Перечень последовательностей
Настоящая заявка содержит перечень последовательностей, который был подан в электронном виде в формате ASCII и тем самым включен в данный документ посредством ссылки во всей своей полноте. Указанная копия ASCII, созданная 31 августа 2020 года, имеет название PAT058332_SL.txt, и ее размер составляет 653193 байта.
Область техники, к которой относится изобретение
Настоящее изобретение относится к слитым белкам, характеризующимся способностью связывать как интегрин, так и фосфатидилсерин. Слитые белки можно применять в качестве терапевтических средств, в частности, для предупреждения или лечения острых или хронических воспалительных нарушений, а также нарушений со стороны органов и микрососудистого русла, обусловленных иммунной системой или коагуляцией.
Предпосылки изобретения
Как показывает опыт, острые воспалительные повреждения органов (AOI) представляют собой сложные заболевания с высокой заболеваемостью, смертностью и значительной неудовлетворенной медицинской потребностью. Типичные AOI включают инфаркт миокарда (MI) и инсульт, которые ежегодно возникают у 32,4 миллионов пациентов во всем мире. Всемирная организация здравоохранения рассматривает пациентов с перенесенными MI и инсультом в качестве группы наиболее высокого риска последующих коронарных и церебральных явлений, которые входят в число основных причин заболеваемости в развитых странах мира. Еще одним AOI является острое повреждение почек (AKI), которое возникает у приблизительно 13,3 миллионов человек в год. В странах с высоким уровнем дохода частота возникновения AKI составляет 3-5/1000, и оно ассоциировано с высокой смертностью (14-46%) (Metha et al., (2015) Lancet, 385(9987): 2616-43). Лица, выжившие после AKI, аналогично таковым в случае MI и инсульта, часто не могут полностью восстановиться и подвергаются повышенному риску развития хронического заболевания почек или терминальной стадии почечной недостаточности. К настоящему времени не существует одобренного FDA лекарственного средства для предупреждения или лечения AKI. Разработка новых видов лечения AKI оказалась сложной, и до настоящего времени успешные результаты клинических испытаний отсутствовали. Вероятно, это связано с многофакторной и комплексной природой патофизиологии AKI, которая предусматривает варианты патомеханизма, связанные с нарушением функции воспаления, микрососудистого русла и нефротоксичностью, вызванными септическими, ишемическими/реперфузионными и/или нефротоксическими поражениями. Эти факторы могут действовать совместно или последовательно, вызывая главным образом повреждение клеток канальцев, но также и клеток клубочков, утрату функционального резерва почек и, в конечном итоге, почечную недостаточность.
Одним общим фактором при AOI является повышенный уровень гибели клеток вследствие повреждения тканей, повышенного образования клеточных фрагментов и протромботических/провоспалительных микрочастиц, которые могут попадать в кровоток и в поврежденную ткань. После инфильтрации ткани нейтрофилами для обеспечения защиты от инфекции нейтрофилы подвергаются апоптозу или другим формам гибели клеток в пораженной ткани. Нейтрофилы содержат вредные соединения, в том числе протеолитические ферменты и молекулярные фрагменты, ассоциированные с опасностью (DAMP), которые могут способствовать повреждению тканей хозяина и распространению воспаления. Эффективный захват погибающих клеток запускает события передачи сигналов, которые приводят к перепрограммированию макрофагов (MΦ) в сторону невоспалительного фенотипа, способствующего разрешению, и высвобождению основных медиаторов для успешного разрешения и восстановления пораженной ткани. Это перепрограммирование недавно объяснялось метаболической передачей сигналов, которая активирует фагоцитарные противовоспалительные ответы в макрофагах (Zhang et al., (2019) Cell Metabolism, 29(2): 443-56). Такое удаление дебриса или состарившихся или погибающих клеток невоспалительным путем называется "эффероцитозом".
Однако в случае задержки эффероцитоза некротические клетки могут накапливаться и вызывать, например, воспалительные ответы, при которых запускается выработка провоспалительных цитокинов (TNF-α) или иммуносупрессивного IL-10 макрофагами (Greenlee-Wacker (2016) Immunol. Reviews, 273: 357-370). Кроме того, если клеточный дебрис и твердые частицы эффективно не удаляются, они могут обуславливать образование скоплений и агрегатов клеток, таких как кластеры фрагментов нейтрофилов и тромбоцитов, микротромбы, и/или обеспечивать высвобождение молекулярных фрагментов, ассоциированных с опасностью (DAMPS), таких как АТФ, ДНК, гистоны или HMGB1. Последствия могут включать окклюзию, нарушение функции и выраженное стерильное воспаление микроциркуляторного русла, приводящие в результате к прогрессированию повреждения тканей, первичной и вторичной органной недостаточности или недостаточно адаптируемой репарации.
В острой фазе AOI для эффероцитотических путей, по-видимому, характерно значительное подавление. Воспаление или острый ответ на повреждение (механические раздражители, гипоксия, окислительный стресс, облучение, воспаление и инфекция) подавляют эффективный эффероцитоз или фагоцитоз посредством снижения содержания специальных белков, связывающих фосфатидилсерин (PS), которые включают мостиковые белки и рецепторы эффероцитоза/клиренса клеточной поверхности. Примером утраты функции рецептора эффероцитоза является протеолитическое отщепление рецепторов семейства ТАМ, таких как тирозинкиназа Mer (MerTK). MerTK представляет собой интегральный мембранный белок, преимущественно экспрессируемый на фагоцитирующих клетках, где он выступает в качестве сигнального белка, а также способствует эффероцитозу (посредством таких белков, как Gas6 или белок S) и ингибирует передачу воспалительных сигналов. Протеолитическое расщепление и высвобождение растворимого эктодомена MerTK индуцируется металлопротеиназой ADAM17. Процесс отщепления может обеспечивать снижение эффероцитоза, осуществляемого фагоцитирующими клетками, вследствие потери поверхностного MerTK. Кроме того, высвобожденный эктодомен также может обеспечивать ингибирование эффероцитоза in vitro (Zhang et al., (2015) J Mol Cell Cardiol., 87:171-9; Miller et al., (2017) Clin Cancer Res., 23(3):623-629). Повышенные количества растворимого Mer в сыворотке/плазме крови обычно наблюдаются при воспалительных, злокачественных или аутоиммунных заболеваниях, таких как диабетическая нефропатия или системная красная волчанка (SLE), и могут служить маркером тяжести заболевания (Ochodnicky P (2017) Am J Pathol., 187(9):1971-1983; Wu et al., (2011) Arthritis Res Ther. 13:R88). Кроме того содержание мостиковых белков, таких как белок жировых глобул молока-EGF-фактор 8 (MFG-E8), также снижается в ходе большинства острых и хронических воспалительных заболеваний. Аналогично растворимому Mer, сниженная концентрация MFG-E8 в сыворотке/плазме крови может быть обнаружена у пациентов с MI или пациентов со стабильной стенокардией (Dai et al., (2016) World J Cardiol., 8(1): 1-23) и может служить маркером тяжести заболевания, как описано для хронического обструктивного заболевания легких (COPD; Zhang et al., (2015) выше).
Представление фосфатидилсерина (PS) на погибающих клетках является эволюционно консервативным противовоспалительным и иммуносупрессивным сигналом для иммунных клеток. Большое количество основных патогенов млекопитающих использует опосредованное PS поглощение в качестве части вирулентной инфекции клеток (Birge et al., (2016) Cell Death Diff., 23(6): 962-78). Вирусы, например, могут связываться с PS-связывающими рецепторами непосредственно или посредством белков, таких как Gas6 (Morizono & Chen (2014) J Virol., 88(8):4275-90). Возможно, что инактивация путей эндогенного клиренса в ответ на повреждение представляет собой эволюционно развитой ответ, направленный на снижение эффективности проникновения возбудителя инфекции в клетки и захвата клеток им после повреждения с обеспечением таким образом ускользания от иммунного ответа и защиты хозяев. Как следствие, модулирование с обеспечением подавления путей клиренса будет обеспечивать повышение эффективности врожденных и адаптивных иммунных эффекторов для борьбы с инфекцией. Как следствие "дружественного огня", эффероцитоз может быть временно нарушен во время острого повреждения органов, и могут возникнуть вышеупомянутые осложнения при AOI. Накопление погибающих клеток, дебриса и провоспалительных и протромботических MP являются отличительными признаками AOI и представляют собой основные триггеры воспаления и повреждения микрососудистого русла. Примечательно, что такое накопление провоспалительных и протромботических микрочастиц характерно для тяжелых заболеваний с высокой потребностью в медицинской помощи и может способствовать их тяжести. Примерами таких показаний являются сепсис и рак (Yang et al., (2016) Tumour Biol., 37(6): 7881-91; Zhao et al., (2016) J Exp Clin Cancer Res., 35: 54; Muhsin-Sharafaldine et al., (2017) Biochim Biophys Acta Gen Subj., 1861(2): 286-295; Ma et al., (2017) Sci Rep., 7(1): 4978; Souza et al., (2015) Kidney Int. 87(6): 1100-8). Предыдущие усилия по обнаружению лекарственных средств в этой области были сосредоточены на PS-связывающих белках, которые могут выступать в качестве основы для разработки лекарств-кандидатов, согласно обзору (Li et al., (2013) Exp Opin Ther Targets, 17(11): 1275-1285).
Подгруппа PS-связывающих белков также распознает интегрины, такие как αvβ3 и αvβ5, которые экспрессируются на многих типах клеток, в том числе фагоцитах, и связывается с ними. Эти белки осуществляют мостиковое соединение апоптотических/погибающих клеток, представляющих PS, с интегринами, что приводит в результате к их эффероцитозу (также называемому фагоцитозом) макрофагами и непрофессиональными фагоцитами. Содержание некоторых мостиковых белков также снижается при большинстве острых и хронических воспалительных заболеваний. Ранее для таких мостиковых белков или их усеченных версий предлагались пути терапевтического применения (WO 2006122327 (сепсис), WO 2009064448 (повреждение органов после ишемии/реперфузии), WO 2012149254 (церебральная ишемия), The Feinstein Institute for Medical Research; WO 2015025959 (инфаркт миокарда) Kyushu University & Tokyo Medical University; WO 20150175512 (резорбция костной ткани) University of Pennsylvania; WO 2017018698 (фиброз тканей), Korea University Research and Business Foundation и US20180334486 (фиброз тканей) Nexel Co., Ltd.; однако применение белков дикого типа или встречающихся в природе белков ограничено рядом проблем. Например, считается, что MFG-E8 дикого типа (wtMFG-E8) характеризуется слабой способностью к проявлению, низкой растворимостью и экспрессией с очень низким выходом при культивировании в клеточных системах экспрессии. В работе Castellanos et al., (2016) было показано, что MFG-E8, экспрессируемый в клетках насекомых или CHO в виде слитого продукта Fc-IgG, является полностью агрегированным и может быть эффективно очищен только посредством добавления детергентов, таких как Triton X-100 или CHAPS (Castellanos et al., (2016) Protein Exp. Pur., 124: 10-22).
Удаление погибающих клеток, дебриса и микрочастиц посредством мостиковых белков, например MFG-E8, EDIL3, Gas6, может обеспечивать устранение основных причин стерильного воспаления и нарушения функции микрососудистого русла и, таким образом, предупреждать прогрессирование повреждения тканей и обеспечивать разрешение воспаления. Таким образом, терапевтический подход, способствующий клиренсу погибающих клеток в ходе AOI, может быть применен для снижения или по меньшей мере ослабления выраженности патологии AOI и может быть значимым при других условиях заболевания, при которых клиренс погибающих клеток или микрочастиц, представляющих PS, является недостаточным. Таким образом, существует потребность в терапевтическом средстве, которое можно применять для снижения повреждения тканей и воспаления и которое характеризуется требуемыми производственными характеристиками для удовлетворения неудовлетворенных медицинских потребностей при AOI.
Сущность изобретения
В настоящем изобретении заявители создали рекомбинантные терапевтические слитые белки на основе структуры встречающихся в природе мостиковых белков (например, MFG-E8) без вышеупомянутых нежелательных свойств и проблем, связанных с продуцированием белка дикого типа. Слитые белки по настоящему изобретению содержат интегрин-связывающий домен, PS-связывающий домен и солюбилизирующий домен. Слитые белки сохраняют основные биологические функции белка дикого типа, т. е. MFG-E8, например, они функционируют с обеспечением связывания PS-представляющих погибающих клеток, дебриса и микрочастиц с фагоцитами и, таким образом, обеспечивают запуск эффероцитоза. Кроме того, терапевтические слитые белки по настоящему изобретению характеризуются улучшенным потенциалом к разработке, в частности, сниженной липкостью и улучшенной растворимостью по сравнению с белком дикого типа MFG-E8 (SEQ ID NO: 1). Кроме того, эти терапевтические слитые белки характеризуются более длительным временем воздействия в плазме крови и характеризуются более высоким выходом при экспрессии в клеточных системах экспрессии по сравнению с белком MFG-E8 дикого типа.
В настоящем документе предусмотрены терапевтические слитые белки для усиления эффероцитоза, содержащие интегрин-связывающий домен, фосфатидилсерин (PS) -связывающий домен и солюбилизирующий домен.
В некоторых вариантах осуществления солюбилизирующий домен слитого белка соединен с интегрин-связывающим доменом. В некоторых вариантах осуществления солюбилизирующий домен соединен с PS-связывающим доменом. В некоторых вариантах осуществления солюбилизирующий домен соединен как с интегрин-связывающим доменом, так и с PS-связывающим доменом, т. е. расположен между интегрин-связывающим доменом и PS-связывающим доменом. В некоторых вариантах осуществления солюбилизирующий домен вставлен в интегрин-связывающий домен или вставлен в PS-связывающий домен. В одном варианте осуществления терапевтический слитый белок от N-конца до С-конца имеет структуру интегрин-связывающий домен - солюбилизирующий домен - PS-связывающий домен.
В некоторых вариантах осуществления интегрин-связывающий домен терапевтического слитого белка содержит мотив связывания аргинин-глицин-аспарагиновая кислота (RGD) и связывается с интегрином(интегринами) αvβ3 и/или αvβ5 или α8β1.
В некоторых вариантах осуществления солюбилизирующий домен терапевтического слитого белка соединен непосредственно с интегрин-связывающим доменом и/или соединен с PS-связывающим доменом, т. е. вставлен между указанными доменами. В альтернативном варианте осуществления солюбилизирующий домен соединен с интегрин-связывающим доменом и/или PS-связывающим доменом опосредованно, с помощью линкера, такого как внешний линкер. В некоторых вариантах осуществления солюбилизирующий домен предусматривает сывороточный альбумин человека (HSA), домен 3 HSA (D3 HSA) или Fc-область IgG (Fc-IgG) или их функциональный вариант.
В некоторых вариантах осуществления терапевтический слитый белок содержит С-конец интегрин-связывающего домена, соединенный с N-концом солюбилизирующего домена, и С-конец солюбилизирующего домена, соединенный с PS-связывающим доменом. В некоторых вариантах осуществления терапевтический слитый белок содержит общую структуру EGF-HSA-C1-C2, где EGF представляет собой интегрин-связывающий, EGF-подобный домен MFG-E8, EDIL3 или другие белки, содержащие интегрин-связывающий домен, как указано в таблице 1, и C1-C2 представляет собой PS-связывающий домен, обнаруженный в MFG-E8, EDIL3 или других белках, содержащих PS-связывающий домен, как указано в таблице 2. Примеры белков, содержащих как интегрин-связывающий домен, так и PS-связывающий домен, например MFG-E8 (SEQ ID NO: 1) и EDIL3 (SEQ ID NO: 11), приведены в таблице 3.
В некоторых вариантах осуществления интегрин-связывающий домен представляет собой EGF-подобный домен, например, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 2, или аминокислотную последовательность, характеризующуюся по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% идентичностью с ней или ее усеченными вариантами. В одном варианте осуществления EGF-подобный домен предусматривает EGF-подобный домен MFG-E8 человека или его функциональный вариант, содержащий одну, две, три, четыре, пять и до 10 аминокислотных модификаций. В одном варианте осуществления EGF-подобный домен предусматривает EGF-подобный домен EDIL3 человека или его функциональный вариант, содержащий одну, две, три, четыре, пять и до 10 аминокислотных модификаций.
В некоторых вариантах осуществления PS-связывающий домен содержит два субдомена дискоидина C1-C2, например, PS-связывающий домен MFG-E8 человека имеет аминокислотную последовательность, представленную под SEQ ID NO: 3, или аминокислотную последовательность, характеризующуюся по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% идентичностью с ней или ее усеченными вариантами. В одном варианте осуществления PS-связывающий домен предусматривает PS-связывающий домен MFG-E8 человека или его функциональный вариант, содержащий одну, две, три, четыре, пять и до 10 аминокислотных модификаций. В одном варианте осуществления PS-связывающий домен предусматривает PS-связывающий домен EDIL3 человека или его функциональный вариант, содержащий одну, две, три, четыре, пять и до 10 аминокислотных модификаций.
В некоторых вариантах осуществления солюбилизирующий домен представляет собой HSA или его функциональный вариант, например, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 4, или аминокислотную последовательность, характеризующуюся по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% идентичностью с ней или ее усеченными вариантами. В одном варианте осуществления HSA содержит аминокислотную замену C34S, функция которой заключается в снижении способности белка к агрегации, и содержит аминокислотную последовательность, представленную под SEQ ID NO: 5. В некоторых вариантах осуществления солюбилизирующий домен предусматривает сывороточный альбумин человека (HSA) или его функциональный вариант, содержащий одну, две, три, четыре, пять и до 10 аминокислотных модификаций, например HSA C34S или усеченный вариант HSA, например домен 3 HSA (D3 HSA) или его функциональный вариант. В предпочтительном варианте осуществления солюбилизирующий домен представляет собой HSA C34S.
В альтернативном варианте осуществления солюбилизирующий домен предусматривает Fc-область IgG (Fc-IgG), например Fc-область IgG1, IgG2, IgG3 или IgG4 человека или их функциональный вариант. В одном варианте осуществления солюбилизирующий домен предусматривает Fc-область Fc-IgG1 человека, содержащую аминокислотную последовательность, представленную под SEQ ID NO: 7, или аминокислотную последовательность, характеризующуюся по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% идентичностью с ней или ее усеченными вариантами. В одном варианте осуществления Fc-IgG1 содержит аминокислотные замены D265A и P329A для снижения эффекторной функции Fc и содержит аминокислотную последовательность, представленную под SEQ ID NO: 8. В другом варианте осуществления Fc-IgG1 содержит аминокислотную замену T366W для создания "выступа" или может содержать аминокислотные замены T366S, L368A, Y407V для создания "впадины". Кроме того, выступ Fc-IgG1 может содержать аминокислотную замену S354C, а впадина Fc-IgG1 может содержать аминокислотную замену Y349C, так что при спаривании образуется цистеиновый мостик. В дополнение к модификациям типа "выступ во впадину" Fc-IgG1 может также содержать замены D265A и P329A для снижения эффекторной функции Fc. В одном варианте осуществления Fc-IgG1 содержит аминокислотную последовательность, представленную под SEQ ID NO: 9 или 10.
В предпочтительном варианте осуществления терапевтический слитый белок содержит белок жировых глобул молока-EGF-фактор 8 (MFG-E8) и солюбилизирующий домен, при этом MFG-E8 содержит интегрин-связывающий EGF-подобный домен (SEQ ID NO: 2) и фосфатидилсерин-связывающие домены C1-C2 (SEQ ID NO: 3 или SEQ ID NO: 76). MFG-E8 может предусматривать встречающийся в природе MFG-E8 или MFG-E8 человека дикого типа (SEQ ID NO: 1) или MFGE-8 с SEQ ID NO: 75 или его функциональный вариант. В одном варианте осуществления солюбилизирующий домен соединен с N- или C-концом MFG-E8. В одном варианте осуществления солюбилизирующий домен вставлен между EGF-подобным доменом и доменом C1 или между доменами C1 и C2. В предпочтительном варианте осуществления солюбилизирующий домен соединен с С-концом EGF-подобного домена и соединен с N-концом домена C1. Солюбилизирующий домен может быть соединен непосредственно или опосредованно с С-концом EGF-подобного домена и соединен непосредственно или опосредованно с N-концом домена С1. В некоторых вариантах осуществления опосредованное соединение осуществляется с помощью внешнего линкера, например линкера на основе глицина-серина.
В одном варианте осуществления терапевтический слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 42 (FP330). В одном варианте осуществления терапевтический слитый белок может содержать гистидиновую метку (His-метка; SEQ ID NO: 67) для облегчения обнаружения и/или очистки в анализах на присутствие и экспрессии белка. В одном варианте осуществления терапевтический слитый белок содержит С-концевую His-метку и содержит аминокислотную последовательность, представленную под SEQ ID NO: 44 (FP278). Терапевтические слитые белки FP278 и FP330 имеют одинаковую аминокислотную последовательность, за исключением добавления His-метки к FP278.
В некоторых вариантах осуществления терапевтический слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 42 (FP330), или аминокислотную последовательность, характеризующуюся по меньшей мере 90%, 95%, 96%, 97%, 98% или 99% идентичностью с ней или ее усеченными вариантами. Например, терапевтический слитый белок FP776 содержит аминокислотную последовательность, представленную под SEQ ID NO: 48, и характеризуется 97,7% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP068 содержит аминокислотную последовательность, представленную под SEQ ID NO: 46, и характеризуется 98,3% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP816 содержит аминокислотную последовательность, представленную под SEQ ID NO: 58, и характеризуется 98,5% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP811 содержит аминокислотную последовательность, представленную под SEQ ID NO: 54, и характеризуется 99,0% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP010 содержит аминокислотную последовательность, представленную под SEQ ID NO: 56, и характеризуется 99,5% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP138 содержит аминокислотную последовательность, представленную под SEQ ID NO: 52, и характеризуется 99,8% идентичностью последовательности с FP330 (SEQ ID NO: 42). Например, терапевтический слитый белок FP284 содержит аминокислотную последовательность, представленную под SEQ ID NO: 50, и характеризуется 99,9% идентичностью последовательности с FP330 (SEQ ID NO: 42).
В некоторых вариантах осуществления и как описано в разделе "Примеры", терапевтические слитые белки по настоящему изобретению функционируют с обеспечением способствования эффероцитозу эндотелиальными клетками в анализе эффероцитоза клеток Jurkat эндотелиальными клетками человека, а также восстановления нарушенного эффероцитоза и усиления исходного уровня эффероцитоза макрофагами в анализе эффероцитоза нейтрофилов макрофагами человека; слитые белки функционируют с обеспечением снижения количества микрочастиц в плазме крови посредством клиренса в анализе эффероцитоза микрочастиц эндотелиальными клетками человека; и/или слитые белки обеспечивают защиту от полиорганного повреждения в модели острой ишемии почек.
В данном документе также раскрыты способы, пути применения, диагностические реагенты, фармацевтические композиции и наборы, в которых применяются или содержатся эти терапевтические слитые белки. В данном документе также предусмотрены нуклеиновые кислоты, кодирующие раскрытые слитые белки, векторы клонирования и экспрессии, содержащие такие нуклеиновые кислоты, клетки-хозяева, содержащие такие нуклеиновые кислоты, и способы получения раскрытых слитых белков посредством культивирования таких клеток-хозяев.
Краткое описание графических материалов
На фигуре 1 представлено схематическое изображение примеров терапевтических слитых белков по настоящему изобретению. Солюбилизирующий домен (обозначенный "SD") был соединен либо с С-концом, либо с N-концом, либо между доменами EGF, C1 или C2 MFG-E8.
На фигуре 2 изображено несколько белковых SDS-PAGE-гелей слитых белков, экспрессированных в клетках HEK. Фиг. 2A: белок EGF-HSA-C1-C2 (FP330; SEQ ID NO: 42); фиг. 2B: белок EGF-HSA-C1-C2 из EDIL3 (FP050; SEQ ID NO: 12); фиг. 2C: белок EGF-Fc(KiH) C1-C2, невосстановленный и восстановленный (этот белок представляет собой гетеродимер FP071 (EGF-Fc(выступ)-C1-C2; SEQ ID NO: 18) с впадиной Fc-IgG1 (SEQ ID NO: 10)); фиг. 2D: белок EGF-HSA-C1 (FP260; SEQ ID NO: 34). Для каждой из фиг. 2A, 2C и 2D в первом столбце изображены стандартные маркеры Precision Plus Protein Unstained, а во втором столбце изображен соответствующий слитый белок. На фиг. 2В в первом столбце изображен слитый белок, а во втором столбце изображены стандартные маркеры Precision Plus Protein Unstained. На фигуре 2Е изображены другие рекомбинантные белки, которые были получены и очищены.
На фигуре 3 представлен пример эффекта потери MFG-E8 дикого типа (wt) по сравнению со слитым белком FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) при работе с ними на практике. На фиг. 3А изображена потеря эффективности wtMFG-E8 в конкурентном анализе связывания L-α-фосфатидилсерина, если разведение белков готовили в полипропиленовых планшетах (символ: □) по сравнению с разведениями, приготовленными в несвязывающих планшетах (символ: ●). В отличие от этого, на фиг. 3B практически не видно потери эффективности слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в конкурентном анализе связывания PS, если разведения белков готовили в полипропиленовых планшетах (символ: □) по сравнению с несвязывающими планшетами (символ: ●).
На фигуре 4 изображено связывание слитых белков с L-α-фосфатидилсерином. На фиг. 4А изображено связывание FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) с иммобилизованным L-α-фосфатидилсерином и в меньшей степени с фосфолипидом кардиолипином зависимым от концентрации образом. На фиг. 4В изображено связывание wtMFG-E8 человека и ряда терапевтических слитых белков: FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44), FP250 (EGF-HSA; SEQ ID NO: 32), FP260 (EGF-HSA-C1; SEQ ID NO: 34) и FP270 (EGF-HSA-C2; SEQ ID NO: 36) с иммобилизованным L-α-фосфатидилсерином зависимым от концентрации образом в формате конкурентного анализа (конкуренция за связывание биотинилированного мышиного wtMFG-E8 с L-α-фосфатидилсерином).
На фигуре 5 изображена зависимая от интегрина αv адгезия клеток к слитым белкам. На фиг. 5А изображено, что адгезия клеток к FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42) полностью блокируется ингибитором интегрина αv циленгитидом или 10 мМ EDTA. Единственная точечная мутация в интегрин-связывающем мотиве RGD (RGD > RGE) EGF-подобного домена (FP280; SEQ ID NO: 38) приводит в результате к полному устранению адгезии клеток, как изображено на фиг. 5В. На фиг. 5C изображено, что иммобилизованный белок EGF-HSA (FP250; SEQ ID NO: 32), несмотря на наличие EGF-подобного домена, не способствует адгезии клеток BW5147.G.1.4 или лишь умеренно способствует ей. Как изображено на фиг. 5D, слитый белок по настоящему изобретению (FP330; SEQ ID NO: 42) способствует зависимой от интегрина αv адгезии клеток подобно wtMFG-E8 при экспрессии в клетках CHO или в клетках HEK.
На фигуре 6 изображен эффект терапевтического слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в отношении способствования эффероцитозу погибающих нейтрофилов макрофагами человека. Концентрация слитого белка изображена на оси x, а уровень эффероцитоза [%] изображен на оси y.
На фигуре 7 изображено, что терапевтический слитый белок FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) может обеспечивать восстановление нарушенного эндотоксином (липополисахаридом) эффероцитоза погибающих нейтрофилов макрофагами человека. На фиг. 7А изображено нарушение эффероцитоза погибающих нейтрофилов человека макрофагами в присутствии 100 мкг/мл липополисахарида (LPS) у трех доноров-людей. На левой панели изображен ответ отдельного донора, на правой панели изображено среднее значение нарушения эффероцитоза (%) для трех доноров. На фиг. 7В изображено восстановление этого нарушенного эндотоксином (LPS) эффероцитоза погибающих нейтрофилов макрофагами человека в присутствии терапевтического слитого белка FP278. Показатели эффероцитоза у 3 различных доноров макрофагов человека нормализовали и наносили на график в виде уровня эффероцитоза (%).
На фигуре 8 изображено восстановление индуцированного частицами S. aureus нарушения эффероцитоза погибающих нейтрофилов макрофагами человека с помощью терапевтического слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44). На фиг. 8А изображен эффект FP278 в концентрации 100 нМ в отношении стимулирования эффероцитоза выше исходного уровня (пунктирная линия; левая часть фигуры), а также эффект 100 нМ FP278 в отношении восстановления нарушения эффероцитоза, вызванного введением S. aureus (правая часть фигуры). На фигуре 8B изображен эффект повышения концентраций слитого белка FP278 (EC50 8 нM) в отношении восстановления нарушенного эффероцитоза, вызванного введением S. aureus, и в отношении способствования эффероцитозу после достижения исходных уровней эффероцитоза.
На фигуре 9 изображен эффект терапевтического слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в отношении способствования эффероцитозу погибающих клеток Jurkat эндотелиальными клетками человека (HUVEC). Эффективность слитого белка в анализе эффероцитоза эндотелиальными клетками зависит от присутствия тандемного домена C1-C2 или C1-C1, поскольку, как изображено на фигуре 9, слитый белок со структурой EGF-HSA-C2 (FP270; SEQ ID NO: 36) является неэффективным в этом анализе.
На фигуре 10 изображено, что расположение домена HSA в терапевтическом слитом белке, а именно в N- или С-концевом положении (FP220 (HSA-EGF-C1-C2; SEQ ID NO: 30) или FP110 (EGF-C1-C2-HSA; SEQ ID NO: 28) соответственно), сообщает слитому белку MFG-E8 HSA функцию блокирования эффероцитоза в анализе эффероцитоза макрофагами. Концентрация слитого белка изображена на оси x, уровень эффероцитоза [%] изображен на оси y.
На фигуре 11 изображено сравнение способствования эффероцитозу различными форматами терапевтических слитых белков, содержащих фрагмент HSA или Fc. Концентрация слитого белка изображена на оси x (нМ), уровень эффероцитоза [MFI] изображен на оси y. На фиг. 11А изображено сравнение слитых белков, содержащих HSA, с HSA, расположенным на С-конце или N-конце, или между EGF-подобным доменом и доменом C1; FP110 (EGF-C1-C2-HSA; SEQ ID NO: 28), FP220 (HSA-EGF-C1-C2; SEQ ID NO: 30) и FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) соответственно. На фиг. 11B изображено сравнение слитых белков, содержащих фрагмент Fc, с Fc, расположенным на С-конце (FP060 (EGF-C1-C2-Fc [S354C, T366W]; SEQ ID NO: 14) и FP080 (EGF-C1-C2-Fc; SEQ ID NO: 22)) или между EGF-подобным доменом и доменом C1 (FP070 (EGF-Fc-C1-C2; SEQ ID NO: 16)) по сравнению с MFG-EG дикого типа (SEQ ID NO: 1). Изображены два формата фрагмента Fc: Fc дикого типа (FP080; SEQ ID NO: 22) и фрагмент Fc с модификациями S354C и T366W (нумерация согласно EU; FP060; SEQ ID NO: 14). На фиг. 11C изображено сравнение трех партий слитого белка FP090 (Fc-EGF-C1-C2; SEQ ID NO: 24), содержащего фрагмент Fc, расположенный на N-конце, при трех различных концентрациях (0,72, 7,2 и 72 нМ) по сравнению с контролем wt-MFG-E8. На фиг. 11D изображено способствование эффероцитозу, обеспечиваемое конструкцией слитого белка FP050, содержащей HSA, вставленный между EGF-подобным доменом и доменом C1-C2 EDIL3 (EGF-HSA-C1-C2 на основе EDIL3; SEQ ID NO: 12). На фигуре 11E изображены дополнительные примеры слитых белков по настоящему изобретению, например химерные варианты (FP145; SEQ ID NO: 80, FP1145; SEQ ID NO: 103, FP146; SEQ ID NO: 82, FP1146) и комбинации интегрин-связывающих доменов MFGE8 или EDIL3 и PS-связывающих доменов, таких как домен V IgSF TIM4 или домен GLA мостикового белка GAS6 (FP1147 и FP1148). На фигуре 11F показана функция стимулирования эффероцитоза рекомбинантных слитых белков, состоящих из химерных белков, объединяющих домены EDIL3 и MFG-E8 со вставкой HSA. Данные демонстрируют, что FP145 (SEQ ID NO: 80) и FP146 (SEQ ID NO: 82) индуцировали эффероцитоз умирающих нейтрофилов макрофагами человека зависимым от концентрации образом. На фигуре 11G показана функция стимулирования эффероцитоза рекомбинантных слитых белков, состоящих из химерных белков, объединяющих домены EDIL3 и MFG-E8 со вставкой HSA. Данные демонстрируют, что FP145 (SEQ ID NO: 80) и FP146 (SEQ ID NO: 82) индуцировали эффероцитоз умирающих клеток Jurkat эндотелиальными клетками человека (HUVEC) зависимым от концентрации образом.
На фигуре 12 изображено способствование эффероцитозу клетками HUVEC, осуществляемое терапевтическим слитым белком FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44), протестированное при 3 различных концентрациях до 30 нМ. Стимуляция эффероцитоза зависела от концентрации, при этом эффероцитоз повышался по мере повышения концентрации слитого белка FP278.
На фигуре 13 изображено, что терапевтические слитые белки FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42; фиг. 13A), FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44; фиг. 13B) и FP776 (EGF-HSA-C1-C2; SEQ ID NO: 48; фиг. 13С) могут обеспечивать восстановление нарушенного эндотоксином (липополисахаридом) эффероцитоза погибающих нейтрофилов макрофагами человека. Концентрация слитого белка изображена на оси x, уровень эффероцитоза [%] изображен на оси y.
На фигуре 14 изображен эффект слитых белков FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42; фиг. 14A), FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44; фиг. 14B) и FP776 (EGF-HSA-C1-C2; SEQ ID NO: 48; фиг. 14С) в отношении способствования эффероцитозу погибающих клеток Jurkat эндотелиальными клетками (HUVEC). Концентрация слитого белка изображена на оси x, уровень эффероцитоза [%] изображен на оси y.
На фигуре 15 изображено, что однократная доза терапевтических слитых белков FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44), FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42) или FP776 (EGF-HSA-C1-C2; SEQ ID NO: 48) обеспечивает защиту функции почек в модели острого повреждения почек, индуцированного ишемией-реперфузией (AKI). На фиг. 15А изображено, что повышение уровня креатинина в сыворотке крови (sCr) (мг/дл; ось y) снижается при внутрибрюшинном (i. p.) введении 0,16 мг/кг или 0,5 мг/кг FP278 (SEQ ID NO: 44) (ось x). Как изображено на фиг. 15В, внутривенное (i. v.) введение 0,5 мг/кг или 1,5 мг/кг слитого белка FP330 (SEQ ID NO: 42) обеспечивало значительное снижение уровня креатинина в сыворотке крови. На фиг. 15C изображено, что i. v. введение слитого белка FP776 (SEQ ID NO: 48) обеспечивало снижение уровня креатинина в сыворотке крови дозозависимым образом.
На фигуре 16 изображено, что однократная доза терапевтического слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в дозе либо 0,16 мг/кг, либо 0,5 мг/кг приводила к снижению уровня азота мочевины в крови (BUN) в мышиной модели острого повреждения почек.
На фигуре 17 изображено, что однократная доза терапевтического слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) обеспечивает защиту отдаленных органов от ответа острой фазы, запускаемого при AKI, индуцированном ишемией-реперфузией, на основе экспрессии генов маркеров повреждения. На фиг. 17А представлен пример такого индуцированного AKI ответа сывороточного амилоидного белка (SAA) в сердце мыши, а на фиг. 17В представлен пример такого индуцированного AKI ответа (SAA) в легком мыши, оба из которых были в значительной степени заблокированы после однократной i.p. инъекции слитого белка FP278, происходящего из MFGE8 (SEQ ID NO: 44) в дозе 0,16 мг/кг или 0,5 мг/кг/i.p.
На фигуре 18 изображено поглощение контрастного вещества на основе суперпарамагнитного оксида железа (SPIO) (Endorem®) печенью в динамике. Endorem® вводили посредством инъекции внутривенно в виде болюса в течение 1,2 с животным с AKI (через 24 часа после индукции заболевания) или после имитационной операции (животным через 24 ч. после нефрэктомии). У животных с AKI было отмечено значительно сниженное поглощение контрастного вещества печенью (мишень=клетки Купфера) по сравнению с животными при имитационном воздействии. Обработка слитым белком FP776 (EGF-HSA-C1-C2; SEQ ID NO: 48), дозу которого вводили профилактически за 30 мин. до индукции AKI, или дозу которого вводили терапевтически через 5 ч. после индукции повреждения по типу ишемия-реперфузия, обеспечивала защиту от прекращения накопления контрастного вещества в печени мышей с AKI.
Подробное описание изобретения
В данном документе раскрыты терапевтические слитые белки, содержащие интегрин-связывающий домен, PS-связывающий домен и солюбилизирующий домен. В данном документе также раскрыты способы лечения с применением слитых белков по настоящему изобретению, а также анализы, такие как анализ эффероцитоза, применимые для определения характеристик слитых белков.
Определения
Для того чтобы настоящее изобретение можно было легче понять, по ходу подробного описания конкретно приведены определения некоторых терминов. Если не указано иное, все технические и научные термины, используемые в данном документе, имеют то же значение, которое обычно понятно специалистам средней квалификации в данной области техники, к которой относится настоящее изобретение.
Во всех случаях, когда термин "содержать", "содержит", "содержащий" или тому подобные используются в отношении последовательности (например, аминокислотной последовательности), следует понимать, что указанная последовательность также может быть ограничена термином "состоять", "состоит", "состоящий" или тому подобное. Используемая в данном документе выражение "состоящий по сути из" относится к родам или видам активных фармацевтических средств, предусмотренных в способе или композиции, а также любым вспомогательным веществам, не проявляющим активность в отношении предполагаемой цели применения способов или композиций. В некоторых аспектах выражение "состоящий по сути из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от мультиспецифической связывающий молекулы по настоящему изобретению. В некоторых аспектах выражение "состоящий по сути из" однозначно исключает включение одного или нескольких дополнительных активных средств, отличных от мультиспецифической связывающий молекулы по настоящему изобретению, и второго средства, вводимого совместно.
Термин "эффероцитоз", используемый в данном документе, относится к процессу в клеточной биологии, при котором погибающие или мертвые клетки, такие как апоптотические, некротические или состарившиеся клетки, или высокоактивированные клетки, или внеклеточные клеточные везикулы (микрочастицы), или клеточный дебрис, совместно называемые "добычей", удаляются посредством фагоцитоза, т. е. поглощаются фагоцитирующей клеткой и расщепляются. Во время эффероцитоза фагоцитирующие клетки активно прикрепляются и поглощают добычу, образуя внутриклеточные крупные заполненные жидкостью везикулы, содержащие добычу, называемые эфферосомами, которые в результате превращаются в лизосомальный компартмент, в котором инициируется разрушение добычи. Во время апоптоза эффероцитоз обеспечивает удаление погибающих клеток до того, как целостность их мембраны будет нарушена и их содержимое сможет просочиться в окружающие ткани, предупреждая воздействие на окружающие ткани DAMP, таких как токсичные ферменты, оксиданты и другие внутриклеточные компоненты, такие как ДНК, гистоны и протеазы. Профессиональные фагоцитирующие клетки включают клетки миелоидного происхождения, такие как макрофаги и дендритные клетки, однако другие клетки, например стромальные клетки, такие как эпителиальные и эндотелиальные клетки и фибробласты, также могут осуществлять эффероцитоз. Была показана связь между нарушением эффероцитоза и аутоиммунными заболеваниями и повреждением тканей, и наличие нарушения эффероцитоза было продемонстрирован при таких заболеваниях, как муковисцидоз, бронхоэктаз, COPD, астма, идиопатический легочный фиброз, ревматоидный артрит, системная красная волчанка, гломерулонефрит и атеросклероз (Vandivier RW et al (2006) Chest, 129(6): 1673-82). К настоящему времени в клинической практике отсутствует терапия, которая специфически способствует эффероцитозу.
Термин "анализ эффероцитоза", используемый в данном документе и описанный в разделе "Примеры", относится к системе анализа, разработанной для профилирования слитых белков, в которой в качестве фагоцитирующих клеток используются макрофаги человека или эндотелиальные клетки человека (HUVEC). Примерами в данном документе являются анализ эффероцитоза нейтрофилов макрофагами, анализ эффероцитоза клеток Jurkat эндотелиальными клетками или анализ эффероцитоза микрочастиц эндотелиальными клетками. Эти анализы, как более подробно описано в разделе "Примеры", можно использовать для демонстрации того, что биотерапевтические средства, происходящие из MFG-E8, такие как слитые белки по настоящему изобретению, эффективно способствуют эффероцитозу погибающих клеток и микрочастиц макрофагами или эндотелиальными клетками. Кроме того, описанный анализ с использованием нейтрофилов и макрофагов подходит для демонстрации того, что такие соединения по настоящему изобретению могут даже приводить к восстановлению нарушенного посредством LPS или S. aureus эффероцитоза погибающих клеток.
Термины "полипептид" и "белок" используются в данном документе взаимозаменяемо для обозначения полимера из аминокислотных остатков. Данные фразы также применимы к полимерам из аминокислот, в которых один или несколько аминокислотных остатков представляют собой искусственный химический миметик соответствующей встречающейся в природе аминокислоты, а также к полимерам из встречающихся в природе аминокислот и к полимеру из не встречающихся в природе аминокислот. Если не указано иное, конкретная полипептидная последовательность также в неявной форме охватывает ее варианты с консервативными модификациями.
Термин "липкость", используемый в данном документе, в отношении белков по настоящему изобретению относится к результату ошибочной укладки белка, которая способствует слипанию или агрегации белка. Эти нежелательные и нефункциональные эффекты являются результатом поверхностных гидрофобных взаимодействий.
Термин "C-конец", используемый в данном документе, относится к карбоксиконцевой аминокислоте полипептидной цепи, содержащей свободную карбоксильную группу (-СООН). Термин "N-конец", используемый в данном документе, относится к аминоконцевой аминокислоте полипептидной цепи, содержащей свободную аминогруппу (-NH2).
Используемый в данном документе термин ‘слитый белок' относится к белку, содержащему ряд доменов, которые могут не представлять собой полностью природный белок или белок дикого типа, но могут быть ограничены активным доменом всего белка, ответственного за связывание с соответствующим рецептором на поверхности клетки. Слитые белки могут быть получены с применением разработки рекомбинантных белков, где термин "рекомбинантный белок" относится к белку, который был получен, экспрессирован, создан или выделен посредством технологии рекомбинантной ДНК. Тандемное слияние, например, относится к методике, посредством которой белки или белковые домены, представляющие интерес, просто соединяются впритык посредством слияния N- или C-концов между белками. Оно обеспечивает гибкую мостиковую структуру, при этом остается достаточно места между партнерами по слиянию для обеспечения надлежащей укладки. Однако N- или C-концы пептида часто являются основными компонентами для получения требуемого паттерна укладки рекомбинантного белка, в результате чего простое соединение доменов впритык может быть неэффективным. В качестве альтернативы процесс вставки домена включает слияние последовательных белковых доменов посредством кодирования требуемых структур в единую полипептидную цепь, а иногда и вставку домена в другой домен. В обоих этих вышеупомянутых процессах домены "непосредственно соединены" или "соединены непосредственно". Вставку домена часто бывает сложнее осуществить, чем тандемное слияние, вследствие сложности поиска подходящего сайта лигирования в гене, представляющем интерес.
В дополнение к вышеупомянутым методикам слияния с непосредственным соединением, для сохранения функциональности белковых доменов в слитом белке можно использовать внешний линкер. Такой линкер относится к участку аминокислот, который соединяет белковый домен с другим белковым доменом, и упоминается в данном документе как "опосредованный линкер". Таким образом, домены являются "опосредованно соединенными" или "соединенными опосредованно". Например, специалистам средней квалификации в данной области техники понятно, что полипептид, структура которого включает два или более функциональных или организационных домена, часто включает участок аминокислот между такими доменами, который соединяет их друг с другом. Линкер обеспечивает взаимодействие доменов, усиливает стабильность и может снижать стерические затруднения, что часто делает их предпочтительными для применения в разработке сконструированных белков, даже если N- и C-концы могут быть слитыми. В некоторых вариантах осуществления линкер характеризуется тем, что ему свойственно не принимать жесткую трехмерную структуру, а придавать полипептиду гибкость. Различные типы встречающихся в природе линкеров применяли в сконструированных белках, например, шарнирную область иммуноглобулина, которая функционирует в качестве линкера во многих рекомбинантных терапевтических белках, в частности в конструкциях сконструированных антител (Pack P et al., (1995) J. Mol. Biol.,246: 28-34). Помимо природных линкеров, было разработано множество искусственных линкеров, которые можно разделить на три категории: гибкие, жесткие и расщепляемые in vivo линкеры (Yu K et al., (2015) Biotech. Advances, 33(1): 155-64; Chen X et al., (2013) Ad. Drug Delivery Reviews, 65(10): 1357-69). Наиболее широко применяемые гибкие линкерные последовательности представляют собой (Gly)n (Sabourin et al., (2007) Yeast, 24: 39-45) и (Gly4Ser)n (SEQ ID NO: 64) (Huston et al., 1988, 85: 5879-83), где длина линкера может регулироваться количеством копий "n". В некоторых вариантах осуществления полипептид, содержащий линкерный элемент, содержит общую структуру общей формы D1-линкер-D2, при этом D1 и D2 могут быть одинаковыми или разными и представлять собой два домена, ассоциированные друг с другом посредством линкера. В некоторых вариантах осуществления полипептидный линкер составляет по меньшей мере 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95, 100 или больше аминокислот в длину.
Термины "модификация" или "мутация" аминокислотного остатка/положения, используемые в данном документе, относятся к изменению первичной аминокислотной последовательности по сравнению с первоначальной аминокислотной последовательностью, при этом изменение представляет собой результат изменения последовательности, содержащей указанные аминокислотные остатки/положения. Например, типичные модификации включают замену остатка (или замену в указанном положении) другой аминокислотой (например, консервативная или неконсервативная замена), вставку одной или нескольких аминокислот, смежных с указанным остатком/положением, и делецию указанного остатка/положения. Аминокислотная "замена" или ее разновидность относится к замене существующего аминокислотного остатка в предварительно определенной (первоначальной) аминокислотной последовательности другим аминокислотным остатком. Обычно и предпочтительно модификация приводит в результате к изменению по меньшей мере одной физико-биохимической активности вариантного полипептида по сравнению с полипептидом, содержащим первоначальную (или "дикого типа") аминокислотную последовательность.
Термин "консервативно модифицированный вариант" используется в отношении как аминокислотных последовательностей, так и последовательностей нуклеиновой кислоты. Применительно к конкретным последовательностям нуклеиновой кислоты консервативно модифицированные варианты относятся к тем нуклеиновым кислотам, которые кодируют идентичные или по сути идентичные аминокислотные последовательности, или в случае, если нуклеиновая кислота не кодирует аминокислотную последовательность, они относятся к по сути идентичным последовательностям. Вследствие вырожденности генетического кода любой заданный белок кодируется большим количеством функционально идентичных нуклеиновых кислот. Например, все кодоны GCA, GCC, GCG и GCU кодируют аминокислоту аланин. Таким образом, в каждом положении, в котором кодоном задан аланин, кодон может быть изменен на любой из соответствующих описанных кодонов без изменения кодируемого полипептида. Такие разновидности нуклеиновой кислоты представляют собой "молчащие разновидности", являющиеся одним из видов разновидностей с консервативными модификациями. Любая последовательность аминокислот в данном документе, которая кодирует полипептид, также описывает каждую возможную молчащую разновидность нуклеиновой кислоты. Специалисту в данной области техники будет понятно, что каждый кодон в нуклеиновой кислоте (за исключением AUG, который обычно является единственным кодоном для метионина, и TGG, который обычно является единственным кодоном для триптофана) может быть модифицирован с получением функционально идентичной молекулы. Соответственно, каждая молчащая разновидность нуклеиновой кислоты, которая кодирует полипептид, неявно определена в каждой описанной последовательности.
Применительно к полипептидным последовательностям варианты с консервативными модификациями включают отдельные замены, делеции или добавления в полипептидную последовательность, которые приводят к замене аминокислоты химически сходной аминокислотой. Таблицы консервативных замен, дающие функционально сходные аминокислоты, известны из уровня техники. Такие варианты с консервативными модификациями дополняют и не исключают полиморфные варианты, межвидовые гомологи и аллели. Следующие восемь групп содержат аминокислоты, которые являются консервативными заменами друг для друга: 1) аланин (A), глицин (G); 2) аспарагиновая кислота (D), глутаминовая кислота (E); 3) аспарагин (N), глутамин (Q); 4) аргинин (R), лизин (K); 5) изолейцин (I), лейцин (L), метионин (M), валин (V); 6) фенилаланин (F), тирозин (Y), триптофан (W); 7) серин (S), треонин (T) и 8) цистеин (C), метионин (M) (см., например, Creighton, Proteins (1984)). В некоторых вариантах осуществления выражение "консервативные модификации последовательности" используется для обозначения аминокислотных модификаций, которые не оказывают значительного влияния на характеристики связывания связывающих доменов сконструированных белков по настоящему изобретению или не изменяют их.
Термин "белковый вариант" или "вариант белка", упоминаемый в данном документе, относится к белку, предусматривающему разновидность, в которой одна или несколько, например 2, 3, 4, 5, 6, 7, 8, 9, 10, аминокислот были модифицированы. Термин "функциональный вариант" белка, упоминаемый в данном документе, относится к белковому варианту, содержащему модификацию, которая приводит в результате к изменению аминокислотной последовательности, но не приводит к изменению общих свойств белка или его функции. Термин "усеченный вариант" белка, упоминаемый в данном документе, относится к укороченной версии белка, однако при этом укороченная версия белка сохраняет функцию родительского белка. Для определения того, не характеризуется ли функциональный вариант или усеченный вариант изменением общего свойства или функции, эти вариантные белки можно протестировать в сравнении с полноразмерным или немодифицированным родительским белком в отношении их эффекта в ряде анализов, как описано в настоящем изобретении. Например, стимуляция эффероцитоза эндотелиальными клетками в анализе эффероцитоза клеток Jurkat эндотелиальными клетками человека, восстановление нарушенного эффероцитоза макрофагами в анализе эффероцитоза нейтрофилов макрофагами человека, снижение количества микрочастиц плазмы крови за счет клиренса в анализе эффероцитоза микрочастиц эндотелиальными клетками человека, и/или обеспечение защиты от полиорганного повреждения в модели острой ишемии почек.
Термины "процентная идентичность" или "процентная идентичность последовательности" в контексте двух или более нуклеиновых кислот или полипептидных последовательностей относятся к двум или более последовательностям или подпоследовательностям, которые являются одинаковыми. Две последовательности являются "по сути идентичными" и характеризуются "идентичностью последовательностей", если две последовательности характеризуются определенной процентной долей аминокислотных остатков или нуклеотидов, которые являются одинаковыми (т. е. характеризуются по меньшей мере 60%, необязательно по меньшей мере 65%, 70%, 75%, 80%, 85%, 90%, 95%, 96%, 97%, 98% или 99% идентичностью по определенной области или, если она не указана, по всей последовательности) при сравнении и выравнивании для достижения максимального соответствия в окне сравнения или определенной области, например, при измерении с применением одного из следующих алгоритмов сравнения последовательностей или с помощью выравнивания в ручном режиме и визуальной оценки. Необязательно, идентичность существует в области, длина которой составляет по меньшей мере приблизительно 50 нуклеотидов (или 10 аминокислот), или в области, длина которой составляет от 100 до 500, или 1000, или 2000, или 3000 или более нуклеотидов, или в качестве альтернативы длина которой составляет от 30 до 200, или 300, или 500, или 700, или 800, или 900, или 1000 или более аминокислот.
При сравнении последовательностей в типичном случае одна последовательность выступает в качестве эталонной последовательности, с которой сравнивают тестируемые последовательности. При использовании алгоритма сравнения последовательностей тестируемую и эталонную последовательности вводят в компьютер, при необходимости устанавливают координаты подпоследовательностей и устанавливают программные параметры алгоритма для анализа последовательностей. Могут применяться программные параметры по умолчанию или можно устанавливать альтернативные параметры. На основании программных параметров алгоритм сравнения последовательностей затем рассчитывает значения процента идентичности последовательностей для тестируемых последовательностей относительно эталонной последовательности.
Термин "окно сравнения", используемый в данном документе, предусматривает ссылку на сегмент из любого количества смежных положений нуклеиновых кислот или аминокислот, выбранного из группы, состоящей из от 20 до 600, обычно от приблизительно 50 до приблизительно 200, чаще от приблизительно 100 до приблизительно 150, в котором последовательность можно сравнивать с эталонной последовательностью с таким же количеством смежных положений, после того как две последовательности подвергли оптимальному выравниванию. Способы выравнивания последовательностей для проведения сравнения известны из уровня техники. Оптимальное выравнивание последовательностей для сравнения можно проводить, например, с помощью алгоритма поиска локальной гомологии Смита-Уотермана (1970) Adv. Appl. Math. 2:482c, с помощью алгоритма выравнивания областей гомологии Нидлмана-Вунша (1970) J. Mol. Biol., 48: 443, с помощью способа поиска сходства Пирсона-Липмана (1988) PNAS USA, 85: 2444, с помощью компьютерных реализаций этих алгоритмов (GAP, BESTFIT, FASTA и TFASTA в составе пакета программного обеспечения Wisconsin Genetics, Genetics Computer Group, 575 Science Dr., Мэдисон, Висконсин) или с помощью выравнивания в ручном режиме и визуальной оценки (см., например, Brent et al., (2003) Current Protocols in Molecular Biology).
Двумя примерами алгоритмов, которые являются подходящими для определения процента идентичности последовательностей и сходства последовательностей, являются алгоритмы BLAST и BLAST 2.0, которые описаны в Altschul et al., (1977) Nuc. Acids Res,. 25: 3389-3402; и Altschul et al., (1990) J. Mol. Biol., 215: 403-410 соответственно. Программное обеспечение для осуществления анализов BLAST доступно для общего пользования в Национальном центре биотехнологической информации.
Алгоритм BLAST также осуществляет статистический анализ сходства между двумя последовательностями (см., например, Karlin & Altschul, (1993) PNAS. USA, 90: 5873-5787). Одной мерой сходства, предусмотренной в алгоритме BLAST, является наименьшая суммарная вероятность (P(N)), которая указывает на вероятность, с которой совпадение между двумя нуклеотидными или аминокислотными последовательностями возникло случайно. Например, нуклеиновая кислота считается сходной с эталонной последовательностью, если наименьшая суммарная вероятность при сравнении тестируемой нуклеиновой кислоты с эталонной нуклеиновой кислотой составляет менее приблизительно 0,2, более предпочтительно менее приблизительно 0,01 и наиболее предпочтительно менее приблизительно 0,001.
Процент идентичности двух аминокислотных последовательностей также можно определить с использованием алгоритма согласно E. Meyers и W. Miller (Comput. Appl. Biosci. 4:11-17 (1988)), который был включен в программу ALIGN (версия 2.0), с применением таблицы весов замен остатков PAM120, штрафа за продление гэпа, составляющего 12, и штрафа за открытие гэпа, составляющего 4. Кроме того, процент идентичности между двумя аминокислотными последовательностями можно определить с применением алгоритма Нидлмана-Вунша (выше), который был включен в программу GAP в пакете программного обеспечения GCG (доступного на www.gcg.com), с использованием либо матрицы Blossom 62, либо матрицы PAM250, а также штрафа за открытие гэпа 16, 14, 12, 10, 8, 6 или 4 и штрафа за продолжение гэпа 1, 2, 3, 4, 5 или 6.
Полипептид, как правило, является по сути идентичным второму полипептиду, например, если два пептида отличаются только консервативными заменами. Другим показателем того, что две последовательности нуклеиновой кислоты являются по сути идентичными, является то, что две молекулы или их комплементарные цепи гибридизируются друг с другом в жестких условиях.
Термин "нуклеиновая кислота" используется в данном документе взаимозаменяемо с термином "полинуклеотид" и относится к дезоксирибонуклеотидам или рибонуклеотидам и их полимерам либо в однонитевой, либо в двухнитевой форме. Термин охватывает нуклеиновые кислоты, содержащие известные аналоги нуклеотидов или модифицированные остатки или связи в остове, которые являются синтетическими, встречающимися в природе и не встречающимися в природе, которые характеризуются свойствами связывания, сходными с таковыми у эталонной нуклеиновой кислоты, и которые метаболизируются способом, сходным с таковым для эталонных нуклеотидов. Примеры таких аналогов включают без ограничения фосфоротиоаты, фосфорамидаты, метилфосфонаты, хиральные метилфосфонаты, 2-O-метилрибонуклеотиды, пептидонуклеиновые кислоты (PNA).
Если не указано иное, конкретная последовательность нуклеиновой кислоты также потенциально охватывает ее варианты с консервативными модификациями (например, замены вырожденными кодонами) и комплементарные последовательности, а также непосредственно указанную последовательность. В частности, замены, приводящие к вырожденным кодонам, могут достигаться посредством образования последовательностей, в которых в третьем положении одного или нескольких выбранных (или всех) кодонов находится замена на смешанное основание и/или остатки дезоксиинозина (Batzer et al., (1991) Nucleic Acid Res., 19: 5081; Ohtsuka et al., (1985) J Biol Chem., 260: 2605-2608; и Rossolini et al., (1994) Mol Cell Probes, 8: 91-98). Термин "оптимизированная нуклеотидная последовательность", используемый в данном документе, означает, что данная нуклеотидная последовательность была изменена таким образом, чтобы она кодировала аминокислотную последовательность с использованием кодонов, которые являются предпочтительными в продуцирующей клетке, например клетке яичника китайского хомячка (CHO). Оптимизированную нуклеотидную последовательность конструируют таким образом, чтобы полностью сохранить аминокислотную последовательность, изначально кодируемую первоначальной нуклеотидной последовательностью, которая также известна как "родительская" последовательность. В конкретных вариантах осуществления оптимизированные последовательности в контексте данного документа были сконструированы так, чтобы они содержали кодоны, которые являются предпочтительными в клетках млекопитающих, представляющих собой CHO.
Терапевтические слитые белки
Интегрин-связывающие домены
Интегрины представляют собой трансмембранные рецепторы, которые способствуют адгезии клеток и внеклеточного матрикса (ECM). После связывания лиганда интегрины активируют пути передачи сигнала, которые опосредуют клеточные сигналы, такие как регуляция клеточного цикла, организация внутриклеточного цитоскелета и перемещение новых рецепторов к клеточной мембране (Giancotti & Ruoslahti (1999) Science, 285 (5430): 1028-32). Присутствие интегринов позволяет осуществлять быстрые и гибкие ответы на события на клеточной поверхности. Существует несколько типов интегринов, и одна клетка может содержать на своей поверхности несколько разных типов. Интегрины содержат две субъединицы: α (альфа) и β (бета), каждая из которых проходит через плазматическую мембрану и содержит несколько цитоплазматических доменов (Nermut MV et al (1988). EMBO J., 7 (13): 4093-9). Кислая аминокислота присутствует в сайте взаимодействия с интегрином многих белков ЕСМ, например, в виде части аминокислотной последовательности аргинин-глицин-аспарагиновая кислота ("RGD" в однобуквенном коде аминокислоты). Мотив RGD был обнаружен во многих белках матрикса, таких как фибронектин, фибриноген, витронектин и остеопонтин, и облегчает адгезию клеток. Мотив RGD обнаружен в ряде белков в консервативном белковом домене, известном как EGF-подобный домен, который получил свое название от эпидермального фактора роста, в котором он был впервые описан. EGF-подобный домен представляет собой один из наиболее распространенных доменов, обнаруживаемых во внеклеточных белках (Hidai C (2018) Open Access J Trans Med Res., 2(2): 67-71), и некоторые примеры EGF-подобных доменов, которые содержат мотив связывания RGD, приведены ниже в таблице 1.
Таблица 1. Примеры белков, содержащих EGF-подобный домен, и белков, содержащих мотив RGD интегрина
Термин "интегрин-связывающий домен", используемый в данном документе, относится к участку аминокислот или белковому домену, которые характеризуются функцией связывания с интегринами. В одном варианте осуществления настоящего изобретения термин "интегрин-связывающий домен", используемый в данном документе, относится к участку аминокислот или белковому домену, который характеризуется функцией связывания с интегринами и содержит мотив RGD. В варианте осуществления настоящего изобретения интегрин-связывающий домен представляет собой EGF-подобный домен из MFG-E8 человека, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 2. В альтернативном варианте осуществления настоящего изобретения интегрин-связывающий домен представляет собой EGF-подобный домен из EDIL3 человека (любую из следующих последовательностей: SEQ ID NO: 11, SEQ ID NO: 77, SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100 или SEQ ID NO: 101); например, где EGF-подобные домены могут находиться в пределах участка аминокислот 1-132 из SEQ ID NO: 11.
Термин "связывается с интегринами", используемый в данном документе, относится к интегрин-связывающей активности. Интегрин-связывающую активность можно определить способами, хорошо известными из уровня техники. Например, анализ опосредованной интегрином адгезии описан в разделе "Примеры", раздел 3.2, в котором определяли адгезию флуоресцентно меченных клеток лимфомы, экспрессирующих интегрин αvβ3, в отношении терапевтических слитых белков по настоящему изобретению. Считается, что интегрин-связывающий домен характеризуется интегрин-связывающей активностью, если он характеризуется наличием по меньшей мере 10%, например по меньшей мере 25%, по меньшей мере 50%, по меньшей мере 75%, более предпочтительно по меньшей мере 80%, например по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% интегрин-связывающей активности, наблюдаемой для белка MFG-E8 человека (SEQ ID NO:1), при тестировании посредством того же способа определения соответствующей активности, предпочтительно при тестировании с применением анализа, описанного в разделе "Примеры", раздел 3.2.
Фосфатидилсерин-связывающие домены
Термин "фосфатидилсерин" (PS), используемый в данном документе, относится к фосфолипиду, который представляет собой компонент клеточной мембраны. PS главным образом приурочен к внутреннему листку клеточной мембраны, в то время как фосфатидилхолин и сфингомиелин локализованы в основном во внешнем листке. Асимметричное распределение фосфолипидов поддерживается действием флиппаз (P4-АТФаз, таких как ATP11A и 11C) в плазматической мембране для активной транслокации PS из наружного листка во внутренний листок. Представление PS на клеточной поверхности наблюдается не только в апоптотических клетках, но и в активированных лимфоцитах, активированных тромбоцитах, состарившихся эритроцитах и некоторых раковых клетках и соответствующих микрочастицах (Sakuragi et al., (2019) PNAS USA, 116(8): 2907-12). Представление PS может представлять собой биомаркер протромботического, воспалительного или ишемического заболевания (Pasalic et al., (2018) J Thromb Haemost., 16(6): 1198-2010; Ma et al., (2017), выше; Zhao et al., (2016), выше. PS функционирует во множестве путей передачи сигналов в клетках и в качестве незаменимого фосфолипида при свертывании крови, где он может выступать в качестве усилителя образования комплексов теназы (факторы IXa, VIIIa и X) и протромбиназы (факторы Xa, Va и протромбин) (Spronk et al., (2014) Thromb Res. 133 (Suppl 1): S54-6). Возможно, наиболее изученной функцией экстернализованного PS по-прежнему является выполнение роли маркера "съешь меня" для фагоцитирующих клеток, таких как макрофаги, для поглощения апоптотических клеток, клеточного дебриса или активированных клеток, представляющих PS. Термин "фосфатидилсерин-связывающий домен" или "PS-связывающий домен", используемый в данном документе, относится к участку аминокислот или белковому домену, который характеризуется функцией связывания с PS. Примеры эндогенных белков с PS-связывающими доменами можно найти в таблице 2 ниже.
Таблица 2. Примеры рецепторов/белков с фосфатидилсерин-связывающими доменами
В варианте осуществления настоящего изобретения PS-связывающий домен получен из MFG-E8 человека, содержащего аминокислотную последовательность, представленную под SEQ ID NO: 3 или SEQ ID NO: 76. В альтернативном варианте осуществления настоящего изобретения интегрин-связывающий домен представляет собой PS-связывающий домен из EDIL3 человека (SEQ ID NO: 11), где PS-связывающий домен содержит аминокислоты 135-453 из SEQ ID NO: 11.
PS-связывающую активность можно определить способами, хорошо известными из уровня техники. Например, анализ связывания PS описан в разделе "Примеры", раздел 3.1, при этом связывание слитых белков по настоящему изобретению с PS, нанесенным на микротитрационные планшеты, оценивали с использованием конкуренции за связывание с биотинилированным мышиным MFG-E8. В соответствии с настоящим изобретением считается, что PS-связывающий домен характеризуется PS-связывающей активностью, если он характеризуется наличием по меньшей мере 10%, например по меньшей мере 25%, по меньшей мере 50%, по меньшей мере 75%, по меньшей мере 80%, предпочтительно по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% PS-связывающей активности, наблюдаемой для белка MFG-E8 человека, представленного под SEQ ID NO:1, при тестировании посредством того же способа определения соответствующей активности, предпочтительно при тестировании с применением анализа, описанного в разделе "Примеры", раздел 3.1.
Мостиковые белки
Существует ряд эндогенных белков, которые содержат как интегрин-связывающий домен, так и PS-связывающий домен. Примеры таких "мостиковых белков" показаны в таблице 3 ниже.
Таблица 3. Мостиковые белки, содержащие как интегрин, так и фосфатидилсерин-связывающие домены
(DEL-1)
(αv- β2)
Для обеспечения терапевтической ценности, полезно, чтобы мостиковый белок содержал интегрин-связывающий домен, который распознает интегрины на фагоцитах, которые обычно не чувствительны к протеолитическому расщеплению или отщеплению, как это наблюдалось у членов семейства ТАМ или других PS-связывающих рецепторов. Было показано, что PS-связывающий домен и интегрин-связывающий домен, например MFG-E8 или его паралог EDIL3/DEL1, индуцирует эффероцитоз in vitro и, таким образом, может характеризоваться терапевтической ценностью в качестве индукторов эффероцитоза при AOI. В отличие от этого, белок GAS6, например, может не быть особенно эффективным в стимулировании эффероцитоза при AOI, поскольку его рецептор на фагоцитах (MerTK) протеолитически расщепляется во время воспаления и инфекции, как указано выше.
Одним из примеров мостикового белка, перечисленных в таблице 3 выше, является MFG-E8, который представляет собой один из основных белков, обнаруживаемых в мембране жировых глобул молока (MFGM). MFG-E8 экспрессируется и секретируется несколькими различными типами клеток (например, эпителиальными клетками молочной железы, клетками сосудов, эпителиальными клетками придатка яичка, гладкомышечными клетками аорты, активированными макрофагами, стимулированным эндометрием и незрелыми дендритными клетками) и тканями (например, сердца, легких, молочных желез, селезенки, кишечника, печени, почек, головного мозга, крови и эндотелия). Белок MFG-E8 также известен под несколькими различными названиями, такими как лактадгерин, BP47, компоненты 15/16, MFGM, MGP57/53, гликопротеин PAS-6/PAS-7, белок клеточной стенки SED1, поверхностный белок спермы SP47, антиген BA46 эпителия молочной железы и О-ацетил-ганглиозид-GD3-синтаза (AGS). Ген MFG-E8 расположен на хромосоме 1 у крыс, на хромосоме 7 у мышей и на хромосоме 15 у людей. Альтернативный сплайсинг пре-мРНК MFG-E8 приводит в результате к образованию трех изоформ белка человека и двух форм мРНК, длинные и короткие варианты экспрессируются в молочных железах мыши. Ген MFG-E8 человека (UniProtKB - Q08431) кодирует белок, длина которого составляет 387 остатков, который процессируется с образованием нескольких белковых продуктов. Аминокислотная последовательность MFG-E8 человека, которая содержит сигнальный пептид (остатки 1-23; подчеркнуты), EGF-подобный домен (остатки 24-67; выделены курсивом), домен C1 (остатки 70-225; выделены жирным) и домен C2 (остатки 230-387; выделены жирным и подчеркнуты), представлена ниже.
MPRPRLLAAL CGALLCAPSL LVA LDICSKN PCHNGGLCEE ISQEVRGDVF PSYTCTCLKG YAGNHCETKC VEPLGLENGN IANSQIAASS VRVTFLGLQH WVPELARLNR AGMVNAWTPS SNDDNPWIQV NLLRRMWVTG VVTQGASRLA SHEYLKAFKV AYSLNGHEFD FIHDVNKKHK EFVGNWNKNA VHVNLFETPV EAQYVRLYPT SCHTACTLRF ELLGCELNGC ANPLGLKNNS IPDKQITASS SYKTWGLHLF SWNPSYARLD KQGNFNAWVA GSYGNDQWLQ VDLGSSKEVT
GIITQGARNF GSVQFVASYK VAYSNDSANW TEYQDPRTGS SKIFPGNWDN
HSHKKNLFET PILARYVRIL PVAWHNRIAL RLELLGC (SEQ ID NO: 1).
MFG-E8 характеризуется отсутствием трансмембранной функции, которая характерна для MFGM, и поэтому выступает в качестве периферического мембранного белка. MFG-E8 человека состоит из одного N-концевого EGF-подобного домена (SEQ ID NO: 2), который связывается с интегринами αvβ3 и αvβ5, экспрессируемыми на фагоцитах, и PS-связывающего домена (SEQ ID NO: 3), содержащего два субдомена F5/8-дискоидина (C1 и C2), которые с высокой аффинностью связываются с анионными фосфолипидами. Связывание интегрина происходит в результате наличия мотива RGD, расположенного в остатках 46-48 MFG-E8 человека (SEQ ID NO: 1). Апоптотические клетки, клеточный дебрис, гиперактивированные клетки и большинство микрочастиц (MP) представляют PS и являются мишенями для MFG-E8, который, действуя в качестве мостиковой молекулы, обеспечивает опсонизацию этих клеток и микрочастиц и их соединение с интегринами αvβ3 и αvβ5 на фагоцитах. Это действие в качестве мостика запускает эффективную программу поглощения, ведущую к интернализации клеток, дебриса и микрочастиц. Белки, обнаруживаемые в MFGM, являются высококонсервативными у всех видов. Структура белка MFG-E8 зависит от вида; все известные в настоящее время виды содержат два домена C, но различаются по количеству EGF-подобных доменов. Например, белок MFG-E8 человека содержит один EGF-подобный домен, в то время как бычий MFG-E8 и мышиный MFG-E8 (SEQ ID NO: 68) содержат два EGF-подобных домена, а у курицы, лягушки и данио-рерио содержится три EGF-подобных домена. Домены MFG-E8 ранее были предложены в качестве составных частей терапевтических средств, в частности описано, что PS-связывающие домены (Kooijmans et al., (2018) Nanoscale, 10(5): 2413-2426) и фрагменты MFG-E8 действуют в моделях фиброза (заявка на патент США US 2018/0334486).
Не связанное с воспалением поглощение погибающих клеток, дебриса и микрочастиц профессиональными и непрофессиональными фагоцитами играет решающую роль в гомеостазе после повреждения ткани (Greenlee-Wacker (2016), выше). Важность надлежащего клиренса стала еще более очевидной в генетических моделях, где мыши с нокаутом MFG-E8 демонстрировали, например, повышенное количество (не подвергшихся клиренсу) погибающих клеток в тканях, повышенный воспалительный ответ в моделях заболеваний, таких как неонатальный сепсис, аутоиммунитет, слабый ангиогенез и нарушение заживления ран (Hanayama et al., (2004) Science, 204(5474): 1147-50; Das et al., (2016) J Immunol., 196(12): 5089-5100; Hansen et al., (2017) J Pediatr Surg., 52(9): 1520-7).
В дополнение, было показано, что MFG-E8 создает толерогенную среду посредством подавления активации и пролиферации Т-клеток, ингибирования субпопуляций Th1, Th2 и Th17 при одновременном обеспечении повышения в отношении субпопуляций регуляторных Т-клеток (Treg). Интересно, что Treg в свою очередь способствуют разрешению воспаления посредством индуцирования эффероцитоза макрофагами (Proto et al., (2018) Immunity, 49(4): 666-77). Было описано, что MFG-E8 способствует аллогенному приживлению тканей, происходящих из эмбриональных стволовых клеток, в обход барьера MHC (Tan et al., (2015) Stem Cell Reports, 5(5): 741-752). Для MFG-E8 также имеется множество путей применения в области питания, которые способствуют развитию тканей и защите от возбудителей инфекций. Гликопротеины, такие как MFG-E8, представляют собой потенциальные нутрицевтики, способствующие улучшению здоровья, для пищевых и фармацевтических областей применения. MFG-E8 также можно комбинировать с другими питательными веществами (например, пробиотиками, мицеллами сывороточного белка, альфа-гидроксиизокапроновой кислотой, цитруллином и жирными кислотами с разветвленной цепью).
Солюбилизирующий домен
Как описано в данном документе, терапевтические слитые белки по настоящему изобретению содержат интегрин-связывающий домен и PS-связывающий домен. Кроме того, слитые белки также содержат дополнительный домен, который сообщает слитому белку ряд требуемых свойств. Этот дополнительный домен, который для целей настоящей заявки был назван "солюбилизирующим доменом", сообщает улучшенные биологические свойства, такие как повышенная растворимость, сниженная агрегация и повышенная биоактивность. В результате слитый белок демонстрирует требуемые фармакокинетические профили. Кроме того, присутствие солюбилизирующего домена улучшает стабильность терапевтического слитого белка и приводит в результате к улучшению экспрессии слитого белка по сравнению с белком дикого типа в клеточных системах экспрессии, о чем свидетельствует увеличение выхода после очистки.
Присутствие солюбилизирующего домена может также сообщать терапевтическому слитому белку продленное время полужизни. Например, многие белковые лекарственные средства соединены с полиэтиленгликолем (PEG), reCODE PEG, каркасом антитела, полисиаловой кислотой (PSA), гидроксиэтилкрахмалом (HES) и сывороточными белками, такими как альбумин, IgG и FcRn, для продления их полужизни в плазме крови и для достижения усиленных терапевтических эффектов (Kim et al., (2010) J Pharmacol Exp Ther., 334: 682-92; Weimer et al., (2008) Thromb Haemost. 99: 659-67; Dumont et al., (2006) BioDrugs, 20: 151-60; Schellenberger et al., (2009) Nat Biotechnol., 27: 1186-90).
В некоторых вариантах осуществления солюбилизирующий домен представляет собой белок альбумин, такой как сывороточный альбумин человека (HSA; SEQ ID NO: 4) или его варианты. Например, HSA, содержащий аминокислотную замену C34S для снижения склонности к агрегации (SEQ ID NO: 5), или домены HSA, такие как D3 HSA; (SEQ ID NO: 6). HSA характеризуется очень длительным временем полужизни в сыворотке крови вследствие ряда факторов, в том числе его относительно большого размера, который обеспечивает снижение уровня почечной фильтрации, и его способности связываться с неонатальным Fc-рецептором (FcRn), что позволяет ему избегать внутриклеточного разрушения. Также было предложено использование N-концевых фрагментов HSA для продуктов слияния с полипептидами (например, патентная заявка EP399666). Соответственно, генетическое или химическое слияние или конъюгирование молекул с альбумином может обеспечить их стабилизацию, или продление срока хранения, и/или сохранение активности молекулы в течение продленных периодов времени в растворе, in vitro и/или in vivo. Дополнительные способы, относящиеся к продуктам слияния с HSA, можно найти, например, в международных патентных заявках WO 2001/077137 и WO 2003/060071.
В некоторых вариантах осуществления солюбилизирующий домен содержит Fc-домен антитела, такой как Fc-иммуноглобулин G1 человека (Fc-IgG1; SEQ ID NO: 7). Fc-домен также можно модифицировать, например, с применением модификаций на основе "выступ во впадину" (KiH) для улучшения гетеродимеризации Fc посредством введения комплементарных аминокислотных замен в CH3-домен Fc. Например, замена T366W для создания "выступа" в одном CH3-домене и замены T366S, L368A и Y407V для создания "впадины" в другом CH3-домене (Merchant et al (1998) Nat. Biotechnol., 16(7): 677-81; нумерация IgG1 согласно EU). Дополнительные модификации, которые могут быть включены в Fc-домен отдельно или в комбинации с модификациями для улучшения гетеродимеризации, могут включать, например, аминокислотные замены на цистеин для создания дополнительной цистеиновой связи, например S354C и/или Y349C, и аминокислотные замены для снижения или устранения связывания с рецепторами Fcγ и белком комплемента C1q для обеспечения "сайленсинга" иммунной эффекторной функции. Так называемая двойная мутация "LALA" (L234A вместе с L235A; нумерация согласно EU) приводит в результате к ослаблению эффекторных функций (Lund et al., (1992) Mol Immunol., 29: 53-9). В качестве альтернативы двойная мутация "DAPA" (D265A вместе с P329A; нумерация согласно ЕС) приводит в результате к ослаблению эффекторных функций. В варианте осуществления настоящего изобретения Fc-домен может содержать аминокислотные замены D265A, P329A для обеспечения сайленсинга Fc и/или аминокислотные замены KiH T366W (выступ) или T366S, L368A и Y407V (впадина). В одном варианте осуществления Fc-домен получен из IgG1 человека и содержит аминокислотные замены D265A, P329A (SEQ ID NO: 8). В другом варианте осуществления Fc-домен получен из IgG1 человека и содержит аминокислотные замены D265A, P329A, S354C и аминокислотную замену T366W (Fc-IgG1-выступ; SEQ ID NO: 9). В другом варианте осуществления Fc-домен получен из IgG1 человека и содержит аминокислотные замены D265A, P329A, Y349C и аминокислотные замены T366S, L368A и Y407V (Fc-IgG1-впадина; SEQ ID NO: 10).
В некоторых вариантах осуществления солюбилизирующий домен содержит Fc-домен антитела, полученный из IgA, IgD, IgE или IgM человека.
В некоторых вариантах осуществления солюбилизирующий домен содержит SUMO (малый убиквитин-подобный модификатор), убиквитин, GST (глутатион-S-трансферазу) или их варианты.
Соединение и ориентация доменов терапевтических слитых белков
Интегрин-связывающий домен, PS-связывающий домен и солюбилизирующий домен слитых белков по настоящему изобретению являются соединенными. Термины "соединенный" или "соединяющий", используемые в данном документе, относится к одному домену слитого белка, присоединенному, непосредственно или опосредованно, к другому домену слитого белка. Непосредственное присоединение представляет собой форму соединения и упоминается в данном документе как "слитый" или "слияние". Используя в качестве примера молекулу, характеризующуюся формой ABC: домен A соединен непосредственно с доменом B и соединен непосредственно с доменом C. Таким образом, домен A также может быть описан как слитый с доменом B, который слит с доменом C. В качестве другого примера, домен A соединен непосредственно с доменом B и соединен опосредованно с доменом C. Таким образом, домен A также может быть описан как слитый с доменом B, который соединен с доменом C опосредованно с помощью внутреннего линкера.
В некоторых вариантах осуществления соединение представляет собой непосредственное соединение, и домены таким образом, являются слитыми друг с другом. В некоторых вариантах осуществления интегрин-связывающий домен слит с PS-связывающим доменом, который слит с солюбилизирующим доменом. В частности, PS-связывающий домен (например, субдомены C1-C2 дискоидина) слит с С-концом интегрин-связывающего домена (например, EGF-подобного домена) и слит с N-концом солюбилизирующего домена (например, HSA). В некоторых вариантах осуществления солюбилизирующий домен слит с интегрин-связывающим доменом, который слит с PS-связывающим доменом. В частности, интегрин-связывающий домен (например, EGF-подобный домен) слит с С-концом солюбилизирующего домена (например, HSA) и слит с N-концом PS-связывающего домена (например, субдомены C1-C2 дискоидина). В некоторых вариантах осуществления интегрин-связывающий домен слит с PS-связывающим доменом, содержащим субдомены C1-C2 дискоидина, и солюбилизирующий домен вставлен между субдоменами C1-C2 дискоидина. В частности, С-конец интегрин-связывающего домена (например, EGF-подобного домена) слит с N-концом субдомена С1 дискоидина, С-конец субдомена С1 дискоидина слит с N-концом солюбилизирующего домена (например, HSA) и С-конец солюбилизирующего домена слит с N-концом субдомена С2 дискоидина. В другом варианте осуществления интегрин-связывающий домен слит с солюбилизирующим доменом, который слит с PS-связывающим доменом. В частности, солюбилизирующий домен (например, HSA) слит с С-концом интегрин-связывающего домена (например, EGF-подобного домена) и с N-концом PS-связывающего домена (например, субдоменов C1-C2 дискоидина). В одном варианте осуществления HSA слит с С-концом EGF-подобного домена и слит с N-концом домена C1 дискоидина.
В некоторых вариантах осуществления слияние солюбилизирующего домена (например, HSA) осуществлено между интегрин-связывающим доменом и PS-связывающим доменом. В некоторых вариантах осуществления интегрин-связывающий домен расположен на N-конце слитого белка, а PS-связывающий домен расположен на С-конце слитого белка.
В некоторых вариантах осуществления слитый белок содержит первую область, содержащую интегрин-связывающий домен, например EGF-подобный домен, вторую область, содержащую солюбилизирующий домен (например, HSA), и третью область, содержащую PS-связывающий домен, например домен С1 и/или С2 дискоидина. В некоторых вариантах осуществления интегрин-связывающий домен расположен на N-конце слитого белка, а PS-связывающий домен расположен на С-конце слитого белка.
В некоторых вариантах осуществления слияние солюбилизирующего домена (например, HSA) осуществлено между интегрин-связывающим доменом и PS-связывающим доменом. В некоторых вариантах осуществления интегрин-связывающий домен расположен на N-конце слитого белка, а PS-связывающий домен расположен на С-конце слитого белка.
В некоторых вариантах осуществления слитый белок содержит первую область, содержащую интегрин-связывающий домен, например EGF-подобный домен, вторую область, содержащую солюбилизирующий домен (например, HSA или Fc), и третью область, содержащую PS-связывающий домен, например домен С1 и/или С2 дискоидина. В некоторых вариантах осуществления интегрин-связывающий домен расположен на N-конце слитого белка, а PS-связывающий домен расположен на С-конце слитого белка.
В некоторых вариантах осуществления солюбилизирующий домен представляет собой HSA.
В некоторых вариантах осуществления солюбилизирующий домен представляет собой Fc антитела, представляющего собой иммуноглобулин G1 (Fc-IgG1; SEQ ID NO: 7).
В некоторых вариантах осуществления солюбилизирующий домен (например, HSA) представляет собой HSA, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 5 или его функциональный вариант.
В предпочтительном варианте осуществления HSA, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 5, слит с С-концом EGF-подобного домена MFG-E8 и слит с N-концом PS-связывающего домена MFG-E8. В одном варианте осуществления слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 46 (FP068). В одном варианте осуществления слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 48 (FP776).
В альтернативном варианте осуществления HSA, содержащий аминокислотную последовательность, представленную под SEQ ID NO: 5, слит с С-концом EGF-подобного домена EDIL3 и слит с N-концом PS-связывающего домена EDIL3. В одном варианте осуществления слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 70 (FP1068). В одном варианте осуществления слитый белок содержит аминокислотную последовательность, представленную под SEQ ID NO: 69 (FP1776).
В некоторых вариантах осуществления соединение осуществляется с помощью полипептидного линкера, и полипептидный линкер, который, например, соединяет солюбилизирующий домен с PS-связывающим доменом в слитом белке по настоящему изобретению, обозначается как "внешний линкер". Эти внешние линкеры обычно содержат глицин (G) и/или серин (S) и могут также содержать глицин и лейцин (GL) или глицин и валин (GL). В некоторых вариантах осуществления линкер содержит несколько остатков G и S, например G2S и их множества, как например (G2S)4, представленный под SEQ ID NO: 62, (GS)4, представленный под SEQ ID NO: 63, G4S, представленный под SEQ ID NO: 64, или (G4S)2, представленный под SEQ ID NO: 65.
В некоторых вариантах осуществления слияние внешнего линкера осуществлено между С-концом интегрин-связывающего домена и N-концом солюбилизирующего домена. В частности, внешний линкер слит с С-концом EGF-подобного домена и N-концом HSA. В некоторых вариантах осуществления слияние внешнего линкера осуществлено между С-концом солюбилизирующего домена и N-концом PS-связывающего домена. В частности, внешний линкер слит с С-концом HSA и N-концом PS-связывающего домена. В некоторых вариантах осуществления слияние внешнего линкера осуществлено между С-концом интегрин-связывающего домена и N-концом солюбилизирующего домена, а слияние дополнительного внешнего линкера осуществлено между С-концом солюбилизирующего домена и N-концом PS-связывающего домена. В частности, внешний линкер слит с С-концом EGF-подобного домена и N-концом HSA, а дополнительный внешний линкер слит с C-концом HSA и N-концом PS-связывающего домена.
В некоторых вариантах осуществления внешний линкер, содержащий GS, слит с С-концом интегрин-связывающего домена и с N-концом солюбилизирующего домена. В некоторых вариантах осуществления внешний линкер, содержащий GL, слит с С-концом солюбилизирующего домена и с N-концом PS-связывающего домена. В некоторых вариантах осуществления внешний линкер, содержащий (G2S)4 (SEQ ID NO: 62), слит с С-концом солюбилизирующего домена и с N-концом PS-связывающего домена. В некоторых вариантах осуществления внешний линкер, содержащий G4S (SEQ ID NO: 64), слит с С-концом солюбилизирующего домена и с N-концом PS-связывающего домена. В некоторых вариантах осуществления внешний линкер, содержащий (G4S)2 (SEQ ID NO: 65), слит с С-концом солюбилизирующего домена и с N-концом PS-связывающего домена.
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA. Слитый белок по настоящему изобретению, содержащий данную структуру, содержит аминокислотную последовательность, представленную под SEQ ID NO: 42 (FP330).
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA, а дополнительный внешний линкер, содержащий (GS)4 (SEQ ID NO: 63), слит с С-концом HSA и с N-концом PS-связывающего домена.
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA, а дополнительный внешний линкер, содержащий (G2S)4 (SEQ ID NO: 62), слит с С-концом HSA и с N-концом PS-связывающего домена. Слитый белок по настоящему изобретению, содержащий данную структуру, содержит аминокислотную последовательность, представленную под SEQ ID NO: 42 (FP330).
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA. С-конец HSA непосредственно слит с N-концом PS-связывающего домена.
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA, а дополнительный внешний линкер, содержащий G4S (SEQ ID NO: 64), слит с С-концом HSA и с N-концом PS-связывающего домена. Слитый белок по настоящему изобретению, содержащий данную структуру, содержит аминокислотную последовательность, представленную под SEQ ID NO: 54 (FP811).
В одном варианте осуществления внешний линкер, содержащий GS, слит с С-концом EGF-подобного домена и с N-концом HSA, а дополнительный внешний линкер, содержащий (G4S)2 (SEQ ID NO: 65), слит с С-концом HSA и с N-концом PS-связывающего домена. Слитый белок по настоящему изобретению, содержащий данную структуру, содержит аминокислотную последовательность, представленную под SEQ ID NO: 56 (FP010).
В некоторых вариантах осуществления His-метка слита с внешним линкером, содержащим GS (GS-6xHis; SEQ ID NO: 66), который слит с С-концом PS-связывающего домена. В одном варианте осуществления слитый белок по настоящему изобретению, содержащий His-метку, содержит аминокислотную последовательность, представленную под SEQ ID NO: 44 (FP278) или SEQ ID NO: 60 (FP114 или FP260).
Функциональные свойства терапевтических слитых белков
В настоящем изобретении предусмотрены слитые белки, полученные из MFG-E8 человека, которые эффективно способствуют эффероцитозу и, следовательно, активны в устранении основных факторов системного воспаления и патологии микрососудистого русла. Как указано в разделе "Примеры", было показано, что слитые белки, имеющие общую структуру EGF-HSA-C1-C2, эффективны в ряде анализов эффероцитоза. Например, слитые белки были эффективны в восстановлении нарушенного липополисахаридом (LPS) или S. aureus эффероцитоза посредством макрофагов и усилении эффероцитоза микрочастиц и погибающих клеток посредством эндотелиальных клеток. Слитые белки также оказались эффективными в защите функции почек и защите от потери веса тела в мышиной модели острого повреждения почек.
Иллюстративные последовательности белка
Аминокислотные последовательности в таблице 4 включают примеры терапевтических слитых белков по настоящему изобретению, а также их частей.
По всему тексту настоящей заявки может быть несоответствие между текстом описания (например таблица 4) и перечнем последовательностей, при этом текст описания имеет преимущественную силу.
Таблица 4. Иллюстративные последовательности белка
MFG-E8
(субдомены C1-C2)
EDIL3 EGF-HSA-C1-C2
Нуклеиновая кислота
EGF-C1-C2-Fc [S354C, T366W]
Нуклеиновая кислота
EGF-Fc-C1-C2
Нуклеиновая кислота
EGF-Fc(выступ)-C1-C2
Нуклеиновая кислота
EGF-Fc(впадина)-C1-C2
Нуклеиновая кислота
EGF-C1-C2-Fc
Нуклеиновая кислота
Fc-EGF-C1-C2
Нуклеиновая кислота
EGF-C2-C2
Нуклеиновая кислота
EGF-C1-C2-HSA
Нуклеиновая кислота
HSA-EGF-C1-C2
Нуклеиновая кислота
EGF-HSA
Нуклеиновая кислота
EGF-HSA-C1
Нуклеиновая кислота
EGF-HSA-C2
Нуклеиновая кислота
EGF(RGE)-HSA-C1-C2
Нуклеиновая кислота
EGF-HSA D3-C1-C2-His
Нуклеиновая кислота
EGF-HSA-C1-C2
Нуклеиновая кислота
EGF-HSA-C1-C2 His-метка
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
Нуклеиновая кислота
=133
=121
=119
Не-M 3163
=147
= 74
= 73
=117
Настоящая заявка также включает варианты каждой из SEQ ID NO: 69, 70 и 72, где EGF-подобный домен последовательности EDIL3, включенный в нее, соответствует любой из следующих последовательностей: SEQ ID NO: 96, SEQ ID NO: 97, SEQ ID NO: 98, SEQ ID NO: 99, SEQ ID NO: 100 или SEQ ID NO: 101.
Настоящая заявка также включает терапевтический слитый белок, содержащий интегрин-связывающие домены из MFGE8 или EDIL3 и PS-связывающие домены, такие как домен IgSF V TIM4 или домен GLA вариантов мостикового белка GAS6 (например, FP1147 и FP1148).
Модификация белков по настоящему изобретению
Настоящая заявка предусматривает варианты белков, описанных в данном документе, и/или их фрагментов, содержащих различные модификации в доменах, а также продукты слияния и конъюгаты раскрытых молекул. Например, домен терапевтического слитого белка может содержать консервативную модификацию аминокислотных остатков, и при этом модифицированные белки сохраняют улучшенные свойства по сравнению с слитым белком, содержащим родительский домен, или характеризуются ими. В качестве альтернативы, домен терапевтического слитого белка может содержать делецию(делеции) аминокислотных остатков, при этом модифицированные слитые белки сохраняют улучшенные свойства по сравнению с белком, содержащим родительский домен, или характеризуются ими. В качестве альтернативы, терапевтические слитые белки могут содержать вставку(вставки) аминокислотных остатков, при этом модифицированные белки сохраняют улучшенные свойства по сравнению с немодифицированным белком, или характеризуются ими. В одном варианте осуществления такая аминокислотная вставка содержит остатки глицина или серина в ряде комбинаций для функционирования в качестве линкера между доменами родительского белка.
Сайт-направленный мутагенез или ПЦР-опосредованный мутагенез, можно осуществлять для введения мутации(мутаций), а эффект в отношении связывания интегрина и/или PS или другое функциональное свойство, представляющее интерес, может быть оценено в анализах in vitro или in vivo. Могут быть введены консервативные модификации (как обсуждалось выше), и/или мутации могут представлять собой аминокислотные замены, добавления или делеции. Более того, как правило, изменяют не более одного, двух, трех, четырех или пяти остатков в пределах связывающего домена.
Варианты аминокислотной последовательности терапевтических слитых белков, которые характеризуются по сути сходными свойствами с немодифицированными вариантами, могут быть получены посредством внесения соответствующих нуклеотидных изменений в кодирующие ДНК или посредством синтеза требуемых вариантов. Такие варианты включают, например, делеции, вставки или замены остатков в аминокислотных последовательностях молекул по настоящему изобретению. В некоторых вариантах осуществления варианты могут включать дополнительные линкерные последовательности, укороченные линкерные последовательности или удаление линкерных последовательностей, и/или аминокислотные мутации или замены и делецию одной или нескольких аминокислот. Для получения конечной конструкции используется любая комбинация делеции, вставки и замены при условии, что конечная конструкция характеризуется требуемыми характеристиками. Изменения аминокислот также могут обеспечивать изменение посттрансляционных процессов для молекул, такое как изменение количества или положения возможных сайтов гликозилирования.
Способы получения рекомбинантных молекул
Нуклеиновые кислоты и системы экспрессии
В одном варианте осуществления в настоящей заявке предусмотрен способ рекомбинантного получения одной или нескольких полипептидных цепей терапевтического слитого белка, включающий: 1) получение одной или нескольких конструкций ДНК, содержащих молекулу нуклеиновой кислоты, кодирующую полипептидную цепь мультиспецифической связывающей молекулы; 2) введение указанной(указанных) ДНК-конструкции(конструкций) в один или несколько векторов экспрессии; 3) осуществление совместной трансфекции указанного(указанных) вектора(векторов) экспрессии в одну или несколько клеток-хозяев; и 4) обеспечение экспрессии и сборки молекулы в клетке-хозяине или в растворе.
В этом отношении в настоящем изобретении предусмотрены выделенные нуклеиновые кислоты, например один или несколько полинуклеотидов, кодирующих терапевтические слитые белки, описанные в данном документе. Молекулы нуклеиновых кислот включают ДНК и РНК как в одноцепочечной, так и в двухцепочечной форме, а также соответствующие комплементарные последовательности. Молекулы нуклеиновых кислот по настоящему изобретению включают полноразмерные гены или молекулы cDNA, а также комбинации их фрагментов. Нуклеиновые кислоты по настоящему изобретению получены из человеческих источников, однако настоящее изобретение предусматривает нуклеиновые кислоты, полученные из видов, отличных от человека.
Термин "выделенная нуклеиновая кислота", представляет собой нуклеиновую кислоту, которая была отделена от смежных генетических последовательностей, присутствующих в геноме организма, из которого нуклеиновая кислота была выделена, в случае нуклеиновых кислот, выделенных из встречающихся в природе источников. В случае нуклеиновых кислот, синтезированных ферментативным путем из матрицы или химическим путем, таких как, например, ПЦР-продукты, молекулы cDNA или олигонуклеотиды, следует понимать, что нуклеиновые кислоты, полученные в результате таких способов, представляют собой выделенные нуклеиновые кислоты. Под выделенной молекулой нуклеиновой кислоты понимается молекула нуклеиновой кислоты в виде отдельного фрагмента или компонента более крупной конструкции нуклеиновой кислоты. В одном предпочтительном варианте осуществления нуклеиновые кислоты по сути не содержат контаминирующего эндогенного материала. Молекулу нуклеиновой кислоты предпочтительно получают из ДНК или РНК, выделенной по меньшей мере один раз в по сути чистой форме и в количестве или концентрации, позволяющих идентифицировать, манипулировать и восстанавливать составляющие ее нуклеотидные последовательности с помощью стандартных биохимических способов (таких как способы, описанные в Sambrook et al., Molecular Cloning: A Laboratory Manual, 2nd ed., Cold Spring Harbor Laboratory, Cold Spring Harbor, NY (1989)). Такие последовательности предпочтительно получают и/или конструируют в форме открытой рамки считывания, не прерываемой внутренними нетранслируемыми последовательностями или интронами, которые, как правило, присутствуют в генах эукариот. Последовательности нетранслируемой ДНК могут находиться в направлении 5'- или 3'-конца от открытой рамки считывания, где они не нарушают манипулирование с кодирующей областью или ее экспрессию.
Настоящее изобретение также предусматривает системы и конструкции экспрессии в форме плазмид, векторов экспрессии, транскрипционных или экспрессионных кассет, которые содержат по меньшей мере один полинуклеотид, как описано выше. Кроме того, настоящее изобретение предусматривает клетки-хозяева, содержащие такие системы или конструкции экспрессии.
В одном варианте осуществления в настоящем изобретении предусмотрен способ получения терапевтического слитого белка, включающий следующие стадии: (а) культивирование клетки-хозяина, содержащей нуклеиновую кислоту, кодирующую слитый белок, где культивируемая клетка-хозяин экспрессирует слитый белок; и (b) извлечение слитого белка из культуры клетки-хозяина.
В настоящем изобретении также предусмотрены векторы экспрессии и клетки-хозяева для получения терапевтических слитых белков, описанных выше. Термин "вектор" означает любую молекулу или объект (например, нуклеиновую кислоту, плазмиду, бактериофаг или вирус), который подходит для трансформации или трансфекции клетки-хозяина и содержит последовательности нуклеиновых кислот, которые направляют и/или контролируют (вместе с клеткой-хозяином) экспрессию одной или нескольких гетерологичных кодирующих областей, функционально связанных с ними. Для экспрессии полинуклеотидов, кодирующих цепи или связывающие домены молекулы, можно применять различные векторы экспрессии. Для получения терапевтического слитого белка в клетке-хозяине млекопитающего можно применять как вирусные, так и невирусные векторы экспрессии. Невирусные векторы и системы включают плазмиды, эписомные векторы, как правило с кассетой экспрессии для экспрессии белка или РНК и искусственные хромосомы человека (см., например, Harrington et al., (1997) Nat Genet. 15: 345). Например, невирусные векторы, применимые для экспрессии полинуклеотидов и полипептидов в клетках млекопитающих (например, человека), включают pThioHis A, B и C, pcDNA3.1/His, pEBVHis A, B и C (Invitrogen, Сан-Диего, штат Калифорния, США), векторы MPSV и многочисленные другие векторы, известные из уровня техники для экспрессии других белков. Применимые вирусные векторы включают векторы на основе ретровирусов, аденовирусов, аденоассоциированных вирусов, вирусов герпеса, векторы на основе SV40, папилломавируса, вируса Эпштейна-Барр HBP, векторы на основе вируса осповакцины и вируса леса Семлики (SFV). См. Brent et al., (1995), выше; Smith, Annu. Rev. Microbiol. 49: 807; и Rosenfeld et al., (1992) Cell 68: 143.
Выбор вектора экспрессии зависит от предполагаемых клеток-хозяев, в которых должен экспрессироваться вектор. Как правило, векторы экспрессии содержат промотор и другие регуляторные последовательности (например, энхансеры), которые функционально соединены с полинуклеотидами, кодирующими терапевтический слитый белок. В некоторых вариантах осуществления индуцируемый промотор используют для предупреждения экспрессии вставленных последовательностей в условиях, отличающихся от индуцирующих. Индуцируемые промоторы включают, например, арабинозный, lacZ, металлотионеиновый промотор или промотор белка теплового шока. Культуры трансформированных организмов можно размножать в неиндуцирующих условиях без смещения популяции в сторону кодирующих последовательностей, продукты экспрессии которых лучше переносятся клетками-хозяевами. Для эффективной экспрессии терапевтических слитых белков кроме промоторов также могут быть необходимы или могут требоваться другие регуляторные элементы. Такие элементы, как правило, включают инициаторный кодон ATG и смежный сайт связывания рибосомы или другие последовательности. В дополнение, эффективность экспрессии можно повысить за счет включения энхансеров, соответствующих используемой клеточной системе (см., например, Scharf et al., (1994) Results Probl. Cell Differ. 20: 125; и Bittner et al., (1987) Meth. Enzymol., 153:516). Например, для повышения экспрессии в клетках-хозяевах, являющихся клетками млекопитающих, можно использовать энхансер SV40 или энхансер CMV.
Векторы экспрессии могут также обеспечивать положение для секреторной сигнальной последовательности для образования слитого белка с полипептидами, кодируемыми посредством вставки описанных выше последовательностей связывающих доменов и/или солюбилизирующих доменов. Чаще вставляемые последовательности соединяют с сигнальными последовательностями перед включением в вектор. Векторы, которые обеспечивают экспрессию связывающих доменов и солюбилизирующего домена в виде слитых белков, тем самым приводят к получению интактных сконструированных белков. При культивировании в соответствующих условиях клетка-хозяин может быть использована для экспрессии сконструированного белка, который впоследствии может быть собран из среды для культивирования (если клетка-хозяин секретирует его в среду) или непосредственно из клетки-хозяина, продуцирующей его (если он не секретируется). Выбор подходящей клетки-хозяина будет зависеть от различных факторов, таких как требуемые уровни экспрессии, модификации полипептидов, которые требуются или необходимы для активности (такие как гликозилирование или фосфорилирование), и простота укладки в биологически активную молекулу. Клетка-хозяин может быть эукариотической или прокариотической.
Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, известны из уровня техники и включают без ограничения иммортализованные линии клеток, доступные из Американской коллекции типовых культур (АТСС), и любые линии клеток, используемые в системе экспрессии, известной из уровня техники, могут быть использованы для получения рекомбинантных слитых белков по настоящему изобретению. Как правило, клетки-хозяева трансформируют рекомбинантным вектором экспрессии, который содержит ДНК, кодирующую требуемый слитый белок. Среди клеток-хозяев, которые можно применять, присутствуют клетки прокариот, дрожжей или высших эукариот. Прокариоты включают грамотрицательные или грамположительные организмы, например E. coli или бациллы. К клеткам высших эукариот относятся клетки насекомых и стабильные линии клеток млекопитающих. Примеры подходящих линий клеток-хозяев млекопитающих включают клетки COS-7, клетки L, клетки Cl27, клетки 3T3, клетки яичника китайского хомячка (CHO) или их производные и соответствующие линии клеток, которые растут в бессывороточной среде, клетки HeLa, линии клеток BHK, линию клеток CV-1 EBNA, клетки почки эмбриона человека (HEK), такие как 293, 293 EBNA или MSR 293, эпидермальные клетки A431 человека, клетки Colo205 человека, другие трансформированные линии клеток приматов, нормальные диплоидные клетки, клеточные линии, полученные из in vitro культуры первичной ткани, первичные эксплантаты, клетки HL-60, U937, HaK или Jurkat. Необязательно линии клеток млекопитающих, такие как HepG2/3B, KB, NIH 3T3 или S49, например, могут быть использованы для экспрессии полипептида, если необходимо использовать полипептид в различных анализах трансдукции сигнала или анализах с использованием репортерного гена. В качестве альтернативы полипептид можно получить в низших эукариотах, таких как дрожжи, или в прокариотах, таких как бактерии. Подходящие виды дрожжей включают P. pastoris, S. cerevisiae, S. pombe, штаммы Kluyveromyces, Candida или любой штамм дрожжей, способный экспрессировать гетерологичные полипептиды. Подходящие бактериальные штаммы включают штаммы E. coli, B. subtilis, S. typhimurium или любой бактериальный штамм, способный экспрессировать гетерологичные полипептиды. Если слитый белок получают в дрожжах или бактериях, для получения функционального продукта может быть необходимым модифицировать продуцируемый ими продукт, например, посредством фосфорилирования или гликозилирования соответствующих сайтов. Такие ковалентные присоединения могут быть выполнены с применением известных химических или ферментативных способов.
Способы введения векторов экспрессии, содержащих полинуклеотидные последовательности, представляющие интерес, варьируют в зависимости от типа клетки-хозяина. Например, трансфекцию с использованием хлорида кальция обычно используют для прокариотических клеток, тогда как обработку фосфатом кальция или электропорацию можно применять для других клеток-хозяев. Другие способы включают, например, электропорацию, обработку фосфатом кальция, трансформацию, опосредованную липосомами, инъекцию и микроинъекцию, баллистические способы, виросомы, иммунолипосомы, конъюгаты поликатион:нуклеиновая кислота, депротеинизированную ДНК, искусственные вирионы, слияние со структурным белком VP22 вируса герпеса, усиленное средством поглощение ДНК и трансдукцию ex vivo. Для долговременного получения рекомбинантных белков с высоким выходом зачастую будет необходима стабильная экспрессия. Например, линии клеток, которые стабильно экспрессируют сконструированные белки, можно получать с помощью векторов экспрессии по настоящему изобретению, которые содержат вирусные точки начала репликации или эндогенные элементы для обеспечения экспрессии и ген селектируемого маркера. После введения вектора клетки могут оставлять для роста на 1-2 дня в обогащенной среде перед их переносом в селективную среду. Задачей селектируемого маркера является сообщение устойчивости к факторам отбора, и его присутствие обеспечивает возможность роста в селективной среде клеток, которые успешно экспрессируют введенные последовательности. Пролиферацию устойчивых стабильно трансфицированных клеток можно обеспечивать с применением методик культуры тканей, соответствующих типу клеток.
Слитые белки, как правило, извлекаются из среды для культивирования в виде секретируемого полипептида, хотя они также могут быть выделены из лизата клетки-хозяина при непосредственном получении без секреторной сигнальной последовательности. Если полипептид является мембраносвязанным, его можно высвободить из мембраны с помощью подходящего раствора детергента (например, Triton-X 100).
При получении слитого белка в рекомбинантной клетке, отличной от клетки человеческого происхождения, он полностью не содержит белков или полипептидов человеческого происхождения. Однако имеется необходимость в очищении слитого белка от рекомбинантных клеточных белков или полипептидов. В качестве первой стадии среду для культивирования или лизат обычно центрифугируют для удаления твердого клеточного дебриса. Полученные молекулы можно легко очистить посредством хроматографии на гидроксилапатите, гель-электрофореза, диализа или аффинной хроматографии, при этом аффинная хроматография является предпочтительной методикой очистки. Также доступны другие методики очистки белка, такие как фракционирование на ионообменной колонке, осаждение этанолом, обращенно-фазовая HPLC, хроматография на диоксиде кремния, хроматография на гепарин-сефарозе, хроматография на анионообменной или катионообменной смоле (такой как колонка с полиаспарагиновой кислотой), хроматофокусирование, SDS-PAGE и осаждение сульфатом аммония.
В определенных аспектах в данном документе предусмотрен вирусный вектор, содержащий полинуклеотид, кодирующий терапевтический слитый белок по настоящему изобретению. В некоторых вариантах осуществления вирусный вектор получен из AAV. В определенных некоторых вариантах осуществления вирусный вектор вводят субъекту, например человеку, при этом терапевтический слитый белок экспрессируется, и его можно применять для лечения и/или предупреждения заболеваний, приведенных в данном документе.
Фармацевтические композиции
В другом аспекте в настоящем изобретении предусмотрена композиция, например фармацевтическая композиция, содержащая терапевтический слитый белок по настоящему изобретению в комбинации с одним или несколькими фармацевтически приемлемыми вспомогательными веществами, разбавителями или носителями. Такие композиции могут включать один терапевтический слитый белок по настоящему изобретению или их комбинацию (например, двух или более разных).
Фармацевтические композиции, описанные в данном документе, также можно вводить в составе комбинированной терапии, т. е. в комбинации с другими средствами. Например, комбинированная терапия может включать слитый белок по настоящему изобретению в комбинации, например, по меньшей мере с одним противовоспалительным, противоинфекционным средством или иммунодепрессантным средством. Примеры терапевтических средств, которые можно применять в комбинированной терапии, более подробно описаны ниже в разделе, посвященном путям применения терапевтических слитых белков по настоящему изобретению.
Для приготовления фармацевтических или стерильных композиций, содержащих слитый белок по настоящему изобретению, слитый белок смешивают с фармацевтически приемлемым носителем или вспомогательным веществом.
Выражение "фармацевтически приемлемый" означает одобренный регулирующим органом федерального правительства или правительства штата или указанный в Фармакопее США или другой общепризнанной фармакопее для применения у животных и, более конкретно, у людей.
Термин "фармацевтическая композиция" относится к смеси по меньшей мере одного активного ингредиента (например, сконструированного белка) и по меньшей мере одного фармацевтически приемлемого вспомогательного вещества, разбавителя или носителя.
"Лекарственный препарат" относится к веществу, применяемому для медицинского лечения.
Используемый в данном документе "фармацевтически приемлемый носитель" предусматривает все возможные растворители, дисперсионные среды, покрытия, антибактериальные и противогрибковые средства, изотонические и замедляющие абсорбцию средства и т.п., которые являются физиологически совместимыми. Носитель должен быть подходящим для внутривенного, внутримышечного, подкожного, парентерального, спинального или эпидермального введения (например, посредством инъекции или вливания). В одном варианте осуществления носитель должен быть подходящим для подкожного пути. В зависимости от пути введения активное соединение, т. е. слитый белок, может быть покрыто материалом для защиты соединения от действия кислот и других естественных условий, которые могут инактивировать соединение.
Фармацевтические композиции, описанные в данном документе, могут содержать одну или несколько фармацевтически приемлемых солей. Фармацевтическая композиция, описанная в данном документе, может также содержать фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; жирорастворимые антиоксиданты, такие как пальмитат аскорбила, бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), лецитин, пропилгаллат, альфа-токоферол и т.п.; и металлические хелатообразующие средства, такие как лимонная кислота, этилендиаминтетрауксусная кислота (EDTA), сорбит, винная кислота, фосфорная кислота и т.п.
Примеры подходящих водных и неводных носителей, которые могут быть использованы в фармацевтических композициях, описанных в данном документе, включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, и органические сложные эфиры для инъекций, такие как этилолеат. Надлежащую текучесть можно поддерживать, например, с помощью использования материалов для покрытия, таких как лецитин, посредством поддержания необходимого размера частиц в случае дисперсий и с помощью использования поверхностно-активных веществ.
Эти композиции также могут содержать вспомогательные средства, такие как консерванты, смачивающие средства, эмульгирующие средства и диспергирующие средства. Предупреждение присутствия микроорганизмов может быть обеспечено процедурами стерилизации и посредством включения различных антибактериальных и противогрибковых средств, например парабена, хлорбутанола, фенолсорбиновой кислоты и т.п. Также может быть необходимым включение изотонических средств, таких как сахара, хлорид натрия и т.п., в состав композиций. Кроме того, пролонгированная абсорбция фармацевтических форм для инъекций может быть обеспечена включением средств, которые замедляют абсорбцию, таких как моностеарат алюминия и желатин.
Фармацевтически приемлемые носители включают стерильные водные растворы или дисперсии и стерильные порошки для немедленного приготовления стерильных растворов или дисперсий для инъекций. Применение такой среды и средств для фармацевтически активных веществ известно из уровня техники. За исключением случаев, когда любые общепринятые среды или средство являются несовместимыми с активным соединением, предполагается их использование в фармацевтических композициях по настоящему изобретению. Дополнительные активные соединения также могут быть включены в состав композиций.
Терапевтические композиции, как правило, должны быть стерильными и стабильными в условиях изготовления и хранения. Композиция может быть составлена в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для обеспечения высокой концентрации лекарственного средства. Носитель может представлять собой растворитель или дисперсионную среду, содержащие, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Надлежащую текучесть можно поддерживать, например, посредством применения покрытия, такого как лецитин, посредством поддержания требуемого размера частиц в случае дисперсии и посредством применения поверхностно-активных веществ. Во многих случаях в составе композиции можно включать изотонические средства, например сахара, полиспирты, такие как маннит, сорбит или хлорид натрия.
Обзоры разработки стабильных составов на основе белка можно найти в Cleland et al., (1993) Crit Reviews Ther Drug Carrier Systems, 10(4): 307-377 и Wei W (1999) Int J Pharmaceutics, 185: 129-88.
Растворы или суспензии, которые применяют для внутрикожного или подкожного применения, как правило, включают один или более из следующих компонентов: стерильный разбавитель, такой как вода для инъекций, физиологический раствор, нелетучие масла, полиэтиленгликоли, глицерин, пропиленгликоль или другие синтетические растворители, антибактериальные средства, такие как бензиловый спирт или метилпарабены, антиоксиданты, такие как аскорбиновая кислота или бисульфит натрия, хелатирующие средства, такие как этилендиаминтетрауксусная кислота, буферы, такие как ацетаты, цитраты или фосфаты, и средства для регулировки тонуса, такие как хлорид натрия или декстроза. Уровень pH можно регулировать посредством кислот или оснований, таких как хлористоводородная кислота или гидроксид натрия. Такие препараты можно заключить в ампулы, одноразовые шприцы или пузырьки многоразового использования, сделанные из стекла или пластика.
Стерильные растворы для инъекций можно приготовить посредством включения активного соединения в требуемом количестве в подходящем растворителе с одним или комбинацией ингредиентов, перечисленных выше, если потребуется, с последующей стерилизационной микрофильтрацией. Как правило, дисперсии получают посредством включения слитых белков по настоящему изобретению в стерильный носитель, который содержит базовую дисперсионную среду и другие требуемые ингредиенты из перечисленных выше. В случае стерильных порошков для приготовления стерильных растворов для инъекций способы получения представляют собой вакуумную сушку и лиофильную сушку (лиофилизацию), которые дают на выходе порошок активного ингредиента плюс любого дополнительного необходимого ингредиента из их предварительно стерильно отфильтрованного раствора.
Количество активного ингредиента, которое можно объединять с материалом носителя для получения стандартной лекарственной формы, будет варьировать в зависимости от субъекта, подлежащего лечению, и конкретного способа введения. Количество активного ингредиента, которое можно объединять с материалом носителя для получения единичной лекарственной формы, обычно будет представлять собой такое количество композиции, которое обеспечивает терапевтический эффект. Как правило, в пересчете на сто процентов данное количество будет находиться в диапазоне от приблизительно 0,01 процента до приблизительно девяноста девяти процентов активного ингредиента, от приблизительно 0,1 процента до приблизительно 70 процентов или от приблизительно 1 процента до приблизительно 30 процентов активного ингредиента в комбинации с фармацевтически приемлемым носителем.
Выбор схемы введения для терапевтического сконструированного белка зависит от нескольких факторов, в том числе скорости метаболизма объекта в сыворотке крови или ткани, уровня симптомов, иммуногенности объекта и доступности клеток-мишеней в биологической матрице. В определенных вариантах осуществления схема введения обеспечивает увеличение до максимума количества терапевтического средства, доставляемого пациенту, в соответствии с приемлемым уровнем побочных эффектов. Соответственно, количество доставляемого белка частично зависит от конкретного объекта и тяжести состояния, подлежащего лечению. Доступны руководства по выбору соответствующих доз биологических средств и малых молекул (см., например, Bach (ed.) (1993) Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, N.Y.; Baert, et al. (2003) New Engl. J. Med. 348:601-608; Milgrom, et al. (1999) New Engl. J. Med. 341:1966-1973; Slamon, et al. (2001) New Engl. J. Med. 344:783-792; Beniaminovitz, et al. (2000) New Engl. J. Med. 342:613-619; Ghosh, et al. (2003) New Engl. J. Med. 348:24-32; Lipsky, et al. (2000) New Engl. J. Med. 343:1594-1602).
Определение подходящей дозы выполняет лечащий врач, например, с использованием параметров или факторов, которые, как известно или предполагается из уровня техники, влияют на лечение или согласно прогнозам влияют на лечение. Введение доз обычно начинают с количества, несколько меньшего, чем оптимальная доза, и после этого его увеличивают с небольшими приращениями до тех пор, пока не достигается требуемый или оптимальный эффект по сравнению с любыми отрицательными побочными эффектами. Важные диагностические показатели включают, например, показатели симптомов воспаления или уровня продуцируемых воспалительных цитокинов.
Фактические уровни доз активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать для того, чтобы получить количество активного ингредиента, которое является эффективным для достижения требуемого терапевтического ответа у конкретного пациента, эффективным в плане композиции и способа введения, не будучи токсичным для пациента. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, в том числе активности конкретных используемых композиций по настоящему изобретению, пути введения, времени введения, скорости выведения конкретного используемого соединения, длительности лечения, других лекарственных средств, соединений и/или материалов, применяемых в комбинации с конкретными используемыми композициями, возраста, пола, веса тела, состояния, общего состояния здоровья и анамнеза пациента, подлежащего лечению, и аналогичных факторов, известных в области техники медицины.
Режимы введения дозы корректируют для обеспечения оптимального требуемого ответа. Например, можно вводить одну болюсную дозу, можно вводить несколько разделенных доз в течение некоторого времени, или дозу можно пропорционально снижать или увеличивать в соответствии с потребностями терапевтической ситуации. Особенно преимущественным является составление композиций для парентерального применения в виде единичной дозированной формы для удобства введения и однородности дозирования. Единичная дозированная форма, применяемая в данном документе, относится к физически дискретным единицам, подходящим в качестве единичных доз для субъектов, подлежащих лечению; каждая единица содержит предварительно определенное количество активного соединения, рассчитанное для получения необходимого терапевтического эффекта, в сочетании с необходимым фармацевтическим носителем. Спецификация единичных дозированных форм данного изобретения определяется и непосредственно зависит от уникальных свойств активного соединения и конкретного терапевтического эффекта, который следует получить, и ограничений, свойственных данной области техники при получении составов такого активного соединения для лечения чувствительности у индивидуумов.
Для введения терапевтического слитого белка доза находится в диапазоне от приблизительно 0,0001 до 150 мг/кг, например, при подкожном введении - 5, 15 и 50 мг/кг и чаще от 0,01 до 5 мг/кг веса тела хозяина. Иллюстративный режим лечения включает введение один раз в неделю, один раз в две недели, один раз в три недели, один раз в четыре недели, один раз в месяц, один раз в 3 месяца или один раз каждые 3-6 месяцев.
Терапевтические слитые белки по настоящему изобретению можно вводить многократно. Интервалы между однократными дозами могут быть, например, недельными, месячными, трехмесячными или ежегодными. Интервалы также могут быть нерегулярными, на что указывает измерение уровня сконструированного белка в крови у пациента. В некоторых способах дозу корректируют для достижения концентрации белка в плазме крови приблизительно 1-1000 мкг/мл, а в некоторых способах приблизительно 25-300 мкг/мл.
В качестве альтернативы, терапевтический слитый белок можно вводить в виде состава с замедленным высвобождением, и в этом случае требуется менее частое введение. Доза и частота варьируют в зависимости от времени полужизни белка у пациента и могут варьировать в зависимости от того, является ли лечение профилактическим или терапевтическим. В профилактических вариантах применения относительно низкую дозу вводят с относительно большими интервалами в течение длительного периода времени. Некоторые пациенты могут продолжать получать лечение до конца своей жизни. В терапевтических применениях иногда необходимо введение относительно высокой дозы с относительно короткими интервалами до тех пор, пока прогрессирование состояния или заболевания не снизится или завершится, или до тех пор, пока у пациента не будет наблюдаться частичное или полное облегчение симптомов состояния или заболевания. После этого пациенту можно осуществлять введение согласно профилактическому режиму.
Фактические уровни дозы активных ингредиентов в фармацевтических композициях по настоящему изобретению можно варьировать для того, чтобы получить количество активного ингредиента, которое является эффективным для достижения необходимого терапевтического ответа у конкретного пациента, эффективным в отношении композиции и способа введения, при этом не токсично для пациента. Выбранный уровень дозы будет зависеть от ряда фармакокинетических факторов, в том числе активности конкретных используемых композиций по настоящему изобретению, пути введения, времени введения, скорости выведения конкретного используемого соединения, длительности лечения, других лекарственных средств, соединений и/или материалов, применяемых в комбинации с конкретными используемыми композициями, возраста, пола, веса тела, состояния, общего состояния здоровья и анамнеза пациента, подлежащего лечению, и аналогичных факторов, хорошо известных в области техники медицины.
"Терапевтически эффективная доза" слитого белка по настоящему изобретению может приводить к снижению тяжести состояния или симптомов заболевания и/или предупреждению ухудшения или инвалидности вследствие состояния.
Композицию по настоящему изобретению можно вводить одним или несколькими путями введения с применением одного или нескольких разнообразных способов, известных из уровня техники. Как следует понимать специалисту в данной области техники, путь и/или способ введения будут варьировать в зависимости от требуемых результатов. Пути введения сконструированных белков по настоящему изобретению включают внутривенный, внутримышечный, внутрикожный, внутрибрюшинный, подкожный, спинальный или другие парентеральные пути введения, например, с помощью инъекции или вливания. Выражение "парентеральное введение", используемое в данном документе, означает способы введения, отличные от энтерального и местного введения, обычно осуществляемые посредством инъекции, и включает без ограничения внутривенные, внутримышечные, внутриартериальные, интратекальные, внутрикапсульные, интраорбитальные, внутрисердечные, внутрикожные, внутрибрюшинные, транстрахеальные, подкожные, субкутикулярные, внутрисуставные, подкапсулярные, субарахноидальные, интраспинальные, эпидуральные и интрастернальные инъекцию и вливание.
В качестве альтернативы, терапевтический слитый белок по настоящему изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или слизистый путь введения.
Терапевтические слитые белки по настоящему изобретению могут быть приготовлены с носителями, которые будут защищать белки от быстрого высвобождения, как, например, состав с контролируемым высвобождением, в том числе имплантаты, трансдермальные пластыри и микрокапсулированные системы доставки. Можно применять биоразлагаемые, биосовместимые полимеры, такие как этиленвинилацетат, полиангидриды, полигликолевая кислота, коллаген, сложные полиортоэфиры и полимолочная кислота. Способы получения таких составов запатентованы или общеизвестны для специалистов в данной области техники. См., например, Sustained and Controlled Release Drug Delivery Systems, J.R. Robinson, ed., Marcel Dekker, Inc., New York, 1978.
В определенных вариантах осуществления терапевтические слитые белки по настоящему изобретению могут составляться так, чтобы гарантировать точное распределение in vivo. Например, гематоэнцефалический барьер (BBB) не пропускает множество высокогидрофильных соединений. Чтобы гарантировать то, что терапевтические соединения по настоящему изобретению пересекают BBB (при необходимости), их можно составлять, например, в липосомах. Способы изготовления липосом см., например, в патентах США 4522811, 5374548 и 5399331. Липосомы могут содержать один или несколько фрагментов, которые селективно транспортируются в специфические клетки или органы, с улучшением тем самым нацеленной доставки лекарственного средства (см., например, Ranade VV (1989) J. Clin. Pharmacol., 29:685).
Пути терапевтического применения и способы по настоящему изобретению
Терапевтические слитые белки по настоящему изобретению характеризуются диагностическими и терапевтическими областями применения in vitro и in vivo. Например, эти молекулы можно вводить в клетки в культуре, например in vitro, или в организме субъекта, например in vivo, для лечения, предупреждения или диагностики различных нарушений. Способы являются особенно подходящими для лечения, предупреждения или диагностики острых или хронических воспалительных нарушений, а также нарушений органов и микрососудистого русла, обусловленных иммунной системой или коагуляцией.
Терапевтические слитые белки по настоящему изобретению без ограничения применимы для лечения, предупреждения или облегчения острых и хронических воспалительных поражений органов, в частности воспалительных поражений, при которых эндогенные механизмы гомеостатического клиренса или пути эффероцитоза для удаления погибающих клеток, клеточных фрагментов и протромботических/провоспалительных микрочастиц значительно подавлены. Примеры острых воспалительных поражений органов включают инфаркт миокарда, острое повреждение почек (AKI), острый инсульт и воспаление, а также повреждения органов в результате ишемии/реперфузии, такой как ишемия/реперфузия желудочно-кишечного тракта, печени, селезенки, легкого, почки, поджелудочной железы, сердца, головного мозга, спинного мозга и/или размозженной конечности.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения ингибирования или замедления свертывания крови, лечения микробиома, воспалительного заболевания кишечника (IBD), поглощения жирных кислот и/или снижения перистальтики желудка, нарушений, зависимых от микротромбов, атеросклероза, ремоделирования сердца, фиброза тканей, острого повреждения печени, хронических заболеваний печени, неалкогольного стеатогепатита (NASH), сосудистых заболеваний, возрастных сосудистых заболеваний, заболеваний кишечника, сепсиса, нарушений со стороны костей, рака, талассемии, панкреатита, гепатита, эндокардита, пневмонии, острого повреждения легкого, остеоартрита, периодонтита, воспаления тканей, индуцированного травмой, колита, сахарного диабета, геморрагического шока, отторжения трансплантата, радиационно-индуцированного повреждения, спленомегалии, индуцированного сепсисом AKI или полиорганной недостаточности, острых ожогов, респираторного дистресс-синдрома взрослых и детей, заживления ран, восстановления сухожилий и неврологических заболеваний.
В одном варианте осуществления неврологические заболевания могут быть выбраны из состояний, характеризующихся нейропсихиатрическим, нейровоспалительным и/или нейродегенеративным компонентом, в том числе такими симптомами, как синдромы, связанные с тошнотой, тошнота, пассивное избегание, подавление поведенческой гибкости, нарушение памяти и дисфункция памяти. Примеры неврологических заболеваний включают неврологические заболевания, связанные с бета-амилоидом, такие как болезнь Альцгеймера, болезнь Паркинсона и депрессия.
В одном варианте осуществления нарушения со стороны костей могут быть выбраны из состояний, включающих остеопороз, остеомаляцию, остеосклероз и остеопетроз. Более конкретно, введение слитого белка по настоящему изобретению может ингибировать экспрессию по меньшей мере одного маркера остеокластов, такого как NFATc1, катепсин К и интегрин αvβ3. В одном варианте осуществления введение ингибирует остеокластогенез. В другом варианте осуществления введение ингибирует RANKL-индуцированный остеокластогенез. В еще одном варианте осуществления введение ингибирует резорбцию костной ткани. В еще одном варианте осуществления введение ингибирует экспрессию по меньшей мере одного стимулятора резорбции костной ткани, такого как стимулятор резорбции костной ткани, содержащий TNF, IL-6, IL-17A, MMP-9, Ptgs2, RANKL, Tnfsf11, CXCL1, CXCL2, CXCL3, CXCL5 и их комбинации. В другом варианте осуществления введение ингибирует экспрессию по меньшей мере одного провоспалительного цитокина, выбранного из группы, состоящей из IL-8 и CCL2/MCP-1.
В одном варианте осуществления фиброз тканей может представлять собой фиброз в печени, легком, диафрагме, почке, головном мозге, сердце, в которых слитый белок по настоящему изобретению снижает экспрессию коллагена. В одном варианте осуществления фиброз легкого представляет собой интерстициальный фиброз легких (IPF). В одном варианте осуществления фиброз печени представляет собой цирроз печени, который может быть обусловлен или не обусловлен NASH.
Для многих респираторных заболеваний свойственно накопление апоптотических клеток. Кроме того, дефектные эффероцитоз и фагоцитоз макрофагами при хроническом обструктивном заболевании легких (COPD) ассоциированы с обострениями и тяжестью. Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения респираторных заболеваний, таких как острый респираторный дистресс-синдром или COPD. Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения острого повреждения легкого (ALI), например, повреждения легкого, индуцированного вдыханием или аспирацией токсичных экзогенных или эндогенных соединений или лекарственных средств; повреждения легкого, вызванного отеком легкого, шоком, панкреатитом, ожогами, травмами грудной клетки или политравмами, облучением, сепсисом, патогенами (бактериями, вирусами или паразитами, такими как плазмодии); хронических заболеваний легочной недостаточности, приводящих к гипоксемии.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести повреждения легкого, вызванного вирусами типа вирусов из семейства Coronaviridae, например SARS-CoV, SARS-CoV-2 или MERS-CoV. В одном варианте осуществления терапевтические слитые белки по настоящему изобретению предназначены для применения при лечении инфекции, обусловленной SARS-CoV-2, у пациентов с COVID-19.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести легочной недостаточности, ассоциированной с переливанием крови (TRALI).
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести хронической легочной недостаточности, приводящей к гипоксемии.
Терапевтические слитые белки по настоящему изобретению, например терапевтические слитые белки, содержащие домен EDIL3 по настоящему изобретению, также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести послеоперационных перитонеальных спаек.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести сердечной недостаточности.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести гемодиализа.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести отсроченной функции трансплантата или реакции "трансплантат против хозяина".
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести тяжелых обморожений, траншейной стопы, гангренозной пиодермии/гангрены.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести патологий, индуцированных бактериями, грибами, вирусами или паразитами (например, сепсиса или других патологий, непосредственно индуцируемых патогенами, такими как сибирская язва, чума, некротизирующие инфекции мягких тканей (NSTI, такие как некротизирующий фасцит), остеомиелит, малярия).
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести травмы/политравмы, вызванной несчастными случаями, приводящими к травмам, такими как несчастные случаи на производстве, падения, дорожно-транспортные происшествия, баллистические и боевые травмы или другие механизмы травмы.
Терапевтические слитые белки по настоящему изобретению также могут быть применимы для диагностики, лечения, предупреждения или облегчения тяжести патологии, опосредованной остеокластами.
Терапевтические слитые белки по настоящему изобретению можно вводить в виде единственного активного ингредиента или в комбинации, например, в качестве вспомогательного средства к или в комбинации с другими лекарственными средствами, например иммуносупрессивными или иммуномодулирующими средствами или другими противовоспалительными средствами, например цитотоксическими или противораковыми средствами, например, для лечения или предупреждения заболеваний, упомянутых выше.
Как используется в данном документе, вводимый "в комбинации", в отношении дополнительного терапевтического средства, означает, что два (или более) различных средства для лечения доставляют субъекту в период, когда субъект страдает нарушением, например, два или более средства для лечения доставляют после того, как у субъекта было диагностировано нарушение, и до того, как нарушение было излечено или устранено или лечение было прекращено по другим причинам. В некоторых вариантах осуществления доставка одного средства для лечения все еще осуществляется, когда начинается доставка второго, так что в отношении введения имеет место перекрывание. Это иногда упоминается в данном документе как "совместная" или "одновременная" доставка. В других вариантах осуществления доставка одного средства для лечения заканчивается до начала доставки другого средства для лечения. В некоторых вариантах осуществления в любом из случаев лечение является более эффективным благодаря комбинированному введению. Например, второе средство для лечения является более эффективным, например, эквивалентный эффект наблюдается при меньшем количестве второго средства для лечения, или второе средство для лечения снижает интенсивность симптомов в большей степени, чем наблюдалось бы при введении второго средства для лечения в отсутствие первого средства для лечения, или аналогичная ситуация наблюдается с первым средством для лечения. В некоторых вариантах осуществления доставка является такой, при которой снижение интенсивности симптома или другого параметра, связанного с нарушением, является большим, чем наблюдалось бы при доставке одного средства для лечения в отсутствие другого. Эффект двух средств для лечения может быть частично аддитивным, полностью аддитивным или превышающим аддитивный. Доставка может быть такой, что эффект от первого доставленного средства для лечения все еще поддается выявлению при доставке второго.
Термин "одновременно" не ограничивается введением средств терапии (например, профилактических или терапевтических средств) точно в одно и то же время, а скорее означает, что фармацевтическую композицию, содержащую терапевтические слитые белки по настоящему изобретению, вводят субъекту в такой последовательности и в течение такого интервала времени, что слитые белки могут действовать вместе с дополнительным(дополнительными) терапевтическим(терапевтическими) средством(средствами) с обеспечением большей пользы, чем если бы их вводили иным способом. Например, каждое средство терапии можно вводить субъекту в одно и то же время или последовательно в любом порядке в разные моменты времени; однако, если их не вводят в одно и то же время, их следует вводить достаточно близко друг к другу по времени, чтобы обеспечить требуемый терапевтический или профилактический эффект. Каждое средство терапии можно вводиться субъекту отдельно в любой подходящей форме и с помощью любого подходящего пути.
Терапевтический слитый белок, описанный в данном документе, и дополнительное(дополнительные) терапевтическое(терапевтические) средство(средства) можно вводить совместно, в той же самой или в отдельной фармацевтической композиции, что и описанный слитый белок, или последовательно. Для последовательного введения слитый белок, описанный в данном документе, можно вводить первым, а дополнительное средство можно вводить вторым, или порядок введения можно изменить на обратный. Дополнительный(дополнительные) терапевтический(терапевтические) средство(средства) можно вводить субъекту с помощью тех же или других путей введения по сравнению со слитым белком.
Терапевтический слитый белок, описанный в данном документе, и/или дополнительное(дополнительные) терапевтическое(терапевтические) средство(средства), процедуры или способы воздействия можно применять в течение периодов активного проявления нарушения или в течение периода ремиссии или менее активного проявления заболевания. Терапевтический слитый белок, описанный в данном документе, можно вводить перед другим средством для лечения, одновременно со средством для лечения, после средства для лечения или во время ремиссии нарушения.
При введении в комбинации терапевтический слитый белок, описанный в данном документе, и дополнительное терапевтическое средство (например, второе или третье средство) или все из них можно вводить в количестве или дозе, которые являются более высокими, более низкими или такими же по сравнению с количеством или дозой каждого средства, применяемого в отдельности, например, в качестве монотерапии. В некоторых вариантах осуществления терапевтический слитый белок, описанный в данном документе, дополнительное средство (например, второе или третье средство) или все из них составляют меньше (например, на по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40% или по меньшей мере 50%), чем количество или доза каждого средства, применяемого в отдельности, например, в качестве монотерапии. В других вариантах осуществления количество или доза терапевтического слитого белка, описанного в данном документе, дополнительного средства (например, второго или третьего средства) или их всех, которые дают требуемый эффект (например, лечение воспалительного заболевания или состояния), являются более низкими (например, на по меньшей мере 20%, по меньшей мере 30%, по меньшей мере 40% или по меньшей мере 50% более низкими), чем количество или доза каждого средства, применяемого в отдельности, например в качестве монотерапии, необходимые для достижения того же терапевтического эффекта.
Например, терапевтические слитые белки по настоящему изобретению можно применять в комбинации с DMARD, например солями золота, сульфасалазином, противомалярийными средствами, метотрексатом, D-пеницилламином, азатиоприном, микофеноловой кислотой, такролимусом, сиролимусум, миноциклином, лефлуномидом, глюкокортикоидами; ингибитором кальциневрина, например циклоспорином А или FK 506; модулятором рециркуляции лимфоцитов, например аналогами FTY720 и FTY720; ингибитором mTOR, например рапамицином, 40-O-(2-гидроксиэтил)-рапамицином, CCI779, ABT578, AP23573 или TAFA-93; аскомицином, характеризующимся иммуносупрессивными свойствами, например АВТ-281, ASM981 и т. д.; кортикостероидами; циклофосфамидом; азатиоприном; лефлуномидом; мизорибином; микофенолата мофетилом; 15-дезоксиспергуалином или его иммуносупрессивным гомологом, аналогом или производным; иммуносупрессивными моноклональными антителами, например моноклональными антителами к рецепторам лейкоцитов, например MHC, CD2, CD3, CD4, CD7, CD8, CD25, CD28, CD40, CD45, CD58, CD80, CD86 или их лигандами; другими иммуномодулирующими соединениями, например рекомбинантной связывающей молекулой, содержащей по меньшей мере часть внеклеточного домена CTLA4 или его мутанта, например по меньшей мере внеклеточную часть CTLA4 или его мутанта, присоединенную к последовательности белка, отличного от CTLA4, например, CTLA4Ig (например, обозначенного ATCC 68629) или его мутанта, например LEA29Y; ингибиторами молекул адгезии, т. е. антагонистами LFA-1, антагонистами ICAM-1 или -3, антагонистами VCAM-4 или антагонистами VLA-4; или химиотерапевтическим средством, например паклитакселом, гемцитабином, цисплатином, доксорубицином или 5-фторурацилом; средствами, направленными против TNF, т. е. моноклональными антитела к TNF, например, инфликсимабом, адалимумабом, CDP870 или конструкциями рецепторов к TNF-RI или TNF-RII, например этанерцептом, PEG-TNF-RI; блокаторами провоспалительных цитокинов, блокаторами IL-1, в т. ч. анакинра или ловушкой IL-1, канакинумабом, блокаторами IL-13, блокаторами IL-4, блокаторами IL-6; блокаторами хемокинов, например ингибиторами или активаторами протеаз, например металлопротеаз, антителами к IL-15, антителами к IL-6, антителами к IL-4, антителами к IL-13, антителами к CD20, NSAID, такими как аспирин или противоинфекционное средство; антагонистами дистресс-ассоциированного молекулярного паттерна (DAMP), или патоген-ассоциированного молекулярного паттерна (PAMP), например конверторами, детоксикантами, средствами для удаления, т. е. АТФ-конверторами, модуляторами HMGB-1, детоксикантами гистонов; ингибиторами иммунных ответов, индуцированных суперантигеном; ингибиторами комплемента и устройствами для экстракорпорального плазмафереза.
Наборы
Также в объем настоящего изобретения входят наборы, состоящие из композиций, например терапевтических слитых белков по настоящему изобретению, и инструкции по применению. Такие наборы содержат терапевтически эффективное количество слитого белка в соответствии с настоящим изобретением. Кроме того, такие наборы могут содержать средства для введения терапевтического слитого белка (например, автодозатор, шприц и флакон, предварительно заполненный шприц, предварительно заполненная ручка) и инструкции по применению. Эти наборы могут содержать дополнительные терапевтические средства (описанные ниже) для лечения пациента с аутоиммунным заболеванием или воспалительным нарушением или AOI. Такие наборы могут также содержать инструкции по введению терапевтического слитого белка для лечения пациента. В таких инструкциях может быть указана доза, способ введения, режим и общая продолжительность лечения для применения прилагаемого слитого белка. Наборы, как правило, включают этикетку, указывающую на предполагаемое применение содержимого набора. Термин "этикетка" включает любой письменный или записанный материал, поставляемый на наборе или с набором, или который иным образом сопровождает набор. Набор может дополнительно содержать инструменты для диагностики того, принадлежит ли пациент к группе, которая будет реагировать на лечение терапевтическим слитым белком по настоящему изобретению, как определено выше.
Варианты осуществления
В настоящем изобретении предусмотрены следующие варианты осуществления.
1. Терапевтический слитый белок для усиления эффероцитоза, содержащий интегрин-связывающий домен, фосфатидилсерин (PS)-связывающий домен и солюбилизирующий домен.
2. Слитый белок по варианту осуществления 1, где солюбилизирующий домен:
(i) соединен с интегрин-связывающим доменом;
(ii) соединен с PS-связывающим доменом;
(iii) вставлен между интегрин-связывающим доменом и PS-связывающим доменом;
(iv) вставлен в интегрин-связывающий домен или
(v) вставлен в PS-связывающий домен.
3. Слитый белок по варианту осуществления 1 или варианту осуществления 2, где интегрин-связывающий домен связывается с одним или несколькими интегринами.
4. Слитый белок по варианту осуществления 3, где интегрин-связывающий домен связывается с интегрином αvβ3, и/или αvβ5, и/или α8β1.
5. Слитый белок по варианту осуществления 3 или варианту осуществления 4, где интегрин-связывающий домен содержит мотив аргинин-глицин-аспарагиновая кислота (RGD).
6. Слитый белок по любому из предыдущих вариантов осуществления, где солюбилизирующий домен соединен непосредственно с интегрин-связывающим доменом, с PS-связывающим доменом или с обоими доменами.
7. Слитый белок по любому из вариантов осуществления 1-6, где солюбилизирующий домен опосредованно соединен с интегрин-связывающим доменом и/или PS-связывающим доменом с помощью линкера.
8. Слитый белок по любому из вариантов осуществления, где солюбилизирующий домен содержит сывороточный альбумин человека (HSA), домен 3 HSA (HSA D3), Fc-IgG или их функциональный вариант.
9. Слитый белок по любому из предыдущих вариантов осуществления, где солюбилизирующий домен содержит сывороточный альбумин человека (HSA) или его функциональный вариант.
7. Слитый белок по любому из предыдущих вариантов осуществления, где интегрин-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 2 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
8. Слитый белок по любому из предыдущих вариантов осуществления, где PS-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 3 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней, или PS-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 76 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
9. Слитый белок по любому из предыдущих вариантов осуществления, где солюбилизирующий домен представляет собой HSA и содержит аминокислотную последовательность под SEQ ID NO: 4 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
10. Слитый белок по любому из предыдущих вариантов осуществления, где интегрин-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 2 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней, и PS-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 78 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
11. Слитый белок по любому из предыдущих вариантов осуществления, где интегрин-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 77 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней, и PS-связывающий домен содержит аминокислотную последовательность под SEQ ID NO: 3 или под SEQ ID NO: 76 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
12. Слитый белок по любому из предыдущих вариантов осуществления, где указанный слитый белок:
a. способствует эффероцитозу эндотелиальными клетками в анализе эффероцитоза клеток Jurkat эндотелиальными клетками человека;
b. восстанавливает нарушенный эффероцитоз макрофагами в анализе эффероцитоза нейтрофилов макрофагами человека;
c. снижает количество микрочастиц в плазме крови за счет клиренса в анализе эффероцитоза микрочастиц эндотелиальными клетками человека; и/или
d. обеспечивает защиту от полиорганного повреждения в модели острого повреждения почек.
13. Слитый белок по любому из предыдущих вариантов осуществления, содержащий в последовательности интегрин-связывающий домен-HSA-PS-связывающий домен.
14. Терапевтический слитый белок, содержащий MFG-E8 и солюбилизирующий домен, где MFG-E8 содержит от N-конца к С-концу: EGF-подобный домен, домен C1 и домен C2, и содержит последовательность MFG-E8 дикого типа человека (SEQ ID NO: 1) или MFG-E8 под SEQ ID NO: 75 или его функциональный вариант.
15. Слитый белок по варианту осуществления 14, где солюбилизирующий домен вставлен между EGF-подобным доменом и доменом C1.
16. Слитый белок по варианту осуществления 14 или варианту осуществления 15, где солюбилизирующий домен представляет собой HSA, HSA D3 или Fc-IgG или их функциональный вариант.
17. Слитый белок по любому из вариантов осуществления 1-16, где сконструированный белок содержит аминокислотную последовательность под SEQ ID NO: 42 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
18. Слитый белок по любому из предыдущих вариантов осуществления, где слитый белок содержит аминокислотную последовательность под SEQ ID NO: 44 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней; или SEQ ID NO: 47 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней; или SEQ ID NO: 48 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
19. Слитый белок по любому из предыдущих вариантов осуществления, где слитый белок содержит аминокислотную последовательность под SEQ ID NO: 80 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
20. Слитый белок по любому из предыдущих вариантов осуществления, где слитый белок содержит аминокислотную последовательность под SEQ ID NO: 82 или последовательность, характеризующуюся по меньшей мере 90% идентичностью последовательности с ней.
21. Выделенная нуклеиновая кислота, кодирующая аминокислотную последовательность по любому из вариантов осуществления 17-20.
22. Вектор клонирования или экспрессии, содержащий нуклеиновую кислоту в соответствии с вариантом осуществления 21.
23. Вирусный вектор, содержащий выделенную нуклеиновую кислоту в соответствии с вариантом осуществления 21, предпочтительно вирусный вектор, содержащий выделенную нуклеиновую кислоту в соответствии с вариантом осуществления 21, получен из AAV.
24. Вирусный вектор в соответствии с вариантом осуществления 23, где вектор вводится субъекту, например человеку, нуждающемуся в этом.
25. Вирусный вектор в соответствии с вариантом осуществления 23 для применения в лечении и/или предупреждении заболеваний, приведенных в данном документе.
26. Рекомбинантная клетка-хозяин, подходящая для продуцирования терапевтического слитого белка, содержащая один или несколько векторов клонирования или экспрессии в соответствии с вариантом осуществления 22 и необязательно секреторные сигнальные последовательности.
27. Рекомбинантная клетка-хозяин по варианту осуществления 26, где клетка-хозяин представляет собой, например, прокариотическую, дрожжевую клетку, клетку насекомого или млекопитающего.
28. Слитый белок по любому из вариантов осуществления 1-20, где экспрессия белка в клетке-хозяине обеспечивает выход, составляющий по меньшей мере 10 мг/л.
29. Слитый белок по любому из вариантов осуществления 1-20, где экспрессия белка в клетке млекопитающего приводит к повышению выхода в по меньшей мере 100 раз по сравнению с MFG-E8 дикого типа (SEQ ID NO: 1).
30. Фармацевтическая композиция, содержащая слитый белок по любому из вариантов осуществления 1-20 и по меньшей мере один фармацевтически приемлемый носитель.
31. Способ лечения или предупреждения воспалительного нарушения или воспалительного поражения органов у индивидуума, нуждающегося в этом, включающий введение индивидууму терапевтически эффективного количества слитого белка по любому из вариантов осуществления 1-20.
32. Слитый белок по любому из вариантов осуществления 1-20 для применения в лечении или предупреждении воспалительного нарушения или воспалительного поражения органов у индивидуума, нуждающегося в этом.
33. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где воспалительное нарушение или воспалительное поражение органов представляет собой острое повреждение почек, сепсис, инфаркт миокарда, острый инсульт, ожоги, травматическое повреждение и воспалительные повреждения и повреждения органов, возникающие в результате ишемии/реперфузии.
34. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где воспалительное нарушение или воспалительное поражение органов представляет собой острое повреждение почек.
35. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где воспалительное нарушение или воспалительное поражение органов представляет собой инфаркт миокарда.
36. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где воспалительное нарушение или воспалительное поражение органов представляет собой инсульт.
37. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где воспалительное нарушение или воспалительное поражение органов представляет собой острое повреждение легкого (например, острый респираторный дистресс-синдром), или повреждение печени, или острое повреждение кишечника.
38. Способ по варианту осуществления 31 или применение по варианту осуществления 32, где слитый белок вводят в комбинации с другим терапевтическим средством.
39. Способ или применение по варианту осуществления 38, где другое терапевтическое средство представляет собой иммуносупрессивное средство, иммуномодулирующее средство, противовоспалительное средство, антиоксидант, противоинфекционное средство, цитотоксическое средство или противораковое средство.
40. Терапевтический слитый белок, содержащий MFG-E8 и солюбилизирующий домен, где MFG-E8 содержит от N-конца к С-концу: EGF-подобный домен, домен C1 и домен C2, и содержит последовательность MFG-E8 дикого типа человека (SEQ ID NO: 1) или SEQ ID NO: 75 или его функциональный вариант.
41. Слитый белок по варианту осуществления 40, где солюбилизирующий домен соединен с N-концом или C-концом MFG-E8 (SEQ ID NO: 1 или SEQ ID NO: 75).
42. Слитый белок по варианту осуществления 40, где солюбилизирующий домен вставлен между EGF-подобным доменом и доменом C1.
43. Слитый белок по варианту осуществления 41, где солюбилизирующий домен вставлен между доменом C1 и доменом C2.
44. Слитый белок по любому из вариантов осуществления 40-43, где солюбилизирующий домен представляет собой HSA, D3 HSA или Fc-IgG или их функциональный вариант.
38. Выделенная нуклеиновая кислота, кодирующая слитый белок по любому из вариантов осуществления 33-37.
39. Вектор клонирования или экспрессии, содержащий нуклеиновую кислоту в соответствии с вариантом осуществления 38.
40. Вирусный вектор, содержащий выделенную нуклеиновую кислоту в соответствии с вариантом осуществления 38, предпочтительно вирусный вектор, содержащий выделенную нуклеиновую кислоту в соответствии с вариантом осуществления 38, получен из AAV.
41. Вирусный вектор в соответствии с вариантом осуществления 40, где вектор вводится субъекту, например человеку, нуждающемуся в этом.
42. Вирусный вектор в соответствии с вариантом осуществления 40 для применения в лечении и/или предупреждении заболеваний, приведенных в данном документе.
43. Рекомбинантная клетка-хозяин, подходящая для продуцирования терапевтического слитого белка, содержащая один или несколько векторов клонирования или экспрессии в соответствии с вариантом осуществления 39 и необязательно секреторные сигнальные последовательности.
44. Рекомбинантная клетка-хозяин по варианту осуществления 43, где клетка-хозяин представляет собой, например, прокариотическую, дрожжевую клетку, клетку насекомого или млекопитающего.
45. Слитый белок по любому из вариантов осуществления 33-37, где экспрессия белка в клетке-хозяине обеспечивает выход, составляющий по меньшей мере 10 мг/л.
46. Слитый белок по любому из вариантов осуществления 33-37, где экспрессия белка в клетке млекопитающего приводит к повышению выхода в по меньшей мере 100 раз по сравнению с MFG-E8 дикого типа.
Следует понимать, что каждый вариант осуществления может быть объединен с одним или несколькими другими вариантами осуществления в той степени, в которой такая комбинация соответствует описанию вариантов осуществления. Кроме того, следует понимать, что представленные выше варианты осуществления понимаются как включающие все варианты осуществления, в том числе такие варианты осуществления, которые являются результатом комбинаций вариантов осуществления.
Все литературные источники, процитированные в данном документе, включая патенты, заявки на патенты, бумажная документация, публикации, учебные пособия и т.п., а также литературные источники, процитированные в них, в тех случаях, когда они еще не упоминались, настоящим включены в данный документ посредством ссылки во всей своей полноте.
Примеры
Следующие примеры предусмотрены для дополнительной иллюстрации настоящего изобретения, но не для ограничения его объема. Другие варианты настоящего изобретения будут полностью очевидны специалисту средней квалификации в данной области техники и охвачены прилагаемой формулой изобретения.
Пример 1. Получение слитых белков
MFG-E8 представляет собой мультидоменный белок, состоящий из N-концевого домена эпидермального фактора роста (EGF-подобного) и двух С-концевых доменов C лектинового типа (C1 и C2). Попытки получить рекомбинантный полноразмерный белок человека, как описано в литературе, продемонстрировали, что белок агрегирует и уровни экспрессии являются очень низкими (Castellanos et al., (2016) Protein Expression Purification 1124: 10-22). Поэтому, для попытки растворения белка и повышения его экспрессии, авторами настоящего изобретения был исследован эффект слияния ряда белков с MFG-E8.
Солюбилизирующий домен (SD), полученный из Fc-IgG1 человека, сывороточного альбумина человека (HSA) и домена 3 HSA (D3 HSA), сливали с MFG-E8 в разных положениях: на N- или С-конце или между доменами EGF и C1 или C1 и C2, как схематически изображено на фигуре 1. Кроме того, продукты слияния с Fc-IgG1 или HSA потенциально могут приводить к продлению времени полужизни молекулы in vivo, поскольку эти белки связываются с FcRn. Слияние MFG-E8 с Fc-IgG1 или HSA также может обеспечивать повышение уровня продуцирования и растворимости (Castellanos et al., (2016), выше) слитого белка, как показано в следующих примерах.
В таблице 5 показано связывание слитого белка FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42), содержащего вставку HSA, с неонатальным Fc-рецептором человека (см. также пример 5.1).
Таблица 5. Аффинность связывания слитого белка FP330 с FcRn человека
(pH 5,8)
(pH 7,4)
Пример 2. Получение продуктов слияния HSA с wtMFG-E8 и MFG-E8; экспрессия и очистка
Способы получения слитых белков описаны ниже; вкратце, MFG-E8 и продукты слияния с MFG-E8 и продукты слияния с EDIL, в частности продукты слияния с HSA, получали в соответствии со следующим способом.
ДНК синтезировали в GeneArt (Регенсбург, Германия) и клонировали в вектор экспрессии млекопитающих с применением методик клонирования, основанных на лигировании с использованием рестрикционных ферментов. Полученной плазмидой трансфицировали клетки HEK293T. Для временной экспрессии белков векторы для цепей дикого типа или сконструированных цепей трансфицировали в адаптированные к культивированию в суспензии клетки HEK293T с использованием полиэтиленимина (PEI; кат. № 24765, Polysciences, Inc.). Как правило, 100 мл клеток в суспензии с плотностью 1-2 млн. клеток на мл трансфицировали с использованием ДНК, содержащей 100мкг векторов экспрессии, кодирующих сконструированные цепи. Затем рекомбинантные векторы экспрессии вводили в клетки-хозяева, а конструкцию получали посредством дальнейшего культивирования клеток в течение 7 дней для обеспечения секреции в среду для культивирования (НЕК, бессывороточная среда) с добавлением 0,1% плюроновой кислоты, 4 мМ глутамина и 0,25 мкг/мл антибиотика.
Затем полученные конструкции очищали из бесклеточного супернатанта с применением аффинной хроматографии с иммобилизованными ионами металлов (IMAC) или хроматографии с использованием захвата белком А или захвата антителом к HSA.
После захвата меченого his белка с помощью IMAC, отфильтрованную кондиционированную среду смешивали со смолой IMAC (GE Healthcare), уравновешенной с помощью 1% тритона и 20 мМ NaPO4, 0,5 Мн NaCl, 20 мМ имидазола, рН 7,0. Смолу трижды промывали 15 колоночными объемами 20 мМ NaPO4, 0,5 Мн NaCl, 20 мМ имидазола, рН 7,0, до того, как белок элюировали 10 колоночными объемами элюирующего буфера (20 мМ NaPO4, 0,5 Мн NaCl, 500 мМ имидазола, рН 7,0).
Когда белок улавливали посредством хроматографии с использованием белка А или антитела к HSA, отфильтрованную кондиционированную среду смешивали со смолой с белком А (CaptivA PriMab™, Repligen) или смолой с антителом к HSA (матрица аффинности для альбумина человека Capture Select, Thermo), уравновешенной с помощью PBS, pH 7,4. Смолу трижды промывали 15 колоночными объемами PBS, pH 7,4, до того, как белок элюировали 10 колоночными объемами элюирующего буфера (50 мМ цитрата, 90 мМ NaCl, pH 2,5) и pH нейтрализовали с использованием 1 M TRIS pH 10,0.
В конечном итоге осуществляли тонкую очистку элюированных фракций путем применения эксклюзионной хроматографии (HiPrep Superdex 200, 16/60, GE Healthcare Life Sciences) и анализировали посредством SDS-PAGE в отношении стандартных маркеров Precision Plus Protein Unstained (Biorad, рег. № 161-0363).
Типичные гели для анализа экспрессии слитых белков изображены на фигуре 2: Фиг. 2A: белок EGF-HSA-C1-C2 (FP330; SEQ ID NO: 42); фиг. 2B: белок EGF-HSA-C1-C2 из EDIL3 (FP050; SEQ ID NO: 12); фиг. 2C: белок EGF-Fc(KiH)-C1-C2, невосстановленный и восстановленный. Этот белок представляет собой гетеродимер FP071 (EGF-Fc(выступ)-C1-C2; SEQ ID NO: 18) с впадиной Fc-IgG1 (SEQ ID NO: 10); фиг. 2D: белок EGF-HSA-C1 (FP260; SEQ ID NO: 34). Белок в восстанавливающих и невосстанавливающих условиях изображен на фиг. 2C, поскольку гетеродимеры подвержены распаду в восстанавливающих условиях, поэтому тестировали оба состояния. Результаты экспрессии и выход после очистки дополнительной совокупности слитых белков показаны в таблице 6; как видно из данных по экспрессии, продукты слияния с HSA, полученные из MFG-E8, даже с HSA в других положениях, демонстрируют по меньшей мере 100-кратное улучшение в отношении экспрессии по сравнению с wtMFG-E8. Как показано в правом столбце таблицы 6, продукты слияния HSA и MFG-E8 также демонстрируют по меньшей мере 100-кратное повышение выхода по сравнению с wtMFG-E8.
Таблица 6. Экспрессия и выход слитых белков, экспрессируемых в линии клеток НЕК
Другие примеры терапевтических слитых белков по настоящему изобретению получали в соответствии с вышеуказанным способом и дополнительно анализировали посредством SDS-PAGE (электрофорез в полиакриламидном геле с додецилсульфатом натрия), где белки разделяли на основе их молекулярной массы. Каждый белок смешивали с буфером Лэммли перед нанесением на полиакриламидный гель (Biorad, 4-20% Mini-PROTEAN TGX Stain free). После 30 мин. миграции при 200 В в подвижном буфере TRIS-Glycine-SDS белки, содержащиеся в геле, выявляли в сканере без окрашивания (Biorad, Gel Doc EZ). Как изображено на фигуре 2E, на SDS-PAGE видны рекомбинантные белки, которые получили и очистили:
Полоса 1, 12: Маркер молекулярной массы (Biorad, Precision Plus Protein)
Полоса 2: His6_EGF[MFG-E8]_C1[MFG-E8] 23,87 кДа
Полоса 3: EGF[MFG-E8]_C1[MFG-E8]_His6 SEQ ID 115 23,87 кДа
Полоса 4: EGF[MFG-E8]_HSA_C1[MFG-E8] SEQ ID 117 90,38 кДа
Полоса 5: EGF[MFG-E8]_HSA_C1[MFG-E8] SEQ ID 74 89,27 кДа
Полоса 6: EGF[MFG-E8]_HSA_C1[MFG-E8] SEQ ID 73 88,72 кДа
Полоса 7: EGF[EDIL3]_HSA_C1[EDIL3] SEQ ID 71 98,22 кДа
Полоса 8: EGF[EDIL3]_HSA_C2[EDIL3] SEQ ID 135 98,20 кДа
Полоса 9: EGF[MFG-E8]_HSA_C2[MFG-E8] SEQ ID 137 88,45 кДа
Полоса 10: EGF[EDIL3]_HSA_C1_C2[MFG-E8] SEQ ID 80 115,67 кДа
Полоса 11: EGF[MFG-E8]_HSA_C1_C2[EDIL3] SEQ ID 82 107,32 кДа
Пример 3. Определение характеристик сконструированных белков MFG-E8-HSA
3.1 Связывание фосфатидилсерина (биохимическое)
L-α-фосфатидилсерин (головной мозг, свиной, Avanti 840032, Алабама, США) растворяли в хлороформе, разбавляли в метаноле и наносили на 384-луночные планшеты для микротитрования (Corning™ 3653, Кеннебанк, Мэн, США) в концентрации 1 мкг/мл. После инкубации в течение ночи при 4°С растворитель выпаривали с использованием системы SpeedVac™ (Thermo Scientific™). Планшеты обрабатывали фосфатно-солевым буфером (PBS), содержащим 3% бычьего сывороточного альбумина (BSA), не содержащего жирных кислот, при к. т. в течение 1,5 ч.
Связывание слитых белков с L-α-фосфатидилсерином оценивали посредством конкуренции со связыванием биотинилированного мышиного MFG-E8/лактадгерина (собственного производства, mMFG-E8:биотин). Белки разбавляли в PBS, содержащем 3% BSA, не содержащем жирные кислоты, pH 7,4, и инкубировали с микротитрационными планшетами, покрытыми L-α-фосфатидилсерином, в течение 30 мин. mMFG-E8:биотин в PBS, содержащем 3% BSA, не содержащем жирные кислоты, pH 7,4, добавляли в количестве 1 нМ и инкубировали в течение дополнительных 30 мин. Несвязанный mMFG-E8:биотин удаляли с помощью трех стадий промывания промывочным буфером для усиленного диссоциацией лантанидного флюоресцентного иммуноанализа (DELFIA™) (Perkin Elmer 1244-114 Массачусетс, США). Меченный европием стрептавидин (Perkin Elmer 1244-360, Wallac Oy, Финляндия) добавляли в аналитический буфер DELFIA™ (Perkin Elmer 1244-111, Массачусетс, США) при к. т. в течение 20 мин. После этого осуществляли три стадии промывания аналитическим буфером DELFIA™. Европий выявляли в соответствии с инструкциями производителя (Perkin Elmer 1244-105, Бостон, штат Массачусетс, США). Флуоресценцию европия с временным разрешением определяли количественно с помощью планшет-ридера Envision™2103 с несколькими метками, Perkin Elmer, штат Коннектикут, США). Анализ данных проводили с применением программного обеспечения MS Excel и GraphPad Prism.
Полипропиленовые планшеты представляют собой микротитрационные планшеты с низким уровнем связывания белка, которые обычно применяются в лабораториях для серийных разведений. По сравнению с полистироловыми, эти планшеты характеризуются тем преимуществом, что приводят к снижению потери белка во время разведения, и, как правило, классифицируются как планшеты с низким уровнем связывания белка. Когда разведения wtMFG-E8 готовили в полипропиленовых планшетах, по сравнению с разведениями, выполненными в несвязывающих планшетах, wtMFG-E8 утрачивал активность в конкурентном анализе связывания L-α-фосфатидилсерина. Эти данные, как изображено на фигуре 3, предполагают, что wtMFG-E8 частично утрачивается во время работы с жидкостью и стадий разведения при использовании полипропиленовых планшетов, которые уже были оптимизированы для обеспечения низкого уровня связывания белка (фиг. 3A). Эти результаты указывают на то, что присущая wtMFG-E8 липкость представляет собой проблему при работе в лаборатории и, вероятнее всего, во время изготовления и производства лекарственных средств, где требуются стадии захвата и тонкой очистки для получения лекарственной субстанции с высоким выходом и очень высокой чистотой. В отличие от этого, липкость сконструированного белка FP278 (метка EGF-HSA-C1-C2-His; SEQ ID NO: 44) была значительно снижена по сравнению с таковой wtMFG-E8, и разницы между разведениями, выполненными в несвязывающих планшетах и полипропиленовых планшетах, практически не наблюдали (фиг. 3B). Эти данные свидетельствуют о том, что вставка солюбилизирующего домена в белки по настоящему изобретению может приводить к улучшению их технической обработки для повышения выхода на стадиях и, таким образом, общего выхода в процессе изготовления.
Оценка связывания слитых белков с L-α-фосфатидилсерином изображена на фигуре 4. Сконструированный белок FP278, происходящий из MFG-E8 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44), связывался с иммобилизованным PS и в меньшей степени с фосфолипидом кардиолипином зависимым от концентрации образом (фиг. 4A). Связывание FP278 с иммобилизованным L-α-фосфатидилсерином или связывание с кардиолипином (1,3-бис(sn-3'-фосфатидил)-sn-глицерин) определяли с использованием антитела к домену EGF-L wtMFG-E8. Сила связывания нескольких рекомбинантных слитых белков с иммобилизованным L-α-фосфатидилсерином изображена на фиг. 4В. wtMFG-E8 человека и слитые белки FP278 (метка EGF-HSA-C1-C2-His; SEQ ID NO: 44) и FP260 (EGF-HSA-C1; SEQ ID NO: 34) эффективно конкурировали за связывание с 1 нМ биотинилированного мышиного MFG-E8 за иммобилизованный L-α-фосфатидилсерин зависимым от концентрации образом. Значения IC50, полученные для слитых белков, означают, что значения силы связывания L-α-фосфатидилсерина с доменами C1-C2 сконструированного белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) являются очень сходными с таковыми для wtMFG-E8 человека. Неожиданно эти данные также свидетельствуют о том, что домен C2 человека не взаимодействует или лишь слабо взаимодействует с L-α-фосфатидилсерином, как продемонстрировано результатом для FP270 (EGF-HSA-C2; SEQ ID NO: 36), который наряду с FP250 (EGF-HSA; SEQ ID NO: 32) не конкурировал в этом формате анализа. FP100, белок EGF-C2-C2 (SEQ ID NO: 26) тестировали, и он не конкурировал в этом формате анализа (не продемонстрировано), следовательно домен C1 является основным PS-связывающим фрагментом в MFG-E8 человека. Этот результат был неожиданным, поскольку большая часть литературных данных предполагает, что домен C2 MFG-E8 представляет собой основной домен, ответственный за связывание PS (Andersen et al., (2000) Biochemistry, 39(20): 6200-6; Shi & Gilbert (2003) Blood, 101: 2628-2636; Shao et al., (2008) J Biol Chem., 283(11): 7230-41). В заключение, эти результаты демонстрируют, что домен C1 представляет собой основной интегральный PS-связывающий домен сконструированных белков MFG-E8 и является важным для зависимых от связывания PS функций. Таким образом, домен C1 может быть применимым для замещения в гетерологичных белках для сообщения PS-связывающей способности; однако наиболее высокую PS-связывающую способность продемонстрировали для слитых белков, содержащих тандемный домен C1-C2 или C1-C1 (последний не изображен).
3.2 Анализ опосредованной интегрином адгезии αv
Слитые белки разбавляли в фосфатно-солевом буфере (PBS), pH 7,4, и 50 мкл 24 нМ раствора иммобилизовали посредством адсорбции (96-луночный планшет, Nunc Maxisorb) в течение ночи (1,2 нМ/лунка). Затем планшеты обрабатывали с помощью PBS, содержащего 3% бычьего сывороточного альбумина (BSA), не содержащего жирные кислоты, при к. т. в течение 1,5 ч. Клетки лимфомы, экспрессирующие интегрин αvβ3 (ATCC-TIB-48 BW5147.G.1.4, ATCC, США), культивировали в среде RPMI 1640 с добавлением GlutaMax, 25 мМ HEPES, 10% FBS, Pen/Strep, 1 мМ пирувата Na, 50 мкМ β-меркаптоэтанола. Клетки отсоединяли за день до эксперимента на адгезию. Клетки метили с помощью 3 мкг/мл сложного ацетоксиметилового эфира 2',7'-бис-(2-карбоксиэтил)-5-(-6-)карбоксифлуоресцеина (BCECF AM) (Thermo Fisher Scientific Inc, США) в течение 30 мин. Клетки BW5147.G.1.4 ресуспендировали в буфере для адгезии (TBS, 0,5% BSA, 1 мМ MnCl2, pH 7,4) и 50000 клеток/лунка оставляли для прикрепления при к. т. в течение 40 мин. Не прикрепившиеся клетки удаляли повторным промыванием буфером для адгезии. Флуоресценцию прикрепившихся клеток количественно определяли с использованием многофункционального планшет-ридера Envision™2103, Perkin Elmer, США. Анализ данных проводили с применением программного обеспечения MS Excel и GraphPad Prism.
Адгезия клеток к иммобилизованному слитому белку FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42) полностью блокировалась ингибитором интегрина αv циленгитидом или 10 мМ EDTA, демонстрируя зависимую от интегрина адгезию клеток к иммобилизованному сконструированному белку (фиг. 5А). Единственная точечная мутация в интегрин-связывающем мотиве RGD (RGD > RGE) EGF-подобного домена (FP280; SEQ ID NO: 38) приводила к полному устранению адгезии клеток, демонстрируя, что функциональный и доступный RGD-связывающий мотив в слитом белке необходим для зависимой от интегрина αv адгезии (фиг. 5B). Иммобилизованный белок EGF-HSA, не содержащий доменов C1-C2, FP250 (SEQ ID NO: 32), не способствовал адгезии клеток BW5147.G.1.4 или способствовал ей лишь в незначительной мере, несмотря на наличие EGF-подобного домена (фиг. 5C). Этот результат позволяет предположить, что в экспериментальных условиях тестирования петля RGD в EGF-подобном домене, слитом с HSA, может быть недостаточно доступной для интегринов клеточной поверхности, возможно, из-за причин, связанных со стерическим эффектом. Это нарушение не проявлялось, когда С1, С2 или С1-С2 сливались с EGF-HSA в С-концевом положении. Рекомбинантные белки по настоящему изобретению, например FP330, способствуют зависимой от интегрина αv адгезии клеток, сходной с таковой в случае wtMFG-E8 при экспрессии в клетках CHO или клетках HEK (фиг. 5D).
В совокупности эти данные демонстрируют, что слитые белки по настоящему изобретению связываются с клеточными интегринами, способствуют зависимой от интегринов адгезии клеток, и указывают на то, что в белках со вставкой домена HSA С-концевой EGF-подобный домен может иметь функциональные преимущества, обусловленные наличием слитого с C-концом белкового домена, для способствования связыванию интегрина.
3.3 Анализ эффероцитоза нейтрофилов макрофагами человека
Мононуклеарные клетки периферической крови человека (PBMC) выделяли из лейкоцитарной пленки посредством центрифугирования в градиенте фиколла (Ficoll®-Paque PLUS, GE Healthcare, Швеция) с последующей отрицательной селекцией моноцитов с использованием набора для выделения стволовых клеток (Stemcell 19059, Ванкувер, Канада). Дифференцирование моноцитов в макрофаги "M0" осуществляли с использованием рекомбинантного M-CSF человека, 40 нг/мл (Macrophage Colony Stimulating Factor, R&D Systems, США) в RPMI 1640, содержащем 25 мМ HEPES, 10% FBS, Pen/Strep, 1 мМ NaPyr, 50 мкМ β-Мерк в течение 5 дней. За день до эффероцитоза макрофаги метили с помощью PKH26 с применением набора Red Fluorescent Dye Linker (Sigma MINI26, США). Клетки ресуспендировали в RPMI 1640, содержащем 25 мМ HEPES, 10% FBS, Pen/Strep, 1 мМ NaPyr, 50 мкМ β-Merc, высевали в 96-луночные планшеты черного цвета (Corning, США) в количестве 40000 клеток/лунка и оставляли для прикрепления на 20 ч.
Нейтрофилы. Нейтрофилы человека выделяли из лейкоцитарных пленок посредством осаждения декстраном в комбинации с градиентом плотности в Ficoll™ следующим образом. Плазму крови из лейкоцитарного слоя удаляли центрифугированием разведенного лейкоцитарного слоя. Собранные клетки разбавляли 1% декстраном (из Leuconostoc spp., MW 450000-650000; Sigma, США) и оставляли для осаждения на льду на 20-30 мин.
Лейкоциты из супернатанта собирали и наносили на слой Ficoll™-Paque (GE Healthcare, Швеция). После центрифугирования осадок собирали, а оставшиеся эритроциты лизировали с использованием буфера для лизиса эритроцитов (RBC) (BioConcept, Швейцария). Нейтрофилы однократно промывали средой (RPMI 1640+GlutaMax, содержащей 25 мМ HEPES, 10% FBS, Pen/Strep, 0,1 мМ NaPyr, 50 мкМ b-Merc) и оставляли на ночь при 15°C. Апоптоз/гибель клеток индуцировали обработкой нейтрофилов с помощью 1 мкг/мл Superfas Ligand (Enzo Life Sciences, Лозанна, Швейцария) при 37°C в течение 3 ч. Нейтрофилы окрашивали как с помощью Hoechst 33342 (Life Technologies, США) в течение 25 мин., так и с помощью DRAQ5 (eBioscience, Великобритания, разведение 1:2000) при 37°С в темноте в течение 5 мин.
Анализ эффероцитоза
Макрофаги М0 инкубировали со слитыми белками в течение 30 мин. Апоптотические меченые нейтрофилы добавляли в соотношении М0/нейтрофил, составляющем 1:4. Эффероцитоз апоптотических нейтрофилов макрофагами визуализировали с использованием увеличения интенсивности флуоресценции DRAQ5 при локализации нейтрофилов в лизосомальном компартменте макрофагов M0 с низким pH.
Эффероцитоз количественно определяли с применением широкопольной системы одновременного многопараметрического анализа ImageXpress Micro XLS (Molecular DEVICES. Калифорния, США). Макрофаги идентифицировали посредством флуоресценции РКН26. Индекс эффероцитоза (EI, отображаемый в %) рассчитывали в виде соотношения количества макрофагов, содержащих по меньшей мере одно событие поглощенного апоптотического нейтрофила (DRAQ5high), к общему количеству макрофагов. Анализ данных проводили с применением программного обеспечения MS Excel и GraphPad Prism.
Эффект слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в отношении способствования эффероцитозу погибающих нейтрофилов макрофагами человека изображен на фигуре 6. Слитые белки обеспечивают повышение уровня интернализации погибающих нейтрофилов человека, меченных pHrodo, в макрофаги по сравнению с уже высокой способностью к эффероцитозу макрофагов M0, изображенной в качестве исходного уровня. На фигуре 7 изображено, что рекомбинантный слитый белок FP278 может обеспечивать восстановление нарушенного эндотоксином (липополисахаридом) эффероцитоза умирающих нейтрофилов макрофагами человека. На фиг. 7А изображено нарушение эффероцитоза погибающих нейтрофилов человека макрофагами в присутствии 100 мкг/мл липополисахарида (LPS) у трех доноров-людей. На левой панели изображен ответ отдельного донора, на правой панели изображено среднее значение нарушения эффероцитоза (%) для трех доноров. На фиг. 7В изображено восстановление этого нарушенного эндотоксином (LPS) эффероцитоза погибающих нейтрофилов макрофагами человека с помощью слитого белка FP278.
Восстановление нарушенного частицами S. Aureus эффероцитоза погибающих нейтрофилов макрофагами человека с помощью слитого белка FP330 изображено на фигуре 8. На фиг. 8А изображен эффект слитого белка в концентрации 100 нМ в отношении стимулирования эффероцитоза выше исходного уровня (пунктирная линия; левая часть фигуры), а также эффект 100 нМ слитого белка в отношении восстановления нарушения эффероцитоза, вызванного добавлением S. aureus (правая часть фигуры). На фиг. 8B изображен эффект повышения концентрации слитого белка FP278 (EC50 8 нМ) в отношении восстановления нарушенного эффероцитоза, вызванного добавлением S. aureus, и в отношении способствования эффероцитозу после достижения исходных уровней эффероцитоза.
3.4 Анализ эффероцитоза клеток Jurkat эндотелиальными клетками человека
Культура клеток
Эндотелиальные клетки пупочной вены человека (HUVEC) получали от Lonza (Базель, Швейцария). Клетки культивировали в колбах, покрытых желатином (из бычьей кожи, конечная концентрация 0,2% в PBS, разбавление 2% исходным раствором, Sigma, Германия). Клетки выращивали на среде для культивирования 199 (Thermo Fischer Scientific, США) с добавлением 10% FBS (GE Healthcare, Великобритания), 1% Pen/Strep (Thermo Fischer Scientific, США), 1% Glutamax (Thermo Fischer Scientific, США) и 1 нг/мл рекомбинантного основного фактора роста фибробластов (Peprotech, Великобритания). Клетки отделяли для сбора или пассирования с использованием Accutase™ (Thermo Fischer Scientific, США).
Клетки Jurkat E6-1 получали из ATCC (Американская коллекция типовых культур, США) и выращивали в среде для культивирования RPMI 1640 (Thermo Fischer Scientific, США) с добавлением 10% FBS (GE Healthcare, Великобритания), 1% Pen/Strep (Thermo Fischer Scientific, США), 10 мМ пирувата натрия (Thermo Fischer Scientific, США) и 10 мМ HEPES (4-(2-гидроксиэтил)-1-пиперазинэтансульфокислоты, Thermo Fischer Scientific, США).
Апоптоз клеток Jurkat E6-1 индуцировали с использованием рекомбинантного TRAIL человека (R&D Systems, США). Апоптотические клетки метили сложноэфирным красителем pHrodo™ Green STP (Thermo Fischer Scientific, США). Буфер для проточной цитометрии готовили с использованием PBS (Thermo Fischer Scientific, США) с добавлением 1% FBS (GE Healthcare, Великобритания), 0,05% вес./об. азида натрия (Merck, Германия) и 0,5 мМ EDTA (этилендиаминтетрауксусной кислоты, Thermo Fischer Scientific, США).
Анализ эффероцитоза
В день 1 HUVEC (конфлюэнтность 70-90%) собирали посредством отделения с помощью Accutase™ в течение 5 минут, промывали с помощью PBS и ресуспендировали в среде для культивирования клеток. Численность и жизнеспособность клеток оценивали с применением проточного цитометра Guava EasyCyte (Merck, Германия) и реагента Guava ViaCount (Merck, Германия) в соответствии с инструкциями производителя. Необходимое количество клеток центрифугировали при 300×g в течение 5 мин. при к. т. и ресуспендировали в среде для культивирования с получением количества клеток, составляющего 6,6×104 клеток/мл. 150 мкл/лунка этой клеточной суспензии добавляли в 96-луночные планшеты для культуры тканей (Corning™, США). HUVEC инкубировали в инкубаторе при 37 °C/5% CO2/95% влажности в течение дополнительных 16-20 часов.
Количество клеток Jurkat E6-1 и статус жизнеспособности/гибели клеток оценивали с применением проточного цитометра Guava EasyCyte (Merck, Германия) и реагента Guava ViaCount (Merck, Германия) в соответствии с инструкциями производителя. Необходимое количество клеток центрифугировали при 300×g в течение 5 мин. при к. т. и ресуспендировали при плотности 1×106 клеток/мл в среде для культивирования с добавлением рекомбинантного TRAIL человека в конечной концентрации 50 нг/мл. Гибель клеток индуцировали при 37 °C/5% CO2/95% влажности в течение ночи.
В день 2 среду удаляли из HUVEC путем аспирации и добавляли 25 мкл свежей, предварительно нагретой (37°C) среды для культивирования с последующим добавлением 25 мкл слитого белка или контролей, разведенных в предварительно нагретой (37°C) среде для культивирования. Для разведения использовали 96-луночные обработанные планшеты с несвязывающей поверхностью (NBS) (Corning™, США). Слитые белки оставляли для взаимодействия с HUVEC на 30 мин. при 37°C/5% CO2/95% влажности перед добавлением погибающих клеток Jurkat.
Количество апоптотических/погибающих клеток Jurkat E6-1 подсчитывали с применением проточного цитометра Guava EasyCyte (Merck, Германия) и реагента Guava ViaCount (Merck, Германия). Необходимое количество апоптотических клеток центрифугировали при 400xg при к. т. в течение 5 мин. и ресуспендировали при плотности 5×106 клеток/мл в среде RPMI 1640 (без FBS) с добавлением сложноэфирного красителя pHrodo™ Green STP в конечной концентрации 5 мкг/мл (окрашивающая среда). После окрашивания в течение 10 мин. при 37°C оставшийся реакционноспособный сложный эфир pHrodo™ Green STP инактивировали окрашивающей средой с добавлением 10% FBS в течение дополнительных 5 мин. при 37°C. Клетки, меченные pHrodo™ Green, промывали один раз и количество клеток доводили до 3×106 клеток/мл в среде для культивирования HUVEC. 1,5×106/лунка клеток Jurkat, меченых pHrodo™ Green добавляли к HUVEC и инкубировали при 37°C/5% CO2/95% влажности в течение 5 ч. Среду удаляли, клетки HUVEC однократно промывали в PBS и отделяли с помощью 40 мкл/лунка раствора Accutase™. Клетки собирали посредством добавления 80 мкл ледяного буфера для проточной цитометрии, переносили в 1,5 мл полипропиленового 96-луночного блока, промывали избытком ледяного буфера для проточной цитометрии и центрифугировали при 400xg (4°C) в течение 5 мин. Супернатанты удаляли путем аспирации, а осадки ресуспендировали в 80 мкл ледяного буфера для проточной цитометрии и переносили в 96-луночный микротитрационный планшет с V-образным дном (BD Biosciences, США). Затем образцы измеряли на проточном цитометре BD LSRFortessa™ (BD Biosciences, США). Регистрировали интенсивность флуоресценции pHrodo™ Green в качестве индикатора лизосомальной локализации поглощенных клеток Jurkat. Анализ данных проточной цитометрии проводили с применением программного обеспечения FlowJo™. Медианные значения интенсивности флуоресценции (MFI) сигнала pHrodo™ Green от синглетно гейтированных HUVEC использовали в качестве регистрируемого показателя. Анализ данных проводили с применением программного обеспечения MS Excel и GraphPad Prism для расчета EC50.
Эффект слитых белков FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) и FP270 (EGF-HSA-C2; SEQ ID NO: 36) в отношении способствования эффероцитозу погибающих клеток Jurkat эндотелиальными клетками HUVEC изображен на фигуре 9. Интернализации меченых pHrodo погибающих Т-клеток Jurkat человека клетками HUVEC в значительной степени способствует слитый белок FP278. Результаты демонстрируют, что эндотелиальные клетки вооружаются слитым белком, становясь таким образом эффективными фагоцитами погибающих клеток. Удивительно, но эффективность слитых белков в этом анализе отчетливо зависит от присутствия тандемного домена C1-C2 или C1-C1. Слитый белок, состоящий из EGF-HSA-C2 (FP270), например, является неактивным в этих экспериментальных условиях, как изображено на фигуре 9. На фигуре 10 продемонстрирован весьма неожиданный результат авторов настоящего изобретения, свидетельствующий о том, что расположение домена HSA в сконструированных белках, а именно в N- или C-концевом положении (HSA-EGF-C1-C2 (FP220; SEQ ID NO: 30) или EGF-C1-C2-HSA (FP110; SEQ ID NO: 28) соответственно), сообщает сконструированным белкам на основе MFG-E8 и HSA способность блокировать эффероцитоз в анализе эффероцитоза посредством макрофагов. Эти данные ясно демонстрируют важность расположения домена HSA между интегрин-связывающим и PS-связывающим доменами для эффективного стимулирования эффероцитоза посредством слитых белков по настоящему изобретению.
На фигуре 11 изображено сравнение различных форматов слитых белков, содержащих комбинации домена EGF, домена C1-C2, HSA или Fc-домена, в отношении способствования эффероцитозу эндотелиальными клетками. На фиг. 11А изображено сравнение слитых белков, содержащих HSA, с HSA, расположенным на С-конце или N-конце, или между EGF-подобным доменом и доменом C1-C2; EGF-C1-C2-HSA (FP110; SEQ ID NO: 28), HSA-EGF-C1-C2 (FP220; SEQ ID NO: 30) и EGF-HSA-C1-C2-His-метка (FP278; SEQ ID NO: 44) соответственно. На фиг. 11В изображено сравнение слитых белков, содержащих Fc-домен, с Fc, расположенным на С-конце или между EGF-подобным доменом и доменом C1. Изображены два формата фрагмента Fc: Fc дикого типа (SEQ ID NO: 7), обнаруживаемый в FP070 (EGF-Fc-C1-C2; SEQ ID NO: 17) и FP080 (EGF-C1-C2-Fc; SEQ ID NO: 22), и фрагменты Fc с модификациями KiH S354C и T366W на одном плече Fc (FP060; EGF-C1-C2-Fc [S354C, T366W]; SEQ ID NO: 14), нумерация согласно EU (Merchant et al (1998), выше). На фиг. 11C изображено сравнение слитых белков FP090 (Fc-EGF-C1-C2; SEQ ID NO: 24), содержащих фрагмент Fc, расположенный на N-конце, для трех партий FP090 в трех различных концентрациях (0,72, 7,2 и 72 нМ) по сравнению с контролем wtMFG-E8. Эффероцитозу погибающих клеток Jurkat, осуществляемому HUVEC, способствовали только сконструированными белками с фрагментом HSA или Fc, вставленным после EGF-подобного домена. На фиг. 11D изображено, что вставка солюбилизирующего домена может приводить к образованию нового биоактивного слитого белка на основе эндогенного мостикового белка EDIL3, паралога MFG-E8. Как изображено на фиг. 11D, HSA был вставлен между EGF-подобным доменом и доменом C1-C2 EDIL3, паралогом MFG-E8. Эта конструкция EDIL3 (FP050 (EGF-HSA-C1-C2 на основе EDIL3; SEQ ID NO: 12) содержит только один (содержащий петлю RGD) из 3 EGF-подобных доменов, которые обнаруживаются в wtEDIL3. В этой конструкции авторы настоящего изобретения неожиданно обнаружили сходную переносимость вставки домена HSA в отношении экспрессии нового рекомбинантного сконструированного белка с очень высокой степенью чистоты (фиг. 2B). В дополнение, неожиданно было обнаружено, что рекомбинантный сконструированный белок FP050, происходящий из EDIL3, способствует эффероцитозу погибающих клеток Jurkat эндотелиальными клетками (HUVEC), демонстрируя основную функциональность мостикового белка и наглядное доказательство того, что домены мостиковых белков можно применять для разработки функциональных новых рекомбинантных сконструированных белков.
Пример 4. Эффероцитоз протромбоцитарных плазматических микрочастиц
4.1 Анализ эффероцитоза микрочастиц эндотелиальными клетками человека
Культура клеток
Клетки HUVEC получали от Lonza (Базель, Швейцария). Клетки культивировали в колбах, покрытых желатином (из бычьей кожи, конечная концентрация 0,2% в PBS, разбавление 2% исходным раствором, Sigma Aldrich/Merck, Германия). Клетки выращивали на среде для культивирования 199 (Thermo Fischer Scientific, США) с добавлением 10% FBS (GE Healthcare, Великобритания), 1% Pen/Strep (Thermo Fischer Scientific, США), 1% Glutamax (Thermo Fischer Scientific, США) и 1 нг/мл рекомбинантного основного фактора роста фибробластов (Peprotech, Великобритания). Клетки отделяли для сбора или пассирования с использованием Accutase™ (Thermo Fischer Scientific, США).
Микрочастицы тромбоцитарного происхождения готовили в соответствии со следующей процедурой: цитратную венозную кровь собирали (Coagulation 9NC Citrate Monovette, Зарштедт, Германия) у здоровых взрослых добровольцев после получения письменного информированного согласия. Обогащенную тромбоцитами плазму крови (PRP) готовили посредством центрифугирования (200xg, 15 минут, без задержки, при комнатной температуре). Микрочастицы/дебрис тромбоцитарного происхождения получали посредством проведения трех циклов мгновенного замораживания PRP с использованием жидкого азота и размораживания при 37°C. Фрагменты/микрочастицы тромбоцитов осаждали центрифугированием при 20000×g в течение 15 мин. при к. т. Осадок ресуспендировали в PBS, готовили аликвоты и хранили при -80°С. Препараты микрочастиц были на 85-100% PS-положительными, как определяли посредством проточной цитометрии с использованием меченого Alexa Fluor™ 488 мышиного MFG-E8/лактадгерина (собственного производства Novartis). Количество микрочастиц определяли с использованием специальных счетных гранул (BioCytex/Stago, Франция). Буфер для проточной цитометрии готовили с использованием PBS (Thermo Fischer Scientific, США) с добавлением 1% FBS (GE Healthcare, Великобритания), 0,05% вес./об. азида натрия (Merck, Германия) и 0,5 мМ EDTA (этилендиаминтетрауксусной кислоты, Thermo Fischer Scientific, США).
4.2 Анализ эффероцитоза
В день 1 клетки HUVEC (конфлюэнтность 70-90%) собирали посредством отделения с помощью Accutase™ в течение 5 мин., промывали с помощью PBS и ресуспендировали в среде для культивирования клеток. Численность и жизнеспособность клеток оценивали с применением проточного цитометра Guava EasyCyte (Merck, Германия) и реагента Guava ViaCount (Merck, Германия) в соответствии с инструкциями производителя. Необходимое количество клеток центрифугировали при 300×g в течение 5 мин. при к. т. и ресуспендировали в среде для культивирования с получением количества клеток, составляющего 6,6×104 клеток/мл. 150 мкл/лунка этой клеточной суспензии добавляли в 96-луночные планшеты для культуры тканей (Corning™, США). Клетки HUVEC инкубировали в инкубаторе при 37°C/5% CO2/95% влажности в течение дополнительных 16-20 часов.
В день 2 среду удаляли из клеток HUVEC путем аспирации и добавляли 25 мкл свежей, предварительно нагретой (37°C) среды для культивирования, а затем добавляли 25 мкл слитого белка FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) в трех различных концентрациях: 0,3 нМ, 3 нМ или 30 нМ, или контроль, разведенный в предварительно нагретой (37°C) среде для культивирования. Для разведения использовали 96-луночные обработанные планшеты с несвязывающей поверхностью (NBS) (Corning™, США). Тестируемые белки оставляли для взаимодействия с клетками HUVEC при 37°C/5% CO2/95% влажности на 30 мин. перед добавлением микрочастиц тромбоцитарного происхождения.
Необходимое количество микрочастиц центрифугировали при 20000xg при 4°C в течение 15 мин. и ресуспендировали при плотности 2×108 частиц/мл в среде RPMI 1640 (без FBS) с добавлением сложноэфирного красителя pHrodo™ Green STP в конечной концентрации 5 мкг/мл (среда для окрашивания). После окрашивания в течение 10 мин. при 37°C оставшийся реакционноспособный сложный эфир pHrodo™ Green STP инактивировали окрашивающей средой с добавлением 10% FBS в течение дополнительных 5 мин. при 37°C. Микрочастицы, меченные pHrodo™ Green, однократно промывали путем центрифугирования при 20000xg при 4°C в течение 15 мин. и количество доводили до 1×108 частиц/мл в среде для культивирования клеток HUVEC. 5×106 частиц/лунка, меченые pHrodo™ Green, добавляли к клеткам HUVEC и инкубировали при 37°C/5% CO2/95% влажности в течение 5 ч. Среду удаляли, клетки HUVEC однократно промывали в PBS и отделяли с помощью 40 мкл/лунка раствора Accutase™. Клетки собирали посредством добавления 80 мкл ледяного буфера для проточной цитометрии, переносили в 1,5 мл полипропиленового 96-луночного блока, промывали избытком ледяного буфера для проточной цитометрии и центрифугировали при 400xg (4°C) в течение 5 мин. Супернатанты удаляли путем аспирации, а осадки ресуспендировали в 80 мкл ледяного буфера для проточной цитометрии и переносили в 96-луночный микротитрационный планшет с V-образным дном (BD Biosciences, США). Образцы измеряли на проточном цитометре BD LSRFortessa™ (BD Biosciences, США). Регистрировали интенсивность флуоресценции pHrodo™ Green в качестве индикатора лизосомальной локализации поглощенных микрочастиц. Анализ данных проточной цитометрии проводили с применением программного обеспечения FlowJo™. Медианные значения интенсивности флуоресценции (MFI) сигнала pHrodo™ Green от синглетно гейтированных клеток HUVEC использовали в качестве регистрируемого показателя. Анализ данных проводили с применением программного обеспечения MS Excel и GraphPad Prism для расчета EC50. Слитый белок FP278 способствовал эффероцитозу микрочастиц тромбоцитарного происхождения эндотелиальными клетками зависимым от концентрации образом, как изображено на фигуре 12. Способствование поглощению было зависимым от концентрации, а также наблюдалось в других типах эндотелиальных клеток (не показано).
Пример 5. Технические свойства слитых белков MFG-E8-HSA
5.1 Анализ связывания слитого белка FP330 с FcRn посредством поверхностного плазмонного
резонанса (SPR)
Проводили анализ непосредственного связывания для характеристики связывания слитого белка FP330 (EGF-HSA-C1-C2; SEQ ID NO: 42) с FcRn. Кинетические константы аффинности связывания (KD) измеряли на захваченном белке с использованием рекомбинантного FcRn человека в качестве аналита. Измерения проводили на BIAcore® T200 (GE Healthcare, Глатбруг, Швейцария) при комнатной температуре и при pH 5,8 и 7,4 соответственно. Для измерения аффинности белки разбавляли в 10 мМ NaP, 150 мМ NaCl, 0,05% Tween 20, pH 5,8 и иммобилизовали на проточных ячейках сенсорного чипа высокой чистоты CM5 (GE Healthcare, BR-1000-14) с применением стандартной процедуры в соответствии с рекомендациями производителя (GE Healthcare). Для целей сравнения в одной проточной ячейке иммобилизовали холостой раствор. Данные по связыванию получали посредством последовательного введения посредством инъекции разведений аналита в проточную ячейку сравнения и измерительную проточную ячейку. Образцы с нулевой концентрацией (только подвижный буфер) включали для обеспечения двойного сравнения во время оценки данных. Для оценки данных использовали сенсограммы, полученные с использованием двойного сравнения, и анализировали константы диссоциации (KD).
Слитый белок FP330 связывается с FcRn при рН 5,8 с аффинностью 1380 нМ, тогда как при рН 7,4 связывание не наблюдали (см. таблицу 5 выше). Эти результаты хорошо согласуются с HSA дикого типа (1000-2000 нМ, при pH 5,8, данные не показаны).
5.2 Дифференциальная сканирующая калориметрия (DSC) MFG-E8 и вариантов
Термостабильность сконструированного белкового варианта MFG-E8 FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) измеряли с применением дифференциальной сканирующей калориметрии. Измерения проводили на дифференциальном сканирующем микрокалориметре (Nano DSC, TA Instruments). Объем ячейки составлял 0,5 мл, скорость нагрева 1°С/мин. Белок использовали в концентрации 1 мг/мл в PBS (рН 7,4). Молярную теплоемкость белка оценивали посредством сравнения с двойными образцами, содержащими идентичный буфер, из которого был исключен белок. Значения парциальной молярной теплоемкости и кривые плавления анализировали с применением стандартной процедуры. Термограммы строили с учетом поправки на исходный уровень и нормализовали по концентрации. Наблюдали два события плавления с первой Tm при 50°С, второй Tm при 64°С.
5.3 Измерения способности к агрегации и растворимости вариантов MFG-E8
Во-первых, способность к агрегации варианта белка FP278 MFG-E8 (EGF-HSA-C1-C2-His-метка; SEQ ID NO: 44) измеряли посредством динамического светорассеяния (DLS, Wyatt). Динамическое светорассеяние применяли для измерения коэффициентов поступательной диффузии FP278 в растворе посредством количественной оценки динамических флуктуаций в рассеянном свете. Распределение белковых вариантов по размерам без фракционирования, обеспечивающее получение оценок полидисперсности, а также значений гидродинамического радиуса, измеряли при концентрации 1 мг/мл. Гидродинамические радиусы слитого белка FP278 определяли с помощью планшетного ридера DynaPro™ (Wyatt Technology Europe GmbH, Дернбах, Германия) в комбинации с программным обеспечением DYNAMICS (версия 7.1.0.25, Wyatt). Параметры 50 мкл неразведенного и отфильтрованного (PVDF-фильтр с размером пор 0,22 мкм (Millex® Syringe-driven Filter Unit, Millipore, Биллерика, США)) раствора белка измеряли в 384-луночном планшете (384-луночный планшет, полистирол, Thermo Scientific, Лангензельбольд, Германия). Агрегаты с более высокой молекулярной массой в образце белка идентифицировать не удалось. Гидродинамический радиус белка составлял около 5-6 нм, что указывало на присутствие мономерного белка в растворе.
Во-вторых, для оценки растворимости белка выполняли измерения гидродинамического радиуса слитого белка FP278 в зависимости от концентрации. Применяли концентрации белка, составляющие до 22 мг/мл. Гидродинамические радиусы определяли, как описано выше. При повышении концентрации слитого белка FP278 повышения значения радиуса (5-7 нм) не наблюдали, тогда как измерение динамического светорассеяния wtMFG-E8 (SEQ ID NO: 1) стало невозможным вследствие высокой степени агрегации при концентрациях около 0,2 мг/мл.
Пример 6. Оптимизация слитых белков MFG-E8
Масс-спектрометрию (MS) применяли для исследования слитого белка FP330 (EGF-HSA-C1-C2) для создания панели вариантных слитых белков на основе MFG-E8, оптимизированных для улучшения экспрессии и выхода. При создании панели вариантных белков использовали линкеры разного размера и структуры, например линкеры, содержащие GS между доменами EGF и HSA и/или множество GS или G4S (SEQ ID NO: 64) между HSA и доменами C1. Кроме того, в некоторые из вариантов были включены аминокислотные модификации (обозначенные как HSA* в таблице 7), содержащие делеции или замены. Сводные данные по панели вариантных слитых белков представлены в таблице 7 ниже.
Таблица 7. Сводные данные по вариантным слитым белкам
1 Положение аминокислотной модификации пронумеровано в соответствии с SEQ ID NO: 42 (FP330).
Пример 7. Вариантные слитые белки MFG-E8; экспрессия и очистка
Способы получения слитых белков в линиях клеток НЕК описаны в примере 2. Для экспрессии в линии клеток CHO нуклеиновые кислоты, кодирующие варианты MFG-E8, синтезировали в Geneart (LifeTechnologies) и клонировали в вектор экспрессии млекопитающих с применением методик клонирования на основе лигирования с использованием рестрикционных ферментов. Полученными в результате плазмидами трансфицировали клетки CHO-S (Thermo). Вкратце, для временной экспрессии слитых белков вектор экспрессии трансфицировали в адаптированные к культивированию в суспензии клетки CHO-S с использованием средства для трансфекции CHO Expifectamine (Thermo). Как правило, 400 мл клеток в суспензии с плотностью 6 млн. клеток на мл трансфицировали с помощью ДНК, содержащей 400 мкг вектора экспрессии, кодирующего сконструированный белок. Затем рекомбинантный вектор экспрессии вводили в клетки-хозяева для дополнительной секреции в течение семи дней в среде для культивирования (среда для экспрессии ExpiCHO с добавлением реагентов ExpiCHO Feed и ExpiCHO Enhancer (Thermo)).
Как видно из данных экспрессии, показанных в таблице 8, вариантные слитые белки FP068 (SEQ ID NO: 46) и FP776 (SEQ ID NO: 48) продемонстрировали примерно двукратное улучшение экспрессии по сравнению со слитым белком FP330 (SEQ ID NO: 42).
Таблица 8. Экспрессия вариантных слитых белков в линиях клеток HEK и CHO*
(мг/л)
* Обозначает слитый белок, продуцируемый в линии клеток CHO.
Пример 8. Определение характеристик вариантных слитых белков
Эффект вариантных слитых белков в отношении эффероцитоза определяли посредством осуществления анализов эффероцитоза, описанных в примере 3.
В первом анализе эффект вариантных слитых белков в анализе эффероцитоза нейтрофилов макрофагами человека определяли в соответствии со способом, описанным в разделе 3.3 выше. Макрофаги M0 инкубировали со слитым белком FP330 (EGF-HSA-C1-C2; SEQ ID No: 42) или вариантами FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID No: 44) или FP776 (EGF-HSA-C1-C2; SEQ ID No: 48) в течение 30 мин. Как показано на фигуре 13, слитые белки FP330, FP278 и FP776 могут обеспечивать восстановление нарушенного эндотоксином (липополисахаридом (LPS)) эффероцитоза погибающих нейтрофилов макрофагами человека. Повышение концентрации слитых белков FP330 (EC50=1,6 нМ; фиг. 13A), FP278 (EC50=1,78 нМ; фиг. 13B) и FP776 (EC50=0,5 нМ; фиг. 13C) обеспечивало восстановление нарушенного эффероцитоза, вызванного добавлением LPS, и даже способствовало эффероцитозу после достижения исходных уровней.
Характеристики слитых белков FP330, FP278 и FP776 дополнительно определяли в анализе эффероцитоза клеток Jurkat эндотелиальными клетками человека (HUVEC) в соответствии со способом, описанным в разделе 3.4 выше. Эффект слитых белков FP330, FP278 и FP776 в отношении способствования эффероцитозу погибающих клеток Jurkat эндотелиальными клетками HUVEC изображен на фигуре 14. Интернализации меченых pHrodo погибающих Т-клеток Jurkat человека клетками HUVEC значительно способствовало повышение концентрации FP330 (EC50=3,4 нМ; фиг. 14A), FP278 (EC50=2,4 нМ; фиг. 14B) и FP776 (EC50=3 нМ; фиг. 14С). Эти результаты демонстрируют, что эндотелиальные клетки вооружаются слитыми белками, становясь таким образом эффективными фагоцитами погибающих клеток.
Пример 9. Защита мышей от AKI и острого органного ответа, вызванного AKI
9.1 Модель острого повреждения почек
Самок мышей C57BL/6 (18-22 г) приобретали в Charles River (Франция) и содержали в помещении с контролируемой температурой в клетках с защитой в виде встроенного в крышку фильтра и 12-часовыми циклами свет/темнота. С животными обращались в строгом соответствии со швейцарскими федеральными законами и принципами NIH по уходу за лабораторными животными. Тестируемый терапевтический слитый белок вводили либо внутрибрюшинно (i.p.), либо внутривенно (i.v.) за два часа до хирургического вмешательства. Бупренорфин (Indivior Schweiz AG) применяли подкожно (s.c.) в дозе 0,1 мг/кг за 60-30 минут до хирургического вмешательства. Ингаляционную анестезию изофлураном индуцировали в наркозной камере (3,5-5 об. %, газ-носитель: кислород) за 5 мин. до хирургического вмешательства. Во время хирургического вмешательства животное поддерживали под наркозом с помощью намордной маски с использованием 1-2 об. % изофлурана/кислорода, скорость потока газа составляла 0,8-1,2 л/мин. Кожу живота выбривали и дезинфицировали с помощью Betaseptic (Mundipharma, Франция). Животных помещали на гомеотермическое одеяло (Rothacher, Швейцария) с системой гомеотермического мониторинга (PhysiTemp, США, Physitemp Instruments LLC, США) и накрывали стерильной марлей. Температуру тела контролировали на протяжении всего хирургического вмешательства посредством ректального датчика (Physitemp Instruments LLC, США) и контролировали для обеспечения температуры тела 36,5-37,5°C. Всем животным, в том числе контрольной имитационной группе, выполняли одностороннюю нефрэктомию правой почки. После срединного разреза/лапаротомии содержимое брюшной полости отводили влево для открытия правой почки. Правый мочеточник и почечные сосуды отсоединяли и перевязывали, затем удаляли правую почку. У животных, подвергшихся AKI, содержимое брюшной полости располагали справа на стерильной марле, а левую почечную артерию и вену рассекали для обеспечения возможности пережатия для индукции ишемии. Зажим для получения микроаневризмы (B Braun, Швейцария) использовали для пережатия почечной ножки (артерии и вены вместе с применением одного зажима) для блокирования кровотока в почке и индуцирования почечной ишемии. Успешное создание ишемии подтверждали по изменению цвета почки с красного на темно-фиолетовый, которое происходило в течение нескольких секунд. После индуцирования ишемии (35-38 минут) зажим для получения микроаневризмы снимали. Теплый стерильный физиологический раствор (~ 2 мл, 37°C) использовали для промывания содержимого брюшной полости для регидратации тканей перед закрытием раны. После промывания добавляли дополнительно 1 мл стерильного физиологического раствора i.p. в качестве заменителя жидкости. При начале реперфузии рану закрывали в два слоя (мышечная ткань и кожа отдельно). Затем животных содержали под красной теплой лампой до полного выздоровления. Бупренорфин повторно вводили через 1 ч. и 4 ч. после хирургического вмешательства в дозе 0,1 мг/кг, а также включали в питьевую воду (9,091 мкг/мл). Через 24 ч. животных подвергали эвтаназии для анализа.
9.2 Введение терапевтических слитых белков
Терапевтические слитые белки FP330 (EGF-HSA-C1-C2; SEQ ID No: 42), FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID No: 44) и FP776 (EGF-HSA-C1-C2; SEQ ID No: 48) тестировали в модели AKI, как описано выше, в дозах, указанных в таблице 9 ниже. Для исследований по обнаружению сывороточных маркеров и экспрессии маркеров qPCR за 2 часа до хирургического вмешательства вводили слитый белок FP278. FP330 и FP776 вводили i.v. за 30 мин. до начала повреждения по типу ишемия-реперфузия. Для исследования измерения поглощения контрастного вещества посредством магнитно-резонансной томографии слитый белок FP776 вводили профилактически за 30 мин. до индукции AKI в дозе 1,26 мг/кг или вводили терапевтически в дозе 2 мг/кг i.v. через 5 ч. после индукции повреждения по типу ишемия-реперфузия.
Таблица 9. Введение доз терапевтических слитых белков
9.3 Регистрируемые показатели/анализ на защиту от AKI
Сывороточные маркеры
Образцы сыворотки отбирали через 24 ч. после индукции ишемии-реперфузии и анализировали в отношении содержания креатинина в сыворотке крови и азота мочевины в крови (BUN) с применением клинического анализатора Hitachi M40 в соответствии с инструкциями производителя (Axonlab, Швейцария).
Определение уровня экспрессии маркера в органах с помощью qPCR
Органы (почка, печень, легкое и сердце) собирали через 24 ч. после индукции AKI, разрезали на фрагменты по 1 см и хранили в буфере RNA Later (Thermo Fisher Scientific Inc, США) при 4°C в течение ночи. Фрагменты органов переносили в буфер RLT (RNeasy Mini Kit, Qiagen, Дания), содержащий 134 мМ бета-меркаптоэтанола (Merck, Дания), в пробирках Lysing Matrix D (MP Biomedicals, Франция) и гомогенизировали с использованием инструмента FastPrep-24 (MP Biomedicals). Затем фиброзную ткань сердца расщепляли протеиназой К (набор RNeasy Mini), а лизаты почек, печени и легкого непосредственно центрифугировали в течение 3 мин. на полной скорости в микроцентрифуге (Eppendorf, Дания). Супернатанты переносили на центрифужную колонку QIAshredder (Qiagen, Дания) и центрифугировали в течение 2 мин. Экстракцию РНК из фильтрата проводили в соответствии с руководством для набора RNeasy Mini, в том числе расщепление ДНКазой. Концентрацию РНК измеряли посредством устройства Nano Drop 1000 (Thermo Fisher Scientific Inc). Образцы по 2 мкг РНК подвергали обратной транскрипции в соответствии с руководством по набору для обратной транскрипции High-Capacity cDNA (Thermo Fisher Scientific Inc) с применением термоциклера SimpliAmp (Applied Biosystems, США). cDNA объединяли с водой, не содержащей нуклеаз (Thermo Fisher Scientific Inc), зондом TaqMan (анализ экспрессии генов TaqMan (FAM), Thermo Fisher Scientific Inc) и мастер-миксом для анализа экспрессии генов TaqMan (Thermo Fisher Scientific Inc) в 384-луночном микропланшете (384-луночный реакционный планшет MicroAmp Optical, Thermo Fisher Scientific Inc). qPCR проводили на системе для ПЦР в реальном времени ViiA 7 (Applied Biosystems, США). Условия были следующие: 1-2 мин., 50°C; 2-10 мин., 95°C; 3-15 с, 95°C; 4-1 мин., 60°C. Стадии 3 и 4 повторяли 45 циклов. Анализ данных проводили с применением программного обеспечения ViiA 7, программное обеспечение для анализа данных qPCR выполняли с применением программного обеспечения MS Excel и GraphPad Prism.
Поглощение контрастного вещества печенью согласно измерениям с помощью магнитно-резонансной томографии (МРТ)
Способы осуществления МРТ были адаптированы на основе публикации Egger et al (Egger et al., (2015) J Magn Reson Imaging, 41: 829-840). Эксперименты проводили в системе МРТ 7-T Bruker Biospec (Bruker Biospin, Эттлинген, Германия). Во время получения сигнала МРТ мышей помещали в положение лежа на спине на платформе из плексигласа. Температуру тела поддерживали на уровне 37±1°С с помощью грелки. После короткого периода индукции анестезию поддерживали с помощью прим. 1,4% изофлурана в смеси O2/N2O (1:2), вводимой через носовой конус. Все измерения проводили на самостоятельно дышащих животных; не применяли стимуляции ни сердца, ни дыхания.
После помещения мыши в сканер с целью локализации получали быстрые ориентировочные изображения. Анализы перфузии проводили с использованием внутрисосудистого средства, содержащего наночастицы суперпарамагнитного оксида железа (SPIO) (Endorem®, Guerbet, Франция). Endorem® вводили посредством инъекции внутривенно в виде болюса в течение 1,2 с животным с AKI (через 24 часа после индукции заболевания) или после имитационной операции (животным через 24 ч. после нефрэктомии). Первый болюс вводили в течение 1,2 с в сочетании с последовательным получением эхо-планарных изображений с разрешением 400 мс/изображение. После получения 25 исходных изображений вводили второй болюс в течение 1,2 с, и после введения болюса получали дополнительные 575 изображений, в результате чего за 4 мин. получали в общей сложности 600 изображений. Суперпарамагнитное контрастное вещество индуцировало локальные изменения чувствительности, что приводило в результате к ослаблению сигнала, пропорциональному степени перфузии почки. Для серии изображений значения интенсивности сигнала оценивали в областях, представляющих интерес (ROI), расположенных в коре/внешней полосе наружного мозгового вещества. Положение, форма и размер ROI тщательно выбирали таким образом, чтобы гарантировать, что они покрывают примерно одну и ту же область, несмотря на движения почек, вызванные дыханием. Средние значения интенсивности сигнала для изображений до инъекции принимали в качестве исходных значений интенсивности (S(0)). Показатели перфузии определяли из средних значений следующих соотношений (Rosen et al., (1990) Magn Reson Med., 14: 249-265):
-ln[S(t)/S(0)] ~ TE.V.cT (t),
где TE представляет собой время появления эхо-сигнала, V представляет собой объем крови, cT представляет собой концентрацию контрастного вещества.
Наночастицы SPIO, применяемые в исследовании, характеризуются средним диаметром приблизительно 150 нм и поглощаются клетками Купфера в печени. Таким образом, в дополнение к перфузии почек, МРТ также позволяла осуществлять мониторинг поглощения наночастиц в печени путем определения изменения контраста, оцениваемого в ROI, располагающихся в печени.
9.4 Результаты
Как изображено на фигуре 15, слитые белки FP330 (EGF-HSA-C1-C2; SEQ ID No: 42), FP278 (EGF-HSA-C1-C2-His-метка; SEQ ID No: 44) и FP776 (EGF-HSA-C1-C2; SEQ ID No: 48) обеспечивали защиту функции почек в этой модели острого повреждения почек (AKI) при введении либо i.p., (FP278), либо i.v. (FP330 и FP776). Эта защита выражается в блокировании повышения уровня креатинина в сыворотке крови (sCr). На фиг. 15А изображено, что слитый белок FP278 в обеих тестируемых дозах обеспечивал значительное снижение уровней креатинина в сыворотке крови (p < 0,0001) по сравнению с таковым у животных, обработанных средой-носителем, и являлся таким же эффективным, как мышиный MFG-E8. Как изображено на фиг. 15В, слитый белок FP330 обеспечивал защиту функции почек зависимым от дозы образом, и аналогичным образом слитый белок FP776 (фиг. 15С), где уровни креатинина в сыворотке крови также блокировались зависимым от дозы образом.
Нарушение функции почек также отражалось на уровнях азота мочевины в крови (BUN) у тестируемых мышей, и эффект слитого белка FP278 в отношении уровней BUN изображен на фигуре 16.
Таким образом, как изображено на фигурах 15 и 16, слитые белки FP278, FP330 и FP776 обеспечивали эффективную защиту от повышения уровня этих маркеров, применяемых для клинической диагностики почечной недостаточности. Наблюдаемую эффективность подтверждали гистологическим анализом (не показано).
Кроме того, как изображено на фиг. 17, однократная доза слитого белка FP278 обеспечивает защиту отдаленных органов от ответа острой фазы, вызванной AKI. AKI индуцирует множество ответов с участием мРНК, которые можно измерить посредством qPCR в лизатах отдаленных органов с высокой перфузией, таких как селезенка, легкое, печень, сердце и головной мозг. Типичные мРНК индуцировали выборочное повреждение (NGAL, KIM-1), индукцию хемокинов (не показано) или индукцию индукции белка ответа острой фазы, такого как сывороточный амилоид A (SAA). На фиг. 17А и 17В приведен пример такого индуцированного AKI ответа (сывороточный амилоид А (SAA)) в сердце и легком мышей, который в значительной степени блокировали и возвращали к уровням имитационного контроля после однократной инъекции слитого белка.
Поглощение контрастного вещества SPIO Endorem® печенью в динамике изображено на фигуре 18. У животных с AKI было отмечено значительно сниженное поглощение контрастного вещества печенью (мишень=клетки Купфера) по сравнению с животными при имитационном воздействии. Обработка с помощью FP776 (профилактически в дозе 1,26 мг/кг, за 30 мин. до индукции AKI или терапевтически в дозе 2 мг/кг, через 5 ч. после индукции повреждения по типу ишемия-реперфузия) обеспечивала защиту от прекращения накопления контрастного вещества в печени мышей с AKI. Эти результаты позволяют предположить, что в этой мышиной модели AKI вызывает значительное нарушение эндогенного клиренса частиц, опосредованного клетками Купфера, и что AKI вызывает микрососудистые нарушения, которые влияют на накопление контрастного вещества на основе частиц железа в печени. Обработка слитым белком FP776 обеспечивала защиту от прекращения осуществления клиренса и возникновения нарушений микрососудистого русла и даже обеспечивала усиление поглощения контрастного вещества при обеих тестируемых дозах по сравнению с животными в группе с имитационным воздействием.
Терапевтические слитые белки, например, по варианту осуществления 19 (например, SEQ ID NO: 80) или по варианту осуществления 20 (например, SEQ ID NO: 82), способствуют адгезии клеток - интегрин av и способствуют эффероцитозу подобно FPJ776 при тестировании в вышеуказанных экспериментах. Следовательно, они подходят для вариантов терапевтического применения, раскрытых в данном документе.
В совокупности эти данные демонстрируют, что слитые белки по настоящему изобретению, например, со вставкой домена HSA, являются функциональными и эффективными и поэтому могут быть применимы в качестве терапевтических средств.
Следует понимать, что примеры и варианты осуществления, описанные в данном документе, предназначены только для иллюстративных целей, и что различные модификации или изменения в их контексте будут предложены специалистам в данной области техники и должны быть включены в сущность и область действия настоящей заявки и объем прилагаемой формулы изобретения. Все публикации, патенты и заявки на патенты, цитируемые в данном документе, включены в данный документ посредством ссылки для всех целей.
--->
ПЕРЕЧЕНЬ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> NOVARTIS AG
<120> Терапевтические слитые белки
<130> PAT058332
<140>
<141>
<150> EP 19196045.9
<151> 2019-09-06
<160> 148
<170> PatentIn версия 3.5
<210> 1
<211> 387
<212> БЕЛОК
<213> Homo sapiens
<400> 1
Met Pro Arg Pro Arg Leu Leu Ala Ala Leu Cys Gly Ala Leu Leu Cys
1 5 10 15
Ala Pro Ser Leu Leu Val Ala Leu Asp Ile Cys Ser Lys Asn Pro Cys
20 25 30
His Asn Gly Gly Leu Cys Glu Glu Ile Ser Gln Glu Val Arg Gly Asp
35 40 45
Val Phe Pro Ser Tyr Thr Cys Thr Cys Leu Lys Gly Tyr Ala Gly Asn
50 55 60
His Cys Glu Thr Lys Cys Val Glu Pro Leu Gly Leu Glu Asn Gly Asn
65 70 75 80
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
85 90 95
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
100 105 110
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
115 120 125
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
130 135 140
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
145 150 155 160
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
165 170 175
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
180 185 190
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
195 200 205
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
210 215 220
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
225 230 235 240
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
245 250 255
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
260 265 270
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
275 280 285
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
290 295 300
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
305 310 315 320
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
325 330 335
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
340 345 350
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
355 360 365
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
370 375 380
Leu Gly Cys
385
<210> 2
<211> 46
<212> БЕЛОК
<213> Homo sapiens
<400> 2
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys
35 40 45
<210> 3
<211> 318
<212> БЕЛОК
<213> Homo sapiens
<400> 3
Cys Val Glu Pro Leu Gly Leu Glu Asn Gly Asn Ile Ala Asn Ser Gln
1 5 10 15
Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp
20 25 30
Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp
35 40 45
Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu
50 55 60
Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu
65 70 75 80
Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn
85 90 95
Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu
100 105 110
Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu
115 120 125
Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His
130 135 140
Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
145 150 155 160
Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln
165 170 175
Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser
180 185 190
Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala
195 200 205
Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu
210 215 220
Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn
225 230 235 240
Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn
245 250 255
Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser
260 265 270
Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu
275 280 285
Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala
290 295 300
Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
305 310 315
<210> 4
<211> 583
<212> БЕЛОК
<213> Homo sapiens
<400> 4
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Cys Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Ala Leu
580
<210> 5
<211> 583
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 5
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Ala Leu
580
<210> 6
<211> 206
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 6
Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe
1 5 10 15
Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr
20 25 30
Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser
35 40 45
Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala
50 55 60
Lys Arg Met Pro Cys Ala Glu Asp Cys Leu Ser Val Phe Leu Asn Gln
65 70 75 80
Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys
85 90 95
Cys Cys Thr Glu Ser Leu Val Asn Gly Arg Pro Cys Phe Ser Ala Leu
100 105 110
Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe
115 120 125
Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile
130 135 140
Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala
145 150 155 160
Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val
165 170 175
Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu
180 185 190
Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu
195 200 205
<210> 7
<211> 217
<212> БЕЛОК
<213> Homo sapiens
<400> 7
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 8
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 8
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 9
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 9
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 10
<211> 217
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 10
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
1 5 10 15
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
20 25 30
Val Val Ala Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
35 40 45
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
50 55 60
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
65 70 75 80
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
85 90 95
Ala Leu Ala Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
100 105 110
Pro Arg Glu Pro Gln Val Cys Thr Leu Pro Pro Ser Arg Glu Glu Met
115 120 125
Thr Lys Asn Gln Val Ser Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro
130 135 140
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
145 150 155 160
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
165 170 175
Val Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
180 185 190
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
195 200 205
Lys Ser Leu Ser Leu Ser Pro Gly Lys
210 215
<210> 11
<211> 457
<212> БЕЛОК
<213> Homo sapiens
<400> 11
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly
130 135 140
Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala Leu
145 150 155 160
Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys
165 170 175
Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp
180 185 190
Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile Thr
195 200 205
Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys
210 215 220
Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val Lys
225 230 235 240
Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn Thr
245 250 255
Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg
260 265 270
Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu Leu
275 280 285
Leu Gly Cys Glu Leu Ser Gly Cys Ser Glu Pro Leu Gly Met Lys Ser
290 295 300
Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr
305 310 315 320
Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp
325 330 335
Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn Asp Gln Ser
340 345 350
Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val Thr Gly Ile
355 360 365
Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe Val Gly Ser
370 375 380
Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr Val Tyr Gln
385 390 395 400
Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn Phe Asp Asn
405 410 415
Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr Ala Arg His
420 425 430
Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser
435 440 445
Glu Leu Leu Gly Cys Thr Glu Glu Glu
450 455
<210> 12
<211> 1052
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 12
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Gly Ser Asp Ala His Lys Ser Glu Val Ala
130 135 140
His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu
145 150 155 160
Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val
165 170 175
Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp
180 185 190
Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp
195 200 205
Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala
210 215 220
Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln
225 230 235 240
His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val
245 250 255
Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys
260 265 270
Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro
275 280 285
Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys
290 295 300
Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu
305 310 315 320
Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys
325 330 335
Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val
340 345 350
Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser
355 360 365
Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly
370 375 380
Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile
385 390 395 400
Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu
405 410 415
Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp
420 425 430
Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser
435 440 445
Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly
450 455 460
Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val
465 470 475 480
Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys
485 490 495
Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu
500 505 510
Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys
515 520 525
Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu
530 535 540
Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val
545 550 555 560
Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His
565 570 575
Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val
580 585 590
Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg
595 600 605
Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe
610 615 620
Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala
625 630 635 640
Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu
645 650 655
Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys
660 665 670
Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala
675 680 685
Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe
690 695 700
Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly
705 710 715 720
Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys Ser Gly
725 730 735
Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala
740 745 750
Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr
755 760 765
Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala
770 775 780
Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys Met
785 790 795 800
Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro
805 810 815
Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr
820 825 830
Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe Arg
835 840 845
Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro
850 855 860
Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg His
865 870 875 880
Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly Cys Ser
885 890 895
Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln Ile Thr
900 905 910
Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr Trp Glu
915 920 925
Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala Trp Thr
930 935 940
Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu Leu Val
945 950 955 960
Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp Phe Gly
965 970 975
His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn Asp Gly
980 985 990
Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp Lys Val
995 1000 1005
Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val Ile
1010 1015 1020
Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser
1025 1030 1035
Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
1040 1045 1050
<210> 13
<211> 3156
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 13
gacatctgcg accccaatcc ttgcgagaat ggcggcattt gtctgcctgg actggccgat 60
ggcagcttct cttgtgaatg ccccgatggc ttcacagacc ccaattgcag ctctgtggtg 120
gaagtggcca gcgacgagga agaacctaca agcgctggcc cctgcacacc caatccatgt 180
cataatggcg gaacctgcga gatcagcgag gcctacagag gcgatacctt catcggctac 240
gtgtgcaagt gccccagagg cttcaatggc atccactgcc agcacaacat caacgagtgc 300
gaggtggaac catgcaagaa cggcggcatc tgtaccgacc tggtggccaa ttactcttgc 360
gagtgccctg gcgagttcat gggcagaaac tgccagtaca agggatccga cgctcacaag 420
tctgaggtgg cccacagatt caaggacctg ggcgaagaga acttcaaggc cctggtgctg 480
atcgccttcg ctcagtatct gcagcagagc cctttcgagg accacgtgaa gctggtcaac 540
gaagtgaccg agttcgccaa gacctgtgtg gccgatgaga gcgccgagaa ctgtgacaag 600
agcctgcaca cactgttcgg cgacaagctg tgtaccgtgg ccacactgag agaaacctac 660
ggcgagatgg ccgactgctg tgccaagcaa gagcccgaga gaaacgagtg cttcctgcag 720
cacaaggacg acaaccccaa cctgcctaga ctcgtgcgac ccgaagtgga tgtgatgtgc 780
accgcctttc acgacaacga ggaaaccttc ctgaagaagt acctgtacga gatcgccaga 840
cggcacccct acttttatgc ccctgagctg ctgttcttcg ccaagcggta taaggccgcc 900
ttcaccgaat gttgccaggc cgctgataag gctgcctgtc tgctgcctaa gctggacgag 960
ctgagagatg agggcaaagc cagctctgcc aagcagagac tgaagtgcgc cagcctgcag 1020
aagttcggcg agagagcttt taaggcctgg gccgttgcca gactgagcca gagatttcct 1080
aaggccgagt ttgccgaggt gtccaagctc gtgaccgatc tgacaaaggt gcacaccgag 1140
tgctgtcacg gcgatctgct ggaatgtgcc gacgatagag ccgacctggc caagtatatc 1200
tgcgagaacc aggacagcat cagcagcaag ctgaaagagt gctgcgagaa gcccctgctg 1260
gaaaagtctc actgtatcgc cgaggtcgag aacgacgaga tgcctgctga tctgcctagc 1320
ctggccgccg atttcgtgga aagcaaggat gtgtgcaaga actacgccga ggccaaagat 1380
gtgtttctgg gcatgtttct gtatgagtac gcccgcagac accccgacta ttctgtggtt 1440
ctgctgctgc ggctggccaa gacatacgag acaaccctgg aaaaatgctg cgccgctgcc 1500
gatcctcacg agtgttatgc caaggtgttc gacgagttca agccactggt ggaagaaccc 1560
cagaacctga tcaagcagaa ctgcgagctg ttcgagcagc tgggcgagta caagttccag 1620
aatgccctgc tcgtgcggta caccaagaaa gtgcctcagg tgtccacacc tacactggtt 1680
gaggtgtccc ggaatctggg caaagtgggc agcaagtgtt gcaagcaccc tgaggccaag 1740
agaatgcctt gcgccgagga ttacctgagc gtggtgctga atcagctgtg cgtgctgcac 1800
gagaaaaccc ctgtgtccga cagagtgacc aagtgctgta ccgagagcct cgtgaacaga 1860
aggccttgct ttagcgccct ggaagtggac gagacatacg tgcccaaaga gttcaacgcc 1920
gagacattca ccttccacgc cgatatctgc accctgtccg agaaagagcg gcagatcaag 1980
aagcagacag ccctggtcga gctggttaag cacaagccca aggccaccaa agaacagctg 2040
aaggccgtga tggacgactt cgccgccttt gtcgagaagt gctgcaaggc cgacgacaaa 2100
gagacatgct tcgccgaaga gggcaagaaa ctggtggctg cctctcaggc tgctctcgga 2160
cttggaggaa gcggaggatc tggcggttcc ggaggaagtt gttctggccc tcttggcatc 2220
gaaggcggca tcatcagcaa tcagcagatc accgccagca gcacccacag agcactgttt 2280
ggactgcaga aatggtatcc ctactacgcc cggctgaaca agaagggcct gattaacgcc 2340
tggacagccg ccgagaatga cagatggccc tggattcaga tcaacctgca gcggaagatg 2400
agagtgaccg gcgttatcac acagggcgcc aaaagaatcg gcagccccga gtacatcaag 2460
agctacaaga tcgcctacag caacgacggc aagacctggg ccatgtacaa agtgaagggc 2520
accaacgagg acatggtgtt ccggggcaac atcgacaaca acacccctta cgccaacagc 2580
ttcacccctc ctatcaaggc ccagtacgtg cggctgtacc ctcaagtgtg cagaaggcac 2640
tgtaccctga gaatggaact gctgggctgc gaactgtctg gctgttctga gccactgggc 2700
atgaagtccg gccacatcca ggattaccag atcacagcct ccagcatctt cagaaccctg 2760
aacatggata tgttcacctg ggagccccgg aaggccagac tggataagca gggaaaagtg 2820
aatgcctgga ccagcggcca caacgaccag tctcaatggc tgcaagtgga cctgctggtg 2880
cccaccaaag tgaccggaat cattactcag ggcgcaaagg acttcggcca cgtgcagttt 2940
gtgggctcct acaagctggc ctactccaac gatggcgagc actggacagt gtaccaggac 3000
gagaagcagc gcaaggataa ggtgttccag ggaaacttcg ataacgatac ccaccggaag 3060
aacgtgatcg accctccaat ctacgccaga cacatcagaa tcctgccttg gtcttggtac 3120
ggcagaatca ccctgagatc cgagctgctg ggatgc 3156
<210> 14
<211> 596
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 14
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys Val
35 40 45
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
50 55 60
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
65 70 75 80
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
85 90 95
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
100 105 110
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
115 120 125
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
130 135 140
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
145 150 155 160
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
165 170 175
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
180 185 190
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala
195 200 205
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
210 215 220
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
225 230 235 240
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
245 250 255
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
260 265 270
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
275 280 285
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
290 295 300
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
305 310 315 320
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
325 330 335
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
340 345 350
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Gly Gly Gly
355 360 365
Thr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
370 375 380
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
385 390 395 400
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
405 410 415
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
420 425 430
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
435 440 445
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
450 455 460
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
465 470 475 480
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
485 490 495
Val Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val
500 505 510
Ser Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
515 520 525
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
530 535 540
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
545 550 555 560
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
565 570 575
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
580 585 590
Ser Pro Gly Lys
595
<210> 15
<211> 1788
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 15
ctggacatct gcagcaagaa cccctgccac aacggcggcc tgtgcgaaga gatcagccag 60
gaagtgcggg gcgacgtgtt ccccagctac acctgtacct gcctgaaggg ctacgccggc 120
aaccactgcg agactaagtg cgtggaaccc ctgggcatgg aaaacggcaa tattgccaac 180
agccagatcg ccgccagctc cgtgcgcgtg acctttctgg gactgcagca ctgggtgccc 240
gagctggcca gactgaacag agccggcatg gtgaacgcct ggacccccag cagcaacgac 300
gacaaccctt ggatccaggt gaacctgctg cggcggatgt gggtgacagg cgtggtgaca 360
cagggcgcca gcagactggc cagccacgag tacctgaagg cctttaaggt ggcctacagc 420
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaaacacaa agaatttgtg 480
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacccgt ggaagcccag 540
tacgtgcggc tgtaccccac cagctgccac accgcctgca ccctgagatt cgagctgctg 600
ggctgcgagc tgaacggctg cgccaacccc ctgggcctga agaacaacag catccccgac 660
aagcagatca ccgcctccag cagctacaag acctggggcc tgcacctgtt cagctggaac 720
cccagctacg cccggctgga caagcagggc aacttcaacg cctgggtggc cggcagctac 780
ggcaacgacc agtggctgca ggtggacctg ggcagcagca aagaagtgac cggcatcatc 840
acccaggggg ccagaaactt cggcagcgtg cagttcgtgg ccagctacaa agtggcctac 900
tccaacgaca gcgccaactg gaccgagtac caggaccccc ggaccggcag ctccaagatc 960
ttccccggca actgggacaa ccacagccac aagaagaatc tgttcgaaac ccccatcctg 1020
gccagatacg tgcggatcct gcccgtggcc tggcacaacc ggatcgccct gagactggaa 1080
ctgctgggat gtgggggagg cggtaccgac aagacccaca cctgcccccc ctgcccagcc 1140
ccagagctgc tgggcggacc ctccgtgttc ctgttccccc ccaagcccaa ggacaccctg 1200
atgatcagca ggacccccga ggtgacctgc gtggtggtgg acgtgagcca cgaggaccca 1260
gaggtgaagt tcaactggta cgtggacggc gtggaggtgc acaacgccaa gaccaagccc 1320
agagaggagc agtacaacag cacctacagg gtggtgtccg tgctgaccgt gctgcaccag 1380
gactggctga acggcaagga atacaagtgc aaggtctcca acaaggccct gccagccccc 1440
atcgaaaaga ccatcagcaa ggccaagggc cagccacggg agccccaggt gtacaccctg 1500
cccccctgcc gggaggagat gaccaagaac caggtgtccc tgtggtgtct ggtgaagggc 1560
ttctacccca gcgacatcgc cgtggagtgg gagagcaacg gccagcccga gaacaactac 1620
aagaccaccc ccccagtgct ggacagcgac ggcagcttct tcctgtacag caagctgacc 1680
gtggacaagt ccaggtggca gcagggcaac gtgttcagct gcagcgtgat gcacgaggcc 1740
ctgcacaacc actacaccca gaagagcctg agcctgtccc ccggcaag 1788
<210> 16
<211> 613
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 16
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
50 55 60
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
65 70 75 80
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
85 90 95
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
100 105 110
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
115 120 125
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
130 135 140
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
145 150 155 160
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
165 170 175
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
180 185 190
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
195 200 205
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
210 215 220
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
225 230 235 240
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
245 250 255
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
260 265 270
Pro Gly Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys
275 280 285
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
290 295 300
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
305 310 315 320
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
325 330 335
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
340 345 350
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
355 360 365
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
370 375 380
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
385 390 395 400
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
405 410 415
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
420 425 430
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
435 440 445
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
450 455 460
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
465 470 475 480
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
485 490 495
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
500 505 510
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
515 520 525
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
530 535 540
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
545 550 555 560
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
565 570 575
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
580 585 590
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Ser His
595 600 605
His His His His His
610
<210> 17
<211> 1839
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 17
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgacaag acccacacct gccccccctg cccagcccca 180
gagctgctgg gcggaccctc cgtgttcctg ttccccccca agcccaagga caccctgatg 240
atcagcagga cccccgaggt gacctgcgtg gtggtggacg tgagccacga ggacccagag 300
gtgaagttca actggtacgt ggacggcgtg gaggtgcaca acgccaagac caagcccaga 360
gaggagcagt acaacagcac ctacagggtg gtgtccgtgc tgaccgtgct gcaccaggac 420
tggctgaacg gcaaggaata caagtgcaag gtctccaaca aggccctgcc agcccccatc 480
gaaaagacca tcagcaaggc caagggccag ccacgggagc cccaggtgta caccctgccc 540
ccctcccggg aggagatgac caagaaccag gtgtccctga cctgtctggt gaagggcttc 600
taccccagcg acatcgccgt ggagtgggag agcaacggcc agcccgagaa caactacaag 660
accacccccc cagtgctgga cagcgacggc agcttcttcc tgtacagcaa gctgaccgtg 720
gacaagtcca ggtggcagca gggcaacgtg ttcagctgca gcgtgatgca cgaggccctg 780
cacaaccact acacccagaa gagcctgagc ctgtcccccg gcaagggagg aagcggagga 840
tctggcggtt ccggaggctc ttgtgtggaa cccctcggca tggaaaacgg caatatcgcc 900
aatagccaga ttgccgccag cagcgtcaga gtgacatttc tgggactgca gcactgggtg 960
cccgagctgg ctagactgaa tagagccggc atggtcaacg cctggacacc cagcagcaac 1020
gacgataacc cttggattca agtgaacctg ctgcggcgta tgtgggtcac aggtgttgtt 1080
acacagggcg cctctagact ggccagccac gagtatctga aggcctttaa ggtggcctac 1140
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaagagttt 1200
gtcggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacacc tgtggaagcc 1260
cagtacgtgc ggctgtaccc tacaagctgt cacaccgcct gcactctgag attcgaactg 1320
ctgggatgcg agctgaacgg ctgtgctaat cctctgggcc tgaagaacaa cagcatcccc 1380
gataagcaga tcaccgccag ctccagctat aagacatggg gcctgcacct gttcagctgg 1440
aacccttctt acgccagact ggacaagcag ggcaacttca atgcttgggt ggccggcagc 1500
tacggcaatg atcagtggct gcaagtggac ctgggcagca gcaaagaagt gacaggcatc 1560
atcacccagg gcgccagaaa tttcggcagc gtgcagtttg tggccagcta caaagtggcc 1620
tactccaacg acagcgccaa ctggaccgag tatcaggacc ctagaaccgg cagctccaag 1680
atcttccccg gcaattggga caaccacagc cacaagaaga atctgttcga aacccctatc 1740
ctggccagat atgtgcgcat tctgcccgtg gcctggcaca acagaattgc cctgagactg 1800
gaactgctcg gctgtggctc tcaccaccac catcaccat 1839
<210> 18
<211> 613
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 18
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
50 55 60
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
65 70 75 80
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
85 90 95
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
100 105 110
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
115 120 125
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
130 135 140
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
145 150 155 160
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
165 170 175
Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser
180 185 190
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
195 200 205
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
210 215 220
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
225 230 235 240
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
245 250 255
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
260 265 270
Pro Gly Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys
275 280 285
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
290 295 300
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
305 310 315 320
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
325 330 335
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
340 345 350
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
355 360 365
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
370 375 380
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
385 390 395 400
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
405 410 415
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
420 425 430
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
435 440 445
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
450 455 460
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
465 470 475 480
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
485 490 495
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
500 505 510
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
515 520 525
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
530 535 540
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
545 550 555 560
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
565 570 575
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
580 585 590
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Ser His
595 600 605
His His His His His
610
<210> 19
<211> 1839
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 19
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgacaag acccacacct gccccccctg cccagcccca 180
gagctgctgg gcggaccctc cgtgttcctg ttccccccca agcccaagga caccctgatg 240
atcagcagga cccccgaggt gacctgcgtg gtggtggacg tgagccacga ggacccagag 300
gtgaagttca actggtacgt ggacggcgtg gaggtgcaca acgccaagac caagcccaga 360
gaggagcagt acaacagcac ctacagggtg gtgtccgtgc tgaccgtgct gcaccaggac 420
tggctgaacg gcaaggaata caagtgcaag gtctccaaca aggccctgcc agcccccatc 480
gaaaagacca tcagcaaggc caagggccag ccacgggagc cccaggtgta caccctgccc 540
ccctgccggg aggagatgac caagaaccag gtgtccctgt ggtgtctggt gaagggcttc 600
taccccagcg acatcgccgt ggagtgggag agcaacggcc agcccgagaa caactacaag 660
accacccccc cagtgctgga cagcgacggc agcttcttcc tgtacagcaa gctgaccgtg 720
gacaagtcca ggtggcagca gggcaacgtg ttcagctgca gcgtgatgca cgaggccctg 780
cacaaccact acacccagaa gagcctgagc ctgtcccccg gcaagggagg aagcggagga 840
tctggcggtt ccggaggctc ttgtgtggaa cccctcggca tggaaaacgg caatatcgcc 900
aatagccaga ttgccgccag cagcgtcaga gtgacatttc tgggactgca gcactgggtg 960
cccgagctgg ctagactgaa tagagccggc atggtcaacg cctggacacc cagcagcaac 1020
gacgataacc cttggattca agtgaacctg ctgcggcgta tgtgggtcac aggtgttgtt 1080
acacagggcg cctctagact ggccagccac gagtatctga aggcctttaa ggtggcctac 1140
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaagagttt 1200
gtcggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacacc tgtggaagcc 1260
cagtacgtgc ggctgtaccc tacaagctgt cacaccgcct gcactctgag attcgaactg 1320
ctgggatgcg agctgaacgg ctgtgctaat cctctgggcc tgaagaacaa cagcatcccc 1380
gataagcaga tcaccgccag ctccagctat aagacatggg gcctgcacct gttcagctgg 1440
aacccttctt acgccagact ggacaagcag ggcaacttca atgcttgggt ggccggcagc 1500
tacggcaatg atcagtggct gcaagtggac ctgggcagca gcaaagaagt gacaggcatc 1560
atcacccagg gcgccagaaa tttcggcagc gtgcagtttg tggccagcta caaagtggcc 1620
tactccaacg acagcgccaa ctggaccgag tatcaggacc ctagaaccgg cagctccaag 1680
atcttccccg gcaattggga caaccacagc cacaagaaga atctgttcga aacccctatc 1740
ctggccagat atgtgcgcat tctgcccgtg gcctggcaca acagaattgc cctgagactg 1800
gaactgctcg gctgtggctc tcaccaccac catcaccat 1839
<210> 20
<211> 613
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 20
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
50 55 60
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
65 70 75 80
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
85 90 95
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
100 105 110
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
115 120 125
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
130 135 140
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
145 150 155 160
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
165 170 175
Cys Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
180 185 190
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
195 200 205
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
210 215 220
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
225 230 235 240
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
245 250 255
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
260 265 270
Pro Gly Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys
275 280 285
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
290 295 300
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
305 310 315 320
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
325 330 335
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
340 345 350
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
355 360 365
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
370 375 380
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
385 390 395 400
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
405 410 415
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
420 425 430
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
435 440 445
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
450 455 460
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
465 470 475 480
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
485 490 495
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
500 505 510
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
515 520 525
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
530 535 540
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
545 550 555 560
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
565 570 575
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
580 585 590
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Ser His
595 600 605
His His His His His
610
<210> 21
<211> 1839
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 21
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgacaag acccacacct gccccccctg cccagcccca 180
gagctgctgg gcggaccctc cgtgttcctg ttccccccca agcccaagga caccctgatg 240
atcagcagga cccccgaggt gacctgcgtg gtggtggacg tgagccacga ggacccagag 300
gtgaagttca actggtacgt ggacggcgtg gaggtgcaca acgccaagac caagcccaga 360
gaggagcagt acaacagcac ctacagggtg gtgtccgtgc tgaccgtgct gcaccaggac 420
tggctgaacg gcaaggaata caagtgcaag gtctccaaca aggccctgcc agcccccatc 480
gaaaagacca tcagcaaggc caagggccag ccacgggagc cccaggtgtg caccctgccc 540
ccctcccggg aggagatgac caagaaccag gtgtccctgt cctgtgcggt gaagggcttc 600
taccccagcg acatcgccgt ggagtgggag agcaacggcc agcccgagaa caactacaag 660
accacccccc cagtgctgga cagcgacggc agcttcttcc tggtcagcaa gctgaccgtg 720
gacaagtcca ggtggcagca gggcaacgtg ttcagctgca gcgtgatgca cgaggccctg 780
cacaaccact acacccagaa gagcctgagc ctgtcccccg gcaagggagg aagcggagga 840
tctggcggtt ccggaggctc ttgtgtggaa cccctcggca tggaaaacgg caatatcgcc 900
aatagccaga ttgccgccag cagcgtcaga gtgacatttc tgggactgca gcactgggtg 960
cccgagctgg ctagactgaa tagagccggc atggtcaacg cctggacacc cagcagcaac 1020
gacgataacc cttggattca agtgaacctg ctgcggcgta tgtgggtcac aggtgttgtt 1080
acacagggcg cctctagact ggccagccac gagtatctga aggcctttaa ggtggcctac 1140
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaagagttt 1200
gtcggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacacc tgtggaagcc 1260
cagtacgtgc ggctgtaccc tacaagctgt cacaccgcct gcactctgag attcgaactg 1320
ctgggatgcg agctgaacgg ctgtgctaat cctctgggcc tgaagaacaa cagcatcccc 1380
gataagcaga tcaccgccag ctccagctat aagacatggg gcctgcacct gttcagctgg 1440
aacccttctt acgccagact ggacaagcag ggcaacttca atgcttgggt ggccggcagc 1500
tacggcaatg atcagtggct gcaagtggac ctgggcagca gcaaagaagt gacaggcatc 1560
atcacccagg gcgccagaaa tttcggcagc gtgcagtttg tggccagcta caaagtggcc 1620
tactccaacg acagcgccaa ctggaccgag tatcaggacc ctagaaccgg cagctccaag 1680
atcttccccg gcaattggga caaccacagc cacaagaaga atctgttcga aacccctatc 1740
ctggccagat atgtgcgcat tctgcccgtg gcctggcaca acagaattgc cctgagactg 1800
gaactgctcg gctgtggctc tcaccaccac catcaccat 1839
<210> 22
<211> 596
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 22
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys Val
35 40 45
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
50 55 60
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
65 70 75 80
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
85 90 95
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
100 105 110
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
115 120 125
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
130 135 140
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
145 150 155 160
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
165 170 175
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
180 185 190
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala
195 200 205
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
210 215 220
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
225 230 235 240
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
245 250 255
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
260 265 270
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
275 280 285
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
290 295 300
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
305 310 315 320
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
325 330 335
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
340 345 350
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Gly Gly Gly
355 360 365
Thr Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu
370 375 380
Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu
385 390 395 400
Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser
405 410 415
His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu
420 425 430
Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr
435 440 445
Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn
450 455 460
Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro
465 470 475 480
Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln
485 490 495
Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val
500 505 510
Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val
515 520 525
Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro
530 535 540
Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr
545 550 555 560
Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val
565 570 575
Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu
580 585 590
Ser Pro Gly Lys
595
<210> 23
<211> 1788
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 23
ctggacatct gcagcaagaa cccctgccac aacggcggcc tgtgcgaaga gatcagccag 60
gaagtgcggg gcgacgtgtt ccccagctac acctgtacct gcctgaaggg ctacgccggc 120
aaccactgcg agactaagtg cgtggaaccc ctgggcatgg aaaacggcaa tatcgccaac 180
agccagatcg ccgccagctc cgtgcgcgtg acctttctgg gactgcagca ctgggtgccc 240
gagctggcca gactgaacag agccggcatg gtgaacgcct ggacccccag cagcaacgac 300
gacaaccctt ggatccaggt gaacctgctg cggcggatgt gggtgacagg cgtggtgaca 360
cagggcgcca gcagactggc cagccacgag tacctgaagg cctttaaggt ggcctacagc 420
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaaacacaa agaatttgtg 480
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacccgt ggaagcccag 540
tacgtgcggc tgtaccccac cagctgccac accgcctgca ccctgagatt cgagctgctg 600
ggctgcgagc tgaacggctg cgccaacccc ctgggcctga agaacaacag catccccgac 660
aagcagatca ccgcctccag cagctacaag acctggggcc tgcacctgtt cagctggaac 720
cccagctacg cccggctgga caagcagggc aacttcaacg cctgggtggc cggcagctac 780
ggcaacgacc agtggctgca ggtggacctg ggcagcagca aagaagtgac cggcatcatc 840
acccaggggg ccagaaactt cggcagcgtg cagttcgtgg ccagctacaa agtggcctac 900
tccaacgaca gcgccaactg gaccgagtac caggaccccc ggaccggcag ctccaagatc 960
ttccccggca actgggacaa ccacagccac aagaagaatc tgttcgaaac ccccatcctg 1020
gccagatacg tgcggatcct gcccgtggcc tggcacaacc ggatcgccct gagactggaa 1080
ctgctgggat gtgggggagg cggtaccgac aagacccaca cctgcccccc ctgcccagcc 1140
ccagagctgc tgggcggacc ctccgtgttc ctgttccccc ccaagcccaa ggacaccctg 1200
atgatcagca ggacccccga ggtgacctgc gtggtggtgg acgtgagcca cgaggaccca 1260
gaggtgaagt tcaactggta cgtggacggc gtggaggtgc acaacgccaa gaccaagccc 1320
agagaggagc agtacaacag cacctacagg gtggtgtccg tgctgaccgt gctgcaccag 1380
gactggctga acggcaagga atacaagtgc aaggtctcca acaaggccct gccagccccc 1440
atcgaaaaga ccatcagcaa ggccaagggc cagccacggg agccccaggt gtacaccctg 1500
cccccctccc gggaggagat gaccaagaac caggtgtccc tgacctgtct ggtgaagggc 1560
ttctacccca gcgacatcgc cgtggagtgg gagagcaacg gccagcccga gaacaactac 1620
aagaccaccc ccccagtgct ggacagcgac ggcagcttct tcctgtacag caagctgacc 1680
gtggacaagt ccaggtggca gcagggcaac gtgttcagct gcagcgtgat gcacgaggcc 1740
ctgcacaacc actacaccca gaagagcctg agcctgtccc ccggcaag 1788
<210> 24
<211> 604
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 24
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys Gly Ser Leu Glu Val Leu Phe Gln Gly Pro Gly Ser Ser
225 230 235 240
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
245 250 255
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
260 265 270
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys Val
275 280 285
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
290 295 300
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
305 310 315 320
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
325 330 335
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
340 345 350
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
355 360 365
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
370 375 380
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
385 390 395 400
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
405 410 415
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
420 425 430
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala
435 440 445
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
450 455 460
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
465 470 475 480
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
485 490 495
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
500 505 510
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
515 520 525
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
530 535 540
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
545 550 555 560
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
565 570 575
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
580 585 590
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
595 600
<210> 25
<211> 1812
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 25
gacaagaccc acacctgtcc cccctgccct gctcctgagc tgctgggagg acccagcgtg 60
ttcctgttcc cccccaagcc caaggacacc ctgatgatca gccggacccc cgaagtgacc 120
tgcgtggtgg tggacgtgtc ccacgaggac cctgaagtga agttcaattg gtacgtggac 180
ggcgtggagg tgcacaacgc caagaccaag ccccgggagg aacagtacaa cagcacctac 240
cgggtggtgt ccgtgctgac cgtgctgcac caggactggc tgaacggcaa agaatacaag 300
tgcaaggtgt ccaacaaggc cctgcctgcc cccatcgaga aaaccatcag caaggccaag 360
ggccagccca gagaacccca ggtgtacaca ctcccaccaa gccgggagga aatgaccaag 420
aaccaggtgt ccctgacctg cctggtgaag ggcttctacc ccagcgacat tgccgtggag 480
tgggagagca acggccagcc tgagaacaac tacaagacca cccctccagt cctcgattct 540
gatggatctt tcttcctgta ctccaagctg accgtggaca agagccggtg gcagcaggga 600
aacgtctttt cctgttccgt catgcatgag gctctccaca atcactacac ccagaagtcc 660
ctgagcctga gccccggcaa gggatccctc gaggtgctgt ttcagggacc aggcagcagc 720
ctggacatct gcagcaagaa cccctgccac aacggcggcc tgtgcgaaga gatcagccag 780
gaagtgcggg gcgacgtgtt ccccagctac acctgtacct gcctgaaggg ctacgccggc 840
aaccactgcg agactaagtg cgtggaaccc ctgggaatgg aaaacggcaa tatcgccaac 900
agccagatcg ccgccagctc cgtcagagtg acctttctgg gactccagca ctgggtgccc 960
gagctggcca gactgaatag agccggcatg gtcaacgcct ggacccccag cagcaacgac 1020
gacaacccct ggattcaagt gaacctgctg cggcgtatgt gggtcaccgg cgtcgtgaca 1080
cagggcgcta gcagactggc cagccacgag tacctgaagg cctttaaggt ggcctacagc 1140
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaaacacaa agaatttgtg 1200
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacccgt ggaagcccag 1260
tacgtgcggc tgtaccctac cagctgtcac accgcctgca ccttaagatt cgagctgctg 1320
ggctgcgagc tgaacggctg cgctaatcct ctgggcctga agaacaacag catccccgac 1380
aagcagatca ccgcctccag cagctacaag acctggggac tgcacctgtt cagctggaac 1440
cctagctacg cccggctgga caagcagggc aacttcaatg cttgggtggc cggcagctac 1500
ggcaacgacc agtggctcca ggtggacctg ggcagcagca aagaagtgac cggcatcatc 1560
acccaggggg ccagaaactt cggcagcgtg cagttcgtgg cctcctacaa agtggcctac 1620
tccaacgaca gcgccaactg gaccgagtac caggacccta gaaccggcag ctccaagatt 1680
ttccccggca actgggataa ccacagccac aagaagaatc tgttcgaaac ccccatcctg 1740
gcccgctacg tgcgcattct accggtcgcc tggcacaacc ggatcgccct gagactggaa 1800
ctgctgggat gc 1812
<210> 26
<211> 364
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 26
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Cys
35 40 45
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
50 55 60
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
65 70 75 80
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
85 90 95
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
100 105 110
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
115 120 125
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
130 135 140
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
145 150 155 160
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
165 170 175
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
180 185 190
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Cys Ala
195 200 205
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
210 215 220
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
225 230 235 240
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
245 250 255
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
260 265 270
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
275 280 285
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
290 295 300
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
305 310 315 320
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
325 330 335
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
340 345 350
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
355 360
<210> 27
<211> 1092
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 27
ctggacatct gcagcaagaa cccctgccac aacggcggcc tgtgcgaaga gatcagccag 60
gaagtgcggg gcgacgtgtt ccccagctac acctgtacct gcctgaaggg ctacgccggc 120
aaccactgcg agacaaaggg ctgcgccaac cccctgggcc tgaagaacaa cagcatcccc 180
gacaagcaga tcaccgccag cagcagctac aagacctggg gcctgcacct gttcagctgg 240
aaccccagct acgcccggct ggacaagcag ggcaacttca acgcctgggt ggccggcagc 300
tacggcaacg accagtggct gcaggtggac ctgggcagca gcaaagaagt gaccggcatc 360
atcacccagg gcgccagaaa cttcggcagc gtgcagttcg tggccagcta caaggtggcc 420
tacagcaacg acagcgccaa ctggaccgag taccaggacc cccggaccgg cagctccaag 480
atcttccccg gcaactggga caaccacagc cacaagaaga acctgttcga gacacccatc 540
ctggccagat acgtgcggat cctgcccgtg gcctggcaca accggatcgc cctgagactg 600
gaactgctgg gctgcggctg tgccaatcct ctgggactga aaaacaattc catccctgat 660
aagcagatta cagcctccag ctcctataag acatgggggc tgcatctgtt ttcttggaac 720
ccctcctacg ctagactgga taagcaggga aatttcaatg cttgggtggc cgggtcctat 780
ggaaatgatc agtggctgca ggtggacctg ggatcctcca aagaagtgac agggattatt 840
acacaggggg ctcggaactt tggctctgtg cagtttgtgg cttcctacaa agtggcttac 900
tccaacgatt ccgccaattg gacagaatat caggatccca gaaccggctc cagcaagatc 960
tttcctggaa attgggataa tcactcccac aagaaaaatc tgtttgaaac ccctattctg 1020
gctcgctatg tgcgcattct gcctgtggct tggcataata gaatcgctct gcggctggaa 1080
ctgctgggat gc 1092
<210> 28
<211> 967
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 28
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys Val
35 40 45
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
50 55 60
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
65 70 75 80
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
85 90 95
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
100 105 110
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
115 120 125
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
130 135 140
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
145 150 155 160
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
165 170 175
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
180 185 190
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala
195 200 205
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
210 215 220
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
225 230 235 240
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
245 250 255
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
260 265 270
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
275 280 285
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
290 295 300
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
305 310 315 320
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
325 330 335
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
340 345 350
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Gly Ser Gly
355 360 365
Gly Ser Gly Gly Ser Gly Gly Ser Asp Ala His Lys Ser Glu Val Ala
370 375 380
His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu
385 390 395 400
Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val
405 410 415
Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp
420 425 430
Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp
435 440 445
Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala
450 455 460
Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln
465 470 475 480
His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val
485 490 495
Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys
500 505 510
Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro
515 520 525
Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys
530 535 540
Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu
545 550 555 560
Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys
565 570 575
Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val
580 585 590
Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser
595 600 605
Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly
610 615 620
Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile
625 630 635 640
Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu
645 650 655
Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp
660 665 670
Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser
675 680 685
Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly
690 695 700
Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val
705 710 715 720
Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys
725 730 735
Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu
740 745 750
Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys
755 760 765
Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu
770 775 780
Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val
785 790 795 800
Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His
805 810 815
Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val
820 825 830
Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg
835 840 845
Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe
850 855 860
Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala
865 870 875 880
Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu
885 890 895
Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys
900 905 910
Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala
915 920 925
Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe
930 935 940
Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly
945 950 955 960
Leu His His His His His His
965
<210> 29
<211> 2901
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 29
ctggacatct gcagcaagaa cccctgccac aacggcggcc tgtgcgaaga gatcagccag 60
gaagtgcggg gcgacgtgtt ccccagctac acctgtacct gcctgaaggg ctacgccggc 120
aaccactgcg agactaagtg cgtggaaccc ctgggcatgg aaaacggcaa tatcgccaac 180
agccagatcg ccgccagctc cgtgcgcgtg acctttctgg gactgcagca ctgggtgccc 240
gagctggcca gactgaacag agccggcatg gtgaacgcct ggacccccag cagcaacgac 300
gacaaccctt ggatccaggt gaacctgctg cggcggatgt gggtgacagg cgtggtgaca 360
cagggcgcca gcagactggc cagccacgag tacctgaagg cctttaaggt ggcctacagc 420
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaaacacaa agaatttgtg 480
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacccgt ggaagcccag 540
tacgtgcggc tgtaccccac cagctgccac accgcctgca ccctgagatt cgagctgctg 600
ggctgcgagc tgaacggctg cgccaacccc ctgggcctga agaacaacag catccccgac 660
aagcagatca ccgcctccag cagctacaag acctggggcc tgcacctgtt cagctggaac 720
cccagctacg cccggctgga caagcagggc aacttcaacg cctgggtggc cggcagctac 780
ggcaacgacc agtggctgca ggtggacctg ggcagcagca aagaagtgac cggcatcatc 840
acccaggggg ccagaaactt cggcagcgtg cagttcgtgg ccagctacaa agtggcctac 900
tccaacgaca gcgccaactg gaccgagtac caggaccccc ggaccggcag ctccaagatc 960
ttccccggca actgggacaa ccacagccac aagaagaatc tgttcgaaac ccccatcctg 1020
gccagatacg tgcggatcct gcccgtggcc tggcacaacc ggatcgccct gagactggaa 1080
ctgctgggat gtggaggaag cggaggatct ggcggttccg gaggctctga cgcccacaag 1140
agcgaggtgg cccaccggtt caaggacctg ggcgaggaaa acttcaaggc cctggtgctg 1200
atcgccttcg cccagtacct gcagcagagc cccttcgaag atcacgtaaa gttagtcaac 1260
gaggttacgg aattcgcaaa gacatgcgtt gctgacgaat ccgctgagaa ttgtgacaag 1320
agtttgcaca ctttattcgg agataagttg tgtactgtag ctactttgag agagacttac 1380
ggtgaaatgg ctgactgctg tgcaaaacag gaaccagaac gtaacgaatg tttccttcag 1440
cataaggatg ataaccctaa ccttccaagg cttgttaggc cagaagtcga cgtgatgtgc 1500
accgccttcc atgataatga agagactttt cttaaaaagt acctatacga gattgcaagg 1560
cgtcatccat atttttacgc cccagagctg ttgtttttcg caaagagata caaagctgca 1620
tttactgagt gttgccaagc tgccgacaag gccgcttgtt tgctaccaaa gttggacgaa 1680
ttgagagacg agggtaaggc atcatctgcc aagcagagat taaaatgtgc atctttgcaa 1740
aaatttggag agagagcttt taaggcatgg gctgttgccc gactaagcca aagattccca 1800
aaagccgaat ttgctgaagt atccaagctg gtgactgatt tgactaaagt acatacagaa 1860
tgttgccatg gcgacctttt agaatgtgct gatgacagag cagatttggc taagtatatc 1920
tgcgaaaatc aagattcaat cagctctaag ctgaaggaat gttgcgagaa accactgtta 1980
gaaaaatcgc attgtattgc tgaagttgaa aatgatgaga tgcctgctga cttgccttct 2040
cttgccgctg attttgttga gtcgaaggat gtctgtaaga attatgctga agctaaagac 2100
gttttcctgg gtatgttctt atatgagtac gcaagacgtc acccagatta ctctgtggtt 2160
ctgctactga gattggctaa aacatacgag acaacgctgg agaagtgctg tgctgccgct 2220
gaccctcatg agtgctatgc aaaggttttt gatgaattca aaccattggt tgaagagcct 2280
caaaacttga taaagcagaa ctgtgagctg tttgagcaat tgggtgagta taagttccaa 2340
aatgccctgt tggtgagata tacaaaaaag gtaccccaag tttcaacgcc cactttagtt 2400
gaagtgtcca gaaatcttgg taaagtgggt agcaaatgtt gcaagcatcc agaagccaag 2460
cgaatgccct gtgctgagga ttatctgtcc gtcgtgttga accaattgtg cgtattacac 2520
gaaaaaaccc cagtctctga tagagtcacc aaatgttgca ctgagtcact agttaataga 2580
aggccttgtt tttccgcttt ggaagttgat gaaacctacg tgcctaagga atttaacgct 2640
gagaccttta cctttcacgc tgacatttgt actttgagtg aaaaagagcg tcaaatcaaa 2700
aagcaaaccg ctcttgttga attggtgaaa cacaagccta aggctacgaa ggagcagctt 2760
aaagccgtca tggacgattt cgccgcattt gttgaaaaat gctgtaaagc tgatgacaag 2820
gaaacatgtt tcgctgaaga gggaaagaaa ttggttgcgg ccagtcaggc cgcacttggt 2880
ttgcaccatc atcaccatca c 2901
<210> 30
<211> 965
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 30
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
1 5 10 15
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
20 25 30
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
35 40 45
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
50 55 60
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
65 70 75 80
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
85 90 95
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
100 105 110
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
115 120 125
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
130 135 140
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
145 150 155 160
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
165 170 175
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
180 185 190
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
195 200 205
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
210 215 220
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
225 230 235 240
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
245 250 255
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
260 265 270
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
275 280 285
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
290 295 300
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
305 310 315 320
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
325 330 335
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
340 345 350
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
355 360 365
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
370 375 380
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
385 390 395 400
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
405 410 415
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
420 425 430
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
435 440 445
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
450 455 460
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
465 470 475 480
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
485 490 495
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
500 505 510
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
515 520 525
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
530 535 540
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
545 550 555 560
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
565 570 575
Ala Ala Ser Gln Ala Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly
580 585 590
Ser Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys
595 600 605
Glu Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr
610 615 620
Cys Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys
625 630 635 640
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
645 650 655
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
660 665 670
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
675 680 685
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
690 695 700
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
705 710 715 720
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
725 730 735
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
740 745 750
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
755 760 765
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
770 775 780
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
785 790 795 800
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
805 810 815
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
820 825 830
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
835 840 845
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
850 855 860
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
865 870 875 880
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
885 890 895
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
900 905 910
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
915 920 925
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
930 935 940
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys Gly Ser His
945 950 955 960
His His His His His
965
<210> 31
<211> 2895
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 31
gacgcccaca agagcgaggt ggcccaccgg ttcaaggacc tgggcgagga aaacttcaag 60
gccctggtgc tgatcgcctt cgcccagtac ctgcagcaga gccccttcga agatcacgta 120
aagttagtca acgaggttac ggaattcgca aagacatgcg ttgctgacga atccgctgag 180
aattgtgaca agagtttgca cactttattc ggagataagt tgtgtactgt agctactttg 240
agagagactt acggtgaaat ggctgactgc tgtgcaaaac aggaaccaga acgtaacgaa 300
tgtttccttc agcataagga tgataaccct aaccttccaa ggcttgttag gccagaagtc 360
gacgtgatgt gcaccgcctt ccatgataat gaagagactt ttcttaaaaa gtacctatac 420
gagattgcaa ggcgtcatcc atatttttac gccccagagc tgttgttttt cgcaaagaga 480
tacaaagctg catttactga gtgttgccaa gctgccgaca aggccgcttg tttgctacca 540
aagttggacg aattgagaga cgagggtaag gcatcatctg ccaagcagag attaaaatgt 600
gcatctttgc aaaaatttgg agagagagct tttaaggcat gggctgttgc ccgactaagc 660
caaagattcc caaaagccga atttgctgaa gtatccaagc tggtgactga tttgactaaa 720
gtacatacag aatgttgcca tggcgacctt ttagaatgtg ctgatgacag agcagatttg 780
gctaagtata tctgcgaaaa tcaagattca atcagctcta agctgaagga atgttgcgag 840
aaaccactgt tagaaaaatc gcattgtatt gctgaagttg aaaatgatga gatgcctgct 900
gacttgcctt ctcttgccgc tgattttgtt gagtcgaagg atgtctgtaa gaattatgct 960
gaagctaaag acgttttcct gggtatgttc ttatatgagt acgcaagacg tcacccagat 1020
tactctgtgg ttctgctact gagattggct aaaacatacg agacaacgct ggagaagtgc 1080
tgtgctgccg ctgaccctca tgagtgctat gcaaaggttt ttgatgaatt caaaccattg 1140
gttgaagagc ctcaaaactt gataaagcag aactgtgagc tgtttgagca attgggtgag 1200
tataagttcc aaaatgccct gttggtgaga tatacaaaaa aggtacccca agtttcaacg 1260
cccactttag ttgaagtgtc cagaaatctt ggtaaagtgg gtagcaaatg ttgcaagcat 1320
ccagaagcca agcgaatgcc ctgtgctgag gattatctgt ccgtcgtgtt gaaccaattg 1380
tgcgtattac acgaaaaaac cccagtctct gatagagtca ccaaatgttg cactgagtca 1440
ctagttaata gaaggccttg tttttccgct ttggaagttg atgaaaccta cgtgcctaag 1500
gaatttaacg ctgagacctt tacctttcac gctgacattt gtactttgag tgaaaaagag 1560
cgtcaaatca aaaagcaaac cgctcttgtt gaattggtga aacacaagcc taaggctacg 1620
aaggagcagc ttaaagccgt catggacgat ttcgccgcat ttgttgaaaa atgctgtaaa 1680
gctgatgaca aggaaacatg tttcgctgaa gagggaaaga aattggttgc ggccagtcag 1740
gccggaggaa gcggaggatc tggcggttcc ggaggctctc tagacatctg cagcaagaac 1800
ccctgccaca acggcggcct gtgcgaagag atcagccagg aagtgcgggg cgacgtgttc 1860
cccagctaca cctgtacctg cctgaagggc tacgccggca accactgcga gactaagtgc 1920
gtggaacccc tgggcatgga aaacggcaat atcgccaaca gccagatcgc cgccagctcc 1980
gtgcgcgtga cctttctggg actgcagcac tgggtgcccg agctggccag actgaacaga 2040
gccggcatgg tgaacgcctg gacccccagc agcaacgacg acaacccttg gatccaggtg 2100
aacctgctgc ggcggatgtg ggtgacaggc gtggtgacac agggcgccag cagactggcc 2160
agccacgagt acctgaaggc ctttaaggtg gcctacagcc tgaacggcca cgagttcgac 2220
ttcatccacg acgtgaacaa gaaacacaaa gaatttgtgg gcaactggaa caagaacgcc 2280
gtgcacgtga acctgttcga gacacccgtg gaagcccagt acgtgcggct gtaccccacc 2340
agctgccaca ccgcctgcac cctgagattc gagctgctgg gctgcgagct gaacggctgc 2400
gccaaccccc tgggcctgaa gaacaacagc atccccgaca agcagatcac cgcctccagc 2460
agctacaaga cctggggcct gcacctgttc agctggaacc ccagctacgc ccggctggac 2520
aagcagggca acttcaacgc ctgggtggcc ggcagctacg gcaacgacca gtggctgcag 2580
gtggacctgg gcagcagcaa agaagtgacc ggcatcatca cccagggggc cagaaacttc 2640
ggcagcgtgc agttcgtggc cagctacaaa gtggcctact ccaacgacag cgccaactgg 2700
accgagtacc aggacccccg gaccggcagc tccaagatct tccccggcaa ctgggacaac 2760
cacagccaca agaagaatct gttcgaaacc cccatcctgg ccagatacgt gcggatcctg 2820
cccgtggcct ggcacaaccg gatcgccctg agactggaac tgctgggatg tggctctcac 2880
caccaccatc accat 2895
<210> 32
<211> 641
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 32
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Ser His His His His His
625 630 635 640
His
<210> 33
<211> 1923
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 33
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gctctcacca ccaccatcac 1920
cat 1923
<210> 34
<211> 809
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 34
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Gly Ser His His His His His His
805
<210> 35
<211> 2427
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 35
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcagcactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aacccttgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcctcta gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcggctctc accaccacca tcaccat 2427
<210> 36
<211> 811
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 36
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
645 650 655
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
660 665 670
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
675 680 685
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
690 695 700
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
705 710 715 720
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
725 730 735
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
740 745 750
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
755 760 765
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
770 775 780
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
785 790 795 800
Leu Gly Cys Gly Ser His His His His His His
805 810
<210> 37
<211> 2433
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 37
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgacgcc cacaagagcg aggtggccca ccggttcaag 180
gacctgggcg aggaaaactt caaggccctg gtgctgatcg ccttcgccca gtacctgcag 240
cagagcccct tcgaagatca cgtaaagtta gtcaacgagg ttacggaatt cgcaaagaca 300
tgcgttgctg acgaatccgc tgagaattgt gacaagagtt tgcacacttt attcggagat 360
aagttgtgta ctgtagctac tttgagagag acttacggtg aaatggctga ctgctgtgca 420
aaacaggaac cagaacgtaa cgaatgtttc cttcagcata aggatgataa ccctaacctt 480
ccaaggcttg ttaggccaga agtcgacgtg atgtgcaccg ccttccatga taatgaagag 540
acttttctta aaaagtacct atacgagatt gcaaggcgtc atccatattt ttacgcccca 600
gagctgttgt ttttcgcaaa gagatacaaa gctgcattta ctgagtgttg ccaagctgcc 660
gacaaggccg cttgtttgct accaaagttg gacgaattga gagacgaggg taaggcatca 720
tctgccaagc agagattaaa atgtgcatct ttgcaaaaat ttggagagag agcttttaag 780
gcatgggctg ttgcccgact aagccaaaga ttcccaaaag ccgaatttgc tgaagtatcc 840
aagctggtga ctgatttgac taaagtacat acagaatgtt gccatggcga ccttttagaa 900
tgtgctgatg acagagcaga tttggctaag tatatctgcg aaaatcaaga ttcaatcagc 960
tctaagctga aggaatgttg cgagaaacca ctgttagaaa aatcgcattg tattgctgaa 1020
gttgaaaatg atgagatgcc tgctgacttg ccttctcttg ccgctgattt tgttgagtcg 1080
aaggatgtct gtaagaatta tgctgaagct aaagacgttt tcctgggtat gttcttatat 1140
gagtacgcaa gacgtcaccc agattactct gtggttctgc tactgagatt ggctaaaaca 1200
tacgagacaa cgctggagaa gtgctgtgct gccgctgacc ctcatgagtg ctatgcaaag 1260
gtttttgatg aattcaaacc attggttgaa gagcctcaaa acttgataaa gcagaactgt 1320
gagctgtttg agcaattggg tgagtataag ttccaaaatg ccctgttggt gagatataca 1380
aaaaaggtac cccaagtttc aacgcccact ttagttgaag tgtccagaaa tcttggtaaa 1440
gtgggtagca aatgttgcaa gcatccagaa gccaagcgaa tgccctgtgc tgaggattat 1500
ctgtccgtcg tgttgaacca attgtgcgta ttacacgaaa aaaccccagt ctctgataga 1560
gtcaccaaat gttgcactga gtcactagtt aatagaaggc cttgtttttc cgctttggaa 1620
gttgatgaaa cctacgtgcc taaggaattt aacgctgaga cctttacctt tcacgctgac 1680
atttgtactt tgagtgaaaa agagcgtcaa atcaaaaagc aaaccgctct tgttgaattg 1740
gtgaaacaca agcctaaggc tacgaaggag cagcttaaag ccgtcatgga cgatttcgcc 1800
gcatttgttg aaaaatgctg taaagctgat gacaaggaaa catgtttcgc tgaagaggga 1860
aagaaattgg ttgcggccag tcaggccgca cttggtttgg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgcgc caaccccctg ggcctgaaga acaacagcat ccccgacaag 1980
cagatcaccg cctccagcag ctacaagacc tggggcctgc acctgttcag ctggaacccc 2040
agctacgccc ggctggacaa gcagggcaac ttcaacgcct gggtggccgg cagctacggc 2100
aacgaccagt ggctgcaggt ggacctgggc agcagcaaag aagtgaccgg catcatcacc 2160
cagggggcca gaaacttcgg cagcgtgcag ttcgtggcca gctacaaagt ggcctactcc 2220
aacgacagcg ccaactggac cgagtaccag gacccccgga ccggcagctc caagatcttc 2280
cccggcaact gggacaacca cagccacaag aagaatctgt tcgaaacccc catcctggcc 2340
agatacgtgc ggatcctgcc cgtggcctgg cacaaccgga tcgccctgag actggaactg 2400
ctgggatgtg gctctcacca ccaccatcac cat 2433
<210> 38
<211> 971
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 38
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Glu Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
805 810 815
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
820 825 830
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
835 840 845
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
850 855 860
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
865 870 875 880
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
885 890 895
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
900 905 910
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
915 920 925
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
930 935 940
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
945 950 955 960
Leu Gly Cys Gly Ser His His His His His His
965 970
<210> 39
<211> 2913
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 39
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgaggtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcagcactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aacccttgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcctcta gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggctgtgc taatcctctg ggcctgaaga acaacagcat ccccgataag 2460
cagatcaccg ccagctccag ctataagaca tggggcctgc acctgttcag ctggaaccct 2520
tcttacgcca gactggacaa gcagggcaac ttcaatgctt gggtggccgg cagctacggc 2580
aatgatcagt ggctgcaagt ggacctgggc agcagcaaag aagtgacagg catcatcacc 2640
cagggcgcca gaaatttcgg cagcgtgcag tttgtggcca gctacaaagt ggcctactcc 2700
aacgacagcg ccaactggac cgagtatcag gaccctagaa ccggcagctc caagatcttc 2760
cccggcaatt gggacaacca cagccacaag aagaatctgt tcgaaacccc tatcctggcc 2820
agatatgtgc gcattctgcc cgtggcctgg cacaacagaa ttgccctgag actggaactg 2880
ctcggctgtg gctctcacca ccaccatcac cat 2913
<210> 40
<211> 592
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 40
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe
50 55 60
Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr
65 70 75 80
Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser
85 90 95
Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala
100 105 110
Lys Arg Met Pro Cys Ala Glu Asp Cys Leu Ser Val Phe Leu Asn Gln
115 120 125
Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys
130 135 140
Cys Cys Thr Glu Ser Leu Val Asn Gly Arg Pro Cys Phe Ser Ala Leu
145 150 155 160
Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe
165 170 175
Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile
180 185 190
Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala
195 200 205
Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val
210 215 220
Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu
225 230 235 240
Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly
245 250 255
Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly
260 265 270
Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val
275 280 285
Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg
290 295 300
Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp
305 310 315 320
Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr
325 330 335
Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu
340 345 350
Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe
355 360 365
Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn
370 375 380
Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln
385 390 395 400
Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg
405 410 415
Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly
420 425 430
Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser
435 440 445
Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala
450 455 460
Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr
465 470 475 480
Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val
485 490 495
Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe
500 505 510
Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr
515 520 525
Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn
530 535 540
Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu
545 550 555 560
Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala
565 570 575
Leu Arg Leu Glu Leu Leu Gly Cys Gly Ser His His His His His His
580 585 590
<210> 41
<211> 1776
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 41
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccctggtg gaagaacccc agaacctgat caagcagaac 180
tgcgagctgt tcgagcagct gggcgagtac aagttccaga atgccctgct cgtgcggtac 240
accaagaaag tgcctcaggt gtccacacct acactggtcg aggtgtccag aaacctgggc 300
aaagtgggca gcaagtgctg caagcaccct gaggccaaaa gaatgccttg cgccgaggat 360
tgcctgagcg tgttcctgaa tcagctgtgc gtgctgcacg agaaaacccc tgtgtccgac 420
agagtgacca agtgctgtac cgagagcctg gtcaacggca gaccttgctt tagcgccctg 480
gaagtggatg agacatacgt gcccaaagag ttcaacgccg agacattcac cttccacgcc 540
gacatctgta ccctgagcga gaaagagcgg cagatcaaga agcagacagc cctggtcgag 600
ctggtcaagc acaagcctaa ggccaccaaa gaacagctga aggccgtgat ggacgacttc 660
gccgccttcg tggaaaagtg ttgcaaggcc gacgacaaag agacatgctt cgccgaagag 720
ggcaagaaac tggtggctgc ctctcaggct gctctcggac ttggaggaag cggaggatct 780
ggcggttccg gaggctcttg tgtggaaccc ctcggcatgg aaaacggcaa tatcgccaat 840
agccagatcg ccgccagcag cgtcagagtg acatttctgg gactgcagca ctgggtgcca 900
gagctggcta gactgaatag agccggcatg gtcaacgcct ggacacccag cagcaacgac 960
gacaacccct ggattcaagt gaacctgctg cggcgtatgt gggtcacagg tgttgttaca 1020
cagggcgcct ctagactggc cagccacgag tatctgaagg cctttaaggt ggcctacagc 1080
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaagcacaa agagtttgtc 1140
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacctgt ggaagcccag 1200
tacgtgcggc tgtaccctac aagctgtcac accgcctgca cactgagatt cgaactgctg 1260
ggatgcgagc tgaacggctg tgctaatcct ctgggcctga agaacaacag catccccgat 1320
aagcagatca ccgcctccag cagctataag acatggggcc tgcacctgtt cagctggaac 1380
cctagctacg ccagactgga caagcagggc aactttaatg cctgggtggc cggcagctac 1440
ggcaatgatc aatggctgca agtggacctg ggcagcagca aagaagtgac cggcatcatt 1500
acccagggcg caagaaattt cggcagcgtg cagttcgtgg ccagctacaa agtggcctac 1560
tccaacgaca gcgccaactg gaccgagtat caggacccta gaaccggcag ctccaagatc 1620
ttccccggca attgggacaa ccacagccac aagaagaatc tgttcgaaac ccctatcctg 1680
gccagatatg tgcgcattct gcccgtggcc tggcacaaca gaattgccct gagactggaa 1740
ctgctcggct gtggctctca ccaccaccat caccat 1776
<210> 42
<211> 963
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 42
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
805 810 815
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
820 825 830
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
835 840 845
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
850 855 860
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
865 870 875 880
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
885 890 895
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
900 905 910
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
915 920 925
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
930 935 940
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
945 950 955 960
Leu Gly Cys
<210> 43
<211> 2889
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 43
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcagcactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aacccttgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcctcta gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggctgtgc taatcctctg ggcctgaaga acaacagcat ccccgataag 2460
cagatcaccg ccagctccag ctataagaca tggggcctgc acctgttcag ctggaaccct 2520
tcttacgcca gactggacaa gcagggcaac ttcaatgctt gggtggccgg cagctacggc 2580
aatgatcagt ggctgcaagt ggacctgggc agcagcaaag aagtgacagg catcatcacc 2640
cagggcgcca gaaatttcgg cagcgtgcag tttgtggcca gctacaaagt ggcctactcc 2700
aacgacagcg ccaactggac cgagtatcag gaccctagaa ccggcagctc caagatcttc 2760
cccggcaatt gggacaacca cagccacaag aagaatctgt tcgaaacccc tatcctggcc 2820
agatatgtgc gcattctgcc cgtggcctgg cacaacagaa ttgccctgag actggaactg 2880
ctcggctgt 2889
<210> 44
<211> 971
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 44
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
805 810 815
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
820 825 830
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
835 840 845
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
850 855 860
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
865 870 875 880
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
885 890 895
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
900 905 910
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
915 920 925
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
930 935 940
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
945 950 955 960
Leu Gly Cys Gly Ser His His His His His His
965 970
<210> 45
<211> 2913
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 45
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcagcactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aacccttgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcctcta gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggctgtgc taatcctctg ggcctgaaga acaacagcat ccccgataag 2460
cagatcaccg ccagctccag ctataagaca tggggcctgc acctgttcag ctggaaccct 2520
tcttacgcca gactggacaa gcagggcaac ttcaatgctt gggtggccgg cagctacggc 2580
aatgatcagt ggctgcaagt ggacctgggc agcagcaaag aagtgacagg catcatcacc 2640
cagggcgcca gaaatttcgg cagcgtgcag tttgtggcca gctacaaagt ggcctactcc 2700
aacgacagcg ccaactggac cgagtatcag gaccctagaa ccggcagctc caagatcttc 2760
cccggcaatt gggacaacca cagccacaag aagaatctgt tcgaaacccc tatcctggcc 2820
agatatgtgc gcattctgcc cgtggcctgg cacaacagaa ttgccctgag actggaactg 2880
ctcggctgtg gctctcacca ccaccatcac cat 2913
<210> 46
<211> 947
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 46
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala
610 615 620
Ser Gln Ala Ala Leu Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
625 630 635 640
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
645 650 655
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
660 665 670
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
675 680 685
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
690 695 700
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
705 710 715 720
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
725 730 735
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
740 745 750
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
755 760 765
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
770 775 780
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
785 790 795 800
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
805 810 815
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
820 825 830
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
835 840 845
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
850 855 860
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
865 870 875 880
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
885 890 895
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
900 905 910
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
915 920 925
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
930 935 940
Leu Gly Cys
945
<210> 47
<211> 2841
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 47
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggctg cttctcaggc cgctctgtgt gtggaacccc tcggcatgga aaacggcaat 1920
atcgccaata gccagattgc cgccagcagc gtcagagtga catttctggg actgcaacac 1980
tgggtgcccg agctggctag actgaataga gccggcatgg tcaacgcctg gacacccagc 2040
agcaacgacg ataatccctg gattcaagtg aacctgctgc ggcgtatgtg ggtcacaggt 2100
gttgttacac agggcgcaag cagactggcc agccacgagt atctgaaggc ctttaaggtg 2160
gcctacagcc tgaacggcca cgagttcgac ttcatccacg acgtgaacaa gaagcacaaa 2220
gagtttgtcg gcaactggaa caagaacgcc gtgcacgtga acctgttcga gacacctgtg 2280
gaagcccagt acgtgcggct gtaccctaca agctgtcaca ccgcctgcac tctgagattc 2340
gaactgctgg gatgcgagct gaacggctgt gctaatcctc tgggcctgaa gaacaacagc 2400
atccccgata agcagatcac cgccagctcc agctataaga catggggcct gcacctgttc 2460
agctggaacc cttcttacgc cagactggac aagcagggca acttcaatgc ttgggtggcc 2520
ggcagctacg gcaatgatca gtggctgcaa gtggacctgg gcagcagcaa agaagtgaca 2580
ggcatcatca cccaaggggc cagaaatttc ggcagcgtgc agttcgtggc cagctacaaa 2640
gtggcctact ccaacgacag cgccaactgg accgagtatc aggaccctag aaccggcagc 2700
tccaagatct tccccggcaa ttgggacaac cacagccaca agaagaatct gttcgaaacc 2760
cctatcctgg ccagatatgt gcgcattctg cccgtggcct ggcacaacag aattgccctg 2820
agactggaac tgctcggctg c 2841
<210> 48
<211> 941
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 48
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
625 630 635 640
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
645 650 655
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
660 665 670
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
675 680 685
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
690 695 700
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
705 710 715 720
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
725 730 735
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
740 745 750
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
755 760 765
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
770 775 780
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
785 790 795 800
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
805 810 815
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
820 825 830
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
835 840 845
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
850 855 860
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
865 870 875 880
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
885 890 895
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
900 905 910
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
915 920 925
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
930 935 940
<210> 49
<211> 2823
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 49
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggcct gtgtggaacc cctcggcatg gaaaacggca atatcgccaa tagccagatt 1920
gccgccagca gcgtcagagt gacatttctg ggactgcaac actgggtgcc cgagctggct 1980
agactgaata gagccggcat ggtcaacgcc tggacaccca gcagcaacga cgataatccc 2040
tggattcaag tgaacctgct gcggcgtatg tgggtcacag gtgttgttac acagggcgca 2100
agcagactgg ccagccacga gtatctgaag gcctttaagg tggcctacag cctgaacggc 2160
cacgagttcg acttcatcca cgacgtgaac aagaagcaca aagagtttgt cggcaactgg 2220
aacaagaacg ccgtgcacgt gaacctgttc gagacacctg tggaagccca gtacgtgcgg 2280
ctgtacccta caagctgtca caccgcctgc actctgagat tcgaactgct gggatgcgag 2340
ctgaacggct gtgctaatcc tctgggcctg aagaacaaca gcatccccga taagcagatc 2400
accgccagct ccagctataa gacatggggc ctgcacctgt tcagctggaa cccttcttac 2460
gccagactgg acaagcaggg caacttcaat gcttgggtgg ccggcagcta cggcaatgat 2520
cagtggctgc aagtggacct gggcagcagc aaagaagtga caggcatcat cacccaaggg 2580
gccagaaatt tcggcagcgt gcagttcgtg gccagctaca aagtggccta ctccaacgac 2640
agcgccaact ggaccgagta tcaggaccct agaaccggca gctccaagat cttccccggc 2700
aattgggaca accacagcca caagaagaat ctgttcgaaa cccctatcct ggccagatat 2760
gtgcgcattc tgcccgtggc ctggcacaac agaattgccc tgagactgga actgctcggc 2820
tgc 2823
<210> 50
<211> 963
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 50
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Val Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
805 810 815
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
820 825 830
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
835 840 845
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
850 855 860
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
865 870 875 880
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
885 890 895
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
900 905 910
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
915 920 925
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
930 935 940
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
945 950 955 960
Leu Gly Cys
<210> 51
<211> 2889
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 51
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggagtgg gaggaagcgg aggatctggc 1920
ggttccggag gctcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcagcactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aacccttgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcctcta gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggctgtgc taatcctctg ggcctgaaga acaacagcat ccccgataag 2460
cagatcaccg ccagctccag ctataagaca tggggcctgc acctgttcag ctggaaccct 2520
tcttacgcca gactggacaa gcagggcaac ttcaatgctt gggtggccgg cagctacggc 2580
aatgatcagt ggctgcaagt ggacctgggc agcagcaaag aagtgacagg catcatcacc 2640
cagggcgcca gaaatttcgg cagcgtgcag tttgtggcca gctacaaagt ggcctactcc 2700
aacgacagcg ccaactggac cgagtatcag gaccctagaa ccggcagctc caagatcttc 2760
cccggcaatt gggacaacca cagccacaag aagaatctgt tcgaaacccc tatcctggcc 2820
agatatgtgc gcattctgcc cgtggcctgg cacaacagaa ttgccctgag actggaactg 2880
ctcggctgt 2889
<210> 52
<211> 961
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 52
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser
625 630 635 640
Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala
645 650 655
Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu
660 665 670
Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val
675 680 685
Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val
690 695 700
Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala
705 710 715 720
Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr
725 730 735
Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys
740 745 750
His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn
755 760 765
Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr
770 775 780
Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu
785 790 795 800
Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro
805 810 815
Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His
820 825 830
Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn
835 840 845
Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln
850 855 860
Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly
865 870 875 880
Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala
885 890 895
Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr
900 905 910
Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys
915 920 925
Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu
930 935 940
Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly
945 950 955 960
Cys
<210> 53
<211> 2883
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 53
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggaggaa gcggaggatc tggcggttcc 1920
ggaggctctt gtgtggaacc cctcggcatg gaaaacggca atatcgccaa tagccagatt 1980
gccgccagca gcgtcagagt gacatttctg ggactgcagc actgggtgcc cgagctggct 2040
agactgaata gagccggcat ggtcaacgcc tggacaccca gcagcaacga cgataaccct 2100
tggattcaag tgaacctgct gcggcgtatg tgggtcacag gtgttgttac acagggcgcc 2160
tctagactgg ccagccacga gtatctgaag gcctttaagg tggcctacag cctgaacggc 2220
cacgagttcg acttcatcca cgacgtgaac aagaagcaca aagagtttgt cggcaactgg 2280
aacaagaacg ccgtgcacgt gaacctgttc gagacacctg tggaagccca gtacgtgcgg 2340
ctgtacccta caagctgtca caccgcctgc actctgagat tcgaactgct gggatgcgag 2400
ctgaacggct gtgctaatcc tctgggcctg aagaacaaca gcatccccga taagcagatc 2460
accgccagct ccagctataa gacatggggc ctgcacctgt tcagctggaa cccttcttac 2520
gccagactgg acaagcaggg caacttcaat gcttgggtgg ccggcagcta cggcaatgat 2580
cagtggctgc aagtggacct gggcagcagc aaagaagtga caggcatcat cacccagggc 2640
gccagaaatt tcggcagcgt gcagtttgtg gccagctaca aagtggccta ctccaacgac 2700
agcgccaact ggaccgagta tcaggaccct agaaccggca gctccaagat cttccccggc 2760
aattgggaca accacagcca caagaagaat ctgttcgaaa cccctatcct ggccagatat 2820
gtgcgcattc tgcccgtggc ctggcacaac agaattgccc tgagactgga actgctcggc 2880
tgt 2883
<210> 54
<211> 954
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 54
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Gly Gly Gly Ser Cys Val Glu Pro
625 630 635 640
Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser
645 650 655
Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu
660 665 670
Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser
675 680 685
Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp
690 695 700
Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu
705 710 715 720
Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe
725 730 735
Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val Gly Asn
740 745 750
Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro Val Glu
755 760 765
Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr
770 775 780
Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro
785 790 795 800
Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser
805 810 815
Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser
820 825 830
Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly
835 840 845
Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys
850 855 860
Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val
865 870 875 880
Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn
885 890 895
Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro
900 905 910
Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu Thr Pro
915 920 925
Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His Asn Arg
930 935 940
Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
945 950
<210> 55
<211> 2862
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 55
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaagctgct ctcggaggcg gaggatcttg tgtggaaccc 1920
ctcggcatgg aaaacggcaa tatcgccaat agccagattg ccgccagcag cgtcagagtg 1980
acatttctgg gactgcagca ctgggtgccc gagctggcta gactgaatag agccggcatg 2040
gtcaacgcct ggacacccag cagcaacgac gataaccctt ggattcaagt gaacctgctg 2100
cggcgtatgt gggtcacagg tgttgttaca cagggcgcct ctagactggc cagccacgag 2160
tatctgaagg cctttaaggt ggcctacagc ctgaacggcc acgagttcga cttcatccac 2220
gacgtgaaca agaagcacaa agagtttgtc ggcaactgga acaagaacgc cgtgcacgtg 2280
aacctgttcg agacacctgt ggaagcccag tacgtgcggc tgtaccctac aagctgtcac 2340
accgcctgca ctctgagatt cgaactgctg ggatgcgagc tgaacggctg tgctaatcct 2400
ctgggcctga agaacaacag catccccgat aagcagatca ccgccagctc cagctataag 2460
acatggggcc tgcacctgtt cagctggaac ccttcttacg ccagactgga caagcagggc 2520
aacttcaatg cttgggtggc cggcagctac ggcaatgatc agtggctgca agtggacctg 2580
ggcagcagca aagaagtgac aggcatcatc acccagggcg ccagaaattt cggcagcgtg 2640
cagtttgtgg ccagctacaa agtggcctac tccaacgaca gcgccaactg gaccgagtat 2700
caggacccta gaaccggcag ctccaagatc ttccccggca attgggacaa ccacagccac 2760
aagaagaatc tgttcgaaac ccctatcctg gccagatatg tgcgcattct gcccgtggcc 2820
tggcacaaca gaattgccct gagactggaa ctgctcggct gc 2862
<210> 56
<211> 959
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 56
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Gly Gly Gly Ser Gly Gly Gly Gly
625 630 635 640
Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser
645 650 655
Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His
660 665 670
Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala
675 680 685
Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu
690 695 700
Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg
705 710 715 720
Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu
725 730 735
Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys
740 745 750
Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe
755 760 765
Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys
770 775 780
His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn
785 790 795 800
Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys
805 810 815
Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe
820 825 830
Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn
835 840 845
Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp
850 855 860
Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg
865 870 875 880
Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser
885 890 895
Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser
900 905 910
Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn
915 920 925
Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val
930 935 940
Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
945 950 955
<210> 57
<211> 2877
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 57
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atccgatgct cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctgcagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa gtgcgccagc ctgcagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgtaccc tgagcgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaagctgct ctcggaggcg gaggctccgg aggcggagga 1920
tcttgtgtgg aacccctcgg catggaaaac ggcaatatcg ccaatagcca gattgccgcc 1980
agcagcgtca gagtgacatt tctgggactg cagcactggg tgcccgagct ggctagactg 2040
aatagagccg gcatggtcaa cgcctggaca cccagcagca acgacgataa cccttggatt 2100
caagtgaacc tgctgcggcg tatgtgggtc acaggtgttg ttacacaggg cgcctctaga 2160
ctggccagcc acgagtatct gaaggccttt aaggtggcct acagcctgaa cggccacgag 2220
ttcgacttca tccacgacgt gaacaagaag cacaaagagt ttgtcggcaa ctggaacaag 2280
aacgccgtgc acgtgaacct gttcgagaca cctgtggaag cccagtacgt gcggctgtac 2340
cctacaagct gtcacaccgc ctgcactctg agattcgaac tgctgggatg cgagctgaac 2400
ggctgtgcta atcctctggg cctgaagaac aacagcatcc ccgataagca gatcaccgcc 2460
agctccagct ataagacatg gggcctgcac ctgttcagct ggaacccttc ttacgccaga 2520
ctggacaagc agggcaactt caatgcttgg gtggccggca gctacggcaa tgatcagtgg 2580
ctgcaagtgg acctgggcag cagcaaagaa gtgacaggca tcatcaccca gggcgccaga 2640
aatttcggca gcgtgcagtt tgtggccagc tacaaagtgg cctactccaa cgacagcgcc 2700
aactggaccg agtatcagga ccctagaacc ggcagctcca agatcttccc cggcaattgg 2760
gacaaccaca gccacaagaa gaatctgttc gaaaccccta tcctggccag atatgtgcgc 2820
attctgcccg tggcctggca caacagaatt gccctgagac tggaactgct cggctgc 2877
<210> 58
<211> 949
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 58
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala
610 615 620
Ser Gln Ala Ala Leu Gly Leu Cys Val Glu Pro Leu Gly Met Glu Asn
625 630 635 640
Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr
645 650 655
Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg
660 665 670
Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro
675 680 685
Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val
690 695 700
Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe
705 710 715 720
Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp
725 730 735
Val Asn Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala
740 745 750
Val His Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg
755 760 765
Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu
770 775 780
Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn
785 790 795 800
Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr
805 810 815
Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp
820 825 830
Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp
835 840 845
Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile
850 855 860
Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser
865 870 875 880
Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln
885 890 895
Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn
900 905 910
His Ser His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr
915 920 925
Val Arg Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu
930 935 940
Glu Leu Leu Gly Cys
945
<210> 59
<211> 2847
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 59
ctggacatct gcagcaagaa tccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctgcag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaagtgcgc cagcctgcag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgt 1680
accctgagcg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggctg cttctcaggc tgccctggga ctgtgtgtgg aacccctcgg catggaaaac 1920
ggcaatatcg ccaatagcca gattgccgcc agcagcgtca gagtgacatt tctgggactg 1980
cagcactggg tgcccgagct ggctagactg aatagagccg gcatggtcaa cgcctggaca 2040
cccagcagca acgacgataa tccctggatc caagtgaacc tgctgcggcg tatgtgggtc 2100
acaggtgttg ttacacaggg cgcctctaga ctggccagcc acgagtatct gaaggccttt 2160
aaggtggcct acagcctgaa cggccacgag ttcgacttca tccacgacgt gaacaagaag 2220
cacaaagagt ttgtcggcaa ctggaacaag aacgccgtgc acgtgaacct gttcgagaca 2280
cctgtggaag cccagtacgt gcggctgtac cctacaagct gtcacaccgc ctgcactctg 2340
agattcgaac tgctgggatg cgagctgaac ggctgtgcta atcctctggg cctgaagaac 2400
aacagcatcc ccgataagca gatcaccgcc agctccagct ataagacatg gggcctgcac 2460
ctgttcagct ggaacccttc ttacgccaga ctggacaagc agggcaactt caatgcttgg 2520
gtggccggca gctacggcaa tgatcagtgg ctgcaagtgg acctgggcag cagcaaagaa 2580
gtgacaggca tcatcaccca gggcgccaga aatttcggca gcgtgcagtt tgtggccagc 2640
tacaaagtgg cctactccaa cgacagcgcc aactggaccg agtaccagga tcctagaacc 2700
ggcagctcca agatcttccc cggcaattgg gacaaccaca gccacaagaa gaatctgttc 2760
gaaaccccta tcctggccag atatgtgcgg attctgcccg tggcctggca caacagaatt 2820
gccctgagac tggaactgct cggctgt 2847
<210> 60
<400> 60
000
<210> 61
<400> 61
000
<210> 62
<211> 12
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 62
Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
1 5 10
<210> 63
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 63
Gly Ser Gly Ser Gly Ser Gly Ser
1 5
<210> 64
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 64
Gly Gly Gly Gly Ser
1 5
<210> 65
<211> 10
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 65
Gly Gly Gly Gly Ser Gly Gly Gly Gly Ser
1 5 10
<210> 66
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 66
Gly Ser His His His His His His
1 5
<210> 67
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
Синтетическая 6xHis метка"
<400> 67
His His His His His His
1 5
<210> 68
<211> 463
<212> БЕЛОК
<213> Mus musculus
<400> 68
Met Gln Val Ser Arg Val Leu Ala Ala Leu Cys Gly Met Leu Leu Cys
1 5 10 15
Ala Ser Gly Leu Phe Ala Ala Ser Gly Asp Phe Cys Asp Ser Ser Leu
20 25 30
Cys Leu Asn Gly Gly Thr Cys Leu Thr Gly Gln Asp Asn Asp Ile Tyr
35 40 45
Cys Leu Cys Pro Glu Gly Phe Thr Gly Leu Val Cys Asn Glu Thr Glu
50 55 60
Arg Gly Pro Cys Ser Pro Asn Pro Cys Tyr Asn Asp Ala Lys Cys Leu
65 70 75 80
Val Thr Leu Asp Thr Gln Arg Gly Asp Ile Phe Thr Glu Tyr Ile Cys
85 90 95
Gln Cys Pro Val Gly Tyr Ser Gly Ile His Cys Glu Thr Glu Thr Asn
100 105 110
Tyr Tyr Asn Leu Asp Gly Glu Tyr Met Phe Thr Thr Ala Val Pro Asn
115 120 125
Thr Ala Val Pro Thr Pro Ala Pro Thr Pro Asp Leu Ser Asn Asn Leu
130 135 140
Ala Ser Arg Cys Ser Thr Gln Leu Gly Met Glu Gly Gly Ala Ile Ala
145 150 155 160
Asp Ser Gln Ile Ser Ala Ser Ser Val Tyr Met Gly Phe Met Gly Leu
165 170 175
Gln Arg Trp Gly Pro Glu Leu Ala Arg Leu Tyr Arg Thr Gly Ile Val
180 185 190
Asn Ala Trp Thr Ala Ser Asn Tyr Asp Ser Lys Pro Trp Ile Gln Val
195 200 205
Asn Leu Leu Arg Lys Met Arg Val Ser Gly Val Met Thr Gln Gly Ala
210 215 220
Ser Arg Ala Gly Arg Ala Glu Tyr Leu Lys Thr Phe Lys Val Ala Tyr
225 230 235 240
Ser Leu Asp Gly Arg Lys Phe Glu Phe Ile Gln Asp Glu Ser Gly Gly
245 250 255
Asp Lys Glu Phe Leu Gly Asn Leu Asp Asn Asn Ser Leu Lys Val Asn
260 265 270
Met Phe Asn Pro Thr Leu Glu Ala Gln Tyr Ile Lys Leu Tyr Pro Val
275 280 285
Ser Cys His Arg Gly Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu
290 295 300
Leu His Gly Cys Ser Glu Pro Leu Gly Leu Lys Asn Asn Thr Ile Pro
305 310 315 320
Asp Ser Gln Met Ser Ala Ser Ser Ser Tyr Lys Thr Trp Asn Leu Arg
325 330 335
Ala Phe Gly Trp Tyr Pro His Leu Gly Arg Leu Asp Asn Gln Gly Lys
340 345 350
Ile Asn Ala Trp Thr Ala Gln Ser Asn Ser Ala Lys Glu Trp Leu Gln
355 360 365
Val Asp Leu Gly Thr Gln Arg Gln Val Thr Gly Ile Ile Thr Gln Gly
370 375 380
Ala Arg Asp Phe Gly His Ile Gln Tyr Val Ala Ser Tyr Lys Val Ala
385 390 395 400
His Ser Asp Asp Gly Val Gln Trp Thr Val Tyr Glu Glu Gln Gly Ser
405 410 415
Ser Lys Val Phe Gln Gly Asn Leu Asp Asn Asn Ser His Lys Lys Asn
420 425 430
Ile Phe Glu Lys Pro Phe Met Ala Arg Tyr Val Arg Val Leu Pro Val
435 440 445
Ser Trp His Asn Arg Ile Thr Leu Arg Leu Glu Leu Leu Gly Cys
450 455 460
<210> 69
<211> 1030
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 69
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Cys Ser Gly Pro Leu Gly Ile Glu Gly
705 710 715 720
Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala
725 730 735
Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys
740 745 750
Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro
755 760 765
Trp Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile
770 775 780
Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr
785 790 795 800
Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val
805 810 815
Lys Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn
820 825 830
Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val
835 840 845
Arg Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu
850 855 860
Leu Leu Gly Cys Glu Leu Ser Gly Cys Ser Glu Pro Leu Gly Met Lys
865 870 875 880
Ser Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile Phe Arg
885 890 895
Thr Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala Arg Leu
900 905 910
Asp Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn Asp Gln
915 920 925
Ser Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val Thr Gly
930 935 940
Ile Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe Val Gly
945 950 955 960
Ser Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr Val Tyr
965 970 975
Gln Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn Phe Asp
980 985 990
Asn Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr Ala Arg
995 1000 1005
His Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr Leu
1010 1015 1020
Arg Ser Glu Leu Leu Gly Cys
1025 1030
<210> 70
<211> 1036
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 70
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Cys Ser Gly
705 710 715 720
Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala
725 730 735
Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr
740 745 750
Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala
755 760 765
Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys Met
770 775 780
Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro
785 790 795 800
Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr
805 810 815
Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe Arg
820 825 830
Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro
835 840 845
Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg His
850 855 860
Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly Cys Ser
865 870 875 880
Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln Ile Thr
885 890 895
Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr Trp Glu
900 905 910
Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala Trp Thr
915 920 925
Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu Leu Val
930 935 940
Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp Phe Gly
945 950 955 960
His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn Asp Gly
965 970 975
Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp Lys Val
980 985 990
Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val Ile Asp
995 1000 1005
Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser Trp
1010 1015 1020
Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
1025 1030 1035
<210> 71
<211> 872
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 71
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Cys Ser Gly Pro Leu Gly Ile Glu Gly
705 710 715 720
Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala
725 730 735
Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys
740 745 750
Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro
755 760 765
Trp Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile
770 775 780
Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr
785 790 795 800
Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val
805 810 815
Lys Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn
820 825 830
Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val
835 840 845
Arg Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu
850 855 860
Leu Leu Gly Cys Glu Leu Ser Gly
865 870
<210> 72
<211> 878
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 72
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Cys Ser Gly
705 710 715 720
Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala
725 730 735
Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr
740 745 750
Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala
755 760 765
Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys Met
770 775 780
Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro
785 790 795 800
Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr
805 810 815
Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe Arg
820 825 830
Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro
835 840 845
Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg His
850 855 860
Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly
865 870 875
<210> 73
<211> 783
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 73
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
625 630 635 640
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
645 650 655
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
660 665 670
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
675 680 685
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
690 695 700
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
705 710 715 720
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
725 730 735
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
740 745 750
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
755 760 765
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
770 775 780
<210> 74
<211> 789
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 74
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala
610 615 620
Ser Gln Ala Ala Leu Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
625 630 635 640
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
645 650 655
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
660 665 670
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
675 680 685
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
690 695 700
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
705 710 715 720
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
725 730 735
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
740 745 750
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
755 760 765
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
770 775 780
Cys Glu Leu Asn Gly
785
<210> 75
<211> 387
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 75
Met Pro Arg Pro Arg Leu Leu Ala Ala Leu Cys Gly Ala Leu Leu Cys
1 5 10 15
Ala Pro Ser Leu Leu Val Ala Leu Asp Ile Cys Ser Lys Asn Pro Cys
20 25 30
His Asn Gly Gly Leu Cys Glu Glu Ile Ser Gln Glu Val Arg Gly Asp
35 40 45
Val Phe Pro Ser Tyr Thr Cys Thr Cys Leu Lys Gly Tyr Ala Gly Asn
50 55 60
His Cys Glu Thr Lys Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
65 70 75 80
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
85 90 95
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
100 105 110
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
115 120 125
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
130 135 140
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
145 150 155 160
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
165 170 175
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
180 185 190
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
195 200 205
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
210 215 220
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
225 230 235 240
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
245 250 255
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
260 265 270
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
275 280 285
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
290 295 300
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
305 310 315 320
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
325 330 335
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
340 345 350
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
355 360 365
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
370 375 380
Leu Gly Cys
385
<210> 76
<211> 318
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 76
Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln
1 5 10 15
Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp
20 25 30
Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp
35 40 45
Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu
50 55 60
Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu
65 70 75 80
Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn
85 90 95
Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu
100 105 110
Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu
115 120 125
Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His
130 135 140
Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
145 150 155 160
Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln
165 170 175
Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser
180 185 190
Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala
195 200 205
Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu
210 215 220
Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn
225 230 235 240
Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn
245 250 255
Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser
260 265 270
Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu
275 280 285
Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala
290 295 300
Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
305 310 315
<210> 77
<211> 134
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 77
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys
130
<210> 78
<211> 323
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 78
Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln
1 5 10 15
Ile Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp
20 25 30
Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp
35 40 45
Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln
50 55 60
Arg Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile
65 70 75 80
Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp
85 90 95
Gly Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met
100 105 110
Val Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe
115 120 125
Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys
130 135 140
Arg Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser
145 150 155 160
Gly Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr
165 170 175
Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe
180 185 190
Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn
195 200 205
Ala Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp
210 215 220
Leu Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys
225 230 235 240
Asp Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser
245 250 255
Asn Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys
260 265 270
Asp Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn
275 280 285
Val Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp
290 295 300
Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys Thr
305 310 315 320
Glu Glu Glu
<210> 79
<211> 319
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 79
Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln
1 5 10 15
Ile Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp
20 25 30
Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp
35 40 45
Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln
50 55 60
Arg Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile
65 70 75 80
Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp
85 90 95
Gly Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met
100 105 110
Val Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe
115 120 125
Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys
130 135 140
Arg Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser
145 150 155 160
Gly Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr
165 170 175
Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe
180 185 190
Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn
195 200 205
Ala Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp
210 215 220
Leu Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys
225 230 235 240
Asp Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser
245 250 255
Asn Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys
260 265 270
Asp Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn
275 280 285
Val Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp
290 295 300
Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
305 310 315
<210> 80
<211> 1029
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 80
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Cys Val Glu Pro Leu Gly Met Glu Asn
705 710 715 720
Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr
725 730 735
Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg
740 745 750
Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro
755 760 765
Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val
770 775 780
Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe
785 790 795 800
Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp
805 810 815
Val Asn Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala
820 825 830
Val His Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg
835 840 845
Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu
850 855 860
Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn
865 870 875 880
Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr
885 890 895
Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp
900 905 910
Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp
915 920 925
Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile
930 935 940
Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser
945 950 955 960
Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln
965 970 975
Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn
980 985 990
His Ser His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr
995 1000 1005
Val Arg Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg
1010 1015 1020
Leu Glu Leu Leu Gly Cys
1025
<210> 81
<211> 3087
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 81
gacatctgcg accccaatcc ttgcgagaat ggcggcattt gtctgcctgg actggccgat 60
ggcagcttct cttgtgaatg ccccgatggc ttcacagacc ccaattgcag ctctgtggtg 120
gaagtggcca gcgacgagga agaacctaca agcgctggcc cctgcacacc caatccatgt 180
cataatggcg gaacctgcga gatcagcgag gcctacagag gcgatacctt catcggctac 240
gtgtgcaagt gccccagagg cttcaatggc atccactgcc agcacaacat caacgagtgc 300
gaggtggaac catgcaagaa cggcggcatc tgtaccgacc tggtggccaa ttactcttgc 360
gagtgccctg gcgagttcat gggcagaaac tgccagtaca aggacgccca caagagcgag 420
gtggcccaca gattcaagga cctgggcgaa gagaacttca aggccctggt gctgatcgcc 480
ttcgctcagt atctccagca gagccctttc gaggaccacg tgaagctggt caacgaagtg 540
accgagttcg ccaagacctg tgtggccgat gagagcgccg agaactgtga caagagcctg 600
cacacactgt tcggcgacaa gctgtgtacc gtggccacac tgagagaaac ctacggcgag 660
atggccgact gctgtgccaa gcaagagccc gagagaaacg agtgcttcct ccagcacaag 720
gatgacaacc ccaacctgcc tagactcgtg cggcctgaag tggatgtgat gtgcaccgcc 780
tttcacgaca acgaggaaac cttcctgaag aagtacctgt acgagatcgc cagacggcac 840
ccctactttt atgcccctga gctgctgttc ttcgccaagc ggtataaggc cgccttcacc 900
gaatgttgcc aggccgctga taaggctgcc tgtctgctgc ctaagctgga cgagctgaga 960
gatgagggca aagccagctc tgccaagcag agactgaaat gcgccagcct ccagaagttc 1020
ggcgagagag cttttaaggc ctgggccgtt gccagactga gccagagatt tcctaaggcc 1080
gagtttgccg aggtgtccaa gctcgtgacc gatctgacaa aggtgcacac cgagtgctgt 1140
cacggcgatc tgctggaatg tgccgacgat agagccgacc tggccaagta tatctgcgag 1200
aaccaggaca gcatcagcag caagctgaaa gagtgctgcg agaagcccct gctggaaaag 1260
tctcactgta tcgccgaagt ggaaaacgac gagatgcccg ccgatctgcc ttctctggct 1320
gccgatttcg tggaaagcaa ggatgtgtgc aagaactacg ccgaggccaa agatgtgttt 1380
ctgggcatgt ttctgtatga gtacgcccgc agacaccccg actattctgt ggttctgctg 1440
ctgcggctgg ccaagacata cgagacaacc ctggaaaaat gctgcgccgc tgccgatcct 1500
cacgagtgtt atgccaaggt gttcgacgag ttcaagccac tggtggaaga accccagaac 1560
ctgatcaagc agaactgcga gctgttcgag cagctgggcg agtacaagtt ccagaatgcc 1620
ctgctcgtgc ggtacaccaa gaaagtgcct caggtgtcca cacctacact ggttgaggtg 1680
tcccggaatc tgggcaaagt gggcagcaag tgttgcaagc accctgaggc caagagaatg 1740
ccttgcgccg aggattacct gagcgtggtg ctgaatcagc tgtgcgtgct gcacgagaaa 1800
acccctgtgt ccgacagagt gaccaagtgc tgtaccgaga gcctcgtgaa cagaaggcct 1860
tgctttagcg ccctggaagt ggacgagaca tacgtgccca aagagttcaa cgccgagaca 1920
ttcaccttcc acgccgatat ctgcaccctg tccgagaaag agcggcagat caagaagcag 1980
acagccctgg tcgagctggt taagcacaag cccaaggcca ccaaagaaca gctgaaggcc 2040
gtgatggacg acttcgccgc ctttgtcgag aagtgctgca aggccgacga caaagagaca 2100
tgcttcgccg aagagggcaa gaaactggtg gcctgtgtgg aacccctcgg catggaaaac 2160
ggcaatatcg ccaatagcca gattgccgcc agcagcgtca gagtgacatt tctgggactg 2220
caacactggg tgcccgagct ggctagactg aatagagccg gcatggtcaa cgcctggaca 2280
cccagcagca acgacgataa cccctggatt caagtgaacc tgctgcggcg tatgtgggtc 2340
acaggtgttg ttacacaggg cgcaagcaga ctggcctctc acgagtacct gaaggccttt 2400
aaggtggcct acagcctgaa cggccacgag ttcgacttca tccacgacgt gaacaagaag 2460
cacaaagagt ttgtcggcaa ctggaacaag aacgccgtgc acgtgaacct gttcgagaca 2520
cctgtggaag cccagtacgt gcggctgtac cctacaagct gtcacaccgc ctgcactctg 2580
agattcgaac tgctgggatg cgagctgaac ggctgtgcta atcctctggg cctgaagaac 2640
aacagcatcc ccgataagca gatcaccgcc agctccagct ataagacatg gggcctgcac 2700
ctgttcagct ggaacccttc ttacgccaga ctggacaagc agggcaactt caatgcttgg 2760
gtggccggca gctacggcaa tgatcagtgg ctgcaagtgg atctgggcag cagcaaagaa 2820
gtgacaggca tcatcaccca aggggccaga aatttcggca gcgtgcagtt cgtggccagc 2880
tacaaagtgg cctactccaa cgacagcgcc aactggaccg agtatcagga ccctagaacc 2940
ggcagctcca agatcttccc cggcaattgg gacaaccaca gccacaagaa gaatctcttc 3000
gagactccca tcctggccag atatgtgcgg attctgcctg tggcctggca caacagaatc 3060
gccctgagac tggaactgct cggctgt 3087
<210> 82
<211> 942
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 82
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile
625 630 635 640
Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr
645 650 655
Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr
660 665 670
Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg
675 680 685
Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly
690 695 700
Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly
705 710 715 720
Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val
725 730 735
Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr
740 745 750
Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg
755 760 765
Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly
770 775 780
Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln
785 790 795 800
Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr
805 810 815
Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala
820 825 830
Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu
835 840 845
Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp
850 855 860
Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn
865 870 875 880
Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp
885 890 895
Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val
900 905 910
Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser
915 920 925
Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
930 935 940
<210> 83
<211> 2826
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 83
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggcct gttctggccc tctgggcatc gaaggcggca tcatcagcaa tcagcagatc 1920
accgccagca gcacccacag agcactgttt ggcctgcaaa agtggtatcc ctactacgcc 1980
cggctgaaca agaagggcct gattaacgcc tggacagccg ccgagaatga cagatggccc 2040
tggattcaga tcaacctcca gcggaagatg agagtgaccg gcgttatcac acagggcgca 2100
aagagaatcg gctcccctga gtacatcaag agctacaaga tcgcctacag caacgacggc 2160
aagacctggg ccatgtacaa agtgaagggc accaacgagg acatggtgtt ccggggcaac 2220
atcgacaaca acacccctta cgccaacagc ttcacccctc ctatcaaggc ccagtacgtg 2280
cggctgtacc ctcaagtgtg cagaaggcac tgtaccctga gaatggaact gctgggctgc 2340
gaactgtctg gctgttctga gccactggga atgaagtccg gccacatcca ggactaccag 2400
attaccgcct ccagcatctt cagaaccctg aacatggata tgttcacctg ggagccccgg 2460
aaggccagac tggataagca gggaaaagtg aatgcctgga ccagcggcca caacgaccag 2520
tctcaatggc tgcaagtgga cctgctggtg cctaccaaag tgaccggaat catcacccaa 2580
ggcgctaagg atttcggcca cgtgcagttc gtgggctcct acaagctggc ctactccaat 2640
gatggcgagc actggaccgt gtaccaggac gagaagcagc ggaaggataa ggtgttccag 2700
ggaaacttcg ataacgatac ccaccggaag aacgtgatcg accctccaat ctacgccaga 2760
cacatcagaa tcctgccttg gtcttggtac ggcagaatca ccctgagatc cgagctgctg 2820
ggatgc 2826
<210> 84
<211> 968
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 84
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Gly Ser Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys
50 55 60
Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala
65 70 75 80
Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn
85 90 95
Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu
100 105 110
Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr
115 120 125
Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala
130 135 140
Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp
145 150 155 160
Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys
165 170 175
Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr
180 185 190
Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe
195 200 205
Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala
210 215 220
Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu
225 230 235 240
Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln
245 250 255
Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser
260 265 270
Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr
275 280 285
Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu
290 295 300
Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln
305 310 315 320
Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu
325 330 335
Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala
340 345 350
Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys
355 360 365
Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr
370 375 380
Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg
385 390 395 400
Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala
405 410 415
Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu
420 425 430
Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu
435 440 445
Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr
450 455 460
Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg
465 470 475 480
Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys
485 490 495
Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu
500 505 510
Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys
515 520 525
Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu
530 535 540
Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr
545 550 555 560
Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys
565 570 575
Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr
580 585 590
Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu
595 600 605
Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly
610 615 620
Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser
625 630 635 640
Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys Ser Gly Pro Leu Gly Ile
645 650 655
Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His
660 665 670
Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu
675 680 685
Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg
690 695 700
Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly
705 710 715 720
Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys
725 730 735
Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr
740 745 750
Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp
755 760 765
Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln
770 775 780
Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg
785 790 795 800
Met Glu Leu Leu Gly Cys Glu Leu Ser Gly Cys Ser Glu Pro Leu Gly
805 810 815
Met Lys Ser Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile
820 825 830
Phe Arg Thr Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala
835 840 845
Arg Leu Asp Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn
850 855 860
Asp Gln Ser Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val
865 870 875 880
Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe
885 890 895
Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr
900 905 910
Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn
915 920 925
Phe Asp Asn Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr
930 935 940
Ala Arg His Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr
945 950 955 960
Leu Arg Ser Glu Leu Leu Gly Cys
965
<210> 85
<211> 963
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 85
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Gly Ser Asp
35 40 45
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
50 55 60
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
65 70 75 80
Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
85 90 95
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
100 105 110
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
115 120 125
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
130 135 140
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
145 150 155 160
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
165 170 175
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
180 185 190
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
195 200 205
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
210 215 220
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
225 230 235 240
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
245 250 255
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
260 265 270
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
275 280 285
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
290 295 300
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
305 310 315 320
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
325 330 335
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
340 345 350
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
355 360 365
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
370 375 380
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
385 390 395 400
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
405 410 415
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
420 425 430
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
435 440 445
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
450 455 460
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
465 470 475 480
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
485 490 495
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
500 505 510
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
515 520 525
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
530 535 540
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
545 550 555 560
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
565 570 575
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
580 585 590
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
595 600 605
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
610 615 620
Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly Gly
625 630 635 640
Ser Gly Gly Ser Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile
645 650 655
Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly
660 665 670
Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu
675 680 685
Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln
690 695 700
Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly
705 710 715 720
Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala
725 730 735
Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr
740 745 750
Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr
755 760 765
Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Ser Gly Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His
805 810 815
Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn
820 825 830
Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln
835 840 845
Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp
850 855 860
Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr
865 870 875 880
Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys
885 890 895
Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu
900 905 910
Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr
915 920 925
His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg
930 935 940
Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu
945 950 955 960
Leu Gly Cys
<210> 86
<211> 957
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 86
Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys
1 5 10 15
Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met
20 25 30
Gly Arg Asn Cys Gln Tyr Lys Gly Ser Asp Ala His Lys Ser Glu Val
35 40 45
Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val
50 55 60
Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His
65 70 75 80
Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala
85 90 95
Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly
100 105 110
Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met
115 120 125
Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu
130 135 140
Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu
145 150 155 160
Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu
165 170 175
Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala
180 185 190
Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu
195 200 205
Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp
210 215 220
Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys
225 230 235 240
Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala
245 250 255
Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val
260 265 270
Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His
275 280 285
Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr
290 295 300
Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys
305 310 315 320
Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn
325 330 335
Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu
340 345 350
Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu
355 360 365
Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val
370 375 380
Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys
385 390 395 400
Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp
405 410 415
Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn
420 425 430
Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu
435 440 445
Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu
450 455 460
Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys
465 470 475 480
His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val
485 490 495
Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp
500 505 510
Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys
515 520 525
Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn
530 535 540
Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys
545 550 555 560
Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His
565 570 575
Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe
580 585 590
Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys
595 600 605
Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu
610 615 620
Gly Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys Ser
625 630 635 640
Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr
645 650 655
Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro
660 665 670
Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala
675 680 685
Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys
690 695 700
Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser
705 710 715 720
Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys
725 730 735
Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe
740 745 750
Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro
755 760 765
Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg
770 775 780
His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly Cys
785 790 795 800
Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln Ile
805 810 815
Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr Trp
820 825 830
Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala Trp
835 840 845
Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu Leu
850 855 860
Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp Phe
865 870 875 880
Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn Asp
885 890 895
Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp Lys
900 905 910
Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val Ile
915 920 925
Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser Trp
930 935 940
Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
945 950 955
<210> 87
<211> 1013
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 87
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Gly
85 90 95
Ser Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly
100 105 110
Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu
115 120 125
Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr
130 135 140
Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp
145 150 155 160
Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr
165 170 175
Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu
180 185 190
Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn
195 200 205
Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe
210 215 220
His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala
225 230 235 240
Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys
245 250 255
Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala
260 265 270
Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala
275 280 285
Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly
290 295 300
Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe
305 310 315 320
Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr
325 330 335
Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp
340 345 350
Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile
355 360 365
Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser
370 375 380
His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro
385 390 395 400
Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr
405 410 415
Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala
420 425 430
Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys
435 440 445
Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His
450 455 460
Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu
465 470 475 480
Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly
485 490 495
Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val
500 505 510
Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly
515 520 525
Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro
530 535 540
Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu
545 550 555 560
His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu
565 570 575
Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu
580 585 590
Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala
595 600 605
Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr
610 615 620
Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln
625 630 635 640
Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys
645 650 655
Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu
660 665 670
Val Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser
675 680 685
Gly Gly Ser Gly Gly Ser Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly
690 695 700
Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala Leu
705 710 715 720
Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys
725 730 735
Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp
740 745 750
Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile Thr
755 760 765
Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys
770 775 780
Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val Lys
785 790 795 800
Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn Thr
805 810 815
Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg
820 825 830
Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu Leu
835 840 845
Leu Gly Cys Glu Leu Ser Gly Cys Ser Glu Pro Leu Gly Met Lys Ser
850 855 860
Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr
865 870 875 880
Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp
885 890 895
Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn Asp Gln Ser
900 905 910
Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val Thr Gly Ile
915 920 925
Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe Val Gly Ser
930 935 940
Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr Val Tyr Gln
945 950 955 960
Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn Phe Asp Asn
965 970 975
Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr Ala Arg His
980 985 990
Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser
995 1000 1005
Glu Leu Leu Gly Cys
1010
<210> 88
<211> 1002
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 88
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn Ile Asn
35 40 45
Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr Asp Leu
50 55 60
Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly Arg Asn
65 70 75 80
Cys Gln Tyr Lys Gly Ser Asp Ala His Lys Ser Glu Val Ala His Arg
85 90 95
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
100 105 110
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
115 120 125
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
130 135 140
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
145 150 155 160
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
165 170 175
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
180 185 190
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
195 200 205
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
210 215 220
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
225 230 235 240
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
245 250 255
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
260 265 270
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
275 280 285
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
290 295 300
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
305 310 315 320
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
325 330 335
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
340 345 350
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
355 360 365
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
370 375 380
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
385 390 395 400
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
405 410 415
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
420 425 430
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
435 440 445
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
450 455 460
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
465 470 475 480
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
485 490 495
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
500 505 510
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
515 520 525
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
530 535 540
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
545 550 555 560
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
565 570 575
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
580 585 590
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
595 600 605
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
610 615 620
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
625 630 635 640
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
645 650 655
Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly
660 665 670
Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys Ser Gly Pro Leu
675 680 685
Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser
690 695 700
Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala
705 710 715 720
Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn
725 730 735
Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val
740 745 750
Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr
755 760 765
Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala
770 775 780
Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn
785 790 795 800
Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys
805 810 815
Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr
820 825 830
Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser Gly Cys Ser Glu Pro
835 840 845
Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser
850 855 860
Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg
865 870 875 880
Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly
885 890 895
His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr
900 905 910
Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val
915 920 925
Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His
930 935 940
Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln
945 950 955 960
Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro
965 970 975
Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg
980 985 990
Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
995 1000
<210> 89
<211> 1007
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 89
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly
50 55 60
Ile Cys Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu
65 70 75 80
Phe Met Gly Arg Asn Cys Gln Tyr Lys Gly Ser Asp Ala His Lys Ser
85 90 95
Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala
100 105 110
Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu
115 120 125
Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys
130 135 140
Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu
145 150 155 160
Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly
165 170 175
Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys
180 185 190
Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg
195 200 205
Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr
210 215 220
Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe
225 230 235 240
Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe
245 250 255
Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys
260 265 270
Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg
275 280 285
Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala
290 295 300
Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala
305 310 315 320
Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys
325 330 335
Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala
340 345 350
Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu
355 360 365
Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val
370 375 380
Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe
385 390 395 400
Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val
405 410 415
Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr
420 425 430
Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu
435 440 445
Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val
450 455 460
Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys
465 470 475 480
Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn
485 490 495
Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro
500 505 510
Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys
515 520 525
Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu
530 535 540
Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val
545 550 555 560
Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg
565 570 575
Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu
580 585 590
Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser
595 600 605
Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val
610 615 620
Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp
625 630 635 640
Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu
645 650 655
Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala Ser Gln Ala
660 665 670
Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser
675 680 685
Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln
690 695 700
Ile Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp
705 710 715 720
Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp
725 730 735
Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln
740 745 750
Arg Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile
755 760 765
Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp
770 775 780
Gly Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met
785 790 795 800
Val Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe
805 810 815
Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys
820 825 830
Arg Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser
835 840 845
Gly Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr
850 855 860
Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe
865 870 875 880
Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn
885 890 895
Ala Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp
900 905 910
Leu Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys
915 920 925
Asp Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser
930 935 940
Asn Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys
945 950 955 960
Asp Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn
965 970 975
Val Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp
980 985 990
Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
995 1000 1005
<210> 90
<211> 2904
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 90
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc ggcagcgacg cccacaagag cgaggtggcc 180
caccggttca aggacctggg cgaggagaac ttcaaggccc tggtgctgat cgccttcgcc 240
cagtacctgc agcagagccc cttcgaggac cacgtgaagc tggtgaacga ggtgaccgag 300
ttcgccaaga cctgcgtggc cgacgagagc gccgagaact gcgacaagag cctgcacacc 360
ctgttcggcg acaagctgtg caccgtggcc accctgcggg agacctacgg cgagatggcc 420
gactgctgcg ccaagcagga gcccgagcgg aacgagtgct tcctgcagca caaggacgac 480
aaccccaacc tgccccggct ggtgcggccc gaggtggacg tgatgtgcac cgccttccac 540
gacaacgagg agaccttcct gaagaagtac ctgtacgaga tcgcccggcg gcacccctac 600
ttctacgccc ccgagctgct gttcttcgcc aagcggtaca aggccgcctt caccgagtgc 660
tgccaggccg ccgacaaggc cgcctgcctg ctgcccaagc tggacgagct gcgggacgag 720
ggcaaggcca gcagcgccaa gcagcggctg aagtgcgcca gcctgcagaa gttcggcgag 780
cgggccttca aggcctgggc cgtggcccgg ctgagccagc ggttccccaa ggccgagttc 840
gccgaggtga gcaagctggt gaccgacctg accaaggtgc acaccgagtg ctgccacggc 900
gacctgctgg agtgcgccga cgaccgggcc gacctggcca agtacatctg cgagaaccag 960
gacagcatca gcagcaagct gaaggagtgc tgcgagaagc ccctgctgga gaagagccac 1020
tgcatcgccg aggtggagaa cgacgagatg cccgccgacc tgcccagcct ggccgccgac 1080
ttcgtggaga gcaaggacgt gtgcaagaac tacgccgagg ccaaggacgt gttcctgggc 1140
atgttcctgt acgagtacgc ccggcggcac cccgactaca gcgtggtgct gctgctgcgg 1200
ctggccaaga cctacgagac caccctggag aagtgctgcg ccgccgccga cccccacgag 1260
tgctacgcca aggtgttcga cgagttcaag cccctggtgg aggagcccca gaacctgatc 1320
aagcagaact gcgagctgtt cgagcagctg ggcgagtaca agttccagaa cgccctgctg 1380
gtgcggtaca ccaagaaggt gccccaggtg agcaccccca ccctggtgga ggtgagccgg 1440
aacctgggca aggtgggcag caagtgctgc aagcaccccg aggccaagcg gatgccctgc 1500
gccgaggact acctgagcgt ggtgctgaac cagctgtgcg tgctgcacga gaagaccccc 1560
gtgagcgacc gggtgaccaa gtgctgcacc gagagcctgg tgaaccggcg gccctgcttc 1620
agcgccctgg aggtggacga gacctacgtg cccaaggagt tcaacgccga gaccttcacc 1680
ttccacgccg acatctgcac cctgagcgag aaggagcggc agatcaagaa gcagaccgcc 1740
ctggtggagc tggtgaagca caagcccaag gccaccaagg agcagctgaa ggccgtgatg 1800
gacgacttcg ccgccttcgt ggagaagtgc tgcaaggccg acgacaagga gacctgcttc 1860
gccgaggagg gcaagaagct ggtggccgcc agccaggccg ccctgggcct gggcggcagc 1920
ggcggcagcg gcggcagcgg cggcagctgc agcggccccc tgggcatcga gggcggcatc 1980
atcagcaacc agcagatcac cgccagcagc acccaccggg ccctgttcgg cctgcagaag 2040
tggtacccct actacgcccg gctgaacaag aagggcctga tcaacgcctg gaccgccgcc 2100
gagaacgacc ggtggccctg gatccagatc aacctgcagc ggaagatgcg ggtgaccggc 2160
gtgatcaccc agggcgccaa gcggatcggc agccccgagt acatcaagag ctacaagatc 2220
gcctacagca acgacggcaa gacctgggcc atgtacaagg tgaagggcac caacgaggac 2280
atggtgttcc ggggcaacat cgacaacaac accccctacg ccaacagctt cacccccccc 2340
atcaaggccc agtacgtgcg gctgtacccc caggtgtgcc ggcggcactg caccctgcgg 2400
atggagctgc tgggctgcga gctgagcggc tgcagcgagc ccctgggcat gaagagcggc 2460
cacatccagg actaccagat caccgccagc agcatcttcc ggaccctgaa catggacatg 2520
ttcacctggg agccccggaa ggcccggctg gacaagcagg gcaaggtgaa cgcctggacc 2580
agcggccaca acgaccagag ccagtggctg caggtggacc tgctggtgcc caccaaggtg 2640
accggcatca tcacccaggg cgccaaggac ttcggccacg tgcagttcgt gggcagctac 2700
aagctggcct acagcaacga cggcgagcac tggaccgtgt accaggacga gaagcagcgg 2760
aaggacaagg tgttccaggg caacttcgac aacgacaccc accggaagaa cgtgatcgac 2820
ccccccatct acgcccggca catccggatc ctgccctgga gctggtacgg ccggatcacc 2880
ctgcggagcg agctgctggg ctgc 2904
<210> 91
<211> 2889
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 91
agcgccggcc cctgcacccc caacccctgc cacaacggcg gcacctgcga gatcagcgag 60
gcctaccggg gcgacacctt catcggctac gtgtgcaagt gcccccgggg cttcaacggc 120
atccactgcc agcacggcag cgacgcccac aagagcgagg tggcccaccg gttcaaggac 180
ctgggcgagg agaacttcaa ggccctggtg ctgatcgcct tcgcccagta cctgcagcag 240
agccccttcg aggaccacgt gaagctggtg aacgaggtga ccgagttcgc caagacctgc 300
gtggccgacg agagcgccga gaactgcgac aagagcctgc acaccctgtt cggcgacaag 360
ctgtgcaccg tggccaccct gcgggagacc tacggcgaga tggccgactg ctgcgccaag 420
caggagcccg agcggaacga gtgcttcctg cagcacaagg acgacaaccc caacctgccc 480
cggctggtgc ggcccgaggt ggacgtgatg tgcaccgcct tccacgacaa cgaggagacc 540
ttcctgaaga agtacctgta cgagatcgcc cggcggcacc cctacttcta cgcccccgag 600
ctgctgttct tcgccaagcg gtacaaggcc gccttcaccg agtgctgcca ggccgccgac 660
aaggccgcct gcctgctgcc caagctggac gagctgcggg acgagggcaa ggccagcagc 720
gccaagcagc ggctgaagtg cgccagcctg cagaagttcg gcgagcgggc cttcaaggcc 780
tgggccgtgg cccggctgag ccagcggttc cccaaggccg agttcgccga ggtgagcaag 840
ctggtgaccg acctgaccaa ggtgcacacc gagtgctgcc acggcgacct gctggagtgc 900
gccgacgacc gggccgacct ggccaagtac atctgcgaga accaggacag catcagcagc 960
aagctgaagg agtgctgcga gaagcccctg ctggagaaga gccactgcat cgccgaggtg 1020
gagaacgacg agatgcccgc cgacctgccc agcctggccg ccgacttcgt ggagagcaag 1080
gacgtgtgca agaactacgc cgaggccaag gacgtgttcc tgggcatgtt cctgtacgag 1140
tacgcccggc ggcaccccga ctacagcgtg gtgctgctgc tgcggctggc caagacctac 1200
gagaccaccc tggagaagtg ctgcgccgcc gccgaccccc acgagtgcta cgccaaggtg 1260
ttcgacgagt tcaagcccct ggtggaggag ccccagaacc tgatcaagca gaactgcgag 1320
ctgttcgagc agctgggcga gtacaagttc cagaacgccc tgctggtgcg gtacaccaag 1380
aaggtgcccc aggtgagcac ccccaccctg gtggaggtga gccggaacct gggcaaggtg 1440
ggcagcaagt gctgcaagca ccccgaggcc aagcggatgc cctgcgccga ggactacctg 1500
agcgtggtgc tgaaccagct gtgcgtgctg cacgagaaga cccccgtgag cgaccgggtg 1560
accaagtgct gcaccgagag cctggtgaac cggcggccct gcttcagcgc cctggaggtg 1620
gacgagacct acgtgcccaa ggagttcaac gccgagacct tcaccttcca cgccgacatc 1680
tgcaccctga gcgagaagga gcggcagatc aagaagcaga ccgccctggt ggagctggtg 1740
aagcacaagc ccaaggccac caaggagcag ctgaaggccg tgatggacga cttcgccgcc 1800
ttcgtggaga agtgctgcaa ggccgacgac aaggagacct gcttcgccga ggagggcaag 1860
aagctggtgg ccgccagcca ggccgccctg ggcctgggcg gcagcggcgg cagcggcggc 1920
agcggcggca gctgcagcgg ccccctgggc atcgagggcg gcatcatcag caaccagcag 1980
atcaccgcca gcagcaccca ccgggccctg ttcggcctgc agaagtggta cccctactac 2040
gcccggctga acaagaaggg cctgatcaac gcctggaccg ccgccgagaa cgaccggtgg 2100
ccctggatcc agatcaacct gcagcggaag atgcgggtga ccggcgtgat cacccagggc 2160
gccaagcgga tcggcagccc cgagtacatc aagagctaca agatcgccta cagcaacgac 2220
ggcaagacct gggccatgta caaggtgaag ggcaccaacg aggacatggt gttccggggc 2280
aacatcgaca acaacacccc ctacgccaac agcttcaccc cccccatcaa ggcccagtac 2340
gtgcggctgt acccccaggt gtgccggcgg cactgcaccc tgcggatgga gctgctgggc 2400
tgcgagctga gcggctgcag cgagcccctg ggcatgaaga gcggccacat ccaggactac 2460
cagatcaccg ccagcagcat cttccggacc ctgaacatgg acatgttcac ctgggagccc 2520
cggaaggccc ggctggacaa gcagggcaag gtgaacgcct ggaccagcgg ccacaacgac 2580
cagagccagt ggctgcaggt ggacctgctg gtgcccacca aggtgaccgg catcatcacc 2640
cagggcgcca aggacttcgg ccacgtgcag ttcgtgggca gctacaagct ggcctacagc 2700
aacgacggcg agcactggac cgtgtaccag gacgagaagc agcggaagga caaggtgttc 2760
cagggcaact tcgacaacga cacccaccgg aagaacgtga tcgacccccc catctacgcc 2820
cggcacatcc ggatcctgcc ctggagctgg tacggccgga tcaccctgcg gagcgagctg 2880
ctgggctgc 2889
<210> 92
<211> 2871
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 92
aacatcaacg agtgcgaggt ggagccctgc aagaacggcg gcatctgcac cgacctggtg 60
gccaactaca gctgcgagtg ccccggcgag ttcatgggcc ggaactgcca gtacaagggc 120
agcgacgccc acaagagcga ggtggcccac cggttcaagg acctgggcga ggagaacttc 180
aaggccctgg tgctgatcgc cttcgcccag tacctgcagc agagcccctt cgaggaccac 240
gtgaagctgg tgaacgaggt gaccgagttc gccaagacct gcgtggccga cgagagcgcc 300
gagaactgcg acaagagcct gcacaccctg ttcggcgaca agctgtgcac cgtggccacc 360
ctgcgggaga cctacggcga gatggccgac tgctgcgcca agcaggagcc cgagcggaac 420
gagtgcttcc tgcagcacaa ggacgacaac cccaacctgc cccggctggt gcggcccgag 480
gtggacgtga tgtgcaccgc cttccacgac aacgaggaga ccttcctgaa gaagtacctg 540
tacgagatcg cccggcggca cccctacttc tacgcccccg agctgctgtt cttcgccaag 600
cggtacaagg ccgccttcac cgagtgctgc caggccgccg acaaggccgc ctgcctgctg 660
cccaagctgg acgagctgcg ggacgagggc aaggccagca gcgccaagca gcggctgaag 720
tgcgccagcc tgcagaagtt cggcgagcgg gccttcaagg cctgggccgt ggcccggctg 780
agccagcggt tccccaaggc cgagttcgcc gaggtgagca agctggtgac cgacctgacc 840
aaggtgcaca ccgagtgctg ccacggcgac ctgctggagt gcgccgacga ccgggccgac 900
ctggccaagt acatctgcga gaaccaggac agcatcagca gcaagctgaa ggagtgctgc 960
gagaagcccc tgctggagaa gagccactgc atcgccgagg tggagaacga cgagatgccc 1020
gccgacctgc ccagcctggc cgccgacttc gtggagagca aggacgtgtg caagaactac 1080
gccgaggcca aggacgtgtt cctgggcatg ttcctgtacg agtacgcccg gcggcacccc 1140
gactacagcg tggtgctgct gctgcggctg gccaagacct acgagaccac cctggagaag 1200
tgctgcgccg ccgccgaccc ccacgagtgc tacgccaagg tgttcgacga gttcaagccc 1260
ctggtggagg agccccagaa cctgatcaag cagaactgcg agctgttcga gcagctgggc 1320
gagtacaagt tccagaacgc cctgctggtg cggtacacca agaaggtgcc ccaggtgagc 1380
acccccaccc tggtggaggt gagccggaac ctgggcaagg tgggcagcaa gtgctgcaag 1440
caccccgagg ccaagcggat gccctgcgcc gaggactacc tgagcgtggt gctgaaccag 1500
ctgtgcgtgc tgcacgagaa gacccccgtg agcgaccggg tgaccaagtg ctgcaccgag 1560
agcctggtga accggcggcc ctgcttcagc gccctggagg tggacgagac ctacgtgccc 1620
aaggagttca acgccgagac cttcaccttc cacgccgaca tctgcaccct gagcgagaag 1680
gagcggcaga tcaagaagca gaccgccctg gtggagctgg tgaagcacaa gcccaaggcc 1740
accaaggagc agctgaaggc cgtgatggac gacttcgccg ccttcgtgga gaagtgctgc 1800
aaggccgacg acaaggagac ctgcttcgcc gaggagggca agaagctggt ggccgccagc 1860
caggccgccc tgggcctggg cggcagcggc ggcagcggcg gcagcggcgg cagctgcagc 1920
ggccccctgg gcatcgaggg cggcatcatc agcaaccagc agatcaccgc cagcagcacc 1980
caccgggccc tgttcggcct gcagaagtgg tacccctact acgcccggct gaacaagaag 2040
ggcctgatca acgcctggac cgccgccgag aacgaccggt ggccctggat ccagatcaac 2100
ctgcagcgga agatgcgggt gaccggcgtg atcacccagg gcgccaagcg gatcggcagc 2160
cccgagtaca tcaagagcta caagatcgcc tacagcaacg acggcaagac ctgggccatg 2220
tacaaggtga agggcaccaa cgaggacatg gtgttccggg gcaacatcga caacaacacc 2280
ccctacgcca acagcttcac cccccccatc aaggcccagt acgtgcggct gtacccccag 2340
gtgtgccggc ggcactgcac cctgcggatg gagctgctgg gctgcgagct gagcggctgc 2400
agcgagcccc tgggcatgaa gagcggccac atccaggact accagatcac cgccagcagc 2460
atcttccgga ccctgaacat ggacatgttc acctgggagc cccggaaggc ccggctggac 2520
aagcagggca aggtgaacgc ctggaccagc ggccacaacg accagagcca gtggctgcag 2580
gtggacctgc tggtgcccac caaggtgacc ggcatcatca cccagggcgc caaggacttc 2640
ggccacgtgc agttcgtggg cagctacaag ctggcctaca gcaacgacgg cgagcactgg 2700
accgtgtacc aggacgagaa gcagcggaag gacaaggtgt tccagggcaa cttcgacaac 2760
gacacccacc ggaagaacgt gatcgacccc cccatctacg cccggcacat ccggatcctg 2820
ccctggagct ggtacggccg gatcaccctg cggagcgagc tgctgggctg c 2871
<210> 93
<211> 3039
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 93
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc agcgccggcc cctgcacccc caacccctgc 180
cacaacggcg gcacctgcga gatcagcgag gcctaccggg gcgacacctt catcggctac 240
gtgtgcaagt gcccccgggg cttcaacggc atccactgcc agcacggcag cgacgcccac 300
aagagcgagg tggcccaccg gttcaaggac ctgggcgagg agaacttcaa ggccctggtg 360
ctgatcgcct tcgcccagta cctgcagcag agccccttcg aggaccacgt gaagctggtg 420
aacgaggtga ccgagttcgc caagacctgc gtggccgacg agagcgccga gaactgcgac 480
aagagcctgc acaccctgtt cggcgacaag ctgtgcaccg tggccaccct gcgggagacc 540
tacggcgaga tggccgactg ctgcgccaag caggagcccg agcggaacga gtgcttcctg 600
cagcacaagg acgacaaccc caacctgccc cggctggtgc ggcccgaggt ggacgtgatg 660
tgcaccgcct tccacgacaa cgaggagacc ttcctgaaga agtacctgta cgagatcgcc 720
cggcggcacc cctacttcta cgcccccgag ctgctgttct tcgccaagcg gtacaaggcc 780
gccttcaccg agtgctgcca ggccgccgac aaggccgcct gcctgctgcc caagctggac 840
gagctgcggg acgagggcaa ggccagcagc gccaagcagc ggctgaagtg cgccagcctg 900
cagaagttcg gcgagcgggc cttcaaggcc tgggccgtgg cccggctgag ccagcggttc 960
cccaaggccg agttcgccga ggtgagcaag ctggtgaccg acctgaccaa ggtgcacacc 1020
gagtgctgcc acggcgacct gctggagtgc gccgacgacc gggccgacct ggccaagtac 1080
atctgcgaga accaggacag catcagcagc aagctgaagg agtgctgcga gaagcccctg 1140
ctggagaaga gccactgcat cgccgaggtg gagaacgacg agatgcccgc cgacctgccc 1200
agcctggccg ccgacttcgt ggagagcaag gacgtgtgca agaactacgc cgaggccaag 1260
gacgtgttcc tgggcatgtt cctgtacgag tacgcccggc ggcaccccga ctacagcgtg 1320
gtgctgctgc tgcggctggc caagacctac gagaccaccc tggagaagtg ctgcgccgcc 1380
gccgaccccc acgagtgcta cgccaaggtg ttcgacgagt tcaagcccct ggtggaggag 1440
ccccagaacc tgatcaagca gaactgcgag ctgttcgagc agctgggcga gtacaagttc 1500
cagaacgccc tgctggtgcg gtacaccaag aaggtgcccc aggtgagcac ccccaccctg 1560
gtggaggtga gccggaacct gggcaaggtg ggcagcaagt gctgcaagca ccccgaggcc 1620
aagcggatgc cctgcgccga ggactacctg agcgtggtgc tgaaccagct gtgcgtgctg 1680
cacgagaaga cccccgtgag cgaccgggtg accaagtgct gcaccgagag cctggtgaac 1740
cggcggccct gcttcagcgc cctggaggtg gacgagacct acgtgcccaa ggagttcaac 1800
gccgagacct tcaccttcca cgccgacatc tgcaccctga gcgagaagga gcggcagatc 1860
aagaagcaga ccgccctggt ggagctggtg aagcacaagc ccaaggccac caaggagcag 1920
ctgaaggccg tgatggacga cttcgccgcc ttcgtggaga agtgctgcaa ggccgacgac 1980
aaggagacct gcttcgccga ggagggcaag aagctggtgg ccgccagcca ggccgccctg 2040
ggcctgggcg gcagcggcgg cagcggcggc agcggcggca gctgcagcgg ccccctgggc 2100
atcgagggcg gcatcatcag caaccagcag atcaccgcca gcagcaccca ccgggccctg 2160
ttcggcctgc agaagtggta cccctactac gcccggctga acaagaaggg cctgatcaac 2220
gcctggaccg ccgccgagaa cgaccggtgg ccctggatcc agatcaacct gcagcggaag 2280
atgcgggtga ccggcgtgat cacccagggc gccaagcgga tcggcagccc cgagtacatc 2340
aagagctaca agatcgccta cagcaacgac ggcaagacct gggccatgta caaggtgaag 2400
ggcaccaacg aggacatggt gttccggggc aacatcgaca acaacacccc ctacgccaac 2460
agcttcaccc cccccatcaa ggcccagtac gtgcggctgt acccccaggt gtgccggcgg 2520
cactgcaccc tgcggatgga gctgctgggc tgcgagctga gcggctgcag cgagcccctg 2580
ggcatgaaga gcggccacat ccaggactac cagatcaccg ccagcagcat cttccggacc 2640
ctgaacatgg acatgttcac ctgggagccc cggaaggccc ggctggacaa gcagggcaag 2700
gtgaacgcct ggaccagcgg ccacaacgac cagagccagt ggctgcaggt ggacctgctg 2760
gtgcccacca aggtgaccgg catcatcacc cagggcgcca aggacttcgg ccacgtgcag 2820
ttcgtgggca gctacaagct ggcctacagc aacgacggcg agcactggac cgtgtaccag 2880
gacgagaagc agcggaagga caaggtgttc cagggcaact tcgacaacga cacccaccgg 2940
aagaacgtga tcgacccccc catctacgcc cggcacatcc ggatcctgcc ctggagctgg 3000
tacggccgga tcaccctgcg gagcgagctg ctgggctgc 3039
<210> 94
<211> 3006
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 94
agcgccggcc cctgcacccc caacccctgc cacaacggcg gcacctgcga gatcagcgag 60
gcctaccggg gcgacacctt catcggctac gtgtgcaagt gcccccgggg cttcaacggc 120
atccactgcc agcacaacat caacgagtgc gaggtggagc cctgcaagaa cggcggcatc 180
tgcaccgacc tggtggccaa ctacagctgc gagtgccccg gcgagttcat gggccggaac 240
tgccagtaca agggcagcga cgcccacaag agcgaggtgg cccaccggtt caaggacctg 300
ggcgaggaga acttcaaggc cctggtgctg atcgccttcg cccagtacct gcagcagagc 360
cccttcgagg accacgtgaa gctggtgaac gaggtgaccg agttcgccaa gacctgcgtg 420
gccgacgaga gcgccgagaa ctgcgacaag agcctgcaca ccctgttcgg cgacaagctg 480
tgcaccgtgg ccaccctgcg ggagacctac ggcgagatgg ccgactgctg cgccaagcag 540
gagcccgagc ggaacgagtg cttcctgcag cacaaggacg acaaccccaa cctgccccgg 600
ctggtgcggc ccgaggtgga cgtgatgtgc accgccttcc acgacaacga ggagaccttc 660
ctgaagaagt acctgtacga gatcgcccgg cggcacccct acttctacgc ccccgagctg 720
ctgttcttcg ccaagcggta caaggccgcc ttcaccgagt gctgccaggc cgccgacaag 780
gccgcctgcc tgctgcccaa gctggacgag ctgcgggacg agggcaaggc cagcagcgcc 840
aagcagcggc tgaagtgcgc cagcctgcag aagttcggcg agcgggcctt caaggcctgg 900
gccgtggccc ggctgagcca gcggttcccc aaggccgagt tcgccgaggt gagcaagctg 960
gtgaccgacc tgaccaaggt gcacaccgag tgctgccacg gcgacctgct ggagtgcgcc 1020
gacgaccggg ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 1080
ctgaaggagt gctgcgagaa gcccctgctg gagaagagcc actgcatcgc cgaggtggag 1140
aacgacgaga tgcccgccga cctgcccagc ctggccgccg acttcgtgga gagcaaggac 1200
gtgtgcaaga actacgccga ggccaaggac gtgttcctgg gcatgttcct gtacgagtac 1260
gcccggcggc accccgacta cagcgtggtg ctgctgctgc ggctggccaa gacctacgag 1320
accaccctgg agaagtgctg cgccgccgcc gacccccacg agtgctacgc caaggtgttc 1380
gacgagttca agcccctggt ggaggagccc cagaacctga tcaagcagaa ctgcgagctg 1440
ttcgagcagc tgggcgagta caagttccag aacgccctgc tggtgcggta caccaagaag 1500
gtgccccagg tgagcacccc caccctggtg gaggtgagcc ggaacctggg caaggtgggc 1560
agcaagtgct gcaagcaccc cgaggccaag cggatgccct gcgccgagga ctacctgagc 1620
gtggtgctga accagctgtg cgtgctgcac gagaagaccc ccgtgagcga ccgggtgacc 1680
aagtgctgca ccgagagcct ggtgaaccgg cggccctgct tcagcgccct ggaggtggac 1740
gagacctacg tgcccaagga gttcaacgcc gagaccttca ccttccacgc cgacatctgc 1800
accctgagcg agaaggagcg gcagatcaag aagcagaccg ccctggtgga gctggtgaag 1860
cacaagccca aggccaccaa ggagcagctg aaggccgtga tggacgactt cgccgccttc 1920
gtggagaagt gctgcaaggc cgacgacaag gagacctgct tcgccgagga gggcaagaag 1980
ctggtggccg ccagccaggc cgccctgggc ctgggcggca gcggcggcag cggcggcagc 2040
ggcggcagct gcagcggccc cctgggcatc gagggcggca tcatcagcaa ccagcagatc 2100
accgccagca gcacccaccg ggccctgttc ggcctgcaga agtggtaccc ctactacgcc 2160
cggctgaaca agaagggcct gatcaacgcc tggaccgccg ccgagaacga ccggtggccc 2220
tggatccaga tcaacctgca gcggaagatg cgggtgaccg gcgtgatcac ccagggcgcc 2280
aagcggatcg gcagccccga gtacatcaag agctacaaga tcgcctacag caacgacggc 2340
aagacctggg ccatgtacaa ggtgaagggc accaacgagg acatggtgtt ccggggcaac 2400
atcgacaaca acacccccta cgccaacagc ttcacccccc ccatcaaggc ccagtacgtg 2460
cggctgtacc cccaggtgtg ccggcggcac tgcaccctgc ggatggagct gctgggctgc 2520
gagctgagcg gctgcagcga gcccctgggc atgaagagcg gccacatcca ggactaccag 2580
atcaccgcca gcagcatctt ccggaccctg aacatggaca tgttcacctg ggagccccgg 2640
aaggcccggc tggacaagca gggcaaggtg aacgcctgga ccagcggcca caacgaccag 2700
agccagtggc tgcaggtgga cctgctggtg cccaccaagg tgaccggcat catcacccag 2760
ggcgccaagg acttcggcca cgtgcagttc gtgggcagct acaagctggc ctacagcaac 2820
gacggcgagc actggaccgt gtaccaggac gagaagcagc ggaaggacaa ggtgttccag 2880
ggcaacttcg acaacgacac ccaccggaag aacgtgatcg acccccccat ctacgcccgg 2940
cacatccgga tcctgccctg gagctggtac ggccggatca ccctgcggag cgagctgctg 3000
ggctgc 3006
<210> 95
<211> 3021
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 95
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc aacatcaacg agtgcgaggt ggagccctgc 180
aagaacggcg gcatctgcac cgacctggtg gccaactaca gctgcgagtg ccccggcgag 240
ttcatgggcc ggaactgcca gtacaagggc agcgacgccc acaagagcga ggtggcccac 300
cggttcaagg acctgggcga ggagaacttc aaggccctgg tgctgatcgc cttcgcccag 360
tacctgcagc agagcccctt cgaggaccac gtgaagctgg tgaacgaggt gaccgagttc 420
gccaagacct gcgtggccga cgagagcgcc gagaactgcg acaagagcct gcacaccctg 480
ttcggcgaca agctgtgcac cgtggccacc ctgcgggaga cctacggcga gatggccgac 540
tgctgcgcca agcaggagcc cgagcggaac gagtgcttcc tgcagcacaa ggacgacaac 600
cccaacctgc cccggctggt gcggcccgag gtggacgtga tgtgcaccgc cttccacgac 660
aacgaggaga ccttcctgaa gaagtacctg tacgagatcg cccggcggca cccctacttc 720
tacgcccccg agctgctgtt cttcgccaag cggtacaagg ccgccttcac cgagtgctgc 780
caggccgccg acaaggccgc ctgcctgctg cccaagctgg acgagctgcg ggacgagggc 840
aaggccagca gcgccaagca gcggctgaag tgcgccagcc tgcagaagtt cggcgagcgg 900
gccttcaagg cctgggccgt ggcccggctg agccagcggt tccccaaggc cgagttcgcc 960
gaggtgagca agctggtgac cgacctgacc aaggtgcaca ccgagtgctg ccacggcgac 1020
ctgctggagt gcgccgacga ccgggccgac ctggccaagt acatctgcga gaaccaggac 1080
agcatcagca gcaagctgaa ggagtgctgc gagaagcccc tgctggagaa gagccactgc 1140
atcgccgagg tggagaacga cgagatgccc gccgacctgc ccagcctggc cgccgacttc 1200
gtggagagca aggacgtgtg caagaactac gccgaggcca aggacgtgtt cctgggcatg 1260
ttcctgtacg agtacgcccg gcggcacccc gactacagcg tggtgctgct gctgcggctg 1320
gccaagacct acgagaccac cctggagaag tgctgcgccg ccgccgaccc ccacgagtgc 1380
tacgccaagg tgttcgacga gttcaagccc ctggtggagg agccccagaa cctgatcaag 1440
cagaactgcg agctgttcga gcagctgggc gagtacaagt tccagaacgc cctgctggtg 1500
cggtacacca agaaggtgcc ccaggtgagc acccccaccc tggtggaggt gagccggaac 1560
ctgggcaagg tgggcagcaa gtgctgcaag caccccgagg ccaagcggat gccctgcgcc 1620
gaggactacc tgagcgtggt gctgaaccag ctgtgcgtgc tgcacgagaa gacccccgtg 1680
agcgaccggg tgaccaagtg ctgcaccgag agcctggtga accggcggcc ctgcttcagc 1740
gccctggagg tggacgagac ctacgtgccc aaggagttca acgccgagac cttcaccttc 1800
cacgccgaca tctgcaccct gagcgagaag gagcggcaga tcaagaagca gaccgccctg 1860
gtggagctgg tgaagcacaa gcccaaggcc accaaggagc agctgaaggc cgtgatggac 1920
gacttcgccg ccttcgtgga gaagtgctgc aaggccgacg acaaggagac ctgcttcgcc 1980
gaggagggca agaagctggt ggccgccagc caggccgccc tgggcctggg cggcagcggc 2040
ggcagcggcg gcagcggcgg cagctgcagc ggccccctgg gcatcgaggg cggcatcatc 2100
agcaaccagc agatcaccgc cagcagcacc caccgggccc tgttcggcct gcagaagtgg 2160
tacccctact acgcccggct gaacaagaag ggcctgatca acgcctggac cgccgccgag 2220
aacgaccggt ggccctggat ccagatcaac ctgcagcgga agatgcgggt gaccggcgtg 2280
atcacccagg gcgccaagcg gatcggcagc cccgagtaca tcaagagcta caagatcgcc 2340
tacagcaacg acggcaagac ctgggccatg tacaaggtga agggcaccaa cgaggacatg 2400
gtgttccggg gcaacatcga caacaacacc ccctacgcca acagcttcac cccccccatc 2460
aaggcccagt acgtgcggct gtacccccag gtgtgccggc ggcactgcac cctgcggatg 2520
gagctgctgg gctgcgagct gagcggctgc agcgagcccc tgggcatgaa gagcggccac 2580
atccaggact accagatcac cgccagcagc atcttccgga ccctgaacat ggacatgttc 2640
acctgggagc cccggaaggc ccggctggac aagcagggca aggtgaacgc ctggaccagc 2700
ggccacaacg accagagcca gtggctgcag gtggacctgc tggtgcccac caaggtgacc 2760
ggcatcatca cccagggcgc caaggacttc ggccacgtgc agttcgtggg cagctacaag 2820
ctggcctaca gcaacgacgg cgagcactgg accgtgtacc aggacgagaa gcagcggaag 2880
gacaaggtgt tccagggcaa cttcgacaac gacacccacc ggaagaacgt gatcgacccc 2940
cccatctacg cccggcacat ccggatcctg ccctggagct ggtacggccg gatcaccctg 3000
cggagcgagc tgctgggctg c 3021
<210> 96
<211> 50
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 96
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr
50
<210> 97
<211> 45
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 97
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His
35 40 45
<210> 98
<211> 39
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 98
Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys
1 5 10 15
Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met
20 25 30
Gly Arg Asn Cys Gln Tyr Lys
35
<210> 99
<211> 95
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 99
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His
85 90 95
<210> 100
<211> 84
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 100
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn Ile Asn
35 40 45
Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr Asp Leu
50 55 60
Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly Arg Asn
65 70 75 80
Cys Gln Tyr Lys
<210> 101
<211> 89
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 101
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly
50 55 60
Ile Cys Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu
65 70 75 80
Phe Met Gly Arg Asn Cys Gln Tyr Lys
85
<210> 102
<211> 945
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 102
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu
50 55 60
Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr
65 70 75 80
Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val
85 90 95
Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys
100 105 110
Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala
115 120 125
Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln
130 135 140
Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro
145 150 155 160
Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala
165 170 175
Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile
180 185 190
Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala
195 200 205
Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys
210 215 220
Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys
225 230 235 240
Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe
245 250 255
Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg
260 265 270
Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu
275 280 285
Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala
290 295 300
Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser
305 310 315 320
Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys
325 330 335
Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu
340 345 350
Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn
355 360 365
Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr
370 375 380
Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala
385 390 395 400
Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro
405 410 415
His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu
420 425 430
Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu
435 440 445
Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys
450 455 460
Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu
465 470 475 480
Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met
485 490 495
Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val
500 505 510
Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr
515 520 525
Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp
530 535 540
Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His
545 550 555 560
Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln
565 570 575
Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu
580 585 590
Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys
595 600 605
Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys
610 615 620
Leu Val Ala Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala
625 630 635 640
Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu
645 650 655
Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val
660 665 670
Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val
675 680 685
Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala
690 695 700
Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr
705 710 715 720
Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys
725 730 735
His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn
740 745 750
Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr
755 760 765
Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu
770 775 780
Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro
785 790 795 800
Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His
805 810 815
Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn
820 825 830
Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln
835 840 845
Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly
850 855 860
Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala
865 870 875 880
Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr
885 890 895
Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys
900 905 910
Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu
915 920 925
Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly
930 935 940
Cys
945
<210> 103
<211> 940
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 103
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asp Ala His
35 40 45
Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe
50 55 60
Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro
65 70 75 80
Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys
85 90 95
Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His
100 105 110
Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr
115 120 125
Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn
130 135 140
Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu
145 150 155 160
Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu
165 170 175
Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro
180 185 190
Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala
195 200 205
Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu
210 215 220
Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys
225 230 235 240
Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe
245 250 255
Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu
260 265 270
Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr
275 280 285
Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp
290 295 300
Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu
305 310 315 320
Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala
325 330 335
Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala
340 345 350
Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys
355 360 365
Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro
370 375 380
Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr
385 390 395 400
Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala
405 410 415
Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu
420 425 430
Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe
435 440 445
Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser
450 455 460
Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser
465 470 475 480
Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp
485 490 495
Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr
500 505 510
Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn
515 520 525
Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro
530 535 540
Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr
545 550 555 560
Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu
565 570 575
Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val
580 585 590
Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp
595 600 605
Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys Val
610 615 620
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
625 630 635 640
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
645 650 655
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
660 665 670
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
675 680 685
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
690 695 700
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
705 710 715 720
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
725 730 735
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
740 745 750
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
755 760 765
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala
770 775 780
Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr
785 790 795 800
Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn
805 810 815
Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val
820 825 830
Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser
835 840 845
Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly
850 855 860
Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser
865 870 875 880
Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile
885 890 895
Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu
900 905 910
Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His
915 920 925
Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
930 935 940
<210> 104
<211> 934
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 104
Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys
1 5 10 15
Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met
20 25 30
Gly Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His
35 40 45
Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile
50 55 60
Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys
65 70 75 80
Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu
85 90 95
Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys
100 105 110
Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp
115 120 125
Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His
130 135 140
Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp
145 150 155 160
Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys
165 170 175
Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu
180 185 190
Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys
195 200 205
Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu
210 215 220
Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala
225 230 235 240
Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala
245 250 255
Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys
260 265 270
Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp
275 280 285
Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys
290 295 300
Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys
305 310 315 320
Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu
325 330 335
Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys
340 345 350
Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met
355 360 365
Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu
370 375 380
Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys
385 390 395 400
Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe
405 410 415
Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu
420 425 430
Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val
435 440 445
Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu
450 455 460
Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro
465 470 475 480
Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu
485 490 495
Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val
500 505 510
Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser
515 520 525
Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu
530 535 540
Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg
545 550 555 560
Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro
565 570 575
Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala
580 585 590
Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala
595 600 605
Glu Glu Gly Lys Lys Leu Val Ala Cys Val Glu Pro Leu Gly Met Glu
610 615 620
Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val
625 630 635 640
Thr Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn
645 650 655
Arg Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn
660 665 670
Pro Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val
675 680 685
Val Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala
690 695 700
Phe Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His
705 710 715 720
Asp Val Asn Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn
725 730 735
Ala Val His Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val
740 745 750
Arg Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu
755 760 765
Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys
770 775 780
Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys
785 790 795 800
Thr Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu
805 810 815
Asp Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn
820 825 830
Asp Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly
835 840 845
Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala
850 855 860
Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr
865 870 875 880
Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp
885 890 895
Asn His Ser His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg
900 905 910
Tyr Val Arg Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg
915 920 925
Leu Glu Leu Leu Gly Cys
930
<210> 105
<211> 990
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 105
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asp
85 90 95
Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu
100 105 110
Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln
115 120 125
Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe
130 135 140
Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser
145 150 155 160
Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg
165 170 175
Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu
180 185 190
Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro
195 200 205
Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp
210 215 220
Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg
225 230 235 240
His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr
245 250 255
Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys
260 265 270
Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser
275 280 285
Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg
290 295 300
Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys
305 310 315 320
Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val
325 330 335
His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg
340 345 350
Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser
355 360 365
Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys
370 375 380
Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu
385 390 395 400
Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu
405 410 415
Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg
420 425 430
His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr
435 440 445
Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys
450 455 460
Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln
465 470 475 480
Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr
485 490 495
Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln
500 505 510
Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val
515 520 525
Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala
530 535 540
Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu
545 550 555 560
Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu
565 570 575
Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr
580 585 590
Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile
595 600 605
Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu
610 615 620
Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys
625 630 635 640
Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala
645 650 655
Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
660 665 670
Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln
675 680 685
Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp
690 695 700
Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp
705 710 715 720
Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu
725 730 735
Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu
740 745 750
Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn
755 760 765
Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu
770 775 780
Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu
785 790 795 800
Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His
805 810 815
Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
820 825 830
Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln
835 840 845
Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser
850 855 860
Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala
865 870 875 880
Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu
885 890 895
Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn
900 905 910
Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn
915 920 925
Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser
930 935 940
Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu
945 950 955 960
Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala
965 970 975
Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
980 985 990
<210> 106
<211> 979
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 106
Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly Thr Cys
1 5 10 15
Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr Val Cys
20 25 30
Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn Ile Asn
35 40 45
Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr Asp Leu
50 55 60
Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly Arg Asn
65 70 75 80
Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys
85 90 95
Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala
100 105 110
Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn
115 120 125
Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu
130 135 140
Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr
145 150 155 160
Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala
165 170 175
Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp
180 185 190
Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys
195 200 205
Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr
210 215 220
Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe
225 230 235 240
Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala
245 250 255
Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu
260 265 270
Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln
275 280 285
Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser
290 295 300
Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr
305 310 315 320
Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu
325 330 335
Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln
340 345 350
Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu
355 360 365
Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala
370 375 380
Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys
385 390 395 400
Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr
405 410 415
Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg
420 425 430
Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala
435 440 445
Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu
450 455 460
Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu
465 470 475 480
Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr
485 490 495
Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg
500 505 510
Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys
515 520 525
Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu
530 535 540
Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys
545 550 555 560
Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu
565 570 575
Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr
580 585 590
Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys
595 600 605
Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr
610 615 620
Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu
625 630 635 640
Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly
645 650 655
Lys Lys Leu Val Ala Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
660 665 670
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
675 680 685
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
690 695 700
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
705 710 715 720
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
725 730 735
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
740 745 750
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
755 760 765
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
770 775 780
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
785 790 795 800
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
805 810 815
Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser
820 825 830
Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly
835 840 845
Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln
850 855 860
Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp
865 870 875 880
Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr
885 890 895
Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys
900 905 910
Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro
915 920 925
Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser
930 935 940
His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg
945 950 955 960
Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu
965 970 975
Leu Gly Cys
<210> 107
<211> 984
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 107
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Asn Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly
50 55 60
Ile Cys Thr Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu
65 70 75 80
Phe Met Gly Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val
85 90 95
Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val
100 105 110
Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His
115 120 125
Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala
130 135 140
Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly
145 150 155 160
Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met
165 170 175
Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu
180 185 190
Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu
195 200 205
Val Asp Val Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu
210 215 220
Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala
225 230 235 240
Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu
245 250 255
Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp
260 265 270
Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys
275 280 285
Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala
290 295 300
Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val
305 310 315 320
Ser Lys Leu Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His
325 330 335
Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr
340 345 350
Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys
355 360 365
Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn
370 375 380
Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu
385 390 395 400
Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu
405 410 415
Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val
420 425 430
Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys
435 440 445
Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp
450 455 460
Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn
465 470 475 480
Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu
485 490 495
Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu
500 505 510
Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys
515 520 525
His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val
530 535 540
Val Leu Asn Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp
545 550 555 560
Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys
565 570 575
Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn
580 585 590
Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys
595 600 605
Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His
610 615 620
Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe
625 630 635 640
Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys
645 650 655
Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys Val Glu Pro Leu Gly
660 665 670
Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val
675 680 685
Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg
690 695 700
Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp
705 710 715 720
Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr
725 730 735
Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu
740 745 750
Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe
755 760 765
Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn
770 775 780
Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln
785 790 795 800
Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg
805 810 815
Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys Ala Asn Pro Leu Gly
820 825 830
Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile Thr Ala Ser Ser Ser
835 840 845
Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp Asn Pro Ser Tyr Ala
850 855 860
Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp Val Ala Gly Ser Tyr
865 870 875 880
Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly Ser Ser Lys Glu Val
885 890 895
Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe Gly Ser Val Gln Phe
900 905 910
Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp Ser Ala Asn Trp Thr
915 920 925
Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys Ile Phe Pro Gly Asn
930 935 940
Trp Asp Asn His Ser His Lys Lys Asn Leu Phe Glu Thr Pro Ile Leu
945 950 955 960
Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp His Asn Arg Ile Ala
965 970 975
Leu Arg Leu Glu Leu Leu Gly Cys
980
<210> 108
<211> 2835
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 108
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc gacgcccaca agagcgaggt ggcccaccgg 180
ttcaaggacc tgggcgagga gaacttcaag gccctggtgc tgatcgcctt cgcccagtac 240
ctgcagcaga gccccttcga ggaccacgtg aagctggtga acgaggtgac cgagttcgcc 300
aagacctgcg tggccgacga gagcgccgag aactgcgaca agagcctgca caccctgttc 360
ggcgacaagc tgtgcaccgt ggccaccctg cgggagacct acggcgagat ggccgactgc 420
tgcgccaagc aggagcccga gcggaacgag tgcttcctgc agcacaagga cgacaacccc 480
aacctgcccc ggctggtgcg gcccgaggtg gacgtgatgt gcaccgcctt ccacgacaac 540
gaggagacct tcctgaagaa gtacctgtac gagatcgccc ggcggcaccc ctacttctac 600
gcccccgagc tgctgttctt cgccaagcgg tacaaggccg ccttcaccga gtgctgccag 660
gccgccgaca aggccgcctg cctgctgccc aagctggacg agctgcggga cgagggcaag 720
gccagcagcg ccaagcagcg gctgaagtgc gccagcctgc agaagttcgg cgagcgggcc 780
ttcaaggcct gggccgtggc ccggctgagc cagcggttcc ccaaggccga gttcgccgag 840
gtgagcaagc tggtgaccga cctgaccaag gtgcacaccg agtgctgcca cggcgacctg 900
ctggagtgcg ccgacgaccg ggccgacctg gccaagtaca tctgcgagaa ccaggacagc 960
atcagcagca agctgaagga gtgctgcgag aagcccctgc tggagaagag ccactgcatc 1020
gccgaggtgg agaacgacga gatgcccgcc gacctgccca gcctggccgc cgacttcgtg 1080
gagagcaagg acgtgtgcaa gaactacgcc gaggccaagg acgtgttcct gggcatgttc 1140
ctgtacgagt acgcccggcg gcaccccgac tacagcgtgg tgctgctgct gcggctggcc 1200
aagacctacg agaccaccct ggagaagtgc tgcgccgccg ccgaccccca cgagtgctac 1260
gccaaggtgt tcgacgagtt caagcccctg gtggaggagc cccagaacct gatcaagcag 1320
aactgcgagc tgttcgagca gctgggcgag tacaagttcc agaacgccct gctggtgcgg 1380
tacaccaaga aggtgcccca ggtgagcacc cccaccctgg tggaggtgag ccggaacctg 1440
ggcaaggtgg gcagcaagtg ctgcaagcac cccgaggcca agcggatgcc ctgcgccgag 1500
gactacctga gcgtggtgct gaaccagctg tgcgtgctgc acgagaagac ccccgtgagc 1560
gaccgggtga ccaagtgctg caccgagagc ctggtgaacc ggcggccctg cttcagcgcc 1620
ctggaggtgg acgagaccta cgtgcccaag gagttcaacg ccgagacctt caccttccac 1680
gccgacatct gcaccctgag cgagaaggag cggcagatca agaagcagac cgccctggtg 1740
gagctggtga agcacaagcc caaggccacc aaggagcagc tgaaggccgt gatggacgac 1800
ttcgccgcct tcgtggagaa gtgctgcaag gccgacgaca aggagacctg cttcgccgag 1860
gagggcaaga agctggtggc ctgcgtggag cccctgggca tggagaacgg caacatcgcc 1920
aacagccaga tcgccgccag cagcgtgcgg gtgaccttcc tgggcctgca gcactgggtg 1980
cccgagctgg cccggctgaa ccgggccggc atggtgaacg cctggacccc cagcagcaac 2040
gacgacaacc cctggatcca ggtgaacctg ctgcggcgga tgtgggtgac cggcgtggtg 2100
acccagggcg ccagccggct ggccagccac gagtacctga aggccttcaa ggtggcctac 2160
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaggagttc 2220
gtgggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacccc cgtggaggcc 2280
cagtacgtgc ggctgtaccc caccagctgc cacaccgcct gcaccctgcg gttcgagctg 2340
ctgggctgcg agctgaacgg ctgcgccaac cccctgggcc tgaagaacaa cagcatcccc 2400
gacaagcaga tcaccgccag cagcagctac aagacctggg gcctgcacct gttcagctgg 2460
aaccccagct acgcccggct ggacaagcag ggcaacttca acgcctgggt ggccggcagc 2520
tacggcaacg accagtggct gcaggtggac ctgggcagca gcaaggaggt gaccggcatc 2580
atcacccagg gcgcccggaa cttcggcagc gtgcagttcg tggccagcta caaggtggcc 2640
tacagcaacg acagcgccaa ctggaccgag taccaggacc cccggaccgg cagcagcaag 2700
atcttccccg gcaactggga caaccacagc cacaagaaga acctgttcga gacccccatc 2760
ctggcccggt acgtgcggat cctgcccgtg gcctggcaca accggatcgc cctgcggctg 2820
gagctgctgg gctgc 2835
<210> 109
<211> 2820
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 109
agcgccggcc cctgcacccc caacccctgc cacaacggcg gcacctgcga gatcagcgag 60
gcctaccggg gcgacacctt catcggctac gtgtgcaagt gcccccgggg cttcaacggc 120
atccactgcc agcacgacgc ccacaagagc gaggtggccc accggttcaa ggacctgggc 180
gaggagaact tcaaggccct ggtgctgatc gccttcgccc agtacctgca gcagagcccc 240
ttcgaggacc acgtgaagct ggtgaacgag gtgaccgagt tcgccaagac ctgcgtggcc 300
gacgagagcg ccgagaactg cgacaagagc ctgcacaccc tgttcggcga caagctgtgc 360
accgtggcca ccctgcggga gacctacggc gagatggccg actgctgcgc caagcaggag 420
cccgagcgga acgagtgctt cctgcagcac aaggacgaca accccaacct gccccggctg 480
gtgcggcccg aggtggacgt gatgtgcacc gccttccacg acaacgagga gaccttcctg 540
aagaagtacc tgtacgagat cgcccggcgg cacccctact tctacgcccc cgagctgctg 600
ttcttcgcca agcggtacaa ggccgccttc accgagtgct gccaggccgc cgacaaggcc 660
gcctgcctgc tgcccaagct ggacgagctg cgggacgagg gcaaggccag cagcgccaag 720
cagcggctga agtgcgccag cctgcagaag ttcggcgagc gggccttcaa ggcctgggcc 780
gtggcccggc tgagccagcg gttccccaag gccgagttcg ccgaggtgag caagctggtg 840
accgacctga ccaaggtgca caccgagtgc tgccacggcg acctgctgga gtgcgccgac 900
gaccgggccg acctggccaa gtacatctgc gagaaccagg acagcatcag cagcaagctg 960
aaggagtgct gcgagaagcc cctgctggag aagagccact gcatcgccga ggtggagaac 1020
gacgagatgc ccgccgacct gcccagcctg gccgccgact tcgtggagag caaggacgtg 1080
tgcaagaact acgccgaggc caaggacgtg ttcctgggca tgttcctgta cgagtacgcc 1140
cggcggcacc ccgactacag cgtggtgctg ctgctgcggc tggccaagac ctacgagacc 1200
accctggaga agtgctgcgc cgccgccgac ccccacgagt gctacgccaa ggtgttcgac 1260
gagttcaagc ccctggtgga ggagccccag aacctgatca agcagaactg cgagctgttc 1320
gagcagctgg gcgagtacaa gttccagaac gccctgctgg tgcggtacac caagaaggtg 1380
ccccaggtga gcacccccac cctggtggag gtgagccgga acctgggcaa ggtgggcagc 1440
aagtgctgca agcaccccga ggccaagcgg atgccctgcg ccgaggacta cctgagcgtg 1500
gtgctgaacc agctgtgcgt gctgcacgag aagacccccg tgagcgaccg ggtgaccaag 1560
tgctgcaccg agagcctggt gaaccggcgg ccctgcttca gcgccctgga ggtggacgag 1620
acctacgtgc ccaaggagtt caacgccgag accttcacct tccacgccga catctgcacc 1680
ctgagcgaga aggagcggca gatcaagaag cagaccgccc tggtggagct ggtgaagcac 1740
aagcccaagg ccaccaagga gcagctgaag gccgtgatgg acgacttcgc cgccttcgtg 1800
gagaagtgct gcaaggccga cgacaaggag acctgcttcg ccgaggaggg caagaagctg 1860
gtggcctgcg tggagcccct gggcatggag aacggcaaca tcgccaacag ccagatcgcc 1920
gccagcagcg tgcgggtgac cttcctgggc ctgcagcact gggtgcccga gctggcccgg 1980
ctgaaccggg ccggcatggt gaacgcctgg acccccagca gcaacgacga caacccctgg 2040
atccaggtga acctgctgcg gcggatgtgg gtgaccggcg tggtgaccca gggcgccagc 2100
cggctggcca gccacgagta cctgaaggcc ttcaaggtgg cctacagcct gaacggccac 2160
gagttcgact tcatccacga cgtgaacaag aagcacaagg agttcgtggg caactggaac 2220
aagaacgccg tgcacgtgaa cctgttcgag acccccgtgg aggcccagta cgtgcggctg 2280
taccccacca gctgccacac cgcctgcacc ctgcggttcg agctgctggg ctgcgagctg 2340
aacggctgcg ccaaccccct gggcctgaag aacaacagca tccccgacaa gcagatcacc 2400
gccagcagca gctacaagac ctggggcctg cacctgttca gctggaaccc cagctacgcc 2460
cggctggaca agcagggcaa cttcaacgcc tgggtggccg gcagctacgg caacgaccag 2520
tggctgcagg tggacctggg cagcagcaag gaggtgaccg gcatcatcac ccagggcgcc 2580
cggaacttcg gcagcgtgca gttcgtggcc agctacaagg tggcctacag caacgacagc 2640
gccaactgga ccgagtacca ggacccccgg accggcagca gcaagatctt ccccggcaac 2700
tgggacaacc acagccacaa gaagaacctg ttcgagaccc ccatcctggc ccggtacgtg 2760
cggatcctgc ccgtggcctg gcacaaccgg atcgccctgc ggctggagct gctgggctgc 2820
<210> 110
<211> 2802
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 110
aacatcaacg agtgcgaggt ggagccctgc aagaacggcg gcatctgcac cgacctggtg 60
gccaactaca gctgcgagtg ccccggcgag ttcatgggcc ggaactgcca gtacaaggac 120
gcccacaaga gcgaggtggc ccaccggttc aaggacctgg gcgaggagaa cttcaaggcc 180
ctggtgctga tcgccttcgc ccagtacctg cagcagagcc ccttcgagga ccacgtgaag 240
ctggtgaacg aggtgaccga gttcgccaag acctgcgtgg ccgacgagag cgccgagaac 300
tgcgacaaga gcctgcacac cctgttcggc gacaagctgt gcaccgtggc caccctgcgg 360
gagacctacg gcgagatggc cgactgctgc gccaagcagg agcccgagcg gaacgagtgc 420
ttcctgcagc acaaggacga caaccccaac ctgccccggc tggtgcggcc cgaggtggac 480
gtgatgtgca ccgccttcca cgacaacgag gagaccttcc tgaagaagta cctgtacgag 540
atcgcccggc ggcaccccta cttctacgcc cccgagctgc tgttcttcgc caagcggtac 600
aaggccgcct tcaccgagtg ctgccaggcc gccgacaagg ccgcctgcct gctgcccaag 660
ctggacgagc tgcgggacga gggcaaggcc agcagcgcca agcagcggct gaagtgcgcc 720
agcctgcaga agttcggcga gcgggccttc aaggcctggg ccgtggcccg gctgagccag 780
cggttcccca aggccgagtt cgccgaggtg agcaagctgg tgaccgacct gaccaaggtg 840
cacaccgagt gctgccacgg cgacctgctg gagtgcgccg acgaccgggc cgacctggcc 900
aagtacatct gcgagaacca ggacagcatc agcagcaagc tgaaggagtg ctgcgagaag 960
cccctgctgg agaagagcca ctgcatcgcc gaggtggaga acgacgagat gcccgccgac 1020
ctgcccagcc tggccgccga cttcgtggag agcaaggacg tgtgcaagaa ctacgccgag 1080
gccaaggacg tgttcctggg catgttcctg tacgagtacg cccggcggca ccccgactac 1140
agcgtggtgc tgctgctgcg gctggccaag acctacgaga ccaccctgga gaagtgctgc 1200
gccgccgccg acccccacga gtgctacgcc aaggtgttcg acgagttcaa gcccctggtg 1260
gaggagcccc agaacctgat caagcagaac tgcgagctgt tcgagcagct gggcgagtac 1320
aagttccaga acgccctgct ggtgcggtac accaagaagg tgccccaggt gagcaccccc 1380
accctggtgg aggtgagccg gaacctgggc aaggtgggca gcaagtgctg caagcacccc 1440
gaggccaagc ggatgccctg cgccgaggac tacctgagcg tggtgctgaa ccagctgtgc 1500
gtgctgcacg agaagacccc cgtgagcgac cgggtgacca agtgctgcac cgagagcctg 1560
gtgaaccggc ggccctgctt cagcgccctg gaggtggacg agacctacgt gcccaaggag 1620
ttcaacgccg agaccttcac cttccacgcc gacatctgca ccctgagcga gaaggagcgg 1680
cagatcaaga agcagaccgc cctggtggag ctggtgaagc acaagcccaa ggccaccaag 1740
gagcagctga aggccgtgat ggacgacttc gccgccttcg tggagaagtg ctgcaaggcc 1800
gacgacaagg agacctgctt cgccgaggag ggcaagaagc tggtggcctg cgtggagccc 1860
ctgggcatgg agaacggcaa catcgccaac agccagatcg ccgccagcag cgtgcgggtg 1920
accttcctgg gcctgcagca ctgggtgccc gagctggccc ggctgaaccg ggccggcatg 1980
gtgaacgcct ggacccccag cagcaacgac gacaacccct ggatccaggt gaacctgctg 2040
cggcggatgt gggtgaccgg cgtggtgacc cagggcgcca gccggctggc cagccacgag 2100
tacctgaagg ccttcaaggt ggcctacagc ctgaacggcc acgagttcga cttcatccac 2160
gacgtgaaca agaagcacaa ggagttcgtg ggcaactgga acaagaacgc cgtgcacgtg 2220
aacctgttcg agacccccgt ggaggcccag tacgtgcggc tgtaccccac cagctgccac 2280
accgcctgca ccctgcggtt cgagctgctg ggctgcgagc tgaacggctg cgccaacccc 2340
ctgggcctga agaacaacag catccccgac aagcagatca ccgccagcag cagctacaag 2400
acctggggcc tgcacctgtt cagctggaac cccagctacg cccggctgga caagcagggc 2460
aacttcaacg cctgggtggc cggcagctac ggcaacgacc agtggctgca ggtggacctg 2520
ggcagcagca aggaggtgac cggcatcatc acccagggcg cccggaactt cggcagcgtg 2580
cagttcgtgg ccagctacaa ggtggcctac agcaacgaca gcgccaactg gaccgagtac 2640
caggaccccc ggaccggcag cagcaagatc ttccccggca actgggacaa ccacagccac 2700
aagaagaacc tgttcgagac ccccatcctg gcccggtacg tgcggatcct gcccgtggcc 2760
tggcacaacc ggatcgccct gcggctggag ctgctgggct gc 2802
<210> 111
<211> 2970
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 111
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc agcgccggcc cctgcacccc caacccctgc 180
cacaacggcg gcacctgcga gatcagcgag gcctaccggg gcgacacctt catcggctac 240
gtgtgcaagt gcccccgggg cttcaacggc atccactgcc agcacgacgc ccacaagagc 300
gaggtggccc accggttcaa ggacctgggc gaggagaact tcaaggccct ggtgctgatc 360
gccttcgccc agtacctgca gcagagcccc ttcgaggacc acgtgaagct ggtgaacgag 420
gtgaccgagt tcgccaagac ctgcgtggcc gacgagagcg ccgagaactg cgacaagagc 480
ctgcacaccc tgttcggcga caagctgtgc accgtggcca ccctgcggga gacctacggc 540
gagatggccg actgctgcgc caagcaggag cccgagcgga acgagtgctt cctgcagcac 600
aaggacgaca accccaacct gccccggctg gtgcggcccg aggtggacgt gatgtgcacc 660
gccttccacg acaacgagga gaccttcctg aagaagtacc tgtacgagat cgcccggcgg 720
cacccctact tctacgcccc cgagctgctg ttcttcgcca agcggtacaa ggccgccttc 780
accgagtgct gccaggccgc cgacaaggcc gcctgcctgc tgcccaagct ggacgagctg 840
cgggacgagg gcaaggccag cagcgccaag cagcggctga agtgcgccag cctgcagaag 900
ttcggcgagc gggccttcaa ggcctgggcc gtggcccggc tgagccagcg gttccccaag 960
gccgagttcg ccgaggtgag caagctggtg accgacctga ccaaggtgca caccgagtgc 1020
tgccacggcg acctgctgga gtgcgccgac gaccgggccg acctggccaa gtacatctgc 1080
gagaaccagg acagcatcag cagcaagctg aaggagtgct gcgagaagcc cctgctggag 1140
aagagccact gcatcgccga ggtggagaac gacgagatgc ccgccgacct gcccagcctg 1200
gccgccgact tcgtggagag caaggacgtg tgcaagaact acgccgaggc caaggacgtg 1260
ttcctgggca tgttcctgta cgagtacgcc cggcggcacc ccgactacag cgtggtgctg 1320
ctgctgcggc tggccaagac ctacgagacc accctggaga agtgctgcgc cgccgccgac 1380
ccccacgagt gctacgccaa ggtgttcgac gagttcaagc ccctggtgga ggagccccag 1440
aacctgatca agcagaactg cgagctgttc gagcagctgg gcgagtacaa gttccagaac 1500
gccctgctgg tgcggtacac caagaaggtg ccccaggtga gcacccccac cctggtggag 1560
gtgagccgga acctgggcaa ggtgggcagc aagtgctgca agcaccccga ggccaagcgg 1620
atgccctgcg ccgaggacta cctgagcgtg gtgctgaacc agctgtgcgt gctgcacgag 1680
aagacccccg tgagcgaccg ggtgaccaag tgctgcaccg agagcctggt gaaccggcgg 1740
ccctgcttca gcgccctgga ggtggacgag acctacgtgc ccaaggagtt caacgccgag 1800
accttcacct tccacgccga catctgcacc ctgagcgaga aggagcggca gatcaagaag 1860
cagaccgccc tggtggagct ggtgaagcac aagcccaagg ccaccaagga gcagctgaag 1920
gccgtgatgg acgacttcgc cgccttcgtg gagaagtgct gcaaggccga cgacaaggag 1980
acctgcttcg ccgaggaggg caagaagctg gtggcctgcg tggagcccct gggcatggag 2040
aacggcaaca tcgccaacag ccagatcgcc gccagcagcg tgcgggtgac cttcctgggc 2100
ctgcagcact gggtgcccga gctggcccgg ctgaaccggg ccggcatggt gaacgcctgg 2160
acccccagca gcaacgacga caacccctgg atccaggtga acctgctgcg gcggatgtgg 2220
gtgaccggcg tggtgaccca gggcgccagc cggctggcca gccacgagta cctgaaggcc 2280
ttcaaggtgg cctacagcct gaacggccac gagttcgact tcatccacga cgtgaacaag 2340
aagcacaagg agttcgtggg caactggaac aagaacgccg tgcacgtgaa cctgttcgag 2400
acccccgtgg aggcccagta cgtgcggctg taccccacca gctgccacac cgcctgcacc 2460
ctgcggttcg agctgctggg ctgcgagctg aacggctgcg ccaaccccct gggcctgaag 2520
aacaacagca tccccgacaa gcagatcacc gccagcagca gctacaagac ctggggcctg 2580
cacctgttca gctggaaccc cagctacgcc cggctggaca agcagggcaa cttcaacgcc 2640
tgggtggccg gcagctacgg caacgaccag tggctgcagg tggacctggg cagcagcaag 2700
gaggtgaccg gcatcatcac ccagggcgcc cggaacttcg gcagcgtgca gttcgtggcc 2760
agctacaagg tggcctacag caacgacagc gccaactgga ccgagtacca ggacccccgg 2820
accggcagca gcaagatctt ccccggcaac tgggacaacc acagccacaa gaagaacctg 2880
ttcgagaccc ccatcctggc ccggtacgtg cggatcctgc ccgtggcctg gcacaaccgg 2940
atcgccctgc ggctggagct gctgggctgc 2970
<210> 112
<211> 2937
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 112
agcgccggcc cctgcacccc caacccctgc cacaacggcg gcacctgcga gatcagcgag 60
gcctaccggg gcgacacctt catcggctac gtgtgcaagt gcccccgggg cttcaacggc 120
atccactgcc agcacaacat caacgagtgc gaggtggagc cctgcaagaa cggcggcatc 180
tgcaccgacc tggtggccaa ctacagctgc gagtgccccg gcgagttcat gggccggaac 240
tgccagtaca aggacgccca caagagcgag gtggcccacc ggttcaagga cctgggcgag 300
gagaacttca aggccctggt gctgatcgcc ttcgcccagt acctgcagca gagccccttc 360
gaggaccacg tgaagctggt gaacgaggtg accgagttcg ccaagacctg cgtggccgac 420
gagagcgccg agaactgcga caagagcctg cacaccctgt tcggcgacaa gctgtgcacc 480
gtggccaccc tgcgggagac ctacggcgag atggccgact gctgcgccaa gcaggagccc 540
gagcggaacg agtgcttcct gcagcacaag gacgacaacc ccaacctgcc ccggctggtg 600
cggcccgagg tggacgtgat gtgcaccgcc ttccacgaca acgaggagac cttcctgaag 660
aagtacctgt acgagatcgc ccggcggcac ccctacttct acgcccccga gctgctgttc 720
ttcgccaagc ggtacaaggc cgccttcacc gagtgctgcc aggccgccga caaggccgcc 780
tgcctgctgc ccaagctgga cgagctgcgg gacgagggca aggccagcag cgccaagcag 840
cggctgaagt gcgccagcct gcagaagttc ggcgagcggg ccttcaaggc ctgggccgtg 900
gcccggctga gccagcggtt ccccaaggcc gagttcgccg aggtgagcaa gctggtgacc 960
gacctgacca aggtgcacac cgagtgctgc cacggcgacc tgctggagtg cgccgacgac 1020
cgggccgacc tggccaagta catctgcgag aaccaggaca gcatcagcag caagctgaag 1080
gagtgctgcg agaagcccct gctggagaag agccactgca tcgccgaggt ggagaacgac 1140
gagatgcccg ccgacctgcc cagcctggcc gccgacttcg tggagagcaa ggacgtgtgc 1200
aagaactacg ccgaggccaa ggacgtgttc ctgggcatgt tcctgtacga gtacgcccgg 1260
cggcaccccg actacagcgt ggtgctgctg ctgcggctgg ccaagaccta cgagaccacc 1320
ctggagaagt gctgcgccgc cgccgacccc cacgagtgct acgccaaggt gttcgacgag 1380
ttcaagcccc tggtggagga gccccagaac ctgatcaagc agaactgcga gctgttcgag 1440
cagctgggcg agtacaagtt ccagaacgcc ctgctggtgc ggtacaccaa gaaggtgccc 1500
caggtgagca cccccaccct ggtggaggtg agccggaacc tgggcaaggt gggcagcaag 1560
tgctgcaagc accccgaggc caagcggatg ccctgcgccg aggactacct gagcgtggtg 1620
ctgaaccagc tgtgcgtgct gcacgagaag acccccgtga gcgaccgggt gaccaagtgc 1680
tgcaccgaga gcctggtgaa ccggcggccc tgcttcagcg ccctggaggt ggacgagacc 1740
tacgtgccca aggagttcaa cgccgagacc ttcaccttcc acgccgacat ctgcaccctg 1800
agcgagaagg agcggcagat caagaagcag accgccctgg tggagctggt gaagcacaag 1860
cccaaggcca ccaaggagca gctgaaggcc gtgatggacg acttcgccgc cttcgtggag 1920
aagtgctgca aggccgacga caaggagacc tgcttcgccg aggagggcaa gaagctggtg 1980
gcctgcgtgg agcccctggg catggagaac ggcaacatcg ccaacagcca gatcgccgcc 2040
agcagcgtgc gggtgacctt cctgggcctg cagcactggg tgcccgagct ggcccggctg 2100
aaccgggccg gcatggtgaa cgcctggacc cccagcagca acgacgacaa cccctggatc 2160
caggtgaacc tgctgcggcg gatgtgggtg accggcgtgg tgacccaggg cgccagccgg 2220
ctggccagcc acgagtacct gaaggccttc aaggtggcct acagcctgaa cggccacgag 2280
ttcgacttca tccacgacgt gaacaagaag cacaaggagt tcgtgggcaa ctggaacaag 2340
aacgccgtgc acgtgaacct gttcgagacc cccgtggagg cccagtacgt gcggctgtac 2400
cccaccagct gccacaccgc ctgcaccctg cggttcgagc tgctgggctg cgagctgaac 2460
ggctgcgcca accccctggg cctgaagaac aacagcatcc ccgacaagca gatcaccgcc 2520
agcagcagct acaagacctg gggcctgcac ctgttcagct ggaaccccag ctacgcccgg 2580
ctggacaagc agggcaactt caacgcctgg gtggccggca gctacggcaa cgaccagtgg 2640
ctgcaggtgg acctgggcag cagcaaggag gtgaccggca tcatcaccca gggcgcccgg 2700
aacttcggca gcgtgcagtt cgtggccagc tacaaggtgg cctacagcaa cgacagcgcc 2760
aactggaccg agtaccagga cccccggacc ggcagcagca agatcttccc cggcaactgg 2820
gacaaccaca gccacaagaa gaacctgttc gagaccccca tcctggcccg gtacgtgcgg 2880
atcctgcccg tggcctggca caaccggatc gccctgcggc tggagctgct gggctgc 2937
<210> 113
<211> 2952
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 113
gacatctgcg accccaaccc ctgcgagaac ggcggcatct gcctgcccgg cctggccgac 60
ggcagcttca gctgcgagtg ccccgacggc ttcaccgacc ccaactgcag cagcgtggtg 120
gaggtggcca gcgacgagga ggagcccacc aacatcaacg agtgcgaggt ggagccctgc 180
aagaacggcg gcatctgcac cgacctggtg gccaactaca gctgcgagtg ccccggcgag 240
ttcatgggcc ggaactgcca gtacaaggac gcccacaaga gcgaggtggc ccaccggttc 300
aaggacctgg gcgaggagaa cttcaaggcc ctggtgctga tcgccttcgc ccagtacctg 360
cagcagagcc ccttcgagga ccacgtgaag ctggtgaacg aggtgaccga gttcgccaag 420
acctgcgtgg ccgacgagag cgccgagaac tgcgacaaga gcctgcacac cctgttcggc 480
gacaagctgt gcaccgtggc caccctgcgg gagacctacg gcgagatggc cgactgctgc 540
gccaagcagg agcccgagcg gaacgagtgc ttcctgcagc acaaggacga caaccccaac 600
ctgccccggc tggtgcggcc cgaggtggac gtgatgtgca ccgccttcca cgacaacgag 660
gagaccttcc tgaagaagta cctgtacgag atcgcccggc ggcaccccta cttctacgcc 720
cccgagctgc tgttcttcgc caagcggtac aaggccgcct tcaccgagtg ctgccaggcc 780
gccgacaagg ccgcctgcct gctgcccaag ctggacgagc tgcgggacga gggcaaggcc 840
agcagcgcca agcagcggct gaagtgcgcc agcctgcaga agttcggcga gcgggccttc 900
aaggcctggg ccgtggcccg gctgagccag cggttcccca aggccgagtt cgccgaggtg 960
agcaagctgg tgaccgacct gaccaaggtg cacaccgagt gctgccacgg cgacctgctg 1020
gagtgcgccg acgaccgggc cgacctggcc aagtacatct gcgagaacca ggacagcatc 1080
agcagcaagc tgaaggagtg ctgcgagaag cccctgctgg agaagagcca ctgcatcgcc 1140
gaggtggaga acgacgagat gcccgccgac ctgcccagcc tggccgccga cttcgtggag 1200
agcaaggacg tgtgcaagaa ctacgccgag gccaaggacg tgttcctggg catgttcctg 1260
tacgagtacg cccggcggca ccccgactac agcgtggtgc tgctgctgcg gctggccaag 1320
acctacgaga ccaccctgga gaagtgctgc gccgccgccg acccccacga gtgctacgcc 1380
aaggtgttcg acgagttcaa gcccctggtg gaggagcccc agaacctgat caagcagaac 1440
tgcgagctgt tcgagcagct gggcgagtac aagttccaga acgccctgct ggtgcggtac 1500
accaagaagg tgccccaggt gagcaccccc accctggtgg aggtgagccg gaacctgggc 1560
aaggtgggca gcaagtgctg caagcacccc gaggccaagc ggatgccctg cgccgaggac 1620
tacctgagcg tggtgctgaa ccagctgtgc gtgctgcacg agaagacccc cgtgagcgac 1680
cgggtgacca agtgctgcac cgagagcctg gtgaaccggc ggccctgctt cagcgccctg 1740
gaggtggacg agacctacgt gcccaaggag ttcaacgccg agaccttcac cttccacgcc 1800
gacatctgca ccctgagcga gaaggagcgg cagatcaaga agcagaccgc cctggtggag 1860
ctggtgaagc acaagcccaa ggccaccaag gagcagctga aggccgtgat ggacgacttc 1920
gccgccttcg tggagaagtg ctgcaaggcc gacgacaagg agacctgctt cgccgaggag 1980
ggcaagaagc tggtggcctg cgtggagccc ctgggcatgg agaacggcaa catcgccaac 2040
agccagatcg ccgccagcag cgtgcgggtg accttcctgg gcctgcagca ctgggtgccc 2100
gagctggccc ggctgaaccg ggccggcatg gtgaacgcct ggacccccag cagcaacgac 2160
gacaacccct ggatccaggt gaacctgctg cggcggatgt gggtgaccgg cgtggtgacc 2220
cagggcgcca gccggctggc cagccacgag tacctgaagg ccttcaaggt ggcctacagc 2280
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaagcacaa ggagttcgtg 2340
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacccccgt ggaggcccag 2400
tacgtgcggc tgtaccccac cagctgccac accgcctgca ccctgcggtt cgagctgctg 2460
ggctgcgagc tgaacggctg cgccaacccc ctgggcctga agaacaacag catccccgac 2520
aagcagatca ccgccagcag cagctacaag acctggggcc tgcacctgtt cagctggaac 2580
cccagctacg cccggctgga caagcagggc aacttcaacg cctgggtggc cggcagctac 2640
ggcaacgacc agtggctgca ggtggacctg ggcagcagca aggaggtgac cggcatcatc 2700
acccagggcg cccggaactt cggcagcgtg cagttcgtgg ccagctacaa ggtggcctac 2760
agcaacgaca gcgccaactg gaccgagtac caggaccccc ggaccggcag cagcaagatc 2820
ttccccggca actgggacaa ccacagccac aagaagaacc tgttcgagac ccccatcctg 2880
gcccggtacg tgcggatcct gcccgtggcc tggcacaacc ggatcgccct gcggctggag 2940
ctgctgggct gc 2952
<210> 114
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический пептид"
<400> 114
Gly Gly Gly Gly Ser
1 5
<210> 115
<211> 212
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 115
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Cys Val
35 40 45
Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile Ala
50 55 60
Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val Pro
65 70 75 80
Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr Pro
85 90 95
Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg Arg
100 105 110
Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala Ser
115 120 125
His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly His
130 135 140
Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe Val
145 150 155 160
Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr Pro
165 170 175
Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr Ala
180 185 190
Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly His His
195 200 205
His His His His
210
<210> 116
<211> 636
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 116
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaatg tgtggaaccc ctcggcatgg aaaacggcaa tatcgccaat 180
agccagatcg ccgccagcag cgtcagagtg acatttctgg gactgcaaca ctgggtgcca 240
gagctggcca gactgaatag agccggcatg gttaacgcct ggacacccag cagcaacgac 300
gacaacccct ggattcaagt gaacctgctg cggcgtatgt gggtcacagg tgttgttaca 360
cagggcgcaa gcagactggc cagccacgag tatctgaagg cctttaaggt ggcctacagc 420
ctgaacggcc acgagttcga cttcatccac gacgtgaaca agaagcacaa agagttcgtc 480
ggcaactgga acaagaacgc cgtgcacgtg aacctgttcg agacacctgt ggaagcccag 540
tacgtgcggc tgtaccctac aagctgtcac accgcctgca cactgagatt cgagctgctg 600
ggctgtgaac tgaatggcca ccaccaccat caccac 636
<210> 117
<211> 805
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 117
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly
805
<210> 118
<211> 2415
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 118
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atctgatgcc cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctccagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa atgcgccagc ctccagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgcaccc tgtccgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gtggaagcgg aggaagtggt 1920
ggatctggcg gatcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcaacactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aatccctgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcaagca gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggc 2415
<210> 119
<211> 789
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 119
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Ala
610 615 620
Ser Gln Ala Ala Leu Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
625 630 635 640
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
645 650 655
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
660 665 670
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
675 680 685
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
690 695 700
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
705 710 715 720
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
725 730 735
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
740 745 750
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
755 760 765
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
770 775 780
Cys Glu Leu Asn Gly
785
<210> 120
<211> 2367
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 120
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggctg cttctcaggc cgctctgtgt gtggaacccc tcggcatgga aaacggcaat 1920
atcgccaata gccagattgc cgccagcagc gtcagagtga catttctggg actgcaacac 1980
tgggtgcccg agctggctag actgaataga gccggcatgg tcaacgcctg gacacccagc 2040
agcaacgacg ataatccctg gattcaagtg aacctgctgc ggcgtatgtg ggtcacaggt 2100
gttgttacac agggcgcaag cagactggcc agccacgagt atctgaaggc ctttaaggtg 2160
gcctacagcc tgaacggcca cgagttcgac ttcatccacg acgtgaacaa gaagcacaaa 2220
gagtttgtcg gcaactggaa caagaacgcc gtgcacgtga acctgttcga gacacctgtg 2280
gaagcccagt acgtgcggct gtaccctaca agctgtcaca ccgcctgcac tctgagattc 2340
gaactgctgg gatgcgagct gaacggc 2367
<210> 121
<211> 783
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 121
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
625 630 635 640
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
645 650 655
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
660 665 670
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
675 680 685
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
690 695 700
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
705 710 715 720
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
725 730 735
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
740 745 750
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
755 760 765
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
770 775 780
<210> 122
<211> 2349
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 122
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggcct gtgtggaacc cctcggcatg gaaaacggca atatcgccaa tagccagatt 1920
gccgccagca gcgtcagagt gacatttctg ggactgcaac actgggtgcc cgagctggct 1980
agactgaata gagccggcat ggtcaacgcc tggacaccca gcagcaacga cgataatccc 2040
tggattcaag tgaacctgct gcggcgtatg tgggtcacag gtgttgttac acagggcgca 2100
agcagactgg ccagccacga gtatctgaag gcctttaagg tggcctacag cctgaacggc 2160
cacgagttcg acttcatcca cgacgtgaac aagaagcaca aagagtttgt cggcaactgg 2220
aacaagaacg ccgtgcacgt gaacctgttc gagacacctg tggaagccca gtacgtgcgg 2280
ctgtacccta caagctgtca caccgcctgc actctgagat tcgaactgct gggatgcgag 2340
ctgaacggc 2349
<210> 123
<211> 227
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 123
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
130 135 140
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 124
<211> 681
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 124
gataagaccc acacctgtcc tccatgtcct gctccagaac tgctcggcgg accctccgtt 60
ttcctgtttc cacctaagcc taaggacacc ctgatgatca gcagaacccc tgaagtgacc 120
tgtgtggtgg tggccgtgtc tcacgaagat cccgaagtga agttcaattg gtacgtggac 180
ggcgtggaag tgcacaacgc caagaccaag cctagagagg aacagtacaa cagcacctac 240
agagtggtgt ccgtgctgac cgtgctgcac caggattggc tgaacggcaa agagtacaag 300
tgcaaggtgt ccaacaaggc cctggccgct cctatcgaga aaaccatctc taaggccaag 360
ggccagcctc gggaacctca agtctgtaca ctgcctccta gccgggacga gctgaccaaa 420
aatcaggtgt ccctgagctg cgccgtgaag ggcttttacc cttccgatat cgccgtggaa 480
tgggagagca atggccagcc tgagaacaac tacaagacca cacctcctgt gctggacagc 540
gacggctcat tctttctggt gtccaagctg acagtggaca agagcagatg gcagcagggc 600
aacgtgttca gctgttctgt gatgcacgag gccctgcaca accactacac ccagaagtct 660
ctgtctctga gccccggcaa a 681
<210> 125
<211> 605
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 125
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
50 55 60
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
65 70 75 80
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
85 90 95
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
100 105 110
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
115 120 125
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
130 135 140
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
145 150 155 160
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
165 170 175
Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser
180 185 190
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
195 200 205
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
210 215 220
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
225 230 235 240
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
245 250 255
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
260 265 270
Pro Gly Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys
275 280 285
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
290 295 300
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
305 310 315 320
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
325 330 335
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
340 345 350
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
355 360 365
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
370 375 380
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
385 390 395 400
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
405 410 415
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
420 425 430
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
435 440 445
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
450 455 460
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
465 470 475 480
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
485 490 495
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
500 505 510
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
515 520 525
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
530 535 540
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
545 550 555 560
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
565 570 575
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
580 585 590
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
595 600 605
<210> 126
<211> 1815
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 126
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaggg cagcgataag acccacacct gtcctccatg tcctgctcca 180
gaactgctcg gcggaccctc cgttttcctg tttccaccta agcctaagga caccctgatg 240
atcagcagaa cccctgaagt gacctgtgtg gtggtggccg tgtctcacga agatcccgaa 300
gtgaagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga 360
gaggaacagt acaacagcac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat 420
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca aggccctggc cgctcctatc 480
gagaaaacca tctctaaggc caagggccag cctcgggaac ctcaggttta caccctgcct 540
ccatgccggg aagagatgac caagaatcag gtgtccctgt ggtgcctggt caagggcttc 600
tacccttccg atatcgccgt ggaatgggag agcaatggcc agcctgagaa caactacaag 660
accacacctc ctgtgctgga cagcgacggc tcattcttcc tgtacagcaa gctgacagtg 720
gacaagagca gatggcagca gggcaacgtg ttcagctgtt ctgtgatgca cgaggccctg 780
cacaaccact acacccagaa gtctctgtct ctgagccctg gcaaaggcgg aagcggtgga 840
agcggaggat ctggcggatc ttgtgtggaa cccctcggca tggaaaacgg caatatcgcc 900
aatagccaga tcgccgccag cagcgtcaga gtgacatttc tgggactgca acactgggtg 960
ccagagctgg ccagactgaa tagagccggc atggttaacg cctggacacc cagcagcaac 1020
gacgacaacc cctggattca agtgaacctg ctgcggcgta tgtgggtcac aggtgttgtt 1080
acacagggcg caagcagact ggccagccac gagtatctga aggcctttaa ggtggcctac 1140
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaagagttc 1200
gtcggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacacc tgtggaagcc 1260
cagtacgtgc ggctgtaccc tacaagctgt cacaccgcct gcacactgag attcgagctg 1320
ctgggctgcg agctgaatgg ctgtgctaat cctctgggcc tgaagaacaa tagcatcccc 1380
gacaagcaga tcaccgcctc cagcagctat aagacatggg gcctgcacct gtttagctgg 1440
aaccctagct acgccagact ggacaagcag ggaaacttca atgcctgggt ggccggcagc 1500
tacggcaatg atcaatggct gcaagtggac ctgggcagca gcaaagaagt gaccggcatc 1560
attacccagg gcgctagaaa tttcggcagc gtgcagttcg tggccagcta caaagtggcc 1620
tactccaacg acagcgccaa ctggaccgag tatcaggacc ctagaaccgg cagctccaag 1680
atcttccccg gcaattggga caaccacagc cacaagaaga atctgttcga aacccctatc 1740
ctggccagat atgtgcgcat tctgcccgtg gcctggcaca acagaattgc cctgagactg 1800
gaactgctgg gatgc 1815
<210> 127
<211> 227
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 127
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
1 5 10 15
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
20 25 30
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
35 40 45
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
50 55 60
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
65 70 75 80
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
85 90 95
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
100 105 110
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
115 120 125
Tyr Thr Leu Pro Pro Cys Arg Glu Glu Met Thr Lys Asn Gln Val Ser
130 135 140
Leu Trp Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
145 150 155 160
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
165 170 175
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val
180 185 190
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
195 200 205
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
210 215 220
Pro Gly Lys
225
<210> 128
<211> 681
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 128
gataagaccc acacctgtcc tccatgtcct gctccagaac tgctcggcgg accctccgtt 60
ttcctgtttc cacctaagcc taaggacacc ctgatgatca gcagaacccc tgaagtgacc 120
tgtgtggtgg tggccgtgtc tcacgaagat cccgaagtga agttcaattg gtacgtggac 180
ggcgtggaag tgcacaacgc caagaccaag cctagagagg aacagtacaa cagcacctac 240
agagtggtgt ccgtgctgac cgtgctgcac caggattggc tgaacggcaa agagtacaag 300
tgcaaggtgt ccaacaaggc cctggccgct cctatcgaga aaaccatctc taaggccaag 360
ggccagcctc gggaacctca ggtttacacc ctgcctccat gccgggaaga gatgaccaag 420
aatcaggtgt ccctgtggtg cctggtcaag ggcttctacc cttccgatat cgccgtggaa 480
tgggagagca atggccagcc tgagaacaac tacaagacca cacctcctgt gctggacagc 540
gacggctcat tcttcctgta cagcaagctg acagtggaca agagcagatg gcagcagggc 600
aacgtgttca gctgttctgt gatgcacgag gccctgcaca accactacac ccagaagtct 660
ctgtctctga gccccggcaa a 681
<210> 129
<211> 605
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 129
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly
50 55 60
Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met
65 70 75 80
Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Ala Val Ser His
85 90 95
Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val
100 105 110
His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr
115 120 125
Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly
130 135 140
Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Ala Ala Pro Ile
145 150 155 160
Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val
165 170 175
Cys Thr Leu Pro Pro Ser Arg Asp Glu Leu Thr Lys Asn Gln Val Ser
180 185 190
Leu Ser Cys Ala Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu
195 200 205
Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro
210 215 220
Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Val Ser Lys Leu Thr Val
225 230 235 240
Asp Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met
245 250 255
His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser
260 265 270
Pro Gly Lys Gly Gly Ser Gly Gly Ser Gly Gly Ser Gly Gly Ser Cys
275 280 285
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
290 295 300
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
305 310 315 320
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
325 330 335
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
340 345 350
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
355 360 365
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
370 375 380
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
385 390 395 400
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
405 410 415
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
420 425 430
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly Cys
435 440 445
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
450 455 460
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
465 470 475 480
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
485 490 495
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
500 505 510
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
515 520 525
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
530 535 540
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
545 550 555 560
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
565 570 575
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
580 585 590
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
595 600 605
<210> 130
<211> 1815
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 130
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaggg cagcgataag acccacacct gtcctccatg tcctgctcca 180
gaactgctcg gcggaccctc cgttttcctg tttccaccta agcctaagga caccctgatg 240
atcagcagaa cccctgaagt gacctgtgtg gtggtggccg tgtctcacga agatcccgaa 300
gtgaagttca attggtacgt ggacggcgtg gaagtgcaca acgccaagac caagcctaga 360
gaggaacagt acaacagcac ctacagagtg gtgtccgtgc tgaccgtgct gcaccaggat 420
tggctgaacg gcaaagagta caagtgcaag gtgtccaaca aggccctggc cgctcctatc 480
gagaaaacca tctctaaggc caagggccag cctcgggaac ctcaagtctg tacactgcct 540
cctagccggg acgagctgac caaaaatcag gtgtccctga gctgcgccgt gaagggcttt 600
tacccttccg atatcgccgt ggaatgggag agcaatggcc agcctgagaa caactacaag 660
accacacctc ctgtgctgga cagcgacggc tcattctttc tggtgtccaa gctgacagtg 720
gacaagagca gatggcagca gggcaacgtg ttcagctgtt ctgtgatgca cgaggccctg 780
cacaaccact acacccagaa gtctctgtct ctgagccctg gcaaaggcgg aagcggtgga 840
agcggaggat ctggcggatc ttgtgtggaa cccctcggca tggaaaacgg caatatcgcc 900
aatagccaga tcgccgccag cagcgtcaga gtgacatttc tgggactgca acactgggtg 960
ccagagctgg ccagactgaa tagagccggc atggttaacg cctggacacc cagcagcaac 1020
gacgacaacc cctggattca agtgaacctg ctgcggcgta tgtgggtcac aggtgttgtt 1080
acacagggcg caagcagact ggccagccac gagtatctga aggcctttaa ggtggcctac 1140
agcctgaacg gccacgagtt cgacttcatc cacgacgtga acaagaagca caaagagttc 1200
gtcggcaact ggaacaagaa cgccgtgcac gtgaacctgt tcgagacacc tgtggaagcc 1260
cagtacgtgc ggctgtaccc tacaagctgt cacaccgcct gcacactgag attcgagctg 1320
ctgggctgcg agctgaatgg ctgtgctaat cctctgggcc tgaagaacaa tagcatcccc 1380
gacaagcaga tcaccgcctc cagcagctat aagacatggg gcctgcacct gtttagctgg 1440
aaccctagct acgccagact ggacaagcag ggaaacttca atgcctgggt ggccggcagc 1500
tacggcaatg atcaatggct gcaagtggac ctgggcagca gcaaagaagt gaccggcatc 1560
attacccagg gcgctagaaa tttcggcagc gtgcagttcg tggccagcta caaagtggcc 1620
tactccaacg acagcgccaa ctggaccgag tatcaggacc ctagaaccgg cagctccaag 1680
atcttccccg gcaattggga caaccacagc cacaagaaga atctgttcga aacccctatc 1740
ctggccagat atgtgcgcat tctgcccgtg gcctggcaca acagaattgc cctgagactg 1800
gaactgctgg gatgc 1815
<210> 131
<211> 783
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 131
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Glu Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln Ile
625 630 635 640
Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp Val
645 650 655
Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp Thr
660 665 670
Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu Arg
675 680 685
Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu Ala
690 695 700
Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn Gly
705 710 715 720
His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu Phe
725 730 735
Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu Thr
740 745 750
Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His Thr
755 760 765
Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
770 775 780
<210> 132
<211> 2349
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 132
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatcagtcaa 60
gaagtgcggg gcgaagtctt tcccagctac acctgtacct gtctgaaggg ctatgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttatctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggcct gtgtggaacc cctcggcatg gaaaacggca atatcgccaa tagccagatt 1920
gccgccagca gcgtcagagt gacatttctg ggactgcaac actgggtgcc cgagctggct 1980
agactgaata gagccggcat ggtcaacgcc tggacaccca gcagcaacga cgataatccc 2040
tggattcaag tgaacctgct gcggcgtatg tgggtcacag gtgttgttac acagggcgca 2100
agcagactgg ccagccacga gtatctgaag gcctttaagg tggcctacag cctgaacggc 2160
cacgagttcg acttcatcca cgacgtgaac aagaagcaca aagagtttgt cggcaactgg 2220
aacaagaacg ccgtgcacgt gaacctgttc gagacacctg tggaagccca gtacgtgcgg 2280
ctgtacccta caagctgtca caccgcctgc actctgagat tcgaactgct gggatgcgag 2340
ctgaacggc 2349
<210> 133
<211> 872
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 133
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Cys Ser Gly Pro Leu Gly Ile Glu Gly
705 710 715 720
Gly Ile Ile Ser Asn Gln Gln Ile Thr Ala Ser Ser Thr His Arg Ala
725 730 735
Leu Phe Gly Leu Gln Lys Trp Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys
740 745 750
Lys Gly Leu Ile Asn Ala Trp Thr Ala Ala Glu Asn Asp Arg Trp Pro
755 760 765
Trp Ile Gln Ile Asn Leu Gln Arg Lys Met Arg Val Thr Gly Val Ile
770 775 780
Thr Gln Gly Ala Lys Arg Ile Gly Ser Pro Glu Tyr Ile Lys Ser Tyr
785 790 795 800
Lys Ile Ala Tyr Ser Asn Asp Gly Lys Thr Trp Ala Met Tyr Lys Val
805 810 815
Lys Gly Thr Asn Glu Asp Met Val Phe Arg Gly Asn Ile Asp Asn Asn
820 825 830
Thr Pro Tyr Ala Asn Ser Phe Thr Pro Pro Ile Lys Ala Gln Tyr Val
835 840 845
Arg Leu Tyr Pro Gln Val Cys Arg Arg His Cys Thr Leu Arg Met Glu
850 855 860
Leu Leu Gly Cys Glu Leu Ser Gly
865 870
<210> 134
<211> 2616
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 134
gacatctgcg accccaatcc ttgcgagaat ggcggcattt gtctgcctgg actggccgat 60
ggcagcttct cttgtgaatg ccccgatggc ttcacagacc ccaattgcag ctctgtggtg 120
gaagtggcca gcgacgagga agaacctaca agcgctggcc cctgcacacc caatccatgt 180
cataatggcg gaacctgcga gatcagcgag gcctacagag gcgatacctt catcggctac 240
gtgtgcaagt gccccagagg cttcaatggc atccactgcc agcacaacat caacgagtgc 300
gaggtggaac catgcaagaa cggcggcatc tgtaccgacc tggtggccaa ttactcttgc 360
gagtgccctg gcgagttcat gggcagaaac tgccagtaca aggacgccca caagagcgag 420
gtggcccaca gattcaagga cctgggcgaa gagaacttca aggccctggt gctgatcgcc 480
ttcgctcagt atctccagca gagccctttc gaggaccacg tgaagctggt caacgaagtg 540
accgagttcg ccaagacctg tgtggccgat gagagcgccg agaactgtga caagagcctg 600
cacacactgt tcggcgacaa gctgtgtacc gtggccacac tgagagaaac ctacggcgag 660
atggccgact gctgtgccaa gcaagagccc gagagaaacg agtgcttcct ccagcacaag 720
gatgacaacc ccaacctgcc tagactcgtg cggcctgaag tggatgtgat gtgcaccgcc 780
tttcacgaca acgaggaaac cttcctgaag aagtacctgt acgagatcgc cagacggcac 840
ccctactttt atgcccctga gctgctgttc ttcgccaagc ggtataaggc cgccttcacc 900
gaatgttgcc aggccgctga taaggctgcc tgtctgctgc ctaagctgga cgagctgaga 960
gatgagggca aagccagctc tgccaagcag agactgaaat gcgccagcct ccagaagttc 1020
ggcgagagag cttttaaggc ctgggccgtt gccagactga gccagagatt tcctaaggcc 1080
gagtttgccg aggtgtccaa gctcgtgacc gatctgacaa aggtgcacac cgagtgctgt 1140
cacggcgatc tgctggaatg tgccgacgat agagccgacc tggccaagta tatctgcgag 1200
aaccaggaca gcatcagcag caagctgaaa gagtgctgcg agaagcccct gctggaaaag 1260
tctcactgta tcgccgaagt ggaaaacgac gagatgcccg ccgatctgcc ttctctggct 1320
gccgatttcg tggaaagcaa ggatgtgtgc aagaactacg ccgaggccaa agatgtgttt 1380
ctgggcatgt ttctgtatga gtacgcccgc agacaccccg actattctgt ggttctgctg 1440
ctgcggctgg ccaagacata cgagacaacc ctggaaaaat gctgcgccgc tgccgatcct 1500
cacgagtgtt atgccaaggt gttcgacgag ttcaagccac tggtggaaga accccagaac 1560
ctgatcaagc agaactgcga gctgttcgag cagctgggcg agtacaagtt ccagaatgcc 1620
ctgctcgtgc ggtacaccaa gaaagtgcct caggtgtcca cacctacact ggttgaggtg 1680
tcccggaatc tgggcaaagt gggcagcaag tgttgcaagc accctgaggc caagagaatg 1740
ccttgcgccg aggattacct gagcgtggtg ctgaatcagc tgtgcgtgct gcacgagaaa 1800
acccctgtgt ccgacagagt gaccaagtgc tgtaccgaga gcctcgtgaa cagaaggcct 1860
tgctttagcg ccctggaagt ggacgagaca tacgtgccca aagagttcaa cgccgagaca 1920
ttcaccttcc acgccgatat ctgcaccctg tccgagaaag agcggcagat caagaagcag 1980
acagccctgg tcgagctggt taagcacaag cccaaggcca ccaaagaaca gctgaaggcc 2040
gtgatggacg acttcgccgc ctttgtcgag aagtgctgca aggccgacga caaagagaca 2100
tgcttcgccg aagagggcaa gaaactggtg gcctgttctg gccctctggg catcgaaggc 2160
ggcatcatca gcaatcagca gatcaccgcc agcagcaccc acagagcact gtttggcctg 2220
caaaagtggt atccctacta cgcccggctg aacaagaagg gcctgattaa cgcctggaca 2280
gccgccgaga atgacagatg gccctggatt cagatcaacc tccagcggaa gatgagagtg 2340
accggcgtta tcacacaggg cgcaaagaga atcggctccc ctgagtacat caagagctac 2400
aagatcgcct acagcaacga cggcaagacc tgggccatgt acaaagtgaa gggcaccaac 2460
gaggacatgg tgttccgggg caacatcgac aacaacaccc cttacgccaa cagcttcacc 2520
cctcctatca aggcccagta cgtgcggctg taccctcaag tgtgcagaag gcactgtacc 2580
ctgagaatgg aactgctggg ctgcgaactg tctggc 2616
<210> 135
<211> 869
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 135
Asp Ile Cys Asp Pro Asn Pro Cys Glu Asn Gly Gly Ile Cys Leu Pro
1 5 10 15
Gly Leu Ala Asp Gly Ser Phe Ser Cys Glu Cys Pro Asp Gly Phe Thr
20 25 30
Asp Pro Asn Cys Ser Ser Val Val Glu Val Ala Ser Asp Glu Glu Glu
35 40 45
Pro Thr Ser Ala Gly Pro Cys Thr Pro Asn Pro Cys His Asn Gly Gly
50 55 60
Thr Cys Glu Ile Ser Glu Ala Tyr Arg Gly Asp Thr Phe Ile Gly Tyr
65 70 75 80
Val Cys Lys Cys Pro Arg Gly Phe Asn Gly Ile His Cys Gln His Asn
85 90 95
Ile Asn Glu Cys Glu Val Glu Pro Cys Lys Asn Gly Gly Ile Cys Thr
100 105 110
Asp Leu Val Ala Asn Tyr Ser Cys Glu Cys Pro Gly Glu Phe Met Gly
115 120 125
Arg Asn Cys Gln Tyr Lys Asp Ala His Lys Ser Glu Val Ala His Arg
130 135 140
Phe Lys Asp Leu Gly Glu Glu Asn Phe Lys Ala Leu Val Leu Ile Ala
145 150 155 160
Phe Ala Gln Tyr Leu Gln Gln Ser Pro Phe Glu Asp His Val Lys Leu
165 170 175
Val Asn Glu Val Thr Glu Phe Ala Lys Thr Cys Val Ala Asp Glu Ser
180 185 190
Ala Glu Asn Cys Asp Lys Ser Leu His Thr Leu Phe Gly Asp Lys Leu
195 200 205
Cys Thr Val Ala Thr Leu Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys
210 215 220
Cys Ala Lys Gln Glu Pro Glu Arg Asn Glu Cys Phe Leu Gln His Lys
225 230 235 240
Asp Asp Asn Pro Asn Leu Pro Arg Leu Val Arg Pro Glu Val Asp Val
245 250 255
Met Cys Thr Ala Phe His Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr
260 265 270
Leu Tyr Glu Ile Ala Arg Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu
275 280 285
Leu Phe Phe Ala Lys Arg Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln
290 295 300
Ala Ala Asp Lys Ala Ala Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg
305 310 315 320
Asp Glu Gly Lys Ala Ser Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser
325 330 335
Leu Gln Lys Phe Gly Glu Arg Ala Phe Lys Ala Trp Ala Val Ala Arg
340 345 350
Leu Ser Gln Arg Phe Pro Lys Ala Glu Phe Ala Glu Val Ser Lys Leu
355 360 365
Val Thr Asp Leu Thr Lys Val His Thr Glu Cys Cys His Gly Asp Leu
370 375 380
Leu Glu Cys Ala Asp Asp Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu
385 390 395 400
Asn Gln Asp Ser Ile Ser Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro
405 410 415
Leu Leu Glu Lys Ser His Cys Ile Ala Glu Val Glu Asn Asp Glu Met
420 425 430
Pro Ala Asp Leu Pro Ser Leu Ala Ala Asp Phe Val Glu Ser Lys Asp
435 440 445
Val Cys Lys Asn Tyr Ala Glu Ala Lys Asp Val Phe Leu Gly Met Phe
450 455 460
Leu Tyr Glu Tyr Ala Arg Arg His Pro Asp Tyr Ser Val Val Leu Leu
465 470 475 480
Leu Arg Leu Ala Lys Thr Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala
485 490 495
Ala Ala Asp Pro His Glu Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys
500 505 510
Pro Leu Val Glu Glu Pro Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu
515 520 525
Phe Glu Gln Leu Gly Glu Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg
530 535 540
Tyr Thr Lys Lys Val Pro Gln Val Ser Thr Pro Thr Leu Val Glu Val
545 550 555 560
Ser Arg Asn Leu Gly Lys Val Gly Ser Lys Cys Cys Lys His Pro Glu
565 570 575
Ala Lys Arg Met Pro Cys Ala Glu Asp Tyr Leu Ser Val Val Leu Asn
580 585 590
Gln Leu Cys Val Leu His Glu Lys Thr Pro Val Ser Asp Arg Val Thr
595 600 605
Lys Cys Cys Thr Glu Ser Leu Val Asn Arg Arg Pro Cys Phe Ser Ala
610 615 620
Leu Glu Val Asp Glu Thr Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr
625 630 635 640
Phe Thr Phe His Ala Asp Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln
645 650 655
Ile Lys Lys Gln Thr Ala Leu Val Glu Leu Val Lys His Lys Pro Lys
660 665 670
Ala Thr Lys Glu Gln Leu Lys Ala Val Met Asp Asp Phe Ala Ala Phe
675 680 685
Val Glu Lys Cys Cys Lys Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu
690 695 700
Glu Gly Lys Lys Leu Val Ala Cys Ser Glu Pro Leu Gly Met Lys Ser
705 710 715 720
Gly His Ile Gln Asp Tyr Gln Ile Thr Ala Ser Ser Ile Phe Arg Thr
725 730 735
Leu Asn Met Asp Met Phe Thr Trp Glu Pro Arg Lys Ala Arg Leu Asp
740 745 750
Lys Gln Gly Lys Val Asn Ala Trp Thr Ser Gly His Asn Asp Gln Ser
755 760 765
Gln Trp Leu Gln Val Asp Leu Leu Val Pro Thr Lys Val Thr Gly Ile
770 775 780
Ile Thr Gln Gly Ala Lys Asp Phe Gly His Val Gln Phe Val Gly Ser
785 790 795 800
Tyr Lys Leu Ala Tyr Ser Asn Asp Gly Glu His Trp Thr Val Tyr Gln
805 810 815
Asp Glu Lys Gln Arg Lys Asp Lys Val Phe Gln Gly Asn Phe Asp Asn
820 825 830
Asp Thr His Arg Lys Asn Val Ile Asp Pro Pro Ile Tyr Ala Arg His
835 840 845
Ile Arg Ile Leu Pro Trp Ser Trp Tyr Gly Arg Ile Thr Leu Arg Ser
850 855 860
Glu Leu Leu Gly Cys
865
<210> 136
<211> 2607
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 136
gacatctgcg accccaatcc ttgcgagaat ggcggcattt gtctgcctgg actggccgat 60
ggcagcttct cttgtgaatg ccccgatggc ttcacagacc ccaattgcag ctctgtggtg 120
gaagtggcca gcgacgagga agaacctaca agcgctggcc cctgcacacc caatccatgt 180
cataatggcg gaacctgcga gatcagcgag gcctacagag gcgatacctt catcggctac 240
gtgtgcaagt gccccagagg cttcaatggc atccactgcc agcacaacat caacgagtgc 300
gaggtggaac catgcaagaa cggcggcatc tgtaccgacc tggtggccaa ttactcttgc 360
gagtgccctg gcgagttcat gggcagaaac tgccagtaca aggacgccca caagagcgag 420
gtggcccaca gattcaagga cctgggcgaa gagaacttca aggccctggt gctgatcgcc 480
ttcgctcagt atctccagca gagccctttc gaggaccacg tgaagctggt caacgaagtg 540
accgagttcg ccaagacctg tgtggccgat gagagcgccg agaactgtga caagagcctg 600
cacacactgt tcggcgacaa gctgtgtacc gtggccacac tgagagaaac ctacggcgag 660
atggccgact gctgtgccaa gcaagagccc gagagaaacg agtgcttcct ccagcacaag 720
gatgacaacc ccaacctgcc tagactcgtg cggcctgaag tggatgtgat gtgcaccgcc 780
tttcacgaca acgaggaaac cttcctgaag aagtacctgt acgagatcgc cagacggcac 840
ccctactttt atgcccctga gctgctgttc ttcgccaagc ggtataaggc cgccttcacc 900
gaatgttgcc aggccgctga taaggctgcc tgtctgctgc ctaagctgga cgagctgaga 960
gatgagggca aagccagctc tgccaagcag agactgaaat gcgccagcct ccagaagttc 1020
ggcgagagag cttttaaggc ctgggccgtt gccagactga gccagagatt tcctaaggcc 1080
gagtttgccg aggtgtccaa gctcgtgacc gatctgacaa aggtgcacac cgagtgctgt 1140
cacggcgatc tgctggaatg tgccgacgat agagccgacc tggccaagta tatctgcgag 1200
aaccaggaca gcatcagcag caagctgaaa gagtgctgcg agaagcccct gctggaaaag 1260
tctcactgta tcgccgaagt ggaaaacgac gagatgcccg ccgatctgcc ttctctggct 1320
gccgatttcg tggaaagcaa ggatgtgtgc aagaactacg ccgaggccaa agatgtgttt 1380
ctgggcatgt ttctgtatga gtacgcccgc agacaccccg actattctgt ggttctgctg 1440
ctgcggctgg ccaagacata cgagacaacc ctggaaaaat gctgcgccgc tgccgatcct 1500
cacgagtgtt atgccaaggt gttcgacgag ttcaagccac tggtggaaga accccagaac 1560
ctgatcaagc agaactgcga gctgttcgag cagctgggcg agtacaagtt ccagaatgcc 1620
ctgctcgtgc ggtacaccaa gaaagtgcct caggtgtcca cacctacact ggttgaggtg 1680
tcccggaatc tgggcaaagt gggcagcaag tgttgcaagc accctgaggc caagagaatg 1740
ccttgcgccg aggattacct gagcgtggtg ctgaatcagc tgtgcgtgct gcacgagaaa 1800
acccctgtgt ccgacagagt gaccaagtgc tgtaccgaga gcctcgtgaa cagaaggcct 1860
tgctttagcg ccctggaagt ggacgagaca tacgtgccca aagagttcaa cgccgagaca 1920
ttcaccttcc acgccgatat ctgcaccctg tccgagaaag agcggcagat caagaagcag 1980
acagccctgg tcgagctggt taagcacaag cccaaggcca ccaaagaaca gctgaaggcc 2040
gtgatggacg acttcgccgc ctttgtcgag aagtgctgca aggccgacga caaagagaca 2100
tgcttcgccg aagagggcaa gaaactggtg gcctgttctg agccactggg catgaagtct 2160
ggccacatcc aggattacca gatcaccgcc agcagcatct tcagaaccct gaacatggat 2220
atgttcacct gggagccccg gaaggccaga ctggataagc agggaaaagt gaacgcctgg 2280
accagcggcc acaatgacca gtctcagtgg ctgcaagtgg acctgctggt gcctaccaaa 2340
gtgaccggca tcatcacaca gggcgcaaag gatttcggcc acgtgcagtt tgtgggcagc 2400
tacaagctgg cctacagcaa cgatggcgag cactggacag tgtaccagga cgagaagcag 2460
cggaaggata aggtgttcca gggcaacttc gacaacgaca cccaccggaa gaacgtgatc 2520
gaccctccta tctacgcccg gcacatcaga atcctgcctt ggtcttggta cggccggatc 2580
accctgagaa gcgagctgct tggatgt 2607
<210> 137
<211> 781
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 137
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala Cys
610 615 620
Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln Ile
625 630 635 640
Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser Trp
645 650 655
Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala Trp
660 665 670
Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu Gly
675 680 685
Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn Phe
690 695 700
Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn Asp
705 710 715 720
Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser Lys
725 730 735
Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu Phe
740 745 750
Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala Trp
755 760 765
His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
770 775 780
<210> 138
<211> 2343
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 138
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggcct gtgctaaccc tctgggcctg aagaacaaca gcatccccga taagcagatc 1920
accgccagca gcagctataa gacatggggc ctgcacctgt tcagctggaa cccttcttac 1980
gccagactgg acaagcaggg caacttcaat gcttgggtgg ccggcagcta cggcaatgat 2040
cagtggctgc aagtggacct gggcagcagc aaagaagtga caggcatcat cacccagggc 2100
gcaagaaatt tcggcagcgt gcagttcgtg gccagctaca aggtggccta cagcaacgat 2160
agcgccaact ggaccgagta tcaggaccct agaaccggca gctccaagat cttccccggc 2220
aactgggaca accacagcca caagaagaat ctgttcgaga cacccatcct ggccagatac 2280
gtgcggattc tgcctgtggc ctggcacaac agaatcgccc tgagactgga actgctgggc 2340
tgt 2343
<210> 139
<211> 623
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 139
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Asp Ala
35 40 45
His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu Glu Asn
50 55 60
Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln Gln Ser
65 70 75 80
Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu Phe Ala
85 90 95
Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys Ser Leu
100 105 110
His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu Arg Glu
115 120 125
Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro Glu Arg
130 135 140
Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu Pro Arg
145 150 155 160
Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His Asp Asn
165 170 175
Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg Arg His
180 185 190
Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg Tyr Lys
195 200 205
Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala Cys Leu
210 215 220
Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser Ser Ala
225 230 235 240
Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu Arg Ala
245 250 255
Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro Lys Ala
260 265 270
Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys Val His
275 280 285
Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp Arg Ala
290 295 300
Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser Ser Lys
305 310 315 320
Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His Cys Ile
325 330 335
Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser Leu Ala
340 345 350
Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala Glu Ala
355 360 365
Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg Arg His
370 375 380
Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr Tyr Glu
385 390 395 400
Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu Cys Tyr
405 410 415
Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro Gln Asn
420 425 430
Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu Tyr Lys
435 440 445
Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro Gln Val
450 455 460
Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys Val Gly
465 470 475 480
Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys Ala Glu
485 490 495
Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His Glu Lys
500 505 510
Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser Leu Val
515 520 525
Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr Tyr Val
530 535 540
Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp Ile Cys
545 550 555 560
Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala Leu Val
565 570 575
Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu Lys Ala
580 585 590
Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys Ala Asp
595 600 605
Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val Ala
610 615 620
<210> 140
<211> 1869
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 140
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaagga tgcccacaag agcgaggtgg cccacagatt caaggatctg 180
ggcgaagaga acttcaaggc cctggtgctg atcgccttcg ctcagtatct ccagcagagc 240
cctttcgagg accacgtgaa gctggtcaac gaagtgaccg agttcgccaa gacctgtgtg 300
gccgatgaga gcgccgagaa ctgtgataag agcctgcaca ccctgttcgg cgacaagctg 360
tgtacagtgg ccacactgag agaaacctac ggcgagatgg ccgactgctg tgccaagcaa 420
gagcccgaga gaaacgagtg cttcctccag cacaaggacg acaaccccaa cctgcctaga 480
ctcgtgcgac ccgaagtgga tgtgatgtgc accgcctttc acgacaacga ggaaaccttc 540
ctgaagaagt acctgtacga gatcgccaga cggcacccct acttttatgc ccctgagctg 600
ctgttcttcg ccaagcggta taaggccgcc ttcaccgaat gttgccaggc cgctgataag 660
gctgcctgtc tgctgcctaa gctggacgag ctgagagatg agggcaaagc cagctctgcc 720
aagcagagac tgaaatgcgc cagcctccag aagttcggcg agagagcttt taaggcctgg 780
gccgttgcca gactgagcca gagatttcct aaggccgagt ttgccgaggt gtccaagctc 840
gtgaccgatc tgacaaaggt gcacaccgag tgctgtcacg gcgatctgct ggaatgtgcc 900
gacgatagag ccgacctggc caagtacatc tgcgagaacc aggacagcat cagcagcaag 960
ctgaaagagt gctgcgagaa gcccctgctg gaaaagtctc actgtatcgc cgaggtggaa 1020
aacgacgaga tgcctgccga tctgcctagc ctggctgccg atttcgtgga aagcaaggac 1080
gtgtgcaaga actacgccga ggccaaggat gtgtttctgg gcatgtttct gtatgagtac 1140
gcccgcagac accccgacta ttctgtggtt ctgctgctgc ggctggccaa aacctacgag 1200
acaaccctgg aaaaatgctg cgccgctgcc gatcctcacg agtgttatgc caaggtgttc 1260
gacgagttca agcctctggt ggaagaaccc cagaacctga tcaagcagaa ctgcgagctg 1320
ttcgagcagc tgggcgagta caagttccag aatgccctgc tcgtgcggta caccaagaaa 1380
gtgcctcagg tgtccacacc tacactggtt gaggtgtccc ggaatctggg caaagtgggc 1440
agcaagtgtt gcaagcaccc tgaggccaag agaatgcctt gcgccgagga ttacctgagc 1500
gtggtgctga atcagctgtg cgtgctgcac gagaaaaccc ctgtgtccga cagagtgacc 1560
aagtgctgta ccgagagcct cgtgaacaga aggccttgct ttagcgccct ggaagtggac 1620
gagacatacg tgcccaaaga gttcaacgcc gagacattca ccttccacgc cgacatctgc 1680
accctgtccg agaaagagcg gcagatcaag aagcagacag ccctggtcga gctggttaag 1740
cacaagccca aggccaccaa agaacagctg aaggccgtga tggacgactt cgccgccttt 1800
gtcgagaagt gctgcaaggc cgacgacaaa gagacatgct tcgccgaaga gggcaagaaa 1860
ctggtggct 1869
<210> 141
<211> 160
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 141
Cys Val Glu Pro Leu Gly Leu Glu Asn Gly Asn Ile Ala Asn Ser Gln
1 5 10 15
Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp
20 25 30
Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp
35 40 45
Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu
50 55 60
Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu
65 70 75 80
Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn
85 90 95
Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu
100 105 110
Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu
115 120 125
Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His
130 135 140
Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
145 150 155 160
<210> 142
<211> 160
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 142
Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn Ile Ala Asn Ser Gln
1 5 10 15
Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu Gly Leu Gln His Trp
20 25 30
Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly Met Val Asn Ala Trp
35 40 45
Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile Gln Val Asn Leu Leu
50 55 60
Arg Arg Met Trp Val Thr Gly Val Val Thr Gln Gly Ala Ser Arg Leu
65 70 75 80
Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val Ala Tyr Ser Leu Asn
85 90 95
Gly His Glu Phe Asp Phe Ile His Asp Val Asn Lys Lys His Lys Glu
100 105 110
Phe Val Gly Asn Trp Asn Lys Asn Ala Val His Val Asn Leu Phe Glu
115 120 125
Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr Pro Thr Ser Cys His
130 135 140
Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly Cys Glu Leu Asn Gly
145 150 155 160
<210> 143
<211> 158
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 143
Cys Ala Asn Pro Leu Gly Leu Lys Asn Asn Ser Ile Pro Asp Lys Gln
1 5 10 15
Ile Thr Ala Ser Ser Ser Tyr Lys Thr Trp Gly Leu His Leu Phe Ser
20 25 30
Trp Asn Pro Ser Tyr Ala Arg Leu Asp Lys Gln Gly Asn Phe Asn Ala
35 40 45
Trp Val Ala Gly Ser Tyr Gly Asn Asp Gln Trp Leu Gln Val Asp Leu
50 55 60
Gly Ser Ser Lys Glu Val Thr Gly Ile Ile Thr Gln Gly Ala Arg Asn
65 70 75 80
Phe Gly Ser Val Gln Phe Val Ala Ser Tyr Lys Val Ala Tyr Ser Asn
85 90 95
Asp Ser Ala Asn Trp Thr Glu Tyr Gln Asp Pro Arg Thr Gly Ser Ser
100 105 110
Lys Ile Phe Pro Gly Asn Trp Asp Asn His Ser His Lys Lys Asn Leu
115 120 125
Phe Glu Thr Pro Ile Leu Ala Arg Tyr Val Arg Ile Leu Pro Val Ala
130 135 140
Trp His Asn Arg Ile Ala Leu Arg Leu Glu Leu Leu Gly Cys
145 150 155
<210> 144
<211> 161
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 144
Cys Ser Gly Pro Leu Gly Ile Glu Gly Gly Ile Ile Ser Asn Gln Gln
1 5 10 15
Ile Thr Ala Ser Ser Thr His Arg Ala Leu Phe Gly Leu Gln Lys Trp
20 25 30
Tyr Pro Tyr Tyr Ala Arg Leu Asn Lys Lys Gly Leu Ile Asn Ala Trp
35 40 45
Thr Ala Ala Glu Asn Asp Arg Trp Pro Trp Ile Gln Ile Asn Leu Gln
50 55 60
Arg Lys Met Arg Val Thr Gly Val Ile Thr Gln Gly Ala Lys Arg Ile
65 70 75 80
Gly Ser Pro Glu Tyr Ile Lys Ser Tyr Lys Ile Ala Tyr Ser Asn Asp
85 90 95
Gly Lys Thr Trp Ala Met Tyr Lys Val Lys Gly Thr Asn Glu Asp Met
100 105 110
Val Phe Arg Gly Asn Ile Asp Asn Asn Thr Pro Tyr Ala Asn Ser Phe
115 120 125
Thr Pro Pro Ile Lys Ala Gln Tyr Val Arg Leu Tyr Pro Gln Val Cys
130 135 140
Arg Arg His Cys Thr Leu Arg Met Glu Leu Leu Gly Cys Glu Leu Ser
145 150 155 160
Gly
<210> 145
<211> 162
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 145
Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln
1 5 10 15
Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr
20 25 30
Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala
35 40 45
Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu
50 55 60
Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp
65 70 75 80
Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn
85 90 95
Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp
100 105 110
Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val
115 120 125
Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser
130 135 140
Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys Thr Glu
145 150 155 160
Glu Glu
<210> 146
<211> 158
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 146
Cys Ser Glu Pro Leu Gly Met Lys Ser Gly His Ile Gln Asp Tyr Gln
1 5 10 15
Ile Thr Ala Ser Ser Ile Phe Arg Thr Leu Asn Met Asp Met Phe Thr
20 25 30
Trp Glu Pro Arg Lys Ala Arg Leu Asp Lys Gln Gly Lys Val Asn Ala
35 40 45
Trp Thr Ser Gly His Asn Asp Gln Ser Gln Trp Leu Gln Val Asp Leu
50 55 60
Leu Val Pro Thr Lys Val Thr Gly Ile Ile Thr Gln Gly Ala Lys Asp
65 70 75 80
Phe Gly His Val Gln Phe Val Gly Ser Tyr Lys Leu Ala Tyr Ser Asn
85 90 95
Asp Gly Glu His Trp Thr Val Tyr Gln Asp Glu Lys Gln Arg Lys Asp
100 105 110
Lys Val Phe Gln Gly Asn Phe Asp Asn Asp Thr His Arg Lys Asn Val
115 120 125
Ile Asp Pro Pro Ile Tyr Ala Arg His Ile Arg Ile Leu Pro Trp Ser
130 135 140
Trp Tyr Gly Arg Ile Thr Leu Arg Ser Glu Leu Leu Gly Cys
145 150 155
<210> 147
<211> 805
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полипептид"
<400> 147
Leu Asp Ile Cys Ser Lys Asn Pro Cys His Asn Gly Gly Leu Cys Glu
1 5 10 15
Glu Ile Ser Gln Glu Val Arg Gly Asp Val Phe Pro Ser Tyr Thr Cys
20 25 30
Thr Cys Leu Lys Gly Tyr Ala Gly Asn His Cys Glu Thr Lys Gly Ser
35 40 45
Asp Ala His Lys Ser Glu Val Ala His Arg Phe Lys Asp Leu Gly Glu
50 55 60
Glu Asn Phe Lys Ala Leu Val Leu Ile Ala Phe Ala Gln Tyr Leu Gln
65 70 75 80
Gln Ser Pro Phe Glu Asp His Val Lys Leu Val Asn Glu Val Thr Glu
85 90 95
Phe Ala Lys Thr Cys Val Ala Asp Glu Ser Ala Glu Asn Cys Asp Lys
100 105 110
Ser Leu His Thr Leu Phe Gly Asp Lys Leu Cys Thr Val Ala Thr Leu
115 120 125
Arg Glu Thr Tyr Gly Glu Met Ala Asp Cys Cys Ala Lys Gln Glu Pro
130 135 140
Glu Arg Asn Glu Cys Phe Leu Gln His Lys Asp Asp Asn Pro Asn Leu
145 150 155 160
Pro Arg Leu Val Arg Pro Glu Val Asp Val Met Cys Thr Ala Phe His
165 170 175
Asp Asn Glu Glu Thr Phe Leu Lys Lys Tyr Leu Tyr Glu Ile Ala Arg
180 185 190
Arg His Pro Tyr Phe Tyr Ala Pro Glu Leu Leu Phe Phe Ala Lys Arg
195 200 205
Tyr Lys Ala Ala Phe Thr Glu Cys Cys Gln Ala Ala Asp Lys Ala Ala
210 215 220
Cys Leu Leu Pro Lys Leu Asp Glu Leu Arg Asp Glu Gly Lys Ala Ser
225 230 235 240
Ser Ala Lys Gln Arg Leu Lys Cys Ala Ser Leu Gln Lys Phe Gly Glu
245 250 255
Arg Ala Phe Lys Ala Trp Ala Val Ala Arg Leu Ser Gln Arg Phe Pro
260 265 270
Lys Ala Glu Phe Ala Glu Val Ser Lys Leu Val Thr Asp Leu Thr Lys
275 280 285
Val His Thr Glu Cys Cys His Gly Asp Leu Leu Glu Cys Ala Asp Asp
290 295 300
Arg Ala Asp Leu Ala Lys Tyr Ile Cys Glu Asn Gln Asp Ser Ile Ser
305 310 315 320
Ser Lys Leu Lys Glu Cys Cys Glu Lys Pro Leu Leu Glu Lys Ser His
325 330 335
Cys Ile Ala Glu Val Glu Asn Asp Glu Met Pro Ala Asp Leu Pro Ser
340 345 350
Leu Ala Ala Asp Phe Val Glu Ser Lys Asp Val Cys Lys Asn Tyr Ala
355 360 365
Glu Ala Lys Asp Val Phe Leu Gly Met Phe Leu Tyr Glu Tyr Ala Arg
370 375 380
Arg His Pro Asp Tyr Ser Val Val Leu Leu Leu Arg Leu Ala Lys Thr
385 390 395 400
Tyr Glu Thr Thr Leu Glu Lys Cys Cys Ala Ala Ala Asp Pro His Glu
405 410 415
Cys Tyr Ala Lys Val Phe Asp Glu Phe Lys Pro Leu Val Glu Glu Pro
420 425 430
Gln Asn Leu Ile Lys Gln Asn Cys Glu Leu Phe Glu Gln Leu Gly Glu
435 440 445
Tyr Lys Phe Gln Asn Ala Leu Leu Val Arg Tyr Thr Lys Lys Val Pro
450 455 460
Gln Val Ser Thr Pro Thr Leu Val Glu Val Ser Arg Asn Leu Gly Lys
465 470 475 480
Val Gly Ser Lys Cys Cys Lys His Pro Glu Ala Lys Arg Met Pro Cys
485 490 495
Ala Glu Asp Tyr Leu Ser Val Val Leu Asn Gln Leu Cys Val Leu His
500 505 510
Glu Lys Thr Pro Val Ser Asp Arg Val Thr Lys Cys Cys Thr Glu Ser
515 520 525
Leu Val Asn Arg Arg Pro Cys Phe Ser Ala Leu Glu Val Asp Glu Thr
530 535 540
Tyr Val Pro Lys Glu Phe Asn Ala Glu Thr Phe Thr Phe His Ala Asp
545 550 555 560
Ile Cys Thr Leu Ser Glu Lys Glu Arg Gln Ile Lys Lys Gln Thr Ala
565 570 575
Leu Val Glu Leu Val Lys His Lys Pro Lys Ala Thr Lys Glu Gln Leu
580 585 590
Lys Ala Val Met Asp Asp Phe Ala Ala Phe Val Glu Lys Cys Cys Lys
595 600 605
Ala Asp Asp Lys Glu Thr Cys Phe Ala Glu Glu Gly Lys Lys Leu Val
610 615 620
Ala Ala Ser Gln Ala Ala Leu Gly Leu Gly Gly Ser Gly Gly Ser Gly
625 630 635 640
Gly Ser Gly Gly Ser Cys Val Glu Pro Leu Gly Met Glu Asn Gly Asn
645 650 655
Ile Ala Asn Ser Gln Ile Ala Ala Ser Ser Val Arg Val Thr Phe Leu
660 665 670
Gly Leu Gln His Trp Val Pro Glu Leu Ala Arg Leu Asn Arg Ala Gly
675 680 685
Met Val Asn Ala Trp Thr Pro Ser Ser Asn Asp Asp Asn Pro Trp Ile
690 695 700
Gln Val Asn Leu Leu Arg Arg Met Trp Val Thr Gly Val Val Thr Gln
705 710 715 720
Gly Ala Ser Arg Leu Ala Ser His Glu Tyr Leu Lys Ala Phe Lys Val
725 730 735
Ala Tyr Ser Leu Asn Gly His Glu Phe Asp Phe Ile His Asp Val Asn
740 745 750
Lys Lys His Lys Glu Phe Val Gly Asn Trp Asn Lys Asn Ala Val His
755 760 765
Val Asn Leu Phe Glu Thr Pro Val Glu Ala Gln Tyr Val Arg Leu Tyr
770 775 780
Pro Thr Ser Cys His Thr Ala Cys Thr Leu Arg Phe Glu Leu Leu Gly
785 790 795 800
Cys Glu Leu Asn Gly
805
<210> 148
<211> 2415
<212> ДНК
<213> Искусственная последовательность
<220>
<221> источник
<223> /примечание="Описание искусственной последовательности:
синтетический полинуклеотид"
<400> 148
ctggacatct gtagcaagaa cccttgccac aacggcggcc tgtgcgaaga gatttctcaa 60
gaagtgcggg gcgacgtttt ccccagctac acctgtacat gtctgaaggg ctacgccggc 120
aaccactgcg agacaaaagg atctgatgcc cacaagagcg aggtggccca cagattcaag 180
gatctgggcg aagagaactt caaggccctg gtgctgatcg ccttcgctca gtatctccag 240
cagagccctt tcgaggacca cgtgaagctg gtcaacgaag tgaccgagtt cgccaagacc 300
tgtgtggccg atgagagcgc cgagaactgt gataagagcc tgcacaccct gttcggcgac 360
aagctgtgta cagtggccac actgagagaa acctacggcg agatggccga ctgctgtgcc 420
aagcaagagc ccgagagaaa cgagtgcttc ctccagcaca aggacgacaa ccccaacctg 480
cctagactcg tgcgacccga agtggatgtg atgtgcaccg cctttcacga caacgaggaa 540
accttcctga agaagtacct gtacgagatc gccagacggc acccctactt ttatgcccct 600
gagctgctgt tcttcgccaa gcggtataag gccgccttca ccgaatgttg ccaggccgct 660
gataaggctg cctgtctgct gcctaagctg gacgagctga gagatgaggg caaagccagc 720
tctgccaagc agagactgaa atgcgccagc ctccagaagt tcggcgagag agcttttaag 780
gcctgggccg ttgccagact gagccagaga tttcctaagg ccgagtttgc cgaggtgtcc 840
aagctcgtga ccgatctgac aaaggtgcac accgagtgct gtcacggcga tctgctggaa 900
tgtgccgacg atagagccga cctggccaag tacatctgcg agaaccagga cagcatcagc 960
agcaagctga aagagtgctg cgagaagccc ctgctggaaa agtctcactg tatcgccgag 1020
gtggaaaacg acgagatgcc tgccgatctg cctagcctgg ctgccgattt cgtggaaagc 1080
aaggacgtgt gcaagaacta cgccgaggcc aaggatgtgt ttctgggcat gtttctgtat 1140
gagtacgccc gcagacaccc cgactattct gtggttctgc tgctgcggct ggccaaaacc 1200
tacgagacaa ccctggaaaa atgctgcgcc gctgccgatc ctcacgagtg ttatgccaag 1260
gtgttcgacg agttcaagcc tctggtggaa gaaccccaga acctgatcaa gcagaactgc 1320
gagctgttcg agcagctggg cgagtacaag ttccagaatg ccctgctcgt gcggtacacc 1380
aagaaagtgc ctcaggtgtc cacacctaca ctggttgagg tgtcccggaa tctgggcaaa 1440
gtgggcagca agtgttgcaa gcaccctgag gccaagagaa tgccttgcgc cgaggattac 1500
ctgagcgtgg tgctgaatca gctgtgcgtg ctgcacgaga aaacccctgt gtccgacaga 1560
gtgaccaagt gctgtaccga gagcctcgtg aacagaaggc cttgctttag cgccctggaa 1620
gtggacgaga catacgtgcc caaagagttc aacgccgaga cattcacctt ccacgccgac 1680
atctgcaccc tgtccgagaa agagcggcag atcaagaagc agacagccct ggtcgagctg 1740
gttaagcaca agcccaaggc caccaaagaa cagctgaagg ccgtgatgga cgacttcgcc 1800
gcctttgtcg agaagtgctg caaggccgac gacaaagaga catgcttcgc cgaagagggc 1860
aagaaactgg tggctgcctc tcaggctgct ctcggacttg gtggaagcgg aggaagtggt 1920
ggatctggcg gatcttgtgt ggaacccctc ggcatggaaa acggcaatat cgccaatagc 1980
cagattgccg ccagcagcgt cagagtgaca tttctgggac tgcaacactg ggtgcccgag 2040
ctggctagac tgaatagagc cggcatggtc aacgcctgga cacccagcag caacgacgat 2100
aatccctgga ttcaagtgaa cctgctgcgg cgtatgtggg tcacaggtgt tgttacacag 2160
ggcgcaagca gactggccag ccacgagtat ctgaaggcct ttaaggtggc ctacagcctg 2220
aacggccacg agttcgactt catccacgac gtgaacaaga agcacaaaga gtttgtcggc 2280
aactggaaca agaacgccgt gcacgtgaac ctgttcgaga cacctgtgga agcccagtac 2340
gtgcggctgt accctacaag ctgtcacacc gcctgcactc tgagattcga actgctggga 2400
tgcgagctga acggc 2415
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| ТЕРАПЕВТИЧЕСКИЕ СЛИТЫЕ БЕЛКИ | 2020 |
|
RU2837545C1 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2019 |
|
RU2728861C1 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2012 |
|
RU2698655C2 |
| СЛИТЫЕ СЕРПИНОВЫЕ ПОЛИПЕПТИДЫ И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2015 |
|
RU2746550C2 |
| ПОЛИПЕПТИДЫ, КОТОРЫЕ СВЯЗЫВАЮТСЯ С КОМПОНЕНТОМ КОМПЛЕМЕНТА C5 ИЛИ СЫВОРОТОЧНЫМ АЛЬБУМИНОМ, И ИХ БЕЛКИ СЛИЯНИЯ | 2018 |
|
RU2794359C2 |
| БИСПЕЦИФИЧНЫЙ СЛИТЫЙ БЕЛОК И ЕГО ПРИМЕНЕНИЕ | 2021 |
|
RU2801528C2 |
| НУКЛЕОТИДНАЯ ПОСЛЕДОВАТЕЛЬНОСТЬ, КОДИРУЮЩАЯ СЛИТЫЙ БЕЛОК, СОСТОЯЩИЙ ИЗ РАСТВОРИМОГО ВНЕКЛЕТОЧНОГО ФРАГМЕНТА ЧЕЛОВЕЧЕСКОГО Dll4 И КОНСТАНТНОЙ ЧАСТИ ТЯЖЕЛОЙ ЦЕПИ ЧЕЛОВЕЧЕСКОГО IgG4 | 2021 |
|
RU2787060C1 |
| АНТИТЕЛА, СОДЕРЖАЩИЕ ПОЛИПЕПТИД, ВСТРОЕННЫЙ В УЧАСТОК КАРКАСНОЙ ОБЛАСТИ 3 | 2019 |
|
RU2796254C2 |
| СЛИТЫЕ БЕЛКИ С АЛЬБУМИН-СВЯЗЫВАЮЩИМИ ДОМЕНАМИ | 2018 |
|
RU2786444C2 |
| МОЛЕКУЛА TGFβR2 С УКОРОЧЕННЫМ ВНЕКЛЕТОЧНЫМ ДОМЕНОМ, СЛИТЫЙ БЕЛОК МОЛЕКУЛЫ TGFβR2 С УКОРОЧЕННЫМ ВНЕКЛЕТОЧНЫМ ДОМЕНОМ, И АНТИТЕЛА ПРОТИВ EGFR, И ПРОТИВООПУХОЛЕВОЕ ПРИМЕНЕНИЕ СЛИТОГО БЕЛКА | 2021 |
|
RU2824199C1 |

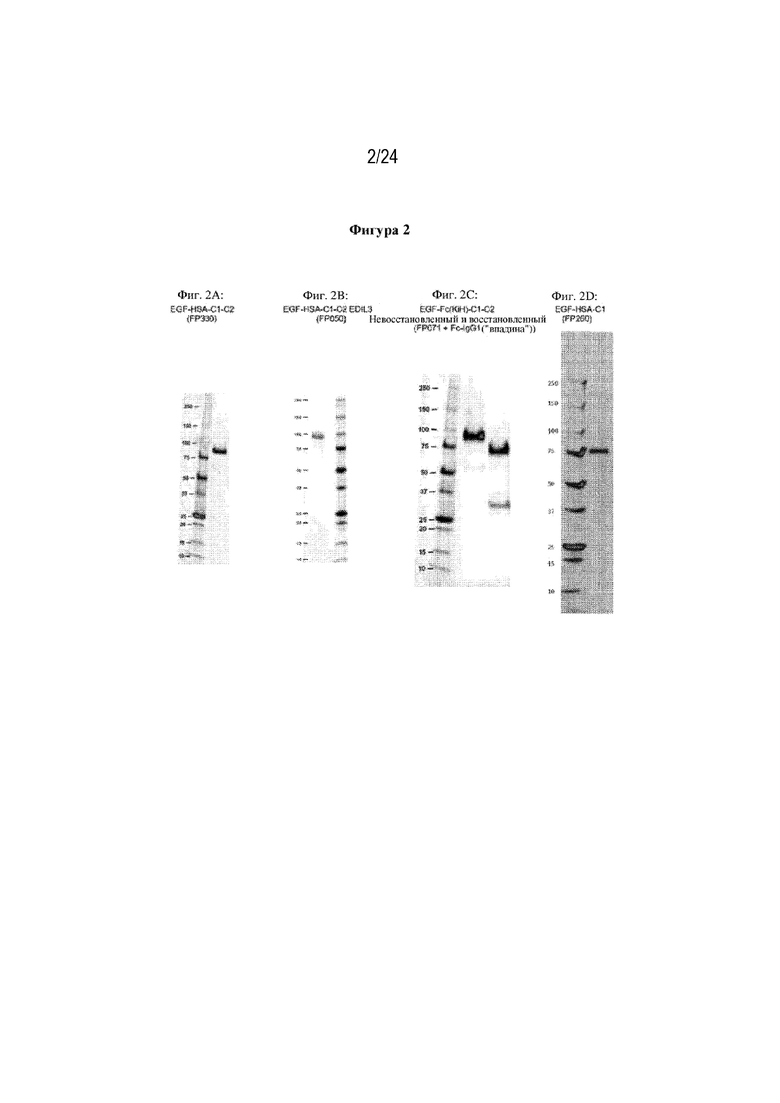

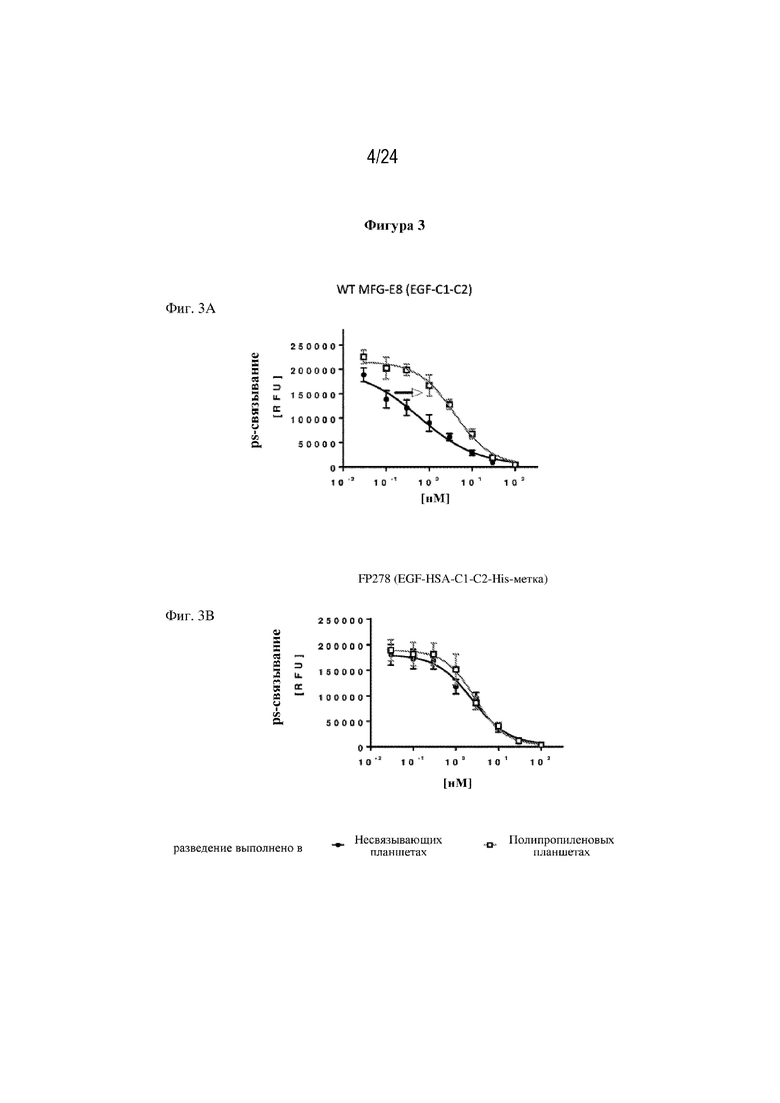
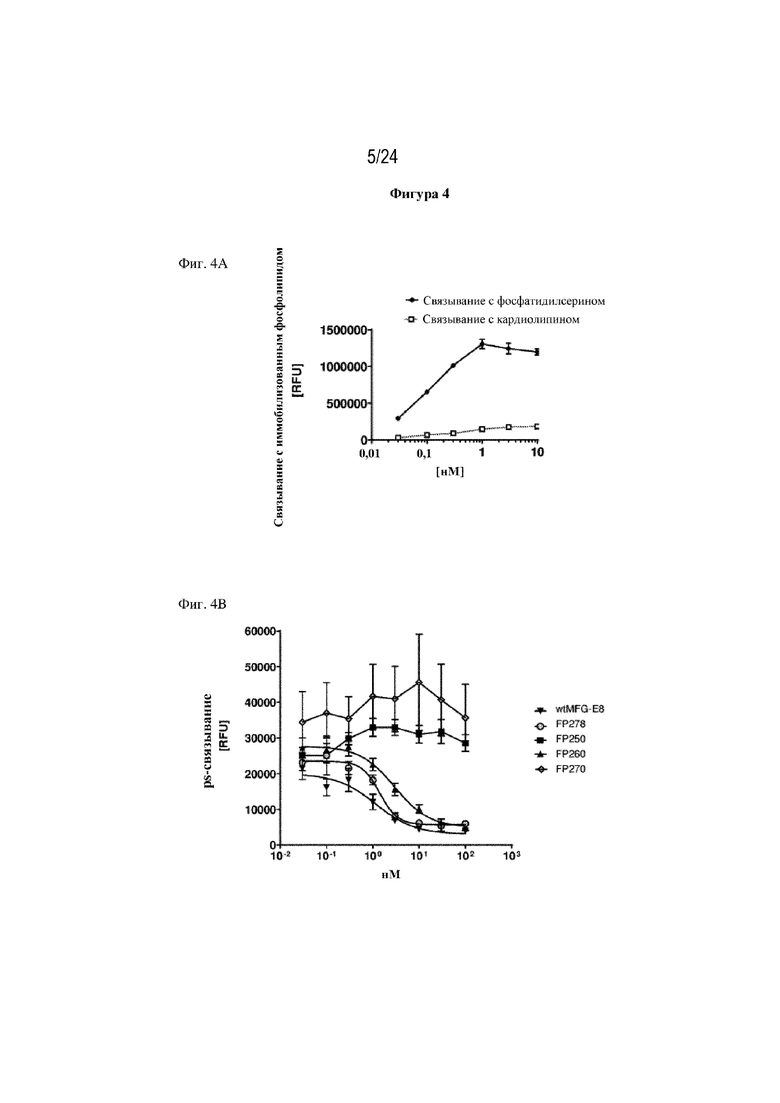
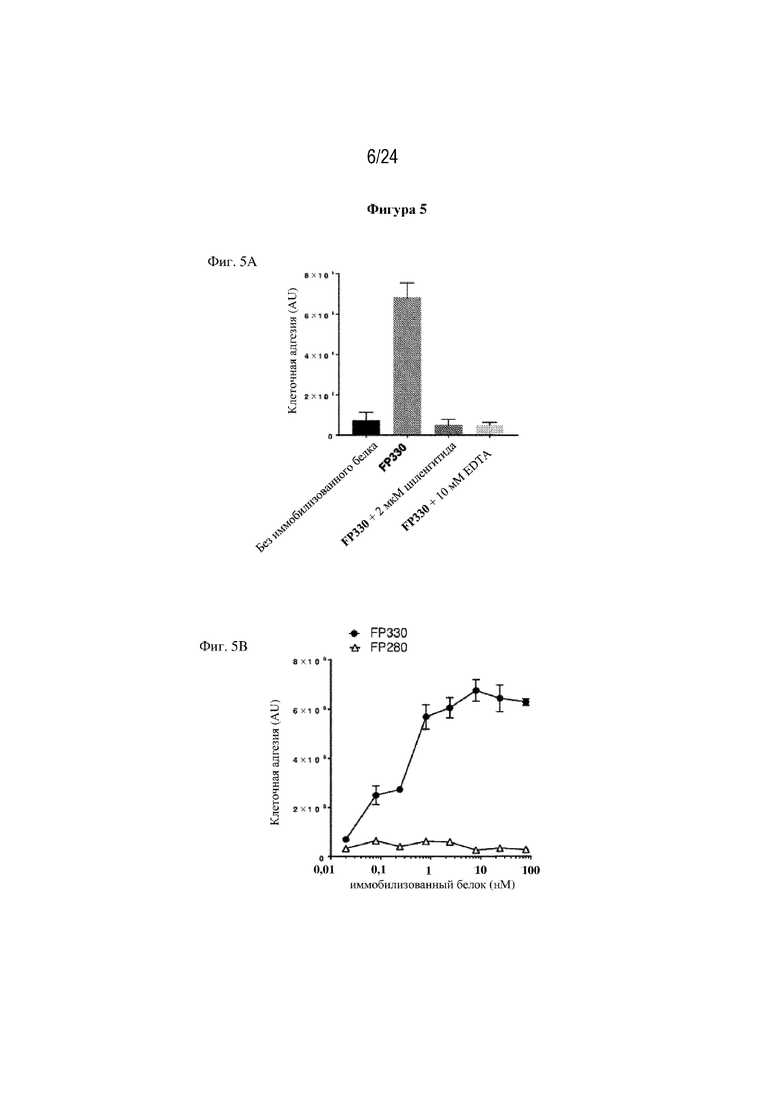
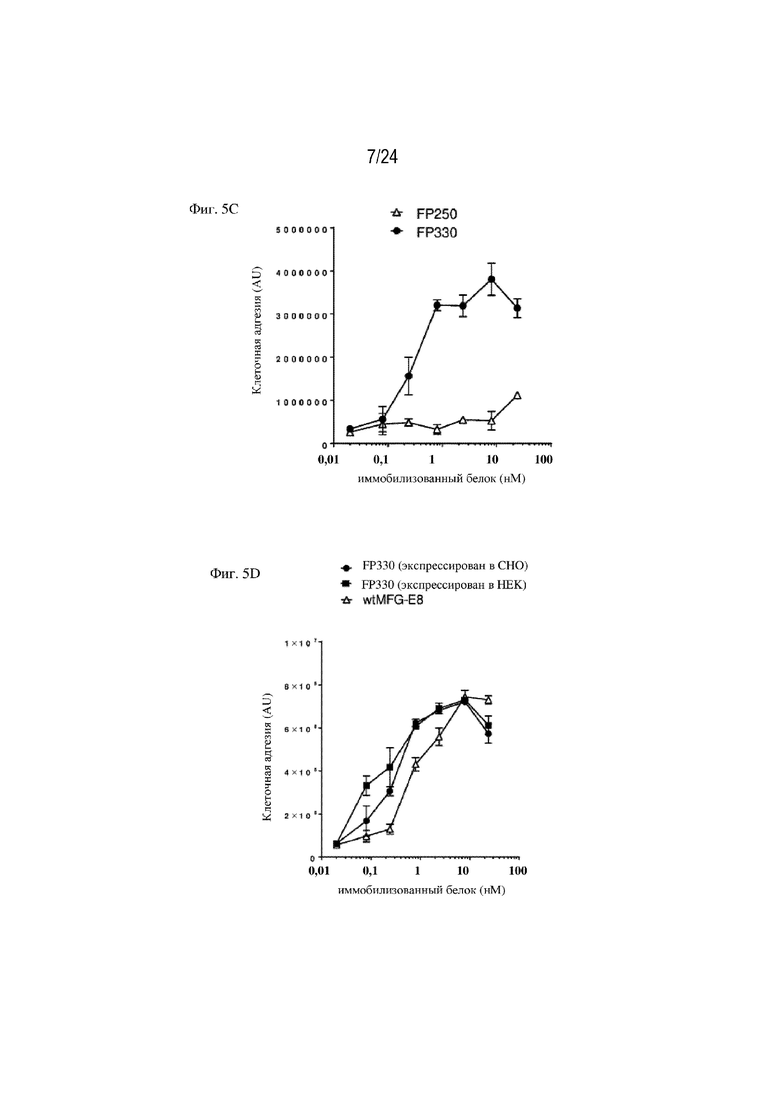
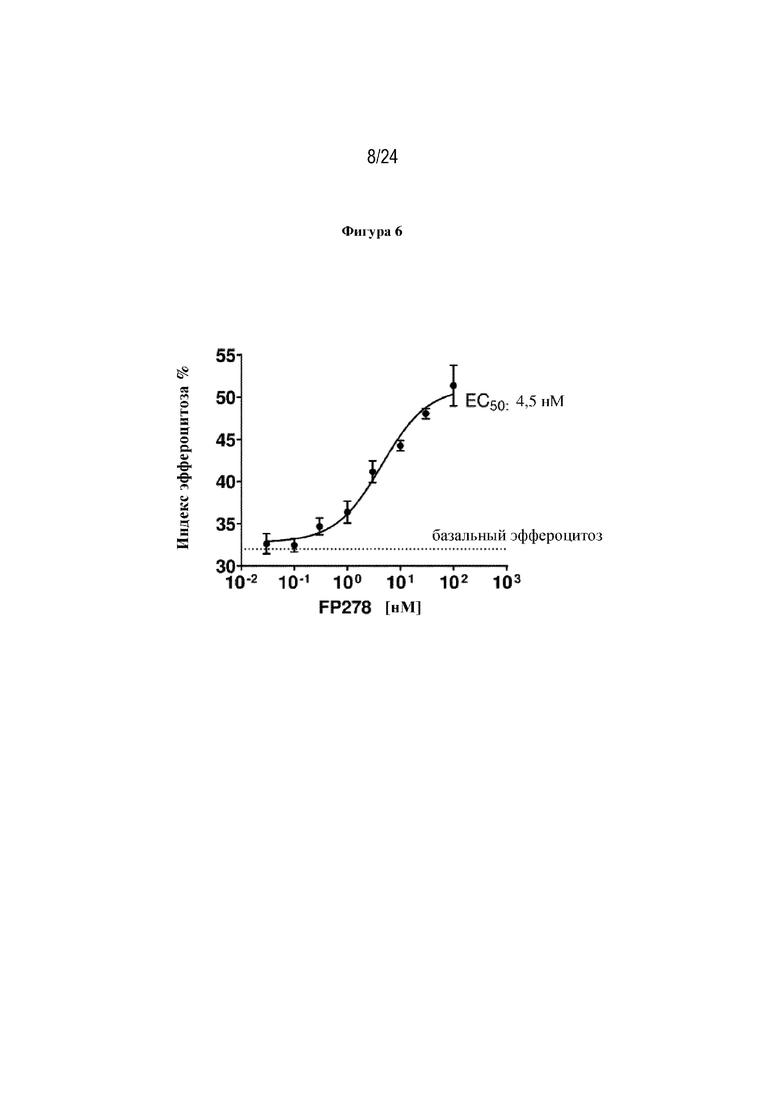
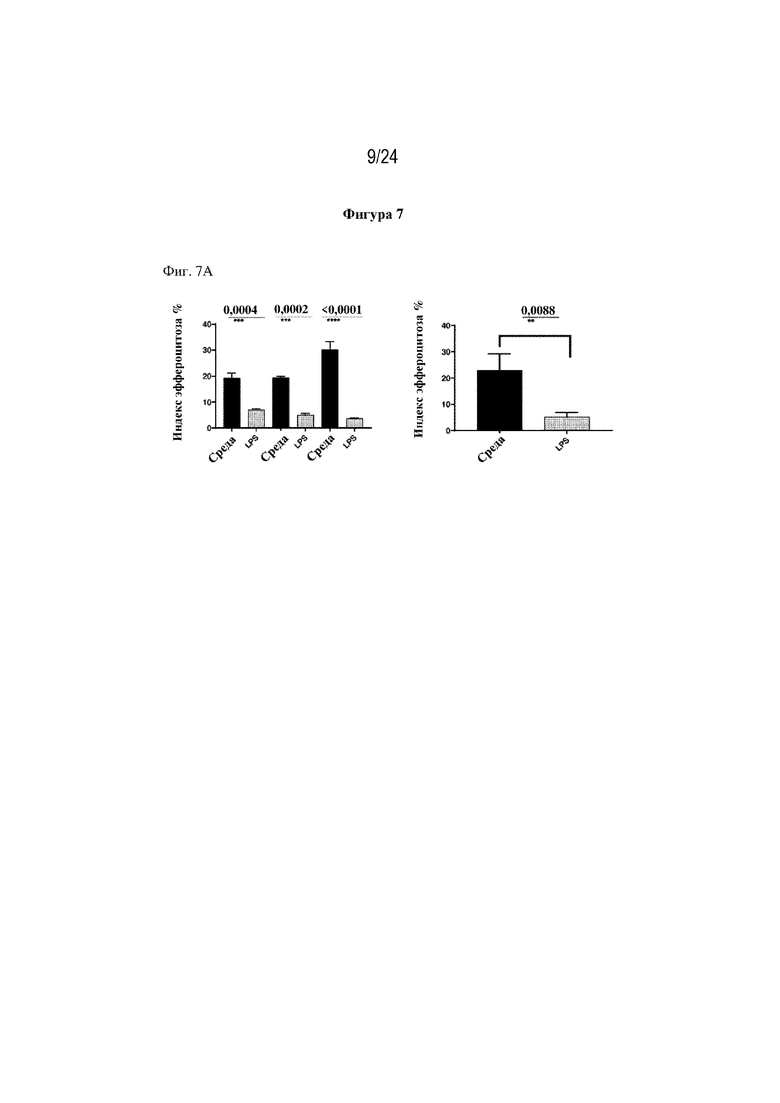
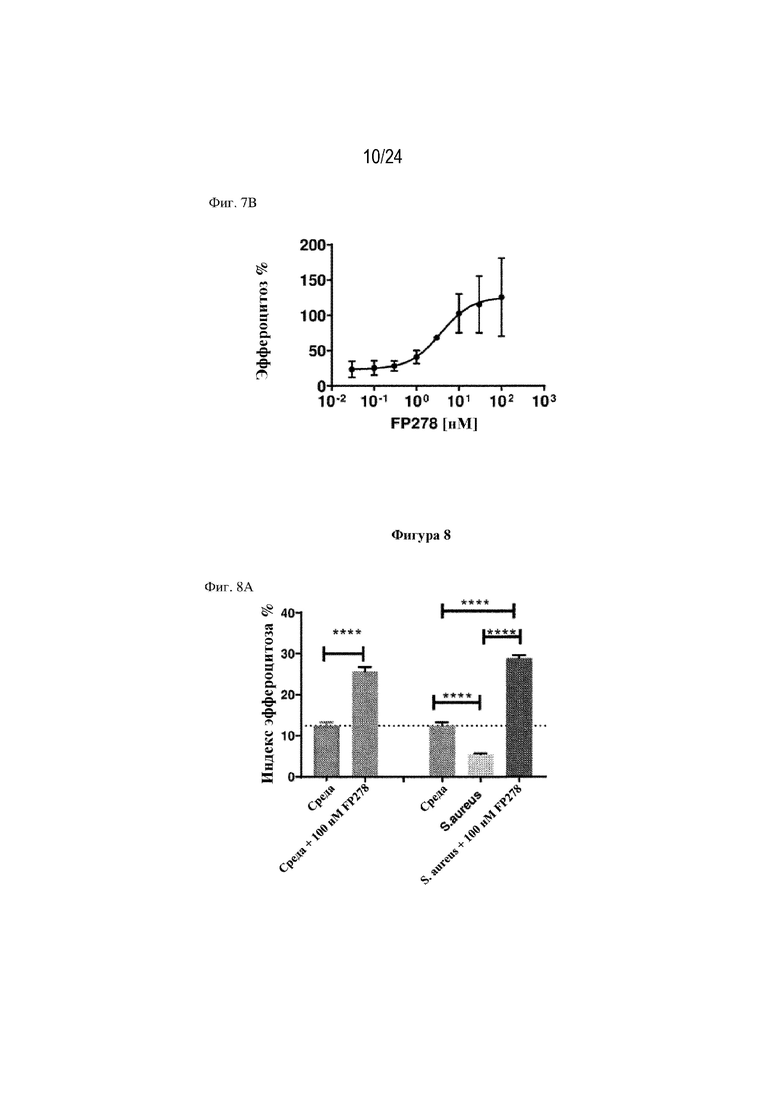
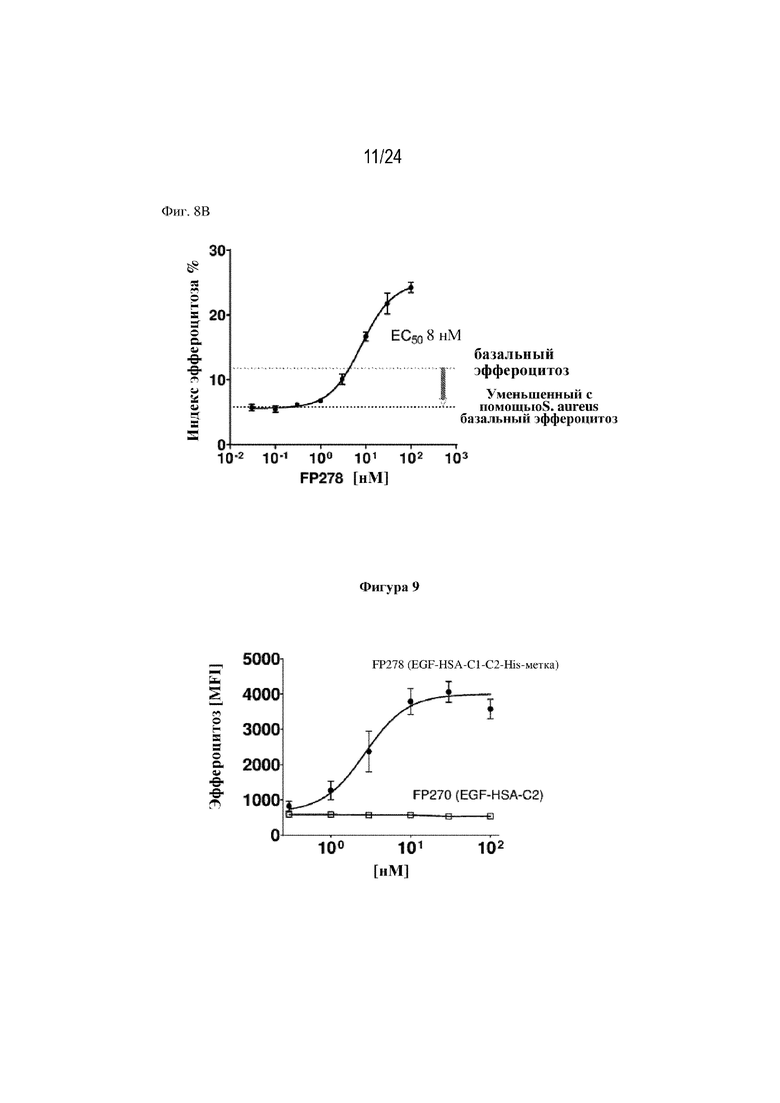
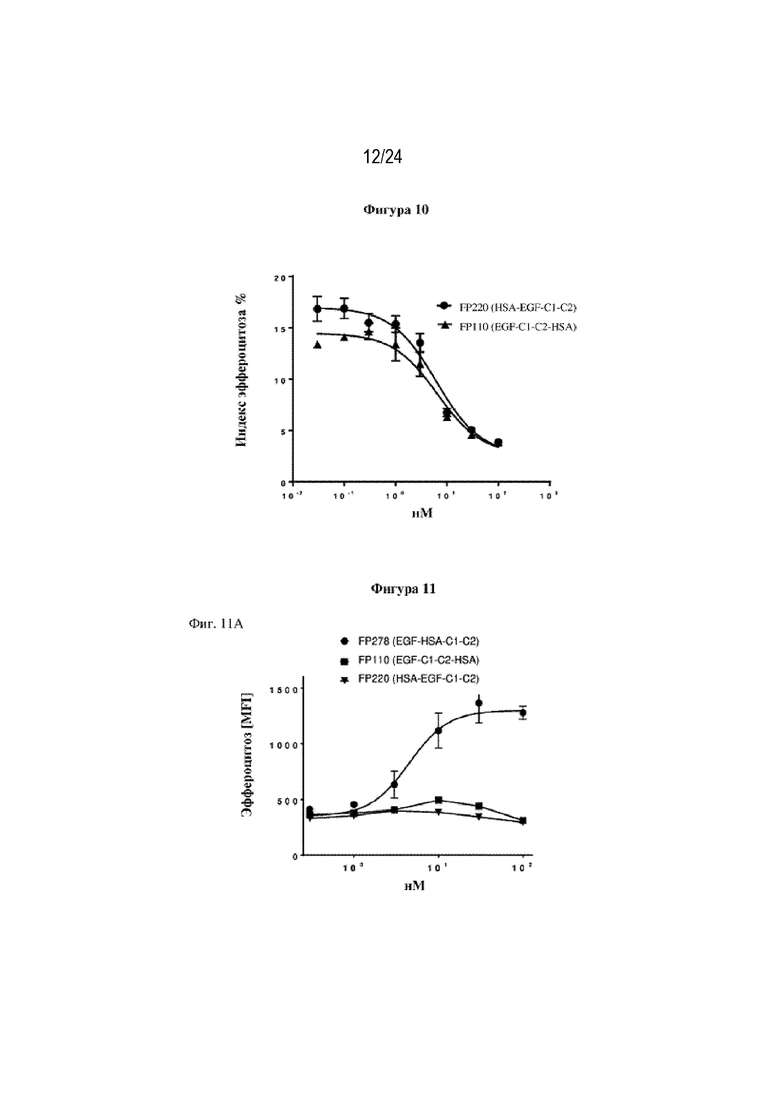
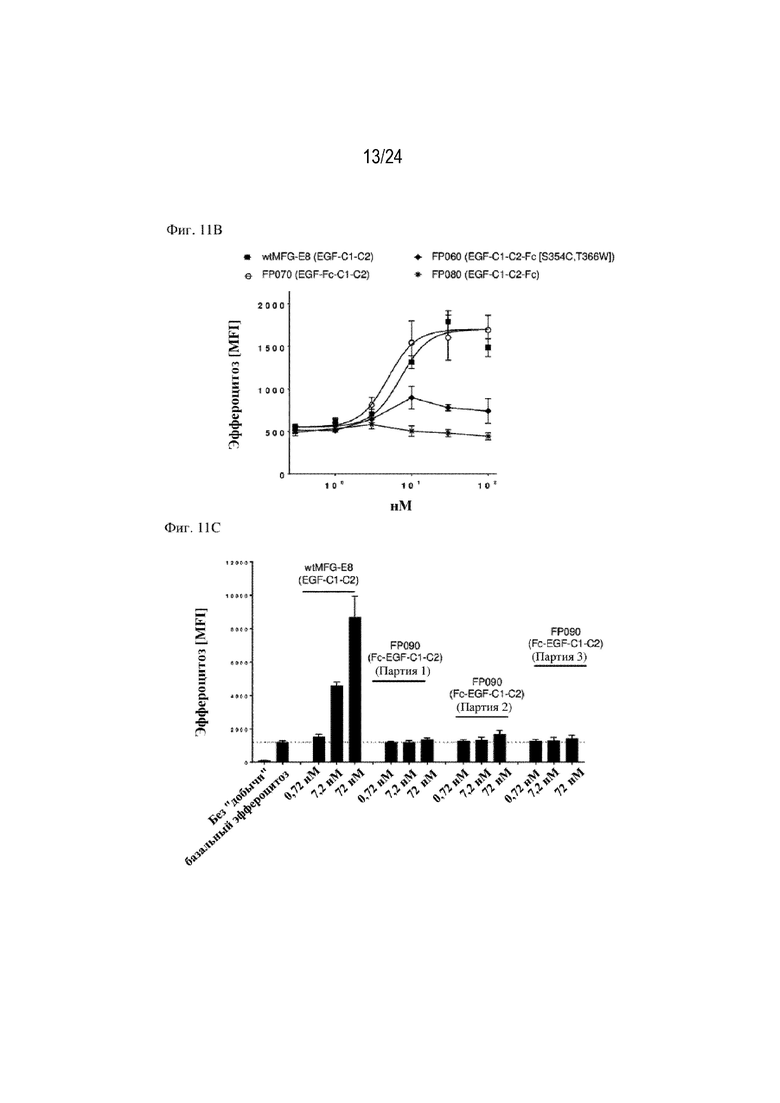
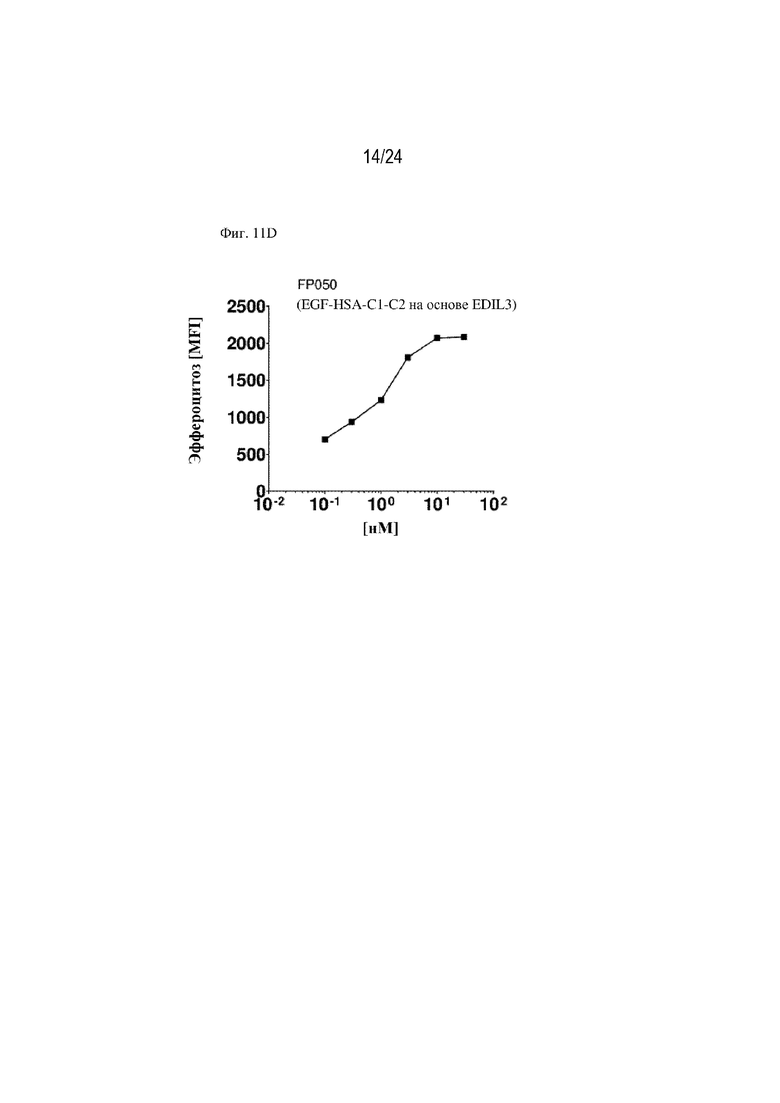
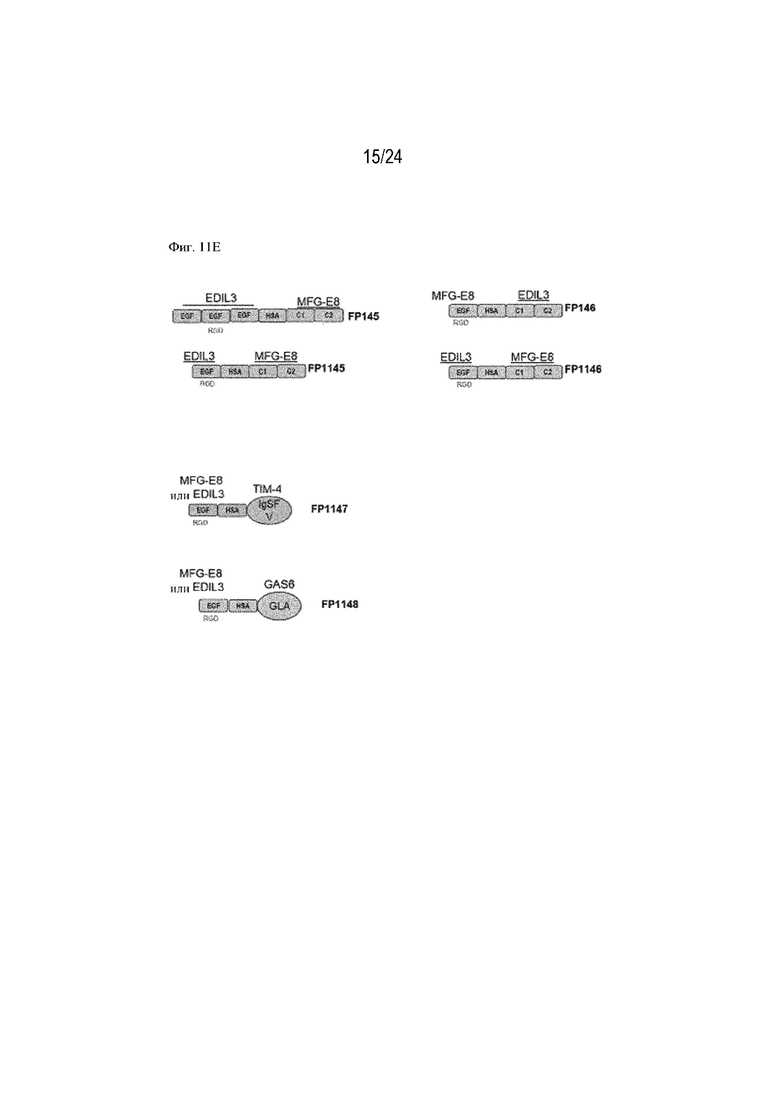
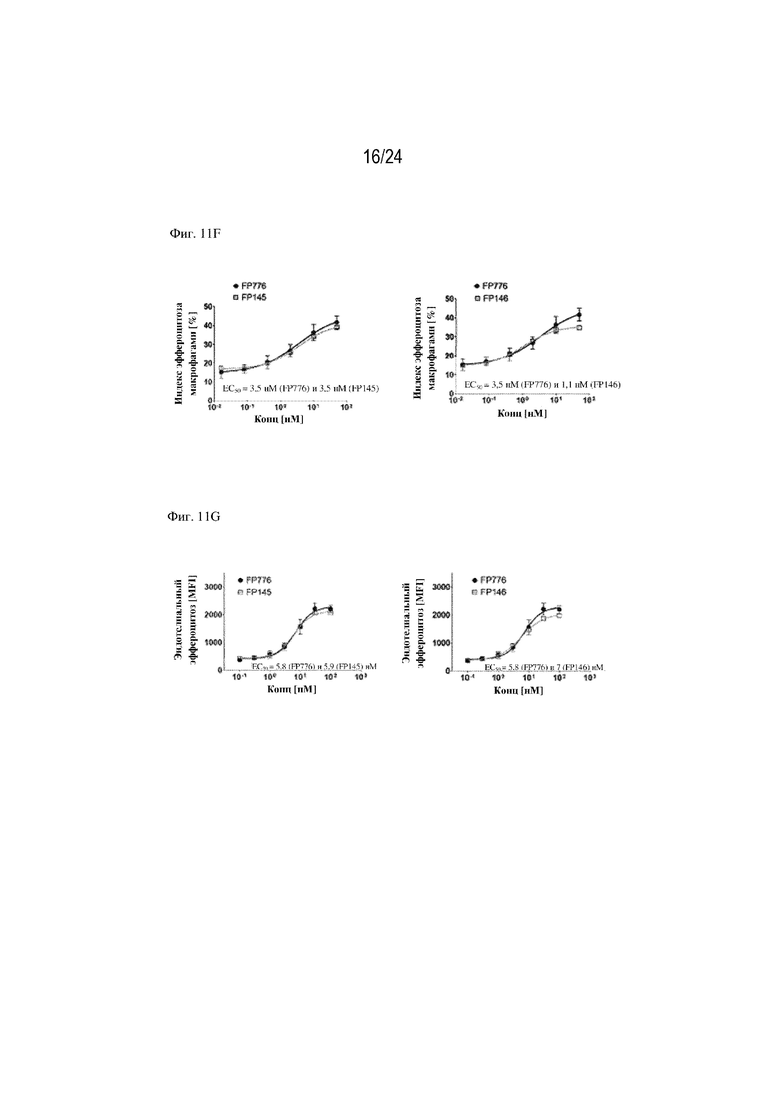
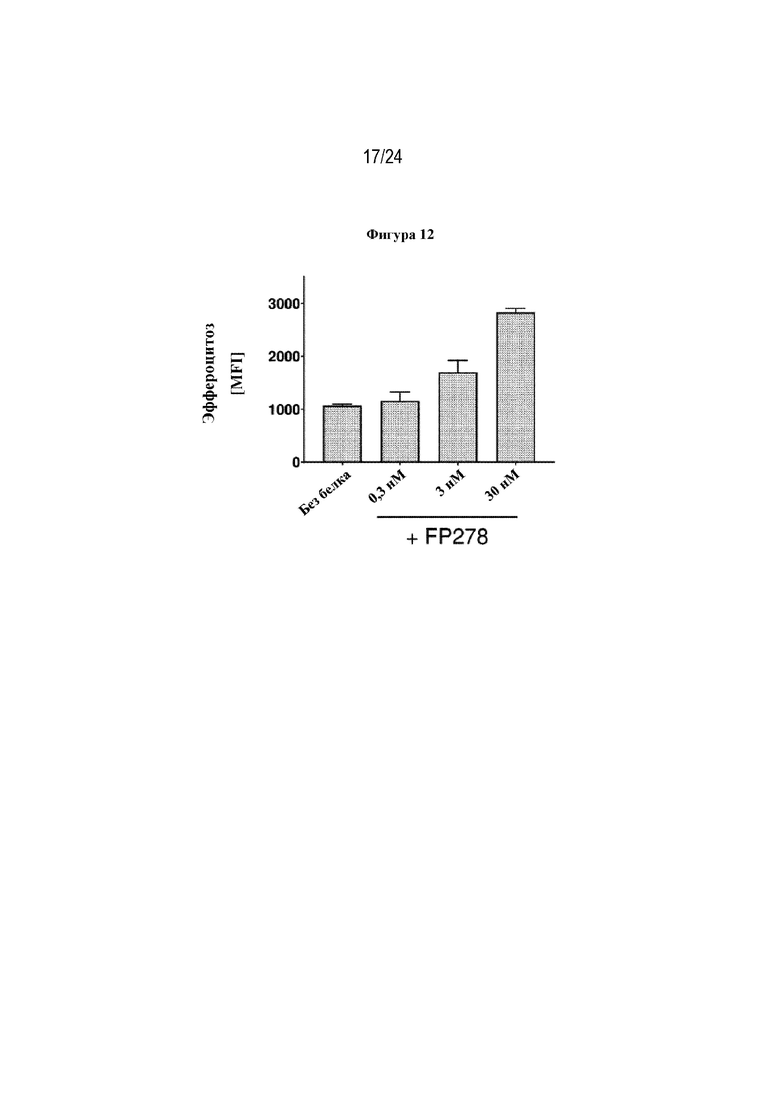
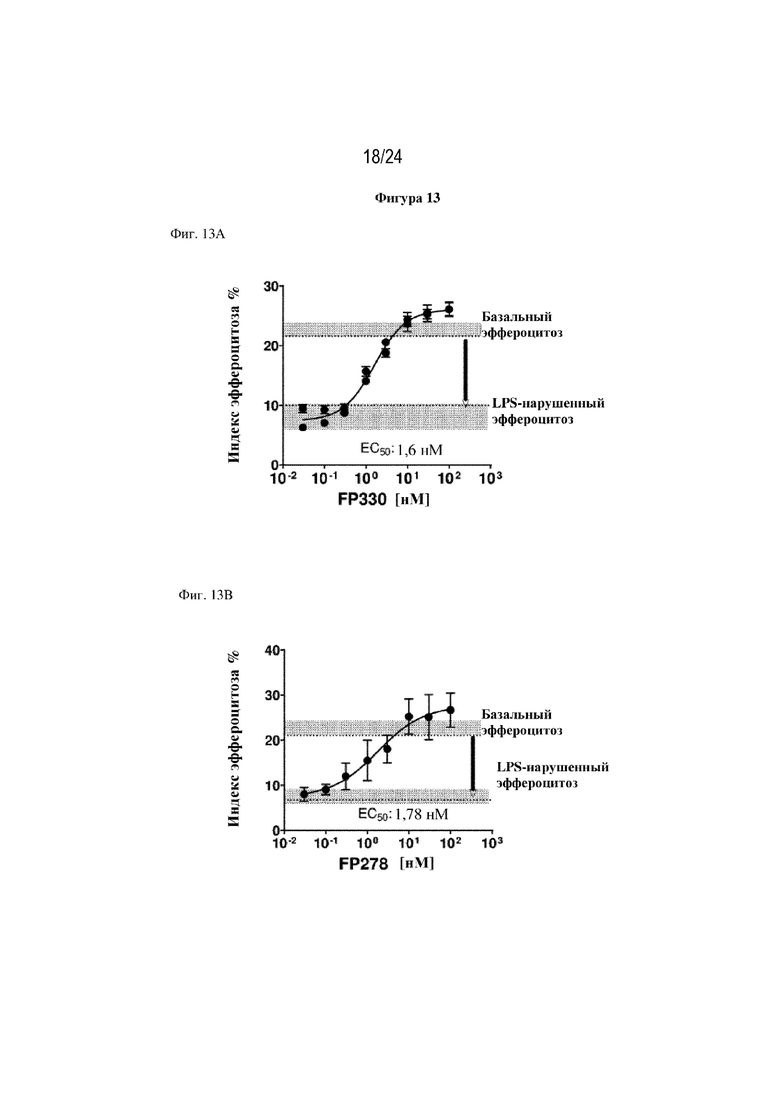
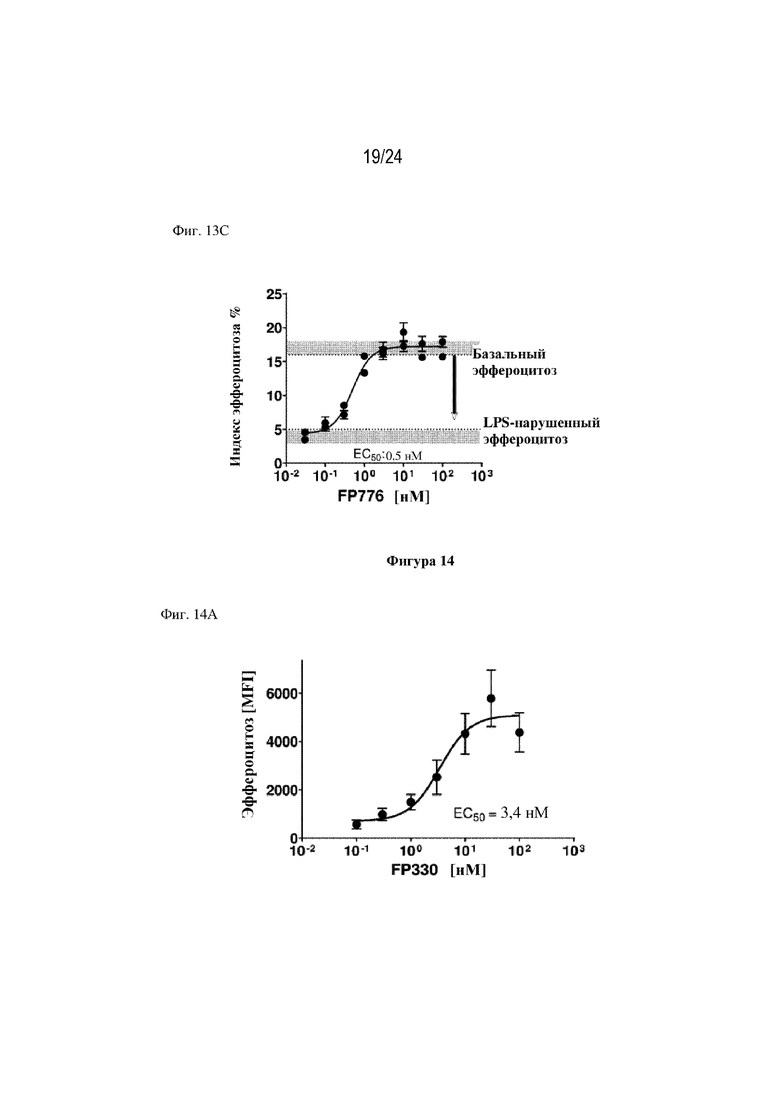
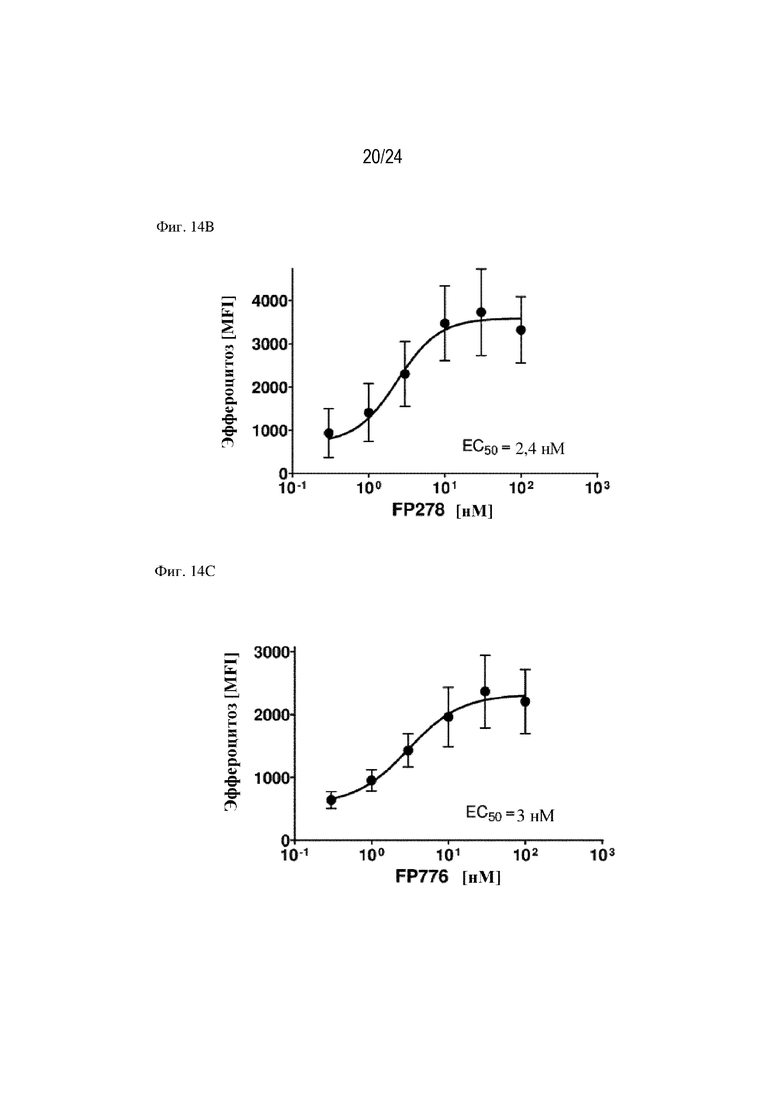
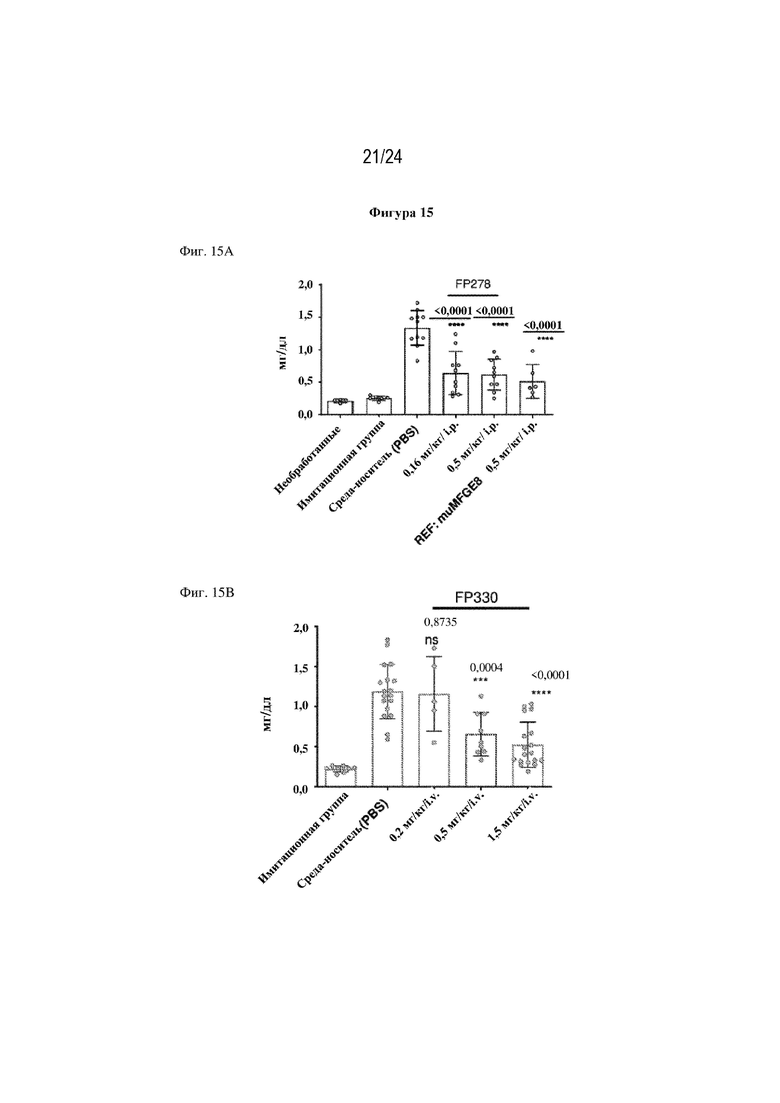
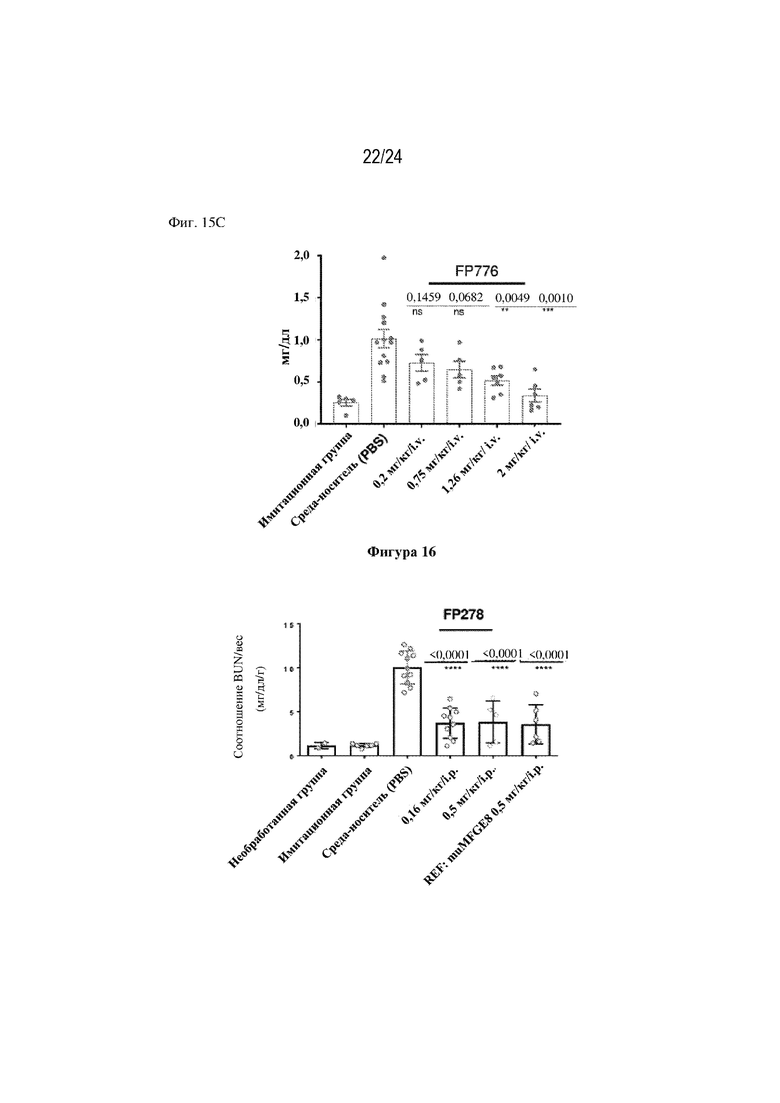
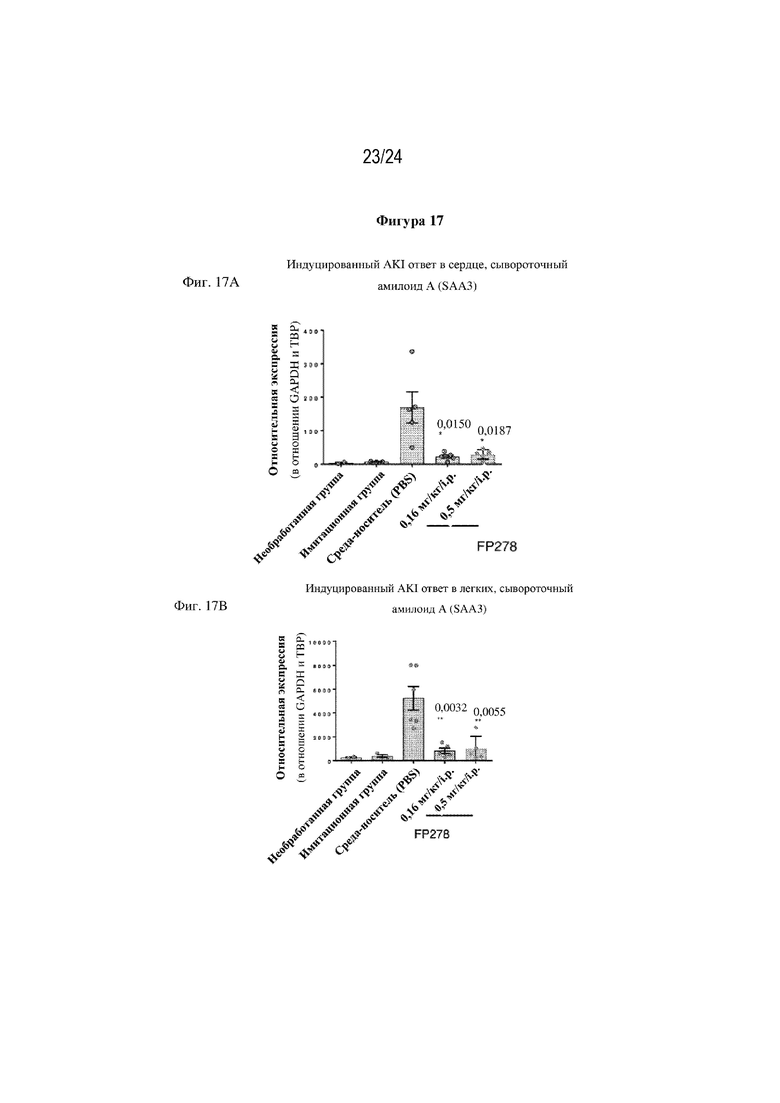
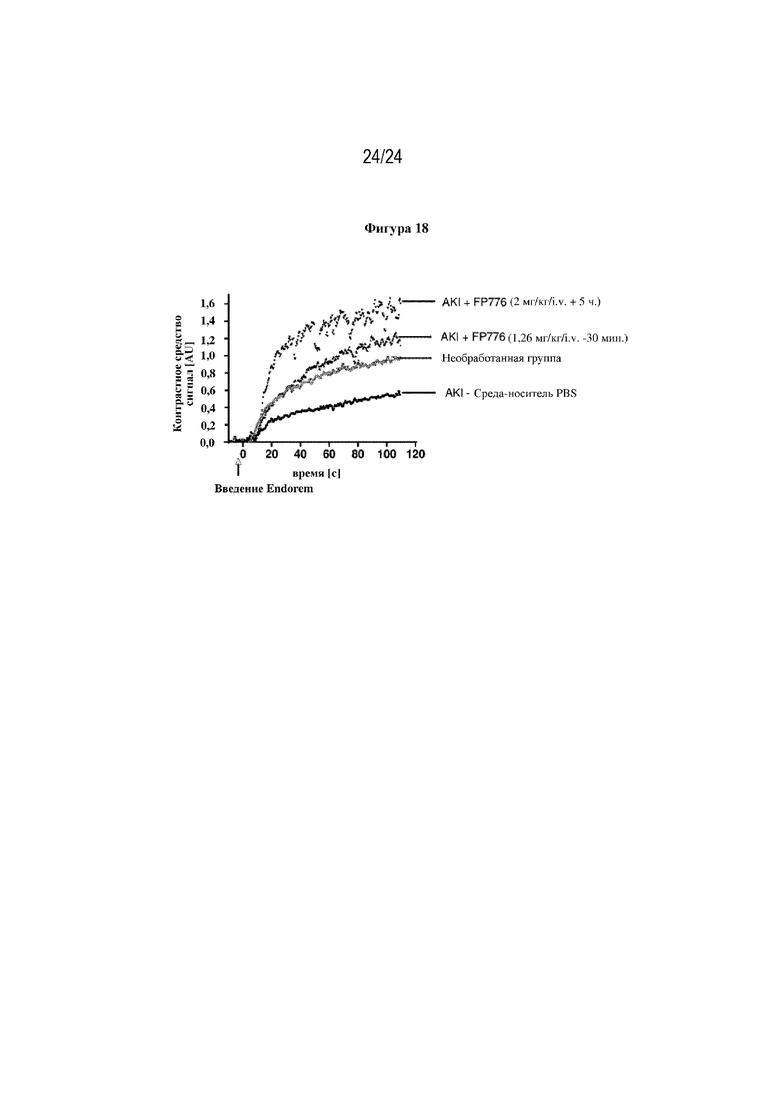
Настоящее изобретение относится к области биотехнологии, конкретно к рекомбинантному получению терапевтических слитых белков, усиливающих эффероцитоз, и может быть использовано в медицине в лечении или предупреждении воспалительного нарушения или воспалительного поражения органов, связанного со сниженным эффероцитозом. Предложены варианты слитых белков на основе интегрин-связывающего домена, фосфатидилсерин (PS)-связывающего домена и солюбилизирующего домена, в которых солюбилизирующий домен вставлен между интегрин-связывающим доменом и PS-связывающим доменом. Изобретение обеспечивает получение слитых белков на основе структуры встречающихся в природе мостиковых белков, а именно MFG-E8, которые функционируют с обеспечением связывания PS-представляющих погибающих клеток, дебриса и микрочастиц с фагоцитами и, таким образом, обеспечивают запуск эффероцитоза, при этом характеризуются сниженной липкостью и улучшенной растворимостью, более длительным временем воздействия в плазме крови и более высоким выходом при экспрессии в клеточных системах экспрессии по сравнению с белком MFG-E8 дикого типа с SEQ ID NO: 1. 8 н.п. ф-лы, 18 ил., 9 табл., 9 пр.
1. Терапевтический слитый белок для усиления эффероцитоза, содержащий интегрин-связывающий домен, фосфатидилсерин (PS)-связывающий домен и солюбилизирующий домен, где солюбилизирующий домен вставлен между интегрин-связывающим доменом и PS-связывающим доменом; и где интегрин-связывающий домен связывается с интегрином,
и где слитый белок имеет аминокислотную последовательность, выбранную из:
- SEQ ID NO: 44 или последовательность по меньшей мере на 90% идентичную ей,
- SEQ ID NO: 46 или последовательность по меньшей мере на 90% идентичную ей,
- SEQ ID NO: 80 или последовательность по меньшей мере на 90% идентичную ей,
- SEQ ID NO: 82 или последовательность по меньшей мере на 90% идентичную ей.
2. Терапевтический слитый белок для усиления эффероцитоза, содержащий интегрин-связывающий домен, фосфатидилсерин (PS)-связывающий домен и солюбилизирующий домен, где солюбилизирующий домен вставлен между интегрин-связывающим доменом и PS-связывающим доменом; и где интегрин-связывающий домен связывается с интегрином,
где слитый белок имеет аминокислотную последовательность SEQ ID NO: 42 или последовательность по меньшей мере на 90% идентичную ей.
3. Терапевтический слитый белок для усиления эффероцитоза, содержащий интегрин-связывающий домен, фосфатидилсерин (PS)-связывающий домен и солюбилизирующий домен, где солюбилизирующий домен вставлен между интегрин-связывающим доменом и PS-связывающим доменом; и где интегрин-связывающий домен связывается с интегрином,
где слитый белок имеет аминокислотную последовательность SEQ ID NO: 48 или последовательность по меньшей мере на 90% идентичную ей.
4. Выделенная нуклеиновая кислота, кодирующая слитый белок по любому из пп. 1-3.
5. Вектор экспрессии, содержащий нуклеиновую кислоту по п. 4.
6. Рекомбинантная клетка-хозяин для продуцирования терапевтического слитого белка по любому из пп. 1-3, содержащая вектор экспрессии по п. 5 и необязательно секреторные сигнальные последовательности.
7. Фармацевтическая композиция для усиления эффероцитоза, содержащая эффективное количество слитого белка по любому из пп. 1-3 и фармацевтически приемлемый носитель.
8. Применение слитого белка по любому из пп. 1-3 в лечении или предупреждении воспалительного нарушения или воспалительного поражения органов, связанного со сниженным эффероцитозом у индивидуума, нуждающегося в этом.
| WO 2013049200 A1, 04.04.2013, KOURTZELIS I | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Прибор для промывания газов | 1922 |
|
SU20A1 |
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Приспособление с иглой для прочистки кухонь типа "Примус" | 1923 |
|
SU40A1 |
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |
| Способ восстановления хромовой кислоты, в частности для получения хромовых квасцов | 1921 |
|
SU7A1 |
| Приспособление для выпечки формового хлеба в механических печах с выдвижным подом без смазки форм жировым веществом | 1921 |
|
SU307A1 |
| Приспособление для выпечки формового хлеба в механических печах с выдвижным подом без смазки форм жировым веществом | 1921 |
|
SU307A1 |

