[0001] Настоящая заявка испрашивает приоритет в соответствии с §120 раздела 35 U.S.С. по предварительной заявке на патент США №62/256902, поданной 18 ноября 2015 г., и предварительной заявке на патент США №62/300507, поданной 26 февраля 2016 г., содержание которых включено в данный документ посредством ссылки.
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] К настоящей заявке прилагается перечень последовательностей под названием 80906-PCT_ST25.txt, созданный 16 ноября 2016 г., размер которого составляет примерно 392 килобайта. Этот перечень последовательностей включен в данный документ посредством ссылки во всей своей полноте. Этот перечень последовательностей подается вместе с настоящей заявкой посредством EFS-Web и соответствует §1.824(а)(2)-(6) и (b) раздела 37 C.F.R.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
[0003] Раскрытый в данном документе объект изобретения относится к диагностическому выявлению индукции гаплоидии ("HI") или ее отсутствия и/или присутствия у растений, которые являются индукторами гаплоидии или не являются таковыми. Более конкретно, раскрытый в данном документе объект изобретения относится к нуклеиновым кислотам, которые можно использовать для индуцирования HI у растений, и/или видам биологической активности, которые можно модифицировать для получения или предупреждения HI у растения, которое в противном случае будет демонстрировать HI, либо у растения, которое в противном случае не будет демонстрировать HI. Еще более конкретно, раскрытый в данном документе объект изобретения относится к молекуле нуклеиновой кислоты, которая кодирует биологически активную молекулу, а также к способам применения ее для регулирования HI у растений.
[0004] В данном документе предусматриваются серия независимых индуцированных человеком мутаций, обнаруживаемых по меньшей мере в одном гене пататин-подобной фосфолипазы А2α ("PLA") маиса; растения маиса, имеющие эти мутации по меньшей мере в одном из их генов PLA; и способ создания и идентификации сходных и/или дополнительных мутаций в гене PLA посредством скрининга объединенных и/или отдельных растений маиса. Растения маиса по настоящему изобретению индуцируют гаплоидию в результате нетрансгенных мутаций по меньшей мере в одном из их генов PLA. Также предусматриваются способы индуцирования de novo индукции гаплоидии посредством нанесения химических веществ на репродуктивные ткани растений. Также предусматриваются способы увеличения уровня завязывания семян и уровня индукции гаплоидии ("HIR", определенного в данном документе как количество выживших гаплоидных зерен по сравнению с общим количеством зерен после опыления початка пыльцой, индуцирующей гаплоидию) посредством нанесения химических веществ на репродуктивные ткани растения во время опыления.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
[0005] Опыление является сложным процессом. Пыльцевые зерна покрытосеменных состоят из крупной вегетативной клетки и двух мужских гамет (спермиев). После попадания на дыхальце из зерна прорастает пыльцевая трубка, которая демонстрирует быстрый концевой рост по мере перемещения вниз по женским передающим путям под воздействием хемоаттрактантов, секретируемых двумя синергидными клетками на микропилярном конце зародышевого мешка. Во время прохождения вниз по трубке спермин соединены друг с другом и с вегетативным ядром посредством тяжистой цитоплазмы, называемой мужской зародышевой единицей. Вскоре после контакта с одной из двух синергид пыльцевая трубка разрывается, и два спермия продвигаются через цитоплазму умирающей синергидной клетки, чтобы независимо слиться с яйцеклеткой и центральными клетками зародышевого мешка, завершая двойное оплодотворение. Даже после первоначального контакта неудачные события оплодотворения могут быть исправлены второй пыльцевой трубкой, которая оплодотворяет зародышевый мешок посредством взаимодействия с персистентной синергидной клеткой.
[0006] Селекционеры скрещивают инбредные родительские линии, при этом одна выступает в качестве мужской особи, а одна - в качестве женской особи, для образования гибридных семян. Процесс разработки инбредных родительских линий, которые являются практически гомозиготными, обычно требует выбора гибридного продукта скрещивания и его самоопыления (самооплодотворения) в течение множества поколений для того, чтобы он стал почти гомозиготным. Этот процесс является времязатратным и дорогостоящим. Для сокращения времени для разработки гомозиготных инбредных растений маиса, риса, пшеницы, ячменя и других сельскохозяйственных культур селекционеры могут предпочесть использовать линию, индуцирующую гаплоидию, для индукции образования гаплоидных семян у гибридной родительской особи. Число хромосом гаплоидных растений затем удваивают, например, посредством средства, удваивающего число хромосом, такого как колхицин, с образованием двойных гаплоидных гомозиготных инбредных линий.
[0007] Индукция гаплоидии ("HI") представляет собой класс явлений у растений, характеризующийся утратой хромосом индуктора в ходе развития зародыша. В WO2012/030893, включенном в данный документ посредством ссылки, описана область хромосомы 1 маиса, которая может отвечать за индукцию гаплоидии. Идентифицированные маркеры в этой области, увеличивающие индукцию гаплоидии, описаны как находящиеся в положениях 48249509-51199249, которые соответствуют имеющемуся в открытом доступе маркеру umc 1169, имеющему физическое положение (60213661). Эта область, по-видимому, не выравнивается с областью индукции гаплоидии у Stock 6. В Dong et al., (2013) Theor. Appl. Genet. 126: 1713-1720 описан QTL, расположенный в секции 1.04, который объясняет до 66% генотипической дисперсии включительно для HIR.
[0008] Индукцию гаплоидии наблюдали у множества видов растений, таких как сорго, ячмень, пшеница и другие злаковые растения. У маиса HI, по-видимому, является результатом реаранжировки, мутаций и/или рекомбинаций, вставки или делеций в пределах области хромосомы 1 (с существенным исключением для индукции гаплоидии ig-типа, которая является результатом мутации в гене INDETERMINATE GAMETOPHYTE1 в хромосоме 3). Предусматриваемые линии HI были изучены и приближенно идентифицированы. Однако не были представлены экспериментальные данные, демонстрирующие генетический причинный фактор HI у маиса. Также ранее не были идентифицированы перечисленные в данном документе маркеры, которые ассоциированы с этим признаком.
[0009] У маиса гаплоидные семена или зародыши, в частности, образуются посредством осуществления скрещиваний между мужской особью, индуцирующей гаплоидию (т.е. "пыльцой, индуцирующей гаплоидию"), и практически любым выбранным початком, при этом початок может быть получен из любой инбредной, гибридной или другой зародышевой плазмы. Гаплоиды образуются, если ДНК пыльцы, индуцирующей гаплоидию, не полностью передается и/или сохраняется на протяжении первых клеточных делений зародышей. Полученный в результате фенотип не является полностью пенетрантным, при этом некоторые семязачатки содержат гаплоидные зародыши, а другие содержат диплоидные зародыши, анеуплоидные зародыши, химерные зародыши или недоразвитые зародыши. Гаплоидные зерна имеют зародыши, которые содержат только материнскую ДНК вместе с нормальным триплоидным эндоспермом. После индукции гаплоидии гаплоидные зародыши или семена, как правило, отделяют от диплоидных и анеуплоидных сибсов с помощью скрининга по фенотипическим или генетическим маркерам, и из них выращивают или культивируют гаплоидные растения. Эти растения затем превращают естественным путем либо посредством химической манипуляции (т.е. с помощью колхицина) в двойные гаплоидные (DH) растения, которые затем образуют инбредные семена.
[0010] Линии HI содержат локус количественного признака ("QTL") в хромосоме 1, отвечающий по меньшей мере за 66% изменчивости индукции гаплоидии. QTL вызывает индукцию гаплоидии на различных уровнях, если он интрогрессирован в различные окружения. Все линии, индуцирующие гаплоидию, применяемые в промышленном семеноводстве, являются производными линии-основателя HI, известной как Stock6, и все они имеют мутацию в QTL хромосомы 1, индуцирующую гаплоидию. В данном документе авторы настоящего изобретения раскрывают ключевую мутацию в QTL, при комплементации которой восстанавливается нормальное размножение. Несмотря на то, что причины этой мутации являются неясными, она возникает во всех индуцирующих линиях, в том числе Stock6.
[0011] Селекция растений облегчается посредством применения двойных гаплоидных (DH) растений. Получение DH-растений позволяет селекционерам-растениеводам получать инбредные линии без инбридинга в нескольких поколениях, сокращая таким образом время, необходимое для получения гомозиготных растений. DH-растения обеспечивают бесценный инструмент для селекционеров-растениеводов, в частности для получения инбредных линий, картирования QTL, цитоплазматических превращений, интрогрессии признаков и скрининга F2 в отношении высокопроизводительного улучшения признака. Экономится большое количество времени, поскольку гомозиготные линии образуются по существу за одно поколение с исключением необходимости в традиционном инбридинге в нескольких поколениях. В частности, поскольку DH-растения являются полностью гомозиготными, они хорошо поддаются количественным генетическим исследованиям. Образование гаплоидных семян является критически важным для процесса селекции двойных гаплоидов. Гаплоидные семена образуются в материнской зародышевой плазме при оплодотворении пыльцой от гиногенетического индуктора, такого как Stock 6.
[0012] Растения маиса, индуцирующие гаплоидию, образуют пыльцу, которая при скрещивании с неиндуцирующей зародышевой плазмой обуславливает гиногенетическое развитие гаплоидных семян. К сожалению, в результате этого процесса часто получают низкую частоту гаплоидных зерен. Неэффективная частота индукции гаплоидии является ограничивающим фактором в программах селекции двойных гаплоидов маиса.
КРАТКОЕ ОПИСАНИЕ
[0013] Высокий HIR обеспечивает большую частоту образования гаплоидных семян родительским растением, представляющим интерес.Родительские растения можно подвергнуть предварительному скринингу с использованием генетических маркеров, ассоциированных с необходимыми признаками, или фенотипически наблюдаемых признаков для обогащения генетического потенциала родительских растений. Если эти желаемые родительские растения опыляют индуктором гаплоидии, который характеризуется более высоким HIR, то получают более высокий потенциал получения желаемых двойных гаплоидов с необходимым генотипом и фенотипом.
[0014] Несмотря на то, что процесс селекции двойных гаплоидов приводил в результате к более быстрому получению гомозиготных инбредных растений, объем получаемых двойных гаплоидных инбредных растений может быть ограничен. Известные индуцирующие линии включают без ограничения Stock 6, MHI (молдавский индуктор гаплоидии), линию с мутацией неопределенного гаметофита ("ig"), KEMS, RWK, ZEM, ZMS и KMS. Все из них характеризуются относительно низким HIR. Stock 6, например, индуцирует образование только 1-3% гаплоидных семян. В силу этого индукция образования гаплоидов представляет собой ограничивающую уровень стадию в процессе получения двойных гаплоидных линий.
[0015] Авторы настоящего изобретения изобрели способ индукции получения гаплоидов и/или увеличения уровня индукции гаплоидии у растений посредством обработки растений липидным соединением, ингибитором фосфолипаз и ингибитором десатураз жирных кислот.Одна такая группа способов включает нанесение специфических химических веществ на репродуктивные ткани при скрещивании с пыльцой дикого типа (не индуцирующей гаплоидию). Авторы настоящего изобретения впервые химическим путем вызвали индукцию гаплоидии de novo. Это осуществляется посредством введения ингибитора фосфолипаз метил-альфа-линоленоилфторфосфоната (MALFP) в определенной концентрации в цветок во время опыления, что приводит к высокому уровню индукции гаплоидии, составляющему до 9% HIR включительно. Кроме того, авторы настоящего изобретения вызвали индукцию гаплоидии de novo посредством введения арахидонилфторфосфоната (MAFP) в определенной концентрации в цветок во время опыления. Кроме того, авторы настоящего изобретения вызвали индукцию гаплоидии de novo посредством введения 1,2-дистеароил-sn-глицеро-3-фосфатидилхолина (также известного как дистеарилфосфатидилхолин; "DSPC") в определенной концентрации в цветок во время опыления. Авторы настоящего изобретения также вызвали индукцию гаплоидии de novo посредством введения альфа-линоленовой кислоты в определенной концентрации в цветок во время опыления. Авторы настоящего изобретения также вызвали индукцию гаплоидии de novo посредством введения легкодоступных соединений, в том числе кукурузного масла и льняного масла, а также химически синтезированных линолевой кислоты, этилового сложного эфира олеиновой кислоты (ОАЕЕ), метилового сложного эфира арахидоновой кислоты (ААМЕ) и ингибитора фосфолипаз маноалида. Авторы настоящего изобретения также увеличили уровень индукции гаплоидии при скрещивании растений с линией, индуцирующей гаплоидию (например, RWK, Stock 6 или ZMS), посредством введения ингибитора фосфолипаз метил-альфа-линоленоилфторфосфоната (MALFP) в определенной концентрации. При концентрациях 2% MALFP+ смесь поверхностно-активных веществ 91, эмульгированных буферным раствором DML, авторы настоящего изобретения удвоили уровень индукции гаплоидии в опылениях с применением RWK в качестве опылителя. Типичный уровень индукции для RWK составляет приблизительно 10-18%. При нанесении MALFP уровень индукции увеличивается до 20-35%. Авторы настоящего изобретения также повышали уровень индукции гаплоидии посредством нанесения линолевой кислоты (LLE), этилового сложного эфира линолевой кислоты (LLAEE) и ингибитора фосфолипаз, называемого метиларахидонилфторфосфонатом (MAFP), в определенных концентрациях на цветок во время опыления. Кроме того, при выполнении этого таким образом также снижается уровень недоразвитости зародышей и увеличивается уровень образования зерен для MALFP, LLAEE, MAFP и LLE. В совокупности эти эффекты приводят к увеличению общего количества гаплоидных зародышей, воспроизводящихся на початке после опыления.
[0016] Авторы настоящего изобретения также клонировали ген, отвечающий за индукцию гаплоидии у маиса, и определили его характеристики. Ген имеет название PLA2 и характеризуется экспрессией, специфической для пыльцы. Белок PLA2, по-видимому, локализуется в цитоплазме спермия, возможно, в эндоплазматическом ретикулуме или тельцах Гольджи. Идентификация гена привела к изобретениям нескольких новых методик улучшения процесса индукции гаплоидии, определяемых в качестве действий по получению гаплоидных зародышей, зерен, семян или растений посредством скрещивания любого початка с пыльцой, индуцирующей гаплоидию. Идентификация гена также привела к изобретениям новых способов индукции образования гаплоидов. Другая группа включает способы создания новых линий, индуцирующих гаплоидию, путем изменения последовательности причинного гена посредством направленного мутагенеза, TILLING или CRISPR/Cas9. Экспрессия белка PLA2 может быть подавлена с помощью RNAi или посредством применения направленного мутагенеза в промоторе, 3'-UTR, 5'-UTR или сайтах сплайсинга.
[0017] С учетом идентификации этой мутации авторы настоящего изобретения изобрели способы модификации и улучшения процесса индукции гаплоидии, и авторы настоящего изобретения впервые раскрывают способы получения гаплоидов de novo посредством химической обработки во время опыления. Авторы настоящего изобретения демонстрируют способы увеличения уровня индукции гаплоидии ("HIR," т.е. процентного количества гаплоидных зародышей, обнаруживаемых на указанном початке, подвергнутом индукции гаплоидии), а также способы увеличения уровня выживаемости зерен в ходе индукции гаплоидии. Авторы настоящего изобретения также обсуждают способы создания новых линий, индуцирующих гаплоидию, с применением стратегий генетической модификации ("GM") или направленного мутагенеза.
[0018] Процесс индукции гаплоидии можно улучшить посредством множества способов. Во-первых, можно стремиться улучшить средний HIR. HIR имеет ограниченный уровень при крупномасштабном получении двойных гаплоидных ("DH") растений, поскольку HIR является относительно низким. См. фигуру 1. За редкими исключениями, HIR обычно составляет менее 25% и наиболее часто находится в диапазоне 10-20%, что означает, что 75-90% зерен являются диплоидными. С некоторой низкой частотой в ходе индукции гаплоидии также образуются анеуплоидные зародыши. Примеры соединений, которые, как было показано, увеличивают количество образующихся гаплоидов и/или уровень образования гаплоидов в ходе скрещиваний с индуктором гаплоидии без увеличения уровня недоразвитости зародышей, включают метил-альфа-линоленоилфторфосфонат (MALFP), метиларахидонилфторфосфонат (MAFP), линолевую кислоту (LLA) и этиловый сложный эфир линолевой кислоты (LLAEE). Во-вторых, можно уменьшить уровень недоразвитости зерен и частоту неудачного оплодотворения в ходе индукции гаплоидии, что приводит к образованию большего количества зерен на один початок. Недоразвитость зерна возникает после того, как происходит успешное оплодотворение, но при этом не могут развиться функциональные зародыш или эндосперм, и в результате вырастает небольшое, бесцветное зерно, не имеющее внутри зрелого зародыша. Уровень недоразвитости зерен является высоким в процессах индукции гаплоидии - 10-50% оплодотворенных семязачатков в початке. Это ограничивает количество гаплоидных зерен, которое может воспроизводиться на один початок. Неудачное оплодотворение происходит, если семязачаток неспособен быть оплодотворен пыльцевым зерном, и характеризуется отсутствием какого-либо развития зерен после опыления. Этот уровень в большинстве случаев оказывается высоким в ходе индукции гаплоидии - 10-70% зерен в некоторых початках, подвергнутых индукции гаплоидии, неспособны быть оплодотворены, в зависимости от синхронности созревания мужского и женского цветков, типа осуществляемого скрещивания и генетических характеристик мужской и женской особей. Примеры соединений, которые, как было показано, увеличивают уровень образования жизнеспособных зерен посредством снижения частоты неудачного оплодотворения и уровня недоразвитости зародышей, включают те же четыре соединения, упомянутые выше: MALFP, MAFP, LLA и LLAEE. Эти четыре молекулы при нанесении на пыльцу, метелки или другие части цветка в ходе скрещиваний для индукции гаплоидии, увеличивают количество образующихся гаплоидов посредством увеличения как уровня индукции гаплоидии, так и числа зерен.
[0019] И наконец, можно улучшить процесс индукции гаплоидии с исключением необходимости в мужских особях, индуцирующих гаплоидию, посредством обеспечения возникновения индукции гаплоидии de novo на початке или в семязачатке при его самоскрещивании или скрещивании с пыльцой от линии, не индуцирующей гаплоидию. Эти способы индукции гаплоидии de novo, в том числе путем нанесения фосфолипидов, таких как DSPC, жирных кислот, таких как линоленовая кислота (LNA), широко известных смесей триглицеридов, таких как кукурузное масло и льняное масло, или ингибиторов фосфолипаз, таких как MALFP, можно применять при любом ауткроссе или самоопылении для индукции образования гаплоидов у кукурузы. В данном документе авторы настоящего изобретения описывают примеры, которые относятся к одной или нескольким из этих категорий, представляющие улучшения процесса индукции гаплоидии или новые изобретения в нем.
[0020] Настоящее изобретение относится к способу индуцирования образования гаплоидных зародышей при скрещивании между двумя родительскими растениями. Это осуществляют путем изменения экспрессии фосфолипазы в одном из родительских растений. Это изменение можно осуществлять несколькими путями: либо вызывая экспрессию мутантной фосфолипазы одним из родительских растений; либо посредством введения малой интерферирующей РНК в одно или оба родительских растения, что вызывает подавление экспрессии фосфолипазы; либо посредством трансформации одного из родительских растений мутантной фосфолипазой; либо посредством редактирования фосфолипазы одного из родительских растений, например, посредством сайт-направленного мутагенеза, такого как технологии на основе CRISPR или TALEN. Если экспрессия фосфолипазы в одном из родительских растения изменена посредством одной из этих методик, то в случае, если это родительское растение применяется в скрещивании, образуется по меньшей мере один гаплоидный зародыш.
[0021] В одном варианте осуществления способа фосфолипаза представляет собой пататин-подобную фосфолипазу. В другом варианте осуществления пататин-подобная фосфолипаза является ортологом pPLAIIα, которая кодируется нуклеотидной последовательностью, содержащей SEQ ID NO: 1, или последовательностью, по меньшей мере на 70% идентичной SEQ ID NO: 1. Нуклеотидная последовательность, кодирующая пататин-подобную фосфолипазу, может быть подвергнута мутации, и в одном варианте осуществления нуклеотидная последовательность имеет мутацию сдвига рамки считывания, при которой создается искусственный стоп-кодон. Последовательность с мутацией сдвига рамки считывания содержит SEQ ID NO: 3 или последовательность, по меньшей мере на 70% идентичную SEQ ID NO: 3.
[0022] Другие мутации являются возможными и находятся в пределах объема настоящего изобретения. Сайт-направленный мутагенез можно применять для создания большего количества мутаций фосфолипазы. Применение CRISPR/Cas9, TALEN, "цинковых пальцев" и мегануклеаз является способами осуществления сайт-направленного мутагенеза в соответствии с вариантами осуществления настоящего изобретения.
[0023] Настоящее изобретение является применимым при многих типах скрещиваний между растениями. В одном варианте осуществления родительские растения, применяемые в скрещивании, являются однодольными растениями, такими как маис, рис, ячмень и пшеница. Родительские растения могут относиться к одному и тому же виду однодольных, или же они могут относиться к разным видам. В другом варианте осуществления родительские растения, применяемые в скрещивании, представляют собой двудольные растения, такие как соя, подсолнечник, томат, перец, сахарная свекла или разновидности брюссельской капусты. В предпочтительном варианте осуществления родительские растения представляют собой растения маиса или риса. В пределах объема настоящего изобретения находятся гаплоидный зародыш, полученный с помощью данного способа, гаплоидное семя, содержащее гаплоидный зародыш, и гаплоидное растение, выращенное из гаплоидного семени. Также в пределах объема настоящего изобретения находится двойной гаплоид, полученный посредством воздействия на гаплоидный зародыш средства, удваивающего число хромосом, такого как колхицин или трифлуралин.
[0024] Настоящее изобретение относится к кДНК, содержащей SEQ ID NO: 3, или последовательность, ортологичную SEQ ID NO: 3, или последовательность, на 70% идентичную SEQ ID NO: 3. В предпочтительных вариантах осуществления последовательность, ортологичная SEQ ID NO: 3, охватывает пататин-подобные фосфолипазы из маиса, риса, пшеницы, сои и подсолнечника. В частности, последовательность, ортологичная SEQ ID NO: 3, содержит ген Os03g27610 риса. В более предпочтительном варианте осуществления последовательность, ортологичная SEQ ID NO: 3, охватывает SEQ ID NO: 23 и 73-81.
[0025] Настоящее изобретение относится к растению, содержащему индуцированную человеком нетрансгенную мутацию в его гене пататин-подобной фосфолипазы. В предпочтительном варианте осуществления ген пататин-подобной фосфолипазы представляет собой pPLAIIα. В другом варианте осуществления мутация вызывает кодирование преждевременного стоп-кодона в гене. В более предпочтительном варианте осуществления растение представляет собой любое однодольное или любое двудольное растение, но особенно предпочтительными являются маис или рис.
[0026] Настоящее изобретение также относится к способу индуцирования образования гаплоидных зародышей и семян посредством обработки репродуктивных тканей растения соединением, включающим в себя липид или ингибитор фосфолипаз. В одном варианте осуществления обработку осуществляют до, во время или непосредственно после опыления. Обрабатываемые растения могут представлять собой любые однодольные или двудольные растения, но в предпочтительных вариантах осуществления растения представляют собой маис, рис, пшеницу, сою, подсолнечник и сахарную свеклу. В другом варианте осуществления липид или ингибитор фосфолипаз выбраны из группы, указанной в таблице 7. В предпочтительных вариантах осуществления соединение для обработки включает в себя метил-альфа-линоленоилфторфосфонат ("MALFP"), этиловый сложный эфир линолевой кислоты ("LLAEE"), линолевую кислоту ("LLA"), кукурузное масло, дистеарилфосфатидилхолин ("DSPC") или метиларахидонилфторфосфонат ("MAFP").
[0027] В другом варианте осуществления липид или ингибитор фосфолипаз соответствуют следующей формуле (I): 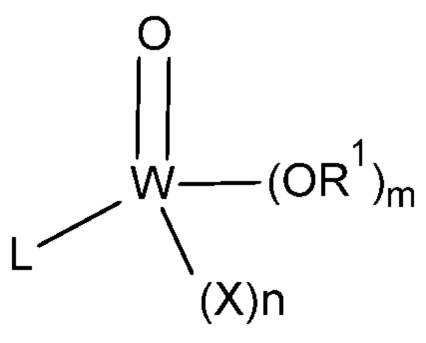 . В пределах объема настоящего изобретения W в формуле (I) представляет собой углерод ("С"), фосфор ("Р") или серу ("S"); m может равняться 0 или 1; n может равняться 0 или 1; X выбран из группы, состоящей из ОН, CN, O(С1-С4алкил), галогена, С1-С4алкила и С1-С4алкила, замещенного одним, двумя или тремя галогенами или карбонилами; R1 выбран из группы, состоящей из Н, C1-С6алкила и C1-С6алкила, замещенного одной или несколькими гидроксильными группами, где одна или несколько из указанных гидроксильных групп необязательно этерифицированы радикалом, независимо выбранным из группы, состоящей из
. В пределах объема настоящего изобретения W в формуле (I) представляет собой углерод ("С"), фосфор ("Р") или серу ("S"); m может равняться 0 или 1; n может равняться 0 или 1; X выбран из группы, состоящей из ОН, CN, O(С1-С4алкил), галогена, С1-С4алкила и С1-С4алкила, замещенного одним, двумя или тремя галогенами или карбонилами; R1 выбран из группы, состоящей из Н, C1-С6алкила и C1-С6алкила, замещенного одной или несколькими гидроксильными группами, где одна или несколько из указанных гидроксильных групп необязательно этерифицированы радикалом, независимо выбранным из группы, состоящей из 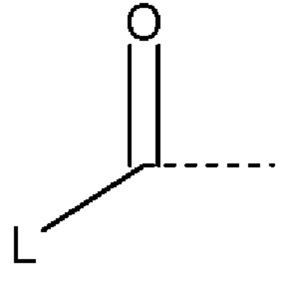 и
и  ; или R1 представляет собой связь с W, если W представляет собой S; каждый L независимо представляет собой углеродную цепь С2-С30, при этом указанная углеродная цепь необязательно содержит одну или несколько групп, независимо выбранных из алкенила, алкинила, фенила и гетероарила, и указанная углеродная цепь необязательно прерывается 1-6 атомами кислорода. Если W представляет собой С и m равняется 1, тогда n равняется 0; однако если W представляет собой С и m равняется 0, тогда n равняется 1.
; или R1 представляет собой связь с W, если W представляет собой S; каждый L независимо представляет собой углеродную цепь С2-С30, при этом указанная углеродная цепь необязательно содержит одну или несколько групп, независимо выбранных из алкенила, алкинила, фенила и гетероарила, и указанная углеродная цепь необязательно прерывается 1-6 атомами кислорода. Если W представляет собой С и m равняется 1, тогда n равняется 0; однако если W представляет собой С и m равняется 0, тогда n равняется 1.
[0028] В предпочтительном варианте осуществления X представляет собой F, Cl, CF3, CCl3, CF2H, CCl2H, CF2CF3, CCl2CCl3, CF2Cl, CF2CH3, С(O)СН3 или CN. В более предпочтительном варианте осуществления галоген представляет собой F или Cl.
[0029] Важно отметить, что в R1 C1-С6алкил включает в себя линейные, разветвленные и циклические алкильные группы. В предпочтительном варианте осуществления R1 представляет собой C1-С6алкил, замещенный одной-шестью гидроксильными группами.
[0030] В одном варианте осуществления каждый L в формуле (I) независимо представляет собой углеродную цепь С2-С30, в том числе разветвленные цепи, которые могут быть насыщенными, ненасыщенными или полиненасыщенными. В предпочтительном варианте осуществления углеродная цепь L содержит от одной до четырех групп, независимо выбранных из алкенила, алкинила, фенила и гетероарила. Ненасыщенность представлена в виде двойных или тройных связей. Алкенил или алкинил могут находиться в пределах углеродной цепи или на конце углеродной цепи. Фенильные и/или гетероарильные кольца могут быть присоединены к углеродной цепи в орто-, мета- или пара-положениях или могут находиться на конце углеродной цепи. Арильные кольца могут быть необязательно замещены. В предпочтительном варианте осуществления углеродная цепь прерывается одним-шестью атомами кислорода. Как используется в данном документе, "прерывается" означает то, что углеродная цепь содержит по меньшей мере два атома углерода в последовательности, за которыми расположен атом кислорода. Например, -СН2-СН2-О-СН2-СН2-СН3 представляет собой углеродную цепь, прерываемую атомом кислорода. В предпочтительном варианте осуществления углеродная цепь прерывается одним-двумя атомами кислорода.
[0031] Примеры подходящих углеродных цепей, соответствующих требованиям L, включают (СН2)8-(СН)2-СН2-(СН)2-СН2-(СН)2-СН2-СН3; (СН2)3-(СН)2-СН2-(СН)2-СН2-(СН)2-СН2-(СН)2-(СН2)4-СН3; (СН2)7-(СН)2-(СН2)7-СН3; (СН2)8-(СН)2-СН2-фенил-СН2-(СН)2-СН2-СН3; (СН2)8-(СН)2-(СН2)2-O-СН2-(СН)2-СН2-СН3 и (СН2)8-(СН)2-СН2-фенил-O-(СН2)3-СН3.
[0032] Обработка этими соединениями осуществляется посредством нанесения соединений с помощью любой из следующих методик: погружения, инъекции, местного применения с использованием пульверизатора, распыления, местного применения с использованием пипетки и местного применения с использованием кисти, а также любого другого вида местного применения. В предпочтительных вариантах осуществления применяют аэрозоль или распылитель.
[0033] Настоящее изобретение дополнительно относится к способу увеличения завязывания семян и снижения недоразвитости зародышей у растений в ходе индукции гаплоидии, включающему обработку репродуктивных тканей растения, таких как рыльца, метелки, пыльца, початки, зерна или другие цветущие ткани, соединением в подходящей концентрации до, во время или после опыления. В одном варианте осуществления соединение выбрано из группы, состоящей из членов из таблицы 7. В другом варианте осуществления соединение представляет собой метил-альфа-линоленоилфторфосфонат (MALFP). В другом варианте осуществления соединение представляет собой линолевую кислоту (LLA), этиловый сложный эфир линолевой кислоты (LLAEE), линоленовую кислоту (LNA), дистеароилфосфатидилхолин (DSPC) или метиларахидонилфторфосфонат (MAFP).
[0034] Настоящее изобретение дополнительно относится к способу увеличения уровня индукции гаплоидии у растения, включающему нанесение липидной композиции на ткани растения непосредственно до, во время или непосредственно после опыления. В одном варианте осуществления растение является однодольным или двудольным растением; или растение представляет собой растение маиса или растение риса. В другом варианте осуществления липид выступает в качестве ингибитора фосфолипаз и/или ингибитора десатураз жирных кислот. В другом варианте осуществления липид представляет собой жирную кислоту (например, LLA) или сложный эфир жирной кислоты (например, LLAEE) с определенной длиной цепи и степенью насыщенности (восемнадцать атомов углерода и две двойные связи), относящиеся к классу длин цепей жирных кислот, не представленному в пыльце, индуцирующей гаплоидию. В целях иллюстрации, а не ограничения, липид представляет собой, например, ингибитор фосфолипаз метил-альфа-линоленоилфторфосфонат (MALFP), растворенный в буферном растворе DMSO при концентрациях MALFP от 0,0001 мг/мл до 1 г/мл или растворенный в составе на основе поверхностно-активных веществ и затем эмульгированный в буферном растворе диметилацетамида (DML) при концентрациях MALF от 0,0001 мг/мл до 1 г/мл. В целях дополнительной иллюстрации, а не ограничения, липидную композицию наносят посредством погружения, инъекции, опрыскивания, разбрызгивания, распыления, полива, нанесения кистью или любого другого способа нанесения на репродуктивные ткани растения. В одном варианте осуществления липидную композицию объединяют с пыльцой в смесь, и эту смесь затем наносят на ткани растения. В другом варианте осуществления смесь наносят на репродуктивные ткани растения, например, на пыльцу или рыльца растения маиса.
[0035] Настоящее изобретение относится к способу индуцирования de novo индукции гаплоидии у растения, включающему введение липидного соединения по меньшей мере в репродуктивную ткань растения во время опыления, перед опылением или после опыления. В одном варианте осуществления растение выбрано из группы, состоящей из однодольных и двудольных растений. В другом варианте осуществления растение выбрано из группы, состоящей из риса, маиса, пшеницы, сорго, томата, сахарной свеклы, проса, ячменя, сои, подсолнечника, хлопчатника, разновидностей овса, табака, овощных культур, плодовых культур и любых других культурных растений.
[0036] В соответствии с одним иллюстративным вариантом осуществления настоящее изобретение включает растение маиса или риса, способное индуцировать гаплоидию в результате индуцированной человеком мутации в гене пататин-подобной фосфолипазы AIIα ("PLA"), а также семена, пыльцу, части растения и потомство этого растения.
[0037] В соответствии с еще одним другим иллюстративным вариантом осуществления настоящее изобретение включает растение маиса или риса, способное индуцировать образование гаплоидов, созданное посредством стадий получения растительного материала из родительского растения маиса или риса, индуцирования по меньшей мере одной мутации по меньшей мере в одной копии гена PLA растительного материала посредством обработки растительного материала мутагеном с созданием растительного материала, подвергнутого мутагенезу, культивирования растительного материала, подвергнутого мутагенезу, с получением растений-потомков риса или маиса, анализа растений-потомков риса или маиса для выявления по меньшей мере одной мутации по меньшей мере в одной копии гена PLA, отбора растений-потомков риса или маиса, которые обладают способностью индуцировать образование гаплоидов, по сравнению с родительским растением риса или маиса; и повторения цикла культивирования растений-потомков риса или маиса с получением дополнительных растений-потомков, обладающих способностью индуцировать образование гаплоидов.
КРАТКОЕ ОПИСАНИЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ В ПЕРЕЧНЕ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0038] SEQ ID NO: 1 представляет собой последовательность кДНК немутантной фосфолипазы, обнаруживаемую в GRMZM2G471240-NIL. Немутантный аллель гена фосфолипазы в данном документе переименован в MATRILINEAL.
[0039] SEQ ID NO: 2 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 1.
[0040] SEQ ID NO: 3 представляет собой нуклеотидную последовательность кДНК мутантной фосфолипазы, обнаруживаемую в GRMZM2G471240-mtl, содержащую вставку из 4 пар оснований. Мутантный аллель гена фосфолипазы в данном документе переименован в matrilineal.
[0041] SEQ ID NO: 4 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 3.
[0042] SEQ ID NO: 5 представляет собой праймер GRMZM2G471240_nil.F1.
[0043] SEQ ID NO: 6 представляет собой праймер GRMZM2G471240_nil.R1.
[0044] SEQ ID NO: 7 представляет собой праймер GRMZM2G471240_rwk.F1.
[0045] SEQ ID NO: 8 представляет собой праймер GRMZM2G471240_rwk.R1.
[0046] SEQ ID NO: 9 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-3954, представляющего собой объект 39А.
[0047] SEQ ID NO: 10 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-3924, представляющего собой объект 23 А.
[0048] SEQ ID NO: 11 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL у индивидуума Т1 с ID 22808-3932, представляющего собой объект 81А, индивидуума с ID 22808-3317, представляющего собой объект 81А, и индивидуума с ID 22808-3303, представляющего собой объект 81 А.
[0049] SEQ ID NO: 12 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 39А с ID 22808-3954.
[0050] SEQ ID NO: 13 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 23А с ID 22808-3924.
[0051] SEQ ID NO: 14 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22808-4108, представляющего собой объект 38А.
[0052] SEQ ID NO: 15 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22807-4016, представляющего собой объект 18А.
[0053] SEQ ID NO: 16 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22807-4073, представляющего собой объект 27А.
[0054] SEQ ID NO: 17 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 1 у индивидуума Т1 с ID 22807-4081, представляющего собой объект 27А.
[0055] SEQ ID NO: 18 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22873-3999, представляющего собой объект 76А.
[0056] SEQ ID NO: 19 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL у индивидуума Т1 с ID 22873-3991, представляющего собой объект 32А.
[0057] SEQ ID NO: 20 представляет собой нуклеотидную последовательность направляющей CRISPR-PHK.
[0058] SEQ ID NO: 21 представляет собой геномную нуклеотидную последовательность Os03g27610, ортолога PLA2 риса.
[0059] SEQ ID NO: 22 представляет собой последовательность кДНК для SEQ ID NO: 21.
[0060] SEQ ID NO: 23 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 22.
[0061] SEQ ID NO: 24 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-B73.
[0062] SEQ ID NO: 25 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-RWK.
[0063] SEQ ID NO: 26 представляет собой нуклеотидную последовательность немутантного GRMZM2G471240-ST6.
[0064] SEQ ID NO: 27 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 24.
[0065] SEQ ID NO: 28 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 25.
[0066] SEQ ID NO: 29 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 26.
[0067] SEQ ID NO: 30 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22466, содержащую MATRILINEAL дикого типа.
[0068] SEQ ID NO: 31 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22467, содержащую ген фосфоглицератмутазы дикого типа.
[0069] SEQ ID NO: 32 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22503, содержащей последовательность, кодирующую структуру типа "стебель-петля", нацеленную на экзон 2MATRILINEAL.
[0070] SEQ ID NO: 33 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22513, содержащую последовательность, кодирующую структуру типа стебель-петля, нацеленную на экзон 4MATRILINEAL.
[0071] SEQ ID NO: 34 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22807, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0072] SEQ ID NO: 35 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22808, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0073] SEQ ID NO: 36 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 22873, содержащую последовательности, кодирующие аппарат редактирования CRISPR/Cas9, нацеленный на MATRILINEAL в NP2222.
[0074] SEQ ID NO: 37 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23123, содержащую последовательности, кодирующие аппарат редактирования TALEN, нацеленный на MATRILINEAL в NP2222.
[0075]SEQ ID NO: 38 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с двумя направляющими последовательностями gRNA, нацеленными на экзон 4 риса.
[0076] SEQ ID NO: 39 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с одной направляющей последовательностью gRNA, нацеленной на экзон 4 риса.
[0077] SEQ ID NO: 40 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с двумя направляющими последовательностями gRNA, нацеленными на экзон 1 риса.
[0078] SEQ ID NO: 41 представляет собой нуклеотидную последовательность кассеты экспрессии конструкции 23501 с одной направляющей последовательностью gRNA, нацеленной на экзон 1 риса.
[0079] SEQ ID NO: 42 представляет собой нуклеотидную последовательность с TALEN-индуцированной мутацией MTL в аллеле 2 у объекта 38А с ID 22808-4108.
[0080] SEQ ID NO: 43 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 2 у объекта 27А с ID 22807-4073.
[0081] SEQ ID NO: 44 представляет собой нуклеотидную последовательность с CRISPR-индуцированной мутацией MTL в аллеле 2 у объекта 27А с ID 22807-4081.
[0082] SEQ ID NO: 45 представляет собой нуклеотидную последовательность линии 1139, полученной с помощью TILLING.
[0083] SEQ ID NO: 46 представляет собой нуклеотидную последовательность линии 3594, полученной с помощью TILLING.
[0084] SEQ ID NO: 47 представляет собой нуклеотидную последовательность линии 0505, полученной с помощью TILLING.
[0085] SEQ ID NO: 48 представляет собой нуклеотидную последовательность линии 2658, полученной с помощью TILLING.
[0086] SEQ ID NO: 49 представляет собой нуклеотидную последовательность линии 1983, полученной с помощью TILLING.
[0087] SEQ ID NO: 50 представляет собой нуклеотидную последовательность линии 2732, полученной с помощью TILLING.
[0088] SEQ ID NO: 51 представляет собой нуклеотидную последовательность линии 2414, полученной с помощью TILLING.
[0089] SEQ ID NO: 52 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 45.
[0090] SEQ ID NO: 53 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 46.
[0091] SEQ ID NO: 54 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 47.
[0092]SEQ ID NO: 55 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 48.
[0093] SEQ ID NO: 56 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 49.
[0094] SEQ ID NO: 57 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 50.
[0095]SEQ ID NO: 58 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 51.
[0096] SEQ ID NO: 59 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 9.
[0097] SEQ ID NO: 60 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 10.
[0098] SEQ ID NO: 61 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 11.
[0099] SEQ ID NO: 62 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 12.
[00100] SEQ ID NO: 63 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 13.
[00101] SEQ ID NO: 64 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 14.
[00102] SEQ ID NO: 65 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 15.
[00103] SEQ ID NO: 66 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 16.
[00104] SEQ ID NO: 67 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 17.
[00105] SEQ ID NO: 68 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 18.
[00106] SEQ ID NO: 69 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 19.
[00107] SEQ ID NO: 70 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 42.
[00108] SEQ ID NO: 71 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 43.
[00109] SEQ ID NO: 72 представляет собой аминокислотную последовательность, кодируемую SEQ ID NO: 44.
[00110] SEQ ID NO: 73 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Sorghum bicolor.
[00111] SEQ ID NO: 74 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Setaria italica.
[00112] SEQ ID NO: 75 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Hordeum vulgare.
[00113] SEQ ID NO: 76 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Brachypodium distachyon.
[00114] SEQ ID NO: 77 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Oryza sativa v. indica.
[00115] SEQ ID NO: 78 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Triticum aestivum.
[00116] SEQ ID NO: 79 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого b Musa acuminata.
[00117] SEQ ID NO: 80 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Elaeis guineensis.
[00118] SEQ ID NO: 81 представляет собой аминокислотную последовательность ортолога MTL, обнаруживаемого в Arabidopsis thaliana.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00119] На фигуре 1 представлена схема картирования, применяемая для картирования признака индукции гаплоидии в RWK.
[00120] На фигуре 2 показано тонкое картирование, ограничивающее главный QTL очень небольшим интервалом в секции 1.04 от 67,85 млн п.о. до 68,42 млн п.о. Эта область имеет семь аннотированных генов. Авторы настоящего изобретения секвенировали и собирали гены в этом интервале в нескольких линиях. Два гена с наиболее значительными мутациями в линиях, индуцирующих гаплоидию, показаны в правой нижней части (GRMZM2G471240 и GRMZM2G062320).
[00121] На фигуре 3 показаны различия в экспрессии GRMZM2G471240 в пыльце, индуцирующей и не индуцирующей гаплоидию, и пыльцевых мешках после цветения (спорофитной ткани с удаленными пыльцевыми зернами). Этот ген, в частности, экспрессируется в мужском гаметофите.
[00122] На фигуре 4а показаны результаты сплайс-специфической qRT-PCR для GRMZM2G471240. Три биологические реплики пыльников на стадии R1 тестировали в трех технических повторностях, и для каждой реакции рассчитывали среднее значение Ct и среднеквадратическое отклонение. Относительное количество каждого типа транскрипта сравнивали с эндогенным контролем с применением log2-регрессии дельта Ct. Два набора праймеров применяли для оценки относительной представленности каждого из двух аннотированных сплайс-вариантов по сравнению с набором праймеров, инвариантным относительно сплайс-вариантов. Более короткий вариант транскрипта по сравнению с длинным транскриптом характеризуется относительно низкой представленностью в генотипах как NP2222 (дикий тип), так и NP2222-HI (индуктор гаплоидии). Экспрессия мутантных копий гена в NP2222-HI была значительно более высокой для всех трех тестируемых пар праймеров.
[00123] На фигуре 4b показано пять биологических реплик свежей пыльцы от растений NP2222 и MTLTAL-FS (растений Т1, которые являются гомозиготными по отредактированным mtl-подобным аллелям), которые тестировали в трех технических повторах с универсальным праймером, и для каждой реакции рассчитывали среднее значение Ct и среднеквадратическое отклонение. Относительное количество каждого типа транскрипта сравнивали с эндогенным контролем с применением log2-регрессии дельта Ct. Пыльца MTLTAL-FS характеризуется более низкой представленностью транскриптов, чем пыльца NP2222 (дикого типа).
[00124] На фигуре 5а показано выравнивание предсказанной аминокислотной последовательности белка В73, кодируемой длинным сплайс-вариантом гена GRMZM2G471240 в В73 и RWK-NIL, с предсказанной последовательностью, кодируемой аллелем mil, обнаруживаемым в RWK и Stock 6 (S6). Аминокислоты, которые отличаются, указаны красным цветом; аминокислоты, которые совпадают, указаны обычным серым текстом, а стоп-кодоны указаны точкой. Две точечные мутации приводят в результате к аминокислотным заменам: гистидина (Н) на тирозин (Y), а лизина (K) на аргинин (N). Эти изменения не являются консервативными; возможно, что одно или оба из них модифицируют фенотип индукции гаплоидии что позволяет предположить, что аллельная серия может быть обнаружена при дополнительном исследовании вариантов.
[00125] На фигуре 5b показано, что MTL дикого типа и мутантный (усеченный) MTL, кодируемый аллелем mil, характеризуются фосфолипазной активностью in vitro. Фосфолипазную активность PLA2 измеряли посредством флуоресцентного липосомного анализа рекомбинантного очищенного белка, полученного с применением кДНК MTL и mtl. Планки погрешностей указывают на среднеквадратическую погрешность, вычисленную по среднему значению для четырех повторностей.
[00126] На фигуре 6 показано, что mtl отвечает за плейотропные фенотипы, ассоциированные с индукцией гаплоидии. 6А: скорость прорастания пыльцевой трубки была сходной у индукторов и неиндукторов (n=200). 6 В: первоначальное удлинение пыльцевой трубки также было сходным (n=25). 6С: RWK, но не RWK-NIL, подвергается нарушению сегрегации (SD), как следует из низкой (25%) степени передачи признаков прорастающему потомству (n=300). 6D: Комплементационные линии MTL/0 также демонстрируют SD в отношении mtl у прорастающего потомства (n=400). 6Е: диаграмма Венна, на которой показаны результаты профилирования двух пар почти изогенных линий, индуцирующих гаплоидию, методом секвенирования РНК (левая часть - RWK по сравнению с RWK-NIL; правая часть - NP2222-HI по сравнению с NP2222; красный текст -активация; зеленый текст - подавление). Было обнаружено, что только 60 генов значительно изменились в одном и том же направлении.
[00127] На фигуре 7 показано выравнивание аминокислотных последовательностей, кодируемых геном MTL маиса и имеющимися в открытом доступе ортологами MTL восьми видов злаковых растений, двух однодольных растений, отличных от злаковых, и Arabidopsis (резуховидки Таля). Это выравнивание охватывает маис (Zea mays), сорго (Sorghum bicolor, 92% идентичность последовательностей с MTL), итальянское просо (Setaria italica, 85% идентичность), ячмень (Hordeum vulgare, 78% идентичность), Brachypodium distachyon (78% идентичность), индийский и японский сорта риса (Oryza sativa v. indica и japonica, Os3g27610, 78% и 79% идентичность соответственно), пшеницу обыкновенную (Triticum aestivum, 55% идентичность), банан (Musa acuminata, 57% идентичность), масличную пальму (Elaeis guineensis, 56% идентичность) и Arabidopsis thaliana (52% идентичность).
[00128] Фигура 8. Профиль экспрессии фосфолипаз риса (по материалам Singh, A., et al., Rice phospholipase A superfamily: organization, phylogenetic and expression analysis during abiotic stresses and development, PLoS One 7: e30947 (2012)). Ближайшим гомологом MTL является ген риса OspPLAIIα Os3g27610).
[00129] Фигура 9. Схема, на которой показан путь редактирования Os3g27610 для получения линий, индуцирующих гаплоидию. Можно целенаправленно воздействовать на любую часть гена (в данном документе показано целенаправленное воздействие на первый и четвертый экзоны) и ожидать создания мутаций сдвига рамки считывания, которые приведут к нокауту и утрате функции гена, что приведет к индукции гаплоидии.
[00130] На фигуре 10 показана атомная структура метил-альфа-линоленоилфторфосфоната (MALFP).
[00131] На фигуре 11 показана атомная структура метиларахидонилфторфосфоната (MAFP).
[00132] На фигуре 12 показана атомная структура пальмитилтрифторметилкетона (PACOCF3).
[00133] На фигуре 13 показана атомная структура арахидонилтрифторметилкетона (AACOCF3).
[00134] На фигуре 14 показана атомная структура маноалида.
[00135] На фигуре 15 показана атомная структура этилового сложного эфира линолевой кислоты (LLAEE).
[00136] На фигуре 16 показана атомная структура этилового сложного эфира линоленовой кислоты (LNAEE).
[00137] На фигуре 17 показана атомная структура метилового сложного эфира арахидоновой кислоты (ААМЕ).
[00138] На фигуре 18 показана атомная структура метилового сложного эфира олеиновой кислоты (ОАМЕ).
[00139] На фигуре 19 показана атомная структура этилового сложного эфира олеиновой кислоты (ОАЕЕ).
[00140] На фигуре 20 показана атомная структура этилового сложного эфира пальмитиновой кислоты (РАЕЕ).
[00141] На фигуре 21 показана атомная структура этилового сложного эфира пальмитолеиновой кислоты (PLAEE).
[00142] На фигуре 22 показана атомная структура альфа-линоленовой кислоты (aLNA).
[00143] На фигуре 23 показана атомная структура гамма-линоленовой кислоты (gLNA).
[00144] На фигуре 24 показана атомная структура олеиновой кислоты.
[00145] На фигуре 25 показана атомная структура линолевой кислоты.
[00146] На фигуре 26 показана атомная структура арахидоновой кислоты.
[00147] На фигуре 27 показана атомная структура стеариновой кислоты.
[00148] На фигуре 28 показана атомная структура 9(Z)-11(Е)-сопряженной линолевой кислоты.
[00149] На фигуре 29 показана атомная структура дистеароилфосфатидилхолина (DSPC).
[00150] На фигуре 30 показана атомная структура 2-олеоил-1-пальмитоил-sn-глицеро-3-фосфоэтаноламина.
[00151] На фигуре 31 показана общая атомная структура молекул, пригодных к использованию в заявленном изобретении.
ОПРЕДЕЛЕНИЯ
[00152] Хотя следующие термины, как предполагается, хорошо понятны среднему специалисту в данной области, следующие определения приведены для облегчения пояснения раскрытого в данном документе объекта изобретения.
[00153] Все используемые в данном документе технические и научные термины, если ниже не указано иное, предполагаются как имеющие такое же значение, которое обычно понятно среднему специалисту в данной области. Ссылки на используемые в данном документе методики предполагаются как относящиеся к методикам, общепринятым в данной области техники, в том числе к изменениям этих методик и/или заменам на эквивалентные методики, которые будут очевидны специалистам в данной области. Хотя следующие термины, как предполагается, хорошо понятны среднему специалисту в данной области, следующие определения приведены для облегчения пояснения раскрытого в данном документе объекта изобретения.
[00154] Согласно устоявшейся традиции в патентном праве формы существительного единственного числа относятся к "одному или нескольким" при использовании в настоящей заявке, в том числе в формуле изобретения. Например, фраза "клетка" относится к одной или нескольким клеткам и в некоторых вариантах осуществления может относиться к ткани и/или органу. Аналогичным образом, фраза "по меньшей мере один" при использовании в данном документе для обозначения объекта относится, например, к 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 15, 20, 25, 30, 35, 40, 45, 50, 75, 100 или большему количеству этих объектов, включая без ограничения все целочисленные значения от 1 до 100, а также целые числа больше 100.
[00155] Если не указано иное, все числа, выражающие количества ингредиентов, условия реакции и т.д., используемые в настоящем описании и формуле изобретения, следует понимать как модифицированные во всех случаях термином "приблизительно". Термин "приблизительно", используемый в данном документе в отношении измеряемого значения, такого как величина массы, веса, времени, объема, концентрации или процентного значения, предназначено для охвата изменений определенного количества, составляющих в некоторых вариантах осуществления ±20%, в некоторых вариантах осуществления ±10%, в некоторых вариантах осуществления ±5%, в некоторых вариантах осуществления ±1%, в некоторых вариантах осуществления ±0,5% и в некоторых вариантах осуществления ±0,1%, поскольку такие изменения подходят для осуществления раскрытых способов и/или использования раскрытых композиций, нуклеиновых кислот, полипептидов и т.д. Соответственно, если не указано обратное, числовые параметры, изложенные в настоящем описании и прилагаемой формуле изобретения, являются приблизительными величинами, которые могут варьироваться в зависимости от желаемых свойств, которых стремятся достичь посредством раскрытого в данном документе объекта изобретения.
[00156] Используемый в данном документе термин "аллель" относится к варианту или альтернативной форме последовательности в генетическом локусе. У диплоидов отдельный аллель наследуется индивидуумом-потомком от каждой родительской особи отдельно в каждом локусе. Два аллеля данного локуса, присутствующие в диплоидном организме, занимают соответствующие места в паре гомологичных хромосом, хотя специалист в данной области понимает, что аллели в любом конкретном индивидууме необязательно представляют все аллели, которые присутствуют у данного вида.
[00157] Используемый в данном документе термин "и/или" при использовании применительно к перечню объектов относится к объектам, присутствующим в отдельности или в комбинации. Таким образом, например, фраза "А, В, С и/или D" включает А, В, С и D отдельно, но также включает все возможные комбинации и подкомбинации А, В, С и D (например, АВ, AC, AD, ВС, BD, CD, ABC, ABD и BCD). В некоторых вариантах осуществления один или несколько элементов, к которым относится "и/или", может также отдельно присутствовать в однократных или многократных случаях в комбинации (комбинациях) и/или подкомбинации(подкомбинациях).
[00158] Используемая в данном документе фраза "ассоциированный с" относится к распознаваемой и/или анализируемой взаимосвязи между двумя объектами. Например, фраза "ассоциированный с HI" относится к признаку, локусу, гену, аллелю, маркеру, фенотипу и т.д. или их экспрессии, присутствие или отсутствие которых может влиять на меру и/или степень, в которой растение или его потомство демонстрируют HI. Соответственно, маркер является "ассоциированным с" признаком, если он сцеплен с ним и если наличие маркера является индикатором того, будут ли и/или в какой степени будут желаемый признак или форма признака встречаться у растения/в зародышевой плазме, содержащих маркер. Аналогичным образом, маркер является "ассоциированным с" аллелем, если он сцеплен с ним и если наличие маркера является индикатором того, присутствует ли аллель у растения/в зародышевой плазме, содержащих маркер. Например, "маркер, ассоциированный с HI", относится к маркеру, наличие или отсутствие которого можно применять для предсказания того, будет ли и/или в какой степени будет растение проявлять индукцию гаплоидии.
[00159] Термин "содержащий", который является синонимом "включающего", "вмещающего" и "характеризующегося", является включающим или открытым и не исключает дополнительных, не упомянутых элементов и/или стадий способа. "Содержащий" является термином из уровня техники, который означает, что названные элементы и/или стадии присутствуют, но могут быть добавлены другие элементы и/или стадии, которые при этом по-прежнему находятся в пределах объема соответствующего объекта изобретения.
[00160] Используемая в данном документе фраза "состоящий из" исключает любой элемент, стадию или ингредиент, не упомянутые конкретно. Если фраза "состоит из" появляется в основной части пункта формулы изобретения, а не сразу после ограничительной части, то она ограничивает только элемент, изложенный в этой части; другие элементы не исключаются из пункта формулы изобретения в целом.
[00161] Используемая в данном документе фраза "состоящий фактически из" ограничивает объем связанного раскрытия или пункта формулы изобретения указанными материалами и/или стадиями, а также материалами и/или стадиями, которые существенно не влияют на основную (основные) и новую (новые) характеристику (характеристики) раскрытого и/или заявленного объекта изобретения.
[00162] Что касается терминов "содержащий", "состоящий фактически из" и "состоящий из", если в данном документе используется один из этих трех терминов, то раскрытый в данном документе и заявленный объект изобретения может включать в некоторых вариантах осуществления применение любого из двух других терминов. Например, это справедливо, если объект изобретения относится в некоторых вариантах осуществления к нуклеиновым кислотам, кодирующим полипептиды, содержащие аминокислотные последовательности, которые по меньшей мере на 95% идентичны SEQ ID NO: 2 или 3. Следует понимать, что раскрытый объект изобретения, таким образом, также охватывает нуклеиновые кислоты, кодирующие полипептиды, которые в некоторых вариантах осуществления состоят фактически из аминокислотных последовательностей, по меньшей мере на 95% идентичных этим SEQ ID NO: 2 или 3, а также нуклеиновые кислоты, кодирующие полипептиды, которые в некоторых вариантах осуществления состоят из аминокислотных последовательностей, по меньшей мере на 95% идентичных этим SEQ ID NO: 2 или 3. Аналогичным образом, также следует понимать, что в некоторых вариантах осуществления способы согласно раскрытому объекту изобретения включают стадии, раскрытые в данном документе, в некоторых вариантах осуществления способы согласно раскрытому в данном документе объекту изобретения состоят фактически из раскрытых стадий, и в некоторых вариантах осуществления способы согласно раскрытому в данном документе объекту изобретения состоят из стадий, раскрытых в данном документе.
[00163] Используемый в данном документе термин "индукция гаплоидии de novo" относится к вызыванию индукции гаплоидии посредством введения средства, индуцирующего спонтанную гаплоидию. Такое введение можно осуществлять посредством местного опрыскивания, опыления ручным способом, мутагенеза или трансгенных способов. Термины "индукция гаплоидии de novo", "HI de novo" и "de novo индукция гаплоидии" используются взаимозаменяемо во всем настоящем описании.
[00164] Используемый в данном документе термин "ген" относится к единице наследственности, содержащей последовательность ДНК, которая занимает определенное местоположение в хромосоме и которая содержит генетическую инструкцию для определенной характеристики или признака, свойственных организму.
[00165] "Генетическая карта" представляет собой описание взаимосвязей при генетическом сцеплении между локусами в одной или нескольких хромосомах у данного вида, обычно изображаемое в форме схемы или таблицы.
[00166] Как используется в данном документе, растение, называемое "гаплоидным", имеет одинарный набор (геном) хромосом, и уменьшенное количество хромосом (n) в гаплоидном растении равно количеству хромосом в гамете. Как используется в данном документе, растение, называемое "двойным гаплоидным", развивается посредством удвоения гаплоидного набора хромосом. Растение или семя, которые получают из двойного гаплоидного растения, самооплодотворяемого в течение любого количества поколений, можно по-прежнему идентифицировать как двойное гаплоидное растение. Двойное гаплоидное растение считается гомозиготным растением. Растение считается двойным гаплоидным, если оно является фертильным, даже если вся вегетативная часть растения состоит из клеток не с двойным набором хромосом; то есть растение будет считаться двойным гаплоидным, если оно содержит жизнеспособные гаметы, даже если оно является химерным.
[00167] Используемый в данном документе термин "индуцированная человеком мутация" относится к любой мутации, появляющейся в результате непосредственного либо опосредованного действия человека. Этот термин включает без ограничения, мутации, получаемые посредством любого способа направленного мутагенеза.
[00168] Используемые в данном документе термины "маркерный зонд" и "зонд" относятся к нуклеотидной последовательности или молекуле нуклеиновой кислоты, которые можно применять для выявления наличия или отсутствия последовательности в пределах большей последовательности посредством гибридизации нуклеиновых кислот, например, к зонду на основе нуклеиновой кислоты, комплементарному всему маркеру или маркерному локусу или его части. Для гибридизации нуклеиновых кислот можно применять маркерные зонды, содержащие приблизительно 8, 10, 15, 20, 30, 40, 50, 60, 70, 80, 90, 100 или более смежных нуклеотидов.
[00169] Используемый в данном документе термин "молекулярный маркер" можно использовать для обозначения генетического маркера, определенного выше, или кодируемого им продукта (например, белка), применяемого в качестве исходной точки при идентификации наличия/отсутствия HI-ассоциированного локуса. Молекулярный маркер может быть получен из геномных нуклеотидных последовательностей или из экспрессируемых нуклеотидных последовательностей (например, из РНК, кДНК и т.д.). Данный термин также относится к нуклеотидным последовательностям, комплементарным маркерным последовательностям или фланкирующим их, таким как нуклеотидные последовательности, применяемые в качестве зондов и/или праймеров, способных амплифицировать маркерную последовательность. Нуклеотидные последовательности являются "комплементарными", если они специфически гибридизируются в растворе (например, согласно правилам спаривания оснований Уотсона-Крика). Данный термин также относится к генетическим маркерам, которые указывают на признак посредством отсутствия нуклеотидных последовательностей, комплементарных маркерным последовательностям или фланкирующих их, таких как нуклеотидные последовательности, применяемые в качестве зондов и/или праймеров, способных амплифицировать маркерную последовательность.
[00170] Используемые в данном документе термины "нуклеотидная последовательность", "полинуклеотид", "последовательность нуклеиновой кислоты", "молекула нуклеиновой кислоты" и "фрагмент нуклеиновой кислоты" относятся к полимеру РНК или ДНК, который является одно- или двухнитевым, необязательно содержащему синтетические, неприродные и/или измененные нуклеотидные основания. "Нуклеотид" представляет собой мономерную единицу, из которой сконструированы полимеры ДНК или РНК и которая состоит из пуринового или пиримидинового основания, пентозы и группы фосфорной кислоты. Нуклеотиды (обычно обнаруживаемые в форме их 5'-монофосфата) называют их однобуквенным обозначением следующим образом: "А" в случае аденилата или дезоксиаденилата (соответственно в случае РНК или ДНК), "С" в случае цитидилата или дезоксицитидилата, "G" в случае гуанилата или дезоксигуанилата, "U" в случае уридилата, "Т" в случае дезокситимидилата, "R" в случае пуринов (А или G), "Y" в случае пиримидинов (С или Т), "K" в случае G или Т, "Н" в случае А, или С, или Т, "I" в случае инозина и "N" в случае любого возможного нуклеотида.
[00171] Используемый в данном документе термин "идентичность нуклеотидных последовательностей" относится к наличию идентичных нуклеотидов в соответствующих положениях двух полинуклеотидов. Полинуклеотиды имеют "идентичные" последовательности в случае, если последовательность нуклеотидов в двух полинуклеотидах является одной и той же при выравнивании с максимальным соответствием (например, в окне сравнения). Сравнение последовательностей двух или более нуклеотидов обычно осуществляют путем сравнения частей двух последовательностей в окне сравнения для выявления и сравнения локальных областей сходства последовательностей. Как правило, окно сравнения составляет от приблизительно 20 до 200 смежных нуклеотидов. "Процентное значение идентичности последовательностей" для полинуклеотидов, как, например, приблизительно 50-, 55-, 60-, 65-, 70-, 75-, 80-, 85-, 90-, 95-, 98-, 99-или 100-процентную идентичность последовательностей, можно определять путем сравнения двух оптимально выровненных последовательностей в окне сравнения, где часть полинуклеотидной последовательности в окне сравнения может содержать добавления или делеции (т.е. гэпы) по сравнению с эталонной последовательностью для оптимального выравнивания двух последовательностей. В некоторых вариантах осуществления процентное значение рассчитывают путем (а) определения числа положений, в которых идентичное основание нуклеиновой кислоты встречается в обеих последовательностях; (b) деления числа совпадающих положений на общее число положений в окне сравнения и (с) умножения результата на 100. Оптимальное выравнивание последовательностей для сравнения можно также проводить с помощью компьютерных реализаций известных алгоритмов или путем визуального осмотра. Общедоступными алгоритмами сравнения последовательностей и множественного выравнивания последовательностей являются соответственно средство поиска основного локального выравнивания (BLAST) и программы ClustalW/ClustalW2/Clustal Omega, доступные в Интернете (например, на веб-сайте EMBL-EBI). Другие подходящие программы включают без ограничений GAP, BestFit, PlotSimilarity и FASTA, которые являются частью пакета программ Accelrys GCG, доступного от Accelrys, Inc. Сан-Диего, Калифорния, Соединенные Штаты Америки. См. также Smith & Waterman, 1981; Needleman & Wunsch, 1970; Pearson & Lipman, 1988; Ausubel et al., 1988 и Sambrook & Russell, 2001.
[00172] Одним примером алгоритма, который подходит для определения процентного значения идентичности последовательностей и сходства последовательностей, является алгоритм BLAST, который описан в Altschul et al., 1990. В некоторых вариантах осуществления процентное значение идентичности последовательностей относится к идентичности последовательностей по всей длине одной из сравниваемых последовательностей gDNA, кДНК или предсказываемого белка в наибольшей ORF SEQ ID NO: 1. В некоторых вариантах осуществления при расчете с целью определения процентного значения идентичности последовательностей нуклеиновых кислот не включаются в расчет какие-либо положения нуклеотидов, в которых любая из сравниваемых нуклеиновых кислот содержит "N" (т.е. где в данном положении может находиться любой нуклеотид).
[00173] Термин "открытая рамка считывания" (ORF) относится к последовательности нуклеиновой кислоты, которая кодирует полипептид. В некоторых вариантах осуществления ORF содержит кодон инициации трансляции, кодон терминации трансляции (т.е. стоп-кодон) и последовательность нуклеиновой кислоты между ними, которая кодирует аминокислоты, присутствующие в полипептиде. Термины "инициирующий кодон" и "терминирующий кодон" относятся к единице из трех прилегающих нуклеотидов (т.е. кодону) в кодирующей последовательности, которая определяет соответственно инициацию и терминацию цепи в синтезе белка (трансляции мРНК).
[00174] Пататин-подобная фосфолипаза А2α может также быть известна как PLA, pPLA, pPLAIIA, pPLAIIα, PbA2-альфа или PLA2 или другие сходные варианты. Пататин-подобную фосфолипазу AIIα также называют MATRILINEAL. Данные термины везде используются взаимозаменяемо. Ген MATRILINEAL, содержащий мутацию сдвига рамки считывания, охватывающую четыре пары оснований, в данном документе называется matrilineal.
[00175] Используемые в данном документе термины "фенотип", "фенотипический признак" или "признак" относятся к одному или нескольким признакам растения или растительной клетки. Фенотип можно наблюдать невооруженным глазом или с помощью любых других средств оценки, известных в данной области техники, например, микроскопии, биохимического анализа или электромеханического анализа. В некоторых случаях фенотип непосредственно контролируется одним геном или генетическим локусом (т.е. соответствует "признаку, определенному одним геном"). В случае индукции гаплоидии применение маркеров цвета, таких как R-Navajo, и других маркеров, в том числе трансгенов, визуализируемых по наличию или отсутствию цвета в семени, дает свидетельство того, является ли семя семенем с индуцированной гаплоидией. Применение R-Navajo в качестве маркера цвета и применение трансгенов хорошо известно из уровня техники в качестве способов выявления индукции образования гаплоидных семян на женском растении. В других случаях фенотип является результатом взаимодействий между несколькими генами, и в некоторых вариантах осуществления он также являются результатом взаимодействия растения и/или растительной клетки с их окружающей средой.
[00176] Используемый в данном документе термин "растение" может относиться к целому растению, любой его части или культуре клеток или тканей, полученным из растения. Таким образом, термин "растение" может относиться к любому из целых растений, компонентов или органов растения (например, листьев, стеблей, корней и т.д.), растительных тканей, семян и/или растительных клеток.
[00177] Растительная клетка представляет собой клетку растения, взятую из растения или полученную посредством культивирования из клетки, взятой из растения. Таким образом, термин "растительная клетка" включает без ограничения клетки в семенах, суспензионных культурах, зародышах, областях меристемы, каллюсной ткани, листьях, побегах, гаметофитах, спорофитах, пыльце и микроспорах. Фраза "часть растения" относится к части растения, в том числе к отдельным клеткам и клеточным тканям, таким как растительные клетки, являющиеся интактными в растениях, скопления клеток и культуры тканей, из которых можно регенерировать растения. Примеры частей растения включают без ограничения отдельные клетки и ткани из пыльцы, семязачатков, листьев, зародышей, корней, кончиков корней, пыльников, цветков, плодов, стеблей, побегов и семян; а также привоев, подвоев, протопластов, каллюсов и т.п.
[00178] Используемый в данном документе термин "праймер" относится к олигонуклеотиду, который способен отжигаться с нуклеиновой кислотой-мишенью (в некоторых вариантах осуществления специфически отжигаться с нуклеиновой кислотой-мишенью), обеспечивая присоединение к ней ДНК-полимер азы и/или обратной транскриптазы и выступая таким образом в качестве точки инициации синтеза ДНК, будучи помещенным в условия, в которых индуцируется синтез продукта удлинения праймера (например, в присутствии нуклеотидов и средства для полимеризации, такого как ДНК-полимераза, и при подходящих значениях температуры и рН). В некоторых вариантах осуществления для амплификации нуклеиновых кислот растения используют одну или несколько совокупностей праймеров (например, с применением полимеразной цепной реакции; ПНР).
[00179] Используемый в данном документе термин "зонд" относится к нуклеиновой кислоте (например, однонитевой нуклеиновой кислоте или нити двухнитевой нуклеиновой кислоты или нуклеиновой кислоты более высокого порядка или их подпоследовательности), которая может образовывать стабилизированный водородными связями дуплекс с комплементарной последовательностью в последовательности нуклеиновой кислоты-мишени. Как правило, зонд имеет достаточную длину для образования стабильной и специфичной для последовательности дуплексной молекулы с комплементарной ему последовательностью, и, следовательно, может использоваться в некоторых вариантах осуществления для выявления последовательности, представляющей интерес, присутствующей во множестве нуклеиновых кислот.
[00180] Используемые в данном документе термины "потомство" и "растение-потомок" относятся к растению, полученному в результате вегетативного или полового размножения от одного или нескольких родительских растений. При индукции гаплоидии семена на женской родительской особи являются гаплоидными и, следовательно, не являются потомством линии, индуцирующей гаплоидию. Потомство гаплоидных семян не является единственным желаемым потомством. Существуют также семена HI и последующие растения- и семена-потомки растения, индуцирующего гаплоидию. Как гаплоидные семена, так и семена HI могут являться потомством. Растение-потомка можно получать путем клонирования или самооплодотворения одного родительского растения или путем скрещивания двух или более родительских растений. Например, растение-потомка можно получать путем клонирования или самооплодотворения одного родительского растения или путем скрещивания двух родительских растений и включает продукты самооплодотворения, а также F1 или F2 или более далекие поколения. F1 представляет собой потомство первого поколения, полученное от родительских особей, по меньшей мере одну из которых впервые используют в качестве донора признака, тогда как потомство второго поколения (F2) или последующих поколений (F3, F4 и т.п.) представляет собой образцы, полученные в результате самооплодотворений, перекрестных опылений, возвратных скрещиваний и/или других скрещиваний F1, F2 и т.п.F1, таким образом, может представлять собой (и в некоторых вариантах осуществления представляет собой) гибрид, полученный в результате скрещивания двух родительских особей из чистых линий (т.е. каждая из родительских особей из чистых линий является гомозиготной по признаку, представляющему интерес, или его аллелю), тогда как F2 может представлять собой (и в некоторых вариантах осуществления представляет собой) потомство, полученное в результате самоопыления гибридов F1.
[00181] Используемая в данном документе фраза "рекомбинация" относится к обмену фрагментами ДНК между двумя молекулами ДНК или хроматидами парных хромосом ("кроссинговеру") в области сходства или идентичности нуклеотидных последовательностей. "Событие рекомбинации" в данном документе понимают как относящееся в некоторых вариантах осуществления к мейотическому кроссинговеру.
[00182] Используемый в данном документе термин "эталонная последовательность" относится к определенной нуклеотидной последовательности, применяемой в качестве основы для сравнения нуклеотидных последовательностей. В некоторых вариантах осуществления любая из SEQ ID NO: 1-4, 22-23 или 73-81 может выступать в качестве эталонной последовательности для сравнения с другими последовательностями, полученными из растений.
[00183] Используемый в данном документе термин "регенерировать" и его грамматические варианты относятся к получению растения из культуры тканей.
[00184] Используемая в данном документе фраза "жесткие условия гибридизации" относится к условиям, при которых полинуклеотид гибридизируется со своей подпоследовательностью-мишенью, как правило, в сложной смеси нуклеиновых кислот, но фактически не делает этого с другими последовательностями. Жесткие условия зависят от последовательности и могут различаться при разных обстоятельствах.
[00185] Более длинные последовательности, как правило, специфически гибридизируются при более высоких температурах. Исчерпывающее руководство по гибридизации нуклеиновых кислот можно найти в Sambrook & Russell, 2001. Как правило, жесткие условия выбирают так, чтобы температура была на приблизительно 5-10°С ниже температурной точки плавления (Tm) для конкретной последовательности при определенных ионной силе и рН. Tm представляет собой температуру (при определенных ионной силе, рН и концентрации нуклеиновой кислоты), при которой 50% зондов, комплементарных мишени, гибридизируются с последовательностью-мишенью в равновесном состоянии (поскольку последовательности-мишени присутствуют в избытке, то при Tm в равновесном состоянии занято 50% зондов). Иллюстративными жесткими условиями являются условия, при которых концентрация солей составляет менее чем приблизительно 1,0 М ионов натрия, как правило, концентрация ионов натрия (или других солей) составляет приблизительно 0,01-1,0 М при рН 7,0-8,3, а температура составляет по меньшей мере приблизительно 30°С для коротких зондов (например, 10-50 нуклеотидов) и по меньшей мере приблизительно 60°С для длинных зондов (например, более 50 нуклеотидов).
[00186] Жестких условий также можно достичь путем добавления дестабилизирующих средств, таких как формамид. Дополнительные иллюстративные жесткие условия гибридизации включают инкубирование в 50% формамиде, 5х SSC и 1% SDS при 42°С; или инкубирование в SSC, 1% SDS при 65°С; с одной или несколькими стадиями отмывки в 0,2х SSC и 0,1% SDS при 65°С. Для ПЦР температура, составляющая приблизительно 36°С, является типичной для амплификации в условиях низкой жесткости, хотя значения температуры отжига могут варьироваться от приблизительно 32°С до 48°С (или выше) в зависимости от длины праймера. Дополнительные руководства по определению параметров гибридизации представлены во множестве литературных источников (см. например, Ausubel et al., 1999).
[00187] Используемый в данном документе термин "признак" относится к фенотипу, представляющему интерес, гену, вносящему вклад в фенотип, представляющий интерес, а также к последовательности нуклеиновой кислоты, ассоциированной с геном, вносящим вклад в фенотип, представляющий интерес. Например, "признак HI" относится к фенотипу индукции гаплоидии, а также к гену, который вносит вклад в индукцию гаплоидии, и последовательности нуклеиновой кислоты (например, продукту гена, ассоциированному с HI), ассоциированной с наличием или отсутствием фенотипа индукции гаплоидии.
[00188] Используемый в данном документе термин "трансген" относится к молекуле нуклеиновой кислоты, введенной в организм или одного или нескольких его предков посредством некоторой формы методики искусственного переноса. Посредством методики искусственного переноса, таким образом, создают "трансгенный организм" или "трансгенную клетку". Следует понимать, что методику искусственного переноса можно осуществлять в организме-предке (или его клетке и/или клетке, из которой может развиться организм-предок), и тем не менее любой индивидуум-потомок, который имеет искусственно перенесенную молекулу нуклеиновой кислоты или ее фрагмент, по-прежнему считается трансгенным объектом, если в результате одного или нескольких естественных и/или вспомогательных скрещиваний искусственно перенесенная молекула нуклеиновой кислоты присутствует в индивидууме-потомке.
[00189] Используемый в данном документе термин "направленный мутагенез" или "стратегия мутагенеза" относится к любому способу мутагенеза, который приводит в результате к преднамеренному мутагенезу выбранного гена. Направленный мутагенез включает способы с использованием CRISPR, TILLING, TALEN и другие способы, которые еще не обнаружены, но которые можно применять для достижения такого же результата.
[00190] Используемый в данном документе термин "уровень индукции гаплоидии" ("HIR") означает количество выживших гаплоидных зерен по сравнению с общим количеством зерен после опыления початка пыльцой, индуцирующей гаплоидию.
[00191] Конкретными проблемами, преследующими эту индукцию гаплоидии, являются увеличенные уровни недоразвитости зародышей и увеличенные частоты неудачного оплодотворения (пониженные уровни завязывания семян). В силу описанных причин существует потребность в успешном определении причины HI и применении этого знания для определения способов создания гаплоидных растений в стабильном или увеличивающемся объеме при одновременном снижении частоты неудачного оплодотворения и недоразвитости зародышей.
[00192] В частности, предполагается, что промотор можно подвергнуть мутагенезу, чтобы потенциально улучшить пригодность элементов для экспрессии трансгенов у растений. Мутагенез этих элементов можно осуществлять случайным образом, и подвергнутые мутагенезу промоторные последовательности можно подвергать скринингу в отношении активности по методу проб и ошибок. В качестве альтернативы, можно идентифицировать определенные последовательности, которые придают промотору желаемые характеристики экспрессии или придают промотору активность усиления экспрессии, и эти или сходные последовательности можно вводить в промотор посредством мутации. Дополнительно предполагается, что эти последовательности можно подвергнуть мутагенезу для усиления экспрессии трансгенов с их помощью у определенного вида. Способы осуществления мутагенеза сегмента ДНК, кодирующего промоторную последовательность по настоящему изобретению, хорошо известны специалистам в данной области. Как указано, модификации промотора или другого регуляторного элемента можно осуществлять посредством процедур случайного или сайт-специфического мутагенеза. Промотор и другой регуляторный элемент можно модифицировать путем изменения их структуры посредством добавления или делеции одного или нескольких нуклеотидов из последовательности, которая кодирует соответствующие немодифицированные последовательности.
[00193] Мутагенез можно осуществлять в соответствии с любыми методиками, известными из уровня техники, такими как без ограничения синтез олигонуклеотида, имеющего одну или несколько мутаций в пределах последовательности в конкретной регуляторной последовательности. В частности, сайт-специфический мутагенез представляет собой методику, применимую в получении мутантных промоторов посредством специфического мутагенеза лежащей в их основе ДНК. Также можно применять РНК-управляемые эндонуклеазы ("RGEN", например, CRISPR/Cas9). Методика дополнительно обеспечивает легкодоступную возможность получения и тестирования вариантов последовательностей, например, с учетом одного или нескольких из вышеуказанных соображений, посредством введения одного или нескольких изменений нуклеотидной последовательности в ДНК. Сайт-специфический мутагенез обеспечивает получение мутантных форм посредством применения специфических олигонуклеотидных последовательностей, которые кодируют последовательность ДНК с желаемой мутацией, а также достаточного количества прилегающих нуклеотидов, для получения последовательности праймера, имеющей достаточные размер и сложность последовательности для образования стабильного дуплекса по обе стороны от пересекаемой границы делеции. Как правило, предпочтительным является праймер, составляющий в длину от приблизительно 17 до приблизительно 75 нуклеотидов или более, имеющий от приблизительно 10 до приблизительно 25 или более остатков по обе стороны от границы в последовательности, которую изменяют.
[00194] В случае, если клон, содержащий промотор, был выделен в соответствии с настоящим изобретением, можно пожелать определить границы существенных областей промотора в пределах клона. Один эффективный направленный способ для получения подвергнутых мутагенезу промоторов основан на идентификации предполагаемых регуляторных элементов в промоторной последовательности. Ее можно начать со сравнения с промоторными последовательностями, которые, как известно, характеризуются сходными тканеспецифическими или уникальными для стадии развития профилями экспрессии. Последовательности, которые являются общими среди промоторов со сходными профилями экспрессии, являются вероятными кандидатами для связывания с факторами транскрипции и, таким образом, являются вероятными элементами, обеспечивающими профили экспрессии. Подтверждение этих предполагаемых регуляторных элементов можно осуществлять посредством делеционного анализа каждой предполагаемой регуляторной последовательности с последующим функциональным анализом каждой делеционной конструкции посредством анализа репортерного гена, функционально присоединенного к каждой конструкции. Таким образом, после получения исходной промоторной последовательности можно легко получить любую из ряда различных делеционных мутантных форм исходного промотора.
[00195] В настоящем изобретении, раскрытом в данном документе, предусматриваются молекулы полинуклеотидов, содержащие фрагменты регуляторных элементов, которые можно применять при конструировании новых химерных регуляторных элементов. Новые комбинации, содержащие фрагменты этих молекул полинуклеотидов и по меньшей мере один другой регуляторный элемент или фрагмент, можно конструировать и тестировать у растений, и они считаются находящимися в пределах объема настоящего изобретения. Таким образом, разработка, конструирование и применение химерных регуляторных элементов составляют один вариант осуществления настоящего изобретения. Промоторы по настоящему изобретению включают в себя гомологи цис-действующих элементов, которые, как известно, влияют на регуляцию экспрессии генов, демонстрирующие гомологию с промоторными последовательностями по настоящему изобретению.
[00196] Функциональные эквивалентные фрагменты одной из нуклеиновых кислот, регулирующих транскрипцию, описанных в данном документе, содержат по меньшей мере 50, 100, 150, 200, 250, 300, 350, 400, 450, 500, 550, 600, 650, 700, 750, 800, 850, 900, 950 или 1000 пар оснований нуклеиновой кислоты, регулирующей транскрипцию. Из эквивалентных фрагментов нуклеиновых кислот, регулирующих транскрипцию, которые получают посредством делеции области, кодирующей 5'-нетранслируемую область мРНК, можно затем получить только (нетранскрибируемую) промоторную область. 5'-Нетранслируемую область можно легко определить с помощью способов, известных из уровня техники (таких как анализ 5'-RACE). Соответственно, некоторые из нуклеиновых кислот, регулирующих транскрипцию, описанных в данном документе, являются эквивалентными фрагментами других последовательностей.
[00197] Как указано выше, также можно случайным образом получать и затем анализировать делеционные мутантные формы промотора по настоящему изобретению. Следуя этой стратегии, получают серию конструкций, каждая из которых содержит отличную от других часть промотора (субклон), и затем эти конструкции подвергают скринингу в отношении активности. Подходящими способами для осуществления скрининга в отношении активности являются присоединение промоторной или интронной конструкции с делецией, которая содержит сегмент, подвергнутый делеции, к селектируемому или подвергаемому скринингу маркеру, и выделение только тех клеток, которые экспрессируют маркерный ген. Таким путем идентифицируют ряд различных промоторных конструкций с делециями, которые по-прежнему сохраняют желаемую или даже повышенную активность. Благодаря этому идентифицируют наименьший сегмент, требуемый для обеспечения активности, посредством сравнения выбранных конструкций. Этот сегмент можно затем применять для конструирования векторов для экспрессии экзогенных генов.
[00198] Кассета экспрессии, описанная в данном документе, может содержать дополнительные регуляторные элементы. Этот термин в данном контексте следует понимать в широком значении, включающем все последовательности, которые могут влиять на конструкцию или функцию кассеты экспрессии. Регуляторные элементы могут, например, модифицировать транскрипцию и/или трансляцию у прокариотических или эукариотических организмов. Кассета экспрессии, описанная в данном документе, может находиться ниже (в 3'-направлении) последовательности нуклеиновой кислоты, подлежащей экспрессии, и необязательно может содержать дополнительные регуляторные элементы, такие как транскрипционные или трансляционные энхансеры. Каждый дополнительный регуляторный элемент может быть функционально связан с последовательностью нуклеиновой кислоты, подлежащей экспрессии (или нуклеотидной последовательностью, регулирующей транскрипцию). Дополнительные регуляторные элементы могут включать в себя дополнительные промоторы, минимальные промоторы, промоторные элементы или транспозонные элементы, которые могут модифицировать или усиливать свойства регуляции экспрессии. Кассета экспрессии может также содержать один или несколько интронов, один или несколько экзонов и один или несколько терминаторов.
[00199] Кроме того, предполагается, что могут быть применимыми промоторы, в которых объединены элементы из более чем одного промотора. Например, в патенте США №5491288 раскрывается объединение промотора вируса мозаики цветной капусты с гистоновым промотором. Таким образом, элементы из промоторов, раскрытых в данном документе, можно объединять с элементами из других промоторов. Промоторы, применимые для экспрессии трансгенов у растений, включают индуцируемые, вирусные, синтетические, конститутивные (Odell Nature 313: 810-812 (1985)), регулируемые во времени, регулируемые в пространстве, тканеспецифические и регулируемые в пространстве и времени промоторы. Применяя регуляторные элементы, описанные в данном документе, можно экспрессировать множество агрономически значимых генов в трансформированных растениях. Более конкретно, растения можно создавать методами генной инженерии для экспрессии различных фенотипов, представляющих агрономический интерес.
[00200] Соединения по настоящему изобретению могут существовать в виде различных геометрических или оптических изомеров (диастереоизомеров и энантиомеров) или таутомерных форм. Настоящее изобретение охватывает все такие изомеры и таутомеры и их смеси во всех количественных соотношениях, а также изотопные формы, такие как дейтерированные соединения. Настоящее изобретение также охватывает все соли, N-оксиды и металлоидные комплексы соединений по настоящему изобретению.
[00201] Каждый алкильный фрагмент в отдельности либо как часть большей группы (такой как алкокси, алкоксикарбонил, алкилкарбонил, алкиламинокарбонил, диалкиламинокарбонил) имеет линейную или разветвленную цепь и представляет собой, например, метил, этил, н-пропил, н-бутил, н-пентил, н-гексил, изопропил, н-бутил, втор-бутил, изобутил, трет-бутил или неопентил. Алкильные группы включают в себя С1-С6алкил, С1- С4алкил и С1-С3алкил.
[00202] Термин "алкенил", используемый в данном документе, представляет собой алкильный фрагмент, имеющий по меньшей мере одну углерод-углеродную двойную связь, например, С2-С6алкенил. Конкретные примеры включают винил и аллил. Алкенильный фрагмент может представлять собой часть большей группы (такой как алкенокси, алкеноксикарбонил, алкенилкарбонил, алкениламинокарбонил, диалкениламинокарбонил).
[00203] Термин "ацетокси" относится к -ОС(=O)СН3.
[00204] Термин "алкинил", используемый в данном документе, представляет собой алкильный фрагмент, имеющий по меньшей мере одну углерод-углеродную тройную связь, например, С2-С6алкинил. Конкретные примеры включают этинил и пропаргил. Алкинильный фрагмент может представлять собой часть большей группы (такой как алкинокси, алкиноксикарбонил, алкинилкарбонил, алкиниламинокарбонил, диалкиниламинокарбонил).
[00205] Галоген представляет собой фтор (F), хлор (Cl), бром (Br) или йод (I).
[00206] Галогеналкильные группы (в отдельности либо как часть большей группы, такой как галогеналкокси или галогеналкилтио) являются алкильными группами, замещенными одним или несколькими одинаковыми или разными атомами галогенов, и представляют собой, например, -CF3, -CF2C1, -CH2CF3 или -CH2CHF2.
[00207] Гидроксиалкильные группы являются алкильными группами, замещенными одной или несколькими гидроксильными группами, и представляют собой, например, -CH2OH, -CH2CH2OH или -СН(ОН)СН3.
[00208] Алкоксиалкильные группы представляют собой алкоксигруппу, связанную с алкилом (R-O-R'), например, -(CH2)rO(CH2)sCH3, где г равняется 1-6, и s равняется 1-5.
[00209] В контексте настоящего описания термин "арил" относится к кольцевой системе, которая может быть моно-, би- или трициклической. Примеры таких колец включают фенил, нафталинил, антраценил, инденил или фенантренил.
[00210] Если не указано иное, алкенил и алкинил сами по себе или как часть другого заместителя могут иметь линейную или разветвленную цепь и могут содержать от 2 до 6 атомов углерода и в соответствующих случаях могут находиться либо в (Е)-, либо (Z)-конфигурации. Примеры включают винил, аллил, этинил и пропаргил.
[00211] Если не указано иное, циклоалкил может быть моно- или бициклическим, может быть необязательно замещен одной или несколькими С1-С6алкильными группами и содержит от 3 до 7 атомов углерода. Примеры циклоалкила включают циклопропил, 1-метилциклопропил, 2-метилциклопропил, циклобутил, циклопентил и циклогексил.
[00212] Термин "гетероциклил" относится к кольцевой системе, содержащей от одного до четырех гетероатомов, выбранных из N, О и S, где атомы азота и серы необязательно окислены, и атом (атомы) азота необязательно кватернизирован (кватернизированы). Гетероциклил включает в себя гетероарил, его насыщенные аналоги и, кроме того, их ненасыщенные или частично ненасыщенные аналоги, такие как 4,5,6,7-тетрагидробензотиофенил, 9Н-флуоренил, 3,4-дигидро-2Н-бензо-1,4-диоксепинил, 2,3-дигидробензофуранил, пиперидинил, 1,3-диоксоланил, 1,3-диоксанил, 4,5-дигидроизоксазолил, тетрагидрофуранил и морфолинил. Кроме того, термин "гетероциклил" включает гетероциклоалкил, неароматическое моноциклическое или полициклическое кольцо, содержащее атомы углерода и водорода и по меньшей мере один гетероатом, выбранный из атомов азота, кислорода и серы, например, азоксетанил или тиетанил. Моноциклический гетероциклоалкил может содержать 3-7 членов.
[00213] Термин "гетероарил" относится к ароматической кольцевой системе, содержащей от одного до четырех гетероатомов, выбранных из N, О и S, где атомы азота и серы необязательно окислены, например, имеющей 5, 6, 9 или 10 членов и состоящей из одного кольца либо из двух или более конденсированных колец. Отдельные кольца могут содержать не более трех гетероатомов, а бициклические системы - не более четырех гетероатомов, которые предпочтительно будут выбраны из атомов азота, кислорода и серы. Примеры таких групп включают пиридил, пиридазинил, пиримидинил, пиразинил, фуранил, тиенил, оксазолил, изоксазолил, оксадиазолил, тиазолил, изотиазолил, тиадиазолил, пирролил, пиразолил, имидазолил, триазолил и тетразолил.
ПРИМЕРЫ
I. Идентификация мутации сдвига рамки считывания в PLA
[00214] В настоящем изобретении определены серия независимых индуцированных человеком мутаций, обнаруживаемых по меньшей мере в одном гене пататин-подобной фосфолипазы AIIα (pPLAIIα) маиса; растения маиса, имеющие эти мутации по меньшей мере в одном из их генов PLA; и способ создания и идентификации сходных и/или дополнительных мутаций в гене PLA посредством скрининга объединенных и/или отдельных растений риса и маиса. Растения риса и маиса по настоящему изобретению индуцируют гаплоидию в результате нетрансгенных мутаций по меньшей мере в одном из их генов PLA.
[00215] Более конкретно, с помощью настоящего изобретения получают новые линии маиса, индуцирующие гаплоидию. Существует ряд известных линий маиса, индуцирующих гаплоидию, в том числе без ограничения Stock 6, MHI (молдавский индуктор гаплоидии), линия с мутацией неопределенного гаметофита ("ig"), KEMS, ZEM, ZMS, KMS, RWS и RWK. Настоящее изобретение относится к способу идентификации и/или отбора зародышевой плазмы, которая способна или не способна индуцировать образование гаплоидов. Настоящее изобретение также относится к увеличению количества и дополнительной разработке отобранной зародышевой плазмы, индуцирующей гаплоидию. Настоящее изобретение дополнительно относится к способу улучшения зародышевой плазмы, индуцирующей гаплоидию, для увеличения индукции образования гаплоидов у родительской особи, образующей семена.
[00216] Первоначальная стадия в получении гаплоидных семян от гибридного или сегрегантного материнского родительского растения начинается с опыления початка растения, образующего семена, пыльцой индуктора гаплоидии. Результатом этого процесса гибридизации является получение диплоидных и материнских гаплоидных (1n) зерен. Между зернами с индуцированной гаплоидией (1n) и диплоидными семенами часто проводят различие посредством применения маркеров цвета, которые указывают на плоидность зародыша. Диплоидные семена обычно отбрасывают, тогда как гаплоидные зерна или зародыши часто подвергают процессу удвоения числа хромосом с получением двойных гаплоидных растений. Более конкретно, гаплоидный генетический материал обрабатывают одним или несколькими средствами, блокирующими митоз, для обеспечения образования гомологичных пар в гаплоидном (1n) наборе хромосом в одной или нескольких клетках. После процедуры химической обработки химическое(химические) вещество(вещества), удваивающее(удваивающие) число хромосом, удаляют. Маису, являющемуся теперь двойным гаплоидом, позволяют созревать, и полученные в результате двойные гаплоидные семена при посадке будут давать гомозиготные растения (также называемые инбредными растениями или линиями). Эти инбредные линии являются материалами, которые селекционеры используют для проведения своих программ выведения гибридов.
[00217] Локус признака индукции гаплоидии был подвергнут тонкому картированию. Хотя главный QTL в хромосоме 1, отвечающий за индукцию гаплоидию, был картирован и опубликован в Dong et al. Theor. Appl. Genet (2013) 126: 1713-1720, точный ген/генетический элемент, отвечающий за процесс индукции, до сегодняшнего дня не был идентифицирован. Чтобы прояснить генетические механизмы развития, лежащие в основе индукции гаплоидии, производную Stock 6 RWK (~13% HIR) получали из Гогенгеймского университета в 2006 г., скрещивали с инбредными линиями NP2460 и NP2391 и впоследствии подвергали возвратному скрещиванию с RWK для образования картирующих популяций. См. фигуру 1.
[00218] Повышенный HIR в обоих популяциях, косегрегировал с маркером SM020SDQ в секции 1.04, что согласуется с недавними сообщениями о QTL, называемом qhirl. См. Prigge, et al., New Insights into the Genetics of in Vivo Induction of Maternal Haploids, the Backbone of Doubled Haploid Technology in Maize, Genetics (2012) 190:781 793 (в которой обсуждается главный QTL для HI в секции 1.04 [qhirl] и минорные QTL для HI в секциях 3.02 [qhir2], 3.06 [qhir3], 4.03 [qhir4], 5.01 [qhir5], 5.04 [qhir6], 7.01 [qhir7] и 9.01 [qhirS]); Liu, et al., Fine mapping of qhir8 affecting in vivo haploid induction in maize, Theor. Appl. Genet. (2015) 128:2507-2515 (в которой описывается тонкое картирование тридцати пяти генов с отнесением их к qhir8); Hu, et al., Genetic Basis of Haploid Induction in Maize Identified with a Novel Genome-Wide Association Method, Genetics (2016) 202:1267-1276 (в которой утверждается, что qhirl представляет собой два QTL: qhirl 1 и qhir12, и описывается тонкое картирование qhir6 с отнесением его к области размером 1,1 млн п.о.). Авторы настоящего изобретения провели несколько циклов тонкого картирования и ограничили QTL областью размером примерно 0,57 млн п о. между 67,85 млн п. о. и 68,42 млн п. о., которая лежит в пределах qhirl 1. Эта область имеет семь аннотированных генов (фигура 2).
[00219] С помощью Illumina HiSeq2000 авторы настоящего изобретения секвенировали RWK, Stock 6 и неиндуцирующую линию BC3F5 "RWK-NIL", которая является почти изогенной RWK, но имеет гаплотипы NP2391 в интервале qhirl 1. Путем сравнения индуцирующей и неиндуцирующей зародышевой плазмы было определено, что вставка из четырех нуклеотидов, присутствующая у индукторов гаплоидии, которая сдвигает рамку считывания для кодирования аминокислот в GRMZM2G471240, не присутствует в неиндуцирующей зародышевой плазме. Таким образом, в настоящем изобретении был идентифицирован ген с мутацией сдвига рамки считывания в индуцирующей зародышевой плазме в качестве отвечающего за индукцию гаплоидии у маиса. Ген-кандидат, соответствующий генной модели GRMZM2G471240, кодирует пататин-подобную фосфолипазу AIIα (pPLAIIα), которую авторы настоящего изобретения переименовали в MATRILINEAL (MTL) для представления аллеля дикого типа, а аллель со сдвигом рамки считывания называется matrilineal (mtl).
[00220] Последовательность ДНК получали для каждого гена-кандидата из двух индуцирующих линий (Stock6 и RWK) и одной неиндуцирующей линии (RWK-NIL). Кроме того, имеющиеся в открытом доступе данные о геноме В73 использовали в качестве второй неиндуцирующей линии. Информацию из генной модели сравнивали с данными о EST/кДНК для подтверждения структуры каждого гена. Аннотированные данные о последовательностях сравнивали с различиями по каталогу между четырьмя аллелями каждого гена.
[00221] Сравнения последовательностей выявили, что аллели В73 и RWK-NIL являлись сходными друг с другом, и аллели RWK и Stock 6 являлись сходными друг с другом. Большинство различий между последовательностями представляли собой однонуклеотидные полиморфизмы, которые не изменяют последовательность, кодирующую белок. Наблюдались некоторые вставки и некоторые делеции, большинство из которых находились в последовательности, не кодирующей белок.
[00222] В таблице 1 ниже перечислены наименования генов в показанном интервале. Эта информация относится к хромосоме 1, см., например, Patrick S. Schnable et al., The В 73 Maize Genome: Complexity, Diversity, and Dynamics, 326 Science 1112-15 (2009), включенный в данный документ посредством ссылки во всей своей полноте, и включает краткое описание других кодируемых белков из генов в пределах локуса, индуцирующего гаплоидию.
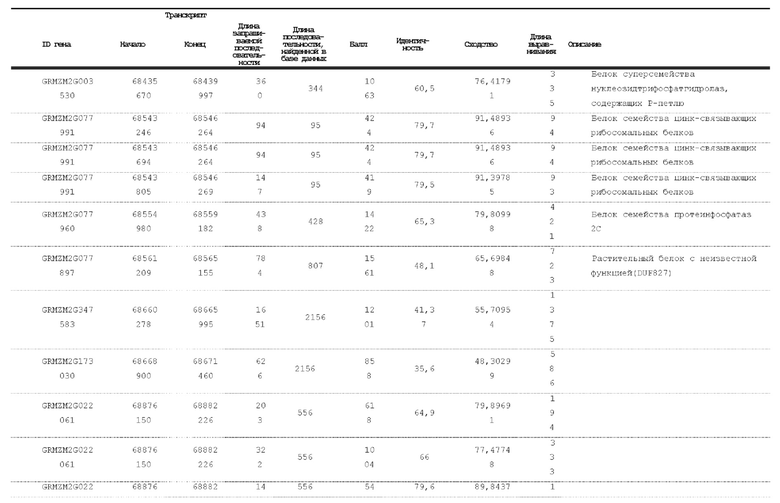
[00223] После завершения тонкого картирования признака индукции гаплоидии с отнесением его к интервалу, содержащему только семь генов, авторы настоящего изобретения сосредоточились на них при сборке и анализе последовательностей. Последовательности семи генов были почти идентичными в сравнении между В73 и RWK-NIL, но в RWK и Stock 6 отсутствовал GRMZM2G062320, ген фосфоглицератмутазы (PGM), и имелась вставка из 4 пар оснований ("п.о.") в четвертом экзоне GRMZM2G471240, гена пататин-подобной фосфолипазы АПа (pPLAIIa) (фигура 2). Авторы настоящего изобретения обнаружили, что как RWK, так и Stock6 имели одну и ту же вставку из 4 п.о. в четвертом экзоне pPLAIIa, и что этот ген специфически экспрессируется в пыльце (см. maizegdb.org/gene_center/gene?id=GRMZM2G471240, включенный в данный документ посредством ссылки). Немутантный GRMZM2G471240 представлен под SEQ ID NO: 1. GRMZM2G471240, содержащий вставку из 4 п.о. в четвертом экзоне, представлен под SEQ ID NO:3.
[00224] Большинство гаплоидов, которые были идентифицированы, были обнаружены с помощью маркерного теста Taqman. В данном маркерном тесте используются преимущества различия в гене pPLAIIa между скрещиваемыми RWK и NP2222. В скрещиваниях, где авторы настоящего изобретения применяли RWK в качестве женской особи, а NP2222 в качестве мужской особи, родительская особь RWK является гомозиготной по аллелю mtl, тогда как NP2222 является гомозиготной по аллелю MTL. Диплоидное потомство представляет собой MTL/mtl, а гиногенетическое гаплоидное потомство представляет собой mtl/0. Таким образом, при выполнении этого теста результаты Taqman показывают 1 копию аллеля mtl и одну копию аллеля MTL у диплоидного потомства, а также 1 копию mtl и отсутствие копий MTL у гаплоидного потомства. При осуществлении этого типа скрещивания початки собирают в период 12-21 дня после опыления, зародыши извлекают, и небольшой образец зародышей отбирают для маркерного анализа Taqman. В качестве альтернативы, зародыши высевают на твердые среды и проращивают в темноте с тем, чтобы можно было отобрать более крупный образец удлиненного побега или корня 2-10 дней спустя для маркерного анализа. В то же время некоторую часть ткани сохраняют для анализа плоидности. В этом последнем случае после применения молекулярного теста более крупные образцы гаплоидов можно анализировать на анализаторе плоидности CyFlow Space и подтверждать в качестве гаплоидов. В большинстве случае это приводит в результате к положительной идентификации гаплоидов. В нескольких более редких случаях это приводит в результате к опровержению ложноположительных результатов маркерного анализа и коррекции сигнала как диплоидного.
[00225] Другим способом, которым авторы настоящего изобретения проводили тестирование на наличие гаплоидов, является анализ доминантных маркеров. В данном случае применяли мужских особей линии Х26. Эта линия является гомозиготной в отношении маркера, который действует доминантным образом. В таком скрещивании в качестве женской особи можно применять растение любой линии, при условии, что оно не имеет маркера или каких-либо генов или аллелей, функционирующих как подавляющие маркерный фенотип. Линия Х26 является неиндуцирующей и гомозиготной по MTL. При применении такой линии потомство срезали в период 12-21 дня после опыления и оценивали в отношении наличия маркера, или его исследовали непосредственно на початке, или высушенные зерна собирали и оценивали в отношении наличия маркера. Диплоидное потомство демонстрирует маркерный фенотип, поскольку оно имеет одну копию маркерного гена от мужской родительской особи Х26, тогда как гиногенетическое гаплоидное потомство не демонстрирует маркерный фенотип. Пенетрантность маркера и уровень спонтанной индукции гаплоидии у Х26 тестировали во множестве контрольных скрещиваний. С применением этой системы авторы настоящего изобретения осуществляли скрининг в отношении гаплоидов и затем тестировали их на анализаторе плоидности для подтверждения того, что они действительно являются гаплоидами.
[00226] Авторы настоящего изобретения разработали ПЦР-тесты для специфического выявления аллелей "дикого типа" и "мутантных" аллелей для скрининга девятнадцати индукторов, происходящих от Stock 6, в том числе индуктора гаплоидии NP2222 (NP2222-HI), линию ВС3, полученную в результате интрогрессии RWK в стандартную трансформируемую инбредную линию Syngenta NP2222. Авторы настоящего изобретения также подвергли скринингу девять неиндуцирующих контрольных линий.
[00227] Для разработки ПЦР-теста, с помощью которого можно было бы провести различие между гаплотипами RWK/Stock6 и RWK-NIL, разрабатывали две пары праймеров: одна пара должна амплифицировать аллель со сдвигом рамки считывания RWK/Stock6, тогда как другая должна амплифицировать аллель B73/RWK-NIL. Эти пары функционировали согласно ожидаемому для ДНК RWK-NIL, RWK и Stock6: gDNA RWK-NIL амплифицировалась только парой праймеров RWK-NIL. gDNA RWK и Stock6 амплифицировалась только парой праймеров RWK/Stock6, которая специфически выявляет аллель со сдвигом рамки считывания. Продукты ПЦР секвенировали, и последовательности были идентичными последовательностям, определенным в результате полногеномного секвенирования. SNP, которые идентифицировали в рамках полногеномного секвенирования, подтверждали в продуктах ПЦР. Ниже, на фигуре 2, ДНК, применяемая в каждой реакции, указана прописными буквами. Праймерами являются "nil.Fl/Rl" и "rwk.Fl/Rl." GRMZM2G471240 nil.Fl: GTACGCCGTGCGCTAACA (SEQ ГО NO: 5). GRMZM2G471240 nil.Rl: TCGTACCTCCCTGTCTCCAC (SEQ ID NO: 6). GRMZM2G471240 rwk.Fl: TACGCCGTGCGCTAACATA (SEQ ID NO: 7). GRMZM2G471240 rwk.Rl: GTACCTCGCTCCCTGTCTCC (SEQ ID NO: 8).
[00228] Пары праймеров "rwk.Fl/Rl" и "nil.Fl/Rl" применяли для генотипирования панели сильных, слабых индукторов и неиндукторов. Авторы настоящего изобретения обнаружили, что все 19 линий, индуцирующих гаплоидию, в том числе, среди прочих, Stock6 (3% уровень индукции гаплоидии ["HIR"]), RWK (линия, полученная из запасов Гогенгеймского университета, 10-15% HIR), RWS и Z22, имели вставку из 4 п.о. В отличие от этого, во всех девяти линиях, не индуцирующих гаплоидию (средний HIR 0,1%), был обнаружен аллель дикого типа. Данные указывают на то, что гомозиготность по аллелю со сдвигом рамки считывания коррелирует с индукционной способностью: 12/12 сильных и 7/7 слабых индукторов амплифицировались в анализе сдвига рамки считывания, но не в анализе дикого типа, тогда как 9/9 неиндукторов амплифицировались в анализе дикого типа, но не сдвига рамки считывания. Это указывает на то, что индукционная способность коррелирует с мутацией GRMZM2G471240, и что pPLAIIα лежит в основе qhir11 и является основной мутацией, отвечающей за индукцию гаплоидии в этих линиях.
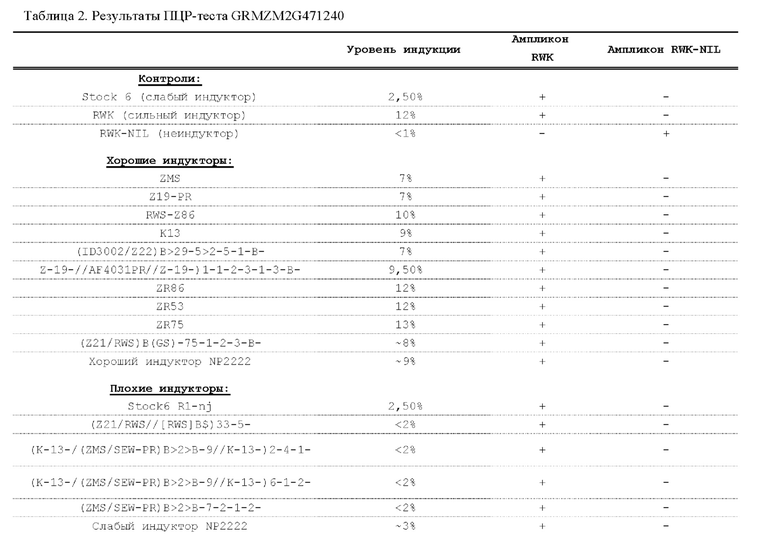
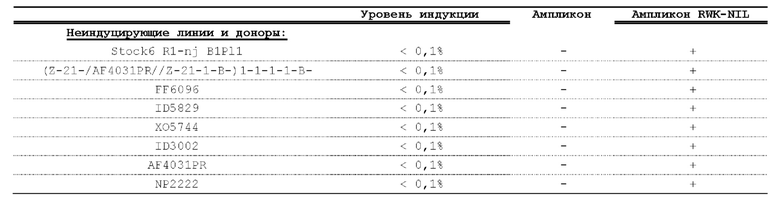
[00229] Авторы настоящего изобретения также идентифицировали ряд однонуклеотидных полиморфизмов ("SNP") между аллелем со сдвигом рамки считывания и аллелем RWK-NIL. Для многих из этих SNP последовательности STOCK6 и RWK согласовывались с другими инбредными растениями, которые авторы настоящего изобретения секвенировали, и, следовательно, по всей вероятности представляют собой естественную изменчивость. Действительно, большинство из этих SNP не изменяло аминокислотную последовательность и, следовательно, по всей вероятности не вносит вклад в фенотип индукции гаплоидии. Два SNP приводили в результате к аминокислотным изменениям (H107Y; K232N), и они не являются высококонсервативными изменениями, поэтому они могут вносить небольшой вклад в фенотип, но в большинстве случаев они не влияют на фенотип, поскольку сдвиг рамки считывания вызывает потерю функции.
[00230] Авторы настоящего изобретения переименовали pPLAIIα в "MATRILINEAL" (MTL; т.е. SEQ ID NO: 1) и нативный аллель со вставкой из 4 п.о. в "matrilineal" (mtl; т.е. SEQ ID NO: 3). В соответствии с предсказанной последовательностью белка вставка из 4 п.о. вызывает сдвиг открытой рамки считывания белка в положении аминокислоты ("АА") 352 из 401. Сдвиг рамки считывания приводит к появлению преждевременного стоп-кодона.
[00231] После обнаружения нокаут-мутации сдвига рамки считывания авторы настоящего изобретения непосредственно тестировали эффект, который она оказывала на индукцию гаплоидии, посредством комплементации линии, индуцирующей гаплоидию, трансгеном pPLAIIα дикого типа. Гетерологичная комплементация NP2222-HI (10,2% HIR) копией MATRILINEAL дикого типа практически устраняла индукцию гаплоидии и недоразвитость зерна. По сравнению с контролями HIR уменьшался в 50 раз, от 10% до 0,23%. Она также уменьшала уровень недоразвитости зародышей до 0,65%. Также получали линии с полноразмерными функциональными репортерами с помощью трансгенных слияний гена MTL дикого типа с GFP, а также мутантного аллеля mtl с GFP для того, чтобы как визуализировать субклеточную локализацию MTL дикого типа, так также и увидеть, правильно ли локализуется мутантная версия белка или вырабатывается ли она вообще. Эти линии также служили в качестве дополнительного материала для тестирования комплементации. Материал, индуцирующий гаплоидию (NP2222-HI), который являлся гомозиготным по трансгену MTL-GFP, также не демонстрировал фенотип индукции гаплоидии. Уровень индукции NP2222-HI падает до 0,60%, если он является гомозиготным по MTL-GFP'. Кроме того, трансген MTL-GFP также снижал недоразвитость зародышей до 4,86%. И наконец, авторы настоящего изобретения тестировали, обеспечивает ли мутантный аллель mtl, слитый с GFP, комплементацию фенотипа индукции гаплоидии, и было обнаружено, что этого не происходит. Уровни индукции гаплоидии и недоразвитости зародышей были весьма сходными у NP2222-HI по сравнению с NP2222-HI, который являлся гомозиготным по мутантному слитому трансгену mtl-GFP. См. таблицу 3. Это представляет убедительное доказательство того, что сдвиг рамки считывания в MATRILINEAL отвечает за индукцию гаплоидии и необходим для этого. Для применения этих знаний авторы настоящего изобретения демонстрируют, что осуществление мутации или модулирование экспрессии pPLAIIa в линии дикого типа приводит к созданию новых линий с индуцированной гаплоидией.

[00232] Несколько mtl-подобных аллелей создавали в инбредном растении NP2222 посредством введения небольших делеций в MTL вблизи сайта вставки из 4 п.о. в mtl, с применением эффекторных нуклеаз, подобных активаторам транскрипции (TALEN) (Boch, J. et al., Breaking the code of DNA binding specificity of TAL-type III effectors, SCIENCE 326:1509-1512 (2009), включенный в данный документ посредством ссылки). Несколько мутантных объектов подвергали самоопылению, и растения Т1, лишенные вставки Т-ДНК, введенной с помощью TALEN, но гомозиготные по небольшим делециям в MTL, подвергали ауткроссингу с NP2222. Отредактированные линии, гомозиготные по делециям со сдвигом рамки считывания в MTL (далее в данном документе называемые MTLTAL-FS), демонстрировали HIR, составляющий 4,0 - 12,5% (в среднем 6,65%) (таблица 4). Статус плоидности 118/127 предполагаемых гаплоидов подтверждали посредством проточной цитометрии, и фенотипические оценки указывали на то, что эти растения являлись гаплоидами. Эти результаты доказывают, что сдвиг рамки считывания в MTL является достаточным для индуцирования высоких уровней индукции гаплоидии. Картировали другие факторы, вносящие вклад в фенотип, в том числе соседний qhirl2 (см. Liu, 2016), который может являться причиной различия в HIR между MTLTAL-FS и NP2222-HI. Разумно сделать вывод о том, что уровни завязывания семян, HIR и недоразвитости зерна устанавливаются благодаря mtl посредством специфических взаимодействий отцовского и материнского генотипа.
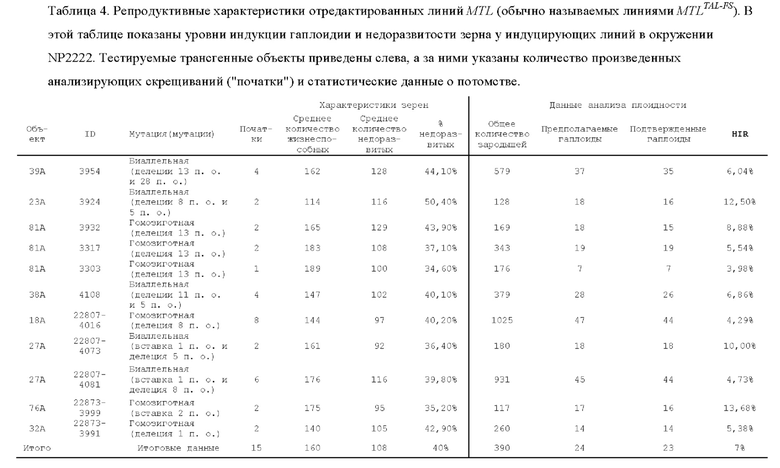
[00233] Образование гаплоидных семян у маиса имеет постзиготический характер и вызывается дефектом в мужском гаметофите. Этот факт отражен в данных об экспрессии MTL. Имеющиеся в открытом доступе профили секвенирования РНК указывают на то, что транскрипт MTL дикого типа является специфическим для пыльников на стадии цветения (см. Sekhon, R. S., et al. Genome-wide atlas of transcription during maize development, PLANT JOURNAL, 66, 553-563 (2011), включенный в данный документ посредством ссылки), что согласуется с профилем развития, обнаруживаемым исключительно в пыльниках до вскрывания (см. Zhai, J., et al. Spatiotemporally dynamic, cell-type-dependentpremeiotic and meiotic phasiRNAs in maize anthers, PNAS 112, 3146-3151 (2015), включенный в данный документ посредством ссылки). Авторы настоящего изобретения обнаружили, что пыльца дикого типа имела в 18 раз больше транскриптов MTL, чем пыльцевые мешки после цветения, что указывало на то, что ген является специфическим для мужского гаметофита (фигура 3). Попытки осуществления нокдауна MTL посредством RNAi привели к повышению уровней образования гаплоидов только для MTLRNAi (таблица 5). Существует два аннотированных сплайс-варианта MTL, отображающие альтернативный 81 нуклеотид сайта сплайсинга перед 3'-концом экзона 2. По сравнению с NP2222 уровень Mtl был повышен в пыльце NP2222-HI, но не MTLTAL-FS (фигура 4), тогда как представленность двух аннотированных сплайс-вариантов была единообразной.
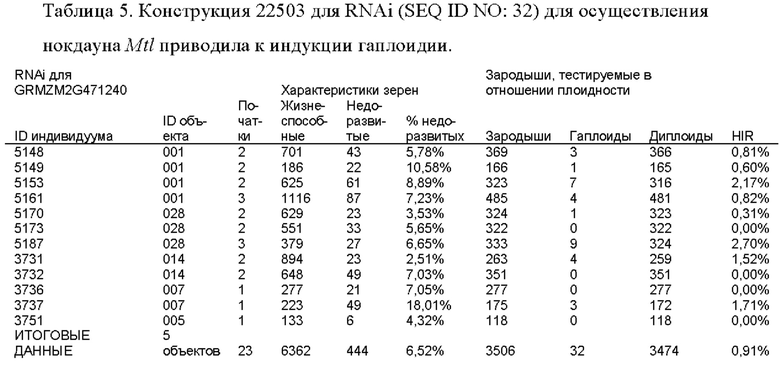
[00234] Сдвиг рамки считывания в mtl появляется в положении аминокислоты 380, что приводит к изменению 20 аминокислот с появлением расположенного после них преждевременного стоп-кодона, который усекает белок на 29 аминокислот (фигура 5а). Белок МТХ дикого типа был обнаружен в профилях LS-MS пыльцы RWK-NIL и NP2222, но его уровень находился ниже предела выявления в трех из трех образцов RWK и 3 из трех 3 образцов NP2222-HI (таблица 6). Это демонстрирует, что, несмотря на наличие мутантного транскрипта mtl, образующегося в пыльце, белок не выявляется, что подтверждает наличие мутации потери функции. Как мутантные формы, так и формы дикого типа рекомбинантного белка MTL проявляют фосфолипазную активность in vitro во флуоресцентных анализах расщепления липосом pPLAIIα-подобным белком (фигура 5b). Это демонстрирует, что функциональная аннотация гена MTL (т.е. то, что он кодирует белок фосфолипазу) является правильной.
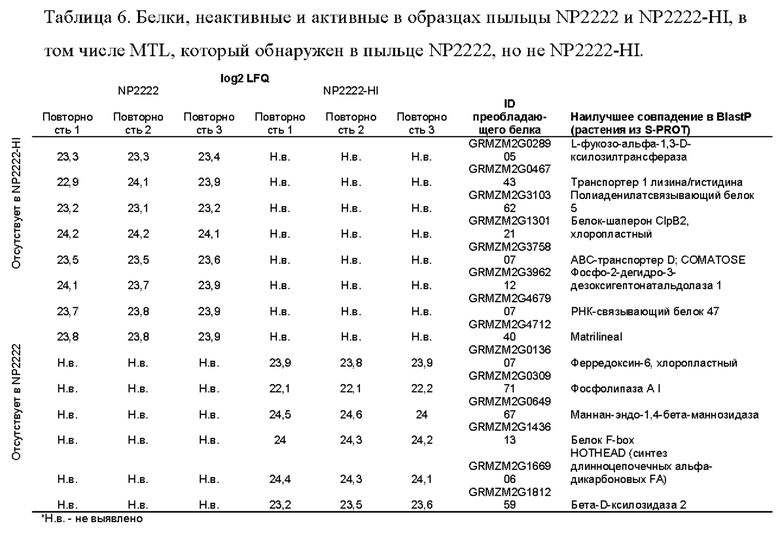
II. Эксперименты по проращиванию пыльцы и определению локализации
[00235] Линии с полноразмерными функциональными репортерами использовали для определения характеристик локализации MTL. Сигнал не был обнаружен в пыльце NP2222 или NP2222+mtl-GFPImtl-GFP. В отличие от этого, пыльца NP2222+MTL-GFP/MTL-GFP демонстрировала сильный сигнал в цитоплазме двух спермиев. Этот сигнал был обнаружен в тяжистой цитоплазме гамет в пределах проросших пыльцевых трубок. Зародышевые мешки NP2222, фиксированные через 18 часов после опыления пыльцой MTL-GFP, давали сигнал в области дегенерирующей синергиды, согласующийся с сигналом SC, доставляемых в ходе оплодотворения. Это указывает на то, что MTL является частью мужской зародышевой единицы, которая откладывается в зародышевом мешке после разрыва пыльцевой трубки. MTL-GFP, но не mtl-GFP устранял индукцию гаплоидии в NP2222-HI (таблица 3). Совместно эти данные указывают на то, что MTL является фосфолипазой, специфической для цитоплазмы SC, и что сдвиг рамки считывания в mtl нарушает локализацию или стабильность MTL в пыльце, индуцирующей гаплоидию.
[00236] Идентификациями в качестве причинного гена при индукции гаплоидии у маиса позволила провести анализ плейотропных фенотипов, традиционно ассоциированных с признаком. Мутации фосфолипазы ассоциированы с замедленным прорастанием пыльцы и ростом трубки (см. Kim, Н. J., et al. Endoplasmic reticulum- and golgi-localized phospholipase A2 plays critical roles in Arabidopsis pollen development and germination, PLANT CELL 23, 94-110 (2011)), но они являлись нормальными в линиях RWK, Stock 6 и MTLTAL-FS (фигура 6А, В). Початки, опыленные пыльцой NP2222-HI и MTLTAL-FS, демонстрируют -10-25% частоту неудачного оплодотворения, и конкурентный анализ пыльцы показал, что RWK подвергается нарушению сегрегации (SD) (фигура 6С), что согласуется с предыдущими сообщениями (см. Xu, X., et al. Gametophytic and zygotic selection leads to segregation distortion through in vivo induction of a maternal haploid in maize, J. EXP. ВОТ. 64, 1083-1096 (2013)). Скрещивания с гемизиготной пыльцой NP2222-HI+MTL/0 давали пропорциональное смещение в сторону потомства MTL+(фигура 6D), что указывало на то, что индукция SD может быть отнесена на счет mtl. Недоразвитость зародышей, неизменный побочный результат индукции гаплоидии, связанный с неудачной пролиферацией эндосперма, имела место на сходных уровнях у нативного индуктора и индуктора MTLTAL-FS (таблица 4). Совместно эти данные подразумевают участие mtl в каждом репродуктивном дефекте, ассоциированном с индукцией гаплоидии. Двумя механизмами, которые, как правило, предлагают для объяснения образования гаплоидов, являются одинарное оплодотворение и постзиготическая элиминация генома (см. Sarkar, K. R. & Сое, Е. Н, A genetic analysis of the origin of maternal haploids in maize, GENETICS 54, 453-464 (1966); Zhang, Z., et al., Chromosome elimination and in vivo haploid production induced by Stock 6-derived inducer line in maize (Zea mays L.J, PLANT CELL REP. 27, 1851-1860 (2008); и Barret, P., Brinkmann, M., & Beckert, M., A major locus expressed in the male gametophyte with incomplete penetrance is responsible for in situ gynogenesis in maize, THEOR. APPL. GENET. 117, 581-594 (2008)). В первом случае гаплоиды образуются в результате оплодотворения центральной клетки, но не яйцеклетки, которая впоследствии развивается посредством партеногенеза. В последнем случае двойное оплодотворение предшествует элиминации мужских хромосом. Выяснение точного механизма потребует тщательных эмбриологических исследований после опылений MTLTAL-FS наряду с получением количественных данных для отслеживания редкой стойкости мужской ДНК у гаплоидов маиса.
[00237] Индукцию гаплоидии недавно осуществляли в Arabidopsis посредством манипуляции с геном центромерного варианта гистона Н3, который вызывает однородительскую элиминацию генома посредством постзиготического дисбаланса центромер между гибридизированными геномами. Попытка повторить это у маиса была успешной (см. Ravi, М. & Chan, S. W. L. Haploid plants produced by centromere-mediated genome elimination, NATURE 464, 615-618 (2010)), но эта публикация является первым случаем описания системы индукции гаплоидии, запускаемой цитоплазматическим белком, который не связывается с хроматином. Таким образом, данная работа подчеркивает важность неядерных компонентов спермия в репродуктивном успехе и точном переносе генома. Консервативность MTL у злаковых растений (см. фиг. 7), в частности у риса, где ближайший гомолог является специфическим для пыльцы и также обнаружен в спермин, позволяет предположить, что эти полученные результаты приведут к разработке новых внутривидовых линий важных культурных растений, индуцирующих гаплоидию.
III. Химические аэрозоли для индукции HI de novo
[00238] После обнаружения того, что фосфолипаза MTL (и в частности, мутация потери функции со сдвигом рамки считывания в mtl) вызывает индукцию гаплоидию, авторы настоящего изобретения тестировали, могут ли ингибиторы фосфолипаз, жирные кислоты или другие липидные соединения выступать в качестве химических индукторов гаплоидии.
[00239] Авторы настоящего изобретения успешно достигли индукции гаплоидии de novo посредством нанесения различных химических веществ на цветки, рыльца, початки, метелки и пыльцу. Все эти химические вещества принадлежат к классу липидов или ингибиторов фосфолипаз. В таблице 7 в общих чертах изложены эти химические вещества и уровни индукции гаплоидии, которые были получены в результате их нанесения. Также в таблице 7 представлена информация о составе и информация о способе нанесения. Из этой таблицы понятно, что множество липидных соединений индуцирует образование гаплоидов de novo при нанесении до, во время или после опыления на рыльца, которые представляют собой рыльцевую ткань, специфичную для растения маиса, пыльцу, метелки (которые содержат мужские цветки, содержащие пыльцу до выбрасывания пыльцы) и початки. Такие соединения включают смешанные масла, жирные кислоты, сложные эфиры жирных кислот, фосфолипиды и ингибиторы фосфолипаз.
[00240] Авторы настоящего изобретения тестировали много ингибиторов фосфолипаз, в том числе маноалид, ингибитор фосфолипаз, не имеющий каких-либо цепей жирных кислот.Это химическое вещество было способно индуцировать образование гаплоидов de novo. Имея этот результат, авторы настоящего изобретения начали тестировать больше соединений. Авторы настоящего изобретения обнаружили, что наиболее активным соединение для индукции de novo является метил-альфа-линоленоилфторфосфонат (MALFP), сильнодействующий ингибитор фосфолипаз и ингибитор других липид-модифицирующих ферментов. Это соединение содержит три цепи жирной кислоты линолената (восемнадцать атомов углерода с тремя тройными связями), а также метальную группу и атом фтора в положении группы головки. Другим высокоактивным соединением, индуцирующим гаплоидию de novo, является метиларахидонилфторфосфонат (MAFP), другой ингибитор фосфолипаз с несколько отличающейся структурой цепи жирной кислоты (20 атомов углерода и четыре двойные связи). Не желая ограничиваться какой-либо теорией, полагают, что MALFP и MAFP ингибируют фосфолипазы, располагаясь в связывающем кармане для цепей жирных кислот и в некоторых случаях катализируя необратимое фосфорилирование аминокислоты серина в активном центре (см. Lio Y.C., Reynolds L.J., Balsinde J., и Dennis E.A. Irreversible inhibition of Ca(2+)-independent phospholipase A2 by methyl arachidonylfluorophosphonate. BIOCHIMBlOPHYS ACTA 1302:55-60 (1996)). Некоторые жирные кислоты, такие как линолевая кислота и линоленовая кислота, могут также неконкурентно ингибировать фосфолипазы, располагаясь в связывающем кармане для цепей жирных кислот без ковалентной модификации активного центра, однако эффект этого ингибирования является более слабым. Ballou, L.R., и Cheung, W.Y. Inhibition of human platelet phospholipase A2 activity by unsaturated fatty acids PROC NATL ACAD SCI 82 (2): 371-375 (1985)). Эти литературные источники могут помочь объяснить основания для полученных авторами настоящего изобретения результатов, заключающихся в том, что индукция гаплоидии de novo вызывается не только сильными ингибиторами фосфолипаз, такими как MALFP и MAFP, но также и сложными эфирами жирных кислот, в том числе метиловым сложным эфиром арахидоновой кислоты (ААМЕ), который имеет 20 атомов углерода и четыре двойные связи (сложный эфир полиненасыщенной жирной кислоты из 20 атомов углерода), а также линолевой кислотой (LLE), которая имеет 18 атомов углерода и две двойные связи (18:2), и линоленовой кислотой (LNA), которая имеет 18 атомов углерода и три двойные связи (18:3), и этиловым сложным эфиром олеиновой кислоты (ОАЕЕ), и арахидоновой кислотой (которая имеет 20 атомов углерода и четыре двойные связи). Опять же, полагают, что эти цепи ненасыщенных жирных кислот, будь то в их форме свободной жирной кислоты, в их форме сложного эфира (которые синтезируются в лаборатории и могут быть способны проникать в ткани) или в некоторой другой форме, могут выступать в качестве ингибиторов фосфолипаз. Несмотря на то, что, как правило, не признается, что эти соединения в форме сложного эфира могут ингибировать фосфолипазы (см. Ballou, выше), авторы настоящего изобретения обнаружили, что такие метиловые и этиловые сложные эфиры жирных кислот могут в действительности приводить к индукции гаплоидии de novo. См. таблицу 7. Следует отметить, что тестировали насыщенные жирные кислоты и их производные, в том числе этиловый сложный эфир стеариновой кислоты и пальмитиновой кислоты. В предшествующем уровне техники указано, что молекулы, которые содержат боковые цепи насыщенных жирных кислот, неспособны или не будут способны конкурентно ингибировать фосфолипазу, находясь в ее активном центре. См. Ballou, выше. Считалось, что такие соединения не обладают способностью ингибировать фосфолипазы. Однако авторы настоящего изобретения неожиданно смогли применять молекулы, содержащие боковые цепи насыщенных жирных кислот, для индукции образования гаплоидов de novo. Например, этиловый сложный эфир пальмитиновой кислоты (РАЕЕ) индуцирует образование гаплоидов. См. таблицу 7.
[00241] Авторы настоящего изобретения также решили протестировать, могут ли определенные масла и фосфолипиды, в том числе широко известные растительные масла, вызывать индукцию гаплоидии de novo. Авторы настоящего изобретения обнаружили, что одна высокоактивная липидная смесь, индуцирующая гаплоидию de novo, представляет собой кукурузное масло, натуральный растительный экстракт, который можно приобрести в продуктовом магазине. Другая высокоактивная липидная смесь представляет собой льняное масло, которое можно приобрести в магазине хозяйственных товаров. Также будет действовать масло канолы, растительное масло, арахисовое масло, кунжутное масло или любое другое масло при нанесении правильным образом. Другое высокоактивное соединение представляет собой 1,2-дистеарил-8п-глицерофосфатидилхолин (DSPC) (также известный как PC 18:0/18:0), фосфолипид с фосфатидилхолиновой группой головки и двумя цепями насыщенной жирной кислоты из 18 атомов углерода. Этот широко известный фосфолипид присутствует во многих биологических материалах, поскольку он является одним из основных и наиболее распространенных компонентов фосфолипидных бислоев, которые составляют клеточные мембраны всех живых существ на планете. Другое соединение, индуцирующее гаплоидию, представляет собой фосфатидилэтаноламин с одной стеарильной цепью и одной олеоильной цепью (также известный как 1-стеарил-2-олеоил-sn-фосфатидилэтаноламин), другой широко известный фосфолипид. Другие фосфолипиды, а также лизофосфолипиды и другие триацилглицериды, диацилглицерины, лизофосфолипиды, сложные эфиры тритерпеноидов или глицеролипиды, будут выступать в качестве индукторов гаплоидии de novo. Эти масла и фосфолипиды не являются общеизвестными ингибиторами фосфолипаз, но они могут являться исходным материалом для получения побочных продуктов фосфолипазной активности, в том числе тех самых жирных кислот, которые ингибируют фосфолипазы. Тот факт, что эти соединения выступают в качестве химических веществ, индуцирующих гаплоидию de novo, таким образом, указывает на то, что эти масла и фосфолипиды могут ингибировать фосфолипазы опосредованно, или, возможно, они вызывают дисбаланс или изменение состава липидов или мембраны пыльцы, что приводит к индукции гаплоидии. В любом случае из данной работы ясно, что огромный ряд липидосодержащих соединений, в дополнение к нелипидным ингибиторам фосфолипаз, при нанесении на цветки может выступать в качестве химических индукторов образования гаплоидных семян.
[00242] Авторы настоящего изобретения также тестировали, могут ли буферы, которые авторы настоящего изобретения использовали для нанесения этих соединений на цветки, индуцировать образование гаплоидов сами по себе. Эти буферы вариативно содержали определенные смеси поверхностно-активных веществ, и/или соль, и/или DMSO (диметилсульфоксид), и/или другие близкие добавки. Эти буферы не могли индуцировать образование гаплоидов, что можно увидеть в строках 121 123 таблицы 7. Кроме того, индукция гаплоидии de novo без добавления каких-либо соединений является чрезвычайно редкой у маиса, что можно увидеть в строке 1. Среди 3073 тестированных потомков было обнаружено два гаплоида (Kelliher, et al. Maternal haploids are preferentially induced by CENH3-tailswap transgenic complementation in maize, FRONTIERS IN PLANT SCIENCE 7: 414 (2016)). Таким образом, было обнаружено, что контрольный уровень индукции составляет примерно 0,065%, что попадает в диапазон гиногенетической индукции гаплоидии, о котором сообщается в предшествующих литературных источниках (0,05-0,1%) (Chang, М. -Т., и Сое, Е. "Doubled haploids", в Molecular Genetic Approaches to Maize Improvement, eds A.L.Kriz и В.А. Larkins (Heidelberg: Springer), 127 142).
[00243] Способ нанесения этих различных липидных соединений и соединений-ингибиторов липаз может являться достаточно вариативным и при этом по-прежнему обеспечивать образование гаплоидов. Один такой способ нанесения заключается в растворении соединений в солевом растворителе с добавлением 1% DMSO. Другой такой растворитель представляет собой смесь поверхностно-активных веществ (таблица 8). Ткань для нанесения может также изменяться, что видно из таблицы 7, где нанесение как на пыльцу, так и на рыльца во время опыления, а также на метелки за несколько часов или дней до опыления может привести в результате к образованию гаплоидов. Кроме того, соединения могут индуцировать образование гаплоидов в большом разнообразии концентраций от 20 мкМ до 100 мМ или от 0,2 мг/мл до 50 мг/мл включительно. На самом деле образование гаплоидов способен индуцировать намного более широкий диапазон, чем этот, что можно увидеть из таблицы 7.
[00244] В двух случаях авторы настоящего изобретения также разработали новые смеси поверхностно-активных веществ, чтобы обеспечить способность конкретных классов липидов к эмульгированию и образованию микроэмульсии (водно-органической смеси с липидными каплями, имеющими размер, меньший определенного размера по меньшей мере меньше 10 микрон в диаметре и во многих случаях меньше 1 микрона в диаметре). См. таблицу 8 для состава двух смесей поверхностно-активных веществ. Смесь "91" в идеальном варианте применяют с жирными кислотами в концентрации 6,5 части смеси поверхностно-активных веществ на 1 часть жирной кислоты. Однако также могут действовать несколько близких концентраций и близких смесей поверхностно-активных веществ. Смесь поверхностно-активных веществ "92" в идеальном варианте применяют со сложными эфирами жирных кислот, а также маслами в чистой форме либо в виде смеси триглицеридов. В идеальном варианте их смешивают в соотношении 10 частей смеси поверхностно-активных веществ на 1 часть сложного эфира или масла. Любая из этих смесей или близких смесей соединений может являться подходящей для надлежащего растворения или образования микроэмульсии с близкими молекулярными классами липидов помимо жирных кислот, сложных эфиров и масел. Они могут включать в себя фосфолипиды, диацилглицерины, лизофосфолипиды, сложные эфиры тритерпеноидов или глицеролипиды.
[00245] Эти смеси поверхностно-активных веществ, как правило, смешивают с активным ингредиентом и затем смешивают в определенном процентном соотношении с водным буфером с получением эмульсии и в идеальном варианте микроэмульсии. При получении эмульсии процентное содержание смеси поверхностно-активных веществ+активного ингредиента в водном буфере может иметь любое значение в пределах 0,01-50%. Водный буфер может состоять из любого количества элементов. Авторы настоящего изобретения применяли 1x PBS+1% DML и PBS+50% DML, но другие составы также действуют."PBS" означает фосфатно-солевой буферный раствор; "DML" означает диметиллактамид.
[00246] Применение этих буферов в отдельности не было способно привести к индукции гаплоидии de novo, что можно увидеть в строках 121-123 таблицы 7.
[00247] Способ нанесения соединений может иметь любое количество форм, включая погружение цветочной почки, инъекцию в цветочную почку, микроинъекцию, замачивание пыльцы, разбрызгивание на пыльцу, опрыскивание пыльцы, разбрызгивание на цветочную почку, опрыскивание цветочной почки, окунание рыльца, опрыскивание рыльца и т.д. Авторы настоящего изобретения чаще всего применяли опрыскивания метелок, пыльцы и рыльца до, во время или после опыления. Авторы настоящего изобретения достигали образования гаплоидов de novo посредством DSPC и линоленовой кислоты в случае опрыскивания метелок за два дня до опыления. Авторы настоящего изобретения также достигали образования гаплоидов de novo посредством DSPC в случае опрыскивания метелок всего за несколько минут до сбора пыльцы и последующего опыления. Аналогичным образом, авторы настоящего изобретения достигали образования гаплоидов de novo посредством DSPC в случае опрыскивания пыльцы непосредственно до или во время опыления, а также в случае опрыскивания рылец до, во время или после опыления, в том числе в период до двух дней включительно до опыления и до шести часов включительно после опыления.
[00248] Авторы настоящего изобретения также продемонстрировали образование гаплоидов de novo при применении различных аппликаторов, в том числе дешевого пластикового пульверизатора, который можно приобрести в магазине товаров повседневного спроса, и распылителя, который представляет собой медицинское устройство, применяемое для пероральной доставки лекарственных препаратов в форме очень маленьких водных капель. При том, что пульверизатор, как правило, образует капли размером 50-150 микрон в диаметре, распылитель способен образовывать капли размером менее 10 микрон. Это является преимущественным для нанесения на пыльцу, поскольку размеры пыльцы у большинства растений находятся в диапазоне 20-200 микрон в диаметре (примерно 70 микрон у маиса; примерно 50 микрон у риса), и если одна из капель из обычного пульверизатора попадет на зерно, то капля будет больше, чем зерно. Пыльца чрезвычайно чувствительна к влажности и осмотическому шоку. Если пыльцевое зерно вступает в контакт с каплей слишком большого размера, то из этого зерна неспособна прорасти успешная растущая пыльцевая трубка. То же самое может быть и в случае, если зерно попадает на дыхальце или рыльце, которое является слишком влажным например, если это рыльце или дыхальце получило слишком много наносимого липида (будь то микроэмульсия или просто липидные капли или мицеллы, образовавшиеся в водном растворе). Если бы авторы настоящего изобретения наносили более 3 мл аэрозольного липида на пыльцу или рыльца с помощью пульверизатора или распылителя, авторы настоящего изобретения часто получали бы очень низкий уровень завязывания семян.
[00249] Таким образом, способ нанесения имеет решающее значение, и, в частности, наиболее важно попытаться нанести как можно меньший объем аэрозоля на пыльцу, или дыхальце, или рыльца, или на цветок в целом, и наилучшим способом распределения активного ингредиента, в частности, липида, в как можно меньшем объеме является получение микроэмульсии этого липида в водном буфере, где размеры липидных капель находятся на субмикронном уровне, а затем дозирование этой микроэмульсии с помощью распылителя или аналогичного устройства, способного образовывать капли, имеющие размер в диапазоне 1-2 микрона. Это приводит в результате к доставке миллионов капель активного ингредиента в "парообразной" форме в соответствующие ткани, при этом размер капли по объему в >100000 раз меньше размера самого пыльцевого зерна (если капля имеет размер 1 микрон, а пыльцевое зерно имеет размер 50 микрон, то диаметр капли в 50 раз меньше диаметра зерна). А 503=125000, таким образом, объем капли в >105000 раз меньше объема пыльцевого зерна. При использовании микроэмульсий размер липидных капель может находиться в диапазоне примерно 20-1000 нм в диаметре, таким образом, в зависимости от концентрации смеси поверхностно-активных веществ вместе с активным ингредиентом в водном растворе в каждой капле, доставляемой к каждому пыльцевому зерну или рыльцу, может находиться одна или несколько связок липидов. Кроме того, принимая во внимание то, что некоторые молекулы липидного типа или ингибиторы фосфолипазы можно растворить в водном растворе вместо доставки в виде микроэмульсии, авторы настоящего изобретения обнаружили, что соединения могут индуцировать образование гаплоидов в большом разнообразии концентраций от 20 мкг/мл до 50 мг/мл включительно. Возможно, в более высоких концентрациях соединения будут индуцировать образование гаплоидов de novo.
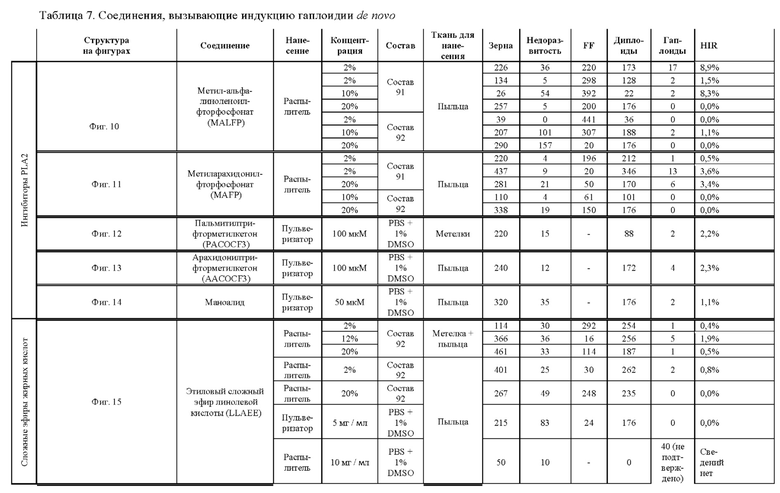
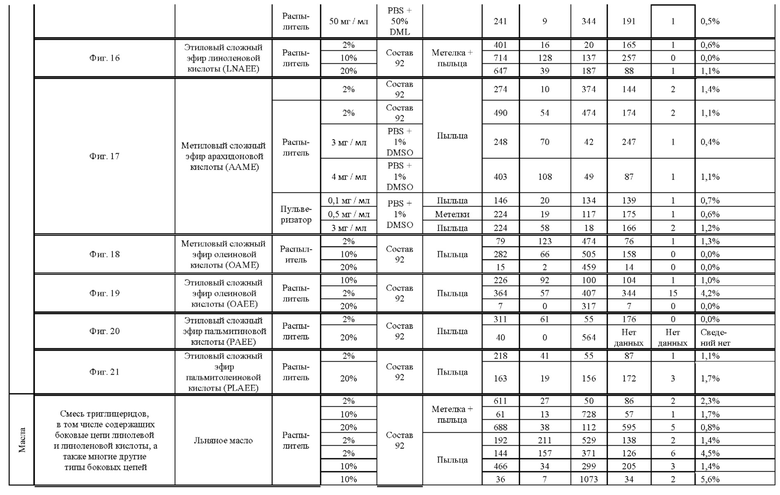
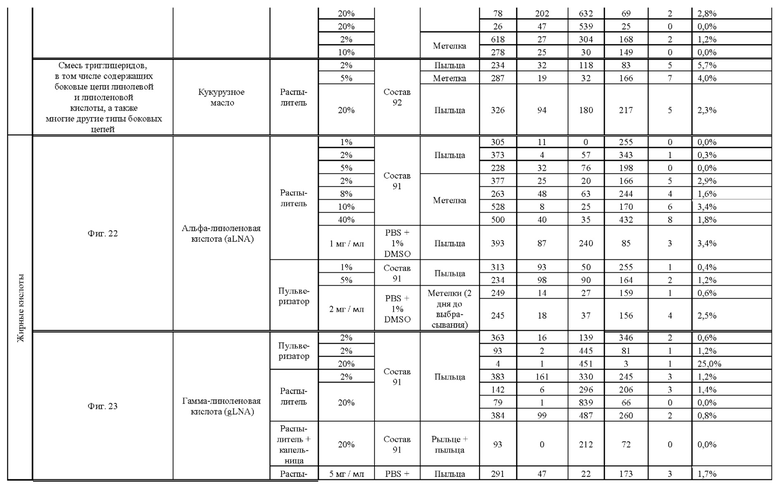
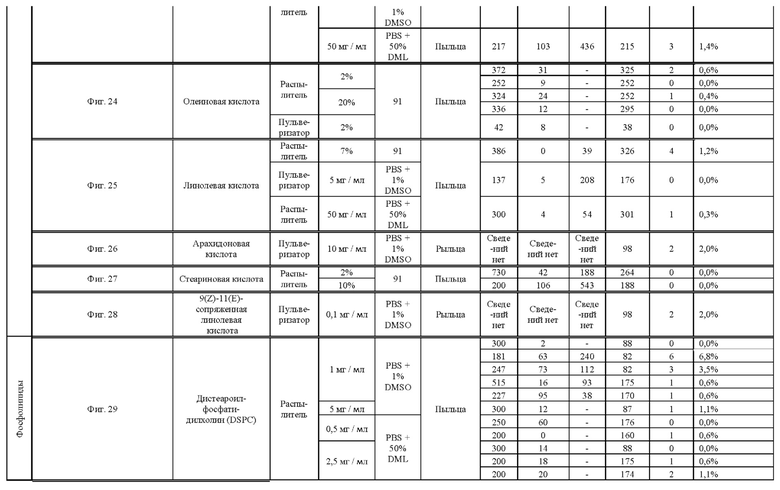
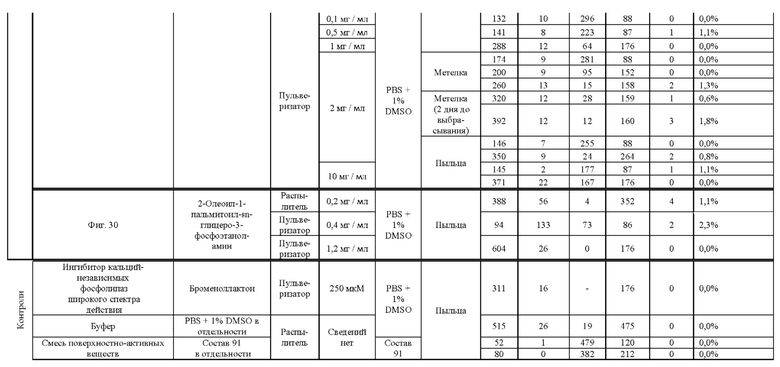

IV. Увеличение HIR и восстановление завязывания семян посредством нанесения липидного соединения
[00250] Молекулярное профилирование, в том числе метаболомическое и липомическое профилирование, пыльцы, индуцирующей и не индуцирующей гаплоидию (сравнительно), показывает, что пыльца, индуцирующая гаплоидию, является, в частности, дефицитной по определенным типам классов липидов и избыточной по другим. Этот дефицит особенно выражен в классе молекул липидов с цепью из 18 атомов углерода, хотя он также наблюдается в классах с цепью из 20 атомов углерода. Дефицит, в частности, выражен для липидов с цепью из 18 атомов углерода с одной и двумя двойными связями (так называемых олеатов и линолеатов), а избыток был обнаружен, в частности, для липидов с цепью из 18 атомов углерода с тремя двойными связями (так называемых линоленатов). Что касается типов липидов, измененных в пыльце, индуцирующей гаплоидию, то их спектр является очень широким и включает триглицериды, диацилглицериды, свободные жирные кислоты, лизофосфолипиды и фосфолипиды. Изменения содержания липидов в пыльце, индуцирующей гаплоидию, также варьируются среди жирных кислот с различными уровнями насыщения. После рассмотрения этих данных авторы настоящего изобретения решили наносить липидные соединения на цветки во время опыления пыльцой, индуцирующей гаплоидию, чтобы увидеть, может ли простое добавление определенных типов липидов повлиять на уровень образования гаплоидов или число зерен. Авторы настоящего изобретения были особенно заинтересованы в нанесении соединений, которые имели в своей молекулярной структуре некоторые насыщенные жирные кислоты с цепями из 18 или 20 атомов углерода, находившиеся среди тех, представленность которых, как было замечено, является более низкой в пыльце, индуцирующей гаплоидию (например, олеат и линолеаты).
[00251] Нанесение липидов может также привести к увеличению уровня образования гаплоидов, завязывания семян и снижению частоты недоразвития зерен, что указано в таблицах 10-13. В таблице 10 показано девять отдельных контролируемых экспериментов с применением соединения метил-альфа-линоленоилфторфосфоната (MALFP). Каждый эксперимент предусматривал скрещивание двух початков с одним и тем же мужским родительским растением, индуцирующим гаплоидию, в один и тот же день почти точно в одно и то же время (в пределах 5 минут друг от друга) из точно одной и той же популяции пыльцы, собранной с одной и той же родительской мужской особи. Эти два початка получали наносимый контрольный буфер либо буфер вместе с активным ингредиентом. Как мужские, так и женские тестируемые растения выращивали рядом друг с другом в одних и тех же условиях в одной и той же теплице. В среднем, и по сравнению с контрольными процедурами нанесения буфера без MALFP, в случае нанесения MALFP уровень индукции гаплоидии увеличивался на 3,6%, количество зерен увеличивалось почти на 42 на один початок, и общее количество воспроизводящихся гаплоидов на один початок увеличивалось на 11,8 гаплоидов на один початок, а частота недоразвития зародышей уменьшалась на 1,1% (таблица 9). Все это представляет улучшения схемы получения индуцированных гаплоидов. Улучшения были особенно очевидными, если состав 91 применяли в качестве смеси поверхностно-активных веществ для растворения MALFP. Механизм, посредством которого эти ингибиторы оказывают аддитивный эффект вместе с mtl, приводящий к более высоким уровням индукции, может заключаться в том, что они разрушают другие фосфолипазы или липид-модифицирующие белки.
[00252] Тот же тип данных собирали для этилового сложного эфира жирной кислоты LLAEE, который дает увеличение уровня индукции гаплоидии на 2,4%, увеличение образования зерен более чем на 43 на один початок, увеличение образования гаплоидов более чем на 8 на один початок и уменьшение частоты недоразвития зародышей на 3,7%. Хотя многие из этих видов обработки предусматривали различные количества активного ингредиента, в большинстве из этих экспериментов LLAEE давал повышение этих статистических показателей образования гаплоидов и завязывания семян. См. таблицу 10.
[00253] Тот же тип данных собирали для жирной кислоты линолевой кислоты (LLA) (таблица 11), которая давала увеличение уровня индукции гаплоидии на 1,5%, увеличение образования жизнеспособных зерен на 41 на один початок и гаплоидов на 7,6 на один початок и уменьшение уровня недоразвитости зародышей на 2,9%. Это является результатом 22 отдельных экспериментов, при этом опять же каждый эксперимент состоит из опыления одного початка контрольным буфером и опыления одного початка буфером вместе с активным ингредиентом (LLE). Интересно, что представленность 18:2 снижена в пыльце, индуцирующей гаплоидию (что показывают липомические данные). Не желая ограничиваться какой-либо теорией, разумно выдвинуть гипотезу, что это может вызывать проблемы оплодотворения и недоразвитость зародышей или вносить вклад в них. Если авторы настоящего изобретения добавляли их обратно посредством нанесения LLA и LLAEE путем опрыскивания, эти молекулы могли ингибировать фосфолипазы, что приводило к более высокому НПЧ, и восполнять запасы 18:2, что приводило к большему количеству зерен.
[00254] И наконец, аналогичные данные собирали для MAFP, и были отмечены аналогичные результаты, в том числе повышение уровня индукции гаплоидии на 1,9%, увеличение количества воспроизводящихся гаплоидов на один початок на 6,6 и общего количества зерен на 28, а также уменьшение уровня недоразвитости зародышей на 2,2% (таблица 12). Эти соединения, а также другие липиды, можно применять для повышения частоты образования гаплоидных зародышей в рассадниках для индукции гаплоидии и также можно применять для увеличения уровня завязывания семян в случае с самоопыляемыми линиями, индуцирующими гаплоидию, для увеличения запаса семян этих линий.

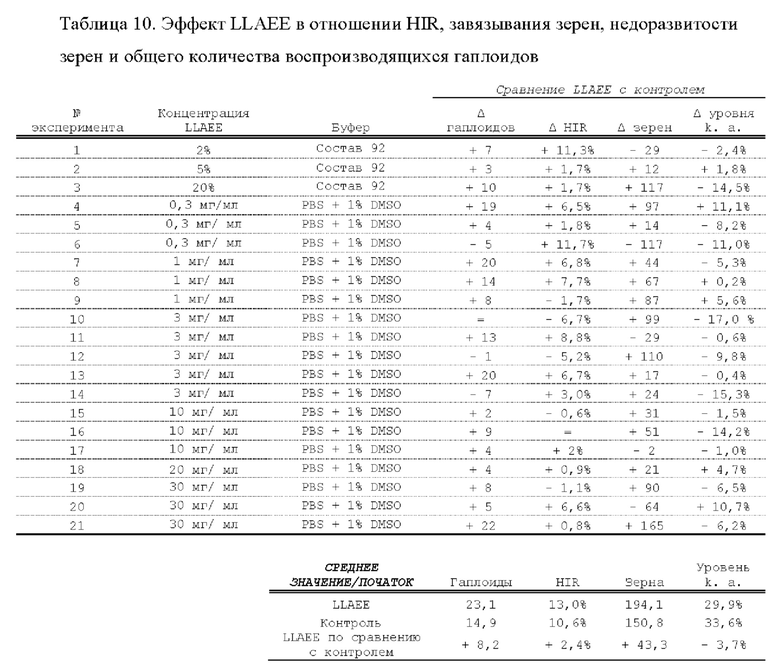
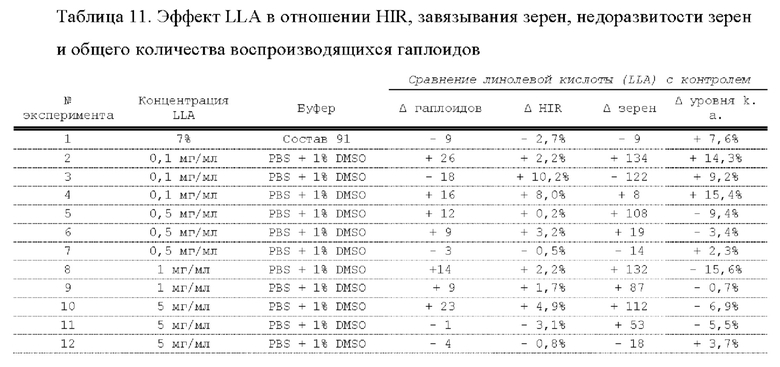
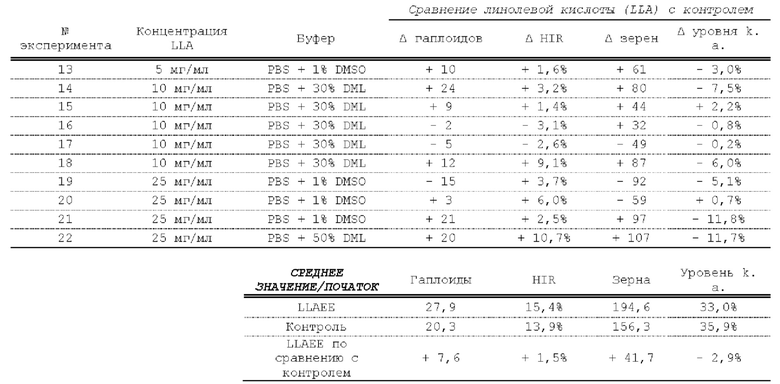
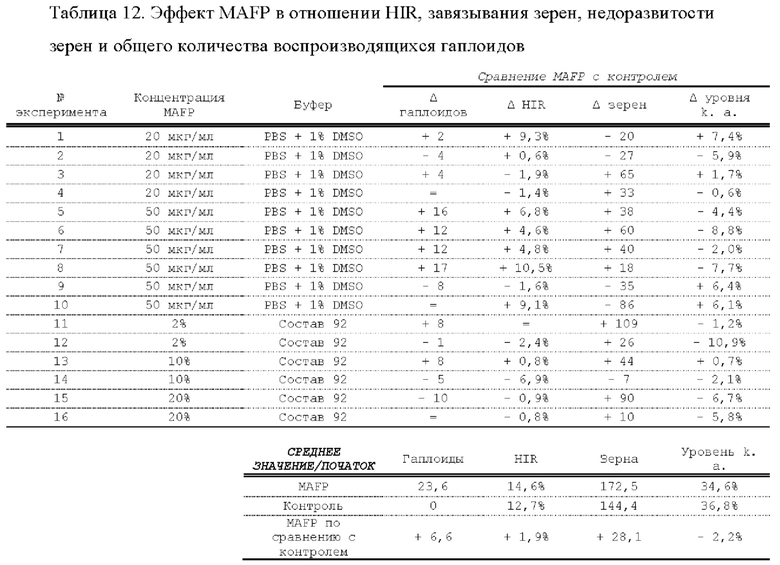
V. Мутагенез/нокаутные варианты PLA
[00255] В попытке изменить уровень индукции гаплоидии или уменьшить уровень недоразвитости зародышей в ходе скрещиваний для индукции гаплоидии авторы настоящего изобретения создавали или получали несколько мутантных линий посредством нескольких способов, в том числе GM-линии, полученные с помощью RNAi, линии, полученные с помощью TILLING, линии, полученные с помощью CRISPR, и линии, полученные с помощью TALEN. Вначале авторы настоящего изобретения искали доказательства того, что направленный мутагенез pPLAIIα является осуществимой стратегией для создания новых линий, индуцирующих гаплоидию. Таким образом, авторы настоящего изобретения тестировали стратегию направленных мутаций маиса с использованием как CRISPR/CAS9, так и TALEN, направленную на одну и ту же последовательность, которая содержит сдвиг рамки считывания в мутантном аллеле, индуцирующем гаплоидию. Это приводило к образованию линий с новыми мутациями, которые авторы настоящего изобретения тестировали в отношении индукции гаплоидии.
[00256] Существует три ключевых компонента для CRISPR-опосредованного процесса. См. патент США №8697359 В1, включенный в данный документ посредством ссылки во всей своей полноте. Первым ключевым компонентом является последовательность-мишень. Вторым является Cas9, которая представляет собой эндонуклеазу. Третьим ключевым компонентом является направляющая РНК ("gRNA"), которая является комплементарной последовательности-мишени и отвечает за привлечение Cas9 в необходимое местоположение. Последовательность-мишень составляет 18-20 п.о. в длину и необязательно должна располагаться сразу после мотива, прилегающего к протоспейсеру ("РАМ"), в 5'-направлении относительно него в геноме растения. Для Cas9 из Streptococcus pyogenes последовательность РАМ должна представлять собой 5'-NGG-3'. Транскрипция gRNA может управляться промотором Pol III U3 (если РНК начинается с А) или U6 (если РНК начинается с G). gRNA должна нести последовательность-мишень на 5'-конце непосредственно после А (U3) или G (U6). Cas9 будет образовывать двухнитевой разрыв ("DSB") в последовательности-мишени на расстоянии в три пары оснований в 5'-направлении относительно последовательности РАМ. Аминокислотная последовательность Cas9 является такой же, как у Cas9 из Streptococcus pyogenes штамма SF370, с двумя аминокислотными изменениями L1164V и I1179V в PI-домене (1099-1368) в NUC-доле. Активность Cas9, как было показано в экспериментах по трансформации, обуславливает примерно 90% частоту мутаций тестируемой последовательности-мишени у кукурузы. Как правило, рекомендуется идентифицировать несколько кандидатов среди РАМ и последовательностей-мишеней в области-мишени, а затем отыскать наилучшую, рассматривая то, какая из последовательностей является уникальной в геноме мишени. Растение-мишень представляет собой растение маиса, риса или любое однодольное растение.
[00257] Этой стратегии следовали для идентификации последовательностей-мишеней для CRISPR, которые перекрываются с существующей мутацией сдвига рамки считывания. Точный сайт разрезания находится всего в двух парах оснований от точки вставки в место сдвига рамки считывания. Конструкции, содержащие как Cas9, так и gRNA, вводили в растения маиса путем трансформации. Как правило, из множества образованных объектов воспроизводимыми являются биаллельные или гомозиготные мутантные растения, но гетерозиготные мутантные растения также являются применимыми. Гетерозиготные растения подвергали самооплодотворению, затем семена Т1 выращивали, подвергали скринингу в отношении гомозиготности мутации и подвергали ауткроссингу. Гомозиготных или биаллельных мутантных трансформантов Т0 просто подвергали самооплодотворению и ауткроссингу с трансформированной NP2222. Всех полученных в результате ауткроссинга зародышей выделяли для анализа плоидности для обнаружения гаплоидов.
[00258] Три различные созданные конструкции для направленного мутагенеза: CRISPR/CAS9 I, CRISPR/CAS9 II и TALEN. Различие между CRISPR/CAS9 I и II является незначительным. Локус сайта-мишени для всех трех конструкций представлял собой ту же область, где был обнаружен сдвиг рамки считывания в линиях, индуцирующих гаплоидию. Для конструкций CRISPR последовательность направляющей РНК начинается с нуклеотида +1560: -GTCAACGTGGAGACAGGG-(т.е. SEQ ID NO: 20). Сайт -AGG- РАМ SEQ ID NO: 20 подчеркнут и выделен курсивом. Вставка четырех пар оснований в линиях, индуцирующих гаплоидию, находится точно в том же сайте в нуклеотиде +1576. После трансформации несколько различных объектов, полученных с помощью CRISPR I (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 34), объектов, полученных с помощью CRISPR II (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 36), и объектов, полученных с помощью TALEN (содержащих экспрессионную конструкцию, находящуюся под SEQ ID NO: 35), были отобраны, выращены до зрелости и завязывали жизнеспособные семена. В поколении Т0 авторы настоящего изобретения осуществляли ПЦР в сайте-мишени и секвенировали продукты ПЦР после субклонирования. Авторы настоящего изобретения идентифицировали много уникальных мутаций среди этих объектов (и многие из объектов являлись химерами или имели несколько аллелей).
[00259] Многие растения являлись химерными, о чем свидетельствует появление нескольких различных последовательностей в поколении Т1. После самоопыления Т0 растения Т1 сегрегировали 1:2:1 по конструкции для направленного мутагенеза, и многие из них имели новые мутации в локусе-мишени в биаллельном либо в гомозиготном состоянии. Авторы настоящего изобретения осуществляли скрининг проростков на уровне ДНК с применением маркеров TAQMAN, идентифицировали биаллельные объекты, у которых отсутствовали трансгены, введенные с помощью Cas9 или TALEN, и осуществляли ПЦР-секвенирование с получением продукта ПЦР, прочитываемого от +1494 до +1691 пары оснований в последовательности гена GRMZM2G471240. Авторы настоящего изобретения затем тестировали гомозиготные мутантные формы в отношении способности индуцировать гаплоидию. См. SEQ ID NO: 9-19 и 42-44 в отношении последовательностей новых растений Т1 в гене mtl.
[00260] HIR измеряли для новых предполагаемых линий. См. таблицу 4, выше. Эти данные HIR получены из скрещиваний, где мужская особь относилась к предполагаемой линии, индуцирующей гаплоидию, а женская особь относилась к собственной стандартной инбредной трансформируемой линии NP2222. Предполагаемые линии, индуцирующие гаплоидию, создавали посредством направленной мутации локуса pPLAIIα, опосредованной TALEN либо CRISPR/CAS9. Среди показанных в данном документе существует одиннадцать различных предполагаемых индуцирующих растений, содержащих восемь различных объектов из трех отдельных трансформационных конструкций. Объект 39А представлял собой объект, полученный с помощью TALEN. Объекты 18А и 27А представляли собой объекты, полученные с помощью CRISPR. Последний представлял собой химеру, используемую в качестве растения Т0, и после его самоопыления в популяции Т1, в том числе в "биаллельных" растениях (под биаллельными авторы настоящего изобретения подразумевают то, что при секвенировании области pPLAIIα, которая была подвергнута мутации, авторы настоящего изобретения обнаружили два различных новых аллеля - так что очевидно, что обе копии гена дикого типа были подвергнуты мутации, но они были подвергнуты мутации различным образом, поэтому они представляют собой два новых аллеля), было обнаружено несколько мутаций. Каждое из этих одиннадцати отдельных растений, таким образом, имело различные комбинации мутаций в pPLAIIα. Общим у них всех было то, что ни одно из одиннадцати растений не имело копии pPLAII дикого типаα. Таким образом, все они являются "гомозиготными мутантными" по гену pPLAIIα. Все мутации представляли собой сдвиги рамки считывания в экзоне 4, имитирующие исходную мутацию в нативных линиях, индуцирующих гаплоидию. Используя эти пять растений в качестве мужских особей, авторы настоящего изобретения скрещивали их с одним либо несколькими женскими початками с образованием нескольких тысяч зародышей. Авторы настоящего изобретения срезали это потомство, осуществляли в его отношении анализ плоидности и обнаружили, что каждый из наборов потомков имел по меньшей мере 3,98% гаплоидов при максимальном содержании 12,5% гаплоидов. Это демонстрирует то, что образование мутаций в pPLAIIα приводит к индукции гаплоидии. Авторы настоящего изобретения думают, что другие типы мутаций, помимо сдвигов рамки считывания, также приводят к индукции гаплоидии. Эти мутации могут находиться в любом месте в гене, и они могут представлять собой точечные мутации, или вставки, или делеции, или другие типы мутаций.
[00261] Для образования линий, индуцирующих гаплоидию, также применяли RNAi. Для RNAi получали две шпилечные конструкции; одну, соответствующую границе между экзоном 1 и 2, и другую, соответствующую экзону 4 (фигуры 8 и 9). Конструкции вводили в растения дикого типа путем трансформации, и растения Т0 подвергали самооплодотворению. Семена Т1 из трех объектов на одну конструкцию выращивали, подвергали скринингу в отношении гомозиготности по трансгену и подвергали ауткроссингу с несколькими початками в качестве тестеров индукции гаплоидии. После исследования более 1500 зерен, полученных в результате этих ауткроссингов, авторы настоящего изобретения обнаружили, что оба объекта индуцировали гаплоидию на уровне, составляющем примерно 1%-2%. Наиболее высокий уровень индукции гаплоидии, полученный на одном початке, составлял 4,3%. Этот початок содержал приблизительно 300 зерен, так что авторы настоящего изобретения могут сделать вывод, что уровень недоразвитости зародышей также был ниже, чем у обычной линии, являющейся сильным индуктором. Эта работа демонстрирует то, что стратегию RNAi+GM можно применять для создания новых линий, индуцирующих гаплоидию, из линий, в иных отношениях обычно являющихся линиями дикого типа, путем изменения экспрессии pPLAIIα.
[00262] Для создания и идентификации мутаций фосфолипазы и маиса по настоящему изобретению также применяли способ мутагенеза TILLING. Публикации, в которых описан TILLING, доступны для культурных растений, таких как рис: Till et al., ВМС Plant Biology 7:19 (2007), томат: Rigola et al. PLOS ONE March 13, 2009, и маис: Till et al. BMC Plant Biol. 2004 Jul 28;4:12 (2004), все из которых включены в данный документ посредством ссылки. В базовой методике TILLING растительный материал, такой как семена, подвергают химическому мутагенезу, в результате которого создается серия мутаций в геномах клеток семян. Из семян, подвергнутых мутагенезу, выращивают взрослые растения M1, и их подвергают самоопылению. Образцы ДНК из полученных в результате этого растений М2 объединяют и затем подвергают скринингу в отношении мутаций в гене, представляющем интерес.После идентификации мутации в гене, представляющем интерес, из семян растений М2, несущих эту мутацию, выращивают взрослые растения М3, и их подвергают скринингу в отношении фенотипических характеристик, ассоциированных с геном, представляющим интерес.
[00263] Любой культивар маиса, имеющий по меньшей мере один ген фосфолипазы со значительной гомологией с SEQ ID NO: 1, можно применять в соответствии с настоящим изобретением. Используемое в данном документе выражение "значительная гомология" означает, что последовательность ДНК гена является достаточно сходной с SEQ ID NO: 1 на нуклеотидном уровне, чтобы кодировать белок, эквивалентный кодируемому SEQ ID NO: 1, при возможном наличии аллельных различий между культиварами. В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения "значительная гомология" может иметь место уже тогда, когда гомология между геном фосфолипазы и SEQ ID NO: 1 составляет приблизительно 85%, при условии, что гомология в консервативных областях гена является более высокой (например, по меньшей мере приблизительно 90%). Предпочтительно, процентная идентичность в кодирующей области составляет 85-90%, более предпочтительно 90-95%, и в оптимальном варианте она составляет более 95%. Специалист в данной области может предпочитать культивар маиса, имеющий коммерческую популярность или имеющий конкретные желаемые характеристики, из которого можно создать маис с мутантной фосфолипазой. В качестве альтернативы, специалист в данной области может предпочитать культивар маиса, имеющий незначительное количество полиморфизмов, такой как инбредный культивар, для облегчения скрининга в отношении мутаций в пределах локусов генов фосфолипаз.
[00264] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения семена риса и маиса подвергали мутагенезу, и затем из них выращивали растения M1. Затем обеспечивали самоопыление растений M1, и из семян растений M1 выращивали растения М2, которые затем подвергали скринингу в отношении мутаций в их локусе гена фосфолипазы. Хотя скринингу в отношении мутаций можно подвергать и растения M1, преимущество осуществления скрининга растений М2 заключается в том, что все соматические мутации соответствуют герминативным мутациям. Специалисту в данной области будет понятно, что множество растительных материалов маиса, в том числе без ограничения семена, пыльцу, растительную ткань или растительные клетки, можно подвергнуть мутагенезу для создания маиса с мутантной фосфолипазой по настоящему изобретению. Однако тип растительного материала, подвергаемого мутагенезу, может оказывать влияние при осуществлении скрининга растительной ДНК в отношении мутаций. Например, если пыльцу подвергают мутагенезу до опыления не подвергнутого мутагенезу растения, то из семян, полученных в результате этого опыления, выращивают растения M1. Каждая клетка растений M1 будет содержать мутаций, созданные в пыльце, поэтому эти растения M1 можно затем подвергать скринингу в отношении мутации фосфолипазы вместо того, чтобы ждать получения поколения М2.
[00265] Для создания мутаций по настоящему изобретению можно применять мутагены, которые создают главным образом точечные мутации и короткие делеции, вставки, трансверсии и или транзиции (от приблизительно 1 до приблизительно 5 нуклеотидов), такие как химические мутагены или радиоактивное излучение. Мутагены, соответствующие способу по настоящему изобретению, включают в себя без ограничения этилметансульфонат (EMS), метилметансульфонат (MMS), N-этил-N-нитрозомочевину (ENU), триэтилмеламин (ТЕМ), N-метил-N-нитрозомочевину (MNU), прокарбазин, хлорамбуцил, циклофосфамид, диэтилсульфат, мономерный акриламид, мелфалан, азотистый иприт, винкристин, диметилнитрозамин, N-метил-N'-нитронитрозогуанидин (MNNG), нитрозогуанидин, 2-аминопурин, 7,12-диметилбенз(а)антрацен (DMBA), этиленоксид, гексаметилфосфорамид, бисульфан, диэпоксиалканы (диэпоксиоктан (DEO), диэпоксибутан (ВЕВ) и т.п.), дигидрохлорид 2-метокси-6-хлор-9[3-(этил-2-хлорэтил)аминопропиламино]акридина (ICR-170) и формальдегид. Спонтанные мутации в области ядрышкового организатора ("NOR"), которые могут не быть непосредственно вызваны мутагеном, можно также идентифицировать в соответствии с различными вариантами осуществления настоящего изобретения.
[00266] Любой подходящий способ получения растительной ДНК, который известен на данный момент или будет разработан в будущем, можно применять для получения ДНК растения маиса для скрининга в отношении мутации фосфолипазы. Например, см. Chen и Ronald, Plant Molecular Biology Reporter 17:53-57, 1999; Stewart и Via, Bio Techniques 14:748-749, 1993. Кроме того, доступно несколько коммерческих наборов, в том числе наборы от Qiagen (Валенсия, Калифорния) и Qbiogene (Карлсбад, Калифорния).
[00267] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения образцы ДНК из отдельных растений маиса получали и затем объединяли для ускорения скрининга в отношении мутаций в фосфолипазе во всей популяции растений, происходящих из растительной ткани, подвергнутой мутагенезу. Размер объединенной группы может зависеть от чувствительности применяемого способа скрининга. В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения отдельные растения маиса объединяли в группы из четырех или более растений.
[00268] В соответствии с другим аспектом иллюстративного варианта осуществления после объединения образцов ДНК пулы подвергали методикам амплификации, специфичным в отношении последовательности фосфолипазы, таким как полимеразная цепная реакция (ПЦР). В отношении общего обзора ПЦР см. PCR Protocols: A Guide to Methods and Applications (Innis, Gelfand, Sninsky, J., и White, eds.), Academic Press, San Diego, 1990, который включен в данный документ посредством ссылки. Любой праймер, специфичный в отношении локуса гена фосфолипазы или последовательностей, непосредственно прилегающих к локусу гена фосфолипазы, можно использовать для амплификации последовательностей фосфолипазы в объединенном образце ДНК. Праймер предпочтительно разрабатывают для амплификации областей локуса гена фосфолипазы, где с наибольшей вероятностью возникнут полезные мутации. Праймер наиболее предпочтительно разрабатывают для выявления мутаций в кодирующей области гена фосфолипазы. Кроме того, предпочтительно, чтобы праймер избегал известных полиморфных сайтов для облегчения скрининга в отношении точечных мутаций. Для облегчения выявления продуктов ПЦР в геле праймер для ПЦР можно метить с помощью традиционного или разработанного в будущем способа мечения.
[00269] В соответствии с одним аспектом иллюстративного варианта осуществления настоящего изобретения продукты ПЦР-амплификации можно подвергнуть скринингу в отношении мутаций фосфолипазы с помощью любого способа, обеспечивающего идентификацию нуклеотидных различий между последовательностью дикого типа и мутантной последовательностью. Он может включать без ограничения секвенирование, денатурирующую жидкостную хроматографию высокого давления (dHPLC), капиллярный электрофорез с постоянной концентрацией денатурирующего средства (CDCE), капиллярный электрофорез с температурным градиентом (TGCE) (см. Li et al., Electrophoresis 23(10): 1499-1511, 2002) или фрагментацию с помощью ферментативного расщепления, как, например, применяемого в высокопроизводительном способе, описанном в Colbert et al., Plant Physiology 126:480-484, 2001. Продукты ПЦР-амплификации предпочтительно инкубируют с эндонуклеазой, которая преимущественно расщепляет участки ошибочного спаривания оснований в гетеродуплексах между последовательностью дикого типа и мутантной последовательностью. В соответствии с другим аспектом иллюстративного варианта осуществления продукты расщепления подвергают электрофорезу с помощью автоматизированного устройства для электрофореза в секвенирующем геле, и изображения геля анализируют с помощью стандартной коммерческой программы для обработки изображений.
[00270] Авторы настоящего изобретения определили, что для достижения индукции гаплоидии у маиса желаемыми являются мутации, которые изменяют функцию фосфолипазы. Предпочтительные мутации включают миссенс-мутации, нонсенс-мутации и изменения границы сплайсинга, в том числе мутации, проявляющиеся в преждевременном завершении трансляции белка фосфолипазы с информационной РНК, как, например, такие мутации, которые создают стоп-кодон в пределах кодирующих областей гена фосфолипазы. Такие мутации включают вставки, повторения последовательностей, модификации открытых рамок считывания (ORF) и, наиболее предпочтительно, точечные мутации.
[00271] В соответствии с еще одним другим аспектом иллюстративного варианта осуществления настоящего изобретения после идентификации растения М2, имеющего мутантную последовательность фосфолипазы, мутации анализируют для определения их эффектов в отношении экспрессии, трансляции и/или активности белка. В соответствии с одним иллюстративным вариантом осуществления фрагмент фосфолипазы, содержащий мутацию, секвенируют с помощью стандартных методик секвенирования для определения точного местоположения мутации относительно всей последовательности фосфолипазы. Каждую мутацию оценивают для того, чтобы предсказать ее влияние на функцию белка (т.е. от полной переносимости до потери функции), с помощью биоинформатических инструментов, таких как SIFT (рассортировка непереносимых и переносимых мутаций; Ng et al., Nucleic Acids Research 31:3812-3814, 2003), PSSM (позиционно-специфическая матрица замен; Henikoff и Henikoff, Computer Applications in the Biosciences 12:135-143, 1996) и PARSESNP (Taylor и Greene, Nucleic Acids Research 31:3808-3811, 2003). Например, балл SIFT, который составляет менее 0,05, и значительное изменение балла PSSM (например, примерно на 10 или более) указывает на мутацию, которая по всей вероятности оказывает пагубный эффект в отношении функции белка.
[00272] В соответствии с дополнительным аспектом иллюстративного варианта осуществления в случае, если первоначальная оценка мутации у растения М2 указывает на то, что она имеет полезную природу и находится в полезном положении в гене фосфолипазы, то проводят дополнительный фенотипический анализ растения маиса, содержащего эту мутацию. Вначале растение М2 дважды подвергают возвратному скрещиванию или ауткроссингу для создания растения ВС1 с целью устранения фоновых мутаций. Затем полученное в результате возвратного скрещивания или ауткроссинга растение ВС1 подвергают самоопылению для создания растения BC1F2, которое является гомозиготным по мутации фосфолипазы.
[00273] Оценивают несколько физических характеристик этих гомозиготных по фосфолипазе мутантных растений для определения того, приводит ли мутация в результате к полезному фенотипическому изменению у маиса. Мутантный по фосфолипазе маис оценивают в отношении индукции гаплоидии в сравнении с нормальным родительским маисом (например, дикого типа) или с контрольным сибсом маиса дикого типа. В таблице 13 показаны новые мутации, полученные с помощью TILLING.
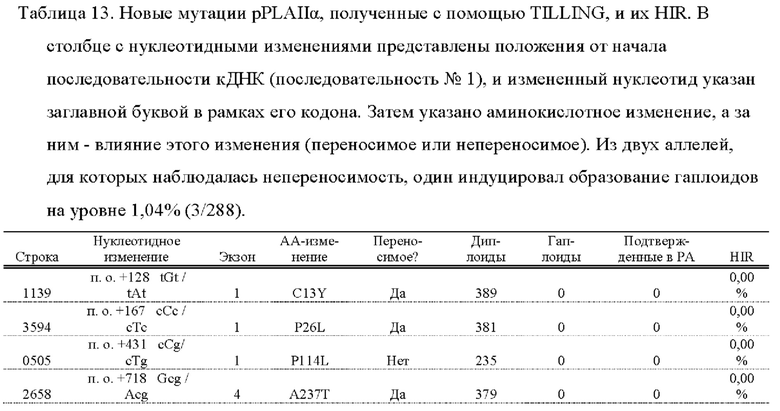
[00274] С помощью номенклатуры, применяемой в таблице 13, указываются нуклеотид или аминокислота дикого типа, за которыми указано их положение в соответствии с эталонной SEQ ID NO: 1, за которыми указаны измененные нуклеотид или аминокислота в этом положении, с применением стандартной терминологии генетического кода.
[00275] В случае с маисом в результате TILLING гена pPLAIIα маиса образуются новые аллели, которые характеризуются низкими уровнями индукции гаплоидии. Это позволяет создавать аллельные серии, в том числе нокаутные варианты, GRMZM2G471240. Последовательность двух сегментов этого гена (максимум 1,5 т.п.о., что равняется 20 ампликонам на один ген) подвергают скринингу в отношении мутаций. Эти последовательности включали в себя геномную последовательность, содержащую интроны, а также предсказанную последовательность кДНК и кодирующие последовательности для двух сплайс-вариантов. На основании этих последовательностей разрабатывают соответствующие и уникальные последовательности ампликонов, и скрининг в отношении мутаций осуществляют в существующей общей популяции кукурузы М2. Идентифицированные мутантные формы характеризуют в категориях последовательности ДНК и последствий в отношении транслируемой последовательности белка. Семена М3 выращивают и подвергают самооплодотворению с получением линий М4 с сегрегирующими предполагаемыми мутантными гомозиготными индивидуумами. Этих индивидуумов идентифицируют посредством ПЦР-секвенирования и подвергают ауткроссингу и самооплодотворению для тестирования способности этих мутантных линий индуцировать образование гаплоидов.
[00276] Для осуществления анализирующих скрещиваний новые линии выращивают рядом с маркерной линией, которая является гомозиготной рецессивной по нелетальному гену маркера цвета. Реципрокные скрещивания применяют для тестирования специфичности индукции в отношении передачи по мужской и женской линии посредством оценки полученных в результате растений в отношении наличия гаплоидов, которые демонстрируют фенотип цвета. Положительные хиты подтверждают посредством анализа плоидности, описанного выше.
[00277] Индивидуумов, которые являются гомозиготными в отношении мутаций SNP, скрещивали в качестве мужской особи с женской особью маркерной линии, что в некоторых случаях приводило к образованию гаплоидов на низком уровне (см. таблицу 18). Положительные хиты подтверждают посредством анализа плоидности, описанного выше. В частности, линия, которая давала образование гаплоидов, имела мутацию замены G на А в паре оснований 1077 последовательности кДНК. Эта мутация вызывает аминокислотную замену метионина (М) на изолейцин (I) в положении аминокислоты 356. Это является не консервативным аминокислотным изменение, которое может нарушать активность белка, что приводит к низкому уровню образования гаплоидов. Среди 288 тестированных потомков авторы настоящего изобретения обнаружили трех гаплоидов при уровне индукции 1% (3/288).
VI. Создание линий риса, индуцирующих гаплоидию
[00278] У риса ближайшим гомологом pPLAIIα маиса является Os03g27610, пататин-подобная фосфолипаза риса (OspPLAIIα) со сходной аннотацией, структурой гена и профилем экспрессии, т.е. она экспрессируется в пыльце и отсутствует в других частях растений (фигура 10). SEQ ID NO: 21 содержит последовательность геномной ДНК Os03g27610, SEQ ID NO: 22 содержит последовательность кДНК, и SEQ ID NO: 23 содержит аминокислотную последовательность. Близкое совпадение этих признаков, наряду с коротким эволюционным расстоянием между этими двумя злаковыми растениями, позволяет предположить, что мутация в гене риса может также способствовать образованию линии, индуцирующей гаплоидию. В недавней публикации белок риса был выявлен в ядрах спермиев пыльцевых зерен (Abiko et al., 2013), что позволяет предположить участие этого белка в оплодотворении и/или развитии зиготы.
[00279] Для улучшения уровня индукции гаплоидии у маиса и создания первых линий риса, индуцирующих гаплоидию, применяли подход "обратной генетики" TILLING для получения новых индивидуумов, мутантных по гену GRMZM2G471240 маиса и гену Os03g27610 риса. См. McCallum CM et al. (2000) Targeting induced local lesions IN genomes (TILLING) for plant functional genomics, PLANTPHYSIOL. 123: 439-42, включенный в данный документ посредством ссылки. TILLING представляет собой беспристрастный подход для образования новых мутантных форм, поскольку исследователь не может контролировать, где мутаген этилметансульфонат (EMS) будет создавать новые мутации. Получали новые аллели в определенном разнообразии и представленности, и их тестировали в отношении уровня индукции гаплоидии.
[00280] С помощью TILLING получали тринадцать различных линий М3. См. таблицу 14. В столбце "Положение в геноме" указано нуклеотидное положение мутации и изменение (например, G803A указывает на то, что пара оснований G в положении 803 была изменена на А). Эффектом является изменение аминокислоты или другое изменение белка в результате мутации (например, А209Т указывает на то, что аланин в положении аминокислоты 209 был изменен на треонин). Балл BLOSUM является предсказательной оценкой силы эффекта, который изменение аминокислоты будет оказывать на конформацию или сворачивание белка (чем она более отрицательная, тем сильнее эффект). "Тип" указывает на тип изменения аминокислоты ("NSM" означает немолчащую мутацию; "PSM" означает частично молчащую мутацию; "молчащая" означает молчащую мутацию; "сплайс" означает мутацию сайта сплайсинга, приводящую в результате к аберрантному сплайсингу; "интрон" означает, что мутация происходит в интроне). Наконец, № в GSOR представляет собой ID линии в коллекции генетического фонда Oryza в USDA.
[00281] Эти тринадцать линий подвергали самооплодотворению с получением М4, и семена М4 выращивали и тестировали в отношении гомозиготности. Гомозиготных мутантных индивидуумов подвергали самооплодотворению и ауткроссингу для тестирования способности индуцировать гаплоидию. Полученное в результате потомство исследовали в отношении содержания ДНК на одну клетку (плоидности) с помощью анализатора плоидности.
[00282] Неконсервативные изменения, такие как изменения в сайте сплайсинга и изменения с наиболее отрицательными баллами BLOSUM, имеют наибольший потенциал индукции гаплоидии. Они должны оказывать более выраженные дестабилизирующие эффекты на белковый продукт и поэтому в сравнении с другими индуцирующими гаплоидию аллелями, полученными с помощью TILLING, превосходят их, способствуя образованию большего количества гаплоидов на одно скрещивание для индукции гаплоидии и по всей вероятности обуславливая частично нарушенное завязывание семян. В действительности авторы настоящего изобретения уже начали наблюдать это в некоторых самоопылениях Т4. Линия с наиболее низким уровнем завязывания семян содержала мутацию G153A в сайте сплайсинга, при этом воспроизводилось только 29 семян на 12 скрещенных гомозиготных мутантных растений М4. Другие линии имели более 100 воспроизводящихся семян.
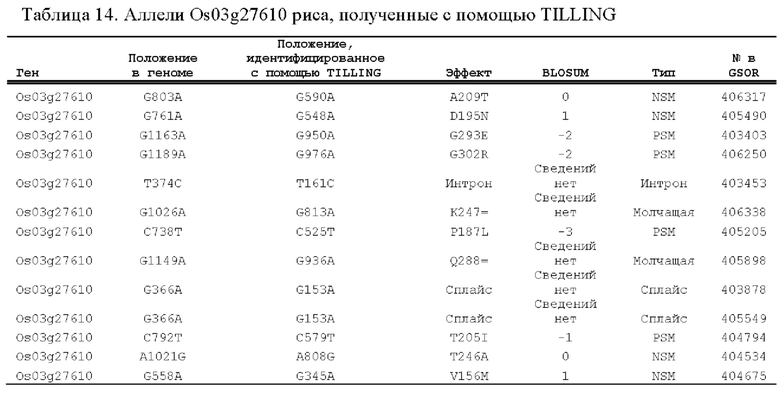
[00283] В качестве альтернативы, ген фосфолипазы риса, обнаруженный в Os03g27610, может быть отредактирован с помощью способов CRISPR/Cas9. Как указано выше, существует три ключевых компонента для CRISPR-опосредованного процесса. Первым ключевым компонентом является последовательность-мишень. Вторым является Cas9, которая представляет собой эндонуклеазу. Третьим ключевым компонентом является направляющая РНК ("gRNA"), которая является комплементарной последовательности-мишени и отвечает за привлечение Cas9 в необходимое местоположение. Направляющие РНК могут находиться в форме одинарной направляющей РНК (sgRNA) или двойной направляющей РНК (dgRNA). Для риса авторы настоящего изобретения создали четыре конструкции, целенаправленно воздействующие на ген фосфолипазы риса. SEQ ID NO: 38 содержит кассету экспрессии, которая обеспечивает нацеливание dgRNA на экзон 4 Os03g27610 очень близко к тому месту, где в гомологе маиса расположена природная мутация, затрагивающая четыре пары оснований. В гене риса сайтом-мишенью для направляющей РНК является GAGACCGGCAGGTACGTCGAGG. SEQ ID NO: 39 содержит кассету экспрессии, которая обеспечивает нацеливание sgRNA на тот же сайт-мишень для gRNA в экзоне 4 Os03g27610, на который нацеливается SEQ ID NO: 38. Ожидается, что в случае как с SEQ ID NO: 38, так и с SEQ ГО NO: 39 мутации сдвига рамки считывания появятся там, где помещена вертикальная черта между G и Т в последовательности CAGGTACG TCGAGG (в паре оснований +1150 последовательности gDNA в SEQ ID NO: 22). Таким образом, как ожидается, обе из этих конструкций будут обеспечивать образование мутаций, индуцирующих гаплоидию, расположенных всего на семь пар оснований ниже места, где у маиса расположена вставка, индуцирующая гаплоидию. Эти мутации в большинстве случаев будут представлять собой мутации сдвига рамки считывания, которые индуцируют небольшие вставки или делеции, например, делецию G или Т в сайте разрезания, или любую другую аналогичную мутацию. SEQ ID NO: 40 содержит кассету экспрессии, которая обеспечивает нацеливание dgRNA на Os03g27610. SEQ ID NO: 41 содержит кассету экспрессии, которая обеспечивает нацеливание sgRNA на Os03g27610. Обе они содержат направляющие РНК, которые нацеливаются на последовательность CCTCGCCGATTACTTCGACTGCA в экзоне 1. В результате этого должен образовываться нокаут большей части кодирующей последовательности гена. Образующаяся мутация должна появиться в сайте разрезания, где помещена вертикальная черта между С и С в последовательности CCTCGC CGATTAC (в паре оснований +215 последовательности кДНК в SEQ ID NO: 22). Таким образом, конструкции как 40, так и 41, как ожидается, будут обеспечивать высокую частоту образования растений, содержащих нокаут-мутации гена, что должно также привести к высоким уровням индукции гаплоидии у риса.
[00284] Растения риса трансформируют с помощью трансформационной конструкции, содержащей последовательность, выбранную из группы, состоящей из SEQ ID NO: 38-41. Посредством аппарата CRISPR/Cas9, закодированного в трансформационной конструкции, у трансформантов, т.е. растений риса Т0, образуются новые аллели гена фосфолипазы. Растения риса Т0 выращивают и скрещивают (т.е. подвергают самоопылению) для создания растений Т1. Растения риса Т1 тестируют в отношении гомозиготности по новому аллелю гена фосфолипазы. Гомозиготные растения риса Т1 скрещивают с линией риса, и полученное в результате потомство тестируют в отношении гаплоидии с помощью анализатора плоидности. Гаплоидные зародыши, не содержащие выявляемой ДНК Т1, идентифицируют и подсчитывают, и измеряют HTR. В результате скрещивания образуется по меньшей мере один гаплоидный зародыш, и HIR является повышенным. HIR предпочтительно составляет по меньшей мере 5%. По меньшей мере одного гаплоидного зародыша обрабатывают средством, удваивающим число хромосом, например, колхицином, и из него выращивают двойное гаплоидное растение.
В нижеприведенной первой таблице показаны различные аллели, полученные путем редактирования матрилинейной последовательности дикого типа (SEQ ID NO: 1). В ней приведены дополнительные данные, подтверждающие другие изменения в SEQ ID NO: 1. Как указывалось ранее, SEQ ID NO: 1 - это последовательность, которая будет изменена путем редактирования гена. SEQ ID NO: 3 - это всего лишь одна отредактированная последовательность, которая может быть получена.

Таким образом, было обнаружено, что 4 из 86 подтвержденных гаплоидов имели мутации в гене MATL. Эти растения являются гаплоидами и не содержат никакой ДНК Cas9. Ясно, что трансген Cas9 был удален вместе с остальной ДНК мужского происхождения во время эмбриогенеза, и что в женском (полученном из яйцеклетки) геноме произошли изменения в процессе эмбриогенеза.
Эти изменения являются новыми и произошли в женском геноме в процессе эмбриогенеза, потому что линия индуктора гаплоидов обычно создает материнские гаплоиды, и было подтверждено, что они действительно являются гаплоидами. Кто-то может попытаться возразить, что есть вероятность, что это на самом деле отцовские гаплоиды, и что изменения, которые мы видим, на самом деле являются изменениями, которые уже присутствовали в отцовской ДНК. Однако это не так. Во-первых, мутации не совпадают с мутациями родителя по отцовской линии. Это хорошо видно из таблиц 3 и 4 (приведены ниже). Отредактированное гаплоидное растение USR01350343-1 было гомозиготным по вставке одного нуклеотида (буква "А"), но родительское растение мужского пола имело делецию 13 нуклеотидов. Аналогично, растение USR01350328-1 было гомозиготным по делеции А, но у родительского мужчины была делеция 13 нуклеотидов. Эти примеры, взятые вместе, доказывают, что в процессе индукции гаплоидов возможно редактирование материнского генома, что приводит к образованию отредактированных материнских гаплоидов. В соответствии с этим и на основе анализа, обнаруживающего присутствие MATL, и подтверждения с помощью анализа плоидности, а также с использованием трансгена Cas9 на мужской стороне под контролем промотора убиквитина кукурузы, скорость редактирования во время процесса индукции гаплоидов составляет около 4/86, или 4,65%.
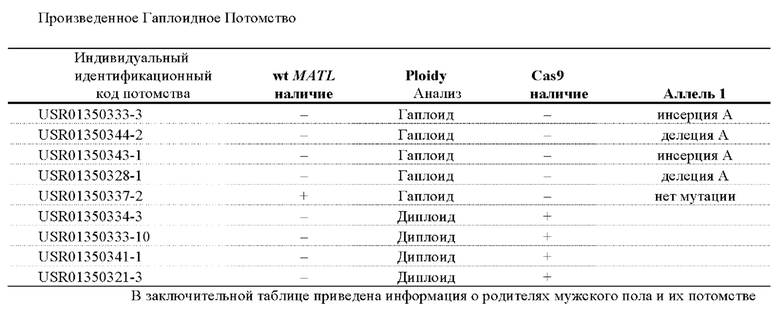
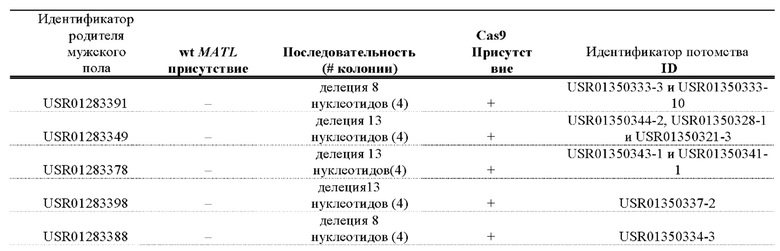
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Syngenta Participations, AG
Kelliher, Tim
Starr, Dakota
Wendeborn, Sebastian
Delzer, Brent
Chintamanani, Satya
Skibbe, David
Chen, Zhongying
<120> КОМПОЗИЦИИ ДЛЯ ИНДУКЦИИ ГАПЛОИДИИ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ
<130> 80906-WO-REG-ORG-P-1
<150> US 62/256,902
<151> 2015-11-15
<150> US 62/300,507
<151> 2016-02-16
<160> 81
<170> PatentIn версия 3.5
<210> 1
<211> 1371
<212> ДНК
<213> Zea mays
<400> 1
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 2
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 2
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 3
<211> 1375
<212> ДНК
<213> Zea mays
<400> 3
agttcatcac taatcacact tattgtgccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctggaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacta cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gcgctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaacccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gcatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct acgcatccag 1020
gacaactcgc tccgtggcgc cgcggcaacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagcggg tgtccagggt caacgtggag 1140
acagggagcg aggtacgaac cggtgaccgg agaaggaagc aatgccgatg ccctcggtgg 1200
gctcgctagg cagctctccg aggagaggag aacaaggctc gcgcgccgcg tctctgccat 1260
caaccccaga agctctagat gtgcgcccta cgatatctaa gacaagtggc tttactgtca 1320
atcacatgct tgtaaataag tagactttat tttaataaaa tataaaaata tatat 1375
<210> 4
<211> 372
<212> БЕЛОК
<213> Zea mays
<400> 4
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn Tyr Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Ser Glu Val Arg Thr Gly Asp Arg Arg Arg Lys Gln Cys Arg Cys Pro
355 360 365
Arg Trp Ala Arg
370
<210> 5
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер GRMZM2G471240_nil.F1
<400> 5
gtacgccgtg cgctaaca 18
<210> 6
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер GRMZM2G471240_nil.R1
<400> 6
tcgtacctcc ctgtctccac 20
<210> 7
<211> 19
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер GRMZM2G471240_rwk.F1
<400> 7
tacgccgtgc gctaacata 19
<210> 8
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> праймер GRMZM2G471240_rwk.R1
<400> 8
gtacctcgct ccctgtctcc 20
<210> 9
<211> 1358
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL в аллеле 1 у индивидуума с ID
22808-3954, представляющего собой объект 39А
<400> 9
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
aaccggtgac tggcgaagga agcaatgccg atgccctcgg tgggctcgct aggcagctct 1200
ccgaggagag gagaacaagg ctcgcgcgcc gcgtctctgc catcaaccca agaggctcta 1260
gatgtgcgtc gtacgatatc taagacaagt ggctttactg tcagtcacat gcttgtaaat 1320
aagtagactt tattttaata aaacataaaa atatatat 1358
<210> 10
<211> 1366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL в аллеле 1 у индивидуума с ID
22808-3924, представляющего собой объект 23А
<400> 10
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggagaa ccggtgactg gcgaaggaag caatgccgat gccctcggtg ggctcgctag 1200
gcagctctcc gaggagagga gaacaaggct cgcgcgccgc gtctctgcca tcaacccaag 1260
aggctctaga tgtgcgtcgt acgatatcta agacaagtgg ctttactgtc agtcacatgc 1320
ttgtaaataa gtagacttta ttttaataaa acataaaaat atatat 1366
<210> 11
<211> 1358
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL у объекта 81A
<400> 11
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
aaccggtgac tggcgaagga agcaatgccg atgccctcgg tgggctcgct aggcagctct 1200
ccgaggagag gagaacaagg ctcgcgcgcc gcgtctctgc catcaaccca agaggctcta 1260
gatgtgcgtc gtacgatatc taagacaagt ggctttactg tcagtcacat gcttgtaaat 1320
aagtagactt tattttaata aaacataaaa atatatat 1358
<210> 12
<211> 1343
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL в аллеле 2 у индивидуума с ID
22808-3954, представляющего собой объект 39А
<400> 12
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acggaagcaa tgccgatgcc ctcggtgggc tcgctaggca gctctccgag gagaggagaa 1200
caaggctcgc gcgccgcgtc tctgccatca acccaagagg ctctagatgt gcgtcgtacg 1260
atatctaaga caagtggctt tactgtcagt cacatgcttg taaataagta gactttattt 1320
taataaaaca taaaaatata tat 1343
<210> 13
<211> 1363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL в аллеле 2 у объекта 23A с ID
22808-3924
<400> 13
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaggaaccg gtgactggcg aaggaagcaa tgccgatgcc ctcggtgggc tcgctaggca 1200
gctctccgag gagaggagaa caaggctcgc gcgccgcgtc tctgccatca acccaagagg 1260
ctctagatgt gcgtcgtacg atatctaaga caagtggctt tactgtcagt cacatgcttg 1320
taaataagta gactttattt taataaaaca taaaaatata tat 1363
<210> 14
<211> 1360
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация MTL в аллеле 1 у индивидуума с ID
22808-4108, представляющего собой объект 38А
<400> 14
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaaccggtg actggcgaag gaagcaatgc cgatgccctc ggtgggctcg ctaggcagct 1200
ctccgaggag aggagaacaa ggctcgcgcg ccgcgtctct gccatcaacc caagaggctc 1260
tagatgtgcg tcgtacgata tctaagacaa gtggctttac tgtcagtcac atgcttgtaa 1320
ataagtagac tttattttaa taaaacataa aaatatatat 1360
<210> 15
<211> 1363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация MTL у объекта 18А с ID 22807-4016
<400> 15
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaggaaccg gtgactggcg aaggaagcaa tgccgatgcc ctcggtgggc tcgctaggca 1200
gctctccgag gagaggagaa caaggctcgc gcgccgcgtc tctgccatca acccaagagg 1260
ctctagatgt gcgtcgtacg atatctaaga caagtggctt tactgtcagt cacatgcttg 1320
taaataagta gactttattt taataaaaca taaaaatata tat 1363
<210> 16
<211> 1372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация MTL в аллеле 1 у индивидуума с
ID 22807-4801, представляющего собой объект 27A
<400> 16
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaagggagg tacgaaccgg tgactggcga aggaagcaat gccgatgccc tcggtgggct 1200
cgctaggcag ctctccgagg agaggagaac aaggctcgcg cgccgcgtct ctgccatcaa 1260
cccaagaggc tctagatgtg cgtcgtacga tatctaagac aagtggcttt actgtcagtc 1320
acatgcttgt aaataagtag actttatttt aataaaacat aaaaatatat at 1372
<210> 17
<211> 1372
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация в аллеле 1 у индивидуума с ID
22807-4081, представляющего собой объект 27A
<400> 17
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaagggagg tacgaaccgg tgactggcga aggaagcaat gccgatgccc tcggtgggct 1200
cgctaggcag ctctccgagg agaggagaac aaggctcgcg cgccgcgtct ctgccatcaa 1260
cccaagaggc tctagatgtg cgtcgtacga tatctaagac aagtggcttt actgtcagtc 1320
acatgcttgt aaataagtag actttatttt aataaaacat aaaaatatat at 1372
<210> 18
<211> 1373
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация MTL у индивидуума с ID 22873-3999,
представляющего собой объект 76A
<400> 18
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaaagggag gtacgaaccg gtgactggcg aaggaagcaa tgccgatgcc ctcggtgggc 1200
tcgctaggca gctctccgag gagaggagaa caaggctcgc gcgccgcgtc tctgccatca 1260
acccaagagg ctctagatgt gcgtcgtacg atatctaaga caagtggctt tactgtcagt 1320
cacatgcttg taaataagta gactttattt taataaaaca taaaaatata tat 1373
<210> 19
<211> 1370
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация у индивидуума с ID 22873-3991,
представляющего собой объект 32A
<400> 19
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acgggaggta cgaaccggtg actggcgaag gaagcaatgc cgatgccctc ggtgggctcg 1200
ctaggcagct ctccgaggag aggagaacaa ggctcgcgcg ccgcgtgtct gccatcaacc 1260
caagaggctc tagatgtgcg tcgtacgata tctaagacaa gtggctttac tgtcagtcac 1320
atgcttgtaa ataagtagac tttattttaa taaaacataa aaatatatat 1370
<210> 20
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Нуклеотидная последовательность, кодирующая направляющую РНК
<400> 20
gtcaacgtgg agacaggg 18
<210> 21
<211> 1823
<212> ДНК
<213> Oryza sativa
<220>
<221> 5'-UTR
<222> (1)..(78)
<220>
<221> экзон
<222> (79)..(444)
<223> Экзон 1
<220>
<221> Интрон
<222> (445)..(536)
<223> Интрон 1
<220>
<221> экзон
<222> (537)..(717)
<223> Экзон 2
<220>
<221> Интрон
<222> (718)..(803)
<223> Интрон 2
<220>
<221> экзон
<222> (804)..(967)
<223> Экзон 3
<220>
<221> Интрон
<222> (968)..(1074)
<223> Интрон 3
<220>
<221> экзон
<222> (1075)..(1662)
<223> Экзон 4
<220>
<221> 3'-UTR
<222> (1663)..(1823)
<400> 21
acagtgacta gtgacaaacg atcgatcgat ccctccatcc acaaaccctc ctcgatctca 60
tcttccttcg tctcgtca atg gcg gcg agc tac tcg tgc cgg cgg aca tgc 111
Met Ala Ala Ser Tyr Ser Cys Arg Arg Thr Cys
1 5 10
gag gcg tgc agc acg agg gcg atg gcc ggg tgc gtg gtg ggc gag ccg 159
Glu Ala Cys Ser Thr Arg Ala Met Ala Gly Cys Val Val Gly Glu Pro
15 20 25
gcg tcg gcg ccg ggg cag cgg gtg acg ttg ctg gcg atc gac ggc ggc 207
Ala Ser Ala Pro Gly Gln Arg Val Thr Leu Leu Ala Ile Asp Gly Gly
30 35 40
ggc atc agg ggc ctc atc ccg ggc acc atc ctc gcc ttc ctc gag gcc 255
Gly Ile Arg Gly Leu Ile Pro Gly Thr Ile Leu Ala Phe Leu Glu Ala
45 50 55
agg ctg cag gag ctg gat ggc ccc gac gcg cgc ctc gcc gat tac ttc 303
Arg Leu Gln Glu Leu Asp Gly Pro Asp Ala Arg Leu Ala Asp Tyr Phe
60 65 70 75
gac tgc atc gcc ggg acc agc acc ggc ggc ctc atc acc gcc atg ctc 351
Asp Cys Ile Ala Gly Thr Ser Thr Gly Gly Leu Ile Thr Ala Met Leu
80 85 90
gcc gcg ccc ggc gac cac ggc cgc ccg ctc ttc gcc gcc agc gac atc 399
Ala Ala Pro Gly Asp His Gly Arg Pro Leu Phe Ala Ala Ser Asp Ile
95 100 105
aac cgc ttc tac ctc gac aac ggc cca ctc atc ttc cca caa aag 444
Asn Arg Phe Tyr Leu Asp Asn Gly Pro Leu Ile Phe Pro Gln Lys
110 115 120
taactgatca cctcgaattc gatctcctct cttcgatctc tgcattattt gatttgattg 504
gggattgtgg gcggcgtggc gtggcgtcca gg agg tgc ggc atg gcg gcg gcc 557
Arg Cys Gly Met Ala Ala Ala
125
atg gcg gcg ctg acg agg ccg agg tac aac ggc aag tac ctg cag ggg 605
Met Ala Ala Leu Thr Arg Pro Arg Tyr Asn Gly Lys Tyr Leu Gln Gly
130 135 140 145
aag atc agg aag atg ctg ggc gag acg agg gtg cgc gac acg ctg acg 653
Lys Ile Arg Lys Met Leu Gly Glu Thr Arg Val Arg Asp Thr Leu Thr
150 155 160
aac gtc gtc atc ccc acg ttc gac gtc agg ctg ctc cag cca acc atc 701
Asn Val Val Ile Pro Thr Phe Asp Val Arg Leu Leu Gln Pro Thr Ile
165 170 175
ttc tcc aca tac gac g tgcgtgcgtt gattccatcc gcattggcgt tggaatcagc 757
Phe Ser Thr Tyr Asp
180
tgattgtttg attgatcgaa caattgatcg gttaaaattt tgcagg cg aag agc 811
Ala Lys Ser
185
atg ccg ctc aag aac gcg ctc ctc tcc gac atc tgc atc agc aca tcc 859
Met Pro Leu Lys Asn Ala Leu Leu Ser Asp Ile Cys Ile Ser Thr Ser
190 195 200
gcg gcg ccg acc tac ctc ccc gcg cac tgc ttc cag acc acc gac gac 907
Ala Ala Pro Thr Tyr Leu Pro Ala His Cys Phe Gln Thr Thr Asp Asp
205 210 215
gcc acc ggc aag gtc cgc gag ttc gac ctc atc gac ggc ggc gtc gcc 955
Ala Thr Gly Lys Val Arg Glu Phe Asp Leu Ile Asp Gly Gly Val Ala
220 225 230
gcc aac aac ccg gtaactaatc aatcaagcaa tccatcaaac gaagatccac 1007
Ala Asn Asn Pro
235
atgtgcattc ctgtggtaca aatgctgatc gatcgatgga tggatcgatt ttcgcgagaa 1067
cgtacag acg atg gtg gcc atg acg cag atc acc aag aag ata atg gtg 1116
Thr Met Val Ala Met Thr Gln Ile Thr Lys Lys Ile Met Val
240 245 250
aag gac aag gag gag ctg tac ccg gta aag ccg tcg gac tgc ggt aag 1164
Lys Asp Lys Glu Glu Leu Tyr Pro Val Lys Pro Ser Asp Cys Gly Lys
255 260 265
ttc ctg gtg ctg tcc gtg ggc acc ggg tcg acg tcg gac cag ggg atg 1212
Phe Leu Val Leu Ser Val Gly Thr Gly Ser Thr Ser Asp Gln Gly Met
270 275 280
tac acg gcg agg cag tgc tcg cgg tgg ggg atc gtc cgg tgg ctg cgc 1260
Tyr Thr Ala Arg Gln Cys Ser Arg Trp Gly Ile Val Arg Trp Leu Arg
285 290 295
aac aag ggg atg gcg ccc atc atc gac atc ttc atg gcg gcc agc tcc 1308
Asn Lys Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser
300 305 310 315
gac ctc gtc gac atc cac gcc gcc gtc atg ttc cag tcg ctg cac agc 1356
Asp Leu Val Asp Ile His Ala Ala Val Met Phe Gln Ser Leu His Ser
320 325 330
gac ggc gac tac ctc cgc atc cag gac aac acg ctc cac ggc gac gcc 1404
Asp Gly Asp Tyr Leu Arg Ile Gln Asp Asn Thr Leu His Gly Asp Ala
335 340 345
gcc acg gtg gac gcc gcc acc agg gac aac atg cgg gcg ctc gtc ggg 1452
Ala Thr Val Asp Ala Ala Thr Arg Asp Asn Met Arg Ala Leu Val Gly
350 355 360
atc ggc gag cgg atg ctg gcg cag cgg gtg tcg agg gtc aac gtc gag 1500
Ile Gly Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu
365 370 375
acc ggc agg tac gtc gag gtg ccc ggc gcc ggc agc aac gcc gac gcg 1548
Thr Gly Arg Tyr Val Glu Val Pro Gly Ala Gly Ser Asn Ala Asp Ala
380 385 390 395
ctg agg ggc ttc gcc agg cag ctc tcc gag gag agg agg gcg agg cta 1596
Leu Arg Gly Phe Ala Arg Gln Leu Ser Glu Glu Arg Arg Ala Arg Leu
400 405 410
ggt cgg cga aac gcc tgc ggc ggc ggc ggc gaa gga gag ccc agc ggc 1644
Gly Arg Arg Asn Ala Cys Gly Gly Gly Gly Glu Gly Glu Pro Ser Gly
415 420 425
gtg gcg tgc aag cgt tag taactgtaca cgcatcatgc tgacgcgatc 1692
Val Ala Cys Lys Arg
430
ttttttattt ttcttttttt ttttttacct ttctagcgga catggggaat aacaagacgt 1752
gacagtagtg caatcggttt gtaacgtgcg tataccaaca ttgatccatt tcttcatcac 1812
agtttcagtt c 1823
<210> 22
<211> 1299
<212> ДНК
<213> Oryza sativa
<400> 22
atggcggcga gctactcgtg ccggcggaca tgcgaggcgt gcagcacgag ggcgatggcc 60
gggtgcgtgg tgggcgagcc ggcgtcggcg ccggggcagc gggtgacgtt gctggcgatc 120
gacggcggcg gcatcagggg cctcatcccg ggcaccatcc tcgccttcct cgaggccagg 180
ctgcaggagc tggatggccc cgacgcgcgc ctcgccgatt acttcgactg catcgccggg 240
accagcaccg gcggcctcat caccgccatg ctcgccgcgc ccggcgacca cggccgcccg 300
ctcttcgccg ccagcgacat caaccgcttc tacctcgaca acggcccact catcttccca 360
caaaagaggt gcggcatggc ggcggccatg gcggcgctga cgaggccgag gtacaacggc 420
aagtacctgc aggggaagat caggaagatg ctgggcgaga cgagggtgcg cgacacgctg 480
acgaacgtcg tcatccccac gttcgacgtc aggctgctcc agccaaccat cttctccaca 540
tacgacgcga agagcatgcc gctcaagaac gcgctcctct ccgacatctg catcagcaca 600
tccgcggcgc cgacctacct ccccgcgcac tgcttccaga ccaccgacga cgccaccggc 660
aaggtccgcg agttcgacct catcgacggc ggcgtcgccg ccaacaaccc gacgatggtg 720
gccatgacgc agatcaccaa gaagataatg gtgaaggaca aggaggagct gtacccggta 780
aagccgtcgg actgcggtaa gttcctggtg ctgtccgtgg gcaccgggtc gacgtcggac 840
caggggatgt acacggcgag gcagtgctcg cggtggggga tcgtccggtg gctgcgcaac 900
aaggggatgg cgcccatcat cgacatcttc atggcggcca gctccgacct cgtcgacatc 960
cacgccgccg tcatgttcca gtcgctgcac agcgacggcg actacctccg catccaggac 1020
aacacgctcc acggcgacgc cgccacggtg gacgccgcca ccagggacaa catgcgggcg 1080
ctcgtcggga tcggcgagcg gatgctggcg cagcgggtgt cgagggtcaa cgtcgagacc 1140
ggcaggtacg tcgaggtgcc cggcgccggc agcaacgccg acgcgctgag gggcttcgcc 1200
aggcagctct ccgaggagag gagggcgagg ctaggtcggc gaaacgcctg cggcggcggc 1260
ggcgaaggag agcccagcgg cgtggcgtgc aagcgttag 1299
<210> 23
<211> 432
<212> БЕЛОК
<213> Oryza sativa
<400> 23
Met Ala Ala Ser Tyr Ser Cys Arg Arg Thr Cys Glu Ala Cys Ser Thr
1 5 10 15
Arg Ala Met Ala Gly Cys Val Val Gly Glu Pro Ala Ser Ala Pro Gly
20 25 30
Gln Arg Val Thr Leu Leu Ala Ile Asp Gly Gly Gly Ile Arg Gly Leu
35 40 45
Ile Pro Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu
50 55 60
Asp Gly Pro Asp Ala Arg Leu Ala Asp Tyr Phe Asp Cys Ile Ala Gly
65 70 75 80
Thr Ser Thr Gly Gly Leu Ile Thr Ala Met Leu Ala Ala Pro Gly Asp
85 90 95
His Gly Arg Pro Leu Phe Ala Ala Ser Asp Ile Asn Arg Phe Tyr Leu
100 105 110
Asp Asn Gly Pro Leu Ile Phe Pro Gln Lys Arg Cys Gly Met Ala Ala
115 120 125
Ala Met Ala Ala Leu Thr Arg Pro Arg Tyr Asn Gly Lys Tyr Leu Gln
130 135 140
Gly Lys Ile Arg Lys Met Leu Gly Glu Thr Arg Val Arg Asp Thr Leu
145 150 155 160
Thr Asn Val Val Ile Pro Thr Phe Asp Val Arg Leu Leu Gln Pro Thr
165 170 175
Ile Phe Ser Thr Tyr Asp Ala Lys Ser Met Pro Leu Lys Asn Ala Leu
180 185 190
Leu Ser Asp Ile Cys Ile Ser Thr Ser Ala Ala Pro Thr Tyr Leu Pro
195 200 205
Ala His Cys Phe Gln Thr Thr Asp Asp Ala Thr Gly Lys Val Arg Glu
210 215 220
Phe Asp Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val
225 230 235 240
Ala Met Thr Gln Ile Thr Lys Lys Ile Met Val Lys Asp Lys Glu Glu
245 250 255
Leu Tyr Pro Val Lys Pro Ser Asp Cys Gly Lys Phe Leu Val Leu Ser
260 265 270
Val Gly Thr Gly Ser Thr Ser Asp Gln Gly Met Tyr Thr Ala Arg Gln
275 280 285
Cys Ser Arg Trp Gly Ile Val Arg Trp Leu Arg Asn Lys Gly Met Ala
290 295 300
Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile
305 310 315 320
His Ala Ala Val Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu
325 330 335
Arg Ile Gln Asp Asn Thr Leu His Gly Asp Ala Ala Thr Val Asp Ala
340 345 350
Ala Thr Arg Asp Asn Met Arg Ala Leu Val Gly Ile Gly Glu Arg Met
355 360 365
Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Tyr Val
370 375 380
Glu Val Pro Gly Ala Gly Ser Asn Ala Asp Ala Leu Arg Gly Phe Ala
385 390 395 400
Arg Gln Leu Ser Glu Glu Arg Arg Ala Arg Leu Gly Arg Arg Asn Ala
405 410 415
Cys Gly Gly Gly Gly Glu Gly Glu Pro Ser Gly Val Ala Cys Lys Arg
420 425 430
<210> 24
<211> 1371
<212> ДНК
<213> Zea mays
<400> 24
agttcatcac taatcacact tattgtgccc tcgacgagta tctatagcta gctcattaat 60
cgattcgggg gtgtgttgtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctggaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
cagaactgcc cgcgcatctt tcctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtctc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 25
<211> 1375
<212> ДНК
<213> Zea mays
<400> 25
agttcatcac taatcacact tattgtgccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctggaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacta cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gcgctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaacccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gcatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct acgcatccag 1020
gacaactcgc tccgtggcgc cgcggcaacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagcggg tgtccagggt caacgtggag 1140
acagggagcg aggtacgaac cggtgaccgg agaaggaagc aatgccgatg ccctcggtgg 1200
gctcgctagg cagctctccg aggagaggag aacaaggctc gcgcgccgcg tctctgccat 1260
caaccccaga agctctagat gtgcgcccta cgatatctaa gacaagtggc tttactgtca 1320
atcacatgct tgtaaataag tagactttat tttaataaaa tataaaaata tatat 1375
<210> 26
<211> 1375
<212> ДНК
<213> Zea mays
<400> 26
agttcatcac taatcacact tattgtgccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctggaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacta cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gcgctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaacccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gcatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct acgcatccag 1020
gacaactcgc tccgtggcgc cgcggcaacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagcggg tgtccagggt caacgtggag 1140
acagggagcg aggtacgaac cggtgaccgg agaaggaagc aatgccgatg ccctcggtgg 1200
gctcgctagg cagctctccg aggagaggag aacaaggctc gcgcgccgcg tctctgccat 1260
caaccccaga agctctagat gtgcgcccta cgatatctaa gacaagtggc tttactgtca 1320
atcacatgct tgtaaataag tagactttat tttaataaaa tataaaaata tatat 1375
<210> 27
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 27
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 28
<211> 372
<212> БЕЛОК
<213> Zea mays
<400> 28
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn Tyr Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Ser Glu Val Arg Thr Gly Asp Arg Arg Arg Lys Gln Cys Arg Cys Pro
355 360 365
Arg Trp Ala Arg
370
<210> 29
<211> 372
<212> БЕЛОК
<213> Zea mays
<400> 29
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn Tyr Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Ser Glu Val Arg Thr Gly Asp Arg Arg Arg Lys Gln Cys Arg Cys Pro
355 360 365
Arg Trp Ala Arg
370
<210> 30
<211> 15463
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22466
<220>
<221> промотор
<222> (214)..(2339)
<220>
<221> ген
<222> (2341)..(3018)
<220>
<221> Терминатор
<222> (3024)..(4036)
<220>
<221> Энхансер
<222> (4044)..(4237)
<223> eFMV
<220>
<221> Энхансер
<222> (4244)..(4536)
<223> e35S
<220>
<221> промотор
<222> (4544)..(8537)
<223> Ubi
<220>
<221> Терминатор
<222> (10009)..(11008)
<223> Ubi
<220>
<221> промотор
<222> (11028)..(12618)
<223> pPLA2
<220>
<221> экзон
<222> (12619)..(12977)
<223> PLA2 exon1
<220>
<221> Интрон
<222> (12978)..(13096)
<223> PLA2 intron1
<220>
<221> экзон
<222> (13097)..(13193)
<223> PLA2 exon2
<220>
<221> Интрон
<222> (13194)..(13388)
<223> PLA2 intron2
<220>
<221> экзон
<222> (13389)..(13550)
<223> PLA2 exon3
<220>
<221> Интрон
<222> (13551)..(13660)
<223> PLA2 intron3
<220>
<221> экзон
<222> (13661)..(14248)
<223> PLA2 exon4
<220>
<221> Терминатор
<222> (14249)..(15244)
<223> tPLA2
<400> 30
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga cccgaaagta gcaaacaaca ggttcatgtg 240
cactataaaa agacaaaatt ctcgagtttc atcttttatt ccacataagc cttatatttt 300
ccattttcat atgattttta gtttaagttt gtgtcttaac tttttcgtta atacgtaatt 360
ctatgcatta tggatgcgtg aagtattttt gtttaaaaaa atgaaatgtc aaaatacgtt 420
ttgtgatcta tttccatgtt ttcacctaac aggtggtttt tactatatat tctgccataa 480
ctctagcctt agatgtaaat cgaaaaaaaa tgagagatga gctggagata gccttagatg 540
aagcgtctga aatataaaag aaagagtaat gttgaacgca gtaggtgtag cagctgtagt 600
tccatctcta ggaaagggaa ctgcaatccg ggctccgggc ctcgcgcaat ctggcctgtc 660
gtgtagatgc agccctgtcc atgacggccc aagcaacgcc cgcggctctc gatccaccac 720
ggaacccact ccgacacaca ctgacacaca catgctggat gtggatgtgc tgtccaatta 780
ttagtagcaa ttcggtaggc acaggcacgt actggccggt gttttagctg taagtaccga 840
accaatcacg gttaagaacc gattaatccg tgcccagccg ccgagtgcgt tcgtacgtgc 900
atcggatgca ctgcatgaat tgagagcatc atcatatcat acgcaggagt agtacgacgc 960
cgctgctgtc ttgtccggct aatgctttgc tcacagatta gtccatcgcc cacggtcggt 1020
gtggtgtgga tcgctgatgc cactgctttt tgtttggttt ttattcccct gataatcctc 1080
cgcgtccctg aatgtatcta tttattttca ttccgaaatc cctttcacga aaaagaaaac 1140
gaataaaaag agagttacga atacgcttcc ggcggcccac atcaccttcc agcgaacatc 1200
gcgccgcgct gacgtgtcgc ccatcgcggc cgtccatatc gccatccgac gaccgtggaa 1260
gctggcagcg gccgctccgt tccgtcgaag gggcaggtca gtcaggtcac ccacacggcc 1320
acacccgcgc gggggatacg cggtggaaaa cccggcgacc acatcaaaac acgaggcgtc 1380
tcccgcagga ctggtcactc ggcacgcagg cagaggcagc acagcagcag ccagctccat 1440
ccatcctctt tcccctcctc gcttcgcttc ctcggcggat tcctcctccc tcggccgtcc 1500
ccgtcccctt cttcgccgcg ccagctcgcc cgagttggta cgcccatcct ctgctgtact 1560
ccccccgttc ctgctgtgcg aaaccgagca cccaaaccct agcctaagct attcgcatcg 1620
caaactctat tgagcgacgg atcgcgaaac gcgcgccgcc gcattcggac gagacctcgt 1680
ggcccgtacc ttcctactct accgtcgtcg ccggagacca gctcaccggt tagggtttcg 1740
ggattagggg cttggggctt ggattcccgg gggactccat acccggtggc cttagaaggg 1800
gaagggggcc cagcggtggt ggtggttcga ctgtcagcga gcgagacggc ggggcaccgc 1860
cgatcgggcg tcgctggaca tttgattgag ctgggcaggc gaaggcgtgg agcttgctct 1920
tcgattggga ttagaggagg gcggaggtgg tatggtgggc ggctcggctg gctcaatgga 1980
gccgctggcc ggtggtgggg caggcgagat cacgctctct catagagcgt aatgggttgc 2040
gtaacacact cctgtttgta tttcgatccg atctaataat taaagtttat ttacgtggtg 2100
cacatgtaga attttcccat gcccactgct aacttggatt ggatttttgc caaaaaaacg 2160
ggtgtgcacc caatcactta tagtgaaacc gttgtgtgtt tgatgtgctt ctaaccaagt 2220
acaaatccag tgaaaacaca cctaccattc aacaatccac attttggttg caattatcag 2280
cattctagaa aggtgcatgt gcccattgat acacttgcta ttggtgcagg caaacccacc 2340
atggcctcct ccgagaacgt catcaccgag ttcatgcgct tcaaggtgcg catggagggc 2400
accgtgaacg gccacgagtt cgagatcgag ggcgagggcg agggccgccc ctacgagggc 2460
cacaacaccg tgaagctgaa ggtgaccaag ggcggccccc tgcccttcgc ctgggacatc 2520
ctgtcccccc agttccagta cggctccaag gtgtacgtga agcaccccgc cgacatcccc 2580
gactacaaga agctgtcctt ccccgagggc ttcaagtggg agcgcgtgat gaacttcgag 2640
gacggcggcg tggcgaccgt gacccaggac tcctccctgc aggacggctg cttcatctac 2700
aaggtgaagt tcatcggcgt gaacttcccc tccgacggcc ccgtgatgca gaagaagacc 2760
atgggctggg aggcctccac cgagcgcctg tacccccgcg acggcgtgct gaagggcgag 2820
acccacaagg ccctgaagct gaaggacggc ggccactacc tggtggagtt caagtccatc 2880
tacatggcca agaagcccgt gcagctgccc ggctactact acgtggacgc caagctggac 2940
atcacctccc acaacgagga ctacaccatc gtggagcagt acgagcgcac cgagggccgc 3000
caccacctgt tcctgtagga gctcgcatca tgatcatgca tcatggactc ggcctactac 3060
tgtggatttg tatgccatta tagacttggt gctgtgaaag actgcttgat gatttgcggg 3120
tttgttgctg tgtaaaaaaa ggtcccttgg ctcccagaag accatgaagg ttcggatcta 3180
tcatgtaatt ccttgttatc tgccaattat gtatggacta tggacatgtg ttgcgctgtt 3240
caacttacta ctacaaataa gtaatcgata tgttcccttc ccatgtctcg gtgacaattg 3300
tctggagaag cttaggggtc gtttgtttgg gattatgtct ggagaaactt attttaaact 3360
aagtgtgagt tcaagttaag ttagattata taatctaggc agattataat tccaagcgaa 3420
caggtcctta gtgtttttgg aaaatcctag gtgttctttt ggctacattg ttgtgtgtgc 3480
agatcccttg ttggtctgta agcgtgggga agtaagaatc gtccgtttct actgaagacc 3540
tgctcgagtt aggcaccgag gatgccggta accaaacaga gcaatagtgt ctctgtgggc 3600
acagtggagt gtgaatctgt gtgatgcaaa tccgtcattt gtttagcaaa atttccagcg 3660
ttgcatgatg cagtttcttt aacacggact taagggaagg gaaaaaaatg ttgagccagg 3720
agatccttca atgtgttaga ctgacgtgat agccaactaa accacgacgc aatgttgtcg 3780
ttaatgacaa aaaaactatt tgttcctaaa tccttggcga cattgcatgg ctgtctcatg 3840
agataatggt ctcatctctt atttatctct tatttatagc cggaagtggt agtgacccct 3900
gcttgattgc tcgtatgcca tctcaagttc tcaaccgtgt cgagcagcca ttttcccatc 3960
tcaagcgcat catcgtttcg tttgacctca tctgctatcc tgctcctagt gcaaatcaca 4020
tgcgacagaa agtgtgcgga cccagctgct tgtggggacc agacaaaaaa ggaatggtgc 4080
agaattgtta ggcgcaccta ccaaaagcaa ctttgccttt attgcaaaga taaagcagat 4140
tcctctagta caagtgggga acaaaataac gtggaaaaga gctgtcctga cagcccactc 4200
actattgcgt ttgacgaacg cagtgacgac cacaaaactc gagacttttc aacaaagggt 4260
attatccgga aacctcctcg gattccattg cccagctatc tgtcacttta ttgtgaagat 4320
agtggaaaag gaaggtggct cctacaaatg ccatcattgc gataaaggaa aggctatcgt 4380
tgaagatgcc tctgccgaca gtggtcccaa agatggaccc ccacccacga ggagcatcgt 4440
ggaaaaagaa gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg atatctccac 4500
tgacgtaagg gttgacgaac aatcccacta tccttcaagc tttggaaaac caaaacaacg 4560
aataagcaaa ctgcaggaaa agtatgcagt ggaaaccaac ccagattcgg acgataggaa 4620
agtatcaagt gaatgatttg ccaggaaaag gagaggggta aaaaggggcg aagatttaga 4680
agatctaaag cacaagaacc agagattaga ttgaacaata gggaacttgg agcatccttt 4740
ttttcttcag ggaaaaactg aaaatccaaa ccatgttgag caaaaccgag tgggattgga 4800
aaccaaaaaa cccgagataa agaaactcga gaaaaagcat gaaatcgaaa ccaacttcag 4860
taaaacaaaa ggaggacaga aaagaaagtc ggaagctata aagaatacat taacattcag 4920
tgaaacagca tgctgtcttc ttcttttttt atgcacaaca gagcatacat atataccttc 4980
ccaggctgag gacttggcgg aggagagccg cggataggtt ggcggtgcag acggtctgga 5040
cgggcccgaa gacggagacg aacagcgggc ccttcctgcc caggcaccac gcttggaacg 5100
ccaagcacgc gccaaccgcg gccccgccga ggacgacgat ccccgcgaca agcgtggcgt 5160
cgatcctggg cgaccccaag ccgaggaacc tcccttccag gacgagccgt aggacggcgg 5220
tgagagaggc acccgtcgcg gaggtggcgc agcacaaggt gagcagcgcg gggaaggcgg 5280
cgagcgtggc ggcctgcagg acggtgacga gcgcgaagac ggtgacgccg gcgacgaggc 5340
agcagcagcc gaggatccag tcgtaggagg aagccggacc aaaccgggca atgcaacctg 5400
cagatgcact agacggaggt aacgaggagg aggagaaaac agagcaagag caggcggaga 5460
gaagatagag caaaacacga gtgaggcaca gcgtaagcac tcggtagaag tctccagagg 5520
cgaggtgcgc acaggagaac agatgagtaa agtcagccaa ggatccacga tccaacggct 5580
acgaattttt ggagtgacgt ggataggctc aaaggcgcca tttccatccg gctttatagt 5640
attttaaaaa aattcatttt cctccctcta gtgtgtgcgg aggcgtgagc ccgtttaacg 5700
gcgttgagaa gtctaacgga caccaaccac aaccaggaac cagcgccggc cgcgccgccg 5760
agtgaagcag actgcatacg gcacggcgcg gcatctctct ggctgcctct cgagagttcc 5820
gcccccacct tcccgcggta gcgtggtggt ttcgctttcc gctgtcggca tccggaagtt 5880
gcgtggcaga gtggacggag acgaggccgg gtcctccagc tcctctcaaa cgtcacggca 5940
ccggcatccg gcagccagcg cggtccttcc caaccactcg ttcccaaccc atcccccttc 6000
ctcgcccgcc gtcataaata gccagcccca tccccagctt ctttccccaa cctcatcttc 6060
tctccttttg ctctgaacgc acacaccgcc cggtctccga tctccgatcc ccgatcccct 6120
cgtcgatcct aggtacggcg accatcctac cccccccccc cccccctctc tctctgcctt 6180
ctctagatcg gcgatccgat ccatgcttac ttggttaggg cctgctaact atgttcatgt 6240
ttgcgttaga tccgtgcatg gacgcgatct gtacacacca gacgcgttct gattgctagc 6300
taactcgcca gtacctggga atcctgggat ggctgtagcc ggccccgcac gcagacggga 6360
ccgatttcat gattctctat ttttttcttt gtttcgttgc ctagggtttc gttcgatcga 6420
tccgcgttat tctttatttc catatattct ggtacgatgt tgatacggtt cgaccgtgct 6480
gcttacgttc tgtgcgcttg tttgccgggt catttttacc ttgccttttt tgtatggttt 6540
ggttgtggcg atgtggtctg gtcgggctgt cgttctagat cggagtagag tgctgtttca 6600
aactgtctag cggatctatt agatttggat ctgcatgtgt gacatatatc ttcgtagtta 6660
agatgatgca tctgtatgtg tgacatgcgg atctattaga tttggatctg tatgtgtgac 6720
atatatcttc gtagttgaga tgatgcatct gtatgtgtga catatatctt cgtagttaag 6780
attatgcatg gaaatatcaa tcctttagat aaggacgggt atacttgttg ctgtgggttt 6840
tactggtact tcgatagatg catatacatg atctaacatg cttagataca tgaagtaaca 6900
tgctgctacg gtttaataat tcttgagttg atttttactg gtacttagat agatgtatat 6960
acatgcttag atacatgaag taacatgctc ctacagttcc tttaatcatt attgagtacc 7020
tatatattct aataaatcag tatgttttaa attattttga ttttactggt acttagatag 7080
atgtatatat acatgctcaa acatgcttag atacatgaag taacatgctg ctacggttta 7140
gtcattattg agtgcctata tattctaata aatcagtatg ttttaaatta ttttgatttt 7200
actggtactt agatagatgt atatatacat gctcaaacat gcttagatac atgaagtaat 7260
atgctactac ggtttaattg ttcttgagta cctatatatt ctaataaatc agtatgtttt 7320
aaattatttc gattttactg gtacttagat agatgtatat atacatgctt agatacatga 7380
agtaacatgc tactacggtt taattgttct tgaataccta tatattctaa taaatcagta 7440
tgttttaaat tatttcgatt ttactggtac ttagatagat gtatatatac atgctcgaac 7500
atgcttagat acatgaagta acatgctaca tatatattat aataaatcag tatgtcttaa 7560
attattttga ttttactggt acttagatag atgtatatac atgctcaaac atgcttagat 7620
acatgaagta acatgctact acggtttaat cattattgag tacctatata ttctaataaa 7680
tcagtatgtt ttcaattgtt ttgattttac tggtacttag atatatgtat atatacatgc 7740
tcgaacatgc ttagatacgt gaagtaacat gctactatgg ttaattgttc ttgagtacct 7800
atatattcta ataaatcagt atgttttaaa ttatttcgat tttactggta cttagataga 7860
tgtatatata catgctcgaa catgcttaga tacatgaagt aacatgctac tacggtttaa 7920
tcgttcttga gtacctatat attctaataa atcagtatgt cttaaattat cttgatttta 7980
ctggtactta gatagatgta tatacatgct tagatacatg aagtaacatg ctactatgat 8040
ttaatcgttc ttgagtacct atatattcta ataaatcagt atgtttttaa ttattttgat 8100
tttactggta cttagataga tgtatatata catgctcgaa catgcttaga tacatgaagt 8160
aacatgctac tacggtttaa tcattcttga gtacctatat attctaataa atcagtatgt 8220
ttttaattat tttgatatta ctggtactta acatgtttag atacatcata tagcatgcac 8280
atgctgctac tgtttaatca ttcgtgaata cctatatatt ctaatatatc agtatgtctt 8340
ctaattatta tgattttgat gtacttgtat ggtggcatat gctgcagcta tgtgtagatt 8400
ttgaataccc agtgtgatga gcatgcatgg cgccttcata gttcatatgc tgtttatttc 8460
ctttgagact gttctttttt gttgatagtc accctgttgt ttggtgattc ttatgcagat 8520
ccagatcttc gtaaaccatg gctctggtgg ctaggccagt gctgtctgct agagtggctg 8580
cttctaggcc aagggtggcc gctagaaagg ctgtgagggt gagcgctgcc agtcacggcg 8640
ctagctctag gccagccacc gctaggaagt ccagcggcct gagcggcacc gtgcgcatcc 8700
caggcgacaa gagcatcagt caccgcagct tcatgttcgg cggcctggcc agcggcgaga 8760
ctaggatcac cggcctgctc gagggcgagg acgtgatcaa caccggcaag gccatgcagg 8820
ctatgggcgc tcgcatccgc aaggagggcg acacctggat catcgacggc gtgggtaatg 8880
gcggcctgct ggccccagag gccccactgg acttcggcaa cgccgccacc ggctgcaggc 8940
tgactatggg cctggtgggc gtgtacgact tcgacagcac ctttatcggc gacgccagcc 9000
tgaccaagag gccgatgggc agggtgctga acccgctgcg cgagatgggc gtgcaggtca 9060
agagcgagga cggcgacagg ctgccggtga ccctgagggg cccaaagacc ccgaccccga 9120
tcacctaccg cgtgccaatg gccagcgccc aggtgaagtc agccgtgctg ctggccggcc 9180
tgaacacccc aggcatcacc accgtgatcg agccgatcat gacccgcgat cacaccgaga 9240
agatgctgca gggcttcggc gccaacctga ccgtcgagac tgacgctgac ggcgtgagga 9300
ccatcaggct cgagggcagg ggcaagctga ccggccaggt gatcgacgtg ccgggcgacc 9360
caagcagcac cgccttccca ctggtggccg ccctgctggt gccaggctct gacgtgacca 9420
tcctgaacgt gctgatgaac ccgagccgca ccggcctgat cctgaccctg caggagatgg 9480
gcgccgatat cgaggtgatc aacccgaggc tggctggcgg cgaggacgtc gccgacctga 9540
gggtgaggtc cagcaccctg aagggcgtga ccgtgccaga ggacagggcc ccgagtatga 9600
ttgacgagta cccgatcctg gccgtggccg ctgccttcgc cgagggcgcc accgtgatga 9660
acggcctcga ggagctgcgc gtgaaggaga gcgacaggct gagcgctgtg gccaacggcc 9720
tgaagctgaa cggcgtggac tgcgacgagg gcgagactag cctggtggtg aggggcaggc 9780
cagacggcaa gggcctgggc aacgcttccg gcgctgccgt ggccacccac ctggatcaca 9840
ggatcgccat gtcgttcctc gtgatgggcc tcgtgagcga gaacccggtg accgtggacg 9900
acgccaccat gatcgccacc agcttccccg agttcatgga cctgatggcc ggcctgggcg 9960
ctaagatcga gctgagcgac accaaggccg cctgatcatc tagagctcgc caaggttcaa 10020
ttaagctgct gctgtacctg ggtatctgcg tcgtctggtg ccctctggtg tacctctata 10080
tggatgtcgt cgtctaataa acatctgtgg tttgtgtgtc atgaatcgtg gttgtggctt 10140
cgttggttta atggacctgt tgtgtcctct gtgttgtacc caaaactctt ctgcagcagt 10200
atggcttgaa tccttatgaa gtttgatatt tgaacttaaa agtctgctca ttatgttttt 10260
ttctggttat atctcctaat taactgcctg ggatcaaatt tgattcgctg gtgtttattg 10320
gacccctccc aggttcttgc tttctaccgt ttcttgctga atgttaactt gattctgtca 10380
ggctcagttt cccactatgg cttacagctt aacgtgtttg gtttgttgaa tgttaacttg 10440
gttttgtcaa gctcagtttt ttactctggc ttacagcata acatgtttga cttttggttt 10500
tgctgctttg ttattgggtt ctgggtagtt cttgatgaat ccaaaagatc atgtgcacag 10560
ccatattatc tatttaagcg atccaggtta ttactatgaa aggatgcctt ctagctaagg 10620
agtagttagg ttttttcttc aaggttaaat tttctcgatg ctctagtgtt cctgtgacca 10680
taatcataat aattcctttg aaagctctat ggtccctgga agcagggcat acaatgcaag 10740
acagcaactt gatcacatca actgaagtat acagggttct cttaactctt ggtgacttcg 10800
gtttaatgga ccggttgtac tcgtgttcta tccgtaaccg ttgtgatgtc ttgtgtgttt 10860
ggttgcggga tagctgggac cacgacgttt ccgtctaatt ctgatggata gctatagacg 10920
gcactgagat ggttatatta taacctctga tcctgaactc tacgagatcg tctcatccgt 10980
cattgccacc aaatacacca ttaaattact aattagctaa cggaccctat ttgtactcat 11040
tccatgtctc ataaactttg ggcaccatcc atccaacaca tccaatctaa acacaccaaa 11100
cgatggggaa tggaaagagc agtattcgat tcaacaatgg caaacaaata tcactgaatt 11160
agaccaagaa taaacctaat tagacaacga cctcccaacc atcattcgtc aggctgtaaa 11220
gaagataaag ctgccatggg gcatggatca agcagaacac cagagatgaa tccaaacaca 11280
cagaaaatca cgcgcgctgt ctacaatgac aacaagcccc acatttcatt gcagtacact 11340
gggctacaaa ggcacgtaca acaaagagct agggaaacat tgcggagggc acgagagagc 11400
agctaacttg acaatatagc agactgagct tgcactgtta gcaggcgagg aagggaatca 11460
tggggacgga gaatggggtc catgcccgcg aaggagaagg cggacgccgc cacggtggca 11520
ccggcgcacg cgcacacagg gaacccgcac aggcagccat ggatgctgcc tcgccattgc 11580
gccggtcgtc tctgccacgc tcctctctct ctcccgctgc atcgccgtgg atggggcaag 11640
cagagagcag ggactgcgac gatctgggcg gaggactcgc cttggagagc gcggacgcag 11700
acgggattct agggagagag cgaagacggg gcgcgcgcgg cgctcgcgcg gcgtggtggc 11760
ggcgagatta gcgggggtgg ggggagggcg gagccgtggt gagggtgtgg acgccctcct 11820
taccctctta agtagtagta gagatataat ccgttccaaa atatccatcc gttcaattta 11880
tatttcgttt gatcttttta ccctaaattt gattgactca tcttattaaa aaagttcata 11940
actattatta atctttattg agatatcatt tagcatataa tatactttaa gtgtggtttt 12000
agattttttt taaaaaaaaa aattcgcaaa aattaaatga aacgacccaa tcaaacttga 12060
aaagtaaaac taattataaa tttgaacgga aggagtaaga ggatgtttga atgtactaga 12120
gctaatagtt ggttgcttta aaatttgcta gtagaattag ctagctaata aatatctaga 12180
taactattag ctaatttgct aaaacagcta atagttgaac tattagctag attgtttgga 12240
tgtattcggc taattttaat ggctaactat tagctatagt acaatattca aacacctcct 12300
aattaaaatg gacaaatatc tcttcttttg gtcccttgcg ttagattttt catatctcct 12360
tatttagtat aaaagaatca tcaaaaagtg gacaacccct agtggaacac cattttagta 12420
gtggttgcat gaaacctttc gcgcaccagt ttctatgtgt cactctaaaa atgggacagc 12480
atgtacgtag tgcctatata tatacaagtc atctatcgtt gcctcctcag ttcatcacta 12540
atcacactta ttgtgccctc gacgagtatc tatagctagc tcattaatcg attcgggggt 12600
gtgttgtcga aggcggca atg gcg agc tac tcg tcg cgg cgt cca tgc aat 12651
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn
1 5 10
acc tgt agc acg aag gcg atg gcc ggg agc gtg gtc ggc gag ccc gtc 12699
Thr Cys Ser Thr Lys Ala Met Ala Gly Ser Val Val Gly Glu Pro Val
15 20 25
gtg ctg ggg cag agg gtg acg gtg ctg acg gtg gac ggc ggc ggc gtc 12747
Val Leu Gly Gln Arg Val Thr Val Leu Thr Val Asp Gly Gly Gly Val
30 35 40
cgg ggt ctc atc ccg gga acc atc ctc gcc ttc ctg gag gcc agg ctg 12795
Arg Gly Leu Ile Pro Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu
45 50 55
cag gag ctg gac ggg ccg gag gcg agg ctg gcg gac tac ttc gac tac 12843
Gln Glu Leu Asp Gly Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr
60 65 70 75
atc gcc gga acc agc acc ggc ggt ctc atc acc gcc atg ctc acc gcg 12891
Ile Ala Gly Thr Ser Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala
80 85 90
ccc ggc aag gac aag cgg cct ctc tac gct gcc aag gac atc aac cac 12939
Pro Gly Lys Asp Lys Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His
95 100 105
ttt tac atg cag aac tgc ccg cgc atc ttt cct cag aa gtgagtccga 12987
Phe Tyr Met Gln Asn Cys Pro Arg Ile Phe Pro Gln Lys
110 115 120
tgctgccgcc attgttcttg catccatcca gcatcgtacg tacgtcctct atacatctgc 13047
ggatcatcat gtgcgcatgt ttgtggcatg catgcatgca tgtgagcag g agc agg 13103
Ser Arg
ctt gcg gcc gcc atg tcc gcg ctg agg aag cca aag tac aac ggc aag 13151
Leu Ala Ala Ala Met Ser Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys
125 130 135
tgc atg cgc agc ctg att agg agc atc ctc ggc gag acg agg 13193
Cys Met Arg Ser Leu Ile Arg Ser Ile Leu Gly Glu Thr Arg
140 145 150
gtaagcgaga cgctgaccaa cgtcatcatc cctgccttcg acatcaggct gctgcagcct 13253
atcatcttct ctacctacga cgtacgtacg tcgtcacgaa tgattcatct gtacgtcgtc 13313
gcatgcgaat ggctgcctac gtacgccgtg cgctaacata ctcagctctt tcctatctgc 13373
tgcgccaatt tgcag gcc aag agc acg cct ctg aag aac gct ctg ctc tcg 13424
Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser
155 160
gac gtg tgc att ggc acg tcc gcc gcg ccg acc tac ctc ccg gcg cac 13472
Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His
165 170 175 180
tac ttc cag act gaa gac gcc aac ggc aag gag cgc gaa tac aac ctc 13520
Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu
185 190 195
atc gac ggc ggt gtg gcg gcc aac aac ccg gtaactgact agctaactgg 13570
Ile Asp Gly Gly Val Ala Ala Asn Asn Pro
200 205
aaaacggacg cacagactcc atgtccatgg cggcccacaa ggtcgatgct aattgttgct 13630
tatgtatgtc gcccgattgc acatgcgtag acg atg gtt gcg atg acg cag atc 13684
Thr Met Val Ala Met Thr Gln Ile
210
acc aaa aag atg ctt gcc agc aag gac aag gcc gag gag ctg tac cca 13732
Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr Pro
215 220 225 230
gtg aag ccg tcg aac tgc cgc agg ttc ctg gtg ctg tcc atc ggg acg 13780
Val Lys Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly Thr
235 240 245
ggg tcg acg tcc gag cag ggc ctc tac acg gcg cgg cag tgc tcc cgg 13828
Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser Arg
250 255 260
tgg ggt atc tgc cgg tgg ctc cgc aac aac ggc atg gcc ccc atc atc 13876
Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile Ile
265 270 275
gac atc ttc atg gcg gcc agc tcg gac ctg gtg gac atc cac gtc gcc 13924
Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val Ala
280 285 290
gcg atg ttc cag tcg ctc cac agc gac ggc gac tac ctg cgc atc cag 13972
Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile Gln
295 300 305 310
gac aac tcg ctc cgt ggc gcc gcg gcc acc gtg gac gcg gcg acg ccg 14020
Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr Pro
315 320 325
gag aac atg cgg acg ctc gtc ggg atc ggg gag cgg atg ctg gca cag 14068
Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala Gln
330 335 340
agg gtg tcc agg gtc aac gtg gag aca ggg agg tac gaa ccg gtg act 14116
Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Tyr Glu Pro Val Thr
345 350 355
ggc gaa gga agc aat gcc gat gcc ctc ggt ggg ctc gct agg cag ctc 14164
Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly Gly Leu Ala Arg Gln Leu
360 365 370
tcc gag gag agg aga aca agg ctc gcg cgc cgc gtc tct gcc atc aac 14212
Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg Arg Val Ser Ala Ile Asn
375 380 385 390
cca aga ggc tct aga tgt gcg tcg tac gat atc taa gacaagtggc 14258
Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp Ile
395 400
tttactgtca gtcacatgct tgtaaataag tagactttat tttaataaaa cataaaaata 14318
tatatatgtt cttgaatata aaattgataa ccaaattaaa attcgaacca tcacttatac 14378
ataattttac tttatttttt ataaaacgtg aacgggaagg actaccgtga atgactatag 14438
aaccaatcat actagtataa aatatatgat gacactacgg gagagacaaa ctttgtctgg 14498
cgctaaatat tttgccgagt gtgaattcac gggcactagg caaagatctt ctttgccgag 14558
tgttacgctg ggcaaagtaa gacactaggt aaatcagtca tttgccgagt gtccgccact 14618
aggcaaagca aaacactggc aaatcaaaag tttacctagt gccagacact aggcaaaaaa 14678
aaaacgctcg gcaaatcgga agtttcccta gtgccagaca ctagacaaag aaaaacactt 14738
gataaactag cgtcgtcagc taacaccatc caccaaccgt taacgttgcc gagtatctga 14798
cttcgacact cggcaaagaa ggtctctttg cctagtgtcg gtctggaaca ctaggcaaag 14858
aggcacttta cctagtgtcg tattttgaca ctcagtaaaa taattttttt tctttctgct 14918
tccaaacttt ttatgatgtg ttcctatagc acctagaact acatgtcaag ttttggtaaa 14978
atttttgaag tttttgctat atttacttaa tttattttat ttaattgaat ttcttttgat 15038
aattcaaatt tgaactcggc aaggtaagaa gcgagggtag cctggaaaca cactttgcct 15098
agtgttacac tcggtacagg agcctcccct gcctagtgct gcactcgaca aaagattcgc 15158
ctttgcctag cgctgcactc ggcacaggag tcgcctttgc ctagtgctgc actaggcaaa 15218
gcctccgtta ccgtgccttc catcgtcgga ccgcctgcag gcccgggggc gcgccctaat 15278
tagctaacgg ccaggatcgc cgcgtgagcc tttagcaact agctagatta attaacgcaa 15338
tctgttatta agttgtctaa gcgtcaattt gtttacacca caatatatcc tgccaccagc 15398
cagccaacag ctccccgacc ggcagctcgg cacaaaatca ccactcgata caggcagccc 15458
atcag 15463
<210> 31
<211> 15757
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22467
<220>
<221> промотор
<222> (214)..(2339)
<220>
<221> ген
<222> (2341)..(3018)
<220>
<221> Терминатор
<222> (3024)..(4036)
<220>
<221> Энхансер
<222> (4044)..(4237)
<223> eFMV
<220>
<221> Энхансер
<222> (4244)..(4536)
<223> e35S
<220>
<221> промотор
<222> (4544)..(8537)
<223> Ubi
<220>
<221> ген
<222> (8538)..(9995)
<223> EPSPS
<220>
<221> Терминатор
<222> (10009)..(11008)
<223> Ubi
<220>
<221> ген
<222> (11028)..(15538)
<223> GRMZM2G062320
<220>
<221> промотор
<222> (11028)..(12596)
<223> pPGM
<220>
<221> экзон
<222> (12597)..(12834)
<223> Экзон 1 PGM
<220>
<221> Интрон
<222> (12835)..(12974)
<223> Интрон 1 PGM
<220>
<221> экзон
<222> (12975)..(13198)
<223> Экзон 2 PGM
<220>
<221> Интрон
<222> (13199)..(13295)
<223> Интрон 2 PGM
<220>
<221> экзон
<222> (13296)..(13382)
<223> Экзон 3 PGM
<220>
<221> Интрон
<222> (13383)..(13718)
<223> Интрон 3 PGM
<220>
<221> экзон
<222> (13719)..(13787)
<223> Экзон 4 PGM
<220>
<221> Интрон
<222> (13788)..(14102)
<223> Интрон 4 PGM
<220>
<221> экзон
<222> (14103)..(14204)
<223> Экзон 5 PGM
<220>
<221> Интрон
<222> (14205)..(14304)
<223> Интрон 5 PGM
<220>
<221> экзон
<222> (14305)..(14424)
<223> Экзон 6 PGM
<220>
<221> Интрон
<222> (14425)..(14492)
<223> Интрон 6 PGM
<220>
<221> экзон
<222> (14493)..(14540)
<223> Экзон 7 PGM
<220>
<221> Терминатор
<222> (14541)..(15538)
<223> tPGM
<400> 31
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga cccgaaagta gcaaacaaca ggttcatgtg 240
cactataaaa agacaaaatt ctcgagtttc atcttttatt ccacataagc cttatatttt 300
ccattttcat atgattttta gtttaagttt gtgtcttaac tttttcgtta atacgtaatt 360
ctatgcatta tggatgcgtg aagtattttt gtttaaaaaa atgaaatgtc aaaatacgtt 420
ttgtgatcta tttccatgtt ttcacctaac aggtggtttt tactatatat tctgccataa 480
ctctagcctt agatgtaaat cgaaaaaaaa tgagagatga gctggagata gccttagatg 540
aagcgtctga aatataaaag aaagagtaat gttgaacgca gtaggtgtag cagctgtagt 600
tccatctcta ggaaagggaa ctgcaatccg ggctccgggc ctcgcgcaat ctggcctgtc 660
gtgtagatgc agccctgtcc atgacggccc aagcaacgcc cgcggctctc gatccaccac 720
ggaacccact ccgacacaca ctgacacaca catgctggat gtggatgtgc tgtccaatta 780
ttagtagcaa ttcggtaggc acaggcacgt actggccggt gttttagctg taagtaccga 840
accaatcacg gttaagaacc gattaatccg tgcccagccg ccgagtgcgt tcgtacgtgc 900
atcggatgca ctgcatgaat tgagagcatc atcatatcat acgcaggagt agtacgacgc 960
cgctgctgtc ttgtccggct aatgctttgc tcacagatta gtccatcgcc cacggtcggt 1020
gtggtgtgga tcgctgatgc cactgctttt tgtttggttt ttattcccct gataatcctc 1080
cgcgtccctg aatgtatcta tttattttca ttccgaaatc cctttcacga aaaagaaaac 1140
gaataaaaag agagttacga atacgcttcc ggcggcccac atcaccttcc agcgaacatc 1200
gcgccgcgct gacgtgtcgc ccatcgcggc cgtccatatc gccatccgac gaccgtggaa 1260
gctggcagcg gccgctccgt tccgtcgaag gggcaggtca gtcaggtcac ccacacggcc 1320
acacccgcgc gggggatacg cggtggaaaa cccggcgacc acatcaaaac acgaggcgtc 1380
tcccgcagga ctggtcactc ggcacgcagg cagaggcagc acagcagcag ccagctccat 1440
ccatcctctt tcccctcctc gcttcgcttc ctcggcggat tcctcctccc tcggccgtcc 1500
ccgtcccctt cttcgccgcg ccagctcgcc cgagttggta cgcccatcct ctgctgtact 1560
ccccccgttc ctgctgtgcg aaaccgagca cccaaaccct agcctaagct attcgcatcg 1620
caaactctat tgagcgacgg atcgcgaaac gcgcgccgcc gcattcggac gagacctcgt 1680
ggcccgtacc ttcctactct accgtcgtcg ccggagacca gctcaccggt tagggtttcg 1740
ggattagggg cttggggctt ggattcccgg gggactccat acccggtggc cttagaaggg 1800
gaagggggcc cagcggtggt ggtggttcga ctgtcagcga gcgagacggc ggggcaccgc 1860
cgatcgggcg tcgctggaca tttgattgag ctgggcaggc gaaggcgtgg agcttgctct 1920
tcgattggga ttagaggagg gcggaggtgg tatggtgggc ggctcggctg gctcaatgga 1980
gccgctggcc ggtggtgggg caggcgagat cacgctctct catagagcgt aatgggttgc 2040
gtaacacact cctgtttgta tttcgatccg atctaataat taaagtttat ttacgtggtg 2100
cacatgtaga attttcccat gcccactgct aacttggatt ggatttttgc caaaaaaacg 2160
ggtgtgcacc caatcactta tagtgaaacc gttgtgtgtt tgatgtgctt ctaaccaagt 2220
acaaatccag tgaaaacaca cctaccattc aacaatccac attttggttg caattatcag 2280
cattctagaa aggtgcatgt gcccattgat acacttgcta ttggtgcagg caaacccacc 2340
atggcctcct ccgagaacgt catcaccgag ttcatgcgct tcaaggtgcg catggagggc 2400
accgtgaacg gccacgagtt cgagatcgag ggcgagggcg agggccgccc ctacgagggc 2460
cacaacaccg tgaagctgaa ggtgaccaag ggcggccccc tgcccttcgc ctgggacatc 2520
ctgtcccccc agttccagta cggctccaag gtgtacgtga agcaccccgc cgacatcccc 2580
gactacaaga agctgtcctt ccccgagggc ttcaagtggg agcgcgtgat gaacttcgag 2640
gacggcggcg tggcgaccgt gacccaggac tcctccctgc aggacggctg cttcatctac 2700
aaggtgaagt tcatcggcgt gaacttcccc tccgacggcc ccgtgatgca gaagaagacc 2760
atgggctggg aggcctccac cgagcgcctg tacccccgcg acggcgtgct gaagggcgag 2820
acccacaagg ccctgaagct gaaggacggc ggccactacc tggtggagtt caagtccatc 2880
tacatggcca agaagcccgt gcagctgccc ggctactact acgtggacgc caagctggac 2940
atcacctccc acaacgagga ctacaccatc gtggagcagt acgagcgcac cgagggccgc 3000
caccacctgt tcctgtagga gctcgcatca tgatcatgca tcatggactc ggcctactac 3060
tgtggatttg tatgccatta tagacttggt gctgtgaaag actgcttgat gatttgcggg 3120
tttgttgctg tgtaaaaaaa ggtcccttgg ctcccagaag accatgaagg ttcggatcta 3180
tcatgtaatt ccttgttatc tgccaattat gtatggacta tggacatgtg ttgcgctgtt 3240
caacttacta ctacaaataa gtaatcgata tgttcccttc ccatgtctcg gtgacaattg 3300
tctggagaag cttaggggtc gtttgtttgg gattatgtct ggagaaactt attttaaact 3360
aagtgtgagt tcaagttaag ttagattata taatctaggc agattataat tccaagcgaa 3420
caggtcctta gtgtttttgg aaaatcctag gtgttctttt ggctacattg ttgtgtgtgc 3480
agatcccttg ttggtctgta agcgtgggga agtaagaatc gtccgtttct actgaagacc 3540
tgctcgagtt aggcaccgag gatgccggta accaaacaga gcaatagtgt ctctgtgggc 3600
acagtggagt gtgaatctgt gtgatgcaaa tccgtcattt gtttagcaaa atttccagcg 3660
ttgcatgatg cagtttcttt aacacggact taagggaagg gaaaaaaatg ttgagccagg 3720
agatccttca atgtgttaga ctgacgtgat agccaactaa accacgacgc aatgttgtcg 3780
ttaatgacaa aaaaactatt tgttcctaaa tccttggcga cattgcatgg ctgtctcatg 3840
agataatggt ctcatctctt atttatctct tatttatagc cggaagtggt agtgacccct 3900
gcttgattgc tcgtatgcca tctcaagttc tcaaccgtgt cgagcagcca ttttcccatc 3960
tcaagcgcat catcgtttcg tttgacctca tctgctatcc tgctcctagt gcaaatcaca 4020
tgcgacagaa agtgtgcgga cccagctgct tgtggggacc agacaaaaaa ggaatggtgc 4080
agaattgtta ggcgcaccta ccaaaagcaa ctttgccttt attgcaaaga taaagcagat 4140
tcctctagta caagtgggga acaaaataac gtggaaaaga gctgtcctga cagcccactc 4200
actattgcgt ttgacgaacg cagtgacgac cacaaaactc gagacttttc aacaaagggt 4260
attatccgga aacctcctcg gattccattg cccagctatc tgtcacttta ttgtgaagat 4320
agtggaaaag gaaggtggct cctacaaatg ccatcattgc gataaaggaa aggctatcgt 4380
tgaagatgcc tctgccgaca gtggtcccaa agatggaccc ccacccacga ggagcatcgt 4440
ggaaaaagaa gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg atatctccac 4500
tgacgtaagg gttgacgaac aatcccacta tccttcaagc tttggaaaac caaaacaacg 4560
aataagcaaa ctgcaggaaa agtatgcagt ggaaaccaac ccagattcgg acgataggaa 4620
agtatcaagt gaatgatttg ccaggaaaag gagaggggta aaaaggggcg aagatttaga 4680
agatctaaag cacaagaacc agagattaga ttgaacaata gggaacttgg agcatccttt 4740
ttttcttcag ggaaaaactg aaaatccaaa ccatgttgag caaaaccgag tgggattgga 4800
aaccaaaaaa cccgagataa agaaactcga gaaaaagcat gaaatcgaaa ccaacttcag 4860
taaaacaaaa ggaggacaga aaagaaagtc ggaagctata aagaatacat taacattcag 4920
tgaaacagca tgctgtcttc ttcttttttt atgcacaaca gagcatacat atataccttc 4980
ccaggctgag gacttggcgg aggagagccg cggataggtt ggcggtgcag acggtctgga 5040
cgggcccgaa gacggagacg aacagcgggc ccttcctgcc caggcaccac gcttggaacg 5100
ccaagcacgc gccaaccgcg gccccgccga ggacgacgat ccccgcgaca agcgtggcgt 5160
cgatcctggg cgaccccaag ccgaggaacc tcccttccag gacgagccgt aggacggcgg 5220
tgagagaggc acccgtcgcg gaggtggcgc agcacaaggt gagcagcgcg gggaaggcgg 5280
cgagcgtggc ggcctgcagg acggtgacga gcgcgaagac ggtgacgccg gcgacgaggc 5340
agcagcagcc gaggatccag tcgtaggagg aagccggacc aaaccgggca atgcaacctg 5400
cagatgcact agacggaggt aacgaggagg aggagaaaac agagcaagag caggcggaga 5460
gaagatagag caaaacacga gtgaggcaca gcgtaagcac tcggtagaag tctccagagg 5520
cgaggtgcgc acaggagaac agatgagtaa agtcagccaa ggatccacga tccaacggct 5580
acgaattttt ggagtgacgt ggataggctc aaaggcgcca tttccatccg gctttatagt 5640
attttaaaaa aattcatttt cctccctcta gtgtgtgcgg aggcgtgagc ccgtttaacg 5700
gcgttgagaa gtctaacgga caccaaccac aaccaggaac cagcgccggc cgcgccgccg 5760
agtgaagcag actgcatacg gcacggcgcg gcatctctct ggctgcctct cgagagttcc 5820
gcccccacct tcccgcggta gcgtggtggt ttcgctttcc gctgtcggca tccggaagtt 5880
gcgtggcaga gtggacggag acgaggccgg gtcctccagc tcctctcaaa cgtcacggca 5940
ccggcatccg gcagccagcg cggtccttcc caaccactcg ttcccaaccc atcccccttc 6000
ctcgcccgcc gtcataaata gccagcccca tccccagctt ctttccccaa cctcatcttc 6060
tctccttttg ctctgaacgc acacaccgcc cggtctccga tctccgatcc ccgatcccct 6120
cgtcgatcct aggtacggcg accatcctac cccccccccc cccccctctc tctctgcctt 6180
ctctagatcg gcgatccgat ccatgcttac ttggttaggg cctgctaact atgttcatgt 6240
ttgcgttaga tccgtgcatg gacgcgatct gtacacacca gacgcgttct gattgctagc 6300
taactcgcca gtacctggga atcctgggat ggctgtagcc ggccccgcac gcagacggga 6360
ccgatttcat gattctctat ttttttcttt gtttcgttgc ctagggtttc gttcgatcga 6420
tccgcgttat tctttatttc catatattct ggtacgatgt tgatacggtt cgaccgtgct 6480
gcttacgttc tgtgcgcttg tttgccgggt catttttacc ttgccttttt tgtatggttt 6540
ggttgtggcg atgtggtctg gtcgggctgt cgttctagat cggagtagag tgctgtttca 6600
aactgtctag cggatctatt agatttggat ctgcatgtgt gacatatatc ttcgtagtta 6660
agatgatgca tctgtatgtg tgacatgcgg atctattaga tttggatctg tatgtgtgac 6720
atatatcttc gtagttgaga tgatgcatct gtatgtgtga catatatctt cgtagttaag 6780
attatgcatg gaaatatcaa tcctttagat aaggacgggt atacttgttg ctgtgggttt 6840
tactggtact tcgatagatg catatacatg atctaacatg cttagataca tgaagtaaca 6900
tgctgctacg gtttaataat tcttgagttg atttttactg gtacttagat agatgtatat 6960
acatgcttag atacatgaag taacatgctc ctacagttcc tttaatcatt attgagtacc 7020
tatatattct aataaatcag tatgttttaa attattttga ttttactggt acttagatag 7080
atgtatatat acatgctcaa acatgcttag atacatgaag taacatgctg ctacggttta 7140
gtcattattg agtgcctata tattctaata aatcagtatg ttttaaatta ttttgatttt 7200
actggtactt agatagatgt atatatacat gctcaaacat gcttagatac atgaagtaat 7260
atgctactac ggtttaattg ttcttgagta cctatatatt ctaataaatc agtatgtttt 7320
aaattatttc gattttactg gtacttagat agatgtatat atacatgctt agatacatga 7380
agtaacatgc tactacggtt taattgttct tgaataccta tatattctaa taaatcagta 7440
tgttttaaat tatttcgatt ttactggtac ttagatagat gtatatatac atgctcgaac 7500
atgcttagat acatgaagta acatgctaca tatatattat aataaatcag tatgtcttaa 7560
attattttga ttttactggt acttagatag atgtatatac atgctcaaac atgcttagat 7620
acatgaagta acatgctact acggtttaat cattattgag tacctatata ttctaataaa 7680
tcagtatgtt ttcaattgtt ttgattttac tggtacttag atatatgtat atatacatgc 7740
tcgaacatgc ttagatacgt gaagtaacat gctactatgg ttaattgttc ttgagtacct 7800
atatattcta ataaatcagt atgttttaaa ttatttcgat tttactggta cttagataga 7860
tgtatatata catgctcgaa catgcttaga tacatgaagt aacatgctac tacggtttaa 7920
tcgttcttga gtacctatat attctaataa atcagtatgt cttaaattat cttgatttta 7980
ctggtactta gatagatgta tatacatgct tagatacatg aagtaacatg ctactatgat 8040
ttaatcgttc ttgagtacct atatattcta ataaatcagt atgtttttaa ttattttgat 8100
tttactggta cttagataga tgtatatata catgctcgaa catgcttaga tacatgaagt 8160
aacatgctac tacggtttaa tcattcttga gtacctatat attctaataa atcagtatgt 8220
ttttaattat tttgatatta ctggtactta acatgtttag atacatcata tagcatgcac 8280
atgctgctac tgtttaatca ttcgtgaata cctatatatt ctaatatatc agtatgtctt 8340
ctaattatta tgattttgat gtacttgtat ggtggcatat gctgcagcta tgtgtagatt 8400
ttgaataccc agtgtgatga gcatgcatgg cgccttcata gttcatatgc tgtttatttc 8460
ctttgagact gttctttttt gttgatagtc accctgttgt ttggtgattc ttatgcagat 8520
ccagatcttc gtaaaccatg gctctggtgg ctaggccagt gctgtctgct agagtggctg 8580
cttctaggcc aagggtggcc gctagaaagg ctgtgagggt gagcgctgcc agtcacggcg 8640
ctagctctag gccagccacc gctaggaagt ccagcggcct gagcggcacc gtgcgcatcc 8700
caggcgacaa gagcatcagt caccgcagct tcatgttcgg cggcctggcc agcggcgaga 8760
ctaggatcac cggcctgctc gagggcgagg acgtgatcaa caccggcaag gccatgcagg 8820
ctatgggcgc tcgcatccgc aaggagggcg acacctggat catcgacggc gtgggtaatg 8880
gcggcctgct ggccccagag gccccactgg acttcggcaa cgccgccacc ggctgcaggc 8940
tgactatggg cctggtgggc gtgtacgact tcgacagcac ctttatcggc gacgccagcc 9000
tgaccaagag gccgatgggc agggtgctga acccgctgcg cgagatgggc gtgcaggtca 9060
agagcgagga cggcgacagg ctgccggtga ccctgagggg cccaaagacc ccgaccccga 9120
tcacctaccg cgtgccaatg gccagcgccc aggtgaagtc agccgtgctg ctggccggcc 9180
tgaacacccc aggcatcacc accgtgatcg agccgatcat gacccgcgat cacaccgaga 9240
agatgctgca gggcttcggc gccaacctga ccgtcgagac tgacgctgac ggcgtgagga 9300
ccatcaggct cgagggcagg ggcaagctga ccggccaggt gatcgacgtg ccgggcgacc 9360
caagcagcac cgccttccca ctggtggccg ccctgctggt gccaggctct gacgtgacca 9420
tcctgaacgt gctgatgaac ccgagccgca ccggcctgat cctgaccctg caggagatgg 9480
gcgccgatat cgaggtgatc aacccgaggc tggctggcgg cgaggacgtc gccgacctga 9540
gggtgaggtc cagcaccctg aagggcgtga ccgtgccaga ggacagggcc ccgagtatga 9600
ttgacgagta cccgatcctg gccgtggccg ctgccttcgc cgagggcgcc accgtgatga 9660
acggcctcga ggagctgcgc gtgaaggaga gcgacaggct gagcgctgtg gccaacggcc 9720
tgaagctgaa cggcgtggac tgcgacgagg gcgagactag cctggtggtg aggggcaggc 9780
cagacggcaa gggcctgggc aacgcttccg gcgctgccgt ggccacccac ctggatcaca 9840
ggatcgccat gtcgttcctc gtgatgggcc tcgtgagcga gaacccggtg accgtggacg 9900
acgccaccat gatcgccacc agcttccccg agttcatgga cctgatggcc ggcctgggcg 9960
ctaagatcga gctgagcgac accaaggccg cctgatcatc tagagctcgc caaggttcaa 10020
ttaagctgct gctgtacctg ggtatctgcg tcgtctggtg ccctctggtg tacctctata 10080
tggatgtcgt cgtctaataa acatctgtgg tttgtgtgtc atgaatcgtg gttgtggctt 10140
cgttggttta atggacctgt tgtgtcctct gtgttgtacc caaaactctt ctgcagcagt 10200
atggcttgaa tccttatgaa gtttgatatt tgaacttaaa agtctgctca ttatgttttt 10260
ttctggttat atctcctaat taactgcctg ggatcaaatt tgattcgctg gtgtttattg 10320
gacccctccc aggttcttgc tttctaccgt ttcttgctga atgttaactt gattctgtca 10380
ggctcagttt cccactatgg cttacagctt aacgtgtttg gtttgttgaa tgttaacttg 10440
gttttgtcaa gctcagtttt ttactctggc ttacagcata acatgtttga cttttggttt 10500
tgctgctttg ttattgggtt ctgggtagtt cttgatgaat ccaaaagatc atgtgcacag 10560
ccatattatc tatttaagcg atccaggtta ttactatgaa aggatgcctt ctagctaagg 10620
agtagttagg ttttttcttc aaggttaaat tttctcgatg ctctagtgtt cctgtgacca 10680
taatcataat aattcctttg aaagctctat ggtccctgga agcagggcat acaatgcaag 10740
acagcaactt gatcacatca actgaagtat acagggttct cttaactctt ggtgacttcg 10800
gtttaatgga ccggttgtac tcgtgttcta tccgtaaccg ttgtgatgtc ttgtgtgttt 10860
ggttgcggga tagctgggac cacgacgttt ccgtctaatt ctgatggata gctatagacg 10920
gcactgagat ggttatatta taacctctga tcctgaactc tacgagatcg tctcatccgt 10980
cattgccacc aaatacacca ttaaattact aattagctaa cggacccctt gcctcggtta 11040
caattgagtt tgtcataaga aagaatgaga aagaaaagaa gcaatccaag cgcaagagct 11100
caaaggaaca caagtcactc tctcactagt cactattgat tggaattgaa ctagggactt 11160
gggagaggat tttatctctt tggtgtgtct tgtattgaat gctatagctc ttgtaatgtg 11220
tagaatgttg gaaacttgga tgccattgaa tgtggggtgg ttggggtatt tatagcccca 11280
accaccacaa tgtggtcgtt ggaagtctgc tgtcgcatgg cgcaccggat agtccgatgc 11340
gccaccggac actgtccggt gcgccagcca cgtcagccag ccgttggggt tcgaccgttg 11400
gagctctgac ttgtggggcc tctgggctgt ccggtggtgc accggacagg tcctgtagac 11460
tgtccggtgc accaactgcg cgtgctctga cctctgcgcg cgcaggcgcg cattaaatgc 11520
gttgcagtcg accgttgcgc ttgaagtagt cgttgctccg ctggcacacc ggacagtgtc 11580
cggtgaatta tagcggagcg gcctccattt tcccgaaggt agcgagttca gcgtcgagtt 11640
ccctggtgca ccggacactg tccggtggca caccggacag tccggtgcgc cagaccaggg 11700
cacacttcgg ttgtctttgg ctctctttgt ttgaaccctt tcttggtctt tttattggtc 11760
tattgtgaac ctttggcacc tgtaaaactt ataatctaga gtaaactagt tagtccaatt 11820
atttgtgttg ggcaattcaa ccaccaaaat caattaggaa aaggtgtaag cctatttccc 11880
tttcaggagg cgtacgtgag agggagaggt gaaaaggaac aacgcgtata ccagataagg 11940
tcccacagcc taagtaggta gccttctaat atctctacta actattaagg agagagtgta 12000
gactgccccc gctccctacc caacgccccc cgctacctgt tcaccgcgcg ccagcgaaac 12060
ctccgcacgc ccactgccca tctgttcccc gtgcgccagc gaaacatccg cacgcccgcg 12120
gcccgcctgt tccccgcgca tcccgctgca cgacttctgc taccgcaacg gccacccacg 12180
cacgcccgcc tgttcaccgc gcatcccgct gacctcccct tcacgctcgc acacgctccg 12240
ttcccccacc ccaccgcaat ccccgacgct ataagagcgg taaccaactc catctccctg 12300
gtgccacgca ttgttgagtt cttaaggtgc gtttcgttga ggacttgttc atttttgttg 12360
gtcatgtatt ccattttact gctctaccat tttgtggaat aaagggagga atgttttcac 12420
tagaagagtt catcaatctt atgttggttt cttggatcag ttttgctcta tggctaaatg 12480
gtcgaattga gcctatttca ttataaagtt agcgagcgaa taattgttca gcctcttcct 12540
agaactcatt accagtagaa tcagttacta actgcttttc tttttcttgg attaga atg 12599
Met
1
gct ggg gct atc tct cac cat gcg cta gca ttt tca caa tcc cac tgg 12647
Ala Gly Ala Ile Ser His His Ala Leu Ala Phe Ser Gln Ser His Trp
5 10 15
tgc agt gcg aag aac tct aga ttc gga aag agg acg ggc aat gct cgc 12695
Cys Ser Ala Lys Asn Ser Arg Phe Gly Lys Arg Thr Gly Asn Ala Arg
20 25 30
ctg gtt tat cta aaa gga aga tgt ggt tca ggc agc aga aaa ctg ggt 12743
Leu Val Tyr Leu Lys Gly Arg Cys Gly Ser Gly Ser Arg Lys Leu Gly
35 40 45
ttg atg tgg gcc tcg agc tcg cag tct tct gtc atg gag ccg acg cac 12791
Leu Met Trp Ala Ser Ser Ser Gln Ser Ser Val Met Glu Pro Thr His
50 55 60 65
cta cca tct gat ggc aac agc agc cac acc cca aaa aaa tca a 12834
Leu Pro Ser Asp Gly Asn Ser Ser His Thr Pro Lys Lys Ser
70 75
gtaattttaa cgacctccta tggtggttat ttgtttttaa tttgagaaaa ctatccattt 12894
gacacattta actttgggct tctcagaatt tgggggcata taataagatc tgctaatctg 12954
ttatctctat gtcgttgtag gt gaa agc gct ctt ata ttg att tgg cat ggt 13006
Ser Glu Ser Ala Leu Ile Leu Ile Trp His Gly
80 85 90
gaa tcc ctg tgg aac gag aaa aat cta ttt cct ggc tgc atc gat gta 13054
Glu Ser Leu Trp Asn Glu Lys Asn Leu Phe Pro Gly Cys Ile Asp Val
95 100 105
ccc ctg aca ccg aag ggt gtt gag gag gcc att gag gca ggt aaa agg 13102
Pro Leu Thr Pro Lys Gly Val Glu Glu Ala Ile Glu Ala Gly Lys Arg
110 115 120
ata tgc aat atc cca atc gat gtg ata tat act tca tca ctg att tgt 13150
Ile Cys Asn Ile Pro Ile Asp Val Ile Tyr Thr Ser Ser Leu Ile Cys
125 130 135
gct cag atg acc gca atg ctt gcc atg atg cag cat cga cgc aag aag 13198
Ala Gln Met Thr Ala Met Leu Ala Met Met Gln His Arg Arg Lys Lys
140 145 150
gtttgtgtct ttcctttgaa attccagtaa tttcttctag catttgtatg aacttgccgg 13258
agaaatcatg ctttgctggt gatatatgta tttatag atc cta gtt atc acg cat 13313
Ile Leu Val Ile Thr His
155 160
aat gag agt gaa caa gct cac agg tgg agt cag ata tac agt gag gag 13361
Asn Glu Ser Glu Gln Ala His Arg Trp Ser Gln Ile Tyr Ser Glu Glu
165 170 175
aca atg aaa cag tcc att cct gtcatcacag cttggcaatt gaatgaacgg 13412
Thr Met Lys Gln Ser Ile Pro
180
atgtaatact ttctccatac tctttgattt gctaattact ccctctgtct caaaatagta 13472
ttaattttag ctcttgattt ttatgtctat attcaaatag atgatgataa atctagattc 13532
tagacacaaa tataaaacat atacatcaag tattatatga atctattaat ttactaagac 13592
caattttaat ttgggacaga gggagtatac gattataata gttgtttgac tgtgcttctc 13652
tttaaatatc ccttgacatt tctaggtatg gtgagctaca aggccttaac aagcaagaaa 13712
ctgtag atc gat ttg gca aag aac aag ttc atg agt ggc gcc gca gtt 13760
Ile Asp Leu Ala Lys Asn Lys Phe Met Ser Gly Ala Ala Val
185 190 195
atg ata ttc ctc cgc caa atg gag aaa gtctagagaa gtgtgctgag 13807
Met Ile Phe Leu Arg Gln Met Glu Lys
200 205
agagctgttg cttatttcaa agatcaggca catctagcaa ggccacttta cactaattga 13867
aagatacact ttttacttgg gttattggtc ttgctgcagt attggtatgc atgctaaagg 13927
ttattcttga atcgatgaat tcctctacta tgggatgcag aaatgcatgt gcttagtttt 13987
ctttctattg tgctagctca tatcaaattt ataacctgaa ttttttattt atgttcgact 14047
ctaaaaaaca gttttttcta gctcgatttg acctatagta atttttccgt aatag att 14105
Ile
att cca caa ctt gtg gct gga aaa cat gtg atg gtt gct gca cat ggg 14153
Ile Pro Gln Leu Val Ala Gly Lys His Val Met Val Ala Ala His Gly
210 215 220
aat tca ctt cgt tca att ata atg cat ctg gac aaa tta act tct cag 14201
Asn Ser Leu Arg Ser Ile Ile Met His Leu Asp Lys Leu Thr Ser Gln
225 230 235
aag gtaattcact gtcgtttttg tctttccatc aaaaaggact cggctaaaca 14254
Lys
240
gaacatgtag cattatgtta agtttgggag tgagcctttc gtcccttcag gta ata 14310
Val Ile
agc ctt gag ctg tct act ggc att ccc atg ctt tac ata ttc aaa gag 14358
Ser Leu Glu Leu Ser Thr Gly Ile Pro Met Leu Tyr Ile Phe Lys Glu
245 250 255
gga aag ttt att cga cgt ggg act cca gta gga cct tcg gag gcc agt 14406
Gly Lys Phe Ile Arg Arg Gly Thr Pro Val Gly Pro Ser Glu Ala Ser
260 265 270
gtt tat gct tat acc agg gtaagattct ttcccccaca tgttctacca 14454
Val Tyr Ala Tyr Thr Arg
275 280
taggacgata ctccagttta caaaccttat ctgtacag acc aaa cga ttt gct gag 14510
Thr Lys Arg Phe Ala Glu
285
cac att aca ttt cag aac aaa ttg gcc tag aagataaggg gtgtttggtt 14560
His Ile Thr Phe Gln Asn Lys Leu Ala
290 295
tgagaaatca ctctattcaa aatgagatgg tgtatcatgg gtccatttct caaatttggt 14620
gggatgaccc tattcctcat attagtacta actaggtgag tgtccgtgcg ttgcaacggg 14680
aacatataat aacatgataa cttatataca aaatgtgtct tatattgtta taagaaaatg 14740
tttcataatc catttgtaat cctggccata cataaatttt gttattttaa tttagctgtt 14800
tcactactac attgcaacca tcagtatcat gcagacttcg atatatgtca cgatttgcat 14860
agtcttatca ttgaagagca cgtgtcacac ctaccggtag aagttccctc gtacattgtc 14920
agggggacct catggtcgtc gctaaagccg gcgactctct ggttgttctg ggcaccgcat 14980
ccgacgacaa caccgtcacg ctgtccagct catcatccac ctgaagatca acctgccacg 15040
taagtcggta gtgaccggcc atgaattctt gtgcgctggg gcaacgggcg ttgcattccg 15100
ttgtgtgagt gttttagaac aagatgtatg taaaggagta gcacatccgg tggtgcaatg 15160
gctaggtgta ctacctcgct gatgagctcg cggtgcactt cgtttagcag cccagccaag 15220
agtcatcggt acttgccatg tcgtgcgtgt tcgacgacta ctatatcaag gactgtggcg 15280
tcctcttggc gccagaggtg aggagtaccg acagcaatga cctctcgcca tcgtcggggt 15340
agcttttgtg ctggcccgcc tttaattctc ttcgtaaaca catacacttg ccttttttgt 15400
ttaagcaagt gagtatagtg gtgcgtacct gatgactgac aatgtacgag gaaacttcta 15460
ctggcaggtg tggcacgtgc tcttcaatga tgagaccatg taaatcgtga catatatcga 15520
agtctgcatg atactgatcg gaccgcctgc aggcccgggg gcgcgcccta attagctaac 15580
ggccaggatc gccgcgtgag cctttagcaa ctagctagat taattaacgc aatctgttat 15640
taagttgtct aagcgtcaat ttgtttacac cacaatatat cctgccacca gccagccaac 15700
agctccccga ccggcagctc ggcacaaaat caccactcga tacaggcagc ccatcag 15757
<210> 32
<211> 15355
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22503
<220>
<221> Энхансер
<222> (214)..(407)
<223> eFMV
<220>
<221> Энхансер
<222> (414)..(706)
<223> e35S
<220>
<221> промотор
<222> (714)..(4707)
<223> Ubi
<220>
<221> ген
<222> (4708)..(6165)
<223> EPSPS
<220>
<221> Терминатор
<222> (6179)..(7178)
<223> Ubi
<220>
<221> промотор
<222> (7198)..(9285)
<223> pPLA2m
<220>
<221> стебель_петля
<222> (9298)..(10212)
<223> нацеливающаяся на экзон 2 PLA2
<220>
<221> Терминатор
<222> (10223)..(11218)
<223> tPLA2
<220>
<221> промотор
<222> (11226)..(13349)
<220>
<221> ген
<222> (13352)..(14131)
<220>
<221> другой_признак
<222> (14039)..(14128)
<220>
<221> Терминатор
<222> (14138)..(15136)
<400> 32
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga cccagctgct tgtggggacc agacaaaaaa 240
ggaatggtgc agaattgtta ggcgcaccta ccaaaagcaa ctttgccttt attgcaaaga 300
taaagcagat tcctctagta caagtgggga acaaaataac gtggaaaaga gctgtcctga 360
cagcccactc actattgcgt ttgacgaacg cagtgacgac cacaaaactc gagacttttc 420
aacaaagggt attatccgga aacctcctcg gattccattg cccagctatc tgtcacttta 480
ttgtgaagat agtggaaaag gaaggtggct cctacaaatg ccatcattgc gataaaggaa 540
aggctatcgt tgaagatgcc tctgccgaca gtggtcccaa agatggaccc ccacccacga 600
ggagcatcgt ggaaaaagaa gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg 660
atatctccac tgacgtaagg gttgacgaac aatcccacta tccttcaagc tttggaaaac 720
caaaacaacg aataagcaaa ctgcaggaaa agtatgcagt ggaaaccaac ccagattcgg 780
acgataggaa agtatcaagt gaatgatttg ccaggaaaag gagaggggta aaaaggggcg 840
aagatttaga agatctaaag cacaagaacc agagattaga ttgaacaata gggaacttgg 900
agcatccttt ttttcttcag ggaaaaactg aaaatccaaa ccatgttgag caaaaccgag 960
tgggattgga aaccaaaaaa cccgagataa agaaactcga gaaaaagcat gaaatcgaaa 1020
ccaacttcag taaaacaaaa ggaggacaga aaagaaagtc ggaagctata aagaatacat 1080
taacattcag tgaaacagca tgctgtcttc ttcttttttt atgcacaaca gagcatacat 1140
atataccttc ccaggctgag gacttggcgg aggagagccg cggataggtt ggcggtgcag 1200
acggtctgga cgggcccgaa gacggagacg aacagcgggc ccttcctgcc caggcaccac 1260
gcttggaacg ccaagcacgc gccaaccgcg gccccgccga ggacgacgat ccccgcgaca 1320
agcgtggcgt cgatcctggg cgaccccaag ccgaggaacc tcccttccag gacgagccgt 1380
aggacggcgg tgagagaggc acccgtcgcg gaggtggcgc agcacaaggt gagcagcgcg 1440
gggaaggcgg cgagcgtggc ggcctgcagg acggtgacga gcgcgaagac ggtgacgccg 1500
gcgacgaggc agcagcagcc gaggatccag tcgtaggagg aagccggacc aaaccgggca 1560
atgcaacctg cagatgcact agacggaggt aacgaggagg aggagaaaac agagcaagag 1620
caggcggaga gaagatagag caaaacacga gtgaggcaca gcgtaagcac tcggtagaag 1680
tctccagagg cgaggtgcgc acaggagaac agatgagtaa agtcagccaa ggatccacga 1740
tccaacggct acgaattttt ggagtgacgt ggataggctc aaaggcgcca tttccatccg 1800
gctttatagt attttaaaaa aattcatttt cctccctcta gtgtgtgcgg aggcgtgagc 1860
ccgtttaacg gcgttgagaa gtctaacgga caccaaccac aaccaggaac cagcgccggc 1920
cgcgccgccg agtgaagcag actgcatacg gcacggcgcg gcatctctct ggctgcctct 1980
cgagagttcc gcccccacct tcccgcggta gcgtggtggt ttcgctttcc gctgtcggca 2040
tccggaagtt gcgtggcaga gtggacggag acgaggccgg gtcctccagc tcctctcaaa 2100
cgtcacggca ccggcatccg gcagccagcg cggtccttcc caaccactcg ttcccaaccc 2160
atcccccttc ctcgcccgcc gtcataaata gccagcccca tccccagctt ctttccccaa 2220
cctcatcttc tctccttttg ctctgaacgc acacaccgcc cggtctccga tctccgatcc 2280
ccgatcccct cgtcgatcct aggtacggcg accatcctac cccccccccc cccccctctc 2340
tctctgcctt ctctagatcg gcgatccgat ccatgcttac ttggttaggg cctgctaact 2400
atgttcatgt ttgcgttaga tccgtgcatg gacgcgatct gtacacacca gacgcgttct 2460
gattgctagc taactcgcca gtacctggga atcctgggat ggctgtagcc ggccccgcac 2520
gcagacggga ccgatttcat gattctctat ttttttcttt gtttcgttgc ctagggtttc 2580
gttcgatcga tccgcgttat tctttatttc catatattct ggtacgatgt tgatacggtt 2640
cgaccgtgct gcttacgttc tgtgcgcttg tttgccgggt catttttacc ttgccttttt 2700
tgtatggttt ggttgtggcg atgtggtctg gtcgggctgt cgttctagat cggagtagag 2760
tgctgtttca aactgtctag cggatctatt agatttggat ctgcatgtgt gacatatatc 2820
ttcgtagtta agatgatgca tctgtatgtg tgacatgcgg atctattaga tttggatctg 2880
tatgtgtgac atatatcttc gtagttgaga tgatgcatct gtatgtgtga catatatctt 2940
cgtagttaag attatgcatg gaaatatcaa tcctttagat aaggacgggt atacttgttg 3000
ctgtgggttt tactggtact tcgatagatg catatacatg atctaacatg cttagataca 3060
tgaagtaaca tgctgctacg gtttaataat tcttgagttg atttttactg gtacttagat 3120
agatgtatat acatgcttag atacatgaag taacatgctc ctacagttcc tttaatcatt 3180
attgagtacc tatatattct aataaatcag tatgttttaa attattttga ttttactggt 3240
acttagatag atgtatatat acatgctcaa acatgcttag atacatgaag taacatgctg 3300
ctacggttta gtcattattg agtgcctata tattctaata aatcagtatg ttttaaatta 3360
ttttgatttt actggtactt agatagatgt atatatacat gctcaaacat gcttagatac 3420
atgaagtaat atgctactac ggtttaattg ttcttgagta cctatatatt ctaataaatc 3480
agtatgtttt aaattatttc gattttactg gtacttagat agatgtatat atacatgctt 3540
agatacatga agtaacatgc tactacggtt taattgttct tgaataccta tatattctaa 3600
taaatcagta tgttttaaat tatttcgatt ttactggtac ttagatagat gtatatatac 3660
atgctcgaac atgcttagat acatgaagta acatgctaca tatatattat aataaatcag 3720
tatgtcttaa attattttga ttttactggt acttagatag atgtatatac atgctcaaac 3780
atgcttagat acatgaagta acatgctact acggtttaat cattattgag tacctatata 3840
ttctaataaa tcagtatgtt ttcaattgtt ttgattttac tggtacttag atatatgtat 3900
atatacatgc tcgaacatgc ttagatacgt gaagtaacat gctactatgg ttaattgttc 3960
ttgagtacct atatattcta ataaatcagt atgttttaaa ttatttcgat tttactggta 4020
cttagataga tgtatatata catgctcgaa catgcttaga tacatgaagt aacatgctac 4080
tacggtttaa tcgttcttga gtacctatat attctaataa atcagtatgt cttaaattat 4140
cttgatttta ctggtactta gatagatgta tatacatgct tagatacatg aagtaacatg 4200
ctactatgat ttaatcgttc ttgagtacct atatattcta ataaatcagt atgtttttaa 4260
ttattttgat tttactggta cttagataga tgtatatata catgctcgaa catgcttaga 4320
tacatgaagt aacatgctac tacggtttaa tcattcttga gtacctatat attctaataa 4380
atcagtatgt ttttaattat tttgatatta ctggtactta acatgtttag atacatcata 4440
tagcatgcac atgctgctac tgtttaatca ttcgtgaata cctatatatt ctaatatatc 4500
agtatgtctt ctaattatta tgattttgat gtacttgtat ggtggcatat gctgcagcta 4560
tgtgtagatt ttgaataccc agtgtgatga gcatgcatgg cgccttcata gttcatatgc 4620
tgtttatttc ctttgagact gttctttttt gttgatagtc accctgttgt ttggtgattc 4680
ttatgcagat ccagatcttc gtaaaccatg gctctggtgg ctaggccagt gctgtctgct 4740
agagtggctg cttctaggcc aagggtggcc gctagaaagg ctgtgagggt gagcgctgcc 4800
agtcacggcg ctagctctag gccagccacc gctaggaagt ccagcggcct gagcggcacc 4860
gtgcgcatcc caggcgacaa gagcatcagt caccgcagct tcatgttcgg cggcctggcc 4920
agcggcgaga ctaggatcac cggcctgctc gagggcgagg acgtgatcaa caccggcaag 4980
gccatgcagg ctatgggcgc tcgcatccgc aaggagggcg acacctggat catcgacggc 5040
gtgggtaatg gcggcctgct ggccccagag gccccactgg acttcggcaa cgccgccacc 5100
ggctgcaggc tgactatggg cctggtgggc gtgtacgact tcgacagcac ctttatcggc 5160
gacgccagcc tgaccaagag gccgatgggc agggtgctga acccgctgcg cgagatgggc 5220
gtgcaggtca agagcgagga cggcgacagg ctgccggtga ccctgagggg cccaaagacc 5280
ccgaccccga tcacctaccg cgtgccaatg gccagcgccc aggtgaagtc agccgtgctg 5340
ctggccggcc tgaacacccc aggcatcacc accgtgatcg agccgatcat gacccgcgat 5400
cacaccgaga agatgctgca gggcttcggc gccaacctga ccgtcgagac tgacgctgac 5460
ggcgtgagga ccatcaggct cgagggcagg ggcaagctga ccggccaggt gatcgacgtg 5520
ccgggcgacc caagcagcac cgccttccca ctggtggccg ccctgctggt gccaggctct 5580
gacgtgacca tcctgaacgt gctgatgaac ccgagccgca ccggcctgat cctgaccctg 5640
caggagatgg gcgccgatat cgaggtgatc aacccgaggc tggctggcgg cgaggacgtc 5700
gccgacctga gggtgaggtc cagcaccctg aagggcgtga ccgtgccaga ggacagggcc 5760
ccgagtatga ttgacgagta cccgatcctg gccgtggccg ctgccttcgc cgagggcgcc 5820
accgtgatga acggcctcga ggagctgcgc gtgaaggaga gcgacaggct gagcgctgtg 5880
gccaacggcc tgaagctgaa cggcgtggac tgcgacgagg gcgagactag cctggtggtg 5940
aggggcaggc cagacggcaa gggcctgggc aacgcttccg gcgctgccgt ggccacccac 6000
ctggatcaca ggatcgccat gtcgttcctc gtgatgggcc tcgtgagcga gaacccggtg 6060
accgtggacg acgccaccat gatcgccacc agcttccccg agttcatgga cctgatggcc 6120
ggcctgggcg ctaagatcga gctgagcgac accaaggccg cctgatcatc tagagctcgc 6180
caaggttcaa ttaagctgct gctgtacctg ggtatctgcg tcgtctggtg ccctctggtg 6240
tacctctata tggatgtcgt cgtctaataa acatctgtgg tttgtgtgtc atgaatcgtg 6300
gttgtggctt cgttggttta atggacctgt tgtgtcctct gtgttgtacc caaaactctt 6360
ctgcagcagt atggcttgaa tccttatgaa gtttgatatt tgaacttaaa agtctgctca 6420
ttatgttttt ttctggttat atctcctaat taactgcctg ggatcaaatt tgattcgctg 6480
gtgtttattg gacccctccc aggttcttgc tttctaccgt ttcttgctga atgttaactt 6540
gattctgtca ggctcagttt cccactatgg cttacagctt aacgtgtttg gtttgttgaa 6600
tgttaacttg gttttgtcaa gctcagtttt ttactctggc ttacagcata acatgtttga 6660
cttttggttt tgctgctttg ttattgggtt ctgggtagtt cttgatgaat ccaaaagatc 6720
atgtgcacag ccatattatc tatttaagcg atccaggtta ttactatgaa aggatgcctt 6780
ctagctaagg agtagttagg ttttttcttc aaggttaaat tttctcgatg ctctagtgtt 6840
cctgtgacca taatcataat aattcctttg aaagctctat ggtccctgga agcagggcat 6900
acaatgcaag acagcaactt gatcacatca actgaagtat acagggttct cttaactctt 6960
ggtgacttcg gtttaatgga ccggttgtac tcgtgttcta tccgtaaccg ttgtgatgtc 7020
ttgtgtgttt ggttgcggga tagctgggac cacgacgttt ccgtctaatt ctgatggata 7080
gctatagacg gcactgagat ggttatatta taacctctga tcctgaactc tacgagatcg 7140
tctcatccgt cattgccacc aaatacacca ttaaattact aattagctaa cggaccctat 7200
ttgtactcat tccatgtctc ataaactttg ggcaccatcc atccaacaca tccaatctaa 7260
acacaccaaa cgatggggaa tggaaagagc agtattcgat tcaacaatgg caaacaaata 7320
tcactgaatt agaccaagaa taaacctaat tagacaacga cctcccaacc atcattcgtc 7380
aggctgtaaa gaagataaag ctgccttggg gcatggatca agcagaacac cagagatgaa 7440
tccaaacaca cagaaaatca cgcgcgctgt ctacaatgac aacaagcccc acatttcatt 7500
gcagtacact gggctacaaa ggcacgtaca acaaagagct agggaaacat tgcggagggc 7560
acgagagagc agctaacttg acaatatagc agactgagct tgcactgtta gcaggcgagg 7620
aagggaatca tggggacgga gaatggggtc catgcccgcg aaggagaagg cggacgccgc 7680
cacggtggca ccggcgcacg cgcacacagg gaacccgcac aggcagccaa ggatgctgcc 7740
tcgccattgc gccggtcgtc tctgccacgc tcctctctct ctcccgctgc atcgccgtgg 7800
atggggcaag cagagagcag ggactgcgac gatctgggcg gaggactcgc cttggagagc 7860
gcggacgcag acgggattct agggagagag cgaagacggg gcgcgcgcgg cgctcgcgcg 7920
gcgtggtggc ggcgagatta gcgggggtgg ggggagggcg gagccgtggt gagggtgtgg 7980
acgccctcct taccctctta agtagtagta gagatataat ccgttccaaa atatccatcc 8040
gttcaattta tatttcgttt gatcttttta ccctaaattt gattgactca tcttattaaa 8100
aaagttcata actattatta atctttattg agatatcatt tagcatataa tatactttaa 8160
gtgtggtttt agattttttt taaaaaaaaa aattcgcaaa aattaaatga aacgacccaa 8220
tcaaacttga aaagtaaaac taattataaa tttgaacgga aggagtaaga ggatgtttga 8280
atgtactaga gctaatagtt ggttgcttta aaatttgcta gtagaattag ctagctaata 8340
aatatctaga taactattag ctaatttgct aaaacagcta atagttgaac tattagctag 8400
attgtttgga tgtattcggc taattttaat ggctaactat tagctatagt acaatattca 8460
aacacctcct aattaaaatg gacaaatatc tcttcttttg gtcccttgcg ttagattttt 8520
catatctcct tatttagtat aaaagaatca tcaaaaagtg gacaacccct agtggaacac 8580
cattttagta gtggttgcat gaaacctttc gcgcaccagt ttctatgtgt cactctaaaa 8640
atgggacagc atgtacgtag tgcctatata tatacaagtc atctatcgtt gcctcctcag 8700
ttcatcacta atcacactta ttgtgccctc gacgagtatc tatagctagc tcattaatcg 8760
attcgggggt gtgttgtcga aggcggcatt ggcgagctac tcgtcgcggc gtccaagcaa 8820
tacctgtagc acgaaggcga tcgccgggag cgtggtcggc gagcccgtcg tgctggggca 8880
gagggtgacg gtgctgacgg tggacggcgg cggcgtccgg ggtctcatcc cgggaaccat 8940
cctcgccttc ctggaggcca ggctgcagga gctggacgca ccggaggcga ggctggcgga 9000
ctacttcgac tacatcgccg gaaccagcac cggcggtctc atcaccgcct tgctgaccgc 9060
gcccggcaag gacaagcggc ctctctaggc tgccaaggac atcaaccact tttacatcca 9120
taactgcccg cgcatctttc ctcagaagtg agtccgatgc tgccgccatt gttcttgcat 9180
ccatccagca tcgtacgtac gtcctctata catctgcgga tcatcatgtg cgcatgtttg 9240
tggcatgcat gcatgcatgt gagcaggagc aggcttgcga aaaccccatg gagatctgaa 9300
caggcttgcg gccgccgtgt ccgcgctgag gaagccaaag tacaacggca agtgcatgcg 9360
cagcctgatt aggagcatcc tcggcgagac gaggggtcga ctcgggagca tgccctttgt 9420
gcttatgcct ccgttctgcc ttctgacgaa tttggtactg gaagcagatg agttttggtt 9480
cactatcatt ctgaatttac acctgcgctt gctgtcagac taggcaacca agtgactttt 9540
gtgactttga tcatgttcag tgtgtttcca agtcctaatc aatcaaaaag aaaaacagtt 9600
tgttaacgat tgtttgccat gtctatataa taaagttgct tttatagtag cttagaattc 9660
aatcggccaa ctttatctcg tacgctgaca gtaaaggtac atttaaaagg tgacaatgga 9720
tagtctaata cttgaactga caatagagac acattacatg tcagttgatt aagtttgtaa 9780
cagaaaaata aacaatacta cataattgca aagtttcttt gatgtctttc tttcaagaaa 9840
cacaaatata tcaatgctac agtattgctg atgaatttat ccatgttgag atgtttttct 9900
ggtttctgat ctgatcagtc tcaattggtg tgctgtttca ttttcatttg ctgatgatcg 9960
tccgagtagt taattcttac taatatttag ataatttggc atacaagcga atcacgtaga 10020
acatgatact tttgaatgaa tttatcaaag ttttatcact tggtgagttg tttcatggtt 10080
ttcctactga tgtctcttct tcagatttct cgagccctcg tctcgccgag gatgctccta 10140
atcaggctgc gcatgcactt gccgttgtac tttggcttcc tcagcgcgga catggcggcc 10200
gcaagcctgc tcggatccat gggacaagtg gctttactgt cagtcacatg cttgtaaata 10260
agtagacttt attttaataa aacataaaaa tatatatatg ttcttgaata taaaattgat 10320
aaccaaatta aaattcgaac catcacttat acataatttt actttatttt ttataaaacg 10380
tgaacgggaa ggactaccgt gaatgactat agaaccaatc atactagtat aaaatatatg 10440
atgacactac gggagagaca aactttgtct ggcgctaaat attttgccga gtgtgaattc 10500
acgggcacta ggcaaagatc ttctttgccg agtgttacgc tgggcaaagt aagacactag 10560
gtaaatcagt catttgccga gtgtccgcca ctaggcaaag caaaacactg gcaaatcaaa 10620
agtttaccta gtgccagaca ctaggcaaaa aaaaaacgct cggcaaatcg gaagtttccc 10680
tagtgccaga cactagacaa agaaaaacac ttgataaact agcgtcgtca gctaacacca 10740
tccaccaacc gttaacgttg ccgagtatct gacttcgaca ctcggcaaag aaggtctctt 10800
tgcctagtgt cggtctggaa cactaggcaa agaggcactt tacctagtgt cgtattttga 10860
cactcagtaa aataattttt tttctttctg cttccaaact ttttatgatg tgttcctata 10920
gcacctagaa ctacatgtca agttttggta aaatttttga agtttttgct atatttactt 10980
aatttatttt atttaattga atttcttttg ataattcaaa tttgaactcg gcaaggtaag 11040
aagcgagggt agcctggaaa cacactttgc ctagtgttac actcggtaca ggagcctccc 11100
ctgcctagtg ctgcactcga caaaagattc gcctttgcct agcgctgcac tcggcacagg 11160
agtcgccttt gcctagtgct gcactaggca aagcctccgt taccgtgcct tccatcgtcg 11220
gaccctggag aggcatgaac ccatcatgta tactcctgga cttggaagag gaaatgtcaa 11280
ccaaagtgaa aggttttctt ctagttgctg ctaagagata aattgaatac tagatctctc 11340
ctaagacatc ggggcattcg tctggactac tacacatgcg aaaattgcat cttacagtgg 11400
gaagaaacta tatctcacct cttcttgcgg tgtaactttg cccgaagatg ttggctcact 11460
gttggaatca ctccgccccg aacgttggat ctagcgcttg cagtgctaca tattagagca 11520
agactgacat tgtcgtggag aatggaaggt attataacca tgcatgtcat ggtacatatg 11580
gaaatgtcaa aataactgga tattcgaaaa cataccgcca acggtggcgg tctgcaagga 11640
aatgttcaag attgaaatga actacatatg caaccaggtt aagcttgaga caggagataa 11700
aagtagaaac tggatacaac actttgtaac atagcgacac tctctccttc ctttctttta 11760
ccttagaact atacatagaa tctacattca ataaaaatac agtaggtacg ccgagagatt 11820
taaaatgagt aagctaacat accaactaag gccctgtttg tttcggatta taatctctcc 11880
agattatata atccagcgta aataattcag cagataaaca aacacctaaa ttatatgttc 11940
agattatata atctatagcg gagattatga taatctcgta atctcctaag agtagcttat 12000
ttgagatttt tttggcaaaa gacccactac ccaaggttat gtaaatagaa ttacaatata 12060
tgacatcctt ctttcttcac ctcaaataaa caaacaaggg tactgttgtc tttatgaata 12120
atctacattt atataatcta gactaacaaa caactacata tagattataa tatgtctaga 12180
ttataatcta gattatataa tttaaattat agtccatatt atataatcta taagctaaaa 12240
caaataggcc ctaattatta gctattagtt gttagctatt taaatctaag ctaaaaccaa 12300
ctaatagctt attagttgaa ttacaattag ctcaacggaa ttctctgttt ttctaaaaaa 12360
aaactgcccc tctcttacag caaattgtcc gctgcccgtc gtccagatac aatgaacgta 12420
cctagtagga actcttttac acgctcggtc gctcgccgcg gatcggagtc cccggaacac 12480
gacaccactg tggaacacga caaagtctgc tcagaggcgg ccacaccctg gcgtgcaccg 12540
agccggagcc cggataagca cggtaaggag agtacggcgg gacgtggcga cccgtgtgtc 12600
tgctgccacg cagccttcct ccacgtagcc gcgcggccgc gccacgtacc agggcccggc 12660
gctggtataa cgcgccacct ccgctttagt tctgcataca gccaacccaa cacacacccg 12720
agcatatcac agtgacagac actacacggt gagcgccaga atcgttgtcc tcctcgccac 12780
cctcctatcc gctgccgccg cagtcgcgtc gtcctgggag gacgacaacc tccaccacca 12840
cgggggccac aagtccgggc gtagcgtgcg gcggtgcgag gaccggccct ggcaccagcg 12900
cccccggtgc ctggagcagt gcagggagga ggagcgggag aagcggcaag agcggagcag 12960
gctagaggcc gacgaccgca gcggcaaggg ttcgtcggag gtagagcgcg agcgagagca 13020
ggaggaggag aagcagaagg accggcggcc gtacgtgttc gaccggcgca gcttccgtcg 13080
cgtggtccgg agcgagcagg ggtctctgag ggtgctccgg ccgttcgacg aggtgtccag 13140
gctcctccgc ggcatccggg actaccgcgt ggcggtccta gaggcgaacc cgcgctcgtt 13200
cgtggtgccc agccataccg tagcgcactg catctgctac gtggtggaag gtacgttcgt 13260
ccggtccctg agaccacata gatcgagggc gacgccggcc gtgcacgtta gctgatctgt 13320
tgttgcctcg tgactaggcg agggataaac catggcctcc tccgagaacg tcatcaccga 13380
gttcatgcgc ttcaaggtgc gcatggaggg caccgtgaac ggccacgagt tcgagatcga 13440
gggcgagggc gagggccgcc cctacgaggg ccacaacacc gtgaagctga aggtgaccaa 13500
gggcggcccc ctgcccttcg cctgggacat cctgtccccc cagttccagt acggctccaa 13560
ggtgtacgtg aagcaccccg ccgacatccc cgactacaag aagctgtcct tccccgaggg 13620
cttcaagtgg gagcgcgtga tgaacttcga ggacggcggc gtggcgaccg tgacccagga 13680
ctcctccctg caggacggct gcttcatcta caaggtgaag ttcatcggcg tgaacttccc 13740
ctccgacggc cccgtgatgc agaagaagac aatgggctgg gaggcctcca ccgagcgcct 13800
gtacccccgc gacggcgtgc tgaagggcga gacccacaag gccctgaagc tgaaggacgg 13860
cggccactac ctggtggagt tcaagtccat ctacatggcc aagaagcccg tgcagctgcc 13920
cggctactac tacgtggacg ccaagctgga catcacctcc cacaacgagg actacaccat 13980
cgtggagcag tacgagcgca ccgagggccg ccaccacctg ttcctgagat ctcgagctga 14040
tccaaaaaag aagagaaagg tagatccaaa aaagaagaga aaggtagatc caaaaaagaa 14100
gagaaaggta ggctccaccg gatctagata agagctcggc cgccgcgctc gccaaaacga 14160
gcaagaagca acaagagggt ggcgcgcgac cgacgtgcgt acgtagctga gcctgagtgg 14220
agacgttgga cgtgtatgta tatacctctc tgcgtgttaa ctatgtacgt aagcggcagg 14280
cagtgcaata agtgtggctc tgtagtatgt acgtgcggat acgatgctgt atgctactga 14340
ggcaagtccc ataaataaat aatgacacgt gcgtgttcta taatctcttc gcttcttcag 14400
tgtccccttg cggagtttgg catccattga tgccgttaca ctgagaacat aacggacaca 14460
gtagacgaac cacttgagtt cttgtatgaa attctgaccc ttttttttga aggatcagga 14520
gggggaaacc cctactggcc tggtcctttt acaatcaata aaaagaaaag aaaaagaaag 14580
aaaaaggaag agctatttct atatttgcca caaatttagc cagttcggga ttgaggtgtc 14640
gaattctccc cgtgctctgt gaatgattag tcgcaattct ctactaaatt cctgagtaca 14700
gcggagcacg gagggatctc tatttgaaaa caaccactcg tttcggcttt tccagatgct 14760
ccaagtcata agaatagtga tctccataga gaagggaacg ttgagctggt gtttgaggtt 14820
cacagatgcc tcttcaaggt tggaaattct gggtggggta atgccaattg agttccagca 14880
ggctttcgcg aagttgcatc tgaggaaaag atgataaagg gtctcctctc tttgccaaag 14940
gcagttttcg caagagtagg aatctagctc catgtttctc cttctcaaca tatttctggt 15000
gtttagcctg ttcttaagcc acagccaata gaagactcta tgcttaggct gacacttact 15060
tttccaaact agtttgtaga taggatgagt ttgcatctgg cccatgaaag atctgtaagc 15120
tttttgggac gagaatcgga ccgcctgcag gcccgggggc gcgccctaat tagctaacgg 15180
ccaggatcgc cgcgtgagcc tttagcaact agctagatta attaacgcaa tctgttatta 15240
agttgtctaa gcgtcaattt gtttacacca caatatatcc tgccaccagc cagccaacag 15300
ctccccgacc ggcagctcgg cacaaaatca ccactcgata caggcagccc atcag 15355
<210> 33
<211> 15441
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22513
<220>
<221> Энхансер
<222> (214)..(407)
<223> eFMV
<220>
<221> Энхансер
<222> (414)..(706)
<223> e35S
<220>
<221> промотор
<222> (714)..(4707)
<223> Ubi
<220>
<221> ген
<222> (4708)..(6165)
<223> EPSPS
<220>
<221> Терминатор
<222> (6179)..(7178)
<223> Ubi
<220>
<221> промотор
<222> (7198)..(9285)
<223> pPLA2
<220>
<221> стебель_петля
<222> (9292)..(10386)
<223> нацеливающаяся на экзон 4 PLA2
<220>
<221> Терминатор
<222> (10397)..(11392)
<223> tPLA2
<220>
<221> промотор
<222> (11400)..(13526)
<220>
<221> ген
<222> (13527)..(14204)
<220>
<221> Терминатор
<222> (14210)..(15222)
<400> 33
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga cccagctgct tgtggggacc agacaaaaaa 240
ggaatggtgc agaattgtta ggcgcaccta ccaaaagcaa ctttgccttt attgcaaaga 300
taaagcagat tcctctagta caagtgggga acaaaataac gtggaaaaga gctgtcctga 360
cagcccactc actattgcgt ttgacgaacg cagtgacgac cacaaaactc gagacttttc 420
aacaaagggt attatccgga aacctcctcg gattccattg cccagctatc tgtcacttta 480
ttgtgaagat agtggaaaag gaaggtggct cctacaaatg ccatcattgc gataaaggaa 540
aggctatcgt tgaagatgcc tctgccgaca gtggtcccaa agatggaccc ccacccacga 600
ggagcatcgt ggaaaaagaa gacgttccaa ccacgtcttc aaagcaagtg gattgatgtg 660
atatctccac tgacgtaagg gttgacgaac aatcccacta tccttcaagc tttggaaaac 720
caaaacaacg aataagcaaa ctgcaggaaa agtatgcagt ggaaaccaac ccagattcgg 780
acgataggaa agtatcaagt gaatgatttg ccaggaaaag gagaggggta aaaaggggcg 840
aagatttaga agatctaaag cacaagaacc agagattaga ttgaacaata gggaacttgg 900
agcatccttt ttttcttcag ggaaaaactg aaaatccaaa ccatgttgag caaaaccgag 960
tgggattgga aaccaaaaaa cccgagataa agaaactcga gaaaaagcat gaaatcgaaa 1020
ccaacttcag taaaacaaaa ggaggacaga aaagaaagtc ggaagctata aagaatacat 1080
taacattcag tgaaacagca tgctgtcttc ttcttttttt atgcacaaca gagcatacat 1140
atataccttc ccaggctgag gacttggcgg aggagagccg cggataggtt ggcggtgcag 1200
acggtctgga cgggcccgaa gacggagacg aacagcgggc ccttcctgcc caggcaccac 1260
gcttggaacg ccaagcacgc gccaaccgcg gccccgccga ggacgacgat ccccgcgaca 1320
agcgtggcgt cgatcctggg cgaccccaag ccgaggaacc tcccttccag gacgagccgt 1380
aggacggcgg tgagagaggc acccgtcgcg gaggtggcgc agcacaaggt gagcagcgcg 1440
gggaaggcgg cgagcgtggc ggcctgcagg acggtgacga gcgcgaagac ggtgacgccg 1500
gcgacgaggc agcagcagcc gaggatccag tcgtaggagg aagccggacc aaaccgggca 1560
atgcaacctg cagatgcact agacggaggt aacgaggagg aggagaaaac agagcaagag 1620
caggcggaga gaagatagag caaaacacga gtgaggcaca gcgtaagcac tcggtagaag 1680
tctccagagg cgaggtgcgc acaggagaac agatgagtaa agtcagccaa ggatccacga 1740
tccaacggct acgaattttt ggagtgacgt ggataggctc aaaggcgcca tttccatccg 1800
gctttatagt attttaaaaa aattcatttt cctccctcta gtgtgtgcgg aggcgtgagc 1860
ccgtttaacg gcgttgagaa gtctaacgga caccaaccac aaccaggaac cagcgccggc 1920
cgcgccgccg agtgaagcag actgcatacg gcacggcgcg gcatctctct ggctgcctct 1980
cgagagttcc gcccccacct tcccgcggta gcgtggtggt ttcgctttcc gctgtcggca 2040
tccggaagtt gcgtggcaga gtggacggag acgaggccgg gtcctccagc tcctctcaaa 2100
cgtcacggca ccggcatccg gcagccagcg cggtccttcc caaccactcg ttcccaaccc 2160
atcccccttc ctcgcccgcc gtcataaata gccagcccca tccccagctt ctttccccaa 2220
cctcatcttc tctccttttg ctctgaacgc acacaccgcc cggtctccga tctccgatcc 2280
ccgatcccct cgtcgatcct aggtacggcg accatcctac cccccccccc cccccctctc 2340
tctctgcctt ctctagatcg gcgatccgat ccatgcttac ttggttaggg cctgctaact 2400
atgttcatgt ttgcgttaga tccgtgcatg gacgcgatct gtacacacca gacgcgttct 2460
gattgctagc taactcgcca gtacctggga atcctgggat ggctgtagcc ggccccgcac 2520
gcagacggga ccgatttcat gattctctat ttttttcttt gtttcgttgc ctagggtttc 2580
gttcgatcga tccgcgttat tctttatttc catatattct ggtacgatgt tgatacggtt 2640
cgaccgtgct gcttacgttc tgtgcgcttg tttgccgggt catttttacc ttgccttttt 2700
tgtatggttt ggttgtggcg atgtggtctg gtcgggctgt cgttctagat cggagtagag 2760
tgctgtttca aactgtctag cggatctatt agatttggat ctgcatgtgt gacatatatc 2820
ttcgtagtta agatgatgca tctgtatgtg tgacatgcgg atctattaga tttggatctg 2880
tatgtgtgac atatatcttc gtagttgaga tgatgcatct gtatgtgtga catatatctt 2940
cgtagttaag attatgcatg gaaatatcaa tcctttagat aaggacgggt atacttgttg 3000
ctgtgggttt tactggtact tcgatagatg catatacatg atctaacatg cttagataca 3060
tgaagtaaca tgctgctacg gtttaataat tcttgagttg atttttactg gtacttagat 3120
agatgtatat acatgcttag atacatgaag taacatgctc ctacagttcc tttaatcatt 3180
attgagtacc tatatattct aataaatcag tatgttttaa attattttga ttttactggt 3240
acttagatag atgtatatat acatgctcaa acatgcttag atacatgaag taacatgctg 3300
ctacggttta gtcattattg agtgcctata tattctaata aatcagtatg ttttaaatta 3360
ttttgatttt actggtactt agatagatgt atatatacat gctcaaacat gcttagatac 3420
atgaagtaat atgctactac ggtttaattg ttcttgagta cctatatatt ctaataaatc 3480
agtatgtttt aaattatttc gattttactg gtacttagat agatgtatat atacatgctt 3540
agatacatga agtaacatgc tactacggtt taattgttct tgaataccta tatattctaa 3600
taaatcagta tgttttaaat tatttcgatt ttactggtac ttagatagat gtatatatac 3660
atgctcgaac atgcttagat acatgaagta acatgctaca tatatattat aataaatcag 3720
tatgtcttaa attattttga ttttactggt acttagatag atgtatatac atgctcaaac 3780
atgcttagat acatgaagta acatgctact acggtttaat cattattgag tacctatata 3840
ttctaataaa tcagtatgtt ttcaattgtt ttgattttac tggtacttag atatatgtat 3900
atatacatgc tcgaacatgc ttagatacgt gaagtaacat gctactatgg ttaattgttc 3960
ttgagtacct atatattcta ataaatcagt atgttttaaa ttatttcgat tttactggta 4020
cttagataga tgtatatata catgctcgaa catgcttaga tacatgaagt aacatgctac 4080
tacggtttaa tcgttcttga gtacctatat attctaataa atcagtatgt cttaaattat 4140
cttgatttta ctggtactta gatagatgta tatacatgct tagatacatg aagtaacatg 4200
ctactatgat ttaatcgttc ttgagtacct atatattcta ataaatcagt atgtttttaa 4260
ttattttgat tttactggta cttagataga tgtatatata catgctcgaa catgcttaga 4320
tacatgaagt aacatgctac tacggtttaa tcattcttga gtacctatat attctaataa 4380
atcagtatgt ttttaattat tttgatatta ctggtactta acatgtttag atacatcata 4440
tagcatgcac atgctgctac tgtttaatca ttcgtgaata cctatatatt ctaatatatc 4500
agtatgtctt ctaattatta tgattttgat gtacttgtat ggtggcatat gctgcagcta 4560
tgtgtagatt ttgaataccc agtgtgatga gcatgcatgg cgccttcata gttcatatgc 4620
tgtttatttc ctttgagact gttctttttt gttgatagtc accctgttgt ttggtgattc 4680
ttatgcagat ccagatcttc gtaaaccatg gctctggtgg ctaggccagt gctgtctgct 4740
agagtggctg cttctaggcc aagggtggcc gctagaaagg ctgtgagggt gagcgctgcc 4800
agtcacggcg ctagctctag gccagccacc gctaggaagt ccagcggcct gagcggcacc 4860
gtgcgcatcc caggcgacaa gagcatcagt caccgcagct tcatgttcgg cggcctggcc 4920
agcggcgaga ctaggatcac cggcctgctc gagggcgagg acgtgatcaa caccggcaag 4980
gccatgcagg ctatgggcgc tcgcatccgc aaggagggcg acacctggat catcgacggc 5040
gtgggtaatg gcggcctgct ggccccagag gccccactgg acttcggcaa cgccgccacc 5100
ggctgcaggc tgactatggg cctggtgggc gtgtacgact tcgacagcac ctttatcggc 5160
gacgccagcc tgaccaagag gccgatgggc agggtgctga acccgctgcg cgagatgggc 5220
gtgcaggtca agagcgagga cggcgacagg ctgccggtga ccctgagggg cccaaagacc 5280
ccgaccccga tcacctaccg cgtgccaatg gccagcgccc aggtgaagtc agccgtgctg 5340
ctggccggcc tgaacacccc aggcatcacc accgtgatcg agccgatcat gacccgcgat 5400
cacaccgaga agatgctgca gggcttcggc gccaacctga ccgtcgagac tgacgctgac 5460
ggcgtgagga ccatcaggct cgagggcagg ggcaagctga ccggccaggt gatcgacgtg 5520
ccgggcgacc caagcagcac cgccttccca ctggtggccg ccctgctggt gccaggctct 5580
gacgtgacca tcctgaacgt gctgatgaac ccgagccgca ccggcctgat cctgaccctg 5640
caggagatgg gcgccgatat cgaggtgatc aacccgaggc tggctggcgg cgaggacgtc 5700
gccgacctga gggtgaggtc cagcaccctg aagggcgtga ccgtgccaga ggacagggcc 5760
ccgagtatga ttgacgagta cccgatcctg gccgtggccg ctgccttcgc cgagggcgcc 5820
accgtgatga acggcctcga ggagctgcgc gtgaaggaga gcgacaggct gagcgctgtg 5880
gccaacggcc tgaagctgaa cggcgtggac tgcgacgagg gcgagactag cctggtggtg 5940
aggggcaggc cagacggcaa gggcctgggc aacgcttccg gcgctgccgt ggccacccac 6000
ctggatcaca ggatcgccat gtcgttcctc gtgatgggcc tcgtgagcga gaacccggtg 6060
accgtggacg acgccaccat gatcgccacc agcttccccg agttcatgga cctgatggcc 6120
ggcctgggcg ctaagatcga gctgagcgac accaaggccg cctgatcatc tagagctcgc 6180
caaggttcaa ttaagctgct gctgtacctg ggtatctgcg tcgtctggtg ccctctggtg 6240
tacctctata tggatgtcgt cgtctaataa acatctgtgg tttgtgtgtc atgaatcgtg 6300
gttgtggctt cgttggttta atggacctgt tgtgtcctct gtgttgtacc caaaactctt 6360
ctgcagcagt atggcttgaa tccttatgaa gtttgatatt tgaacttaaa agtctgctca 6420
ttatgttttt ttctggttat atctcctaat taactgcctg ggatcaaatt tgattcgctg 6480
gtgtttattg gacccctccc aggttcttgc tttctaccgt ttcttgctga atgttaactt 6540
gattctgtca ggctcagttt cccactatgg cttacagctt aacgtgtttg gtttgttgaa 6600
tgttaacttg gttttgtcaa gctcagtttt ttactctggc ttacagcata acatgtttga 6660
cttttggttt tgctgctttg ttattgggtt ctgggtagtt cttgatgaat ccaaaagatc 6720
atgtgcacag ccatattatc tatttaagcg atccaggtta ttactatgaa aggatgcctt 6780
ctagctaagg agtagttagg ttttttcttc aaggttaaat tttctcgatg ctctagtgtt 6840
cctgtgacca taatcataat aattcctttg aaagctctat ggtccctgga agcagggcat 6900
acaatgcaag acagcaactt gatcacatca actgaagtat acagggttct cttaactctt 6960
ggtgacttcg gtttaatgga ccggttgtac tcgtgttcta tccgtaaccg ttgtgatgtc 7020
ttgtgtgttt ggttgcggga tagctgggac cacgacgttt ccgtctaatt ctgatggata 7080
gctatagacg gcactgagat ggttatatta taacctctga tcctgaactc tacgagatcg 7140
tctcatccgt cattgccacc aaatacacca ttaaattact aattagctaa cggaccctat 7200
ttgtactcat tccatgtctc ataaactttg ggcaccatcc atccaacaca tccaatctaa 7260
acacaccaaa cgatggggaa tggaaagagc agtattcgat tcaacaatgg caaacaaata 7320
tcactgaatt agaccaagaa taaacctaat tagacaacga cctcccaacc atcattcgtc 7380
aggctgtaaa gaagataaag ctgccttggg gcatggatca agcagaacac cagagatgaa 7440
tccaaacaca cagaaaatca cgcgcgctgt ctacaatgac aacaagcccc acatttcatt 7500
gcagtacact gggctacaaa ggcacgtaca acaaagagct agggaaacat tgcggagggc 7560
acgagagagc agctaacttg acaatatagc agactgagct tgcactgtta gcaggcgagg 7620
aagggaatca tggggacgga gaatggggtc catgcccgcg aaggagaagg cggacgccgc 7680
cacggtggca ccggcgcacg cgcacacagg gaacccgcac aggcagccaa ggatgctgcc 7740
tcgccattgc gccggtcgtc tctgccacgc tcctctctct ctcccgctgc atcgccgtgg 7800
atggggcaag cagagagcag ggactgcgac gatctgggcg gaggactcgc cttggagagc 7860
gcggacgcag acgggattct agggagagag cgaagacggg gcgcgcgcgg cgctcgcgcg 7920
gcgtggtggc ggcgagatta gcgggggtgg ggggagggcg gagccgtggt gagggtgtgg 7980
acgccctcct taccctctta agtagtagta gagatataat ccgttccaaa atatccatcc 8040
gttcaattta tatttcgttt gatcttttta ccctaaattt gattgactca tcttattaaa 8100
aaagttcata actattatta atctttattg agatatcatt tagcatataa tatactttaa 8160
gtgtggtttt agattttttt taaaaaaaaa aattcgcaaa aattaaatga aacgacccaa 8220
tcaaacttga aaagtaaaac taattataaa tttgaacgga aggagtaaga ggatgtttga 8280
atgtactaga gctaatagtt ggttgcttta aaatttgcta gtagaattag ctagctaata 8340
aatatctaga taactattag ctaatttgct aaaacagcta atagttgaac tattagctag 8400
attgtttgga tgtattcggc taattttaat ggctaactat tagctatagt acaatattca 8460
aacacctcct aattaaaatg gacaaatatc tcttcttttg gtcccttgcg ttagattttt 8520
catatctcct tatttagtat aaaagaatca tcaaaaagtg gacaacccct agtggaacac 8580
cattttagta gtggttgcat gaaacctttc gcgcaccagt ttctatgtgt cactctaaaa 8640
atgggacagc atgtacgtag tgcctatata tatacaagtc atctatcgtt gcctcctcag 8700
ttcatcacta atcacactta ttgtgccctc gacgagtatc tatagctagc tcattaatcg 8760
attcgggggt gtgttgtcga aggcggcatt ggcgagctac tcgtcgcggc gtccaagcaa 8820
tacctgtagc acgaaggcga tcgccgggag cgtggtcggc gagcccgtcg tgctggggca 8880
gagggtgacg gtgctgacgg tggacggcgg cggcgtccgg ggtctcatcc cgggaaccat 8940
cctcgccttc ctggaggcca ggctgcagga gctggacgca ccggaggcga ggctggcgga 9000
ctacttcgac tacatcgccg gaaccagcac cggcggtctc atcaccgcct tgctgaccgc 9060
gcccggcaag gacaagcggc ctctctaggc tgccaaggac atcaaccact tttacatcca 9120
taactgcccg cgcatctttc ctcagaagtg agtccgatgc tgccgccatt gttcttgcat 9180
ccatccagca tcgtacgtac gtcctctata catctgcgga tcatcatgtg cgcatgtttg 9240
tggcatgcat gcatgcatgt gagcaggagc aggcttgcga aaaccccatg gagatctttg 9300
cgatggcgca gatcaccaaa aagatgcttg ccaacaagga caaggccgag gagctgtacc 9360
cagtgaagcc gtcgaactgc cgcaggttcc tggtgctgtc catcgggacg gggtcgacgt 9420
ccgagcaggg cctctacacg gcgcggcagt gctcccggtg gggtatctgc cggtggctcc 9480
gcgtcgactc gggagcatgc cctttgtgct tatgcctccg ttctgccttc tgacgaattt 9540
ggtactggaa gcagatgagt tttggttcac tatcattctg aatttacacc tgcgcttgct 9600
gtcagactag gcaaccaagt gacttttgtg actttgatca tgttcagtgt gtttccaagt 9660
cctaatcaat caaaaagaaa aacagtttgt taacgattgt ttgccatgtc tatataataa 9720
agttgctttt atagtagctt agaattcaat cggccaactt tatctcgtac gctgacagta 9780
aaggtacatt taaaaggtga caatggatag tctaatactt gaactgacaa tagagacaca 9840
ttacatgtca gttgattaag tttgtaacag aaaaataaac aatactacat aattgcaaag 9900
tttctttgat gtctttcttt caagaaacac aaatatatca atgctacagt attgctgatg 9960
aatttatcca tgttgagatg tttttctggt ttctgatctg atcagtctca attggtgtgc 10020
tgtttcattt tcatttgctg atgatcgtcc gagtagttaa ttcttactaa tatttagata 10080
atttggcata caagcgaatc acgtagaaca tgatactttt gaatgaattt atcaaagttt 10140
tatcacttgg tgagttgttt catggttttc ctactgatgt ctcttcttca gatttctcga 10200
ggcggagcca ccggcagata ccccaccggg agcactgccg cgccgtgtag aggccctgct 10260
cggacgtcga ccccgtcccg atggacagca ccaggaacct gcggcagttc gacggcttca 10320
ctgggtacag ctcctcggcc ttgtccttgc tggcaagcat ctttttggtg atctgcgtca 10380
tcgcaaggat ccatgggaca agtggcttta ctgtcagtca catgcttgta aataagtaga 10440
ctttatttta ataaaacata aaaatatata tatgttcttg aatataaaat tgataaccaa 10500
attaaaattc gaaccatcac ttatacataa ttttacttta ttttttataa aacgtgaacg 10560
ggaaggacta ccgtgaatga ctatagaacc aatcatacta gtataaaata tatgatgaca 10620
ctacgggaga gacaaacttt gtctggcgct aaatattttg ccgagtgtga attcacgggc 10680
actaggcaaa gatcttcttt gccgagtgtt acgctgggca aagtaagaca ctaggtaaat 10740
cagtcatttg ccgagtgtcc gccactaggc aaagcaaaac actggcaaat caaaagttta 10800
cctagtgcca gacactaggc aaaaaaaaaa cgctcggcaa atcggaagtt tccctagtgc 10860
cagacactag acaaagaaaa acacttgata aactagcgtc gtcagctaac accatccacc 10920
aaccgttaac gttgccgagt atctgacttc gacactcggc aaagaaggtc tctttgccta 10980
gtgtcggtct ggaacactag gcaaagaggc actttaccta gtgtcgtatt ttgacactca 11040
gtaaaataat tttttttctt tctgcttcca aactttttat gatgtgttcc tatagcacct 11100
agaactacat gtcaagtttt ggtaaaattt ttgaagtttt tgctatattt acttaattta 11160
ttttatttaa ttgaatttct tttgataatt caaatttgaa ctcggcaagg taagaagcga 11220
gggtagcctg gaaacacact ttgcctagtg ttacactcgg tacaggagcc tcccctgcct 11280
agtgctgcac tcgacaaaag attcgccttt gcctagcgct gcactcggca caggagtcgc 11340
ctttgcctag tgctgcacta ggcaaagcct ccgttaccgt gccttccatc gtcggacccg 11400
aaagtagcaa acaacaggtt catgtgcact ataaaaagac aaaattctcg agtttcatct 11460
tttattccac ataagcctta tattttccat tttcatatga tttttagttt aagtttgtgt 11520
cttaactttt tcgttaatac gtaattctat gcattatgga tgcgtgaagt atttttgttt 11580
aaaaaaatga aatgtcaaaa tacgttttgt gatctatttc catgttttca cctaacaggt 11640
ggtttttact atatattctg ccataactct agccttagat gtaaatcgaa aaaaaatgag 11700
agatgagctg gagatagcct tagatgaagc gtctgaaata taaaagaaag agtaatgttg 11760
aacgcagtag gtgtagcagc tgtagttcca tctctaggaa agggaactgc aatccgggct 11820
ccgggcctcg cgcaatctgg cctgtcgtgt agatgcagcc ctgtccatga cggcccaagc 11880
aacgcccgcg gctctcgatc caccacggaa cccactccga cacacactga cacacacatg 11940
ctggatgtgg atgtgctgtc caattattag tagcaattcg gtaggcacag gcacgtactg 12000
gccggtgttt tagctgtaag taccgaacca atcacggtta agaaccgatt aatccgtgcc 12060
cagccgccga gtgcgttcgt acgtgcatcg gatgcactgc atgaattgag agcatcatca 12120
tatcatacgc aggagtagta cgacgccgct gctgtcttgt ccggctaatg ctttgctcac 12180
agattagtcc atcgcccacg gtcggtgtgg tgtggatcgc tgatgccact gctttttgtt 12240
tggtttttat tcccctgata atcctccgcg tccctgaatg tatctattta ttttcattcc 12300
gaaatccctt tcacgaaaaa gaaaacgaat aaaaagagag ttacgaatac gcttccggcg 12360
gcccacatca ccttccagcg aacatcgcgc cgcgctgacg tgtcgcccat cgcggccgtc 12420
catatcgcca tccgacgacc gtggaagctg gcagcggccg ctccgttccg tcgaaggggc 12480
aggtcagtca ggtcacccac acggccacac ccgcgcgggg gatacgcggt ggaaaacccg 12540
gcgaccacat caaaacacga ggcgtctccc gcaggactgg tcactcggca cgcaggcaga 12600
ggcagcacag cagcagccag ctccatccat cctctttccc ctcctcgctt cgcttcctcg 12660
gcggattcct cctccctcgg ccgtccccgt ccccttcttc gccgcgccag ctcgcccgag 12720
ttggtacgcc catcctctgc tgtactcccc ccgttcctgc tgtgcgaaac cgagcaccca 12780
aaccctagcc taagctattc gcatcgcaaa ctctattgag cgacggatcg cgaaacgcgc 12840
gccgccgcat tcggacgaga cctcgtggcc cgtaccttcc tactctaccg tcgtcgccgg 12900
agaccagctc accggttagg gtttcgggat taggggcttg gggcttggat tcccggggga 12960
ctccataccc ggtggcctta gaaggggaag ggggcccagc ggtggtggtg gttcgactgt 13020
cagcgagcga gacggcgggg caccgccgat cgggcgtcgc tggacatttg attgagctgg 13080
gcaggcgaag gcgtggagct tgctcttcga ttgggattag aggagggcgg aggtggtatg 13140
gtgggcggct cggctggctc aatggagccg ctggccggtg gtggggcagg cgagatcacg 13200
ctctctcata gagcgtaatg ggttgcgtaa cacactcctg tttgtatttc gatccgatct 13260
aataattaaa gtttatttac gtggtgcaca tgtagaattt tcccatgccc actgctaact 13320
tggattggat ttttgccaaa aaaacgggtg tgcacccaat cacttatagt gaaaccgttg 13380
tgtgtttgat gtgcttctaa ccaagtacaa atccagtgaa aacacaccta ccattcaaca 13440
atccacattt tggttgcaat tatcagcatt ctagaaaggt gcatgtgccc attgatacac 13500
ttgctattgg tgcaggcaaa cccaccatgg cctcctccga gaacgtcatc accgagttca 13560
tgcgcttcaa ggtgcgcatg gagggcaccg tgaacggcca cgagttcgag atcgagggcg 13620
agggcgaggg ccgcccctac gagggccaca acaccgtgaa gctgaaggtg accaagggcg 13680
gccccctgcc cttcgcctgg gacatcctgt ccccccagtt ccagtacggc tccaaggtgt 13740
acgtgaagca ccccgccgac atccccgact acaagaagct gtccttcccc gagggcttca 13800
agtgggagcg cgtgatgaac ttcgaggacg gcggcgtggc gaccgtgacc caggactcct 13860
ccctgcagga cggctgcttc atctacaagg tgaagttcat cggcgtgaac ttcccctccg 13920
acggccccgt gatgcagaag aagaccatgg gctgggaggc ctccaccgag cgcctgtacc 13980
cccgcgacgg cgtgctgaag ggcgagaccc acaaggccct gaagctgaag gacggcggcc 14040
actacctggt ggagttcaag tccatctaca tggccaagaa gcccgtgcag ctgcccggct 14100
actactacgt ggacgccaag ctggacatca cctcccacaa cgaggactac accatcgtgg 14160
agcagtacga gcgcaccgag ggccgccacc acctgttcct gtaggagctc gcatcatgat 14220
catgcatcat ggactcggcc tactactgtg gatttgtatg ccattataga cttggtgctg 14280
tgaaagactg cttgatgatt tgcgggtttg ttgctgtgta aaaaaaggtc ccttggctcc 14340
cagaagacca tgaaggttcg gatctatcat gtaattcctt gttatctgcc aattatgtat 14400
ggactatgga catgtgttgc gctgttcaac ttactactac aaataagtaa tcgatatgtt 14460
cccttcccat gtctcggtga caattgtctg gagaagctta ggggtcgttt gtttgggatt 14520
atgtctggag aaacttattt taaactaagt gtgagttcaa gttaagttag attatataat 14580
ctaggcagat tataattcca agcgaacagg tccttagtgt ttttggaaaa tcctaggtgt 14640
tcttttggct acattgttgt gtgtgcagat cccttgttgg tctgtaagcg tggggaagta 14700
agaatcgtcc gtttctactg aagacctgct cgagttaggc accgaggatg ccggtaacca 14760
aacagagcaa tagtgtctct gtgggcacag tggagtgtga atctgtgtga tgcaaatccg 14820
tcatttgttt agcaaaattt ccagcgttgc atgatgcagt ttctttaaca cggacttaag 14880
ggaagggaaa aaaatgttga gccaggagat ccttcaatgt gttagactga cgtgatagcc 14940
aactaaacca cgacgcaatg ttgtcgttaa tgacaaaaaa actatttgtt cctaaatcct 15000
tggcgacatt gcatggctgt ctcatgagat aatggtctca tctcttattt atctcttatt 15060
tatagccgga agtggtagtg acccctgctt gattgctcgt atgccatctc aagttctcaa 15120
ccgtgtcgag cagccatttt cccatctcaa gcgcatcatc gtttcgtttg acctcatctg 15180
ctatcctgct cctagtgcaa atcacatgcg acagaaagtg tgcggaccgc ctgcaggccc 15240
gggggcgcgc cctaattagc taacggccag gatcgccgcg tgagccttta gcaactagct 15300
agattaatta acgcaatctg ttattaagtt gtctaagcgt caatttgttt acaccacaat 15360
atatcctgcc accagccagc caacagctcc ccgaccggca gctcggcaca aaatcaccac 15420
tcgatacagg cagcccatca g 15441
<210> 34
<211> 10671
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22807
<220>
<221> Энхансер
<222> (165)..(256)
<223> eNOS
<220>
<221> промотор
<222> (301)..(2097)
<223> Ubi
<220>
<221> ген
<222> (2114)..(6283)
<223> Cas9
<220>
<221> другой_признак
<222> (6218)..(6274)
<223> NLS-метка
<220>
<221> Терминатор
<222> (6289)..(6541)
<223> tNOS
<220>
<221> промотор
<222> (6548)..(6922)
<223> pU3
<220>
<221> другой_признак
<222> (6924)..(6941)
<220>
<221> другой_признак
<222> (6924)..(7026)
<223> sgRNA
<220>
<221> промотор
<222> (7037)..(9028)
<223> Ubi
<220>
<221> ген
<222> (9045)..(10223)
<223> PMI
<220>
<221> Терминатор
<222> (10246)..(10498)
<223> tNOS
<400> 34
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tgaaggcggg aaacgacaat ctgatcatga gcggagaatt 180
aagggagtca cgttatgacc cccgccgatg acgcgggaca agccgtttta cgtttggaac 240
tgacagaacc gcaacgctgc aggaattggc cgcagcggcc atttaaacaa agcttggtac 300
cattatgtgg tctaggtagg ttctatatat aagaaaactt gaaatgttct aaaaaaaaat 360
tcaagcccat gcatgattga agcaaacggt atagcaacgg tgttaacctg atctagtgat 420
ctcttgcaat ccttaacggc cacctaccgc aggtagcaaa cggcgtcccc ctcctcgata 480
tctccgcggc gacctctggc tttttccgcg gaattgcgcg gtggggacgg attccacgag 540
accgcgacgc aaccgcctct cgccgctggg ccccacaccg ctcggtgccg tagcctcacg 600
ggactctttc tccctcctcc cccgttataa attggcttca tcccctcctt gcctcatcca 660
tccaaatccc agtccccaat cccatccctt cgtaggagaa attcatcgaa gctaagcgaa 720
tcctcgcgat cctctcaagg tactgcgagt tttcgatccc cctctcgacc cctcgtatgt 780
ttgtgtttgt cgtagcgttt gattaggtat gctttccctg tttgtgttcg tcgtagcgtt 840
tgattaggta tgctttccct gttcgtgttc atcgtagtgt ttgattaggt cgtgtgaggc 900
gatggcctgc tcgcgtcctt cgatctgtag tcgatttgcg ggtcgtggtg tagatctgcg 960
ggctgtgatg aagttatttg gtgtgatctg ctcgcctgat tctgcgggtt ggctcgagta 1020
gatatgatgg ttggaccggt tggttcgttt accgcgctag ggttgggctg ggatgatgtt 1080
gcatgcgccg ttgcgcgtga tcccgcagca ggacttgcgt ttgattgcca gatctcgtta 1140
cgattatgtg atttggtttg gactttttag atctgtagct tctgcttatg tgccagatgc 1200
gcctactgct catatgcctg atgataatca taaatggctg tggaactaac tagttgattg 1260
cggagtcatg tatcagctac aggtgtaggg actagctaca ggtgtaggga cttgcgtcta 1320
attgtttggt cctttactca tgttgcaatt atgcaattta gtttagattg tttgttccac 1380
tcatctaggc tgtaaaaggg acactgctta gattgctgtt taatcttttt agtagattat 1440
attatattgg taacttatta cccctattac atgccatacg tgacttctgc tcatgcctga 1500
tgataatcat agatcactgt ggaattaatt agttgattgt tgaatcatgt ttcatgtaca 1560
taccacggca caattgctta gttccttaac aaatgcaaat tttactgatc catgtatgat 1620
ttgcgtggtt ctctaatgtg aaatactata gctacttgtt agtaagaatc aggttcgtat 1680
gcttaatgct gtatgtgcct tctgctcatg cctgatgata atcatatatc actggaatta 1740
attagttgat cgtttaatca tatatcaagt acataccatg ccacaatttt tagtcactta 1800
acccatgcag attgaactgg tccctgcatg ttttgctaaa ttgttctatt ctgattagac 1860
catatatcat gtattttttt ttggtaatgg ttctcttatt ttaaatgcta tatagttctg 1920
gtacttgtta gaaagatctg cttcatagtt tagttgccta tccctcgaat taggatgctg 1980
agcagctgat cctatagctt tgtttcatgt atcaattctt ttgtgttcaa cagtcagttt 2040
ttgttagatt cattgtaact tatggtcgct tactcttctg gtcctcaatg cttgcaggat 2100
cgcggccgct catatggaca agaagtacag catcggcctg gacatcggca ccaacagcgt 2160
gggctgggcc gtgatcaccg acgagtacaa ggtgccgagc aagaagttca aggtgctggg 2220
caacaccgac aggcacagca tcaagaagaa cctgatcggc gccctgctgt tcgacagcgg 2280
cgagaccgcc gaggccacca ggctgaagag gaccgccagg aggaggtaca ccaggaggaa 2340
gaacaggatc tgctacctgc aggagatctt cagcaacgag atggccaagg tggacgacag 2400
cttcttccac aggctggagg agagcttcct ggtggaggag gacaagaagc acgagaggca 2460
cccgatcttc ggcaacatcg tggacgaggt ggcctaccac gagaagtacc cgaccatcta 2520
ccacctgagg aagaagctgg tggacagcac cgacaaggcc gacctgaggc tgatctacct 2580
ggccctggcc cacatgatca agttcagggg ccacttcctg atcgagggcg acctgaaccc 2640
ggacaacagc gacgtggaca agctgttcat ccagctggtg cagacctaca accagctgtt 2700
cgaggagaac ccgatcaacg ccagcggcgt ggacgccaag gccatcctga gcgccaggct 2760
gagcaagagc aggaggctgg agaacctgat cgcccagctg ccgggcgaga agaagaacgg 2820
cctgttcggc aacctgatcg ccctgagcct gggcctgacc ccgaacttca agagcaactt 2880
cgacctggcc gaggacgcca agctgcagct gagcaaggac acctacgacg acgacctgga 2940
caacctgctg gcccagatcg gcgaccagta cgccgacctg ttcctggccg ccaagaacct 3000
gagcgacgcc atcctgctga gcgacatcct gagggtgaac accgagatca ccaaggcccc 3060
gctgagcgcc agcatgatca agaggtacga cgagcaccac caggacctga ccctgctgaa 3120
ggccctggtg aggcagcagc tgccggagaa gtacaaggag atcttcttcg accagagcaa 3180
gaacggctac gccggctaca tcgacggcgg cgccagccag gaggagttct acaagttcat 3240
caagccgatc ctggagaaga tggacggcac cgaggagctg ctggtgaagc tgaacaggga 3300
ggacctgctg aggaagcaga ggaccttcga caacggcagc atcccgcacc agatccacct 3360
gggcgagctg cacgccatcc tgaggaggca ggaggacttc tacccgttcc tgaaggacaa 3420
cagggagaag atcgagaaga tcctgacctt ccgcatcccg tactacgtgg gcccgctggc 3480
caggggcaac agcaggttcg cctggatgac caggaagagc gaggagacca tcaccccgtg 3540
gaacttcgag gaggtggtgg acaagggcgc cagcgcccag agcttcatcg agaggatgac 3600
caacttcgac aagaacctgc cgaacgagaa ggtgctgccg aagcacagcc tgctgtacga 3660
gtacttcacc gtgtacaacg agctgaccaa ggtgaagtac gtgaccgagg gcatgaggaa 3720
gccggccttc ctgagcggcg agcagaagaa ggccatcgtg gacctgctgt tcaagaccaa 3780
caggaaggtg accgtgaagc agctgaagga ggactacttc aagaagatcg agtgcttcga 3840
cagcgtggag atcagcggcg tggaggacag gttcaacgcc agcctgggca cctaccacga 3900
cctgctgaag atcatcaagg acaaggactt cctggacaac gaggagaacg aggacatcct 3960
ggaggacatc gtgctgaccc tgaccctgtt cgaggacagg gagatgatcg aggagaggct 4020
gaagacctac gcccacctgt tcgacgacaa ggtgatgaag cagctgaaga ggaggaggta 4080
caccggctgg ggcaggctga gcaggaagct gatcaacggc atcagggaca agcagagcgg 4140
caagaccatc ctggacttcc tgaagagcga cggcttcgcc aacaggaact tcatgcagct 4200
gatccacgac gacagcctga ccttcaagga ggacatccag aaggcccagg tgagcggcca 4260
gggcgacagc ctgcacgagc acatcgccaa cctggccggc agcccggcca tcaagaaggg 4320
catcctgcag accgtgaagg tggtggacga gctggtgaag gtgatgggca ggcacaagcc 4380
ggagaacatc gtgatcgaga tggccaggga gaaccagacc acccagaagg gccagaagaa 4440
cagcagggag aggatgaaga ggatcgagga gggcatcaag gagctgggca gccagatcct 4500
gaaggagcac ccggtggaga acacccagct gcagaacgag aagctgtacc tgtactacct 4560
gcagaacggc agggacatgt acgtggacca ggagctggac atcaacaggc tgagcgacta 4620
cgacgtggac cacatcgtgc cgcagagctt cctgaaggac gacagcatcg acaacaaggt 4680
gctgaccagg agcgacaaga acaggggcaa gagcgacaac gtgccgagcg aggaggtggt 4740
gaagaagatg aaaaactact ggaggcagct gctgaacgcc aagctgatca cccagaggaa 4800
gttcgacaac ctgaccaagg ccgagagggg cggcctgagc gagctggaca aggccggctt 4860
cattaaaagg cagctggtgg agaccaggca gatcaccaag cacgtggccc agatcctgga 4920
cagcaggatg aacaccaagt acgacgagaa cgacaagctg atcagggagg tgaaggtgat 4980
caccctgaag agcaagctgg tgagcgactt caggaaggac ttccagttct acaaggtgag 5040
ggagatcaat aattaccacc acgcccacga cgcctacctg aacgccgtgg tgggcaccgc 5100
cctgattaaa aagtacccga agctggagag cgagttcgtg tacggcgact acaaggtgta 5160
cgacgtgagg aagatgatcg ccaagagcga gcaggagatc ggcaaggcca ccgccaagta 5220
cttcttctac agcaacatca tgaacttctt caagaccgag atcaccctgg ccaacggcga 5280
gatcaggaag aggccgctga tcgagaccaa cggcgagacc ggcgagatcg tgtgggacaa 5340
gggcagggac ttcgccaccg tgaggaaggt gctgtccatg ccgcaggtga acatcgtgaa 5400
gaagaccgag gtgcagaccg gcggcttcag caaggagagc atcctgccga agaggaacag 5460
cgacaagctg atcgccagga agaaggactg ggacccgaag aagtacggcg gcttcgacag 5520
cccgaccgtg gcctacagcg tgctggtggt ggccaaggtg gagaagggca agagcaagaa 5580
gctgaagagc gtgaaggagc tggtgggcat caccatcatg gagaggagca gcttcgagaa 5640
gaacccagtg gacttcctgg aggccaaggg ctacaaggag gtgaagaagg acctgatcat 5700
taaactgccg aagtacagcc tgttcgagct ggagaacggc aggaagagga tgctggccag 5760
cgccggcgag ctgcagaagg gcaacgagct ggccctgccg agcaagtacg tgaacttcct 5820
gtacctggcc agccactacg agaagctgaa gggcagcccg gaggacaacg agcagaagca 5880
gctgttcgtg gagcagcaca agcactacct ggacgagatc atcgagcaga tcagcgagtt 5940
cagcaagagg gtgatcctgg ccgacgccaa cctggacaag gtgctgagcg cctacaacaa 6000
gcacagggac aagccgatca gggagcaggc cgagaacatc atccacctgt tcaccctgac 6060
caacctgggc gccccggccg ccttcaagta cttcgacacc accatcgaca ggaagaggta 6120
caccagcacc aaggaggtgc tggacgccac cctgatccac cagagcatca ccggcctgta 6180
cgagaccagg atcgacctga gccagctggg cggcgacagc agcccgccga agaagaagag 6240
gaaggtgagc tggaaggacg ccagcggctg gagcaggatg tgaagcttga tcgttcaaac 6300
atttggcaat aaagtttctt aagattgaat cctgttgccg gtcttgcgat gattatcata 6360
taatttctgt tgaattacgt taagcatgta ataattaaca tgtaatgcat gacgttattt 6420
atgagatggg tttttatgat tagagtcccg caattataca tttaatacgc gatagaaaac 6480
aaaatatagc gcgcaaacta ggataaatta tcgcgcgcgg tgtcatctat gttactagat 6540
cttcgaaggg atctttaaac atacgaacag atcacttaaa gttcttctga agcaacttaa 6600
agttatcagg catgcatgga tcttggagga atcagatgtg cagtcaggga ccatagcaca 6660
ggacaggcgt cttctactgg tgctaccagc aaatgctgga agccgggaac actgggtacg 6720
ttggaaacca cgtgatgtgg agtaagataa actgtaggag aaaagcattt cgtagtgggc 6780
catgaagcct ttcaggacat gtattgcagt atgggccggc ccattacgca attggacgac 6840
aacaaagact agtattagta ccacctcggc tatccacata gatcaaagct ggtttaaaag 6900
agttgtgcag atgatccgtg gcagtcaacg tggagacagg ggttttagag ctagaaatag 6960
caagttaaaa taaggctagt ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt 7020
ttttttcgga ccgcgcctgc agtgcagcgt gacccggtcg tgcccctctc tagagataat 7080
gagcattgca tgtctaagtt ataaaaaatt accacatatt ttttttgtca cacttgtttg 7140
aagtgcagtt tatctatctt tatacatata tttaaacttt actctacgaa taatataatc 7200
tatagtacta caataatatc agtgttttag agaatcatat aaatgaacag ttagacatgg 7260
tctaaaggac aattgagtat tttgacaaca ggactctaca gttttatctt tttagtgtgc 7320
atgtgttctc cttttttttt gcaaatagct tcacctatat aatacttcat ccattttatt 7380
agtacatcca tttagggttt agggttaatg gtttttatag actaattttt ttagtacatc 7440
tattttattc tattttagcc tctaaattaa gaaaactaaa actctatttt agttttttta 7500
tttaataatt tagatataaa atagaataaa ataaagtgac taaaaattaa acaaataccc 7560
tttaagaaat taaaaaaact aaggaaacat ttttcttgtt tcgagtagat aatgccagcc 7620
tgttaaacgc cgtcgacgag tctaacggac accaaccagc gaaccagcag cgtcgcgtcg 7680
ggccaagcga agcagacggc acggcatctc tgtcgctgcc tctggacccc tctcgagagt 7740
tccgctccac cgttggactt gctccgctgt cggcatccag aaattgcgtg gcggagcggc 7800
agacgtgagc cggcacggca ggcggcctcc tcctcctctc acggcaccgg cagctacggg 7860
ggattccttt cccaccgctc cttcgctttc ccttcctcgc ccgccgtaat aaatagacac 7920
cccctccaca ccctctttcc ccaacctcgt gttgttcgga gcgcacacac acacaaccag 7980
atctccccca aatccacccg tcggcacctc cgcttcaagg tacgccgctc gtcctccccc 8040
ccccccctct ctaccttctc tagatcggcg ttccggtcca tggttagggc ccggtagttc 8100
tacttctgtt catgtttgtg ttagatccgt gtttgtgtta gatccgtgct gctagcgttc 8160
gtacacggat gcgacctgta cgtcagacac gttctgattg ctaacttgcc agtgtttctc 8220
tttggggaat cctgggatgg ctctagccgt tccgcagacg ggatcgattt catgattttt 8280
tttgtttcgt tgcatagggt ttggtttgcc cttttccttt atttcaatat atgccgtgca 8340
cttgtttgtc gggtcatctt ttcatgcttt tttttgtctt ggttgtgatg atgtggtctg 8400
gttgggcggt cgttctagat cggagtagaa ttctgtttca aactacctgg tggatttatt 8460
aattttggat ctgtatgtgt gtgccataca tattcatagt tacgaattga agatgatgga 8520
tggaaatatc gatctaggat aggtatacat gttgatgcgg gttttactga tgcatataca 8580
gagatgcttt ttgttcgctt ggttgtgatg atgtggtgtg gttgggcggt cgttcattcg 8640
ttctagatcg gagtagaata ctgtttcaaa ctacctggtg tatttattaa ttttggaact 8700
gtatgtgtgt gtcatacatc ttcatagtta cgagtttaag atggatggaa atatcgatct 8760
aggataggta tacatgttga tgtgggtttt actgatgcat atacatgatg gcatatgcag 8820
catctattca tatgctctaa ccttgagtac ctatctatta taataaacaa gtatgtttta 8880
taattatttt gatcttgata tacttggatg atggcatatg cagcagctat atgtggattt 8940
ttttagccct gccttcatac gctatttatt tgcttggtac tgtttctttt gtcgatgctc 9000
accctgttgt ttggtgttac ttctgcaggg atccggcagc agccatgcag aagctgatca 9060
acagcgtgca gaactacgcc tggggcagca agaccgccct gaccgagctg tacggcatgg 9120
agaaccccag cagccagccc atggccgagc tgtggatggg cgcccacccc aagagcagca 9180
gccgcgtgca gaacgccgcc ggcgacatcg tgagcctgcg cgacgtgatc gagagcgaca 9240
agagcaccct gctgggcgag gccgtggcca agcgcttcgg cgagctgccc ttcctgttca 9300
aggtgctgtg cgccgcccag cccctgagca tccaggtgca ccccaacaag cacaacagcg 9360
agatcggctt cgccaaggag aacgccgccg gcatccccat ggacgccgcc gagcgcaact 9420
acaaggaccc caaccacaag cccgagctgg tgttcgccct gacccccttc ctggccatga 9480
acgccttccg cgagttcagc gagatcgtga gcctgctgca gcccgtggcc ggcgcccacc 9540
ccgccatcgc ccacttcctg cagcagcccg acgccgagcg cctgagcgag ctgttcgcca 9600
gcctgctgaa catgcagggc gaggagaaga gccgcgccct ggccatcctg aagagcgccc 9660
tggacagcca gcagggcgag ccctggcaga ccatccgcct gatcagcgag ttctaccccg 9720
aggacagcgg cctgttcagc cccctgctgc tgaacgtggt gaagctgaac cccggcgagg 9780
ccatgttcct gttcgccgag accccccacg cctacctgca gggcgtggcc ctggaggtga 9840
tggccaacag cgacaacgtg ctgcgcgccg gcctgacccc caagtacatc gacatccccg 9900
agctggtggc caacgtgaag ttcgaggcca agcccgccaa ccagctgctg acccagcccg 9960
tgaagcaggg cgccgagctg gacttcccca tccccgtgga cgacttcgcc ttcagcctgc 10020
acgacctgag cgacaaggag accaccatca gccagcagag cgccgccatc ctgttctgcg 10080
tggagggcga cgccaccctg tggaagggca gccagcagct gcagctgaag cccggcgaga 10140
gcgccttcat cgccgccaac gagagccccg tgaccgtgaa gggccacggc cgcctggccc 10200
gcgtgtacaa caagctgtga taggagctcg atccgtcgac ctgcagatcg ttcaaacatt 10260
tggcaataaa gtttcttaag attgaatcct gttgccggtc ttgcgatgat tatcatataa 10320
tttctgttga attacgttaa gcatgtaata attaacatgt aatgcatgac gttatttatg 10380
agatgggttt ttatgattag agtcccgcaa ttatacattt aatacgcgat agaaaacaaa 10440
atatagcgcg caaactagga taaattatcg cgcgcggtgt catctatgtt actagatcgg 10500
cgcgccgcaa ttgaagtttg ggcggccagc atggccgtat ccgcaatgtg ttattaagtt 10560
gtctaagcgt caatttgttt acaccacaat atatcctgcc accagccagc caacagctcc 10620
ccgaccggca gctcggcaca aaatcaccac tcgatacagg cagcccatca g 10671
<210> 35
<211> 14475
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22808
<220>
<221> Энхансер
<222> (165)..(256)
<223> eNOS
<220>
<221> промотор
<222> (301)..(2097)
<223> Ubi
<220>
<221> ген
<222> (2116)..(5190)
<223> TNFw
<220>
<221> Терминатор
<222> (2116)..(5190)
<223> tNOS
<220>
<221> промотор
<222> (5483)..(7475)
<223> Ubi
<220>
<221> ген
<222> (7489)..(10563)
<223> TNRv
<220>
<221> Терминатор
<222> (10570)..(10822)
<223> tNOS
<220>
<221> промотор
<222> (10841)..(12832)
<223> Ubi
<220>
<221> ген
<222> (12849)..(14027)
<223> PMI
<220>
<221> Терминатор
<222> (14050)..(14302)
<223> tNOS
<400> 35
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tgaaggcggg aaacgacaat ctgatcatga gcggagaatt 180
aagggagtca cgttatgacc cccgccgatg acgcgggaca agccgtttta cgtttggaac 240
tgacagaacc gcaacgctgc aggaattggc cgcagcggcc atttaaacaa agcttggtac 300
cattatgtgg tctaggtagg ttctatatat aagaaaactt gaaatgttct aaaaaaaaat 360
tcaagcccat gcatgattga agcaaacggt atagcaacgg tgttaacctg atctagtgat 420
ctcttgcaat ccttaacggc cacctaccgc aggtagcaaa cggcgtcccc ctcctcgata 480
tctccgcggc gacctctggc tttttccgcg gaattgcgcg gtggggacgg attccacgag 540
accgcgacgc aaccgcctct cgccgctggg ccccacaccg ctcggtgccg tagcctcacg 600
ggactctttc tccctcctcc cccgttataa attggcttca tcccctcctt gcctcatcca 660
tccaaatccc agtccccaat cccatccctt cgtaggagaa attcatcgaa gctaagcgaa 720
tcctcgcgat cctctcaagg tactgcgagt tttcgatccc cctctcgacc cctcgtatgt 780
ttgtgtttgt cgtagcgttt gattaggtat gctttccctg tttgtgttcg tcgtagcgtt 840
tgattaggta tgctttccct gttcgtgttc atcgtagtgt ttgattaggt cgtgtgaggc 900
gatggcctgc tcgcgtcctt cgatctgtag tcgatttgcg ggtcgtggtg tagatctgcg 960
ggctgtgatg aagttatttg gtgtgatctg ctcgcctgat tctgcgggtt ggctcgagta 1020
gatatgatgg ttggaccggt tggttcgttt accgcgctag ggttgggctg ggatgatgtt 1080
gcatgcgccg ttgcgcgtga tcccgcagca ggacttgcgt ttgattgcca gatctcgtta 1140
cgattatgtg atttggtttg gactttttag atctgtagct tctgcttatg tgccagatgc 1200
gcctactgct catatgcctg atgataatca taaatggctg tggaactaac tagttgattg 1260
cggagtcatg tatcagctac aggtgtaggg actagctaca ggtgtaggga cttgcgtcta 1320
attgtttggt cctttactca tgttgcaatt atgcaattta gtttagattg tttgttccac 1380
tcatctaggc tgtaaaaggg acactgctta gattgctgtt taatcttttt agtagattat 1440
attatattgg taacttatta cccctattac atgccatacg tgacttctgc tcatgcctga 1500
tgataatcat agatcactgt ggaattaatt agttgattgt tgaatcatgt ttcatgtaca 1560
taccacggca caattgctta gttccttaac aaatgcaaat tttactgatc catgtatgat 1620
ttgcgtggtt ctctaatgtg aaatactata gctacttgtt agtaagaatc aggttcgtat 1680
gcttaatgct gtatgtgcct tctgctcatg cctgatgata atcatatatc actggaatta 1740
attagttgat cgtttaatca tatatcaagt acataccatg ccacaatttt tagtcactta 1800
acccatgcag attgaactgg tccctgcatg ttttgctaaa ttgttctatt ctgattagac 1860
catatatcat gtattttttt ttggtaatgg ttctcttatt ttaaatgcta tatagttctg 1920
gtacttgtta gaaagatctg cttcatagtt tagttgccta tccctcgaat taggatgctg 1980
agcagctgat cctatagctt tgtttcatgt atcaattctt ttgtgttcaa cagtcagttt 2040
ttgttagatt cattgtaact tatggtcgct tactcttctg gtcctcaatg cttgcaggat 2100
cgcggccgcg ccaccatggg aaaacctatt cctaatcctc tgctgggcct ggattctacc 2160
ggaggcatgg cccctaagaa aaagcggaag gtggacggcg gagtggacct gagaacactg 2220
ggatattctc agcagcagca ggagaagatc aagcccaagg tgagatctac agtggcccag 2280
caccacgaag ccctggtggg acacggattt acacacgccc acattgtggc cctgtctcag 2340
caccctgccg ccctgggaac agtggccgtg aaatatcagg atatgattgc cgccctgcct 2400
gaggccacac acgaagccat tgtgggagtg ggaaaacagt ggtctggagc cagagccctg 2460
gaagccctgc tgacagtggc cggagaactg agaggacctc ctctgcagct ggatacagga 2520
cagctgctga agattgccaa aaggggcgga gtgaccgcgg tggaagccgt gcacgcctgg 2580
agaaatgccc tgacaggagc ccctctgaac ctgacccccg aacaggtggt ggccattgcc 2640
agccacgacg gcggcaagca ggccctggaa accgtgcaga gactgctgcc cgtgctgtgc 2700
caggcccatg gcctgacacc tgaacaggtg gtggctatcg cctctcacga cggaggaaaa 2760
caggctctgg aaacagtgca gcggctgctg cctgtgctgt gtcaggctca cggcttgact 2820
ccagaacagg tggtggctat tgcttccaat attgggggga aacaggccct ggaaactgtg 2880
cagcgcctgc tgccagtgct gtgccaggct cacggactga cccccgaaca ggtggtggcc 2940
attgccagca acaacggcgg caagcaggcc ctggaaaccg tgcagagact gctgcccgtg 3000
ctgtgccagg cccatggcct gacacctgaa caggtggtgg ctatcgcctc taacaacgga 3060
ggaaaacagg ctctggaaac agtgcagcgg ctgctgcctg tgctgtgtca ggctcacggc 3120
ttgactccag aacaggtggt ggctattgct tccaacaacg gggggaaaca ggccctggaa 3180
actgtgcagc gcctgctgcc agtgctgtgc caggctcacg ggctgacccc cgaacaggtg 3240
gtggccattg ccagcaacgg cggcggcaag caggccctgg aaaccgtgca gagactgctg 3300
cccgtgctgt gccaggccca tggcctgaca cctgaacagg tggtggctat cgcctctcac 3360
gacggaggaa aacaggctct ggaaacagtg cagcggctgc tgcctgtgct gtgtcaggct 3420
cacggcttga ctccagaaca ggtggtggct attgcttcca atattggggg gaaacaggcc 3480
ctggaaactg tgcagcgcct gctgccagtg ctgtgccagg ctcacggcct cactcccgaa 3540
caggtggtgg ccattgccag caacatcggc ggcaagcagg ccctggaaac cgtgcagaga 3600
ctgctgcccg tgctgtgcca ggcccatggc ctgacacctg aacaggtggt ggctatcgcc 3660
tctcacgacg gaggaaaaca ggctctggaa acagtgcagc ggctgctgcc tgtgctgtgt 3720
caggctcacg gcttgactcc agaacaggtg gtggctattg cttccaacaa cggggggaaa 3780
caggccctgg aaactgtgca gcgcctgctg ccagtgctgt gccaggctca cggactgacc 3840
cccgaacagg tggtggccat tgccagcaac ggcggcggca agcaggccct ggaaaccgtg 3900
cagagactgc tgcccgtgct gtgccaggcc catggcctga cacctgaaca ggtggtggct 3960
atcgcctcta acaacggagg aaaacaagca ctcgagacag tgcagcggct gctgcctgtg 4020
ctgtgtcagg ctcacggctt gactccagaa caggtggtgg ctattgcttc caacaacggg 4080
gggaaacagg ccctggaaac tgtgcagcgc ctgctgccag tgctgtgcca ggctcacggg 4140
ctgacccccg aacaggtggt ggccattgcc agcaacatcg gcggcaagca ggccctggaa 4200
accgtgcaga gactgctgcc cgtgctgtgc caggcccatg gcctgacacc tgaacaggtg 4260
gtggctatcg cctctaacaa cggaggaaaa caggctctgg aaacagtgca gcggctgctg 4320
cctgtgctgt gtcaggctca cggcttgact ccacagcagg tcgtggcaat tgctagcaat 4380
atcggcggac ggcccgccct ggagagcatt gtggcccagc tgtctagacc tgatcctgcc 4440
ctggccgccc tgacaaatga tcacctggtg gccctggcct gtctgggagg cagacctgcc 4500
ctggatgccg tgaaaaaagg actgcctcac gcccctgccc tgattaaaag aacaaataga 4560
agaatccccg agcggacctc tcacagagtg gccggatccc agctggtgaa atctgagctg 4620
gaggagaaga agtctgagct gagacacaag ctgaagtacg tgcctcacga gtacatcgag 4680
ctgatcgaga tcgccagaaa tagcacccag gatagaatcc tggagatgaa ggtgatggag 4740
ttcttcatga aagtgtacgg ctacagagga aagcatctgg gaggaagcag aaaacctgac 4800
ggagccattt atacagtggg cagccctatc gattatggcg tgatcgtgga tacaaaggcc 4860
tacagcggag gctacaatct gcctattgga caggccgatg agatgcagag atacgtggag 4920
gagaaccaaa ccaggaacaa gcatatcaac cctaacgagt ggtggaaggt gtacccttct 4980
agcgtgaccg agttcaagtt cctgtttgtg agcggccact tcaagggcaa ttataaggcc 5040
cagctgacca ggctgaacca catcacaaat tgtaatggcg ccgtgctgtc tgtggaggaa 5100
ctgctgattg gaggagagat gattaaggcc ggaacactga cactggagga ggtgagaaga 5160
aagttcaaca acggcgagat caacttctga aagcttgatc gttcaaacat ttggcaataa 5220
agtttcttaa gattgaatcc tgttgccggt cttgcgatga ttatcatata atttctgttg 5280
aattacgtta agcatgtaat aattaacatg taatgcatga cgttatttat gagatgggtt 5340
tttatgatta gagtcccgca attatacatt taatacgcga tagaaaacaa aatatagcgc 5400
gcaaactagg ataaattatc gcgcgcggtg tcatctatgt tactagatct tcgaacccta 5460
gtcgaagaca accggtgcat gcctgcagtg cagcgtgacc cggtcgtgcc cctctctaga 5520
gataatgagc attgcatgtc taagttataa aaaattacca catatttttt ttgtcacact 5580
tgtttgaagt gcagtttatc tatctttata catatattta aactttactc tacgaataat 5640
ataatctata gtactacaat aatatcagtg ttttagagaa tcatataaat gaacagttag 5700
acatggtcta aaggacaatt gagtattttg acaacaggac tctacagttt tatcttttta 5760
gtgtgcatgt gttctccttt ttttttgcaa atagcttcac ctatataata cttcatccat 5820
tttattagta catccattta gggtttaggg ttaatggttt ttatagacta atttttttag 5880
tacatctatt ttattctatt ttagcctcta aattaagaaa actaaaactc tattttagtt 5940
tttttattta ataatttaga tataaaatag aataaaataa agtgactaaa aattaaacaa 6000
atacccttta agaaattaaa aaaactaagg aaacattttt cttgtttcga gtagataatg 6060
ccagcctgtt aaacgccgtc gacgagtcta acggacacca accagcgaac cagcagcgtc 6120
gcgtcgggcc aagcgaagca gacggcacgg catctctgtc gctgcctctg gacccctctc 6180
gagagttccg ctccaccgtt ggacttgctc cgctgtcggc atccagaaat tgcgtggcgg 6240
agcggcagac gtgagccggc acggcaggcg gcctcctcct cctctcacgg caccggcagc 6300
tacgggggat tcctttccca ccgctccttc gctttccctt cctcgcccgc cgtaataaat 6360
agacaccccc tccacaccct ctttccccaa cctcgtgttg ttcggagcgc acacacacac 6420
aaccagatct cccccaaatc cacccgtcgg cacctccgct tcaaggtacg ccgctcgtcc 6480
tccccccccc cccctctcta ccttctctag atcggcgttc cggtccatgg ttagggcccg 6540
gtagttctac ttctgttcat gtttgtgtta gatccgtgtt tgtgttagat ccgtgctgct 6600
agcgttcgta cacggatgcg acctgtacgt cagacacgtt ctgattgcta acttgccagt 6660
gtttctcttt ggggaatcct gggatggctc tagccgttcc gcagacggga tcgatttcat 6720
gatttttttt gtttcgttgc atagggtttg gtttgccctt ttcctttatt tcaatatatg 6780
ccgtgcactt gtttgtcggg tcatcttttc atgctttttt ttgtcttggt tgtgatgatg 6840
tggtctggtt gggcggtcgt tctagatcgg agtagaattc tgtttcaaac tacctggtgg 6900
atttattaat tttggatctg tatgtgtgtg ccatacatat tcatagttac gaattgaaga 6960
tgatggatgg aaatatcgat ctaggatagg tatacatgtt gatgcgggtt ttactgatgc 7020
atatacagag atgctttttg ttcgcttggt tgtgatgatg tggtgtggtt gggcggtcgt 7080
tcattcgttc tagatcggag tagaatactg tttcaaacta cctggtgtat ttattaattt 7140
tggaactgta tgtgtgtgtc atacatcttc atagttacga gtttaagatg gatggaaata 7200
tcgatctagg ataggtatac atgttgatgt gggttttact gatgcatata catgatggca 7260
tatgcagcat ctattcatat gctctaacct tgagtaccta tctattataa taaacaagta 7320
tgttttataa ttattttgat cttgatatac ttggatgatg gcatatgcag cagctatatg 7380
tggatttttt tagccctgcc ttcatacgct atttatttgc ttggtactgt ttcttttgtc 7440
gatgctcacc ctgttgtttg gtgttacttc tgcagcggcc gcgccaccat gggaaaacct 7500
attcctaatc ctctgctggg cctggattct accggaggca tggcccctaa gaaaaagcgg 7560
aaggtggacg gcggagtgga cctgagaaca ctgggatatt ctcagcagca gcaggagaag 7620
atcaagccca aggtgagatc tacagtggcc cagcaccacg aagccctggt gggacacgga 7680
tttacacacg cccacattgt ggccctgtct cagcaccctg ccgccctggg aacagtggcc 7740
gtgaaatatc aggatatgat tgccgccctg cctgaggcca cacacgaagc cattgtggga 7800
gtgggaaaac agtggtctgg agccagagcc ctggaagccc tgctgacagt ggccggagaa 7860
ctgagaggac ctcctctgca gctggataca ggacagctgc tgaagattgc caaaaggggc 7920
ggagtgaccg cggtggaagc cgtgcacgcc tggagaaatg ccctgacagg agcccctctg 7980
aacctgaccc ccgaacaggt ggtggccatt gccagcaaca acggcggcaa gcaggccctg 8040
gaaaccgtgc agagactgct gcccgtgctg tgccaggccc atggcctgac acctgaacag 8100
gtggtggcta tcgcctctca cgacggagga aaacaggctc tggaaacagt gcagcggctg 8160
ctgcctgtgc tgtgtcaggc tcacggcttg actccagaac aggtggtggc tattgcttcc 8220
aacggcgggg ggaaacaggc cctggaaact gtgcagcgcc tgctgccagt gctgtgccag 8280
gctcacggac tgacccccga acaggtggtg gccattgcca gcaacggcgg cggcaagcag 8340
gccctggaaa ccgtgcagag actgctgccc gtgctgtgcc aggcccatgg cctgacacct 8400
gaacaggtgg tggctatcgc ctctcacgac ggaggaaaac aggctctgga aacagtgcag 8460
cggctgctgc ctgtgctgtg tcaggctcac ggcttgactc cagaacaggt ggtggctatt 8520
gcttcccacg acggggggaa acaggccctg gaaactgtgc agcgcctgct gccagtgctg 8580
tgccaggctc acgggctgac ccccgaacag gtggtggcca ttgccagcaa cggcggcggc 8640
aagcaggccc tggaaaccgt gcagagactg ctgcccgtgc tgtgccaggc ccatggcctg 8700
acacctgaac aggtggtggc tatcgcctct aacggcggag gaaaacaggc tctggaaaca 8760
gtgcagcggc tgctgcctgt gctgtgtcag gctcacggct tgactccaga acaggtggtg 8820
gctattgctt cccacgacgg ggggaaacag gccctggaaa ctgtgcagcg cctgctgcca 8880
gtgctgtgcc aggctcacgg cctcactccc gaacaggtgg tggccattgc cagcaacaac 8940
ggcggcaagc aggccctgga aaccgtgcag agactgctgc ccgtgctgtg ccaggcccat 9000
ggcctgacac ctgaacaggt ggtggctatc gcctctcacg acggaggaaa acaggctctg 9060
gaaacagtgc agcggctgct gcctgtgctg tgtcaggctc acggcttgac tccagaacag 9120
gtggtggcta ttgcttccca cgacgggggg aaacaggccc tggaaactgt gcagcgcctg 9180
ctgccagtgc tgtgccaggc tcacggactg acccccgaac aggtggtggc cattgccagc 9240
aacatcggcg gcaagcaggc cctggaaacc gtgcagagac tgctgcccgt gctgtgccag 9300
gcccatggcc tgacacctga acaggtggtg gctatcgcct ctaacaacgg aggaaaacaa 9360
gcactcgaga cagtgcagcg gctgctgcct gtgctgtgtc aggctcacgg cttgactcca 9420
gaacaggtgg tggctattgc ttccaacggc ggggggaaac aggccctgga aactgtgcag 9480
cgcctgctgc cagtgctgtg ccaggctcac gggctgaccc ccgaacaggt ggtggccatt 9540
gccagccacg acggcggcaa gcaggccctg gaaaccgtgc agagactgct gcccgtgctg 9600
tgccaggccc atggcctgac acctgaacag gtggtggcta tcgcctctaa tatcggagga 9660
aaacaggctc tggaaacagt gcagcggctg ctgcctgtgc tgtgtcaggc tcacggcttg 9720
actccacagc aggtcgtggc aattgctagc cacgacggcg gacggcccgc cctggagagc 9780
attgtggccc agctgtctag acctgatcct gccctggccg ccctgacaaa tgatcacctg 9840
gtggccctgg cctgtctggg aggcagacct gccctggatg ccgtgaaaaa aggactgcct 9900
cacgcccctg ccctgattaa aagaacaaat agaagaatcc ccgagcggac ctctcacaga 9960
gtggccggat cccagctggt gaaatctgag ctggaggaga agaagtctga gctgagacac 10020
aagctgaagt acgtgcctca cgagtacatc gagctgatcg agatcgccag aaatagcacc 10080
caggatagaa tcctggagat gaaggtgatg gagttcttca tgaaagtgta cggctacaga 10140
ggaaagcatc tgggaggaag cagaaaacct gacggagcca tttatacagt gggcagccct 10200
atcgattatg gcgtgatcgt ggatacaaag gcctacagcg gaggctacaa tctgcctatt 10260
ggacaggccg atgagatgca gagatacgtg gaggagaacc aaaccaggaa caagcatatc 10320
aaccctaacg agtggtggaa ggtgtaccct tctagcgtga ccgagttcaa gttcctgttt 10380
gtgagcggcc acttcaaggg caattataag gcccagctga ccaggctgaa ccacatcaca 10440
aattgtaatg gcgccgtgct gtctgtggag gaactgctga ttggaggaga gatgattaag 10500
gccggaacac tgacactgga ggaggtgaga agaaagttca acaacggcga gatcaacttc 10560
tgaaagcttg atcgttcaaa catttggcaa taaagtttct taagattgaa tcctgttgcc 10620
ggtcttgcga tgattatcat ataatttctg ttgaattacg ttaagcatgt aataattaac 10680
atgtaatgca tgacgttatt tatgagatgg gtttttatga ttagagtccc gcaattatac 10740
atttaatacg cgatagaaaa caaaatatag cgcgcaaact aggataaatt atcgcgcgcg 10800
gtgtcatcta tgttactaga tcttcgaaga cggaccgcgc ctgcagtgca gcgtgacccg 10860
gtcgtgcccc tctctagaga taatgagcat tgcatgtcta agttataaaa aattaccaca 10920
tatttttttt gtcacacttg tttgaagtgc agtttatcta tctttataca tatatttaaa 10980
ctttactcta cgaataatat aatctatagt actacaataa tatcagtgtt ttagagaatc 11040
atataaatga acagttagac atggtctaaa ggacaattga gtattttgac aacaggactc 11100
tacagtttta tctttttagt gtgcatgtgt tctccttttt ttttgcaaat agcttcacct 11160
atataatact tcatccattt tattagtaca tccatttagg gtttagggtt aatggttttt 11220
atagactaat ttttttagta catctatttt attctatttt agcctctaaa ttaagaaaac 11280
taaaactcta ttttagtttt tttatttaat aatttagata taaaatagaa taaaataaag 11340
tgactaaaaa ttaaacaaat accctttaag aaattaaaaa aactaaggaa acatttttct 11400
tgtttcgagt agataatgcc agcctgttaa acgccgtcga cgagtctaac ggacaccaac 11460
cagcgaacca gcagcgtcgc gtcgggccaa gcgaagcaga cggcacggca tctctgtcgc 11520
tgcctctgga cccctctcga gagttccgct ccaccgttgg acttgctccg ctgtcggcat 11580
ccagaaattg cgtggcggag cggcagacgt gagccggcac ggcaggcggc ctcctcctcc 11640
tctcacggca ccggcagcta cgggggattc ctttcccacc gctccttcgc tttcccttcc 11700
tcgcccgccg taataaatag acaccccctc cacaccctct ttccccaacc tcgtgttgtt 11760
cggagcgcac acacacacaa ccagatctcc cccaaatcca cccgtcggca cctccgcttc 11820
aaggtacgcc gctcgtcctc cccccccccc ctctctacct tctctagatc ggcgttccgg 11880
tccatggtta gggcccggta gttctacttc tgttcatgtt tgtgttagat ccgtgtttgt 11940
gttagatccg tgctgctagc gttcgtacac ggatgcgacc tgtacgtcag acacgttctg 12000
attgctaact tgccagtgtt tctctttggg gaatcctggg atggctctag ccgttccgca 12060
gacgggatcg atttcatgat tttttttgtt tcgttgcata gggtttggtt tgcccttttc 12120
ctttatttca atatatgccg tgcacttgtt tgtcgggtca tcttttcatg cttttttttg 12180
tcttggttgt gatgatgtgg tctggttggg cggtcgttct agatcggagt agaattctgt 12240
ttcaaactac ctggtggatt tattaatttt ggatctgtat gtgtgtgcca tacatattca 12300
tagttacgaa ttgaagatga tggatggaaa tatcgatcta ggataggtat acatgttgat 12360
gcgggtttta ctgatgcata tacagagatg ctttttgttc gcttggttgt gatgatgtgg 12420
tgtggttggg cggtcgttca ttcgttctag atcggagtag aatactgttt caaactacct 12480
ggtgtattta ttaattttgg aactgtatgt gtgtgtcata catcttcata gttacgagtt 12540
taagatggat ggaaatatcg atctaggata ggtatacatg ttgatgtggg ttttactgat 12600
gcatatacat gatggcatat gcagcatcta ttcatatgct ctaaccttga gtacctatct 12660
attataataa acaagtatgt tttataatta ttttgatctt gatatacttg gatgatggca 12720
tatgcagcag ctatatgtgg atttttttag ccctgccttc atacgctatt tatttgcttg 12780
gtactgtttc ttttgtcgat gctcaccctg ttgtttggtg ttacttctgc agggatccgg 12840
cagcagccat gcagaagctg atcaacagcg tgcagaacta cgcctggggc agcaagaccg 12900
ccctgaccga gctgtacggc atggagaacc ccagcagcca gcccatggcc gagctgtgga 12960
tgggcgccca ccccaagagc agcagccgcg tgcagaacgc cgccggcgac atcgtgagcc 13020
tgcgcgacgt gatcgagagc gacaagagca ccctgctggg cgaggccgtg gccaagcgct 13080
tcggcgagct gcccttcctg ttcaaggtgc tgtgcgccgc ccagcccctg agcatccagg 13140
tgcaccccaa caagcacaac agcgagatcg gcttcgccaa ggagaacgcc gccggcatcc 13200
ccatggacgc cgccgagcgc aactacaagg accccaacca caagcccgag ctggtgttcg 13260
ccctgacccc cttcctggcc atgaacgcct tccgcgagtt cagcgagatc gtgagcctgc 13320
tgcagcccgt ggccggcgcc caccccgcca tcgcccactt cctgcagcag cccgacgccg 13380
agcgcctgag cgagctgttc gccagcctgc tgaacatgca gggcgaggag aagagccgcg 13440
ccctggccat cctgaagagc gccctggaca gccagcaggg cgagccctgg cagaccatcc 13500
gcctgatcag cgagttctac cccgaggaca gcggcctgtt cagccccctg ctgctgaacg 13560
tggtgaagct gaaccccggc gaggccatgt tcctgttcgc cgagaccccc cacgcctacc 13620
tgcagggcgt ggccctggag gtgatggcca acagcgacaa cgtgctgcgc gccggcctga 13680
cccccaagta catcgacatc cccgagctgg tggccaacgt gaagttcgag gccaagcccg 13740
ccaaccagct gctgacccag cccgtgaagc agggcgccga gctggacttc cccatccccg 13800
tggacgactt cgccttcagc ctgcacgacc tgagcgacaa ggagaccacc atcagccagc 13860
agagcgccgc catcctgttc tgcgtggagg gcgacgccac cctgtggaag ggcagccagc 13920
agctgcagct gaagcccggc gagagcgcct tcatcgccgc caacgagagc cccgtgaccg 13980
tgaagggcca cggccgcctg gcccgcgtgt acaacaagct gtgataggag ctcgatccgt 14040
cgacctgcag atcgttcaaa catttggcaa taaagtttct taagattgaa tcctgttgcc 14100
ggtcttgcga tgattatcat ataatttctg ttgaattacg ttaagcatgt aataattaac 14160
atgtaatgca tgacgttatt tatgagatgg gtttttatga ttagagtccc gcaattatac 14220
atttaatacg cgatagaaaa caaaatatag cgcgcaaact aggataaatt atcgcgcgcg 14280
gtgtcatcta tgttactaga tcggcgcgcc gcaattgaag tttgggcggc cagcatggcc 14340
gtatccgcaa tgtgttatta agttgtctaa gcgtcaattt gtttacacca caatatatcc 14400
tgccaccagc cagccaacag ctccccgacc ggcagctcgg cacaaaatca ccactcgata 14460
caggcagccc atcag 14475
<210> 36
<211> 10668
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 22873
<220>
<221> Энхансер
<222> (165)..(256)
<223> eNOS
<220>
<221> промотор
<222> (301)..(2097)
<223> Ubi
<220>
<221> ген
<222> (2114)..(6280)
<223> Cas9m
<220>
<221> другой_признак
<222> (6215)..(6271)
<223> NLS-метка
<220>
<221> Терминатор
<222> (6286)..(6538)
<223> tNOS
<220>
<221> промотор
<222> (6545)..(6919)
<223> pU3
<220>
<221> другой_признак
<222> (6921)..(6938)
<220>
<221> другой_признак
<222> (6921)..(7023)
<223> sgRNA
<220>
<221> промотор
<222> (7034)..(9025)
<223> Ubi
<220>
<221> ген
<222> (9042)..(10220)
<223> PMI
<220>
<221> Терминатор
<222> (10243)..(10495)
<223> tNOS
<400> 36
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tgaaggcggg aaacgacaat ctgatcatga gcggagaatt 180
aagggagtca cgttatgacc cccgccgatg acgcgggaca agccgtttta cgtttggaac 240
tgacagaacc gcaacgctgc aggaattggc cgcagcggcc atttaaacaa agcttggtac 300
cattatgtgg tctaggtagg ttctatatat aagaaaactt gaaatgttct aaaaaaaaat 360
tcaagcccat gcatgattga agcaaacggt atagcaacgg tgttaacctg atctagtgat 420
ctcttgcaat ccttaacggc cacctaccgc aggtagcaaa cggcgtcccc ctcctcgata 480
tctccgcggc gacctctggc tttttccgcg gaattgcgcg gtggggacgg attccacgag 540
accgcgacgc aaccgcctct cgccgctggg ccccacaccg ctcggtgccg tagcctcacg 600
ggactctttc tccctcctcc cccgttataa attggcttca tcccctcctt gcctcatcca 660
tccaaatccc agtccccaat cccatccctt cgtaggagaa attcatcgaa gctaagcgaa 720
tcctcgcgat cctctcaagg tactgcgagt tttcgatccc cctctcgacc cctcgtatgt 780
ttgtgtttgt cgtagcgttt gattaggtat gctttccctg tttgtgttcg tcgtagcgtt 840
tgattaggta tgctttccct gttcgtgttc atcgtagtgt ttgattaggt cgtgtgaggc 900
gatggcctgc tcgcgtcctt cgatctgtag tcgatttgcg ggtcgtggtg tagatctgcg 960
ggctgtgatg aagttatttg gtgtgatctg ctcgcctgat tctgcgggtt ggctcgagta 1020
gatatgatgg ttggaccggt tggttcgttt accgcgctag ggttgggctg ggatgatgtt 1080
gcatgcgccg ttgcgcgtga tcccgcagca ggacttgcgt ttgattgcca gatctcgtta 1140
cgattatgtg atttggtttg gactttttag atctgtagct tctgcttatg tgccagatgc 1200
gcctactgct catatgcctg atgataatca taaatggctg tggaactaac tagttgattg 1260
cggagtcatg tatcagctac aggtgtaggg actagctaca ggtgtaggga cttgcgtcta 1320
attgtttggt cctttactca tgttgcaatt atgcaattta gtttagattg tttgttccac 1380
tcatctaggc tgtaaaaggg acactgctta gattgctgtt taatcttttt agtagattat 1440
attatattgg taacttatta cccctattac atgccatacg tgacttctgc tcatgcctga 1500
tgataatcat agatcactgt ggaattaatt agttgattgt tgaatcatgt ttcatgtaca 1560
taccacggca caattgctta gttccttaac aaatgcaaat tttactgatc catgtatgat 1620
ttgcgtggtt ctctaatgtg aaatactata gctacttgtt agtaagaatc aggttcgtat 1680
gcttaatgct gtatgtgcct tctgctcatg cctgatgata atcatatatc actggaatta 1740
attagttgat cgtttaatca tatatcaagt acataccatg ccacaatttt tagtcactta 1800
acccatgcag attgaactgg tccctgcatg ttttgctaaa ttgttctatt ctgattagac 1860
catatatcat gtattttttt ttggtaatgg ttctcttatt ttaaatgcta tatagttctg 1920
gtacttgtta gaaagatctg cttcatagtt tagttgccta tccctcgaat taggatgctg 1980
agcagctgat cctatagctt tgtttcatgt atcaattctt ttgtgttcaa cagtcagttt 2040
ttgttagatt cattgtaact tatggtcgct tactcttctg gtcctcaatg cttgcaggat 2100
cgcggccgct aaaatggata agaagtactc tattggcctc gatattggaa ccaactctgt 2160
gggctgggcc gtgatcaccg atgagtacaa ggtgccatct aagaagttca aggtgctcgg 2220
caacaccgat aggcactcta tcaagaagaa cctcatcggt gccctcctct tcgattctgg 2280
cgagaccgcc gaggccacca ggctcaagag gaccgccagg aggaggtaca ccaggaggaa 2340
gaacaggatc tgctacctcc aggagatctt ctctaacgag atggccaagg tggatgattc 2400
tttcttccac aggctcgagg agtctttcct cgtggaggag gataagaagc acgagaggca 2460
cccaatcttc ggcaacatcg tggatgaggt ggcctaccac gagaagtacc caaccatcta 2520
ccacctcagg aagaagctcg tggattctac cgataaggcc gatctcaggc tcatctacct 2580
cgccctcgcc cacatgatca agttcagggg ccacttcctc atcgagggcg atctcaaccc 2640
agataactct gatgtggata agctcttcat ccagctcgtg cagacctaca accagctctt 2700
cgaggagaac ccaatcaacg cctctggcgt ggatgccaag gccatcctct ctgccaggct 2760
ctctaagtct aggaggctcg agaacctcat cgcccagctc ccaggcgaga agaagaacgg 2820
cctcttcggc aacctcatcg ccctctctct cggcctcacc ccaaacttca agtctaactt 2880
cgatctcgcc gaggatgcca agctccagct ctctaaggat acctacgatg atgatctcga 2940
taacctcctc gcccagatcg gcgatcagta cgccgatctc ttcctcgccg ccaagaacct 3000
ctctgatgcc atcctcctct ctgatatcct cagggtgaac accgagatca ccaaggcccc 3060
actctctgcc tctatgatca agaggtacga tgagcaccac caggatctca ccctcctcaa 3120
ggccctcgtg aggcagcagc tcccagagaa gtacaaggag atcttcttcg atcagtctaa 3180
gaacggctac gccggctaca tcgatggcgg cgcctctcag gaggagttct acaagttcat 3240
caagccaatc ctcgagaaga tggatggcac cgaggagctc ctcgtgaagc tcaacaggga 3300
ggatctcctc aggaagcaga ggaccttcga taacggctct atcccacacc agatccacct 3360
cggcgagctc cacgccatcc tcaggaggca ggaggatttc tacccattcc tcaaggataa 3420
cagggagaag atcgagaaga tcctcacctt ccgcatccca tactacgtgg gcccactcgc 3480
caggggcaac tctaggttcg cctggatgac caggaagtct gaggagacca tcaccccttg 3540
gaacttcgag gaggtggtgg ataagggcgc ctctgcccag tctttcatcg agaggatgac 3600
caacttcgat aagaacctcc caaacgagaa ggtgctccca aagcactctc tcctctacga 3660
gtacttcacc gtgtacaacg agctcaccaa ggtgaagtac gtgaccgagg gcatgaggaa 3720
gccagccttc ctctctggcg agcagaagaa ggccatcgtg gatctcctct tcaagaccaa 3780
caggaaggtg accgtgaagc agctcaagga ggattacttc aagaagatcg agtgcttcga 3840
ttctgtggag atctctggcg tggaggatag gttcaacgcc tctctcggca cctaccacga 3900
tctcctcaag atcatcaagg ataaggattt cctcgataac gaggagaacg aggatatcct 3960
cgaggatatc gtgctcaccc tcaccctctt cgaggatagg gagatgatcg aggagaggct 4020
caagacctac gcccacctct tcgatgataa ggtgatgaag cagctcaaga ggaggaggta 4080
caccggctgg ggcaggctct ctaggaagct catcaacggc atcagggata agcagtctgg 4140
caagaccatc ctcgatttcc tcaagtctga tggcttcgcc aacaggaact tcatgcagct 4200
catccacgat gattctctca ccttcaagga ggatatccag aaggctcagg tgtctggcca 4260
gggccactct ctccacgagc agatcgccaa cctcgccggc tctccagcca tcaagaaggg 4320
catcctccag accgtgaagg tggtggatga gctcgtgaag gtgatgggcc acaagccaga 4380
gaacatcgtg atcgagatgg ccagggagaa ccagaccacc cagaagggcc agaagaactc 4440
tagggagagg atgaagagga tcgaggaggg catcaaggag ctcggctctc agatcctcaa 4500
ggagcaccca gtggagaaca cccagctcca gaacgagaag ctctacctct actacctcca 4560
gaacggcagg gatatgtacg tggatcagga gctcgatatc aacaggctct ctgattacga 4620
tgtggatcac atcgtgccac agtctttcct caaggatgat tctatcgata acaaggtgct 4680
caccaggtct gataagaaca ggggcaagtc tgataacgtg ccatctgagg aggtggttaa 4740
gaagatgaag aactactgga ggcagctcct caacgccaag ctcatcaccc agaggaagtt 4800
cgataacctc accaaggccg agaggggcgg cctctctgag ctcgataagg ccggcttcat 4860
caagaggcag ctcgtggaga ccaggcagat cactaagcac gtggcccaga tcctcgattc 4920
taggatgaac accaagtacg atgagaacga taagctcatc agggaggtga aggtgatcac 4980
cctcaagtct aagctcgtgt ctgatttcag gaaggatttc cagttctaca aggtgaggga 5040
gatcaacaac taccaccacg cccacgatgc ctacctcaac gccgtggtgg gcaccgccct 5100
catcaagaag tacccaaaac tcgagtctga gttcgtgtac ggcgattaca aggtgtacga 5160
tgtgaggaag atgatcgcca agtctgagca ggagatcggc aaggccaccg ccaagtactt 5220
cttctactct aacatcatga acttcttcaa gaccgagatc accctcgcca acggcgagat 5280
caggaagagg ccactcatcg agaccaacgg cgagaccggc gagatcgtgt gggataaggg 5340
cagggatttc gccaccgtga ggaaggtgct ctctatgcca caggtgaaca tcgtgaagaa 5400
gaccgaggtg cagaccggcg gcttctctaa ggagtctatc ctcccaaaga ggaactctga 5460
taagctcatc gccaggaaga aggattggga cccgaagaag tacggcggct tcgacagccc 5520
gaccgtggcc tacagcgtgc tggtggtggc caaggtggag aagggcaaga gcaagaagct 5580
gaagagcgtg aaggagctgg tgggcatcac catcatggag aggagcagct tcgagaagaa 5640
cccagtggac ttcctggagg ccaagggcta caaggaggtg aagaaggacc tgatcattaa 5700
actgccgaag tacagcctgt tcgagctgga gaacggcagg aagaggatgc tggccagcgc 5760
cggcgagctg cagaagggca acgagctggc cctgccgagc aagtacgtga acttcctgta 5820
cctggccagc cactacgaga agctgaaggg cagcccggag gacaacgagc agaagcagct 5880
gttcgtggag cagcacaagc actacctgga cgagatcatc gagcagatca gcgagttcag 5940
caagagggtg atcctggccg acgccaacct ggacaaggtg ctgagcgcct acaacaagca 6000
cagggacaag ccgatcaggg agcaggccga gaacatcatc cacctgttca ccctgaccaa 6060
cctgggcgcc ccggccgcct tcaagtactt cgacaccacc atcgacagga agaggtacac 6120
cagcaccaag gaggtgctgg acgccaccct gatccaccag agcatcaccg gcctgtacga 6180
gaccaggatc gacctgagcc agctgggcgg cgacagcagc ccgccgaaga agaagaggaa 6240
ggtgagctgg aaggacgcca gcggctggag caggatgtga agcttgatcg ttcaaacatt 6300
tggcaataaa gtttcttaag attgaatcct gttgccggtc ttgcgatgat tatcatataa 6360
tttctgttga attacgttaa gcatgtaata attaacatgt aatgcatgac gttatttatg 6420
agatgggttt ttatgattag agtcccgcaa ttatacattt aatacgcgat agaaaacaaa 6480
atatagcgcg caaactagga taaattatcg cgcgcggtgt catctatgtt actagatctt 6540
cgaagggatc tttaaacata cgaacagatc acttaaagtt cttctgaagc aacttaaagt 6600
tatcaggcat gcatggatct tggaggaatc agatgtgcag tcagggacca tagcacagga 6660
caggcgtctt ctactggtgc taccagcaaa tgctggaagc cgggaacact gggtacgttg 6720
gaaaccacgt gatgtggagt aagataaact gtaggagaaa agcatttcgt agtgggccat 6780
gaagcctttc aggacatgta ttgcagtatg ggccggccca ttacgcaatt ggacgacaac 6840
aaagactagt attagtacca cctcggctat ccacatagat caaagctggt ttaaaagagt 6900
tgtgcagatg atccgtggca gtcaacgtgg agacaggggt tttagagcta gaaatagcaa 6960
gttaaaataa ggctagtccg ttatcaactt gaaaaagtgg caccgagtcg gtgctttttt 7020
tttcggaccg cgcctgcagt gcagcgtgac ccggtcgtgc ccctctctag agataatgag 7080
cattgcatgt ctaagttata aaaaattacc acatattttt tttgtcacac ttgtttgaag 7140
tgcagtttat ctatctttat acatatattt aaactttact ctacgaataa tataatctat 7200
agtactacaa taatatcagt gttttagaga atcatataaa tgaacagtta gacatggtct 7260
aaaggacaat tgagtatttt gacaacagga ctctacagtt ttatcttttt agtgtgcatg 7320
tgttctcctt tttttttgca aatagcttca cctatataat acttcatcca ttttattagt 7380
acatccattt agggtttagg gttaatggtt tttatagact aattttttta gtacatctat 7440
tttattctat tttagcctct aaattaagaa aactaaaact ctattttagt ttttttattt 7500
aataatttag atataaaata gaataaaata aagtgactaa aaattaaaca aatacccttt 7560
aagaaattaa aaaaactaag gaaacatttt tcttgtttcg agtagataat gccagcctgt 7620
taaacgccgt cgacgagtct aacggacacc aaccagcgaa ccagcagcgt cgcgtcgggc 7680
caagcgaagc agacggcacg gcatctctgt cgctgcctct ggacccctct cgagagttcc 7740
gctccaccgt tggacttgct ccgctgtcgg catccagaaa ttgcgtggcg gagcggcaga 7800
cgtgagccgg cacggcaggc ggcctcctcc tcctctcacg gcaccggcag ctacggggga 7860
ttcctttccc accgctcctt cgctttccct tcctcgcccg ccgtaataaa tagacacccc 7920
ctccacaccc tctttcccca acctcgtgtt gttcggagcg cacacacaca caaccagatc 7980
tcccccaaat ccacccgtcg gcacctccgc ttcaaggtac gccgctcgtc ctcccccccc 8040
cccctctcta ccttctctag atcggcgttc cggtccatgg ttagggcccg gtagttctac 8100
ttctgttcat gtttgtgtta gatccgtgtt tgtgttagat ccgtgctgct agcgttcgta 8160
cacggatgcg acctgtacgt cagacacgtt ctgattgcta acttgccagt gtttctcttt 8220
ggggaatcct gggatggctc tagccgttcc gcagacggga tcgatttcat gatttttttt 8280
gtttcgttgc atagggtttg gtttgccctt ttcctttatt tcaatatatg ccgtgcactt 8340
gtttgtcggg tcatcttttc atgctttttt ttgtcttggt tgtgatgatg tggtctggtt 8400
gggcggtcgt tctagatcgg agtagaattc tgtttcaaac tacctggtgg atttattaat 8460
tttggatctg tatgtgtgtg ccatacatat tcatagttac gaattgaaga tgatggatgg 8520
aaatatcgat ctaggatagg tatacatgtt gatgcgggtt ttactgatgc atatacagag 8580
atgctttttg ttcgcttggt tgtgatgatg tggtgtggtt gggcggtcgt tcattcgttc 8640
tagatcggag tagaatactg tttcaaacta cctggtgtat ttattaattt tggaactgta 8700
tgtgtgtgtc atacatcttc atagttacga gtttaagatg gatggaaata tcgatctagg 8760
ataggtatac atgttgatgt gggttttact gatgcatata catgatggca tatgcagcat 8820
ctattcatat gctctaacct tgagtaccta tctattataa taaacaagta tgttttataa 8880
ttattttgat cttgatatac ttggatgatg gcatatgcag cagctatatg tggatttttt 8940
tagccctgcc ttcatacgct atttatttgc ttggtactgt ttcttttgtc gatgctcacc 9000
ctgttgtttg gtgttacttc tgcagggatc cggcagcagc catgcagaag ctgatcaaca 9060
gcgtgcagaa ctacgcctgg ggcagcaaga ccgccctgac cgagctgtac ggcatggaga 9120
accccagcag ccagcccatg gccgagctgt ggatgggcgc ccaccccaag agcagcagcc 9180
gcgtgcagaa cgccgccggc gacatcgtga gcctgcgcga cgtgatcgag agcgacaaga 9240
gcaccctgct gggcgaggcc gtggccaagc gcttcggcga gctgcccttc ctgttcaagg 9300
tgctgtgcgc cgcccagccc ctgagcatcc aggtgcaccc caacaagcac aacagcgaga 9360
tcggcttcgc caaggagaac gccgccggca tccccatgga cgccgccgag cgcaactaca 9420
aggaccccaa ccacaagccc gagctggtgt tcgccctgac ccccttcctg gccatgaacg 9480
ccttccgcga gttcagcgag atcgtgagcc tgctgcagcc cgtggccggc gcccaccccg 9540
ccatcgccca cttcctgcag cagcccgacg ccgagcgcct gagcgagctg ttcgccagcc 9600
tgctgaacat gcagggcgag gagaagagcc gcgccctggc catcctgaag agcgccctgg 9660
acagccagca gggcgagccc tggcagacca tccgcctgat cagcgagttc taccccgagg 9720
acagcggcct gttcagcccc ctgctgctga acgtggtgaa gctgaacccc ggcgaggcca 9780
tgttcctgtt cgccgagacc ccccacgcct acctgcaggg cgtggccctg gaggtgatgg 9840
ccaacagcga caacgtgctg cgcgccggcc tgacccccaa gtacatcgac atccccgagc 9900
tggtggccaa cgtgaagttc gaggccaagc ccgccaacca gctgctgacc cagcccgtga 9960
agcagggcgc cgagctggac ttccccatcc ccgtggacga cttcgccttc agcctgcacg 10020
acctgagcga caaggagacc accatcagcc agcagagcgc cgccatcctg ttctgcgtgg 10080
agggcgacgc caccctgtgg aagggcagcc agcagctgca gctgaagccc ggcgagagcg 10140
ccttcatcgc cgccaacgag agccccgtga ccgtgaaggg ccacggccgc ctggcccgcg 10200
tgtacaacaa gctgtgatag gagctcgatc cgtcgacctg cagatcgttc aaacatttgg 10260
caataaagtt tcttaagatt gaatcctgtt gccggtcttg cgatgattat catataattt 10320
ctgttgaatt acgttaagca tgtaataatt aacatgtaat gcatgacgtt atttatgaga 10380
tgggttttta tgattagagt cccgcaatta tacatttaat acgcgataga aaacaaaata 10440
tagcgcgcaa actaggataa attatcgcgc gcggtgtcat ctatgttact agatcggcgc 10500
gccgcaattg aagtttgggc ggccagcatg gccgtatccg caatgtgtta ttaagttgtc 10560
taagcgtcaa tttgtttaca ccacaatata tcctgccacc agccagccaa cagctccccg 10620
accggcagct cggcacaaaa tcaccactcg atacaggcag cccatcag 10668
<210> 37
<211> 10673
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 23123
<220>
<221> Энхансер
<222> (165)..(256)
<223> eNOS
<220>
<221> промотор
<222> (301)..(2097)
<223> Ubi
<220>
<221> ген
<222> (2114)..(6283)
<223> Cas9
<220>
<221> другой_признак
<222> (6218)..(6274)
<223> NLS-метка
<220>
<221> Терминатор
<222> (6289)..(6541)
<223> tNOS
<220>
<221> промотор
<222> (6548)..(6922)
<223> pU3
<220>
<221> другой_признак
<222> (6924)..(6943)
<220>
<221> другой_признак
<222> (6924)..(7028)
<223> sgRNA
<220>
<221> промотор
<222> (7039)..(9030)
<223> Ubi
<220>
<221> ген
<222> (9047)..(10225)
<223> PMI
<220>
<221> Терминатор
<222> (10248)..(10500)
<223> tNOS
<400> 37
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tgaaggcggg aaacgacaat ctgatcatga gcggagaatt 180
aagggagtca cgttatgacc cccgccgatg acgcgggaca agccgtttta cgtttggaac 240
tgacagaacc gcaacgctgc aggaattggc cgcagcggcc atttaaacaa agcttggtac 300
cattatgtgg tctaggtagg ttctatatat aagaaaactt gaaatgttct aaaaaaaaat 360
tcaagcccat gcatgattga agcaaacggt atagcaacgg tgttaacctg atctagtgat 420
ctcttgcaat ccttaacggc cacctaccgc aggtagcaaa cggcgtcccc ctcctcgata 480
tctccgcggc gacctctggc tttttccgcg gaattgcgcg gtggggacgg attccacgag 540
accgcgacgc aaccgcctct cgccgctggg ccccacaccg ctcggtgccg tagcctcacg 600
ggactctttc tccctcctcc cccgttataa attggcttca tcccctcctt gcctcatcca 660
tccaaatccc agtccccaat cccatccctt cgtaggagaa attcatcgaa gctaagcgaa 720
tcctcgcgat cctctcaagg tactgcgagt tttcgatccc cctctcgacc cctcgtatgt 780
ttgtgtttgt cgtagcgttt gattaggtat gctttccctg tttgtgttcg tcgtagcgtt 840
tgattaggta tgctttccct gttcgtgttc atcgtagtgt ttgattaggt cgtgtgaggc 900
gatggcctgc tcgcgtcctt cgatctgtag tcgatttgcg ggtcgtggtg tagatctgcg 960
ggctgtgatg aagttatttg gtgtgatctg ctcgcctgat tctgcgggtt ggctcgagta 1020
gatatgatgg ttggaccggt tggttcgttt accgcgctag ggttgggctg ggatgatgtt 1080
gcatgcgccg ttgcgcgtga tcccgcagca ggacttgcgt ttgattgcca gatctcgtta 1140
cgattatgtg atttggtttg gactttttag atctgtagct tctgcttatg tgccagatgc 1200
gcctactgct catatgcctg atgataatca taaatggctg tggaactaac tagttgattg 1260
cggagtcatg tatcagctac aggtgtaggg actagctaca ggtgtaggga cttgcgtcta 1320
attgtttggt cctttactca tgttgcaatt atgcaattta gtttagattg tttgttccac 1380
tcatctaggc tgtaaaaggg acactgctta gattgctgtt taatcttttt agtagattat 1440
attatattgg taacttatta cccctattac atgccatacg tgacttctgc tcatgcctga 1500
tgataatcat agatcactgt ggaattaatt agttgattgt tgaatcatgt ttcatgtaca 1560
taccacggca caattgctta gttccttaac aaatgcaaat tttactgatc catgtatgat 1620
ttgcgtggtt ctctaatgtg aaatactata gctacttgtt agtaagaatc aggttcgtat 1680
gcttaatgct gtatgtgcct tctgctcatg cctgatgata atcatatatc actggaatta 1740
attagttgat cgtttaatca tatatcaagt acataccatg ccacaatttt tagtcactta 1800
acccatgcag attgaactgg tccctgcatg ttttgctaaa ttgttctatt ctgattagac 1860
catatatcat gtattttttt ttggtaatgg ttctcttatt ttaaatgcta tatagttctg 1920
gtacttgtta gaaagatctg cttcatagtt tagttgccta tccctcgaat taggatgctg 1980
agcagctgat cctatagctt tgtttcatgt atcaattctt ttgtgttcaa cagtcagttt 2040
ttgttagatt cattgtaact tatggtcgct tactcttctg gtcctcaatg cttgcaggat 2100
cgcggccgct catatggaca agaagtacag catcggcctg gacatcggca ccaacagcgt 2160
gggctgggcc gtgatcaccg acgagtacaa ggtgccgagc aagaagttca aggtgctggg 2220
caacaccgac aggcacagca tcaagaagaa cctgatcggc gccctgctgt tcgacagcgg 2280
cgagaccgcc gaggccacca ggctgaagag gaccgccagg aggaggtaca ccaggaggaa 2340
gaacaggatc tgctacctgc aggagatctt cagcaacgag atggccaagg tggacgacag 2400
cttcttccac aggctggagg agagcttcct ggtggaggag gacaagaagc acgagaggca 2460
cccgatcttc ggcaacatcg tggacgaggt ggcctaccac gagaagtacc cgaccatcta 2520
ccacctgagg aagaagctgg tggacagcac cgacaaggcc gacctgaggc tgatctacct 2580
ggccctggcc cacatgatca agttcagggg ccacttcctg atcgagggcg acctgaaccc 2640
ggacaacagc gacgtggaca agctgttcat ccagctggtg cagacctaca accagctgtt 2700
cgaggagaac ccgatcaacg ccagcggcgt ggacgccaag gccatcctga gcgccaggct 2760
gagcaagagc aggaggctgg agaacctgat cgcccagctg ccgggcgaga agaagaacgg 2820
cctgttcggc aacctgatcg ccctgagcct gggcctgacc ccgaacttca agagcaactt 2880
cgacctggcc gaggacgcca agctgcagct gagcaaggac acctacgacg acgacctgga 2940
caacctgctg gcccagatcg gcgaccagta cgccgacctg ttcctggccg ccaagaacct 3000
gagcgacgcc atcctgctga gcgacatcct gagggtgaac accgagatca ccaaggcccc 3060
gctgagcgcc agcatgatca agaggtacga cgagcaccac caggacctga ccctgctgaa 3120
ggccctggtg aggcagcagc tgccggagaa gtacaaggag atcttcttcg accagagcaa 3180
gaacggctac gccggctaca tcgacggcgg cgccagccag gaggagttct acaagttcat 3240
caagccgatc ctggagaaga tggacggcac cgaggagctg ctggtgaagc tgaacaggga 3300
ggacctgctg aggaagcaga ggaccttcga caacggcagc atcccgcacc agatccacct 3360
gggcgagctg cacgccatcc tgaggaggca ggaggacttc tacccgttcc tgaaggacaa 3420
cagggagaag atcgagaaga tcctgacctt ccgcatcccg tactacgtgg gcccgctggc 3480
caggggcaac agcaggttcg cctggatgac caggaagagc gaggagacca tcaccccgtg 3540
gaacttcgag gaggtggtgg acaagggcgc cagcgcccag agcttcatcg agaggatgac 3600
caacttcgac aagaacctgc cgaacgagaa ggtgctgccg aagcacagcc tgctgtacga 3660
gtacttcacc gtgtacaacg agctgaccaa ggtgaagtac gtgaccgagg gcatgaggaa 3720
gccggccttc ctgagcggcg agcagaagaa ggccatcgtg gacctgctgt tcaagaccaa 3780
caggaaggtg accgtgaagc agctgaagga ggactacttc aagaagatcg agtgcttcga 3840
cagcgtggag atcagcggcg tggaggacag gttcaacgcc agcctgggca cctaccacga 3900
cctgctgaag atcatcaagg acaaggactt cctggacaac gaggagaacg aggacatcct 3960
ggaggacatc gtgctgaccc tgaccctgtt cgaggacagg gagatgatcg aggagaggct 4020
gaagacctac gcccacctgt tcgacgacaa ggtgatgaag cagctgaaga ggaggaggta 4080
caccggctgg ggcaggctga gcaggaagct gatcaacggc atcagggaca agcagagcgg 4140
caagaccatc ctggacttcc tgaagagcga cggcttcgcc aacaggaact tcatgcagct 4200
gatccacgac gacagcctga ccttcaagga ggacatccag aaggcccagg tgagcggcca 4260
gggcgacagc ctgcacgagc acatcgccaa cctggccggc agcccggcca tcaagaaggg 4320
catcctgcag accgtgaagg tggtggacga gctggtgaag gtgatgggca ggcacaagcc 4380
ggagaacatc gtgatcgaga tggccaggga gaaccagacc acccagaagg gccagaagaa 4440
cagcagggag aggatgaaga ggatcgagga gggcatcaag gagctgggca gccagatcct 4500
gaaggagcac ccggtggaga acacccagct gcagaacgag aagctgtacc tgtactacct 4560
gcagaacggc agggacatgt acgtggacca ggagctggac atcaacaggc tgagcgacta 4620
cgacgtggac cacatcgtgc cgcagagctt cctgaaggac gacagcatcg acaacaaggt 4680
gctgaccagg agcgacaaga acaggggcaa gagcgacaac gtgccgagcg aggaggtggt 4740
gaagaagatg aaaaactact ggaggcagct gctgaacgcc aagctgatca cccagaggaa 4800
gttcgacaac ctgaccaagg ccgagagggg cggcctgagc gagctggaca aggccggctt 4860
cattaaaagg cagctggtgg agaccaggca gatcaccaag cacgtggccc agatcctgga 4920
cagcaggatg aacaccaagt acgacgagaa cgacaagctg atcagggagg tgaaggtgat 4980
caccctgaag agcaagctgg tgagcgactt caggaaggac ttccagttct acaaggtgag 5040
ggagatcaat aattaccacc acgcccacga cgcctacctg aacgccgtgg tgggcaccgc 5100
cctgattaaa aagtacccga agctggagag cgagttcgtg tacggcgact acaaggtgta 5160
cgacgtgagg aagatgatcg ccaagagcga gcaggagatc ggcaaggcca ccgccaagta 5220
cttcttctac agcaacatca tgaacttctt caagaccgag atcaccctgg ccaacggcga 5280
gatcaggaag aggccgctga tcgagaccaa cggcgagacc ggcgagatcg tgtgggacaa 5340
gggcagggac ttcgccaccg tgaggaaggt gctgtccatg ccgcaggtga acatcgtgaa 5400
gaagaccgag gtgcagaccg gcggcttcag caaggagagc atcctgccga agaggaacag 5460
cgacaagctg atcgccagga agaaggactg ggacccgaag aagtacggcg gcttcgacag 5520
cccgaccgtg gcctacagcg tgctggtggt ggccaaggtg gagaagggca agagcaagaa 5580
gctgaagagc gtgaaggagc tggtgggcat caccatcatg gagaggagca gcttcgagaa 5640
gaacccagtg gacttcctgg aggccaaggg ctacaaggag gtgaagaagg acctgatcat 5700
taaactgccg aagtacagcc tgttcgagct ggagaacggc aggaagagga tgctggccag 5760
cgccggcgag ctgcagaagg gcaacgagct ggccctgccg agcaagtacg tgaacttcct 5820
gtacctggcc agccactacg agaagctgaa gggcagcccg gaggacaacg agcagaagca 5880
gctgttcgtg gagcagcaca agcactacct ggacgagatc atcgagcaga tcagcgagtt 5940
cagcaagagg gtgatcctgg ccgacgccaa cctggacaag gtgctgagcg cctacaacaa 6000
gcacagggac aagccgatca gggagcaggc cgagaacatc atccacctgt tcaccctgac 6060
caacctgggc gccccggccg ccttcaagta cttcgacacc accatcgaca ggaagaggta 6120
caccagcacc aaggaggtgc tggacgccac cctgatccac cagagcatca ccggcctgta 6180
cgagaccagg atcgacctga gccagctggg cggcgacagc agcccgccga agaagaagag 6240
gaaggtgagc tggaaggacg ccagcggctg gagcaggatg tgaagcttga tcgttcaaac 6300
atttggcaat aaagtttctt aagattgaat cctgttgccg gtcttgcgat gattatcata 6360
taatttctgt tgaattacgt taagcatgta ataattaaca tgtaatgcat gacgttattt 6420
atgagatggg tttttatgat tagagtcccg caattataca tttaatacgc gatagaaaac 6480
aaaatatagc gcgcaaacta ggataaatta tcgcgcgcgg tgtcatctat gttactagat 6540
cttcgaaggg atctttaaac atacgaacag atcacttaaa gttcttctga agcaacttaa 6600
agttatcagg catgcatgga tcttggagga atcagatgtg cagtcaggga ccatagcaca 6660
ggacaggcgt cttctactgg tgctaccagc aaatgctgga agccgggaac actgggtacg 6720
ttggaaacca cgtgatgtgg agtaagataa actgtaggag aaaagcattt cgtagtgggc 6780
catgaagcct ttcaggacat gtattgcagt atgggccggc ccattacgca attggacgac 6840
aacaaagact agtattagta ccacctcggc tatccacata gatcaaagct ggtttaaaag 6900
agttgtgcag atgatccgtg gcagggtcaa cgtggagaca ggggttttag agctagaaat 6960
agcaagttaa aataaggcta gtccgttatc aacttgaaaa agtggcaccg agtcggtgct 7020
ttttttttcg gaccgcgcct gcagtgcagc gtgacccggt cgtgcccctc tctagagata 7080
atgagcattg catgtctaag ttataaaaaa ttaccacata ttttttttgt cacacttgtt 7140
tgaagtgcag tttatctatc tttatacata tatttaaact ttactctacg aataatataa 7200
tctatagtac tacaataata tcagtgtttt agagaatcat ataaatgaac agttagacat 7260
ggtctaaagg acaattgagt attttgacaa caggactcta cagttttatc tttttagtgt 7320
gcatgtgttc tccttttttt ttgcaaatag cttcacctat ataatacttc atccatttta 7380
ttagtacatc catttagggt ttagggttaa tggtttttat agactaattt ttttagtaca 7440
tctattttat tctattttag cctctaaatt aagaaaacta aaactctatt ttagtttttt 7500
tatttaataa tttagatata aaatagaata aaataaagtg actaaaaatt aaacaaatac 7560
cctttaagaa attaaaaaaa ctaaggaaac atttttcttg tttcgagtag ataatgccag 7620
cctgttaaac gccgtcgacg agtctaacgg acaccaacca gcgaaccagc agcgtcgcgt 7680
cgggccaagc gaagcagacg gcacggcatc tctgtcgctg cctctggacc cctctcgaga 7740
gttccgctcc accgttggac ttgctccgct gtcggcatcc agaaattgcg tggcggagcg 7800
gcagacgtga gccggcacgg caggcggcct cctcctcctc tcacggcacc ggcagctacg 7860
ggggattcct ttcccaccgc tccttcgctt tcccttcctc gcccgccgta ataaatagac 7920
accccctcca caccctcttt ccccaacctc gtgttgttcg gagcgcacac acacacaacc 7980
agatctcccc caaatccacc cgtcggcacc tccgcttcaa ggtacgccgc tcgtcctccc 8040
ccccccccct ctctaccttc tctagatcgg cgttccggtc catggttagg gcccggtagt 8100
tctacttctg ttcatgtttg tgttagatcc gtgtttgtgt tagatccgtg ctgctagcgt 8160
tcgtacacgg atgcgacctg tacgtcagac acgttctgat tgctaacttg ccagtgtttc 8220
tctttgggga atcctgggat ggctctagcc gttccgcaga cgggatcgat ttcatgattt 8280
tttttgtttc gttgcatagg gtttggtttg cccttttcct ttatttcaat atatgccgtg 8340
cacttgtttg tcgggtcatc ttttcatgct tttttttgtc ttggttgtga tgatgtggtc 8400
tggttgggcg gtcgttctag atcggagtag aattctgttt caaactacct ggtggattta 8460
ttaattttgg atctgtatgt gtgtgccata catattcata gttacgaatt gaagatgatg 8520
gatggaaata tcgatctagg ataggtatac atgttgatgc gggttttact gatgcatata 8580
cagagatgct ttttgttcgc ttggttgtga tgatgtggtg tggttgggcg gtcgttcatt 8640
cgttctagat cggagtagaa tactgtttca aactacctgg tgtatttatt aattttggaa 8700
ctgtatgtgt gtgtcataca tcttcatagt tacgagttta agatggatgg aaatatcgat 8760
ctaggatagg tatacatgtt gatgtgggtt ttactgatgc atatacatga tggcatatgc 8820
agcatctatt catatgctct aaccttgagt acctatctat tataataaac aagtatgttt 8880
tataattatt ttgatcttga tatacttgga tgatggcata tgcagcagct atatgtggat 8940
ttttttagcc ctgccttcat acgctattta tttgcttggt actgtttctt ttgtcgatgc 9000
tcaccctgtt gtttggtgtt acttctgcag ggatccggca gcagccatgc agaagctgat 9060
caacagcgtg cagaactacg cctggggcag caagaccgcc ctgaccgagc tgtacggcat 9120
ggagaacccc agcagccagc ccatggccga gctgtggatg ggcgcccacc ccaagagcag 9180
cagccgcgtg cagaacgccg ccggcgacat cgtgagcctg cgcgacgtga tcgagagcga 9240
caagagcacc ctgctgggcg aggccgtggc caagcgcttc ggcgagctgc ccttcctgtt 9300
caaggtgctg tgcgccgccc agcccctgag catccaggtg caccccaaca agcacaacag 9360
cgagatcggc ttcgccaagg agaacgccgc cggcatcccc atggacgccg ccgagcgcaa 9420
ctacaaggac cccaaccaca agcccgagct ggtgttcgcc ctgaccccct tcctggccat 9480
gaacgccttc cgcgagttca gcgagatcgt gagcctgctg cagcccgtgg ccggcgccca 9540
ccccgccatc gcccacttcc tgcagcagcc cgacgccgag cgcctgagcg agctgttcgc 9600
cagcctgctg aacatgcagg gcgaggagaa gagccgcgcc ctggccatcc tgaagagcgc 9660
cctggacagc cagcagggcg agccctggca gaccatccgc ctgatcagcg agttctaccc 9720
cgaggacagc ggcctgttca gccccctgct gctgaacgtg gtgaagctga accccggcga 9780
ggccatgttc ctgttcgccg agacccccca cgcctacctg cagggcgtgg ccctggaggt 9840
gatggccaac agcgacaacg tgctgcgcgc cggcctgacc cccaagtaca tcgacatccc 9900
cgagctggtg gccaacgtga agttcgaggc caagcccgcc aaccagctgc tgacccagcc 9960
cgtgaagcag ggcgccgagc tggacttccc catccccgtg gacgacttcg ccttcagcct 10020
gcacgacctg agcgacaagg agaccaccat cagccagcag agcgccgcca tcctgttctg 10080
cgtggagggc gacgccaccc tgtggaaggg cagccagcag ctgcagctga agcccggcga 10140
gagcgccttc atcgccgcca acgagagccc cgtgaccgtg aagggccacg gccgcctggc 10200
ccgcgtgtac aacaagctgt gataggagct cgatccgtcg acctgcagat cgttcaaaca 10260
tttggcaata aagtttctta agattgaatc ctgttgccgg tcttgcgatg attatcatat 10320
aatttctgtt gaattacgtt aagcatgtaa taattaacat gtaatgcatg acgttattta 10380
tgagatgggt ttttatgatt agagtcccgc aattatacat ttaatacgcg atagaaaaca 10440
aaatatagcg cgcaaactag gataaattat cgcgcgcggt gtcatctatg ttactagatc 10500
ggcgcgccgc aattgaagtt tgggcggcca gcatggccgt atccgcaatg tgttattaag 10560
ttgtctaagc gtcaatttgt ttacaccaca atatatcctg ccaccagcca gccaacagct 10620
ccccgaccgg cagctcggca caaaatcacc actcgataca ggcagcccat cag 10673
<210> 38
<211> 10693
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 23501, нацеливающаяся gRNA в виде 1
двойной направляющей последовательности
<220>
<221> промотор
<222> (214)..(734)
<223> 35S
<220>
<221> ген
<222> (748)..(4917)
<223> Cas9
<220>
<221> другой_признак
<222> (4852)..(4908)
<223> NLS-метка
<220>
<221> Терминатор
<222> (4923)..(5175)
<223> tNOS
<220>
<221> промотор
<222> (5196)..(5570)
<223> pU3
<220>
<221> другой_признак
<222> (5572)..(5591)
<220>
<221> другой_признак
<222> (5592)..(5636)
<223> crRNA
<220>
<221> промотор
<222> (5643)..(6158)
<223> pU6
<220>
<221> другой_признак
<222> (6159)..(6246)
<223> tracrRNA
<220>
<221> промотор
<222> (6253)..(8245)
<223> Ubi
<220>
<221> ген
<222> (8257)..(9435)
<223> PMI
<220>
<221> Терминатор
<222> (9440)..(10474)
<223> Ubi
<400> 38
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga ccgagtcaaa gattcaaata gaggacctaa 240
cagaactcgc cgtaaagact ggcgaacagt tcatacagag tctcttacga ctcaatgaca 300
agaagaaaat cttcgtcaac ttggtggagc acgacacgct agtctactcc aaaaatatca 360
aagatacagt ctcagaagac caaagggcaa ttgagacttt tcaacaaagg gtaatatccg 420
gaaacctcct cggattccat tgcccagcta tctgtcactt aattgtgaag atagtggaaa 480
aggaaggtgg ctcctacaaa tgccatcatt gcgataaagg aaaggccatc gttgaagatg 540
cctctgccga cagtggtccc aaagatggac ccccacccac gaggagcatc gtggtaaaag 600
aagacgttcc aaccacgtct tcaaagcaag tggattgatg tgatatctcc actgacgtaa 660
gggatgacgc acaatcccac tatccttcgc aagacccttc ctctatataa ggaagttcat 720
ttcatttgga gaggataatt atccaccatg gacaagaagt acagcatcgg cctggacatc 780
ggcaccaaca gcgtgggctg ggccgtgatc accgacgagt acaaggtgcc gagcaagaag 840
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 900
ctgttcgaca gcggcgagac cgccgaggcc accaggctga agaggaccgc caggaggagg 960
tacaccagga ggaagaacag gatctgctac ctgcaggaga tcttcagcaa cgagatggcc 1020
aaggtggacg acagcttctt ccacaggctg gaggagagct tcctggtgga ggaggacaag 1080
aagcacgaga ggcacccgat cttcggcaac atcgtggacg aggtggccta ccacgagaag 1140
tacccgacca tctaccacct gaggaagaag ctggtggaca gcaccgacaa ggccgacctg 1200
aggctgatct acctggccct ggcccacatg atcaagttca ggggccactt cctgatcgag 1260
ggcgacctga acccggacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 1320
tacaaccagc tgttcgagga gaacccgatc aacgccagcg gcgtggacgc caaggccatc 1380
ctgagcgcca ggctgagcaa gagcaggagg ctggagaacc tgatcgccca gctgccgggc 1440
gagaagaaga acggcctgtt cggcaacctg atcgccctga gcctgggcct gaccccgaac 1500
ttcaagagca acttcgacct ggccgaggac gccaagctgc agctgagcaa ggacacctac 1560
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 1620
gccgccaaga acctgagcga cgccatcctg ctgagcgaca tcctgagggt gaacaccgag 1680
atcaccaagg ccccgctgag cgccagcatg atcaagaggt acgacgagca ccaccaggac 1740
ctgaccctgc tgaaggccct ggtgaggcag cagctgccgg agaagtacaa ggagatcttc 1800
ttcgaccaga gcaagaacgg ctacgccggc tacatcgacg gcggcgccag ccaggaggag 1860
ttctacaagt tcatcaagcc gatcctggag aagatggacg gcaccgagga gctgctggtg 1920
aagctgaaca gggaggacct gctgaggaag cagaggacct tcgacaacgg cagcatcccg 1980
caccagatcc acctgggcga gctgcacgcc atcctgagga ggcaggagga cttctacccg 2040
ttcctgaagg acaacaggga gaagatcgag aagatcctga ccttccgcat cccgtactac 2100
gtgggcccgc tggccagggg caacagcagg ttcgcctgga tgaccaggaa gagcgaggag 2160
accatcaccc cgtggaactt cgaggaggtg gtggacaagg gcgccagcgc ccagagcttc 2220
atcgagagga tgaccaactt cgacaagaac ctgccgaacg agaaggtgct gccgaagcac 2280
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaggtgaa gtacgtgacc 2340
gagggcatga ggaagccggc cttcctgagc ggcgagcaga agaaggccat cgtggacctg 2400
ctgttcaaga ccaacaggaa ggtgaccgtg aagcagctga aggaggacta cttcaagaag 2460
atcgagtgct tcgacagcgt ggagatcagc ggcgtggagg acaggttcaa cgccagcctg 2520
ggcacctacc acgacctgct gaagatcatc aaggacaagg acttcctgga caacgaggag 2580
aacgaggaca tcctggagga catcgtgctg accctgaccc tgttcgagga cagggagatg 2640
atcgaggaga ggctgaagac ctacgcccac ctgttcgacg acaaggtgat gaagcagctg 2700
aagaggagga ggtacaccgg ctggggcagg ctgagcagga agctgatcaa cggcatcagg 2760
gacaagcaga gcggcaagac catcctggac ttcctgaaga gcgacggctt cgccaacagg 2820
aacttcatgc agctgatcca cgacgacagc ctgaccttca aggaggacat ccagaaggcc 2880
caggtgagcg gccagggcga cagcctgcac gagcacatcg ccaacctggc cggcagcccg 2940
gccatcaaga agggcatcct gcagaccgtg aaggtggtgg acgagctggt gaaggtgatg 3000
ggcaggcaca agccggagaa catcgtgatc gagatggcca gggagaacca gaccacccag 3060
aagggccaga agaacagcag ggagaggatg aagaggatcg aggagggcat caaggagctg 3120
ggcagccaga tcctgaagga gcacccggtg gagaacaccc agctgcagaa cgagaagctg 3180
tacctgtact acctgcagaa cggcagggac atgtacgtgg accaggagct ggacatcaac 3240
aggctgagcg actacgacgt ggaccacatc gtgccgcaga gcttcctgaa ggacgacagc 3300
atcgacaaca aggtgctgac caggagcgac aagaacaggg gcaagagcga caacgtgccg 3360
agcgaggagg tggtgaagaa gatgaaaaac tactggaggc agctgctgaa cgccaagctg 3420
atcacccaga ggaagttcga caacctgacc aaggccgaga ggggcggcct gagcgagctg 3480
gacaaggccg gcttcattaa aaggcagctg gtggagacca ggcagatcac caagcacgtg 3540
gcccagatcc tggacagcag gatgaacacc aagtacgacg agaacgacaa gctgatcagg 3600
gaggtgaagg tgatcaccct gaagagcaag ctggtgagcg acttcaggaa ggacttccag 3660
ttctacaagg tgagggagat caataattac caccacgccc acgacgccta cctgaacgcc 3720
gtggtgggca ccgccctgat taaaaagtac ccgaagctgg agagcgagtt cgtgtacggc 3780
gactacaagg tgtacgacgt gaggaagatg atcgccaaga gcgagcagga gatcggcaag 3840
gccaccgcca agtacttctt ctacagcaac atcatgaact tcttcaagac cgagatcacc 3900
ctggccaacg gcgagatcag gaagaggccg ctgatcgaga ccaacggcga gaccggcgag 3960
atcgtgtggg acaagggcag ggacttcgcc accgtgagga aggtgctgtc catgccgcag 4020
gtgaacatcg tgaagaagac cgaggtgcag accggcggct tcagcaagga gagcatcctg 4080
ccgaagagga acagcgacaa gctgatcgcc aggaagaagg actgggatcc gaagaagtac 4140
ggcggcttcg acagcccgac cgtggcctac agcgtgctgg tggtggccaa ggtggagaag 4200
ggcaagagca agaagctgaa gagcgtgaag gagctggtgg gcatcaccat catggagagg 4260
agcagcttcg agaagaaccc agtggacttc ctggaggcca agggctacaa ggaggtgaag 4320
aaggacctga tcattaaact gccgaagtac agcctgttcg agctggagaa cggcaggaag 4380
aggatgctgg ccagcgccgg cgagctgcag aagggcaacg agctggccct gccgagcaag 4440
tacgtgaact tcctgtacct ggccagccac tacgagaagc tgaagggcag cccggaggac 4500
aacgagcaga agcagctgtt cgtggagcag cacaagcact acctggacga gatcatcgag 4560
cagatcagcg agttcagcaa gagggtgatc ctggccgacg ccaacctgga caaggtgctg 4620
agcgcctaca acaagcacag ggacaagccg atcagggagc aggccgagaa catcatccac 4680
ctgttcaccc tgaccaacct gggcgccccg gccgccttca agtacttcga caccaccatc 4740
gacaggaaga ggtacaccag caccaaggag gtgctggacg ccaccctgat ccaccagagc 4800
atcaccggcc tgtacgagac caggatcgac ctgagccagc tgggcggcga cagcagcccg 4860
ccgaagaaga agaggaaggt gagctggaag gacgccagcg gctggagcag gatgtgaagc 4920
ttgatcgttc aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg 4980
cgatgattat catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat 5040
gcatgacgtt atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat 5100
acgcgataga aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat 5160
ctatgttact agatcgggac cggaattcgg gacccgggat ctttaaacat acgaacagat 5220
cacttaaagt tcttctgaag caacttaaag ttatcaggca tgcatggatc ttggaggaat 5280
cagatgtgca gtcagggacc atagcacagg acaggcgtct tctactggtg ctaccagcaa 5340
atgctggaag ccgggaacac tgggtacgtt ggaaaccacg tgatgtggag taagataaac 5400
tgtaggagaa aagcatttcg tagtgggcca tgaagccttt caggacatgt attgcagtat 5460
gggccggccc attacgcaat tggacgacaa caaagactag tattagtacc acctcggcta 5520
tccacataga tcaaagctgg tttaaaagag ttgtgcagat gatccgtggc atgcagtcga 5580
agtaatcggc ggttttagag ctatgctgtt ttgaatggtc ccaaaacttt ttttttacta 5640
gttttgtgaa agttgaatta cggcatagcc gaaggaataa cagaatcgtt tcacactttc 5700
gtaacaaagg tcttcttatc atgtttcaga cgatggaggc aaggctgatc aaagtgatca 5760
agcacataaa cgcatttttt taccatgttt cactccataa gcgtctgaga ttatcacaag 5820
tcacgtctag tagtttgatg gtacactagt gacaatcagt tcgtgcagac agagctcata 5880
cttgactact tgagcgatta caggcgaaag tgtgaaacgc atgtgatgtg ggctgggagg 5940
aggagaatat atactaatgg gccgtatcct gatttgggct gcgtcggaag gtgcagccca 6000
cgcgcgccgt accgcgcggg tggcgctgct acccacttta gtccgttgga tggggatccg 6060
atggtttgcg cggtggcgtt gcgggggatg tttagtacca catcggaaac cgaaagacga 6120
tggaaccagc ttataaaccc gcgcgctgta gtcagcttgg aaccattcaa aacagcatag 6180
caagttaaaa taaggctagt ccgttatcaa cttgaaaaag tggcaccgag tcggtgcttt 6240
ttttttccta ggctgcagtg cagcgtgacc cggtcgtgcc cctctctaga gataatgagc 6300
attgcatgtc taagttataa aaaattacca catatttttt ttgtcacact tgtttgaagt 6360
gcagtttatc tatctttata catatattta aactttactc tacgaataat ataatctata 6420
gtactacaat aatatcagtg ttttagagaa tcatataaat gaacagttag acatggtcta 6480
aaggacaatt gagtattttg acaacaggac tctacagttt tatcttttta gtgtgcatgt 6540
gttctccttt ttttttgcaa atagcttcac ctatataata cttcatccat tttattagta 6600
catccattta gggtttaggg ttaatggttt ttatagacta atttttttag tacatctatt 6660
ttattctatt ttagcctcta aattaagaaa actaaaactc tattttagtt tttttattta 6720
ataatttaga tataaaatag aataaaataa agtgactaaa aattaaacaa atacccttta 6780
agaaattaaa aaaactaagg aaacattttt cttgtttcga gtagataatg ccagcctgtt 6840
aaacgccgtc gacgagtcta acggacacca accagcgaac cagcagcgtc gcgtcgggcc 6900
aagcgaagca gacggcacgg catctctgtc gctgcctctg gacccctctc gagagttccg 6960
ctccaccgtt ggacttgctc cgctgtcggc atccagaaat tgcgtggcgg agcggcagac 7020
gtgagccggc acggcaggcg gcctcctcct cctctcacgg caccggcagc tacgggggat 7080
tcctttccca ccgctccttc gctttccctt cctcgcccgc cgtaataaat agacaccccc 7140
tccacaccct ctttccccaa cctcgtgttg ttcggagcgc acacacacac aaccagatct 7200
cccccaaatc cacccgtcgg cacctccgct tcaaggtacg ccgctcgtcc tccccccccc 7260
cccctctcta ccttctctag atcggcgttc cggtccatgg ttagggcccg gtagttctac 7320
ttctgttcat gtttgtgtta gatccgtgtt tgtgttagat ccgtgctgct agcgttcgta 7380
cacggatgcg acctgtacgt cagacacgtt ctgattgcta acttgccagt gtttctcttt 7440
ggggaatcct gggatggctc tagccgttcc gcagacggga tcgatttcat gatttttttt 7500
gtttcgttgc atagggtttg gtttgccctt ttcctttatt tcaatatatg ccgtgcactt 7560
gtttgtcggg tcatcttttc atgctttttt ttgtcttggt tgtgatgatg tggtctggtt 7620
gggcggtcgt tctagatcgg agtagaattc tgtttcaaac tacctggtgg atttattaat 7680
tttggatctg tatgtgtgtg ccatacatat tcatagttac gaattgaaga tgatggatgg 7740
aaatatcgat ctaggatagg tatacatgtt gatgcgggtt ttactgatgc atatacagag 7800
atgctttttg ttcgcttggt tgtgatgatg tggtgtggtt gggcggtcgt tcattcgttc 7860
tagatcggag tagaatactg tttcaaacta cctggtgtat ttattaattt tggaactgta 7920
tgtgtgtgtc atacatcttc atagttacga gtttaagatg gatggaaata tcgatctagg 7980
ataggtatac atgttgatgt gggttttact gatgcatata catgatggca tatgcagcat 8040
ctattcatat gctctaacct tgagtaccta tctattataa taaacaagta tgttttataa 8100
ttattttgat cttgatatac ttggatgatg gcatatgcag cagctatatg tggatttttt 8160
tagccctgcc ttcatacgct atttatttgc ttggtactgt ttcttttgtc gatgctcacc 8220
ctgttgtttg gtgttacttc tgcagtgact aaatagatgc agaagctgat caacagcgtg 8280
cagaactacg cctggggcag caagaccgcc ctgaccgagc tgtacggcat ggagaacccc 8340
agcagccagc ccatggccga gctgtggatg ggcgcccacc ccaagagctc aagccgcgtg 8400
cagaacgccg ccggcgatat cgttagcctg cgcgacgtga tcgagagcga caagagcacc 8460
ctgctgggcg aggccgtggc caagcgcttc ggcgagctgc ccttcctgtt caaggtgctg 8520
tgcgccgctc agcccctgag catccaggtg caccctaaca agcacaacag cgagatcggc 8580
ttcgccaagg agaacgccgc cggcatcccc atggacgccg ccgagcgcaa ctacaaggac 8640
cccaaccaca agcccgagct ggtgttcgcc ctgaccccct tcctggccat gaacgccttc 8700
cgcgagttca gcgagatcgt tagcctgctg cagcccgtgg ccggcgccca ccccgctatc 8760
gcccacttcc ttcagcagcc cgacgccgag cgcctgagcg agctgttcgc cagcctgctg 8820
aacatgcagg gtgaggagaa gtcacgcgcc ctggccatcc tgaagagcgc cctggacagc 8880
cagcagggcg agccctggca gacaatccgc ctgatcagcg agttctaccc cgaggatagc 8940
ggcctgttca gccccctgct gctgaacgtg gtgaagctga accccggcga ggccatgttc 9000
ctgttcgccg agacccccca cgcctacctg cagggcgtgg ccctggaggt gatggccaac 9060
agcgacaacg tgctgcgcgc cggcctgacc cccaagtaca tcgacatccc cgagctggtg 9120
gccaacgtga agttcgaggc taagcccgcc aaccagctgc tgacccagcc cgtgaagcag 9180
ggcgccgagc tggacttccc tatccccgtt gacgacttcg ccttcagcct gcacgacctg 9240
agcgacaagg agaccactat cagccagcag agcgccgcga tcctgttctg cgtggagggc 9300
gacgccaccc tgtggaaggg cagccagcag ctgcagctga agcccggcga gagcgccttt 9360
atcgccgcca acgagagccc cgtgaccgtg aagggccacg gccgcctggc ccgcgtgtac 9420
aacaagctgt gatagctacg tcatgggtcg tttaagctgc cgatgtgcct gcgtcgtctg 9480
gtgccctctc tccatatgga ggttgtcaaa gtatctgctg ttcgtgtcat gagtcgtgtc 9540
agtgttggtt taataatgga ccggttgtgt tgtgtgtgcg tactacccag aactatgaca 9600
aatcatgaat aagtttgatg tttgaaatta aagcctgtgc tcattatgtt ctgtctttca 9660
gttgtctcct aatatttgcc tgcaggtact ggctatctac cgtttcttac ttaggaggtg 9720
tttgaatgca ctaaaactaa tagttagtgg ctaaaattag ttaaaacatc caaacaccat 9780
agctaatagt tgaactatta gctatttttg gaaaattagt taatagtgag gtagttattt 9840
gttagctagc taattcaact aacaattttt agccaactaa caattagttt cagtgcattc 9900
aaacaccccc ttaatgttaa cgtggttcta tctaccgtct cctaatatat ggttgattgt 9960
tcggtttgtt gctatgctat tgggttctga ttgctgctag ttcttgctga atccagaagt 10020
tctcgtagta tagctcagat tcatattatt tatttgagtg ataagtgatc caggttatta 10080
ctatgttagc taggtttttt ttacaaggat aaattatctg tgatcataat tcttatgaaa 10140
gctttatgtt tcctggaggc agtggcatgc aatgcatgac agcaacttga tcacaccagc 10200
tgaggtagat acggtaacaa ggttcttaaa tctgttcacc aaatcattgg agaacacaca 10260
tacacattct tgccagtctt ggttagagaa atttcatgac aaaatgccaa agctgtcttg 10320
actcttcact tttggccatg agtcgtgact tagtttggtt taatggaccg gttctcctag 10380
cttgttctac tcaaaactgt tgttgatgcg aataagttgt gatggttgat ctctggattt 10440
tgttttgctc tcaatagtgg acgagattag atagcggacc gcctgcaggc ccgggggcgc 10500
gccctaatta gctaacggcc aggatcgccg cgtgagcctt tagcaactag ctagattaat 10560
taacgcaatc tgttattaag ttgtctaagc gtcaatttgt ttacaccaca atatatcctg 10620
ccaccagcca gccaacagct ccccgaccgg cagctcggca caaaatcacc actcgataca 10680
ggcagcccat cag 10693
<210> 39
<211> 10264
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 23501, нацеливающаяся gRNA в виде
1 одинарной направляющей последовательности
<220>
<221> промотор
<222> (214)..(734)
<223> 35S
<220>
<221> ген
<222> (748)..(4917)
<223> Cas9
<220>
<221> другой_признак
<222> (4852)..(4908)
<223> NLS-метка
<220>
<221> Терминатор
<222> (4923)..(5175)
<223> tNOS
<220>
<221> промотор
<222> (5196)..(5711)
<223> pU6
<220>
<221> другой_признак
<222> (5712)..(5817)
<223> sgRNA
<220>
<221> другой_признак
<222> (5713)..(5732)
<220>
<221> промотор
<222> (5824)..(7816)
<223> Промотор Ubi
<220>
<221> ген
<222> (7828)..(9006)
<223> PMI
<220>
<221> Терминатор
<222> (9011)..(10045)
<223> Терминатор Ubi
<400> 39
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga ccgagtcaaa gattcaaata gaggacctaa 240
cagaactcgc cgtaaagact ggcgaacagt tcatacagag tctcttacga ctcaatgaca 300
agaagaaaat cttcgtcaac ttggtggagc acgacacgct agtctactcc aaaaatatca 360
aagatacagt ctcagaagac caaagggcaa ttgagacttt tcaacaaagg gtaatatccg 420
gaaacctcct cggattccat tgcccagcta tctgtcactt aattgtgaag atagtggaaa 480
aggaaggtgg ctcctacaaa tgccatcatt gcgataaagg aaaggccatc gttgaagatg 540
cctctgccga cagtggtccc aaagatggac ccccacccac gaggagcatc gtggtaaaag 600
aagacgttcc aaccacgtct tcaaagcaag tggattgatg tgatatctcc actgacgtaa 660
gggatgacgc acaatcccac tatccttcgc aagacccttc ctctatataa ggaagttcat 720
ttcatttgga gaggataatt atccaccatg gacaagaagt acagcatcgg cctggacatc 780
ggcaccaaca gcgtgggctg ggccgtgatc accgacgagt acaaggtgcc gagcaagaag 840
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 900
ctgttcgaca gcggcgagac cgccgaggcc accaggctga agaggaccgc caggaggagg 960
tacaccagga ggaagaacag gatctgctac ctgcaggaga tcttcagcaa cgagatggcc 1020
aaggtggacg acagcttctt ccacaggctg gaggagagct tcctggtgga ggaggacaag 1080
aagcacgaga ggcacccgat cttcggcaac atcgtggacg aggtggccta ccacgagaag 1140
tacccgacca tctaccacct gaggaagaag ctggtggaca gcaccgacaa ggccgacctg 1200
aggctgatct acctggccct ggcccacatg atcaagttca ggggccactt cctgatcgag 1260
ggcgacctga acccggacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 1320
tacaaccagc tgttcgagga gaacccgatc aacgccagcg gcgtggacgc caaggccatc 1380
ctgagcgcca ggctgagcaa gagcaggagg ctggagaacc tgatcgccca gctgccgggc 1440
gagaagaaga acggcctgtt cggcaacctg atcgccctga gcctgggcct gaccccgaac 1500
ttcaagagca acttcgacct ggccgaggac gccaagctgc agctgagcaa ggacacctac 1560
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 1620
gccgccaaga acctgagcga cgccatcctg ctgagcgaca tcctgagggt gaacaccgag 1680
atcaccaagg ccccgctgag cgccagcatg atcaagaggt acgacgagca ccaccaggac 1740
ctgaccctgc tgaaggccct ggtgaggcag cagctgccgg agaagtacaa ggagatcttc 1800
ttcgaccaga gcaagaacgg ctacgccggc tacatcgacg gcggcgccag ccaggaggag 1860
ttctacaagt tcatcaagcc gatcctggag aagatggacg gcaccgagga gctgctggtg 1920
aagctgaaca gggaggacct gctgaggaag cagaggacct tcgacaacgg cagcatcccg 1980
caccagatcc acctgggcga gctgcacgcc atcctgagga ggcaggagga cttctacccg 2040
ttcctgaagg acaacaggga gaagatcgag aagatcctga ccttccgcat cccgtactac 2100
gtgggcccgc tggccagggg caacagcagg ttcgcctgga tgaccaggaa gagcgaggag 2160
accatcaccc cgtggaactt cgaggaggtg gtggacaagg gcgccagcgc ccagagcttc 2220
atcgagagga tgaccaactt cgacaagaac ctgccgaacg agaaggtgct gccgaagcac 2280
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaggtgaa gtacgtgacc 2340
gagggcatga ggaagccggc cttcctgagc ggcgagcaga agaaggccat cgtggacctg 2400
ctgttcaaga ccaacaggaa ggtgaccgtg aagcagctga aggaggacta cttcaagaag 2460
atcgagtgct tcgacagcgt ggagatcagc ggcgtggagg acaggttcaa cgccagcctg 2520
ggcacctacc acgacctgct gaagatcatc aaggacaagg acttcctgga caacgaggag 2580
aacgaggaca tcctggagga catcgtgctg accctgaccc tgttcgagga cagggagatg 2640
atcgaggaga ggctgaagac ctacgcccac ctgttcgacg acaaggtgat gaagcagctg 2700
aagaggagga ggtacaccgg ctggggcagg ctgagcagga agctgatcaa cggcatcagg 2760
gacaagcaga gcggcaagac catcctggac ttcctgaaga gcgacggctt cgccaacagg 2820
aacttcatgc agctgatcca cgacgacagc ctgaccttca aggaggacat ccagaaggcc 2880
caggtgagcg gccagggcga cagcctgcac gagcacatcg ccaacctggc cggcagcccg 2940
gccatcaaga agggcatcct gcagaccgtg aaggtggtgg acgagctggt gaaggtgatg 3000
ggcaggcaca agccggagaa catcgtgatc gagatggcca gggagaacca gaccacccag 3060
aagggccaga agaacagcag ggagaggatg aagaggatcg aggagggcat caaggagctg 3120
ggcagccaga tcctgaagga gcacccggtg gagaacaccc agctgcagaa cgagaagctg 3180
tacctgtact acctgcagaa cggcagggac atgtacgtgg accaggagct ggacatcaac 3240
aggctgagcg actacgacgt ggaccacatc gtgccgcaga gcttcctgaa ggacgacagc 3300
atcgacaaca aggtgctgac caggagcgac aagaacaggg gcaagagcga caacgtgccg 3360
agcgaggagg tggtgaagaa gatgaaaaac tactggaggc agctgctgaa cgccaagctg 3420
atcacccaga ggaagttcga caacctgacc aaggccgaga ggggcggcct gagcgagctg 3480
gacaaggccg gcttcattaa aaggcagctg gtggagacca ggcagatcac caagcacgtg 3540
gcccagatcc tggacagcag gatgaacacc aagtacgacg agaacgacaa gctgatcagg 3600
gaggtgaagg tgatcaccct gaagagcaag ctggtgagcg acttcaggaa ggacttccag 3660
ttctacaagg tgagggagat caataattac caccacgccc acgacgccta cctgaacgcc 3720
gtggtgggca ccgccctgat taaaaagtac ccgaagctgg agagcgagtt cgtgtacggc 3780
gactacaagg tgtacgacgt gaggaagatg atcgccaaga gcgagcagga gatcggcaag 3840
gccaccgcca agtacttctt ctacagcaac atcatgaact tcttcaagac cgagatcacc 3900
ctggccaacg gcgagatcag gaagaggccg ctgatcgaga ccaacggcga gaccggcgag 3960
atcgtgtggg acaagggcag ggacttcgcc accgtgagga aggtgctgtc catgccgcag 4020
gtgaacatcg tgaagaagac cgaggtgcag accggcggct tcagcaagga gagcatcctg 4080
ccgaagagga acagcgacaa gctgatcgcc aggaagaagg actgggatcc gaagaagtac 4140
ggcggcttcg acagcccgac cgtggcctac agcgtgctgg tggtggccaa ggtggagaag 4200
ggcaagagca agaagctgaa gagcgtgaag gagctggtgg gcatcaccat catggagagg 4260
agcagcttcg agaagaaccc agtggacttc ctggaggcca agggctacaa ggaggtgaag 4320
aaggacctga tcattaaact gccgaagtac agcctgttcg agctggagaa cggcaggaag 4380
aggatgctgg ccagcgccgg cgagctgcag aagggcaacg agctggccct gccgagcaag 4440
tacgtgaact tcctgtacct ggccagccac tacgagaagc tgaagggcag cccggaggac 4500
aacgagcaga agcagctgtt cgtggagcag cacaagcact acctggacga gatcatcgag 4560
cagatcagcg agttcagcaa gagggtgatc ctggccgacg ccaacctgga caaggtgctg 4620
agcgcctaca acaagcacag ggacaagccg atcagggagc aggccgagaa catcatccac 4680
ctgttcaccc tgaccaacct gggcgccccg gccgccttca agtacttcga caccaccatc 4740
gacaggaaga ggtacaccag caccaaggag gtgctggacg ccaccctgat ccaccagagc 4800
atcaccggcc tgtacgagac caggatcgac ctgagccagc tgggcggcga cagcagcccg 4860
ccgaagaaga agaggaaggt gagctggaag gacgccagcg gctggagcag gatgtgaagc 4920
ttgatcgttc aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg 4980
cgatgattat catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat 5040
gcatgacgtt atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat 5100
acgcgataga aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat 5160
ctatgttact agatcgggac cggaattcgg gaccctttgt gaaagttgaa ttacggcata 5220
gccgaaggaa taacagaatc gtttcacact ttcgtaacaa aggtcttctt atcatgtttc 5280
agacgatgga ggcaaggctg atcaaagtga tcaagcacat aaacgcattt ttttaccatg 5340
tttcactcca taagcgtctg agattatcac aagtcacgtc tagtagtttg atggtacact 5400
agtgacaatc agttcgtgca gacagagctc atacttgact acttgagcga ttacaggcga 5460
aagtgtgaaa cgcatgtgat gtgggctggg aggaggagaa tatatactaa tgggccgtat 5520
cctgatttgg gctgcgtcgg aaggtgcagc ccacgcgcgc cgtaccgcgc gggtggcgct 5580
gctacccact ttagtccgtt ggatggggat ccgatggttt gcgcggtggc gttgcggggg 5640
atgtttagta ccacatcgga aaccgaaaga cgatggaacc agcttataaa cccgcgcgct 5700
gtagtcagct tgtgcagtcg aagtaatcgg cggttttaga gctagaaata gcaagttaaa 5760
ataaggctag tccgttatca acttgaaaaa gtggcaccga gtcggtgctt tttttttcct 5820
aggctgcagt gcagcgtgac ccggtcgtgc ccctctctag agataatgag cattgcatgt 5880
ctaagttata aaaaattacc acatattttt tttgtcacac ttgtttgaag tgcagtttat 5940
ctatctttat acatatattt aaactttact ctacgaataa tataatctat agtactacaa 6000
taatatcagt gttttagaga atcatataaa tgaacagtta gacatggtct aaaggacaat 6060
tgagtatttt gacaacagga ctctacagtt ttatcttttt agtgtgcatg tgttctcctt 6120
tttttttgca aatagcttca cctatataat acttcatcca ttttattagt acatccattt 6180
agggtttagg gttaatggtt tttatagact aattttttta gtacatctat tttattctat 6240
tttagcctct aaattaagaa aactaaaact ctattttagt ttttttattt aataatttag 6300
atataaaata gaataaaata aagtgactaa aaattaaaca aatacccttt aagaaattaa 6360
aaaaactaag gaaacatttt tcttgtttcg agtagataat gccagcctgt taaacgccgt 6420
cgacgagtct aacggacacc aaccagcgaa ccagcagcgt cgcgtcgggc caagcgaagc 6480
agacggcacg gcatctctgt cgctgcctct ggacccctct cgagagttcc gctccaccgt 6540
tggacttgct ccgctgtcgg catccagaaa ttgcgtggcg gagcggcaga cgtgagccgg 6600
cacggcaggc ggcctcctcc tcctctcacg gcaccggcag ctacggggga ttcctttccc 6660
accgctcctt cgctttccct tcctcgcccg ccgtaataaa tagacacccc ctccacaccc 6720
tctttcccca acctcgtgtt gttcggagcg cacacacaca caaccagatc tcccccaaat 6780
ccacccgtcg gcacctccgc ttcaaggtac gccgctcgtc ctcccccccc ccccctctct 6840
accttctcta gatcggcgtt ccggtccatg gttagggccc ggtagttcta cttctgttca 6900
tgtttgtgtt agatccgtgt ttgtgttaga tccgtgctgc tagcgttcgt acacggatgc 6960
gacctgtacg tcagacacgt tctgattgct aacttgccag tgtttctctt tggggaatcc 7020
tgggatggct ctagccgttc cgcagacggg atcgatttca tgattttttt tgtttcgttg 7080
catagggttt ggtttgccct tttcctttat ttcaatatat gccgtgcact tgtttgtcgg 7140
gtcatctttt catgcttttt tttgtcttgg ttgtgatgat gtggtctggt tgggcggtcg 7200
ttctagatcg gagtagaatt ctgtttcaaa ctacctggtg gatttattaa ttttggatct 7260
gtatgtgtgt gccatacata ttcatagtta cgaattgaag atgatggatg gaaatatcga 7320
tctaggatag gtatacatgt tgatgcgggt tttactgatg catatacaga gatgcttttt 7380
gttcgcttgg ttgtgatgat gtggtgtggt tgggcggtcg ttcattcgtt ctagatcgga 7440
gtagaatact gtttcaaact acctggtgta tttattaatt ttggaactgt atgtgtgtgt 7500
catacatctt catagttacg agtttaagat ggatggaaat atcgatctag gataggtata 7560
catgttgatg tgggttttac tgatgcatat acatgatggc atatgcagca tctattcata 7620
tgctctaacc ttgagtacct atctattata ataaacaagt atgttttata attattttga 7680
tcttgatata cttggatgat ggcatatgca gcagctatat gtggattttt ttagccctgc 7740
cttcatacgc tatttatttg cttggtactg tttcttttgt cgatgctcac cctgttgttt 7800
ggtgttactt ctgcagtgac taaatagatg cagaagctga tcaacagcgt gcagaactac 7860
gcctggggca gcaagaccgc cctgaccgag ctgtacggca tggagaaccc cagcagccag 7920
cccatggccg agctgtggat gggcgcccac cccaagagct caagccgcgt gcagaacgcc 7980
gccggcgata tcgttagcct gcgcgacgtg atcgagagcg acaagagcac cctgctgggc 8040
gaggccgtgg ccaagcgctt cggcgagctg cccttcctgt tcaaggtgct gtgcgccgct 8100
cagcccctga gcatccaggt gcaccctaac aagcacaaca gcgagatcgg cttcgccaag 8160
gagaacgccg ccggcatccc catggacgcc gccgagcgca actacaagga ccccaaccac 8220
aagcccgagc tggtgttcgc cctgaccccc ttcctggcca tgaacgcctt ccgcgagttc 8280
agcgagatcg ttagcctgct gcagcccgtg gccggcgccc accccgctat cgcccacttc 8340
cttcagcagc ccgacgccga gcgcctgagc gagctgttcg ccagcctgct gaacatgcag 8400
ggtgaggaga agtcacgcgc cctggccatc ctgaagagcg ccctggacag ccagcagggc 8460
gagccctggc agacaatccg cctgatcagc gagttctacc ccgaggatag cggcctgttc 8520
agccccctgc tgctgaacgt ggtgaagctg aaccccggcg aggccatgtt cctgttcgcc 8580
gagacccccc acgcctacct gcagggcgtg gccctggagg tgatggccaa cagcgacaac 8640
gtgctgcgcg ccggcctgac ccccaagtac atcgacatcc ccgagctggt ggccaacgtg 8700
aagttcgagg ctaagcccgc caaccagctg ctgacccagc ccgtgaagca gggcgccgag 8760
ctggacttcc ctatccccgt tgacgacttc gccttcagcc tgcacgacct gagcgacaag 8820
gagaccacta tcagccagca gagcgccgcg atcctgttct gcgtggaggg cgacgccacc 8880
ctgtggaagg gcagccagca gctgcagctg aagcccggcg agagcgcctt tatcgccgcc 8940
aacgagagcc ccgtgaccgt gaagggccac ggccgcctgg cccgcgtgta caacaagctg 9000
tgatagctac gtcatgggtc gtttaagctg ccgatgtgcc tgcgtcgtct ggtgccctct 9060
ctccatatgg aggttgtcaa agtatctgct gttcgtgtca tgagtcgtgt cagtgttggt 9120
ttaataatgg accggttgtg ttgtgtgtgc gtactaccca gaactatgac aaatcatgaa 9180
taagtttgat gtttgaaatt aaagcctgtg ctcattatgt tctgtctttc agttgtctcc 9240
taatatttgc ctgcaggtac tggctatcta ccgtttctta cttaggaggt gtttgaatgc 9300
actaaaacta atagttagtg gctaaaatta gttaaaacat ccaaacacca tagctaatag 9360
ttgaactatt agctattttt ggaaaattag ttaatagtga ggtagttatt tgttagctag 9420
ctaattcaac taacaatttt tagccaacta acaattagtt tcagtgcatt caaacacccc 9480
cttaatgtta acgtggttct atctaccgtc tcctaatata tggttgattg ttcggtttgt 9540
tgctatgcta ttgggttctg attgctgcta gttcttgctg aatccagaag ttctcgtagt 9600
atagctcaga ttcatattat ttatttgagt gataagtgat ccaggttatt actatgttag 9660
ctaggttttt tttacaagga taaattatct gtgatcataa ttcttatgaa agctttatgt 9720
ttcctggagg cagtggcatg caatgcatga cagcaacttg atcacaccag ctgaggtaga 9780
tacggtaaca aggttcttaa atctgttcac caaatcattg gagaacacac atacacattc 9840
ttgccagtct tggttagaga aatttcatga caaaatgcca aagctgtctt gactcttcac 9900
ttttggccat gagtcgtgac ttagtttggt ttaatggacc ggttctccta gcttgttcta 9960
ctcaaaactg ttgttgatgc gaataagttg tgatggttga tctctggatt ttgttttgct 10020
ctcaatagtg gacgagatta gatagcggac cgcctgcagg cccgggggcg cgccctaatt 10080
agctaacggc caggatcgcc gcgtgagcct ttagcaacta gctagattaa ttaacgcaat 10140
ctgttattaa gttgtctaag cgtcaatttg tttacaccac aatatatcct gccaccagcc 10200
agccaacagc tccccgaccg gcagctcggc acaaaatcac cactcgatac aggcagccca 10260
tcag 10264
<210> 40
<211> 10692
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 23501, нацеливающаяся gRNA в виде 2
двойной направляющей последовательности
<220>
<221> промотор
<222> (214)..(734)
<223> 35S
<220>
<221> ген
<222> (748)..(4917)
<223> Cas9
<220>
<221> другой_признак
<222> (4852)..(4908)
<223> NLS-метка
<220>
<221> Терминатор
<222> (4923)..(5175)
<223> tNOS
<220>
<221> промотор
<222> (5196)..(5570)
<223> pU3
<220>
<221> другой_признак
<222> (5572)..(5590)
<220>
<221> другой_признак
<222> (5591)..(5635)
<223> crRNA
<220>
<221> промотор
<222> (5642)..(6157)
<223> pU6
<220>
<221> другой_признак
<222> (6158)..(6245)
<223> tracrRNA
<220>
<221> промотор
<222> (6252)..(8244)
<223> Промотор Ubi
<220>
<221> ген
<222> (8256)..(9434)
<223> PMI
<220>
<221> Терминатор
<222> (9439)..(10473)
<223> Терминатор Ubi
<400> 40
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga ccgagtcaaa gattcaaata gaggacctaa 240
cagaactcgc cgtaaagact ggcgaacagt tcatacagag tctcttacga ctcaatgaca 300
agaagaaaat cttcgtcaac ttggtggagc acgacacgct agtctactcc aaaaatatca 360
aagatacagt ctcagaagac caaagggcaa ttgagacttt tcaacaaagg gtaatatccg 420
gaaacctcct cggattccat tgcccagcta tctgtcactt aattgtgaag atagtggaaa 480
aggaaggtgg ctcctacaaa tgccatcatt gcgataaagg aaaggccatc gttgaagatg 540
cctctgccga cagtggtccc aaagatggac ccccacccac gaggagcatc gtggtaaaag 600
aagacgttcc aaccacgtct tcaaagcaag tggattgatg tgatatctcc actgacgtaa 660
gggatgacgc acaatcccac tatccttcgc aagacccttc ctctatataa ggaagttcat 720
ttcatttgga gaggataatt atccaccatg gacaagaagt acagcatcgg cctggacatc 780
ggcaccaaca gcgtgggctg ggccgtgatc accgacgagt acaaggtgcc gagcaagaag 840
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 900
ctgttcgaca gcggcgagac cgccgaggcc accaggctga agaggaccgc caggaggagg 960
tacaccagga ggaagaacag gatctgctac ctgcaggaga tcttcagcaa cgagatggcc 1020
aaggtggacg acagcttctt ccacaggctg gaggagagct tcctggtgga ggaggacaag 1080
aagcacgaga ggcacccgat cttcggcaac atcgtggacg aggtggccta ccacgagaag 1140
tacccgacca tctaccacct gaggaagaag ctggtggaca gcaccgacaa ggccgacctg 1200
aggctgatct acctggccct ggcccacatg atcaagttca ggggccactt cctgatcgag 1260
ggcgacctga acccggacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 1320
tacaaccagc tgttcgagga gaacccgatc aacgccagcg gcgtggacgc caaggccatc 1380
ctgagcgcca ggctgagcaa gagcaggagg ctggagaacc tgatcgccca gctgccgggc 1440
gagaagaaga acggcctgtt cggcaacctg atcgccctga gcctgggcct gaccccgaac 1500
ttcaagagca acttcgacct ggccgaggac gccaagctgc agctgagcaa ggacacctac 1560
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 1620
gccgccaaga acctgagcga cgccatcctg ctgagcgaca tcctgagggt gaacaccgag 1680
atcaccaagg ccccgctgag cgccagcatg atcaagaggt acgacgagca ccaccaggac 1740
ctgaccctgc tgaaggccct ggtgaggcag cagctgccgg agaagtacaa ggagatcttc 1800
ttcgaccaga gcaagaacgg ctacgccggc tacatcgacg gcggcgccag ccaggaggag 1860
ttctacaagt tcatcaagcc gatcctggag aagatggacg gcaccgagga gctgctggtg 1920
aagctgaaca gggaggacct gctgaggaag cagaggacct tcgacaacgg cagcatcccg 1980
caccagatcc acctgggcga gctgcacgcc atcctgagga ggcaggagga cttctacccg 2040
ttcctgaagg acaacaggga gaagatcgag aagatcctga ccttccgcat cccgtactac 2100
gtgggcccgc tggccagggg caacagcagg ttcgcctgga tgaccaggaa gagcgaggag 2160
accatcaccc cgtggaactt cgaggaggtg gtggacaagg gcgccagcgc ccagagcttc 2220
atcgagagga tgaccaactt cgacaagaac ctgccgaacg agaaggtgct gccgaagcac 2280
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaggtgaa gtacgtgacc 2340
gagggcatga ggaagccggc cttcctgagc ggcgagcaga agaaggccat cgtggacctg 2400
ctgttcaaga ccaacaggaa ggtgaccgtg aagcagctga aggaggacta cttcaagaag 2460
atcgagtgct tcgacagcgt ggagatcagc ggcgtggagg acaggttcaa cgccagcctg 2520
ggcacctacc acgacctgct gaagatcatc aaggacaagg acttcctgga caacgaggag 2580
aacgaggaca tcctggagga catcgtgctg accctgaccc tgttcgagga cagggagatg 2640
atcgaggaga ggctgaagac ctacgcccac ctgttcgacg acaaggtgat gaagcagctg 2700
aagaggagga ggtacaccgg ctggggcagg ctgagcagga agctgatcaa cggcatcagg 2760
gacaagcaga gcggcaagac catcctggac ttcctgaaga gcgacggctt cgccaacagg 2820
aacttcatgc agctgatcca cgacgacagc ctgaccttca aggaggacat ccagaaggcc 2880
caggtgagcg gccagggcga cagcctgcac gagcacatcg ccaacctggc cggcagcccg 2940
gccatcaaga agggcatcct gcagaccgtg aaggtggtgg acgagctggt gaaggtgatg 3000
ggcaggcaca agccggagaa catcgtgatc gagatggcca gggagaacca gaccacccag 3060
aagggccaga agaacagcag ggagaggatg aagaggatcg aggagggcat caaggagctg 3120
ggcagccaga tcctgaagga gcacccggtg gagaacaccc agctgcagaa cgagaagctg 3180
tacctgtact acctgcagaa cggcagggac atgtacgtgg accaggagct ggacatcaac 3240
aggctgagcg actacgacgt ggaccacatc gtgccgcaga gcttcctgaa ggacgacagc 3300
atcgacaaca aggtgctgac caggagcgac aagaacaggg gcaagagcga caacgtgccg 3360
agcgaggagg tggtgaagaa gatgaaaaac tactggaggc agctgctgaa cgccaagctg 3420
atcacccaga ggaagttcga caacctgacc aaggccgaga ggggcggcct gagcgagctg 3480
gacaaggccg gcttcattaa aaggcagctg gtggagacca ggcagatcac caagcacgtg 3540
gcccagatcc tggacagcag gatgaacacc aagtacgacg agaacgacaa gctgatcagg 3600
gaggtgaagg tgatcaccct gaagagcaag ctggtgagcg acttcaggaa ggacttccag 3660
ttctacaagg tgagggagat caataattac caccacgccc acgacgccta cctgaacgcc 3720
gtggtgggca ccgccctgat taaaaagtac ccgaagctgg agagcgagtt cgtgtacggc 3780
gactacaagg tgtacgacgt gaggaagatg atcgccaaga gcgagcagga gatcggcaag 3840
gccaccgcca agtacttctt ctacagcaac atcatgaact tcttcaagac cgagatcacc 3900
ctggccaacg gcgagatcag gaagaggccg ctgatcgaga ccaacggcga gaccggcgag 3960
atcgtgtggg acaagggcag ggacttcgcc accgtgagga aggtgctgtc catgccgcag 4020
gtgaacatcg tgaagaagac cgaggtgcag accggcggct tcagcaagga gagcatcctg 4080
ccgaagagga acagcgacaa gctgatcgcc aggaagaagg actgggatcc gaagaagtac 4140
ggcggcttcg acagcccgac cgtggcctac agcgtgctgg tggtggccaa ggtggagaag 4200
ggcaagagca agaagctgaa gagcgtgaag gagctggtgg gcatcaccat catggagagg 4260
agcagcttcg agaagaaccc agtggacttc ctggaggcca agggctacaa ggaggtgaag 4320
aaggacctga tcattaaact gccgaagtac agcctgttcg agctggagaa cggcaggaag 4380
aggatgctgg ccagcgccgg cgagctgcag aagggcaacg agctggccct gccgagcaag 4440
tacgtgaact tcctgtacct ggccagccac tacgagaagc tgaagggcag cccggaggac 4500
aacgagcaga agcagctgtt cgtggagcag cacaagcact acctggacga gatcatcgag 4560
cagatcagcg agttcagcaa gagggtgatc ctggccgacg ccaacctgga caaggtgctg 4620
agcgcctaca acaagcacag ggacaagccg atcagggagc aggccgagaa catcatccac 4680
ctgttcaccc tgaccaacct gggcgccccg gccgccttca agtacttcga caccaccatc 4740
gacaggaaga ggtacaccag caccaaggag gtgctggacg ccaccctgat ccaccagagc 4800
atcaccggcc tgtacgagac caggatcgac ctgagccagc tgggcggcga cagcagcccg 4860
ccgaagaaga agaggaaggt gagctggaag gacgccagcg gctggagcag gatgtgaagc 4920
ttgatcgttc aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg 4980
cgatgattat catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat 5040
gcatgacgtt atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat 5100
acgcgataga aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat 5160
ctatgttact agatcgggac cggaattcgg gacccgggat ctttaaacat acgaacagat 5220
cacttaaagt tcttctgaag caacttaaag ttatcaggca tgcatggatc ttggaggaat 5280
cagatgtgca gtcagggacc atagcacagg acaggcgtct tctactggtg ctaccagcaa 5340
atgctggaag ccgggaacac tgggtacgtt ggaaaccacg tgatgtggag taagataaac 5400
tgtaggagaa aagcatttcg tagtgggcca tgaagccttt caggacatgt attgcagtat 5460
gggccggccc attacgcaat tggacgacaa caaagactag tattagtacc acctcggcta 5520
tccacataga tcaaagctgg tttaaaagag ttgtgcagat gatccgtggc agagaccggc 5580
aggtacgtcg gttttagagc tatgctgttt tgaatggtcc caaaactttt tttttactag 5640
ttttgtgaaa gttgaattac ggcatagccg aaggaataac agaatcgttt cacactttcg 5700
taacaaaggt cttcttatca tgtttcagac gatggaggca aggctgatca aagtgatcaa 5760
gcacataaac gcattttttt accatgtttc actccataag cgtctgagat tatcacaagt 5820
cacgtctagt agtttgatgg tacactagtg acaatcagtt cgtgcagaca gagctcatac 5880
ttgactactt gagcgattac aggcgaaagt gtgaaacgca tgtgatgtgg gctgggagga 5940
ggagaatata tactaatggg ccgtatcctg atttgggctg cgtcggaagg tgcagcccac 6000
gcgcgccgta ccgcgcgggt ggcgctgcta cccactttag tccgttggat ggggatccga 6060
tggtttgcgc ggtggcgttg cgggggatgt ttagtaccac atcggaaacc gaaagacgat 6120
ggaaccagct tataaacccg cgcgctgtag tcagcttgga accattcaaa acagcatagc 6180
aagttaaaat aaggctagtc cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt 6240
tttttcctag gctgcagtgc agcgtgaccc ggtcgtgccc ctctctagag ataatgagca 6300
ttgcatgtct aagttataaa aaattaccac atattttttt tgtcacactt gtttgaagtg 6360
cagtttatct atctttatac atatatttaa actttactct acgaataata taatctatag 6420
tactacaata atatcagtgt tttagagaat catataaatg aacagttaga catggtctaa 6480
aggacaattg agtattttga caacaggact ctacagtttt atctttttag tgtgcatgtg 6540
ttctcctttt tttttgcaaa tagcttcacc tatataatac ttcatccatt ttattagtac 6600
atccatttag ggtttagggt taatggtttt tatagactaa tttttttagt acatctattt 6660
tattctattt tagcctctaa attaagaaaa ctaaaactct attttagttt ttttatttaa 6720
taatttagat ataaaataga ataaaataaa gtgactaaaa attaaacaaa taccctttaa 6780
gaaattaaaa aaactaagga aacatttttc ttgtttcgag tagataatgc cagcctgtta 6840
aacgccgtcg acgagtctaa cggacaccaa ccagcgaacc agcagcgtcg cgtcgggcca 6900
agcgaagcag acggcacggc atctctgtcg ctgcctctgg acccctctcg agagttccgc 6960
tccaccgttg gacttgctcc gctgtcggca tccagaaatt gcgtggcgga gcggcagacg 7020
tgagccggca cggcaggcgg cctcctcctc ctctcacggc accggcagct acgggggatt 7080
cctttcccac cgctccttcg ctttcccttc ctcgcccgcc gtaataaata gacaccccct 7140
ccacaccctc tttccccaac ctcgtgttgt tcggagcgca cacacacaca accagatctc 7200
ccccaaatcc acccgtcggc acctccgctt caaggtacgc cgctcgtcct cccccccccc 7260
ccctctctac cttctctaga tcggcgttcc ggtccatggt tagggcccgg tagttctact 7320
tctgttcatg tttgtgttag atccgtgttt gtgttagatc cgtgctgcta gcgttcgtac 7380
acggatgcga cctgtacgtc agacacgttc tgattgctaa cttgccagtg tttctctttg 7440
gggaatcctg ggatggctct agccgttccg cagacgggat cgatttcatg attttttttg 7500
tttcgttgca tagggtttgg tttgcccttt tcctttattt caatatatgc cgtgcacttg 7560
tttgtcgggt catcttttca tgcttttttt tgtcttggtt gtgatgatgt ggtctggttg 7620
ggcggtcgtt ctagatcgga gtagaattct gtttcaaact acctggtgga tttattaatt 7680
ttggatctgt atgtgtgtgc catacatatt catagttacg aattgaagat gatggatgga 7740
aatatcgatc taggataggt atacatgttg atgcgggttt tactgatgca tatacagaga 7800
tgctttttgt tcgcttggtt gtgatgatgt ggtgtggttg ggcggtcgtt cattcgttct 7860
agatcggagt agaatactgt ttcaaactac ctggtgtatt tattaatttt ggaactgtat 7920
gtgtgtgtca tacatcttca tagttacgag tttaagatgg atggaaatat cgatctagga 7980
taggtataca tgttgatgtg ggttttactg atgcatatac atgatggcat atgcagcatc 8040
tattcatatg ctctaacctt gagtacctat ctattataat aaacaagtat gttttataat 8100
tattttgatc ttgatatact tggatgatgg catatgcagc agctatatgt ggattttttt 8160
agccctgcct tcatacgcta tttatttgct tggtactgtt tcttttgtcg atgctcaccc 8220
tgttgtttgg tgttacttct gcagtgacta aatagatgca gaagctgatc aacagcgtgc 8280
agaactacgc ctggggcagc aagaccgccc tgaccgagct gtacggcatg gagaacccca 8340
gcagccagcc catggccgag ctgtggatgg gcgcccaccc caagagctca agccgcgtgc 8400
agaacgccgc cggcgatatc gttagcctgc gcgacgtgat cgagagcgac aagagcaccc 8460
tgctgggcga ggccgtggcc aagcgcttcg gcgagctgcc cttcctgttc aaggtgctgt 8520
gcgccgctca gcccctgagc atccaggtgc accctaacaa gcacaacagc gagatcggct 8580
tcgccaagga gaacgccgcc ggcatcccca tggacgccgc cgagcgcaac tacaaggacc 8640
ccaaccacaa gcccgagctg gtgttcgccc tgaccccctt cctggccatg aacgccttcc 8700
gcgagttcag cgagatcgtt agcctgctgc agcccgtggc cggcgcccac cccgctatcg 8760
cccacttcct tcagcagccc gacgccgagc gcctgagcga gctgttcgcc agcctgctga 8820
acatgcaggg tgaggagaag tcacgcgccc tggccatcct gaagagcgcc ctggacagcc 8880
agcagggcga gccctggcag acaatccgcc tgatcagcga gttctacccc gaggatagcg 8940
gcctgttcag ccccctgctg ctgaacgtgg tgaagctgaa ccccggcgag gccatgttcc 9000
tgttcgccga gaccccccac gcctacctgc agggcgtggc cctggaggtg atggccaaca 9060
gcgacaacgt gctgcgcgcc ggcctgaccc ccaagtacat cgacatcccc gagctggtgg 9120
ccaacgtgaa gttcgaggct aagcccgcca accagctgct gacccagccc gtgaagcagg 9180
gcgccgagct ggacttccct atccccgttg acgacttcgc cttcagcctg cacgacctga 9240
gcgacaagga gaccactatc agccagcaga gcgccgcgat cctgttctgc gtggagggcg 9300
acgccaccct gtggaagggc agccagcagc tgcagctgaa gcccggcgag agcgccttta 9360
tcgccgccaa cgagagcccc gtgaccgtga agggccacgg ccgcctggcc cgcgtgtaca 9420
acaagctgtg atagctacgt catgggtcgt ttaagctgcc gatgtgcctg cgtcgtctgg 9480
tgccctctct ccatatggag gttgtcaaag tatctgctgt tcgtgtcatg agtcgtgtca 9540
gtgttggttt aataatggac cggttgtgtt gtgtgtgcgt actacccaga actatgacaa 9600
atcatgaata agtttgatgt ttgaaattaa agcctgtgct cattatgttc tgtctttcag 9660
ttgtctccta atatttgcct gcaggtactg gctatctacc gtttcttact taggaggtgt 9720
ttgaatgcac taaaactaat agttagtggc taaaattagt taaaacatcc aaacaccata 9780
gctaatagtt gaactattag ctatttttgg aaaattagtt aatagtgagg tagttatttg 9840
ttagctagct aattcaacta acaattttta gccaactaac aattagtttc agtgcattca 9900
aacaccccct taatgttaac gtggttctat ctaccgtctc ctaatatatg gttgattgtt 9960
cggtttgttg ctatgctatt gggttctgat tgctgctagt tcttgctgaa tccagaagtt 10020
ctcgtagtat agctcagatt catattattt atttgagtga taagtgatcc aggttattac 10080
tatgttagct aggttttttt tacaaggata aattatctgt gatcataatt cttatgaaag 10140
ctttatgttt cctggaggca gtggcatgca atgcatgaca gcaacttgat cacaccagct 10200
gaggtagata cggtaacaag gttcttaaat ctgttcacca aatcattgga gaacacacat 10260
acacattctt gccagtcttg gttagagaaa tttcatgaca aaatgccaaa gctgtcttga 10320
ctcttcactt ttggccatga gtcgtgactt agtttggttt aatggaccgg ttctcctagc 10380
ttgttctact caaaactgtt gttgatgcga ataagttgtg atggttgatc tctggatttt 10440
gttttgctct caatagtgga cgagattaga tagcggaccg cctgcaggcc cgggggcgcg 10500
ccctaattag ctaacggcca ggatcgccgc gtgagccttt agcaactagc tagattaatt 10560
aacgcaatct gttattaagt tgtctaagcg tcaatttgtt tacaccacaa tatatcctgc 10620
caccagccag ccaacagctc cccgaccggc agctcggcac aaaatcacca ctcgatacag 10680
gcagcccatc ag 10692
<210> 41
<211> 10262
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Кассета экспрессии конструкции 23501, нацеливающаяся gRNA в
виде 2 одинарной направляющей последовательности
<220>
<221> промотор
<222> (214)..(734)
<223> Промотор 35S
<220>
<221> ген
<222> (748)..(4917)
<223> Cas9
<220>
<221> Терминатор
<222> (4923)..(5175)
<223> tNOS
<220>
<221> промотор
<222> (5196)..(5711)
<220>
<221> другой_признак
<222> (5712)..(5730)
<220>
<221> другой_признак
<222> (5712)..(5815)
<223> sgRNA
<220>
<221> промотор
<222> (5822)..(7814)
<223> Промотор Ubi
<220>
<221> ген
<222> (7826)..(9004)
<223> PMI
<220>
<221> Терминатор
<222> (9009)..(10043)
<223> Терминатор Ubi
<400> 41
cctgtggttg gcatgcacat acaaatggac gaacggataa accttttcac gcccttttaa 60
atatccgatt attctaataa acgctctttt ctcttaggtt tacccgccaa tatatcctgt 120
caaacactga tagtttaaac tggcactagc ctaacggtgt tgactaacta ggccgcttcc 180
ctaattagct aacccggggg cgcgccggga ccgagtcaaa gattcaaata gaggacctaa 240
cagaactcgc cgtaaagact ggcgaacagt tcatacagag tctcttacga ctcaatgaca 300
agaagaaaat cttcgtcaac ttggtggagc acgacacgct agtctactcc aaaaatatca 360
aagatacagt ctcagaagac caaagggcaa ttgagacttt tcaacaaagg gtaatatccg 420
gaaacctcct cggattccat tgcccagcta tctgtcactt aattgtgaag atagtggaaa 480
aggaaggtgg ctcctacaaa tgccatcatt gcgataaagg aaaggccatc gttgaagatg 540
cctctgccga cagtggtccc aaagatggac ccccacccac gaggagcatc gtggtaaaag 600
aagacgttcc aaccacgtct tcaaagcaag tggattgatg tgatatctcc actgacgtaa 660
gggatgacgc acaatcccac tatccttcgc aagacccttc ctctatataa ggaagttcat 720
ttcatttgga gaggataatt atccaccatg gacaagaagt acagcatcgg cctggacatc 780
ggcaccaaca gcgtgggctg ggccgtgatc accgacgagt acaaggtgcc gagcaagaag 840
ttcaaggtgc tgggcaacac cgacaggcac agcatcaaga agaacctgat cggcgccctg 900
ctgttcgaca gcggcgagac cgccgaggcc accaggctga agaggaccgc caggaggagg 960
tacaccagga ggaagaacag gatctgctac ctgcaggaga tcttcagcaa cgagatggcc 1020
aaggtggacg acagcttctt ccacaggctg gaggagagct tcctggtgga ggaggacaag 1080
aagcacgaga ggcacccgat cttcggcaac atcgtggacg aggtggccta ccacgagaag 1140
tacccgacca tctaccacct gaggaagaag ctggtggaca gcaccgacaa ggccgacctg 1200
aggctgatct acctggccct ggcccacatg atcaagttca ggggccactt cctgatcgag 1260
ggcgacctga acccggacaa cagcgacgtg gacaagctgt tcatccagct ggtgcagacc 1320
tacaaccagc tgttcgagga gaacccgatc aacgccagcg gcgtggacgc caaggccatc 1380
ctgagcgcca ggctgagcaa gagcaggagg ctggagaacc tgatcgccca gctgccgggc 1440
gagaagaaga acggcctgtt cggcaacctg atcgccctga gcctgggcct gaccccgaac 1500
ttcaagagca acttcgacct ggccgaggac gccaagctgc agctgagcaa ggacacctac 1560
gacgacgacc tggacaacct gctggcccag atcggcgacc agtacgccga cctgttcctg 1620
gccgccaaga acctgagcga cgccatcctg ctgagcgaca tcctgagggt gaacaccgag 1680
atcaccaagg ccccgctgag cgccagcatg atcaagaggt acgacgagca ccaccaggac 1740
ctgaccctgc tgaaggccct ggtgaggcag cagctgccgg agaagtacaa ggagatcttc 1800
ttcgaccaga gcaagaacgg ctacgccggc tacatcgacg gcggcgccag ccaggaggag 1860
ttctacaagt tcatcaagcc gatcctggag aagatggacg gcaccgagga gctgctggtg 1920
aagctgaaca gggaggacct gctgaggaag cagaggacct tcgacaacgg cagcatcccg 1980
caccagatcc acctgggcga gctgcacgcc atcctgagga ggcaggagga cttctacccg 2040
ttcctgaagg acaacaggga gaagatcgag aagatcctga ccttccgcat cccgtactac 2100
gtgggcccgc tggccagggg caacagcagg ttcgcctgga tgaccaggaa gagcgaggag 2160
accatcaccc cgtggaactt cgaggaggtg gtggacaagg gcgccagcgc ccagagcttc 2220
atcgagagga tgaccaactt cgacaagaac ctgccgaacg agaaggtgct gccgaagcac 2280
agcctgctgt acgagtactt caccgtgtac aacgagctga ccaaggtgaa gtacgtgacc 2340
gagggcatga ggaagccggc cttcctgagc ggcgagcaga agaaggccat cgtggacctg 2400
ctgttcaaga ccaacaggaa ggtgaccgtg aagcagctga aggaggacta cttcaagaag 2460
atcgagtgct tcgacagcgt ggagatcagc ggcgtggagg acaggttcaa cgccagcctg 2520
ggcacctacc acgacctgct gaagatcatc aaggacaagg acttcctgga caacgaggag 2580
aacgaggaca tcctggagga catcgtgctg accctgaccc tgttcgagga cagggagatg 2640
atcgaggaga ggctgaagac ctacgcccac ctgttcgacg acaaggtgat gaagcagctg 2700
aagaggagga ggtacaccgg ctggggcagg ctgagcagga agctgatcaa cggcatcagg 2760
gacaagcaga gcggcaagac catcctggac ttcctgaaga gcgacggctt cgccaacagg 2820
aacttcatgc agctgatcca cgacgacagc ctgaccttca aggaggacat ccagaaggcc 2880
caggtgagcg gccagggcga cagcctgcac gagcacatcg ccaacctggc cggcagcccg 2940
gccatcaaga agggcatcct gcagaccgtg aaggtggtgg acgagctggt gaaggtgatg 3000
ggcaggcaca agccggagaa catcgtgatc gagatggcca gggagaacca gaccacccag 3060
aagggccaga agaacagcag ggagaggatg aagaggatcg aggagggcat caaggagctg 3120
ggcagccaga tcctgaagga gcacccggtg gagaacaccc agctgcagaa cgagaagctg 3180
tacctgtact acctgcagaa cggcagggac atgtacgtgg accaggagct ggacatcaac 3240
aggctgagcg actacgacgt ggaccacatc gtgccgcaga gcttcctgaa ggacgacagc 3300
atcgacaaca aggtgctgac caggagcgac aagaacaggg gcaagagcga caacgtgccg 3360
agcgaggagg tggtgaagaa gatgaaaaac tactggaggc agctgctgaa cgccaagctg 3420
atcacccaga ggaagttcga caacctgacc aaggccgaga ggggcggcct gagcgagctg 3480
gacaaggccg gcttcattaa aaggcagctg gtggagacca ggcagatcac caagcacgtg 3540
gcccagatcc tggacagcag gatgaacacc aagtacgacg agaacgacaa gctgatcagg 3600
gaggtgaagg tgatcaccct gaagagcaag ctggtgagcg acttcaggaa ggacttccag 3660
ttctacaagg tgagggagat caataattac caccacgccc acgacgccta cctgaacgcc 3720
gtggtgggca ccgccctgat taaaaagtac ccgaagctgg agagcgagtt cgtgtacggc 3780
gactacaagg tgtacgacgt gaggaagatg atcgccaaga gcgagcagga gatcggcaag 3840
gccaccgcca agtacttctt ctacagcaac atcatgaact tcttcaagac cgagatcacc 3900
ctggccaacg gcgagatcag gaagaggccg ctgatcgaga ccaacggcga gaccggcgag 3960
atcgtgtggg acaagggcag ggacttcgcc accgtgagga aggtgctgtc catgccgcag 4020
gtgaacatcg tgaagaagac cgaggtgcag accggcggct tcagcaagga gagcatcctg 4080
ccgaagagga acagcgacaa gctgatcgcc aggaagaagg actgggatcc gaagaagtac 4140
ggcggcttcg acagcccgac cgtggcctac agcgtgctgg tggtggccaa ggtggagaag 4200
ggcaagagca agaagctgaa gagcgtgaag gagctggtgg gcatcaccat catggagagg 4260
agcagcttcg agaagaaccc agtggacttc ctggaggcca agggctacaa ggaggtgaag 4320
aaggacctga tcattaaact gccgaagtac agcctgttcg agctggagaa cggcaggaag 4380
aggatgctgg ccagcgccgg cgagctgcag aagggcaacg agctggccct gccgagcaag 4440
tacgtgaact tcctgtacct ggccagccac tacgagaagc tgaagggcag cccggaggac 4500
aacgagcaga agcagctgtt cgtggagcag cacaagcact acctggacga gatcatcgag 4560
cagatcagcg agttcagcaa gagggtgatc ctggccgacg ccaacctgga caaggtgctg 4620
agcgcctaca acaagcacag ggacaagccg atcagggagc aggccgagaa catcatccac 4680
ctgttcaccc tgaccaacct gggcgccccg gccgccttca agtacttcga caccaccatc 4740
gacaggaaga ggtacaccag caccaaggag gtgctggacg ccaccctgat ccaccagagc 4800
atcaccggcc tgtacgagac caggatcgac ctgagccagc tgggcggcga cagcagcccg 4860
ccgaagaaga agaggaaggt gagctggaag gacgccagcg gctggagcag gatgtgaagc 4920
ttgatcgttc aaacatttgg caataaagtt tcttaagatt gaatcctgtt gccggtcttg 4980
cgatgattat catataattt ctgttgaatt acgttaagca tgtaataatt aacatgtaat 5040
gcatgacgtt atttatgaga tgggttttta tgattagagt cccgcaatta tacatttaat 5100
acgcgataga aaacaaaata tagcgcgcaa actaggataa attatcgcgc gcggtgtcat 5160
ctatgttact agatcgggac cggaattcgg gaccctttgt gaaagttgaa ttacggcata 5220
gccgaaggaa taacagaatc gtttcacact ttcgtaacaa aggtcttctt atcatgtttc 5280
agacgatgga ggcaaggctg atcaaagtga tcaagcacat aaacgcattt ttttaccatg 5340
tttcactcca taagcgtctg agattatcac aagtcacgtc tagtagtttg atggtacact 5400
agtgacaatc agttcgtgca gacagagctc atacttgact acttgagcga ttacaggcga 5460
aagtgtgaaa cgcatgtgat gtgggctggg aggaggagaa tatatactaa tgggccgtat 5520
cctgatttgg gctgcgtcgg aaggtgcagc ccacgcgcgc cgtaccgcgc gggtggcgct 5580
gctacccact ttagtccgtt ggatggggat ccgatggttt gcgcggtggc gttgcggggg 5640
atgtttagta ccacatcgga aaccgaaaga cgatggaacc agcttataaa cccgcgcgct 5700
gtagtcagct tgagaccggc aggtacgtcg gttttagagc tagaaatagc aagttaaaat 5760
aaggctagtc cgttatcaac ttgaaaaagt ggcaccgagt cggtgctttt tttttcctag 5820
gctgcagtgc agcgtgaccc ggtcgtgccc ctctctagag ataatgagca ttgcatgtct 5880
aagttataaa aaattaccac atattttttt tgtcacactt gtttgaagtg cagtttatct 5940
atctttatac atatatttaa actttactct acgaataata taatctatag tactacaata 6000
atatcagtgt tttagagaat catataaatg aacagttaga catggtctaa aggacaattg 6060
agtattttga caacaggact ctacagtttt atctttttag tgtgcatgtg ttctcctttt 6120
tttttgcaaa tagcttcacc tatataatac ttcatccatt ttattagtac atccatttag 6180
ggtttagggt taatggtttt tatagactaa tttttttagt acatctattt tattctattt 6240
tagcctctaa attaagaaaa ctaaaactct attttagttt ttttatttaa taatttagat 6300
ataaaataga ataaaataaa gtgactaaaa attaaacaaa taccctttaa gaaattaaaa 6360
aaactaagga aacatttttc ttgtttcgag tagataatgc cagcctgtta aacgccgtcg 6420
acgagtctaa cggacaccaa ccagcgaacc agcagcgtcg cgtcgggcca agcgaagcag 6480
acggcacggc atctctgtcg ctgcctctgg acccctctcg agagttccgc tccaccgttg 6540
gacttgctcc gctgtcggca tccagaaatt gcgtggcgga gcggcagacg tgagccggca 6600
cggcaggcgg cctcctcctc ctctcacggc accggcagct acgggggatt cctttcccac 6660
cgctccttcg ctttcccttc ctcgcccgcc gtaataaata gacaccccct ccacaccctc 6720
tttccccaac ctcgtgttgt tcggagcgca cacacacaca accagatctc ccccaaatcc 6780
acccgtcggc acctccgctt caaggtacgc cgctcgtcct cccccccccc ccctctctac 6840
cttctctaga tcggcgttcc ggtccatggt tagggcccgg tagttctact tctgttcatg 6900
tttgtgttag atccgtgttt gtgttagatc cgtgctgcta gcgttcgtac acggatgcga 6960
cctgtacgtc agacacgttc tgattgctaa cttgccagtg tttctctttg gggaatcctg 7020
ggatggctct agccgttccg cagacgggat cgatttcatg attttttttg tttcgttgca 7080
tagggtttgg tttgcccttt tcctttattt caatatatgc cgtgcacttg tttgtcgggt 7140
catcttttca tgcttttttt tgtcttggtt gtgatgatgt ggtctggttg ggcggtcgtt 7200
ctagatcgga gtagaattct gtttcaaact acctggtgga tttattaatt ttggatctgt 7260
atgtgtgtgc catacatatt catagttacg aattgaagat gatggatgga aatatcgatc 7320
taggataggt atacatgttg atgcgggttt tactgatgca tatacagaga tgctttttgt 7380
tcgcttggtt gtgatgatgt ggtgtggttg ggcggtcgtt cattcgttct agatcggagt 7440
agaatactgt ttcaaactac ctggtgtatt tattaatttt ggaactgtat gtgtgtgtca 7500
tacatcttca tagttacgag tttaagatgg atggaaatat cgatctagga taggtataca 7560
tgttgatgtg ggttttactg atgcatatac atgatggcat atgcagcatc tattcatatg 7620
ctctaacctt gagtacctat ctattataat aaacaagtat gttttataat tattttgatc 7680
ttgatatact tggatgatgg catatgcagc agctatatgt ggattttttt agccctgcct 7740
tcatacgcta tttatttgct tggtactgtt tcttttgtcg atgctcaccc tgttgtttgg 7800
tgttacttct gcagtgacta aatagatgca gaagctgatc aacagcgtgc agaactacgc 7860
ctggggcagc aagaccgccc tgaccgagct gtacggcatg gagaacccca gcagccagcc 7920
catggccgag ctgtggatgg gcgcccaccc caagagctca agccgcgtgc agaacgccgc 7980
cggcgatatc gttagcctgc gcgacgtgat cgagagcgac aagagcaccc tgctgggcga 8040
ggccgtggcc aagcgcttcg gcgagctgcc cttcctgttc aaggtgctgt gcgccgctca 8100
gcccctgagc atccaggtgc accctaacaa gcacaacagc gagatcggct tcgccaagga 8160
gaacgccgcc ggcatcccca tggacgccgc cgagcgcaac tacaaggacc ccaaccacaa 8220
gcccgagctg gtgttcgccc tgaccccctt cctggccatg aacgccttcc gcgagttcag 8280
cgagatcgtt agcctgctgc agcccgtggc cggcgcccac cccgctatcg cccacttcct 8340
tcagcagccc gacgccgagc gcctgagcga gctgttcgcc agcctgctga acatgcaggg 8400
tgaggagaag tcacgcgccc tggccatcct gaagagcgcc ctggacagcc agcagggcga 8460
gccctggcag acaatccgcc tgatcagcga gttctacccc gaggatagcg gcctgttcag 8520
ccccctgctg ctgaacgtgg tgaagctgaa ccccggcgag gccatgttcc tgttcgccga 8580
gaccccccac gcctacctgc agggcgtggc cctggaggtg atggccaaca gcgacaacgt 8640
gctgcgcgcc ggcctgaccc ccaagtacat cgacatcccc gagctggtgg ccaacgtgaa 8700
gttcgaggct aagcccgcca accagctgct gacccagccc gtgaagcagg gcgccgagct 8760
ggacttccct atccccgttg acgacttcgc cttcagcctg cacgacctga gcgacaagga 8820
gaccactatc agccagcaga gcgccgcgat cctgttctgc gtggagggcg acgccaccct 8880
gtggaagggc agccagcagc tgcagctgaa gcccggcgag agcgccttta tcgccgccaa 8940
cgagagcccc gtgaccgtga agggccacgg ccgcctggcc cgcgtgtaca acaagctgtg 9000
atagctacgt catgggtcgt ttaagctgcc gatgtgcctg cgtcgtctgg tgccctctct 9060
ccatatggag gttgtcaaag tatctgctgt tcgtgtcatg agtcgtgtca gtgttggttt 9120
aataatggac cggttgtgtt gtgtgtgcgt actacccaga actatgacaa atcatgaata 9180
agtttgatgt ttgaaattaa agcctgtgct cattatgttc tgtctttcag ttgtctccta 9240
atatttgcct gcaggtactg gctatctacc gtttcttact taggaggtgt ttgaatgcac 9300
taaaactaat agttagtggc taaaattagt taaaacatcc aaacaccata gctaatagtt 9360
gaactattag ctatttttgg aaaattagtt aatagtgagg tagttatttg ttagctagct 9420
aattcaacta acaattttta gccaactaac aattagtttc agtgcattca aacaccccct 9480
taatgttaac gtggttctat ctaccgtctc ctaatatatg gttgattgtt cggtttgttg 9540
ctatgctatt gggttctgat tgctgctagt tcttgctgaa tccagaagtt ctcgtagtat 9600
agctcagatt catattattt atttgagtga taagtgatcc aggttattac tatgttagct 9660
aggttttttt tacaaggata aattatctgt gatcataatt cttatgaaag ctttatgttt 9720
cctggaggca gtggcatgca atgcatgaca gcaacttgat cacaccagct gaggtagata 9780
cggtaacaag gttcttaaat ctgttcacca aatcattgga gaacacacat acacattctt 9840
gccagtcttg gttagagaaa tttcatgaca aaatgccaaa gctgtcttga ctcttcactt 9900
ttggccatga gtcgtgactt agtttggttt aatggaccgg ttctcctagc ttgttctact 9960
caaaactgtt gttgatgcga ataagttgtg atggttgatc tctggatttt gttttgctct 10020
caatagtgga cgagattaga tagcggaccg cctgcaggcc cgggggcgcg ccctaattag 10080
ctaacggcca ggatcgccgc gtgagccttt agcaactagc tagattaatt aacgcaatct 10140
gttattaagt tgtctaagcg tcaatttgtt tacaccacaa tatatcctgc caccagccag 10200
ccaacagctc cccgaccggc agctcggcac aaaatcacca ctcgatacag gcagcccatc 10260
ag 10262
<210> 42
<211> 1366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> TALEN-индуцированная мутация в аллеле 2 у объекта 38А с ID 22808-4108
<400> 42
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaggacgaa ccggtgactg gcgaaggaag caatgccgat gccctcggtg ggctcgctag 1200
gcagctctcc gaggagagga gaacaaggct cgcgcgccgc gtctctgcca tcaacccaag 1260
aggctctaga tgtgcgtcgt acgatatcta agacaagtgg ctttactgtc agtcacatgc 1320
ttgtaaataa gtagacttta ttttaataaa acataaaaat atatat 1366
<210> 43
<211> 1366
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация MTL в аллеле 2 у объекта 27А с
ID 22807-4073
<400> 43
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggagaa ccggtgactg gcgaaggaag caatgccgat gccctcggtg ggctcgctag 1200
gcagctctcc gaggagagga gaacaaggct cgcgcgccgc gtctctgcca tcaacccaag 1260
aggctctaga tgtgcgtcgt acgatatcta agacaagtgg ctttactgtc agtcacatgc 1320
ttgtaaataa gtagacttta ttttaataaa acataaaaat atatat 1366
<210> 44
<211> 1363
<212> ДНК
<213> Искусственная последовательность
<220>
<223> CRISPR-индуцированная мутация MTL в аллеле 2 у объекта 27А
с ID 22807-4081
<400> 44
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acaggaaccg gtgactggcg aaggaagcaa tgccgatgcc ctcggtgggc tcgctaggca 1200
gctctccgag gagaggagaa caaggctcgc gcgccgcgtc tctgccatca acccaagagg 1260
ctctagatgt gcgtcgtacg atatctaaga caagtggctt tactgtcagt cacatgcttg 1320
taaataagta gactttattt taataaaaca taaaaatata tat 1363
<210> 45
<211> 1371
<212> ДНК
<213> Zea mays
<400> 45
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctata gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 46
<211> 1371
<212> ДНК
<213> Zea mays
<400> 46
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagctcgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 47
<211> 1371
<212> ДНК
<213> Zea mays
<400> 47
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc tgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 48
<211> 1371
<212> ДНК
<213> Zea mays
<400> 48
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttacg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 49
<211> 1371
<212> ДНК
<213> Zea mays
<400> 49
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatacgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 50
<211> 1371
<212> ДНК
<213> Zea mays
<400> 50
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gattggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggagaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 51
<211> 1371
<212> ДНК
<213> Zea mays
<400> 51
agttcatcac taatcacact tattgttccc tcgacgagta tctagctagc tcattaatcg 60
atcaatcggg gtgtgcggtc gaaggcggca atggcgagct actcgtcgcg gcgtccatgc 120
aatacctgta gcacgaaggc gatggccggg agcgtggtcg gcgagcccgt cgtgctgggg 180
cagagggtga cggtgctgac ggtggacggc ggcggcgtcc ggggtctcat cccgggaacc 240
atcctcgcct tcctcgaggc caggctgcag gagctggacg gaccggaggc gaggctggcg 300
gactacttcg actacatcgc cggaaccagc accggcggtc tcatcaccgc catgctcacc 360
gcgcccggca aggacaagcg gcctctctac gctgccaagg acatcaacca cttttacatg 420
gagaactgcc cgcgcatctt ccctcagaag agcaggcttg cggccgccat gtccgcgctg 480
aggaagccaa agtacaacgg caagtgcatg cgcagcctga ttaggagcat cctcggcgag 540
acgagggcca agagcacgcc tctgaagaac gctctgctct cggacgtgtg cattggcacg 600
tccgccgcgc cgacctacct cccggcgcac tacttccaga ctgaagacgc caacggcaag 660
gagcgcgaat acaacctcat cgacggcggt gtggcggcca acaacccgac gatggttgcg 720
atgacgcaga tcaccaaaaa gatgcttgcc agcaaggaca aggccgagga gctgtaccca 780
gtgaagccgt cgaactgccg caggttcctg gtgctgtcca tcgggacggg gtcgacgtcc 840
gagcagggcc tctacacggc gcggcagtgc tcccggtggg gtatctgccg gtggctccgc 900
aacaacggca tggcccccat catcgacatc ttcatggcgg ccagctcgga cctggtggac 960
atccacgtcg ccgcgatgtt ccagtcgctc cacagcgacg gcgactacct gcgcatccag 1020
gacaactcgc tccgtggcgc cgcggccacc gtggacgcgg cgacgccgga gaacatgcgg 1080
acgctcgtcg ggatcgggga gcggatgctg gcacagaggg tgtccagggt caacgtggag 1140
acagggaggt acgaaccggt gactggcgaa ggaagcaatg ccgatgccct cggtgggctc 1200
gctaggcagc tctccgagga gaggaaaaca aggctcgcgc gccgcgtgtc tgccatcaac 1260
ccaagaggct ctagatgtgc gtcgtacgat atctaagaca agtggcttta ctgtcagtca 1320
catgcttgta aataagtaga ctttatttta ataaaacata aaaatatata t 1371
<210> 52
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 52
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Tyr Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 53
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 53
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Leu Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 54
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 54
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Leu Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 55
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 55
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Thr Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 56
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 56
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Ile Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 57
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 57
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Ile Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 58
<211> 401
<212> БЕЛОК
<213> Zea mays
<400> 58
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Glu Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Ala Lys Ser Thr Pro Leu Lys Asn
145 150 155 160
Ala Leu Leu Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr
165 170 175
Leu Pro Ala His Tyr Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg
180 185 190
Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met
195 200 205
Val Ala Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys
210 215 220
Ala Glu Glu Leu Tyr Pro Val Lys Pro Ser Asn Cys Arg Arg Phe Leu
225 230 235 240
Val Leu Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr
245 250 255
Ala Arg Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn
260 265 270
Gly Met Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu
275 280 285
Val Asp Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly
290 295 300
Asp Tyr Leu Arg Ile Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr
305 310 315 320
Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly
325 330 335
Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly
340 345 350
Arg Tyr Glu Pro Val Thr Gly Glu Gly Ser Asn Ala Asp Ala Leu Gly
355 360 365
Gly Leu Ala Arg Gln Leu Ser Glu Glu Arg Lys Thr Arg Leu Ala Arg
370 375 380
Arg Val Ser Ala Ile Asn Pro Arg Gly Ser Arg Cys Ala Ser Tyr Asp
385 390 395 400
Ile
<210> 59
<211> 379
<212> БЕЛОК
<213> Zea mays
<400> 59
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Asn Arg
370 375
<210> 60
<211> 396
<212> БЕЛОК
<213> Zea mays
<400> 60
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Thr Gly Asp Trp
370 375 380
Arg Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 61
<211> 379
<212> БЕЛОК
<213> Zea mays
<400> 61
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Asn Arg
370 375
<210> 62
<211> 394
<212> БЕЛОК
<213> Zea mays
<400> 62
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Thr Gly Asp Trp Arg Arg
370 375 380
Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390
<210> 63
<211> 395
<212> БЕЛОК
<213> Zea mays
<400> 63
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Thr Gly Asp Trp Arg
370 375 380
Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 64
<211> 393
<212> БЕЛОК
<213> Zea mays
<400> 64
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Thr Gly Asp Trp Arg Arg
370 375 380
Lys Gln Cys Arg Cys Pro Arg Trp Ala
385 390
<210> 65
<211> 395
<212> БЕЛОК
<213> Zea mays
<400> 65
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Thr Gly Asp Trp Arg
370 375 380
Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 66
<211> 398
<212> БЕЛОК
<213> Zea mays
<400> 66
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Arg Glu Val Arg Thr Gly
370 375 380
Asp Trp Arg Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 67
<211> 398
<212> БЕЛОК
<213> Zea mays
<400> 67
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Arg Glu Val Arg Thr Gly
370 375 380
Asp Trp Arg Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 68
<211> 384
<212> БЕЛОК
<213> Zea mays
<400> 68
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Lys Gly Gly Thr Asn Arg
370 375 380
<210> 69
<211> 383
<212> БЕЛОК
<213> Zea mays
<400> 69
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Gly Thr Asn Arg
370 375 380
<210> 70
<211> 396
<212> БЕЛОК
<213> Zea mays
<400> 70
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Thr Gly Asp Trp
370 375 380
Arg Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 71
<211> 396
<212> БЕЛОК
<213> Zea mays
<400> 71
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Thr Gly Asp Trp
370 375 380
Arg Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 72
<211> 395
<212> БЕЛОК
<213> Zea mays
<400> 72
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Thr Cys Ser Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Val Asp Gly Gly Gly Val Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp Lys
85 90 95
Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn His Phe Tyr Met Gln Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Lys Ser Arg Leu Ala Ala Ala Met Ser
115 120 125
Ala Leu Arg Lys Pro Lys Tyr Asn Gly Lys Cys Met Arg Ser Leu Ile
130 135 140
Arg Ser Ile Leu Gly Glu Thr Arg Val Ser Glu Thr Leu Thr Asn Val
145 150 155 160
Ile Ile Pro Ala Phe Asp Ile Arg Leu Leu Gln Pro Ile Ile Phe Ser
165 170 175
Thr Tyr Asp Ala Lys Ser Thr Pro Leu Lys Asn Ala Leu Leu Ser Asp
180 185 190
Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His Tyr
195 200 205
Phe Gln Thr Glu Asp Ala Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile
210 215 220
Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln
225 230 235 240
Ile Thr Lys Lys Met Leu Ala Ser Lys Asp Lys Ala Glu Glu Leu Tyr
245 250 255
Pro Val Asn Pro Ser Asn Cys Arg Arg Phe Leu Val Leu Ser Ile Gly
260 265 270
Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg Gln Cys Ser
275 280 285
Arg Trp Gly Ile Cys Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile
290 295 300
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Val
305 310 315 320
Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Ala Ala Thr
340 345 350
Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Thr Gly Asp Trp Arg
370 375 380
Arg Lys Gln Cys Arg Cys Pro Arg Trp Ala Arg
385 390 395
<210> 73
<211> 437
<212> БЕЛОК
<213> Sorghum bicolor
<400> 73
Met Ala Thr Tyr Tyr Ser Ser Arg Arg Pro Cys Asn Ala Cys Ser Thr
1 5 10 15
Lys Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Leu Gly Gln
20 25 30
Arg Val Thr Val Leu Thr Val Asp Gly Gly Gly Ile Arg Gly Leu Ile
35 40 45
Pro Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp
50 55 60
Gly Pro Glu Val Arg Leu Ala Asp Tyr Phe Asp Tyr Ile Ala Gly Thr
65 70 75 80
Ser Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Lys Asp
85 90 95
Arg Arg Pro Leu Tyr Ala Ala Lys Asp Ile Asn Gln Phe Tyr Met Glu
100 105 110
Asn Cys Pro Arg Ile Phe Pro Gln Lys Ser Ser Arg Leu Ala Ala Ala
115 120 125
Met Ser Ala Leu Arg Lys Pro Arg Tyr Asn Gly Lys Cys Leu Arg Asn
130 135 140
Leu Ile Met Ser Met Leu Gly Glu Thr Arg Val Ser Asp Thr Leu Thr
145 150 155 160
Asn Val Ile Ile Pro Thr Phe Asp Val Arg Leu Leu Gln Pro Ile Ile
165 170 175
Phe Ser Thr Tyr Asp Ala Lys Ser Met Pro Leu Lys Asn Ala Leu Leu
180 185 190
Ser Asp Val Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala
195 200 205
His Tyr Phe Gln Thr Lys Asp Ala Gly Ser Gly Lys Glu Arg Glu Tyr
210 215 220
Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala
225 230 235 240
Met Thr Gln Ile Thr Lys Lys Met Leu Ala Ser Lys Glu Lys Ala Glu
245 250 255
Glu Leu Tyr Pro Val Lys Pro Trp Asn Cys Arg Lys Phe Leu Val Leu
260 265 270
Ser Ile Gly Thr Gly Ser Thr Ser Glu Gln Gly Leu Tyr Thr Ala Arg
275 280 285
Gln Cys Ser Arg Trp Gly Ile Cys Arg Trp Ile Arg Asn Asn Gly Met
290 295 300
Ala Pro Ile Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp
305 310 315 320
Ile His Val Ala Ala Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr
325 330 335
Leu Arg Ile Gln Asp Asn Ser Leu His Gly Ala Ala Ala Thr Val Asp
340 345 350
Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val Gly Ile Gly Glu Arg
355 360 365
Met Leu Ala Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Tyr
370 375 380
Glu Pro Val Pro Gly Glu Gly Ser Asn Ala Asp Ala Leu Ala Gly Ile
385 390 395 400
Ala Arg Gln Leu Ser Glu Glu Arg Arg Thr Arg Leu Ala Arg Arg Thr
405 410 415
Ser Ala Ile Val Ser Ser Gly Gly Ala Ser Arg Arg Thr Cys Ala Ser
420 425 430
Lys Val Ser Asn Val
435
<210> 74
<211> 421
<212> БЕЛОК
<213> Setaria italica
<400> 74
Met Ala Ser Tyr Ser Ser Arg Arg Pro Cys Asn Ala Cys Arg Thr Lys
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Glu Pro Val Val Pro Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Ile Asp Gly Gly Gly Ile Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Glu Ala Arg Leu Ala Asp Tyr Phe Asp Cys Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Ile Thr Thr Pro Gly Glu Asp Lys
85 90 95
Arg Pro Leu Phe Ala Ala Arg Asp Ile Asn Arg Phe Tyr Phe Asp Asn
100 105 110
Cys Pro Arg Ile Phe Pro Gln Ser Arg Ser Ser Leu Ala Ala Ala Met
115 120 125
Ser Ala Leu Arg Lys Pro Arg Tyr Asn Gly Lys Tyr Leu Arg Ser Thr
130 135 140
Ile Arg Ser Met Leu Gly Glu Thr Arg Val Ser Asp Ala Leu Thr Asn
145 150 155 160
Val Val Ile Pro Thr Phe Asp Ile Lys Leu Ile Gln Pro Ile Ile Phe
165 170 175
Ser Thr Tyr Asp Val Lys Asn Met Pro Leu Lys Asn Ala Leu Leu Ser
180 185 190
Asp Val Cys Ile Ser Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His
195 200 205
Tyr Phe Gln Ile Gln Asp Ala Gly Gly Lys Thr Arg Glu Tyr Asn Leu
210 215 220
Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr
225 230 235 240
Gln Ile Thr Lys Met Met Leu Ala Lys Asp Lys Glu Glu Leu Tyr Pro
245 250 255
Val Lys Pro Glu Asp Cys Arg Lys Phe Leu Val Leu Ser Ile Gly Thr
260 265 270
Gly Ser Thr Ser Asp Glu Gly Leu Phe Thr Ala Arg Gln Cys Ser Arg
275 280 285
Trp Gly Val Val Arg Trp Leu Arg Asn Asn Gly Met Ala Pro Ile Ile
290 295 300
Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Ala Ala
305 310 315 320
Val Leu Phe Gln Ser Leu His Ser Asp Gly His Ser Leu Arg Gly Ala
325 330 335
Ala Ala Thr Val Asp Ala Ala Thr Pro Glu Asn Met Arg Thr Leu Val
340 345 350
Gly Ile Gly Glu Arg Met Leu Ala Gln Arg Val Ser Arg Val Asn Val
355 360 365
Glu Thr Gly Arg Tyr Glu Pro Val Pro Gly Glu Gly Ser Asn Ala Asp
370 375 380
Ala Leu Val Ala Leu Ala Arg Gln Leu Ser Asp Glu Arg Arg Ala Arg
385 390 395 400
Ile Ala Arg Arg Ala Ala Ala Ala Cys Ala Gly Gly Ser Arg Cys Cys
405 410 415
Ser Pro Val Lys Thr
420
<210> 75
<211> 422
<212> БЕЛОК
<213> Hordeum vulgare
<400> 75
Met Ala Ser Tyr Trp Cys Arg Arg Pro Cys Glu Ser Cys Ser Thr Arg
1 5 10 15
Ala Met Ala Gly Ser Val Val Gly Gln Pro Val Ala Pro Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Ile Asp Gly Gly Gly Ile Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Asp Ala Arg Leu Ala Asp Tyr Phe Asp Cys Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Leu Thr Ala Pro Gly Gln Asp Gly
85 90 95
Arg Pro Leu Phe Ala Ala Lys Asp Val Asn Arg Phe Tyr Leu Asp Asn
100 105 110
Gly Pro Tyr Ile Phe Pro Gln Arg Arg Cys Ala Leu Ala Ala Val Thr
115 120 125
Ala Ser Leu Arg Arg Pro Arg Tyr Ser Gly Lys Tyr Leu His Gly Lys
130 135 140
Ile Arg Ser Met Leu Gly Glu Thr Arg Leu Cys Asp Ala Leu Thr Asp
145 150 155 160
Val Val Ile Pro Thr Phe Asp Val Lys Leu Leu Gln Pro Ile Ile Phe
165 170 175
Ser Thr Tyr Asp Ala Arg Asn Met Pro Leu Lys Asn Ala Arg Leu Ala
180 185 190
Asp Ile Cys Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His
195 200 205
His Phe His Thr Gln Asp Asp Asn Gly Lys Glu Arg Glu Tyr Asn Leu
210 215 220
Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Thr Met Thr
225 230 235 240
Gln Ile Thr Lys Lys Met Met Val Lys Asp Arg Glu Glu Leu Tyr Pro
245 250 255
Val Lys Pro Ser Asp Cys Gly Lys Phe Leu Val Leu Ser Ile Gly Thr
260 265 270
Gly Ser Thr Ser Asp Gln Gly Leu Tyr Thr Ala Lys Gln Cys Ser Gln
275 280 285
Trp Gly Ile Ile Arg Trp Leu Arg Asn Lys Gly Met Ala Pro Ile Ile
290 295 300
Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Ala Ala
305 310 315 320
Val Leu Phe Gln Ser Leu His Ser Asp Gly Asn Tyr Leu Arg Ile Gln
325 330 335
Asp Asn Ser Leu His Gly Pro Ala Ala Thr Val Asp Ala Ala Thr Pro
340 345 350
Glu Asn Met Ala Glu Leu Leu Arg Ile Gly Glu Arg Met Leu Ala Gln
355 360 365
Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Tyr Glu Glu Ile Arg
370 375 380
Gly Ala Gly Ser Asn Ala Asp Ala Leu Ala Gly Phe Ala Lys Gln Leu
385 390 395 400
Ser Asp Glu Arg Arg Thr Arg Leu Gly Arg Arg Arg Val Gly Ala Gly
405 410 415
Arg Leu Lys Ser Arg Arg
420
<210> 76
<211> 423
<212> БЕЛОК
<213> Brachypodium distachyon
<400> 76
Met Ala Ser Tyr Ala Cys Arg Arg Pro Cys Glu Ser Cys Arg Thr Arg
1 5 10 15
Ala Met Ala Gly Gly Val Val Gly Glu Pro Thr Thr Pro Gly Gln Arg
20 25 30
Val Thr Val Leu Thr Ile Asp Gly Gly Gly Ile Arg Gly Leu Ile Pro
35 40 45
Gly Thr Ile Leu Ala Phe Leu Glu Asp Arg Leu Gln Glu Leu Asp Gly
50 55 60
Pro Asp Ala Arg Leu Ala Asp Tyr Phe Asp Cys Ile Ala Gly Thr Ser
65 70 75 80
Thr Gly Gly Leu Ile Thr Ala Met Ile Thr Ala Pro Gly Glu Glu Gly
85 90 95
Arg Pro Leu Phe Ala Ala Glu Asp Ile Asn Arg Phe Tyr Leu Asp Asn
100 105 110
Gly Pro Gln Ile Phe Pro Gln Lys Arg Ser Ser Leu Met Ser Val Leu
115 120 125
Ala Ser Leu Thr Arg Pro Arg Tyr Asn Gly Lys Phe Leu His Gly Lys
130 135 140
Ile Arg Ser Met Leu Gly Glu Thr Arg Val Cys Asp Thr Leu Thr Asp
145 150 155 160
Val Val Ile Pro Thr Phe Asp Val Arg Leu Leu Gln Pro Ile Ile Phe
165 170 175
Ser Thr Tyr Asp Ala Lys Ser Met Pro Leu Lys Asn Ala Leu Leu Ser
180 185 190
Asp Val Cys Ile Ser Thr Ser Ala Ala Pro Thr Phe Leu Pro Ala His
195 200 205
Tyr Phe Gln Thr Glu Asp Asp Asn Gly Lys Val Arg Glu Tyr Asn Leu
210 215 220
Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr
225 230 235 240
Gln Ile Thr Lys Lys Ile Met Ala Lys Asp Lys Glu Glu Leu Tyr Pro
245 250 255
Val Lys Pro Ser Asp Cys Gly Lys Phe Leu Val Leu Ser Ile Gly Thr
260 265 270
Gly Ser Thr Ser Asp Gln Gly Leu Tyr Thr Ala Lys Gln Cys Ser Arg
275 280 285
Trp Gly Ile Ile Arg Trp Leu Arg Asn Lys Gly Met Ala Pro Ile Ile
290 295 300
Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Ala Ala
305 310 315 320
Val Leu Phe Gln Ser Leu His Ser Asp Gly Asp Cys Tyr Leu Arg Ile
325 330 335
Gln Asp Asn Ser Leu Arg Gly Ala Ala Ala Thr Val Asp Thr Ala Thr
340 345 350
Pro Asp Asn Met Arg Glu Leu Val Arg Ile Gly Glu Arg Met Leu Ala
355 360 365
Gln Arg Val Ser Lys Val Asn Val Glu Thr Gly Arg Tyr Glu Glu Met
370 375 380
Gln Gly Ala Gly Thr Asn Ala Asp Ala Leu Ala Gly Phe Ala Arg Gln
385 390 395 400
Leu Ser Asp Glu Arg Arg Ala Arg Phe Gly Pro Arg Asp Gly Ala Pro
405 410 415
Ala Asn Gly Lys Ser Arg Cys
420
<210> 77
<211> 414
<212> БЕЛОК
<213> Oryza sativa
<400> 77
Met Ala Gly Cys Val Val Gly Glu Pro Ala Ser Ala Pro Gly Gln Arg
1 5 10 15
Val Thr Leu Leu Ala Ile Asp Gly Gly Gly Ile Arg Gly Leu Ile Pro
20 25 30
Gly Thr Ile Leu Ala Phe Leu Glu Ala Arg Leu Gln Glu Leu Asp Gly
35 40 45
Pro Asp Ala Arg Leu Ala Asp Tyr Phe Asp Cys Ile Ala Gly Thr Ser
50 55 60
Thr Gly Gly Leu Ile Thr Ala Met Leu Ala Ala Pro Gly Asp His Gly
65 70 75 80
Arg Pro Leu Phe Ala Ala Ser Asp Ile Asn Arg Phe Tyr Leu Asp Asn
85 90 95
Gly Pro Arg Ile Phe Pro Gln Lys Arg Cys Gly Met Ala Ala Ala Met
100 105 110
Ala Ala Leu Thr Arg Pro Arg Tyr Asn Gly Lys Tyr Leu Gln Gly Lys
115 120 125
Ile Arg Lys Met Leu Gly Glu Thr Arg Val Arg Asp Thr Leu Thr Asn
130 135 140
Val Val Ile Pro Thr Phe Asp Val Arg Leu Leu Gln Pro Thr Ile Phe
145 150 155 160
Ser Thr Tyr Asp Ala Lys Ser Met Pro Leu Lys Asn Ala Leu Leu Ser
165 170 175
Asp Ile Cys Ile Ser Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His
180 185 190
Cys Phe Gln Thr Thr Asp Asp Ala Thr Gly Lys Val Arg Glu Phe Asp
195 200 205
Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met
210 215 220
Thr Gln Ile Thr Lys Lys Ile Met Val Lys Asp Lys Glu Glu Leu Tyr
225 230 235 240
Pro Val Lys Pro Ser Asp Cys Gly Lys Phe Leu Val Leu Ser Leu Gly
245 250 255
Thr Gly Ser Thr Ser Asp Gln Gly Met Tyr Thr Ala Arg Gln Cys Ser
260 265 270
Arg Trp Gly Ile Val Arg Trp Leu Arg Asn Lys Gly Met Ala Pro Ile
275 280 285
Ile Asp Ile Phe Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Ala
290 295 300
Ala Val Met Phe Gln Ser Leu His Ser Asp Gly Asp Tyr Leu Arg Ile
305 310 315 320
Gln Asp Asn Thr Leu His Gly Asp Ala Ala Thr Val Asp Ala Ala Thr
325 330 335
Arg Asp Asn Met Arg Ala Leu Val Gly Ile Gly Glu Arg Met Leu Ala
340 345 350
Gln Arg Val Ser Arg Val Asn Val Glu Thr Gly Arg Tyr Val Glu Val
355 360 365
Pro Gly Ala Gly Ser Asn Ala Asp Ala Leu Arg Gly Phe Ala Arg Gln
370 375 380
Leu Ser Glu Glu Arg Arg Ala Arg Leu Gly Arg Arg Asn Ala Cys Gly
385 390 395 400
Gly Gly Gly Glu Gly Glu Pro Ser Gly Val Ala Cys Lys Arg
405 410
<210> 78
<211> 275
<212> БЕЛОК
<213> Triticum aestivum
<400> 78
Met Leu Gly Glu Thr Arg Leu Ser Asp Ala Leu Thr Asp Val Val Ile
1 5 10 15
Pro Thr Phe Asp Val Lys Leu Leu Gln Pro Ile Ile Phe Ser Thr Tyr
20 25 30
Asp Ala Lys Ser Met Pro Leu Lys Asn Ala Arg Leu Ala Asp Val Cys
35 40 45
Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Ala His His Phe His
50 55 60
Thr His Asp Gly Asn Gly Lys Glu Arg Glu Tyr Asn Leu Ile Asp Gly
65 70 75 80
Gly Val Ala Ala Asn Asn Pro Thr Met Val Ala Met Thr Gln Ile Thr
85 90 95
Lys Lys Met Met Gly Lys Asp Arg Glu Glu Leu Tyr Pro Val Glu Pro
100 105 110
Ser Asp Cys Gly Lys Phe Leu Val Leu Ser Val Gly Thr Gly Ser Thr
115 120 125
Ser Asp Gln Gly Leu Tyr Thr Ala Lys Gln Cys Ser Gln Trp Gly Ile
130 135 140
Ile Ser Trp Leu Arg Asn Lys Gly Met Ala Pro Ile Ile Asp Ile Phe
145 150 155 160
Met Ala Ala Ser Ser Asp Leu Val Asp Ile His Ala Ala Val Leu Phe
165 170 175
Gln Ser Leu His Ser Asp Ala Asn Tyr Leu Arg Ile Gln Asp Asn Ser
180 185 190
Leu His Gly Pro Ala Ala Thr Val Asp Ala Ala Thr Pro Glu Asn Met
195 200 205
Ala Glu Leu Leu Arg Ile Gly Glu Arg Met Leu Ala Gln Arg Val Ser
210 215 220
Arg Val Asn Val Glu Thr Gly Arg Tyr Glu Glu Val Lys Gly Ala Gly
225 230 235 240
Asn Asn Ala Asp Ala Leu Ala Gly Phe Ala Arg Gln Leu Ser Asp Glu
245 250 255
Arg Arg Thr Arg Leu Gly Ser Arg Arg Gly Gly Ala Gly Arg Leu Lys
260 265 270
Ser Ser Arg
275
<210> 79
<211> 416
<212> БЕЛОК
<213> Musa acuminata
<400> 79
Met Ala Ser Asp Gln Thr Pro Pro Ala Thr Ala Asp Ala Pro Ile Ser
1 5 10 15
Thr Pro Pro Pro Ser Phe Gly Lys Arg Val Thr Val Leu Cys Ile Asp
20 25 30
Gly Gly Gly Val Arg Gly Leu Ile Pro Ala Thr Ile Ile Ala Phe Leu
35 40 45
Glu Ala Glu Leu Gln Lys Leu Asp Gly Pro Glu Ala Arg Ile Ala Asp
50 55 60
Tyr Phe Asp Val Ile Ala Gly Thr Ser Thr Gly Gly Leu Val Thr Ala
65 70 75 80
Met Leu Thr Ala Pro Asn Lys Glu Lys Arg Pro Leu Phe Ala Ala Lys
85 90 95
Asp Ile Val Gln Phe Tyr Leu Asp Asn Ser Pro Lys Ile Phe Pro Gln
100 105 110
Lys Asn Ala Gly Leu Phe Asn Ser Ala Leu Asn Leu Val Gly Ala Val
115 120 125
Ser Gly Pro Lys Tyr Asp Gly Lys Tyr Leu His Ala Ile Ile Arg Gln
130 135 140
Leu Leu Gly Asp Thr Arg Leu Ser Gln Thr Leu Thr Asn Val Val Ile
145 150 155 160
Pro Thr Phe Asp Ile Lys Phe Leu Gln Pro Thr Ile Phe Ser Thr Tyr
165 170 175
Gln Thr Lys Ser Thr Pro Leu Lys Asp Ala Leu Leu Ser Asp Ile Cys
180 185 190
Ile Gly Thr Ser Ala Ala Pro Thr Tyr Leu Pro Gly His Tyr Phe Glu
195 200 205
Thr Lys Asp Asp Asn Gly Asn Lys Arg Ser Phe Asn Leu Val Asp Gly
210 215 220
Gly Val Ala Ala Asn Asn Pro Thr Leu Thr Ala Met Thr Glu Val Ser
225 230 235 240
Lys Glu Ile Leu Leu Ser Asn Pro Asp Phe Phe Ser Tyr Gln Pro Val
245 250 255
Glu Tyr Asp Arg Phe Leu Val Ile Ser Leu Gly Thr Gly Ala Pro Lys
260 265 270
Gln Glu Glu Lys Phe Thr Ala Gln Glu Ser Ala Lys Trp Gly Val Leu
275 280 285
Gly Trp Leu Leu Asn Lys Gly Thr Thr Pro Leu Ile Asp Ile Phe Thr
290 295 300
Gln Ala Ser Ala Asp Met Val Asp Ile His Ala Ser Val Leu Phe Gln
305 310 315 320
Ala Leu Asn Lys Gly Lys Asn Tyr Leu Arg Ile Glu Asp Asp Thr Leu
325 330 335
Thr Gly Gln Thr Ser Ser Val Asp Val Ser Thr Lys Lys Asn Leu Gln
340 345 350
Asp Leu Val Asp Ile Gly Asn Ser Leu Leu Lys Lys Pro Val Ser Arg
355 360 365
Val Asn Ile Glu Thr Gly His Ser Glu Ala Val Asp Gly Glu Gly Thr
370 375 380
Asn Glu Ala Ala Leu Thr Gly Phe Ala Lys Lys Leu Ser Asp Glu Lys
385 390 395 400
Arg Arg Arg Gln Ser Lys Gln Leu Thr Ser Ser Asp Ala Thr Gln His
405 410 415
<210> 80
<211> 402
<212> БЕЛОК
<213> Elaeis guineensis
<400> 80
Met Gly Ser Asn Glu Ser Pro Val Ala Asn Pro Pro Pro Cys Lys Gly
1 5 10 15
Lys Val Val Thr Ile Leu Ser Ile Asp Gly Gly Gly Val Arg Gly Ile
20 25 30
Ile Pro Gly Thr Ile Leu Glu Phe Leu Glu Ala Lys Leu Gln Glu Leu
35 40 45
Asp Gly Pro Asp Ala Arg Ile Ala Asp Tyr Phe Asp Ile Ile Ala Gly
50 55 60
Thr Ser Thr Gly Gly Leu Val Thr Thr Met Leu Thr Ala Pro Asn Lys
65 70 75 80
Asp Asn Arg Pro Leu Phe Ser Ala Lys Asp Ile Ile Gln Phe Tyr Leu
85 90 95
Glu Asn Cys Pro Lys Ile Phe Pro Gln Arg Thr Gly Leu Leu Ala Gly
100 105 110
Ala Leu Asn Leu Phe Gly Ala Val Ser Gly Pro Lys Tyr Asp Gly Lys
115 120 125
Phe Leu His Ser Lys Val Lys Glu Leu Leu Gly Asp Thr Lys Leu His
130 135 140
Gln Thr Leu Thr Asn Ile Val Ile Pro Ala Phe Asp Ile Lys Leu Leu
145 150 155 160
Gln Pro Val Ile Phe Ser Thr Phe Glu Thr Lys Thr Asp Pro Ser Lys
165 170 175
Asp Ala Leu Leu Ser Asp Ile Cys Ile Ser Thr Ser Ala Ala Pro Thr
180 185 190
Tyr Leu Pro Ala His Tyr Phe Glu Thr Lys Asp Ser Gln Gly Lys Thr
195 200 205
Arg Ser Phe Asn Leu Val Asp Gly Gly Val Ala Ala Asn Asn Pro Met
210 215 220
Leu Ile Ala Thr Ser Gln Ile Thr Lys Gln Ile Phe Trp Asn Asn Glu
225 230 235 240
Asp Phe Ser Lys Phe Lys Pro Thr Asp Phe Ala Lys Phe Leu Val Val
245 250 255
Ser Leu Gly Thr Gly Ser Pro Lys Gln Glu Gln Lys Phe Thr Ala Pro
260 265 270
Glu Ser Ala Lys Trp Gly Leu Leu Gly Trp Leu Gln Asn Lys Gly Ser
275 280 285
Thr Pro Ile Ile Asp Ile Phe Ser Gln Ser Ser Ala Asp Met Val Asp
290 295 300
Ile His Ala Ser Ile Leu Phe Gln Ala Leu Arg Ser Glu Lys Asn Tyr
305 310 315 320
Leu Arg Ile Gln Asp Asp Thr Leu Glu Gly Asp Thr Ala Ser Val Asp
325 330 335
Val Ser Thr Ser Glu Asn Leu Arg Lys Leu Val Gln Val Gly Gln Asp
340 345 350
Leu Leu Lys Lys Pro Val Ser Arg Val Asn Leu Glu Thr Gly Val Ser
355 360 365
Glu Ala Cys Asp Val Glu Gly Thr Asn Glu Asp Ala Leu Ile Arg Phe
370 375 380
Ala Lys Met Leu Ser Asn Glu Arg Lys Ser Arg Asn Ala Lys Met Ser
385 390 395 400
Ala Ala
<210> 81
<211> 407
<212> БЕЛОК
<213> Arabidopsis thaliana
<400> 81
Met Gln Met Asp Ser Pro Lys Ser Pro Leu Gln Pro Pro Thr Tyr Gly
1 5 10 15
Asn Leu Val Thr Ile Leu Ser Ile Asp Gly Gly Gly Ile Arg Gly Leu
20 25 30
Ile Pro Ala Val Ile Leu Gly Phe Leu Glu Ser Glu Leu Gln Lys Leu
35 40 45
Asp Gly Glu Glu Ala Arg Leu Ala Asp Tyr Phe Asp Val Ile Ala Gly
50 55 60
Thr Ser Thr Gly Gly Leu Val Thr Ala Met Leu Thr Ala Pro Asn Lys
65 70 75 80
Glu Gly Arg Pro Leu Phe Ala Ala Ser Glu Ile Lys Asp Phe Tyr Leu
85 90 95
Glu Gln Cys Pro Lys Ile Phe Pro Gln Asp His Phe Pro Phe Ser Ala
100 105 110
Ala Lys Lys Leu Val Lys Ser Leu Thr Gly Pro Lys Tyr Asp Gly Lys
115 120 125
Tyr Leu His Gln Leu Ile His Ala Lys Leu Gly Asp Thr Lys Leu Ser
130 135 140
Gln Thr Leu Thr Asn Val Val Ile Pro Thr Phe Asp Ile Lys His Leu
145 150 155 160
Gln Pro Thr Ile Phe Ser Ser Tyr Glu Val Lys Asn His Pro Leu Lys
165 170 175
Asp Ala Ala Leu Ala Asp Ile Ala Ile Ser Thr Ser Ala Ala Pro Thr
180 185 190
Tyr Leu Pro Ala His Phe Phe Lys Val Glu Asp Leu Asn Gly Asn Ala
195 200 205
Lys Glu Tyr Asn Leu Ile Asp Gly Gly Val Ala Ala Asn Asn Pro Ala
210 215 220
Leu Leu Ala Ile Gly Glu Val Thr Asn Glu Ile Ser Gly Gly Ser Ser
225 230 235 240
Asp Phe Phe Pro Ile Arg Pro Asn Asp Tyr Gly Arg Phe Leu Val Leu
245 250 255
Ser Leu Gly Thr Gly Asn His Lys Ala Glu Glu Lys Phe Asn Ala Lys
260 265 270
Glu Val Ala Gly Trp Gly Leu Leu Asn Trp Leu Thr His Asp Asn Ser
275 280 285
Thr Pro Ile Ile Asp Ala Phe Ser Gln Ala Ser Ser Asp Met Val Asp
290 295 300
Phe His Leu Ser Ala Val Phe Arg Ala Leu His Ser Glu Ala Asn Tyr
305 310 315 320
Ile Arg Ile Gln Asp Asp Thr Leu Thr Gly Asp Ala Ala Ser Val Asp
325 330 335
Ile Ala Thr Val Glu Asn Leu Asp Ile Leu Ala Lys Thr Gly Asp Glu
340 345 350
Leu Leu Lys Lys Pro Val Ala Arg Val Asn Leu Asp Ser Gly Cys Asn
355 360 365
Glu Asn Ala Tyr Glu Thr Thr Asn Glu His Ala Leu Ile Lys Leu Ala
370 375 380
Gly Ile Leu Ser Lys Glu Lys Lys Ile Arg Asp Ile Arg Ser Pro His
385 390 395 400
Ala Lys Ala Pro Ile Arg Ile
405
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| HPV-СПЕЦИФИЧЕСКИЕ СВЯЗЫВАЮЩИЕ МОЛЕКУЛЫ | 2018 |
|
RU2804664C2 |
| СОЧЕТАНИЕ КЛЕТОЧНОЙ ТЕРАПИИ И ИММУНОМОДУЛЯТОРНОГО СОЕДИНЕНИЯ | 2018 |
|
RU2777911C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ИНТЕРЛЕЙКИНА-33 (IL-33) | 2015 |
|
RU2829846C2 |
| МОДИФИЦИРОВАННЫЕ НЕПРИРОДНЫЕ ЛИГАНДЫ NKG2D, КОТОРЫЕ ИЗБИРАТЕЛЬНО ДОСТАВЛЯЮТ ПРИСОЕДИНЕННЫЕ ГЕТЕРОЛОГИЧНЫЕ МОЛЕКУЛЫ К НЕПРИРОДНЫМ РЕЦЕПТОРАМ NKG2D НА CAR-КЛЕТКАХ | 2020 |
|
RU2823728C2 |
| СИНЦИТИАЛЬНЫЕ ОНКОЛИТИЧЕСКИЕ МУТАНТЫ HERPES SIMPLEX В КАЧЕСТВЕ МОЩНЫХ ТЕРАПЕВТИЧЕСКИХ СРЕДСТВ ДЛЯ ЛЕЧЕНИЯ РАКА | 2020 |
|
RU2821999C2 |
| НЕПРИРОДНЫЕ РЕЦЕПТОРЫ NKG2D, КОТОРЫЕ НЕПОСРЕДСТВЕННО НЕ ПЕРЕДАЮТ СИГНАЛ КЛЕТКАМ, С КОТОРЫМИ ОНИ СВЯЗАНЫ | 2019 |
|
RU2832538C2 |
| НОВЫЕ БЕЛКИ, ИМЕЮЩИЕ ИНГИБИРУЮЩЕЕ ДЕЙСТВИЕ В ОТНОШЕНИИ НАСЕКОМЫХ | 2017 |
|
RU2781075C2 |
| Антитела против CXCR2 и их применение | 2019 |
|
RU2807067C2 |
| ГИБРИДНЫЕ НЕЙРОТОКСИНЫ | 2017 |
|
RU2782382C2 |
| МУТАНТЫ БЕЛКА F RSV | 2016 |
|
RU2788403C2 |

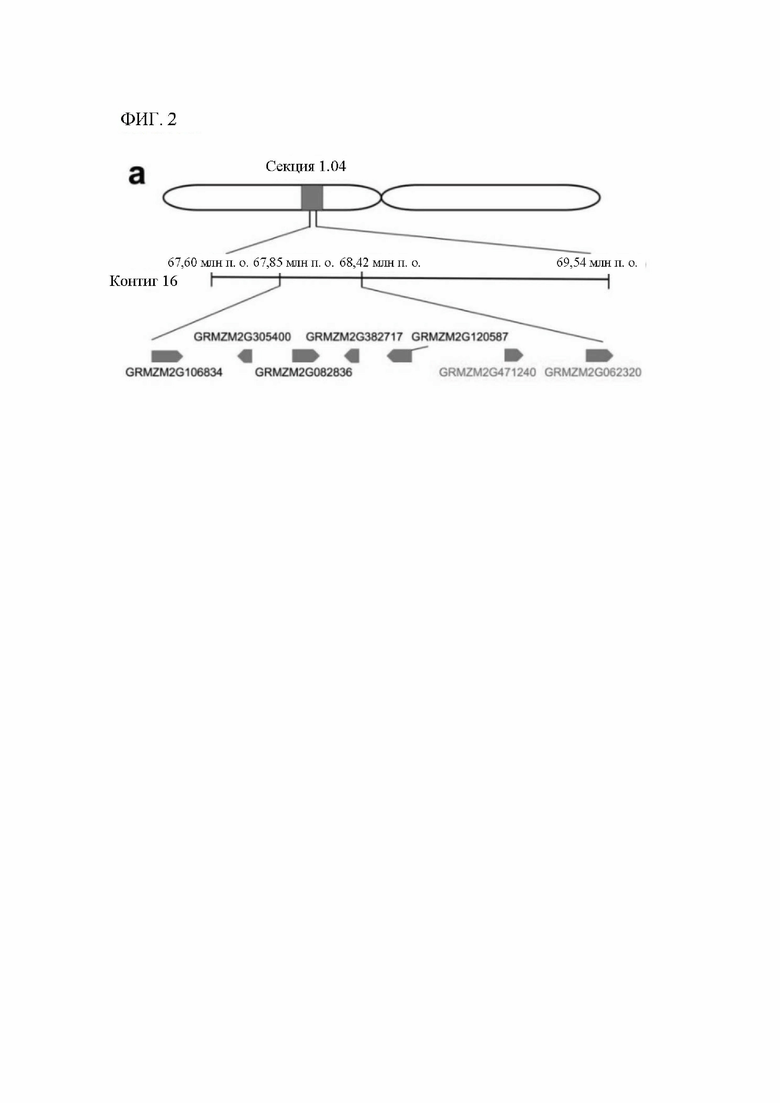
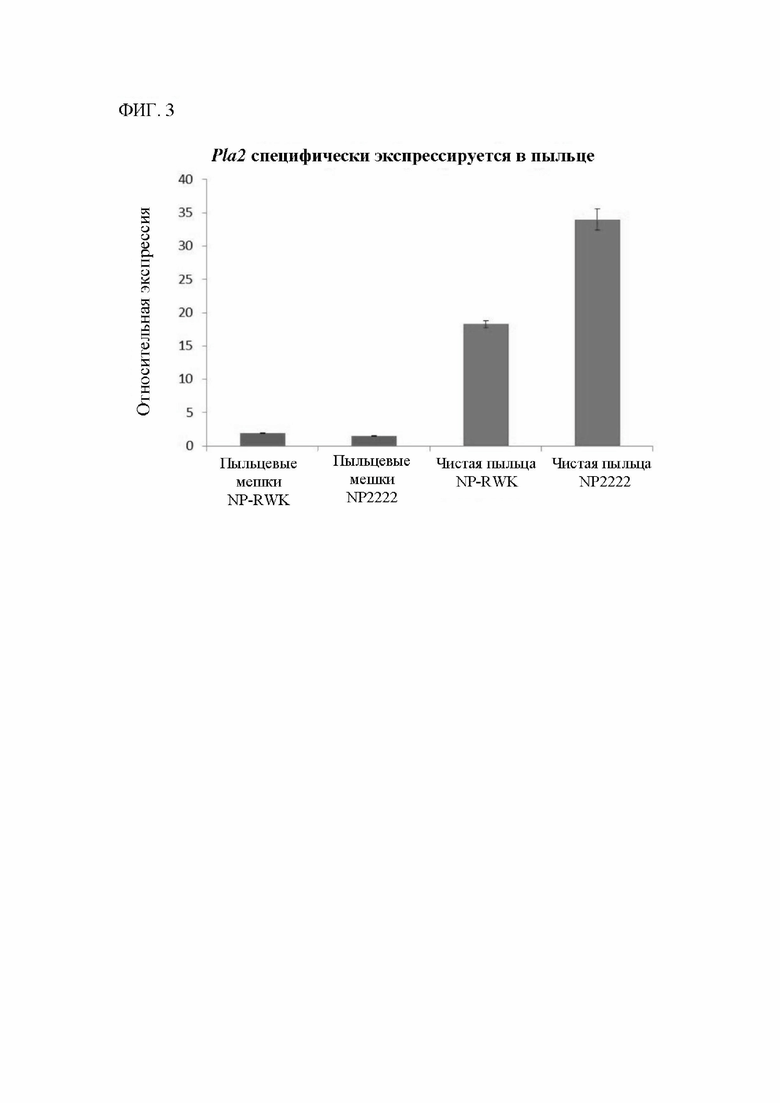
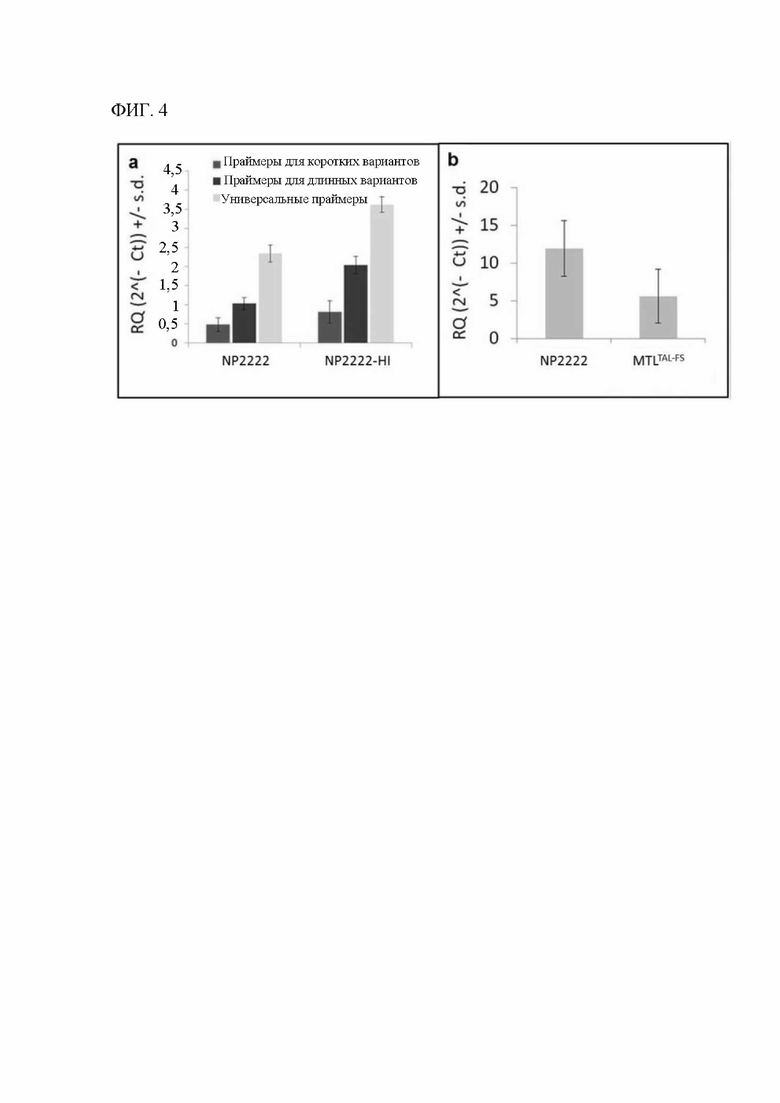
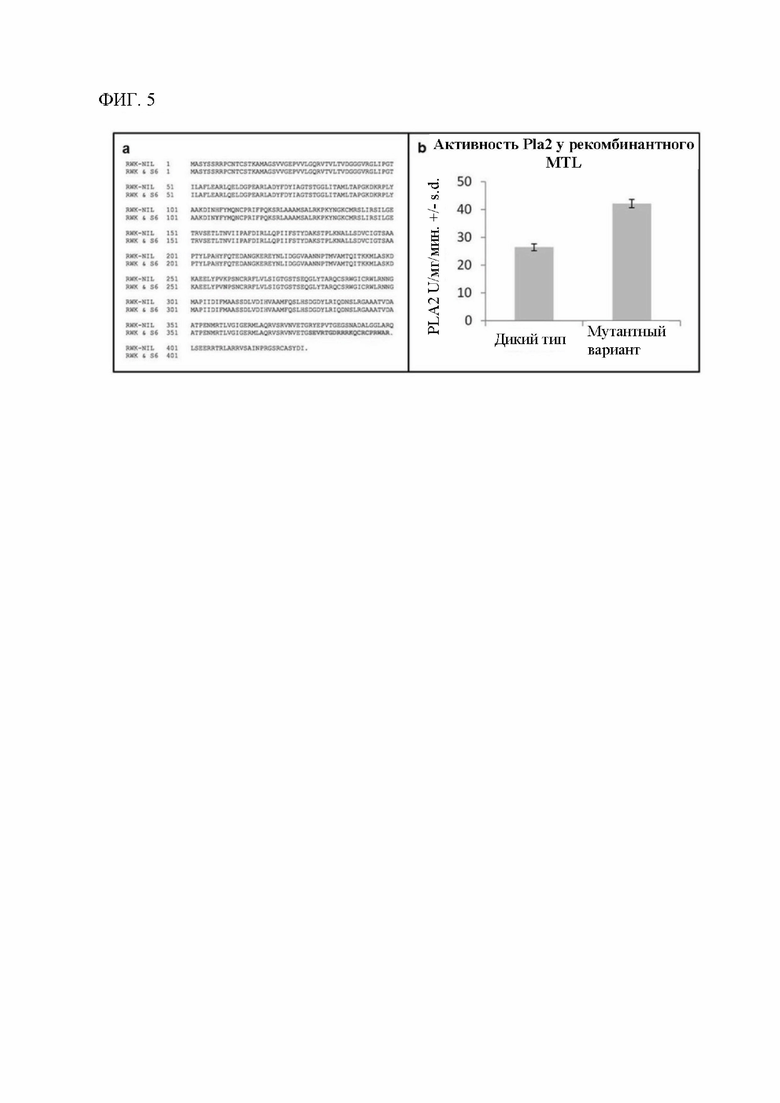
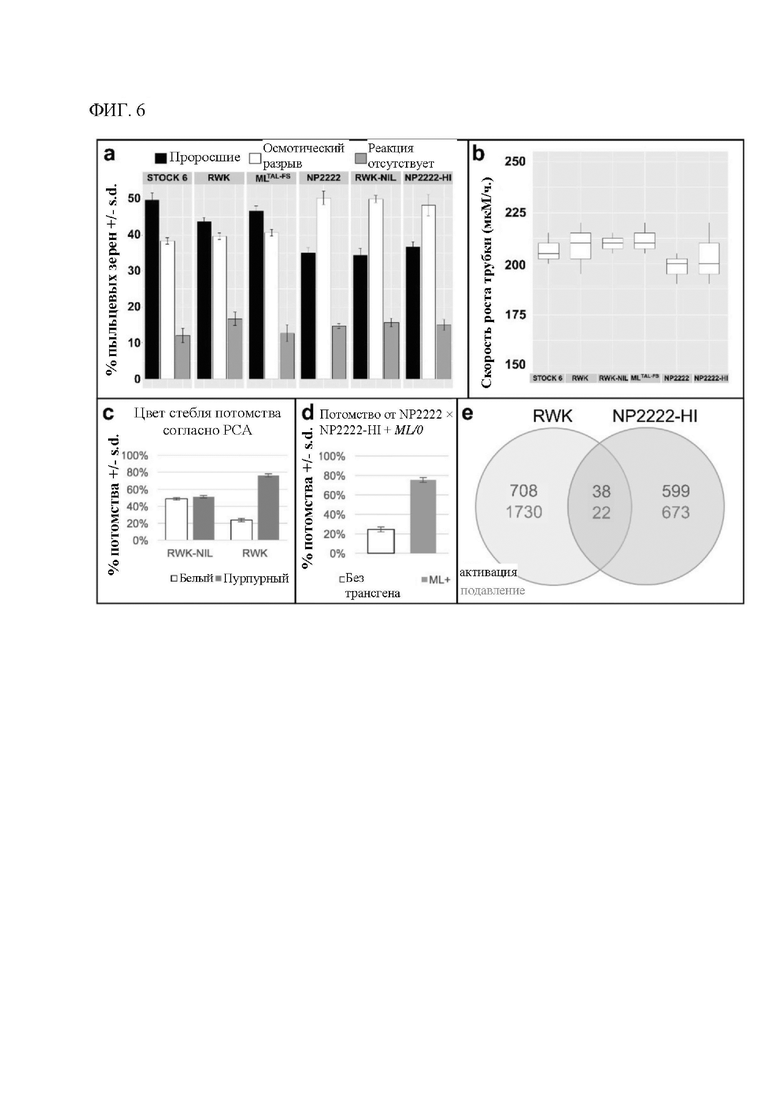
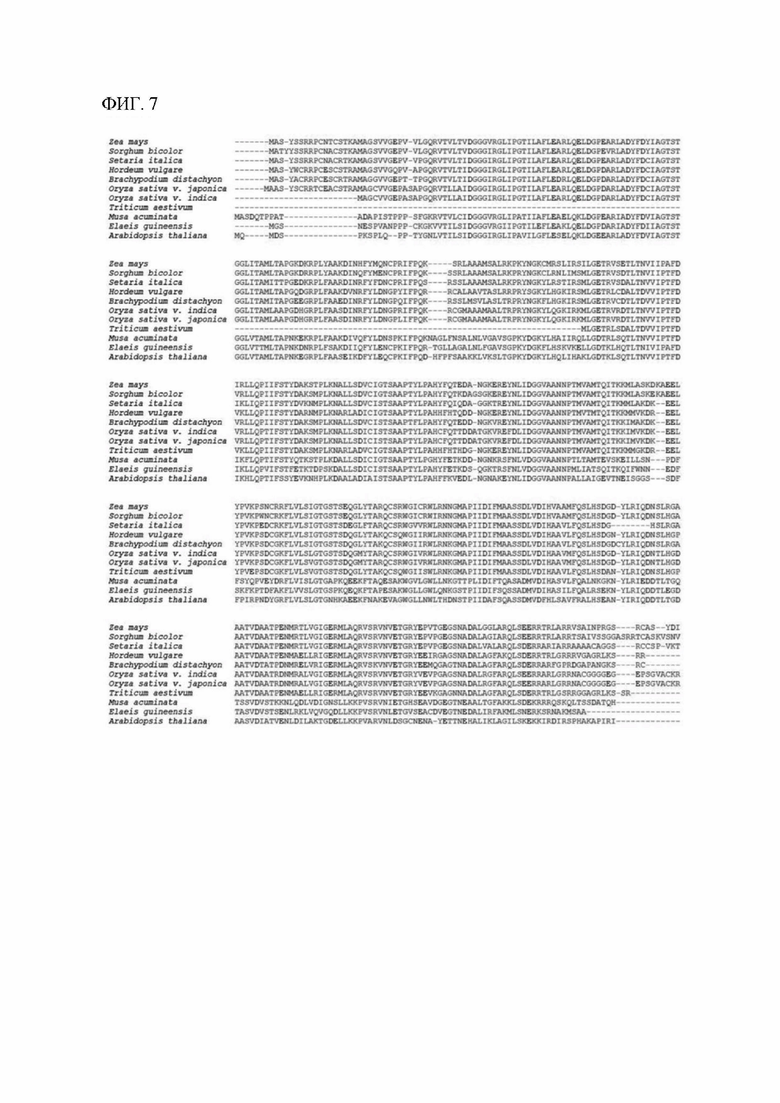


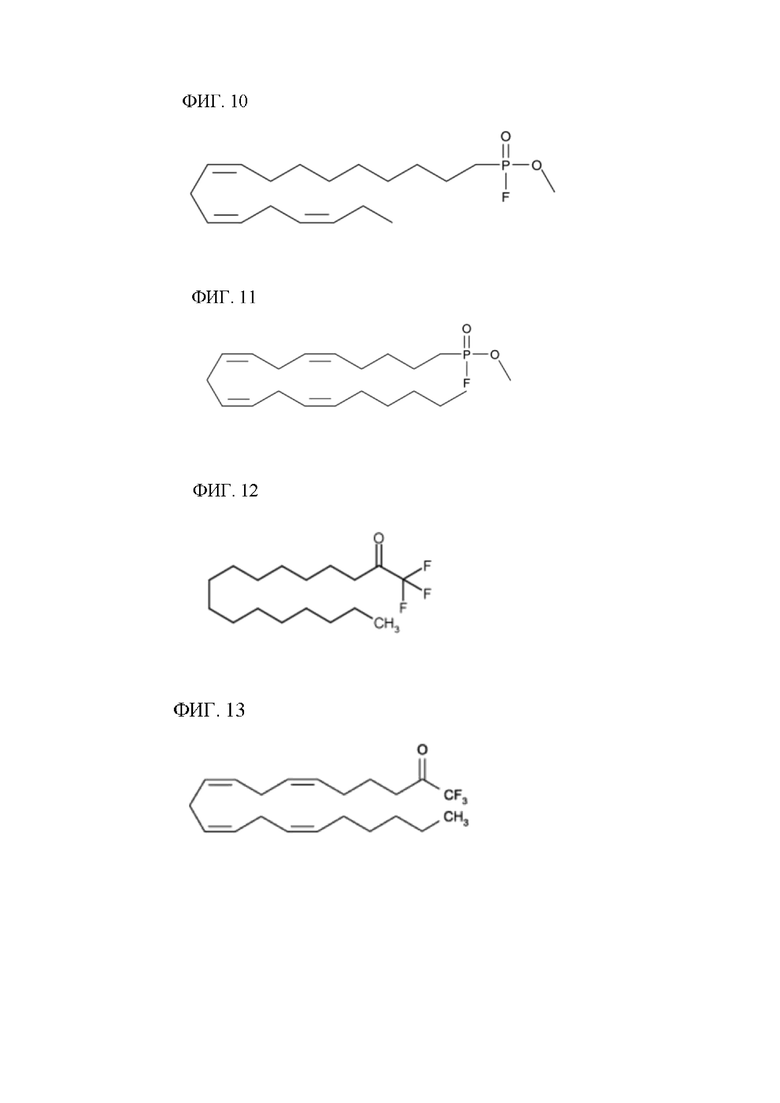
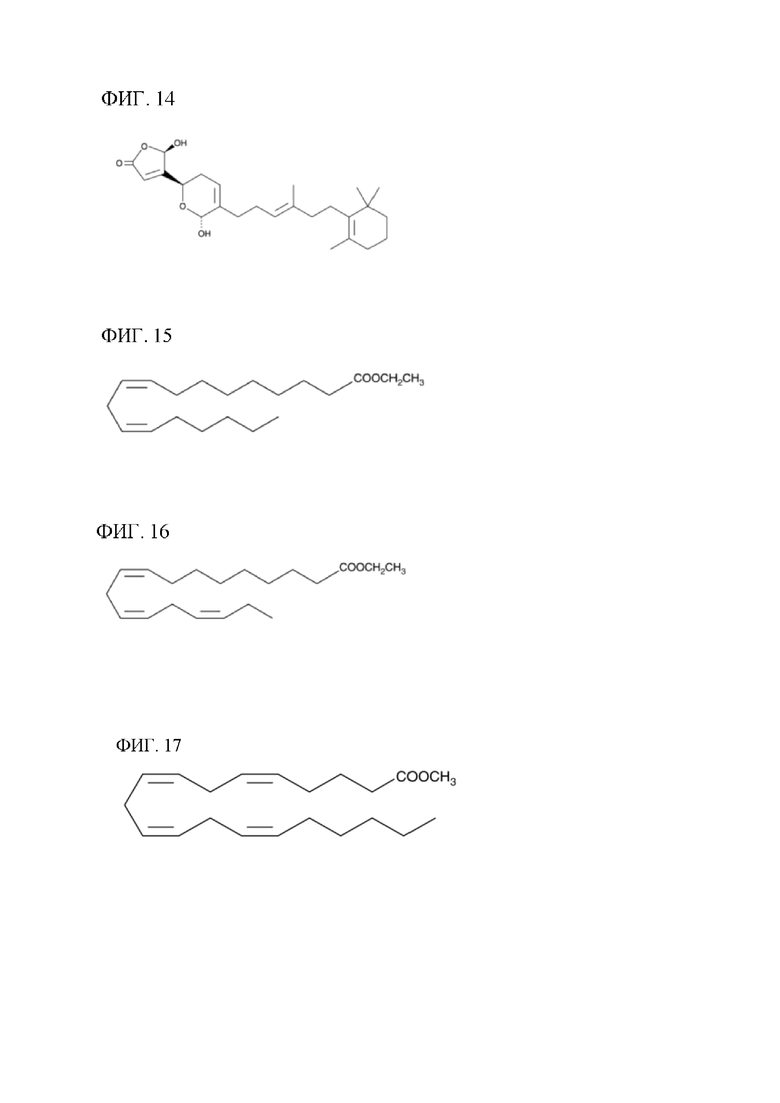
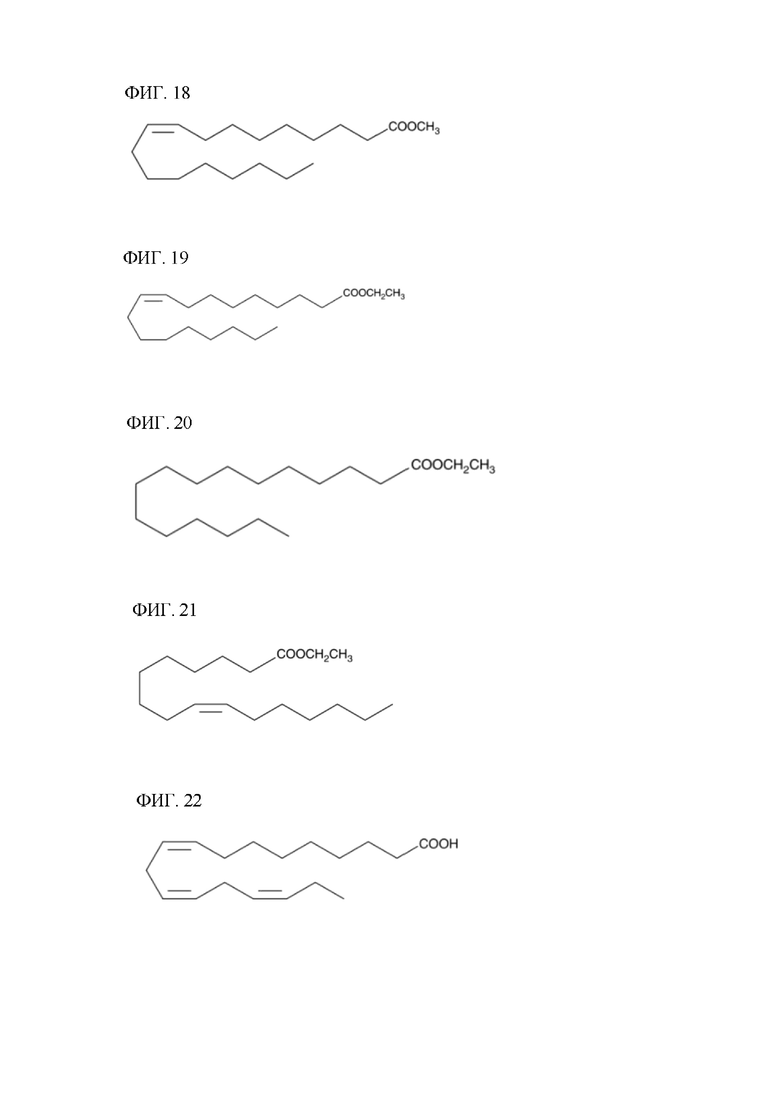
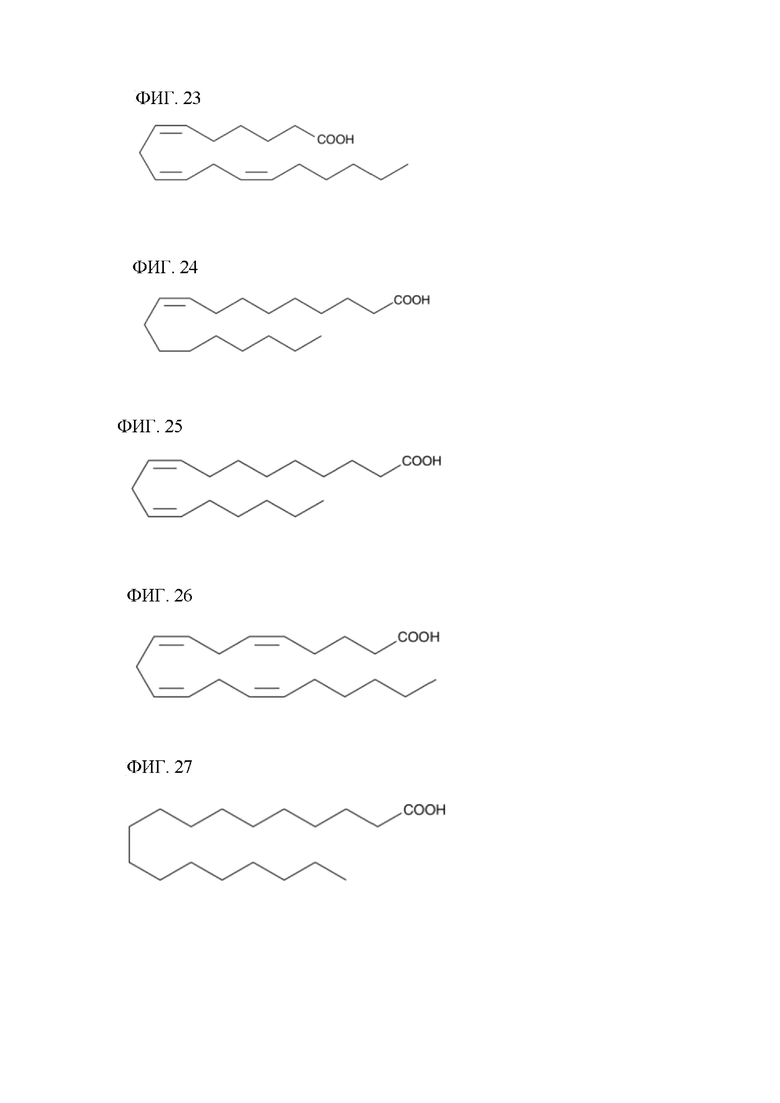
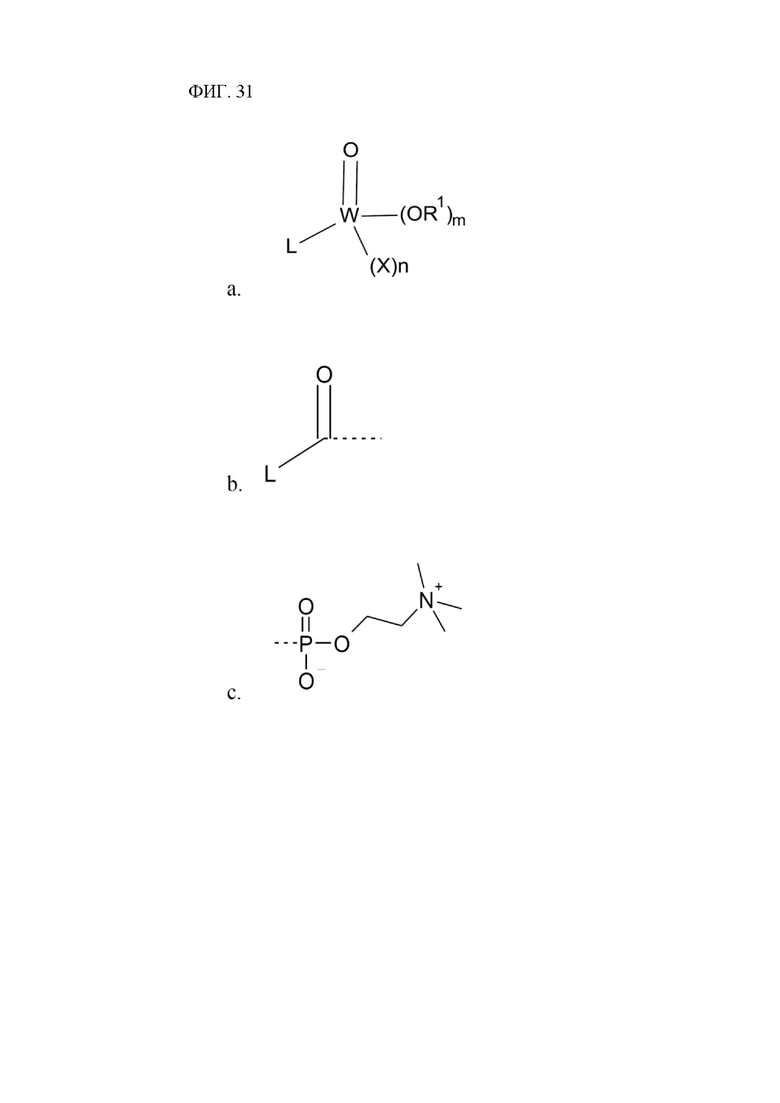
Изобретение относится к области биохимии, в частности к способу индукции образования гаплоидных зародышей. Изобретение позволяет эффективно получать гаплоидные зародыши при скрещивании двух растений. 3 з.п. ф-лы, 31 ил., 12 табл., 1 пр.
1. Способ индукции образования гаплоидных зародышей при скрещивании двух растений, при этом способ включает: осуществление мутации в последовательности пататин-подобной фосфолипазы растения посредством редактирования гена,
где пататин-подобная фосфолипаза кодируется последовательностью нуклеиновой кислоты, содержащей SEQ ID NO: 1, или последовательностью, на 90% идентичной SEQ ID NO: 1; где мутация представляет собой нокаут-мутацию, и
где растение-мутант применяют в качестве родительского растения при скрещивании между двумя растениями, так что в результате скрещивания образуется по меньшей мере один гаплоидный зародыш, где скрещивание двух растений представляет собой скрещивание двух растений маиса.
2. Способ по п. 1, где мутантная пататин-подобная фосфолипаза кодируется нуклеиновой кислотой, содержащей SEQ ID NO: 3.
3. Способ по п. 1, где редактирование гена осуществляют посредством сайт-направленного мутагенеза.
4. Способ по п. 3, где сайт-направленный мутагенез осуществляют посредством методики, выбранной из группы, состоящей из CRISPR/Cas9, TALEN, "цинковых пальцев" и мегануклеаз.
| WO 2013169802 A1, 14.11.2013 | |||
| WO 2014093718 A1, 19.06.2014 | |||
| CN 104726449 A, 24.06.2015 | |||
| RU 2013119981 A, 20.11.2014. |

