует критериям, перечисленным выше, анализ считается недействительным.
6. Окончательное значение:
1) Для клинического образца окончательное значение для образца определяется как среднее из двух или трех действительных и независимых результатов анализа, как описано ниже:
Разница в% рассчитывается следующим образом:
Абсолютное значение (результат анализа 1 - результат анализа 2) / среднее значение (результат анализа 1, результат анализа 2) × 100%
2) Если % Разницы результатов двух анализов <= 20%, укажите средние результаты двух анализов.
3) Если % Разницы результатов двух анализов > 20%, выполните 1 дополнительный соответствующий анализ.
4) Если CV результатов анализа трех образцов <= 25%, укажите средние результаты трех анализов.
5) Если CV анализа трех образцов составляет > 25%, окончательное значение отсутствует. Установите несоответствие в отношении подтверждающего тестирования.
6) Если окончательное значение для образца не соответствует спецификациям, перечисленным в СОА (сертификате исследования), установите несоответствие в отношении подтверждающего тестирования.
7. Подтверждающее тестирование.
Выполните повторную проверку образца следующим образом:
1) Повторно протестируйте образец с тремя действительными и независимыми анализами.
2) Если CV результатов анализа трех образцов <= 25%, укажите средние результаты трех анализов.
3) Если CV анализа трех образцов составляем 25%, окончательное значение отсутствует.
4) Если результат повторного тестирования не соответствует спецификации (OOS), указанной в СОА, заключение считается неудачным.
Пример 7.
Определение специфического связывания иммобилизованного IMP321 с клетками Raji в растворе в анализе BLI
Анализ BLI, как описано в Примере 6, использовали для определения связывания иммобилизованного IMP321 в растворе с клетками Raji в различных концентрациях (8Е6/мл, 4Е6/мл, 2Е6/мл, 1Е6/мл). Клетки Jurket использовали в качестве отрицательного контроля. Полученные кривые ассоциации и диссоциации показаны на Фиг. 11(a). На Фиг. 11(b) показан график сигнала связывания, полученный для различных концентраций клеток Raji. Результаты показывают, что сигнал связывания зависел от концентрации клеток Raji, то есть чем выше концентрация клеток Raji, тем выше скорость связывания и полученное верхнее плато. В том же анализе не наблюдалось никакого специфического связывания клеток Jurket.
Дальнейший анализ BLI проводили, как описано в Примере 6, но для сравнения связывания иммобилизованного IMP321 с клетками Раджи со связыванием иммобилизованного Хумира или Авастина. Полученные кривые ассоциации и диссоциации показаны на Фиг. 12(a). На Фиг. 12 (b) показан график сигнала связывания, полученный для различных иммобилизованных белков. Результаты показывают, что IMP321, но не Хумира или Авастин, связывается с клетками Raji.
Из этих результатов был сделан вывод, что анализ BLI способен определять специфическое связывание иммобилизованного IMP321 с клетками Raji в растворе.
Пример 8
Корреляция активности связывания IMP321, измеренной с помощью анализа BLI, с известной эффективностью связывания
Образцы IMP321, разведенные из эталонного стандарта с различными уровнями эффективности связывания клеток Raji, использовали в анализе BLI, чтобы определить, коррелирует ли активность связывания, измеренная анализом, с известной эффективностью связывания образцов. Результаты показаны в таблице ниже. На Фиг. 13 показан график процентной активности связывания, измеренной анализом BLI, в сравнении с их ожидаемой активностью;
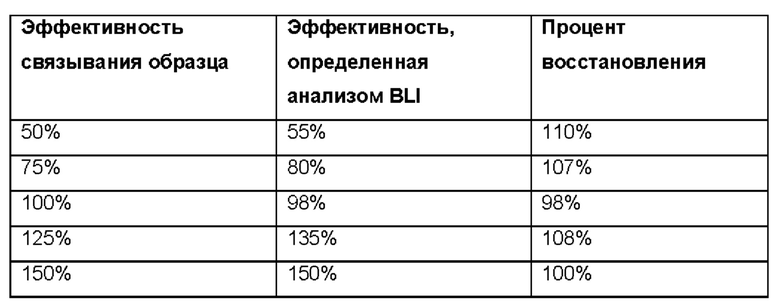
Результаты показывают хорошую корреляцию между эффективностью связывания, измеренной анализом BLI, и ожидаемой эффективностью связывания. Средние значения восстановления для каждого образца составляли от 90% до 110% с хорошим параллелизмом кривых связывания (то есть приемлемое отношение углового коэффициента и объединения плато).
Пример 9
Использование замороженных клеток в анализе BLI для определения активности связывания ГКГС класса II
Анализ ВЫ, как описано в Примере 6, проводили для сравнения связывания иммобилизованного IMP321 с клетками Raji в растворе, полученном из культуры или из замороженного исходного раствора. График сигнала связывания, полученный в отношении связывания различных концентраций иммобилизованного IMP321 с культивируемыми клетками Raji в растворе, показан на Фиг. 14(a). График сигнала связывания, полученный в отношении связывания различных концентраций иммобилизованного IMP321 с ранее замороженными клетками Raji в растворе, показан на Фиг. 14(b).
Результаты показывают, что замороженные клетки Raji ведут себя очень сходно с культивируемыми клетками Raji, и поэтому замороженный исходный раствор можно использовать вместо свежего культурального раствора, для обеспечения лучшей надежности анализа и применимости.
Пример 10
Тестирования образца без остановки процесса
Анализы BLI, как описано в Примере 6, были проведены для определения активности связывания ГКГС класса II различных отличающихся друг от друга препаратов IMP321 и для сравнения биологической активности препаратов, определенной с помощью анализа высвобождения CCL4.
ТНР-1 представляет собой единичную клеточную линию лейкоза человека. При индукции белком LAG-3 или стресс-образцами клетки ТНР-1 секретируют цитокин CCL4, который можно количественно определить с помощью набора ELISA для CCL4. Уровень высвобождения CCL4 можно использовать для измерения биологической активности препарата белка LAG-3 или его фрагмента, производного или его аналога.

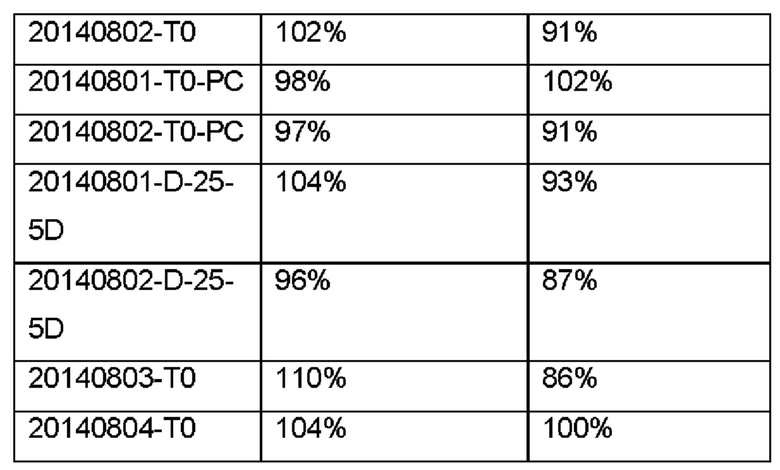
Был сделан вывод, что биологическая активность различных образцов IMP321 коррелировала с биологической активностью, определенной с помощью анализа высвобождения CCL4.
Пример 11
Анализ BLI для тестируемых образцов IMP321 и корреляции с клеточным анализом высвобождения CCL4
Анализы BLI, как описано в Примере 6, использовали для определения активности связывания ГКГС класса II в образцах IMP321, которые подвергались различным обработкам (дегликозилированию посредством обработки ПНГазой, хранению при 37°C, оксидации путем обработки 1% или 0,1% перекисью водорода, обработке кислотой при рН 3,0, 3,6 или 3,1, обработке щелочью при рН 9,2, 9,75 или воздействию света). Результаты показаны на Фиг. 15-24.
На Фиг. 15(a) показан график снижения высвобождения CCL4, полученного с помощью клеточного анализа при связывании различных концентраций иммобилизованного IMP321 или дегликозилированного IMP321 с клетками Raji;
На Фиг. 15(b) показан график сигнала связывания, полученный с помощью анализа BLI при связывании различных концентраций иммобилизованного IMP321 или дегликозилированного IMP321 с клетками Raji;
На Фиг. 16 показаны графики сигнала связывания иммобилизованных IMP321 или IMP321 различных концентраций, которые хранятся ненадлежащим образом (при 37°C в течение 12 дней), с клетками Raji. Результаты, показанные на Фиг. 16(a), были получены с помощью клеточного анализа, измеряющего высвобождение CCL4, а результаты, показанные на Фиг. 16(b), были получены с помощью анализа BLI;
На Фиг. 17 показаны графики сигнала связывания различных концентраций иммобилизованного IMP321 или IMP321, хранящихся ненадлежащим образом (при 37°C в течение 1 месяца), с клетками Raji. Результаты, показанные на Фиг. 17(a), были получены с помощью клеточного анализа, измеряющего высвобождение CCL4, а результаты, показанные на Фиг. 17(b), были получены с помощью анализа BLI;
На Фиг. 18 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 18а), или с помощью анализа BLI (Фиг. 18b), при связывании различных концентраций иммобилизованного необработанного IMP321 или окисленного IMP321 (с 1%-ой перекисью водорода) с клетками Raji;
На Фиг. 19 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 19а), или с помощью анализа BLI (Фиг. 19b), при связывании различных концентраций иммобилизованного необработанного IMP321 или окисленного IMP321 (с 0,1%-ой перекисью водорода) с клетками Раджи; На Фиг. 20 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (фигура 20а), или с помощью анализа BLI (фигура 20b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного кислотой (при рН 3,0), с клетками Raji;
На Фиг. 21 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 21а), или с помощью анализа BLI (Фиг. 21b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного кислотой (при рН 3,1, или рН 3,6), с клетками Raji;
На Фиг. 22 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 22а) или с помощью анализа BLI (Фиг. 22b), при связывании различных концентраций иммобилизованного IMP321, необработанного или обработанного щелочами (при рН 9,2 или рН). 9,75), с клетками Raji;
На Фиг. 23 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 23а), или с помощью анализа BLI (Фиг. 23b), при связывании различных концентраций иммобилизованного IMP321, необработанного или находившегося на свету (при 25°C в течение 5 дней) с клетками Raji; и
На Фиг. 24 показаны графики сигнала, полученного с помощью клеточного анализа, измеряющего высвобождение CCL4 (Фиг. 24а), или с помощью анализа BLI (Фиг. 24b), при связывании различных концентраций иммобилизованного IMP321, необработанного или находившегося на свету (при 25°C в течение 10 дней).
Биологическая активность (определенная по высвобождению CCL4 различных образцов IMP321 по сравнению с их активностью связывания ГКГС класса II (определенная способом, описанным в Примере 6) показана в таблице ниже:
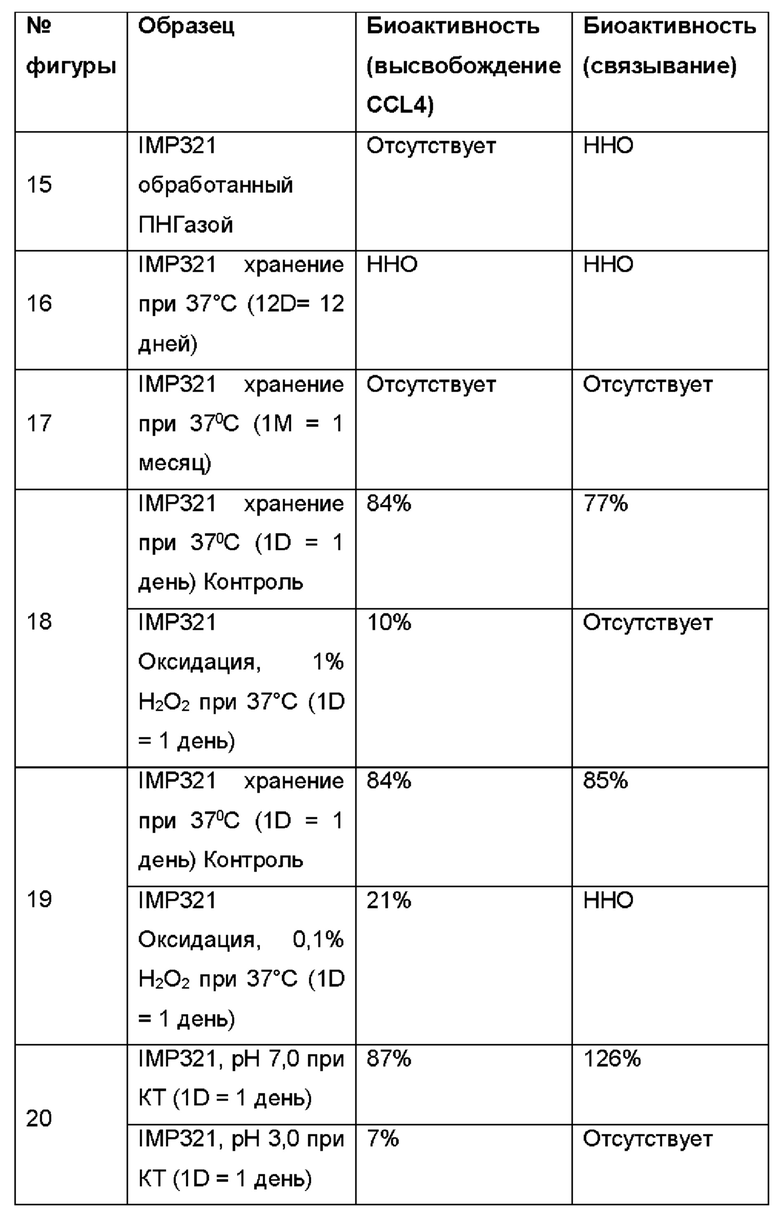
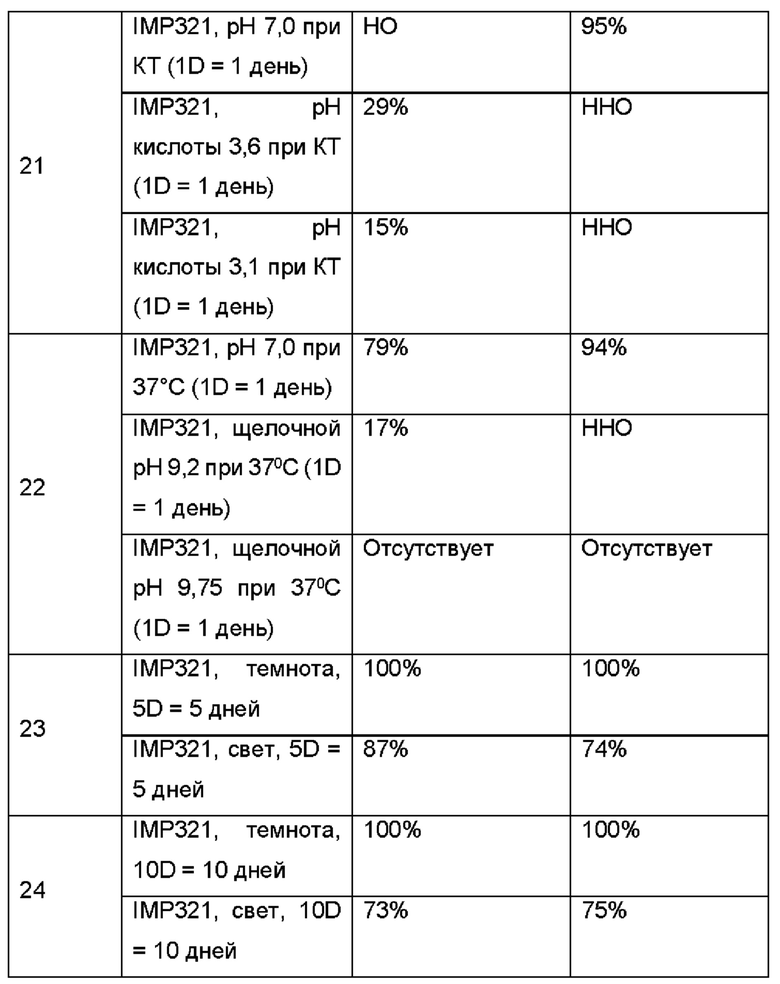
Результаты показывают хорошую корреляцию между биологической активностью каждого обработанного образца IMP321, как определено высвобождением CCL4, и его активностью связывания ГКГС класса II, как определено анализом BLI согласно изобретению. Был сделан вывод, что определение связывающей активности ГКГС класса II с помощью анализа BLI можно использовать для определения биологической активности препаратов IMP321.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
10 20 30 40 50 60
LQPGAEVPVV WAQEGAPAQL PCSPTIPLQD LSLLRRAGVT WQHQPDSGPP AAAPGHPLAP
70 80 90 100 110 120
GPHPAAPSSW GPRPRRYTVL SVGPGGLRSG RLPLQPRVQL DERGRQRGDF SLWLRPARRA
130 140 150 160 170 180
DAGEYRAAVH LRDRALSCRL RLRLGQASMT ASPPGSLRAS DWVILNCSFS RPDRPASVHW
190 200 210 220 230 240
FRNRGQGRVP VRESPHHHLA ESFLFLPQVS PMDSGPWGCI LTYRDGFNVS IMYNLTVLGL
250 260 270 280 290 300
EPPTPLTVYA GAGSRVGLPC RLPAGVGTRS FLTAKWTPPG GGPDLLVTGD NGDFTLRLED
310 320 330 340 350 360
VSQAQAGTYT CHIHLQEQQL NATVTLAIIT VTPKSFGSPG SLGKLLCEVT PVSGQERFVW
370 380 390 400 410 420
SSLDTPSQRS FSGPWLEAQE AQLLSQPWQC QLYQGERLLG AAVYFTELSS PGAQRSGRAP
430 440 450 460 470 480
GALPAGHLLL FLTLGVLSLL LLVTGAFGFH LWRRQWRPRR FSALEQGIHP QAQSKIEELE
490 500
QEPEPEPEPE PEPEPEPEPE QL
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНАЛИЗЫ | 2020 |
|
RU2834240C2 |
| КОМБИНИРОВАННЫЕ ПРЕПАРАТЫ ДЛЯ ЛЕЧЕНИЯ РАКА ИЛИ ИНФЕКЦИИ | 2016 |
|
RU2777945C2 |
| ЧЕЛОВЕЧЕСКИЕ АНТИТЕЛА И КОНЪЮГАТЫ АНТИТЕЛО-ПРЕПАРАТ ПРОТИВ CD74 | 2012 |
|
RU2636029C2 |
| Анти-LAG-3 антитела | 2016 |
|
RU2760582C2 |
| РАСТВОРИМЫЕ ПОЛИПЕПТИДНЫЕ ФРАКЦИИ ПРОТЕИНА LAG-3, СПОСОБ ПОЛУЧЕНИЯ, ТЕРАПЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, АНТИТЕЛО | 1995 |
|
RU2178306C2 |
| МОНОКЛОНАЛЬНОЕ АНТИ-CD20-АНТИТЕЛО | 2006 |
|
RU2415871C2 |
| ТВЕРДОФАЗНЫЙ ИММУНОФЕРМЕНТНЫЙ АНАЛИЗ (ELISA) ДЛЯ ФАКТОРА РОСТА ЭНДОТЕЛИЯ СОСУДОВ (VEGF) | 2007 |
|
RU2517301C2 |
| АНТИТЕЛО ПРОТИВ LAG-3, ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И ИХ ФАРМАЦЕВТИЧЕСКОЕ ПРИМЕНЕНИЕ | 2017 |
|
RU2757813C2 |
| ОПРЕДЕЛЯЮЩИЕ КОМПЛЕМЕНТАРНОСТЬ УЧАСТКИ ДЛЯ СВЯЗЫВАНИЯ CD3 И СОДЕРЖАЩАЯ ИХ БИСПЕЦИФИЧЕСКАЯ АНТИГЕНСВЯЗЫВАЮЩАЯ МОЛЕКУЛА | 2019 |
|
RU2738802C1 |
| ИЗГОТОВЛЕНИЕ АКТИВНЫХ ВЫСОКОФОСФОРИЛИРОВАННЫХ ЛИЗОСОМАЛЬНЫХ ФЕРМЕНТОВ СУЛЬФАТАЗ ЧЕЛОВЕКА И ИХ ПРИМЕНЕНИЕ | 2009 |
|
RU2510820C2 |

Изобретение относится к биотехнологии. Описан способ определения связывающей активности средства, содержащего белок гена активации лимфоцитов 3 (LAG-3) или его фрагмент, производное или аналог. Определяют связывание белка LAG-3, его фрагмента, производного или аналога с молекулами Главного Комплекса Гистосовместимости (ГКГС) класса II с использованием биослойной интерферометрии (BLI). Причем указанный способ включает в себя определение связывания белка LAG-3, его фрагмента, производного или аналога с молекулами ГКГС класса II, присутствующими на клетках, экспрессирующих ГКГС класса II; и белок LAG-3, его фрагмент, производное или аналог иммобилизованы на слое реагента зонда BLI, и клетки, экспрессирующие ГКГС класса II, находятся в растворе. Представлен сенсор для BLI для определения связывающей активности фрагмента, производного или аналога белка LAG-3с молекулами ГКГС класса II, который содержит слой реагента, на котором иммобилизированы фрагмент, производное или аналог белка LAG-3. Представлен набор для определения связывающей активности белка LAG-3 или его фрагмента, производного или аналога с молекулами ГКГС класса II, который включает зонд BLI, имеющий слой реагента, на котором белок LAG-3 или его фрагмент, производное или аналог иммобилизованы, а также клетки, экспрессирующие ГКГС класса II. Изобретение позволяет определить специфическое связывание производного LAG-3 IMP321 с молекулами ГКГС класса II, экспрессируемыми на поверхности иммунных клеток. 3 н. и 19 з.п. ф-лы, 25 ил., 17 табл., 11 пр.
1. Способ определения связывающей активности средства, содержащего белок гена активации лимфоцитов 3 (LAG-3) или его фрагмент, производное или аналог, при этом указанный способ включает в себя определение связывания белка LAG-3, его фрагмента, производного или аналога с молекулами Главного Комплекса Гистосовместимости (ГКГС) класса II с использованием биослойной интерферометрии (BLI), причем указанный способ включает в себя определение связывания белка LAG-3, его фрагмента, производного или аналога с молекулами ГКГС класса II, присутствующими на клетках, экспрессирующих ГКГС класса II; и белок LAG-3, его фрагмент, производное или аналог иммобилизованы на слое реагента зонда BLI, и клетки, экспрессирующие ГКГС класса II, находятся в растворе.
2. Способ по п. 1, отличающийся тем, что клетки, экспрессирующие ГКГС класса II, присутствуют при плотности, по меньшей мере, 1 × 106/мл, предпочтительно, по меньшей мере, 4 × 106/мл или 8 × 106/мл.
3. Способ по п. 1 или 2, отличающийся тем, что слой реагента предварительно обрабатывают блокирующим реагентом, чтобы минимизировать неспецифическое связывание клеток, экспрессирующих ГКГС класса II, со слоем реагента.
4. Способ по п. 3, отличающийся тем, что блокирующий реагент содержит альбумин, предпочтительно бычий сывороточный альбумин (БСА).
5. Способ по любому из пп. 1-4, отличающийся тем, что клетки, экспрессирующие ГКГС класса II, представляют собой клетки Raji.
6. Способ по любому из пп. 1-5, отличающийся тем, что клетки, экспрессирующие ГКГС класса II, представляют собой оттаявшие, готовые к использованию клетки, полученные из замороженного исходного раствора.
7. Способ по любому из предыдущих пунктов, который включает определение степени связывания белка LAG-3, фрагмента, производного или аналога с молекулами ГКГС класса II для множества различных концентраций белка LAG-3, его фрагмента, производного или аналога, и генерирование кривой доза-ответ для скоростей связывания.
8. Способ по любому из предыдущих пунктов, который дополнительно включает определение связывающей активности контрольного образца белка LAG-3 или его фрагмента, производного или аналога с молекулами ГКГС класса II путем определения связывания белка, фрагмента, производного или аналога эталонного образца LAG-3 с молекулами ГКГС класса II с использованием BLI в тех же условиях, которые используются для определения связывания средства белка LAG-3, его фрагмента, производного или аналога, и сравнение активности связывания ГКГС класса II, определенной для эталонного образца, с активностью связывания ГКГС класса II, определенной для средства.
9. Способ по п. 8, отличающийся тем, что связывающая активность с молекулами ГКГС класса II контрольного образца установлена как 100%.
10. Способ по п. 8 или 9, отличающийся тем, что контрольный образец содержит белок LAG-3 или его фрагмент, производное или аналог, который был обработан для снижения его активности связывания с ГКГС класса II.
11. Способ по п. 10, отличающийся тем, что белок LAG-3, его фрагмент, производное или аналог эталонного образца дегликозилирован, хранится при 37°С в течение по меньшей мере 12 дней, окисляется, денатурируется кислотным или щелочным воздействием, или подвергается воздействию света в течение не менее 5 дней.
12. Сенсор для BLI для определения связывающей активности фрагмента, производного или аналога белка LAG-3с молекулами ГКГС класса II, который содержит слой реагента, на котором иммобилизированы фрагмент, производное или аналог белка LAG-3.
13. Сенсор по п. 12, отличающийся тем, что указанный слой реагента предварительно обработан блокирующим реагентом для минимизации неспецифического связывания клеток, экспрессирующих ГКГС класса II, со слоем реагента.
14. Сенсор по п. 13, отличающийся тем, что указанный блокирующий реагент содержит альбумин, предпочтительно БСА.
15. Набор для определения связывающей активности белка LAG-3 или его фрагмента, производного или аналога с молекулами ГКГС класса II, который включает зонд BLI, имеющий слой реагента, на котором белок LAG-3 или его фрагмент, производное или аналог иммобилизованы, а также клетки, экспрессирующие ГКГС класса II.
16. Набор по п. 15, отличающийся тем, что слой реагента зонда BLI предварительно обработан блокирующим реагентом, чтобы минимизировать неспецифическое связывание клеток, экспрессирующих ГКГС класса II, со слоем реагента.
17. Набор по п. 16, отличающийся тем, что блокирующий реагент содержит альбумин, предпочтительно БСА.
18. Набор по любому из пп. 15-17, отличающийся тем, что клетки, экспрессирующие ГКГС класса II, представляют собой замороженные клетки.
19. Набор по любому из пп. 15-18, отличающийся тем, что клетки представляют собой клетки Raji.
20. Набор по любому из пп. 15-20, отличающийся тем, что клетки присутствуют при плотности, по меньшей мере, 1 × 106/мл, предпочтительно, по меньшей мере, 4 × 106/мл или 8 × 106/мл.
21. Набор по любому из пп. 15-20, который дополнительно включает эталонный образец, содержащий белок LAG-3 или его фрагмент, производное или аналог.
22. Набор по п. 21, отличающийся тем, что связывающая активность эталонного образца c молекулами ГКГС класса II известна.
| Приспособление для суммирования отрезков прямых линий | 1923 |
|
SU2010A1 |
| Токарный резец | 1924 |
|
SU2016A1 |
| Способ обработки целлюлозных материалов, с целью тонкого измельчения или переведения в коллоидальный раствор | 1923 |
|
SU2005A1 |
| Пломбировальные щипцы | 1923 |
|
SU2006A1 |
| Колосоуборка | 1923 |
|
SU2009A1 |

