Настоящее изобретение относится к фармацевтическим препаратам для лечения нарушений свертываемости крови.
Уровень техники
Фактор VIII («FVIII») представляет собой гликопротеин плазмы крови с молекулярной массой, составляющей приблизительно 280 кДа. Он участвует в каскаде реакций коагуляции, которые приводят к свертываемости крови. Наиболее распространенное расстройство свертываемости крови, гемофилия А, обусловлено дефицитом функционального Фактора VIII. Лечится путем замены Фактора VIII либо на основе плазмы, либо рекомбинантным. Фактор VIII используется для неотложного и профилактического лечения кровотечений у пациентов с гемофилией А.
Аминокислотная последовательность Фактора VIII организована в три структурных домена: утроенный домен А из 330 аминокислот, один B-домен из 980 аминокислот и дублированной домен С из 150 аминокислот. B-домен не имеет гомологии с другими белками и обеспечивает 18 из 25 потенциальных аспарагин(N)-связанных сайтов гликозилирования этого белка. B-домен не имеет по-видимому никакой функции в коагуляции. Молекулы Фактора VIII с делетированным В-доменом обладают неизменной прокоагулянтной активностью по сравнению с полноразмерным Фактором VIII. Некоторые препараты рекомбинантного Фактора VIII (rFVIII) являются делетированными по B-домену.
В плазме Фактор VIII стабилизирован ассоциацией с белком Фактора фон Виллебранда («vWF»), который, как представляется, ингибирует клиренс Фактора VIII, например, с помощью протеолиза или рецептор-опосредованного клиренса через LRP-рецептор. В кровотоке, Фактор фон Виллебранда присутствует в 50-кратном молярном избытке по отношению к Фактору VIII в нормальных физиологических условиях.
Фактор фон Виллебранда является мультимерным адгезивным гликопротеином, присутствующим в плазме у млекопитающих, который обладает многочисленными физиологическими функциями. Во время первичного гемостаза, Фактор фон Виллебранда выступает в качестве посредника между специфическими рецепторами на поверхности тромбоцитов и компонентами внеклеточного матрикса, такими как коллаген. Кроме того, Фактор фон Виллебранда служит в качестве носителя и стабилизирующего белка для прокоагулянтного фактора VIII. Фактор фон Виллебранда синтезируется в эндотелиальных клетках и мегакариоцитах в виде молекулы-предшественника из 2813 аминокислот. Полипептид-предшественник, пре-про-Фактор фон Виллебранда, состоит из сигнального пептида из 22 остатков, про-пептида из 741 остатка и полипептида из 2050 остатков, обнаруженного в зрелом факторе фон Виллебранда в плазме (Fischer et al., FEBS Lett. 351: 345-348, 1994). После секреции в плазме Фактор фон Виллебранда циркулирует в форме различных видов с различными молекулярными размерами. Эти молекулы Фактора фон Виллебранда состоят из олиго- и мультимеров зрелой субъединицы, содержащей 2050 аминокислотных остатков. Фактор фон Виллебранда может, как правило, быть обнаружен в плазме в виде мультимеров с размером в диапазоне приблизительно от 500 до 20000 кДа (Furlan, Ann Hematol. 1996 Jun;72(6):341-8).
Средний период полужизни человеческого Фактора VIII в кровотоке человека in vivo составляет приблизительно 12 часов. Фактор фон Виллебранда может уменьшить возможные иммунные реакции против Фактора VIII, будучи в комплексе с Фактором VIII, экранируя FVIII от известных потенциальных сайтов ингибирующих антител на тяжелой цепи (домен А2) и легкой цепи (A3/C2 домен) (Ragni, J Thromb. Haemost. 10: 2324-2327, 2012) или на других потенциальных ингибирующих сайтах антител на молекуле фактора VIII.
Другое нарушение свертываемости крови у человека представляет собой болезнь фон Виллебранда (БВ). В зависимости от тяжести симптомов кровотечения, БВ можно лечить с помощью заместительной терапии с использованием концентратов, содержащих Фактор фон Виллебранда, в основном, полученного из плазмы, но рекомбинантный Фактор фон Виллебранда, также находится на стадии разработки. Фактор фон Виллебранда, как известно, стабилизирует Фактор VIII in vivo, и, таким образом, играет ключевую роль в регуляции уровня Фактора VIII в плазме и, как следствие, является центральным фактором для контроля первичного и вторичного гемостаза.
До сегодняшнего дня, стандартное лечение гемофилии А и БВ включало в себя частые внутривенные инфузии препаратов Фактора VIII и концентратов Фактора VIII/Фактора Виллебранда. Эти заместительные терапии, как правило, эффективны, однако, например, пациентам с тяжелой формой гемофилии А, подвергающихся профилактическому лечению, Фактор VIII необходимо вводить внутривенно (i.v.) примерно 3 раза в неделю из-за короткого периода полужизни Фактора VIII в плазме, составляющего примерно 12 часов. Уже при достижении уровня Фактора VIII выше 1% в нормальной человеческой плазмы, соответствующему повышению уровня Фактора VIII на 0,01 ед/мл, тяжелая гемофилия А превращается в умеренную гемофилию А. При профилактической терапии, схема дозирования разрабатывается таким образом, чтобы уровень активности Фактора VIII не опускается ниже уровня 2-3% от активности Фактора VIII у не страдающих гемофилией.
Введение Фактора VIII с помощью внутривенного введения (i.v.) является трудоемким, связанным с болью и влечет за собой риск инфекции, особенно если это делается в домашних условиях самими пациентами или родителями детей с диагнозом гемофилии А. Кроме того, частые внутривенные инъекции неизбежно приводят к образованию рубцов, препятствующих будущим инфузиям. Кроме того, i.v. лечение может потребоваться в чрезвычайной ситуации или при хирургическом вмешательстве, то есть когда высокий уровень Фактора VIII будет необходим немедленно.
Подкожное введение (s.c.) был предложено для Фактора VIII, например, в WO 95/01804 А1 и WO 95/026750 А1. Однако требовалось введение очень высоких доз Фактора VIII для достижения приемлемой биодоступности.
Другой подход к улучшению биодоступности при не внутривенном введении состоял в использовании альбумин-слитого Фактора VIII (WO 2011/020866 А2).
В WO 2013/057167 А1 предложено введение Фактора VIII в комбинации с сульфатированными гликозаминогликанами посредством не внутривенного введения, необязательно вместе с Фактором фон Виллебранда.
В WO 2008/151817 А1 описано общее применение нерасщепленных мультимеров Фактора фон Виллебранда для стабилизации Фактора VIII, полученного из плазмы или рекомбинантного (полноразмерного и делеционных мутантов), предназначенных для экстраваскулярного лечения.
В WO 2013/160005 А1 описано общее применение рекомбинантного Фактора фон Виллебранда или рекомбинантных фрагментов Фактора фон Виллебранда для улучшения биодоступности после s.c.-лечения для очень специфичных молекул Фактора VIII, где указанные молекулы Фактора VIII содержат укороченный B-домен размером 100-400 аминокислот. В соответствии с WO 2013/160005 A1 молекулы Фактора VIII с укороченными доменами B от 100 до 400 аминокислот, обладают более высокой биодоступностью Фактора VIII по сравнению с Фактором VIII, содержащим полный домен B, или с молекулами Фактора VIII, содержащими укороченный домен В, в котором нет или содержится несколько аминокислот.
Все еще существует потребность в препаратах Фактора VIII, демонстрирующих улучшенную биодоступность, стабильность и/или меньший риск для выработки антител, избегая таким образом недостатки предшествующего уровня техники.
Целью настоящего изобретения является предложение альтернативных препаратов Фактора VIII. Предпочтительно, эти препараты должны демонстрировать повышенную стабильность, улучшенную биодоступность и/или уменьшенный риск для иммунологических реакций.
В одном воплощении, эту задачу решают с помощью композиции, содержащей комплекс Фактора VIII и одного или нескольких пептидов Фактора фон Виллебранда, где пептиды Фактора фон Виллебранда содержат, по меньшей мере аминокислоты 764-1035 и 1691-1905 из SEQ ID No. 1, но не содержат аминокислоты 2255-2645 из SEQ ID NO 1.
Согласно настоящему изобретению, предлагается препарат Фактора VIII, содержащий пептиды Фактор фон Виллебранда. Фактор VIII образует комплекс с содержанием пептидов Фактора фон Виллебранда.
Используемый в настоящем изобретении Фактор VIII охватывает полноразмерный Фактор VIII, где B-домен заменен на искусственный линкер или фрагмент естественного B-домена или их комбинацию, то есть B-домен имеет отличный размер по сравнению с полноразмерным Фактором VIII. Он охватывает также Фактор VIII с ограниченным числом модификаций, включающих вставки, делеции или замены, особенно Фактор VIII, адаптированный к гаплотипам, как описано у K.R. Viel, et al. New England J Med 2009; 360: 1618-1627. Предпочтительно, гомология последовательности с Фактором VIII (как определено в аминокислотах 20-2351 P00451 из SwissProt 21 июля, 1986), но без учета гомологии в B-домене, составляет 99% согласно FASTA, реализованной с помощью FASTA версии 36 на основе WR Pearson (1996) «Effective protein sequence comparison» Meth. Enzymol. 266:227-258. Другими словами, при вычислении гомологии последовательностей, В-домен не включают в сравнение обоих белков. Также охвачены модифицированный Фактор VIII, типа сшитых белков HES-Фактор VIII или ПЭГ-Фактор VIII или Фактор VIII Fc, и сшитые белки Фактора VIII с альбумином, как это описано в Oldenburg, Haemophilia (2014), 20 (Suppl. 4), 23-28.
Фактор VIII по настоящему изобретению может быть получен из плазмы или может быть рекомбинантным Фактором VIII. Когда используют рекомбинантный Фактор VIII, то предпочтительно он экспрессируется в человеческой клеточной линии, чтобы имитировать человеческий профиль гликозилирования (Casademunt, Eur J Haematol 2012; 89:. 165-76), или, как описано в публикации WO 2010/020690.
Пептиды Фактора фон Виллебранда, используемые в настоящем документе, представляют собой пептиды, содержащие, по меньшей мере, аминокислоты 764-1035 из SEQ ID NO:1 и 1691-1905 из SEQ ID NO:1 в одной аминокислотной цепи. Эти аминокислоты могут быть частью более длинной последовательности, содержащей обе эти последовательности вместе. Другими словами, пептиды фон Виллебранда согласно изобретению содержат как SEQ ID NO: 5, так и SEQ ID NO: 6. Они могут содержать дополнительные части Фактора фон Виллебранда, за исключением всех аминокислот 2255-2645 (SEQ ID NO: 7). Пептиды фон Виллебранда могут содержать другие последовательности, которые являются частью последовательности SEQ ID NO: 1, или последовательности, которые не являются частью SEQ ID NO: 1, например, аминокислотные линкеры или тому подобное. Предпочтительно, общее количество аминокислот, которые не являются частью SEQ ID NO: 1, составляет не более чем 50, не более чем 20 или не более чем 10 аминокислот.
Один из важных аспектов изобретения состоит в том, что аминокислоты 2255-2645 из SEQ ID No. 1 не являются частью пептидов Фактора фон Виллебранда. Другими словами, пептиды Фактора фон Виллебранда не содержат какой-либо последовательности, которая имеет, по меньшей мере, 90% гомологии с SEQ ID NO: 7, в соответствии с FASTA, описанной ниже.
SEQ ID NO: 1 представляет собой последовательность P04275 базы данных Swiss Prot по состоянию на 11 января 2011 года.
Пептиды Фактор фон Виллебранда в композиции по настоящему изобретению могут представлять собой пептиды, имеющие ту же последовательность, или могут представлять собой смесь пептидов, имеющих последовательности, как это определено выше.
Обычно молекулярное соотношение Фактора VIII и пептидов Фактора фон Виллебранда будет находиться в диапазоне от 1:1 до 1:20, предпочтительно от 1:2 до 1:10. Если пептиды Фактора Виллебранда представлены в форме димеров или мультимеров, то молекулярное соотношение рассчитывается на одной аминокислотной цепи, т.е. комплекс молекулы Фактора VIII с димером пептидов Фактора фон Виллебранда будет иметь соотношение 1:2.
Комплекс, используемый в данном документе, относится к нековалентному связыванию Фактора VIII с одним или несколькими пептидами Фактора фон Виллебранда.
В предпочтительном воплощении настоящего изобретения, пептиды фактора фон Виллебранда являются фрагментами фактора фон Виллебранда, т.е. N-концевыми и/или С-концевыми укороченными формами фактора фон Виллебранда.
В одном воплощении, фрагменты содержат аминокислоты 764-1905 SEQ ID NO: 1.
Следующее воплощение изобретения представляет собой композицию, содержащую комплекс фактора VIII и один или несколько пептидов Фактора Виллебранда, которые являются фрагментами фактора фон Виллебранда и имеют аминокислотную последовательность, которая соответствует аминокислотной последовательности SEQ ID NO: 1, начиная от аминокислоты 764 и заканчивая между аминокислотами 1905 и 2153 при наличии до 20 или до 10 модификаций, выбранных из аминокислотных делеций, вставок или замен.
Предпочтительные пептиды Фактор фон Виллебранда представляют собой:
Пептиды, имеющие последовательность 764-1905 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1906 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1907 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1908 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1909 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1910 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1911 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1912 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1913 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1914 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1915 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1916 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1917 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1918 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1919 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1920 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1921 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1922 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1923 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1924 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1925 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1926 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1927 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1928 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1929 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1930 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1931 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1932 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1933 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1934 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1935 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1936 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1937 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1938 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1939 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1940 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1941 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1942 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1943 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1944 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1945 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1946 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1947 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1948 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1949 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1950 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1951 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1952 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1953 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1954 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764 -1955 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1956 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1957 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1958 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1959 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1960 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1961 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1962 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1963 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1964 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1965 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1966 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1967 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1968 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1969 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1970 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1971 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1972 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1973 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1974 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1975 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1976 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1977 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1978 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1979 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1980 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1981 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1982 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1983 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1984 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1985 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1986 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1987 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1988 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1989 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1990 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1991 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1992 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1993 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1994 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1995 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1996 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1997 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1998 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-1999 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2000 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2001 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2002 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2003 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2004 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2005 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2006 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2007 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2008 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2009 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2010 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2011 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2012 в SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2013 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2014 в SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2015 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2016 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2017 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2018 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2019 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2020 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2021 из SEQ ID NO: 1
Пептиды, имеющие последовательность с 764-2022 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2023 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2024 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2025 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2026 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2027 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2028 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2029 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2030 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2031 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2032 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2033 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2034 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2035 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2036 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2037 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2038 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2039 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2040 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2041 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2042 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2043 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2044 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2045 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2046 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2047 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2048 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2049 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2050 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2051 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2052 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2053 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2054 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2055 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2056 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2057 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2058 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2059 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2060 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2061 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2062 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2063 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2064 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2065 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2066 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2067 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2068 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2069 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2070 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2071 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2072 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2073 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2074 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2075 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2076 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2077 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2078 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2079 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2080 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2081 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2082 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2083 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2084 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2085 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2086 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2087 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2088 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2089 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2090 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2091 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2092 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2093 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2094 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2095 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2096 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2097 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2098 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2099 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2100 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2101 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2102 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2103 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2104 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2105 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2106 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2107 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2108 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2109 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2110 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2111 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2112 SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2113 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2114 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2115 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2116 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2117 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2118 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2119 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2120 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2121 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2122 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2123 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2124 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2125 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2126 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2127 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2128 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2129 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2130 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2131 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2132 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2133 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2134 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2135 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2136 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2137 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2138 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2139 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2140 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2141 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2142 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2143 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2144 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2145 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2146 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2147 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2148 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2149 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2150 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2151 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2152 из SEQ ID NO: 1
Пептиды, имеющие последовательность 764-2153 из SEQ ID NO: 1
Следующее воплощение изобретения представляет собой композицию, содержащую комплекс Фактора VIII с одним или несколькими пептидами Фактора фон Виллебранда, где
- пептиды Фактора фон Виллебранда являются фрагментами фактора фон Виллебранда
- Комплекс фактора VIII и фрагментов Фактора фон Виллебранда демонстрирует уменьшенное связывание с фосфолипидами мембран по сравнению с Фактором VIII индивидуально
- Комплекс Фактора VIII и фрагментов Фактора фон Виллебранда демонстрирует уменьшенное связывание с коллагеном III по сравнению с комплексом фактора VIII и полноразмерного Фактора фон Виллебранда
- Комплекс Фактора VIII и фрагментов Фактора фон Виллебранда демонстрирует уменьшенное связывание с гепарином по сравнению с комплексом Фактора VIII и полноразмерного Фактора фон Виллебранда.
Предпочтительно, пептиды Фактора фон Виллебранда имеют молекулярную массу <500 кДа, предпочтительно <400 кДа. Поскольку Фактор фон Виллебранда часто образует олигомеры или мультимеры, то пептиды по настоящему изобретению также могут быть представлены в виде мультимеров или олигомеров.
В предпочтительном воплощении, пептиды по настоящему изобретению обладают, по меньшей мере, одним свойством, выбранным из группы, состоящей из
(i) константа аффинного связывания для гепарина KD> 1 нМ, предпочтительно ≥ 2,43 нМ
(ii) константа аффинного связывания для коллагена III в KD> 5 нМ, предпочтительно ≥ 17,02 нМ
(iii) константа аффинного связывания для Фактора VIII KD <100 нМ или <10 нМ, предпочтительно ≤ 6,19 нМ и
(iv) ингибирование связывания Фактора VIII с фосфолипидами составляет, по меньшей мере, 70%, предпочтительно по меньшей мере, 80% или, по меньшей мере, 90%.
Пептиды Фактора фон Виллебранда по изобретению предпочтительно демонстрируют уменьшенное связывание с гепарином, более низкую аффинность к коллагену (например, коллагену III), более низкую аффинность к фосфолипидам, но при этом сильное связывание с Фактором VIII.
Удивительно, но слабое связывание с фосфолипидами и коллагеном повышает скорость высвобождения в случае не внутривенного введения, особенно подкожного.
Измерение соответствующих констант аффинности связывания описано в экспериментальной части.
В одном воплощении, пептиды фактора фон Виллебранда являются производными от фактора фон Виллебранда с помощью протеолитического или химического расщепления. Если используется протеолитическое расщепление, то особенно предпочтительна протеаза V-8 S. aureus.
Предпочтительно, композиция по настоящему изобретению включает, по меньшей мере, одно из следующих свойств:
(i) пептиды Фактор фон Виллебранда экранируют Фактор VIII от связывания с антителами с минимизацией образования ингибитора у пациента
(ii) стабилизирует Фактор VIII для обеспечения остаточной активности Фактора VIII, составляющей, по меньшей мере, 90% после хранения в течение 12 месяцев в замороженном виде в жидком состоянии при-70 °C
(iii) стабилизирует Фактор VIII для обеспечения остаточной активности Фактора VIII, составляющей, по меньшей мере, 90% после хранения в течение 24 месяцев в лиофилизированной форме при температуре 5 °C
(iv) стабилизирует Фактор VIII для обеспечения остаточной активности Фактора VIII, составляющей, по меньшей мере, 90% после хранения в течение 12 месяцев в лиофилизированной форме при 25 °C
(v) пролонгирует период полужизни Фактора VIII в естественных условиях, по меньшей мере, на 20% и
(vi) уменьшает образование ингибитора у ранее не получавших лечение пациентов до менее чем 20%, предпочтительно до менее чем 10% после обработки композицией в течение 6 месяцев.
Удивительно, что пептиды Фактора фон Виллебранда, оказывается, увеличивают стабильность Фактора при хранении (срок годности), и/или уменьшают образование ингибиторов у пациентов. Образование ингибиторов является одной из основных проблем при лечении хронических нарушений свертываемости.
Композиция по настоящему изобретению является особенно полезной при лечении или профилактике нарушения свертываемости крови.
Таким образом, еще одним воплощением изобретения является способ лечения нарушения свертываемости крови, включающий введение пациенту, нуждающемуся в этом, эффективного количества композиции по настоящему изобретению.
Количество зависит от заболевания или состояния, подлежащего лечению, и может быть выбрано специалистом в данной области техники. Для длительного лечения, как правило, подходит количество, составляющее 20-40 МЕ/кг массы тела на применение. В чрезвычайной ситуации, количество может составлять примерно 10-50 МЕ/кг массы тела.
Композиция по изобретению может применяться путем внутривенного или не внутривенного введения. Не внутривенное введение может представлять собой подкожную инъекцию, внутрикожную инъекцию или внутримышечное введение.
Одним из преимуществ способа по настоящему изобретению является возможность использования нанофильтрации для удаления вирусов. Фактор фон Виллебранда из-за его размера не может быть подвергнут нанофильтрации с использованием нанофильтров с малым размером пор для удаления вирусов. Поскольку размер пептидов Фактора фон Виллебранда намного меньше полноразмерной молекулы Фактора Виллебранда, то нанофильтрация с использованием малого размера пор становится возможной. Нанофильтрацию осуществляют при таком размере пор и при таких условиях, которые приводят к уменьшению концентрации одного из самых маленьких известных вирусов - парвовируса свиней, по меньшей мере, в 100 раз (2 log), предпочтительно по меньшей мере в 1000 (3 log) и, наиболее предпочтительно, до концентрация ниже предела обнаружения теста на парвовирус, необязательно с использованием одного или нескольких нанофильтров последовательно. Для этого теста, парвовирус свиней выкалывали в образец и анализировали после фильтрации.
Таким образом, следующее воплощение изобретения представляет собой способ уменьшения количества вируса, включающий стадию нанофильтрации пептидов Фактора фон Виллебранда до или после комбинации с Фактором VIII, в результате чего количество парвовируса свиней будет уменьшено, по меньшей мере, в 2 log.
Предпочтительный буфер для введения композиции по изобретению содержит мелизитозу, предпочтительно в количестве до 1000 мМ, конкретно, примерно от 10 мМ примерно до 200 мМ, конкретно, примерно от 10 мМ примерно до 100 мМ.
Следующее воплощение изобретения представляет собой способ получения пептидов Фактора фон Виллебранда, включающий следующие стадии:
Инкубация Фактора фон Виллебранда с протеазой V-8 S. aureus в течение 2-16 часов при массовом соотношении фермента к Фактору фон Виллебранда масс./масс., составляющем 1:5-1:100
Связывание и очищение на анионообменной смоле и сбор целевых очищенных пептидов VWF во фракцию, поступающую из анионообменной смолы с применения увеличенного количества концентрации соли.
Краткое описание чертежей
На Фигуре 1 представлена очистка фрагмента III (SPIII) из pdVWF, гидролизованных с помощью протеазы V8 S. aureus. A - MonoQ Хроматограмма профиля фрагмента III (показан стрелкой). B - SDS-PAGE-гель очищенного фрагмента; сниж.- снижается; н.с.- не снижается.
На Фигуре 2 представлена очистка фрагмента I (III-Т4) из фрагмента III, гидролизованного трипсином. MonoQ хроматограмма профиля элюирования фрагмента I (показано стрелкой). Изображение SDS-PAGE очищенного фрагмента в не восстанавливающих условиях показано на вставке.
На Фигуре 3 представлена очистка фрагмента II от фрагмента III после второго гидролиза протеазой V8 S. aureus. A - MonoQ хроматограмма профиля элюирования фрагмента II (показано стрелкой), второго продукта расщепления, а также протеазы V8. B - хроматограмма второй MonoQ хроматографии, необходимой для полного удаления протеазы. Изображение SDS-PAGE очищенного фрагмента в восстанавливающих условиях показано на вставке.
На Фигуре 4 представлено связывание pdVWF фрагментов II и III с rFVIII. A, B, C Сенсограммы связывания (серые кривые), а также выравнивание кривой (черные кривые), репрезентативное для взаимодействия между иммобилизованным rFVIII и pdVWF/очищенными фрагментами II и III. Концентрации и тип образца указаны на диаграмме. C- константы диссоциации (KD) выражены как среднее и SEM; n=8.
На Фигуре 5 представлено связывание rFVIII с фосфолипидным монослоем в SPR и ингибирование с помощью pdVWF. А - сенсограмма связывания rFVIII и rFVIII в присутствии либо 108 нМ БСА (бычий сывороточный альбумин), либо 47,6 нМ pdVWF; каждый образец в трех экземплярах. B - среднее и SD уровня связывания, измеренного через 120 сек после окончания инъекции аналита, выраженное в процентах от связывания rFVIII; n=3.
На Фигуре 6 представлено ингибирование взаимодействия rFVIII-фосфолипид с помощью фрагментов, производных от Фактора фон Виллебранда, по измерениям в SPR. Связывание rFVIII с фосфолипидным монослоем проводили в присутствии трех различных концентраций трех фрагментов, производных от Фактора фон Виллебранда (концентрации и тип фрагмента обозначены на графике). График представляет собой среднее и SD уровней связывания, измеренных через 120 сек после окончания инъекции аналита, выраженные в процентах от связывания rFVIII; n=3.
На Фигуре 7 представлено зависимое от концентрации ингибирование связывания rFVIII с фосфолипидным монослоем фрагмента III. А - сенсограммы связывания rFVIII с фосфолипидным монослоем в присутствии различных концентраций фрагмента III (концентрации указаны на графике), каждый образец в трех экземплярах. B - среднее и SD уровней связывания, измеренных через 120 сек после окончания инъекции аналита, выраженные в процентах от связывания rFVIII; n=3.
На Фигуре 8 показаны связывания из pdVWF и фрагмента III с коллагеном типа III. A, B - сенсограммы связывания (серые кривые), а также выравнивание кривой (черные кривые), репрезентативное для взаимодействия между иммобилизованным коллагеном типа III и pdVWF/очищенного фрагмента III. Концентрации и тип образца указаны на диаграмме. C- константы диссоциации (KD) выражены как среднее и SEM; n=9.
На Фигуре 9 представлено связывание pdVWF и фрагмента III с гепарином. A, B - сенсограммы связывания (серые кривые) и выравнивание кривой (черные кривые), репрезентативное для взаимодействия между иммобилизованным гепарином и pdVWF/очищенным фрагментом III. Концентрации и тип образца указаны на диаграмме. C - константы диссоциации (KD) выражены как среднее и SEM; n=6.
На Фигуре 10 представлено сравнение значений времени свертывания цельной крови (WBCT), измеренных в образцах крови собак с гемофилией А, подвергнутых лечению подкожно с использованием FVIII индивидуально или в комбинации с фрагментом III VWF. WBCT, полученные после подкожного применения FVIII в комбинации с пятикратным молярным избытком фрагмента III VWF, применяли при 200 МЕ FVIII/кг массы тела. Горизонтальная пунктирная линия отмечает верхний предел времени свертывания крови у здоровых собак (12 минут).
На Фигуре 11 показана активность FVIII, измеренная с помощью теста хромогенной активности FVIII на образцах плазмы собак с гемофилией А, полученных после применения FVIII или FVIII в комбинации с фрагментом III VWF. А- FVIII, или FVIII с пятикратным молярным избытком фрагмента III VWF применяли подкожно при 200 МЕ FVIII/кг массы тела; площадь под кривой (AUC) для образца FVIII индивидуально составила 2,867, а для FVIII в комбинации с фрагментом III VWF-4,917. B- FVIII, или FVIII с пятикратным молярным избытком фрагмента III VWF применяли внутривенно в дозе 200 МЕ FVIII/кг массы тела. AUC для FVIII образца индивидуально составила 27,69, а для FVIII в комбинации с фрагментом III VWF-45,72.
На Фигуре 12 представлено связывание рекомбинантного мономера фрагмента III, рекомбинантного димера фрагмента III и VWF (flVWF) из плазмы с rFVIII. A, B, C - сенсограммы связывания (серые кривые), и выравнивание кривой (черные кривые), репрезентативное для взаимодействия между иммобилизованным VWF или рекомбинантными VWF-фрагментами и FVIII. Тип образца указан на диаграмме. Концентрация применяемого FVIII составили 0, 0,2, 0,6, 1,7, 5, 15, 45 и 135 нМ. D - константы диссоциации (KD) выражены как среднее и SD; n=4.
На Фигуре 13 представлена стабилизация FVIII с помощью фрагмента III VWF. FVIII активность индивидуально или FVIII, в комплексе с фрагментом III VWF, инкубированном при 40 °С, измеряли в различные моменты времени.
На Фигуре14 представлено связывание с гепарином с использованием аффинной хроматографии с гепарином двух фрагментов VWF, как описано в Примере 9.
Примеры
Изобретение дополнительно поясняется следующими, не ограничивающими примерами.
Пример 1
Получение и очистка фрагментов, производных плазматического Фактора фон Виллебранда.
Получение и очистка фрагмента III (SPIII, ост. 764-2128) (Согласно Marti et al. Identification of disulfide-bridged substructures within human von Willebrand factor. Biochemistry 1987; 26:8099-8109 с модификациями) (SEQ. ID. No. 2):
SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVA
LERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKY
LFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNV
KRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQ
NNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRIL
TSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTA
TLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKIL
DELLQTCVDPEDCPVCEVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGL
VVPPTDAPVSPTTLYVEDISEPPLHDFYCSRLLDLVFLLDGSSRLSEAEFEVLKAFVVDM
MERLRISQKWVRVAVVEYHDGSHAYIGLKDRKRPSELRRIASQVKYAGSQVASTSEVLKY
TLFQIFSKIDRPEASRITLLLMASQEPQRMSRNFVRYVQGLKKKKVIVIPVGIGPHANLK
QIRLIEKQAPENKAFVLSSVDELEQQRDEIVSYLCDLAPEAPPPTLPPDMAQVTVGPGLL
GVSTLGPKRNSMVLDVAFVLEGSDKIGEADFNRSKEFMEEVIQRMDVGQDSIHVTVLQYS
YMVTVEYPFSEAQSKGDILQRVREIRYQGGNRTNTGLALRYLSDHSFLVSQGDREQAPNL
VYMVTGNPASDEIKRLPGDIQVVPIGVGPNANVQELERIGWPNAPILIQDFETLPREAPD
LVLQRCCSGEGLQIPTLSPAPDCSQPLDVILLLDGSSSFPASYFDEMKSFAKAFISKANI
GPRLTQVSVLQYGSITTIDVPWNVVPEKAHLLSLVDVMQREGGPSQIGDALGFAVRYLTS
EMHGARPGASKAVVILVTDVSVDSVDAAADAARSNRVTVFPIGIGDRYDAAQLRILAGPA
GDSNVVKLQRIEDLPTMVTLGNSFLHKLCSGFVRICMDEDGNEKRPGDVWTLPDQCHTVT
CQPDGQTLLKSHRVNCDRGLRPSCPNSQSPVKVEETCGCRWTCPCVCTGSSTRHIVTFDG
QNFKLTGSCSYVLFQNKEQDLEVILHNGACSPGARQGCMKSIEVKHSALSVELHSDMEVT
VNGRLVSVPYVGGNMEVNVYGAIMHEVRFNHLGHIFTFTPQNNEFQLQLSPKTFASKTYG
LCGICDENGANDFMLRDGTVTTDWKTLVQEWTVQRPGQTCQPILE
Фрагмент III получали путем гидролиза выделенного из плазмы Фактора фон Виллебранда (pdVWF) с помощью протеазы V-8 S. aureus. Гидролиз проводили в течение 3 часов при температуре 37 °С в буфере 50 мМ Трис-HCl, 150 мМ NaCl, рН 7,8 при массовом соотношении фермента к белку, составляющем 1:40.
Очистку фрагмента проводили с использованием колонки с сильной анионообменной смолой (MonoQ). Рабочий буфер представляет собой 20 мМ Трис-HCl, рН 7,4, и элюирующий буфер (буфер В) 20 мМ Трис-HCl, 500 мМ NaCl, рН 7,4. V-8 протеаза S. aureus элюируется из анионообменной колонки при приблизительно 22 мСм/см (около 40% буфера В), поэтому длинная стадия промывки при 42% до элюирования фрагмента требуется для вымывания протеазы. В качестве альтернативы может быть проведена стадия SEC на Superose 6 10/300 GL для удаления протеазы.
Очистка фрагмента III и полученный продукт изображены на Фиг. 1. Последовательность, определенная Marti et al. 1987, была подтверждена с помощью анализа Масс-спектрометрии.
Получение и очистка фрагмента I (III-T4, ост. 764-1035) (Согласно Marti et al. 1987 с модификациями) (SEQ. ID. No. 3):
SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVA
LERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKY
LFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNV
KRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQ
NNDLTSSNLQVEEDPVDFGNSWKVSSQCADTR
Фрагмент I получали из фрагмента III (SPIII) с помощью гидролиза трипсином (обработанный ТРСК от крупного рогатого скота). Гидролиз проводили в течение 1,5 ч в буфере 100 мМ NH4HCO3 рН 8 при массовом соотношении фермента к белку 1:100.Гидролиз завершали путем добавления соевого ингибитора трипсина.
Очистку фрагмента I проводили с использованием колонки с сильной анионообменной смолой (MonoQ) с последующей SEC на Superose 6, 10/300 GL. Рабочий буфер для анионообменной колонки представляет собой 20 мМ Трис-HCl, рН 7,4, и элюирующий буфер (буфер В) 20 мМ Трис-HCl, 500 мМ NaCl, рН 7,4.Рабочий буфер для SEC представляет собой PBS (фосфатный буферный раствор) pH 7.
Очистка фрагмента I и полученный продукт изображены на Фиг. 2. Последовательность, определенная Marti et al. 1987, была подтверждена с помощью анализа MS.
Получение и очистка фрагмента II (ост. 764-1673) (SEQ. ID. No. 4):
SLSCRPPMVKLVCPADNLRAEGLECTKTCQNYDLECMSMGCVSGCLCPPGMVRHENRCVA
LERCPCFHQGKEYAPGETVKIGCNTCVCQDRKWNCTDHVCDATCSTIGMAHYLTFDGLKY
LFPGECQYVLVQDYCGSNPGTFRILVGNKGCSHPSVKCKKRVTILVEGGEIELFDGEVNV
KRPMKDETHFEVVESGRYIILLLGKALSVVWDRHLSISVVLKQTYQEKVCGLCGNFDGIQ
NNDLTSSNLQVEEDPVDFGNSWKVSSQCADTRKVPLDSSPATCHNNIMKQTMVDSSCRIL
TSDVFQDCNKLVDPEPYLDVCIYDTCSCESIGDCACFCDTIAAYAHVCAQHGKVVTWRTA
TLCPQSCEERNLRENGYECEWRYNSCAPACQVTCQHPEPLACPVQCVEGCHAHCPPGKIL
DELLQTCVDPEDCPVCEVAGRRFASGKKVTLNPSDPEHCQICHCDVVNLTCEACQEPGGL
VVPPTDAPVSPTTLYVEDISEPPLHDFYCSRLLDLVFLLDGSSRLSEAEFEVLKAFVVDM
MERLRISQKWVRVAVVEYHDGSHAYIGLKDRKRPSELRRIASQVKYAGSQVASTSEVLKY
TLFQIFSKIDRPEASRITLLLMASQEPQRMSRNFVRYVQGLKKKKVIVIPVGIGPHANLK
QIRLIEKQAPENKAFVLSSVDELEQQRDEIVSYLCDLAPEAPPPTLPPDMAQVTVGPGLL
GVSTLGPKRNSMVLDVAFVLEGSDKIGEADFNRSKEFMEEVIQRMDVGQDSIHVTVLQYS
YMVTVEYPFSEAQSKGDILQRVREIRYQGGNRTNTGLALRYLSDHSFLVSQGDREQAPNL
VYMVTGNPASDEIKRLPGDIQVVPIGVGPNANVQELERIGWPNAPILIQDFETLPREAPD
LVLQRCCSGE
Фрагмент II получали из фрагмента III с помощью второго гидролиза протеазой V8 S. aureus. Гидролиз проводили в течение 21 ч в буфере 50 мМ Трис-HCl, 150 мМ NaCl, рН 7,8 с массовым соотношением фермента к белку, составляющим 1:10.
Очистку фрагмента II проводили с использованием колонки с сильной анионообменной смолой (MonoQ). Рабочий буфер представляет собой 20 мМ Трис-HCl, рН 7,4, и элюирующий буфер (буфер В) 20 мМ Трис-HCl, 500 мМ NaCl, рН 7,4.Вторая очистка MonoQ с длинной стадией промывки при 42% B требуется для удаления протеазы.
Очистка фрагмента II и полученный продукт изображены на Фиг. 3. Второй сайт расщепления V8 между Glu1673-Gly1674 был определен Fretto et al. 1986 и подтвержден анализом масс-спектрометрии.
Пример 2
Определение аффинности связывания Фактора VIII
Анализ проводили с использованием прибора BIAcore 2000 (GE Healthcare) в соответствии с McCormick et al. 2004 с модификациями. Вкратце, rFVIII ковалентно связывали с сенсорным чипом CM5, что приводило к получению уровня покрытия ~ 200RU. Затем фрагменты Фактора фон Виллебранда, а также полноразмерный Фактор фон Виллебранда (flvWF) инъецировали на поверхность сенсорного чипа. Рабочий буфер представляет собой 20 мМ Hepes, 150 мМ NaCl, 5 мМ CaCl2, 0,02% твин-20. Константы аффинности диссоциации определяли для flVWF, а также для фрагментов II и III, при этом не было никакого значительного связывания фрагмента I с Фактором VIII, следовательно, KD не определяли. Сенсограммы связывания и расчетные значения KD изображены на Фиг. 4. flVWF связывался с rFVIII с KD 0,67 нМ, фрагмент III связывался с более низкой аффинностью (KD 6,18 нМ), аффинность дополнительно снижалась для фрагмента II (KD 154,6 нМ).
Пример 3
Определение связывания Фактора VIII с фосфолипидным монослоем и ингибирование с помощью Фактора фон Виллебранда и фрагментов, производных Фактора фон Виллебранда.
Анализ проводили с использованием прибора BIAcore 2000 (GE Healthcare) в соответствии с Saenko et al.1999 с модификациями. Вкратце, фосфолипидные везикулы готовили из DOPC (1,2-диолеоил-sn-глицерин-3-фосфохолин) и DOPS (1,2-диолеоил-sn-глицерин-3-фосфо-L-серин). Однослойные везикулы готовили в соответствии с MacDonal et al. 1991 с использованием экструдера и наносили в виде покрытия на сенсорный чип HPA. Затем соединения, представляющие интерес, инъецировали на поверхность PCPS и оценивали уровень связывания через 120 с после инъекции.
Отрицательные контроли; Фактор фон Виллебранда и БСА не связывались с поверхностью PSPC (не показано), в отличие от этого, был продемонстрирован высокий уровень связывания rFVIII. Это связывание может быть полностью ингибировано Фактором фон Виллебранда, в отличие от добавления высокой концентрации БСА, которое не оказывало влияния на связывание (Фиг. 5).
Для того чтобы оценить, способны ли фрагменты, полученные с помощью ограниченного гидролиза, ингибировать связывание PSPC аналогично flVWF, фрагменты I, II и III инъецировали на поверхность сенсорного чипа. Только фрагмент III обладает способностью ингибировать взаимодействие между rFVIII и фосфолипидным монослоем (Фиг. 6). Этот эффект имел дозозависимый характер при почти полном ингибировании при 2,5х избытке фрагмента III по отношению к rFVIII (Фиг. 7).
Пример 4
Определение аффинности связывания коллагена III с flVWF и с фрагментом III.
Анализ проводили с использованием прибора BIAcore 2000 (GE Healthcare) в соответствии с Romjin et al. 2003 с модификациями. Вкратце, человеческий коллаген типа III, гидролизованный пепсином, ковалентно связывали с поверхностью сенсорного чипа CM5. Затем образцы инъецировали на поверхность сенсорного чипа. Рабочий буфер представлял собой 10 мМ Hepes, 150 мМ NaCl, 3,4 мМ ЭДТА, 0,005% Tween-20. FlVWF связывался с коллагеном III с очень высокой аффинностью (0,75 нМ), связывание фрагмента III значительно снижалось до 17,02 нм (Фиг. 8).
Пример 5
Определение аффинности связывания гепарина с flVWF и фрагментом III.
Анализ проводили с использованием прибора Biacore T200 (GE Healthcare) в соответствии с Sarafanov et al. 2001. Вкратце, гепарин из слизистой оболочки кишечника свиньи биотинилировали с использованием набора реагентов NHS-биотин и связывали с поверхностью сенсорного чипа SA. Эталонную проточную ячейку покрывали биотином. Затем образцы инъецировали на поверхность сенсорного чипа. Рабочий буфер представлял собой 150 мМ Hepes, 150 мМ NaCl, 5 мМ CaCl2, 0,05% Tween-20. FlVWF связывался с гепарином с аффинностью 0,65 нМ, аффинность связывания фрагмента III была значительно снижена до 2,43 нм (Фиг. 9).
Пример 6
Определение восстановления FVIII или комплекса FVIII/Фрагмент III VWF и периода полужизни в кровотоке у собак с гемофилией А.
Две собаки с гемофилией А были подвергнуты подкожной и затем внутривенной инъекции рекомбинантного FVIII с делетированным В-доменом индивидуально или в комбинации с пятикратным молярным избытком Фрагмент III VWF. Собака 1 получала 200 МЕ/кг массы BW FVIII индивидуально, и собака 2 получала 200 МЕ/кг BW FVIII в комплексе с Фрагментом III VWF. Образцы крови собирали через 0,5, 1, 2, 4, 8, 12, 24, 32, 48, 72 и 96 часов после каждого подкожного или внутривенного введения лекарственного средства. Образцы анализировали на время свертывания цельной крови (WBCT) и на активность в хромогенном тесте FVIII активности. Подкожное введение Фрагмента III VWF в комплексе с FVIII приводило к 1,4-кратному увеличению времени, необходимому для превышения времени свертывания крови здоровой собаки по сравнению с подкожным введением FVIII индивидуально (Фиг. 10). Введение Фрагмента III VWF вместе с FVIII приводило также к увеличению активности FVIII в плазме собак с течением времени и почти удваивались значения площади под кривой (AUC) для обоих применений, подкожного и внутривенного, по сравнению с введением FVIII индивидуально (Фиг. 11).
Пример 7
Определение аффинности связывания FVIII с рекомбинантным мономером и димером фрагмента III.
Рекомбинантный фрагмент III транзиторно экспрессировали в клеточной линии HEK293, с С-концевым Strep-маркером и очищали с помощью аффинной хроматографии с «Strep-tactin». Мономеры и димеры фрагмента III разделяли с помощью эксклюзионной хроматографии (SEC). Анализ проводили с использованием прибора Biacore 2000. Мономеры и димеры Фрагмента III иммобилизовали на CM5, и серии концентраций FVIII инъецировали на поверхность сенсорного чипа. В качестве контроля использовали полноразмерный VWF, полученный из плазмы. Рабочий буфер представлял собой 150 мМ Hepes, 150 мМ NaCl, 5 мМ CaCl2, 0,05% Tween-20. FVIII связывался с димером фрагмента III с константой аффинности 1,9 нМ. Аффинность FVIII к мономерному Фрагменту III была значительно ниже (KD=14,3 нМ) (Фиг. 12).
Пример 8
Стабилизация rFVIII в растворе с помощью Фрагмента III VWF.
2000 МЕ рекомбинантного FVIII (Nuwiq®) восстанавливали в 2,5 мл воды, с добавлением или без добавления пятикратного молярного избытка Фрагмента III VWF. Оба препарата инкубировали при 40 °С, и аликвоты отбирали через 48, 96, 192, 384, 408 и 672 часа. Образцы анализировали на активность FVIII в хромогенном анализе активности FVIII. Фрагмент III VWF способствовал значительно более продолжительной активности FVIII при 40 °С (Фиг. 13).
Пример 9
Сравнение связывания гепарина между рекомбинантным фрагментом III и фрагментом NovoSeq21.
Рекомбинантный фрагмент III и фрагмент NovoSeq21 (SEQ ID N 21 из WO2013/160005A1) транзиторно экспрессировали в клеточной линии HEK293, с С-концевым Strep-маркером и очищали с помощью Strep-tactin аффинной хроматографии. Связывание гепарина тестировали с использованием аффинной хроматографии с гепарином. Оба рекомбинантных фрагмента связывали с гепариновой колонкой (HiTrap Гепарин HP 1мл, GE Healthcare) и элюировали в линейном градиенте соли в диапазоне концентраций от 0 до 500 мМ NaCl. Оба фрагмента прогоняли в трех экземплярах, см. Фиг. 14. Средний пик элюции для фрагмента NovoSeq21 выходил при 15,57 ± 0,04 мин, что соответствует 285,381 мМ NaCl, и для фрагмента III при 15,47 ± 0,02 мин, что соответствует 282,051 мМ NaCl. Это указывает на более высокую аффинность к гепарину фрагмента NovoSeq21.
Аналитические методы
Описание аналитических методов
FVIII:C, Метод скрининга на основе Coatest
Метод основан на двухстадийном принципе, и его осуществляли с использованием метода микропланшетов. На первой стадии, активированный фактор X (Xa) генерируется посредством внутреннего пути, где FVIII:C выступает в качестве кофактора. На второй стадии, фактор Ха затем определяют с использованием синтетического хромогенного субстрата, S-2222 в присутствии ингибитора тромбина I-2581 для предотвращения гидролиза субстрата тромбином. Реакцию останавливают с помощью кислоты, и активность VIII:C, которая пропорциональна высвобождения pNA (пара-нитроанилина), определяют фотометрически при 405 нм против пустого реагента.
Метод соответствует требованиям, изложенным в Европейской Фармакопее. Единицы FVIII: C выражаются в международных единицах (МЕ), как это определено в настоящее время Международным Стандартом Концентратов (IS), установленным Всемирной организацией здравоохранения (ВОЗ). Была подтверждена процедура с использованием буфера, содержащего 1% BSA вместо тяжелой гемофильной плазмы для предварительных разведений. Смотрите также ссылки на литературу (European Pharmacopoeia Supplement 2000, general Methods, 2.7. Assay of Blood Coagulation FVIII; Rosén S (1984) Assay of FVIII: C with a Chromogenic Substrate. J, Haematol, Suppl 40, vol 33, 139-145, 1984; Carlebjörk G, Oswaldsson U, Rosén S (1987) A simple and accurate micro plate assay for the determination of FVIII activity. Thrombosis Research 47; 5-14, 1987; Mire-Sluis AR, Gerrard T, Gaines das R, Padilla A and Thorpe R. Biological assays: Their Role in the development and quality Control of Recombinant Biological Medicinal Products. Biological, 24, 351-362 (1996)).
Определение общего содержания белка в соответствии с методом Брэдфорда
Определение белка по методу Брэдфорда основано на наблюдении того, что максимальное оптическое поглощение для кислотного раствора Кумасси бриллиантового синего G-250 сдвигается от 465 нм до 595 нм при связывании с белком. Оба гидрофобных и ионных взаимодействия стабилизируют анионную форму красителя, вызывая видимое изменение цвета. Анализ полезен, так как коэффициент экстинкции комплексного раствора красителя-альбумина является постоянным в диапазоне 10-ти кратного изменения концентраций. См. также для дополнительной информации ссылку Bradford, MM. A rapid and sensitive method for the quantisation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254. 1976.
Определение общего содержания белка в соответствии с аминокислотным анализом (ААА)
Перед AAA все белки гидролизуют с помощью 6 М HCl в течение 24 ч при 110 °С. Аминокислоты разделяют с помощью катионообменной хроматографии на сульфированных полистирольных смолах и непрерывно детектируют в элюенте. Детектирование основано на постколоночной модификации нингидрина с использованием двойного фотометра для одновременного измерения при 440 нм для пролина и гидроксипролина и при 570 нм для всех остальных аминокислот. Обе аминокислоты, аспарагин и глутамин, дезамидируются в течение AAA и определяются как аспарагиновая кислота и глутаминовая кислота, соответственно. Таким образом, результаты по аспарагиновой кислоте и глутаминовой кислоте в исходном образце представляют собой сумму аспарагиновой кислоты/аспарагина (ASX) и глутаминовой кислоты/глутамина (GLX), соответственно. Триптофан не создает четкого ответа при использовании этого метода, и, таким образом, не определяется количественно с помощью ААА. Цистеин разрушается во время гидролиза и не определяется количественно. ААА дополнительно описан в ссылке: Total protein AAA analytical method. Spackman, D. H., Stein, W. H., and Moore, S. (1958) Anal. Biochem. 30: 1190-1206.
Чистота или удельная активность (FVIII: C/общий белок)
Чистоту (или так называемую удельную активность) для образца, рассчитывают с использованием значения, полученного из анализа FVIII:C, поделенного на значение, полученного из анализа общего белка.
SDS-PAGE (Распределение по молекулярной массе)
Электрофорез в SDS-полиакриламидном геле (SDS-PAGE) включает разделение белков в зависимости от их размера. Этот метод описывает SDS-PAGE белков, которые прогоняют на геле в восстанавливающих условиях. При нагревании образца в денатурирующих и восстанавливающих условиях, белки становятся развернутыми и покрываются анионным детергентом додецилсульфатом натрия (SDS), приобретая высокий чистый отрицательный заряд, который пропорционален длине полипептидной цепи. При загрузке на матрикс полиакриламидного геля и при помещении в электрическое поле отрицательно заряженные молекулы белка мигрируют в сторону положительно заряженного электрода и разделяются с помощью эффекта молекулярного просеивания, т.е. по их молекулярной массе. Полиакриламидные гели сдерживают более крупные молекулы от настолько быстрой миграции, как у более мелких молекул. Поскольку отношение заряда к массе почти одинаковое среди SDS-денатурированных полипептидов, окончательное разделение белков почти полностью зависит от различий в относительной молекулярной массе полипептидов. В геле однородной плотности расстояние относительной миграции белка (Rf) отрицательно пропорционально логарифму его массы. Если белки известной массы прогоняются одновременно с неизвестными, то связь между Rf и массой может быть определена графически, а массы неизвестных белков можно оценить. Белковые полосы, разделенные электрофорезом, визуализируют путем окрашивания серебром. Оценку осуществляют визуально путем определения проявления стандартов, эталона (контрольный образец) и анализируемых образцов.
Содержание антигена Фактора VIII (FVIII: Ag)
Количество содержания антигена Фактора VIII (FVIII: Ag) измеряют с помощью набора тИФА (ASSERACHROM® VIII: Ag, иммуноферментный анализ для Фактора VIII, набор, Diagnostica Stago (Франция), как дополнительно описано (18) с заменой предоставляемого в наборе буфера на Трис-NaCl буфер+1% бычий сывороточный альбумин для разбавлений образца.
Эксклюзионная хроматография (SEC)
Мономер, агрегат и фрагмент определяют с помощью аналитической колонки (Superdex 200, 10/300 GL, GE Healthcare) эксклюзионной хроматографии (SEC-HPLC), разработанной в условиях нативного буфера (25 мМ HEPES, 0,5 М NaCl, 0,3 М аргинина, 50 мМ CaCl2, 0,02% полисорбат 80, рН 7,5). Загрузка образца составляет приблизительно 1% от колонки эксклюзионной хроматографии, и концентрация Фактора VIII:С составляет приблизительно 1000 МЕ/мл.
Вестерн-блот против Фактора VIII
Продукт вырождения Фактора VIII на основе размера измеряют с помощью Вестерн-блота FVIII. Распределение по молекулярной массе. Белки и пептиды FVIII в препаратах Фактора VIII разделяются по молекулярной массе с помощью электрофореза в додецилсульфат натрия (SDS)-полиакриламидном геле (ПААГ) при восстанавливающих условиях. Затем белки переносят электрофоретически из гелевого матрикса на нитроцеллюлозную мембрану, которую затем инкубируют с блокирующим агентом. Коммерчески доступные поликлональные антитела овец, направленные против цельной молекулы человеческого фактора VIII затем добавляют с последующим добавлением вторичного антитела, меченного ферментом, в качестве зонда. В качестве третьей стадии добавляют хемилюминесцентный субстрат и в сочетании с ферментом излучается свет в качестве побочного продукта. Световой поток захватывается в качестве изображения в реальном времени с использованием охлажденной камеры с устройством с зарядовой связью. Интенсивность сигнала коррелирует с представленностью антигена (FVIII) на мембране блота.
2D-PAGE
2D-электрофорез с окрашиванием серебром проводили с целью изучения профиля электрофоретического разделения белковой цепи Фактора VIII. Изоэлектрическое фокусирование осуществляли в качестве прогона в первом направлении с использованием линейного градиента рН от рН 3 до 10. Второе направление SDS-PAGE прогоняли с использованием Tris-ацетатных гелей (3-8%). Гели окрашивали серебром после прогона во втором направлении.
Общий белок (по Брэдфорду)
Определение белка по Брэдфорду основано на том наблюдении, что максимальное оптическое поглощение для кислотного раствора Кумасси бриллиантового синего G-250 сдвигается от 465 нм до 595 нм, когда происходит связывание с белком. Оба взаимодействия, гидрофобное и ионное, стабилизируют анионную форму красителя, вызывая видимое изменение цвета. Этот анализ полезен, так как коэффициент экстинкции комплексного раствора краситель-альбумин является постоянным в 10-ти кратном диапазоне концентраций. См. также для дополнительной информации ссылку Bradford, MM. A rapid and sensitive method for the quantisation of microgram quantities of protein utilizing the principle of protein-dye binding. Analytical Biochemistry 72: 248-254. 1976.
Все ссылки, упомянутые в настоящем документе, включены в качестве ссылки в полном объеме, в котором включение не противоречит ясно выраженному изложению настоящего описания.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Octapharma AG
<120> Препарат, содержащий Фактор VIII и пептиды Фактора фон Виллебранда
<130> 151432WO
<160> 7
<170> PatentIn version 3.5
<210> 1
<211> 2813
<212> Белок
<213> Homo sapiens
<400> 1
Met Ile Pro Ala Arg Phe Ala Gly Val Leu Leu Ala Leu Ala Leu Ile
1 5 10 15
Leu Pro Gly Thr Leu Cys Ala Glu Gly Thr Arg Gly Arg Ser Ser Thr
20 25 30
Ala Arg Cys Ser Leu Phe Gly Ser Asp Phe Val Asn Thr Phe Asp Gly
35 40 45
Ser Met Tyr Ser Phe Ala Gly Tyr Cys Ser Tyr Leu Leu Ala Gly Gly
50 55 60
Cys Gln Lys Arg Ser Phe Ser Ile Ile Gly Asp Phe Gln Asn Gly Lys
65 70 75 80
Arg Val Ser Leu Ser Val Tyr Leu Gly Glu Phe Phe Asp Ile His Leu
85 90 95
Phe Val Asn Gly Thr Val Thr Gln Gly Asp Gln Arg Val Ser Met Pro
100 105 110
Tyr Ala Ser Lys Gly Leu Tyr Leu Glu Thr Glu Ala Gly Tyr Tyr Lys
115 120 125
Leu Ser Gly Glu Ala Tyr Gly Phe Val Ala Arg Ile Asp Gly Ser Gly
130 135 140
Asn Phe Gln Val Leu Leu Ser Asp Arg Tyr Phe Asn Lys Thr Cys Gly
145 150 155 160
Leu Cys Gly Asn Phe Asn Ile Phe Ala Glu Asp Asp Phe Met Thr Gln
165 170 175
Glu Gly Thr Leu Thr Ser Asp Pro Tyr Asp Phe Ala Asn Ser Trp Ala
180 185 190
Leu Ser Ser Gly Glu Gln Trp Cys Glu Arg Ala Ser Pro Pro Ser Ser
195 200 205
Ser Cys Asn Ile Ser Ser Gly Glu Met Gln Lys Gly Leu Trp Glu Gln
210 215 220
Cys Gln Leu Leu Lys Ser Thr Ser Val Phe Ala Arg Cys His Pro Leu
225 230 235 240
Val Asp Pro Glu Pro Phe Val Ala Leu Cys Glu Lys Thr Leu Cys Glu
245 250 255
Cys Ala Gly Gly Leu Glu Cys Ala Cys Pro Ala Leu Leu Glu Tyr Ala
260 265 270
Arg Thr Cys Ala Gln Glu Gly Met Val Leu Tyr Gly Trp Thr Asp His
275 280 285
Ser Ala Cys Ser Pro Val Cys Pro Ala Gly Met Glu Tyr Arg Gln Cys
290 295 300
Val Ser Pro Cys Ala Arg Thr Cys Gln Ser Leu His Ile Asn Glu Met
305 310 315 320
Cys Gln Glu Arg Cys Val Asp Gly Cys Ser Cys Pro Glu Gly Gln Leu
325 330 335
Leu Asp Glu Gly Leu Cys Val Glu Ser Thr Glu Cys Pro Cys Val His
340 345 350
Ser Gly Lys Arg Tyr Pro Pro Gly Thr Ser Leu Ser Arg Asp Cys Asn
355 360 365
Thr Cys Ile Cys Arg Asn Ser Gln Trp Ile Cys Ser Asn Glu Glu Cys
370 375 380
Pro Gly Glu Cys Leu Val Thr Gly Gln Ser His Phe Lys Ser Phe Asp
385 390 395 400
Asn Arg Tyr Phe Thr Phe Ser Gly Ile Cys Gln Tyr Leu Leu Ala Arg
405 410 415
Asp Cys Gln Asp His Ser Phe Ser Ile Val Ile Glu Thr Val Gln Cys
420 425 430
Ala Asp Asp Arg Asp Ala Val Cys Thr Arg Ser Val Thr Val Arg Leu
435 440 445
Pro Gly Leu His Asn Ser Leu Val Lys Leu Lys His Gly Ala Gly Val
450 455 460
Ala Met Asp Gly Gln Asp Val Gln Leu Pro Leu Leu Lys Gly Asp Leu
465 470 475 480
Arg Ile Gln His Thr Val Thr Ala Ser Val Arg Leu Ser Tyr Gly Glu
485 490 495
Asp Leu Gln Met Asp Trp Asp Gly Arg Gly Arg Leu Leu Val Lys Leu
500 505 510
Ser Pro Val Tyr Ala Gly Lys Thr Cys Gly Leu Cys Gly Asn Tyr Asn
515 520 525
Gly Asn Gln Gly Asp Asp Phe Leu Thr Pro Ser Gly Leu Ala Glu Pro
530 535 540
Arg Val Glu Asp Phe Gly Asn Ala Trp Lys Leu His Gly Asp Cys Gln
545 550 555 560
Asp Leu Gln Lys Gln His Ser Asp Pro Cys Ala Leu Asn Pro Arg Met
565 570 575
Thr Arg Phe Ser Glu Glu Ala Cys Ala Val Leu Thr Ser Pro Thr Phe
580 585 590
Glu Ala Cys His Arg Ala Val Ser Pro Leu Pro Tyr Leu Arg Asn Cys
595 600 605
Arg Tyr Asp Val Cys Ser Cys Ser Asp Gly Arg Glu Cys Leu Cys Gly
610 615 620
Ala Leu Ala Ser Tyr Ala Ala Ala Cys Ala Gly Arg Gly Val Arg Val
625 630 635 640
Ala Trp Arg Glu Pro Gly Arg Cys Glu Leu Asn Cys Pro Lys Gly Gln
645 650 655
Val Tyr Leu Gln Cys Gly Thr Pro Cys Asn Leu Thr Cys Arg Ser Leu
660 665 670
Ser Tyr Pro Asp Glu Glu Cys Asn Glu Ala Cys Leu Glu Gly Cys Phe
675 680 685
Cys Pro Pro Gly Leu Tyr Met Asp Glu Arg Gly Asp Cys Val Pro Lys
690 695 700
Ala Gln Cys Pro Cys Tyr Tyr Asp Gly Glu Ile Phe Gln Pro Glu Asp
705 710 715 720
Ile Phe Ser Asp His His Thr Met Cys Tyr Cys Glu Asp Gly Phe Met
725 730 735
His Cys Thr Met Ser Gly Val Pro Gly Ser Leu Leu Pro Asp Ala Val
740 745 750
Leu Ser Ser Pro Leu Ser His Arg Ser Lys Arg Ser Leu Ser Cys Arg
755 760 765
Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp Asn Leu Arg Ala Glu
770 775 780
Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr Asp Leu Glu Cys Met
785 790 795 800
Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro Pro Gly Met Val Arg
805 810 815
His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys Pro Cys Phe His Gln
820 825 830
Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys Ile Gly Cys Asn Thr
835 840 845
Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr Asp His Val Cys Asp
850 855 860
Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr Leu Thr Phe Asp Gly
865 870 875 880
Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr Val Leu Val Gln Asp
885 890 895
Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile Leu Val Gly Asn Lys
900 905 910
Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys Arg Val Thr Ile Leu
915 920 925
Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly Glu Val Asn Val Lys
930 935 940
Arg Pro Met Lys Asp Glu Thr His Phe Glu Val Val Glu Ser Gly Arg
945 950 955 960
Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser Val Val Trp Asp Arg
965 970 975
His Leu Ser Ile Ser Val Val Leu Lys Gln Thr Tyr Gln Glu Lys Val
980 985 990
Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln Asn Asn Asp Leu Thr
995 1000 1005
Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val Asp Phe Gly Asn
1010 1015 1020
Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg Lys Val Pro
1025 1030 1035
Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met Lys Gln
1040 1045 1050
Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val Phe
1055 1060 1065
Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
1070 1075 1080
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala
1085 1090 1095
Cys Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln
1100 1105 1110
His Gly Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln
1115 1120 1125
Ser Cys Glu Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu
1130 1135 1140
Trp Arg Tyr Asn Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln
1145 1150 1155
His Pro Glu Pro Leu Ala Cys Pro Val Gln Cys Val Glu Gly Cys
1160 1165 1170
His Ala His Cys Pro Pro Gly Lys Ile Leu Asp Glu Leu Leu Gln
1175 1180 1185
Thr Cys Val Asp Pro Glu Asp Cys Pro Val Cys Glu Val Ala Gly
1190 1195 1200
Arg Arg Phe Ala Ser Gly Lys Lys Val Thr Leu Asn Pro Ser Asp
1205 1210 1215
Pro Glu His Cys Gln Ile Cys His Cys Asp Val Val Asn Leu Thr
1220 1225 1230
Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu Val Val Pro Pro Thr
1235 1240 1245
Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val Glu Asp Ile Ser
1250 1255 1260
Glu Pro Pro Leu His Asp Phe Tyr Cys Ser Arg Leu Leu Asp Leu
1265 1270 1275
Val Phe Leu Leu Asp Gly Ser Ser Arg Leu Ser Glu Ala Glu Phe
1280 1285 1290
Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu Arg
1295 1300 1305
Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr His Asp
1310 1315 1320
Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser
1325 1330 1335
Glu Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln
1340 1345 1350
Val Ala Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile
1355 1360 1365
Phe Ser Lys Ile Asp Arg Pro Glu Ala Ser Arg Ile Thr Leu Leu
1370 1375 1380
Leu Met Ala Ser Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val
1385 1390 1395
Arg Tyr Val Gln Gly Leu Lys Lys Lys Lys Val Ile Val Ile Pro
1400 1405 1410
Val Gly Ile Gly Pro His Ala Asn Leu Lys Gln Ile Arg Leu Ile
1415 1420 1425
Glu Lys Gln Ala Pro Glu Asn Lys Ala Phe Val Leu Ser Ser Val
1430 1435 1440
Asp Glu Leu Glu Gln Gln Arg Asp Glu Ile Val Ser Tyr Leu Cys
1445 1450 1455
Asp Leu Ala Pro Glu Ala Pro Pro Pro Thr Leu Pro Pro Asp Met
1460 1465 1470
Ala Gln Val Thr Val Gly Pro Gly Leu Leu Gly Val Ser Thr Leu
1475 1480 1485
Gly Pro Lys Arg Asn Ser Met Val Leu Asp Val Ala Phe Val Leu
1490 1495 1500
Glu Gly Ser Asp Lys Ile Gly Glu Ala Asp Phe Asn Arg Ser Lys
1505 1510 1515
Glu Phe Met Glu Glu Val Ile Gln Arg Met Asp Val Gly Gln Asp
1520 1525 1530
Ser Ile His Val Thr Val Leu Gln Tyr Ser Tyr Met Val Thr Val
1535 1540 1545
Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly Asp Ile Leu Gln
1550 1555 1560
Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg Thr Asn Thr
1565 1570 1575
Gly Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu Val Ser
1580 1585 1590
Gln Gly Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val Thr
1595 1600 1605
Gly Asn Pro Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile
1610 1615 1620
Gln Val Val Pro Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu
1625 1630 1635
Leu Glu Arg Ile Gly Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp
1640 1645 1650
Phe Glu Thr Leu Pro Arg Glu Ala Pro Asp Leu Val Leu Gln Arg
1655 1660 1665
Cys Cys Ser Gly Glu Gly Leu Gln Ile Pro Thr Leu Ser Pro Ala
1670 1675 1680
Pro Asp Cys Ser Gln Pro Leu Asp Val Ile Leu Leu Leu Asp Gly
1685 1690 1695
Ser Ser Ser Phe Pro Ala Ser Tyr Phe Asp Glu Met Lys Ser Phe
1700 1705 1710
Ala Lys Ala Phe Ile Ser Lys Ala Asn Ile Gly Pro Arg Leu Thr
1715 1720 1725
Gln Val Ser Val Leu Gln Tyr Gly Ser Ile Thr Thr Ile Asp Val
1730 1735 1740
Pro Trp Asn Val Val Pro Glu Lys Ala His Leu Leu Ser Leu Val
1745 1750 1755
Asp Val Met Gln Arg Glu Gly Gly Pro Ser Gln Ile Gly Asp Ala
1760 1765 1770
Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser Glu Met His Gly Ala
1775 1780 1785
Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu Val Thr Asp Val
1790 1795 1800
Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala Arg Ser Asn
1805 1810 1815
Arg Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr Asp Ala
1820 1825 1830
Ala Gln Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser Asn Val
1835 1840 1845
Val Lys Leu Gln Arg Ile Glu Asp Leu Pro Thr Met Val Thr Leu
1850 1855 1860
Gly Asn Ser Phe Leu His Lys Leu Cys Ser Gly Phe Val Arg Ile
1865 1870 1875
Cys Met Asp Glu Asp Gly Asn Glu Lys Arg Pro Gly Asp Val Trp
1880 1885 1890
Thr Leu Pro Asp Gln Cys His Thr Val Thr Cys Gln Pro Asp Gly
1895 1900 1905
Gln Thr Leu Leu Lys Ser His Arg Val Asn Cys Asp Arg Gly Leu
1910 1915 1920
Arg Pro Ser Cys Pro Asn Ser Gln Ser Pro Val Lys Val Glu Glu
1925 1930 1935
Thr Cys Gly Cys Arg Trp Thr Cys Pro Cys Val Cys Thr Gly Ser
1940 1945 1950
Ser Thr Arg His Ile Val Thr Phe Asp Gly Gln Asn Phe Lys Leu
1955 1960 1965
Thr Gly Ser Cys Ser Tyr Val Leu Phe Gln Asn Lys Glu Gln Asp
1970 1975 1980
Leu Glu Val Ile Leu His Asn Gly Ala Cys Ser Pro Gly Ala Arg
1985 1990 1995
Gln Gly Cys Met Lys Ser Ile Glu Val Lys His Ser Ala Leu Ser
2000 2005 2010
Val Glu Leu His Ser Asp Met Glu Val Thr Val Asn Gly Arg Leu
2015 2020 2025
Val Ser Val Pro Tyr Val Gly Gly Asn Met Glu Val Asn Val Tyr
2030 2035 2040
Gly Ala Ile Met His Glu Val Arg Phe Asn His Leu Gly His Ile
2045 2050 2055
Phe Thr Phe Thr Pro Gln Asn Asn Glu Phe Gln Leu Gln Leu Ser
2060 2065 2070
Pro Lys Thr Phe Ala Ser Lys Thr Tyr Gly Leu Cys Gly Ile Cys
2075 2080 2085
Asp Glu Asn Gly Ala Asn Asp Phe Met Leu Arg Asp Gly Thr Val
2090 2095 2100
Thr Thr Asp Trp Lys Thr Leu Val Gln Glu Trp Thr Val Gln Arg
2105 2110 2115
Pro Gly Gln Thr Cys Gln Pro Ile Leu Glu Glu Gln Cys Leu Val
2120 2125 2130
Pro Asp Ser Ser His Cys Gln Val Leu Leu Leu Pro Leu Phe Ala
2135 2140 2145
Glu Cys His Lys Val Leu Ala Pro Ala Thr Phe Tyr Ala Ile Cys
2150 2155 2160
Gln Gln Asp Ser Cys His Gln Glu Gln Val Cys Glu Val Ile Ala
2165 2170 2175
Ser Tyr Ala His Leu Cys Arg Thr Asn Gly Val Cys Val Asp Trp
2180 2185 2190
Arg Thr Pro Asp Phe Cys Ala Met Ser Cys Pro Pro Ser Leu Val
2195 2200 2205
Tyr Asn His Cys Glu His Gly Cys Pro Arg His Cys Asp Gly Asn
2210 2215 2220
Val Ser Ser Cys Gly Asp His Pro Ser Glu Gly Cys Phe Cys Pro
2225 2230 2235
Pro Asp Lys Val Met Leu Glu Gly Ser Cys Val Pro Glu Glu Ala
2240 2245 2250
Cys Thr Gln Cys Ile Gly Glu Asp Gly Val Gln His Gln Phe Leu
2255 2260 2265
Glu Ala Trp Val Pro Asp His Gln Pro Cys Gln Ile Cys Thr Cys
2270 2275 2280
Leu Ser Gly Arg Lys Val Asn Cys Thr Thr Gln Pro Cys Pro Thr
2285 2290 2295
Ala Lys Ala Pro Thr Cys Gly Leu Cys Glu Val Ala Arg Leu Arg
2300 2305 2310
Gln Asn Ala Asp Gln Cys Cys Pro Glu Tyr Glu Cys Val Cys Asp
2315 2320 2325
Pro Val Ser Cys Asp Leu Pro Pro Val Pro His Cys Glu Arg Gly
2330 2335 2340
Leu Gln Pro Thr Leu Thr Asn Pro Gly Glu Cys Arg Pro Asn Phe
2345 2350 2355
Thr Cys Ala Cys Arg Lys Glu Glu Cys Lys Arg Val Ser Pro Pro
2360 2365 2370
Ser Cys Pro Pro His Arg Leu Pro Thr Leu Arg Lys Thr Gln Cys
2375 2380 2385
Cys Asp Glu Tyr Glu Cys Ala Cys Asn Cys Val Asn Ser Thr Val
2390 2395 2400
Ser Cys Pro Leu Gly Tyr Leu Ala Ser Thr Ala Thr Asn Asp Cys
2405 2410 2415
Gly Cys Thr Thr Thr Thr Cys Leu Pro Asp Lys Val Cys Val His
2420 2425 2430
Arg Ser Thr Ile Tyr Pro Val Gly Gln Phe Trp Glu Glu Gly Cys
2435 2440 2445
Asp Val Cys Thr Cys Thr Asp Met Glu Asp Ala Val Met Gly Leu
2450 2455 2460
Arg Val Ala Gln Cys Ser Gln Lys Pro Cys Glu Asp Ser Cys Arg
2465 2470 2475
Ser Gly Phe Thr Tyr Val Leu His Glu Gly Glu Cys Cys Gly Arg
2480 2485 2490
Cys Leu Pro Ser Ala Cys Glu Val Val Thr Gly Ser Pro Arg Gly
2495 2500 2505
Asp Ser Gln Ser Ser Trp Lys Ser Val Gly Ser Gln Trp Ala Ser
2510 2515 2520
Pro Glu Asn Pro Cys Leu Ile Asn Glu Cys Val Arg Val Lys Glu
2525 2530 2535
Glu Val Phe Ile Gln Gln Arg Asn Val Ser Cys Pro Gln Leu Glu
2540 2545 2550
Val Pro Val Cys Pro Ser Gly Phe Gln Leu Ser Cys Lys Thr Ser
2555 2560 2565
Ala Cys Cys Pro Ser Cys Arg Cys Glu Arg Met Glu Ala Cys Met
2570 2575 2580
Leu Asn Gly Thr Val Ile Gly Pro Gly Lys Thr Val Met Ile Asp
2585 2590 2595
Val Cys Thr Thr Cys Arg Cys Met Val Gln Val Gly Val Ile Ser
2600 2605 2610
Gly Phe Lys Leu Glu Cys Arg Lys Thr Thr Cys Asn Pro Cys Pro
2615 2620 2625
Leu Gly Tyr Lys Glu Glu Asn Asn Thr Gly Glu Cys Cys Gly Arg
2630 2635 2640
Cys Leu Pro Thr Ala Cys Thr Ile Gln Leu Arg Gly Gly Gln Ile
2645 2650 2655
Met Thr Leu Lys Arg Asp Glu Thr Leu Gln Asp Gly Cys Asp Thr
2660 2665 2670
His Phe Cys Lys Val Asn Glu Arg Gly Glu Tyr Phe Trp Glu Lys
2675 2680 2685
Arg Val Thr Gly Cys Pro Pro Phe Asp Glu His Lys Cys Leu Ala
2690 2695 2700
Glu Gly Gly Lys Ile Met Lys Ile Pro Gly Thr Cys Cys Asp Thr
2705 2710 2715
Cys Glu Glu Pro Glu Cys Asn Asp Ile Thr Ala Arg Leu Gln Tyr
2720 2725 2730
Val Lys Val Gly Ser Cys Lys Ser Glu Val Glu Val Asp Ile His
2735 2740 2745
Tyr Cys Gln Gly Lys Cys Ala Ser Lys Ala Met Tyr Ser Ile Asp
2750 2755 2760
Ile Asn Asp Val Gln Asp Gln Cys Ser Cys Cys Ser Pro Thr Arg
2765 2770 2775
Thr Glu Pro Met Gln Val Ala Leu His Cys Thr Asn Gly Ser Val
2780 2785 2790
Val Tyr His Glu Val Leu Asn Ala Met Glu Cys Lys Cys Ser Pro
2795 2800 2805
Arg Lys Cys Ser Lys
2810
<210> 2
<211> 1365
<212> Белок
<213> Homo sapiens
<400> 2
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Cys
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu
465 470 475 480
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
485 490 495
Glu Asp Ile Ser Glu Pro Pro Leu His Asp Phe Tyr Cys Ser Arg Leu
500 505 510
Leu Asp Leu Val Phe Leu Leu Asp Gly Ser Ser Arg Leu Ser Glu Ala
515 520 525
Glu Phe Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu
530 535 540
Arg Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr His Asp
545 550 555 560
Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser Glu
565 570 575
Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln Val Ala
580 585 590
Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile Phe Ser Lys
595 600 605
Ile Asp Arg Pro Glu Ala Ser Arg Ile Thr Leu Leu Leu Met Ala Ser
610 615 620
Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val Arg Tyr Val Gln Gly
625 630 635 640
Leu Lys Lys Lys Lys Val Ile Val Ile Pro Val Gly Ile Gly Pro His
645 650 655
Ala Asn Leu Lys Gln Ile Arg Leu Ile Glu Lys Gln Ala Pro Glu Asn
660 665 670
Lys Ala Phe Val Leu Ser Ser Val Asp Glu Leu Glu Gln Gln Arg Asp
675 680 685
Glu Ile Val Ser Tyr Leu Cys Asp Leu Ala Pro Glu Ala Pro Pro Pro
690 695 700
Thr Leu Pro Pro Asp Met Ala Gln Val Thr Val Gly Pro Gly Leu Leu
705 710 715 720
Gly Val Ser Thr Leu Gly Pro Lys Arg Asn Ser Met Val Leu Asp Val
725 730 735
Ala Phe Val Leu Glu Gly Ser Asp Lys Ile Gly Glu Ala Asp Phe Asn
740 745 750
Arg Ser Lys Glu Phe Met Glu Glu Val Ile Gln Arg Met Asp Val Gly
755 760 765
Gln Asp Ser Ile His Val Thr Val Leu Gln Tyr Ser Tyr Met Val Thr
770 775 780
Val Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly Asp Ile Leu Gln
785 790 795 800
Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg Thr Asn Thr Gly
805 810 815
Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu Val Ser Gln Gly
820 825 830
Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val Thr Gly Asn Pro
835 840 845
Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile Gln Val Val Pro
850 855 860
Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu Leu Glu Arg Ile Gly
865 870 875 880
Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp Phe Glu Thr Leu Pro Arg
885 890 895
Glu Ala Pro Asp Leu Val Leu Gln Arg Cys Cys Ser Gly Glu Gly Leu
900 905 910
Gln Ile Pro Thr Leu Ser Pro Ala Pro Asp Cys Ser Gln Pro Leu Asp
915 920 925
Val Ile Leu Leu Leu Asp Gly Ser Ser Ser Phe Pro Ala Ser Tyr Phe
930 935 940
Asp Glu Met Lys Ser Phe Ala Lys Ala Phe Ile Ser Lys Ala Asn Ile
945 950 955 960
Gly Pro Arg Leu Thr Gln Val Ser Val Leu Gln Tyr Gly Ser Ile Thr
965 970 975
Thr Ile Asp Val Pro Trp Asn Val Val Pro Glu Lys Ala His Leu Leu
980 985 990
Ser Leu Val Asp Val Met Gln Arg Glu Gly Gly Pro Ser Gln Ile Gly
995 1000 1005
Asp Ala Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser Glu Met His
1010 1015 1020
Gly Ala Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu Val Thr
1025 1030 1035
Asp Val Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala Arg
1040 1045 1050
Ser Asn Arg Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr
1055 1060 1065
Asp Ala Ala Gln Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser
1070 1075 1080
Asn Val Val Lys Leu Gln Arg Ile Glu Asp Leu Pro Thr Met Val
1085 1090 1095
Thr Leu Gly Asn Ser Phe Leu His Lys Leu Cys Ser Gly Phe Val
1100 1105 1110
Arg Ile Cys Met Asp Glu Asp Gly Asn Glu Lys Arg Pro Gly Asp
1115 1120 1125
Val Trp Thr Leu Pro Asp Gln Cys His Thr Val Thr Cys Gln Pro
1130 1135 1140
Asp Gly Gln Thr Leu Leu Lys Ser His Arg Val Asn Cys Asp Arg
1145 1150 1155
Gly Leu Arg Pro Ser Cys Pro Asn Ser Gln Ser Pro Val Lys Val
1160 1165 1170
Glu Glu Thr Cys Gly Cys Arg Trp Thr Cys Pro Cys Val Cys Thr
1175 1180 1185
Gly Ser Ser Thr Arg His Ile Val Thr Phe Asp Gly Gln Asn Phe
1190 1195 1200
Lys Leu Thr Gly Ser Cys Ser Tyr Val Leu Phe Gln Asn Lys Glu
1205 1210 1215
Gln Asp Leu Glu Val Ile Leu His Asn Gly Ala Cys Ser Pro Gly
1220 1225 1230
Ala Arg Gln Gly Cys Met Lys Ser Ile Glu Val Lys His Ser Ala
1235 1240 1245
Leu Ser Val Glu Leu His Ser Asp Met Glu Val Thr Val Asn Gly
1250 1255 1260
Arg Leu Val Ser Val Pro Tyr Val Gly Gly Asn Met Glu Val Asn
1265 1270 1275
Val Tyr Gly Ala Ile Met His Glu Val Arg Phe Asn His Leu Gly
1280 1285 1290
His Ile Phe Thr Phe Thr Pro Gln Asn Asn Glu Phe Gln Leu Gln
1295 1300 1305
Leu Ser Pro Lys Thr Phe Ala Ser Lys Thr Tyr Gly Leu Cys Gly
1310 1315 1320
Ile Cys Asp Glu Asn Gly Ala Asn Asp Phe Met Leu Arg Asp Gly
1325 1330 1335
Thr Val Thr Thr Asp Trp Lys Thr Leu Val Gln Glu Trp Thr Val
1340 1345 1350
Gln Arg Pro Gly Gln Thr Cys Gln Pro Ile Leu Glu
1355 1360 1365
<210> 3
<211> 272
<212> Белок
<213> Homo sapiens
<400> 3
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
<210> 4
<211> 910
<212> Белок
<213> Homo sapiens
<400> 4
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
Lys Val Pro Leu Asp Ser Ser Pro Ala Thr Cys His Asn Asn Ile Met
275 280 285
Lys Gln Thr Met Val Asp Ser Ser Cys Arg Ile Leu Thr Ser Asp Val
290 295 300
Phe Gln Asp Cys Asn Lys Leu Val Asp Pro Glu Pro Tyr Leu Asp Val
305 310 315 320
Cys Ile Tyr Asp Thr Cys Ser Cys Glu Ser Ile Gly Asp Cys Ala Cys
325 330 335
Phe Cys Asp Thr Ile Ala Ala Tyr Ala His Val Cys Ala Gln His Gly
340 345 350
Lys Val Val Thr Trp Arg Thr Ala Thr Leu Cys Pro Gln Ser Cys Glu
355 360 365
Glu Arg Asn Leu Arg Glu Asn Gly Tyr Glu Cys Glu Trp Arg Tyr Asn
370 375 380
Ser Cys Ala Pro Ala Cys Gln Val Thr Cys Gln His Pro Glu Pro Leu
385 390 395 400
Ala Cys Pro Val Gln Cys Val Glu Gly Cys His Ala His Cys Pro Pro
405 410 415
Gly Lys Ile Leu Asp Glu Leu Leu Gln Thr Cys Val Asp Pro Glu Asp
420 425 430
Cys Pro Val Cys Glu Val Ala Gly Arg Arg Phe Ala Ser Gly Lys Lys
435 440 445
Val Thr Leu Asn Pro Ser Asp Pro Glu His Cys Gln Ile Cys His Cys
450 455 460
Asp Val Val Asn Leu Thr Cys Glu Ala Cys Gln Glu Pro Gly Gly Leu
465 470 475 480
Val Val Pro Pro Thr Asp Ala Pro Val Ser Pro Thr Thr Leu Tyr Val
485 490 495
Glu Asp Ile Ser Glu Pro Pro Leu His Asp Phe Tyr Cys Ser Arg Leu
500 505 510
Leu Asp Leu Val Phe Leu Leu Asp Gly Ser Ser Arg Leu Ser Glu Ala
515 520 525
Glu Phe Glu Val Leu Lys Ala Phe Val Val Asp Met Met Glu Arg Leu
530 535 540
Arg Ile Ser Gln Lys Trp Val Arg Val Ala Val Val Glu Tyr His Asp
545 550 555 560
Gly Ser His Ala Tyr Ile Gly Leu Lys Asp Arg Lys Arg Pro Ser Glu
565 570 575
Leu Arg Arg Ile Ala Ser Gln Val Lys Tyr Ala Gly Ser Gln Val Ala
580 585 590
Ser Thr Ser Glu Val Leu Lys Tyr Thr Leu Phe Gln Ile Phe Ser Lys
595 600 605
Ile Asp Arg Pro Glu Ala Ser Arg Ile Thr Leu Leu Leu Met Ala Ser
610 615 620
Gln Glu Pro Gln Arg Met Ser Arg Asn Phe Val Arg Tyr Val Gln Gly
625 630 635 640
Leu Lys Lys Lys Lys Val Ile Val Ile Pro Val Gly Ile Gly Pro His
645 650 655
Ala Asn Leu Lys Gln Ile Arg Leu Ile Glu Lys Gln Ala Pro Glu Asn
660 665 670
Lys Ala Phe Val Leu Ser Ser Val Asp Glu Leu Glu Gln Gln Arg Asp
675 680 685
Glu Ile Val Ser Tyr Leu Cys Asp Leu Ala Pro Glu Ala Pro Pro Pro
690 695 700
Thr Leu Pro Pro Asp Met Ala Gln Val Thr Val Gly Pro Gly Leu Leu
705 710 715 720
Gly Val Ser Thr Leu Gly Pro Lys Arg Asn Ser Met Val Leu Asp Val
725 730 735
Ala Phe Val Leu Glu Gly Ser Asp Lys Ile Gly Glu Ala Asp Phe Asn
740 745 750
Arg Ser Lys Glu Phe Met Glu Glu Val Ile Gln Arg Met Asp Val Gly
755 760 765
Gln Asp Ser Ile His Val Thr Val Leu Gln Tyr Ser Tyr Met Val Thr
770 775 780
Val Glu Tyr Pro Phe Ser Glu Ala Gln Ser Lys Gly Asp Ile Leu Gln
785 790 795 800
Arg Val Arg Glu Ile Arg Tyr Gln Gly Gly Asn Arg Thr Asn Thr Gly
805 810 815
Leu Ala Leu Arg Tyr Leu Ser Asp His Ser Phe Leu Val Ser Gln Gly
820 825 830
Asp Arg Glu Gln Ala Pro Asn Leu Val Tyr Met Val Thr Gly Asn Pro
835 840 845
Ala Ser Asp Glu Ile Lys Arg Leu Pro Gly Asp Ile Gln Val Val Pro
850 855 860
Ile Gly Val Gly Pro Asn Ala Asn Val Gln Glu Leu Glu Arg Ile Gly
865 870 875 880
Trp Pro Asn Ala Pro Ile Leu Ile Gln Asp Phe Glu Thr Leu Pro Arg
885 890 895
Glu Ala Pro Asp Leu Val Leu Gln Arg Cys Cys Ser Gly Glu
900 905 910
<210> 5
<211> 272
<212> Белок
<213> Homo sapiens
<400> 5
Ser Leu Ser Cys Arg Pro Pro Met Val Lys Leu Val Cys Pro Ala Asp
1 5 10 15
Asn Leu Arg Ala Glu Gly Leu Glu Cys Thr Lys Thr Cys Gln Asn Tyr
20 25 30
Asp Leu Glu Cys Met Ser Met Gly Cys Val Ser Gly Cys Leu Cys Pro
35 40 45
Pro Gly Met Val Arg His Glu Asn Arg Cys Val Ala Leu Glu Arg Cys
50 55 60
Pro Cys Phe His Gln Gly Lys Glu Tyr Ala Pro Gly Glu Thr Val Lys
65 70 75 80
Ile Gly Cys Asn Thr Cys Val Cys Gln Asp Arg Lys Trp Asn Cys Thr
85 90 95
Asp His Val Cys Asp Ala Thr Cys Ser Thr Ile Gly Met Ala His Tyr
100 105 110
Leu Thr Phe Asp Gly Leu Lys Tyr Leu Phe Pro Gly Glu Cys Gln Tyr
115 120 125
Val Leu Val Gln Asp Tyr Cys Gly Ser Asn Pro Gly Thr Phe Arg Ile
130 135 140
Leu Val Gly Asn Lys Gly Cys Ser His Pro Ser Val Lys Cys Lys Lys
145 150 155 160
Arg Val Thr Ile Leu Val Glu Gly Gly Glu Ile Glu Leu Phe Asp Gly
165 170 175
Glu Val Asn Val Lys Arg Pro Met Lys Asp Glu Thr His Phe Glu Val
180 185 190
Val Glu Ser Gly Arg Tyr Ile Ile Leu Leu Leu Gly Lys Ala Leu Ser
195 200 205
Val Val Trp Asp Arg His Leu Ser Ile Ser Val Val Leu Lys Gln Thr
210 215 220
Tyr Gln Glu Lys Val Cys Gly Leu Cys Gly Asn Phe Asp Gly Ile Gln
225 230 235 240
Asn Asn Asp Leu Thr Ser Ser Asn Leu Gln Val Glu Glu Asp Pro Val
245 250 255
Asp Phe Gly Asn Ser Trp Lys Val Ser Ser Gln Cys Ala Asp Thr Arg
260 265 270
<210> 6
<211> 174
<212> Белок
<213> Homo sapiens
<400> 6
Val Ile Leu Leu Leu Asp Gly Ser Ser Ser Phe Pro Ala Ser Tyr Phe
1 5 10 15
Asp Glu Met Lys Ser Phe Ala Lys Ala Phe Ile Ser Lys Ala Asn Ile
20 25 30
Gly Pro Arg Leu Thr Gln Val Ser Val Leu Gln Tyr Gly Ser Ile Thr
35 40 45
Thr Ile Asp Val Pro Trp Asn Val Val Pro Glu Lys Ala His Leu Leu
50 55 60
Ser Leu Val Asp Val Met Gln Arg Glu Gly Gly Pro Ser Gln Ile Gly
65 70 75 80
Asp Ala Leu Gly Phe Ala Val Arg Tyr Leu Thr Ser Glu Met His Gly
85 90 95
Ala Arg Pro Gly Ala Ser Lys Ala Val Val Ile Leu Val Thr Asp Val
100 105 110
Ser Val Asp Ser Val Asp Ala Ala Ala Asp Ala Ala Arg Ser Asn Arg
115 120 125
Val Thr Val Phe Pro Ile Gly Ile Gly Asp Arg Tyr Asp Ala Ala Gln
130 135 140
Leu Arg Ile Leu Ala Gly Pro Ala Gly Asp Ser Asn Val Val Lys Leu
145 150 155 160
Gln Arg Ile Glu Asp Leu Pro Thr Met Val Thr Leu Gly Asn
165 170
<210> 7
<211> 390
<212> Белок
<213> Homo sapiens
<400> 7
Gln Cys Ile Gly Glu Asp Gly Val Gln His Gln Phe Leu Glu Ala Trp
1 5 10 15
Val Pro Asp His Gln Pro Cys Gln Ile Cys Thr Cys Leu Ser Gly Arg
20 25 30
Lys Val Asn Cys Thr Thr Gln Pro Cys Pro Thr Ala Lys Ala Pro Thr
35 40 45
Cys Gly Leu Cys Glu Val Ala Arg Leu Arg Gln Asn Ala Asp Gln Cys
50 55 60
Cys Pro Glu Tyr Glu Cys Val Cys Asp Pro Val Ser Cys Asp Leu Pro
65 70 75 80
Pro Val Pro His Cys Glu Arg Gly Leu Gln Pro Thr Leu Thr Asn Pro
85 90 95
Gly Glu Cys Arg Pro Asn Phe Thr Cys Ala Cys Arg Lys Glu Glu Cys
100 105 110
Lys Arg Val Ser Pro Pro Ser Cys Pro Pro His Arg Leu Pro Thr Leu
115 120 125
Arg Lys Thr Gln Cys Cys Asp Glu Tyr Glu Cys Ala Cys Asn Cys Val
130 135 140
Asn Ser Thr Val Ser Cys Pro Leu Gly Tyr Leu Ala Ser Thr Ala Thr
145 150 155 160
Asn Asp Cys Gly Cys Thr Thr Thr Thr Cys Leu Pro Asp Lys Val Cys
165 170 175
Val His Arg Ser Thr Ile Tyr Pro Val Gly Gln Phe Trp Glu Glu Gly
180 185 190
Cys Asp Val Cys Thr Cys Thr Asp Met Glu Asp Ala Val Met Gly Leu
195 200 205
Arg Val Ala Gln Cys Ser Gln Lys Pro Cys Glu Asp Ser Cys Arg Ser
210 215 220
Gly Phe Thr Tyr Val Leu His Glu Gly Glu Cys Cys Gly Arg Cys Leu
225 230 235 240
Pro Ser Ala Cys Glu Val Val Thr Gly Ser Pro Arg Gly Asp Ser Gln
245 250 255
Ser Ser Trp Lys Ser Val Gly Ser Gln Trp Ala Ser Pro Glu Asn Pro
260 265 270
Cys Leu Ile Asn Glu Cys Val Arg Val Lys Glu Glu Val Phe Ile Gln
275 280 285
Gln Arg Asn Val Ser Cys Pro Gln Leu Glu Val Pro Val Cys Pro Ser
290 295 300
Gly Phe Gln Leu Ser Cys Lys Thr Ser Ala Cys Cys Pro Ser Cys Arg
305 310 315 320
Cys Glu Arg Met Glu Ala Cys Met Leu Asn Gly Thr Val Ile Gly Pro
325 330 335
Gly Lys Thr Val Met Ile Asp Val Cys Thr Thr Cys Arg Cys Met Val
340 345 350
Gln Val Gly Val Ile Ser Gly Phe Lys Leu Glu Cys Arg Lys Thr Thr
355 360 365
Cys Asn Pro Cys Pro Leu Gly Tyr Lys Glu Glu Asn Asn Thr Gly Glu
370 375 380
Cys Cys Gly Arg Cys Leu
385 390
<---
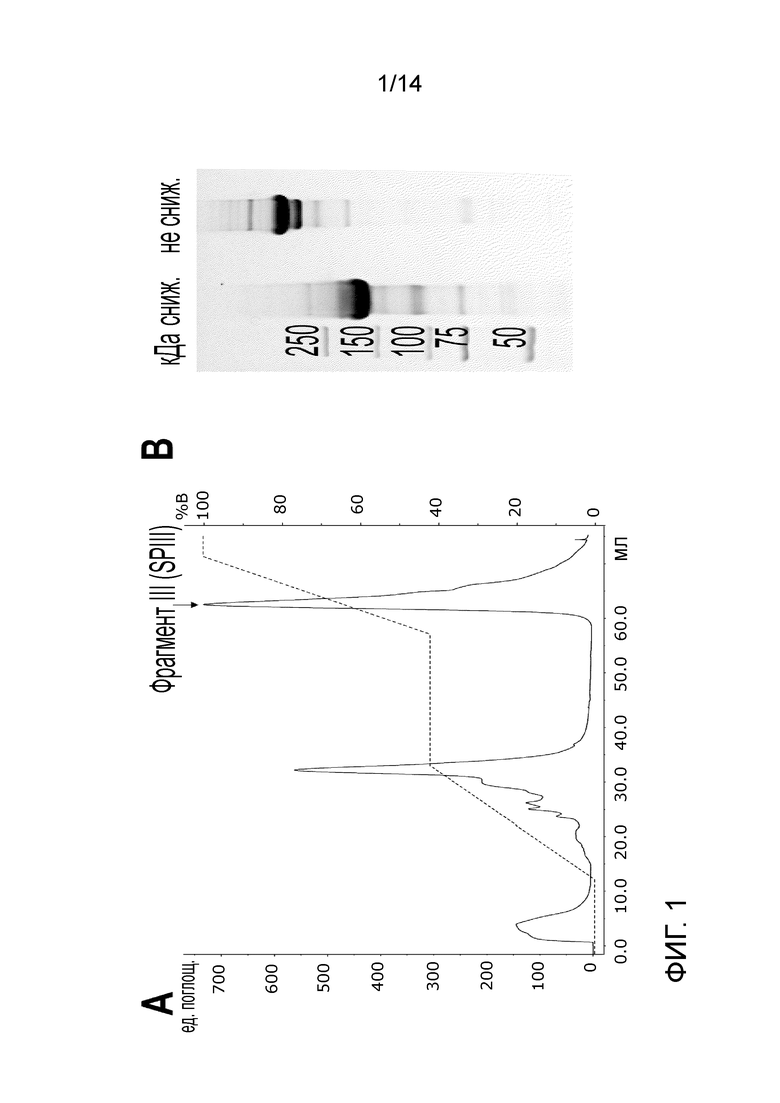
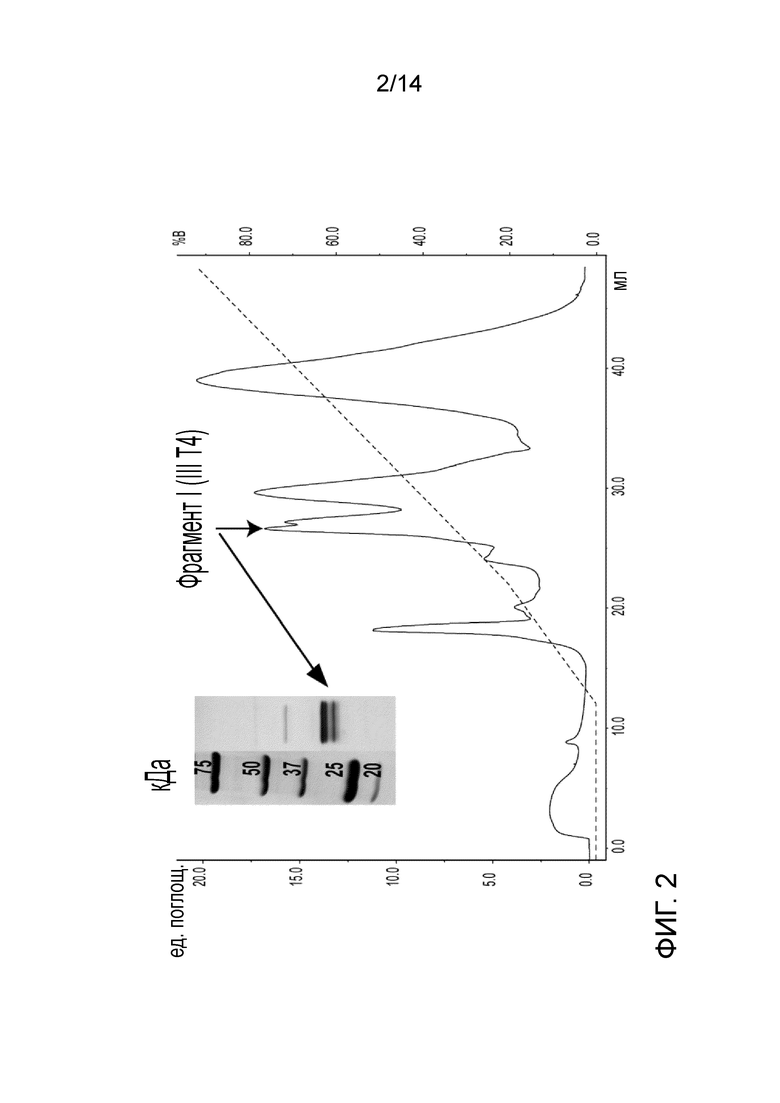
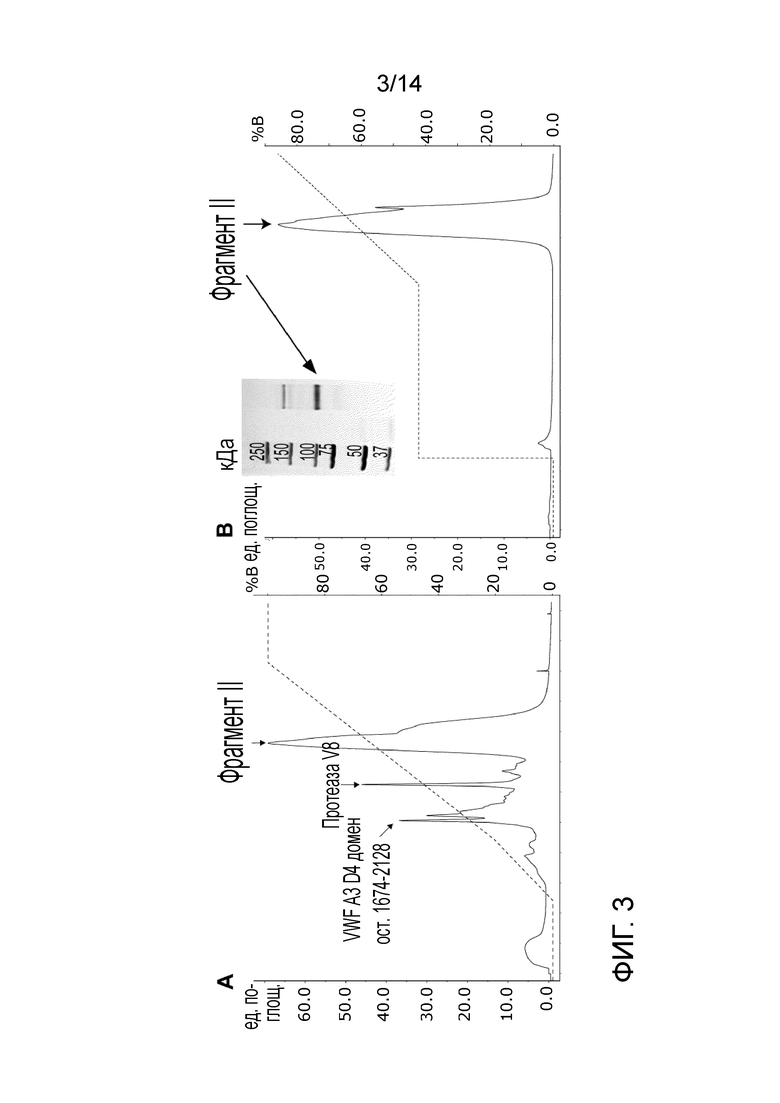
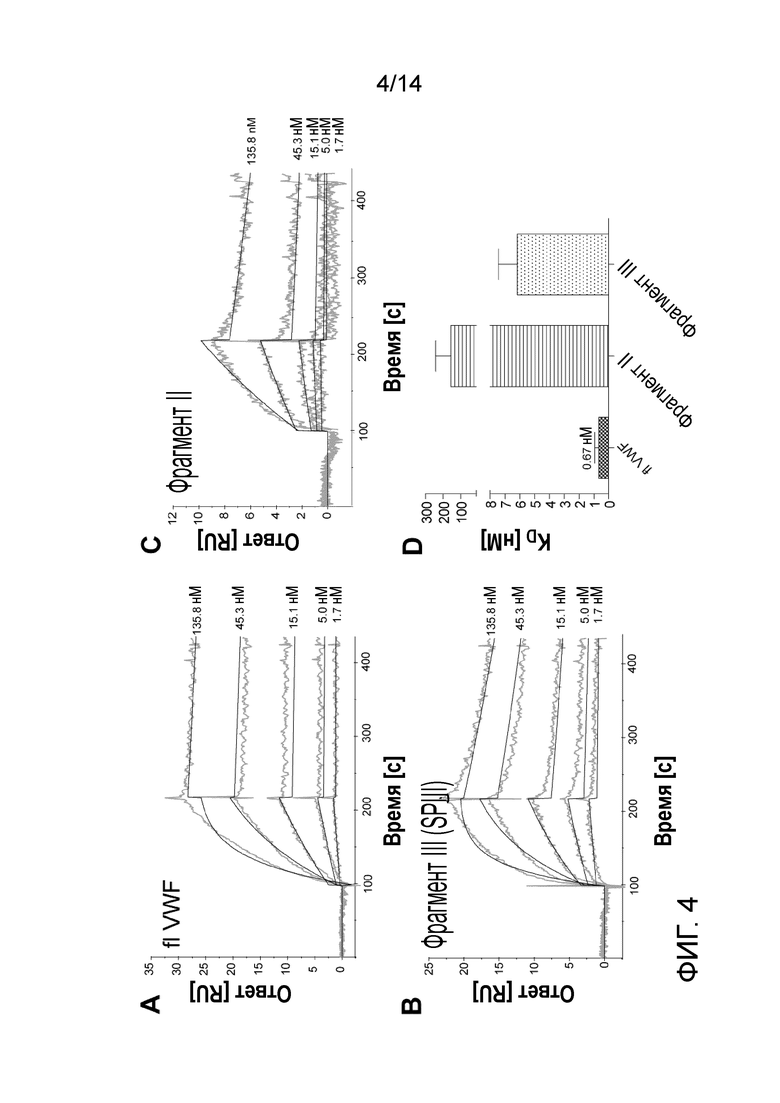
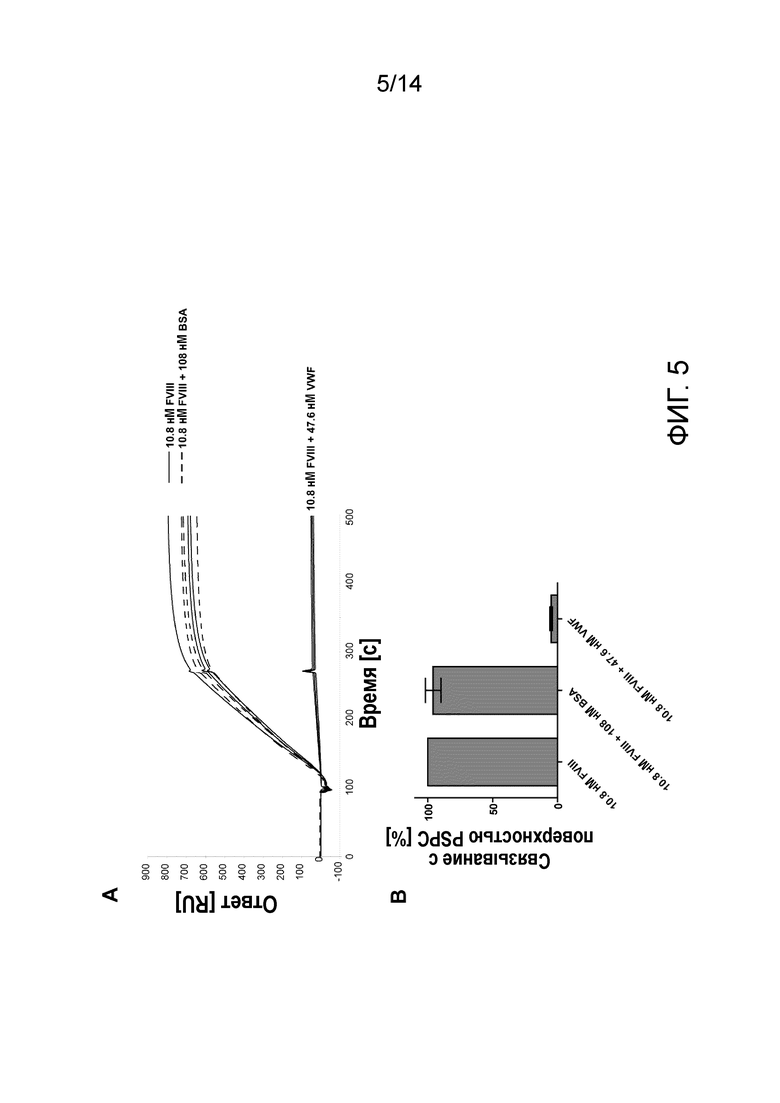
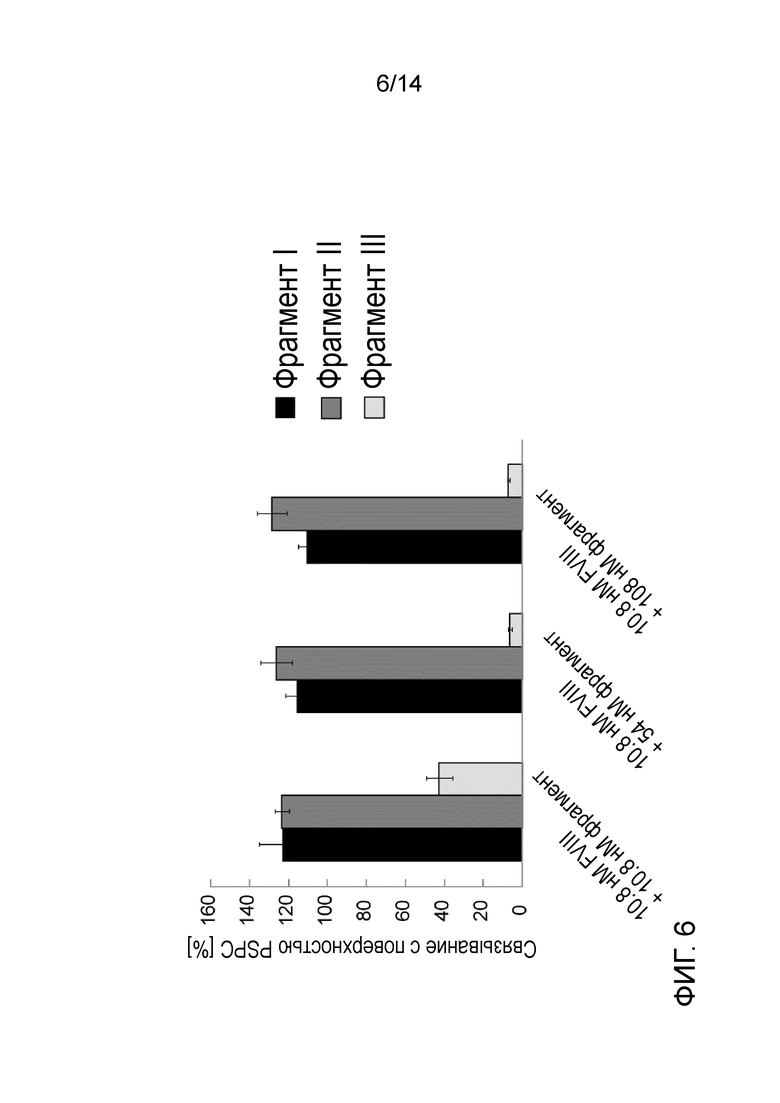
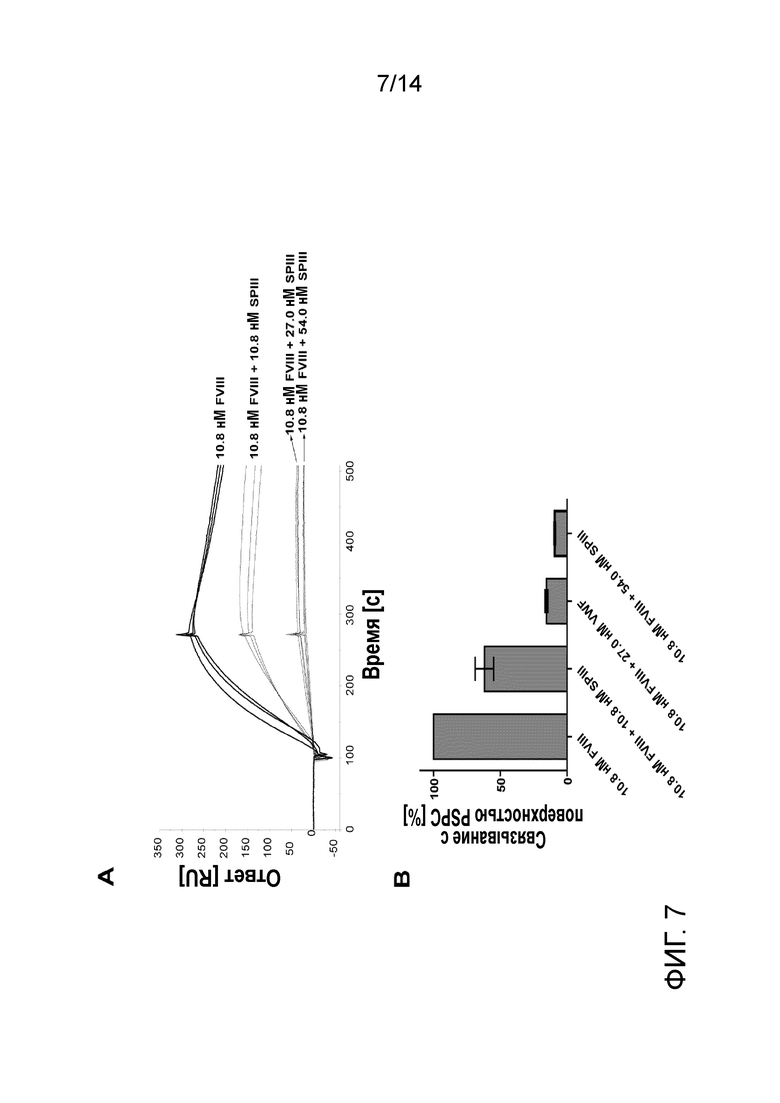
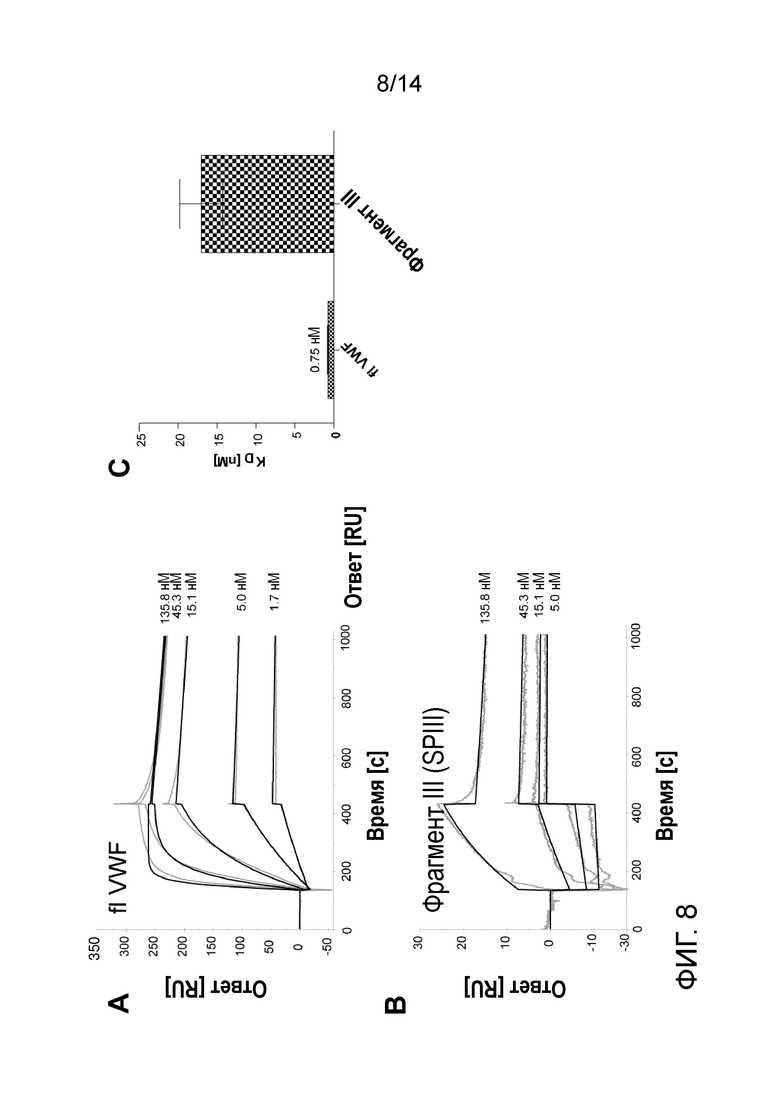
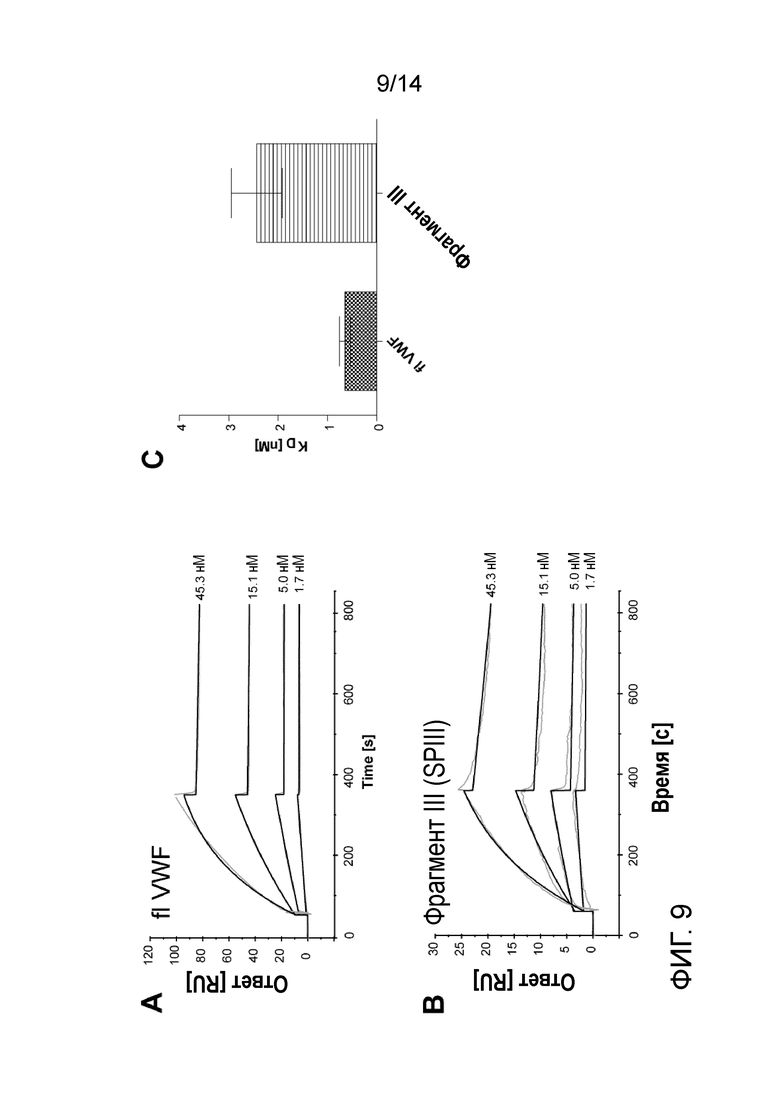
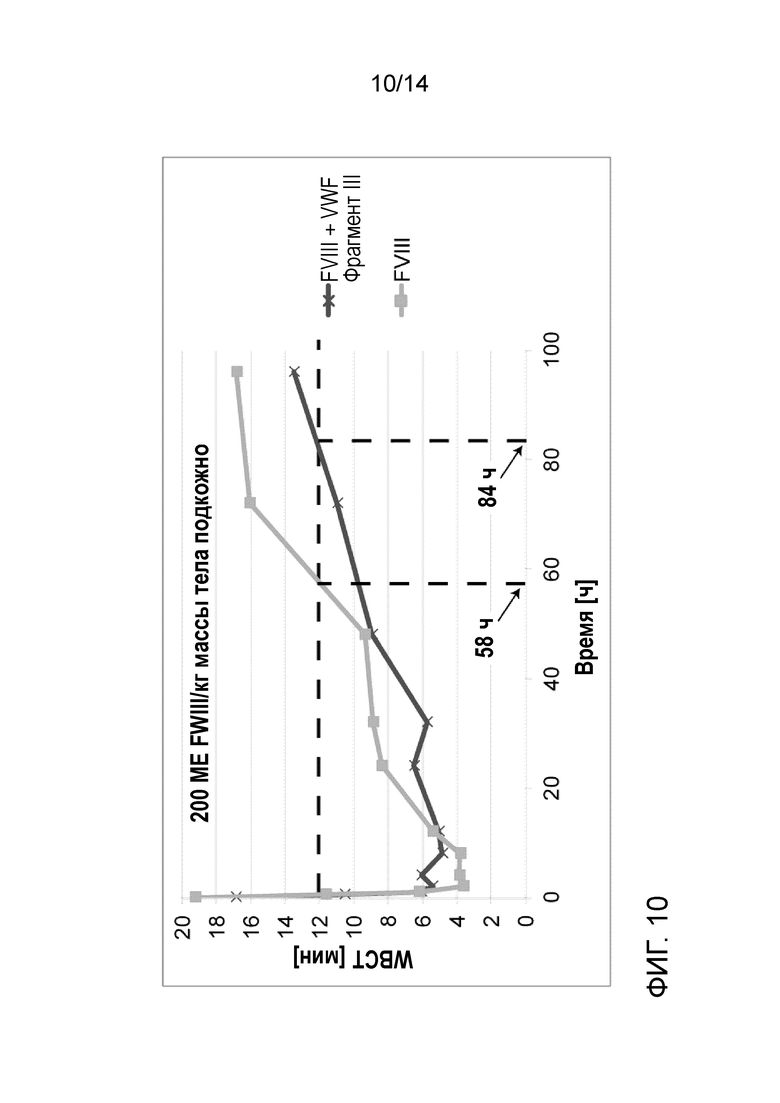
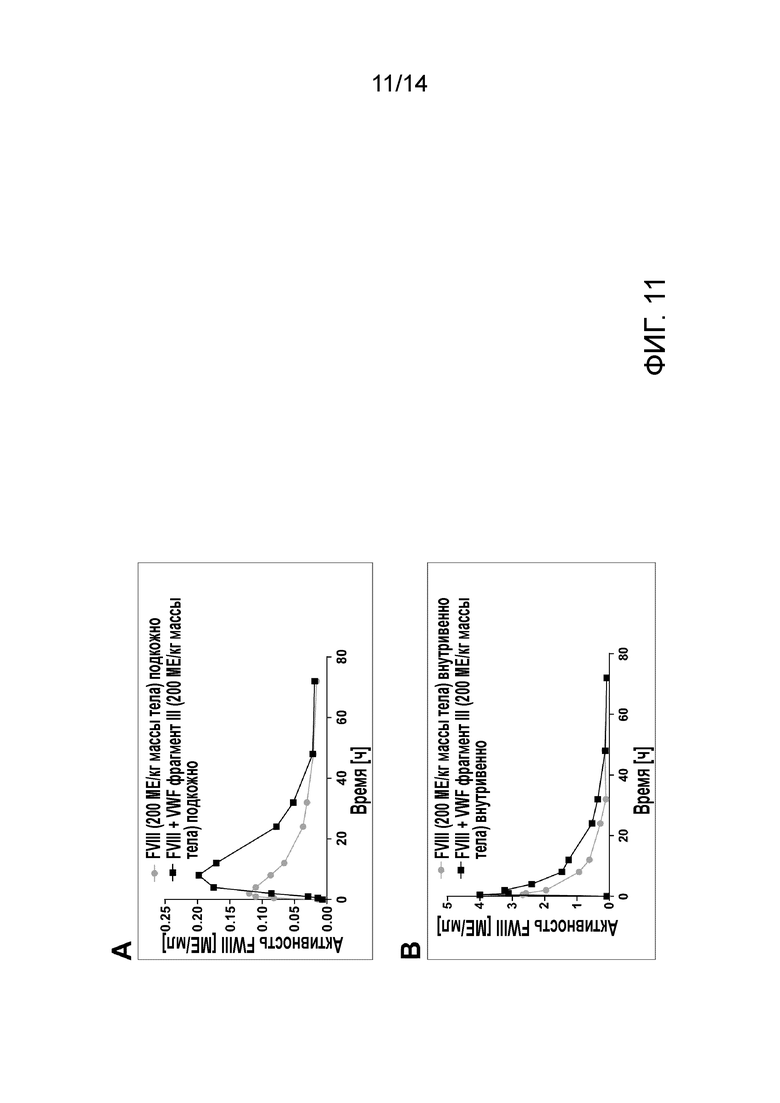
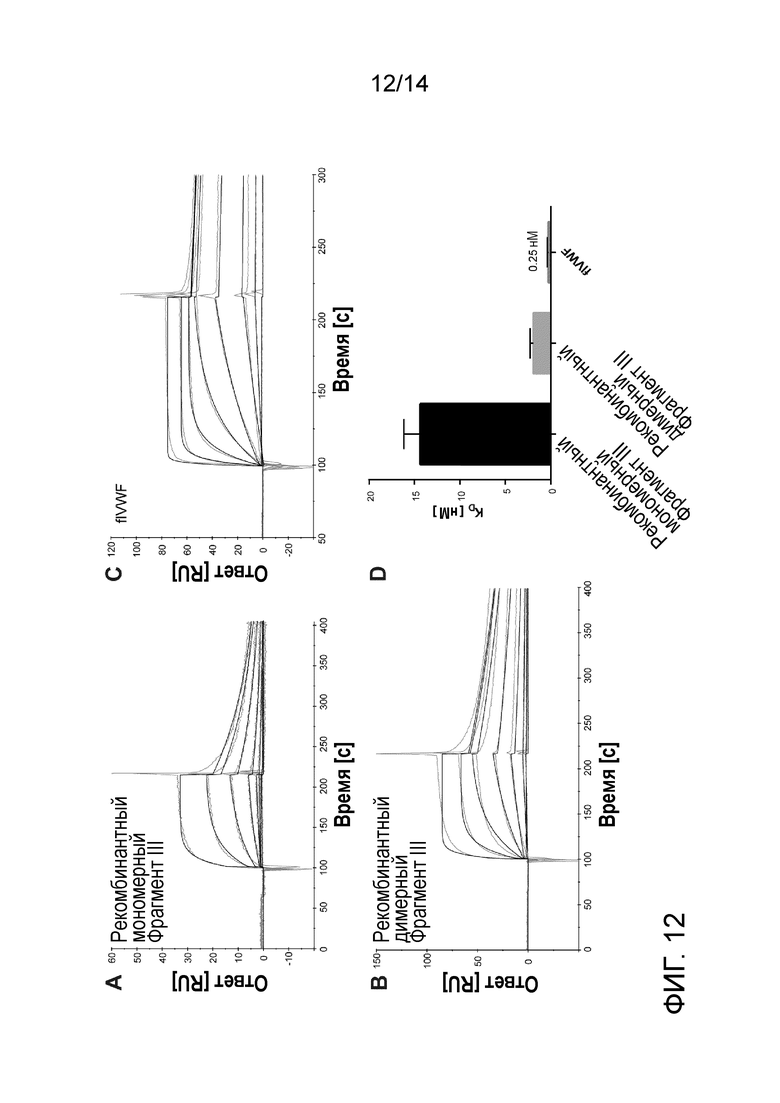
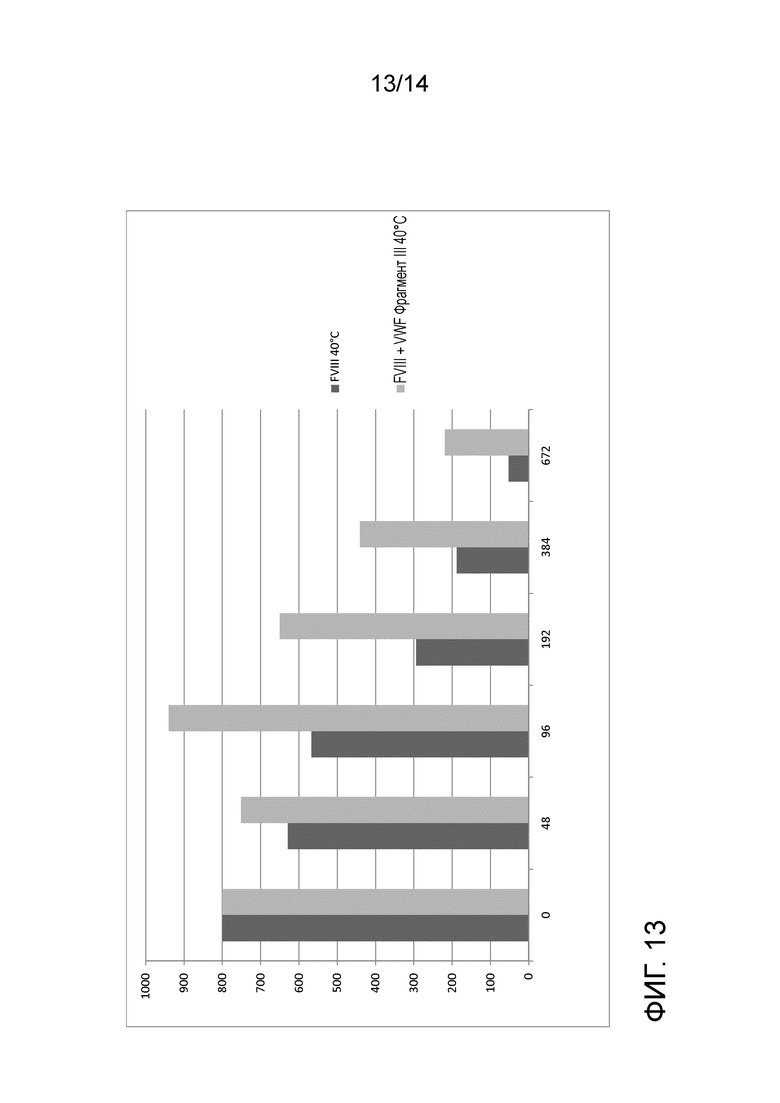
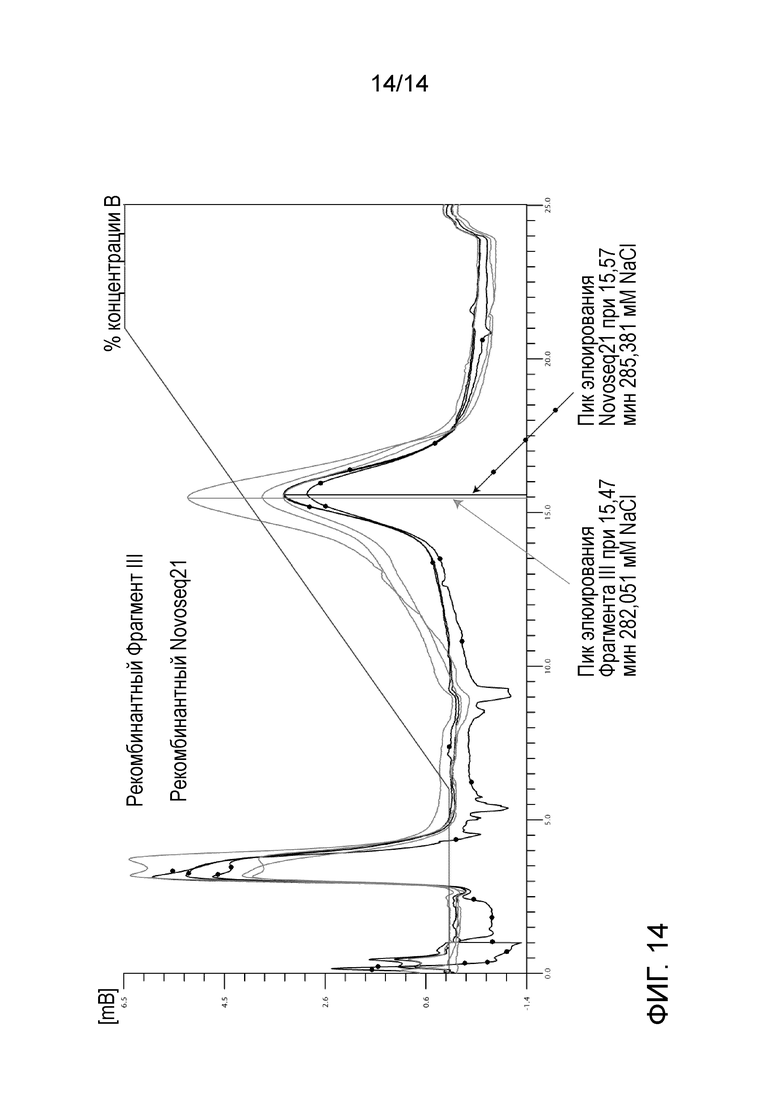
Изобретение относится к области биотехнологии, конкретно к композиции, содержащей комплекс Фактора VIII и одного или более пептидов Фактора фон Виллебранда, где пептиды Фактора фон Виллебранда содержат по меньшей мере аминокислоты 764-1035 и 1691-1905 из SEQ ID NO: 1, но не содержат аминокислоты 2255-2645 из SEQ ID NO: 1, и может быть использовано в медицине. Указанная композиция используется для эффективного лечения гемофилии. 2 н. и 10 з.п. ф-лы, 14 ил., 9 пр.
1. Композиция, содержащая комплекс эффективного количества Фактора VIII и эффективного количества одного или более пептидов Фактора фон Виллебранда, для лечения гемофилии, где пептиды Фактора фон Виллебранда содержат по меньшей мере аминокислоты 764-1035 и 1691-1905 из SEQ ID NO: 1, но не содержат аминокислоты 2255-2645 из SEQ ID NO: 1, где пептиды Фактора фон Виллебранда имеют константу аффинности связывания для гепарина KD>1 нМ, предпочтительно ≥ 2,43 нМ.
2. Композиция по п.1, отличающаяся тем, что пептиды Фактора фон Виллебранда являются фрагментами Фактора фон Виллебранда.
3. Композиция по п.2, отличающаяся тем, что фрагменты Фактора фон Виллебранда содержат аминокислоты 764-1905 из SEQ ID NO: 1.
4. Композиция по п.2, отличающаяся тем, что фрагменты Фактора фон Виллебранда имеют аминокислотную последовательность, которая соответствует аминокислотной последовательности SEQ ID NO: 1, начиная с аминокислоты 764 и заканчивая между аминокислотами 1905 и 2153.
5. Композиция по п.4, отличающаяся тем, что пептид Фактора фон Виллебранда имеет молекулярную массу <500 кДа, предпочтительно <400 кДа.
6. Композиция по любому из пп.1-5, отличающаяся тем, что пептиды Фактора фон Виллебранда обладают по меньшей мере одним из свойств, выбранных из группы, состоящей из
(i) константа аффинности связывания для коллагена III KD>5 нМ, предпочтительно ≥ 17,02 нМ,
(ii) константа аффинности связывания для Фактора VIII KD<100 нМ, или <10 нМ, предпочтительно ≤ 6,19 нМ и
(iii) ингибирование связывания Фактора VIII с фосфолипидами составляет по меньшей мере 70%, предпочтительно по меньшей мере 80%.
7. Композиция по п.1, отличающаяся тем, что пептиды Фактора фон Виллебранда получены из Фактора фон Виллебранда путем протеолитического расщепления или химического расщепления, предпочтительно путем протеолитического расщепления с помощью протеазы V-8 S. aureus.
8. Композиция по п.1, отличающаяся тем, что Фактор VIII представляет собой полноразмерный Фактор VIII, Фактор VIII с делетированным B-доменом или Фактор VIII, где B-домен заменен на искусственный линкер или фрагмент природного B-домена или их комбинацию.
9. Композиция по п.1, отличающаяся тем, что Фактор VIII представляет собой Фактор VIII, выделенный из плазмы, или рекомбинантный Фактор VIII.
10. Композиция по п.1, отличающаяся тем, что композиция обладает по меньшей мере одним из свойств, выбранных из группы, состоящей из
(i) пептиды Фактора фон Виллебранда экранируют Фактор VIII от связывания с антителами с минимизацией образования ингибитора у пациента,
(ii) стабилизирует Фактор VIII, обеспечивая остаточную активность Фактора VIII, составляющую по меньшей мере 90% после хранения в течение 12 месяцев в виде замороженной жидкости при -70°C,
(iii) стабилизирует Фактор VIII, обеспечивая остаточную активность Фактора VIII, составляющую по меньшей мере 90% после хранения в течение 24 месяцев в лиофилизированной форме при 5°C,
(iv) стабилизирует Фактор VIII, обеспечивая остаточную активность Фактора VIII, составляющую по меньшей мере 90% после хранения в течение 12 месяцев в лиофилизированной форме при 25°C,
(v) продлевает период полужизни Фактора VIII in vivo по меньшей мере на 20% и
(vi) уменьшает образование ингибитора у ранее не получавших лечение пациентов до менее чем 20% после лечения с использованием композиции в течение 6 месяцев.
11. Применение композиции по любому из пп.1-10 для лечения или предотвращения гемофилии.
12. Применение по п.11, включающее невнутривенное введение композиции, предпочтительно, где невнутривенное введение является подкожной инъекцией.
| WO 2013083858, 13.06.2013 | |||
| WO 2013160005, 31.10.2013 | |||
| ROMIJN R.A | |||
| et al | |||
| Переносная печь для варки пищи и отопления в окопах, походных помещениях и т.п. | 1921 |
|
SU3A1 |
| ПАРОВАЯ ИЛИ ГАЗОВАЯ ТУРБИНА | 1914 |
|
SU278A1 |
| Печь для сжигания твердых и жидких нечистот | 1920 |
|
SU17A1 |
| ЛИХАЧЕВА Е.А | |||
| и др | |||
| Международный опыт диагностики и лечения болезни Виллебранда, Гематология и трансфузиология, 2013, V | |||
| Способ окисления боковых цепей ароматических углеводородов и их производных в кислоты и альдегиды | 1921 |
|
SU58A1 |
| Очаг для массовой варки пищи, выпечки хлеба и кипячения воды | 1921 |
|
SU4A1 |

