Область техники
Изобретение относится к области молекулярной биотехнологии и экспериментальной медицины, в частности к применению аденоассоциированного вирусного вектора, кодирующего hAIPL1.
Уровень техники
Моногенные заболевания сетчатки представляют собой группу из порядка 300 разнородных по клиническим проявлениям изолированных или синдромальных заболеваний, которые объединены схожими механизмами развития повреждением одного из звеньев фототранс дукции или целостности клеток фоторецепторов/вспомогательных клеток. Частота встречаемости моногенных заболеваний сетчатки колеблется от 1:5000 (болезнь Штаргардта) до 1:100000 (синдром Ашера, ахроматопсия). На сегодняшний день в широкой клинической практике отсутствует патогенетически направленное эффективное лечение данной группы заболеваний, однако на разных стадиях клинических и доклинических исследований находятся перспективные методы лечения.
Амавроз Лебера (Leber Congenital Amaurosis - LCA), самая быстрая и тяжелая форма наследственной дистрофии сетчатки, обычно наследуется по аутосомно-рецессивному типу и характеризуется ранней потерей зрения, нистагмом и отсутствующей или существенно сниженной электроретинограммой (ЭРГ) [den Hollander AI, Roepman R, Koenekoop RK, Cremers FP. Leber congenital amaurosis: genes, proteins and disease mechanisms. Prog Retin Eye Res. 2008 Jul;27(4):391-419]. Встречается с частотой приблизительно 1:200000 человек. На сегодняшний день мутации в 26 различных генах, кодирующих белки, играющие важную роль в развитии и физиологической функции сетчатки, вызывают клинически различные типы LCA [RetNet https://sph.uth.edu/retnet, дата обращения 12.12.2023]. Среди них мутации в гене белка-подобного 1, взаимодействующего с арилуглеводородными рецепторами (aryl hydrocarbon receptor interacting protein like 1 - AIPL1), связаны с LCA типа IV (LCA4).
Несмотря на то, что на него приходится всего 5 10% случаев LCA, клинический фенотип, вызванный дефицитом AIPL1, находится в спектре LCA с тяжелым течением. Такие серьезные симптомы вызваны обширной и необратимой дегенерацией фоторецепторов палочек и колбочек, критических для зрительной фототрансдукции, в которой AIPL1 играет косвенную, но важную роль для поддержания функциональной целостности [Yadav RP, Artemyev NO. AIPL1: A specialized chaperone for the phototransduction effector. Cell Signal. 2017 Dec;40:183-189].
Экспрессия AIPL1 происходит исключительно в фоторецепторах сетчатки и эпифизе [van der Spuy J, Chappie JP, Clark BJ, Luthert PJ, Sethi CS, Cheetham ME. The Leber congenital amaurosis gene product AIPL1 is localized exclusively in rod photoreceptors of the adult human retina. Hum Mol Genet. 2002 Apr 1;11(7):823-31]. В сетчатке AIPL1 действует как специализированный ко-шаперон циклической нуклеотидной фосфодиэстеразы шестого семейства (PDE6), важного эффектора фермента в пути фототрансдукции [Sacristan-Reviriego A, van der Spuy J. The Leber Congenital Amaurosis-Linked Protein AIPL1 and Its Critical Role in Photoreceptors. Adv Exp Med Biol. 2018;1074:381-386]. После стимуляции светом активированный PDE6 гидролизует циклический ГМФ (цГМФ), запуская закрытие цГМФ-зависимых ионных каналов Са2+ и распространение «светового» электрического сигнала за счет гиперполяризации плазматической мембраны [Yadav RP, Artemyev NO. AIPL1: A specialized chaperone for the phototransduction effector. Cell Signal. 2017 Dec;40:183-189]. Стабилизация PDE6 обеспечивается шаперонным гетерокомплексом, включающим HSP90 и родственный РОЕб-специфический ко-шаперон AIPL1 [Hidalgo-de-Quintana J, Evans RJ, Cheetham ME, van der Spuy J. The Leber congenital amaurosis protein AIPL1 functions as part of a chaperone heterocomplex. Invest Ophthalmol Vis Sci. 2008 Jul;49(7):2878-87], которые необходимы для стабильной сборки холофермента PDE6 [Kolandaivelu S, Huang J, Hurley JB, Ramamurthy V. AIPL1, a protein associated with childhood blindness, interacts with alpha-subunit of rod phosphodiesterase (PDE6) and is essential for its proper assembly. J Biol Chem. 2009 Nov 6;284(45):30853-61].
Мутации AIPL1 влияют на функциональные домены транслируемого белка, нарушая взаимодействие AIPL1 с изопренилированным PDE6 или HSP90, что предотвращает сборку гетерокомплекса шаперона PDE6 [Sacristan-Reviriego A, van der Spuy J. The Leber Congenital Amaurosis-Linked Protein AIPL1 and Its Critical Role in Photoreceptors. Adv Exp Med Biol. 2018;1074:381-386]. У мышей с нокаутом AIPL1 и у гипоморфных мышей потеря функции AIPL1 вызывает неправильную сборку холофермента PDE6, вызывая дестабилизацию и быструю протеасомную деградацию субъединиц PDE6. Следовательно, происходит быстрая дегенерация палочковых фоторецепторов из-за увеличения внутриклеточного цГМФ, что приводит к длительному открытию циклических нуклеотид-управляемых каналов и избыточному притоку Са2+ [Wang Т, Tsang SH, Chen J. Two pathways of rod photoreceptor cell death induced by elevated cGMP. Hum Mol Genet. 2017 Jun 15;26(12):2299-2306].
Учитывая уникальные особенности глаза, наследственные дистрофии сетчатки являются одной из самых привлекательных мишеней для генной терапии. За последние годы количество клинических испытаний генной терапии наследственных дистрофий сетчатки увеличилось, и кульминацией стала регистрация препарата Luxturna - рекомбинантного аденоассоциированного вируса (rAAV), экспрессирующего ген RPE65 для лечения амавроза Лебера II типа (LCA II) [Auricchio F, Scavone С, Cimmaruta D, Di Mauro G, Capuano A, Sportiello L, Rafaniello C. Drugs approved for the treatment of multiple sclerosis: review of their safety profile. Expert Opin Drug Saf. 2017 Dec; 16(12): 1359-1371]. AAV являются перспективными и наиболее успешными векторами для доставки терапевтических генов и стандартным выбором для трансдукции фоторецепторов. Было показано, что несколько природных или рекомбинантных серотипов AAV эффективно трансдуцируют фоторецепторы в моделях на животных [Day TP, Byrne LC, Schaffer DV, Flannery JG. Advances in AAV vector development for gene therapy in the retina. Adv Exp Med Biol. 2014;801:687-93].
В заявке на патент US 20030022165 A1 раскрывается нормальный и мутантный ген AIPL1 и его роль в развитии LCA4, а также способ лечения заболевания сетчатки, включающий, в частности, введение животному эффективного количества полинуклеотидной последовательности tquhAIPLI дикого типа.
Иммунный ответ является одним из наиболее важных препятствий, ограничивающих эффективную и долгосрочную экспрессию трансгена [Muhuri М, Maeda Y, Ma Н, et al. Overcoming innate immune barriers that impede AAV gene therapy vectors. J Clin Invest. 2021;131(1):e143780. doi:10.1172/JCI143780]. Кроме того, до настоящего времени наблюдаются проблемы низкой эффективности генной терапии, которые связаны с гиперактивацией иммунного ответа, в том числе активацией, связанной с двухцепочечными РНК [Shao W, Earley LF, Chai Z, et al. Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction. JCI Insight. 2018;3(12):e120474. Published 2018 Jun 21. doi: 10.1172/jci.insight. 120474].
Ранее для решения проблемы повышенной иммуногенности препаратов аденоассоциированных вирусных векторов использовали подход, включающий изменение последовательности капсидных белков для снижения иммуногенности вирусных капсидов. Так, например, патент US 10081659 раскрывает модифицированный капсидный белок аденоассоциированного вируса (AAV), содержащий по меньшей мере одну ненативную аминокислоту в аминокислотном положении, участвующем в связывании капсидного белка с клеточным рецептором. За счет этого модифицированные частицы AAV приобретали новые свойства, такие как повышенная эффективность трансдукции и пониженная иммуногенность.
В заявке на изобретение US 20090317417 A1 раскрыты композиции, в которых функциональность гепарин-связывающих доменов на поверхности капсидных белков AAV изменена с целью снижения иммуногенности и, в частности, Т-клеточного иммунного ответа против AAV. Это может достигаться удалением или маскированием мотива RXXR на поверхности капсидных белков AAV.
В случае аденоассоциированных вирусных векторов исследования показали, что геном вектора активирует Toll-подобный рецептор 9 (TLR9), так называемый рецептор распознавания паттерна, который воспринимает чужеродную ДНК.
Чан с соавторами разработали менее иммуногенные векторы AAV путем включения непосредственно в геном вектора коротких ДНК-олигонуклеотидов, которые препятствуют активации TLR9. Сконструированные векторы вызывали заметное снижение врожденного иммунного ответа и Т-клеточных ответов. Было показано, что связывание специфических иммуномодулирующих некодирующих
последовательностей с гораздо более длинными терапевтическими нуклеиновыми кислотами может «скрыть» вектор от индукции нежелательного иммунного ответа во многих, но не во всех моделях [Chan YK, Wang SK, Chu CJ, et al. Engineering adeno-associated viral vectors to evade innate immune and inflammatory responses. Sci Transl Med. 2021;13(580):eabd3438. doi:10.1126/scitranslmed.abd3438].
Все разработанные стратегии снижения иммуногенности AAV не учитывали возможность активации врожденного иммунного ответа за счет образования двухцепочечных РНК в результате трансдукции AAV, которое наблюдается в ограниченном наборе клеточных линий, что подчеркивает роль специфических внутриклеточных условий в регуляции активации врожденного иммунного ответа.
В клинических исследованиях у пациентов с наследственными формами слепоты в некоторых исследованиях сообщалось, что терапевтический эффект постепенно улучшался, достигал пика, а затем впоследствии снижался у пациентов с врожденным амаврозом Лебера после субретинального введения AAV векторов, кодирующих терапевтические трансгены. Остается невыясненным, происходит ли снижение экспрессии трансгена в результате активации врожденного иммунитета в более поздние моменты времени после применения генной терапии или другие факторы влияют на экспрессию трансгена у пациентов со слепотой. Поскольку число пациентов, участвующих в исследованиях генной терапии амавроза Лебера с использованием AAV, ограниченно, потребуется продолжительное наблюдение за пациентами, чтобы оценить важность врожденного иммунного ответа в различных тканях/органах после применения. Тем не менее было продемонстрировано, что врожденный иммунный ответ, опосредованный двухцепочечными РНК, со временем снижает эффективность генной терапии [Shao W, Earley LF, Chai Z, et al. Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction. JCI Insight. 2018;3(12):e120474. Published 2018 Jun 21. doi: 10.1172/jci.insight. 120474].
Технической задачей, на решение которой направлено настоящее изобретение, является решение как минимум одной из вышеуказанных в уровне техники проблем.
Сущность изобретения
Техническим решением является использование описанного признаками в пунктах формулы изобретения.
Одной из возможных технических задач, на решение которой может быть направлено настоящее изобретение, являлось создание экспрессионного вектора для доставки рекомбинантного гена hAIPL1 в клетки сетчатки, позволяющего снизить внутриклеточный иммунный ответ на введение чужеродного вирусного вектора. Изобретение может быть использовано при генной терапии офтальмологических заболеваний.
Техническим результатом настоящего изобретения является снижение экспрессии провоспалительных генов (IL6, IL1RAPL) и повышение экспрессии противовоспалительных генов (IL1RN), при устойчивой экспрессии гена hAIPL1, введенного в клетки сетчатки человека с помощью вирусного экспрессионного вектора на основе AAV.
Задача может решаться применением экспрессионного вектора на основе аденоассоциированного вируса, содержащего последовательность нуклеотидов Seq Id No.:2 по новому назначению, а именно для снижения внутриклеточного иммунного ответа при введении в клетки сетчатки человека.
Техническое решение основано на неожиданном эффекте, полученном при применении рекомбинантного AAV, экспрессирующего кодонно оптимизированную последовательность гена hAIPL1 Seq Id No.:2. Анализ экспрессии генов на уровне транскриптома показал, что применение экспрессионного вектора на основе аденоассоциированного вируса, содержащего кодонно оптимизированную последовательность гена hAIPL1 Seq Id No.:2, для снижения внутриклеточного иммунного ответа при введении в клетки сетчатки человека, приводит к значительному изменению паттерна экспрессии генов, связанных с внутриклеточным иммунитетом, а именно к снижению экспрессии провоспалительных генов IL6, IL1RAPL и повышению экспрессии противовоспалительного гена IL1RN, таким образом обеспечивая достижение технического результата. Также показано снижение экспрессии провоспалительных генов семейства OAS, активируемых двухцепочечными РНК.
Фигуры
Изобретение иллюстрируются следующими графическими материалами:
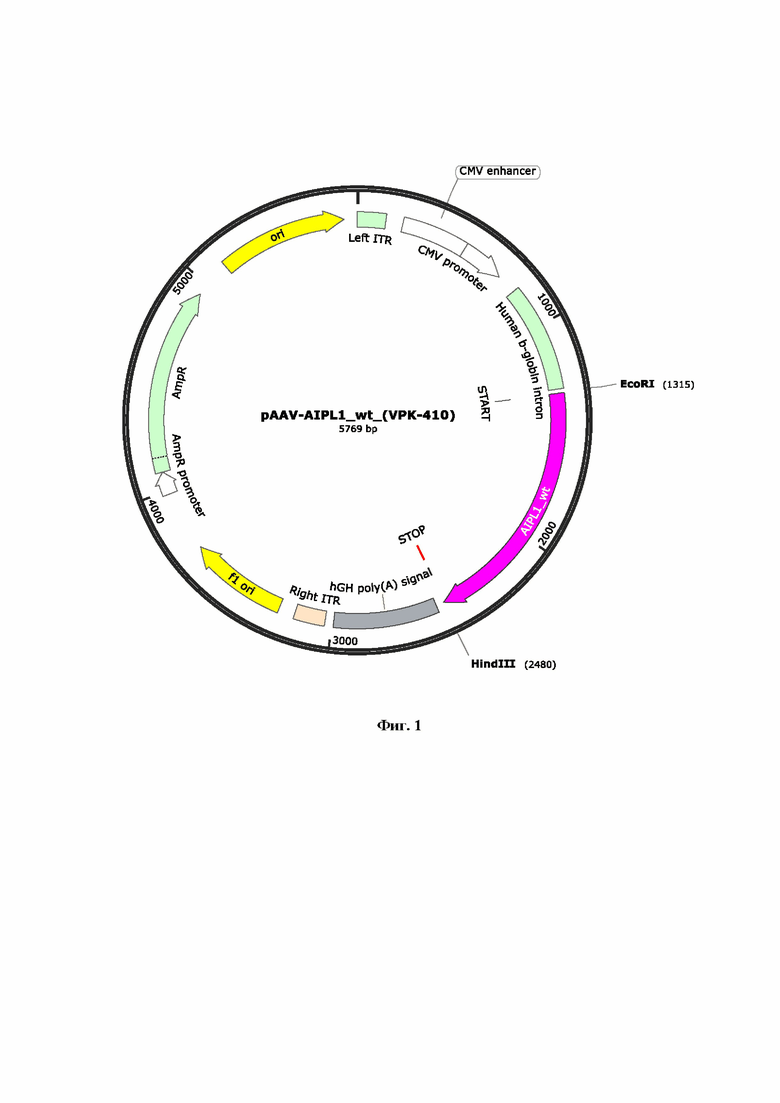
На Фиг. 1 представлен рекомбинантный плазмидный экспрессионный вектор pAAV-AIPL1 wt, кодирующий ген hAIPL1 дикого типа. Ген hAIPL1 был синтезирован и клонирован в плазмиду pAAV-MCS по сайтам рестрикции (EcoRI HindIII) под контролем CMV промотора.
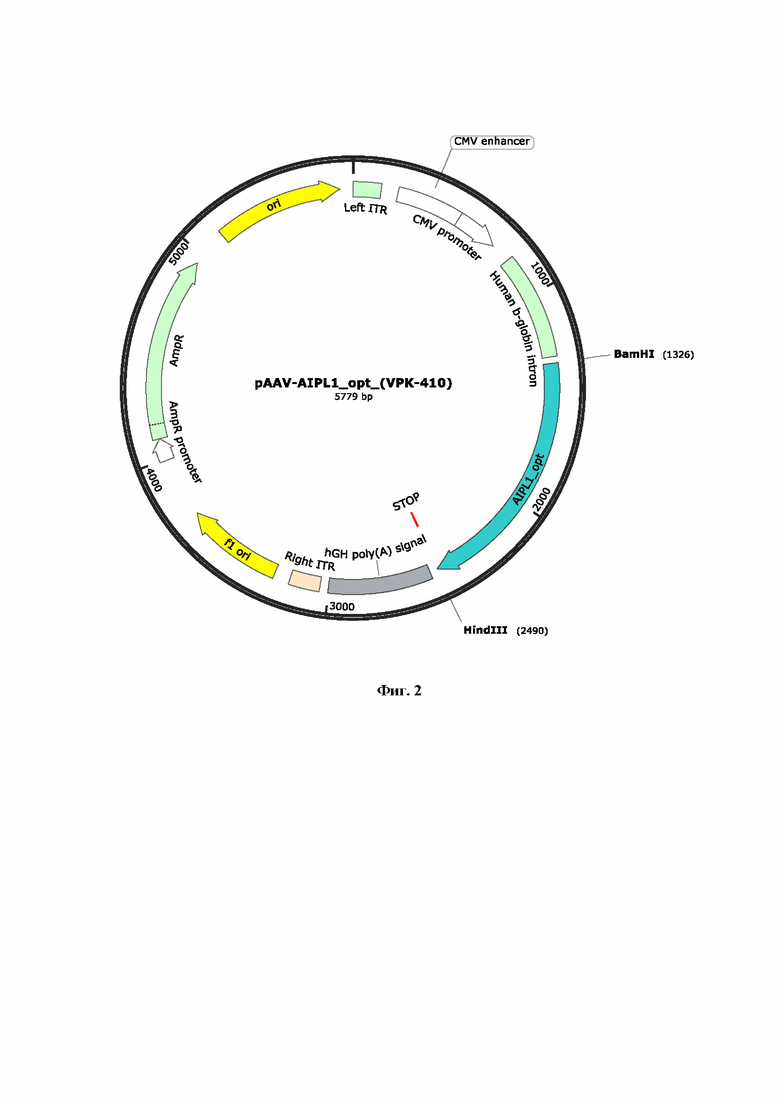
На Фиг. 2 представлен рекомбинантный плазмидный экспрессионный вектор pAAV-AIPL1opt, кодирующий кодонно оптимизированный вариант гена hAIPL1. Ген hAIPL1 opt был синтезирован и клонирован в плазмиду pAAV-MCS по сайтам рестрикции (EcoRI Hindlll) под контролем CMV промотора.
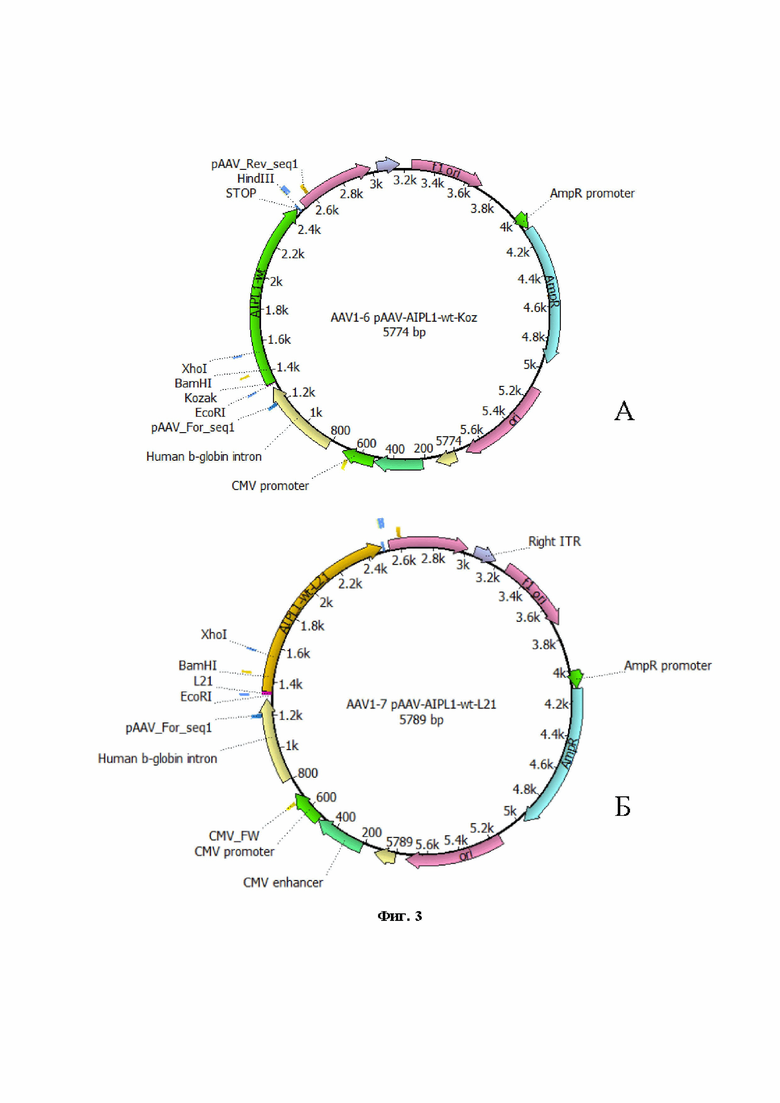
На Фиг. 3 представлен рекомбинантный плазмидный экспрессионный вектор pAAV-AIPL1 wt, кодирующий ген hAIPL1 дикого типа, дополнительно содержащий регуляторный элемент Kozak (А) и пептид L21 (Б).
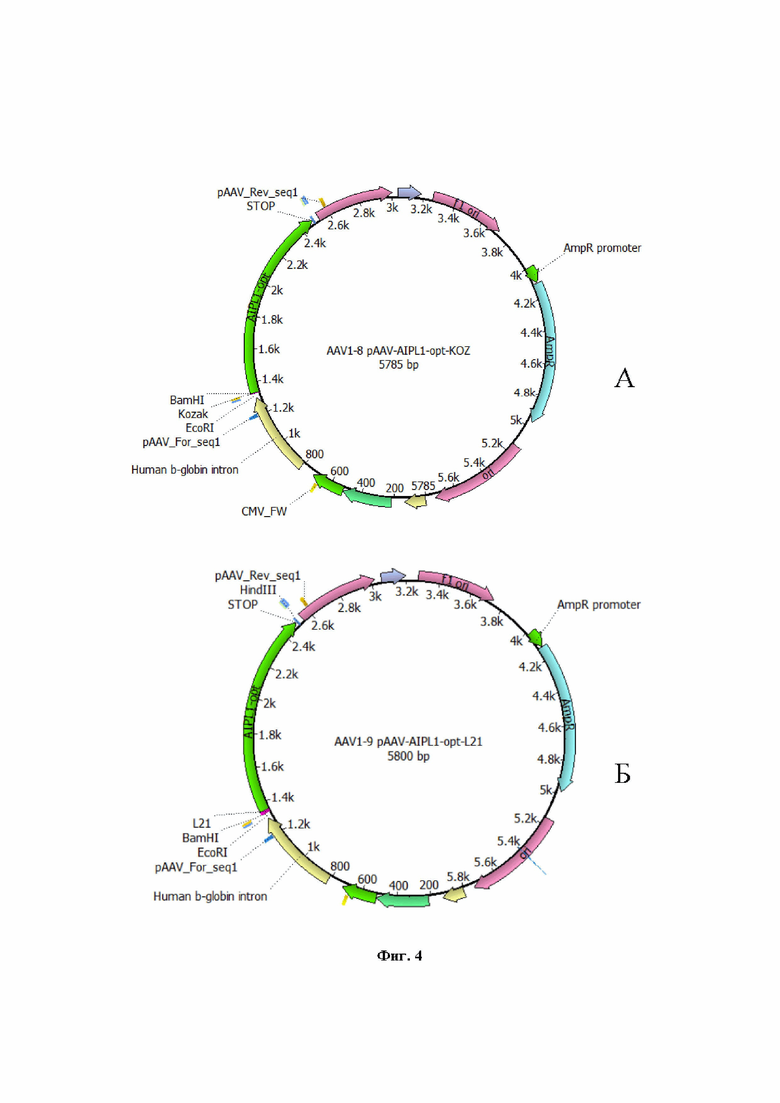
На Фиг. 4 представлен рекомбинантный плазмидный экспрессионный вектор pAAV-AIPL1 opt, кодирующий кодонно оптимизированный вариант гена hAIPL1, дополнительно содержащий регуляторный элемент Kozak (А) и L21 (Б).
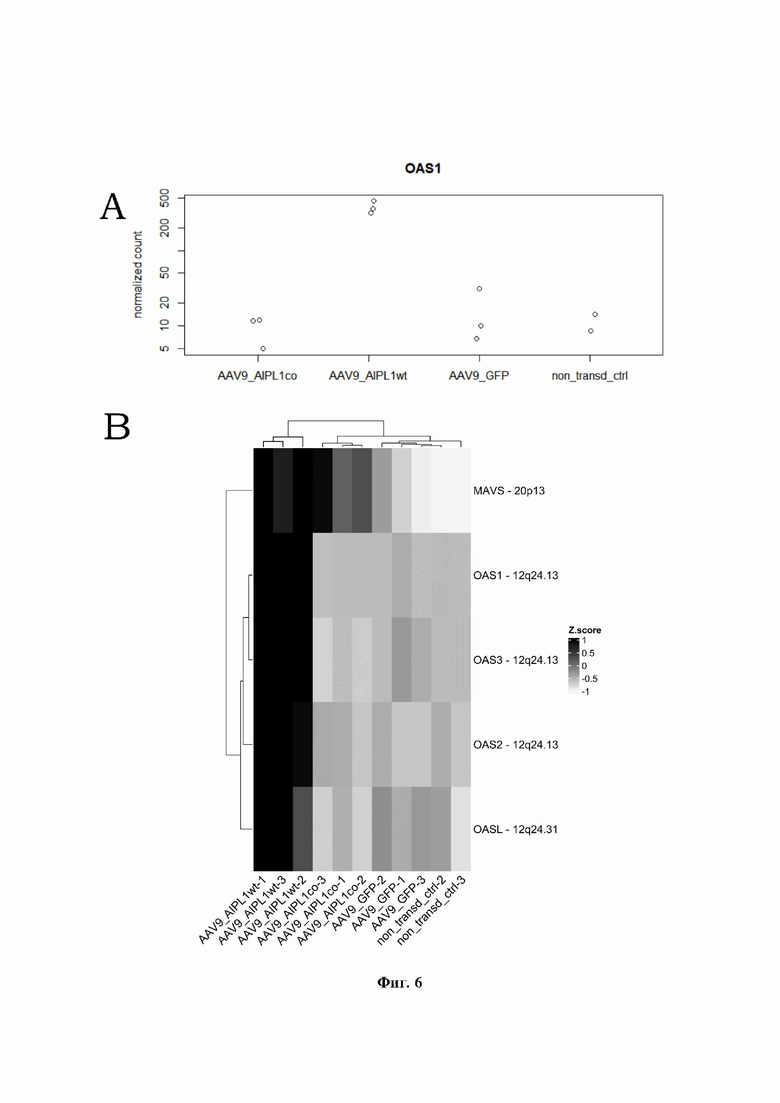
На Фиг. 5 представлена карта экспрессии генов, транскрипция которых стимулируется интерфероном. Z-оценка и оттенки представляют собой масштабированный диапазон нормализованного количества прочтений.
На Фиг. 6 представлена карта экспрессии генов, транскрипция которых модулируется дцРНК, Z-оценка и оттенки представляют собой масштабированный диапазон нормализованного количества прочтений.
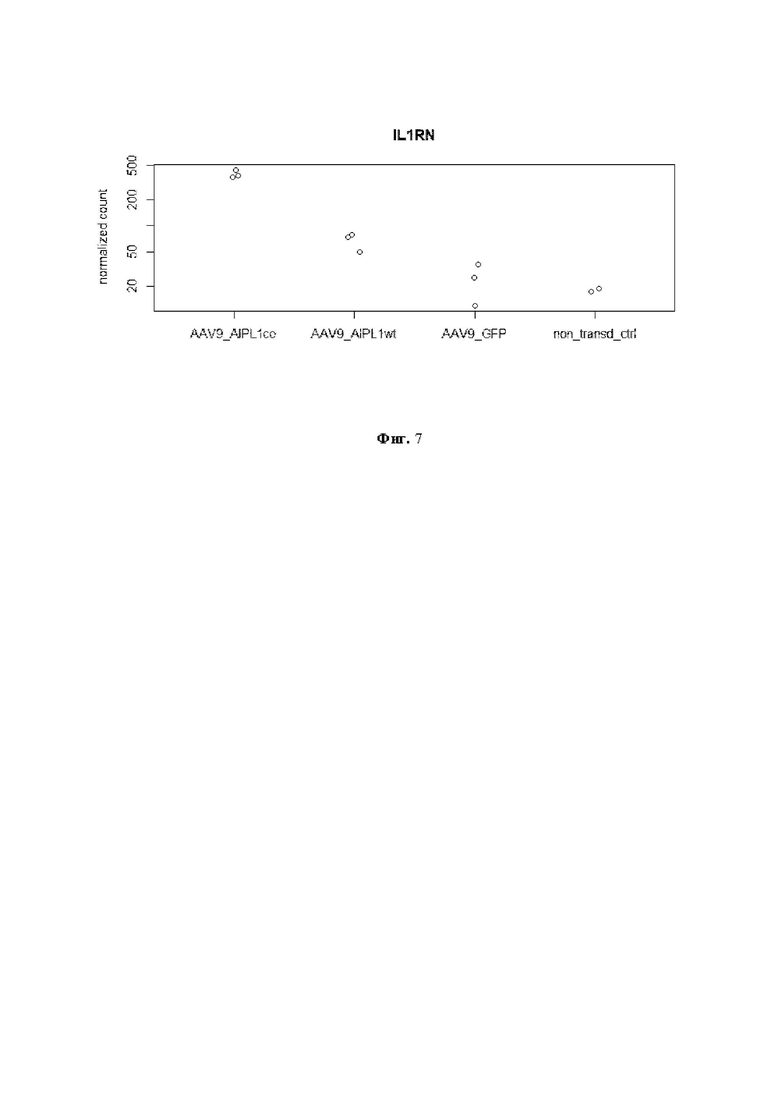
На Фиг. 7 демонстрируется нормализованное количество прочтений гена IL1RN в разных образцах. По оси ординат - количество прочтений.
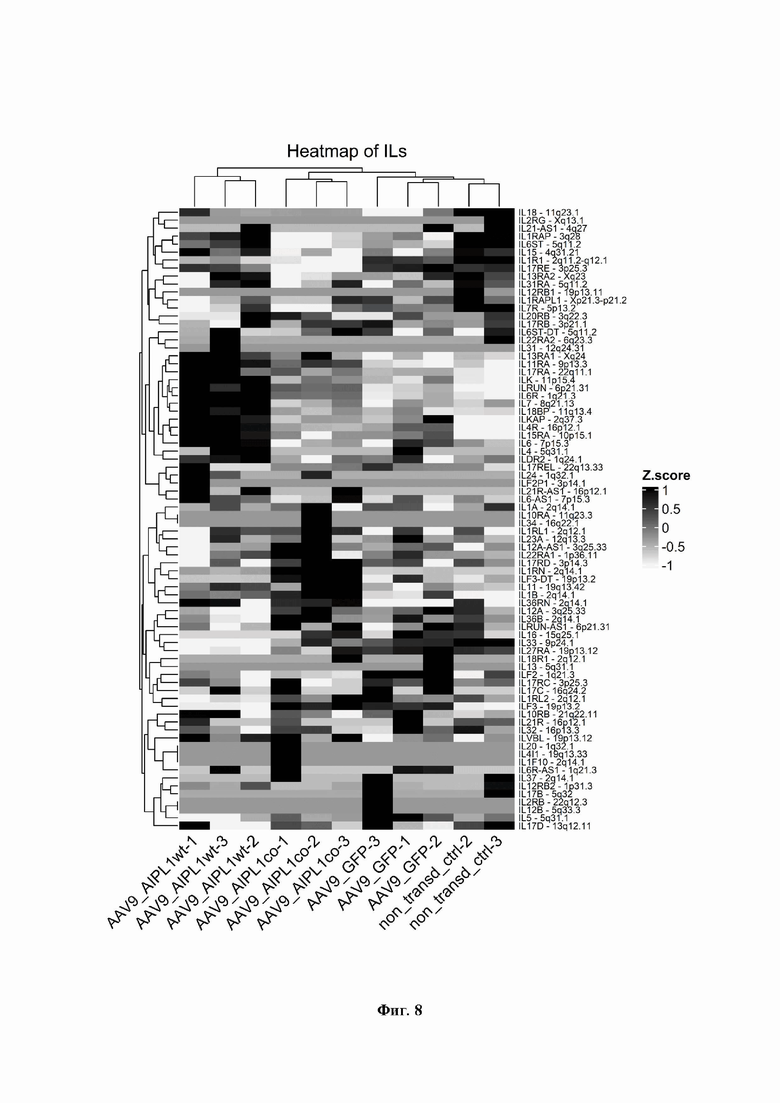
На Фиг. 8 демонстрируется нормализованное количество прочтений обоих вариантов гена hAIPL1 в образцах. Планки погрешности представляют собой стандартное отклонение между повторами в группах образцов.
На Фиг. 9 представлена карта экспрессии генов, транскрипция которых регулируется NF-kBP. Z-оценка и оттенки представляют собой масштабированный диапазон нормализованного количества прочтений.
Детальное описание изобретения
Если не указано иначе, предполагается, что все термины, обозначения и другие научные термины, используемые в данной заявке, имеют значения, которые обычно понимают специалисты в области, к которой относится настоящее изобретение. В некоторых случаях определения терминов с общепринятыми значениями приведены в данной заявке для ясности и/или для быстрой справки и понимания, и включение таких определений в настоящее описание не должно истолковываться как наличие существенного отличия значения термина от обычно подразумеваемого в данной области.
Кроме того, если по контексту не требуется иное, термины в единственном числе включают в себя термины во множественном числе, и термины во множественном числе включают в себя термины в единственном числе. Как правило, используемая классификация и методы культивирования клеток, молекулярной биологии, иммунологии, микробиологии, генетики, аналитической химии, химии органического синтеза, медицинской и фармацевтической химии, а также гибридизации и химии белка и нуклеиновых кислот, описанные в настоящем документе, хорошо известны специалистам и широко применяются в данной области. Ферментативные реакции и способы очистки осуществляют в соответствии с инструкциями производителя, как это обычно осуществляется в данной области, или как описано в настоящем документе.
В настоящем изобретении предлагается применение экспрессионного вектора на основе аденоассоциированного вируса, содержащего кодонно оптимизированную последовательность гена hAIPL1 Seq Id No.:2, для снижения внутриклеточного иммунного ответа при введении в клетки сетчатки человека.
Последовательность нуклеотидов Seq Id No.: 2, кодирующая aipl1 человека, была создана на основе последовательности гена hAIPL1 дикого типа (NP 055151.3, Gene ID: 23746, Seq Id No.: 1) посредством оптимизации кодонов для того, чтобы увеличить стабильность синтезируемой на основе этого гена матричной РНК и уровень экспрессии гетерологичного гена hAIPL1 при введении в клетки в составе плазмидного и/или вирусного экспрессионного вектора [патент РФ №2785621].
Применение экспрессионного вектора на основе аденоассоциированного вируса по настоящему изобретению не ограничивает серотип AAV. Примеры таких векторов на основе AAV можно найти в статье: Peters, С.W., Maguire, С.А., & Hanlon, К. S. (2021). Delivering AAV to the Central Nervous and Sensory Systems. Trends in Pharmacological Sciences, 42(6), 461^74. Векторы на AAV признаются эффективными для генной терапии производных нервной ткани, в том числе тканей сенсорных органов, таких как глаз. AAV представляет собой небольшой, неспособный к самостоятельной репликации, вирус, не имеющий оболочки размером около 20 нм. У человека и приматов описано множество различных серотипов AAV. Геном всех известных серотипов AAV организован сходно: он представляет собой линейную одноцепочечную молекулу ДНК размером менее чем примерно 5000 нуклеотидов (нт). Инвертированные концевые повторы (англ. inverted terminal repeats, ITR) ограничивают внутри себя уникальные нуклеотидные последовательности, кодирующие капсидные белки (Сар) и белки репликации (Rep). Ген Сар кодирует белки VP (VP1, VP2 и VP3), которые образуют капсид. При образовании рекомбинантного вектора AAV кассету экспрессии, не содержащую гены Rep и ограниченную ITR, помещают в капсид AAV. Рекомбинантный AAV не способен к репликации и в настоящее время признается одним из самых безопасных и широко используемых вирусных экспрессионных векторов для переноса генов in vivo. Векторы на основе AAV могут проникать в клетки различных тканей, обеспечивая стабильную экспрессию гетерологичного гена. Эти вирусы непатогенны и обладают относительно низкой иммуногенностью [High КА, Aubourg P. rAAV human trial experience. Methods Mol Biol. 2011;807:429-57].
Термин «внутриклеточный иммунный ответ» в контексте описания настоящего изобретения означает комплекс сигнальных каскадов реакций, активирующихся при внутриклеточной локализации патогена и обеспечивающий врожденный противовирусный иммунитет на уровне клетки. Часто внутриклеточный иммунитет активируется в ответ на появление в цитоплазме клетки чужеродных нуклеиновых кислот и приводит к выработке интерферонов. Примером чужеродных нуклеиновых кислот могут быть двухцепочечные РНК, которые отсутствуют в здоровой клетке и могут образовываться в процессе вирусного заражения. В защитной системе клетки существуют специфические механизмы, детектирующие проникновение чужеродных молекул, так называемых патоген-ассоциированных молекулярных паттернов, в ее внутреннее пространство [Алямкина Е.А., Долгова Е.В., Проскурина А.С., и др. Внутриклеточные системы обнаружения экзогенных нуклеиновых кислот и механизмы запуска иммунных реакций в ответ на интернализацию экзогенной ДНК. Медицинская иммунология. 2013;15(5):413-430. doi:10.15789/1563-0625-2013-5-413-430]. Результатом активации каскада событий, запускаемых цитозольной двухцепочечной ДНК или РНК, является индукция ядерных транскрипционных факторов NF-κВ и IRF3/IRF7 и усиление продукции интерферона-бета, а также процессинг предшественников провоспалительных цитокинов IL-1β и IL-18.
В фоторецепторных клетках сетчатки, которые являются мишенью для доставки терапевтических векторов, эффективность долговременных эффектов генной терапии может снижаться из-за развития противовирусного иммунного ответа, опосредованного двухцепочечными РНК [Shao W, Earley LF, Chai Z, et al. Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction. JCI Insight. 2018;3(12):e120474. Published 2018 Jun 21. doi: 10.1172/jci.insight. 120474].
Анализ дифференциальной экспрессии генов клеток пигментного эпителия сетчатки линии ARPE-19 после трансдукции векторами AAV9-AIPL1wt и AAV9-AIPL1co, содержащими последовательность гена hAIPL1 дикого типа (Seq Id No.: 1) и кодонно оптимизированного варианта (Seq Id No.: 2) соответственно, показал, что применение аденоассоциированных вирусных векторов, содержащих кодонно оптимизированную последовательность гена hAIPL1 Seq Id No.: 2, приводит к значительному снижению внутриклеточного противовирусного иммунного ответа, выражающемуся в снижении экспрессии генов, кодирующих провоспалительные белки, такие как IL6, IL1RAPL, а также в повышении экспрессии гена, кодирующего противовоспалительный белок IL1RN. Также было показано, что применение по настоящему изобретению не приводит к активации олигоаденилатсинтаз (OAS), которые активируются в ответ на появление двухцепочечных РНК в клетке [Schwartz SL, Park EN, Vachon VK, et al. Human OAS1 activation is highly dependent on both RNA sequence and context of activating RNA motifs. Nucleic Acids Res. 2020;48(13):7520-7531. doi:10.1093/nar/gkaa513; Kristiansen H, Gad HH, Eskildsen-Larsen S, et al. The oligoadenylate synthetase family: an ancient protein family with multiple antiviral activities. J Interferon Cytokine Res. 2011;31(1):41-47. doi:10.1089/jir.2010.0107].
Сущность и промышленная применимость изобретения поясняются следующими примерами:
Пример 1. Создание вирусного экспрессионного вектора.
Пример 1.1. Синтез последовательностей нуклеотидов, кодирующих ген hAIPL1wt и hAIPL1opt и подготовка плазмидных экспрессионных векторов.
Нуклеиновую кислоту с последовательностью гена hAIPL1 дикого типа (hAIPL1wt, Seq Id No: 1) и с оптимизированной последовательностью нуклеотидов по настоящему изобретению (hAIPL1opt, Seq Id No: 2) синтезировали на заказ при помощи сервиса TopGenetech (Канада). Для дальнейшего получения вирусных векторов использовали экспрессионные плазмиды, а также коммерчески доступные хелперную плазмиду (pHelper) и упаковочную плазмиду pRC2/8/9 для разных серотипов вируса: AAV2, AAV8, AAV9 соответственно (Cell Biolabs Inc, США).
Для получения вирусных векторов использовали экспрессионную плазмиду AAV-2 в которую методом клонирования по сайтам рестрикции BamHI и HindIII доставляли нуклеотидную последовательность гена hAIPL1wt или hAIPL1opt. Были созданы и наработаны, проверены рестрикционно и секвенированием, заложены в банк плазмидные экспрессионные векторы с экспрессионными кассетами AAV с генами hAIPL1 дикого типа (Фиг. 1) и оптимизированными вариантами, включая hAIPL1opt (Фиг. 2), плазмиды с His-tag, необходимые для оценки уровня трансляции а также плазмидные экспрессионные векторы, дополнительно несущие консенсусную последовательность Kozak или лидерный пептид L21 (Фиг. 3А, 4А) и L21 (Фиг. 3Б, 4Б).
Пример 1.2. Получение и контроль качества AAV
Для наработки векторов использовали линию НЕК293, адаптированную к росту на суспензионной среде. Для получения вирусных векторов использовали экспрессионную плазмиду AAV-2, получение которой описано в примере 1.1. Хелперная плазмида (pHelper) и упаковочная плазмида (pRC9) для вируса серотипа AAV9 для сборки вирусных экспрессионных векторов использовались коммерчески доступные, например, производства Cell Biolabs, США. Суспензионную культуру клеток линии НЕК293 трансфицировали композицией плазмид pAAV-RC2/9, pHelper и плазмидным экспрессионным вектором pAAV-AIPL1co, pAAV-AIPL1wt или pAAV-GFP в молярном соотношении 5:2:1 соответственно с использованием трансфекционного реагента PEI МАХ (линейный полимер, MW 40'000, Polysciences inc.), смешанного с плазмидной ДНК в пропорции 5:1. Рекомбинантные AAV получали путем культивирования в колбах Эрленмейера объемом 250 мл (Corning, США). Через 96 часов клетки собирали и лизировали 0,05% Твин-20 в течение 1 часа и дополнительно обрабатывали бензоназой (30 Ед.) в течение 2 часов. Затем лизаты осветляли с помощью диатомовой земли (HyFloSuperCel) путем фильтрации образца через фильтр с размером пор 0,22 мкм. Тангенциальную фильтрацию осветленных лизатов AAV9-AIPL1co, AAV9-AIPL1wt и AAV9-GFP проводили на системе Labscale TFF с использованием кассеты 100 кДа (Millipore Pellicon® XL100, Merck). Хроматографическую очистку проводили с помощью аффинной смолы CaptureSelect POROS AAVX (ThermoFischer Scientific, США) с системой Bio-Rad Quest 10 Plus. Наконец, рекомбинантные векторы AAV концентрировали до объема 1 мл посредством диализа в 1× PBS (0,37 М NaCl) с использованием системы тангенциальной фильтрации VivaFlow200 (Sartorius, США) и добавляли 0,001% Pluronic F-68 (Sigma-Aldrich, Великобритания). Очищенный вирусный препарат подвергали аналитическому тестированию с использованием метода количественной ПЦР (определение копий генома), малоуглового динамического рассеяния света (агрегаты, концентрация вирусных частиц), размерно-исключающей хроматографии (агрегаты, низкомолекулярные фракции), просвечивающей электронной микроскопии (идентичность, чистота, отношение пустого к полному); Денатурирующий электрофорез полиакрил амид но го геля (идентичность, чистота).
Пример 1.3. Контроль экспрессии hAIPL1.
Для подтверждения экспрессии hAIPL1co и hAIPL1wt использовали ПЦР с обратной транскрипцией (ОТ-ПЦР). Разработана универсальная пара праймеров для дикого типа и оптимизированной последовательности гена hAIPL1 (прямой: 5'-GTGGCTGAAGCTGGAGAAG-3', обратный: 5'-CTCCAGCTCCAGCACTTTC-3') с флуоресцентными зондами, специфичными для каждого гена (WT: 5'-FAM-AGTGCCTGCTGAAGAAGAG-BHQ1-3', ОРТ: 5'-FAM-CAACACCCTGATCCTGAAC-BHQ1-3'). Ген PPIA использовали в качестве эталона для нормализации экспрессии генов [Lemma S, Avnet S, Meade MJ, et al. Validation of Suitable Housekeeping Genes for the Normalization of mRNA Expression for Studying Tumor Acidosis, hit J Mol Sci. 2018;19(10):2930. doi: 10.3390/ijmsl9102930 8]. Для количественной оценки уровня мРНК hAIPL1co и hAIPL1wt в трансфицированных или трансдуцированных клетках проводили количественная ПЦР в режиме реального времени (кПЦР). Статистический анализ экспрессии генов проводили методом ддКТ [Samuel W, Jaworski С, Postnikova OA, et al. Appropriately differentiated ARPE-19 cells regain phenotype and gene expression profiles similar to those of native RPE cells. Mol Vis. 2017;23:60-89]. Экспрессию генов фоторецептор-ассоциированных маркеров также измеряли в клетках пигментного эпителия сетчатки линии ARPE-19. Для этого клетки ARPE-19 инкубировали в течение 5 дней при 37°С с 5% СО2 без смены среды. Клетки лизировали с помощью набора Lyra+ (Биолабмикс, Россия) по протоколу выделения РНК в соответствии с рекомендациями производителя. Синтезировали кДНК с помощью набора M-MuLV-RH RT (Биолабмикс, Россия) и 2 мкг общей РНК с полиА-праймерами. Амплифицировали гены-мишени (PPIA, NUB1, AHR, LCA5, PDE6B, RDH12, ND4, KLF4, AIPL1, TUBB3) со 100 нг кДНК на реакцию с использованием набора HS-qPCR Hi-ROX (Биолабмикс, Россия) и специфических праймеров. Кроме того, экспрессию генов (PPIA, OPN3, KLF4, TUBB3) измеряли в клетках ARPE-19, обработанных основным фактором роста фибробластов (bFGF) в концентрации 20 нг/мл в течение 5 дней.
Пример 1.4. Вестерн-блоттинг hAIPL1.
Наличие hAIPL1 подтверждали методом вестерн-блоттинга. Вкратце, НЕК293Т клетки трансфицировали плазмидами pAAV-AIPL1co и pAAV-AIPL1wt и Lipofectamine 3000 (ThermoFisher Scientific, США). Через 24 часа после трансфекции клетки собирали и промывали в холодном PBS и лизировали в буфере RIP А. Лизат белка разделяли с помощью электрофореза в 12% полиакриламидном геле в денатурирующих условиях. Затем белки переносили на нитроцеллюлозную мембрану размером 0,1 мкм (GE, США). Свободные места связывания блокировали 5% сухим молоком в PBS-T и обрабатывали козьими анти-His антителами (Abeam, США) в течение 1 часа при комнатной температуре. Для нормализации использовали анти-тубулиновые антитела (Abeam, США). Для визуализации с помощью системы визуализации Chemidoc MP (BioRad, США) использовали конъюгированные с пероксидазой хрена анти-кроличьи антитела (Abeam, США) и хемилюминесцентный субстрат Clarity ECL (BioRad, США). В клетках НЕК-293Т, трансфицированных AAV-AIPL1co, количество hAIPL1 в 1,39 раза выше по сравнению с клетками, трансфицированными pAAV-AIPL1wt.
Пример 2. Анализ иммуногенности вирусных векторов
Пример 2.1. Трансдукция клеток пигментного эпителия сетчатки линии ARPE-19 Клетки ARPE-19 из коллекции клеточных культур ИБР РАН культивировали в среде Дюльбекко-Игла (1:1) с добавлением 10% фетальной бычьей сыворотки при 37°С в атмосфере влажного воздуха с 5% СО2. Для индуцирования дифференцировки клетки по достижении конфлюэнтности обрабатывали в течение 120 часов 3 мкМ раствором N-(4-гидроксифенил)ретинамида (фенретинид, Sigma-Aldrich, США).
Клетки ARPE-19, дифференцированные 3 мкМ фенретинидом в течение 5 дней, трансдуцировали либо AAV9-AIPL1co, либо AAV9-AIPL1wt, либо AAV9-GFP в количестве 100000 вирусных геномов на клетку (MOI). Для каждого образца: AAV9-AIPL1co, AAV9-AIPL1wt, AAV9-GFP, нетрансдуцированный контроль - использовали три повторности. Лизировали клетки через 18 часов после трансдукции с использованием препарата Лира+ (Биолабмикс, Россия). Клеточные лизаты замораживали в жидком азоте и хранили при -80°С.
Пример 2.2. Секвенирование транскриптома
Выделение РНК, подготовку библиотек и секвенирование заказывали в ООО «Геноаналитика». Общую РНК экстрагировали с помощью PureLink RNA Micro Kit в соответствии с инструкцией к набору. Анализ фрагментов РНК проводили с помощью прибора Agilent Bioanalyzer 2100 с набором Bioanalyzer RNA Analysis kit. Библиотеки РНК были подготовлены с помощью набора New England Biolabs NEBNext® Ultra™ RNA II Library Prep Kit для Illumina® с модулем протокола магнитного выделения polyA мРНК в соответствии с инструкциями производителя. Контроль качества полученных библиотек фрагментов ДНК проводили в Agilent Bioanalyzer 2100 с использованием High Sensitivity Kit в соответствии с протоколом производителя. Секвенирование проводили с помощью HiSeq 1500 (Illumina, США) по стандартным протоколам Illumina. Было сгенерировано более 10,1-12,8 миллиона прочтений на выборку, и уникально сопоставленные чтения использовались для последующего анализа.
Пример 2.3. Обработка результатов секвенирования
На высокопроизводительном вычислительном кластере «Сириус #2» проводили контроль качества прочтений, построение геномного индекса, картирование, суммирование прочитываний по генам и контроль качества выравнивания. Все необходимое программное обеспечение было установлено с помощью пакетного менеджера Conda [anaconda.org, дата обращения 13.12.2023]. Контроль качества сырых считываний осуществляли с помощью инструмента FastQC [https://www.bioinformatics.babraharn.ac.uk/projects/fastqc/, дата обращения 13.12.2023], суммирование данных и формирование сводного отчета осуществляли с помощью инструмента MultiQC [Ewels Р, Magnusson М, Lundin S, Kaller M. MultiQC: summarize analysis results for multiple tools and samples in a single report. Bioinformatics. 2016;32(19):3047-3048. doi:10.1093/bioinformatics/btw354]. Геномные индексы строили для референсного генома человека GRCh38 первичной сборки [Frankish A, Diekhans М, Jungreis I, et al. GENCODE 2021. Nucleic Acids Res. 2021;49(D1):D916-D923. doi:10.1093/nar/gkaa1087] и аннотации v43 из базы данных Gencode [Frankish A, Diekhans M, Ferreira AM, et al. GENCODE reference annotation for the human and mouse genomes. Nucleic Acids Res. 2019;47(D1):D766-D773. doi:10.1093/nar/gky955] с помощью инструмента STAR [Dobin A, Davis CA, Schlesinger F, et al. STAR: ultrafast universal RNA-seq aligner. Bioinformatics. 2013;29(1):15-21. doi:10.1093/bioinformatics/bts635] версии 2.7.10b с параметрами по умолчанию. Считывание секвенирования было сопоставлено с референсным геномом также с помощью STAR в квантовом режиме "GeneCounts". Суммирование прочтений по генам проводили с помощью HTSeq-count [Anders S, Pyl PT, Huber W. HTSeq--a Python framework to work with high-throughput sequencing data. Bioinformatics. 2015;31(2): 166-169. doi:10.1093/bioinformatics/btu638] в режиме «объединения экзонов», что рекомендовано для большинства случаев использования в дифференциальном анализе экспрессии генов. Контролировали качество выравнивания с помощью пакета RseQC [Wang L, Wang S, Li W. RSeQC: quality control of RNA-seq experiments. Bioinformatics. 2012;28(16):2184-2185. doi:10.1093/bioinformatics/bts356], суммировали данные и формировали сводный отчет с помощью инструмента MultiQC. Последующий анализ проводили в интегрированной среде разработки RStudio [https://posit.co/download/rstudio-desktop/, дата обращения 13.12.2023] настольной версии для персонального компьютера. Нормализация подсчетов, контроль качества на уровне выборки и отдельных генов, анализ дифференциальной экспрессии проводили с использованием пакета DEseq2 [Love MI, Huber W, Anders S. Moderated estimation of fold change and dispersion for RNA-seq data with DESeq2. Genome Biol. 2014;15(12):550. doi:10.1186/sl3059-014-0550-8] версии 1.28.1 для языка программирования R [https://www.r-project.org/, дата обращения 13.12.2023]. Для модели DEseq2 был задан дизайн формулы, включающий только тип выборки. Контроль качества на уровне выборки с использованием методов главного компонентного анализа (РСА) и иерархической кластеризации осуществляли с помощью встроенных функций пакета DEseq2. Попарные сравнения между различными типами выборок проводили с использованием отрицательной биномиальной обобщенной линейной модели в DEseq2. Статистические (р-значение с поправкой = 0,005) и биологические (log2FoldChange±1,58) пороги были установлены как минимальные (менее консервативные), при которых мы перестаем наблюдать изменения числа дифференциально экспрессируемых генов. Сгенерированные списки дифференциально экспрессируемых генов (с р-значением, настроенным менее 0,005 и log2FoldChange = 1,58 или выше) были запущены для терминов Gene Ontology с использованием базы данных GO через функцию API из пакета clusterProfiler R [https://rdrr.io/bioc/clusterProfiler/, дата обращения 13.12.2023]. Визуализацию и иллюстрации выполняли с помощью пакета ggplot2 R [https://link.springer.com/book/10.1007/978-3-319-24277-4, дата обращения 13.12.2023]. Для сравнения выборок и обработки отдельных генов было построено пересечение наборов генов. Для функционального анализа были взяты списки генов из пересечений и исключений, а также соответствующие наборы генов из других баз данных.
Для оценки влияния самого эффекта трансдукции результаты секвенирования транскриптома клеток, трансдуцированных векторами, экспрессирующими природную или кодонно оптимизированную последовательность гена hAIPL1 сравнивали с результатами, полученными на клеточной линии, трансдуцированной AAV9-GFP. В качестве контроля использовали нетрансдуцированные клетки.
Ранее было показано, что интерфероновый ответ может быть вызван некодирующими РНК, синтезируемыми с повторов гетерохроматиновых регионов или эндогенных ретровирусов [Zhao Н, Boije Н, Granberg F, et al. Activation of the interferon-induced STAT pathway during an adenovirus type 12 infection. Virology. 2009;392(2): 186-195. doi: 10.1016/j.virol.2009.07.006]. В ходе нашего исследования мы обнаружили, что трансдукция клеток ARPE-19 вектором AAV9-AIPL1co вызывает меньший внутриклеточный иммунный ответ, чем трансдукция AAV9-AIPL1wt. Например, экспрессия интерферон-стимулируемых генов (ISG), в частности генов OAS1, IFIT1, ISG15, IFI6, была ниже, чем в клетках, трансдуцированных AAV9-AIPL1wt. Также было отмечено, что меньшее количество ISG активируется в клетках после трансдукции AAV9-AIPL1co по сравнению с клетками после трансдукции AAV9-AIPL1wt (Фиг. 5).
OAS1 принадлежит к семейству генов, кодирующих ферменты 2'-5'-олигоаденилатсинтетаз (OAS): OAS1, OAS2, OAS3 и OASL. Все 4 гена сосредоточены в геноме человека в локусе 12q24.1, и OASL в 12q24.2, и все они индуцируются IFN, но OASL активируется непосредственно при вирусной инфекции. Эти ферменты нуждаются в дцРНК для своей каталитической активности. Таким образом, обычно функция OAS1 связана с ограничением инфицирования и прогрессирования РНК-содержащего вируса [Schwartz SL, Park EN, Vachon VK, et al. Human OAS1 activation is highly dependent on both RNA sequence and context of activating RNA motifs. Nucleic Acids Res. 2020;48(13):7520-7531. doi:10.1093/nar/gkaa513]. Сообщалось, что трансдукция AAV может индуцировать активацию врожденного иммунного ответа, опосредованную дцРНК, причем этот процесс может зависеть как от серотипа, так и от структуры трансгена [Shao W, Earley LF, Chai Z, et al. Double-stranded RNA innate immune response activation from long-term adeno-associated virus vector transduction. JCI Insight. 2018;3(12):e120474. Published 2018 Jun 21. doi: 10.1172/jci.insight. 120474]. Наши результаты демонстрируют скоординированную сверхэкспрессию OAS1 и других дцРНК-чувствительных OAS-генов в образцах, трансдуцированных AAV9-AIPL1wt, что характеризует повышенную реактивность клеток в ответ на вирусную доставку hAIPL1 дикого типа. При введении в клетки AAV9-AIPL1co уровень экспрессии дцРНК-чувствительных генов семейства OAS оставался на уровне экспрессии в контрольных нетрансдуцированных клетках (Фиг. 6) Кодонно оптимизированная последовательность гена hAIPL1_opt имеет уникальную комбинацию нуклеотидов обеспечивающую точную передачу генетического кода для трансляции AIPL1 и энергетически выгодной структуры РНК.
Противовоспалительный антагонист рецептора ILIRa, кодируемый геном IL1RN, принадлежит к семейству интерлейкинов-1 и способен модулировать иммунный ответ, блокируя рецепторы интерлейкина-1. По данным анализа секвенирования транскриптома, (Фиг. 7) значительное повышение уровня экспрессии IL1RN происходит в результате трансдукции AAV9-AIPL1wt (в 3 раза по сравнению с контрольным образцом), а также трансдукции AAV9-AIPL1co (в 16 раз). Это говорит о том, что применение вектора по настоящему изобретению ингибирует провоспалительный сигнальный каскад IL-1.
Мы также наблюдаем значительные изменения в экспрессии и других генов, связанных с IL-1 (Фиг. 8). Большинство генов семейства IL-1 расположены в одном локусе на хромосоме 2. Следовательно, согласованное изменение локализованных близко друг к другу генов возможно благодаря непосредственной близости регуляторных областей внутри кластера генов [Sharaf N, Nicklin MJ, di Giovine FS. Long-range DNA interactions at the IL-1/IL-36/IL-37 gene cluster (2ql3) are induced by activation of monocytes. Cytokine. 2014;68(1):16-22. doi:10.1016/j.cyto.2014.03.002]. Повышенная экспрессия гена IL1RN в клетках, трансдуцированных AAV9-AIPL1co, может влиять на местный иммунитет, в том числе на провоспалительные цитокины, интерфероновый ответ и активацию Toll-подобного рецептора (TLR). IL-IRa широко известен своей способностью подавлять активность интерлейкина-1, провоспалительного цитокина, участвующего в иммунном ответе, путем избирательного связывания его рецептора [Dripps DJ, Brandhuber BJ, Thompson RC, et al. Interleukin-1 (IL-1) receptor antagonist binds to the 80-kDa IL-1 receptor but does not initiate IL-1 signal transduction. J Biol Chem. 1991;266(16):10331-10336.].
В целом, представленные данные свидетельствуют о том, что сигнальный путь IL-1 ингибируется в клетках, трансдуцированных AAV9-AIPL1co. Уровень IL-6 был также снижен в клетках после трансдукции AAV9-AIPL1co по сравнению с AAV9-AIPL1wt, и находится почти на таком же уровне, как и в нетрансдуцированном контроле (Фиг. 8.
Транскрипционный фактор NF-кВ, играющий важную роль во врожденном и адаптативном иммунном ответе, может быть активирован интерлейкином-1. Мы показали обширную активацию генов, ассоциированных с NF-κВ, в транскриптоме клеток, трансдуцированных AAV9-AILPlwt (Фиг. 9) по сравнению с нетрансдуцированными клетками. При этом клетки, трансдуцированные AAV9-AIPL1co, имеют значительно более низкие уровни экспрессии генов того же набора.
Взаимодействие между интерферонами и IL-IRa, включая активацию наименьшего количества генов в ответ на IFN [Mayer-Barber KD, Yan В. Clash of the Cytokine Titans: counter-regulation of interleukin-1 and type I interferon-mediated inflammatory responses. Cell Mol Immunol. 2017;14(1):22-35. doi:10.1038/cmi.2016.25; Gander-Bui HTT, Schlafli J, Baumgartner J, et al. Targeted removal of macrophage-secreted interleukin-1 receptor antagonist protects against lethal Candida albicans sepsis. Immunity. 2023;56(8): 1743-1760.e9. doi:10.1016/j.immuni.2023.06.023], может привести к изменениям в передаче иммунных сигналов.
Суммируя полученные данные, показано, что применение экспрессионного вектора на основе аденоассоциированного вируса, содержащего кодонно оптимизированную последовательность гена hAIPL1 Seq Id No.: 2, приводит к снижению экспрессии генов, ответственных за развитие иммунного ответа и повышению экспрессии генов противовоспалительных факторов по сравнению с экспрессией генов в ответ на введение AAV, содержащего последовательность дикого типа, что говорит о снижении внутриклеточного иммунного ответа при вирусной доставке hAIPL1.
Все публикации, патенты и заявки на патенты включены в настоящий документ посредством ссылки. Хотя в вышеприведенном описании это изобретение было описано в отношении некоторых предпочтительных вариантов его осуществления, и многие детали были изложены в целях иллюстрации, для специалистов в данной области техники будет очевидно, что изобретение допускает дополнительные варианты осуществления и что некоторые детали, описанные в данном документе, могут значительно изменяться без отклонения от сущности изобретения.
Использование терминов в единственном числе в контексте описания изобретения должно толковаться как охватывающее как единственное, так и множественное число, если иное не указано в данном документе или явно не противоречит контексту. Термины «состоящий из», «имеющий», «включающий» и «содержащий» следует толковать как неограничивающие термины, т.е. означающие «включая, но не ограничиваясь», если не указано иное. Перечисление диапазонов значений в данном документе просто предназначено для использования в качестве сокращенного способа индивидуальной ссылки на каждое отдельное значение, попадающее в этот диапазон, если здесь не указано иное, и каждое отдельное значение включено в спецификацию, как если бы оно было отдельно изложено в данном документе. Все способы, описанные в данном документе, могут выполняться в любом подходящем порядке, если иное не указано в данном документе или иным образом явно не противоречит контексту. Использование любых и всех примеров или иллюстративного языка (например, «такой как»), представленных в данном документе, предназначено просто для лучшего описания изобретения и не налагает ограничения на объем изобретения, если иное не заявлено. Никакие формулировки в описании не следует истолковывать как указывающие на какой-либо не заявленный элемент как существенный для практического применения изобретения.
Здесь описаны варианты осуществления этого изобретения, включая лучший из известных изобретателям способа осуществления изобретения. Разновидности этих вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения предшествующего описания. Авторы ожидают, что квалифицированные специалисты будут использовать такие варианты в зависимости от обстоятельств, и авторы предполагают, что изобретение будет реализовано на практике иначе, чем конкретно описано в данном документе. Соответственно, это изобретение включает в себя все модификации и эквиваленты признаков, изложенных в прилагаемой формуле изобретения, как это разрешено действующим законодательством. Более того, любая комбинация вышеописанных признаков во всех их возможных вариациях охватывается изобретением, если иное не указано в данном документе или иным образом явно не противоречит контексту.
Заявитель просит рассмотреть представленные материалы заявки «Применение вектора для снижения иммунного ответа при вирусной доставке А1РЫ» на предмет выдачи патента на изобретение.
--->
<?xml version="1.0" encoding="UTF-8"?>
<!DOCTYPE ST26SequenceListing PUBLIC "-//WIPO//DTD Sequence Listing
1.3//EN" "ST26SequenceListing_V1_3.dtd">
<ST26SequenceListing dtdVersion="V1_3"
fileName="AIPL1new_перечень.xml" softwareName="WIPO Sequence"
softwareVersion="2.3.0" productionDate="2023-12-22">
<ApplicationIdentification>
<IPOfficeCode>RU</IPOfficeCode>
<ApplicationNumberText>2023134691</ApplicationNumberText>
<FilingDate>2023-12-22</FilingDate>
</ApplicationIdentification>
<ApplicantName languageCode="ru">Автономная некоммерческая
образовательная организация высшего образования
"Научно-Технологический Университет
"Сириус"</ApplicantName>
<ApplicantNameLatin>Sirius University of Science and
Technology</ApplicantNameLatin>
<InventionTitle languageCode="ru">Применение вектора для снижения
иммунного ответа при вирусной доставке AIPL1</InventionTitle>
<SequenceTotalQuantity>2</SequenceTotalQuantity>
<SequenceData sequenceIDNumber="1">
<INSDSeq>
<INSDSeq_length>1158</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1158</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>genomic DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q2">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>Homo sapiens</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggatgccgctctgctcctgaacgtggaaggggtcaagaaaaccattc
tgcacgggggcacgggcgagctcccaaacttcatcaccggatcccgagtgatctttcatttccgcaccat
gaaatgtgatgaggagcggacagtcattgacgacagtcggcaggtgggccagcccatgcacatcatcatc
ggaaacatgttcaagctcgaggtctgggagatcctgcttacctccatgcgggtgcacgaggtggccgagt
tctggtgcgacaccatccacacgggggtctaccccatcctatcccggagcctgaggcagatggcccaggg
caaggaccccacagagtggcacgtgcacacgtgcgggctggccaacatgttcgcctaccacacgctgggc
tacgaggacctggacgagctgcagaaggagcctcagcctctggtctttgtgatcgagctgctgcaggttg
atgccccgagtgattaccagagggagacctggaacctgagcaatcatgagaagatgaaggcggtgcccgt
cctccacggagagggaaatcggctcttcaagctgggccgctacgaggaggcctcttccaagtaccaggag
gccatcatctgcctaaggaacctgcagaccaaggagaagccatgggaggtgcagtggctgaagctggaga
agatgatcaatactctgatcctcaactactgccagtgcctgctgaagaaggaggagtactatgaggtgct
ggagcacaccagtgatattctccggcaccacccaggcatcgtgaaggcctactacgtgcgtgcccgggct
cacgcagaggtgtggaatgaggccgaggccaaggcggacctccagaaagtgctggagctggagccgtcca
tgcagaaggcggtgcgcagggagctgaggctgctggagaaccgcatggcggagaagcaggaggaggagcg
gctgcgctgccggaacatgctgagccagggtgccacgcagcctcccgcagagccacccacagagccaccc
gcacagtcatccacagagccacctgcagagccacccacagcaccatctgcagagctgtccgcagggcccc
ctgcagagccagccacagagccacccccgtccccagggcactcgctgcagcactaatga</INSDSeq_s
equence>
</INSDSeq>
</SequenceData>
<SequenceData sequenceIDNumber="2">
<INSDSeq>
<INSDSeq_length>1158</INSDSeq_length>
<INSDSeq_moltype>DNA</INSDSeq_moltype>
<INSDSeq_division>PAT</INSDSeq_division>
<INSDSeq_feature-table>
<INSDFeature>
<INSDFeature_key>source</INSDFeature_key>
<INSDFeature_location>1..1158</INSDFeature_location>
<INSDFeature_quals>
<INSDQualifier>
<INSDQualifier_name>mol_type</INSDQualifier_name>
<INSDQualifier_value>other DNA</INSDQualifier_value>
</INSDQualifier>
<INSDQualifier id="q4">
<INSDQualifier_name>organism</INSDQualifier_name>
<INSDQualifier_value>synthetic construct</INSDQualifier_value>
</INSDQualifier>
</INSDFeature_quals>
</INSDFeature>
</INSDSeq_feature-table>
<INSDSeq_sequence>atggacgccgctctgctgctgaatgtggaaggcgtgaaaaagaccatcc
tgcacggtggaacaggcgagctgcctaacttcatcaccggctccagagtgatcttccacttccggaccat
gaagtgcgacgaagagagaaccgtgattgacgatagcagacaggtgggccagcctatgcacatcatcatc
ggcaacatgtttaagctggaggtctgggaaatcctgctgaccagcatgagagtgcatgaagttgccgagt
tctggtgcgatacaatccacacaggagtgtaccctatcctgtctagaagcctgagacagatggcccaggg
aaaggaccccacagaatggcacgtgcatacctgcggcctggccaatatgttcgcttaccacaccctggga
tacgaggacctggatgagcttcagaaggagccacagcctctggtctttgtgatcgagctgcttcaagtgg
acgcccctagcgactaccagcgggaaacctggaacctgagcaaccacgaaaagatgaaagctgtgcctgt
gctgcacggcgaaggcaatcggctgttcaaactgggcagatacgaggaggccagcagcaagtaccaggag
gccatcatttgtttgcgcaacctgcagaccaaggaaaaaccttgggaggtgcagtggctgaagctggaga
agatgatcaacaccctgatcctgaactactgccagtgtctgctgaagaaagaggagtactacgaggtcct
ggagcacacatctgacatcctgagacaccaccccggcatcgtgaaggcctattacgtgcgggcccgggcc
cacgccgaagtgtggaacgaggccgaggccaaggcggacctccagaaagtgctggagctggagccgtcca
tgcagaaggcggtgcgcagggagctgaggctgctggagaaccgcatggcggagaagcaggaggaggagcg
gctgcgctgccggaacatgctgagccagggtgccacgcagcctcccgcagagccacccacagagccaccc
gcacagtcatccacagagccacctgcagagccacccacagcaccatctgcagagctgtccgcagggcccc
ctgcagagccagccacagagccacccccgtccccagggcactcgctgcagcactaatga</INSDSeq_s
equence>
</INSDSeq>
</SequenceData>
</ST26SequenceListing>
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| МИНИ-БЕЛОК USH2A, НУКЛЕИНОВАЯ КИСЛОТА, КОДИРУЮЩАЯ МИНИБЕЛОК USH2A, И СОДЕРЖАЩИЙ ЕЕ ЭКСПРЕССИОННЫЙ ВЕКТОР ДЛЯ ГЕННОЙ ТЕРАПИИ | 2023 |
|
RU2822884C1 |
| Пептид митохондриальной локализации, нуклеиновая кислота для аллотопической экспрессии гена MT-ND4, содержащий ее экспрессионный вектор и его применение | 2023 |
|
RU2817420C1 |
| Нуклеиновая кислота для аллотопической экспрессии гена MT-ND4 | 2023 |
|
RU2809065C1 |
| Кодон-оптимизированная нуклеиновая кислота, которая кодирует белок SMN1, и ее применение | 2020 |
|
RU2742837C1 |
| ВЕКТОРЫ И ПОСЛЕДОВАТЕЛЬНОСТИ ДЛЯ ЛЕЧЕНИЯ ЗАБОЛЕВАНИЙ | 2011 |
|
RU2588667C2 |
| Способ терапии ламеллярного ихтиоза с использованием рекомбинантного аденоассоциированного вируса на основе генетической конструкции, кодирующей нуклеотидную последовательность гена трансглутаминазы 1 | 2023 |
|
RU2821567C1 |
| Препарат для стимулирования регенерации и нейропротекции нервной ткани и способ его использования | 2023 |
|
RU2828544C1 |
| Нуклеиновая кислота, предназначенная для снижения массы тела млекопитающего, экспрессионный вектор для экспрессии в клетках млекопитающего, способ его доставки и способ снижения массы тела млекопитающего | 2023 |
|
RU2810191C1 |
| Препарат для лечения метахроматической лейкодистрофии и способ ее лечения | 2021 |
|
RU2769577C1 |


Изобретение относится к области биотехнологии. Описано применение экспрессионного вектора на основе аденоассоциированного вируса, содержащего кодонно-оптимизированную последовательность гена AIPL1 SEQ ID NO: 2, для снижения внутриклеточного иммунного ответа при введении в клетки сетчатки человека. Техническим результатом настоящего изобретения является снижение экспрессии провоспалительных генов (IL6, IL1RAPL) и повышение экспрессии противовоспалительных генов (IL1RN), при устойчивой экспрессии гена hAIPL1, введенного в клетки сетчатки человека с помощью вирусного экспрессионного вектора на основе AAV. 1 з.п. ф-лы, 9 ил., 2 пр.
1. Применение экспрессионного вектора на основе аденоассоциированного вируса, содержащего кодонно-оптимизированную последовательность гена AIPL1 SEQ ID NO: 2, для снижения внутриклеточного иммунного ответа при введении в клетки сетчатки человека.
2. Применение по п. 1, в котором в качестве аденоассоциированного вируса используют вирус серотипа AAV-9.
| Кодон-оптимизированная последовательность нуклеотидов, кодирующая hAIPL1, и её содержащий экспрессионный вектор | 2021 |
|
RU2785621C1 |
| US 20030022165 A1, 30.01.2003 | |||
| Sun, X., Pawlyk, B | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Gene Therapy, 2010, 17(1), p.117-131. | |||

