ОБЛАСТЬ ТЕХНИКИ
Для настоящей заявки испрашивается приоритет по заявке на корейский патент №10-2020-0066030, поданной 1 июня 2020 года, описание которой включено в настоящую заявку в своем полном объеме.
Настоящее изобретение относится к биспецифическому антителу, характеризующемуся высоким показателем образования гетеродимера, или к его антигенсвязывающему фрагменту и способу его получения.
УРОВЕНЬ ТЕХНИКИ
Природный иммуноглобулин человека (Ig) состоит из двух идентичных тяжелых цепей и двух легких цепей, соединенных вместе. Кроме того, иммуноглобулины функционально могут быть разделены на область антигенсвязывающего фрагмента (Fab), которая образует связывание антиген-антитело, и область кристаллизуемого фрагмента (Fc), в которой две тяжелые цепи образуют димер. Эти природные иммуноглобулины, т.е. моноклональные антитела (mAb), обычно нацеливаются на один и тот же сайт (эпитоп) связывания одного и того же антигена, поскольку две Fab-области идентичны друг другу, и они характеризуются бивалентными и моноспецифическими связывающими свойствами.
Иммуноглобулины широко используются в качестве терапевтического средства при различных заболеваниях из-за антигенсвязывающей функции Fab-области, специфической в отношении мишени, а также функции Fc-области, заключающейся в индуцировании иммунных клеток и увеличении периода полужизни in vivo. Однако в последнее время при трудноизлечимых заболеваниях, таких как рак или аутоиммунные заболевания, синергическое лечение с использованием двух лекарственных средств, действующих на разные мишени, показало больший терапевтический эффект, чем использование одного лекарственного средства, поэтому разрабатываются различные биспецифические антитела, которые могут действовать на два вещества-мишени.
Среди биспецифических антител, разрабатываемых таким образом, биспецифические антитела типа scFv-scFv, в которых фрагменты вариабельной области разных антител связаны полипептидной цепью, обладают недостатком, заключающимся в том, что они не способны находиться в организме в течение длительного времени и сохранять эффективность из-за их низкой молекулярной массы и низкой стабильности. Кроме того, биспецифические антитела типа scFv-Fab-Fc или Fab-Fc-scFv, в которых конец полипептида природного иммуноглобулина связан с фрагментом вариабельной области другого антитела, характеризуются недостатками, связанными с возможностью образования агрегата из-за низкой структурной стабильности, при этом они проявляют иммуногенность in vivo. Поэтому для решения упомянутых проблем необходимо разработать биспецифические антитела, содержащие разные вариабельные области, максимально повторяющие форму природного иммуноглобулина.
Для образования такого типа биспецифического антитела между Fc-областями двух разных комплексов Fab-Fc должен образоваться гетеродимер. Поскольку аминокислоты в положениях, где СН3-домены взаимодействуют друг с другом, играют важную роль в образовании гетеродимера Fc, технология гетеродимеров Fc разрабатывается таким образом, что эти аминокислоты заменяются другими аминокислотами (патент США №5731168 А (1998.03.24), патент Кореи №10-2098919 (2020.04.02)).
Однако существующая технология гетеродимеров Fc, разработанная таким образом, также обладает ограничением, заключающимся в том, что она не может обеспечивать получение всех структур Fc в форме 100% гетеродимеров, то есть образуется смесь гетеродимеров и гомодимеров.
Поэтому существует необходимость в разработке белкового комплекса с улучшенной эффективностью образования гетеродимеров, биспецифического антитела, содержащего его, или его антигеневязывающего фрагмента, рецепторов и рецептор-связывающих агонистов, антагонистов, лигандов, биконъюгатов на основе рецептора-ловушки и т.п.
ОПИСАНИЕ ВАРИАНТОВ ОСУЩЕСТВЛЕНИЯ ТЕХНИЧЕСКАЯ ЗАДАЧА
В одном аспекте представлен белковый комплекс, характеризующийся высоким показателем образования гетеродимера.
В другом аспекте представлен способ получения белкового комплекса, характеризующегося высоким показателем образования гетеродимера.
В еще одном аспекте представлена фармацевтическая композиция для профилактики или лечения заболевания, включающая белковый комплекс, характеризующийся высоким показателем образования гетеродимера.
В еще одном аспекте представлен способ профилактики или лечения заболевания с применением белкового комплекса, характеризующегося высоким показателем образования гетеродимера.
В еще одном аспекте представлены варианты применения белкового комплекса, характеризующегося высоким показателем образования гетеродимера, для получения профилактического или терапевтического средства для профилактики или лечения заболевания.
РЕШЕНИЕ ЗАДАЧИ
В одном аспекте представлен белковый комплекс, включающий первый полипептид, включающий первый СН3 константной области антитела, и второй полипептид, включающий второй СН3 константной области антитела, где первый полипептид и второй полипептид образуют гетеродимер.
В одном варианте осуществления первый СН3 константной области антитела включает триптофан (W) в положении 366, и второй СН3 константной области антитела включает серии (S) в положении 366, аланин (А) в положении 368 и валин (V) в положении 407; и по меньшей мере один из первого СН3 константной области антитела и второго СН3 константной области антитела может включать по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (Н), глицина (G), валина (V), метионина (М) и аланина (А), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394.
Белковый комплекс, включает первый полипептид, включающий первый СН3 константной области антитела, и второй полипептид, включающий второй СН3 константной области антитела, где первый полипептид и второй полипептид образуют гетеродимер.
Термин "антитело" используется взаимозаменяемо с термином "иммуноглобулин (Ig)". Полное антитело характеризуется структурой, состоящей из двух полноразмерных легких цепей и двух полноразмерных тяжелых цепей, при этом каждая легкая цепь связана с тяжелой цепью посредством дисульфидной связи (SS-связью). Существует два типа легких цепей: λ и κ, при этом каждая состоит из примерно 211-217 аминокислот. В каждом антителе человека имеется только один тип легкой цепи. Легкая цепь состоит из непрерывно соединенных константной области и вариабельной области. Существует пять типов тяжелых цепей (γ, δ, α, μ, ε), при этом тяжелая цепь определяет тип антитела, α и γ состоят из 450 аминокислот, а μ и ε состоят из 550 аминокислот. Тяжелая цепь имеет две области: вариабельную область и константную область. Вариабельная область относится к области, с которой антиген связывается в антителе. Вариабельная область может включать определяющую комплементарность область (CDR), придающую антигенсвязывающую специфичность.
Антитело может включать область антигенсвязывающего фрагмента (Fab), которая связывается с антигеном, и область кристаллизуемого фрагмента (Fc), которая связывается с рецептором клеточной поверхности. При расщеплении папаином полное антитело может быть расщеплено на две Fab-области и одну Fc-область. Fab-область может быть областью, в которой полипептид, включающий домен вариабельной области тяжелой цепи (VH) и домен 1 константной области тяжелой цепи (СН1), и полипептид, включающий домен вариабельной области легкой цепи (VL) и домен константной области легкой цепи (CL), связаны дисульфидной связью. Fc-область может представлять собой такую область, в которой связаны два полипептида, включающие домен 2 константной области тяжелой цепи (СН2) и домен 3 константной области тяжелой цепи (СН3). Fc-область может образовывать шарнирную область.
СН3 константной области антитела относится к домену 3 константной области тяжелой цепи антитела.
Первый полипептид и второй полипептид могут образовывать Fc-область антитела.
Гетеродимер относится к комбинации двух полипептидов, характеризующихся разными последовательностями, количествами или типами аминокислотных остатков. Белковый комплекс может представлять собой биспецифическое антитело или его антигенсвязывающий фрагмент, образованный путем объединения двух полипептидов, которые специфически связываются с мишенями, отличными друг от друга.
Белковый комплекс может представлять собой биспецифическое антитело или его антигенсвязывающий фрагмент, конъюгат на основе рецептора и агониста, конъюгат на основе рецептора и антагониста, конъюгат на основе рецептора и лиганда или конъюгат на основе лиганда и рецептора-ловушки. Кроме того, белковый комплекс может включать любое, выбранное из группы, состоящей из антигенсвязывающего фрагмента (Fab), одноцепочечного вариабельного фрагмента (scFv), внеклеточного домена мембранного рецептора, агониста, антагониста, лиганда, рецептора-ловушки, цитокина, фактора коагуляции и аффинной метки.
Антитело может представлять собой, например, IgA, IgD, IgE, IgG или IgM. Антитело может представлять собой моноклональное антитело или поликлональное антитело. Антитело может быть антителом, полученным от животного, химерным антителом мыши-человека, гуманизированным антителом или антителом человека.
Термин "антигенсвязывающий фрагмент" относится к фрагменту структуры полного иммуноглобулина и относится к части полипептида, включающей часть, способную к связыванию с антигеном. Например, антигенсвязывающий фрагмент может представлять собой scFv, (scFv)2, Fv, Fab, Fab', Fv F(ab')2 или их комбинацию.
Термин "биспецифический" относится к специфическому распознаванию белков-мишеней, отличающихся друг от друга. Биспецифическое антитело или его антигенсвязывающий фрагмент относится к антителу или его антигенсвязывающему фрагменту, имеющему два антигенсвязывающих сайта, которые распознают разные антигены-мишени. Биспецифическое антитело или его антигенсвязывающий фрагмент также может называться биспецифическим антителом (BsAb).
Биспецифическое антитело или его антигенсвязывающий фрагмент может представлять собой IgG, полученный посредством похода "выступ-во-впадину" (kih), scFv-Fc, scFv2-Fc, TrioMab, IgG-подобное антитело, CrossMab, 2:1 CrossMab, 2:2 CrossMab, DuoBody, DVD-Ig (иммуноглобулин с двойным вариабельным доменом), scFv-IgG, IgG-IgG, Fab-scFv-Fc, ADPTIR, BiTE (биспецифический рекрутер Т-клеток)-Fc, DART (переориентирующееся антитело с двойной аффинностью)-Fc, четырехвалентное DART-Fc, LP-DART, CODV (антитело с перекрестным двойным вариабельным доменом)-Ig, CODV-Fab-TL, HLE (с удлиненным периодом полужизни)-Bite, Tandem VHH (тяжелая цепь-единственный вариабельный домен)-Fc или их комбинацию.
Термин "рецептор" относится к веществу, которое принимает или передает сигнал, который может быть передан в биологическую систему. Рецептор может быть белковым рецептором. Рецептор может связываться с агонистом, антагонистом, лигандом или цитокином. Агонист может представлять собой вещество, которое связывается с рецептором и активирует рецептор, инициируя биологический ответ. Антагонист может представлять собой вещество, которое связывается с рецептором и ингибирует рецептор, подавляя биологический ответ. Лиганд может представлять собой вещество, которое связывается с рецептором. Лиганд может связываться с рецептором-ловушкой. Рецептор-ловушка относится к рецептору, который специфически связывается с лигандом, подавляя за счет этого передачу сигнала действительным рецептором. Цитокин относится к небольшому белку, который действует на клеточную передачу сигнала, регуляцию и поддержание воспалительных процессов.
Белковый комплекс может быть модифицирован. Например, белковый комплекс может быть модифицирован путем конъюгации или связывания, гликозилирования, присоединения метки или их комбинации. Антитело может быть конъюгировано с другими лекарственными средствами, такими как противораковые лекарственные средства. Например, белковый комплекс может быть объединен с пероксидазой хрена (HRP), щелочной фосфатазой, гаптеном, биотином, стрептавидином, флуоресцентным материалом, радиоактивным материалом, квантовыми точками, полиэтиленгликолем (PEG), гистидиновой меткой или их комбинацией. Флуоресцентное вещество может представлять собой Alexa Fluor®532, Alexa Fluor®546, Alexa Fluor®568, Alexa Fluor®680, Alexa Fluor®750, Alexa Fluor®790 или Alexa Fluor®350.
Положение аминокислоты соответствует индексу EU согласно Кабату (EU-индекс, как описано в публикации 'Kabat et al., Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. (1991)'). Положение аминокислоты в СН3-домене и соответствующий тип аминокислоты основаны на IgG1 человека.
В белковом комплексе первый СН3 константной области антитела может включать триптофан (W) в положении 366. В первом СН3 константной области антитела положение 366 может быть положением (T366W), в котором треонин (Т) заменен триптофаном (W). Второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368 и валин (V) в положении 407. Во втором СН3 константной области антитела положение 366 может быть положением (T366S), в котором треонин (Т) заменен серином (S). Во втором СН3 константной области антитела положение 368 может быть положением (L368A), в котором лейцин (L) заменен аланином (А). Во втором СН3 константной области антитела положение 407 может быть положением (Y407V), в котором тирозин (Y) заменен валином (V).
В белковом комплексе по меньшей мере один из первого СН3 константной области антитела и второго СН3 константной области антитела может включать по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (Н), глицина (G), валина (V), метионина (М), аланина (А), изолейцина (I) и серина (S), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394.
Например, первый СН3 константной области антитела может включать по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (Н), глицина (G), валина (V), метионина (М), аланина (А) и изолейцина (I), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394. Кроме того, второй СН3 константной области антитела может включать по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (Н), глицина (G), валина (V), метионина (М), аланина (А) и серина (S), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394.
В одном варианте осуществления первый СН3 константной области антитела может включать триптофан (W) в положении 366 и фенилаланин (F), гистидин (Н) или триптофан (W) в положении 394. Положение 394 может представлять собой положение, в котором треонин (Т) заменен фенилаланином (F), гистидином (Н) или триптофаном (W) (T394F, Т394Н или T394W).
В другом варианте осуществления первый СН3 константной области антитела может включать триптофан (W) в положении 366 и может включать триптофан (W), валин (V), аланин (А) или фенилаланин (F) в положении 351. Положение 351 может представлять собой положение, в котором лейцин (L) заменен триптофаном (W), валином (V), аланином (А) или фенилаланином (F) (L351W, L351V, L351A или L351F).
В другом варианте осуществления второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и аланин (А), глицин (G), валин (V), метионин (М) или фенилаланин (F) в положении 351. Положение 351 может представлять собой положение, в котором лейцин (L) заменен аланином (А), глицином (G), валином (V), метионином (М) или фенилаланином (F) (L351A, L351G, L351V, L351M или L351F).
Первый СН3 константной области антитела может включать одно, выбранное из группы, состоящей из:
триптофана (W) в положении 366;
триптофана (W) в положении 366 и гистидина (Н) в положении 394;
триптофана (W) в положении 366 и фенилаланина (F) в положении 394;
триптофана (W) в положении 366 и триптофана (W) в положении 394;
триптофана (W) в положении 366 и фенилаланина (F) в положении 351;
триптофана (W) в положении 366 и триптофана (W) в положении 351;
триптофана (W) в положении 366 и валина (V) в положении 351;
триптофана (W) в положении 366 и аланина (А) в положении 351; и
триптофана (W) в положении 366, гистидина в положении 394 (Н) и фенилаланина в положении 351 (F).
Второй СН3 константной области антитела может включать одно, выбранное из группы, состоящей из:
серина (S) в положении 366, аланина (А) в положении 368 и валина (V) в положении 407;
серина (S) в положении 366, аланина (А) в положении 368, валина (V) в положении 407 и аланина (А) в положении 351;
серина (S) в положении 366, аланина (А) в положении 368, валина (V) в положении 407 и глицина (G) в положении 351;
серина (S) в положении 366, аланина (А) в положении 368, валина (V) в положении 407 и валина (V) в положении 351;
серина (S) в положении 366, аланина (А) в положении 368, валина (V) в положении 407 и метионина (М) в положении 351; и
серина (S) в положении 366, аланина (А) в положении 368, валина (V) в положении 407 и фенилаланина (F) в положении 351.
Первый СН3 константной области антитела может включать триптофан (W) в положении 366 и фенилаланин (F), гистидин (Н) или триптофан (W) в положении 394; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368 и валин (V) в положении 407.
Первый СН3 константной области антитела может включать триптофан (W) в положении 366 и фенилаланин (F) в положении 351; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и аланин (А) в положении 351. Первый СН3 константной области антитела может дополнительно включать гистидин (Н) в положении 394.
Первый СН3 константной области антитела может включать триптофан (W) в положении 366; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и глицин (G) в положении 351. Первый СН3 константной области антитела может дополнительно включать фенилаланин (F) или триптофан (W) в положении 351.
Первый СН3 константной области антитела может включать триптофан (W) в положении 366; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и аланин (А), фенилаланин (F), валин (V) или метионин (М) в положении 351. Первый СН3 константной области антитела может дополнительно включать валин (V) или аланин (А) в положении 351. Например, первый СН3 константной области антитела может включать триптофан (W) в положении 366 и валин (V) в положении 351; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и аланин (А) в положении 351. Кроме того, первый СН3 константной области антитела может включать триптофан (W) в положении 366 и аланин (А) в положении 351; и второй СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и фенилаланин (F) в положении 351.
Белковый комплекс согласно одному аспекту может быть вариантом Fc-варианта, замещенным обратной последовательностью. Используемый в данном документе термин "вариант с замещением обратной последовательностью" относится к структуре, в которой левая и правая константные области тяжелой цепи белкового комплекса симметричны относительно оси Y на координатной плоскости. Например, вариант с замещением обратной последовательностью, имеет одинаковые конкретное положение и тип аминокислотной замены в белковом комплексе согласно варианту осуществления, но первый СН3 константной области антитела и второй СН3 константной области антитела могут иметь структуру, симметричную относительно оси Y на координатной плоскости. Поскольку вариант с замещением обратной последовательностью образует гетеродимер с показателем, подобным показателю до замены конкретной аминокислоты, возможно образование Fc-варианта с высокой эффективностью. Конкретные детали, касающиеся белкового комплекса, описаны выше.
В одном варианте осуществления первый СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368 и валин (V) в положении 407; и второй СН3 константной области антитела может включать триптофан (W) в положении 366; и по меньшей мере один из первого СН3 константной области антитела и второго СН3 константной области антитела может включать по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (Н), глицина (G), валина (V), метионина (М) и аланина (А), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394. Конкретные подробности, касающиеся первого СН3 константной области антитела и второго СН3 константной области антитела, описаны выше.
Кроме того, первый СН3 константной области антитела может включать серии (S) в положении 366, аланин (А) в положении 368, валин (V) в положении 407 и глицин (G) в положении 351; и второй СН3 константной области антитела может включать триптофан (W) в положении 366. Второй СН3 константной области антитела может дополнительно включать фенилаланин (F) или триптофан (W) в положении 351.
В одном варианте осуществления в результате сравнения показателей образования гетеродимеров у биспецифического антитела IgG1 дикого типа и биспецифического антитела от Genentech, полученного посредством подхода "выступы-во-впадины" (KiH), с показателем образования у Fc-вариантов, выбранных после замены аминокислот в определенных положениях в СН3 константной области антитела, было подтверждено, что показатель образования гетеродимера у Fc-вариантов был заметно выше, при этом термодинамическая стабильность также была превосходной. Кроме того, было подтверждено, что показатель образования гетеродимера также заметно выше у вариантов из Fc-вариантов, замещенных обратной последовательностью, по сравнению с IgG1 дикого типа и биспецифическим антителом от Genentech, полученным посредством подхода "выступы-во-впадины" (KiH). Таким образом, белковый комплекс согласно одному аспекту характеризуется улучшенной эффективностью образования гетеродимера, и, следовательно, белковый комплекс может быть использован в качестве биспецифического антитела, включающего его же или его антигенсвязывающий фрагмент, рецептор и рецептор-связывающий агонист, антагонист, лиганд, цитокин, биконъюгат на основе рецептора-ловушки и т.п.
В другом аспекте представлен способ получения белкового комплекса, предусматривающий трансформацию одной или нескольких клеток одним или несколькими векторами экспрессии, кодирующими первый полипептид согласно одному аспекту, второй полипептид согласно одному аспекту или их комбинацию; и обеспечение экспрессии первого полипептида, второго полипептида или их комбинации.
Конкретные подробности, касающиеся первого полипептида, второго полипептида и белкового комплекса, описаны выше.
"Вектор экспрессии" относится к вектору экспрессии, способному к экспрессированию белка-мишени в соответствующих клетках-хозяевах, и он относится к вектору, включающему ключевые регуляторные элементы, функционально связанные таким образом, что может экспрессироваться вставленная последовательность нуклеиновой кислоты. "Функционально связанный" означает, что регуляторная последовательность для контроля экспрессии нуклеиновой кислоты и нуклеиновая кислота, кодирующая целевой белок, функционально связаны для выполнения общей функции. Вектор экспрессии может включать полинуклеотид, кодирующий первый полипептид, второй полипептид или их комбинацию. Вектор экспрессии может включать регуляторную область, необходимую для экспрессии гена, например, энхансер, промотор, последовательность поли(А) и т.п.
Клетки могут представлять собой раковые клетки. Клетки могут представлять собой клетки in vitro. Клетки могут представлять собой бактерии, дрожжи, растительные клетки или клетки млекопитающих. Бактерии могут представлять собой Е. coli. Клетки млекопитающих относятся к клеткам, полученным от мышей, крыс, кроликов, собак, кошек, овец, коров, лошадей, обезьян, шимпанзе или людей. Клетки могут представлять собой клеточную линию. Клетки могут быть выбраны, например, из группы, состоящей из клеток яичника китайского хомячка (СНО), клеток почки эмбриона человека (HEK), клеток почки детеныша хомячка (BHK), клеток NS0, клеток PER.C6, клеток HeLa, клеток почки собаки Мадина-Дарби (MDCK), клеток миеломы мыши SP2/0, клеток COS-7 и клеток миеломы крысы YB2/0. Клетки СНО могут представлять собой клетки СНО DG44, СНО-K1, CHO-S, GS-CHO или СНО DUKX (DXB11). Клетки HEK могут представлять собой клетки HEK 293.
"Трансформация" относится к способу вставки определенного фрагмента нуклеиновой кислоты в геном клетки таким образом, что вставленная нуклеиновая кислота экспрессируется.
Векторами экспрессии, кодирующими первый полипептид, и векторами экспрессии, кодирующими второй полипептид, могут быть совместно трансфицированы клетки, или векторами экспрессии, кодирующими первый полипептид, и векторами экспрессии, кодирующими второй полипептид, могут быть трансформированы по меньшей мере два типа клеток, при этом одним вектором для каждого типа клеток.
Клетки могут культивироваться в культуральной клеточной среде. Культуральная клеточная среда относится к раствору, содержащему питательные вещества, необходимые для культивирования клеток. Среда включает коммерческую или приготовленную среду, используемую для культивирования клеток. Культуральная клеточная среда может содержать антибиотик. Культуральная клеточная среда может включать G418 (генетицин), пуромицин, бластицидин, зеоцин или их комбинацию. Культуральная клеточная среда может включать химически определенную среду.
Клетка может культивироваться в условиях, которые обеспечивают выживание или пролиферацию клеток. Условия, обеспечивающие выживание или пролиферацию клеток, могут варьироваться в зависимости от типа клеток. Клетки можно культивировать при температуре от приблизительно 25°С до приблизительно 42°С, от приблизительно 25°С до приблизительно 40°С, от приблизительно 30°С до приблизительно 40°С, от приблизительно 30°С до приблизительно 37°С или при приблизительно 37°С. Клетки можно культивировать в присутствии воздуха, содержащего от приблизительно 1% СО2 до приблизительно 10% СО2 или от приблизительно 5% СО2 до приблизительно 10% СО2. Клетки можно культивировать в среде с показателем рН, составляющим от приблизительно рН 6 до приблизительно рН 8, от приблизительно рН 6,2 до приблизительно рН 7,8, от приблизительно рН 6,4 до приблизительно рН 7,6, от приблизительно рН 6,6 до приблизительно рН 7,4 или от приблизительно рН 6,8 до приблизительно рН 7,2. Клетки можно культивировать в атмосфере, содержащей от приблизительно 10% до приблизительно 80%, от приблизительно 15% до приблизительно 70% от или приблизительно 20% до приблизительно 60% растворенного кислорода.
Культура может варьироваться в зависимости от типа клеток. Для культивирования можно использовать известный способ. Культивирование может быть выполнено на планшете, в колбе или т.п. Культивирование может быть выполнено путем прикрепления клеток к субстрату или в виде плавающих в культуральной среде клеток. Культура может представлять собой субкультуру, периодическую культуру, подпитываемую культуру, перфузионную культуру или их комбинацию. В ходе культивирования культуральную клеточную среду можно периодически заменять свежей средой. Клетки можно культивировать в течение приблизительно 1 дня или больше, приблизительно 2 дней или больше, приблизительно 3 дней или больше, приблизительно 4 дней или больше, приблизительно 5 дней или больше, приблизительно 6 дней или больше, приблизительно 1 недели или больше, приблизительно 10 дней или больше, приблизительно 2 недель или больше, приблизительно 3 недель или больше, приблизительно 1 месяца или больше, от приблизительно 1 дня до приблизительно 1 месяца, от приблизительно 1 дня до приблизительно 3 недель, от приблизительно 1 дня до приблизительно 2 недель, от приблизительно 2 дней до приблизительно 2 недель, от приблизительно 3 дней до приблизительно 2 недель, от приблизительно 4 дней до приблизительно 2 недель, от приблизительно 5 дней до приблизительно 2 недель, от приблизительно 6 дней до приблизительно 2 недель или от приблизительно 1 недели до приблизительно 2 недель.
Может быть включено получение белкового комплекса на основе первого полипептида, второго полипептида или первого полипептида и второго полипептида из клеток или культуральной клеточной среды.
Культуральная клеточная среда может представлять собой культуральную среду без клеток.
При совместной трансформации вектором экспрессии клеток белковый комплекс на основе первого полипептида и второго полипептида может быть получен из клеток или культуральной клеточной среды. Если по меньшей мере два типа клеток трансформируют вектором экспрессии, кодирующим первый полипептид, и вектором экспрессии, кодирующим второй полипептид, при этом один вектор предназначен для каждого типа клеток, то первый полипептид и второй полипептид могут быть получены из клеток или культуральных клеточных сред.
Получение белкового комплекса может включать инкубацию полученного первого полипептида и полученного второго полипептида с образованием белкового комплекса. Инкубацию можно выполнять в восстановительном условии. Восстановительное условие может быть обеспечено в присутствии 2-меркаптоэтанола (2-МЕ), дитиотреитола (DTT) или их комбинации.
Получение белкового комплекса может включать очистку белкового комплекса. Очистка может быть выполнена посредством фильтрации, центрифугирования, хроматографии, диализа, иммунопреципитации или их комбинации.
В другом аспекте представлена фармацевтическая композиция для профилактики или лечения рака, включающая белковый комплекс согласно одному аспекту.
Конкретные детали, касающиеся белкового комплекса, описаны выше.
Рак может представлять солидный рак или рак, не относящийся к солидному. Солидным раком называют злокачественные опухоли, возникшие в таких органах, как печень, легкое, молочная железа, кожа и т.д. Виды рака, не относящиеся к солидным, представляют собой виды рака, возникшие в крови, которые также называются гемобластозами. Рак может представлять собой карциному, саркому, рак, происходящий из кроветворных клеток, герминогенную опухоль или бластому. Рак может быть выбран из группы, состоящей из рака молочной железы, рака кожи, рака головы и шеи, рака поджелудочной железы, рака легкого, колоректального рака, рака желудка, рака яичника, рака предстательной железы, рака мочевого пузыря, рака уретры, рака печени, рака почки, светлоклеточной саркомы, меланомы, цереброспинальной опухоли, рака головного мозга, тимомы, мезотелиомы, рака пищевода, рака желчевыводящих путей, рака яичка, герминогенной опухоли, рака щитовидной железы, рака паращитовидной железы, рака шейки матки, рака эндометрия, лимфомы, миелодиспластических синдромов (MDS), миелофиброза, острого лейкоза, хронического лейкоза, множественной миеломы, болезни Ходжкина, эндокринного рака и саркомы.
Термин "профилактика" относится к любому действию, которое подавляет заболевание или задерживает проявление заболевания посредством введения фармацевтической композиции. Термин "лечение" относится к любому действию, которое улучшает или благоприятно изменяет симптомы заболевания за счет введения фармацевтической композиции.
Фармацевтическая композиция может включать фармацевтически приемлемый носитель. Термин "носитель" используют для охвата вспомогательных средств, разбавителей или вспомогательных веществ. Например, носитель может быть выбран из группы, состоящей из лактозы, декстрозы, сахарозы, сорбита, маннита, ксилита, эритрита, мальтита, крахмала, гуммиарабика, альгината, желатина, фосфата кальция, силиката кальция, целлюлозы, метилцеллюлозы, поливинилпирролидона, воды, физиологического солевого раствора, буферов, таких как PBS, метилгидроксибензоата, пропилгидроксибензоата, талька, стеарата магния и минерального масла. Композиция может включать наполнитель, антиагрегирующее средство, смазывающее средство, смачивающее средство, вкусоароматическое средство, эмульгатор, консервант или их комбинацию.
Фармацевтическая композиция может быть получена в любом составе согласно способу из уровня техники. Композиция может быть составлена, например, в виде состава для перорального введения (например, порошка, таблетки, капсулы, сиропа, пилюли или гранулы) или состава для парентерального введения (например, путем инъекции). Кроме того, композиция может быть получена в виде состава для системного введения или в виде состава для местного введения.
Фармацевтическая композиция может дополнительно включать другие противораковые средства. Противораковое средство может представлять собой цетуксимаб, панитумумаб, эрлотиниб, гефитиниб, трастузумаб, T-DM1, перьету, лапатиниб, паклитаксел, таксол, тамоксифен, цисплатин или их комбинацию. Фармацевтическая композиция может представлять собой одну композицию или отдельные композиции. Например, композиция на основе антитела или его антигенсвязывающего фрагмента может представлять собой композицию для парентерального введения, а противораковое средство может представлять собой композицию для перорального введения.
Фармацевтическая композиция может включать белковый комплекс в эффективном количестве. Термин "эффективное количество" означает количество, достаточное для проявления эффекта профилактики или лечения при введении субъекту, нуждающемуся в профилактике или лечении заболевания. Эффективное количество может быть соответствующим образом выбрано специалистами в данной области техники в зависимости от клетки или субъекта. Эффективное количество может быть определено на основании тяжести заболевания, возраста, массы тела, состояния здоровья, пола пациента, чувствительности пациента к лекарственному средству, времени введения, пути введения и скорости выведения, продолжительности лечения, лекарственного средства, используемого в комбинации или одновременно с используемой композицией, и других факторов, хорошо известных в области медицины. Эффективное количество может составлять от приблизительно 0,5 мкг до приблизительно 2 г, от приблизительно 1 мкг до приблизительно 1 г, от приблизительно 10 мкг до приблизительно 500 мг, от приблизительно 100 мкг до приблизительно 100 мг или от приблизительно 1 мг до приблизительно 50 мг на фармацевтическую композицию.
Дозировка фармацевтической композиции для взрослых может находиться, например, в диапазоне от приблизительно 0,001 мг/кг до приблизительно 100 мг/кг, от приблизительно 0,01 мг/кг до приблизительно 10 мг/кг или от приблизительно 0,1 мг/кг до приблизительно 1 мг/кг. Введение можно осуществлять один раз в день, несколько раз в день, один раз в неделю, один раз в 2 недели, один раз в 3 недели или от одного раза в 4 недели до одного раза в год.
В другом аспекте представлен способ профилактики или лечения рака, предусматривающий введение белкового комплекса согласно одному аспекту в клетку или субъекту.
Конкретные подробности, касающиеся белкового комплекса, клеток, рака, профилактики или лечения, описаны выше.
Субъектом может быть млекопитающее, например, человек, корова, лошадь, свинья, собака, овца, коза или кошка. Субъект может быть субъектом, у которого имеется рак или имеется риск заболевания раком.
Способ может дополнительно предусматривать введение второго активного ингредиента субъекту. Второй активный ингредиент может представлять собой активный ингредиент для профилактики или лечения рака. Активный ингредиент можно вводить одновременно, отдельно или последовательно с белковым комплексом.
Белковый комплекс может быть, например, введен непосредственно субъекту любым способом, таким как пероральное, внутривенное, внутримышечное, трансдермальное, чресслизистое, интраназальное, интратрахеальное или подкожное введение. Белковый комплекс можно вводить системно или местно, отдельно или в комбинации с другими фармацевтически активными соединениями.
Эффективное количество белкового комплекса может варьироваться в зависимости от состояния пациента и массы тела, тяжести заболевания, состава лекарственного средства, пути и продолжительности введения и т.д., и может быть соответствующим образом подобрано специалистами в данной области техники. Дозировка может находиться, например, в диапазоне от приблизительно 0,001 мг/кг до приблизительно 100 мг/кг, от приблизительно 0,01 мг/кг до приблизительно 10 мг/кг или от приблизительно 0,1 мг/кг до приблизительно 1 мг/кг для взрослых. Введение белкового комплекса можно осуществлять один раз в день, несколько раз в день, один раз в неделю, один раз в 2 недели, один раз в 3 недели или от одного раза в 4 недели до одного раза в год.
ПОЛЕЗНЫЕ ЭФФЕКТЫ ИЗОБРЕТЕНИЯ
Согласно белковому комплексу, характеризующемуся высоким показателем образования гетеродимера, способу его получения, фармацевтической композиции для профилактики или лечения рака, включающей белковый комплекс, и способу профилактики или лечения рака посредством ее применения, биспецифические антитела или антигенсвязывающие фрагменты с повышенной стабильностью, рецепторы и рецептор-связывающие агонисты, антагонисты, лиганды, цитокины или биконъюгаты на основе рецептора-ловушки могут быть легко получены и использованы в различных областях, таких как профилактика или лечение заболеваний и диагностика заболеваний.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 представлено изображение, показывающее результаты анализа посредством SDS-PAGE Fc-вариантов (М: маркер размера, WT: PTCWT (группа отрицательного контроля), 019: РТС019 (группаположительного контроля), 039: РТС039, 040: РТС040, 074: РТС074, 111: РТС111).
На фиг. 2 представлен график, показывающий результаты анализа Fc-вариантов посредством капиллярного электрофореза-SDS (CE-SDS).
На фиг. 3А-3F представлены графики, показывающие результаты анализа посредством эксклюзионной высокоэффективной жидкостной хроматографии (SE-HPLC) PTCWT, РТС019, РТС039, РТС040, РТС074 и РТС111 соответственно.
На фиг. 4 представлено изображение, показывающее результаты анализа посредством SDS-PAGE Fc-вариантов с замещением обратной последовательностью (М: маркер размера, WT: PTCWT (группа отрицательного контроля), 019: РТС019 (группа положительного контроля), 088: РТС088, 089: РТС089, 090: РТС090).
На фиг. 5 представлено изображение, показывающее результаты анализа посредством SDS-PAGE Fc-вариантов, в которых часть последовательности замещена (М: маркер размера, WT: PTCWT (группа отрицательного контроля), 019: РТС019 (группа положительного контроля), 074: РТС074, 097: РТС097, 098: РТС098, 099: РТС099, 111: РТС111, 113: РТС113).
На фиг. 6 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × scFv-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × scFv-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × scFv-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × scFv-Fc с последовательностью РТС074, встроенной в СН3-домен).
На фиг. 7 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × scFv-scFv-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью РТС074, встроенной в СН3-домен).
На фиг. 8 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × цитокин-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × цитокин-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × цитокин-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × цитокин-Fc с последовательностью РТС074, встроенной в СН3-домен).
На фиг. 9 представлен график оценки способности биспецифических антител к PD-L1 × scFv к CD3 индуцировать гибель раковых клеток in vitro (авелумаб: группа положительного контроля, BsAb к PD-L1 × scFv к CD3: биспецифическое антитело к PD-L1 × scFv к CD3, имеющее последовательность РТС074 в СН3-домене).
На фиг. 10 представлен график оценки способности биспецифических антител к PD-L1 × scFv к CD3 подавлять рост раковых клеток in vivo (авелумаб: группа положительного контроля, BsAb к PD-L1 × scFv к CD3: биспецифическое антитело к PD-L1 × scFv к CD3, имеющее последовательность РТС074 в СН3-домене).
ПРИНЦИП ИЗОБРЕТЕНИЯ
Далее представлены предпочтительные примеры для лучшего понимания настоящего изобретения. Тем не менее, следующие примеры приведены исключительно для упрощения понимания настоящего изобретения, при этом содержание настоящего изобретения не ограничивается следующими примерами.
Примеры
Пример 1. Получение гетеродимерных Fc-вариантов
Для оценки гетеродимер-образующей способности Fc, которая изменяется в результате аминокислотной замены в СН3-домене антитела, конструировали систему экспрессии для каждого сконструированного Fc-варианта.
Чтобы облегчить различение и анализ двух полипептидов Fc, составляющих гетеродимер, первый полипептид конструировали для экспрессии цепи IgG (Fc1) интактной формы, в которой тяжелая цепь и легкая цепи связаны, и второй полипептид конструировали для экспрессии только тяжелой цепи Fc-области (Fc2). В Fc1 аминокислоты в СН3-домене заменяли на основе антитела авелумаб (Bavencio®, Pfizer), включающего Fc-область IgG1 дикого типа, и в Fc2 аминокислоты в СН3-домене заменяли на основе Fc-области антитела IgG1 дикого типа. Легкая цепь Fc1 имеет ту же аминокислотную последовательность, что и легкая цепь антитела авелумаб (SEQ ID NO: 2).
Для сравнения IgG1 дикого типа (соответствующий "PTCWT" в таблице 1) использовали в качестве группы отрицательного контроля, и биспецифическое антитело, полученное посредством подхода "выступы-во-впадины" (KiH) от Genentech (соответствующее "РТС019" в таблице 1), использовали в качестве группы положительного контроля.
В частности, векторы экспрессии получали путем введения нуклеотидной открытой рамки считывания (ORF), кодирующей каждый полипептид, в векторы pCHO1.O. Клеточную линию ExpiCHO-S™ (Thermo Fisher) культивировали в среде для экспрессии ExpiCHO™. Векторы для экспрессии Fc1 и векторы для экспрессии Fc2 смешивали в соотношении 1:1 и ими трансфицировали клеточную линию ExpiCHO-S™ с использованием набора ExpiFectamine™ СНО Transfection Kit (Thermo Fisher). После культивирования трансфицированных клеток в среде для экспрессии культуральную среду отделяли и извлекали. Извлеченную культуральную среду очищали с использованием колонки Protein A HP SpinTrap™ (GE Healthcare). Для очищенного белка буфер заменяли на PBS (рН 7,4) и измеряли концентрацию белка.
Экспрессированные аминокислотные последовательности были такими, как описано ниже.
[PTCWT]
Тяжелая цепь из Fc1:

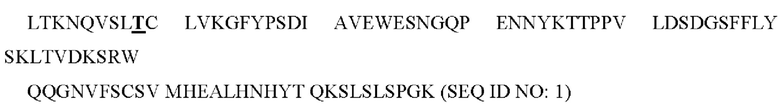
(жирным шрифтом: L351, жирным шрифтом и подчеркнуто: Т366).
Легкая цепь из Fc1:

Fc-цепь из Fc2:

(жирным шрифтом: L351, жирным шрифтом и подчеркнуто: Т366, подчеркнуто: L368, курсивом: Y407).
[РТС019]
Тяжелая цепь из Fc1:


(жирным шрифтом: L351, жирным шрифтом и подчеркнуто: T366W).
Легкая цепь из Fc1 (SEQ ID NO: 2)
Fc-цепь из Fc2:

(жирным шрифтом: L351, жирным шрифтом и подчеркнуто: T366S, подчеркнуто: L368A, курсивом: Y407V).
[РТС039]
Тяжелая цепь из Fc1:


(жирным шрифтом: L351F, жирным шрифтом и подчеркнуто: T366W).
Легкая цепь из Fc1 (SEQ ID NO: 2)
Fc-цепь из Fc2:

(жирным шрифтом: L351G, жирным шрифтом и подчеркнуто: T366S, подчеркнуто: L368A, курсивом: Y407V).
[РТС040]
Тяжелая цепь из Fc1:

(жирным шрифтом: L351W, жирным шрифтом и подчеркнуто: T366W).
Легкая цепь из Fc1 (SEQ ID NO: 2)
Fc-цепь из Fc2 (SEQ ID NO: 7)
[PTC074]
Тяжелая цепь из Fc1 (SEQ ID NO: 4)
Легкая цепь из Fc1 (SEQ ID NO: 2)
Fc-цепь из Fc2 (SEQ ID NO: 7)
[PTC111]
Тяжелая цепь из Fc1 (SEQ ID NO: 4)
Легкая цепь из Fc1 (SEQ ID NO: 2)
Fc-цепь из Fc2:

(жирным шрифтом: L351F, жирным шрифтом и подчеркнуто: T366S, подчеркнуто: L368A, курсивом: Y407V).
Пример 2. Сравнение гетеродимер-образующих способностей у Fc-вариантов
Используя систему транзиентной экспрессии, сконструированной в примере 1 выше, сравнивали способности к образованию гетеродимеров Fc, которые изменены посредством аминокислотной замены в СН3-домене, с отбором тем самым высокоэффективных гетеродимерных Fc-вариантов.
В качестве способа оценки, после выполнения SDS-PAGE в невосстанавливающих условиях, измеряли и сравнивали интенсивность полос, полученных в PAGE, соответствующих гетеродимерам.
В частности, белок, очищенный в примере 1, восстанавливали посредством 2-меркаптоэтанола или образец готовили без обработки 2-меркаптоэтанолом. Восстановленный или невосстановленный образец подвергали электрофорезу посредством способа SDS-PAGE и интенсивность электрофоретической полосы измеряли с использованием системы ChemiDoc™ Imaging System (Bio-Rad) и Image Lab™ Software (Bio-Rad).
Способность к образованию гетеродимеров Fc согласно аминокислотной замене в СН3-домене рассчитывали с учетом измеренных интенсивностей полос, и результаты представлены в таблице 1 ниже.

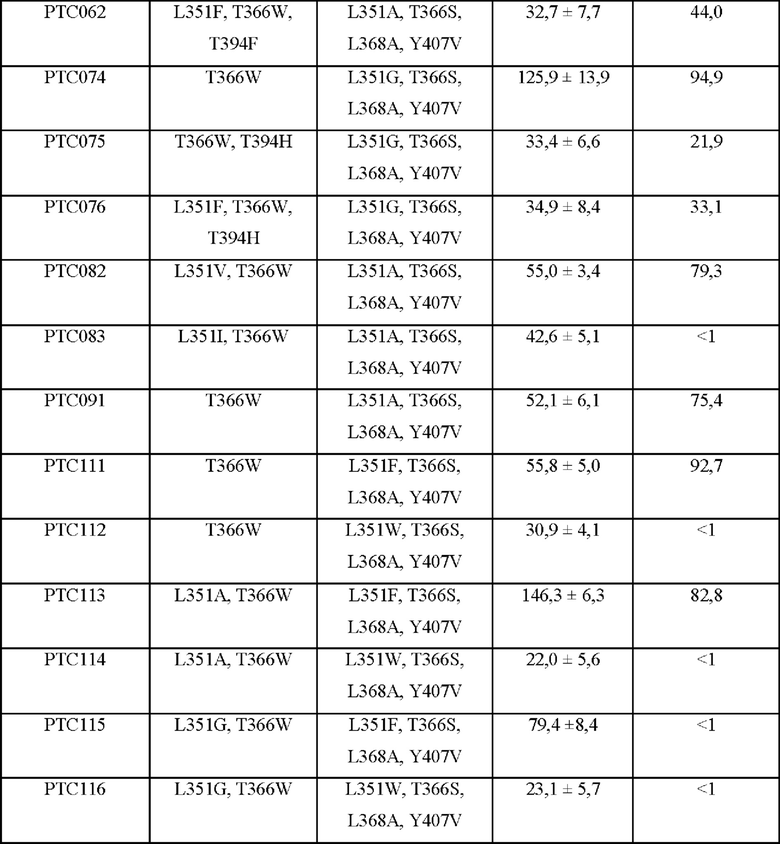
В результате, как показано в таблице 1, показатели образования гетеродимеров в группе отрицательного контроля (PTCWT) и группе положительного контроля (РТС019) составляли 41,5% и 64,6% соответственно, тогда как было подтверждено, что показатели образования гетеродимеров РТС032, РТС033, РТС034, РТС037, РТС039, РТС040, РТС061, РТС074, РТС082, РТС091, РТС111 и РТС113 составляли 78,7%, 76,4%, 76,1%, 85,8%, 93,8%, 94,6%, 70,9%, 94,9%, 79,3%, 75,4%, 92,7% и 82,8% соответственно. В частности, подтвердили, что у РТС039, РТС040, РТС074 и РТС111 соотношение гетеродимеров Fc превышало 90%.
То есть Fc-вариант согласно одному аспекту может проявлять превосходную способность к образованию гетеродимеров Fc.
Пример 3. Оценка гетеродимер-образующих способностей у высокоэффективных Fc-вариантов
3-1. Анализ посредством SDS-PAGE
Для оценки гетеродимер-образующей способности у высокоэффективных Fc-вариантов, отобранных в примере 2, выполняли анализ посредством SDS-PAGE таким же образом, как и в примере 2.
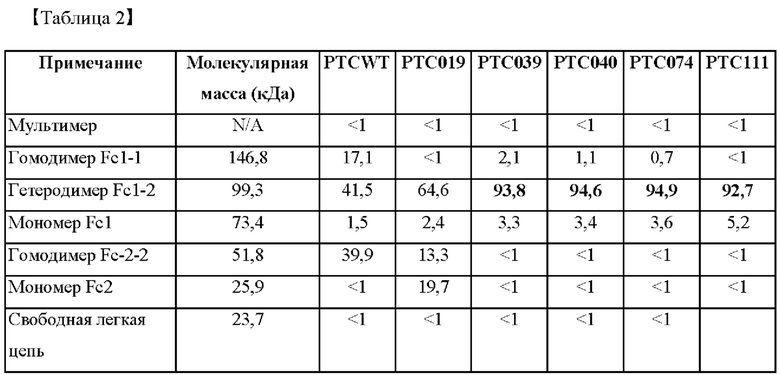
В частности, измеряли интенсивности электрофоретических полос, полученных при анализе посредством SDS-PAGE. После этого показатели образования гетеродимеров Fc согласно аминокислотной замене в СН3-домене подтверждали по интенсивности измеренных полос, и результаты представлены в таблице 2 ниже (N/A: не применимо).
На фиг. 1 представлено изображение, показывающее результаты анализа Fc-вариантов посредством SDS-PAGE.
В результате, как показано на фигуре 1, было подтверждено, что интенсивность полосы, соответствующей гомодимеру Fc1-1 (146,8 кДа), у выбранных РТС039, РТС040, РТС074 и РТС111 была заметно снижена по сравнению с группой отрицательного контроля (PTCWT), а интенсивность полосы, соответствующей мономеру Fc2 (25,9 кДа), была заметно снижена по сравнению с группой положительного контроля (РТС019). С другой стороны, было подтверждено, что гетеродимер Fc1-2 образовывался с высокой эффективностью при 99,3 кДа.
Кроме того, как показано в таблице 1, в случаях группы отрицательного контроля (PTCWT) и группы положительного контроля (РТС019) было подтверждено, что не только гетеродимер Fc1-2, но и гомодимер Fc1-1 образовывался с показателем 17,1% в группе отрицательного контроля (PTCWT), и подтверждено, что гомодимер Fc-2-2 образовывался с показателем 39,9% в группе отрицательного контроля (PTCWT) и 13,3% в группе положительного контроля (РТС019). С другой стороны, в случаях выбранных Fc-вариантов РТС039, РТС040, РТС074 и РТС111 было подтверждено, что показатель образования гомодимеров Fc1-1 и гомодимеров Fc-2-2 был низким, а показатель образования гетеродимеров Fc1-2 был заметно выше.
То есть можно видеть, что Fc-вариант согласно одному аспекту способен избирательно образовывать гетеродимеры Fc1-2.
3-2. Анализ посредством CE-SDS
Для того чтобы проверить гетеродимер-образующие способности у высокоэффективных Fc-вариантов, выбранных в примере 2, выполняли анализ посредством капиллярного электрофореза-SDS (CE-SDS).
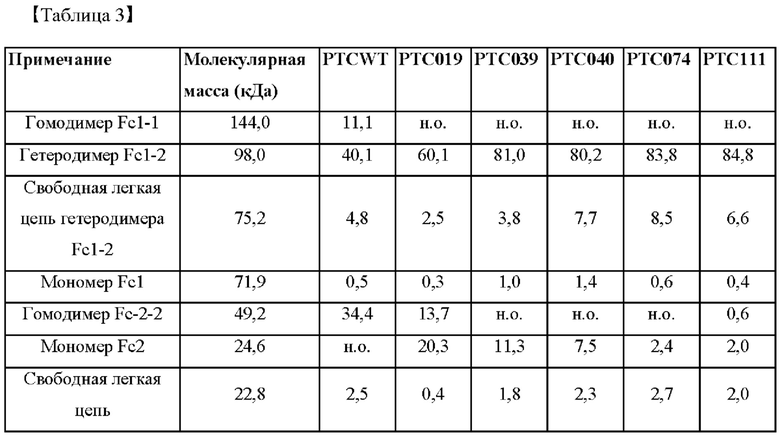
В частности, капиллярный электрофорез выполняли с использованием фармацевтической аналитической системы РА 800 plus™ (SCIEX) для проверки показателей образования гетеродимеров Fc у Fc-вариантов, и его результаты показаны в таблице 3 ниже.
На фиг. 2 представлен график, показывающий результаты анализа Fc-вариантов посредством капиллярного электрофореза-SDS (CE-SDS).
В результате, как показано на фиг. 2, при времени удерживания около 23 минут показатель образования гомодимеров Fc-2-2 в группе отрицательного контроля (PTCWT) составил 34,44%, а показатель образования гомодимеров Fc-2-2 в группе положительного контроля (РТС019) составил 13,71%. С другой стороны, в случаях РСТ039, РТС040 и РТС074 гомодимер Fc-2-2 не образовывался, а в случае РСТ111 показатель образования гомодимеров Fc-2-2 составлял 0,57%, и этот показатель был заметно ниже по сравнению с группой отрицательного контроля (PTCWT) и группой положительного контроля (РСТ019).
Кроме того, как показано в таблице 2, показатели образования гетеродимеров у РТС039, РТС040, РТС074 и РТС111 составляли 81,0%, 80,2%, 83,8% и 84,8% соответственно, и они были значительно выше, чем у группы отрицательного контроля (PCTWT) и группы положительного контроля (РТС019).
То есть можно видеть, что Fc-вариант согласно одному аспекту обладает превосходной гетеродимер-образующей способностью.
3-3. Анализ посредством SE-HPLC
Для того чтобы проверить гетеродимер-образующие способности у высокоэффективных Fc-вариантов, выбранных в примере 2, выполняли анализ посредством эксклюзионной высокоэффективной жидкостной хроматографии (SE-HPLC), и его результаты показаны в таблице 4 ниже.
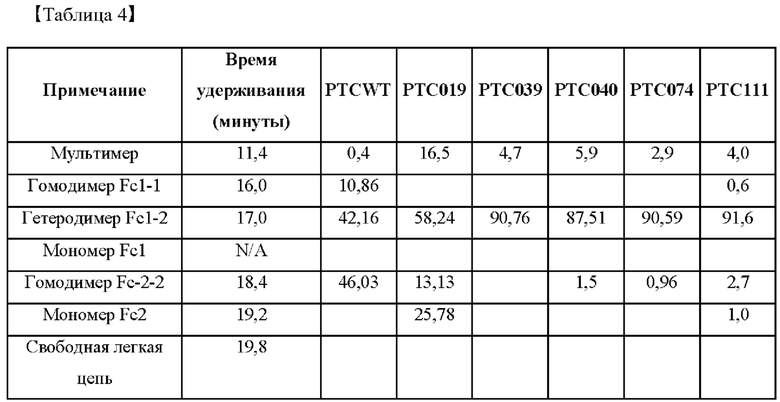
На фиг. 3А-3F представлены графики, показывающие результаты анализа посредством эксклюзионной высокоэффективной жидкостной хроматографии (SE-HPLC) PTCWT, РТС019, РТС039, РТС040, РТС074 и РТС111 соответственно.
В результате, как показано на фиг. 3А-3F, в анализе белка, очищенного с помощью белка А, посредством SE-HPLC основные пики гетеродимера наблюдали при времени удерживания от 16,989 минуты до 17,197 минуты в Fc-вариантах, включая группу отрицательного контроля (PTCWT) и группу положительного контроля (РТС019). В частности, в случае группы отрицательного контроля (PTCWT) пик гомодимера Fc-1-1 наблюдали при времени удерживания 16,079 минуты, а пик гомодимера Fc2-2 наблюдали при 18,377 минуты (фиг. 3А). Кроме того, в случае группы положительного контроля (РТС019) пик гомодимера Fc-2-2 наблюдали при времени удерживания 18,343 минуты, а пик мономера Fc-2 наблюдали при 19,14 минуты (фиг. 3В). То есть в случае группы отрицательного контроля (PTCWT) и группы положительного контроля (РСТ019) показатель образования гетеродимеров подтверждали как низкий, так как было показано несколько пиков, включающих гетеродимер Fc1-2, с другой стороны, в случае Fc-вариантов РТС039, РТС040, РТС074 и РТС111 основной пик наблюдали в виде отдельного пика. То есть в случае выбранных Fc-вариантов показатель образования гетеродимеров подтверждали как превосходный по сравнению с группой отрицательного контроля (PTCWT) и группой положительного контроля (РТС019).
Кроме того, как показано в таблице 4, подтверждали, что выбранные Fc-варианты РТС039, РТС040, РТС074 и РТС111 обладали лучшими способностями к образованию гетеродимеров по сравнению с двумя группами контроля PTCWT и РТС019.
Таким образом, Fc-вариант согласно одному аспекту может образовывать гетеродимер с высокой эффективностью, и поэтому легко могут быть получены биспецифические антитела.
Пример 4. Оценка термодинамической стабильности гетеродимеров высокоэффективных Fc-вариантов
Для косвенного подтверждения структурных стабильностей гетеродимеров высокоэффективных Fc-вариантов, идентифицированных в примере 3, выполняли оценку термодинамической стабильности с использованием дифференциальной сканирующей калориметрии (DSC).
В частности, термические стабильности выбранных Fc-вариантов измеряли с использованием Nano DSC™ (ТА instruments) и рассчитывали температуры плавления (Tm,°С). Рассчитанные точки плавления Fc-вариантов показаны в таблице 5 ниже.
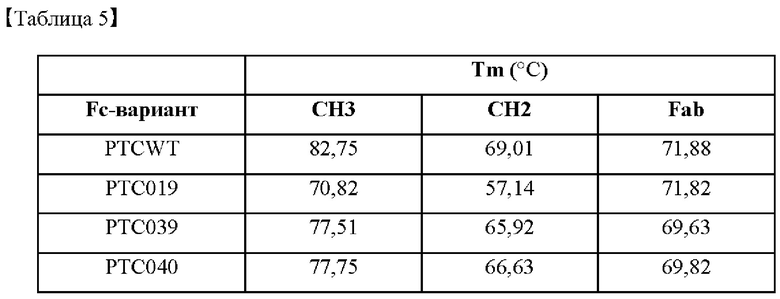
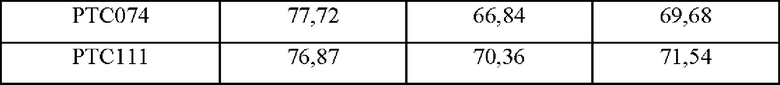
В результате, как показано в таблице 5, было подтверждено, что выбранные Fc-варианты РТС039, РТС040, РТС074 и РТС111 характеризовались несколько нестабильной термической стабильностью по сравнению с гетеродимером Fc дикого типа (PTCWT), но характеризовались относительно высокой термодинамической стабильностью по сравнению с гетеродимером Fc KiH (РТС019) от Genentech.
То есть Fc-вариант согласно одному аспекту характеризуется превосходной гетеродимер-образующей способностью, а также отличной термодинамической стабильностью, поэтому возможно образование стабильных биспецифических антител.
Пример 5. Сравнение гетеродимер-образующих способностей у вариантов, замещенных обратной последовательностью в СН3-домене, среди высокоэффективных Fc-вариантов
Для оценки гетеродимер-образующих способностей у вариантов, замещенных обратной последовательностью в СН3-домене, среди высокоэффективных Fc-вариантов, выбранных в примере 3 выше, выполняли анализ посредством SDS-PAGE таким же образом, как и в примере 2.
Способности к образованию гетеродимера Fc согласно аминокислотной замене в СН3-домене рассчитывали с учетом интенсивностей полос, измеренных в анализе посредством SDS-PAGE, и результаты представлены в таблице 6 ниже.
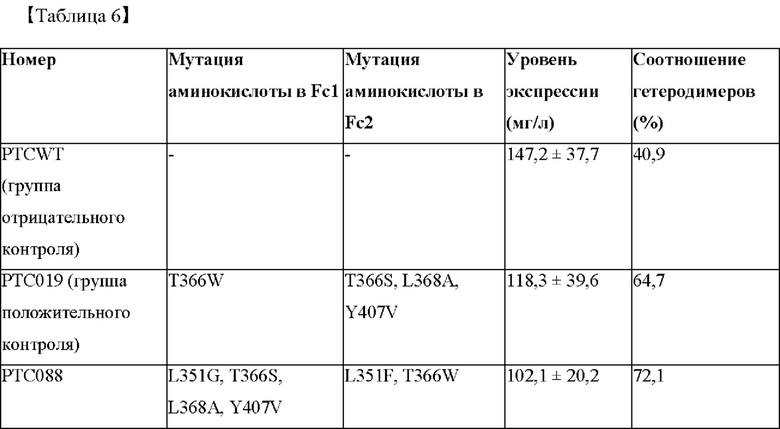
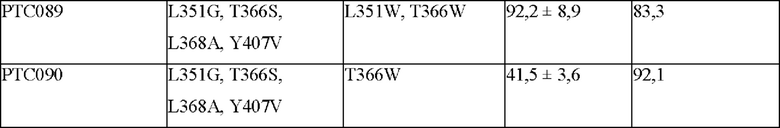
На фиг. 4 представлено изображение, показывающее результаты анализа посредством SDS-PAGE Fc-вариантов с замещением обратной последовательностью (М: маркер размера, WT: PTCWT (группа отрицательного контроля), 019: РТС019 (группа положительного контроля), 088: РТС088, 089: РТС089, 090: РТС090).
В результате, как показано на фиг. 4, было подтверждено, что Fc-варианты с замещением обратной последовательностью не образовывали полосу размером около 150 кДа по сравнению с группой отрицательного контроля (PTCWT), а полосы были слабо сформированы при 49,2 кДа. То есть подтвердили, что Fc-варианты с замещением обратной последовательностью не образовывали гомодимер Fc-1-l, а образование гомодимеров Fc-2-2 было снижено. Кроме того, было подтверждено, что по сравнению с группой положительного контроля (РСТ019) слабо формировалась полоса размером около 25 кДа. То есть можно видеть, что Fc-варианты с заменой обратной последовательностью характеризуются сниженным образованием мономеров Fc-2 и демонстрируют показатели образования гетеродимеров, аналогичные РТС039, РТС040 и РТС074 (см. фиг. 1).
Кроме того, как показано в таблице 6, показатели образования гетеродимеров в группе отрицательного контроля (PTCWT) и группе положительного контроля (РТС019) составляли 40,9% и 64,7% соответственно, тогда как было подтверждено, что РТС088, РТС089 и РТС090, которые являются Fc-вариантами с замещением обратной последовательностью РТС039, РТС040 и РТС074, демонстрировали показатели образования гетеродимеров 72,1%, 83,3% и 92,1% соответственно.
Таким образом, можно видеть, что варианты с замещение обратной последовательностью Fc-варианта согласно одному аспекту образуют гетеродимер при аналогичном соотношении до и после замещения. Кроме того, поскольку варианты с замещением обратной последовательностью Fc-варианта характеризуются превосходным показателем образования гетеродимера, Fc-варианты могут быть образованы с высокой эффективностью независимо от того, замещена ли последовательность, вставленная в Fc1 или Fc2, или нет.
Пример 6. Сравнение гетеродимер-образующих способностей у Fc-вариантов с последовательностью, измененной в положении 351 СН3-домена
Для оценки гетеродимер-образующей способности у вариантов, у которых положение 351 в СН3-домене заменено аминокислотой, имеющей аналогичные свойства, подобно РТС074 и РТС111, которые являются высокоэффективными Fc-вариантами, выбранными в примере 3, выполняли анализ посредством SDS-PAGE таким же образом, как и в примере 2.
Гетеродимер-образующие способности у Fc согласно аминокислотной замене в СН3-домене рассчитывали с учетом интенсивностей полос, измеренных посредством анализа SDS-PAGE, и результаты представлены в таблице 7 ниже.

На фиг. 5 представлено изображение, показывающее результаты анализа Fc-вариантов посредством SDS-PAGE, в которых часть последовательности заменена (М: маркер размера, WT: PTCWT (группа отрицательного контроля), 019: РТС019 (группа положительного контроля), 074: РТС074, 097: РТС097, 098: РТС098, 099: РТС099, 111: РТС111, 113: РТС113).
В результате, как показано на фиг. 5, в случаях РТС074 и РТС111 подтвердили, что не образовывалась полоса около 150 кДа по сравнению с группой отрицательного контроля (PTCWT), а по сравнению с группой положительного контроля (РТС019) интенсивность полос при 49,2 кДа и 24,6 кДа была заметно снижена. Кроме того, подтвердили, что интенсивность полос при 24,6 кДа была заметно снижена по сравнению с РТС097, РТС098 и РТС099, в которых последовательность заменена в положении 351 в Fc-2. С другой стороны, подтвердили, что интенсивность полосы увеличивалась при 98 кДа, что является расчетной молекулярной массой гетеродимера Fc-1-2.
Кроме того, как показано в таблице 7, показатели образования гетеродимеров у РТС074, РТС097, РТС099 и РТС111 составляли 93,5%, 82,7%, 86,5% и 92,7% соответственно, и подтвердили, что они были значительно выше, чем у группы отрицательного контроля (PCTWT) и группы положительного контроля (РТС019). В частности, в случаях РТС074 и РТС111, в которых положение 351 в СН3-домене было заменено глицином (G) или фенилаланином (F), подтвердили, что показатель образования гетеродимеров Fc превышал 90%.
То есть можно видеть, что Fc-вариант согласно одному аспекту обладает превосходной способностью к образованию гетеродимеров Fc за счет замены положения 351 в СН3-домене определенной аминокислотой.
Пример 7. Оценка гетеродимер-образующей способности у биспецифических антител, включающих высокоэффективные Fc-варианты
7-1. Биспецифическое антитело Fab-Fc × scFv-Fc
Для оценки показателя образования гетеродимеров у гетеробиспецифического антитела, включающего высокоэффективные Fc-варианты, выбранные в примере 3, первый полипептид конструировали для экспрессии цепи IgG (Fc1) интактной формы, в которой тяжелая цепь и легкая цепи связаны, и второй полипептид конструировали для экспрессии формы scFV-Fc (Fc2). После этого осуществляли экспрессию Fc1 на основе антитела авелумаб (Bavencio, Pfizer), включающего Fc-область IgG1, а экспрессию Fc2 осуществляли в форме scFv, включающей Fc-область IgG1, на основе последовательности scFv к CD3 блинатумомаба.
Для сравнения в качестве группы отрицательного контроля использовали гетеробиспецифическое антитело Fab-Fc × scFv-Fc, в котором scFv к CD3 блинатумомаба связан с Fc2 из Fc-варианта, имеющего последовательность СН3-домена дикого типа (соответствующую "PTCWT" в таблице 1), и в качестве группы положительного контроля использовали гетеробиспецифическое антитело Fab-Fc × scFv-Fc, в котором scFv к CD3 блинатумомаба связан с Fc2 из Fc-варианта (соответствующего "РТС019" в таблице 1), имеющего в СН3-домене последовательность, полученную посредством подхода "выступы-во-впадины" (KiH) от Genentech.
Экспрессию каждого тестируемого вещества осуществляли с использованием системы транзиентной экспрессии, сконструированной в примере 1, и после выполнения SDS-PAGE в невосстанавливающих условиях измеряли интенсивности полос PAGE, соответствующих гетеродимерам, для сравнения показателей образования гетеродимеров.
Способность к образованию гетеродимера Fc согласно аминокислотной замене в СН3-домене рассчитывали с учетом измеренной интенсивности полос, и результаты представлены в таблице 8 ниже.
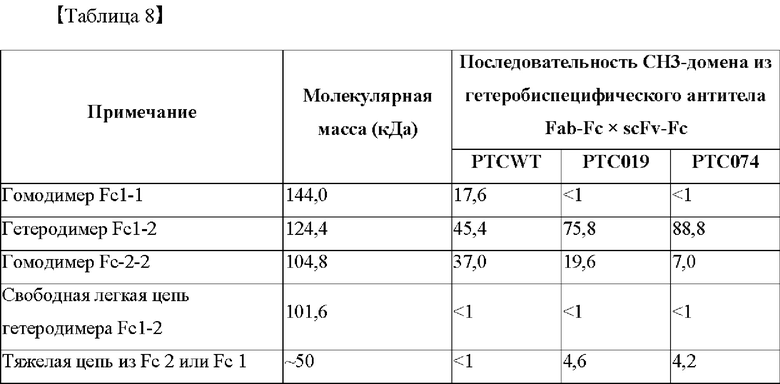
На фиг. 6 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × scFv-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × scFv-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × scFv-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × scFv-Fc с последовательностью РТС074, встроенной в домен СН3).
В результате, как показано на фиг. 6, подтвердили, что гетеробиспецифическое антитело Fab-Fc × scFv-Fc, в которое была встроена последовательность РТС074, не образовывало полосу на уровне 144 кДа в отличие от гетеробиспецифического антитела, в которое была встроена последовательность из группы отрицательного контроля (PTCWT). Кроме того, было подтверждено, что интенсивность полосы была снижена при 104,8 кДа по сравнению с гетеробиспецифическим антителом, в которое была встроена последовательность РТС019. С другой стороны, при 124,4 кДа, что является расчетной молекулярной массой полного гетеробиспецифического антитела Fab-Fc × scFv-Fc, было подтверждено, что интенсивность полосы была увеличена по сравнению с группой отрицательного контроля (PCTWT) и группой положительного контроля (РТС019).
Кроме того, как показано в таблице 8, показатель образования гетеродимера Fc1-2 у гетеробиспецифического антитела Fab-Fc × scFv-Fc, которое содержит последовательность РТС074 СН3-домена, составлял 88,8%, что подтвердило, что показатель образования гетеродимеров был значительно выше, чем у группы отрицательного контроля (PTCWT) и группы положительного контроля (РТС019).
То есть можно видеть, что Fc-вариант согласно одному аспекту может применяться в качестве структуры для гетеробиспецифического антитела, которая образует антитела разных структур и обладает превосходной способностью к образованию гетеродимеров.
7-2. Биспецифическое антитело Fab-Fc × scFv-scFv-Fc
Показатели образования гетеродимеров оценивали таким же образом, как и в примере 7-1, за исключением того, что использовали биспецифические антитела Fab-Fc × scFv-scFv-Fc, сконструированные для экспрессии второго полипептида в форме scFv-scFv-Fc (Fc2), a Fc2 экспрессировали в форме двойного scFv (scFv × scFv), включающего Fc-область IgG1, на основе последовательности scFv к CD3 блинатумомаба и последовательности VH-VL бевацизумаба (Avastin, Roche).
Для сравнения в качестве группы отрицательного контроля использовали гетеробиспецифическое антитело Fab-Fc × scFv-scFv-Fc, в котором двойной scFv связан с Fc2 из Fc-варианта, имеющего последовательность СН3-домена дикого типа (соответствующую "PTCWT" в таблице 1), и в качестве группы положительного контроля использовали гетеробиспецифическое антитело Fab-Fc × scFv-scFv-Fc, в котором двойной scFv связан с Fc2 из Fc-варианта (соответствующего "РТС019" в таблице 1), имеющего в СН3-домене последовательность, полученную посредством подхода "выступы-во-впадины" (KiH) от Genentech.

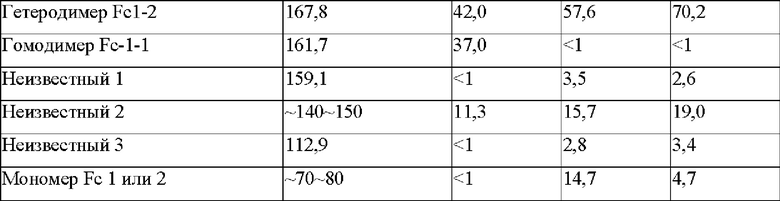
На фиг. 7 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × scFv-scFv-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × scFv-scFv-Fc с последовательностью РТС074, встроенной в СН3-домен).
В результате, как показано на фиг. 7, было подтверждено, что биконъюгат Fab-Fc × scFv × scFv-Fc, в который была встроена последовательность РТС074, имел заметно сниженную интенсивность полосы, соответствующую размеру 196 кДа, по сравнению с конъюгатом, в который были встроены последовательности группы положительного контроля (PTCWT) и группы положительного контроля (РТС019). Кроме того, поскольку интенсивности полос были снижены за исключением полосы гетеродимера, показанного в биконъюгате, в который была встроена последовательность группы положительного контроля (РТС019), было подтверждено, что был получен с высокой эффективностью целевой гетеродимер Fc 1-2 размером 167,8 кДа.
Кроме того, как показано в таблице 9, показатель образования гетеродимера Fc1-2 у гетеробиспецифического антитела Fab-Fc × scFv-scFv-Fc, содержащего последовательность СН3-домена РТС074, составлял 70,2%, и было подтверждено, что показатель образования гетеродимера был значительно выше, чем у группы отрицательного контроля (PTCWT) и группы положительного контроля (РТС019).
То есть можно видеть, что Fc-вариант согласно одному аспекту не только с легкостью можно использовать в качестве структуры гетеробиспецифического антитела, характеризующегося структурой с большой молекулярной массой, поскольку несколько структур соединены, но он также характеризуется превосходным образованием гетеродимеров при использовании в качестве структуры.
7-3. Биспецифическое антитело Fab-Fc × цитокин-Fc
Показатели образования гетеродимеров оценивали таким же образом, как и в примере 7-1, за исключением того, что использовали биспецифические антитела Fab-Fc × цитокин-Fc, сконструированные для экспрессии второго полипептида в форме цитокин-Fc (Fc2), и экспрессию Fc2 осуществляли в форме цитокина, включающего Fc-область IgG1, которая основана на последовательности интерлейкина-2 (IL-2, альдеслейкина).
Для сравнения в качестве группы отрицательного контроля использовали гетеробиспецифическое антитело Fab-Fc × IL-2v-Fc, в котором вариант IL-2 (IL-2v) связан с Fc2 из Fc-варианта, имеющего последовательность СН3-домена дикого типа (соответствующую "PTCWT" в таблице 1), и в качестве группы положительного контроля использовали гетеробиспецифическое антитело Fab-Fc × IL-2v-Fc, в котором вариант IL-2 (IL-2v) связан с Fc2 из Fc-варианта (соответствующего "РТС019" в таблице 1), имеющего последовательность в СН3-домене, полученную посредством подхода "выступы-во-впадины" (KiH) от Genentech.
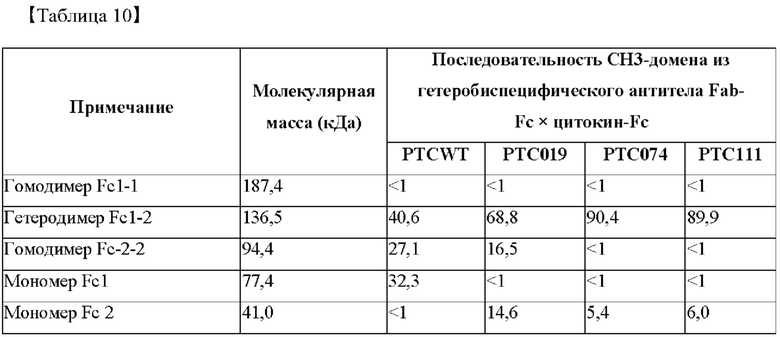
На фиг. 8 представлено изображение, показывающее результаты анализа посредством SDS-PAGE биконъюгатов Fab-Fc × цитокин-Fc (M: маркер размера, WT: биконъюгат Fab-Fc × цитокин-Fc с последовательностью PTCWT, встроенной в СН3-домен (группа отрицательного контроля), 019: биконъюгат Fab-Fc × цитокин-Fc с последовательностью РТС019, встроенной в СН3-домен (группа положительного контроля), 074: биконъюгат Fab-Fc × цитокин-Fc с последовательностью РТС074, встроенной в СН3-домен).
В результате, как показано на фиг. 8, было подтверждено, что биконъюгат Fab-Fc × цитокин-Fc, в который были встроены последовательности РТС074 и РТС111, характеризовался пониженной интенсивностью полосы, соответствующей размеру 94,4 кДа (гомодимер Fc-2-2) и 77,4 кДа (мономер Fc1), и повышенной интенсивностью, соответствующей размеру 136,5 кДа (гетеродимер Fc-1-2), по сравнению с группой отрицательного контроля (PTCWT) или группой положительного контроля (РТС019). Таким образом, в биконъюгате Fab-Fc × цитокин-Fc, в который были встроены последовательности РТС074 и РТС111, образование полосы, соответствующей гомодимеру Fc-2-2 и мономерам Fc-1, было снижено, и, таким образом, можно видеть, что гетеродимеры Fc-1-2 образовывались с высокой эффективностью.
Кроме того, как показано в таблице 10, согласно структуре, в которой последовательности группы отрицательного контроля (PTCWT), группы положительного контроля (РТС019), РТС074 и РТС111 присутствуют в биконъюгате Fab-Fc × цитокин-Fc, соотношение гетеродимеров Fc-1-2 составляло 40,6%, 68,8%, 90,4% и 89,9% соответственно, и было подтверждено, что биконъюгаты, демонстрирующие РТС074 и РТС111, характеризовались значительно более высоким показателем образования гетеродимеров по сравнению с группой отрицательного контроля (PTCWT) и группой положительного контроля (РТС019).
То есть можно видеть, что Fc-вариант согласно одному аспекту может применяться в качестве структуры, которая образует антитело, и при применении белка цитокина способность к образованию гетеродимера является превосходной.
7-4. Биспецифическое антитело Fab-Fc × Fab-Fc
Для оценки показателей образования гетеродимеров у гетеробиспецифических антител, включающих высокоэффективные Fc-варианты, выбранные в примере 3, как первый полипептид, так и второй полипептид конструировали для экспрессии биспецифических антител интактной формы, в которой тяжелая и легкая цепи связаны. В частности, Fc1 экспрессировали на основе антитела авелумаб (Bavencio, Pfizer), включающего Fc-область IgG1 дикого типа, и экспрессию Fc2 осуществляли на основе антитела бевацизумаб (Avastin, Roche), включающего Fc-область IgG1.
Для сравнения в качестве группы отрицательного контроля использовали гетеробиспецифическое антитело Fab-Fc × Fab-Fc, включающего Fc-вариант, имеющий последовательность СН3-домена дикого типа (соответствующую "PTCWT" в таблице 1), и в качестве группы положительного контроля использовали гетеробиспецифическое антитело Fab-Fc × Fab-Fc, включающее Fc-вариант (соответствующий "РТС019" в таблице 1), имеющий последовательность в СН3-домене, полученную посредством подхода "выступы-во-впадины" (KiH) от Genentech.
В качестве способа оценки использовали способ анализа посредством Intact MASS (ESI-LC-MS). В частности, выполняли жидкостную хроматографию-масс-спектрометрию (LC-MS) с использованием Thermo Scientific Dionex™ UHPLC Ultimate 3000 и TripleTOF 5600+(AB sciex), и программное обеспечение Analyst® и PeakView® Software использовали в качестве программ анализа для расчета гетеродимер-образующей способности, и результаты представлены в таблице 11 ниже.

Как показано в таблице 11, было подтверждено, что показатели образования гетеродимера Fc-1-2 у гетеробиспецифических антител Fab-Fc × Fab-Fc, в которые были встроены последовательности группы отрицательного контроля (PTCWT) и группы положительного контроля (РТС019), составляли 25,4% и 32,4% соответственно, с другой стороны, показатели образования гетеродимера Fc-1-2 у гетеробиспецифических антител, имеющих последовательности РТС039, РТС040 и РТС074 СН3-домена, составляли 58,1%, 40,5% и 46,8% соответственно.
То есть, можно видеть, что Fc-вариант согласно одному аспекту не только с легкостью можно использовать в качестве биспецифического антитела, но он также характеризуется превосходным образованием гетеродимеров при использовании в качестве упомянутой выше структуры.
Пример 8. Оценка противораковой активности биспецифических антител Fab-Fc × scFv-Fc, включающих высокоэффективные Fc-варианты
8-1. Подтверждение гибели раковых клеток и подавления роста раковых клеток
Биспецифическое антитело к PD-L1 × scFv к CD3, полученное в примере 7-1, оценивали в отношении его способности к уничтожению раковых клеток и подавлению роста раковых клеток.
В частности, 10% (об./об.) FBS и 1% (об./об.) смеси пенициллин-стрептомицин добавляли в среду RPMI1640 (№11875-093, Gibco), и клетки рака молочной железы (MDA-МВ-231) культивировали при 37°С в атмосфере с 5% СО2. После деления клеток в 96-луночном планшете при концентрации 10000 клеток/лунка клетки культивировали в течение ночи при 37°С в атмосфере с 5% СО2, а мононуклеарные клетки периферической крови человека (РВМС) (эффекторные клетки) совместно культивировали после деления в соотношении 20:1 (эффекторные клетки:клетки-мишени (MDA-MB-231)). Затем в лунках клетки обрабатывали биспецифическими антителами к PD-L1 × scFv к CD3, а затем их культивировали в течение 48 часов. Затем после извлечения надосадочной жидкости цитотоксичность анализировали в соответствии с уравнением 1 ниже с использованием набора для анализа цитотоксичности LDH (№С20301, Invitrogen). Авелумаб (антитело к PD-L1, Bavencio®) использовали в качестве группы положительного контроля.


На фиг. 9 представлен график оценки способности биспецифических антител к PD-L1 × scFv к CD3 индуцировать гибель раковых клеток in vitro (авелумаб: группа положительного контроля, BsAb к PD-L1 × scFv к CD3: биспецифическое антитело к PD-L1 × scFv к CD3, имеющее последовательность РТС074 в СН3-домене).
В результате, как показано на фиг. 9, Emax (максимальный цитотоксический эффект) в группе положительного контроля составил 26% при максимальной концентрации вещества, тогда как Emax для гетеробиспецифического антитела Fab-Fc × scFv-Fc составил 76,3%, что подтвердило, что гетеробиспецифическое антитело характеризуется заметно высоким цитотоксическим эффектом в отношении раковых клеток. Кроме того, группа положительного контроля демонстрировала насыщение без дальнейшего увеличения лизиса клеток при концентрации вещества 2log нг/мл или больше, что подтвердило, что эффективное количество образовывалось в диапазоне от 0log нг/мл до 2log нг/мл. С другой стороны, гетеробиспецифическое антитело Fab-Fc × scFv-Fc демонстрировало насыщение без дальнейшего увеличения лизиса клеток при концентрации вещества 1log нг/мл или больше, что подтвердило, что эффективное количество образовалось в диапазоне от -1log нг/мл до 1log нг/мл.
Кроме того, как показано в таблице 12, в случае группы положительного контроля эффект подавления роста раковых клеток MDA-MB-231 был представлен показателем ЕС50 11,8 нг/мл, тогда как гетеробиспецифическое антитело Fab-Fc × scFc-Fc характеризовалось ЕС50 1,3 нг/мл, что подтвердило, что гетеробиспецифическое антитело более эффективно в подавлении роста раковых клеток.
То есть гетеробиспецифическое антитело Fab-Fc × scFv-Fc, образованное посредством Fc-варианта согласно одному аспекту, демонстрирует превосходный противораковый эффект при более низкой концентрации по сравнению с известными иммунотерапевтическими лекарственными средствами, при этом могут быть уменьшены побочные эффекты существующих терапевтических средств, такие как разрушение здоровых клеток или резистентность.
8-2. Подтверждение противораковой эффективности
Противораковую эффективность in vivo биспецифического антитела к PD-L1 × scFv к CD3 оценивали с использованием гуманизированной мышиной модели опухолевого ксенотрансплантата. В частности, смесь клеток MDA-MB-231 (5×106) и очищенных Т-клеток человека (2,5×106) смешивали с таким же количеством Matrigel и подкожно вводили в бок мышам NOD/SCID (самкам, возрастом 6 недель, Jabio) в дозе 0,2 мл/мышь. Через один час после инокуляции внутривенно вводили 2,5 мг/кг биспецифического антитела к PD-L1 × scFv к CD3, полученного в примере 7-1 (5 раз/неделя). Авелумаб, положительный контроль, вводили внутривенно (3 раза/неделя) в дозе 20 мг/кг. Через 7 дней после инокуляции измеряли объемы опухоли (3 раза/неделя) для сравнения и оценки противораковой эффективности двух веществ.
На фиг. 10 представлен график оценки способности биспецифических антител к PD-L1 × scFv к CD3 подавлять рост раковых клеток in vivo (авелумаб: группа положительного контроля, BsAb к PD-L1 × scFv к CD3: биспецифическое антитело к PD-L1 × scFv к CD3, имеющее последовательность РТС074 в СН3-домене).
В результате, как показано на фиг. 10, в случае группы отрицательного контроля средний объем опухоли постепенно увеличивался в течение 28 дней, пока не достиг 600 мм3 или больше, что подтвердило отсутствие эффекта подавления роста опухоли. Кроме того, в случае группы положительного контроля объем опухоли продолжал увеличиваться, что подтвердило, что эффект подавления роста опухоли составил 59% в день 28. С другой стороны, в случае Fab-Fc × scFv-Fc опухолевая регрессия за счет подавления ее роста сохранялась такой же с дня 9 по 21, при этом эффект подавления роста опухоли составил 80% в день 28, что подтвердило, что биспецифическое антитело эффективно снижает рост опухоли в гуманизированных мышиных моделях опухолевого ксенотрансплантата.
То есть гетеробиспецифическое антитело Fab-Fc × scFv-Fc, образованное посредством Fc-варианта согласно одному аспекту, обладает превосходным эффектом подавления роста опухоли, и, таким образом, может быть использовано для профилактики или лечения различных видов рака, включая рак молочной железы.
Приведенное выше описание настоящего изобретения представлено в иллюстративных целях, и специалистам в области, к которой относится настоящее изобретение, будет понятно, что примеры и варианты осуществления могут быть легко модифицированы без изменения технической идеи или существенных признаков настоящего изобретения. Поэтому следует учитывать, что приведенные выше примеры не являются ограничивающими, а иллюстрируют все аспекты.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> MUSTBIO Co., Ltd.
<120> БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ИЛИ ЕГО АНТИГЕНСВЯЗЫВАЮЩИЙ ФРАГМЕНТ И СПОСОБ ИХ ПОЛУЧЕНИЯ
<130> PX064553PCT
<150> KR10-2020-0066030
<151> 2020-06-04
<160> 9
<170> PatentIn версия 3.2
<210> 1
<211> 450
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь из Fc1 в PTCWT
<400> 1
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 2
<211> 216
<212> БЕЛОК
<213> Искусственная
<220>
<223> Легкая цепь из Fc1 в PTCWT
<400> 2
Gln Ser Ala Leu Thr Gln Pro Ala Ser Val Ser Gly Ser Pro Gly Gln
1 5 10 15
Ser Ile Thr Ile Ser Cys Thr Gly Thr Ser Ser Asp Val Gly Gly Tyr
20 25 30
Asn Tyr Val Ser Trp Tyr Gln Gln His Pro Gly Lys Ala Pro Lys Leu
35 40 45
Met Ile Tyr Asp Val Ser Asn Arg Pro Ser Gly Val Ser Asn Arg Phe
50 55 60
Ser Gly Ser Lys Ser Gly Asn Thr Ala Ser Leu Thr Ile Ser Gly Leu
65 70 75 80
Gln Ala Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ser Tyr Thr Ser Ser
85 90 95
Ser Thr Arg Val Phe Gly Thr Gly Thr Lys Val Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Asn Pro Thr Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Gly Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Lys Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 3
<211> 232
<212> БЕЛОК
<213> Искусственная
<220>
<223> Fc-цепь из Fc2 в PTCWT
<400> 3
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 4
<211> 450
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь из Fc1 в PTC019
<400> 4
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 5
<211> 232
<212> БЕЛОК
<213> Искусственная
<220>
<223> Fc-цепь из Fc2 в PTC019
<400> 5
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 6
<211> 450
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь из Fc1 в PTC039
<400> 6
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Phe Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 7
<211> 232
<212> БЕЛОК
<213> Искусственная
<220>
<223> Fc-цепь из Fc2 в PTC039
<400> 7
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Gly Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<210> 8
<211> 450
<212> БЕЛОК
<213> Искусственная
<220>
<223> Тяжелая цепь из Fc1 в PTC040
<400> 8
Glu Val Gln Leu Leu Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Ile Met Met Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Ser Ile Tyr Pro Ser Gly Gly Ile Thr Phe Tyr Ala Asp Thr Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Ile Lys Leu Gly Thr Val Thr Thr Val Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Trp Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser Leu
355 360 365
Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly Lys
450
<210> 9
<211> 232
<212> БЕЛОК
<213> Искусственная
<220>
<223> Fc-цепь из Fc2 в PTC111
<400> 9
Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala
1 5 10 15
Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro
20 25 30
Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val
35 40 45
Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val
50 55 60
Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln
65 70 75 80
Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln
85 90 95
Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala
100 105 110
Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro
115 120 125
Arg Glu Pro Gln Val Tyr Thr Phe Pro Pro Ser Arg Asp Glu Leu Thr
130 135 140
Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser
145 150 155 160
Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr
165 170 175
Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val
180 185 190
Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe
195 200 205
Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys
210 215 220
Ser Leu Ser Leu Ser Pro Gly Lys
225 230
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО ПРОТИВ CD3E/BCMA И ЕГО ПРИМЕНЕНИЕ | 2019 |
|
RU2800164C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CD19 ЧЕЛОВЕКА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2016 |
|
RU2731156C1 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2839153C2 |
| БИСПЕЦИФИЧЕСКОЕ АНТИТЕЛО | 2021 |
|
RU2840407C1 |
| АНТАГОНИСТЫ | 2019 |
|
RU2839429C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2810924C2 |
| Антитела к TSLP человека и их применение | 2021 |
|
RU2825460C1 |
| ИНДУЦИРУЮЩИЙ ЦИТОТОКСИЧНОСТЬ ТЕРАПЕВТИЧЕСКИЙ АГЕНТ | 2022 |
|
RU2824285C2 |
| АНТИТЕЛА, СВЯЗЫВАЮЩИЕСЯ С CD3 | 2019 |
|
RU2832669C2 |
| Двухвалентное биспецифическое химерное антитело, включающее гетеродимер на основе MHC или MHC-подобных белков | 2021 |
|
RU2820683C2 |
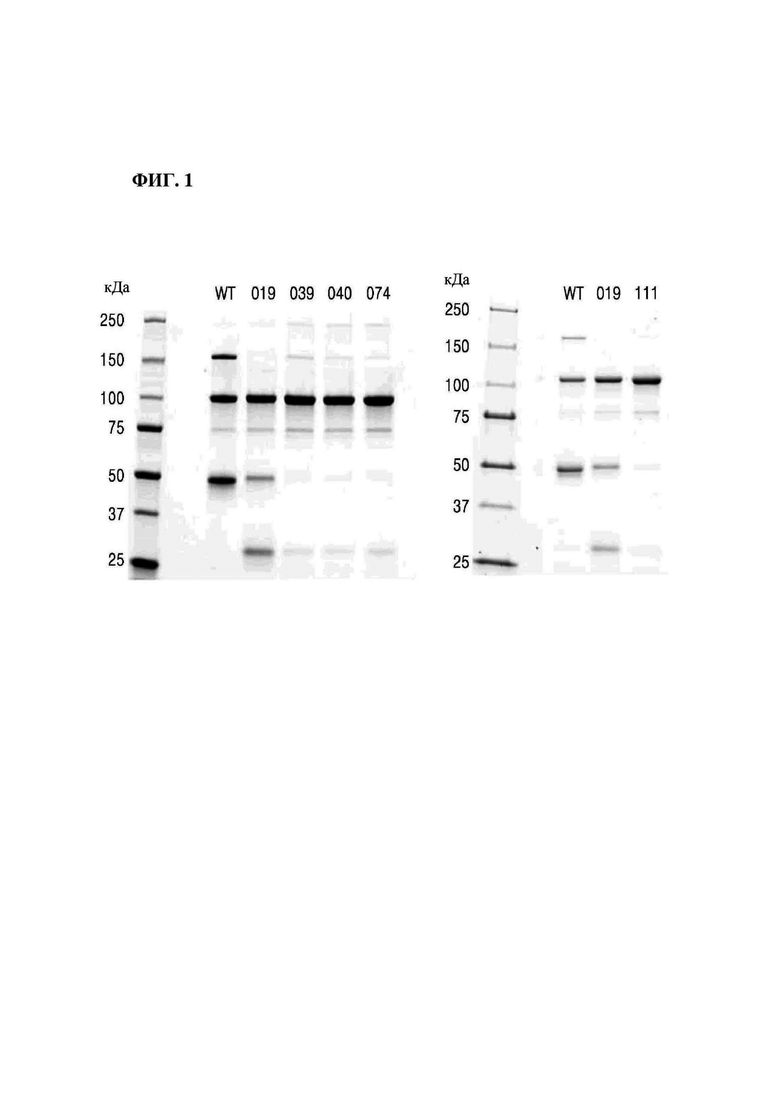
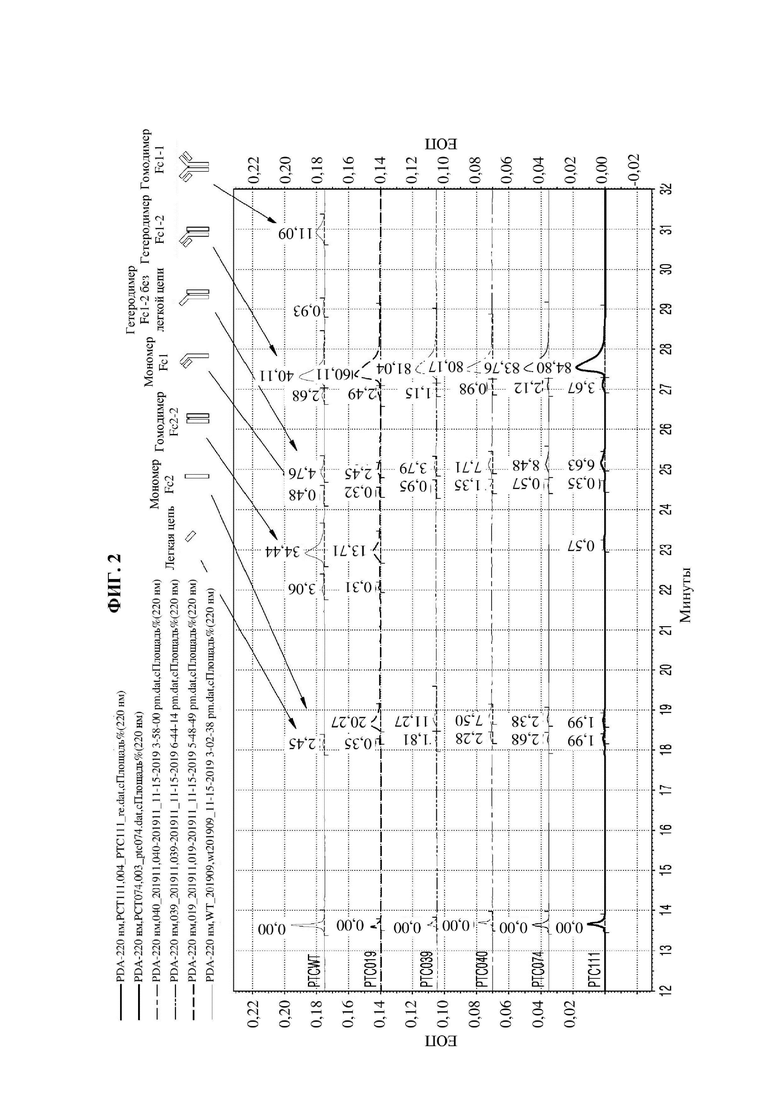
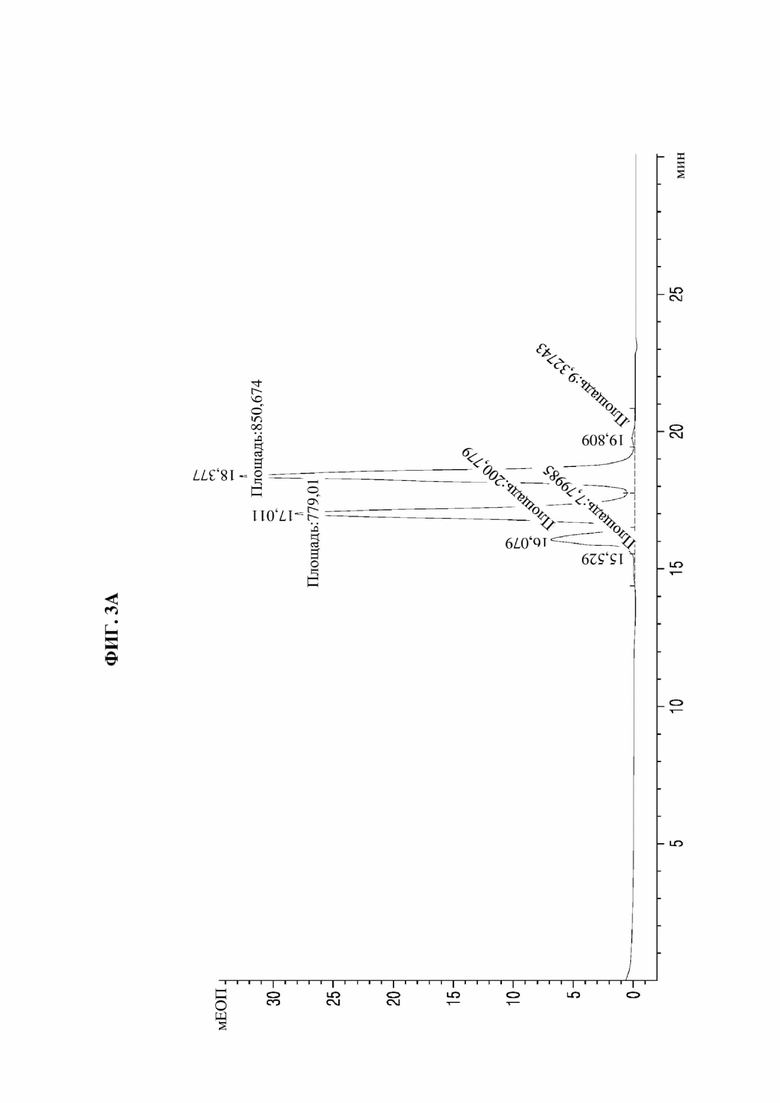
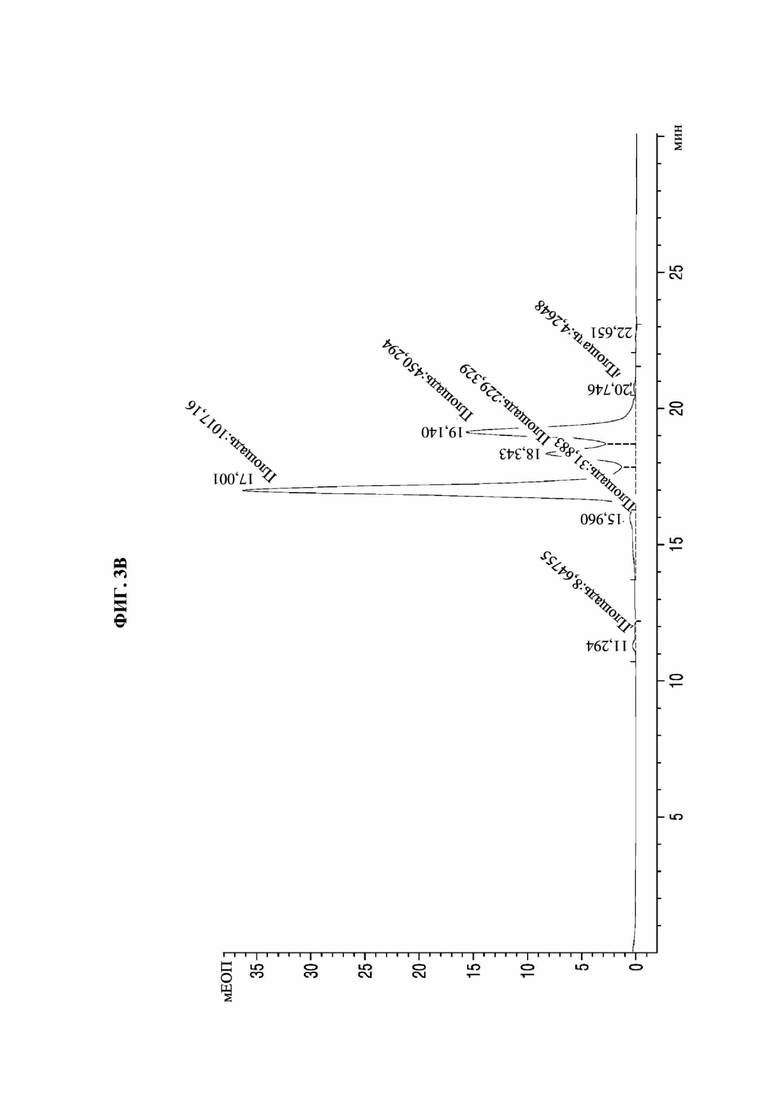
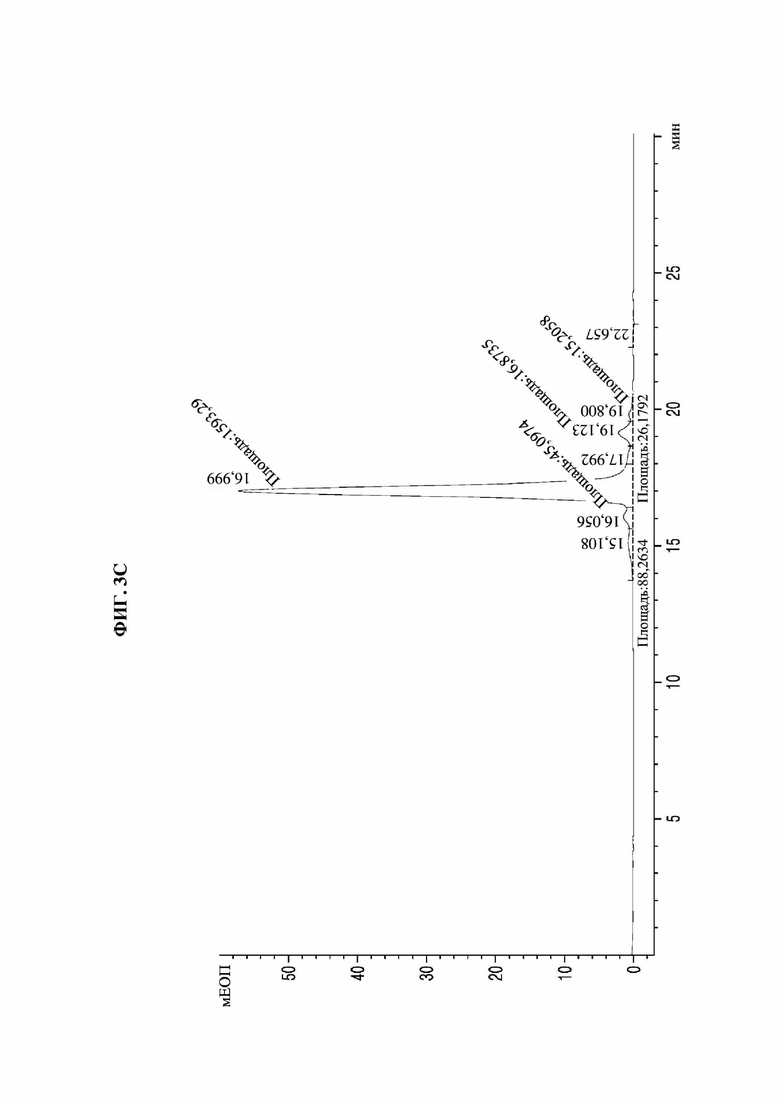
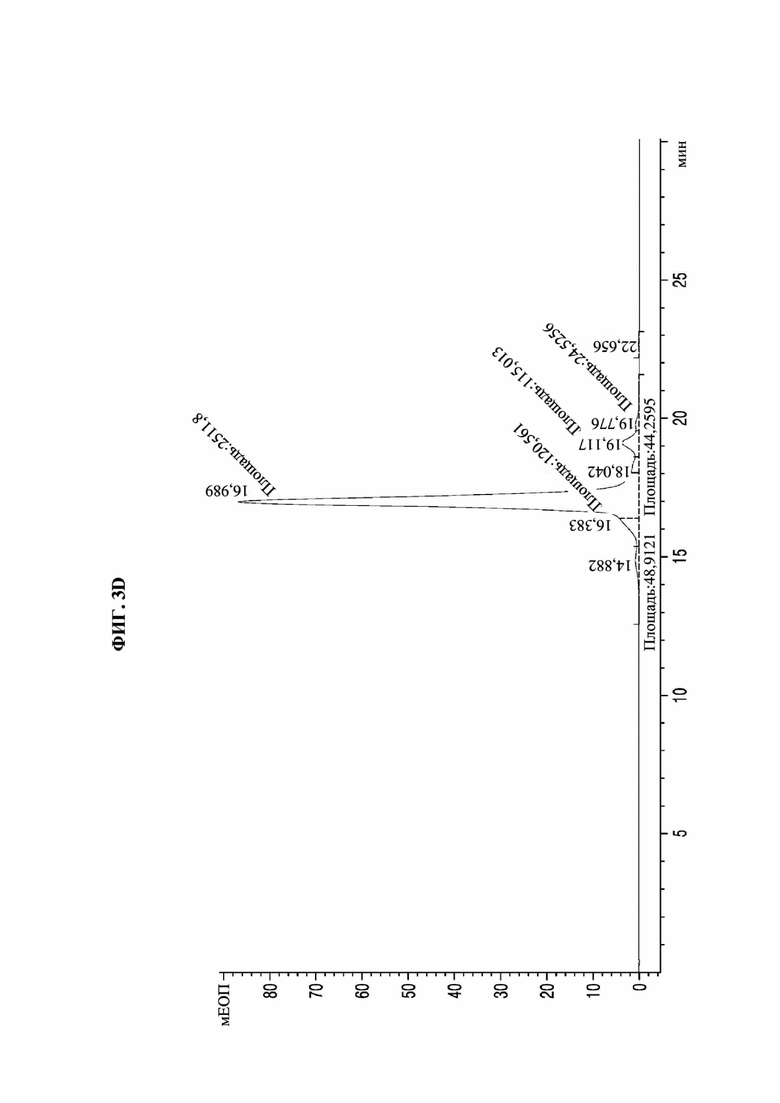
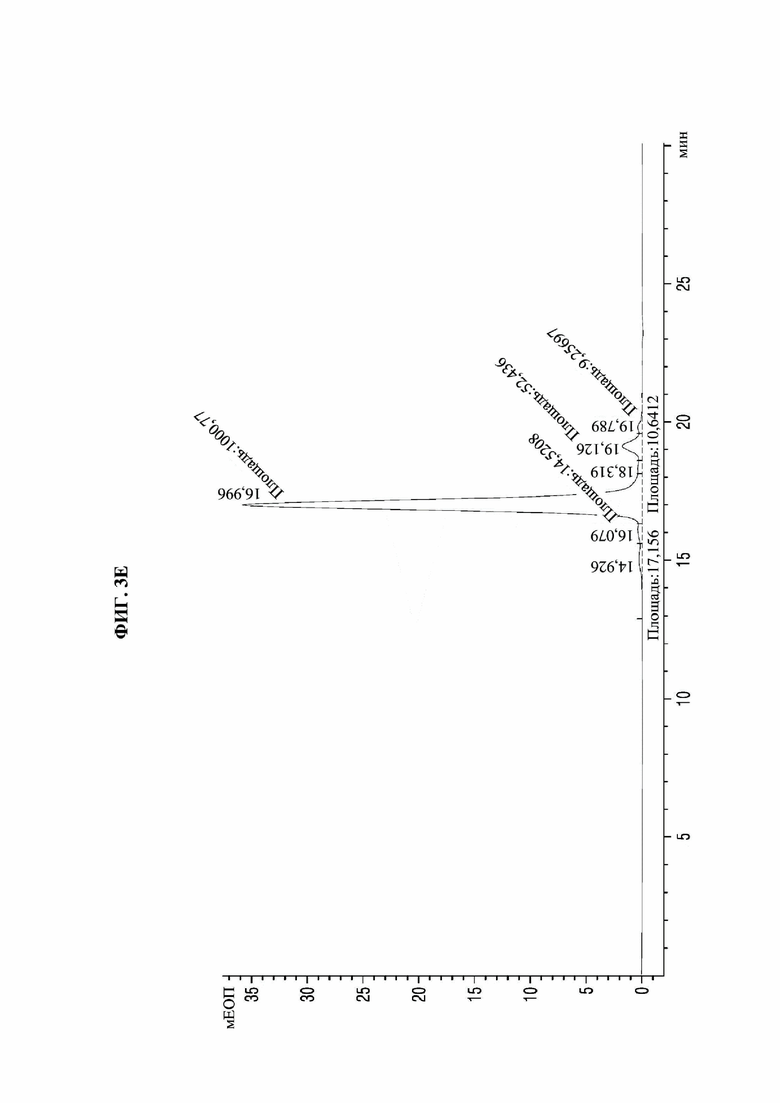
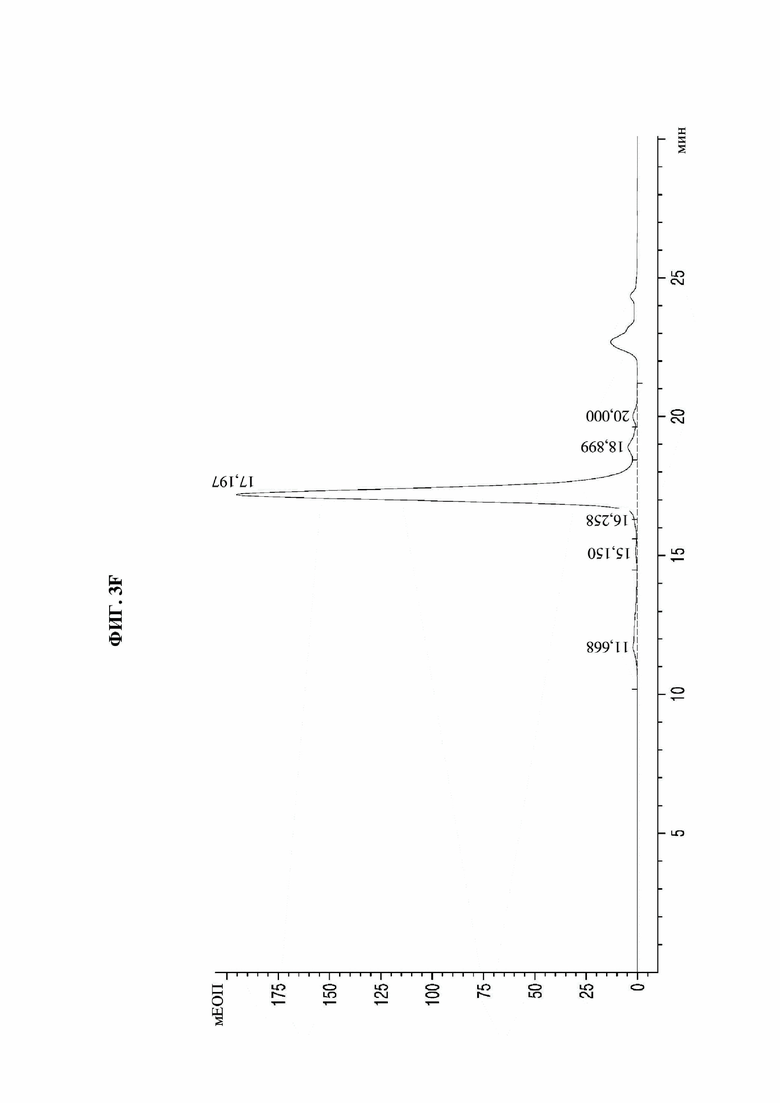
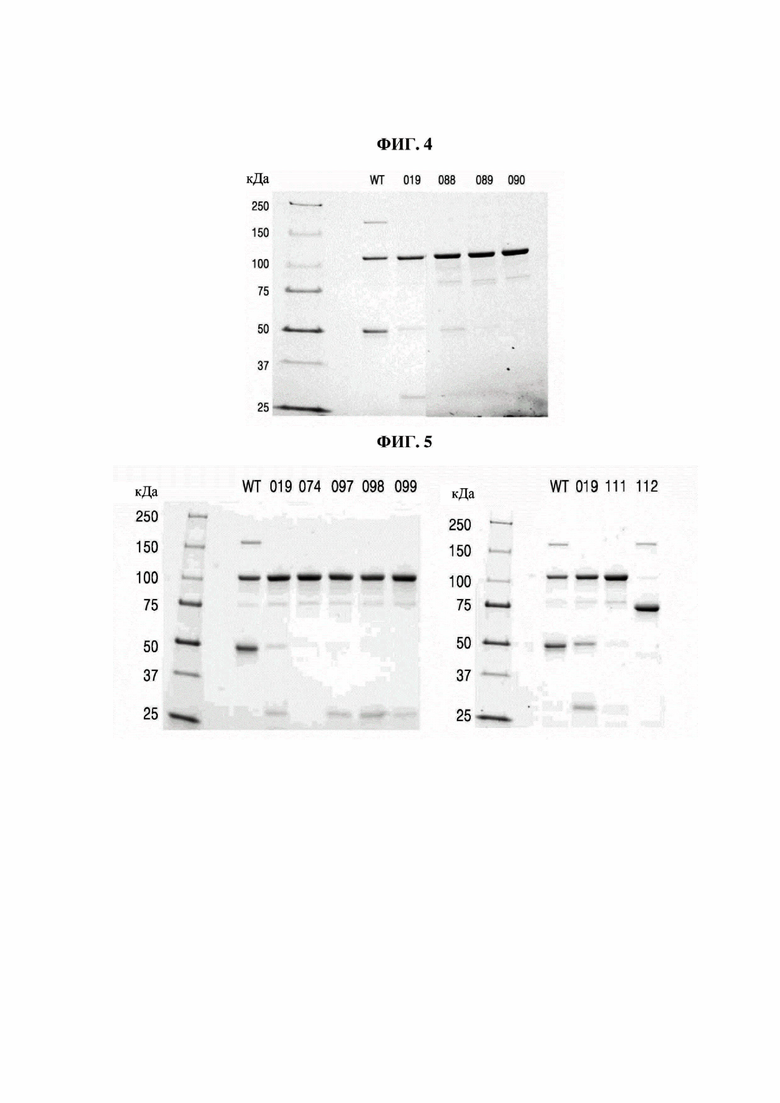

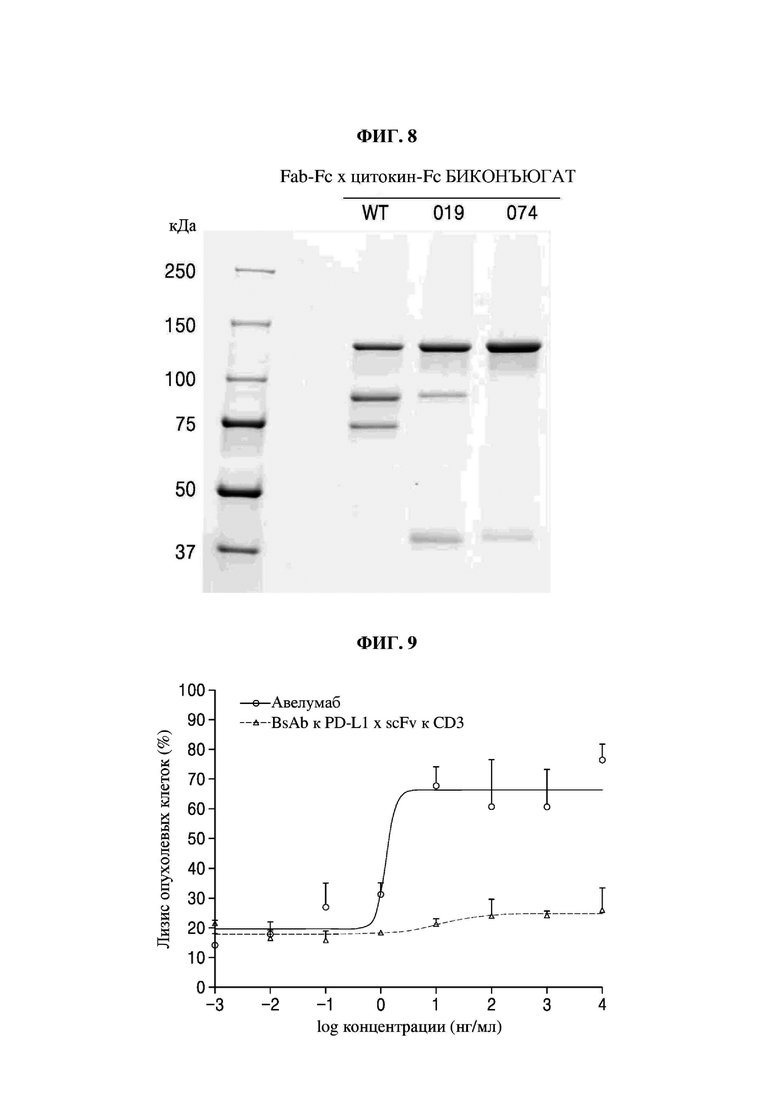
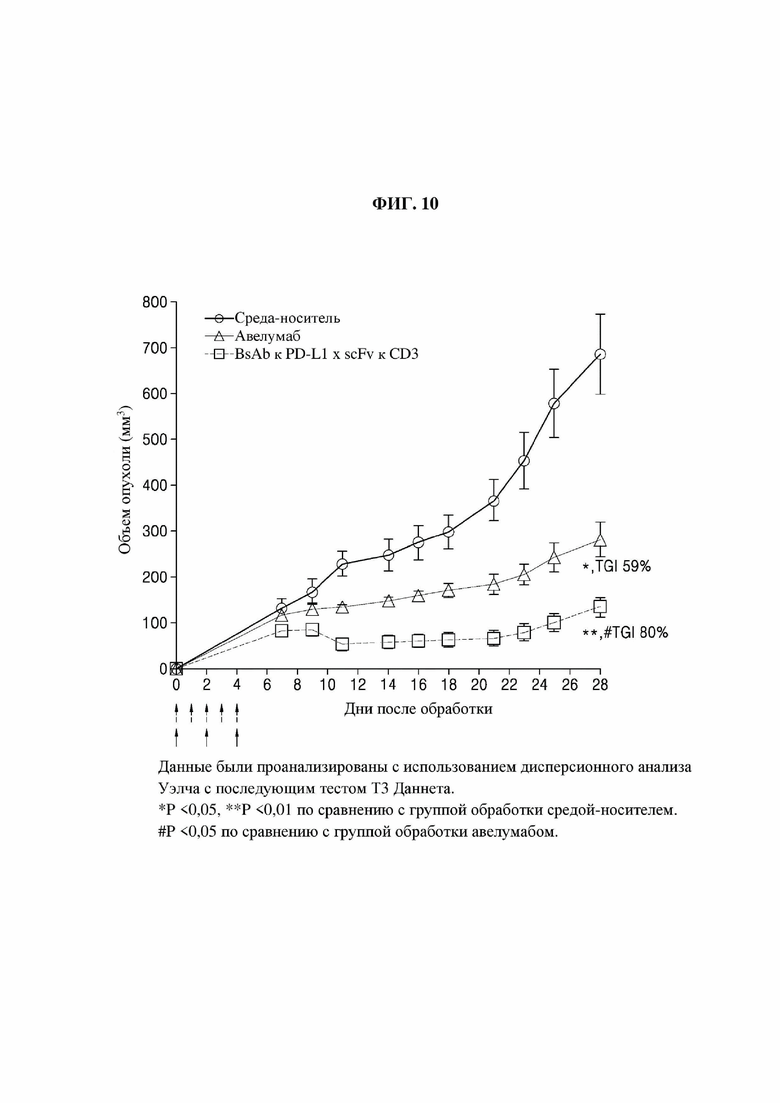
Изобретение относится к биотехнологии и медицине, в частности к белковому комплексу, характеризующемуся высоким показателем образования гетеродимера, и к способу его получения. Изобретение позволяет получить белковые комплексы с улучшенной эффективностью образования гетеродимеров для использования в различных областях, таких как профилактика или лечение заболеваний и диагностика заболеваний. 3 н. и 22 з.п. ф-лы, 10 ил., 12 табл., 8 пр.
1. Белковый комплекс, обладающий повышенной способностью к образованию гетеродимера Fc по сравнению с его диким типом или с типом "выступ-во-впадину" (KiH), содержащий первый полипептид, включающий первый CH3-домен константной области человеческого IgG1 антитела; и второй полипептид, включающий второй CH3-домен константной области человеческого IgG1 антитела, где первый полипептид и второй полипептид образуют гетеродимер;
при этом первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366, и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368 и валин (V) в положении 407; и
по меньшей мере один из первого CH3-домена константной области человеческого IgG1 антитела и второго CH3-домена константной области человеческого IgG1 антитела содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (H), глицина (G), валина (V), метионина (M) и аланина (A), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394, где положение аминокислоты соответствует индексу EU согласно 20 Кабату.
2. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (H), глицина (G), валина (V), метионина (M) и аланина (A), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394.
3. Белковый комплекс по п. 1, где второй CH3-домен константной области человеческого IgG1 антитела содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (H), глицина (G), валина (V), метионина (M) и аланина (A), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394.
4. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и фенилаланин (F), гистидин (H) или триптофан (W) в положении 394.
5. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и валин (V), триптофан (W), аланин (A) или фенилаланин (F) в положении 351.
6. Белковый комплекс по п. 1, где второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и аланин (A), глицин (G), валин (V), метионин (M) или фенилаланин (F) в положении 351.
7. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит одно, выбранное из группы, состоящей из:
триптофана (W) в положении 366;
триптофана (W) в положении 366 и гистидина (H) в положении 394;
триптофана (W) в положении 366 и фенилаланина (F) в положении 394;
триптофана (W) в положении 366 и триптофана (W) в положении 394;
триптофана (W) в положении 366 и фенилаланина (F) в положении 351;
триптофана (W) в положении 366 и триптофана (W) в положении 351;
триптофана (W) в положении 366 и валина (V) в положении 351;
триптофана (W) в положении 366 и аланина (A) в положении 351; и
триптофана (W) в положении 366, гистидина в положении 394 (H) и фенилаланина в положении 351 (F).
8. Белковый комплекс по п. 1, где второй CH3-домен константной области человеческого IgG1 антитела содержит одно, выбранное из группы, состоящей из:
серина (S) в положении 366, аланина (A) в положении 368 и валина (V) в положении 407;
серина (S) в положении 366, аланина (A) в положении 368, валина (V) в положении 407 и аланина (A) в положении 351;
серина (S) в положении 366, аланина (A) в положении 368, валина (V) в положении 407 и глицина (G) в положении 351;
серина (S) в положении 366, аланина (A) в положении 368, валина (V) в положении 407 и валина (V) в положении 351;
серина (S) в положении 366, аланина (A) в положении 368, валина (V) в положении 407 и метионина (M) в положении 351; и
серина (S) в положении 366, аланина (A) в положении 368, валина (V) в положении 407 и фенилаланина (F) в положении 351.
9. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и фенилаланин (F), гистидин (H) или триптофан (W) в положении 394; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368 и валин (V) в положении 407.
10. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и фенилаланин (F) в положении 351; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и аланин (A) в положении 351.
11. Белковый комплекс по п. 10, где первый CH3-домен константной области человеческого IgG1 антитела дополнительно содержит гистидин (H) в положении 394.
12. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и глицин (G) в положении 351.
13. Белковый комплекс по п. 12, где первый CH3-домен константной области человеческого IgG1 антитела дополнительно содержит фенилаланин (F) или триптофан (W) в положении 351.
14. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и аланин (A), валин (V) или метионин (M) в положении 351.
15. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и фенилаланин (F) в положении 351.
16. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и валин (V) в положении 351; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и аланин (A) в положении 351.
17. Белковый комплекс по п. 1, где первый CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366 и аланин (A) в положении 351; и второй CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и фенилаланин (F) в положении 351.
18. Белковый комплекс по п. 1, где первый полипептид и/или второй полипептид дополнительно содержат любое, выбранное из группы, состоящей из антигенсвязывающего фрагмента (Fab), одноцепочечного вариабельного фрагмента (scFv), внеклеточного домена мембранного рецептора, агониста, антагониста, лиганда, рецептора-ловушки, цитокина, фактора коагуляции и аффинной метки.
19. Белковый комплекс, обладающий повышенной способностью к образованию гетеродимера Fc по сравнению с его диким типом или с типом "выступ-во-впадину” (KiH), содержащий первый полипептид, включающий первый CH3-домен константной области человеческого IgG1 антитела; и второй полипептид, включающий второй CH3-домен константной области человеческого IgG1 антитела, где первый полипептид и второй полипептид образуют гетеродимер,
при этом первый CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368 и валин (V) в положении 407, второй CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366, и
по меньшей мере один из первого CH3-домен константной области человеческого IgG1 антитела и второго CH3-домен константной области человеческого IgG1 антитела содержит по меньшей мере одну аминокислоту, выбранную из группы, состоящей из фенилаланина (F), триптофана (W), гистидина (H), глицина (G), валина (V), метионина (M) и аланина (A), в одном или нескольких положениях, выбранных из группы, состоящей из положений 351 и 394, где положение аминокислоты соответствует индексу EU согласно Кабату.
20. Белковый комплекс по п. 19, где первый CH3-домен константной области человеческого IgG1 антитела содержит серин (S) в положении 366, аланин (A) в положении 368, валин (V) в положении 407 и глицин (G) в положении 351; и второй CH3-домен константной области человеческого IgG1 антитела содержит триптофан (W) в положении 366.
21. Белковый комплекс по п. 20, где второй CH3-домен константной области человеческого IgG1 антитела дополнительно содержит фенилаланин (F) или триптофан (W) в положении 351.
22. Способ получения белкового комплекса, предусматривающий трансформацию одной или нескольких клеток посредством одного или нескольких векторов экспрессии, кодирующих первый полипептид по п. 1 или 19, второй полипептид по п. 1 или 19 или их комбинацию; и обеспечение экспрессии первого полипептида, второго полипептида или их комбинации.
23. Способ по п. 22, где вектором экспрессии, кодирующим первый полипептид, и вектором экспрессии, кодирующим второй полипептид, совместно трансфицируют клетки, или вектором экспрессии, кодирующим первый полипептид, и вектором экспрессии, кодирующим второй полипептид, соответственно трансформируют по меньшей мере два типа клеток.
24. Способ по п. 22, предусматривающий получение белкового комплекса на основе первого полипептида, второго полипептида или первого полипептида и второго полипептида из клеток или культуральной клеточной среды.
25. Способ по п. 22, где первый полипептид и/или второй полипептид дополнительно содержат одно, выбранное из группы, состоящей из его антигенсвязывающего фрагмента (Fab), одноцепочечного вариабельного фрагмента (scFv), внеклеточного домена мембранного рецептора, агониста, антагониста, лиганда, рецептора-ловушки, цитокина, фактора коагуляции и аффинной метки.
| WO 2015017822 A1, 05.02.2015 | |||
| CHOI H.J | |||
| et al | |||
| Engineering of Immunoglobulin Fc Heterodimers Using Yeast Surface-Displayed Combinatorial Fc Library Screening, PLoS One, 2015, vol | |||
| Печь-кухня, могущая работать, как самостоятельно, так и в комбинации с разного рода нагревательными приборами | 1921 |
|
SU10A1 |
| Способ гальванического снятия позолоты с серебряных изделий без заметного изменения их формы | 1923 |
|
SU12A1 |
| XU Y | |||
| et al | |||
| Production of bispecific antibodies in "knobs-into-holes" using a cell-free expression system, MAbs, 2015, vol | |||
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Машина для удаления камней из почвы | 1922 |
|
SU231A1 |

