Ссылка на список последовательностей, таблицу или компьютерную программу
[0001] Список последовательностей, одновременно представленный в соответствии с § 1.821 раздела 37 Свода федеральных нормативных актов США в машиночитаемой форме (CRF) через EFS-Web, как файл с именем KALSEC_76_PCT_Sequence_Listing_26_Sept_2019.txt, включен в настоящее описание путем ссылки. Электронная копия Списка последовательностей была создана 26 сентября 2019 года.
Область техники, к которой относится изобретение
Настоящее изобретение относится к способу получения горького компонента пива посредством катализируемой ферментами биоконверсии изо-альфа-кислот, полученных из хмеля, в дигидро-(rho)-изо-альфа-кислоты, а также к новым ферментным катализаторам, которые могут быть использованы в таком способе. Дигидро-(rho)-изо-альфа-кислоты обладают превосходными характеристиками, улучшающими их применимость в качестве добавки к напитку. Потребители могут предпочесть дигидро-(rho)-изо-альфа-кислоты, полученные с помощью данного способа, который не требует использования агрессивных химических реагентов и использует ферменты, которые могут встречаться в природе.
Уровень техники изобретения
[0003] Традиционные способы придания горечи пиву используют цельный свежий хмель, цельный сушеный хмель или гранулы хмеля, добавленные во время кипячения в котле. Экстракты хмеля, полученные путем экстрагирования хмеля сверхкритическим диоксидом углерода, или изомеризованные гранулы хмеля, полученные путем нагревания хмеля в присутствии катализатора, являются более поздними инновациями в получении горького компонента, которые также были приняты пивоварами. Гранулы хмеля также могут быть добавлены позже в процессе пивоварения, а в случае сухого охмеления хмель добавляется в готовое пиво перед фильтрацией. Эти способы имеют в качестве недостатков низкий уровень использования горьких соединений, присутствующих в хмеле, что неблагоприятно сказывается на стоимости. Пиво или другие солодовые напитки, произведенные таким образом, нестабильны на свету и должны быть упакованы в темно-коричневые бутылки или банки или помещены в темное место во избежание индуцированного светом образования 3-метил-2-бутен-1-тиола (3-МБТ), который дает ярко выраженный «засвеченный» или выдохшийся аромат. Помещение бутылок в картонные коробки или полное оборачивание их светонепроницаемой или светофильтрующей бумагой, фольгой или пластиковыми покрытиями является еще одним дорогостоящим способом защиты этих напитков от выдохшегося на свету вкуса и аромата.
[0004] Горечь в пиве, сваренном традиционным способом, в первую очередь связана с изо-альфа-кислотами. Эти соединения образуются в процессе пивоварения в результате изомеризации гумулонов, которые являются естественными соединениями в лупулиновых железах хмеля. Следствием этого, учитывая естественную нестабильность изо-альфа-кислот по отношению к фотохимическим реакциям в пиве, является то, что напиток склонен к образованию характерного привкуса или неприятного запаха.
[0005] Полностью светлое стабильное пиво или другие солодовые напитки можно приготовить с использованием так называемых усовершенствованных или модифицированных кислот хмеля. Пиво, приготовленное с использованием этих горьких компонентов, может быть упаковано в бутылки из бесцветного стекла, не опасаясь образования неприятного запаха. Дигидро-(rho)-изо-альфа-кислоты являются продуктами восстановления изо-альфа-кислот, которые светостабильны. На сегодняшний день эти соединения в природе не обнаружены. Традиционно часть изо-альфа-кислот, которая отвечает за фотохимические процессы, изменялась восстановлением карбонильной группы с использованием боргидрида натрия.
[0006] Боргидрид натрия представляет собой неорганическое соединение, которое можно использовать для восстановления кетонов. Оно чрезвычайно опасно при попадании на кожу, в глаза, вдыхании или проглатывании, при пероральной LD50 160 мг/кг (для крыс). Боргидрид натрия также легко воспламеняется, вызывает коррозию и чрезвычайно активен по отношению к окислителям, кислотам, щелочам и влаге (боргидрид натрия; MSDS No.S9125; Sigma-Aldrich Co.: Saint Louis, MO, 01 ноября 2015 г.).
[0007] Потребители все чаще отдают предпочтение натуральным материалам перед синтетическими или полусинтетическими. Таким образом, существует потребность не только в создании композиций, использующих натуральные материалы в качестве горьких компонентов для пива и других напитков, но также в способах более естественного производства указанных материалов.
[0008] Биокаталитическое производство представляет собой новую технологию, которая обеспечивает высокоселективное, безопасное, чистое и масштабируемое производство ценных химических соединений. Биокаталитическое производство основано на природных ферментах, которые заменяют химические катализаторы.
[0009] Ферменты представляют собой встречающиеся в природе белки, способные катализировать определенные химические реакции. В природе существуют ферменты, которые в настоящее время способны заменять химические катализаторы при получении модифицированных соединений горечи хмеля (Robinson, P.K., Enzymes: principles and biotechnological applications. Essays Biochem 2015, 59, 1-41.).
[0010] Гумулон представляет собой естественный вторичный метаболит, который может подвергаться воздействию грибов и бактерий, сосуществующих с растением Humulus lupulus. Возможно, что обитающие в почве и растениях грибы и бактерии обладают ферментами, способными модифицировать гумулон в целях детоксикации или удаления. Кроме того, в организмах могли возникнуть ферменты для модификации гумулоноподобных молекул, но из-за неизбирательной активности эти ферменты обладают активностью в отношении представляющих интерес соединений, изо-альфа-кислот (Hult, K.; Berglund, P., Enzyme promiscuity: mechanism and applications. Trends Biotechnol. 2007, 25 (5), 231-238; Nobeli, I.; Favia, A. D.; Thornton, J. M., Protein promiscuity and its implications for biotechnology. Nat. Biotechnol. 2009, 27 (2), 157-167.).
[0011] Ферменты, которые катализируют реакции окисления/восстановления, то есть перенос атомов или электронов водорода и кислорода от одного вещества к другому, в целом классифицируются как оксидоредуктазы. Более конкретно, ферменты, восстанавливающие кетонные группы до гидроксильных групп, известны как кеторедуктазы или карбонилредуктазы и зависят от добавления экзогенного источника восстанавливающих эквивалентов (например, кофакторов НАДН, НАДФН). В соответствии с существующим названием охарактеризованных здесь ферментов, эти ферменты будут называться «кеторедуктазами».
[0012] Затраты по использованию дорогих кофакторов (НАДН, НАДФН) могут быть снижены путем включения дополнительных ферментов и субстратов для рециклирования кофакторов, например, глюкозодегидрогеназы и глюкозы, или путем использования кеторедуктазы, которая также способна окислять дешевое и естественный сырье, такое как этанол.
[0013] Существует много примеров применения ферментов в пивоварении и их благоприятного влияния на конечные свойства пива (Pozen, M., Enzymes in Brewing. Ind. Eng. Chem, 1934, 26 (11), 1127-1133.). Известно, что присутствие дрожжевых ферментов при естественном брожении пива приводит к образованию соединений, влияющих на вкус и аромат конечного напитка (Praet, T.; Opstaele, F.; Jaskula-Goiris, B.; Aerts, G.; De Cooman, L., Biotransformations of hop-derived aroma compounds by Saccharomyces cerevisiae upon fermentation. Cerevisia, 2012, 36, 125-132.). Экзогенно добавленные ферменты обеспечивают различные улучшения процесса пивоварения, такие как снижение вязкости, увеличение количества сбраживаемых сахаров, защита от холода и осветление (Wallerstein, L. (1947) Bentonite and Proteolytic Enzyme Treatment of Beer, US Patent 2,433,411.; Ghionno, L.; Marconi, O.; Sileoni, V.; De Francesco, G.; Perretti, G., Brewing with prolyl endopeptidase from Aspergillus niger: the impact of enzymatic treatment on gluten levels, quality attributes, and sensory profile. Int. J. Food Sci. Technol, 2017, 52 (6), 1367-1374.). Кроме того, экстракты хмеля специально предварительно обрабатывали ферментами для модификации ароматических соединений, полученных из хмеля (Gros, J.; Tran, T.T. H.; Collin, S., Enzymatic release of odourant polyfunctional thiols from cysteine conjugates in hop. J. Inst. Brew. 2013, 119 (4), 221-227.).
[0014] Однако до настоящего изобретения ферменты, способные катализировать восстановление изо-альфа-кислот до дигидро-(rho)-изо-альфа-кислот, не наблюдались в природе и, таким образом, не были описаны в литературе. Раскрытый в настоящем изобретении способ представляет собой новую ферментативную реакцию.
Задача изобретения
[0015] Задачей настоящего изобретения является создание способа ферментативного получения дигидро-(rho)-изо-альфа-кислот, модифицированного варианта природных горьких веществ, полученных из растения хмеля. Настоящий способ разработан для замены существующих производственных процессов, в которых используется химический реагент, боргидрид натрия. Еще одной задачей настоящего изобретения является получение новых ферментных катализаторов, которые можно использовать в таком способе.
Сущность изобретения
[0016] Настоящее изобретение относится к способу, который можно масштабировать до промышленных уровней для биоконверсии изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты, которые затем можно использовать в качестве природного и светостабильного горького компонента для напитков.
[0017] В одном аспекте настоящее изобретение относится к способу высокопроизводительной биоконверсии изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты с использованием фермента кеторедуктазы или микроорганизма, экспрессирующего ген, кодирующий указанную кеторедуктазу.
[0018] В другом аспекте настоящее изобретения относится к такому способу получения дигидро-(rho)-изо-альфа-кислот, где способ проводят в водной системе с мягкими условиями температуры и pH, что делает его экологически безопасным производственным процессом.
[0019] В одном варианте осуществления настоящего изобретения биоконверсия изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает добавление к смеси изо-альфа-кислот очищенного фермента кеторедуктазы и НАДФН или НАДФ с последующей инкубацией до получения желаемого выхода.
[0020] В другом варианте осуществления настоящего изобретения биоконверсия изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает добавление к смеси изо-альфа-кислот очищенного фермента кеторедуктазы и НАДФН или НАДФ в присутствии изопропанола для рециклирования кофактора с последующей инкубацией до получения желаемого выхода.
[0021] В другом варианте осуществления настоящего изобретения концентрация изо-альфа-кислот, то есть субстрата, является максимальной для увеличения объемной производительности биоконверсии.
[0022] В другом варианте осуществления настоящего изобретения концентрация кофактора НАДФН или НАДФ в смеси минимизирована для улучшения экономических показателей биоконверсии.
[0023] В одном варианте осуществления настоящего изобретения биоконверсия изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает добавление к смеси изоальфа-кислот очищенного фермента кеторедуктазы и НАДФН или НАДФ в присутствии другого фермента (такого как глюкозодегидрогеназа) для рециклирования кофактора с последующей инкубацией до получения желаемого выхода.
[0024] В другом варианте осуществления настоящего изобретения биоконверсия изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает добавление к смеси изо-альфа-кислот цельноклеточного биокатализатора с последующей инкубацией до получения желаемого выхода, где цельноклеточный биокатализатор представляет собой иммобилизованный микроорганизм, экспрессирующий ген, кодирующий кеторедуктазу.
[0025] В другом варианте осуществления настоящего изобретения биоконверсия изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает обеспечение растущего микроорганизма, экспрессирующего ген, кодирующий кеторедуктазу, изо-альфа-кислотой.
[0026] В другом варианте осуществления настоящего изобретения биоконверсия альфа-кислот в дигидро-(rho)-изо-альфа-кислоты включает добавление к экстракту альфа-кислот термостабильного фермента кеторедуктазы, где смесь нагревают и инкубируют до желаемого выхода дигидро-(rho)-изо-альфа кислот.
[0027] Настоящее изобретение также относится к новым ферментным катализаторам, которые можно использовать в способе настоящего изобретения, как определено выше.
[0028] Редуктаза настоящего изобретения необязательно проявляет активность по восстановлению карбонильной группы в боковой цепи у C(4) изо-альфа-кислот, превращая светочувствительную ацилоиновую группу во вторичный спирт с получением светостабильного производного изо-альфа-кислоты (Фигура 1).
[0029] В другом варианте осуществления настоящего изобретения кеторедуктаза, используемая в способе настоящего изобретения, катализирует предпочтительно в минимальной степени или вообще не катализирует восстановление любого конкретного члена из шести основных изо-альфа-кислот: цис-изогумулона, транс-изогумулона, цис-изокогумулона, транс-изокогумулона, цис-изоадгумулона и транс-изоадгумулона.
[0030] В другом варианте осуществления изобретения кеторедуктаза, используемая в способе настоящего изобретения, специфично восстанавливает цис-изогумулон, цис-изокогумулон и цис-изоадгумулон.
[0031] В другом варианте осуществления настоящего изобретения кеторедуктаза, используемая в способе настоящего изобретения, специфично восстанавливает транс-изогумулон, транс-изокогумулон и транс-изоадгумулон.
[0032] В другом варианте осуществления настоящего изобретения смесь 2 или более ферментов кеторедуктазы, проявляющих указанную выше субстратную специфичность, используется в способе настоящего изобретения для восстановления смеси цис- и транс-изо-альфа-кислот до их соответствующих дигидро-изо-альфа-кислот.
[0033] В другом варианте осуществления настоящего изобретения смесь 2 или более ферментов кеторедуктазы, проявляющих субстратную специфичность, может быть добавлена к реакционной смеси для получения уникальной смеси дигидро-изо-альфа-кислот, которая отличается от смеси, производимой химическими восстановителями, такими как борогидрид натрия.
[0034] В другом варианте осуществления настоящее изобретение относится к способу, как определено выше, где фермент редуктаза представляет собой кеторедуктазу.
[0035] В другом варианте осуществления настоящее изобретение относится к способу, как определено выше, где фермент кеторедуктаза или микроорганизм, экспрессирующий ген, кодирующий фермент кеторедуктазу, содержит аминокислотную последовательность SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
[0036] В другом варианте осуществления настоящее изобретение относится к способу, как определено выше, где фермент кеторедуктаза или микроорганизм, экспрессирующий ген, кодирующий фермент кеторедуктазу, может необязательно иметь одно или несколько различий в аминокислотных остатках по сравнению с ферментом кеторедуктаза, выбранным из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22.
[0037] В еще одном варианте осуществления настоящее изобретение относится к ферменту кеторедуктаза или микроорганизму, экспрессирующему ген, который кодирует фермент кеторедуктаза, который содержит аминокислотную последовательность SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
[0038] В другом варианте осуществления настоящее изобретение относится к ферменту кеторедуктаза или микроорганизму, экспрессирующему ген, кодирующий редуктазу, который может необязательно иметь одно или несколько отличий в аминокислотных остатках по сравнению с последовательностью фермента редуктазы SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
[0039] В еще одном варианте осуществления настоящее изобретение относится к ферменту кеторедуктаза или микроорганизму, экспрессирующему ген, кодирующий фермент кеторедуктазу, который на 99, 95, 90, 85, 80, 75 или 70 процентов гомологичен ферменту кеторедуктаза, выбранному из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20. и SEQ ID NO: 22.
[0040] В другом аспект настоящее изобретение относится к вектору, содержащему полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
[0041] Настоящее изобретение также относится к такому вектору, который дополнительно содержит, по меньшей мере, одну регуляторную последовательность.
[0042] Настоящее изобретение также относится к клетке-хозяину, содержащей такой вектор, содержащий полинуклеотид, кодирующий аминокислотную последовательность SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
[0043] Настоящее изобретение также относится к способу получения кеторедуктазы SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22, включающему культивирование указанной клетки-хозяина в условиях, при которых кеторедуктаза продуцируется указанной клеткой-хозяином.
[0044] Настоящее изобретение также относится к способу получения кеторедуктазы SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22, дополнительно включающему этап выделения кеторедуктазы, продуцируемой указанной клеткой-хозяином.
Краткое описание чертежей
[0045] На Фигуре 1 показано катализируемое ферментом восстановление репрезентативного эпимера изо-альфа-кислот.
[0046] На Фигуре 2 показан анализ очищенных редуктаз с помощью ДСН-ПААГ.
[0047] На Фигуре 3 показаны СВЭЖХ-хроматограммы для изо-альфа-кислот, инкубированных с редуктазой R20 (два верхних изображения) и без редуктазы R20 (два нижних изображения) в течение 24 часов при температуре 30°C. Указаны пики, соответствующие продукту, дигидро-(rho)-изо-альфа-кислотам.
[0048] На Фигуре 4 показана структурная модель редуктазы R17 (темно-серый, визуализация поверхности) с репрезентативным субстратом (транс-изогумулон, черный) и кофактором (НАДФН, светло-серый), связанными с полостью активного сайта.
[0049] На Фигуре 5 показаны СВЭЖХ-хроматограммы для (A) кеторедуктазы, которая продуцирует только один диастереомер дигидро-(rho)-изо-альфа-кислот, и (B) кеторедуктазы, которая продуцирует оба диастереомера дигидро-(rho)-изо-альфа-кислот. Указаны пики, соответствующие продукту, дигидро-(rho)-изо-альфа-кислотам.
[0050] На Фигуре 6 показано выравнивание аминокислотной последовательности, полученное с помощью Clustal Omega (www.ebi.ac.uk/Tools/msa/clustalo/) активных гомологов кеторедуктазы: R4, R17, R20, R21 и R23.
Подробное описание изобретения
[0051] В настоящем изобретении фермент кеторедуктаза заменяет функцию боргидрида натрия и позволяет использовать более естественный способ получения добавки к напитку, дигидро-(rho)-изо-альфа кислоты. Фермент может представлять собой любую кеторедуктазу, специфично восстанавливающую кетонную группу до гидроксигруппы любого или всех изомеров изо-альфа-кислоты (ко-, н-ад- и цис/транс-). Фермент может иметь бактериальное, грибное или растительное происхождение, но не ограничивается ими. Фермент может быть кофактор-зависимым (НАДН или НАДФН) или независимым.
[0052] Используемые здесь термины «изо-альфа-кислоты», «изо-альфа-кислоты хмеля» и «изо-альфа-кислоты из хмеля» могут использоваться взаимозаменяемо.
[0053] В соответствии с настоящим изобретением раствор изо-альфа-кислоты подвергают ферментативной обработке с использованием одного или нескольких очищенных ферментов или смеси, содержащей фермент(ы) и, при необходимости, дополнительные ферменты для рециклирования кофактора. Количество фермента зависит от параметров инкубации, включая продолжительность, температуру, количество и концентрацию субстрата.
[0054] Альтернативно раствор изо-альфа кислоты подвергают ферментативной обработке с использованием смеси, содержащей микроорганизм, экспрессирующий указанный фермент(ы). Кроме того, настоящее изобретение относится к способу восстановления изо-альфа-кислот в соответствии с настоящим изобретением, который включает культивирование микроорганизма, продуцирующего кеторедуктазу, при необходимости индуцируя экспрессию кеторедуктазы. Интактные клетки можно собирать и добавлять непосредственно в реакцию вместо выделенного фермента для восстановления изо-альфа-кислот, как описано выше. Кроме того, собранные клетки можно иммобилизовать перед добавлением в реакцию восстановления. Микроорганизм можно культивировать и ферментировать известными способами. Микроорганизм может представлять собой бактерии или грибы.
[0055] Смесь цис- и транс-изо-альфа-кислот можно инкубировать с одной кеторедуктазой, проявляющей способность восстанавливать оба изомера. Альтернативно, смесь цис- и транс-изо-альфа-кислот можно инкубировать с 2 или более кеторедуктазами, проявляющими разную специфичность, если полученный продукт представляет собой смесь цис- и транс-дигидро-изо-альфа-кислот.
[0056] Альтернативно, раствор, содержащий только цис-изо-альфа-кислоты, можно инкубировать с кеторедуктазой(ами), специфичной для цис-изомера, и полученный продукт представляет собой раствор цис-дигидро-изо-альфа-кислот. Раствор, содержащий только цис-дигидро-изо-альфа-кислоты, может обладать желательными свойствами горечи и/или термостабильности.
[0057] Альтернативно, раствор, содержащий только транс-изо-альфа-кислоты, можно инкубировать с кеторедуктазой(ами), специфичной для транс-изомера, и полученный продукт представляет собой раствор транс-дигидро-изо-альфа-кислот. Раствор, содержащий только транс-дигидро-изо-альфа-кислоты, может обладать желательными свойствами горечи.
[0058] Создаваемые для конкретного случая смеси транс- и цис-изо-альфа-кислот можно инкубировать с 1 или несколькими кеторедуктазами, проявляющими переменную субстратную специфичность, для получения уникальных смесей дигидро-изо-альфа-кислот, которые иначе недостижимы.
[0059] Смесь изо-альфа-кислот может быть подвергнута ферментативной реакции с использованием фермента кеторедуктазы(з) в дополнение к ферментам для катализа дополнительных желаемых модификаций, таким как, но без ограничений, дегидрогеназы, изомеразы, гидратазы и лиазы. Ферменты с различной активностью можно комбинировать в реакции в одной емкости или добавлять последовательно.
[0060] Подходящий растворитель для применения при инкубации ферментов включает воду и смеси воды с другим совместимым с ферментом растворителем, таким как этанол или изопропанол. Ферментативная активность улучшается за счет буферизации водных растворов. Буферные агенты включают без ограничений трис(гидроксиметил)аминометан (также известный как Трис), 4-(2-гидроксиэтил) пиперазин-1-этансульфоновую кислоту (также известную как HEPES), фосфат натрия и фосфат калия.
[0061] Фермент(ы) и изо-альфа-кислоты инкубируют в подходящем диапазоне значений pH, например, pH от 6 до 10, и диапазоне температур, например, от 10 до 90°C, и выдерживают при этой температуре в течение достаточного времени для превращения изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты с желаемым выходом. Непрерывное перемешивание обеспечивает постоянную температуру и воздействие фермента на субстрат. Продолжительность реакции, обычно составляет от 24 до 48 часов, и она будет зависеть от количества и концентрации фермента и субстрата, присутствующего растворителя и выбранной температуры.
[0062] Фермент(ы) может быть свободным в растворе, иммобилизованным на гранулах или подобных смешиваемых основах или иммобилизованным на пленке или смоле, через которые пропускают раствор изо-альфа-кислот. Уровень чистоты фермента может варьировать от 30 до 90+% в зависимости от способа очистки.
[0063] Фермент(ы) может быть удален из конечного продукта путем физической фильтрации или центрифугирования. Фермент(ы) также можно инактивировать путем воздействия экстремальной температуры или pH, и он может оставаться в конечном продукте.
[0064] Ферменты редуктазы, охватываемые настоящим изобретением, включают ферменты кеторедуктазы.
[0065] Информация о 23 успешно очищенных ферментах приведена в Таблице 1, включая сокращенное обозначение, идентификационный номер последовательности и аминокислотную последовательность.
Таблица 1. Очищенные редуктазы
[0066] Почти все кандидаты были в значительной степени чистыми (целевой белок составляет >80%) после одностадийной очистки (см. Фигуру 2).
[0067] Ферменты редуктазы, охватываемые настоящим изобретением, включают ферменты, содержащие следующие аминокислотные последовательности: SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22.
[0068] Ферменты редуктазы, охватываемые настоящим изобретением, также включают ферменты, имеющие одно или несколько различий в аминокислотных остатках по сравнению со следующими аминокислотными последовательностями: SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22.
[0069] Ферменты редуктазы, охватываемые настоящим изобретением, также включают ферменты, содержащие аминокислотную последовательность, которая идентична, по меньшей мере, на 40% (включая, по меньшей мере, на 50%, по меньшей мере, на 60%, по меньшей мере, на 70%, по меньшей мере, на 80%, по меньшей мере, на 85%, по меньшей мере, на 90% и, по меньшей мере, на 95%) следующим аминокислотным последовательностям: SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22.
[0070] Используемые здесь термины «процент гомологии последовательностей», «процент гомологии» и «процент идентичности» относятся к сравнениям между полинуклеотидными последовательностями или полипептидными последовательностями и определяются путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где для оптимального выравнивания двух последовательностей часть полинуклеотидной или полипептидной последовательности в окне сравнения может содержать вставки или делеции (т.е. пробелы) по сравнению с эталонной последовательностью. Процент рассчитывается путем определения количества положений, в которых или идентичное основание нуклеиновой кислоты, или аминокислотный остаток встречается в обеих последовательностях, либо основание нуклеиновой кислоты или аминокислотный остаток выравниваются с пробелом, чтобы получить количество совпадающих положений, деления количества совпадающих положений на общее количество положений в окне сравнения и умножения результата на 100, чтобы получить процент идентичности последовательностей. Определение оптимального выравнивания и процента гомологии последовательностей выполняется с использованием алгоритмов BLAST и BLAST 2.0 (см., Например, Altschul et al., J. Mol. Biol. 215: 403-410 [1990]; и Altschul et al., Nucleic Acids Res. 3389-3402 [1977]). Программное обеспечение для проведения анализа алгоритмом BLAST общедоступно на веб-сайте Национального центра биотехнологической информации.
[0071] Неизбирательные ферменты могут катализировать одну и ту же химическую реакцию, несмотря на низкую идентичность общих аминокислот. Кеторедуктаза R4 (SEQ ID NO: 3) изначально была выбрана для скрининга из-за ее неизбирательной природы [Guo et al. Biochim. Biophys. Acta 2014, 1844]. Пять дополнительных кеторедуктаз (R17 (SEQ ID NO: 16), R20 (SEQ ID NO: 19), R21 (SEQ ID NO: 20), R22 (SEQ ID NO: 21) и R23 (SEQ ID NO: 22)), которые содержат один и тот же домен фермента (IPR001509: НАД-зависимая эпимераза/дегидратаза) и имеют общую аминокислотную идентичность с R4 (SEQ ID NO: 3), были получены как синтетические гены, очищены и охарактеризованы. Редуктазы специально отбирали с все более низкой идентичностью последовательностей, чтобы установить предел по идентичности последовательностей.
[0072] Несмотря на относительно низкую процентную идентичность (34-39% по всей длине фермента) с R4 (SEQ ID NO: 3), ферменты R17 (SEQ ID NO: 16), R20 (SEQ ID NO: 19) , R21 (SEQ ID NO: 20) и R23 (SEQ ID NO: 22) катализируют превращение изо-альфа-кислот в дигидро-(rho)-изоальфа-кислоты. R22 (SEQ ID NO: 21), который на 33% идентичен R4 (SEQ ID NO: 3), не катализирует превращение изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты, но в остальном является активным ферментом в очищенном состоянии (установлено путем измерения ферментативно-катализируемой окислительной активности изопропанола).
[0073] Признак, который отделяет функциональные редуктазы от нефункциональных для получения дигидро-(rho)-изо-альфа кислот, иллюстрируется множественным выравниванием последовательностей (Фигура 6). Кеторедуктаза R4 (SEQ ID NO: 3) и все гомологи R4 (SEQ ID NO: 3), охарактеризованные здесь как способные превращать изо-альфа-кислоты в дигидро-(rho)-изо-альфа-кислоты, обладают доменом, находящимся между остатками 100 и 200, состоящим из 13 аминокислот с хорошей гомологией (>53% идентичности) и 9 аминокислот с высокой гомологией (>55%), разделенных 36-39 аминокислотами с низкой гомологией (38-46%). Этот домен отсутствует в нефункциональном полипептиде R22 (SEQ ID NO: 21). Таким образом, домен считается признаком активности кеторедуктазы для получения дигидро-(rho)-изо-альфа-кислот.
[0074] Раскрытые здесь конкретные варианты осуществления изобретения могут быть дополнительно ограничены в формуле изобретения использованием терминов «состоящий из» или «в основном состоящий из». При использовании в формуле изобретения, независимо от того, является ли она первоначально поданной или измененной в ходе экспертизы, переходный термин «состоящий из» исключает любой элемент, этап или ингредиент, не указанные в формуле изобретения. Переходный термин «в основном состоящий из» ограничивает объем формулы изобретения указанными материалами или этапами, а также теми, которые существенно не влияют на основные и новые характеристики. Заявленные таким образом варианты осуществления настоящего изобретения по умолчанию или явно описаны и доступны здесь.
[0075] Используемый здесь термин «содержащий» или «содержит» предназначен для обозначения того, что композиции и способы включают перечисленные элементы, но не исключают другие.
[0076] Термин «эффективное количество» относится к тому количеству редуктазы, которое достаточно для превращения изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты. Определение эффективного количества для данного применения хорошо известно среднему специалисту в области фармацевтики.
[0077] В способе получения дигидро-(rho)-изо-альфа-кислот раствор изо-альфа-кислоты подвергают ферментативной обработке с использованием одного или нескольких очищенных ферментов редуктазы или смеси, содержащей фермент(ы) редуктазы и, необязательно, дополнительные ферменты для рециклирования кофактора в количестве, эффективном для превращения изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты. Количество фермента зависит от параметров инкубации, включая продолжительность, температуру, количество и концентрацию субстрата.
[0078] Альтернативно раствор изо-альфа кислоты подвергают ферментативной обработке с использованием смеси, содержащей микроорганизм, экспрессирующий указанный фермент.
[0079] Смесь цис- и транс-изо-альфа-кислот можно инкубировать с одной редуктазой/кеторедуктазой, проявляющей способность восстанавливать оба изомера. Альтернативно, смесь цис- и транс-изоальфа кислот можно инкубировать с 2 или более кеторедуктазами, проявляющими различную специфичность, если полученный продукт представляет собой смесь цис- и транс-дигидро-изо-альфа кислот.
[0080] Индивидуальные смеси транс- и цис-изо-альфа кислот можно инкубировать с 1 или более редуктазами/ кеторедуктазами, проявляющими переменную субстратную специфичность, для получения уникальных смесей дигидро-изо-альфа-кислот, которые иначе недостижимы.
[0081] Смесь изо-альфа-кислот может быть подвергнута ферментативной реакции с использованием фермента редуктазы в дополнение к ферментам для катализа дополнительных желаемых модификаций, таких как, но без ограничений, дегидрогеназы, изомеразы, гидратазы и лиазы. Ферменты с различной активностью можно комбинировать в реакции в одной емкости или добавлять последовательно.
[0082] Подходящий растворитель для применения при инкубации ферментов включает воду и смеси воды с другим совместимым с ферментом растворителем, таким как этанол или изопропанол. Ферментативная активность улучшается за счет буферизации водных растворов. Буферные агенты включают без ограничений трис(гидроксиметил)аминометан (также известный как Трис), 4-(2-гидроксиэтил) пиперазин-1-этансульфоновую кислоту (также известную как HEPES), фосфат натрия и фосфат калия.
[0083] Фермент и изо-альфа-кислоты инкубируют в подходящем диапазоне значений pH, например, pH от 6 до 10, и диапазоне температур, например, от 10 до 90°C, и выдерживают при этой температуре в течение достаточного времени для превращения изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты с желаемым выходом. Непрерывное перемешивание обеспечивает постоянную температуру и воздействие фермента на субстрат. Продолжительность реакции, обычно составляет от 24 до 48 часов, и она будет зависеть от количества и концентрации фермента и субстрата, присутствующего растворителя и выбранной температуры.
[0084] Фермент редуктаза может быть свободным в растворе, иммобилизованным на гранулах или подобных смешиваемых основах или иммобилизованным на пленке или смоле, через которые пропускают раствор изо-альфа-кислот. Уровень чистоты фермента может варьировать от 30 до 90+% в зависимости от способа очистки.
[0085] Редуктаза может быть удалена из конечного продукта путем физической фильтрации или центрифугирования. Фермент также можно инактивировать путем воздействия экстремальной температуры или pH, и он может оставаться в конечном продукте.
[0086] Настоящее изобретение представляет собой новый способ использования редуктаз для преобразования изо-альфа-кислот в дигидро-(rho)-изоальфа-кислоты. Оптимизированные по кодонам гены редуктазы обеспечили выход более 100 мг очищенного фермента на L-клеточную культуру в E. coli BL21 (DE3). Все ферменты использовали НАДФН в качестве кофактора. Редуктазы, охарактеризованные в данном исследовании, обладают ферментативной активностью, которая ранее не описывалась. Эти ферменты образуют основу для новых биокатализаторов, которые могут быть использованы в новой биотрансформации для замены существующих способов с использованием боргидрида натрия.
Примеры
[0087] Следующие ниже примеры иллюстрируют изобретение, не ограничивая его объем.
Пример 1
Получение редуктазы и скрининг
Методы
Идентификация кандидата
[0088] Кандидаты в редуктазы отбирали после обширного поиска в литературе охарактеризованных ферментативных реакций, аналогичных желаемой реакции, с последующим биоинформатическим анализом трех общедоступных баз данных последовательностей белков: UniProt (www.uniprot.org/), Pfam ( //pfam.xfam.org/) и InterPro (www.ebi.ac.uk/interpro/E. coli). Биоинформатический анализ основывался на выравнивании последовательностей охарактеризованных ферментов и кандидатов в редуктазы при помощи алгоритма BLASTP (//blast.ncbi.nlm.nih.gov/Blast.cgi).
Экспрессия и очистка фермента
[0089] Плазмидную ДНК получали несколькими способами: 1) в экспрессионном векторе из репозитория плазмид DNASU (www.dnasu.org), 2) в векторе для клонирования из репозитория плазмид DNASU и последующего клонировали в собственный экспрессионный вектор, 3) как синтетический ген в экспрессионном векторе от Atum (www.atum.bio) или 4) как синтетический ген в экспрессионном векторе от General Biosystems (www.generalbiosystems.com). Синтетические гены оптимизировали по кодонам для экспрессии в E. coli.
[0090] 5 мл бульона Луриа с соответствующими антибиотиками инокулировали штаммом E. coli BL21 (DE3), содержащим целевой экспрессионный вектор, с чашки с агаром, и инкубировали при температуре 30°C при встряхивании в течение ночи. На следующий день ночную культуру разводили 1:100 в свежем 0,5 л бульона Луриа с антибиотиками и инкубировали при температуре 37°C в течение 2-3 часов со встряхиванием 220 об./мин до достижения оптической плотности 0,5. Культуры индуцировали конечной концентрацией 0,2 мМ изопропил-β-D-1-тиогалактопиранозида (IPTG) и инкубировали при температуре 25°C со встряхиванием 180 об./мин в течение 16 часов. Клетки собирали центрифугированием при 4800 об./мин в течение 15 мин. Осадок клеток ресуспендировали в 12 мл связывающего буфера (10 мМ HEPES, 50 мМ NaCl, pH 7,5), и клетки лизировали ультразвуком в течение 15 минут (5 секунд в режиме включено, 5 секунд паузы). Лизат клеток осветляли центрифугированием при 10000 об./мин в течение 20 мин. Меченый белок очищали из осветленного клеточного лизата с помощью сродства к кобальту, сродства к мальтозе или аффинной хроматографии с глутатионом. Растворы белков заменяли на буфер для хранения белка (20 мМ Трис-HCl, 50 мМ NaCl, pH 7,0) посредством центрифугирования. Концентрацию белка измеряли по оптической плотности при 280 нм, используя коэффициенты экстинкции, рассчитанные с использованием соответствующей аминокислотной последовательности. Добавляли глицерин до конечной концентрации 20% и замораживают растворы ферментов при -20 или -80°C.
Анализ восстановления изо-альфа-кислот
[0091] Очищенные кандидаты в ферменты тестировали на их способность восстанавливать изо-альфа-кислоты. Конкретная реакция приводит к восстановление определенной кетонной группы до гидроксигруппы любого или всех изомеров и родственных соединений изо-альфа-кислоты (со-, н-, ад- и цис/транс-). В микроцентрифужной пробирке на 2 мл 100 мкл раствора фермента (конечная концентрация 0,15-1,8 мг/мл фермента) добавляли к 900 мкл забуференного водного раствора с рециклированием кофактора глюкозодегидрогеназой (263 мМ фосфата натрия, 1,7 мМ сульфата магния, 1,1 мМ НАДФ+, 1,1 мМ НАД+, 80 мМ D-глюкозы, 4,3 ед/мл глюкозодегидрогеназы, pH 7,0). Добавляли 5 мкл щелочного раствора изо-альфа-кислоты (ISOLONE®, 29% изо-альфа-кислот) до конечной концентрации 0,29% изо-альфа-кислот. Реакционную смесь инкубировали при температуре 30°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь фильтровали для удаления фермента. Изо-альфа-кислоты и дигидро-(rho)-изо-альфа-кислоты определяли с помощью СВЭЖХ-МС/МС. Образец отрицательного контроля содержал все вышеуказанные компоненты реакции, в которых раствор фермента был заменен буфером для хранения белка.
Результаты
Выбор кандидата
[0092] На основе общедоступных функциональных аннотаций и сходства аминокислотных последовательностей 60 уникальных ферментных последовательностей были идентифицированы как кандидаты на редуктазу.
Экспрессия и очистка ферментов
[0093] 30 кандидатов были отобраны для экспрессии и очистки на основании доступности ДНК и достаточной выборки разнообразия аминокислотных последовательностей, представленных в начальной группе из 60 кандидатов. Большинство кандидатов показали хорошие уровни экспрессии и растворимости в E. coli BL21 (DE3) с выходами, варьирующими от 5 до 100 мг очищенного белка на литр культуры. От нескольких кандидатов отказались из-за плохой растворимости в организме хозяина.
Характеристика редуктаз
[0094] Было определено, что ферменты восстанавливают изо-альфа-кислоты, если пики, соответствующие цис/транс- со/ад/н-дигидро-(rho)-изо-альфа-кислоте, детектируются с помощью СВЭЖХ с большей интенсивностью, чем контрольный образец, в котором отсутствует фермент. Десять уникальных ферментов были определены как редуктазы изо-альфа-кислот (см. Фигуру 3). Информация об этих ферментах представлена в Таблице 2. Из-за растворимости и выхода ферментов конечная концентрация собственных ферментов в анализе варьировала от 0,15 до 1,8 мг/мл. Более низкая концентрация фермента способствует выходу дигидро-(rho)-изо-альфа кислот.
[0095] Ферменты первоначально тестировали на активность редуктазы в присутствии глюкозы, глюкозодегидрогеназы и НАД, чтобы рециклировать НАДФ, необходимый для восстановления изо-альфа кислот. После определения активности редуктазы ферменты были охарактеризованы по их способности окислять изопропанол, более экономичную альтернативу рециклирования кофакторов. Способность эффективно окислять изопропанол указана в Таблице 2.
Таблица 2. Охарактеризованные новые редуктазы изо-альфа-кислот.
Субстратная специфичность
[0096] Идеальная кеторедуктаза для целей биотрансформации не проявляет субстратной специфичности в отношении конгенеров изогумулона, которые варьируются в зависимости от состава боковой цепи (н-, ад- и ко-изогумулон). Кроме того, кеторедуктаза не проявляет специфичности в отношении цис- и транс-изомеров изогумулона, которые пространственно варьируются в группе третичного спирта C4, проксимальной к участку ферментативного восстановления. Субстратная специфичность определяется аминокислотной последовательностью и, следовательно, геометрией субстрат-связывающего кармана фермента. Более крупные связывающие карманы подходят для более крупных субстратов, а также для большего разнообразия субстратов по сравнению с более ограниченными связывающими карманами. (См. Фигуру 4).
[0097] Среди охарактеризованных редуктаз наблюдали два варианта стереоспецифичности восстановления (см. Фигуру 5).
[0098] Несмотря на присутствие двух дополнительных кетонных групп в молекуле изо-альфа-кислоты, для всех охарактеризованных кеторедуктаз наблюдалось только желаемое восстановление боковой цепи С4.
Пример 2
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием кофактора путем окисления изопропанола
[0060] В микроцентрифужной пробирке на 1,5 мл 10 мг редуктазы ресуспендируют в 700 мкл забуференного водного раствора (например, фосфата натрия, pH 7,5). Добавляют 290 мкл изопропанола. Добавляют 10 мкл щелочного раствора изо-альфа кислот (29% изо-альфа кислот) до конечной концентрации 0,29% изо-альфа кислот. Реакционную смесь инкубируют при температуре 30°C с орбитальным встряхиванием при 180 об./мин в течение 48 часов. Полученную реакционную смесь фильтруют для удаления фермента. Изо-альфа-кислоты и дигидро-(rho)-изо-альфа-кислоты количественно определяют с помощью ВЭЖХ.
Пример 3
Ферментативная обработка подкисленных изо-альфа-кислот хмеля с рециклированием кофактора путем окисления изопропанола
[00100] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2, где источником изо-альфа-кислот является высококонцентрированный материал (68,9% изо-альфа-кислот), имеющий значение pH <7.
Пример 4
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием кофактора глюкозодегидрогеназой
[00101] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2, за исключением того, что изопропанол заменяют на 4,3 ед./мл глюкозодегидрогеназы, 0,7 г/л мМ НАД и 14,4 г/л D-глюкозы.
Пример 5
Ферментативная обработка изо-альфа-кислот хмеля без рециклирования кофактора
[00102] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2, за исключением того, что изопропанол заменяют эквимолярным количеством НАДФН в качестве субстрата.
Пример 6
Ферментативная обработка изо-альфа-кислот хмеля термостабильной редуктазой
[00103] Природные термостабильные редуктазы получают из термофильных бактериальных и архейных организмов, таких как Thermotoga maritima. В микроцентрифужной пробирке на 1,5 мл 100 мкл раствора фермента (1,5-15,0 мг/мл фермента) добавляют к 900 мкл забуференного водного раствора (263 мМ фосфата натрия, pH 7,0, 1,7 мМ сульфата магния, 4,3 ед/мл глюкозодегидрогеназы, 1,1 мМ НАДФ+, 1,1 мМ НАД+, 80 мМ D-глюкоза). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакцию инкубируют при температуре 60-80°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь фильтруют для удаления фермента.
Пример 7
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием кофактора путем окисления этанола
[00104] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2, за исключением того, что изопропанол заменяют этанолом.
Пример 8
Ферментативная обработка изо-альфа-кислот хмеля кеторедуктазой, иммобилизованной на SiO2
[00105] Кеторедуктазу адсорбируют на SiO2 и сшивают с глутаральдегидом с получением материала иммобилизованной кеторедуктазы. Изо-альфа-кислоты обрабатывают иммобилизованной кеторедуктазой способом, описанным в Примере 2. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 9
Ферментативная обработка изо-альфа-кислот хмеля кеторедуктазой, иммобилизованной на ДЭАЭ-целлюлозе
[00106] Кеторедуктазу сшивают с глутаральдегидом и адсорбируют на ДЭАЭ-целлюлозе с получением материала иммобилизованной кеторедуктазы. Изо-альфа-кислоты обрабатывают иммобилизованной кеторедуктазой способом, описанным в Примере 2. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 10
Ферментативная обработка изо-альфа-кислот хмеля кеторедуктазой, иммобилизованной на оксиде алюминия, обработанном PEI
[00107] Кеторедуктазу сшивают с глутаральдегидом и адсорбируют на оксиде алюминия, обработанном полиэтилимином (PEI), с получением материала иммобилизованной кеторедуктазы. Изо-альфа-кислоты обрабатывают иммобилизованной кеторедуктазой способом, описанным в Примере 2. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 11
Ферментативная обработка изо-альфа-кислот хмеля коиммобилизованными ферментами
[00108] Редуктазу и фермент рециклирования кофактора, такой как глюкозодегидрогеназа, иммобилизуют последовательно или вместе в единой композиции с использованием любого из вышеупомянутых способов с получением коиммобилизованного материала. Коиммобилизованный материал добавляют до концентрации 0,1-10 мг/мл в забуференный водный раствор (50-250 мМ фосфата натрия, 0,1-1,0 мМ НАДФН, 10-40% изопропанола, pH 7-9). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакционную смесь инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./ мин в течение 24 часов. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 12
Ферментативная обработка изо-альфа-кислот хмеля сшитыми/сферонизированными клетками
[00109] Клетку (бактериальную, грибковую, растительную), экспрессирующую редуктазу, сшивают полиамином/глутаральдегидом, экструдируют и сферонизируют с получением материала иммобилизованной редуктазы. Иммобилизованную редуктазу добавляют до концентрации 0,1-10 мг/мл в забуференный водный раствор (50-250 мМ фосфата натрия, 0,1-1,0 мМ НАДФН, 10-40% изопропанола, pH 7-9). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакционную смесь инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 13
Ферментативная обработка изо-альфа-кислот хмеля, сшитыми/захваченными клетками
[00110] Клетку (бактериальную, грибковую, растительную), экспрессирующую редуктазу, сшивают с глутаральдегидом и захватывют желатиновыми или полимерными гранулами с получением иммобилизованного материала редуктазы. Иммобилизованную редуктазу добавляют до концентрации 0,1-10 мг/мл в забуференный водный раствор (50-250 мМ фосфата натрия, 0,1-1,0 мМ НАДФН, 10-40% изопропанола, pH 7-9). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакционную смесь инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь центрифугируют при 10000 g для удаления иммобилизованного фермента.
Пример 14
Ферментативная обработка изо-альфа-кислот хмеля живыми клетками
[00111] Микроорганизм (бактерии, грибок), экспрессирующий редуктазу, выращивают путем ферментации до высокой плотности, собирают, промывают и гранулируют с образованием клеточной пасты. Клеточную пасту ресуспендируют в свежей питательной среде, содержащей 0,145-16% изо-альфа-кислот. Культуру клеток инкубируют при температуре 25-37°C при перемешивании в течение 24-72 часов. Культуру клеток центрифугируют при 10000 g для удаления клеток из отработанной питательной среды. Дигидро-(rho)-изо-альфа кислоты экстрагируют этанолом из отработанной питательной среды.
Пример 15
Ферментативная обработка изо-альфа-кислот хмеля клеточным лизатом
[00112] Микроорганизм (бактерии, грибок), экспрессирующий редуктазу, выращивают путем ферментации до высокой плотности, собирают, промывают и лизируют с получением неочищенного клеточного лизата. К неочищенному клеточному лизату добавляют изо-альфа-кислоты до конечной концентрации 0,145-16% изо-альфа-кислот. Культуру клеток инкубируют при температуре 25-40°C при перемешивании в течение 24 часов. Реакционную смесь центрифугируют при 10000 g или фильтруют для удаления клеточного материала из лизата. Дигидро-(rho)-изо-альфа кислоты экстрагируют из осветленного лизата этанолом.
Пример 16
Ферментативная обработка изо-альфа-кислот хмеля психрофильной редуктазой
[00113] Ферментативная обработка, где редуктаза является гомологом из психрофильного (толерантного к холоду) микроорганизма. Редуктазу добавляют до концентрации 0,1-10 мг/мл в забуференный водный раствор (50-250 мМ фосфата натрия, 0,1-1,0 мМ НАДФН, 10-40% изопропанола, pH 7-9). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакцию инкубируют при температуре 0-20°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь фильтруют для удаления фермента.
Пример 17
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием кофактора НАДН
[00114] Обработка ферментом, при которой кофактор НАДФН заменяется НАДН. Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2, но НАДФН заменяют на НАД.
Пример 18
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием кофактора путем окисления этанола
[00115] Ферментативная обработка, при которой исходный изопропаноловый материал заменяется этанолом, при этом редуктаза добавляется до концентрации 0,1-10 мг/мл в забуференный водном растворе (50-250 мМ фосфата натрия, 0,1-1,0 мМ НАДН, 10-40% этанол, pH 7-9). Раствор изомеризованного экстракта хмеля ISOLONE® (29% изо-альфа-кислот) добавляют до конечной концентрации 0,145-16% изо-альфа-кислот. Реакционную смесь инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь фильтруют для удаления фермента.
Пример 19
Ферментативная обработка изо-альфа-кислот хмеля с последующей экстракцией
[00116] Ферментативную обработку выполняют с последующей экстракцией для увеличения конечной концентрации дигидро-(rho)-изо-альфа-кислот. Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2. Полученную реакционную смесь фильтруют для удаления фермента и экстрагируют пищевым растворителем для достижения желаемой концентрации дигидро-(rho)-изо-альфа-кислот.
Пример 20
Ферментативная обработка изо-альфа-кислот хмеля с последующей термической инактивацией
[00117] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2. Реакцию инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./мин в течение 24 часов. Полученную реакционную смесь нагревают при температуре 80-100°C в течение 10-30 минут для инактивации фермента.
Пример 21
Ферментативная обработка изо-альфа-кислот хмеля с последующей химической инактивацией
[00118] Изо-альфа-кислоты обрабатывают способом, описанным в Примере 2. Реакцию инкубируют при температуре 30-40°C с орбитальным встряхиванием при 180 об./ мин в течение 24 часов. Пищевой этанол добавляют до конечной концентрации >50% для инактивации фермента.
Пример 22
Ферментативная обработка изо-альфа-кислот хмеля с рециклированием иммобилизованной кеторедуктазы
[00119] Кеторедуктазу сшивают с глутаральдегидом и адсорбируют на ДЭАЭ-целлюлозе с получением материала иммобилизованной кеторедуктазы. Затем изо-альфа-кислоты обрабатывают иммобилизованной кеторедуктазой способом, описанным в Примере 2. Полученную реакционную смесь центрифугируют при 10000 g для отделения иммобилизованной кеторедуктазы от реакционной смеси. Иммобилизованную кеторедуктазу восстанавливают, промывается водой или водным буфером и повторно используется в новой реакционной смеси.
Заключение
[00120] 23 кеторедуктазы были охарактеризованы как превращающие изо-альфа-кислоты в дигидро-(rho)-изо-альфа-кислоты. Кеторедуктазы, охарактеризованные в этом исследовании, обладают ферментативной активностью, которая ранее не описывалась. Кеторедуктазы, охарактеризованные в этом исследовании, восстанавливают кетонную группу до спирта и, таким образом, являются кеторедуктазами. Эти результаты демонстрируют, что биокатализатор кеторедуктаза можно использовать для превращения изо-альфа-кислот в дигидро-(rho)-изо-альфа-кислоты в новом процессе биотрансформации. Настоящее изобретение предназначено для замены существующих процессов, в которых используется боргидрид натрия.
[00121] Настоящее изобретение не должно ограничиваться в объеме описанными здесь конкретными вариантами его осуществления. А именно, различные модификации настоящего изобретения в дополнение к описанным здесь станут очевидными для специалистов в данной области техники из предшествующего описания. Предполагается, что такие модификации входят в объем прилагаемой формулы изобретения.
[00122] Все патенты, заявки, публикации, способы тестирования, литература и другие материалы, цитируемые в описании настоящего изобретения, включены в него путем ссылки.
Цитируемые ссылки
1. Sodium Borohydride; MSDS No. S9125; Sigma-Aldrich Co.: Saint Louis, MO November 01, 2015. (accessed 06/08/17).
2. Robinson, P. K., Enzymes: principles and biotechnological applications. Essays Biochem 2015, 59, 1-41.
3. Hult, K.; Berglund, P., Enzyme promiscuity: mechanism and applications. Trends Biotechnol. 2007, 25 (5), 231-238.
4. Nobeli, I.; Favia, A. D.; Thornton, J. M., Protein promiscuity and its implications for biotechnology. Nat. Biotechnol. 2009, 27 (2), 157-167.
5. Pozen, M., Enzymes in Brewing. Ind. Eng. Chem, 1934, 26 (11), 1127-1133.
6. Praet, T.; Opstaele, F.; Jaskula-Goiris, B.; Aerts, G.; De Cooman, L., Biotransformations of hop-derived aroma compounds by Saccharomyces cerevisiae upon fermentation. Cerevisia, 2012, 36, 125-132.
7. Wallerstein, L. (1947) Bentonite and Proteolytic Enzyme Treatment of Beer, US Patent 2,433,411.
8. Ghionno, L.; Marconi, O.; Sileoni, V.; De Francesco, G.; Perretti, G., Brewing with prolyl endopeptidase from Aspergillus niger: the impact of enzymatic treatment on gluten levels, quality attributes, and sensory profile. Int. J. Food Sci. Technol, 2017, 52 (6), 1367-1374.
9. Gros, J.; Tran, T. T. H.; Collin, S., Enzymatic release of odourant polyfunctional thiols from cysteine conjugates in hop. J. Inst. Brew. 2013, 119 (4), 221-227.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> KALAMAZOO HOLDINGS, INC.
<120> ФЕРМЕНТАТИВНЫЙ СПОСОБ ПОЛУЧЕНИЯ МОДИФИЦИРОВАННЫХ
ХМЕЛЕПРОДУКТОВ
<130> KALSEC 76 PCT SEQ
<140>
<141> 2019-09-26
<150> US 62/736,558
<151> 2018-09-26
<160> 22
<170> Патентная версия 3.5
<210> 1
<211> 277
<212> Белок
<213> Homo sapiens
<400> 1
Met Ser Ser Gly Ile His Val Ala Leu Val Thr Gly Gly Asn Lys Gly
1 5 10 15
Ile Gly Leu Ala Ile Val Arg Asp Leu Cys Arg Leu Phe Ser Gly Asp
20 25 30
Val Val Leu Thr Ala Arg Asp Val Thr Arg Gly Gln Ala Ala Val Gln
35 40 45
Gln Leu Gln Ala Glu Gly Leu Ser Pro Arg Phe His Gln Leu Asp Ile
50 55 60
Asp Asp Leu Gln Ser Ile Arg Ala Leu Arg Asp Phe Leu Arg Lys Glu
65 70 75 80
Tyr Gly Gly Leu Asp Val Leu Val Asn Asn Ala Gly Ile Ala Phe Lys
85 90 95
Val Ala Asp Pro Thr Pro Phe His Ile Gln Ala Glu Val Thr Met Lys
100 105 110
Thr Asn Phe Phe Gly Thr Arg Asp Val Cys Thr Glu Leu Leu Pro Leu
115 120 125
Ile Lys Pro Gln Gly Arg Val Val Asn Val Ser Ser Ile Met Ser Val
130 135 140
Arg Ala Leu Lys Ser Cys Ser Pro Glu Leu Gln Gln Lys Phe Arg Ser
145 150 155 160
Glu Thr Ile Thr Glu Glu Glu Leu Val Gly Leu Met Asn Lys Phe Val
165 170 175
Glu Asp Thr Lys Lys Gly Val His Gln Lys Glu Gly Trp Pro Ser Ser
180 185 190
Ala Tyr Gly Val Thr Lys Ile Gly Val Thr Val Leu Ser Arg Ile His
195 200 205
Ala Arg Lys Leu Ser Glu Gln Arg Lys Gly Asp Lys Ile Leu Leu Asn
210 215 220
Ala Cys Cys Pro Gly Trp Val Arg Thr Asp Met Ala Gly Pro Lys Ala
225 230 235 240
Thr Lys Ser Pro Glu Glu Gly Ala Glu Thr Pro Val Tyr Leu Ala Leu
245 250 255
Leu Pro Pro Asp Ala Glu Gly Pro His Gly Gln Phe Val Ser Glu Lys
260 265 270
Arg Val Glu Gln Trp
275
<210> 2
<211> 246
<212> Белок
<213> Thermotoga maritima
<400> 2
Met Arg Leu Glu Gly Lys Val Cys Leu Ile Thr Gly Ala Ala Ser Gly
1 5 10 15
Ile Gly Lys Ala Thr Thr Leu Leu Phe Ala Gln Glu Gly Ala Thr Val
20 25 30
Ile Ala Gly Asp Ile Ser Lys Glu Asn Leu Asp Ser Leu Val Lys Glu
35 40 45
Ala Glu Gly Leu Pro Gly Lys Val Asp Pro Tyr Val Leu Asn Val Thr
50 55 60
Asp Arg Asp Gln Ile Lys Glu Val Val Glu Lys Val Val Gln Lys Tyr
65 70 75 80
Gly Arg Ile Asp Val Leu Val Asn Asn Ala Gly Ile Thr Arg Asp Ala
85 90 95
Leu Leu Val Arg Met Lys Glu Glu Asp Trp Asp Ala Val Ile Asn Val
100 105 110
Asn Leu Lys Gly Val Phe Asn Val Thr Gln Met Val Val Pro Tyr Met
115 120 125
Ile Lys Gln Arg Asn Gly Ser Ile Val Asn Val Ser Ser Val Val Gly
130 135 140
Ile Tyr Gly Asn Pro Gly Gln Thr Asn Tyr Ala Ala Ser Lys Ala Gly
145 150 155 160
Val Ile Gly Met Thr Lys Thr Trp Ala Lys Glu Leu Ala Gly Arg Asn
165 170 175
Ile Arg Val Asn Ala Val Ala Pro Gly Phe Ile Glu Thr Pro Met Thr
180 185 190
Glu Lys Leu Pro Glu Lys Ala Arg Glu Thr Ala Leu Ser Arg Ile Pro
195 200 205
Leu Gly Arg Phe Gly Lys Pro Glu Glu Val Ala Gln Val Ile Leu Phe
210 215 220
Leu Ala Ser Asp Glu Ser Ser Tyr Val Thr Gly Gln Val Ile Gly Ile
225 230 235 240
Asp Gly Gly Leu Val Ile
245
<210> 3
<211> 342
<212> Белок
<213> Saccharomyces cerevisiae
<400> 3
Met Ser Val Phe Val Ser Gly Ala Asn Gly Phe Ile Ala Gln His Ile
1 5 10 15
Val Asp Leu Leu Leu Lys Glu Asp Tyr Lys Val Ile Gly Ser Ala Arg
20 25 30
Ser Gln Glu Lys Ala Glu Asn Leu Thr Glu Ala Phe Gly Asn Asn Pro
35 40 45
Lys Phe Ser Met Glu Val Val Pro Asp Ile Ser Lys Leu Asp Ala Phe
50 55 60
Asp His Val Phe Gln Lys His Gly Lys Asp Ile Lys Ile Val Leu His
65 70 75 80
Thr Ala Ser Pro Phe Cys Phe Asp Ile Thr Asp Ser Glu Arg Asp Leu
85 90 95
Leu Ile Pro Ala Val Asn Gly Val Lys Gly Ile Leu His Ser Ile Lys
100 105 110
Lys Tyr Ala Ala Asp Ser Val Glu Arg Val Val Leu Thr Ser Ser Tyr
115 120 125
Ala Ala Val Phe Asp Met Ala Lys Glu Asn Asp Lys Ser Leu Thr Phe
130 135 140
Asn Glu Glu Ser Trp Asn Pro Ala Thr Trp Glu Ser Cys Gln Ser Asp
145 150 155 160
Pro Val Asn Ala Tyr Cys Gly Ser Lys Lys Phe Ala Glu Lys Ala Ala
165 170 175
Trp Glu Phe Leu Glu Glu Asn Arg Asp Ser Val Lys Phe Glu Leu Thr
180 185 190
Ala Val Asn Pro Val Tyr Val Phe Gly Pro Gln Met Phe Asp Lys Asp
195 200 205
Val Lys Lys His Leu Asn Thr Ser Cys Glu Leu Val Asn Ser Leu Met
210 215 220
His Leu Ser Pro Glu Asp Lys Ile Pro Glu Leu Phe Gly Gly Tyr Ile
225 230 235 240
Asp Val Arg Asp Val Ala Lys Ala His Leu Val Ala Phe Gln Lys Arg
245 250 255
Glu Thr Ile Gly Gln Arg Leu Ile Val Ser Glu Ala Arg Phe Thr Met
260 265 270
Gln Asp Val Leu Asp Ile Leu Asn Glu Asp Phe Pro Val Leu Lys Gly
275 280 285
Asn Ile Pro Val Gly Lys Pro Gly Ser Gly Ala Thr His Asn Thr Leu
290 295 300
Gly Ala Thr Leu Asp Asn Lys Lys Ser Lys Lys Leu Leu Gly Phe Lys
305 310 315 320
Phe Arg Asn Leu Lys Glu Thr Ile Asp Asp Thr Ala Ser Gln Ile Leu
325 330 335
Lys Phe Glu Gly Arg Ile
340
<210> 4
<211> 272
<212> Белок
<213> Cytophaga hutchinsonii
<400> 4
Met Asn Gln Val Val Leu Val Thr Gly Gly Ser Ser Gly Ile Gly Lys
1 5 10 15
Ser Ile Cys Leu Tyr Leu His Glu Lys Gly Tyr Ile Val Tyr Gly Thr
20 25 30
Ser Arg Asn Pro Ala Arg Tyr Ala His Glu Val Pro Phe Lys Leu Ile
35 40 45
Ala Leu Asp Val Leu Asp Asp Thr Thr Ile Thr Pro Ala Leu Lys Thr
50 55 60
Ile Ile Asp Ala Glu Gly Lys Leu Asp Val Leu Val Asn Asn Ala Gly
65 70 75 80
Ile Gly Met Leu Gly Ser Ile Glu Asp Ser Thr Ala Glu Glu Val Lys
85 90 95
Glu Val Phe Glu Thr Asn Val Tyr Gly Ile Leu Arg Thr Cys Gln Ala
100 105 110
Val Leu Pro His Met Arg Glu Arg Lys Met Gly Leu Ile Ile Asn Val
115 120 125
Ser Ser Ile Ala Gly Tyr Met Gly Leu Pro Tyr Arg Gly Ile Tyr Ser
130 135 140
Ala Thr Lys Ala Ser Val His Met Ile Thr Glu Ala Met Arg Met Glu
145 150 155 160
Leu Lys Pro Tyr Gly Val His Ala Cys Val Val Asp Pro Gly Asp Phe
165 170 175
Ala Thr Asn Ile Ser Asp Asn Arg Lys Val Ala His Ala Gly Arg Ser
180 185 190
Gly Ser Val Tyr Met Glu Glu Ile Asn Arg Ile Glu Ala Met Ile Asn
195 200 205
Ala Glu Val Ala His Ser Ser Asp Pro Leu Leu Met Gly Lys Ala Ile
210 215 220
Glu Lys Ile Ile Arg Ser Ser Asn Pro Asp Ile Asn Tyr Leu Val Gly
225 230 235 240
Lys Pro Met Gln Lys Leu Ser Ile Leu Val Arg Arg Leu Val Pro Lys
245 250 255
Lys Trp Phe Glu Lys Ile Ile Ala Ser His Tyr Asn Met Pro Val Lys
260 265 270
<210> 5
<211> 326
<212> Белок
<213> Arabidopsis thaliana
<400> 5
Met Ala Asn Ser Gly Glu Gly Lys Val Val Cys Val Thr Gly Ala Ser
1 5 10 15
Gly Tyr Ile Ala Ser Trp Leu Val Lys Phe Leu Leu Ser Arg Gly Tyr
20 25 30
Thr Val Lys Ala Ser Val Arg Asp Pro Ser Asp Pro Lys Lys Thr Gln
35 40 45
His Leu Val Ser Leu Glu Gly Ala Lys Glu Arg Leu His Leu Phe Lys
50 55 60
Ala Asp Leu Leu Glu Gln Gly Ser Phe Asp Ser Ala Ile Asp Gly Cys
65 70 75 80
His Gly Val Phe His Thr Ala Ser Pro Phe Phe Asn Asp Ala Lys Asp
85 90 95
Pro Gln Ala Glu Leu Ile Asp Pro Ala Val Lys Gly Thr Leu Asn Val
100 105 110
Leu Asn Ser Cys Ala Lys Ala Ser Ser Val Lys Arg Val Val Val Thr
115 120 125
Ser Ser Met Ala Ala Val Gly Tyr Asn Gly Lys Pro Arg Thr Pro Asp
130 135 140
Val Thr Val Asp Glu Thr Trp Phe Ser Asp Pro Glu Leu Cys Glu Ala
145 150 155 160
Ser Lys Met Trp Tyr Val Leu Ser Lys Thr Leu Ala Glu Asp Ala Ala
165 170 175
Trp Lys Leu Ala Lys Glu Lys Gly Leu Asp Ile Val Thr Ile Asn Pro
180 185 190
Ala Met Val Ile Gly Pro Leu Leu Gln Pro Thr Leu Asn Thr Ser Ala
195 200 205
Ala Ala Ile Leu Asn Leu Ile Asn Gly Ala Lys Thr Phe Pro Asn Leu
210 215 220
Ser Phe Gly Trp Val Asn Val Lys Asp Val Ala Asn Ala His Ile Gln
225 230 235 240
Ala Phe Glu Val Pro Ser Ala Asn Gly Arg Tyr Cys Leu Val Glu Arg
245 250 255
Val Val His His Ser Glu Ile Val Asn Ile Leu Arg Glu Leu Tyr Pro
260 265 270
Asn Leu Pro Leu Pro Glu Arg Cys Val Asp Glu Asn Pro Tyr Val Pro
275 280 285
Thr Tyr Gln Val Ser Lys Asp Lys Thr Arg Ser Leu Gly Ile Asp Tyr
290 295 300
Ile Pro Leu Lys Val Ser Ile Lys Glu Thr Val Glu Ser Leu Lys Glu
305 310 315 320
Lys Gly Phe Ala Gln Phe
325
<210> 6
<211> 236
<212> Белок
<213> Pseudomonas savastanoi
<400> 6
Met Thr Leu Ser Ser Ala Pro Ile Leu Ile Thr Gly Ala Ser Gln Arg
1 5 10 15
Val Gly Leu His Cys Ala Leu Arg Leu Leu Glu His Gly His Arg Val
20 25 30
Ile Ile Ser Tyr Arg Thr Glu His Ala Ser Val Thr Glu Leu Arg Gln
35 40 45
Ala Gly Ala Val Ala Leu Tyr Gly Asp Phe Ser Cys Glu Thr Gly Ile
50 55 60
Met Ala Phe Ile Asp Leu Leu Lys Thr Gln Thr Ser Ser Leu Arg Ala
65 70 75 80
Val Val His Asn Ala Ser Glu Trp Leu Ala Glu Thr Pro Gly Glu Glu
85 90 95
Ala Asp Asn Phe Thr Arg Met Phe Ser Val His Met Leu Ala Pro Tyr
100 105 110
Leu Ile Asn Leu His Cys Glu Pro Leu Leu Thr Ala Ser Glu Val Ala
115 120 125
Asp Ile Val His Ile Ser Asp Asp Val Thr Arg Lys Gly Ser Ser Lys
130 135 140
His Ile Ala Tyr Cys Ala Thr Lys Ala Gly Leu Glu Ser Leu Thr Leu
145 150 155 160
Ser Phe Ala Ala Arg Phe Ala Pro Leu Val Lys Val Asn Gly Ile Ala
165 170 175
Pro Ala Leu Leu Met Phe Gln Pro Lys Asp Asp Ala Ala Tyr Arg Ala
180 185 190
Asn Ala Leu Ala Lys Ser Ala Leu Gly Ile Glu Pro Gly Ala Glu Val
195 200 205
Ile Tyr Gln Ser Leu Arg Tyr Leu Leu Asp Ser Thr Tyr Val Thr Gly
210 215 220
Thr Thr Leu Thr Val Asn Gly Gly Arg His Val Lys
225 230 235
<210> 7
<211> 246
<212> Белок
<213> Pseudomonas putida
<400> 7
Met Ser Leu Gln Gly Lys Val Ala Leu Val Thr Gly Ala Ser Arg Gly
1 5 10 15
Ile Gly Gln Ala Ile Ala Leu Glu Leu Gly Arg Gln Gly Ala Thr Val
20 25 30
Ile Gly Thr Ala Thr Ser Ala Ser Gly Ala Glu Arg Ile Ala Ala Thr
35 40 45
Leu Lys Glu His Gly Ile Thr Gly Thr Gly Met Glu Leu Asn Val Thr
50 55 60
Ser Ala Glu Ser Val Glu Ala Val Leu Ala Ala Ile Gly Glu Gln Phe
65 70 75 80
Gly Ala Pro Ala Ile Leu Val Asn Asn Ala Gly Ile Thr Arg Asp Asn
85 90 95
Leu Met Leu Arg Met Lys Asp Asp Glu Trp Phe Asp Val Ile Asp Thr
100 105 110
Asn Leu Asn Ser Leu Tyr Arg Leu Ser Lys Gly Val Leu Arg Gly Met
115 120 125
Thr Lys Ala Arg Trp Gly Arg Ile Ile Ser Ile Gly Ser Val Val Gly
130 135 140
Ala Met Gly Asn Ala Gly Gln Ala Asn Tyr Ala Ala Ala Lys Ala Gly
145 150 155 160
Leu Glu Gly Phe Ser Arg Ala Leu Ala Arg Glu Val Gly Ser Arg Gly
165 170 175
Ile Thr Val Asn Ser Val Thr Pro Gly Phe Ile Asp Thr Asp Met Thr
180 185 190
Arg Glu Leu Pro Glu Ala Gln Arg Glu Ala Leu Gln Thr Gln Ile Pro
195 200 205
Leu Gly Arg Leu Gly Gln Ala Asp Glu Ile Ala Lys Val Val Ser Phe
210 215 220
Leu Ala Ser Asp Gly Ala Ala Tyr Val Thr Gly Ala Thr Val Pro Val
225 230 235 240
Asn Gly Gly Met Tyr Met
245
<210> 8
<211> 262
<212> Белок
<213> Enterococcus faecalis
<400> 8
Met Asp Leu Thr Asn Lys Val Val Val Val Thr Gly Gly Ser Ala Gly
1 5 10 15
Leu Gly Glu Gln Ile Cys Tyr Glu Ala Ala Lys Gln Gly Ala Val Val
20 25 30
Val Val Cys Ala Arg Arg Ile Asn Leu Ile Gly Lys Val Arg Glu Gln
35 40 45
Cys Ala Val Leu Ser Gly Arg Glu Ala Phe Ser Tyr Gln Leu Asp Ile
50 55 60
Ala Asp Pro Glu Ser Val Glu Arg Val Val Glu Ala Ile Ser Ala Glu
65 70 75 80
Val Gly Pro Ile Asp Val Leu Val Asn Asn Ala Gly Phe Gly Leu Phe
85 90 95
Glu Asn Phe Val Glu Ile Asp Leu Ala Val Ala Arg Gln Met Phe Asp
100 105 110
Val Asn Val Leu Gly Met Met Thr Phe Thr Gln Lys Val Ala Ile Lys
115 120 125
Met Ile Glu Ala Gly Gln Gly His Ile Ile Asn Val Ala Ser Met Ala
130 135 140
Gly Lys Met Ala Thr Ala Lys Ser Thr Val Tyr Ser Ala Thr Lys Phe
145 150 155 160
Ala Val Leu Gly Phe Ser Asn Ala Leu Arg Leu Glu Leu Lys Pro Leu
165 170 175
Gly Val Ala Val Thr Thr Val Asn Pro Gly Pro Ile Gln Thr Glu Phe
180 185 190
Phe Asp Lys Ala Asp Pro Thr Gly Thr Tyr Leu Ala Ala Val Asp Lys
195 200 205
Ile Val Leu Asp Pro Thr Lys Leu Ala Lys Glu Val Val Gly Ser Met
210 215 220
Gly Thr Ser Arg Arg Glu Ile Asn Arg Pro Phe Val Met Glu Ala Ala
225 230 235 240
Ala Arg Phe Tyr Thr Leu Phe Pro His Leu Gly Asp Phe Ile Ala Gly
245 250 255
Asn Ile Leu Asn Lys Lys
260
<210> 9
<211> 319
<212> Белок
<213> Pseudomonas syringae
<400> 9
Met Arg Arg Ile Leu Ile Thr Gly Ala Asn Gly Phe Val Gly Gln Ile
1 5 10 15
Leu Cys Ser Met Leu Arg Gln Ala Gly His His Val Ile Ala Leu Val
20 25 30
Gly Ala Glu Ser Ala Leu Ser Ser His Ala Asp Glu Ser Val Arg Cys
35 40 45
Asp Ile Arg Asp Ala Ser Gly Leu Glu Gln Ala Leu Cys Arg Ala Ala
50 55 60
Pro Thr His Val Val His Leu Ala Ala Ile Thr His Val Pro Thr Ser
65 70 75 80
Phe Asn Asn Pro Val Leu Thr Trp Gln Thr Asn Val Met Gly Ser Val
85 90 95
Asn Leu Leu Gln Ala Leu Gln Arg Ser Ala Pro Glu Ala Phe Val Leu
100 105 110
Phe Val Ser Ser Ser Glu Val Tyr Gly Glu Thr Phe Lys Gln Gly Thr
115 120 125
Ala Leu Gly Glu Asp Ser Ala Cys Lys Pro Met Asn Pro Tyr Ala Ala
130 135 140
Ser Lys Leu Ala Ala Glu Ala Ala Phe Asn Glu Tyr Phe Arg Gln Gly
145 150 155 160
Arg Lys Gly Ile Val Val Arg Pro Phe Asn His Ile Gly Ala Arg Gln
165 170 175
Ser Pro Asp Phe Ala Thr Ala Ser Phe Ala Arg Gln Ile Ala Leu Ile
180 185 190
Glu Ala Gly Lys Gln Ala Pro Gln Leu Lys Val Gly Asn Leu Gln Ala
195 200 205
Ala Arg Asp Phe Leu Asp Val His Asp Val Cys Asp Ala Tyr Val Ala
210 215 220
Leu Leu Gln Leu Ala Asp Glu Gln Glu Arg Tyr Pro Gly Cys Leu Asn
225 230 235 240
Ile Cys Arg Gly Glu Pro Thr Ser Leu Gln Thr Leu Leu Thr Gln Leu
245 250 255
Met Ala Leu Ser Ser Ser Val Ile Glu Val Thr Ile Asp Pro Asp Arg
260 265 270
Met Arg Pro Ser Asp Ile Pro Ser Ala Phe Gly Asn Asn Ser Ala Met
275 280 285
Arg Cys Ala Thr Gly Trp Lys Pro Lys Thr Lys Leu Asp Asp Thr Leu
290 295 300
Glu Ala Leu Leu Asn Tyr Trp Arg His Glu Val Ile Ser Ala Val
305 310 315
<210> 10
<211> 368
<212> Белок
<213> Pseudomonas cannabina
<400> 10
Met Ser Leu Leu Leu Glu Pro Tyr Thr Leu Arg Gln Leu Thr Leu Arg
1 5 10 15
Asn Arg Ile Ala Val Ser Pro Met Cys Gln Tyr Ser Ser Val Asp Gly
20 25 30
Leu Ala Asn Asp Trp His Leu Val His Leu Gly Ser Arg Ala Val Gly
35 40 45
Gly Ala Gly Leu Val Ile Ser Glu Ala Met Ala Val Thr Pro Asp Gly
50 55 60
Arg Ile Thr Pro Glu Asp Leu Gly Leu Trp Asn Asp Glu Gln Ile Glu
65 70 75 80
Pro Leu Gln Arg Ile Thr Arg Phe Ile Asn Thr Gln Gly Ala Val Ala
85 90 95
Gly Ile Gln Leu Ala His Ala Gly Arg Lys Ala Ser Thr Trp Arg Pro
100 105 110
Trp Leu Gly Lys His Gly Ser Val Pro Leu Thr Glu Gly Gly Trp Thr
115 120 125
Pro Val Gly Pro Ser Ala Ile Ala Phe Asp Pro Gln His Thr Ala Pro
130 135 140
Leu Gln Leu Ser Glu Thr Gln Ile Gln Glu Leu Ile Lys Ala Phe Val
145 150 155 160
Asp Ser Ala Arg Arg Ala Leu Thr Ala Gly Phe Lys Val Val Glu Ile
165 170 175
His Ala Ala His Gly Tyr Leu Leu His Gln Phe Leu Ser Pro Leu Ser
180 185 190
Asn Gln Arg Thr Asp Gln Tyr Gly Gly Ser Phe Glu Asn Arg Ile Arg
195 200 205
Leu Thr Leu Gln Val Thr Glu Ala Val Arg Ala Val Trp Pro Gln Glu
210 215 220
Leu Pro Leu Phe Val Arg Val Ser Ala Thr Asp Trp Val Glu Asp Gly
225 230 235 240
Trp Asn Ala Glu Glu Thr Val Glu Leu Ala Arg Arg Leu Lys Ala Leu
245 250 255
Gly Thr Asp Leu Ile Asp Val Ser Ser Gly Gly Thr Ser Ala Asn Ala
260 265 270
Glu Ile Pro Val Gly Pro Gly Tyr Gln Thr Arg Phe Ala Glu Gln Val
275 280 285
Arg Lys Glu Ala Asp Ile Ala Thr Gly Thr Val Gly Met Ile Thr Asp
290 295 300
Pro Ala Gln Ala Glu His Ile Leu Arg Thr Gly Gln Ala Asp Ile Ile
305 310 315 320
Leu Leu Ala Arg Glu Leu Leu Arg Asp Pro Tyr Trp Pro Leu Arg Ala
325 330 335
Asp Glu Asp Leu Gly Gly Arg Gln Ala Thr Trp Pro Ala Gln Tyr Gln
340 345 350
Arg Ala Thr His Arg Asp Gln Pro Ile His Glu Ser Asp Leu Arg Asp
355 360 365
<210> 11
<211> 344
<212> Белок
<213> Saccharomyces cerevisiae
<400> 11
Met Ser Ser Ser Ser Leu Arg Val Leu Ala Ile Gly Asn Asn Pro Asn
1 5 10 15
Ile Leu Phe Tyr Thr Ser Arg Phe Gln Leu Ala Lys Asn Ile Asp Leu
20 25 30
Tyr His Val Asn Asp Ser Lys Ser Cys Gln Phe Glu Ile Glu Thr Glu
35 40 45
Tyr Tyr Gly Lys Asp Arg Phe Glu Leu Glu Asn His Phe Thr Ser Ile
50 55 60
Glu His Leu Thr Glu Ala Leu Ser Ser Lys Ser Ser Glu Ala Val Phe
65 70 75 80
Asp Ile Ile Ile Met Ser Ala Pro Ser Leu Gln Glu Leu Ser Ser Leu
85 90 95
Ala Ser Lys Leu Thr Ser Ile Ile Asp Ser Asn Thr Lys Ile Phe Leu
100 105 110
Glu Ser Ser Gly Phe Ile Gln Leu Glu Pro Phe Val Lys Leu Ser Met
115 120 125
Glu Ser Pro His Val Asn Val Phe Ser Ile Leu Thr Asp Leu Asp Ile
130 135 140
Arg Gln Ile Gly Pro Asn His Phe Lys His Phe Pro Ser Thr Ala Lys
145 150 155 160
Glu Asn Thr Ile Tyr Leu Gly Glu Ser Lys Ser Ser Thr Glu Lys Tyr
165 170 175
Ser Ser Gly Val Ile Thr Leu Leu Thr Thr Phe Glu Lys Leu Phe Ala
180 185 190
Lys Leu Phe Ser Asn Ile Lys Ile Asn Leu Cys Asn Phe Ser Ser Ile
195 200 205
Glu Phe Leu Ser Gln Gln Trp Lys Leu Ala Ile Ser Arg Ile Cys Phe
210 215 220
Asp Pro Leu Leu Ile Met Phe Glu Gln Glu Asn Pro Ser Asp Leu Asp
225 230 235 240
Gln Gln Ile Ile Ala Lys Pro Leu Ile Ser Gly Leu Val Thr Glu Ile
245 250 255
Ile Thr Val Ala Lys Thr Met Gly Ala Arg Leu Asn Ser Ser His Asp
260 265 270
Asn Glu Asn Ser Leu Leu Ser Leu Trp Lys Asn Ser Tyr His Ser Thr
275 280 285
Asn Lys Pro Pro Ala Leu Val Tyr His Phe Ile His Gln Thr Thr Pro
290 295 300
Leu Asn Ile Asp Ile Leu Leu Leu Gln Thr Ile Leu Leu Ala Asp Asp
305 310 315 320
Phe Gly Ile Lys Thr Pro Tyr Leu Glu Phe Leu Tyr Ser Val Leu Ser
325 330 335
Gln Phe Glu Arg Leu Asn Ser Gly
340
<210> 12
<211> 319
<212> Белок
<213> Thermotoga maritima
<400> 12
Met Glu Tyr Arg Lys Val Gly Lys Trp Gly Val Lys Ile Ser Glu Leu
1 5 10 15
Ser Leu Gly Ser Trp Leu Thr Phe Gly Lys Gln Leu Asp Leu Asp Thr
20 25 30
Ala Thr Glu Val Val Lys Lys Ala Phe Asn Ser Gly Ile Asn Phe Phe
35 40 45
Asp Thr Ala Glu Ala Tyr Ala Gly Gly Ile Ala Glu Ala Met Leu Gly
50 55 60
Lys Ile Leu Lys Asn Phe Arg Arg Glu Asp Leu Val Val Ser Thr Lys
65 70 75 80
Ile Phe Trp Gly Gly Ser Gly Pro Asn Asp Leu Gly Leu Ser Lys Lys
85 90 95
His Leu Leu Glu Gly Thr Trp Asn Ser Leu Lys Arg Leu Gln Met Asp
100 105 110
Tyr Val Asp Ile Leu Tyr Cys His Arg Pro Asp Pro Asn Val Pro Met
115 120 125
Glu Glu Val Val Phe Ala Met Asp Tyr Ile Leu Arg Glu Gly Leu Ala
130 135 140
Leu Tyr Trp Gly Thr Ser Glu Trp Ser Ala Lys Glu Ile Glu Glu Ala
145 150 155 160
His Arg Val Cys Lys Glu Leu Gly Val Met Pro Pro Ile Val Glu Gln
165 170 175
Pro Gln Tyr Asn Met Phe Val Arg Glu Arg Val Glu Lys Glu Tyr Ala
180 185 190
Pro Leu Tyr Glu Lys Tyr Gly Met Gly Leu Thr Thr Tyr Ser Pro Leu
195 200 205
Ala Ser Gly Leu Leu Ser Gly Lys Tyr Asn Asn Gly Ile Pro Glu Gly
210 215 220
Ser Arg Leu Ala Thr Phe Pro Gln Val Arg Lys Trp Leu Glu Glu Gly
225 230 235 240
Gly Leu Leu Asn Glu Lys Thr Phe Lys Lys Leu Arg Lys Leu Gln Asn
245 250 255
Ile Ala Asp Gln Leu Gly Ala Ser Leu Pro Gln Leu Ala Ile Ala Trp
260 265 270
Ile Leu Lys Asn Lys Asn Val Ser Ser Val Ile Leu Gly Val Ser Arg
275 280 285
Pro Glu Gln Leu Glu Glu Asn Leu Lys Ala Val Glu Ile Lys Glu Lys
290 295 300
Leu Thr Glu Asp Val Met Glu Glu Ile Glu Lys Ile Leu Asn Glu
305 310 315
<210> 13
<211> 326
<212> Белок
<213> Agrobacterium fabrum
<400> 13
Met Thr Leu Ala Asn Leu Pro Pro Leu Val Thr Val Phe Gly Gly Ser
1 5 10 15
Gly Phe Val Gly Arg His Val Val Arg Met Leu Ala Lys Arg Gly Tyr
20 25 30
Arg Ile Arg Val Ala Val Arg Arg Pro Asp Leu Ala Gly Phe Leu Gln
35 40 45
Pro Leu Gly Asn Val Gly Gln Ile Ser Phe Ala Gln Ala Asn Leu Arg
50 55 60
Tyr Arg Asp Ser Ile Ile Lys Ala Val Glu Asp Ala Asp His Val Val
65 70 75 80
Asn Cys Val Gly Ile Leu Ala Glu Ser Gly Arg Asn Thr Phe Asp Ala
85 90 95
Val Gln Glu Phe Gly Ala Lys Ala Ile Ala Glu Ala Ala Arg Asp Thr
100 105 110
Gly Ala Thr Leu Thr His Ile Ser Ala Ile Gly Ala Asp Ala Asn Ser
115 120 125
Gln Thr Gly Tyr Gly Arg Thr Lys Gly Arg Ala Glu Ala Ala Ile His
130 135 140
Ser Val Leu Pro Gly Ala Val Ile Leu Arg Pro Ser Ile Ile Phe Gly
145 150 155 160
Pro Glu Asp Asp Phe Phe Asn Lys Phe Ala Lys Met Ala Arg Asn Leu
165 170 175
Pro Phe Leu Pro Leu Ile Gly Gly Gly Lys Thr Lys Phe Gln Pro Val
180 185 190
Tyr Val Glu Asp Val Ala Glu Ala Val Ala Arg Ser Val Asp Gly Lys
195 200 205
Leu Lys Pro Gly Ala Ile Tyr Glu Leu Gly Gly Pro Asp Val Met Thr
210 215 220
Phe Arg Asp Cys Leu Glu Ala Val Leu Ala Ala Thr Tyr Arg Glu Arg
225 230 235 240
Ser Phe Val Asn Leu Pro Phe Gly Val Ala Ser Met Ile Gly Lys Leu
245 250 255
Ala Ser Leu Val Pro Leu Ile Thr Pro Pro Leu Thr Pro Asp Gln Val
260 265 270
Thr Met Leu Lys Lys Asp Asn Val Val Ser Ala Glu Ala Glu Lys Lys
275 280 285
Gly Leu Thr Leu Glu Gly Ile Gly Ile Thr Pro Val Arg Val Ala Ser
290 295 300
Val Leu Pro Ser Tyr Met Val Gln Tyr Arg Gln His Gly Gln Phe Ser
305 310 315 320
Asn Ala Gly Lys Ala Ala
325
<210> 14
<211> 311
<212> Белок
<213> Rhizobium meliloti
<400> 14
Met Thr Ala Glu Val Phe Asp Pro Arg Ala Leu Arg Asp Ala Phe Gly
1 5 10 15
Ala Phe Ala Thr Gly Val Thr Val Val Thr Ala Ser Asp Ala Ala Gly
20 25 30
Lys Pro Ile Gly Phe Thr Ala Asn Ser Phe Thr Ser Val Ser Leu Asp
35 40 45
Pro Pro Leu Leu Leu Val Cys Leu Ala Lys Ser Ser Arg Asn Tyr Glu
50 55 60
Ser Met Thr Ser Ala Gly Arg Phe Ala Ile Asn Val Leu Ser Glu Thr
65 70 75 80
Gln Lys Asp Val Ser Asn Thr Phe Ala Arg Pro Val Glu Asp Arg Phe
85 90 95
Ala Ala Val Asp Trp Arg Leu Gly Arg Asp Gly Cys Pro Ile Phe Ser
100 105 110
Asp Val Ala Ala Trp Phe Glu Cys Ser Met Gln Asp Ile Ile Glu Ala
115 120 125
Gly Asp His Val Ile Ile Ile Gly Arg Val Thr Ala Phe Glu Asn Ser
130 135 140
Gly Leu Asn Gly Leu Gly Tyr Ala Arg Gly Gly Tyr Phe Thr Pro Arg
145 150 155 160
Leu Ala Gly Lys Ala Val Ser Ala Ala Val Glu Gly Glu Ile Arg Leu
165 170 175
Gly Ala Val Leu Glu Gln Gln Gly Ala Val Phe Leu Ala Gly Asn Glu
180 185 190
Thr Leu Ser Leu Pro Asn Cys Thr Val Glu Gly Gly Asp Pro Ala Arg
195 200 205
Thr Leu Ala Ala Tyr Leu Glu Gln Leu Thr Gly Leu Asn Val Thr Ile
210 215 220
Gly Phe Leu Tyr Ser Val Tyr Glu Asp Lys Ser Asp Gly Arg Gln Asn
225 230 235 240
Ile Val Tyr His Ala Leu Ala Ser Asp Gly Ala Pro Arg Gln Gly Arg
245 250 255
Phe Leu Arg Pro Ala Glu Leu Ala Ala Ala Lys Phe Ser Ser Ser Ala
260 265 270
Thr Ala Asp Ile Ile Asn Arg Phe Val Leu Glu Ser Ser Ile Gly Asn
275 280 285
Phe Gly Ile Tyr Phe Gly Asp Glu Thr Gly Gly Thr Val His Pro Ile
290 295 300
Ala Asn Lys Asp Ala His Ser
305 310
<210> 15
<211> 240
<212> Белок
<213> Lactobacillus paraplantarum
<400> 15
Met Asp Glu Val Ile Leu Val Thr Gly Ala Ala Lys Gly Ile Gly Leu
1 5 10 15
Ala Thr Val Lys Arg Leu Ser Ser Gln Gly Ala Arg Val Ile Leu Asn
20 25 30
Val His His Glu Ile Glu Ala Thr Asp Trp Gln Ala Leu Thr Ala Glu
35 40 45
Tyr Pro Arg Leu Thr Gln Leu Val Gly Asp Val Ser Asp Asp Gln Ser
50 55 60
Ala Ala Asn Leu Ile Asp Thr Val Met Thr Asn Phe Gly Arg Leu Asp
65 70 75 80
Gly Leu Val Asn Asn Ala Gly Val Thr His Asp Gln Leu Leu Thr Arg
85 90 95
Leu His Ala Glu Asp Phe Met Ser Val Ile Gln Thr Asn Leu Leu Gly
100 105 110
Thr Phe Asn Met Thr Lys Tyr Ala Leu Lys Val Met Gln Arg Gln Arg
115 120 125
Gln Gly Ala Ile Val Asn Val Ala Ser Val Val Gly Leu His Gly Asn
130 135 140
Val Gly Gln Ala Asn Tyr Ala Ala Ser Lys Ala Gly Ile Ile Gly Leu
145 150 155 160
Thr Lys Thr Thr Ala Lys Glu Ala Ala Arg Arg Gln Val Arg Cys Asn
165 170 175
Ala Val Ala Pro Gly Met Ile Thr Thr Ala Met Thr Ala Gln Leu Asn
180 185 190
Asp Arg Val Thr Ala Ala Ala Leu Ser Asp Ile Pro Leu Lys Arg Phe
195 200 205
Gly Thr Pro Asp Glu Ile Ala Gln Ala Ile Asp Phe Leu Leu His Gln
210 215 220
Pro Tyr Leu Thr Gly Gln Val Leu Thr Val Asp Gly Gly Met Thr Ile
225 230 235 240
<210> 16
<211> 340
<212> Белок
<213> Botryontinia fuckeliana
<400> 16
Met Arg Val Leu Leu Thr Gly Gly Ser Gly Phe Ile Ala Ala His Ile
1 5 10 15
Leu Asp Ile Leu Leu Ser Arg Gly His Thr Val Ile Thr Thr Val Arg
20 25 30
Ser Gln Gln Lys Ile Asp Ala Ile Lys Ala Ala His Pro Asp Val Pro
35 40 45
Ala Ser Lys Leu Asp Phe Phe Ile Val Glu Asp Ile Ala Lys Glu Asn
50 55 60
Ala Phe Asp Glu Cys Leu Lys Lys Phe Gly Glu Gly Leu Glu Ala Val
65 70 75 80
Leu His Thr Ala Ser Pro Phe His Phe Asn Val Thr Asp Thr Lys Lys
85 90 95
Asp Leu Leu Asp Pro Ala Ile Ile Gly Thr Thr Ala Ile Leu His Ala
100 105 110
Ile Lys Lys Phe Ala Pro Ser Val Thr Arg Val Val Val Thr Ser Ser
115 120 125
Phe Ala Ser Ile Ile Asp Ala Ser Lys Gly Asn Trp Pro Asp His Thr
130 135 140
Tyr Thr Glu Glu Asp Trp Asn Pro Ile Thr Leu Ser Glu Ala Val Glu
145 150 155 160
Asn Pro Ser Asn Gly Tyr Arg Ala Ser Lys Thr Phe Ala Glu Lys Ala
165 170 175
Ala Trp Glu Phe Val Glu Lys Glu Asn Pro Asn Phe Thr Leu Ser Thr
180 185 190
Met Asn Pro Pro Leu Val Leu Gly Pro Ile Val His Tyr Leu Asn Ser
195 200 205
Leu Asp Ala Leu Asn Thr Ser Asn Gln Arg Val Arg Asp Val Leu Gln
210 215 220
Gly Lys Trp Lys Glu Glu Ile Pro Gly Thr Gly Thr Phe Ile Trp Ile
225 230 235 240
Asp Val Arg Asp Leu Ala Leu Ala His Val Lys Ala Ile Glu Ile Ala
245 250 255
Glu Ala Ala Gly Lys Arg Phe Phe Ile Thr Glu Gly Tyr Phe Ser Asn
260 265 270
Lys Glu Ile Cys Glu Ile Ile Arg Lys Asn Phe Pro Glu Asp Gly Gly
275 280 285
Glu Leu Pro Gly Lys Glu Val Lys Gly Gly Gly Tyr Pro Glu Gly Gly
290 295 300
Ile Tyr Lys Phe Asp Asn Ala Arg Thr Arg Ser Val Leu Gly Leu Glu
305 310 315 320
Phe Arg Gly Leu Glu Glu Ser Ile Val Asp Leu Val Lys Ser Leu Lys
325 330 335
Glu Val Gly Val
340
<210> 17
<211> 243
<212> Белок
<213> Fusarium oxysporum
<400> 17
Met Ser Arg Asn Leu Ala Leu Val Thr Gly Ser Thr Gln Gly Ile Gly
1 5 10 15
Leu Ala Val Ala Lys Glu Leu Ala Ile Lys His Asn Tyr Gln Val Leu
20 25 30
Leu Gly Val Arg Asn Thr Lys Ala Gly Glu Glu Ile Ala Ser Asp Leu
35 40 45
Arg Lys Glu Gly His Glu Ala Ser Val Val Glu Leu Asp Leu Thr Ser
50 55 60
Ala Asp Ser Ile Asp Lys Ala Val Lys His Ile Asp Glu Lys Tyr Gly
65 70 75 80
Tyr Leu Asp Val Leu Ile Asn Asn Ala Gly Val Leu Leu Asp Arg Gln
85 90 95
Glu Gly Leu Ser Thr Trp Asp Leu Phe Ser Lys Thr Phe Thr Thr Asn
100 105 110
Val Phe Gly Thr Gly Cys Leu Thr Gln Ser Leu Leu Pro Leu Leu Arg
115 120 125
Lys Ala Lys Asn Ser Pro Pro Arg Ile Val Phe Val Thr Ser Val Met
130 135 140
Gly Ser Leu Thr Lys Ala Thr Asp Glu Thr Thr Thr Tyr Tyr Asn Ile
145 150 155 160
Asp Tyr Lys Ala Tyr Asp Ala Ser Lys Ala Ala Val Asn Met Leu Met
165 170 175
Phe Asn Phe Ala Arg Glu Leu Asp Ala Val Gly Gly Lys Val Asn Ser
180 185 190
Val Cys Pro Gly Leu Val Lys Thr Gly Leu Thr Asn Tyr His Glu Trp
195 200 205
Gly Thr Ser Pro Glu Thr Gly Ala Glu Arg Ile Val Glu Met Ala Thr
210 215 220
Ile Gly Glu Asp Gly Pro Thr Lys Thr Ile Ser Asp Arg Asn Gly Glu
225 230 235 240
Leu Pro Leu
<210> 18
<211> 240
<212> Белок
<213> Lactobacillus delbrueckii
<400> 18
Met Asp Leu Gln Asn Lys Arg Val Leu Val Thr Gly Ser Thr Gln Gly
1 5 10 15
Ile Gly Ala Ala Thr Ala Leu Ala Phe Ala Gln Lys Gly Cys Gln Val
20 25 30
Leu Leu Asn Gly Arg Arg Pro Glu Leu Pro Glu Glu Ile Ala Asp Gln
35 40 45
Leu Glu Lys Ile Gly Ala Asp Tyr Gln Tyr Phe Ser Ala Asp Val Ser
50 55 60
Asp Glu Gly Ala Ile Lys Gln Leu Phe Lys Glu Ile Gly Glu Ile Asp
65 70 75 80
Ile Leu Val Asn Asn Ala Gly Ile Thr Lys Asp Gln Ile Met Ile Gly
85 90 95
Met Lys Leu Ala Asp Phe Asp Gln Val Ile Lys Val Asn Leu Arg Ser
100 105 110
Ser Phe Met Leu Thr Gln Lys Ala Leu Lys Lys Met Leu Lys Lys Arg
115 120 125
Ser Gly Ala Ile Ile Asn Met Ala Ser Ile Val Gly Gln His Gly Asn
130 135 140
Ala Gly Gln Ala Asn Tyr Ala Ala Ser Lys Ala Gly Val Ile Ala Leu
145 150 155 160
Thr Gln Thr Ala Ala Lys Glu Ala Ala Gly Arg Gly Val Arg Val Asn
165 170 175
Ala Ile Ala Pro Gly Met Ile Ala Ser Gln Met Thr Ala Val Leu Pro
180 185 190
Asp Glu Val Lys Glu Gln Ala Leu Ser Gln Ile Pro Leu Ala Arg Phe
195 200 205
Gly Lys Ala Glu Glu Val Ala Gln Ala Ala Val Phe Leu Ala Glu Asn
210 215 220
Asp Tyr Val Thr Gly Gln Thr Leu Val Val Asp Gly Gly Met Thr Ile
225 230 235 240
<210> 19
<211> 338
<212> Белок
<213> Acremonium
<400> 19
Met Thr Lys Val Leu Val Ala Gly Gly Ser Gly Phe Ile Gly Ala His
1 5 10 15
Ile Leu Glu Gln Leu Leu Ala Lys Gly His Ser Val Val Thr Thr Val
20 25 30
Arg Ser Lys Glu Lys Ala Gln Lys Ile Leu Asp Ala His Lys Ala Glu
35 40 45
Ala Asp Arg Leu Glu Val Ala Ile Val Pro Glu Ile Ala Arg Glu Asp
50 55 60
Ala Phe Asp Glu Val Val Lys Thr Pro Gly Ile Glu Val Val Ile His
65 70 75 80
Pro Ala Ser Pro Cys His Leu Asn Phe Thr Asp Pro Gln Lys Glu Leu
85 90 95
Ile Asp Pro Ala Val Leu Gly Thr Thr Asn Ile Leu Arg Ala Ile Lys
100 105 110
Arg Asp Ala Pro Gln Val Arg Arg Val Ile Ile Thr Ser Ser Val Ala
115 120 125
Ala Ile Phe Asn Thr Lys Asp Pro Val Ser Thr Leu Thr Glu Gln Ser
130 135 140
Trp Asn Pro Asn Asp Leu Ser Asn Ile His Asp Ser Arg Ala Val Ala
145 150 155 160
Tyr Cys Val Ser Lys Thr Leu Ala Glu Arg Ala Ala Trp Asp Tyr Val
165 170 175
Asp Gln Glu Lys Pro Asn Phe Asp Leu Val Thr Val Asn Pro Pro Leu
180 185 190
Val Leu Gly Pro Val Val Gly His Phe Ser Asn Val Asp Ser Ile Asn
195 200 205
Ala Ser Asn Glu Cys Leu Ala Asn Leu Val Arg Gly Lys Trp Arg Asp
210 215 220
Glu Ile Pro Pro Thr Gly Pro Val Asn Ile Trp Ile Asp Val Arg Asp
225 230 235 240
Val Ala Ala Ala His Val Arg Ala Met Glu Arg Gln Glu Ala Gly Gly
245 250 255
Lys Arg Leu Phe Thr Val Gly Gly Arg Phe Ser Tyr Thr Lys Ile Ala
260 265 270
Glu Ile Val Arg Glu His Gly Pro Asp Arg Phe Lys Asp Lys Met Pro
275 280 285
Arg Ala Glu Ala Arg Ser Gly Asp Ala Asn Tyr Thr Gly Pro Val Leu
290 295 300
Lys Phe Asp Asn Gly Glu Thr Asn Arg Ile Leu Gly Ile Glu Trp Thr
305 310 315 320
Pro Leu Glu Lys Ser Val Leu Asp Phe Val Glu Ser Ile Lys Glu Phe
325 330 335
Asp Leu
<210> 20
<211> 348
<212> Белок
<213> Tricoderma
<400> 20
Met Thr Lys Val Leu Leu Thr Gly Gly Ser Gly Phe Ile Ala Ala His
1 5 10 15
Ile Leu Glu Gln Leu Leu Ala Lys Asn Tyr Thr Val Ile Thr Thr Val
20 25 30
Arg Thr Lys Ser Lys Ala Asp Leu Ile Lys Glu Ala His Ala Asp Leu
35 40 45
Val Lys Ser Gly Arg Leu Ser Val Ala Ile Val Pro Asp Ile Ala Val
50 55 60
Leu Ser Ala Phe Asp Asp Leu Val Ala Lys Ile Ala Ser Gly Pro Asp
65 70 75 80
Gly Asp Leu Glu Tyr Val Val His Thr Ala Ser Pro Leu Phe Phe Thr
85 90 95
Phe Thr Asp Ala Gln Lys Glu Ile Ile Thr Pro Ala Leu Asn Gly Thr
100 105 110
Arg Gly Ile Leu Glu Ala Val Lys Arg Ser Ala Pro Lys Val Lys Arg
115 120 125
Val Val Ile Thr Ser Ser Phe Ala Ala Ile Leu Ser Glu Asp Asp Phe
130 135 140
Thr Asn Pro Asn Ala Thr Phe Ser Glu Ser Ser Trp Asn Pro Asp Thr
145 150 155 160
Val Lys Asp Ala Asn Arg Ser Ile Ala Thr Gly Tyr His Val Ser Lys
165 170 175
Val Glu Ser Glu Arg Leu Ala Trp Asp Phe Ile Lys Asn Glu Lys Pro
180 185 190
Asn Phe Asp Leu Val Thr Val Asn Pro Pro Leu Val Leu Gly Pro Val
195 200 205
Ala His Ser Leu Ala Ser Val Asp Ala Ile Asn Ala Ser Asn Glu Arg
210 215 220
Ile Ala Asp Leu Leu Arg Gly Lys Trp Lys Ala Glu Ile Pro Glu Thr
225 230 235 240
Gly Ala Val Asp Leu Tyr Ile Asp Val Arg Asp Thr Ala Lys Ala His
245 250 255
Ile Lys Ala Leu Glu Leu Pro Glu Ala Ser Gly His Arg Leu Phe Pro
260 265 270
Val Ala Ser Arg Thr Ser Asn His Glu Ile Ala Lys Ile Ile Arg Asp
275 280 285
Asn Phe Pro Glu Phe Ala Glu Arg Leu Pro Gly Pro Glu Val Lys Gly
290 295 300
Gly Glu His Val Asp Glu Asn Lys Ala Tyr Lys Trp Asn Cys Asp Glu
305 310 315 320
Thr Asn Lys Leu Leu Lys Ile Asp Trp Ile Pro Ile Glu Gln Ser Met
325 330 335
Ile Asp Thr Val Asn Ser Leu Lys Asp Lys Gly Ile
340 345
<210> 21
<211> 302
<212> Белок
<213> Coprinopsis
<400> 21
Met Pro Thr Val Ser Pro Gly Ser Lys Val Leu Val Thr Gly Ala Asn
1 5 10 15
Gly Phe Ile Ala Ile Trp Val Val Arg Arg Leu Leu Glu Glu Gly Tyr
20 25 30
Ser Val Arg Gly Thr Val Arg Ala Ala Ser Lys Ala Ser His Leu Lys
35 40 45
Asp Ile Phe Lys Ser Tyr Gly Glu Lys Leu Glu Val Val Val Val Pro
50 55 60
Asp Phe Thr Lys Glu Gly Ala Phe Asp Glu Leu Ile Lys Gly Met Asp
65 70 75 80
Ala Ile Gln His Ile Ala Ser Pro Gly Pro Ala Asn Thr Asp Asp Leu
85 90 95
Tyr Glu Ile Val Asn Pro Ala Val Asp Gly Thr Leu Asn Leu Leu Asn
100 105 110
Thr Ala Leu Lys His Gly Ser Gly Leu Lys Arg Ile Val Ile Thr Ser
115 120 125
Gly Ala Gly Ala Ile Ile Asp Thr Thr Thr Ala Trp Lys Phe Tyr Asn
130 135 140
Asp His Lys Asn Val Ile Lys Trp Asp Leu Thr Val Leu Asn Pro Val
145 150 155 160
Phe Val Phe Gly Pro Pro Ile His Glu Ile Gly Ala Ser Pro Met Thr
165 170 175
Leu Asn Ser Ser Met Val His Phe Trp Val Asn Val Ile Ser Thr Asp
180 185 190
Thr Pro Lys Thr Lys Glu Gly Leu Ser Phe Ala Ala Ser Trp Val Asp
195 200 205
Val Arg Asp Val Ala Gln Gly His Val Leu Ala Leu Gln Lys Glu Ala
210 215 220
Ala Gly Gly Glu Arg Ile Ile Leu Ser Glu Gly Ser Phe Val Trp Gln
225 230 235 240
Asp Trp Val Asp Val Ala Asn Lys Phe Lys Ser Lys Arg Glu Leu Pro
245 250 255
Lys Gly Met Pro Glu Ile Glu Arg Val Tyr Lys Phe Gln Met Asp Ala
260 265 270
Ser Lys Ala Thr Arg Ile Leu Gly Ile Thr Tyr Arg Ser Lys Glu Asp
275 280 285
Thr Met Lys Asp Leu Leu Glu Asp Phe Glu Arg Arg Gly Trp
290 295 300
<210> 22
<211> 239
<212> Белок
<213> Exophiala
<400> 22
Met Lys Val Leu Leu Thr Gly Gly Ser Gly Phe Ile Ala Thr His Cys
1 5 10 15
Leu Asp Ala Leu Leu Lys His Gly His Glu Val Val Ile Thr Val Arg
20 25 30
Ser Ala Glu Lys Gly Gln Ala Leu Val Asp Leu Phe Lys Gly Gln Lys
35 40 45
Val Ser Tyr Thr Ile Val Lys Asp Ile Ser Val Pro Gly Ala Phe Asp
50 55 60
Gln Ala Val Ile Ser Asp Pro Pro Phe Asp Ala Val Val His Thr Ala
65 70 75 80
Ser Pro Phe His Tyr Asp Val Gln Asp Asn Lys Arg Asp Leu Leu Asp
85 90 95
Pro Ala Ile Ile Gly Thr Thr Gly Ile Leu Glu Ser Ile Gln Lys Gly
100 105 110
Ala Pro Ser Val Lys Lys Val Val Val Thr Ser Ser Phe Ala Ala Ile
115 120 125
Ser Asn Pro Thr Ala Pro Pro Lys Val Tyr Asp Glu Thr Val Trp Asn
130 135 140
Gln Met Thr Met Glu Glu Ala Leu Thr Thr Lys Asp Pro Gln Ala Val
145 150 155 160
Tyr Arg Gly Ser Lys Thr Phe Ala Glu Lys Ala Ala Trp Glu Phe Val
165 170 175
Glu Arg Glu Lys Pro Asn Phe Thr Leu Thr Val Leu Asn Pro Pro Val
180 185 190
Ser His Phe Leu Phe Ser Arg His Lys Asp Val Ala Val Thr Phe Phe
195 200 205
Ser Asp Ser Phe Gln His Cys Arg Trp Ser Thr Ala Arg Ser Cys Thr
210 215 220
Pro Trp His His Trp Thr Ile Ser Thr Pro Arg Ala Ser Glu Ser
225 230 235
<---
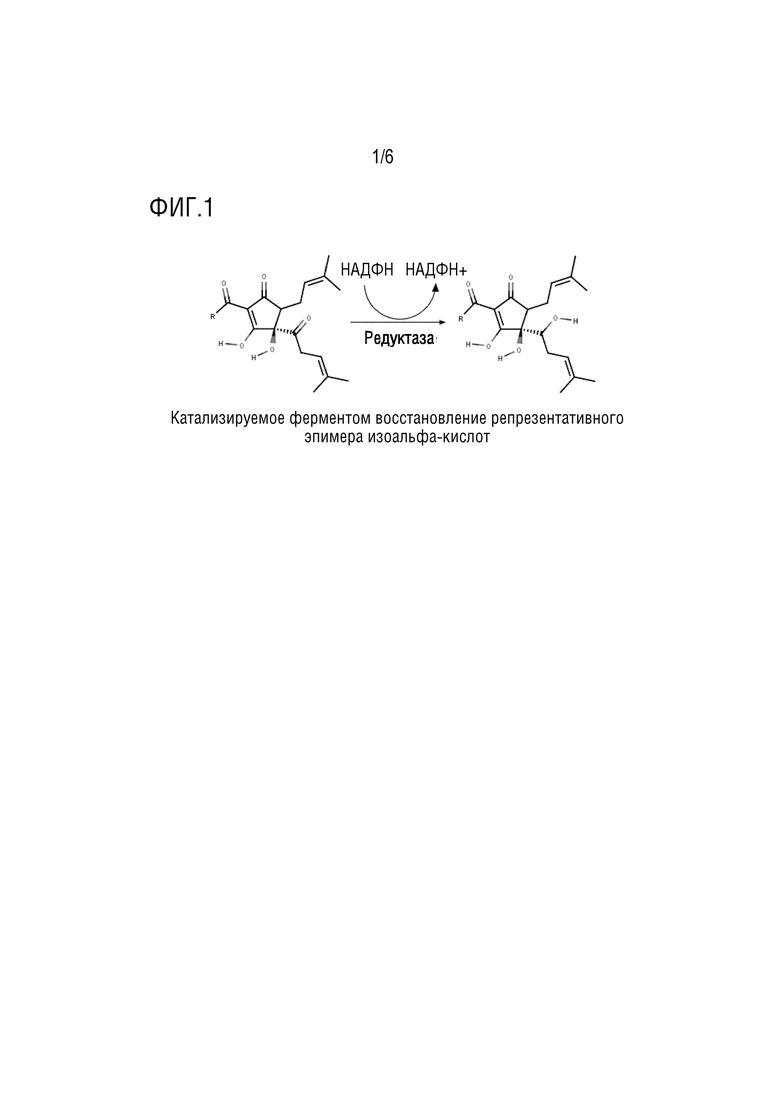
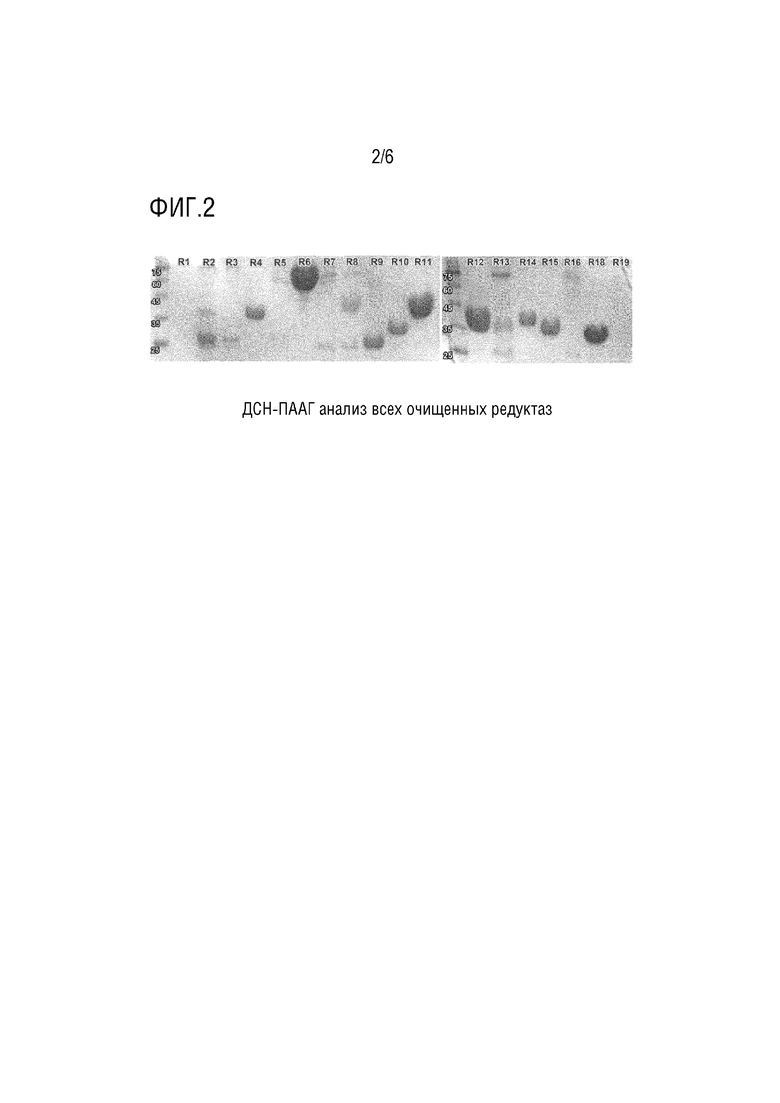
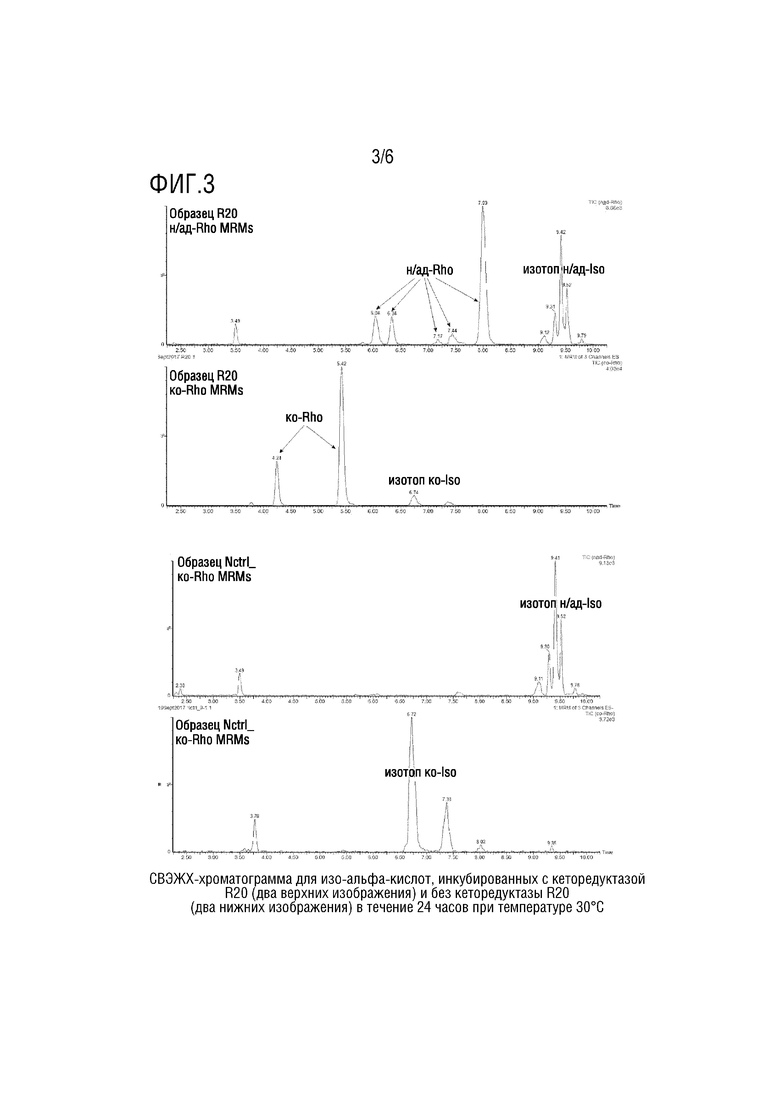



Изобретение относится к биотехнологии и представляет собой способ получения дигидро-изо-альфа кислот или rho-изо-альфа-кислот, включающий обработку изо-альфа-кислот ферментом кеторедуктазой или микроорганизмом, экспрессирующим ген, кодирующий кеторедуктазу, где фермент кеторедуктаза содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22, или где фермент кеторедуктаза на 99, 95, 90, 85, 80, 75 или 70 процентов гомологичен ферменту кеторедуктазе, выбранному из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22. Изобретение позволяет получать дигидро-изо-альфа кислоты с высокой степенью эффективности. 15 з.п. ф-лы, 6 ил., 2 табл., 22 пр.
1. Способ получения дигидро-изо-альфа кислот (или rho-изо-альфа-кислот), включающий обработку изо-альфа-кислот ферментом кеторедуктазой или микроорганизмом, экспрессирующим ген, кодирующий кеторедуктазу, где фермент кеторедуктаза содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22, или где фермент кеторедуктаза на 99, 95, 90, 85, 80, 75 или 70 процентов гомологичен ферменту кеторедуктазе, выбранному из группы, состоящей из SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 и SEQ ID NO: 22.
2. Способ по п.1, где способ проводят в водной системе.
3. Способ по п.2, где способ проводят в условиях умеренной температуры и pH.
4. Способ по п.1, включающий добавление к смеси изо-альфа-кислот фермента кеторедуктазы и НАДФН или НАДФ с последующей инкубацией.
5. Способ по п.1, включающий добавление к смеси изо-альфа-кислот фермента кеторедуктазы и НАДФН или НАДФ в присутствии изопропанола для рециклирования кофактора с последующей инкубацией.
6. Способ по п.5, где концентрация изо-альфа-кислот, то есть субстрата, является максимальной для увеличения объемной производительности биоконверсии.
7. Способ по п.5, где концентрация кофактора НАДФН или НАДФ в смеси минимизирована для улучшения экономических показателей биоконверсии.
8. Способ по п.1, включающий добавление к смеси изоальфа-кислот фермента кеторедуктазы и НАДФН или НАДФ в присутствии другого фермента для рециклирования кофактора с последующей инкубацией.
9. Способ по п.1, включающий добавление к смеси изоальфа-кислот цельноклеточного биокатализатора, где цельноклеточный биокатализатор представляет собой иммобилизованный микроорганизм, экспрессирующий ген, кодирующий кеторедуктазу, с последующей инкубацией.
10. Способ по п.1, включающий культивирование микроорганизма, экспрессирующего ген, кодирующий кеторедуктазу, и добавление изо-альфа-кислот в культуру.
11. Способ по п.1, включающий добавление к экстракту изо-альфа-кислот фермента кеторедуктазы, где кеторедуктаза является термостабильной и где смесь нагревают и инкубируют.
12. Способ по п.1, где кеторедуктаза специфично восстанавливает цис-изогумулон, цис-изокогумулон и цис-изоадгумулон.
13. Способ по п.1, где кеторедуктаза специфично восстанавливает транс-изогумулон, транс-изокогумулон и транс-изоадгумулон.
14. Способ по п.1, включающий добавление смеси 2 или более ферментов кеторедуктазы в количестве, эффективном для восстановления смеси цис- и транс-изо-альфа-кислот до их соответствующих дигидро-изо-альфа-кислот.
15. Способ по п.14, где смесь 2 или более ферментов кеторедуктазы обеспечивает получение уникальной смеси дигидро-изо-альфа-кислот, которая отличается от смеси, производимой химическими восстановителями, такими как боргидрид натрия.
16. Способ по п.1, где кеторедуктаза на 99, 95, 90, 85, 80, 75 или 70 процентов гомологична кеторедуктазе SEQ ID NO: 2, SEQ ID NO: 3, SEQ ID NO: 6, SEQ ID NO: 8, SEQ ID NO: 12, SEQ ID NO: 13, SEQ ID NO: 16, SEQ ID NO: 19, SEQ ID NO: 20 или SEQ ID NO: 22.
| WO 2009077611 A2, 25.06.2009 | |||
| WO 2010025238 A3, 04.03.2010 | |||
| WO 2009029554 A3, 02.07.2009 | |||
| СПОСОБ ПОЛУЧЕНИЯ ГИДРОКСИЛИРОВАННЫХ ЦИКЛОПЕНТАПИРИМИДИНОВЫХ СОЕДИНЕНИЙ И ИХ СОЛЕЙ | 2013 |
|
RU2642311C2 |

