ССЫЛКА НА ПРИОРИТЕТНЫЕ ЗАЯВКИ
[01] Настоящая заявка испрашивает приоритет согласно предварительной заявке на патент США № 62/673670, поданной 18 мая 2018 года, предварительной заявке на патент США № 62/712880, поданной 31 июля 2018 года, предварительной заявке на патент США № 62/773785, поданной 30 ноября 2018 года, и предварительной заявке на патент США № 62/801576, поданной 5 февраля 2019 года, каждая из которых включена в данный документ посредством ссылки во всей своей полноте.
ССЫЛКА НА ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ, ПОДАННЫЙ В ЭЛЕКТРОННОМ ВИДЕ
[02] Содержание перечня последовательностей, поданного в электронном виде в виде текстового файла в формате ASCII (название: SA9-461PC_SeqListing.txt; размер: 922 килобайт; дата создания: 14 мая 2019 г.), включено в данный документ посредством ссылки во всей своей полноте.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[03] Гемофилия A представляет собой нарушение свертываемости крови, вызванное дефектами гена, кодирующего фактор коагуляции VIII (FVIII), и поражает 1-2 из 10000 новорожденных мальчиков. Graw et al., Nat. Rev. Genet. 6(6): 488-501 (2005). Пациентов, страдающих гемофилией А, можно подвергать лечению посредством инфузий очищенного или полученного рекомбинантным путем FVIII. Известно, что многие коммерчески доступные продукты FVIII характеризуются периодом полужизни приблизительно 8-12 часов, что требует частого внутривенного введения пациентам. См. Weiner M.A. and Cairo, M.S., Pediatric Hematology Secrets, Lee, M.T., 12. Disorders of Coagulation, Elsevier Health Sciences, 2001; Lillicrap, D. Thromb. Res. 122 Suppl 4:S2-8 (2008). Кроме того, был опробован ряд подходов для продления периода полужизни FVIII. Например, разрабатываемые подходы для продления периода полужизни факторов свертывания крови включают пегилирование, гликопегилирование и конъюгирование с альбумином. См. Dumont et al., Blood. 119(13): 3024-3030 (2012). Достоверные результаты были продемонстрированы у людей, например, сообщалось, что rFVIIIFc улучшает период полужизни в до ~1,7 раза по сравнению с ADVATE® у пациентов с гемофилией А. См. Powell et al., Blood. 119(13): 3031-3037 (2012). Следовательно, увеличение периода полужизни, несмотря на незначительные улучшения, указывает на присутствие других факторов, ограничивающих период полужизни. См. Liu, T. et al., 2007 ISTH meeting, abstract #P-M-035; Henrik, A. et al., 2011 ISTH meeting, abstract #P=MO-181; Liu, T. et al., 2011 ISTH meeting abstract #P-WE-131.
[04] Текущий рекомендуемый стандарт лечения включает регулярное введение FVIII (обычную профилактику) для минимизации количества эпизодов кровотечения. Было установлено, что обычная профилактика связана с улучшением долгосрочных исходов, но она представляет собой сложный режим, ограниченный необходимостью частого внутривенного (IV) введения. См. Manco-Johnson et al., N Engl J Med. 357(6):535-44 (2007). Продукты с продленным периодом полужизни FVIII обеспечивали снижение частоты введения FVIII для профилактики; однако все они взаимодействуют с фактором фон Виллебранда (VWF) и имеют сравнимые периоды полужизни в кровотоке, что соответствует верхнему пределу периода полужизни вариантов rFVIII, обусловленному периодом полужизни эндогенного VWF. См., например, Pipe et al., Blood. 128(16):2007-16 (2016). Профилактическое введение доз этих продуктов FVIII осуществляется каждые 3-5 дней.
[05] Продукты с продленным периодом полужизни FVIII следующего поколения, которые обеспечивают предупреждение и контроль эпизодов кровотечения в течение более длительных периодов времени, что приводит к менее частому введению, потенциально могут решить проблемы соблюдения сложных профилактических режимов, что, в свою очередь, может улучшить качество жизни пациентов с гемофилией.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[06] Определенные аспекты настоящего изобретения относятся к способу лечения гемофилии А у субъекта-человека, нуждающегося в этом, предусматривающему введение субъекту нескольких доз химерного полипептида, содержащего (i) белковый фактор VIII (FVIII) и (ii) фрагмент фактора фон Виллебранда (VWF), содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней.
[07] В некоторых вариантах осуществления несколько доз предусматривают по меньшей мере две дозы, по меньшей мере три дозы, по меньшей мере четыре дозы, по меньшей мере пять доз, по меньшей мере шесть доз, по меньшей мере семь доз, по меньшей мере восемь доз, по меньшей мере девять доз, по меньшей мере десять доз, по меньшей мере одиннадцать доз, по меньшей мере двенадцать доз, по меньшей мере тринадцать доз, по меньшей мере четырнадцать доз, по меньшей мере пятнадцать доз, по меньшей мере шестнадцать доз, по меньшей мере семнадцать доз, по меньшей мере восемнадцать доз, по меньшей мере девятнадцать доз, по меньшей мере двадцать доз или больше.
[08] В некоторых вариантах осуществления лечение гемофилии А предусматривает контроль или уменьшение встречаемости или частоты эпизодов кровотечения у субъекта-человека, нуждающегося в этом. В некоторых вариантах осуществления лечение гемофилии А предусматривает предупреждение или лечение эпизода кровотечения у субъекта-человека, нуждающегося в этом.
[09] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 20 МЕ/кг до приблизительно 95 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 90 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 85 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 75 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 70 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 65 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 60 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 55 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 50 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 45 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 40 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 35 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 30 МЕ/кг или от приблизительно 20 МЕ/кг до приблизительно 25 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 20 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 25 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 30 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 35 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 40 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 45 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 55 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 70 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 75 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 80 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 85 МЕ/кг до приблизительно 100 МЕ/кг или от приблизительно 90 МЕ/кг до приблизительно 100 МЕ/кг.
[010] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 20 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 25 МЕ/кг до приблизительно 75 МЕ/кг, от приблизительно 30 МЕ/кг до приблизительно 70 МЕ/кг, от приблизительно 35 МЕ/кг до приблизительно 65 МЕ/кг, от приблизительно 40 МЕ/кг до приблизительно 60 МЕ/кг или от приблизительно 45 МЕ/кг до приблизительно 55 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг.
[011] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 20 МЕ/кг, приблизительно 25 МЕ/кг, приблизительно 30 МЕ/кг, приблизительно 35 МЕ/кг, приблизительно 40 МЕ/кг, приблизительно 45 МЕ/кг, приблизительно 50 МЕ/кг, приблизительно 55 МЕ/кг, приблизительно 60 МЕ/кг, приблизительно 65 МЕ/кг, приблизительно 70 МЕ/кг, приблизительно 75 МЕ/кг, приблизительно 80 МЕ/кг, приблизительно 85 МЕ/кг, приблизительно 90 МЕ/кг, приблизительно 95 МЕ/кг или приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг.
[012] В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 5 дней, по меньшей мере приблизительно 6 дней, по меньшей мере приблизительно 7 дней, по меньшей мере приблизительно 8 дней, по меньшей мере приблизительно 9 дней, по меньшей мере приблизительно 10 дней, по меньшей мере приблизительно 11 дней, по меньшей мере приблизительно 12 дней, по меньшей мере приблизительно 13 дней, по меньшей мере приблизительно 14 дней, по меньшей мере приблизительно 15 дней, по меньшей мере приблизительно 16 дней, по меньшей мере приблизительно 17 дней, по меньшей мере приблизительно 18 дней, по меньшей мере приблизительно 19 дней, по меньшей мере приблизительно 20 дней, по меньшей мере приблизительно 21 день, по меньшей мере приблизительно 22 дня, по меньшей мере приблизительно 23 дня, по меньшей мере приблизительно 24 дня, по меньшей мере приблизительно 25 дней, по меньшей мере приблизительно 26 дней, по меньшей мере приблизительно 27 дней, по меньшей мере приблизительно 28 дней, по меньшей мере приблизительно 29 дней, по меньшей мере приблизительно 30 дней или по меньшей мере приблизительно 31 день.
[013] В некоторых вариантах осуществления частота введения доз составляет по меньшей мере один раз в неделю, по меньшей мере один раз в 2 недели, по меньшей мере один раз в 3 недели или по меньшей мере один раз в 4 недели. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере одну неделю. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере две недели.
[014] В некоторых вариантах осуществления химерный полипептид вводят для профилактического лечения.
[015] В некоторых вариантах осуществления белок FVIII связан с фрагментом VWF с помощью ковалентной связи. В некоторых вариантах осуществления ковалентная связь представляет собой пептидную связь или дисульфидную связь.
[016] В некоторых вариантах осуществления белок FVIII содержит полипептид FVIII и первый компонент, обеспечивающий продление периода полужизни. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, слит с С-концом или N-концом полипептида FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в полипептид FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в домен B полипептида FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в полипептид FVIII непосредственно ниже аминокислоты, соответствующей аминокислотному остатку 745 в SEQ ID NO: 65. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, слит с полипептидом FVIII с помощью линкера.
[017] В некоторых вариантах осуществления фрагмент VWF содержит второй компонент, обеспечивающий продление периода полужизни. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с С-концом или N-концом фрагмента VWF. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, вставлен во фрагмент VWF. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с С-концом фрагмента VWF. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с фрагментом VWF с помощью линкера.
[018] В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, второй компонент, обеспечивающий продление периода полужизни, или они оба выбраны из группы, состоящей из альбумина, Fc-области иммуноглобулина, последовательности XTEN, C-концевого пептида (CTP) β-субъединицы хорионического гонадотропина человека, последовательности PAS, последовательности HAP, трансферрина, альбуминсвязывающих компонентов или любых их фрагментов, производных, вариантов и любой их комбинации.
[019] В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, содержит первый XTEN.
[020] В некоторых вариантах осуществления первый XTEN вставлен в полипептид FVIII непосредственно ниже аминокислоты, соответствующей аминокислотному остатку 745 в SEQ ID NO: 65.
[021] В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, содержит второй XTEN. В некоторых вариантах осуществления второй XTEN слит с С-концом фрагмента VWF.
[022] В некоторых вариантах осуществления белок FVIII содержит константную область первого иммуноглобулина (Ig) или ее часть. В некоторых вариантах осуществления первая константная область Ig или ее часть слита с С-концом или N-концом полипептида FVIII. В некоторых вариантах осуществления первая константная область Ig или ее часть вставлена в полипептид FVIII. В некоторых вариантах осуществления первая константная область Ig или ее часть слита с С-концом полипептида FVIII. В некоторых вариантах осуществления первая константная область Ig или ее часть слита с полипептидом FVIII с помощью линкера. В некоторых вариантах осуществления первая константная область Ig или ее часть содержит первый Fc-домен или его часть.
[023] В некоторых вариантах осуществления фрагмент VWF содержит вторую константную область Ig или ее часть. В некоторых вариантах осуществления вторая константная область Ig или ее часть слита с С-концом или N-концом фрагмента VWF. В некоторых вариантах осуществления вторая константная область Ig или ее часть вставлена во фрагмент VWF. В некоторых вариантах осуществления вторая константная область Ig или ее часть слита с С-концом фрагмента VWF. В некоторых вариантах осуществления вторая константная область Ig или ее часть слита с фрагментом VWF с помощью линкера. В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. В некоторых вариантах осуществления вторая константная область Ig или ее часть содержит первый Fc-домен или его часть.
[024] В некоторых вариантах осуществления белок FVIII и фрагмент VWF связаны друг с другом посредством ковалентной связи между первым Fc-доменом и вторым Fc-доменом. В некоторых вариантах осуществления белок FVIII и фрагмент VWF дополнительно связаны друг с другом посредством нековалентного взаимодействия между белком FVIII и фрагментом VWF.
[025] В одном аспекте в данном документе раскрыт способ лечения гемофилии А у субъекта-человека, предусматривающий введение субъекту, нуждающемуся в этом, нескольких доз химерного полипептида с интервалом между введениями доз, где химерный полипептид содержит: (i) белок FVIII, содержащий первый фрагмент полипептида FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 215; первую последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 8 (AE288); второй фрагмент полипептида FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 216; и первую Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217; и (ii) белок VWF, содержащий: домен D' VWF, который содержит аминокислотную последовательность под SEQ ID NO: 210; домен D3 VWF, который содержит аминокислотную последовательность под SEQ ID NO: 214; вторую последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 58 (AE144_5A); линкер а2, который содержит аминокислотную последовательность под SEQ ID NO: 88; и вторую Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217, и где первая Fc-область ковалентно соединена со второй Fc-областью с помощью дисульфидной связи.
[026] В некоторых вариантах осуществления химерный полипептид содержит белок FVIII, содержащий полипептид FVIII, первую последовательность XTEN, первую Fc-область, и белок VWF, содержащий домен D' VWF, домен D3 VWF, вторую последовательность XTEN, линкер a2 FVIII и вторую Fc-область, где полипептид FVIII содержит аминокислотную последовательность под SEQ ID NO: 215, первая последовательность XTEN содержит аминокислотную последовательность AE288 (SEQ ID NO: 8) и слита с С-концом SEQ ID NO: 215, указанный полипептид FVIII дополнительно содержит аминокислотную последовательность под SEQ ID NO: 216; первая Fc-область содержит аминокислотную последовательность под SEQ ID NO: 217 и слита с С-концом SEQ ID NO: 216; домен D' VWF содержит аминокислотную последовательность под SEQ ID NO: 210; домен D3 VWF содержит аминокислотную последовательность под SEQ ID NO: 214, вторая последовательность XTEN содержит аминокислотную последовательность AE144_5A (SEQ ID NO: 58) и слита с С-концом домена D3 VWF; линкер a2 содержит аминокислотную последовательность под SEQ ID NO: 88 и слит с С-концом второй последовательности XTEN; вторая Fc-область содержит аминокислотную последовательность под SEQ ID NO: 217 и слита с С-концом линкера a2; и где первая Fc-область ковалентно соединена со второй Fc-областью с помощью дисульфидной связи.
[027] В некоторых вариантах осуществления химерный полипептид содержит белок FVIII, содержащий сигнальный пептид FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 64. В некоторых вариантах осуществления химерный полипептид содержит белок VWF, содержащий сигнальный пептид VWF, который содержит аминокислотную последовательность под SEQ ID NO: 208. В некоторых вариантах осуществления химерный полипептид содержит белок VWF, содержащий домен D1D2 VWF, который содержит аминокислотную последовательность под SEQ ID NO: 209.
[028] В некоторых вариантах осуществления химерный полипептид содержит белок FVIII, содержащий аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную SEQ ID NO: 201, SEQ ID NO: 203 или SEQ ID NO: 207; и белок VWF, содержащий аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную SEQ ID NO: 202 или SEQ ID NO: 205.
[029] В одном варианте осуществления химерный полипептид содержит белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 203, и белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 205. В другом варианте осуществления химерный полипептид содержит белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 201, и белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 202. В другом варианте осуществления химерный полипептид содержит белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 207, и белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 202.
[030] В некоторых вариантах осуществления химерный полипептид содержит белок FVIII, содержащий аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную последовательности, выбранной из FVIII-161 (SEQ ID NO: 69), FVIII-169 (SEQ ID NO: 70), FVIII-170 (SEQ ID NO: 71), FVIII-173 (SEQ ID NO: 72); FVIII-195 (SEQ ID NO: 73); FVIII-196 (SEQ ID NO: 74), FVIII199 (SEQ ID NO: 75), FVIII-201 (SEQ ID NO: 76); FVIII-203 (SEQ ID NO: 77), FVIII-204 (SEQ ID NO: 78), FVIII-205 (SEQ ID NO: 79), FVIII-266 (SEQ ID NO: 80), FVIII-267 (SEQ ID NO: 81), FVIII-268 (SEQ ID NO: 82), FVIII-269 (SEQ ID NO: 83), FVIII-271 (SEQ ID NO: 84), FVIII-272 (SEQ ID NO: 85), FVIII-312 (SEQ ID NO: 173) или FVIII-312A (SEQ ID NO: 203); и белок VWF, содержащий аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную последовательности, выбранной из VWF031 (SEQ ID NO: 86), VWF034 (SEQ ID NO: 87), VWF059 (SEQ ID NO: 197), VWF059A (SEQ ID NO: 202) или VWF036.
[031] В некоторых вариантах осуществления химерный полипептид вводят посредством пути, выбранного из группы, состоящей из внутривенной инъекции, внутривенной инфузии, подкожного введения, внутримышечного введения, перорального введения, назального введения и легочного введения.
[032] В некоторых вариантах осуществления химерный полипептид после введения приводит к уровню активности FVIII в плазме крови, составляющему по меньшей мере приблизительно 1%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 9% или по меньшей мере приблизительно 10%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 3%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5%.
[033] В некоторых вариантах осуществления химерный полипептид после введения приводит к уровню активности FVIII в плазме крови, составляющему по меньшей мере приблизительно 1 МЕ/дл, по меньшей мере приблизительно 2 МЕ/дл, по меньшей мере приблизительно 3 МЕ/дл, по меньшей мере приблизительно 4 МЕ/дл, по меньшей мере приблизительно 5 МЕ/дл, по меньшей мере приблизительно 6 МЕ/дл, по меньшей мере приблизительно 7 МЕ/дл, по меньшей мере приблизительно 8 МЕ/дл, по меньшей мере приблизительно 9 МЕ/дл или по меньшей мере приблизительно 10 МЕ/дл. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 3 МЕ/дл. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5 МЕ/дл.
[034] В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 10 МЕ/дл через по меньшей мере приблизительно 5 дней после введения химерного полипептида. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5 МЕ/дл через по меньшей мере приблизительно 7 дней после введения химерного полипептида. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 3 МЕ/дл через по меньшей мере приблизительно 8 дней после введения химерного полипептида. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 1 МЕ/дл через по меньшей мере приблизительно 10 дней после введения химерного полипептида.
[035] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней.
[036] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг. В некоторых вариантах осуществления несколько доз составляют приблизительно 50 МЕ/кг, а интервал между введениями доз составляет приблизительно 5 дней. В некоторых вариантах осуществления несколько доз составляют приблизительно 50 МЕ/кг, а интервал между введениями доз составляет приблизительно 7 дней. В некоторых вариантах осуществления несколько доз составляют приблизительно 50 МЕ/кг, а интервал между введениями доз составляет приблизительно 14 дней.
[037] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней.
[038] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней.
[039] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней.
[040] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 1 неделю. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 1 неделю. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 1 неделю.
[041] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 2 недели. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 2 недели. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 2 недели.
[042] В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В некоторых вариантах осуществления интервал между введениями доз составляет от приблизительно 5 дней до приблизительно 14 дней. В некоторых вариантах осуществления интервал между введениями доз составляет от приблизительно 7 дней до приблизительно 14 дней. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В некоторых вариантах осуществления интервал между введениями доз составляет от приблизительно 10 дней до приблизительно 21 дня. В некоторых вариантах осуществления интервал между введениями доз составляет от приблизительно 14 дней до приблизительно 21 дня. В некоторых вариантах осуществления интервал между введениями доз составляет приблизительно 14 дней.
[043] В некоторых вариантах осуществления субъект-человек является женщиной. В некоторых вариантах осуществления субъект-человек является ребенком. В некоторых вариантах осуществления субъект-человек является ребенком в возрасте, равном приблизительно 12 лет или меньше, в возрасте менее чем приблизительно 11 лет, в возрасте менее чем приблизительно 10 лет, в возрасте менее чем приблизительно 9 лет, в возрасте менее чем приблизительно 8 лет, в возрасте менее чем приблизительно 7 лет, в возрасте менее чем приблизительно 6 лет, в возрасте менее чем приблизительно 5 лет, в возрасте менее чем приблизительно 4 года, в возрасте менее чем приблизительно 3 года, в возрасте менее чем приблизительно 2 года или в возрасте менее чем приблизительно 1 год.
[044] В некоторых вариантах осуществления введение приводит к индукции иммунной толерантности к FVIII у субъекта-человека. В некоторых вариантах осуществления введение приводит к ослаблению ингибирующего иммунного ответа на FVIII у человека. В некоторых вариантах осуществления ингибирующий иммунный ответ на FVIII предусматривает высокий титр антител к FVIII у субъекта-человека.
[045] В некоторых вариантах осуществления введение химерного полипептида не приводит к индукции образования ингибитора FVIII после приблизительно 7 дней, приблизительно 10 дней, приблизительно 11 дней, приблизительно 12 дней, приблизительно 13 дней, приблизительно 14 дней, приблизительно 15 дней, приблизительно 20 дней, приблизительно 24 дней, приблизительно 25 дней, приблизительно 28 дней, приблизительно 30 дней или приблизительно 35 дней введения. В некоторых вариантах осуществления введение химерного полипептида не приводит к индукции образования ингибитора FVIII после приблизительно 28 дней введения.
КРАТКОЕ ОПИСАНИЕ ГРАФИЧЕСКИХ МАТЕРИАЛОВ/ФИГУР
[046] Фиг. 1 представляет собой схематическое изображение rFVIIIFc-VWF-XTEN. FVIII: фактор VIII; VWF: фактор фон Виллебранда; A1, A2, A3, C1, C2: домены FVIII; D'D3: домены VWF; Fc: Fc-область константной области иммуноглобулина.
[047] На фиг. 2A и 2B показан протокол тестирования безопасности и эффективности rFVIIIFc-VWF-XTEN у пациентов-людей в когорте с низкой дозой, которым вводили 25 МЕ/кг rFVIIIFc-VWF-XTEN (фиг. 2A), и в когорте с высокой дозой, которым вводили 65 МЕ/кг rFVIIIFc-VWF-XTEN (фиг. 2B).
[048] Фиг. 3A-3B представляют собой графические изображения уровней активности FVIII с поправкой на исходный уровень на основании теста активированного частичного тромбопластинового времени (aPTT) у субъектов-людей с тяжелой формой гемофилии А, которым вводили 25 МЕ/кг rFVIII с последующим периодом отмывки, а затем 25 МЕ/кг rFVIIIFc-VWF-XTEN (фиг. 3A; когорта с низкой дозой) или 65 МЕ/кг rFVIII с последующим периодом отмывки и затем 65 МЕ/кг rFVIIIFc-VWF-XTEN (фиг. 3B; когорта с высокой дозой). Горизонтальные пунктирные линии указывают на 3%, 5%, 10% и 20% активность FVIII.
[049] Фиг. 4 представляет собой схематическое изображение плана клинического исследования для оценки безопасности и переносимости в общей сложности четырех еженедельных доз rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг или 65 МЕ/кг у взрослых пациентов-мужчин, ранее получавших лечение (PTP), в возрасте от 18 до 65 лет (включительно) с тяжелой формой гемофилии A. EOS=окончание исследования; ET=досрочное прекращение; PK=фармакокинетические показатели.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[050] Настоящее изобретение относится к способу лечения заболевания или состояния, связанного с кровотечением, например, гемофилии A, у субъекта-человека, нуждающегося в этом, предусматривающему введение субъекту нескольких доз химерного полипептида, содержащего (i) полипептидный фактор VIII (FVIII) и (ii) фрагмент фактора фон Виллебранда (VWF), содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 7 дней.
I. Определения
[051] Следует отметить, что форма единственного числа объекта относится к одному или нескольким таким объектам; например, под "нуклеотидной последовательностью" понимают одну или несколько нуклеотидных последовательностей. В связи с этим формы единственного числа, термины "один или несколько" и "по меньшей мере один" могут использоваться в данном документе взаимозаменяемо.
[052] Кроме того, "и/или" при использовании в данном документе следует понимать как конкретное раскрытие каждого из двух указанных признаков или компонентов с другим или без него. Таким образом, подразумевается, что термин "и/или", используемый в данном документе в такой фразе, как "А и/или В", включает "А и В", "А или В", "А" (отдельно) и "В" (отдельно). Аналогично, подразумевается, что термин "и/или", используемый в такой фразе, как "А, В и/или С", охватывает каждый из следующих аспектов: A, B и C; A, B или C; A или C; A или B; B или C; A и C; A и B; B и C; A (отдельно); B (отдельно) и C (отдельно).
[053] Следует понимать, что во всех случаях, когда аспекты описываются в данном документе с формулировкой "содержащий", также предусмотрены другие аналогичные аспекты, описываемые терминами "состоящий из" и/или "состоящий по сути из".
[054] Если не определено иное, то все используемые в данном документе технические и научные термины имеют то же значение, которое обычно понимает средний специалист в области техники, к которой относится настоящее изобретение. Например, Concise Dictionary of Biomedicine and Molecular Biology, Juo, Pei-Show, 2nd ed., 2002, CRC Press; The Dictionary of Cell and Molecular Biology, 3rd ed., 1999, Academic Press и Oxford Dictionary Of Biochemistry And Molecular Biology, Revised, 2000, Oxford University Press обеспечивают специалиста общим словарем многих терминов, используемых в настоящем раскрытии.
[055] Единицы измерения, приставки и символы обозначены в их форме, принятой согласно Международной системе единиц (SI). Числовые диапазоны включают числа, определяющие диапазон. Если не указано иное, аминокислотные последовательности записаны слева направо в направлении от амино- к карбокси-концу. Приведенные в данном документе заголовки не являются ограничением различных аспектов настоящего изобретения. Соответственно, термины, определенные непосредственно ниже, более полно определяются посредством ссылки на настоящее описание во всей его полноте.
[056] Термин "приблизительно" используется в данном документе в значении примерно, порядка, около или ориентировочно. Если термин "приблизительно" используется в сочетании с числовым диапазоном, то он модифицирует данный диапазон, расширяя границы выше и ниже изложенных числовых значений. В целом, термин "приблизительно" может модифицировать числовое значение, делая его большим или меньшим заявленного значения, с отклонением, например, на 10 процентов, вверх или вниз (выше или ниже).
[057] Подразумевается, что термин "полинуклеотид" или "нуклеотид" охватывает нуклеиновую кислоту в единственном числе, а также нуклеиновые кислоты во множественном числе и относится к выделенной молекуле или конструкции нуклеиновой кислоты, например, матричной РНК (мРНК) или плазмидной ДНК (пДНК). В определенных вариантах осуществления полинуклеотид содержит традиционную фосфодиэфирную связь или нетрадиционную связь (например, амидную связь, такую как встречающаяся в пептидных нуклеиновых кислотах (PNA)). Термин "нуклеиновая кислота" относится к любому одному или нескольким сегментам нуклеиновой кислоты, например, фрагментам ДНК или РНК, присутствующим в полинуклеотиде. Под "выделенными" нуклеиновой кислотой или полинуклеотидом подразумевается молекула нуклеиновой кислоты, ДНК или РНК, которая была извлечена из ее нативного окружения. Например, рекомбинантный полинуклеотид, кодирующий полипептидный фактор VIII, который содержится в векторе, в рамках настоящего изобретения считают выделенным. Дополнительные примеры выделенного полинуклеотида включают рекомбинантные полинуклеотиды, содержащиеся в гетерологичных клетках-хозяевах или очищенные (частично или по существу) от других полинуклеотидов в растворе. Выделенные молекулы РНК включают РНК-транскрипты полинуклеотидов по настоящему изобретению, полученные in vivo или in vitro. Выделенные полинуклеотиды или нуклеиновые кислоты в соответствии с настоящим изобретением дополнительно включают такие молекулы, полученные путем синтеза. Кроме того, полинуклеотид или нуклеиновая кислота могут содержать регуляторные элементы, такие как промоторы, энхансеры, сайты связывания рибосом или сигналы терминации транскрипции.
[058] Определенные белки, секретируемые клетками млекопитающих, связаны с секреторным сигнальным пептидом, отщепляющимся от зрелого белка после начала экспорта растущей белковой цепи через гранулярный эндоплазматический ретикулум. Средним специалистам в данной области известно, что сигнальные полипептиды обычно слиты с N-концом полипептида и отщепляются от полного или "полноразмерного" полипептида с образованием секретируемой или "зрелой" формы полипептида. В определенных вариантах осуществления применяют нативный сигнальный пептид или функциональное производное такой последовательности, которое сохраняет способность к управлению секрецией полипептида, функционально связанного с ним. В качестве альтернативы можно применять гетерологичный сигнальный пептид млекопитающего, например, тканевой активатор плазминогена (ТРА) человека или сигнальный пептид ß-глюкуронидазы мыши или его функциональное производное.
[059] Подразумевается, что используемый в данном документе термин "полипептид" охватывает "полипептид" в единственном числе, а также "полипептиды" во множественном числе и относится к молекуле, состоящей из мономеров (аминокислот), линейно соединенных амидными связями (также известными как пептидные связи). Термин "полипептид" относится к любой цепи или цепям из двух или более аминокислот и не относится к конкретной длине продукта. Таким образом, пептиды, дипептиды, трипептиды, олигопептиды, "белок", "аминокислотная цепь" или любой другой термин, используемый для обозначения цепи или цепей из двух или более аминокислот, включены в определение "полипептида", и термин "полипептид" можно использовать вместо любого из этих терминов или взаимозаменяемо с любым из них. Также подразумевается, что термин "полипептид" относится к продуктам постэкспрессионных модификаций полипептида, включая без ограничения гликозилирование, ацетилирование, фосфорилирование, амидирование, дериватизацию с помощью известных защитных/блокирующих групп, протеолитическое расщепление или модификацию с помощью аминокислот, не встречающихся в природе. Полипептид может происходить из природного биологического источника или быть получен с помощью рекомбинантной технологии, но не обязательно быть получен в результате трансляции определенной последовательности нуклеиновой кислоты. Он может быть получен любым способом, в том числе путем химического синтеза.
[060] "Выделенный" полипептид или его фрагмент, вариант или производное относятся к полипептиду, который не находится в своем естественном окружении. Никакой конкретный уровень очистки не требуется. Например, выделенный полипептид может быть просто извлечен из его нативной или природной среды. Полученные рекомбинантным путем полипептиды и белки, экспрессирующиеся в клетках-хозяевах, считаются выделенными для целей настоящего изобретения, как и нативные или рекомбинантные полипептиды, которые были отделены, фракционированы или частично или в значительной степени очищены с помощью любой подходящей методики.
[061] Также в настоящее изобретение включены фрагменты или варианты полипептидов и любая их комбинация. Термины "фрагмент" или "вариант" в отношении связывающих доменов полипептидов или связывающих молекул по настоящему изобретению включают любые полипептиды, которые сохраняют по меньшей мере некоторые из свойств эталонного полипептида (например, аффинность связывания с FcRn для FcRn-связывающего домена или варианта Fc, активность коагуляции для варианта FVIII или активность связывания FVIII для фрагмента VWF). Фрагменты полипептидов включают фрагменты, полученные посредством протеолиза, а также фрагменты, полученные посредством делеции, в дополнение к конкретным фрагментам антитела, обсуждаемым в данном документе в другом месте, но не включают встречающийся в природе полноразмерный полипептид (или зрелый полипептид). Варианты связывающих доменов полипептидов или связывающих молекул по настоящему изобретению включают фрагменты, описанные выше, а также полипептиды с аминокислотными последовательностями, измененными в результате аминокислотных замен, делеций или вставок. Варианты могут быть встречающимися в природе или не встречающимися в природе. Не встречающиеся в природе варианты можно получить с помощью известных из уровня техники методик мутагенеза. Варианты полипептидов могут содержать консервативные или неконсервативные аминокислотные замены, делеции или добавления.
[062] Термин "белок VWF" или "белки VWF", используемый в данном документе, означает любые фрагменты VWF, которые взаимодействуют с FVIII и сохраняют по меньшей мере одно или несколько свойств, которые обычно обеспечиваются у FVIII благодаря полноразмерному VWF, например, предотвращение преждевременной активации до FVIIIa, предотвращение преждевременного протеолиза, предотвращение ассоциации с фосфолипидными мембранами, которая может привести к преждевременному выведению, предотвращение связывания с рецепторами, опосредующими выведение FVIII, которые могут связываться с "голым" FVIII, но не с FVIII, связанным с VWF, и/или стабилизацию взаимодействий тяжелой цепи и легкой цепи FVIII. Фрагмент VWF, упоминаемый в данном документе, представляет собой полипептид VWF, имеющий меньший размер, чем полноразмерный белок VWF, при этом фрагмент VWF сохраняет способность взаимодействовать и/или связываться с FVIII.
[063] "Консервативная аминокислотная замена" представляет собой замену, при которой аминокислотный остаток замещается аминокислотным остатком, имеющим сходную боковую цепь. Семейства аминокислотных остатков, имеющих сходные боковые цепи, были определены в уровне техники, включая основные боковые цепи (например, лизин, аргинин, гистидин), кислые боковые цепи (например, аспарагиновая кислота, глутаминовая кислота), незаряженные полярные боковые цепи (например, глицин, аспарагин, глутамин, серин, треонин, тирозин, цистеин), неполярные боковые цепи (например, аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан), бета-разветвленные боковые цепи (например, треонин, валин, изолейцин) и ароматические боковые цепи (например, тирозин, фенилаланин, триптофан, гистидин). Таким образом, если аминокислота в полипептиде замещается другой аминокислотой из того же семейства боковых цепей, то замена считается консервативной. В других вариантах осуществления нить из аминокислот можно подвергнуть консервативному замещению сходной в структурном отношении нитью, которая отличается порядком расположения и/или составом представителей семейства боковых цепей.
[064] Как известно из уровня техники, "идентичность последовательностей" между двумя полипептидами определяется путем сравнения аминокислотной последовательности одного полипептида с последовательностью второго полипептида. При обсуждении в данном документе можно определить, является ли какой-либо конкретный полипептид на по меньшей мере 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 99% или 100% идентичным другому полипептиду, с применением способов и компьютерных программ/программного обеспечения, известных в данной области техники, таких как, без ограничения, программа BESTFIT (Wisconsin Sequence Analysis Package, версия 8 для Unix, Genetics Computer Group, University Research Park, 575 Science Drive, Мэдисон, Висконсин 53711). В BESTFIT используется алгоритм поиска локальной гомологии Смита-Уотермана, Advances in Applied Mathematics 2: 482-489 (1981), для нахождения наилучшего сегмента гомологии между двумя последовательностями. При использовании BESTFIT или любой другой программы выравнивания последовательностей для определения того, является ли конкретная последовательность, например, на 95% идентичной эталонной последовательности согласно настоящему изобретению, параметры, разумеется, устанавливаются так, что процент идентичности рассчитывается по полной длине эталонной полипептидной последовательности, и допускаются гэпы в гомологии в количестве до 5% от общего числа аминокислот в эталонной последовательности.
[065] Используемая в данном документе "соответствующая аминокислота" или "эквивалентная аминокислота" в последовательности VWF или последовательности белка FVIII идентифицируются путем выравнивания для максимального увеличения идентичности или сходства между первой последовательностью VWF или FVIII и второй последовательностью VWF или FVIII. Номер, используемый для идентификации эквивалентной аминокислоты во второй последовательности VWF или FVIII, соответствует номеру, используемому для идентификации соответствующей аминокислоты в первой последовательности VWF или FVIII.
[066] Используемый в данном документе термин "сайт вставки" относится к положению в полипептиде FVIII или его фрагменте, варианте или производном, которое находится непосредственно ниже положения, в которое может быть вставлен компонент, обеспечивающий продление периода полужизни, или гетерологичный компонент. "Сайт вставки" указан в виде числа, при этом число является номером аминокислоты в зрелом нативном FVIII (SEQ ID NO: 65), которому соответствует сайт вставки, который располагается непосредственно за положением вставки в направлении С-конца. Например, фраза "a3 содержит XTEN в сайте вставки, который соответствует аминокислоте 1656 в SEQ ID NO: 65" указывает на то, что гетерологичный компонент расположен между двумя аминокислотами, соответствующими аминокислоте 1656 и аминокислоте 1657 в SEQ ID NO: 65.
[067] Используемая в данном документе фраза "непосредственно ниже аминокислоты" относится к положению сразу возле концевой карбоксильной группы аминокислоты. Например, сайт вставки непосредственно ниже аминокислоты 745, соответствующей зрелому белку FVIII дикого типа, означает, что сайт вставки находится между аминокислотой 745 и аминокислотой 746, соответствующей зрелому белку FVIII дикого типа. Аналогично, фраза "непосредственно выше аминокислоты" относится к положению сразу возле концевой аминогруппы аминокислоты.
[068] Используемая в данном документе фраза "между двумя аминокислотами в сайте вставки" относится к положению, в котором XTEN или любой другой полипептид вставлен между двумя соседними аминокислотами. Таким образом, фразы "вставлен непосредственно ниже аминокислоты" и "вставлен между двумя аминокислотами в сайте вставки" используются как синонимы фразы "вставлен в сайт вставки".
[069] Термины "вставленный", "вставлен", "вставленный в" или грамматически родственные термины, используемые в данном документе, относятся к положению XTEN в химерном полипептиде относительно аналогичного положения в нативном зрелом FVIII человека. Используемые в данном документе термины относятся к характеристикам рекомбинантного полипептида FVIII относительно нативного зрелого FVIII человека и не указывают, не подразумевают или не предполагают никаких способов или процесса, посредством которых был получен химерный полипептид. Например, по отношению к химерному полипептиду, предусмотренному в данном документе, фраза "XTEN вставлен непосредственно ниже остатка 745 полипептида FVIII" означает, что химерный полипептид содержит XTEN непосредственно ниже аминокислоты, соответствующей аминокислоте 745 в природном зрелом FVIII человека, например, ограничен аминокислотами, соответствующими аминокислотам 745 и 746 природного зрелого FVIII человека.
[070] "Слитый" или "химерный" белок содержит первую аминокислотную последовательность, соединенную со второй аминокислотной последовательностью, с которой она в естественных условиях не соединена в природе. Аминокислотные последовательности, которые в обычных условиях существуют в отдельных белках, могут быть объединены в слитом полипептиде, или аминокислотные последовательности, которые в обычных условиях существуют в одном и том же белке, могут быть размещены в новом порядке в слитом полипептиде, например, при слиянии домена фактора VIII по настоящему изобретению с Fc-доменом Ig. Слитый белок создают, например, путем химического синтеза или путем создания и трансляции полинуклеотида, в котором области пептида кодируются в необходимом взаиморасположении. Химерный полипептид может дополнительно содержать вторую аминокислотную последовательность, связанную с первой аминокислотной последовательностью с помощью ковалентной непептидной связи или нековалентной связи.
[071] Термин "соединенный", используемый в данном документе, относится к первой аминокислотной последовательности или нуклеотидной последовательности, ковалентно или нековалентно присоединенной соответственно ко второй аминокислотной последовательности или нуклеотидной последовательности. Первая аминокислотная или нуклеотидная последовательность может быть непосредственно присоединена ко второй аминокислотной или нуклеотидной последовательности или объединена с ней, или, в качестве альтернативы, промежуточная последовательность может ковалентно присоединять первую последовательность ко второй последовательности. Термин "соединенный" означает не только слияние первой аминокислотной последовательности со второй аминокислотной последовательностью на С-конце или N-конце, но также включает вставку всей первой аминокислотной последовательности (или второй аминокислотной последовательности) между любыми двумя аминокислотами во второй аминокислотной последовательности (или соответственно в первой аминокислотной последовательности). В некоторых вариантах осуществления первая аминокислотная последовательность может быть соединена со второй аминокислотной последовательностью с помощью пептидной связи или линкера. Первая нуклеотидная последовательность может быть соединена со второй нуклеотидной последовательностью с помощью фосфодиэфирной связи или линкера. Линкер может представлять собой пептид или полипептид (в случае полипептидных цепей), или нуклеотид или цепь нуклеотидов (в случае цепей нуклеотидов), или любой химический компонент (как в случае полипептидных, так и полинуклеотидных цепей). Термин "соединенный" также обозначается дефисом (-).
[072] Используемый в данном документе термин "связанный с" относится к ковалентной или нековалентной связи, образованной между первой цепью аминокислот и второй цепью аминокислот. В некоторых вариантах осуществления термин "связанный с" означает ковалентную непептидную связь или нековалентную связь. Эта связь может быть обозначена двоеточием, т. e. (:). В других вариантах осуществления это означает любую ковалентную связь, за исключением пептидной связи. Например, аминокислота цистеин содержит тиольную группу, которая может образовывать дисульфидную связь или мостик с тиольной группой во втором цистеиновом остатке. В большинстве встречающихся в природе молекул IgG области CH1 и CL связаны дисульфидной связью, а две тяжелые цепи связаны двумя дисульфидными связями в положениях, соответствующих 239 и 242 согласно системе нумерации по Kabat (положения 226 или 229, система нумерации EU). Примеры ковалентных связей включают без ограничения пептидную связь, металлическую связь, водородную связь, дисульфидную связь, сигма-связь, пи-связь, дельта-связь, гликозидную связь, агостическую связь, банановую связь, диполярную связь, обратную донорно-акцепторную Pi-связь, двойную связь, тройную связь, четверную связь, пятерную связь, шестерную связь, конъюгирование, гиперконъюгирование, ароматичность, гаптичность или антисвязывание. Неограничивающие примеры нековалентной связи включают ионную связь (например, катионную пи-связь или солевую связь), металлическую связь, водородную связь (например, диводородную связь, диводородный комплекс, низкобарьерную водородную связь или симметричную водородную связь), силу Ван-дер-Ваальса, лондоновскую дисперсионную силу, механическую связь, галогенную связь, аурофильность, интеркаляцию, стэкинг, энтропийную силу или химическую полярность.
[073] Используемый в данном документе термин "сайт расщепления" или "сайт ферментативного расщепления" относится к сайту, распознаваемому ферментом. Определенные сайты ферментативного расщепления включают сайт внутриклеточного процессинга. В других вариантах осуществления полипептид содержит сайт ферментативного расщепления, расщепляемый ферментом, который активируется во время каскада свертывания крови, так что расщепление таких сайтов происходит в месте образования сгустка. Примеры таких сайтов включают, например, сайты, распознаваемые тромбином, фактором XIa или фактором Xa. Иллюстративные сайты расщепления для FXIa включают, например, TQSFNDFTR (SEQ ID NO: 1) и SVSQTSKLTR (SEQ ID NO: 3). Иллюстративные сайты расщепления для тромбина включают, например, DFLAEGGGVR (SEQ ID NO: 4), TTKIKPR (SEQ ID NO: 5), LVPRG (SEQ ID NO: 6) , ALRPR (SEQ ID NO: 7), ISDKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 106), DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88) и IEPRSFS (SEQ ID NO: 194). Другие сайты ферментативного расщепления известны из уровня техники и описаны в данном документе в другом месте.
[074] Используемый в данном документе термин "сайт процессинга" или "сайт внутриклеточного процессинга" относится к типу сайта ферментативного расщепления в полипептиде, который является мишенью для ферментов, функционирующих после трансляции полипептида. В других вариантах осуществления такие ферменты функционируют во время транспорта из полости аппарата Гольджи в транс-отдел аппарата Гольджи. Ферменты внутриклеточного процессинга расщепляют полипептиды перед секрецией белка из клетки. Примеры таких сайтов процессинга включают, например, сайты, на которые нацеливается семейство PACE/фуриновых (где PACE является аббревиатурой для фермента, расщепляющего белок в месте спаренных основных аминокислот) эндопептидаз. Эти ферменты локализованы на мембране аппарата Гольджи и расщепляют белки с карбоксиконцевой стороны от мотива последовательности Arg-[любой остаток]-(Lys или Arg)-Arg. Используемое в данном документе семейство "фуриновых" ферментов включает, например, PCSK1 (также известный как PC1/PC3), PCSK2 (также известный как PC2), PCSK3 (также известный как фурин или PACE), PCSK4 (также известный как PC4), PCSK5 (также известный как PC5 или PC6), PCSK6 (также известный как PACE4) или PCSK7 (также известный как PC7/LPC, PC8 или SPC7). Другие сайты процессинга известны из уровня техники.
[075] Следует понимать, что в конструкциях, которые содержат более одного сайта процессинга или расщепления, такие сайты могут быть одинаковыми или разными.
[076] "Процессируемый линкер", как используется в данном документе, относится к линкеру, содержащему по меньшей мере один сайт внутриклеточного процессинга, который описан в данном документе в другом месте.
[077] Используемый в данном документе термин "период полужизни" относится к биологическому периоду полужизни конкретного полипептида in vivo. Период полужизни можно выразить в виде времени, требуемого для выведения из кровотока и/или других тканей животного половины количества, введенного субъекту. Если кривую выведения данного полипептида строят в виде функции времени, кривая обычно является двухфазной с быстрой α-фазой и более длинной β-фазой. Обычно α-фаза отображает уравновешивание содержания введенного полипептида Fc между внутри- и внесосудистым пространством и частично определяется размером полипептида. Обычно β-фаза отображает катаболизм полипептида во внутрисосудистом пространстве. В некоторых вариантах осуществления FVIII и химерные полипептиды, содержащие FVIII, являются монофазными и, таким образом, характеризуются отсутствием альфа-фазы и наличием одной только бета-фазы. Следовательно, в определенных вариантах осуществления термин "период полужизни", используемый в данном документе, относится к периоду полужизни полипептида в β-фазе. У человека типичный период полужизни в бета-фазе для антитела человека составляет 21 день. В определенных вариантах осуществления период полужизни выражается как период полужизни в конечной фазе.
[078] Гемостатическое нарушение, как используется в данном документе, означает генетически унаследованное или приобретенное состояние, характеризующееся склонностью к кровоизлиянию, спонтанно либо в результате травмы, из-за нарушенной способности или неспособности образовывать фибриновый сгусток. Примеры таких нарушений включают формы гемофилии. Тремя основными формами являются гемофилия A (дефицит фактора VIII), гемофилия B (дефицит фактора IX или "болезнь Кристмаса") и гемофилия C (дефицит фактора XI, легкая склонность к кровотечению). Другие гемостатические нарушения включают, например, болезнь фон Виллебранда, дефицит фактора XI (дефицит PTA), дефицит фактора XII, дефициты или аномалии структуры фибриногена, протромбина, фактора V, фактора VII, фактора X или фактора XIII, синдром Бернара-Сулье, который представляет собой дефект или дефицит GPIb. GPIb, рецептор VWF, может быть дефектным и приводить к невозможности образования первичного сгустка (первичного гемостаза) и увеличенной склонности к кровотечению, а также к тромбастении Гланцманна-Негели (тромбастении Гланцманна). При печеночной недостаточности (острой и хронической формах) имеет место недостаточная выработка печенью факторов коагуляции; это может увеличивать риск кровотечения.
[079] "Вводить" или "введение", как используется в данном документе, относится к доставке субъекту композиции, описанной в данном документе, например, химерного полипептида. Композицию, например, химерный полипептид, можно вводить субъекту с применением способов, известных из уровня техники. В частности, композицию можно вводить внутривенно, подкожно, внутримышечно, внутрикожно или через любую поверхность слизистой оболочки, например, перорально, сублингвально, трансбуккально, назально, ректально, вагинально или легочным путем - химерный полипептид. В некоторых вариантах осуществления введение представляет собой самостоятельное введение. В некоторых вариантах осуществления родитель вводит химерный полипептид ребенку. В некоторых вариантах осуществления химерный полипептид вводит субъекту медицинский работник, такой как врач, санитар или медсестра.
[080] Используемый в данном документе термин "доза" относится к однократному введению композиции субъекту. Однократная доза может быть введена сразу, например, в виде болюса, или в течение определенного периода времени, например, путем внутривенной инфузии. Термин "несколько доз" означает более одной дозы, например, более чем одно введение.
[081] Если речь идет о совместном введении более чем одной композиции, доза композиции A может вводиться одновременно с дозой композиции B. В качестве альтернативы дозу композиции A можно вводить до или после введения дозы композиции B. В некоторых вариантах осуществления композиция A и композиция B объединены в один состав.
[082] Используемый в данном документе термин "интервал" или "интервал между введениями доз" относится к количеству времени, которое проходит между первой дозой композиции A и последующей дозой той же композиции, вводимой субъекту. Интервал между введениями доз может относиться к времени, которое проходит между первой дозой и второй дозой, или интервал между введениями доз может относиться к количеству времени, которое проходит между несколькими дозами.
[083] Термин "частота введения доз", используемый в данном документе, относится к количеству доз, вводимых в расчете на конкретный интервал между введениями доз. Например, частота введения доз может быть записана как один раз в неделю, один раз в две недели и т. д. Следовательно, интервал между введениями доз, составляющий 7 дней, также может быть записан как интервал между введениями доз один раз в 7 дней, или каждую неделю, или один раз в неделю.
[084] Используемый в данном документе термин "профилактическое лечение" относится к применению терапии для лечения гемофилии, где такое лечение предназначено для предупреждения или снижения тяжести одного или нескольких симптомов гемофилии, например, эпизодов кровотечения, например, одного или нескольких эпизодов спонтанного кровотечения, и/или повреждения суставов. См. Jimenez-Yuste et al., Blood Transfus. 12(3):314-19 (2014). Для предупреждения или снижения тяжести таких симптомов, например, эпизодов кровотечения и прогрессирования заболевания суставов, пациенты с гемофилией А могут получать регулярные инфузии фактора свертывания крови в рамках режима профилактического лечения. Основой такого профилактического лечения является наблюдение, что пациенты с гемофилией с уровнем фактора свертывания крови, например, FVIII, составляющим 1% или больше, редко испытывают эпизоды спонтанного кровотечения и имеют меньше сопутствующих заболеваний, связанных с гемофилией, по сравнению с пациентами с тяжелой формой гемофилии. См., например, Coppola A. et al, Semin. Thromb. Hemost. 38(1): 79-94 (2012) . Практикующие врачи, лечащие таких пациентов с гемофилией, предположили, что поддержание уровней фактора при около 1% с помощью регулярных инфузий потенциально может снизить риск симптомов гемофилии, в том числе эпизодов кровотечения и повреждения суставов. См. там же. Последующие исследования подтвердили эти преимущества у педиатрических пациентов с гемофилией, получающих профилактическое лечение с помощью фактора свертывания крови, что делает профилактическое лечение целью для людей с тяжелой формой гемофилии. См. там же.
[085] "Профилактическое" лечение также может относиться к упреждающему введению композиции, описанной в данном документе, например, химерного полипептида, субъекту с целью контроля частоты возникновения или тяжести одного или нескольких симптомов гемофилии А, например, эпизодов кровотечения, управления ими, их предупреждения или снижения. Профилактическое лечение с помощью фактора свертывания крови, например, FVIII, является стандартом лечения субъектов с тяжелой формой гемофилии A. См., например, Oldenburg, Blood 125:2038-44 (2015). В некоторых вариантах осуществления профилактическое лечение относится к введению композиции, раскрытой в данном документе, субъекту, нуждающемуся в этом, для снижения частоты возникновения одного или нескольких симптомов гемофилии А. Профилактическое лечение может включать введение нескольких доз. Несколько доз, используемых при профилактическом лечении, обычно вводятся с определенными интервалами между введениями доз. В определенных вариантах осуществления частота кровотечений в годовом исчислении может быть снижена до менее 10, менее 9, менее 8, менее 7, менее 6, менее 5, менее 4, менее 3, менее 2 или менее 1.
[086] Термин "лечение по необходимости" или "эпизодическое лечение" относится к введению "по мере необходимости" химерной молекулы в ответ на симптомы гемофилии А, например, эпизод кровотечения, или перед началом деятельности, которая может вызвать кровотечение. В одном аспекте лечение по необходимости может быть назначено субъекту, когда начинается кровотечение, например, после травмы, или когда ожидается кровотечение, например, перед хирургическим вмешательством. В другом аспекте лечение по необходимости может быть назначено перед началом действий, которые увеличивают риск кровотечения, таких как контактные виды спорта. В некоторых вариантах осуществления лечение по необходимости назначается в виде однократной дозы. В других вариантах осуществления лечение по необходимости назначают в виде первой дозы, за которой следуют одна или несколько дополнительных доз. Если химерный полипептид вводят по необходимости, то одну или несколько дополнительных доз можно вводить через по меньшей мере приблизительно 12 часов, по меньшей мере приблизительно 24 часа, по меньшей мере приблизительно 36 часов, по меньшей мере приблизительно 48 часов, по меньшей мере приблизительно 60 часов, по меньшей мере приблизительно 72 часа, по меньшей мере приблизительно 84 часа, по меньшей мере приблизительно 96 часов, по меньшей мере приблизительно 108 часов или по меньшей мере приблизительно 120 часов после первой дозы. Однако следует отметить, что интервал между введениями доз, связанный с лечением по необходимости, не совпадает с интервалом между введениями доз, используемым для профилактического лечения.
[087] В некоторых вариантах осуществления субъект, нуждающийся в общем гемостатическом средстве, переносит или вскоре перенесет хирургическое вмешательство. Химерный полипептид по настоящему изобретению можно вводить до или после хирургического вмешательства. Химерный полипептид по настоящему изобретению можно также вводить во время или после хирургического вмешательства для контроля эпизода острого кровотечения. Если химерный полипептид вводят до хирургического вмешательства, то введение можно выполнять за по меньшей мере приблизительно 1 час, по меньшей мере приблизительно 2 часа, по меньшей мере приблизительно 4 часа, по меньшей мере приблизительно 8 часов, по меньшей мере приблизительно 12 часов, по меньшей мере приблизительно 24 часа, по меньшей мере приблизительно 36 часов, по меньшей мере приблизительно 48 часов или по меньшей мере приблизительно 72 часа до хирургического вмешательства. Если химерный полипептид вводят после хирургического вмешательства, то введение можно выполнять через по меньшей мере приблизительно 1 час, по меньшей мере приблизительно 2 часа, по меньшей мере приблизительно 4 часа, по меньшей мере приблизительно 8 часов, по меньшей мере приблизительно 12 часов, по меньшей мере приблизительно 24 часа, по меньшей мере приблизительно 36 часов, по меньшей мере приблизительно 48 часов или по меньшей мере приблизительно 72 часа после хирургического вмешательства. Хирургическое вмешательство может включать без ограничения трансплантацию печени, резекцию печени, стоматологические процедуры или трансплантацию стволовых клеток.
[088] Используемый в данном документе термин "острое кровотечение" относится к эпизоду кровотечения независимо от первопричины. Например, у субъекта может быть травма, уремия, наследственное нарушение свертываемости крови (например, дефицит фактора VII), тромбоцитарное нарушение или устойчивость вследствие развития антител к факторам свертывания крови.
[089] "Лечить", "лечение", "осуществление лечения", как используется в данном документе, относится, например, к снижению тяжести заболевания или состояния; снижению продолжительности течения заболевания; уменьшению интенсивности одного или нескольких симптомов, ассоциированных с заболеванием или состоянием; обеспечению благоприятных эффектов у субъекта с заболеванием или состоянием, при этом не обязательно с излечением заболевания или состояния, или профилактике одного или нескольких симптомов, ассоциированных с заболеванием или состоянием. В некоторых вариантах осуществления термин "осуществление лечения" или "лечение" означает поддержание у субъекта остаточного уровня FVIII, составляющего по меньшей мере приблизительно 1 МЕ/дл, 2 МЕ/дл, 3 МЕ/дл, 4 МЕ/дл, 5 МЕ/дл, 6 МЕ/дл, 7 МЕ/дл, 8 МЕ/дл, 9 МЕ/дл, 10 МЕ/дл, 11 МЕ/дл, 12 МЕ/дл, 13 МЕ/дл, 14 МЕ/дл, 15 МЕ/дл, 16 МЕ/дл, 17 МЕ/дл, 18 МЕ/дл, 19 МЕ/дл или 20 МЕ/дл, путем введения химерного полипептида или фрагмента VWF по настоящему изобретению. Используемый в данном документе "остаточный уровень" у пациента с гемофилией представляет собой измеренное значение наиболее низкой концентрации, достигаемой с помощью терапии фактором свертывания крови, например, терапии посредством FVIII, перед введением следующей дозы. В других вариантах осуществления осуществление лечения или лечение означает поддержание остаточного уровня FVIII, составляющего по меньшей мере приблизительно 1 МЕ/дл, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание остаточного уровня FVIII, составляющего по меньшей мере приблизительно 3 МЕ/дл, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание остаточного уровня FVIII, составляющего по меньшей мере приблизительно 5 МЕ/дл, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание остаточного уровня FVIII, составляющего от приблизительно 1 до приблизительно 20 МЕ/дл, от приблизительно 2 до приблизительно 20 МЕ/дл, от приблизительно 3 до приблизительно 20 МЕ/дл, от приблизительно 4 до приблизительно 20 МЕ/дл, от приблизительно 5 до приблизительно 20 МЕ/дл, от приблизительно 6 до приблизительно 20 МЕ/дл, от приблизительно 7 до приблизительно 20 МЕ/дл, от приблизительно 8 до приблизительно 20 МЕ/дл, от приблизительно 9 до приблизительно 20 МЕ/дл или от приблизительно 10 до приблизительно 20 МЕ/дл, в течение интервала между введениями доз.
[090] "Лечение" или "осуществление лечения" заболевания или состояния может также включать поддержание активности FVIII у субъекта на уровне, сравнимом с по меньшей мере приблизительно 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19% или 20% активности FVIII у субъекта, не страдающего гемофилией, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 1%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 2%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 3%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 4%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 5%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 6%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 7%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 8%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 9%, в интервале между введениями доз. В других вариантах осуществления осуществление лечения или лечение означает поддержание уровня активности FVIII, составляющего по меньшей мере приблизительно 10%, в интервале между введениями доз. Минимальный остаточный уровень, требуемый для лечения, может быть измерен с помощью одного или нескольких известных способов (например, с помощью анализов aPTT или хромогенных анализов, описанных в данном документе) и может быть скорректирован (увеличен или уменьшен) для каждого человека.
II. Способы по настоящему изобретению
[091] Определенные аспекты настоящего изобретения относятся к способам лечения гемофилии A у субъекта, нуждающегося в этом, предусматривающим введение субъекту химерного полипептида, содержащего белок FVIII и фрагмент VWF, с интервалом между введениями доз. В некоторых вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, например, rFVIIIFc-VWF-XTEN, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз. В других аспектах настоящее изобретение относится к способам лечения заболевания или состояния, связанного с кровотечением, например, гемофилии A, у субъекта, нуждающегося в этом, предусматривающим введение субъекту нескольких доз белка FVIII и нескольких доз фрагмента VWF с интервалом между введениями доз.
[092] В некоторых вариантах осуществления несколько доз предусматривают по меньшей мере две дозы, по меньшей мере три дозы, по меньшей мере четыре дозы, по меньшей мере пять доз, по меньшей мере шесть доз, по меньшей мере семь доз, по меньшей мере восемь доз, по меньшей мере девять доз, по меньшей мере десять доз, по меньшей мере одиннадцать доз, по меньшей мере двенадцать доз, по меньшей мере тринадцать доз, по меньшей мере четырнадцать доз, по меньшей мере пятнадцать доз, по меньшей мере шестнадцать доз, по меньшей мере семнадцать доз, по меньшей мере восемнадцать доз, по меньшей мере девятнадцать доз, по меньшей мере двадцать доз или больше. В некоторых вариантах осуществления несколько доз вводят в течение по меньшей мере приблизительно 1 месяца, по меньшей мере приблизительно 2 месяцев, по меньшей мере приблизительно 3 месяцев, по меньшей мере приблизительно 4 месяцев, по меньшей мере приблизительно 5 месяцев, по меньшей мере приблизительно 6 месяцев, по меньшей мере приблизительно 12 месяцев, по меньшей мере приблизительно 18 месяцев, по меньшей мере приблизительно 2 лет, по меньшей мере приблизительно 3 лет, по меньшей мере приблизительно 4 лет, по меньшей мере приблизительно 5 лет, по меньшей мере приблизительно 10 лет, по меньшей мере приблизительно 15 лет, по меньшей мере приблизительно 20 лет или в течение по меньшей мере приблизительно 25 лет.
[093] В определенных вариантах осуществления способы по настоящему изобретению относятся к лечению гемофилии А. В некоторых вариантах осуществления лечение гемофилии А включает предупреждение эпизода кровотечения у субъекта-человека, нуждающегося в этом. В некоторых вариантах осуществления лечение гемофилии А предусматривает лечение эпизода кровотечения у субъекта-человека, нуждающегося в этом. В некоторых вариантах осуществления лечение гемофилии А предусматривает контроль встречаемости или частоты эпизодов кровотечения у субъекта-человека, нуждающегося в этом. В некоторых вариантах осуществления лечение гемофилии А предусматривает уменьшение встречаемости или частоты эпизодов кровотечения у субъекта-человека, нуждающегося в этом.
A. Дозы
[094] В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят в виде однократной дозы или в виде нескольких доз. В некоторых вариантах осуществления количества каждой из нескольких доз являются одинаковыми. В других вариантах осуществления одна или несколько из нескольких доз отличаются от одной или нескольких из других нескольких доз. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 5 МЕ/кг до приблизительно 200 МЕ/кг или от приблизительно 10 МЕ/кг до приблизительно 150 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 20 МЕ/кг до приблизительно 95 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 90 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 85 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 75 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 70 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 65 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 60 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 55 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 50 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 45 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 40 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 35 МЕ/кг, от приблизительно 20 МЕ/кг до приблизительно 30 МЕ/кг или от приблизительно 20 МЕ/кг до приблизительно 25 МЕ/кг.
[095] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 20 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 25 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 30 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 35 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 40 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 45 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 55 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 70 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 75 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 80 МЕ/кг до приблизительно 100 МЕ/кг, от приблизительно 85 МЕ/кг до приблизительно 100 МЕ/кг или от приблизительно 90 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг.
[096] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 50 МЕ/кг до приблизительно 150 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 140 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 130 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 120 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 110 МЕ/кг или от приблизительно 50 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 50 МЕ/кг до приблизительно 95 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 90 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 85 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 75 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 70 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 60 МЕ/кг, от приблизительно 50 МЕ/кг до приблизительно 55 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг.
[097] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 60 МЕ/кг до приблизительно 150 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 140 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 130 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 120 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 110 МЕ/кг или от приблизительно 60 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 60 МЕ/кг до приблизительно 95 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 90 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 85 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 75 МЕ/кг, от приблизительно 60 МЕ/кг до приблизительно 70 МЕ/кг или от приблизительно 60 МЕ/кг до приблизительно 65 МЕ/кг.
[098] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 65 МЕ/кг до приблизительно 150 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 140 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 130 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 120 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 110 МЕ/кг или от приблизительно 65 МЕ/кг до приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 65 МЕ/кг до приблизительно 95 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 90 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 85 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг, от приблизительно 65 МЕ/кг до приблизительно 75 МЕ/кг или от приблизительно 65 МЕ/кг до приблизительно 70 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз химерного полипептида составляет от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг.
[099] В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 5 МЕ/кг, приблизительно 10 МЕ/кг, приблизительно 15 МЕ/кг, приблизительно 20 МЕ/кг, приблизительно 25 МЕ/кг, приблизительно 30 МЕ/кг, приблизительно 35 МЕ/кг, приблизительно 40 МЕ/кг, приблизительно 45 МЕ/кг, приблизительно 50 МЕ/кг, приблизительно 55 МЕ/кг, приблизительно 60 МЕ/кг, приблизительно 65 МЕ/кг, приблизительно 70 МЕ/кг, приблизительно 75 МЕ/кг, приблизительно 80 МЕ/кг, приблизительно 85 МЕ/кг, приблизительно 90 МЕ/кг, приблизительно 95 МЕ/кг, приблизительно 100 МЕ/кг, приблизительно 125 МЕ/кг, приблизительно 150 МЕ/кг, приблизительно 175 МЕ/кг или приблизительно 200 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 30 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 35 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 40 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 45 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 55 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 60 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 70 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 75 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 85 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 90 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 95 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 100 МЕ/кг.
[0100] В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 25 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 30 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 35 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 40 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 45 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 50 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 55 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 60 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 65 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 70 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 75 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 80 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 85 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 90 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 95 МЕ/кг. В некоторых вариантах осуществления каждая доза из нескольких доз составляет приблизительно 100 МЕ/кг.
[0101] В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят в целях профилактики. При профилактическом введении по меньшей мере одна из нескольких доз может составлять от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг. В определенных вариантах осуществления по меньшей мере одна из нескольких доз, вводимых в целях профилактики, составляет приблизительно 25 МЕ/кг, приблизительно 30 МЕ/кг, приблизительно 35 МЕ/кг, приблизительно 40 МЕ/кг, приблизительно 45 МЕ/кг, приблизительно 50 МЕ/кг, приблизительно 55 МЕ/кг, приблизительно 60 МЕ/кг, приблизительно 65 МЕ/кг, приблизительно 70 МЕ/кг, приблизительно 75 МЕ/кг, приблизительно 80 МЕ/кг, приблизительно 85 МЕ/кг, приблизительно 90 МЕ/кг, приблизительно 95 МЕ/кг или приблизительно 100 МЕ/кг. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз, вводимых в целях профилактики, составляет приблизительно 25 МЕ/кг. В других вариантах осуществления по меньшей мере одна из нескольких доз, вводимых в целях профилактики, составляет приблизительно 50 МЕ/кг. В других вариантах осуществления по меньшей мере одна из нескольких доз, вводимых в целях профилактики, составляет приблизительно 65 МЕ/кг. В других вариантах осуществления по меньшей мере одна из нескольких доз, вводимых в целях профилактики, составляет приблизительно 80 МЕ/кг.
[0102] В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят по необходимости. При введении по необходимости химерный полипептид можно вводить в виде однократной дозы или в виде нескольких доз. В некоторых вариантах осуществления химерный полипептид вводят в виде одной или нескольких доз, составляющих от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг. В определенных вариантах осуществления химерный полипептид вводят по необходимости в виде одной или нескольких доз, составляющих приблизительно 25 МЕ/кг, приблизительно 30 МЕ/кг, приблизительно 35 МЕ/кг, приблизительно 40 МЕ/кг, приблизительно 45 МЕ/кг, приблизительно 50 МЕ/кг, приблизительно 55 МЕ/кг, приблизительно 60 МЕ/кг, приблизительно 65 МЕ/кг, приблизительно 70 МЕ/кг, приблизительно 75 МЕ/кг, приблизительно 80 МЕ/кг, приблизительно 85 МЕ/кг, приблизительно 90 МЕ/кг, приблизительно 95 МЕ/кг или приблизительно 100 МЕ/кг. В некоторых вариантах осуществления химерный полипептид вводят по необходимости в виде одной или нескольких доз, составляющих приблизительно 25 МЕ/кг. В других вариантах осуществления химерный полипептид вводят по необходимости в виде одной или нескольких доз, составляющих приблизительно 50 МЕ/кг. В других вариантах осуществления химерный полипептид вводят по необходимости в виде одной или нескольких доз, составляющих приблизительно 65 МЕ/кг. В других вариантах осуществления химерный полипептид вводят по необходимости в виде одной или нескольких доз, составляющих приблизительно 80 МЕ/кг.
[0103] В некоторых вариантах осуществления введение химерного полипептида в соответствии со способами по настоящему изобретению не приводит к индукции образования ингибитора FVIII после приблизительно 7 дней, приблизительно 10 дней, приблизительно 11 дней, приблизительно 12 дней, приблизительно 13 дней, приблизительно 14 дней, приблизительно 15 дней, приблизительно 20 дней, приблизительно 24 дней, приблизительно 25 дней, приблизительно 28 дней, приблизительно 30 дней или приблизительно 35 дней введения. В некоторых вариантах осуществления введение химерного полипептида не приводит к индукции образования ингибитора FVIII после приблизительно 28 дней введения.
B. Интервал между введениями доз
[0104] В определенных вариантах осуществления, в частности для профилактического лечения, химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят в виде нескольких доз с интервалом между введениями доз. В некоторых вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 3 дня, по меньшей мере приблизительно 4 дня, по меньшей мере приблизительно 5 дней, по меньшей мере приблизительно 6 дней, по меньшей мере приблизительно 7 дней, по меньшей мере приблизительно 8 дней, по меньшей мере приблизительно 9 дней, по меньшей мере приблизительно 10 дней, по меньшей мере приблизительно 11 дней, по меньшей мере приблизительно 12 дней, по меньшей мере приблизительно 13 дней, по меньшей мере приблизительно 14 дней, по меньшей мере приблизительно 15 дней, по меньшей мере приблизительно 16 дней, по меньшей мере приблизительно 17 дней, по меньшей мере приблизительно 18 дней, по меньшей мере приблизительно 19 дней, по меньшей мере приблизительно 20 дней, по меньшей мере приблизительно 21 день, по меньшей мере приблизительно 22 дня, по меньшей мере приблизительно 23 дня, по меньшей мере приблизительно 24 дня, по меньшей мере приблизительно 25 дней, по меньшей мере приблизительно 26 дней, по меньшей мере приблизительно 27 дней, по меньшей мере приблизительно 28 дней, по меньшей мере приблизительно 29 дней, по меньшей мере приблизительно 30 дней или по меньшей мере приблизительно 31 день.
[0105] В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 6 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 21 день. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 27 дней. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 30 дней.
[0106] В некоторых вариантах осуществления частота введения доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере два раза в неделю, по меньшей мере один раз в неделю, по меньшей мере один раз в 2 недели, по меньшей мере один раз в 3 недели, по меньшей мере один раз в 4 недели, по меньшей мере один раз в 5 недель или по меньшей мере один раз в 6 недель. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере одну неделю. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере 2 недели. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере 3 недели. В определенных вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере 4 недели.
[0107] В некоторых вариантах осуществления частота введения доз, например, для профилактического лечения гемофилии A, составляет приблизительно 2 раза в три месяца, приблизительно 1 раз в месяц, приблизительно 2 раза в месяц, приблизительно 3 раза в месяц, приблизительно 4 раза в месяц, приблизительно 5 раз в месяц, приблизительно 6 раз в месяц, приблизительно 7 раз в месяц или приблизительно 8 раз в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 1 раз в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 2 раза в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 3 раза в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 4 раза в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 5 раз в месяц. В определенных вариантах осуществления частота введения доз составляет приблизительно 6 раз в месяц.
[0108] В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 3-5 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 4-6 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 4-7 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 4-8 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 4-9 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 4-10 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 5-7 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 5-8 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 5-9 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 5-10 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления интервал между введениями доз, например, для профилактического лечения гемофилии A, составляет по меньшей мере приблизительно 14-21 день.
[0109] В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в соответствии с дозой и частотой введения доз, описанными в таблице 1.
Таблица 1A. Количества дозы и соответствующая частота введения доз, например, для профилактического лечения гемофилии
Таблица 1B. Количества дозы и соответствующие интервалы между введениями доз, например, для профилактического лечения гемофилии
[0110] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0111] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 25 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0112] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 50 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0113] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 65 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0114] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе, составляющей приблизительно 80 МЕ/кг, с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0115] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0116] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 50 МЕ/кг до приблизительно 65 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0117] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0118] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-8 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-9 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-10 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-11 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-12 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-13 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8-14 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10-21 день. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13-15 дней. В некоторых вариантах осуществления по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14-21 день.
[0119] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 7 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 6 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 5 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 4 дня. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 3 дня. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 2 дня. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 8 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 9 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 10 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 11 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 13 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 14 дней. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 21 день.
[0120] В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с интервалом между введениями доз, составляющим по меньшей мере приблизительно 1 неделю. В некоторых вариантах осуществления, как, например, для профилактического лечения гемофилии A у педиатрического субъекта, по меньшей мере одну из нескольких доз вводят в дозе от приблизительно 65 МЕ/кг до приблизительно 80 МЕ/кг с частотой введения доз, составляющей по меньшей мере приблизительно два раза в 1 неделю.
[0121] В определенных вариантах осуществления, как, например, при профилактическом лечении гемофилии A, способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, например, rFVIIIFc-VWF-XTEN, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления, как, например, при профилактическом лечении гемофилии A, способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет от приблизительно 15 МЕ/кг до приблизительно 100 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 15 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 16 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 17 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 18 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 19 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 20 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг до приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 21 день.
[0122] В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, например, rFVIIIFc-VWF-XTEN, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 6 дней. В некоторых вариантах осуществления способ предназначен для профилактического лечения гемофилии А. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 15 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 16 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 17 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 18 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 19 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 20 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 25 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 21 день.
[0123] В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, например, rFVIIIFc-VWF-XTEN, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 6 дней. В некоторых вариантах осуществления способ предназначен для профилактического лечения гемофилии А. В некоторых вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 15 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 16 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 17 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 18 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 19 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 20 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 50 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 21 день.
[0124] В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, например, rFVIIIFc-VWF-XTEN, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 6 дней. В некоторых вариантах осуществления способ предназначен для профилактического лечения гемофилии А. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 15 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 16 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 17 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 18 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 19 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 20 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 65 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 21 день.
[0125] В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, например, rFVIIIFc-VWF-XTEN, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 5 дней. В определенных вариантах осуществления способ предусматривает введение субъекту нескольких доз химерного полипептида, содержащего (i) белок FVIII и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, с интервалом между введениями доз, где по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 6 дней. В некоторых вариантах осуществления способ предназначен для профилактического лечения гемофилии А. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 7 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 8 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 9 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 10 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 11 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 12 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 13 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 14 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 15 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 16 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 17 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 18 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 19 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 20 дней. В определенных вариантах осуществления по меньшей мере одна из нескольких доз составляет от приблизительно 80 МЕ/кг, а интервал между введениями доз составляет по меньшей мере приблизительно 21 день.
[0126] В некоторых вариантах осуществления химерный полипептид вводят для профилактического лечения гемофилии A. Профилактическое лечение гемофилии A включает облегчение или снижение тяжести симптомов гемофилии A на непрерывной или почти непрерывной основе. В некоторых вариантах осуществления профилактическое лечение проводят до возникновения симптома гемофилии А, например, до случая кровотечения. В других вариантах осуществления профилактическое лечение проводят на регулярной основе, например, с интервалом между введениями доз, описанным в данном документе, для предупреждения появления или уменьшения тяжести симптома до появления симптома. Любой интервал между введениями доз, раскрытый в данном документе, можно использовать для профилактического лечения гемофилии А.
[0127] В других вариантах осуществления химерный полипептид вводят перед началом деятельности, которая может вызвать один или несколько симптомов гемофилии A, например, в качестве лечения по необходимости. Например, химерный полипептид по настоящему изобретению можно вводить субъекту, страдающему гемофилией A, до того, как субъект перенесет хирургическое вмешательство или начнет деятельность, которая в ином случае увеличивает риск физической травмы и/или случая кровотечения. При введении по необходимости химерный полипептид можно вводить в виде однократной дозы или в виде нескольких доз. В некоторых вариантах осуществления лечение по необходимости включает введение нескольких доз химерного полипептида с интервалом между введениями доз, составляющим по меньшей мере приблизительно 12 часов, по меньшей мере приблизительно 24 часа, по меньшей мере приблизительно 36 часов, по меньшей мере приблизительно 48 часов, по меньшей мере приблизительно 60 часов, по меньшей мере приблизительно 72 часа, по меньшей мере приблизительно 84 часа, по меньшей мере приблизительно 96 часов, по меньшей мере приблизительно 108 часов или по меньшей мере приблизительно 120 часов. В определенных вариантах осуществления лечение по необходимости включает введение по меньшей мере 2 доз, по меньшей мере 3 доз, по меньшей мере 4 доз или по меньшей мере 5 доз химерного полипептида.
[0128] В некоторых вариантах осуществления субъект ранее получал лечение с помощью одного или нескольких средств заместительной терапии на основе FVIII. В определенных вариантах осуществления субъект не отвечал на предшествующую заместительную терапию с помощью FVIII. В некоторых вариантах осуществления средство заместительной терапии на основе FVIII представляет собой ELOCTATE® или ADVATE®. В некоторых вариантах осуществления субъект является взрослым. В некоторых вариантах осуществления субъект является взрослым мужчиной. В других вариантах осуществления субъект является взрослой женщиной. В других вариантах осуществления субъект является ребенком, например, в возрасте, равном приблизительно 12 лет или меньше, в возрасте, равном приблизительно 11 лет или меньше, в возрасте, равном приблизительно 10 лет или меньше, в возрасте, равном приблизительно 9 лет или меньше, в возрасте, равном приблизительно 8 лет или меньше, в возрасте, равном приблизительно 7 лет или меньше, в возрасте, равном приблизительно 6 лет или меньше, в возрасте, равном приблизительно 5 лет или меньше, в возрасте, равном приблизительно 4 года или меньше, в возрасте, равном приблизительно 3 года или меньше, в возрасте, равном приблизительно 2 года или меньше, или в возрасте, равном приблизительно 1 год или меньше. В некоторых вариантах осуществления субъект является женщиной. В некоторых вариантах осуществления субъект является мужчиной. В некоторых вариантах осуществления субъект является женщиной в возрасте, равном приблизительно 12 лет или меньше. В некоторых вариантах осуществления субъект является женщиной в возрасте, равном приблизительно 11 лет или меньше. В некоторых вариантах осуществления субъект является женщиной в возрасте, равном приблизительно 10 лет или меньше.
[0129] Химерный полипептид, описанный в данном документе, можно вводить любыми способами, известными из уровня техники. В некоторых вариантах осуществления химерный полипептид вводят посредством пути, выбранного из группы, состоящей из внутривенной инъекции, внутривенной инфузии, подкожного введения, внутримышечного введения, перорального введения, назального введения и легочного введения. В некоторых вариантах осуществления химерный полипептид вводят внутривенно. В других вариантах осуществления химерный полипептид вводят подкожно.
[0130] В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, после введения приводит к уровню активности FVIII в плазме крови, составляющему по меньшей мере приблизительно 1%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 9% или по меньшей мере приблизительно 10%. В определенных вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 3%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 4%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 4%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5,6%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 7%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 10%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 12%. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 12,95%. Используемый в данном документе уровень активности в плазме крови выражается в процентах (%). В качестве альтернативы активность в плазме крови может быть выражена в единицах МЕ/дл, где 1% равен 1 МЕ/дл.
[0131] В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 10% через по меньшей мере приблизительно 5 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 12% через по меньшей мере приблизительно 5 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 12,95% через по меньшей мере приблизительно 5 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5% через по меньшей мере приблизительно 7 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 5,6% через по меньшей мере приблизительно 7 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 3% через по меньшей мере приблизительно 8 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг. В некоторых вариантах осуществления уровень активности FVIII в плазме крови составляет по меньшей мере приблизительно 1% через по меньшей мере приблизительно 10 дней после введения химерного полипептида, например, в дозе 25 МЕ/кг.
[0132] Настоящее изобретение также включает способ лечения гемофилии A, предусматривающий введение субъекту композиции, содержащей (i) белок FVIII и (ii) фрагмент VWF. В некоторых вариантах осуществления белок FVIII присутствует в первом составе, а фрагмент VWF присутствует во втором составе. В других вариантах осуществления белок FVIII и фрагмент VWF присутствуют в одном и том же составе. В конкретных вариантах осуществления белок FVIII и фрагмент VWF являются отдельными компонентами одного состава. В определенных вариантах осуществления белок FVIII, например, одноцепочечный FVIII, FVIII-Fc, пегилированный FVIII, полноразмерный зрелый FVIII или FVIII с делецией домена B, вводят в дозе, раскрытой в данном документе, например, составляющей 25 МЕ/кг, 30 МЕ/кг, 35 МЕ/кг, 40 МЕ/кг, 45 МЕ/кг, 50 МЕ/кг, 55 МЕ/кг, 60 МЕ/кг или 65 МЕ/кг, и с интервалом между введениями доз, составляющим по меньшей мере 6 дней, по меньшей мере 7 дней, по меньшей мере 8 дней, по меньшей мере 9 дней, по меньшей мере 10 дней, по меньшей мере 11 дней, по меньшей мере 12 дней, по меньшей мере 13 дней или по меньшей мере 14 дней, а фрагмент VWF вводят в дозе, соответствующей соотношению вводимых фрагмента VWF и молекулы FVIII, составляющему по меньшей мере приблизительно 0,25:1, 0,5:1, 1:1, 2:1, 3:1, 4:1, 5:1, 10:1, 15:1, 20:1, 25:1, 30:1, 35:1, 40:1, 45:1, 50:1, 100:1, 200:1, 300:1, 400:1, 500:1, 1000:1, 2000:1, 3000:1, 4000:1, 5000:1, 10000:1, 50000:1, 100000:1 или 500000:1. В других вариантах осуществления фрагмент VWF вводят субъекту одновременно с белком FVIII, сразу после него или непосредственно перед ним. В определенных вариантах осуществления фрагмент VWF вводят за приблизительно 0,5 часа, приблизительно 1 час, приблизительно 1,5 часа, приблизительно 2 часа, приблизительно 2,5 часа, приблизительно 3 часа, приблизительно 3,5 часа, приблизительно 4 часа, приблизительно 4,5 часа, приблизительно 5 часов, приблизительно 5,5 часа или приблизительно 6 часов до белка FVIII. В других вариантах осуществления белок FVIII вводят за приблизительно 0,5 часа, приблизительно 1 час, приблизительно 1,5 часа, приблизительно 2 часа, приблизительно 2,5 часа, приблизительно 3 часа, приблизительно 3,5 часа, приблизительно 4 часа, приблизительно 4,5 часа, приблизительно 5 часов, приблизительно 5,5 часа или приблизительно 6 часов до фрагмента VWF.
C. Индукция иммунной толерантности
[0133] Определенные аспекты настоящего изобретения относятся к способам лечения гемофилии A у субъекта, нуждающегося в этом, предусматривающим введение субъекту химерного полипептида, содержащего белок FVIII и фрагмент VWF, с интервалом между введениями доз, где химерный полипептид индуцирует иммунную толерантность к FVIII у субъекта. Некоторые аспекты настоящего изобретения относятся к способам лечения гемофилии A у субъекта, нуждающегося в этом, предусматривающим введение субъекту химерного полипептида, содержащего белок FVIII и фрагмент VWF, с интервалом между введениями доз, где химерный полипептид снижает ингибирующий ответ на FVIII у субъекта. Некоторые аспекты настоящего изобретения относятся к способам лечения гемофилии A у субъекта, нуждающегося в этом, предусматривающим введение субъекту химерного полипептида, содержащего белок FVIII и фрагмент VWF, с интервалом между введениями доз, где химерный полипептид не индуцирует иммунный ответ на химерный полипептид после введения. Другой аспект настоящего изобретения относится к способу индукции иммунной толерантности у человека с гемофилией, предусматривающему (1) введение человеку эффективного количества химерного полипептида, описанного в данном документе, например, rFVIIIFc-VWF-XTEN, где эффективное количество химерного полипептида индуцирует иммунную толерантность у человека. Различные способы индукции иммунной толерантности с применением химерных полипептидов, содержащих FVIII и Fc, раскрыты в международной публикации № WO 2018/102760 A1, которая включена в данный документ посредством ссылки во всей своей полноте.
[0134] В определенных вариантах осуществления способ дополнительно предусматривает (2) после индукции иммунной толерантности применение в отношении человека режима с постепенным снижением дозы композиции или химерного полипептида. В определенных вариантах осуществления индукция иммунной толерантности происходит, если титр ингибирующих антител у человека составляет менее чем приблизительно 0,6 БЕ. В определенных вариантах осуществления индукция иммунной толерантности происходит, если титр ингибирующих антител у человека составляет менее чем приблизительно 0,6 БЕ и имеет место 60% восстановление активности фактора свертывания крови, отслеживаемое в плазме крови. В некоторых вариантах осуществления настоящего изобретения способ дополнительно включает (3) после применения режима с постепенным снижением дозы введение человеку профилактической дозы фактора свертывания крови, например, химерного полипептида, описанного в данного документе, например, rFVIIIFc-VWF-XTEN.
[0135] В определенных аспектах человек не получал лечение с помощью предшествующей терапии для формирования иммунной толерантности к фактору свертывания крови, например, к FVIII. Химерный полипептид, например, rFVIIIFc-VWF-XTEN, можно вводить человеку в любое время после того, как было определено, что у человека развился ингибирующий иммунный ответ, например, после измерения уровня ингибирующего иммунного ответа у человека. В других вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, можно вводить человеку, у которого еще не развились один или несколько ингибирующих иммунных ответов, для предупреждения развития ингибирующего иммунного ответа. В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку, у которого имеется высокая вероятность развития ингибирующего иммунного ответа (например, семейный анамнез, генетическая предрасположенность или наличие биомаркера). В некоторых вариантах осуществления способ дополнительно предусматривает измерение уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа перед введением.
[0136] В некоторых вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку через менее чем приблизительно 1 день, менее чем приблизительно 2 дня, менее чем приблизительно 3 дня, менее чем приблизительно 4 дня, менее чем приблизительно 5 дней, менее чем приблизительно 6 дней, менее чем приблизительно 7 дней, менее чем приблизительно 2 недели, менее чем приблизительно 3 недели, менее чем приблизительно 4 недели, менее чем приблизительно 2 месяца, менее чем приблизительно 3 месяца, менее чем приблизительно 4 месяца, менее чем приблизительно 5 месяцев, менее чем приблизительно 6 месяцев, менее чем приблизительно 1 год, менее чем приблизительно 2 года, менее чем приблизительно 3 года, менее чем приблизительно 4 года или менее чем приблизительно 5 лет после определения того, что у человека развился ингибирующий иммунный ответ или что у человека имеется вероятность развития ингибирующего иммунного ответа, например, после измерения уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа у человека. В определенных вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку непосредственно после определения того, что у человека развился ингибирующий иммунный ответ или что у человека имеется вероятность развития ингибирующего иммунного ответа, например, после измерения уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа у человека. В конкретных вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку через менее чем приблизительно 5 минут, менее чем приблизительно 10 минут, менее чем приблизительно 15 минут, менее чем приблизительно 20 минут, менее чем приблизительно 30 минут, менее чем приблизительно 45 минут, менее чем приблизительно 1 час, менее чем приблизительно 2 часа, менее чем приблизительно 3 часа, менее чем приблизительно 4 часа, менее чем приблизительно 5 часов, менее чем приблизительно 6 часов, менее чем приблизительно 7 часов, менее чем приблизительно 8 часов, менее чем приблизительно 9 часов, менее чем приблизительно 10 часов, менее чем приблизительно 11 часов, менее чем приблизительно 12 часов, приблизительно 18 часов или менее чем приблизительно 24 часа после определения того, что у человека развился ингибирующий иммунный ответ или что у человека имеется вероятность развития ингибирующего иммунного ответа, например, после измерения уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа у человека. В конкретных вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку через приблизительно 5 минут, приблизительно 10 минут, приблизительно 15 минут, приблизительно 20 минут, приблизительно 30 минут, приблизительно 45 минут, приблизительно 1 час, приблизительно 2 часа, приблизительно 3 часа, приблизительно 4 часа, приблизительно 5 часов, приблизительно 6 часов, приблизительно 7 часов, приблизительно 8 часов, приблизительно 9 часов, приблизительно 10 часов, приблизительно 11 часов, приблизительно 12 часов, приблизительно 18 часов или приблизительно 24 часа после определения того, что у человека развился ингибирующий иммунный ответ или что у человека имеется вероятность развития ингибирующего иммунного ответа, например, после измерения уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа у человека. В определенных вариантах осуществления химерный полипептид, например, rFVIIIFc-VWF-XTEN, вводят человеку через менее чем приблизительно 1 день после определения того, что у человека развился ингибирующий иммунный ответ или что у человека имеется вероятность развития ингибирующего иммунного ответа, например, после измерения уровня ингибирующего иммунного ответа или вероятности развития ингибирующего иммунного ответа у человека.
[0137] Индукцию иммунного ответа можно продолжать до тех пор, пока уровень ингибитора не станет ниже определенного уровня или пока ингибиторы не будут выявляться. В определенных вариантах осуществления период индукции может продолжаться в течение по меньшей мере приблизительно 24 недель, по меньшей мере приблизительно 26 недель, по меньшей мере приблизительно 28 недель, по меньшей мере приблизительно 30 недель, по меньшей мере приблизительно 32 недель, по меньшей мере приблизительно 34 недель, по меньшей мере приблизительно 36 недель, по меньшей мере приблизительно 38 недель, по меньшей мере приблизительно 40 недель, по меньшей мере приблизительно 42 недель, по меньшей мере приблизительно 44 недель, по меньшей мере приблизительно 46 недель, по меньшей мере приблизительно 48 недель, по меньшей мере приблизительно 50 недель, по меньшей мере приблизительно 52 недель, по меньшей мере приблизительно 54 недель, по меньшей мере приблизительно 56 недель, по меньшей мере приблизительно 58 недель, по меньшей мере приблизительно 60 недель, по меньшей мере приблизительно 62 недель, по меньшей мере приблизительно 64 недель, по меньшей мере приблизительно 66 недель, по меньшей мере приблизительно 68 недель, по меньшей мере приблизительно 70 недель. В конкретном варианте осуществления период индукции составляет менее 60 недель.
[0138] Ингибирующий иммунный ответ, лечение которого осуществляют с помощью способов по настоящему изобретению, может включать любой ответ в организме человека, который отрицательно влияет на один или несколько эффектов лечения с помощью фактора свертывания крови. В некоторых вариантах осуществления ингибирующий иммунный ответ включает выработку ингибирующих антител к фактору свертывания крови, например, ингибирующих антител к FVIII. В определенных вариантах осуществления способ по настоящему изобретению дополнительно включает измерение титра одного или нескольких ингибирующих антител у человека до (например, на исходном уровне) и после введения эффективного количества химерного полипептида, описанного в данном документе, например, rFVIIIFc-VWF-XTEN, или полинуклеотида, кодирующего его. В некоторых вариантах осуществления титр ингибирующих антител перед введением (например, на исходном уровне) составляет по меньшей мере приблизительно 0,6 единицы Бетезда (БЕ). В определенных вариантах осуществления титр ингибирующих антител перед введением (например, на исходном уровне) составляет по меньшей мере приблизительно 1 БЕ, по меньшей мере приблизительно 2 БЕ, по меньшей мере приблизительно 3 БЕ, по меньшей мере приблизительно 4 БЕ, по меньшей мере приблизительно 5 БЕ, по меньшей мере приблизительно 6 БЕ, по меньшей мере приблизительно 7 БЕ, по меньшей мере приблизительно 10 БЕ, по меньшей мере приблизительно 20 БЕ, по меньшей мере приблизительно 30 БЕ, по меньшей мере приблизительно 40 БЕ, по меньшей мере приблизительно 50 БЕ, по меньшей мере приблизительно 100 БЕ, по меньшей мере приблизительно 150 БЕ или по меньшей мере приблизительно 200 БЕ. В одном конкретном варианте осуществления титр ингибирующих антител перед введением (например, на исходном уровне) составляет по меньшей мере приблизительно 5 БЕ.
[0139] В некоторых вариантах осуществления способы по настоящему изобретению обеспечивают снижение титра ингибирующих антител у субъекта-человека по сравнению с титром ингибирующих антител перед введением. В определенных вариантах осуществления титр ингибирующих антител после введения составляет менее чем приблизительно 0,6 БЕ. В некоторых вариантах осуществления титр ингибирующих антител после введения составляет менее чем приблизительно 0,5 БЕ, менее чем приблизительно 0,4 БЕ, менее чем приблизительно 0,3 БЕ, менее чем приблизительно 0,2 БЕ или менее чем приблизительно 0,1 БЕ. В одном конкретном варианте осуществления титр ингибирующих антител после введения составляет 0 БЕ. В других вариантах осуществления титр ингибирующих антител после введения составляет менее 5 БЕ, менее 4 БЕ, менее 3 БЕ, менее 2 БЕ, менее 1 БЕ, менее 0,9 БЕ, менее 0,8 БЕ, менее 0,7 БЕ или менее 0,6 БЕ.
[0140] В некоторых вариантах осуществления введение химерного полипептида, описанного в данном документе, например, rFVIIIFc-VWF-XTEN, обеспечивает увеличение интенсивности дифференцировки макрофагов у человека в направлении М2-подобного фенотипа по сравнению с дифференцировкой макрофагов у не получавших лечение контрольных лиц и у людей, получавших лечение с помощью фактора свертывания крови в отдельности. В некоторых вариантах осуществления M2-подобный фенотип предусматривает повышение экспрессии генов сигнального пути NRF2, сигнального пути PPAR-гамма или как сигнального пути NRF2, так и сигнального пути PPAR-гамма. В некоторых вариантах осуществления M2-подобный фенотип предусматривает повышение экспрессии CD206 (MRC1). В некоторых вариантах осуществления M2-подобный фенотип предусматривает повышение экспрессии ARG1. В некоторых вариантах осуществления M2-подобный фенотип предусматривает повышение экспрессии CD206 (MRC1) и ARG1.
[0141] В некоторых вариантах осуществления введение химерного полипептида, описанного в данном документе, например, rFVIIIFc-VWF-XTEN, приводит к большей экспрессии одного или нескольких генов у человека по сравнению с экспрессией одного или нескольких генов у не получавшего лечение субъекта или у субъекта, получавшего лечение с помощью фактора свертывания крови в отдельности. В некоторых вариантах осуществления введение приводит к большей экспрессии одного или нескольких генов, выбранных из группы, состоящей из генов Hmox1, PPAR-гамма, LPL, EGR2, SLCO4A1, гемоксигеназы 1 (HO-1), индуцируемого окислительным стрессом ингибитора роста 1 (OSGIN1), супероксиддисмутазы 1 (SOD1), глутатиондисульфидредуктазы (GSR), каталитической субъединицы глутаматцистеинлигазы (GCLC), модифицирующей субъединицы глутаматцистеинлигазы (GCLM), NAD(P)H-хинондегидрогеназы 1 (NQO1), белка 5, связывающего жирные кислоты (FABP5), B7-H3 (CD276), представителя 3 семейства SLAM (SLAMF3; лимфоцитарный антиген 9; LY9), представителя 7 семейства SLAM (SLAMF7), маннозного рецептора C-типа 1 (MRC1), представителя 4 семейства 12 переносчиков растворенных веществ (SLC12A), нейропилина 1 (NRP1) и любой их комбинации. В некоторых вариантах осуществления введение приводит к большей экспрессии одного или нескольких генов сигнального пути NRF2. В определенных вариантах осуществления один или несколько генов сигнального пути NRF2 выбраны из группы, состоящей из генов HO-1, OSGIN1, SOD1, GSR, GCLC, GCLM, NQO1 и любой их комбинации. В некоторых вариантах осуществления введение приводит к большей экспрессии одного или нескольких генов сигнального пути PPAR-гамма. В некоторых вариантах осуществления один или несколько генов сигнального пути PPAR-гамма выбраны из группы, состоящей из генов PPAR-гамма, LPL, FABP5, EGR2 и любой их комбинации. В некоторых вариантах осуществления введение приводит к большей экспрессии одного или нескольких генов, выбранных из группы, состоящей из генов B7-H3 (CD276), SLAMF3, SLAMF7, MRC1, SLC12A, NRP1 и любой их комбинации. В конкретных вариантах осуществления введение приводит к большей экспрессии одного или нескольких генов по сравнению с экспрессией одного или нескольких генов у не получавшего лечение человека или у человека, которому вводили фактор свертывания крови в отдельности, где экспрессия является в по меньшей мере приблизительно 1,5 раза большей, в по меньшей мере приблизительно 2 раза большей, в по меньшей мере приблизительно 2,5 раза большей, в по меньшей мере приблизительно 3 раза большей, в по меньшей мере приблизительно 3,5 раза большей, в по меньшей мере приблизительно 4 раза большей, в по меньшей мере приблизительно 4,5 раза большей или в по меньшей мере приблизительно 5 раз большей.
[0142] В некоторых вариантах осуществления дифференциальная экспрессия одного или нескольких генов наблюдается через менее чем 6 часов после введения химерного полипептида, описанного в данном документе, например, rFVIIIFc-VWF-XTEN. В некоторых вариантах осуществления дифференциальная экспрессия наблюдается через менее чем 12 часов после введения. В некоторых вариантах осуществления дифференциальная экспрессия наблюдается через менее чем 18 часов после введения. В некоторых вариантах осуществления дифференциальная экспрессия наблюдается через менее чем 24 часа после введения.
[0143] В некоторых вариантах осуществления ингибирующий иммунный ответ включает клеточноопосредованный иммунный ответ. В определенных вариантах осуществления клеточноопосредованный иммунный ответ включает высвобождение цитокина. В некоторых вариантах осуществления цитокин представляет собой любой цитокин, ассоциированный с усиленным иммунным ответом. В некоторых вариантах осуществления цитокин выбран из группы, состоящей из IL-1, IL-6, IL-16, IL-12, IL-4, IL-17, фактора некроза опухоли α (TNF-α), интерферона α, интерферона γ и любой их комбинации. В одном варианте осуществления клеточноопосредованный иммунный ответ предусматривает увеличенные уровни IL-12 в сыворотке крови. В другом варианте осуществления клеточноопосредованный иммунный ответ предусматривает увеличенные уровни IL-4 в сыворотке крови. В другом варианте осуществления клеточноопосредованный иммунный ответ предусматривает увеличенные уровни IL-17 в сыворотке крови. В другом варианте осуществления клеточноопосредованный иммунный ответ предусматривает увеличенные уровни TNF-α в сыворотке крови.
[0144] Было установлено, что различные генные мутации связаны с увеличенным риском развития ингибирующего иммунного ответа. Например, полиморфизм TNF-α -308G>A в Hap2, который ассоциирован с увеличенными уровнями конститутивной и индуцируемой транскрипции TNF, был связан с увеличенным риском развития ингибирующего иммунного ответа. См. Astermark et al., Blood 108: 3739-3745 (2006), которая включена в данный документ посредством ссылки во всей своей полноте. Таким образом, в некоторых вариантах осуществления у человека имеется генетический полиморфизм, ассоциированный с увеличенным уровнем TNF-α. В некоторых вариантах осуществления полиморфизм представляет собой полиморфизм TNF-α -308G>A. В некоторых вариантах осуществления у человека имеется полиморфизм в гене IL10, например, полиморфизм, ассоциированный с увеличенной секрецией IL10. В некоторых вариантах осуществления FVIII-Fc вводят субъекту с аллелем 134 микросателлита IL10G в промоторной области гена IL10. См. Astermark et al. Hemostatis, Thrombosis, and Vascular Biology 108: 3739-3745 (2006), которая включена в данный документ посредством ссылки во всей своей полноте.
[0145] В некоторых вариантах осуществления у человека имеется генетический полиморфизм, ассоциированный с уменьшенной экспрессией CTLA-4 (антигена 4 цитотоксических Т-лимфоцитов). В некоторых вариантах осуществления у человека имеется мутация в молекулах DR15 (HLA-DR15) или DQB0602 MHC (главного комплекса гистосовместимости) класса II. Другими молекулами MHC класса II, ассоциированными с развитием ингибирующего иммунного ответа у субъектов с гемофилией, являются A3, B7, C7, DQA0102, C2, DQA0103, DQB0603 и DR13 (см. Inhibitors in Patients with Hemophilia, E.C. Rodriguez-Merchan & C.A. Lee, Eds., Blackwell Science, Ltd., 2002).
[0146] В некоторых вариантах осуществления способы по настоящему изобретению обеспечивают снижение уровня одного или нескольких цитокинов у субъекта по сравнению с уровнем одного или нескольких цитокинов у субъекта после предшествующего лечения с помощью полипептида, состоящего из белка FVIII. В другом варианте осуществления способы по настоящему изобретению обеспечивают снижение уровня одного или нескольких цитокинов у субъекта по сравнению с уровнем одного или нескольких цитокинов у субъекта до введения. В других вариантах осуществления экспрессия одной или нескольких толерогенных молекул увеличивается после применения способов по настоящему изобретению по сравнению с уровнем экспрессии одной или нескольких толерогенных молекул до введения. В определенных вариантах осуществления одна или несколько толерогенных молекул выбраны из IL-10, TGF-β, IL-35, IDO-1 и любой их комбинации.
[0147] В других вариантах осуществления иммунный ответ включает клинический симптом, выбранный из группы, состоящей из увеличенной склонности к кровотечению, высокого потребления фактора свертывания крови, отсутствия ответа на терапию фактором свертывания крови, уменьшенной эффективности терапии фактором свертывания крови, сокращенного периода полужизни фактора свертывания крови и любой их комбинации. В определенных вариантах осуществления иммунный ответ включает клинический симптом, выбранный из группы, состоящей из увеличенной склонности к кровотечению, высокого потребления фактора свертывания крови, отсутствия ответа на терапию фактором свертывания крови, уменьшенной эффективности терапии фактором свертывания крови, уменьшенного восстановления активности фактора свертывания крови, отслеживаемого в плазме крови, сокращенного периода полужизни фактора свертывания крови и любой их комбинации.
[0148] В определенных вариантах осуществления у человека ранее было диагностировано наличие ингибирующего иммунного ответа. Такой диагноз можно поставить с помощью любых способов, известных из уровня техники. Например, человек может характеризоваться наличием иммунного ответа на фактор свертывания крови, например, FVIII, если у человека имеется одно или несколько из следующего: (a) титра ингибирующих антител к фактору свертывания крови, превышающего или равного 0,6 БЕ; (b) увеличенных уровней одного или нескольких цитокинов, выбранных из группы, состоящей из IL-12, IL-4, IL-17 и TNF-α, в сыворотке крови; (c) увеличенной склонности к кровотечению; (d) высокого потребления фактора свертывания крови; (e) отсутствия ответа на терапию фактором свертывания крови; (f) уменьшенной эффективности терапии фактором свертывания крови; (g) сокращенного периода полужизни фактора свертывания крови и любой их комбинации. В одном конкретном варианте осуществления человек характеризуется наличием иммунного ответа на фактор свертывания крови, если у человека титр ингибирующих антител к фактору свертывания крови превышает или равен 0,6 БЕ.
[0149] В некоторых вариантах осуществления у человека ранее было диагностировано наличие развившегося ингибирующего иммунного ответа на фактор свертывания крови за по меньшей мере приблизительно 1 месяц, по меньшей мере приблизительно 2 месяца, по меньшей мере приблизительно 3 месяца, по меньшей мере приблизительно 4 месяца, по меньшей мере приблизительно 5 месяцев, по меньшей мере приблизительно 6 месяцев, по меньшей мере приблизительно 7 месяцев, по меньшей мере приблизительно 8 месяцев, по меньшей мере приблизительно 9 месяцев, по меньшей мере приблизительно 10 месяцев, по меньшей мере приблизительно 11 месяцев, по меньшей мере приблизительно 12 месяцев, по меньшей мере приблизительно 13 месяцев, по меньшей мере приблизительно 14 месяцев, по меньшей мере приблизительно 15 месяцев, по меньшей мере приблизительно 16 месяцев, по меньшей мере приблизительно 17 месяцев, по меньшей мере приблизительно 18 месяцев, по меньшей мере приблизительно 19 месяцев, по меньшей мере приблизительно 20 месяцев, по меньшей мере приблизительно 21 месяц, по меньшей мере приблизительно 22 месяца, по меньшей мере приблизительно 23 месяца, по меньшей мере приблизительно 24 месяца, по меньшей мере приблизительно 27 месяцев, по меньшей мере приблизительно 30 месяцев, по меньшей мере приблизительно 33 месяца, по меньшей мере приблизительно 36 месяцев, по меньшей мере приблизительно 39 месяцев, по меньшей мере приблизительно 42 месяца, по меньшей мере приблизительно 45 месяцев, по меньшей мере приблизительно 48 лет, по меньшей мере приблизительно 51 месяц, по меньшей мере приблизительно 54 месяца, по меньшей мере приблизительно 57 месяцев, по меньшей мере приблизительно 60 месяцев, по меньшей мере приблизительно 6 лет, по меньшей мере приблизительно 7 лет, по меньшей мере приблизительно 8 лет, по меньшей мере приблизительно 10 лет, по меньшей мере приблизительно 15 лет или по меньшей мере приблизительно 20 лет до введения. В одном варианте осуществления у человека ранее было диагностировано наличие развившегося ингибирующего иммунного ответа на фактор свертывания крови за по меньшей мере приблизительно 5 лет до введения.
[0150] В некоторых вариантах осуществления способы по настоящему изобретению обеспечивают улучшенное время достижения толерантности по сравнению со стандартными способами индукции иммунной толерантности. Используемый в данном документе термин "время достижения толерантности" относится к количеству времени между введением первой дозы композиции или химерного белка, содержащих фактор свертывания крови и Fc-область, и развитием иммунной толерантности у человека. Уменьшение времени достижения толерантности может иметь значительные преимущества для человека, в том числе без ограничения уменьшать общее финансовое бремя, требуемое для достижения толерантности. В некоторых вариантах осуществления время достижения толерантности составляет от приблизительно 1 до приблизительно 24 недель, от приблизительно 1 до приблизительно 23 недель, от приблизительно 1 до приблизительно 22 недель, от приблизительно 1 до приблизительно 21 недели, от приблизительно 2 до приблизительно 20 недель, от приблизительно 2 до приблизительно 19 недель, от приблизительно 2 до приблизительно 18 недель, от приблизительно 2 до приблизительно 17 недель, от приблизительно 3 до приблизительно 16 недель, от приблизительно 3 до приблизительно 15 недель, от приблизительно 3 до приблизительно 14 недель, от приблизительно 3 до приблизительно 13 недель, от приблизительно 4 до приблизительно 12 недель, от приблизительно 4 до приблизительно 11 недель, от приблизительно 4 до приблизительно 10 недель, от приблизительно 4 до приблизительно 9 недель, от приблизительно 5 до приблизительно 8 недель, от приблизительно 5 до приблизительно 7 недель, от приблизительно 5 до приблизительно 6 недель, от приблизительно 1 до приблизительно 12 недель, от приблизительно 1 до приблизительно 11 недель, от приблизительно 1 до приблизительно 10 недель, от приблизительно 1 до приблизительно 9 недель, от приблизительно 1 до приблизительно 8 недель, от приблизительно 1 до приблизительно 7 недель, от приблизительно 1 до приблизительно 6 недель, от приблизительно 1 до приблизительно 5 недель или от приблизительно 1 до приблизительно 4 недель. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 70 недель, менее чем приблизительно 65 недель, менее чем приблизительно 60 недель, менее чем приблизительно 58 недель, менее чем приблизительно 56 недель, менее чем приблизительно 54 недели, менее чем приблизительно 52 недели, менее чем приблизительно 50 недель, менее чем приблизительно 48 недель, менее чем приблизительно 46 недель, менее чем приблизительно 44 недели, менее чем приблизительно 42 недели, менее чем приблизительно 40 недель, менее чем приблизительно 38 недель, менее чем приблизительно 36 недель, менее чем приблизительно 34 недели, менее чем приблизительно 32 недели, менее чем приблизительно 30 недель, менее чем приблизительно 28 недель, менее чем приблизительно 26 недель, менее чем приблизительно 24 недели, менее чем приблизительно 23 недели, менее чем приблизительно 22 недели, менее чем приблизительно 21 неделю, менее чем приблизительно 20 недель, менее чем приблизительно 19 недель, менее чем приблизительно 18 недель, менее чем приблизительно 17 недель, менее чем приблизительно 16 недель, менее чем приблизительно 15 недель, менее чем приблизительно 14 недель, менее чем приблизительно 13 недель, менее чем приблизительно 12 недель, менее чем приблизительно 11 недель, менее чем приблизительно 10 недель, менее чем приблизительно 9 недель, менее чем приблизительно 8 недель, менее чем приблизительно 7 недель, менее чем приблизительно 6 недель, менее чем приблизительно 5 недель, менее чем приблизительно 4 недели, менее чем приблизительно 3 недели, менее чем приблизительно 2 недели или менее чем приблизительно 1 неделю. В определенных вариантах осуществления время достижения толерантности составляет от приблизительно 4 до приблизительно 12 недель. В одном варианте осуществления время достижения толерантности составляет приблизительно 4 недели. В другом варианте осуществления время достижения толерантности составляет приблизительно 12 недель. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 10 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 9 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 8 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 7 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 6 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 5 месяцев. В некоторых вариантах осуществления время достижения толерантности составляет менее чем приблизительно 4 месяца. В некоторых вариантах осуществления способы согласно настоящему изобретению приводят к более короткому времени достижения толерантности у человека после лечения с помощью композиции или химерного белка, содержащих фактор свертывания крови и Fc-область, по сравнению со временем достижения толерантности после лечения с помощью фактора свертывания крови в отдельности.
[0151] В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим менее чем приблизительно 0,6 БЕ. В некоторых вариантах осуществления формирование иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим приблизительно 0,5 БЕ. В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим менее чем приблизительно 0,4 БЕ. В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим менее чем приблизительно 0,3 БЕ. В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим менее чем приблизительно 0,2 БЕ. В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим менее чем приблизительно 0,1 БЕ. В некоторых вариантах осуществления развитие иммунной толерантности характеризуется титром ингибирующего антитела к фактору свертывания крови, составляющим 0,0 БЕ. В определенных вариантах осуществления титр ингибирующих иммунных антител наблюдают при двух последовательных измерениях, например, в две последовательные недели в течение четырехнедельного периода.
[0152] В некоторых вариантах осуществления развитие иммунной толерантности характеризуется постепенным восстановлением до > 66% (например, постепенным восстановлением до приблизительно 67%, приблизительно 68%, приблизительно 69%, приблизительно 70%, приблизительно 71%, приблизительно 72%, приблизительно 73%, приблизительно 74%, приблизительно 75%, приблизительно 76%, приблизительно 77%, приблизительно 78%, приблизительно 79%, приблизительно 80%, приблизительно 81%, приблизительно 82%, приблизительно 83%, приблизительно 84%, приблизительно 85%, приблизительно 86%, приблизительно 87%, приблизительно 88%, приблизительно 89%, приблизительно 90%, приблизительно 91%, приблизительно 92%, приблизительно 93%, приблизительно 94%, приблизительно 95%, приблизительно 96%, приблизительно 97%, приблизительно 98%, приблизительно 99% или приблизительно 100%). Используемый в данном документе термин "постепенное восстановление" относится к пиковым уровням FVIII через 15-30 минут после инфузии.
[0153] После завершения периода индукции и периода постепенного снижения дозы субъект затем может получать профилактическое лечение с помощью химерного белка. Профилактический режим введения доз может представлять собой любой режим введения доз, раскрытый в данном документе.
[0154] В некоторых вариантах осуществления человек, получающий лечение с помощью способов по настоящему изобретению, получает или недавно получил иммуностимулирующую терапию. Например, о наличии ингибиторов также сообщалось у HCV-положительных пациентов с гемофилией А, проходящих лечение интерфероном, а также у HIV-положительных пациентов с гемофилией А, у которых имеется воспалительный синдром восстановления иммунитета, ассоциированный с антиретровирусной терапией. См. Report of Expert Meeting on Factor VIII Products and Inhibitor Development, European Medicines Agency (February 28, 2006 - March 2, 2006). Таким образом, в некоторых вариантах осуществления человек получает терапию интерфероном. В некоторых вариантах осуществления человек получает противовирусную терапию. В некоторых вариантах осуществления человек получает антиретровирусную терапию, и у него имеется воспалительный синдром восстановления иммунитета.
[0155] В определенных вариантах осуществления человек принимал фактор свертывания крови, например, FVIII, в течение менее 150 дней воздействия (ED). В одном варианте осуществления человек принимал его в течение менее 50 ED. В другом варианте осуществления человек принимал его в течение менее 20 ED.
[0156] Некоторые аспекты настоящего изобретения относятся к способам снижения тяжести или частоты возникновения аллергической или анафилактической реакции на фактор свертывания крови у субъекта, нуждающегося в этом, предусматривающим введение субъекту композиции или химерного белка, содержащих фактор свертывания крови и Fc-область. В некоторых вариантах осуществления введение композиции или химерного белка обеспечивает снижение тяжести анафилактоидной реакции на фактор свертывания крови. В некоторых вариантах осуществления введение композиции или химерного белка обеспечивает снижение тяжести аллергической реакции на фактор свертывания крови.
III. Химерные полипептиды
[0157] Химерные полипептиды, применимые для настоящего изобретения, содержат белок FVIII, содержащий полипептид FVIII, и фрагмент VWF, содержащий домен D' и домен D3 VWF. Фрагмент VWF используется для предотвращения или блокирования взаимодействия белка FVIII с эндогенным VWF, за счет чего обеспечивается предотвращение выведения белка FVIII через путь выведения VWF. Для предотвращения диссоциации белка FVIII и фрагмента VWF в некоторых вариантах осуществления белок FVIII и фрагмент VWF связаны непосредственно или опосредованно друг с другом с помощью связи, более сильной, чем естественное взаимодействие между FVIII и VWF.
[0158] В определенных вариантах осуществления белок FVIII связан с фрагментом VWF с помощью ковалентной связи. Ковалентная связь может представлять собой любую ковалентную связь, известную из уровня техники. В некоторых вариантах осуществления ковалентная связь представляет собой дисульфидную связь. В некоторых вариантах осуществления ковалентная связь представляет собой пептидную связь. В других вариантах осуществления белок FVIII модифицирован для увеличения силы взаимодействия между белком FVIII и фрагментом VWF. В других вариантах осуществления фрагмент VWF модифицирован для увеличения силы взаимодействия между белком FVIII и фрагментом VWF. В других вариантах осуществления как белок FVIII, так и фрагмент VWF модифицированы для увеличения силы взаимодействия между белком FVIII и фрагментом VWF.
[0159] В конкретных вариантах осуществления белок FVIII, содержащий полипептид FVIII, и фрагмент VWF, содержащий домен D' и домен D3 VWF, связаны непосредственно с помощью по меньшей мере одной ковалентной связи между по меньшей мере одной аминокислотой в последовательности полипептида FVIII и по меньшей мере одной аминокислотой в доменах D' и D3 VWF. В других вариантах осуществления белок FVIII и фрагмент VWF связаны опосредованно с помощью по меньшей мере одной ковалентной связи между по меньшей мере одной аминокислотой в последовательности полипептида FVIII и по меньшей мере одной аминокислотой в гетерологичной последовательности, непосредственно или опосредованно слитой с доменами D' и D3 VWF. В других вариантах осуществления белок FVIII и фрагмент VWF связаны опосредованно с помощью по меньшей мере одной ковалентной связи между по меньшей мере одной аминокислотой в последовательности фрагмента VWF и по меньшей мере одной аминокислотой в гетерологичной последовательности, непосредственно или опосредованно слитой с полипептидом FVIII. В еще нескольких других вариантах осуществления белок FVIII и фрагмент VWF связаны опосредованно с помощью по меньшей мере одной ковалентной связи между по меньшей мере одной аминокислотой в гетерологичной последовательности, непосредственно или опосредованно слитой с доменами D' и D3 VWF, и по меньшей мере одной аминокислотой в гетерологичной последовательности, непосредственно или опосредованно слитой с полипептидом FVIII.
[0160] Белок FVIII по настоящему изобретению может содержать полипептид FVIII и один или несколько гетерологичных компонентов, например, компонентов, обеспечивающих продление периода полужизни, непосредственно или опосредованно слитых с полипептидом FVIII. Фрагмент VWF по настоящему изобретению также может содержать домен D' и домен D3 VWF и один или несколько гетерологичных компонентов, например, компонентов, обеспечивающих продление периода полужизни, непосредственно или опосредованно слитых с доменами D' и D3 VWF. В некоторых вариантах осуществления белок FVIII, применимый для настоящего изобретения, состоит по сути из полипептида FVIII или состоит из него, а фрагмент VWF, применимый для настоящего изобретения, содержит домен D' и домен D3 VWF и один или несколько гетерологичных компонентов, например, компонентов, обеспечивающих продление периода полужизни, непосредственно или опосредованно слитых с доменами D' и D3 VWF. В некоторых вариантах осуществления белок FVIII содержит полипептид FVIII и один или несколько гетерологичных компонентов, например, компонентов, обеспечивающих продление периода полужизни, непосредственно или опосредованно слитых с полипептидом FVIII, а фрагмент VWF, применимый для настоящего изобретения, состоит по сути из домена D' и домена D3 VWF или состоит из них. В некоторых вариантах осуществления химерный полипептид содержит белок FVIII, состоящий по сути из полипептида FVIII или состоящий из него, и фрагмент VWF, состоящий по сути из домена D' и домена D3 VWF или состоящий из них.
[0161] В некоторых вариантах осуществления химерный полипептид или белок, раскрытый в данном документе, представляет собой гетеродимер FVIII-XTEN-Fc/D'D3-XTEN-Fc. В одном варианте осуществления гетеродимерный химерный полипептид FVIII-XTEN-Fc/D'D3-XTEN-Fc содержит: (i) белок FVIII, содержащий полипептид FVIII, XTEN, вставленный в домен B полипептида FVIII, и первую Fc-область; и (ii) белок VWF, содержащий фрагмент VWF, вторую последовательность XTEN, линкер a2 и вторую Fc-область. Схематическое изображение иллюстративного гетеродимера FVIII-XTEN-Fc/D'D3-XTEN-Fc, который представляет собой rFVIIIFc-VWF-XTEN, представлено на фиг. 1.
[0162] В конкретном варианте осуществления химерный полипептид представляет собой rFVIIIFc-VWF-XTEN. В другом конкретном варианте осуществления rFVIIIFc-VWF-XTEN содержит (i) белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 203, и (ii) белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 205. В еще одном конкретном варианте осуществления rFVIIIFc-VWF-XTEN содержит (i) белок FVIII и (ii) белок VWF, которые ковалентно соединены друг с другом посредством дисульфидной связи.
III.A. Полипептидные факторы VIII
[0163] "Фактор VIII", сокращенно называемый во всей настоящей заявке как "FVIII", как используется в данном документе, означает функциональный полипептид FVIII с его нормальной ролью в коагуляции, если не указано иное. Таким образом, термин "FVIII" включает варианты полипептидов, которые являются функциональными. "Белок FVIII" используется для обозначения полипептида (или белка) FVIII в отдельности, полипептида FVIII, слитого с дополнительными полипептидами, и полипептида FVIII, связанного с одним или несколькими дополнительными полипептидами, при условии, что белок FVIII демонстрирует функцию/активность FVIII. Термины "полипептид FVIII", "часть FVIII" и "FVIII" относятся к последовательности полипептида FVIII в отдельности. Примеры функций/видов активности FVIII включают без ограничения способность активировать коагуляцию, способность действовать в качестве кофактора для фактора IX или способность образовывать теназный комплекс с фактором IX в присутствии Ca2+ и фосфолипидов, который затем обеспечивает превращение фактора X в активированную форму Xa. Полипептид FVIII может представлять собой полипептид FVIII человека, свиньи, собаки, крысы или мыши. Кроме того, путем сравнения FVIII от людей и FVIII от других видов были идентифицированы консервативные остатки, которые, вероятно, требуются для функционирования (Cameron et al., Thromb. Haemost. 79:317-22 (1998); US 6251632). Известны полноразмерные полипептидные и полинуклеотидные последовательности, а также многие функциональные фрагменты, мутантные и модифицированные варианты. Различные аминокислотные и нуклеотидные последовательности FVIII раскрыты, например, в публикациях заявок на патент США №№ 2015/0158929 A1, 2014/0308280 A1 и 2014/0370035 A1 и в международной публикации № WO 2015/106052 A1. Полипептиды FVIII включают в себя, например, полноразмерный FVIII, полноразмерный FVIII без Met на N-конце, зрелый полипептид FVIII (без сигнальной последовательности), зрелый полипептид FVIII с дополнительным Met на N-конце и/или полипептид FVIII с полной или частичной делецией домена B. Варианты FVIII содержат делеции домена B, будь то частичные или полные делеции.
[0164] Полипептид FVIII химерного полипептида, используемого в данном документе, обладает активностью FVIII в плазме крови. Активность FVIII можно измерять с помощью любых способов, известных из уровня техники. Доступен ряд тестов для оценки функции системы коагуляции: тест активированного частичного тромбопластинового времени (aPTT), хромогенный анализ, анализ ROTEM, тест протромбинового времени (PT) (также применяется для определения INR), тестирование содержания фибриногена (зачастую с помощью способа Клаусса), подсчет тромбоцитов, тестирование функции тромбоцитов (зачастую с помощью PFA-100), TCT, определение времени свертывания крови, тест смешивания (устраняется ли аномалия при смешивании плазмы крови пациента с нормальной плазмой крови), анализы факторов коагуляции, антител к фосфолипидам, D-димера, генетические тесты (например, лейденовской мутации фактора V, мутации протромбина G20210A), определение времени свертывания крови с разбавленным ядом гадюки Рассела (dRVVT), различные тесты функции тромбоцитов, тромбоэластография (TEG или Sonoclot), тромбоэластометрия (TEM®, например, ROTEM®) или определение времени лизиса эуглобулинов (ELT).
[0165] Тест aPTT является показателем функционирования, измеряющим эффективность как "внутреннего" (также называемого контактным путем активации), так и общего путей коагуляции. Этот тест обычно применяют для измерения свертывающей активности коммерчески доступных рекомбинантных факторов свертывания крови, например, FVIII. Его, как правило, применяют в сочетании с протромбиновым временем (PT), с помощью которого измеряют внешний путь. (См., например, Kamal et al., Mayo Clin Proc., 82(7):864-873 (2007)). В одном варианте осуществления aPTT тестируют с помощью анализа, в котором активность FVIII измеряют с использованием реагента для определения активированного PTT Dade® Actin® FSL (Siemens Health Care Diagnostics) на анализаторе BCS® XP (Siemens Healthcare Diagnostics). В определенных вариантах осуществления химерный полипептид характеризуется активностью FVIII в плазме крови, составляющей по меньшей мере приблизительно 1%, по меньшей мере приблизительно 2%, по меньшей мере приблизительно 3%, по меньшей мере приблизительно 4%, по меньшей мере приблизительно 5%, по меньшей мере приблизительно 6%, по меньшей мере приблизительно 7%, по меньшей мере приблизительно 8%, по меньшей мере приблизительно 9%, по меньшей мере приблизительно 10%, по меньшей мере приблизительно 11%, по меньшей мере приблизительно 12%, по меньшей мере приблизительно 13%, по меньшей мере приблизительно 14%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 15%, по меньшей мере приблизительно 16%, по меньшей мере приблизительно 17%, по меньшей мере приблизительно 18%, по меньшей мере приблизительно 19% или по меньшей мере приблизительно 20% согласно измерению с помощью теста aPTT.
[0166] Анализ aPTT также можно применять для оценки активности химерного полипептида перед введением пациенту или субъекту. (Hubbard AR, et al. J Thromb Haemost 11: 988-9 (2013)). Анализ aPTT можно дополнительно применять в сочетании с любым из анализов, описанных в данном документе, перед введением либо после введения пациенту или субъекту.
[0167] Анализ ROTEM предоставляет информацию обо всех кинетических показателях гемостаза: времени свертывания крови, образовании сгустка, стабильности и лизисе сгустка. Различные параметры тромбоэластометрии зависят от активности системы коагуляции плазмы крови, функции тромбоцитов, фибринолиза или многих факторов, которые влияют на эти взаимодействия. Этот анализ может дать полное представление о вторичном гемостазе.
[0168] Механизм хромогенного анализа основан на принципах каскада коагуляции крови, где активированный FVIII ускоряет превращение фактора X в фактор Xa в присутствии активированного фактора IX, фосфолипидов и ионов кальция. Активность фактора Ха оценивают путем гидролиза п-нитроанилидного (pNA) субстрата, специфичного для фактора Ха. Начальная скорость высвобождения п-нитроанилина, измеренная при 405 нМ, прямо пропорциональна активности фактора Xa и, следовательно, активности FVIII в образце. В одном варианте осуществления хромогенный анализ представляет собой анализ BIOPHEN FVIII: C (Hyphen Biomed, Невиль-сюр-Уаз, Франция).
[0169] Хромогенный анализ рекомендован подкомитетом по FVIII и фактору IX научного и стандартизационного комитета (SSC) Международного общества по тромбозу и гемостазу (ISTH). С 1994 года хромогенный анализ также являлся эталонным способом определения активности концентрата FVIII согласно Европейской фармакопее. Таким образом, в некоторых вариантах осуществления химерный полипептид, содержащий полипептид FVIII, обладает активностью FVIII, сравнимой с таковой у химерного полипептида, содержащего зрелый полипептид FVIII или полипептид FVIII BDD (например, RECOMBINATE®, KOGENATE FS®, HELIXATE FS®, XYNTHA/REFACTO AB®, HEMOFIL-M®, MONARC-M®, MONOCLATE-P®, HUMATE-P®, ALPHANATE®, KOATE-DVI®, AFSTYLA® и HYATE:C®).
[0170] Хромогенный анализ также можно применять для оценки активности химерного полипептида перед введением пациенту или субъекту (Hubbard AR, et al. J Thromb Haemost 11: 988-9 (2013)). Хромогенный анализ можно дополнительно применять в сочетании с любым из анализов, описанных в данном документе, перед введением либо после введения пациенту или субъекту.
[0171] В других вариантах осуществления химерный полипептид, содержащий полипептид FVIII по настоящему изобретению, характеризуется скоростью образования фактора Ха, сравнимой с таковой у химерного полипептида, содержащего зрелый полипептид FVIII или полипептид FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®).
[0172] Для активации фактора X до фактора Xa активированный фактор IX (фактор IXa) гидролизует одну связь между аргинином и изолейцином в факторе X с образованием фактора Xa в присутствии Ca2+, мембранных фосфолипидов и кофактора FVIII. Следовательно, взаимодействие FVIII с фактором IX является критически важным в пути коагуляции. В определенных вариантах осуществления химерный полипептид, содержащий полипептид FVIII, может взаимодействовать с фактором IXa со скоростью, сравнимой с таковой у химерного полипептида, содержащего последовательность зрелого полипептида FVIII или полипептид FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®).
[0173] Кроме того, FVIII связывается с фактором фон Виллебранда, находясь в неактивной форме в кровотоке. FVIII быстро разрушается, если он не связан с VWF, и высвобождается от VWF под действием тромбина. В некоторых вариантах осуществления химерный полипептид, содержащий полипептид FVIII, связывается с фактором фон Виллебранда, например, фрагментом VWF, раскрытым в данном документе, на уровне, сравнимом с таковым у химерного полипептида, содержащего последовательность зрелого полипептида FVIII или полипептид FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®).
[0174] FVIII можно инактивировать с помощью активированного белка С в присутствии кальция и фосфолипидов. Активированный белок C производит расщепление тяжелой цепи FVIII после аргинина 336 в домене A1, что разрушает сайт взаимодействия с субстратом, представляющим собой фактор X, и производит расщепление после аргинина 562 в домене A2, что усиливает диссоциацию домена A2, а также разрушает сайт взаимодействия с фактором IXa. Это расщепление также приводит к разделению пополам домена A2 (43 кДа) и образованию доменов A2-N (18 кДа) и A2-C (25 кДа). Таким образом, активированный белок C может катализировать расщепление в нескольких сайтах тяжелой цепи. В некоторых вариантах осуществления химерный полипептид, содержащий полипептид FVIII, инактивируется под действием активированного белка С на уровне, сравнимом с таковым у химерного полипептида, содержащего последовательность зрелого полипептида FVIII или FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®).
[0175] В других вариантах осуществления химерный полипептид, содержащий полипептид FVIII, обладает активностью FVIII in vivo, сравнимой с таковой у химерного полипептида, содержащего последовательность зрелого полипептида FVIII или полипептид FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®). В конкретном варианте осуществления химерный полипептид, содержащий полипептид FVIII, способен обеспечивать защиту у мыши HemA на уровне, сравнимом с таковым у химерного полипептида, содержащего последовательность зрелого полипептида FVIII или полипептид FVIII BDD (например, ADVATE®, REFACTO® или ELOCTATE®), в модели рассечения хвостовой вены на мышах HemA.
[0176] Примеры последовательностей FVIII человека (полноразмерных) показаны ниже.
домен B выделен курсивом; и легкая цепь FVIII представлена в виде обычного текста)
*Подчеркнутые нуклеиновые кислоты кодируют сигнальный пептид.
721 CTTCTTTCTG TGCCTTTTGC GATTCTGCTT TAGTGCCACC AGAAGATACT ACCTGGGTGC
781 AGTGGAACTG TCATGGGACT ATATGCAAAG TGATCTCGGT GAGCTGCCTG TGGACGCAAG
841 ATTTCCTCCT AGAGTGCCAA AATCTTTTCC ATTCAACACC TCAGTCGTGT ACAAAAAGAC
901 TCTGTTTGTA GAATTCACGG ATCACCTTTT CAACATCGCT AAGCCAAGGC CACCCTGGAT
961 GGGTCTGCTA GGTCCTACCA TCCAGGCTGA GGTTTATGAT ACAGTGGTCA TTACACTTAA
1021 GAACATGGCT TCCCATCCTG TCAGTCTTCA TGCTGTTGGT GTATCCTACT GGAAAGCTTC
1081 TGAGGGAGCT GAATATGATG ATCAGACCAG TCAAAGGGAG AAAGAAGATG ATAAAGTCTT
1141 CCCTGGTGGA AGCCATACAT ATGTCTGGCA GGTCCTGAAA GAGAATGGTC CAATGGCCTC
1201 TGACCCACTG TGCCTTACCT ACTCATATCT TTCTCATGTG GACCTGGTAA AAGACTTGAA
1261 TTCAGGCCTC ATTGGAGCCC TACTAGTATG TAGAGAAGGG AGTCTGGCCA AGGAAAAGAC
1321 ACAGACCTTG CACAAATTTA TACTACTTTT TGCTGTATTT GATGAAGGGA AAAGTTGGCA
1381 CTCAGAAACA AAGAACTCCT TGATGCAGGA TAGGGATGCT GCATCTGCTC GGGCCTGGCC
1441 TAAAATGCAC ACAGTCAATG GTTATGTAAA CAGGTCTCTG CCAGGTCTGA TTGGATGCCA
1501 CAGGAAATCA GTCTATTGGC ATGTGATTGG AATGGGCACC ACTCCTGAAG TGCACTCAAT
1561 ATTCCTCGAA GGTCACACAT TTCTTGTGAG GAACCATCGC CAGGCGTCCT TGGAAATCTC
1621 GCCAATAACT TTCCTTACTG CTCAAACACT CTTGATGGAC CTTGGACAGT TTCTACTGTT
1681 TTGTCATATC TCTTCCCACC AACATGATGG CATGGAAGCT TATGTCAAAG TAGACAGCTG
1741 TCCAGAGGAA CCCCAACTAC GAATGAAAAA TAATGAAGAA GCGGAAGACT ATGATGATGA
1801 TCTTACTGAT TCTGAAATGG ATGTGGTCAG GTTTGATGAT GACAACTCTC CTTCCTTTAT
1861 CCAAATTCGC TCAGTTGCCA AGAAGCATCC TAAAACTTGG GTACATTACA TTGCTGCTGA
1921 AGAGGAGGAC TGGGACTATG CTCCCTTAGT CCTCGCCCCC GATGACAGAA GTTATAAAAG
1981 TCAATATTTG AACAATGGCC CTCAGCGGAT TGGTAGGAAG TACAAAAAAG TCCGATTTAT
2041 GGCATACACA GATGAAACCT TTAAGACTCG TGAAGCTATT CAGCATGAAT CAGGAATCTT
2101 GGGACCTTTA CTTTATGGGG AAGTTGGAGA CACACTGTTG ATTATATTTA AGAATCAAGC
2161 AAGCAGACCA TATAACATCT ACCCTCACGG AATCACTGAT GTCCGTCCTT TGTATTCAAG
2221 GAGATTACCA AAAGGTGTAA AACATTTGAA GGATTTTCCA ATTCTGCCAG GAGAAATATT
2281 CAAATATAAA TGGACAGTGA CTGTAGAAGA TGGGCCAACT AAATCAGATC CTCGGTGCCT
2341 GACCCGCTAT TACTCTAGTT TCGTTAATAT GGAGAGAGAT CTAGCTTCAG GACTCATTGG
2401 CCCTCTCCTC ATCTGCTACA AAGAATCTGT AGATCAAAGA GGAAACCAGA TAATGTCAGA
2461 CAAGAGGAAT GTCATCCTGT TTTCTGTATT TGATGAGAAC CGAAGCTGGT ACCTCACAGA
2521 GAATATACAA CGCTTTCTCC CCAATCCAGC TGGAGTGCAG CTTGAGGATC CAGAGTTCCA
2581 AGCCTCCAAC ATCATGCACA GCATCAATGG CTATGTTTTT GATAGTTTGC AGTTGTCAGT
2641 TTGTTTGCAT GAGGTGGCAT ACTGGTACAT TCTAAGCATT GGAGCACAGA CTGACTTCCT
2701 TTCTGTCTTC TTCTCTGGAT ATACCTTCAA ACACAAAATG GTCTATGAAG ACACACTCAC
2761 CCTATTCCCA TTCTCAGGAG AAACTGTCTT CATGTCGATG GAAAACCCAG GTCTATGGAT
2821 TCTGGGGTGC CACAACTCAG ACTTTCGGAA CAGAGGCATG ACCGCCTTAC TGAAGGTTTC
2881 TAGTTGTGAC AAGAACACTG GTGATTATTA CGAGGACAGT TATGAAGATA TTTCAGCATA
2941 CTTGCTGAGT AAAAACAATG CCATTGAACC AAGAAGCTTC TCCCAGAATT CAAGACACCC
3001 TAGCACTAGG CAAAAGCAAT TTAATGCCAC CACAATTCCA GAAAATGACA TAGAGAAGAC
3061 TGACCCTTGG TTTGCACACA GAACACCTAT GCCTAAAATA CAAAATGTCT CCTCTAGTGA
3121 TTTGTTGATG CTCTTGCGAC AGAGTCCTAC TCCACATGGG CTATCCTTAT CTGATCTCCA
3181 AGAAGCCAAA TATGAGACTT TTTCTGATGA TCCATCACCT GGAGCAATAG ACAGTAATAA
3241 CAGCCTGTCT GAAATGACAC ACTTCAGGCC ACAGCTCCAT CACAGTGGGG ACATGGTATT
3301 TACCCCTGAG TCAGGCCTCC AATTAAGATT AAATGAGAAA CTGGGGACAA CTGCAGCAAC
3361 AGAGTTGAAG AAACTTGATT TCAAAGTTTC TAGTACATCA AATAATCTGA TTTCAACAAT
3421 TCCATCAGAC AATTTGGCAG CAGGTACTGA TAATACAAGT TCCTTAGGAC CCCCAAGTAT
3481 GCCAGTTCAT TATGATAGTC AATTAGATAC CACTCTATTT GGCAAAAAGT CATCTCCCCT
3541 TACTGAGTCT GGTGGACCTC TGAGCTTGAG TGAAGAAAAT AATGATTCAA AGTTGTTAGA
3601 ATCAGGTTTA ATGAATAGCC AAGAAAGTTC ATGGGGAAAA AATGTATCGT CAACAGAGAG
3661 TGGTAGGTTA TTTAAAGGGA AAAGAGCTCA TGGACCTGCT TTGTTGACTA AAGATAATGC
3721 CTTATTCAAA GTTAGCATCT CTTTGTTAAA GACAAACAAA ACTTCCAATA ATTCAGCAAC
3781 TAATAGAAAG ACTCACATTG ATGGCCCATC ATTATTAATT GAGAATAGTC CATCAGTCTG
3841 GCAAAATATA TTAGAAAGTG ACACTGAGTT TAAAAAAGTG ACACCTTTGA TTCATGACAG
3901 AATGCTTATG GACAAAAATG CTACAGCTTT GAGGCTAAAT CATATGTCAA ATAAAACTAC
3961 TTCATCAAAA AACATGGAAA TGGTCCAACA GAAAAAAGAG GGCCCCATTC CACCAGATGC
4021 ACAAAATCCA GATATGTCGT TCTTTAAGAT GCTATTCTTG CCAGAATCAG CAAGGTGGAT
4081 ACAAAGGACT CATGGAAAGA ACTCTCTGAA CTCTGGGCAA GGCCCCAGTC CAAAGCAATT
4141 AGTATCCTTA GGACCAGAAA AATCTGTGGA AGGTCAGAAT TTCTTGTCTG AGAAAAACAA
4201 AGTGGTAGTA GGAAAGGGTG AATTTACAAA GGACGTAGGA CTCAAAGAGA TGGTTTTTCC
4261 AAGCAGCAGA AACCTATTTC TTACTAACTT GGATAATTTA CATGAAAATA ATACACACAA
4321 TCAAGAAAAA AAAATTCAGG AAGAAATAGA AAAGAAGGAA ACATTAATCC AAGAGAATGT
4381 AGTTTTGCCT CAGATACATA CAGTGACTGG CACTAAGAAT TTCATGAAGA ACCTTTTCTT
4441 ACTGAGCACT AGGCAAAATG TAGAAGGTTC ATATGACGGG GCATATGCTC CAGTACTTCA
4501 AGATTTTAGG TCATTAAATG ATTCAACAAA TAGAACAAAG AAACACACAG CTCATTTCTC
4561 AAAAAAAGGG GAGGAAGAAA ACTTGGAAGG CTTGGGAAAT CAAACCAAGC AAATTGTAGA
4621 GAAATATGCA TGCACCACAA GGATATCTCC TAATACAAGC CAGCAGAATT TTGTCACGCA
4681 ACGTAGTAAG AGAGCTTTGA AACAATTCAG ACTCCCACTA GAAGAAACAG AACTTGAAAA
4741 AAGGATAATT GTGGATGACA CCTCAACCCA GTGGTCCAAA AACATGAAAC ATTTGACCCC
4801 GAGCACCCTC ACACAGATAG ACTACAATGA GAAGGAGAAA GGGGCCATTA CTCAGTCTCC
4861 CTTATCAGAT TGCCTTACGA GGAGTCATAG CATCCCTCAA GCAAATAGAT CTCCATTACC
4921 CATTGCAAAG GTATCATCAT TTCCATCTAT TAGACCTATA TATCTGACCA GGGTCCTATT
4981 CCAAGACAAC TCTTCTCATC TTCCAGCAGC ATCTTATAGA AAGAAAGATT CTGGGGTCCA
5041 AGAAAGCAGT CATTTCTTAC AAGGAGCCAA AAAAAATAAC CTTTCTTTAG CCATTCTAAC
5101 CTTGGAGATG ACTGGTGATC AAAGAGAGGT TGGCTCCCTG GGGACAAGTG CCACAAATTC
5161 AGTCACATAC AAGAAAGTTG AGAACACTGT TCTCCCGAAA CCAGACTTGC CCAAAACATC
5221 TGGCAAAGTT GAATTGCTTC CAAAAGTTCA CATTTATCAG AAGGACCTAT TCCCTACGGA
5281 AACTAGCAAT GGGTCTCCTG GCCATCTGGA TCTCGTGGAA GGGAGCCTTC TTCAGGGAAC
5341 AGAGGGAGCG ATTAAGTGGA ATGAAGCAAA CAGACCTGGA AAAGTTCCCT TTCTGAGAGT
5401 AGCAACAGAA AGCTCTGCAA AGACTCCCTC CAAGCTATTG GATCCTCTTG CTTGGGATAA
5461 CCACTATGGT ACTCAGATAC CAAAAGAAGA GTGGAAATCC CAAGAGAAGT CACCAGAAAA
5521 AACAGCTTTT AAGAAAAAGG ATACCATTTT GTCCCTGAAC GCTTGTGAAA GCAATCATGC
5581 AATAGCAGCA ATAAATGAGG GACAAAATAA GCCCGAAATA GAAGTCACCT GGGCAAAGCA
5641 AGGTAGGACT GAAAGGCTGT GCTCTCAAAA CCCACCAGTC TTGAAACGCC ATCAACGGGA
5701 AATAACTCGT ACTACTCTTC AGTCAGATCA AGAGGAAATT GACTATGATG ATACCATATC
5761 AGTTGAAATG AAGAAGGAAG ATTTTGACAT TTATGATGAG GATGAAAATC AGAGCCCCCG
5821 CAGCTTTCAA AAGAAAACAC GACACTATTT TATTGCTGCA GTGGAGAGGC TCTGGGATTA
5881 TGGGATGAGT AGCTCCCCAC ATGTTCTAAG AAACAGGGCT CAGAGTGGCA GTGTCCCTCA
5941 GTTCAAGAAA GTTGTTTTCC AGGAATTTAC TGATGGCTCC TTTACTCAGC CCTTATACCG
6001 TGGAGAACTA AATGAACATT TGGGACTCCT GGGGCCATAT ATAAGAGCAG AAGTTGAAGA
6061 TAATATCATG GTAACTTTCA GAAATCAGGC CTCTCGTCCC TATTCCTTCT ATTCTAGCCT
6121 TATTTCTTAT GAGGAAGATC AGAGGCAAGG AGCAGAACCT AGAAAAAACT TTGTCAAGCC
6181 TAATGAAACC AAAACTTACT TTTGGAAAGT GCAACATCAT ATGGCACCCA CTAAAGATGA
6241 GTTTGACTGC AAAGCCTGGG CTTATTTCTC TGATGTTGAC CTGGAAAAAG ATGTGCACTC
6301 AGGCCTGATT GGACCCCTTC TGGTCTGCCA CACTAACACA CTGAACCCTG CTCATGGGAG
6361 ACAAGTGACA GTACAGGAAT TTGCTCTGTT TTTCACCATC TTTGATGAGA CCAAAAGCTG
6421 GTACTTCACT GAAAATATGG AAAGAAACTG CAGGGCTCCC TGCAATATCC AGATGGAAGA
6481 TCCCACTTTT AAAGAGAATT ATCGCTTCCA TGCAATCAAT GGCTACATAA TGGATACACT
6541 ACCTGGCTTA GTAATGGCTC AGGATCAAAG GATTCGATGG TATCTGCTCA GCATGGGCAG
6601 CAATGAAAAC ATCCATTCTA TTCATTTCAG TGGACATGTG TTCACTGTAC GAAAAAAAGA
6661 GGAGTATAAA ATGGCACTGT ACAATCTCTA TCCAGGTGTT TTTGAGACAG TGGAAATGTT
6721 ACCATCCAAA GCTGGAATTT GGCGGGTGGA ATGCCTTATT GGCGAGCATC TACATGCTGG
6781 GATGAGCACA CTTTTTCTGG TGTACAGCAA TAAGTGTCAG ACTCCCCTGG GAATGGCTTC
6841 TGGACACATT AGAGATTTTC AGATTACAGC TTCAGGACAA TATGGACAGT GGGCCCCAAA
6901 GCTGGCCAGA CTTCATTATT CCGGATCAAT CAATGCCTGG AGCACCAAGG AGCCCTTTTC
6961 TTGGATCAAG GTGGATCTGT TGGCACCAAT GATTATTCAC GGCATCAAGA CCCAGGGTGC
7021 CCGTCAGAAG TTCTCCAGCC TCTACATCTC TCAGTTTATC ATCATGTATA GTCTTGATGG
7081 GAAGAAGTGG CAGACTTATC GAGGAAATTC CACTGGAACC TTAATGGTCT TCTTTGGCAA
7141 TGTGGATTCA TCTGGGATAA AACACAATAT TTTTAACCCT CCAATTATTG CTCGATACAT
7201 CCGTTTGCAC CCAACTCATT ATAGCATTCG CAGCACTCTT CGCATGGAGT TGATGGGCTG
7261 TGATTTAAAT AGTTGCAGCA TGCCATTGGG AATGGAGAGT AAAGCAATAT CAGATGCACA
7321 GATTACTGCT TCATCCTACT TTACCAATAT GTTTGCCACC TGGTCTCCTT CAAAAGCTCG
7381 ACTTCACCTC CAAGGGAGGA GTAATGCCTG GAGACCTCAG GTGAATAATC CAAAAGAGTG
7441 GCTGCAAGTG GACTTCCAGA AGACAATGAA AGTCACAGGA GTAACTACTC AGGGAGTAAA
7501 ATCTCTGCTT ACCAGCATGT ATGTGAAGGA GTTCCTCATC TCCAGCAGTC AAGATGGCCA
7561 TCAGTGGACT CTCTTTTTTC AGAATGGCAA AGTAAAGGTT TTTCAGGGAA ATCAAGACTC
7621 CTTCACACCT GTGGTGAACT CTCTAGACCC ACCGTTACTG ACTCGCTACC TTCGAATTCA
7681 CCCCCAGAGT TGGGTGCACC AGATTGCCCT GAGGATGGAG GTTCTGGGCT GCGAGGCACA
7741 GGACCTCTAC
[0177] Полипептиды FVIII включают в себя полноразмерный FVIII, полноразмерный FVIII без Met на N-конце, зрелый FVIII (без сигнальной последовательности), зрелый FVIII с дополнительным Met на N-конце и/или FVIII с полной или частичной делецией домена B. В определенных вариантах осуществления варианты FVIII содержат делеции домена B, будь то частичные или полные делеции.
[0178] Последовательность нативного зрелого полипептида FVIII человека представлена под SEQ ID NO: 65. Нативный полипептид FVIII имеет следующую формулу: A1-a1-A2-a2-B-a3-A3-C1-C2, где A1, A2 и A3 представляют собой структурно родственные "домены A", B представляет собой "домен B", C1 и C2 представляют собой структурно родственные "домены C", а а1, а2 и а3 представляют собой кислые спейсерные области. Что касается положения первичной аминокислотной последовательности в SEQ ID NO: 65, домен A1 FVIII человека простирается от Ala1 до приблизительно Arg336, спейсерная область a1 простирается от приблизительно Met337 до приблизительно Val374, домен A2 простирается от приблизительно Ala375 до приблизительно Tyr719, спейсерная область a2 простирается от приблизительно Glu720 до приблизительно Arg740, домен B простирается от приблизительно Ser741 до приблизительно Arg 1648, спейсерная область a3 простирается от приблизительно Glu1649 до приблизительно Arg1689, домен A3 простирается от приблизительно Ser1690 до приблизительно Leu2025, C1 домен простирается от приблизительно Gly2026 до приблизительно Asn2072, а домен C2 простирается от приблизительно Ser2073 до Tyr2332. За исключением конкретных сайтов протеолитического расщепления, обозначение местоположений границ между доменами и областями FVIII может варьироваться в различных литературных источниках. Поэтому указанные в данном документе границы обозначены как примерные с использованием термина "приблизительно".
[0179] Ген FVIII человека был выделен и экспрессирован в клетках млекопитающих (Toole, J. J., et al., Nature 312:342-347 (1984); Gitschier, J., et al., Nature 312:326-330 (1984); Wood, W. I., et al., Nature 312:330-337 (1984); Vehar, G. A., et al., Nature 312:337-342 (1984); WO 87/04187; WO 88/08035; WO 88/03558; а также патент США № 4757006). Аминокислотная последовательность FVIII была выведена из кДНК, как показано в патенте США № 4965199. Кроме того, FVIII с частичной или полной делецией домена B показан в патентах США №№ 4994371 и 4868112. В некоторых вариантах осуществления домен B фактора FVIII человека замещен доменом B фактора V человека, как показано в патенте США № 5004803. Последовательность кДНК, кодирующая фактор VIII человека, и аминокислотная последовательность показаны под SEQ ID NO: 1 и 2 соответственно в публикации заявки на патент США № 2005/0100990.
[0180] Последовательность FVIII свиньи опубликована в Toole, J. J., et al., Proc. Natl. Acad. Sci. USA 83:5939-5942 (1986). Кроме того, полная последовательность кДНК свиньи, полученная в результате ПЦР-амплификации последовательностей FVIII из библиотеки кДНК селезенки свиньи, описана в Healey, J. F., et al., Blood 88:4209-4214 (1996). Гибридный FVIII человека/свиньи, имеющий замены всех доменов, всех субъединиц и конкретных аминокислотных последовательностей, раскрыт в патенте США № 5364771 авторства Lollar и Runge, а также в WO 93/20093. Совсем недавно нуклеотидные и соответствующие аминокислотные последовательности доменов A1 и A2 FVIII свиньи и химерного FVIII с доменами A1 и/или A2 свиньи, замещенными соответствующими доменами человека, были описаны в WO 94/11503. В патенте США № 5859204, Lollar, J. S., также раскрыты кДНК свиньи и выведенные аминокислотные последовательности. В патенте США № 6458563 описан FVIII свиньи с делецией домена B.
[0181] В патенте США № 5859204, выданном Lollar, J. S., сообщается о функциональных мутантных формах FVIII, характеризующихся пониженной антигенностью и пониженной иммунореактивностью. В патенте США № 6376463, выданном Lollar, J. S., также сообщается о мутантных формах FVIII, характеризующихся пониженной иммунореактивностью. В публикации заявки на патент США № 2005/0100990 авторства Saenko et al. сообщается о функциональных мутациях в домене A2 FVIII.
[0182] В некоторых вариантах осуществления полипептид FVIII (или часть FVIII химерного полипептида) может быть на по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичным аминокислотной последовательности FVIII из аминокислот 1-1438 SEQ ID NO: 67 или аминокислот 1-2332 SEQ ID NO: 65 (без сигнальной последовательности) или аминокислотной последовательности FVIII из аминокислот 1-19 SEQ ID NO: 64 и 1-1438 SEQ ID NO: 67 или аминокислот 1-19 SEQ ID NO: 64 и аминокислот 1-2332 SEQ ID NO: 65 (с сигнальной последовательностью), где FVIII обладает свертывающей активностью, например, активирует фактор IX в качестве кофактора для превращения фактора X в активированный фактор X. Полипептид FVIII (или часть химерного полипептида, представляющая собой FVIII) может быть идентичен аминокислотной последовательность FVIII из аминокислот 1-1438 SEQ ID NO: 67 или аминокислот 1-2332 SEQ ID NO: 65 (без сигнальной последовательности). Полипептид FVIII может дополнительно содержать сигнальную последовательность.
[0183] "Домен B" в FVIII, как используется в данном документе, является тем же доменом В, известным из уровня техники, который определяется по идентичности внутренней аминокислотной последовательности и сайтам протеолитического расщепления, например, остаткам Ser741-Arg1648 полноразмерного FVIII человека. Другие домены FVIII человека определяются по следующим аминокислотным остаткам: A1 - остатки Ala1-Arg372; A2 - остатки Ser373-Arg740; A3 - остатки Ser1690-Asn2019; C1 - остатки Lys2020-Asn2172; C2 - остатки Ser2173-Tyr2332. Последовательность A3-C1-C2 содержит остатки Ser1690-Tyr2332. Оставшаяся последовательность, остатки Glu1649-Arg1689, обычно называется кислой областью а3. Из уровня техники также известны местоположения границ всех доменов, в том числе доменов B, для FVIII свиньи, мыши и собаки. В некоторых вариантах осуществления домен B FVIII подвергнут делеции (как в "факторе VIII с делецией домена В" или "FVIII BDD"). Примером FVIII BDD является REFACTO® (рекомбинантный FVIII BDD), который имеет ту же последовательность, что и часть последовательности фактора VIII в таблице 4. (Тяжелая цепь FVIII BDD выделена двойным подчеркиванием; домен B выделен курсивом; легкая цепь FVIII BDD представлена в виде обычного текста). В некоторых вариантах осуществления FVIII с доменом B имеет делецию всего домена B, за исключением 5 аминокислот, как показано в таблице 5 (SEQ ID NO: 68) (домен B выделен курсивом).
[0184] "FVIII с делецией домена B" может иметь полную или частичную делеции, раскрытые в патентах США №№ 6316226, 6346513, 7041635, 5789203, 6060447, 5595886, 6228620, 5972885, 6048720, 5543502, 5610278, 5171844, 5112950, 4868112 и 6458563, публикациях заявок на патент США №№ 2017/0073393 A1 и 2012/308641 A1, а также международных публикациях №№ WO 2011/041770 A1, WO 2015/106052 A1 (PCT/US2015/010738) и WO 2016/025764. В некоторых вариантах осуществления последовательность FVIII с делецией домена B, применяемая в способах по настоящему изобретению, содержит любую из делеций, раскрытых от столбца 4, строки 4 до столбца 5, строки 28 и в примерах 1-5 патента США № 6316226 (также в US 6346513). В других вариантах осуществления фактор VIII с делецией домена В представляет собой фактор VIII с делецией S743/Q1638 домена В (FVIII BDD SQ) (например, фактор VIII, имеющий делецию от аминокислоты 744 до аминокислоты 1637, например, фактор VIII, содержащий аминокислоты 1-743 и аминокислоты 1638-2332 зрелого FVIII). В некоторых вариантах осуществления FVIII с делецией домена B, применяемый в способах по настоящему изобретению, имеет делецию, раскрытую в столбце 2, строках 26-51 и в примерах 5-8 патента США № 5789203 (также в US 6060447, US 5595886 и US 6228620). В некоторых вариантах осуществления фактор VIII с делецией домена В имеет делецию, раскрытую от столбца 1, строки 25 до столбца 2, строки 40 патента США № 5972885; в столбце 6, строках 1-22 и в примере 1 патента США № 6048720; в столбце 2, строках 17-46 патента США № 5543502; от столбца 4, строки 22 до столбца 5, строки 36 патента США № 5171844; в столбце 2, строках 55-68, на фигуре 2 и в примере 1 патента США № 5112950; от столбца 2, строки 2 до столбца 19, строки 21 и в таблице 2 патента США № 4868112; от столбца 2, строки 1 до столбца 3, строки 19, от столбца 3, строки 40 до столбца 4, строки 67, от столбца 7, строки 43 до столбца 8, строки 26 и от столбца 11, строки 5 до столбца 13, строки 39 патента США № 7041635 или в столбце 4, строках 25-53 патента США № 6458563. В некоторых вариантах осуществления полипептид FVIII с делецией домена B имеет делецию большей части домена В, но по-прежнему содержит аминоконцевые последовательности домена В, существенно важные для протеолитического процессинга первичного продукта трансляции in vivo до двух полипептидных цепей, как раскрыто в WO 91/09122. В некоторых вариантах осуществления полипептид FVIII с делецией домена B сконструирован с делецией аминокислот 747-1638, т. е. с практически полной делецией домена В. Hoeben R.C., et al. J. Biol. Chem. 265 (13): 7318-7323 (1990). Полипептидный фактор VIII с делецией домена В также может содержать делецию аминокислот 771-1666 или аминокислот 868-1562 FVIII. Meulien P., et al. Protein Eng. 2(4): 301-6 (1988). Дополнительные варианты делеции В-домена, которые являются частью настоящего изобретения, включают делецию аминокислот 982-1562 или 760-1639 (Toole et al., Proc. Natl. Acad. Sci. U.S.A. (1986) 83, 5939-5942)), 797-1562 (Eaton, et al. Biochemistry (1986) 25:8343-8347)), 741-1646 (Kaufman (опубликованная заявка согласно PCT № WO 87/04187)), 747-1560 (Sarver, et al., DNA (1987) 6:553-564)), 741-1648 (Pasek (заявка согласно PCT № 88/00831)), или 816-1598, или 741-1648 (Lagner (Behring Inst. Mitt. (1988) No 82:16-25, EP 295597)). В конкретных вариантах осуществления полипептид FVIII с делецией домена B содержит делецию аминокислотных остатков 746-1648 зрелого FVIII (что соответствует делеции 765-1665 полноразмерного FVIII). В других вариантах осуществления полипептид FVIII с делецией домена B содержит делецию аминокислотных остатков 745-1648 зрелого FVIII (что соответствует делеции 764-1665 полноразмерного FVIII).
[0185] В других вариантах осуществления FVIII BDD включает в себя полипептид FVIII, содержащий фрагменты домена В, в которых сохранены один или несколько сайтов N-связанного гликозилирования, например, остатки 757, 784, 828, 900, 963 или необязательно 943, которые соответствуют аминокислотной последовательности полноразмерной последовательности FVIII. Примеры фрагментов домена В включают 226 аминокислот или 163 аминокислоты из домена В, как раскрыто в Miao, H.Z., et al., Blood 103(a): 3412-3419 (2004), Kasuda, A, et al., J. Thromb. Haemost. 6. 1352-1359 (2008) и Pipe, S.W., et al., J. Thromb. Haemost. 9. 2235-2242 (2011) (т. е. сохранены первые 226 аминокислот или 163 аминокислоты из домена В). В еще нескольких других вариантах осуществления FVIII BDD дополнительно содержит точечную мутацию в остатке 309 (по типу замены Phe на Ser) для улучшения экспрессии полипептида FVIII BDD. См. Miao, H.Z., et al., Blood 103(a): 3412-3419 (2004). В еще нескольких других вариантах осуществления FVIII BDD включает в себя полипептид FVIII, содержащий часть домена В, но не содержащий один или несколько сайтов расщепления фурином (например, Arg1313 и Arg 1648). См. Pipe, S.W., et al., J. Thromb. Haemost. 9. 2235-2242 (2011). В некоторых вариантах осуществления FVIII BDD включает в себя одноцепочечный FVIII, который содержит делецию аминокислот 765-1652, соответствующих зрелому полноразмерному FVIII (также известный как одноцепочечный rFVIII и AFSTYLA®). См. патент США № 7041635. Каждый из вышеизложенных вариантов делеции может быть осуществлен в любой последовательности FVIII.
[0186] В некоторых вариантах осуществления FVIII имеет частичный домен B. В некоторых вариантах осуществления полипептид FVIII с частичным доменом B представляет собой FVIII198. FVIII198 представляет собой одноцепочечную молекулу FVIIIFc-226N6, содержащую частичный домен B. Число 226 представляет N-концевые 226 аминокислот домена B FVIII, а N6 представляет шесть сайтов N-гликозилирования в домене B.
[0187] В определенных вариантах осуществления полипептид FVIII выбран из полипептида FVIII, раскрытого в публикациях №№ WO 2017/117630 A1, WO 2018/087271 A1, US 9878017 B2, US 8575104 B2, US 8754194 B2, US 7939632 B2, US 2018/0161402 A1, US 9956269 B2, US 9107902 B2, US 2017/209546 A1.
[0188] В некоторых вариантах осуществления FVIII расщепляется сразу после аргинина по аминокислоте 1648 (в полноразмерном факторе VIII или SEQ ID NO: 65), аминокислоте 754 (в факторе VIII с делецией S743/Q1638 домена B или в SEQ ID NO: 67) или соответствующему аргининовому остатку (в других вариантах), в результате чего образуются тяжелая цепь и легкая цепь. В других вариантах осуществления полипептид FVIII содержит тяжелую цепь и легкую цепь, которые соединены или связаны нековалентной связью, опосредованной ионами металлов.
[0189] В других вариантах осуществления FVIII представляет собой одноцепочечный FVIII, который не был расщеплен сразу после аргинина по аминокислоте 1648 (в полноразмерном FVIII или SEQ ID NO: 65), аминокислоте 754 (в FVIII с делецией S743/Q1638 домена B или в SEQ ID NO: 67) или соответствующему аргининовому остатку (в других вариантах). Одноцепочечный FVIII может содержать одну или несколько аминокислотных замен. В некоторых вариантах осуществления аминокислотная замена происходит по остатку, соответствующему остатку 1648, остатку 1645 или им обоим в полноразмерном зрелом полипептидном факторе VIII (SEQ ID NO: 65) или остатку 754, остатку 751 или им обоим в факторе VIII BDD SQ (SEQ ID NO: 67). Заменяющая аминокислота может представлять собой любую аминокислоту, отличную от аргинина, например, изолейцин, лейцин, лизин, метионин, фенилаланин, треонин, триптофан, валин, аланин, аспарагин, аспарагиновую кислоту, цистеин, глутаминовую кислоту, глутамин, глицин, пролин, селеноцистеин, серин, тирозин, гистидин, орнитин, пирролизин или таурин.
[0190] FVIII может дополнительно расщепляться тромбином и затем активироваться в виде FVIIIa, выступая в качестве кофактора для активированного фактора IX (FIXa). А активированный FIX вместе с активированным FVIII образуют X-азный комплекс и превращают фактор X в активированный фактор X (FXa). Для активации FVIII расщепляется тромбином после трех аргининовых остатков по аминокислотам 372, 740 и 1689 (которые соответствуют аминокислотам 372, 740 и 795 в последовательности FVIII с делецией домена B), при этом при расщеплении образуется FVIIIa, имеющий цепи A1 размером 50 кДа, A2 размером 43 кДа и A3-C1-C2 размером 73 кДа. В некоторых вариантах осуществления полипептид FVIII, применимый для настоящего изобретения, представляет собой неактивный FVIII. В других вариантах осуществления полипептид FVIII представляет собой активированный FVIII.
[0191] Белок, имеющий полипептид FVIII, соединенный или связанный с белком VWF, может содержать последовательность, на по меньшей мере 50%, 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO: 65 или 67, где последовательность обладает свертывающей активностью FVIII, например, активируя фактор IX в качестве кофактора для превращения фактора X в активированный фактор X (FXa).
[0192] "Гибридные" или "химерные" полипептиды и белки, как используется в данном документе, содержат комбинацию первого полипептида и второго полипептида. В некоторых вариантах осуществления гибридный или химерный полипептид содержит одну полипептидную цепь, например, в случае с химерным полипептидом, содержащим полипептид FVIII и XTEN. См., например, US 2015/0158929 A1, включенный в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления гибридный или химерный полипептид содержит комбинацию первой полипептидной цепи, например, фрагмента VWF, слитого с последовательностью XTEN и первой константной областью Ig или ее частью, и второй полипептидной цепи, например, полипептида FVIII, слитого со второй константной областью Ig или ее частью, с образованием тем самым гетеродимера. См., например, US 2015/0266943 A1, US 2016/0251408 A1, US 2017/0073393 A1, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления первый полипептид и второй полипептид в гибридной молекуле могут быть связаны друг с другом посредством белок-белковых взаимодействий, таких как взаимодействия между зарядами или гидрофобные взаимодействия. В других вариантах осуществления первый полипептид содержит слитый белок белок VWF-XTEN-Fc, а второй полипептид содержит слитый белок FVIII-Fc, что делает гибрид гетеродимером, в котором XTEN содержит менее 288 аминокислот. В других вариантах осуществления первый полипептид содержит слитый белок белок VWF-XTEN-Fc, а второй полипептид содержит слитый белок FVIII(X)-Fc, что делает гибрид гетеродимером, в котором XTEN содержит менее 288 аминокислот. Первый полипептид и второй полипептид могут быть связаны посредством ковалентной связи, например, дисульфидной связи, между первой Fc-областью и второй Fc-областью. Первый полипептид и второй полипептид могут быть дополнительно связаны друг с другом посредством образования связи между фрагментом VWF и полипептидом FVIII.
[0193] В определенных вариантах осуществления химерный полипептид, раскрытый в данном документе, содержит белок FVIII, содержащий первый фрагмент полипептида FVIII, последовательность XTEN, второй фрагмент полипептида FVIII и Fc-область. В некоторых вариантах осуществления белок FVIII содержит в порядке от N-конца к C-концу первый фрагмент полипептида FVIII, слитый с последовательностью XTEN, слитой со вторым фрагментом полипептида FVIII, слитым с Fc-областью. В конкретном варианте осуществления белок FVIII содержит первый фрагмент полипептида FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 215; последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 8; второй полипептид FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 216; и/или первую Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217.
[0194] В одном конкретном варианте осуществления химерный полипептид представляет собой rFVIIIFc-VWF-XTEN и содержит белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 201. В другом конкретном варианте осуществления rFVIIIFc-VWF-XTEN содержит аминокислотную последовательность под SEQ ID NO: 207.
[0195] В некоторых вариантах осуществления белок FVIII дополнительно содержит последовательность сигнального пептида FVIII. В одном конкретном варианте осуществления белок FVIII содержит сигнальный пептид FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 64, первый фрагмент полипептида FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 215; последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 8; второй полипептид FVIII, который содержит аминокислотную последовательность под SEQ ID NO: 216; и/или первую Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217.
[0196] В одном конкретном варианте осуществления химерный полипептид представляет собой rFVIIIFc-VWF-XTEN и содержит белок FVIII, содержащий аминокислотную последовательность под SEQ ID NO: 203. В другом конкретном варианте осуществления rFVIIIFc-VWF-XTEN содержит белок FVIII, кодируемый последовательностью нуклеиновой кислоты под SEQ ID NO: 204, или его фрагмент. Дополнительные иллюстративные полипептидные последовательности, относящиеся к белкам FVIII химерных полипептидов, раскрытых в данном документе, представлены в таблицах 18-19.
[0197] Известно множество функциональных вариантов FVIII, обсуждаемых выше и ниже. Кроме того, у пациентов с гемофилией были идентифицированы сотни нефункциональных мутаций в FVIII, и было определено, что эффект данных мутаций в отношении функции FVIII обусловлен в большей степени их местоположением в 3-мерной структуре FVIII, а не природой замены (Cutler et al., Hum. Mutat. 19:274-8 (2002), включенная в данный документ посредством ссылки во всей своей полноте). Кроме того, путем сравнения FVIII от людей и FVIII от других видов были идентифицированы консервативные остатки, которые, вероятно, требуются для функционирования (Cameron et al., Thromb. Haemost. 79:317-22 (1998); US 6251632, включенные в данный документ посредством ссылки во всей своей полноте).
III.B. Фрагменты фактора фон Виллебранда (VWF)
[0198] VWF (также известный как F8VWF) представляет собой крупный мультимерный гликопротеин, присутствующий в плазме крови и вырабатываемый конститутивно в эндотелии (в тельцах Вайбеля-Паладе), мегакариоцитах (α-гранулах тромбоцитов) и субэндотелиальной соединительной ткани. Основным мономером VWF является белок из 2813 аминокислот. Каждый мономер содержит ряд специфических доменов с конкретной функцией - домен D'/D3 (который связывается с фактором VIII), домен A1 (который связывается с тромбоцитарным рецептором GPIb, гепарином и/или, возможно, коллагеном), домен A3 (который связывается с коллагеном), домен C1 (в котором домен RGD связывается с тромбоцитарным интегрином αIIbβ3, когда он активирован) и домен "цистеиновый узел" на C-конце белка (который является общим для VWF и тромбоцитарного фактора роста (PDGF), трансформирующего фактора роста β (TGFβ) и β-субъединицы хорионического гонадотропина человека (βHCG)).
[0199] Термин "фрагмент VWF", используемый в данном документе, включает без ограничения функциональные фрагменты VWF, содержащие домен D' и домен D3, которые способны ингибировать связывание эндогенного VWF с FVIII. В некоторых вариантах осуществления фрагмент VWF связывается с белком FVIII. В других вариантах осуществления фрагмент VWF блокирует сайт связывания VWF в белке FVIII, тем самым ингибируя взаимодействие белка FVIII с эндогенным VWF. В других вариантах осуществления фрагмент VWF блокирует связывание FVIII с эндогенным VWF, тем самым предотвращая выведение белка FVIII через путь выведения VWF. Фрагменты VWF включают производные, варианты, мутантные формы или аналоги, у которых сохраняются эти виды активности VWF.
[0200] Последовательность мономерного VWF человека из 2813 аминокислот приведена в Genbank под номером доступа NP000543.2. Нуклеотидная последовательность, кодирующая VWF человека, приведена в Genbank под номером доступа NM000552.3. Нуклеотидная последовательность VWF человека обозначена как SEQ ID NO: 20. SEQ ID NO: 21 представляет собой аминокислотную последовательность полноразмерного VWF. Каждый домен VWF приведен в таблице 6.
ТАБЛИЦА 6. Последовательности VWF (человека)
SEQ ID NO: 208
(Аминокислоты 1-22 из SEQ ID NO: 21)
SEQ ID NO: 209
(Аминокислоты 23-763 из SEQ ID NO: 21)
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKR 763
SEQ ID NO: 210
(Аминокислоты 764-866 из SEQ ID NO: 21)
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDAT 866
SEQ ID NO: 211
(Аминокислоты 867-1240 из SEQ ID NO: 21)
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCACF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY ECEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP 1240
SEQ ID NO: 212
(Аминокислоты 1241-1479 из SEQ ID NO: 21)
1251 PVSPTTLYVE DISEPPLHDF YCSRLLDLVF LLDGSSRLSE AEFEVLKAFV
1301 VDMMERLRIS QKWVRVAVVE YHDGSHAYIG LKDRKRPSEL RRIASQVKYA
1351 GSQVASTSEV LKYTLFQIFS KIDRPEASRI ALLLMASQEP QRMSRNFVRY
1401 VQGLKKKKVI VIPVGIGPHA NLKQIRLIEK QAPENKAFVL SSVDELEQQR
1451 DEIVSYLCDL APEAPPPTLP PDMAQVTVG 1479
C-конца
SEQ ID NO: 213
(Аминокислоты 1480-2813 из SEQ ID NO: 21)
1501 FVLEGSDKIG EADFNRSKEF MEEVIQRMDV GQDSIHVTVL QYSYMVTVEY
1551 PFSEAQSKGD ILQRVREIRY QGGNRTNTGL ALRYLSDHSF LVSQGDREQA
1601 PNLVYMVTGN PASDEIKRLP GDIQVVPIGV GPNANVQELE RIGWPNAPIL
1651 IQDFETLPRE APDLVLQRCC SGEGLQIPTL SPAPDCSQPL DVILLLDGSS
1701 SFPASYFDEM KSFAKAFISK ANIGPRLTQV SVLQYGSITT IDVPWNVVPE
1751 KAHLLSLVDV MQREGGPSQI GDALGFAVRY LTSEMHGARP GASKAVVILV
1801 TDVSVDSVDA AADAARSNRV TVFPIGIGDR YDAAQLRILA GPAGDSNVVK
1851 LQRIEDLPTM VTLGNSFLHK LCSGFVRICM DEDGNEKRPG DVWTLPDQCH
1901 TVTCQPDGQT LLKSHRVNCD RGLRPSCPNS QSPVKVEETC GCRWTCPCVC
1951 TGSSTRHIVT FDGQNFKLTG SCSYVLFQNK EQDLEVILHN GACSPGARQG
2001 CMKSIEVKHS ALSVEXHSDM EVTVNGRLVS VPYVGGNMEV NVYGAIMHEV
2051 RFNHLGHIFT FTPQNNEFQL QLSPKTFASK TYGLCGICDE NGANDFMLRD
2101 GTVTTDWKTL VQEWTVQRPG QTCQPILEEQ CLVPDSSHCQ VLLLPLFAEC
2151 HKVLAPATFY AICQQDSCHQ EQVCEVIASY AHLCRTNGVC VDWRTPDFCA
2201 MSCPPSLVYN HCEHGCPRHC DGNVSSCGDH PSEGCFCPPD KVMLEGSCVP
2251 EEACTQCIGE DGVQHQFLEA WVPDHQPCQI CTCLSGRKVN CTTQPCPTAK
2301 APTCGLCEVA RLRQNADQCC PEYECVCDPV SCDLPPVPHC ERGLQPTLTN
2351 PGECRPNFTC ACRKEECKRV SPPSCPPHRL PTLRKTQCCD EYECACNCVN
2401 STVSCPLGYL ASTATNDCGC TTTTCLPDKV CVHRSTIYPV GQFWEEGCDV
2451 CTCTDMEDAV MGLRVAQCSQ KPCEDSCRSG FTYVLHEGEC CGRCLPSACE
2501 VVTGSPRGDS QSSWKSVGSQ WASPENPCLI NECVRVKEEV FIQQRNVSCP
2551 QLEVPVCPSG FQLSCKTSAC CPSCRCERME ACMLNGTVIG PGKTVMIDVC
2601 TTCRCMVQVG VISGFKLECR KTTCNPCPLG YKEENNTGEC CGRCLPTACT
2651 IQLRGGQIMT LKRDETLQDG CDTHFCKVNE RGEYFWEKRV TGCPPFDEHK
2701 CLAEGGKIMK IPGTCCDTCE EPECNDITAR LQYVKVGSCK SEVEVDIHYC
2751 QGKCASKAMY SIDINDVQDQ CSCCSPTRTE PMQVALHCTN GSVVYHEVLN
2801 AMECKCSPRK CSK 2813
[0201] Белок VWF, используемый в данном документе, может представлять собой фрагмент VWF, содержащий домен D' и домен D3 VWF, где фрагмент VWF связывается с фактором VIII (FVIII) и ингибирует связывание эндогенного VWF (полноразмерного VWF) с FVIII. Фрагмент VWF, содержащий домен D' и домен D3, может дополнительно содержать домен VWF, выбранный из группы, состоящей из домена A1, домена A2, домена A3, домена D1, домена D2, домена D4, домена B1, домена B2, домена B3, домена C1, домена C2, домена CK, одного или нескольких их фрагментов и любых их комбинаций. В некоторых вариантах осуществления фрагмент VWF содержит: (1) домены D' и D3 VWF или их фрагменты; (2) домены D1, D' и D3 VWF или их фрагменты; (3) домены D2, D' и D3 VWF или их фрагменты; (4) домены D1, D2, D' и D3 VWF или их фрагменты или (5) домены D1, D2, D', D3 и A1 VWF или их фрагменты, состоит по сути из них или состоит из них. Фрагмент VWF, описанный в данном документе, не содержит сайт связывания с рецептором, опосредующим выведение VWF. В других вариантах осуществления фрагмент VWF, описанный в данном документе, не представляет собой аминокислоты 764-1274 из SEQ ID NO: 21. Фрагмент VWF по настоящему изобретению может содержать любые другие последовательности, соединенные или слитые с фрагментом VWF. Например, фрагмент VWF, описанный в данном документе, может дополнительно содержать сигнальный пептид.
[0202] В некоторых вариантах осуществления фрагмент VWF, содержащий домен D' и домен D3, связывается с белком FVIII или соединен с ним. См., например, US 2015/0023959 A1, US 2015/0266943 A1, US 2016/0251408 A1, US 2017/0073393 A1, US 2018/185455 A1, US 2018/0051067 A1, US 2017/0152300 A1, US 9878017 B2, US 9458223 B2, US 8575104 B2, WO 2017/117630 A1, US 2018/0161402 A1, WO 2017/117631 A1, WO 2018/087271 A1, US 9107902 B2, WO 2017/222337 A1, WO 2015/185758 A1, каждая из которых включена в данный документ посредством ссылки во всей своей полноте. Связываясь или соединяясь с белком FVIII, фрагмент VWF по настоящему изобретению защищает FVIII от расщепления протеазой и активации FVIII, стабилизирует тяжелую цепь и легкую цепь FVIII и предотвращает выведение FVIII, опосредованное фагоцитарными рецепторами. В других вариантах осуществления фрагмент VWF связывается или соединяется с белком FVIII и блокирует или предотвращает связывание белка FVIII с фосфолипидом и активированным белком C. Посредством предотвращения или ингибирования связывания белка FVIII с эндогенным полноразмерным VWF фрагмент VWF по настоящему изобретению снижает выведение FVIII с помощью рецепторов, опосредующих выведение VWF и таким образом продлевает период полужизни химерного полипептида. Продление периода полужизни химерного полипептида, таким образом, происходит вследствие связывания или соединения с фрагментом VWF, в котором отсутствует сайт связывания рецептора, опосредующего выведение VWF, с белком FVIII, и экранирования или защиты белка FVIII от эндогенного VWF, который содержит сайт связывания рецептора, опосредующего выведение VWF, с помощью фрагмента VWF. Белок FVIII, связанный с фрагментом VWF или защищенный им, также может обеспечивать рециркуляцию белка FVIII. Посредством устранения сайтов связывания рецепторов пути выведения VWF, содержащихся в полноразмерной молекуле VWF, гетеродимеры FVIII/VWF по настоящему изобретению экранируются от пути выведения VWF с дополнительным продлением периода полужизни FVIII.
[0203] В некоторых вариантах осуществления белок VWF, применимый для настоящего изобретения, содержит домен D' и домен D3 VWF, где домен D' на по меньшей мере 60%, 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичен аминокислотам 764-866 из SEQ ID NO: 21, где белок VWF предотвращает или ингибирует связывание эндогенного VWF с FVIII. В других вариантах осуществления белок VWF содержит домен D' и домен D3 VWF, где домен D3 на по меньшей мере 60%, 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичен аминокислотам 867-1240 из SEQ ID NO: 21, где белок VWF предотвращает или ингибирует связывание эндогенного VWF с FVIII. В некоторых вариантах осуществления белок VWF, описанный в данном документе, содержит домен D' и домен D3 VWF, которые на по меньшей мере 60%, 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичны аминокислотам 764-1240 из SEQ ID NO: 21, состоит по сути из них или состоит из них, где белок VWF предотвращает или ингибирует связывание эндогенного VWF с FVIII. В других вариантах осуществления белок VWF содержит домены D1, D2, D' и D3, на по меньшей мере 60%, 70%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичные аминокислотам 23-1240 из SEQ ID NO: 21, состоит по сути из них или состоит из них, где белок VWF предотвращает или ингибирует связывание эндогенного VWF с FVIII. В еще нескольких других вариантах осуществления белок VWF дополнительно содержит сигнальный пептид, функционально связанный с ним.
[0204] В некоторых вариантах осуществления белок VWF, применимый для настоящего изобретения, состоит по сути из (1) домена D'D3, домена D1D'D3, домена D2D'D3 или домена D1D2D'D3 и (2) дополнительной последовательности VWF размером до приблизительно 10 аминокислот (например, любых последовательностей от аминокислот 764-1240 из SEQ ID NO: 21 до аминокислот 764-1250 из SEQ ID NO: 21), до приблизительно 15 аминокислот (например, любых последовательностей от аминокислот 764-1240 из SEQ ID NO: 21 до аминокислот 764-1255 из SEQ ID NO: 21), до приблизительно 20 аминокислот (например, любых последовательностей от аминокислот 764-1240 из SEQ ID NO: 21 до аминокислот 764-1260 из SEQ ID NO: 21), до приблизительно 25 аминокислот (например, любых последовательностей от аминокислот 764-1240 из SEQ ID NO: 21 до аминокислот 764-1265 из SEQ ID NO: 21) или до приблизительно 30 аминокислот (например, любых последовательностей от аминокислот 764-1240 из SEQ ID NO: 21 до аминокислот 764-1260 из SEQ ID NO: 21) или состоит по сути из них. В некоторых вариантах осуществления белок VWF, содержащий домен D' и домен D3 или состоящий по сути из них, не представляет собой ни аминокислоты 764-1274 из SEQ ID NO: 21, ни полноразмерный зрелый VWF. В некоторых вариантах осуществления домен D1D2 экспрессируется в транс-положении относительно домена D'D3. В некоторых вариантах осуществления домен D1D2 экспрессируется в цис-положении относительно домена D'D3.
[0205] В других вариантах осуществления белок VWF, содержащий домены D'D3, соединенные с доменами D1D2, дополнительно содержит сайт внутриклеточного расщепления (например, сайт расщепления PACE (фурином) или PC5), позволяющий отщеплять домены D1D2 от доменов D'D3 при экспрессии. Неограничивающие примеры сайта внутриклеточного расщепления раскрыты в данном документе в другом месте.
[0206] В еще некоторых других вариантах осуществления белок VWF содержит домен D' и домен D3, но не содержит аминокислотную последовательность, выбранную из группы, состоящей из последовательностей (1) аминокислот 1241-2813, соответствующих SEQ ID NO: 21, (2) от аминокислоты 1270 до аминокислоты 2813, соответствующих SEQ ID NO: 21, (3) от аминокислоты 1271 до аминокислоты 2813, соответствующих SEQ ID NO: 21, (4) от аминокислоты 1272 до аминокислоты 2813, соответствующих SEQ ID NO: 21, (5) от аминокислоты 1273 до аминокислоты 2813, соответствующих SEQ ID NO: 21, (6) от аминокислоты 1274 до аминокислоты 2813, соответствующих SEQ ID NO: 21, и любых их комбинаций.
[0207] В еще нескольких других вариантах осуществления белок VWF по настоящему изобретению содержит аминокислотную последовательность, соответствующую домену D', домену D3 и домену A1, состоит по сути из нее или состоит из нее, где аминокислотная последовательность на по меньшей мере 60%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотам 764-1479 из SEQ ID NO: 21, где белок VWF предотвращает связывание эндогенного VWF с FVIII. В конкретных вариантах осуществления белок VWF не представляет собой аминокислоты 764-1274 из SEQ ID NO: 21.
[0208] В некоторых вариантах осуществления белок VWF по настоящему изобретению содержит домен D' и домен D3, но не содержит по меньшей мере один домен VWF, выбранный из группы, состоящей из (1) домена A1, (2) домена A2, (3) домена A3, (4) домена D4, (5) домена B1, (6) домена B2, (7) домена B3, (8) домена C1, (9) домена C2, (10) домена CK, (11) домена CK и домена C2, (12) домена CK, домена C2 и домена C1, (13) домена CK, домена C2, домена C1, домена B3, (14) домена CK, домена C2, домена C1, домена B3, домена B2, (15) домена CK, домена C2, домена C1, домена B3, домена B2 и домена B1, (16) домена CK, домена C2, домена C1, домена B3, домена B2, домена B1 и домена D4, (17) домена CK, домена C2, домена C1, домена B3, домена B2, домена B1, домена D4 и домена A3, (18) домена CK, домена C2, домена C1, домена B3, домена B2, домена B1, домена D4, домена A3 и домена A2, (19) домена CK, домена C2, домена C1, домена B3, домена B2, домена B1, домена D4, домена A3, домена A2 и домена A1, а также (20) любых их комбинаций.
[0209] В еще некоторых других вариантах осуществления белок VWF содержит домены D'D3 и один или несколько доменов или модулей. Примеры таких доменов или модулей включают без ограничения домены и модули, раскрытые в Zhour et al., Blood, опубликованной в Интернете 6 апреля 2012 г.: DOI 10.1182/blood-2012-01-405134, включенной в данный документ посредством ссылки во всей своей полноте. Например, белок VWF может содержать домен D'D3 и один или несколько доменов или модулей, выбранных из группы, состоящей из домена A1, домена A2, домена A3, модуля D4N, модуля VWD4, модуля C8-4, модуля TIL-4, модуля C1, модуля C2, модуля C3, модуля C4, модуля C5, модуля C5, модуля C6 и любых их комбинаций.
[0210] В еще нескольких других вариантах осуществления белок VWF соединен с гетерологичным компонентом, где гетерологичный компонент соединен с N-концом или C-концом белка VWF или вставлен непосредственно ниже одной или нескольких аминокислот (например, в одном или нескольких сайтах вставки XTEN) в белке VWF. Например, сайты вставки гетерологичного компонента в белке VWF могут находиться в домене D', домене D3 или в них обоих. Гетерологичный компонент может представлять собой компонент, обеспечивающий продление периода полужизни.
[0211] В определенных вариантах осуществления белок VWF, применимый для настоящего изобретения, образует мультимер, например, димер, тример, тетрамер, пентамер, гексамер, гептамер или мультимеры более высокого порядка. В других вариантах осуществления белок VWF представляет собой мономер, содержащий только один белок VWF. В некоторых вариантах осуществления белок VWF по настоящему изобретению может иметь одну или несколько аминокислотных замен, делеций, добавлений или модификаций. В некоторых вариантах осуществления белок VWF может содержать аминокислотные замены, делеции, добавления или модификации, при которых белок VWF не способен образовывать дисульфидную связь или образовывать димер или мультимер. В других вариантах осуществления аминокислотная замена находится в домене D' и домене D3. В конкретном варианте осуществления белок VWF, применимый для настоящего изобретения, содержит по меньшей мере одну аминокислотную замену по остатку, соответствующему остатку 1099, остатку 1142 или как остатку 1099, так и остатку 1142, соответствующим SEQ ID NO: 21. По меньшей мере одна заменяющая аминокислота может представлять собой любую аминокислоту, которая не встречается в природе в VWF дикого типа. Например, заменяющая аминокислота может представлять собой любую аминокислоту, отличную от цистеина, например, изолейцин, аланин, лейцин, аспарагин, лизин, аспарагиновую кислоту, метионин, фенилаланин, глутаминовую кислоту, треонин, глутамин, триптофан, глицин, валин, пролин, серин, тирозин, аргинин или гистидин. В другом примере заменяющая аминокислота включает в себя одну или несколько аминокислот, которые предотвращают или ингибируют образование мультимеров белками VWF.
[0212] В определенных вариантах осуществления белок VWF, применимый в данном документе, может быть дополнительно модифицирован для улучшения его взаимодействия с FVIII, например, для улучшения аффинности связывания с FVIII. В качестве неограничивающего примера белок VWF содержит сериновый остаток на месте остатка, соответствующего аминокислоте 764 в SEQ ID NO: 21, и лизиновый остаток на месте остатка, соответствующего аминокислоте 773 в SEQ ID NO: 21. Остатки 764 и/или 773 могут вносить вклад в аффинность связывания белков VWF с FVIII. В других вариантах осуществления белки VWF, применимые для настоящего изобретения, могут иметь другие модификации, например, белок может быть пегилированным, гликозилированным, гезилированным или полисиалилированным.
[0213] В определенных вариантах осуществления химерный полипептид, раскрытый в данном документе, содержит белок VWF, содержащий фрагмент VWF, последовательность XTEN, линкер a2 FVIII и Fc-область. В некоторых вариантах осуществления белок VWF содержит в порядке от N-конца к C-концу фрагмент VWF, слитый с последовательностью XTEN, слитой с линкером a2, слитым с Fc-областью. В определенных вариантах осуществления белок VWF содержит домен D' VWF, который содержит аминокислотную последовательность под SEQ ID NO: 210, домен D3 VWF, который содержит аминокислотную последовательность под SEQ ID NO: 214; последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 58 (AE144_5A); линкер а2, который содержит аминокислотную последовательность под SEQ ID NO: 88; и/или Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217.
[0214] В одном конкретном варианте осуществления химерный полипептид представляет собой rFVIIIFc-VWF-XTEN и содержит белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 202.
[0215] В некоторых вариантах осуществления белок VWF содержит фрагмент VWF, содержащий домен D1, D2, D' и/или D3 VWF. В одном варианте осуществления фрагмент VWF содержит домен D1D2 VWF, содержащий аминокислотную последовательность под SEQ ID NO: 209. В некоторых вариантах осуществления белок VWF дополнительно содержит последовательность сигнального пептида VWF. В одном варианте осуществления сигнальный пептид VWF содержит аминокислотную последовательность под SEQ ID NO: 208. В одном конкретном варианте осуществления белок VWF содержит сигнальный пептид VWF, который содержит аминокислотную последовательность под SEQ ID NO: 208, область D1D2 VWF, которая содержит аминокислотную последовательность под SEQ ID NO: 209, домен D' VWF, который содержит аминокислотную последовательность под SEQ ID NO: 210, домен D3 VWF, который содержит аминокислотную последовательность под SEQ ID NO: 214; последовательность XTEN, которая содержит аминокислотную последовательность под SEQ ID NO: 58 (AE144_5A); линкер а2, который содержит аминокислотную последовательность под SEQ ID NO: 88; и/или Fc-область, которая содержит аминокислотную последовательность под SEQ ID NO: 217.
[0216] В одном конкретном варианте осуществления химерный полипептид представляет собой rFVIIIFc-VWF-XTEN и содержит белок VWF, содержащий аминокислотную последовательность под SEQ ID NO: 205. В другом конкретном варианте осуществления rFVIIIFc-VWF-XTEN содержит белок VWF, кодируемый последовательностью нуклеиновой кислоты под SEQ ID NO: 206, или его фрагмент. Дополнительные иллюстративные полипептидные последовательности, относящиеся к белкам VWF химерных полипептидов, раскрытых в данном документе, представлены в таблицах 18-19.
III.C. Компоненты, обеспечивающие продление периода полужизни
[0217] В некоторых вариантах осуществления химерный полипептид по настоящему изобретению содержит один или несколько компонентов, обеспечивающих продление периода полужизни. В некоторых вариантах осуществления полипептид FVIII химерного полипептида слит или связан с одним или несколькими компонентами, обеспечивающими продление периода полужизни (т. е. в белке FVIII). В других вариантах осуществления химерный полипептид содержит по меньшей мере два компонента, обеспечивающих продление периода полужизни, при этом первый компонент, обеспечивающий продление периода полужизни, слит с полипептидом FVIII, и второй компонент, обеспечивающий продление периода полужизни, слит с доменами D'D3 VWF. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, слит с С-концом или N-концом полипептида FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в полипептид FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в домен B полипептида FVIII. В некоторых вариантах осуществления первый компонент, обеспечивающий продление периода полужизни, вставлен в полипептид FVIII непосредственно ниже аминокислоты, соответствующей аминокислотному остатку 745 в SEQ ID NO: 65. В некоторых вариантах осуществления первый гетерологичный компонент слит с полипептидом FVIII с помощью линкера.
[0218] В определенных вариантах осуществления домены D'D3 VWF во фрагменте VWF могут быть слиты или связаны со вторым компонентом, обеспечивающим продление периода полужизни. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с С-концом или N-концом фрагмента VWF. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, вставлен во фрагмент VWF. В некоторых вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с С-концом фрагмента VWF. В определенных вариантах осуществления второй компонент, обеспечивающий продление периода полужизни, слит с фрагментом VWF с помощью линкера.
[0219] В других вариантах осуществления химерный полипептид содержит полипептид FVIII и фрагмент VWF, содержащий домен D' и домен D3 VWF, где домен D'D3 VWF слит с одним или несколькими фрагментами, обеспечивающими продление периода полужизни, и где домен D'D3 VWF и полипептид FVIII связаны более сильной связью, чем встречающаяся в природе связь между FVIII и VWF.
[0220] В некоторых вариантах осуществления химерный полипептид содержит полипептид FVIII и фрагмент VWF, содержащий домен D' и домен D3 VWF, где домен D'D3 VWF слит с полипептидом FVIII посредством линкера. В некоторых вариантах осуществления линкер представляет собой расщепляемый линкер. В некоторых вариантах осуществления домен D' и домен D3 VWF дополнительно связаны с полипептидом FVIII с помощью по меньшей мере одной нековалентной связи. В определенных вариантах осуществления домен D' и домен D3 VWF дополнительно слиты с Fc. В некоторых вариантах осуществления домен D' и домен D3 VWF слиты с Fc с помощью линкера. В некоторых вариантах осуществления Fc слит со вторым Fc с помощью дополнительного линкера. В определенных вариантах осуществления Fc и второй Fc связаны друг с другом с помощью ковалентной связи, например, дисульфидной связи. В конкретных вариантах осуществления химерный полипептид содержит полипептид FVIII и фрагмент VWF, содержащий домен D' и домен D3 VWF, как раскрыто в международной публикации № WO 2017/222337 A1, которая включена в данный документ посредством ссылки во всей своей полноте.
[0221] Первый компонент, обеспечивающий продление периода полужизни, второй компонент, обеспечивающий продление периода полужизни, или они оба могут быть выбраны из группы, состоящей из партнера по связыванию с FcRn, например, альбумина или Fc-области иммуноглобулина, последовательности XTEN, C-концевого пептида (CTP) β-субъединицы хорионического гонадотропина человека, последовательности PAS, последовательности HAP, трансферрина, альбуминсвязывающих компонентов или любых их фрагментов, производных, вариантов и любой их комбинации.
III.C.1. Константная область Ig или ее часть
[0222] В некоторых вариантах осуществления химерный полипептид по настоящему изобретению также содержит первую константную область Ig или ее часть, слитую с полипептидом FVIII с помощью необязательного линкера. Первая константная область Ig или ее часть может быть вставлена в полипептид FVIII или слита с С-концом или N-концом полипептида FVIII. В некоторых вариантах осуществления химерный полипептид дополнительно содержит вторую константную область Ig или ее часть, слитую с белком VWF. Первая константная область Ig или ее часть может быть вставлена во фрагмент VWF или слита с С-концом или N-концом фрагмента VWF. В конкретных вариантах осуществления первая константная область Ig соединена или связана со второй константной областью Ig с помощью ковалентной связи, например, дисульфидной связи.
[0223] Константная область Ig или ее часть могут улучшать фармакокинетические или фармакодинамические свойства химерного полипептида в соединении с дополнительным гетерологичным компонентом, например, последовательностью XTEN, и белком VWF. В определенных вариантах осуществления константная область Ig или ее часть обеспечивает продление периода полужизни молекулы, слитой с константной областью Ig или ее частью.
[0224] Константная область Ig состоит из доменов, обозначенных как домены СН (константной области тяжелой цепи) (СН1, СН2 и т. д.). В зависимости от изотипа (т. е. IgG, IgM, IgA, IgD или IgE) константная область может состоять из трех или четырех доменов СН. Константные области некоторых изотипов (например, IgG) также содержат шарнирную область. См. Janeway et al. 2001, Immunobiology, Garland Publishing, N.Y., N.Y.
[0225] Константная область Ig или ее часть для получения химерного полипептида по настоящему изобретению могут быть получены из ряда различных источников. В некоторых вариантах осуществления константная область Ig или ее часть получены из Ig человека. Однако следует понимать, что константная область Ig или ее часть могут быть получены из Ig другого вида млекопитающего, в том числе, например, вида грызуна (например, мыши, крысы, кролика, морской свинки) или примата, отличного от человека (например, шимпанзе, макака). Более того, константная область Ig или ее часть могут быть получены из любого класса Ig, в том числе IgM, IgG, IgD, IgA и IgE, и любого изотипа Ig, в том числе IgG1, IgG2, IgG3 и IgG4. В некоторых вариантах осуществления применяют изотип IgG1 человека.
[0226] Различные последовательности генов константных областей Ig (например, последовательности генов константных областей человека) доступны в форме общедоступных депонирований. Можно выбрать последовательность домена константной области, обладающую конкретной эффекторной функцией (или не обладающую конкретной эффекторной функцией) или имеющую конкретную модификацию для снижения иммуногенности. Было опубликовано большое количество последовательностей антител и генов, кодирующих антитела, и подходящие последовательности константных областей Ig (например, последовательности шарнирных областей, СН2 и/или СН3 или их части) могут быть получены из этих последовательностей с применением методик, принятых в данной области техники. Генетический материал, полученный с применением любого из вышеуказанных способов, можно затем подвергать изменению или синтезу с получением полипептидов по настоящему изобретению. Дополнительно следует понимать, что объем настоящего изобретения охватывает аллели, варианты и мутации последовательностей ДНК константных областей.
[0227] Последовательности константной области Ig или ее части могут быть клонированы, например, с применением полимеразной цепной реакции и праймеров, выбранных для амплификации домена, представляющего интерес. Для клонирования последовательности константной области Ig или ее части из антитела можно выделить мРНК из клеток гибридомы, селезенки или лимфатических клеток, подвергнуть ее обратной транскрипции с образованием ДНК и амплифицировать гены антитела с помощью ПЦР. Способы ПЦР-амплификации подробно описаны в патентах США №№ 4683195; 4683202; 4800159; 4965188 и, например, в "PCR Protocols: A Guide to Methods and Applications" Innis et al. eds., Academic Press, San Diego, CA (1990); Ho et al. 1989. Gene 77:51; Horton et al. 1993. Methods Enzymol. 217:270. ПЦР можно инициировать с помощью консенсусных праймеров для константной области или с помощью более специфических праймеров на основе опубликованных последовательностей ДНК и аминокислот, соответствующих тяжелым и легким цепям. Как обсуждалось выше, ПЦР также можно применять для выделения клонов ДНК, кодирующих легкую и тяжелую цепи антитела. В этом случае можно осуществлять скрининг библиотек с помощью консенсусных праймеров или более крупных гомологичных зондов, таких как зонды для константных областей мыши. Из уровня техники известны многочисленные наборы праймеров, подходящие для амплификации генов антител (например, 5'-праймеры на основе N-концевой последовательности очищенных антител (Benhar and Pastan. 1994. Protein Engineering 7:1509); праймеры для быстрой амплификации концов кДНК (Ruberti, F. et al. 1994. J. Immunol. Methods 173:33); праймеры для лидерных последовательностей антител (Larrick et al. 1989 Biochem. Biophys. Res. Commun. 160:1250)). Клонирование последовательностей антител дополнительно описано в выданном Newman et al. патенте США № 5658570, заявка на который была подана 25 января 1995 г., который включен в данный документ посредством ссылки.
[0228] Используемая в данном документе константная область Ig может содержать все домены и шарнирную область или их части. В некоторых вариантах осуществления константная область Ig или ее часть содержит домен СН2, домен СН3 и шарнирную область, т. е. Fc-область или партнера по связыванию с FcRn.
[0229] Используемый в данном документе термин "Fc-область" определяется как часть полипептида, которая соответствует Fc-области нативного Ig, т. e. образована путем димерной ассоциации соответствующих Fc-доменов двух его тяжелых цепей. Нативная Fc-область образует гомодимер с другой Fc-областью. В отличие от этого, термины "генетически слитая Fc-область" или "одноцепочечная Fc-область" (scFc-область), используемые в данном документе, относятся к синтетической димерной Fc-области, состоящей из Fc-доменов, генетически соединенных в одну полипептидную цепь (т. е. кодируемых одной непрерывной генетической последовательностью).
[0230] В некоторых вариантах осуществления "Fc-область" относится к части одной тяжелой цепи Ig, начинающейся в шарнирной области непосредственно выше сайта расщепления папаином (т. е. остатка 216 в IgG, если принять первый остаток константной области тяжелой цепи за 114) и заканчивающейся на С-конце антитела. Соответственно, полный Fc-домен содержит по меньшей мере шарнирный домен, домен CH2 и домен CH3.
[0231] Fc-область константной области Ig в зависимости от изотипа Ig может содержать домены CH2, CH3 и CH4, а также шарнирную область. Химерные полипептиды, содержащие Fc-область Ig, наделяют химерный полипептид несколькими необходимыми свойствами, включая увеличенную стабильность, увеличенный период полужизни в сыворотке крови (см. Capon et al., 1989, Nature 337:525), а также связывание с Fc-рецепторами, такими как неонатальный Fc-рецептор (FcRn) (патенты США №№ 6086875, 6485726, 6030613; WO 03/077834; US2003-0235536A1, которые включены в данный документ посредством ссылки во всей своей полноте).
[0232] Константная область Ig или ее часть может являться партнером по связыванию с FcRn. FcRn активен в зрелых эпителиальных тканях и экспрессируется в просвете кишечника, легочных дыхательных путях, поверхностях носа, поверхностях влагалища, поверхностях толстой и прямой кишки (патент США № 6485726). Партнер по связыванию с FcRn представляет собой часть Ig, которая связывается с FcRn. Другим примером партнера по связыванию с FcRn является альбумин, который дополнительно описан ниже.
[0233] Рецептор FcRn был выделен у некоторых видов млекопитающих, в том числе у людей. Известны последовательности FcRn человека, FcRn обезьяны, FcRn крысы и FcRn мыши (Story et al. 1994, J. Exp. Med. 180:2377). Рецептор FcRn связывается с IgG (но не с Ig других классов, таких как IgA, IgM, IgD и IgE) при относительно низком значении pH, осуществляет активный транспорт IgG через клетки по направлению от просвета к серозной оболочке, а затем высвобождает IgG при относительно более высоком значении pH, обнаруживаемом в интерстициальных жидкостях. Он экспрессируется в зрелой эпителиальной ткани (патенты США №№ 6485726, 6030613, 6086875; WO 03/077834; US2003-0235536A1), в том числе в эпителии легких и кишечника (Israel et al. 1997, Immunology 92:69), эпителии почечных проксимальных канальцев (Kobayashi et al. 2002, Am. J. Physiol. Renal Physiol. 282: F358), а также эпителии полости носа, на поверхностях влагалища и поверхностях системы желчных протоков.
[0234] Партнеры по связыванию с FcRn, применимые в настоящем изобретении, охватывают молекулы, которые могут специфично связываться с рецептором FcRn, в том числе целый IgG, Fc-фрагмент IgG и другие фрагменты, которые содержат полную связывающую область для рецептора FcRn. Область Fc-части IgG, которая связывается с рецептором FcRn, была описана с использованием рентгеноструктурной кристаллографии (Burmeister et al. 1994, Nature 372:379). Основная область контакта Fc с FcRn находится вблизи стыка доменов CH2 и CH3. Все области контакта Fc-FcRn находятся в пределах одной тяжелой цепи Ig. Партнеры по связыванию с FcRn включают в себя целый IgG, Fc-фрагмент IgG и другие фрагменты IgG, которые содержат полную связывающую область для FcRn. Основные сайты контакта включают аминокислотные остатки 248, 250-257, 272, 285, 288, 290-291, 308-311 и 314 из домена CH2 и аминокислотные остатки 385-387, 428 и 433-436 из домена CH3. Все из ссылок на нумерацию аминокислот Ig или фрагментов или областей Ig делаются на основе Kabat et al. 1991, Sequences of Proteins of Immunological Interest, U.S. Department of Public Health, Bethesda, Md.
[0235] Fc-области или партнеры по связыванию с FcRn, связанные с FcRn, могут эффективно переноситься через эпителиальные барьеры с помощью FcRn, за счет чего обеспечивается неинвазивный способ системного введения необходимой терапевтической молекулы. Кроме того, слитые белки, содержащие Fc-область или партнера по связыванию с FcRn, подвергаются эндоцитозу клетками, экспрессирующими FcRn. Но вместо маркировки для разрушения эти слитые белки подвергаются рециркуляции в кровоток, за счет чего увеличивается период полужизни этих белков in vivo. В определенных вариантах осуществления части константных областей Ig представляют собой Fc-область или партнера по связыванию с FcRn, которые обычно связываются посредством дисульфидных связей и других неспецифических взаимодействий с другой Fc-областью или другим партнером по связыванию с FcRn с образованием димеров и мультимеров более высокого порядка.
[0236] Два рецептора FcRn могут связываться с одной молекулой Fc. Кристаллографические данные позволяют предположить, что каждая молекула FcRn связывается с одним полипептидом гомодимера Fc. В некоторых вариантах осуществления связывание партнера по связыванию с FcRn, например Fc-фрагмента IgG, с биологически активной молекулой обеспечивает способ доставки биологически активной молекулы пероральным, трансбуккальным, сублингвальным, ректальным, вагинальным путем, в виде аэрозоля, вводимого назально или посредством легочного пути, или посредством глазного пути. В других вариантах осуществления химерный полипептид можно вводить инвазивным способом, например, подкожно, внутривенно.
[0237] Область партнера по связыванию с FcRn представляет собой молекулу или ее часть, которая может специфично связываться с рецептором FcRn с последующим активным транспортом Fc-области с помощью рецептора FcRn. "Специфично связанный" относится к двум молекулам, образующим комплекс, который является относительно стабильным в физиологических условиях. Специфичное связывание характеризуется высокой аффинностью и емкостью от низкой до умеренной, в отличие от неспецифичного связывания, которое обычно характеризуется низкой аффинностью и емкостью от умеренной до высокой. Как правило, связывание считается специфичным, если константа аффинности KA превышает 106 M-1 или превышает 108 M-1. При необходимости неспецифичное связывание можно снизить без существенного влияния на специфичное связывание путем изменения условий связывания. Соответствующие условия связывания, такие как концентрация молекул, ионная сила раствора, температура, допустимое время связывания, концентрация блокирующего средства (например, сывороточного альбумина, казеина молока) и т. д., могут быть оптимизированы специалистом в данной области с применением обычных методик.
[0238] В определенных вариантах осуществления химерный полипептид по настоящему изобретению содержит одну или несколько усеченных Fc-областей, которых, тем не менее, достаточно для придания Fc-области свойств связывания с Fc-рецептором (FcR). Например, часть Fc-области, которая связывается с FcRn (т. e. FcRn-связывающая часть), содержит приблизительно аминокислоты 282-438 из IgG1 согласно нумерации EU, при этом основными сайтами контакта являются аминокислоты 248, 250-257, 272, 285, 288, 290-291, 308-311 и 314 в домене CH2 и аминокислотные остатки 385-387, 428 и 433-436 в домене CH3. Таким образом, Fc-область по настоящему изобретению может содержать FcRn-связывающую часть или состоять из нее. FcRn-связывающие части могут быть получены из тяжелых цепей любого изотипа, в том числе IgG1, IgG2, IgG3 и IgG4. В некоторых вариантах осуществления применяют FcRn-связывающую часть из антитела изотипа IgG1 человека. В других вариантах осуществления применяют FcRn-связывающую часть из антитела изотипа IgG4 человека.
[0239] В других вариантах осуществления "Fc-область" содержит аминокислотную последовательность Fc-домена или аминокислотную последовательность, полученную из Fc-домена. В определенных вариантах осуществления Fc-область содержит по меньшей мере одно из шарнирного домена (например, верхней, средней и/или нижней шарнирной области) (приблизительно аминокислоты 216-230 Fc-области антитела в соответствии с нумерацией EU), домена CH2 (приблизительно аминокислоты 231-340 Fc-области антитела в соответствии с нумерацией EU), домена CH3 (приблизительно аминокислоты 341-438 Fc-области антитела в соответствии с нумерацией EU), домена CH4 или их варианта, части или фрагмента. В других вариантах осуществления Fc-область содержит полный Fc-домен (т. е. шарнирный домен, домен CH2 и домен CH3). В некоторых вариантах осуществления Fc-область содержит шарнирный домен (или его часть), слитый с доменом CH3 (или его частью), шарнирный домен (или его часть), слитый с доменом CH2 (или его частью), домен CH2 (или его часть), слитый с доменом CH3 (или его частью), домен CH2 (или его часть), слитый как с шарнирным доменом (или его частью), так и доменом CH3 (или его частью), состоит по сути из них или состоит из них. В еще нескольких других вариантах осуществления Fc-область не содержит по меньшей мере часть домена CH2 (например, весь домен CH2 или его часть). В конкретном варианте осуществления Fc-область содержит аминокислоты 221-447 в соответствии с нумерацией EU или состоит из них.
[0240] Fc-области, обозначенные в данном документе как F, F1 или F2, могут быть получены из ряда различных источников. В некоторых вариантах осуществления Fc-область полипептида получена из Ig человека. Однако следует понимать, что Fc-область может быть получена из Ig другого вида млекопитающего, в том числе, например, вида грызуна (например, мыши, крысы, кролика или морской свинки) или отличного от человека примата (например, шимпанзе, макака). Более того, полипептидные Fc-домены или их части могут быть получены из любого класса Ig, в том числе IgM, IgG, IgD, IgA и IgE, и любого изотипа Ig, в том числе IgG1, IgG2, IgG3 и IgG4. В других вариантах осуществления применяют изотип IgG1 человека.
[0241] В определенных вариантах осуществления вариант Fc обеспечивает изменение по меньшей мере одной эффекторной функции, придаваемой Fc-областью, содержащей указанный Fc-домен дикого типа (например, улучшение или снижение способности Fc-области к связыванию с Fc-рецепторами (например, FcγRI, FcγRII или FcγRIII) или белками системы комплемента (например, C1q) или инициированию антителозависимой цитотоксичности (ADCC), фагоцитоза или комплементзависимой цитотоксичности (CDCC)). В других вариантах осуществления в варианте Fc представлен сконструированный цистеиновый остаток.
[0242] В качестве Fc-областей по настоящему изобретению можно использовать известные из уровня техники варианты Fc, которые, как известно, придают изменение (например, усиление или снижение) эффекторной функции и/или связывания с FcR или FcRn. В частности, связывающая молекула по настоящему изобретению может содержать, например, изменение (например, замену) в одном или нескольких аминокислотных положениях, раскрытых в международных публикациях согласно PCT WO88/07089A1, WO96/14339A1, WO98/05787A1, WO98/23289A1, WO99/51642A1, WO99/58572A1, WO00/09560A2, WO00/32767A1, WO00/42072A2, WO02/44215A2, WO02/060919A2, WO03/074569A2, WO04/016750A2, WO04/029207A2, WO04/035752A2, WO04/063351A2, WO04/074455A2, WO04/099249A2, WO05/040217A2, WO04/044859, WO05/070963A1, WO05/077981A2, WO05/092925A2, WO05/123780A2, WO06/019447A1, WO06/047350A2 и WO06/085967A2; публикациях заявок на патент США №№ US2007/0231329, US2007/0231329, US2007/0237765, US2007/0237766, US2007/0237767, US2007/0243188, US20070248603, US20070286859, US20080057056 или патентах США №№ 5648260; 5739277; 5834250; 5869046; 6096871; 6121022; 6194551; 6242195; 6277375; 6528624; 6538124; 6737056; 6821505; 6998253; 7083784; 7404956 и 7317091, каждый из которых включен в данный документ посредством ссылки. В некоторых вариантах осуществления конкретное изменение (например, конкретная замена одной или нескольких аминокислот, раскрытых в уровне техники) может быть осуществлено в одном или нескольких раскрытых аминокислотных положениях. В других вариантах осуществления может быть осуществлено другое изменение в одном или нескольких раскрытых аминокислотных положениях (например, другая замена в одном или нескольких аминокислотных положениях, раскрытых в уровне техники).
[0243] Fc-область IgG или его партнер по связыванию с FcRn могут быть модифицированы в соответствии с хорошо известными процедурами, такими как сайт-направленный мутагенез и т. п., с получением модифицированных IgG или Fc-фрагментов или их частей, которые будут связываться с FcRn. Такие модификации включают модификации в отдалении от сайтов контакта с FcRn, а также модификации в пределах сайтов контакта, которые обеспечивают сохранение или даже усиление связывания с FcRn. Например, следующие отдельные аминокислотные остатки в Fc IgG1 человека (Fcγ1) могут быть заменены без значительной потери аффинности связывания Fc с FcRn: P238A, S239A, K246A, K248A, D249A, M252A, T256A, E258A, T260A, D265A, S267A, H268A, E269A, D270A, E272A, L274A, N276A, Y278A, D280A, V282A, E283A, H285A, N286A, T289A, K290A, R292A, E293A, E294A, Q295A, Y296F, N297A, S298A, Y300F, R301A, V303A, V305A, T307A, L309A, Q311A, D312A, N315A, K317A, E318A, K320A, K322A, S324A, K326A, A327Q, P329A, A330Q, P331A, E333A, K334A, T335A, S337A, K338A, K340A, Q342A, R344A, E345A, Q347A, R355A, E356A, M358A, T359A, K360A, N361A, Q362A, Y373A, S375A, D376A, A378Q, E380A, E382A, S383A, N384A, Q386A, E388A, N389A, N390A, Y391F, K392A, L398A, S400A, D401A, D413A, K414A, R416A, Q418A, Q419A, N421A, V422A, S424A, E430A, N434A, T437A, Q438A, K439A, S440A, S444A, и K447A, где, например, P238A представляет собой пролин дикого типа, замененный аланином в положении номер 238. В качестве примера некоторые варианты осуществления включают мутацию N297A, обеспечивающую удаление высококонсервативного сайта N-гликозилирования. Аминокислоты дикого типа в положениях, указанных выше, могут быть заменены другими аминокислотами в дополнение к аланину. Мутации могут быть введены в Fc по отдельности, что дает более ста Fc-областей, отличных от нативного Fc. Кроме того, комбинации из двух, трех или более из этих отдельных мутаций могут быть введены совместно, что дает еще несколько сотен Fc-областей. Более того, одна из Fc-областей конструкции по настоящему изобретению может быть подвергнута мутации, а другая Fc-область конструкции вообще не подвергнута мутации, или они обе могут быть подвергнуты мутации, но с использованием разных мутаций.
[0244] Определенные из вышеуказанных мутаций могут придавать новое функциональное свойство Fc-области или партнеру по связыванию с FcRn. Например, некоторые варианты осуществления включают N297A, обеспечивающую удаление высококонсервативного сайта N-гликозилирования. Эффект этой мутации заключается в снижении иммуногенности, за счет чего увеличивается период полужизни Fc-области в кровотоке, и лишении Fc-области способности к связыванию с Fc-гамма-RI, Fc-гамма-RIIA, Fc-гамма-RIIB и Fc-гамма-RIIIA без нарушения аффинности в отношении FcRn (Routledge et al. 1995, Transplantation 60:847; Friend et al. 1999, Transplantation 68:1632; Shields et al. 1995, J. Biol. Chem. 276:6591). В качестве дополнительного примера нового функционального свойства, появляющегося в результате вышеописанных мутаций, аффинность в отношении FcRn в некоторых случаях может увеличиваться, превышая таковую у дикого типа. Эту увеличенную аффинность могут отражать увеличенная скорость ассоциации, уменьшенная скорость диссоциации или как увеличенная скорость ассоциации, так и уменьшенная скорость диссоциации. Примеры мутаций, которые, как полагают, придают увеличенную аффинность в отношении FcRn, включают без ограничения T256A, T307A, E380A и N434A (Shields et al. 2001, J. Biol. Chem. 276:6591).
[0245] Кроме того, по меньшей мере три Fc-гамма-рецептора человека, по-видимому, распознают сайт связывания в IgG в нижней шарнирной области, обычно аминокислоты 234-237. Следовательно, другой пример нового функционального свойства и потенциальной уменьшенной иммуногенности может появляться в результате мутаций в этой области, как, например, путем замещения аминокислот 233-236 "ELLG" из IgG1 человека соответствующей последовательностью "PVA" из IgG2 (с делецией одной аминокислоты). Было показано, что FcγRI, FcγRII и FcγRIII, которые опосредуют различные эффекторные функции, не будут связываться с IgG1 после введения таких мутаций. Ward and Ghetie 1995, Therapeutic Immunology 2:77, и Armour et al. 1999, Eur. J. Immunol. 29:2613.
[0246] В некоторых вариантах осуществления константная область Ig или ее часть, например, Fc-область, представляет собой полипептид, содержащий последовательность PKNSSMISNTP (SEQ ID NO: 89 или SEQ ID NO: 3 из патента США № 5739277) и необязательно дополнительно содержащий последовательность, выбранную из HQSLGTQ (SEQ ID NO: 90), HQNLSDGK (SEQ ID NO: 91), HQNISDGK (SEQ ID NO: 92) или VISSHLGQ (SEQ ID NO: 93) (или SEQ ID NO: 11, 1, 2 и 31 соответственно из патента США № 5739277).
[0247] В других вариантах осуществления константная область иммуноглобулина или ее часть содержит аминокислотную последовательность в шарнирной области или ее части, которая образует одну или несколько дисульфидных связей со другой константной областью иммуноглобулина или ее частью. Дисульфидная связь, образуемая константной областью иммуноглобулина или ее частью, объединяет первый полипептид, содержащий полипептид FVIII, и второй полипептид, содержащий фрагмент VWF, так что эндогенный VWF не замещает фрагмент VWF и не связывается с полипептидом FVIII. Следовательно, дисульфидная связь между первой константной областью иммуноглобулина или ее частью и второй константной областью иммуноглобулина или ее частью предотвращает взаимодействие между эндогенным VWF и полипептидом FVIII. Это ингибирование взаимодействия между VWF и полипептидом FVIII обеспечивает продление периода полужизни химерного полипептида свыше двукратного предела. Шарнирная область или ее часть могут дополнительно быть соединены с одним или несколькими доменами СН1, СН2, СН3, их фрагментом и любыми их комбинациями. В конкретном варианте осуществления константная область иммуноглобулина или ее часть представляет собой шарнирную область и CH2.
[0248] В определенных вариантах осуществления константная область Ig или ее часть является полугликозилированной. Например, химерный полипептид, содержащий две Fc-области или два партнера по связыванию с FcRn, может содержать гликозилированные первые Fc-область (например, гликозилированную область CH2) или партнера по связыванию с FcRn и агликозилированные вторые Fc-область (например, агликозилированную область CH2) или партнера по связыванию с FcRn. В некоторых вариантах осуществления линкер может быть помещен между гликозилированной и агликозилированной Fc-областями. В других вариантах осуществления Fc-область или партнер по связыванию с FcRn являются полностью гликозилированными, т. е. все Fc-области являются гликозилированными. В других вариантах осуществления Fc-область может быть агликозилированной, т. е. ни один из Fc-компонентов не является гликозилированным.
[0249] В определенных вариантах осуществления химерный полипептид по настоящему изобретению содержит аминокислотную замену в константной области Ig или ее части (например, варианты Fc), которая изменяет антигеннезависимые эффекторные функции константной области Ig, в частности, период полужизни белка в кровотоке.
[0250] Такие белки демонстрируют увеличенное либо уменьшенное связывание с FcRn по сравнению с белками, не содержащими таких замен, и, следовательно, характеризуются соответственно увеличенным или уменьшенным периодом полужизни в сыворотке крови. Предполагается, что варианты Fc с улучшенной аффинностью в отношении FcRn характеризуются более длительными периодами полужизни в сыворотке крови, и такие молекулы имеют полезную применимость в способах лечения млекопитающих, при которых необходим длительный период полужизни введенного полипептида, например, для лечения хронического заболевания или нарушения (см., например, патенты США 7348004, 7404956 и 7862820). В отличие от этого, ожидается, что варианты Fc с уменьшенной аффинностью связывания с FcRn будут характеризоваться более короткими периодами полужизни, и такие молекулы также применимы, например, для введения млекопитающему, при котором целесообразным может быть сокращенное время циркуляции, например, для диагностической визуализации in vivo или в ситуациях, когда исходный полипептид оказывает токсические побочные эффекты, если он присутствует в кровотоке в течение длительных периодов. Варианты Fc с уменьшенной аффинностью связывания с FcRn также с меньшей долей вероятности пересекают плацентарный барьер и, таким образом, также являются применимыми в лечении заболеваний или нарушений у беременных женщин. Кроме того, другие пути применения, при которых может быть необходимой сниженная аффинность связывания с FcRn, включают такие пути применения, при которых необходима локализация в головном мозге, почке и/или печени. В одном иллюстративном варианте осуществления химерные полипептиды по настоящему изобретению демонстрируют сниженный транспорт через эпителий почечных клубочков из сосудистой сети. В других вариантах осуществления химерные полипептиды по настоящему изобретению демонстрируют сниженный транспорт через гематоэнцефалический барьер (ВВВ) из головного мозга в сосудистое пространство. В некоторых вариантах осуществления белок с измененным связыванием с FcRn содержит по меньшей мере одну Fc-область или партнера по связыванию с FcRn (например, одну или две Fc-области или один или два партнера по связыванию с FcRn), имеющих одну или несколько аминокислотных замен в "FcRn-связывающей петле" в константной области Ig. FcRn-связывающая петля состоит из аминокислотных остатков 280-299 (в соответствии с нумерацией EU) полноразмерной Fc-области дикого типа. В других вариантах осуществления константная область Ig или ее часть в химерном полипептиде по настоящему изобретению, характеризующемся измененной аффинностью связывания c FcRn, содержит по меньшей мере одну Fc-область или партнера по связыванию с FcRn, имеющих одну или несколько аминокислотных замен в пределах "зоны контакта" с FcRn размером 15 Ǻ. Используемый в данном документе термин "зона контакта" с FcRn размером 15 Ǻ включает остатки в следующих положениях полноразмерного Fc-компонента дикого типа: 243-261, 275-280, 282-293, 302-319, 336-348, 367, 369, 372-389, 391, 393, 408, 424, 425-440 (нумерация EU). В других вариантах осуществления константная область Ig или ее часть по настоящему изобретению, характеризующиеся измененной аффинностью связывания с FcRn, содержат по меньшей мере одну Fc-область или партнера по связыванию с FcRn, имеющих одну или несколько аминокислотных замен в аминокислотном положении, соответствующем любому из следующих EU-положений: 256, 277-281, 283-288, 303-309, 313, 338, 342, 376, 381, 384, 385, 387, 434 (например, N434A или N434K) и 438. Иллюстративные аминокислотные замены, обеспечивающие изменение активности связывания с FcRn, раскрыты в международной публикации согласно PCT № WO05/047327, которая включена в данный документ посредством ссылки.
[0251] Fc-область или партнер по связыванию с FcRn, применяемые в настоящем изобретении, также могут содержать известную из уровня техники аминокислотную замену, обеспечивающую изменение гликозилирования химерного полипептида. Например, Fc-область или партнер по связыванию с FcRn химерного полипептида, соединенные с доменом D'D3 VWF или полипептидом FVIII, может включать в себя Fc-область, имеющую мутацию, приводящую к пониженному гликозилированию (например, N- или O-связанному гликозилированию), или может включать в себя измененную гликоформу Fc-компонента дикого типа (например, гликан с низким содержанием фукозы или гликан, не содержащий фукозу).
[0252] В некоторых вариантах осуществления непроцессированный химерный полипептид по настоящему изобретению может содержать генетически слитую Fc-область (т. e. scFc-область), имеющую две или более составляющих ее константных областей Ig или их частей, независимо выбранных из константной области Ig или ее части, описанных в данном документе. В некоторых вариантах осуществления Fc-области димерной Fc-области являются одинаковыми. В некоторых вариантах осуществления по меньшей мере две Fc-области являются разными. Например, Fc-области или партнеры по связыванию с FcRn белков по настоящему изобретению содержат одинаковое количество аминокислотных остатков, или они могут отличаться по длине одним или несколькими аминокислотными остатками (например, приблизительно 5 аминокислотными остатками (например, 1, 2, 3, 4 или 5 аминокислотными остатками), приблизительно 10 остатками, приблизительно 15 остатками, приблизительно 20 остатками, приблизительно 30 остатками, приблизительно 40 остатками или приблизительно 50 остатками). В еще некоторых других вариантах осуществления Fc-области или партнеры по связыванию с FcRn белка по настоящему изобретению могут отличаться по последовательности в одном или нескольких аминокислотных положениях. Например, по меньшей мере две Fc-области или два партнера по связыванию с FcRn могут отличаться по приблизительно 5 аминокислотным положениям (например, 1, 2, 3, 4 или 5 аминокислотным положениям), приблизительно 10 положениям, приблизительно 15 положениям, приблизительно 20 положениям, приблизительно 30 положениям, приблизительно 40 положениям или приблизительно 50 положениям.
III.C.2. Последовательности XTEN
[0253] Используемое в данном документе выражение "последовательность XTEN" относится к полипептидам увеличенной длины с не встречающимися в природе по существу неповторяющимися последовательностями, которые состоят главным образом из небольших гидрофильных аминокислот, при этом последовательность характеризуется низкой степенью образования вторичной или третичной структуры в физиологических условиях или отсутствием таковой. Являясь партнером для химерного полипептида, XTEN могут служить в качестве носителя, придавая определенные необходимые фармакокинетические, физико-химические и фармацевтические свойства, будучи соединенными с белком VWF или последовательностью FVIII по настоящему изобретению с образованием химерного полипептида. Такие необходимые свойства включают без ограничения улучшенные фармакокинетические параметры и характеристики растворимости. Используемый в данном документе термин "XTEN" конкретным образом исключает антитела или фрагменты антител, такие как одноцепочечные антитела или Fc-фрагменты легкой цепи или тяжелой цепи.
[0254] В некоторых вариантах осуществления более короткая последовательность XTEN обеспечивает улучшенное свойство продления периода полужизни по сравнению с более длинной последовательностью XTEN, если последовательность XTEN слита с белком VWF и/или второй константной областью Ig или ее частью. Следовательно, в некоторых вариантах осуществления последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, имеет длину менее 288 аминокислот, т. е. короче 288 аминокислот. В некоторых вариантах осуществления последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, состоит из аминокислотной последовательности, имеющей длину от 12 аминокислот до 287 аминокислот. В других вариантах осуществления последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, содержит по меньшей мере приблизительно 36 аминокислот, по меньшей мере приблизительно 42 аминокислоты, по меньшей мере приблизительно 72 аминокислоты или по меньшей мере приблизительно 144 аминокислоты, но менее 288 аминокислот. В других вариантах осуществления последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, выбрана из AE36, AG36, AE42, AG42, AE72, AG72, AE144 или AG144. В некоторых вариантах осуществления последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, представляет собой аминокислотную последовательность, на по меньшей мере 80%, 85%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентичную SEQ ID NO: 9, SEQ ID NO: 10, SEQ ID NO: 11 или SEQ ID NO: 14, где химерный полипептид демонстрирует улучшенный период полужизни по сравнению с химерным полипептидом без последовательности XTEN.
[0255] Химерный полипептид по настоящему изобретению может дополнительно содержать дополнительные (вторую, третью или больше) последовательности XTEN. Дополнительная последовательность XTEN дополнительно может быть слита с полипептидом FVIII или первой константной областью Ig или ее частью. Дополнительные последовательности XTEN могут иметь любую длину. Например, дополнительная последовательность XTEN, слитая с полипептидом FVIII или первой константной областью Ig или ее частью, представляет собой пептид или полипептид, содержащий более чем приблизительно 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950, 1000, 1200, 1400, 1600, 1800 или 2000 аминокислотных остатков. В определенных вариантах осуществления дополнительная последовательность XTEN представляет собой пептид или полипептид, содержащий более чем от приблизительно 20 до приблизительно 3000 аминокислотных остатков, более чем от приблизительно 30 до приблизительно 2500 остатков, более чем от приблизительно 40 до приблизительно 2000 остатков, более чем от приблизительно 50 до приблизительно 1500 остатков, более чем от приблизительно 60 до приблизительно 1000 остатков, более чем от приблизительно 70 до приблизительно 900 остатков, более чем от приблизительно 80 до приблизительно 800 остатков, более чем от приблизительно 90 до приблизительно 700 остатков, более чем от приблизительно 100 до приблизительно 600 остатков, более чем от приблизительно 110 до приблизительно 500 остатков или более чем от приблизительно 120 до приблизительно 400 остатков. В определенных вариантах осуществления дополнительная последовательность XTEN, слитая с полипептидом FVIII, содержит по меньшей мере приблизительно 288 аминокислот. В конкретных вариантах осуществления дополнительная последовательность XTEN, слитая с полипептидом FVIII, содержит приблизительно 288 аминокислот. В конкретных вариантах осуществления дополнительная последовательность XTEN, слитая с полипептидом FVIII, содержит аминокислотную последовательность, которая на по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или приблизительно 100% идентична аминокислотной последовательности AE288 (SEQ ID NO: 8).
[0256] Последовательности XTEN (т. е. последовательность XTEN, слитая с белком VWF и/или второй константной областью Ig или ее частью, или последовательность XTEN, слитая с полипептидом FVIII и/или первой константной областью Ig или ее частью или вставленная в один или несколько сайтов вставки в полипептиде FVIII) могут содержать один или несколько мотивов последовательности из 9-14 аминокислотных остатков или аминокислотную последовательность, на по меньшей мере 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную мотиву последовательности, где мотив содержит 4-6 типов аминокислот, выбранных из группы, состоящей из глицина (G), аланина (A), серина (S), треонина (T), глутамата (E) и пролина (P), состоит по сути из них или состоит из них. См. US 2010-0239554 A1.
[0257] В некоторых вариантах осуществления последовательность XTEN содержит неперекрывающиеся мотивы последовательностей, в которых по меньшей мере приблизительно 80%, или по меньшей мере приблизительно 85%, или по меньшей мере приблизительно 90%, или по меньшей мере приблизительно 91%, или по меньшей мере приблизительно 92%, или по меньшей мере приблизительно 93%, или по меньшей мере приблизительно 94%, или по меньшей мере приблизительно 95%, или по меньшей мере приблизительно 96%, или по меньшей мере приблизительно 97%, или по меньшей мере приблизительно 98%, или по меньшей мере приблизительно 99%, или приблизительно 100% последовательности состоит из нескольких звеньев неперекрывающихся последовательностей, выбранных из одного семейства мотивов, выбранного из таблицы 7, что в результате дает последовательность, характерную для определенного семейства. Используемый в данном документе термин "семейство" означает, что XTEN содержит мотивы, выбранные только из одной категории мотивов из таблицы 7, т. е. XTEN на основе AD, AE, AF, AG, AM, AQ, BC или BD, и что любые другие аминокислоты в XTEN, не относящиеся к мотиву из определенного семейства, выбраны для достижения необходимого свойства, как, например, для обеспечения встраивания сайта рестрикции рядом с кодирующими нуклеотидами, встраивания последовательности для расщепления или для достижения лучшего соединения с FVIII или VWF. В некоторых вариантах осуществления семейств XTEN последовательность XTEN содержит несколько звеньев неперекрывающихся мотивов последовательностей из семейства мотивов AD, или семейства мотивов AE, или семейства мотивов AF, или семейства мотивов AG, или семейства мотивов AM, или семейства мотивов AQ, или семейства BC, или семейства BD, в результате чего XTEN демонстрирует диапазон гомологии, описанный выше. В других вариантах осуществления XTEN содержит несколько звеньев последовательностей мотивов из двух или более семейств мотивов из таблицы 7. Эти последовательности могут быть выбраны для достижения необходимых физических/химических характеристик, в том числе таких свойств, как суммарный заряд, гидрофильность, отсутствие вторичной структуры или отсутствие повторяемости, которые определяются аминокислотным составом мотивов, более полно описанным ниже. В вариантах осуществления, описанных выше в данном абзаце данного документа, мотивы, встраиваемые в XTEN, могут быть отобраны и собраны с применением способов, описанных в данном документе, для получения XTEN из от приблизительно 36 до приблизительно 3000 аминокислотных остатков.
Таблица 7. Мотивы последовательностей XTEN из 12 аминокислот и семейства мотивов
* Обозначает отдельные последовательности мотивов, которые при совместном применении в различных перестановках дают в результате "последовательность, характерную для определенного семейства"
[0258] В некоторых вариантах осуществления последовательность XTEN, применяемая в настоящем изобретении, на по меньшей мере 60%, 70%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 100% идентична последовательности, выбранной из группы, состоящей из AE42, AG42, AE48, AM48, AE72, AG72, AE108, AG108, AE144, AF144, AG144, AE180, AG180, AE216, AG216, AE252, AG252, AE288, AG288, AE324, AG324, AE360, AG360, AE396, AG396, AE432, AG432, AE468, AG468, AE504, AG504, AF504, AE540, AG540, AF540, AD576, AE576, AF576, AG576, AE612, AG612, AE624, AE648, AG648, AG684, AE720, AG720, AE756, AG756, AE792, AG792, AE828, AG828, AD836, AE864, AF864, AG864, AM875, AE912, AM923, AM1318, BC864, BD864, AE948, AE1044, AE1140, AE1236, AE1332, AE1428, AE1524, AE1620, AE1716, AE1812, AE1908, AE2004A, AG948, AG1044, AG1140, AG1236, AG1332, AG1428, AG1524, AG1620, AG1716, AG1812, AG1908 и AG2004. См. US 2010-0239554 A1.
[0259] В некоторых вариантах осуществления последовательность XTEN на по меньшей мере 60%, 70%, 80%, 90%, 95%, 96%, 97%, 98%, 99% или 100% идентична аминокислотной последовательности, выбранной из группы, состоящей из AE42 (SEQ ID NO: 9), AE72 (SEQ ID NO: 10), AE144_2A (SEQ IDNO: 55), AE144_3B (SEQ ID NO: 56), AE144_4A (SEQ ID NO: 57), AE144_5A (SEQ ID NO: 58), AE144_6B (SEQ ID NO: 59), AG144_A (SEQ ID NO: 60), AG144_B (SEQ ID NO: 61), AG144_C (SEQ ID NO: 62), AG144_F (SEQ IDNO: 63), AE864 (SEQ ID NO: 15), AE576 (SEQ ID NO: 16), AE288 (SEQ ID NO: 8), AE288_2 (SEQ ID NO: 54), AE144 (SEQ ID NO: 11), AG864 (SEQ ID NO: 17), AG576 (SEQ ID NO: 18), AG288 (SEQ ID NO: 19), AG144 (SEQ ID NO: 14) и любых их комбинаций. В других вариантах осуществления последовательность XTEN выбрана из группы, состоящей из AE42 (SEQ ID NO: 9), AE72 (SEQ ID NO: 10), AE144_2A (SEQ IDNO: 55), AE144_3B (SEQ ID NO: 56), AE144_4A (SEQ ID NO: 57), AE144_5A (SEQ IDNO: 58), AE144_6B (SEQ ID NO: 59), AG144_A (SEQ ID NO: 60), AG144_B (SEQ ID NO: 61), AG144_C (SEQ ID NO: 62), AG144_F (SEQ IDNO: 63), AE864 (SEQ ID NO: 15), AE576 (SEQ ID NO: 16), AE288 (SEQ ID NO: 8), AE288_2 (SEQ ID NO: 54), AE144 (SEQ ID NO: 11), AG864 (SEQ ID NO: 17), AG576 (SEQ ID NO: 18), AG288 (SEQ ID NO: 19), AG144 (SEQ ID NO: 14) и любых их комбинаций. В некоторых вариантах осуществления последовательность XTEN представляет собой AE288. Аминокислотные последовательности для определенных последовательностей XTEN по настоящему изобретению показаны в таблице 8.
ТАБЛИЦА 8. Последовательности XTEN
SEQ ID NO: 9
(SEQ ID NO: 10)
TSGSETPGTS ESATPESGPG TSTEPSEGSA PGASS
(SEQ ID NO: 11)
SAPGSEPATSGSETPGSEPATSGSETPGSEPATSGSETPGTSTEPSEGSAPGTSESA
PESGPGSEPATSGSETPGTSTEPSEGSAP
(SEQ ID NO: 55)
(SEQ ID NO: 56)
(SEQ ID NO: 57)
(SEQ ID NO: 58)
(SEQ ID NO: 59)
SEQ ID NO: 14
GSPGASPGTSSTGSPGSSTPSGATGSPGSSPSASTGTGPGASPGTSSTGSPGSSPSA
STGTGPGTPGSGTASSSPGSSTPSGATGSP
(SEQ ID NO: 60)
(SEQ ID NO: 61)
(SEQ ID NO: 62)
(SEQ ID NO: 63)
SEQ ID NO: 8
PGTSTEPSEGSAPGSPAGSPTSTEEGTSESATPESGPGSEPATSGSETPGTSESATPES
GPGSPAGSPTSTEEGSPAGSPTSTEEGTSTEPSEGSAPGTSESATPESGPGTSESATPE
SGPGTSESATPESGPGSEPATSGSETPGSEPATSGSETPGSPAGSPTSTEEGTSTEPSE
GSAPGTSTEPSEGSAPGSEPATSGSETPGTSESATPESGPGTSTEPSEGSAP
(SEQ ID NO: 54)
SEQ ID NO: 19
SPGSSTPSGATGSPGTPGSGTASSSPGSSTPSGATGSPGSSTPSGATGSPGSSPSASTG
TGPGSSPSASTGTGPGASPGTSSTGSPGTPGSGTASSSPGSSTPSGATGSPGSSPSAST
GTGPGSSPSASTGTGPGASPGTSSTGSPGASPGTSSTGSPGSSTPSGATGSPGSSPSAS
TGTGPGASPGTSSTGSPGSSPSASTGTGPGTPGSGTASSSPGSSTPSGATGS
SEQ ID NO: 16
PGTSTEPSEGSAPGTSESATPESGPGSEPATSGSETPGSEPATSGSETPGSPAGSPTST
EEGTSESATPESGPGTSTEPSEGSAPGTSTEPSEGSAPGSPAGSPTSTEEGTSTEPSEG
SAPGTSTEPSEGSAPGTSESATPESGPGTSTEPSEGSAPGTSESATPESGPGSEPATSG
SETPGTSTEPSEGSAPGTSTEPSEGSAPGTSESATPESGPGTSESATPESGPGSPAGSP
TSTEEGTSESATPESGPGSEPATSGSETPGTSESATPESGPGTSTEPSEGSAPGTSTEP
SEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGSPAG
SPTSTEEGTSTEPSEGSAPGTSESATPESGPGSEPATSGSETPGTSESATPESGPGSEP
ATSGSETPGTSESATPESGPGTSTEPSEGSAPGTSESATPESGPGSPAGSPTSTEEGSP
AGSPTSTEEGSPAGSPTSTEEGTSESATPESGPGTSTEPSEGSAP
SEQ ID NO: 18
SPGSSTPSGATGSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTAS
SSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGSSPSASTGTGPGTPGSGTA
SSSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGSSTPSGATGSPGSSTPSG
ATGSPGASPGTSSTGSPGTPGSGTASSSPGSSTPSGATGSPGSSTPSGATGSPGSSTPS
GATGSPGSSPSASTGTGPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTASSSPGASPG
TSSTGSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTASSSPGSST
PSGATGSPGTPGSGTASSSPGSSTPSGATGSPGTPGSGTASSSPGSSTPSGATGSPGSS
TPSGATGSPGSSPSASTGTGPGSSPSASTGTGPGASPGTSSTGSPGTPGSGTASSSPGS
STPSGATGSPGSSPSASTGTGPGSSPSASTGTGPGASPGTSSTGS
SEQ ID NO: 15
PGTSTEPSEGSAPGTSESATPESGPGSEPATSGSETPGSEPATSGSETPGSPAGSPTST
EEGTSESATPESGPGTSTEPSEGSAPGTSTEPSEGSAPGSPAGSPTSTEEGTSTEPSEG
SAPGTSTEPSEGSAPGTSESATPESGPGTSTEPSEGSAPGTSESATPESGPGSEPATSG
SETPGTSTEPSEGSAPGTSTEPSEGSAPGTSESATPESGPGTSESATPESGPGSPAGSP
TSTEEGTSESATPESGPGSEPATSGSETPGTSESATPESGPGTSTEPSEGSAPGTSTEP
SEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGTSTEPSEGSAPGSPAG
SPTSTEEGTSTEPSEGSAPGTSESATPESGPGSEPATSGSETPGTSESATPESGPGSEP
ATSGSETPGTSESATPESGPGTSTEPSEGSAPGTSESATPESGPGSPAGSPTSTEEGSP
AGSPTSTEEGSPAGSPTSTEEGTSESATPESGPGTSTEPSEGSAPGTSESATPESGPGS
EPATSGSETPGTSESATPESGPGSEPATSGSETPGTSESATPESGPGTSTEPSEGSAPG
SPAGSPTSTEEGTSESATPESGPGSEPATSGSETPGTSESATPESGPGSPAGSPTSTEE
GSPAGSPTSTEEGTSTEPSEGSAPGTSESATPESGPGTSESATPESGPGTSESATPESG
PGSEPATSGSETPGSEPATSGSETPGSPAGSPTSTEEGTSTEPSEGSAPGTSTEPSEGS
APGSEPATSGSETPGTSESATPESGPGTSTEPSEGSAP
SEQ ID NO: 17
PGSSPSASTGTGPGASPGTSSTGSPGTPGSGTASSSPGSSTPSGATGSPGTPGSGTASS
SPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTASSSPGSSTPSGATGSPGASPGTSST
GSPGTPGSGTASSSPGSSTPSGATGSPGSSPSASTGTGPGSSPSASTGTGPGSSTPSGA
TGSPGSSTPSGATGSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGTPGSGT
ASSSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGSSPSASTGTGPGTPGSG
TASSSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGSSTPSGATGSPGSSTP
SGATGSPGASPGTSSTGSPGTPGSGTASSSPGSSTPSGATGSPGSSTPSGATGSPGSST
PSGATGSPGSSPSASTGTGPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTASSSPGAS
PGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGASPGTSSTGSPGTPGSGTASSSPGS
STPSGATGSPGTPGSGTASSSPGSSTPSGATGSPGTPGSGTASSSPGSSTPSGATGSPG
SSTPSGATGSPGSSPSASTGTGPGSSPSASTGTGPGASPGTSSTGSPGTPGSGTASSSP
GSSTPSGATGSPGSSPSASTGTGPGSSPSASTGTGPGASPGTSSTGSPGASPGTSSTGS
PGSSTPSGATGSPGSSPSASTGTGPGASPGTSSTGSPGSSPSASTGTGPGTPGSGTASS
SPGSSTPSGATGSPGSSTPSGATGSPGASPGTSSTGSP
[0260] В тех вариантах осуществления, где компонент(компоненты) XTEN содержит(содержат) менее 100% аминокислот, состоящих из 4, 5 или 6 типов аминокислот, выбранных из глицина (G), аланина (A), серина (S), треонина (T), глутамата (E) и пролина (P), или менее 100% последовательности, состоящей из мотивов последовательностей из таблицы 7 или последовательностей XTEN из таблицы 8, другие аминокислотные остатки XTEN выбраны из любых других 14 природных L-аминокислот, но предпочтительно выбраны из гидрофильных аминокислот, так что последовательность XTEN содержит по меньшей мере приблизительно 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или по меньшей мере приблизительно 99% гидрофильных аминокислот. Аминокислоты XTEN, которые не являются глицином (G), аланином (A), серином (S), треонином (T), глутаматом (E) и пролином (P), распределены по всей последовательности XTEN, расположены в пределах мотивов последовательностей или между ними либо сконцентрированы в одном или нескольких коротких отрезках последовательности XTEN, например, для создания линкера между компонентами XTEN и FVIII или VWF. В таких случаях, когда компонент XTEN содержит аминокислоты, отличные от глицина (G), аланина (A), серина (S), треонина (T), глутамата (E) и пролина (P), предпочтительно, чтобы менее чем приблизительно 2% или менее чем приблизительно 1% аминокислот представляли собой гидрофобные остатки, так что полученные последовательности обычно не имеют вторичной структуры, например, имеют не более 2% альфа-спиралей или 2% бета-складчатых слоев, как определено посредством способов, раскрытых в данном документе. Гидрофобные остатки, которые являются менее предпочтительными при конструировании XTEN, включают триптофан, фенилаланин, тирозин, лейцин, изолейцин, валин и метионин. Кроме того, можно разработать последовательности XTEN так, чтобы они содержали менее 5%, или менее 4%, или менее 3%, или менее 2%, или менее 1%, или ни одной из следующих аминокислот: цистеина (во избежание образования дисульфидных связей и окисления), метионина (во избежание окисления), аспарагина и глутамина (во избежание дезамидирования). Таким образом, в некоторых вариантах осуществления компонент XTEN, содержащий другие аминокислоты в дополнение к глицину (G), аланину (A), серину (S), треонину (T), глутамату (E) и пролину (P), имеет последовательность с менее чем 5% остатков, вносящих вклад в образование альфа-спиралей и бета-складчатых слоев, как измерено с помощью алгоритма Чоу-Фасмана, и характеризуются по меньшей мере 90% или по меньшей мере приблизительно 95% или большей вероятностью образования случайного клубка, как измерено с помощью алгоритма GOR.
[0261] В дополнительных вариантах осуществления последовательность XTEN, применяемая в настоящем изобретении, влияет на физические или химические свойства, например, фармакокинетические показатели, химерного полипептида по настоящему изобретению. Последовательность XTEN, применяемая в настоящем изобретении, может демонстрировать одно или несколько из следующих преимущественных свойств: конформационную гибкость, повышенную растворимость в воде, высокую степень устойчивости к протеазам, низкую иммуногенность, слабое связывание с рецепторами млекопитающих или увеличенные значения гидродинамического радиуса (или радиуса Стокса). В некоторых вариантах осуществления последовательность XTEN, соединенная с белком FVIII согласно настоящему изобретению, улучшает фармакокинетические свойства, как, например, обеспечивает более длительный конечный период полужизни или увеличенную площадь под кривой (AUC), так что описанный в данном документе химерный полипептид сохраняется in vivo в течение увеличенного периода времени по сравнению с FVIII дикого типа. В дополнительных вариантах осуществления последовательность XTEN, применяемая в настоящем изобретении, улучшает фармакокинетические свойства, как, например, обеспечивает более длительный конечный период полужизни или увеличенную площадь под кривой (AUC), так что белок FVIII сохраняется in vivo в течение увеличенного периода времени по сравнению с FVIII дикого типа.
[0262] Для определения физических/химических свойств белков, содержащих последовательность XTEN, можно использовать различные способы и анализы. Такие способы включают без ограничения аналитическое центрифугирование, EPR, ионообменную HPLC, эксклюзионную HPLC, обращенно-фазовую HPLC, рассеяние света, капиллярный электрофорез, круговой дихроизм, дифференциальную сканирующую калориметрию, флуоресценцию, ионообменную HPLC, эксклюзионную HPLC, IR-спектроскопию, NMR, рамановскую спектрометрию, рефрактометрию и спектроскопию в UV- и видимой области спектра. Дополнительные способы раскрыты в Amau et al., Prot Expr and Purif 48, 1-13 (2006).
[0263] Дополнительные примеры последовательностей XTEN, которые можно применять согласно настоящему изобретению, раскрыты в публикациях заявок на патент США №№ 2010/0239554 A1, 2010/0323956 A1, 2011/0046060 A1, 2011/0046061 A1, 2011/0077199 A1 или 2011/0172146 A1 или публикациях международных заявок на патент №№ WO 2010091122 A1, WO 2010144502 A2, WO 2010144508 A1, WO 2011028228 A1, WO 2011028229 A1, WO 2011028344 A2 или WO 20130122617 A1.
III.C.2. Альбумины
[0264] Альбумин или его часть может являться партнером по связыванию с FcRn. В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один полипептид альбумин или его фрагмент, вариант или производное. Сывороточный альбумин человека (HSA или HA), представляющий собой белок из 609 аминокислот в своей полноразмерной форме, ответственен за значительную долю осмотического давления сыворотки крови, а также выполняет функцию носителя эндогенных и экзогенных лигандов. Термин "альбумин", используемый в данном документе, включает полноразмерный альбумин или его функциональный фрагмент, вариант, производное или аналог. Примеры альбумина или его фрагментов или вариантов раскрыты в публикациях заявок на патент США №№ 2008/0194481A1, 2008/0004206 A1, 2008/0161243 A1, 2008/0261877 A1 или 2008/0153751 A1 или в публикациях заявок согласно PCT №№ 2008/033413 A2, 2009/058322 A1 или 2007/021494 A2, которые включены в данный документ посредством ссылки во всей своей полноте.
[0265] Альбуминсвязывающие полипептиды (ABP) могут включать без ограничения бактериальные альбуминсвязывающие домены, альбуминсвязывающие пептиды или фрагменты альбуминсвязывающих антител, которые могут связываться с альбумином. Домен 3 из стрептококкового белка G, раскрытого в Kraulis et al., FEBS Lett. 378:190-194 (1996) и Linhult et al., Protein Sci. 11:206-213 (2002), является примером бактериального альбуминсвязывающего домена. Примеры альбуминсвязывающих пептидов раскрыты в Dennis et al., J. Biol. Chem. 2002, 277: 35035-35043 (2002). Примеры фрагментов альбуминсвязывающих антител раскрыты в Muller and Kontermann, Curr. Opin. Mol. Ther. 9:319-326 (2007); Roovers et al., Cancer Immunol. Immunother. 56:303-317 (2007), и Holt et al., Prot. Eng. Design Sci., 21:283-288 (2008), которые включены в данный документ посредством ссылки во всей своей полноте.
[0266] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один сайт присоединения неполипептидной малой молекулы, ее варианта или производного, которые могут связываться с альбумином. Например, химерный полипептид может содержать один или несколько органических альбуминсвязывающих компонентов. Примером таких альбуминсвязывающих компонентов является 2-(3-малеимидопропанамидо)-6-(4-(4-йодфенил)бутанамидо)гексаноат (метка "Albu"), раскрытый в Trussel et al., Bioconjugate Chem. 20:2286-2292 (2009).
[0267] В определенных вариантах осуществления фрагмент VWF, описанный в данном документе, слит с альбумином. В некоторых вариантах осуществления конструкции фрагмент VWF-альбумин образуют гомодимеры. В некоторых вариантах осуществления домен D'D3 VWF, который слит с альбумином, связывается с полипептидом FVIII. В некоторых вариантах осуществления домен D'D3 VWF, слитый с альбумином, связывается с полипептидом FVIII посредством взаимодействия, более сильного, чем природные нековалентные взаимодействия между FVIII дикого типа и VWF. В конкретных вариантах осуществления домен D'D3 VWF и/или полипептид FVIII содержат одну или несколько мутаций, которые увеличивают аффинность между доменом D'D3 VWF и полипептидом FVIII. В определенных вариантах осуществления домен D'D3 VWF и/или полипептид FVIII содержат одну или несколько мутаций, которые обеспечивают образование дисульфидной связи между доменом D'D3 VWF и/или полипептидом FVIII.
III.C.3. CTP
[0268] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один C-концевой пептид (CTP) β-субъединицы хорионического гонадотропина человека или его фрагмент, вариант или производное. Известно, что CTP-пептиды обеспечивают увеличение периода полужизни данного белка. См., например, патент США № 5712122, включенный в данный документ посредством ссылки во всей своей полноте. Неограничивающие иллюстративные CTP-пептиды раскрыты в публикации заявки на патент США № US 2009/0087411 A1, включенной посредством ссылки.
III.C.4. PAS
[0269] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один пептид PAS или его фрагмент, вариант или производное. Пептид PAS или последовательность PAS, как используется в данном документе, означают аминокислотную последовательность, содержащую главным образом аланиновые и сериновые остатки или содержащую главным образом аланиновые, сериновые и пролиновые остатки, при этом аминокислотная последовательность образует конформацию случайного клубка в физиологических условиях. Соответственно, последовательность PAS представляет собой структурный блок, полимер из аминокислот или последовательность-кассету, которые содержат аланин, серин и пролин, состоят по сути из них или состоят из них, которые можно применять в качестве части гетерологичного компонента в химерном полипептиде. Полимер из аминокислот также может образовывать конформацию случайного клубка при добавлении в последовательность PAS остатков, отличных от аланина, серина и пролина, в качестве вспомогательной составляющей. Под "вспомогательной составляющей" подразумевается, что аминокислоты, отличные от аланина, серина и пролина, могут быть добавлены в последовательность PAS в определенной мере, например, в количестве до приблизительно 12%, т. e. приблизительно 12 из 100 аминокислот последовательности PAS, до приблизительно 10%, до приблизительно 9%, до приблизительно 8%, приблизительно 6%, приблизительно 5%, приблизительно 4%, приблизительно 3%, т. e. приблизительно 2%, или приблизительно 1% аминокислот. Аминокислоты, отличные от аланина, серина и пролина, могут быть выбраны из группы, состоящей из Arg, Asn, Asp, Cys, Gln, Glu, Gly, His, Ile, Leu, Lys, Met, Phe, Thr, Trp, Tyr и Val. В физиологических условиях пептид PAS образует конформацию случайного клубка и за счет этого может опосредовать увеличенную стабильность рекомбинантного белка по настоящему изобретению in vivo и/или in vitro и обладает прокоагулянтной активностью.
[0270] Неограничивающие примеры пептидов PAS раскрыты, например, в публикации заявки на патент США № 2010/0292130 A1; публикации заявки согласно PCT № WO 2008/155134 A1 и выданном европейском патенте № EP2173890.
III.C.5. HAP
[0271] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один пептид, являющийся гомополимером из аминокислот (HAP), или его фрагмент, вариант или производное. Пептид HAP может содержать последовательность из повторяющихся глициновых остатков, которая имеет длину по меньшей мере 50 аминокислот, по меньшей мере 100 аминокислот, 120 аминокислот, 140 аминокислот, 160 аминокислот, 180 аминокислот, 200 аминокислот, 250 аминокислот, 300 аминокислот, 350 аминокислот, 400 аминокислот, 450 аминокислот или 500 аминокислот. Последовательность HAP способна обеспечивать продление периода полужизни компонента, слитого или связанного с последовательностью HAP. Неограничивающие примеры последовательности HAP включают без ограничения (Gly)n, (Gly4Ser)n или S(Gly4Ser)n, где n равняется 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, или 20. В некоторых вариантах осуществления n равняется 20, 21, 22, 23, 24, 25, 26, 26, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, или 40. В других вариантах осуществления n равняется 50, 60, 70, 80, 90, 100, 110, 120, 130, 140, 150, 160, 170, 180, 190, или 200. См., например, Schlapschy M et al., Protein Eng. Design Selection, 20: 273-284 (2007).
III.C.6. Трансферрин
[0272] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один пептид трансферрин или его фрагмент, вариант или производное. Любой трансферрин можно сливать с химерным полипептидом, применяемым в способах по настоящему изобретению. В качестве примера, Tf человека дикого типа (Tf) представляет собой белок из 679 аминокислот размером примерно 75 кДа (без учета гликозилирования) с двумя основными доменами N (приблизительно 330 аминокислот) и C (приблизительно 340 аминокислот), которые, по-видимому, образуются в результате дупликации гена. См. номера доступа в GenBank NM001063, XM002793, M12530, XM039845, XM 039847 и S95936 (www.ncbi.nlm.nih.gov), все из которых включены в данный документ посредством ссылки во всей своей полноте.
[0273] Трансферрин транспортирует железо путем эндоцитоза, опосредованного рецепторами трансферрина (TfR). После высвобождения железа в эндосомальный компартмент и рециркуляции комплекса Tf-TfR на клеточную поверхность Tf высвобождается обратно во внеклеточное пространство для следующего цикла транспорта железа. Tf обладает длительным периодом полужизни, превышающим 14-17 дней (Li et al., Trends Pharmacol. Sci. 23:206-209 (2002)). Слитые белки на основе трансферрина исследовались в отношении продления периода полужизни, целенаправленной доставки для терапии рака, пероральной доставки и устойчивой активации проинсулина (Brandsma et al., Biotechnol. Adv., 29: 230-238 (2011); Bai et al., Proc. Natl. Acad. Sci. USA 102:7292-7296 (2005); Kim et al., J. Pharmacol. Exp. Ther., 334:682-692 (2010); Wang et al., J. Controlled Release 155:386-392 (2011)).
III.C.7. PEG
[0274] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один сайт присоединения неполипептидного гетерологичного компонента или его фрагмента, варианта или производного. Например, химерный полипептид, применяемый в способах по настоящему изобретению, может содержать один или несколько полиэтиленгликолевых (PEG) компонентов, присоединенных к одному или нескольким аминокислотным остаткам в факторе свертывания крови и/или Fc-области.
[0275] Пегилирование белка может относиться к конъюгату, образуемому между белком и по меньшей мере одной молекулой полиэтиленгликоля (PEG). PEG коммерчески доступен в широком ассортименте с различными значениями молекулярной массы и диапазонами средней молекулярной массы. Типичные примеры диапазонов средней молекулярной массы PEG включают без ограничения приблизительно 200, приблизительно 300, приблизительно 400, приблизительно 600, приблизительно 1000, приблизительно 1300-1600, приблизительно 1450, приблизительно 2000, приблизительно 3000, приблизительно 3000-3750, приблизительно 3350, приблизительно 3000-7000, приблизительно 3500-4500, приблизительно 5000-7000, приблизительно 7000-9000, приблизительно 8000, приблизительно 10000, приблизительно 8500-11500, приблизительно 16000-24000, приблизительно 35000, приблизительно 40000, приблизительно 60000 и приблизительно 80000 дальтонов. Эти значения средней молекулярной массы приведены исключительно в качестве примеров и не подразумеваются как ограничивающие каким-либо образом.
[0276] Химерный полипептид, применяемый в способах по настоящему изобретению, может быть пегилирован со включением одного или нескольких (например, 2-4) PEG-компонентов. Пегилирование можно выполнять с помощью любой из реакций пегилирования, известных из уровня техники. Способы получения продукта на основе пегилированного белка обычно будут включать (i) осуществление реакции полипептида с полиэтиленгликолем (таким как реакционноспособное сложноэфирное или альдегидное производное PEG) в условиях, при которых происходит присоединение пептида по настоящему изобретению к одной или нескольким группам PEG; и (ii) получение продукта(продуктов) реакции. В целом, оптимальные реакционные условия для реакций будут определяться в каждом конкретном случае, исходя из известных параметров и необходимого результата.
[0277] Существует ряд способов присоединения PEG, которые доступны специалистам в данной области, например, в Malik F et al., Exp. Hematol. 20:1028-35 (1992); Francis, Focus on Growth Factors 3(2):4-10 (1992); публикациях заявок на европейский патент №№ EP0401384, EP0154316 и EP0401384 и публикациях международных заявок на патент №№ WO92/16221 и WO95/34326. В качестве неограничивающего примера, варианты FVIII могут содержать замены на цистеин, и цистеиновые остатки могут быть дополнительно конъюгированы с полимером PEG. См. Mei et al., Blood 116:270-279 (2010) и патент США № 7632921, которые включены в данный документ посредством ссылки во всей своей полноте.
[0278] В определенных вариантах осуществления химерный полипептид содержит одноцепочечный полипептид FVIII, ковалентно соединенный с фрагментом VWF, содержащим домен D' и домен D3 VWF, где полипептид FVIII содержит делецию всего домена B или его части, где полипептид FVIII является пегилированным, и где фрагмент VWF непосредственно или опосредованно слит с N-концом полипептида FVIII (например, в PEG-scFVIII-D'D3, "MG1121" или LAFATE). В других вариантах осуществления химерный полипептид содержит одноцепочечный полипептид FVIII, ковалентно соединенный с фрагментом VWF, содержащим домен D' и домен D3 VWF, где полипептид FVIII содержит делецию всего домена B или его части, где фрагмент VWF является пегилированным, и где фрагмент VWF непосредственно или опосредованно слит с N-концом полипептида FVIII (например, в scFVIII-D'D3-PEG). В других вариантах осуществления химерный полипептид содержит одноцепочечный полипептид FVIII, ковалентно соединенный с фрагментом VWF, содержащим домен D' и домен D3 VWF, где полипептид FVIII содержит делецию всего домена B или его части, где полипептид FVIII и фрагмент VWF являются пегилированными, и где фрагмент VWF непосредственно или опосредованно слит с N-концом полипептида FVIII (например, в scFVIII-D'D3-PEG).
III.C.8. HES
[0279] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один полимер гидроксиэтилкрахмал (HES). HES является производным встречающегося в природе амилопектина и разрушается под действием альфа-амилазы в организме. HES демонстрирует преимущественные биологические свойства и применяется в качестве средства для восполнения объема крови и в гемодилюционной терапии в клиниках. См., например, Sommermeyer et al., Krankenhauspharmazie 8:271-278 (1987); и Weidler et al., Arzneim.-Forschung/Drug Res. 41: 494-498 (1991).
[0280] HES главным образом характеризуют по молекулярно-массовому распределению и степени замещения. HES характеризуется средней молекулярной массой (средневзвешенной) от 1 до 300 кДа, от 2 до 200 кДа, от 3 до 100 кДа или от 4 до 70 кДа. Гидроксиэтилкрахмал может дополнительно характеризоваться степенью молярного замещения от 0,1 до 3, от 0,1 до 2, от 0,1 до 0,9 или от 0,1 до 0,8 и соотношением замещения С2:С6 в диапазоне от 2 до 20 применительно к гидроксиэтильным группам. HES со средней молекулярной массой приблизительно 130 кДа представляет собой Voluven® от Fresenius. Voluven® представляет собой искусственный коллоид, используемый, например, для восполнения объема, который применяют при терапевтическом показании для терапии и профилактики гиповолемии. Существует ряд способов присоединения HES, доступных специалистам в данной области, например, аналогичные способам присоединения PEG, которые описаны выше.
III.C.9. PSA
[0281] В определенных аспектах химерный полипептид, применяемый в способах по настоящему изобретению, содержит по меньшей мере один полимер полисиаловую кислоту (PSA). PSA представляют собой встречающиеся в природе неразветвленные полимеры сиаловой кислоты, вырабатываемые определенными штаммами бактерий и у млекопитающих в определенных клетках. См., например, Roth J. et al. (1993) в Polysialic Acid: From Microbes to Man, eds. Roth J., Rutishauser U., Troy F. A. (BirkhäuserVerlag, Basel, Switzerland), pp. 335-348. PSA могут быть получены с различной степенью полимеризации от n=приблизительно 80 или больше остатков сиаловой кислоты до n=2 путем неполного кислотного гидролиза, или путем расщепления нейраминидазами, или путем фракционирования природных форм полимера бактериального происхождения. Существует ряд способов присоединения PSA, доступных специалистам в данной области, например, аналогичные способам присоединения PEG, которые описаны выше. В определенных аспектах активированная PSA также может быть присоединена к цистеиновому аминокислотному остатку в факторе свертывания крови, например, в FVIII, или в Fc-области. См., например, патент США № 5846951.
III.C.10. Рецепторы, опосредующие выведение
[0282] В определенных аспектах период полужизни химерного полипептида, применяемого в способах согласно настоящему изобретению, может быть продлен, если фактор свертывания крови в составе химерного полипептида содержит полипептид FVIII и по меньшей мере один фрагмент рецептора, опосредующего выведение FVIII, или его FVIII-связывающий фрагмент, вариант или производное. Посредством вставки растворимых форм рецепторов, опосредующих выведение, таких как белок LRP1, родственный рецепторам липопротеинов низкой плотности, или их фрагментов можно блокировать связывание FVIII с рецепторами, опосредующими выведение, и за счет этого продлевать его период полужизни, например, период полужизни in vivo. LRP1 представляет собой интегральный мембранный белок размером 600 кДа, который участвует в рецепторно-опосредованном выведении различных белков, в том числе FVIII. См., например, Lenting et al., Haemophilia 16:6-16 (2010). Другими подходящими рецепторами, опосредующими выведение FVIII, являются, например, LDLR (рецептор липопротеинов низкой плотности), VLDLR (рецептор липопротеинов очень низкой плотности) и мегалин (LRP-2) или их фрагменты. См., например, Bovenschen et al., Blood 106:906-912 (2005); Bovenschen, Blood 116:5439-5440 (2010); Martinelli et al., Blood 116:5688-5697 (2010).
III.D. Сайты вставки
[0283] В некоторых аспектах сайт вставки в полипептиде FVIII расположен в одном или нескольких доменах полипептида FVIII, которые представляют собой N-конец, домен A1, домен A2, домен A3, домен B, домен C1, домен C2, C-конец или две или более их комбинации, или между двумя доменами полипептида FVIII, которые представляют собой домен A1 и кислую область a1, и кислую область a1 и домен A2, домен A2 и кислую область a2, кислую область a2 и домен B, домен B и домен A3, и домен A3 и домен C1, домен C1 и домен C2 или любые их комбинации. Например, сайты вставки, в которые может быть вставлена последовательность XTEN, выбраны из группы, состоящей из N-конца и домена A1, N-конца и домена A2, N-конца и домена A3, N-конца и домена В, N-конца и домена C1, N-конца и домена C2, N-конца и C-конца, доменов A1 и A2, доменов A1 и A3, доменов A1 и B, доменов A1 и C1, доменов A1 и C2, домена A1 и C-конца, доменов A2 и A3, доменов A2 и B, доменов A2 и C1, доменов A2 и C2, домена A2 и C-конца, доменов A3 и B, доменов A3 и C1, доменов A3 и C2, домена A3 и C-конца, доменов B и C1, доменов B и C2, домена B и C-конца, доменов C1 и C2, домена C1 и C-конца, домена C2 и C-конца, а также двух или более их комбинаций.
[0284] Полипептид FVIII, в котором последовательность XTEN вставлена непосредственно ниже одной или нескольких аминокислот (например, в одном или нескольких сайтах вставки XTEN) в полипептиде FVIII или соединена с С-концом или N-концом, сохраняет активность FVIII после соединения с последовательностью XTEN или ее вставки. Последовательность XTEN может быть вставлена в полипептид FVIII один или более одного раза, два раза, три раза, четыре раза, пять раз или шесть раз, таким образом, что вставки не влияют на активность FVIII (т. е. белок FVIII по-прежнему сохраняет свойство коагуляции).
[0285] Полипептид FVIII, применимый в настоящем изобретении, может быть соединен с одним или несколькими полипептидами XTEN на N-конце или C-конце полипептида FVIII с помощью необязательного линкера, или они могут быть вставлены непосредственно ниже одной или нескольких аминокислот (например, в одном или нескольких сайтах вставки XTEN) в полипептиде FVIII с помощью одного или нескольких необязательных линкеров.
[0286] В еще некоторых других вариантах осуществления один или несколько XTEN вставлены в домен B FVIII. В одном примере XTEN вставлен между аминокислотами 740 и 1640, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 740 и 1640 необязательно отсутствует. В другом примере XTEN вставлен между аминокислотами 741 и 1690, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 740 и 1690 необязательно отсутствует. В других примерах XTEN вставлен между аминокислотами 741 и 1648, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 741 и 1648 необязательно отсутствует. В еще некоторых других примерах XTEN вставлен между аминокислотами 743 и 1638, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 743 и 1638 необязательно отсутствует. В еще нескольких других примерах XTEN вставлен между аминокислотами 745 и 1656, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 745 и 1656 необязательно отсутствует. В некоторых примерах XTEN вставлен между аминокислотами 745 и 1657, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 745 и 1657 необязательно отсутствует. В определенных примерах XTEN вставлен между аминокислотами 745 и 1667, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 745 и 1667 необязательно отсутствует. В еще нескольких других примерах XTEN вставлен между аминокислотами 745 и 1686, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 745 и 1686 необязательно отсутствует. В некоторых других примерах XTEN вставлен между аминокислотами 747 и 1642, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 747 и 1642 необязательно отсутствует. В еще нескольких других примерах XTEN вставлен между аминокислотами 751 и 1667, соответствующими SEQ ID NO: 65, где последовательность FVIII между аминокислотами 751 и 1667 необязательно отсутствует. В некоторых вариантах осуществления последовательность XTEN вставлена между аминокислотами 745 и 746 зрелого полипептида FVIII или в соответствующем сайте вставки полипептида FVIII с делецией домена B. В конкретном варианте осуществления последовательность XTEN, содержащая аминокислотную последовательность, которая характеризуется по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или приблизительно 100% идентичностью последовательности с аминокислотной последовательностью под SEQ AE288 (SEQ ID NO: 8), вставлена между аминокислотами 745 и 746 зрелого FVIII, где зрелый FVIII имеет делецию аминокислотных остатков 746-1648. В конкретных вариантах осуществления XTEN вставлен непосредственно ниже аминокислоты 745 в FVIII с делецией домена B из таблицы 5 (SEQ ID NO: 68).
III.E. Линкеры
[0287] В определенных вариантах осуществления химерный полипептид по настоящему изобретению дополнительно содержит один или несколько линкеров в белке FVIII и/или во фрагменте VWF. Одним из типов линкеров является расщепляемый линкер, который может расщепляться различными протеазами при введении субъекту in vivo, например, в участке коагуляции. В некоторых вариантах осуществления расщепляемый линкер обеспечивает отщепление компонента, например, белка VWF, от последовательности XTEN и, таким образом, от химерного полипептида в участке каскада коагуляции, за счет чего обеспечивается наличие активности FVIIIa у активированного FVIII (FVIIIa). Другим типом линкеров является процессируемый линкер, который содержит сайт внутриклеточного расщепления и, таким образом, может расщепляться ферментом внутриклеточного процессинга в клетке-хозяине, обеспечивая удобную экспрессию полипептида и образование химерного полипептида.
[0288] Один или несколько линкеров могут присутствовать между любыми двумя белками химерного полипептида. В некоторых вариантах осуществления химерный полипептид содержит первый полипептид, который содержит (i) полипептид FVIII и (ii) первую константную область Ig или ее часть, и второй полипептид, который содержит (iii) белок VWF, (iv) линкер (например, расщепляемый линкер), (v) последовательность XTEN и (vi) вторую константную область Ig или ее часть. В других вариантах осуществления химерный полипептид содержит первый полипептид, который содержит (i) полипептид FVIII и (ii) первую константную область Ig или ее часть, и второй полипептид, который содержит (iii) белок VWF, (iv) последовательность XTEN, (v) линкер (например, расщепляемый линкер), а также (vi) вторую константную область Ig или ее часть. В других вариантах осуществления химерный полипептид содержит первый полипептид, который содержит (i) полипептид FVIII и (ii) первую константную область Ig или ее часть, и второй полипептид, который содержит (iii) белок VWF, (iv) первый линкер (например, расщепляемый линкер), (v) последовательность XTEN, (vi) второй линкер (например, расщепляемый линкер), а также (vii) вторую константную область Ig или ее часть. В некоторых вариантах осуществления первый полипептид дополнительно содержит линкер, например, расщепляемый линкер между полипептидом FVIII и первой константной областью Ig.
[0289] В определенных вариантах осуществления химерный полипептид содержит одну цепь, содержащую (i) полипептид FVIII, (ii) первую константную область Ig или ее часть, (iii) линкер (например, процессируемый линкер), (iv) белок VWF, (v) последовательность XTEN и (vi) вторую константную область Ig или ее часть. В других вариантах осуществления химерный полипептид содержит одну цепь, содержащую (i) полипептид FVIII, (ii) первую константную область Ig или ее часть, (iii) первый линкер (например, процессируемый линкер), (iv) белок VWF, (v) второй линкер (например, расщепляемый линкер), (vi) последовательность XTEN и (vii) вторую константную область Ig или ее часть. Процессируемый линкер может подвергаться процессингу после экспрессии химерного полипептида в клетке-хозяине; таким образом, химерный полипептид, вырабатываемый в клетке-хозяине, может быть представлен в конечной форме, содержащей две или три полипептидные цепи.
[0290] Линкер, применимый в настоящем изобретении, может включать в себя любую органическую молекулу. Примеры линкеров, которые можно применять в химерных полипептидах по настоящему изобретению, раскрыты, например, в US 2011/0183907 A1, US 2016/0229903 A1 и US 2016/0251408 A1, каждый из которых включен в данный документ посредством ссылки во всей своей полноте. В некоторых вариантах осуществления линкер содержит полимер, например, полиэтиленгликоль (PEG) или гидроксиэтилкрахмал (HES). В других вариантах осуществления линкер содержит аминокислотную последовательность. Линкер может содержать по меньшей мере приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300, 400,500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900 или 2000 аминокислот. Линкер может содержать 1-5 аминокислот, 1-10 аминокислот, 1-20 аминокислот, 10-50 аминокислот, 50-100 аминокислот, 100-200 аминокислот, 200-300 аминокислот, 300-400 аминокислот, 400-500 аминокислот, 500-600 аминокислот, 600-700 аминокислот, 700-800 аминокислот, 800-900 аминокислот или 900-1000 аминокислот. В некоторых вариантах осуществления линкер содержит последовательность XTEN. Дополнительные примеры XTEN, которые можно применять согласно настоящему изобретению, раскрыты в публикациях заявок на патент США №№ 2010/0239554 A1, 2010/0323956 A1, 2011/0046060 A1, 2011/0046061 A1, 2011/0077199 A1 или 2011/0172146 A1 или публикациях международных заявок на патент №№ WO 2010091122 A1, WO 2010144502 A2, WO 2010144508 A1, WO 2011028228 A1, WO 2011028229 A1 или WO 2011028344 A2. В других вариантах осуществления линкер представляет собой последовательность PAS.
[0291] В определенных вариантах осуществления линкер представляет собой полимер, например, полиэтиленгликоль (PEG) или гидроксиэтилкрахмал (HES). В других вариантах осуществления линкер представляет собой аминокислотную последовательность. Линкер может содержать по меньшей мере приблизительно 10, 20, 30, 40, 50, 60, 70, 80, 90, 100, 150, 200, 300, 400, 500, 600, 700, 800, 900, 1000, 1100, 1200, 1300, 1400, 1500, 1600, 1700, 1800, 1900, или 2000 аминокислот. Линкер может содержать 1-5 аминокислот, 1-10 аминокислот, 1-20 аминокислот, 10-50 аминокислот, 50-100 аминокислот, 100-200 аминокислот, 200-300 аминокислот, 300-400 аминокислот, 400-500 аминокислот, 500-600 аминокислот, 600-700 аминокислот, 700-800 аминокислот, 800-900 аминокислот или 900-1000 аминокислот.
[0292] Примеры линкеров хорошо известны из уровня техники. В определенных вариантах осуществления линкер содержит последовательность Gn. Линкер может содержать последовательность (GA)n. Линкер может содержать последовательность (GGS)n. В других вариантах осуществления линкер содержит (GGGS)n (SEQ ID NO: 101). В еще нескольких других вариантах осуществления линкер содержит последовательность (GGS)n(GGGGS)n (SEQ ID NO: 95). В этих случаях n может являться целым числом от 1 до 100. В других случаях n может являться целым числом от 1 до 20, т. е.1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, или 20. Примеры линкеров включают без ограничения GGG, SGGSGGS (SEQ ID NO: 96), GGSGGSGGSGGSGGG (SEQ ID NO: 97), GGSGGSGGGGSGGGGS (SEQ ID NO: 98), GGSGGSGGSGGSGGSGGS (SEQ ID NO: 99), или GGGGSGGGGSGGGGS (SEQ ID NO: 100). Линкер не устраняет и не снижает активность белка VWF или свертывающую активность фактора VIII. Линкер необязательно усиливает активность белка VWF или свертывающую активность белкового фактора VIII, например, за счет дополнительного снижения эффектов стерического несоответствия и обеспечения большей доступности белка VWF или части фактора VIII для их сайта связывания в мишени.
[0293] В определенных вариантах осуществления линкер, применимый для химерного полипептида, имеет длину 15-25 аминокислот. В других вариантах осуществления линкер, применимый для химерного полипептида, имеет длину 15-20 аминокислот. В некоторых вариантах осуществления линкер для химерного полипептида имеет длину 10-25 аминокислот. В других вариантах осуществления линкер для химерного полипептида имеет длину 15 аминокислот. В еще нескольких других вариантах осуществления линкер для химерного полипептида представляет собой (GGGGS)n (SEQ ID NO: 94), где G представляет собой глицин, S представляет собой серин, и n представляет собой целое число от 1 до 20.
III.F. Сайты расщепления
[0294] Расщепляемый линкер может содержать в своем составе компонент, способный расщепляться химическим (например, путем гидролиза сложноэфирной связи), ферментативным (т. е. при встраивании последовательности для расщепления протеазой) или фотолитическим путем (например, в случае с хромофором, таким как 3-амино-3-(2-нитрофенил)пропионовая кислота (ANP)) для высвобождения одной молекулы от другой. Примеры расщепляемых линкеров, подходящих для химерных полипептидов по настоящему изобретению, можно найти, например, в US 2016/0229903 A1 и US 2016/0251408 A1, каждый из которых включен в данный документ посредством ссылки во всей своей полноте.
[0295] В определенных вариантах осуществления расщепляемый линкер содержит один или несколько сайтов расщепления на N-конце или C-конце или на них обоих. В других вариантах осуществления расщепляемый линкер состоит по сути из одного или нескольких расщепляемых сайтов или состоит из них. В других вариантах осуществления расщепляемый линкер содержит гетерологичные аминокислотные линкерные последовательности, описанные в данном документе, или полимеры и один или несколько расщепляемых сайтов.
[0296] В определенных вариантах осуществления расщепляемый линкер содержит один или несколько сайтов расщепления, которые могут расщепляться в клетке-хозяине (т. е. сайты внутриклеточного процессинга). Неограничивающие примеры сайта расщепления включают RRRR (SEQ ID NO: 102), RKRRKR (SEQ ID NO: 103), а также RRRRS (SEQ ID NO: 104).
[0297] В некоторых вариантах осуществления расщепляемый линкер содержит область a1 из FVIII, область a2 из FVIII, область a3 из FVIII, сайт расщепления тромбином, который содержит X-V-P-R (SEQ ID NO: 105) и мотив PAR1, взаимодействующий с экзосайтами, где X представляет собой алифатическую аминокислоту, или любые их комбинации. Он содержит область a2, которая содержит аминокислотную последовательность, на по меньшей мере приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% или на 100% идентичную последовательности от Glu720 до Arg740, соответствующей полноразмерному FVIII, при этом область a2 способна расщепляться тромбином. В конкретных вариантах осуществления расщепляемый линкер, применимый для настоящего изобретения, содержит область a2, которая содержит ISDKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 106) или DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88).. В других вариантах осуществления расщепляемый линкер по настоящему изобретению содержит область a1, которая содержит аминокислотную последовательность, на по меньшей мере приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% или на 100% идентичную последовательности от Met337 до Arg372, соответствующей полноразмерному FVIII, при этом область a1 способна расщепляться тромбином. В конкретных вариантах осуществления область a1 содержит ISMKNNEEAEDYDDDLTDSEMDVVRFDDDNSPSFIQIRSV (SEQ ID NO: 107). В некоторых вариантах осуществления расщепляемый линкер по настоящему изобретению содержит область a3, которая содержит аминокислотную последовательность, на по меньшей мере приблизительно 80%, приблизительно 85%, приблизительно 90%, приблизительно 95% или на 100% идентичную последовательности от Glu1649 до Arg1689, соответствующей полноразмерному FVIII, при этом область a3 способна расщепляться тромбином. В определенных вариантах осуществления расщепляемый линкер по настоящему изобретению содержит область a3, содержащую ISEITRTTLQSDQEEIDYDDTISVEMKKEDFDIYDEDENQSPRSFQ (SEQ ID NO: 108).
[0298] В других вариантах осуществления расщепляемый линкер содержит сайт расщепления для тромбина, который содержит X-V-P-R (SEQ ID NO: 105) и мотив PAR1, взаимодействующий с экзосайтами, и где мотив PAR1, взаимодействующий с экзосайтами, содержит S-F-L-L-R-N (SEQ ID NO: 109). Мотив PAR1, взаимодействующий с экзосайтами, может дополнительно содержать аминокислотную последовательность, выбранную из P, P-N, P-N-D, P-N-D-K (SEQ ID NO: 110), P-N-D-K-Y (SEQ ID NO: 111), P-N-D-K-Y-E (SEQ ID NO: 112), P-N-D-K-Y-E-P (SEQ ID NO: 113), P-N-D-K-Y-E-P-F (SEQ ID NO: 114), P-N-D-K-Y-E-P-F-W (SEQ ID NO: 115), P-N-D-K-Y-E-P-F-W-E (SEQ ID NO: 116), P-N-D-K-Y-E-P-F-W-E-D (SEQ ID NO: 117), P-N-D-K-Y-E-P-F-W-E-D-E (SEQ ID NO: 118), P-N-D-K-Y-E-P-F-W-E-D-E-E (SEQ ID NO: 119), P-N-D-K-Y-E-P-F-W-E-D-E-E-S (SEQ ID NO: 120), или любую их комбинацию. В некоторых вариантах осуществления алифатическая аминокислота выбрана из глицина, аланина, валина, лейцина или изолейцина.
[0299] В других вариантах осуществления расщепляемый линкер содержит один или несколько сайтов расщепления, которые расщепляются протеазой после введения субъекту химерного полипептида, содержащего расщепляемый линкер. В некоторых вариантах осуществления сайт расщепления расщепляется протеазой, выбранной из группы, состоящей из фактора XIa, фактора XIIa, калликреина, фактора VIIa, фактора IXa, фактора Xa, фактора IIa (тромбина), эластазы-2, MMP-12, MMP-13, MMP-17 и MMP-20. В других вариантах осуществления сайт расщепления выбран из группы, состоящей из сайта расщепления для FXIa (например, KLTR↓AET (SEQ ID NO: 121)), сайта расщепления для FXIa (например, DFTR↓VVG (SEQ ID NO: 122)), сайта расщепления для FXIIa (например, TMTR↓IVGG (SEQ ID NO: 123)), сайта расщепления для калликреина (например, SPFR↓STGG (SEQ ID NO: 124)), сайта расщепления для FVIIa (например, LQVR↓IVGG (SEQ ID NO: 125)), сайта расщепления для FIXa (например, PLGR↓IVGG (SEQ ID NO: 126)), сайта расщепления для FXa (например, IEGR↓TVGG (SEQ ID NO: 127)), сайта расщепления для FIIa (тромбина) (например, LTPR↓SLLV (SEQ ID NO: 128)), сайта расщепления для эластазы-2 (например, LGPV↓SGVP (SEQ ID NO: 129)), сайта расщепления для гранзима В (например, VAGD↓SLEE (SEQ ID NO: 130)), сайта расщепления для MMP-12 (например, GPAG↓LGGA (SEQ ID NO: 131)), сайта расщепления для MMP-13 (например, GPAG↓LRGA (SEQ ID NO: 132)), сайта расщепления для MMP-17 (например, APLG↓LRLR (SEQ ID NO: 133)), сайта расщепления для MMP-20 (например, PALP↓LVAQ (SEQ ID NO: 134)), сайта расщепления для TEV (например, ENLYFQ↓G (SEQ ID NO: 135)), сайта расщепления для энтерокиназы (например, DDDK↓IVGG (SEQ ID NO: 136)), сайта расщепления для протеазы 3C (PreScission™) (например, LEVLFQ↓GP (SEQ ID NO: 137)), а также сайта расщепления для сортазы A (например, LPKT↓GSES) (SEQ ID NO: 138). В определенных вариантах осуществления сайты расщепления для FXIa включают без ограничения, например, TQSFNDFTR (SEQ ID NO: 1) и SVSQTSKLTR (SEQ ID NO: 3). Неограничивающие иллюстративные примеры сайтов расщепления для тромбина включают, например, DFLAEGGGVR (SEQ ID NO: 4), TTKIKPR (SEQ ID NO: 5), LVPRG (SEQ ID NO: 6), DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88) или IEPRSFS (SEQ ID NO: 194), а также последовательность, содержащую ALRPR (SEQ ID NO: 7) (например, ALRPRVVGGA (SEQ ID NO: 145)), состоящую по сути из нее или состоящую из нее.
[0300] В некоторых вариантах осуществления сайт расщепления представляет собой TLDPRSFLLRNPNDKYEPFWEDEEK (SEQ ID NO: 146). В других вариантах осуществления сайт расщепления содержит DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88) или его фрагмент. В конкретных вариантах осуществления сайт расщепления содержит IEPRSFS (SEQ ID NO: 194). В других вариантах осуществления сайт расщепления содержит EPRSFS (SEQ ID NO: 195), где сайт расщепления не представляет собой полноразмерную область а2 FVIII. В еще нескольких других вариантах осуществления сайт расщепления содержит IEPR (SEQ ID NO: 200). В еще некоторых других вариантах осуществления сайт расщепления содержит IEPR (SEQ ID NO: 200), где сайт расщепления не представляет собой полноразмерную область а2 FVIII или не содержит полноразмерную область а2 FVIII. В других вариантах осуществления сайт расщепления содержит DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88), KNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 139), NTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 140), TGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 141), GDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 142), DYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 143)¸ YYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 144)¸ YEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 176), EDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 177), DSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 178), SYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 179), YEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 180), EDISAYLLSKNNAIEPRSFS (SEQ ID NO: 181), DISAYLLSKNNAIEPRSFS (SEQ ID NO: 182)¸ ISAYLLSKNNAIEPRSFS (SEQ ID NO: 183), SAYLLSKNNAIEPRSFS (SEQ ID NO: 184), AYLLSKNNAIEPRSFS (SEQ ID NO: 185), YLLSKNNAIEPRSFS (SEQ ID NO: 186), LLSKNNAIEPRSFS (SEQ ID NO: 187), LSKNNAIEPRSFS (SEQ ID NO: 188), SKNNAIEPRSFS (SEQ ID NO: 189), KNNAIEPRSFS (SEQ ID NO: 190), NNAIEPRSFS (SEQ ID NO: 191), NAIEPRSFS (SEQ ID NO: 192), AIEPRSFS (SEQ ID NO: 193), или IEPRSFS (SEQ ID NO: 194). В других вариантах осуществления сайт расщепления содержит DKNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 88), KNTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 139), NTGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 140), TGDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 141), GDYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 142), DYYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 143)¸ YYEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 144)¸ YEDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 176), EDSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 177), DSYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 178), SYEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 179), YEDISAYLLSKNNAIEPRSFS (SEQ ID NO: 180), EDISAYLLSKNNAIEPRSFS (SEQ ID NO: 181), DISAYLLSKNNAIEPRSFS (SEQ ID NO: 182)¸ ISAYLLSKNNAIEPRSFS (SEQ ID NO: 183), SAYLLSKNNAIEPRSFS (SEQ ID NO: 184), AYLLSKNNAIEPRSFS (SEQ ID NO: 185), YLLSKNNAIEPRSFS (SEQ ID NO: 186), LLSKNNAIEPRSFS (SEQ ID NO: 187), LSKNNAIEPRSFS (SEQ ID NO: 188), SKNNAIEPRSFS (SEQ ID NO: 189), KNNAIEPRSFS (SEQ ID NO: 190), NNAIEPRSFS (SEQ ID NO: 191), NAIEPRSFS (SEQ ID NO: 192), AIEPRSFS (SEQ ID NO: 193), или IEPRSFS (SEQ ID NO:194), где сайт расщепления не представляет собой полноразмерную область a2 FVIII. В определенных вариантах осуществления расщепляемый линкер расщепляется в анализе расщепления тромбином, как предусмотрено в данном документе или как известно из уровня техники.
[0301] В конкретных вариантах осуществления химерный полипептид по настоящему изобретению содержит: (i) белок FVIII, содержащий полипептид FVIII, первую последовательность XTEN и первую Fc-область; и (ii) фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, вторую последовательность XTEN, линкер a2 FVIII и вторую Fc-область; где:
(a) белок FVIII имеет делецию аминокислот 746-1648, соответствующих зрелому FVIII;
(b) первая последовательность XTEN вставлена в полипептид FVIII непосредственно ниже аминокислоты 745, соответствующей зрелому FVIII;
(с) первая последовательность XTEN содержит аминокислотную последовательность, которая характеризуется по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или приблизительно 100% идентичностью последовательности с аминокислотной последовательностью под SEQ AE288 (SEQ ID NO: 8);
(d) первая Fc-область слита с С-концом полипептида FVIII;
(e) вторая последовательность XTEN слита с С-концом фрагмента VWF;
(f) вторая последовательность XTEN содержит аминокислотную последовательность, которая характеризуется по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или приблизительно 100% идентичностью последовательности с аминокислотной последовательностью под SEQ AE144_5A (SEQ ID NO: 58);
(g) линкер a2 слит с C-концом XTEN;
(h) линкер а2 содержит аминокислотную последовательность, которая характеризуется по меньшей мере приблизительно 70%, по меньшей мере приблизительно 75%, по меньшей мере приблизительно 80%, по меньшей мере приблизительно 85%, по меньшей мере приблизительно 90%, по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% или приблизительно 100% идентичностью последовательности с аминокислотной последовательностью под SEQ ID NO: 88;
(i) вторая Fc-область слита с С-концом линкера a2; и
(j) первая Fc-область ковалентно соединена со второй Fc-областью с помощью дисульфидной связи.
[0302] В конкретных вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную последовательности, выбранной из FVIII-161 (SEQ ID NO: 69), FVIII-169 (SEQ ID NO: 70), FVIII-170 (SEQ ID NO: 71), FVIII-173 (SEQ ID NO: 72); FVIII-195 (SEQ ID NO: 73); FVIII-196 (SEQ ID NO: 74), FVIII199 (SEQ ID NO: 75), FVIII-201 (SEQ ID NO: 76); FVIII-203 (SEQ ID NO: 77), FVIII-204 (SEQ ID NO: 78), FVIII-205 (SEQ ID NO: 79), FVIII-266 (SEQ ID NO: 80), FVIII-267 (SEQ ID NO: 81), FVIII-268 (SEQ ID NO: 82), FVIII-269 (SEQ ID NO: 83), FVIII-271 (SEQ ID NO: 84) или FVIII-272 (SEQ ID NO: 85), и вторую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную последовательности, выбранной из VWF031 (SEQ ID NO: 86), VWF034 (SEQ ID NO: 87) или VWF-036.
[0303] В некоторых вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную аминокислотной последовательности, приведенной под SEQ ID NO: 201; и вторую полипептидную последовательность, содержащую фрагмент VWF, содержащий домен D' VWF и домен D3 VWF, и Fc-область. В некоторых вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую полипептид FVIII и Fc-область; и вторую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную аминокислотной последовательности, приведенной под SEQ ID NO: 202. В определенных вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную аминокислотной последовательности, приведенной под SEQ ID NO: 201, и вторую полипептидную последовательность, содержащую аминокислотную последовательность, на по меньшей мере приблизительно 80%, 90%, 95% или 100% идентичную аминокислотной последовательности, приведенной под SEQ ID NO: 202. В конкретных вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую аминокислотную последовательность, приведенную под SEQ ID NO: 201, и вторую полипептидную последовательность, содержащую аминокислотную последовательность, приведенную под SEQ ID NO: 202. В других конкретных вариантах осуществления химерный полипептид по настоящему изобретению содержит две полипептидные последовательности - первую полипептидную последовательность, содержащую аминокислотную последовательность, приведенную под SEQ ID NO: 201, и вторую полипептидную последовательность, содержащую аминокислотную последовательность, приведенную под SEQ ID NO: 202, где первая полипептидная последовательность и вторая полипептидная последовательность соединены друг с другом с помощью дисульфидной связи.
IV. Фармацевтические композиции
[0304] Композиции, содержащие химерный полипептид по настоящему изобретению, могут содержать подходящий фармацевтически приемлемый носитель. Например, они могут содержать вспомогательные вещества и/или вспомогательные средства, которые способствуют переработке активных соединений в препараты, предназначенные для доставки в место действия.
[0305] Фармацевтическая композиция может быть составлена для парентерального введения (т. е. внутривенного, подкожного или внутримышечного) путем болюсной инъекции. Составы для инъекций могут быть представлены в единичной лекарственной форме, например, в ампулах или в многодозовых контейнерах с добавленным консервантом. Композиции могут иметь такие формы, как суспензии, растворы или эмульсии в масляных или водных средах-носителях, и содержать средства для составления, такие как суспендирующие, стабилизирующие и/или диспергирующие средства. В качестве альтернативы активный ингредиент может быть представлен в форме порошка для разбавления подходящей средой-носителем, например, апирогенной водой.
[0306] Подходящие составы для парентерального введения также включают водные растворы активных соединений в водорастворимой форме, например, водорастворимые соли. Кроме того, можно вводить суспензии активных соединений в виде подходящих масляных инъекционных суспензий. Подходящие липофильные растворители или среды-носители включают жирные масла, например, кунжутное масло, или синтетические сложные эфиры жирных кислот, например, этилолеат или триглицериды. Водные инъекционные суспензии могут содержать вещества, которые увеличивают вязкость суспензии, в том числе, например, натрий-карбоксиметилцеллюлозу, сорбит и декстран. Суспензия также необязательно может содержать стабилизаторы. Также можно применять липосомы для инкапсулирования молекул по настоящему изобретению для доставки в клетки или интерстициальные пространства. Иллюстративными фармацевтически приемлемыми носителями являются физиологически совместимые растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические средства и средства, замедляющие всасывание, вода, солевой раствор, фосфатно-солевой буферный раствор, декстроза, глицерин, этанол и т. п. В некоторых вариантах осуществления композиция содержит изотонические средства, например, сахара, многоатомные спирты, такие как маннит, сорбит, или хлорид натрия. В других вариантах осуществления композиции содержат фармацевтически приемлемые вещества, такие как смачивающие средства или незначительные количества вспомогательных веществ, таких как смачивающие или эмульгирующие средства, консерванты или буферы, которые увеличивают срок годности или эффективность активных ингредиентов.
[0307] Композиции по настоящему изобретению могут быть представлены в различных формах, в том числе, например, в форме жидкости (например, инъекционных и инфузионных растворов), дисперсий, суспензий, полутвердых и твердых лекарственных форм. Предпочтительная форма зависит от способа введения и терапевтического применения.
[0308] Композиция может быть составлена в виде раствора, микроэмульсии, дисперсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Стерильные инъекционные растворы можно получить путем помещения активного ингредиента в требуемом количестве в соответствующий растворитель с одним ингредиентом или комбинацией ингредиентов, перечисленных выше, если это требуется, с последующей стерилизующей фильтрацией. Обычно дисперсии получают путем помещения активного ингредиента в стерильную среду-носитель, которая содержит основную дисперсионную среду и требуемые другие ингредиенты из перечисленных выше. В случае со стерильными порошками для получения стерильных инъекционных растворов предпочтительные способы получения представляют собой вакуумную сушку и сублимационную сушку, которые приводят к получению порошка активного ингредиента и любого дополнительного необходимого ингредиента из их раствора, предварительно подвергнутого стерилизующей фильтрации. Надлежащую текучесть раствора можно поддерживать, например, путем применения покрытия, такого как лецитин, путем поддержания требуемого размера частиц в случае с дисперсией и путем применения поверхностно-активных веществ. Длительное всасывание инъекционных композиций может обеспечиваться включением в композицию средств, которые замедляют всасывание, например, моностеаратных солей и желатина.
[0309] Активный ингредиент может быть составлен в составе или устройстве с контролируемым высвобождением. Примеры таких составов и устройств включают имплантаты, трансдермальные пластыри и микроинкапсулированные системы доставки. Можно применять биоразлагаемые, биосовместимые полимеры, например, этиленвинилацетат, полиангидриды, полигликолевую кислоту, коллаген, сложные полиортоэфиры и полимолочную кислоту. Способы получения таких составов и устройств известны из уровня техники. См., например, Sustained and Controlled Release Drug Delivery Systems, J. R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
[0310] Инъекционные депо-составы можно получать путем образования микроинкапсулированных матриц с лекарственным средством в биоразлагаемых полимерах, таких как сополимер лактида и гликолида. В зависимости от соотношения лекарственного средства и полимера и природы используемого полимера можно контролировать скорость высвобождения лекарственного средства. Другими иллюстративными биоразлагаемыми полимерами являются сложные полиортоэфиры и полиангидриды. Инъекционные депо-составы также можно получать путем захвата лекарственного средства в липосомы или микроэмульсии.
[0311] В композиции могут быть включены дополнительные активные соединения. В некоторых вариантах осуществления химерный полипептид по настоящему изобретению составлен с другим фактором свертывания крови или его вариантом, фрагментом, аналогом или производным. Например, фактор свертывания крови включает без ограничения фактор V, фактор VII, фактор VIII, фактор IX, фактор X, фактор XI, фактор XII, фактор XIII, протромбин, фибриноген, фактор фон Виллебранда или рекомбинантный растворимый тканевой фактор (rsTF) или активированные формы любого из предшествующих. Фактор свертывания крови в гемостатическом средстве также может включать антифибринолитические лекарственные средства, например, эпсилон-аминокапроновую кислоту, транексамовую кислоту.
[0312] Режимы введения доз могут быть скорректированы для обеспечения оптимального необходимого ответа. Например, можно вводить однократную болюсную дозу, можно вводить несколько разделенных доз на протяжении некоторого периода времени, или дозу можно пропорционально снизить или увеличить, как указывают потребности терапевтической ситуации. Преимущественным является составление композиций для парентерального применения в виде единичной лекарственной формы для удобства введения и однородности дозирования. См., например, Remington's Pharmaceutical Sciences (Mack Pub. Co., Easton, Pa. 1980).
[0313] В дополнение к активному соединению жидкая лекарственная форма может содержать инертные ингредиенты, такие как вода, этиловый спирт, этилкарбонат, этилацетат, бензиловый спирт, бензилбензоат, пропиленгликоль, 1,3-бутиленгликоль, диметилформамид, масла, глицерин, тетрагидрофурфуриловый спирт, полиэтиленгликоли и сложные эфиры жирных кислот и сорбитана.
[0314] Неограничивающие примеры подходящих фармацевтических носителей также описаны в Remington's Pharmaceutical Sciences, E.W. Martin. Некоторые примеры вспомогательных веществ включают крахмал, глюкозу, лактозу, сахарозу, желатин, солод, рис, муку, мел, силикагель, стеарат натрия, моностеарат глицерина, тальк, хлорид натрия, сухое обезжиренное молоко, глицерин, пропиленгликоль, воду, этанол и т. п. Композиция также может содержать рН-буферные реагенты, а также смачивающие или эмульгирующие средства.
[0315] В некоторых вариантах осуществления фармацевтическая композиция содержит химерный полипептид и фармацевтически приемлемый носитель. Белок FVIII в химерном полипептиде имеет продленный период полужизни по сравнению с белком FVIII дикого типа или соответствующим белком FVIII без фрагмента VWF. В некоторых вариантах осуществления продленный период полужизни химерного полипептида является в по меньшей мере приблизительно 1,5 раза, по меньшей мере приблизительно 2 раза, по меньшей мере приблизительно 2,5 раза, по меньшей мере приблизительно 3 раза, по меньшей мере приблизительно 4 раза, по меньшей мере приблизительно 5 раз, по меньшей мере приблизительно 6 раз, по меньшей мере приблизительно 7 раз, по меньшей мере приблизительно 8 раз, по меньшей мере приблизительно 9 раз, по меньшей мере приблизительно 10 раз, по меньшей мере приблизительно 11 раз или по меньшей мере приблизительно 12 раз более длительным по сравнению с FVIII дикого типа. В других вариантах осуществления период полужизни фактора VIII составляет по меньшей мере приблизительно 17 часов, по меньшей мере приблизительно 18 часов, по меньшей мере приблизительно 19 часов, по меньшей мере приблизительно 20 часов, по меньшей мере приблизительно 21 час, по меньшей мере приблизительно 22 часа, по меньшей мере приблизительно 23 часа, по меньшей мере приблизительно 24 часа, по меньшей мере приблизительно 25 часов, по меньшей мере приблизительно 26 часов, по меньшей мере приблизительно 27 часов, по меньшей мере приблизительно 28 часов, по меньшей мере приблизительно 29 часов, по меньшей мере приблизительно 30 часов, по меньшей мере приблизительно 31 час, по меньшей мере приблизительно 32 часа, по меньшей мере приблизительно 33 часа, по меньшей мере приблизительно 34 часа, по меньшей мере приблизительно 35 часов, по меньшей мере приблизительно 36 часов, по меньшей мере приблизительно 48 часов, по меньшей мере приблизительно 60 часов, по меньшей мере приблизительно 72 часа, по меньшей мере приблизительно 84 часа, по меньшей мере приблизительно 96 часов или по меньшей мере приблизительно 108 часов. В некоторых вариантах осуществления период полужизни химерного полипептида продлен приблизительно в 3 раза по сравнению с FVIII дикого типа.
[0316] В некоторых вариантах осуществления композицию вводят путем парентерального введения, которое может представлять собой внутривенное или подкожное введение.
[0317] В других вариантах осуществления композицию применяют для лечения заболевания или состояния, связанного с кровотечением, у субъекта, нуждающегося в этом. Заболевание или состояние, связанное с кровотечением, выбрано из группы, состоящей из коагуляционного нарушения свертываемости крови, гемартроза, мышечного кровотечения, кровотечения из полости рта, кровоизлияния, кровоизлияния в мышцы, кровоизлияния в полости рта, травмы, черепно-мозговой травмы, желудочно-кишечного кровотечения, внутричерепного кровоизлияния, внутрибрюшного кровоизлияния, внутригрудного кровоизлияния, перелома кости, кровотечения в центральной нервной системе, кровотечения в заглоточном пространстве, кровотечения в забрюшинном пространстве, кровотечения во влагалище подвздошно-поясничной мышцы и любых их комбинаций. В еще нескольких других вариантах осуществления у субъекта запланировано проведение хирургического вмешательства. В еще нескольких других вариантах осуществления лечение проводят в режиме профилактики или по необходимости.
[0318] Теперь при наличии подробного описания настоящего изобретения таковое будет более четко понятным благодаря ссылке на следующие примеры, которые включены в данный документ только в целях иллюстрации и не предназначены для ограничения настоящего изобретения. Все патенты, публикации и статьи, упомянутые в данном документе, в явной форме и конкретным образом включены в него посредством ссылки.
ПРИМЕРЫ
Пример 1. Исследование гетеродимеров FVIII-XTEN-Fc/D'D3-XTEN-Fc, впервые проводящееся с участием людей
[0319] Заместительная терапия с использованием факторов свертывания крови является фундаментальным компонентом медицинской помощи при гемофилии. Традиционные рекомбинантные продукты FVIII (rFVIII) имеют короткий период полужизни, что требует частого введения. Значительные успехи в уходе за пациентами были достигнуты с помощью rFVIIIFc (Eloctate®), в том числе наиболее длительный интервал между введениями доз при гемофилии А; надлежащий контроль кровотечения, в том числе профилактический, в острых, периоперационных и экстренных ситуациях; защита, выходящая за рамки контроля кровотечения, включая защиту суставов и улучшение качества жизни; и долгосрочный последовательный и хорошо изученный профиль безопасности. Однако профилактическое введение доз фактора VIII с частотой один раз в неделю или реже все еще представляет собой неудовлетворенную потребность для большинства людей с гемофилией А.
[0320] В настоящем исследовании изучаются безопасность и фармакокинетические показатели (PK) rFVIIIFc-VWF-XTEN - химерного полипептида, содержащего (i) полипептид FVIII, XTEN, вставленный в домен B полипептида FVIII, и первую Fc-область; и (ii) фрагмент VWF, вторую последовательность XTEN, линкер a2 и вторую Fc-область (фиг. 1).
[0321] План исследования
[0322] Целью настоящего исследования была оценка безопасности, переносимости и активности фактора VIII (PK) однократной дозы rFVIIIFc-VWF-XTEN по сравнению с rFVIII. Популяция пациентов включала взрослых мужчин с тяжелой формой гемофилии A (уровень эндогенной активности FVIII < 1 МЕ/дл [< 1%]), ранее получавших лечение продуктами FVIII в течение по меньшей мере 150 дней воздействия и при отсутствии положительного результата теста на ингибиторы или клинических признаков ослабленного ответа на введение FVIII в анамнезе. Первичными конечными точками являлись нежелательные явления и клинические аномалии в лабораторных тестах, в том числе выработка ингибиторов. Ключевые вторичные конечные точки включали фармакокинетические параметры, в том числе без ограничения период полужизни (t1/2), клиренс (CL), площадь под кривой от 0 до бесконечности (AUC0-∞), среднее время удержания (MRT) и постепенное восстановление (IR).
[0323] Пациентов включали в когорту с низкой дозой (7 пациентов; фиг. 2A) или в когорту с высокой дозой (9 пациентов; фиг. 2B). Каждому пациенту вводили дозу rFVIII (ADVATE®) с последующим периодом отбора образцов для PK-анализа. После короткого периода отмывки (≥ 3 дня для когорты с низкой дозой (фиг. 2A) и ≥ 4 дня для когорты с высокой дозой (фиг. 2B)) каждому субъекту затем вводили rFVIIIFc-VWF-XTEN с последующим периодом отбора образцов для PK-анализа. Субъекты подвергались наблюдению в отношении безопасности в течение 28 дней после инъекции rFVIIIFc-VWF-XTEN, включающему сбор образцов для оценки ингибиторов через 14 и 28 дней после инъекции rFVIIIFc-VWF-XTEN. Пациенты в когорте с низкой дозой получали 25 МЕ/кг rFVIII и 25 МЕ/кг rFVIIIFc-VWF-XTEN. Пациенты в когорте с высокой дозой получали 65 МЕ/кг rFVIII и 65 МЕ/кг rFVIIIFc-VWF-XTEN.
[0324] Способы
[0325] Фармакокинетические параметры измеряли с помощью модифицированного анализа свертывания крови по активированному частичному тромбопластиновому времени (aPTT) ("одностадийного" анализа). Вкратце, активность FVIII измеряют в модифицированном анализе aPTT с использованием реагента для определения активированного PTT Dade® Actin® FSL (Siemens Health Care Diagnostics) на анализаторе BCS® XP (Siemens Healthcare Diagnostics). Калибровочные кривые по 7 точкам (высокий уровень: 1,500-0,050 МЕ/мл; низкий уровень: 0,150-0,0025 МЕ/мл) получали с использованием специфического калибровочного стандарта для rFVIIIFc-VWF-XTEN. Для построения калибровочной кривой высокого уровня калибровочный стандарт rFVIIIFc-VWF-XTEN [1,000 МЕ/мл] разбавляют в буфере Tris/BSA (50 мМ, 150 мМ хлорида натрия, pH 7,4 +/- 0,2, содержащем 1% BSA). Это разбавление выполняется с помощью анализатора BCS® XP для получения предварительно отобранных калибровочных уровней активности FVIII. Затем их тестируют в рабочем разведении 1:2, приготовленном в буфере Tris/BSA. Для построения калибровочной кривой низкого уровня калибровочный стандарт для rFVIIIFc-VWF-XTEN [1,000 МЕ/мл] разбавляют в иммуноистощенной плазме крови с дефицитом FVIII (Siemens Healthcare Diagnostics) с помощью анализатора BCS® XP и тестируют в неразбавленном виде (при отсутствии рабочего разведения). Калибровку выполняют в двух повторностях. Кривые строят по активности FVIII в МЕ/мл (ось x) и времени свертывания крови в секундах (ось y). Для калибровочных кривых как высокого уровня, так и низкого уровня используется аппроксимация кривой методом логарифмически-линейной регрессии с линейной зависимостью.
[0326] Образцы тестируют в двух повторностях с использованием метатеста с калибровочной кривой высокого уровня (в рабочих разведениях 1:2, 1:4 и 1:8 в Tris/BSA) или метатеста с калибровочной кривой низкого уровня (в неразбавленном виде и в рабочих разведениях 1:2 и 1:4 в иммуноистощенной плазме крови с дефицитом FVIII). После получения соответствующего рабочего разведения все калибровочные стандарты, образцы от пациентов и образцы для контроля качества смешивают в соотношении 1:1 с иммуноистощенной плазмой крови с дефицитом FVIII, содержащей молекулу с активностью фактора VIII в количестве, равном 0,010 МЕ/мл или меньше, и по меньшей мере 0,400 МЕ/мл других факторов свертывания крови. После смешивания образцов с указанной плазмой крови добавляют реагент для APTT, и смесь инкубируют. После инкубирования добавляют хлорид кальция и оптическим путем измеряют время образования сгустка. Посредством алгоритма затем проверяют, находятся ли результаты для отдельных рабочих разведений в пределах +/- 15,0% от среднего значения, для определения регистрируемой активности FVIII. Для определения регистрируемой активности FVIII необходимы результаты как минимум двух метатестов в рабочих разведениях, за исключением значений, близких к нижнему пределу количественного определения (LLOQ).
[0327] Результаты: когорта с низкой дозой
[0328] Семь субъектов были включены в когорту с низкой дозой; один субъект отказался от участия по причинам, не связанным с исследованием, до получения rFVIIIFc-VWF-XTEN. Шесть субъектов получили однократную низкую дозу rFVIIIFc-VWF-XTEN (25 МЕ/кг). Характеристики первых четырех субъектов, получавших лечение в когорте с низкой дозой, включали следующие: ранее получавшие лечение взрослые субъекты-мужчины с тяжелой формой гемофилии A (уровень эндогенной активности FVIII < 1 МЕ/дл [< 1%]); все субъекты относились к европеоидной расе; возраст субъектов находился в диапазоне от 19 до 60 лет, при этом средний возраст составлял 33 года; а средняя масса тела составляла 85 кг (в диапазоне от 67,2 до 100,6 кг).
[0329] У всех пациентов rFVIIIFc-VWF-XTEN обычно хорошо переносился, и в день 14 или день 28 после введения дозы ингибиторы не наблюдались.
[0330] Средняя активность FVIII после инфузии у первых шести субъектов, получавших однократную низкую дозу rFVIIIFc-VWF-XTEN (25 МЕ/кг), через 5 дней составляла приблизительно 12,0%, а через 7 дней составляла приблизительно 5,3% (фиг. 3A). Средняя активность FVIII оставалась на уровне выше 1% в течение 10 дней (фиг. 3A). Фармакокинетические параметры, измеренные путем измерения активированного частичного тромбопластинового времени (aPTT), для rFVIIIFc-VWF-XTEN (25 МЕ/кг) и rFVIII (25 МЕ/кг) показаны в таблице 9. Наблюдалось, что rFVIIIFc-VWF-XTEN у субъектов-людей имеет средний период полужизни, составляющий приблизительно 37,6 часа, что более чем в 4 раза превышает средний период полужизни rFVIII (приблизительно 9,12 часа), вводимого в той же дозе.
Таблица 9. Фармакокинетические (PK) параметры для когорты с низкой дозой
среднее геометрическое значение
[диапазон] (N=6)
среднее геометрическое значение
[диапазон] (N=7)
[95% CI]
[49,7-98,9]
[43,3-61,9]
P=0,032
[0331] Результаты: когорта с высокой дозой
[0332] Девять субъектов были включены в когорту с высокой дозой. Субъектам вводили однократную высокую дозу rFVIIIFc-VWF-XTEN (65 МЕ/кг). Один субъект не мог быть подвергнут тестированию в отношении PK-параметров по причинам, не связанным с исследованием. Характеристики субъектов, получавших лечение в когорте с высокой дозой, включали следующие: ранее получавшие лечение взрослые субъекты-мужчины с тяжелой формой гемофилии A (уровень эндогенной активности FVIII < 1 МЕ/дл [< 1%]); 7 из 9 субъектов относились к европеоидной расе; возраст субъектов находился в диапазоне от 32 до 63 лет, при этом средний возраст составлял 44 года; а средняя масса тела составляла 81,1 кг (в диапазоне от 61,5 до 100,7 кг).
[0333] Средняя активность FVIII после инфузии у восьми оцениваемых субъектов, получавших однократную высокую дозу rFVIIIFc-VWF-XTEN (65 МЕ/кг), через 5 дней составляла приблизительно 38,0%, а через 7 дней составляла приблизительно 17% (фиг. 3B). Средняя активность FVIII оставалась на уровне выше 1% в течение 14 дней (фиг. 3B). Фармакокинетические параметры, измеренные путем измерения активированного частичного тромбопластинового времени (aPTT), для rFVIIIFc-VWF-XTEN (65 МЕ/кг) и rFVIII (65 МЕ/кг) показаны в таблице 10. Наблюдалось, что rFVIIIFc-VWF-XTEN у субъектов-людей имеет средний период полужизни, составляющий приблизительно 42,5 часа, что более чем в 3 раза превышает средний период полужизни rFVIII (приблизительно 13 часов), вводимого в той же дозе.
Таблица 10. Фармакокинетические (PK) параметры для когорты с высокой дозой
среднее геометрическое значение
[диапазон] (N=8)
среднее геометрическое значение
[диапазон] (N=9)
[95% CI]
[39,72-45,56]
[10,89-15,87]
P < 0,001
[142-183]
[117-162]
P < 0,001
[11,100-14,900]
[1670-2310]
P < 0,001
[62,59-73,14]
[14,06-17,45]
P < 0,001
[0,44-0,59]
[2,81-3,88]
P < 0,001
[2,18-2,82]
[1,79-2,49]
P < 0,001
[0334] Выводы
[0335] rFVIIIFc-VWF-XTEN хорошо переносился всеми пациентами с тяжелой формой гемофилии А, которые получали лечение с помощью однократной низкой дозы (25 МЕ/кг) либо однократной высокой дозы (65 МЕ/кг). Ни у одного пациента не вырабатывался ингибитор FVIII. rFVIIIFc-VWF-XTEN продемонстрировал период полужизни, который был в от 3 раз до 4 раз выше, чем у rFVIII, что свидетельствует о прорыве в направлении достижения верхнего предела периода полужизни, налагаемого VWF. Устойчивая высокая активность фактора VIII может обеспечить улучшенную защиту от всех типов кровотечений.
Пример 2. Исследование с повторными дозами гетеродимеров FVIII-XTEN-Fc/D'D3-XTEN-Fc
[0336] rFVIIIFc-VWF-XTEN хорошо переносился и обеспечивал устойчивые уровни FVIII в когорте субъектов, получивших однократную дозу rFVIIIFc-VWF-XTEN. (Пример 1). В настоящем исследовании будут оцениваться безопасность и переносимость, а также определяться фармакокинетические характеристики rFVIIIFc-VWF-XTEN при повторном введении доз (фиг. 1).
[0337] План исследования
[0338] В общей сложности 4 еженедельные дозы rFVIIIFc-VWF-XTEN будут введены взрослым пациентам-мужчинам с тяжелой формой гемофилии A, которые ранее получали лечение посредством лечения с помощью FVIII и получали такое лечение с помощью FVIII в течение по меньшей мере 150 дней воздействия. Будут включены две когорты, каждая из которых состоит из примерно десяти взрослых мужчин в возрасте от 18 до 65 лет. Каждый субъект в когорте 1 получит в общей сложности четыре еженедельные дозы по 50 МЕ/кг rFVIIIFc-VWF-XTEN в дни 1, 8, 15 и 22. Каждый субъект в когорте 2 получит в общей сложности четыре еженедельные дозы по 65 МЕ/кг rFVIIIFc-VWF-XTEN в дни 1, 8, 15 и 22.
[0339] Схематическое изображение плана клинического исследования изображено на фиг. 4. Образец для фармакокинетического анализа до введения дозы будет взят в день 1. Кроме того, после введения дозы в дни 1 и 22 будут взяты несколько образцов для PK-анализа, и в дни 8, 15 и 22 перед введением дозы будут взяты образцы с остаточным уровнем (168 ч.). Субъекты будут проходить период наблюдения в отношении безопасности в течение 28 дней после последней дозы rFVIIIFc-VWF-XTEN, в том числе подвергаться оценке ингибиторов через 14 и 28 дней после последней дозы rFVIIIFc-VWF-XTEN. Для субъектов, завершивших прием всех четырех еженедельных доз rFVIIIFc-VWF-XTEN, период наблюдения в отношении безопасности заканчивается визитом окончания исследования (EOS) в день 50 (+3 дня). Для субъектов, которые досрочно прекращают лечение, период наблюдения в отношении безопасности заканчивается визитом досрочного прекращения исследования (ЕТ) через 28 дней после последней дозы rFVIIIFc-VWF-XTEN.
[0340] Первичной целью исследования является оценка безопасности и переносимости в общей сложности четырех еженедельных внутривенно вводимых (IV) доз rFVIIIFc-VWF-XTEN. Первичными конечными точками являются возникновение нежелательных явлений (AE) и возникновение клинически значимых аномалий в лабораторных тестах, в том числе выработка ингибиторов (нейтрализующих антител, направленных против FVIII), что определяется с помощью анализа Бетезда в модификации Неймегена (Verbruggen et al, Thrombosis and Haemostasis, 73: 247-251 (1995)).
[0341] Вторичной целью исследования является определение фармакокинетических характеристик rFVIIIFc-VWF-XTEN после в общей сложности четырех еженедельных IV доз, при этом активность FVIII определяется с помощью одностадийного анализа свертывания крови по активированному частичному тромбопластиновому времени (aPTT), как описано в примере 1. Вторичными конечными точками являются фармакокинетические показатели, оцениваемые посредством оценки параметров, включающих без ограничения максимальную активность (Cmax), период полужизни (t½), общий клиренс в равновесном состоянии (CLss), индекс накопления (AI), площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз (AUC0-tau), объем распределения в равновесном состоянии (Vss), среднее время удержания (MRT), IR, наиболее низкую (остаточную) концентрацию, которой лекарственное средство достигает до введения следующей дозы (Ctrough), и время до достижения активности FVIII, на 1% превышающей исходный уровень.
[0342] Поисковой целью исследования является определение фармакокинетических характеристик rFVIIIFc-VWF-XTEN после в общей сложности четырех еженедельных IV доз, при этом активность FVIII определяется с помощью двухстадийного хромогенного анализа коагуляции. Поисковыми конечными точками являются PK-показатели, оцениваемые посредством оценки параметров, включающих без ограничения Cmax, t½, CLss, AI, AUC0-tau, Vss, MRT, IR, Ctrough и время до достижения активности FVIII, на 1% превышающей исходный уровень.
[0343] Способы
[0344] Активность FVIII определяли с помощью модифицированного анализа свертывания крови по активированному частичному тромбопластиновому времени (aPTT) ("одностадийного" анализа), как подробно описано выше в примере 1.
[0345] Посредством двухстадийного хромогенного анализа коагуляции измеряют активность FVIII с применением анализа BIOPHEN FVIII:C (Hyphen Biomed 221402-RUO) на анализаторе BCS® XP (Siemens Healthcare Diagnostics). Вкратце, фосфолипиды, кальций и FVIII:C, активированный тромбином, образуют ферментный комплекс в присутствии постоянного количества фактора IXa. Этот ферментный комплекс, в свою очередь, активирует фактор X (FX), который предоставляется в постоянной концентрации и в избытке, до фактора Xa (FXa). Активация FX до FXa напрямую связана с количеством FVIII:C, которое является лимитирующим фактором в анализе в присутствии постоянного и избыточного количества фактора IXa. Затем активность образованного FXa измеряют с использованием специфического хромогенного субстрата FXa, представляющего собой SXa-11 (Hyphen Biomed). Фактор Ха расщепляет SXa-11 и высвобождает паранитроанилин (pNA). Количество высвободившегося pNA прямо пропорционально количеству образованного FXa. Существует прямая зависимость между количеством FVIII:C в образце, количеством образованного FXa и количеством высвободившегося pNA. Высвобождение pNA определяется по проявлению окрашивания при длине волны 405 нм.
[0346] Калибровочные кривые по 8 точкам (высокий уровень: 1,500-0,150 МЕ/мл; низкий уровень: 0,200-0,008 МЕ/мл) получали с использованием специфического калибровочного стандарта для rFVIIIFc-VWF-XTEN. Для построения калибровочной кривой высокого уровня rFVIIIFc-VWF-XTEN при 1 МЕ/мл разбавляют в буфере Tris/BSA для получения предварительно отобранных калибровочных уровней активности FVIII. Это разбавление выполняется с помощью анализатора BCS® XP. Предварительно отобранные калибровочные уровни активности FVIII тестируют в рабочем разведении 1:4 в буфере Tris/BSA. Для построения калибровочной кривой низкого уровня rFVIIIFc-VWF-XTEN в концентрации 1 МЕ/мл предварительно разбавляют 1:4 в плазме крови с врожденным дефицитом FVIII (Helena Laboratories), а затем дополнительно разбавляют с помощью анализатора BCS ® XP в буфере Tris/BSA для получения предварительно отобранных калибровочных уровней активности FVIII, которые тестируют при отсутствии дополнительного разбавления в реакционной установке.
[0347] Калибровку выполняют в двух повторностях. Кривые строят по активности FVIII в МЕ/мл (ось x) и изменению оптической плотности/мин (ось y). Для калибровочной кривой высокого уровня используется аппроксимация кривой методом двойной логарифмической регрессии с линейной зависимостью. Для калибровочной кривой низкого уровня используется аппроксимация кривой методом двойной линейной регрессии с линейной зависимостью. Образцы тестируют в двух повторностях по калибровочной кривой высокого уровня в рабочем разведении 1:4 либо по калибровочной кривой низкого уровня в разведении 1:4 в плазме крови с врожденным дефицитом FVIII или в неразбавленном виде (при отсутствии рабочего разведения).
[0348] Результаты
[0349] Демографические данные и характеристики исходного уровня
[0350] 10 взрослых субъектов-мужчин в возрасте от 18 до 65 лет (включительно) были включены в когорту 1 (50 МЕ/кг), и 5 взрослых субъектов-мужчин в возрасте от 18 до 65 лет (включительно) были включены в когорту 2 (65 МЕ/кг). Все 10 субъектов в дозовой когорте 50 МЕ/кг получили 4 дозы rFVIIIFc-VWF-XTEN. Трое из 5 субъектов, включенных в дозовую когорту 65 МЕ/кг, получили по меньшей мере одну дозу rFVIIIFc-VWF-XTEN. Все 15 субъектов, включенных в исследование, были из одного исследовательского центра в Софии, Болгария. Из 15 субъектов, включенных в исследование, 13 получили по меньшей мере одну дозу BIVV001 и были включены в сводный обзор демографических данных и характеристик исходного уровня. Все субъекты были мужчинами. Медианный возраст составлял 35 лет (диапазон: от 25 до 55 лет), и все субъекты были белыми. Медианная масса тела составляла 86,7 кг (диапазон: от 51,0 до 130,9 кг), а медианный индекс массы тела составлял 25,9 кг/м2 (диапазон: от 16,4 до 40,9 кг/м2).
[0351] Анализ безопасности
[0352] Для промежуточного анализа в выборку для анализа безопасности были включены в общей сложности 13 субъектов (n=10 из когорты 50 МЕ/кг и n=3 из когорты 65 МЕ/кг). Все анализы безопасности проводились на основе выборки для анализа безопасности и включали всех субъектов, получивших по меньшей мере одну дозу rFVIIIFc-VWF-XTEN.
[0353] AE, возникшее после начала лечения (TEAE), определяется как любое нежелательное явление, которое началось во время или после первого введения rFVIIIFc-VWF-XTEN и в течение 28 дней после последнего введения rFVIIIFc-VWF-XTEN. Предполагается, что AE с отсутствующими датами начала возникли после начала лечения, если только дата прекращения не была раньше даты введения дозы. В целом, AE были обобщены по встречаемости у субъектов по когортам и в целом, а также представлены в перечне данных по когортам и субъектам. Серьезные нежелательные явления, возникшие после начала лечения (TESAE), также оценивались для каждого субъекта, но ни одно из них не наблюдалось у субъектов, получавших лечение (таблица 11).
[0354] Из 15 субъектов, включенных в исследование, 13 получили по меньшей мере одну дозу rFVIIIFc-VWF-XTEN и были включены в анализ безопасности. У 8 из 13 субъектов (61,5%) сообщалось о 19 TEAE (таблица 11). Не сообщалось о серьезных или связанных TEAE, и ни один субъект не прекратил исследование из-за TEAE. Наиболее частыми TEAE были артралгия, инфекция верхних дыхательных путей, головная боль и ринит (в каждом случае у 2/13 пациентов; 15,4%). Не была выявлена выработка ингибиторов FVIII, и не было сообщений о серьезной гиперчувствительности или анафилаксии. На каждом уровне обобщения у субъектов субъекта подсчитывают один раз, если он сообщил об одном или нескольких явлениях. Процентные значения, указанные в таблице 11, приводятся на основании количества субъектов, получивших по меньшей мере одну дозу rFVIIIFc-VWF-XTEN в каждой когорте и в целом.
Таблица 11. Сводный обзор нежелательных явлений, возникших после начала лечения (TEAE), в выборке для анализа безопасности
50 МЕ/кг
65 МЕ/кг
общее количество получивших лечение
[0355] Анализ иммуногенности
[0356] Всех субъектов оценивали в отношении выработки ингибиторов и антител к rFVIIIFc-VWF-XTEN при скрининге, в день 1, день 8, день 15, день 22, день 36 и день 50 (во время визита EOS или ET). Тестирование на ингибиторы проводилось центральной лабораторией с использованием анализа Бетезда в модификации Неймегена. Если выдаваемый результат анализа Бетезда в модификации Неймегена был ≥ 0,6 БЕ/мл, то собирали отдельный образец, и его тестировали для подтверждения выработки ингибиторов в течение 2-4 недель. Тестирование на потенциальное образование антител проводили в центральной лаборатории с использованием валидированного анализа rFVIIIFc-VWF-XTEN-специфичных антител к лекарственному средству (ADA). Для подтвержденных положительных образцов определяли дополнительные характеристики относительно антител, специфичных к FVIII, D'D3 или XTEN. Количество и процентную долю субъектов с положительным результатом тестирования в отношении выработки ингибиторов, а также количество и процентную долю субъектов с положительным результатом тестирования в отношении антител к rFVIIIFc-VWF-XTEN обобщали по когортам с использованием выборки для анализа безопасности.
[0357] После введения rFVIIIFc-VWF-XTEN ни у одного субъекта не проявлялся положительный результат анализа на антитела к лекарственному средству.
[0358] Фармакокинетический анализ и результаты
[0359] В целом, данные о безопасности и PK-параметрах обобщали с использованием стандартных показателей сводной статистики для непрерывных и категориальных данных. Данные обобщали по когортам. Непрерывные переменные обобщали с использованием показателей описательной статистики, включающих количество непропущенных значений (n), среднее значение, стандартное отклонение (SD), медиану, минимум и максимум. Фармакокинетические параметры также обобщали с использованием среднего геометрического значения и процентного коэффициента вариации (% CV). Категориальные переменные обобщали по количеству и процентным значениям.
[0360] Все PK-анализы проводили на основе выборки для PK-анализов (PKAS) и включали 9 субъектов из дозовой когорты 50 МЕ/кг, у которых были надлежащим образом собраны образцы крови после введения rFVIIIFc-VWF-XTEN.
[0361] Активность FVIII
[0362] Для определения фармакокинетических характеристик rFVIIIFc-VWF-XTEN после в общей сложности четырех еженедельных IV доз активность FVIII определяли у каждого субъекта в заранее определенные моменты времени. Активность FVIII обобщали по когортам, запланированным визитам для PK-анализа и моментам времени как для одностадийного анализа свертывания крови (по активированному частичному тромбопластиновому времени [aPTT]), так и для хромогенного анализа коагуляции. Профили активности FVIII в зависимости от времени строили как в исходном, так и в логарифмическом масштабе для обоих анализов активности FVIII. Индивидуальная активность FVIII была указана для каждого субъекта по когортам, запланированным визитам для PK-анализа и моментам времени для обоих анализов активности FVIII. Результаты относительно активности FVIII согласно одностадийному анализу [aPTT] представлены в таблице 12. Результаты относительно активности FVIII согласно хромогенному анализу представлены в таблице 13.
[0363] Вкратце, после введения дозы в день 1 средние значения активности FVIII согласно одностадийному и хромогенному анализам через 72 ч. составляли 45,53% и 29,37%, через 120 ч. составляли 21,56% и 13,49%, а через 168 ч. составляли 7,91% и 5,97% соответственно. После введения дозы в день 22 средние значения активности FVIII согласно одностадийному и хромогенному анализам через 72 ч. составляли 46,28% и 30,46%, через 120 ч. составляли 22,30% и 14,48%, а через 168 ч. составляли 9,83% и 6,74% соответственно. Обычно для всех пациентов, моментов времени и для обоих анализов Ctrough для rFVIIIFc-VWF-XTEN при 50 МЕ/кг QW в этом промежуточном анализе находилась в диапазоне 7-10%.
[0364] PK-параметры
[0365] Индивидуальные оценки PK-параметров были перечислены для каждого субъекта и обобщены описательно по когортам, запланированным визитам для PK-анализа и моментам времени для обоих анализов активности FVIII.
[0366] Среднее геометрическое значение периода полувыведения (t1/2) для всех субъектов, получавших rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг QW в этом промежуточном анализе, составляло 41,24 часа согласно одностадийному анализу (таблица 16B) и 43,87 часа согласно хромогенному анализу (таблица 17B).
[0367] После введения дозы в день 1 среднее геометрическое значение максимальной активности FVIII (Cmax) составляло 113 МЕ/дл в одностадийном анализе (таблица 14) и 108 МЕ/дл в хромогенном анализе (таблица 15). После введения дозы в день 22 среднее геометрическое значение активности FVIII в равновесном состоянии (Cmaxss) составляло 127 МЕ/дл согласно одностадийному анализу (таблица 16A) и 115 МЕ/дл согласно хромогенному анализу (таблица 17A).
[0368] После введения дозы в день 1 среднее геометрическое значение совокупной активности FVIII (AUC0-tau) составляло 7650 ч.*МЕ/дл согласно одностадийному анализу (таблица 14) и 5630 ч.*МЕ/дл согласно хромогенному анализу (таблица 15). После введения дозы в день 22 среднее геометрическое значение AUC0-tau составляло 8270 ч.*МЕ/дл согласно одностадийному анализу (таблица 16А) и 5880 ч.*МЕ/дл согласно хромогенному анализу (таблица 17А).
[0369] Данные о Cmax и AUC0-tau в день 1 и 22 введения доз указывали на минимальное накопление, что подтверждается индексом накопления (AI), составляющим 1,07 (одностадийный анализ, таблица 16B) и 1,08 (хромогенный анализ, таблица 17B). Для всех данных о PK-параметрах, представленных в таблицах 12-17, значения ниже предела количественного определения (BLQ) обрабатывались как 0 для целей анализа. При необходимости для расчета PK-параметров, поскольку время окончания инфузии не было зафиксировано, продолжительность инфузии была условно рассчитана как 8 минут для каждого субъекта в соответствии с руководством по введению в документе "Указания по обращению и введению" исследования.
Таблица 12. Сводный обзор уровней активности FVIII с поправкой на исходный уровень (МЕ/дл) согласно одностадийному анализу свертывания крови по aPTT: выборка для PK-анализа (уровень дозы: 50 МЕ/кг)
Сокращения: SD=стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, мин. = минимальное значение, макс. = максимальное значение, SOI=начало инъекции
Таблица 13. Сводный обзор уровней активности FVIII с поправкой на исходный уровень (МЕ/дл) согласно хромогенному анализу: выборка для PK-анализа (уровень дозы: 50 МЕ/кг)
Сокращения: SD=стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, мин. = минимальное значение, макс. = максимальное значение, SOI=начало инъекции
Таблица 14. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно одностадийному анализу свертывания крови по aPTT после введения дозы в день 1: выборка для PK-анализа
(МЕ/кг)
(МЕ/дл)
(ч.)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
(МЕ/дл на МЕ/кг)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
Таблица 15. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно хромогенному анализу в день 1: выборка для PK-анализа
(МЕ/кг)
(МЕ/дл)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
(МЕ/дл на МЕ/кг)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
Таблица 16A. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно одностадийному анализу свертывания крови по aPTT в день 22: выборка для PK-анализа (часть 1)
(МЕ/кг)
(МЕ/дл)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
Таблица 16B. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно одностадийному анализу свертывания крови по aPTT в день 22: выборка для PK-анализа (часть 2)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
Таблица 17A. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно хромогенному анализу в день 22: выборка для PK-анализа (часть 1)
(МЕ/кг)
(МЕ/дл)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
(ч.*МЕ/дл)
(ч.*кг*МЕ/дл/МЕ)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
Таблица 17B. Сводный обзор фармакокинетических параметров активности FVIII с поправкой на исходный уровень согласно хромогенному анализу в день 22: выборка для PK-анализа (часть 2)
Сокращения: станд. отклон. = стандартное отклонение, ср. геом. = среднее геометрическое значение, % CV=процентный коэффициент вариации, CMAX=максимальная концентрация в равновесном состоянии, TMAX=время достижения максимальной концентрации, CMAXSS=максимальная концентрация в равновесном состоянии, IVR=восстановление in vivo, AUC0-inf=площадь под кривой зависимости активности от времени от момента времени 0 до бесконечности, AUC0-tau=площадь под кривой зависимости активности от времени от часа 0 в течение интервала между введениями доз, MRTINF=среднее время удержания, экстраполированное на бесконечность, AI=индекс накопления, IR=постепенное восстановление, Ctrough=наиболее низкая концентрация, которой лекарственное средство достигает до введения следующей дозы, Cavg=средняя концентрация после достижения равновесного состояния, t1/2=период полувыведения, CLss=клиренс в равновесном состоянии, Vss=объем распределения в равновесном состоянии.
[0370] Выводы
[0371] В соответствии с результатами, наблюдаемыми в исследовании с однократной дозой (пример 1), еженедельное введение доз rFVIIIFc-VWF-XTEN в целом хорошо переносилось, и ингибиторы не выявлялись в течение 28 дней после последнего введения rFVIIIFc-VWF-XTEN. rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг продемонстрировал средний период полувыведения (t1/2), составляющий 41,24 и 43,87 часа в одностадийном и хромогенном анализах соответственно. Эти промежуточные результаты демонстрируют, что увеличенный период полужизни, наблюдаемый при однократном введении дозы rFVIIIFc-VWF-XTEN, сохраняется при повторном введении доз QW, что убедительно свидетельствует о том, что преимущества rFVIIIFc-VWF-XTEN могут быть реализованы у пациентов с гемофилией A, получающих лечение в соответствии с профилактической схемой введения доз FVIII.
Пример 3. Исследование rFVIIIFc-VWF-XTEN фазы 3 у пациентов с тяжелой формой гемофилии A, ранее получавших лечение
[0372] В настоящем исследовании будут оцениваться безопасность и переносимость, а также определяться фармакокинетические характеристики rFVIIIFc-VWF-XTEN при еженедельном введении 50 МЕ/кг (фиг. 1). Первичной целью исследования является оценивание эффективности профилактического лечения с помощью rFVIIIFc-VWF-XTEN по сравнению с лечением по необходимости с помощью rFVIIIFc-VWF-XTEN. Вторичной целью исследования является оценивание rFVIIIFc-VWF-XTEN в качестве средства профилактического лечения, его эффекта в отношении эпизодов кровотечения и потребления rFVIIIFc-VWF-XTEN, функционального состояния суставов и качества жизни, что измеряют по исходам, сообщаемым пациентами (PRO), и оценивание rFVIIIFc-VWF-XTEN для лечения по необходимости.
[0373] Кроме того, целью фармакокинетического анализа (PK) является определение PK-показателей rFVIIIFc-VWF-XTEN при профилактическом применении на исходном уровне для всех субъектов и повторно через три месяца для заранее определенной подгруппы субъектов согласно анализу активности FVIII по aPTT. Дополнительными целями являются оценивание эффективности rFVIIIFc-VWF-XTEN у субъектов, переносящих хирургическое вмешательство, а также оценивание безопасности и переносимости rFVIIIFc-VWF-XTEN. Кроме того, структурные исходы в отношении функционального состояния суставов будут оцениваться с помощью ультразвукового исследования, и будет оцениваться влияние лечения на другие измеряемые показатели качества жизни и на использование ресурсов здравоохранения.
[0374] План исследования
[0375] Это опорное открытое многоцентровое исследование фазы 3 в отношении безопасности, эффективности и PK-показателей rFVIIIFc-VWF-XTEN при внутривенном введении у пациентов в возрасте ≥ 12 лет с тяжелой формой гемофилии А (определяемой как уровень эндогенного фактора VIII < 1 МЕ/дл [< 1%]). Примерно двадцать пять субъектов, в настоящее время проходящих режим лечения по необходимости, будут получать rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг IV по необходимости в течение двадцати шести недель, затем будут получать rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг IV один раз в неделю в режиме профилактического лечения в течение двадцати шести недель. Субъекты, которые в настоящее время проходят режим профилактического лечения, будут получать rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг IV один раз в неделю в режиме профилактического лечения в течение пятидесяти двух недель.
[0376] В исследование будут включены примерно 140 субъектов в возрасте ≥ 12 лет; повторные оценки PK-показателей будут проведены в подгруппе из примерно шестнадцати субъектов через 12 недель.
[0377] Субъекты будут подвергнуты скринингу и после определения соответствия критериям включения перейдут к одному из двух режимов лечения (т. е. по необходимости или для профилактики) с помощью rFVIIIFc-VWF-XTEN в дозе 50 МЕ/кг IV. Все субъекты будут подвергнуты оценкам PK-показателей на исходном уровне после периода отмывки (по меньшей мере 4-5 дней в зависимости от текущей терапии). На протяжении всего исследования субъекты будут подвергаться оценкам эффективности и безопасности. Эти оценки безопасности будут включать тестирование в отношении образования потенциальных ингибиторов. Визит для оценки безопасности или телефонный звонок в период последующего наблюдения будет происходить через двадцать восемь дней после последней дозы rFVIIIFc-VWF-XTEN, если только субъект не будет включен в открытое расширенное исследование.
[0378] Окончание исследования (EOS) может произойти, если соблюдены оба критерия: (i) по меньшей мере 104 субъекта достигли 50 дней воздействия (ED) и выполнили достоверный тест на ингибиторы после 50-го ED; и (ii) по меньшей мере двадцать пять субъектов завершили исследование в группе лечения по необходимости, в том числе 26-недельный период профилактического лечения, или досрочно прекратили участие в исследовании. Кроме того, в подгруппу переносящих хирургическое вмешательство будут включены субъекты из любой группы, которые переносят хирургическое вмешательство во время исследования. Для оценки контроля и предупреждения кровотечения в хирургической ситуации будет намечено не менее десяти обширных хирургических вмешательств у по меньшей мере пяти субъектов.
[0379] Субъекты будут посещать клинику в течение примерно 28-дневного периода скрининга для определения соответствия критериям участия в исследовании. В дополнение к скрининговому(скрининговым) визиту(визитам) субъекты будут повторно посещать клинику для визитов на исходном уровне, в неделю 4, неделю 12, неделю 26, неделю 36, неделю 52 и при EOS/ET. Период введения доз составляет более 52 недель. Субъекты, входящие в группу профилактического лечения, будут получать 52 еженедельные дозы rFVIIIFc-VWF-XTEN и дополнительные дозы, необходимые для лечения эпизодов кровотечения. Субъекты, которые входят в группу лечения по необходимости, будут получать rFVIIIFc-VWF-XTEN по необходимости в течение 26-недельного периода лечения по необходимости, а также 26 еженедельных доз rFVIIIFc-VWF-XTEN и дополнительные дозы, необходимые для лечения эпизодов кровотечения, в течение последующего 26-недельного периода профилактического лечения. Безопасность будет оцениваться на протяжении всего исследования. Визит для оценки безопасности или телефонный звонок в период последующего наблюдения будет происходить через 28 дней после последней дозы rFVIIIFc-VWF-XTEN, если только субъект не будет включен в открытое расширенное исследование.
[0380] Первичной конечной точкой исследования будет интрасубъектное сравнение частоты кровотечения в годовом исчислении (ABR) в случае, когда субъекты проходят лечение с помощью rFVIIIFc-VWF-XTEN по необходимости (26 недель), и в случае, когда субъекты проходят профилактическое лечение один раз в неделю (26 недель).
[0381] Вторичные конечные точки исследования будут включать (i) интрасубъектное сравнение ABR при профилактическом лечении с помощью rFVIIIFc-VWF-XTEN и исторической ABR (для субъектов, проходящих профилактическое лечение, которые участвовали в предшествующем наблюдательном исследовании); (ii) частоту возникновения эпизодов кровотечения и потребление (общую ABR, ABR по типу и местоположению, а также общее потребление rFVIIIFc-VWF-XTEN на одного субъекта в годовом исчислении); (iii) ответ на лечение; (iv) индекс эффективности (процентную долю субъектов, у которых сохраняется остаточный уровень L3% в день 7; (v) функциональное состояние суставов и (vi) изменения в измеряемых показателях качества жизни (физической функции согласно опросникам Haem-A-QoL и PROMIS-SF для субъектов в возрасте по меньшей мере 18 лет, а также физической активности согласно опросникам Haemo-QoL и педиатрическому PROMIS-SF для субъектов в возрасте менее 18 лет).
[0382] Ответ на лечение будет включать (i) количество инфузий и дозу rFVIIIFc-VWF-XTEN, требуемые для устранения эпизода кровотечения; (ii) процентную долю эпизодов кровотечения, для устранения которых требуется однократная инфузия; (iii) оценку субъектом ответа на лечение отдельных эпизодов кровотечения с помощью rFVIIIFc-VWF-XTEN по 4-балльной шкале ответа и (iv) общую оценку исследователем ответа субъекта на лечение с помощью rFVIIIFc-VWF-XTEN по 4-балльной шкале ответа.
[0383] Оценка функционального состояния суставов будет включать (i) ABR для эпизодов кровотечения в суставах; (ii) восстановление суставов-мишеней через 52 недели на основании критериев Международного общества по изучению тромбозов и гемостаза (ISTH); (iii) функциональные исходы на основании абсолютного изменения от исходного уровня до 52 недель, оцениваемого по модифицированной шкале оценки функционального состояния суставов при гемофилии (mHJHS или HJHS); (iv) процентную долю субъектов без ухудшения состояния суставов через 52 недели, оцениваемого по mHJHS или HJHS.
[0384] Дополнительные конечные точки включают (i) максимальную (пиковую) концентрацию лекарственного средства в плазме крови (Cmax), период полувыведения (t½), индекс накопления (AI), площадь под кривой зависимости концентрации в плазме крови от времени (AUC), кажущийся объем распределения в равновесном состоянии (Vss), среднее время удержания (MRT), постепенное восстановление (IR), остаточную концентрацию в плазме крови (Ctrough) и время превышения предварительно определенных уровней активности FVIII; (ii) оценки исследователями или хирургами гемостатического ответа субъектов на лечение с помощью rFVIIIFc-VWF-XTEN по 4-балльной шкале ответа, количество и дозу инфузий, требуемых для поддержания гемостаза во время хирургического вмешательства, общее потребление rFVIIIFc-VWF-XTEN во время хирургического периода, ожидаемую кровопотерю во время хирургического вмешательства, количество и тип переливаний компонентов крови, требуемых во время хирургического вмешательства, и эпизоды кровотечений после хирургического вмешательства во время хирургического периода; (iii) возникновение нежелательных явлений (AE) и серьезных нежелательных явлений (SAE), клинически значимые аномалии при физикальном обследовании, в показателях жизненно важных функций и в лабораторных тестах, а также выработку ингибиторов (нейтрализующих антител, направленных против фактора VIII, как определено с помощью анализа Бетезда в модификации Неймегена); и (iv) анатомические структурные исходы в отношении функционального состояния суставов, полученные с помощью ультразвуковой визуализации в субпопуляции, изменения в других измеряемых показателях качества жизни (баллов EQ-5D-5L, PGIC (только неделя 52), интенсивности боли согласно опроснику PROMIS, влияния боли согласно опроснику PROMIS-SF (субъекты ≥ 18 лет), влияния боли согласно педиатрическому опроснику PROMIS-SF (субъекты < 18 лет)) и HRU.
[0385] Критерии включения/исключения
[0386] Пригодные кандидаты должны соответствовать следующим критериям участия в исследовании: (1) возраст ≥ 12 лет на момент подписания информированного согласия; (2) тяжелая форма гемофилии A, определяемая как уровень эндогенного фактора VIII < 1 МЕ/дл (< 1%), что документально подтверждено сертифицированной клинической лабораторией; (3) предшествующее лечение гемофилии А (профилактическое или по необходимости) с помощью любого рекомбинантного и/или полученного из плазмы крови FVIII или криопреципитата в течение по меньшей мере 150 ED; (4) в настоящее время проходят режим профилактического лечения с помощью представленного на рынке продукта FVIII и имели по меньшей мере четыре эпизода кровотечения за двенадцать месяцев до включения в исследование или в настоящее время проходят режим лечения по необходимости с помощью представленного на рынке продукта FVIII и имели по меньшей мере двенадцать эпизодов кровотечения за двенадцать месяцев до включения в исследование; и (5) количество тромбоцитов ≥ 100000 клеток/мкл при скрининге (в тесте, проведенном центральной лабораторией и проанализированном до введения дозы в день 1). Субъекты будут исключены по любой из следующих причин: (1) другое(другие) известное(известные) нарушение(нарушения) коагуляции в дополнение к гемофилии А; (2) гиперчувствительность или анафилаксия в анамнезе, связанная с любым продуктом FVIII; (3) положительный результат теста на ингибиторы или клинические признаки ослабленного ответа на введение FVIII в анамнезе (при наличии ингибиторов в семейном анамнезе субъект не исключается); (4) положительный результат теста на ингибиторы, полученный при скрининге, определенный как ≥ 0,6 БЕ/мл; (5) аномалия почечной функции, определяемая как уровень креатинина в сыворотке крови > 2,0 мг/дл, полученный при скрининге; (6) уровень аланинаминотрансферазы (ALT) или аспартатаминотрансферазы (AST) в сыворотке крови > 5 × верхний предел нормы (ULN), полученный при скрининге; (7) уровень общего билирубина в сыворотке крови > 3 × ULN, полученный при скрининге; (8) применение фитусирана или эмицизумаба в течение 12 недель до скрининга; (9) лечение в течение 12 недель до скрининга с помощью терапевтического моноклонального антитела, слитого белка на основе Fc, отличного от rFVIIIFc, или иммуноглобулина IV; (10) лечение с помощью ацетилсалициловой кислоты (ASA) в течение 2 недель до скрининга или лечение с помощью нестероидных противовоспалительных лекарственных средств (NSAID) в максимальной дозе, указанной в региональной инструкции по применению для каждого продукта, или в дозе, превышающей ее; (11) системное лечение в течение 12 недель до скрининга с помощью химиотерапии и/или других иммунодепрессивных лекарственных средств (за исключением лечения инфекции, вызываемой вирусом гепатита С [HCV] или HIV; применение кортикостероидов допускается, за исключением лечения системными кортикостероидами, проводимого ежедневно или через день в дозе 20 мг/день преднизона или его эквивалента в течение > 14 дней; местное, наружное и/или ингаляционное применение стероидов разрешено) и (12) обширное хирургическое вмешательство в течение 8 недель до скрининга (обширное хирургическое вмешательство определяется как любая хирургическая процедура (плановая или экстренная), которая, как правило, но не всегда, включает общую анестезию и/или искусственную вентиляцию легких, при которой проникают в большую полость тела и обнажают ее или производят существенное нарушение физических или физиологических функций (например, лапаротомия, торакотомия, краниотомия, замена сустава или ампутация конечности)).
ПОСЛЕДОВАТЕЛЬНОСТИ
Нуклеотидная последовательность VWF031 (SEQ ID NO: 147)
1 ATGAT TCCTG CCAGA TTTGC CGGGG TGCTG CTTGC TCTGG CCCTC ATTTT
51 GCCAG GGACC CTTTG TGCAG AAGGA ACTCG CGGCA GGTCA TCCAC GGCCC
101 GATGC AGCCT TTTCG GAAGT GACTT CGTCA ACACC TTTGA TGGGA GCATG
151 TACAG CTTTG CGGGA TACTG CAGTT ACCTC CTGGC AGGGG GCTGC CAGAA
201 ACGCT CCTTC TCGAT TATTG GGGAC TTCCA GAATG GCAAG AGAGT GAGCC
251 TCTCC GTGTA TCTTG GGGAA TTTTT TGACA TCCAT TTGTT TGTCA ATGGT
301 ACCGT GACAC AGGGG GACCA AAGAG TCTCC ATGCC CTATG CCTCC AAAGG
351 GCTGT ATCTA GAAAC TGAGG CTGGG TACTA CAAGC TGTCC GGTGA GGCCT
401 ATGGC TTTGT GGCCA GGATC GATGG CAGCG GCAAC TTTCA AGTCC TGCTG
451 TCAGA CAGAT ACTTC AACAA GACCT GCGGG CTGTG TGGCA ACTTT AACAT
501 CTTTG CTGAA GATGA CTTTA TGACC CAAGA AGGGA CCTTG ACCTC GGACC
551 CTTAT GACTT TGCCA ACTCA TGGGC TCTGA GCAGT GGAGA ACAGT GGTGT
601 GAACG GGCAT CTCCT CCCAG CAGCT CATGC AACAT CTCCT CTGGG GAAAT
651 GCAGA AGGGC CTGTG GGAGC AGTGC CAGCT TCTGA AGAGC ACCTC GGTGT
701 TTGCC CGCTG CCACC CTCTG GTGGA CCCCG AGCCT TTTGT GGCCC TGTGT
751 GAGAA GACTT TGTGT GAGTG TGCTG GGGGG CTGGA GTGCG CCTGC CCTGC
801 CCTCC TGGAG TACGC CCGGA CCTGT GCCCA GGAGG GAATG GTGCT GTACG
851 GCTGG ACCGA CCACA GCGCG TGCAG CCCAG TGTGC CCTGC TGGTA TGGAG
901 TATAG GCAGT GTGTG TCCCC TTGCG CCAGG ACCTG CCAGA GCCTG CACAT
951 CAATG AAATG TGTCA GGAGC GATGC GTGGA TGGCT GCAGC TGCCC TGAGG
1001 GACAG CTCCT GGATG AAGGC CTCTG CGTGG AGAGC ACCGA GTGTC CCTGC
1051 GTGCA TTCCG GAAAG CGCTA CCCTC CCGGC ACCTC CCTCT CTCGA GACTG
1101 CAACA CCTGC ATTTG CCGAA ACAGC CAGTG GATCT GCAGC AATGA AGAAT
1151 GTCCA GGGGA GTGCC TTGTC ACTGG TCAAT CCCAC TTCAA GAGCT TTGAC
1201 AACAG ATACT TCACC TTCAG TGGGA TCTGC CAGTA CCTGC TGGCC CGGGA
1251 TTGCC AGGAC CACTC CTTCT CCATT GTCAT TGAGA CTGTC CAGTG TGCTG
1301 ATGAC CGCGA CGCTG TGTGC ACCCG CTCCG TCACC GTCCG GCTGC CTGGC
1351 CTGCA CAACA GCCTT GTGAA ACTGA AGCAT GGGGC AGGAG TTGCC ATGGA
1401 TGGCC AGGAC ATCCA GCTCC CCCTC CTGAA AGGTG ACCTC CGCAT CCAGC
1451 ATACA GTGAC GGCCT CCGTG CGCCT CAGCT ACGGG GAGGA CCTGC AGATG
1501 GACTG GGATG GCCGC GGGAG GCTGC TGGTG AAGCT GTCCC CCGTC TATGC
1551 CGGGA AGACC TGCGG CCTGT GTGGG AATTA CAATG GCAAC CAGGG CGACG
1601 ACTTC CTTAC CCCCT CTGGG CTGGC GGAGC CCCGG GTGGA GGACT TCGGG
1651 AACGC CTGGA AGCTG CACGG GGACT GCCAG GACCT GCAGA AGCAG CACAG
1701 CGATC CCTGC GCCCT CAACC CGCGC ATGAC CAGGT TCTCC GAGGA GGCGT
1751 GCGCG GTCCT GACGT CCCCC ACATT CGAGG CCTGC CATCG TGCCG TCAGC
1801 CCGCT GCCCT ACCTG CGGAA CTGCC GCTAC GACGT GTGCT CCTGC TCGGA
1851 CGGCC GCGAG TGCCT GTGCG GCGCC CTGGC CAGCT ATGCC GCGGC CTGCG
1901 CGGGG AGAGG CGTGC GCGTC GCGTG GCGCG AGCCA GGCCG CTGTG AGCTG
1951 AACTG CCCGA AAGGC CAGGT GTACC TGCAG TGCGG GACCC CCTGC AACCT
2001 GACCT GCCGC TCTCT CTCTT ACCCG GATGA GGAAT GCAAT GAGGC CTGCC
2051 TGGAG GGCTG CTTCT GCCCC CCAGG GCTCT ACATG GATGA GAGGG GGGAC
2101 TGCGT GCCCA AGGCC CAGTG CCCCT GTTAC TATGA CGGTG AGATC TTCCA
2151 GCCAG AAGAC ATCTT CTCAG ACCAT CACAC CATGT GCTAC TGTGA GGATG
2201 GCTTC ATGCA CTGTA CCATG AGTGG AGTCC CCGGA AGCTT GCTGC CTGAC
2251 GCTGT CCTCA GCAGT CCCCT GTCTC ATCGC AGCAA AAGGA GCCTA TCCTG
2301 TCGGC CCCCC ATGGT CAAGC TGGTG TGTCC CGCTG ACAAC CTGCG GGCTG
2351 AAGGG CTCGA GTGTA CCAAA ACGTG CCAGA ACTAT GACCT GGAGT GCATG
2401 AGCAT GGGCT GTGTC TCTGG CTGCC TCTGC CCCCC GGGCA TGGTC CGGCA
2451 TGAGA ACAGA TGTGT GGCCC TGGAA AGGTG TCCCT GCTTC CATCA GGGCA
2501 AGGAG TATGC CCCTG GAGAA ACAGT GAAGA TTGGC TGCAA CACTT GTGTC
2551 TGTCG GGACC GGAAG TGGAA CTGCA CAGAC CATGT GTGTG ATGCC ACGTG
2601 CTCCA CGATC GGCAT GGCCC ACTAC CTCAC CTTCG ACGGG CTCAA ATACC
2651 TGTTC CCCGG GGAGT GCCAG TACGT TCTGG TGCAG GATTA CTGCG GCAGT
2701 AACCC TGGGA CCTTT CGGAT CCTAG TGGGG AATAA GGGAT GCAGC CACCC
2751 CTCAG TGAAA TGCAA GAAAC GGGTC ACCAT CCTGG TGGAG GGAGG AGAGA
2801 TTGAG CTGTT TGACG GGGAG GTGAA TGTGA AGAGG CCCAT GAAGG ATGAG
2851 ACTCA CTTTG AGGTG GTGGA GTCTG GCCGG TACAT CATTC TGCTG CTGGG
2901 CAAAG CCCTC TCCGT GGTCT GGGAC CGCCA CCTGA GCATC TCCGT GGTCC
2951 TGAAG CAGAC ATACC AGGAG AAAGT GTGTG GCCTG TGTGG GAATT TTGAT
3001 GGCAT CCAGA ACAAT GACCT CACCA GCAGC AACCT CCAAG TGGAG GAAGA
3051 CCCTG TGGAC TTTGG GAACT CCTGG AAAGT GAGCT CGCAG TGTGC TGACA
3101 CCAGA AAAGT GCCTC TGGAC TCATC CCCTG CCACC TGCCA TAACA ACATC
3151 ATGAA GCAGA CGATG GTGGA TTCCT CCTGT AGAAT CCTTA CCAGT GACGT
3201 CTTCC AGGAC TGCAA CAAGC TGGTG GACCC CGAGC CATAT CTGGA TGTCT
3251 GCATT TACGA CACCT GCTCC TGTGA GTCCA TTGGG GACTG CGCCG CATTC
3301 TGCGA CACCA TTGCT GCCTA TGCCC ACGTG TGTGC CCAGC ATGGC AAGGT
3351 GGTGA CCTGG AGGAC GGCCA CATTG TGCCC CCAGA GCTGC GAGGA GAGGA
3401 ATCTC CGGGA GAACG GGTAT GAGGC TGAGT GGCGC TATAA CAGCT GTGCA
3451 CCTGC CTGTC AAGTC ACGTG TCAGC ACCCT GAGCC ACTGG CCTGC CCTGT
3501 GCAGT GTGTG GAGGG CTGCC ATGCC CACTG CCCTC CAGGG AAAAT CCTGG
3551 ATGAG CTTTT GCAGA CCTGC GTTGA CCCTG AAGAC TGTCC AGTGT GTGAG
3601 GTGGC TGGCC GGCGT TTTGC CTCAG GAAAG AAAGT CACCT TGAAT CCCAG
3651 TGACC CTGAG CACTG CCAGA TTTGC CACTG TGATG TTGTC AACCT CACCT
3701 GTGAA GCCTG CCAGG AGCCG ATATC TGGCG GTGGA GGTTC CGGTG GCGGG
3751 GGATC CGGCG GTGGA GGTTC CGGCG GTGGA GGTTC CGGTG GCGGG GGATC
3801 CGGTG GCGGG GGATC CCTGG TCCCC CGGGG CAGCG GCGGT GGAGG TTCCG
3851 GTGGC GGGGG ATCCG ACAAA ACTCA CACAT GCCCA CCGTG CCCAG CTCCA
3901 GAACT CCTGG GCGGA CCGTC AGTCT TCCTC TTCCC CCCAA AACCC AAGGA
3951 CACCC TCATG ATCTC CCGGA CCCCT GAGGT CACAT GCGTG GTGGT GGACG
4001 TGAGC CACGA AGACC CTGAG GTCAA GTTCA ACTGG TACGT GGACG GCGTG
4051 GAGGT GCATA ATGCC AAGAC AAAGC CGCGG GAGGA GCAGT ACAAC AGCAC
4101 GTACC GTGTG GTCAG CGTCC TCACC GTCCT GCACC AGGAC TGGCT GAATG
4151 GCAAG GAGTA CAAGT GCAAG GTCTC CAACA AAGCC CTCCC AGCCC CCATC
4201 GAGAA AACCA TCTCC AAAGC CAAAG GGCAG CCCCG AGAAC CACAG GTGTA
4251 CACCC TGCCC CCATC CCGCG ATGAG CTGAC CAAGA ACCAG GTCAG CCTGA
4301 CCTGC CTGGT CAAAG GCTTC TATCC CAGCG ACATC GCCGT GGAGT GGGAG
4351 AGCAA TGGGC AGCCG GAGAA CAACT ACAAG ACCAC GCCTC CCGTG TTGGA
4401 CTCCG ACGGC TCCTT CTTCC TCTAC AGCAA GCTCA CCGTG GACAA GAGCA
4451 GGTGG CAGCA GGGGA ACGTC TTCTC ATGCT CCGTG ATGCA TGAGG CTCTG
4501 CACAA CCACT ACACG CAGAA GAGCC TCTCC CTGTC TCCGG GTAAA TGA
Последовательность белка VWF031 (SEQ ID NO: 86)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGGGGSGGG
1251 GSGGGGSGGG GSGGGGSGGG GSLVPRGSGG GGSGGGGSDK THTCPPCPAP
1301 ELLGGPSVFL FPPKPKDTLM ISRTPEVTCV VVDVSHEDPE VKFNWYVDGV
1351 EVHNAKTKPR EEQYNSTYRV VSVLTVLHQD WLNGKEYKCK VSNKALPAPI
1401 EKTISKAKGQ PREPQVYTLP PSRDELTKNQ VSLTCLVKGF YPSDIAVEWE
1451 SNGQPENNYK TTPPVLDSDG SFFLYSKLTV DKSRWQQGNV FSCSVMHEAL
1501 HNHYTQKSLS LSPGK*
Нуклеотидная последовательность VWF034 (SEQ ID NO: 148)
1 ATGAT TCCTG CCAGA TTTGC CGGGG TGCTG CTTGC TCTGG CCCTC ATTTT
51 GCCAG GGACC CTTTG TGCAG AAGGA ACTCG CGGCA GGTCA TCCAC GGCCC
101 GATGC AGCCT TTTCG GAAGT GACTT CGTCA ACACC TTTGA TGGGA GCATG
151 TACAG CTTTG CGGGA TACTG CAGTT ACCTC CTGGC AGGGG GCTGC CAGAA
201 ACGCT CCTTC TCGAT TATTG GGGAC TTCCA GAATG GCAAG AGAGT GAGCC
251 TCTCC GTGTA TCTTG GGGAA TTTTT TGACA TCCAT TTGTT TGTCA ATGGT
301 ACCGT GACAC AGGGG GACCA AAGAG TCTCC ATGCC CTATG CCTCC AAAGG
351 GCTGT ATCTA GAAAC TGAGG CTGGG TACTA CAAGC TGTCC GGTGA GGCCT
401 ATGGC TTTGT GGCCA GGATC GATGG CAGCG GCAAC TTTCA AGTCC TGCTG
451 TCAGA CAGAT ACTTC AACAA GACCT GCGGG CTGTG TGGCA ACTTT AACAT
501 CTTTG CTGAA GATGA CTTTA TGACC CAAGA AGGGA CCTTG ACCTC GGACC
551 CTTAT GACTT TGCCA ACTCA TGGGC TCTGA GCAGT GGAGA ACAGT GGTGT
601 GAACG GGCAT CTCCT CCCAG CAGCT CATGC AACAT CTCCT CTGGG GAAAT
651 GCAGA AGGGC CTGTG GGAGC AGTGC CAGCT TCTGA AGAGC ACCTC GGTGT
701 TTGCC CGCTG CCACC CTCTG GTGGA CCCCG AGCCT TTTGT GGCCC TGTGT
751 GAGAA GACTT TGTGT GAGTG TGCTG GGGGG CTGGA GTGCG CCTGC CCTGC
801 CCTCC TGGAG TACGC CCGGA CCTGT GCCCA GGAGG GAATG GTGCT GTACG
851 GCTGG ACCGA CCACA GCGCG TGCAG CCCAG TGTGC CCTGC TGGTA TGGAG
901 TATAG GCAGT GTGTG TCCCC TTGCG CCAGG ACCTG CCAGA GCCTG CACAT
951 CAATG AAATG TGTCA GGAGC GATGC GTGGA TGGCT GCAGC TGCCC TGAGG
1001 GACAG CTCCT GGATG AAGGC CTCTG CGTGG AGAGC ACCGA GTGTC CCTGC
1051 GTGCA TTCCG GAAAG CGCTA CCCTC CCGGC ACCTC CCTCT CTCGA GACTG
1101 CAACA CCTGC ATTTG CCGAA ACAGC CAGTG GATCT GCAGC AATGA AGAAT
1151 GTCCA GGGGA GTGCC TTGTC ACTGG TCAAT CCCAC TTCAA GAGCT TTGAC
1201 AACAG ATACT TCACC TTCAG TGGGA TCTGC CAGTA CCTGC TGGCC CGGGA
1251 TTGCC AGGAC CACTC CTTCT CCATT GTCAT TGAGA CTGTC CAGTG TGCTG
1301 ATGAC CGCGA CGCTG TGTGC ACCCG CTCCG TCACC GTCCG GCTGC CTGGC
1351 CTGCA CAACA GCCTT GTGAA ACTGA AGCAT GGGGC AGGAG TTGCC ATGGA
1401 TGGCC AGGAC ATCCA GCTCC CCCTC CTGAA AGGTG ACCTC CGCAT CCAGC
1451 ATACA GTGAC GGCCT CCGTG CGCCT CAGCT ACGGG GAGGA CCTGC AGATG
1501 GACTG GGATG GCCGC GGGAG GCTGC TGGTG AAGCT GTCCC CCGTC TATGC
1551 CGGGA AGACC TGCGG CCTGT GTGGG AATTA CAATG GCAAC CAGGG CGACG
1601 ACTTC CTTAC CCCCT CTGGG CTGGC GGAGC CCCGG GTGGA GGACT TCGGG
1651 AACGC CTGGA AGCTG CACGG GGACT GCCAG GACCT GCAGA AGCAG CACAG
1701 CGATC CCTGC GCCCT CAACC CGCGC ATGAC CAGGT TCTCC GAGGA GGCGT
1751 GCGCG GTCCT GACGT CCCCC ACATT CGAGG CCTGC CATCG TGCCG TCAGC
1801 CCGCT GCCCT ACCTG CGGAA CTGCC GCTAC GACGT GTGCT CCTGC TCGGA
1851 CGGCC GCGAG TGCCT GTGCG GCGCC CTGGC CAGCT ATGCC GCGGC CTGCG
1901 CGGGG AGAGG CGTGC GCGTC GCGTG GCGCG AGCCA GGCCG CTGTG AGCTG
1951 AACTG CCCGA AAGGC CAGGT GTACC TGCAG TGCGG GACCC CCTGC AACCT
2001 GACCT GCCGC TCTCT CTCTT ACCCG GATGA GGAAT GCAAT GAGGC CTGCC
2051 TGGAG GGCTG CTTCT GCCCC CCAGG GCTCT ACATG GATGA GAGGG GGGAC
2101 TGCGT GCCCA AGGCC CAGTG CCCCT GTTAC TATGA CGGTG AGATC TTCCA
2151 GCCAG AAGAC ATCTT CTCAG ACCAT CACAC CATGT GCTAC TGTGA GGATG
2201 GCTTC ATGCA CTGTA CCATG AGTGG AGTCC CCGGA AGCTT GCTGC CTGAC
2251 GCTGT CCTCA GCAGT CCCCT GTCTC ATCGC AGCAA AAGGA GCCTA TCCTG
2301 TCGGC CCCCC ATGGT CAAGC TGGTG TGTCC CGCTG ACAAC CTGCG GGCTG
2351 AAGGG CTCGA GTGTA CCAAA ACGTG CCAGA ACTAT GACCT GGAGT GCATG
2401 AGCAT GGGCT GTGTC TCTGG CTGCC TCTGC CCCCC GGGCA TGGTC CGGCA
2451 TGAGA ACAGA TGTGT GGCCC TGGAA AGGTG TCCCT GCTTC CATCA GGGCA
2501 AGGAG TATGC CCCTG GAGAA ACAGT GAAGA TTGGC TGCAA CACTT GTGTC
2551 TGTCG GGACC GGAAG TGGAA CTGCA CAGAC CATGT GTGTG ATGCC ACGTG
2601 CTCCA CGATC GGCAT GGCCC ACTAC CTCAC CTTCG ACGGG CTCAA ATACC
2651 TGTTC CCCGG GGAGT GCCAG TACGT TCTGG TGCAG GATTA CTGCG GCAGT
2701 AACCC TGGGA CCTTT CGGAT CCTAG TGGGG AATAA GGGAT GCAGC CACCC
2751 CTCAG TGAAA TGCAA GAAAC GGGTC ACCAT CCTGG TGGAG GGAGG AGAGA
2801 TTGAG CTGTT TGACG GGGAG GTGAA TGTGA AGAGG CCCAT GAAGG ATGAG
2851 ACTCA CTTTG AGGTG GTGGA GTCTG GCCGG TACAT CATTC TGCTG CTGGG
2901 CAAAG CCCTC TCCGT GGTCT GGGAC CGCCA CCTGA GCATC TCCGT GGTCC
2951 TGAAG CAGAC ATACC AGGAG AAAGT GTGTG GCCTG TGTGG GAATT TTGAT
3001 GGCAT CCAGA ACAAT GACCT CACCA GCAGC AACCT CCAAG TGGAG GAAGA
3051 CCCTG TGGAC TTTGG GAACT CCTGG AAAGT GAGCT CGCAG TGTGC TGACA
3101 CCAGA AAAGT GCCTC TGGAC TCATC CCCTG CCACC TGCCA TAACA ACATC
3151 ATGAA GCAGA CGATG GTGGA TTCCT CCTGT AGAAT CCTTA CCAGT GACGT
3201 CTTCC AGGAC TGCAA CAAGC TGGTG GACCC CGAGC CATAT CTGGA TGTCT
3251 GCATT TACGA CACCT GCTCC TGTGA GTCCA TTGGG GACTG CGCCG CATTC
3301 TGCGA CACCA TTGCT GCCTA TGCCC ACGTG TGTGC CCAGC ATGGC AAGGT
3351 GGTGA CCTGG AGGAC GGCCA CATTG TGCCC CCAGA GCTGC GAGGA GAGGA
3401 ATCTC CGGGA GAACG GGTAT GAGGC TGAGT GGCGC TATAA CAGCT GTGCA
3451 CCTGC CTGTC AAGTC ACGTG TCAGC ACCCT GAGCC ACTGG CCTGC CCTGT
3501 GCAGT GTGTG GAGGG CTGCC ATGCC CACTG CCCTC CAGGG AAAAT CCTGG
3551 ATGAG CTTTT GCAGA CCTGC GTTGA CCCTG AAGAC TGTCC AGTGT GTGAG
3601 GTGGC TGGCC GGCGT TTTGC CTCAG GAAAG AAAGT CACCT TGAAT CCCAG
3651 TGACC CTGAG CACTG CCAGA TTTGC CACTG TGATG TTGTC AACCT CACCT
3701 GTGAA GCCTG CCAGG AGCCG ATATC GGGTA CCTCA GAGTC TGCTA CCCCC
3751 GAGTC AGGGC CAGGA TCAGA GCCAG CCACC TCCGG GTCTG AGACA CCCGG
3801 GACTT CCGAG AGTGC CACCC CTGAG TCCGG ACCCG GGTCC GAGCC CGCCA
3851 CTTCC GGCTC CGAAA CTCCC GGCAC AAGCG AGAGC GCTAC CCCAG AGTCA
3901 GGACC AGGAA CATCT ACAGA GCCCT CTGAA GGCTC CGCTC CAGGG TCCCC
3951 AGCCG GCAGT CCCAC TAGCA CCGAG GAGGG AACCT CTGAA AGCGC CACAC
4001 CCGAA TCAGG GCCAG GGTCT GAGCC TGCTA CCAGC GGCAG CGAGA CACCA
4051 GGCAC CTCTG AGTCC GCCAC ACCAG AGTCC GGACC CGGAT CTCCC GCTGG
4101 GAGCC CCACC TCCAC TGAGG AGGGA TCTCC TGCTG GCTCT CCAAC ATCTA
4151 CTGAG GAAGG TACCT CAACC GAGCC ATCCG AGGGA TCAGC TCCCG GCACC
4201 TCAGA GTCGG CAACC CCGGA GTCTG GACCC GGAAC TTCCG AAAGT GCCAC
4251 ACCAG AGTCC GGTCC CGGGA CTTCA GAATC AGCAA CACCC GAGTC CGGCC
4301 CTGGG TCTGA ACCCG CCACA AGTGG TAGTG AGACA CCAGG ATCAG AACCT
4351 GCTAC CTCAG GGTCA GAGAC ACCCG GATCT CCGGC AGGCT CACCA ACCTC
4401 CACTG AGGAG GGCAC CAGCA CAGAA CCAAG CGAGG GCTCC GCACC CGGAA
4451 CAAGC ACTGA ACCCA GTGAG GGTTC AGCAC CCGGC TCTGA GCCGG CCACA
4501 AGTGG CAGTG AGACA CCCGG CACTT CAGAG AGTGC CACCC CCGAG AGTGG
4551 CCCAG GCACT AGTAC CGAGC CCTCT GAAGG CAGTG CGCCA GATTC TGGCG
4601 GTGGA GGTTC CGGTG GCGGG GGATC CGGTG GCGGG GGATC CGGTG GCGGG
4651 GGATC CGGTG GCGGG GGATC CCTGG TCCCC CGGGG CAGCG GAGGC GACAA
4701 AACTC ACACA TGCCC ACCGT GCCCA GCTCC AGAAC TCCTG GGCGG ACCGT
4751 CAGTC TTCCT CTTCC CCCCA AAACC CAAGG ACACC CTCAT GATCT CCCGG
4801 ACCCC TGAGG TCACA TGCGT GGTGG TGGAC GTGAG CCACG AAGAC CCTGA
4851 GGTCA AGTTC AACTG GTACG TGGAC GGCGT GGAGG TGCAT AATGC CAAGA
4901 CAAAG CCGCG GGAGG AGCAG TACAA CAGCA CGTAC CGTGT GGTCA GCGTC
4951 CTCAC CGTCC TGCAC CAGGA CTGGC TGAAT GGCAA GGAGT ACAAG TGCAA
5001 GGTCT CCAAC AAAGC CCTCC CAGCC CCCAT CGAGA AAACC ATCTC CAAAG
5051 CCAAA GGGCA GCCCC GAGAA CCACA GGTGT ACACC CTGCC CCCAT CCCGG
5101 GATGA GCTGA CCAAG AACCA GGTCA GCCTG ACCTG CCTGG TCAAA GGCTT
5151 CTATC CCAGC GACAT CGCCG TGGAG TGGGA GAGCA ATGGG CAGCC GGAGA
5201 ACAAC TACAA GACCA CGCCT CCCGT GTTGG ACTCC GACGG CTCCT TCTTC
5251 CTCTA CAGCA AGCTC ACCGT GGACA AGAGC AGGTG GCAGC AGGGG AACGT
5301 CTTCT CATGC TCCGT GATGC ATGAG GCTCT GCACA ACCAC TACAC GCAGA
5351 AGAGC CTCTC CCTGT CTCCG GGTAA ATGA
Последовательность белка VWF034 (SEQ ID NO: 87)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGTSESATP
1251 ESGPGSEPAT SGSETPGTSE SATPESGPGS EPATSGSETP GTSESATPES
1301 GPGTSTEPSE GSAPGSPAGS PTSTEEGTSE SATPESGPGS EPATSGSETP
1351 GTSESATPES GPGSPAGSPT STEEGSPAGS PTSTEEGTST EPSEGSAPGT
1401 SESATPESGP GTSESATPES GPGTSESATP ESGPGSEPAT SGSETPGSEP
1451 ATSGSETPGS PAGSPTSTEE GTSTEPSEGS APGTSTEPSE GSAPGSEPAT
1501 SGSETPGTSE SATPESGPGT STEPSEGSAP DIGGGGGSGG GGSLVPRGSG
1551 GDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE
1601 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY
1651 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
1701 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
1751 GNVFSCSVMH EALHNHYTQK SLSLSPGK*
Нуклеотидная последовательность VWF050 (с тройной мутацией IHH) (SEQ ID NO: 149)
1 ATGAT TCCTG CCAGA TTTGC CGGGG TGCTG CTTGC TCTGG CCCTC ATTTT
51 GCCAG GGACC CTTTG TGCAG AAGGA ACTCG CGGCA GGTCA TCCAC GGCCC
101 GATGC AGCCT TTTCG GAAGT GACTT CGTCA ACACC TTTGA TGGGA GCATG
151 TACAG CTTTG CGGGA TACTG CAGTT ACCTC CTGGC AGGGG GCTGC CAGAA
201 ACGCT CCTTC TCGAT TATTG GGGAC TTCCA GAATG GCAAG AGAGT GAGCC
251 TCTCC GTGTA TCTTG GGGAA TTTTT TGACA TCCAT TTGTT TGTCA ATGGT
301 ACCGT GACAC AGGGG GACCA AAGAG TCTCC ATGCC CTATG CCTCC AAAGG
351 GCTGT ATCTA GAAAC TGAGG CTGGG TACTA CAAGC TGTCC GGTGA GGCCT
401 ATGGC TTTGT GGCCA GGATC GATGG CAGCG GCAAC TTTCA AGTCC TGCTG
451 TCAGA CAGAT ACTTC AACAA GACCT GCGGG CTGTG TGGCA ACTTT AACAT
501 CTTTG CTGAA GATGA CTTTA TGACC CAAGA AGGGA CCTTG ACCTC GGACC
551 CTTAT GACTT TGCCA ACTCA TGGGC TCTGA GCAGT GGAGA ACAGT GGTGT
601 GAACG GGCAT CTCCT CCCAG CAGCT CATGC AACAT CTCCT CTGGG GAAAT
651 GCAGA AGGGC CTGTG GGAGC AGTGC CAGCT TCTGA AGAGC ACCTC GGTGT
701 TTGCC CGCTG CCACC CTCTG GTGGA CCCCG AGCCT TTTGT GGCCC TGTGT
751 GAGAA GACTT TGTGT GAGTG TGCTG GGGGG CTGGA GTGCG CCTGC CCTGC
801 CCTCC TGGAG TACGC CCGGA CCTGT GCCCA GGAGG GAATG GTGCT GTACG
851 GCTGG ACCGA CCACA GCGCG TGCAG CCCAG TGTGC CCTGC TGGTA TGGAG
901 TATAG GCAGT GTGTG TCCCC TTGCG CCAGG ACCTG CCAGA GCCTG CACAT
951 CAATG AAATG TGTCA GGAGC GATGC GTGGA TGGCT GCAGC TGCCC TGAGG
1001 GACAG CTCCT GGATG AAGGC CTCTG CGTGG AGAGC ACCGA GTGTC CCTGC
1051 GTGCA TTCCG GAAAG CGCTA CCCTC CCGGC ACCTC CCTCT CTCGA GACTG
1101 CAACA CCTGC ATTTG CCGAA ACAGC CAGTG GATCT GCAGC AATGA AGAAT
1151 GTCCA GGGGA GTGCC TTGTC ACTGG TCAAT CCCAC TTCAA GAGCT TTGAC
1201 AACAG ATACT TCACC TTCAG TGGGA TCTGC CAGTA CCTGC TGGCC CGGGA
1251 TTGCC AGGAC CACTC CTTCT CCATT GTCAT TGAGA CTGTC CAGTG TGCTG
1301 ATGAC CGCGA CGCTG TGTGC ACCCG CTCCG TCACC GTCCG GCTGC CTGGC
1351 CTGCA CAACA GCCTT GTGAA ACTGA AGCAT GGGGC AGGAG TTGCC ATGGA
1401 TGGCC AGGAC ATCCA GCTCC CCCTC CTGAA AGGTG ACCTC CGCAT CCAGC
1451 ATACA GTGAC GGCCT CCGTG CGCCT CAGCT ACGGG GAGGA CCTGC AGATG
1501 GACTG GGATG GCCGC GGGAG GCTGC TGGTG AAGCT GTCCC CCGTC TATGC
1551 CGGGA AGACC TGCGG CCTGT GTGGG AATTA CAATG GCAAC CAGGG CGACG
1601 ACTTC CTTAC CCCCT CTGGG CTGGC GGAGC CCCGG GTGGA GGACT TCGGG
1651 AACGC CTGGA AGCTG CACGG GGACT GCCAG GACCT GCAGA AGCAG CACAG
1701 CGATC CCTGC GCCCT CAACC CGCGC ATGAC CAGGT TCTCC GAGGA GGCGT
1751 GCGCG GTCCT GACGT CCCCC ACATT CGAGG CCTGC CATCG TGCCG TCAGC
1801 CCGCT GCCCT ACCTG CGGAA CTGCC GCTAC GACGT GTGCT CCTGC TCGGA
1851 CGGCC GCGAG TGCCT GTGCG GCGCC CTGGC CAGCT ATGCC GCGGC CTGCG
1901 CGGGG AGAGG CGTGC GCGTC GCGTG GCGCG AGCCA GGCCG CTGTG AGCTG
1951 AACTG CCCGA AAGGC CAGGT GTACC TGCAG TGCGG GACCC CCTGC AACCT
2001 GACCT GCCGC TCTCT CTCTT ACCCG GATGA GGAAT GCAAT GAGGC CTGCC
2051 TGGAG GGCTG CTTCT GCCCC CCAGG GCTCT ACATG GATGA GAGGG GGGAC
2101 TGCGT GCCCA AGGCC CAGTG CCCCT GTTAC TATGA CGGTG AGATC TTCCA
2151 GCCAG AAGAC ATCTT CTCAG ACCAT CACAC CATGT GCTAC TGTGA GGATG
2201 GCTTC ATGCA CTGTA CCATG AGTGG AGTCC CCGGA AGCTT GCTGC CTGAC
2251 GCTGT CCTCA GCAGT CCCCT GTCTC ATCGC AGCAA AAGGA GCCTA TCCTG
2301 TCGGC CCCCC ATGGT CAAGC TGGTG TGTCC CGCTG ACAAC CTGCG GGCTG
2351 AAGGG CTCGA GTGTA CCAAA ACGTG CCAGA ACTAT GACCT GGAGT GCATG
2401 AGCAT GGGCT GTGTC TCTGG CTGCC TCTGC CCCCC GGGCA TGGTC CGGCA
2451 TGAGA ACAGA TGTGT GGCCC TGGAA AGGTG TCCCT GCTTC CATCA GGGCA
2501 AGGAG TATGC CCCTG GAGAA ACAGT GAAGA TTGGC TGCAA CACTT GTGTC
2551 TGTCG GGACC GGAAG TGGAA CTGCA CAGAC CATGT GTGTG ATGCC ACGTG
2601 CTCCA CGATC GGCAT GGCCC ACTAC CTCAC CTTCG ACGGG CTCAA ATACC
2651 TGTTC CCCGG GGAGT GCCAG TACGT TCTGG TGCAG GATTA CTGCG GCAGT
2701 AACCC TGGGA CCTTT CGGAT CCTAG TGGGG AATAA GGGAT GCAGC CACCC
2751 CTCAG TGAAA TGCAA GAAAC GGGTC ACCAT CCTGG TGGAG GGAGG AGAGA
2801 TTGAG CTGTT TGACG GGGAG GTGAA TGTGA AGAGG CCCAT GAAGG ATGAG
2851 ACTCA CTTTG AGGTG GTGGA GTCTG GCCGG TACAT CATTC TGCTG CTGGG
2901 CAAAG CCCTC TCCGT GGTCT GGGAC CGCCA CCTGA GCATC TCCGT GGTCC
2951 TGAAG CAGAC ATACC AGGAG AAAGT GTGTG GCCTG TGTGG GAATT TTGAT
3001 GGCAT CCAGA ACAAT GACCT CACCA GCAGC AACCT CCAAG TGGAG GAAGA
3051 CCCTG TGGAC TTTGG GAACT CCTGG AAAGT GAGCT CGCAG TGTGC TGACA
3101 CCAGA AAAGT GCCTC TGGAC TCATC CCCTG CCACC TGCCA TAACA ACATC
3151 ATGAA GCAGA CGATG GTGGA TTCCT CCTGT AGAAT CCTTA CCAGT GACGT
3201 CTTCC AGGAC TGCAA CAAGC TGGTG GACCC CGAGC CATAT CTGGA TGTCT
3251 GCATT TACGA CACCT GCTCC TGTGA GTCCA TTGGG GACTG CGCCG CATTC
3301 TGCGA CACCA TTGCT GCCTA TGCCC ACGTG TGTGC CCAGC ATGGC AAGGT
3351 GGTGA CCTGG AGGAC GGCCA CATTG TGCCC CCAGA GCTGC GAGGA GAGGA
3401 ATCTC CGGGA GAACG GGTAT GAGGC TGAGT GGCGC TATAA CAGCT GTGCA
3451 CCTGC CTGTC AAGTC ACGTG TCAGC ACCCT GAGCC ACTGG CCTGC CCTGT
3501 GCAGT GTGTG GAGGG CTGCC ATGCC CACTG CCCTC CAGGG AAAAT CCTGG
3551 ATGAG CTTTT GCAGA CCTGC GTTGA CCCTG AAGAC TGTCC AGTGT GTGAG
3601 GTGGC TGGCC GGCGT TTTGC CTCAG GAAAG AAAGT CACCT TGAAT CCCAG
3651 TGACC CTGAG CACTG CCAGA TTTGC CACTG TGATG TTGTC AACCT CACCT
3701 GTGAA GCCTG CCAGG AGCCG ATATC TGGCG GTGGA GGTTC CGGTG GCGGG
3751 GGATC CGGCG GTGGA GGTTC CGGCG GTGGA GGTTC CGGTG GCGGG GGATC
3801 CGGTG GCGGG GGATC CCTGG TCCCC CGGGG CAGCG GCGGT GGAGG TTCCG
3851 GTGGC GGGGG ATCCG ACAAA ACTCA CACAT GCCCA CCGTG CCCAG CTCCA
3901 GAACT CCTGG GCGGA CCGTC AGTCT TCCTC TTCCC CCCAA AACCC AAGGA
3951 CACCC TCATG GCCTC CCGGA CCCCT GAGGT CACAT GCGTG GTGGT GGACG
4001 TGAGC CACGA AGACC CTGAG GTCAA GTTCA ACTGG TACGT GGACG GCGTG
4051 GAGGT GCATA ATGCC AAGAC AAAGC CGCGG GAGGA GCAGT ACAAC AGCAC
4101 GTACC GTGTG GTCAG CGTCC TCACC GTCCT GGCCC AGGAC TGGCT GAATG
4151 GCAAG GAGTA CAAGT GCAAG GTCTC CAACA AAGCC CTCCC AGCCC CCATC
4201 GAGAA AACCA TCTCC AAAGC CAAAG GGCAG CCCCG AGAAC CACAG GTGTA
4251 CACCC TGCCC CCATC CCGCG ATGAG CTGAC CAAGA ACCAG GTCAG CCTGA
4301 CCTGC CTGGT CAAAG GCTTC TATCC CAGCG ACATC GCCGT GGAGT GGGAG
4351 AGCAA TGGGC AGCCG GAGAA CAACT ACAAG ACCAC GCCTC CCGTG TTGGA
4401 CTCCG ACGGC TCCTT CTTCC TCTAC AGCAA GCTCA CCGTG GACAA GAGCA
4451 GGTGG CAGCA GGGGA ACGTC TTCTC ATGCT CCGTG ATGCA TGAGG CTCTG
4501 CACAA CGCCT ACACG CAGAA GAGCC TCTCC CTGTC TCCGG GTAAA TGA
Последовательность белка VWF050 (с тройной мутацией IHH) (SEQ ID NO: 150)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGGGGSGGG
1251 GSGGGGSGGG GSGGGGSGGG GSLVPRGSGG GGSGGGGSDK THTCPPCPAP
1301 ELLGGPSVFL FPPKPKDTLM ASRTPEVTCV VVDVSHEDPE VKFNWYVDGV
1351 EVHNAKTKPR EEQYNSTYRV VSVLTVLAQD WLNGKEYKCK VSNKALPAPI
1401 EKTISKAKGQ PREPQVYTLP PSRDELTKNQ VSLTCLVKGF YPSDIAVEWE
1451 SNGQPENNYK TTPPVLDSDG SFFLYSKLTV DKSRWQQGNV FSCSVMHEAL
1501 HNAYTQKSLS LSPGK*
Нуклеотидная последовательность VWF057 (SEQ ID NO: 151)
1 ATGAT TCCTG CCAGA TTTGC CGGGG TGCTG CTTGC TCTGG CCCTC ATTTT
51 GCCAG GGACC CTTTG TGCAG AAGGA ACTCG CGGCA GGTCA TCCAC GGCCC
101 GATGC AGCCT TTTCG GAAGT GACTT CGTCA ACACC TTTGA TGGGA GCATG
151 TACAG CTTTG CGGGA TACTG CAGTT ACCTC CTGGC AGGGG GCTGC CAGAA
201 ACGCT CCTTC TCGAT TATTG GGGAC TTCCA GAATG GCAAG AGAGT GAGCC
251 TCTCC GTGTA TCTTG GGGAA TTTTT TGACA TCCAT TTGTT TGTCA ATGGT
301 ACCGT GACAC AGGGG GACCA AAGAG TCTCC ATGCC CTATG CCTCC AAAGG
351 GCTGT ATCTA GAAAC TGAGG CTGGG TACTA CAAGC TGTCC GGTGA GGCCT
401 ATGGC TTTGT GGCCA GGATC GATGG CAGCG GCAAC TTTCA AGTCC TGCTG
451 TCAGA CAGAT ACTTC AACAA GACCT GCGGG CTGTG TGGCA ACTTT AACAT
501 CTTTG CTGAA GATGA CTTTA TGACC CAAGA AGGGA CCTTG ACCTC GGACC
551 CTTAT GACTT TGCCA ACTCA TGGGC TCTGA GCAGT GGAGA ACAGT GGTGT
601 GAACG GGCAT CTCCT CCCAG CAGCT CATGC AACAT CTCCT CTGGG GAAAT
651 GCAGA AGGGC CTGTG GGAGC AGTGC CAGCT TCTGA AGAGC ACCTC GGTGT
701 TTGCC CGCTG CCACC CTCTG GTGGA CCCCG AGCCT TTTGT GGCCC TGTGT
751 GAGAA GACTT TGTGT GAGTG TGCTG GGGGG CTGGA GTGCG CCTGC CCTGC
801 CCTCC TGGAG TACGC CCGGA CCTGT GCCCA GGAGG GAATG GTGCT GTACG
851 GCTGG ACCGA CCACA GCGCG TGCAG CCCAG TGTGC CCTGC TGGTA TGGAG
901 TATAG GCAGT GTGTG TCCCC TTGCG CCAGG ACCTG CCAGA GCCTG CACAT
951 CAATG AAATG TGTCA GGAGC GATGC GTGGA TGGCT GCAGC TGCCC TGAGG
1001 GACAG CTCCT GGATG AAGGC CTCTG CGTGG AGAGC ACCGA GTGTC CCTGC
1051 GTGCA TTCCG GAAAG CGCTA CCCTC CCGGC ACCTC CCTCT CTCGA GACTG
1101 CAACA CCTGC ATTTG CCGAA ACAGC CAGTG GATCT GCAGC AATGA AGAAT
1151 GTCCA GGGGA GTGCC TTGTC ACTGG TCAAT CCCAC TTCAA GAGCT TTGAC
1201 AACAG ATACT TCACC TTCAG TGGGA TCTGC CAGTA CCTGC TGGCC CGGGA
1251 TTGCC AGGAC CACTC CTTCT CCATT GTCAT TGAGA CTGTC CAGTG TGCTG
1301 ATGAC CGCGA CGCTG TGTGC ACCCG CTCCG TCACC GTCCG GCTGC CTGGC
1351 CTGCA CAACA GCCTT GTGAA ACTGA AGCAT GGGGC AGGAG TTGCC ATGGA
1401 TGGCC AGGAC ATCCA GCTCC CCCTC CTGAA AGGTG ACCTC CGCAT CCAGC
1451 ATACA GTGAC GGCCT CCGTG CGCCT CAGCT ACGGG GAGGA CCTGC AGATG
1501 GACTG GGATG GCCGC GGGAG GCTGC TGGTG AAGCT GTCCC CCGTC TATGC
1551 CGGGA AGACC TGCGG CCTGT GTGGG AATTA CAATG GCAAC CAGGG CGACG
1601 ACTTC CTTAC CCCCT CTGGG CTGGC GGAGC CCCGG GTGGA GGACT TCGGG
1651 AACGC CTGGA AGCTG CACGG GGACT GCCAG GACCT GCAGA AGCAG CACAG
1701 CGATC CCTGC GCCCT CAACC CGCGC ATGAC CAGGT TCTCC GAGGA GGCGT
1751 GCGCG GTCCT GACGT CCCCC ACATT CGAGG CCTGC CATCG TGCCG TCAGC
1801 CCGCT GCCCT ACCTG CGGAA CTGCC GCTAC GACGT GTGCT CCTGC TCGGA
1851 CGGCC GCGAG TGCCT GTGCG GCGCC CTGGC CAGCT ATGCC GCGGC CTGCG
1901 CGGGG AGAGG CGTGC GCGTC GCGTG GCGCG AGCCA GGCCG CTGTG AGCTG
1951 AACTG CCCGA AAGGC CAGGT GTACC TGCAG TGCGG GACCC CCTGC AACCT
2001 GACCT GCCGC TCTCT CTCTT ACCCG GATGA GGAAT GCAAT GAGGC CTGCC
2051 TGGAG GGCTG CTTCT GCCCC CCAGG GCTCT ACATG GATGA GAGGG GGGAC
2101 TGCGT GCCCA AGGCC CAGTG CCCCT GTTAC TATGA CGGTG AGATC TTCCA
2151 GCCAG AAGAC ATCTT CTCAG ACCAT CACAC CATGT GCTAC TGTGA GGATG
2201 GCTTC ATGCA CTGTA CCATG AGTGG AGTCC CCGGA AGCTT GCTGC CTGAC
2251 GCTGT CCTCA GCAGT CCCCT GTCTC ATCGC AGCAA AAGGA GCCTA TCCTG
2301 TCGGC CCCCC ATGGT CAAGC TGGTG TGTCC CGCTG ACAAC CTGCG GGCTG
2351 AAGGG CTCGA GTGTA CCAAA ACGTG CCAGA ACTAT GACCT GGAGT GCATG
2401 AGCAT GGGCT GTGTC TCTGG CTGCC TCTGC CCCCC GGGCA TGGTC CGGCA
2451 TGAGA ACAGA TGTGT GGCCC TGGAA AGGTG TCCCT GCTTC CATCA GGGCA
2501 AGGAG TATGC CCCTG GAGAA ACAGT GAAGA TTGGC TGCAA CACTT GTGTC
2551 TGTCG GGACC GGAAG TGGAA CTGCA CAGAC CATGT GTGTG ATGCC ACGTG
2601 CTCCA CGATC GGCAT GGCCC ACTAC CTCAC CTTCG ACGGG CTCAA ATACC
2651 TGTTC CCCGG GGAGT GCCAG TACGT TCTGG TGCAG GATTA CTGCG GCAGT
2701 AACCC TGGGA CCTTT CGGAT CCTAG TGGGG AATAA GGGAT GCAGC CACCC
2751 CTCAG TGAAA TGCAA GAAAC GGGTC ACCAT CCTGG TGGAG GGAGG AGAGA
2801 TTGAG CTGTT TGACG GGGAG GTGAA TGTGA AGAGG CCCAT GAAGG ATGAG
2851 ACTCA CTTTG AGGTG GTGGA GTCTG GCCGG TACAT CATTC TGCTG CTGGG
2901 CAAAG CCCTC TCCGT GGTCT GGGAC CGCCA CCTGA GCATC TCCGT GGTCC
2951 TGAAG CAGAC ATACC AGGAG AAAGT GTGTG GCCTG TGTGG GAATT TTGAT
3001 GGCAT CCAGA ACAAT GACCT CACCA GCAGC AACCT CCAAG TGGAG GAAGA
3051 CCCTG TGGAC TTTGG GAACT CCTGG AAAGT GAGCT CGCAG TGTGC TGACA
3101 CCAGA AAAGT GCCTC TGGAC TCATC CCCTG CCACC TGCCA TAACA ACATC
3151 ATGAA GCAGA CGATG GTGGA TTCCT CCTGT AGAAT CCTTA CCAGT GACGT
3201 CTTCC AGGAC TGCAA CAAGC TGGTG GACCC CGAGC CATAT CTGGA TGTCT
3251 GCATT TACGA CACCT GCTCC TGTGA GTCCA TTGGG GACTG CGCCG CATTC
3301 TGCGA CACCA TTGCT GCCTA TGCCC ACGTG TGTGC CCAGC ATGGC AAGGT
3351 GGTGA CCTGG AGGAC GGCCA CATTG TGCCC CCAGA GCTGC GAGGA GAGGA
3401 ATCTC CGGGA GAACG GGTAT GAGGC TGAGT GGCGC TATAA CAGCT GTGCA
3451 CCTGC CTGTC AAGTC ACGTG TCAGC ACCCT GAGCC ACTGG CCTGC CCTGT
3501 GCAGT GTGTG GAGGG CTGCC ATGCC CACTG CCCTC CAGGG AAAAT CCTGG
3551 ATGAG CTTTT GCAGA CCTGC GTTGA CCCTG AAGAC TGTCC AGTGT GTGAG
3601 GTGGC TGGCC GGCGT TTTGC CTCAG GAAAG AAAGT CACCT TGAAT CCCAG
3651 TGACC CTGAG CACTG CCAGA TTTGC CACTG TGATG TTGTC AACCT CACCT
3701 GTGAA GCCTG CCAGG AGCCG ATATC GGGCG CGCCA ACATC AGAGA GCGCC
3751 ACCCC TGAAA GTGGT CCCGG GAGCG AGCCA GCCAC ATCTG GGTCG GAAAC
3801 GCCAG GCACA AGTGA GTCTG CAACT CCCGA GTCCG GACCT GGCTC CGAGC
3851 CTGCC ACTAG CGGCT CCGAG ACTCC GGGAA CTTCC GAGAG CGCTA CACCA
3901 GAAAG CGGAC CCGGA ACCAG TACCG AACCT AGCGA GGGCT CTGCT CCGGG
3951 CAGCC CAGCC GGCTC TCCTA CATCC ACGGA GGAGG GCACT TCCGA ATCCG
4001 CCACC CCGGA GTCAG GGCCA GGATC TGAAC CCGCT ACCTC AGGCA GTGAG
4051 ACGCC AGGAA CGAGC GAGTC CGCTA CACCG GAGAG TGGGC CAGGG AGCCC
4101 TGCTG GATCT CCTAC GTCCA CTGAG GAAGG GTCAC CAGCG GGCTC GCCCA
4151 CCAGC ACTGA AGAAG GTGCC TCGAG CGGCG GTGGA GGTTC CGGTG GCGGG
4201 GGATC CGGTG GCGGG GGATC CGGTG GCGGG GGATC CGGTG GCGGG GGATC
4251 CCTGG TCCCC CGGGG CAGCG GAGGC GACAA AACTC ACACA TGCCC ACCGT
4301 GCCCA GCTCC AGAAC TCCTG GGCGG ACCGT CAGTC TTCCT CTTCC CCCCA
4351 AAACC CAAGG ACACC CTCAT GATCT CCCGG ACCCC TGAGG TCACA TGCGT
4401 GGTGG TGGAC GTGAG CCACG AAGAC CCTGA GGTCA AGTTC AACTG GTACG
4451 TGGAC GGCGT GGAGG TGCAT AATGC CAAGA CAAAG CCGCG GGAGG AGCAG
4501 TACAA CAGCA CGTAC CGTGT GGTCA GCGTC CTCAC CGTCC TGCAC CAGGA
4551 CTGGC TGAAT GGCAA GGAGT ACAAG TGCAA GGTCT CCAAC AAAGC CCTCC
4601 CAGCC CCCAT CGAGA AAACC ATCTC CAAAG CCAAA GGGCA GCCCC GAGAA
4651 CCACA GGTGT ACACC CTGCC CCCAT CCCGG GATGA GCTGA CCAAG AACCA
4701 GGTCA GCCTG ACCTG CCTGG TCAAA GGCTT CTATC CCAGC GACAT CGCCG
4751 TGGAG TGGGA GAGCA ATGGG CAGCC GGAGA ACAAC TACAA GACCA CGCCT
4801 CCCGT GTTGG ACTCC GACGG CTCCT TCTTC CTCTA CAGCA AGCTC ACCGT
4851 GGACA AGAGC AGGTG GCAGC AGGGG AACGT CTTCT CATGC TCCGT GATGC
4901 ATGAG GCTCT GCACA ACCAC TACAC GCAGA AGAGC CTCTC CCTGT CTCCG
4951 GGTAA ATGA
Последовательность белка VWF057 (SEQ ID NO: 152)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGAPTSESA
1251 TPESGPGSEP ATSGSETPGT SESATPESGP GSEPATSGSE TPGTSESATP
1301 ESGPGTSTEP SEGSAPGSPA GSPTSTEEGT SESATPESGP GSEPATSGSE
1351 TPGTSESATP ESGPGSPAGS PTSTEEGSPA GSPTSTEEGA SSGGGGSGGG
1401 GSGGGGSGGG GSGGGGSLVP RGSGGDKTHT CPPCPAPELL GGPSVFLFPP
1451 KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
1501 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE
1551 PQVYTLPPSR DELTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP
1601 PVLDSDGSFF LYSKLTVDKS RWQQGNVFSC SVMHEALHNH YTQKSLSLSP
1651 GK*
Нуклеотидная последовательность VWF058 (VWF034 с мутацией IHH) (SEQ ID NO: 153)
1 ATGAT TCCTG CCAGA TTTGC CGGGG TGCTG CTTGC TCTGG CCCTC ATTTT
51 GCCAG GGACC CTTTG TGCAG AAGGA ACTCG CGGCA GGTCA TCCAC GGCCC
101 GATGC AGCCT TTTCG GAAGT GACTT CGTCA ACACC TTTGA TGGGA GCATG
151 TACAG CTTTG CGGGA TACTG CAGTT ACCTC CTGGC AGGGG GCTGC CAGAA
201 ACGCT CCTTC TCGAT TATTG GGGAC TTCCA GAATG GCAAG AGAGT GAGCC
251 TCTCC GTGTA TCTTG GGGAA TTTTT TGACA TCCAT TTGTT TGTCA ATGGT
301 ACCGT GACAC AGGGG GACCA AAGAG TCTCC ATGCC CTATG CCTCC AAAGG
351 GCTGT ATCTA GAAAC TGAGG CTGGG TACTA CAAGC TGTCC GGTGA GGCCT
401 ATGGC TTTGT GGCCA GGATC GATGG CAGCG GCAAC TTTCA AGTCC TGCTG
451 TCAGA CAGAT ACTTC AACAA GACCT GCGGG CTGTG TGGCA ACTTT AACAT
501 CTTTG CTGAA GATGA CTTTA TGACC CAAGA AGGGA CCTTG ACCTC GGACC
551 CTTAT GACTT TGCCA ACTCA TGGGC TCTGA GCAGT GGAGA ACAGT GGTGT
601 GAACG GGCAT CTCCT CCCAG CAGCT CATGC AACAT CTCCT CTGGG GAAAT
651 GCAGA AGGGC CTGTG GGAGC AGTGC CAGCT TCTGA AGAGC ACCTC GGTGT
701 TTGCC CGCTG CCACC CTCTG GTGGA CCCCG AGCCT TTTGT GGCCC TGTGT
751 GAGAA GACTT TGTGT GAGTG TGCTG GGGGG CTGGA GTGCG CCTGC CCTGC
801 CCTCC TGGAG TACGC CCGGA CCTGT GCCCA GGAGG GAATG GTGCT GTACG
851 GCTGG ACCGA CCACA GCGCG TGCAG CCCAG TGTGC CCTGC TGGTA TGGAG
901 TATAG GCAGT GTGTG TCCCC TTGCG CCAGG ACCTG CCAGA GCCTG CACAT
951 CAATG AAATG TGTCA GGAGC GATGC GTGGA TGGCT GCAGC TGCCC TGAGG
1001 GACAG CTCCT GGATG AAGGC CTCTG CGTGG AGAGC ACCGA GTGTC CCTGC
1051 GTGCA TTCCG GAAAG CGCTA CCCTC CCGGC ACCTC CCTCT CTCGA GACTG
1101 CAACA CCTGC ATTTG CCGAA ACAGC CAGTG GATCT GCAGC AATGA AGAAT
1151 GTCCA GGGGA GTGCC TTGTC ACTGG TCAAT CCCAC TTCAA GAGCT TTGAC
1201 AACAG ATACT TCACC TTCAG TGGGA TCTGC CAGTA CCTGC TGGCC CGGGA
1251 TTGCC AGGAC CACTC CTTCT CCATT GTCAT TGAGA CTGTC CAGTG TGCTG
1301 ATGAC CGCGA CGCTG TGTGC ACCCG CTCCG TCACC GTCCG GCTGC CTGGC
1351 CTGCA CAACA GCCTT GTGAA ACTGA AGCAT GGGGC AGGAG TTGCC ATGGA
1401 TGGCC AGGAC ATCCA GCTCC CCCTC CTGAA AGGTG ACCTC CGCAT CCAGC
1451 ATACA GTGAC GGCCT CCGTG CGCCT CAGCT ACGGG GAGGA CCTGC AGATG
1501 GACTG GGATG GCCGC GGGAG GCTGC TGGTG AAGCT GTCCC CCGTC TATGC
1551 CGGGA AGACC TGCGG CCTGT GTGGG AATTA CAATG GCAAC CAGGG CGACG
1601 ACTTC CTTAC CCCCT CTGGG CTGGC GGAGC CCCGG GTGGA GGACT TCGGG
1651 AACGC CTGGA AGCTG CACGG GGACT GCCAG GACCT GCAGA AGCAG CACAG
1701 CGATC CCTGC GCCCT CAACC CGCGC ATGAC CAGGT TCTCC GAGGA GGCGT
1751 GCGCG GTCCT GACGT CCCCC ACATT CGAGG CCTGC CATCG TGCCG TCAGC
1801 CCGCT GCCCT ACCTG CGGAA CTGCC GCTAC GACGT GTGCT CCTGC TCGGA
1851 CGGCC GCGAG TGCCT GTGCG GCGCC CTGGC CAGCT ATGCC GCGGC CTGCG
1901 CGGGG AGAGG CGTGC GCGTC GCGTG GCGCG AGCCA GGCCG CTGTG AGCTG
1951 AACTG CCCGA AAGGC CAGGT GTACC TGCAG TGCGG GACCC CCTGC AACCT
2001 GACCT GCCGC TCTCT CTCTT ACCCG GATGA GGAAT GCAAT GAGGC CTGCC
2051 TGGAG GGCTG CTTCT GCCCC CCAGG GCTCT ACATG GATGA GAGGG GGGAC
2101 TGCGT GCCCA AGGCC CAGTG CCCCT GTTAC TATGA CGGTG AGATC TTCCA
2151 GCCAG AAGAC ATCTT CTCAG ACCAT CACAC CATGT GCTAC TGTGA GGATG
2201 GCTTC ATGCA CTGTA CCATG AGTGG AGTCC CCGGA AGCTT GCTGC CTGAC
2251 GCTGT CCTCA GCAGT CCCCT GTCTC ATCGC AGCAA AAGGA GCCTA TCCTG
2301 TCGGC CCCCC ATGGT CAAGC TGGTG TGTCC CGCTG ACAAC CTGCG GGCTG
2351 AAGGG CTCGA GTGTA CCAAA ACGTG CCAGA ACTAT GACCT GGAGT GCATG
2401 AGCAT GGGCT GTGTC TCTGG CTGCC TCTGC CCCCC GGGCA TGGTC CGGCA
2451 TGAGA ACAGA TGTGT GGCCC TGGAA AGGTG TCCCT GCTTC CATCA GGGCA
2501 AGGAG TATGC CCCTG GAGAA ACAGT GAAGA TTGGC TGCAA CACTT GTGTC
2551 TGTCG GGACC GGAAG TGGAA CTGCA CAGAC CATGT GTGTG ATGCC ACGTG
2601 CTCCA CGATC GGCAT GGCCC ACTAC CTCAC CTTCG ACGGG CTCAA ATACC
2651 TGTTC CCCGG GGAGT GCCAG TACGT TCTGG TGCAG GATTA CTGCG GCAGT
2701 AACCC TGGGA CCTTT CGGAT CCTAG TGGGG AATAA GGGAT GCAGC CACCC
2751 CTCAG TGAAA TGCAA GAAAC GGGTC ACCAT CCTGG TGGAG GGAGG AGAGA
2801 TTGAG CTGTT TGACG GGGAG GTGAA TGTGA AGAGG CCCAT GAAGG ATGAG
2851 ACTCA CTTTG AGGTG GTGGA GTCTG GCCGG TACAT CATTC TGCTG CTGGG
2901 CAAAG CCCTC TCCGT GGTCT GGGAC CGCCA CCTGA GCATC TCCGT GGTCC
2951 TGAAG CAGAC ATACC AGGAG AAAGT GTGTG GCCTG TGTGG GAATT TTGAT
3001 GGCAT CCAGA ACAAT GACCT CACCA GCAGC AACCT CCAAG TGGAG GAAGA
3051 CCCTG TGGAC TTTGG GAACT CCTGG AAAGT GAGCT CGCAG TGTGC TGACA
3101 CCAGA AAAGT GCCTC TGGAC TCATC CCCTG CCACC TGCCA TAACA ACATC
3151 ATGAA GCAGA CGATG GTGGA TTCCT CCTGT AGAAT CCTTA CCAGT GACGT
3201 CTTCC AGGAC TGCAA CAAGC TGGTG GACCC CGAGC CATAT CTGGA TGTCT
3251 GCATT TACGA CACCT GCTCC TGTGA GTCCA TTGGG GACTG CGCCG CATTC
3301 TGCGA CACCA TTGCT GCCTA TGCCC ACGTG TGTGC CCAGC ATGGC AAGGT
3351 GGTGA CCTGG AGGAC GGCCA CATTG TGCCC CCAGA GCTGC GAGGA GAGGA
3401 ATCTC CGGGA GAACG GGTAT GAGGC TGAGT GGCGC TATAA CAGCT GTGCA
3451 CCTGC CTGTC AAGTC ACGTG TCAGC ACCCT GAGCC ACTGG CCTGC CCTGT
3501 GCAGT GTGTG GAGGG CTGCC ATGCC CACTG CCCTC CAGGG AAAAT CCTGG
3551 ATGAG CTTTT GCAGA CCTGC GTTGA CCCTG AAGAC TGTCC AGTGT GTGAG
3601 GTGGC TGGCC GGCGT TTTGC CTCAG GAAAG AAAGT CACCT TGAAT CCCAG
3651 TGACC CTGAG CACTG CCAGA TTTGC CACTG TGATG TTGTC AACCT CACCT
3701 GTGAA GCCTG CCAGG AGCCG ATATC GGGTA CCTCA GAGTC TGCTA CCCCC
3751 GAGTC AGGGC CAGGA TCAGA GCCAG CCACC TCCGG GTCTG AGACA CCCGG
3801 GACTT CCGAG AGTGC CACCC CTGAG TCCGG ACCCG GGTCC GAGCC CGCCA
3851 CTTCC GGCTC CGAAA CTCCC GGCAC AAGCG AGAGC GCTAC CCCAG AGTCA
3901 GGACC AGGAA CATCT ACAGA GCCCT CTGAA GGCTC CGCTC CAGGG TCCCC
3951 AGCCG GCAGT CCCAC TAGCA CCGAG GAGGG AACCT CTGAA AGCGC CACAC
4001 CCGAA TCAGG GCCAG GGTCT GAGCC TGCTA CCAGC GGCAG CGAGA CACCA
4051 GGCAC CTCTG AGTCC GCCAC ACCAG AGTCC GGACC CGGAT CTCCC GCTGG
4101 GAGCC CCACC TCCAC TGAGG AGGGA TCTCC TGCTG GCTCT CCAAC ATCTA
4151 CTGAG GAAGG TACCT CAACC GAGCC ATCCG AGGGA TCAGC TCCCG GCACC
4201 TCAGA GTCGG CAACC CCGGA GTCTG GACCC GGAAC TTCCG AAAGT GCCAC
4251 ACCAG AGTCC GGTCC CGGGA CTTCA GAATC AGCAA CACCC GAGTC CGGCC
4301 CTGGG TCTGA ACCCG CCACA AGTGG TAGTG AGACA CCAGG ATCAG AACCT
4351 GCTAC CTCAG GGTCA GAGAC ACCCG GATCT CCGGC AGGCT CACCA ACCTC
4401 CACTG AGGAG GGCAC CAGCA CAGAA CCAAG CGAGG GCTCC GCACC CGGAA
4451 CAAGC ACTGA ACCCA GTGAG GGTTC AGCAC CCGGC TCTGA GCCGG CCACA
4501 AGTGG CAGTG AGACA CCCGG CACTT CAGAG AGTGC CACCC CCGAG AGTGG
4551 CCCAG GCACT AGTAC CGAGC CCTCT GAAGG CAGTG CGCCA GATTC TGGCG
4601 GTGGA GGTTC CGGTG GCGGG GGATC CGGTG GCGGG GGATC CGGTG GCGGG
4651 GGATC CGGTG GCGGG GGATC CCTGG TCCCC CGGGG CAGCG GAGGC GACAA
4701 AACTC ACACA TGCCC ACCGT GCCCA GCTCC AGAAC TCCTG GGCGG ACCGT
4751 CAGTC TTCCT CTTCC CCCCA AAACC CAAGG ACACC CTCAT GGCCT CCCGG
4801 ACCCC TGAGG TCACA TGCGT GGTGG TGGAC GTGAG CCACG AAGAC CCTGA
4851 GGTCA AGTTC AACTG GTACG TGGAC GGCGT GGAGG TGCAT AATGC CAAGA
4901 CAAAG CCGCG GGAGG AGCAG TACAA CAGCA CGTAC CGTGT GGTCA GCGTC
4951 CTCAC CGTCC TGGCC CAGGA CTGGC TGAAT GGCAA GGAGT ACAAG TGCAA
5001 GGTCT CCAAC AAAGC CCTCC CAGCC CCCAT CGAGA AAACC ATCTC CAAAG
5051 CCAAA GGGCA GCCCC GAGAA CCACA GGTGT ACACC CTGCC CCCAT CCCGC
5101 GATGA GCTGA CCAAG AACCA GGTCA GCCTG ACCTG CCTGG TCAAA GGCTT
5151 CTATC CCAGC GACAT CGCCG TGGAG TGGGA GAGCA ATGGG CAGCC GGAGA
5201 ACAAC TACAA GACCA CGCCT CCCGT GTTGG ACTCC GACGG CTCCT TCTTC
5251 CTCTA CAGCA AGCTC ACCGT GGACA AGAGC AGGTG GCAGC AGGGG AACGT
5301 CTTCT CATGC TCCGT GATGC ATGAG GCTCT GCACA ACGCC TACAC GCAGA
5351 AGAGC CTCTC CCTGT CTCCG GGTAA ATGA
Последовательность белка VWF058 (VWF034 с мутацией IHH) (SEQ ID NO: 154)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGTSESATP
1251 ESGPGSEPAT SGSETPGTSE SATPESGPGS EPATSGSETP GTSESATPES
1301 GPGTSTEPSE GSAPGSPAGS PTSTEEGTSE SATPESGPGS EPATSGSETP
1351 GTSESATPES GPGSPAGSPT STEEGSPAGS PTSTEEGTST EPSEGSAPGT
1401 SESATPESGP GTSESATPES GPGTSESATP ESGPGSEPAT SGSETPGSEP
1451 ATSGSETPGS PAGSPTSTEE GTSTEPSEGS APGTSTEPSE GSAPGSEPAT
1501 SGSETPGTSE SATPESGPGT STEPSEGSAP DSGGGGSGGG GSGGGGSGGG
1551 GSGGGGSLVP RGSGGDKTHT CPPCPAPELL GGPSVFLFPP KPKDTLMASR
1601 TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ YNSTYRVVSV
1651 LTVLAQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE PQVYTLPPSR
1701 DELTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP PVLDSDGSFF
1751 LYSKLTVDKS RWQQGNVFSC SVMHEALHNA YTQKSLSLSP GK*
Нуклеотидная последовательность FVIII 169 (SEQ ID NO: 155)
1 ATGCA AATAG AGCTC TCCAC CTGCT TCTTT CTGTG CCTTT TGCGA TTCTG
51 CTTTA GTGCC ACCAG AAGAT ACTAC CTGGG TGCAG TGGAA CTGTC ATGGG
101 ACTAT ATGCA AAGTG ATCTC GGTGA GCTGC CTGTG GACGC AAGAT TTCCT
151 CCTAG AGTGC CAAAA TCTTT TCCAT TCAAC ACCTC AGTCG TGTAC AAAAA
201 GACTC TGTTT GTAGA ATTCA CGGAT CACCT TTTCA ACATC GCTAA GCCAA
251 GGCCA CCCTG GATGG GTCTG CTAGG TCCTA CCATC CAGGC TGAGG TTTAT
301 GATAC AGTGG TCATT ACACT TAAGA ACATG GCTTC CCATC CTGTC AGTCT
351 TCATG CTGTT GGTGT ATCCT ACTGG AAAGC TTCTG AGGGA GCTGA ATATG
401 ATGAT CAGAC CAGTC AAAGG GAGAA AGAAG ATGAT AAAGT CTTCC CTGGT
451 GGAAG CCATA CATAT GTCTG GCAGG TCCTG AAAGA GAATG GTCCA ATGGC
501 CTCTG ACCCA CTGTG CCTTA CCTAC TCATA TCTTT CTCAT GTGGA CCTGG
551 TAAAA GACTT GAATT CAGGC CTCAT TGGAG CCCTA CTAGT ATGTA GAGAA
601 GGGAG TCTGG CCAAG GAAAA GACAC AGACC TTGCA CAAAT TTATA CTACT
651 TTTTG CTGTA TTTGA TGAAG GGAAA AGTTG GCACT CAGAA ACAAA GAACT
701 CCTTG ATGCA GGATA GGGAT GCTGC ATCTG CTCGG GCCTG GCCTA AAATG
751 CACAC AGTCA ATGGT TATGT AAACA GGTCT CTGCC AGGTC TGATT GGATG
801 CCACA GGAAA TCAGT CTATT GGCAT GTGAT TGGAA TGGGC ACCAC TCCTG
851 AAGTG CACTC AATAT TCCTC GAAGG TCACA CATTT CTTGT GAGGA ACCAT
901 CGCCA GGCTA GCTTG GAAAT CTCGC CAATA ACTTT CCTTA CTGCT CAAAC
951 ACTCT TGATG GACCT TGGAC AGTTT CTACT GTTTT GTCAT ATCTC TTCCC
1001 ACCAA CATGA TGGCA TGGAA GCTTA TGTCA AAGTA GACAG CTGTC CAGAG
1051 GAACC CCAAC TACGA ATGAA AAATA ATGAA GAAGC GGAAG ACTAT GATGA
1101 TGATC TTACT GATTC TGAAA TGGAT GTGGT CAGGT TTGAT GATGA CAACT
1151 CTCCT TCCTT TATCC AAATT CGCTC AGTTG CCAAG AAGCA TCCTA AAACT
1201 TGGGT ACATT ACATT GCTGC TGAAG AGGAG GACTG GGACT ATGCT CCCTT
1251 AGTCC TCGCC CCCGA TGACA GAAGT TATAA AAGTC AATAT TTGAA CAATG
1301 GCCCT CAGCG GATTG GTAGG AAGTA CAAAA AAGTC CGATT TATGG CATAC
1351 ACAGA TGAAA CCTTT AAGAC TCGTG AAGCT ATTCA GCATG AATCA GGAAT
1401 CTTGG GACCT TTACT TTATG GGGAA GTTGG AGACA CACTG TTGAT TATAT
1451 TTAAG AATCA AGCAA GCAGA CCATA TAACA TCTAC CCTCA CGGAA TCACT
1501 GATGT CCGTC CTTTG TATTC AAGGA GATTA CCAAA AGGTG TAAAA CATTT
1551 GAAGG ATTTT CCAAT TCTGC CAGGA GAAAT ATTCA AATAT AAATG GACAG
1601 TGACT GTAGA AGATG GGCCA ACTAA ATCAG ATCCT CGGTG CCTGA CCCGC
1651 TATTA CTCTA GTTTC GTTAA TATGG AGAGA GATCT AGCTT CAGGA CTCAT
1701 TGGCC CTCTC CTCAT CTGCT ACAAA GAATC TGTAG ATCAA AGAGG AAACC
1751 AGATA ATGTC AGACA AGAGG AATGT CATCC TGTTT TCTGT ATTTG ATGAG
1801 AACCG AAGCT GGTAC CTCAC AGAGA ATATA CAACG CTTTC TCCCC AATCC
1851 AGCTG GAGTG CAGCT TGAGG ATCCA GAGTT CCAAG CCTCC AACAT CATGC
1901 ACAGC ATCAA TGGCT ATGTT TTTGA TAGTT TGCAG TTGTC AGTTT GTTTG
1951 CATGA GGTGG CATAC TGGTA CATTC TAAGC ATTGG AGCAC AGACT GACTT
2001 CCTTT CTGTC TTCTT CTCTG GATAT ACCTT CAAAC ACAAA ATGGT CTATG
2051 AAGAC ACACT CACCC TATTC CCATT CTCAG GAGAA ACTGT CTTCA TGTCG
2101 ATGGA AAACC CAGGT CTATG GATTC TGGGG TGCCA CAACT CAGAC TTTCG
2151 GAACA GAGGC ATGAC CGCCT TACTG AAGGT TTCTA GTTGT GACAA GAACA
2201 CTGGT GATTA TTACG AGGAC AGTTA TGAAG ATATT TCAGC ATACT TGCTG
2251 AGTAA AAACA ATGCC ATTGA ACCAA GAAGC TTCTC TCAAA ACGGC GCGCC
2301 AGGTA CCTCA GAGTC TGCTA CCCCC GAGTC AGGGC CAGGA TCAGA GCCAG
2351 CCACC TCCGG GTCTG AGACA CCCGG GACTT CCGAG AGTGC CACCC CTGAG
2401 TCCGG ACCCG GGTCC GAGCC CGCCA CTTCC GGCTC CGAAA CTCCC GGCAC
2451 AAGCG AGAGC GCTAC CCCAG AGTCA GGACC AGGAA CATCT ACAGA GCCCT
2501 CTGAA GGCTC CGCTC CAGGG TCCCC AGCCG GCAGT CCCAC TAGCA CCGAG
2551 GAGGG AACCT CTGAA AGCGC CACAC CCGAA TCAGG GCCAG GGTCT GAGCC
2601 TGCTA CCAGC GGCAG CGAGA CACCA GGCAC CTCTG AGTCC GCCAC ACCAG
2651 AGTCC GGACC CGGAT CTCCC GCTGG GAGCC CCACC TCCAC TGAGG AGGGA
2701 TCTCC TGCTG GCTCT CCAAC ATCTA CTGAG GAAGG TACCT CAACC GAGCC
2751 ATCCG AGGGA TCAGC TCCCG GCACC TCAGA GTCGG CAACC CCGGA GTCTG
2801 GACCC GGAAC TTCCG AAAGT GCCAC ACCAG AGTCC GGTCC CGGGA CTTCA
2851 GAATC AGCAA CACCC GAGTC CGGCC CTGGG TCTGA ACCCG CCACA AGTGG
2901 TAGTG AGACA CCAGG ATCAG AACCT GCTAC CTCAG GGTCA GAGAC ACCCG
2951 GATCT CCGGC AGGCT CACCA ACCTC CACTG AGGAG GGCAC CAGCA CAGAA
3001 CCAAG CGAGG GCTCC GCACC CGGAA CAAGC ACTGA ACCCA GTGAG GGTTC
3051 AGCAC CCGGC TCTGA GCCGG CCACA AGTGG CAGTG AGACA CCCGG CACTT
3101 CAGAG AGTGC CACCC CCGAG AGTGG CCCAG GCACT AGTAC CGAGC CCTCT
3151 GAAGG CAGTG CGCCA GCCTC GAGCC CACCA GTCTT GAAAC GCCAT CAAGC
3201 TGAAA TAACT CGTAC TACTC TTCAG TCAGA TCAAG AGGAA ATCGA TTATG
3251 ATGAT ACCAT ATCAG TTGAA ATGAA GAAGG AAGAT TTTGA CATTT ATGAT
3301 GAGGA TGAAA ATCAG AGCCC CCGCA GCTTT CAAAA GAAAA CACGA CACTA
3351 TTTTA TTGCT GCAGT GGAGA GGCTC TGGGA TTATG GGATG AGTAG CTCCC
3401 CACAT GTTCT AAGAA ACAGG GCTCA GAGTG GCAGT GTCCC TCAGT TCAAG
3451 AAAGT TGTTT TCCAG GAATT TACTG ATGGC TCCTT TACTC AGCCC TTATA
3501 CCGTG GAGAA CTAAA TGAAC ATTTG GGACT CCTGG GGCCA TATAT AAGAG
3551 CAGAA GTTGA AGATA ATATC ATGGT AACTT TCAGA AATCA GGCCT CTCGT
3601 CCCTA TTCCT TCTAT TCTAG CCTTA TTTCT TATGA GGAAG ATCAG AGGCA
3651 AGGAG CAGAA CCTAG AAAAA ACTTT GTCAA GCCTA ATGAA ACCAA AACTT
3701 ACTTT TGGAA AGTGC AACAT CATAT GGCAC CCACT AAAGA TGAGT TTGAC
3751 TGCAA AGCCT GGGCT TATTT CTCTG ATGTT GACCT GGAAA AAGAT GTGCA
3801 CTCAG GCCTG ATTGG ACCCC TTCTG GTCTG CCACA CTAAC ACACT GAACC
3851 CTGCT CATGG GAGAC AAGTG ACAGT ACAGG AATTT GCTCT GTTTT TCACC
3901 ATCTT TGATG AGACC AAAAG CTGGT ACTTC ACTGA AAATA TGGAA AGAAA
3951 CTGCA GGGCT CCCTG CAATA TCCAG ATGGA AGATC CCACT TTTAA AGAGA
4001 ATTAT CGCTT CCATG CAATC AATGG CTACA TAATG GATAC ACTAC CTGGC
4051 TTAGT AATGG CTCAG GATCA AAGGA TTCGA TGGTA TCTGC TCAGC ATGGG
4101 CAGCA ATGAA AACAT CCATT CTATT CATTT CAGTG GACAT GTGTT CACTG
4151 TACGA AAAAA AGAGG AGTAT AAAAT GGCAC TGTAC AATCT CTATC CAGGT
4201 GTTTT TGAGA CAGTG GAAAT GTTAC CATCC AAAGC TGGAA TTTGG CGGGT
4251 GGAAT GCCTT ATTGG CGAGC ATCTA CATGC TGGGA TGAGC ACACT TTTTC
4301 TGGTG TACAG CAATA AGTGT CAGAC TCCCC TGGGA ATGGC TTCTG GACAC
4351 ATTAG AGATT TTCAG ATTAC AGCTT CAGGA CAATA TGGAC AGTGG GCCCC
4401 AAAGC TGGCC AGACT TCATT ATTCC GGATC AATCA ATGCC TGGAG CACCA
4451 AGGAG CCCTT TTCTT GGATC AAGGT GGATC TGTTG GCACC AATGA TTATT
4501 CACGG CATCA AGACC CAGGG TGCCC GTCAG AAGTT CTCCA GCCTC TACAT
4551 CTCTC AGTTT ATCAT CATGT ATAGT CTTGA TGGGA AGAAG TGGCA GACTT
4601 ATCGA GGAAA TTCCA CTGGA ACCTT AATGG TCTTC TTTGG CAATG TGGAT
4651 TCATC TGGGA TAAAA CACAA TATTT TTAAC CCTCC AATTA TTGCT CGATA
4701 CATCC GTTTG CACCC AACTC ATTAT AGCAT TCGCA GCACT CTTCG CATGG
4751 AGTTG ATGGG CTGTG ATTTA AATAG TTGCA GCATG CCATT GGGAA TGGAG
4801 AGTAA AGCAA TATCA GATGC ACAGA TTACT GCTTC ATCCT ACTTT ACCAA
4851 TATGT TTGCC ACCTG GTCTC CTTCA AAAGC TCGAC TTCAC CTCCA AGGGA
4901 GGAGT AATGC CTGGA GACCT CAGGT GAATA ATCCA AAAGA GTGGC TGCAA
4951 GTGGA CTTCC AGAAG ACAAT GAAAG TCACA GGAGT AACTA CTCAG GGAGT
5001 AAAAT CTCTG CTTAC CAGCA TGTAT GTGAA GGAGT TCCTC ATCTC CAGCA
5051 GTCAA GATGG CCATC AGTGG ACTCT CTTTT TTCAG AATGG CAAAG TAAAG
5101 GTTTT TCAGG GAAAT CAAGA CTCCT TCACA CCTGT GGTGA ACTCT CTAGA
5151 CCCAC CGTTA CTGAC TCGCT ACCTT CGAAT TCACC CCCAG AGTTG GGTGC
5201 ACCAG ATTGC CCTGA GGATG GAGGT TCTGG GCTGC GAGGC ACAGG ACCTC
5251 TACGA CAAAA CTCAC ACATG CCCAC CGTGC CCAGC TCCAG AACTC CTGGG
5301 CGGAC CGTCA GTCTT CCTCT TCCCC CCAAA ACCCA AGGAC ACCCT CATGA
5351 TCTCC CGGAC CCCTG AGGTC ACATG CGTGG TGGTG GACGT GAGCC ACGAA
5401 GACCC TGAGG TCAAG TTCAA CTGGT ACGTG GACGG CGTGG AGGTG CATAA
5451 TGCCA AGACA AAGCC GCGGG AGGAG CAGTA CAACA GCACG TACCG TGTGG
5501 TCAGC GTCCT CACCG TCCTG CACCA GGACT GGCTG AATGG CAAGG AGTAC
5551 AAGTG CAAGG TCTCC AACAA AGCCC TCCCA GCCCC CATCG AGAAA ACCAT
5601 CTCCA AAGCC AAAGG GCAGC CCCGA GAACC ACAGG TGTAC ACCCT GCCCC
5651 CATCC CGGGA TGAGC TGACC AAGAA CCAGG TCAGC CTGAC CTGCC TGGTC
5701 AAAGG CTTCT ATCCC AGCGA CATCG CCGTG GAGTG GGAGA GCAAT GGGCA
5751 GCCGG AGAAC AACTA CAAGA CCACG CCTCC CGTGT TGGAC TCCGA CGGCT
5801 CCTTC TTCCT CTACA GCAAG CTCAC CGTGG ACAAG AGCAG GTGGC AGCAG
5851 GGGAA CGTCT TCTCA TGCTC CGTGA TGCAT GAGGC TCTGC ACAAC CACTA
5901 CACGC AGAAG AGCCT CTCCC TGTCT CCGGG TAAAT GA
Последовательность белка FVIII 169 (SEQ ID NO: 70)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
801 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
851 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
901 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
951 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1001 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1051 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1101 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1151 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1201 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1251 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1301 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1351 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1401 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1451 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1501 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1551 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1601 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1651 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1701 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1751 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE
1801 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY
1851 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
1901 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
1951 GNVFSCSVMH EALHNHYTQK SLSLSPGK*
Нуклеотидная последовательность FVIII 263 (с тройной мутацией IHH) (SEQ ID NO: 156)
1 ATGCA AATAG AGCTC TCCAC CTGCT TCTTT CTGTG CCTTT TGCGA TTCTG
51 CTTTA GTGCC ACCAG AAGAT ACTAC CTGGG TGCAG TGGAA CTGTC ATGGG
101 ACTAT ATGCA AGGCG CGCCA ACATC AGAGA GCGCC ACCCC TGAAA GTGGT
151 CCCGG GAGCG AGCCA GCCAC ATCTG GGTCG GAAAC GCCAG GCACA AGTGA
201 GTCTG CAACT CCCGA GTCCG GACCT GGCTC CGAGC CTGCC ACTAG CGGCT
251 CCGAG ACTCC GGGAA CTTCC GAGAG CGCTA CACCA GAAAG CGGAC CCGGA
301 ACCAG TACCG AACCT AGCGA GGGCT CTGCT CCGGG CAGCC CAGCC GGCTC
351 TCCTA CATCC ACGGA GGAGG GCACT TCCGA ATCCG CCACC CCGGA GTCAG
401 GGCCA GGATC TGAAC CCGCT ACCTC AGGCA GTGAG ACGCC AGGAA CGAGC
451 GAGTC CGCTA CACCG GAGAG TGGGC CAGGG AGCCC TGCTG GATCT CCTAC
501 GTCCA CTGAG GAAGG GTCAC CAGCG GGCTC GCCCA CCAGC ACTGA AGAAG
551 GTGCC TCGAG CAGTG ATCTC GGTGA GCTGC CTGTG GACGC AAGAT TTCCT
601 CCTAG AGTGC CAAAA TCTTT TCCAT TCAAC ACCTC AGTCG TGTAC AAAAA
651 GACTC TGTTT GTAGA ATTCA CGGAT CACCT TTTCA ACATC GCTAA GCCAA
701 GGCCA CCCTG GATGG GTCTG CTAGG TCCTA CCATC CAGGC TGAGG TTTAT
751 GATAC AGTGG TCATT ACACT TAAGA ACATG GCTTC CCATC CTGTC AGTCT
801 TCATG CTGTT GGTGT ATCCT ACTGG AAAGC TTCTG AGGGA GCTGA ATATG
851 ATGAT CAGAC CAGTC AAAGG GAGAA AGAAG ATGAT AAAGT CTTCC CTGGT
901 GGAAG CCATA CATAT GTCTG GCAGG TCCTG AAAGA GAATG GTCCA ATGGC
951 CTCTG ACCCA CTGTG CCTTA CCTAC TCATA TCTTT CTCAT GTGGA CCTGG
1001 TAAAA GACTT GAATT CAGGC CTCAT TGGAG CCCTA CTAGT ATGTA GAGAA
1051 GGGAG TCTGG CCAAG GAAAA GACAC AGACC TTGCA CAAAT TTATA CTACT
1101 TTTTG CTGTA TTTGA TGAAG GGAAA AGTTG GCACT CAGAA ACAAA GAACT
1151 CCTTG ATGCA GGATA GGGAT GCTGC ATCTG CTCGG GCCTG GCCTA AAATG
1201 CACAC AGTCA ATGGT TATGT AAACA GGTCT CTGCC AGGTC TGATT GGATG
1251 CCACA GGAAA TCAGT CTATT GGCAT GTGAT TGGAA TGGGC ACCAC TCCTG
1301 AAGTG CACTC AATAT TCCTC GAAGG TCACA CATTT CTTGT GAGGA ACCAT
1351 CGCCA GGCTA GCTTG GAAAT CTCGC CAATA ACTTT CCTTA CTGCT CAAAC
1401 ACTCT TGATG GACCT TGGAC AGTTT CTACT GTTTT GTCAT ATCTC TTCCC
1451 ACCAA CATGA TGGCA TGGAA GCTTA TGTCA AAGTA GACAG CTGTC CAGAG
1501 GAACC CCAAC TACGA ATGAA AAATA ATGAA GAAGC GGAAG ACTAT GATGA
1551 TGATC TTACT GATTC TGAAA TGGAT GTGGT CAGGT TTGAT GATGA CAACT
1601 CTCCT TCCTT TATCC AAATT CGCTC AGTTG CCAAG AAGCA TCCTA AAACT
1651 TGGGT ACATT ACATT GCTGC TGAAG AGGAG GACTG GGACT ATGCT CCCTT
1701 AGTCC TCGCC CCCGA TGACA GAAGT TATAA AAGTC AATAT TTGAA CAATG
1751 GCCCT CAGCG GATTG GTAGG AAGTA CAAAA AAGTC CGATT TATGG CATAC
1801 ACAGA TGAAA CCTTT AAGAC TCGTG AAGCT ATTCA GCATG AATCA GGAAT
1851 CTTGG GACCT TTACT TTATG GGGAA GTTGG AGACA CACTG TTGAT TATAT
1901 TTAAG AATCA AGCAA GCAGA CCATA TAACA TCTAC CCTCA CGGAA TCACT
1951 GATGT CCGTC CTTTG TATTC AAGGA GATTA CCAAA AGGTG TAAAA CATTT
2001 GAAGG ATTTT CCAAT TCTGC CAGGA GAAAT ATTCA AATAT AAATG GACAG
2051 TGACT GTAGA AGATG GGCCA ACTAA ATCAG ATCCT CGGTG CCTGA CCCGC
2101 TATTA CTCTA GTTTC GTTAA TATGG AGAGA GATCT AGCTT CAGGA CTCAT
2151 TGGCC CTCTC CTCAT CTGCT ACAAA GAATC TGTAG ATCAA AGAGG AAACC
2201 AGATA ATGTC AGACA AGAGG AATGT CATCC TGTTT TCTGT ATTTG ATGAG
2251 AACCG AAGCT GGTAC CTCAC AGAGA ATATA CAACG CTTTC TCCCC AATCC
2301 AGCTG GAGTG CAGCT TGAGG ATCCA GAGTT CCAAG CCTCC AACAT CATGC
2351 ACAGC ATCAA TGGCT ATGTT TTTGA TAGTT TGCAG TTGTC AGTTT GTTTG
2401 CATGA GGTGG CATAC TGGTA CATTC TAAGC ATTGG AGCAC AGACT GACTT
2451 CCTTT CTGTC TTCTT CTCTG GATAT ACCTT CAAAC ACAAA ATGGT CTATG
2501 AAGAC ACACT CACCC TATTC CCATT CTCAG GAGAA ACTGT CTTCA TGTCG
2551 ATGGA AAACC CAGGT CTATG GATTC TGGGG TGCCA CAACT CAGAC TTTCG
2601 GAACA GAGGC ATGAC CGCCT TACTG AAGGT TTCTA GTTGT GACAA GAACA
2651 CTGGT GATTA TTACG AGGAC AGTTA TGAAG ATATT TCAGC ATACT TGCTG
2701 AGTAA AAACA ATGCC ATTGA ACCAA GAAGC TTCTC TCAAA ACGGC GCGCC
2751 AGGTA CCTCA GAGTC TGCTA CCCCC GAGTC AGGGC CAGGA TCAGA GCCAG
2801 CCACC TCCGG GTCTG AGACA CCCGG GACTT CCGAG AGTGC CACCC CTGAG
2851 TCCGG ACCCG GGTCC GAGCC CGCCA CTTCC GGCTC CGAAA CTCCC GGCAC
2901 AAGCG AGAGC GCTAC CCCAG AGTCA GGACC AGGAA CATCT ACAGA GCCCT
2951 CTGAA GGCTC CGCTC CAGGG TCCCC AGCCG GCAGT CCCAC TAGCA CCGAG
3001 GAGGG AACCT CTGAA AGCGC CACAC CCGAA TCAGG GCCAG GGTCT GAGCC
3051 TGCTA CCAGC GGCAG CGAGA CACCA GGCAC CTCTG AGTCC GCCAC ACCAG
3101 AGTCC GGACC CGGAT CTCCC GCTGG GAGCC CCACC TCCAC TGAGG AGGGA
3151 TCTCC TGCTG GCTCT CCAAC ATCTA CTGAG GAAGG TACCT CAACC GAGCC
3201 ATCCG AGGGA TCAGC TCCCG GCACC TCAGA GTCGG CAACC CCGGA GTCTG
3251 GACCC GGAAC TTCCG AAAGT GCCAC ACCAG AGTCC GGTCC CGGGA CTTCA
3301 GAATC AGCAA CACCC GAGTC CGGCC CTGGG TCTGA ACCCG CCACA AGTGG
3351 TAGTG AGACA CCAGG ATCAG AACCT GCTAC CTCAG GGTCA GAGAC ACCCG
3401 GATCT CCGGC AGGCT CACCA ACCTC CACTG AGGAG GGCAC CAGCA CAGAA
3451 CCAAG CGAGG GCTCC GCACC CGGAA CAAGC ACTGA ACCCA GTGAG GGTTC
3501 AGCAC CCGGC TCTGA GCCGG CCACA AGTGG CAGTG AGACA CCCGG CACTT
3551 CAGAG AGTGC CACCC CCGAG AGTGG CCCAG GCACT AGTAC CGAGC CCTCT
3601 GAAGG CAGTG CGCCA GCCTC GAGCC CACCA GTCTT GAAAC GCCAT CAAGC
3651 TGAAA TAACT CGTAC TACTC TTCAG TCAGA TCAAG AGGAA ATCGA TTATG
3701 ATGAT ACCAT ATCAG TTGAA ATGAA GAAGG AAGAT TTTGA CATTT ATGAT
3751 GAGGA TGAAA ATCAG AGCCC CCGCA GCTTT CAAAA GAAAA CACGA CACTA
3801 TTTTA TTGCT GCAGT GGAGA GGCTC TGGGA TTATG GGATG AGTAG CTCCC
3851 CACAT GTTCT AAGAA ACAGG GCTCA GAGTG GCAGT GTCCC TCAGT TCAAG
3901 AAAGT TGTTT TCCAG GAATT TACTG ATGGC TCCTT TACTC AGCCC TTATA
3951 CCGTG GAGAA CTAAA TGAAC ATTTG GGACT CCTGG GGCCA TATAT AAGAG
4001 CAGAA GTTGA AGATA ATATC ATGGT AACTT TCAGA AATCA GGCCT CTCGT
4051 CCCTA TTCCT TCTAT TCTAG CCTTA TTTCT TATGA GGAAG ATCAG AGGCA
4101 AGGAG CAGAA CCTAG AAAAA ACTTT GTCAA GCCTA ATGAA ACCAA AACTT
4151 ACTTT TGGAA AGTGC AACAT CATAT GGCAC CCACT AAAGA TGAGT TTGAC
4201 TGCAA AGCCT GGGCT TATTT CTCTG ATGTT GACCT GGAAA AAGAT GTGCA
4251 CTCAG GCCTG ATTGG ACCCC TTCTG GTCTG CCACA CTAAC ACACT GAACC
4301 CTGCT CATGG GAGAC AAGTG ACAGT ACAGG AATTT GCTCT GTTTT TCACC
4351 ATCTT TGATG AGACC AAAAG CTGGT ACTTC ACTGA AAATA TGGAA AGAAA
4401 CTGCA GGGCT CCCTG CAATA TCCAG ATGGA AGATC CCACT TTTAA AGAGA
4451 ATTAT CGCTT CCATG CAATC AATGG CTACA TAATG GATAC ACTAC CTGGC
4501 TTAGT AATGG CTCAG GATCA AAGGA TTCGA TGGTA TCTGC TCAGC ATGGG
4551 CAGCA ATGAA AACAT CCATT CTATT CATTT CAGTG GACAT GTGTT CACTG
4601 TACGA AAAAA AGAGG AGTAT AAAAT GGCAC TGTAC AATCT CTATC CAGGT
4651 GTTTT TGAGA CAGTG GAAAT GTTAC CATCC AAAGC TGGAA TTTGG CGGGT
4701 GGAAT GCCTT ATTGG CGAGC ATCTA CATGC TGGGA TGAGC ACACT TTTTC
4751 TGGTG TACAG CAATA AGTGT CAGAC TCCCC TGGGA ATGGC TTCTG GACAC
4801 ATTAG AGATT TTCAG ATTAC AGCTT CAGGA CAATA TGGAC AGTGG GCCCC
4851 AAAGC TGGCC AGACT TCATT ATTCC GGATC AATCA ATGCC TGGAG CACCA
4901 AGGAG CCCTT TTCTT GGATC AAGGT GGATC TGTTG GCACC AATGA TTATT
4951 CACGG CATCA AGACC CAGGG TGCCC GTCAG AAGTT CTCCA GCCTC TACAT
5001 CTCTC AGTTT ATCAT CATGT ATAGT CTTGA TGGGA AGAAG TGGCA GACTT
5051 ATCGA GGAAA TTCCA CTGGA ACCTT AATGG TCTTC TTTGG CAATG TGGAT
5101 TCATC TGGGA TAAAA CACAA TATTT TTAAC CCTCC AATTA TTGCT CGATA
5151 CATCC GTTTG CACCC AACTC ATTAT AGCAT TCGCA GCACT CTTCG CATGG
5201 AGTTG ATGGG CTGTG ATTTA AATAG TTGCA GCATG CCATT GGGAA TGGAG
5251 AGTAA AGCAA TATCA GATGC ACAGA TTACT GCTTC ATCCT ACTTT ACCAA
5301 TATGT TTGCC ACCTG GTCTC CTTCA AAAGC TCGAC TTCAC CTCCA AGGGA
5351 GGAGT AATGC CTGGA GACCT CAGGT GAATA ATCCA AAAGA GTGGC TGCAA
5401 GTGGA CTTCC AGAAG ACAAT GAAAG TCACA GGAGT AACTA CTCAG GGAGT
5451 AAAAT CTCTG CTTAC CAGCA TGTAT GTGAA GGAGT TCCTC ATCTC CAGCA
5501 GTCAA GATGG CCATC AGTGG ACTCT CTTTT TTCAG AATGG CAAAG TAAAG
5551 GTTTT TCAGG GAAAT CAAGA CTCCT TCACA CCTGT GGTGA ACTCT CTAGA
5601 CCCAC CGTTA CTGAC TCGCT ACCTT CGAAT TCACC CCCAG AGTTG GGTGC
5651 ACCAG ATTGC CCTGA GGATG GAGGT TCTGG GCTGC GAGGC ACAGG ACCTC
5701 TACGA CAAAA CTCAC ACATG CCCAC CGTGC CCAGC TCCAG AACTC CTGGG
5751 CGGAC CGTCA GTCTT CCTCT TCCCC CCAAA ACCCA AGGAC ACCCT CATGG
5801 CCTCC CGGAC CCCTG AGGTC ACATG CGTGG TGGTG GACGT GAGCC ACGAA
5851 GACCC TGAGG TCAAG TTCAA CTGGT ACGTG GACGG CGTGG AGGTG CATAA
5901 TGCCA AGACA AAGCC GCGGG AGGAG CAGTA CAACA GCACG TACCG TGTGG
5951 TCAGC GTCCT CACCG TCCTG GCCCA GGACT GGCTG AATGG CAAGG AGTAC
6001 AAGTG CAAGG TCTCC AACAA AGCCC TCCCA GCCCC CATCG AGAAA ACCAT
6051 CTCCA AAGCC AAAGG GCAGC CCCGA GAACC ACAGG TGTAC ACCCT GCCCC
6101 CATCC CGCGA TGAGC TGACC AAGAA CCAGG TCAGC CTGAC CTGCC TGGTC
6151 AAAGG CTTCT ATCCC AGCGA CATCG CCGTG GAGTG GGAGA GCAAT GGGCA
6201 GCCGG AGAAC AACTA CAAGA CCACG CCTCC CGTGT TGGAC TCCGA CGGCT
6251 CCTTC TTCCT CTACA GCAAG CTCAC CGTGG ACAAG AGCAG GTGGC AGCAG
6301 GGGAA CGTCT TCTCA TGCTC CGTGA TGCAT GAGGC TCTGC ACAAC GCCTA
6351 CACGC AGAAG AGCCT CTCCC TGTCT CCGGG TAAAT GA
Последовательность белка FVIII 263 (с тройной мутацией IHH) (SEQ ID NO: 157)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS
151 ESATPESGPG SPAGSPTSTE EGSPAGSPTS TEEGASSSDL GELPVDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
951 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
1001 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
1051 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
1101 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1151 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1201 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1251 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1301 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1351 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1401 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1451 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1501 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1551 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1601 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1651 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1701 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1751 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1801 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1851 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1901 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMASRTPEV TCVVVDVSHE
1951 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL AQDWLNGKEY
2001 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
2051 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
2101 GNVFSCSVMH EALHNAYTQK SLSLSPGK*
Нуклеотидная последовательность FVIII 282 (SEQ ID NO: 158)
1 ATGCA AATAG AGCTC TCCAC CTGCT TCTTT CTGTG CCTTT TGCGA TTCTG
51 CTTTA GTGCC ACCAG AAGAT ACTAC CTGGG TGCAG TGGAA CTGTC ATGGG
101 ACTAT ATGCA AAGTG ATCTC GGTGA GCTGC CTGTG GACGC AAGAT TTCCT
151 CCTAG AGTGC CAAAA TCTTT TCCAT TCAAC ACCTC AGTCG TGTAC AAAAA
201 GACTC TGTTT GTAGA ATTCA CGGAT CACCT TTTCA ACATC GCTAA GCCAA
251 GGCCA CCCTG GATGG GTCTG CTAGG TCCTA CCATC CAGGC TGAGG TTTAT
301 GATAC AGTGG TCATT ACACT TAAGA ACATG GCTTC CCATC CTGTC AGTCT
351 TCATG CTGTT GGTGT ATCCT ACTGG AAAGC TTCTG AGGGA GCTGA ATATG
401 ATGAT CAGAC CAGTC AAAGG GAGAA AGAAG ATGAT AAAGT CTTCC CTGGT
451 GGAAG CCATA CATAT GTCTG GCAGG TCCTG AAAGA GAATG GTCCA ATGGC
501 CTCTG ACCCA CTGTG CCTTA CCTAC TCATA TCTTT CTCAT GTGGA CCTGG
551 TAAAA GACTT GAATT CAGGC CTCAT TGGAG CCCTA CTAGT ATGTA GAGAA
601 GGGAG TCTGG CCAAG GAAAA GACAC AGACC TTGCA CAAAT TTATA CTACT
651 TTTTG CTGTA TTTGA TGAAG GGAAA AGTTG GCACT CAGAA ACAAA GAACT
701 CCTTG ATGCA GGATA GGGAT GCTGC ATCTG CTCGG GCCTG GCCTA AAATG
751 CACAC AGTCA ATGGT TATGT AAACA GGTCT CTGCC AGGTC TGATT GGATG
801 CCACA GGAAA TCAGT CTATT GGCAT GTGAT TGGAA TGGGC ACCAC TCCTG
851 AAGTG CACTC AATAT TCCTC GAAGG TCACA CATTT CTTGT GAGGA ACCAT
901 CGCCA GGCTA GCTTG GAAAT CTCGC CAATA ACTTT CCTTA CTGCT CAAAC
951 ACTCT TGATG GACCT TGGAC AGTTT CTACT GTTTT GTCAT ATCTC TTCCC
1001 ACCAA CATGA TGGCA TGGAA GCTTA TGTCA AAGTA GACAG CTGTC CAGAG
1051 GAACC CCAAC TACGA ATGAA AAATA ATGAA GAAGC GGAAG ACTAT GATGA
1101 TGATC TTACT GATTC TGAAA TGGAT GTGGT CAGGT TTGAT GATGA CAACT
1151 CTCCT TCCTT TATCC AAATT CGCTC AGTTG CCAAG AAGCA TCCTA AAACT
1201 TGGGT ACATT ACATT GCTGC TGAAG AGGAG GACTG GGACT ATGCT CCCTT
1251 AGTCC TCGCC CCCGA TGACA GAAGT TATAA AAGTC AATAT TTGAA CAATG
1301 GCCCT CAGCG GATTG GTAGG AAGTA CAAAA AAGTC CGATT TATGG CATAC
1351 ACAGA TGAAA CCTTT AAGAC TCGTG AAGCT ATTCA GCATG AATCA GGAAT
1401 CTTGG GACCT TTACT TTATG GGGAA GTTGG AGACA CACTG TTGAT TATAT
1451 TTAAG AATCA AGCAA GCAGA CCATA TAACA TCTAC CCTCA CGGAA TCACT
1501 GATGT CCGTC CTTTG TATTC AAGGA GATTA CCAAA AGGTG TAAAA CATTT
1551 GAAGG ATTTT CCAAT TCTGC CAGGA GAAAT ATTCA AATAT AAATG GACAG
1601 TGACT GTAGA AGATG GGCCA ACTAA ATCAG ATCCT CGGTG CCTGA CCCGC
1651 TATTA CTCTA GTTTC GTTAA TATGG AGAGA GATCT AGCTT CAGGA CTCAT
1701 TGGCC CTCTC CTCAT CTGCT ACAAA GAATC TGTAG ATCAA AGAGG AAACC
1751 AGATA ATGTC AGACA AGAGG AATGT CATCC TGTTT TCTGT ATTTG ATGAG
1801 AACCG AAGCT GGTAC CTCAC AGAGA ATATA CAACG CTTTC TCCCC AATCC
1851 AGCTG GAGTG CAGCT TGAGG ATCCA GAGTT CCAAG CCTCC AACAT CATGC
1901 ACAGC ATCAA TGGCT ATGTT TTTGA TAGTT TGCAG TTGTC AGTTT GTTTG
1951 CATGA GGTGG CATAC TGGTA CATTC TAAGC ATTGG AGCAC AGACT GACTT
2001 CCTTT CTGTC TTCTT CTCTG GATAT ACCTT CAAAC ACAAA ATGGT CTATG
2051 AAGAC ACACT CACCC TATTC CCATT CTCAG GAGAA ACTGT CTTCA TGTCG
2101 ATGGA AAACC CAGGT CTATG GATTC TGGGG TGCCA CAACT CAGAC TTTCG
2151 GAACA GAGGC ATGAC CGCCT TACTG AAGGT TTCTA GTTGT GACAA GAACA
2201 CTGGT GATTA TTACG AGGAC AGTTA TGAAG ATATT TCAGC ATACT TGCTG
2251 AGTAA AAACA ATGCC ATTGA ACCAA GAAGC TTCTC TCAAA ACGGC GCGCC
2301 AACAT CAGAG AGCGC CACCC CTGAA AGTGG TCCCG GGAGC GAGCC AGCCA
2351 CATCT GGGTC GGAAA CGCCA GGCAC AAGTG AGTCT GCAAC TCCCG AGTCC
2401 GGACC TGGCT CCGAG CCTGC CACTA GCGGC TCCGA GACTC CGGGA ACTTC
2451 CGAGA GCGCT ACACC AGAAA GCGGA CCCGG AACCA GTACC GAACC TAGCG
2501 AGGGC TCTGC TCCGG GCAGC CCAGC CGGCT CTCCT ACATC CACGG AGGAG
2551 GGCAC TTCCG AATCC GCCAC CCCGG AGTCA GGGCC AGGAT CTGAA CCCGC
2601 TACCT CAGGC AGTGA GACGC CAGGA ACGAG CGAGT CCGCT ACACC GGAGA
2651 GTGGG CCAGG GAGCC CTGCT GGATC TCCTA CGTCC ACTGA GGAAG GGTCA
2701 CCAGC GGGCT CGCCC ACCAG CACTG AAGAA GGTGC CTCGA GCCCA CCAGT
2751 CTTGA AACGC CATCA AGCTG AAATA ACTCG TACTA CTCTT CAGTC AGATC
2801 AAGAG GAAAT CGATT ATGAT GATAC CATAT CAGTT GAAAT GAAGA AGGAA
2851 GATTT TGACA TTTAT GATGA GGATG AAAAT CAGAG CCCCC GCAGC TTTCA
2901 AAAGA AAACA CGACA CTATT TTATT GCTGC AGTGG AGAGG CTCTG GGATT
2951 ATGGG ATGAG TAGCT CCCCA CATGT TCTAA GAAAC AGGGC TCAGA GTGGC
3001 AGTGT CCCTC AGTTC AAGAA AGTTG TTTTC CAGGA ATTTA CTGAT GGCTC
3051 CTTTA CTCAG CCCTT ATACC GTGGA GAACT AAATG AACAT TTGGG ACTCC
3101 TGGGG CCATA TATAA GAGCA GAAGT TGAAG ATAAT ATCAT GGTAA CTTTC
3151 AGAAA TCAGG CCTCT CGTCC CTATT CCTTC TATTC TAGCC TTATT TCTTA
3201 TGAGG AAGAT CAGAG GCAAG GAGCA GAACC TAGAA AAAAC TTTGT CAAGC
3251 CTAAT GAAAC CAAAA CTTAC TTTTG GAAAG TGCAA CATCA TATGG CACCC
3301 ACTAA AGATG AGTTT GACTG CAAAG CCTGG GCTTA TTTCT CTGAT GTTGA
3351 CCTGG AAAAA GATGT GCACT CAGGC CTGAT TGGAC CCCTT CTGGT CTGCC
3401 ACACT AACAC ACTGA ACCCT GCTCA TGGGA GACAA GTGAC AGTAC AGGAA
3451 TTTGC TCTGT TTTTC ACCAT CTTTG ATGAG ACCAA AAGCT GGTAC TTCAC
3501 TGAAA ATATG GAAAG AAACT GCAGG GCTCC CTGCA ATATC CAGAT GGAAG
3551 ATCCC ACTTT TAAAG AGAAT TATCG CTTCC ATGCA ATCAA TGGCT ACATA
3601 ATGGA TACAC TACCT GGCTT AGTAA TGGCT CAGGA TCAAA GGATT CGATG
3651 GTATC TGCTC AGCAT GGGCA GCAAT GAAAA CATCC ATTCT ATTCA TTTCA
3701 GTGGA CATGT GTTCA CTGTA CGAAA AAAAG AGGAG TATAA AATGG CACTG
3751 TACAA TCTCT ATCCA GGTGT TTTTG AGACA GTGGA AATGT TACCA TCCAA
3801 AGCTG GAATT TGGCG GGTGG AATGC CTTAT TGGCG AGCAT CTACA TGCTG
3851 GGATG AGCAC ACTTT TTCTG GTGTA CAGCA ATAAG TGTCA GACTC CCCTG
3901 GGAAT GGCTT CTGGA CACAT TAGAG ATTTT CAGAT TACAG CTTCA GGACA
3951 ATATG GACAG TGGGC CCCAA AGCTG GCCAG ACTTC ATTAT TCCGG ATCAA
4001 TCAAT GCCTG GAGCA CCAAG GAGCC CTTTT CTTGG ATCAA GGTGG ATCTG
4051 TTGGC ACCAA TGATT ATTCA CGGCA TCAAG ACCCA GGGTG CCCGT CAGAA
4101 GTTCT CCAGC CTCTA CATCT CTCAG TTTAT CATCA TGTAT AGTCT TGATG
4151 GGAAG AAGTG GCAGA CTTAT CGAGG AAATT CCACT GGAAC CTTAA TGGTC
4201 TTCTT TGGCA ATGTG GATTC ATCTG GGATA AAACA CAATA TTTTT AACCC
4251 TCCAA TTATT GCTCG ATACA TCCGT TTGCA CCCAA CTCAT TATAG CATTC
4301 GCAGC ACTCT TCGCA TGGAG TTGAT GGGCT GTGAT TTAAA TAGTT GCAGC
4351 ATGCC ATTGG GAATG GAGAG TAAAG CAATA TCAGA TGCAC AGATT ACTGC
4401 TTCAT CCTAC TTTAC CAATA TGTTT GCCAC CTGGT CTCCT TCAAA AGCTC
4451 GACTT CACCT CCAAG GGAGG AGTAA TGCCT GGAGA CCTCA GGTGA ATAAT
4501 CCAAA AGAGT GGCTG CAAGT GGACT TCCAG AAGAC AATGA AAGTC ACAGG
4551 AGTAA CTACT CAGGG AGTAA AATCT CTGCT TACCA GCATG TATGT GAAGG
4601 AGTTC CTCAT CTCCA GCAGT CAAGA TGGCC ATCAG TGGAC TCTCT TTTTT
4651 CAGAA TGGCA AAGTA AAGGT TTTTC AGGGA AATCA AGACT CCTTC ACACC
4701 TGTGG TGAAC TCTCT AGACC CACCG TTACT GACTC GCTAC CTTCG AATTC
4751 ACCCC CAGAG TTGGG TGCAC CAGAT TGCCC TGAGG ATGGA GGTTC TGGGC
4801 TGCGA GGCAC AGGAC CTCTA CGACA AAACT CACAC ATGCC CACCG TGCCC
4851 AGCTC CAGAA CTCCT GGGCG GACCG TCAGT CTTCC TCTTC CCCCC AAAAC
4901 CCAAG GACAC CCTCA TGATC TCCCG GACCC CTGAG GTCAC ATGCG TGGTG
4951 GTGGA CGTGA GCCAC GAAGA CCCTG AGGTC AAGTT CAACT GGTAC GTGGA
5001 CGGCG TGGAG GTGCA TAATG CCAAG ACAAA GCCGC GGGAG GAGCA GTACA
5051 ACAGC ACGTA CCGTG TGGTC AGCGT CCTCA CCGTC CTGCA CCAGG ACTGG
5101 CTGAA TGGCA AGGAG TACAA GTGCA AGGTC TCCAA CAAAG CCCTC CCAGC
5151 CCCCA TCGAG AAAAC CATCT CCAAA GCCAA AGGGC AGCCC CGAGA ACCAC
5201 AGGTG TACAC CCTGC CCCCA TCCCG GGATG AGCTG ACCAA GAACC AGGTC
5251 AGCCT GACCT GCCTG GTCAA AGGCT TCTAT CCCAG CGACA TCGCC GTGGA
5301 GTGGG AGAGC AATGG GCAGC CGGAG AACAA CTACA AGACC ACGCC TCCCG
5351 TGTTG GACTC CGACG GCTCC TTCTT CCTCT ACAGC AAGCT CACCG TGGAC
5401 AAGAG CAGGT GGCAG CAGGG GAACG TCTTC TCATG CTCCG TGATG CATGA
5451 GGCTC TGCAC AACCA CTACA CGCAG AAGAG CCTCT CCCTG TCTCC GGGTA
5501 AATGA
Последовательность белка FVIII 282 (SEQ ID NO: 159)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNGAPTSE SATPESGPGS EPATSGSETP GTSESATPES
801 GPGSEPATSG SETPGTSESA TPESGPGTST EPSEGSAPGS PAGSPTSTEE
851 GTSESATPES GPGSEPATSG SETPGTSESA TPESGPGSPA GSPTSTEEGS
901 PAGSPTSTEE GASSPPVLKR HQAEITRTTL QSDQEEIDYD DTISVEMKKE
951 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1001 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1051 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1101 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1151 FALFFTIFDE TKSWYFTENM ERNCRAPCNI QMEDPTFKEN YRFHAINGYI
1201 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1251 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1301 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1351 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1401 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1451 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1501 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1551 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1601 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1651 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1701 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1751 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1801 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Нуклеотидная последовательность FVIII 283 (FVIII 169 с тройной мутацией IHH) (SEQ ID NO: 160)
1 ATGCA AATAG AGCTC TCCAC CTGCT TCTTT CTGTG CCTTT TGCGA TTCTG
51 CTTTA GTGCC ACCAG AAGAT ACTAC CTGGG TGCAG TGGAA CTGTC ATGGG
101 ACTAT ATGCA AAGTG ATCTC GGTGA GCTGC CTGTG GACGC AAGAT TTCCT
151 CCTAG AGTGC CAAAA TCTTT TCCAT TCAAC ACCTC AGTCG TGTAC AAAAA
201 GACTC TGTTT GTAGA ATTCA CGGAT CACCT TTTCA ACATC GCTAA GCCAA
251 GGCCA CCCTG GATGG GTCTG CTAGG TCCTA CCATC CAGGC TGAGG TTTAT
301 GATAC AGTGG TCATT ACACT TAAGA ACATG GCTTC CCATC CTGTC AGTCT
351 TCATG CTGTT GGTGT ATCCT ACTGG AAAGC TTCTG AGGGA GCTGA ATATG
401 ATGAT CAGAC CAGTC AAAGG GAGAA AGAAG ATGAT AAAGT CTTCC CTGGT
451 GGAAG CCATA CATAT GTCTG GCAGG TCCTG AAAGA GAATG GTCCA ATGGC
501 CTCTG ACCCA CTGTG CCTTA CCTAC TCATA TCTTT CTCAT GTGGA CCTGG
551 TAAAA GACTT GAATT CAGGC CTCAT TGGAG CCCTA CTAGT ATGTA GAGAA
601 GGGAG TCTGG CCAAG GAAAA GACAC AGACC TTGCA CAAAT TTATA CTACT
651 TTTTG CTGTA TTTGA TGAAG GGAAA AGTTG GCACT CAGAA ACAAA GAACT
701 CCTTG ATGCA GGATA GGGAT GCTGC ATCTG CTCGG GCCTG GCCTA AAATG
751 CACAC AGTCA ATGGT TATGT AAACA GGTCT CTGCC AGGTC TGATT GGATG
801 CCACA GGAAA TCAGT CTATT GGCAT GTGAT TGGAA TGGGC ACCAC TCCTG
851 AAGTG CACTC AATAT TCCTC GAAGG TCACA CATTT CTTGT GAGGA ACCAT
901 CGCCA GGCTA GCTTG GAAAT CTCGC CAATA ACTTT CCTTA CTGCT CAAAC
951 ACTCT TGATG GACCT TGGAC AGTTT CTACT GTTTT GTCAT ATCTC TTCCC
1001 ACCAA CATGA TGGCA TGGAA GCTTA TGTCA AAGTA GACAG CTGTC CAGAG
1051 GAACC CCAAC TACGA ATGAA AAATA ATGAA GAAGC GGAAG ACTAT GATGA
1101 TGATC TTACT GATTC TGAAA TGGAT GTGGT CAGGT TTGAT GATGA CAACT
1151 CTCCT TCCTT TATCC AAATT CGCTC AGTTG CCAAG AAGCA TCCTA AAACT
1201 TGGGT ACATT ACATT GCTGC TGAAG AGGAG GACTG GGACT ATGCT CCCTT
1251 AGTCC TCGCC CCCGA TGACA GAAGT TATAA AAGTC AATAT TTGAA CAATG
1301 GCCCT CAGCG GATTG GTAGG AAGTA CAAAA AAGTC CGATT TATGG CATAC
1351 ACAGA TGAAA CCTTT AAGAC TCGTG AAGCT ATTCA GCATG AATCA GGAAT
1401 CTTGG GACCT TTACT TTATG GGGAA GTTGG AGACA CACTG TTGAT TATAT
1451 TTAAG AATCA AGCAA GCAGA CCATA TAACA TCTAC CCTCA CGGAA TCACT
1501 GATGT CCGTC CTTTG TATTC AAGGA GATTA CCAAA AGGTG TAAAA CATTT
1551 GAAGG ATTTT CCAAT TCTGC CAGGA GAAAT ATTCA AATAT AAATG GACAG
1601 TGACT GTAGA AGATG GGCCA ACTAA ATCAG ATCCT CGGTG CCTGA CCCGC
1651 TATTA CTCTA GTTTC GTTAA TATGG AGAGA GATCT AGCTT CAGGA CTCAT
1701 TGGCC CTCTC CTCAT CTGCT ACAAA GAATC TGTAG ATCAA AGAGG AAACC
1751 AGATA ATGTC AGACA AGAGG AATGT CATCC TGTTT TCTGT ATTTG ATGAG
1801 AACCG AAGCT GGTAC CTCAC AGAGA ATATA CAACG CTTTC TCCCC AATCC
1851 AGCTG GAGTG CAGCT TGAGG ATCCA GAGTT CCAAG CCTCC AACAT CATGC
1901 ACAGC ATCAA TGGCT ATGTT TTTGA TAGTT TGCAG TTGTC AGTTT GTTTG
1951 CATGA GGTGG CATAC TGGTA CATTC TAAGC ATTGG AGCAC AGACT GACTT
2001 CCTTT CTGTC TTCTT CTCTG GATAT ACCTT CAAAC ACAAA ATGGT CTATG
2051 AAGAC ACACT CACCC TATTC CCATT CTCAG GAGAA ACTGT CTTCA TGTCG
2101 ATGGA AAACC CAGGT CTATG GATTC TGGGG TGCCA CAACT CAGAC TTTCG
2151 GAACA GAGGC ATGAC CGCCT TACTG AAGGT TTCTA GTTGT GACAA GAACA
2201 CTGGT GATTA TTACG AGGAC AGTTA TGAAG ATATT TCAGC ATACT TGCTG
2251 AGTAA AAACA ATGCC ATTGA ACCAA GAAGC TTCTC TCAAA ACGGC GCGCC
2301 AGGTA CCTCA GAGTC TGCTA CCCCC GAGTC AGGGC CAGGA TCAGA GCCAG
2351 CCACC TCCGG GTCTG AGACA CCCGG GACTT CCGAG AGTGC CACCC CTGAG
2401 TCCGG ACCCG GGTCC GAGCC CGCCA CTTCC GGCTC CGAAA CTCCC GGCAC
2451 AAGCG AGAGC GCTAC CCCAG AGTCA GGACC AGGAA CATCT ACAGA GCCCT
2501 CTGAA GGCTC CGCTC CAGGG TCCCC AGCCG GCAGT CCCAC TAGCA CCGAG
2551 GAGGG AACCT CTGAA AGCGC CACAC CCGAA TCAGG GCCAG GGTCT GAGCC
2601 TGCTA CCAGC GGCAG CGAGA CACCA GGCAC CTCTG AGTCC GCCAC ACCAG
2651 AGTCC GGACC CGGAT CTCCC GCTGG GAGCC CCACC TCCAC TGAGG AGGGA
2701 TCTCC TGCTG GCTCT CCAAC ATCTA CTGAG GAAGG TACCT CAACC GAGCC
2751 ATCCG AGGGA TCAGC TCCCG GCACC TCAGA GTCGG CAACC CCGGA GTCTG
2801 GACCC GGAAC TTCCG AAAGT GCCAC ACCAG AGTCC GGTCC CGGGA CTTCA
2851 GAATC AGCAA CACCC GAGTC CGGCC CTGGG TCTGA ACCCG CCACA AGTGG
2901 TAGTG AGACA CCAGG ATCAG AACCT GCTAC CTCAG GGTCA GAGAC ACCCG
2951 GATCT CCGGC AGGCT CACCA ACCTC CACTG AGGAG GGCAC CAGCA CAGAA
3001 CCAAG CGAGG GCTCC GCACC CGGAA CAAGC ACTGA ACCCA GTGAG GGTTC
3051 AGCAC CCGGC TCTGA GCCGG CCACA AGTGG CAGTG AGACA CCCGG CACTT
3101 CAGAG AGTGC CACCC CCGAG AGTGG CCCAG GCACT AGTAC CGAGC CCTCT
3151 GAAGG CAGTG CGCCA GCCTC GAGCC CACCA GTCTT GAAAC GCCAT CAAGC
3201 TGAAA TAACT CGTAC TACTC TTCAG TCAGA TCAAG AGGAA ATCGA TTATG
3251 ATGAT ACCAT ATCAG TTGAA ATGAA GAAGG AAGAT TTTGA CATTT ATGAT
3301 GAGGA TGAAA ATCAG AGCCC CCGCA GCTTT CAAAA GAAAA CACGA CACTA
3351 TTTTA TTGCT GCAGT GGAGA GGCTC TGGGA TTATG GGATG AGTAG CTCCC
3401 CACAT GTTCT AAGAA ACAGG GCTCA GAGTG GCAGT GTCCC TCAGT TCAAG
3451 AAAGT TGTTT TCCAG GAATT TACTG ATGGC TCCTT TACTC AGCCC TTATA
3501 CCGTG GAGAA CTAAA TGAAC ATTTG GGACT CCTGG GGCCA TATAT AAGAG
3551 CAGAA GTTGA AGATA ATATC ATGGT AACTT TCAGA AATCA GGCCT CTCGT
3601 CCCTA TTCCT TCTAT TCTAG CCTTA TTTCT TATGA GGAAG ATCAG AGGCA
3651 AGGAG CAGAA CCTAG AAAAA ACTTT GTCAA GCCTA ATGAA ACCAA AACTT
3701 ACTTT TGGAA AGTGC AACAT CATAT GGCAC CCACT AAAGA TGAGT TTGAC
3751 TGCAA AGCCT GGGCT TATTT CTCTG ATGTT GACCT GGAAA AAGAT GTGCA
3801 CTCAG GCCTG ATTGG ACCCC TTCTG GTCTG CCACA CTAAC ACACT GAACC
3851 CTGCT CATGG GAGAC AAGTG ACAGT ACAGG AATTT GCTCT GTTTT TCACC
3901 ATCTT TGATG AGACC AAAAG CTGGT ACTTC ACTGA AAATA TGGAA AGAAA
3951 CTGCA GGGCT CCCTG CAATA TCCAG ATGGA AGATC CCACT TTTAA AGAGA
4001 ATTAT CGCTT CCATG CAATC AATGG CTACA TAATG GATAC ACTAC CTGGC
4051 TTAGT AATGG CTCAG GATCA AAGGA TTCGA TGGTA TCTGC TCAGC ATGGG
4101 CAGCA ATGAA AACAT CCATT CTATT CATTT CAGTG GACAT GTGTT CACTG
4151 TACGA AAAAA AGAGG AGTAT AAAAT GGCAC TGTAC AATCT CTATC CAGGT
4201 GTTTT TGAGA CAGTG GAAAT GTTAC CATCC AAAGC TGGAA TTTGG CGGGT
4251 GGAAT GCCTT ATTGG CGAGC ATCTA CATGC TGGGA TGAGC ACACT TTTTC
4301 TGGTG TACAG CAATA AGTGT CAGAC TCCCC TGGGA ATGGC TTCTG GACAC
4351 ATTAG AGATT TTCAG ATTAC AGCTT CAGGA CAATA TGGAC AGTGG GCCCC
4401 AAAGC TGGCC AGACT TCATT ATTCC GGATC AATCA ATGCC TGGAG CACCA
4451 AGGAG CCCTT TTCTT GGATC AAGGT GGATC TGTTG GCACC AATGA TTATT
4501 CACGG CATCA AGACC CAGGG TGCCC GTCAG AAGTT CTCCA GCCTC TACAT
4551 CTCTC AGTTT ATCAT CATGT ATAGT CTTGA TGGGA AGAAG TGGCA GACTT
4601 ATCGA GGAAA TTCCA CTGGA ACCTT AATGG TCTTC TTTGG CAATG TGGAT
4651 TCATC TGGGA TAAAA CACAA TATTT TTAAC CCTCC AATTA TTGCT CGATA
4701 CATCC GTTTG CACCC AACTC ATTAT AGCAT TCGCA GCACT CTTCG CATGG
4751 AGTTG ATGGG CTGTG ATTTA AATAG TTGCA GCATG CCATT GGGAA TGGAG
4801 AGTAA AGCAA TATCA GATGC ACAGA TTACT GCTTC ATCCT ACTTT ACCAA
4851 TATGT TTGCC ACCTG GTCTC CTTCA AAAGC TCGAC TTCAC CTCCA AGGGA
4901 GGAGT AATGC CTGGA GACCT CAGGT GAATA ATCCA AAAGA GTGGC TGCAA
4951 GTGGA CTTCC AGAAG ACAAT GAAAG TCACA GGAGT AACTA CTCAG GGAGT
5001 AAAAT CTCTG CTTAC CAGCA TGTAT GTGAA GGAGT TCCTC ATCTC CAGCA
5051 GTCAA GATGG CCATC AGTGG ACTCT CTTTT TTCAG AATGG CAAAG TAAAG
5101 GTTTT TCAGG GAAAT CAAGA CTCCT TCACA CCTGT GGTGA ACTCT CTAGA
5151 CCCAC CGTTA CTGAC TCGCT ACCTT CGAAT TCACC CCCAG AGTTG GGTGC
5201 ACCAG ATTGC CCTGA GGATG GAGGT TCTGG GCTGC GAGGC ACAGG ACCTC
5251 TACGA CAAAA CTCAC ACATG CCCAC CGTGC CCAGC TCCAG AACTC CTGGG
5301 CGGAC CGTCA GTCTT CCTCT TCCCC CCAAA ACCCA AGGAC ACCCT CATGG
5351 CCTCC CGGAC CCCTG AGGTC ACATG CGTGG TGGTG GACGT GAGCC ACGAA
5401 GACCC TGAGG TCAAG TTCAA CTGGT ACGTG GACGG CGTGG AGGTG CATAA
5451 TGCCA AGACA AAGCC GCGGG AGGAG CAGTA CAACA GCACG TACCG TGTGG
5501 TCAGC GTCCT CACCG TCCTG GCCCA GGACT GGCTG AATGG CAAGG AGTAC
5551 AAGTG CAAGG TCTCC AACAA AGCCC TCCCA GCCCC CATCG AGAAA ACCAT
5601 CTCCA AAGCC AAAGG GCAGC CCCGA GAACC ACAGG TGTAC ACCCT GCCCC
5651 CATCC CGGGA TGAGC TGACC AAGAA CCAGG TCAGC CTGAC CTGCC TGGTC
5701 AAAGG CTTCT ATCCC AGCGA CATCG CCGTG GAGTG GGAGA GCAAT GGGCA
5751 GCCGG AGAAC AACTA CAAGA CCACG CCTCC CGTGT TGGAC TCCGA CGGCT
5801 CCTTC TTCCT CTACA GCAAG CTCAC CGTGG ACAAG AGCAG GTGGC AGCAG
5851 GGGAA CGTCT TCTCA TGCTC CGTGA TGCAT GAGGC TCTGC ACAAC GCCTA
5901 CACGC AGAAG AGCCT CTCCC TGTCT CCGGG TAAAT GA
Последовательность белка FVIII 283 (FVIII 169 с тройной мутацией IHH) (SEQ ID NO: 161)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
801 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
851 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
901 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
951 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1001 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1051 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1101 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1151 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1201 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1251 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1301 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1351 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1401 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1451 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1501 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1551 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1601 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1651 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1701 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1751 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMASRTPEV TCVVVDVSHE
1801 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL AQDWLNGKEY
1851 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
1901 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
1951 GNVFSCSVMH EALHNAYTQK SLSLSPGK*
Нуклеотидная последовательность pSYNFVIII 010 (двухцепочечный FVIIIFc) (SEQ ID NO: 162)
1 ATGCAAATAG AGCTCTCCAC CTGCTTCTTT CTGTGCCTTT TGCGATTCTG
51 CTTTAGTGCC ACCAGAAGAT ACTACCTGGG TGCAGTGGAA CTGTCATGGG
101 ACTATATGCA AAGTGATCTC GGTGAGCTGC CTGTGGACGC AAGATTTCCT
151 CCTAGAGTGC CAAAATCTTT TCCATTCAAC ACCTCAGTCG TGTACAAAAA
201 GACTCTGTTT GTAGAATTCA CGGATCACCT TTTCAACATC GCTAAGCCAA
251 GGCCACCCTG GATGGGTCTG CTAGGTCCTA CCATCCAGGC TGAGGTTTAT
301 GATACAGTGG TCATTACACT TAAGAACATG GCTTCCCATC CTGTCAGTCT
351 TCATGCTGTT GGTGTATCCT ACTGGAAAGC TTCTGAGGGA GCTGAATATG
401 ATGATCAGAC CAGTCAAAGG GAGAAAGAAG ATGATAAAGT CTTCCCTGGT
451 GGAAGCCATA CATATGTCTG GCAGGTCCTG AAAGAGAATG GTCCAATGGC
501 CTCTGACCCA CTGTGCCTTA CCTACTCATA TCTTTCTCAT GTGGACCTGG
551 TAAAAGACTT GAATTCAGGC CTCATTGGAG CCCTACTAGT ATGTAGAGAA
601 GGGAGTCTGG CCAAGGAAAA GACACAGACC TTGCACAAAT TTATACTACT
651 TTTTGCTGTA TTTGATGAAG GGAAAAGTTG GCACTCAGAA ACAAAGAACT
701 CCTTGATGCA GGATAGGGAT GCTGCATCTG CTCGGGCCTG GCCTAAAATG
751 CACACAGTCA ATGGTTATGT AAACAGGTCT CTGCCAGGTC TGATTGGATG
801 CCACAGGAAA TCAGTCTATT GGCATGTGAT TGGAATGGGC ACCACTCCTG
851 AAGTGCACTC AATATTCCTC GAAGGTCACA CATTTCTTGT GAGGAACCAT
901 CGCCAGGCGT CCTTGGAAAT CTCGCCAATA ACTTTCCTTA CTGCTCAAAC
951 ACTCTTGATG GACCTTGGAC AGTTTCTACT GTTTTGTCAT ATCTCTTCCC
1001 ACCAACATGA TGGCATGGAA GCTTATGTCA AAGTAGACAG CTGTCCAGAG
1051 GAACCCCAAC TACGAATGAA AAATAATGAA GAAGCGGAAG ACTATGATGA
1101 TGATCTTACT GATTCTGAAA TGGATGTGGT CAGGTTTGAT GATGACAACT
1151 CTCCTTCCTT TATCCAAATT CGCTCAGTTG CCAAGAAGCA TCCTAAAACT
1201 TGGGTACATT ACATTGCTGC TGAAGAGGAG GACTGGGACT ATGCTCCCTT
1251 AGTCCTCGCC CCCGATGACA GAAGTTATAA AAGTCAATAT TTGAACAATG
1301 GCCCTCAGCG GATTGGTAGG AAGTACAAAA AAGTCCGATT TATGGCATAC
1351 ACAGATGAAA CCTTTAAGAC TCGTGAAGCT ATTCAGCATG AATCAGGAAT
1401 CTTGGGACCT TTACTTTATG GGGAAGTTGG AGACACACTG TTGATTATAT
1451 TTAAGAATCA AGCAAGCAGA CCATATAACA TCTACCCTCA CGGAATCACT
1501 GATGTCCGTC CTTTGTATTC AAGGAGATTA CCAAAAGGTG TAAAACATTT
1551 GAAGGATTTT CCAATTCTGC CAGGAGAAAT ATTCAAATAT AAATGGACAG
1601 TGACTGTAGA AGATGGGCCA ACTAAATCAG ATCCTCGGTG CCTGACCCGC
1651 TATTACTCTA GTTTCGTTAA TATGGAGAGA GATCTAGCTT CAGGACTCAT
1701 TGGCCCTCTC CTCATCTGCT ACAAAGAATC TGTAGATCAA AGAGGAAACC
1751 AGATAATGTC AGACAAGAGG AATGTCATCC TGTTTTCTGT ATTTGATGAG
1801 AACCGAAGCT GGTACCTCAC AGAGAATATA CAACGCTTTC TCCCCAATCC
1851 AGCTGGAGTG CAGCTTGAGG ATCCAGAGTT CCAAGCCTCC AACATCATGC
1901 ACAGCATCAA TGGCTATGTT TTTGATAGTT TGCAGTTGTC AGTTTGTTTG
1951 CATGAGGTGG CATACTGGTA CATTCTAAGC ATTGGAGCAC AGACTGACTT
2001 CCTTTCTGTC TTCTTCTCTG GATATACCTT CAAACACAAA ATGGTCTATG
2051 AAGACACACT CACCCTATTC CCATTCTCAG GAGAAACTGT CTTCATGTCG
2101 ATGGAAAACC CAGGTCTATG GATTCTGGGG TGCCACAACT CAGACTTTCG
2151 GAACAGAGGC ATGACCGCCT TACTGAAGGT TTCTAGTTGT GACAAGAACA
2201 CTGGTGATTA TTACGAGGAC AGTTATGAAG ATATTTCAGC ATACTTGCTG
2251 AGTAAAAACA ATGCCATTGA ACCAAGAAGC TTCTCTCAAA ACCCACCAGT
2301 CTTGAAACGC CATCAACGGG AAATAACTCG TACTACTCTT CAGTCAGATC
2351 AAGAGGAAAT TGACTATGAT GATACCATAT CAGTTGAAAT GAAGAAGGAA
2401 GATTTTGACA TTTATGATGA GGATGAAAAT CAGAGCCCCC GCAGCTTTCA
2451 AAAGAAAACA CGACACTATT TTATTGCTGC AGTGGAGAGG CTCTGGGATT
2501 ATGGGATGAG TAGCTCCCCA CATGTTCTAA GAAACAGGGC TCAGAGTGGC
2551 AGTGTCCCTC AGTTCAAGAA AGTTGTTTTC CAGGAATTTA CTGATGGCTC
2601 CTTTACTCAG CCCTTATACC GTGGAGAACT AAATGAACAT TTGGGACTCC
2651 TGGGGCCATA TATAAGAGCA GAAGTTGAAG ATAATATCAT GGTAACTTTC
2701 AGAAATCAGG CCTCTCGTCC CTATTCCTTC TATTCTAGCC TTATTTCTTA
2751 TGAGGAAGAT CAGAGGCAAG GAGCAGAACC TAGAAAAAAC TTTGTCAAGC
2801 CTAATGAAAC CAAAACTTAC TTTTGGAAAG TGCAACATCA TATGGCACCC
2851 ACTAAAGATG AGTTTGACTG CAAAGCCTGG GCTTATTTCT CTGATGTTGA
2901 CCTGGAAAAA GATGTGCACT CAGGCCTGAT TGGACCCCTT CTGGTCTGCC
2951 ACACTAACAC ACTGAACCCT GCTCATGGGA GACAAGTGAC AGTACAGGAA
3001 TTTGCTCTGT TTTTCACCAT CTTTGATGAG ACCAAAAGCT GGTACTTCAC
3051 TGAAAATATG GAAAGAAACT GCAGGGCTCC CTGCAATATC CAGATGGAAG
3101 ATCCCACTTT TAAAGAGAAT TATCGCTTCC ATGCAATCAA TGGCTACATA
3151 ATGGATACAC TACCTGGCTT AGTAATGGCT CAGGATCAAA GGATTCGATG
3201 GTATCTGCTC AGCATGGGCA GCAATGAAAA CATCCATTCT ATTCATTTCA
3251 GTGGACATGT GTTCACTGTA CGAAAAAAAG AGGAGTATAA AATGGCACTG
3301 TACAATCTCT ATCCAGGTGT TTTTGAGACA GTGGAAATGT TACCATCCAA
3351 AGCTGGAATT TGGCGGGTGG AATGCCTTAT TGGCGAGCAT CTACATGCTG
3401 GGATGAGCAC ACTTTTTCTG GTGTACAGCA ATAAGTGTCA GACTCCCCTG
3451 GGAATGGCTT CTGGACACAT TAGAGATTTT CAGATTACAG CTTCAGGACA
3501 ATATGGACAG TGGGCCCCAA AGCTGGCCAG ACTTCATTAT TCCGGATCAA
3551 TCAATGCCTG GAGCACCAAG GAGCCCTTTT CTTGGATCAA GGTGGATCTG
3601 TTGGCACCAA TGATTATTCA CGGCATCAAG ACCCAGGGTG CCCGTCAGAA
3651 GTTCTCCAGC CTCTACATCT CTCAGTTTAT CATCATGTAT AGTCTTGATG
3701 GGAAGAAGTG GCAGACTTAT CGAGGAAATT CCACTGGAAC CTTAATGGTC
3751 TTCTTTGGCA ATGTGGATTC ATCTGGGATA AAACACAATA TTTTTAACCC
3801 TCCAATTATT GCTCGATACA TCCGTTTGCA CCCAACTCAT TATAGCATTC
3851 GCAGCACTCT TCGCATGGAG TTGATGGGCT GTGATTTAAA TAGTTGCAGC
3901 ATGCCATTGG GAATGGAGAG TAAAGCAATA TCAGATGCAC AGATTACTGC
3951 TTCATCCTAC TTTACCAATA TGTTTGCCAC CTGGTCTCCT TCAAAAGCTC
4001 GACTTCACCT CCAAGGGAGG AGTAATGCCT GGAGACCTCA GGTGAATAAT
4051 CCAAAAGAGT GGCTGCAAGT GGACTTCCAG AAGACAATGA AAGTCACAGG
4101 AGTAACTACT CAGGGAGTAA AATCTCTGCT TACCAGCATG TATGTGAAGG
4151 AGTTCCTCAT CTCCAGCAGT CAAGATGGCC ATCAGTGGAC TCTCTTTTTT
4201 CAGAATGGCA AAGTAAAGGT TTTTCAGGGA AATCAAGACT CCTTCACACC
4251 TGTGGTGAAC TCTCTAGACC CACCGTTACT GACTCGCTAC CTTCGAATTC
4301 ACCCCCAGAG TTGGGTGCAC CAGATTGCCC TGAGGATGGA GGTTCTGGGC
4351 TGCGAGGCAC AGGACCTCTA CGACAAAACT CACACATGCC CACCGTGCCC
4401 AGCTCCAGAA CTCCTGGGCG GACCGTCAGT CTTCCTCTTC CCCCCAAAAC
4451 CCAAGGACAC CCTCATGATC TCCCGGACCC CTGAGGTCAC ATGCGTGGTG
4501 GTGGACGTGA GCCACGAAGA CCCTGAGGTC AAGTTCAACT GGTACGTGGA
4551 CGGCGTGGAG GTGCATAATG CCAAGACAAA GCCGCGGGAG GAGCAGTACA
4601 ACAGCACGTA CCGTGTGGTC AGCGTCCTCA CCGTCCTGCA CCAGGACTGG
4651 CTGAATGGCA AGGAGTACAA GTGCAAGGTC TCCAACAAAG CCCTCCCAGC
4701 CCCCATCGAG AAAACCATCT CCAAAGCCAA AGGGCAGCCC CGAGAACCAC
4751 AGGTGTACAC CCTGCCCCCA TCCCGGGATG AGCTGACCAA GAACCAGGTC
4801 AGCCTGACCT GCCTGGTCAA AGGCTTCTAT CCCAGCGACA TCGCCGTGGA
4851 GTGGGAGAGC AATGGGCAGC CGGAGAACAA CTACAAGACC ACGCCTCCCG
4901 TGTTGGACTC CGACGGCTCC TTCTTCCTCT ACAGCAAGCT CACCGTGGAC
4951 AAGAGCAGGT GGCAGCAGGG GAACGTCTTC TCATGCTCCG TGATGCATGA
5001 GGCTCTGCAC AACCACTACA CGCAGAAGAG CCTCTCCCTG TCTCCGGGTA
5051 AATGA
Последовательность белка pSYNFVIII 010 (двухцепочечный FVIIIFc) (SEQ ID NO: 163)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNPPVLKR HQREITRTTL QSDQEEIDYD DTISVEMKKE
801 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
851 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
901 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
951 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1001 FALFFTIFDE TKSWYFTENM ERNCRAPCNI QMEDPTFKEN YRFHAINGYI
1051 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1101 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1151 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1201 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1251 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1301 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1351 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1401 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1451 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1501 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1551 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1601 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1651 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность белка FVIII 195 (двухцепочечный FVIIIFc с двумя XTEN AE-144 в аминокислотных положениях 1656 и 1900) (SEQ ID NO: 73)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNPPVLKR HQREITRTTL QGAPGTPGSG TASSSPGASP
801 GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
851 TASSSPGASP GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSTPSGA
901 TGSPGSSTPS GATGSPGASP GTSSTGSPAS SSDQEEIDYD DTISVEMKKE
951 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1001 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1051 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1101 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1151 FALFFTIFDE TKSWYFTENM ERNCRGAPTS ESATPESGPG SEPATSGSET
1201 PGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG
1251 TSESATPESG PGSPAGSPTS TEEGSPAGSP TSTEEGSPAG SPTSTEEGTS
1301 ESATPESGPG TSTEPSEGSA PGASSAPCNI QMEDPTFKEN YRFHAINGYI
1351 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1401 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1451 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1501 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1551 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1601 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1651 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1701 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1751 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1801 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1851 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1901 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1951 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность зрелого белка pSYN-FVIII-173 (SEQ ID NO: 72):
1 ATRRYYLGAV ELSWDYMQSD LGELPVDARF PPRVPKSFPF NTSVVYKKTL
51 FVEFTDHLFN IAKPRPPWMG LLGPTIQAEV YDTVVITLKN MASHPVSLHA
101 VGVSYWKASE GAEYDDQTSQ REKEDDKVFP GGSHTYVWQV LKENGPMASD
151 PLCLTYSYLS HVDLVKDLNS GLIGALLVCR EGSLAKEKTQ TLHKFILLFA
201 VFDEGKSWHS ETKNSLMQDR DAASARAWPK MHTVNGYVNR SLPGLIGCHR
251 KSVYWHVIGM GTTPEVHSIF LEGHTFLVRN HRQASLEISP ITFLTAQTLL
301 MDLGQFLLFC HISSHQHDGM EAYVKVDSCP EEPQLRMKNN EEAEDYDDDL
351 TDSEMDVVRF DDDNSPSFIQ IRSVAKKHPK TWVHYIAAEE EDWDYAPLVL
401 APDDRSYKSQ YLNNGPQRIG RKYKKVRFMA YTDETFKTRE AIQHESGILG
451 PLLYGEVGDT LLIIFKNQAS RPYNIYPHGI TDVRPLYSRR LPKGVKHLKD
501 FPILPGEIFK YKWTVTVEDG PTKSDPRCLT RYYSSFVNME RDLASGLIGP
551 LLICYKESVD QRGNQIMSDK RNVILFSVFD ENRSWYLTEN IQRFLPNPAG
601 VQLEDPEFQA SNIMHSINGY VFDSLQLSVC LHEVAYWYIL SIGAQTDFLS
651 VFFSGYTFKH KMVYEDTLTL FPFSGETVFM SMENPGLWIL GCHNSDFRNR
701 GMTALLKVSS CDKNTGDYYE DSYEDISAYL LSKNNAIEPR SFSQNGAPGT
751 SESATPESGP GSEPATSGSE TPGTSESATP ESGPGSEPAT SGSETPGTSE
801 SATPESGPGT STEPSEGSAP GSPAGSPTST EEGTSESATP ESGPGSEPAT
851 SGSETPGTSE SATPESGPGS PAGSPTSTEE GSPAGSPTST EEGTSTEPSE
901 GSAPGTSESA TPESGPGTSE SATPESGPGT SESATPESGP GSEPATSGSE
951 TPGSEPATSG SETPGSPAGS PTSTEEGTST EPSEGSAPGT STEPSEGSAP
1001 GSEPATSGSE TPGTSESATP ESGPGTSTEP SEGSAPASSP PVLKRHQREI
1051 TRTTLQSDQE EIDYDDTISV EMKKEDFDIY DEDENQSPRS FQKKTRHYFI
1101 AAVERLWDYG MSSSPHVLRN RAQSGSVPQF KKVVFQEFTD GSFTQPLYRG
1151 ELNEHLGLLG PYIRAEVEDN IMVTFRNQAS RPYSFYSSLI SYEEDQRQGA
1201 EPRKNFVKPN ETKTYFWKVQ HHMAPTKDEF DCKAWAYFSD VDLEKDVHSG
1251 LIGPLLVCHT NTLNPAHGRQ VTVQEFALFF TIFDETKSWY FTENMERNCR
1301 APCNIQMEDP TFKENYRFHA INGYIMDTLP GLVMAQDQRI RWYLLSMGSN
1351 ENIHSIHFSG HVFTVRKKEE YKMALYNLYP GVFETVEMLP SKAGIWRVEC
1401 LIGEHLHAGM STLFLVYSNK CQTPLGMASG HIRDFQITAS GQYGQWAPKL
1451 ARLHYSGSIN AWSTKEPFSW IKVDLLAPMI IHGIKTQGAR QKFSSLYISQ
1501 FIIMYSLDGK KWQTYRGNST GTLMVFFGNV DSSGIKHNIF NPPIIARYIR
1551 LHPTHYSIRS TLRMELMGCD LNSCSMPLGM ESKAISDAQI TASSYFTNMF
1601 ATWSPSKARL HLQGRSNAWR PQVNNPKEWL QVDFQKTMKV TGVTTQGVKS
1651 LLTSMYVKEF LISSSQDGHQ WTLFFQNGKV KVFQGNQDSF TPVVNSLDPP
1701 LLTRYLRIHP QSWVHQIALR MEVLGCEAQD LYDKTHTCPP CPAPELLGGP
1751 SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY VDGVEVHNAK
1801 TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL PAPIEKTISK
1851 AKGQPREPQV YTLPPSRDEL TKNQVSLTCL VKGFYPSDIA VEWESNGQPE
1901 NNYKTTPPVL DSDGSFFLYS KLTVDKSRWQ QGNVFSCSVM HEALHNHYTQ
1951 KSLSLSPGK
Последовательность белка FVIII 196 (двухцепочечный FVIIIFc с тремя XTEN AE-144 в аминокислотных положениях 26, 1656 и 1900) (SEQ ID NO: 74)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVGAPGS
51 SPSASTGTGP GSSPSASTGT GPGASPGTSS TGSPGASPGT SSTGSPGSST
101 PSGATGSPGS SPSASTGTGP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
151 TASSSPGSST PSGATGSPGS STPSGATGSP GASPGTSSTG SPASSDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNPPVLKR HQREITRTTL QGAPGTPGSG TASSSPGASP
951 GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
1001 TASSSPGASP GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSTPSGA
1051 TGSPGSSTPS GATGSPGASP GTSSTGSPAS SSDQEEIDYD DTISVEMKKE
1101 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1151 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1201 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1251 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1301 FALFFTIFDE TKSWYFTENM ERNCRGAPTS ESATPESGPG SEPATSGSET
1351 PGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG
1401 TSESATPESG PGSPAGSPTS TEEGSPAGSP TSTEEGSPAG SPTSTEEGTS
1451 ESATPESGPG TSTEPSEGSA PGASSAPCNI QMEDPTFKEN YRFHAINGYI
1501 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1551 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1601 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1651 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1701 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1751 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1801 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1851 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1901 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1951 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
2001 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
2051 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
2101 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность белка FVIII 199 (одноцепочечный FVIIIFc с тремя XTEN AE-144
в аминокислотных положениях 1656 и 1900) (SEQ ID NO: 75)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNPPVLKR HQAEITRTTL QGAPGTPGSG TASSSPGASP
801 GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
851 TASSSPGASP GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSTPSGA
901 TGSPGSSTPS GATGSPGASP GTSSTGSPAS SSDQEEIDYD DTISVEMKKE
951 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1001 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1051 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1101 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1151 FALFFTIFDE TKSWYFTENM ERNCRGAPTS ESATPESGPG SEPATSGSET
1201 PGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG
1251 TSESATPESG PGSPAGSPTS TEEGSPAGSP TSTEEGSPAG SPTSTEEGTS
1301 ESATPESGPG TSTEPSEGSA PGASSAPCNI QMEDPTFKEN YRFHAINGYI
1351 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1401 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1451 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1501 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1551 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1601 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1651 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1701 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1751 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1801 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1851 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1901 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1951 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность белка FVIII 201 (одноцепочечный FVIIIFc с тремя XTEN AE-144
в аминокислотных положениях 26, 1656 и 1900) (SEQ ID NO: 76)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVGAPGS
51 SPSASTGTGP GSSPSASTGT GPGASPGTSS TGSPGASPGT SSTGSPGSST
101 PSGATGSPGS SPSASTGTGP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
151 TASSSPGSST PSGATGSPGS STPSGATGSP GASPGTSSTG SPASSDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNPPVLKR HQAEITRTTL QGAPGTPGSG TASSSPGASP
951 GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSPSAST GTGPGTPGSG
1001 TASSSPGASP GTSSTGSPGA SPGTSSTGSP GASPGTSSTG SPGSSTPSGA
1051 TGSPGSSTPS GATGSPGASP GTSSTGSPAS SSDQEEIDYD DTISVEMKKE
1101 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1151 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1201 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1251 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1301 FALFFTIFDE TKSWYFTENM ERNCRGAPTS ESATPESGPG SEPATSGSET
1351 PGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG
1401 TSESATPESG PGSPAGSPTS TEEGSPAGSP TSTEEGSPAG SPTSTEEGTS
1451 ESATPESGPG TSTEPSEGSA PGASSAPCNI QMEDPTFKEN YRFHAINGYI
1501 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1551 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1601 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1651 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1701 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1751 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1801 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1851 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1901 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1951 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
2001 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
2051 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
2101 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность белка FVIII 203 (одноцепочечный FVIIIFc с двумя XTEN AE - одним XTEN AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 1900) (SEQ ID NO: 77)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
801 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
851 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
901 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
951 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1001 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1051 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1101 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1151 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1201 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1251 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1301 IFDETKSWYF TENMERNCRG APTSESATPE SGPGSEPATS GSETPGTSES
1351 ATPESGPGSE PATSGSETPG TSESATPESG PGTSTEPSEG SAPGTSESAT
1401 PESGPGSPAG SPTSTEEGSP AGSPTSTEEG SPAGSPTSTE EGTSESATPE
1451 SGPGTSTEPS EGSAPGASSA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1501 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1551 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1601 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1651 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1701 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1751 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1801 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1851 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1901 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE
1951 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY
2001 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
2051 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
2101 GNVFSCSVMH EALHNHYTQK SLSLSPGK*
Последовательность белка FVIII 204 (одноцепочечный FVIIIFc с двумя XTEN AE - одним XTEN AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 403) (SEQ ID NO: 78)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDGAPTSTEP SEGSAPGSPA GSPTSTEEGT
451 STEPSEGSAP GTSTEPSEGS APGTSESATP ESGPGTSTEP SEGSAPGTSE
501 SATPESGPGS EPATSGSETP GTSTEPSEGS APGTSTEPSE GSAPGTSESA
551 TPESGPGTSE SATPESGPGA SSDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
951 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
1001 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
1051 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
1101 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1151 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1201 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1251 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1301 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1351 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1401 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1451 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1501 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1551 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1601 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1651 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1701 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1751 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1801 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1851 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1901 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE
1951 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY
2001 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
2051 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
2101 GNVFSCSVMH EALHNHYTQK SLSLSPGK*
Последовательность белка FVIII 205 (одноцепочечный FVIIIFc с двумя XTEN AE - одним XTEN AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 18) (SEQ ID NO: 79)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS
151 ESATPESGPG SPAGSPTSTE EGSPAGSPTS TEEGASSSDL GELPVDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
951 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
1001 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
1051 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
1101 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1151 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1201 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1251 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1301 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1351 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1401 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1451 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1501 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1551 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1601 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1651 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1701 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1751 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1801 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1851 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1901 YDKTHTCPPC PAPELLGGPS VFLFPPKPKD TLMISRTPEV TCVVVDVSHE
1951 DPEVKFNWYV DGVEVHNAKT KPREEQYNST YRVVSVLTVL HQDWLNGKEY
2001 KCKVSNKALP APIEKTISKA KGQPREPQVY TLPPSRDELT KNQVSLTCLV
2051 KGFYPSDIAV EWESNGQPEN NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ
2101 GNVFSCSVMH EALHNHYTQK SLSLSPGK*
Последовательность белка pSYN FVIII 266 (FVIII Fc с XTEN AE-42 в аминокислотном положении 18 и XTEN AE-288 в домене B) (SEQ ID NO: 80)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP GSPAGSPTST
51 EEGTSESATP ESGPGSEPAT SGSETPASSS DLGELPVDAR FPPRVPKSFP
101 FNTSVVYKKT LFVEFTDHLF NIAKPRPPWM GLLGPTIQAE VYDTVVITLK
151 NMASHPVSLH AVGVSYWKAS EGAEYDDQTS QREKEDDKVF PGGSHTYVWQ
201 VLKENGPMAS DPLCLTYSYL SHVDLVKDLN SGLIGALLVC REGSLAKEKT
251 QTLHKFILLF AVFDEGKSWH SETKNSLMQD RDAASARAWP KMHTVNGYVN
301 RSLPGLIGCH RKSVYWHVIG MGTTPEVHSI FLEGHTFLVR NHRQASLEIS
351 PITFLTAQTL LMDLGQFLLF CHISSHQHDG MEAYVKVDSC PEEPQLRMKN
401 NEEAEDYDDD LTDSEMDVVR FDDDNSPSFI QIRSVAKKHP KTWVHYIAAE
451 EEDWDYAPLV LAPDDRSYKS QYLNNGPQRI GRKYKKVRFM AYTDETFKTR
501 EAIQHESGIL GPLLYGEVGD TLLIIFKNQA SRPYNIYPHG ITDVRPLYSR
551 RLPKGVKHLK DFPILPGEIF KYKWTVTVED GPTKSDPRCL TRYYSSFVNM
601 ERDLASGLIG PLLICYKESV DQRGNQIMSD KRNVILFSVF DENRSWYLTE
651 NIQRFLPNPA GVQLEDPEFQ ASNIMHSING YVFDSLQLSV CLHEVAYWYI
701 LSIGAQTDFL SVFFSGYTFK HKMVYEDTLT LFPFSGETVF MSMENPGLWI
751 LGCHNSDFRN RGMTALLKVS SCDKNTGDYY EDSYEDISAY LLSKNNAIEP
801 RSFSQNGAPG TSESATPESG PGSEPATSGS ETPGTSESAT PESGPGSEPA
851 TSGSETPGTS ESATPESGPG TSTEPSEGSA PGSPAGSPTS TEEGTSESAT
901 PESGPGSEPA TSGSETPGTS ESATPESGPG SPAGSPTSTE EGSPAGSPTS
951 TEEGTSTEPS EGSAPGTSES ATPESGPGTS ESATPESGPG TSESATPESG
1001 PGSEPATSGS ETPGSEPATS GSETPGSPAG SPTSTEEGTS TEPSEGSAPG
1051 TSTEPSEGSA PGSEPATSGS ETPGTSESAT PESGPGTSTE PSEGSAPASS
1101 PPVLKRHQAE ITRTTLQSDQ EEIDYDDTIS VEMKKEDFDI YDEDENQSPR
1151 SFQKKTRHYF IAAVERLWDY GMSSSPHVLR NRAQSGSVPQ FKKVVFQEFT
1201 DGSFTQPLYR GELNEHLGLL GPYIRAEVED NIMVTFRNQA SRPYSFYSSL
1251 ISYEEDQRQG AEPRKNFVKP NETKTYFWKV QHHMAPTKDE FDCKAWAYFS
1301 DVDLEKDVHS GLIGPLLVCH TNTLNPAHGR QVTVQEFALF FTIFDETKSW
1351 YFTENMERNC RAPCNIQMED PTFKENYRFH AINGYIMDTL PGLVMAQDQR
1401 IRWYLLSMGS NENIHSIHFS GHVFTVRKKE EYKMALYNLY PGVFETVEML
1451 PSKAGIWRVE CLIGEHLHAG MSTLFLVYSN KCQTPLGMAS GHIRDFQITA
1501 SGQYGQWAPK LARLHYSGSI NAWSTKEPFS WIKVDLLAPM IIHGIKTQGA
1551 RQKFSSLYIS QFIIMYSLDG KKWQTYRGNS TGTLMVFFGN VDSSGIKHNI
1601 FNPPIIARYI RLHPTHYSIR STLRMELMGC DLNSCSMPLG MESKAISDAQ
1651 ITASSYFTNM FATWSPSKAR LHLQGRSNAW RPQVNNPKEW LQVDFQKTMK
1701 VTGVTTQGVK SLLTSMYVKE FLISSSQDGH QWTLFFQNGK VKVFQGNQDS
1751 FTPVVNSLDP PLLTRYLRIH PQSWVHQIAL RMEVLGCEAQ DLYDKTHTCP
1801 PCPAPELLGG PSVFLFPPKP KDTLMISRTP EVTCVVVDVS HEDPEVKFNW
1851 YVDGVEVHNA KTKPREEQYN STYRVVSVLT VLHQDWLNGK EYKCKVSNKA
1901 LPAPIEKTIS KAKGQPREPQ VYTLPPSRDE LTKNQVSLTC LVKGFYPSDI
1951 AVEWESNGQP ENNYKTTPPV LDSDGSFFLY SKLTVDKSRW QQGNVFSCSV
2001 MHEALHNHYT QKSLSLSPGK *
Последовательность белка pSYN FVIII 267 (FVIII Fc с XTEN AE-72 в аминокислотном положении 18 и XTEN AE-288 в домене B) (SEQ ID NO: 81)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGASSSDLGE LPVDARFPPR VPKSFPFNTS VVYKKTLFVE
151 FTDHLFNIAK PRPPWMGLLG PTIQAEVYDT VVITLKNMAS HPVSLHAVGV
201 SYWKASEGAE YDDQTSQREK EDDKVFPGGS HTYVWQVLKE NGPMASDPLC
251 LTYSYLSHVD LVKDLNSGLI GALLVCREGS LAKEKTQTLH KFILLFAVFD
301 EGKSWHSETK NSLMQDRDAA SARAWPKMHT VNGYVNRSLP GLIGCHRKSV
351 YWHVIGMGTT PEVHSIFLEG HTFLVRNHRQ ASLEISPITF LTAQTLLMDL
401 GQFLLFCHIS SHQHDGMEAY VKVDSCPEEP QLRMKNNEEA EDYDDDLTDS
451 EMDVVRFDDD NSPSFIQIRS VAKKHPKTWV HYIAAEEEDW DYAPLVLAPD
501 DRSYKSQYLN NGPQRIGRKY KKVRFMAYTD ETFKTREAIQ HESGILGPLL
551 YGEVGDTLLI IFKNQASRPY NIYPHGITDV RPLYSRRLPK GVKHLKDFPI
601 LPGEIFKYKW TVTVEDGPTK SDPRCLTRYY SSFVNMERDL ASGLIGPLLI
651 CYKESVDQRG NQIMSDKRNV ILFSVFDENR SWYLTENIQR FLPNPAGVQL
701 EDPEFQASNI MHSINGYVFD SLQLSVCLHE VAYWYILSIG AQTDFLSVFF
751 SGYTFKHKMV YEDTLTLFPF SGETVFMSME NPGLWILGCH NSDFRNRGMT
801 ALLKVSSCDK NTGDYYEDSY EDISAYLLSK NNAIEPRSFS QNGAPGTSES
851 ATPESGPGSE PATSGSETPG TSESATPESG PGSEPATSGS ETPGTSESAT
901 PESGPGTSTE PSEGSAPGSP AGSPTSTEEG TSESATPESG PGSEPATSGS
951 ETPGTSESAT PESGPGSPAG SPTSTEEGSP AGSPTSTEEG TSTEPSEGSA
1001 PGTSESATPE SGPGTSESAT PESGPGTSES ATPESGPGSE PATSGSETPG
1051 SEPATSGSET PGSPAGSPTS TEEGTSTEPS EGSAPGTSTE PSEGSAPGSE
1101 PATSGSETPG TSESATPESG PGTSTEPSEG SAPASSPPVL KRHQAEITRT
1151 TLQSDQEEID YDDTISVEMK KEDFDIYDED ENQSPRSFQK KTRHYFIAAV
1201 ERLWDYGMSS SPHVLRNRAQ SGSVPQFKKV VFQEFTDGSF TQPLYRGELN
1251 EHLGLLGPYI RAEVEDNIMV TFRNQASRPY SFYSSLISYE EDQRQGAEPR
1301 KNFVKPNETK TYFWKVQHHM APTKDEFDCK AWAYFSDVDL EKDVHSGLIG
1351 PLLVCHTNTL NPAHGRQVTV QEFALFFTIF DETKSWYFTE NMERNCRAPC
1401 NIQMEDPTFK ENYRFHAING YIMDTLPGLV MAQDQRIRWY LLSMGSNENI
1451 HSIHFSGHVF TVRKKEEYKM ALYNLYPGVF ETVEMLPSKA GIWRVECLIG
1501 EHLHAGMSTL FLVYSNKCQT PLGMASGHIR DFQITASGQY GQWAPKLARL
1551 HYSGSINAWS TKEPFSWIKV DLLAPMIIHG IKTQGARQKF SSLYISQFII
1601 MYSLDGKKWQ TYRGNSTGTL MVFFGNVDSS GIKHNIFNPP IIARYIRLHP
1651 THYSIRSTLR MELMGCDLNS CSMPLGMESK AISDAQITAS SYFTNMFATW
1701 SPSKARLHLQ GRSNAWRPQV NNPKEWLQVD FQKTMKVTGV TTQGVKSLLT
1751 SMYVKEFLIS SSQDGHQWTL FFQNGKVKVF QGNQDSFTPV VNSLDPPLLT
1801 RYLRIHPQSW VHQIALRMEV LGCEAQDLYD KTHTCPPCPA PELLGGPSVF
1851 LFPPKPKDTL MISRTPEVTC VVVDVSHEDP EVKFNWYVDG VEVHNAKTKP
1901 REEQYNSTYR VVSVLTVLHQ DWLNGKEYKC KVSNKALPAP IEKTISKAKG
1951 QPREPQVYTL PPSRDELTKN QVSLTCLVKG FYPSDIAVEW ESNGQPENNY
2001 KTTPPVLDSD GSFFLYSKLT VDKSRWQQGN VFSCSVMHEA LHNHYTQKSL
2051 SLSPGK*
Последовательность белка pSYN FVIII 268 (FVIII Fc с XTEN AE-144 в аминокислотном положении 18) (SEQ ID NO: 82)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS
151 ESATPESGPG SPAGSPTSTE EGSPAGSPTS TEEGASSSDL GELPVDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNPPVLKR HQAEITRTTL QSDQEEIDYD DTISVEMKKE
951 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
1001 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
1051 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
1101 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1151 FALFFTIFDE TKSWYFTENM ERNCRAPCNI QMEDPTFKEN YRFHAINGYI
1201 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1251 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1301 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1351 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1401 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1451 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1501 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1551 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1601 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1651 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1701 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1751 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1801 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Последовательность белка pSYN FVIII 269 (FVIII Fc с XTEN AE-72 в аминокислотном положении 18) (SEQ ID NO: 83)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGASSSDLGE LPVDARFPPR VPKSFPFNTS VVYKKTLFVE
151 FTDHLFNIAK PRPPWMGLLG PTIQAEVYDT VVITLKNMAS HPVSLHAVGV
201 SYWKASEGAE YDDQTSQREK EDDKVFPGGS HTYVWQVLKE NGPMASDPLC
251 LTYSYLSHVD LVKDLNSGLI GALLVCREGS LAKEKTQTLH KFILLFAVFD
301 EGKSWHSETK NSLMQDRDAA SARAWPKMHT VNGYVNRSLP GLIGCHRKSV
351 YWHVIGMGTT PEVHSIFLEG HTFLVRNHRQ ASLEISPITF LTAQTLLMDL
401 GQFLLFCHIS SHQHDGMEAY VKVDSCPEEP QLRMKNNEEA EDYDDDLTDS
451 EMDVVRFDDD NSPSFIQIRS VAKKHPKTWV HYIAAEEEDW DYAPLVLAPD
501 DRSYKSQYLN NGPQRIGRKY KKVRFMAYTD ETFKTREAIQ HESGILGPLL
551 YGEVGDTLLI IFKNQASRPY NIYPHGITDV RPLYSRRLPK GVKHLKDFPI
601 LPGEIFKYKW TVTVEDGPTK SDPRCLTRYY SSFVNMERDL ASGLIGPLLI
651 CYKESVDQRG NQIMSDKRNV ILFSVFDENR SWYLTENIQR FLPNPAGVQL
701 EDPEFQASNI MHSINGYVFD SLQLSVCLHE VAYWYILSIG AQTDFLSVFF
751 SGYTFKHKMV YEDTLTLFPF SGETVFMSME NPGLWILGCH NSDFRNRGMT
801 ALLKVSSCDK NTGDYYEDSY EDISAYLLSK NNAIEPRSFS QNPPVLKRHQ
851 AEITRTTLQS DQEEIDYDDT ISVEMKKEDF DIYDEDENQS PRSFQKKTRH
901 YFIAAVERLW DYGMSSSPHV LRNRAQSGSV PQFKKVVFQE FTDGSFTQPL
951 YRGELNEHLG LLGPYIRAEV EDNIMVTFRN QASRPYSFYS SLISYEEDQR
1001 QGAEPRKNFV KPNETKTYFW KVQHHMAPTK DEFDCKAWAY FSDVDLEKDV
1051 HSGLIGPLLV CHTNTLNPAH GRQVTVQEFA LFFTIFDETK SWYFTENMER
1101 NCRAPCNIQM EDPTFKENYR FHAINGYIMD TLPGLVMAQD QRIRWYLLSM
1151 GSNENIHSIH FSGHVFTVRK KEEYKMALYN LYPGVFETVE MLPSKAGIWR
1201 VECLIGEHLH AGMSTLFLVY SNKCQTPLGM ASGHIRDFQI TASGQYGQWA
1251 PKLARLHYSG SINAWSTKEP FSWIKVDLLA PMIIHGIKTQ GARQKFSSLY
1301 ISQFIIMYSL DGKKWQTYRG NSTGTLMVFF GNVDSSGIKH NIFNPPIIAR
1351 YIRLHPTHYS IRSTLRMELM GCDLNSCSMP LGMESKAISD AQITASSYFT
1401 NMFATWSPSK ARLHLQGRSN AWRPQVNNPK EWLQVDFQKT MKVTGVTTQG
1451 VKSLLTSMYV KEFLISSSQD GHQWTLFFQN GKVKVFQGNQ DSFTPVVNSL
1501 DPPLLTRYLR IHPQSWVHQI ALRMEVLGCE AQDLYDKTHT CPPCPAPELL
1551 GGPSVFLFPP KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH
1601 NAKTKPREEQ YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT
1651 ISKAKGQPRE PQVYTLPPSR DELTKNQVSL TCLVKGFYPS DIAVEWESNG
1701 QPENNYKTTP PVLDSDGSFF LYSKLTVDKS RWQQGNVFSC SVMHEALHNH
1751 YTQKSLSLSP GK*
Последовательность белка pSYNFVIII 271 (FVIII Fc с XTEN AE-42 в аминокислотном положении 18) (SEQ ID NO: 84)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP GSPAGSPTST
51 EEGTSESATP ESGPGSEPAT SGSETPASSS DLGELPVDAR FPPRVPKSFP
101 FNTSVVYKKT LFVEFTDHLF NIAKPRPPWM GLLGPTIQAE VYDTVVITLK
151 NMASHPVSLH AVGVSYWKAS EGAEYDDQTS QREKEDDKVF PGGSHTYVWQ
201 VLKENGPMAS DPLCLTYSYL SHVDLVKDLN SGLIGALLVC REGSLAKEKT
251 QTLHKFILLF AVFDEGKSWH SETKNSLMQD RDAASARAWP KMHTVNGYVN
301 RSLPGLIGCH RKSVYWHVIG MGTTPEVHSI FLEGHTFLVR NHRQASLEIS
351 PITFLTAQTL LMDLGQFLLF CHISSHQHDG MEAYVKVDSC PEEPQLRMKN
401 NEEAEDYDDD LTDSEMDVVR FDDDNSPSFI QIRSVAKKHP KTWVHYIAAE
451 EEDWDYAPLV LAPDDRSYKS QYLNNGPQRI GRKYKKVRFM AYTDETFKTR
501 EAIQHESGIL GPLLYGEVGD TLLIIFKNQA SRPYNIYPHG ITDVRPLYSR
551 RLPKGVKHLK DFPILPGEIF KYKWTVTVED GPTKSDPRCL TRYYSSFVNM
601 ERDLASGLIG PLLICYKESV DQRGNQIMSD KRNVILFSVF DENRSWYLTE
651 NIQRFLPNPA GVQLEDPEFQ ASNIMHSING YVFDSLQLSV CLHEVAYWYI
701 LSIGAQTDFL SVFFSGYTFK HKMVYEDTLT LFPFSGETVF MSMENPGLWI
751 LGCHNSDFRN RGMTALLKVS SCDKNTGDYY EDSYEDISAY LLSKNNAIEP
801 RSFSQNPPVL KRHQAEITRT TLQSDQEEID YDDTISVEMK KEDFDIYDED
851 ENQSPRSFQK KTRHYFIAAV ERLWDYGMSS SPHVLRNRAQ SGSVPQFKKV
901 VFQEFTDGSF TQPLYRGELN EHLGLLGPYI RAEVEDNIMV TFRNQASRPY
951 SFYSSLISYE EDQRQGAEPR KNFVKPNETK TYFWKVQHHM APTKDEFDCK
1001 AWAYFSDVDL EKDVHSGLIG PLLVCHTNTL NPAHGRQVTV QEFALFFTIF
1051 DETKSWYFTE NMERNCRAPC NIQMEDPTFK ENYRFHAING YIMDTLPGLV
1101 MAQDQRIRWY LLSMGSNENI HSIHFSGHVF TVRKKEEYKM ALYNLYPGVF
1151 ETVEMLPSKA GIWRVECLIG EHLHAGMSTL FLVYSNKCQT PLGMASGHIR
1201 DFQITASGQY GQWAPKLARL HYSGSINAWS TKEPFSWIKV DLLAPMIIHG
1251 IKTQGARQKF SSLYISQFII MYSLDGKKWQ TYRGNSTGTL MVFFGNVDSS
1301 GIKHNIFNPP IIARYIRLHP THYSIRSTLR MELMGCDLNS CSMPLGMESK
1351 AISDAQITAS SYFTNMFATW SPSKARLHLQ GRSNAWRPQV NNPKEWLQVD
1401 FQKTMKVTGV TTQGVKSLLT SMYVKEFLIS SSQDGHQWTL FFQNGKVKVF
1451 QGNQDSFTPV VNSLDPPLLT RYLRIHPQSW VHQIALRMEV LGCEAQDLYD
1501 KTHTCPPCPA PELLGGPSVF LFPPKPKDTL MISRTPEVTC VVVDVSHEDP
1551 EVKFNWYVDG VEVHNAKTKP REEQYNSTYR VVSVLTVLHQ DWLNGKEYKC
1601 KVSNKALPAP IEKTISKAKG QPREPQVYTL PPSRDELTKN QVSLTCLVKG
1651 FYPSDIAVEW ESNGQPENNY KTTPPVLDSD GSFFLYSKLT VDKSRWQQGN
1701 VFSCSVMHEA LHNHYTQKSL SLSPGK*
Последовательность белка pSYN FVIII 272 (FVIII с XTEN AE-144 в аминокислотном положении 18 и XTEN AE-244 в домене B - без Fc) (SEQ ID NO: 85)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQGAP TSESATPESG
51 PGSEPATSGS ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
101 TSTEPSEGSA PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS
151 ESATPESGPG SPAGSPTSTE EGSPAGSPTS TEEGASSSDL GELPVDARFP
201 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
251 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
301 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
351 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
401 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
451 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
501 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
551 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
601 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
651 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
701 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
751 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
801 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
851 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
901 SKNNAIEPRS FSQNGAPGTS ESATPESGPG SEPATSGSET PGTSESATPE
951 SGPGSEPATS GSETPGTSES ATPESGPGTS TEPSEGSAPG SPAGSPTSTE
1001 EGTSESATPE SGPGSEPATS GSETPGTSES ATPESGPGSP AGSPTSTEEG
1051 SPAGSPTSTE EGTSTEPSEG SAPGTSESAT PESGPGTSES ATPESGPGTS
1101 ESATPESGPG SEPATSGSET PGSEPATSGS ETPGSPAGSP TSTEEGTSTE
1151 PSEGSAPGTS TEPSEGSAPG SEPATSGSET PGTSESATPE SGPGTSTEPS
1201 EGSAPASSPP VLKRHQAEIT RTTLQSDQEE IDYDDTISVE MKKEDFDIYD
1251 EDENQSPRSF QKKTRHYFIA AVERLWDYGM SSSPHVLRNR AQSGSVPQFK
1301 KVVFQEFTDG SFTQPLYRGE LNEHLGLLGP YIRAEVEDNI MVTFRNQASR
1351 PYSFYSSLIS YEEDQRQGAE PRKNFVKPNE TKTYFWKVQH HMAPTKDEFD
1401 CKAWAYFSDV DLEKDVHSGL IGPLLVCHTN TLNPAHGRQV TVQEFALFFT
1451 IFDETKSWYF TENMERNCRA PCNIQMEDPT FKENYRFHAI NGYIMDTLPG
1501 LVMAQDQRIR WYLLSMGSNE NIHSIHFSGH VFTVRKKEEY KMALYNLYPG
1551 VFETVEMLPS KAGIWRVECL IGEHLHAGMS TLFLVYSNKC QTPLGMASGH
1601 IRDFQITASG QYGQWAPKLA RLHYSGSINA WSTKEPFSWI KVDLLAPMII
1651 HGIKTQGARQ KFSSLYISQF IIMYSLDGKK WQTYRGNSTG TLMVFFGNVD
1701 SSGIKHNIFN PPIIARYIRL HPTHYSIRST LRMELMGCDL NSCSMPLGME
1751 SKAISDAQIT ASSYFTNMFA TWSPSKARLH LQGRSNAWRP QVNNPKEWLQ
1801 VDFQKTMKVT GVTTQGVKSL LTSMYVKEFL ISSSQDGHQW TLFFQNGKVK
1851 VFQGNQDSFT PVVNSLDPPL LTRYLRIHPQ SWVHQIALRM EVLGCEAQDL
1901 Y*
Последовательность белка pSYN-FVIII-161 (SEQ ID NO: 69) (аминокислотные положения 1-1457 последовательности FVIII; область, выделенная подчеркиванием, представляет Fc-область; область, выделенная волнистым подчеркиванием, представляет расщепляемый линкер между первым Fc и фрагментом VWF; область, выделенная двойным подчеркиванием, представляет фрагмент VWF; область, выделенная жиирным шрифтом, представляет расщепляемый линкер между фрагментом VWF и Fc)
1 MQIELSTCFF LCLLRFCFSA TRRYYLGAVE LSWDYMQSDL GELPVDARFP
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNPPVLKR HQREITRTTL QSDQEEIDYD DTISVEMKKE
801 DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP HVLRNRAQSG
851 SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA EVEDNIMVTF
901 RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY FWKVQHHMAP
951 TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP AHGRQVTVQE
1001 FALFFTIFDE TKSWYFTENM ERNCRAPCNI QMEDPTFKEN YRFHAINGYI
1051 MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV RKKEEYKMAL
1101 YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL VYSNKCQTPL
1151 GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK EPFSWIKVDL
1201 LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY RGNSTGTLMV
1251 FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME LMGCDLNSCS
1301 MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR SNAWRPQVNN
1351 PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS QDGHQWTLFF
1401 QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH QIALRMEVLG
1451 CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI SRTPEVTCVV
1501 VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV SVLTVLHQDW
1551 LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP SRDELTKNQV
1601 SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS FFLYSKLTVD
1651 KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGKRRRRSG GGGSGGGGSG
1701 GGGSGGGGSG GGGSGGGGSR KRRKRSLSCR PPMVKLVCPA DNLRAEGLEC
1751 TKTCQNYDLE CMSMGCVSGC LCPPGMVRHE NRCVALERCP CFHQGKEYAP
1801 GETVKIGCNT CVCRDRKWNC TDHVCDATCS TIGMAHYLTF DGLKYLFPGE
1851 CQYVLVQDYC GSNPGTFRIL VGNKGCSHPS VKCKKRVTIL VEGGEIELFD
1901 GEVNVKRPMK DETHFEVVES GRYIILLLGK ALSVVWDRHL SISVVLKQTY
1951 QEKVCGLCGN FDGIQNNDLT SSNLQVEEDP VDFGNSWKVS SQCADTRKVP
2001 LDSSPATCHN NIMKQTMVDS SCRILTSDVF QDCNKLVDPE PYLDVCIYDT
2051 CSCESIGDCA AFCDTIAAYA HVCAQHGKVV TWRTATLCPQ SCEERNLREN
2101 GYEAEWRYNS CAPACQVTCQ HPEPLACPVQ CVEGCHAHCP PGKILDELLQ
2151 TCVDPEDCPV CEVAGRRFAS GKKVTLNPSD PEHCQICHCD VVNLTCEACQ
2201 EPISGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP GSEPATSGSE
2251 TPGTSESATP ESGPGTSTEP SEGSAPGSPA GSPTSTEEGT SESATPESGP
2301 GSEPATSGSE TPGTSESATP ESGPGSPAGS PTSTEEGSPA GSPTSTEEGT
2351 STEPSEGSAP GTSESATPES GPGTSESATP ESGPGTSESA TPESGPGSEP
2401 ATSGSETPGS EPATSGSETP GSPAGSPTST EEGTSTEPSE GSAPGTSTEP
2451 SEGSAPGSEP ATSGSETPGT SESATPESGP GTSTEPSEGS APDSGGGGSG
2501 GGGSGGGGSG GGGSGGGGSL VPRGSGG DKT HTCPPCPAPE LLGGPSVFLF
2551 PPKPKDTLMI SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE
2601 EQYNSTYRVV SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP
2651 REPQVYTLPP SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT
2701 TPPVLDSDGS FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL
2751 SPGK
Последовательность белка pSYN-FVIII-170 (SEQ ID NO: 71)
1 SLSCRPPMVK LVCPADNLRA EGLECTKTCQ NYDLECMSMG CVSGCLCPPG
51 MVRHENRCVA LERCPCFHQG KEYAPGETVK IGCNTCVCRD RKWNCTDHVC
101 DATCSTIGMA HYLTFDGLKY LFPGECQYVL VQDYCGSNPG TFRILVGNKG
151 CSHPSVKCKK RVTILVEGGE IELFDGEVNV KRPMKDETHF EVVESGRYII
201 LLLGKALSVV WDRHLSISVV LKQTYQEKVC GLCGNFDGIQ NNDLTSSNLQ
251 VEEDPVDFGN SWKVSSQCAD TRKVPLDSSP ATCHNNIMKQ TMVDSSCRIL
301 TSDVFQDCNK LVDPEPYLDV CIYDTCSCES IGDCAAFCDT IAAYAHVCAQ
351 HGKVVTWRTA TLCPQSCEER NLRENGYEAE WRYNSCAPAC QVTCQHPEPL
401 ACPVQCVEGC HAHCPPGKIL DELLQTCVDP EDCPVCEVAG RRFASGKKVT
451 LNPSDPEHCQ ICHCDVVNLT CEACQEPISG TSESATPESG PGSEPATSGS
501 ETPGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG TSTEPSEGSA
551 PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG
601 SPAGSPTSTE EGSPAGSPTS TEEGTSTEPS EGSAPGTSES ATPESGPGTS
651 ESATPESGPG TSESATPESG PGSEPATSGS ETPGSEPATS GSETPGSPAG
701 SPTSTEEGTS TEPSEGSAPG TSTEPSEGSA PGSEPATSGS ETPGTSESAT
751 PESGPGTSTE PSEGSAPDSG GGGSGGGGSG GGGSGGGGSG GGGSLVPRGS
801 GGASATRRYY LGAVELSWDY MQSDLGELPV DARFPPRVPK SFPFNTSVVY
851 KKTLFVEFTD HLFNIAKPRP PWMGLLGPTI QAEVYDTVVI TLKNMASHPV
901 SLHAVGVSYW KASEGAEYDD QTSQREKEDD KVFPGGSHTY VWQVLKENGP
951 MASDPLCLTY SYLSHVDLVK DLNSGLIGAL LVCREGSLAK EKTQTLHKFI
1001 LLFAVFDEGK SWHSETKNSL MQDRDAASAR AWPKMHTVNG YVNRSLPGLI
1051 GCHRKSVYWH VIGMGTTPEV HSIFLEGHTF LVRNHRQASL EISPITFLTA
1101 QTLLMDLGQF LLFCHISSHQ HDGMEAYVKV DSCPEEPQLR MKNNEEAEDY
1151 DDDLTDSEMD VVRFDDDNSP SFIQIRSVAK KHPKTWVHYI AAEEEDWDYA
1201 PLVLAPDDRS YKSQYLNNGP QRIGRKYKKV RFMAYTDETF KTREAIQHES
1251 GILGPLLYGE VGDTLLIIFK NQASRPYNIY PHGITDVRPL YSRRLPKGVK
1301 HLKDFPILPG EIFKYKWTVT VEDGPTKSDP RCLTRYYSSF VNMERDLASG
1351 LIGPLLICYK ESVDQRGNQI MSDKRNVILF SVFDENRSWY LTENIQRFLP
1401 NPAGVQLEDP EFQASNIMHS INGYVFDSLQ LSVCLHEVAY WYILSIGAQT
1451 DFLSVFFSGY TFKHKMVYED TLTLFPFSGE TVFMSMENPG LWILGCHNSD
1501 FRNRGMTALL KVSSCDKNTG DYYEDSYEDI SAYLLSKNNA IEPRSFSQNP
1551 PVLKRHQREI TRTTLQSDQE EIDYDDTISV EMKKEDFDIY DEDENQSPRS
1601 FQKKTRHYFI AAVERLWDYG MSSSPHVLRN RAQSGSVPQF KKVVFQEFTD
1651 GSFTQPLYRG ELNEHLGLLG PYIRAEVEDN IMVTFRNQAS RPYSFYSSLI
1701 SYEEDQRQGA EPRKNFVKPN ETKTYFWKVQ HHMAPTKDEF DCKAWAYFSD
1751 VDLEKDVHSG LIGPLLVCHT NTLNPAHGRQ VTVQEFALFF TIFDETKSWY
1801 FTENMERNCR APCNIQMEDP TFKENYRFHA INGYIMDTLP GLVMAQDQRI
1851 RWYLLSMGSN ENIHSIHFSG HVFTVRKKEE YKMALYNLYP GVFETVEMLP
1901 SKAGIWRVEC LIGEHLHAGM STLFLVYSNK CQTPLGMASG HIRDFQITAS
1951 GQYGQWAPKL ARLHYSGSIN AWSTKEPFSW IKVDLLAPMI IHGIKTQGAR
2001 QKFSSLYISQ FIIMYSLDGK KWQTYRGNST GTLMVFFGNV DSSGIKHNIF
2051 NPPIIARYIR LHPTHYSIRS TLRMELMGCD LNSCSMPLGM ESKAISDAQI
2101 TASSYFTNMF ATWSPSKARL HLQGRSNAWR PQVNNPKEWL QVDFQKTMKV
2151 TGVTTQGVKS LLTSMYVKEF LISSSQDGHQ WTLFFQNGKV KVFQGNQDSF
2201 TPVVNSLDPP LLTRYLRIHP QSWVHQIALR MEVLGCEAQD LY
Нуклеотидная последовательность pSYN FVIII 310 (кодирующая FVIII с полной делецией домена B, за исключением 2 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 742) (SEQ ID NO: 170)
1 ATGCAAATAG AGCTCTCCAC CTGCTTCTTT CTGTGCCTTT TGCGATTCTG
51 CTTTAGTGCC ACCAGAAGAT ACTACCTGGG TGCAGTGGAA CTGTCATGGG
101 ACTATATGCA AAGTGATCTC GGTGAGCTGC CTGTGGACGC AAGATTTCCT
151 CCTAGAGTGC CAAAATCTTT TCCATTCAAC ACCTCAGTCG TGTACAAAAA
201 GACTCTGTTT GTAGAATTCA CGGATCACCT TTTCAACATC GCTAAGCCAA
251 GGCCACCCTG GATGGGTCTG CTAGGTCCTA CCATCCAGGC TGAGGTTTAT
301 GATACAGTGG TCATTACACT TAAGAACATG GCTTCCCATC CTGTCAGTCT
351 TCATGCTGTT GGTGTATCCT ACTGGAAAGC TTCTGAGGGA GCTGAATATG
401 ATGATCAGAC CAGTCAAAGG GAGAAAGAAG ATGATAAAGT CTTCCCTGGT
451 GGAAGCCATA CATATGTCTG GCAGGTCCTG AAAGAGAATG GTCCAATGGC
501 CTCTGACCCA CTGTGCCTTA CCTACTCATA TCTTTCTCAT GTGGACCTGG
551 TAAAAGACTT GAATTCAGGC CTCATTGGAG CCCTACTAGT ATGTAGAGAA
601 GGGAGTCTGG CCAAGGAAAA GACACAGACC TTGCACAAAT TTATACTACT
651 TTTTGCTGTA TTTGATGAAG GGAAAAGTTG GCACTCAGAA ACAAAGAACT
701 CCTTGATGCA GGATAGGGAT GCTGCATCTG CTCGGGCCTG GCCTAAAATG
751 CACACAGTCA ATGGTTATGT AAACAGGTCT CTGCCAGGTC TGATTGGATG
801 CCACAGGAAA TCAGTCTATT GGCATGTGAT TGGAATGGGC ACCACTCCTG
851 AAGTGCACTC AATATTCCTC GAAGGTCACA CATTTCTTGT GAGGAACCAT
901 CGCCAGGCGT CCTTGGAAAT CTCGCCAATA ACTTTCCTTA CTGCTCAAAC
951 ACTCTTGATG GACCTTGGAC AGTTTCTACT GTTTTGTCAT ATCTCTTCCC
1001 ACCAACATGA TGGCATGGAA GCTTATGTCA AAGTAGACAG CTGTCCAGAG
1051 GAACCCCAAC TACGAATGAA AAATAATGAA GAAGCGGAAG ACTATGATGA
1101 TGATCTTACT GATTCTGAAA TGGATGTGGT CAGGTTTGAT GATGACAACT
1151 CTCCTTCCTT TATCCAAATT CGCTCAGTTG CCAAGAAGCA TCCTAAAACT
1201 TGGGTACATT ACATTGCTGC TGAAGAGGAG GACTGGGACT ATGCTCCCTT
1251 AGTCCTCGCC CCCGATGACA GAAGTTATAA AAGTCAATAT TTGAACAATG
1301 GCCCTCAGCG GATTGGTAGG AAGTACAAAA AAGTCCGATT TATGGCATAC
1351 ACAGATGAAA CCTTTAAGAC TCGTGAAGCT ATTCAGCATG AATCAGGAAT
1401 CTTGGGACCT TTACTTTATG GGGAAGTTGG AGACACACTG TTGATTATAT
1451 TTAAGAATCA AGCAAGCAGA CCATATAACA TCTACCCTCA CGGAATCACT
1501 GATGTCCGTC CTTTGTATTC AAGGAGATTA CCAAAAGGTG TAAAACATTT
1551 GAAGGATTTT CCAATTCTGC CAGGAGAAAT ATTCAAATAT AAATGGACAG
1601 TGACTGTAGA AGATGGGCCA ACTAAATCAG ATCCTCGGTG CCTGACCCGC
1651 TATTACTCTA GTTTCGTTAA TATGGAGAGA GATCTAGCTT CAGGACTCAT
1701 TGGCCCTCTC CTCATCTGCT ACAAAGAATC TGTAGATCAA AGAGGAAACC
1751 AGATAATGTC AGACAAGAGG AATGTCATCC TGTTTTCTGT ATTTGATGAG
1801 AACCGAAGCT GGTACCTCAC AGAGAATATA CAACGCTTTC TCCCCAATCC
1851 AGCTGGAGTG CAGCTTGAGG ATCCAGAGTT CCAAGCCTCC AACATCATGC
1901 ACAGCATCAA TGGCTATGTT TTTGATAGTT TGCAGTTGTC AGTTTGTTTG
1951 CATGAGGTGG CATACTGGTA CATTCTAAGC ATTGGAGCAC AGACTGACTT
2001 CCTTTCTGTC TTCTTCTCTG GATATACCTT CAAACACAAA ATGGTCTATG
2051 AAGACACACT CACCCTATTC CCATTCTCAG GAGAAACTGT CTTCATGTCG
2101 ATGGAAAACC CAGGTCTATG GATTCTGGGG TGCCACAACT CAGACTTTCG
2151 GAACAGAGGC ATGACCGCCT TACTGAAGGT TTCTAGTTGT GACAAGAACA
2201 CTGGTGATTA TTACGAGGAC AGTTATGAAG ATATTTCAGC ATACTTGCTG
2251 AGTAAAAACA ATGCCATTGA ACCAAGAAGC TTCGGTACCT CAGAGTCTGC
2301 TACCCCCGAG TCAGGGCCAG GATCAGAGCC AGCCACCTCC GGGTCTGAGA
2351 CACCCGGGAC TTCCGAGAGT GCCACCCCTG AGTCCGGACC CGGGTCCGAG
2401 CCCGCCACTT CCGGCTCCGA AACTCCCGGC ACAAGCGAGA GCGCTACCCC
2451 AGAGTCAGGA CCAGGAACAT CTACAGAGCC CTCTGAAGGC TCCGCTCCAG
2501 GGTCCCCAGC CGGCAGTCCC ACTAGCACCG AGGAGGGAAC CTCTGAAAGC
2551 GCCACACCCG AATCAGGGCC AGGGTCTGAG CCTGCTACCA GCGGCAGCGA
2601 GACACCAGGC ACCTCTGAGT CCGCCACACC AGAGTCCGGA CCCGGATCTC
2651 CCGCTGGGAG CCCCACCTCC ACTGAGGAGG GATCTCCTGC TGGCTCTCCA
2701 ACATCTACTG AGGAAGGTAC CTCAACCGAG CCATCCGAGG GATCAGCTCC
2751 CGGCACCTCA GAGTCGGCAA CCCCGGAGTC TGGACCCGGA ACTTCCGAAA
2801 GTGCCACACC AGAGTCCGGT CCCGGGACTT CAGAATCAGC AACACCCGAG
2851 TCCGGCCCTG GGTCTGAACC CGCCACAAGT GGTAGTGAGA CACCAGGATC
2901 AGAACCTGCT ACCTCAGGGT CAGAGACACC CGGATCTCCG GCAGGCTCAC
2951 CAACCTCCAC TGAGGAGGGC ACCAGCACAG AACCAAGCGA GGGCTCCGCA
3001 CCCGGAACAA GCACTGAACC CAGTGAGGGT TCAGCACCCG GCTCTGAGCC
3051 GGCCACAAGT GGCAGTGAGA CACCCGGCAC TTCAGAGAGT GCCACCCCCG
3101 AGAGTGGCCC AGGCACTAGT ACCGAGCCCT CTGAAGGCAG TGCGCCAGCC
3151 TCGAGCGAAA TAACTCGTAC TACTCTTCAG TCAGATCAAG AGGAAATCGA
3201 TTATGATGAT ACCATATCAG TTGAAATGAA GAAGGAAGAT TTTGACATTT
3251 ATGATGAGGA TGAAAATCAG AGCCCCCGCA GCTTTCAAAA GAAAACACGA
3301 CACTATTTTA TTGCTGCAGT GGAGAGGCTC TGGGATTATG GGATGAGTAG
3351 CTCCCCACAT GTTCTAAGAA ACAGGGCTCA GAGTGGCAGT GTCCCTCAGT
3401 TCAAGAAAGT TGTTTTCCAG GAATTTACTG ATGGCTCCTT TACTCAGCCC
3451 TTATACCGTG GAGAACTAAA TGAACATTTG GGACTCCTGG GGCCATATAT
3501 AAGAGCAGAA GTTGAAGATA ATATCATGGT AACTTTCAGA AATCAGGCCT
3551 CTCGTCCCTA TTCCTTCTAT TCTAGCCTTA TTTCTTATGA GGAAGATCAG
3601 AGGCAAGGAG CAGAACCTAG AAAAAACTTT GTCAAGCCTA ATGAAACCAA
3651 AACTTACTTT TGGAAAGTGC AACATCATAT GGCACCCACT AAAGATGAGT
3701 TTGACTGCAA AGCCTGGGCT TATTTCTCTG ATGTTGACCT GGAAAAAGAT
3751 GTGCACTCAG GCCTGATTGG ACCCCTTCTG GTCTGCCACA CTAACACACT
3801 GAACCCTGCT CATGGGAGAC AAGTGACAGT ACAGGAATTT GCTCTGTTTT
3851 TCACCATCTT TGATGAGACC AAAAGCTGGT ACTTCACTGA AAATATGGAA
3901 AGAAACTGCA GGGCTCCCTG CAATATCCAG ATGGAAGATC CCACTTTTAA
3951 AGAGAATTAT CGCTTCCATG CAATCAATGG CTACATAATG GATACACTAC
4001 CTGGCTTAGT AATGGCTCAG GATCAAAGGA TTCGATGGTA TCTGCTCAGC
4051 ATGGGCAGCA ATGAAAACAT CCATTCTATT CATTTCAGTG GACATGTGTT
4101 CACTGTACGA AAAAAAGAGG AGTATAAAAT GGCACTGTAC AATCTCTATC
4151 CAGGTGTTTT TGAGACAGTG GAAATGTTAC CATCCAAAGC TGGAATTTGG
4201 CGGGTGGAAT GCCTTATTGG CGAGCATCTA CATGCTGGGA TGAGCACACT
4251 TTTTCTGGTG TACAGCAATA AGTGTCAGAC TCCCCTGGGA ATGGCTTCTG
4301 GACACATTAG AGATTTTCAG ATTACAGCTT CAGGACAATA TGGACAGTGG
4351 GCCCCAAAGC TGGCCAGACT TCATTATTCC GGATCAATCA ATGCCTGGAG
4401 CACCAAGGAG CCCTTTTCTT GGATCAAGGT GGATCTGTTG GCACCAATGA
4451 TTATTCACGG CATCAAGACC CAGGGTGCCC GTCAGAAGTT CTCCAGCCTC
4501 TACATCTCTC AGTTTATCAT CATGTATAGT CTTGATGGGA AGAAGTGGCA
4551 GACTTATCGA GGAAATTCCA CTGGAACCTT AATGGTCTTC TTTGGCAATG
4601 TGGATTCATC TGGGATAAAA CACAATATTT TTAACCCTCC AATTATTGCT
4651 CGATACATCC GTTTGCACCC AACTCATTAT AGCATTCGCA GCACTCTTCG
4701 CATGGAGTTG ATGGGCTGTG ATTTAAATAG TTGCAGCATG CCATTGGGAA
4751 TGGAGAGTAA AGCAATATCA GATGCACAGA TTACTGCTTC ATCCTACTTT
4801 ACCAATATGT TTGCCACCTG GTCTCCTTCA AAAGCTCGAC TTCACCTCCA
4851 AGGGAGGAGT AATGCCTGGA GACCTCAGGT GAATAATCCA AAAGAGTGGC
4901 TGCAAGTGGA CTTCCAGAAG ACAATGAAAG TCACAGGAGT AACTACTCAG
4951 GGAGTAAAAT CTCTGCTTAC CAGCATGTAT GTGAAGGAGT TCCTCATCTC
5001 CAGCAGTCAA GATGGCCATC AGTGGACTCT CTTTTTTCAG AATGGCAAAG
5051 TAAAGGTTTT TCAGGGAAAT CAAGACTCCT TCACACCTGT GGTGAACTCT
5101 CTAGACCCAC CGTTACTGAC TCGCTACCTT CGAATTCACC CCCAGAGTTG
5151 GGTGCACCAG ATTGCCCTGA GGATGGAGGT TCTGGGCTGC GAGGCACAGG
5201 ACCTCTACGA CAAAACTCAC ACATGCCCAC CGTGCCCAGC TCCAGAACTC
5251 CTGGGCGGAC CGTCAGTCTT CCTCTTCCCC CCAAAACCCA AGGACACCCT
5301 CATGATCTCC CGGACCCCTG AGGTCACATG CGTGGTGGTG GACGTGAGCC
5351 ACGAAGACCC TGAGGTCAAG TTCAACTGGT ACGTGGACGG CGTGGAGGTG
5401 CATAATGCCA AGACAAAGCC GCGGGAGGAG CAGTACAACA GCACGTACCG
5451 TGTGGTCAGC GTCCTCACCG TCCTGCACCA GGACTGGCTG AATGGCAAGG
5501 AGTACAAGTG CAAGGTCTCC AACAAAGCCC TCCCAGCCCC CATCGAGAAA
5551 ACCATCTCCA AAGCCAAAGG GCAGCCCCGA GAACCACAGG TGTACACCCT
5601 GCCCCCATCC CGGGATGAGC TGACCAAGAA CCAGGTCAGC CTGACCTGCC
5651 TGGTCAAAGG CTTCTATCCC AGCGACATCG CCGTGGAGTG GGAGAGCAAT
5701 GGGCAGCCGG AGAACAACTA CAAGACCACG CCTCCCGTGT TGGACTCCGA
5751 CGGCTCCTTC TTCCTCTACA GCAAGCTCAC CGTGGACAAG AGCAGGTGGC
5801 AGCAGGGGAA CGTCTTCTCA TGCTCCGTGA TGCATGAGGC TCTGCACAAC
5851 CACTACACGC AGAAGAGCCT CTCCCTGTCT CCGGGTAAAT GA
Последовательность белка pSYN FVIII 310 (FVIII с полной делецией домена B, за исключением 2 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 742) (SEQ ID NO: 171)
1 ATRRYYLGAV ELSWDYMQSD LGELPVDARF PPRVPKSFPF NTSVVYKKTL
51 FVEFTDHLFN IAKPRPPWMG LLGPTIQAEV YDTVVITLKN MASHPVSLHA
101 VGVSYWKASE GAEYDDQTSQ REKEDDKVFP GGSHTYVWQV LKENGPMASD
151 PLCLTYSYLS HVDLVKDLNS GLIGALLVCR EGSLAKEKTQ TLHKFILLFA
201 VFDEGKSWHS ETKNSLMQDR DAASARAWPK MHTVNGYVNR SLPGLIGCHR
251 KSVYWHVIGM GTTPEVHSIF LEGHTFLVRN HRQASLEISP ITFLTAQTLL
301 MDLGQFLLFC HISSHQHDGM EAYVKVDSCP EEPQLRMKNN EEAEDYDDDL
351 TDSEMDVVRF DDDNSPSFIQ IRSVAKKHPK TWVHYIAAEE EDWDYAPLVL
401 APDDRSYKSQ YLNNGPQRIG RKYKKVRFMA YTDETFKTRE AIQHESGILG
451 PLLYGEVGDT LLIIFKNQAS RPYNIYPHGI TDVRPLYSRR LPKGVKHLKD
501 FPILPGEIFK YKWTVTVEDG PTKSDPRCLT RYYSSFVNME RDLASGLIGP
551 LLICYKESVD QRGNQIMSDK RNVILFSVFD ENRSWYLTEN IQRFLPNPAG
601 VQLEDPEFQA SNIMHSINGY VFDSLQLSVC LHEVAYWYIL SIGAQTDFLS
651 VFFSGYTFKH KMVYEDTLTL FPFSGETVFM SMENPGLWIL GCHNSDFRNR
701 GMTALLKVSS CDKNTGDYYE DSYEDISAYL LSKNNAIEPR SFGTSESATP
751 ESGPGSEPAT SGSETPGTSE SATPESGPGS EPATSGSETP GTSESATPES
801 GPGTSTEPSE GSAPGSPAGS PTSTEEGTSE SATPESGPGS EPATSGSETP
851 GTSESATPES GPGSPAGSPT STEEGSPAGS PTSTEEGTST EPSEGSAPGT
901 SESATPESGP GTSESATPES GPGTSESATP ESGPGSEPAT SGSETPGSEP
951 ATSGSETPGS PAGSPTSTEE GTSTEPSEGS APGTSTEPSE GSAPGSEPAT
1001 SGSETPGTSE SATPESGPGT STEPSEGSAP ASSEITRTTL QSDQEEIDYD
1051 DTISVEMKKE DFDIYDEDEN QSPRSFQKKT RHYFIAAVER LWDYGMSSSP
1101 HVLRNRAQSG SVPQFKKVVF QEFTDGSFTQ PLYRGELNEH LGLLGPYIRA
1151 EVEDNIMVTF RNQASRPYSF YSSLISYEED QRQGAEPRKN FVKPNETKTY
1201 FWKVQHHMAP TKDEFDCKAW AYFSDVDLEK DVHSGLIGPL LVCHTNTLNP
1251 AHGRQVTVQE FALFFTIFDE TKSWYFTENM ERNCRAPCNI QMEDPTFKEN
1301 YRFHAINGYI MDTLPGLVMA QDQRIRWYLL SMGSNENIHS IHFSGHVFTV
1351 RKKEEYKMAL YNLYPGVFET VEMLPSKAGI WRVECLIGEH LHAGMSTLFL
1401 VYSNKCQTPL GMASGHIRDF QITASGQYGQ WAPKLARLHY SGSINAWSTK
1451 EPFSWIKVDL LAPMIIHGIK TQGARQKFSS LYISQFIIMY SLDGKKWQTY
1501 RGNSTGTLMV FFGNVDSSGI KHNIFNPPII ARYIRLHPTH YSIRSTLRME
1551 LMGCDLNSCS MPLGMESKAI SDAQITASSY FTNMFATWSP SKARLHLQGR
1601 SNAWRPQVNN PKEWLQVDFQ KTMKVTGVTT QGVKSLLTSM YVKEFLISSS
1651 QDGHQWTLFF QNGKVKVFQG NQDSFTPVVN SLDPPLLTRY LRIHPQSWVH
1701 QIALRMEVLG CEAQDLYDKT HTCPPCPAPE LLGGPSVFLF PPKPKDTLMI
1751 SRTPEVTCVV VDVSHEDPEV KFNWYVDGVE VHNAKTKPRE EQYNSTYRVV
1801 SVLTVLHQDW LNGKEYKCKV SNKALPAPIE KTISKAKGQP REPQVYTLPP
1851 SRDELTKNQV SLTCLVKGFY PSDIAVEWES NGQPENNYKT TPPVLDSDGS
1901 FFLYSKLTVD KSRWQQGNVF SCSVMHEALH NHYTQKSLSL SPGK*
Нуклеотидная последовательность pSYN FVIII 312 (кодирующая FVIII с полной делецией домена B, за исключением 5 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 745 - вариант B5) (SEQ ID NO: 172)
1 ATGCAAATAG AGCTCTCCAC CTGCTTCTTT CTGTGCCTTT TGCGATTCTG
51 CTTTAGTGCC ACCAGAAGAT ACTACCTGGG TGCAGTGGAA CTGTCATGGG
101 ACTATATGCA AAGTGATCTC GGTGAGCTGC CTGTGGACGC AAGATTTCCT
151 CCTAGAGTGC CAAAATCTTT TCCATTCAAC ACCTCAGTCG TGTACAAAAA
201 GACTCTGTTT GTAGAATTCA CGGATCACCT TTTCAACATC GCTAAGCCAA
251 GGCCACCCTG GATGGGTCTG CTAGGTCCTA CCATCCAGGC TGAGGTTTAT
301 GATACAGTGG TCATTACACT TAAGAACATG GCTTCCCATC CTGTCAGTCT
351 TCATGCTGTT GGTGTATCCT ACTGGAAAGC TTCTGAGGGA GCTGAATATG
401 ATGATCAGAC CAGTCAAAGG GAGAAAGAAG ATGATAAAGT CTTCCCTGGT
451 GGAAGCCATA CATATGTCTG GCAGGTCCTG AAAGAGAATG GTCCAATGGC
501 CTCTGACCCA CTGTGCCTTA CCTACTCATA TCTTTCTCAT GTGGACCTGG
551 TAAAAGACTT GAATTCAGGC CTCATTGGAG CCCTACTAGT ATGTAGAGAA
601 GGGAGTCTGG CCAAGGAAAA GACACAGACC TTGCACAAAT TTATACTACT
651 TTTTGCTGTA TTTGATGAAG GGAAAAGTTG GCACTCAGAA ACAAAGAACT
701 CCTTGATGCA GGATAGGGAT GCTGCATCTG CTCGGGCCTG GCCTAAAATG
751 CACACAGTCA ATGGTTATGT AAACAGGTCT CTGCCAGGTC TGATTGGATG
801 CCACAGGAAA TCAGTCTATT GGCATGTGAT TGGAATGGGC ACCACTCCTG
851 AAGTGCACTC AATATTCCTC GAAGGTCACA CATTTCTTGT GAGGAACCAT
901 CGCCAGGCGT CCTTGGAAAT CTCGCCAATA ACTTTCCTTA CTGCTCAAAC
951 ACTCTTGATG GACCTTGGAC AGTTTCTACT GTTTTGTCAT ATCTCTTCCC
1001 ACCAACATGA TGGCATGGAA GCTTATGTCA AAGTAGACAG CTGTCCAGAG
1051 GAACCCCAAC TACGAATGAA AAATAATGAA GAAGCGGAAG ACTATGATGA
1101 TGATCTTACT GATTCTGAAA TGGATGTGGT CAGGTTTGAT GATGACAACT
1151 CTCCTTCCTT TATCCAAATT CGCTCAGTTG CCAAGAAGCA TCCTAAAACT
1201 TGGGTACATT ACATTGCTGC TGAAGAGGAG GACTGGGACT ATGCTCCCTT
1251 AGTCCTCGCC CCCGATGACA GAAGTTATAA AAGTCAATAT TTGAACAATG
1301 GCCCTCAGCG GATTGGTAGG AAGTACAAAA AAGTCCGATT TATGGCATAC
1351 ACAGATGAAA CCTTTAAGAC TCGTGAAGCT ATTCAGCATG AATCAGGAAT
1401 CTTGGGACCT TTACTTTATG GGGAAGTTGG AGACACACTG TTGATTATAT
1451 TTAAGAATCA AGCAAGCAGA CCATATAACA TCTACCCTCA CGGAATCACT
1501 GATGTCCGTC CTTTGTATTC AAGGAGATTA CCAAAAGGTG TAAAACATTT
1551 GAAGGATTTT CCAATTCTGC CAGGAGAAAT ATTCAAATAT AAATGGACAG
1601 TGACTGTAGA AGATGGGCCA ACTAAATCAG ATCCTCGGTG CCTGACCCGC
1651 TATTACTCTA GTTTCGTTAA TATGGAGAGA GATCTAGCTT CAGGACTCAT
1701 TGGCCCTCTC CTCATCTGCT ACAAAGAATC TGTAGATCAA AGAGGAAACC
1751 AGATAATGTC AGACAAGAGG AATGTCATCC TGTTTTCTGT ATTTGATGAG
1801 AACCGAAGCT GGTACCTCAC AGAGAATATA CAACGCTTTC TCCCCAATCC
1851 AGCTGGAGTG CAGCTTGAGG ATCCAGAGTT CCAAGCCTCC AACATCATGC
1901 ACAGCATCAA TGGCTATGTT TTTGATAGTT TGCAGTTGTC AGTTTGTTTG
1951 CATGAGGTGG CATACTGGTA CATTCTAAGC ATTGGAGCAC AGACTGACTT
2001 CCTTTCTGTC TTCTTCTCTG GATATACCTT CAAACACAAA ATGGTCTATG
2051 AAGACACACT CACCCTATTC CCATTCTCAG GAGAAACTGT CTTCATGTCG
2101 ATGGAAAACC CAGGTCTATG GATTCTGGGG TGCCACAACT CAGACTTTCG
2151 GAACAGAGGC ATGACCGCCT TACTGAAGGT TTCTAGTTGT GACAAGAACA
2201 CTGGTGATTA TTACGAGGAC AGTTATGAAG ATATTTCAGC ATACTTGCTG
2251 AGTAAAAACA ATGCCATTGA ACCAAGAAGC TTCTCTCAAA ACGGTACCTC
2301 AGAGTCTGCT ACCCCCGAGT CAGGGCCAGG ATCAGAGCCA GCCACCTCCG
2351 GGTCTGAGAC ACCCGGGACT TCCGAGAGTG CCACCCCTGA GTCCGGACCC
2401 GGGTCCGAGC CCGCCACTTC CGGCTCCGAA ACTCCCGGCA CAAGCGAGAG
2451 CGCTACCCCA GAGTCAGGAC CAGGAACATC TACAGAGCCC TCTGAAGGCT
2501 CCGCTCCAGG GTCCCCAGCC GGCAGTCCCA CTAGCACCGA GGAGGGAACC
2551 TCTGAAAGCG CCACACCCGA ATCAGGGCCA GGGTCTGAGC CTGCTACCAG
2601 CGGCAGCGAG ACACCAGGCA CCTCTGAGTC CGCCACACCA GAGTCCGGAC
2651 CCGGATCTCC CGCTGGGAGC CCCACCTCCA CTGAGGAGGG ATCTCCTGCT
2701 GGCTCTCCAA CATCTACTGA GGAAGGTACC TCAACCGAGC CATCCGAGGG
2751 ATCAGCTCCC GGCACCTCAG AGTCGGCAAC CCCGGAGTCT GGACCCGGAA
2801 CTTCCGAAAG TGCCACACCA GAGTCCGGTC CCGGGACTTC AGAATCAGCA
2851 ACACCCGAGT CCGGCCCTGG GTCTGAACCC GCCACAAGTG GTAGTGAGAC
2901 ACCAGGATCA GAACCTGCTA CCTCAGGGTC AGAGACACCC GGATCTCCGG
2951 CAGGCTCACC AACCTCCACT GAGGAGGGCA CCAGCACAGA ACCAAGCGAG
3001 GGCTCCGCAC CCGGAACAAG CACTGAACCC AGTGAGGGTT CAGCACCCGG
3051 CTCTGAGCCG GCCACAAGTG GCAGTGAGAC ACCCGGCACT TCAGAGAGTG
3101 CCACCCCCGA GAGTGGCCCA GGCACTAGTA CCGAGCCCTC TGAAGGCAGT
3151 GCGCCAGCCT CGAGCGAAAT AACTCGTACT ACTCTTCAGT CAGATCAAGA
3201 GGAAATCGAT TATGATGATA CCATATCAGT TGAAATGAAG AAGGAAGATT
3251 TTGACATTTA TGATGAGGAT GAAAATCAGA GCCCCCGCAG CTTTCAAAAG
3301 AAAACACGAC ACTATTTTAT TGCTGCAGTG GAGAGGCTCT GGGATTATGG
3351 GATGAGTAGC TCCCCACATG TTCTAAGAAA CAGGGCTCAG AGTGGCAGTG
3401 TCCCTCAGTT CAAGAAAGTT GTTTTCCAGG AATTTACTGA TGGCTCCTTT
3451 ACTCAGCCCT TATACCGTGG AGAACTAAAT GAACATTTGG GACTCCTGGG
3501 GCCATATATA AGAGCAGAAG TTGAAGATAA TATCATGGTA ACTTTCAGAA
3551 ATCAGGCCTC TCGTCCCTAT TCCTTCTATT CTAGCCTTAT TTCTTATGAG
3601 GAAGATCAGA GGCAAGGAGC AGAACCTAGA AAAAACTTTG TCAAGCCTAA
3651 TGAAACCAAA ACTTACTTTT GGAAAGTGCA ACATCATATG GCACCCACTA
3701 AAGATGAGTT TGACTGCAAA GCCTGGGCTT ATTTCTCTGA TGTTGACCTG
3751 GAAAAAGATG TGCACTCAGG CCTGATTGGA CCCCTTCTGG TCTGCCACAC
3801 TAACACACTG AACCCTGCTC ATGGGAGACA AGTGACAGTA CAGGAATTTG
3851 CTCTGTTTTT CACCATCTTT GATGAGACCA AAAGCTGGTA CTTCACTGAA
3901 AATATGGAAA GAAACTGCAG GGCTCCCTGC AATATCCAGA TGGAAGATCC
3951 CACTTTTAAA GAGAATTATC GCTTCCATGC AATCAATGGC TACATAATGG
4001 ATACACTACC TGGCTTAGTA ATGGCTCAGG ATCAAAGGAT TCGATGGTAT
4051 CTGCTCAGCA TGGGCAGCAA TGAAAACATC CATTCTATTC ATTTCAGTGG
4101 ACATGTGTTC ACTGTACGAA AAAAAGAGGA GTATAAAATG GCACTGTACA
4151 ATCTCTATCC AGGTGTTTTT GAGACAGTGG AAATGTTACC ATCCAAAGCT
4201 GGAATTTGGC GGGTGGAATG CCTTATTGGC GAGCATCTAC ATGCTGGGAT
4251 GAGCACACTT TTTCTGGTGT ACAGCAATAA GTGTCAGACT CCCCTGGGAA
4301 TGGCTTCTGG ACACATTAGA GATTTTCAGA TTACAGCTTC AGGACAATAT
4351 GGACAGTGGG CCCCAAAGCT GGCCAGACTT CATTATTCCG GATCAATCAA
4401 TGCCTGGAGC ACCAAGGAGC CCTTTTCTTG GATCAAGGTG GATCTGTTGG
4451 CACCAATGAT TATTCACGGC ATCAAGACCC AGGGTGCCCG TCAGAAGTTC
4501 TCCAGCCTCT ACATCTCTCA GTTTATCATC ATGTATAGTC TTGATGGGAA
4551 GAAGTGGCAG ACTTATCGAG GAAATTCCAC TGGAACCTTA ATGGTCTTCT
4601 TTGGCAATGT GGATTCATCT GGGATAAAAC ACAATATTTT TAACCCTCCA
4651 ATTATTGCTC GATACATCCG TTTGCACCCA ACTCATTATA GCATTCGCAG
4701 CACTCTTCGC ATGGAGTTGA TGGGCTGTGA TTTAAATAGT TGCAGCATGC
4751 CATTGGGAAT GGAGAGTAAA GCAATATCAG ATGCACAGAT TACTGCTTCA
4801 TCCTACTTTA CCAATATGTT TGCCACCTGG TCTCCTTCAA AAGCTCGACT
4851 TCACCTCCAA GGGAGGAGTA ATGCCTGGAG ACCTCAGGTG AATAATCCAA
4901 AAGAGTGGCT GCAAGTGGAC TTCCAGAAGA CAATGAAAGT CACAGGAGTA
4951 ACTACTCAGG GAGTAAAATC TCTGCTTACC AGCATGTATG TGAAGGAGTT
5001 CCTCATCTCC AGCAGTCAAG ATGGCCATCA GTGGACTCTC TTTTTTCAGA
5051 ATGGCAAAGT AAAGGTTTTT CAGGGAAATC AAGACTCCTT CACACCTGTG
5101 GTGAACTCTC TAGACCCACC GTTACTGACT CGCTACCTTC GAATTCACCC
5151 CCAGAGTTGG GTGCACCAGA TTGCCCTGAG GATGGAGGTT CTGGGCTGCG
5201 AGGCACAGGA CCTCTACGAC AAAACTCACA CATGCCCACC GTGCCCAGCT
5251 CCAGAACTCC TGGGCGGACC GTCAGTCTTC CTCTTCCCCC CAAAACCCAA
5301 GGACACCCTC ATGATCTCCC GGACCCCTGA GGTCACATGC GTGGTGGTGG
5351 ACGTGAGCCA CGAAGACCCT GAGGTCAAGT TCAACTGGTA CGTGGACGGC
5401 GTGGAGGTGC ATAATGCCAA GACAAAGCCG CGGGAGGAGC AGTACAACAG
5451 CACGTACCGT GTGGTCAGCG TCCTCACCGT CCTGCACCAG GACTGGCTGA
5501 ATGGCAAGGA GTACAAGTGC AAGGTCTCCA ACAAAGCCCT CCCAGCCCCC
5551 ATCGAGAAAA CCATCTCCAA AGCCAAAGGG CAGCCCCGAG AACCACAGGT
5601 GTACACCCTG CCCCCATCCC GGGATGAGCT GACCAAGAAC CAGGTCAGCC
5651 TGACCTGCCT GGTCAAAGGC TTCTATCCCA GCGACATCGC CGTGGAGTGG
5701 GAGAGCAATG GGCAGCCGGA GAACAACTAC AAGACCACGC CTCCCGTGTT
5751 GGACTCCGAC GGCTCCTTCT TCCTCTACAG CAAGCTCACC GTGGACAAGA
5801 GCAGGTGGCA GCAGGGGAAC GTCTTCTCAT GCTCCGTGAT GCATGAGGCT
5851 CTGCACAACC ACTACACGCA GAAGAGCCTC TCCCTGTCTC CGGGTAAATG
Последовательность белка pSYN FVIII 312 (FVIII с полной делецией домена B, за исключением 5 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 745 - вариант B5) (SEQ ID NO: 173)
1 ATRRYYLGAV ELSWDYMQSD LGELPVDARF PPRVPKSFPF NTSVVYKKTL
51 FVEFTDHLFN IAKPRPPWMG LLGPTIQAEV YDTVVITLKN MASHPVSLHA
101 VGVSYWKASE GAEYDDQTSQ REKEDDKVFP GGSHTYVWQV LKENGPMASD
151 PLCLTYSYLS HVDLVKDLNS GLIGALLVCR EGSLAKEKTQ TLHKFILLFA
201 VFDEGKSWHS ETKNSLMQDR DAASARAWPK MHTVNGYVNR SLPGLIGCHR
251 KSVYWHVIGM GTTPEVHSIF LEGHTFLVRN HRQASLEISP ITFLTAQTLL
301 MDLGQFLLFC HISSHQHDGM EAYVKVDSCP EEPQLRMKNN EEAEDYDDDL
351 TDSEMDVVRF DDDNSPSFIQ IRSVAKKHPK TWVHYIAAEE EDWDYAPLVL
401 APDDRSYKSQ YLNNGPQRIG RKYKKVRFMA YTDETFKTRE AIQHESGILG
451 PLLYGEVGDT LLIIFKNQAS RPYNIYPHGI TDVRPLYSRR LPKGVKHLKD
501 FPILPGEIFK YKWTVTVEDG PTKSDPRCLT RYYSSFVNME RDLASGLIGP
551 LLICYKESVD QRGNQIMSDK RNVILFSVFD ENRSWYLTEN IQRFLPNPAG
601 VQLEDPEFQA SNIMHSINGY VFDSLQLSVC LHEVAYWYIL SIGAQTDFLS
651 VFFSGYTFKH KMVYEDTLTL FPFSGETVFM SMENPGLWIL GCHNSDFRNR
701 GMTALLKVSS CDKNTGDYYE DSYEDISAYL LSKNNAIEPR SFSQNGTSES
751 ATPESGPGSE PATSGSETPG TSESATPESG PGSEPATSGS ETPGTSESAT
801 PESGPGTSTE PSEGSAPGSP AGSPTSTEEG TSESATPESG PGSEPATSGS
851 ETPGTSESAT PESGPGSPAG SPTSTEEGSP AGSPTSTEEG TSTEPSEGSA
901 PGTSESATPE SGPGTSESAT PESGPGTSES ATPESGPGSE PATSGSETPG
951 SEPATSGSET PGSPAGSPTS TEEGTSTEPS EGSAPGTSTE PSEGSAPGSE
1001 PATSGSETPG TSESATPESG PGTSTEPSEG SAPASSEITR TTLQSDQEEI
1051 DYDDTISVEM KKEDFDIYDE DENQSPRSFQ KKTRHYFIAA VERLWDYGMS
1101 SSPHVLRNRA QSGSVPQFKK VVFQEFTDGS FTQPLYRGEL NEHLGLLGPY
1151 IRAEVEDNIM VTFRNQASRP YSFYSSLISY EEDQRQGAEP RKNFVKPNET
1201 KTYFWKVQHH MAPTKDEFDC KAWAYFSDVD LEKDVHSGLI GPLLVCHTNT
1251 LNPAHGRQVT VQEFALFFTI FDETKSWYFT ENMERNCRAP CNIQMEDPTF
1301 KENYRFHAIN GYIMDTLPGL VMAQDQRIRW YLLSMGSNEN IHSIHFSGHV
1351 FTVRKKEEYK MALYNLYPGV FETVEMLPSK AGIWRVECLI GEHLHAGMST
1401 LFLVYSNKCQ TPLGMASGHI RDFQITASGQ YGQWAPKLAR LHYSGSINAW
1451 STKEPFSWIK VDLLAPMIIH GIKTQGARQK FSSLYISQFI IMYSLDGKKW
1501 QTYRGNSTGT LMVFFGNVDS SGIKHNIFNP PIIARYIRLH PTHYSIRSTL
1551 RMELMGCDLN SCSMPLGMES KAISDAQITA SSYFTNMFAT WSPSKARLHL
1601 QGRSNAWRPQ VNNPKEWLQV DFQKTMKVTG VTTQGVKSLL TSMYVKEFLI
1651 SSSQDGHQWT LFFQNGKVKV FQGNQDSFTP VVNSLDPPLL TRYLRIHPQS
1701 WVHQIALRME VLGCEAQDLY DKTHTCPPCP APELLGGPSV FLFPPKPKDT
1751 LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY
1801 RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT
1851 LPPSRDELTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
1901 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPGK*
Нуклеотидная последовательность pSYN VWF059 (кодирующая D'D3 VWF-Fc с сайтом расщепления тромбином в кислой области 2 (а2) в линкере) (SEQ ID NO: 196)
1 ATGATTCCTG CCAGATTTGC CGGGGTGCTG CTTGCTCTGG CCCTCATTTT
51 GCCAGGGACC CTTTGTGCAG AAGGAACTCG CGGCAGGTCA TCCACGGCCC
101 GATGCAGCCT TTTCGGAAGT GACTTCGTCA ACACCTTTGA TGGGAGCATG
151 TACAGCTTTG CGGGATACTG CAGTTACCTC CTGGCAGGGG GCTGCCAGAA
201 ACGCTCCTTC TCGATTATTG GGGACTTCCA GAATGGCAAG AGAGTGAGCC
251 TCTCCGTGTA TCTTGGGGAA TTTTTTGACA TCCATTTGTT TGTCAATGGT
301 ACCGTGACAC AGGGGGACCA AAGAGTCTCC ATGCCCTATG CCTCCAAAGG
351 GCTGTATCTA GAAACTGAGG CTGGGTACTA CAAGCTGTCC GGTGAGGCCT
401 ATGGCTTTGT GGCCAGGATC GATGGCAGCG GCAACTTTCA AGTCCTGCTG
451 TCAGACAGAT ACTTCAACAA GACCTGCGGG CTGTGTGGCA ACTTTAACAT
501 CTTTGCTGAA GATGACTTTA TGACCCAAGA AGGGACCTTG ACCTCGGACC
551 CTTATGACTT TGCCAACTCA TGGGCTCTGA GCAGTGGAGA ACAGTGGTGT
601 GAACGGGCAT CTCCTCCCAG CAGCTCATGC AACATCTCCT CTGGGGAAAT
651 GCAGAAGGGC CTGTGGGAGC AGTGCCAGCT TCTGAAGAGC ACCTCGGTGT
701 TTGCCCGCTG CCACCCTCTG GTGGACCCCG AGCCTTTTGT GGCCCTGTGT
751 GAGAAGACTT TGTGTGAGTG TGCTGGGGGG CTGGAGTGCG CCTGCCCTGC
801 CCTCCTGGAG TACGCCCGGA CCTGTGCCCA GGAGGGAATG GTGCTGTACG
851 GCTGGACCGA CCACAGCGCG TGCAGCCCAG TGTGCCCTGC TGGTATGGAG
901 TATAGGCAGT GTGTGTCCCC TTGCGCCAGG ACCTGCCAGA GCCTGCACAT
951 CAATGAAATG TGTCAGGAGC GATGCGTGGA TGGCTGCAGC TGCCCTGAGG
1001 GACAGCTCCT GGATGAAGGC CTCTGCGTGG AGAGCACCGA GTGTCCCTGC
1051 GTGCATTCCG GAAAGCGCTA CCCTCCCGGC ACCTCCCTCT CTCGAGACTG
1101 CAACACCTGC ATTTGCCGAA ACAGCCAGTG GATCTGCAGC AATGAAGAAT
1151 GTCCAGGGGA GTGCCTTGTC ACTGGTCAAT CCCACTTCAA GAGCTTTGAC
1201 AACAGATACT TCACCTTCAG TGGGATCTGC CAGTACCTGC TGGCCCGGGA
1251 TTGCCAGGAC CACTCCTTCT CCATTGTCAT TGAGACTGTC CAGTGTGCTG
1301 ATGACCGCGA CGCTGTGTGC ACCCGCTCCG TCACCGTCCG GCTGCCTGGC
1351 CTGCACAACA GCCTTGTGAA ACTGAAGCAT GGGGCAGGAG TTGCCATGGA
1401 TGGCCAGGAC ATCCAGCTCC CCCTCCTGAA AGGTGACCTC CGCATCCAGC
1451 ATACAGTGAC GGCCTCCGTG CGCCTCAGCT ACGGGGAGGA CCTGCAGATG
1501 GACTGGGATG GCCGCGGGAG GCTGCTGGTG AAGCTGTCCC CCGTCTATGC
1551 CGGGAAGACC TGCGGCCTGT GTGGGAATTA CAATGGCAAC CAGGGCGACG
1601 ACTTCCTTAC CCCCTCTGGG CTGGCGGAGC CCCGGGTGGA GGACTTCGGG
1651 AACGCCTGGA AGCTGCACGG GGACTGCCAG GACCTGCAGA AGCAGCACAG
1701 CGATCCCTGC GCCCTCAACC CGCGCATGAC CAGGTTCTCC GAGGAGGCGT
1751 GCGCGGTCCT GACGTCCCCC ACATTCGAGG CCTGCCATCG TGCCGTCAGC
1801 CCGCTGCCCT ACCTGCGGAA CTGCCGCTAC GACGTGTGCT CCTGCTCGGA
1851 CGGCCGCGAG TGCCTGTGCG GCGCCCTGGC CAGCTATGCC GCGGCCTGCG
1901 CGGGGAGAGG CGTGCGCGTC GCGTGGCGCG AGCCAGGCCG CTGTGAGCTG
1951 AACTGCCCGA AAGGCCAGGT GTACCTGCAG TGCGGGACCC CCTGCAACCT
2001 GACCTGCCGC TCTCTCTCTT ACCCGGATGA GGAATGCAAT GAGGCCTGCC
2051 TGGAGGGCTG CTTCTGCCCC CCAGGGCTCT ACATGGATGA GAGGGGGGAC
2101 TGCGTGCCCA AGGCCCAGTG CCCCTGTTAC TATGACGGTG AGATCTTCCA
2151 GCCAGAAGAC ATCTTCTCAG ACCATCACAC CATGTGCTAC TGTGAGGATG
2201 GCTTCATGCA CTGTACCATG AGTGGAGTCC CCGGAAGCTT GCTGCCTGAC
2251 GCTGTCCTCA GCAGTCCCCT GTCTCATCGC AGCAAAAGGA GCCTATCCTG
2301 TCGGCCCCCC ATGGTCAAGC TGGTGTGTCC CGCTGACAAC CTGCGGGCTG
2351 AAGGGCTCGA GTGTACCAAA ACGTGCCAGA ACTATGACCT GGAGTGCATG
2401 AGCATGGGCT GTGTCTCTGG CTGCCTCTGC CCCCCGGGCA TGGTCCGGCA
2451 TGAGAACAGA TGTGTGGCCC TGGAAAGGTG TCCCTGCTTC CATCAGGGCA
2501 AGGAGTATGC CCCTGGAGAA ACAGTGAAGA TTGGCTGCAA CACTTGTGTC
2551 TGTCGGGACC GGAAGTGGAA CTGCACAGAC CATGTGTGTG ATGCCACGTG
2601 CTCCACGATC GGCATGGCCC ACTACCTCAC CTTCGACGGG CTCAAATACC
2651 TGTTCCCCGG GGAGTGCCAG TACGTTCTGG TGCAGGATTA CTGCGGCAGT
2701 AACCCTGGGA CCTTTCGGAT CCTAGTGGGG AATAAGGGAT GCAGCCACCC
2751 CTCAGTGAAA TGCAAGAAAC GGGTCACCAT CCTGGTGGAG GGAGGAGAGA
2801 TTGAGCTGTT TGACGGGGAG GTGAATGTGA AGAGGCCCAT GAAGGATGAG
2851 ACTCACTTTG AGGTGGTGGA GTCTGGCCGG TACATCATTC TGCTGCTGGG
2901 CAAAGCCCTC TCCGTGGTCT GGGACCGCCA CCTGAGCATC TCCGTGGTCC
2951 TGAAGCAGAC ATACCAGGAG AAAGTGTGTG GCCTGTGTGG GAATTTTGAT
3001 GGCATCCAGA ACAATGACCT CACCAGCAGC AACCTCCAAG TGGAGGAAGA
3051 CCCTGTGGAC TTTGGGAACT CCTGGAAAGT GAGCTCGCAG TGTGCTGACA
3101 CCAGAAAAGT GCCTCTGGAC TCATCCCCTG CCACCTGCCA TAACAACATC
3151 ATGAAGCAGA CGATGGTGGA TTCCTCCTGT AGAATCCTTA CCAGTGACGT
3201 CTTCCAGGAC TGCAACAAGC TGGTGGACCC CGAGCCATAT CTGGATGTCT
3251 GCATTTACGA CACCTGCTCC TGTGAGTCCA TTGGGGACTG CGCCGCATTC
3301 TGCGACACCA TTGCTGCCTA TGCCCACGTG TGTGCCCAGC ATGGCAAGGT
3351 GGTGACCTGG AGGACGGCCA CATTGTGCCC CCAGAGCTGC GAGGAGAGGA
3401 ATCTCCGGGA GAACGGGTAT GAGGCTGAGT GGCGCTATAA CAGCTGTGCA
3451 CCTGCCTGTC AAGTCACGTG TCAGCACCCT GAGCCACTGG CCTGCCCTGT
3501 GCAGTGTGTG GAGGGCTGCC ATGCCCACTG CCCTCCAGGG AAAATCCTGG
3551 ATGAGCTTTT GCAGACCTGC GTTGACCCTG AAGACTGTCC AGTGTGTGAG
3601 GTGGCTGGCC GGCGTTTTGC CTCAGGAAAG AAAGTCACCT TGAATCCCAG
3651 TGACCCTGAG CACTGCCAGA TTTGCCACTG TGATGTTGTC AACCTCACCT
3701 GTGAAGCCTG CCAGGAGCCG ATATCGGGCG CGCCAACATC AGAGAGCGCC
3751 ACCCCTGAAA GTGGTCCCGG GAGCGAGCCA GCCACATCTG GGTCGGAAAC
3801 GCCAGGCACA AGTGAGTCTG CAACTCCCGA GTCCGGACCT GGCTCCGAGC
3851 CTGCCACTAG CGGCTCCGAG ACTCCGGGAA CTTCCGAGAG CGCTACACCA
3901 GAAAGCGGAC CCGGAACCAG TACCGAACCT AGCGAGGGCT CTGCTCCGGG
3951 CAGCCCAGCC GGCTCTCCTA CATCCACGGA GGAGGGCACT TCCGAATCCG
4001 CCACCCCGGA GTCAGGGCCA GGATCTGAAC CCGCTACCTC AGGCAGTGAG
4051 ACGCCAGGAA CGAGCGAGTC CGCTACACCG GAGAGTGGGC CAGGGAGCCC
4101 TGCTGGATCT CCTACGTCCA CTGAGGAAGG GTCACCAGCG GGCTCGCCCA
4151 CCAGCACTGA AGAAGGTGCC TCGATATCTG ACAAGAACAC TGGTGATTAT
4201 TACGAGGACA GTTATGAAGA TATTTCAGCA TACTTGCTGA GTAAAAACAA
4251 TGCCATTGAA CCAAGAAGCT TCTCTGACAA AACTCACACA TGCCCACCGT
4301 GCCCAGCTCC AGAACTCCTG GGCGGACCGT CAGTCTTCCT CTTCCCCCCA
4351 AAACCCAAGG ACACCCTCAT GATCTCCCGG ACCCCTGAGG TCACATGCGT
4401 GGTGGTGGAC GTGAGCCACG AAGACCCTGA GGTCAAGTTC AACTGGTACG
4451 TGGACGGCGT GGAGGTGCAT AATGCCAAGA CAAAGCCGCG GGAGGAGCAG
4501 TACAACAGCA CGTACCGTGT GGTCAGCGTC CTCACCGTCC TGCACCAGGA
4551 CTGGCTGAAT GGCAAGGAGT ACAAGTGCAA GGTCTCCAAC AAAGCCCTCC
4601 CAGCCCCCAT CGAGAAAACC ATCTCCAAAG CCAAAGGGCA GCCCCGAGAA
4651 CCACAGGTGT ACACCCTGCC CCCATCCCGG GATGAGCTGA CCAAGAACCA
4701 GGTCAGCCTG ACCTGCCTGG TCAAAGGCTT CTATCCCAGC GACATCGCCG
4751 TGGAGTGGGA GAGCAATGGG CAGCCGGAGA ACAACTACAA GACCACGCCT
4801 CCCGTGTTGG ACTCCGACGG CTCCTTCTTC CTCTACAGCA AGCTCACCGT
4851 GGACAAGAGC AGGTGGCAGC AGGGGAACGT CTTCTCATGC TCCGTGATGC
4901 ATGAGGCTCT GCACAACCAC TACACGCAGA AGAGCCTCTC CCTGTCTCCG
4951 GGTAAATGA
Последовательность белка pSYN VWF059 (D'D3 VWF-Fc с сайтом расщепления FVIII тромбином в области a2 в линкере) - зона, выделенная жирным шрифтом и подчеркиванием, показывает область a2 (SEQ ID NO: 197)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGAPTSESA
1251 TPESGPGSEP ATSGSETPGT SESATPESGP GSEPATSGSE TPGTSESATP
1301 ESGPGTSTEP SEGSAPGSPA GSPTSTEEGT SESATPESGP GSEPATSGSE
1351 TPGTSESATP ESGPGSPAGS PTSTEEGSPA GSPTSTEEGA SISDKNTGDY
1401 YEDSYEDISA YLLSKNNAIE PRSFSDKTHT CPPCPAPELL GGPSVFLFPP
1451 KPKDTLMISR TPEVTCVVVD VSHEDPEVKF NWYVDGVEVH NAKTKPREEQ
1501 YNSTYRVVSV LTVLHQDWLN GKEYKCKVSN KALPAPIEKT ISKAKGQPRE
1551 PQVYTLPPSR DELTKNQVSL TCLVKGFYPS DIAVEWESNG QPENNYKTTP
1601 PVLDSDGSFF LYSKLTVDKS RWQQGNVFSC SVMHEALHNH YTQKSLSLSP
1651 GK*
Нуклеотидная последовательность pSYN VWF062 (кодирующая D'D3 VWF-Fc без сайта расщепления тромбином в линкере) (SEQ ID NO: 198)
1 ATGATTCCTG CCAGATTTGC CGGGGTGCTG CTTGCTCTGG CCCTCATTTT
51 GCCAGGGACC CTTTGTGCAG AAGGAACTCG CGGCAGGTCA TCCACGGCCC
101 GATGCAGCCT TTTCGGAAGT GACTTCGTCA ACACCTTTGA TGGGAGCATG
151 TACAGCTTTG CGGGATACTG CAGTTACCTC CTGGCAGGGG GCTGCCAGAA
201 ACGCTCCTTC TCGATTATTG GGGACTTCCA GAATGGCAAG AGAGTGAGCC
251 TCTCCGTGTA TCTTGGGGAA TTTTTTGACA TCCATTTGTT TGTCAATGGT
301 ACCGTGACAC AGGGGGACCA AAGAGTCTCC ATGCCCTATG CCTCCAAAGG
351 GCTGTATCTA GAAACTGAGG CTGGGTACTA CAAGCTGTCC GGTGAGGCCT
401 ATGGCTTTGT GGCCAGGATC GATGGCAGCG GCAACTTTCA AGTCCTGCTG
451 TCAGACAGAT ACTTCAACAA GACCTGCGGG CTGTGTGGCA ACTTTAACAT
501 CTTTGCTGAA GATGACTTTA TGACCCAAGA AGGGACCTTG ACCTCGGACC
551 CTTATGACTT TGCCAACTCA TGGGCTCTGA GCAGTGGAGA ACAGTGGTGT
601 GAACGGGCAT CTCCTCCCAG CAGCTCATGC AACATCTCCT CTGGGGAAAT
651 GCAGAAGGGC CTGTGGGAGC AGTGCCAGCT TCTGAAGAGC ACCTCGGTGT
701 TTGCCCGCTG CCACCCTCTG GTGGACCCCG AGCCTTTTGT GGCCCTGTGT
751 GAGAAGACTT TGTGTGAGTG TGCTGGGGGG CTGGAGTGCG CCTGCCCTGC
801 CCTCCTGGAG TACGCCCGGA CCTGTGCCCA GGAGGGAATG GTGCTGTACG
851 GCTGGACCGA CCACAGCGCG TGCAGCCCAG TGTGCCCTGC TGGTATGGAG
901 TATAGGCAGT GTGTGTCCCC TTGCGCCAGG ACCTGCCAGA GCCTGCACAT
951 CAATGAAATG TGTCAGGAGC GATGCGTGGA TGGCTGCAGC TGCCCTGAGG
1001 GACAGCTCCT GGATGAAGGC CTCTGCGTGG AGAGCACCGA GTGTCCCTGC
1051 GTGCATTCCG GAAAGCGCTA CCCTCCCGGC ACCTCCCTCT CTCGAGACTG
1101 CAACACCTGC ATTTGCCGAA ACAGCCAGTG GATCTGCAGC AATGAAGAAT
1151 GTCCAGGGGA GTGCCTTGTC ACTGGTCAAT CCCACTTCAA GAGCTTTGAC
1201 AACAGATACT TCACCTTCAG TGGGATCTGC CAGTACCTGC TGGCCCGGGA
1251 TTGCCAGGAC CACTCCTTCT CCATTGTCAT TGAGACTGTC CAGTGTGCTG
1301 ATGACCGCGA CGCTGTGTGC ACCCGCTCCG TCACCGTCCG GCTGCCTGGC
1351 CTGCACAACA GCCTTGTGAA ACTGAAGCAT GGGGCAGGAG TTGCCATGGA
1401 TGGCCAGGAC ATCCAGCTCC CCCTCCTGAA AGGTGACCTC CGCATCCAGC
1451 ATACAGTGAC GGCCTCCGTG CGCCTCAGCT ACGGGGAGGA CCTGCAGATG
1501 GACTGGGATG GCCGCGGGAG GCTGCTGGTG AAGCTGTCCC CCGTCTATGC
1551 CGGGAAGACC TGCGGCCTGT GTGGGAATTA CAATGGCAAC CAGGGCGACG
1601 ACTTCCTTAC CCCCTCTGGG CTGGCGGAGC CCCGGGTGGA GGACTTCGGG
1651 AACGCCTGGA AGCTGCACGG GGACTGCCAG GACCTGCAGA AGCAGCACAG
1701 CGATCCCTGC GCCCTCAACC CGCGCATGAC CAGGTTCTCC GAGGAGGCGT
1751 GCGCGGTCCT GACGTCCCCC ACATTCGAGG CCTGCCATCG TGCCGTCAGC
1801 CCGCTGCCCT ACCTGCGGAA CTGCCGCTAC GACGTGTGCT CCTGCTCGGA
1851 CGGCCGCGAG TGCCTGTGCG GCGCCCTGGC CAGCTATGCC GCGGCCTGCG
1901 CGGGGAGAGG CGTGCGCGTC GCGTGGCGCG AGCCAGGCCG CTGTGAGCTG
1951 AACTGCCCGA AAGGCCAGGT GTACCTGCAG TGCGGGACCC CCTGCAACCT
2001 GACCTGCCGC TCTCTCTCTT ACCCGGATGA GGAATGCAAT GAGGCCTGCC
2051 TGGAGGGCTG CTTCTGCCCC CCAGGGCTCT ACATGGATGA GAGGGGGGAC
2101 TGCGTGCCCA AGGCCCAGTG CCCCTGTTAC TATGACGGTG AGATCTTCCA
2151 GCCAGAAGAC ATCTTCTCAG ACCATCACAC CATGTGCTAC TGTGAGGATG
2201 GCTTCATGCA CTGTACCATG AGTGGAGTCC CCGGAAGCTT GCTGCCTGAC
2251 GCTGTCCTCA GCAGTCCCCT GTCTCATCGC AGCAAAAGGA GCCTATCCTG
2301 TCGGCCCCCC ATGGTCAAGC TGGTGTGTCC CGCTGACAAC CTGCGGGCTG
2351 AAGGGCTCGA GTGTACCAAA ACGTGCCAGA ACTATGACCT GGAGTGCATG
2401 AGCATGGGCT GTGTCTCTGG CTGCCTCTGC CCCCCGGGCA TGGTCCGGCA
2451 TGAGAACAGA TGTGTGGCCC TGGAAAGGTG TCCCTGCTTC CATCAGGGCA
2501 AGGAGTATGC CCCTGGAGAA ACAGTGAAGA TTGGCTGCAA CACTTGTGTC
2551 TGTCGGGACC GGAAGTGGAA CTGCACAGAC CATGTGTGTG ATGCCACGTG
2601 CTCCACGATC GGCATGGCCC ACTACCTCAC CTTCGACGGG CTCAAATACC
2651 TGTTCCCCGG GGAGTGCCAG TACGTTCTGG TGCAGGATTA CTGCGGCAGT
2701 AACCCTGGGA CCTTTCGGAT CCTAGTGGGG AATAAGGGAT GCAGCCACCC
2751 CTCAGTGAAA TGCAAGAAAC GGGTCACCAT CCTGGTGGAG GGAGGAGAGA
2801 TTGAGCTGTT TGACGGGGAG GTGAATGTGA AGAGGCCCAT GAAGGATGAG
2851 ACTCACTTTG AGGTGGTGGA GTCTGGCCGG TACATCATTC TGCTGCTGGG
2901 CAAAGCCCTC TCCGTGGTCT GGGACCGCCA CCTGAGCATC TCCGTGGTCC
2951 TGAAGCAGAC ATACCAGGAG AAAGTGTGTG GCCTGTGTGG GAATTTTGAT
3001 GGCATCCAGA ACAATGACCT CACCAGCAGC AACCTCCAAG TGGAGGAAGA
3051 CCCTGTGGAC TTTGGGAACT CCTGGAAAGT GAGCTCGCAG TGTGCTGACA
3101 CCAGAAAAGT GCCTCTGGAC TCATCCCCTG CCACCTGCCA TAACAACATC
3151 ATGAAGCAGA CGATGGTGGA TTCCTCCTGT AGAATCCTTA CCAGTGACGT
3201 CTTCCAGGAC TGCAACAAGC TGGTGGACCC CGAGCCATAT CTGGATGTCT
3251 GCATTTACGA CACCTGCTCC TGTGAGTCCA TTGGGGACTG CGCCGCATTC
3301 TGCGACACCA TTGCTGCCTA TGCCCACGTG TGTGCCCAGC ATGGCAAGGT
3351 GGTGACCTGG AGGACGGCCA CATTGTGCCC CCAGAGCTGC GAGGAGAGGA
3401 ATCTCCGGGA GAACGGGTAT GAGGCTGAGT GGCGCTATAA CAGCTGTGCA
3451 CCTGCCTGTC AAGTCACGTG TCAGCACCCT GAGCCACTGG CCTGCCCTGT
3501 GCAGTGTGTG GAGGGCTGCC ATGCCCACTG CCCTCCAGGG AAAATCCTGG
3551 ATGAGCTTTT GCAGACCTGC GTTGACCCTG AAGACTGTCC AGTGTGTGAG
3601 GTGGCTGGCC GGCGTTTTGC CTCAGGAAAG AAAGTCACCT TGAATCCCAG
3651 TGACCCTGAG CACTGCCAGA TTTGCCACTG TGATGTTGTC AACCTCACCT
3701 GTGAAGCCTG CCAGGAGCCG ATATCGGGCG CGCCAACATC AGAGAGCGCC
3751 ACCCCTGAAA GTGGTCCCGG GAGCGAGCCA GCCACATCTG GGTCGGAAAC
3801 GCCAGGCACA AGTGAGTCTG CAACTCCCGA GTCCGGACCT GGCTCCGAGC
3851 CTGCCACTAG CGGCTCCGAG ACTCCGGGAA CTTCCGAGAG CGCTACACCA
3901 GAAAGCGGAC CCGGAACCAG TACCGAACCT AGCGAGGGCT CTGCTCCGGG
3951 CAGCCCAGCC GGCTCTCCTA CATCCACGGA GGAGGGCACT TCCGAATCCG
4001 CCACCCCGGA GTCAGGGCCA GGATCTGAAC CCGCTACCTC AGGCAGTGAG
4051 ACGCCAGGAA CGAGCGAGTC CGCTACACCG GAGAGTGGGC CAGGGAGCCC
4101 TGCTGGATCT CCTACGTCCA CTGAGGAAGG GTCACCAGCG GGCTCGCCCA
4151 CCAGCACTGA AGAAGGTGCC TCGAGCGACA AAACTCACAC ATGCCCACCG
4201 TGCCCAGCTC CAGAACTCCT GGGCGGACCG TCAGTCTTCC TCTTCCCCCC
4251 AAAACCCAAG GACACCCTCA TGATCTCCCG GACCCCTGAG GTCACATGCG
4301 TGGTGGTGGA CGTGAGCCAC GAAGACCCTG AGGTCAAGTT CAACTGGTAC
4351 GTGGACGGCG TGGAGGTGCA TAATGCCAAG ACAAAGCCGC GGGAGGAGCA
4401 GTACAACAGC ACGTACCGTG TGGTCAGCGT CCTCACCGTC CTGCACCAGG
4451 ACTGGCTGAA TGGCAAGGAG TACAAGTGCA AGGTCTCCAA CAAAGCCCTC
4501 CCAGCCCCCA TCGAGAAAAC CATCTCCAAA GCCAAAGGGC AGCCCCGAGA
4551 ACCACAGGTG TACACCCTGC CCCCATCCCG GGATGAGCTG ACCAAGAACC
4601 AGGTCAGCCT GACCTGCCTG GTCAAAGGCT TCTATCCCAG CGACATCGCC
4651 GTGGAGTGGG AGAGCAATGG GCAGCCGGAG AACAACTACA AGACCACGCC
4701 TCCCGTGTTG GACTCCGACG GCTCCTTCTT CCTCTACAGC AAGCTCACCG
4751 TGGACAAGAG CAGGTGGCAG CAGGGGAACG TCTTCTCATG CTCCGTGATG
4801 CATGAGGCTC TGCACAACCA CTACACGCAG AAGAGCCTCT CCCTGTCTCC
4851 GGGTAAATGA
Последовательность белка pSYN VWF062 (D'D3 VWF-Fc без сайта расщепления тромбином в линкере) (SEQ ID NO: 199)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP ISGAPTSESA
1251 TPESGPGSEP ATSGSETPGT SESATPESGP GSEPATSGSE TPGTSESATP
1301 ESGPGTSTEP SEGSAPGSPA GSPTSTEEGT SESATPESGP GSEPATSGSE
1351 TPGTSESATP ESGPGSPAGS PTSTEEGSPA GSPTSTEEGA SSDKTHTCPP
1401 CPAPELLGGP SVFLFPPKPK DTLMISRTPE VTCVVVDVSH EDPEVKFNWY
1451 VDGVEVHNAK TKPREEQYNS TYRVVSVLTV LHQDWLNGKE YKCKVSNKAL
1501 PAPIEKTISK AKGQPREPQV YTLPPSRDEL TKNQVSLTCL VKGFYPSDIA
1551 VEWESNGQPE NNYKTTPPVL DSDGSFFLYS KLTVDKSRWQ QGNVFSCSVM
1601 HEALHNHYTQ KSLSLSPGK*
Нуклеотидная последовательность pSYN VWF073 (кодирующая D1D2D'D3 VWF-XTEN AE-144-FVIII с усеченным сайтом расщепления тромбина в a2-Fc) (SEQ ID NO: 174)
1 ATGATTCCTG CCAGATTTGC CGGGGTGCTG CTTGCTCTGG CCCTCATTTT
51 GCCAGGGACC CTTTGTGCAG AAGGAACTCG CGGCAGGTCA TCCACGGCCC
101 GATGCAGCCT TTTCGGAAGT GACTTCGTCA ACACCTTTGA TGGGAGCATG
151 TACAGCTTTG CGGGATACTG CAGTTACCTC CTGGCAGGGG GCTGCCAGAA
201 ACGCTCCTTC TCGATTATTG GGGACTTCCA GAATGGCAAG AGAGTGAGCC
251 TCTCCGTGTA TCTTGGGGAA TTTTTTGACA TCCATTTGTT TGTCAATGGT
301 ACCGTGACAC AGGGGGACCA AAGAGTCTCC ATGCCCTATG CCTCCAAAGG
351 GCTGTATCTA GAAACTGAGG CTGGGTACTA CAAGCTGTCC GGTGAGGCCT
401 ATGGCTTTGT GGCCAGGATC GATGGCAGCG GCAACTTTCA AGTCCTGCTG
451 TCAGACAGAT ACTTCAACAA GACCTGCGGG CTGTGTGGCA ACTTTAACAT
501 CTTTGCTGAA GATGACTTTA TGACCCAAGA AGGGACCTTG ACCTCGGACC
551 CTTATGACTT TGCCAACTCA TGGGCTCTGA GCAGTGGAGA ACAGTGGTGT
601 GAACGGGCAT CTCCTCCCAG CAGCTCATGC AACATCTCCT CTGGGGAAAT
651 GCAGAAGGGC CTGTGGGAGC AGTGCCAGCT TCTGAAGAGC ACCTCGGTGT
701 TTGCCCGCTG CCACCCTCTG GTGGACCCCG AGCCTTTTGT GGCCCTGTGT
751 GAGAAGACTT TGTGTGAGTG TGCTGGGGGG CTGGAGTGCG CCTGCCCTGC
801 CCTCCTGGAG TACGCCCGGA CCTGTGCCCA GGAGGGAATG GTGCTGTACG
851 GCTGGACCGA CCACAGCGCG TGCAGCCCAG TGTGCCCTGC TGGTATGGAG
901 TATAGGCAGT GTGTGTCCCC TTGCGCCAGG ACCTGCCAGA GCCTGCACAT
951 CAATGAAATG TGTCAGGAGC GATGCGTGGA TGGCTGCAGC TGCCCTGAGG
1001 GACAGCTCCT GGATGAAGGC CTCTGCGTGG AGAGCACCGA GTGTCCCTGC
1051 GTGCATTCCG GAAAGCGCTA CCCTCCCGGC ACCTCCCTCT CTCGAGACTG
1101 CAACACCTGC ATTTGCCGAA ACAGCCAGTG GATCTGCAGC AATGAAGAAT
1151 GTCCAGGGGA GTGCCTTGTC ACTGGTCAAT CCCACTTCAA GAGCTTTGAC
1201 AACAGATACT TCACCTTCAG TGGGATCTGC CAGTACCTGC TGGCCCGGGA
1251 TTGCCAGGAC CACTCCTTCT CCATTGTCAT TGAGACTGTC CAGTGTGCTG
1301 ATGACCGCGA CGCTGTGTGC ACCCGCTCCG TCACCGTCCG GCTGCCTGGC
1351 CTGCACAACA GCCTTGTGAA ACTGAAGCAT GGGGCAGGAG TTGCCATGGA
1401 TGGCCAGGAC ATCCAGCTCC CCCTCCTGAA AGGTGACCTC CGCATCCAGC
1451 ATACAGTGAC GGCCTCCGTG CGCCTCAGCT ACGGGGAGGA CCTGCAGATG
1501 GACTGGGATG GCCGCGGGAG GCTGCTGGTG AAGCTGTCCC CCGTCTATGC
1551 CGGGAAGACC TGCGGCCTGT GTGGGAATTA CAATGGCAAC CAGGGCGACG
1601 ACTTCCTTAC CCCCTCTGGG CTGGCGGAGC CCCGGGTGGA GGACTTCGGG
1651 AACGCCTGGA AGCTGCACGG GGACTGCCAG GACCTGCAGA AGCAGCACAG
1701 CGATCCCTGC GCCCTCAACC CGCGCATGAC CAGGTTCTCC GAGGAGGCGT
1751 GCGCGGTCCT GACGTCCCCC ACATTCGAGG CCTGCCATCG TGCCGTCAGC
1801 CCGCTGCCCT ACCTGCGGAA CTGCCGCTAC GACGTGTGCT CCTGCTCGGA
1851 CGGCCGCGAG TGCCTGTGCG GCGCCCTGGC CAGCTATGCC GCGGCCTGCG
1901 CGGGGAGAGG CGTGCGCGTC GCGTGGCGCG AGCCAGGCCG CTGTGAGCTG
1951 AACTGCCCGA AAGGCCAGGT GTACCTGCAG TGCGGGACCC CCTGCAACCT
2001 GACCTGCCGC TCTCTCTCTT ACCCGGATGA GGAATGCAAT GAGGCCTGCC
2051 TGGAGGGCTG CTTCTGCCCC CCAGGGCTCT ACATGGATGA GAGGGGGGAC
2101 TGCGTGCCCA AGGCCCAGTG CCCCTGTTAC TATGACGGTG AGATCTTCCA
2151 GCCAGAAGAC ATCTTCTCAG ACCATCACAC CATGTGCTAC TGTGAGGATG
2201 GCTTCATGCA CTGTACCATG AGTGGAGTCC CCGGAAGCTT GCTGCCTGAC
2251 GCTGTCCTCA GCAGTCCCCT GTCTCATCGC AGCAAAAGGA GCCTATCCTG
2301 TCGGCCCCCC ATGGTCAAGC TGGTGTGTCC CGCTGACAAC CTGCGGGCTG
2351 AAGGGCTCGA GTGTACCAAA ACGTGCCAGA ACTATGACCT GGAGTGCATG
2401 AGCATGGGCT GTGTCTCTGG CTGCCTCTGC CCCCCGGGCA TGGTCCGGCA
2451 TGAGAACAGA TGTGTGGCCC TGGAAAGGTG TCCCTGCTTC CATCAGGGCA
2501 AGGAGTATGC CCCTGGAGAA ACAGTGAAGA TTGGCTGCAA CACTTGTGTC
2551 TGTCGGGACC GGAAGTGGAA CTGCACAGAC CATGTGTGTG ATGCCACGTG
2601 CTCCACGATC GGCATGGCCC ACTACCTCAC CTTCGACGGG CTCAAATACC
2651 TGTTCCCCGG GGAGTGCCAG TACGTTCTGG TGCAGGATTA CTGCGGCAGT
2701 AACCCTGGGA CCTTTCGGAT CCTAGTGGGG AATAAGGGAT GCAGCCACCC
2751 CTCAGTGAAA TGCAAGAAAC GGGTCACCAT CCTGGTGGAG GGAGGAGAGA
2801 TTGAGCTGTT TGACGGGGAG GTGAATGTGA AGAGGCCCAT GAAGGATGAG
2851 ACTCACTTTG AGGTGGTGGA GTCTGGCCGG TACATCATTC TGCTGCTGGG
2901 CAAAGCCCTC TCCGTGGTCT GGGACCGCCA CCTGAGCATC TCCGTGGTCC
2951 TGAAGCAGAC ATACCAGGAG AAAGTGTGTG GCCTGTGTGG GAATTTTGAT
3001 GGCATCCAGA ACAATGACCT CACCAGCAGC AACCTCCAAG TGGAGGAAGA
3051 CCCTGTGGAC TTTGGGAACT CCTGGAAAGT GAGCTCGCAG TGTGCTGACA
3101 CCAGAAAAGT GCCTCTGGAC TCATCCCCTG CCACCTGCCA TAACAACATC
3151 ATGAAGCAGA CGATGGTGGA TTCCTCCTGT AGAATCCTTA CCAGTGACGT
3201 CTTCCAGGAC TGCAACAAGC TGGTGGACCC CGAGCCATAT CTGGATGTCT
3251 GCATTTACGA CACCTGCTCC TGTGAGTCCA TTGGGGACTG CGCCGCATTC
3301 TGCGACACCA TTGCTGCCTA TGCCCACGTG TGTGCCCAGC ATGGCAAGGT
3351 GGTGACCTGG AGGACGGCCA CATTGTGCCC CCAGAGCTGC GAGGAGAGGA
3401 ATCTCCGGGA GAACGGGTAT GAGGCTGAGT GGCGCTATAA CAGCTGTGCA
3451 CCTGCCTGTC AAGTCACGTG TCAGCACCCT GAGCCACTGG CCTGCCCTGT
3501 GCAGTGTGTG GAGGGCTGCC ATGCCCACTG CCCTCCAGGG AAAATCCTGG
3551 ATGAGCTTTT GCAGACCTGC GTTGACCCTG AAGACTGTCC AGTGTGTGAG
3601 GTGGCTGGCC GGCGTTTTGC CTCAGGAAAG AAAGTCACCT TGAATCCCAG
3651 TGACCCTGAG CACTGCCAGA TTTGCCACTG TGATGTTGTC AACCTCACCT
3701 GTGAAGCCTG CCAGGAGCCG GGCGCGCCAA CATCAGAGAG CGCCACCCCT
3751 GAAAGTGGTC CCGGGAGCGA GCCAGCCACA TCTGGGTCGG AAACGCCAGG
3801 CACAAGTGAG TCTGCAACTC CCGAGTCCGG ACCTGGCTCC GAGCCTGCCA
3851 CTAGCGGCTC CGAGACTCCG GGAACTTCCG AGAGCGCTAC ACCAGAAAGC
3901 GGACCCGGAA CCAGTACCGA ACCTAGCGAG GGCTCTGCTC CGGGCAGCCC
3951 AGCCGGCTCT CCTACATCCA CGGAGGAGGG CACTTCCGAA TCCGCCACCC
4001 CGGAGTCAGG GCCAGGATCT GAACCCGCTA CCTCAGGCAG TGAGACGCCA
4051 GGAACGAGCG AGTCCGCTAC ACCGGAGAGT GGGCCAGGGA GCCCTGCTGG
4101 ATCTCCTACG TCCACTGAGG AAGGGTCACC AGCGGGCTCG CCCACCAGCA
4151 CTGAAGAAGG TGCCTCGAGC GGCGGTGGAG GATCCGGTGG CGGGGGATCC
4201 GGTGGCGGGG GATCCGGTGG CGGGGGATCC GGTGGCGGGG GATCCGGTGG
4251 CGGGGGATCC ATTGAACCAA GAAGCTTCTC TGGCAGCGGA GGCGACAAAA
4301 CTCACACATG CCCACCGTGC CCAGCTCCAG AACTCCTGGG CGGACCGTCA
4351 GTCTTCCTCT TCCCCCCAAA ACCCAAGGAC ACCCTCATGA TCTCCCGGAC
4401 CCCTGAGGTC ACATGCGTGG TGGTGGACGT GAGCCACGAA GACCCTGAGG
4451 TCAAGTTCAA CTGGTACGTG GACGGCGTGG AGGTGCATAA TGCCAAGACA
4501 AAGCCGCGGG AGGAGCAGTA CAACAGCACG TACCGTGTGG TCAGCGTCCT
4551 CACCGTCCTG CACCAGGACT GGCTGAATGG CAAGGAGTAC AAGTGCAAGG
4601 TCTCCAACAA AGCCCTCCCA GCCCCCATCG AGAAAACCAT CTCCAAAGCC
4651 AAAGGGCAGC CCCGAGAACC ACAGGTGTAC ACCCTGCCCC CATCCCGGGA
4701 TGAGCTGACC AAGAACCAGG TCAGCCTGAC CTGCCTGGTC AAAGGCTTCT
4751 ATCCCAGCGA CATCGCCGTG GAGTGGGAGA GCAATGGGCA GCCGGAGAAC
4801 AACTACAAGA CCACGCCTCC CGTGTTGGAC TCCGACGGCT CCTTCTTCCT
4851 CTACAGCAAG CTCACCGTGG ACAAGAGCAG GTGGCAGCAG GGGAACGTCT
4901 TCTCATGCTC CGTGATGCAT GAGGCTCTGC ACAACCACTA CACGCAGAAG
4951 AGCCTCTCCC TGTCTCCGGG TAAATGA
Последовательность белка pSYN VWF073 (D1D2D'D3 VWF-XTEN AE-144-усеченный сайт расщепления тромбином в a2-Fc) (SEQ ID NO: 175)
1 MIPARFAGVL LALALILPGT LCAEGTRGRS STARCSLFGS DFVNTFDGSM
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP GAPTSESATP
1251 ESGPGSEPAT SGSETPGTSE SATPESGPGS EPATSGSETP GTSESATPES
1301 GPGTSTEPSE GSAPGSPAGS PTSTEEGTSE SATPESGPGS EPATSGSETP
1351 GTSESATPES GPGSPAGSPT STEEGSPAGS PTSTEEGASS GGGGSGGGGS
1401 GGGGSGGGGS GGGGSGGGGS IEPRSFSGSG GDKTHTCPPC PAPELLGGPS
1451 VFLFPPKPKD TLMISRTPEV TCVVVDVSHE DPEVKFNWYV DGVEVHNAKT
1501 KPREEQYNST YRVVSVLTVL HQDWLNGKEY KCKVSNKALP APIEKTISKA
1551 KGQPREPQVY TLPPSRDELT KNQVSLTCLV KGFYPSDIAV EWESNGQPEN
1601 NYKTTPPVLD SDGSFFLYSK LTVDKSRWQQ GNVFSCSVMH EALHNHYTQK
1651 SLSLSPGK*
Таблица 18. Иллюстративные последовательности химерных полипептидов
51 PRVPKSFPFN TSVVYKKTLF VEFTDHLFNI AKPRPPWMGL LGPTIQAEVY
101 DTVVITLKNM ASHPVSLHAV GVSYWKASEG AEYDDQTSQR EKEDDKVFPG
151 GSHTYVWQVL KENGPMASDP LCLTYSYLSH VDLVKDLNSG LIGALLVCRE
201 GSLAKEKTQT LHKFILLFAV FDEGKSWHSE TKNSLMQDRD AASARAWPKM
251 HTVNGYVNRS LPGLIGCHRK SVYWHVIGMG TTPEVHSIFL EGHTFLVRNH
301 RQASLEISPI TFLTAQTLLM DLGQFLLFCH ISSHQHDGME AYVKVDSCPE
351 EPQLRMKNNE EAEDYDDDLT DSEMDVVRFD DDNSPSFIQI RSVAKKHPKT
401 WVHYIAAEEE DWDYAPLVLA PDDRSYKSQY LNNGPQRIGR KYKKVRFMAY
451 TDETFKTREA IQHESGILGP LLYGEVGDTL LIIFKNQASR PYNIYPHGIT
501 DVRPLYSRRL PKGVKHLKDF PILPGEIFKY KWTVTVEDGP TKSDPRCLTR
551 YYSSFVNMER DLASGLIGPL LICYKESVDQ RGNQIMSDKR NVILFSVFDE
601 NRSWYLTENI QRFLPNPAGV QLEDPEFQAS NIMHSINGYV FDSLQLSVCL
651 HEVAYWYILS IGAQTDFLSV FFSGYTFKHK MVYEDTLTLF PFSGETVFMS
701 MENPGLWILG CHNSDFRNRG MTALLKVSSC DKNTGDYYED SYEDISAYLL
751 SKNNAIEPRS FSQNGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP
801 GSEPATSGSE TPGTSESATP ESGPGTSTEP SEGSAPGSPA GSPTSTEEGT
851 SESATPESGP GSEPATSGSE TPGTSESATP ESGPGSPAGS PTSTEEGSPA
901 GSPTSTEEGT STEPSEGSAP GTSESATPES GPGTSESATP ESGPGTSESA
951 TPESGPGSEP ATSGSETPGS EPATSGSETP GSPAGSPTST EEGTSTEPSE
1001 GSAPGTSTEP SEGSAPGSEP ATSGSETPGT SESATPESGP GTSTEPSEGS
1051 APASSEITRT TLQSDQEEID YDDTISVEMK KEDFDIYDED ENQSPRSFQK
1101 KTRHYFIAAV ERLWDYGMSS SPHVLRNRAQ SGSVPQFKKV VFQEFTDGSF
1151 TQPLYRGELN EHLGLLGPYI RAEVEDNIMV TFRNQASRPY SFYSSLISYE
1201 EDQRQGAEPR KNFVKPNETK TYFWKVQHHM APTKDEFDCK AWAYFSDVDL
1251 EKDVHSGLIG PLLVCHTNTL NPAHGRQVTV QEFALFFTIF DETKSWYFTE
1301 NMERNCRAPC NIQMEDPTFK ENYRFHAING YIMDTLPGLV MAQDQRIRWY
1351 LLSMGSNENI HSIHFSGHVF TVRKKEEYKM ALYNLYPGVF ETVEMLPSKA
1401 GIWRVECLIG EHLHAGMSTL FLVYSNKCQT PLGMASGHIR DFQITASGQY
1451 GQWAPKLARL HYSGSINAWS TKEPFSWIKV DLLAPMIIHG IKTQGARQKF
1501 SSLYISQFII MYSLDGKKWQ TYRGNSTGTL MVFFGNVDSS GIKHNIFNPP
1551 IIARYIRLHP THYSIRSTLR MELMGCDLNS CSMPLGMESK AISDAQITAS
1601 SYFTNMFATW SPSKARLHLQ GRSNAWRPQV NNPKEWLQVD FQKTMKVTGV
1651 TTQGVKSLLT SMYVKEFLIS SSQDGHQWTL FFQNGKVKVF QGNQDSFTPV
1701 VNSLDPPLLT RYLRIHPQSW VHQIALRMEV LGCEAQDLYD KTHTCPPCPA
1751 PELLGGPSVF LFPPKPKDTL MISRTPEVTC VVVDVSHEDP EVKFNWYVDG
1801 VEVHNAKTKP REEQYNSTYR VVSVLTVLHQ DWLNGKEYKC KVSNKALPAP
1851 IEKTISKAKG QPREPQVYTL PPSRDELTKN QVSLTCLVKG FYPSDIAVEW
1901 ESNGQPENNY KTTPPVLDSD GSFFLYSKLT VDKSRWQQGN VFSCSVMHEA
1951 LHNHYTQKSL SLSPG*
51 CTTTAGTGCC ACCAGAAGAT ACTACCTGGG TGCAGTGGAA CTGTCATGGG
101 ACTATATGCA AAGTGATCTC GGTGAGCTGC CTGTGGACGC AAGATTTCCT
151 CCTAGAGTGC CAAAATCTTT TCCATTCAAC ACCTCAGTCG TGTACAAAAA
201 GACTCTGTTT GTAGAATTCA CGGATCACCT TTTCAACATC GCTAAGCCAA
251 GGCCACCCTG GATGGGTCTG CTAGGTCCTA CCATCCAGGC TGAGGTTTAT
301 GATACAGTGG TCATTACACT TAAGAACATG GCTTCCCATC CTGTCAGTCT
351 TCATGCTGTT GGTGTATCCT ACTGGAAAGC TTCTGAGGGA GCTGAATATG
401 ATGATCAGAC CAGTCAAAGG GAGAAAGAAG ATGATAAAGT CTTCCCTGGT
451 GGAAGCCATA CATATGTCTG GCAGGTCCTG AAAGAGAATG GTCCAATGGC
501 CTCTGACCCA CTGTGCCTTA CCTACTCATA TCTTTCTCAT GTGGACCTGG
551 TAAAAGACTT GAATTCAGGC CTCATTGGAG CCCTACTAGT ATGTAGAGAA
601 GGGAGTCTGG CCAAGGAAAA GACACAGACC TTGCACAAAT TTATACTACT
651 TTTTGCTGTA TTTGATGAAG GGAAAAGTTG GCACTCAGAA ACAAAGAACT
701 CCTTGATGCA GGATAGGGAT GCTGCATCTG CTCGGGCCTG GCCTAAAATG
751 CACACAGTCA ATGGTTATGT AAACAGGTCT CTGCCAGGTC TGATTGGATG
801 CCACAGGAAA TCAGTCTATT GGCATGTGAT TGGAATGGGC ACCACTCCTG
851 AAGTGCACTC AATATTCCTC GAAGGTCACA CATTTCTTGT GAGGAACCAT
901 CGCCAGGCGT CCTTGGAAAT CTCGCCAATA ACTTTCCTTA CTGCTCAAAC
951 ACTCTTGATG GACCTTGGAC AGTTTCTACT GTTTTGTCAT ATCTCTTCCC
1001 ACCAACATGA TGGCATGGAA GCTTATGTCA AAGTAGACAG CTGTCCAGAG
1051 GAACCCCAAC TACGAATGAA AAATAATGAA GAAGCGGAAG ACTATGATGA
1101 TGATCTTACT GATTCTGAAA TGGATGTGGT CAGGTTTGAT GATGACAACT
1151 CTCCTTCCTT TATCCAAATT CGCTCAGTTG CCAAGAAGCA TCCTAAAACT
1201 TGGGTACATT ACATTGCTGC TGAAGAGGAG GACTGGGACT ATGCTCCCTT
1251 AGTCCTCGCC CCCGATGACA GAAGTTATAA AAGTCAATAT TTGAACAATG
1301 GCCCTCAGCG GATTGGTAGG AAGTACAAAA AAGTCCGATT TATGGCATAC
1351 ACAGATGAAA CCTTTAAGAC TCGTGAAGCT ATTCAGCATG AATCAGGAAT
1401 CTTGGGACCT TTACTTTATG GGGAAGTTGG AGACACACTG TTGATTATAT
1451 TTAAGAATCA AGCAAGCAGA CCATATAACA TCTACCCTCA CGGAATCACT
1501 GATGTCCGTC CTTTGTATTC AAGGAGATTA CCAAAAGGTG TAAAACATTT
1551 GAAGGATTTT CCAATTCTGC CAGGAGAAAT ATTCAAATAT AAATGGACAG
1601 TGACTGTAGA AGATGGGCCA ACTAAATCAG ATCCTCGGTG CCTGACCCGC
1651 TATTACTCTA GTTTCGTTAA TATGGAGAGA GATCTAGCTT CAGGACTCAT
1701 TGGCCCTCTC CTCATCTGCT ACAAAGAATC TGTAGATCAA AGAGGAAACC
1751 AGATAATGTC AGACAAGAGG AATGTCATCC TGTTTTCTGT ATTTGATGAG
1801 AACCGAAGCT GGTACCTCAC AGAGAATATA CAACGCTTTC TCCCCAATCC
1851 AGCTGGAGTG CAGCTTGAGG ATCCAGAGTT CCAAGCCTCC AACATCATGC
1901 ACAGCATCAA TGGCTATGTT TTTGATAGTT TGCAGTTGTC AGTTTGTTTG
1951 CATGAGGTGG CATACTGGTA CATTCTAAGC ATTGGAGCAC AGACTGACTT
2001 CCTTTCTGTC TTCTTCTCTG GATATACCTT CAAACACAAA ATGGTCTATG
2051 AAGACACACT CACCCTATTC CCATTCTCAG GAGAAACTGT CTTCATGTCG
2101 ATGGAAAACC CAGGTCTATG GATTCTGGGG TGCCACAACT CAGACTTTCG
2151 GAACAGAGGC ATGACCGCCT TACTGAAGGT TTCTAGTTGT GACAAGAACA
2201 CTGGTGATTA TTACGAGGAC AGTTATGAAG ATATTTCAGC ATACTTGCTG
2251 AGTAAAAACA ATGCCATTGA ACCAAGAAGC TTCTCTCAAA ACGGTACCTC
2301 AGAGTCTGCT ACCCCCGAGT CAGGGCCAGG ATCAGAGCCA GCCACCTCCG
2351 GGTCTGAGAC ACCCGGGACT TCCGAGAGTG CCACCCCTGA GTCCGGACCC
2401 GGGTCCGAGC CCGCCACTTC CGGCTCCGAA ACTCCCGGCA CAAGCGAGAG
2451 CGCTACCCCA GAGTCAGGAC CAGGAACATC TACAGAGCCC TCTGAAGGCT
2501 CCGCTCCAGG GTCCCCAGCC GGCAGTCCCA CTAGCACCGA GGAGGGAACC
2551 TCTGAAAGCG CCACACCCGA ATCAGGGCCA GGGTCTGAGC CTGCTACCAG
2601 CGGCAGCGAG ACACCAGGCA CCTCTGAGTC CGCCACACCA GAGTCCGGAC
2651 CCGGATCTCC CGCTGGGAGC CCCACCTCCA CTGAGGAGGG ATCTCCTGCT
2701 GGCTCTCCAA CATCTACTGA GGAAGGAACC TCAACCGAGC CATCCGAGGG
2751 ATCAGCTCCC GGCACCTCAG AGTCGGCAAC CCCGGAGTCT GGACCCGGAA
2801 CTTCCGAAAG TGCCACACCA GAGTCCGGTC CCGGGACTTC AGAATCAGCA
2851 ACACCCGAGT CCGGCCCTGG GTCTGAACCC GCCACAAGTG GTAGTGAGAC
2901 ACCAGGATCA GAACCTGCTA CCTCAGGGTC AGAGACACCC GGATCTCCGG
2951 CAGGCTCACC AACCTCCACT GAGGAGGGCA CCAGCACAGA ACCAAGCGAG
3001 GGCTCCGCAC CCGGAACAAG CACTGAACCC AGTGAGGGTT CAGCACCCGG
3051 CTCTGAGCCG GCCACAAGTG GCAGTGAGAC ACCCGGCACT TCAGAGAGTG
3101 CCACCCCCGA GAGTGGCCCA GGCACTAGTA CCGAGCCCTC TGAAGGCAGT
3151 GCGCCAGCCT CGAGCGAAAT AACTCGTACT ACTCTTCAGT CAGATCAAGA
3201 GGAAATTGAC TATGATGATA CCATATCAGT TGAAATGAAG AAGGAAGATT
3251 TTGACATTTA TGATGAGGAT GAAAATCAGA GCCCCCGCAG CTTTCAAAAG
3301 AAAACACGAC ACTATTTTAT TGCTGCAGTG GAGAGGCTCT GGGATTATGG
3351 GATGAGTAGC TCCCCACATG TTCTAAGAAA CAGGGCTCAG AGTGGCAGTG
3401 TCCCTCAGTT CAAGAAAGTT GTTTTCCAGG AATTTACTGA TGGCTCCTTT
3451 ACTCAGCCCT TATACCGTGG AGAACTAAAT GAACATTTGG GACTCCTGGG
3501 GCCATATATA AGAGCAGAAG TTGAAGATAA TATCATGGTA ACTTTCAGAA
3551 ATCAGGCCTC TCGTCCCTAT TCCTTCTATT CTAGCCTTAT TTCTTATGAG
3601 GAAGATCAGA GGCAAGGAGC AGAACCTAGA AAAAACTTTG TCAAGCCTAA
3651 TGAAACCAAA ACTTACTTTT GGAAAGTGCA ACATCATATG GCACCCACTA
3701 AAGATGAGTT TGACTGCAAA GCCTGGGCTT ATTTCTCTGA TGTTGACCTG
3751 GAAAAAGATG TGCACTCAGG CCTGATTGGA CCCCTTCTGG TCTGCCACAC
3801 TAACACACTG AACCCTGCTC ATGGGAGACA AGTGACAGTA CAGGAATTTG
3851 CTCTGTTTTT CACCATCTTT GATGAGACCA AAAGCTGGTA CTTCACTGAA
3901 AATATGGAAA GAAACTGCAG GGCTCCCTGC AATATCCAGA TGGAAGATCC
3951 CACTTTTAAA GAGAATTATC GCTTCCATGC AATCAATGGC TACATAATGG
4001 ATACACTACC TGGCTTAGTA ATGGCTCAGG ATCAAAGGAT TCGATGGTAT
4051 CTGCTCAGCA TGGGCAGCAA TGAAAACATC CATTCTATTC ATTTCAGTGG
4101 ACATGTGTTC ACTGTACGAA AAAAAGAGGA GTATAAAATG GCACTGTACA
4151 ATCTCTATCC AGGTGTTTTT GAGACAGTGG AAATGTTACC ATCCAAAGCT
4201 GGAATTTGGC GGGTGGAATG CCTTATTGGC GAGCATCTAC ATGCTGGGAT
4251 GAGCACACTT TTTCTGGTGT ACAGCAATAA GTGTCAGACT CCCCTGGGAA
4301 TGGCTTCTGG ACACATTAGA GATTTTCAGA TTACAGCTTC AGGACAATAT
4351 GGACAGTGGG CCCCAAAGCT GGCCAGACTT CATTATTCCG GATCAATCAA
4401 TGCCTGGAGC ACCAAGGAGC CCTTTTCTTG GATCAAGGTG GATCTGTTGG
4451 CACCAATGAT TATTCACGGC ATCAAGACCC AGGGTGCCCG TCAGAAGTTC
4501 TCCAGCCTCT ACATCTCTCA GTTTATCATC ATGTATAGTC TTGATGGGAA
4551 GAAGTGGCAG ACTTATCGAG GAAATTCCAC TGGAACCTTA ATGGTCTTCT
4601 TTGGCAATGT GGATTCATCT GGGATAAAAC ACAATATTTT TAACCCTCCA
4651 ATTATTGCTC GATACATCCG TTTGCACCCA ACTCATTATA GCATTCGCAG
4701 CACTCTTCGC ATGGAGTTGA TGGGCTGTGA TTTAAATAGT TGCAGCATGC
4751 CATTGGGAAT GGAGAGTAAA GCAATATCAG ATGCACAGAT TACTGCTTCA
4801 TCCTACTTTA CCAATATGTT TGCCACCTGG TCTCCTTCAA AAGCTCGACT
4851 TCACCTCCAA GGGAGGAGTA ATGCCTGGAG ACCTCAGGTG AATAATCCAA
4901 AAGAGTGGCT GCAAGTGGAC TTCCAGAAGA CAATGAAAGT CACAGGAGTA
4951 ACTACTCAGG GAGTAAAATC TCTGCTTACC AGCATGTATG TGAAGGAGTT
5001 CCTCATCTCC AGCAGTCAAG ATGGCCATCA GTGGACTCTC TTTTTTCAGA
5051 ATGGCAAAGT AAAGGTTTTT CAGGGAAATC AAGACTCCTT CACACCTGTG
5101 GTGAACTCTC TAGACCCACC GTTACTGACT CGCTACCTTC GAATTCACCC
5151 CCAGAGTTGG GTGCACCAGA TTGCCCTGAG GATGGAGGTT CTGGGCTGCG
5201 AGGCACAGGA CCTCTACGAC AAAACTCACA CATGCCCACC GTGCCCAGCT
5251 CCAGAACTCC TGGGCGGACC GTCAGTCTTC CTCTTCCCCC CAAAACCCAA
5301 GGACACCCTC ATGATCTCCC GGACCCCTGA GGTCACATGC GTGGTGGTGG
5351 ACGTGAGCCA CGAAGACCCT GAGGTCAAGT TCAACTGGTA TGTGGACGGC
5401 GTGGAAGTGC ATAATGCCAA GACAAAGCCG CGGGAGGAGC AGTACAACAG
5451 CACGTACCGT GTGGTCAGCG TCCTCACCGT CCTGCACCAA GACTGGCTGA
5501 ATGGCAAGGA GTACAAGTGC AAGGTCTCCA ACAAAGCCCT CCCAGCCCCC
5551 ATCGAGAAAA CCATCTCCAA AGCCAAAGGG CAGCCCCGAG AACCACAGGT
5601 GTACACCCTG CCCCCATCCC GGGATGAGCT GACCAAGAAC CAAGTTAGCC
5651 TGACCTGCCT GGTCAAAGGC TTCTATCCCA GCGACATCGC CGTGGAGTGG
5701 GAGAGCAATG GGCAGCCGGA GAACAACTAC AAGACCACGC CTCCCGTGTT
5751 GGACTCCGAC GGCTCCTTCT TCCTCTACTC CAAGCTCACC GTGGACAAGA
5801 GCAGGTGGCA GCAGGGGAAC GTCTTCTCAT GCTCCGTGAT GCATGAGGCT
5851 CTGCACAACC ACTACACGCA GAAGAGCCTC TCCCTGTCTC CGGGTTGA
51 YSFAGYCSYL LAGGCQKRSF SIIGDFQNGK RVSLSVYLGE FFDIHLFVNG
101 TVTQGDQRVS MPYASKGLYL ETEAGYYKLS GEAYGFVARI DGSGNFQVLL
151 SDRYFNKTCG LCGNFNIFAE DDFMTQEGTL TSDPYDFANS WALSSGEQWC
201 ERASPPSSSC NISSGEMQKG LWEQCQLLKS TSVFARCHPL VDPEPFVALC
251 EKTLCECAGG LECACPALLE YARTCAQEGM VLYGWTDHSA CSPVCPAGME
301 YRQCVSPCAR TCQSLHINEM CQERCVDGCS CPEGQLLDEG LCVESTECPC
351 VHSGKRYPPG TSLSRDCNTC ICRNSQWICS NEECPGECLV TGQSHFKSFD
401 NRYFTFSGIC QYLLARDCQD HSFSIVIETV QCADDRDAVC TRSVTVRLPG
451 LHNSLVKLKH GAGVAMDGQD IQLPLLKGDL RIQHTVTASV RLSYGEDLQM
501 DWDGRGRLLV KLSPVYAGKT CGLCGNYNGN QGDDFLTPSG LAEPRVEDFG
551 NAWKLHGDCQ DLQKQHSDPC ALNPRMTRFS EEACAVLTSP TFEACHRAVS
601 PLPYLRNCRY DVCSCSDGRE CLCGALASYA AACAGRGVRV AWREPGRCEL
651 NCPKGQVYLQ CGTPCNLTCR SLSYPDEECN EACLEGCFCP PGLYMDERGD
701 CVPKAQCPCY YDGEIFQPED IFSDHHTMCY CEDGFMHCTM SGVPGSLLPD
751 AVLSSPLSHR SKRSLSCRPP MVKLVCPADN LRAEGLECTK TCQNYDLECM
801 SMGCVSGCLC PPGMVRHENR CVALERCPCF HQGKEYAPGE TVKIGCNTCV
851 CRDRKWNCTD HVCDATCSTI GMAHYLTFDG LKYLFPGECQ YVLVQDYCGS
901 NPGTFRILVG NKGCSHPSVK CKKRVTILVE GGEIELFDGE VNVKRPMKDE
951 THFEVVESGR YIILLLGKAL SVVWDRHLSI SVVLKQTYQE KVCGLCGNFD
1001 GIQNNDLTSS NLQVEEDPVD FGNSWKVSSQ CADTRKVPLD SSPATCHNNI
1051 MKQTMVDSSC RILTSDVFQD CNKLVDPEPY LDVCIYDTCS CESIGDCAAF
1101 CDTIAAYAHV CAQHGKVVTW RTATLCPQSC EERNLRENGY EAEWRYNSCA
1151 PACQVTCQHP EPLACPVQCV EGCHAHCPPG KILDELLQTC VDPEDCPVCE
1201 VAGRRFASGK KVTLNPSDPE HCQICHCDVV NLTCEACQEP GTSESATPES
1251 GPGSEPATSG SETPGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP
1301 GTSTEPSEGS APGSPAGSPT STEEGTSESA TPESGPGSEP ATSGSETPGT
1351 SESATPESGP GSPAGSPTST EEGSPAGSPT STEEGASSDK NTGDYYEDSY
1401 EDISAYLLSK NNAIEPRSFS DKTHTCPPCP APELLGGPSV FLFPPKPKDT
1451 LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY
1501 RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT
1551 LPPSRDELTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS
1601 DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPG*
51 GCCAGGGACC CTTTGTGCAG AAGGAACTCG CGGCAGGTCA TCCACGGCCC
101 GATGCAGCCT TTTCGGAAGT GACTTCGTCA ACACCTTTGA TGGGAGCATG
151 TACAGCTTTG CGGGATACTG CAGTTACCTC CTGGCAGGGG GCTGCCAGAA
201 ACGCTCCTTC TCGATTATTG GGGACTTCCA GAATGGCAAG AGAGTGAGCC
251 TCTCCGTGTA TCTTGGGGAA TTTTTTGACA TCCATTTGTT TGTCAATGGT
301 ACCGTGACAC AGGGGGACCA AAGAGTCTCC ATGCCCTATG CCTCCAAAGG
351 GCTGTATCTA GAAACTGAGG CTGGGTACTA CAAGCTGTCC GGTGAGGCCT
401 ATGGCTTTGT GGCCAGGATC GATGGCAGCG GCAACTTTCA AGTCCTGCTG
451 TCAGACAGAT ACTTCAACAA GACCTGCGGG CTGTGTGGCA ACTTTAACAT
501 CTTTGCTGAA GATGACTTTA TGACCCAAGA AGGGACCTTG ACCTCGGACC
551 CTTATGACTT TGCCAACTCA TGGGCTCTGA GCAGTGGAGA ACAGTGGTGT
601 GAACGGGCAT CTCCTCCCAG CAGCTCATGC AACATCTCCT CTGGGGAAAT
651 GCAGAAGGGC CTGTGGGAGC AGTGCCAGCT TCTGAAGAGC ACCTCGGTGT
701 TTGCCCGCTG CCACCCTCTG GTGGACCCCG AGCCTTTTGT GGCCCTGTGT
751 GAGAAGACTT TGTGTGAGTG TGCTGGGGGG CTGGAGTGCG CCTGCCCTGC
801 CCTCCTGGAG TACGCCCGGA CCTGTGCCCA GGAGGGAATG GTGCTGTACG
851 GCTGGACCGA CCACAGCGCG TGCAGCCCAG TGTGCCCTGC TGGTATGGAG
901 TATAGGCAGT GTGTGTCCCC TTGCGCCAGG ACCTGCCAGA GCCTGCACAT
951 CAATGAAATG TGTCAGGAGC GATGCGTGGA TGGCTGCAGC TGCCCTGAGG
1001 GACAGCTCCT GGATGAAGGC CTCTGCGTGG AGAGCACCGA GTGTCCCTGC
1051 GTGCATTCCG GAAAGCGCTA CCCTCCCGGC ACCTCCCTCT CTCGAGACTG
1101 CAACACCTGC ATTTGCCGAA ACAGCCAGTG GATCTGCAGC AATGAAGAAT
1151 GTCCAGGGGA GTGCCTTGTC ACTGGTCAAT CCCACTTCAA GAGCTTTGAC
1201 AACAGATACT TCACCTTCAG TGGGATCTGC CAGTACCTGC TGGCCCGGGA
1251 TTGCCAGGAC CACTCCTTCT CCATTGTCAT TGAGACTGTC CAGTGTGCTG
1301 ATGACCGCGA CGCTGTGTGC ACCCGCTCCG TCACCGTCCG GCTGCCTGGC
1351 CTGCACAACA GCCTTGTGAA ACTGAAGCAT GGGGCAGGAG TTGCCATGGA
1401 TGGCCAGGAC ATCCAGCTCC CCCTCCTGAA AGGTGACCTC CGCATCCAGC
1451 ATACAGTGAC GGCCTCCGTG CGCCTCAGCT ACGGGGAGGA CCTGCAGATG
1501 GACTGGGATG GCCGCGGGAG GCTGCTGGTG AAGCTGTCCC CCGTCTATGC
1551 CGGGAAGACC TGCGGCCTGT GTGGGAATTA CAATGGCAAC CAGGGCGACG
1601 ACTTCCTTAC CCCCTCTGGG CTGGCGGAGC CCCGGGTGGA GGACTTCGGG
1651 AACGCCTGGA AGCTGCACGG GGACTGCCAG GACCTGCAGA AGCAGCACAG
1701 CGATCCCTGC GCCCTCAACC CGCGCATGAC CAGGTTCTCC GAGGAGGCGT
1751 GCGCGGTCCT GACGTCCCCC ACATTCGAGG CCTGCCATCG TGCCGTCAGC
1801 CCGCTGCCCT ACCTGCGGAA CTGCCGCTAC GACGTGTGCT CCTGCTCGGA
1851 CGGCCGCGAG TGCCTGTGCG GCGCCCTGGC CAGCTATGCC GCGGCCTGCG
1901 CGGGGAGAGG CGTGCGCGTC GCGTGGCGCG AGCCAGGCCG CTGTGAGCTG
1951 AACTGCCCGA AAGGCCAGGT GTACCTGCAG TGCGGGACCC CCTGCAACCT
2001 GACCTGCCGC TCTCTCTCTT ACCCGGATGA GGAATGCAAT GAGGCCTGCC
2051 TGGAGGGCTG CTTCTGCCCC CCAGGGCTCT ACATGGATGA GAGGGGGGAC
2101 TGCGTGCCCA AGGCCCAGTG CCCCTGTTAC TATGACGGTG AGATCTTCCA
2151 GCCAGAAGAC ATCTTCTCAG ACCATCACAC CATGTGCTAC TGTGAGGATG
2201 GCTTCATGCA CTGTACCATG AGTGGAGTCC CCGGAAGCTT GCTGCCTGAC
2251 GCTGTCCTCA GCAGTCCCCT GTCTCATCGC AGCAAAAGGA GCCTATCCTG
2301 TCGGCCCCCC ATGGTCAAGC TGGTGTGTCC CGCTGACAAC CTGCGGGCTG
2351 AAGGGCTCGA GTGTACCAAA ACGTGCCAGA ACTATGACCT GGAGTGCATG
2401 AGCATGGGCT GTGTCTCTGG CTGCCTCTGC CCCCCGGGCA TGGTCCGGCA
2451 TGAGAACAGA TGTGTGGCCC TGGAAAGGTG TCCCTGCTTC CATCAGGGCA
2501 AGGAGTATGC CCCTGGAGAA ACAGTGAAGA TTGGCTGCAA CACTTGTGTC
2551 TGTCGGGACC GGAAGTGGAA CTGCACAGAC CATGTGTGTG ATGCCACGTG
2601 CTCCACGATC GGCATGGCCC ACTACCTCAC CTTCGACGGG CTCAAATACC
2651 TGTTCCCCGG GGAGTGCCAG TACGTTCTGG TGCAGGATTA CTGCGGCAGT
2701 AACCCTGGGA CCTTTCGGAT CCTAGTGGGG AATAAGGGAT GCAGCCACCC
2751 CTCAGTGAAA TGCAAGAAAC GGGTCACCAT CCTGGTGGAG GGAGGAGAGA
2801 TTGAGCTGTT TGACGGGGAG GTGAATGTGA AGAGGCCCAT GAAGGATGAG
2851 ACTCACTTTG AGGTGGTGGA GTCTGGCCGG TACATCATTC TGCTGCTGGG
2901 CAAAGCCCTC TCCGTGGTCT GGGACCGCCA CCTGAGCATC TCCGTGGTCC
2951 TGAAGCAGAC ATACCAGGAG AAAGTGTGTG GCCTGTGTGG GAATTTTGAT
3001 GGCATCCAGA ACAATGACCT CACCAGCAGC AACCTCCAAG TGGAGGAAGA
3051 CCCTGTGGAC TTTGGGAACT CCTGGAAAGT GAGCTCGCAG TGTGCTGACA
3101 CCAGAAAAGT GCCTCTGGAC TCATCCCCTG CCACCTGCCA TAACAACATC
3151 ATGAAGCAGA CGATGGTGGA TTCCTCCTGT AGAATCCTTA CCAGTGACGT
3201 CTTCCAGGAC TGCAACAAGC TGGTGGACCC CGAGCCATAT CTGGATGTCT
3251 GCATTTACGA CACCTGCTCC TGTGAGTCCA TTGGGGACTG CGCCGCATTC
3301 TGCGACACCA TTGCTGCCTA TGCCCACGTG TGTGCCCAGC ATGGCAAGGT
3351 GGTGACCTGG AGGACGGCCA CATTGTGCCC CCAGAGCTGC GAGGAGAGGA
3401 ATCTCCGGGA GAACGGGTAT GAGGCTGAGT GGCGCTATAA CAGCTGTGCA
3451 CCTGCCTGTC AAGTCACGTG TCAGCACCCT GAGCCACTGG CCTGCCCTGT
3501 GCAGTGTGTG GAGGGCTGCC ATGCCCACTG CCCTCCAGGG AAAATCCTGG
3551 ATGAGCTTTT GCAGACCTGC GTTGACCCTG AAGACTGTCC AGTGTGTGAG
3601 GTGGCTGGCC GGCGTTTTGC CTCAGGAAAG AAAGTCACCT TGAATCCCAG
3651 TGACCCTGAG CACTGCCAGA TTTGCCACTG TGATGTTGTC AACCTCACCT
3701 GTGAAGCCTG CCAGGAGCCG GGTACATCAG AGAGCGCCAC CCCTGAAAGT
3751 GGTCCCGGGA GCGAGCCAGC CACATCTGGG TCGGAAACGC CAGGCACATC
3801 CGAGTCTGCA ACTCCCGAGT CCGGACCTGG CTCCGAGCCT GCCACTAGCG
3851 GCTCCGAGAC TCCGGGAACT TCCGAGAGCG CTACACCAGA AAGCGGACCC
3901 GGAACCAGTA CCGAACCTAG CGAGGGCTCT GCTCCGGGCA GCCCAGCCGG
3951 CTCTCCTACA TCCACGGAGG AGGGCACTTC CGAATCCGCC ACCCCGGAGT
4001 CAGGGCCAGG ATCTGAACCC GCTACCTCAG GCAGTGAGAC GCCAGGAACG
4051 AGCGAGTCCG CTACACCGGA GAGTGGGCCA GGGAGCCCTG CTGGATCTCC
4101 TACGTCCACT GAGGAAGGGT CACCAGCGGG CTCGCCCACC AGCACTGAAG
4151 AAGGTGCCTC GTCTGACAAG AACACTGGTG ATTATTACGA GGACAGTTAT
4201 GAAGATATTT CAGCATACTT GCTGAGTAAA AACAATGCCA TTGAACCAAG
4251 AAGCTTCTCT GACAAAACTC ACACATGCCC ACCGTGCCCA GCTCCAGAAC
4301 TCCTGGGCGG ACCGTCAGTC TTCCTCTTCC CCCCAAAACC CAAGGACACC
4351 CTCATGATCT CCCGGACCCC TGAGGTCACA TGCGTGGTGG TGGACGTGAG
4401 CCACGAAGAC CCTGAGGTCA AGTTCAACTG GTATGTGGAC GGCGTGGAAG
4451 TGCATAATGC CAAGACAAAG CCGCGGGAGG AGCAGTACAA CAGCACGTAC
4501 CGTGTGGTCA GCGTCCTCAC CGTCCTGCAC CAAGACTGGC TGAATGGCAA
4551 GGAGTACAAG TGCAAGGTCT CCAACAAAGC CCTCCCAGCC CCCATCGAGA
4601 AAACCATCTC CAAAGCCAAA GGGCAGCCCC GAGAACCACA GGTGTACACC
4651 CTGCCCCCAT CCCGGGATGA GCTGACCAAG AACCAAGTTA GCCTGACCTG
4701 CCTGGTCAAA GGCTTCTATC CCAGCGACAT CGCCGTGGAG TGGGAGAGCA
4751 ATGGGCAGCC GGAGAACAAC TACAAGACCA CGCCTCCCGT GTTGGACTCC
4801 GACGGCTCCT TCTTCCTCTA CTCCAAGCTC ACCGTGGACA AGAGCAGGTG
4851 GCAGCAGGGG AACGTCTTCT CATGCTCCGT GATGCATGAG GCTCTGCACA
4901 ACCACTACAC GCAGAAGAGC CTCTCCCTGT CTCCGGGTTG A
Таблица 19. Дополнительные последовательности химерных полипептидов
SEQ ID NO:
SEQ ID NO: 8
GTSTEPSEGS APGSPAGSPT STEEGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP 120
GSPAGSPTST EEGSPAGSPT STEEGTSTEP SEGSAPGTSE SATPESGPGT SESATPESGP 180
GTSESATPES GPGSEPATSG SETPGSEPAT SGSETPGSPA GSPTSTEEGT STEPSEGSAP 240
GTSTEPSEGS APGSEPATSG SETPGTSESA TPESGPGTST EPSEGSAP 288
SEQ ID NO: 13
SEQ ID NO: 22
SEQ ID NO: 173
IAKPRPPWMG LLGPTIQAEV YDTVVITLKN MASHPVSLHA VGVSYWKASE GAEYDDQTSQ 120
REKEDDKVFP GGSHTYVWQV LKENGPMASD PLCLTYSYLS HVDLVKDLNS GLIGALLVCR 180
EGSLAKEKTQ TLHKFILLFA VFDEGKSWHS ETKNSLMQDR DAASARAWPK MHTVNGYVNR 240
SLPGLIGCHR KSVYWHVIGM GTTPEVHSIF LEGHTFLVRN HRQASLEISP ITFLTAQTLL 300
MDLGQFLLFC HISSHQHDGM EAYVKVDSCP EEPQLRMKNN EEAEDYDDDL TDSEMDVVRF 360
DDDNSPSFIQ IRSVAKKHPK TWVHYIAAEE EDWDYAPLVL APDDRSYKSQ YLNNGPQRIG 420
RKYKKVRFMA YTDETFKTRE AIQHESGILG PLLYGEVGDT LLIIFKNQAS RPYNIYPHGI 480
TDVRPLYSRR LPKGVKHLKD FPILPGEIFK YKWTVTVEDG PTKSDPRCLT RYYSSFVNME 540
RDLASGLIGP LLICYKESVD QRGNQIMSDK RNVILFSVFD ENRSWYLTEN IQRFLPNPAG 600
VQLEDPEFQA SNIMHSINGY VFDSLQLSVC LHEVAYWYIL SIGAQTDFLS VFFSGYTFKH 660
KMVYEDTLTL FPFSGETVFM SMENPGLWIL GCHNSDFRNR GMTALLKVSS CDKNTGDYYE 720
DSYEDISAYL LSKNNAIEPR SFSQNGTSES ATPESGPGSE PATSGSETPG TSESATPESG 780
PGSEPATSGS ETPGTSESAT PESGPGTSTE PSEGSAPGSP AGSPTSTEEG TSESATPESG 840
PGSEPATSGS ETPGTSESAT PESGPGSPAG SPTSTEEGSP AGSPTSTEEG TSTEPSEGSA 900
PGTSESATPE SGPGTSESAT PESGPGTSES ATPESGPGSE PATSGSETPG SEPATSGSET 960
PGSPAGSPTS TEEGTSTEPS EGSAPGTSTE PSEGSAPGSE PATSGSETPG TSESATPESG 1020
PGTSTEPSEG SAPASSEITR TTLQSDQEEI DYDDTISVEM KKEDFDIYDE DENQSPRSFQ 1080
KKTRHYFIAA VERLWDYGMS SSPHVLRNRA QSGSVPQFKK VVFQEFTDGS FTQPLYRGEL 1140
NEHLGLLGPY IRAEVEDNIM VTFRNQASRP YSFYSSLISY EEDQRQGAEP RKNFVKPNET 1200
KTYFWKVQHH MAPTKDEFDC KAWAYFSDVD LEKDVHSGLI GPLLVCHTNT LNPAHGRQVT 1260
VQEFALFFTI FDETKSWYFT ENMERNCRAP CNIQMEDPTF KENYRFHAIN GYIMDTLPGL 1320
VMAQDQRIRW YLLSMGSNEN IHSIHFSGHV FTVRKKEEYK MALYNLYPGV FETVEMLPSK 1380
AGIWRVECLI GEHLHAGMST LFLVYSNKCQ TPLGMASGHI RDFQITASGQ YGQWAPKLAR 1440
LHYSGSINAW STKEPFSWIK VDLLAPMIIH GIKTQGARQK FSSLYISQFI IMYSLDGKKW 1500
QTYRGNSTGT LMVFFGNVDS SGIKHNIFNP PIIARYIRLH PTHYSIRSTL RMELMGCDLN 1560
SCSMPLGMES KAISDAQITA SSYFTNMFAT WSPSKARLHL QGRSNAWRPQ VNNPKEWLQV 1620
DFQKTMKVTG VTTQGVKSLL TSMYVKEFLI SSSQDGHQWT LFFQNGKVKV FQGNQDSFTP 1680
VVNSLDPPLL TRYLRIHPQS WVHQIALRME VLGCEAQDLY DKTHTCPPCP APELLGGPSV 1740
FLFPPKPKDT LMISRTPEVT CVVVDVSHED PEVKFNWYVD GVEVHNAKTK PREEQYNSTY 1800
RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK GQPREPQVYT LPPSRDELTK 1860
NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS DGSFFLYSKL TVDKSRWQQG 1920
NVFSCSVMHE ALHNHYTQKS LSLSPGK
SEQ ID NO: 54
GTSTEPSEGS APGTSTEPSE GSAPGTSTEP SEGSAPGTST EPSEGSAPGT STEPSEGSAP 120
GSPAGSPTST EEGTSTEPSE GSAPGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP 180
GSEPATSGSE TPGTSESATP ESGPGTSTEP SEGSAPGTSE SATPESGPGS PAGSPTSTEE 240
GSPAGSPTST EEGSPAGSPT STEEGTSESA TPESGPGTST EPSEGSAP 288
SEQ ID NO: 58
TSTEPSEGSA PGSPAGSPTS TEEGTSESAT PESGPGSEPA TSGSETPGTS ESATPESGPG 120
SPAGSPTSTE EGSPAGSPTS TEEG
SEQ ID NO: 88
SEQ ID NO: 64
SEQ ID NO: 215
IAKPRPPWMG LLGPTIQAEV YDTVVITLKN MASHPVSLHA VGVSYWKASE GAEYDDQTSQ 120
REKEDDKVFP GGSHTYVWQV LKENGPMASD PLCLTYSYLS HVDLVKDLNS GLIGALLVCR 180
EGSLAKEKTQ TLHKFILLFA VFDEGKSWHS ETKNSLMQDR DAASARAWPK MHTVNGYVNR 240
SLPGLIGCHR KSVYWHVIGM GTTPEVHSIF LEGHTFLVRN HRQASLEISP ITFLTAQTLL 300
MDLGQFLLFC HISSHQHDGM EAYVKVDSCP EEPQLRMKNN EEAEDYDDDL TDSEMDVVRF 360
DDDNSPSFIQ IRSVAKKHPK TWVHYIAAEE EDWDYAPLVL APDDRSYKSQ YLNNGPQRIG 420
RKYKKVRFMA YTDETFKTRE AIQHESGILG PLLYGEVGDT LLIIFKNQAS RPYNIYPHGI 480
TDVRPLYSRR LPKGVKHLKD FPILPGEIFK YKWTVTVEDG PTKSDPRCLT RYYSSFVNME 540
RDLASGLIGP LLICYKESVD QRGNQIMSDK RNVILFSVFD ENRSWYLTEN IQRFLPNPAG 600
VQLEDPEFQA SNIMHSINGY VFDSLQLSVC LHEVAYWYIL SIGAQTDFLS VFFSGYTFKH 660
KMVYEDTLTL FPFSGETVFM SMENPGLWIL GCHNSDFRNR GMTALLKVSS CDKNTGDYYE 720
DSYEDISAYL LSKNNAIEPR SFSQN
химерного полипептида
SEQ ID NO: 216
YGMSSSPHVL RNRAQSGSVP QFKKVVFQEF TDGSFTQPLY RGELNEHLGL LGPYIRAEVE 120
DNIMVTFRNQ ASRPYSFYSS LISYEEDQRQ GAEPRKNFVK PNETKTYFWK VQHHMAPTKD 180
EFDCKAWAYF SDVDLEKDVH SGLIGPLLVC HTNTLNPAHG RQVTVQEFAL FFTIFDETKS 240
WYFTENMERN CRAPCNIQME DPTFKENYRF HAINGYIMDT LPGLVMAQDQ RIRWYLLSMG 300
SNENIHSIHF SGHVFTVRKK EEYKMALYNL YPGVFETVEM LPSKAGIWRV ECLIGEHLHA 360
GMSTLFLVYS NKCQTPLGMA SGHIRDFQIT ASGQYGQWAP KLARLHYSGS INAWSTKEPF 420
SWIKVDLLAP MIIHGIKTQG ARQKFSSLYI SQFIIMYSLD GKKWQTYRGN STGTLMVFFG 480
NVDSSGIKHN IFNPPIIARY IRLHPTHYSI RSTLRMELMG CDLNSCSMPL GMESKAISDA 540
QITASSYFTN MFATWSPSKA RLHLQGRSNA WRPQVNNPKE WLQVDFQKTM KVTGVTTQGV 600
KSLLTSMYVK EFLISSSQDG HQWTLFFQNG KVKVFQGNQD SFTPVVNSLD PPLLTRYLRI 660
HPQSWVHQIA LRMEVLGCEA QDLY
SEQ ID NO: 208
SEQ ID NO: 209
SLSVYLGEFF DIHLFVNGTV TQGDQRVSMP YASKGLYLET EAGYYKLSGE AYGFVARIDG 120
SGNFQVLLSD RYFNKTCGLC GNFNIFAEDD FMTQEGTLTS DPYDFANSWA LSSGEQWCER 180
ASPPSSSCNI SSGEMQKGLW EQCQLLKSTS VFARCHPLVD PEPFVALCEK TLCECAGGLE 240
CACPALLEYA RTCAQEGMVL YGWTDHSACS PVCPAGMEYR QCVSPCARTC QSLHINEMCQ 300
ERCVDGCSCP EGQLLDEGLC VESTECPCVH SGKRYPPGTS LSRDCNTCIC RNSQWICSNE 360
ECPGECLVTG QSHFKSFDNR YFTFSGICQY LLARDCQDHS FSIVIETVQC ADDRDAVCTR 420
SVTVRLPGLH NSLVKLKHGA GVAMDGQDIQ LPLLKGDLRI QHTVTASVRL SYGEDLQMDW 480
DGRGRLLVKL SPVYAGKTCG LCGNYNGNQG DDFLTPSGLA EPRVEDFGNA WKLHGDCQDL 540
QKQHSDPCAL NPRMTRFSEE ACAVLTSPTF EACHRAVSPL PYLRNCRYDV CSCSDGRECL 600
CGALASYAAA CAGRGVRVAW REPGRCELNC PKGQVYLQCG TPCNLTCRSL SYPDEECNEA 660
CLEGCFCPPG LYMDERGDCV PKAQCPCYYD GEIFQPEDIF SDHHTMCYCE DGFMHCTMSG 720
VPGSLLPDAV LSSPLSHRSK R
SEQ ID NO: 210
LERCPCFHQG KEYAPGETVK IGCNTCVCRD RKWNCTDHVC DAT 103
SEQ ID NO: 214
ILVEGGEIEL FDGEVNVKRP MKDETHFEVV ESGRYIILLL GKALSVVWDR HLSISVVLKQ 120
TYQEKVCGLC GNFDGIQNND LTSSNLQVEE DPVDFGNSWK VSSQCADTRK VPLDSSPATC 180
HNNIMKQTMV DSSCRILTSD VFQDCNKLVD PEPYLDVCIY DTCSCESIGD CAAFCDTIAA 240
YAHVCAQHGK VVTWRTATLC PQSCEERNLR ENGYEAEWRY NSCAPACQVT CQHPEPLACP 300
VQCVEGCHAH CPPGKILDEL LQTCVDPEDC PVCEVAGRRF ASGKKVTLNP SDPEHCQICH 360
CDVVNLTCEA CQEP 374
SEQ ID NO: 217
GVEVHNAKTK PREEQYNSTY RVVSVLTVLH QDWLNGKEYK CKVSNKALPA PIEKTISKAK 120
GQPREPQVYT LPPSRDELTK NQVSLTCLVK GFYPSDIAVE WESNGQPENN YKTTPPVLDS 180
DGSFFLYSKL TVDKSRWQQG NVFSCSVMHE ALHNHYTQKS LSLSPG
SEQ ID NO: 218
GTSTEPSEGS APGSPAGSPT STEEGTSESA TPESGPGSEP ATSGSETPGT SESATPESGP 120
GSPAGSPTST EEGSPAGSPT STEEGTSTEP SEGSAPGTSE SATPESGPGT SESATPESGP 180
GTSESATPES GPGSEPATSG SETPGSEPAT SGSETPGSPA GSPTSTEEGT STEPSEGSAP 240
GTSTEPSEGS APGSEPATSG SETPGTSESA TPESGPGTST EPSEGSAPAS S 291
[0387] Предшествующее описание конкретных вариантов осуществления настолько полно раскрывает общий характер настоящего изобретения, что другие могут, используя знания в пределах квалификации в данной области техники, легко модифицировать и/или адаптировать такие конкретные варианты осуществления для различных вариантов применения без проведения дополнительных экспериментов, не отступая от общей концепции настоящего изобретения. Следовательно, такие адаптации и модификации предназначены для того, чтобы находиться в пределах значения и диапазона эквивалентов раскрытых вариантов осуществления, основанных на идее и принципе, представленных в данном документе. Следует понимать, что формулировки или терминология в данном документе предназначены для целей описания, а не ограничения, и вследствие этого терминологию или формулировки в настоящем описании квалифицированному специалисту следует интерпретировать в свете данных идей и принципов.
[0388] Другие варианты осуществления настоящего изобретения будут очевидны специалистам в данной области при рассмотрении описания и практическом применении настоящего изобретения, раскрытого в данном документе. Предполагается, что описание и примеры должны рассматриваться только в качестве иллюстративных, а действительные объем и сущность настоящего изобретения указаны в нижеследующей формуле изобретения.
Все патенты и публикации, цитируемые в данном документе, включены в данный документ посредством ссылки во всей своей полноте.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Bioverativ Therapeutics Inc.
<120> СПОСОБЫ ЛЕЧЕНИЯ ГЕМОФИЛИИ A
<130> 612297: SA9-461PC
<150> US 62/673,670
<151> 2018-05-18
<150> US 62/712,880
<151> 2018-07-31
<150> US 62/773,785
<151> 2018-11-30
<150> US 62/801,576
<151> 2019-02-05
<160> 218
<170> PatentIn версия 3.5
<210> 1
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 1
Thr Gln Ser Phe Asn Asp Phe Thr Arg
1 5
<210> 2
<211> 302
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 312A
<400> 2
Pro Arg Ser Phe Ser Gln Asn Gly Thr Ser Glu Ser Ala Thr Pro Glu
1 5 10 15
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
20 25 30
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
35 40 45
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
50 55 60
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
65 70 75 80
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
85 90 95
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
100 105 110
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
115 120 125
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
130 135 140
Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly
145 150 155 160
Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
165 170 175
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser
180 185 190
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
195 200 205
Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
210 215 220
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu
225 230 235 240
Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly
245 250 255
Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
260 265 270
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
275 280 285
Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile Thr Arg
290 295 300
<210> 3
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 3
Ser Val Ser Gln Thr Ser Lys Leu Thr Arg
1 5 10
<210> 4
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 4
Asp Phe Leu Ala Glu Gly Gly Gly Val Arg
1 5 10
<210> 5
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 5
Thr Thr Lys Ile Lys Pro Arg
1 5
<210> 6
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 6
Leu Val Pro Arg Gly
1 5
<210> 7
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 7
Ala Leu Arg Pro Arg
1 5
<210> 8
<211> 288
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE288
<400> 8
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1 5 10 15
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
20 25 30
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
35 40 45
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
50 55 60
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
65 70 75 80
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
85 90 95
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
100 105 110
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr
115 120 125
Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
130 135 140
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu
145 150 155 160
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro
165 170 175
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
180 185 190
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro
195 200 205
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr
210 215 220
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
225 230 235 240
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
245 250 255
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
260 265 270
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
275 280 285
<210> 9
<211> 42
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE42
<400> 9
Gly Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
1 5 10 15
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
20 25 30
Thr Ser Gly Ser Glu Thr Pro Ala Ser Ser
35 40
<210> 10
<211> 78
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE72
<400> 10
Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1 5 10 15
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala
20 25 30
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
35 40 45
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
50 55 60
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ala Ser Ser
65 70 75
<210> 11
<211> 143
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144
<400> 11
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
1 5 10 15
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
20 25 30
Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
35 40 45
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
50 55 60
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly
65 70 75 80
Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
85 90 95
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu
100 105 110
Ser Ala Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
115 120 125
Glu Thr Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
130 135 140
<210> 12
<211> 56
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF057
<400> 12
Thr Ser Thr Glu Glu Gly Ala Ser Ser Gly Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
20 25 30
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg
35 40 45
Gly Ser Gly Gly Asp Lys Thr His
50 55
<210> 13
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF059
<400> 13
Thr Ser Thr Glu Glu Gly Ala Ser Ile Ser Asp Lys Asn Thr Gly Asp
1 5 10 15
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
20 25 30
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Asp Lys Thr His
35 40 45
<210> 14
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG144
<400> 14
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
1 5 10 15
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr
20 25 30
Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
35 40 45
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro
50 55 60
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
65 70 75 80
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
85 90 95
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Pro
100 105 110
Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly Ser Gly Thr Ala
115 120 125
Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
130 135 140
<210> 15
<211> 864
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE864
<400> 15
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
1 5 10 15
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu
20 25 30
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
35 40 45
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
50 55 60
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
65 70 75 80
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
85 90 95
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala
100 105 110
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro
115 120 125
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
130 135 140
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala
145 150 155 160
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu
165 170 175
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
180 185 190
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
195 200 205
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
210 215 220
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
225 230 235 240
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
245 250 255
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
260 265 270
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
275 280 285
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
290 295 300
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
305 310 315 320
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
325 330 335
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
340 345 350
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu
355 360 365
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
370 375 380
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
385 390 395 400
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
405 410 415
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
420 425 430
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
435 440 445
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
450 455 460
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
465 470 475 480
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
485 490 495
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
500 505 510
Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
515 520 525
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
530 535 540
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro
545 550 555 560
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
565 570 575
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
580 585 590
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
595 600 605
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
610 615 620
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
625 630 635 640
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
645 650 655
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
660 665 670
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
675 680 685
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr
690 695 700
Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
705 710 715 720
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu
725 730 735
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro
740 745 750
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
755 760 765
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro
770 775 780
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr
785 790 795 800
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
805 810 815
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
820 825 830
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
835 840 845
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
850 855 860
<210> 16
<211> 576
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE576
<400> 16
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
1 5 10 15
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu
20 25 30
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
35 40 45
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
50 55 60
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
65 70 75 80
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
85 90 95
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala
100 105 110
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro
115 120 125
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
130 135 140
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala
145 150 155 160
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu
165 170 175
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
180 185 190
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
195 200 205
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
210 215 220
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
225 230 235 240
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
245 250 255
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
260 265 270
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
275 280 285
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
290 295 300
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
305 310 315 320
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
325 330 335
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
340 345 350
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu
355 360 365
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
370 375 380
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
385 390 395 400
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
405 410 415
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
420 425 430
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
435 440 445
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
450 455 460
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
465 470 475 480
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
485 490 495
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
500 505 510
Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
515 520 525
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
530 535 540
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro
545 550 555 560
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
565 570 575
<210> 17
<211> 864
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG864
<400> 17
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Pro
1 5 10 15
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr
20 25 30
Gly Thr Gly Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
35 40 45
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
85 90 95
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Thr Pro Gly
100 105 110
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
115 120 125
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
130 135 140
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
145 150 155 160
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
165 170 175
Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
180 185 190
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
195 200 205
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr
210 215 220
Gly Thr Gly Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
225 230 235 240
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala Ser Pro
245 250 255
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
260 265 270
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
275 280 285
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro
290 295 300
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
305 310 315 320
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
325 330 335
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly
340 345 350
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
355 360 365
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
370 375 380
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr
385 390 395 400
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
405 410 415
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
420 425 430
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
435 440 445
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
450 455 460
Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
465 470 475 480
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro
485 490 495
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
500 505 510
Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
515 520 525
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro
530 535 540
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
545 550 555 560
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
565 570 575
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
580 585 590
Pro Ser Gly Ala Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala
595 600 605
Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
610 615 620
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
625 630 635 640
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
645 650 655
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
660 665 670
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro
675 680 685
Gly Thr Ser Ser Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala
690 695 700
Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
705 710 715 720
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro
725 730 735
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
740 745 750
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
755 760 765
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
770 775 780
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
785 790 795 800
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
805 810 815
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
820 825 830
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
835 840 845
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
850 855 860
<210> 18
<211> 576
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG576
<400> 18
Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser
1 5 10 15
Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser
20 25 30
Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly
35 40 45
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser
50 55 60
Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser
65 70 75 80
Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
85 90 95
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Thr Pro
100 105 110
Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser
115 120 125
Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
130 135 140
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser
145 150 155 160
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly Ser Gly Thr
165 170 175
Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
180 185 190
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser
195 200 205
Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly
210 215 220
Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
225 230 235 240
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Thr Pro
245 250 255
Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly
260 265 270
Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
275 280 285
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser
290 295 300
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser
305 310 315 320
Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
325 330 335
Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser
340 345 350
Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser
355 360 365
Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
370 375 380
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Thr Pro
385 390 395 400
Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly
405 410 415
Ala Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser
420 425 430
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Thr Pro
435 440 445
Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly
450 455 460
Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
465 470 475 480
Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser
485 490 495
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser
500 505 510
Ser Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser
515 520 525
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser
530 535 540
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser
545 550 555 560
Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
565 570 575
<210> 19
<211> 288
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG288
<400> 19
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser
1 5 10 15
Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr
20 25 30
Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
35 40 45
Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser
50 55 60
Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr
65 70 75 80
Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
85 90 95
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser
100 105 110
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser
115 120 125
Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser
130 135 140
Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser
145 150 155 160
Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser
165 170 175
Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly
180 185 190
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser
195 200 205
Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly
210 215 220
Ala Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly
225 230 235 240
Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser
245 250 255
Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly Ser Gly Thr
260 265 270
Ala Ser Ser Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
275 280 285
<210> 20
<211> 8442
<212> ДНК
<213> Homo sapiens
<400> 20
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcrgagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgcctgcttc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gagtgtgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
ggaggcctgg tggtgcctcc cacagatgcc ccggtgagcc ccaccactct gtatgtggag 3780
gacatctcgg aaccgccgtt gcacgatttc tactgcagca ggctactgga cctggtcttc 3840
ctgctggatg gctcctccag gctgtccgag gctgagtttg aagtgctgaa ggcctttgtg 3900
gtggacatga tggagcggct gcgcatctcc cagaagtggg tccgcgtggc cgtggtggag 3960
taccacgacg gctcccacgc ctacatcggg ctcaaggacc ggaagcgacc gtcagagctg 4020
cggcgcattg ccagccaggt gaagtatgcg ggcagccagg tggcctccac cagcgaggtc 4080
ttgaaataca cactgttcca aatcttcagc aagatcgacc gccctgaagc ctcccgcatc 4140
gccctgctcc tgatggccag ccaggagccc caacggatgt cccggaactt tgtccgctac 4200
gtccagggcc tgaagaagaa gaaggtcatt gtgatcccgg tgggcattgg gccccatgcc 4260
aacctcaagc agatccgcct catcgagaag caggcccctg agaacaaggc cttcgtgctg 4320
agcagtgtgg atgagctgga gcagcaaagg gacgagatcg ttagctacct ctgtgacctt 4380
gcccctgaag cccctcctcc tactctgccc cccgacatgg cacaagtcac tgtgggcccg 4440
gggctcttgg gggtttcgac cctggggccc aagaggaact ccatggttct ggatgtggcg 4500
ttcgtcctgg aaggatcgga caaaattggt gaagccgact tcaacaggag caaggagttc 4560
atggaggagg tgattcagcg gatggatgtg ggccaggaca gcatccacgt cacggtgctg 4620
cagtactcct acatggtgac cgtggagtac cccttcagcg aggcacagtc caaaggggac 4680
atcctgcagc gggtgcgaga gatccgctac cagggcggca acaggaccaa cactgggctg 4740
gccctgcggt acctctctga ccacagcttc ttggtcagcc agggtgaccg ggagcaggcg 4800
cccaacctgg tctacatggt caccggaaat cctgcctctg atgagatcaa gaggctgcct 4860
ggagacatcc aggtggtgcc cattggagtg ggccctaatg ccaacgtgca ggagctggag 4920
aggattggct ggcccaatgc ccctatcctc atccaggact ttgagacgct cccccgagag 4980
gctcctgacc tggtgctgca gaggtgctgc tccggagagg ggctgcagat ccccaccctc 5040
tcccctgcac ctgactgcag ccagcccctg gacgtgatcc ttctcctgga tggctcctcc 5100
agtttcccag cttcttattt tgatgaaatg aagagtttcg ccaaggcttt catttcaaaa 5160
gccaatatag ggcctcgtct cactcaggtg tcagtgctgc agtatggaag catcaccacc 5220
attgacgtgc catggaacgt ggtcccggag aaagcccatt tgctgagcct tgtggacgtc 5280
atgcagcggg agggaggccc cagccaaatc ggggatgcct tgggctttgc tgtgcgatac 5340
ttgacttcag aaatgcatgg tgccaggccg ggagcctcaa aggcggtggt catcctggtc 5400
acggacgtct ctgtggattc agtggatgca gcagctgatg ccgccaggtc caacagagtg 5460
acagtgttcc ctattggaat tggagatcgc tacgatgcag cccagctacg gatcttggca 5520
ggcccagcag gcgactccaa cgtggtgaag ctccagcgaa tcgaagacct ccctaccatg 5580
gtcaccttgg gcaattcctt cctccacaaa ctgtgctctg gatttgttag gatttgcatg 5640
gatgaggatg ggaatgagaa gaggcccggg gacgtctgga ccttgccaga ccagtgccac 5700
accgtgactt gccagccaga tggccagacc ttgctgaaga gtcatcgggt caactgtgac 5760
cgggggctga ggccttcgtg ccctaacagc cagtcccctg ttaaagtgga agagacctgt 5820
ggctgccgct ggacctgccc ctgygtgtgc acaggcagct ccactcggca catcgtgacc 5880
tttgatgggc agaatttcaa gctgactggc agctgttctt atgtcctatt tcaaaacaag 5940
gagcaggacc tggaggtgat tctccataat ggtgcctgca gccctggagc aaggcagggc 6000
tgcatgaaat ccatcgaggt gaagcacagt gccctctccg tcgagstgca cagtgacatg 6060
gaggtgacgg tgaatgggag actggtctct gttccttacg tgggtgggaa catggaagtc 6120
aacgtttatg gtgccatcat gcatgaggtc agattcaatc accttggtca catcttcaca 6180
ttcactccac aaaacaatga gttccaactg cagctcagcc ccaagacttt tgcttcaaag 6240
acgtatggtc tgtgtgggat ctgtgatgag aacggagcca atgacttcat gctgagggat 6300
ggcacagtca ccacagactg gaaaacactt gttcaggaat ggactgtgca gcggccaggg 6360
cagacgtgcc agcccatcct ggaggagcag tgtcttgtcc ccgacagctc ccactgccag 6420
gtcctcctct taccactgtt tgctgaatgc cacaaggtcc tggctccagc cacattctat 6480
gccatctgcc agcaggacag ttgccaccag gagcaagtgt gtgaggtgat cgcctcttat 6540
gcccacctct gtcggaccaa cggggtctgc gttgactgga ggacacctga tttctgtgct 6600
atgtcatgcc caccatctct ggtctacaac cactgtgagc atggctgtcc ccggcactgt 6660
gatggcaacg tgagctcctg tggggaccat ccctccgaag gctgtttctg ccctccagat 6720
aaagtcatgt tggaaggcag ctgtgtccct gaagaggcct gcactcagtg cattggtgag 6780
gatggagtcc agcaccagtt cctggaagcc tgggtcccgg accaccagcc ctgtcagatc 6840
tgcacatgcc tcagcgggcg gaaggtcaac tgcacaacgc agccctgccc cacggccaaa 6900
gctcccacgt gtggcctgtg tgaagtagcc cgcctccgcc agaatgcaga ccagtgctgc 6960
cccgagtatg agtgtgtgtg tgacccagtg agctgtgacc tgcccccagt gcctcactgt 7020
gaacgtggcc tccagcccac actgaccaac cctggcgagt gcagacccaa cttcacctgc 7080
gcctgcagga aggaggagtg caaaagagtg tccccaccct cctgcccccc gcaccgtttg 7140
cccacccttc ggaagaccca gtgctgtgat gagtatgagt gtgcctgcaa ctgtgtcaac 7200
tccacagtga gctgtcccct tgggtacttg gcctcaaccg ccaccaatga ctgtggctgt 7260
accacaacca cctgccttcc cgacaaggtg tgtgtccacc gaagcaccat ctaccctgtg 7320
ggccagttct gggaggaggg ctgcgatgtg tgcacctgca ccgacatgga ggatgccgtg 7380
atgggcctcc gcgtggccca gtgctcccag aagccctgtg aggacagctg tcggtcgggc 7440
ttcacttacg ttctgcatga aggcgagtgc tgtggaaggt gcctgccatc tgcctgtgag 7500
gtggtgactg gctcaccgcg gggggactcc cagtcttcct ggaagagtgt cggctcccag 7560
tgggcctccc cggagaaccc ctgcctcatc aatgagtgtg tccgagtgaa ggaggaggtc 7620
tttatacaac aaaggaacgt ctcctgcccc cagctggagg tccctgtctg cccctcgggc 7680
tttcagctga gctgtaagac ctcagcgtgc tgcccaagct gtcgctgtga gcgcatggag 7740
gcctgcatgc tcaatggcac tgtcattggg cccgggaaga ctgtgatgat cgatgtgtgc 7800
acgacctgcc gctgcatggt gcaggtgggg gtcatctctg gattcaagct ggagtgcagg 7860
aagaccacct gcaacccctg ccccctgggt tacaaggaag aaaataacac aggtgaatgt 7920
tgtgggagat gtttgcctac ggcttgcacc attcagctaa gaggaggaca gatcatgaca 7980
ctgaagcgtg atgagacgct ccaggatggc tgtgatactc acttctgcaa ggtcaatgag 8040
agaggagagt acttctggga gaagagggtc acaggctgcc caccctttga tgaacacaag 8100
tgtcttgctg agggaggtaa aattatgaaa attccaggca cctgctgtga cacatgtgag 8160
gagcctgagt gcaacgacat cactgccagg ctgcagtatg tcaaggtggg aagctgtaag 8220
tctgaagtag aggtggatat ccactactgc cagggcaaat gtgccagcaa agccatgtac 8280
tccattgaca tcaacgatgt gcaggaccag tgctcctgct gctctccgac acggacggag 8340
cccatgcagg tggccctgca ctgcaccaat ggctctgttg tgtaccatga ggttctcaat 8400
gccatggagt gcaaatgctc ccccaggaag tgcagcaagt ga 8442
<210> 21
<211> 2813
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> другой_признак
<222> (2016)..(2016)
<223> Xaa может представлять собой любую втречающуюся в природе аминокислоту
<400> 21
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Cys Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu Val Val Pro Pro Thr
1235 1240 1245
Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile Ser
1250 1255 1260
Glu Pro Pro Leu His Asp Phe Tyr Cys Ser Arg Leu Leu Asp Leu
1265 1270 1275
Val Phe Leu Leu Asp Gly Ser Ser Arg Leu Ser Glu Ala Glu Phe
1280 1285 1290
Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu Arg
1295 1300 1305
Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr His Asp
1310 1315 1320
Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser
1325 1330 1335
Glu Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln
1340 1345 1350
Val Ala Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile
1355 1360 1365
Phe Ser Lys Ile Asp Arg Pro Glu Ala Ser Arg Ile Ala Leu Leu
1370 1375 1380
Leu Met Ala Ser Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val
1385 1390 1395
Arg Tyr Val Gln Gly Leu Lys Lys Lys Lys Val Ile Val Ile Pro
1400 1405 1410
Val Gly Ile Gly Pro His Ala Asn Leu Lys Gln Ile Arg Leu Ile
1415 1420 1425
Glu Lys Gln Ala Pro Glu Asn Lys Ala Phe Val Leu Ser Ser Val
1430 1435 1440
Asp Glu Leu Glu Gln Gln Arg Asp Glu Ile Val Ser Tyr Leu Cys
1445 1450 1455
Asp Leu Ala Pro Glu Ala Pro Pro Pro Thr Leu Pro Pro Asp Met
1460 1465 1470
Ala Gln Val Thr Val Gly Pro Gly Leu Leu Gly Val Ser Thr Leu
1475 1480 1485
Gly Pro Lys Arg Asn Ser Met Val Leu Asp Val Ala Phe Val Leu
1490 1495 1500
Glu Gly Ser Asp Lys Ile Gly Glu Ala Asp Phe Asn Arg Ser Lys
1505 1510 1515
Glu Phe Met Glu Glu Val Ile Gln Arg Met Asp Val Gly Gln Asp
1520 1525 1530
Ser Ile His Val Thr Val Leu Gln Tyr Ser Tyr Met Val Thr Val
1535 1540 1545
Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly Asp Ile Leu Gln
1550 1555 1560
Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg Thr Asn Thr
1565 1570 1575
Gly Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu Val Ser
1580 1585 1590
Gln Gly Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val Thr
1595 1600 1605
Gly Asn Pro Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile
1610 1615 1620
Gln Val Val Pro Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu
1625 1630 1635
Leu Glu Arg Ile Gly Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp
1640 1645 1650
Phe Glu Thr Leu Pro Arg Glu Ala Pro Asp Leu Val Leu Gln Arg
1655 1660 1665
Cys Cys Ser Gly Glu Gly Leu Gln Ile Pro Thr Leu Ser Pro Ala
1670 1675 1680
Pro Asp Cys Ser Gln Pro Leu Asp Val Ile Leu Leu Leu Asp Gly
1685 1690 1695
Ser Ser Ser Phe Pro Ala Ser Tyr Phe Asp Glu Met Lys Ser Phe
1700 1705 1710
Ala Lys Ala Phe Ile Ser Lys Ala Asn Ile Gly Pro Arg Leu Thr
1715 1720 1725
Gln Val Ser Val Leu Gln Tyr Gly Ser Ile Thr Thr Ile Asp Val
1730 1735 1740
Pro Trp Asn Val Val Pro Glu Lys Ala His Leu Leu Ser Leu Val
1745 1750 1755
Asp Val Met Gln Arg Glu Gly Gly Pro Ser Gln Ile Gly Asp Ala
1760 1765 1770
Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser Glu Met His Gly Ala
1775 1780 1785
Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu Val Thr Asp Val
1790 1795 1800
Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala Arg Ser Asn
1805 1810 1815
Arg Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr Asp Ala
1820 1825 1830
Ala Gln Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser Asn Val
1835 1840 1845
Val Lys Leu Gln Arg Ile Glu Asp Leu Pro Thr Met Val Thr Leu
1850 1855 1860
Gly Asn Ser Phe Leu His Lys Leu Cys Ser Gly Phe Val Arg Ile
1865 1870 1875
Cys Met Asp Glu Asp Gly Asn Glu Lys Arg Pro Gly Asp Val Trp
1880 1885 1890
Thr Leu Pro Asp Gln Cys His Thr Val Thr Cys Gln Pro Asp Gly
1895 1900 1905
Gln Thr Leu Leu Lys Ser His Arg Val Asn Cys Asp Arg Gly Leu
1910 1915 1920
Arg Pro Ser Cys Pro Asn Ser Gln Ser Pro Val Lys Val Glu Glu
1925 1930 1935
Thr Cys Gly Cys Arg Trp Thr Cys Pro Cys Val Cys Thr Gly Ser
1940 1945 1950
Ser Thr Arg His Ile Val Thr Phe Asp Gly Gln Asn Phe Lys Leu
1955 1960 1965
Thr Gly Ser Cys Ser Tyr Val Leu Phe Gln Asn Lys Glu Gln Asp
1970 1975 1980
Leu Glu Val Ile Leu His Asn Gly Ala Cys Ser Pro Gly Ala Arg
1985 1990 1995
Gln Gly Cys Met Lys Ser Ile Glu Val Lys His Ser Ala Leu Ser
2000 2005 2010
Val Glu Xaa His Ser Asp Met Glu Val Thr Val Asn Gly Arg Leu
2015 2020 2025
Val Ser Val Pro Tyr Val Gly Gly Asn Met Glu Val Asn Val Tyr
2030 2035 2040
Gly Ala Ile Met His Glu Val Arg Phe Asn His Leu Gly His Ile
2045 2050 2055
Phe Thr Phe Thr Pro Gln Asn Asn Glu Phe Gln Leu Gln Leu Ser
2060 2065 2070
Pro Lys Thr Phe Ala Ser Lys Thr Tyr Gly Leu Cys Gly Ile Cys
2075 2080 2085
Asp Glu Asn Gly Ala Asn Asp Phe Met Leu Arg Asp Gly Thr Val
2090 2095 2100
Thr Thr Asp Trp Lys Thr Leu Val Gln Glu Trp Thr Val Gln Arg
2105 2110 2115
Pro Gly Gln Thr Cys Gln Pro Ile Leu Glu Glu Gln Cys Leu Val
2120 2125 2130
Pro Asp Ser Ser His Cys Gln Val Leu Leu Leu Pro Leu Phe Ala
2135 2140 2145
Glu Cys His Lys Val Leu Ala Pro Ala Thr Phe Tyr Ala Ile Cys
2150 2155 2160
Gln Gln Asp Ser Cys His Gln Glu Gln Val Cys Glu Val Ile Ala
2165 2170 2175
Ser Tyr Ala His Leu Cys Arg Thr Asn Gly Val Cys Val Asp Trp
2180 2185 2190
Arg Thr Pro Asp Phe Cys Ala Met Ser Cys Pro Pro Ser Leu Val
2195 2200 2205
Tyr Asn His Cys Glu His Gly Cys Pro Arg His Cys Asp Gly Asn
2210 2215 2220
Val Ser Ser Cys Gly Asp His Pro Ser Glu Gly Cys Phe Cys Pro
2225 2230 2235
Pro Asp Lys Val Met Leu Glu Gly Ser Cys Val Pro Glu Glu Ala
2240 2245 2250
Cys Thr Gln Cys Ile Gly Glu Asp Gly Val Gln His Gln Phe Leu
2255 2260 2265
Glu Ala Trp Val Pro Asp His Gln Pro Cys Gln Ile Cys Thr Cys
2270 2275 2280
Leu Ser Gly Arg Lys Val Asn Cys Thr Thr Gln Pro Cys Pro Thr
2285 2290 2295
Ala Lys Ala Pro Thr Cys Gly Leu Cys Glu Val Ala Arg Leu Arg
2300 2305 2310
Gln Asn Ala Asp Gln Cys Cys Pro Glu Tyr Glu Cys Val Cys Asp
2315 2320 2325
Pro Val Ser Cys Asp Leu Pro Pro Val Pro His Cys Glu Arg Gly
2330 2335 2340
Leu Gln Pro Thr Leu Thr Asn Pro Gly Glu Cys Arg Pro Asn Phe
2345 2350 2355
Thr Cys Ala Cys Arg Lys Glu Glu Cys Lys Arg Val Ser Pro Pro
2360 2365 2370
Ser Cys Pro Pro His Arg Leu Pro Thr Leu Arg Lys Thr Gln Cys
2375 2380 2385
Cys Asp Glu Tyr Glu Cys Ala Cys Asn Cys Val Asn Ser Thr Val
2390 2395 2400
Ser Cys Pro Leu Gly Tyr Leu Ala Ser Thr Ala Thr Asn Asp Cys
2405 2410 2415
Gly Cys Thr Thr Thr Thr Cys Leu Pro Asp Lys Val Cys Val His
2420 2425 2430
Arg Ser Thr Ile Tyr Pro Val Gly Gln Phe Trp Glu Glu Gly Cys
2435 2440 2445
Asp Val Cys Thr Cys Thr Asp Met Glu Asp Ala Val Met Gly Leu
2450 2455 2460
Arg Val Ala Gln Cys Ser Gln Lys Pro Cys Glu Asp Ser Cys Arg
2465 2470 2475
Ser Gly Phe Thr Tyr Val Leu His Glu Gly Glu Cys Cys Gly Arg
2480 2485 2490
Cys Leu Pro Ser Ala Cys Glu Val Val Thr Gly Ser Pro Arg Gly
2495 2500 2505
Asp Ser Gln Ser Ser Trp Lys Ser Val Gly Ser Gln Trp Ala Ser
2510 2515 2520
Pro Glu Asn Pro Cys Leu Ile Asn Glu Cys Val Arg Val Lys Glu
2525 2530 2535
Glu Val Phe Ile Gln Gln Arg Asn Val Ser Cys Pro Gln Leu Glu
2540 2545 2550
Val Pro Val Cys Pro Ser Gly Phe Gln Leu Ser Cys Lys Thr Ser
2555 2560 2565
Ala Cys Cys Pro Ser Cys Arg Cys Glu Arg Met Glu Ala Cys Met
2570 2575 2580
Leu Asn Gly Thr Val Ile Gly Pro Gly Lys Thr Val Met Ile Asp
2585 2590 2595
Val Cys Thr Thr Cys Arg Cys Met Val Gln Val Gly Val Ile Ser
2600 2605 2610
Gly Phe Lys Leu Glu Cys Arg Lys Thr Thr Cys Asn Pro Cys Pro
2615 2620 2625
Leu Gly Tyr Lys Glu Glu Asn Asn Thr Gly Glu Cys Cys Gly Arg
2630 2635 2640
Cys Leu Pro Thr Ala Cys Thr Ile Gln Leu Arg Gly Gly Gln Ile
2645 2650 2655
Met Thr Leu Lys Arg Asp Glu Thr Leu Gln Asp Gly Cys Asp Thr
2660 2665 2670
His Phe Cys Lys Val Asn Glu Arg Gly Glu Tyr Phe Trp Glu Lys
2675 2680 2685
Arg Val Thr Gly Cys Pro Pro Phe Asp Glu His Lys Cys Leu Ala
2690 2695 2700
Glu Gly Gly Lys Ile Met Lys Ile Pro Gly Thr Cys Cys Asp Thr
2705 2710 2715
Cys Glu Glu Pro Glu Cys Asn Asp Ile Thr Ala Arg Leu Gln Tyr
2720 2725 2730
Val Lys Val Gly Ser Cys Lys Ser Glu Val Glu Val Asp Ile His
2735 2740 2745
Tyr Cys Gln Gly Lys Cys Ala Ser Lys Ala Met Tyr Ser Ile Asp
2750 2755 2760
Ile Asn Asp Val Gln Asp Gln Cys Ser Cys Cys Ser Pro Thr Arg
2765 2770 2775
Thr Glu Pro Met Gln Val Ala Leu His Cys Thr Asn Gly Ser Val
2780 2785 2790
Val Tyr His Glu Val Leu Asn Ala Met Glu Cys Lys Cys Ser Pro
2795 2800 2805
Arg Lys Cys Ser Lys
2810
<210> 22
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF059A
<400> 22
Thr Ser Thr Glu Glu Gly Ala Ser Ser Asp Lys Asn Thr Gly Asp Tyr
1 5 10 15
Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn
20 25 30
Asn Ala Ile Glu Pro Arg Ser Phe Ser Asp Lys Thr His
35 40 45
<210> 23
<211> 54
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF073
<400> 23
Thr Ser Thr Glu Glu Gly Ala Ser Ser Gly Gly Gly Gly Ser Gly Gly
1 5 10 15
Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
20 25 30
Gly Ser Gly Gly Gly Gly Ser Ile Glu Pro Arg Ser Phe Ser Gly Ser
35 40 45
Gly Gly Asp Lys Thr His
50
<210> 24
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AD XTEN
<400> 24
Gly Glu Ser Pro Gly Gly Ser Ser Gly Ser Glu Ser
1 5 10
<210> 25
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AD XTEN
<400> 25
Gly Ser Glu Gly Ser Ser Gly Pro Gly Glu Ser Ser
1 5 10
<210> 26
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AD XTEN
<400> 26
Gly Ser Ser Glu Ser Gly Ser Ser Glu Gly Gly Pro
1 5 10
<210> 27
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AD XTEN
<400> 27
Gly Ser Gly Gly Glu Pro Ser Glu Ser Gly Ser Ser
1 5 10
<210> 28
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AE, AM XTEN
<400> 28
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
1 5 10
<210> 29
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AE, AM, AQ XTEN
<400> 29
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
1 5 10
<210> 30
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AE, AM, AQ XTEN
<400> 30
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1 5 10
<210> 31
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AE, AM, AQ XTEN
<400> 31
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
1 5 10
<210> 32
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AF, AM XTEN
<400> 32
Gly Ser Thr Ser Glu Ser Pro Ser Gly Thr Ala Pro
1 5 10
<210> 33
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AF, AM XTEN
<400> 33
Gly Thr Ser Thr Pro Glu Ser Gly Ser Ala Ser Pro
1 5 10
<210> 34
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AF, AM
<400> 34
Gly Thr Ser Pro Ser Gly Glu Ser Ser Thr Ala Pro
1 5 10
<210> 35
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AF, AM XTEN
<400> 35
Gly Ser Thr Ser Ser Thr Ala Glu Ser Pro Gly Pro
1 5 10
<210> 36
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AG, AM XTEN
<400> 36
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
1 5 10
<210> 37
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AG, AM XTEN
<400> 37
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
1 5 10
<210> 38
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AG, AM XTEN
<400> 38
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
1 5 10
<210> 39
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейства мотивов AG, AM XTEN
<400> 39
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
1 5 10
<210> 40
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 40
Gly Glu Pro Ala Gly Ser Pro Thr Ser Thr Ser Glu
1 5 10
<210> 41
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 41
Gly Thr Gly Glu Pro Ser Ser Thr Pro Ala Ser Glu
1 5 10
<210> 42
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 42
Gly Ser Gly Pro Ser Thr Glu Ser Ala Pro Thr Glu
1 5 10
<210> 43
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 43
Gly Ser Glu Thr Pro Ser Gly Pro Ser Glu Thr Ala
1 5 10
<210> 44
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 44
Gly Pro Ser Glu Thr Ser Thr Ser Glu Pro Gly Ala
1 5 10
<210> 45
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов AQ XTEN
<400> 45
Gly Ser Pro Ser Glu Pro Thr Glu Gly Thr Ser Ala
1 5 10
<210> 46
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BC XTEN
<400> 46
Gly Ser Gly Ala Ser Glu Pro Thr Ser Thr Glu Pro
1 5 10
<210> 47
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BC XTEN
<400> 47
Gly Ser Glu Pro Ala Thr Ser Gly Thr Glu Pro Ser
1 5 10
<210> 48
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BC XTEN
<400> 48
Gly Thr Ser Glu Pro Ser Thr Ser Glu Pro Gly Ala
1 5 10
<210> 49
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BC XTEN
<400> 49
Gly Thr Ser Thr Glu Pro Ser Glu Pro Gly Ser Ala
1 5 10
<210> 50
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BD XTEN
<400> 50
Gly Ser Thr Ala Gly Ser Glu Thr Ser Thr Glu Ala
1 5 10
<210> 51
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BD XTEN
<400> 51
Gly Ser Glu Thr Ala Thr Ser Gly Ser Glu Thr Ala
1 5 10
<210> 52
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BD XTEN
<400> 52
Gly Thr Ser Glu Ser Ala Thr Ser Glu Ser Gly Ala
1 5 10
<210> 53
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Семейство мотивов BD XTEN
<400> 53
Gly Thr Ser Thr Glu Ala Ser Glu Gly Ser Ala Ser
1 5 10
<210> 54
<211> 288
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE288_2
<400> 54
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
1 5 10 15
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
20 25 30
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
35 40 45
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
50 55 60
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu
65 70 75 80
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
85 90 95
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
100 105 110
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
115 120 125
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
130 135 140
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
145 150 155 160
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
165 170 175
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
180 185 190
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
195 200 205
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
210 215 220
Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
225 230 235 240
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
245 250 255
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro
260 265 270
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
275 280 285
<210> 55
<211> 143
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144_2A
<400> 55
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser
1 5 10 15
Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
20 25 30
Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr
35 40 45
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro
50 55 60
Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
65 70 75 80
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
85 90 95
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro
100 105 110
Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
115 120 125
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
130 135 140
<210> 56
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144_3B
<400> 56
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
1 5 10 15
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
20 25 30
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
35 40 45
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu
50 55 60
Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly
65 70 75 80
Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
85 90 95
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu
100 105 110
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
115 120 125
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
130 135 140
<210> 57
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144_4A
<400> 57
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1 5 10 15
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
20 25 30
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
35 40 45
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
50 55 60
Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
65 70 75 80
Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
85 90 95
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
100 105 110
Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu
115 120 125
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
130 135 140
<210> 58
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144_5A
<400> 58
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1 5 10 15
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
20 25 30
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
35 40 45
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
50 55 60
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
65 70 75 80
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
85 90 95
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
100 105 110
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
115 120 125
Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
130 135 140
<210> 59
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE144_6B
<400> 59
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
1 5 10 15
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
20 25 30
Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
35 40 45
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala
50 55 60
Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
65 70 75 80
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
85 90 95
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala
100 105 110
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
115 120 125
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
130 135 140
<210> 60
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG144_A
<400> 60
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Pro
1 5 10 15
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr
20 25 30
Gly Thr Gly Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
35 40 45
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
85 90 95
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Thr Pro Gly
100 105 110
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
115 120 125
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
130 135 140
<210> 61
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG144_B
<400> 61
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
1 5 10 15
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
20 25 30
Thr Gly Ser Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro
35 40 45
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro Ser Ala Ser Thr
65 70 75 80
Gly Thr Gly Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro
85 90 95
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala Ser Pro
100 105 110
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
115 120 125
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
130 135 140
<210> 62
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AG144_C
<400> 62
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro
1 5 10 15
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
20 25 30
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
35 40 45
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly
50 55 60
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
85 90 95
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr
100 105 110
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
115 120 125
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
130 135 140
<210> 63
<211> 144
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> AG144_F
<400> 63
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro
1 5 10 15
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
20 25 30
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
35 40 45
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
85 90 95
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
100 105 110
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
115 120 125
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
130 135 140
<210> 64
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сигнальный пептид FVIII
<400> 64
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser
<210> 65
<211> 2332
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Зрелый фактор VIII
<400> 65
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Ser Arg His Pro Ser Thr Arg
740 745 750
Gln Lys Gln Phe Asn Ala Thr Thr Ile Pro Glu Asn Asp Ile Glu Lys
755 760 765
Thr Asp Pro Trp Phe Ala His Arg Thr Pro Met Pro Lys Ile Gln Asn
770 775 780
Val Ser Ser Ser Asp Leu Leu Met Leu Leu Arg Gln Ser Pro Thr Pro
785 790 795 800
His Gly Leu Ser Leu Ser Asp Leu Gln Glu Ala Lys Tyr Glu Thr Phe
805 810 815
Ser Asp Asp Pro Ser Pro Gly Ala Ile Asp Ser Asn Asn Ser Leu Ser
820 825 830
Glu Met Thr His Phe Arg Pro Gln Leu His His Ser Gly Asp Met Val
835 840 845
Phe Thr Pro Glu Ser Gly Leu Gln Leu Arg Leu Asn Glu Lys Leu Gly
850 855 860
Thr Thr Ala Ala Thr Glu Leu Lys Lys Leu Asp Phe Lys Val Ser Ser
865 870 875 880
Thr Ser Asn Asn Leu Ile Ser Thr Ile Pro Ser Asp Asn Leu Ala Ala
885 890 895
Gly Thr Asp Asn Thr Ser Ser Leu Gly Pro Pro Ser Met Pro Val His
900 905 910
Tyr Asp Ser Gln Leu Asp Thr Thr Leu Phe Gly Lys Lys Ser Ser Pro
915 920 925
Leu Thr Glu Ser Gly Gly Pro Leu Ser Leu Ser Glu Glu Asn Asn Asp
930 935 940
Ser Lys Leu Leu Glu Ser Gly Leu Met Asn Ser Gln Glu Ser Ser Trp
945 950 955 960
Gly Lys Asn Val Ser Ser Thr Glu Ser Gly Arg Leu Phe Lys Gly Lys
965 970 975
Arg Ala His Gly Pro Ala Leu Leu Thr Lys Asp Asn Ala Leu Phe Lys
980 985 990
Val Ser Ile Ser Leu Leu Lys Thr Asn Lys Thr Ser Asn Asn Ser Ala
995 1000 1005
Thr Asn Arg Lys Thr His Ile Asp Gly Pro Ser Leu Leu Ile Glu
1010 1015 1020
Asn Ser Pro Ser Val Trp Gln Asn Ile Leu Glu Ser Asp Thr Glu
1025 1030 1035
Phe Lys Lys Val Thr Pro Leu Ile His Asp Arg Met Leu Met Asp
1040 1045 1050
Lys Asn Ala Thr Ala Leu Arg Leu Asn His Met Ser Asn Lys Thr
1055 1060 1065
Thr Ser Ser Lys Asn Met Glu Met Val Gln Gln Lys Lys Glu Gly
1070 1075 1080
Pro Ile Pro Pro Asp Ala Gln Asn Pro Asp Met Ser Phe Phe Lys
1085 1090 1095
Met Leu Phe Leu Pro Glu Ser Ala Arg Trp Ile Gln Arg Thr His
1100 1105 1110
Gly Lys Asn Ser Leu Asn Ser Gly Gln Gly Pro Ser Pro Lys Gln
1115 1120 1125
Leu Val Ser Leu Gly Pro Glu Lys Ser Val Glu Gly Gln Asn Phe
1130 1135 1140
Leu Ser Glu Lys Asn Lys Val Val Val Gly Lys Gly Glu Phe Thr
1145 1150 1155
Lys Asp Val Gly Leu Lys Glu Met Val Phe Pro Ser Ser Arg Asn
1160 1165 1170
Leu Phe Leu Thr Asn Leu Asp Asn Leu His Glu Asn Asn Thr His
1175 1180 1185
Asn Gln Glu Lys Lys Ile Gln Glu Glu Ile Glu Lys Lys Glu Thr
1190 1195 1200
Leu Ile Gln Glu Asn Val Val Leu Pro Gln Ile His Thr Val Thr
1205 1210 1215
Gly Thr Lys Asn Phe Met Lys Asn Leu Phe Leu Leu Ser Thr Arg
1220 1225 1230
Gln Asn Val Glu Gly Ser Tyr Asp Gly Ala Tyr Ala Pro Val Leu
1235 1240 1245
Gln Asp Phe Arg Ser Leu Asn Asp Ser Thr Asn Arg Thr Lys Lys
1250 1255 1260
His Thr Ala His Phe Ser Lys Lys Gly Glu Glu Glu Asn Leu Glu
1265 1270 1275
Gly Leu Gly Asn Gln Thr Lys Gln Ile Val Glu Lys Tyr Ala Cys
1280 1285 1290
Thr Thr Arg Ile Ser Pro Asn Thr Ser Gln Gln Asn Phe Val Thr
1295 1300 1305
Gln Arg Ser Lys Arg Ala Leu Lys Gln Phe Arg Leu Pro Leu Glu
1310 1315 1320
Glu Thr Glu Leu Glu Lys Arg Ile Ile Val Asp Asp Thr Ser Thr
1325 1330 1335
Gln Trp Ser Lys Asn Met Lys His Leu Thr Pro Ser Thr Leu Thr
1340 1345 1350
Gln Ile Asp Tyr Asn Glu Lys Glu Lys Gly Ala Ile Thr Gln Ser
1355 1360 1365
Pro Leu Ser Asp Cys Leu Thr Arg Ser His Ser Ile Pro Gln Ala
1370 1375 1380
Asn Arg Ser Pro Leu Pro Ile Ala Lys Val Ser Ser Phe Pro Ser
1385 1390 1395
Ile Arg Pro Ile Tyr Leu Thr Arg Val Leu Phe Gln Asp Asn Ser
1400 1405 1410
Ser His Leu Pro Ala Ala Ser Tyr Arg Lys Lys Asp Ser Gly Val
1415 1420 1425
Gln Glu Ser Ser His Phe Leu Gln Gly Ala Lys Lys Asn Asn Leu
1430 1435 1440
Ser Leu Ala Ile Leu Thr Leu Glu Met Thr Gly Asp Gln Arg Glu
1445 1450 1455
Val Gly Ser Leu Gly Thr Ser Ala Thr Asn Ser Val Thr Tyr Lys
1460 1465 1470
Lys Val Glu Asn Thr Val Leu Pro Lys Pro Asp Leu Pro Lys Thr
1475 1480 1485
Ser Gly Lys Val Glu Leu Leu Pro Lys Val His Ile Tyr Gln Lys
1490 1495 1500
Asp Leu Phe Pro Thr Glu Thr Ser Asn Gly Ser Pro Gly His Leu
1505 1510 1515
Asp Leu Val Glu Gly Ser Leu Leu Gln Gly Thr Glu Gly Ala Ile
1520 1525 1530
Lys Trp Asn Glu Ala Asn Arg Pro Gly Lys Val Pro Phe Leu Arg
1535 1540 1545
Val Ala Thr Glu Ser Ser Ala Lys Thr Pro Ser Lys Leu Leu Asp
1550 1555 1560
Pro Leu Ala Trp Asp Asn His Tyr Gly Thr Gln Ile Pro Lys Glu
1565 1570 1575
Glu Trp Lys Ser Gln Glu Lys Ser Pro Glu Lys Thr Ala Phe Lys
1580 1585 1590
Lys Lys Asp Thr Ile Leu Ser Leu Asn Ala Cys Glu Ser Asn His
1595 1600 1605
Ala Ile Ala Ala Ile Asn Glu Gly Gln Asn Lys Pro Glu Ile Glu
1610 1615 1620
Val Thr Trp Ala Lys Gln Gly Arg Thr Glu Arg Leu Cys Ser Gln
1625 1630 1635
Asn Pro Pro Val Leu Lys Arg His Gln Arg Glu Ile Thr Arg Thr
1640 1645 1650
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile
1655 1660 1665
Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp
1670 1675 1680
Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr
1685 1690 1695
Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser
1700 1705 1710
Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro
1715 1720 1725
Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe
1730 1735 1740
Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
1745 1750 1755
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val
1760 1765 1770
Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser
1775 1780 1785
Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
1790 1795 1800
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys
1805 1810 1815
Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys
1820 1825 1830
Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His
1835 1840 1845
Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu
1850 1855 1860
Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu
1865 1870 1875
Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu
1880 1885 1890
Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu
1895 1900 1905
Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly
1910 1915 1920
Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln
1925 1930 1935
Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile
1940 1945 1950
His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys
1955 1960 1965
Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe
1970 1975 1980
Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val
1985 1990 1995
Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu
2000 2005 2010
Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala
2015 2020 2025
Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr
2030 2035 2040
Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser
2045 2050 2055
Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val
2060 2065 2070
Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly
2075 2080 2085
Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile
2090 2095 2100
Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn
2105 2110 2115
Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser
2120 2125 2130
Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr
2135 2140 2145
Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg
2150 2155 2160
Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu
2165 2170 2175
Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser
2180 2185 2190
Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala
2195 2200 2205
Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val
2210 2215 2220
Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met
2225 2230 2235
Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr
2240 2245 2250
Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly
2255 2260 2265
His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe
2270 2275 2280
Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp
2285 2290 2295
Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp
2300 2305 2310
Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala
2315 2320 2325
Gln Asp Leu Tyr
2330
<210> 66
<211> 7053
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Полноразмерный FVIII
<400> 66
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctcccaga attcaagaca ccctagcact aggcaaaagc aatttaatgc caccacaatt 2340
ccagaaaatg acatagagaa gactgaccct tggtttgcac acagaacacc tatgcctaaa 2400
atacaaaatg tctcctctag tgatttgttg atgctcttgc gacagagtcc tactccacat 2460
gggctatcct tatctgatct ccaagaagcc aaatatgaga ctttttctga tgatccatca 2520
cctggagcaa tagacagtaa taacagcctg tctgaaatga cacacttcag gccacagctc 2580
catcacagtg gggacatggt atttacccct gagtcaggcc tccaattaag attaaatgag 2640
aaactgggga caactgcagc aacagagttg aagaaacttg atttcaaagt ttctagtaca 2700
tcaaataatc tgatttcaac aattccatca gacaatttgg cagcaggtac tgataataca 2760
agttccttag gacccccaag tatgccagtt cattatgata gtcaattaga taccactcta 2820
tttggcaaaa agtcatctcc ccttactgag tctggtggac ctctgagctt gagtgaagaa 2880
aataatgatt caaagttgtt agaatcaggt ttaatgaata gccaagaaag ttcatgggga 2940
aaaaatgtat cgtcaacaga gagtggtagg ttatttaaag ggaaaagagc tcatggacct 3000
gctttgttga ctaaagataa tgccttattc aaagttagca tctctttgtt aaagacaaac 3060
aaaacttcca ataattcagc aactaataga aagactcaca ttgatggccc atcattatta 3120
attgagaata gtccatcagt ctggcaaaat atattagaaa gtgacactga gtttaaaaaa 3180
gtgacacctt tgattcatga cagaatgctt atggacaaaa atgctacagc tttgaggcta 3240
aatcatatgt caaataaaac tacttcatca aaaaacatgg aaatggtcca acagaaaaaa 3300
gagggcccca ttccaccaga tgcacaaaat ccagatatgt cgttctttaa gatgctattc 3360
ttgccagaat cagcaaggtg gatacaaagg actcatggaa agaactctct gaactctggg 3420
caaggcccca gtccaaagca attagtatcc ttaggaccag aaaaatctgt ggaaggtcag 3480
aatttcttgt ctgagaaaaa caaagtggta gtaggaaagg gtgaatttac aaaggacgta 3540
ggactcaaag agatggtttt tccaagcagc agaaacctat ttcttactaa cttggataat 3600
ttacatgaaa ataatacaca caatcaagaa aaaaaaattc aggaagaaat agaaaagaag 3660
gaaacattaa tccaagagaa tgtagttttg cctcagatac atacagtgac tggcactaag 3720
aatttcatga agaacctttt cttactgagc actaggcaaa atgtagaagg ttcatatgac 3780
ggggcatatg ctccagtact tcaagatttt aggtcattaa atgattcaac aaatagaaca 3840
aagaaacaca cagctcattt ctcaaaaaaa ggggaggaag aaaacttgga aggcttggga 3900
aatcaaacca agcaaattgt agagaaatat gcatgcacca caaggatatc tcctaataca 3960
agccagcaga attttgtcac gcaacgtagt aagagagctt tgaaacaatt cagactccca 4020
ctagaagaaa cagaacttga aaaaaggata attgtggatg acacctcaac ccagtggtcc 4080
aaaaacatga aacatttgac cccgagcacc ctcacacaga tagactacaa tgagaaggag 4140
aaaggggcca ttactcagtc tcccttatca gattgcctta cgaggagtca tagcatccct 4200
caagcaaata gatctccatt acccattgca aaggtatcat catttccatc tattagacct 4260
atatatctga ccagggtcct attccaagac aactcttctc atcttccagc agcatcttat 4320
agaaagaaag attctggggt ccaagaaagc agtcatttct tacaaggagc caaaaaaaat 4380
aacctttctt tagccattct aaccttggag atgactggtg atcaaagaga ggttggctcc 4440
ctggggacaa gtgccacaaa ttcagtcaca tacaagaaag ttgagaacac tgttctcccg 4500
aaaccagact tgcccaaaac atctggcaaa gttgaattgc ttccaaaagt tcacatttat 4560
cagaaggacc tattccctac ggaaactagc aatgggtctc ctggccatct ggatctcgtg 4620
gaagggagcc ttcttcaggg aacagaggga gcgattaagt ggaatgaagc aaacagacct 4680
ggaaaagttc cctttctgag agtagcaaca gaaagctctg caaagactcc ctccaagcta 4740
ttggatcctc ttgcttggga taaccactat ggtactcaga taccaaaaga agagtggaaa 4800
tcccaagaga agtcaccaga aaaaacagct tttaagaaaa aggataccat tttgtccctg 4860
aacgcttgtg aaagcaatca tgcaatagca gcaataaatg agggacaaaa taagcccgaa 4920
atagaagtca cctgggcaaa gcaaggtagg actgaaaggc tgtgctctca aaacccacca 4980
gtcttgaaac gccatcaacg ggaaataact cgtactactc ttcagtcaga tcaagaggaa 5040
attgactatg atgataccat atcagttgaa atgaagaagg aagattttga catttatgat 5100
gaggatgaaa atcagagccc ccgcagcttt caaaagaaaa cacgacacta ttttattgct 5160
gcagtggaga ggctctggga ttatgggatg agtagctccc cacatgttct aagaaacagg 5220
gctcagagtg gcagtgtccc tcagttcaag aaagttgttt tccaggaatt tactgatggc 5280
tcctttactc agcccttata ccgtggagaa ctaaatgaac atttgggact cctggggcca 5340
tatataagag cagaagttga agataatatc atggtaactt tcagaaatca ggcctctcgt 5400
ccctattcct tctattctag ccttatttct tatgaggaag atcagaggca aggagcagaa 5460
cctagaaaaa actttgtcaa gcctaatgaa accaaaactt acttttggaa agtgcaacat 5520
catatggcac ccactaaaga tgagtttgac tgcaaagcct gggcttattt ctctgatgtt 5580
gacctggaaa aagatgtgca ctcaggcctg attggacccc ttctggtctg ccacactaac 5640
acactgaacc ctgctcatgg gagacaagtg acagtacagg aatttgctct gtttttcacc 5700
atctttgatg agaccaaaag ctggtacttc actgaaaata tggaaagaaa ctgcagggct 5760
ccctgcaata tccagatgga agatcccact tttaaagaga attatcgctt ccatgcaatc 5820
aatggctaca taatggatac actacctggc ttagtaatgg ctcaggatca aaggattcga 5880
tggtatctgc tcagcatggg cagcaatgaa aacatccatt ctattcattt cagtggacat 5940
gtgttcactg tacgaaaaaa agaggagtat aaaatggcac tgtacaatct ctatccaggt 6000
gtttttgaga cagtggaaat gttaccatcc aaagctggaa tttggcgggt ggaatgcctt 6060
attggcgagc atctacatgc tgggatgagc acactttttc tggtgtacag caataagtgt 6120
cagactcccc tgggaatggc ttctggacac attagagatt ttcagattac agcttcagga 6180
caatatggac agtgggcccc aaagctggcc agacttcatt attccggatc aatcaatgcc 6240
tggagcacca aggagccctt ttcttggatc aaggtggatc tgttggcacc aatgattatt 6300
cacggcatca agacccaggg tgcccgtcag aagttctcca gcctctacat ctctcagttt 6360
atcatcatgt atagtcttga tgggaagaag tggcagactt atcgaggaaa ttccactgga 6420
accttaatgg tcttctttgg caatgtggat tcatctggga taaaacacaa tatttttaac 6480
cctccaatta ttgctcgata catccgtttg cacccaactc attatagcat tcgcagcact 6540
cttcgcatgg agttgatggg ctgtgattta aatagttgca gcatgccatt gggaatggag 6600
agtaaagcaa tatcagatgc acagattact gcttcatcct actttaccaa tatgtttgcc 6660
acctggtctc cttcaaaagc tcgacttcac ctccaaggga ggagtaatgc ctggagacct 6720
caggtgaata atccaaaaga gtggctgcaa gtggacttcc agaagacaat gaaagtcaca 6780
ggagtaacta ctcagggagt aaaatctctg cttaccagca tgtatgtgaa ggagttcctc 6840
atctccagca gtcaagatgg ccatcagtgg actctctttt ttcagaatgg caaagtaaag 6900
gtttttcagg gaaatcaaga ctccttcaca cctgtggtga actctctaga cccaccgtta 6960
ctgactcgct accttcgaat tcacccccag agttgggtgc accagattgc cctgaggatg 7020
gaggttctgg gctgcgaggc acaggacctc tac 7053
<210> 67
<211> 1438
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII BDD
<400> 67
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu Lys Arg His
740 745 750
Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile
755 760 765
Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp
770 775 780
Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
785 790 795 800
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly
805 810 815
Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser
820 825 830
Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser
835 840 845
Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
850 855 860
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr
865 870 875 880
Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile
885 890 895
Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe
900 905 910
Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His
915 920 925
Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
930 935 940
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro
945 950 955 960
Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln
965 970 975
Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
980 985 990
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro
995 1000 1005
Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg
1010 1015 1020
Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu
1025 1030 1035
Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met
1040 1045 1050
Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val
1055 1060 1065
Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn
1070 1075 1080
Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys
1085 1090 1095
Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His
1100 1105 1110
Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln
1115 1120 1125
Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile
1130 1135 1140
Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg
1145 1150 1155
Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro
1160 1165 1170
Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His
1175 1180 1185
Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr
1190 1195 1200
Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp
1205 1210 1215
Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe
1220 1225 1230
Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro
1235 1240 1245
Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser
1250 1255 1260
Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn
1265 1270 1275
Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp
1280 1285 1290
Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr
1295 1300 1305
Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn
1310 1315 1320
Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val
1325 1330 1335
Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly
1340 1345 1350
Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile
1355 1360 1365
Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn
1370 1375 1380
Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro
1385 1390 1395
Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg
1400 1405 1410
Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu
1415 1420 1425
Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr
1430 1435
<210> 68
<211> 1429
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Аминокислотная последовательность фактора VIII с делецией домена B
(делеция всего домена B, за исключением 5 аминокислотных остатков
<400> 68
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Glu Ile Thr Arg Thr Thr Leu
740 745 750
Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu
755 760 765
Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser
770 775 780
Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val
785 790 795 800
Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg
805 810 815
Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe
820 825 830
Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu
835 840 845
Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val
850 855 860
Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr
865 870 875 880
Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
885 890 895
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr
900 905 910
Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp
915 920 925
Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val
930 935 940
His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu
945 950 955 960
Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe
965 970 975
Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met
980 985 990
Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr
995 1000 1005
Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met
1010 1015 1020
Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg
1025 1030 1035
Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile
1040 1045 1050
His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr
1055 1060 1065
Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val
1070 1075 1080
Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu
1085 1090 1095
Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val
1100 1105 1110
Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His
1115 1120 1125
Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp
1130 1135 1140
Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala
1145 1150 1155
Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu
1160 1165 1170
Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln
1175 1180 1185
Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser
1190 1195 1200
Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly
1205 1210 1215
Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys
1220 1225 1230
His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu
1235 1240 1245
His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu
1250 1255 1260
Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu
1265 1270 1275
Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe
1280 1285 1290
Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His
1295 1300 1305
Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro
1310 1315 1320
Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr
1325 1330 1335
Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr
1340 1345 1350
Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp
1355 1360 1365
Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn
1370 1375 1380
Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu
1385 1390 1395
Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln
1400 1405 1410
Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu
1415 1420 1425
Tyr
<210> 69
<211> 2754
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN-FVIII-161
<400> 69
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu
755 760 765
Lys Arg His Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln
770 775 780
Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu
785 790 795 800
Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe
805 810 815
Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp
820 825 830
Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln
835 840 845
Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
850 855 860
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His
865 870 875 880
Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile
885 890 895
Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser
900 905 910
Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
915 920 925
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val
930 935 940
Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp
945 950 955 960
Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu
965 970 975
Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His
980 985 990
Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
995 1000 1005
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1010 1015 1020
Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1025 1030 1035
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1040 1045 1050
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1055 1060 1065
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1070 1075 1080
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1085 1090 1095
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1100 1105 1110
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1115 1120 1125
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1130 1135 1140
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1145 1150 1155
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1160 1165 1170
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1175 1180 1185
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1190 1195 1200
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1205 1210 1215
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1220 1225 1230
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1235 1240 1245
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1250 1255 1260
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1265 1270 1275
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1280 1285 1290
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1295 1300 1305
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1310 1315 1320
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1325 1330 1335
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1340 1345 1350
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1355 1360 1365
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1370 1375 1380
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1385 1390 1395
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1400 1405 1410
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1415 1420 1425
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1430 1435 1440
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1445 1450 1455
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1460 1465 1470
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1475 1480 1485
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1490 1495 1500
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1505 1510 1515
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1520 1525 1530
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1535 1540 1545
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1550 1555 1560
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1565 1570 1575
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1580 1585 1590
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1595 1600 1605
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1610 1615 1620
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1625 1630 1635
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1640 1645 1650
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1655 1660 1665
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1670 1675 1680
Lys Arg Arg Arg Arg Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1685 1690 1695
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
1700 1705 1710
Ser Gly Gly Gly Gly Ser Arg Lys Arg Arg Lys Arg Ser Leu Ser
1715 1720 1725
Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu
1730 1735 1740
Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp
1745 1750 1755
Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
1760 1765 1770
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg
1775 1780 1785
Cys Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr
1790 1795 1800
Val Lys Ile Gly Cys Asn Thr Cys Val Cys Arg Asp Arg Lys Trp
1805 1810 1815
Asn Cys Thr Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly
1820 1825 1830
Met Ala His Tyr Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro
1835 1840 1845
Gly Glu Cys Gln Tyr Val Leu Val Gln Asp Tyr Cys Gly Ser Asn
1850 1855 1860
Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys Gly Cys Ser His
1865 1870 1875
Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu Val Glu Gly
1880 1885 1890
Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys Arg Pro
1895 1900 1905
Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg Tyr
1910 1915 1920
Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
1925 1930 1935
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys
1940 1945 1950
Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp
1955 1960 1965
Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe
1970 1975 1980
Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys
1985 1990 1995
Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
2000 2005 2010
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp
2015 2020 2025
Val Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu
2030 2035 2040
Asp Val Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp
2045 2050 2055
Cys Ala Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys
2060 2065 2070
Ala Gln His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys
2075 2080 2085
Pro Gln Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu
2090 2095 2100
Ala Glu Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr
2105 2110 2115
Cys Gln His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu
2120 2125 2130
Gly Cys His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu
2135 2140 2145
Leu Gln Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val
2150 2155 2160
Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro
2165 2170 2175
Ser Asp Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn
2180 2185 2190
Leu Thr Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Thr Ser Glu
2195 2200 2205
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
2210 2215 2220
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
2225 2230 2235
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
2240 2245 2250
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
2255 2260 2265
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro
2270 2275 2280
Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
2285 2290 2295
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
2300 2305 2310
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala
2315 2320 2325
Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro
2330 2335 2340
Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
2345 2350 2355
Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
2360 2365 2370
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu
2375 2380 2385
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
2390 2395 2400
Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
2405 2410 2415
Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
2420 2425 2430
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
2435 2440 2445
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser
2450 2455 2460
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
2465 2470 2475
Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Asp
2480 2485 2490
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly
2495 2500 2505
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg
2510 2515 2520
Gly Ser Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
2525 2530 2535
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
2540 2545 2550
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
2555 2560 2565
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
2570 2575 2580
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
2585 2590 2595
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
2600 2605 2610
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
2615 2620 2625
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
2630 2635 2640
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
2645 2650 2655
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
2660 2665 2670
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
2675 2680 2685
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
2690 2695 2700
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
2705 2710 2715
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
2720 2725 2730
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
2735 2740 2745
Ser Leu Ser Pro Gly Lys
2750
<210> 70
<211> 1978
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 169
<400> 70
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly
755 760 765
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
770 775 780
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
785 790 795 800
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
805 810 815
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
820 825 830
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
835 840 845
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
850 855 860
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
865 870 875 880
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
885 890 895
Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
900 905 910
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
915 920 925
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
930 935 940
Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
945 950 955 960
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala
965 970 975
Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
980 985 990
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
995 1000 1005
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1010 1015 1020
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1025 1030 1035
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1040 1045 1050
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1055 1060 1065
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1070 1075 1080
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1085 1090 1095
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1100 1105 1110
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1115 1120 1125
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1130 1135 1140
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1145 1150 1155
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1160 1165 1170
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1175 1180 1185
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1190 1195 1200
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1205 1210 1215
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1220 1225 1230
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1235 1240 1245
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1250 1255 1260
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1265 1270 1275
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1280 1285 1290
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1295 1300 1305
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1310 1315 1320
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1325 1330 1335
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1340 1345 1350
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1355 1360 1365
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1370 1375 1380
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1385 1390 1395
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1400 1405 1410
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1415 1420 1425
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1430 1435 1440
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1445 1450 1455
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1460 1465 1470
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1475 1480 1485
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1490 1495 1500
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1505 1510 1515
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1520 1525 1530
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1535 1540 1545
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1550 1555 1560
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1565 1570 1575
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1580 1585 1590
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1595 1600 1605
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1610 1615 1620
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1625 1630 1635
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1640 1645 1650
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1655 1660 1665
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1670 1675 1680
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1685 1690 1695
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1700 1705 1710
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1715 1720 1725
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1730 1735 1740
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1745 1750 1755
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1760 1765 1770
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
1775 1780 1785
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1790 1795 1800
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1805 1810 1815
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1820 1825 1830
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
1835 1840 1845
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
1850 1855 1860
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
1865 1870 1875
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
1880 1885 1890
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
1895 1900 1905
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
1910 1915 1920
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
1925 1930 1935
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
1940 1945 1950
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
1955 1960 1965
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1970 1975
<210> 71
<211> 2242
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN-FVIII-170
<400> 71
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Ala
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly
465 470 475 480
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
485 490 495
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
500 505 510
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
515 520 525
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
530 535 540
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
545 550 555 560
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
565 570 575
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
580 585 590
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
595 600 605
Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
610 615 620
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
625 630 635 640
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
645 650 655
Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
660 665 670
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala
675 680 685
Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
690 695 700
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
705 710 715 720
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala
725 730 735
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
740 745 750
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Asp
755 760 765
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
770 775 780
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly Ser
785 790 795 800
Gly Gly Ala Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu
805 810 815
Ser Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala
820 825 830
Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val
835 840 845
Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn
850 855 860
Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile
865 870 875 880
Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala
885 890 895
Ser His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala
900 905 910
Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu
915 920 925
Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val
930 935 940
Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr
945 950 955 960
Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu
965 970 975
Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys
980 985 990
Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu
995 1000 1005
Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp
1010 1015 1020
Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val
1025 1030 1035
Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His
1040 1045 1050
Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro
1055 1060 1065
Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
1070 1075 1080
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu
1085 1090 1095
Thr Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe
1100 1105 1110
Cys His Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val
1115 1120 1125
Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn
1130 1135 1140
Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu
1145 1150 1155
Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile
1160 1165 1170
Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
1175 1180 1185
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val
1190 1195 1200
Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn
1205 1210 1215
Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
1220 1225 1230
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His
1235 1240 1245
Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp
1250 1255 1260
Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn
1265 1270 1275
Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg
1280 1285 1290
Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu
1295 1300 1305
Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
1310 1315 1320
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser
1325 1330 1335
Ser Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly
1340 1345 1350
Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn
1355 1360 1365
Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe
1370 1375 1380
Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe
1385 1390 1395
Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe Gln
1400 1405 1410
Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
1415 1420 1425
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile
1430 1435 1440
Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser
1445 1450 1455
Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
1460 1465 1470
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn
1475 1480 1485
Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn
1490 1495 1500
Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn
1505 1510 1515
Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr
1520 1525 1530
Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln
1535 1540 1545
Asn Pro Pro Val Leu Lys Arg His Gln Arg Glu Ile Thr Arg Thr
1550 1555 1560
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile
1565 1570 1575
Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp
1580 1585 1590
Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr
1595 1600 1605
Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser
1610 1615 1620
Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro
1625 1630 1635
Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe
1640 1645 1650
Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
1655 1660 1665
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val
1670 1675 1680
Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser
1685 1690 1695
Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
1700 1705 1710
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys
1715 1720 1725
Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys
1730 1735 1740
Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His
1745 1750 1755
Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu
1760 1765 1770
Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu
1775 1780 1785
Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu
1790 1795 1800
Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu
1805 1810 1815
Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly
1820 1825 1830
Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln
1835 1840 1845
Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile
1850 1855 1860
His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys
1865 1870 1875
Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe
1880 1885 1890
Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val
1895 1900 1905
Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu
1910 1915 1920
Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala
1925 1930 1935
Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr
1940 1945 1950
Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser
1955 1960 1965
Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val
1970 1975 1980
Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly
1985 1990 1995
Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile
2000 2005 2010
Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn
2015 2020 2025
Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser
2030 2035 2040
Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr
2045 2050 2055
Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg
2060 2065 2070
Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu
2075 2080 2085
Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser
2090 2095 2100
Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala
2105 2110 2115
Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val
2120 2125 2130
Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met
2135 2140 2145
Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr
2150 2155 2160
Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly
2165 2170 2175
His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe
2180 2185 2190
Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp
2195 2200 2205
Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp
2210 2215 2220
Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala
2225 2230 2235
Gln Asp Leu Tyr
2240
<210> 72
<211> 1959
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN-FVIII-173
<400> 72
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly Thr Ser Glu
740 745 750
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
755 760 765
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
770 775 780
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
785 790 795 800
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu
805 810 815
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
820 825 830
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
835 840 845
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
850 855 860
Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
865 870 875 880
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr
885 890 895
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
900 905 910
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
915 920 925
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
930 935 940
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly
945 950 955 960
Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
965 970 975
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
980 985 990
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly
995 1000 1005
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1010 1015 1020
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser
1025 1030 1035
Ser Pro Pro Val Leu Lys Arg His Gln Arg Glu Ile Thr Arg Thr
1040 1045 1050
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile
1055 1060 1065
Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp
1070 1075 1080
Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr
1085 1090 1095
Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser
1100 1105 1110
Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro
1115 1120 1125
Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe
1130 1135 1140
Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
1145 1150 1155
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val
1160 1165 1170
Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser
1175 1180 1185
Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
1190 1195 1200
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys
1205 1210 1215
Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys
1220 1225 1230
Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His
1235 1240 1245
Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu
1250 1255 1260
Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu
1265 1270 1275
Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu
1280 1285 1290
Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu
1295 1300 1305
Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly
1310 1315 1320
Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln
1325 1330 1335
Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile
1340 1345 1350
His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys
1355 1360 1365
Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe
1370 1375 1380
Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val
1385 1390 1395
Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu
1400 1405 1410
Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala
1415 1420 1425
Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr
1430 1435 1440
Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser
1445 1450 1455
Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val
1460 1465 1470
Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly
1475 1480 1485
Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile
1490 1495 1500
Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn
1505 1510 1515
Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser
1520 1525 1530
Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr
1535 1540 1545
Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg
1550 1555 1560
Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu
1565 1570 1575
Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser
1580 1585 1590
Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala
1595 1600 1605
Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val
1610 1615 1620
Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met
1625 1630 1635
Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr
1640 1645 1650
Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly
1655 1660 1665
His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe
1670 1675 1680
Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp
1685 1690 1695
Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp
1700 1705 1710
Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala
1715 1720 1725
Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1730 1735 1740
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1745 1750 1755
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
1760 1765 1770
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
1775 1780 1785
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
1790 1795 1800
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
1805 1810 1815
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
1820 1825 1830
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
1835 1840 1845
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
1850 1855 1860
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
1865 1870 1875
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
1880 1885 1890
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
1895 1900 1905
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
1910 1915 1920
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
1925 1930 1935
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
1940 1945 1950
Ser Leu Ser Pro Gly Lys
1955
<210> 73
<211> 1984
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 195 (двухцепочечный FVIIIFc с двумя XTEN AE-144 в
аминокислотных положениях 1656 и 1900)
<400> 73
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu
755 760 765
Lys Arg His Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Gly Ala Pro
770 775 780
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro
785 790 795 800
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
805 810 815
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
820 825 830
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly
835 840 845
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
850 855 860
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
865 870 875 880
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr
885 890 895
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
900 905 910
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
915 920 925
Ala Ser Ser Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser
930 935 940
Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn
945 950 955 960
Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala
965 970 975
Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val
980 985 990
Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
995 1000 1005
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1010 1015 1020
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1025 1030 1035
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1040 1045 1050
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1055 1060 1065
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1070 1075 1080
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1085 1090 1095
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1100 1105 1110
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1115 1120 1125
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1130 1135 1140
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1145 1150 1155
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1160 1165 1170
Cys Arg Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1175 1180 1185
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
1190 1195 1200
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1205 1210 1215
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
1220 1225 1230
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
1235 1240 1245
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1250 1255 1260
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
1265 1270 1275
Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr
1280 1285 1290
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1295 1300 1305
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ala
1310 1315 1320
Ser Ser Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1325 1330 1335
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1340 1345 1350
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1355 1360 1365
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1370 1375 1380
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1385 1390 1395
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1400 1405 1410
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1415 1420 1425
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1430 1435 1440
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1445 1450 1455
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1460 1465 1470
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1475 1480 1485
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1490 1495 1500
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1505 1510 1515
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1520 1525 1530
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1535 1540 1545
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1550 1555 1560
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1565 1570 1575
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1580 1585 1590
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1595 1600 1605
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1610 1615 1620
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1625 1630 1635
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1640 1645 1650
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1655 1660 1665
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1670 1675 1680
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1685 1690 1695
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1700 1705 1710
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1715 1720 1725
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1730 1735 1740
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1745 1750 1755
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1760 1765 1770
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1775 1780 1785
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1790 1795 1800
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1805 1810 1815
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1820 1825 1830
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1835 1840 1845
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1850 1855 1860
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1865 1870 1875
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1880 1885 1890
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1895 1900 1905
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1910 1915 1920
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1925 1930 1935
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1940 1945 1950
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1955 1960 1965
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1970 1975 1980
Lys
<210> 74
<211> 2134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 196 (двухцепочечный FVIIIFc с тремя XTEN AE-144 в
аминокислотных положениях 26, 1656 и 1900)
<400> 74
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Gly Ala Pro
35 40 45
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
85 90 95
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
100 105 110
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
115 120 125
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
130 135 140
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
145 150 155 160
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
165 170 175
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
180 185 190
Ala Ser Ser Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Pro Pro Val Leu Lys Arg His Gln Arg Glu Ile Thr Arg Thr
915 920 925
Thr Leu Gln Gly Ala Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser
930 935 940
Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala
945 950 955 960
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr
965 970 975
Ser Ser Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr
980 985 990
Gly Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala
995 1000 1005
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly
1010 1015 1020
Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
1025 1030 1035
Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
1040 1045 1050
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala
1055 1060 1065
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Ala Ser Ser Ser Asp
1070 1075 1080
Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys
1085 1090 1095
Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro
1100 1105 1110
Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val
1115 1120 1125
Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu
1130 1135 1140
Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
1145 1150 1155
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1160 1165 1170
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1175 1180 1185
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1190 1195 1200
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1205 1210 1215
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1220 1225 1230
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1235 1240 1245
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1250 1255 1260
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1265 1270 1275
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1280 1285 1290
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1295 1300 1305
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1310 1315 1320
Cys Arg Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1325 1330 1335
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
1340 1345 1350
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1355 1360 1365
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
1370 1375 1380
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
1385 1390 1395
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1400 1405 1410
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
1415 1420 1425
Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr
1430 1435 1440
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1445 1450 1455
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ala
1460 1465 1470
Ser Ser Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1475 1480 1485
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1490 1495 1500
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1505 1510 1515
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1520 1525 1530
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1535 1540 1545
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1550 1555 1560
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1565 1570 1575
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1580 1585 1590
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1595 1600 1605
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1610 1615 1620
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1625 1630 1635
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1640 1645 1650
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1655 1660 1665
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1670 1675 1680
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1685 1690 1695
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1700 1705 1710
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1715 1720 1725
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1730 1735 1740
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1745 1750 1755
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1760 1765 1770
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1775 1780 1785
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1790 1795 1800
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1805 1810 1815
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1820 1825 1830
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1835 1840 1845
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1850 1855 1860
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1865 1870 1875
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1880 1885 1890
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1895 1900 1905
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1910 1915 1920
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1925 1930 1935
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1940 1945 1950
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1955 1960 1965
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1970 1975 1980
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1985 1990 1995
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
2000 2005 2010
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
2015 2020 2025
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
2030 2035 2040
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
2045 2050 2055
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
2060 2065 2070
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
2075 2080 2085
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
2090 2095 2100
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
2105 2110 2115
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
2120 2125 2130
Lys
<210> 75
<211> 1984
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 199 (одноцепочечный FVIIIFc с тремя XTEN AE-144 в
аминокислотных положениях 1656 и 1900)
<400> 75
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu
755 760 765
Lys Arg His Gln Ala Glu Ile Thr Arg Thr Thr Leu Gln Gly Ala Pro
770 775 780
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro
785 790 795 800
Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
805 810 815
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
820 825 830
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Thr Pro Gly
835 840 845
Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
850 855 860
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
865 870 875 880
Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ser Ser Thr
885 890 895
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
900 905 910
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
915 920 925
Ala Ser Ser Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser
930 935 940
Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn
945 950 955 960
Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala
965 970 975
Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val
980 985 990
Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
995 1000 1005
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1010 1015 1020
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1025 1030 1035
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1040 1045 1050
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1055 1060 1065
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1070 1075 1080
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1085 1090 1095
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1100 1105 1110
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1115 1120 1125
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1130 1135 1140
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1145 1150 1155
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1160 1165 1170
Cys Arg Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1175 1180 1185
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
1190 1195 1200
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1205 1210 1215
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
1220 1225 1230
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
1235 1240 1245
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1250 1255 1260
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
1265 1270 1275
Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr
1280 1285 1290
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1295 1300 1305
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ala
1310 1315 1320
Ser Ser Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1325 1330 1335
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1340 1345 1350
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1355 1360 1365
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1370 1375 1380
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1385 1390 1395
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1400 1405 1410
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1415 1420 1425
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1430 1435 1440
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1445 1450 1455
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1460 1465 1470
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1475 1480 1485
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1490 1495 1500
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1505 1510 1515
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1520 1525 1530
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1535 1540 1545
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1550 1555 1560
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1565 1570 1575
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1580 1585 1590
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1595 1600 1605
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1610 1615 1620
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1625 1630 1635
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1640 1645 1650
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1655 1660 1665
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1670 1675 1680
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1685 1690 1695
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1700 1705 1710
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1715 1720 1725
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1730 1735 1740
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1745 1750 1755
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1760 1765 1770
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1775 1780 1785
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1790 1795 1800
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1805 1810 1815
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1820 1825 1830
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1835 1840 1845
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1850 1855 1860
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1865 1870 1875
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1880 1885 1890
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1895 1900 1905
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1910 1915 1920
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1925 1930 1935
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1940 1945 1950
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1955 1960 1965
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1970 1975 1980
Lys
<210> 76
<211> 2134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 201 (одноцепочечный FVIIIFc с тремя XTEN AE-144 в
аминокислотных положениях 26, 1656 и 1900)
<400> 76
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Gly Ala Pro
35 40 45
Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro Gly Ser Ser Pro
50 55 60
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
65 70 75 80
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
85 90 95
Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Pro
100 105 110
Ser Ala Ser Thr Gly Thr Gly Pro Gly Ala Ser Pro Gly Thr Ser Ser
115 120 125
Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr Gly Pro
130 135 140
Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ser Ser Thr
145 150 155 160
Pro Ser Gly Ala Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala
165 170 175
Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro
180 185 190
Ala Ser Ser Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Pro Pro Val Leu Lys Arg His Gln Ala Glu Ile Thr Arg Thr
915 920 925
Thr Leu Gln Gly Ala Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser
930 935 940
Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala
945 950 955 960
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr
965 970 975
Ser Ser Thr Gly Ser Pro Gly Ser Ser Pro Ser Ala Ser Thr Gly Thr
980 985 990
Gly Pro Gly Thr Pro Gly Ser Gly Thr Ala Ser Ser Ser Pro Gly Ala
995 1000 1005
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly
1010 1015 1020
Thr Ser Ser Thr Gly Ser Pro Gly Ala Ser Pro Gly Thr Ser Ser
1025 1030 1035
Thr Gly Ser Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser
1040 1045 1050
Pro Gly Ser Ser Thr Pro Ser Gly Ala Thr Gly Ser Pro Gly Ala
1055 1060 1065
Ser Pro Gly Thr Ser Ser Thr Gly Ser Pro Ala Ser Ser Ser Asp
1070 1075 1080
Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys
1085 1090 1095
Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro
1100 1105 1110
Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val
1115 1120 1125
Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu
1130 1135 1140
Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
1145 1150 1155
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1160 1165 1170
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1175 1180 1185
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1190 1195 1200
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1205 1210 1215
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1220 1225 1230
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1235 1240 1245
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1250 1255 1260
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1265 1270 1275
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1280 1285 1290
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1295 1300 1305
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1310 1315 1320
Cys Arg Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1325 1330 1335
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
1340 1345 1350
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1355 1360 1365
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
1370 1375 1380
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
1385 1390 1395
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1400 1405 1410
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
1415 1420 1425
Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr
1430 1435 1440
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
1445 1450 1455
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ala
1460 1465 1470
Ser Ser Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1475 1480 1485
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1490 1495 1500
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1505 1510 1515
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1520 1525 1530
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1535 1540 1545
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1550 1555 1560
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1565 1570 1575
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1580 1585 1590
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1595 1600 1605
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1610 1615 1620
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1625 1630 1635
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1640 1645 1650
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1655 1660 1665
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1670 1675 1680
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1685 1690 1695
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1700 1705 1710
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1715 1720 1725
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1730 1735 1740
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1745 1750 1755
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1760 1765 1770
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1775 1780 1785
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1790 1795 1800
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1805 1810 1815
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1820 1825 1830
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1835 1840 1845
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1850 1855 1860
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1865 1870 1875
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1880 1885 1890
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1895 1900 1905
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1910 1915 1920
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1925 1930 1935
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1940 1945 1950
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1955 1960 1965
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1970 1975 1980
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1985 1990 1995
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
2000 2005 2010
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
2015 2020 2025
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
2030 2035 2040
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
2045 2050 2055
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
2060 2065 2070
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
2075 2080 2085
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
2090 2095 2100
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
2105 2110 2115
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
2120 2125 2130
Lys
<210> 77
<211> 2128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 203 (одноцепочечный FVIIIFc с двумя XTEN AE — одним XTEN
AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 1900)
<400> 77
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly
755 760 765
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
770 775 780
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
785 790 795 800
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
805 810 815
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
820 825 830
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
835 840 845
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
850 855 860
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
865 870 875 880
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
885 890 895
Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
900 905 910
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
915 920 925
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
930 935 940
Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
945 950 955 960
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala
965 970 975
Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
980 985 990
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
995 1000 1005
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1010 1015 1020
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1025 1030 1035
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1040 1045 1050
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1055 1060 1065
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1070 1075 1080
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1085 1090 1095
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1100 1105 1110
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1115 1120 1125
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1130 1135 1140
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1145 1150 1155
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1160 1165 1170
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1175 1180 1185
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1190 1195 1200
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1205 1210 1215
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1220 1225 1230
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1235 1240 1245
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1250 1255 1260
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1265 1270 1275
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1280 1285 1290
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1295 1300 1305
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Gly Ala Pro Thr
1310 1315 1320
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
1325 1330 1335
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
1340 1345 1350
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
1355 1360 1365
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
1370 1375 1380
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
1385 1390 1395
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr
1400 1405 1410
Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
1415 1420 1425
Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr
1430 1435 1440
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
1445 1450 1455
Pro Ser Glu Gly Ser Ala Pro Gly Ala Ser Ser Ala Pro Cys Asn
1460 1465 1470
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1475 1480 1485
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1490 1495 1500
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1505 1510 1515
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1520 1525 1530
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1535 1540 1545
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1550 1555 1560
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1565 1570 1575
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1580 1585 1590
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1595 1600 1605
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1610 1615 1620
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1625 1630 1635
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1640 1645 1650
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1655 1660 1665
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1670 1675 1680
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1685 1690 1695
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1700 1705 1710
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1715 1720 1725
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1730 1735 1740
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1745 1750 1755
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1760 1765 1770
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1775 1780 1785
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1790 1795 1800
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1805 1810 1815
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1820 1825 1830
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1835 1840 1845
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1850 1855 1860
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1865 1870 1875
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1880 1885 1890
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1895 1900 1905
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1910 1915 1920
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
1925 1930 1935
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1940 1945 1950
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1955 1960 1965
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1970 1975 1980
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
1985 1990 1995
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
2000 2005 2010
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
2015 2020 2025
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
2030 2035 2040
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
2045 2050 2055
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
2060 2065 2070
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
2075 2080 2085
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
2090 2095 2100
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
2105 2110 2115
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
2120 2125
<210> 78
<211> 2128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 204 (одноцепочечный FVIIIFc с двумя XTEN AE — одним XTEN
AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 403)
<400> 78
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Gly Ala Pro Thr Ser Thr Glu Pro Ser Glu
420 425 430
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
435 440 445
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
450 455 460
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
465 470 475 480
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
485 490 495
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
500 505 510
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Thr Glu Pro Ser Glu
515 520 525
Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
530 535 540
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu
545 550 555 560
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ala Ser Ser Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
915 920 925
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
930 935 940
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
945 950 955 960
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
965 970 975
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro
980 985 990
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr
995 1000 1005
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
1010 1015 1020
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1025 1030 1035
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
1040 1045 1050
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
1055 1060 1065
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1070 1075 1080
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1085 1090 1095
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1100 1105 1110
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser
1115 1120 1125
Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
1130 1135 1140
Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
1145 1150 1155
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1160 1165 1170
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1175 1180 1185
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1190 1195 1200
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1205 1210 1215
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1220 1225 1230
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1235 1240 1245
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1250 1255 1260
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1265 1270 1275
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1280 1285 1290
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1295 1300 1305
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1310 1315 1320
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1325 1330 1335
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1340 1345 1350
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1355 1360 1365
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1370 1375 1380
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1385 1390 1395
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1400 1405 1410
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1415 1420 1425
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1430 1435 1440
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1445 1450 1455
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1460 1465 1470
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1475 1480 1485
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1490 1495 1500
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1505 1510 1515
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1520 1525 1530
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1535 1540 1545
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1550 1555 1560
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1565 1570 1575
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1580 1585 1590
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1595 1600 1605
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1610 1615 1620
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1625 1630 1635
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1640 1645 1650
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1655 1660 1665
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1670 1675 1680
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1685 1690 1695
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1700 1705 1710
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1715 1720 1725
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1730 1735 1740
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1745 1750 1755
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1760 1765 1770
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1775 1780 1785
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1790 1795 1800
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1805 1810 1815
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1820 1825 1830
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1835 1840 1845
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1850 1855 1860
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1865 1870 1875
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1880 1885 1890
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1895 1900 1905
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1910 1915 1920
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
1925 1930 1935
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1940 1945 1950
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1955 1960 1965
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1970 1975 1980
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
1985 1990 1995
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
2000 2005 2010
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
2015 2020 2025
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
2030 2035 2040
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
2045 2050 2055
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
2060 2065 2070
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
2075 2080 2085
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
2090 2095 2100
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
2105 2110 2115
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
2120 2125
<210> 79
<211> 2128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 205 (одноцепочечный FVIIIFc с двумя XTEN AE — одним XTEN
AE-288 в домене B и одним XTEN AE-144 в аминокислотном положении 18)
<400> 79
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
115 120 125
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
130 135 140
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
145 150 155 160
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
165 170 175
Ser Pro Thr Ser Thr Glu Glu Gly Ala Ser Ser Ser Asp Leu Gly Glu
180 185 190
Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
915 920 925
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
930 935 940
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
945 950 955 960
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
965 970 975
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro
980 985 990
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr
995 1000 1005
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
1010 1015 1020
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1025 1030 1035
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
1040 1045 1050
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
1055 1060 1065
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1070 1075 1080
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1085 1090 1095
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1100 1105 1110
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser
1115 1120 1125
Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
1130 1135 1140
Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
1145 1150 1155
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1160 1165 1170
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1175 1180 1185
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1190 1195 1200
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1205 1210 1215
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1220 1225 1230
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1235 1240 1245
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1250 1255 1260
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1265 1270 1275
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1280 1285 1290
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1295 1300 1305
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1310 1315 1320
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1325 1330 1335
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1340 1345 1350
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1355 1360 1365
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1370 1375 1380
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1385 1390 1395
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1400 1405 1410
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1415 1420 1425
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1430 1435 1440
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1445 1450 1455
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1460 1465 1470
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1475 1480 1485
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1490 1495 1500
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1505 1510 1515
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1520 1525 1530
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1535 1540 1545
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1550 1555 1560
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1565 1570 1575
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1580 1585 1590
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1595 1600 1605
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1610 1615 1620
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1625 1630 1635
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1640 1645 1650
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1655 1660 1665
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1670 1675 1680
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1685 1690 1695
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1700 1705 1710
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1715 1720 1725
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1730 1735 1740
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1745 1750 1755
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1760 1765 1770
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1775 1780 1785
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1790 1795 1800
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1805 1810 1815
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1820 1825 1830
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1835 1840 1845
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1850 1855 1860
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1865 1870 1875
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1880 1885 1890
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1895 1900 1905
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1910 1915 1920
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro
1925 1930 1935
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1940 1945 1950
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1955 1960 1965
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1970 1975 1980
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
1985 1990 1995
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
2000 2005 2010
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
2015 2020 2025
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
2030 2035 2040
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
2045 2050 2055
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
2060 2065 2070
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
2075 2080 2085
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
2090 2095 2100
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
2105 2110 2115
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
2120 2125
<210> 80
<211> 2020
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 266 (FVIII Fc с XTEN AE-42 в аминокислотном положении
18 и XTEN AE-288 в домене B)
<400> 80
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
35 40 45
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
50 55 60
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Ala Ser Ser Ser
65 70 75 80
Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro
85 90 95
Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe
100 105 110
Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro
115 120 125
Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr
130 135 140
Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His
145 150 155 160
Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp
165 170 175
Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly
180 185 190
Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met
195 200 205
Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp
210 215 220
Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys
225 230 235 240
Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe
245 250 255
Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu
260 265 270
Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala
275 280 285
Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro
290 295 300
Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly
305 310 315 320
Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr
325 330 335
Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile
340 345 350
Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu
355 360 365
Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr
370 375 380
Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn
385 390 395 400
Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met
405 410 415
Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile
420 425 430
Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala
435 440 445
Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp
450 455 460
Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile
465 470 475 480
Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr
485 490 495
Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro
500 505 510
Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn
515 520 525
Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val
530 535 540
Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys
545 550 555 560
Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val
565 570 575
Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg
580 585 590
Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu
595 600 605
Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly
610 615 620
Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe
625 630 635 640
Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu
645 650 655
Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser
660 665 670
Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu
675 680 685
Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly
690 695 700
Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys
705 710 715 720
His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly
725 730 735
Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly
740 745 750
Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys
755 760 765
Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr
770 775 780
Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro
785 790 795 800
Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr
805 810 815
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
820 825 830
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu
835 840 845
Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
850 855 860
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
865 870 875 880
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
885 890 895
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
900 905 910
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
915 920 925
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro
930 935 940
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
945 950 955 960
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
965 970 975
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
980 985 990
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
995 1000 1005
Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
1010 1015 1020
Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
1025 1030 1035
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
1040 1045 1050
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser
1055 1060 1065
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1070 1075 1080
Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala
1085 1090 1095
Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu Ile Thr Arg
1100 1105 1110
Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr
1115 1120 1125
Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu
1130 1135 1140
Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His
1145 1150 1155
Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser
1160 1165 1170
Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val
1175 1180 1185
Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser
1190 1195 1200
Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly
1205 1210 1215
Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met
1220 1225 1230
Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser
1235 1240 1245
Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro
1250 1255 1260
Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp
1265 1270 1275
Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys
1280 1285 1290
Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val
1295 1300 1305
His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr
1310 1315 1320
Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala
1325 1330 1335
Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr
1340 1345 1350
Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met
1355 1360 1365
Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn
1370 1375 1380
Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp
1385 1390 1395
Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn
1400 1405 1410
Ile His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys
1415 1420 1425
Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val
1430 1435 1440
Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg
1445 1450 1455
Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr
1460 1465 1470
Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met
1475 1480 1485
Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln
1490 1495 1500
Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly
1505 1510 1515
Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys
1520 1525 1530
Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln
1535 1540 1545
Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile
1550 1555 1560
Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly
1565 1570 1575
Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser
1580 1585 1590
Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg
1595 1600 1605
Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu
1610 1615 1620
Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro
1625 1630 1635
Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala
1640 1645 1650
Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys
1655 1660 1665
Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln
1670 1675 1680
Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr
1685 1690 1695
Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu
1700 1705 1710
Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp
1715 1720 1725
Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val
1730 1735 1740
Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu
1745 1750 1755
Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser
1760 1765 1770
Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu
1775 1780 1785
Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro Cys Pro
1790 1795 1800
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
1805 1810 1815
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr
1820 1825 1830
Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe
1835 1840 1845
Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys
1850 1855 1860
Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val
1865 1870 1875
Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
1880 1885 1890
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr
1895 1900 1905
Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
1910 1915 1920
Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
1925 1930 1935
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
1940 1945 1950
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
1955 1960 1965
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu
1970 1975 1980
Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys
1985 1990 1995
Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
2000 2005 2010
Leu Ser Leu Ser Pro Gly Lys
2015 2020
<210> 81
<211> 2056
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 267 (FVIII Fc с XTEN AE-72 в аминокислотном положении
18 и XTEN AE-288 в домене B)
<400> 81
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ala Ser Ser Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro
115 120 125
Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys
130 135 140
Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys
145 150 155 160
Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu
165 170 175
Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro
180 185 190
Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly
195 200 205
Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys
210 215 220
Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu
225 230 235 240
Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu
245 250 255
Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala
260 265 270
Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr
275 280 285
Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser
290 295 300
Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala
305 310 315 320
Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn
325 330 335
Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp
340 345 350
His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu
355 360 365
Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu
370 375 380
Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu
385 390 395 400
Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly
405 410 415
Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu
420 425 430
Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr
435 440 445
Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser
450 455 460
Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val
465 470 475 480
His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val
485 490 495
Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly
500 505 510
Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr
515 520 525
Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly
530 535 540
Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile
545 550 555 560
Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly
565 570 575
Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val
580 585 590
Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr
595 600 605
Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg
610 615 620
Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu
625 630 635 640
Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val
645 650 655
Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu
660 665 670
Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile
675 680 685
Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu
690 695 700
Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp
705 710 715 720
Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile
725 730 735
Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly
740 745 750
Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe
755 760 765
Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu
770 775 780
Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr
785 790 795 800
Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr
805 810 815
Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn
820 825 830
Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly Thr Ser
835 840 845
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
850 855 860
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
865 870 875 880
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
885 890 895
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser
900 905 910
Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
915 920 925
Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu
930 935 940
Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
945 950 955 960
Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
965 970 975
Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
980 985 990
Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr
995 1000 1005
Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1010 1015 1020
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1025 1030 1035
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro
1040 1045 1050
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro
1055 1060 1065
Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1070 1075 1080
Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
1085 1090 1095
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
1100 1105 1110
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser
1115 1120 1125
Glu Gly Ser Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His
1130 1135 1140
Gln Ala Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu
1145 1150 1155
Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp
1160 1165 1170
Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe
1175 1180 1185
Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu
1190 1195 1200
Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg
1205 1210 1215
Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln
1220 1225 1230
Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu
1235 1240 1245
Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu
1250 1255 1260
Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg
1265 1270 1275
Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln
1280 1285 1290
Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu
1295 1300 1305
Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr
1310 1315 1320
Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val
1325 1330 1335
Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu
1340 1345 1350
Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val
1355 1360 1365
Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
1370 1375 1380
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala
1385 1390 1395
Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr
1400 1405 1410
Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly
1415 1420 1425
Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser
1430 1435 1440
Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His
1445 1450 1455
Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr
1460 1465 1470
Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser
1475 1480 1485
Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu
1490 1495 1500
His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys
1505 1510 1515
Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln
1520 1525 1530
Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala
1535 1540 1545
Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu
1550 1555 1560
Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile
1565 1570 1575
His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu
1580 1585 1590
Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys
1595 1600 1605
Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe
1610 1615 1620
Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn
1625 1630 1635
Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr
1640 1645 1650
Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu
1655 1660 1665
Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser
1670 1675 1680
Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala
1685 1690 1695
Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser
1700 1705 1710
Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln
1715 1720 1725
Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln
1730 1735 1740
Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu
1745 1750 1755
Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln
1760 1765 1770
Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr
1775 1780 1785
Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu
1790 1795 1800
Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met
1805 1810 1815
Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His
1820 1825 1830
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
1835 1840 1845
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
1850 1855 1860
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
1865 1870 1875
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
1880 1885 1890
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
1895 1900 1905
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
1910 1915 1920
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
1925 1930 1935
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
1940 1945 1950
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
1955 1960 1965
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
1970 1975 1980
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
1985 1990 1995
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
2000 2005 2010
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
2015 2020 2025
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
2030 2035 2040
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
2045 2050 2055
<210> 82
<211> 1834
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 268 (FVIII Fc с XTEN AE-144 в аминокислотном
положении 18)
<400> 82
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
115 120 125
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
130 135 140
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
145 150 155 160
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
165 170 175
Ser Pro Thr Ser Thr Glu Glu Gly Ala Ser Ser Ser Asp Leu Gly Glu
180 185 190
Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Pro Pro Val Leu Lys Arg His Gln Ala Glu Ile Thr Arg Thr
915 920 925
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser
930 935 940
Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn
945 950 955 960
Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala
965 970 975
Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val
980 985 990
Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
995 1000 1005
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1010 1015 1020
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1025 1030 1035
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1040 1045 1050
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1055 1060 1065
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1070 1075 1080
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1085 1090 1095
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1100 1105 1110
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1115 1120 1125
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1130 1135 1140
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1145 1150 1155
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1160 1165 1170
Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1175 1180 1185
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1190 1195 1200
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1205 1210 1215
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1220 1225 1230
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1235 1240 1245
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1250 1255 1260
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1265 1270 1275
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1280 1285 1290
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1295 1300 1305
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1310 1315 1320
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1325 1330 1335
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1340 1345 1350
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1355 1360 1365
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1370 1375 1380
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1385 1390 1395
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1400 1405 1410
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1415 1420 1425
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1430 1435 1440
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1445 1450 1455
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1460 1465 1470
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1475 1480 1485
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1490 1495 1500
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1505 1510 1515
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1520 1525 1530
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1535 1540 1545
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1550 1555 1560
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1565 1570 1575
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1580 1585 1590
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1595 1600 1605
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1610 1615 1620
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1625 1630 1635
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1640 1645 1650
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1655 1660 1665
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1670 1675 1680
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1685 1690 1695
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1700 1705 1710
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1715 1720 1725
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1730 1735 1740
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1745 1750 1755
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1760 1765 1770
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1775 1780 1785
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1790 1795 1800
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1805 1810 1815
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1820 1825 1830
Lys
<210> 83
<211> 1762
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 269 (FVIII Fc with 72 AE-XTEN at amino acid 18)
<400> 83
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ala Ser Ser Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro
115 120 125
Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys
130 135 140
Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys
145 150 155 160
Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu
165 170 175
Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro
180 185 190
Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly
195 200 205
Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys
210 215 220
Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu
225 230 235 240
Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu
245 250 255
Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala
260 265 270
Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr
275 280 285
Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser
290 295 300
Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala
305 310 315 320
Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn
325 330 335
Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp
340 345 350
His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu
355 360 365
Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu
370 375 380
Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu
385 390 395 400
Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly
405 410 415
Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu
420 425 430
Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr
435 440 445
Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser
450 455 460
Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val
465 470 475 480
His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val
485 490 495
Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly
500 505 510
Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr
515 520 525
Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly
530 535 540
Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile
545 550 555 560
Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly
565 570 575
Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val
580 585 590
Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr
595 600 605
Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg
610 615 620
Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu
625 630 635 640
Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val
645 650 655
Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu
660 665 670
Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile
675 680 685
Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu
690 695 700
Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp
705 710 715 720
Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile
725 730 735
Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly
740 745 750
Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe
755 760 765
Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu
770 775 780
Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr
785 790 795 800
Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr
805 810 815
Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn
820 825 830
Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu Lys Arg
835 840 845
His Gln Ala Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu
850 855 860
Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe
865 870 875 880
Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys
885 890 895
Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
900 905 910
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly
915 920 925
Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly
930 935 940
Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly
945 950 955 960
Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val
965 970 975
Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu
980 985 990
Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn
995 1000 1005
Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln
1010 1015 1020
His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp
1025 1030 1035
Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly
1040 1045 1050
Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro
1055 1060 1065
Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe
1070 1075 1080
Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met
1085 1090 1095
Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro
1100 1105 1110
Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile
1115 1120 1125
Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile
1130 1135 1140
Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser
1145 1150 1155
Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu
1160 1165 1170
Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr
1175 1180 1185
Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys
1190 1195 1200
Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu
1205 1210 1215
Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly
1220 1225 1230
His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln
1235 1240 1245
Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn
1250 1255 1260
Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu
1265 1270 1275
Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg
1280 1285 1290
Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr
1295 1300 1305
Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr
1310 1315 1320
Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile
1325 1330 1335
Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg
1340 1345 1350
Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu
1355 1360 1365
Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met
1370 1375 1380
Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr
1385 1390 1395
Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu
1400 1405 1410
His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn
1415 1420 1425
Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val
1430 1435 1440
Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met
1445 1450 1455
Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln
1460 1465 1470
Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly
1475 1480 1485
Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro
1490 1495 1500
Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His
1505 1510 1515
Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp
1520 1525 1530
Leu Tyr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
1535 1540 1545
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
1550 1555 1560
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
1565 1570 1575
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
1580 1585 1590
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
1595 1600 1605
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
1610 1615 1620
Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
1625 1630 1635
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
1640 1645 1650
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
1655 1660 1665
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
1670 1675 1680
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
1685 1690 1695
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
1700 1705 1710
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
1715 1720 1725
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
1730 1735 1740
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
1745 1750 1755
Ser Pro Gly Lys
1760
<210> 84
<211> 1726
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 271 (FVIII Fc с XTEN AE-42 в аминокислотном
положении 18)
<400> 84
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
35 40 45
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
50 55 60
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Ala Ser Ser Ser
65 70 75 80
Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro
85 90 95
Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe
100 105 110
Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro
115 120 125
Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr
130 135 140
Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His
145 150 155 160
Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp
165 170 175
Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly
180 185 190
Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met
195 200 205
Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp
210 215 220
Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys
225 230 235 240
Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe
245 250 255
Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu
260 265 270
Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala
275 280 285
Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro
290 295 300
Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly
305 310 315 320
Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr
325 330 335
Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile
340 345 350
Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu
355 360 365
Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr
370 375 380
Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn
385 390 395 400
Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met
405 410 415
Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile
420 425 430
Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala
435 440 445
Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp
450 455 460
Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile
465 470 475 480
Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr
485 490 495
Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro
500 505 510
Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn
515 520 525
Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val
530 535 540
Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys
545 550 555 560
Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val
565 570 575
Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg
580 585 590
Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu
595 600 605
Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly
610 615 620
Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe
625 630 635 640
Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu
645 650 655
Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser
660 665 670
Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu
675 680 685
Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly
690 695 700
Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys
705 710 715 720
His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly
725 730 735
Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly
740 745 750
Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys
755 760 765
Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr
770 775 780
Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro
785 790 795 800
Arg Ser Phe Ser Gln Asn Pro Pro Val Leu Lys Arg His Gln Ala Glu
805 810 815
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp
820 825 830
Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp
835 840 845
Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His
850 855 860
Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser
865 870 875 880
Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln
885 890 895
Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln
900 905 910
Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro
915 920 925
Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn
930 935 940
Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
945 950 955 960
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys Pro
965 970 975
Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met Ala Pro
980 985 990
Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val
995 1000 1005
Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu
1010 1015 1020
Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val
1025 1030 1035
Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
1040 1045 1050
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala
1055 1060 1065
Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr
1070 1075 1080
Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly
1085 1090 1095
Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser
1100 1105 1110
Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His
1115 1120 1125
Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr
1130 1135 1140
Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser
1145 1150 1155
Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu
1160 1165 1170
His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys
1175 1180 1185
Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln
1190 1195 1200
Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala
1205 1210 1215
Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu
1220 1225 1230
Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile
1235 1240 1245
His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu
1250 1255 1260
Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys
1265 1270 1275
Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe
1280 1285 1290
Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn
1295 1300 1305
Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr
1310 1315 1320
Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu
1325 1330 1335
Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser
1340 1345 1350
Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala
1355 1360 1365
Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser
1370 1375 1380
Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln
1385 1390 1395
Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln
1400 1405 1410
Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu
1415 1420 1425
Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln
1430 1435 1440
Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr
1445 1450 1455
Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu
1460 1465 1470
Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met
1475 1480 1485
Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His
1490 1495 1500
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
1505 1510 1515
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
1520 1525 1530
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
1535 1540 1545
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
1550 1555 1560
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
1565 1570 1575
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
1580 1585 1590
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
1595 1600 1605
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
1610 1615 1620
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
1625 1630 1635
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
1640 1645 1650
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
1655 1660 1665
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
1670 1675 1680
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
1685 1690 1695
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
1700 1705 1710
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1715 1720 1725
<210> 85
<211> 1901
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 272 (FVIII с XTEN AE-144 в аминокислотном положении
18 и XTEN AE-244 в домене B - без Fc)
<400> 85
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
115 120 125
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
130 135 140
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
145 150 155 160
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
165 170 175
Ser Pro Thr Ser Thr Glu Glu Gly Ala Ser Ser Ser Asp Leu Gly Glu
180 185 190
Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
915 920 925
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
930 935 940
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
945 950 955 960
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
965 970 975
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro
980 985 990
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr
995 1000 1005
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
1010 1015 1020
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1025 1030 1035
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
1040 1045 1050
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
1055 1060 1065
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1070 1075 1080
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1085 1090 1095
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1100 1105 1110
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser
1115 1120 1125
Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
1130 1135 1140
Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
1145 1150 1155
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1160 1165 1170
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1175 1180 1185
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1190 1195 1200
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1205 1210 1215
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1220 1225 1230
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1235 1240 1245
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1250 1255 1260
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1265 1270 1275
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1280 1285 1290
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1295 1300 1305
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1310 1315 1320
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1325 1330 1335
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1340 1345 1350
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1355 1360 1365
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1370 1375 1380
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1385 1390 1395
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1400 1405 1410
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1415 1420 1425
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1430 1435 1440
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1445 1450 1455
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1460 1465 1470
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1475 1480 1485
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1490 1495 1500
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1505 1510 1515
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1520 1525 1530
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1535 1540 1545
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1550 1555 1560
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1565 1570 1575
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1580 1585 1590
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1595 1600 1605
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1610 1615 1620
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1625 1630 1635
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1640 1645 1650
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1655 1660 1665
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1670 1675 1680
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1685 1690 1695
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1700 1705 1710
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1715 1720 1725
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1730 1735 1740
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1745 1750 1755
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1760 1765 1770
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1775 1780 1785
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1790 1795 1800
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1805 1810 1815
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1820 1825 1830
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1835 1840 1845
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1850 1855 1860
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1865 1870 1875
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1880 1885 1890
Gly Cys Glu Ala Gln Asp Leu Tyr
1895 1900
<210> 86
<211> 1515
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF031
<400> 86
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Gly Gly Gly Ser Gly
1235 1240 1245
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1250 1255 1260
Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly Ser
1265 1270 1275
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr
1280 1285 1290
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
1295 1300 1305
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg
1310 1315 1320
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
1325 1330 1335
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
1340 1345 1350
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
1355 1360 1365
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
1370 1375 1380
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
1385 1390 1395
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
1400 1405 1410
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
1415 1420 1425
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
1430 1435 1440
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
1445 1450 1455
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
1460 1465 1470
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
1475 1480 1485
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His
1490 1495 1500
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1505 1510 1515
<210> 87
<211> 1778
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF034
<400> 87
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Thr Ser Glu Ser Ala
1235 1240 1245
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1250 1255 1260
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1265 1270 1275
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1280 1285 1290
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro
1295 1300 1305
Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
1310 1315 1320
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1325 1330 1335
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1340 1345 1350
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser
1355 1360 1365
Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser
1370 1375 1380
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
1385 1390 1395
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
1400 1405 1410
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala
1415 1420 1425
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1430 1435 1440
Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
1445 1450 1455
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
1460 1465 1470
Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro
1475 1480 1485
Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1490 1495 1500
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1505 1510 1515
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Asp Ile Gly
1520 1525 1530
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly
1535 1540 1545
Ser Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
1550 1555 1560
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
1565 1570 1575
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
1580 1585 1590
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
1595 1600 1605
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
1610 1615 1620
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
1625 1630 1635
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
1640 1645 1650
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
1655 1660 1665
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
1670 1675 1680
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
1685 1690 1695
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
1700 1705 1710
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
1715 1720 1725
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
1730 1735 1740
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
1745 1750 1755
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
1760 1765 1770
Leu Ser Pro Gly Lys
1775
<210> 88
<211> 32
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 88
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
1 5 10 15
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25 30
<210> 89
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Часть константной области Ig
<400> 89
Pro Lys Asn Ser Ser Met Ile Ser Asn Thr Pro
1 5 10
<210> 90
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Часть константной области Ig
<400> 90
His Gln Ser Leu Gly Thr Gln
1 5
<210> 91
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Часть константной области Ig
<400> 91
His Gln Asn Leu Ser Asp Gly Lys
1 5
<210> 92
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Часть константной области Ig
<400> 92
His Gln Asn Ile Ser Asp Gly Lys
1 5
<210> 93
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Часть константной области Ig
<400> 93
Val Ile Ser Ser His Leu Gly Gln
1 5
<210> 94
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер, в котором последовательность может повторяться 1-20 раз
<400> 94
Gly Gly Gly Gly Ser
1 5
<210> 95
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер, в котором последовательность может повторяться 1-100 раз
или 1-20 раз
<400> 95
Gly Gly Ser Gly Gly Gly Gly Ser
1 5
<210> 96
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 96
Ser Gly Gly Ser Gly Gly Ser
1 5
<210> 97
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 97
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Gly
1 5 10 15
<210> 98
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 98
Gly Gly Ser Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 99
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 99
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly
1 5 10 15
Gly Ser
<210> 100
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 100
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
<210> 101
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер, в котором последовательность может повторяться 1-00 раз
или 1-20 раз
<400> 101
Gly Gly Gly Ser
1
<210> 102
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления
<400> 102
Arg Arg Arg Arg
1
<210> 103
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления
<400> 103
Arg Lys Arg Arg Lys Arg
1 5
<210> 104
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления
<400> 104
Arg Arg Arg Arg Ser
1 5
<210> 105
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления тромбином
<220>
<221> другой_признак
<222> (1)..(1)
<223> Xaa представляет собой алифатическую аминокислоту
<400> 105
Xaa Val Pro Arg
1
<210> 106
<211> 34
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 106
Ile Ser Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp
1 5 10 15
Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser
20 25 30
Phe Ser
<210> 107
<211> 40
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 107
Ile Ser Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu
1 5 10 15
Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro
20 25 30
Ser Phe Ile Gln Ile Arg Ser Val
35 40
<210> 108
<211> 46
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 108
Ile Ser Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile
1 5 10 15
Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp
20 25 30
Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln
35 40 45
<210> 109
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 109
Ser Phe Leu Leu Arg Asn
1 5
<210> 110
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 110
Pro Asn Asp Lys
1
<210> 111
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 111
Pro Asn Asp Lys Tyr
1 5
<210> 112
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 112
Pro Asn Asp Lys Tyr Glu
1 5
<210> 113
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 113
Pro Asn Asp Lys Tyr Glu Pro
1 5
<210> 114
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 114
Pro Asn Asp Lys Tyr Glu Pro Phe
1 5
<210> 115
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 115
Pro Asn Asp Lys Tyr Glu Pro Phe Trp
1 5
<210> 116
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 116
Pro Asn Asp Lys Tyr Glu Pro Phe Trp Glu
1 5 10
<210> 117
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 117
Pro Asn Asp Lys Tyr Glu Pro Phe Trp Glu Asp
1 5 10
<210> 118
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 118
Pro Asn Asp Lys Tyr Glu Pro Phe Trp Glu Asp Glu
1 5 10
<210> 119
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 119
Pro Asn Asp Lys Tyr Glu Pro Phe Trp Glu Asp Glu Glu
1 5 10
<210> 120
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Мотив PAR1, взаимодействующий с экзосайтами
<400> 120
Pro Asn Asp Lys Tyr Glu Pro Phe Trp Glu Asp Glu Glu Ser
1 5 10
<210> 121
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления FXIa
<400> 121
Lys Leu Thr Arg Ala Glu Thr
1 5
<210> 122
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления FXIa
<400> 122
Asp Phe Thr Arg Val Val Gly
1 5
<210> 123
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для FXIIa
<400> 123
Thr Met Thr Arg Ile Val Gly Gly
1 5
<210> 124
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для калликреина
<400> 124
Ser Pro Phe Arg Ser Thr Gly Gly
1 5
<210> 125
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для FVIIa
<400> 125
Leu Gln Val Arg Ile Val Gly Gly
1 5
<210> 126
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для FIXa
<400> 126
Pro Leu Gly Arg Ile Val Gly Gly
1 5
<210> 127
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для FXa
<400> 127
Ile Glu Gly Arg Thr Val Gly Gly
1 5
<210> 128
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для FIIa (тромбина)
<400> 128
Leu Thr Pro Arg Ser Leu Leu Val
1 5
<210> 129
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для эластазы-2
<400> 129
Leu Gly Pro Val Ser Gly Val Pro
1 5
<210> 130
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для гранзима B
<400> 130
Val Ala Gly Asp Ser Leu Glu Glu
1 5
<210> 131
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для MMP-12
<400> 131
Gly Pro Ala Gly Leu Gly Gly Ala
1 5
<210> 132
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для MMP-13
<400> 132
Gly Pro Ala Gly Leu Arg Gly Ala
1 5
<210> 133
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для MMP-17
<400> 133
Ala Pro Leu Gly Leu Arg Leu Arg
1 5
<210> 134
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для MMP-20
<400> 134
Pro Ala Leu Pro Leu Val Ala Gln
1 5
<210> 135
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для TEV
<400> 135
Glu Asn Leu Tyr Phe Gln Gly
1 5
<210> 136
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для энтерокиназы
<400> 136
Asp Asp Asp Lys Ile Val Gly Gly
1 5
<210> 137
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для протеазы 3C (PRESCISSION)
<400> 137
Leu Glu Val Leu Phe Gln Gly Pro
1 5
<210> 138
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для сортазы A
<400> 138
Leu Pro Lys Thr Gly Ser Glu Ser
1 5
<210> 139
<211> 31
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 139
Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala
1 5 10 15
Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25 30
<210> 140
<211> 30
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 140
Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr
1 5 10 15
Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25 30
<210> 141
<211> 29
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 141
Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu
1 5 10 15
Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25
<210> 142
<211> 28
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 142
Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu
1 5 10 15
Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25
<210> 143
<211> 27
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 143
Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser
1 5 10 15
Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25
<210> 144
<211> 26
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 144
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
1 5 10 15
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25
<210> 145
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Сайт расщепления для тромбина
<400> 145
Ala Leu Arg Pro Arg Val Val Gly Gly Ala
1 5 10
<210> 146
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 146
Thr Leu Asp Pro Arg Ser Phe Leu Leu Arg Asn Pro Asn Asp Lys Tyr
1 5 10 15
Glu Pro Phe Trp Glu Asp Glu Glu Lys
20 25
<210> 147
<211> 4548
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VWF031
<400> 147
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatctggcg gtggaggttc cggtggcggg ggatccggcg gtggaggttc cggcggtgga 3780
ggttccggtg gcgggggatc cggtggcggg ggatccctgg tcccccgggg cagcggcggt 3840
ggaggttccg gtggcggggg atccgacaaa actcacacat gcccaccgtg cccagctcca 3900
gaactcctgg gcggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 3960
atctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 4020
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 4080
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct gcaccaggac 4140
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 4200
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 4260
ccatcccgcg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 4320
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 4380
accacgcctc ccgtgttgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 4440
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 4500
cacaaccact acacgcagaa gagcctctcc ctgtctccgg gtaaatga 4548
<210> 148
<211> 5379
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VWF034
<400> 148
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatcgggta cctcagagtc tgctaccccc gagtcagggc caggatcaga gccagccacc 3780
tccgggtctg agacacccgg gacttccgag agtgccaccc ctgagtccgg acccgggtcc 3840
gagcccgcca cttccggctc cgaaactccc ggcacaagcg agagcgctac cccagagtca 3900
ggaccaggaa catctacaga gccctctgaa ggctccgctc cagggtcccc agccggcagt 3960
cccactagca ccgaggaggg aacctctgaa agcgccacac ccgaatcagg gccagggtct 4020
gagcctgcta ccagcggcag cgagacacca ggcacctctg agtccgccac accagagtcc 4080
ggacccggat ctcccgctgg gagccccacc tccactgagg agggatctcc tgctggctct 4140
ccaacatcta ctgaggaagg tacctcaacc gagccatccg agggatcagc tcccggcacc 4200
tcagagtcgg caaccccgga gtctggaccc ggaacttccg aaagtgccac accagagtcc 4260
ggtcccggga cttcagaatc agcaacaccc gagtccggcc ctgggtctga acccgccaca 4320
agtggtagtg agacaccagg atcagaacct gctacctcag ggtcagagac acccggatct 4380
ccggcaggct caccaacctc cactgaggag ggcaccagca cagaaccaag cgagggctcc 4440
gcacccggaa caagcactga acccagtgag ggttcagcac ccggctctga gccggccaca 4500
agtggcagtg agacacccgg cacttcagag agtgccaccc ccgagagtgg cccaggcact 4560
agtaccgagc cctctgaagg cagtgcgcca gattctggcg gtggaggttc cggtggcggg 4620
ggatccggtg gcgggggatc cggtggcggg ggatccggtg gcgggggatc cctggtcccc 4680
cggggcagcg gaggcgacaa aactcacaca tgcccaccgt gcccagctcc agaactcctg 4740
ggcggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 4800
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 4860
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 4920
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 4980
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 5040
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 5100
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 5160
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 5220
cccgtgttgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 5280
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 5340
tacacgcaga agagcctctc cctgtctccg ggtaaatga 5379
<210> 149
<211> 4548
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VWF050 (с тройной мутацией IHH)
<400> 149
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatctggcg gtggaggttc cggtggcggg ggatccggcg gtggaggttc cggcggtgga 3780
ggttccggtg gcgggggatc cggtggcggg ggatccctgg tcccccgggg cagcggcggt 3840
ggaggttccg gtggcggggg atccgacaaa actcacacat gcccaccgtg cccagctcca 3900
gaactcctgg gcggaccgtc agtcttcctc ttccccccaa aacccaagga caccctcatg 3960
gcctcccgga cccctgaggt cacatgcgtg gtggtggacg tgagccacga agaccctgag 4020
gtcaagttca actggtacgt ggacggcgtg gaggtgcata atgccaagac aaagccgcgg 4080
gaggagcagt acaacagcac gtaccgtgtg gtcagcgtcc tcaccgtcct ggcccaggac 4140
tggctgaatg gcaaggagta caagtgcaag gtctccaaca aagccctccc agcccccatc 4200
gagaaaacca tctccaaagc caaagggcag ccccgagaac cacaggtgta caccctgccc 4260
ccatcccgcg atgagctgac caagaaccag gtcagcctga cctgcctggt caaaggcttc 4320
tatcccagcg acatcgccgt ggagtgggag agcaatgggc agccggagaa caactacaag 4380
accacgcctc ccgtgttgga ctccgacggc tccttcttcc tctacagcaa gctcaccgtg 4440
gacaagagca ggtggcagca ggggaacgtc ttctcatgct ccgtgatgca tgaggctctg 4500
cacaacgcct acacgcagaa gagcctctcc ctgtctccgg gtaaatga 4548
<210> 150
<211> 1515
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF050 (с тройной мутацией IHH)
<400> 150
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Gly Gly Gly Ser Gly
1235 1240 1245
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1250 1255 1260
Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly Ser
1265 1270 1275
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Asp Lys Thr His Thr
1280 1285 1290
Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
1295 1300 1305
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ala Ser Arg
1310 1315 1320
Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
1325 1330 1335
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
1340 1345 1350
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
1355 1360 1365
Arg Val Val Ser Val Leu Thr Val Leu Ala Gln Asp Trp Leu Asn
1370 1375 1380
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala
1385 1390 1395
Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu
1400 1405 1410
Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys
1415 1420 1425
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
1430 1435 1440
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
1445 1450 1455
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
1460 1465 1470
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly
1475 1480 1485
Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Ala
1490 1495 1500
Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1505 1510 1515
<210> 151
<211> 4959
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VWF057
<400> 151
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatcgggcg cgccaacatc agagagcgcc acccctgaaa gtggtcccgg gagcgagcca 3780
gccacatctg ggtcggaaac gccaggcaca agtgagtctg caactcccga gtccggacct 3840
ggctccgagc ctgccactag cggctccgag actccgggaa cttccgagag cgctacacca 3900
gaaagcggac ccggaaccag taccgaacct agcgagggct ctgctccggg cagcccagcc 3960
ggctctccta catccacgga ggagggcact tccgaatccg ccaccccgga gtcagggcca 4020
ggatctgaac ccgctacctc aggcagtgag acgccaggaa cgagcgagtc cgctacaccg 4080
gagagtgggc cagggagccc tgctggatct cctacgtcca ctgaggaagg gtcaccagcg 4140
ggctcgccca ccagcactga agaaggtgcc tcgagcggcg gtggaggttc cggtggcggg 4200
ggatccggtg gcgggggatc cggtggcggg ggatccggtg gcgggggatc cctggtcccc 4260
cggggcagcg gaggcgacaa aactcacaca tgcccaccgt gcccagctcc agaactcctg 4320
ggcggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 4380
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 4440
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 4500
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 4560
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 4620
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 4680
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 4740
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 4800
cccgtgttgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 4860
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 4920
tacacgcaga agagcctctc cctgtctccg ggtaaatga 4959
<210> 152
<211> 1652
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF057
<400> 152
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Ala Pro Thr Ser Glu
1235 1240 1245
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
1250 1255 1260
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1265 1270 1275
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1280 1285 1290
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
1295 1300 1305
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro
1310 1315 1320
Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1325 1330 1335
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1340 1345 1350
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala
1355 1360 1365
Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro
1370 1375 1380
Thr Ser Thr Glu Glu Gly Ala Ser Ser Gly Gly Gly Gly Ser Gly
1385 1390 1395
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1400 1405 1410
Gly Gly Gly Ser Leu Val Pro Arg Gly Ser Gly Gly Asp Lys Thr
1415 1420 1425
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
1430 1435 1440
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
1445 1450 1455
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
1460 1465 1470
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
1475 1480 1485
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
1490 1495 1500
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
1505 1510 1515
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
1520 1525 1530
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
1535 1540 1545
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
1550 1555 1560
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
1565 1570 1575
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
1580 1585 1590
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
1595 1600 1605
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
1610 1615 1620
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
1625 1630 1635
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1640 1645 1650
<210> 153
<211> 5379
<212> ДНК
<213> Искусственная последовательность
<220>
<223> VWF058 (VWF034 с мутацией IHH)
<400> 153
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatcgggta cctcagagtc tgctaccccc gagtcagggc caggatcaga gccagccacc 3780
tccgggtctg agacacccgg gacttccgag agtgccaccc ctgagtccgg acccgggtcc 3840
gagcccgcca cttccggctc cgaaactccc ggcacaagcg agagcgctac cccagagtca 3900
ggaccaggaa catctacaga gccctctgaa ggctccgctc cagggtcccc agccggcagt 3960
cccactagca ccgaggaggg aacctctgaa agcgccacac ccgaatcagg gccagggtct 4020
gagcctgcta ccagcggcag cgagacacca ggcacctctg agtccgccac accagagtcc 4080
ggacccggat ctcccgctgg gagccccacc tccactgagg agggatctcc tgctggctct 4140
ccaacatcta ctgaggaagg tacctcaacc gagccatccg agggatcagc tcccggcacc 4200
tcagagtcgg caaccccgga gtctggaccc ggaacttccg aaagtgccac accagagtcc 4260
ggtcccggga cttcagaatc agcaacaccc gagtccggcc ctgggtctga acccgccaca 4320
agtggtagtg agacaccagg atcagaacct gctacctcag ggtcagagac acccggatct 4380
ccggcaggct caccaacctc cactgaggag ggcaccagca cagaaccaag cgagggctcc 4440
gcacccggaa caagcactga acccagtgag ggttcagcac ccggctctga gccggccaca 4500
agtggcagtg agacacccgg cacttcagag agtgccaccc ccgagagtgg cccaggcact 4560
agtaccgagc cctctgaagg cagtgcgcca gattctggcg gtggaggttc cggtggcggg 4620
ggatccggtg gcgggggatc cggtggcggg ggatccggtg gcgggggatc cctggtcccc 4680
cggggcagcg gaggcgacaa aactcacaca tgcccaccgt gcccagctcc agaactcctg 4740
ggcggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat ggcctcccgg 4800
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 4860
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 4920
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tggcccagga ctggctgaat 4980
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 5040
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgc 5100
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 5160
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 5220
cccgtgttgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 5280
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaacgcc 5340
tacacgcaga agagcctctc cctgtctccg ggtaaatga 5379
<210> 154
<211> 1792
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF058 (VWF034 с мутацией IHH)
<400> 154
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Thr Ser Glu Ser Ala
1235 1240 1245
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1250 1255 1260
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1265 1270 1275
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1280 1285 1290
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro
1295 1300 1305
Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
1310 1315 1320
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1325 1330 1335
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1340 1345 1350
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser
1355 1360 1365
Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser
1370 1375 1380
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
1385 1390 1395
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
1400 1405 1410
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala
1415 1420 1425
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1430 1435 1440
Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
1445 1450 1455
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
1460 1465 1470
Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro
1475 1480 1485
Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1490 1495 1500
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1505 1510 1515
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Asp Ser Gly
1520 1525 1530
Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1535 1540 1545
Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly Ser
1550 1555 1560
Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu
1565 1570 1575
Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
1580 1585 1590
Asp Thr Leu Met Ala Ser Arg Thr Pro Glu Val Thr Cys Val Val
1595 1600 1605
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
1610 1615 1620
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
1625 1630 1635
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val
1640 1645 1650
Leu Ala Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val
1655 1660 1665
Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys
1670 1675 1680
Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro
1685 1690 1695
Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
1700 1705 1710
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser
1715 1720 1725
Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
1730 1735 1740
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
1745 1750 1755
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
1760 1765 1770
His Glu Ala Leu His Asn Ala Tyr Thr Gln Lys Ser Leu Ser Leu
1775 1780 1785
Ser Pro Gly Lys
1790
<210> 155
<211> 5937
<212> ДНК
<213> Искусственная последовательность
<220>
<223> FVIII 169
<400> 155
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcta gcttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acggcgcgcc aggtacctca gagtctgcta cccccgagtc agggccagga 2340
tcagagccag ccacctccgg gtctgagaca cccgggactt ccgagagtgc cacccctgag 2400
tccggacccg ggtccgagcc cgccacttcc ggctccgaaa ctcccggcac aagcgagagc 2460
gctaccccag agtcaggacc aggaacatct acagagccct ctgaaggctc cgctccaggg 2520
tccccagccg gcagtcccac tagcaccgag gagggaacct ctgaaagcgc cacacccgaa 2580
tcagggccag ggtctgagcc tgctaccagc ggcagcgaga caccaggcac ctctgagtcc 2640
gccacaccag agtccggacc cggatctccc gctgggagcc ccacctccac tgaggaggga 2700
tctcctgctg gctctccaac atctactgag gaaggtacct caaccgagcc atccgaggga 2760
tcagctcccg gcacctcaga gtcggcaacc ccggagtctg gacccggaac ttccgaaagt 2820
gccacaccag agtccggtcc cgggacttca gaatcagcaa cacccgagtc cggccctggg 2880
tctgaacccg ccacaagtgg tagtgagaca ccaggatcag aacctgctac ctcagggtca 2940
gagacacccg gatctccggc aggctcacca acctccactg aggagggcac cagcacagaa 3000
ccaagcgagg gctccgcacc cggaacaagc actgaaccca gtgagggttc agcacccggc 3060
tctgagccgg ccacaagtgg cagtgagaca cccggcactt cagagagtgc cacccccgag 3120
agtggcccag gcactagtac cgagccctct gaaggcagtg cgccagcctc gagcccacca 3180
gtcttgaaac gccatcaagc tgaaataact cgtactactc ttcagtcaga tcaagaggaa 3240
atcgattatg atgataccat atcagttgaa atgaagaagg aagattttga catttatgat 3300
gaggatgaaa atcagagccc ccgcagcttt caaaagaaaa cacgacacta ttttattgct 3360
gcagtggaga ggctctggga ttatgggatg agtagctccc cacatgttct aagaaacagg 3420
gctcagagtg gcagtgtccc tcagttcaag aaagttgttt tccaggaatt tactgatggc 3480
tcctttactc agcccttata ccgtggagaa ctaaatgaac atttgggact cctggggcca 3540
tatataagag cagaagttga agataatatc atggtaactt tcagaaatca ggcctctcgt 3600
ccctattcct tctattctag ccttatttct tatgaggaag atcagaggca aggagcagaa 3660
cctagaaaaa actttgtcaa gcctaatgaa accaaaactt acttttggaa agtgcaacat 3720
catatggcac ccactaaaga tgagtttgac tgcaaagcct gggcttattt ctctgatgtt 3780
gacctggaaa aagatgtgca ctcaggcctg attggacccc ttctggtctg ccacactaac 3840
acactgaacc ctgctcatgg gagacaagtg acagtacagg aatttgctct gtttttcacc 3900
atctttgatg agaccaaaag ctggtacttc actgaaaata tggaaagaaa ctgcagggct 3960
ccctgcaata tccagatgga agatcccact tttaaagaga attatcgctt ccatgcaatc 4020
aatggctaca taatggatac actacctggc ttagtaatgg ctcaggatca aaggattcga 4080
tggtatctgc tcagcatggg cagcaatgaa aacatccatt ctattcattt cagtggacat 4140
gtgttcactg tacgaaaaaa agaggagtat aaaatggcac tgtacaatct ctatccaggt 4200
gtttttgaga cagtggaaat gttaccatcc aaagctggaa tttggcgggt ggaatgcctt 4260
attggcgagc atctacatgc tgggatgagc acactttttc tggtgtacag caataagtgt 4320
cagactcccc tgggaatggc ttctggacac attagagatt ttcagattac agcttcagga 4380
caatatggac agtgggcccc aaagctggcc agacttcatt attccggatc aatcaatgcc 4440
tggagcacca aggagccctt ttcttggatc aaggtggatc tgttggcacc aatgattatt 4500
cacggcatca agacccaggg tgcccgtcag aagttctcca gcctctacat ctctcagttt 4560
atcatcatgt atagtcttga tgggaagaag tggcagactt atcgaggaaa ttccactgga 4620
accttaatgg tcttctttgg caatgtggat tcatctggga taaaacacaa tatttttaac 4680
cctccaatta ttgctcgata catccgtttg cacccaactc attatagcat tcgcagcact 4740
cttcgcatgg agttgatggg ctgtgattta aatagttgca gcatgccatt gggaatggag 4800
agtaaagcaa tatcagatgc acagattact gcttcatcct actttaccaa tatgtttgcc 4860
acctggtctc cttcaaaagc tcgacttcac ctccaaggga ggagtaatgc ctggagacct 4920
caggtgaata atccaaaaga gtggctgcaa gtggacttcc agaagacaat gaaagtcaca 4980
ggagtaacta ctcagggagt aaaatctctg cttaccagca tgtatgtgaa ggagttcctc 5040
atctccagca gtcaagatgg ccatcagtgg actctctttt ttcagaatgg caaagtaaag 5100
gtttttcagg gaaatcaaga ctccttcaca cctgtggtga actctctaga cccaccgtta 5160
ctgactcgct accttcgaat tcacccccag agttgggtgc accagattgc cctgaggatg 5220
gaggttctgg gctgcgaggc acaggacctc tacgacaaaa ctcacacatg cccaccgtgc 5280
ccagctccag aactcctggg cggaccgtca gtcttcctct tccccccaaa acccaaggac 5340
accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa 5400
gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca 5460
aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg 5520
caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca 5580
gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac 5640
accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc 5700
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 5760
aactacaaga ccacgcctcc cgtgttggac tccgacggct ccttcttcct ctacagcaag 5820
ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat 5880
gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaatga 5937
<210> 156
<211> 6387
<212> ДНК
<213> Искусственная последовательность
<220>
<223> FVIII 263 (с тройной мутацией IHH)
<400> 156
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aggcgcgcca 120
acatcagaga gcgccacccc tgaaagtggt cccgggagcg agccagccac atctgggtcg 180
gaaacgccag gcacaagtga gtctgcaact cccgagtccg gacctggctc cgagcctgcc 240
actagcggct ccgagactcc gggaacttcc gagagcgcta caccagaaag cggacccgga 300
accagtaccg aacctagcga gggctctgct ccgggcagcc cagccggctc tcctacatcc 360
acggaggagg gcacttccga atccgccacc ccggagtcag ggccaggatc tgaacccgct 420
acctcaggca gtgagacgcc aggaacgagc gagtccgcta caccggagag tgggccaggg 480
agccctgctg gatctcctac gtccactgag gaagggtcac cagcgggctc gcccaccagc 540
actgaagaag gtgcctcgag cagtgatctc ggtgagctgc ctgtggacgc aagatttcct 600
cctagagtgc caaaatcttt tccattcaac acctcagtcg tgtacaaaaa gactctgttt 660
gtagaattca cggatcacct tttcaacatc gctaagccaa ggccaccctg gatgggtctg 720
ctaggtccta ccatccaggc tgaggtttat gatacagtgg tcattacact taagaacatg 780
gcttcccatc ctgtcagtct tcatgctgtt ggtgtatcct actggaaagc ttctgaggga 840
gctgaatatg atgatcagac cagtcaaagg gagaaagaag atgataaagt cttccctggt 900
ggaagccata catatgtctg gcaggtcctg aaagagaatg gtccaatggc ctctgaccca 960
ctgtgcctta cctactcata tctttctcat gtggacctgg taaaagactt gaattcaggc 1020
ctcattggag ccctactagt atgtagagaa gggagtctgg ccaaggaaaa gacacagacc 1080
ttgcacaaat ttatactact ttttgctgta tttgatgaag ggaaaagttg gcactcagaa 1140
acaaagaact ccttgatgca ggatagggat gctgcatctg ctcgggcctg gcctaaaatg 1200
cacacagtca atggttatgt aaacaggtct ctgccaggtc tgattggatg ccacaggaaa 1260
tcagtctatt ggcatgtgat tggaatgggc accactcctg aagtgcactc aatattcctc 1320
gaaggtcaca catttcttgt gaggaaccat cgccaggcta gcttggaaat ctcgccaata 1380
actttcctta ctgctcaaac actcttgatg gaccttggac agtttctact gttttgtcat 1440
atctcttccc accaacatga tggcatggaa gcttatgtca aagtagacag ctgtccagag 1500
gaaccccaac tacgaatgaa aaataatgaa gaagcggaag actatgatga tgatcttact 1560
gattctgaaa tggatgtggt caggtttgat gatgacaact ctccttcctt tatccaaatt 1620
cgctcagttg ccaagaagca tcctaaaact tgggtacatt acattgctgc tgaagaggag 1680
gactgggact atgctccctt agtcctcgcc cccgatgaca gaagttataa aagtcaatat 1740
ttgaacaatg gccctcagcg gattggtagg aagtacaaaa aagtccgatt tatggcatac 1800
acagatgaaa cctttaagac tcgtgaagct attcagcatg aatcaggaat cttgggacct 1860
ttactttatg gggaagttgg agacacactg ttgattatat ttaagaatca agcaagcaga 1920
ccatataaca tctaccctca cggaatcact gatgtccgtc ctttgtattc aaggagatta 1980
ccaaaaggtg taaaacattt gaaggatttt ccaattctgc caggagaaat attcaaatat 2040
aaatggacag tgactgtaga agatgggcca actaaatcag atcctcggtg cctgacccgc 2100
tattactcta gtttcgttaa tatggagaga gatctagctt caggactcat tggccctctc 2160
ctcatctgct acaaagaatc tgtagatcaa agaggaaacc agataatgtc agacaagagg 2220
aatgtcatcc tgttttctgt atttgatgag aaccgaagct ggtacctcac agagaatata 2280
caacgctttc tccccaatcc agctggagtg cagcttgagg atccagagtt ccaagcctcc 2340
aacatcatgc acagcatcaa tggctatgtt tttgatagtt tgcagttgtc agtttgtttg 2400
catgaggtgg catactggta cattctaagc attggagcac agactgactt cctttctgtc 2460
ttcttctctg gatatacctt caaacacaaa atggtctatg aagacacact caccctattc 2520
ccattctcag gagaaactgt cttcatgtcg atggaaaacc caggtctatg gattctgggg 2580
tgccacaact cagactttcg gaacagaggc atgaccgcct tactgaaggt ttctagttgt 2640
gacaagaaca ctggtgatta ttacgaggac agttatgaag atatttcagc atacttgctg 2700
agtaaaaaca atgccattga accaagaagc ttctctcaaa acggcgcgcc aggtacctca 2760
gagtctgcta cccccgagtc agggccagga tcagagccag ccacctccgg gtctgagaca 2820
cccgggactt ccgagagtgc cacccctgag tccggacccg ggtccgagcc cgccacttcc 2880
ggctccgaaa ctcccggcac aagcgagagc gctaccccag agtcaggacc aggaacatct 2940
acagagccct ctgaaggctc cgctccaggg tccccagccg gcagtcccac tagcaccgag 3000
gagggaacct ctgaaagcgc cacacccgaa tcagggccag ggtctgagcc tgctaccagc 3060
ggcagcgaga caccaggcac ctctgagtcc gccacaccag agtccggacc cggatctccc 3120
gctgggagcc ccacctccac tgaggaggga tctcctgctg gctctccaac atctactgag 3180
gaaggtacct caaccgagcc atccgaggga tcagctcccg gcacctcaga gtcggcaacc 3240
ccggagtctg gacccggaac ttccgaaagt gccacaccag agtccggtcc cgggacttca 3300
gaatcagcaa cacccgagtc cggccctggg tctgaacccg ccacaagtgg tagtgagaca 3360
ccaggatcag aacctgctac ctcagggtca gagacacccg gatctccggc aggctcacca 3420
acctccactg aggagggcac cagcacagaa ccaagcgagg gctccgcacc cggaacaagc 3480
actgaaccca gtgagggttc agcacccggc tctgagccgg ccacaagtgg cagtgagaca 3540
cccggcactt cagagagtgc cacccccgag agtggcccag gcactagtac cgagccctct 3600
gaaggcagtg cgccagcctc gagcccacca gtcttgaaac gccatcaagc tgaaataact 3660
cgtactactc ttcagtcaga tcaagaggaa atcgattatg atgataccat atcagttgaa 3720
atgaagaagg aagattttga catttatgat gaggatgaaa atcagagccc ccgcagcttt 3780
caaaagaaaa cacgacacta ttttattgct gcagtggaga ggctctggga ttatgggatg 3840
agtagctccc cacatgttct aagaaacagg gctcagagtg gcagtgtccc tcagttcaag 3900
aaagttgttt tccaggaatt tactgatggc tcctttactc agcccttata ccgtggagaa 3960
ctaaatgaac atttgggact cctggggcca tatataagag cagaagttga agataatatc 4020
atggtaactt tcagaaatca ggcctctcgt ccctattcct tctattctag ccttatttct 4080
tatgaggaag atcagaggca aggagcagaa cctagaaaaa actttgtcaa gcctaatgaa 4140
accaaaactt acttttggaa agtgcaacat catatggcac ccactaaaga tgagtttgac 4200
tgcaaagcct gggcttattt ctctgatgtt gacctggaaa aagatgtgca ctcaggcctg 4260
attggacccc ttctggtctg ccacactaac acactgaacc ctgctcatgg gagacaagtg 4320
acagtacagg aatttgctct gtttttcacc atctttgatg agaccaaaag ctggtacttc 4380
actgaaaata tggaaagaaa ctgcagggct ccctgcaata tccagatgga agatcccact 4440
tttaaagaga attatcgctt ccatgcaatc aatggctaca taatggatac actacctggc 4500
ttagtaatgg ctcaggatca aaggattcga tggtatctgc tcagcatggg cagcaatgaa 4560
aacatccatt ctattcattt cagtggacat gtgttcactg tacgaaaaaa agaggagtat 4620
aaaatggcac tgtacaatct ctatccaggt gtttttgaga cagtggaaat gttaccatcc 4680
aaagctggaa tttggcgggt ggaatgcctt attggcgagc atctacatgc tgggatgagc 4740
acactttttc tggtgtacag caataagtgt cagactcccc tgggaatggc ttctggacac 4800
attagagatt ttcagattac agcttcagga caatatggac agtgggcccc aaagctggcc 4860
agacttcatt attccggatc aatcaatgcc tggagcacca aggagccctt ttcttggatc 4920
aaggtggatc tgttggcacc aatgattatt cacggcatca agacccaggg tgcccgtcag 4980
aagttctcca gcctctacat ctctcagttt atcatcatgt atagtcttga tgggaagaag 5040
tggcagactt atcgaggaaa ttccactgga accttaatgg tcttctttgg caatgtggat 5100
tcatctggga taaaacacaa tatttttaac cctccaatta ttgctcgata catccgtttg 5160
cacccaactc attatagcat tcgcagcact cttcgcatgg agttgatggg ctgtgattta 5220
aatagttgca gcatgccatt gggaatggag agtaaagcaa tatcagatgc acagattact 5280
gcttcatcct actttaccaa tatgtttgcc acctggtctc cttcaaaagc tcgacttcac 5340
ctccaaggga ggagtaatgc ctggagacct caggtgaata atccaaaaga gtggctgcaa 5400
gtggacttcc agaagacaat gaaagtcaca ggagtaacta ctcagggagt aaaatctctg 5460
cttaccagca tgtatgtgaa ggagttcctc atctccagca gtcaagatgg ccatcagtgg 5520
actctctttt ttcagaatgg caaagtaaag gtttttcagg gaaatcaaga ctccttcaca 5580
cctgtggtga actctctaga cccaccgtta ctgactcgct accttcgaat tcacccccag 5640
agttgggtgc accagattgc cctgaggatg gaggttctgg gctgcgaggc acaggacctc 5700
tacgacaaaa ctcacacatg cccaccgtgc ccagctccag aactcctggg cggaccgtca 5760
gtcttcctct tccccccaaa acccaaggac accctcatgg cctcccggac ccctgaggtc 5820
acatgcgtgg tggtggacgt gagccacgaa gaccctgagg tcaagttcaa ctggtacgtg 5880
gacggcgtgg aggtgcataa tgccaagaca aagccgcggg aggagcagta caacagcacg 5940
taccgtgtgg tcagcgtcct caccgtcctg gcccaggact ggctgaatgg caaggagtac 6000
aagtgcaagg tctccaacaa agccctccca gcccccatcg agaaaaccat ctccaaagcc 6060
aaagggcagc cccgagaacc acaggtgtac accctgcccc catcccgcga tgagctgacc 6120
aagaaccagg tcagcctgac ctgcctggtc aaaggcttct atcccagcga catcgccgtg 6180
gagtgggaga gcaatgggca gccggagaac aactacaaga ccacgcctcc cgtgttggac 6240
tccgacggct ccttcttcct ctacagcaag ctcaccgtgg acaagagcag gtggcagcag 6300
gggaacgtct tctcatgctc cgtgatgcat gaggctctgc acaacgccta cacgcagaag 6360
agcctctccc tgtctccggg taaatga 6387
<210> 157
<211> 2128
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 263 (с тройной мутацией IHH)
<400> 157
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Gly Ala Pro Thr Ser Glu Ser Ala Thr Pro Glu
35 40 45
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
50 55 60
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
65 70 75 80
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
85 90 95
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
100 105 110
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
115 120 125
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
130 135 140
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
145 150 155 160
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
165 170 175
Ser Pro Thr Ser Thr Glu Glu Gly Ala Ser Ser Ser Asp Leu Gly Glu
180 185 190
Leu Pro Val Asp Ala Arg Phe Pro Pro Arg Val Pro Lys Ser Phe Pro
195 200 205
Phe Asn Thr Ser Val Val Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr
210 215 220
Asp His Leu Phe Asn Ile Ala Lys Pro Arg Pro Pro Trp Met Gly Leu
225 230 235 240
Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr Asp Thr Val Val Ile Thr
245 250 255
Leu Lys Asn Met Ala Ser His Pro Val Ser Leu His Ala Val Gly Val
260 265 270
Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser
275 280 285
Gln Arg Glu Lys Glu Asp Asp Lys Val Phe Pro Gly Gly Ser His Thr
290 295 300
Tyr Val Trp Gln Val Leu Lys Glu Asn Gly Pro Met Ala Ser Asp Pro
305 310 315 320
Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His Val Asp Leu Val Lys Asp
325 330 335
Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu Val Cys Arg Glu Gly Ser
340 345 350
Leu Ala Lys Glu Lys Thr Gln Thr Leu His Lys Phe Ile Leu Leu Phe
355 360 365
Ala Val Phe Asp Glu Gly Lys Ser Trp His Ser Glu Thr Lys Asn Ser
370 375 380
Leu Met Gln Asp Arg Asp Ala Ala Ser Ala Arg Ala Trp Pro Lys Met
385 390 395 400
His Thr Val Asn Gly Tyr Val Asn Arg Ser Leu Pro Gly Leu Ile Gly
405 410 415
Cys His Arg Lys Ser Val Tyr Trp His Val Ile Gly Met Gly Thr Thr
420 425 430
Pro Glu Val His Ser Ile Phe Leu Glu Gly His Thr Phe Leu Val Arg
435 440 445
Asn His Arg Gln Ala Ser Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr
450 455 460
Ala Gln Thr Leu Leu Met Asp Leu Gly Gln Phe Leu Leu Phe Cys His
465 470 475 480
Ile Ser Ser His Gln His Asp Gly Met Glu Ala Tyr Val Lys Val Asp
485 490 495
Ser Cys Pro Glu Glu Pro Gln Leu Arg Met Lys Asn Asn Glu Glu Ala
500 505 510
Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser Glu Met Asp Val Val Arg
515 520 525
Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile Gln Ile Arg Ser Val Ala
530 535 540
Lys Lys His Pro Lys Thr Trp Val His Tyr Ile Ala Ala Glu Glu Glu
545 550 555 560
Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr
565 570 575
Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr
580 585 590
Lys Lys Val Arg Phe Met Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg
595 600 605
Glu Ala Ile Gln His Glu Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly
610 615 620
Glu Val Gly Asp Thr Leu Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg
625 630 635 640
Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr Asp Val Arg Pro Leu Tyr
645 650 655
Ser Arg Arg Leu Pro Lys Gly Val Lys His Leu Lys Asp Phe Pro Ile
660 665 670
Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp Thr Val Thr Val Glu Asp
675 680 685
Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser
690 695 700
Phe Val Asn Met Glu Arg Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu
705 710 715 720
Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln Arg Gly Asn Gln Ile Met
725 730 735
Ser Asp Lys Arg Asn Val Ile Leu Phe Ser Val Phe Asp Glu Asn Arg
740 745 750
Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala
755 760 765
Gly Val Gln Leu Glu Asp Pro Glu Phe Gln Ala Ser Asn Ile Met His
770 775 780
Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu Gln Leu Ser Val Cys Leu
785 790 795 800
His Glu Val Ala Tyr Trp Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp
805 810 815
Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr Phe Lys His Lys Met Val
820 825 830
Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe Ser Gly Glu Thr Val Phe
835 840 845
Met Ser Met Glu Asn Pro Gly Leu Trp Ile Leu Gly Cys His Asn Ser
850 855 860
Asp Phe Arg Asn Arg Gly Met Thr Ala Leu Leu Lys Val Ser Ser Cys
865 870 875 880
Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser
885 890 895
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
900 905 910
Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
915 920 925
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
930 935 940
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
945 950 955 960
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
965 970 975
Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro
980 985 990
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr
995 1000 1005
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
1010 1015 1020
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1025 1030 1035
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
1040 1045 1050
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
1055 1060 1065
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1070 1075 1080
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
1085 1090 1095
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1100 1105 1110
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser
1115 1120 1125
Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
1130 1135 1140
Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
1145 1150 1155
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1160 1165 1170
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1175 1180 1185
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1190 1195 1200
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1205 1210 1215
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1220 1225 1230
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1235 1240 1245
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1250 1255 1260
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1265 1270 1275
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1280 1285 1290
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1295 1300 1305
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1310 1315 1320
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1325 1330 1335
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1340 1345 1350
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1355 1360 1365
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1370 1375 1380
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1385 1390 1395
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1400 1405 1410
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1415 1420 1425
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1430 1435 1440
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1445 1450 1455
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1460 1465 1470
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1475 1480 1485
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1490 1495 1500
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1505 1510 1515
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1520 1525 1530
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1535 1540 1545
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1550 1555 1560
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1565 1570 1575
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1580 1585 1590
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1595 1600 1605
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1610 1615 1620
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1625 1630 1635
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1640 1645 1650
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1655 1660 1665
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1670 1675 1680
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1685 1690 1695
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1700 1705 1710
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1715 1720 1725
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1730 1735 1740
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1745 1750 1755
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1760 1765 1770
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1775 1780 1785
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1790 1795 1800
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1805 1810 1815
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1820 1825 1830
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1835 1840 1845
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1850 1855 1860
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1865 1870 1875
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1880 1885 1890
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1895 1900 1905
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1910 1915 1920
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ala Ser Arg Thr Pro
1925 1930 1935
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1940 1945 1950
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1955 1960 1965
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1970 1975 1980
Val Ser Val Leu Thr Val Leu Ala Gln Asp Trp Leu Asn Gly Lys
1985 1990 1995
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
2000 2005 2010
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
2015 2020 2025
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
2030 2035 2040
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
2045 2050 2055
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
2060 2065 2070
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
2075 2080 2085
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
2090 2095 2100
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Ala Tyr Thr
2105 2110 2115
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
2120 2125
<210> 158
<211> 5505
<212> ДНК
<213> Искусственная последовательность
<220>
<223> FVIII 282
<400> 158
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcta gcttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acggcgcgcc aacatcagag agcgccaccc ctgaaagtgg tcccgggagc 2340
gagccagcca catctgggtc ggaaacgcca ggcacaagtg agtctgcaac tcccgagtcc 2400
ggacctggct ccgagcctgc cactagcggc tccgagactc cgggaacttc cgagagcgct 2460
acaccagaaa gcggacccgg aaccagtacc gaacctagcg agggctctgc tccgggcagc 2520
ccagccggct ctcctacatc cacggaggag ggcacttccg aatccgccac cccggagtca 2580
gggccaggat ctgaacccgc tacctcaggc agtgagacgc caggaacgag cgagtccgct 2640
acaccggaga gtgggccagg gagccctgct ggatctccta cgtccactga ggaagggtca 2700
ccagcgggct cgcccaccag cactgaagaa ggtgcctcga gcccaccagt cttgaaacgc 2760
catcaagctg aaataactcg tactactctt cagtcagatc aagaggaaat cgattatgat 2820
gataccatat cagttgaaat gaagaaggaa gattttgaca tttatgatga ggatgaaaat 2880
cagagccccc gcagctttca aaagaaaaca cgacactatt ttattgctgc agtggagagg 2940
ctctgggatt atgggatgag tagctcccca catgttctaa gaaacagggc tcagagtggc 3000
agtgtccctc agttcaagaa agttgttttc caggaattta ctgatggctc ctttactcag 3060
cccttatacc gtggagaact aaatgaacat ttgggactcc tggggccata tataagagca 3120
gaagttgaag ataatatcat ggtaactttc agaaatcagg cctctcgtcc ctattccttc 3180
tattctagcc ttatttctta tgaggaagat cagaggcaag gagcagaacc tagaaaaaac 3240
tttgtcaagc ctaatgaaac caaaacttac ttttggaaag tgcaacatca tatggcaccc 3300
actaaagatg agtttgactg caaagcctgg gcttatttct ctgatgttga cctggaaaaa 3360
gatgtgcact caggcctgat tggacccctt ctggtctgcc acactaacac actgaaccct 3420
gctcatggga gacaagtgac agtacaggaa tttgctctgt ttttcaccat ctttgatgag 3480
accaaaagct ggtacttcac tgaaaatatg gaaagaaact gcagggctcc ctgcaatatc 3540
cagatggaag atcccacttt taaagagaat tatcgcttcc atgcaatcaa tggctacata 3600
atggatacac tacctggctt agtaatggct caggatcaaa ggattcgatg gtatctgctc 3660
agcatgggca gcaatgaaaa catccattct attcatttca gtggacatgt gttcactgta 3720
cgaaaaaaag aggagtataa aatggcactg tacaatctct atccaggtgt ttttgagaca 3780
gtggaaatgt taccatccaa agctggaatt tggcgggtgg aatgccttat tggcgagcat 3840
ctacatgctg ggatgagcac actttttctg gtgtacagca ataagtgtca gactcccctg 3900
ggaatggctt ctggacacat tagagatttt cagattacag cttcaggaca atatggacag 3960
tgggccccaa agctggccag acttcattat tccggatcaa tcaatgcctg gagcaccaag 4020
gagccctttt cttggatcaa ggtggatctg ttggcaccaa tgattattca cggcatcaag 4080
acccagggtg cccgtcagaa gttctccagc ctctacatct ctcagtttat catcatgtat 4140
agtcttgatg ggaagaagtg gcagacttat cgaggaaatt ccactggaac cttaatggtc 4200
ttctttggca atgtggattc atctgggata aaacacaata tttttaaccc tccaattatt 4260
gctcgataca tccgtttgca cccaactcat tatagcattc gcagcactct tcgcatggag 4320
ttgatgggct gtgatttaaa tagttgcagc atgccattgg gaatggagag taaagcaata 4380
tcagatgcac agattactgc ttcatcctac tttaccaata tgtttgccac ctggtctcct 4440
tcaaaagctc gacttcacct ccaagggagg agtaatgcct ggagacctca ggtgaataat 4500
ccaaaagagt ggctgcaagt ggacttccag aagacaatga aagtcacagg agtaactact 4560
cagggagtaa aatctctgct taccagcatg tatgtgaagg agttcctcat ctccagcagt 4620
caagatggcc atcagtggac tctctttttt cagaatggca aagtaaaggt ttttcaggga 4680
aatcaagact ccttcacacc tgtggtgaac tctctagacc caccgttact gactcgctac 4740
cttcgaattc acccccagag ttgggtgcac cagattgccc tgaggatgga ggttctgggc 4800
tgcgaggcac aggacctcta cgacaaaact cacacatgcc caccgtgccc agctccagaa 4860
ctcctgggcg gaccgtcagt cttcctcttc cccccaaaac ccaaggacac cctcatgatc 4920
tcccggaccc ctgaggtcac atgcgtggtg gtggacgtga gccacgaaga ccctgaggtc 4980
aagttcaact ggtacgtgga cggcgtggag gtgcataatg ccaagacaaa gccgcgggag 5040
gagcagtaca acagcacgta ccgtgtggtc agcgtcctca ccgtcctgca ccaggactgg 5100
ctgaatggca aggagtacaa gtgcaaggtc tccaacaaag ccctcccagc ccccatcgag 5160
aaaaccatct ccaaagccaa agggcagccc cgagaaccac aggtgtacac cctgccccca 5220
tcccgggatg agctgaccaa gaaccaggtc agcctgacct gcctggtcaa aggcttctat 5280
cccagcgaca tcgccgtgga gtgggagagc aatgggcagc cggagaacaa ctacaagacc 5340
acgcctcccg tgttggactc cgacggctcc ttcttcctct acagcaagct caccgtggac 5400
aagagcaggt ggcagcaggg gaacgtcttc tcatgctccg tgatgcatga ggctctgcac 5460
aaccactaca cgcagaagag cctctccctg tctccgggta aatga 5505
<210> 159
<211> 1834
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 282
<400> 159
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Thr
755 760 765
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
770 775 780
Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
785 790 795 800
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
805 810 815
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro
820 825 830
Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
835 840 845
Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
850 855 860
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala
865 870 875 880
Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr
885 890 895
Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ala
900 905 910
Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu Ile Thr Arg Thr
915 920 925
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser
930 935 940
Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn
945 950 955 960
Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr Phe Ile Ala
965 970 975
Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val
980 985 990
Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val
995 1000 1005
Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr
1010 1015 1020
Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile
1025 1030 1035
Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln
1040 1045 1050
Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu
1055 1060 1065
Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
1070 1075 1080
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met
1085 1090 1095
Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe
1100 1105 1110
Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly
1115 1120 1125
Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly
1130 1135 1140
Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
1145 1150 1155
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1160 1165 1170
Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1175 1180 1185
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1190 1195 1200
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1205 1210 1215
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1220 1225 1230
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1235 1240 1245
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1250 1255 1260
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1265 1270 1275
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1280 1285 1290
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1295 1300 1305
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1310 1315 1320
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1325 1330 1335
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1340 1345 1350
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1355 1360 1365
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1370 1375 1380
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1385 1390 1395
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1400 1405 1410
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1415 1420 1425
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1430 1435 1440
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1445 1450 1455
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1460 1465 1470
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1475 1480 1485
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1490 1495 1500
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1505 1510 1515
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1520 1525 1530
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1535 1540 1545
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1550 1555 1560
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1565 1570 1575
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1580 1585 1590
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1595 1600 1605
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1610 1615 1620
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1625 1630 1635
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1640 1645 1650
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1655 1660 1665
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1670 1675 1680
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1685 1690 1695
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1700 1705 1710
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1715 1720 1725
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1730 1735 1740
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1745 1750 1755
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1760 1765 1770
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1775 1780 1785
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1790 1795 1800
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1805 1810 1815
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1820 1825 1830
Lys
<210> 160
<211> 5937
<212> ДНК
<213> Искусственная последовательность
<220>
<223> FVIII 283 (FVIII 169 с тройной мутацией IHH)
<400> 160
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcta gcttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acggcgcgcc aggtacctca gagtctgcta cccccgagtc agggccagga 2340
tcagagccag ccacctccgg gtctgagaca cccgggactt ccgagagtgc cacccctgag 2400
tccggacccg ggtccgagcc cgccacttcc ggctccgaaa ctcccggcac aagcgagagc 2460
gctaccccag agtcaggacc aggaacatct acagagccct ctgaaggctc cgctccaggg 2520
tccccagccg gcagtcccac tagcaccgag gagggaacct ctgaaagcgc cacacccgaa 2580
tcagggccag ggtctgagcc tgctaccagc ggcagcgaga caccaggcac ctctgagtcc 2640
gccacaccag agtccggacc cggatctccc gctgggagcc ccacctccac tgaggaggga 2700
tctcctgctg gctctccaac atctactgag gaaggtacct caaccgagcc atccgaggga 2760
tcagctcccg gcacctcaga gtcggcaacc ccggagtctg gacccggaac ttccgaaagt 2820
gccacaccag agtccggtcc cgggacttca gaatcagcaa cacccgagtc cggccctggg 2880
tctgaacccg ccacaagtgg tagtgagaca ccaggatcag aacctgctac ctcagggtca 2940
gagacacccg gatctccggc aggctcacca acctccactg aggagggcac cagcacagaa 3000
ccaagcgagg gctccgcacc cggaacaagc actgaaccca gtgagggttc agcacccggc 3060
tctgagccgg ccacaagtgg cagtgagaca cccggcactt cagagagtgc cacccccgag 3120
agtggcccag gcactagtac cgagccctct gaaggcagtg cgccagcctc gagcccacca 3180
gtcttgaaac gccatcaagc tgaaataact cgtactactc ttcagtcaga tcaagaggaa 3240
atcgattatg atgataccat atcagttgaa atgaagaagg aagattttga catttatgat 3300
gaggatgaaa atcagagccc ccgcagcttt caaaagaaaa cacgacacta ttttattgct 3360
gcagtggaga ggctctggga ttatgggatg agtagctccc cacatgttct aagaaacagg 3420
gctcagagtg gcagtgtccc tcagttcaag aaagttgttt tccaggaatt tactgatggc 3480
tcctttactc agcccttata ccgtggagaa ctaaatgaac atttgggact cctggggcca 3540
tatataagag cagaagttga agataatatc atggtaactt tcagaaatca ggcctctcgt 3600
ccctattcct tctattctag ccttatttct tatgaggaag atcagaggca aggagcagaa 3660
cctagaaaaa actttgtcaa gcctaatgaa accaaaactt acttttggaa agtgcaacat 3720
catatggcac ccactaaaga tgagtttgac tgcaaagcct gggcttattt ctctgatgtt 3780
gacctggaaa aagatgtgca ctcaggcctg attggacccc ttctggtctg ccacactaac 3840
acactgaacc ctgctcatgg gagacaagtg acagtacagg aatttgctct gtttttcacc 3900
atctttgatg agaccaaaag ctggtacttc actgaaaata tggaaagaaa ctgcagggct 3960
ccctgcaata tccagatgga agatcccact tttaaagaga attatcgctt ccatgcaatc 4020
aatggctaca taatggatac actacctggc ttagtaatgg ctcaggatca aaggattcga 4080
tggtatctgc tcagcatggg cagcaatgaa aacatccatt ctattcattt cagtggacat 4140
gtgttcactg tacgaaaaaa agaggagtat aaaatggcac tgtacaatct ctatccaggt 4200
gtttttgaga cagtggaaat gttaccatcc aaagctggaa tttggcgggt ggaatgcctt 4260
attggcgagc atctacatgc tgggatgagc acactttttc tggtgtacag caataagtgt 4320
cagactcccc tgggaatggc ttctggacac attagagatt ttcagattac agcttcagga 4380
caatatggac agtgggcccc aaagctggcc agacttcatt attccggatc aatcaatgcc 4440
tggagcacca aggagccctt ttcttggatc aaggtggatc tgttggcacc aatgattatt 4500
cacggcatca agacccaggg tgcccgtcag aagttctcca gcctctacat ctctcagttt 4560
atcatcatgt atagtcttga tgggaagaag tggcagactt atcgaggaaa ttccactgga 4620
accttaatgg tcttctttgg caatgtggat tcatctggga taaaacacaa tatttttaac 4680
cctccaatta ttgctcgata catccgtttg cacccaactc attatagcat tcgcagcact 4740
cttcgcatgg agttgatggg ctgtgattta aatagttgca gcatgccatt gggaatggag 4800
agtaaagcaa tatcagatgc acagattact gcttcatcct actttaccaa tatgtttgcc 4860
acctggtctc cttcaaaagc tcgacttcac ctccaaggga ggagtaatgc ctggagacct 4920
caggtgaata atccaaaaga gtggctgcaa gtggacttcc agaagacaat gaaagtcaca 4980
ggagtaacta ctcagggagt aaaatctctg cttaccagca tgtatgtgaa ggagttcctc 5040
atctccagca gtcaagatgg ccatcagtgg actctctttt ttcagaatgg caaagtaaag 5100
gtttttcagg gaaatcaaga ctccttcaca cctgtggtga actctctaga cccaccgtta 5160
ctgactcgct accttcgaat tcacccccag agttgggtgc accagattgc cctgaggatg 5220
gaggttctgg gctgcgaggc acaggacctc tacgacaaaa ctcacacatg cccaccgtgc 5280
ccagctccag aactcctggg cggaccgtca gtcttcctct tccccccaaa acccaaggac 5340
accctcatgg cctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa 5400
gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca 5460
aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg 5520
gcccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca 5580
gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac 5640
accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc 5700
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 5760
aactacaaga ccacgcctcc cgtgttggac tccgacggct ccttcttcct ctacagcaag 5820
ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat 5880
gaggctctgc acaacgccta cacgcagaag agcctctccc tgtctccggg taaatga 5937
<210> 161
<211> 1978
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 283 (FVIII 169 с тройной мутацией IHH)
<400> 161
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly
755 760 765
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
770 775 780
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
785 790 795 800
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
805 810 815
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
820 825 830
Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
835 840 845
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
850 855 860
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
865 870 875 880
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
885 890 895
Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly
900 905 910
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser
915 920 925
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
930 935 940
Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
945 950 955 960
Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala
965 970 975
Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
980 985 990
Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
995 1000 1005
Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
1010 1015 1020
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
1025 1030 1035
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
1040 1045 1050
Ala Pro Ala Ser Ser Pro Pro Val Leu Lys Arg His Gln Ala Glu
1055 1060 1065
Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1070 1075 1080
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile
1085 1090 1095
Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys
1100 1105 1110
Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr
1115 1120 1125
Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser
1130 1135 1140
Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
1145 1150 1155
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu
1160 1165 1170
His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp
1175 1180 1185
Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser
1190 1195 1200
Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly
1205 1210 1215
Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr
1220 1225 1230
Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu
1235 1240 1245
Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu
1250 1255 1260
Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His
1265 1270 1275
Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln
1280 1285 1290
Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp
1295 1300 1305
Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
1310 1315 1320
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His
1325 1330 1335
Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met
1340 1345 1350
Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser
1355 1360 1365
Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr
1370 1375 1380
Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr
1385 1390 1395
Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly
1400 1405 1410
Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly
1415 1420 1425
Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro
1430 1435 1440
Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala
1445 1450 1455
Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His
1460 1465 1470
Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser
1475 1480 1485
Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile
1490 1495 1500
Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser
1505 1510 1515
Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr
1520 1525 1530
Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn
1535 1540 1545
Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
1550 1555 1560
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg
1565 1570 1575
Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys
1580 1585 1590
Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln
1595 1600 1605
Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser
1610 1615 1620
Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp
1625 1630 1635
Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe
1640 1645 1650
Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys
1655 1660 1665
Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser
1670 1675 1680
Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys
1685 1690 1695
Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val
1700 1705 1710
Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His
1715 1720 1725
Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu
1730 1735 1740
Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro
1745 1750 1755
Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu
1760 1765 1770
Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ala Ser Arg Thr Pro
1775 1780 1785
Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
1790 1795 1800
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala
1805 1810 1815
Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
1820 1825 1830
Val Ser Val Leu Thr Val Leu Ala Gln Asp Trp Leu Asn Gly Lys
1835 1840 1845
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
1850 1855 1860
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
1865 1870 1875
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln
1880 1885 1890
Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile
1895 1900 1905
Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
1910 1915 1920
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
1925 1930 1935
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
1940 1945 1950
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn Ala Tyr Thr
1955 1960 1965
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1970 1975
<210> 162
<211> 5055
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 010 (двухцепочечный FVIIIFc)
<400> 162
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acccaccagt cttgaaacgc catcaacggg aaataactcg tactactctt 2340
cagtcagatc aagaggaaat tgactatgat gataccatat cagttgaaat gaagaaggaa 2400
gattttgaca tttatgatga ggatgaaaat cagagccccc gcagctttca aaagaaaaca 2460
cgacactatt ttattgctgc agtggagagg ctctgggatt atgggatgag tagctcccca 2520
catgttctaa gaaacagggc tcagagtggc agtgtccctc agttcaagaa agttgttttc 2580
caggaattta ctgatggctc ctttactcag cccttatacc gtggagaact aaatgaacat 2640
ttgggactcc tggggccata tataagagca gaagttgaag ataatatcat ggtaactttc 2700
agaaatcagg cctctcgtcc ctattccttc tattctagcc ttatttctta tgaggaagat 2760
cagaggcaag gagcagaacc tagaaaaaac tttgtcaagc ctaatgaaac caaaacttac 2820
ttttggaaag tgcaacatca tatggcaccc actaaagatg agtttgactg caaagcctgg 2880
gcttatttct ctgatgttga cctggaaaaa gatgtgcact caggcctgat tggacccctt 2940
ctggtctgcc acactaacac actgaaccct gctcatggga gacaagtgac agtacaggaa 3000
tttgctctgt ttttcaccat ctttgatgag accaaaagct ggtacttcac tgaaaatatg 3060
gaaagaaact gcagggctcc ctgcaatatc cagatggaag atcccacttt taaagagaat 3120
tatcgcttcc atgcaatcaa tggctacata atggatacac tacctggctt agtaatggct 3180
caggatcaaa ggattcgatg gtatctgctc agcatgggca gcaatgaaaa catccattct 3240
attcatttca gtggacatgt gttcactgta cgaaaaaaag aggagtataa aatggcactg 3300
tacaatctct atccaggtgt ttttgagaca gtggaaatgt taccatccaa agctggaatt 3360
tggcgggtgg aatgccttat tggcgagcat ctacatgctg ggatgagcac actttttctg 3420
gtgtacagca ataagtgtca gactcccctg ggaatggctt ctggacacat tagagatttt 3480
cagattacag cttcaggaca atatggacag tgggccccaa agctggccag acttcattat 3540
tccggatcaa tcaatgcctg gagcaccaag gagccctttt cttggatcaa ggtggatctg 3600
ttggcaccaa tgattattca cggcatcaag acccagggtg cccgtcagaa gttctccagc 3660
ctctacatct ctcagtttat catcatgtat agtcttgatg ggaagaagtg gcagacttat 3720
cgaggaaatt ccactggaac cttaatggtc ttctttggca atgtggattc atctgggata 3780
aaacacaata tttttaaccc tccaattatt gctcgataca tccgtttgca cccaactcat 3840
tatagcattc gcagcactct tcgcatggag ttgatgggct gtgatttaaa tagttgcagc 3900
atgccattgg gaatggagag taaagcaata tcagatgcac agattactgc ttcatcctac 3960
tttaccaata tgtttgccac ctggtctcct tcaaaagctc gacttcacct ccaagggagg 4020
agtaatgcct ggagacctca ggtgaataat ccaaaagagt ggctgcaagt ggacttccag 4080
aagacaatga aagtcacagg agtaactact cagggagtaa aatctctgct taccagcatg 4140
tatgtgaagg agttcctcat ctccagcagt caagatggcc atcagtggac tctctttttt 4200
cagaatggca aagtaaaggt ttttcaggga aatcaagact ccttcacacc tgtggtgaac 4260
tctctagacc caccgttact gactcgctac cttcgaattc acccccagag ttgggtgcac 4320
cagattgccc tgaggatgga ggttctgggc tgcgaggcac aggacctcta cgacaaaact 4380
cacacatgcc caccgtgccc agctccagaa ctcctgggcg gaccgtcagt cttcctcttc 4440
cccccaaaac ccaaggacac cctcatgatc tcccggaccc ctgaggtcac atgcgtggtg 4500
gtggacgtga gccacgaaga ccctgaggtc aagttcaact ggtacgtgga cggcgtggag 4560
gtgcataatg ccaagacaaa gccgcgggag gagcagtaca acagcacgta ccgtgtggtc 4620
agcgtcctca ccgtcctgca ccaggactgg ctgaatggca aggagtacaa gtgcaaggtc 4680
tccaacaaag ccctcccagc ccccatcgag aaaaccatct ccaaagccaa agggcagccc 4740
cgagaaccac aggtgtacac cctgccccca tcccgggatg agctgaccaa gaaccaggtc 4800
agcctgacct gcctggtcaa aggcttctat cccagcgaca tcgccgtgga gtgggagagc 4860
aatgggcagc cggagaacaa ctacaagacc acgcctcccg tgttggactc cgacggctcc 4920
ttcttcctct acagcaagct caccgtggac aagagcaggt ggcagcaggg gaacgtcttc 4980
tcatgctccg tgatgcatga ggctctgcac aaccactaca cgcagaagag cctctccctg 5040
tctccgggta aatga 5055
<210> 163
<211> 1684
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 010 (двухцепочечный FVIIIFc)
<400> 163
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Pro Pro Val Leu
755 760 765
Lys Arg His Gln Arg Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln
770 775 780
Glu Glu Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu
785 790 795 800
Asp Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe
805 810 815
Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp
820 825 830
Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln
835 840 845
Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr
850 855 860
Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His
865 870 875 880
Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile
885 890 895
Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser
900 905 910
Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
915 920 925
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val
930 935 940
Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp
945 950 955 960
Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His Ser Gly Leu
965 970 975
Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu Asn Pro Ala His
980 985 990
Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe
995 1000 1005
Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn
1010 1015 1020
Cys Arg Ala Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys
1025 1030 1035
Glu Asn Tyr Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr
1040 1045 1050
Leu Pro Gly Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr
1055 1060 1065
Leu Leu Ser Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe
1070 1075 1080
Ser Gly His Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met
1085 1090 1095
Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met
1100 1105 1110
Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly
1115 1120 1125
Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser
1130 1135 1140
Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg
1145 1150 1155
Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
1160 1165 1170
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser
1175 1180 1185
Thr Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro
1190 1195 1200
Met Ile Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe
1205 1210 1215
Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp
1220 1225 1230
Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu
1235 1240 1245
Met Val Phe Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn
1250 1255 1260
Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro
1265 1270 1275
Thr His Tyr Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly
1280 1285 1290
Cys Asp Leu Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys
1295 1300 1305
Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn
1310 1315 1320
Met Phe Ala Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln
1325 1330 1335
Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu
1340 1345 1350
Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val
1355 1360 1365
Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys
1370 1375 1380
Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu
1385 1390 1395
Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
1400 1405 1410
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr
1415 1420 1425
Arg Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala
1430 1435 1440
Leu Arg Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp
1445 1450 1455
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1460 1465 1470
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
1475 1480 1485
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val
1490 1495 1500
Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly
1505 1510 1515
Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr
1520 1525 1530
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
1535 1540 1545
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
1550 1555 1560
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
1565 1570 1575
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp
1580 1585 1590
Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly
1595 1600 1605
Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln
1610 1615 1620
Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp
1625 1630 1635
Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
1640 1645 1650
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala
1655 1660 1665
Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1670 1675 1680
Lys
<210> 164
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер, в котором последовательность может повторяться 1-10 раз
<400> 164
Ser Gly Gly Gly Gly Ser
1 5
<210> 165
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 165
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 166
<211> 34
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Линкер
<400> 166
Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10 15
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Leu Val Pro Arg Gly Ser
20 25 30
Gly Gly
<210> 167
<211> 314
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 169
<400> 167
Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala
1 5 10 15
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
20 25 30
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
35 40 45
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala
50 55 60
Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
65 70 75 80
Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr
85 90 95
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
100 105 110
Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
115 120 125
Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser
130 135 140
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro
145 150 155 160
Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
165 170 175
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
180 185 190
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
195 200 205
Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
210 215 220
Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr
225 230 235 240
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro
245 250 255
Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
260 265 270
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
275 280 285
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Pro Pro Val
290 295 300
Leu Lys Arg His Gln Ala Glu Ile Thr Arg
305 310
<210> 168
<211> 302
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 310
<400> 168
Pro Arg Ser Phe Gly Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
1 5 10 15
Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
20 25 30
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala
35 40 45
Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu
50 55 60
Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly
65 70 75 80
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser
85 90 95
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
100 105 110
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
115 120 125
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly
130 135 140
Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly
145 150 155 160
Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly
165 170 175
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser
180 185 190
Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
195 200 205
Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
210 215 220
Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu
225 230 235 240
Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly
245 250 255
Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
260 265 270
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
275 280 285
Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile Thr Arg
290 295 300
<210> 169
<211> 305
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 312
<400> 169
Pro Arg Ser Phe Ser Gln Asn Gly Ala Pro Gly Thr Ser Glu Ser Ala
1 5 10 15
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
20 25 30
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
35 40 45
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala
50 55 60
Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
65 70 75 80
Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr
85 90 95
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
100 105 110
Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
115 120 125
Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser
130 135 140
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro
145 150 155 160
Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
165 170 175
Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
180 185 190
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
195 200 205
Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
210 215 220
Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr
225 230 235 240
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro
245 250 255
Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
260 265 270
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
275 280 285
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile Thr
290 295 300
Arg
305
<210> 170
<211> 5892
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 310 (кодирующая FVIII с полной делецией домена B,
за исключением 2 аминокислотных остатков, и XTEN AE-288, вставленным
после а. к. 742)
<400> 170
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttcggtacct cagagtctgc tacccccgag tcagggccag gatcagagcc agccacctcc 2340
gggtctgaga cacccgggac ttccgagagt gccacccctg agtccggacc cgggtccgag 2400
cccgccactt ccggctccga aactcccggc acaagcgaga gcgctacccc agagtcagga 2460
ccaggaacat ctacagagcc ctctgaaggc tccgctccag ggtccccagc cggcagtccc 2520
actagcaccg aggagggaac ctctgaaagc gccacacccg aatcagggcc agggtctgag 2580
cctgctacca gcggcagcga gacaccaggc acctctgagt ccgccacacc agagtccgga 2640
cccggatctc ccgctgggag ccccacctcc actgaggagg gatctcctgc tggctctcca 2700
acatctactg aggaaggtac ctcaaccgag ccatccgagg gatcagctcc cggcacctca 2760
gagtcggcaa ccccggagtc tggacccgga acttccgaaa gtgccacacc agagtccggt 2820
cccgggactt cagaatcagc aacacccgag tccggccctg ggtctgaacc cgccacaagt 2880
ggtagtgaga caccaggatc agaacctgct acctcagggt cagagacacc cggatctccg 2940
gcaggctcac caacctccac tgaggagggc accagcacag aaccaagcga gggctccgca 3000
cccggaacaa gcactgaacc cagtgagggt tcagcacccg gctctgagcc ggccacaagt 3060
ggcagtgaga cacccggcac ttcagagagt gccacccccg agagtggccc aggcactagt 3120
accgagccct ctgaaggcag tgcgccagcc tcgagcgaaa taactcgtac tactcttcag 3180
tcagatcaag aggaaatcga ttatgatgat accatatcag ttgaaatgaa gaaggaagat 3240
tttgacattt atgatgagga tgaaaatcag agcccccgca gctttcaaaa gaaaacacga 3300
cactatttta ttgctgcagt ggagaggctc tgggattatg ggatgagtag ctccccacat 3360
gttctaagaa acagggctca gagtggcagt gtccctcagt tcaagaaagt tgttttccag 3420
gaatttactg atggctcctt tactcagccc ttataccgtg gagaactaaa tgaacatttg 3480
ggactcctgg ggccatatat aagagcagaa gttgaagata atatcatggt aactttcaga 3540
aatcaggcct ctcgtcccta ttccttctat tctagcctta tttcttatga ggaagatcag 3600
aggcaaggag cagaacctag aaaaaacttt gtcaagccta atgaaaccaa aacttacttt 3660
tggaaagtgc aacatcatat ggcacccact aaagatgagt ttgactgcaa agcctgggct 3720
tatttctctg atgttgacct ggaaaaagat gtgcactcag gcctgattgg accccttctg 3780
gtctgccaca ctaacacact gaaccctgct catgggagac aagtgacagt acaggaattt 3840
gctctgtttt tcaccatctt tgatgagacc aaaagctggt acttcactga aaatatggaa 3900
agaaactgca gggctccctg caatatccag atggaagatc ccacttttaa agagaattat 3960
cgcttccatg caatcaatgg ctacataatg gatacactac ctggcttagt aatggctcag 4020
gatcaaagga ttcgatggta tctgctcagc atgggcagca atgaaaacat ccattctatt 4080
catttcagtg gacatgtgtt cactgtacga aaaaaagagg agtataaaat ggcactgtac 4140
aatctctatc caggtgtttt tgagacagtg gaaatgttac catccaaagc tggaatttgg 4200
cgggtggaat gccttattgg cgagcatcta catgctggga tgagcacact ttttctggtg 4260
tacagcaata agtgtcagac tcccctggga atggcttctg gacacattag agattttcag 4320
attacagctt caggacaata tggacagtgg gccccaaagc tggccagact tcattattcc 4380
ggatcaatca atgcctggag caccaaggag cccttttctt ggatcaaggt ggatctgttg 4440
gcaccaatga ttattcacgg catcaagacc cagggtgccc gtcagaagtt ctccagcctc 4500
tacatctctc agtttatcat catgtatagt cttgatggga agaagtggca gacttatcga 4560
ggaaattcca ctggaacctt aatggtcttc tttggcaatg tggattcatc tgggataaaa 4620
cacaatattt ttaaccctcc aattattgct cgatacatcc gtttgcaccc aactcattat 4680
agcattcgca gcactcttcg catggagttg atgggctgtg atttaaatag ttgcagcatg 4740
ccattgggaa tggagagtaa agcaatatca gatgcacaga ttactgcttc atcctacttt 4800
accaatatgt ttgccacctg gtctccttca aaagctcgac ttcacctcca agggaggagt 4860
aatgcctgga gacctcaggt gaataatcca aaagagtggc tgcaagtgga cttccagaag 4920
acaatgaaag tcacaggagt aactactcag ggagtaaaat ctctgcttac cagcatgtat 4980
gtgaaggagt tcctcatctc cagcagtcaa gatggccatc agtggactct cttttttcag 5040
aatggcaaag taaaggtttt tcagggaaat caagactcct tcacacctgt ggtgaactct 5100
ctagacccac cgttactgac tcgctacctt cgaattcacc cccagagttg ggtgcaccag 5160
attgccctga ggatggaggt tctgggctgc gaggcacagg acctctacga caaaactcac 5220
acatgcccac cgtgcccagc tccagaactc ctgggcggac cgtcagtctt cctcttcccc 5280
ccaaaaccca aggacaccct catgatctcc cggacccctg aggtcacatg cgtggtggtg 5340
gacgtgagcc acgaagaccc tgaggtcaag ttcaactggt acgtggacgg cgtggaggtg 5400
cataatgcca agacaaagcc gcgggaggag cagtacaaca gcacgtaccg tgtggtcagc 5460
gtcctcaccg tcctgcacca ggactggctg aatggcaagg agtacaagtg caaggtctcc 5520
aacaaagccc tcccagcccc catcgagaaa accatctcca aagccaaagg gcagccccga 5580
gaaccacagg tgtacaccct gcccccatcc cgggatgagc tgaccaagaa ccaggtcagc 5640
ctgacctgcc tggtcaaagg cttctatccc agcgacatcg ccgtggagtg ggagagcaat 5700
gggcagccgg agaacaacta caagaccacg cctcccgtgt tggactccga cggctccttc 5760
ttcctctaca gcaagctcac cgtggacaag agcaggtggc agcaggggaa cgtcttctca 5820
tgctccgtga tgcatgaggc tctgcacaac cactacacgc agaagagcct ctccctgtct 5880
ccgggtaaat ga 5892
<210> 171
<211> 1944
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 310 (FVIII с полной делецией домена B, за исключением
2 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 742)
<400> 171
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
740 745 750
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
755 760 765
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr
770 775 780
Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
785 790 795 800
Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser
805 810 815
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala
820 825 830
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
835 840 845
Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
850 855 860
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser
865 870 875 880
Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
885 890 895
Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
900 905 910
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala
915 920 925
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu
930 935 940
Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser
945 950 955 960
Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro
965 970 975
Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser
980 985 990
Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr
995 1000 1005
Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu
1010 1015 1020
Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile Thr Arg Thr
1025 1030 1035
Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp Thr Ile
1040 1045 1050
Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp Glu Asp
1055 1060 1065
Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg His Tyr
1070 1075 1080
Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser Ser
1085 1090 1095
Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro
1100 1105 1110
Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe
1115 1120 1125
Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu
1130 1135 1140
Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val
1145 1150 1155
Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser
1160 1165 1170
Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg
1175 1180 1185
Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys
1190 1195 1200
Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp Cys Lys
1205 1210 1215
Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp Val His
1220 1225 1230
Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn Thr Leu
1235 1240 1245
Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe Ala Leu
1250 1255 1260
Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe Thr Glu
1265 1270 1275
Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln Met Glu
1280 1285 1290
Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile Asn Gly
1295 1300 1305
Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln Asp Gln
1310 1315 1320
Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn Ile
1325 1330 1335
His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys
1340 1345 1350
Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe
1355 1360 1365
Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val
1370 1375 1380
Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu
1385 1390 1395
Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala
1400 1405 1410
Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr
1415 1420 1425
Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser Gly Ser
1430 1435 1440
Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile Lys Val
1445 1450 1455
Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr Gln Gly
1460 1465 1470
Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe Ile Ile
1475 1480 1485
Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg Gly Asn
1490 1495 1500
Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp Ser Ser
1505 1510 1515
Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala Arg Tyr
1520 1525 1530
Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr Leu Arg
1535 1540 1545
Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met Pro Leu
1550 1555 1560
Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala Ser
1565 1570 1575
Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala
1580 1585 1590
Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val
1595 1600 1605
Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met
1610 1615 1620
Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr
1625 1630 1635
Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly
1640 1645 1650
His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe
1655 1660 1665
Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser Leu Asp
1670 1675 1680
Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln Ser Trp
1685 1690 1695
Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys Glu Ala
1700 1705 1710
Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1715 1720 1725
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1730 1735 1740
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
1745 1750 1755
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn
1760 1765 1770
Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro
1775 1780 1785
Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu
1790 1795 1800
Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys
1805 1810 1815
Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
1820 1825 1830
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu
1835 1840 1845
Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr
1850 1855 1860
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
1865 1870 1875
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
1880 1885 1890
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
1895 1900 1905
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser
1910 1915 1920
Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu
1925 1930 1935
Ser Leu Ser Pro Gly Lys
1940
<210> 172
<211> 5900
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 312 (FVIII с полной делецией домена B, за исключением
5 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 745 –
вариант B5 FVIII)
<400> 172
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acggtacctc agagtctgct acccccgagt cagggccagg atcagagcca 2340
gccacctccg ggtctgagac acccgggact tccgagagtg ccacccctga gtccggaccc 2400
gggtccgagc ccgccacttc cggctccgaa actcccggca caagcgagag cgctacccca 2460
gagtcaggac caggaacatc tacagagccc tctgaaggct ccgctccagg gtccccagcc 2520
ggcagtccca ctagcaccga ggagggaacc tctgaaagcg ccacacccga atcagggcca 2580
gggtctgagc ctgctaccag cggcagcgag acaccaggca cctctgagtc cgccacacca 2640
gagtccggac ccggatctcc cgctgggagc cccacctcca ctgaggaggg atctcctgct 2700
ggctctccaa catctactga ggaaggtacc tcaaccgagc catccgaggg atcagctccc 2760
ggcacctcag agtcggcaac cccggagtct ggacccggaa cttccgaaag tgccacacca 2820
gagtccggtc ccgggacttc agaatcagca acacccgagt ccggccctgg gtctgaaccc 2880
gccacaagtg gtagtgagac accaggatca gaacctgcta cctcagggtc agagacaccc 2940
ggatctccgg caggctcacc aacctccact gaggagggca ccagcacaga accaagcgag 3000
ggctccgcac ccggaacaag cactgaaccc agtgagggtt cagcacccgg ctctgagccg 3060
gccacaagtg gcagtgagac acccggcact tcagagagtg ccacccccga gagtggccca 3120
ggcactagta ccgagccctc tgaaggcagt gcgccagcct cgagcgaaat aactcgtact 3180
actcttcagt cagatcaaga ggaaatcgat tatgatgata ccatatcagt tgaaatgaag 3240
aaggaagatt ttgacattta tgatgaggat gaaaatcaga gcccccgcag ctttcaaaag 3300
aaaacacgac actattttat tgctgcagtg gagaggctct gggattatgg gatgagtagc 3360
tccccacatg ttctaagaaa cagggctcag agtggcagtg tccctcagtt caagaaagtt 3420
gttttccagg aatttactga tggctccttt actcagccct tataccgtgg agaactaaat 3480
gaacatttgg gactcctggg gccatatata agagcagaag ttgaagataa tatcatggta 3540
actttcagaa atcaggcctc tcgtccctat tccttctatt ctagccttat ttcttatgag 3600
gaagatcaga ggcaaggagc agaacctaga aaaaactttg tcaagcctaa tgaaaccaaa 3660
acttactttt ggaaagtgca acatcatatg gcacccacta aagatgagtt tgactgcaaa 3720
gcctgggctt atttctctga tgttgacctg gaaaaagatg tgcactcagg cctgattgga 3780
ccccttctgg tctgccacac taacacactg aaccctgctc atgggagaca agtgacagta 3840
caggaatttg ctctgttttt caccatcttt gatgagacca aaagctggta cttcactgaa 3900
aatatggaaa gaaactgcag ggctccctgc aatatccaga tggaagatcc cacttttaaa 3960
gagaattatc gcttccatgc aatcaatggc tacataatgg atacactacc tggcttagta 4020
atggctcagg atcaaaggat tcgatggtat ctgctcagca tgggcagcaa tgaaaacatc 4080
cattctattc atttcagtgg acatgtgttc actgtacgaa aaaaagagga gtataaaatg 4140
gcactgtaca atctctatcc aggtgttttt gagacagtgg aaatgttacc atccaaagct 4200
ggaatttggc gggtggaatg ccttattggc gagcatctac atgctgggat gagcacactt 4260
tttctggtgt acagcaataa gtgtcagact cccctgggaa tggcttctgg acacattaga 4320
gattttcaga ttacagcttc aggacaatat ggacagtggg ccccaaagct ggccagactt 4380
cattattccg gatcaatcaa tgcctggagc accaaggagc ccttttcttg gatcaaggtg 4440
gatctgttgg caccaatgat tattcacggc atcaagaccc agggtgcccg tcagaagttc 4500
tccagcctct acatctctca gtttatcatc atgtatagtc ttgatgggaa gaagtggcag 4560
acttatcgag gaaattccac tggaacctta atggtcttct ttggcaatgt ggattcatct 4620
gggataaaac acaatatttt taaccctcca attattgctc gatacatccg tttgcaccca 4680
actcattata gcattcgcag cactcttcgc atggagttga tgggctgtga tttaaatagt 4740
tgcagcatgc cattgggaat ggagagtaaa gcaatatcag atgcacagat tactgcttca 4800
tcctacttta ccaatatgtt tgccacctgg tctccttcaa aagctcgact tcacctccaa 4860
gggaggagta atgcctggag acctcaggtg aataatccaa aagagtggct gcaagtggac 4920
ttccagaaga caatgaaagt cacaggagta actactcagg gagtaaaatc tctgcttacc 4980
agcatgtatg tgaaggagtt cctcatctcc agcagtcaag atggccatca gtggactctc 5040
ttttttcaga atggcaaagt aaaggttttt cagggaaatc aagactcctt cacacctgtg 5100
gtgaactctc tagacccacc gttactgact cgctaccttc gaattcaccc ccagagttgg 5160
gtgcaccaga ttgccctgag gatggaggtt ctgggctgcg aggcacagga cctctacgac 5220
aaaactcaca catgcccacc gtgcccagct ccagaactcc tgggcggacc gtcagtcttc 5280
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 5340
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc 5400
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 5460
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc 5520
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 5580
cagccccgag aaccacaggt gtacaccctg cccccatccc gggatgagct gaccaagaac 5640
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 5700
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgtt ggactccgac 5760
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac 5820
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 5880
tccctgtctc cgggtaaatg 5900
<210> 173
<211> 1947
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN FVIII 312 (FVIII с полной делецией домена B, за исключением
5 аминокислотных остатков, и XTEN AE-288, вставленным после а. к. 745 –
вариант B5 FVIII)
<400> 173
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Thr Ser Glu Ser Ala Thr
740 745 750
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
755 760 765
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu
770 775 780
Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
785 790 795 800
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
805 810 815
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
820 825 830
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
835 840 845
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
850 855 860
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro
865 870 875 880
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
885 890 895
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
900 905 910
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
915 920 925
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
930 935 940
Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
945 950 955 960
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
965 970 975
Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser
980 985 990
Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
995 1000 1005
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
1010 1015 1020
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile
1025 1030 1035
Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp
1040 1045 1050
Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr
1055 1060 1065
Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr
1070 1075 1080
Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly
1085 1090 1095
Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly
1100 1105 1110
Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp
1115 1120 1125
Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His
1130 1135 1140
Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn
1145 1150 1155
Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe
1160 1165 1170
Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala
1175 1180 1185
Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr
1190 1195 1200
Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe
1205 1210 1215
Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys
1220 1225 1230
Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr
1235 1240 1245
Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu
1250 1255 1260
Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr
1265 1270 1275
Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile
1280 1285 1290
Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala
1295 1300 1305
Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala
1310 1315 1320
Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn
1325 1330 1335
Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr Val
1340 1345 1350
Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro
1355 1360 1365
Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile
1370 1375 1380
Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met
1385 1390 1395
Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu
1400 1405 1410
Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser
1415 1420 1425
Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr
1430 1435 1440
Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp
1445 1450 1455
Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys
1460 1465 1470
Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln
1475 1480 1485
Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr
1490 1495 1500
Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val
1505 1510 1515
Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile
1520 1525 1530
Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser
1535 1540 1545
Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser
1550 1555 1560
Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile
1565 1570 1575
Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro
1580 1585 1590
Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg
1595 1600 1605
Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln
1610 1615 1620
Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser
1625 1630 1635
Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser
1640 1645 1650
Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val
1655 1660 1665
Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn
1670 1675 1680
Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro
1685 1690 1695
Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly
1700 1705 1710
Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro
1715 1720 1725
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
1730 1735 1740
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
1745 1750 1755
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
1760 1765 1770
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
1775 1780 1785
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
1790 1795 1800
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
1805 1810 1815
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
1820 1825 1830
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
1835 1840 1845
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
1850 1855 1860
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
1865 1870 1875
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
1880 1885 1890
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
1895 1900 1905
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
1910 1915 1920
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
1925 1930 1935
Lys Ser Leu Ser Leu Ser Pro Gly Lys
1940 1945
<210> 174
<211> 4977
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN VWF073 (D1D2D'D3 VWF-XTEN AE-144-FVIII с усеченным сайтом
расщепления тромбина в a2-Fc)
<400> 174
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
ggcgcgccaa catcagagag cgccacccct gaaagtggtc ccgggagcga gccagccaca 3780
tctgggtcgg aaacgccagg cacaagtgag tctgcaactc ccgagtccgg acctggctcc 3840
gagcctgcca ctagcggctc cgagactccg ggaacttccg agagcgctac accagaaagc 3900
ggacccggaa ccagtaccga acctagcgag ggctctgctc cgggcagccc agccggctct 3960
cctacatcca cggaggaggg cacttccgaa tccgccaccc cggagtcagg gccaggatct 4020
gaacccgcta cctcaggcag tgagacgcca ggaacgagcg agtccgctac accggagagt 4080
gggccaggga gccctgctgg atctcctacg tccactgagg aagggtcacc agcgggctcg 4140
cccaccagca ctgaagaagg tgcctcgagc ggcggtggag gatccggtgg cgggggatcc 4200
ggtggcgggg gatccggtgg cgggggatcc ggtggcgggg gatccggtgg cgggggatcc 4260
attgaaccaa gaagcttctc tggcagcgga ggcgacaaaa ctcacacatg cccaccgtgc 4320
ccagctccag aactcctggg cggaccgtca gtcttcctct tccccccaaa acccaaggac 4380
accctcatga tctcccggac ccctgaggtc acatgcgtgg tggtggacgt gagccacgaa 4440
gaccctgagg tcaagttcaa ctggtacgtg gacggcgtgg aggtgcataa tgccaagaca 4500
aagccgcggg aggagcagta caacagcacg taccgtgtgg tcagcgtcct caccgtcctg 4560
caccaggact ggctgaatgg caaggagtac aagtgcaagg tctccaacaa agccctccca 4620
gcccccatcg agaaaaccat ctccaaagcc aaagggcagc cccgagaacc acaggtgtac 4680
accctgcccc catcccggga tgagctgacc aagaaccagg tcagcctgac ctgcctggtc 4740
aaaggcttct atcccagcga catcgccgtg gagtgggaga gcaatgggca gccggagaac 4800
aactacaaga ccacgcctcc cgtgttggac tccgacggct ccttcttcct ctacagcaag 4860
ctcaccgtgg acaagagcag gtggcagcag gggaacgtct tctcatgctc cgtgatgcat 4920
gaggctctgc acaaccacta cacgcagaag agcctctccc tgtctccggg taaatga 4977
<210> 175
<211> 1658
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF073 (D1D2D'D3 VWF-XTEN AE-144-усеченный сайт расщепления
тромбина в a2-Fc)
<400> 175
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Ala Pro Thr Ser Glu Ser Ala
1235 1240 1245
Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser
1250 1255 1260
Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1265 1270 1275
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1280 1285 1290
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro
1295 1300 1305
Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser
1310 1315 1320
Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
1325 1330 1335
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
1340 1345 1350
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser
1355 1360 1365
Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser
1370 1375 1380
Thr Glu Glu Gly Ala Ser Ser Gly Gly Gly Gly Ser Gly Gly Gly
1385 1390 1395
Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly
1400 1405 1410
Gly Ser Gly Gly Gly Gly Ser Ile Glu Pro Arg Ser Phe Ser Gly
1415 1420 1425
Ser Gly Gly Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro
1430 1435 1440
Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
1445 1450 1455
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
1460 1465 1470
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
1475 1480 1485
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg
1490 1495 1500
Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr
1505 1510 1515
Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys
1520 1525 1530
Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser
1535 1540 1545
Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
1550 1555 1560
Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu Thr Cys
1565 1570 1575
Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
1580 1585 1590
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
1595 1600 1605
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
1610 1615 1620
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
1625 1630 1635
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser
1640 1645 1650
Leu Ser Pro Gly Lys
1655
<210> 176
<211> 25
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 176
Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn
1 5 10 15
Asn Ala Ile Glu Pro Arg Ser Phe Ser
20 25
<210> 177
<211> 24
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 177
Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn
1 5 10 15
Ala Ile Glu Pro Arg Ser Phe Ser
20
<210> 178
<211> 23
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 178
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
1 5 10 15
Ile Glu Pro Arg Ser Phe Ser
20
<210> 179
<211> 22
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 179
Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile
1 5 10 15
Glu Pro Arg Ser Phe Ser
20
<210> 180
<211> 21
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 180
Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu
1 5 10 15
Pro Arg Ser Phe Ser
20
<210> 181
<211> 20
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 181
Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro
1 5 10 15
Arg Ser Phe Ser
20
<210> 182
<211> 19
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 182
Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg
1 5 10 15
Ser Phe Ser
<210> 183
<211> 18
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 183
Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser
1 5 10 15
Phe Ser
<210> 184
<211> 17
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 184
Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe
1 5 10 15
Ser
<210> 185
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 185
Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10 15
<210> 186
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 186
Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10 15
<210> 187
<211> 14
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 187
Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10
<210> 188
<211> 13
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 188
Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10
<210> 189
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 189
Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10
<210> 190
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 190
Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10
<210> 191
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 191
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5 10
<210> 192
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 192
Asn Ala Ile Glu Pro Arg Ser Phe Ser
1 5
<210> 193
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 193
Ala Ile Glu Pro Arg Ser Phe Ser
1 5
<210> 194
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 194
Ile Glu Pro Arg Ser Phe Ser
1 5
<210> 195
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 195
Glu Pro Arg Ser Phe Ser
1 5
<210> 196
<211> 4959
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN VWF059 (D'D3 VWF-Fc с сайтом расщепления тромбином в кислой
области 2 (а2) в линкере)
<400> 196
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatcgggcg cgccaacatc agagagcgcc acccctgaaa gtggtcccgg gagcgagcca 3780
gccacatctg ggtcggaaac gccaggcaca agtgagtctg caactcccga gtccggacct 3840
ggctccgagc ctgccactag cggctccgag actccgggaa cttccgagag cgctacacca 3900
gaaagcggac ccggaaccag taccgaacct agcgagggct ctgctccggg cagcccagcc 3960
ggctctccta catccacgga ggagggcact tccgaatccg ccaccccgga gtcagggcca 4020
ggatctgaac ccgctacctc aggcagtgag acgccaggaa cgagcgagtc cgctacaccg 4080
gagagtgggc cagggagccc tgctggatct cctacgtcca ctgaggaagg gtcaccagcg 4140
ggctcgccca ccagcactga agaaggtgcc tcgatatctg acaagaacac tggtgattat 4200
tacgaggaca gttatgaaga tatttcagca tacttgctga gtaaaaacaa tgccattgaa 4260
ccaagaagct tctctgacaa aactcacaca tgcccaccgt gcccagctcc agaactcctg 4320
ggcggaccgt cagtcttcct cttcccccca aaacccaagg acaccctcat gatctcccgg 4380
acccctgagg tcacatgcgt ggtggtggac gtgagccacg aagaccctga ggtcaagttc 4440
aactggtacg tggacggcgt ggaggtgcat aatgccaaga caaagccgcg ggaggagcag 4500
tacaacagca cgtaccgtgt ggtcagcgtc ctcaccgtcc tgcaccagga ctggctgaat 4560
ggcaaggagt acaagtgcaa ggtctccaac aaagccctcc cagcccccat cgagaaaacc 4620
atctccaaag ccaaagggca gccccgagaa ccacaggtgt acaccctgcc cccatcccgg 4680
gatgagctga ccaagaacca ggtcagcctg acctgcctgg tcaaaggctt ctatcccagc 4740
gacatcgccg tggagtggga gagcaatggg cagccggaga acaactacaa gaccacgcct 4800
cccgtgttgg actccgacgg ctccttcttc ctctacagca agctcaccgt ggacaagagc 4860
aggtggcagc aggggaacgt cttctcatgc tccgtgatgc atgaggctct gcacaaccac 4920
tacacgcaga agagcctctc cctgtctccg ggtaaatga 4959
<210> 197
<211> 1652
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF059 (D'D3 VWF-Fc с сайтом расщепления тромбином LVPR в
линкере)
<400> 197
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Ala Pro Thr Ser Glu
1235 1240 1245
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
1250 1255 1260
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1265 1270 1275
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1280 1285 1290
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
1295 1300 1305
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro
1310 1315 1320
Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1325 1330 1335
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1340 1345 1350
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala
1355 1360 1365
Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro
1370 1375 1380
Thr Ser Thr Glu Glu Gly Ala Ser Ile Ser Asp Lys Asn Thr Gly
1385 1390 1395
Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu
1400 1405 1410
Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Asp Lys Thr
1415 1420 1425
His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
1430 1435 1440
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
1445 1450 1455
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
1460 1465 1470
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
1475 1480 1485
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser
1490 1495 1500
Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp
1505 1510 1515
Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu
1520 1525 1530
Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
1535 1540 1545
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu
1550 1555 1560
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
1565 1570 1575
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu
1580 1585 1590
Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser
1595 1600 1605
Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln
1610 1615 1620
Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His
1625 1630 1635
Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1640 1645 1650
<210> 198
<211> 4860
<212> ДНК
<213> Искусственная последовательность
<220>
<223> pSYN VWF062 (D'D3 VWF-Fc без сайта расщепления тромбином в линкере)
<400> 198
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
atatcgggcg cgccaacatc agagagcgcc acccctgaaa gtggtcccgg gagcgagcca 3780
gccacatctg ggtcggaaac gccaggcaca agtgagtctg caactcccga gtccggacct 3840
ggctccgagc ctgccactag cggctccgag actccgggaa cttccgagag cgctacacca 3900
gaaagcggac ccggaaccag taccgaacct agcgagggct ctgctccggg cagcccagcc 3960
ggctctccta catccacgga ggagggcact tccgaatccg ccaccccgga gtcagggcca 4020
ggatctgaac ccgctacctc aggcagtgag acgccaggaa cgagcgagtc cgctacaccg 4080
gagagtgggc cagggagccc tgctggatct cctacgtcca ctgaggaagg gtcaccagcg 4140
ggctcgccca ccagcactga agaaggtgcc tcgagcgaca aaactcacac atgcccaccg 4200
tgcccagctc cagaactcct gggcggaccg tcagtcttcc tcttcccccc aaaacccaag 4260
gacaccctca tgatctcccg gacccctgag gtcacatgcg tggtggtgga cgtgagccac 4320
gaagaccctg aggtcaagtt caactggtac gtggacggcg tggaggtgca taatgccaag 4380
acaaagccgc gggaggagca gtacaacagc acgtaccgtg tggtcagcgt cctcaccgtc 4440
ctgcaccagg actggctgaa tggcaaggag tacaagtgca aggtctccaa caaagccctc 4500
ccagccccca tcgagaaaac catctccaaa gccaaagggc agccccgaga accacaggtg 4560
tacaccctgc ccccatcccg ggatgagctg accaagaacc aggtcagcct gacctgcctg 4620
gtcaaaggct tctatcccag cgacatcgcc gtggagtggg agagcaatgg gcagccggag 4680
aacaactaca agaccacgcc tcccgtgttg gactccgacg gctccttctt cctctacagc 4740
aagctcaccg tggacaagag caggtggcag caggggaacg tcttctcatg ctccgtgatg 4800
catgaggctc tgcacaacca ctacacgcag aagagcctct ccctgtctcc gggtaaatga 4860
<210> 199
<211> 1619
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> pSYN VWF062 (D'D3 VWF-Fc без сайта расщепления тромбином в линкере)
<400> 199
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Ile Ser Gly Ala Pro Thr Ser Glu
1235 1240 1245
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
1250 1255 1260
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1265 1270 1275
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1280 1285 1290
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
1295 1300 1305
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro
1310 1315 1320
Thr Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1325 1330 1335
Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly
1340 1345 1350
Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala
1355 1360 1365
Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro
1370 1375 1380
Thr Ser Thr Glu Glu Gly Ala Ser Ser Asp Lys Thr His Thr Cys
1385 1390 1395
Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe
1400 1405 1410
Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
1415 1420 1425
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro
1430 1435 1440
Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
1445 1450 1455
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
1460 1465 1470
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
1475 1480 1485
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
1490 1495 1500
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro
1505 1510 1515
Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn
1520 1525 1530
Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
1535 1540 1545
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
1550 1555 1560
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
1565 1570 1575
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
1580 1585 1590
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr
1595 1600 1605
Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
1610 1615
<210> 200
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Расщепляемый линкер
<400> 200
Ile Glu Pro Arg
1
<210> 201
<211> 1945
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII(XTEN)-Fc
<400> 201
Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr Met
1 5 10 15
Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro Arg
20 25 30
Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys Thr
35 40 45
Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro Arg
50 55 60
Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val Tyr
65 70 75 80
Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val Ser
85 90 95
Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala Glu
100 105 110
Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val Phe
115 120 125
Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn Gly
130 135 140
Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser His
145 150 155 160
Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu Leu
165 170 175
Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu His
180 185 190
Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp His
195 200 205
Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser Ala
210 215 220
Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg Ser
225 230 235 240
Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His Val
245 250 255
Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu Gly
260 265 270
His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile Ser
275 280 285
Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly Gln
290 295 300
Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met Glu
305 310 315 320
Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg Met
325 330 335
Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp Ser
340 345 350
Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe Ile
355 360 365
Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His Tyr
370 375 380
Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu Ala
385 390 395 400
Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro Gln
405 410 415
Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr Asp
420 425 430
Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile Leu
435 440 445
Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile Phe
450 455 460
Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile Thr
465 470 475 480
Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys His
485 490 495
Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys Trp
500 505 510
Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys Leu
515 520 525
Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala Ser
530 535 540
Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp Gln
545 550 555 560
Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe Ser
565 570 575
Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln Arg
580 585 590
Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe Gln
595 600 605
Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser Leu
610 615 620
Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu Ser
625 630 635 640
Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr Thr
645 650 655
Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro Phe
660 665 670
Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp Ile
675 680 685
Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala Leu
690 695 700
Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp
705 710 715 720
Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile
725 730 735
Glu Pro Arg Ser Phe Ser Gln Asn Gly Thr Ser Glu Ser Ala Thr Pro
740 745 750
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
755 760 765
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
770 775 780
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
785 790 795 800
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
805 810 815
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Glu
820 825 830
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
835 840 845
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
850 855 860
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro Ala
865 870 875 880
Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu
885 890 895
Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
900 905 910
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu
915 920 925
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
930 935 940
Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
945 950 955 960
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr
965 970 975
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser Glu
980 985 990
Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
995 1000 1005
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
1010 1015 1020
Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile Thr
1025 1030 1035
Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp Asp
1040 1045 1050
Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr Asp
1055 1060 1065
Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg
1070 1075 1080
His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met
1085 1090 1095
Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser
1100 1105 1110
Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly
1115 1120 1125
Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu
1130 1135 1140
Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile
1145 1150 1155
Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr
1160 1165 1170
Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala Glu
1175 1180 1185
Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr Phe
1190 1195 1200
Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe Asp
1205 1210 1215
Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys Asp
1220 1225 1230
Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr Asn
1235 1240 1245
Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu Phe
1250 1255 1260
Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr Phe
1265 1270 1275
Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile Gln
1280 1285 1290
Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala Ile
1295 1300 1305
Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln
1310 1315 1320
Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu
1325 1330 1335
Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg
1340 1345 1350
Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly
1355 1360 1365
Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp
1370 1375 1380
Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met Ser
1385 1390 1395
Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly
1400 1405 1410
Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser Gly
1415 1420 1425
Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr Ser
1430 1435 1440
Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp Ile
1445 1450 1455
Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys Thr
1460 1465 1470
Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln Phe
1475 1480 1485
Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr Arg
1490 1495 1500
Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val Asp
1505 1510 1515
Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile Ala
1520 1525 1530
Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser Thr
1535 1540 1545
Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met
1550 1555 1560
Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr
1565 1570 1575
Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser
1580 1585 1590
Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro
1595 1600 1605
Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys
1610 1615 1620
Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu
1625 1630 1635
Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser Gln
1640 1645 1650
Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val Lys
1655 1660 1665
Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn Ser
1670 1675 1680
Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro Gln
1685 1690 1695
Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly Cys
1700 1705 1710
Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro Cys
1715 1720 1725
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro
1730 1735 1740
Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val
1745 1750 1755
Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys
1760 1765 1770
Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr
1775 1780 1785
Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
1790 1795 1800
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr
1805 1810 1815
Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
1820 1825 1830
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
1835 1840 1845
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
1850 1855 1860
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
1865 1870 1875
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr
1880 1885 1890
Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys
1895 1900 1905
Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser
1910 1915 1920
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
1925 1930 1935
Ser Leu Ser Leu Ser Pro Gly
1940 1945
<210> 202
<211> 883
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> VWF(D'D3)-XTEN-линкер a2-Fc
<400> 202
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Ala
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Thr Ser
465 470 475 480
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
485 490 495
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
500 505 510
Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser
515 520 525
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser
530 535 540
Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
545 550 555 560
Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu
565 570 575
Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
580 585 590
Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
595 600 605
Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ala Ser
610 615 620
Ser Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile
625 630 635 640
Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala Ile Glu Pro Arg Ser Phe
645 650 655
Ser Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
660 665 670
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
675 680 685
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
690 695 700
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
705 710 715 720
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
725 730 735
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
740 745 750
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
755 760 765
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
770 775 780
Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
785 790 795 800
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
805 810 815
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
820 825 830
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
835 840 845
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
850 855 860
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
865 870 875 880
Ser Pro Gly
<210> 203
<211> 1965
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Конечная аминокислотная последовательность FVIII 312A
<400> 203
Met Gln Ile Glu Leu Ser Thr Cys Phe Phe Leu Cys Leu Leu Arg Phe
1 5 10 15
Cys Phe Ser Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser
20 25 30
Trp Asp Tyr Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg
35 40 45
Phe Pro Pro Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val
50 55 60
Tyr Lys Lys Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile
65 70 75 80
Ala Lys Pro Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln
85 90 95
Ala Glu Val Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser
100 105 110
His Pro Val Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser
115 120 125
Glu Gly Ala Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp
130 135 140
Asp Lys Val Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu
145 150 155 160
Lys Glu Asn Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser
165 170 175
Tyr Leu Ser His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile
180 185 190
Gly Ala Leu Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr
195 200 205
Gln Thr Leu His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly
210 215 220
Lys Ser Trp His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp
225 230 235 240
Ala Ala Ser Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr
245 250 255
Val Asn Arg Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val
260 265 270
Tyr Trp His Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile
275 280 285
Phe Leu Glu Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser
290 295 300
Leu Glu Ile Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met
305 310 315 320
Asp Leu Gly Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His
325 330 335
Asp Gly Met Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro
340 345 350
Gln Leu Arg Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp
355 360 365
Leu Thr Asp Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser
370 375 380
Pro Ser Phe Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr
385 390 395 400
Trp Val His Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro
405 410 415
Leu Val Leu Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn
420 425 430
Asn Gly Pro Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met
435 440 445
Ala Tyr Thr Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu
450 455 460
Ser Gly Ile Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu
465 470 475 480
Leu Ile Ile Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro
485 490 495
His Gly Ile Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys
500 505 510
Gly Val Lys His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe
515 520 525
Lys Tyr Lys Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp
530 535 540
Pro Arg Cys Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg
545 550 555 560
Asp Leu Ala Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu
565 570 575
Ser Val Asp Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val
580 585 590
Ile Leu Phe Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu
595 600 605
Asn Ile Gln Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp
610 615 620
Pro Glu Phe Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val
625 630 635 640
Phe Asp Ser Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp
645 650 655
Tyr Ile Leu Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe
660 665 670
Ser Gly Tyr Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr
675 680 685
Leu Phe Pro Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro
690 695 700
Gly Leu Trp Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly
705 710 715 720
Met Thr Ala Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp
725 730 735
Tyr Tyr Glu Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys
740 745 750
Asn Asn Ala Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Thr Ser Glu
755 760 765
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly
770 775 780
Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
785 790 795 800
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
805 810 815
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu
820 825 830
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
835 840 845
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
850 855 860
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
865 870 875 880
Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
885 890 895
Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr
900 905 910
Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro
915 920 925
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
930 935 940
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
945 950 955 960
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly
965 970 975
Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
980 985 990
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr
995 1000 1005
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser
1010 1015 1020
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser
1025 1030 1035
Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala
1040 1045 1050
Ser Ser Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu
1055 1060 1065
Ile Asp Tyr Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp
1070 1075 1080
Phe Asp Ile Tyr Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe
1085 1090 1095
Gln Lys Lys Thr Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu
1100 1105 1110
Trp Asp Tyr Gly Met Ser Ser Ser Pro His Val Leu Arg Asn Arg
1115 1120 1125
Ala Gln Ser Gly Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln
1130 1135 1140
Glu Phe Thr Asp Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu
1145 1150 1155
Leu Asn Glu His Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu
1160 1165 1170
Val Glu Asp Asn Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg
1175 1180 1185
Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln
1190 1195 1200
Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu
1205 1210 1215
Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met Ala Pro Thr
1220 1225 1230
Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val
1235 1240 1245
Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu
1250 1255 1260
Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val
1265 1270 1275
Thr Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr
1280 1285 1290
Lys Ser Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala
1295 1300 1305
Pro Cys Asn Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr
1310 1315 1320
Arg Phe His Ala Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly
1325 1330 1335
Leu Val Met Ala Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser
1340 1345 1350
Met Gly Ser Asn Glu Asn Ile His Ser Ile His Phe Ser Gly His
1355 1360 1365
Val Phe Thr Val Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr
1370 1375 1380
Asn Leu Tyr Pro Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser
1385 1390 1395
Lys Ala Gly Ile Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu
1400 1405 1410
His Ala Gly Met Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys
1415 1420 1425
Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln
1430 1435 1440
Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala
1445 1450 1455
Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu
1460 1465 1470
Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile
1475 1480 1485
His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu
1490 1495 1500
Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys
1505 1510 1515
Trp Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe
1520 1525 1530
Phe Gly Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn
1535 1540 1545
Pro Pro Ile Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr
1550 1555 1560
Ser Ile Arg Ser Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu
1565 1570 1575
Asn Ser Cys Ser Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser
1580 1585 1590
Asp Ala Gln Ile Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala
1595 1600 1605
Thr Trp Ser Pro Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser
1610 1615 1620
Asn Ala Trp Arg Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln
1625 1630 1635
Val Asp Phe Gln Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln
1640 1645 1650
Gly Val Lys Ser Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu
1655 1660 1665
Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln
1670 1675 1680
Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr
1685 1690 1695
Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu
1700 1705 1710
Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg Met
1715 1720 1725
Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His
1730 1735 1740
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser
1745 1750 1755
Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
1760 1765 1770
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
1775 1780 1785
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
1790 1795 1800
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
1805 1810 1815
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu
1820 1825 1830
Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro
1835 1840 1845
Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
1850 1855 1860
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
1865 1870 1875
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
1880 1885 1890
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
1895 1900 1905
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe
1910 1915 1920
Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln
1925 1930 1935
Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn
1940 1945 1950
His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
1955 1960 1965
<210> 204
<211> 5898
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Конечная нуклеотидная последовательность FVIII 312A (SEQ ID NO: 204)
<400> 204
atgcaaatag agctctccac ctgcttcttt ctgtgccttt tgcgattctg ctttagtgcc 60
accagaagat actacctggg tgcagtggaa ctgtcatggg actatatgca aagtgatctc 120
ggtgagctgc ctgtggacgc aagatttcct cctagagtgc caaaatcttt tccattcaac 180
acctcagtcg tgtacaaaaa gactctgttt gtagaattca cggatcacct tttcaacatc 240
gctaagccaa ggccaccctg gatgggtctg ctaggtccta ccatccaggc tgaggtttat 300
gatacagtgg tcattacact taagaacatg gcttcccatc ctgtcagtct tcatgctgtt 360
ggtgtatcct actggaaagc ttctgaggga gctgaatatg atgatcagac cagtcaaagg 420
gagaaagaag atgataaagt cttccctggt ggaagccata catatgtctg gcaggtcctg 480
aaagagaatg gtccaatggc ctctgaccca ctgtgcctta cctactcata tctttctcat 540
gtggacctgg taaaagactt gaattcaggc ctcattggag ccctactagt atgtagagaa 600
gggagtctgg ccaaggaaaa gacacagacc ttgcacaaat ttatactact ttttgctgta 660
tttgatgaag ggaaaagttg gcactcagaa acaaagaact ccttgatgca ggatagggat 720
gctgcatctg ctcgggcctg gcctaaaatg cacacagtca atggttatgt aaacaggtct 780
ctgccaggtc tgattggatg ccacaggaaa tcagtctatt ggcatgtgat tggaatgggc 840
accactcctg aagtgcactc aatattcctc gaaggtcaca catttcttgt gaggaaccat 900
cgccaggcgt ccttggaaat ctcgccaata actttcctta ctgctcaaac actcttgatg 960
gaccttggac agtttctact gttttgtcat atctcttccc accaacatga tggcatggaa 1020
gcttatgtca aagtagacag ctgtccagag gaaccccaac tacgaatgaa aaataatgaa 1080
gaagcggaag actatgatga tgatcttact gattctgaaa tggatgtggt caggtttgat 1140
gatgacaact ctccttcctt tatccaaatt cgctcagttg ccaagaagca tcctaaaact 1200
tgggtacatt acattgctgc tgaagaggag gactgggact atgctccctt agtcctcgcc 1260
cccgatgaca gaagttataa aagtcaatat ttgaacaatg gccctcagcg gattggtagg 1320
aagtacaaaa aagtccgatt tatggcatac acagatgaaa cctttaagac tcgtgaagct 1380
attcagcatg aatcaggaat cttgggacct ttactttatg gggaagttgg agacacactg 1440
ttgattatat ttaagaatca agcaagcaga ccatataaca tctaccctca cggaatcact 1500
gatgtccgtc ctttgtattc aaggagatta ccaaaaggtg taaaacattt gaaggatttt 1560
ccaattctgc caggagaaat attcaaatat aaatggacag tgactgtaga agatgggcca 1620
actaaatcag atcctcggtg cctgacccgc tattactcta gtttcgttaa tatggagaga 1680
gatctagctt caggactcat tggccctctc ctcatctgct acaaagaatc tgtagatcaa 1740
agaggaaacc agataatgtc agacaagagg aatgtcatcc tgttttctgt atttgatgag 1800
aaccgaagct ggtacctcac agagaatata caacgctttc tccccaatcc agctggagtg 1860
cagcttgagg atccagagtt ccaagcctcc aacatcatgc acagcatcaa tggctatgtt 1920
tttgatagtt tgcagttgtc agtttgtttg catgaggtgg catactggta cattctaagc 1980
attggagcac agactgactt cctttctgtc ttcttctctg gatatacctt caaacacaaa 2040
atggtctatg aagacacact caccctattc ccattctcag gagaaactgt cttcatgtcg 2100
atggaaaacc caggtctatg gattctgggg tgccacaact cagactttcg gaacagaggc 2160
atgaccgcct tactgaaggt ttctagttgt gacaagaaca ctggtgatta ttacgaggac 2220
agttatgaag atatttcagc atacttgctg agtaaaaaca atgccattga accaagaagc 2280
ttctctcaaa acggtacctc agagtctgct acccccgagt cagggccagg atcagagcca 2340
gccacctccg ggtctgagac acccgggact tccgagagtg ccacccctga gtccggaccc 2400
gggtccgagc ccgccacttc cggctccgaa actcccggca caagcgagag cgctacccca 2460
gagtcaggac caggaacatc tacagagccc tctgaaggct ccgctccagg gtccccagcc 2520
ggcagtccca ctagcaccga ggagggaacc tctgaaagcg ccacacccga atcagggcca 2580
gggtctgagc ctgctaccag cggcagcgag acaccaggca cctctgagtc cgccacacca 2640
gagtccggac ccggatctcc cgctgggagc cccacctcca ctgaggaggg atctcctgct 2700
ggctctccaa catctactga ggaaggaacc tcaaccgagc catccgaggg atcagctccc 2760
ggcacctcag agtcggcaac cccggagtct ggacccggaa cttccgaaag tgccacacca 2820
gagtccggtc ccgggacttc agaatcagca acacccgagt ccggccctgg gtctgaaccc 2880
gccacaagtg gtagtgagac accaggatca gaacctgcta cctcagggtc agagacaccc 2940
ggatctccgg caggctcacc aacctccact gaggagggca ccagcacaga accaagcgag 3000
ggctccgcac ccggaacaag cactgaaccc agtgagggtt cagcacccgg ctctgagccg 3060
gccacaagtg gcagtgagac acccggcact tcagagagtg ccacccccga gagtggccca 3120
ggcactagta ccgagccctc tgaaggcagt gcgccagcct cgagcgaaat aactcgtact 3180
actcttcagt cagatcaaga ggaaattgac tatgatgata ccatatcagt tgaaatgaag 3240
aaggaagatt ttgacattta tgatgaggat gaaaatcaga gcccccgcag ctttcaaaag 3300
aaaacacgac actattttat tgctgcagtg gagaggctct gggattatgg gatgagtagc 3360
tccccacatg ttctaagaaa cagggctcag agtggcagtg tccctcagtt caagaaagtt 3420
gttttccagg aatttactga tggctccttt actcagccct tataccgtgg agaactaaat 3480
gaacatttgg gactcctggg gccatatata agagcagaag ttgaagataa tatcatggta 3540
actttcagaa atcaggcctc tcgtccctat tccttctatt ctagccttat ttcttatgag 3600
gaagatcaga ggcaaggagc agaacctaga aaaaactttg tcaagcctaa tgaaaccaaa 3660
acttactttt ggaaagtgca acatcatatg gcacccacta aagatgagtt tgactgcaaa 3720
gcctgggctt atttctctga tgttgacctg gaaaaagatg tgcactcagg cctgattgga 3780
ccccttctgg tctgccacac taacacactg aaccctgctc atgggagaca agtgacagta 3840
caggaatttg ctctgttttt caccatcttt gatgagacca aaagctggta cttcactgaa 3900
aatatggaaa gaaactgcag ggctccctgc aatatccaga tggaagatcc cacttttaaa 3960
gagaattatc gcttccatgc aatcaatggc tacataatgg atacactacc tggcttagta 4020
atggctcagg atcaaaggat tcgatggtat ctgctcagca tgggcagcaa tgaaaacatc 4080
cattctattc atttcagtgg acatgtgttc actgtacgaa aaaaagagga gtataaaatg 4140
gcactgtaca atctctatcc aggtgttttt gagacagtgg aaatgttacc atccaaagct 4200
ggaatttggc gggtggaatg ccttattggc gagcatctac atgctgggat gagcacactt 4260
tttctggtgt acagcaataa gtgtcagact cccctgggaa tggcttctgg acacattaga 4320
gattttcaga ttacagcttc aggacaatat ggacagtggg ccccaaagct ggccagactt 4380
cattattccg gatcaatcaa tgcctggagc accaaggagc ccttttcttg gatcaaggtg 4440
gatctgttgg caccaatgat tattcacggc atcaagaccc agggtgcccg tcagaagttc 4500
tccagcctct acatctctca gtttatcatc atgtatagtc ttgatgggaa gaagtggcag 4560
acttatcgag gaaattccac tggaacctta atggtcttct ttggcaatgt ggattcatct 4620
gggataaaac acaatatttt taaccctcca attattgctc gatacatccg tttgcaccca 4680
actcattata gcattcgcag cactcttcgc atggagttga tgggctgtga tttaaatagt 4740
tgcagcatgc cattgggaat ggagagtaaa gcaatatcag atgcacagat tactgcttca 4800
tcctacttta ccaatatgtt tgccacctgg tctccttcaa aagctcgact tcacctccaa 4860
gggaggagta atgcctggag acctcaggtg aataatccaa aagagtggct gcaagtggac 4920
ttccagaaga caatgaaagt cacaggagta actactcagg gagtaaaatc tctgcttacc 4980
agcatgtatg tgaaggagtt cctcatctcc agcagtcaag atggccatca gtggactctc 5040
ttttttcaga atggcaaagt aaaggttttt cagggaaatc aagactcctt cacacctgtg 5100
gtgaactctc tagacccacc gttactgact cgctaccttc gaattcaccc ccagagttgg 5160
gtgcaccaga ttgccctgag gatggaggtt ctgggctgcg aggcacagga cctctacgac 5220
aaaactcaca catgcccacc gtgcccagct ccagaactcc tgggcggacc gtcagtcttc 5280
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc 5340
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta tgtggacggc 5400
gtggaagtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt 5460
gtggtcagcg tcctcaccgt cctgcaccaa gactggctga atggcaagga gtacaagtgc 5520
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg 5580
cagccccgag aaccacaggt gtacaccctg cccccatccc gggatgagct gaccaagaac 5640
caagttagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg 5700
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgtt ggactccgac 5760
ggctccttct tcctctactc caagctcacc gtggacaaga gcaggtggca gcaggggaac 5820
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc 5880
tccctgtctc cgggttga 5898
<210> 205
<211> 1646
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Конечная аминокислотная последовательность полипептида VWF059A
<400> 205
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Ala Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Ala Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Thr Ser Glu Ser Ala Thr Pro
1235 1240 1245
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
1250 1255 1260
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1265 1270 1275
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
1280 1285 1290
Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu
1295 1300 1305
Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
1310 1315 1320
Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser
1325 1330 1335
Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser
1340 1345 1350
Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr
1355 1360 1365
Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu
1370 1375 1380
Glu Gly Ala Ser Ser Asp Lys Asn Thr Gly Asp Tyr Tyr Glu Asp
1385 1390 1395
Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
1400 1405 1410
Ile Glu Pro Arg Ser Phe Ser Asp Lys Thr His Thr Cys Pro Pro
1415 1420 1425
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
1430 1435 1440
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
1445 1450 1455
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
1460 1465 1470
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
1475 1480 1485
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
1490 1495 1500
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
1505 1510 1515
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
1520 1525 1530
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
1535 1540 1545
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
1550 1555 1560
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
1565 1570 1575
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
1580 1585 1590
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
1595 1600 1605
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
1610 1615 1620
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
1625 1630 1635
Lys Ser Leu Ser Leu Ser Pro Gly
1640 1645
<210> 206
<211> 4941
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Конечная нуклеотидная последовательность VWF059A
<400> 206
atgattcctg ccagatttgc cggggtgctg cttgctctgg ccctcatttt gccagggacc 60
ctttgtgcag aaggaactcg cggcaggtca tccacggccc gatgcagcct tttcggaagt 120
gacttcgtca acacctttga tgggagcatg tacagctttg cgggatactg cagttacctc 180
ctggcagggg gctgccagaa acgctccttc tcgattattg gggacttcca gaatggcaag 240
agagtgagcc tctccgtgta tcttggggaa ttttttgaca tccatttgtt tgtcaatggt 300
accgtgacac agggggacca aagagtctcc atgccctatg cctccaaagg gctgtatcta 360
gaaactgagg ctgggtacta caagctgtcc ggtgaggcct atggctttgt ggccaggatc 420
gatggcagcg gcaactttca agtcctgctg tcagacagat acttcaacaa gacctgcggg 480
ctgtgtggca actttaacat ctttgctgaa gatgacttta tgacccaaga agggaccttg 540
acctcggacc cttatgactt tgccaactca tgggctctga gcagtggaga acagtggtgt 600
gaacgggcat ctcctcccag cagctcatgc aacatctcct ctggggaaat gcagaagggc 660
ctgtgggagc agtgccagct tctgaagagc acctcggtgt ttgcccgctg ccaccctctg 720
gtggaccccg agccttttgt ggccctgtgt gagaagactt tgtgtgagtg tgctgggggg 780
ctggagtgcg cctgccctgc cctcctggag tacgcccgga cctgtgccca ggagggaatg 840
gtgctgtacg gctggaccga ccacagcgcg tgcagcccag tgtgccctgc tggtatggag 900
tataggcagt gtgtgtcccc ttgcgccagg acctgccaga gcctgcacat caatgaaatg 960
tgtcaggagc gatgcgtgga tggctgcagc tgccctgagg gacagctcct ggatgaaggc 1020
ctctgcgtgg agagcaccga gtgtccctgc gtgcattccg gaaagcgcta ccctcccggc 1080
acctccctct ctcgagactg caacacctgc atttgccgaa acagccagtg gatctgcagc 1140
aatgaagaat gtccagggga gtgccttgtc actggtcaat cccacttcaa gagctttgac 1200
aacagatact tcaccttcag tgggatctgc cagtacctgc tggcccggga ttgccaggac 1260
cactccttct ccattgtcat tgagactgtc cagtgtgctg atgaccgcga cgctgtgtgc 1320
acccgctccg tcaccgtccg gctgcctggc ctgcacaaca gccttgtgaa actgaagcat 1380
ggggcaggag ttgccatgga tggccaggac atccagctcc ccctcctgaa aggtgacctc 1440
cgcatccagc atacagtgac ggcctccgtg cgcctcagct acggggagga cctgcagatg 1500
gactgggatg gccgcgggag gctgctggtg aagctgtccc ccgtctatgc cgggaagacc 1560
tgcggcctgt gtgggaatta caatggcaac cagggcgacg acttccttac cccctctggg 1620
ctggcggagc cccgggtgga ggacttcggg aacgcctgga agctgcacgg ggactgccag 1680
gacctgcaga agcagcacag cgatccctgc gccctcaacc cgcgcatgac caggttctcc 1740
gaggaggcgt gcgcggtcct gacgtccccc acattcgagg cctgccatcg tgccgtcagc 1800
ccgctgccct acctgcggaa ctgccgctac gacgtgtgct cctgctcgga cggccgcgag 1860
tgcctgtgcg gcgccctggc cagctatgcc gcggcctgcg cggggagagg cgtgcgcgtc 1920
gcgtggcgcg agccaggccg ctgtgagctg aactgcccga aaggccaggt gtacctgcag 1980
tgcgggaccc cctgcaacct gacctgccgc tctctctctt acccggatga ggaatgcaat 2040
gaggcctgcc tggagggctg cttctgcccc ccagggctct acatggatga gaggggggac 2100
tgcgtgccca aggcccagtg cccctgttac tatgacggtg agatcttcca gccagaagac 2160
atcttctcag accatcacac catgtgctac tgtgaggatg gcttcatgca ctgtaccatg 2220
agtggagtcc ccggaagctt gctgcctgac gctgtcctca gcagtcccct gtctcatcgc 2280
agcaaaagga gcctatcctg tcggcccccc atggtcaagc tggtgtgtcc cgctgacaac 2340
ctgcgggctg aagggctcga gtgtaccaaa acgtgccaga actatgacct ggagtgcatg 2400
agcatgggct gtgtctctgg ctgcctctgc cccccgggca tggtccggca tgagaacaga 2460
tgtgtggccc tggaaaggtg tccctgcttc catcagggca aggagtatgc ccctggagaa 2520
acagtgaaga ttggctgcaa cacttgtgtc tgtcgggacc ggaagtggaa ctgcacagac 2580
catgtgtgtg atgccacgtg ctccacgatc ggcatggccc actacctcac cttcgacggg 2640
ctcaaatacc tgttccccgg ggagtgccag tacgttctgg tgcaggatta ctgcggcagt 2700
aaccctggga cctttcggat cctagtgggg aataagggat gcagccaccc ctcagtgaaa 2760
tgcaagaaac gggtcaccat cctggtggag ggaggagaga ttgagctgtt tgacggggag 2820
gtgaatgtga agaggcccat gaaggatgag actcactttg aggtggtgga gtctggccgg 2880
tacatcattc tgctgctggg caaagccctc tccgtggtct gggaccgcca cctgagcatc 2940
tccgtggtcc tgaagcagac ataccaggag aaagtgtgtg gcctgtgtgg gaattttgat 3000
ggcatccaga acaatgacct caccagcagc aacctccaag tggaggaaga ccctgtggac 3060
tttgggaact cctggaaagt gagctcgcag tgtgctgaca ccagaaaagt gcctctggac 3120
tcatcccctg ccacctgcca taacaacatc atgaagcaga cgatggtgga ttcctcctgt 3180
agaatcctta ccagtgacgt cttccaggac tgcaacaagc tggtggaccc cgagccatat 3240
ctggatgtct gcatttacga cacctgctcc tgtgagtcca ttggggactg cgccgcattc 3300
tgcgacacca ttgctgccta tgcccacgtg tgtgcccagc atggcaaggt ggtgacctgg 3360
aggacggcca cattgtgccc ccagagctgc gaggagagga atctccggga gaacgggtat 3420
gaggctgagt ggcgctataa cagctgtgca cctgcctgtc aagtcacgtg tcagcaccct 3480
gagccactgg cctgccctgt gcagtgtgtg gagggctgcc atgcccactg ccctccaggg 3540
aaaatcctgg atgagctttt gcagacctgc gttgaccctg aagactgtcc agtgtgtgag 3600
gtggctggcc ggcgttttgc ctcaggaaag aaagtcacct tgaatcccag tgaccctgag 3660
cactgccaga tttgccactg tgatgttgtc aacctcacct gtgaagcctg ccaggagccg 3720
ggtacatcag agagcgccac ccctgaaagt ggtcccggga gcgagccagc cacatctggg 3780
tcggaaacgc caggcacatc cgagtctgca actcccgagt ccggacctgg ctccgagcct 3840
gccactagcg gctccgagac tccgggaact tccgagagcg ctacaccaga aagcggaccc 3900
ggaaccagta ccgaacctag cgagggctct gctccgggca gcccagccgg ctctcctaca 3960
tccacggagg agggcacttc cgaatccgcc accccggagt cagggccagg atctgaaccc 4020
gctacctcag gcagtgagac gccaggaacg agcgagtccg ctacaccgga gagtgggcca 4080
gggagccctg ctggatctcc tacgtccact gaggaagggt caccagcggg ctcgcccacc 4140
agcactgaag aaggtgcctc gtctgacaag aacactggtg attattacga ggacagttat 4200
gaagatattt cagcatactt gctgagtaaa aacaatgcca ttgaaccaag aagcttctct 4260
gacaaaactc acacatgccc accgtgccca gctccagaac tcctgggcgg accgtcagtc 4320
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca 4380
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtatgtggac 4440
ggcgtggaag tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac 4500
cgtgtggtca gcgtcctcac cgtcctgcac caagactggc tgaatggcaa ggagtacaag 4560
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa 4620
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgggatga gctgaccaag 4680
aaccaagtta gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag 4740
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gttggactcc 4800
gacggctcct tcttcctcta ctccaagctc accgtggaca agagcaggtg gcagcagggg 4860
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc 4920
ctctccctgt ctccgggttg a 4941
<210> 207
<211> 1946
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII(XTEN)-Fc с лидерной последовательностью A
<400> 207
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn Gly Thr Ser Glu Ser Ala Thr
740 745 750
Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
755 760 765
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu
770 775 780
Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr
785 790 795 800
Pro Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala
805 810 815
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
820 825 830
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
835 840 845
Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
850 855 860
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Ser Pro
865 870 875 880
Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser
885 890 895
Glu Gly Ser Ala Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly
900 905 910
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser
915 920 925
Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser
930 935 940
Gly Ser Glu Thr Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
945 950 955 960
Pro Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu Gly Thr Ser
965 970 975
Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Thr Glu Pro Ser
980 985 990
Glu Gly Ser Ala Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr
995 1000 1005
Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr
1010 1015 1020
Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Ala Ser Ser Glu Ile
1025 1030 1035
Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr Asp
1040 1045 1050
Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr
1055 1060 1065
Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr
1070 1075 1080
Arg His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly
1085 1090 1095
Met Ser Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly
1100 1105 1110
Ser Val Pro Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp
1115 1120 1125
Gly Ser Phe Thr Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His
1130 1135 1140
Leu Gly Leu Leu Gly Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn
1145 1150 1155
Ile Met Val Thr Phe Arg Asn Gln Ala Ser Arg Pro Tyr Ser Phe
1160 1165 1170
Tyr Ser Ser Leu Ile Ser Tyr Glu Glu Asp Gln Arg Gln Gly Ala
1175 1180 1185
Glu Pro Arg Lys Asn Phe Val Lys Pro Asn Glu Thr Lys Thr Tyr
1190 1195 1200
Phe Trp Lys Val Gln His His Met Ala Pro Thr Lys Asp Glu Phe
1205 1210 1215
Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp Val Asp Leu Glu Lys
1220 1225 1230
Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu Val Cys His Thr
1235 1240 1245
Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr Val Gln Glu
1250 1255 1260
Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser Trp Tyr
1265 1270 1275
Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn Ile
1280 1285 1290
Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala
1295 1300 1305
Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala
1310 1315 1320
Gln Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn
1325 1330 1335
Glu Asn Ile His Ser Ile His Phe Ser Gly His Val Phe Thr Val
1340 1345 1350
Arg Lys Lys Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro
1355 1360 1365
Gly Val Phe Glu Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile
1370 1375 1380
Trp Arg Val Glu Cys Leu Ile Gly Glu His Leu His Ala Gly Met
1385 1390 1395
Ser Thr Leu Phe Leu Val Tyr Ser Asn Lys Cys Gln Thr Pro Leu
1400 1405 1410
Gly Met Ala Ser Gly His Ile Arg Asp Phe Gln Ile Thr Ala Ser
1415 1420 1425
Gly Gln Tyr Gly Gln Trp Ala Pro Lys Leu Ala Arg Leu His Tyr
1430 1435 1440
Ser Gly Ser Ile Asn Ala Trp Ser Thr Lys Glu Pro Phe Ser Trp
1445 1450 1455
Ile Lys Val Asp Leu Leu Ala Pro Met Ile Ile His Gly Ile Lys
1460 1465 1470
Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu Tyr Ile Ser Gln
1475 1480 1485
Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp Gln Thr Tyr
1490 1495 1500
Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly Asn Val
1505 1510 1515
Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile Ile
1520 1525 1530
Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser
1535 1540 1545
Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser
1550 1555 1560
Met Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile
1565 1570 1575
Thr Ala Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro
1580 1585 1590
Ser Lys Ala Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg
1595 1600 1605
Pro Gln Val Asn Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln
1610 1615 1620
Lys Thr Met Lys Val Thr Gly Val Thr Thr Gln Gly Val Lys Ser
1625 1630 1635
Leu Leu Thr Ser Met Tyr Val Lys Glu Phe Leu Ile Ser Ser Ser
1640 1645 1650
Gln Asp Gly His Gln Trp Thr Leu Phe Phe Gln Asn Gly Lys Val
1655 1660 1665
Lys Val Phe Gln Gly Asn Gln Asp Ser Phe Thr Pro Val Val Asn
1670 1675 1680
Ser Leu Asp Pro Pro Leu Leu Thr Arg Tyr Leu Arg Ile His Pro
1685 1690 1695
Gln Ser Trp Val His Gln Ile Ala Leu Arg Met Glu Val Leu Gly
1700 1705 1710
Cys Glu Ala Gln Asp Leu Tyr Asp Lys Thr His Thr Cys Pro Pro
1715 1720 1725
Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe
1730 1735 1740
Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu
1745 1750 1755
Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu Val
1760 1765 1770
Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
1775 1780 1785
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val
1790 1795 1800
Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
1805 1810 1815
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
1820 1825 1830
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
1835 1840 1845
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val
1850 1855 1860
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala
1865 1870 1875
Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr
1880 1885 1890
Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
1895 1900 1905
Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
1910 1915 1920
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
1925 1930 1935
Lys Ser Leu Ser Leu Ser Pro Gly
1940 1945
<210> 208
<211> 22
<212> БЕЛОК
<213> Homo sapiens
<400> 208
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys
20
<210> 209
<211> 741
<212> БЕЛОК
<213> Homo sapiens
<400> 209
Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr Ala Arg Cys Ser Leu Phe
1 5 10 15
Gly Ser Asp Phe Val Asn Thr Phe Asp Gly Ser Met Tyr Ser Phe Ala
20 25 30
Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly Cys Gln Lys Arg Ser Phe
35 40 45
Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys Arg Val Ser Leu Ser Val
50 55 60
Tyr Leu Gly Glu Phe Phe Asp Ile His Leu Phe Val Asn Gly Thr Val
65 70 75 80
Thr Gln Gly Asp Gln Arg Val Ser Met Pro Tyr Ala Ser Lys Gly Leu
85 90 95
Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys Leu Ser Gly Glu Ala Tyr
100 105 110
Gly Phe Val Ala Arg Ile Asp Gly Ser Gly Asn Phe Gln Val Leu Leu
115 120 125
Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly Leu Cys Gly Asn Phe Asn
130 135 140
Ile Phe Ala Glu Asp Asp Phe Met Thr Gln Glu Gly Thr Leu Thr Ser
145 150 155 160
Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala Leu Ser Ser Gly Glu Gln
165 170 175
Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser Ser Cys Asn Ile Ser Ser
180 185 190
Gly Glu Met Gln Lys Gly Leu Trp Glu Gln Cys Gln Leu Leu Lys Ser
195 200 205
Thr Ser Val Phe Ala Arg Cys His Pro Leu Val Asp Pro Glu Pro Phe
210 215 220
Val Ala Leu Cys Glu Lys Thr Leu Cys Glu Cys Ala Gly Gly Leu Glu
225 230 235 240
Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala Arg Thr Cys Ala Gln Glu
245 250 255
Gly Met Val Leu Tyr Gly Trp Thr Asp His Ser Ala Cys Ser Pro Val
260 265 270
Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys Val Ser Pro Cys Ala Arg
275 280 285
Thr Cys Gln Ser Leu His Ile Asn Glu Met Cys Gln Glu Arg Cys Val
290 295 300
Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu Leu Asp Glu Gly Leu Cys
305 310 315 320
Val Glu Ser Thr Glu Cys Pro Cys Val His Ser Gly Lys Arg Tyr Pro
325 330 335
Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn Thr Cys Ile Cys Arg Asn
340 345 350
Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys Pro Gly Glu Cys Leu Val
355 360 365
Thr Gly Gln Ser His Phe Lys Ser Phe Asp Asn Arg Tyr Phe Thr Phe
370 375 380
Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg Asp Cys Gln Asp His Ser
385 390 395 400
Phe Ser Ile Val Ile Glu Thr Val Gln Cys Ala Asp Asp Arg Asp Ala
405 410 415
Val Cys Thr Arg Ser Val Thr Val Arg Leu Pro Gly Leu His Asn Ser
420 425 430
Leu Val Lys Leu Lys His Gly Ala Gly Val Ala Met Asp Gly Gln Asp
435 440 445
Ile Gln Leu Pro Leu Leu Lys Gly Asp Leu Arg Ile Gln His Thr Val
450 455 460
Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu Asp Leu Gln Met Asp Trp
465 470 475 480
Asp Gly Arg Gly Arg Leu Leu Val Lys Leu Ser Pro Val Tyr Ala Gly
485 490 495
Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn Gly Asn Gln Gly Asp Asp
500 505 510
Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro Arg Val Glu Asp Phe Gly
515 520 525
Asn Ala Trp Lys Leu His Gly Asp Cys Gln Asp Leu Gln Lys Gln His
530 535 540
Ser Asp Pro Cys Ala Leu Asn Pro Arg Met Thr Arg Phe Ser Glu Glu
545 550 555 560
Ala Cys Ala Val Leu Thr Ser Pro Thr Phe Glu Ala Cys His Arg Ala
565 570 575
Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys Arg Tyr Asp Val Cys Ser
580 585 590
Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly Ala Leu Ala Ser Tyr Ala
595 600 605
Ala Ala Cys Ala Gly Arg Gly Val Arg Val Ala Trp Arg Glu Pro Gly
610 615 620
Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln Val Tyr Leu Gln Cys Gly
625 630 635 640
Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu Ser Tyr Pro Asp Glu Glu
645 650 655
Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe Cys Pro Pro Gly Leu Tyr
660 665 670
Met Asp Glu Arg Gly Asp Cys Val Pro Lys Ala Gln Cys Pro Cys Tyr
675 680 685
Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp Ile Phe Ser Asp His His
690 695 700
Thr Met Cys Tyr Cys Glu Asp Gly Phe Met His Cys Thr Met Ser Gly
705 710 715 720
Val Pro Gly Ser Leu Leu Pro Asp Ala Val Leu Ser Ser Pro Leu Ser
725 730 735
His Arg Ser Lys Arg
740
<210> 210
<211> 103
<212> БЕЛОК
<213> Homo sapiens
<400> 210
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Arg Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr
100
<210> 211
<211> 374
<212> БЕЛОК
<213> Homo sapiens
<400> 211
Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly Leu Lys
1 5 10 15
Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp Tyr Cys
20 25 30
Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys Gly Cys
35 40 45
Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu Val Glu
50 55 60
Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys Arg Pro
65 70 75 80
Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg Tyr Ile
85 90 95
Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg His Leu
100 105 110
Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val Cys Gly
115 120 125
Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr Ser Ser
130 135 140
Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn Ser Trp Lys
145 150 155 160
Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro Leu Asp Ser Ser
165 170 175
Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln Thr Met Val Asp Ser
180 185 190
Ser Cys Arg Ile Leu Thr Ser Asp Val Phe Gln Asp Cys Asn Lys Leu
195 200 205
Val Asp Pro Glu Pro Tyr Leu Asp Val Cys Ile Tyr Asp Thr Cys Ser
210 215 220
Cys Glu Ser Ile Gly Asp Cys Ala Cys Phe Cys Asp Thr Ile Ala Ala
225 230 235 240
Tyr Ala His Val Cys Ala Gln His Gly Lys Val Val Thr Trp Arg Thr
245 250 255
Ala Thr Leu Cys Pro Gln Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn
260 265 270
Gly Tyr Glu Cys Glu Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln
275 280 285
Val Thr Cys Gln His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val
290 295 300
Glu Gly Cys His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu
305 310 315 320
Leu Gln Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala
325 330 335
Gly Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
340 345 350
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr Cys
355 360 365
Glu Ala Cys Gln Glu Pro
370
<210> 212
<211> 239
<212> БЕЛОК
<213> Homo sapiens
<400> 212
Gly Gly Leu Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr
1 5 10 15
Leu Tyr Val Glu Asp Ile Ser Glu Pro Pro Leu His Asp Phe Tyr Cys
20 25 30
Ser Arg Leu Leu Asp Leu Val Phe Leu Leu Asp Gly Ser Ser Arg Leu
35 40 45
Ser Glu Ala Glu Phe Glu Val Leu Lys Ala Phe Val Val Asp Met Met
50 55 60
Glu Arg Leu Arg Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu
65 70 75 80
Tyr His Asp Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg
85 90 95
Pro Ser Glu Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser
100 105 110
Gln Val Ala Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile
115 120 125
Phe Ser Lys Ile Asp Arg Pro Glu Ala Ser Arg Ile Ala Leu Leu Leu
130 135 140
Met Ala Ser Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val Arg Tyr
145 150 155 160
Val Gln Gly Leu Lys Lys Lys Lys Val Ile Val Ile Pro Val Gly Ile
165 170 175
Gly Pro His Ala Asn Leu Lys Gln Ile Arg Leu Ile Glu Lys Gln Ala
180 185 190
Pro Glu Asn Lys Ala Phe Val Leu Ser Ser Val Asp Glu Leu Glu Gln
195 200 205
Gln Arg Asp Glu Ile Val Ser Tyr Leu Cys Asp Leu Ala Pro Glu Ala
210 215 220
Pro Pro Pro Thr Leu Pro Pro Asp Met Ala Gln Val Thr Val Gly
225 230 235
<210> 213
<211> 1334
<212> БЕЛОК
<213> Homo sapiens
<220>
<221> другой_признак
<222> (537)..(537)
<223> Xaa может представлять собой любую втречающуюся в природе
аминокислоту
<400> 213
Pro Gly Leu Leu Gly Val Ser Thr Leu Gly Pro Lys Arg Asn Ser Met
1 5 10 15
Val Leu Asp Val Ala Phe Val Leu Glu Gly Ser Asp Lys Ile Gly Glu
20 25 30
Ala Asp Phe Asn Arg Ser Lys Glu Phe Met Glu Glu Val Ile Gln Arg
35 40 45
Met Asp Val Gly Gln Asp Ser Ile His Val Thr Val Leu Gln Tyr Ser
50 55 60
Tyr Met Val Thr Val Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly
65 70 75 80
Asp Ile Leu Gln Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg
85 90 95
Thr Asn Thr Gly Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu
100 105 110
Val Ser Gln Gly Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val
115 120 125
Thr Gly Asn Pro Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile
130 135 140
Gln Val Val Pro Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu Leu
145 150 155 160
Glu Arg Ile Gly Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp Phe Glu
165 170 175
Thr Leu Pro Arg Glu Ala Pro Asp Leu Val Leu Gln Arg Cys Cys Ser
180 185 190
Gly Glu Gly Leu Gln Ile Pro Thr Leu Ser Pro Ala Pro Asp Cys Ser
195 200 205
Gln Pro Leu Asp Val Ile Leu Leu Leu Asp Gly Ser Ser Ser Phe Pro
210 215 220
Ala Ser Tyr Phe Asp Glu Met Lys Ser Phe Ala Lys Ala Phe Ile Ser
225 230 235 240
Lys Ala Asn Ile Gly Pro Arg Leu Thr Gln Val Ser Val Leu Gln Tyr
245 250 255
Gly Ser Ile Thr Thr Ile Asp Val Pro Trp Asn Val Val Pro Glu Lys
260 265 270
Ala His Leu Leu Ser Leu Val Asp Val Met Gln Arg Glu Gly Gly Pro
275 280 285
Ser Gln Ile Gly Asp Ala Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser
290 295 300
Glu Met His Gly Ala Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu
305 310 315 320
Val Thr Asp Val Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala
325 330 335
Arg Ser Asn Arg Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr
340 345 350
Asp Ala Ala Gln Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser Asn
355 360 365
Val Val Lys Leu Gln Arg Ile Glu Asp Leu Pro Thr Met Val Thr Leu
370 375 380
Gly Asn Ser Phe Leu His Lys Leu Cys Ser Gly Phe Val Arg Ile Cys
385 390 395 400
Met Asp Glu Asp Gly Asn Glu Lys Arg Pro Gly Asp Val Trp Thr Leu
405 410 415
Pro Asp Gln Cys His Thr Val Thr Cys Gln Pro Asp Gly Gln Thr Leu
420 425 430
Leu Lys Ser His Arg Val Asn Cys Asp Arg Gly Leu Arg Pro Ser Cys
435 440 445
Pro Asn Ser Gln Ser Pro Val Lys Val Glu Glu Thr Cys Gly Cys Arg
450 455 460
Trp Thr Cys Pro Cys Val Cys Thr Gly Ser Ser Thr Arg His Ile Val
465 470 475 480
Thr Phe Asp Gly Gln Asn Phe Lys Leu Thr Gly Ser Cys Ser Tyr Val
485 490 495
Leu Phe Gln Asn Lys Glu Gln Asp Leu Glu Val Ile Leu His Asn Gly
500 505 510
Ala Cys Ser Pro Gly Ala Arg Gln Gly Cys Met Lys Ser Ile Glu Val
515 520 525
Lys His Ser Ala Leu Ser Val Glu Xaa His Ser Asp Met Glu Val Thr
530 535 540
Val Asn Gly Arg Leu Val Ser Val Pro Tyr Val Gly Gly Asn Met Glu
545 550 555 560
Val Asn Val Tyr Gly Ala Ile Met His Glu Val Arg Phe Asn His Leu
565 570 575
Gly His Ile Phe Thr Phe Thr Pro Gln Asn Asn Glu Phe Gln Leu Gln
580 585 590
Leu Ser Pro Lys Thr Phe Ala Ser Lys Thr Tyr Gly Leu Cys Gly Ile
595 600 605
Cys Asp Glu Asn Gly Ala Asn Asp Phe Met Leu Arg Asp Gly Thr Val
610 615 620
Thr Thr Asp Trp Lys Thr Leu Val Gln Glu Trp Thr Val Gln Arg Pro
625 630 635 640
Gly Gln Thr Cys Gln Pro Ile Leu Glu Glu Gln Cys Leu Val Pro Asp
645 650 655
Ser Ser His Cys Gln Val Leu Leu Leu Pro Leu Phe Ala Glu Cys His
660 665 670
Lys Val Leu Ala Pro Ala Thr Phe Tyr Ala Ile Cys Gln Gln Asp Ser
675 680 685
Cys His Gln Glu Gln Val Cys Glu Val Ile Ala Ser Tyr Ala His Leu
690 695 700
Cys Arg Thr Asn Gly Val Cys Val Asp Trp Arg Thr Pro Asp Phe Cys
705 710 715 720
Ala Met Ser Cys Pro Pro Ser Leu Val Tyr Asn His Cys Glu His Gly
725 730 735
Cys Pro Arg His Cys Asp Gly Asn Val Ser Ser Cys Gly Asp His Pro
740 745 750
Ser Glu Gly Cys Phe Cys Pro Pro Asp Lys Val Met Leu Glu Gly Ser
755 760 765
Cys Val Pro Glu Glu Ala Cys Thr Gln Cys Ile Gly Glu Asp Gly Val
770 775 780
Gln His Gln Phe Leu Glu Ala Trp Val Pro Asp His Gln Pro Cys Gln
785 790 795 800
Ile Cys Thr Cys Leu Ser Gly Arg Lys Val Asn Cys Thr Thr Gln Pro
805 810 815
Cys Pro Thr Ala Lys Ala Pro Thr Cys Gly Leu Cys Glu Val Ala Arg
820 825 830
Leu Arg Gln Asn Ala Asp Gln Cys Cys Pro Glu Tyr Glu Cys Val Cys
835 840 845
Asp Pro Val Ser Cys Asp Leu Pro Pro Val Pro His Cys Glu Arg Gly
850 855 860
Leu Gln Pro Thr Leu Thr Asn Pro Gly Glu Cys Arg Pro Asn Phe Thr
865 870 875 880
Cys Ala Cys Arg Lys Glu Glu Cys Lys Arg Val Ser Pro Pro Ser Cys
885 890 895
Pro Pro His Arg Leu Pro Thr Leu Arg Lys Thr Gln Cys Cys Asp Glu
900 905 910
Tyr Glu Cys Ala Cys Asn Cys Val Asn Ser Thr Val Ser Cys Pro Leu
915 920 925
Gly Tyr Leu Ala Ser Thr Ala Thr Asn Asp Cys Gly Cys Thr Thr Thr
930 935 940
Thr Cys Leu Pro Asp Lys Val Cys Val His Arg Ser Thr Ile Tyr Pro
945 950 955 960
Val Gly Gln Phe Trp Glu Glu Gly Cys Asp Val Cys Thr Cys Thr Asp
965 970 975
Met Glu Asp Ala Val Met Gly Leu Arg Val Ala Gln Cys Ser Gln Lys
980 985 990
Pro Cys Glu Asp Ser Cys Arg Ser Gly Phe Thr Tyr Val Leu His Glu
995 1000 1005
Gly Glu Cys Cys Gly Arg Cys Leu Pro Ser Ala Cys Glu Val Val
1010 1015 1020
Thr Gly Ser Pro Arg Gly Asp Ser Gln Ser Ser Trp Lys Ser Val
1025 1030 1035
Gly Ser Gln Trp Ala Ser Pro Glu Asn Pro Cys Leu Ile Asn Glu
1040 1045 1050
Cys Val Arg Val Lys Glu Glu Val Phe Ile Gln Gln Arg Asn Val
1055 1060 1065
Ser Cys Pro Gln Leu Glu Val Pro Val Cys Pro Ser Gly Phe Gln
1070 1075 1080
Leu Ser Cys Lys Thr Ser Ala Cys Cys Pro Ser Cys Arg Cys Glu
1085 1090 1095
Arg Met Glu Ala Cys Met Leu Asn Gly Thr Val Ile Gly Pro Gly
1100 1105 1110
Lys Thr Val Met Ile Asp Val Cys Thr Thr Cys Arg Cys Met Val
1115 1120 1125
Gln Val Gly Val Ile Ser Gly Phe Lys Leu Glu Cys Arg Lys Thr
1130 1135 1140
Thr Cys Asn Pro Cys Pro Leu Gly Tyr Lys Glu Glu Asn Asn Thr
1145 1150 1155
Gly Glu Cys Cys Gly Arg Cys Leu Pro Thr Ala Cys Thr Ile Gln
1160 1165 1170
Leu Arg Gly Gly Gln Ile Met Thr Leu Lys Arg Asp Glu Thr Leu
1175 1180 1185
Gln Asp Gly Cys Asp Thr His Phe Cys Lys Val Asn Glu Arg Gly
1190 1195 1200
Glu Tyr Phe Trp Glu Lys Arg Val Thr Gly Cys Pro Pro Phe Asp
1205 1210 1215
Glu His Lys Cys Leu Ala Glu Gly Gly Lys Ile Met Lys Ile Pro
1220 1225 1230
Gly Thr Cys Cys Asp Thr Cys Glu Glu Pro Glu Cys Asn Asp Ile
1235 1240 1245
Thr Ala Arg Leu Gln Tyr Val Lys Val Gly Ser Cys Lys Ser Glu
1250 1255 1260
Val Glu Val Asp Ile His Tyr Cys Gln Gly Lys Cys Ala Ser Lys
1265 1270 1275
Ala Met Tyr Ser Ile Asp Ile Asn Asp Val Gln Asp Gln Cys Ser
1280 1285 1290
Cys Cys Ser Pro Thr Arg Thr Glu Pro Met Gln Val Ala Leu His
1295 1300 1305
Cys Thr Asn Gly Ser Val Val Tyr His Glu Val Leu Asn Ala Met
1310 1315 1320
Glu Cys Lys Cys Ser Pro Arg Lys Cys Ser Lys
1325 1330
<210> 214
<211> 374
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Область D3 VWF с EAE
<400> 214
Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly Leu Lys
1 5 10 15
Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp Tyr Cys
20 25 30
Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys Gly Cys
35 40 45
Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu Val Glu
50 55 60
Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys Arg Pro
65 70 75 80
Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg Tyr Ile
85 90 95
Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg His Leu
100 105 110
Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val Cys Gly
115 120 125
Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr Ser Ser
130 135 140
Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn Ser Trp Lys
145 150 155 160
Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro Leu Asp Ser Ser
165 170 175
Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln Thr Met Val Asp Ser
180 185 190
Ser Cys Arg Ile Leu Thr Ser Asp Val Phe Gln Asp Cys Asn Lys Leu
195 200 205
Val Asp Pro Glu Pro Tyr Leu Asp Val Cys Ile Tyr Asp Thr Cys Ser
210 215 220
Cys Glu Ser Ile Gly Asp Cys Ala Ala Phe Cys Asp Thr Ile Ala Ala
225 230 235 240
Tyr Ala His Val Cys Ala Gln His Gly Lys Val Val Thr Trp Arg Thr
245 250 255
Ala Thr Leu Cys Pro Gln Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn
260 265 270
Gly Tyr Glu Ala Glu Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln
275 280 285
Val Thr Cys Gln His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val
290 295 300
Glu Gly Cys His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu
305 310 315 320
Leu Gln Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala
325 330 335
Gly Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
340 345 350
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr Cys
355 360 365
Glu Ala Cys Gln Glu Pro
370
<210> 215
<211> 745
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> 1-745 в FVIII
<400> 215
Ala Thr Arg Arg Tyr Tyr Leu Gly Ala Val Glu Leu Ser Trp Asp Tyr
1 5 10 15
Met Gln Ser Asp Leu Gly Glu Leu Pro Val Asp Ala Arg Phe Pro Pro
20 25 30
Arg Val Pro Lys Ser Phe Pro Phe Asn Thr Ser Val Val Tyr Lys Lys
35 40 45
Thr Leu Phe Val Glu Phe Thr Asp His Leu Phe Asn Ile Ala Lys Pro
50 55 60
Arg Pro Pro Trp Met Gly Leu Leu Gly Pro Thr Ile Gln Ala Glu Val
65 70 75 80
Tyr Asp Thr Val Val Ile Thr Leu Lys Asn Met Ala Ser His Pro Val
85 90 95
Ser Leu His Ala Val Gly Val Ser Tyr Trp Lys Ala Ser Glu Gly Ala
100 105 110
Glu Tyr Asp Asp Gln Thr Ser Gln Arg Glu Lys Glu Asp Asp Lys Val
115 120 125
Phe Pro Gly Gly Ser His Thr Tyr Val Trp Gln Val Leu Lys Glu Asn
130 135 140
Gly Pro Met Ala Ser Asp Pro Leu Cys Leu Thr Tyr Ser Tyr Leu Ser
145 150 155 160
His Val Asp Leu Val Lys Asp Leu Asn Ser Gly Leu Ile Gly Ala Leu
165 170 175
Leu Val Cys Arg Glu Gly Ser Leu Ala Lys Glu Lys Thr Gln Thr Leu
180 185 190
His Lys Phe Ile Leu Leu Phe Ala Val Phe Asp Glu Gly Lys Ser Trp
195 200 205
His Ser Glu Thr Lys Asn Ser Leu Met Gln Asp Arg Asp Ala Ala Ser
210 215 220
Ala Arg Ala Trp Pro Lys Met His Thr Val Asn Gly Tyr Val Asn Arg
225 230 235 240
Ser Leu Pro Gly Leu Ile Gly Cys His Arg Lys Ser Val Tyr Trp His
245 250 255
Val Ile Gly Met Gly Thr Thr Pro Glu Val His Ser Ile Phe Leu Glu
260 265 270
Gly His Thr Phe Leu Val Arg Asn His Arg Gln Ala Ser Leu Glu Ile
275 280 285
Ser Pro Ile Thr Phe Leu Thr Ala Gln Thr Leu Leu Met Asp Leu Gly
290 295 300
Gln Phe Leu Leu Phe Cys His Ile Ser Ser His Gln His Asp Gly Met
305 310 315 320
Glu Ala Tyr Val Lys Val Asp Ser Cys Pro Glu Glu Pro Gln Leu Arg
325 330 335
Met Lys Asn Asn Glu Glu Ala Glu Asp Tyr Asp Asp Asp Leu Thr Asp
340 345 350
Ser Glu Met Asp Val Val Arg Phe Asp Asp Asp Asn Ser Pro Ser Phe
355 360 365
Ile Gln Ile Arg Ser Val Ala Lys Lys His Pro Lys Thr Trp Val His
370 375 380
Tyr Ile Ala Ala Glu Glu Glu Asp Trp Asp Tyr Ala Pro Leu Val Leu
385 390 395 400
Ala Pro Asp Asp Arg Ser Tyr Lys Ser Gln Tyr Leu Asn Asn Gly Pro
405 410 415
Gln Arg Ile Gly Arg Lys Tyr Lys Lys Val Arg Phe Met Ala Tyr Thr
420 425 430
Asp Glu Thr Phe Lys Thr Arg Glu Ala Ile Gln His Glu Ser Gly Ile
435 440 445
Leu Gly Pro Leu Leu Tyr Gly Glu Val Gly Asp Thr Leu Leu Ile Ile
450 455 460
Phe Lys Asn Gln Ala Ser Arg Pro Tyr Asn Ile Tyr Pro His Gly Ile
465 470 475 480
Thr Asp Val Arg Pro Leu Tyr Ser Arg Arg Leu Pro Lys Gly Val Lys
485 490 495
His Leu Lys Asp Phe Pro Ile Leu Pro Gly Glu Ile Phe Lys Tyr Lys
500 505 510
Trp Thr Val Thr Val Glu Asp Gly Pro Thr Lys Ser Asp Pro Arg Cys
515 520 525
Leu Thr Arg Tyr Tyr Ser Ser Phe Val Asn Met Glu Arg Asp Leu Ala
530 535 540
Ser Gly Leu Ile Gly Pro Leu Leu Ile Cys Tyr Lys Glu Ser Val Asp
545 550 555 560
Gln Arg Gly Asn Gln Ile Met Ser Asp Lys Arg Asn Val Ile Leu Phe
565 570 575
Ser Val Phe Asp Glu Asn Arg Ser Trp Tyr Leu Thr Glu Asn Ile Gln
580 585 590
Arg Phe Leu Pro Asn Pro Ala Gly Val Gln Leu Glu Asp Pro Glu Phe
595 600 605
Gln Ala Ser Asn Ile Met His Ser Ile Asn Gly Tyr Val Phe Asp Ser
610 615 620
Leu Gln Leu Ser Val Cys Leu His Glu Val Ala Tyr Trp Tyr Ile Leu
625 630 635 640
Ser Ile Gly Ala Gln Thr Asp Phe Leu Ser Val Phe Phe Ser Gly Tyr
645 650 655
Thr Phe Lys His Lys Met Val Tyr Glu Asp Thr Leu Thr Leu Phe Pro
660 665 670
Phe Ser Gly Glu Thr Val Phe Met Ser Met Glu Asn Pro Gly Leu Trp
675 680 685
Ile Leu Gly Cys His Asn Ser Asp Phe Arg Asn Arg Gly Met Thr Ala
690 695 700
Leu Leu Lys Val Ser Ser Cys Asp Lys Asn Thr Gly Asp Tyr Tyr Glu
705 710 715 720
Asp Ser Tyr Glu Asp Ile Ser Ala Tyr Leu Leu Ser Lys Asn Asn Ala
725 730 735
Ile Glu Pro Arg Ser Phe Ser Gln Asn
740 745
<210> 216
<211> 684
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> FVIII 1649-2332
<400> 216
Glu Ile Thr Arg Thr Thr Leu Gln Ser Asp Gln Glu Glu Ile Asp Tyr
1 5 10 15
Asp Asp Thr Ile Ser Val Glu Met Lys Lys Glu Asp Phe Asp Ile Tyr
20 25 30
Asp Glu Asp Glu Asn Gln Ser Pro Arg Ser Phe Gln Lys Lys Thr Arg
35 40 45
His Tyr Phe Ile Ala Ala Val Glu Arg Leu Trp Asp Tyr Gly Met Ser
50 55 60
Ser Ser Pro His Val Leu Arg Asn Arg Ala Gln Ser Gly Ser Val Pro
65 70 75 80
Gln Phe Lys Lys Val Val Phe Gln Glu Phe Thr Asp Gly Ser Phe Thr
85 90 95
Gln Pro Leu Tyr Arg Gly Glu Leu Asn Glu His Leu Gly Leu Leu Gly
100 105 110
Pro Tyr Ile Arg Ala Glu Val Glu Asp Asn Ile Met Val Thr Phe Arg
115 120 125
Asn Gln Ala Ser Arg Pro Tyr Ser Phe Tyr Ser Ser Leu Ile Ser Tyr
130 135 140
Glu Glu Asp Gln Arg Gln Gly Ala Glu Pro Arg Lys Asn Phe Val Lys
145 150 155 160
Pro Asn Glu Thr Lys Thr Tyr Phe Trp Lys Val Gln His His Met Ala
165 170 175
Pro Thr Lys Asp Glu Phe Asp Cys Lys Ala Trp Ala Tyr Phe Ser Asp
180 185 190
Val Asp Leu Glu Lys Asp Val His Ser Gly Leu Ile Gly Pro Leu Leu
195 200 205
Val Cys His Thr Asn Thr Leu Asn Pro Ala His Gly Arg Gln Val Thr
210 215 220
Val Gln Glu Phe Ala Leu Phe Phe Thr Ile Phe Asp Glu Thr Lys Ser
225 230 235 240
Trp Tyr Phe Thr Glu Asn Met Glu Arg Asn Cys Arg Ala Pro Cys Asn
245 250 255
Ile Gln Met Glu Asp Pro Thr Phe Lys Glu Asn Tyr Arg Phe His Ala
260 265 270
Ile Asn Gly Tyr Ile Met Asp Thr Leu Pro Gly Leu Val Met Ala Gln
275 280 285
Asp Gln Arg Ile Arg Trp Tyr Leu Leu Ser Met Gly Ser Asn Glu Asn
290 295 300
Ile His Ser Ile His Phe Ser Gly His Val Phe Thr Val Arg Lys Lys
305 310 315 320
Glu Glu Tyr Lys Met Ala Leu Tyr Asn Leu Tyr Pro Gly Val Phe Glu
325 330 335
Thr Val Glu Met Leu Pro Ser Lys Ala Gly Ile Trp Arg Val Glu Cys
340 345 350
Leu Ile Gly Glu His Leu His Ala Gly Met Ser Thr Leu Phe Leu Val
355 360 365
Tyr Ser Asn Lys Cys Gln Thr Pro Leu Gly Met Ala Ser Gly His Ile
370 375 380
Arg Asp Phe Gln Ile Thr Ala Ser Gly Gln Tyr Gly Gln Trp Ala Pro
385 390 395 400
Lys Leu Ala Arg Leu His Tyr Ser Gly Ser Ile Asn Ala Trp Ser Thr
405 410 415
Lys Glu Pro Phe Ser Trp Ile Lys Val Asp Leu Leu Ala Pro Met Ile
420 425 430
Ile His Gly Ile Lys Thr Gln Gly Ala Arg Gln Lys Phe Ser Ser Leu
435 440 445
Tyr Ile Ser Gln Phe Ile Ile Met Tyr Ser Leu Asp Gly Lys Lys Trp
450 455 460
Gln Thr Tyr Arg Gly Asn Ser Thr Gly Thr Leu Met Val Phe Phe Gly
465 470 475 480
Asn Val Asp Ser Ser Gly Ile Lys His Asn Ile Phe Asn Pro Pro Ile
485 490 495
Ile Ala Arg Tyr Ile Arg Leu His Pro Thr His Tyr Ser Ile Arg Ser
500 505 510
Thr Leu Arg Met Glu Leu Met Gly Cys Asp Leu Asn Ser Cys Ser Met
515 520 525
Pro Leu Gly Met Glu Ser Lys Ala Ile Ser Asp Ala Gln Ile Thr Ala
530 535 540
Ser Ser Tyr Phe Thr Asn Met Phe Ala Thr Trp Ser Pro Ser Lys Ala
545 550 555 560
Arg Leu His Leu Gln Gly Arg Ser Asn Ala Trp Arg Pro Gln Val Asn
565 570 575
Asn Pro Lys Glu Trp Leu Gln Val Asp Phe Gln Lys Thr Met Lys Val
580 585 590
Thr Gly Val Thr Thr Gln Gly Val Lys Ser Leu Leu Thr Ser Met Tyr
595 600 605
Val Lys Glu Phe Leu Ile Ser Ser Ser Gln Asp Gly His Gln Trp Thr
610 615 620
Leu Phe Phe Gln Asn Gly Lys Val Lys Val Phe Gln Gly Asn Gln Asp
625 630 635 640
Ser Phe Thr Pro Val Val Asn Ser Leu Asp Pro Pro Leu Leu Thr Arg
645 650 655
Tyr Leu Arg Ile His Pro Gln Ser Trp Val His Gln Ile Ala Leu Arg
660 665 670
Met Glu Val Leu Gly Cys Glu Ala Gln Asp Leu Tyr
675 680
<210> 217
<211> 226
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Fc-область
<400> 217
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly
225
<210> 218
<211> 291
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> XTEN AE288-3
<400> 218
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Glu Pro
1 5 10 15
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
20 25 30
Glu Ser Gly Pro Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro
35 40 45
Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Thr
50 55 60
Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Pro Ala Gly Ser Pro Thr
65 70 75 80
Ser Thr Glu Glu Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
85 90 95
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu
100 105 110
Ser Ala Thr Pro Glu Ser Gly Pro Gly Ser Pro Ala Gly Ser Pro Thr
115 120 125
Ser Thr Glu Glu Gly Ser Pro Ala Gly Ser Pro Thr Ser Thr Glu Glu
130 135 140
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Thr Ser Glu
145 150 155 160
Ser Ala Thr Pro Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro
165 170 175
Glu Ser Gly Pro Gly Thr Ser Glu Ser Ala Thr Pro Glu Ser Gly Pro
180 185 190
Gly Ser Glu Pro Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Glu Pro
195 200 205
Ala Thr Ser Gly Ser Glu Thr Pro Gly Ser Pro Ala Gly Ser Pro Thr
210 215 220
Ser Thr Glu Glu Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
225 230 235 240
Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro Gly Ser Glu Pro
245 250 255
Ala Thr Ser Gly Ser Glu Thr Pro Gly Thr Ser Glu Ser Ala Thr Pro
260 265 270
Glu Ser Gly Pro Gly Thr Ser Thr Glu Pro Ser Glu Gly Ser Ala Pro
275 280 285
Ala Ser Ser
290
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ВЫСОКОГЛИКОЗИЛИРОВАННЫЙ СЛИТЫЙ БЕЛОК НА ОСНОВЕ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ ЧЕЛОВЕКА VIII, СПОСОБ ЕГО ПОЛУЧЕНИЯ И ЕГО ПРИМЕНЕНИЕ | 2016 |
|
RU2722374C1 |
| ФАРМАЦЕВТИЧЕСКИЕ СОСТАВЫ ПЕГИЛИРОВАННЫХ ЛИПОСОМ И ФАКТОРОВ СВЕРТЫВАНИЯ КРОВИ | 2015 |
|
RU2737291C2 |
| ПРЕПАРАТ, СОДЕРЖАЩИЙ ФАКТОР VIII И ПЕПТИДЫ ФАКТОРА ФОН ВИЛЛЕБРАНДА | 2015 |
|
RU2714154C2 |
| ГЛИКОЗИЛИРОВАННЫЕ СЛИТЫЕ БЕЛКИ VWF С УЛУЧШЕННОЙ ФАРМАКОКИНЕТИКОЙ | 2017 |
|
RU2782212C2 |
| ФАРМАЦЕВТИЧЕСКАЯ КОМПОЗИЦИЯ, ПРЕДНАЗНАЧЕННАЯ ДЛЯ ПРИМЕНЕНИЯ ДЛЯ ПРЕДУПРЕЖДЕНИЯ И/ИЛИ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЯ, КОТОРОЕ РАЗВИВАЕТСЯ ИЛИ ПРОГРЕССИРУЕТ ВСЛЕДСТВИЕ СНИЖЕНИЯ ИЛИ УТРАТЫ АКТИВНОСТИ ФАКТОРА СВЕРТЫВАНИЯ КРОВИ VIII И/ИЛИ АКТИВИРОВАННОГО ФАКТОРА СВЕРТЫВАНИЯ КРОВИ VIII | 2015 |
|
RU2721910C2 |
| УЛУЧШЕННЫЙ БЕЛОК СЛИЯНИЯ FVIII И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2789085C2 |
| Выделенная нуклеиновая кислота, которая кодирует слитый белок на основе FVIII-BDD и гетерологичного сигнального пептида, и ее применение | 2022 |
|
RU2818229C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2819144C2 |
| ПРИМЕНЕНИЕ ЛЕНТИВИРУСНЫХ ВЕКТОРОВ, ЭКСПРЕССИРУЮЩИХ ФАКТОР VIII | 2019 |
|
RU2803163C2 |
| МОЛЕКУЛЫ НУКЛЕИНОВОЙ КИСЛОТЫ И ПУТИ ИХ ПРИМЕНЕНИЯ ДЛЯ НЕВИРУСНОЙ ГЕННОЙ ТЕРАПИИ | 2019 |
|
RU2834625C2 |
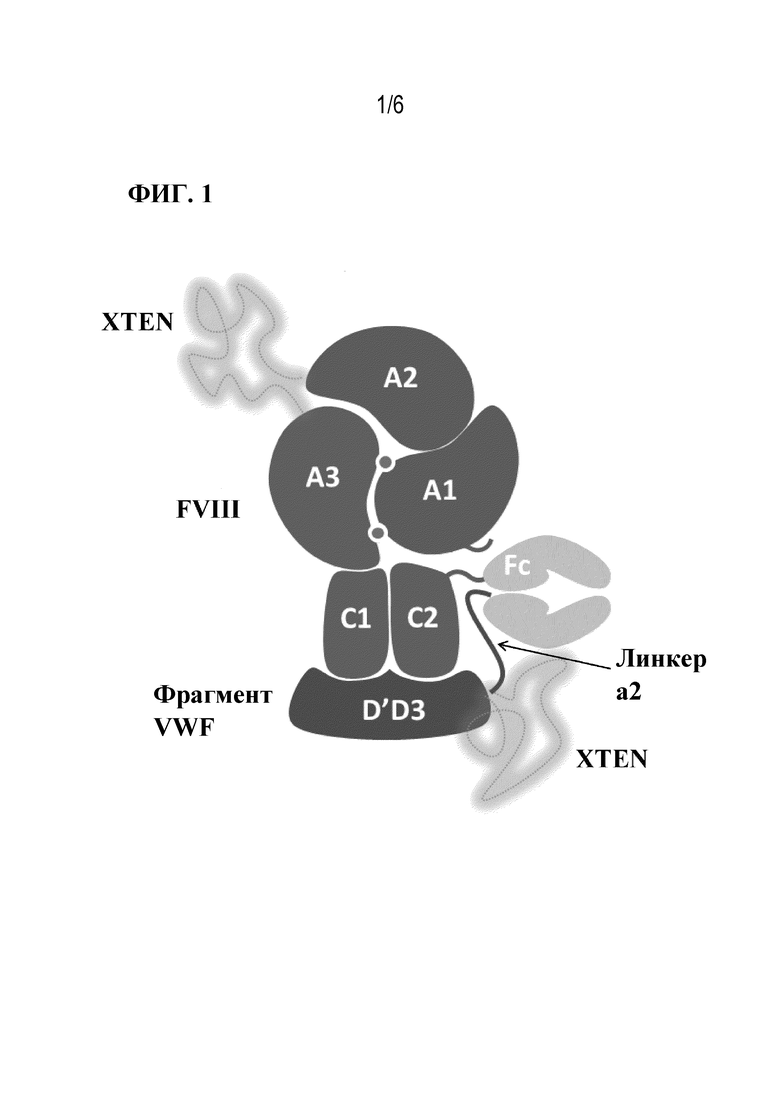
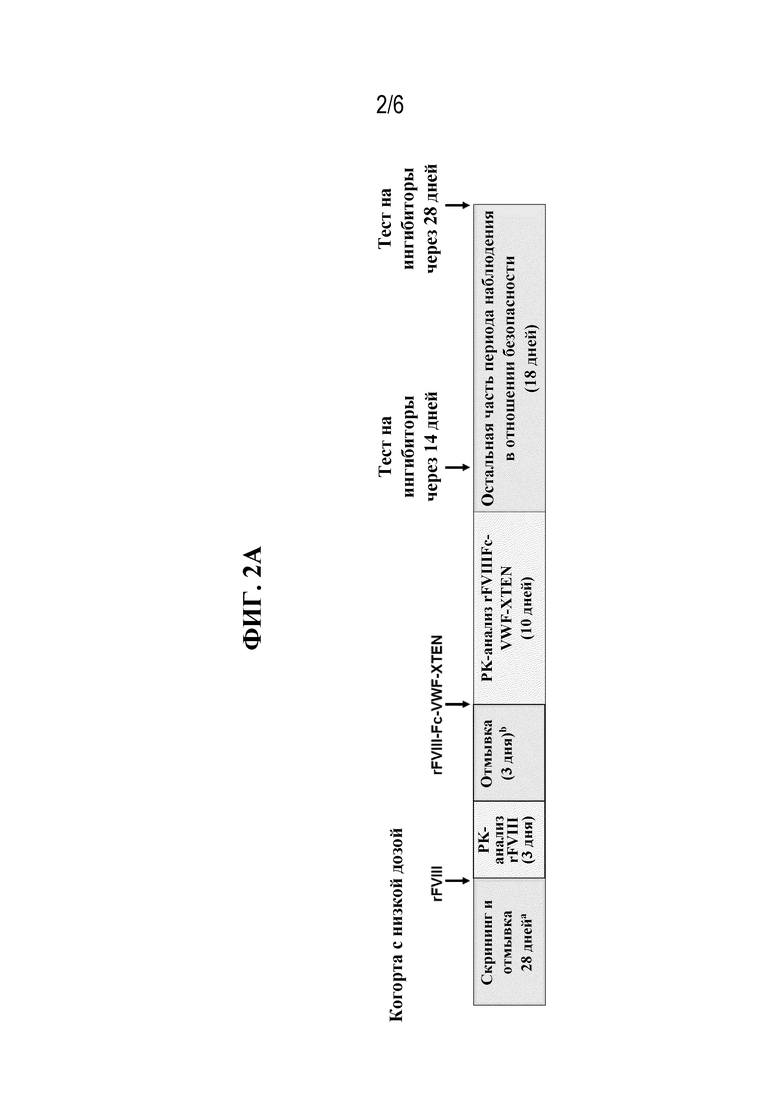
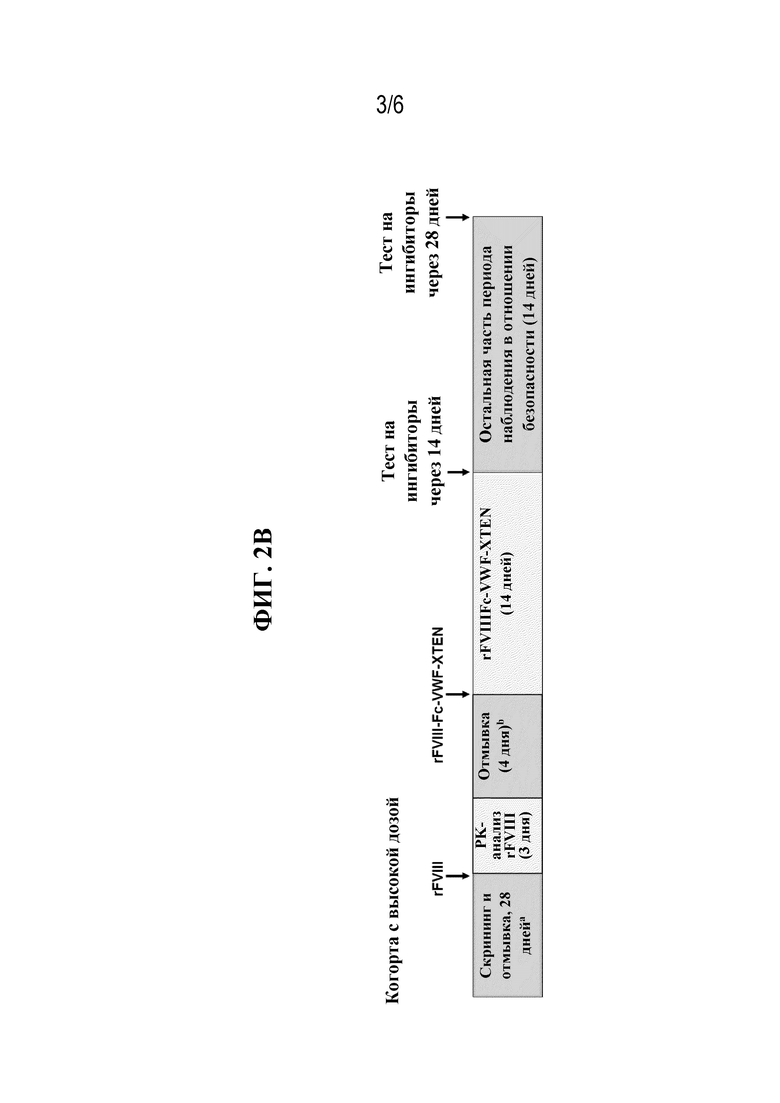
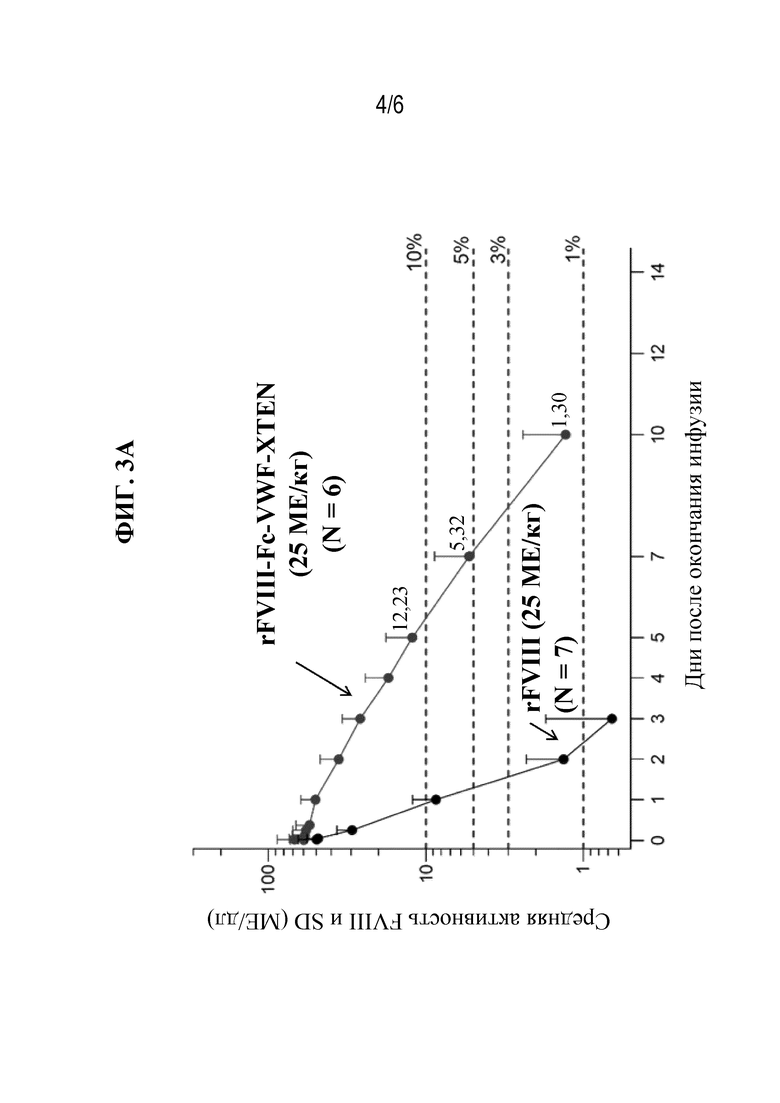
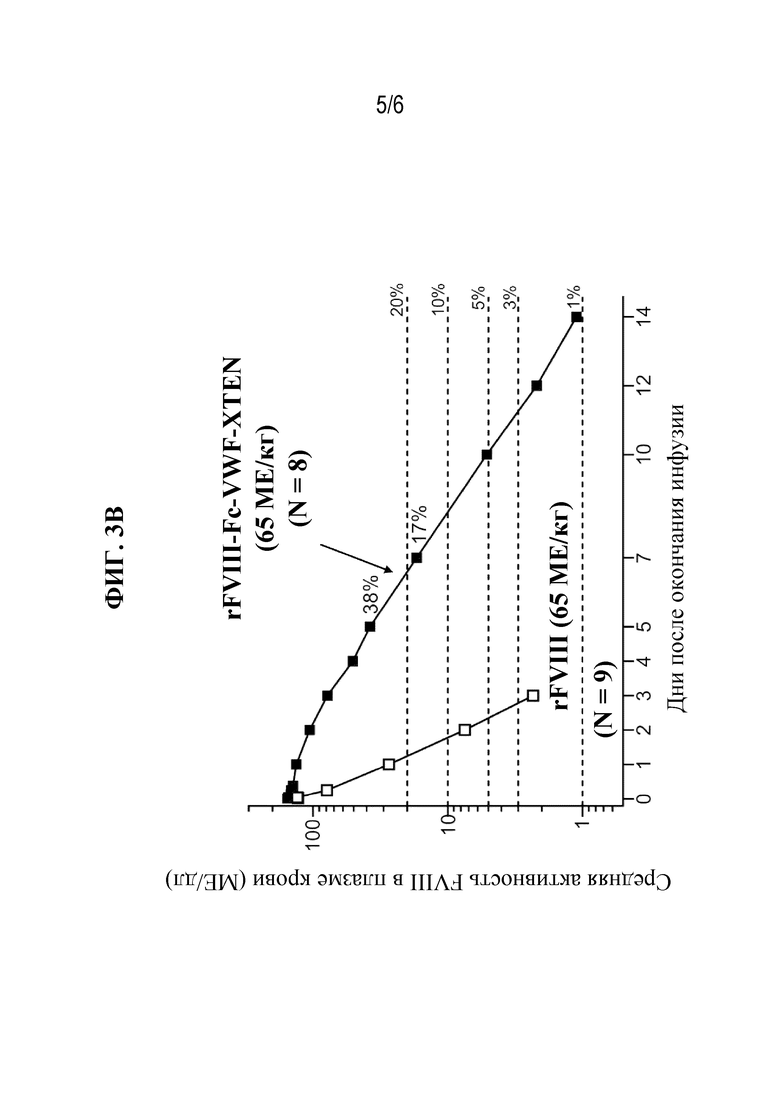
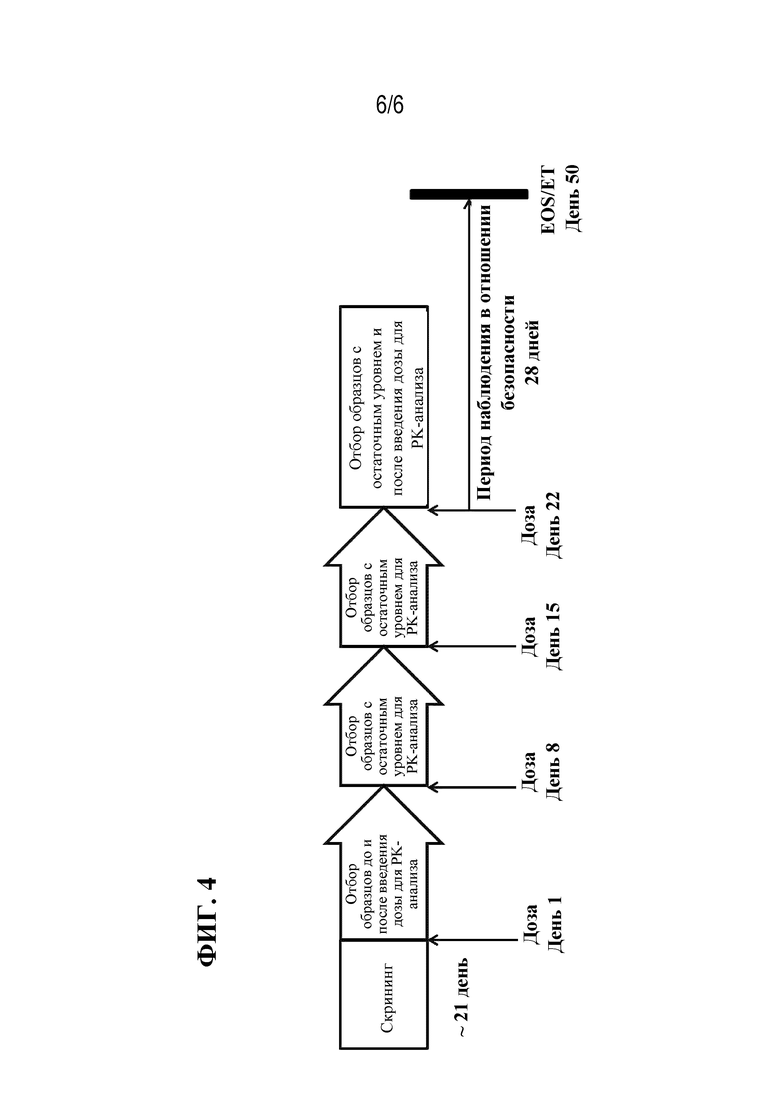
Группа изобретений относится к биотехнологии. Представлены способ лечения и способ профилактического лечения гемофилии A у субъекта, предусматривающий введение субъекту химерного полипептида, содержащего белок фактора VIII (FVIII) и белок фактора фон Виллебранда (VWF). При этом химерный полипептид вводится внутривенно в одной или более дозах, составляющих от 45 МЕ/кг до 55 МЕ/кг, а интервал между введениями доз составляет по меньшей мере 7 дней, где полипептид FVIII связан с фрагментом VWF с помощью ковалентной связи. Изобретение используется для лечения тяжелой формы гемофилии A у субъекта. 2 н. и 12 з.п. ф-лы, 6 ил., 19 табл., 3 пр.
1. Способ лечения гемофилии A у субъекта-человека, нуждающегося в этом, предусматривающий введение субъекту химерного полипептида, содержащего:
(i) белок фактора VIII (FVIII), содержащий аминокислотную последовательность SEQ ID NO: 207; и (ii) белок фактора фон Виллебранда (VWF), содержащий аминокислотную последовательность SEQ ID NO: 202,
где химерный полипептид вводится внутривенно в одной или более доз, составляющих от 45 МЕ/кг до 55 МЕ/кг, а интервал между введениями доз составляет по меньшей мере 7 дней, где белок FVIII связан с белком VWF с помощью ковалентной связи.
2. Способ по п. 1, где ковалентная связь представляет собой дисульфидную связь.
3. Способ по п. 1 или 2, где каждая из нескольких доз составляет 50 МЕ/кг, а интервал между введениями доз составляет приблизительно 7 дней.
4. Способ по любому из пп. 1-3, где химерный полипептид предназначен для профилактического лечения.
5. Способ по любому из пп. 1-4, где субъект имеет тяжелую форму гемофилии A.
6. Способ по любому из пп. 1-5, где субъект ранее получал лечение гемофилии А с помощью любого рекомбинантного FVIII, полученного из плазмы крови FVIII или криопреципитата в течение по меньшей мере 150 дней воздействия (ED).
7. Способ по любому из пп. 1-6, где субъект или
(a) в настоящее время проходит режим профилактического лечения с помощью представленного на рынке продукта FVIII и имеет по меньшей мере четыре эпизода кровотечения за двенадцать месяцев до введения химерного полипептида; или
(b) в настоящее время проходит режим лечения по необходимости с помощью представленного на рынке продукта FVIII и имеет по меньшей мере двенадцать эпизодов кровотечения за двенадцать месяцев до введения химерного полипептида.
8. Способ по любому из пп. 1-7, где субъект имеет количество тромбоцитов по меньшей мере 100000 клеток/мкл.
9. Способ по любому из пп. 1-8, где у субъекта отсутствуют другие нарушения коагуляции.
10. Способ по любому из пп. 1-9, где у субъекта отсутствует в анамнезе выработка ингибиторов к продукту FVIII.
11. Способ по любому из пп. 1-10, где одну или более доз вводят в течение по меньшей мере 1 месяца, по меньшей мере 2 месяцев, по меньшей мере 3 месяцев, по меньшей мере 4 месяцев, по меньшей мере 5 месяцев, по меньшей мере 6 месяцев, по меньшей мере 12 месяцев, по меньшей мере 18 месяцев, по меньшей мере 2 лет, по меньшей мере 3 лет, по меньшей мере 4 лет, по меньшей мере 5 лет, по меньшей мере 10 лет, по меньшей мере 15 лет, по меньшей мере 20 лет или в течение по меньшей мере 25 лет.
12. Способ профилактического лечения тяжелой формы гемофилии A у субъекта, нуждающегося в этом, предусматривающий введение субъекту химерного полипептида, содержащего:
(i) белок FVIII, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 207; и
(ii) белок VWF, содержащий аминокислотную последовательность, представленную в SEQ ID NO: 202,
где химерный полипептид вводится внутривенно в дозе 50 МЕ/кг один раз в неделю.
13. Способ по п. 12, где одна или более доз вводятся в течение по меньшей мере 6 месяцев.
14. Способ по п. 12, где одна или более доз вводятся в течение по меньшей мере 52 недель.
| WO 2015106052 А1, 16.07.2015 | |||
| WO 2015021423 A2, 12.02.2015 | |||
| CHEN, X | |||
| et al | |||
| Fusion protein linkers: Property, design and functionality | |||
| ADVANCED DRUG DELIVERY REVIEWS | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| ЗМАЧИНСКИЙ В.А | |||
| Современные подходы к лечению гемофилии | |||
| Медицинские новости | |||
| Многоступенчатая активно-реактивная турбина | 1924 |
|
SU2013A1 |
| СПОСОБ ПОЛУЧЕНИЯ ПРЕПАРАТА ФАКТОРА VIII | 2004 |
|
RU2253475C1 |
