ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] Настоящая заявка испрашивает приоритет и преимущество на основании одновременно рассматриваемых предварительных заявок на патент США: №62/752293, поданной 29 октября 2018, и № 62/811116, поданной 27 февраля 2019, каждая из которых включена в данный документ в полном объеме посредством ссылки.
УРОВЕНЬ ТЕХНИКИ
ИЛ2
[0002] Интерлейкин-2, ИЛ2 (IL2) представляет собой цитокин, который играет центральную роль как в состоянии покоя, так и в активированном состоянии иммунной системы. Во время стадии покоя ИЛ2 в первую очередь обеспечивает развитие и выживание CD4+ Foxp3+ регуляторных Т-клеток (Treg). Однако во время иммунного ответа он способствует пролиферации и размножению эффекторных Т-клеток, Т-клеток памяти и естественных киллеров (NK). Он также усиливает эффекторную функцию указанных клеток. ИЛ2 секретируется разнообразными клетками при активации (CD8+ T-клетками, NK- и NKT-клетками, DC и тучными клетками), но большая часть его поступает из CD4+ T-клеток; и он может работать аутокринным или паракринным образом.
[0003] В образовании рецептора ИЛ2 участвуют три полипептидные цепи, которые связываются с ИЛ2 с различной аффинностью, когда присутствуют в различных комбинациях (таблица 1). Биологически важные варианты рецептора можно различать по их аффинности к ИЛ2: низкая аффинность (только CD25), промежуточная аффинность (βγ) и высокая аффинность (αβγ). CD122 и общая γ-цепь необходимы для передачи сигнала при связывании ИЛ2, тогда как CD25 увеличивает аффинность к рецептору, но, по-видимому, не передает сигнал.
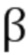 )
)
[0004] Разнообразные функции ИЛ2 в значительной степени являются результатом дифференциальной экспрессии различных субъединиц рецептора ИЛ2 в разных типах клеток. В покоящихся иммунных клетках CD25 в основном ограничен регуляторными Т-клетками, которые также экспрессируют другие субъединицы рецептора ИЛ2, что делает их основной мишенью низких уровней ИЛ2 в состоянии покоя. Что касается эффекторных Т-клеток, на недавно активированных Т-клетках наблюдается стимулирующая регуляция CD25 наряду с незначительным повышением уровней CD122, в то время как экспрессия общей γ-цепи относительно постоянна, что приводит к повышенной чувствительности и высокой зависимости от ИЛ2 во время фазы размножения. На более поздних стадиях иммунного ответа CD8+ Т-клетки памяти и естественные киллеры экспрессируют очень высокие уровни CD122 наряду с γ-цепью, что позволяет им конкурировать с регуляторными Т-клетками за ИЛ2.
[0005] Благодаря роли ИЛ2 как общего фактора роста Т-клеток его уже несколько десятилетий применяют в клиниках в качестве средства для иммунотерапии рака. Однако, учитывая паттерн экспрессии рецепторов, для достаточной активации эффекторных T-клеток и естественных киллеров, несущих рецептор с промежуточной аффинностью, необходима высокая доза ИЛ2, чтобы противодействовать его действию на регуляторные Т-клетки, несущие высокоаффинный рецептор. Другая проблема, связанная с введением ИЛ2, заключается в том, что период его полужизни в крови составляет менее 30 минут, соответственно, необходима его постоянная инфузия или многократные инъекции для поддержания достаточно высокого титра для достижения терапевтического эффекта. Высокодозный режим приводит к множеству побочных эффектов, включая отек легких (из-за экспрессии CD25 в легочных эпителиальных клетках), гипотонию, синдром повышенной проницаемости сосудов и т. п. Кроме того, размножение регуляторных Т-клеток у пациентов представляет собой постоянную угрозу противоопухолевому ответу.
[0006] Чтобы обойти эти ограничения терапии ИЛ2, применяли химическую модификацию, например, пегилирование, для снижения или изменения селективности ИЛ2 в отношении рецепторов и увеличения периода его полужизни. Другой подход заключался в применении специфических антител против ИЛ2 для получения комплекса с ИЛ2, обеспечивающего преимущественное нацеливание на клетки, экспрессирующие CD122. Однако оба этих подхода к настоящему времени показали ограниченный клинический успех, помимо сложностей при производстве. Как вариант, существуют мутанты ИЛ2, которые преимущественно связывают CD122 или больше не связывают CD25, что позволяет эффекторным Т-клеткам более успешно конкурировать с регуляторными Т-клетками за ИЛ2. Однако эти молекулы также имеют короткий период полужизни, как и ИЛ2 дикого типа, и, соответственно, требуются их многократные инфузии.
Антитела контрольных точек
[0007] В нормальных физиологических условиях иммунные контрольные точки представляют собой молекулярные пути, которые развились для поддержания аутотолерантности и защиты тканей от повреждений при ответах иммунной системы на инфекции. Опухоли используют эти пути, неправильно экспрессируя белки иммунных контрольных точек, создавая иммуносупрессивную среду и уклоняясь от иммунной системы. За последнее десятилетие было показано, что антитела, блокирующие пути CTLA-4 и PD1/PDL1, обращают иммуносупрессию, связанную с опухолью, и их применение в клинике оказалось весьма успешным. Однако, несмотря на перспективность, указанные антитела действуют только на небольшую часть пациентов по причинам, которые еще не до конца ясны. Таким образом, существует потребность в лучшем понимании прогностических биомаркеров и терапевтических средств, которые можно использовать в комбинации с антителами контрольных точек.
КРАТКОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[0008] Настоящее изобретение решает описанные выше проблемы за счет применения полученных путем рационального дизайна слитых белковых композиций, которые могут одновременно усиливать противоопухолевый иммунитет или подавлять связанную с опухолью иммуносупрессию наряду с прямой активацией эффекторных клеток ИЛ2 без активации регуляторных Т-клеток. Слитые белки по настоящему изобретению содержат часть антитела или целое антитело к иммунной контрольной точке и мутантный полипептид интерлейкина-2 (ИЛ2). Слитые белки по настоящему изобретению можно использовать для лечения опухолей в качестве монотерапевтических средств или в комбинации с (а) антителами, нацеленными на по меньшей мере другой иммуносупрессивный путь; (б) химиотерапией, таргетной терапией или лучевой терапией; (c) другим механизмом блокирования иммуносупрессивного пути, например, аптамерами или РНК-интерференцией; или (d) другим иммунотерапевтическим агентом, например, цитокином, таргетным терапевтическим средством и т. д.
[0009] В одном аспекте согласно настоящему изобретению предложен слитый белок, в котором антитело к иммунной контрольной точке (также называемое в дальнейшем антителом против контрольной точки или антителом против CP) объединено с мутантным ИЛ2, менее селективным в отношении высокоаффинного рецептора (или, взаимозаменяемо, селективным мутантом рецептора с промежуточной аффинностью, MutIL2). Указанный слитый белок состоит из полного антитела против CP, соединенного с MutIL2 на С-конце тяжелой цепи (фиг. 1А). При указанном подходе добавляют линкерную последовательность между С-концом тяжелой цепи и MutIL2. Кроме того, присутствие Fc-части антитела позволяет использовать путь неонатального Fc-рецептора для увеличения периода полужизни слитого белка на несколько порядков. Кроме того, присутствие нацеленного на опухоль антитела помогает доставлять ИЛ2 преимущественно в опухоль и снижать системные побочные эффекты.
[00010] Согласно одному признаку изобретения антитело против CP представляет собой антитело к PDL1. Предпочтительно, антитело к PDL1 обладает одним или более из следующих признаков, которые могут повышать противоопухолевую способность слитого белка: (а) оно связывает PDL1 с Кd <100 нМ, как для очищенного белка, так и для экспрессируемого на поверхности клетки белка; (б) он блокирует взаимодействие PDL1-PD1 in vitro; и (c) он устраняет репрессию PDL1-опосредованной репрессии эффекторных Т-клеток в активированных культурах мононуклеарных клеток периферической крови (МКПК).
[00011] В одном варианте осуществления антитело к PDL1 представляет собой антитело любого доступного изотипа или любые известные мутантные формы области Fc, которое инактивирует, снижает или усиливает функцию антитело-зависимой клеточной цитотоксичности (АЗКЦ) и/или комплемент-зависимой цитотоксичности (КЗЦ). В предпочтительном варианте осуществления антитело к PDL1 представляет собой антитело изотипа IgG1. В некоторых вариантах осуществления антитело к PDL1 включает пару вариабельной области тяжелой цепи и вариабельной области легкой цепи, соответствующие последовательности которых состоят по существу из следующих: (а) SEQ ID NO: 3 и SEQ ID NO: 6; (b) SEQ ID NO: 9 и SEQ ID NO: 12; или (c) SEQ ID NO: 15 и SEQ ID NO: 18. В некоторых вариантах осуществления антитело к PDL1 включает дополнительно вторую пару вариабельных областей тяжелой и легкой цепей, по существу идентичных вариабельным областям из первой пары. В других вариантах осуществления антитело к PDL1 включает пару тяжелой цепи и легкой цепи, соответствующие последовательности которых состоят по существу из следующих: (а) SEQ ID NO: 2 и SEQ ID NO: 5; (b) SEQ ID NO: 8 и SEQ ID NO: 11; или (c) SEQ ID NO: 14 и SEQ ID NO: 17.
[00012] Согласно одному признаку изобретения MutIL2 обладает одним или обоими из следующих признаков, которые могут повышать противоопухолевую способность слитого белка: (а) сниженная способность активировать CTLL2 и регуляторные Т-клетки человека и мыши при сохранении способности активировать эффекторные Т-клетки или NK-клетки; и (b) увеличение относительной способности активировать эффекторные/регуляторные Т-клетки по сравнению с ИЛ2 дикого типа до 10-кратного - до более чем 1000-кратного. В дополнительном варианте осуществления MutIL2 включает последовательности аминокислот, по существу идентичные последовательностям аминокислот, выбранным из группы, состоящей из SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 и их комбинаций.
[00013] Согласно одному признаку изобретения линкерная последовательность представляет собой гибкий серин-глициновый линкер или другие известные варианты линкеров. В одном варианте осуществления линкерная последовательность представляет собой SEQ ID NO: 19.
[00014] Согласно одному признаку изобретения слитый белок содержит: связывающую антиген часть, содержащую часть антитела или целое антитело к PDL1 и мутантный полипептид интерлейкина-2 (ИЛ2), при этом связывающая антиген часть включает пару тяжелой цепи и легкой цепи, соответствующие последовательности которых состоят по существу из следующих: (а) SEQ ID NO: 2 и SEQ ID NO: 5; (b) SEQ ID NO: 8 и SEQ ID NO: 11; мутантный полипептид ИЛ2 включает последовательность аминокислот, по существу идентичную последовательности аминокислот, выбранной из группы, состоящей из SEQ ID NO: 25, SEQ ID NO: 27; и мутантный полипептид ИЛ2 соединен с С-концами тяжелых цепей. Согласно одному признаку изобретения мутантный полипептид ИЛ2 связан с C-концами тяжелых цепей линкерной последовательностью, содержащей SEQ ID NO: 19.
[00015] В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов протеолитического расщепления для улучшения продуцирования. В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов гликозилирования, без изменения функции белка иным образом, для получения более гомогенного продукта. В другом варианте осуществления альтернативная молекула слитого белка содержит альтернативную секреторную последовательность вместо естественной секреторной последовательности для улучшения секреции.
[00016] В другом аспекте согласно настоящему изобретению предложен слитый белок, где антитело к иммунной контрольной точке (антитело против CP) объединено с мутантом ИЛ2, который является менее селективным в отношении высокоаффинного рецептора (или, взаимозаменяемо, селективный мутант рецептора с промежуточной аффинностью MutIL2), при этом слитый белок представляет собой биспецифическую конструкцию с первым плечом, соответствующим антителу против CP, и вторым плечом, состоящим из MutIL2, слитого с частью Fc (фиг. 1B). Кроме того, присутствие Fc-части белка позволяет использовать путь неонатального Fc-рецептора для увеличения периода полужизни гибридного белка на несколько порядков.
[00017] Согласно одному признаку изобретения антитело против CP представляет собой антитело к PDL1. Антитело к PDL1 обладает одним или более из следующих признаков, которые могут повышать противоопухолевую способность слитого белка: (а) оно связывает PDL1 с Kd <100 нМ, как для очищенного белка, так и для экспрессируемого на поверхности клетки белка; (б) оно блокирует взаимодействие PDL1-PD1 in vitro; и (c) оно устраняет репрессию PDL1-опосредованной репрессии эффекторных Т-клеток в активированных культурах мононуклеарных клеток периферической крови (МКПК).
[00018] В одном варианте осуществления указанное биспецифическое антитело представляет собой антитело любого доступного изотипа или любые известные мутантные формы области Fc, которое инактивирует, снижает или усиливает функции АЗКЦ и/или КЗЦ. В идеальном варианте осуществления биспецифическое антитело представляет собой антитело изотипа IgG1. В некоторых вариантах осуществления первое плечо биспецифического антитела включает пару вариабельной области тяжелой цепи и вариабельной области легкой цепи, соответствующие последовательности которых состоят по существу из следующих: (а) SEQ ID NO: 3 и SEQ ID NO: 6; (b) SEQ ID NO: 9 и SEQ ID NO: 12; или (c) SEQ ID NO: 15 и SEQ ID NO: 18. В других вариантах осуществления антитело к PDL1 включает пару тяжелой цепи и легкой цепи, соответствующие последовательности которых состоят по существу из следующих: (а) SEQ ID NO: 2 и SEQ ID NO: 5; (b) SEQ ID NO: 8 и SEQ ID NO: 11; или (c) SEQ ID NO: 14 и SEQ ID NO: 17.
[00019] Согласно одному признаку изобретения MutIL2 обладает одним или обоими из следующих характеристик, которые могут повышать противоопухолевую способность слитого белка: (а) снижение способности активировать CTLL2 и регуляторные Т-клетки человека и мыши при сохранении способности активировать эффекторные Т-клетки или NK-клетки; и (b) увеличение относительной способности активировать эффекторные/регуляторные Т-клетки по сравнению с ИЛ2 дикого типа до 10-кратного - до более чем 1000-кратного. В дополнительном варианте осуществления MutIL2 включает последовательность аминокислот, по существу идентичную последовательностям аминокислот, выбранным из группы, состоящей из SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 и их комбинаций.
[00020] В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов протеолитического расщепления для улучшения продуцирования. В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов гликозилирования, без изменения функции белка иным образом, для получения более гомогенного продукта. В другом варианте осуществления альтернативная молекула слитого белка содержит альтернативную секреторную последовательность вместо естественной секреторной последовательности для улучшения секреции.
[00021] В другом аспекте настоящего изобретения также предложен слитый белок, который представляет собой полипептид, состоящий из антигенсвязывающего полипептида и мутанта ИЛ2, который менее селективен в отношении высокоаффинного рецептора (или взаимозаменяемо, селективный мутант рецептора с промежуточной аффинностью, MutIL2) (фиг. 2). При указанном подходе добавляют, например, линкерную последовательность между двумя указанными частями. Указанный полипептид представляет собой молекулу меньшего размера, которая может экспрессироваться в нацеленных на опухоль бактериях. В некоторых вариантах осуществления антигенсвязывающий полипептид выбран из группы, состоящей из scFv против CP, лиганда или части лиганда. В некоторых вариантах осуществления scFv против CP представляет собой scFv к PDL1. В некоторых вариантах осуществления лиганд представляет собой PD1 или CTLA4. В некоторых вариантах осуществления антигенсвязывающий полипептид выбран из группы, состоящей из иммунотерапевтических антител, таких как антитела против GD2, против ТФР-бета, против CD47, против OX40, против IBB, или иммунотерапевтического антитела, которое может работать совместно с MutIL2 по настоящему изобретению.
[00022] Согласно одному признаку изобретения, MutIL2 обладает одним или обоими из следующих признаков, которые могут повышать противоопухолевую способность слитого белка: (а) снижение способности активировать CTLL2 и регуляторных Т-клеток человека и мыши при сохранении способности активировать эффекторные Т-клетки или NK-клетки; и (b) увеличение относительной способности активировать эффекторные /регуляторные Т-клетки по сравнению с ИЛ2 дикого типа до 10-кратного - до более чем 1000-кратного. В дополнительном варианте осуществления MutIL2 включает последовательность аминокислот, по существу идентичную последовательностям аминокислот, выбранным из группы, состоящей из SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 и их комбинаций.
[00023] Согласно одному признаку изобретения, линкерная последовательность представляет собой гибкий серин-глициновый линкер или другие известные варианты линкеров. В одном варианте осуществления линкерная последовательность представляет собой SEQ ID NO: 19.
[00024] В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов протеолитического расщепления для улучшения продуцирования. В другом варианте осуществления альтернативная молекула слитого белка содержит одну или более мутированных аминокислот для удаления сайтов гликозилирования, без изменения функции белка иным образом, для получения более гомогенного продукта. В другом варианте осуществления альтернативная молекула слитого белка содержит альтернативную секреторную последовательность вместо естественной секреторной последовательности для улучшения секреции.
[00025] В другом аспекте согласно настоящему изобретению предложен сконструированный белок, содержащий терапевтический белок, например, нацеленный на опухоль агент, который слит или иным образом связан с мутантом полипептида ИЛ2, селективным в отношении рецептора с промежуточной аффинностью. В некоторых вариантах осуществления терапевтический белок представляет собой терапевтическое антитело, нацеленное на опухоль антитело, связывающий антиген опухоли полипептид или нацеленные на опухоль олигонуклеотиды, например, аптамеры или небольшие молекулы. В некоторых вариантах осуществления антитело, нацеленное на опухоль, представляет собой антитело к иммунной контрольной точке. В некоторых вариантах осуществления согласно настоящему изобретению предложен сконструированный белок, содержащий терапевтический белок и мутантный полипептид интерлейкина-2 (ИЛ2). В некоторых вариантах осуществления терапевтический белок представляет собой терапевтическое антитело или связывающий антиген опухоли полипептид. В некоторых вариантах осуществления антитело, направленное на опухоль, или терапевтическое антитело представляет собой антитело против PD-L1, антитело против CD19, антитело против MUC1, антитело против CD22, антитело против HER2, антитело против CD20, антитело, антитело против CD80, антитело против антигена созревания В-клеток (ВСМА), антитело против рецептора эпидермального фактора роста (EGFR) или антитело против мезотелина. Агенты, нацеленные на опухоль, могут помогать доставлять ИЛ2 в опухоль и уменьшать системные побочные эффекты, а мутантный ИЛ2 может непосредственно активировать эффекторные клетки для усиления противоопухолевой активности агентов, нацеленных на опухоль.
[00026] Согласно одному признаку изобретения MutIL2 обладает одним или более из следующих признаков, которые могут повышать противоопухолевую способность слитого белка:(а) снижение способности активировать CTLL2 и регуляторные Т-клетки человека и мыши при сохранении способности активировать эффекторные Т-клетки или NK-клетки; и (b) увеличение относительной способности активировать эффекторные/регуляторные Т-клетки по сравнению с ИЛ2 дикого типа до 10-кратного - до более чем 1000-кратного. В дополнительном варианте осуществления MutIL2 включает последовательности аминокислот, по существу идентичные последовательностям аминокислот, выбранным из группы, состоящей из SEQ ID NO: 23, SEQ ID NO: 25, SEQ ID NO: 27, SEQ ID NO: 29, SEQ ID NO: 31, SEQ ID NO: 33, SEQ ID NO: 35, SEQ ID NO: 37, SEQ ID NO: 39, SEQ ID NO: 41, SEQ ID NO: 43, SEQ ID NO: 45, SEQ ID NO: 47, SEQ ID NO: 49, SEQ ID NO: 51, SEQ ID NO: 53, SEQ ID NO: 55, SEQ ID NO: 57 и их комбинаций.
[00027] В другом аспекте согласно настоящему изобретению предложена фармацевтическая композиция, которая включает слитый белок по любому из вышеперечисленных аспектов. Указанная фармацевтическая композиция дополнительно включает фармацевтически приемлемое вспомогательное вещество, фармацевтически приемлемый носитель или разбавитель.
[00028] В связанном аспекте согласно настоящему изобретению предложен способ терапевтического лечения патологического состояния у нуждающегося в этом субъекта, при этом указанный способ включает введение указанному субъекту терапевтически эффективного количества слитого белка, описанного в данном документе. Указанный способ может дополнительно включать этап введения второго, другого терапевтического антитела против по меньшей мере одного антигена клеточной поверхности, характерного для указанного состояния. Состояние, которое лечат, может быть раком млекопитающих, инфекцией и так далее.
[00029] Предпочтительно, чтобы спектр подлежащих лечению видов рака млекопитающих был выбран из группы, состоящей из рака яичников, рака толстой кишки, рака молочной железы, рака легкого, миелом, опухолей ЦНС, происходящих из нейробластов, моноцитарных лейкозов, В-клеточных лейкозов, Т-клеточных лейкозов, B-клеточных лимфом, T-клеточных лимфом, опухолей, происходящих из тучных клеток, меланом, рака мочевого пузыря, рака желудка, рака печени, уротелиальной карциномы, кожной карциномы, рака почек, рака головы и шеи, рака поджелудочной железы и их комбинаций. В некоторых вариантах осуществления спектр подлежащих лечению видов рака млекопитающих выбран из группы, состоящей из меланомы, рака легкого, рака почек, рака головы и шеи, рака желудка, лимфомы, рака яичников, рака толстой кишки, рака молочной железы, рака легкого, миелом, опухолей головного мозга, лейкозов, лимфом, рака мочевого пузыря, рака печени, уротелиальной карциномы, кожной карциномы, рака поджелудочной железы и их комбинаций.
[00030] В еще одном аспекте согласно настоящему изобретению предложен способ профилактического лечения аналогичных состояний у нуждающегося в этом субъекта, при этом указанный способ включает введение указанному субъекту профилактически эффективного количества фармацевтической композиции по настоящему изобретению. Указанный способ может дополнительно включать этап введения вакцины против указанного состояния. В одном варианте осуществления указанное состояние представляет собой рак.
[00031] В еще одном аспекте согласно настоящему изобретению предложена система экспрессии у млекопитающих, которая продуцирует описанный выше слитый белок.
КРАТКОЕ ОПИСАНИЕ ФИГУР
[00032] На фигурах 1А и 1В схематически изображен дизайн слитого белка по настоящему изобретению.
[00033] Фигура 2 схематически изображает альтернативный дизайн слитого белка по настоящему изобретению.
[00034] На фигурах 3A и 3B показаны репрезентативные данные, сравнивающие связывание PDL1 клеточной поверхности исходными антителами и связывание слитыми белками дикого типа и мутантными слитыми белками ИЛ2 по настоящему изобретению.
[00035] На фигуре 4 показана репрезентативная АЗКЦ-активность различных слитых белков.
[00036] На фигуре 5 представлены репрезентативные данные, сравнивающие связывание тримерного рецептора (высокоаффинного рецептора) ИЛ2 клеточной поверхности слитым белком ИЛ2 дикого типа и связывание двумя мутантными формами слитого белка ИЛ2 по настоящему изобретению.
[00037] На фигуре 6 показаны репрезентативные данные для сравнения способности слитых белков ИЛ2 дикого типа и способности различных мутантных форм слитых белков ИЛ2 по настоящему изобретению запускать и поддерживать пролиферацию клеток CTLL2 (суррогат регуляторных Т-клеток), измеренную с помощью анализа с аламаровым синим.
[00038] На фигуре 7 показаны репрезентативные данные для сравнения способности слитых белков ИЛ2 дикого типа и способности различных мутантных форм слитых белков ИЛ2 поддерживать выживание и активацию регуляторных Т-клеток мыши по настоящему изобретению.
[00039] На фигуре 8 представлены репрезентативные данные для сравнения способности слитых белков ИЛ2 дикого типа и способности различных мутантных форм слитых белков ИЛ2 поддерживать выживание и пролиферацию CD8+ T-клеток мыши (примера эффекторных Т-клеток) по настоящему изобретению.
[00040] На фигурах 9 и 10 показаны репрезентативные данные для сравнения способности слитых белков рекомбинантного ИЛ2 и ИЛ2 дикого типа и способности различных мутантных форм слитого белка ИЛ2 по настоящему изобретению индуцировать фосфорилирование Stat5 в регуляторных и не-регуляторных CD4+ Т-клетках.
[00041] На фигуре 11 показаны репрезентативные данные для сравнения способности слитых белков рекомбинантного ИЛ2 и ИЛ2 дикого типа и способности различных мутантных форм слитого белка ИЛ2 по настоящему изобретению индуцировать фосфорилирование Stat5 в эффекторных CD8+ Т-клетках.
[00042] На фигуре 12 показан фармакокинетический профиль различных мутантных форм слитого белка ИЛ2 по настоящему изобретению.
[00043] На фигуре 13 показана способность различных мутантных форм слитого белка ИЛ2 по настоящему изобретению дифференциально индуцировать преимущественную пролиферацию CD8+ Т-клеток и NK-клеток, а не регуляторных Т-клеток.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
[00044] Если не указано иное, технические термины используют в соответствии с общепринятым значением.
[00045] Используемые в настоящем документе термины в единственном числе могут означать «один или более». При использовании в настоящем документе в сочетании с термином «содержащий» термины в единственном числе могут означать «один или более одного». Используемые в настоящем документе термины «другой», «еще один» могут означать «по меньшей мере второй или более». Также, если иное не продиктовано контекстом, термины в единственном числе включают термины во множественном числа, а термины во множественном числе включают термины в единственном числе.
[00046] Используемый здесь термин «примерно» относится к числовому значению, включая, например, целые числа, дроби и проценты, независимо от того, указаны они явным образом или нет. Термин «примерно» в общем случае относится к диапазону числовых значений (например, ± 5-10% от приведенного значения), которые средний специалист в данной области техники счел бы эквивалентными указанному значению (например, имеющим ту же функцию или результат). В некоторых случаях термин «примерно» может включать числовые значения, округленные до ближайшего значащего числа. Если не указано иное, «примерно» составляет ±10% от приведенных значений.
[00047] «Антигенсвязывающий полипептид» («связывающий антиген полипептид») представляет собой полипептид, содержащий часть, которая связывается с антигеном. Примеры антигенсвязывающих полипептидов включают антитела, фрагменты антител (например, антигенсвязывающую часть антитела), производные антител и аналоги антител.
[00048] Термины «по существу состоит из» или «по существу идентичный» в настоящем документе относятся по меньшей мере к 60%, или 80%, или более предпочтительно - к 85%, 90%, 95% или даже 100% идентичности, например, идентичности выбранной последовательности аминокислот.
[00049] Антигенсвязывающий полипептид или белок может иметь, например, структуру встречающегося в природе антитела (также известного как «иммуноглобулин»). Каждое встречающееся в природе антитело состоит из двух идентичных пар полипептидных цепей, каждая пара имеет одну «легкую» (около 25 кДа) и одну «тяжелую» цепь (около 50-70 кДа). Вариабельные области каждой пары легкой/тяжелой цепи образуют сайт связывания антитела, так что интактное антитело имеет два сайта связывания.
[00050] Вариабельные области встречающихся в природе цепей антител демонстрируют одинаковую общую структуру относительно консервативных каркасных областей (FR), соединенных тремя гипервариабельными областями, также называемыми определяющими комплементарность областями, или CDR. Как легкие, так и тяжелые цепи содержат в направлении от N-конца к C-концу домены FR1, CDR1, FR2, CDR2, FR3, CDR3 и FR4. Назначение аминокислот каждому домену соответствует определениям, приведенным в источнике: Kabat et al. Sequences of Proteins of Immunological Interest, 5th Ed., US Dept. of Health and Human Services, PHS, NIH, NIH Publication no. 91-3242, 1991. Другие системы нумерации аминокислот в цепях иммуноглобулинов включают IMGT (международная информационная система ImMunoGeneTics; Lefranc et al., Dev. Comp. Immunol. 29:185-203; 2005) и AHo (Honegger and Pluckthun, J. Mol. Biol. 309(3):657-670; 2001). В вариабельных областях важное значение имеют области CDR 1, CDR 2 и CDR 3, при этом область CDR3 наиболее важна. Они могут быть идентифицированы с помощью стандартных экспериментов с использованием биоинформатики и мутагенеза.
[00051] Антитела можно получить из таких источников, как сыворотка или плазма, которые содержат иммуноглобулины с различной антигенной специфичностью. Если такие антитела подвергают аффинной очистке, они могут быть обогащены по определенной антигенной специфичности. Такие обогащенные составы с антителами обычно состоят из менее чем примерно 10% антител, обладающих специфической связывающей активностью в отношении конкретного антигена. При проведении нескольких этапов аффинной очистки указанных составов с антителами доля антитела, обладающего специфической связывающей активностью в отношении антигена, может увеличиваться. Антитела, полученные таким образом, часто называют «моноспецифическими». Составы с моноспецифическими антителами могут состоять примерно на 10%, 20%, 30%, 40%, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 99% или 99,9% из антитела, обладающего специфической связывающей активностью в отношении конкретного антигена.
[00052] Термин «антитело» или «Ab» (и их множественные формы) в настоящем документе в широком смысле относится к любой молекуле иммуноглобулина (Ig), состоящей из четырех полипептидных цепей, двух тяжелых (H) цепей и двух легких (L) цепей, или любому ее функциональному фрагменту, мутанту, варианту, производному или аналогу, сохраняющему существенные и специфические особенности связывания эпитопа молекулы Ig. Такие форматы фрагментов, мутантов, вариантов, производных или аналогов антител известны в данной области техники и включают, среди прочего, Fab, F(ab'), F(ab')2, Fv, одноцепочечные антитела (scFv), одноцепочечные антитела (sdAb), фрагменты определяющей комплементарность области (CDR), химерные антитела, диатела, триатела, тетратела и полипептиды, которые содержат по меньшей мере часть иммуноглобулина, достаточную для обеспечения специфического связывания антигена с полипептидом. Фрагменты, производные и аналоги антител могут быть получены методами рекомбинантной ДНК, или путем ферментативного или химического расщепления интактных антител.
[00053] Fab-фрагмент представляет собой моновалентный фрагмент, содержащий домены VL, VH, CL и CH1; фрагмент F(ab')2 представляет собой бивалентный фрагмент, содержащий два Fab-фрагмента, связанных дисульфидным мостиком в шарнирной области; фрагмент Fd содержит домены VH и CH1; фрагмент Fv содержит домены VL и VH одного плеча антитела; и фрагмент dAb содержит домен VH, домен VL или антигенсвязывающий фрагмент домена VH или VL (патенты США №6846634; №6696245, патентные публикации США 20/0202512; 2004/0202995; 2004/0038291; 2004/0009507; 2003/0039958, и Ward et al., Nature 341:544-546, 1989).
[00054] Одноцепочечное антитело (scFv) представляет собой антитело, в котором область VL и область VH соединены линкером (например, синтетической последовательностью остатков аминокислот) с образованием непрерывной белковой цепи, при этом указанный линкер имеет достаточную длину для того, чтобы позволить белковой цепи сворачиваться вовнутрь и формировать моновалентный сайт связывания антигена (см., например, Bird et al., 1988, Science 242:423-26 и Huston et al., 1988, Proc. Natl. Acad. Sci. USA 85:5879-83). Диатела представляют собой бивалентные антитела, содержащие две полипептидные цепи, где каждая полипептидная цепь включает домены VH и VL, соединенные линкером, слишком коротким, чтобы позволять спаривание двух доменов на одной цепи, и таким образом позволяющим каждому домену спариваться с комплементарным доменом другой полипептидной цепи (см, например, Holliger et al., 1993, Proc. Natl. Acad. Sci. USA 90:6444-48, и Poljak et al., 1994, Structure 2:1121-23). Если две полипептидные цепи диатела идентичны, то диатело, полученное в результате их спаривания, будет иметь два идентичных антигенсвязывающих сайта. Полипептидные цепи, имеющие разные последовательности, можно использовать для получения диатела с двумя разными антигенсвязывающими сайтами. Аналогичным образом, триатела и тетратела представляют собой антитела, содержащие три и четыре полипептидных цепи, соответственно, и образующие соответственно три и четыре антигенсвязывающих сайта, которые могут быть одинаковыми или разными.
[00055] Определяющие комплементарность области (CDR) и каркасные области (FR) определенного антитела могут быть идентифицированы с использованием системы, описанной в источниках: Kabat et al. выше; Lefranc et al., выше, и/или Honegger and Pluckthun, выше. Одна или более CDR могут быть включены в молекулу ковалентным или нековалентным образом, с получением антигенсвязывающего белка. Антигенсвязывающий полипептид может включать CDR как часть более крупной полипептидной цепи, может ковалентно связывать CDR с другой полипептидной цепью или может нековалентным образом включать CDR. Указанные CDR позволяют антигенсвязывающему белку специфически связываться с конкретным представляющим интерес антигеном.
[00056] Слитый белок может содержать один или более сайтов связывания. Если имеется более одного сайта связывания, указанные сайты связывания могут быть идентичны друг другу или могут быть разными. Например, встречающийся в природе иммуноглобулин человека обычно содержит два идентичных сайта связывания, в то время как «биспецифическое» или «бифункциональное» антитело содержит два разных сайта связывания.
[00057] Термин «человеческое антитело» или «гуманизированное антитело» включает все антитела, которые содержат одну или более вариабельных и константных областей, происходящих из последовательностей иммуноглобулина человека. В одном варианте осуществления все вариабельные и константные домены происходят из последовательностей иммуноглобулина человека (полностью человеческое или гуманизированное антитело). Указанные антитела могут быть получены различными способами, в том числе путем иммунизации представляющим интерес антигеном мыши, генетически модифицированной для экспрессии антител, происходящих из генов человека, кодирующих тяжелые и/или легкие цепи. Гуманизированное антитело имеет последовательность, которая отличается от последовательности антитела, происходящего от вида, не являющегося человеком, одной или более заменами, делециями и/или добавлениями аминокислот, такими, что указанное гуманизированное антитело при введении субъекту-человеку с меньшей вероятностью вызывает иммунный ответ и/или вызывает менее тяжелый иммунный ответ по сравнению с антителом вида, не являющегося человеком. В одном варианте осуществления определенные аминокислоты в каркасных и константных доменах тяжелых и/или легких цепей антитела вида, не являющегося человеком, мутированы для получения гуманизированного антитела. В другом варианте осуществления константный домен (или домены) из человеческого антитела сливают с вариабельным доменом (или доменами) не являющегося человеком вида. В другом варианте осуществления один или более остатков аминокислот в одной или нескольких последовательностях CDR антитела вида, не являющегося человеком, изменены для снижения вероятной иммуногенности указанного антитела вида, не являющегося человеком, при его введении субъекту-человеку, причем измененные остатки аминокислот либо не являются критическими для иммуноспецифического связывания антитела с его антигеном, либо внесенные в последовательность аминокислот изменения являются консервативными, так что связывание гуманизированного антитела с антигеном существенно не ухудшается по сравнению со связыванием с указанным антигеном антитела вида, не являющегося человеком. Примеры способов получения гуманизированных антител можно найти в патентах США №6054297, №5886152 и №5877293.
[00058] Активированные Т-клетки экспрессируют PD1 на поверхности. Связывание PD-L1 с PD1 активирует PD1 и подавляет PD1+ Т-клетки. «Нейтрализующее антитело» или «ингибирующее антитело» представляет собой антитело, которое блокирует активацию PD1, при этом избыток антитела против PD-L1 снижает уровень указанной активации по меньшей мере примерно на 20% по оценке с использованием такого анализа, как описанные в примерах в настоящем документе. В различных вариантах осуществления антигенсвязывающий белок снижает уровень активации PD1 по меньшей мере на 30%, 40%, 50%, 60%, 70%, 75%, 80%, 85%, 90%, 95%, 97%, 99% и 99,9%.
[00059] Специалисты в данной области техники смогут легко получить фрагменты или аналоги антител, следуя принципам, изложенным в настоящем документе, с использованием методик, известных в данной области техники. Предпочтительные амино- и карбоксиконцы фрагментов или аналогов расположены вблизи границ функциональных доменов. Структурные и функциональные домены могут быть идентифицированы путем сравнения данных последовательностей нуклеотидов и/или аминокислот с общедоступными или собственными базами данных последовательностей. Компьютеризированные методы сравнения могут использоваться для идентификации мотивов последовательностей или доменов предсказанной конформации белка, которые встречаются в других белках с известной структурой и/или функцией. Известны способы идентификации белковых последовательностей, которые укладываются в известную трехмерную структуру. См. Bowie et al., 1991, Science 253:164.
[00060] Слитый белок «специфически связывается» с антигеном (например, PD-L1 человека), если он связывается с антигеном с константой диссоциации, равной 100 нмоль или менее.
[00061] «Антигенсвязывающий домен», «антигенсвязывающая область» или «антигенсвязывающий сайт» представляет собой часть антигенсвязывающего белка, которая содержит остатки аминокислот (или другие фрагменты), которые взаимодействуют с антигеном и вносят вклад в антигенную специфичность связывающего белка и аффинность к антигену. Для того чтобы антитело специфически связывалось со своим антигеном, оно должно включать по меньшей мере часть по меньшей мере одного из его доменов CDR.
[00062] «Эпитоп» представляет собой часть молекулы, которую связывает антигенсвязывающий белок (например, антитело). Эпитоп может включать несмежные части молекулы (например, в случае полипептида, остатки аминокислот, которые не являются смежными в первичной последовательности полипептида, но в контексте третичной и четвертичной структуры полипептида находятся достаточно близко друг к другу, чтобы их связывал антигенсвязывающий белок.
[00063] В контексте настоящего описания термины «полинуклеотид», «олигонуклеотид» и «нуклеиновая кислота» используются взаимозаменяемо и включают молекулы ДНК (например, кДНК или геномной ДНК), молекулы РНК (например, мРНК), аналоги ДНК или РНК, полученные с использованием нуклеотидных аналогов (например, пептидные нуклеиновые кислоты и не встречающиеся в природе аналоги нуклеотидов) и их гибриды. Молекула нуклеиновой кислоты может быть одноцепочечной или двухцепочечной. В одном варианте осуществления молекулы нуклеиновой кислоты по изобретению содержат непрерывную открытую рамку считывания, кодирующую слитый белок, антитело или его фрагмент, производное, мутант или вариант.
[00064] «Вектор» представляет собой нуклеиновую кислоту, которую можно использовать для введения другой, соединенной с ней нуклеиновой кислоты в клетку. Одним из типов вектора является «плазмида», которая представляет собой линейную или кольцевую двухцепочечную молекулу ДНК, с которой могут быть лигированы дополнительные сегменты нуклеиновой кислоты. Другим типом вектора является вирусный вектор (например, репликативно-дефектные ретровирусы, аденовирусы и аденоассоциированные вирусы), при использовании которого дополнительные сегменты ДНК могут быть введены в вирусный геном. Некоторые векторы способны к автономной репликации в клетке-хозяине, в которую их вводят (например, бактериальные векторы, содержащие бактериальную точку начала репликации, и эписомальные векторы млекопитающих). Другие векторы (например, неэписомальные векторы млекопитающих) интегрируются в геном клетки-хозяина после введения в клетку-хозяина и, таким образом, реплицируются вместе с геномом хозяина. «Экспрессионный вектор» представляет собой тип вектора, который может направлять экспрессию выбранного полинуклеотида.
[00065] Нуклеотидная последовательность «функционально связана» с регуляторной последовательностью, если указанная регуляторная последовательность влияет на экспрессию (например, уровень, время или место экспрессии) нуклеотидной последовательности. «Регуляторная последовательность» представляет собой нуклеиновую кислоту, которая влияет на экспрессию (например, уровень, временные характеристики или место экспрессии) нуклеиновой кислоты, с которой она функционально связана. Регуляторная последовательность может, например, оказывать влияние на регулируемую нуклеиновую кислоту непосредственным образом или через действие одной или нескольких других молекул (например, полипептидов, которые связываются с указанной регуляторной последовательностью и/или нуклеиновой кислотой). Примеры регуляторных последовательностей включают промоторы, энхансеры и другие элементы контроля экспрессии (например, сигналы полиаденилирования). Дополнительные примеры регуляторных последовательностей описаны, например, в Goeddel, 1990, Gene Expression Technology: Methods in Enzymology 185, Academic Press, San Diego, Calif.; и Baron et al., 1995, Nucleic Acids Res. 23:3605-06.
[00066] В настоящем документе термины, такие как «терапевтический», относятся к свойству и способности излечивать, замедлять, уменьшать симптомы и/или приостанавливать прогрессирование диагностированного патологического состояния или расстройства. Таким образом, терапевтический белок или полипептид представляет собой белок или полипептид, обладающий указанными выше свойством и способностью. «Лечение» субъекта способами, описанными в настоящем изобретении, проходит успешно, например, если у указанного пациента наблюдается одно или более из следующего: уменьшение количества или полное отсутствие раковых клеток; уменьшение размера опухоли; ингибирование или отсутствие инфильтрации раковых клеток в периферические органы, в том числе распространения рака в мягкие ткани и кости; ингибирование или отсутствие метастазирования опухоли; ингибирование или отсутствие роста опухоли; облегчение одного или нескольких симптомов, связанных с конкретным раком; снижение заболеваемости и смертности; и улучшение качества жизни.
[00067] Предпочтительно, широкий спектр раковых заболеваний млекопитающих, подлежащих лечению композициями по настоящему изобретению, выбран из группы, состоящей из рака яичников, рака толстой кишки, рака молочной железы, рака легкого, миелом, опухолей ЦНС, происходящих из нейробластов, моноцитарных лейкозов, B-клеточных лейкозов, Т-клеточных лейкозов, В-клеточных лимфом, Т-клеточных лимфом, опухолей, происходящих из тучных клеток, меланомы, рака мочевого пузыря, рака желудка, рака печени, уротелиальной карциномы, кожной карциномы, рака почек, рака головы и шеи, рака поджелудочной железы и их комбинаций. В более широком смысле, любой рак, при котором по меньшей мере часть опухолевых клеток экспрессирует детектируемое количество PD-L1, потенциально можно лечить композицией по настоящему изобретению.
[00068] Слитые белки или полипептиды по настоящему изобретению могут быть получены с использованием любых стандартных методов, известных в данной области. В одном примере указанные полипептиды получают методами рекомбинантной ДНК, путем инсерции последовательности нуклеиновой кислоты (например, кДНК), кодирующей полипептид, в рекомбинантный экспрессионный вектор, и экспрессии указанной последовательности ДНК в условиях, способствующих экспрессии.
[00069] Нуклеиновые кислоты, кодирующие любые из различных слитых белков или полипептидов, описанных в настоящем документе, можно синтезировать химическим путем. Частоты использования кодонов могут быть выбраны так, чтобы улучшить экспрессию в клетке. Такие частоты использования кодонов будут зависеть от выбранного типа клетки. Специализированные схемы использования кодонов были разработаны для E. coli и других бактерий, а также для клеток млекопитающих, клеток растений, дрожжевых клеток и клеток насекомых. См., например: Mayfield et al., Proc. Natl. Acad. Sci. USA. 2003 100(2):438-42; Sinclair et al. Protein Expr. Purif. 2002 (1):96-105; Connell N D. Curr. Opin. Biotechnol. 2001 12(5):446-9; Makrides et al. Microbiol. Rev. 1996 60(3):512-38; и Sharp et al. Yeast. 1991 7(7):657-78.
[00070] Общие методы манипуляций с нуклеиновыми кислотами описаны, например, в источниках: Sambrook et al., Molecular Cloning: A Laboratory Manual, Vols. 1-3, Cold Spring Harbor Laboratory Press, 2 ed., 1989, или F. Ausubel et al., Current Protocols in Molecular Biology (Green Publishing and Wiley-Interscience: New York, 1987) с периодическими обновлениями, включенных в настоящий документ посредством ссылки. ДНК, кодирующая полипептид, функционально связана с подходящими регуляторными элементами транскрипции или трансляции, происходящими из генов млекопитающих, вирусов или насекомых. Такие регуляторные элементы включают промотор транскрипции, необязательную последовательность оператора для контроля транскрипции, последовательность, кодирующую подходящие сайты связывания рибосомы с мРНК, и последовательности, которые контролируют терминацию транскрипции и трансляции. Дополнительно включены способность к репликации у хозяина, которую обычно обеспечивает точка начала репликации, и селективный ген для облегчения распознавания трансформантов.
[00071] Рекомбинантная ДНК может также включать любой тип последовательности белковой метки, которая может подходить для очистки белка. Примеры белковых меток включают гистидиновую метку, метку FLAG, метку myc, HA-метку или метку GST, но не ограничиваются перечисленными. Описание подходящих клонирующих и экспрессионных векторов для применения с бактериальными, грибными, дрожжевыми клетками-хозяевами и клетками-хозяевами млекопитающих можно найти в источнике: Cloning Vectors: A Laboratory Manual, (Elsevier, N.Y., 1985).
[00072] Экспрессионную конструкцию вводят в клетку-хозяина с применением способа, подходящего для указанной клетки-хозяина. В данной области известны разнообразные способы введения нуклеиновых кислот в клетки-хозяева, включая, но не ограничиваясь перечисленными, электропорацию; трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию; и инфекцию (если вектор является инфекционным агентом). Подходящие клетки-хозяева включают прокариот, клетки дрожжей, млекопитающих или бактерий.
[00073] Описанные здесь белки также могут быть получены с использованием систем клеточной трансляции. Для этих целей нуклеиновые кислоты, кодирующие полипептид, должны быть модифицированы, чтобы обеспечивать транскрипцию in vitro для продуцирования мРНК и бесклеточную трансляцию указанной мРНК в конкретной используемой бесклеточной системе (эукариотической, такой как система трансляции без клеток млекопитающих или дрожжей, или прокариотической, такой как система трансляции без клеток бактерий).
[00074] Описанная здесь биспецифическая молекула также может быть получена с использованием систем клеточной трансляции. Чтобы упростить получение, существует несколько способов подтверждения преимущественного продуцирования гетеродимерной биспецифической молекулы в клетках за счет использования комплементарных мутаций в двух молекулах тяжелых цепей.
[00075] Слитый белок или полипептид, по настоящему изобретению также можно получить химическим синтезом (например, способами, описанными в источнике: Solid Phase Peptide Synthesis, 2nd ed., 1984, The Pierce Chemical Co., Rockford, Ill.). Модификации белка также могут быть получены путем химического синтеза.
[00076] Другой вариант молекулы слитого белка имеет мутации в одной или более аминокислотах для удаления сайтов протеолитического расщепления для улучшения продуцирования.
[00077] Другой вариант молекулы слитого белка имеет мутации в одной или более аминокислотах для удаления сайтов гликозилирования, без какого-либо иного изменения функции белка, для получения более гомогенного продукта.
[00078] В другом варианте молекулы слитого белка используется альтернативная секреторная последовательность вместо естественной секреторной последовательности антитела для улучшения секреции.
[00079] Слитые белки или полипептиды по настоящему изобретению могут быть очищены методами выделения/очистки белков, общеизвестными в области химии белков. Неограничивающие примеры включают экстракцию, перекристаллизацию, высаливание (например, сульфатом аммония или сульфатом натрия), центрифугирование, диализ, ультрафильтрацию, адсорбционную хроматографию, ионообменную хроматографию, гидрофобную хроматографию, нормально-фазовую хроматографию, обращенно-фазовую хроматографию, гель-фильтрацию, гель-проникающую хроматографию, аффинную хроматографию, электрофорез, противоточное распределение или любую их комбинацию. После очистки полипептидов может быть проведена замена буфера на различные буферы и/или полипептиды могут быть сконцентрированы любым из множества способов, известных в данной области, включая, но не ограничиваясь перечисленными, фильтрацию и диализ.
[00080] Очищенный слитый белок или полипептид преимущественно имеет по меньшей мере 85% чистоту, более предпочтительно по меньшей мере 95% чистоту, и наиболее предпочтительно по меньшей мере 98% чистоту. Независимо от точного числового значения чистоты полипептид является достаточно чистым для использования в качестве фармацевтического продукта.
Посттрансляционные модификации полипептидов
[00081] В некоторых вариантах осуществления слитый белок или полипептиды по изобретению могут дополнительно включать посттрансляционные модификации. Примеры посттрансляционных модификаций белков включают фосфорилирование, ацетилирование, метилирование, АДФ-рибозилирование, убиквитинирование, гликозилирование, карбонилирование, сумоилирование, биотинилирование или добавление боковой цепи полипептида или гидрофобной группы. В результате модифицированные растворимые полипептиды могут содержать неаминокислотные элементы, такие как липиды, поли- или моносахарид и фосфаты. Предпочтительной формой гликозилирования является сиалирование, при котором один или более фрагментов сиаловой кислоты конъюгируют с полипептидом. Фрагменты сиаловой кислоты улучшают растворимость и период полужизни в сыворотке, а также снижают возможную иммуногенность белка. См. Raju et al. Biochemistry. 2001 31; 40(30):8868-76. Эффекты таких неаминокислотных элементов на функциональность полипептида могут быть протестированы на антагонизирующую роль в функции PD-L1 или PD-1, например, на ингибирующее действие на ангиогенез или на рост опухоли.
[00082] В одном варианте осуществления биологическая активность относится к его способности связываться с PD-L1, по оценке на основании коэффициентов диссоциации (KD), скорости ассоциации (Kon) или скорости диссоциации (Koff). В одном конкретном варианте осуществления пегилированный полипептидный белок демонстрирует увеличение связывания с PD-L1 человека по сравнению с непегилированным аналогом. В другом варианте осуществления биологическая активность относится к блокаде взаимодействия PD-L1/PD1.
Терапевтические средства, вакцины и введение
[00083] Настоящее изобретение дополнительно предусматривает способы лечения состояний или предотвращения при предпосылках состояний. Предпочтительными примерами являются состояния, которые характеризуются клеточной гиперпролиферацией и продолжительной инфекцией. Способы и дозировки для введения варьируют в зависимости от типа конкретного полипептида и конкретного состояния, которое лечат. Поскольку регулирующие органы требуют, чтобы состав с белковым реагентом, который применяют в качестве терапевтического средства, содержал приемлемо низкие уровни пирогенов, терапевтические составы по настоящему изобретению можно отличить от других составов на основании того, что они по существу не содержат пирогенов или по меньшей мере содержат не более допустимые уровни пирогена, как определено соответствующим регулирующим органом (например, Управлением по санитарному надзору за качеством пищевых продуктов и медикаментов (FDA)).
[00084] Фармацевтические составы по настоящему изобретению могут включать по меньшей мере один фармацевтически приемлемый разбавитель, носитель или вспомогательное вещество. Вспомогательные вещества, включенные в указанные составы, служат разным целям в зависимости, например, от типа используемой генной конструкции или эффекторных клеток, а также способа введения. Примеры обычно используемых вспомогательных веществ включают, без ограничения: физиологический раствор, забуференный физиологический раствор, декстрозу, воду для инфекций, глицерин, этанол и их комбинации, стабилизирующие агенты, солюбилизирующие агенты и поверхностно-активные вещества, буферы и консерванты, регулирующие тоничность агенты, наполнители и смазывающие агенты.
[00085] В другом варианте осуществления изобретения фармацевтический состав по изобретению вводят пациенту. Примеры способов введения включают внутривенные инъекции, но не ограничиваются ими. Другие способы включают, без ограничения, внутриопухолевый, внутрикожный, подкожный (п/к, п.к., sub-Q, Hypo), внутримышечный (в/м), внутрибрюшинный (в/б), внутриартериальный, интрамедуллярный, внутрисердечный, внутрисуставной (суставной), внутрисиновиальный (в область суставной жидкости), внутричерепной, интраспинальный и интратекальный (в спинномозговую жидкость). Любое известное устройство, подходящее для парентеральных инъекций или инфузии составов, может быть использовано для осуществления такого введения. В настоящем документе термины «лечить» и «лечение» имеют обычные, общепринятые значения и включают одно или более из следующего: блокирование, облегчение или уменьшение тяжести и/или частоты симптома заболевания (например, рака) у субъекта, и/или ингибирование роста, деления, распространения или пролиферации раковых клеток или прогрессирования рака (например, появления новых опухолей) у субъекта. Лечение означает блокирование, облегчение, уменьшение или ингибирование примерно на 5% - примерно на 100% по сравнению с субъектом, у которого не применяли способы по настоящему изобретению. Предпочтительно указанное блокирование, облечение, уменьшение или ингибирование составляет примерно 100%, 99%, 95%, 90%, 80%, 70%, 60%, 50%, 40%, 30%, 20%, 10% или 5% по сравнению с субъектом, у которого не применяли способы по настоящему изобретению.
[00086] Согласно настоящему изобретению также предложен набор, включающий один или более контейнеров, заполненных некоторым количеством генных конструкций, кодирующих слитый белок или полипептиды по изобретению, с фармацевтически приемлемыми вспомогательными веществами. В указанный набор также может входить инструкция по применению. Кроме того, с указанным набором может быть ассоциировано уведомление в форме, предписанной государственным органом, регулирующим производство, применение или продажу фармацевтических или биологических продуктов, отражающее одобрение указанным органом производства, применения или продажи для введения человеку.
ПРИМЕРЫ
[00087] Пример 1
[00088] Легкую цепь к PDL1 (IgL) и тяжелую цепь против PD-L1 (IgH), слитые с MutIL2, клонировали в один эукариотический экспрессионный вектор с двумя кассетами экспрессии - вектор pCHO 1.0. Другие аналогичные векторы коммерчески доступны. Кроме того, указанные два гена можно клонировать в отдельные векторы. Вектором трансфицировали клетки Expi293 в соответствии с рекомендациями производителя. Белок секретировался в супернатант и его очищали до чистоты> 95% на колонке HiTrap с белком A с использованием стандартных процедур. Очищенные белки сравнивали с ИЛ2 дикого типа на предмет способности активировать преимущественно эффекторные Т-клетки, а не регуляторные Т-клетки. В указанном варианте слитые формы названы с использованием названий как антитела к PDL1, так и специфического MutIL2. Например, E1M1 содержит антитело E1 и MutIL2 IL2M1.
[00089] Пример 2
[00090] Чтобы протестировать способность белков связывать PDL1 на клеточной поверхности, авторы изобретения использовали клетки DLD1, экспрессирующие трансгенно экспрессируемый PDL1 человека. Аналогичные результаты могут быть получены либо при трансгенной экспрессии в других клеточных линиях, либо при использовании клеток, которые естественным образом экспрессируют PDL1, или клеток, экспрессия PDL1 в которых индуцирована обработкой интерфероном γ. Вкратце, 105 клеток инкубировали с различными концентрациями белка в течение 30 минут при 4°C. Клетки промывали ФСБ и инкубировали с флуоресцентным антителом против человеческого IgG в течение 30 минут при 4°C. Клетки снова промывали ФСБ и определяли связывание с помощью FACS. На фиг. 3 показано, что использование для слияния ИЛ2 с разными мутациями не влияет на связывание с PDL1 (таблица 2).
[00091] Аналогичным образом, для тестирования способности части антитела блокировать взаимодействия PD1-PDL1 проводили анализы блокирования рецептора в двух разных форматах ИФА ELISA - либо со связыванием биотинилированного Fc-PDL1 с планшетами, покрытыми PD1, либо со связыванием биотинилированного Fc-PD1 с покрытым PDL1 планшетами. В обоих случаях связывание было детектировано с использованием связанной со стрептавидином пероксидазы хрена. В таблице 3 показано, что примеры антител PDL1 демонстрируют высокую эффективность блокирования взаимодействия PD1-PDL1.
[00092] Пример 3
[00093] Для тестирования АЗКЦ-активности авторы изобретения использовали коммерческий набор для анализа АЗКЦ (Promega) на основе клеток Jurkat с геном люциферазы, чувствительным к рецептору Fc. Совместная инкубация этих клеток с клетками PDL1+ MDA-MB-231 и антителом АЗКЦ + запускает активацию рецептора Fc, которую можно измерить по активности люциферазы. Фиг. 4 показывает, что различные слитые белки по изобретению проявляют АЗКЦ-активность.
[00094] Пример 4
[00095] Для тестирования способности мутантных белков связывать рецепторы ИЛ2 на клетках CTLL2 105 клеток инкубировали с различными концентрациями белка в течение 30 минут при 4°C. Клетки промывали ФСБ и инкубировали с флуоресцентным антителом против IgG человека в течение 30 минут при 4°C. Клетки снова промывали ФСБ и определяли связывание с помощью FACS. Фиг. 5 показывает, что различные слитые мутантные ИЛ2 демонстрируют резкое снижение связывания с высокоаффинным рецептором ИЛ2 на клетках CTLL2 по сравнению со слитым ИЛ2 дикого типа.
[00096] Пример 5
[00097] Для тестирования способности мутантных белков активировать клетки CTLL2 (суррогат регуляторных Т-клеток) 104 клеток культивировали в присутствии различных концентраций слитого белка мутантного или дикого типа ИЛ2 в течение 44 часов. Относительное количество жизнеспособных клеток определяли путем инкубации культур с с аламаровым синим в течение 1-4 часов и измерения интенсивности флуоресценции с использованием ридера для планшетов со считыванием флуоресценции. Фиг. 6 показывает, что различные слитые мутантные ИЛ2 демонстрируют более чем 100-кратное снижение способности поддерживать пролиферацию клеток CTLL2 по сравнению со слитым ИЛ2 дикого типа (таблица 4). Это верно для слитых форм, содержащих любое антитело к PDL1. Слитая форма дикого типа сравнимо с рекомбинантным ИЛ2.
[00098] Пример 6
[00099] Для тестирования способности мутантных белков активировать различные подгруппы Т-клеток мыши авторы изобретения использовали клетки из различных источников. Селезенку или лимфатические узлы мышей использовали для выделения CD4, CD8 или регуляторных Т-клеток c использованием набора для выделения клеток на основе магнитных гранул от Stem Cell Technologies в соответствии с рекомендациями производителя. Анализы NK-клеток мыши проводили на тотальных спленоцитах. Коммерчески доступны другие процедуры выделения. Подгруппы Т-клеток человека и NK-клетки человека приобретали у коммерческого поставщика, однако они могут быть выделены из МКПК с использованием наборов, аналогичных тем, которые используют для клеток мыши. Различные типы клеток инкубировали с различными количествами слитого белка мутантного или дикого типа ИЛ2. Например, 2,5×104-105 Т-клеток мышей использовали для экспериментов с Т-клетками мышей. Для NK-клеток мыши использовали 1-5×106 спленоцитов. Культуры регуляторных Т-клеток активировали связанными на планшете антителами против CD3 и также инкубировали с нейтрализующим антителом мыши к ИЛ2. Через 4 дня культивирования пролиферацию клеток измеряли прямым подсчетом (с исключением или без исключения мертвых клеток с использованием йодида пропидия) или методом разведения CFSE с использованием FACS. Кроме того, также оценивали экспрессию маркеров активации, например, CD44 для клеток CD8 мыши. Фиг. 7 показывает, что различные слитые мутантные ИЛ2 демонстрируют значительное снижение способности активировать и поддерживать выживание регуляторных Т-клеток по сравнению со слитым ИЛ2 дикого типа. На фиг. 8 и в таблице 5 показано, что слитые мутантные ИЛ2 сравнимы со слитым белком ИЛ2 дикого типа по способности поддерживать пролиферацию CD8+ Т-клеток мыши. Слитый ИЛ2 дикого типа сравнимо с рекомбинантным ИЛ2 в обоих анализах.
[000100] На фиг. 6, фиг. 7 и фиг. 8 показано, что различные слитые мутантные ИЛ2 демонстрируют более чем 100-кратное увеличение относительной способности активировать эффекторные/регуляторные Т-клетки по сравнению с слитым ИЛ2 дикого типа.
[000101] Пример 7
[000102] Для тестирования способности мутантных белков активировать различные подгруппы Т-клеток человека авторы изобретения использовали клетки периферической крови здоровых доноров-людей. МКПК получали с помощью стандартных процедур и инкубировали с рекомбинантным ИЛ2 или различными слитыми ИЛ2 в течение 15 минут. Фосфорилирование Stat5, запускаемое взаимодействием рецептора ИЛ2 в различных субпопуляциях Т-клеток, оценивали путем окрашивания на соответствующие клеточные маркеры и анализа с помощью FACS. Фиг. 9 и 10 показывают, что по сравнению со слитым ИЛ2 дикого типа различные слитые мутантные ИЛ2 демонстрируют значительное (примерно 100-кратное) увеличение относительной способности активировать не-регуляторные/регуляторные Т-клетки (данные также обобщены в таблицах 6-8). Фиг. 11 показывает, что слитые мутанты ИЛ2 сравнимы со слитым белком ИЛ2 дикого типа по способности индуцировать фосфорилирование Stat5 в CD8 + Т-клетках (данные также обобщены в таблице 9). По сравнению со слитым ИЛ2 дикого типа различные слитые мутантные ИЛ2 демонстрируют значительное (примерно в 1000 раз) увеличение относительной способности «активировать» CD8 + Т-клетки/регуляторные CD4 +Т-клетки (см. таблицу 10). Слитый ИЛ2 дикого типа сравнимо с рекомбинантным ИЛ2 во всех анализах. Эти данные показывают, что различные слитые мутантные ИЛ2 обладают пониженной способностью активировать регуляторные Т-клетки человека, сохраняя при этом способность активировать эффекторные Т-клетки, и демонстрируют более чем 100-кратное увеличение относительной способности активировать эффекторные/регуляторные Т-клетки по сравнению со слитым ИЛ2 дикого типа.
[000103] Пример 8
[000104] Для исследований фармакокинетики (PK) самкам гомозиготных мышей Tg32 (в возрасте 6-8 недель) внутривенно вводили тестируемый слитый белок через хвостовую вену в дозе 0,1-10 мг/кг 5 животным на группу. Кровь брали в нескольких точках времени и получали сыворотку центрифугированием. Количество слитого белка оценивали с помощью сэндвич-анализа ELISA. Титр нормировали по значениям для 1 дня после инъекции.
[000105] На фиг. 12 приведена фармакокинетика для четырех слитых MutIL2. Периоды полужизни слитых MutIL2 приведены в таблице 11. Указанные периоды значительно, примерно на два часа, продолжительнее зарегистрированного периода полужизни рекомбинантного ИЛ2.
[000106] Пример 9
[000107] Для фармакодинамических и токсикологических исследований мышам дикого типа B6 внутривенно вводили тестируемое антитело через хвостовую вену в дозе 0,1-10 мг/кг и наблюдали в течение нескольких дней на предмет активации иммунной системы и побочных реакций. Кровь брали в течение нескольких дней для оценки увеличения компартментов Т-клеток и естественных киллерных клеток. В конце наблюдения мышей умерщвляли и аналогичным образом анализировали селезенку, а в легких и печени оценивали инфильтрацию лимфоцитов и другие иммунные реакции.
[000108] На фиг. 13 показано значительное размножение компартментов естественных киллерных (NK) клеток и CD8+ Т-клеток по сравнению с компартментом CD4+. Способность различных мутантных форм слитых белков ИЛ2 согласно настоящему изобретению к дифференциальному индуцированию преимущественной пролиферации CD8+ Т-клеток и NK-клеток по сравнению с регуляторными Т-клетками указывает на большой потенциал указанных слитых белков в области улучшения противоопухолевой активности антител к контрольным точкам и их клинического применения в лечении рака.
[000109] Пример 10
[000110] Для детекции нацеливания на опухоль слитых белков использовали либо сингенные опухоли (например, MC38 для мышей B6), либо сингенные опухоли, экспрессирующие PDL1 человека. Обычно 1 × 106 опухолевых клеток имплантировали подкожно и позволяли расти, пока они не достигли 100 мм2. Мышам вводили 0,1-10 мг / кг слитого белка. Ткань опухоли и другие органы собирали в различные моменты времени, и накопление слитого белка определяли с помощью сэндвич-анализа ELISA или иммуногистохимии. В качестве альтернативы использовали радиоактивно меченые белки и детектировали их стандартными методами.
Последовательности
[0001111] Примеры последовательностей, которые образуют часть слитого белка согласно представленным изобретениям, перечислены ниже.
[000112] Примеры полноразмерных последовательностей и последовательностей вариабельной области для антител к PDL1, которые блокируют взаимодействие PD1-PDL1, описаны ниже. Последовательности ДНК в качестве примеров также описаны ниже. Другие варианты, основанные на использовании альтернативных кодонов последовательностей ДНК, являются альтернативными вариантами, предусмотренными настоящим изобретением.
[000113] Антитело к PDL1
[000114] Тяжелая цепь B2
[000115] Последовательность ДНК (SEQ ID NO: 1)
ATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTCGCGATTCTTAAGGGTGTCCAGTGCGAAGTGCAGCTGGTGGAGTCTGGGGGAGGCTTGGTACAGCCTGGAGGGTCCCTGAGACTCTCCTGTGCAGCCTCTGGATTCACTTTTAGTGACTATGACATGATCTGGGTCCGCCAGGCTCCAGGCAAGGGGCTGGAGTGGGTGGCAGTTATATCATATGATGGAAGTAATAAATACTATGCAGACTCCGTGAAGGGCCGATTCACCATCTCCAGAGACAATTCCAAGAACACGCTGTATCTGCAAATGAACAGCCTGAGAGCTGAGGACACGGCTGTGTATTACTGTGCGAAAGAGTTCTTTGGTGCTTTTGATATCTGGGGCCAAGGGACAATGGTCACCGTCTCTTCAGCTTCGACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGCGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA
[000116] Последовательность белка
[000117] Полная длина (SEQ ID NO: 2)
MEFGLSWLFLVAILKGVQCEVQLVESGGGLVQPGGSLRLSCAASGFTFSDYDMIWVRQAPGKGLEWVAVISYDGSNKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKEFFGAFDIWGQGTMVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
[000118] Вариабельная область (SEQ ID NO: 3)
EVQLVESGGGLVQPGGSLRLSCAASGFTFSDYDMIWVRQAPGKGLEWVAVISYDGSNKYYADSVKGRFTISRDNSKNTLYLQMNSLRAEDTAVYYCAKEFFGAFDIWGQGTMVTVSS
[000119] Легкая цепь B2
[000120] Последовательность ДНК (SEQ ID NO: 4)
ATGACTTGGACCCCACTCCTCTTCCTCACCCTCCTCCTCCACTGCACAGGAAGCTTATCCTCTTCTGAGCTGACTCAGGACCCTGCTGTGTCGGTGGCCTTGGGACAGACAGTCACGATCACATGCCAAGGAGACAGCCTCAATTACTATTATGCAAACTGGTTCCAGCTGAAGCCAGGGCAGGCCCCTGTACTTGTCCTCTTTGGTAAAAACAACCGGCCCTCAGGGATCCCAGACCGATTCTCTGGCTCCTACTCGGGAAGCACAGCTTCCTTGACCATCACTGGGGCTCAGGCGGAAGATGACGCTGACTATTACTGTAATTCGCGGGACAGCGGTGGTAATCCTTGGGTGTTCGGCGGAGGGACCAAGCTGACCGTCCTAGGTCAACCCAAGGCTGCCCCCTCGGTCACTCTGTTCCCGCCCTCCTCTGAGGAGCTTCAAGCCAACAAGGCCACACTGGTGTGTCTCATAAGTGACTTCTACCCGGGAGCCGTGACAGTGGCCTGGAAGGCAGATAGCAGCCCCGTCAAGGCGGGAGTGGAGACCACCACACCCTCCAAACAAAGCAACAACAAGTACGCGGCCAGCAGCTACCTGAGCCTGACGCCTGAGCAGTGGAAGTCCCACAGAAGCTACAGCTGCCAGGTCACGCATGAAGGGAGCACCGTGGAGAAGACAGTGGCCCCTACAGAATGTTCATGA
[000121] Последовательность белка
[000122] Полная длина (SEQ ID NO: 5)
MTWTPLLFLTLLLHCTGSLSSSELTQDPAVSVALGQTVTITCQGDSLNYYYANWFQLKPGQAPVLVLFGKNNRPSGIPDRFSGSYSGSTASLTITGAQAEDDADYYCNSRDSGGNPWVFGGGTKLTVLGQPKAAPSVTLFPPSSEELQANKATLVCLISDFYPGAVTVAWKADSSPVKAGVETTTPSKQSNNKYAASSYLSLTPEQWKSHRSYSCQVTHEGSTVEKTVAPTECS
[000123] Вариабельная область (SEQ ID NO: 6)
SSELTQDPAVSVALGQTVTITCQGDSLNYYYANWFQLKPGQAPVLVLFGKNNRPSGIPDRFSGSYSGSTASLTITGAQAEDDADYYCNSRDSGGNPWVFGGGTKLTVL
[000124] Тяжелая цепь E1
[000125] Последовательность ДНК (SEQ ID NO: 7)
ATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTCGCGATTCTTAAGGGTGTCCAGTGCCAAATCCAGCTGGTACAATCTGGGGCTGAGGTGAAGATGCCTGGGGCCTCAGTGACGATTTCCTGCGAGGCGTCTGGATACAACTTCATCAGCTACTATATACACTGGGTGCGACAGGCCCCTGGACAAGGCCTTGAGTGGATGGGATTCGTCGTCCCTAGTGGTGGTGCCGCAGGCTACACACAGAAGTTCCAGGGCAGACTCACCGTGACCAGGGACACGTCCACGAGCACAGTCTACATGGACCTGAACAGCCTGACATCTGACGACACGGCCGTGTATTACTGTGTGCGAGAAATGAGTGGTGGCTGGTTTGATTTCTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCGGCTTCGACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGCGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA
[000126] Последовательность белка
[000127] Полная длина (SEQ ID NO: 8)
MEFGLSWLFLVAILKGVQCQIQLVQSGAEVKMPGASVTISCEASGYNFISYYIHWVRQAPGQGLEWMGFVVPSGGAAGYTQKFQGRLTVTRDTSTSTVYMDLNSLTSDDTAVYYCVREMSGGWFDFWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
[000128] Вариабельная область (SEQ ID NO: 9)
SCEASGYNFISYYIHWVRQAPGQGLEWMGFVVPSGGAAGYTQKFQGRLTVTRDTSTSTVYMDLNSLTSDDTAVYYCVREMSGGWFDFWGQGTLVTVSS
[000129] Легкая цепь E1
[000130] Последовательность ДНК (SEQ ID NO: 10)
ATGGACATGCGCGTGCCCGCCCAGCTGCTGGGCCTGCTGCTGCTGTGGTTCCCCGGCTCGCGATGCGACATCGTGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGGGCATTAGAAATGATTTAGGCTGGTATCAGCAAAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCACTTTGCAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGGACAGATTTCACTCTCACCATCAGCAGCCTGCAGGCTGAAGATGTGGCAGTTTATTACTGTCAGCAATATTATAGTACTCCTCTCACTTTCGGCCCTGGGACCAAAGTGGATATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTGA
[000131] Последовательность белка
[000132] Полная длина (SEQ ID NO: 11)
MDMRVPAQLLGLLLLWFPGSRCDIVMTQSPSSLSASVGDRVTITCRASQGIRNDLGWYQQKPGKAPKLLIYAASTLQSGVPSRFSGSGSGTDFTLTISSLQAEDVAVYYCQQYYSTPLTFGPGTKVDIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[000133] Вариабельная область (SEQ ID NO: 12)
DIVMTQSPSSLSASVGDRVTITCRASQGIRNDLGWYQQKPGKAPKLLIYAASTLQSGVPSRFSGSGSGTDFTLTISSLQAEDVAVYYCQQYYSTPLTFGPGTKVDIK
[000134] Тяжелая цепь E3
[000135] Последовательность ДНК (SEQ ID NO: 13)
ATGGAGTTTGGGCTGAGCTGGCTTTTTCTTGTCGCGATTCTTAAGGGTGTCCAGTGCCAAATCCAGCTGGTACAATCTGGGGCTGAGGTGAAGAAGCCTGGGTCCTCGGTGAAGGTCTCCTGCAAGGCTTCTGGAGGCACCTTCAGCAGCTATGCTATCAGCTGGGTGCGACAGGCCCCTGGACAAGGGCTTGAGTGGATGGGAGGGATCATCCCTATCTTTGGTACAGCAAACTACGCACAGAAGTTCCAGGGCAGAGTCACGATTACCGCGGACAAATCCACGAGCACAGCCTACATGGAGCTGAGCAGCCTGAGATCTGAGGACACGGCCGTGTATTACTGTGCCGGAGGGGGAGCAGTGGCGGACAATAGTTACTGGGGCCAGGGAACCCTGGTCACCGTCTCCTCAGCTTCGACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGCGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA
[000136] Последовательность белка
[000137] Полная длина (SEQ ID NO: 14)
MEFGLSWLFLVAILKGVQCQIQLVQSGAEVKKPGSSVKVSCKASGGTFSSYAISWVRQAPGQGLEWMGGIIPIFGTANYAQKFQGRVTITADKSTSTAYMELSSLRSEDTAVYYCAGGGAVADNSYWGQGTLVTVSSASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
[000138] Вариабельная область (SEQ ID NO: 15)
QIQLVQSGAEVKKPGSSVKVSCKASGGTFSSYAISWVRQAPGQGLEWMGGIIPIFGTANYAQKFQGRVTITADKSTSTAYMELSSLRSEDTAVYYCAGGGAVADNSYWGQGTLVTVSS
[000139] Легкая цепь E3
[000140] Последовательность ДНК (SEQ ID NO: 16)
ATGGACATGCGCGTGCCCGCCCAGCTGCTGGGCCTGCTGCTGCTGTGGTTCCCCGGCTCGCGATGCGACATCCGGATGACCCAGTCTCCATCCTCCCTGTCTGCATCTGTAGGAGACAGAGTCACCATCACTTGCCGGGCAAGTCAGGGCATTAGAAATGATTTAGGCTGGTATCAGCAGAAACCAGGGAAAGCCCCTAAGCTCCTGATCTATGCTGCATCCAGTTTACAAAGTGGGGTCCCATCAAGGTTCAGCGGCAGTGGATCTGGCACAGATTTCACTCTCACCATCAGCAGCCTGCAGCCTGAAGATTTTGCAACTTATTACTGTCTACAAGATTACAATTACCCTCGAACGTTCGGCCAAGGGACCAAGGTGGAAATCAAACGTACGGTGGCTGCACCATCTGTCTTCATCTTCCCGCCATCTGATGAGCAGTTGAAATCTGGAACTGCCTCTGTTGTGTGCCTGCTGAATAACTTCTATCCCAGAGAGGCCAAAGTACAGTGGAAGGTGGATAACGCCCTCCAATCGGGTAACTCCCAGGAGAGTGTCACAGAGCAGGACAGCAAGGACAGCACCTACAGCCTCAGCAGCACCCTGACGCTGAGCAAAGCAGACTACGAGAAACACAAAGTCTACGCCTGCGAAGTCACCCATCAGGGCCTGAGCTCGCCCGTCACAAAGAGCTTCAACAGGGGAGAGTGTTGA
[000141] Последовательность белка
[000142] Полная длина (SEQ ID NO: 17)
MDMRVPAQLLGLLLLWFPGSRCDIRMTQSPSSLSASVGDRVTITCRASQGIRNDLGWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQDYNYPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
[000143] Вариабельная область (SEQ ID NO: 18)
DIRMTQSPSSLSASVGDRVTITCRASQGIRNDLGWYQQKPGKAPKLLIYAASSLQSGVPSRFSGSGSGTDFTLTISSLQPEDFATYYCLQDYNYPRTFGQGTKVEIK
[000144] Пример последовательности линкера приведен ниже:
[000145] Линкер Ser-Gly
[000146] Последовательность белка (SEQ ID NO: 19)
GGGGSGGGGSGGGGSGGGGS
[000147] Пример последовательности (а) IL2WT (последовательность ИЛ2 дикого типа); (b) IL2Del (делеция 20 аминокислот); и (c) IL2M1-MNew (MutIL2: мутанты с желаемыми характеристиками, комбинации этих мутаций также могут использоваться) перечислены ниже:
[000148] Мутанты ИЛ2
[000149] IL2WT
[000150] Последовательность ДНК (SEQ ID NO: 20)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCTTCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000151] Последовательность белка (SEQ ID NO: 21)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTFKFYMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000152] IL2Del
[000153] Последовательность ДНК (SEQ ID NO: 22)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000154] Последовательность белка (SEQ ID NO:23)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000155] IL2M1 F42K, Y45R
[000156] Последовательность ДНК (SEQ ID NO: 24)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCAAGAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000157] Последовательность белка (SEQ ID NO:25)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTKKFRMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000158] IL2M2: F42A, Y45R
[000159] Последовательность ДНК (SEQ ID NO: 26)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCGCCAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000160] Последовательность белка (SEQ ID NO:27)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTAKFRMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000161] IL2M3: E62R
[000162] Последовательность ДНК (SEQ ID NO: 28)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCTTCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000163] Последовательность белка (SEQ ID NO:29)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTFKFYMPKKATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000164] IL2M4: R38A, F42A
[000165] Последовательность ДНК (SEQ ID NO: 30)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCGCCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000166] Последовательность белка (SEQ ID NO:31)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTAKFYMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000167] IL2M5: R38A, F42A, Y45A, E62A, C125S
[000168] Последовательность ДНК (SEQ ID NO: 32)
GGAGCTCCTACCTCCTCCTCCACCAAGAAGACCCAGCTCCAGCTGGAACATTTACTGCTGGATTTACAGATGATTTTAAACGGCATCAACAATTACAAGAACCCCAAGCTGACCGCCATGCTGACCGCCAAGTTTGCCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTTTAGAGGAGGCTTTAAAGCCTTTAGAGGAGGTGCTGAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCCCGGGATTTAATCTCCAACATCAACGTGATCGTGCTGGAACTGAAGGGCAGCGAGACCACCTTTATGAGCGAATACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGTCCATCATCTCCACTTTAACCTGA
[000169] Последовательность белка (SEQ ID NO:33)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTAKFAMPKKATELKHLQCLEEALKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMSEYADETATIVEFLNRWITFSQSIISTLT
[000170] IL2M6: R38A, F42K
[000171] Последовательность ДНК (SEQ ID NO:34)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCAAGAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000172] Последовательность белка (SEQ ID NO:35)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTKKFYMPKKATELKHLQCLEEELKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000174] IL2M7: R38A, F42A, E62R
[000174] Последовательность ДНК (SEQ ID NO:36)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCGCCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000175] Последовательность белка (SEQ ID NO:37)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTAKFYMPKKATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000176] IL2M8: F42K, Y45R, E62R
[000177] Последовательность ДНК (SEQ ID NO:38)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCAAGAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000178] Последовательность белка (SEQ ID NO:39)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTKKFRMPKKATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000179] IL2M9: F42A, Y45R, E62R
[000180] Последовательность ДНК (SEQ ID NO:40)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCGCCAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000181] Последовательность белка (SEQ ID NO:41)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTAKFRMPKKATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000182] IL2M10: R38A, F42K, E62R
[000183] Последовательность ДНК (SEQ ID NO:42)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCAAGAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000184] Последовательность белка (SEQ ID NO:43)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTKKFYMPKKATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000185] IL2DelM3: Del, E62R
[000186] Последовательность ДНК (SEQ ID NO:44)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAGGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000187] Последовательность белка (SEQ ID NO: 45)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINATELKHLQCLEERLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000188] L2M3K: E62K
[000189] Последовательность ДНК (SEQ ID NO:46)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCTTCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000190] Последовательность белка (SEQ ID NO:47)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTFKFYMPKKATELKHLQCLEEKLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000191] IL2M6K: R38A, F42K, E62K
[000192] Последовательность ДНК (SEQ ID NO:48)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCAAGAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000193] Последовательность белка (SEQ ID NO:49)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTKKFYMPKKATELKHLQCLEEKLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000194] IL2M7K: R38A, F42A, E62K
[000195] Последовательность ДНК (SEQ ID NO:50)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCGCCATGCTGACCGCCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000196] Последовательность белка (SEQ ID NO:51)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTAMLTAKFYMPKKATELKHLQCLEEKLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000198] IL2M8K: F42K, Y45R, E62K
[000198[ Последовательность ДНК (SEQ ID NO:52)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCAAGAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000199] Последовательность белка (SEQ ID NO:53)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTKKFRMPKKATELKHLQCLEEKLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000200] IL2M9K: F42A, Y45R, E62K
[000201] Последовательность ДНК (SEQ ID NO:54)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCAAGCTGACCCGGATGCTGACCGCCAAGTTCCGCATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGGAAAAGCTGAAGCCTTTAGAGGAAGTGCTCAATTTAGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000202] Последовательность белка (SEQ ID NO:55)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPKLTRMLTAKFRMPKKATELKHLQCLEEKLKPLEEVLNLAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000203] IL2MNew: K35E, R38A, F42A, E61K, L72G
[000204] Последовательность ДНК (SEQ ID NO:56)
GGCGCCCCCACATCCTCCAGCACCAAGAAGACCCAGCTGCAGCTGGAGCATCTGCTGCTGGATCTGCAGATGATTTTAAACGGCATCAACAACTACAAGAACCCCGAGCTGACCGCCATGCTGACCGCCAAGTTCTACATGCCCAAGAAGGCCACCGAGCTGAAGCATTTACAGTGTCTGGAGAAAGAGCTGAAGCCTTTAGAGGAAGTGCTCAATGGCGCCCAGTCCAAGAACTTCCATTTAAGGCCACGTGATTTAATCTCCAACATCAACGTGATCGTGCTGGAGCTGAAGGGCTCCGAGACCACCTTCATGTGCGAGTACGCCGACGAGACCGCCACCATCGTGGAGTTTTTAAATCGTTGGATCACCTTCTGCCAGAGCATCATCTCCACTTTAACCTGA
[000205] Последовательность белка (SEQ ID NO:57)
GAPTSSSTKKTQLQLEHLLLDLQMILNGINNYKNPELTAMLTAKFYMPKKATELKHLQCLEKELKPLEEVLNGAQSKNFHLRPRDLISNINVIVLELKGSETTFMCEYADETATIVEFLNRWITFCQSIISTLT
[000206] Специалистам в данной области техники будет очевидно, что в настоящее изобретение могут быть внесены различные модификации и изменения, не выходящие за рамки объема или сущности изобретения. Другие варианты осуществления изобретения будут очевидны специалистам в данной области техники после изучения настоящего документа и и практической реализации раскрытого здесь изобретения. Предполагается, что представленные описание и примеры являются исключительно иллюстративными, тогда как истинные объем и сущность изобретения раскрыты в приведенной ниже формуле изобретения.
[000207] В настоящей заявке приведены ссылки на различные публикации, патенты и/или патентные заявки для более полного описания уровня техники, к которой относится настоящее изобретение. Содержание указанных публикаций, патентов и/или патентных заявок включено в настоящий документ посредством ссылок во всей полноте в той же степени, как если бы все независимые публикации, патенты и/или патентные заявки были включены посредством ссылок конкретным и индивидуальным образом.
--->
Последовательность ДНК ЛИ, Чиан Дж.
УННИРАМАН, Шиам
БАДЕР, Ханна
ЛАУ, Алан
<120> ПОЛУЧЕННЫЕ ПУТЕМ РАЦИОНАЛЬНОГО ДИЗАЙНА НОВЫЕ БЕЛКОВЫЕ
КОМПОЗИЦИИ
<130> 0902
<150> US 62/752,293
<151> 2018-10-29
<150> US 62/811,116
<151> 2019-02-27
<160> 57
<170> PatentIn, версия 3.5
<210> 1
<211> 1401
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК тяжелой цепи антитела к PDL1 B2
<400> 1
atggagtttg ggctgagctg gctttttctt gtcgcgattc ttaagggtgt ccagtgcgaa
60
gtgcagctgg tggagtctgg gggaggcttg gtacagcctg gagggtccct gagactctcc
120
tgtgcagcct ctggattcac ttttagtgac tatgacatga tctgggtccg ccaggctcca
180
ggcaaggggc tggagtgggt ggcagttata tcatatgatg gaagtaataa atactatgca
240
gactccgtga agggccgatt caccatctcc agagacaatt ccaagaacac gctgtatctg
300
caaatgaaca gcctgagagc tgaggacacg gctgtgtatt actgtgcgaa agagttcttt
360
ggtgcttttg atatctgggg ccaagggaca atggtcaccg tctcttcagc ttcgaccaag
420
ggcccatcgg tcttccccct ggcaccctcc tccaagagca cctctggggg cacagcggcc
480
ctgggctgcc tggtcaagga ctacttcccc gaaccggtga cggtgtcgtg gaactcaggc
540
gccctgacca gcggcgtgca caccttcccg gctgtcctac agtcctcagg actctactcc
600
ctcagcagcg tggtgaccgt gccctccagc agcttgggca cccagaccta catctgcaac
660
gtgaatcaca agcccagcaa caccaaggtg gacaagaaag ttgagcccaa atcttgtgac
720
aaaactcaca catgcccacc gtgcccagca cctgaactcc tggggggacc gtcagtcttc
780
ctcttccccc caaaacccaa ggacaccctc atgatctccc ggacccctga ggtcacatgc
840
gtggtggtgg acgtgagcca cgaagaccct gaggtcaagt tcaactggta cgtggacggc
900
gtggaggtgc ataatgccaa gacaaagccg cgggaggagc agtacaacag cacgtaccgt
960
gtggtcagcg tcctcaccgt cctgcaccag gactggctga atggcaagga gtacaagtgc
1020
aaggtctcca acaaagccct cccagccccc atcgagaaaa ccatctccaa agccaaaggg
1080
cagccccgag aaccacaggt gtacaccctg cccccatccc gcgaggagat gaccaagaac
1140
caggtcagcc tgacctgcct ggtcaaaggc ttctatccca gcgacatcgc cgtggagtgg
1200
gagagcaatg ggcagccgga gaacaactac aagaccacgc ctcccgtgct ggactccgac
1260
ggctccttct tcctctacag caagctcacc gtggacaaga gcaggtggca gcaggggaac
1320
gtcttctcat gctccgtgat gcatgaggct ctgcacaacc actacacgca gaagagcctc
1380
tccctgtctc cgggtaaatg a
1401
<210> 2
<211> 466
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность тяжелой цепи
антитела к PDL1 В2
<400> 2
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln
20 25 30
Pro Gly Gly Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe
35 40 45
Ser Asp Tyr Asp Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu
50 55 60
Glu Trp Val Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala
65 70 75 80
Asp Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn
85 90 95
Thr Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Lys Glu Phe Phe Gly Ala Phe Asp Ile Trp Gly Gln
115 120 125
Gly Thr Met Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
130 135 140
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
145 150 155 160
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
165 170 175
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
180 185 190
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
195 200 205
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
210 215 220
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
225 230 235 240
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
245 250 255
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
260 265 270
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
275 280 285
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
290 295 300
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
305 310 315 320
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
325 330 335
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
340 345 350
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
355 360 365
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
370 375 380
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
385 390 395 400
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
405 410 415
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
420 425 430
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
435 440 445
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
450 455 460
Gly Lys
465
<210> 3
<211> 117
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области тяжелой цепи
антитела к PDL1 В2
<400> 3
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Asp Tyr
20 25 30
Asp Met Ile Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Ser Tyr Asp Gly Ser Asn Lys Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Lys Glu Phe Phe Gly Ala Phe Asp Ile Trp Gly Gln Gly Thr Met
100 105 110
Val Thr Val Ser Ser
115
<210> 4
<211> 705
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК легкой цепи антитела к PDL1 В2
<400> 4
atgacttgga ccccactcct cttcctcacc ctcctcctcc actgcacagg aagcttatcc
60
tcttctgagc tgactcagga ccctgctgtg tcggtggcct tgggacagac agtcacgatc
120
acatgccaag gagacagcct caattactat tatgcaaact ggttccagct gaagccaggg
180
caggcccctg tacttgtcct ctttggtaaa aacaaccggc cctcagggat cccagaccga
240
ttctctggct cctactcggg aagcacagct tccttgacca tcactggggc tcaggcggaa
300
gatgacgctg actattactg taattcgcgg gacagcggtg gtaatccttg ggtgttcggc
360
ggagggacca agctgaccgt cctaggtcaa cccaaggctg ccccctcggt cactctgttc
420
ccgccctcct ctgaggagct tcaagccaac aaggccacac tggtgtgtct cataagtgac
480
ttctacccgg gagccgtgac agtggcctgg aaggcagata gcagccccgt caaggcggga
540
gtggagacca ccacaccctc caaacaaagc aacaacaagt acgcggccag cagctacctg
600
agcctgacgc ctgagcagtg gaagtcccac agaagctaca gctgccaggt cacgcatgaa
660
gggagcaccg tggagaagac agtggcccct acagaatgtt catga
705
<210> 5
<211> 234
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность легкой цепи
антитела к PDL1 В2
<400> 5
Met Thr Trp Thr Pro Leu Leu Phe Leu Thr Leu Leu Leu His Cys Thr
1 5 10 15
Gly Ser Leu Ser Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val
20 25 30
Ala Leu Gly Gln Thr Val Thr Ile Thr Cys Gln Gly Asp Ser Leu Asn
35 40 45
Tyr Tyr Tyr Ala Asn Trp Phe Gln Leu Lys Pro Gly Gln Ala Pro Val
50 55 60
Leu Val Leu Phe Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg
65 70 75 80
Phe Ser Gly Ser Tyr Ser Gly Ser Thr Ala Ser Leu Thr Ile Thr Gly
85 90 95
Ala Gln Ala Glu Asp Asp Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser
100 105 110
Gly Gly Asn Pro Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
115 120 125
Gly Gln Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser
130 135 140
Glu Glu Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp
145 150 155 160
Phe Tyr Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro
165 170 175
Val Lys Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn
180 185 190
Lys Tyr Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys
195 200 205
Ser His Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val
210 215 220
Glu Lys Thr Val Ala Pro Thr Glu Cys Ser
225 230
<210> 6
<211> 108
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области легкой цепи
антитела к PDL1 В2
<400> 6
Ser Ser Glu Leu Thr Gln Asp Pro Ala Val Ser Val Ala Leu Gly Gln
1 5 10 15
Thr Val Thr Ile Thr Cys Gln Gly Asp Ser Leu Asn Tyr Tyr Tyr Ala
20 25 30
Asn Trp Phe Gln Leu Lys Pro Gly Gln Ala Pro Val Leu Val Leu Phe
35 40 45
Gly Lys Asn Asn Arg Pro Ser Gly Ile Pro Asp Arg Phe Ser Gly Ser
50 55 60
Tyr Ser Gly Ser Thr Ala Ser Leu Thr Ile Thr Gly Ala Gln Ala Glu
65 70 75 80
Asp Asp Ala Asp Tyr Tyr Cys Asn Ser Arg Asp Ser Gly Gly Asn Pro
85 90 95
Trp Val Phe Gly Gly Gly Thr Lys Leu Thr Val Leu
100 105
<210> 7
<211> 1404
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК тяжелой цепи антитела к PDL1 Е1
<400> 7
atggagtttg ggctgagctg gctttttctt gtcgcgattc ttaagggtgt ccagtgccaa
60
atccagctgg tacaatctgg ggctgaggtg aagatgcctg gggcctcagt gacgatttcc
120
tgcgaggcgt ctggatacaa cttcatcagc tactatatac actgggtgcg acaggcccct
180
ggacaaggcc ttgagtggat gggattcgtc gtccctagtg gtggtgccgc aggctacaca
240
cagaagttcc agggcagact caccgtgacc agggacacgt ccacgagcac agtctacatg
300
gacctgaaca gcctgacatc tgacgacacg gccgtgtatt actgtgtgcg agaaatgagt
360
ggtggctggt ttgatttctg gggccaggga accctggtca ccgtctcctc ggcttcgacc
420
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg
480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca
540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac
600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc
660
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt
720
gacaaaactc acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc
780
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca
840
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac
900
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac
960
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag
1020
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa
1080
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgcgagga gatgaccaag
1140
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
1200
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc
1260
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg
1320
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc
1380
ctctccctgt ctccgggtaa atga
1404
<210> 8
<211> 467
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность тяжелой цепи
антитела к PDL1 Е1
<400> 8
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Met
20 25 30
Pro Gly Ala Ser Val Thr Ile Ser Cys Glu Ala Ser Gly Tyr Asn Phe
35 40 45
Ile Ser Tyr Tyr Ile His Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Phe Val Val Pro Ser Gly Gly Ala Ala Gly Tyr Thr
65 70 75 80
Gln Lys Phe Gln Gly Arg Leu Thr Val Thr Arg Asp Thr Ser Thr Ser
85 90 95
Thr Val Tyr Met Asp Leu Asn Ser Leu Thr Ser Asp Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Val Arg Glu Met Ser Gly Gly Trp Phe Asp Phe Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Lys
465
<210> 9
<211> 98
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области тяжелой
цепи антитела к PDL1 Е1
<400> 9
Ser Cys Glu Ala Ser Gly Tyr Asn Phe Ile Ser Tyr Tyr Ile His Trp
1 5 10 15
Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met Gly Phe Val Val
20 25 30
Pro Ser Gly Gly Ala Ala Gly Tyr Thr Gln Lys Phe Gln Gly Arg Leu
35 40 45
Thr Val Thr Arg Asp Thr Ser Thr Ser Thr Val Tyr Met Asp Leu Asn
50 55 60
Ser Leu Thr Ser Asp Asp Thr Ala Val Tyr Tyr Cys Val Arg Glu Met
65 70 75 80
Ser Gly Gly Trp Phe Asp Phe Trp Gly Gln Gly Thr Leu Val Thr Val
85 90 95
Ser Ser
<210> 10
<211> 711
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК легкой цепи антитела к PDL1 Е1
<400> 10
atggacatgc gcgtgcccgc ccagctgctg ggcctgctgc tgctgtggtt ccccggctcg
60
cgatgcgaca tcgtgatgac ccagtctcca tcctccctgt ctgcatctgt aggagacaga
120
gtcaccatca cttgccgggc aagtcagggc attagaaatg atttaggctg gtatcagcaa
180
aaaccaggga aagcccctaa gctcctgatc tatgctgcat ccactttgca aagtggggtc
240
ccatcaaggt tcagcggcag tggatctggg acagatttca ctctcaccat cagcagcctg
300
caggctgaag atgtggcagt ttattactgt cagcaatatt atagtactcc tctcactttc
360
ggccctggga ccaaagtgga tatcaaacgt acggtggctg caccatctgt cttcatcttc
420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac
480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac
540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc
600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat
660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgttg a
711
<210> 11
<211> 236
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность легкой цепи
антитела к PDL1 Е1
<400> 11
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Phe Pro Gly Ser Arg Cys Asp Ile Val Met Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
35 40 45
Gln Gly Ile Arg Asn Asp Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln
100 105 110
Tyr Tyr Ser Thr Pro Leu Thr Phe Gly Pro Gly Thr Lys Val Asp Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 12
<211> 107
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области легкой цепи
антитела к PDL1 Е1
<400> 12
Asp Ile Val Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Thr Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Ala
65 70 75 80
Glu Asp Val Ala Val Tyr Tyr Cys Gln Gln Tyr Tyr Ser Thr Pro Leu
85 90 95
Thr Phe Gly Pro Gly Thr Lys Val Asp Ile Lys
100 105
<210> 13
<211> 1404
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК тяжелой цепи антитела к PDL1 E3
<400> 13
atggagtttg ggctgagctg gctttttctt gtcgcgattc ttaagggtgt ccagtgccaa
60
atccagctgg tacaatctgg ggctgaggtg aagaagcctg ggtcctcggt gaaggtctcc
120
tgcaaggctt ctggaggcac cttcagcagc tatgctatca gctgggtgcg acaggcccct
180
ggacaagggc ttgagtggat gggagggatc atccctatct ttggtacagc aaactacgca
240
cagaagttcc agggcagagt cacgattacc gcggacaaat ccacgagcac agcctacatg
300
gagctgagca gcctgagatc tgaggacacg gccgtgtatt actgtgccgg agggggagca
360
gtggcggaca atagttactg gggccaggga accctggtca ccgtctcctc agcttcgacc
420
aagggcccat cggtcttccc cctggcaccc tcctccaaga gcacctctgg gggcacagcg
480
gccctgggct gcctggtcaa ggactacttc cccgaaccgg tgacggtgtc gtggaactca
540
ggcgccctga ccagcggcgt gcacaccttc ccggctgtcc tacagtcctc aggactctac
600
tccctcagca gcgtggtgac cgtgccctcc agcagcttgg gcacccagac ctacatctgc
660
aacgtgaatc acaagcccag caacaccaag gtggacaaga aagttgagcc caaatcttgt
720
gacaaaactc acacatgccc accgtgccca gcacctgaac tcctgggggg accgtcagtc
780
ttcctcttcc ccccaaaacc caaggacacc ctcatgatct cccggacccc tgaggtcaca
840
tgcgtggtgg tggacgtgag ccacgaagac cctgaggtca agttcaactg gtacgtggac
900
ggcgtggagg tgcataatgc caagacaaag ccgcgggagg agcagtacaa cagcacgtac
960
cgtgtggtca gcgtcctcac cgtcctgcac caggactggc tgaatggcaa ggagtacaag
1020
tgcaaggtct ccaacaaagc cctcccagcc cccatcgaga aaaccatctc caaagccaaa
1080
gggcagcccc gagaaccaca ggtgtacacc ctgcccccat cccgcgagga gatgaccaag
1140
aaccaggtca gcctgacctg cctggtcaaa ggcttctatc ccagcgacat cgccgtggag
1200
tgggagagca atgggcagcc ggagaacaac tacaagacca cgcctcccgt gctggactcc
1260
gacggctcct tcttcctcta cagcaagctc accgtggaca agagcaggtg gcagcagggg
1320
aacgtcttct catgctccgt gatgcatgag gctctgcaca accactacac gcagaagagc
1380
ctctccctgt ctccgggtaa atga
1404
<210> 14
<211> 467
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность тяжелой цепи
антитела к PDL1 E3
<400> 14
Met Glu Phe Gly Leu Ser Trp Leu Phe Leu Val Ala Ile Leu Lys Gly
1 5 10 15
Val Gln Cys Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys
20 25 30
Pro Gly Ser Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe
35 40 45
Ser Ser Tyr Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu
50 55 60
Glu Trp Met Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala
65 70 75 80
Gln Lys Phe Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser
85 90 95
Thr Ala Tyr Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val
100 105 110
Tyr Tyr Cys Ala Gly Gly Gly Ala Val Ala Asp Asn Ser Tyr Trp Gly
115 120 125
Gln Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser
130 135 140
Val Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala
145 150 155 160
Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val
165 170 175
Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala
180 185 190
Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val
195 200 205
Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His
210 215 220
Lys Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys
225 230 235 240
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
245 250 255
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
260 265 270
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
275 280 285
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
290 295 300
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
305 310 315 320
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
325 330 335
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
340 345 350
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
355 360 365
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
370 375 380
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
385 390 395 400
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
405 410 415
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
420 425 430
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
435 440 445
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
450 455 460
Pro Gly Lys
465
<210> 15
<211> 118
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области тяжелой цепи
антитела к PDL1 E3
<400> 15
Gln Ile Gln Leu Val Gln Ser Gly Ala Glu Val Lys Lys Pro Gly Ser
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Gly Thr Phe Ser Ser Tyr
20 25 30
Ala Ile Ser Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Ile Ile Pro Ile Phe Gly Thr Ala Asn Tyr Ala Gln Lys Phe
50 55 60
Gln Gly Arg Val Thr Ile Thr Ala Asp Lys Ser Thr Ser Thr Ala Tyr
65 70 75 80
Met Glu Leu Ser Ser Leu Arg Ser Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Gly Gly Gly Ala Val Ala Asp Asn Ser Tyr Trp Gly Gln Gly Thr
100 105 110
Leu Val Thr Val Ser Ser
115
<210> 16
<211> 711
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК легкой цепи антитела к PDL1 E3
<400> 16
atggacatgc gcgtgcccgc ccagctgctg ggcctgctgc tgctgtggtt ccccggctcg
60
cgatgcgaca tccggatgac ccagtctcca tcctccctgt ctgcatctgt aggagacaga
120
gtcaccatca cttgccgggc aagtcagggc attagaaatg atttaggctg gtatcagcag
180
aaaccaggga aagcccctaa gctcctgatc tatgctgcat ccagtttaca aagtggggtc
240
ccatcaaggt tcagcggcag tggatctggc acagatttca ctctcaccat cagcagcctg
300
cagcctgaag attttgcaac ttattactgt ctacaagatt acaattaccc tcgaacgttc
360
ggccaaggga ccaaggtgga aatcaaacgt acggtggctg caccatctgt cttcatcttc
420
ccgccatctg atgagcagtt gaaatctgga actgcctctg ttgtgtgcct gctgaataac
480
ttctatccca gagaggccaa agtacagtgg aaggtggata acgccctcca atcgggtaac
540
tcccaggaga gtgtcacaga gcaggacagc aaggacagca cctacagcct cagcagcacc
600
ctgacgctga gcaaagcaga ctacgagaaa cacaaagtct acgcctgcga agtcacccat
660
cagggcctga gctcgcccgt cacaaagagc ttcaacaggg gagagtgttg a
711
<210> 17
<211> 236
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> полноразмерная белковая последовательность легкой цепи
антитела к PDL1 E3
<400> 17
Met Asp Met Arg Val Pro Ala Gln Leu Leu Gly Leu Leu Leu Leu Trp
1 5 10 15
Phe Pro Gly Ser Arg Cys Asp Ile Arg Met Thr Gln Ser Pro Ser Ser
20 25 30
Leu Ser Ala Ser Val Gly Asp Arg Val Thr Ile Thr Cys Arg Ala Ser
35 40 45
Gln Gly Ile Arg Asn Asp Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys
50 55 60
Ala Pro Lys Leu Leu Ile Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val
65 70 75 80
Pro Ser Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
85 90 95
Ile Ser Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln
100 105 110
Asp Tyr Asn Tyr Pro Arg Thr Phe Gly Gln Gly Thr Lys Val Glu Ile
115 120 125
Lys Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp
130 135 140
Glu Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn
145 150 155 160
Phe Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu
165 170 175
Gln Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp
180 185 190
Ser Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr
195 200 205
Glu Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser
210 215 220
Ser Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
225 230 235
<210> 18
<211> 107
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность вариабельной области легкой цепи
антитела к PDL1 E3
<400> 18
Asp Ile Arg Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Gly Ile Arg Asn Asp
20 25 30
Leu Gly Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Leu Gln Asp Tyr Asn Tyr Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 19
<211> 20
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> линкер Ser-Gly
<400> 19
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser Gly
1 5 10 15
Gly Gly Gly Ser
20
<210> 20
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2WT
<400> 20
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccttca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 21
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2WT
<400> 21
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 22
<211> 345
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2Del
<400> 22
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac gccaccgagc tgaagcattt acagtgtctg
120
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
180
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
240
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
300
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
345
<210> 23
<211> 114
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2Del
<400> 23
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Ala Thr
20 25 30
Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu Lys Pro Leu Glu
35 40 45
Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg
50 55 60
Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser
65 70 75 80
Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val
85 90 95
Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr
100 105 110
Leu Thr
<210> 24
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M1: F42K, Y45R
<400> 24
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccaaga agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 25
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M1: F42K, Y45R
<400> 25
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Lys Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 26
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M2: F42A, Y45R
<400> 26
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccgcca agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 27
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M2: F42A, Y45R
<400> 27
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Ala Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 28
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M3: E62R
<400> 28
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccttca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 29
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M3: E62R
<400> 29
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 30
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M4: R38A, F42A
<400> 30
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccgcca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 31
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M4: R38A, F42A
<400> 31
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 32
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M5: R38A, F42A, Y45A, E62A, C125S
<400> 32
ggagctccta cctcctcctc caccaagaag acccagctcc agctggaaca tttactgctg
60
gatttacaga tgattttaaa cggcatcaac aattacaaga accccaagct gaccgccatg
120
ctgaccgcca agtttgccat gcccaagaag gccaccgagc tgaagcattt acagtgttta
180
gaggaggctt taaagccttt agaggaggtg ctgaatttag cccagtccaa gaacttccat
240
ttaaggcccc gggatttaat ctccaacatc aacgtgatcg tgctggaact gaagggcagc
300
gagaccacct ttatgagcga atacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gtccatcatc tccactttaa cctga
405
<210> 33
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M5: R38A, F42A, Y45A, E62A,
C125S
<400> 33
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Ala Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Ala Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Ser Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Ser Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 34
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M6: R38A, F42K
<400> 34
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccaaga agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaagagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 35
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M6: R38A, F42K
<400> 35
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Lys Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 36
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M7: R38A, F42A, E62R
<400> 36
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccgcca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 37
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Белоквая последовательность IL2M7: R38A, F42A, E62R
<400> 37
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 38
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M8: F42K, Y45R, E62R
<400> 38
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccaaga agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 39
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> Белковая последовательность IL2M8: F42K, Y45R, E62R
<400> 39
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Lys Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 40
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M9: F42A, Y45R, E62R
<400> 40
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccgcca agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 41
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M9: F42A, Y45R, E62R
<400> 41
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Ala Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 42
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> IL2M10: R38A, F42K, E62R последовательность ДНК
<400> 42
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccaaga agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 43
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M10: R38A, F42K, E62R
<400> 43
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Lys Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 44
<211> 345
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> IL2DelM3: Del, E62R последовательность ДНК
<400> 44
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac gccaccgagc tgaagcattt acagtgtctg
120
gaggaaaggc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
180
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
240
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
300
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
345
<210> 45
<211> 114
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2DelM3: Del, E62R
<400> 45
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Ala Thr
20 25 30
Glu Leu Lys His Leu Gln Cys Leu Glu Glu Arg Leu Lys Pro Leu Glu
35 40 45
Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His Leu Arg Pro Arg
50 55 60
Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu Leu Lys Gly Ser
65 70 75 80
Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr Ala Thr Ile Val
85 90 95
Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser Ile Ile Ser Thr
100 105 110
Leu Thr
<210> 46
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M3K: E62K
<400> 46
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccttca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 47
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M3K: E62K
<400> 47
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Phe Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Lys Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 48
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M6K: R38A, F42K, E62K
<400> 48
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccaaga agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 49
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M6K: R38A, F42K, E62K
<400> 49
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Lys Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Lys Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 50
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M7K: R38A, F42A, E62K
<400> 50
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gaccgccatg
120
ctgaccgcca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 51
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M7K: R38A, F42A, E62K
<400> 51
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Lys Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 52
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M8K: F42K, Y45R, E62K
<400> 52
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccaaga agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 53
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M8K: F42K, Y45R, E62K
<400> 53
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Lys Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Lys Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 54
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2M9K: F42A, Y45R, E62K
<400> 54
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccaagct gacccggatg
120
ctgaccgcca agttccgcat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gaggaaaagc tgaagccttt agaggaagtg ctcaatttag cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 55
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2M9K: F42A, Y45R, E62K
<400> 55
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Lys Leu Thr Arg Met Leu Thr Ala Lys Phe Arg Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Glu Lys Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Leu Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<210> 56
<211> 405
<212> DNA/ДНК
<213> Искусственная последовательность
<220>
<223> последовательность ДНК IL2MNew: K35E R38A F42A E61K L72G
<400> 56
ggcgccccca catcctccag caccaagaag acccagctgc agctggagca tctgctgctg
60
gatctgcaga tgattttaaa cggcatcaac aactacaaga accccgagct gaccgccatg
120
ctgaccgcca agttctacat gcccaagaag gccaccgagc tgaagcattt acagtgtctg
180
gagaaagagc tgaagccttt agaggaagtg ctcaatggcg cccagtccaa gaacttccat
240
ttaaggccac gtgatttaat ctccaacatc aacgtgatcg tgctggagct gaagggctcc
300
gagaccacct tcatgtgcga gtacgccgac gagaccgcca ccatcgtgga gtttttaaat
360
cgttggatca ccttctgcca gagcatcatc tccactttaa cctga
405
<210> 57
<211> 134
<212> PRT/БЕЛОК
<213> Искусственная последовательность
<220>
<223> белковая последовательность IL2MNew: K35E R38A F42A E61K L72G
<400> 57
Gly Ala Pro Thr Ser Ser Ser Thr Lys Lys Thr Gln Leu Gln Leu Glu
1 5 10 15
His Leu Leu Leu Asp Leu Gln Met Ile Leu Asn Gly Ile Asn Asn Tyr
20 25 30
Lys Asn Pro Glu Leu Thr Ala Met Leu Thr Ala Lys Phe Tyr Met Pro
35 40 45
Lys Lys Ala Thr Glu Leu Lys His Leu Gln Cys Leu Glu Lys Glu Leu
50 55 60
Lys Pro Leu Glu Glu Val Leu Asn Gly Ala Gln Ser Lys Asn Phe His
65 70 75 80
Leu Arg Pro Arg Asp Leu Ile Ser Asn Ile Asn Val Ile Val Leu Glu
85 90 95
Leu Lys Gly Ser Glu Thr Thr Phe Met Cys Glu Tyr Ala Asp Glu Thr
100 105 110
Ala Thr Ile Val Glu Phe Leu Asn Arg Trp Ile Thr Phe Cys Gln Ser
115 120 125
Ile Ile Ser Thr Leu Thr
130
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ПОЛИВАЛЕНТНЫЕ И ПОЛИСПЕЦИФИЧЕСКИЕ ОХ40-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2773052C2 |
| МУЛЬТИВАЛЕНТНЫЕ И МУЛЬТИСПЕЦИФИЧЕСКИЕ 41BB-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2017 |
|
RU2789648C2 |
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| НОВЫЕ КОМПОЗИЦИИ АНТИТЕЛ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2019 |
|
RU2804490C2 |
| ПОЛИВАЛЕТНЫЕ И ПОЛИСПЕЦИФИЧНЫЕ GITR-СВЯЗЫВАЮЩИЕ СЛИТЫЕ БЕЛКИ | 2016 |
|
RU2753439C2 |
| ДОСТАВКА ТЕРАПЕВТИЧЕСКИХ ПОЛИПЕПТИДОВ ПОСРЕДСТВОМ ПСЕВДОТИПИРОВАННЫХ ОНКОЛИТИЧЕСКИХ ВИРУСОВ | 2017 |
|
RU2758007C2 |
| КОНСТРУКЦИИ СЛИТОГО БЕЛКА ДЛЯ ЗАБОЛЕВАНИЯ, СВЯЗАННОГО С КОМПЛЕМЕНТОМ | 2019 |
|
RU2824402C2 |
| НОВЫЕ СЛИТЫЕ БЕЛКИ, СПЕЦИФИЧЕСКИЕ В ОТНОШЕНИИ CD137 И GPC3 | 2020 |
|
RU2814653C2 |
| СЛИТЫЙ БЕЛОК, ВКЛЮЧАЮЩИЙ IL-12 И АНТИТЕЛО ПРОТИВ FAP, И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2831612C1 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
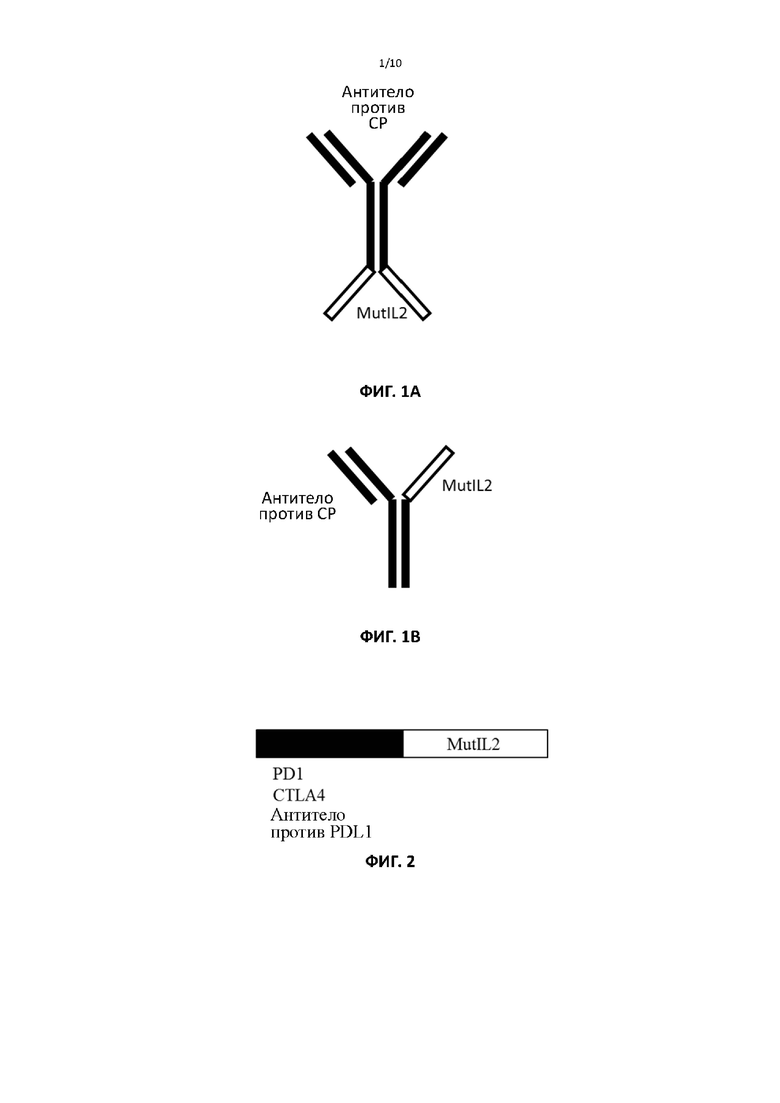
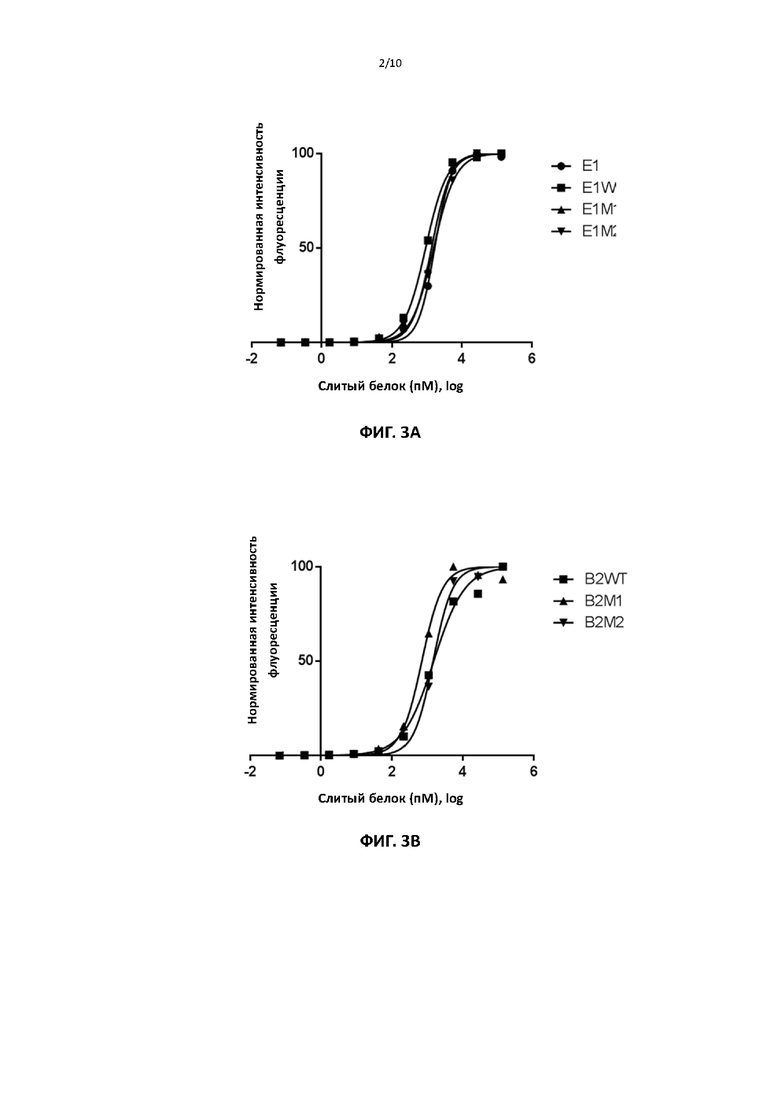
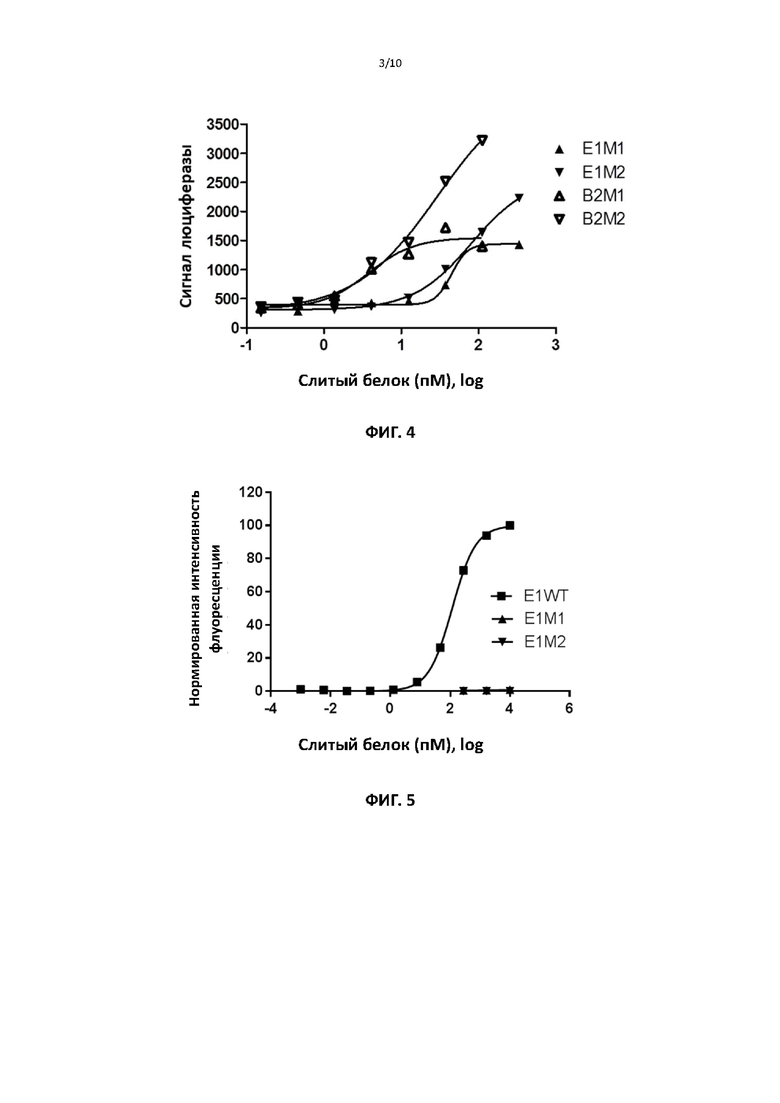
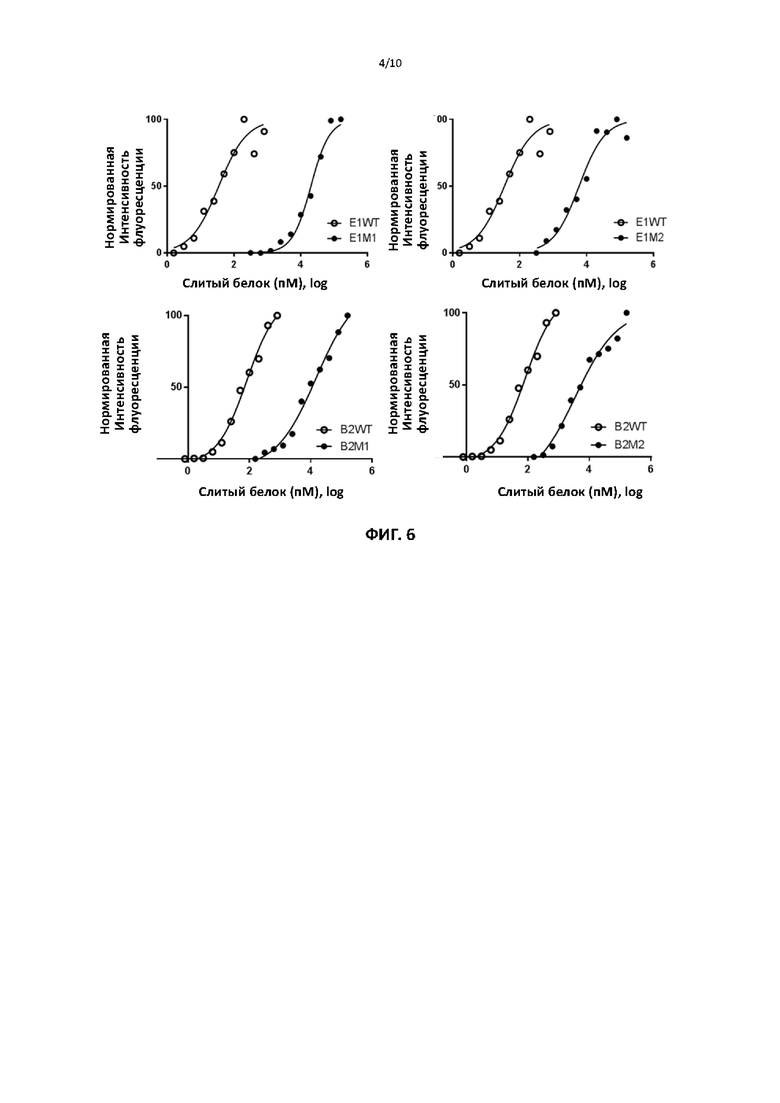
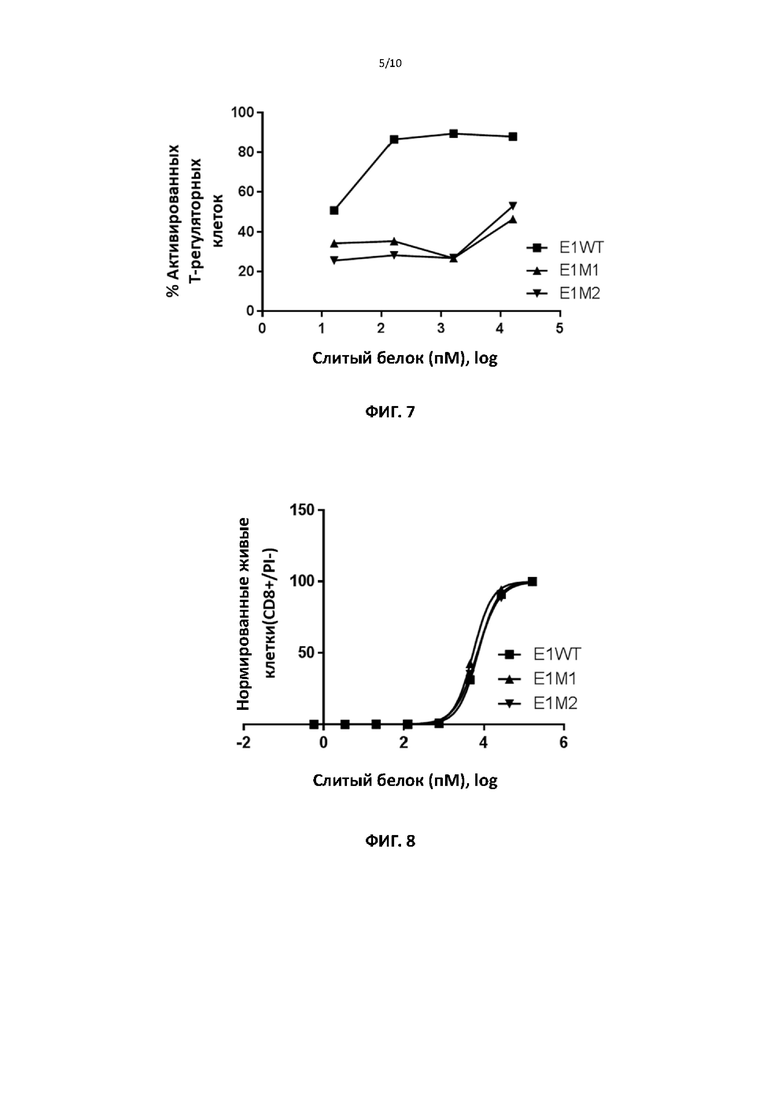
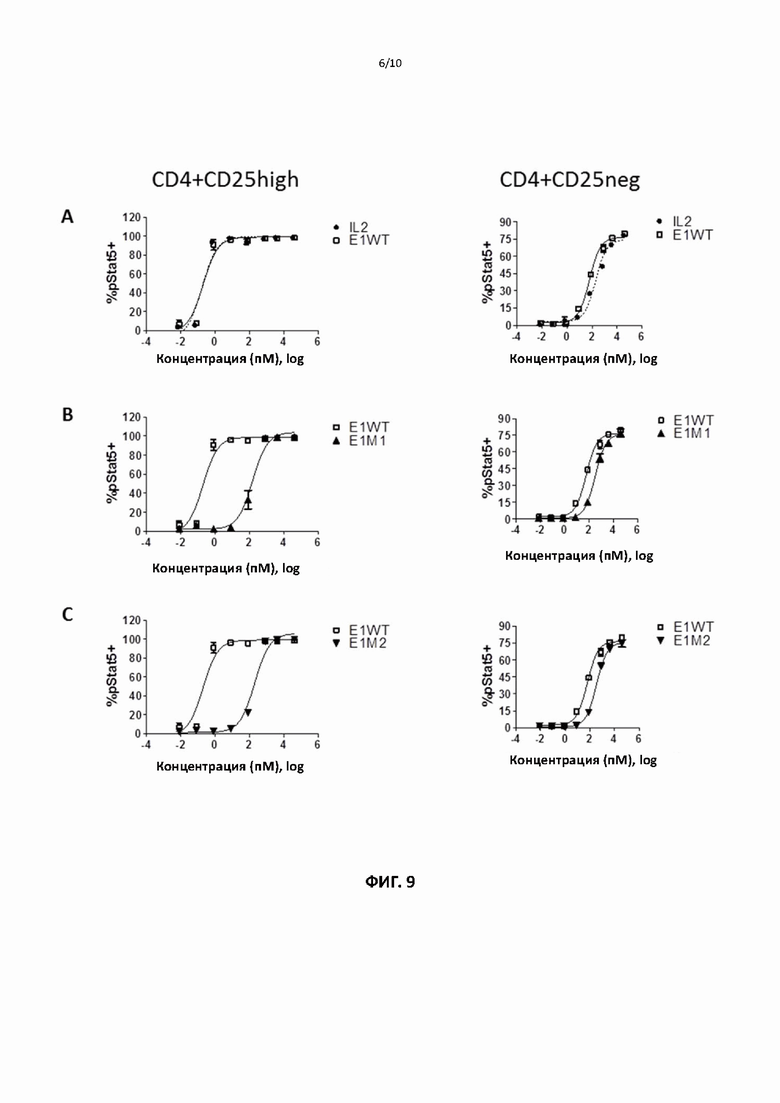
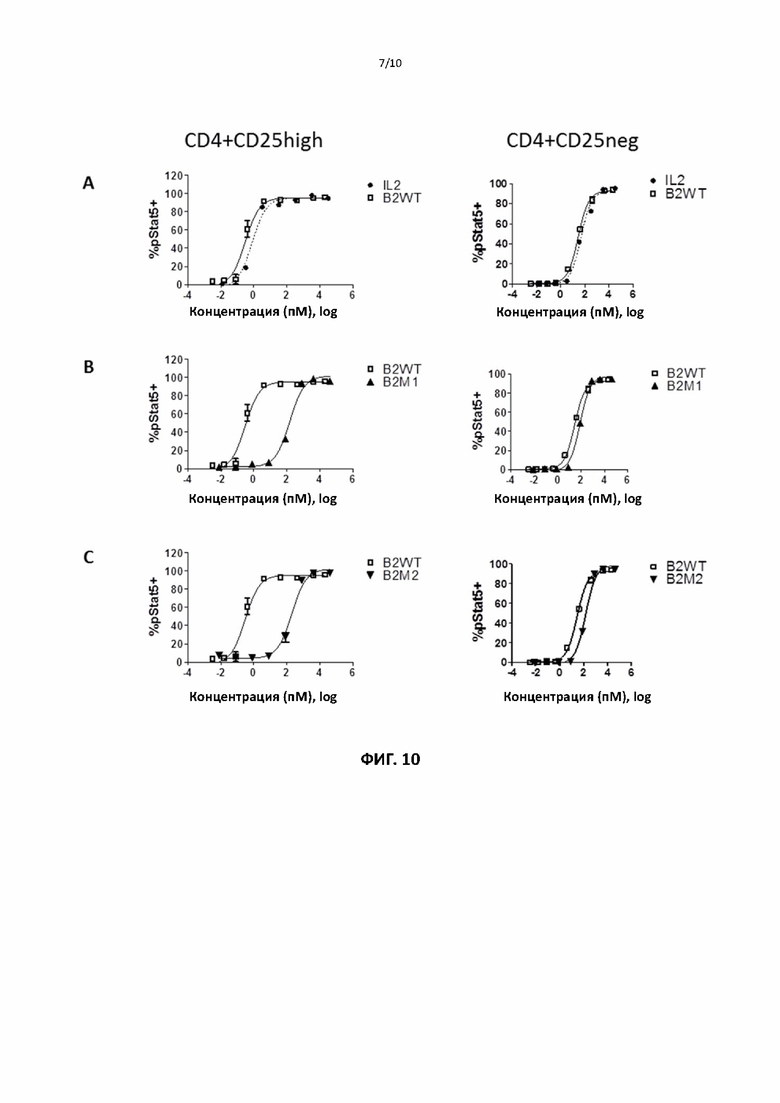
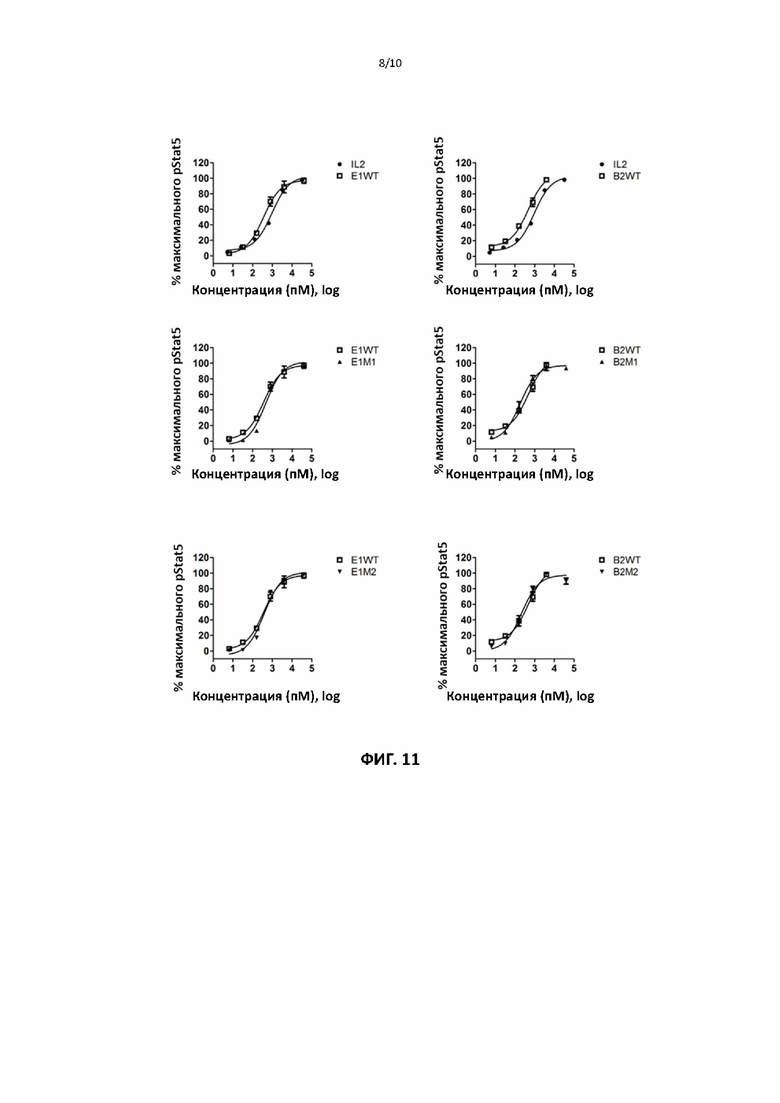
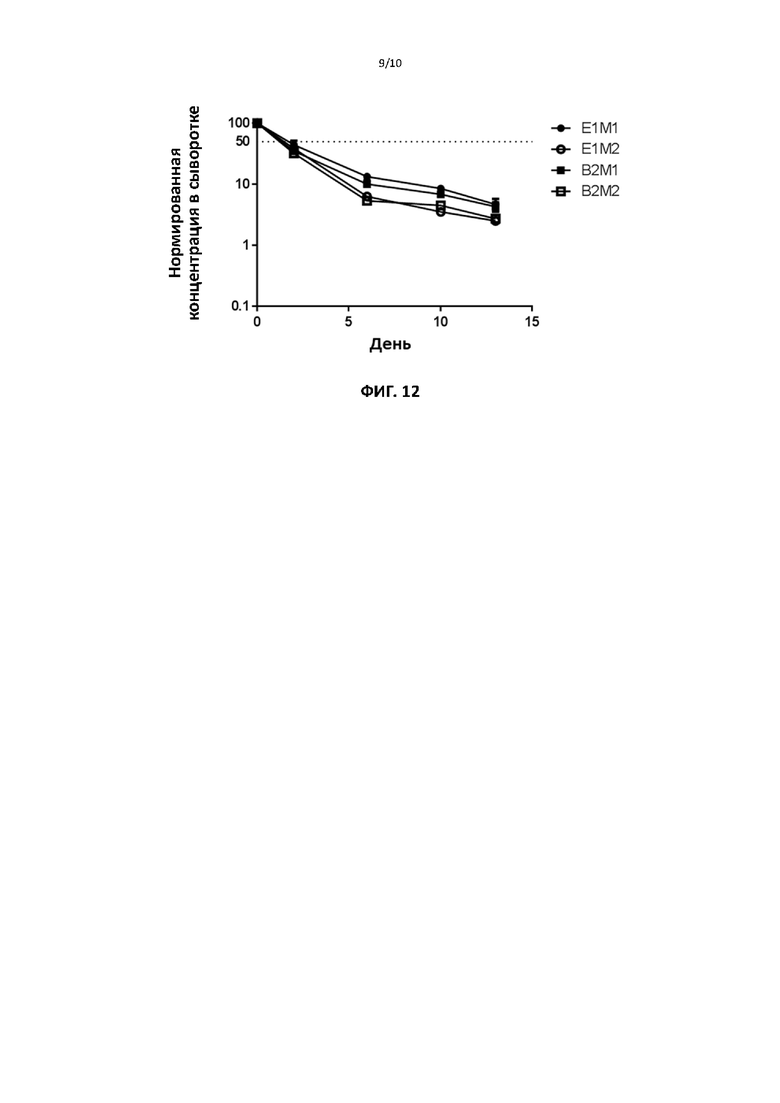
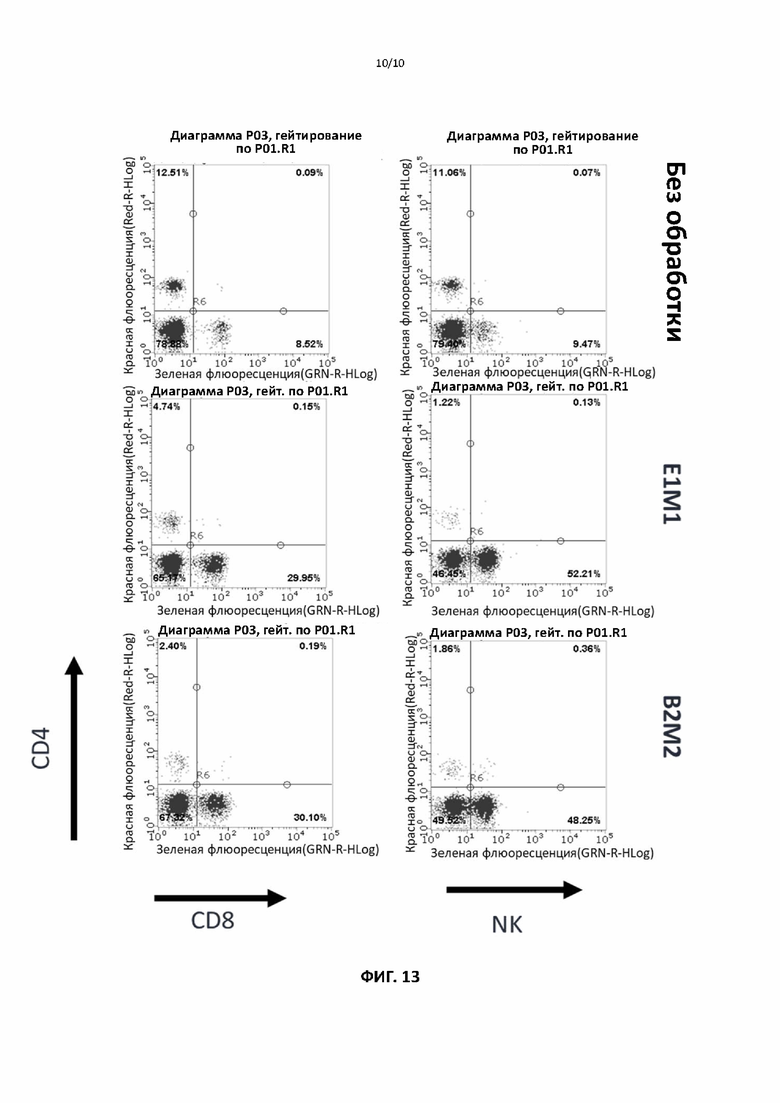
Изобретение относится к области биотехнологии, конкретно к получению иммуноцитокинов с интерлейкином-2, и может быть использовано в медицине для лечения рака. Предложен противоопухолевой слитый белок, содержащий в качестве терапевтического белка целое антитело к PDL1, соединенное с мутантным полипептидом интерлейкина-2 (ИЛ2). При этом мутантный полипептид IL2 соединен с C-концами каждой тяжелой цепи антитела серин-глициновой линкерной последовательностью. Изобретение обеспечивает получение иммуноцитокина с высоким противоопухолевым потенциалом, способного выборочно индуцировать преимущественную пролиферацию CD8+T-клеток и NK-клеток относительно Т-рег клеток. 3 н. и 4 з.п. ф-лы, 13 ил., 11 табл., 10 пр.
1. Противоопухолевый слитый белок, содержащий терапевтический белок, соединенный с мутантным полипептидом интерлейкина-2 (ИЛ2), причем указанный терапевтический белок представляет собой целое антитело к PDL1, содержащее пару тяжелых цепей и пару легких цепей, причем их соответствующие последовательности состоят из аминокислотных последовательностей, идентичных аминокислотным последовательностям: (a) SEQ ID NO: 2 и SEQ ID NO: 5; или (b) SEQ ID NO: 8 и SEQ ID NO: 11, причем указанный мутантный полипептид IL2 соединен с C-концами каждой тяжелой цепи, где указанный мутантный полипептид IL2 содержит аминокислотную последовательность, идентичную аминокислотным последовательностям SEQ ID NO: 25 или SEQ ID NO: 27, причем мутантный полипептид IL2 соединен с C-концом тяжелой цепи серин-глициновой линкерной последовательностью.
2. Слитый белок по п. 1, отличающийся тем, что указанное антитело к PDL1 представляет собой антитело изотипа IgG1, которое опосредует или усиливает функцию АЗКЦ и/или КЗЦ.
3. Противоопухолевая фармацевтическая композиция, содержащая слитый белок по п. 1 или 2 в эффективном количестве и фармацевтически приемлемое вспомогательное вещество, носитель или разбавитель.
4. Способ терапевтического лечения рака у субъекта, включающий введение указанному субъекту терапевтически эффективного количества фармацевтической композиции по п. 3.
5. Способ по п. 4, дополнительно включающий применение (а) химиотерапии, таргетной терапии или лучевой терапии; (b) аптамеров или РНКи; или (с) цитокина.
6. Способ по любому из пп. 4 или 5, отличающийся тем, что указанный рак представляет собой рак млекопитающих.
7. Способ по п. 6, отличающийся тем, что указанный рак выбран из группы, состоящей из меланомы, рака легкого, рака почек, рака головы и шеи, рака желудка, лимфомы, рака яичников, рака толстой кишки, рака молочной железы, миелом, опухолей головного мозга, лейкозов, рака мочевого пузыря, рака печени, уротелиальной карциномы, кожной карциномы, рака поджелудочной железы и их комбинаций.
| WO 2018184964 A1, 11.10.2018. EA 26022 B1, 28.02.2017. CN 107915777 A, 17.04.2018. EA 201300896 A1, 28.02.2014. HUTMACHER C | |||
| ET AL., Antibody-cytokine fusion proteins: Biopharmaceuticals with immunomodulatory properties for cancer therapy, ADVANCED DRUG DELIVERY REVIEWS, 2018, v | |||
| Топливник с глухим подом | 1918 |
|
SU141A1 |
| Приспособление для получения кинематографических стерео снимков | 1919 |
|
SU67A1 |
| E | |||
| et al., Bispecific antibodies, | |||

