ПЕРЕКРЕСТНАЯ ССЫЛКА НА РОДСТВЕННЫЕ ЗАЯВКИ
[0001] По настоящей заявке испрашивается приоритет патентной заявки США № 62/593830, поданной 1 декабря 2017 года, и патентной заявки США № 62/732985, поданной 18 сентября 2018 года, содержание которых включено в настоящее описание в качестве ссылки в полном объеме.
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
[0002] Настоящее описание в качестве ссылки дополнительно включает список последовательностей, поданный с ней 28 ноября 2018 года. В соответствии с 37 C.F.R. § 1.52(e)(5), текстовый файл списка последовательностей, названный PC72320A_Seq_Listing_ST25.txt, имеет размер 112891 байт и создан 15 ноября 2018 года. Список последовательностей, поданный в электронном виде, не выходит за рамки настоящего описания и, таким образом, не содержит новые признаки изобретения.
СТОРОНЫ СОВМЕСТНОГО ИССЛЕДОВАТЕЛЬСКОГО СОГЛАШЕНИЯ
[0003] Описываемое в настоящей заявке изобретение создано указанными ниже сторонами совместного исследовательского соглашения или от их имени. Совместное исследовательское соглашение действовало на день или до дня создания описываемого в заявке изобретения, и описываемое в заявке изобретение было создано в результате действий, предпринятых в объеме совместного исследовательского соглашения. Сторонами совместного исследовательского соглашения являются Попечительский совет Калифорнийского университета от имени его кампуса в г. Сан-Диего и PFIZER INC.
ОБЛАСТЬ ТЕХНИКИ
[0004] Настоящее изобретение относится к антителам и их антигенсвязывающим фрагментам, специфически связывающимся с C-X-C-хемокиновым рецептором типа 5 (CXCR5), и их композициям, способам и применению. Антитела являются фукозилированными и/или афукозилированными и могут демонстрировать измененную эффекторную функцию.
УРОВЕНЬ ТЕХНИКИ
[0005] C-X-C-хемокиновый рецептор типа 5 (CXCR5), также известный как CD185 или рецептор лимфомы Беркитта 1 (BLR1), является природным, сопряженным с G-белком рецептором, экспрессируемым B-клетками, истинными фолликулярными T-хелперными клетками (Tfh) и циркулирующими Tfh-подобными клетками (взаимозаменяемо обозначаемыми в настоящем описании как "cTfh" и "Tfh-подобные" клетки). CXCR5 играет важную роль в иммунном ответе и является потенциальной мишенью для лечения аутоиммунных заболеваний, таких как системная красная волчанка (SLE).
[0006] Герминативные центры (GC), критический компонент гуморального иммунного ответа, являются местами, в которых активированные антигеном B-клетки пролиферируют, дифференцируют и подвергаются соматической гипермутации и переключению классов иммуноглобулинов (Ig) антител, которые они продуцируют. Истинные Tfh-клетки обеспечивают инструктивные сигналы для облегчения этого процесса, кульминацией которого является продукция аффинно зрелых антител. Гуморальная "память" реакции GC поддерживается долгоживущими плазматическими клетками, поддерживающими уровни антител, и B-клетками памяти, которые после повторной стимуляции антигеном запускают вторичные реакции GC с помощью Tfh-клеток памяти, что приводит к продукции большего количества высокоаффинных плазматических клеток (McHeyzer-Williams et al., 2011, Nature Rev. Immunol. 12(1):24-34). Считают, что циркулирующие Tfh-подобные клетки (далее в настоящем описании обозначаемые как "Tfh-подобные" или "cTfh"-клетки) также дифференцируются в истинные Tfh-клетки после повторной стимуляции антигеном для поддержки этих анамнестических реакций (Crotty et al., 2011, Annu. Rev. Immmunol. 29:621-663).
[0007] Важно, что образование аутореактивных B-клеток также может являться результатом реакций GC с нежелательными последствиями. Фактически, многочисленные хронические системные аутоиммунные заболевания, такие как SLE, RA, миозит, синдром Шегрена, ANCA-ассоциированный васкулит и склеродермия, демонстрируют признаки аутореактивных гуморальных ответов. Например, многие из аутоантител, являющих характерными признаками этих заболеваний, являются высокоаффинными, соматически мутировавшими антителами с переключенными изотипами Ig, что позволяет предполагать, что они являются результатом аутореактивных реакций GC (Vinuesa et al., 2009, Nature Rev. Immunol. 9(12):845-857). Кроме того, повышенное количество циркулирующих Tfh-подобных клеток определяют в периферической крови пациентов со многими из этих аутоиммунных заболеваний, уровни которых зачастую коррелируют с титрами аутоантител и/или тяжестью заболевания (Tangye et al., 2013, Nature Rev. Immunol. 13(6):412-426). В совокупности, эти данные свидетельствуют о том, что B-клетки, истинные Tfh-клетки и циркулирующие Tfh-подобные клетки являются потенциальными терапевтическими мишенями в случае многих системных аутоиммунных заболеваний.
[0008] CXCR5 экспрессируется B-клетками, истинными Tfh-клетками и циркулирующими Tfh-подобными клетками и опосредует их миграцию в GC и участие в реакциях GC вместе с градиентом лиганда CXCR5, CXCL13 (Vinuesa and Cyster, 2011, Immunity 35(5):671-680; Ansel et al., 1999, J. Exp. Med. 190(8):1123-1134; Ansel et al., 2000, Nature 406(6793):309-314; Cyster et al., 1999, Curr. Top. Microbiol. Immunol. 246:87-92; Hardtke et al., 2005, Blood 106(6):1924-1931; Haynes et al., 2007, J. Immunol. 179(8):5099-5108). В связи с этим, направленное воздействие на CXCR5 может принести терапевтическую пользу при лечении аутоиммунных заболеваний.
[0009] Кроме того, направленное воздействие на CXCR5 также может принести терапевтическую пользу при злокачественных новообразованиях, отличающихся пролиферацией клеток, экспрессирующих CXCR5, таких как рак поджелудочной железы, рак толстого кишечника, рак мочевого пузыря, T-клеточный лейкоз и B-клеточный лейкоз.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
[00010] В настоящей заявке описаны выделенные антитела и их антигенсвязывающие фрагменты, специфически связывающиеся с CXCR5 (C-X-C-хемокиновым рецептором типа 5). В некоторых аспектах антитела и их антигенсвязывающие фрагменты связываются с CXCR5 и снижают связывание CXCR5 с CXCL13. В других аспектах антитела могут являться фукозилированными, но более предпочтительно они являются афукозилированными. В некоторых аспектах антитела и их антигенсвязывающие фрагменты проявляют измененную эффекторную функцию. В некоторых аспектах антитела и их антигенсвязывающие фрагменты являются афукозилированными и проявляют повышенную антителозависимую клеточную цитотоксичность (ADCC) по сравнению с в остальном идентичными, но фукозилированными, антителами и их антигенсвязывающими фрагментами.
[00011] В некоторых аспектах настоящее изобретение относится к выделенным антителам и их антигенсвязывающим фрагментам, специфически связывающимся с CXCR5, где антитело и его антигенсвязывающий фрагмент связываются с эпитопом, содержащим лейцин (Leu; L) в положении аминокислотного остатка 11 (L11) в соответствии с нумерацией SEQ ID NO:32.
[00012] В другом аспекте настоящее изобретение относится к выделенным антителам или их антигенсвязывающим фрагментам, специфически связывающимся с CXCR5, где антитело и его антигенсвязывающий фрагмент связываются с эпитопом, содержащим аспартат (Asp; D) в положении аминокислотного остатка 22 в соответствии с нумерацией SEQ ID NO:32.
[00013] Специалистам в этой области будут понятны многие эквиваленты конкретных вариантов осуществления, представленных в настоящем описании, или они могут установить их с использованием не более чем рутинного экспериментирования. Такие эквиваленты предназначены для включения в следующие варианты осуществления (E).
E1. Выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с C-X-C-хемокиновым рецептором 5 (CXCR5), где антитело или его антигенсвязывающий фрагмент связывается с эпитопом в N-домене CXCR5 человека (hCXCR5) или CXCR5 яванского макака (cynoCXCR5).
E2. Антитело или его антигенсвязывающий фрагмент по п.E1, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом в аминокислотных остатках 1-50 hCXCR5 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32 или связывается с эпитопом в аминокислотных остатках 1-50 cynoCXCR5 в соответствии с нумерацией SEQ ID NO:33.
E3. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E2, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
E4. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E3, где антитело или его антигенсвязывающий фрагмент, связывается с эпитопом, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
E5. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E4, где антитело или его антигенсвязывающий фрагмент, связывается с эпитопом, содержащим лейцин в положении аминокислотного остатка 11 и аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
E6. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E5, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывается с hCXCR5, если указанный остаток не является лейцином.
E7. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E6, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывается с hCXCR5, если указанный остаток является треонином.
E8. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E7, где антитело или его антигенсвязывающий фрагмент, связывается с эпитопом, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывается с hCXCR5, если указанный остаток не является аспартатом.
E9. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E8, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом, содержащим аспартат в положении аминокислотного остатка 22, в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывается с hCXCR5, если указанный остаток является аланином.
E10. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E9, где антитело или его антигенсвязывающий фрагмент связывается с эпитопом, содержащим лейцин в положении аминокислотного остатка 11 и аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывается с указанным эпитопом, если лейцин замещен треонином, и/или аспартат замещен аланином.
E11. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E10, где антитело или его антигенсвязывающий фрагмент связывается с hCXCR5, экспрессирующимся на B-клетках человека, с кажущейся аффинностью EC50 приблизительно 6,60 пМ со стандартным отклонением приблизительно ±2,33 пМ.
E12. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E11, где антитело или его антигенсвязывающий фрагмент связывается с hCXCR5, экспрессирующимся на циркулирующих фолликулярных T-хелпер-подобных (Tfh-подобных) клетках человека с кажущейся аффинностью EC50 приблизительно 5,89 пМ со стандартным отклонением приблизительно ±1,40 пМ.
E13. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E12, где антитело или его антигенсвязывающий фрагмент связывается с hCXCR5, экспрессирующимся на фолликулярных T-хелперных (Tfh) клетках человека, с кажущейся аффинностью EC50 приблизительно 10,6 пМ.
E14. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E13, где антитело или его антигенсвязывающий фрагмент связывается с cynoCXCR5, экспрессирующимся на B-клетках яванского макака, с кажущейся аффинностью EC50 приблизительно 1,32 пМ.
E15. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E14, где антитело или его антигенсвязывающий фрагмент связывается с cynoCXCR5, экспрессирующимся на Tfh-подобных клетках яванского макака, с кажущейся аффинностью EC50 приблизительно 10,5 пМ.
E16. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E15, где антитело или его антигенсвязывающий фрагмент антагонистически действует на передачу сигнала CXCR5-CXCL13 в репортерном анализе цАМФ с EC50 приблизительно 961 пМ.
E17. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E16, где антитело или его антигенсвязывающий фрагмент проявляет активность ADCC против B-клеток человека, экспрессирующих hCXCR5, с EC50 приблизительно 2,01 пМ со стандартным отклонением приблизительно ±2,28 пМ.
E18. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E17, где антитело или его антигенсвязывающий фрагмент проявляет активность ADCC против Tfh-подобных клеток человека, экспрессирующих hCXCR5, с EC50 приблизительно 4,28 пМ со стандартным отклонением приблизительно ±2,88 пМ.
E19. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E18, где антитело или его антигенсвязывающий фрагмент, проявляет активность ADCC против Tfh-клеток человека, экспрессирующих hCXCR5, с EC50 приблизительно 0,11 пМ.
E20. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E19, где антитело или его антигенсвязывающий фрагмент проявляет активность ADCC против B-клеток яванского макака, экспрессирующих cynoCXCR5, с EC50 приблизительно 15,3 пМ со стандартным отклонением приблизительно ±11,7 пМ.
E21. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E20, где антитело или его антигенсвязывающий фрагмент связывается с hCXCR5, но не связывается детектируемо с хемокиновыми рецепторами человека CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 и XCR1.
E22. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E21, где антитело или его антигенсвязывающий фрагмент истощает B-клетки в периферической крови.
E23. Антитело или его антигенсвязывающий фрагмент по п.E22, где истощение B-клеток в периферической крови является обратимым.
E24. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E23, где антитело или его антигенсвязывающий фрагмент истощает Tfh-подобные клетки в периферической крови.
E25. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E24, где антитело или его антигенсвязывающий фрагмент истощает Tfh-клетки в селезенке.
E26. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E24, где антитело или его антигенсвязывающий фрагмент нарушает гуморальный анамнестический иммунный ответ.
E27. Антитело или его антигенсвязывающий фрагмент по п.E26, где нарушенный гуморальный анамнестический иммунный ответ является ответом на столбнячный анатоксин у яванских макаков.
E28. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E27, где антитело или его антигенсвязывающий фрагмент, ингибирует связывание CXCR5 с CXCL13.
E29. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E28, где антитело или его антигенсвязывающий фрагмент, ингибирует ингибирование CXCL13 продукции цАМФ в клетке, в ином случае запускаемой форсколином, что приводит к повышению уровня цАМФ по сравнению с уровнем цАМФ в отсутствие антитела или его антигенсвязывающего фрагмента.
E30. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E29, где антитело или его антигенсвязывающий фрагмент ингибирует ингибирование CXCL13 продукции цАМФ с EC50 приблизительно 961 пМ.
E31. Антитело или его антигенсвязывающий фрагмент по п.E30, где максимальное ингибирование ингибирования CXCL13 составляет по меньшей мере приблизительно 60%, 70% или 80%.
E32. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E31, где антитело или его антигенсвязывающий фрагмент связывается с CXCR5-экспрессирующими B-клетками человека (например, CXCR5+ B-клетками человека) с кажущейся аффинностью EC50 менее приблизительно 26 пМ, но не связывается с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика.
E33. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E32, где антитело или его антигенсвязывающий фрагмент запускает ADCC CXCR5-экспрессирующих клеток в мононуклеарных клетках периферической крови (PBMC) донора-человека и яванского макака и тонзиллярных мононуклеарных клетках (TMC) донора-человека.
E34. Выделенное антитело, специфически связывающееся с hCXCR5 и конкурирующее с антителом или его антигенсвязывающим фрагментом по любому из пп.E1-E33.
E35. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E34, где антитело или его антигенсвязывающий фрагмент связывается с CXCR5+ B-клетками человека с кажущейся аффинностью EC50 менее приблизительно 26 пМ, но, по существу, не связывается с клетками, экспрессирующими ортологи CXCR5. В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент демонстрирует недетектируемое связывание с клетками, экспрессирующими ортологи CXCR5, например, с учетом любого из анализов, представленных в настоящем описании; или связывается с клетками, экспрессирующими ортологи CXCR5, с кажущейся аффинностью EC50 по меньшей мере в 10000 раз выше, чем при связывании с hCXCR5.
E36. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, где антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:9.
E37. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36, где антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:11.
E38. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E37, где антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:16; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:19; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21.
E39. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E38, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:1, 5, 13, 35, 37, 48-51, 39 и 58-62, и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:6, 10, 12, 17, 18, 36, 40, 53-57 и 63.
E40. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E39, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:1, 5, 13, 35, 37, 48-51, 39 и 58-62, и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:6, 10, 12, 17, 18, 36, 40, 53-57 и 63.
E41. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E40, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324, и VH, содержащую аминокислотную последовательность, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
E42. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35 и E39-E40, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:13, и VH, содержащую аминокислотную последовательность SEQ ID NO:17.
E43. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35 и E38-E40, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:13, и VH, содержащую аминокислотную последовательность SEQ ID NO:18.
E44. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35 и E38-E40, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:37, и VH, содержащую аминокислотную последовательность SEQ ID NO:38.
E45. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36 и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:35, и VH, содержащую аминокислотную последовательность SEQ ID NO:36.
E46. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36 и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:47, и VH, содержащую аминокислотную последовательность SEQ ID NO:52.
E47. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36 и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:5, и VH, содержащую аминокислотную последовательность SEQ ID NO:6.
E48. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36 и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:5, и VH, содержащую аминокислотную последовательность SEQ ID NO:10.
E49. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E36 и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:1, и VH, содержащую аминокислотную последовательность SEQ ID NO:6.
E50. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:1, и VH, содержащую аминокислотную последовательность SEQ ID NO:12.
E51. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, содержащую аминокислотную последовательность SEQ ID NO:39, и VH, содержащую аминокислотную последовательность SEQ ID NO:40.
E52. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит LC, содержащую аминокислотную последовательность SEQ ID NO:22, и HC, содержащую аминокислотную последовательность SEQ ID NO:23.
E53. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35 и E38-E40, где антитело или его антигенсвязывающий фрагмент содержит LC, содержащую аминокислотную последовательность SEQ ID NO:24, и HC, содержащую аминокислотную последовательность SEQ ID NO:25.
E54. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит LC, содержащую аминокислотную последовательность SEQ ID NO:26, и HC, содержащую аминокислотную последовательность SEQ ID NO:27.
E55. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит LC, содержащую аминокислотную последовательность SEQ ID NO:28, и HC, содержащую аминокислотную последовательность SEQ ID NO:29.
E56. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит VL, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:95, и VH, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:96.
E57. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E35, E37, и E39-E41, где антитело или его антигенсвязывающий фрагмент содержит LC, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:97, и HC, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:98.
E58. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E57, где антитело или его антигенсвязывающий фрагмент содержит каркасную последовательность VL и каркасную последовательность VH, и где одна или обе из каркасной последовательности VL или каркасной последовательности VH являются по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичными последовательности зародышевой линии человека, из которой они получены, и где последовательность зародышевой линии VL человека, из которой получена каркасная последовательность VL, выбрана из группы, состоящей из DPK9, DPK12, DPK18, DPK24, HK102_V1, DPK1, DPK8, DPK3, DPK21, Vg_38K, DPK22, DPK15, DPL16, DPL8, V1-22, консенсусной Vλ, консенсусной Vλ1, консенсусной Vλ3, консенсусной Vκ, консенсусной Vκ1, консенсусной Vκ2 и Vκ3, и где последовательность VH зародышевой линии человека, из которой получена каркасная последовательность VH, выбрана из группы, состоящей из DP54, DP47, DP50, DP31, DP46, DP71, DP75, DP10, DP7, DP49, DP51, DP38, DP79, DP78, DP73, VH3, VH5, VH1 и VH4.
E59. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E58, где антитело или его антигенсвязывающий фрагмент является афукозилированным.
E60. Антитело или его антигенсвязывающий фрагмент по п.E59, где антитело или его антигенсвязывающий фрагмент демонстрирует повышенную ADCC.
E61. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие последовательность CDR-H1, CDR-H2 и CDR-H3 VH, содержащей аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:6, 7, 8, 25, 17, 18, 23, 27, 52, 53, 54, 55, 56, 57 и 63.
E62. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие последовательность CDR-L1, CDR-L2 и CDR-L3 VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из аминокислотных последовательностей SEQ ID NO:1, 2, 13, 22, 24, 26, 47, 48, 49, 50, 51, 58, 59, 60, 61 и 62.
E63. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E62, содержащие последовательности CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:6.
E64. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E62, содержащие последовательности CDR-L1, CDR-L2 и CDR-L3 из аминокислотной последовательности SEQ ID NO:1.
E65. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E64, содержащие одну или более из следующих замен аминокислот:
1, 2, 3, 4, 5 или 6 замен в CDR-L1 в соответствующем остатке последовательности VL зародышевой линии человека,
1, 2, 3, 4 или 5 замен в CDR-L2 в соответствующем остатке последовательности VL зародышевой линии человека,
1, 2, 3, 4, 5 или 6 замен в CDR L3 в соответствующем остатке последовательности VL зародышевой линии человека,
1, 2, 3, 4, 5 или 6 замен в CDR-H1 в соответствующем остатке последовательности VH зародышевой линии человека,
1, 2, 3, 4, 5, 6, 7 или 8 замен в CDR-H2 в соответствующем остатке последовательности VH зародышевой линии человека,
где последовательность VL зародышевой линии человека выбрана из группы, состоящей из DPK9, DPK12, DPK18, DPK24, HK102_V1, DPK1, DPK8, DPK3, DPK21, Vg_38K, DPK22, DPK15, DPL16, DPL8, V1-22, консенсусной Vκ1, консенсусной Vκ2 и консенсусной Vκ3, и последовательность VH зародышевой линии человека выбрана, состоящей из DP54, DP47, DP50, DP31, DP46, DP71, DP75, DP10, DP7, DP49, DP51, DP38, DP79, DP78, DP73, VH3, VH5, VH1 и VH4.
E66. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E65, содержащие каркасную последовательность VH, полученную из последовательности VH зародышевой линии человека, выбранной из группы, состоящей из DP54, DP47, DP50, DP31, DP46, DP71, DP75, DP10, DP7, DP49, DP51, DP38, DP79, DP78, DP73, VH3, VH5, VH1 и VH4.
E67. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E66, содержащие каркасную последовательность VH, полученную из последовательности VH3 зародышевой линии человека.
E68. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E67, содержащие каркасную последовательность VH, полученную из последовательности VH зародышевой линии человека, выбранной из группы, состоящей из DP54, DP47, DP50, DP31, DP46, DP49 и DP51.
E69. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E68, содержащие каркасную последовательность VH, полученную из последовательности VH зародышевой линии человека, выбранной из группы, состоящей из DP54, DP47, DP50 и DP31.
E70. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E69, содержащие каркасную последовательность VH, полученную из последовательности DP54 зародышевой линии человека.
E71. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E70, содержащие каркасную последовательность VL, полученную из последовательности VL зародышевой линии человека, выбранной из группы, состоящей из DPK9, DPK12, DPK18, DPK24, HK102_V1, DPK1, DPK8, DPK3, DPK21, Vg_38K, DPK22, DPK15, DPL16, DPL8, V1-22, консенсусной Vκ, консенсусной Vκ1, консенсусной Vκ2 и консенсусной Vκ3.
E72. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E71, содержащие каркасную последовательность VL, полученную из последовательности VL зародышевой линии человека, выбранной из группы, состоящей из DPK9, DPK12, DPK18, DPK24, HK102_V1, DPK1, DPK8, DPK3, DPK21, Vg_38K, DPK22, DPK15, консенсусной Vκ, консенсусной Vκ1, консенсусной Vκ2 и Vκ3.
E73. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E72, содержащие каркасную последовательность VL, полученную из последовательности Vκ1 зародышевой линии человека.
E74. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E73, содержащие каркасную последовательность VL, полученную из последовательности VL зародышевой линии человека, выбранной из группы, состоящей из DPK9, HK102_V1, DPK1 и DPK8.
E75. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E74, содержащие каркасную последовательность VL, полученную из последовательности DPK9 зародышевой линии человека.
E76. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E75, содержащие каркасную последовательность VL и каркасную последовательность VH, и где одна или обе из каркасной последовательности VL или каркасной последовательности VH являются по меньшей мере на 90% идентичными последовательности зародышевой линии человека, из которой они получены.
E77. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E76, содержащие каркасную последовательность VL и каркасную последовательность VH, и где одна или обе из каркасной последовательности VL или каркасной последовательности VH являются по меньшей мере на 66%, 76%, 80%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичными последовательности зародышевой линии человека, из которой они получены.
E78. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E77, содержащие каркасную последовательность VL и каркасную последовательность VH, и где одна или обе из каркасной последовательности VL или каркасной последовательности VH являются идентичными последовательности зародышевой линии человека, из которой они получены.
E79. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E78, содержащие VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO:6.
E80. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E79, содержащие VH, содержащую аминокислотную последовательность, по меньшей мере на 92% идентичную SEQ ID NO:6.
E81. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E80, содержащие VH, содержащую аминокислотную последовательность SEQ ID NO:6.
E82. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E81, содержащие VL, содержащую аминокислотную последовательность, по меньшей мере на 66% идентичную SEQ ID NO:1.
E83. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E82, содержащие VL, содержащую аминокислотную последовательность, по меньшей мере на 66%, 76%, 80%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную SEQ ID NO:1.
E84. Антитело или его антигенсвязывающий фрагмент по любому из пп.E61-E83, содержащие VL, содержащую аминокислотную последовательность SEQ ID NO:1.
E85. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E84, содержащие Fc-домен.
E86. Антитело или его антигенсвязывающий фрагмент по п.E85, где Fc-домен является Fc-доменом IgA (например, IgA1 или IgA2), IgD, IgE, IgM или IgG (например, IgG1, IgG2, IgG3 или IgG4).
E87. Антитело или его антигенсвязывающий фрагмент по п.E86, где Fc-домен является Fc-доменом IgG.
E88. Антитело или его антигенсвязывающий фрагмент по п.E87, где IgG выбран из группы, состоящей из IgG1, IgG2, IgG3 или IgG4.
E89. Антитело или его антигенсвязывающий фрагмент по п.E88, где IgG является IgG1.
E90. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E89, содержащие тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO:29.
E91. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E89, содержащие тяжелую цепь, содержащую аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную SEQ ID NO:29.
E92. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E89, содержащие тяжелую цепь, содержащую аминокислотную последовательность SEQ ID NO:29.
E93. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E93, содержащие LC, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную SEQ ID NO:28.
E94. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E93, содержащие LC, содержащую аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную SEQ ID NO:28.
E95. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E94, содержащие LC, содержащую аминокислотную последовательность SEQ ID NO:28.
E96. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие последовательность VH, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер ATCC PTA-124323.
E97. Выделенное антитело или его антигенсвязывающий фрагмент, содержащие последовательность VL, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер ATCC PTA-124324.
E98. Антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с CXCR5 человека с одним или более из 11G2 мыши, химерного 11G2, VH h11G2 (XC152)/VL (XC151), VH h11G2 (XC152)/VL (XC153), VH h11G2 (XC152)/VL (XC154), VH h11G2 (XC152)/VL (XC346), VH h11G2 (XC152)/VL (XC347), VH h11G2 (XC152)/VL (XC348), VH h11G2 (XC152)/VL (XC349), VH h11G2 (XC155)/VL (XC151), VH h11G2 (XC155)/VL (XC153), VH h11G2 (XC155)/VL (XC154), VH h11G2 (XC155)/VL (XC346), VH h11G2 (XC155)/VL (XC347), VH h11G2 (XC155)/VL (XC3484), VH h11G2 (XC155)/VL (XC349), VH h11G2 (XC156)/VL (XC151), VH h11G2 (XC156)/VL (XC153), VH h11G2 (XC156)/VL (XC154), VH h11G2 (XC156)/VL (XC346), VH h11G2 (XC156)/VL (XC347), VH h11G2 (XC156)/VL (XC348), VH h11G2 (XC156)/VL (XC349), VH h11G2 (XC157)/VL (XC151), VH h11G2 (XC157)/VL (XC153), VH h11G2 (XC157)/VL (XC154), VH h11G2 (XC157)/VL (XC346), VH h11G2 (XC157)/VL (XC347), VH h11G2 (XC157)/VL (XC348), VH h11G2 (XC157)/VL (XC349), VH h11G2 (XC350)/VL (XC151), VH h11G2 (XC350)/VL (XC153), VH h11G2 (XC350)/VL (XC154), VH h11G2 (XC350)/VL (XC346), VH h11G2 (XC350)/VL (XC347), VH h11G2 (XC350)/VL (XC348), VH h11G2 (XC350)/VL (XC349), VH h11G2 (XC351)/VL (XC151), VH h11G2 (XC351)/VL (XC153), VH h11G2 (XC351)/VL (XC154), VH h11G2 (XC351)/VL (XC346), VH h11G2 (XC351)/VL (XC347), VH h11G2 (XC351)/VL (XC348) и VH h11G2 (XC351)/VL (XC349), VH h11G2 (XC352)/VL (XC151), VH h11G2 (XC352)/VL (XC153), VH h11G2 (XC352)/VL (XC154), VH h11G2 (XC352)/VL (XC346), VH h11G2 (XC352)/VL (XC347), VH h11G2 (XC352)/VL (XC348), VH h11G2 (XC352)/VL (XC349), VH h11G2 (XC353)/VL (XC151), VH h11G2 (XC353)/VL (XC153), VH h11G2 (XC353)/VL (XC154), VH h11G2 (XC353)/VL (XC346), VH h11G2 (XC353)/VL (XC347), VH h11G2 (XC353)/VL (XC348), VH h11G2 (XC353)/VL (XC349), VH h11G2 (XC354)/VL (XC151), VH h11G2 (XC354)/VL (XC153), VH h11G2 (XC354)/VL (XC154), VH h11G2 (XC354)/VL (XC346), VH h11G2 (XC354)/VL (XC347), VH h11G2 (XC354)/VL (XC348), VH h11G2 (XC354)/VL (XC349), 41A10 мыши, химерного 41A10, VH h41A10 (XC147)/VL (XC142), VH h41A10 (XC147)/VL (XC143), VH h41A10 (XC147)/VL (XC144), VH h41A10 (XC147)/VL (XC145), VH h41A10 (XC147)/VL (XC146), VH h41A10 (XC147)/VL (XC149), VH h41A10 (XC148)/VL (XC142), VH h41A10 (XC148)/VL (XC143), VH h41A10 (XC148)/VL (XC144), VH h41A10 (XC148)/VL (XC145), VH h41A10 (XC148)/VL (XC146), VH h41A10 (XC148)/VL (XC149), VH h41A10 (XC150)/VL (XC142), VH h41A10 (XC150)/VL (XC143), VH h41A10 (XC150)/VL (XC144), VH h41A10 (XC150)/VL (XC145), VH h41A10 (XC150)/VL (XC146), VH h41A10 (XC150)/VL (XC149), 5H7 мыши и химерного 5H7.
E99. Антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с CXCR5 человека с CXCL13 и антителом по любому из пп.E1-E98.
E100. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E99, где антитело или антигенсвязывающий фрагмент является Fc-слитым белком, монотелом, максителом, бифункциональным антителом, scFab, scFv, пептителом.
E101. Антитело или его антигенсвязывающий фрагмент по п.E1-E100, где антитело или его антигенсвязывающий фрагмент связывается с CXCR5 человека с KD, приблизительно равным или меньшим, чем значение, выбранное из группы, состоящей из приблизительно 10 нМ, 5 нМ, 2 нМ, 1 нМ, 900 пМ, 800 пМ, 700 пМ, 600 пМ, 500 пМ, 400 пМ, 300 пМ, 250 пМ, 200 пМ, 150 пМ, 100 пМ, 50 пМ, 40 пМ, 30 пМ, 25 пМ, 20 пМ, 15 пМ, 10 пМ, 5 пМ и 1 пМ.
E102. Антитело или его антигенсвязывающий фрагмент по п.E1-E101, где антитело или его антигенсвязывающий фрагмент, связывается с CXCR5 яванского макака с KD, приблизительно равным или меньшим, чем значение, выбранное из группы, состоящей из приблизительно 10 нМ, 5 нМ, 2 нМ, 1 нМ, 900 пМ, 800 пМ, 700 пМ, 600 пМ, 500 пМ, 400 пМ, 300 пМ, 250 пМ, 200 пМ, 150 пМ, 100 пМ, 50 пМ, 40 пМ, 30 пМ, 25 пМ, 20 пМ, 15 пМ, 13 пМ, 10 пМ, 5 пМ и 1 пМ.
E103. Антитело или его антигенсвязывающий фрагмент по п.E1-E102, где KD связывания антитела или антигенсвязывающего фрагмента с CXCR5 яванского макака составляет в пределах 1 порядка от KD связывания антитела или его антигенсвязывающего фрагмента с CXCR5 человека.
E104. Антитело или его антигенсвязывающий фрагмент по п.E1-E103, где соотношение KD связывания антитела или антигенсвязывающего фрагмента с CXCR5 человека и связывания с CXCR5 яванского макака составляет от 5:1 до 1:5.
E105. Антитело или его антигенсвязывающий фрагмент по п.E1-E104, где соотношение KD связывания антитела или антигенсвязывающего фрагмента с CXCR5 человека и связывания с CXCR5 яванского макака составляет от 2:1 до 1:2.
E106. Антитело или его антигенсвязывающий фрагмент по п.E1-E105, где соотношение KD связывания антитела или антигенсвязывающего фрагмента с CXCR5 яванского макака и связывания с CXCR5 человека находится в диапазоне, нижний предел которого выбран из группы, состоящей из 0,5, 0,6, 0,7, 0,8, 0,9, 1,0, 1,1, 1,2, 1,3, 1,4, 1,5, 1,6, 1,7,1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9, 3,0, 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,1, 4,2, 4,3, 4,4, 4,5 и 6,2, и верхний предел которого выбран из группы, состоящей из 1,4, 1,5, 1,6, 1,7, 1,8, 1,9, 2,0, 2,1, 2,2, 2,3, 2,4, 2,5, 2,6, 2,7, 2,8, 2,9, 3,0 3,1, 3,2, 3,3, 3,4, 3,5, 3,6, 3,7, 3,8, 3,9, 4,0, 4,1, 4,2, 4,3, 4,4, 4,5, 6,2, 9, 9,2 и 10.
E107. Антитело или его антигенсвязывающий фрагмент по п.E1-E106, где соотношение KD связывания антитела или антигенсвязывающего фрагмента с CXCR5 яванского макака SEQ ID NO:33 и связывания с CXCR5 человека SEQ ID NO:32 составляет от приблизительно 1,0 до приблизительно 10,0.
E108. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-107, где прогнозируемое время полужизни у человека находится в диапазоне от приблизительно одного (1) дня до двадцати одного (21) дня.
E109. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-108, где прогнозируемое время полужизни у человека составляет приблизительно семнадцать (17) дней.
E110. Антитело или его антигенсвязывающий фрагмент, содержащий CDR антитела или его антигенсвязывающего фрагмента, выбранного из группы, состоящей из VH 11G2 мыши, VL 11G2 мыши, химерной VH 11G2, химерной VL 11G2, гуманизированной VH 11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353), VH h11G2 (XC354), VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348), VL h11G2 (XC349), VH 41A10 мыши, VL 41A10 мыши, химерной VH 41A10, химерной VL 41A10, гуманизированной VH 41A10 (XC147), VH h41A10 (XC148), VH h41A10 (XC150), VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146), VL h41A10 (XC149), VH 5H7 мыши, VL 5H7 мыши, химерной VH 5H7 и химерной VL 5H7.
E111. Антитело или его антигенсвязывающий фрагмент, содержащие VL и VH антитела или его антигенсвязывающего фрагмента, выбранную из группы, состоящей из VH 11G2 мыши, VL 11G2 мыши, химерной VH 11G2, химерной VL 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353), VH h11G2 (XC354), VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348), VL h11G2 (XC349), VH 41A10 мыши, VL 41A10 мыши, химерной VH 41A10, химерной VL 41A10, гуманизированной VH 41A10 (XC147), VH h41A10 (XC148), VH h41A10 (XC150), VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146), VL h41A10 (XC149), VH 5H7 мыши, VL 5H7 мыши, химерной VH 5H7 и химерной VL 5H7.
E112. Антитело или его антигенсвязывающий фрагмент, выбранные из группы, состоящей из
a. антитела, содержащего VH 11G2 мыши и VL 11G2 мыши;
b. антитела, содержащего химерную VH 11G2 и химерную VL 11G2;
c. антитела, содержащего гуманизированную VH 11G2 (XC152) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
d. антитела, содержащего гуманизированную VH 11G2 (XC155) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
e. антитела, содержащего гуманизированную VH 11G2 (XC156) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
f. антитела, содержащего гуманизированную VH 11G2 (XC157) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
g. антитела, содержащего гуманизированную VH 11G2 (XC350) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
h. антител, содержащего VH h11G2 (XC351) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
i. антитела, содержащего VH h11G2 (XC352) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
j. антитела, содержащего VH h11G2 (XC353) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
k. антитела, содержащего VH h11G2 (XC354) и VL, выбранную из группы, состоящей из VL 11G2 мыши, химерной VL 11G2, VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348) и VL h11G2 (XC349);
l. антитела, содержащего VL h11G2 (XC151) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
m. антитела, содержащего VL h11G2 (XC153) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
n. антител, содержащего VL h11G2 (XC154) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
o. антитела, содержащего VL h11G2 (XC346) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
p. антитела, содержащего VL h11G2 (XC347) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
q. антитела, содержащего VL h11G2 (XC348) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
r. антитела, содержащего VL h11G2 (XC349) и VH, выбранную из группы, состоящей из VH 11G2 мыши, химерной VH 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353) и VH h11G2 (XC354);
s. антитела, содержащего VH 41A10 мыши и VL, выбранную из группы, состоящей из VL 41A10 мыши, химерной VL 41A10, VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146) и VL h41A10 (XC149);
t. антитела, содержащего химерную VH 41A10 и VL, выбранную из группы, состоящей из VL 41A10 мыши, химерной VL 41A10, VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146) и VL h41A10 (XC149);
u. антитела, содержащего гуманизированную VH 41A10 (XC147) и VL, выбранную из группы, состоящей из VL 41A10 мыши, химерной VL 41A10, VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146) и VL h41A10 (XC149);
v. антитела, содержащего VH h41A10 (XC148) и VL, выбранную из группы, состоящей из VL 41A10 мыши, химерной VL 41A10, VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146) и VL h41A10 (XC149);
w. VH h41A10 (XC150) и VL, выбранную из группы, состоящей из VL 41A10 мыши, химерной VL 41A10, VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146), VL h41A10 (XC149) и VL h41A10 (XC142);
x. антитела, содержащего VL 41A10 мыши и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
y. антитела, содержащего химерную VL 41A10 и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
z. антитела, содержащего VL h41A10 (XC143) и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
aa. антитела, содержащего VL h41A10 (XC144) и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
bb. антитела, содержащего VL h41A10 (XC145) и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
cc. антитела, содержащего VL h41A10 (XC146) и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
dd. антитела, содержащего VL h41A10 (XC149) и VH, выбранную из группы, состоящей из VH 41A10 мыши, химерной VH 41A10, VH h41A10 (XC147), VL h41A10 (XC148) и VH h41A10 (XC150);
ee. антитела, содержащего VH 5H7 мыши и VL, выбранную из группы, состоящей из 5H7 мыши и химерной VL 5H7;
ff. VL 5H7 мыши и VH, выбранную из группы, состоящей из VH 5H7 мыши и химерной VH 5H7;
gg. антитела, содержащего VH h11G2 (XC152) и VL h11G2 (XC151);
hh. антитела, содержащего VH h11G2 (XC155) и VL h11G2 (XC153);
ii. антитела, содержащего VH h11G2 (XC155) и VL h11G2 (XC154);
jj. антитела, содержащего VH h11G2 (XC156) и VL h11G2 (XC153);
kk. антитела, содержащего VH h11G2 (XC157) и h11G2 (XC154);
ll. антитела, содержащего VH 11G2 мыши и VL 11G2 мыши;
mm. антитела, содержащего химерную HC 11G2 и химерную LC 11G2;
nn. антитела, содержащего VH h11G2 (XC152) и VL h11G2 (XC151);
oo. антитела, содержащего VH h11G2 (XC155) и VL h11G2 (XC154);
pp. антитела, содержащего VH h11G2 (XC156) и VL h11G2 (XC153);
qq. антитела, содержащего VH h11G2 (XC157) и VL h11G2 (XC154);
rr. антитела, содержащего VH 41A10 мыши и VL 41A10 мыши;
ss. антитела, содержащего химерную HC 41A10 и химерную LC 41A10;
tt. антитела, содержащего VH h41A10 (XC147) и VL h41A10 (XC142);
uu. антитела, содержащего VH h41A10 (XC147) и VL h41A10 (XC143);
vv. антитела, содержащего VH h41A10 (XC147) и VL h41A10 (XC144);
ww. антитела, содержащего VH h41A10 (XC147) и VL h41A10 (XC145);
xx. антитела, содержащего VH h41A10 (XC148) и VL h41A10 (XC142);
yy. антитела, содержащего VH h41A10 (XC148) и VL h41A10 (XC143);
zz. антитела, содержащего VH h41A10 (XC148) и h41A10VL (XC144);
aaa. антитела, содержащего VH 5H7 мыши и VL 5H7 мыши; и
bbb. антитела, содержащего химерную HC 5H7 и химерную LC 5H7.
E113. Антитело или его антигенсвязывающий фрагмент, выбранные из группы, состоящей из:
(a) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:36, и VL, содержащую аминокислотную последовательность SEQ ID NO:35;
(b) антитела, содержащего CDR-H1, содержащую последовательность SEQ ID NO:7, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8, CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:9, CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3, и CDR-L3, содержащую последовательность SEQ ID NO:4;
(c) антитела, содержащего CDR-H1, содержащую последовательность SEQ ID:7, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8, CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:11, CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3, и CDR-L3, содержащую последовательность SEQ ID NO:4;
(d) антитела, содержащего CDR-H1, содержащую последовательность SEQ ID:19, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21, CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15, и CDR-L3, содержащую последовательность SEQ ID NO:16;
(e) антитела, содержащего CDR-H1, CDR-H2, CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:6, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:1;
(f) антитела, содержащего CDR-H1, CDR-H2, CDR-H3, кодируемые вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323, и CDR-L1, CDR-L2 и CDR-L3, кодируемые вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324;
(g) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:52 (XC152), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(h) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:6 (XC155), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(i) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:10 (XC156), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(j) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:12 (XC157), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(k) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:53 (XC350), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(l) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:54 (XC351), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(m) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:55 (XC352), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(n) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:56 (XC353), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(o) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:55 (XC354), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51;
(p) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:47 (XC151), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(q) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:5 (XC153), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(r) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:1 (XC154), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(s) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:48 (XC346), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(t) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:49 (XC347), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(u) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:50 (XC348), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(v) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:51 (XC349), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57;
(w) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:38 (VH m41A10), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:13, 37, 58, 59, 60, 61 и 62;
(x) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:17 (XC148), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:13, 37, 58, 59, 60, 61 и 62;
(y) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:18 (XC147), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:13, 37, 58, 59, 60, 61 и 62;
(z) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:63 (XC150), и VL, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:13, 37, 58, 59, 60, 61 и 62;
(aa) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:37 (VL h41A10 мыши), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(bb) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:13 (VL h41A10 XC142), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(cc) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:58 (VL h41A10 XC143), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(dd) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:59 (VL h41A10 XC144), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(ee) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:60 (VL h41A10 XC145), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(ff) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:61 (VL h41A10 XC1446), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(gg) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:62 (VL h41A10 XC149), и VH, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:17, 18, 38 и 63;
(hh) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:40 (VH 5H7 мыши), и VL, содержащую аминокислотную последовательность SEQ ID NO:39;
(ii) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:52, и VL, содержащую аминокислотную последовательность SEQ ID NO:47;
(jj) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:6, и VL, содержащую аминокислотную последовательность SEQ ID NO:5;
(kk) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:6, и VL, содержащую аминокислотную последовательность SEQ ID NO:1;
(ll) (ii) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:10, и VL, содержащую аминокислотную последовательность SEQ ID NO:5;
(mm) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:10, и VL, содержащую аминокислотную последовательность SEQ ID NO:5; и
(nn) антитела, содержащего VH, содержащую аминокислотную последовательность SEQ ID NO:12, и VL, содержащую аминокислотную последовательность SEQ ID NO:1.
E114. Антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с CXCR5 человека с антителом или его антигенсвязывающим фрагментом по любому из пп.E1-E113.
E115. Выделенная нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114.
E116. Выделенная молекула нуклеиновой кислоты, содержащая по меньшей мере одну последовательность нуклеиновой кислоты, кодирующую антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E109.
E117. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, выбранную из группы, состоящей из последовательностей, приведенных как SEQ ID NO:95, 96, 97 и 98.
E118. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:95.
E119. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:96.
E120. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:95, и последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:96.
E121. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:97.
E122. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:98.
E123. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:97, и последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:98.
E124. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
E125. Выделенная молекула нуклеиновой кислоты, содержащая последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324.
E126. Выделенная нуклеиновая кислота, кодирующая VH, VL или и ту, и другую антитела или его антигенсвязывающего фрагмента, специфически связывающихся с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты SEQ ID NO:95, последовательность нуклеиновой кислоты SEQ ID NO:107 или и ту, и другую.
E127. Выделенная нуклеиновая кислота, кодирующая тяжелую цепь, легкую цепь или и ту, и другую антитела или его антигенсвязывающего фрагмента, специфически связывающихся с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты SEQ ID NO:97, последовательность нуклеиновой кислоты SEQ ID NO:98 или и ту, и другую.
E128. Выделенная нуклеиновая кислота, кодирующая VH антитела или его антигенсвязывающего фрагмента, специфически связывающихся с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
E129. Выделенная нуклеиновая кислота, кодирующая VL антитела или его антигенсвязывающего фрагмента, специфически связывающихся с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324.
E130. Выделенная нуклеиновая кислота, кодирующая VL и VH антитела или его антигенсвязывающего фрагмента, специфически связывающихся с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324, и последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
E131. Вектор, содержащий молекулу нуклеиновой кислоты по любому из пп.E115-E130.
E132. Клетка-хозяин, содержащая молекулу нуклеиновой кислоты по любому из пп.E115-E130 или вектор по п.E131.
E133. Клетка-хозяин по п.E132, где указанная клетка является клеткой млекопитающего.
E134. Клетка-хозяин по п.E133, где указанная клетка-хозяин является клеткой CHO, клеткой COS, клеткой HEK-293, клеткой NS0, клеткой PER.C6® или клеткой Sp2.0.
E135. Клетка-хозяин по любому из пп.E132-134, где в указанной клетке-хозяине отсутствует функциональная альфа-1,6-фукозилтрансфераза (FUT8).
E136. Клетка-хозяин по любому из пп.E132-135, где указанная клетка не экспрессирует функциональный фермент альфа-1,6-фукозилтрансферазу.
E137. Клетка-хозяин по любому из пп.E132-136, где в указанной клетке отсутствует ген FUT8, кодирующий функциональный фермент.
E138. Клетка-хозяин по любому из пп.E132-137, где в указанной клетке отсутствует ген, кодирующий функциональный ген FUT8.
E139. Клетка-хозяин по любому из пп.E132-139, где указанная клетка является клеткой Potelligent® CHOK1SV или клеткой Lec13 CHO.
E140. Клетка-хозяин по п.E139, где указанная клетка является клеткой Potelligent® CHOK1SV.
E141. Способ получения антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина по любому из пп.E1-E114 в условиях, в которых указанное антитело или антигенсвязывающий фрагмент экспрессируются указанной клеткой-хозяином.
E142. Способ по п.E141, дополнительно включающий выделение указанного антитела или его антигенсвязывающего фрагмента.
E143. Способ получения афукозилированного антитела против CXCR5 или его антигенсвязывающего фрагмента, включающий культивирование клетки-хозяина, содержащей молекулу нуклеиновой кислоты по любому из пп.E115-E130 или вектор по п.E131, где в указанной клетке-хозяине отсутствует функциональный FUT8.
E144. Способ по п.E143, где клетка-хозяин является клеткой Potelligent® CHOK1SV.
E145. Выделенное антитело или его антигенсвязывающий фрагмент, получаемые способом по любому из пп.E141-E114.
E146. Выделенное антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114, где указанное антитело или его антигенсвязывающий фрагмент являются афукозилированными.
E147. Афукозилированное антитело или его антигенсвязывающий фрагмент по п.E146, где указанное афукозилированное антитело или его антигенсвязывающий фрагмент демонстрирует повышенную антителозависимую клеточную цитотоксичность (ADCC) по сравнению с в остальном идентичным антителом или его антигенсвязывающим фрагментом, являющимся фукозилированным.
E148. Афукозилированное антитело или его антигенсвязывающий фрагмент по п.E146, где указанное афукозилированное антитело или его антигенсвязывающий фрагмент демонстрирует в приблизительно 2 раза, приблизительно 5 раз, приблизительно 7 раз, приблизительно 10 раз, приблизительно 20 раз, приблизительно 30 раз, приблизительно 40 раз, приблизительно 50 раз, приблизительно 60 раз, приблизительно 70 раз, приблизительно 80 раз, приблизительно 90 раз, приблизительно 100 раз, приблизительно 110 раз, приблизительно 120 раз, приблизительно 130 раз, приблизительно 140 раз и приблизительно 143 раза более высокую ADCC по сравнению с в остальном идентичным антителом или его антигенсвязывающим фрагментом, являющимся фукозилированным.
E149. Фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114 и E145-E148 и фармацевтически приемлемый носитель или эксципиент.
E150. Фармацевтическая композиция по п.E149, где указанное антитело или его антигенсвязывающий фрагмент являются афукозилированными.
E151. Способ снижения активности CXCR5, включающий введение нуждающемуся в этом индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150.
E152. Способ лечения воспалительного заболевания, включающий введение нуждающемуся в этом индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149-E150.
E153. Способ лечения индивидуума, нуждающегося в иммуносупрессии, включающий введение нуждающемуся в этом индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150.
E154. Способ лечения аутоиммунного заболевания, нарушения или состояния, включающий введение нуждающемуся в этом индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150.
E155. Способ снижения количества клеток, экспрессирующих CXCR5, у больного, включающий введение индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150.
E156. Способ по п.E155, где клетки экспрессируют CXCR5 на своей поверхности.
E157. Способ по п.E156, где клетки являются B-клетками и Tfh-подобными клетками.
E158. Способ по любому из пп.E151-E157, где указанный индивидуум является человеком.
E159. Способ по любому из пп.E151-E158, включающий внутривенное введение указанного антитела или его антигенсвязывающего фрагмента или фармацевтической композиции.
E160. Способ по любому из пп.E151-E158, включающий подкожное введение указанного антитела или его антигенсвязывающего фрагмента или фармацевтической композиции.
E161. Способ по любому из пп.E151-E160, где указанное антитело или его антигенсвязывающий фрагмент или фармацевтическую композицию вводят приблизительно два раза в неделю, один раз в неделю, один раз каждые две недели, один раз каждые три недели, один раз каждые четыре недели, один раз каждые пять недель, один раз каждые шесть недель, один раз каждые семь недель, один раз каждые восемь недель, один раз каждые девять недель, один раз каждые десять недель, дважды в месяц, один раз в месяц, один раз каждые два месяца, один раз каждые три месяца, один раз каждые четыре месяца, один раз каждые пять месяцев, один раз каждые шесть месяцев, один раз каждые семь месяцев, один раз каждые восемь месяцев, один раз каждые девять месяцев, один раз каждые десять месяцев, один раз каждые одиннадцать месяцев или один раз каждые двенадцать месяцев.
E162. Способ снижения количества CXCR5+ клеток в образце, включающий приведение указанной клетки в контакт с антителом или его антигенсвязывающим фрагментом по любому из пп.E1-E114 и E145-E148 или фармацевтической композицией по любому из пп.E149 и E150.
E163. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114 и E145-E148 или фармацевтическая композиция по любому из пп.E149 и E150 для применения в качестве лекарственного средства.
E164. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114 и E145-E148 или фармацевтическая композиция по любому из пп.E149 и E150 для применения в снижении активности CXCR5 у индивидуума.
E165. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114 и E145-E148 или фармацевтическая композиция по любому из пп.E149 и E150 для применения в лечении индивидуума, нуждающегося в иммуносупрессии.
E166. Антитело или его антигенсвязывающий фрагмент по любому из пп.E1-E114 и E145-E148 или фармацевтическая композиция по любому из пп.E149 и E150 для применения в лечении аутоиммунного заболевания, нарушения или состояния у индивидуума.
E167. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 в производстве лекарственного средства для лечения иммунологического заболевания, нарушения или состояния.
E168. Применение фармацевтической композиции по любому из пп.E149 и E150 в производстве лекарственного средства для лечения иммунологического заболевания, нарушения или состояния.
E169. Способ лечения медицинского состояния, включающий введение нуждающемуся в этом индивидууму терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150.
E170. Способ по п.E169, где состояние выбрано из группы, состоящей из воспалительных ответов, таких как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); дерматомиозита; системной склеродермии и склероза; ответов, ассоциированных с воспалительным заболеванием кишечника (таким как болезнь Крона и язвенный колит); респираторного дистресс-синдрома (включая, респираторный дистресс-синдром взрослых; ARDS); дерматита; менингита; энцефалита; увеита; колита; гастрита; гломерулонефрита; аллергических состояний, таких как экзема и астма и другие состояния, включающие инфильтрацию T-клетками и хронические воспалительные ответы; атеросклероза; дефицита адгезии лейкоцитов; ревматоидного артрита; системной красной волчанки (SLE); сахарного диабета (например, сахарного диабета I типа или инсулинозависимого сахарного диабета); рассеянного склероза; синдрома Рейно; аутоиммунного тиреоидита; аллергического энцефаломиелита; синдрома Шегрена; ювенильного диабета; и иммунных ответов, ассоциированных с острой и замедленной реакцией гиперчувствительности, опосредованных цитокинами и T-лимфоцитами, как правило, обнаруживаемых при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; гранулематоза Вегенера; пернициозной анемии (болезни Аддисона); заболеваний, включающих диапедез лейкоцитов; воспалительного нарушения центральной нервной системы (ЦНС); синдрома полиорганной недостаточности; гемолитической анемии (включая, в качестве неограничивающих примеров, криоглобулинемию или аутоиммунную гемолитическую анемию с неполными тепловыми агглютининами); миастении; заболеваний, опосредованных комплексами антиген-антитело; болезни анти-БМК-антител; антифосфолипидного синдрома; аллергического неврита; болезни Грейвса; миастенического синдрома Ламберта-Итона; буллезного пемфигоида; пемфигуса; аутоиммунных полиэндокринопатий; витилиго; болезни Рейтера; синдрома мышечной скованности; болезни Бехчета; гигантоклеточного артериита; иммунокомплексного нефрита; IgA-нефропатии; IgM-полинейропатий; иммунной тромбоцитопенической пурпуры (ITP) или аутоиммунной тромбоцитопении и аутоиммунных гемолитических заболеваний; тиреоидита Хашимото; аутоиммунного гепатита; аутоиммунной гемофилии; аутоиммунного лимфопролиферативного синдрома (ALPS); аутоиммунного увеоретинита; синдрома Гийена-Барре; синдрома Гудпасчера; смешанного заболевания соединительной ткани; аутоиммунно-ассоциированного бесплодия; нодозного полиартериита; гнездной алопеции; идиопатической микседемы; реакции "трансплантат против хозяина"; мышечной дистрофии (дистрофии Дюшена, дистрофии Беккера, миотонической дистрофии, конечностно-поясной дистрофии, плече-лопаточно-лицевой дистрофии, врожденной дистрофии, окулофарингеальной дистрофии, дистальной дистрофии, дистрофии Эмери-Дрейфуса) и контроля пролиферации злокачественных клеток, экспрессирующих CXCR5, такой как рак поджелудочной железы, рак толстого кишечника, рак мочевого пузыря, T-клеточный лейкоз и B-клеточный лейкоз.
E171. Способ детекции CXCR5 в образце, ткани или клетке с использованием антитела или его антигенсвязывающего фрагмента по любому из пп.E1-E114 и E145-E148 или фармацевтической композиции по любому из пп.E149 и E150, включающий приведение образца, ткани или клетки в контакт с антителом и детекторным антителом.
E172. Выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, где антитело является по меньшей мере одним антителом, выбранным из группы, состоящей из:
(a) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:9;
(b) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:11;
(c) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:16; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:19; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21;
(d) антитела, содержащего VL, содержащую аминокислотную последовательность, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324, и VH, содержащую аминокислотную последовательность, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323;
(e) антитела, содержащую VL, содержащую аминокислотную последовательность SEQ ID NO:1, и VH, содержащую аминокислотную последовательность SEQ ID NO:6;
(f) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:13, и VH, содержащую аминокислотную последовательность SEQ ID NO:18;
(g) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:47, и VH, содержащую аминокислотную последовательность SEQ ID NO:52;
(h) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:5, и VH, содержащую аминокислотную последовательность SEQ ID NO:6;
(i) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:5, и VH, содержащую аминокислотную последовательность SEQ ID NO:10;
(j) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:13, и VH, содержащую аминокислотную последовательность SEQ ID NO:17;
(k) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO:1, и VH, содержащую аминокислотную последовательность SEQ ID NO:12;
(l) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO:22, и HC, содержащую аминокислотную последовательность SEQ ID NO:23;
(m) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO:24, и HC, содержащую аминокислотную последовательность SEQ ID NO:25;
(n) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO:26, и HC, содержащую аминокислотную последовательность SEQ ID NO:27;
(o) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO:28, и HC, содержащую аминокислотную последовательность SEQ ID NO:29;
(p) антитела, содержащего VL, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:95, и VH, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:96; и
(q) антитела, содержащего LC, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:97, и HC, кодируемую последовательностью нуклеиновой кислоты SEQ ID NO:98.
E173. Выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с C-X-C-хемокиновым рецептором 5 (CXCR5), где антитело или его антигенсвязывающий фрагмент является по меньшей мере одним антителом, выбранным из группы, состоящей из:
(a) антитела или его антигенсвязывающего фрагмента, связывающегося с эпитопом hCXCR5, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с указанным эпитопом, если указанный остаток не является лейцином;
(b) антитела или его антигенсвязывающего фрагмента, связывающегося с эпитопом hCXCR5, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с указанным эпитопом, если указанный остаток является треонином;
(c) антитела или его антигенсвязывающего фрагмента, связывающегося с эпитопом hCXCR5, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с указанным эпитопом, если указанный остаток не является аспартатом;
(d) антитела или его антигенсвязывающего фрагмента, связывающегося с эпитопом hCXCR5, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с указанным эпитопом, если указанный остаток является аланином;
(e) антитела или его антигенсвязывающего фрагмента, связывающегося с эпитопом hCXCR5, содержащим лейцин в положении аминокислотного остатка 11 и аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с указанным эпитопом, если указанный лейцин замещен треонином и/или указанный аспартат замещен аланином;
(f) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5 или его фрагментом, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с hCXCR5 или его фрагментом, если указанный остаток не является лейцином;
(g) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5 или его фрагментом, содержащим лейцин в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с hCXCR5 или его фрагментом, если указанный остаток является треонином;
(h) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5 или его фрагментом, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с hCXCR5 или его фрагментом, если указанный остаток не является аспартатом;
(i) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5 или его фрагментом, содержащим аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с hCXCR5 или его фрагментом, если указанный остаток является аланином; и
(j) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5 или его фрагментом, содержащим лейцин в положении аминокислотного остатка 11 и аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но не связывающегося с hCXCR5 или его фрагментом, если указанный лейцин замещен треонином и/или указанный аспартат замещен аланином.
E174. Выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с C-X-C-хемокиновым рецептором 5 (CXCR5), где антитело или его антигенсвязывающий фрагмент является по меньшей мере одним антителом, выбранным из группы, состоящей из:
(a) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5, экспрессирующимся на B-клетках человека, с кажущейся аффинностью EC50 приблизительно 6,60 пМ со стандартным отклонением приблизительно ±2,33 пМ;
(b) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5, экспрессирующимся на циркулирующих фолликулярных T-хелпер-подобных клетках человека, с кажущейся аффинностью EC50 приблизительно 5,89 пМ со стандартным отклонением приблизительно ±1,40 пМ;
(c) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5, экспрессирующимся на фолликулярных T-хелперных (Tfh) клетках человека, с кажущейся аффинностью EC50 приблизительно 10,6 пМ;
(d) антитела или его антигенсвязывающего фрагмента, связывающегося с cynoCXCR5, экспрессирующимся на B-клетках яванского макака, с кажущейся аффинностью EC50 приблизительно 1,32 пМ;
(e) антитела или его антигенсвязывающего фрагмента, связывающегося с cynoCXCR5, экспрессирующимся на Tfh-подобных клетках яванского макака, с кажущейся аффинностью EC50 приблизительно 10,5 пМ;
(f) антитела или его антигенсвязывающего фрагмента, антагонистически действующего на передачу сигнала CXCR5-CXCL13 в репортерном анализе цАМФ с EC50 приблизительно 961 пМ;
(g) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против B-клеток человека, экспрессирующих hCXCR5, с EC50 приблизительно 2,01 пМ со стандартным отклонением приблизительно ±2,28 пМ;
(h) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против человек Tfh-подобных клеток, экспрессирующих hCXCR5, с EC50 приблизительно 4,28 пМ со стандартным отклонением приблизительно ±2,88 пМ;
(i) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против Tfh-клеток человека, экспрессирующих hCXCR5, с EC50 приблизительно 0,11 пМ;
(j) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против B-клеток яванского макака, экспрессирующих cynoCXCR5, с EC50 приблизительно 15,3 пМ со стандартным отклонением приблизительно ±11,7 пМ;
(k) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5, но не связывающегося детектируемо с хемокиновыми рецепторами человека CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 и XCR1;
(l) антитела или его антигенсвязывающего фрагмента, ингибирующего связывание CXCR5 с CXCL13.
(m) антитела или его антигенсвязывающего фрагмента, связывающегося с CXCR5+ B-клетками человека с кажущейся аффинностью EC50 менее приблизительно 26 пМ, но не связывающегося с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика;
(n) антитела или его антигенсвязывающего фрагмента, антагонистически действующего на ингибирование CXCL13 высвобождения цАМФ, запускаемого форсколином;
(o) антитела или его антигенсвязывающего фрагмента, запускающего ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и TMC человека-донора;
(p) антитела или его антигенсвязывающего фрагмента, связывающегося с CXCR5 человека, но не связывающегося с хемокиновыми рецепторами человека CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1;
(q) антитела или его антигенсвязывающего фрагмента, истощающего B-клетки в периферической крови;
(r) антитела или его антигенсвязывающего фрагмента, истощающего Tfh-подобные клетки в периферической крови;
(s) антитела или его антигенсвязывающего фрагмента, истощающего истинные Tfh-клетки в селезенке; и
(t) антитела или его антигенсвязывающего фрагмента, нарушающего гуморальный анамнестический иммунный ответ.
E175. Антитело по любому из пп.E172-E174, где указанное антитело или его антигенсвязывающий фрагмент демонстрирует по меньшей мере один из следующих видов биологической активности:
(a) связывается с CXCR5+ B-клетками человека с кажущейся аффинностью EC50 менее приблизительно 26 пМ, но не связывается с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика;
(b) антагонистически действует на ингибирование CXCL13 высвобождения цАМФ, запускаемого форсколином;
(c) запускает ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и TMC человека-донора;
(d) связывается с CXCR5 человека, но не связывается с хемокиновыми рецепторами человека CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1;
(e) истощает B-клетки в периферической крови;
(f) истощает Tfh-подобные клетки в периферической крови;
(g) истощает истинные Tfh-клетки в селезенке; или
(h) нарушает гуморальный анамнестический иммунный ответ.
E176. Антитело по любому из пп.E172-E175, где указанное антитело или его антигенсвязывающий фрагмент является афукозилированным.
E177. Выделенная нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп.E172-E176.
E178. Выделенная нуклеиновая кислота, кодирующая VH, VL или и ту, и другую антитела или его антигенсвязывающего фрагмента, специфически связывающегося с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты SEQ ID NO:95, последовательность нуклеиновой кислоты SEQ ID NO:96 или и ту, и другую.
E179. Выделенная нуклеиновая кислота, кодирующая тяжелую цепь, легкую цепь или и ту, и другую антитела или его антигенсвязывающего фрагмента, специфически связывающегося с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты SEQ ID NO:97, последовательность нуклеиновой кислоты SEQ ID NO:98 или и ту, и другую.
E180. Выделенная нуклеиновая кислота, кодирующая VH, VL или и ту, и другую антитела или его антигенсвязывающего фрагмента, специфически связывающегося с CXCR5, где указанная нуклеиновая кислота содержит последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323, последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324, или и ту, и другую.
E181. Вектор, содержащий нуклеиновую кислоту по любому из пп.E177-E180.
E182. Клетка-хозяин, содержащая вектор по п.E181.
E183. Клетка-хозяин по п.E182, где указанная клетка-хозяин является клеткой млекопитающего, выбранной из группы, состоящей из клетки CHO, клетки COS, клетки HEK-293, клетки NS0, клетки PER.C6® или клетки Sp2.0.
E184. Клетка-хозяин по п.E183, где в указанной клетке отсутствует функциональная альфа-1,6-фукозилтрансфераза (FUT8).
E185. Клетка-хозяин по п.E184, где указанная клетка является клеткой Potelligent® CHOK1SV или клеткой Lec13 CHO.
E186. Способ получения антитела или его антигенсвязывающего фрагмента, включающий использование клетки Potelligent® CHOK1SV по п.E185 в условиях, в которых указанное антитело или его антигенсвязывающий фрагмент экспрессируются указанной клеткой-хозяином и является афукозилированным.
E187. Способ по п.E186, дополнительно включающий выделение указанного антитела или его антигенсвязывающего фрагмента.
E188. Афукозилированное антитело или его антигенсвязывающий фрагмент по п.E186, где указанное антитело демонстрирует повышенную активность ADCC по сравнению с в остальном идентичным антителом или его антигенсвязывающим фрагментом, являющимся фукозилированным.
E189. Фармацевтическая композиция, содержащая антитело или его антигенсвязывающий фрагмент по любому из пп.E172-E176 и E188 и фармацевтически приемлемый носитель или эксципиент.
E190. Антитело или его антигенсвязывающий фрагмент по любому из пп.E172-E176, E188 и фармацевтическая композиция по п.E189 для применения в лечении иммунологического заболевания, нарушения или состояния.
E191. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.E172-E176, E188 или фармацевтической композиции по п.E189 для лечения иммунологического заболевания, нарушения или состояния.
E192. Способ лечения или профилактики иммунологического заболевания, нарушения или состояния, опосредованного CXCR5, у больного, включающий введение индивидууму эффективного количества фармацевтической композиции по п.E189, где указанное заболевание, нарушение или состояние выбрано из группы, состоящей из воспалительных ответов, таких как воспалительные заболевания кожи, включая псориаз и дерматит (например, атопический дерматит); дерматомиозита; системной склеродермии и склероза; ответов, ассоциированных с воспалительным заболеванием кишечника (таким как болезнь Крона и язвенный колит); респираторного дистресс-синдрома (включая респираторный дистресс-синдром взрослых; ARDS); дерматита; менингита; энцефалита; увеита; колита; гастрита; гломерулонефрита; аллергических состояний, таких как экзема и астма, и других состояний, включающих инфильтрацию T-клетками и хронические воспалительные ответы; атеросклероза; дефицита адгезии лейкоцитов; ревматоидного артрита (RA); системной красной волчанки (SLE); сахарного диабета (например, сахарного диабета I типа или инсулинозависимого сахарного диабета); рассеянного склероза; синдрома Рейно; аутоиммунного тиреоидита; аллергического энцефаломиелита; синдрома Шегрена; ювенильного диабета; и иммунных ответов, ассоциированных с острой и замедленной реакцией гиперчувствительности, опосредованных цитокинами и T-лимфоцитами, как правило, обнаруживаемых при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; гранулематоза Вегенера; пернициозной анемии (болезни Аддисона); заболеваний, включающих диапедез лейкоцитов; воспалительного нарушения центральной нервной системы (ЦНС); синдрома полиорганной недостаточности; гемолитической анемии (включая, в качестве неограничивающих примеров, криоглобулинемию или аутоиммунную гемолитическую анемию с неполными тепловыми агглютининами); миастении; заболеваний, опосредованных комплексами антиген-антитело; болезни анти-БМК-антител; антифосфолипидного синдрома; аллергического неврита; болезни Грейвса; миастенического синдрома Ламберта-Итона; буллезного пемфигоида; пемфигуса; аутоиммунных полиэндокринопатий; витилиго; болезни Рейтера; синдрома мышечной скованности; болезни Бехчета; гигантоклеточного артериита; иммунокомплексного нефрита; IgA-нефропатии; IgM-полинейропатий; иммунной тромбоцитопенической пурпуры (ITP) или аутоиммунной тромбоцитопении и аутоиммунных гемолитических заболеваний; тиреоидита Хашимото; аутоиммунного гепатита; аутоиммунной гемофилии; аутоиммунного лимфопролиферативного синдрома (ALPS); аутоиммунного увеоретинита; синдрома Гийена-Барре; синдрома Гудпасчера; смешанного заболевания соединительной ткани; аутоиммунно-ассоциированного бесплодия; нодозного полиартериита; гнездной алопеции; идиопатической микседемы; реакции "трансплантат против хозяина"; мышечной дистрофии (дистрофии Дюшена, дистрофии Беккера, миотонической дистрофии, конечностно-поясной дистрофии, плече-лопаточно-лицевой дистрофии, врожденной дистрофии, окулофарингеальной дистрофии, дистальной дистрофии, дистрофии Эмери-Дрейфуса) и контроля пролиферации злокачественных клеток, экспрессирующих CXCR5, такой как рак поджелудочной железы, рак толстого кишечника, рак мочевого пузыря, T-клеточный лейкоз и B-клеточный лейкоз.
E193. Способ по п.E192, где указанное заболевание является SLE или ревматоидным артритом.
E194. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.E172-E176 или E188 в производстве лекарственного средства для лечения иммунологического заболевания, нарушения или состояния.
E195. Способ детекции CXCR5 в образце, ткани или клетке с использованием антитела или его антигенсвязывающего фрагмента по любому из пп.E172-E176, включающий приведение образца, ткани или клетки в контакт с антителом и детекторным антителом.
E196. Способ снижения биологической активности CXCR5 у больного, включающий введение терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E172-E176 или E188 или фармацевтической композиции по п.E189.
E197. Способ по п.E196, где антитело опосредует истощение по меньшей мере одной клетки, экспрессирующей CXCR5, выбранной из группы, состоящей из Tfh-клетки в селезенке, B-клетки в периферической крови и Tfh-подобной клетки в периферической крови.
E198. Способ ингибирования гуморального иммунного ответа у больного, включающий введение терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.E172-E176 или E188 или фармацевтической композиции по п.E189.
E199. Способ по п.E198, где антитело опосредует истощение по меньшей мере одной клетки, экспрессирующей CXCR5, выбранной из группы, состоящей из Tfh-клетки в селезенке, B-клетки в периферической крови и Tfh-подобной клетки в периферической крови.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
[00014] Описанная выше сущность изобретения, а также следующее подробное описание изобретения будут более понятными при прочтении в сочетании с прилагаемыми чертежами. В иллюстративных целях варианты осуществления представлены на чертежах. Однако следует понимать, что изобретение не ограничено конкретными представленными вариантами и устройствами.
[00015] Фиг. 1 является диаграммой, на которой показана каноническая биантенарная гликоформа, как правило, присутствующая в константных областях тяжелой цепи иммуноглобулина. На диаграмме показана аспарагин-связанная (N-связанная) гликоформа на константном домене IgG в положении аминокислотного остатка 297 (аспарагин 297; Asn297) в соответствии с нумерацией Eu по Edelman et al., 1969, Proc. Natl. Acad. Sci. USA 63(1):78-85, как описано в Kabat et al., 1991.
[00016] Фиг. 2A-2G являются диаграммами, на которых показаны примеры биантенарных гликоформ, как правило, присутствующих в константных областях тяжелой цепи иммуноглобулина. Обозначение "G0" относится к биантенарной структуре, где нет концевых сиаловых кислот (NeuAc) или Gal, обозначение "G1" относится к биантенарной структуре, содержащей одну Gal и несодержащей NeuAc, и обозначение "G2" относится к биантенарной структуре с двумя концевыми Gal и без NeuAc. В каждой гликоформе фукоза, как правило, присутствует в антителах, продуцирующихся в клетках млекопитающих, например, клетках CHO. На фиг. 2A показана G2S2, на фиг. 2B показана G2S1, на фиг. 2C показана G2, на фиг. 2D показана G1, на фиг. 2E показана G0, на фиг. 2F показана G -1, и на фиг. 2G показана G -2.
[00017] На фиг. 3 представлена столбиковая диаграмма, на которой показано связывание различных моноклональных антител (указанных на оси x) с клетками HEK 293, экспрессирующими CXCR5 человека (hCXCR5 293) (более светлые столбики), и/или клетками HEK 293, экспрессирующими CXCR5 яванского макака (cynoCXCR5 293) (более темные столбики). Моноклональные антитела, показанные на фиг. 3, представляют собой, слева направо, 11B9, 18C7, 21A3, 23F1, 30C3, 31F3, 39H4, 47D11, 51D11, 52E7, 56A9, 56G2, 57H5, 58C12, 59F3, 4F1, 6E9, 12E11, 16D10, 18G4, 19D9, 19F8, 20G4, 26E2, 41A10, 48A10, 13F4, 17D5, 18H9, 23C6, 1A11, 1D10, 1C11, 3A1, 5H3, 6D6, 9G11, 10G2, 12C1, 12C3, 15A11, 15C10, 15D5, 18A11, 19C11, 20D10, 21B2, 21G3, 25D4, 25E1, 5H7, 11G2, 18B11, 23D5, 23F7, 31E5, 19G2 и 31G7.
[00018] На фиг. 4a представлена диаграмма, на которой показано отсутствие детектируемого связывания различных антител против CXCR5 с клетками, экспрессирующими CXCR1.
[00019] На фиг. 4B представлена диаграмма, на которой показано отсутствие связывания различных антител против CXCR5 с клетками, экспрессирующими CXCR2.
[00020] На фиг. 4C представлена диаграмма, на которой показано отсутствие связывания различных антител против CXCR5 с клетками, экспрессирующими CXCR3.
[00021] На фиг. 4D представлена диаграмма, на которой показано связывание различных антител против CXCR5 с клетками Jurkat, от природы экспрессирующими CXCR4.
[00022] На фиг. 5 представлен график, на котором показана аффинность связывания с CXCR5-экспрессирующими клетками антител: афукозилированного h11G2 XC154/XC155 (незакрашенные круги), фукозилированного h11G2 XC154/XC155 (закрашенные круги), 2C9 (закрашенные треугольники), 16D7 (закрашенные квадраты) и 11A7 (закрашенные овалы).
[00023] На фиг. 6 показано выравнивание аминокислотных последовательностей CXCR5 человека и мыши. На фигуре также показаны различные внеклеточные домены CXCR5, помеченные как "N", "L1", "L2" и "L3", которые подчеркнуты.
[00024] На фиг. 7 представлена столбиковая диаграмма, на которой показано, что некоторые аминокислотные остатки являлись критичными для связывания антитела с hCXCR5. Т.е. изменение D10G или встраивание SI в последовательность человека устраняло связывание антитела 2C9, но не влияло на связывание другими тремя антителами. Что более важно, замена L11T или D22A устраняла связывание 11G2, но не влияла на связывание 2C9, 16D7 или 11A7. Замена W на K в положении аминокислотного остатка 19 (W19K) устраняла связывание антителами 16D7 и 11A7, но не влияла на связывание 11G2 и 2C9. Эти данные свидетельствуют о том, что эти четыре антитела не имеют общего эпитопа на hCXCR5.
[00025] На фиг. 8A представлен график, на котором показано истощение и восстановление B-клеток периферической крови у самцов яванского макака. Показаны количества B-клеток в периферической крови на мкл крови у самцов до дня 352 поискового исследования токсичности. B-клетки определяли как CD3-CD20+. Представлены данные для отдельных животных. Архивный диапазон B-клеток у самцов обезьян (272-2503 клеток на мкл) указан пунктирными линиями.
[00026] На фиг. 8B представлен график, на котором показано истощение и восстановление B-клеток периферической крови у самок яванского макака. Показаны количества B-клеток в периферической крови на мкл крови у самок до дня 352 поискового исследования токсичности. B-клетки определяли как CD3-CD20+. Представлены данные для отдельных животных. Архивные диапазоны B-клеток у самок обезьян (233-1700 клеток на мкл) указаны пунктирными линиями.
[00027] На фиг. 8C представлен график, на котором показано истощение и восстановление Tfh-подобных (также обозначаемых как "cTfh" или "циркулирующие Tfh") клеток у самцов яванского макака. Показаны количества Tfh-подобных клеток в периферической крови на мкл крови у самцов до дня 352 поискового исследования токсичности. Tfh-подобные клетки определяли как сумму CD3+CD4+CD95+CXCR5+ клеток и CD3+CD4+CD95+hIgG+ клеток, т.к. тестируемый препарат препятствует антителу против CXCR5, используемому для иммунофенотипирования. Представлены данные для отдельных животных.
[00028] На фиг. 8D представлен график, на котором показано истощение и восстановление Tfh-подобных (также обозначаемых как "cTfh" или "циркулирующие Tfh") клеток у самок яванского макака. Показаны количества Tfh-подобных клеток в периферической крови на мкл крови у самок до дня 352 поискового исследования токсичности. Tfh-подобные клетки определяли как сумму CD3+CD4+CD95+CXCR5+ клеток и CD3+CD4+CD95+hIgG+ клеток, т.к. тестируемый препарат препятствует антителу против CXCR5, используемому для иммунофенотипирования. Представлены данные для отдельных животных.
[00029] На фиг. 9A представлен график, на котором показано истощение и восстановление B-клеток периферической крови у яванского макака. Показаны количества B-клеток в периферической крови на мкл крови у обезьян до дня 393 опорного исследования токсичности, соответствующего требованиям GLP. B-клетки определяли как CD3-CD20+ HLA-DR+ клетки. Показаны средние для группы данные (самцы и самки вместе) ± стандартное отклонение.
[00030] На фиг. 9B представлен график, на котором показана истощение и восстановление CXCR5+ B-клеток периферической крови у яванских макаков. Показаны количества CXCR5+ B-клеток периферической крови на мкл крови у обезьян до дня 393 опорного исследования токсичности, соответствующего требованиям GLP. B-клетки определяли как CD3-CD20+ HLA-DR+ клетки. Показаны средние для группы данные (самцы и самки вместе) ± стандартное отклонение.
[00031] На фиг. 9C представлен график, на котором показано истощение и восстановление циркулирующих фолликулярных T-хелперных клеток периферической крови (cTfh; также обозначаемых в других местах настоящего описания как "Tfh-подобные клетки") у яванских макаков. Показаны количества cTfh-клеток периферической крови на мкл крови у обезьян до дня 393 опорного исследования токсичности, соответствующего требованиям GLP. cTfh-клетки определяли как CD3+CD4+CD95+ клетки. Показаны средние для группы данные (самцы и самки вместе) ± стандартное отклонение.
[00032] На фиг. 9D представлен график, на котором показано истощение и восстановление CXCR5+ cTfh-клеток периферической крови (также обозначаемых в других местах настоящего описания как "Tfh-подобные клетки") с детектируемым поверхностным CXCR5 у яванских макаков. Показаны количества CXCR5+ cTfh-клеток периферической крови на мкл крови у обезьян до дня 393 опорного исследования токсичности, соответствующего требованиям GLP. cTfh-клетки определяли как CD3+CD4+CD95+ клетки. Показаны средние для группы данные (самцы и самки вместе) ± стандартное отклонение.
ПОДРОБНОЕ ОПИСАНИЕ
[00033] Настоящее изобретение относится к антителам, специфически связывающимся с CXCR5, и, кроме того, антителам, антагонистически действующим на активность CXCR5 или его взаимодействие с CXCL13. Настоящее изобретение относится к способам получения антител против CXCR5, композициям, содержащим эти антитела, и способам применения этих антител.
[00034] Настоящее изобретение относится к фукозилированным и афукозилированным антителам, связывающимся с CXCR5. В некоторых вариантах осуществления настоящее изобретение также относится к тяжелым цепям и легким цепям афукозилированного антитела, способным образовывать антитела, связывающиеся с CXCR5. В некоторых вариантах осуществления настоящее изобретение относится к афукозилированным антителам, тяжелым цепям и легким цепям, содержащим одну или более конкретных определяющих комплементарность областей (CDR). В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют измененную эффекторную функцию. В некоторых вариантах осуществления антитела по изобретению имеют повышенную активность ADCC относительно в остальном идентичных фукозилированных антител против CXCR5 по изобретению.
[00035] Настоящее изобретение относится к полинуклеотидам, кодирующим антитела, связывающиеся с CXCR5, или их антигенсвязывающие фрагменты. Настоящее изобретение также относится к полинуклеотидам, кодирующим тяжелые цепи или легкие цепи антитела. Настоящее изобретение относится к клеткам-хозяевам, экспрессирующим фукозилированные и/или афукозилированные антитела против CXCR5. Настоящее изобретение относится к способам лечения с использованием афукозилированных и фукозилированных антител против CXCR5. Такие способы включают, в качестве неограничивающих примеров, способы лечения заболевания, ассоциированного или опосредованного экспрессией CXCR5 и/или связыванием с CXCL13, включая, в качестве неограничивающих примеров, воспалительные и иммунологические заболевания.
[00036] Заголовки разделов, используемые в настоящем описании, представлены исключительно в организационных целях, и не следует истолковывать их как ограничение настоящего изобретения.
[00037] Все цитируемые в настоящем описании ссылки, включая патентные заявки, патентные публикации и регистрационные номера Genbank, включены в настоящее описание в качестве ссылки так, как если бы каждая отдельная ссылка была конкретно и отдельно указана как включенная в качестве ссылки в полном объеме.
[00038] Способы, описанные или упомянутые в настоящем описании, как правило, хорошо понятны и общепринято используются специалистами в этой области с использованием общепринятой методологии, например, широко используемыми способами, описанными в Sambrook et al, Molecular Cloning: A Laboratory Manual 3rd. edition (2001) Cold Spring Harbor Laboratory Press, Cold Spring Harbor, N.Y. CURRENT PROTOCOLS IN MOLECULAR BIOLOGY (F. M. Ausubel, et al. eds., (2003)); серии METHODS IN ENZYMOLOGY (Academic Press, Inc.): OCR 2: A PRACTICAL APPROACH (M. J. MacPherson, B. D. Hames и G. R. Taylor eds. (1995)), Harlow and Lane, eds. (1988) ANTIBODIES, A LABORATORY MANUAL, и ANIMAL CELL CULTURE (R. I. Freshney, ed. (1987)); Oligonucleotide Synthesis (M. J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J. E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R. I. Freshney), ed., 1987); Introduction to Cell and Tissue Culture (J. P. Mather и P. E. Roberts, 1998) Plenum Press; Cell and Tissue Culture Laboratory Procedures (A. Doyle, J. B. Griffiths, and D. G. Newell, eds., 1993-8) J. Wiley and Sons; Handbook of Experimental Immunology (D. M. Weir and C. C. Blackwell, eds); Gene Transfer Vectors for Mammalian Cells (J. M. Miller and M. P. Calos, eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al, eds., 1994); Current Protocols in Immunology (J. E. Coligan et al, eds., 1991); Short Protocols in Molecular Biology (Wiley and Sons, 1999); Immunobiology (C. A. Janeway и P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: A Practical Approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal Antibodies: A Practical Approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using Antibodies: A Laboratory Manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999)); Antibodies (M. Zanetti and J. D. Capra, eds., Harwood Academic Publishers, 1995); и Cancer: Principles and Practice of Oncology (V. T. DeVita et al., eds., J.B. Lippincott Company, 1993); и их обновленных версиях.
[00039] Антитела против CXCR5, фукозилированные или афукозилированные, можно использовать в профилактике, лечении и/или улучшении заболеваний, нарушений или состояний, вызванных и/или ассоциированных с активностью CXCR5. Такие заболевания, нарушения или состояния включают, в качестве неограничивающих примеров, воспалительные ответы, такие как системная красная волчанка (SLE); хронические воспалительные ответы; атеросклероз; дефицит адгезии лейкоцитов; ревматоидный артрит; сахарный диабет (например, сахарный диабет I типа или инсулинозависимый сахарный диабет); рассеянный склероз; синдром Рейно; аутоиммунный тиреоидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные ответы, ассоциированные с острой и замедленной реакцией гиперчувствительности, опосредованные цитокинами и T-лимфоцитами, как правило, обнаруживаемые при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; гранулематоз Вегенера; пернициозную анемию (болезнь Аддисона); заболевания, включающие диапедез лейкоцитов; воспалительное нарушение центральной нервной системы (ЦНС); синдром полиорганной недостаточности; гемолитическую анемию (включая, в качестве неограничивающих примеров, криоглобулинемию или аутоиммунную гемолитическую анемию с неполными тепловыми агглютининами); миастению; заболевания, опосредованные комплексами антиген-антитело; болезнь анти-БМК-антител; антифосфолипидный синдром; аллергический неврит; болезнь Грейвса; миастенический синдром Ламберта-Итона; буллезный пемфигоид; пемфигус; аутоиммунные полиэндокринопатии; витилиго; болезнь Рейтера; синдром мышечной скованности; болезнь Бехчета; гигантоклеточный артериит; иммунокомплексный нефрит; IgA-нефропатию; IgM-полинейропатии; иммунную тромбоцитопеническую пурпуру (ITP) или аутоиммунную тромбоцитопению и аутоиммунные гемолитические заболевания; тиреоидит Хашимото; аутоиммунный гепатит; аутоиммунную гемофилию; аутоиммунный лимфопролиферативный синдром (ALPS); аутоиммунный увеоретинит; синдром Гийена-Барре; синдром Гудпасчера; смешанное заболевание соединительной ткани; аутоиммунно-ассоциированное бесплодие; нодозный полиартериит; гнездную алопецию; идиопатическую микседему; реакцию "трансплантат против хозяина"; мышечную дистрофию (дистрофию Дюшена, дистрофию Беккера, миотоническую дистрофию, конечностно-поясную дистрофию, плече-лопаточно-лицевую дистрофию, врожденную дистрофию, окулофарингеальную дистрофию, дистальную дистрофию, дистрофию Эмери-Дрейфуса) и контроль пролиферации злокачественных клеток, экспрессирующих CXCR5, такой как рак поджелудочной железы, рак толстого кишечника, рак мочевого пузыря, T-клеточный лейкоз и B-клеточный лейкоз, как будет понятно специалисту в этой области с учетом идей, представленных в настоящем описании.
I. ОПРЕДЕЛЕНИЯ
[00040] Настоящее изобретение будет более понятным со ссылкой на следующее подробное описание примеров вариантов осуществления изобретения и примеров, включенных в них.
[00041] Если не указано иначе, все технические и научные термины, используемые в настоящем описании, обладают тем же значением, которое общепринято понятно специалисту в области, к которой принадлежит настоящее изобретение. В случае конфликта, настоящее описание, включая определение, будет обладать приоритетом.
[00042] Кроме того, если контекст не требует иного или иное не указано конкретно, термины в единственном числе должны включать множественное число, и термины во множественном числе должны включать единственное число.
[00043] Следует понимать, что аспекты и варианты осуществления изобретения, представленные в настоящем описании, включают "состоящие из" и/или "по существу, состоящие из" аспекты и варианты осуществления. В рамках изобретения, если не указано иначе, термины в единственном числе включают ссылку на множественное число.
[00044] В настоящей заявке использование терминов "или" означает "и/или", если иное не указано конкретно или понятно специалисту в этой области. Что касается пунктов формулы, зависящих от других зависимых пунктов формулы, использование термина "или" включает ссылку на несколько предшествующих независимых или зависимых пунктов.
[00045] Термин "приблизительно" при использовании по отношению к измеримым числовым переменным относится к указанному значению переменной и всем значениям переменной, находящихся в пределах ошибки наблюдения от указанного значения (например, в пределах 95% доверительного интервала для среднего) или в пределах 10 процентов от указанного значения, в зависимости от того что больше. Числовые диапазоны являются инклюзивными для чисел, определяющих диапазон.
[00046] Несмотря на то, что числовые диапазоны и параметры, отражающие широкий объем изобретения, являются приблизительными, числовые значения, приведенные в конкретных примерах, указаны настолько точно насколько возможно. Однако любое числовое значение по определению содержит некоторые ошибки, неизбежно являющиеся результатом стандартного отклонения, обнаруживаемого при соответствующих тестовых измерениях. Кроме того, все диапазоны, представленные в настоящем описании, следует понимать как включающие любые и все поддиапазоны, включенные в них. Например, указанный диапазон "от 1 до 10" следует считать включающим любые и все поддиапазоны от (и включая) минимального значения 1 до максимального значения 10; т.е. все поддиапазоны, начинающиеся с минимального значения 1 или более, например, от 1 до 6,1, и заканчивающиеся максимальным значением 10 или менее, например, от 5,5 до 10.
[00047] На всем протяжении настоящего описания и формулы изобретения термин "содержат" или его варианты, такие как "содержит" или "содержащий", следует понимать как подразумевающие включение указанного целого числа или группы целых чисел, но не исключение любого другого целого числа или группы целых чисел. Если контекст не требует иного, термины в единственном числе должны включать множественное число, и термины во множественном числе должны включать единственное число. Любые примеры, следующие после термина "например", не должны быть исчерпывающими или ограничивающими.
[00048] Следует понимать, что в любом случае, когда варианты осуществления представлены в настоящем описании с термином "содержащий", они также включают в остальном аналогичные варианты осуществления, описанные в терминах "состоящий из" и/или "состоящий, по существу, из".
[00049] Если аспекты или варианты осуществления изобретения описаны в терминах группы Маркуша или другого группирования альтернатив, настоящее изобретение относится не только ко всей указанной группе в целом, но и к каждому члену группы отдельно и всем возможным подгруппам основной группы, а также к основной группе, в которой отсутствуют один или более членов группы. Настоящее изобретение также предусматривают явное исключение одного или более из любых членов группы из описываемого в заявке изобретения.
[00050] Следует понимать, что терминология, используемая в настоящем описании, предназначена исключительно для описания конкретных вариантов осуществления и не является ограничивающей. В настоящем описании и формуле изобретения сделана ссылка на ряд терминов, которые должны обладать следующим значением.
[00051] Термин "выделенная молекула" (если молекула является, например, полипептидом, полинуклеотидом или антителом или его фрагментом) представляет собой молекулу, которая благодаря своему происхождению или источнику получения (1) не ассоциирована с природно ассоциированными компонентами, сопутствующими ей в нативном состоянии, (2) по существу, не содержит другие молекулы того же биологического вида, (3) экспрессируется клеткой другого биологического вида, или (4) не встречается в природе. Таким образом, молекула, являющаяся химически синтезированной или экспрессирующейся в клеточной системе, отличающейся от клетки, из которой она происходит в природе, будет "выделенной" из своих природно ассоциированных компонентов. Молекулу также можно делать, по существу, несодержащей природно ассоциированные компоненты посредством выделения с использованием способов очистки, хорошо известных в этой области. Чистоту или гомогенность молекулы можно анализировать рядом способов, хорошо известных в этой области. Например, чистоту анализа полипептида можно анализировать с использованием электрофореза в полиакриламидном геле и окрашивания геля для визуализации полипептида с использованием способов, хорошо известных в этой области. Для некоторых целей можно достигать высокого разрешения с использованием ВЭЖХ или других способов очистки, хорошо известных в этой области.
[00052] В рамках изобретения термин "по существу, чистый" означает, что целевой вид является преобладающим присутствующим видом (т.е. в пересчете на моли он является более многочисленным, чем любой другой вид в композиции), и, предпочтительно, в значительной степени очищенная фракция представляет собой композицию, где целевой вид (например, гликопротеин, включая антитело или рецептор) составляет по меньшей мере приблизительно 50 процентов (в пересчете на моли) всех присутствующих макромолекулярных видов. Как правило, по существу, чистая композиция будет содержать более приблизительно 80 процентов всех макромолекулярных видов, присутствующих в композиции, более предпочтительно - более приблизительно 85%, 90%, 95% и 99%. Наиболее предпочтительно, целевой вид очищают до значительной гомогенности (в композиции невозможно определить загрязнения общепринятыми способами детекции), где композиция состоит, по существу, из одного вида макромолекул. В некоторых вариантах осуществления, по существу, чистый материал является по меньшей мере на 50% чистым (т.е. несодержащим загрязнения), более предпочтительно - по меньшей мере на 90% чистым, более предпочтительно - по меньшей мере на 95% чистым, еще более предпочтительно - по меньшей мере на 98% чистым, и наиболее предпочтительно - по меньшей мере на 99% чистым.
[00053] Как известно в этой области, термин "идентичность" относится к взаимосвязи между последовательностями двух или более полипептидных молекул или двух или более молекул нуклеиновой кислоты, что определяют посредством сравнения последовательностей. В этой области термин "идентичность" также означает степень родства между последовательностями полипептидных молекул или молекул нуклеиновой кислоты, соответственно, что определяют по совпадению между цепями нуклеотидных или аминокислотных последовательностей. По "идентичности" измеряют процент идентичных совпадений между двумя или более последовательностями с использованием выравнивания с включением пропусков, осуществляемого с помощью конкретной математической модели в компьютерных программах (т.е. "алгоритмах").
[00054] Термин "сходство" представляет собой связанную концепцию, но, в отличие от термина "идентичность", он относится к мере сходства, включающей идентичные совпадения и консервативные замены. Т.к. консервативные замены относятся к полипептидам, а не молекулам нуклеиновой кислоты, термин "сходство" относится только к сравнению полипептидных последовательностей. Если две полипептидные последовательности имеют, например, 10 идентичных аминокислот из 20, и все остальные являются неконсервативными заменами, то каждый из процента идентичности и сходства будет составлять 50%. Если в том же примере есть еще 5 положений, в которых есть консервативные замены, то процент идентичности остается 50%, но процент сходства будет составлять 75% (15 из 20). Таким образом, в случаях, когда есть консервативные замены, степень сходства между двумя полипептидными последовательностями будет выше процента идентичности между этими двумя последовательностями.
[00055] "Фрагменты" или "части" полипептида или антитела по изобретению можно получать посредством укорачивания, например, посредством удаления одной или более аминокислот с N- и/или C-концов полипептида. Таким образом, можно удалять до 10, до 20, до 30, до 40 или более аминокислот с N- и/или C-конца. Фрагменты или части также можно получать посредством одной или более внутренних делеций.
[00056] Вариант антитела может содержать 1, 2, 3, 4, 5, до 10, до 20, до 30 или более замен аминокислот, и/или делеций, и/или инсерций в конкретных последовательностях и фрагментах, описанных выше. Варианты с "делецией" могут содержать делецию отдельных аминокислот, делецию небольших групп аминокислот, таких как 2, 3, 4 или 5 аминокислот, или делецию более крупных аминокислотных областей, такую как делеция конкретных аминокислотных доменов или других признаков. Варианты с "инсерцией" могут содержат инсерцию отдельных аминокислот, инсерцию небольших групп аминокислот, таких как 2, 3, 4 или 5 аминокислот, или инсерцию более крупных аминокислотных областей, такую как инсерция конкретных аминокислотных доменов или других признаков. Варианты с "заменой", предпочтительно, включают замену одной или более аминокислот тем же количеством аминокислот и осуществление консервативных замен аминокислот. Например, аминокислоту можно заменять альтернативной аминокислотой, имеющей схожие свойства, например, другой основной аминокислотой, другой кислой аминокислотой, другой нейтральной аминокислотой, другой заряженной аминокислотой, другой гидрофильной аминокислотой, другой гидрофобной аминокислотой, другой полярной аминокислотой, другой ароматической аминокислотой или другой алифатической аминокислотой. Некоторые свойства 20 главных аминокислот, которые можно использовать для выбора подходящих заместителей, описаны ниже.
[00057] Варианты с заменой содержат по меньшей мере один удаленный аминокислотный остаток в молекуле антитела и другой остаток, встроенный вместо него. Участки, представляющие наибольший интерес в отношении заместительного мутагенеза, включают гипервариабельные области, но также предусмотрены изменения в каркасе. Консервативные замены приведены в таблице 1 под заголовком "консервативные замены". Если такие замены приводят к изменению биологической активности, то можно включать более существенные замены, обозначенные как "примеры замен", представленные ниже или описанные ниже по отношению к классам аминокислот, и подвергать продукты скринингу.
ТАБЛИЦА 1
Аминокислоты и замены
[00058] Значительные модификации биологических свойств антитела осуществляют посредством выбора замен, значительно отличающихся по своему эффекту в отношении поддержания (a) структуры полипептидного остова в области замены, например, в виде бета-листа или спиральной конформации, (b) заряда или гидрофобности молекулы в целевом участке, или (c) объема боковой цепи. Природные остатки разделяют на группы с учетом общих свойств боковой цепи:
i. Неполярные: норлейцин, Met, Ala, Val, Leu, Ile;
ii. Полярные незаряженные: Cys, Ser, Thr, Asn, Gln;
iii. Кислые (отрицательно заряженные): Asp, Glu;
iv. Основные (положительно заряженные): Lys, Arg;
v. Остатки, влияющие на ориентацию цепи: Gly, Pro; и
vi. Ароматические: Trp, Tyr, Phe, His.
[00059] Неконсервативные замены осуществляют посредством замены члена одного из этих классов членом другого класса.
[00060] Одним из типов замен, например, которые можно осуществлять, является замена одного или более цистеинов в антителе, которые могут являться химически реакционноспособными, другим остатком, таким как, в качестве неограничивающих примеров, аланин или серин. Например, можно осуществлять замену неканонического цистеина. Замену можно осуществлять в CDR или каркасной области вариабельного домена или в константной области антитела. В некоторых вариантах осуществления цистеин является каноническим. Любой остаток цистеина, невовлеченный в поддержание правильной конформации антитела, также можно заменять, как правило, серином, для улучшения окислительной стабильности молекулы и предотвращения аномальной перекрестной сшивки. И наоборот, цистеиновые связи можно добавлять в антитело для улучшения его стабильности, в частности, если антитело является фрагментом антитела, таким как Fv-фрагмент.
[00061] "Антитело" является молекулой иммуноглобулина, способного специфически связываться с мишенью, такой как углевод, полинуклеотид, липид, полипептид и т.д., с помощью по меньшей мере одного участка распознавания антигена, локализованного в вариабельной области молекулы иммуноглобулина. В рамках изобретения термин включает не только интактные поликлональные или моноклональные антитела, но также, если не указано иначе, любой его антигенсвязывающий фрагмент, конкурирующий с интактным антителом за специфическое связывание, слитые белки, содержащие антигенсвязывающий фрагмент, и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащей участок распознавания антигена. Антигенсвязывающие фрагменты включают, например, Fab, Fab’, F(ab’)2, Fd, Fv, доменные антитела (dAb, например, антитела акул и Верблюжьих), фрагменты, включающие определяющие комплементарность области (CDR), одноцепочечные антитела из вариабельного фрагмента (scFv), макситела, минитела, интратела, диатела, триатела, тетратела, v-NAR и бис-scFv, и полипептиды, содержащие по меньшей мере фрагмент иммуноглобулина, достаточный для придания полипептиду специфического связывания с антигеном. Антитело включает антитело любого класса, такое как IgG, IgA или IgM (или их подкласс), и антитело может не принадлежать к какому-либо конкретному классу. В зависимости от аминокислотной последовательности константной области антитела в их тяжелых цепях, иммуноглобулины можно приписывать к разным классам. Существует пять основных классов иммуноглобулинов: IgA, IgD, IgE, IgG и IgM, и несколько из них можно дополнительно разделять на подклассы (изотипы), например, IgG1, IgG2, IgG3, IgG4, IgA1 и IgA2. Константные области тяжелой цепи, соответствующие разным классам иммуноглобулинов, называют альфа, дельта, эпсилон, гамма и мю, соответственно. Хорошо известны структуры субъединиц и трехмерные конфигурации разных классов иммуноглобулинов.
[00062] Термины "антигенсвязывающая часть" или "антигенсвязывающий фрагмент" антитела (или просто "часть антитела"), как взаимозаменяемо используют в настоящем описании, относятся к одному или более фрагментам антитела, сохраняющим способность специфически связываются с антигеном (например, CXCR5). Показано, что антигенсвязывающую функцию антитела могут выполнять фрагменты полноразмерного антитела. Примеры связывающих фрагментов, включенных в термин "антигенсвязывающий фрагмент" антитела, включают (i) Fab-фрагмент, моновалентный фрагмент, состоящий из доменов VL, VH, CL и CH1; (ii) F(ab')2-фрагмент, бивалентный фрагмент, содержащий два Fab-фрагмента, соединенных дисульфидным мостиком в шарнирной области; (iii) Fd-фрагмент, состоящий из доменов VH и CH1; (iv) Fv-фрагмент, состоящий из доменов VL и VH одного плеча антитела, (v) dAb-фрагмент (Ward et al., (1989) Nature 341:544-546), состоящий из домена VH; и (vi) выделенную определяющую комплементарность область (CDR), Fv, соединенные дисульфидной связью (dsFv), и антиидиотипические антитела (анти-Id) и интратела. Кроме того, хотя два домена Fv-фрагмента, VL и VH, кодируются отдельными генами, их можно соединять рекомбинантными способами с помощью синтетического линкера, позволяющего получать их в виде единой белковой цепи, в которой области VL и VH спариваются с образованием моновалентных молекул (известных как одноцепочечные Fv (scFv)); см., например, Bird et al. Science 242:423-426 (1988) и Huston et al. Proc. Natl. Acad. Sci. USA 85:5879-5883 (1988)). Такие одноцепочечные антитела также предназначены для включения в термин "антигенсвязывающий фрагмент" антитела. Также включены другие формы одноцепочечных антител, такие как диатела. Диатела являются бивалентными, биспецифическими антителами, в которых домены VH и VL экспрессируются на одной полипептидной цепи, но в них используют линкер, являющийся слишком коротким, чтобы было возможным спаривание между двумя доменами на одной цепи, таким образом, заставляя домены спариваться с комплементарными доменами другой цепи и получая два антигенсвязывающих участка (см. например, Holliger et al. Proc. Natl. Acad. Sci. USA 90:6444-6448 (1993); Poljak et al., 1994, Structure 2:1121-1123).
[00063] Антитела можно получать из любого млекопитающего, включая, в качестве неограничивающих примеров, людей, обезьян, свиней, лошадей, кроликов, собак, кошек, мышей и т.д., или других животных, таких как птицы (например, куры), рыбы (например, акулы) и Верблюжьи (например, ламы).
[00064] Термин "вариабельная область" антитела относится к вариабельной области легкой цепи антитела (VL) или вариабельной области тяжелой цепи антитела (VH) в отдельности или в комбинации. Как известно в этой области, каждая из вариабельных областей тяжелых и легких цепей состоит из четырех каркасных областей (FR), соединенных тремя "определяющими комплементарность областями (CDR)", также известными как гипервариабельные области (HVR), и участвует в образовании антигенсвязывающего участка антитела. Если желательными являются варианты целевой вариабельной области, в частности, с заменой в аминокислотных остатках вне области CDR (т.е. в каркасной области), подходящую замену аминокислоты, предпочтительно, консервативную аминокислотную замену, можно идентифицировать посредством сравнения целевой вариабельной области с вариабельными областями других антител, содержащих последовательности CDR1 и CDR2, в том же каноническом классе в качестве целевой вариабельной области (Chothia and Lesk, J. Mol. Biol. 196(4): 901-917, 1987).
[00065] В некоторых вариантах осуществления точное разграничение CDR и идентификацию остатков, содержащих связывающий участок антитела, осуществляют посредством разрешения структуры антитела и/или разрешения структуры комплекса антитело-лиганд. В некоторых вариантах осуществления, которые можно получать любыми из множества способов, известных специалистам в этой области, такими как рентгеновская кристаллография. В некоторых вариантах осуществления различные способы анализа можно использовать для идентификации или приблизительного определения областей CDR. В некоторых вариантах осуществления различные способы анализа можно использовать для идентификации или приблизительного определения областей CDR. Неограничивающие примеры таких способов включают определение по Kabat, определение по Chothia, определение AbM, определение по системе Contact и конформационное определение.
[00066] Определение по Kabat является стандартом нумерации остатков в антителе, и, как правило, его используют для идентификации областей CDR. См., например, Johnson & Wu, 2000, Nucleic Acids Res., 28: 214-8. Определение по Chothia схоже с определением по Kabat, но в определении по Chothia учитывают положения конкретных областей структурных петель. См., например, Chothia et al., 1986, J. Mol. Biol., 196: 901-17; Chothia et al., 1989, Nature, 342: 877-83. В определении AbM используют встроенный пакет компьютерных программ, производимый Oxford Molecular Group, с помощью которого моделируют структуру антитела. См., например, Martin et al., 1989, Proc Natl Acad Sci (USA), 86:9268-9272; "AbM™, A Computer Program for Modeling Variable Regions of Antibodies," Oxford, UK; Oxford Molecular, Ltd. С помощью определения AbM моделируют третичную структуру антитела с учетом первичной последовательности с использованием комбинации информационных баз данных и способов ab initio, таких как описанные в Samudrala et al., 1999, "Ab Initio Protein Structure Prediction Using a Combined Hierarchical Approach," в PROTEINS, Structure, Function and Genetics Suppl., 3:194-198.
[00067] Определение по системе Contact основано на анализе доступных кристаллических структур комплексов. См., например, MacCallum et al., 1996, J. Mol. Biol., 5:732-45. В другом подходе, обозначенном в настоящем описании как "конформационное определение" CDR, положения CDR можно идентифицировать как остатки, делающие энтальпийный вклад в связывание антигена. См., например, Makabe et al., 2008, Journal of Biological Chemistry, 283:1156-1166. Хотя другие определения границ CDR могут не следовать точно одному из указанных выше подходов, но, тем не менее, будут перекрываться по меньшей мере с частью CDR по Kabat, хотя они могут быть укороченными или удлиненными с учетом прогнозирования или экспериментальных данных о том, что конкретные остатки или группы остатков не влияют значительно на связывание антигена. В рамках изобретения термин "CDR" может относиться к CDR, определяемым с помощью любого подхода, известного в этой области, включая комбинации подходов. В способах, используемых в настоящем описании, можно использовать CDR, определяемые в соответствии с любым из этих подходов. В случае любого указанного варианта осуществления, содержащего несколько CDR, CDR можно определять в соответствии с любым из определения по Kabat, определения по Chothia, расширенного определения, определения AbM, определения по системе Contact и/или конформационного определения.
[00068] В рамках изобретения термин "контактный остаток" в отношении антитела или специфически связанного им антигена относится к аминокислотному остатку, присутствующему на антителе/антигене, содержащему по меньшей мере один тяжелый атом (т.е. не водород), находящийся в пределах 4 Å или менее от тяжелого атома аминокислотного остатка, присутствующего на когнатном антителе/антигене.
[00069] "Каркасные" (FR) остатки являются остатками вариабельного домена антитела, иными, чем остатки CDR. Каркас домена VH или VL содержит четыре каркасных подобласти, FR1, FR2, FR3 и FR4, перемежающихся с CDR в следующей структуре: FR1-CDR1-FR2-CDR2-FR3-CDR3-FR4.
[00070] Как указано выше в настоящем описании, остатки в вариабельном домене, как правило, нумеруют по Kabat, что представляет собой систему нумерации, используемую для вариабельных доменов тяжелой цепи или вариабельных доменов легкой цепи совокупности антител. См., Kabat et al., 1991, Sequences of Proteins of Immunological Interest, 5th Ed. Public Health Service, National Institutes of Health, Bethesda, MD. При использовании этой системы нумерации точная линейная аминокислотная последовательность может содержать меньше аминокислот или дополнительные аминокислоты, соответствующие укорочению или инсерции в FR или CDR вариабельного домена. Например, вариабельный домен тяжелой цепи может включать инсерцию отдельной аминокислоты (остаток 52a по Kabat) после остатка 52 H2 и встроенные остатки (например, остатки 82a, 82b, и 82c по Kabat) после остатка 82 FR тяжелой цепи. Нумерацию остатков по Kabat можно определять для указанного антитела посредством выравнивания в областях гомологии последовательности антитела со "стандартной" пронумерованной по Kabat последовательностью. Доступны различные алгоритмы для приписывания нумерации по Kabat. Алгоритм, воплощенный в версии 2.3.3 выпуска Abysis (www.abysis.org), можно использовать для приписывания нумерации по Kabat вариабельным областям CDR-L1, CDR-L2, CDR-L3, CDR-H2 и CDR-H3, а затем можно использовать определение AbM для CDR-H1.
[00071] В рамках изобретения термин "моноклональное антитело" относится к антителу, получаемому из популяции, по существу, гомогенных антител, т.е. отдельные антитела, составляющие популяцию, являются идентичными, за исключением возможных природных мутаций, которые могут присутствовать в незначительных количествах. Моноклональные антитела являются высокоспецифическими, направленными против одного участка антигена. Кроме того, в отличие от препаратов поликлональных антител, которые, как правило, включают разные антитела против разных детерминант (эпитопов), каждое моноклональное антитело направлено против отдельной детерминанты на антигене. Определение "моноклональное" означает характер антитела как получаемого из, по существу, гомогенной популяции антител, и его не следует истолковывать как требующие получения антитела любым конкретным способом. Например, моноклональные антитела для использования в соответствии с настоящим изобретением можно получать гибридомным способом, впервые описанным в Kohler and Milstein, 1975, Nature 256:495, или можно получать способами рекомбинантной ДНК, такими как описанные в патенте США № 4816567. Моноклональные антитела также можно выделять из фаговых библиотек, полученных способами, описанными, например, в McCafferty et al., 1990, Nature 348:552-554. В рамках изобретения термин "гуманизированное" антитело относится к формам непринадлежащих человеку (например, мышиных) антител, являющихся химерными иммуноглобулинами, цепями иммуноглобулинов или их фрагментами (такими как Fv, Fab, Fab', F(ab')2 или другие антигенсвязывающие подпоследовательности антител), содержащими минимальную последовательность, полученную из непринадлежащего человеку иммуноглобулина. Предпочтительно, гуманизированные антитела являются иммуноглобулинами человека (реципиентным антителом), в которых остатки из CDR реципиентного антитела заменяют остатками из CDR неявляющихся человеком видов (донорного антитела), таких как мышь, крыса или кролик, имеющими желаемую специфичность, аффинность и емкость. Гуманизированное антитело может содержать остатки, необнаруживаемые ни в реципиентном антителе, ни в пересаженной CDR или каркасных последовательностях, но включенные для дополнительной коррекции и оптимизации свойств антитела.
[00072] Антитело или его антигенсвязывающий фрагмент по изобретению могут являться аффинно зрелыми. Например, аффинно зрелое антитело можно получать известными в этой области способами (Marks et al., 1992, Bio/Technology, 10:779-783; Barbas et al., 1994, Proc Nat. Acad. Sci, USA 91:3809-3813; Schier et al., 1995, Gene, 169:147-155; Yelton et al., 1995, J. Immunol., 155:1994-2004; Jackson et al., 1995, J. Immunol., 154(7):3310-9; Hawkins et al., 1992, J. Mol. Biol., 226:889-896 и WO2004/058184).
[00073] "Антитело человека" является антителом, обладающим аминокислотной последовательностью, соответствующей последовательности антитела, продуцируемого человеком и/или полученного любыми из способов получения антител человека, представленных в настоящем описании. Это определение антитела человека конкретно исключает гуманизированное антитело, содержащее антигенсвязывающие остатки, не принадлежащие человеку.
[00074] Термин "химерное антитело" предназначен для обозначения антитела, в котором последовательности вариабельной области, получают из одного вида, а последовательности константной области получают из другого вида, такого как антитело, в котором последовательности вариабельной области получают из антитела мыши, и последовательности константной области получают из антитела человека, или наоборот. Термин также включает антитело, содержащее V-область одного индивидуума одного биологического вида (например, первой мыши) и константную область другого индивидуума того же биологического вида (например, второй мыши).
[00075] Термин "антиген (Ag)" относится к молекуле, используемой для иммунизации иммунокомпетентного позвоночного для получения антитела (Ab), распознающего Ag, или для скрининга экспрессионной библиотеки (например, помимо прочего, библиотеки фагового, дрожжевого или рибосомного дисплея). В настоящем описании термин "Ag" определяют в более широком смысле, и, как правило, он предназначен для включения молекул-мишеней, специфически распознаваемых Ab, таким образом, включая фрагменты или миметики молекулы, используемые для иммунизации для получения Ab или при скрининге библиотеки для селекции Ab. Таким образом, в случае антител по изобретению, связывающихся с CXCR5, полноразмерный CXCR5 млекопитающих (например, CXCR5 человека, обезьяны, мыши и крысы), включая их мономеры и мультимеры, такие как димеры, тримеры и т.д., а также укороченные и другие варианты CXCR5, обозначают как антиген.
[00076] Как правило, термин "эпитоп" относится к области антигена (например, белка, нуклеиновой кислоты, углевода или липида и т.д.), с которым антитело специфически связывается, т.е. области, физически контактирующей с антителом. Таким образом, термин "эпитоп" относится к части молекулы, которая может распознаваться и связываться антителом в одной или более антигенсвязывающих областей антитела. Как правило, эпитоп определяют в контексте молекулярного взаимодействия между "антителом или его антигенсвязывающим фрагментом" (Ab) и соответствующим ему антигеном. Эпитопы зачастую состоят из поверхностной группировки молекул, таких как аминокислоты или сахарные боковые цепи, и имеют специфические трехмерные структурные характеристики, а также специфические характеристики заряда. В некоторых вариантах осуществления эпитоп может являться белковым эпитопом. Белковые эпитопы могут являться линейными или конформационными. В линейном эпитопе все точки взаимодействия между белком и взаимодействующей молекулой (такой как антитело) расположены линейно вдоль первичной аминокислотной последовательности белка. "Нелинейный эпитоп" или "конформационный эпитоп" содержит несмежные полипептиды (или аминокислоты) в антигенном белке, с которым связывается антитело, специфическое для эпитопа. В рамках изобретения термин "антигенный эпитоп" определяют как часть антигена, с которой антитело может специфически связываться, что определяют любым способом, хорошо известным в этой области, например, с помощью общепринятых иммунологических анализов. Альтернативно, во время открытия при получении и характеризации антител можно выяснять информацию о желаемых эпитопах. С помощью этой информации можно осуществлять конкурентный скрининг антител на связывание с тем же эпитопом. Этого можно достигать с помощью анализов конкуренции и перекрестной конкуренции для обнаружения антител, конкурирующих или перекрестно конкурирующих друг с другом за связывание с CXCR5, например, антител, конкурирующих за связывание с антигеном.
[00077] Антитело, "преимущественно связывающееся" или "специфически связывающееся" (термины, используемые взаимозаменяемо в настоящем описании) с эпитопом, представляет собой термин, хорошо известный в этой области, и способы определения такого специфического или преимущественного связывания также хорошо известны в этой области. Указывают, что молекула демонстрирует "специфическое связывание" или "преимущественное связывание", если она реагирует или ассоциируется чаще, быстрее, с большей длительностью и/или с более высокой аффинностью с конкретной клеткой или веществом, чем с альтернативными клетками или веществами. Антитело "специфически связывается" или "преимущественно связывается" с мишенью, если оно связывается с более высокой аффинностью, авидностью, легче и/или с большей длительностью, чем с другими веществами. Кроме того, антитело "специфически связывается" или "преимущественно связывается" с мишенью, если оно связывается с более высокой аффинностью, авидностью, легче и/или с большей длительностью с этой мишенью в образце, чем с другими веществами, присутствующими в образце. Например, антитело, специфически или преимущественно связывающееся с эпитопом CXCR5, является антителом, связывающимся с этим эпитопом в более высокой аффинностью, авидностью, легче и/или с большей длительностью, чем с другими эпитопами CXCR5 или эпитопами, не принадлежащими CXCR5. Также из этого определения следует понимать, например, что антитело (или вещество или эпитоп), специфически или преимущественно связывающееся с первой мишенью может специфически или преимущественно связываться или не связываться со второй мишенью. В связи с этим, "специфическое связывание" или "преимущественное связывание" может не требовать исключительного связывания (хотя может включать его). Как правило, но необязательно, ссылка на связывание означает преимущественное связывание. "Специфическое связывание" или "преимущественное связывание" включает соединение, например, белок, нуклеиновую кислоту, антитело и т.п., распознающее и связывающееся со специфической молекулой, но не обязательно распознающее или связывающее другие молекулы в образце. Например, антитело или пептидный рецептор, распознающее и связывающееся с когнатным лигандом или партнером по связыванию (например, антителом против CXCR5, связывающимся с CXCR5) в образце, но, по существу, не распознающее и не связывающееся с другими молекулами в образце, специфически связывается с этим когнатным лигандом или партнером по связыванию. Таким образом, в условиях описанного анализа указанное связывающее вещество (например, антитело или его антигенсвязывающий фрагмент или рецептор или его лигандсвязывающий фрагмент) связывается преимущественно с конкретной молекулой-мишенью и не связывается в значительной степени с другими компонентами, присутствующими в тестируемом образце.
[00078] Для выбора антитела или пептида, специфически связывающегося с интересующей молекулой, можно использовать множество форматов анализов. Например, твердофазный иммунологический анализ ELISA, иммунопреципитация, Biacore™ (GE Healthcare, Piscataway, NJ), KinExA, активируемая флуоресценцией сортировка клеток (FACS), Octet™ (FortéBio, Inc., Menlo Park, CA) и анализ вестерн-блоттинга являются некоторыми из анализов, которые можно использовать для идентификации антитела, специфически реагирующего с антигеном, или рецептором, или его лигандсвязывающего фрагмента, специфически связывающегося с когнатным лигандом или партнером по связыванию. Как правило, специфическая или селективная реакция будет по меньшей мере в два раза превышать фоновый сигнал или шум, более типично - более чем в 10 раз, даже более типично - более чем в 50 раз, более типично - более чем в 100 раз, еще более типично - более чем в 500 раз, даже более типично - более чем в 1000 раз, и даже более типично - более чем в 10000 раз. Кроме того, указывают, что антитело "специфически связывается" с антигеном, если равновесная константа диссоциации (KD) составляет ≤1 мкМ, предпочтительно - ≤100 нМ, более предпочтительно - ≤10 нМ, даже более предпочтительно - ≤100 пМ, еще более предпочтительно - ≤10 пМ, и даже более предпочтительно - ≤1 пМ. В некоторых вариантах осуществления указывают, что антитело "специфически связывается" с антигеном, если равновесная константа диссоциации (KD) составляет ≤7 нМ.
[00079] В настоящем описании термин "аффинность связывания" используют в качестве меры силы нековалентного взаимодействия между двумя молекулами, например, антителом или его фрагментом и антигеном. Термин "аффинность связывания" используют для описания моновалентных взаимодействий (характерной активности).
[00080] Кроме того, для определения аффинности связывания антител против CXCR5 с CXCR5-экспрессирующими клетками можно осуществлять эксперименты по связыванию с клетками для определения кажущейся аффинности. Кажущаяся аффинность связывания антитела с клетками, экспрессирующими мишень, можно вычислять как EC50 кривых титрования равновесного связывания, в которых геометрическое среднее интенсивности флуоресценции (gMFI) антигенсвязывающей популяции количественно анализируют посредством проточной цитометрии.
[00081] Аффинность связывания между двумя молекулами, например, антителом или его фрагментом и антигеном, посредством моновалентного взаимодействия можно количественно анализировать посредством определения константы диссоциации (KD). В свою очередь, KD можно определять посредством измерения кинетики образования и диссоциации комплексов с использованием, например, способа поверхностного плазмонного резонанса (SPR) (Biacore). Константы скорости, соответствующие ассоциации и диссоциации моновалентного комплекса, обозначают как константу скорости ассоциации ka (или kon) и константу скорости диссоциации kd (или koff), соответственно. KD связана с ka и kd через уравнение KD=kd/ka. Значение константы диссоциации можно определять напрямую хорошо известными способами и можно вычислять даже для сложных смесей способами, такими как способы, например, описанные в Caceci et al. (1984, Byte 9: 340-362). Например, KD можно определять с использованием анализа связывания с двойным нитроцеллюлозным фильтром, такого как анализ, описываемый в Wong & Lohman (1993, Proc. Natl. Acad. Sci. USA 90: 5428-5432). В этой области известны другие стандартные анализы для оценки связывающей способности лигандов, таких как антитела против антигенов-мишеней, включая например, ELISA, вестерн-блоттинг, RIA и проточную цитометрию и другие анализы, примеры которых приведены в других местах настоящего описания. Кинетику связывания и аффинность связывания антитела также можно анализировать с помощью стандартных анализов, известных в этой области, таких как поверхностный плазмонный резонанс (SPR), например, с использованием системы Biacore™ или KinExA.
[00082] Можно осуществлять анализ конкурентного связывания, в котором связывание антитела с антигеном сравнивают со связыванием мишени с другим лигандом этой мишени, таким как другое антитело или растворимый рецептор, иным образом связывающимся с мишенью. Концентрация, при которой происходит 50% ингибирование, известна как Ki. В идеальных условиях Ki эквивалентна KD. Значение Ki никогда не бывает меньше KD, таким образом, измерение Ki можно заменять для получения верхнего предела KD.
[00083] Следуя указанному выше определению, аффинности связывания, ассоциированные с разными молекулярными взаимодействиями, например, аффинность связывания разных антител с указанным антигеном, можно сравнивать посредством сравнения значений KD отдельных комплексов антитело/антиген. Значения KD антител или других партнеров по связыванию можно определять способами, хорошо известными в этой области. Одним из способов определения KD является использование поверхностного плазмонного резонанса, как правило, с помощью биосенсорной системы, такой как система Biacore®.
[00084] Аналогично, специфичность взаимодействия можно анализировать посредством определения и сравнения значения KD интересующего взаимодействия, например, специфического взаимодействия между антителом и антигеном, со значением KD взаимодействия, не представляющего интерес, например, с контрольным антителом, о котором известно, что оно не связывается с CXCR5.
[00085] Антитело, специфически связывающееся с мишенью, может связываться с мишенью с высокой аффинностью, т.е. имеет низкую KD, как указано выше, и может связываться с другими, нецелевыми молекулами с более низкой аффинностью. Например, антитело может связываться с нецелевыми молекулами с KD 1×10-6M или более, более предпочтительно - 1×10-5 M или более, более предпочтительно - 1×10-4 M или более, более предпочтительно - 1×10-3 M или более, даже более предпочтительно - 1×10-2 M или более. Антитело по изобретению, предпочтительно, может связываться с мишенью с аффинностью, по меньшей мере в два раза, 10 раз, 50 раз, 100 раз, 200 раз, 500 раз, 1000 раз или 10000 раз или более превышающей ее аффинность связывания с другой молекулой, неявляющейся CXCR5.
[00086] В рамках изобретения термин "конкурирует" в отношении антитела означает, что первое антитело или его антигенсвязывающий фрагмент связывается с эпитопом, по существу, схожим образом со связыванием второго антитела или его антигенсвязывающего фрагмента таким образом, что результат связывания первого антитела с его когнатным эпитопом детектируемо снижается в присутствии второго антитела по сравнению со связыванием первого антитела в отсутствие второго антитела. Необязательно, может иметь место альтернативный вариант, где связывание второго антитела с его эпитопом также детектируемо снижается в присутствии первого антитела. Т.е. первое антитело может ингибировать связывание второго антитела с его эпитопом без ингибирования вторым антителом связывания первого антитела с соответствующим эпитопом. Однако если каждое антитело детектируемо ингибирует связывание другого антитела с его когнатным эпитопом или лигандом, в той же, большей или меньшей степени, указывают, что антитела "перекрестно конкурируют" друг с другом за связывание с соответствующими эпитопами. Конкурирующие и перекрестно конкурирующие антитела входят в объем настоящего изобретения. Независимо от механизма, посредством которого происходит такая конкуренция или перекрестная конкуренция (например, стерическое препятствие, конформационное изменение или связывание с общим эпитопом или его частью), специалистам в этой области с учетом идей, представленных в настоящем описании, будет известно, что предусмотрены такие конкурирующие и/или перекрестно конкурирующие антитела, и их можно использовать в способах, представленных в настоящем описании.
[00087] Для определения того, конкурируют ли два антитела друг с другом, можно использовать стандартные конкурентные анализы. Один из подходящих анализов конкуренции антител включает использование технологии Biacore, с помощью которого можно измерять степень взаимодействий с использованием поверхностного плазмонного резонанса (SPR), как правило, с помощью биосенсорной системы (такой как система BIACORE®). Например, SPR можно использовать в анализе ингибирования конкурентного связывания in vitro для определения способности одного антитела ингибировать связывание второго антитела. В другом анализе для измерения конкуренции антител используют подход на основе ELISA.
[00088] Кроме того, в международной патентной заявке № WO2003/48731 описывают высокопроизводительный способ "биннинга" антител в зависимости от их конкуренции. Конкуренция имеет место, если одно антитело (или фрагмент) снижает связывание другого антитела (или фрагмента) с CXCR5. Например, можно использовать последовательный анализ конкуренции за связывание с использованием разных антител, добавляемых последовательно. Первое антитело можно добавлять для достижения связывания, близкого к насыщению. Затем добавляют второе антитело. Если связывание второго антитела с CXCR5 не определено или значительно снижено (например, снижение по меньшей мере на приблизительно 10%, по меньшей мере на приблизительно 20%, по меньшей мере на приблизительно 30%, по меньшей мере на приблизительно 40%, по меньшей мере на приблизительно 50%, по меньшей мере на приблизительно 60%, по меньшей мере на приблизительно 70%, по меньшей мере на приблизительно 80% или по меньшей мере на приблизительно 90%) по сравнению с параллельным анализом в отсутствие первого антитела (значение в котором можно принимать за 100%), два антитела считают конкурирующими друг с другом.
[00089] Кроме того, пример анализа биннинга эпитопа антитела с использованием перестановки доменов между белками CXCR5 человека и мыши для оценки потенциальных эпитопов среди нескольких антител представлен в примере 8. Специалистам в этой области с учетом идей, представленных в настоящем описании, будет понятно, что существует широкий спектр известных в этой области анализов, которые можно использовать для определения связывания с мишенью по меньшей мере двух антител относительно друг с другом, и такие анализы включены в настоящем описании.
[00090] Антитела против CXCR5 можно охарактеризовывать хорошо известными в этой области способами. Например, одним из способов является идентификация эпитопа, с которым оно связывается, или "эпитопное картирование". Существует множество известных в этой области способов картирования и характеризации локализации эпитопов на белках, включая разрешение кристаллической структуры комплекса антитело-антиген, конкурентные анализы, анализы экспрессии фрагментов генов и анализы с использованием синтетических пептидов, как описано, например, в главе 11 Harlow and Lane, Using Antibodies, a Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, New York, 1999. В дополнительном примере эпитопное картирование можно использовать для определения последовательности, с которой связывается антитело против CXCR5. Эпитопное картирование является коммерчески доступным в различных источниках, например, Pepscan Systems (15 Edelhertweg 8219 PH Lelystad, The Netherlands). Эпитоп может являться линейным эпитопом, т.е. содержащимся в одном аминокислотном фрагменте, или конформационным эпитопом, образованным посредством трехмерного взаимодействия аминокислот, которые могут не содержаться в одном фрагменте. Пептиды разной длины (например, по меньшей мере 4-6 аминокислот) можно выделять или синтезировать (например, рекомбинантно) и использовать для анализов связывания с использованием антитела против CXCR5.
[00091] Кроме того, эпитоп, с которым связывается антитело против CXCR5, можно определять при системном скрининге с использованием перекрывающихся пептидов, полученных из последовательности CXCR5, и определении связывания антителом. В соответствии с анализами экспрессии фрагментов генов, открытую рамку считывания, кодирующую CXCR5, можно фрагментировать случайным образом или с помощью специфических генетических конструкций и определять реактивность экспрессируемых фрагментов CXCR5 с антителом, подлежащим тестированию. Фрагменты генов можно получать, например, с помощью ПЦР, а затем транскрибировать и транслировать в белок in vitro в присутствии радиоактивно меченых аминокислот. Затем связывание антитела с радиоактивно мечеными фрагментами CXCR5 определяют с помощью иммунопреципитации и электрофореза в геле.
[00092] Некоторые эпитопы также можно идентифицировать с использованием крупных библиотек случайных пептидных последовательностей, экспонируемых на поверхности фаговых частиц (фаговые библиотеки) или дрожжей (дрожжевой дисплей). Альтернативно, определенную библиотеку перекрывающихся пептидных фрагментов можно тестировать на связывание с тестируемым антителом в простых анализах связывания. В дополнительном примере для идентификации остатков, необходимых, достаточных и/или необходимых для связывания эпитопа можно осуществлять мутагенез антигена, эксперименты по перестановке доменов и мутагенез с аланиновым сканированием. Например, можно осуществлять эксперименты по мутагенезу с аланиновым сканированием с использованием мутантного CXCR5, в которых различные остатки полипептида CXCR5 заменяют аланином. Анализируя связывание антитела с мутантным CXCR5, можно оценивать важность конкретных остатков CXCR5 для связывания антитела.
[00093] Еще одним способом, который можно использовать для характеризации антитела CXCR5, является использование конкурентных анализов с помощью других антител, о которых известно, что они связываются с тем же антигеном, т.е. различными фрагментами на CXCR5, для определения того, связывается ли антитело против CXCR5 с тем же эпитопом, что и другие антитела. Конкурентные анализы хорошо известны специалистам в этой области.
[00094] Кроме того, эпитоп для указанной связывающей пары антитело/антиген можно определять и охарактеризовывать на различных уровнях с использованием различных экспериментальных и вычислительных способов эпитопного картирования. Экспериментальные способы включают мутагенез, рентгеновскую кристаллографию, спектроскопию ядерного магнитного резонанса (ЯМР), масс-спектрометрию водородно-дейтериевого обмена (H/D-MS) и различные способы конкурентного связывания, хорошо известные в этой области. Т.к. каждый способ основан на уникальном принципе, описание эпитопа непосредственно связано со способом, посредством которого его определяют. Таким образом, эпитоп для указанной пары антитело/антиген будут определять по-разному в зависимости от используемого способа эпитопного картирования.
[00095] На наиболее детальном уровне эпитоп для взаимодействия между Ag и Ab можно определять по пространственным координатам, определяющим атомные контакты, имеющие место при взаимодействии Ag-Ab, а также информации об их относительном вкладе в термодинамику связывания. На менее детальном уровне эпитоп можно охарактеризовывать по пространственным координатам, определяющим атомные контакты между Ag и Ab. На еще менее детальном уровне эпитоп можно охарактеризовывать по аминокислотным остаткам, которые он содержит, что определяют с помощью конкретного критерия, например, расстояния между атомами (например, тяжелыми, т.е. неводородными атомами) в Ab и Ag. На еще менее детальном уровне эпитоп можно охарактеризовывать по функции, например, по конкурентному связыванию с другими Ab. Эпитоп также можно определять в более общем плане как содержащий аминокислотные остатки, в случае которых замена другой аминокислотой будет изменять характеристики взаимодействия между Ab и Ag (например, с использованием аланинового сканирования).
[00096] Из того, что описание и определения эпитопов в зависимости от используемого способа эпитопного картирования получают на разных уровнях детальности, следует, что сравнение эпитопов для разных Ab на одном и том же Ag аналогичным образом можно осуществлять на разных уровнях детальности.
[00097] Указывают, что эпитопы, описываемые на уровне аминокислот, например, определяемые с помощью рентгеновской структуры, являются идентичными, если они содержат один и тот же набор аминокислотных остатков. Указывают, что эпитопы перекрываются, если по меньшей мере одна аминокислота является общей для эпитопов. Указывают, что эпитопы являются отдельными (уникальными), если у эпитопов нет общих аминокислотных остатков.
[00098] Указывают, что эпитопы, характеризуемые по конкурентному связыванию, перекрываются, если связывание соответствующих антител является взаимоисключающим, т.е. связывание одного антитела исключает одновременное или последовательное связывание другого антитела. Указывают, что эпитопы являются отдельными (уникальными), если антиген может приспосабливаться к одновременному связыванию обоих соответствующих антител.
[00099] Определение термина "паратоп" следует из указанного выше определения "эпитопа" в обратном смысле. Таким образом, термин "паратоп" относится к области на антителе, специфически связывающейся с антигеном, т.е. аминокислотным остаткам на антителе, контактирующим с антигеном (CXCR5) в соответствии с определением "контакта" в других местах настоящего описания.
[000100] Эпитоп и паратоп для указанной пары антитело/антиген можно идентифицировать общепринятыми способами. Например, общую локализацию эпитопа можно определять посредством оценки способности антитела связываться с разными фрагментами или вариантами полипептидов CXCR5. Конкретные аминокислоты в CXCR5, контактирующие с антителом (эпитоп), и конкретные аминокислоты в антителе, контактирующие с CXCR5 (паратоп), также можно определять общепринятыми способами, такими как способы, описанные в примерах. Например, можно комбинировать антитело и молекулу-мишень и кристаллизовать комплекс антитело/антиген. Кристаллическую структуру комплекса можно определять и использовать для идентификации специфических участков взаимодействия между антителом и его мишенью.
[000101] Антитело по изобретению может связываться с тем же эпитопом или доменом CXCR5, что и антитела по изобретению, конкретно представленные в настоящем описании. Анализы, которые можно использовать с целью такой идентификации, включают анализы, в которых оценивают конкуренцию за связывание CXCR5 между интересующим антителом и рецептором CXCR5 в рамках анализов биологической активности, как описано в примерах 1-10, и анализов кристаллической структуры антитела.
[000102] Антитело по изобретению может обладать способностью конкурировать или перекрестно конкурировать с другим антителом по изобретению за связывание с CXCR5, как представлено в настоящем описании. Например, антитело по изобретению может конкурировать или перекрестно конкурировать с антителами, представленными в настоящем описании, за связывание с CXCR5 или с подходящим фрагментом или вариантом CXCR5, связанным антителами, представленными в настоящем описании.
[000103] Т.е., если первое антитело конкурирует со вторым антителом за связывание с CXCR5, но не конкурирует, если второе антитело первым связывается с CXCR5, считают, что оно "конкурирует" со вторым антителом (также обозначаемым как однонаправленная конкуренция). Если антитело конкурирует с другим антителом независимо от того, какое антитело первым связывается с CXCR5, то антитело "перекрестно конкурирует" за связывание с CXCR5 с другим антителом. Такие конкурирующие или перекрестно конкурирующие антитела можно идентифицировать с учетом их способности конкурировать/перекрестно конкурировать с известным антителом по изобретению в стандартных анализах связывания. Например, для демонстрации конкуренции/перекрестной конкуренции можно использовать SPR, например, с использованием системы Biacore™, анализы ELISA или проточную цитометрию. Такая конкуренция/перекрестная конкуренция может позволить предположить, что два антитела связываются с идентичными, перекрывающимися или схожими эпитопами.
[000104] Таким образом, антитело по изобретению можно идентифицировать способом, включающим анализ связывания, в котором оценивают то, может ли тестируемое антитело конкурировать/перекрестно конкурировать с референсным антителом за участок связывания на молекуле-мишени. Способы осуществления анализов конкурентного связывания представлены в настоящем описании и/или хорошо известны в этой области. Например, они могут включать связывание референсного антитела по изобретению с молекулой-мишенью с использованием условий, в которых антитело может связываться с молекулой-мишенью. Затем комплекс антитело/мишень можно подвергать воздействию тестируемого/второго антитела и оценивать степень, с которой тестируемое антитело может вытеснять референсное антитело по изобретению из комплексов антитело/мишень. Альтернативный способ может включать приведение тестируемого антитела в контакт с молекулой-мишенью в условиях, делающих возможным связывание антитела, а затем добавление референсного антитела по изобретению, способного связываться с молекулой-мишенью, и оценку степени, с которой референсное антитело по изобретению может вытеснять тестируемое антитело из комплексов антитело/мишень или одновременно связываться с мишенью (т.е. неконкурирующее антитело).
[000105] Способность тестируемого антитела ингибировать связывание референсного антитела по изобретению с мишенью свидетельствует о том, что тестируемое антитело может конкурировать с референсным антителом по изобретению за связывание с мишенью, и, таким образом, что тестируемое антитело связывается с тем же или, по существу, тем же эпитопом или областью на белке CXCR5 в качестве референсного антитела по изобретению. Тестируемое антитело, идентифицируемое как конкурирующее с референсным антителом по изобретению в таком способе, также является антителом по настоящему изобретению. То, что тестируемое антитело может связываться с CXCR5 в той же области, что и референсное антитело по изобретению, и может конкурировать с референсным антителом по изобретению, позволяет предполагать, что тестируемое антитело может действовать в качестве лиганда в том же участке связывания, что и антитело по изобретению, и что тестируемое антитело, таким образом, может имитировать действие референсного антитела и, таким образом, является антителом по изобретению. Это можно подтверждать посредством сравнения активности CXCR5 в присутствии тестируемого антитела с активностью CXCR5 в присутствии референсного антитела в в остальном идентичных условиях с использованием анализа, что более подробно представлено в других местах настоящего описания.
[000106] Референсное антитело по изобретению может являться антителом, представленным в настоящем описании, таким как 11G2, 41A10, 5H7, и любым его вариантом или частью, представленными в настоящем описании, сохраняющими способность связываться с CXCR5. Антитело по изобретению может связываться с тем же эпитопом, что и референсные антитела, представленные в настоящем описании, или любой их вариант или часть, как представлено в настоящем описании, сохраняющие способность связываться с CXCR5.
[000107] Как указано выше в других местах настоящего описания, специфическое связывание можно анализировать с учетом связывания антитела с молекулой, не являющейся мишенью. Это сравнение можно осуществлять посредством сравнения способности антитела связываться с мишенью и другой молекулой. Это сравнение можно осуществлять, как описано выше для оценки KD или Ki. Другой молекулой, используемой в таком сравнении, может являться любая молекула, не являющаяся молекулой-мишенью. Предпочтительно, другая молекула не является идентичной молекуле-мишени. Предпочтительно, молекула-мишень не является фрагментом молекулы-мишени.
[000108] Другая молекула, используемая для определения специфического связывания, может являться неродственной мишени по структуре или функции. Например, другая молекула может представлять собой неродственный материал или материал, сопутствующий в окружающей среде.
[000109] Другая молекула, используемая для определения специфического связывания, может являться другой молекулой, входящей в тот же путь in vivo, что и молекула-мишень, т.е. CXCR5. Обеспечивая наличие специфичности к CXCR5 у антитела по изобретению по сравнению с другой такой молекулой, можно избегать нежелательной перекрестной реактивности in vivo.
[000110] Антитело по изобретению может сохранять способность связываться с некоторыми молекулами, являющимися родственными для молекулы-мишени.
[000111] Альтернативно, антитело по изобретению может иметь специфичность к конкретной молекуле-мишени. Например, оно может связываться с молекулой-мишенью, представленной в настоящем описании, но может не связываться или может связываться со значительно сниженной аффинностью с другой молекулой-мишенью, представленной в настоящем описании. Например, полноразмерный зрелый CXCR5 человека можно использовать в качестве мишени, но антитело, связывающееся с этой мишенью, может быть неспособным связываться или может связываться с меньшей аффинностью, например, с другими белками CXCR5 других биологических видов, такими как CXCR5 других млекопитающих. В некоторых вариантах осуществления антитело связывается с CXCR5 человека и мыши.
[000112] "Fc-слитый" белок является белком, где один или более полипептидов функционально связаны с Fc-полипептидом. В Fc-слитом белке комбинируют Fc-область иммуноглобулина с партнером по слиянию.
[000113] "Нативная последовательность Fc-области" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. "Вариант Fc-области" содержит аминокислотную последовательность, отличающуюся от нативной последовательности Fc-области в результате по меньшей мере одной модификации аминокислоты, но сохраняющую по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. Предпочтительно, вариант Fc-области содержит по меньшей мере одну замену аминокислоты по сравнению с нативной последовательностью Fc-области или Fc-областью родительского полипептида, например, от приблизительно одной до приблизительно десяти замен аминокислот и, предпочтительно, от приблизительно одной до приблизительно пяти замен аминокислот в нативной последовательности Fc-области или в Fc-области родительского полипептида. Вариант Fc-области в настоящем описании, предпочтительно, будет иметь по меньшей мере приблизительно 80% идентичности последовательности по отношению к нативной последовательности Fc-области и/или Fc-области родительского полипептида, и наиболее предпочтительно - по меньшей мере приблизительно 90% идентичности последовательности по отношению к ней, более предпочтительно - по меньшей мере приблизительно 95%, по меньшей мере приблизительно 96%, по меньшей мере приблизительно 97%, по меньшей мере приблизительно 98%, по меньшей мере приблизительно 99% идентичности последовательности по отношению к ней.
[000114] Как известно в этой области, термин "константная область" антитела относится к константной области легкой цепи антитела или константной области тяжелой цепи антитела в отдельности или в комбинации.
[000115] Термины "Fc-область IgG", "Fc-область", "Fc-домен" и "Fc", как взаимозаменяемо используют в настоящем описании, относятся к части молекулы IgG, соотносимой с кристаллизуемым фрагментом, получаемым посредством расщепления папаином молекулы IgG. В рамках изобретения термины относятся к константной области антитела, за исключением первого домена константной области иммуноглобулина, и дополнительно относится к частям этой области. Таким образом, термин "Fc" относится к последним двум доменам константной области иммуноглобулина IgA, IgD и IgG, последним трем доменам константной области иммуноглобулина IgE и IgM и шибкой шарнирной области, N-концевой относительно этих доменов или их частям. В случае IgA и IgM Fc может включать J-цепь. В случае IgG Fc содержит иммуноглобулиновые домены Cγ2 и Cγ3 (C гамма 2 и C гамма 3) и шарнирную область между Cγ1 (C гамма 1) и Cγ2 (C гамма 2). Хотя границы Fc-области могут варьироваться, Fc-область тяжелой цепи IgG человека, как правило, определяют как содержащую остатки C226 или P230 на карбоксильном конце, где нумерация соответствует индексу EU по Edelman et al., 1969, Proc. Natl. Acad. Sci. USA 63(1):78-85, как описано в Kabat et al., 1991. Как правило, Fc-домен содержит аминокислотные остатки с приблизительно 236 по приблизительно 447 константного домена IgG1 человека. Пример аминокислотной последовательности Fc-домена IgG1 человека дикого типа приведен в SEQ ID NO:31. Термин "Fc-полипептид" может относиться к этой области при выделении или этой области в контексте антитела, или его антигенсвязывающего фрагмента, или Fc-слитого белка.
[000116] Константный домен тяжелой цепи содержит Fc-область и дополнительно содержит домен CH1 и шарнирную область, а также домены CH2 и CH3 (и, необязательно, CH4 из IgA и IgE) тяжелой цепи IgG.
[000117] "Функциональная Fc-область" имеет по меньшей мере одну эффекторную функцию нативной последовательности Fc-области. Примеры "эффекторных функций" включают связывание C1q; обусловленную комплементом цитотоксичность; связывание Fc-рецептора; антителозависимую клеточную цитотоксичность; фагоцитоз; отрицательную регуляцию рецепторов поверхности клеток (например, B-клеточного рецептора) и т.д. Для таких эффекторных функций, как правило, необходимо комбинирование Fc-области со связывающим доменом (например, вариабельным доменом антитела или его антигенсвязывающим фрагментом), и их можно оценивать с использованием различных анализов, известных в этой области для оценки таких эффекторных функций антитела.
[000118] "Нативная последовательность Fc-области" содержит аминокислотную последовательность, идентичную аминокислотной последовательности Fc-области, обнаруживаемой в природе. Нативная последовательность Fc-области человека включает нативную последовательность Fc-области IgG1 человека (аллотипы не-A и A), нативную последовательность Fc-области IgG2 человека, нативную последовательность Fc-области IgG3 человека и нативную последовательность Fc-область IgG4 человека, а также их природные варианты.
[000119] "Вариант Fc-области" содержит аминокислотную последовательность, отличающуюся от нативной последовательности Fc-области в результате по меньшей мере одной модификации аминокислоты.
[000120] С помощью термина "Fc-рецептор" или "FcR" описывают рецептор, связывающийся с Fc-областью антитела. В некоторых вариантах осуществления FcyR является нативным FcR человека. В некоторых вариантах осуществления FcR является FcR, связывающимся с антителом IgG (гамма-рецептор), и включает рецепторы подклассов FcyRI, FcyRII и FcyRIII, включая аллельные варианты и альтернативно сплайсированные формы этих рецепторов. Рецепторы FcyRII включают FcyRIIA ("активирующий рецептор") и FcyRIIB ("ингибирующий рецептор"), имеющие схожие аминокислотные последовательности, отличающиеся, главным образом, своими цитоплазматическими доменами. Активирующий рецептор FcyRIIA содержит иммунорецепторный тирозиновый активирующий мотив (ITAM) в своем цитоплазматическом домене. Ингибирующий рецептор FcyRIIB содержит иммунорецепторный тирозиновый ингибирующий мотив (ITIM) в своем цитоплазматическом домене (см., например, Daeron, Annu. Rev. Immunol. 15:203-234 (1997)). Обзоры FcR приведены, например, в Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991); Capel et ah, Immunomethods 4:25-34 (1994); и de Haas et ah, J. Lab. Clin. Med. 126:330-41 (1995). Другие FcR, включая FcR, которые будут идентифицированы в будущем, включены в термин "FcR" в настоящем описании.
[000121] Термин "Fc-рецептор" или "FcR" также включает неонатальный рецептор FcRn, отвечающий за транспорт материнских IgG в плод (Guyer et ah, J. Immunol. 117:587 (1976) и Kim et ah, J. Immunol. 24:249 (1994)) и регуляцию гомеостаза иммуноглобулинов. Известны способы измерения связывания с FcRn (см., например, Ghetie and Ward., Immunol. Today 18(12):592-598 (1997); Ghetie et al., Nature Biotechnology, 15(7):637-640 (1997); Hinton et al., J. Biol. Chem. 279(8):6213-6216 (2004); WO 2004/92219 (Hinton et al.)).
[000122] Термин "эффекторные функции" относится к видам биологической активности, приписываемым Fc-области антитела и варьирующимся в зависимости от изотипа антитела. Примеры эффекторных функций антитела включают: связывание C1q и обусловленную комплементом цитотоксичность (CDC); связывание Fc-рецептора; антителозависимую клеточную цитотоксичность (ADCC); фагоцитоз; негативную регуляцию рецепторов поверхности клеток (например, B-клеточного рецептора) и активацию B-клеток.
[000123] "Эффекторные клетки человека" являются лейкоцитами, экспрессирующими один или более FcR и выполняющими эффекторные функции. В некоторых вариантах осуществления клетки экспрессируют по меньшей мере FcyRIII и выполняют эффекторные функции ADCC. Примеры лейкоцитов человека, опосредующих ADCC, включают мононуклеарные клетки периферической крови (PBMC), естественные киллеры (NK), моноциты, макрофаги, цитотоксические T-клетки и нейтрофилы. Эффекторные клетки можно выделять из нативного источника, например, крови.
[000124] Термин "антителозависимая клеточная цитотоксичность" или "ADCC" относится к форме цитотоксичности, при которой секретируемые Ig, связанные с Fc-рецепторами (FcR) и присутствующие на некоторых цитотоксических клетках (например, NK-клетках, нейтрофилах и макрофагах), позволяют этим цитотоксическим эффекторным клеткам специфически связываться с несущей антиген клеткой-мишенью, а затем уничтожать клетку-мишень с помощью цитотоксинов. Первичные клетки для опосредования ADCC, NK-клетки, экспрессируют только FcyRIII, в то время как моноциты экспрессируют FcyRI, FcyRII и FcyRIII. Экспрессия FcR на гематопоэтических клетках приведена в таблице 3 на стр. 464 Ravetch and Kinet, Annu. Rev. Immunol 9:457-92 (1991). Для оценки ADCC-активности интересующей молекулы можно проводить анализ ADCC in vitro, такой как описано в патентах США №№ 5500362 или 5821337 или патенте США № 6737056 (Presta). Эффекторные клетки, которые можно использовать для таких анализов, включают PBMC и NK-клетки. Альтернативно или дополнительно, ADCC-активность интересующей молекулы можно анализировать in vivo, например, в модели на животных, такой как описанная в Clynes et al. Proc. Natl. Acad. Sci. (USA) 95:652-656 (1998). Дополнительные антитела с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной активностью ADCC описаны, например, в патенте США № 7923538 и патенте США № 7994290.
[000125] Термин антитело, имеющее "повышенную активность ADCC", относится к антителу, являющемуся более эффективным в опосредовании ADCC in vitro или in vivo по сравнению с родительским антителом, где антитело и родительское антитело отличаются по меньшей мере одним структурным аспектом, и если количества такого антитела и родительского антитела, используемых в анализе, являются, по существу, одинаковыми. В некоторых вариантах осуществления антитело и родительское антитело имеют одинаковую аминокислотную последовательность, но антитело является афукозилированным, в то время как родительское антитело является фукозилированным. В некоторых вариантах осуществления активность ADCC будут определять с использованием анализа ADCC in vitro, как представлено в настоящем описании, но предусмотрены и другие анализы или способы определения активности ADCC, например, в модели на животных и т.д. В некоторых вариантах осуществления антитело с повышенной активностью ADCC имеет повышенную аффинность к Fc гамма RIIIA. В некоторых вариантах осуществления антитело с повышенной активностью ADCC имеет повышенную аффинность к Fc гамма RIIIA (V158). В некоторых вариантах осуществления антитело с повышенной активностью ADCC имеет повышенную аффинность к Fc гамма RIIIA (F158).
[000126] Антитело с "измененной" аффинностью связывания FcR или активностью ADCC является антителом, имеющим повышенную или сниженную активность связывания FcR и/или активность ADCC по сравнению с родительским антителом, где антитело и родительское антитело отличаются по меньшей мере одним структурным аспектом. Антитело, "демонстрирующее повышенное связывание" с FcR, связывается по меньшей мере с одним FcR с лучшей аффинностью, чем родительское антитело. Антитело, "демонстрирующее сниженное связывание" с FcR, связывается по меньшей мере с одним FcR с более низкой аффинностью, чем родительское антитело. Такие антитела, демонстрирующие сниженное связывание с FcR, могут иметь невысокое связывание с FcR или не иметь его, например, иметь 0-20 процентов связывания с FcR по сравнению с нативной последовательностью Fc-области IgG.
[000127] Термин "повышенная аффинность к Fc гамма RIIIA" относится к антителу, имеющему более высокую аффинность к Fc гамма RIIIA (также обозначаемого в некоторых случаях как CD16a), чем родительское антитело, где антитело и родительское антитело отличаются по меньшей мере одним структурным аспектом. В некоторых вариантах осуществления антитело и родительское антитело имеют одинаковую аминокислотную последовательность, но антитело является афукозилированным, в то время как родительское антитело является фукозилированным. Можно использовать любой подходящий способ определения аффинности к Fc гамма RIIIA. В некоторых вариантах осуществления аффинность к Fc гамма RIIIA определяют способом, представленным в настоящем описании. В некоторых вариантах осуществления антитело с повышенной аффинностью к Fc гамма RIIIA имеет повышенную активность ADCC. В некоторых вариантах осуществления антитело с повышенной аффинностью к Fc гамма RIIIA имеет повышенную аффинность к Fc гамма RIIIA(V158). В некоторых вариантах осуществления антитело с повышенной аффинностью к Fc гамма RIIIA имеет повышенную аффинность к Fc гамма RIIIA(F158).
[000128] L-фукоза, также обозначаемая как 6-дезокси-L-гплактоза, является моносахаридом, являющимся компонентом некоторых N- и O-связанных гликанов и гликолипидов у животных. См. Becker and Lowe, Glycobiology 13:41R-51R (2003). Фукоза, как правило, добавляется в качестве концевой модификации к гликанам, включая гликаны, присоединенные к антигенам группы крови, селектинам и антителам. Фукоза может присоединяться к гликанам через α(1,2)-, α(1,3)-, α(1,4)- и α(1,6)-связи с помощью специфических фукозилтрансфераз. α(1,2)-фукозные связи, как правило, ассоциированы с H-группой антигенов крови. α(1,3)- и α(1,4)-фукозные связи ассоциированы с модификацией антигенов Льюса X. α(1,6)-фукозные связи ассоциированы с молекулами N-связанного GlcNAc, такими как молекулы на антителах.
[000129] Молекулы углеводов по настоящему изобретению будут описаны со ссылкой на общеупотребительную номенклатуру для описания олигосахаридов. Обзор химии углеводов, в котором используют эту номенклатуру, можно найти в Hubbard and Ivatt (1981) Ann. Rev. Biochem. 50:555-583. Эта номенклатура включает, например, Man, что соответствует маннозе; GIcNAc, что соответствует 2-N-ацетилглюкозамину; Gal, что соответствует галактозе; Fuc, что соответствует фукозе; и Glc, что соответствует глюкозе. Сиаловые кислоты описывают с помощью сокращенного обозначения NeuNAc для 5-N-ацетилнейраминовой кислоты и NeuNGc для 5-гликолилнейраминовой кислоты (IUB-IUPAC Joint Commission on Biochemical Nomenclature, 1982, J. Biol. Chem. 257: 3347-3351; (1982) J. Biol. Chem. 257: 3352).
[000130] Углеводные структуры по настоящему изобретению находятся на белке, экспрессирующемся в виде N-связанных олигосахаридов. Термин "N-связанное гликозилирование" относится к присоединению молекулы углевода чрез GlcNAc к остатку аспарагина в полипептидной цепи. Все N-связанные углеводы содержат общую коровую структуру Man 1-6(Man1-3)Manβ1-4GlcNAcβ1-4GlcNAcβ-R. Таким образом, в описанной коровой структуре R соответствует остатку аспарагина продуцируемого гликопротеина. Последовательность продуцируемого белка будет содержать аспарагин-X-серин, аспарагин-X-треонин и аспарагин-X-цистеин, где X является любой аминокислотой, за исключением пролина (Asn-Xaa-Ser/Thr). В отличие от этого, "O-связанные" углеводы отличаются общей коровой структурой, представляющей собой GalNAc, присоединенную к гидроксильной группе треонина или серина, но консенсусная последовательность не является необходимой. Из N-связанных углеводов наиболее важными являются "сложные" N-связанные углеводы, такие как "биантенарные" структуры, представленные в настоящем описании.
[000131] Специалистам в этой области понятно, что гликопротеин иммуноглобулина G (IgG) ассоциирован с тремя типами сложных биантенарных структур, содержащих ноль, один или два остатка галактозы (Wormland et al., 1997, Biochemistry 36:1370-1380), общеизвестных как G0, G1 и G2, соответственно. Что касается молекул антитела человека класса IgG, каждая из них содержит N-связанный олигосахарид, присоединенный к амидной боковой цепи Asn 297 β-4-изгиба внутренней поверхности домена CH2 Fc-области (Beale and Feinstein, 1976, Q. Rev. Biophys. 9:253-259; Jefferis et al., 1995, Immunol. Letts. 44:111-117). Олигосахаридный остаток, присоединенный к Asn 297 домена CH2 IgG, принадлежит к сложному биантенарному типу, имеющему идентифицированную гексасахаридную коровую структуру и вариабельные внешние остатки сахара (см. Jefferis et al., 1997, выше; Wyss and Wagner, 1996, Current Opinions in Biotech. 7:409-416). Коровая структура (GIcNAc2Man3GIcNAc) является типичной для биантенарных олигосахаридов и схематически показана на фиг. 1.
[000132] Т.к. каждая коровая структура может содержать бисектный N-ацетилглюкозамин, коровую фукозу и внешние сахариды галактозу или сиаловую кислоту, всего существует 36 структурно уникальных олигосахаридов, которые могут занимать участок Asn 297 (Jefferis and Lund, выше). Также следует понимать, что в конкретном домене CH2 гликозилирование по Asn 297 может быть асимметричным благодаря разным олигосахаридным цепям, присоединенным к остатку Asn 297 в двухцепочечном Fc-домене. Например, хотя тяжелая цепь, синтезируемая в одной антитело-секретирующей клетке, может быть гомогенной по своей аминокислотной последовательности, она, как правило, дифференциально гликозилирована, что приводит к образованию большого количества структурно уникальных гликоформ Ig.
[000133] Основные типы сложных олигосахаридных структур, также обозначаемых как "гликоформы", обнаруживаемых в домене CH2 IgG, описаны в международной патентной публикации № WO 99/22764 на стр. 7.
[000134] В рамках изобретения термин "G0" относится к биантенарной структуре, в которой нет концевых сиаловых кислот (NeuAc) или Gal, термин "G1" относится к биантенарной структуре, содержащей одну Gal и несодержащей NeuAc, и термин "G2" относится к биантенарной структуре с двумя концевыми Gal и без NeuAc. См., например, фиг. 2A-2G, на которых показаны примеры структур G0, G1, G-1 и G2.
[000135] Термины "афукозилированное" антитело или антитело, "в котором отсутствует фукоза", относятся к антителу изотипа IgG1 или IgG3, в котором в гликозилировании константной области отсутствует фукоза. Гликозилирование IgG1 или IgG3 человека происходит в Asn297, т.к. гликозилирование корового фукозилированного биантенарного сложного олигосахарида заканчивается до 2 остатками Gal. В некоторых вариантах осуществления в афукозилированном антителе отсутствует фукоза в Asn297. Эти структуры обозначены как гликановые остатки G0, G1 (1,6 или 1,3) или G2 в зависимости от количества концевых остатков Gal. См., например, Raju, T. S., BioProcess Int. 1: 44-53 (2003). Гликозилирование Fc антитела типа CHO описано, например, в Routier, F. FL, Glycoconjugate J. 14: 201-207 (1997). Различные гликоформы антител показаны на фиг. 2A-2G.
[000136] В некоторых вариантах осуществления термины "фукозил" или "афукозилированное" антитело, как взаимозаменяемо используют в настоящем описании, относятся к антителу, гликоинженерно модифицированных так, чтобы не содержать коровую фукозу. Антитела со сниженным содержанием фукозы в гликановых остатках имеют повышенную аффинность к FcγRIIIa (CD16) и в результате имеют повышенную активность антителозависимой клеточной цитотоксичности (ADCC). Афукозилированные антитела можно получать с использованием линии клеток Potelligent® CHOK1SV (Lonza Biologics), в которой отсутствуют оба аллеля гена, ответственного за добавление фукозы (α1,6-фукозилтрансферазы). Афукозилированные антитела или антитела со сниженной фукозой также можно получать посредством модификации активности биосинтеза олигосахаридов различными способами. Например, гиперэкспрессия N-ацетилглюкозаминтрансферазы III (GnTIII) в аппарате Гольджи линии продуцирующих клеток приводит к образованию бисектных олигосахаридных структур, ассоциированных с константной областью Fc антитела, и супрессирует фукозилирование. В таких системах экспрессии уровень экспрессии GnTIII коррелирует с образованием афукозилированных гликоформ IgG1 и приводит к повышенной активности ADCC. Фукозилирование также может снижаться в культуре клеток при использовании аналогов сахаров, в качестве неограничивающих примеров, таких как аналоги фукозы, как описано в WO 2012/019165. Таким образом, афукозилированные антитела или антитела со сниженным содержанием фукозы можно получать с использованием широкого спектра способов, хорошо известных в этой области.
[000137] В некоторых вариантах осуществления афукозилированное антитело имеет повышенную аффинность к Fc гамма RIIIA. В некоторых вариантах осуществления афукозилированное антитело имеет повышенную аффинность к Fc гамма RIIIA (V158). В некоторых вариантах осуществления афукозилированное антитело имеет повышенную аффинность к Fc гамма RIIIA (F158).
[000138] Термин "гликоформа" относится к сложной олигосахаридной структуре, содержащей связи различных углеводных единиц. Такие структуры описаны, например, в Essentials of Glycobiology Varki et al., eds., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1999), где также представлен обзор стандартной номенклатуры гликобиологии. Такие гликоформы включают, в качестве неограничивающих примеров, G2, G1, G0, G-1 и G-2 (см., например, международную патентную публикацию № WO 99/22764).
[000139] Термин "профиль гликозилирования" определяют как профиль углеводных единиц, ковалентно присоединенных к белку (например, гликоформе), а также к участкам, в которых гликоформы ковалентно присоединяются к пептидному остову белка, более конкретно - к иммуноглобулиновому белку.
[000140] В некоторых вариантах осуществления по меньшей мере 85 процентов партии антител, рекомбинантно экспрессирующихся в негликомодифицированных клетках-хозяевах CHO, являются фукозилированными по Asn297. Что касается композиции, содержащей множество антител, антитела считают афукозилированными, если менее приблизительно 5 процентов антител в композиции содержат фукозу по меньшей мере в одном Asn297. Даже более предпочтительно, уровень афукозилирования составляет приблизительно 100%, т.е. не определяют фукозу на любой гликоформе тяжелой цепи Asn297 с использованием стандартных способов измерения фукозилирования антитела. Способы измерения фукозы включают любые известные в этой области способы, включая способы, представленные в настоящем описании. В некоторых вариантах осуществления фукоза неопределима в композиции, содержащей множество афукозилированных антител. В некоторых вариантах осуществления афукозилированное антитело имеет повышенную активность ADCC.
[000141] Термин "обусловленная комплементом цитотоксичность" или "CDC" относится к лизису клетки-мишени в присутствии комплемента. Активация классического пути комплемента инициируется связыванием первого компонента системы комплемента (C1q) с антителами (подходящего подкласса), связанными с их когнатным антигеном. Для оценки активации комплемента можно проводить анализ CDC, например, как описано в Gazzano-Santoro et al., J. Immunol. Methods 202: 163 (1996). Антитела с измененными аминокислотными последовательностями Fc-области и повышенной или сниженной способностью к связыванию C1q описаны, например, в патенте США № 6194551 B1, патенте США № 7923538, патенте США № 7994290 и WO 1999/51642. См. также, например, Idusogie et al, J.
[000142] В рамках изобретения термины "аминокислота дикого типа", "IgG дикого типа", "антитело дикого типа" или "mAb дикого типа" относятся к последовательности аминокислот или нуклеиновых кислот, встречающихся в природе в некоторой популяции (например, людей, мышей, крыс, клеток и т.д.).
[000143] Термины "C-X-C-хемокиновый рецептор типа 5" или "CXCR5", используемые в настоящем описании взаимозаменяемо, также обозначаемые в этой области как "CD185" и "рецептор лимфомы Беркитта 1 (BLR1)", означают сопряженный с G-белком рецептор, экспрессирующийся в некоторых клетках. Термин "CXCR5" включает гомологи и ортологи CXCR5, включая, помимо прочего, гомологи и ортологи человека, яванского макака, крысы, кролика и мыши. В рамках изобретения термин "CXCR5" относится к CXCR5 млекопитающего, такому как CXCR5 человека, крысы или мыши, а также не являющегося человеком примата, коровы, овцы или свиньи. Неограничивающие примеры CXCR5 включают CXCR5 человека (см., например, регистрационный номер Genbank P60568, SEQ ID NO:32), яванского макака (см., например, регистрационный номер Genbank Q29615, SEQ ID NO:33) и мыши (SEQ ID NO:34). Термин "CXCR5" также включает фрагменты, варианты, изоформы и другие гомологи таких молекул CXCR5. Варианты молекул CXCR5, как правило, будут отличаться наличием того же типа активности, что и природный CXCR5, таким как способность связываться с рецептором CXCR5, способностью индуцировать опосредованную рецепторную активность и способностью связываться или не связываться с антителом или его антигенсвязывающим фрагментом по изобретению.
[000144] CXCR5 может содержать один или более, два или более, три или более, четыре или более, пять или более, шесть или более, семь или более, восемь или более, девять или более, десять или более, двенадцать или более или пятнадцать или более доступных поверхностных остатков CXCR5. На фиг. 6 показана аминокислотная последовательность CXCR5 мыши и человека дикого типа, где подчеркнуты примеры доступных поверхностных остатков. Если CXCR5 содержит гомомультимерную форму CXCR5, мишень может содержать один или более, два или более, три или более, четыре или более, пять или более, шесть или более, семь или более, восемь или более, девять или более, десять или более, двенадцать или более или пятнадцать или более доступных поверхностных остатков первой субъединицы CXCR5, и один или более, два или более, три или более, четыре или более, пять или более, шесть или более, семь или более, восемь или более, девять или более, десять или более, двенадцать или более или пятнадцать или более доступных поверхностных остатков второй субъединицы CXCR5. Молекула-мишень может содержать известный эпитоп из CXCR5.
[000145] Как указано выше, выровненные аминокислотные последовательности CXCR5 человека (hCXCR5) и CXCR5 мыши (mCXCR5) дикого типа, где указаны (подчеркнуты) различные примеры внеклеточных областей (помеченные "N", "L1", "L2" и "3") белков показаны на фиг. 5.
[000146] Как указано в других местах настоящего описания, можно изменять некоторые положения молекулы антитела. В рамках изобретения термин "положение" означает место в последовательности белка. Положения можно нумеровать последовательно или в соответствии с установленным форматом, например, для нумерации аминокислотных остатков антитела можно использовать индекс EU и индекс Kabat. Например, положение 297 является положением в антителе IgG1 человека. Соответствующие положения определяют, как указано выше, как правило, посредством выравнивания с другими родительскими последовательностями.
[000147] В рамках изобретения термин "остаток" означает положение в белке и связанный с ним тип аминокислоты. Например, аспарагин 297 (также обозначаемый как Asn297, также обозначаемый как N297) представляет собой остаток в антителе IgG1 человека.
[000148] Термины "Tfh-клетка", или "Tfh", или "истинная Tfh", или "Tfh-клетка герминативного центра", или " Tfh-клетка GC", как взаимозаменяемо используют в настоящем описании, относятся к фолликулярным хелперным T-клеткам, обнаруживаемым в герминативном центре (GC), являющемся структурой, состоящий из Tfh-клеток GC, B-клеток GC, фолликулярных дендритных клеток (FDC), макрофагов и стромы. См. Crotty, 2014, Immunity 41(4):529-542. Tfh-клетки идентифицируют по конститутивной экспрессии хоминг-рецептора B-клеточного фолликула CXCR5. Функционально Tfh-клетки обеспечивают инструктивные сигналы для B-клеток, направляющие переключения изотипа, соматические гипермутации и быстрое деление клеток для заселения герминативных центров.
[000149] Термины "Tfh-подобные клетки", "циркулирующие Tfh-клетки" и "cTfh", используемые в настоящем описании взаимозаменяемо, относятся к Tfh-клеткам, покинувшим герминативный центр. После выхода из GC клетки приобретают менее активированный, менее поляризованный фенотип и их обозначают как "циркулирующие фолликулярные хелперные T-клетки" (cTfh) или "Tfh-подобные клетки". См. Crotty, 2014, Immunity 41(4):529-542. Эти клетки экспрессируют CXCR5. По сравнению с Tfh-клетками герминативного центра (т.е. "истинными Tfh-клетками", или "Tfh-клетками GC", или "Tfh"), cTfh-клетки (т.е. Tfh-подобные клетки) экспрессируют сниженные уровни ICOS, Bcl-6 и маркеров клеточной активации, таких как CD69 и HLA-DR, но сохраняют способность стимулировать продукцию Ab и переключение классов Ig в B-клетках in vitro после реактивации когнатными антигенами.
[000150] Как известно в этой области, термины "полинуклеотид" или "нуклеиновая кислота", как взаимозаменяемо используют в настоящем описании, относятся к цепям нуклеотидов любой длины и включают ДНК и РНК. Нуклеотиды могут являться дезоксирибонуклеотидами, рибонуклеотидами, модифицированными нуклеотидами или основаниями, и/или их аналогами, или любым субстратом, который может встраиваться в цепь ДНК- или РНК-полимеразами. Полинуклеотид может содержать модифицированные нуклеотиды, такие как метилированные нуклеотиды и их аналоги. Если присутствует модификация в структуру нуклеотида, ее можно включать до или после сборки цепи. Последовательность нуклеотидов может прерываться ненуклеотидными компонентами. Полинуклеотид можно дополнительно модифицировать после полимеризации, например, посредством конъюгации с меченым компонентом. Другие типы модификаций включают, например, "кэп", замену одного или более природных нуклеотидов аналогом, межнуклеотидные модификации, такие как, например, модификации с незаряженными связями (например, метилфосфонатами, фосфотриэфирами, фосфоамидатами, карбаматами и т.д.) и заряженными связями (например, фосфоротиоаты, фосфородитиоаты и т.д.), модификации, содержащие боковые остатки, такие как, например, белки (например, нуклеазы, токсины, антитела, сигнальные пептиды, поли-L-лизин и т.д.), модификации с интеркаляторами (например, акридином, псораленом и т.д.), модификации, содержащие хелаторы (например, металлы, радиоактивные металлы, бор, металлы-окислители и т.д.), модификации, содержащие алкиляторы, модификации с модифицированными связями (например, альфа-аномерные нуклеиновые кислоты и т.д.), а также немодифицированные формы полинуклеотидов. Кроме того, любые из гидроксильных групп, как правило, присутствующие в сахарах, можно заменять, например, фосфонатными группами, фосфатными группами, защищать стандартными защитными группами или активировать для получения дополнительных связей с дополнительными нуклеотидами или их можно конъюгировать с твердыми подложками. 5’- и 3’-концевой OH можно фосфорилировать или замещать аминами или органическими кэп-группировками из 1-20 атомов углерода. Другие гидроксилы также можно дериватизировать до стандартных защитных групп. Полинуклеотиды также могут содержать аналогичные формы сахаров рибозы или дезоксирибозы, как правило, известные в этой области, включая, например, 2’-O-метил-, 2’-O-аллил, 2’-фтор- или 2’-азидо-рибозу, карбоциклические аналоги сахаров, альфа- или бета-аномерные сахара, эпимерные сахара, такие как арабиноза, ксилозы или ликсозы, пиранозные сахара, фуранозные сахара, седогептулозы, ациклические аналоги и нуклеозидные аналоги без азотистого основания, такие как метилрибозид. Одну или более фосфодиэфирных связей можно заменять альтернативными линкерными группами. Эти альтернативные линкерные группы включают, в качестве неограничивающих примеров, варианты осуществления, где фосфат заменяют P(O)S ("тиоатом"), P(S)S ("дитиоатом"), (O)NR2 ("амидатом"), P(O)R, P(O)OR’, CO или CH2 ("формацеталем"), в котором каждый из R или R’ независимо представляет собой H или замещенный или незамещенный алкил (1-20 C), необязательно содержащий эфирную связь (-O-), арил, алкенил, циклоалкил, циклоалкенил или апалдил. Не все связи в полинуклеотиде должны быть идентичными. Предшествующее описание относится ко всем полинуклеотидам, упомянутым в настоящем описании, включая РНК и ДНК.
[000151] В рамках изобретения термин "вектор" означает конструкцию, с помощью которой можно доставлять и, предпочтительно, экспрессировать один или более интересующих генов или последовательностей в клетке-хозяине. Неограничивающие примеры векторов включают вирусные векторы, экспрессирующие векторы депротеинизированной ДНК или РНК, плазмиду, космиду или фаговые векторы, ДНК- или РНК-экспрессирующие векторы, связанные с катионными конденсирующими средствами, ДНК- или РНК-экспрессирующими векторами, инкапсулированными в липосомах, и некоторые эукариотические клетки, такие как клетки-продуценты.
[000152] Термин "клетка-хозяин" включает отдельную клетку или культуру клеток, которые могут являться или являются реципиентом для векторов для встраивания полинуклеотидных вставок. Клетки-хозяева включают потомство одной клетки-хозяина, и потомство может не являться полностью идентичным (по морфологии или геномной ДНК) исходной родительской клетке по причине природной, случайной или умышленной мутации. Клетка-хозяин включает клетки, трансфицированные и/или трансформированные in vivo с использованием полинуклеотида по изобретению.
[000153] Клетки-хозяева могут являться прокариотическими клетками или эукариотическими клетками. Примеры эукариотических клеток включают клетки млекопитающих, такие как клетки примата или клетки животного, не являющегося приматом; клетки грибов, таких как дрожжи; растительные клетки и клетки насекомых.
[000154] Любую клетку-хозяина, восприимчивую к культивированию клеток и экспрессии белка или полипептидов, можно использовать в соответствии с настоящим изобретением. В некоторых вариантах осуществления клетка-хозяин принадлежит млекопитающему. Линии клеток млекопитающих, доступные в качестве хозяев для экспрессии, хорошо известны в этой области и включают многие иммортализованные линии клеток, доступные в American Type Culture Collection (ATCC). Неограничивающие примеры клеток млекопитающих включают клетки NS0, клетки HEK 293 и клетки яичника китайского хомяка (CHO) и их производные, такие как клетки 293-6E и CHO DG44, CHO DXB11 и клетки Potelligent® CHOK1SV (BioWa/Lonza, Allendale, NJ). Клетки-хозяева млекопитающих также включают, в качестве неограничивающих примеров, клетки карциномы шейки матки человека (HeLa, ATCC CCL 2), клетки почки новорожденного хомяка (BHK, ATCC CCL 10), клетки почки обезьяны (COS) и клетки печеночноклеточной карциномы человека (например, Hep G2). Другие неограничивающие примеры клеток млекопитающих, которые можно использовать в соответствии с настоящим изобретением, включают ретинобласты человека (PER.C6®; CruCell, Leiden, The Netherlands); линию клеток печени обезьяны CV1, трансформированную с использованием SV40 (COS-7, ATCC CRL 1651); линию клеток эмбриональной почки человека 293 (HEK 293) или клетки 293, субклонированные для выращивания в суспензионной культуре (Graham et al., 1977, J. Gen Virol. 36:59); клетки Сертоли мыши (TM4, Mather, 1980, Biol. Reprod. 23:243-251); клетки почки обезьяны (CV1 ATCC CCL 70); клетки почки африканской зеленой мартышки (VERO-76, ATCC CRL-1 587); клетки почки собаки (MDCK, ATCC CCL 34); клетки печени крыс линии Buffalo (BRL 3A, ATCC CRL 1442); клетки легких человека (W138, ATCC CCL 75); клетки печени человека (Hep G2, HB 8065); клетки опухоли молочной железы мыши (MMT 060562, ATCC CCL51); клетки TR1 (Mather et al., 1982, Annals N.Y. Acad. Sci. 383:44-68); клетки MRC 5; клетки FS4; линию клеток гепатомы человека (Hep G2); и многочисленные линии клеток миеломы, включая, в качестве неограничивающих примеров, линию клеток миеломы мыши BALB/c (NS0/1, ECACC № 85110503), клетки NS0 и клетки Sp2/0.
[000155] Кроме того, в настоящем изобретении можно использовать любое количество коммерчески и некоммерчески доступных линий клеток, экспрессирующих полипептиды или белки. Специалисту в этой области будет понятно, что разные линии клеток могут иметь разные требования к питательным веществам, и/или им могут требоваться разные условия культивирования для оптимального роста и экспрессии полипептида или белка, и специалист в этой области сможет модифицировать условия по мере необходимости.
[000156] Изобретение включает любую эукариотическую систему экспрессии, известную в этой области или представленную в настоящем описании для продукции интересующих белков, такую как система экспрессии в клетках насекомых, дрожжевая система экспрессии или система клеток млекопитающего, в качестве неограничивающих примеров, такая как клетки CHO.
[000157] В рамках изобретения термин "последовательность контроля экспрессии" означает последовательность нуклеиновой кислоты, направляющую транскрипцию нуклеиновой кислоты. Последовательность контроля экспрессии может являться промотором, таким как конститутивный или индуцибельный промотор, или энхансером. Последовательность контроля экспрессии функционально связана с последовательностью нуклеиновой кислоты, подлежащей транскрипции.
[000158] В рамках изобретения "лечение" представляет собой подход для получения благоприятных или желаемых клинических результатов. В целях по настоящему изобретению, благоприятные или желаемые результаты включают, в качестве неограничивающих примеров, одно или более из следующего: улучшенный коэффициент выживаемости (сниженную смертность), снижение воспалительного ответа на заболевание, снижение степени фиброза тканей, улучшение внешнего вида очагов заболевания, ограничение патологических очагов до фокальных очагов, снижение степени повреждения в результате заболевания, снижение длительности заболевания и/или снижение количества, степени или длительности симптомов, связанных с заболеванием. Термин включает введение соединений или средств по настоящему изобретению для профилактики или задержки дебюта симптомов, осложнений или биохимических показателей заболевания, облегчения симптомов или прекращения или ингибирования дальнейшего развития заболевания, состояния или нарушения. Лечение может представлять собой профилактическую (для профилактики или задержки дебюта заболевания или для профилактики манифестации его клинических или субклинических симптомов) или терапевтическую супрессию или облегчение симптомов после манифестации заболевания.
[000159] Термин "улучшение" означает уменьшение или улучшение одного или более симптомов по сравнению с отсутствием введения антитела CXCR5. Термин "улучшение" также включает уменьшение длительности симптома.
[000160] В рамках изобретения "эффективная доза" или "эффективное количество" лекарственного средства, соединения или фармацевтической композиции является количеством, достаточным для того, чтобы влиять на любые один или более благоприятных или желаемых результатов. В более конкретных аспектах эффективное количество позволяет осуществлять профилактику, облегчать или улучшать симптомы заболевания или инфекции и/или пролонгировать выживаемость индивидуума, подвергаемого лечению. В случае профилактического использования благоприятные или желаемые результаты включают устранение или снижение риска, снижение тяжести или задержку дебюта заболевания, включая биохимические, гистологические и/или поведенческие симптомы заболевания, его осложнения и промежуточные патологические фенотипы, присутствующие во время развития заболевания. В случае терапевтического использования благоприятные или желаемые результаты включают клинические результаты, такие как снижение одного или более симптомов CXCR5-опосредованного заболевания, нарушения или состояния, снижение дозы других лекарственных средств, необходимых для лечения заболевания, усиление эффекта другого лекарственного средства и/или задержку прогрессирования заболевания у пациентов. Эффективную дозу можно вводить за одно или более введений. В целях по настоящему изобретению эффективная доза лекарственного средства, соединения или фармацевтической композиции является количеством, достаточным для прямого или косвенного осуществления профилактического или терапевтического лечения. Как следует понимать в клиническом контексте, эффективной дозы лекарственного средства, соединения или фармацевтической композиции можно достигать или не достигать в комбинации с другим лекарственным средством, соединением или фармацевтической композицией. Таким образом, термин "эффективная доза" можно рассматривать в контексте введения одного или более терапевтических средств, и можно считать, что отдельное средство вводят в эффективном количестве, если при комбинировании с одним или более другими средствами можно достигать или достигают желаемого результата.
[000161] "Индивидуум" представляет собой млекопитающего, более предпочтительно, человека. Млекопитающие также включают, в качестве неограничивающих примеров, сельскохозяйственных животных (например, коров, свиней, лошадей, кур и т.д.), спортивных животных, домашних питомцев, приматов, лошадей, собак, кошек, мышей и крыс. В некоторых вариантах осуществления индивидуум считают имеющим риск развития заболевания, нарушения или состояния, опосредованного или ассоциированного со связыванием CXCR5 с его рецептором и опосредованной им передачи сигнала. В некоторых вариантах осуществления индивидуум имеет аутоиммунное заболевание, нарушение или состояние, такое как диабет 1 типа. В некоторых вариантах осуществления индивидуум нуждается в иммуносупрессорной терапии.
[000162] В рамках изобретения "фармацевтически приемлемый носитель" или "фармацевтически приемлемый эксципиент" включает любой материал, который при комбинировании с активным ингредиентом позволяет ингредиенту сохранять биологическую активность и не реагирует с иммунной системой индивидуума. Неограничивающие примеры включают любые из стандартных фармацевтических носителей, такие как фосфатно-солевой раствор, воду, эмульсии, такие как эмульсия "масло-в-воде" и различные типы увлажнителей. Предпочтительными дилюентами для аэрозольного или парентерального введения являются фосфатно-солевой буфер (PBS) или нормальный (0,9%) физиологический раствор. Композиции, содержащие такие носители, составляют хорошо известными, общепринятыми способами (см., например, Remington's Pharmaceutical Sciences, 18th edition, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy 20th Ed. Mack Publishing, 2000).
[000163] В настоящем описании представлены примеры способов и материалов, хотя в практическом осуществлении или тестировании настоящего изобретения также можно использовать способы и материалы, схожие или эквивалентные способам и материалам, представленным в настоящем описании. Материалы, способы и примеры являются исключительно иллюстративными и не предназначены для ограничения изобретения.
II. АНТИТЕЛА ПРОТИВ CXCR5
[000164] Настоящее изобретение относится к антителам, связывающимся с CXCR5. Предпочтительно, антитела специфически связываются с CXCR5, т.е. они связываются с CXCR5, но они не связываются детектируемо или связываются с более низкой аффинностью с другими молекулами. Изобретение дополнительно относится к антителам против CXCR5, демонстрирующим измененную эффекторную функцию. В некоторых вариантах осуществления измененная эффекторная функция является повышенной ADCC. В некоторых вариантах осуществления в антителах отсутствует фукоза, или они содержат детектируемо сниженные уровни фукозы (т.е. они являются афукозилированными). Изобретение также относится к композициям, содержащим такие антитела, а также к применению таких антител, включая терапевтическое и фармацевтическое применение.
[000165] В одном из вариантов осуществления настоящее изобретение относится к любому из следующих антител или композициям (включая фармацевтические композиции), содержащим антитело, содержащее последовательность легкой цепи или ее фрагмент и последовательность тяжелой цепи или ее фрагмент, полученные из любого из следующих антител: 11G2, 41A10 и 5H7.
[000166] Антитела, которые можно использовать в настоящем изобретении, могут включать моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab’, F(ab’)2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгированные антитела, одноцепочечные антитела (ScFv), их мутанты, слитые белки, содержащие фрагменты антител (например, доменное антитело), гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащей участок распознавания антигена необходимой специфичности, включая варианты гликозилирования антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела могут принадлежать мыши, крысе, человеку или любому другому источнику (включая химерные или гуманизированные антитела). В некоторых вариантах осуществления антитело против CXCR5 является моноклональным антителом. В некоторых вариантах осуществления антитело является антителом человека или гуманизированным антителом.
[000167] Антитела против CXCR5 по изобретению можно получать любым известным в этой области способом. Общие способы получения антител человека и мыши известны в этой области и/или представлены в настоящем описании.
[000168] После исходной идентификации, активность антитела-кандидата против CXCR5 можно дополнительно подтверждать и уточнять посредством известных биологических анализов для тестирования целевых видов биологической активности. В некоторых вариантах осуществления клеточный анализ in vitro используют для дополнительной характеризации антитела-кандидата против CXCR5. Например, биологические анализы можно использовать для прямого скрининга кандидатов. Некоторые из способов идентификации и характеризации антитела против CXCR5 подробно описаны в примерах.
[000169] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит по меньшей мере одну аминокислотную последовательность, выбранную из группы, состоящей из последовательности VH 11G2 мыши, VL 11G2 мыши, VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353), VH h11G2 (XC354), VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348), VL h11G2 (XC349), VH 41A10 мыши, VL 41A10 мыши, VH h41A10 (XC147), VH h41A10 (XC148), VH h41A10 (XC150), VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146), VL h41A10 (XC149), VH 5H7 мыши и VL 5H7 мыши.
[000170] В некоторых аспектах, антитело или его антигенсвязывающий фрагмент конкурирует с любыми из указанных выше антител за связывание с CXCR5.
[000171] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность VH и аминокислотную последовательность VL, включая следующие комбинации: антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с CXCR5 человека с одним или более из 11G2 мыши, химерного 11G2, VH h11G2 (XC152)/VL (XC151), VH h11G2 (XC152)/VL (XC153), VH h11G2 (XC152)/VL (XC154), VH h11G2 (XC152)/VL (XC346), VH h11G2 (XC152)/VL (XC347), VH h11G2 (XC152)/VL (XC348), VH h11G2 (XC152)/VL (XC349), VH h11G2 (XC155)/VL (XC151), VH h11G2 (XC155)/VL (XC153), VH h11G2 (XC155)/VL (XC154), VH h11G2 (XC155)/VL (XC346), VH h11G2 (XC155)/VL (XC347), VH h11G2 (XC155)/VL (XC3484), VH h11G2 (XC155)/VL (XC349), VH h11G2 (XC156)/VL (XC151), VH h11G2 (XC156)/VL (XC153), VH h11G2 (XC156)/VL (XC154), VH h11G2 (XC156)/VL (XC346), VH h11G2 (XC156)/VL (XC347), VH h11G2 (XC156)/VL (XC348), VH h11G2 (XC156)/VL (XC349), VH h11G2 (XC157)/VL (XC151), VH h11G2 (XC157)/VL (XC153), VH h11G2 (XC157)/VL (XC154), VH h11G2 (XC157)/VL (XC346), VH h11G2 (XC157)/VL (XC347), VH h11G2 (XC157)/VL (XC348), VH h11G2 (XC157)/VL (XC349), VH h11G2 (XC350)/VL (XC151), VH h11G2 (XC350)/VL (XC153), VH h11G2 (XC350)/VL (XC154), VH h11G2 (XC350)/VL (XC346), VH h11G2 (XC350)/VL (XC347), VH h11G2 (XC350)/VL (XC348), VH h11G2 (XC350)/VL (XC349), VH h11G2 (XC351)/VL (XC151), VH h11G2 (XC351)/VL (XC153), VH h11G2 (XC351)/VL (XC154), VH h11G2 (XC351)/VL (XC346), VH h11G2 (XC351)/VL (XC347), VH h11G2 (XC351)/VL (XC348) и VH h11G2 (XC351)/VL (XC349), VH h11G2 (XC352)/VL (XC151), VH h11G2 (XC352)/VL (XC153), VH h11G2 (XC352)/VL (XC154), VH h11G2 (XC352)/VL (XC346), VH h11G2 (XC352)/VL (XC347), VH h11G2 (XC352)/VL (XC348), VH h11G2 (XC352)/VL (XC349), VH h11G2 (XC353)/VL (XC151), VH h11G2 (XC353)/VL (XC153), VH h11G2 (XC353)/VL (XC154), VH h11G2 (XC353)/VL (XC346), VH h11G2 (XC353)/VL (XC347), VH h11G2 (XC353)/VL (XC348), VH h11G2 (XC353)/VL (XC349), VH h11G2 (XC354)/VL (XC151), VH h11G2 (XC354)/VL (XC153), VH h11G2 (XC354)/VL (XC154), VH h11G2 (XC354)/VL (XC346), VH h11G2 (XC354)/VL (XC347), VH h11G2 (XC354)/VL (XC348), VH h11G2 (XC354)/VL (XC349), 41A10 мыши, химерного 41A10, VH h41A10 (XC147)/VL (XC142), VH h41A10 (XC147)/VL (XC143), VH h41A10 (XC147)/VL (XC144), VH h41A10 (XC147)/VL (XC145), VH h41A10 (XC147)/VL (XC146), VH h41A10 (XC147)/VL (XC149), VH h41A10 (XC148)/VL (XC142), VH h41A10 (XC148)/VL (XC143), VH h41A10 (XC148)/VL (XC144), VH h41A10 (XC148)/VL (XC145), VH h41A10 (XC148)/VL (XC146), VH h41A10 (XC148)/VL (XC149), VH h41A10 (XC150)/VL (XC142), VH h41A10 (XC150)/VL (XC143), VH h41A10 (XC150)/VL (XC144), VH h41A10 (XC150)/VL (XC145), VH h41A10 (XC150)/VL (XC146), VH h41A10 (XC150)/VL (XC149), 5H7 мыши и химерного 5H7.
[000172] В некоторых аспектах антитело или его антигенсвязывающий фрагмент конкурирует с любыми из указанных выше антител за связывание с CXCR5.
[000173] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000174] В некоторых аспектах антитело или его антигенсвязывающий фрагмент дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности по меньшей мере одной SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56, 57.
[000175] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2, CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:1, и CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:6.
[000176] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2, CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:1, и CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:10.
[000177] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2, CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:1, и CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:12.
[000178] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2, CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:1, и CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:52.
[000179] В некоторых аспектах антитело или его антигенсвязывающий фрагмент дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:6, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000180] В некоторых аспектах антитело или его антигенсвязывающий фрагмент, дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:10, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000181] В некоторых аспектах антитело или его антигенсвязывающий фрагмент, дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:12, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000182] В некоторых аспектах антитело или его антигенсвязывающий фрагмент, дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:52, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности по меньшей мере одной из SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000183] В некоторых аспектах антитело или его антигенсвязывающий фрагмент дополнительно содержит CDR-H1, CDR-H2 и CDR-H3, приведенные в аминокислотной последовательности SEQ ID NO:27, и CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности SEQ ID NO:26.
[000184] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, CDR-L2 и CDR-L3, приведенные в аминокислотной последовательности, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324.
[000185] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-H1, CDR-H2 и CDR-H3, приведенный в аминокислотной последовательности, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
[000186] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит аминокислотную последовательность CDR-L1, CDR-L2 и CDR-L3, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324, и аминокислотную последовательность CDR-H1, CDR-H2 и CDR-H3, кодируемую вставкой плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
[000187] В некоторых аспектах антитело или его антигенсвязывающий фрагмент конкурирует с любым из указанных выше антител за связывание с CXCR5.
[000188] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:9.
[000189] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:11.
[000190] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:2, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:3, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:4, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:19, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21.
[000191] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:16, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:19, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:20, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:21.
[000192] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:16, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:7, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:8, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:9.
[000193] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO:14, CDR-L2, содержащую аминокислотную последовательность SEQ ID NO:15, CDR-L3, содержащую аминокислотную последовательность SEQ ID NO:16, CDR-H1, содержащую аминокислотную последовательность SEQ ID NO:8, CDR-H2, содержащую аминокислотную последовательность SEQ ID NO:9, и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO:10.
[000194] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит аминокислотные последовательности CDR-H1, CDR-H2 и CDR-H3, приведенные по меньшей мере в одной из последовательностей SEQ ID NO:6, 10, 12, 36, 52, 53, 54, 55, 56 и 57.
[000195] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит аминокислотные последовательности CDR-L1, CDR-L2 и CDR-L3, приведенные по меньшей мере в одной из последовательностей SEQ ID NO:1, 5, 35, 47, 48, 49, 50 и 51.
[000196] В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит аминокислотные последовательности CDR-L1, CDR-L2 и CDR-L3, приведенные в SEQ ID NO:5.
[000197] Антитело или его антигенсвязывающий фрагмент может содержать VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:6. VH может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:6. VH может содержать аминокислотную последовательность SEQ ID NO:6.
[000198] Антитело или его антигенсвязывающий фрагмент может содержать VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:10. VH может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:10. VH может содержать аминокислотную последовательность SEQ ID NO:10.
[000199] Антитело или его антигенсвязывающий фрагмент, может содержать VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:12. VH может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:12. VH может содержать аминокислотную последовательность SEQ ID NO:12.
[000200] Антитело или антигенсвязывающий фрагмент может содержать VL, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:1. VL может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:1. VL может содержать аминокислотную последовательность SEQ ID NO:1.
[000201] Антитело или антигенсвязывающий фрагмент может содержать VL, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:5. VL может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:5. VL может содержать аминокислотную последовательность SEQ ID NO:5.
[000202] Антитело или его антигенсвязывающий фрагмент может содержать VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:17. VH может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:17. VH может содержать аминокислотную последовательность SEQ ID NO:17.
[000203] Антитело или его антигенсвязывающий фрагмент, может содержать VH, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:18. VH может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:18. VH может содержать аминокислотную последовательность SEQ ID NO:18.
[000204] Антитело или антигенсвязывающий фрагмент может содержать VL, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:13. VL может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:13. VL может содержать аминокислотную последовательность SEQ ID NO:13.
[000205] Антитело или антигенсвязывающий фрагмент может содержать VL, содержащую аминокислотную последовательность, по меньшей мере на 90% идентичную аминокислотной последовательности SEQ ID NO:58. VL может содержать аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:58. VL может содержать аминокислотную последовательность SEQ ID NO:58.
[000206] В одном из аспектов афукозилированное антитело или его антигенсвязывающий фрагмент может содержать тяжелую цепь, содержащую VH, выбранную из VH, содержащей аминокислотную последовательность SEQ ID NO:10, SEQ ID NO:12, SEQ ID NO:36, SEQ ID NO:52, SEQ ID NO:53, SEQ ID NO:54, SEQ ID NO:55, SEQ ID NO:56 или SEQ ID NO:57 и дополнительно содержащей константный домен IgG1 (SEQ ID NO:31). В одном из аспектов указанный вариант антитела содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 консервативных или неконсервативных замен и/или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 добавлений и/или делеций в полноразмерной тяжелой цепи. В дополнительном аспекте указанный вариант имеет по меньшей мере 65%, по меньшей мере 75%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к полноразмерной тяжелой цепи, и где указанное антитело или антигенсвязывающий фрагмент специфически связываются с CXCR5.
[000207] Афукозилированное антитело по изобретению может содержать легкую цепь, содержащую VL, содержащую аминокислотную последовательность SEQ ID NO:1, SEQ ID NO:5, SEQ ID NO:35, SEQ ID NO:47, SEQ ID NO:48, SEQ ID NO:49, SEQ ID NO:50 или SEQ ID NO:51, где антитело дополнительно содержит константный домен легкой цепи. Как подробно представлено в других местах настоящего описания, константный домен легкой цепи афукозилированного антитела можно выбирать из константной области Cκ или Cλ, например, константной области Cκ SEQ ID NO:30. В одном из аспектов указанный вариант антитела содержит 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 консервативных или неконсервативных замен и/или 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14 или 15 добавлений и/или делеций в полноразмерной легкой цепи. В дополнительном аспекте указанный вариант имеет по меньшей мере 65%, по меньшей мере 75%, по меньшей мере 85%, по меньшей мере 90%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к полноразмерной легкой цепи, и где указанное антитело или антигенсвязывающий фрагмент специфически связывается с CXCR5.
[000208] Антитело или антигенсвязывающий фрагмент может содержать HC, содержащую аминокислотную последовательность, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную аминокислотной последовательности SEQ ID NO:29. HC может содержать аминокислотную последовательность SEQ ID NO:29. Предпочтительно, антитело является афукозилированным.
[000209] Антитело или антигенсвязывающий фрагмент может содержать LC, содержащую аминокислотную последовательность, по меньшей мере на 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% идентичную SEQ ID NO:28. LC может содержать аминокислотную последовательность SEQ ID NO:28. Предпочтительно, антитело является афукозилированным.
[000210] Предпочтительно, антитело или его антигенсвязывающий фрагмент по изобретению является афукозилированным. Даже более предпочтительно, антитело или его антигенсвязывающий фрагмент является афукозилированным и демонстрирует повышенную эффекторную функцию ADCC по сравнению с в остальном идентичным антителом, являющимся фукозилированным.
Замены зародышевой линии
[000211] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент содержит следующие последовательности CDR тяжелой цепи: (i) CDR-H1, содержащую SEQ ID NO:7, CDR-H2, содержащую SEQ ID NO:8, и CDR-H3, содержащую SEQ ID NO:9 или 11; и/или (ii) следующие последовательности CDR легкой цепи: CDR-L1, содержащую SEQ ID NO:2, CDR-L2, содержащую SEQ ID NO:3, и CDR-L3, содержащую SEQ ID NO:4. Они являются CDR мыши и, предпочтительно, являются привитыми или иным образом добавленными в контекст доменов VH и VL человека. Доступен широкий спектр акцепторных последовательностей зародышевой линии человека, и способы "гуманизации" антител не являющихся человеком видов для использования на людях хорошо известны в этой области, а также представлены в других местах настоящего описания. Таким образом, специалистам в этой области будет понятно, что указанные выше последовательности CDR мыши можно помещать в контекст аминокислотных последовательностей V-домена человека. При этом, для сохранения связывания антитела и других желаемых характеристик исходного родительского (т.е. донорного) антитела, как правило, можно делать изменения в акцепторных последовательностях зародышевой линии человека. И CDR, и каркасные области (FW) можно конструировать следующим образом.
[000212] В некоторых вариантах осуществления в CDR-L1 делают не более 11, или не более 10, не более 9, не более 8, не более 7, не более 6, не более 5, не более 4, не более 3, не более 2 или не более 1 замены относительно аминокислотной последовательности SEQ ID NO:2. В некоторых вариантах осуществления в CDR-L2 делают не более 6, не более 5, не более 4, не более 3, не более 3, не более 2 или не более одной замены относительно аминокислотной последовательности SEQ ID NO:3. В некоторых вариантах осуществления в CDR-L3 делают не более 8, не более 7, не более 6, не более 5, не более 4, не более 3, не более 3, не более 2 или не более одной замены относительно аминокислотной последовательности SEQ ID NO:4. В некоторых вариантах осуществления в CDR-H1 делают не более 10, не более 9, не более 8, не более 7, не более 6, не более 5, не более 4, не более 3, не более 2 или не более 1 замены относительно аминокислотной последовательности SEQ ID NO:7. В некоторых вариантах осуществления в CDR-H2 делают не более не более 17, не более 16, не более 15, не более 14, не более 13, не более 12, не более 11 или не более 10, не более 9, не более 8, не более 7, не более 6, не более 5, не более 4, не более 3, не более 2 или не более 1 замены относительно аминокислотной последовательности SEQ ID NO:8. В некоторых вариантах осуществления в CDR-H3 делают не более 12, не более 11, или не более 10, не более 9, не более 8, не более 7, не более 6, не более 5, не более 4, не более 3, не более 2 или не более 1 замены относительно аминокислотной последовательности SEQ ID NO:9 или относительно аминокислотной последовательности SEQ ID NO:11. В некоторых вариантах осуществления замены не изменяют значение аффинности связывания (KD) более чем в 1000 раз, более чем в 100 раз или 10 раз. В некоторых вариантах осуществления замена является консервативной заменой по таблице 1.
[000213] В некоторых вариантах осуществления замена является заменой зародышевой линии человека, в которой (донорный) остаток CDR заменяют соответствующим (акцепторным) остатком зародышевой линии человека для повышения содержания аминокислот человека и потенциального снижения иммуногенности антитела, как описано, например, в публикации патентной заявки США № 2017/0073395 и Townsend et al., 2015, Proc. Nat. Acad. Sci. USA 112(50):15354-15359). Например, если используют каркас DPK9 зародышевой линии человека и сравнивают пример VL 11G2 (XC154) антитела мыши или гуманизированной VL 11G2 (XC154), то выравнивание CDR-L1 антитела VL 11G2 (XC154 мыши и гуманизированного XC154) (SEQ ID NO:2) и DPK9 зародышевой линии человека является следующим:
[000214] В случае положения аминокислоты 28 (выделено курсивом), остаток зародышевой линии человека (акцепторный) и соответствующие остатки VL 11G2 (XC154) (донорные) являются одинаковыми, и замена зародышевой линии невозможна. В случае положений 27, 29, 30, 31, 32, 33, 34 и 35 (выделены полужирным шрифтом и подчеркнуты), остаток зародышевой линии человека (акцепторный) и соответствующий остаток VL 11G2 (XC154) (донорный) отличаются. Остатки VL 11G2 (XC154) в этих положениях можно заменять соответствующим остатком DPK9 зародышевой линии человека для дополнительного повышения содержания остатков человека. То же можно использовать для каждой CDR тяжелой и легкой цепи для повышения содержания аминокислотных остатков человека при сохранении связывающих характеристик, например, связывания эпитопа, аффинности и т.п., и минимизации содержания остатков мыши, таким образом, снижая потенциальную иммуногенность, например, иммунный ответ с образованием антител человека против антитела мыши (HAMA), на антитело у человека.
[000215] Способы и библиотеки для встраивания остатков зародышевой линии человека в CDR антитела подробно описаны в публикации патентной заявки США № 2017/0073395, и Townsend et al., 2015, Proc. Natl. Acad. Sci. USA. 112(50):15354-15359, включенных в настоящее описание в качестве ссылки в полном объеме.
[000216] Антитело или его антигенсвязывающий фрагмент может содержать каркас VH, содержащий каркасную последовательность VH зародышевой линии человека. Каркасную последовательность VH можно получать из VH3 зародышевой линии, VH1 зародышевой линии, VH5 зародышевой линии или VH4 зародышевой линии человека. Предпочтительные каркасы тяжелой цепи зародышевой линии человека являются каркасами, полученными из зародышевых линий VH1, VH3 или VH5. Например, можно использовать каркасы VH из следующих зародышевых линий: IGHV3-23, IGHV3-7 или IGHV1-69 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Предпочтительные каркасы легкой цепи зародышевой линии человека являются каркасами, полученными из зародышевых линий Vκ или Vλ. Например, можно использовать каркасы VL из следующих зародышевых линий: IGKV1-39 или IGKV3-20 (названия зародышевых линий основаны на определении зародышевой линии IMGT). Альтернативно или дополнительно, каркасная последовательность может являться консенсусной каркасной последовательностью зародышевой линии человека, такой как каркас консенсусной последовательности Vλ1человека, консенсусной последовательности Vκ1, консенсусной последовательности Vκ2, консенсусной последовательности Vκ3, консенсусной последовательности зародышевой линии VH3, консенсусной последовательности зародышевой линии VH1, консенсусной последовательности зародышевой линии VH5 или консенсусной последовательности зародышевой линии VH4. Последовательности каркасов зародышевой линии человека доступны в различных общедоступных базах данных, таких как V-base, IMGT, NCBI или Abysis.
[000217] Антитело или его антигенсвязывающий фрагмент может содержать каркас VL, содержащий каркасную последовательность VL зародышевой линии человека. Каркас VL может содержать одну или более замен, добавлений или делеций аминокислот при сохранении функционального и структурного сходства с зародышевой линией, из которой его получают. В некоторых аспектах каркас VL является по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичным каркасной последовательности VL зародышевой линии человека. В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит каркас VL, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 замен, добавлений или делеций аминокислот относительно каркасной последовательности VL зародышевой линии человека. В некоторых аспектах 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 замен, добавлений или делеций аминокислот являются единственными в каркасных областях. В некоторых аспектах % идентичности основан на сходстве с VL, за исключением фрагментов, определяемых в настоящем описании как CDR.
[000218] Каркас VL зародышевой линии человека может являться каркасом DPK9 (название IMGT: IGKV1-39). Каркас VL зародышевой линии человека может являться каркасом DPK12 (название IMGT: IGKV2D-29). Каркас VL зародышевой линии человека может являться каркасом DPK18 (название IMGT: IGKV2-30). Каркас VL зародышевой линии человека может являться каркасом DPK24 (название IMGT: IGKV4-1). Каркас VL зародышевой линии человека может являться каркасом HK102_V1 (название IMGT: IGKV1-5). Каркас VL зародышевой линии человека может являться каркасом DPK1 (название IMGT: IGKV1-33). Каркас VL зародышевой линии человека может являться каркасом DPK8 (название IMGT: IGKV1-9). Каркас VL зародышевой линии человека может являться каркасом DPK3 (название IMGT: IGKV1-6). Каркас VL зародышевой линии человека может являться каркасом DPK21 (название IMGT: IGKV3-15). Каркас VL зародышевой линии человека может являться каркасом Vg_38K (название IMGT: IGKV3-11). Каркас VL зародышевой линии человека может являться каркасом DPK22 (название IMGT: IGKV3-20). Каркас VL зародышевой линии человека может являться каркасом DPK15 (название IMGT: IGKV2-28). Каркас VL зародышевой линии человека может являться каркасом DPL16 (название IMGT: IGLV3-19). Каркас VL зародышевой линии человека может являться каркасом DPL8 (название IMGT: IGLV1-40). Каркас VL зародышевой линии человека может являться каркасом V1-22 (название IMGT: IGLV6-57). Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vλ человека. Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vλ1 человека. Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vλ3 человек. Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vκ человека. Каркас VL зародышевой линии человека может являться каркасом Vκ1 консенсусной последовательности человека. Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vκ2 человека. Каркас VL зародышевой линии человека может являться каркасом консенсусной последовательности Vκ3 человека.
[000219] В некоторых аспектах каркас VL представляет собой DPK9. Прогнозируют, что с помощью других схожих каркасных областей будут получать предпочтительные антитела по изобретению, содержащие CDR SEQ ID NO:2, 3, 4, 7, 8, 9, 11, 14, 15, 16; и CDR, определяемые следующими аминокислотными последовательностями VL: 1, 5, 13, 28, 35, 37, 39, 47, 48, 48, 50, 51, 58, 59, 60, 61, 62, 97, 98, включая DPK5, DPK4, DPK1, IGKV1-5*01, DPK24, DPK21, DPK15, IGKV1-13*02, IGKV1-17*01, DPK8, IGKV3-11*01 и DPK22, имеющие 99, 97, 97, 96, 80, 76, 66, 97, 97, 96, 76 и 74% идентичности, соответственно, по отношению к области FW DPK-9 и одно или меньше отличий аминокислот в общих структурных признаках (нумерация по Kabat): (A) остатках непосредственно под CDR (зоне Вернье), L2, L4, L35, L36, L46, L47, L48, L49, L64, L66, L68, L69, L71, (B) остатках для упаковки цепей VH/VL: L36, L38, L44, L46, L87 и (C) канонических структурных поддерживающих остатках CDR L2, L48, L64, L71 (см. Lo, "Antibody Humanization by CDR Grafting", (2004) Antibody Engineering, Vol. 248, Methods in Molecular biology pp 135-159 и O’Brien and Jones, "Humanization of Monoclonal antibodies by CDR Grafting", (2003) Recombinant Antibodies for Cancer Therapy, Vol. 207, Methods in Molecular biology pp 81-100). Особенно предпочтительными являются каркасные области DPK5, DPK4, DPK1, IGKV1-5*01, DPK24, DPK21 и DPK15, имеющие 99, 97, 97, 96, 80, 76, 66% идентичности по отношению к DPK9, соответственно, и не имеющие отличий аминокислот в этих общих структурных признаках. В некоторых аспектах % идентичности основан на сходстве с VL, за исключением фрагментов, определяемых в настоящем описании как CDR.
[000220] Антитело или его антигенсвязывающий фрагмент может содержать каркас VH, содержащий каркасную последовательность VH зародышевой линии человека. Каркас VH может содержать одну или более замен, добавлений или делеций аминокислот с сохранением функционального и структурного сходства с зародышевой линией, из которых его получают. В некоторых аспектах каркас VH является по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичным каркасной последовательности VH зародышевой линии человека. В некоторых аспектах антитело или его антигенсвязывающий фрагмент содержит каркас VH, содержащий 1, 2, 3, 4, 5, 6, 7, 8, 9, 10 замен, добавлений или делеций аминокислот относительно каркасной последовательности VH зародышевой линии человека. В некоторых аспектах 1, 2, 3, 4, 5, 6, 7, 8, 9 или 10 замен, добавлений или делеций аминокислот являются единственными в каркасных областях. В некоторых аспектах % идентичности основан на сходстве с VH, за исключением фрагментов, определяемых в настоящем описании как CDR.
[000221] Каркас VH зародышевой линии человека может являться каркасом DP54 или IGHV3-7. Каркас VH зародышевой линии человека может являться каркасом DP47 или IGHV3-23. Каркас VH зародышевой линии человека может являться каркасом DP71 или IGHV4-59. Каркас VH зародышевой линии человека может являться каркасом DP75 или IGHV1-2_02. Каркас VH зародышевой линии человека может являться каркасом DP10 или IGHV1-69. Каркас VH зародышевой линии человека может являться каркасом DP7 или IGHV1-46. Каркас VH зародышевой линии человека может являться каркасом DP49 или IGHV3-30. Каркас VH зародышевой линии человека может являться каркасом DP51 или IGHV3-48. Каркас VH зародышевой линии человека может являться каркасом DP38 или IGHV3-15. Каркас VH зародышевой линии человека может являться каркасом DP79 или IGHV4-39. Каркас VH зародышевой линии человека может являться каркасом DP78 или IGHV4-30-4. Каркас VH зародышевой линии человека может являться каркасом DP73 или IGHV5-51. Каркас VH зародышевой линии человека может являться каркасом DP50 или IGHV3-33. Каркас VH зародышевой линии человека может являться каркасом DP46 или IGHV3-30-3. Каркас VH зародышевой линии человека может являться каркасом DP31 или IGHV3-9. Каркас VH зародышевой линии человека может являться каркасом консенсусной последовательности VH зародышевой линии человека. Каркас VH зародышевой линии человека может являться каркасом консенсусной последовательности зародышевой линии VH3 человека. Каркас VH зародышевой линии человека может являться каркасом консенсусной последовательности VH5 зародышевой линии человека. Каркас VH зародышевой линии человека может являться каркасом консенсусной последовательности зародышевой линии VH1 человека. Каркас VH зародышевой линии человека может являться каркасом консенсусной последовательности VH4 зародышевой линии человека.
[000222] В некоторых аспектах каркас VH представляет собой DP-54. Прогнозируют, что с помощью других схожих каркасных областей будут получать предпочтительные антитела по изобретению, содержащие CDR SEQ ID NO:7, 8, 9, 11,19, 20, 21 и CDR, определенные следующими аминокислотными последовательностями VH: 6, 10, 12, 17, 18, 36, 38, 40, 52, 53, 54, 55, 56, 57, 63, 96, включая DP-50, IGHV3-30*09, IGHV3-30*15, IGHV3-48*01, DP-77, DP-51, IGHV3-66*01, DP-53, DP-48, IGHV3-53*01, IGHV3-30*02 и DP-49, имеющие 93, 92, 92, 99, 97, 97, 96, 96, 94, 94, 93, 92% идентичности, соответственно, по отношению к области FW DP-54 и одно или несколько отличий аминокислот в общих структурных признаках (нумерация Kabat): (A) остатках непосредственно под CDR (зоне Вернье), H2, H47, H48, и H49, H67, H69, H71, H73, H93, H94, (B) остатках для упаковки цепей VH/VL: H37, H39, H45, H47, H91, H93 и (C) канонических структурных поддерживающих остатках CDR H24, H71, H94 (см. Lo 2004, и O’Brien and Jones 2003). Особенно предпочтительными являются каркасные области DP-50, IGHV3-30*09, IGHV3-30*15, имеющие 93, 92 и 92% идентичности по отношению к DP-54, соответственно, и неимеющие отличий аминокислот в этих общих структурных признаках. В некоторых аспектах % идентичности основан на сходстве с VH, за исключением фрагментов, определяемых в настоящем описании как CDR.
[000223] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, представленные в настоящем описании, содержит (i) VH, содержащую аминокислотную последовательность, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную аминокислотной последовательности SEQ ID NO:8, и/или (ii) VL, содержащую аминокислотную последовательность, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 66%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 76%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную аминокислотной последовательности SEQ ID NO:1. Изобретение также включает любую комбинацию этих последовательностей VL и VH.
[000224] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, представленный в настоящем описании, содержит (i) HC, содержащую аминокислотную последовательность, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную аминокислотной последовательности SEQ ID NO:29; и/или (ii) LC, содержащую аминокислотную последовательность, по меньшей мере на 50%, по меньшей мере на 60%, по меньшей мере на 70%, по меньшей мере на 75%, по меньшей мере на 80%, по меньшей мере на 85%, по меньшей мере на 90%, по меньшей мере на 91%, по меньшей мере на 92%, по меньшей мере на 93%, по меньшей мере на 94%, по меньшей мере на 95%, по меньшей мере на 96%, по меньшей мере на 97%, по меньшей мере на 98%, по меньшей мере на 99% или на 100% идентичную аминокислотной последовательности SEQ ID NO:28. Изобретение также включает любую комбинацию этих последовательностей HC и LC.
[000225] В некоторых вариантах осуществления антитело или его антигенсвязывающий фрагмент, представленные в настоящем описании, содержат Fc-домен. Fc-домен можно получать из IgA (например, IgA1 или IgA2), IgG, IgE или IgG (например, IgG1, IgG2, IgG3 или IgG4).
[000226] Настоящее изобретение также относится к антителу или его антигенсвязывающему фрагменту, конкурирующему за связывание с CXCR5 человека с любыми антителами или их антигенсвязывающими фрагментами, представленными в настоящем описании, такими как любое из антител, представленных в настоящем описании (или его антигенсвязывающий фрагмент). Например, если связывание антитела или его антигенсвязывающего фрагмента с CXCR5 человека препятствует последующему связыванию CXCL13 с CXCR5 человека, антитело или его антигенсвязывающий фрагмент конкурируют с CXCL13 за связывание с CXCR5 человека.
[000227] Антитела и антигенсвязывающие фрагменты по изобретению включают моноклональные антитела, поликлональные антитела, фрагменты антител (например, Fab, Fab’, F(ab’)2, Fv, Fc и т.д.), химерные антитела, биспецифические антитела, гетероконъюгированные антитела, одноцепочечные антитела (ScFv), их мутанты, слитые белки, содержащие часть антитела, доменные антитела (dAb), гуманизированные антитела и любую другую модифицированную конфигурацию молекулы иммуноглобулина, содержащую участок распознавания антигена необходимой специфичности, включая варианты гликозилирования антител, варианты аминокислотной последовательности антител и ковалентно модифицированные антитела. Антитела и антигенсвязывающие фрагменты могут принадлежать мыши, крысе, человеку или любому другому источнику (включая химерные или гуманизированные антитела). В некоторых вариантах осуществления антитело является моноклональным антителом. В некоторых вариантах осуществления антитело является химерным антителом, гуманизированным антителом или антителом человека. В некоторых вариантах осуществления антитело является антителом человека. В некоторых вариантах осуществления антитело является гуманизированным антителом.
Эпитопное картирование
[000228] Настоящее изобретение также относится к антителу или его антигенсвязывающему фрагменту, связывающемуся с тем же эпитопом CXCR5 человека, что и антитело или его антигенсвязывающий фрагмент, представленные в настоящем описании. Например, конкурентные анализы антител (и анализ перекрывающихся эпитопов) можно осуществлять посредством SPR, проточной цитометрии и любого другого анализа, известного в этой области.
[000229] В одном из аспектов настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, связывающемуся с CXCR5 человека и CXCR5 яванского макака, но не связывающемуся с CXCR5 мыши.
[000230] Настоящее изобретение также включает антитело или его антигенсвязывающий фрагмент, связывающийся с N-концевой областью ("N") CXCR5 человека. N-конец CXCR5 содержит аминокислотные остатки 1-51 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:31.
[000231] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 в эпитопе, содержащем лейцин (L) в положении аминокислотного остатка 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000232] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 в эпитопе, содержащем аспартат (D) в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000233] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 в эпитопе, содержащем лейцин в положении аминокислотного остатка 11 и аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000234] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, если аминокислотный остаток в положении 11 является лейцином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000235] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, если аминокислотный остаток в положении 11 является лейцином, но не связывающиеся с CXCR5, если аминокислотный остаток в положении 11 является треонином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000236] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с дикого типа CXCR5 человека, но не связывающиеся, если аминокислотный остаток в положении 11 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32 не является лейцином.
[000237] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, если аминокислотный остаток в положении 22 является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000238] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с дикого типа CXCR5 человека, но не связывающиеся, если аминокислотный остаток в положении 22 не является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000239] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с дикого типа CXCR5 человека, но не связывающиеся, если аминокислотный остаток в положении 22 является аланином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000240] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, если аминокислотный остаток в положении 11 является лейцином, и аминокислотный остаток в положении 22 является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000241] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 человека, но не связывающиеся с CXCR5, если аминокислотный остаток в положении 11 не является лейцином, и аминокислотный остаток в положении 22 не является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000242] В одном из аспектов настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 человека, но не связывающиеся, если аминокислотный остаток в положении 11 является треонином, и аминокислотный остаток в положении 22 является аланином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000243] Специалистам в этой области с учетом идей, представленных в настоящем описании, будет понятно, что аминокислотный остаток в положении 11 и/или в положении 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32 являются критическими для связывания антитела по изобретению. Более конкретно, эти аминокислотные остатки являются критическими для связывания с CXCR5 антителом 11G2 или его антигенсвязывающим фрагментом. Таким образом, специалистам в этой области будет понятно, что настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5, если аминокислотный остаток в положении 11 является лейцином, и аминокислотный остаток в положении 22 является аспартатом. Замена или делеция этих аминокислотных остатков могут приводить к потере связывания с CXCR5. Некоторые замены аминокислотного остатка в положении 11 могут приводить к сохранению связывания, но не замена этого аминокислотного остатка (лейцина) треонином. Аналогично, некоторые замены или делеция аминокислотного остатка в положении 22 могут приводить к сохранению связывания, но не замена аспартата в этом положении аминокислотного остатка аланином. Таким образом, признаком антитела 11G2, или его антигенсвязывающего фрагмента, или антитела, конкурирующего с ними за связывание, является то, что антитело связывается с CXCR5 человека, если присутствуют лейцин 11 и аспартат 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32, но антитело не связывается, если лейцин 11 заменяют треонином или другим аминокислотным остатком, и где антитело не связывается, если аспартат 22 заменяют аланином или другим аминокислотным остатком. Тестирование на потерю связывания после замены аминокислоты в положении аминокислотного остатка 11 и/или 22 можно осуществлять с использованием широкого спектра способов, хорошо известных в этой области, включая анализ связывания с использованием полипептидов с точечными мутациями, как в случае способов, примеры которых приведены в настоящем описании.
[000244] С учетом идей, представленных в настоящем описании, специалисту в этой области будет понятно, что антитело по изобретению может конкурировать, например, с 11G2 и все равно не содержать эпитоп, содержащий лейцин в положении аминокислотного остатка 11 и/или аспартат в положении аминокислотного остатка 22 в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32. Т.е. антитело может конкурировать с антителом по изобретению за связывание с CXCR5, но связывание конкурирующего антитела не затронуто, если лейцин в положении аминокислотного остатка 11 или аспартат в положении аминокислотного остатка 22 заменяют другой аминокислотой (например, треонином в случае лейцина и/или аланином в случае аспартата), например, 2C9, 11A7 и 16D7. Таким образом, антитело по изобретению конкурирует за связывание с CXCR5, а также не связывается с CXCR5, если аминокислотный остаток в положении 11 не является лейцином, и более конкретно, он является треонином, и/или если аминокислотный остаток в положении 22 не является аспартатом, и более конкретно, он является аланином. Как указано выше в других местах настоящего описания, получение мутантных белков CXCR5 и анализы для оценки конкурентного связывания антитела хорошо известны в этой области, включая способы, представленные в настоящем описании. Таким образом, с учетом идей, представленных в настоящем описании, специалисты в этой области легко могут идентифицировать антитела, связывающиеся с CXCR5, конкурирующие за связывание с антителом по изобретению и потерявшие способность связываться с мутеинами CXCR5, если некоторые аминокислоты заменены, например, лейцин 11, аспартат 22 или и тот и другой в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000245] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 11 не является лейцином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000246] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 22 не является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000247] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 11 не является лейцином, и аминокислотный остаток в положении 22 не является аспартатом в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000248] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 11 является треонином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000249] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 22 является аланином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
[000250] В другом аспекте настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, конкурирующие за связывание с антителом 11G2 или его антигенсвязывающим фрагментом, где антитело не связывается с CXCR5, если аминокислотный остаток в положении 11 является треонином, и аминокислотный остаток в положении 22 является аланином в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32.
Биологическая активность антител против CXCR5
[000251] В дополнение к связыванию эпитопа на CXCR5, антитело или его антигенсвязывающий фрагмент по изобретению могут опосредовать биологическую активность. Т.е. настоящее изобретение включает выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 и опосредующее по меньшей мере одну детектируемую активность, выбранную из следующего: (a) связывается с CXCR5+ клетками с высокой кажущейся аффинностью, но не связывается с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика; (b) антагонистически действует на ингибирование CXCL13 высвобождения цАМФ, запускаемого форсколином; (c) запускает ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и TMC человека-донора; (d) связывается с CXCR5 человека, но не связывается с хемокиновыми рецепторами человека CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1; (e) истощает B-клетки в периферической крови и/или вторичных лимфоидных органах (т.е. лимфоузлах, селезенке, пейеровых бляшках и лимфоидной ткани слизистых оболочек); (f) истощает Tfh-подобные клетки в периферической крови; (g) истощает истинные Tfh-клетки во вторичных лимфоидных органах и (h) нарушает гуморальный анамнестический иммунный ответ.
[000252] В одном из аспектов настоящее изобретение включает антитело, связывающееся с CXCR5+ клетками с высокой кажущейся аффинностью, но не связывающееся с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика. Кажущуюся аффинность связывания можно оценивать с использованием проточной цитометрии для детекции связывания антитела с клетками, экспрессирующими белок-мишень (например, CXCR5). Клетки можно транзиторно или стабильно трансфицировать с использованием нуклеиновой кислоты, кодирующей CXCR5. Альтернативно, клетки могут являться клетками, от природы экспрессирующими CXCR5 на своей поверхности. Независимо от источников CXCR5+ клеток, связывание антитела с клетками легко можно анализировать с использованием различных известных в этой области способов. Антитело или его антигенсвязывающий фрагмент связываются с CXCR5 человека или CXCR5 яванского макака, но не связываются детектируемо или связываются с гораздо меньшей степенью с CXCR5 мыши, крысы или кролика.
[000253] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 и антагонистически действующие на активность, опосредуемую связыванием CXCL13 с CXCR5. Существует множество анализов, известных в этой области для определения ингибирования активности, опосредованной передачей сигнала CXCR5-CXCL13. Одним из таких анализов является репортерный анализ цАМФ. В таком анализе форсколин индуцирует продукцию цАМФ, ингибируемую передачей сигнала CXCR5-CXCL13 в клетке, стабильно экспрессирующей CXCR5. Способность антитела против CXCR5 связываться с CXCR5 и антагонистически действовать на эффект передачи сигнала CXCR5-CXCL13, таким образом, оценивают посредством измерения уровня цАМФ, продуцируемого в присутствии или отсутствие антитела. Предпочтительно, антитело может опосредовать дозозависимое повышение уровней цАМФ с EC50 приблизительно 50 пМ, приблизительно 100 пМ, приблизительно 200, пМ, приблизительно 400 пМ, приблизительно 600 пМ, приблизительно 700 пМ, приблизительно 750 пМ, приблизительно 790 пМ, приблизительно 800 пМ, приблизительно 850 пМ, приблизительно 900 пМ, приблизительно 950 пМ, приблизительно 960 пМ, приблизительно 970 пМ, приблизительно 980 пМ, приблизительно 990 пМ или приблизительно 1000 пМ. Более предпочтительно, антитело или его антигенсвязывающий фрагмент ингибирует ингибирование CXCL13 высвобождения цАМФ, запускаемого форсколином, с EC50 приблизительно 961 пМ.
[000254] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 и запускающие ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и тонзиллярных мононуклеарных клетках (TMC) человека-донора. Для оценки активности ADCC антитела можно использовать многие анализы ADCC. Один из примеров таких анализов представлен в настоящем описании в примерах 6 и 7, но настоящее изобретение не ограничено этим конкретным анализом ADCC. Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, демонстрирующие активность ADCC в отношении B-клеток человека, Tfh-подобных клеток человека, Tfh-клеток человека и B-клеток яванского макака с EC50 приблизительно 0,11 пМ, 0,2 пМ, 0,5 пМ, 1 пМ. 1,5 пМ, 2,0 пМ, 2,5 пМ, 3,0 пМ, 4,5 пМ, 4,8 пМ, 5,0 пМ, 6,0 пМ, 7,0 пМ, 8,0 пМ, 9,0 пМ, 10 пМ, 11 пМ, 12 пМ, 15 пМ, 20, пМ, 25 пМ, 30 пМ, 35 пМ или 40 пМ. Более предпочтительно, антитело или его антигенсвязывающий фрагмент демонстрирует активность ADCC с EC50 приблизительно 2,01±2,28 пМ против B-клеток человека, приблизительно 4,82±2,88 пМ против Tfh-подобных клеток человека, приблизительно 0,11 пМ против Thf-клеток человека и приблизительно 15,3±11,7 пМ против B-клеток яванского макака. Даже более предпочтительно, антитело или его антигенсвязывающий фрагмент является афукозилированным.
[000255] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, связывающемуся с CXCR5 человека, но не связывающемуся детектируемо с белками CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1 человека.
[000256] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, специфически связывающемуся с CXCR5 и демонстрирующему дозозависимое истощение B-клеток в периферической крови. Истощение может являться постоянным или, более предпочтительно, истощение является транзиторным и/или обратимым. В одном из аспектов доза антитела, достаточная для опосредования истощения B-клеток, может находиться в диапазоне от 0,001 до приблизительно 0,2 мг, где 0,0001 мг/кг могут опосредовать истощение B-клеток и Tfh-подобных клеток в периферической крови яванского макака на приблизительно 50% по сравнению с процентной долей B-клеток или Tfh-подобных клеток в периферической крови, если антитело не вводят. Кроме того, антитело может опосредовать максимальное истощение B-клеток и Tfh-подобных клеток в периферической крови яванского макака при введении в дозе менее 5 мг/кг внутривенно (IV). Предпочтительно, после введения антитела происходит частичное или полное восстановление B-клеток, Tfh-подобных клеток и Tfh-клеток.
[000257] Настоящее изобретение включает антитело или его антигенсвязывающий фрагмент, специфически связывающиеся с CXCR5 и нарушающие гуморальный анамнестический иммунный ответ. В одном из аспектов гуморальный анамнестический ответ оценивают с использованием анализа вторичного ответа на вакцину. Т.е. антитело вводят индивидууму, которого ранее вакцинировали с использованием антигена, например, столбнячного анатоксина (TT). Индивидуума подвергают второму введению антигена и иммунный ответ, например, титры IgM и IgG, сравнивают с иммунным ответом у в остальном идентичного индивидуума, которому вводят антитело. Специалистам в этой области с учетом идей, представленных в настоящем описании, будет понятно, что для оценки способности антитела или его антигенсвязывающего фрагмента нарушать гуморальный анамнестический ответ можно использовать многие анализы для определения гуморального анамнестического ответа. Кроме того, специалистам в этой области будет понятно, что способность нарушать гуморальный анамнестический ответ является желаемой характеристикой для антитела или его антигенсвязывающий фрагмент, предназначенных для использования в качестве терапевтического средства для лечения или профилактики иммунологического заболевания у больного.
[000258] Настоящее изобретение относится к антителу или его антигенсвязывающему фрагменту, демонстрирующему по меньшей мере один, предпочтительно - два, даже более предпочтительно - три, даже более предпочтительно - четыре, даже более предпочтительно - пять, даже более предпочтительно - шесть, более предпочтительно - семь, и даже более предпочтительно - все из указанных выше видов биологической активности.
III. АФУКОЗИЛИРОВАННЫЕ АНТИТЕЛА ПРОТИВ CXCR5
[000259] В одном из вариантов осуществления настоящее изобретение относится к антителам, имеющим углеводную структуру, в которой отсутствует фукоза, присоединенная (прямо или косвенно) к Fc-области (т.е. афукозилированным антителам). Например, количество фукозы в композиции, содержащей множество таких антител, может составлять от 0 процентов до приблизительно 30 процентов, от 0 процентов до приблизительно 20 процентов, от 0 процент до приблизительно 15 процентов, от 0 процентов до приблизительно 10 процентов, и более предпочтительно - от 0 процентов до приблизительно 5 процентов. В некоторых вариантах осуществления композиция, содержащая множество таких антител, содержит по меньшей мере 80 процентов, более предпочтительно - по меньшей мере 85 процентов, еще более предпочтительно - по меньшей мере 90 процентов, даже более предпочтительно - по меньшей мере 95 процентов афукозилированных антител, еще более предпочтительно - по меньшей мере 99 процентов, и наиболее предпочтительно - по меньшей мере 99,5 процентов афукозилированных антител. В некоторых вариантах осуществления антитела являются на 100 процентов афукозилированными; т.е. фукозу не определяют на Asn297 с использованием известного в этой области способа детекции фукозы в антителе. Количество фукозы определяют посредством вычисления среднего количества фукозы в сахарной цепи в Asn297 относительно суммы всех гликоструктур, присоединенных к Asn 297 (например, сложных, гибридных и высокоманнозных структур).
[000260] В некоторых вариантах осуществления уровень фукозилирования составляет не более 0,5%, что основано на пределе количественного определения (LOQ) способа тестирования. Таким образом, в некоторых вариантах осуществления уровень афукозилирования составляет 99,5% или более. Для определения уровня фукозилирования, сиалирования, маннозилирования и концевого гликозилирования в образце можно использовать способ профилирования N-связанных олигосахаридов. Способ профилирования N-связанных олигосахаридов можно использовать для оценки N-связанных гликанов. В кратком изложении, N-связанные гликаны ферментативно высвобождают из белка с помощью пептид-N-гликозидазы F. Затем гликаны дериватизируют с помощью флуоресцентного средства и анализируют с использованием жидкостной хроматографии гидрофильных взаимодействий и детекции флуоресценции. Затем хроматографический профиль сравнивают с профилем референсного материала. Этот способ и многие другие способы известны в этой области для анализа фукозилирования антитела, и их можно использовать для определения уровня фукозилирования антитела по настоящему изобретению.
[000261] Неограничивающие примеры способов детекции фукозы в антителе включают масс-спектрометрию MALDI-TOF (см., например, WO 2008/077546), измерение ВЭЖХ высвобождаемых флуоресцентно меченых олигосахаридов (см., например, Schneider et al, "N-Glycan analysis of monoclonal antibodies and other glycoproteins using UHPLC with fluorescence detection," Agilent Technologies, Inc. (2012); Lines, J. Pharm. Biomed. Analysis, 14: 601-608 (1996); Takahasi, J. Chrom., 720: 217-225 (1996)), капиллярный электрофорез высвобождаемых флуоресцентно меченых олигосахаридов (см., например, Ma et al., Anal. Chem., 71: 5185-5192 (1999)) и ВЭЖХ с импульсной амперометрической детекцией для измерения композиции моносахаридов (см., например, Hardy, et al, Analytical Biochem., 170: 54-62 (1988)). Обозначение "Asn297" относится к остатку аспарагина, локализованному в положении приблизительно 297 в Fc-области (нумерация EU остатков Fc-области); однако, Asn297 также может находиться приблизительно на ±3 аминокислоты выше или ниже положения 297, т.е. между положениями 294 и 300, в результате небольших изменений последовательности антител. В антителе против CXCR5, представленном в настоящем описании, Asn297 обнаруживают в последовательности QYNST, и он выделен полужирным шрифтом и подчеркнут в таблице 16 (например, SEQ ID NO:31 Fc-домена IgG1 человека дикого типа).
[000262] Варианты фукозилирования могут иметь улучшенную функцию ADCC. См., например, патентные публикации США №№ 2003/0157108 (Presta, L.); 2004/0093621 (Kyowa Hakko Kogyo Co., Ltd). Примеры публикаций, связанных с "афукозилированными" или "лишенными фукозы" антителами, включают: US 2003/0157108; WO 2000/61739; WO 2001/29246; US 2003/0115614; US 2002/0164328; US 2004/0093621; US 2004/0132140; US 2004/0110704; US 2004/0110282; US 2004/0109865; WO 2003/085119; WO 2003/084570; WO 2005/035586; WO 2005/035778; WO2005/053742; WO2002/031140; Okazaki et al. J. Mol. Biol. 336: 1239-1249 (2004); Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004). Примеры линий клеток, которые могут продуцировать афукозилированные антитела, включают клетки CHO Lec 13, лишенные фукозилирования белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США № 2003/0157108 A1, Presta, L; и WO 2004/056312, Adams et al., особенно в примере 11), и нокаутные линии клеток, такие как линии клеток, в которых отсутствует функциональный ген альфа-1,6-фукозилтрансферазы, FUT8, например, нокаутные клетки CHO (см., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda, Y. et al., Biotechnol. Bioeng, 94(4):680-688 (2006); и WO2003/085107).
[000263] Настоящее изобретение также относится к антителам с бисектными олигосахаридами, например, в которых биантенарный олигосахарид, присоединенный к Fc-области антитела, является бисектным по GlcNAc. Такие антитела могут иметь сниженное фукозилирование и/или улучшенную функцию ADCC. Примеры таких антител описаны, например, в WO 2003/011878 (Jean-Mairet et al.); патенте США № 6602684 (Umaña et al.) и US 2005/0123546 (Umaña et al.). Настоящее изобретение относится к антителам по меньшей мере с одним остатком галактозы в олигосахариде, присоединенным к Fc-области. Такие антитела могут иметь улучшенную функцию CDC. Такие антитела описаны, например, в WO 1997/30087 (Patel et al.); WO 1998/58964 (Raju, S.) и WO 1999/22764 (Raju, S.).
[000264] В некоторых вариантах осуществления изобретения афукозилированное антитело опосредует ADCC в присутствии эффекторных клеток человека более эффективно, чем родительское антитело, содержащее фукозу. В основном, активность ADCC можно определять с использованием анализа ADCC in vitro, как представлено в настоящем описании, но предусмотрены и другие анализы или способы определения активности ADCC, например, в модели на животных и т.д.
[000265] В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют повышенную активность ADCC in vitro и/или in vivo. В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют повышенную активность ADCC in vitro. В некоторых вариантах осуществления активность ADCC in vitro определяют способом, представленным в настоящем описании. В кратком изложении, серийные разведения антител против CXCR5 или изотипического контроля инкубируют с мононуклеарными клетками периферической крови (PBMC) здоровых людей-доноров или яванских макаков. В этом анализе PBMC являются источником эффекторных клеток естественных киллеров (NK) и целевых CXCR5+ B-клеток и Tfh-подобных клеток. Проточную цитометрию используют для анализа количества B-клеток и Tfh-подобных клеток, остающихся через приблизительно 20 ч. Кривые титрования цитотоксичности получали посредством построения графика процента цитотоксичности антигенсвязывающей популяции против логарифма концентрации антитела PF-06835375. Значения EC50 определяли с использованием GraphPad Prism® (версии 6.0, GraphPad Software, Inc, San Diego, CA) с помощью аппроксимации нелинейной регрессионной кривой и логарифма сигмоиды в модели доза агониста-ответ в соответствии со следующим уравнением:
Log (агонист) vs. ответ - переменный угловой коэффициент (четыре параметра)
Y=Минимум+(Максимум-Минимум/(1+10^((LogEC50-X)*угловой коэффициент Хилла))
Где Y является процентом цитотоксичности, X является концентрацией антитела, Максимум является максимальным значением Y, соответствующим верхнему плато сигмоидной кривой, Минимум является минимальным значением Y, соответствующим нижнему плато сигмоидной кривой (ограниченной 0), и LogEC50 является логарифмом концентрации антитела в точке перегиба кривой.
[000266] Значения EC50 суммированы по экспериментам с использованием среднего и стандартного отклонения (STDEV). В некоторых вариантах осуществления способность гуманизированных mAb индуцировать ADCC истинных Tfh-клеток из миндалин человека анализировали аналогичным образом с использованием CD4+ T-клеток, выделенных из тонзиллярных мононуклеарных клеток, с добавлением NK-клеток, выделенных из PBMC. В некоторых вариантах осуществления клетки Ba/F3, экспрессирующие CXCR5, используют в качестве клеток-мишеней. В некоторых вариантах осуществления цитотоксичность определяют посредством количественного анализа LDH с использованием нерадиоактивного анализа цитотоксичности CytoTox (Promega, Madison, WI).
[000267] В некоторых вариантах осуществления максимальный лизис определяют с использованием 5-процентного Triton X-100 и определяют спонтанное высвобождение в отсутствие антитела. В некоторых вариантах осуществления процент специфического лизиса можно определять с использованием формулы: (экспериментальное - спонтанное высвобождение)/(максимальное - спонтанное высвобождение)×100=процент специфического лизиса. В некоторых вариантах осуществления афукозилированное антитело против CXCR5, имеющее повышенную активность ADCC, приводит к специфическому лизису, являющемуся по меньшей мере на 10, по меньшей мере на 15, по меньшей мере на 20, по меньшей мере на 25, по меньшей мере на 30, по меньшей мере на 35, по меньшей мере на 40, по меньшей мере на 45, по меньшей мере на 50, по меньшей мере на 60, по меньшей мере на 65, по меньшей мере на 70 или по меньшей мере на 75 процентных точек более высоким, чем специфический лизис при использовании того же количества фукозилированного антитела по меньшей мере в одной концентрации тестируемого антитела. В некоторых вариантах осуществления афукозилированное антитело против CXCR5, имеющее повышенную активность ADCC, приводит к специфическому лизису, являющемуся по меньшей мере на 10, по меньшей мере на 15, по меньшей мере на 20, по меньшей мере на 25, по меньшей мере на 30, по меньшей мере на 35, по меньшей мере на 40, по меньшей мере на 45, по меньшей мере на 50, по меньшей мере на 60, по меньшей мере на 65, по меньшей мере на 70 или по меньшей мере на 75 процентных точек более высоким, чем специфический лизис при использовании фукозилированного антитела, где каждое антитело находится в концентрации от 0,01 до 1 мкг/мл, и клетки-мишени являются клетками Ba/F3, экспрессирующими CXCR5. В некоторых вариантах осуществления антитела тестируют в концентрациях в диапазоне от 0,000005 мкг/мл до 5 мкг/мл.
[000268] В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют повышенную аффинность к Fc гамма RIIIA. В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют повышенную аффинность к Fc гамма RIIIA(V158). В некоторых вариантах осуществления афукозилированные антитела против CXCR5 имеют повышенную аффинность к Fc гамма RIIIA(F158). В некоторых вариантах осуществления аффинность антитела к Fc гамма RIIIA определяют с использованием поверхностного плазмонного резонанса и/или следующим образом, что описано в отношении Fc гамма RIIIA(V158), но что также подходит для определения аффинности к Fc гамма RIIIA(F158). В кратком изложении, в некоторых вариантах осуществления фукозилированное или афукозилированное антитело против CXCR5 фиксируют на покрытом протеином A декстрановом чипе. Fc гамма RIIIA (V158) (доступный, например, в R & D Systems) инъецируют в различных концентрациях. Константу ассоциации, константу диссоциации и аффинность к Fc гамма RIIIA (V158) фукозилированного и афукозилированного антитела против CXCR5 можно определять, например, с использованием программного обеспечения, поставляемого вместе с системой поверхностного плазмонного резонанса (например, модели связывания 1:1 в программном обеспечении Biacore T200 Evaluation Software). В некоторых вариантах осуществления афукозилированное антитело против CXCR5 может иметь повышенную аффинность к Fc гамма RIIIA (такому как Fc гамма RIIIA(V158) или Fc гамма RIIIA(F158)) и может связываться с Fc гамма RIIIA с по меньшей мере в 2 раза, по меньшей мере 3 раза, по меньшей мере 4 раза, по меньшей мере 5 раз, по меньшей мере 7 раз, по меньшей мере 10 раз, по меньшей мере 12 раз, по меньшей мере 15 раз, по меньшей мере 17 раз, 20 раз, 30 раз, 50 раз, 100 раз, 500 раз или по меньшей мере 1000 раз более высокой аффинностью, чем фукозилированное антитело против CXCR5.
IV. ЭКСПРЕССИЯ И ПРОДУКЦИЯ АНТИТЕЛА ПРОТИВ CXCR5
НУКЛЕИНОВЫЕ КИСЛОТЫ, КОДИРУЮЩИЕ АНТИТЕЛА ПРОТИВ CXCR5
[000269] Изобретение также относится к полинуклеотидам, кодирующим любые из антител, включая фрагменты антител и модифицированные антитела, представленные в настоящем описании. Изобретение также относится к способу получения любых из полинуклеотидов, представленных в настоящем описании. Полинуклеотиды можно получать и экспрессировать известными в этой области способами.
[000270] Последовательность желаемого антитела, определенного фрагмента антитела или его антигенсвязывающего фрагмента и нуклеиновой кислоты, кодирующей такое антитело или его фрагмент, можно определять стандартными способами секвенирования. Последовательность нуклеиновой кислоты, кодирующую желаемое антитело, определенный фрагмент антитела или его антигенсвязывающий фрагмент, можно встраивать в различные векторы (такие как клонирующие и экспрессирующие векторы) для рекомбинантной продукции и характеризации. Нуклеиновую кислоту, кодирующую тяжелую цепь, определенный фрагмент антитела или антигенсвязывающий фрагмент тяжелой цепи, и нуклеиновую кислоту, кодирующую легкую цепь, определенный фрагмент антитела или антигенсвязывающий фрагмент легкой цепи, можно клонировать в один и тот же вектор или разные векторы.
[000271] В одном из аспектов изобретение относится к полинуклеотидам, кодирующим аминокислотные последовательности любого из следующих антител против CXCR5 и их антигенсвязывающих фрагментов: VH 11G2 мыши, VL 11G2 мыши, химерной VH 11G2, химерной VL 11G2, VH h11G2 (XC152), VH h11G2 (XC155), VH h11G2 (XC156), VH h11G2 (XC157), VH h11G2 (XC350), VH h11G2 (XC351), VH h11G2 (XC352), VH h11G2 (XC353), VH h11G2 (XC354), VL h11G2 (XC151), VL h11G2 (XC153), VL h11G2 (XC154), VL h11G2 (XC346), VL h11G2 (XC347), VL h11G2 (XC348), VL h11G2 (XC349), VH 41A10 мыши, VL 41A10 мыши, химерной VH 41A10, химерной VL 41A10, гуманизированной VH 41A10 (XC147), VH h41A10 (XC148), VH h41A10 (XC150), VL h41A10 (XC142), VL h41A10 (XC143), VL h41A10 (XC144), VL h41A10 (XC145), VL h41A10 (XC146), VL h41A10 (XC149), VH 5H7 мыши, VL 5H7 мыши, химерной VH 5H7 и химерной VL 5H7. Полинуклеотид, кодирующий указанные выше аминокислотные последовательности, кодирует аминокислотную последовательность, по меньшей мере на 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99% и, предпочтительно, более идентичную аминокислотной последовательности антитела или его антигенсвязывающего фрагмента по настоящему изобретению, представленного в настоящем описании.
[000272] Изобретение относится к полинуклеотидам, кодирующим аминокислотные последовательности антитела или его антигенсвязывающего фрагмента, связывающегося с, по существу, тем же эпитопом, что и антитело, выбранное из группы, состоящей из: химерного 41A10, гуманизированного 41A10 (142/147), гуманизированного 41A10 (142/148), химерного 11G2, афукозилированного химерного 11G2, гуманизированного 11G2 (151/152), гуманизированного 11G2 (153/155), гуманизированного 11G2 (153/156), гуманизированного 11G2 (154/155), гуманизированного 11G2 (154/157) и афукозилированного гуманизированного 11G2 (154/155).
[000273] Изобретение относится к полинуклеотидам, кодирующим аминокислотные последовательности антитела или его антигенсвязывающего фрагмента, конкурирующего за связывание с CXCR5 с антителом, выбранным из группы, состоящей из: химерного 41A10, гуманизированного 41A10 (142/147), гуманизированного 41A10 (142/148), химерного 11G2, афукозилированного химерного 11G2, гуманизированного 11G2 (151/152), гуманизированного 11G2 (153/155), гуманизированного 11G2 (153/156), гуманизированного 11G2 (154/155), гуманизированного 11G2 (154/157) и афукозилированного гуманизированного 11G2 (154/155).
[000274] Изобретение относится к полинуклеотидам, кодирующим один или более белков, содержащих аминокислотную последовательность, выбранную из группы, состоящей из: SEQ ID NO:1-29, SEQ ID NO:35-40, и SEQ ID NO:47-63.
[000275] Изобретение относится к полинуклеотидам, содержащим последовательность нуклеиновой кислоты, приведенную как одна или более из SEQ ID NO:106, 107, 108 и 109. Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:95. Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:96. Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:97. Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты, приведенную как SEQ ID NO:98.
[000276] Изобретение относится к полинуклеотиду, содержащему одну или обе из последовательностей нуклеиновой кислоты вставки ДНК плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323 и регистрационный номер PTA-124324.
[000277] Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты вставки в плазмиду, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
[000278] Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты вставки в плазмиду, депонируемой в ATCC и имеющей регистрационный номер PTA-124324.
[000279] Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323 и регистрационный номер PTA-124324.
[000280] Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124323.
[000281] Изобретение относится к полинуклеотиду, содержащему последовательность нуклеиновой кислоты вставки плазмиды, депонируемой в ATCC и имеющей регистрационный номер PTA-124324.
[000282] Изобретение относится к клеткам, содержащим одну или более молекул нуклеиновой кислоты, приведенных в одной или более из SEQ ID NO:106, 107, 108 и 109. Изобретение относится к клеткам, содержащим одну или более молекул нуклеиновой кислоты, приведенных в SEQ ID NO:106 и 107. Изобретение относится к клеткам, содержащим одну или более молекул нуклеиновой кислоты, приведенных в SEQ ID NO:108 и 109.
[000283] В другом аспекте изобретение относится к полинуклеотидам и их вариантам, кодирующим антитело против CXCR5, где такие варианты полинуклеотидов имеют по меньшей мере 70%, по меньшей мере 75%, по меньшей мере 80%, по меньшей мере 85%, по меньшей мере 87%, по меньшей мере 89%, по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичности последовательности по отношению к любой из конкретных последовательностей нуклеиновой кислоты, представленных в настоящем описании. Эти количества не должны быть ограничивающими, и приращения между перечисленными процентами конкретно представляют собой часть настоящего изобретения.
[000284] Изобретение относится к полипептидам, кодируемым молекулами нуклеиновой кислоты, представленными в настоящем описании.
[000285] В одном из вариантов осуществления домены VH и VL, или их антигенсвязывающие фрагменты, или полноразмерные HC или LC кодируются отдельными полинуклеотидами. Альтернативно, и VH, и VL, или их антигенсвязывающие фрагменты, или HC и LC кодируются одним полинуклеотидом.
[000286] Полинуклеотиды, комплементарные любым таким последовательностям, также включены в настоящее изобретение. Полинуклеотиды могут являться одноцепочечными (кодирующими или антисмысловыми) или двухцепочечными и могут являться молекулами ДНК (геномной ДНК, кДНК или синтетической ДНК) или РНК. Молекулы РНК включают молекулы HnRNA, содержащие интроны и соответствующие молекуле ДНК один к одному, и молекулы мРНК, не содержащие интроны. Дополнительные кодирующие или некодирующие последовательности, необязательно, могут присутствовать в полинуклеотиде по настоящему изобретению, и полинуклеотид, необязательно, можно соединять с другими молекулами и/или материалами подложки.
[000287] Полинуклеотиды могут содержать нативную последовательность (т.е. эндогенную последовательность, кодирующую антитело или его часть) или могут содержать вариант такой последовательности. Варианты полинуклеотидов содержат одну или более замен, добавлений, делеций и/или инсерций таким образом, что иммунореактивность кодируемого полипептида не снижается относительно нативной иммунореактивной молекулы. Эффект иммунореактивности кодируемого полипептида, как правило, можно анализировать, как представлено в настоящем описании. В некоторых вариантах осуществления варианты имеют по меньшей мере приблизительно 70% идентичности, в некоторых вариантах осуществления - по меньшей мере приблизительно 80% идентичности, в некоторых вариантах осуществления - по меньшей мере приблизительно 90% идентичности, и в некоторых вариантах осуществления - по меньшей мере приблизительно 95% идентичности по отношению к полинуклеотидной последовательности, кодирующей нативное антитело или его часть. Эти количества не должны быть ограничивающими, и приращения между перечисленными процентами конкретно представляют собой часть настоящего изобретения.
[000288] Указывают, что две полинуклеотидные или полипептидные последовательности являются "идентичными", если последовательность нуклеотидов или аминокислот в двух последовательностях является одинаковой при выравнивании с максимальным соответствием, как описано ниже. Сравнение двух последовательностей, как правило, осуществляют посредством сравнения последовательностей в окне сравнения для идентификации и сравнения локальных областей сходства последовательностей. В рамках изобретения термин "окно сравнения" относится к сегменту по меньшей мере из приблизительно 20 смежных положений, как правило, от 30 до приблизительно 75 или от 40 до приблизительно 50, в которых последовательность можно сравнивать с референсной последовательностью того же количества смежных положений после оптимального выравнивания двух последовательностей.
[000289] оптимальное выравнивание последовательностей для сравнения можно осуществлять с использованием программы MegAlign® в пакете биоинформатического программного обеспечения Lasergene® (DNASTAR®, Inc., Madison, WI) с использованием параметров по умолчанию. Этой программе воплощены несколько схем выравнивания, описанных в следующих источниках: Dayhoff, M.O., 1978, A model of evolutionary change in proteins - Matrices for detecting distant relationships. в Dayhoff, M.O. (ed.) Atlas of Protein Sequence and Structure, National Biomedical Research Foundation, Washington DC Vol. 5, Suppl. 3, pp. 345-358; Hein J., 1990, Unified Approach to Alignment and Phylogenes pp. 626-645 Methods in Enzymology vol. 183, Academic Press, Inc., San Diego, CA; Higgins, D.G. и Sharp, P.M., 1989, CABIOS 5:151-153; Myers, E.W. и Muller W., 1988, CABIOS 4:11-17; Robinson, E.D., 1971, Comb. Theor. 11:105; Santou, N., Nes, M., 1987, Mol. Biol. Evol. 4:406-425; Sneath, P.H.A. and Sokal, R.R., 1973, Numerical Taxonomy the Principles and Practice of Numerical Taxonomy, Freeman Press, San Francisco, CA; Wilbur, W.J. and Lipman, D.J., 1983, Proc. Natl. Acad. Sci. USA 80:726-730.
[000290] В некоторых вариантах осуществления "процент идентичности последовательности" определяют посредством сравнения двух оптимально выровненных последовательностей в окне сравнения из по меньшей мере 20 положений, где часть полинуклеотидной или полипептидной последовательности в окне сравнения может содержать добавления или делеции (т.е. пропуски) 20 процентов или менее, как правило, от 5 до 15 процентов или от 10 до 12 процентов по сравнению с референсными последовательностями (не содержащими добавления или делеции) для оптимального выравнивания двух последовательностей. Процент вычисляют посредством определения количества положений, в которых идентичные основания нуклеиновой кислоты или аминокислотный остаток встречаются в обеих последовательностях, для получения количества совпадающих положений, разделяя количество совпадающих положений на общее количество положений в референсной последовательности (т.е. на размер окна сравнения) и умножая результат на 100 для получения процента идентичности последовательности.
[000291] Дополнительно или альтернативно, варианты могут являться, по существу, гомологичными нативному гену или его части или комплементарной цепи. Такие варианты полинуклеотида могут гибридизоваться в условиях умеренной строгости с природной последовательностью ДНК, кодирующей нативное антитело (или комплементарной последовательностью).
[000292] Подходящие "условия умеренной строгости" включают предварительную промывку в растворе 5-кратного SSC, 0,5% SDS, 1,0 мМ ЭДТА (pH 8,0), гибридизацию при 50°C-65°C в 5-кратном SSC в течение ночи с последующей двукратной промывкой при 65°C в течение 20 минут каждым из 2-кратного, 0,5-кратного и 0,2-кратного SSC, содержащего 0,1% SDS.
[000293] В рамках изобретения "условия высокой строгости" или "условия с высокой строгостью" являются условиями, в которых: (1) для промывки используют низкую ионную силу и высокую температуру, например, 0,015 M хлорид натрия/0,0015 M цитрат натрия/0,1% додецилсульфат натрия при 50°C; (2) во время гибридизации используют денатурирующее средство, такое как формамид, например, 50% (об./об.) формамид с 0,1% бычьим сывороточным альбумином/0,1% фиколлом/0,1% поливинилпирролидоном/50 мМ буфером фосфата натрия при pH 6,5 с 750 мМ хлорида натрия, 75 мМ цитрата натрия при 42°C; или (3) используют 50% формамид, 5-кратный SSC (0,75 M NaCl, 0,075 M цитрат натрия), 50 мМ фосфат натрия (pH 6,8), 0,1% пирофосфат натрия, 5-кратный раствор Денхардта, обработанную ультразвуком ДНК молок лососевых (50 мкг/мл), 0,1% SDS и 10% декстран сульфат при 42°C с промывками при 42°C в 0,2-кратном SSC (хлорид натрия/цитрат натрия) и 50% формамиде при 55°C с последующей промывкой в условиях высокой строгости, состоящих из 0,1-кратного SSC, содержащего ЭДТА, при 55°C. Специалистам в этой области будет понятно, как корректировать температуру, ионную силу и т.д. по мере необходимости для адаптации к таким факторам, как длина зонда и т.п.
[000294] Специалистам в этой области следует понимать, что в результате вырожденности генетического кода существует множество нуклеотидных последовательностей, кодирующих полипептид, представленный в настоящем описании. Некоторые из этих полинуклеотидов имеют минимальную гомологию по отношению к нуклеотидной последовательности любого нативного гена. Несмотря на это, полинуклеотиды, которые могут варьироваться благодаря различиям в использовании кодонов, конкретно включены в настоящее изобретение. Кроме того, аллели генов, содержащие полинуклеотидные последовательности, представленные в настоящем описании, входят в объем настоящего изобретения. Аллели представляют собой эндогенные гены, измененные в результате одной или более мутаций, таких как делеции, добавления и/или замены нуклеотидов. Получаемые мРНК и белок, необязательно, могут иметь измененную структуру или функцию. Аллели можно идентифицировать стандартными способами (такими как гибридизация, амплификация и/или сравнение с базами данных последовательностей).
[000295] Полинуклеотиды по изобретению можно получать с использованием химического синтеза, рекомбинантных способов или ПЦР. Способы химического синтеза полинуклеотидов хорошо известны в этой области и не требуют подробного описания в настоящем описании. Для получения желаемой последовательности ДНК специалист в этой области может использовать последовательности, представленные в настоящем описании, и коммерческий ДНК-синтезатор.
[000296] Для получения полинуклеотидов рекомбинантными способами полинуклеотид, содержащий желаемую последовательность, можно встраивать в подходящий вектор, а вектор, в свою очередь, можно встраивать в подходящую клетку-хозяина для репликации и амплификации, как представлено в настоящем описании далее. Полинуклеотиды можно встраивать в клетки-хозяева любыми известными в этой области способами. Клетки трансформируют посредством встраивания экзогенного полинуклеотида посредством прямого захвата, эндоцитоза, трансфекции, F-спаривания или электропорации. После встраивания экзогенный полинуклеотид можно поддерживать в клетке в качестве неинтегрированного вектора (такого как плазмида) или встраивать в геном клетки-хозяина. Амплифицированный таким образом полинуклеотид можно выделять из клетки-хозяина хорошо известными этой области способами. См., например, Sambrook et al., 1989.
[000297] Альтернативно, ПЦР позволяет воспроизводить последовательности ДНК. Технология ПЦР хорошо известна в этой области и описана в патентах США №№ 4683195, 4800159, 4754065 и 4683202, а также в ПЦР: Polymerase chain reaction, Mullis et al. eds., Birkauswer Press, Boston, 1994.
[000298] РНК можно получать с использованием выделенной ДНК в соответствующем векторе и встраивать ее в подходящую клетку-хозяина. Если клетка реплицируется, и ДНК транскрибируется в РНК, РНК можно выделять способами, хорошо известными специалистам в этой области, как описано, например, в Sambrook et al., 1989.
[000299] В некоторых вариантах осуществления первый вектор содержит полинуклеотид, кодирующий тяжелую цепь, и второй вектор содержит полинуклеотид, кодирующий легкую цепь. В некоторых вариантах осуществления с помощью первого вектора и второго вектора трансфицируют клетки-хозяева в схожих количествах (таких как схожие молярные количества или схожие массовые количества). В некоторых вариантах осуществления с помощью первого вектора и второго вектора в молярном или массовом соотношении от 5:1 до 1:5 трансфицируют клетки-хозяева. В некоторых вариантах осуществления используют массовое соотношение от 1:1 до 1:5 вектора, кодирующего тяжелую цепь, и вектора, кодирующего легкую цепь. В некоторых вариантах осуществления используют массовое соотношение 1:2 вектора, кодирующего тяжелую цепь, и вектора, кодирующего легкую цепь.
Векторы
[000300] В некоторых вариантах осуществления выбирают вектор, оптимизированный для экспрессии полипептидов в клетках CHO или клетках, полученных из CHO, или в клетках NSO. Примеры таких векторов описаны, например, в Running Deer et al, Biotechnol. Prog. 20:880-889 (2004).
[000301] Подходящие клонирующие и экспрессирующие векторы могут включать различные компоненты, такие как промотор, энхансер и другие транскрипционные регуляторные последовательности. Также можно конструировать вектор, чтобы сделать возможным последующее клонирование вариабельного домена антитела в другие векторы. Подходящие клонирующие векторы можно конструировать стандартными способами или их можно выбирать из большого количества клонирующих векторов, доступных в этой области. Хотя выбранный клонирующий вектор может варьироваться в зависимости от используемой клетки-хозяина, клонирующие векторы, которые можно использовать, как правило, будут иметь способность к саморепликации, могут иметь одну мишень для конкретной эндонуклеазы рестрикции и/или могут нести гены маркера, который можно использовать в селекции клонов, содержащих вектор. Подходящие примеры включают плазмиды и бактериальные вирусы, например, pUC18, pUC19, Bluescript (например, pBS SK+) и их производные, mp18, mp19, pBR322, пМB9, ColE1, pCR1, RP4, фаговую ДНК и челночные векторы, такие как pSA3 и pAT28. Эти и многие другие клонирующие векторы доступны у коммерческих поставщиков, таких как BioRad, Strategene и Invitrogen. Настоящее изобретение также относится к экспрессирующим векторам. Экспрессирующие векторы, как правило, являются реплицируемыми полинуклеотидными конструкциями, содержащими полинуклеотид по изобретению. Предполагают, что экспрессирующий вектор должен быть реплицируемым в клетках-хозяевах в виде эписом или интегральной части хромосомной ДНК. Подходящие экспрессирующие векторы включают, в качестве неограничивающих примеров, плазмиды, вирусные векторы, включая аденовирусы, аденоассоциированные вирусы, ретровирусы, космиды и экспрессирующие векторы, описанные в публикации PCT № WO 87/04462. Компоненты векторов, как правило, могут включать, в качестве неограничивающих примеров, одно или более из следующего: сигнальную последовательность; участок начала репликации; один или более маркерных генов; подходящие элементы транскрипционного контроля (такие как промоторы, энхансеры и терминатор). Для экспрессии (т.е. трансляции), как правило, также необходимы один или более элементов транскрипционного контроля, такие как участки связывания рибосомы, участки инициации трансляции и стоп-кодоны.
[000302] Векторы, содержащие интересующие полинуклеотиды, и/или сами полинуклеотиды можно встраивать в клетку-хозяина любым из ряда подходящих способов, включая электропорацию, трансфекцию с использованием хлорида кальция, хлорида рубидия, фосфата кальция, DEAE-декстрана или других веществ; бомбардировку микрочастицами; липофекцию и инфекцию (например, если вектор является возбудителем инфекции, таким как вирус осповакцины). Выбор встраиваемых векторов или полинуклеотидов зачастую будет зависеть от признаков клетки-хозяина.
Клетки-хозяева
[000303] Антитело или его антигенсвязывающий фрагмент можно получать рекомбинантно с использованием подходящей клетки-хозяина. Нуклеиновую кислоту, кодирующую антитело или его антигенсвязывающий фрагмент, можно клонировать в экспрессирующий вектор, который затем можно встраивать в клетку-хозяина, такую как клетка E. coli, дрожжевая клетка, клетка насекомого, клетка обезьяны COS, клетка яичника китайского хомяка (CHO) или миеломная клетка, где клетка не продуцирует иммуноглобулиновый белок иным образом, для достижения синтеза антитела в рекомбинантной клетке-хозяине. Предпочтительные клетки-хозяева включают клетку CHO, клетку эмбриональной почки человека HEK-293 или клетку Sp2.0 помимо многих клеток, хорошо известных в этой области. Фрагмент антитела можно получать посредством протеолитической или иной деградации полноразмерного антитела, рекомбинантными способами или посредством химического синтеза. Полипептидный фрагмент антитела, в частности, более короткие полипептиды длиной до приблизительно 50 аминокислот, можно получать посредством химического синтеза. Способы химического синтеза белков и пептидов известны в этой области и коммерчески доступны.
[000304] В различных вариантах осуществления тяжелые цепи антитела против CXCR5 и/или легкие цепи антитела против CXCR5 можно экспрессировать в прокариотических клетках, таких как бактериальные клетки; или в эукариотических клетках, таких как клетки грибов (таких как дрожжи), растительные клетки, клетки насекомых и клетки млекопитающих. Такую экспрессию можно осуществлять, например, способами, известными в этой области. Неограничивающие примеры эукариотических клеток, которые можно использовать для экспрессии полипептидов включают клетки COS, включая клетки COS 7; клетки 293, включая клетки 293-6E; клетки CHO, включая клетки CHO-S, CHO DG44, CHO Lecl3 и клетки FUT8 CHO; клетки PER.C6® (Crucell) и клетки NSO. В некоторых вариантах осуществления тяжелые цепи антитела против CXCR5 и/или легкие цепи антитела против CXCR5 можно экспрессировать в дрожжах. См., например, патентную публикацию США № 2006/0270045 A1. В некоторых вариантах осуществления конкретную эукариотическую клетку-хозяина выбирают с учетом ее способности осуществлять желаемые посттрансляционные модификации в тяжелых цепях антитела против CXCR5 и/или легких цепях антитела против CXCR5. Например, в некоторых вариантах осуществления клетки CHO продуцируют полипептиды, имеющие более высокий уровень сиалирования, чем тот же полипептид, продуцируемый в клетках 293.
[000305] Встраивание одной или более нуклеиновых кислот в желаемую клетку-хозяина можно осуществлять любым способом, включая, в качестве неограничивающих примеров, трансфекцию с фосфатом кальция, трансфекцию, опосредованную DEAE-декстраном, трансфекцию, опосредованную катионными липидами, электропорацию, трансдукцию, инфекцию и т.д. Неограничивающие примеры способов описаны, например, в Sambrook et al., Molecular Cloning, A Laboratory Manual, 3rd ed. Cold Spring Harbor Laboratory Press (2001). С помощью нуклеиновых кислот можно транзиторно или стабильно трансфицировать желаемые клетки-хозяева любым подходящим способом.
[000306] В некоторых вариантах осуществления афукозилированные антитела против CXCR5 получают в клетках, способных продуцировать афукозилированные антитела, таких как клетки Lec3 CHO, лишенные фукозилирования белков (Ripka et al. Arch. Biochem. Biophys. 249:533-545 (1986); патентная заявка США № 2003/0157108; и WO 2004/056312, особенно в примере 11), и нокаутных линиях клеток, таких как линии клеток, в которых отсутствует функциональный ген альфа-1,6-фукозилтрансферазы FUT8, например, нокаутные клетки CHO (см., например, Yamane-Ohnuki et al. Biotech. Bioeng. 87: 614 (2004); Kanda et al, Biotechnol. Bioeng, 94(4):680-688 (2006); и WO2003/085107). В некоторых вариантах осуществления афукозилированные антитела против CXCR5 получают в клетках CHO, в которых отсутствует функциональный ген FUT8. В некоторых вариантах осуществления афукозилированные антитела против CXCR5 получают в клетках Potelligent® CHOK1SV (BioWa/Lonza, Allendale, NJ).
[000307] Антитела против CXCR5 можно очищать любым подходящим способом. Такие способы включают, в качестве неограничивающих примеров, использование аффинных матриц или хроматографии гидрофобных взаимодействий. Подходящие аффинные лиганды включают ECD CXCR5 и лиганды, связывающиеся с константными областями антитела. Например, для связывания константной области и очистки антитела против CXCR5 можно использовать аффинную колонку с протеином A, протеином G, протеином A/G или антителом. Хроматография гидрофобных взаимодействий, например, с использованием бутиловой или фениловой колонки, также может подходить для очистки некоторых полипептидов. В этой области известно множество способов очистки полипептидов.
[000308] В некоторых вариантах осуществления антитело против CXCR5 получают в бесклеточной системе. Неограничивающие примеры бесклеточных систем описаны, например, в Sitaraman et al., Способы Mol. Biol. 498: 229-44 (2009); Spirin, Trends Biotechnol. 22: 538-45 (2004); Endo et al, Biotechnol. Adv. 21: 695-713 (2003).
V. ПРИМЕНЕНИЕ И МЕДИЦИНСКАЯ ТЕРАПИЯ
[000309] В некоторых аспектах настоящее изобретение относится к терапевтическим способам удаления, ингибирования или снижения активности или передачи сигнала CXCR5 с использованием антитела против CXCR5 или его антигенсвязывающего фрагмента, где терапевтические способы включают введение терапевтически эффективного количества фармацевтической композиции, содержащей антитело против CXCR5 или его антигенсвязывающий фрагмент. Подвергаемое лечению нарушение является любым заболеванием или состоянием, улучшаемым, ингибируемым или подвергаемым профилактике посредством устранения, ингибирования или снижения активности или передачи сигнала CXCR5.
[000310] В некоторых аспектах, настоящее изобретение относится к антителу против CXCR5 или его антигенсвязывающему фрагменту для применения в устранении, ингибировании или снижении активности или передачи сигнала CXCR5. В некоторых вариантах осуществления применение может включать введение терапевтически эффективного количества фармацевтической композиции, содержащей антитело против CXCR5 или его антигенсвязывающий фрагмент. В некоторых вариантах осуществления с помощью ингибирования или снижения активности или передачи сигнала CXCR5 можно лечить любое заболевание или состояние, улучшаемое, ингибируемое или подвергаемое профилактике посредством устранения, ингибирования или снижения активности или передачи сигнала CXCR5.
[000311] Такие заболевания, нарушения или состояния включают, в качестве неограничивающих примеров, воспалительные ответы, такие как системная красная волчанка (SLE); хронические воспалительные ответы; атеросклероз; дефицит адгезии лейкоцитов; ревматоидный артрит; сахарный диабет (например, сахарный диабет I типа или инсулинозависимый сахарный диабет); рассеянный склероз; синдром Рейно; аутоиммунный тиреоидит; аллергический энцефаломиелит; синдром Шегрена; ювенильный диабет; и иммунные ответы, ассоциированные с острой и замедленной реакцией гиперчувствительности, опосредованные цитокинами и T-лимфоцитами, как правило, обнаруживаемые при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; гранулематоз Вегенера; пернициозную анемию (болезнь Аддисона); заболевания, включающие диапедез лейкоцитов; воспалительное нарушение центральной нервной системы (ЦНС); синдром полиорганной недостаточности; гемолитическую анемию (включая в качестве неограничивающих примеров криоглобулинемию или аутоиммунную гемолитическую анемию с неполными тепловыми агглютининами); миастению; заболевания, опосредованные комплексами антиген-антитело; болезнь анти-БМК-антител; антифосфолипидный синдром; аллергический неврит; болезнь Грейвса; миастенический синдром Ламберта-Итона; буллезный пемфигоид; пемфигус; аутоиммунные полиэндокринопатии; витилиго; болезнь Рейтера; синдром мышечной скованности; болезнь Бехчета; гигантоклеточный артериит; иммунокомплексный нефрит; IgA-нефропатию; IgM-полинейропатии; иммунную тромбоцитопеническую пурпуру (ITP) или аутоиммунную тромбоцитопению и аутоиммунные гемолитические заболевания; тиреоидит Хашимото; аутоиммунный гепатит; аутоиммунную гемофилию; аутоиммунный лимфопролиферативный синдром (ALPS); аутоиммунный увеоретинит; синдром Гийена-Барре; синдром Гудпасчера; смешанное заболевание соединительной ткани; аутоиммунно-ассоциированное бесплодие; нодозный полиартериит; гнездную алопецию; идиопатическую микседему; реакцию "трансплантат против хозяина"; мышечную дистрофию (дистрофию Дюшена, дистрофию Беккера, миотоническую дистрофию, конечностно-поясную дистрофию, плече-лопаточно-лицевую дистрофию, врожденную дистрофию, окулофарингеальную дистрофию, дистальную дистрофию, дистрофию Эмери-Дрейфуса) и контроль пролиферации злокачественных клеток, экспрессирующих CXCR5, такой как рак поджелудочной железы, рак толстого кишечника, рак мочевого пузыря, T-клеточный лейкоз и B-клеточный лейкоз, как будет понятно специалисту в этой области с учетом идей, представленных в настоящем описании.
[000312] Антитела против CXCR5 или их антигенсвязывающие фрагменты по настоящему изобретению также можно использовать для детекции и/или измерения CXCR5 или CXCR5-экспрессирующих клеток в образце, например, в диагностических целях. Например, антитело против CXCR5 или его фрагмент можно использовать для диагностики состояния или заболевания, отличающегося аномальной экспрессией (например, гиперэкспрессией, недостаточной экспрессией, отсутствием экспрессии и т.д.) CXCR5. Примеры диагностических анализов CXCR5 могут включать, например, приведение образца, полученного от пациента, в контакт с антителом против CXCR5 по изобретению, где антитело против CXCR5 мечено детектируемой меткой или репортерной молекулой.
[000313] В рамках изобретения термин "фармацевтически приемлемый носитель" или "фармацевтически приемлемый эксципиент" включает любой материал, который при комбинировании с активным ингредиентом позволяет ингредиенту сохранять биологическую активность и не реагирует с иммунной системой индивидуума. Неограничивающие примеры включают любые из стандартных фармацевтических носителей, такие как фосфатно-солевой буферный раствор, вода, эмульсии, такие как эмульсия "масло-в-воде", и различные типы увлажнителей. Предпочтительными дилюентами для аэрозольного или парентерального введения являются фосфатно-солевой буфер (PBS) или нормальный (0,9%) физиологический раствор. Композиции, содержащие такие носители, составляют хорошо известными, общепринятыми способами (см., например, Remington's Pharmaceutical Sciences, 18th edition, A. Gennaro, ed., Mack Publishing Co., Easton, PA, 1990; и Remington, The Science and Practice of Pharmacy, 20th Ed., Mack Publishing, 2000).
VI. КОМПОЗИЦИИ
[000314] Настоящее изобретение также относится к фармацевтическим композициям, содержащим эффективное количество антитела против CXCR5 или его антигенсвязывающего фрагмента, представленного в настоящем описании. Примеры таких композиций, а также способы составления также представлены в настоящем описании. В некоторых вариантах осуществления композиция содержит одно или более антител против CXCR5. В других вариантах осуществления антитело против CXCR5 распознает CXCR5. В других вариантах осуществления антитело против CXCR5 является антителом человека. В других вариантах осуществления антитело против CXCR5 является гуманизированным антителом. В некоторых вариантах осуществления антитело против CXCR5 содержит константную область, способную запускать желаемый иммунный ответ, такой как антителоопосредованный лизис или ADCC. В других вариантах осуществления антитело против CXCR5 содержит константную область, являющуюся афукозилированной и обеспечивающей повышенную ADCC по сравнению с в остальном идентичном антителом, являющимся фукозилированным. В других вариантах осуществления антитело против CXCR5 содержит одну или более CDR антитела (например, одну, две, три, четыре, пять или, в некоторых вариантах осуществления, все шесть CDR).
[000315] Следует понимать, что композиции могут содержать несколько антител против CXCR5 или его антигенсвязывающих фрагментов (например, смесь антител против CXCR5, распознающих разные эпитопы CXCR5). Другие примеры композиций содержат несколько антител против CXCR5, распознающих одни и те же эпитопы, или разные типы антител против CXCR5, связывающихся с разными эпитопами CXCR5. В некоторых вариантах осуществления композиции содержат смесь антител против CXCR5, распознающих разные варианты CXCR5.
[000316] Композиция, используемые в настоящем изобретении, может дополнительно содержать фармацевтически приемлемые носители, эксципиенты или стабилизаторы (Remington: The Science and practice of Pharmacy 20th Ed., 2000, Lippincott Williams and Wilkins, Ed. K. E. Hoover) в форме лиофилизированных составов или водных растворов. Приемлемые носители, эксципиенты или стабилизаторы являются нетоксичными для реципиентов в дозах и концентрациях и могут содержать буферы, такие как фосфат, цитрат и другие органические кислоты; антиоксиданты, включая аскорбиновую кислоту и метионин; консерванты (такие как хлорид октадецилдиметилбензиламмония; хлорид гексаметония; хлорид бензалкония, хлорид бензетония; фенол, бутиловый или бензиловый спирт; алкилпарабены, такие как метил- или пропилпарабен; катехол; резорцин; циклогексанол; 3-пентанол и m-крезол); низкомолекулярные (менее приблизительно 10 остатков) полипептиды; белки, такие как сывороточный альбумин, желатин или иммуноглобулины; гидрофильные полимеры, такие как поливинилпирролидон; аминокислоты, такие как глицин, глутамин, аспарагин, гистидин, аргинин или лизин; моносахариды, дисахариды и другие углеводы, включая глюкозу, маннозу или декстраны; хелатирующие средства, такие как ЭДТА; сахара, такие как сахароза, маннит, трегалоза или сорбит; солеобразующие противоионы, такие как натрий; комплексные соединения с металлами (например, комплексы Zn-белок); и/или неионные поверхностно-активные средства, такие как TWEEN™, PLURONIC™ или полиэтиленгликоль (PEG). Фармацевтически приемлемые эксципиенты дополнительно представлены в настоящем описании.
[000317] Антитело против CXCR5, его антигенсвязывающий фрагмент и их композиции также можно использовать в комбинации с другими средствами, служащими для повышения и/или дополнения эффективности средств.
[000318] В некоторых вариантах осуществления антитело против CXCR5 или его антигенсвязывающий фрагмент комплексируют с CXCR5 перед введением. В некоторых вариантах осуществления антитело против CXCR5 не комплексируют с CXCR5 перед введением.
[000319] Настоящее изобретение также относится к композициям, включающим фармацевтические композиции, содержащие любые из полинуклеотидов по изобретению. В некоторых вариантах осуществления композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий антитело, представленное в настоящем описании. В других вариантах осуществления композиция содержит экспрессирующий вектор, содержащий полинуклеотид, кодирующий любые из антител, представленных в настоящем описании. В других вариантах осуществления композиция содержит любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:6, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:10, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:12, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:5 и SEQ ID NO:6, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:5 и SEQ ID NO:10, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:5 и SEQ ID NO:12, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:13 и SEQ ID NO:17, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:13 и SEQ ID NO:18, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:13 и SEQ ID NO:63, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:52, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:53, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:54, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:55, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:56, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:1 и SEQ ID NO:57, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:6 и SEQ ID NO:48, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:6 и SEQ ID NO:49, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:6 и SEQ ID NO:50, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:6 и SEQ ID NO:51, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:17 и SEQ ID NO:58, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:17 и SEQ ID NO:59, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:17 и SEQ ID NO:60, любой или оба из полинуклеотидов, содержащих последовательность, приведенную в SEQ ID NO:17 и SEQ ID NO:61, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:17 и SEQ ID NO:62, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:1, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:5, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:47, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:48, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:49, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:50, любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:52 и SEQ ID NO:51, или любой или оба из полинуклеотидов, кодирующих последовательность, приведенную в SEQ ID NO:35 и SEQ ID NO:36.
[000320] В другом аспекте полинуклеотид может кодировать VH, VL и/или и VH, и VL антитела по изобретению. Т.е. композиция содержит один полинуклеотид или несколько полинуклеотидов, кодирующих антитело или его антигенсвязывающий фрагмент по изобретению.
[000321] Фармацевтические композиции по изобретению также можно вводить в качестве комбинированного лечения, такого как комбинация с другими средствами. Например, комбинированное лечение может включать антитело против CXCR5 или его антигенсвязывающий фрагмент по изобретению в комбинации по меньшей мере с одним другим способом лечения, где лечение может представлять собой хирургическое вмешательство, иммунотерапию или медикаментозную терапию.
[000322] Фармацевтические соединения по изобретению могут включать одну или более фармацевтически приемлемых солей. Примеры таких солей включают кислые соли присоединения и основные соли присоединения. Кислые соли присоединения включают соли, полученные из нетоксичных неорганических кислот, таких как соляная, азотная, фосфорная, серная, бромистоводородная, йодистоводородная, фосфористая и т.п., а также из нетоксичных органических кислот, таких как алифатические моно- и дикарбоновые кислоты, фенил-замещенные алкановые кислоты, гидроксиалкановые кислоты, ароматические кислоты, алифатические и ароматические сульфоновые кислоты и т.п. Основные соли присоединения включают соли, полученные из щелочноземельных металлов, таких как натрий, калий, магний, кальций и т.п., а также из нетоксичных органических аминов, таких как N, N'-дибензилэтилендиамин, N-метилглюкамин, хлорпрокаин, холин, диэтаноламин, этилендиамин, прокаин и т.п.
[000323] Фармацевтическая композиция по изобретению также может включать фармацевтически приемлемый антиоксидант. Примеры фармацевтически приемлемых антиоксидантов включают: (1) водорастворимые антиоксиданты, такие как аскорбиновая кислота, гидрохлорид цистеина, бисульфат натрия, метабисульфит натрия, сульфит натрия и т.п.; (2) жирорастворимые антиоксиданты, такие как аскорбилпальмитат, бутилированный гидроксианизол (BHA), бутилированный гидрокситолуол (BHT), лецитин, пропилгаллат, альфа-токоферол и т.п.; и (3) металл-хелатирующие средства, такие как лимонная кислота, этилендиаминтетрауксусная кислота (ЭДТА), сорбит, винная кислота, фосфорная кислота и т.п.
[000324] Примеры подходящих водных и неводных носителей, которые можно использовать в фармацевтических композициях по изобретению включают воду, этанол, полиолы (такие как глицерин, пропиленгликоль, полиэтиленгликоль и т.п.) и их подходящие смеси, растительные масла, такие как оливковое масло, и инъецируемые органические сложные эфиры, такие как этилолеат. Правильную текучесть можно поддерживать, например, с использованием материалов покрытий, таких как лецитин, посредством поддержания необходимого размера частиц в случае дисперсий, и с использованием поверхностно-активных веществ.
[000325] Эти композиции также могут содержать вспомогательные вещества, такие как консерванты, увлажнители, эмульгаторы и диспергирующие средства. Предотвращения присутствия микроорганизмов можно достигать посредством стерилизации и включения различных антибактериальных и противогрибковых средств, например, парабена, хлорбутанола, фенолсорбиновой кислоты и т.п. Также желательным может являться включение в композиции изотонических средств, таких как сахара, хлорид натрия и т.п. Кроме того, пролонгированной абсорбции инъецируемой фармацевтической формы можно достигать посредством включения средств, замедляющих абсорбцию, таких как моностеарат алюминия и желатин.
[000326] Фармацевтические композиции, как правило, должны быть стерильными и стабильными в условиях производства и хранения. Композицию можно составлять в виде раствора, микроэмульсии, липосомы или другой упорядоченной структуры, подходящей для высокой концентрации лекарственного средства. Носитель может являться растворителем или дисперсионной средой, содержащей, например, воду, этанол, полиол (например, глицерин, пропиленгликоль и жидкий полиэтиленгликоль и т.п.) и их подходящие смеси. Правильную текучесть можно поддерживать, например, с использованием покрытия, такого как лецитин, посредством поддержания необходимого размера частиц в случае дисперсии и с использованием поверхностно-активных веществ. Во многих случаях подходящим может быть включение в композиции изотонических средств, например, сахаров, полиспиртов, таких как маннит, сорбит, или хлорида натрия. Пролонгированной абсорбции инъецируемых композиций можно достигать посредством включения в композицию средства, замедляющего абсорбцию, например, моностеаратных солей и желатина.
[000327] Стерильные инъецируемые растворы можно получать посредством включения активного соединения в необходимом количестве в подходящем растворителе с одним или комбинацией перечисленных выше ингредиентов, при необходимости, с последующей стерилизацией посредством микрофильтрации.
[000328] Как правило, дисперсии получают посредством включения активного соединения в стерильный носитель, содержащий основную дисперсионную среду и другие необходимые ингредиенты из перечисленных выше. В случае стерильных порошков для получения стерильных инъецируемых растворов предпочтительными способами получения являются вакуумная сушка и лиофилизация, посредством которых получают порошок активного ингредиента и любого дополнительного желаемого ингредиента из их ранее стерильного фильтрованного раствора.
[000329] Фармацевтическую композицию по изобретению можно получать, упаковывать или продавать в составе, подходящем для офтальмологического введения. Такие составы, например, могут находиться в форме глазных капель, включающих, например, 0,1%-1,0% (масс./масс.) раствор или суспензию активного ингредиента в водном или жидком масляном носителе. Такие капли могут дополнительно содержать буферные средства, соли или один или более других дополнительных ингредиентов, представленных в настоящем описании. Другие офтальмологически вводимые составы, которые можно использовать, включают составы, содержащие активный ингредиент в микрокристаллической форме или липосомальном препарате.
[000330] В рамках изобретения "дополнительные ингредиенты" включают, в качестве неограничивающих примеров, одно или более из следующего: эксципиенты; поверхностно-активные средства; диспергирующие средства; инертные дилюенты; гранулирующие средства и средства для повышения распадаемости; связывающие средства; смазки; подсластители; ароматизаторы; красители; консерванты; физиологически деградируемые композиции, такие как желатин; водные носители и растворители; масляные носители и растворители; суспендирующие средства; диспергирующие средства или увлажнители; эмульгаторы, смягчающие средства; буферы; соли; загустители; наполнители; эмульгаторы; антиоксиданты; антибиотики; противогрибковые средства; стабилизирующие средства и фармацевтически приемлемые полимерные или гидрофобные материалы. Другие "дополнительные ингредиенты", которые можно включать в фармацевтические композиции по изобретению, известны в этой области и описаны, например в Remington's Pharmaceutical Sciences, Genaro, ed., Mack Publishing Co., Easton, PA (1985), включенный в настоящее описание в качестве ссылки.
[000331] В одном из вариантов осуществления антитело против CXCR5 или его антигенсвязывающий фрагмент вводят во внутривенном составе в виде стерильного водного раствора, содержащего 5 мг/мл, или в некоторых вариантах осуществления приблизительно 10 мг/мл, или в некоторых вариантах осуществления приблизительно 15 мг/мл, или в некоторых вариантах осуществления приблизительно 20 мг/мл антитела, или в некоторых вариантах осуществления приблизительно 25 мг/мл, или в некоторых вариантах осуществления приблизительно 50 мг/мл с ацетатом натрия, полисорбатом 80 и хлоридом натрия при pH в диапазоне приблизительно от 5 до 6. В некоторых вариантах осуществления внутривенный состав является стерильным водным раствором, содержащим 5 или 10 мг/мл антитела с 20 мМ ацетата натрия, 0,2 мг/мл полисорбата 80 и 140 мМ хлорида натрия при pH 5,5. Кроме того, раствор, содержащий антитело или его антигенсвязывающий фрагмент, помимо многих других соединений, может содержать гистидин, маннит, сахарозу, трегалозу, глицин, полиэтиленгликоль, ЭДТА, метионин и любую их комбинацию и многие другие соединения, известные в этой области.
[000332] В одном из вариантов осуществления фармацевтическая композиция по изобретению содержит следующие компоненты: 50 мг/мл антитела против CXCR5 или антигенсвязывающего фрагмента по изобретению, 20 мМ гистидин, 8,5% сахарозу и 0,02% полисорбат 80, 0,005% ЭДТА при pH 5,8; в другом варианте осуществления фармацевтическая композиция по настоящему изобретению содержит следующие компоненты: 100 мг/мл антитела против CXCR5 или антигенсвязывающего фрагмента по изобретению, 10 мМ гистидин, 5% сахарозы и 0,01% полисорбат 80 при pH 5,8. Эту композицию можно получать в виде жидкого состава или лиофилизированного порошка. Когда порошок восстанавливают в полном составе, композиция сохраняет тот же состав. Альтернативно, порошок можно восстанавливать в половине объема, в случае чего композиция содержит 100 мг антитела против CXCR5 или его антигенсвязывающего фрагмента по изобретению, 20 мМ гистидина, 10% сахарозы и 0,02% полисорбата 80 при pH 5,8.
[000333] В одном из вариантов осуществления часть дозы вводят посредством внутривенного болюса, а остальное посредством инфузии состава антитела. Например, 0,01 мг/кг внутривенной инъекции антитела против CXCR5 или его антигенсвязывающего фрагмента можно вводить в виде болюса, и а остальную часть дозы антитела можно вводить посредством внутривенной инъекции. Заранее определенную дозу антитела против CXCR5 или его антигенсвязывающего фрагмента можно вводить, например, в течение часа и от получаса до двух часов или пяти часов.
[000334] Что касается терапевтического средства, если средство является, например, низкомолекулярным соединением, оно может находиться в фармацевтической композиции в форме физиологически приемлемого сложного эфира или соли, например, в комбинации с физиологически приемлемым катионом или анионом, что хорошо известно в этой области.
[000335] Составы фармацевтических композиций, представленные в настоящем описании, можно получать любым способом, известным в области фармакологии, или способом, который будет разработан в будущем. В основном, такие способы получения включают стадию приведения активного ингредиента в контакт с носителем или одним или более другими вспомогательными ингредиентами, а затем, если необходимо или желательно, придания формы или упаковки продукта в желаемый однодозовый или многодозовый контейнер.
[000336] В одном из вариантов осуществления композиции по изобретению являются несодержащими пирогены составами, по существу, не содержащими эндотоксины и/или родственные пирогенные вещества. Эндотоксины включают токсины, локализующиеся внутри микроорганизма и высвобождаемые, когда микроорганизмы разрушаются или погибают. Пирогенные вещества также включают вызывающие лихорадку, термостабильные вещества (гликопротеины) из внешней мембраны бактерий и других микроорганизмов. И те, и другие вещества могут вызывать лихорадку, гипотензию и шок при введении людям. Из-за потенциальных неблагоприятных эффектов предпочтительно удалять даже небольшие количества эндотоксинов из вводимых внутривенно фармацевтических растворов лекарственных средств. Управление по санитарному надзору за качеством пищевых продуктов и медикаментов ("FDA") установило верхний предел 5 единиц эндотоксина (ЕЭ) на дозу на килограмм массы тела за один одночасовой период в случае внутривенного введения лекарственных средств (The United States Pharmacopeial Convention, Pharmacopeial Forum 26 (1):223 (2000)). Если терапевтические белки вводят в количествах несколько сотен или тысяч миллиграммов на килограмм массы тела, предпочтительно удалять даже незначительные количества эндотоксина. В одном из вариантов осуществления уровни эндотоксина и пирогенов в композиции составляют менее 10 ЕЭ/мг, или менее 5 ЕЭ/мг, или менее 1 ЕЭ/мг, или менее 0,1 ЕЭ/мг, или менее 0,01 ЕЭ/мг, или менее 0,001 ЕЭ/мг. В другом варианте осуществления уровни эндотоксина и пирогенов в композиции составляют менее приблизительно 10 ЕЭ/мг, или менее приблизительно 5 ЕЭ/мг, или менее приблизительно 1 ЕЭ/мг, или менее приблизительно 0,1 ЕЭ/мг, или менее приблизительно 0,01 ЕЭ/мг, или менее приблизительно 0,001 ЕЭ/мг.
[000337] В одном из вариантов осуществления настоящее изобретение включает введение композиции, где указанное введение является пероральным, парентеральным, внутримышечным, интраназальным, вагинальным, ректальным, лингвальным, сублингвальным, буккальным, интрабуккальным, внутривенным, кожным, подкожным или трансдермальным.
[000338] В другом варианте осуществления настоящее изобретение дополнительно включает введение композиции в комбинации с другими способами лечения, такими как хирургическое вмешательство, химиотерапия, гормональная терапия, биологическая терапия, иммунотерапия или лучевая терапия.
VII. ДОЗИРОВАНИЕ/ВВЕДЕНИЕ
[000339] Для получения фармацевтических или стерильных композиций, включающих антитело против CXCR5 или его антигенсвязывающий фрагмент по изобретению, антитело смешивают с фармацевтически приемлемым носителем или эксципиентом. Составы терапевтических и диагностических средств можно получать посредством смешивания с физиологически приемлемыми носителями, эксципиентами или стабилизаторами в форме, например, лиофилизированных порошков, суспензий, водных растворов, или лосьонов (см., например, Hardman, et al. (2001) Goodman and Gilman's Pharmaceutical Basis of Therapeutics, McGraw-Hill, New York, N.Y.; Gennaro (2000) Remington: The Science and Practice of Pharmacy, Lippincott, Williams, and Wilkins, New York, N. Y.; Avis, et al. (eds.) (1993) Pharmaceutical Dosage Forms: Parenteral Medications, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Tablets, Marcel Dekker, NY; Lieberman, et al. (eds.) (1990) Pharmaceutical Dosage Forms: Disperse Systems, Marcel Dekker, NY; Weiner and Kotkoskie (2000) Excipient Toxicity and Safety, Marcel Dekker, Inc., New York, N.Y.).
[000340] Выбор схемы введения терапевтического средства зависит от нескольких факторов, включая скорость циркуляции вещества в сыворотке или ткани, уровень симптомов, иммуногенность вещества и доступность клеток-мишеней в биологическом матриксе. В некоторых вариантах осуществления с помощью схемы введения максимизируют количество терапевтического средства, доставляемого пациенту в соответствии с приемлемым уровнем побочных эффектов. Таким образом, количество доставляемого биологического средства частично зависит от конкретного вещества и тяжести состояния, подвергаемого лечению. Доступно руководство по выбору подходящих доз антител, цитокинов и низкомолекулярных соединений (см., например, Wawrzynczak, 1996, Antibody Therapy, Bios Scientific Pub. Ltd, Oxfordshire, UK; Kresina (ed.), 1991, Monoclonal Antibodies, Cytokines and Arthritis, Marcel Dekker, New York, N.Y.; Bach (ed.),1993, Monoclonal Antibodies and Peptide Therapy in Autoimmune Diseases, Marcel Dekker, New York, N. Y.; Baert, et al., 2003, New Engl. J. Med. 348:601-608; Milgrom, et al., 1999, New Engl. J. Med. 341:1966-1973; Slamon, et al., 2001, New Engl. J. Med. 344:783-792; Beniaminovitz, et al., 2000, New Engl. J. Med. 342:613-619; Ghosh, et al., 2003, New Engl. J. Med. 348:24-32; Lipsky, et al., 2000, New Engl. J. Med. 343:1594-1602).
[000341] Определение подходящей дозы осуществляет клиницист, например, с использованием параметров или факторов, которые, как известно или предполагают в этой области, влияют на лечение или, как предполагают, будут влиять на лечение. Как правило, доза начинается с количества, немного меньшего, чем оптимальная доза, и затем ее повышают с небольшими приращениями до достижения желаемого или оптимального эффекта относительно любых отрицательных побочных эффектов. Важные диагностические измерения включают измерение симптомов, например, воспаления или уровня продуцируемых воспалительных цитокинов.
[000342] Точные уровни доз активных ингредиентов в фармацевтических композиций по изобретению можно варьировать так, чтобы получать количество активного ингредиента, являющееся эффективным для достижения желаемого терапевтического ответа в случае конкретного пациента, композиции и способа введения, без токсичности для пациента. Выбранный уровень дозы будет зависеть от различных фармакокинетических факторов, включая активность конкретных используемых композиций по изобретению или их сложного эфира, соли или амида, путь введения, время введение, скорость экскреции конкретного используемого соединения, длительность лечения, другие лекарственные средства, соединения и/или материалы, используемые в комбинации с конкретными используемыми композициями, возраст, пол, массу тела, состояние, общее состояние здоровья и медицинский анамнез пациента, подвергаемого лечению, и подобные факторы, хорошо известные в области медицины.
[000343] Композиции, содержащие антитела против CXCR5 или их антигенсвязывающие фрагменты по изобретению можно вводить посредством непрерывной инфузии или в дозах с интервалами, например, один день, одна неделя или 1-7 раз в неделю. Дозы можно вводить внутривенно, подкожно, местно, перорально, назально, ректально, внутримышечно, интрацеребрально или посредством ингаляции. Конкретный протокол введения является протоколом, включающим максимальную дозу или частоту введения, позволяющие избегать значительных нежелательных побочных эффектов. Общая недельная доза может составлять по меньшей мере 0,05 мкг/кг массы тела, по меньшей мере 0,2 мкг/кг, по меньшей мере 0,5 мкг/кг, по меньшей мере 1 мкг/кг, по меньшей мере 10 мкг/кг, по меньшей мере 100 мкг/кг, по меньшей мере 0,2 мг/кг, по меньшей мере 1,0 мг/кг, по меньшей мере 2,0 мг/кг, по меньшей мере 10 мг/кг, по меньшей мере 15 мг/кг, по меньшей мере 20 мг/кг, по меньшей мере 25 мг/кг или по меньшей мере 50 мг/кг (см., например, Yang, et al., 2003, New Engl. J. Med. 349:427-434; Herold, et al., 2002, New Engl. J. Med. 346:1692-1698; Liu, et al., 1999, J. Neurol. Neurosurg. Psych. 67:451-456; Portielji, et al., 2003, Cancer. Immunol. Immunother. 52: 133-144). Доза может составлять по меньшей мере 15 мкг, по меньшей мере 20 мкг, по меньшей мере 25 мкг, по меньшей мере 30 мкг, по меньшей мере 35 мкг, по меньшей мере 40 мкг, по меньшей мере 45 мкг, по меньшей мере 50 мкг, по меньшей мере 55 мкг, по меньшей мере 60 мкг, по меньшей мере 65 мкг, по меньшей мере 70 мкг, по меньшей мере 75 мкг, по меньшей мере 80 мкг, по меньшей мере 85 мкг, по меньшей мере 90 мкг, по меньшей мере 95 мкг или по меньшей мере 100 мкг. Количество доз, вводимых индивидууму, может составлять по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или более.
[000344] В случае антител против CXCR5 или их антигенсвязывающих фрагментов по изобретению доза, вводимая пациенту, может составлять от 0,0001 мг/кг 100 мг/кг массы тела пациента. Доза может составлять от 0,0001 мг/кг до 20 мг/кг, от 0,0001 мг/кг до 10 мг/кг, от 0,0001 мг/кг до 5 мг/кг, от 0,0001 до 2 мг/кг, от 0,0001 до 1 мг/кг, от 0,0001 мг/кг до 0,75 мг/кг, от 0,0001 мг/кг до 0,5 мг/кг, от 0,0001 мг/кг до 0,25 мг/кг, от 0,0001 до 0,15 мг/кг, от 0,0001 до 0,10 мг/кг, от 0,001 до 0,5 мг/кг, от 0,01 до 0,25 мг/кг или от 0,01 до 0,10 мг/кг массы тела пациента.
[000345] Дозу антитела против CXCR5 или его антигенсвязывающего фрагмента можно вычислять с использованием массы тела пациента в килограммах (кг), умноженной на вводимую дозу в мг/кг. Доза антител по изобретению может составлять 150 мкг/кг или менее, 125 мкг/кг или менее, 100 мкг/кг или менее, 95 мкг/кг или менее, 90 мкг/кг или менее, 85 мкг/кг или менее, 80 мкг/кг или менее, 75 мкг/кг или менее, 70 мкг/кг или менее, 65 мкг/кг или менее, 60 мкг/кг или менее, 55 мкг/кг или менее, 50 мкг/кг или менее, 45 мкг/кг или менее, 40 мкг/кг или менее, 35 мкг/кг или менее, 30 мкг/кг или менее, 25 мкг/кг или менее, 20 мкг/кг или менее, 15 мкг/кг или менее, 10 мкг/кг или менее, 5 мкг/кг или менее, 2,5 мкг/кг или менее, 2 мкг/кг или менее, 1,5 мкг/кг или менее, 1 мкг/кг или менее, 0,5 мкг/кг или менее или 0,1 мкг/кг массы тела пациента или менее.
[000346] Однократная доза антител против CXCR5 или их антигенсвязывающих фрагментов по изобретению может составлять от 0,1 мг до 200 мг, от 0,1 мг до 175 мг, от 0,1 мг до 150 мг, от 0,1 мг до 125 мг, от 0,1 мг до 100 мг, от 0,1 мг до 75 мг, от 0,1 мг до 50 мг, от 0,1 мг до 30 мг, от 0,1 мг до 20 мг, от 0,1 мг до 15 мг, от 0,1 мг до 12 мг, от 0,1 мг до 10 мг, от 0,1 мг до 8 мг, от 0,1 мг до 7 мг, от 0,1 мг до 5 мг, от 0,1 до 2,5 мг, от 0,25 мг до 20 мг, от 0,25 до 15 мг, от 0,25 до 12 мг, от 0,25 до 10 мг, от 0,25 до 8 мг, от 0,25 мг до 7 мг, от 0,25 мг до 5 мг, от 0,5 мг до 2,5 мг, от 1 мг до 20 мг, от 1 мг до 15 мг, от 1 мг до 12 мг, от 1 мг до 10 мг, от 1 мг до 8 мг, от 1 мг до 7 мг, от 1 мг до 5 мг или от 1 мг до 2,5 мг.
[000347] С помощью дозы антител против CXCR5 или их антигенсвязывающих фрагментов по изобретению можно достигать сывороточного титра по меньшей мере 0,1 мкг/мл, по меньшей мере 0,5 мкг/мл, по меньшей мере 1 мкг/мл, по меньшей мере 2 мкг/мл, по меньшей мере 5 мкг/мл, по меньшей мере,6 мкг/мл, по меньшей мере 10 мкг/мл, по меньшей мере 15 мкг/мл, по меньшей мере 20 мкг/мл, по меньшей мере 25 мкг/мл, по меньшей мере 50 мкг/мл, по меньшей мере 100 мкг/мл, по меньшей мере 125 мкг/мл, по меньшей мере 150 мкг/мл, по меньшей мере 175 мкг/мл, по меньшей мере 200 мкг/мл, по меньшей мере 225 мкг/мл, по меньшей мере 250 мкг/мл, по меньшей мере 275 мкг/мл, по меньшей мере 300 мкг/мл, по меньшей мере 325 мкг/мл, по меньшей мере 350 мкг/мл, по меньшей мере 375 мкг/мл или по меньшей мере 400 мкг/мл у индивидуума. Альтернативно, с помощью дозы антител по изобретению можно достигать сывороточного титра по меньшей мере 0,1 мкг/мл, по меньшей мере 0,5 мкг/мл, по меньшей мере 1 мкг/мл, по меньшей мере 2 мкг/мл, по меньшей мере 5 мкг/мл, по меньшей мере 6 мкг/мл, по меньшей мере 10 мкг/мл, по меньшей мере 15 мкг/мл, по меньшей мере 20 мкг/мл, по меньшей мере 25 мкг/мл, по меньшей мере 50 мкг/мл, по меньшей мере 100 мкг/мл, по меньшей мере 125 мкг/мл, по меньшей мере 150 мкг/мл, по меньшей мере 175 мкг/мл, по меньшей мере 200 мкг/мл, по меньшей мере 225 мкг/мл, по меньшей мере 250 мкг/мл, по меньшей мере 275 мкг/мл, по меньшей мере 300 мкг/мл, по меньшей мере 325 мкг/мл, по меньшей мере 350 мкг/мл, по меньшей мере 375 мкг/мл или по меньшей мере 400 мкг/мл у индивидуума.
[000348] Дозы антител против CXCR5 или их антигенсвязывающих фрагментов по изобретению можно повторять и введения можно разделять интервалами по меньшей мере 1 день, 2 дня, 3 дня, 5 дней, 10 дней, 15 дней, 30 дней, 45 дней, 2 месяца, 75 дней, 3 месяца или по меньшей мере 6 месяцев.
[000349] Эффективное количество в случае конкретного пациента может варьироваться в зависимости от таких факторов, как состояние, подвергаемое лечению, общее состояние здоровья пациента, способ, путь введения и доза и тяжесть побочных эффектов (см., например, Maynard, et al., 1996, A Handbook of SOPs for Good Clinical Practice, Interpharm Press, Boca Raton, Fla.; Dent, 2001, Good Laboratory and Good Clinical Practice, Urch Publ, London, UK).
[000350] Путь введения может представлять собой, например, местное или кожное нанесение, инъекцию или инфузию посредством внутривенного, интраперитонеального, интрацеребрального, внутримышечного, внутриглазного, интраартериального, внутриспинномозгового, внутриочагового пути или с помощью систем замедленного высвобождения или имплантата (см., например, Sidman et al., 1983, Biopolymers 22:547-556; Langer, et al., 1981, J. Biomed. Mater. Res. 15: 167-277; Langer, 1982, Chem. Tech. 12:98-105; Epstein, et al., 1985, Proc. Natl. Acad. Sci. USA 82:3688-3692; Hwang, et al., 1980, Proc. Natl. Acad. Sci. USA 77:4030-4034; патенты США №№ 6350466 и 6316024). При необходимости, композиция также может включать солюбилизатор и местный анестетик, такой как лидокаин, для облегчения боли в месте инъекции. Кроме того, также можно использовать легочное введение, например, с помощью ингалятора или небулайзера, и состав с аэрозольным средством. См., например, патенты США №№ 6019968, 5985320, 5985309, 5934272, 5874064, 5855913, 5290540 и 4880078 и публикации PCT №№ WO 92/19244, WO 97/32572, WO 97/44013, WO 98/31346 и WO 99/66903, каждая из которых включена в настоящее описание в качестве ссылки в полном объеме. В одном из вариантов осуществления антитело против CXCR5, или его антигенсвязывающий фрагмент, или композицию по изобретению вводят с использованием технологии легочного введения лекарственных средств Alkermes AIR™ (Alkermes, Inc., Cambridge, Mass.).
[000351] Композицию по изобретению также можно вводить одним или более путями введения одним или более из различных известных в этой области способов. Как будет понятно специалистам в этой области, путь и/или способ введения будет варьироваться в зависимости от желаемых результатов. Выбранные пути введения антител по изобретению включают внутривенный, внутримышечный, внутрикожный, интраперитонеальный, подкожный, спинальный или другие парентеральные пути введения, например, посредством инъекции или инфузии. Парентеральное введение может представлять собой способы введения, иные, чем энтеральное и местное введение, как правило, посредством инъекции, и оно включает, в качестве неограничивающих примеров, внутривенную, внутримышечную, внутриартериальную, интратекальную, интракапсулярную, интраорбитальную, внутрисердечную, внутридермальную, интраперитонеальную, транстрахеальную, подкожную, субкутикулярную, внутрисуставную, подкапсулярную, субарахноидальную, интраспинальную, эпидуральную и внутригрудинную инъекцию и инфузию. Альтернативно, композицию по изобретению можно вводить непарентеральным путем, таким как местный, эпидермальный или слизистый путь введения, например, интраназальный, пероральный, вагинальный, ректальный, сублингвальный или местный муть введения.
[000352] Если антитела против CXCR5 или их антигенсвязывающие фрагменты по изобретению вводят в системе с контролируемым высвобождением или замедленным высвобождением, можно использовать насос для достижения контролируемого или замедленного высвобождения (см. Langer, выше; Sefton, 1987, CRC Crit. Ref. Biomed. Eng. 14:20; Buchwald et al., 1980, Surgery 88:501; Saudek et al., 1989, N. Engl. J. Med. 321:514).
[000353] Для достижения контролируемого или замедленного высвобождения терапевтических средств по изобретению можно использовать полимерные материалы (см. например, Medical Applications of Controlled Release, Langer and Wise (eds.), CRC Pres., Boca Raton, Fla. (1974); Controlled Drug Bioavailability, Drug Product Design and Performance, Smolen and Ball (eds.), Wiley, New York (1984); Ranger and Peppas, 1983, J., Macromol. ScL Rev. Macromol. Chem. 23:61; также см. Levy et al, 1985, Science 11 225:190; During et al., 19Z9, Ann. Neurol. 25:351; Howard et al, 1989, J. Neurosurg. 71: 105; патент США № 5679377; патент США № 5916597; патент США № 5912015; патент США № 5989463; патент США № 5128326; публикация PCT № WO 99/15154; и публикацию PCT № WO 99/20253). Неограничивающие примеры полимеров, используемых в составах с замедленным высвобождением, включают поли(2-гидроксиэтилметакрилат), поли(метилметакрилат), поли(акриловую кислоту), поли(этилен-со-винилацетат), поли(метакриловую кислоту), полигликолиды (PLG), полиангидриды, поли(N-винилпирролидон), поли(виниловый спирт), полиакриламид, поли(этиленгликоль), полилактиды (PLA), поли(лактид-со-гликолиды) (PLGA) и полиортоэфиры. В одном из вариантов осуществления полимер, используемый в составе с замедленным высвобождением, является инертным, несодержащим вымываемые примеси, стабильным при хранении, стерильным и биодеградируемым. Систему с контролируемым или замедленным высвобождением можно помещать вблизи профилактической или терапевтической мишени, и, таким образом, необходима лишь часть системной дозы (см., например, Goodson, в Medical Applications of Controlled Release, выше, vol. 2, pp. 115-138 (1984)).
[000354] Системы с контролируемым высвобождением описаны в обзоре Langer, 1990, Science 249:1527-1533. Для получения составов с замедленным высвобождением, содержащих одно или более антител по изобретению или их конъюгатов, можно использовать любой способ, известный специалисту в этой области. См., например, патент США № 4526938, международные патентные публикации №№ WO 91/05548, WO 96/20698, Ning et al., 1996, "Intratumoral Radioimmunotheraphy of Human Colon Cancer Xenograft Using a Sustained-Release Gel," Radiotherapy and Oncology 59:179-189, Song et al., 1995, " Antibody Mediated Lung Targeting of Long-Circulating Emulsions," PDA Journal of Pharmaceutical Science and Technology 50:372-397, Cleek et ah, 1997, "Biodegradable Polymeric Carriers for a bFGF Antibody for Cardiovascular Application," Pro. Ml. Symp. Control. Rel. Bioact. Mater. 24:853-854, и Lam et al., 1997, "Microencapsulation of Recombinant Humanized Monoclonal Antibody for Local Delivery," Proc. Мл. Symp. Control Rel. Bioact. Mater. 24:759-160, каждый из которых включен в настоящее описание в качестве ссылки в полном объеме.
[000355] Если антитело против CXCR5 или его антигенсвязывающий фрагмент по изобретению вводят местно, его можно составлять в форме мази, крема, трансдермального пластыря, лосьона, геля, шампуня, спрея, аэрозоля, раствора, эмульсии или другой формы, хорошо известной специалисту в этой области. См., например, Remington's Pharmaceutical Sciences and Introduction to Pharmaceutical Dosage Forms, 19th ed., Mack Pub. Co., Easton, Pa. (1995). В случае нераспыляемых местных лекарственных форм, как правило, используют вязкие и полутвердые или твердые формы, содержащие носитель или один или более эксципиентов, совместимых с местным использованием и имеющих динамическую вязкость, в некоторых случаях больше, чем у воды. Подходящие составы включают, в качестве неограничивающих примеров, растворы, суспензии, эмульсии, кремы, мази, порошки, линименты и т.п., которые, при желании, стерилизуют или смешивают с вспомогательными средствами (например, консервантами, стабилизаторами, увлажнителями, буферами или солями) для влияния на различные свойства, такие как, например, осмотическое давление. Другие подходящие местные лекарственные формы включают распыляемые аэрозольные препараты, где активный ингредиент, в некоторых случаях в комбинации с твердым или жидким инертным носителем, упаковывают в смеси с летучим веществом под давлением (например, газообразным пропеллентом, таким как фреон) или в мягкой бутылке. При желании, в фармацевтические композиции и лекарственные формы также можно добавлять увлажнители. Примеры таких дополнительных ингредиентов хорошо известны в этой области.
[000356] Если композиции, содержащие антитела против CXCR5 или их антигенсвязывающие фрагменты вводят интраназально, их можно составлять в аэрозольной форме, спрее или в форме капель. В частности, профилактические или терапевтические средства для применения по изобретению можно вводить в форме аэрозольного спрея в упаковке под давлением или небулайзере с использованием подходящего пропеллента (например, дихлордифторметана, трихлорфторметана, дихлортетрафторэтана, диоксида углерода или другого подходящего газа). В случае аэрозоля под давлением единицу дозирования можно определять с помощью клапана для введения отмеренного количества. Капсулы и картриджи (состоящие, например, из желатина) для использования в ингаляторе или инсуффляторе можно составлять содержащим порошковую смесь соединения и подходящей порошковой основы, такой как лактоза или крахмал.
[000357] Способы совместного введения или лечения с использованием второго терапевтического средства, например, цитокина, стероида, химиотерапевтического средства, антибиотика или лучевой терапии, хорошо известны в этой области (см., например, Hardman, et al. (eds.) (2001) Goodman and Gilman's Pharmacological Basis of Therapeutics, 10 th ed., McGraw-Hill, New York, N.Y.; Poole and Peterson (eds.) (2001) Pharmacotherapeutics for Advanced Practice: A Practical Approach, Lippincott, Williams and Wilkins, Phila., Pa.; Chabner and Longo (eds.) (2001) Cancer Chemotherapy and Biotherapy, Lippincott, Williams and Wilkins, Phila., Pa.). С помощью эффективного количества терапевтического средства можно уменьшать симптомы по меньшей мере на 10 процентов, по меньшей мере на 20 процентов; по меньшей мере на приблизительно 30 процент; по меньшей мере на 40 процентов или по меньшей мере на 50 процентов.
[000358] Дополнительные средства (например, профилактические или терапевтические средства), которые можно вводить в комбинации с антителами против CXCR5 или антигенсвязывающими фрагментами по изобретению, можно вводить с интервалом менее 5 минут, менее 30 минут, 1 час, приблизительно 1 час, от приблизительно 1 до приблизительно 2 часов, от приблизительно 2 часов до приблизительно 3 часов, от приблизительно 3 часов до приблизительно 4 часов, от приблизительно 4 часов до приблизительно 5 часов, от приблизительно 5 часов до приблизительно 6 часов, от приблизительно 6 часов до приблизительно 7 часов, от приблизительно 7 часов до приблизительно 8 часов, от приблизительно 8 часов до приблизительно 9 часов, от приблизительно 9 часов до приблизительно 10 часов, от приблизительно 10 часов до приблизительно 11 часов, от приблизительно 11 часов до приблизительно 12 часов, от приблизительно 12 часов до 18 часов, от 18 часов до 24 часов, от 24 часов до 36 часов, от 36 часов до 48 часов, от 48 часов до 52 часов, от 52 часов до 60 часов, от 60 часов до 72 часов, от 72 часов до 84 часов, от 84 часов до 96 часов или от 96 часов до 120 часов относительно антител по изобретению. Два или более средства можно вводить в рамках одного посещения пациентом лечебного учреждения.
[000359] Антитела CXCR5 или их антигенсвязывающие фрагменты по изобретению и другие средства можно вводить циклически. Циклическая терапия включает введение первого средства (например, первого профилактического или терапевтического средства) в течение периода времени с последующим введением второго средства (например, второго профилактического или терапевтического средства) в течение периода времени, необязательно, с последующим введением третьего средства (например, профилактического или терапевтического средства) в течение периода времени и т.д., и повторение этого последовательного введения, т.е. цикла, для снижения развития резистентности к одному из средств, во избежание или для снижения побочных эффектов одного из средств и/или для улучшения эффективности средств.
[000360] В одном из вариантов осуществления антитела против CXCR5 по изобретению можно вводить совместно с композициями для лечения аутоиммунных заболеваний и нарушений, включая, в качестве неограничивающих примеров, адриамицин, азатиоприн, бусульфан, циклофосфамид, циклоспорин A, цитоксан, флударабин, 5-фторурацил, метотрексат, микофенолат мофетил, 6-меркаптопурин, кортикостероид, нестероидные противовоспалительные средства, сиролимус (рапамицин) и такролимус (FK-506). В альтернативных вариантах осуществления иммуномодулирующее или иммуносупрессирующее средство является антителом, выбранным из группы, состоящей из муромонаба-CD3, алемтузумаба (Campath®), базиликсимаба, даклизумаба, муромонаба (OKT3®), ритуксимаба, антитимоцитарного глобулина и IVIg и др., известных специалисту в этой области.
[000361] В одном из вариантов осуществления антитела против CXCR5 по изобретению можно вводить совместно с композициями для лечения диабета, включая, в качестве неограничивающих примеров, бигуаниды (например, буформин, метформин и фенформин), гормоны и их аналоги (амилин, инсулин, инсулин аспарт, инсулин детемир, инсулин гларгин, инсулин глулизин, инсулин лизпро, лираглутид и прамлинтид), производные сульфонилкарбамида (ацетогексамид, карбутамид, хлорпропамид, глиборнурид, гликлазид, глимепирид, глипизид, гликуидон, глизоксепид, глибурид, глибутиазол, глибузол, глигексамид, глимидин, толазамид, толбутамид, и толцикламид), тиазолидиндионы (пиоглитазон, росиглитазон и троглитазон), акарбозу, эксенатид, миглитол, митиглинид, мураглитазар, натеглинид, репаглинид, ситаглиптин, тезаглитазар, вилдаглиптин и воглибоз.
[000362] В некоторых вариантах осуществления антитела против CXCR5 или их антигенсвязывающие фрагменты по изобретению можно составлять для обеспечения правильного распределения in vivo. Например, гематоэнцефалический барьер (BBB) исключает многие высокогидрофильные соединения. Для обеспечения проникновения терапевтических соединений по изобретению через BBB (при желании), их можно составлять, например, в липосомах. Способы производства липосом см., например, в патентах США №№ 4522811, 5374548 и 5399331. Липосомы могут содержать одно или более веществ, селективно транспортируемых в конкретные клетки или органы, таким образом, повышая направленную доставку лекарственных средств (см., например, V.V. Ranade, 1989, J. Clin. Pharmacol. 29:685). Примеры направляющих веществ включают фолат или биотин (см., например, патент США № 5416016); маннозиды (Umezawa et al., Biochem. Biophys. Res. Commun. 153: 1038); антитела (P. G. Bloeman et al., 1995, FEBS Lett. 357: 140; M. Owais et al., 1995, Antimicrob. Agents Chemother. 39: 180); рецептор белка сурфактанта A (Briscoe et al. (1995) Am. J. Physiol. 1233: 134); p120 (Schreier et al. (1994) J. Biol. Chem. 269:9090); также см. K. Keinanen; M.L. Laukkanen, 1994, FEBS Lett. 346:123; Killion; Fidler, 1994; Immunomethods 4:273.
[000363] Настоящее изобретение относится к способам введения нуждающемуся в этом индивидууму фармацевтической композиции, содержащей антитела против CXCR5 или их антигенсвязывающих фрагментов по изобретению, в отдельности или в комбинации с другими средствами. Средства (например, профилактические или терапевтические средства) в рамках комбинированного лечения по изобретению можно вводить индивидууму одновременно или последовательно. Средство (например, профилактические или терапевтические средства) в рамках комбинированного лечения по изобретения также можно вводить циклически. Циклическая терапия включает введение первого средства (например, первого профилактического или терапевтического средства) в течение периода времени с последующим введением второго средства (например, второго профилактического или терапевтического средства) в течение периода времени и повторением этого последовательного введения, т.е. цикла, для уменьшения развития резистентности к одному из средств во избежание или для снижения побочных эффектов одного из средств и/или для улучшения эффективности средств.
[000364] Средства (например, профилактические или терапевтические средства) в рамках комбинированного лечения по изобретению можно вводить индивидууму одновременно. Термин "одновременно" не органичен введением средства (например, профилактических или терапевтических средств) точно в одно и то же время, но скорее он означает, что фармацевтическую композицию, содержащую антитела против CXCR5 или их антигенсвязывающие фрагменты по изобретению, вводят индивидууму в такой последовательности и в таких пределах временного интервала, что антитела по изобретению или их конъюгаты могут действовать вместе с другими средствами для обеспечения благоприятного эффекта, повышенного относительно их использования иным образом. Например, каждое средство можно вводить индивидууму одновременно или последовательно в любом порядке в разные моменты времени; однако, если их не вводят одновременно, их следует вводить достаточно близко по времени так, чтобы обеспечить желаемый терапевтический или профилактический ответ. Каждое средство можно вводить индивидууму раздельно, в любой подходящей форме и любым подходящим путем. В различных вариантах осуществления средства (например, профилактические или терапевтические средства) вводят индивидууму с интервалом менее 15 минут, менее 30 минут, менее 1 часа, приблизительно 1 час, от приблизительно 1 часа до приблизительно 2 часов, от приблизительно 2 часов до приблизительно 3 часов, от приблизительно 3 часов до приблизительно 4 часов, от приблизительно 4 часов до приблизительно 5 часов, от приблизительно 5 часов до приблизительно 6 часов, от приблизительно 6 часов до приблизительно 7 часов, от приблизительно 7 часов до приблизительно 8 часов, от приблизительно 8 часов до приблизительно 9 часов, от приблизительно 9 часов до приблизительно 10 часов, от приблизительно 10 часов до приблизительно 11 часов, от приблизительно 11 часов до приблизительно 12 часов, 24 часа, 48 часов, 72 часа или 1 неделя. В других вариантах осуществления два или более средства (например, профилактические или терапевтические средства) вводят в рамках одного посещения пациентом лечебного учреждения.
[000365] Профилактические или терапевтические средства в рамках комбинированного лечения можно вводить индивидууму в одной фармацевтической композиции. Альтернативно, профилактические или терапевтические средства в рамках комбинированного лечения можно вводить индивидууму одновременно в отдельных фармацевтических композициях. Профилактические или терапевтические средства можно вводить индивидууму одним и тем же или разными путями введения.
VIII. НАБОРЫ
[000366] Настоящее изобретение также относится к наборам, содержащим любые или все из антител, представленных в настоящем описании. Наборы по изобретению включают один или более контейнеров, содержащих антитело против CXCR5, представленное в настоящем описании, и инструкции по использованию в соответствии с любыми из способов по изобретению, представленных в настоящем описании. Как правило, эти инструкции содержат описание введения антитела для описанного выше терапевтического лечения. В некоторых вариантах осуществления наборы предназначены для получения единицы дозирования с однократной дозой. В некоторых вариантах осуществления набор может содержать первый контейнер, содержащий высушенный белок, и второй контейнер, содержащий водный состав. В некоторых вариантах осуществления предусмотрены наборы, содержащие аппликатор, например, однокамерные и многокамерные предварительно наполненные шприцы (например, шприцы для жидкостей и шприцы с лиофилизатом).
[000367] Инструкции по использованию антитела против CXCR5, как правило, включают информацию о дозировке, схеме введения и пути введения для предполагаемого лечения. Контейнеры могут включать унифицированные дозы, многодозовые упаковки или подъединицы дозирования. Инструкции, поставляемые в наборах по изобретению, как правило, являются письменными инструкциями на ярлыке или вкладыше в упаковку (например, бумажном листе, включенном в набор), но приемлемыми также являются машиночитаемые инструкции (например, инструкции, находящиеся на магнитном или оптическом диске).
[000368] Наборы по изобретению находятся в подходящей упаковке. Подходящая упаковка включает, в качестве неограничивающих примеров, флаконы, бутыли, банки, гибкую упаковку (например, запаянные майларовые или пластиковые пакеты) и т.п. Также предусмотрены упаковки для использования в комбинации с конкретным устройством, таким как ингалятор, устройство для назального введения (например, аэрозольный ингалятор) или инфузионное устройство, такое как мининасос. Набор может содержать стерильный входной порт (например, контейнер может являться мешком для внутривенного раствора или флаконом с пробкой, прокалываемой иглой для подкожных инъекций). Контейнер также может содержать стерильный входной порт (например, контейнер может являться мешком для внутривенного раствора или флаконом с пробкой, прокалываемой иглой для подкожных инъекций). По меньшей мере одно активное средство в композиции является антителом против CXCR5 по изобретению. Контейнер может дополнительно содержать второе фармацевтически активное средство.
[000369] Наборы, необязательно, могут включать дополнительные компоненты, такие как буферы и интерпретируемая информация. Как правило, набор содержит контейнер и ярлык или вкладыш в упаковку на контейнере или вместе с контейнером.
[000370] Настоящее изобретение также относится к диагностическим наборам, содержащим любые или все из антител, представленных в настоящем описании. Диагностические наборы можно использовать, например, для детекции наличия CXCR5 в образце. В некоторых вариантах осуществления диагностический набор можно использовать для идентификации индивидуума с латентным заболеванием, нарушением или состоянием, которое может вызывать риск развития CXCR5-опосредованного заболевания, нарушения или состояния или заболевания, нарушения или состояния с недостаточностью CXCR5. В некоторых вариантах осуществления диагностический набор можно использовать для детекции наличия и/или уровня CXCR5 у индивидуума, как предполагают, имеющего CXCR5-опосредованное заболевание или заболевание, нарушение или состояние с недостаточностью CXCR5.
[000371] Диагностические наборы по изобретению включают один или более контейнеров, содержащих антитело против CXCR5, представленное в настоящем описании, и инструкции по использованию в соответствии с любыми из способов по изобретению, представленных в настоящем описании. Как правило, эти инструкции включают описание использования антитела против CXCR5 для детекции наличия CXCR5 у индивидуумов, имеющих риск развития или, как предполагают, имеющих CXCR5-опосредованное заболевание или заболевание, нарушение или состояние с недостаточностью CXCR5. В некоторых вариантах осуществления пример диагностического набора можно конфигурировать так, чтобы он содержал реагенты, такие как, например, антитело против CXCR5, образец отрицательного контроля, образец положительного контроля и инструкции по использованию набора.
IX. ЭКВИВАЛЕНТЫ
[000372] В изложенном выше описании и следующих примерах подробно описаны некоторые конкретные варианты осуществления изобретения и лучший способ осуществления, предусмотренный авторами настоящего изобретения. Однако следует понимать, что независимо от того, насколько подробно это описано в тексте, настоящее изобретение можно осуществлять на практике множеством способов, и настоящее изобретение должно быть истолковано в соответствии с формулой изобретения и любыми ее эквивалентами.
[000373] Хотя идеи описаны со ссылкой на различное применение, способы, наборы и композиции, следует понимать, что можно осуществлять различные изменения и модификации без отклонения от идей, представленных в настоящем описании и формуле изобретения ниже. Следующие примеры представлены для лучшего иллюстрирования описанных идей и не предназначены для ограничения объема настоящего изобретения. Хотя настоящее изобретения описано в терминах этих примеров вариантов осуществления, специалистам в этой области будет понятно, что возможны многочисленные изменения и модификации этих примеров вариантов осуществления без излишнего экспериментирования. Все такие изменения и модификации входят в объем настоящего изобретения.
[000374] Все цитируемые в настоящем описании ссылки, включая патенты, патентные заявки, статьи, пособия и т.п., и цитируемые в них ссылки, включены, таким образом, в настоящее описание в качестве ссылки в полном объеме. В случае если один или более из включенных литературных источников и схожих материалов отличается или противоречит настоящей заявке, включая, в качестве неограничивающих примеров, определенные термины, использование терминов, описанные способы или т.д., настоящая заявка обладает приоритетом.
X. ОБЩИЕ СПОСОБЫ
[000375] Следует понимать, что настоящее изобретение не ограничено конкретными синтетическими способами получения, которые, разумеется, можно варьировать. Если в настоящем описании не указано иначе, научные и технические термины, используемые в отношении настоящего изобретения, должны обладать значением, общепринято понятным специалистам в этой области. Кроме того, если контекст не требует иного, термины в единственном числе должны включать множественное число, и термины во множественном числе должны включать единственное число. Как правило, используемая номенклатура и способы культивирования клеток и тканей, молекулярной биологии, иммунологии, микробиологии, генетики и химии белков и нуклеиновых кислот и гибридизации, представленные в настоящем описании, хорошо известны и общеупотребительны в этой области.
[000376] Если не указано иначе, в практическом осуществлении настоящего изобретения используют общепринятые способы молекулярной биологии (включая рекомбинантные способы), микробиологии, клеточной биологии, биохимии и иммунологии, известные специалистам в этой области. Такие способы полностью описаны в литературе, например, в Molecular Cloning: A Laboratory Manual, второе издание (Sambrook et al., 1989) Cold Spring Harbor Press; Oligonucleotide Synthesis (M.J. Gait, ed., 1984); Methods in Molecular Biology, Humana Press; Cell Biology: A Laboratory Notebook (J.E. Cellis, ed., 1998) Academic Press; Animal Cell Culture (R.I. Freshney, ed., 1987); Introduction to Cell and Tissue Culture (J.P. Mather and P.E. Roberts, 1998) Plenum Press; Cell and Tissue Culture: Laboratory Procedures (A. Doyle, J.B. Griffiths, and D.G. Newell, eds., 1993-1998) J. Wiley and Sons; Methods in Enzymology (Academic Press, Inc.); Handbook of Experimental Immunology (D.M. Weir and C.C. Blackwell, eds.); Gene Transfer Vectors for Mammalian Cells (J.M. Miller and M.P. Calos, eds., 1987); Current Protocols in Molecular Biology (F.M. Ausubel et al., eds., 1987); PCR: The Polymerase Chain Reaction, (Mullis et al., eds., 1994); Current Protocols in Immunology (J.E. Coligan et al., eds., 1991); Sambrook and Russell, Molecular Cloning: A Laboratory Manual, 3rd. ed., Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (2001); Ausubel et al., Current Protocols in Molecular Biology, John Wiley & Sons, NY (2002); Harlow and Lane, Using Antibodies: A Laboratory Manual, Cold Spring Harbor Laboratory Press, Cold Spring Harbor, NY (1998); Coligan et al., Short Protocols in Protein Science, John Wiley & Sons, NY (2003); Short Protocols in Molecular Biology (Wiley и Sons, 1999); Immunobiology (C.A. Janeway and P. Travers, 1997); Antibodies (P. Finch, 1997); Antibodies: a practical approach (D. Catty., ed., IRL Press, 1988-1989); Monoclonal antibodies: a practical approach (P. Shepherd and C. Dean, eds., Oxford University Press, 2000); Using antibodies: a laboratory manual (E. Harlow and D. Lane (Cold Spring Harbor Laboratory Press, 1999); Antibodies (M. Zanetti and J.D. Capra, eds., Harwood Academic Publishers, 1995).
[000377] Ферментативные реакции и способы очистки осуществляют по инструкциям производителей, как принято в этой области или представлено в настоящем описании. Используемая номенклатура и лабораторные способы аналитической химии, биохимии, иммунологии, молекулярной биологии, синтетической органической химии и медицинской и фармацевтической химии, представленные в настоящем описании, хорошо известны и общеупотребительны в этой области. Для химического синтеза, химических анализов, получения, составления и введения фармацевтических препаратов и лечения пациентов используют стандартные способы.
XI. БИОЛОГИЧЕСКИЙ ДЕПОЗИТ
[000378] Основные материалы по настоящему изобретению депонированы в American Type Culture Collection, 10801 University Boulevard, Manassas, Va. 20110-2209, USA, на 26 июля 2017 года. Вектор h11G2-VH (XC155), имеющей регистрационный номер ATCC PTA-124323, содержит вставку ДНК, кодирующую вариабельную область тяжелой цепи антитела h11G2 (XC155), и вектор h11G2-VL (XC154), имеющей регистрационный номер ATCC PTA-124324, содержит вставку ДНК, кодирующую вариабельную область легкой цепи антитела h11G2 (XC154). Депозиты были сделаны в соответствии с положениями Будапештского договора о международном признании депонирования микроорганизмов для целей патентной процедуры и нормативными актами (Будапештский договор). Это обеспечивает поддержание живых культур депозита в течение 30 лет от даты депонирования. Депозит будет сделан доступным ATCC в рамках Будапештского договора и будет предметом соглашения между Pfizer Inc. и ATCC, обеспечивающим постоянный и неограниченный доступ к потомству культуры депозита общественности после выдачи пертинентного патента США или после опубликования любой патентной заявки США или иной страны, в зависимости от того, что произойдет первым, и обеспечивает доступность потомства лицу, уполномоченному Ведомством по патентам и товарным знакам США в соответствии с 35 U.S.C. разделом 122 и соответствующими правилами Ведомства (включая 37 C.F.R. раздел 1.14 со ссылкой на 886 OG 638).
[000379] Владелец настоящей заявки согласен на то, что, если культура материалов депозита погибает, или утрачена, или разрушена при культивировании в подходящих условиях, материалы будут немедленно заменены по уведомлении другим таким же материалом. Доступность депонируемого материала не следует истолковывать как лицензию на практическое осуществление настоящего изобретения в нарушение прав, предоставленных с разрешения любого правительства в соответствии с патентным законодательством.
ПРИМЕРЫ
[000380] Настоящее изобретение дополнительно подробно описано со ссылкой на следующие экспериментальные примеры. Эти примеры представлены исключительно в иллюстративных целях, и не следует истолковывать их в качестве ограничения, если не указано иначе. Таким образом, настоящее изобретение ни в коем случае не следует истолковывать как ограниченное следующими примерами, а скорее как включающее любые и все варианты, становящиеся очевидными из идей, представленных в настоящем описании.
Пример 1: Гибридомы, связывающиеся с CXCR5 человека и CXCR5 яванского макака
[000381] Самок мышей BALB/c иммунизировали три раза смесью, содержащей 1×106 клеток BaF3, гиперэкспрессирующих CXCR5 человека (SEQ ID NO:32), или клеток 300.19, экспрессирующих CXCR5 человека (SEQ ID NO:32), с 5 мкг адъюванта CPG (ODN1826) (Invivogen) в серии с интервалами по три недели. При конечной бустерной стимуляции мышей иммунизировали 20 мкг вирусоподобных частиц (VLP), полученных из клеток HEK-293, гиперэкспрессирующих CXCR5 человека (SEQ ID NO:32), с использованием системы экспрессии функциональных белков MembranePro™ (Thermo Fisher). Осуществляли пять раундов слияния и получали восемьдесят восемь клонов, связывающих клетки HEK-293, экспрессирующие CXCR5 человека (hCXCR5-293). Эти моноклональные антитела очищали с помощью протеина A и тестировали на связывание с клетками HEK-293, экспрессирующими hCXCR5, и клетками HEK-293, экспрессирующими CXCR5 яванского макака (SEQ ID NO:33), т.е. cynoCXCR5-293. Клетки hCXCR5-293 или cynoCXCR5-293 окрашивали с использованием моноклональных антител в количестве 10 мкг/мл перед окрашиванием с помощью PE-конъюгированных антител против IgG мыши (Southern Biotech). Клетки анализировали посредством проточной цитометрии с использованием анализатора FACSVerse Analyzer (BD Biosciences). При скрининге на связывание с клетками hCXCR5 HEK-293 и cynoCXCR5 HEK-293 идентифицировали тридцать четыре антитела, связывающихся с CXCR5 человека и яванского макака (фиг. 3).
Пример 2: Антитела против CXCR5, связывающиеся с клетками Raji, и антагонистическая активность в анализе переноса кальция
[000382] Антитела, связывающиеся с клетками hCXCR5-293 и cynoCXCR5-293, дополнительно анализировали на связывание с клетками Raji посредством проточной цитометрии с использованием анализатора FACSVerse Analyzer (BD Biosciences, Franklin Lakes, NJ). Кажущуюся аффинность связывания антитела с клетками Raji вычисляли как EC50 кривых титрования равновесного связывания, на которых геометрическое среднее интенсивности флуоресценции (gMFI) антигенсвязывающей популяции количественно анализировали посредством проточной цитометрии, и результаты представлены в таблице 2.
[000383] Данные свидетельствуют о том, что некоторые антитела связываются с клетками Raji с большей/меньшей степенью, чем другие антитела. Эти данные свидетельствуют о том, что антитела распознают эндогенный CXCR5 на клетках Raji, полученных из пациента с лимфомой Беркитта, что позволяет предполагать, что антитела могут связываться с B-клетками in vivo. Кроме того, аффинность антитела 11G2 к CXCR5 на клетках Raji является более высокой, чем у референсного антитела против CXCR5 16D7 (WO 2009/032661), что позволяет предполагать, что 11G2 может быть более активным, чем 16D7.
[000384] Антитела, связывающиеся и с hCXCR5-293, и с cynoCXCR5-293, также тестировали в анализе переноса кальция на антагонистическую активность. Кроме того, в качестве контроля включали референсные антитела против CXCR5 16D7 (WO 2009/032661). В кратком изложении, клетки hCXCR5-HEK-293 высевали на 96-луночный планшет в среды DMEM с высоким содержанием глюкозы. После инкубации в течение ночи среды удаляли и к клеткам добавляли 100 мкл 1-кратного Fluo-4 NW (Thermo Fisher Scientific, Waltham, MA). Затем клетки инкубировали при 37°C в течение одного часа. К клеткам добавляли серийные разведения моноклональных антител и инкубировали при комнатной температуре в течение одного часа. Перенос кальция измеряли с помощью Flexstation (Molecular Devices, Sunnyvale, CA) при длине волны возбуждения 485 нМ и длине волны излучения 525 нМ посредством добавления 111 нМ рекомбинантного hCXCL13 (BPS Bioscience, San Diego, CA). Данные регистрировали с интервалами 1,52 секунд. С помощью устройства определяли перенос кальция посредством вычитания наименьшего считывания из наибольшего считывания. Значения IC50 ингибирования лиганд-индуцируемого переноса кальция определяли с использованием программного обеспечения GraphPad Prism® (версии 6.0, GraphPad Software, Inc, San Diego, CA) с помощью аппроксимации нелинейной регрессионной кривой и логарифма сигмоиды в модели доза агониста-ответ (таблица 2). Ингибирование переноса кальция является свидетельством функционального антагонизма. Таким образом, антитело 11G2 не только является мощным истощающим средством посредством ADCC, но также является антагонистом, что представляет собой второй механизм действия. Антагонизм антитела 11G2 сравним с компараторными антителами, например, 2C9 (референсным антителом, см., например, WO 2012/010582) и 16D7 (референсным антителом, см., например, WO 2009/032661). Данные репортерного анализа цАМФ также свидетельствуют о функциональном антагонизме (см. ниже).
ТАБЛИЦА 2
Пример 3: Клонирование вариабельных областей тяжелой и легкой цепей антитела против CXCR5
[000385] Гибридомы, продуцирующие антитела, анализируемые в примере 2, лизировали с использованием мини-набора RNeasy (Qiagen, Hilden, Germany), а затем первую цепь кДНК синтезировали с использованием системы синтеза первой цепи Superscript III (Thermo Fisher Scientific, Waltham, MA). Вариабельные области легкой и тяжелой цепи (VL и VH) амплифицировали посредством ПЦР с использованием вырожденных праймеров для легкой и тяжелой цепей мыши, содержащих последовательности нуклеиновой кислоты SEQ ID NO:64-88, представленные в таблице 16. После ПЦР VL и VH клонировали в векторы Zero Blunt TA (Thermo Fisher Scientific, Waltham, MA), а затем их секвенировали. Аминокислотные последовательности VL и VH мыши из антител 11G2, 5H7 и 41A10 представлены в таблице 16 как SEQ ID NO:35-40. CDR подчеркнуты.
Пример 4: Получение афукозилированных и фукозилированных химерных антител
[000386] Для оценки эффекта афукозилирования и фукозилирования в отношении активности антитела получали фукозилированные и афукозилированные версии антитела из примера 3 в клетках млекопитающих.
[000387] VL и VH интересующих антител дополнительно клонировали в векторы, содержащие константные области каппа-цепи человека и константные области IgG1 человека, соответственно. Вариабельные области тяжелой цепи клонировали в экспрессирующий вектор млекопитающих pSMED2, содержащий константную область IgG1 человека (SEQ ID NO:89), получая химерные полноразмерные тяжелые цепи мыши-человека. Вариабельные области легкой цепи клонировали в экспрессирующий вектор млекопитающих pSMEN3, содержащий константные области каппа-цепи человека (Cκ) (SEQ ID NO:90), для получения химерных полноразмерных легких цепей мыши-человека.
[000388] С помощью векторов, содержащих гены химерного антитела, транзиторно трансфицировали клетки HEK-293F (Thermo Fisher Scientific, Waltham, MA) для получения фукозилированного химерного антитела. Затем фукозилированные химерные антитела очищали с использованием колонок с протеином A. Кажущуюся аффинность (EC50 связывания клеток) связывания химерного антитела с клетками Raji определяли посредством проточной цитометрии с использованием анализатора FACSVerse Analyzer (BD Bioscience, Franklin Lakes, NJ), и она представлена в таблице 3.
ТАБЛИЦА 3
[000389] Афукозилированные химерные антитела получали с использованием линии клеток Potelligent® CHOK1SV (BioWa/Lonza, Allendale, NJ), в которых отсутствуют оба аллеля гена, отвечающего за добавление фукозы (α-1,6-фукозилтрансферазы, FUT8). Химерную легкую цепь, содержащую константную область каппа-цепи человека, клонировали в вектор Lonza pEE12.4 GS (Lonza Biologics, Basel, Switzerland) и химерную тяжелую цепь, содержащую константную область IgG1 человека, клонировали в вектор Lonza pEE6.4 GS (Lonza Biologics, Basel, Switzerland). Экспрессирующую кассету тяжелой цепи из pEE6.4 очищали посредством расщепления с помощью NotI и PvuI, а затем клонировали по участкам NotI и PvuI в pEE12.4, содержащий химерную легкую цепь. Конечный вектор DGV, содержащий экспрессирующие кассеты тяжелой и легкой цепи, линеаризовали с помощью PvuI, а затем подвергали электропорации в клетки Potelligent® CHOK1SV. Через 24 часа после трансфекции клетки Potelligent® высевали в 96-луночные планшеты в количестве 3-5K клеток/лунку в среды CDCHO (Thermo Fisher Scientific, Waltham, MA), дополненные 1-кратным HT (Thermo Fisher Scientific, Waltham, MA), 1 мМ уридина (Sigma, St. Louis, MO) и 50 мкМ MSX (EMD Millipore, Billerica, MA). После 3-4 недель инкубации супернатанты клонов анализировали на титр антитела с помощью Octet (Pall Fortebio, Fremont, CA). Высокоэкспрессирующие клоны дополнительно выращивали для продукции антител. Затем афукозилированные химерные антитела очищали на колонках с протеином A.
[000390] Для определения специфичности антител к CXCR5 анализировали связывание антител с клетками, экспрессирующими другие цитокины. В кратком изложении, линии клеток с хемокиновыми рецепторами FlowCellect, экспрессирующие CXCR1, CXCR2 или CXCR3 (EMD Millipore, Billerica, MA), и клетки Jurkat (ATCC, Manassas, VA), экспрессирующие CXCR4, окрашивали с помощью 5 мкг/мл или 10 мкг/мл каждого химерного антитела против CXCR5. Затем клетки инкубировали с антителом козы против человека, конъюгированным с PE (Southern Biotech, Birmingham, AL), перед анализом посредством проточной цитометрии с использованием анализатора FACSVerse Analyzer (BD Biosciences, Franklin Lakes, NJ). Как показано на фиг. 4A-4D, все химерные антитела демонстрировали очень низкое связывание с клетками, экспрессирующими CXCR1 (фиг. 4A), CXCR2 (фиг. 4B), CXCR3 (фиг. 4C) или CXCR4/клетками Jurkat (фиг. 4D) по сравнению с положительными контролями.
Пример 5: Гуманизация моноклональных антител мыши
[000391] Во избежание любой потенциальной иммуногенности с образованием HAMA (антител человека против антител мыши) химерные антитела 41A10 (C-41A10) и 11G2 (C-11G2) гуманизировали с использованием каркасных последовательностей зародышевой линии человека из IGKV1-39 (вариабельного домена легкой цепи DPK9, регистрационный номер GenBank X93627.1, SEQ ID NO:93) и каркасной последовательности зародышевой линии человека из IGHV3 (вариабельного домена тяжелой цепи DP54, регистрационный номер GenBank AB019440, SEQ ID NO:91).
[000392] Гуманизированные версии химерных антител 11G2 (C-11G2) и химерных антител 41A10 (C-41A10) получали посредством прививки определяющих комплементарность областей (CDR) (обозначая их как "антитела с привитыми CDR"). Т.е. CDR тяжелой цепи прививали на каркасную область DP-54 человека (подгруппа VH3; SEQ ID NO:91) с сегментом JH4 (SEQ ID NO:92), в то время как CDR легкой цепи прививали на каркас DPK9 человека (подгруппа VKI; SEQ ID NO:93) с сегментом JK4 (SEQ ID NO:94).
[000393] Гуманизированные области VH соединяли с константной областью IgG1 человека (SEQ ID NO:89), а затем субклонировали с собственным экспрессирующим вектором для получения тяжелых цепей с привитыми CDR, включая, в качестве неограничивающих примеров, SEQ ID NO:96 (VH 11G2 с привитыми CDR). Гуманизированные области VL подвергали слиянию с константной области каппа-цепи человека (SEQ ID NO:90), а затем субклонировали в собственный экспрессирующий вектор для получения легких цепей с привитыми CDR, включая, в качестве неограничивающих примеров, SEQ ID NO:97 (VL 11G2 с привитыми CDR).
Пример 6: Аффинность связывания афукозилированных или фукозилированных антител против CXCR5
[000394] Анализировали кажущуюся аффинность химерных и гуманизированных фукозилированных и афукозилированных антител против CXCR5 к CXCR5 на поверхности клеток. Более конкретно, для определения кажущейся аффинности химерных и гуманизированных антител против CXCR5 к CXCR5-экспрессирующим клетками осуществляли эксперименты по связыванию с клетками с использованием мононуклеарных клеток периферической крови (PBMC) человека и яванского макака и тонзиллярных мононуклеарных клеток (TMC) человека. Кажущуюся аффинность связывания mAb против CXCR5 с CXCR5+ клетками (B-клетками, истинными Tfh-клетками и циркулирующими Tfh-подобными клетками) вычисляли как EC50 кривых титрования равновесного связывания, где геометрическое среднее интенсивности флуоресценции (gMFI) антигенсвязывающей популяции количественно анализировали посредством проточной цитометрии.
[000395] Клетки здоровых людей-доноров, полученные с помощью системы для афереза Trima® и обогащенные PBMC, получали из Blood Centers of the Pacific (San Francisco, CA). PBMC выделяли посредством центрифугирования в градиенте плотности с использованием пробирок SepMateTM и среды LymphoprepTM по инструкциям производителя (STEMCELL Technologies, Vancouver, BC, Canada).
[000396] Тонзиллярные мононуклеарные клетки (TMC) выделяли из миндалин человека, полученных из BioOptions (Brea, CA). В кратком изложении, миндалины рассекали на небольшие фрагменты (3-4 мм) в холодной среде RPMI 1640 с использованием стерильного скальпеля. Затем ткань миндалин расщепляли в течение 30 минут при 37°C в среде для расщепления (3 мл 10-кратной коллагеназы, 300 мкл 100-кратной дезоксирибонуклеазы (ДНКазы), 6,7 мл RPMI 1640). Для нейтрализации ферментативной активности добавляли RPMI 1640, дополненную 10% эмбриональной телячьей сывороткой (FBS), а затем ткань фильтровали последовательно через клеточные сита с нейлоновыми фильтрами (нейлон с размером ячеек 70 мкм и 40 мкм). После центрифугирования осуществляли лизис эритроцитов в осадке с использованием лизирующего буфера с хлоридом аммония. Тромбоциты удаляли с использованием низкоскоростного центрифугирования (200×g) в течение 10 минут в фосфатно-солевом буфере (PBS)/2% FBS/2 мМ этилендиаминтетрауксусной кислоте (ЭДТА). CD4+ T-клетки очищали от смеси TMC с использованием набора для обогащения CD4+ T-клеток человека EasySepTM по инструкциям производителя (STEMCELL Technologies, Vancouver, BC, Canada).
[000397] После выделения PBMC высевали при плотности 1,0×105 клеток на лунку и CD4+ TMC высевали при плотности 2,0×105 клеток на лунку в буфере для выделения (PBS, содержащем 1% FBS и 1 мМ ЭДТА) в 96-луночные планшеты с U-образным дном. Затем планшеты центрифугировали при 1500 об./мин. в течение 5 минут при комнатной температуре. PBMC и CD4+ TMC ресуспендировали в буфере для активируемой флуоресценцией сортировки клеток (FACS) (PBS, содержащем 1% FBS), содержащем блокатор кристаллизуемого фрагмента Fc человека (2 мкл на лунку; Biolegend) и 4-кратные серийные разведения антител (серия разведения с 11 точками, начинающаяся с 5000 нг/мл в случае PBMC и 312,5 нг/мл в случае CD4+ TMC) и инкубировали на льду в течение 2 часов. Затем PBMC и CD4+ TMC 3 раза промывали буфером для FACS и ресуспендировали в 50-100 мкл буфера для FACS, содержащего конъюгированные с флуоресцентной меткой антитела для окрашивания субпопуляций лимфоцитов и конъюгированное с флуоресцентной меткой вторичное антитело против Ig человека для детекции моноклональных антител (mAb) против CXCR5. После инкубации в течение 30 минут при 4°C PBMC и CD4+ TMC 2 раза промывали буфером для FACS и ресуспендировали в 125 мкл 0,5% параформальдегида (PFA) в PBS. Планшеты хранили при 4°C до анализа посредством проточной цитометрии (анализатор клеток BD LSRFortessa™, BD Biosciences, Franklin Lakes, NJ).
[000398] Кривые титрования связывания клеток получали посредством построения графика gMFI антигенсвязывающей популяции относительно логарифма концентрации антитела против CXCR5. Значения EC50 определяли с использованием программного обеспечения GraphPad Prism® (версии 6.0, GraphPad Software, Inc, San Diego, CA) с помощью аппроксимации нелинейной регрессионной кривой и логарифма сигмоиды в модели доза агониста-ответ в соответствии со следующим уравнением:
Log (агонист) vs. ответ - переменный угловой коэффициент (четыре параметра)
Y=Минимум+(Максимум-Минимум/(1+10^((LogEC50-X)*угловой коэффициент Хилла))
[000399] Где Y является gMFI, X является концентрацией антитела, Максимум является максимальным значением Y, соответствующим верхнему плато сигмоидной кривой, Минимум является минимальным значением Y, соответствующим нижнему плато сигмоидной кривой, и LogEC50 является логарифмом концентрации антитела в точке перегиба кривой.
[000400] Значения EC50 суммированы по экспериментам с использованием среднего и стандартного отклонения (STDEV), при этом осуществляли множество повторений. Результаты (т.е. средняя кажущаяся аффинность химерных и гуманизированных mAb против CXCR5, являющихся фукозилированными или афукозилированными, к CXCR5-экспрессирующими клетками) представлены в таблице 4. Результаты также включают данные об аффинности связывания, полученные с использованием контрольных антител против CXCR5 2C9 (WO 2012/010582), 16D7 (WO 2009/032661) и 11A7 (WO 2016/028573) которые включали в целях сравнения. Необходимо отметить, что фукозилированные и афукозилированные химерные и гуманизированные антитела 11G2, содержащие различные комбинации VL и VH (т.е. h11G2 XC51/XC152, h11G2 XC153/XC155, h11Gh11G2 XC153/XC156, h11G2 XC154/XC155 и h11G2 XC154/XC157), имеют кажущуюся аффинность к B-клеткам человека, являющуюся приблизительно равной. Т.е. данные свидетельствуют о том, что афукозилирование, по-видимому, не влияет на аффинность антитела к CXCR5-экспрессирующим клеткам. Кроме того, данные свидетельствуют о том, что антитела h11G2 XC154/XC155 (фукозилированные и афукозилированные) демонстрировали приблизительно в 10 раз более высокую аффинность, чем контрольное антитело против CXCR5 2C9, и приблизительно в 100 раз более высокую аффинность, чем 11A7. Контрольное антитело против CXC5 16D7 не демонстрировало насыщаемое связывание. Эти данные свидетельствуют о том, что афукозилирование не влияло на аффинность антител 11G2 в случае связывания с CXCR5-экспрессирующими клетками. Т.к. аффинность является мерой силы взаимодействия между эпитопом и антигенсвязывающим участком антитела (т.е. паратопом), эти данные свидетельствуют о том, что даже если антитела связываются со схожим эпитопом, они не связываются с эпитопом с той же силой, что и 11G2.
ТАБЛИЦА 4
Средняя EC50 ± STDEV (пМ)
Пример 7: Зависящее от концентрации связывание афукозилированных или фукозилированных гуманизированных антител 11G2 по сравнению с фукозилированными референсными антителами
[000401] Зависящее от концентрации связывание фукозилированного гуманизированного 11G2 (VL h11G2 XC154/VH XC155, также обозначаемого как h11G2 154/155 или h11G2 XC154/XC155) и афукозилированного гуманизированного 11G2 CXCR5 (афукозилированного h11G2 154/155) с B-клетками из PBMC человека сравнивали со связыванием компараторных mAb 2C9, 16D7 и 11A7. Кривые титрования связывания клеток получали посредством построения графика gMFI антигенсвязывающей популяции относительно логарифма концентрации антитела против CXCR5, и они показаны на фиг. 5.
[000402] Данные свидетельствуют о том, что фукозилированные и афукозилированные h11G2 154/155 имеют идентичную кривую, что свидетельствует о том, что наличие или отсутствие фукозы на N-связанной гликоформе в положении Asn297 на константных областях по меньшей мере одной, но предпочтительно обеих цепей антитела не влияет на аффинность антитела к CXCR5, находящемуся на клетках.
[000403] Кроме того, данные дополнительно свидетельствуют о том, что EC50 фукозилированных (закрашенные круги, 9,630×10-12 M) и афукозилированных (незакрашенные круги, 1,188×10-11 M) антител h11G2 154/155 была гораздо ниже, чем EC50 компараторных антител: 2C9 (закрашенные треугольники, 6,24×10-11), 11A7 (закрашенные ромбы, 9,265×10-10) и 16D7 (закрашенные квадраты, приблизительно 0,004945, и связывание не было насыщаемым). Эти данные свидетельствуют о том, что антитела h11G2 154/155 имеют иные характеристики связывания, чем компараторные антитела. Таким образом, эти данные свидетельствуют о том, что антитела 11G2 и 2C9, 11A7 и 16D7 не связываются с одним и тем же эпитопом на CXCR5.
Пример 8: Активность ADCC химерных и гуманизированных антител против CXCR5
[000404] Для определения мощности и эффективности, с которыми когорта химерных и гуманизированных антител против CXCR5 стимулирует антителозависимую клеточную цитотоксичность (ADCC), серийные разведения антител против CXCR5 или изотипического контроля инкубировали с мононуклеарными клетками периферической крови (PBMC) здоровых людей-доноров или яванских макаков. В этом анализе PBMC являются источником эффекторных клеток естественных киллеров (NK) и целевых B-клеток и Tfh-подобных клеток. Проточную цитометрию использовали для анализа количества B-клеток и Tfh-подобных клеток, оставшихся через приблизительно 20 ч. Аналогично, способность гуманизированных антител индуцировать ADCC истинных Tfh-клеток из миндалин человека оценивали с использованием CD4+ T-клеток, выделенных из тонзиллярных мононуклеарных клеток, с добавлением NK-клеток, выделенных из PBMC.
[000405] Как описано в примере 6, клетки здоровых людей-доноров, полученные с помощью системы для афереза Trima® и обогащенные на PBMC, получали из Blood Centers of the Pacific (San Francisco, CA). PBMC выделяли посредством центрифугирования в градиенте плотности с использованием пробирок SepMateTM и среды LymphoprepTM по инструкциям производителя (STEMCELL Technologies, Vancouver, BC, Canada). После выделения PBMC высевали в полную среду RPMI при плотности 2,0×105 клеток на лунку в 96-луночные планшеты с U-образным дном.
[000406] В этом анализе PBMC являются источником эффекторных клеток естественных киллеров (NK) и целевых B-клеток и Tfh-подобных клеток. В лунки добавляли серийные разведения mAb против CXCR5 или изотипические контроли и инкубировали клетки при 37°C, 5% CO2 в течение приблизительно 20 часов. Затем планшеты центрифугировали при 1800 об./мин. в течение 5 минут при комнатной температуре (RT). Затем PBMC промывали ледяным буфером для FACS и ресуспендировали в 50 мкл ледяного буфера для FACS, содержащего конъюгированные с флуоресцентной меткой антитела для окрашивания субпопуляций лимфоцитов. После инкубации в течение 15-30 минут при 4°C PBMC 2 раза промывали ледяным буфером для FACS и ресуспендировали в 100 мкл 0,5% параформальдегида (PFA) в PBS. В каждую лунку добавляли бусины CountBrightTM Absolute Counting Beads (Thermo Fisher Scientific) (15 мкл на лунку). Планшеты хранили при 4°C до анализа посредством проточной цитометрии (анализатор клеток BD LSRFortessaTM).
[000407] Тонзиллярные мононуклеарные клетки (TMC) также выделяли, как описано выше в примере 6.
[000408] Кривые титрования цитотоксичности получали способами, описанными в примере 6. Значения EC50 суммированы по экспериментам с использованием среднего и стандартного отклонения (STDEV), при этом осуществляли множество повторений. Результаты представлены в таблице 5. Данные, полученные с использованием референсных антител против CXCR5 2C9, 16D7 и 11A7, включены в целях сравнения.
ТАБЛИЦА 5
Средняя цитотоксичность химерных и гуманизированных mAb против CXCR5 в отношении CXCR5-экспрессирующих клеток
Средняя EC50±STDEV (пМ)
[000409] Данные, представленные в настоящем описании, свидетельствуют о том, что даже при фукозилировании антитело h11G2 154/155 имеет более высокую активность ADCC (253,72±672,14), чем компараторные антитела 2C9 (380,31±757,04) и 16D7 (2590,61±6343,88). Фукозилированное антитело 11A7 демонстрировало более высокую активность ADCC, чем любое тестируемое фукозилированное антитело. Однако данные свидетельствуют о том, что афукозилированное h11G2 154/155 имело более высокую активность ADCC, чем любое тестируемое антитело, включая фукозилированное 11A7. Исследования характеристик, представленные в настоящем описании, показали, что химерные mAb против CXCR5 41A10 и 11G2 запускали ADCC B-клеток человека с EC50 11,73 нМ и 0,56 нМ, соответственно. Афукозилированные версии химерных mAb против CXCR5 41A10 и 11G2 повышали мощность активности ADCC по меньшей мере приблизительно в 100 раз. Гуманизированные варианты запускали ADCC B-клеток человека с мощностями, сравнимыми или превосходящими активность соответствующих химерных антител. Как и в случае химерных mAb против CXCR5, афукозилированный гуманизированный вариант повышал мощность ADCC по сравнению с нормальным соответствующим фукозилированным антителом по меньшей мере приблизительно в 100 раз. Афукозилированное h11G2 являлось более мощным в отношении ADCC, чем фукозилированные версии 2C9 и 16D7, и афукозилированное h11G2 являлось по меньшей мере сравнимым с фукозилированным 11A7.
Пример 9: Антитела 11G2 антагонистически действуют на передачу сигнала CXCL13
[000410] Для определения того, могут ли антитела против CXCR5 функционально антагонистически действовать на передачу сигнала CXCL13, использовали репортерный анализ циклического аденозинмонофосфата (цАМФ) в сконструированной линии клеток, стабильно экспрессирующей CXCR5 человека. В этих клетках CXCL13 ингибирует продукцию цАМФ, запускаемую форсколином, в зависимости от концентрации.
[000411] Для определения того, могут ли антитела против CXCR5 функционально антагонистически действовать на передачу сигнала CXCL13, осуществляли анализ цАМФ Hit Hunter® (DiscoveRx Corporation, Fremont, CA) в клетках CHO-K1, стабильно экспрессирующих CXCR5 человека. В этом анализе активная β-галактозидаза (β-gal) образуется при комплементации фрагментов фермента, когда образуется цАМФ. Затем β-gal может превращать хемилюминесцентный субстрат, генерируя выходной сигнал, детектируемый с помощью стандартного спектрофотометра для чтения микропланшетов. Лиганд CXCR5, CXCL13, ингибирует продукцию цАМФ, запускаемую форсколином (20 мкМ) в зависимости от концентрации. Для тестирования мощности и эффективности антител против CXCR5 при ингибировании CXCL13-опосредованной форсколин-индуцированной продукции цАМФ серийные разведения каждого антитела против CXCR5 добавляли к CXCR5-экспрессирующим клеткам CHO-K1 в присутствии форсколина (20 мкМ) и CXCL13 (используемого при его IC80 600 пМ).
[000412] Кривые титрования функционального антагонизма получали в соответствии с описанным в примере 6, включая определение значения EC50. Результаты представлены в таблице 6.
ТАБЛИЦА 6
Стимуляция CXCL13-ингибируемой продукции цАМФ in vitro в CXCR5+ клетках CHO-K1
[000413] В отдельном исследовании афукозилированное гуманизированное 11G2 154/155 тестировали на функциональный антагонизм, и оно действовало аналогично гуманизированному фукозилированному 11G2 154/155 (т.е. EC50=961,4 пМ).
[000414] Данные свидетельствуют о том, что h11G2 154/155 ингибировало передачу сигнала CXCL13 по меньшей мере так же, как референсные антитела 2C9, 16D7, 11A7. Фактически, h11G2 ингибировало передачу сигнала CXCL13 (120,6%) в значительно большей степени, чем 16D7 (102,4%) и 11A7 (105,9%). Эти данные позволяют предполагать, что антитела 11G2 могут являться полезным новым терапевтическим средством для лечения заболевания, нарушения или состояния, опосредованного или ассоциированного с CXCR5-опосредованной передачей сигнала CXCL13.
Пример 10: Картирование участков связывания антитела 11G2 на hCXCR5
[000415] Участок связывания афукозилированного гуманизированного 11G2 154/155 на CXCR5 идентифицировали посредством прививки аминокислот из внеклеточного N-конца CXCR5 мыши на CXCR5 человека.
[000416] Более конкретно, в стратегии картирования использовали преимущество, состоящее в том, что 11G2 связывается с CXCR5 человека (SEQ ID NO:32), но не связывается с CXCR5 мыши (SEQ ID NO:34). Выравнивание аминокислотных последовательностей CXCR5 человека (hCXCR5) и CXCR5 мыши (mCXCR5) показано на фиг. 6. Аминокислотные остатки внеклеточных доменов CXCR5 подчеркнуты. Внеклеточные домены (ECD) белков CXCR5 мыши и человека указаны полужирным шрифтом и помечены "N", "L1", "L2" и "L3" под последовательностью каждой области на фиг. 6. Эти ECD переставляли между белками мыши и человека для идентификации областей, отвечающих за специфичность 11G2 в отношении CXCR5 человека и за отсутствия связывания с mCXCR5. Каждую из этих областей из CXCR5 мыши заменяли той же областью в CXCR5 человека для получения химерных белков, обозначенных как XC251, XC252, XC253, XC254, XC255 и XC256, где белок CXCR5 человека содержит соответствующий определенный домен CXCR5 мыши. В таблице 7 показано, какие области белка мыши (m) переставлены в CXCR5 человека.
ТАБЛИЦА 7
(химерные белки CXCR5 мыши/человека с переставленными ECD)
[000417] Более конкретно, домен L3 мыши меняли местами с соответствующим доменом L3 человека для получения химерного белка XC251, содержащего домен L3 мыши в контексте белка CXCR5 человека (hCXCR5-mL3). Аналогично, L2 мыши меняли местами с соответствующим доменом человека для получения химерного белка XC252 (hCXCR5-mL2); N-домен мыши меняли местами с соответствующим N-доменом человека для получения химерного белка XC253 (hCXCR5-mN); домены L2 и L3 мыши меняли местами с соответствующими доменами человека для получения химерного белка XC254 (hCXCR5-mL2-mL3); домены mN и L3 мыши меняли местами с соответствующими доменами человека для получения химерного белка XC255 (hCXCR5-mN-mL3); и домены mN и L2 мыши меняли местами с соответствующими доменами человека для получения химерного белка XC255 (hCXCR5-mN-mL2).
[000418] Связывание 11G2 с CXCR5 мыши, CXCR5 человека и различными химерными белками CXCR5 мыши-человека с переставленными ECD (XC251-XC256) анализировали и сравнивали со связыванием различных антител, т.е. антителом крысы против CXCR5 мыши (антителом крысы) (кат. № MAB6198, R&D systems, Minneapolis, MN), антителом кролика против CXCR5 человека (Rb) (кат. № ab46218, Abcam, Cambridge, MA), 2C9, 16D7. Результаты показаны в таблице 8. Числами, выделенные полужирным шрифтом, указаны антитела, связывающиеся с белком.
ТАБЛИЦА 8
(Анализ FACS связывания антитела с химерными белками CXCR5 мыши-человека с переставленными ECD)
(GMFI)
(GMFI)
(GMFI)
(GMFI)
(GMFI)
[000419] Данные свидетельствуют о том, что, если антитело являлось специфическим к CXCR5 мыши, оно связывается с химерным CXCR5 мыши-человека с переставленными ECD при условии, что химерный белок содержит N-домен мыши. Аналогично, в случае антител, связывающихся с CXCR5 человека (11G2, 2C9, 16D7 и Rb), антитела связываются с химерным CXCR5, если N-домен человека присутствовал в химерном белке. Таким образом, видоспецифичность антител, по-видимому, регулируется участком связывания, находящимся в N-домене. Что более важно, эти данные свидетельствуют о том, что N-домен CXCR5 человека был необходим для связывания антитела 11G2, 2C9 и 16D7.
Пример 11: Идентификация аминокислотных остатков, уникальных для антитела 11G2, связывающегося с hCXCR5
[000420] Для более точного определения специфических аминокислотных остатков, ключевых для связывания антитела с hCXCR5 и для дальнейшего дифференцирования антител против CXCR5 человека, осуществляли более подробные исследования N-домена CXCR5. Более конкретно, осуществляли точечные мутации во фрагменте N-домена CXCR5 человека (аминокислоты 1-58 в соответствии с нумерацией SEQ ID NO:32) для получения белков с точечными мутациями, каждый из которых содержит соответствующий аминокислотный остаток мыши из CXCR5. Получали стабильные трансфектанты, экспрессирующие каждый мутеин, и клетки анализировали на связывание антитела с использованием проточной цитометрии. Т.е. получали девятнадцать (19) белков CXCR5 человека с мутантным N-доменом, обозначаемых как XC276-XC294, каждый из которых содержит замену отдельной аминокислоты, где остаток человека из CXCR5 заменяли соответствующим остатком мыши, как показано в таблице 9. Мутеины CXCR5 человека дополнительно содержали эпитоп FLAG (DYKDDDDK), показанный в таблице 9 с использованием строчных букв, на N-конце, чтобы сделать возможной нормализацию с учетом уровня экспрессии; метка FLAG не влияет на связывание антитела с мутеинами. Как правило, замены не осуществляли там, где замена аминокислотного остатка человека аминокислотным остатком мыши будет консервативной заменой, и эти остатки указаны курсивом.
[000421] В таблице 9 показана аминокислотная последовательность N-области hCXCR5 (показаны первые 58 аминокислот) по сравнению с аминокислотной последовательностью N-области CXCR5 мыши. Аминокислоты, отличающиеся между двумя последовательностями, подчеркнуты. На фигуре также показаны различные полученные конструкции (XC276-XC294), где аминокислотный остаток мыши заменяли соответствующей аминокислотой человека в hCXC5. Например, XC276 представляет собой CXCR5 человека с заменой отдельной аминокислоты D (человека) на G (мыши). Остальная часть аминокислотной последовательности hCXCR5 (SEQ ID NO:32) не показана, т.к. ее не изменяли. Мутеины получали для определения того, какие аминокислотные остатки являются критическими для связывания антитела с hCXCR5, т.к. тестируемые антитела не связывались с CXCR5 мыши.
[000422] С помощью нуклеиновых кислот, кодирующих каждый мутантный пептид, транзиторно трансфицировали клетки HEK-293T. Через 2 дня после трансфекции, клетки собирали и окрашивали с помощью 1 мкг/мл PE-конъюгированного антитела против FLAG (Biolegend) или 1 мкг/мл или 3 мкг/мл афукозилированного гуманизированного 11G2 154/155; 2C9; S16D7 или 11A7, а затем PE-конъюгированного антитела против IgG человека (Southern Biotech, Birmingham, AL). Клетки анализировали посредством проточной цитометрии с использованием Accuri (BD Biosciences Franklin Lakes, NJ). Определяли геометрическое среднее интенсивности флуоресценции (gMFI) и вычисляли соотношение gMFI каждого из антител против CXCR5 и антитела против FLAG.
[000423] Как показано на фиг. 7, все четыре антитела связывались с CXCR5 человека дикого типа (XC275; WT). Затем антитела демонстрировали разные профили связывания с пептидами с различными точечными мутациями. Т.е. 2C9 не связывалось с XC276 (содержащим мутацию D10G0) или XC277 (содержащим инсерцию аминокислотных остатков S и I в N-домене CXCR5 человека), в то время как 11G2, 16D7 и 11A7 связывались с этими белками. Эти данные позволяют предполагать, что инсерция этих двух аминокислот в последовательность hCXCR5 устраняла связывание 2C9, но не влияла на связывание трех других антител, что позволяет предполагать, что эпитоп для 2C9 отличается от эпитопа для 11G2, 16D7 и 11A7. Что более важно, мутация лейцина (L) в треонин (T) в положении аминокислотного остатка 11 (с учетом нумерации последовательности SEQ ID NO:32) полностью устраняла связывание только 11G2 с XC278, но не влияла на связывание антител 2C9, 16D7 или 11A7 с этим белком. Таким образом, остаток лейцина, находящийся в положении 11 (относительно аминокислотной последовательности, приведенной в SEQ ID NO:32), по-видимому, является критическим для связывания 11G2 с CXCR5 человека, но неважен для связывания 2C9, 16D7 или 11A7 с hCXCR5. Эти данные свидетельствуют о том, что эпитоп 11G2 не является тем же, что и эпитоп для тех антител. Кроме того, на фиг. 7 показано, что все четыре антитела связывались с XC279 таким образом, что замена E на Y в положении аминокислотного остатка 12 (относительно последовательности SEQ ID NO:32) не являлась критической для связывания любого из этих антител с hCXCR5.
[000424] Связывание этих четырех антител с XC280, содержащим замену W лизином (K) в положении аминокислотного остатка 19 (в соответствии с нумерацией последовательности SEQ ID NO:32), приводило к утрате связывания с XC280 антител 16D7 и 11A7, но не влияло на связывание антител 11G2 и 2C9 с мутеином (фиг. 7). Эти результаты дополнительно свидетельствуют о том, что эти четыре антитела не связываются с одним и тем же эпитопом на hCXCR5.
[000425] На фиг. 7 также показано, что замена D в положении 22 (в соответствии с нумерацией последовательности SEQ ID NO:32) аланином (A) полностью устраняет связывание 11G2 с мутеином XC281, но не влияет на связывание антител 2C9, 16D7, и 11A7. Это еще раз подчеркивает, что эпитоп для 11G2 не является тем же, что эпитопы для антител 2C9, 16D7 и 11A7.
[000426] Все четыре антитела связывались с XC285-294 в равной степени, и на их связывание не влияли какие-либо замены аминокислот в этих мутеинах. Эти данные позволяют предполагать, что аминокислотные остатки в этих положениях (27, 28, 30, 31, 33, 35, 36, 37, 39 и 40; см. таблицу 9) не участвуют в связывании антитела с CXCR5 в случае антител 11G2, 2C9, 11A7 и 16D7.
[000427] Таким образом, эти данные свидетельствуют о том, что остаток лейцина (L) в положении аминокислотного остатка 11 и остаток аспартата (D) в положении аминокислотного остатка 22 (относительно нумерации аминокислотной последовательности SEQ ID NO:32) являются критическими для связывания антитела 11G2 с hCXCR5, т.к. замена любого из этих остатков (или их обоих) соответствующим аминокислотным остатком мыши в этих положениях устраняет связывание антитела с hCXCR5. Эти данные также свидетельствуют о том, что антитело 11G2 не имеет того же эпитопа, что и антитело 2C9, 16D7 или 11A7, т.к. замена L11 и/или W22 не влияет на связывание этих антител с hCXCR5.
[000428] На фиг. 7 также показано, что все четыре антитела связывались с мутантными белками CXCR5 XC282-XC284. Все четыре антитела также связывались с белками XC285-XC294. Эти данные позволяют предполагать, что эпитоп для антител 11G2, 2C9, 16D7 и 11A7 не находится в области N-домена hCXCR5, показанного как мутеины XC282-XC294.
Пример 12: 11G2 является селективным для CXCR5 и не связывается с другими членами семейства хемокиновых GPCR
[000429] Селективность афукозилированного гуманизированного 11G2 154/155 анализировали относительно двадцати (20) членов семейства хемокиновых GPCR с использованием анализа связывания β-аррестина (DiscoveRx). В этом анализе используют цельные клетки, стабильно трансфицированные с использованием каждого рецептора, и напрямую измеряют активность GPCR посредством детекции взаимодействия β-аррестина с активированным GPCR. Т.к. рекрутирование аррестина не зависит от передачи сигнала через G-белок, эти анализы представляют собой универсальную платформу для скрининга и профилирования, которую можно использовать, по существу, для любого Gi-, Gs- или Gq-сопряженного рецептора, обеспечивая стандартизованное и эффективное сравнение среди рецепторов или семейств рецепторов (независимо от сопряжения с G-белком). Анализ осуществляли в агонистическом (без лиганда) и антагонистическом (в присутствии лиганда при EC90) режиме. В панель для изучения селективности включали следующие хемокиновые рецепторы: CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR5, CXCR6, CXCR7, XCR1.
[000430] Не наблюдали значимой активности афукозилированного гуманизированного 11G2 154/155 (тестируемого при 200 нМ) в отношении любого другого хемокинового рецептора при тестировании в агонистическом режиме, и оно было значимо активным только в качестве антагониста CXCR5 (92% ингибирования лиганд-индуцированного связывания β-аррестина) при сравнении в антагонистическом режиме со всеми другими оцениваемыми хемокиновыми рецепторами, т.е. не наблюдали значимого ингибирования в случае любого другого тестируемого хемокинового рецептора.
Пример 13: h11G2 не демонстрирует полиреактивность против ДНК, инсулина или LPS
[000431] Анализ полиреактивности используют для определения того, могут ли антитела потенциально реагировать с различными неродственными антигенами in vivo. Отсутствие полиреактивности позволяет предполагать, что 11G2 является CXCR5-специфическим антителом и не связывается с нецелевыми мишенями in vivo.
[000432] Черные планшеты DELFIA (Thermoscientific) покрывали 10 мкг/мл двухцепочечной ДНК (dsDNA; Millipore), 5 мкг/мл липополисахарида (LPS; Sigma), 10 мкг/мл инсулина (Sigma, St. Louis, MO) или фосфатно-солевым буфером (PBS) и инкубировали при 4°C в течение ночи. Затем планшеты промывали водой с использованием устройства для промывки микропланшетов Biotek ELx405 (Biotek, Winooski, VT), а затем планшет блокировали 200 мкл буфера для анализа, содержащего 1-кратный PBS, 0,05% Tween 20 и 1 мМ ЭДТА при комнатной температуре в течение одного часа. После дополнительной стадии промывки планшеты инкубировали при комнатной температуре в течение одного часа с серийными разведениями антител против CXCR5 (все антитела являлись фукозилированными). Для детекции 100 мкл DELFIA-Eu-N1-антител против IgG человека (50 мкг/мл; Perkin Elmer, Waltham, MA) добавляли на планшеты и инкубировали при комнатной температуре в течение одного часа. И наконец, в каждую лунку добавляли 100 мкл усиливающего раствора DELFIA и планшеты встряхивали при комнатной температуре в течение 15 минут перед считыванием с помощью многоканального спектрофотометра для чтения планшетов VictorX4 (PerkinElmer, Waltham, MA) в условиях флуоресценции европия. Данные представлены в таблице 10.
ТАБЛИЦА 10
Флуоресценция (отсчеты) в случае полиреактивности mAb против CXCR5
[000433] В отдельном исследовании афукозилированное гуманизированное 11G2 154/155 тестировали на полиреактивность и не наблюдали связывание с инсулином, ДНК, LPS или PBS, что соответствует результатам для фукозилированного гуманизированного 11G2 154/155, представленным в таблице 10 выше.
[000434] Эти данные свидетельствуют о том, что 11G2 не реагировало с инсулином, ДНК или LPS и является менее реакционноспособным, чем компараторные антитела 2C9, 16D7 и 11A7. Эти данные позволяют предполагать, что 11G2 является потенциальным полезным новым терапевтическим средством. Полиреактивные антитела, по определению, связываются неспецифическим образом с диапазоном биологических молекул. Тот факт, что 11G2 не реагирует в общем анализе полиреактивности, дополнительно подтверждает то, что 11G2 является специфическим для CXCR5. Данные анализа PK, TK и токсичности также подтверждают, что 11G2 имеет специфичность к CXCR5.
Пример 14: Обобщение различий между 11G2 и компараторными антителами
[000435] В таблице 11 ниже приведены характеристики 11G2 и различных компараторных антител, представленных выше в других местах настоящего описания. Средние ND не определяли.
[000436] Эти данные свидетельствуют о том, что антитело 11G2, и фукозилированное, и афукозилированное, сильно отличается от компараторных антител 2C9, 16D7 и 11A7. Т.е. связывание 11G2 устраняют посредством замены аминокислотного остатка мыши в положениях 11 и 22, в то время как на связывание компараторных антител замена этих остатков (пронумерованных в соответствии с аминокислотной последовательностью SEQ ID NO:32) не влияет. Кроме того, для связывания 2C9 с hCXCR5 необходимо наличие аминокислотных остатков W19, D22 и R23, но ни для одного из антител не требуются все три из них, даже при том, что связывание 11A7 с hCXCR5 устраняется заменой аланина на W в положении 19 (в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32). В отличие от всех остальных антител, связывание 16D7 с hCXCR5 устраняется инсерцией двух остатков мыши между D10 и L11 CXCR5 человека (в соответствии с нумерацией аминокислотной последовательности SEQ ID NO:32) (см. фиг. 7). Таким образом, данные, представленные в настоящем описании, позволяют представлять, что 11G2 не имеет того же эпитопа, что и антитела 2C9, 16D7 и 11A7, и что эти антитела также имеют разные эпитопы относительно друг друга.
[000437] Кроме того, данные, представленные в настоящем описании, свидетельствуют о том, что 11G2 имеет аффинность к hCXCR5 (выраженную как EC50), составляющую по меньшей мере в 10 раз выше (7,08 пМ), чем у 2C9 (65,59 пМ), и по меньшей мере в 100 раз выше, чем у 11A7 (563,63 пМ), и очень отличающуюся от 16D7, которые нельзя сравнить, т.к. в этом случае связывание не являлось насыщаемым.
ТАБЛИЦА 11
(EC50, пМ)
[000438] Эти данные позволяют предполагать, что 11G2 значительно отличается от компараторных антител 2C9, 16D7 и 11A7 и является потенциальным новым полезным терапевтическим средством для лечения или профилактики заболевания, нарушения или состояния, опосредованного или ассоциированного с биологической активностью CXCR5, включая, в качестве неограничивающих примеров, CXCR5-опосредованную передачу сигнала CXCL13. Это особенно актуально в случае афукозилированной версии 11G2, демонстрирующей значительно повышенную активность ADCC, таким образом, повышая его потенциальную полезность в качестве терапевтического средства для лечения, например, иммунологических заболеваний, опосредованных или ассоциированных с экспрессией CXCR5 в клетках.
Пример 15: Фармакодинамика афукозилированного h11G2 XC154/XC155 in vivo
[000439] In vivo дозы в диапазоне от 0,001 до 400 мг/кг/дозу IV или SC изучали в многочисленных исследованиях афукозилированного гуманизированного 11G2 154/155 на яванских макаках. В рамках этого эксперимента составы IV и SC содержали следующие компоненты: 50 мг/мл антитела против CXCR5 или антигенсвязывающего фрагмента по изобретению, 20 мМ гистидина, 8,5% сахарозы и 0,02% полисорбата 80, 0,005% ЭДТА при pH 5,8. В таблице 12 ниже приведен дизайн исследования in vivo афукозилированного 11G2 XC154/XC155 на яванских макаках.
ТАБЛИЦА 12
[000440] У яванских макаков наблюдали дозозависимое истощение B-клеток и Tfh-подобных клеток в периферической крови при введении однократных доз афукозилированного VL h11G2 XC154/VH XC155 (афукозилированного h11G2 XC154/XC155) в диапазоне от 0,001 до 0,2 мг/кг; наименьшая тестируемая доза (0,001 мг/кг) приводила к приблизительно 50% истощению B-клеток и Tfh-подобных (т.е. cTfh) клеток в периферической крови.
[000441] В поисковых и базовых исследованиях токсичности, соответствующих требованиям GLP, на яванских макаках дозы афукозилированного h11G2 XC154/XC155 ≥5 мг/кг IV приводили к значительному истощению B-клеток и Tfh-подобных клеток в периферической крови и снижали клеточность лимфоидных фолликулов в селезенке и других лимфоидных тканях (подмышечных и брыжеечных лимфоузлах и лимфоидной ткани кишечника); эти эффекты коррелировали с фармакологически-опосредованной сниженной клеточностью B-клеточных областей, наблюдаемой иммуногистохимически. В поисковых и базовых исследованиях наблюдали частичное или полное восстановление B-клеток, Tfh-подобных клеток и Tfh-клеток. Кинетика истощения и последующего восстановления связана с уровнями воздействия афукозилированного h11G2 XC154/XC155.
[000442] Для определения того, нарушает ли афукозилированное h11G2 XC154/XC155 гуморальные анамнестические ответы, проводили исследование вторичного ответа на вакцину на яванских макаках. В кратком изложении, афукозилированное h11G2 XC154/XC155 вводили посредством болюсной инъекции IV в дни 1 и 8 на уровнях 2 и 10 мг/кг/дозу яванским макакам (N=6 на группу), вакцинированным против TT (столбнячного анатоксина) перед исследованием. Введение афукозилированного h11G2 XC154/XC155 хорошо переносилось и приводило к ожидаемым фармакологическим результатам (т.е. снижению циркулирующих B-клеток и Tfh-подобных клеток). Повышенные анамнестические ответы с образованием иммуноглобулина M (IgM) и IgG по сравнению с днем 15 наблюдали у всех животных во всех дозовых группах в день 22, через 7 дней после вторичной иммунизации TT. Эти ответы были снижены у животных, которым вводили афукозилированное H11G2 XC154/XC155 (10 мг/кг/дозу), с пиком среднего титра IgG против TT 42667 по сравнению с 72000 в контрольной группе и пиком среднего титра IgM против TT 1583 по сравнению с 3500 в контрольной группе. Ритуксан® (ритуксимаб), истощающее B-клетки антитело, используемое в качестве компаратора, не влияло на анамнестические ответы с образованием IgG против TT. В заключение, эти данные свидетельствуют о том, что афукозилированное h11G2 XC154/XC155 нарушает гуморальные анамнестические ответы in vivo.
Фармакологическое исследование безопасности
[000443] В дизайн базового исследования токсичности включали электрокардиографию и измерения частоты сердечных сокращений. Не наблюдали отклонений результатов электрокардиографии, приписываемых введению афукозилированного h11G2 XC154/XC155. Все электрокардиограммы качественно и количественно находились в пределах нормы, и не наблюдали аритмию.
Пример 16: Фармакокинетика и метаболизм афукозилированного h11G2 XC154/XC155 in vivo
[000444] Для детекции наличия ADA в сыворотке яванского макака осуществляли электрохемилюминесцентный анализ (ECL) с помощью аналитической платформы Meso Scale Discovery® (MSD). Для мониторинга проведения анализа в каждый планшет включали положительный контроль, антиидиотипическое антитело против CXCR5, добавленное в сыворотку яванского макака, и отрицательный контроль, состоящий из объединенной нормальной сыворотки яванского макака. Меченое биотином афукозилированное h11G2 XC154/XC155 и меченое рутением афукозилированное h11G2 XC154/XC155 инкубировали совместно с исследуемыми образцами, положительным контролем и отрицательным контролем. Антитела против афукозилированного h11G2 XC154/XC155, присутствующие в образцах, должны связываться с мечеными биотином и рутением версиями афукозилированного h11G2 XC154/XC155, детектируемыми в этом анализе. Комплексы фиксировали через биотинилированное афукозилированное h11G2 XC154/XC155 на покрытых стрептавидином многоканальных планшетах MSD, считываемых с помощью устройства MSD Sector Imager. Конечную детекцию осуществляли с использованием меченого рутением афукозилированного h11G2 XC154/XC155 и трипропиламина для получения сигнала ECL (RLU) в устройстве MSD Sector Imager.
[000445] Исследуемые образцы тестировали на ADA с использованием многоуровневой стратегии. Исходно образцы тестируют в скрининговом анализе при коэффициенте разведения 75. Образцы, генерирующие RLU ниже порогового значения анализа, регистрировали как отрицательные (<1,88). Образцы, генерирующие RLU на уровне порогового значения анализа или выше, анализировали повторно в полной серии разведения для определения титра антител. Титр антител определяли как кратность разбавления образца, который будет генерировать RLU, равный пороговому значению RLU. Регистрировали логарифм (основание 10) этого титра.
[000446] Выводы, касающиеся индуцирования ADA, делали на основе сравнения образцов, собранных перед введением дозы в день исследования 1 и образцов после введения доз. Если для образца перед введением дозы получали отрицательный результат анализа на ADA, и соответствующий образец после введения дозы являлся положительным, животное считали положительным по индуцированию ADA. Если образцы и до введения дозы, и после введения дозы являлись положительными на ADA, животное считали положительным по индуцированию ADA, только если титр в образце после введения дозы составлял по меньшей мере 0,48 (log 3, коэффициент серийного разведения) или превышал титр в образце перед введением дозы.
Фармакокинетика однократной дозы
[000447] PK афукозилированного h11G2 XC154/XC155 охарактеризовывали после однократного введения IV в дозе 0,001, 0,002, 0,005, 0,1 или 0,2 мг/кг афукозилированного h11G2 XC154/XC155 как части исследования для оценки истощения и восстановления B-клеток и фолликулярных T-хелпер (Tfh)-подобных (Tfh-подобных) клеток периферической крови у самцов и самок яванского макака (n=1/пол/дозовую группу). В рамках этого эксперимента составы IV и SC содержали следующие компоненты: 50 мг/мл антитела против CXCR5 или антигенсвязывающего фрагмента по изобретению, 20 мМ гистидина, 8,5% сахарозы и 0,02% полисорбата 80, 0,005% ЭДТА при pH 5,8.
[000448] После однократного введения IV системное воздействие повышалось с повышением дозы, и PK афукозилированного h11G2 XC154/XC155 отличалась низким CL и низким Vss со средними значениями, находящимися в диапазоне в зависимости от уровней доз приблизительно от 0,2 до 2,6 мл/ч/кг и 0,03 до 0,1 л/кг, соответственно. Средние значения t½ в зависимости от уровней доз находились в диапазоне приблизительно от 1 до 4 дней. При низких дозах (0,001-0,005 мг/кг) наблюдали признаки более высокого CL, вероятно, в результате опосредуемого антителозависимой клеточной цитотоксичностью (ADCC) истощения клеток, которое также может приводить к клиренсу комплекса афукозилированного h11G2 XC154/XC155/CXCR5 (Kryzanski et al., 2016, J. Pharmacokinet. Pharmacodyn. 43(5):513-527; Wang et al., 2010, AAPS J. 12(4):729-740). Схожий механизм клиренса/нелинейности можно наблюдать у людей при низких дозах. Как описано выше в других местах настоящего описания, наблюдали дозозависимое истощение B-клеток и Tfh-подобных клеток в периферической крови, и истощение циркулирующих B-клеток было более устойчивым, чем истощение Tfh-подобных клеток.
Токсикокинетика (TK) при повторном введении у яванского макака
[000449] Оценку TK и ADA проводили после введения IV или SC афукозилированного h11G2 XC154/XC155 в дозе 5 (IV), 20 (SC) или 200 (IV) мг/кг каждые 2 недели (всего 5 доз) самцам и самкам яванского макака (n=3 или 4/пол/дозовую группу) как часть исследования токсичности при повторном введении, соответствующего требованиям GLP.
[000450] Не наблюдали измеримых концентраций афукозилированного h11G2 XC154/XC155 в образцах, собранных и проанализированных в группе контрольного носителя. Не наблюдали очевидных половых различий системного воздействия в дозовых группах; таким образом, средние для группы параметры TK представлены с использованием комбинированных данных для самцов и самок яванского макака (таблица 13).
[000451] После введения раз в две недели в дозе 5 (IV), 20 (SC) или 200 (IV) мг/кг системное воздействие было выше после повторного введения с коэффициентами накопления (день исследования 43/день исследования 1) в диапазоне приблизительно от 1,4 до 1,7. Кроме того, системное воздействие повышалось пропорционально дозе после введения IV. Последние измеримые концентрации афукозилированного h11G2 XC154/XC155 у животных из группы восстановления из дозовой группы 5 мг/кг (IV) наблюдали в дни исследования 148-281 11-месячного периода восстановления. После введения SC биодоступность афукозилированного h11G2 XC154/XC155 была оценена как по меньшей мере приблизительно 50%.
[000452] Частота возникновения ADA против афукозилированного h11G2 XC154/XC155 составляла 19% (3/16 животных), 17% (1/6 животных) и 0% (0/6 животных) после введения афукозилированного h11G2 XC154/XC155 в дозе 5 (IV), 20 (SC) или 200 (IV) мг/кг, соответственно, и концентрации в сыворотке в день исследования 43, как правило, являлись более низкими у ADA-положительных животных по сравнению с ADA-отрицательными животными. Следует отметить, что концентрации циркулирующего афукозилированного h11G2 XC154/XC155 могут мешать детекции ADA.
ТАБЛИЦА 13
(мкг/мл)
(часы)
(мкг·ч/мл)
Распределение
[000453] Исследования связывания белков и распределения в тканях не проводили для афукозилированного h11G2 XC154/XC155 у неклинических видов. Vss афукозилированного h11G2 XC154/XC155 у яванских макаков находился в диапазоне приблизительно от 0,03 до 0,1 л/кг после однократного введения IV, что соответствует ограниченному распределению, ожидаемому для IgG (Lin et al., 1999, J. Pharmacol. Exp. Ther. 288(1):371-378; Mascelli et al., 2007, J. Clin. Pharmacol. 47(5):553-565).
Метаболизм
[000454] Исследования метаболизма не проводили для афукозилированного h11G2 XC154/XC155. Аналогично другим терапевтическим белкам с молекулярными массами выше порога клубочковой фильтрации, ожидают, что афукозилированное h11G2 XC154/XC155 будет метаболизироваться, главным образом, посредством катаболической деградации (Lobo et al., 2004, J. Pharm. Sci. 93(11):2645-2668; Mascelli et al., 2007, выше; Vugmeyster et al., 2012, World J. Biol. Chem. 3(4):73-95).
Фармакокинетические взаимодействия лекарственных средств
[000455] Не проводили исследования фармакокинетического взаимодействия лекарственных средств in vitro или in vivo. Афукозилированное h11G2 XC154/XC155 является гуманизированным моноклональным антителом (mAb) против CXCR5, и показано, что оно модулирует высвобождение цитокинов in vitro, но не in vivo. Показано, что цитокины модулируют экспрессию ферментов цитохрома P450 (CYP) и транспортеров (Lee et al., 2010, Clin. Pharmacokinet. 49(5):295-310; Mahmood & Green, 2007, J. Clin. Pharmacol. 47(12):1540-1554). Таким образом, лечение с использованием афукозилированного h11G2 XC154/XC155 потенциально может влиять на уровни фермента CYP и транспортера и, таким образом, модулировать клиренс сопутствующих лекарственных средств, являющихся субстратами для этих ферментов или транспортеров. Однако, цитокин-опосредованные взаимодействия лекарственных средств, наблюдаемые в клинических условиях для других лекарственных средств, были умеренными, что приводило к менее чем 2-кратным изменениям воздействия совместно вводимых низкомолекулярных лекарственных средств (Huang et al., 2010, Clin. Pharmacol. Ther. 87(4):497-503; Evers et al., 2013, Drug Metab. Dispos. 41(9):1598-1609). Таким образом, не желая быть связанными какой-либо конкретной теорией, полагают, что, если сопутствующее лекарственное средство изменяет экспрессию мишени, оно потенциально может влиять на PK афукозилированного h11G2 XC154/XC155.
Прогнозирование фармакокинетики у людей
[000456] Профили PK афукозилированного h11G2 XC154/XC155 у яванского макака были типичными для mAb IgG1 человека у обезьяны. Таким образом, ожидают, что прогнозируемые значения параметров 2-компартментной PK у человека будут такими же, как у типичного терапевтического mAb IgG1, и предполагают, что они будут линейными в тестируемом диапазоне доз. Используемые значения параметров PK были схожи со значениями, описанными ранее (Singh et al., 2015, In: Developability of Biotherapeutics: Computational Approaches, S. Kumar, Singh S. Kumar, Eds., CRC Press). Эти параметры являлись следующими: 3,2 л в случае центрального объема (Vc), 2,2 л в случае периферического объема (Vp), 0,25 л/сутки в случае центрального клиренса (CLc), 0,45 л/сутки в случае распределительного клиренса (Q), 0,26 л/сутки в случае константы скорости всасывания (ka) при введении SC и 60% в случае биодоступности при введении SC. Учитывая данные, представленные в настоящем описании, прогнозируемое время полужизни афукозилированного h11G2 XC154/XC155 в сыворотке человека составляет приблизительно 17 дней.
Прогнозирование эффективной дозы у людей
[000457] Основанный на модели подход адаптировали для характеризации взаимосвязи между дозой, концентрациями афукозилированного h11G2 XC154/XC155 и модуляцией B-клеток и Tfh-подобных клеток в сыворотке яванского макака. Затем для преобразования параметров истощения B-клеток для афукозилированного h11G2 XC154/XC155 от обезьяны к человеку использовали опубликованные данные о фармакокинетике/фармакодинамике (PK/PD) SBI-087, гуманизированного иммунофармацевтического средства на основе модульного белка малого размера (SMIP), которое, подобно ритуксимабу, связывается с CD20 на B-клетках и истощает их, у обезьяны и человека (Cohen et al., 2016, Clin. Ther. 38(6):1417-1434; Dunussi-Joannopoulos et al., 2008, Ann. Rheum. Dis. 64(Suppl II):190 (Abstr THU0171). В отличие от B-клеток, недоступны данные об истощении Tfh-подобных клеток у человека; таким образом, преобразование данных об истощении Tfh-подобных клеток считают схожим с таковым в случае истощения B-клеток. Используя исходные количества B-клеток и Tfh-подобных клеток у пациентов SLE (Belouski et al., 2010, Cytometry B Clin. Cytom. 78(1):49-58) и прогнозируемые параметры истощения клеток человека, симулировали кинетику истощения B-клеток и Tfh-подобных клеток после введения афукозилированного h11G2 XC154/XC155 людям.
[000458] Прогнозируемая эффективная доза афукозилированного h11G2 XC154/XC155 у человека составляет приблизительно от 10 до 30 мг (IV) и основана на предположении о том, что для клинической эффективности B-клетки в крови необходимо истощать до ≤1 клетки/мл в течение приблизительно 8 недель. Концентрации афукозилированного h11G2 XC154/XC155 в сыворотке после второй дозы от 10 до 30 мг IV (дозы в день 1 и день 29) анализировали как указано выше, и прогнозируемая Cmax составляла приблизительно от 5 до 15 мкг/мл, прогнозируемая площадь под кривой концентрация-время за интервал дозирования tau (AUCtau) составляла приблизительно от 38 до 114 мкг/сутки/мл, и прогнозируемая средняя концентрация (Cav; вычисленная как AUCtau/28) составляла от 1,36 до 4,06 мкг/мл.
Пример 17: Токсикологические исследования афукозилированного h11G2 XC154/XC155
[000459] Афукозилированное h11G2 XC154/XC155 анализировали в серии неклинических исследований токсичности в обзоре исследований тестирования токсичности, как представлено в таблице 14. Выбирали внутривенный (IV) или подкожный (SC) пути введения, т.к. они являются предполагаемыми путями при клиническом введении. В рамках этого эксперимента составы IV и SC содержали следующие компоненты: 50 мг/мл антитела против CXCR5 или антигенсвязывающего фрагмента по изобретению, 20 мМ гистидина, 8,5% сахарозы и 0,02% полисорбата 80, 0,005% ЭДТА при pH 5,8. Как представлено в других местах настоящего описания, исследования с использованием мононуклеарных клеток периферической крови человека или яванского макака показали, что афукозилированное h11G2 XC154/XC155 связывается с CXCR5-экспрессирующими клетками со сравнимой аффинностью и запускает антителозависимую клеточную цитотоксичность (ADCC) схожим образом для клеток человека и обезьяны. Как представлено в других местах настоящего описания, афукозилированное h11G2 XC154/XC155 не связывается с ортологами CXCR5 мыши, крысы или кролика. Таким образом, неклинические исследования токсичности проводили только на яванских макаках.
ТАБЛИЦА 14
[000460] Сокращения в таблице 14 представляют собой следующее: GLP=Надлежащая лабораторная практика; IV=внутривенное введение; OECD=Организация экономического сотрудничества и развития; PBMC=мононуклеарные клетки периферической крови; SC=подкожное введение.
[000461] Все исследования, соответствующие требованиям GLP, проводили в государстве-члене соглашения по взаимному признанию данных OECD. Все исследования in vivo проводили на самцах и самках животных. Если не указано иначе, все дозы выражены как мг белка на кг массы тела на дозу. Цельную кровь человека анализировали при уровне 1, 10, 100 или 1000 мкг/мл (растворимая фаза) или PBMC человека анализировали при уровне 1, 10, или 100 мкг/лунку (твердая фаза).
[000462] Афукозилированное h11G2 XC154/XC155 вводили обезьянам в исследованиях IV и SC с еженедельным введением до 400 мг/кг/дозу (IV) в течение 17 дней (всего 3 дозы) или каждые 2 недели до 200 мг/кг/дозу (IV) в течение 2 месяцев (всего 5 доз). 17-дневное поисковое исследование на обезьянах включало фазу восстановления до дня 352. Базовое 2-месячное исследование на обезьянах (GLP) включало 11-месячную фазу восстановления (день 401). Афукозилированное h11G2 XC154/XC155 индуцировало высвобождение цитокинов в анализе in vitro. Целевым органом, идентифицированным в этих исследованиях, являлась гематолимфопоэтическая система. Уровень без видимых нежелательных явлений (NOAEL) в 2-месячном исследовании на обезьянах представлял собой наибольшую дозу 200 мг/кг/дозу (IV) с Cmax 5220 мкг/мл и AUC168 438,000 мкг·ч/мл в день 43.
Токсичность при повторном введении
[000463] Поисковые и базовые исследования токсичности при повторном введении проводили с использованием афукозилированного h11G2 XC154/XC155 на яванских макаках.
Поисковое исследование токсичности
[000464] В рамках поискового исследования, несоответствующего требованиям GLP, афукозилированное h11G2 XC154/XC155 вводили посредством инъекции IV или SC обезьянам (1-2/пол/дозу) раз в неделю в количестве 0 (IV, носитель SC), 40 (IV), 260 (SC) или 400 (IV) мг/кг/дозу (всего 3 дозы) с последующей фазой восстановления в группах носителя, 40(IV) и 400 (IV) (1/пол/группу) до дня 352. Все животные доживали до запланированной некропсии в день 17 или день 352. Не наблюдали связанных с тестируемым препаратом клинических признаков и эффектов в отношении массы тела, потребления пищи или цитокинов в сыворотке.
[000465] В фазу введения наблюдали снижение общего количества B-клеток, CXCR5+ B-клеток и циркулирующих Tfh-подобных клеток в периферической крови относительно исходного уровня (в 0,00-0,02 раз, 0,00-0,02 раз и 0,03-0,09 раз, соответственно), начиная в дни 2 или 3, с наибольшим снижением относительно исходного уровня, наблюдаемым в день 17. Величина истощения была схожей для всех доз. В селезенке наблюдали значительно более низкие количества B-клеток, CXCR5+ B-клеток и истинных Tfh-клеток (0,018-0,144 раз, 0,003-0,033 раз и 0,037-0,245 раз, соответственно) для всех доз по сравнению с контролями. В периферической крови наблюдали снижение (в 0,33-0,67 раз относительно исходного уровня) абсолютного количества лимфоцитов у всех животных, кроме одного, при ≥40 мг/кг/дозу ко дню 2 и повышение (в 1,11-1,81 раз относительно исходного уровня) иммуноглобулина (Ig) G у животных при ≥40 мг/кг/дозу в день 17. Наблюдали транзиторное снижение количества естественных киллеров (NK) в день 2 (в 0,05-0,14 раз относительно исходного уровня) с частичным или полным восстановлением ко дню 6 у большинства животных. Наблюдали умеренно или значительно сниженную клеточность в лимфоидных фолликулах селезенки; минимально сниженную клеточность в лимфоидных фолликулах подчелюстных, подмышечных и паховых лимфоузлов, и при ≥40 мг/кг/дозу и 260 мг/кг/дозу (SC) в миндалинах наблюдали смешанную клеточную инфильтрацию по результатам микроскопии участков инъекций. При иммуногистохимии определяли независящее от дозы снижение CD20-положительных клеток и CXCR5-положительных клеток в селезенке и снижение CD20-положительных клеток в подчелюстных и паховых лимфоузлах. Эффекты в отношении лимфоидных тканей коррелировали со снижением B-клеток и Tfh-подобных клеток, а также общего количества лимфоцитов в периферической крови, и это соответствовало фармакологическим данным для афукозилированного h11G2 XC154/XC155, являющегося истощающим антителом, направленно воздействующим на CXCR5-экспрессирующие клетки.
[000466] В фазу восстановления сниженное количество B-клеток, CXCR5+ B-клеток и Tfh-подобных клеток периферическая кровь повышалось (медленно) ко дню 199, и к концу фазы восстановления в день 352 все животные имели частичное или полное восстановление этих типов клеток. Во время фазы восстановления NK-клетки и IgG восстанавливали до диапазона исходного уровня (фиг. 8A-8D). Не наблюдали связанных с тестируемым препаратом результатов микроскопии (включая иммуногистохимическую оценку CD20 и CXCR5 в селезенке и подчелюстных и паховых лимфоузлах), и в селезенке количества B-клеток, CXCR5+ B-клеток и истинных Tfh-клеток были схожими или превышали значения для животных в контрольной группе носителя, что позволяет предполагать полное восстановление.
[000467] Не наблюдали эффектов в отношении цитокинов сыворотки или T-клеток, T-хелперных клеток или цитотоксических T-клеток в периферической крови или селезенке при любой дозе. В этом исследовании не определяли очевидного наличия антител против лекарственного средства (ADA).
[000468] Фиг. 8A-8D представляют собой графики, на которых показаны истощение и восстановление B-клеток и Tfh-подобных клеток периферической крови у яванских макаков. Уровни B-клеток (фиг. 8A и ФИГ. 8B) и Tfh-подобных клеток (фиг. 8C и фиг. 8D) периферической крови на мкл крови у самцов (фиг. 8A и фиг. 8C) и самок (фиг. 8B и фиг. 8D) приведены до дня 352 поискового исследования токсичности. B-клетки определяли как CD3-CD20+ клетки. Tfh-подобные клетки определяли как сумму CD3+CD4+CD95+CXCR5+ клеток и CD3+CD4+CD95+hIgG+ клеток, т.к. тестируемый препарат противодействует антителу против CXCR5, используемому для иммунофенотипирования. Архивные диапазоны B-клеток у самцов обезьян (фиг. 8A, 272-2503 клеток на мкл) и самок обезьян (фиг. 8B, 233-1700 клеток на мкл) указаны пунктирными линиями.
Базовое исследование токсичности (GLP)
[000469] В рамках базового исследования афукозилированное h11G2 XC154/XC155 вводили посредством инъекции IV или SC обезьянам (3-8/пол/группу) каждые 2 недели в количестве 0 (IV, SC), 5 (IV), 20 (SC), или 200 (IV) мг/кг/дозу (всего 5 доз) с последующей 11-недельной фазой восстановления (день 401). Для оценки первичного и вторичного T-клеточно-зависимого гуморального ответа (TDAR) обезьянам в день 22 (фаза введения) и в день 253 (фаза восстановления) инъецировали гемоцианин морского блюдца (KLH) и столбнячный анатоксин (TT), соответственно. Животных иммунизировали TT, но они были KLH-наивными перед началом исследования. Образцы крови собирали перед началом исследования (для оценки исходного уровня) в дни 22, 25, 29, 36, 43, 58, 253 (перед иммунизацией в фазу восстановления), 256, 260, 267, 274 и 281 и оценивали на продукцию IgM против KLH, IgG против KLH, IgM против TT и IgG против TT.
[000470] Все животные в фазу введения доживали до запланированной некропсии в день 58. Не наблюдали связанных с тестируемым препаратом клинических признаков и эффектов в отношении массы тела, потребления пищи, цитокинов плазмы, свертывания, клинической биохимии, параметров TDAR-KLH или параметров электрокардиограммы. Все связанные с афукозилированным h11G2 XC154/XC155 результаты в этом исследовании включали ненеблагоприятные и обратимые изменения гематологических параметров, параметров иммунофенотипирования в периферической крови, независящее от дозы снижение значений IgG TDAR-TT и изменение результатов микроскопии и иммунофенотипирования в селезенке и/или лимфоузлах у самцов и самок при 5 и 200 мг/кг/дозу IV и 20 мг/кг/дозу SC. В фазу восстановления одну самку из контрольной группы умерщвляли в день 330 из-за осложнений при заборе крови. Все остальные животные в группе для оценки восстановления доживали до некропсии в день 401.
[000471] Гематологические изменения, связанные с афукозилированным h11G2 XC154/XC155, включали, в целом, независящее от дозы, слабое или умеренное снижение лимфоцитов (в 0,34-0,60 раз относительно исходного уровня) при всех дозах с наибольшей частотой в день 2 и более низкой частотой в последующие моменты времени, вносящее вклад в общее снижение количества лейкоцитов (в 0,25-0,59 раз относительно исходного уровня) в случае отдельных животных при всех дозах. Лимфоциты приближались к исходным и/или контрольным значениям в течение первых 1-2 месяцев фазы восстановления. Также наблюдали небольшое снижение базофилов (в 0,17-0,50 раз) при 200 мг/кг/дозу IV в день 2 и в случае отдельных животных при 5 мг/кг/дозу IV и 20 мг/кг/дозу SC в последующие моменты времени с последующим восстановлением в течением первых 1-3 месяцев фазы восстановления и транзиторное минимальное снижение (в 0,79-0,82 раз) массы эритроцитов (гемоглобина, количества эритроцитов и гематокрита) у самок при 200 мг/кг/дозу IV в день 6.
[000472] Наблюдали значительное связанное с афукозилированным h11G2 XC154/XC155 снижение общего количества B-клеток, CXCR5+ B-клеток и Tfh-подобных клеток (в 0,00-0,56 раз) в периферической крови относительно исходного уровня у всех животных и во всех дозовых группах уже в день 2, и оно сохранялось до конца фазы введения (день 58) с возвращением к исходному уровню или имело абсолютные значения для каждой субпопуляции животных, которым вводили 5 мг/кг/дозу IV в фазу восстановления ко дню 393, находящиеся в диапазоне значений, наблюдаемых у животных в контрольной группе носителя (фиг. 9A-9D). Также наблюдали значительное снижение количества NK-клеток в периферической крови в день 2 у всех животных относительно исходного уровня (в 0,04-0,46 раз); однако, у большинства животных наблюдали частичное или полное восстановление ко дню 6, хотя NK-клетки оставались ниже исходных уровней у некоторых животных в один или более последующих моментов времени с возвращением к исходному уровню к концу фазы восстановления. Также у некоторых животных наблюдали транзиторное снижение общего количества T-клеток, T-хелперных клеток и цитотоксических T-клеток в день 2 (0,45-0,66 раз); однако, эти популяции клеток возвращались к исходным уровням ко дню 6 у большинства животных и ко дню 36 у всех животных. Не наблюдали связанных с афукозилированным h11G2 XC154/XC155 изменений интерлейкина (ИЛ)-2, ИЛ-6, ИЛ-10, ИЛ-13, интерферона (ИФН)-γ и фактора некроза опухоли (ФНО)-α при любой дозе.
[000473] Наблюдали связанное с афукозилированным h11G2 XC154/XC155, независящее от дозы снижение клеточности лимфоидных фолликулов селезенки и лимфоидных фолликулов подмышечных и брыжеечных лимфоузлов и лимфоидной ткани кишечника (GALT) при ≥5 мг/кг/дозу IV и 20 мг/кг/дозу SC. Это было связано со снижением клеточности лимфоидных фолликулов в отношении CD20+ и CXCR5+ клеток в селезенке, оцениваемым посредством иммуногистохимии, за исключением 1 самца при 20 мг/кг/дозу SC; однако, воздействие афукозилированного h11G2 XC154/XC155 у этого животного могло подвергаться влиянию из-за распространения ADA, начиная со дня 15. Кроме того, при некропсии в день 58 определяли более низкие количества B-клеток, CXCR5+ B-клеток и Tfh-клеток в селезенке по сравнению с контролями, которым вводили носитель. У некоторых животных определяли более низкие количества NK-клеток в селезенке по сравнению с контролями. К концу восстановления в день 401 абсолютные значения для каждой исследуемой популяции у всех животных, которым вводили афукозилированное h11G2 XC154/XC155, находились в диапазоне значений для животных в контрольной группе, которой вводили носитель, что свидетельствует о восстановлении после связанного с афукозилированным h11G2 XC154/XC155 снижением количеств клеток, наблюдаемым в фазу введения этого исследования.
[000474] Сниженная лимфоидная клеточность в селезенке и лимфоидных фолликулах в лимфоузлах вместе с более низкими количествами B-клеток, CXCR5+ B-клеток и Tfh-клеток в селезенке коррелировала со снижением общего количества лимфоцитов и B-клеток, CXCR5+ B-клеток и Tfh-подобных клеток в периферической крови при ≥5 мг/кг/дозу IV и 20 мг/кг/дозу SC. К концу фазы восстановления (день 401) не наблюдали связанных с афукозилированным h11G2 XC154/XC155 результатов микроскопии а дозовой группе 5 мг/кг IV, что свидетельствует о полном восстановлении. Эти результаты согласуются с фармакологическими данными о афукозилированном h11G2 XC154/XC155, которое, как ожидают, будет истощать CXCR5-экспрессирующие клетки. Более низкие количества NK-клеток в периферической крови и/или селезенке могут являться результатом гибели эффекторных NK-клеток, что описано для NK-клеток после антителозависимой клеточной цитотоксичности (Warren et al., 2011, J. Immunol. Meth. 370:86-92), и/или перераспределения в ткани; однако, нельзя исключать и другие механизмы.
[000475] Не наблюдали связанных с афукозилированным h11G2 XC154/XC155 эффектов в отношении первичного T-клеточно-зависимого гуморального ответа (TDAR) на гемоцианин морского блюдца (KLH). Ответы с образованием иммуноглобулинов (Ig) M и IgG против KLH наблюдали во всех группах. Всех обезьян вакцинировали против столбнячного анатоксина (TT) перед включением в исследование. Повышенные анамнестические ответы с образованием IgM и IgG наблюдали у всех животных во всех дозовых группах. Связанное с афукозилированным h11G2 XC154/XC155, независящее от дозы снижение вторичного TDAR на TT (значения титра центральной точки IgG) наблюдали у самцов и самок животных в группах 5 IV, 20 SC и/или 200 IV мг/кг/дозу через 7, 14, 21 и 36 дней после иммунизации. Статистически значимое снижение средних для группы значений антител IgG против TT наблюдали в группах 5 и 200 мг/кг/дозу IV через 7 дней после иммунизации.
[000476] После иммунизации в фазу восстановления наблюдали анамнестические ответы с образованием IgM и IgG против KLH у всех животных в дозовой группе 5 мг/кг афукозилированного h11G2 XC154/XC155 в 1 или более моментов времени, находящиеся в пределах диапазона контрольной группы, которой вводили носитель. Во время восстановления наблюдали связанное с афукозилированным h11G2 XC154/XC155 статистически значимое снижение значений титра центральной точки (CPT) IgG против TT в группе, наблюдаемое у животных в дозовой группе 5 мг/кг через 7 дней после иммунизации в фазу восстановления. Значения CPT всех животных в фазу восстановления находились в пределах диапазона контрольной группы, за исключением 2 из 8 животных в дозовой группе 5 мг/кг, имевших значения CPT ниже контрольного диапазона во всем моменты времени в фазу восстановления до 28 дней после иммунизации TT в фазу восстановления. Не наблюдали изменений значений IgM против TT, приписываемых введению афукозилированного h11G2 XC154/XC155, в фазу введения или фазу восстановления.
[000477] Наблюдали первичные ответы с образованием IgM и IgG против KLH во всех группах; некоторые животные в группах 5 и 200 мг/кг/дозу имели CPT выше контрольного диапазона в 1 или более моментов времени после первичной иммунизации. Не наблюдали статистически значимых различий средних по группе значений антител IgM или IgG против KLH в любой из групп в любой момент времени. Т.к. изменения у отдельных животных являлись спорадическими, не зависели от дозы, и значения CPT в группе находили в диапазоне контрольных животных, их не считали связанными с афукозилированным h11G2 XC154/XC155. В фазу восстановления значения у всех животных находились в пределах контрольного диапазона, за исключением 1 из 8 животных в дозовой группе 5 мг/кг, имевшего значения CPT немного ниже контрольного диапазона через 14, 21 и 28 дней после иммунизации KLH. Эти данные (фаза введения или восстановления) свидетельствуют о том, что все животные, которым вводили афукозилированное h11G2 XC154/XC155 (самцы и самки), могли генерировать первичные ответы с образованием IgM и IgG на иммунизацию KLH, схожие с контрольными животными.
[000478] Несмотря на значительное снижение общего количества B-клеток, CXCR5+ B-клеток, Tfh-подобных клеток и NK-клеток в периферической крови и значительно более низкие количества B-клеток, CXCR5+ B-клеток и Tfh-клеток в селезенке, определяемые посредством иммунофенотипирования, снижение количества лимфоцитов и снижение лимфоидной клеточности в селезенке, лимфоузлах и GALT, все результаты не являлись неблагоприятными из-за отсутствия ассоциированных клинических признаков и отсутствия эффекта в отношении первичного TDAR. Другие гематологические результаты, включая снижение параметров общего количества лейкоцитов, базофилов и эритроцитов не являлись неблагоприятными из-за ограниченной тяжести и отсутствия корреляции с результатами микроскопии. Не отмечали связанных с афукозилированным h11G2 XC154/XC155 результатом микроскопии и иммуногистохимии в день 401, что свидетельствует о полном восстановлении.
[000479] NOAEL в случае афукозилированного h11G2 XC154/XC155 у обезьян после SC или IV введения каждую следующую неделю в течение 2 месяцев составлял 200 мг/кг/дозу. Системное воздействие (Cmax и AUC168) при NOAEL составляло 5220 мкг/мл и 438000 мкг·ч/мл, соответственно. В этом исследовании определяли антитела против афукозилированного h11G2 XC154/XC155, и данные приведены в других местах настоящего описания. В других местах настоящего описания представлена оценка риска токсичности для органов-мишеней в исследовании токсичности при повторном введении. В других местах настоящего описания можно найти пороговые концентрации афукозилированного h11G2 XC154/XC155, ассоциированные с ключевыми ответами у обезьян.
Репродуктивная и эмбриональная токсичность
[000480] Не наблюдали связанных с тестируемым препаратом эффектов в отношении репродуктивных тканей самцов или самок, оцениваемых в исследовании токсичности при повторном введении на яванских макаках.
Местная переносимость
[000481] Не проводили независимые исследования местной переносимости с использованием афукозилированного h11G2 XC154/XC155, хотя места инъекций обследовали макроскопически и микроскопически в исследованиях токсичности in vivo.
[000482] В поисковом исследовании на обезьянах в месте SC инъекции при 260 мг/кг/дозу наблюдали умеренную периваскулярную смешанную лейкоцитарную клеточную инфильтрацию (мононуклеарные лейкоциты, нейтрофилы и меньшее количество эозинофилов) по сравнению с минимальными и слабыми инфильтратами в местах SC инъекций у контрольных животных. У самца, которому вводили 260 мг/кг/дозу SC, инфильтрат мультифокально распространялся в мышечную оболочку небольших сосудов. У самки, которой вводили 260 мг/кг/дозу SC, несколько многоядерных гигантских клеток инфильтрировали подкожный коллаген, и в этих клетках определяли внутрицитоплазматические базофильные или эозинофильные фрагменты фибриллярного материала. В базовом 2-месячном исследовании на обезьянах результаты микроскопии в местах введения IV и SC не считали связанными с тестируемым препаратом.
Антигенность
[000483] Иммуногенность оценивали посредством измерения уровней ADA в исследованиях токсичности при повторном введении яванским макакам; эти данные представлены выше в настоящем описании.
Иммунотоксичность
[000484] В исследованиях in vitro и in vivo охарактеризовывали эффект афукозилированного h11G2 XC154/XC155 в отношении иммунной системы. Возможность индуцирования афукозилированным h11G2 XC154/XC155 высвобождения цитокинов (ФНОα, ИЛ-6 и ИФНγ) определяли в анализах высвобождения цитокинов человека с использованием 2 разных форматов in vitro: гомогенного анализа с использованием цельной крови и твердофазного анализа с использованием иммобилизованного афукозилированного h11G2 XC154/XC155 и мононуклеарных клеток периферической крови (PBMC). В каждом из этих форматов оценивали образцы 8 здоровых людей-доноров. В обоих форматах анализа высвобождения цитокинов наблюдали индуцируемое афукозилированным h11G2 XC154/XC155 высвобождением цитокинов (ФНОα, ИЛ-6 и ИФНγ). In vivo в поисковых и базовых исследованиях токсичности при повторном введении на обезьянах охарактеризовывали профиль высвобождения цитокинов in vivo, а также эффекты афукозилированного h11G2 XC154/XC155 в отношении клеток и тканей иммунной системы. Афукозилированное h11G2 XC154/XC155 не индуцировало цитокины in vivo. Истощение лимфоцитов, B-клеток, CXR5+ B-клеток и Tfh/Tfh-подобных клеток наблюдали в периферической крови и селезенке обезьян, которым вводили афукозилированное h11G2 XC154/XC155, как представлено в других местах настоящего описания, эффекты соответствовали ожидаемому механизму действия CXCR5-истощающего антитела. После фазы восстановления в поисковых и базовых исследованиях токсичности параметры лимфоцитов в селезенке и лимфоузлах были сравнимы с контролями, которым вводили носитель.
[000485] Первичные и вторичные ответы TDAR на KLH и TT, соответственно, оценивали в базовом 2-месячном исследовании токсичности, как представлено в другом месте настоящего описания. Не наблюдали связанных с афукозилированным h11G2 XC154/XC155 эффектов в отношении первичного TDAR на KLH. У всех животных развивался вторичный ответ TDAR на TT, но наблюдали связанное с афукозилированным h11G2 XC154/XC155 снижение титров центральных точек в группах 5 и 200 мг/кг/дозу IV через 7 дней после иммунизации во время фазы введения и фазы восстановления в рамках исследования.
Тканевая перекрестная реактивность (TCR)
[000486] В предварительном исследовании способа окрашивания с использованием афукозилированного h11G2 XC154/XC155 проводили обширную предварительную работу по разработке способа, включая использование многочисленных анализов и способов. Ни в одних из условий этого анализа не наблюдали устойчивого окрашивания в ожидаемых клетках и тканях. В соответствующем требованиям GLP исследовании тканевой перекрестной реактивности оценивали связывание биотинилированного афукозилированного h11G2 XC154/XC155 (афукозилированного h11G2 XC154/XC155-Bio; 1 и 5 мкг/мл) с криосрезами тканей яванского макака и человека. Профили окрашивания перекрывались между тканями яванского макака и человека. Окрашивание, общее для человека и обезьяны, наблюдали в мононуклеарных клетках, ретикулоэндотелиальных клетках, различных типах эпителия, глиальных клетках и/или питуицитах, что соответствует ожидаемой реактивности афукозилированного h11G2 XC154/XC155 (Breitfeld et al., 2000, J Exp Med 192(11):1545-1552; Carlsen et al., 2002, Gut 51(3):364-371;Flynn et al., 2003, J. Neuroimmunol. 136(1-2):84-93; Schaerli et al., 2000, J. Exp. Med. 192(11):1553-1562; Schmutz et al., 2005, Arthritis Res. Ther. 7(2):R217-R229), а также коллоида щитовидной железы. Положительное окрашивание с помощью афукозилированного h11G2 XC154/XC155 наблюдали в других тканях обезьяны или человека. В случае тканей человека, окрашивание наблюдали в нервных волокнах зрительного нерва и нейронах корешков спинномозговых нервов, гладкомышечных клетках предстательной железы, интерстициальных клетках яичка и волокнах хрусталика в глазу.
[000487] Тканями, окрашиваемыми только у яванского макака, являлись гематопоэтические клетки-предшественники в костном мозге, тучные клетки кожи и шейки матки, нейропиль в головном мозге и спинном мозге, кардиомиоциты, веретенообразные клетки в плаценте и оболочка зрительного нерва. Хотя это может представлять собой неожиданную перекрестную реактивность тестируемого препарата, окрашивание являлось цитоплазматическим по своей природе, не имело гистологической специфичности и четкого окрашивания плазматической мембраны. С помощью такого цитоплазматического окрашивания нельзя дифференцировать CXCR5-экспрессирующие и неэкспрессирующие клетки и ткани в достаточной степени. Не ожидают, что цитоплазматическое окрашивание будет доступным для тестируемого препарата in vivo, и, как правило, считают, что оно будет иметь небольшое токсикологическое значение или не будет его иметь (Hall et al., 2008, In: Preclinical Safety Evaluation of Biopharmaceuticals: A Science-Based Approach to Facilitating Clinical Trials, p. 208-240, Cavagnaro, J.A., ed., Wiley-Interscience; Leach et al., 2010, Toxicol. Pathol. 38(7):1138-1166). Отсутствие четкого окрашивания мембран позволяет предполагать, что нельзя получить надежный способ для достижения полной оценки. Отсутствие нежелательных результатов в 2-месячном исследовании токсичности при повторном введении подтверждает, что окрашивание, наблюдаемое в исследовании тканевой перекрестной реактивности ex vivo, не преобразуется в эффекты in vivo. Хотя предпринималось множество попыток получить надежный способ TCR, наблюдаемое связывание не приводило к проблемам безопасности, превышающим ожидаемое с учетом предполагаемых фармакологических свойств.
Взаимосвязь результатов и фармакокинетики
[000488] Концентрации афукозилированного h11G2 XC154/XC155 в сыворотке определяли после введения SC или IV самцам и самкам обезьян в дозах 5 (IV), 20 (SC) или 200 (IV) мг/кг/дозу каждые 2 недели в течение 2 месяцев (всего 5 доз). Не наблюдали очевидных половых различий системного воздействия между дозовыми группами; таким образом, средние параметры TK в группе представляли с использованием комбинированных данных для самцов и самок яванского макака. Системное воздействие было более высоким после повторного введения с коэффициентами накопления (день исследования 43/день исследования 1) в диапазоне от приблизительно от 1,4 до 1,7. Кроме того, системное воздействие повышалось пропорционально дозе после введения IV. После введения SC биодоступность афукозилированного h11G2 XC154/XC155 оценивали как >50%.
[000489] Пороговые значения концентраций афукозилированного h11G2 XC154/XC155 в сыворотке, ассоциированные с ключевыми ответами и пределами воздействия, вычисленными относительно этих ключевых ответов, представлены в таблице 15.
ТАБЛИЦА 15
NOAEL
[000490] Сокращения, используемые в таблице 15, являлись следующими: AUC=площадь под кривой концентрация-время; Cav=средняя концентрация; Cmax=максимальная (средняя) концентрация в плазме; Tfh-подобные клетки (альтернативно, cTfh); CXCR5=хемокиновый рецептор типа 5; GALT=лимфоидная ткань кишечника; IgG=иммуноглобулин G; IV=внутривенное введение; NK=естественные киллеры; NOAEL=уровень без видимых нежелательных явлений; SC=подкожное введение; TDAR=T-клеточно-зависимый гуморальный ответ; Tfh=фолликулярные T-хелперные клетки; TT=столбнячный анатоксин; WBC=лейкоциты.
[000491] В исследования повторного введения значения Cmax соответствуют средним концентрациям в плазме. Значения Cav вычисляют, разделяя AUC на интервал забора образцов (48 или 168 часов). Приведенные значения получали ближе к концу фазы введения.
[000492] Пределы воздействия вычисляли, разделяя значения Cav в исследованиях токсичности на животных на прогнозируемую Cav у человека 4,06 мкг/мл при прогнозируемой эффективной дозе у человека 30 мг, IV.
Токсичность для целевых органов
[000493] Учитывая проведенные неклинические исследования, гематолимфопоэтическую систему (селезенку, лимфоузлы, GALT, миндалины, параметры циркулирующих лимфоцитов, лейкоцитов, эритроцитов, высвобождение цитокинов in vitro) идентифицировали в качестве целевых органов/тканей. В небазовом 17-дневном исследовании на обезьянах, проведенном с использованием афукозилированного h11G2 XC154/XC155, связанные с тестируемым препаратом изменения в этих тканях соответствовали наблюдаемому в базовом 2-месячном исследовании токсичности. Все связанные с афукозилированным h11G2 XC154/XC155 результаты полностью или частично возвращались к исходному уровню или значениям в контрольной группе, которой вводили носитель, в фазу восстановления в небазовом и базовом исследованиях токсичности. Ни один из связанных с тестируемым препаратом эффектов не считали нежелательным по причине отсутствия каких-либо клинических результатов или оппортунистических инфекций и по причине минимального влияния на иммунную функцию, измеряемого с помощью анализа TDAR. Таким образом, определяли NOAEL 200 мг/кг/дозу IV у обезьян для связанных с афукозилированным h11G2 XC154/XC155 эффектов.
Гематолимфопоэтическая система
[000494] В исследованиях токсичности при повторном введении на яванских макаках наблюдали связанное с афукозилированным h11G2 XC154/XC155 снижение лимфоцитов в периферической крови при дозах ≥5 мг/кг/дозу (IV) и 20 мг/кг/дозу (SC), начиная со дня исследования 2. Это снижение коррелировало с наблюдаемым снижением общего количества B-клеток, CXCR5+ B-клеток и Tfh-подобных клеток, сохраняющимся до дня 57. Наблюдали полное или частичное возвращение этих сниженных субпопуляций лимфоцитов к исходным значениям к концу фазы восстановления в поисковом исследовании токсичности. Также наблюдали связанное с афукозилированным h11G2 XC154/XC155 снижение NK-клеток, общего количества T-клеток, T-хелперных клеток и цитотоксических T-клеток при ≥5 мг/кг/дозу, начиная со дня 2, но достигали частичного или полного возвращения к исходным значениям к концу фазы введения. В дополнение к снижению популяций лимфоцитов в периферической крови, наблюдали транзиторное минимальное снижение массы эритроцитов у самок обезьян, которым вводили 200 мг/кг/дозу в день 6.
[000495] Сниженные популяции лимфоцитов в периферической крови коррелировали со связанной с афукозилированным h11G2 XC154/XC155 более низкой лимфоидной клеточностью в селезенке и лимфоузлах при ≥5 мг/кг/дозу. Более низкая лимфоидная клеточность отражалась в более низком общем количестве B-клеток, более низких количествах CXCR5+ B-клеток, истинных Tfh-клеток и NK-клеток и более низким количеством фолликулярных CD20+ и CXCR5+ клеток в селезенке. Более низкую фолликулярную лимфоидную клеточность наблюдали в лимфоузлах и GALT при ≥5 мг/кг/дозу и в миндалинах при ≥40 мг/кг/дозу.
[000496] После фазы восстановления в поисковых и базовых исследованиях токсичности, параметры лимфоцитов в селезенке и лимфоузлах были сравнимыми с контролями, которым вводили носитель.
[000497] Снижение общего количества B-клеток, CXCR5+ B-клеток и Tfh/Tfh-подобных клеток соответствует ожидаемому механизму действия афукозилированного h11G2 XC154/XC155, которое, как ожидают, будет истощать CXCR5-экспрессирующие клетки. Более низкие количества NK-клеток в периферической крови и селезенке, вероятно, являются результатом гибели эффекторных клеток, которая, как известно, происходит в NK-клетках после ADCC.
[000498] Хотя наблюдали снижение параметров лимфоцитов в периферической крови и селезенке обезьян, которым вводили афукозилированное h11G2 XC154/XC155, не наблюдали связанного с афукозилированным h11G2 XC154/XC155 эффекта в отношении первичного TDAR на KLH. У всех животных развивался вторичный TDAR но TT, но статистически значимое снижение наблюдали при дозе ≥5 мг/кг/дозу.
[000499] Все связанные с афукозилированным h11G2 XC154/XC155 результаты не считали нежелательными по причине отсутствия соответствующих клинических признаков или оппортунистических инфекций. Эффекты афукозилированного h11G2 XC154/XC155 в отношении лимфоцитов периферической крови, общего количества B-клеток, CXCR5+ B-клеток, Tfh-подобных клеток и NK-клеток, а также первичных и вторичных ответов TDAR можно подвергать мониторингу у людей в клинических условиях.
[000500] В гомогенном и твердофазном форматах анализа высвобождения цитокинов in vitro наблюдали индуцируемое афукозилированным h11G2 XC154/XC155 высвобождение цитокинов (ФНОα, ИЛ-6 и ИФНγ). Индуцируемое афукозилированным h11G2 XC154/XC155 высвобождение цитокинов in vitro являлось ожидаемым в связи с ожидаемыми фармакологическими свойствами этой молекулы. Афукозилированное h11G2 XC154/XC155 не индуцировало высвобождение цитокинов в поисковых или базовых исследованиях in vivo на обезьянах. Можно легко осуществлять мониторинг цитокинов сыворотки и клинических показателей системного высвобождения цитокинов в клинических условиях.
ТАБЛИЦА 16
(ПОСЛЕДОВАТЕЛЬНОСТИ)
(Аминокислотные остатки CDR подчеркнуты)
* указан стоп-кодон
* указан стоп-кодон
* указан стоп-кодон
(VH мыши указана прописными буквами, а константная область IgG1 человека указана строчными буквами)
* указан стоп-кодон
* указан стоп-кодон
* указан стоп-кодон
Последовательность эффекторной функции дикого типа LLGG указана курсивом
N-связанный участок гликозилирования Asn297 NST
Константный домен указан строчными буквами
Константный домен указан строчными буквами
[000501] Хотя описанные идеи представлены со ссылкой на различное применение, способы, наборы и композиции, следует понимать, что можно осуществлять различные изменения и модификации без отклонения от сущности настоящего изобретения и приведенной ниже формулы изобретения. Изложенные выше примеры представлены для лучшего иллюстрирования описываемых идей и не предназначены для их ограничения. Хотя идеи по настоящему изобретению описаны в терминах этих примеров вариантов осуществления, специалистам в этой области будет понятно, что возможны многочисленные варианты и модификации этих примеров вариантов осуществления без излишнего экспериментирования. Все такие варианты и модификации входят в объем идей по настоящему изобретению.
[000502] Все цитируемые в настоящем описании ссылки, включая патенты, патентные заявки, статьи, пособия и т.п., и цитируемые в них ссылки, включены, таким образом, в настоящее описание в качестве ссылки в полном объеме для всех целей. В случае если один или более включенных литературных источников и схожих материалов отличаются или противоречат настоящей заявке, включая, в качестве неограничивающих примеров, определенные термины, использование терминов, описанные способы или т.п., настоящая заявка обладает приоритетом.
[000503] Специалистам в этой области очевидно, что можно осуществлять различные модификации и изменения настоящего изобретения без отклонения от объема или сущности изобретения. Другие варианты осуществления изобретения будут очевидны специалистам в этой области при рассмотрении настоящего описания и практическом осуществлении представленного в настоящем описании изобретения. Следует понимать, что настоящее описание и примеры следует считать исключительно иллюстративными, при этом истинный объем и сущность изобретения указаны в следующей формуле изобретения.
--->
SEQUENCE LISTING
<110> ПФАЙЗЕР ИНК.
ДЗЕ РИДЖЕНТС ОФ ДЗЕ ЮНИВЕРСИТИ ОФ КАЛИФОРНИА
ГРОТ, Рэйчел
СНАЙДЕР, Уилльям, Брайан.
ЦАО, Сяньцзюнь
ДАНН, Роберт, Джозеф
ДАЛ ПОРТО, Джозеф
КАРИН, Майкл
<120> АНТИТЕЛА ПРОТИВ CXCR5 И ИХ КОМПОЗИЦИИ И ПРИМЕНЕНИЕ
<130> 2420-562964RU
<140> RU 2020121591
<141> 2018-11-29
<150> PCT/US2018/062829
<151> 2018-11-28
<150> 62/732,985
<151> 2018-09-18
<150> 62/593,830
<151> 2017-12-01
<160> 100
<170> PatentIn version 3.5
<210> 1
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 1
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 2
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 2
Glu Ser Val Glu Tyr His Gly Thr Ser
1. 5
<210> 3
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 3
Ala Ala Ser Asn Val Glu Ser Gly
1. 5
<210> 4
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 4
Gln Gln Ser Arg Lys Val Pro Trp Thr
1. 5
<210> 5
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 5
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 6
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 6
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 7
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 7
Gly Phe Thr Phe Thr Asp Phe Tyr
1. 5
<210> 8
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 8
Ile Arg Asn Lys Ala Asn Gly Tyr Thr
1. 5
<210> 9
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 9
Ala Arg Val Tyr Gly Ser Thr Leu His Tyr
1. 5 10
<210> 10
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 10
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 11
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 11
Val Arg Val Tyr Gly Ser Thr Leu His Tyr
1. 5 10
<210> 12
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 12
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Ser Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 13
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 13
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 14
<211> 5
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 14
Ser Ser Val Asn Tyr
1. 5
<210> 15
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 15
Glu Thr Ser Arg Leu Ala Ser Gly
1. 5
<210> 16
<211> 9
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 16
Gln Gln Trp Ser Ser Asn Pro Leu Thr
1. 5
<210> 17
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 17
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Thr Tyr Thr Phe Tyr Pro Asp Ile Leu
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala
115 120
<210> 18
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 18
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Thr Tyr Thr Phe Tyr Pro Asp Ile Leu
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala
115 120
<210> 19
<211> 8
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 19
Gly Phe Thr Phe Arg Ser Tyr Gly
1. 5
<210> 20
<211> 7
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 20
Ile Ser Ser Gly Gly Thr Tyr
1. 5
<210> 21
<211> 13
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 21
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His
1. 5 10
<210> 22
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 22
Asp Val Val Val Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Gly Asp Ile Ala Met Tyr Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 23
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 23
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Arg Ala Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Thr Leu Arg Ala Gly Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Ile Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 24
<211> 213
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 24
Gln Ile Val Leu Thr Gln Ser Pro Ala Val Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Thr Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys Arg Thr Val Ala Ala Pro
100 105 110
Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly Thr
115 120 125
Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala Lys
130 135 140
Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln Glu
145 150 155 160
Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser Ser
165 170 175
Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr Ala
180 185 190
Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser Phe
195 200 205
Asn Arg Gly Glu Cys
210
<210> 25
<211> 449
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 25
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Thr Tyr Thr Phe Tyr Pro Asp Ile Leu
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Asn Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val
115 120 125
Phe Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala
130 135 140
Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser
145 150 155 160
Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val
165 170 175
Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro
180 185 190
Ser Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys
195 200 205
Pro Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp
210 215 220
Lys Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly
225 230 235 240
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
245 250 255
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu
260 265 270
Asp Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
275 280 285
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg
290 295 300
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
305 310 315 320
Glu Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu
325 330 335
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
340 345 350
Thr Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu
355 360 365
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
370 375 380
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
385 390 395 400
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp
405 410 415
Lys Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His
420 425 430
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro
435 440 445
Gly
<210> 26
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 26
Asp Val Val Val Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Gly Asp Val Ser Met Phe Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 27
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 27
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asp
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Arg Ala Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Thr Leu Arg Ala Gly Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Ile Leu Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 28
<211> 218
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 28
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg
100 105 110
Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln
115 120 125
Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr
130 135 140
Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser
145 150 155 160
Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr
165 170 175
Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys
180 185 190
His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro
195 200 205
Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 29
<211> 448
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 29
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe
115 120 125
Pro Leu Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu
130 135 140
Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp
145 150 155 160
Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu
165 170 175
Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser
180 185 190
Ser Ser Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro
195 200 205
Ser Asn Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys
210 215 220
Thr His Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro
225 230 235 240
Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser
245 250 255
Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp
260 265 270
Pro Glu Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn
275 280 285
Ala Lys Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val
290 295 300
Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu
305 310 315 320
Tyr Lys Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys
325 330 335
Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr
340 345 350
Leu Pro Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr
355 360 365
Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu
370 375 380
Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu
385 390 395 400
Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys
405 410 415
Ser Arg Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu
420 425 430
Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 30
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 30
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1. 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 31
<211> 329
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 31
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly
325
<210> 32
<211> 372
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 32
Met Asn Tyr Pro Leu Thr Leu Glu Met Asp Leu Glu Asn Leu Glu Asp
1. 5 10 15
Leu Phe Trp Glu Leu Asp Arg Leu Asp Asn Tyr Asn Asp Thr Ser Leu
20 25 30
Val Glu Asn His Leu Cys Pro Ala Thr Glu Gly Pro Leu Met Ala Ser
35 40 45
Phe Lys Ala Val Phe Val Pro Val Ala Tyr Ser Leu Ile Phe Leu Leu
50 55 60
Gly Val Ile Gly Asn Val Leu Val Leu Val Ile Leu Glu Arg His Arg
65 70 75 80
Gln Thr Arg Ser Ser Thr Glu Thr Phe Leu Phe His Leu Ala Val Ala
85 90 95
Asp Leu Leu Leu Val Phe Ile Leu Pro Phe Ala Val Ala Glu Gly Ser
100 105 110
Val Gly Trp Val Leu Gly Thr Phe Leu Cys Lys Thr Val Ile Ala Leu
115 120 125
His Lys Val Asn Phe Tyr Cys Ser Ser Leu Leu Leu Ala Cys Ile Ala
130 135 140
Val Asp Arg Tyr Leu Ala Ile Val His Ala Val His Ala Tyr Arg His
145 150 155 160
Arg Arg Leu Leu Ser Ile His Ile Thr Cys Gly Thr Ile Trp Leu Val
165 170 175
Gly Phe Leu Leu Ala Leu Pro Glu Ile Leu Phe Ala Lys Val Ser Gln
180 185 190
Gly His His Asn Asn Ser Leu Pro Arg Cys Thr Phe Ser Gln Glu Asn
195 200 205
Gln Ala Glu Thr His Ala Trp Phe Thr Ser Arg Phe Leu Tyr His Val
210 215 220
Ala Gly Phe Leu Leu Pro Met Leu Val Met Gly Trp Cys Tyr Val Gly
225 230 235 240
Val Val His Arg Leu Arg Gln Ala Gln Arg Arg Pro Gln Arg Gln Lys
245 250 255
Ala Val Arg Val Ala Ile Leu Val Thr Ser Ile Phe Phe Leu Cys Trp
260 265 270
Ser Pro Tyr His Ile Val Ile Phe Leu Asp Thr Leu Ala Arg Leu Lys
275 280 285
Ala Val Asp Asn Thr Cys Lys Leu Asn Gly Ser Leu Pro Val Ala Ile
290 295 300
Thr Met Cys Glu Phe Leu Gly Leu Ala His Cys Cys Leu Asn Pro Met
305 310 315 320
Leu Tyr Thr Phe Ala Gly Val Lys Phe Arg Ser Asp Leu Ser Arg Leu
325 330 335
Leu Thr Lys Leu Gly Cys Thr Gly Pro Ala Ser Leu Cys Gln Leu Phe
340 345 350
Pro Ser Trp Arg Arg Ser Ser Leu Ser Glu Ser Glu Asn Ala Thr Ser
355 360 365
Leu Thr Thr Phe
370
<210> 33
<211> 372
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 33
Met Asn Tyr Pro Leu Met Leu Glu Met Asp Leu Glu Asn Leu Glu Asp
1. 5 10 15
Leu Phe Leu Glu Phe Asp Lys Phe Asp Asn Tyr Asn Asp Thr Ser Leu
20 25 30
Val Glu Asn His Leu Cys Pro Ala Thr Glu Gly Pro Leu Met Ala Ser
35 40 45
Phe Lys Ala Val Phe Val Pro Val Ala Tyr Ser Leu Ile Phe Leu Leu
50 55 60
Gly Val Ile Gly Asn Val Leu Val Leu Val Ile Leu Glu Arg His Arg
65 70 75 80
Gln Thr Arg Ser Ser Thr Glu Thr Phe Leu Phe His Leu Ala Val Ala
85 90 95
Asp Leu Leu Leu Val Phe Ile Leu Pro Phe Ala Val Ala Glu Gly Ser
100 105 110
Val Gly Trp Val Leu Gly Thr Phe Leu Cys Lys Thr Val Ile Ala Leu
115 120 125
His Lys Val Asn Phe Tyr Cys Ser Ser Leu Leu Leu Ala Cys Ile Ala
130 135 140
Val Asp Arg Tyr Leu Ala Ile Val His Ala Val His Ala Tyr Arg His
145 150 155 160
Arg Arg Leu Leu Ser Ile His Ile Thr Cys Gly Thr Ile Trp Leu Val
165 170 175
Gly Phe Leu Phe Ala Leu Pro Glu Ile Leu Phe Ala Lys Val Ser Gln
180 185 190
Ala His Pro Asn Asn Ser Leu Pro Arg Cys Thr Phe Ser Gln Glu Asn
195 200 205
Gln Ala Glu Thr His Ala Trp Phe Thr Ser Arg Phe Leu Tyr His Val
210 215 220
Ala Gly Phe Leu Leu Pro Met Leu Val Met Gly Trp Cys Tyr Val Gly
225 230 235 240
Val Val His Arg Leu Arg Gln Ala Gln Arg Arg Pro Gln Arg Gln Lys
245 250 255
Ala Val Arg Val Ala Ile Leu Val Thr Ser Ile Phe Phe Leu Cys Trp
260 265 270
Ser Pro Tyr His Ile Val Ile Phe Leu Asp Thr Leu Val Arg Leu Lys
275 280 285
Ala Val Asp Asn Thr Cys Glu Leu Asn Gly Ser Leu Pro Val Ala Ile
290 295 300
Thr Met Cys Glu Phe Leu Gly Leu Ala His Cys Cys Leu Asn Pro Met
305 310 315 320
Leu Tyr Thr Phe Ala Gly Val Lys Phe Arg Ser Asp Leu Ser Arg Leu
325 330 335
Leu Thr Lys Leu Gly Cys Thr Gly Pro Ala Ser Leu Cys Gln Leu Phe
340 345 350
Pro Ser Trp Arg Lys Ser Ser Leu Ser Glu Ser Glu Asn Ala Thr Ser
355 360 365
Leu Thr Thr Phe
370
<210> 34
<211> 374
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 34
Met Asn Tyr Pro Leu Thr Leu Asp Met Gly Ser Ile Thr Tyr Asn Met
1. 5 10 15
Asp Asp Leu Tyr Lys Glu Leu Ala Phe Tyr Ser Asn Ser Thr Glu Ile
20 25 30
Pro Leu Gln Asp Ser Asn Phe Cys Ser Thr Val Glu Gly Pro Leu Leu
35 40 45
Thr Ser Phe Lys Ala Val Phe Met Pro Val Ala Tyr Ser Leu Ile Phe
50 55 60
Leu Leu Gly Met Met Gly Asn Ile Leu Val Leu Val Ile Leu Glu Arg
65 70 75 80
His Arg His Thr Arg Ser Ser Thr Glu Thr Phe Leu Phe His Leu Ala
85 90 95
Val Ala Asp Leu Leu Leu Val Phe Ile Leu Pro Phe Ala Val Ala Glu
100 105 110
Gly Ser Val Gly Trp Val Leu Gly Thr Phe Leu Cys Lys Thr Val Ile
115 120 125
Ala Leu His Lys Ile Asn Phe Tyr Cys Ser Ser Leu Leu Leu Ala Cys
130 135 140
Ile Ala Val Asp Arg Tyr Leu Ala Ile Val His Ala Val His Ala Tyr
145 150 155 160
Arg Arg Arg Arg Leu Leu Ser Ile His Ile Thr Cys Thr Ala Ile Trp
165 170 175
Leu Ala Gly Phe Leu Phe Ala Leu Pro Glu Leu Leu Phe Ala Lys Val
180 185 190
Gly Gln Pro His Asn Asn Asp Ser Leu Pro Gln Cys Thr Phe Ser Gln
195 200 205
Glu Asn Glu Ala Glu Thr Arg Ala Trp Phe Thr Ser Arg Phe Leu Tyr
210 215 220
His Ile Gly Gly Phe Leu Leu Pro Met Leu Val Met Gly Trp Cys Tyr
225 230 235 240
Val Gly Val Val His Arg Leu Leu Gln Ala Gln Arg Arg Pro Gln Arg
245 250 255
Gln Lys Ala Val Arg Val Ala Ile Leu Val Thr Ser Ile Phe Phe Leu
260 265 270
Cys Trp Ser Pro Tyr His Ile Val Ile Phe Leu Asp Thr Leu Glu Arg
275 280 285
Leu Lys Ala Val Asn Ser Ser Cys Glu Leu Ser Gly Tyr Leu Ser Val
290 295 300
Ala Ile Thr Leu Cys Glu Phe Leu Gly Leu Ala His Cys Cys Leu Asn
305 310 315 320
Pro Met Leu Tyr Thr Phe Ala Gly Val Lys Phe Arg Ser Asp Leu Ser
325 330 335
Arg Leu Leu Thr Lys Leu Gly Cys Ala Gly Pro Ala Ser Leu Cys Gln
340 345 350
Leu Phe Pro Asn Trp Arg Lys Ser Ser Leu Ser Glu Ser Glu Asn Ala
355 360 365
Thr Ser Leu Thr Thr Phe
370
<210> 35
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 35
Asp Val Val Val Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Gly Asp Ile Ala Met Tyr Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 36
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 36
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Arg Ala Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Thr Leu Arg Ala Gly Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Ile Leu Thr Val Ser Ser Ala
115 120
<210> 37
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 37
Gln Ile Val Leu Thr Gln Ser Pro Ala Val Met Ser Ala Ser Pro Gly
1. 5 10 15
Glu Lys Val Thr Met Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Ser Gly Thr Ser Pro Lys Arg Trp Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Val Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Ser Tyr Ser Leu Thr Ile Ser Thr Met Glu Ala Glu
65 70 75 80
Asp Ala Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Ala Gly Thr Lys Leu Glu Leu Lys
100 105
<210> 38
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 38
Glu Val Gln Leu Val Glu Ser Gly Gly Asp Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Lys Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Thr Pro Asp Lys Arg Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Thr Tyr Thr Phe Tyr Pro Asp Ile Leu
50 55 60
Lys Gly Arg Ile Thr Ile Ser Arg Asp Asn Ala Lys Asn Thr Leu Tyr
65 70 75 80
Leu Gln Met Ser Asn Leu Lys Ser Glu Asp Thr Ala Met Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His Trp Gly Gln
100 105 110
Gly Thr Ser Val Thr Val Ser Ser Ala
115 120
<210> 39
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 39
Asp Val Val Val Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Gln Arg Ala Thr Ile Ser Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Gln Pro Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Ser Leu Asn Ile His
65 70 75 80
Pro Val Glu Glu Gly Asp Val Ser Met Phe Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
<210> 40
<211> 120
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 40
Glu Val Lys Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Asp
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Pro Pro Gly Arg Ala Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Gln Ser Ile
65 70 75 80
Leu Tyr Leu Gln Met Asn Thr Leu Arg Ala Gly Asp Ser Ala Thr Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Ile Leu Thr Val Ser Ser Ala
115 120
<210> 41
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 41
Asp Ile Val Met Thr Gln Ser Pro Asp Ser Leu Ala Val Ser Leu Gly
1. 5 10 15
Glu Arg Ala Thr Ile Asn Cys Lys Ser Ser Gln Ser Val Leu Tyr Ser
20 25 30
Ser Asn Asn Lys Asn Tyr Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln
35 40 45
Pro Pro Lys Leu Leu Ile Tyr Trp Ala Ser Thr Arg Asp Ser Gly Val
50 55 60
Pro Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr
65 70 75 80
Ile Ser Ser Leu Gln Ala Glu Asp Val Ala Val Tyr Tyr Cys His Gln
85 90 95
Tyr Leu Ser Ser Tyr Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 42
<211> 446
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 42
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Thr Asn
20 25 30
Ala Met Asn Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Arg Val
35 40 45
Ala Arg Ile Arg Ser Lys Ser Asn Asn Tyr Ala Thr Tyr Tyr Ala Asp
50 55 60
Ser Val Lys Asp Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Thr Met Ile Pro Phe Ala Tyr Trp Gly Gln Gly Thr Leu
100 105 110
Val Thr Val Ser Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu
115 120 125
Ala Pro Ser Ser Lys Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys
130 135 140
Leu Val Lys Asp Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser
145 150 155 160
Gly Ala Leu Thr Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser
165 170 175
Ser Gly Leu Tyr Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser
180 185 190
Leu Gly Thr Gln Thr Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn
195 200 205
Thr Lys Val Asp Lys Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His
210 215 220
Thr Cys Pro Pro Cys Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val
225 230 235 240
Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr
245 250 255
Pro Glu Val Thr Cys Val Val Val Asp Val Ser His Glu Asp Pro Glu
260 265 270
Val Lys Phe Asn Trp Tyr Val Asp Gly Val Glu Val His Asn Ala Lys
275 280 285
Thr Lys Pro Arg Glu Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser
290 295 300
Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys
305 310 315 320
Cys Lys Val Ser Asn Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile
325 330 335
Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro
340 345 350
Pro Ser Arg Glu Glu Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu
355 360 365
Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn
370 375 380
Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser
385 390 395 400
Asp Gly Ser Phe Phe Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg
405 410 415
Trp Gln Gln Gly Asn Val Phe Ser Cys Ser Val Met His Glu Ala Leu
420 425 430
His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440 445
<210> 43
<211> 219
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 43
Asp Ile Val Met Thr Gln Ala Ala Pro Ser Val Ala Val Thr Pro Gly
1. 5 10 15
Ala Ser Val Ser Ile Ser Cys Arg Ser Ser Lys Ser Leu Leu His Ser
20 25 30
Ser Gly Lys Thr Tyr Leu Tyr Trp Phe Leu Gln Arg Pro Gly Gln Ser
35 40 45
Pro Gln Leu Leu Ile Tyr Arg Leu Ser Ser Leu Ala Ser Gly Val Pro
50 55 60
Asp Arg Phe Ser Gly Ser Gly Ser Gly Thr Ala Phe Thr Leu Arg Ile
65 70 75 80
Ser Arg Val Glu Ala Glu Asp Val Gly Val Tyr Tyr Cys Met Gln His
85 90 95
Leu Glu Tyr Pro Tyr Thr Phe Gly Gly Gly Thr Lys Leu Glu Ile Lys
100 105 110
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
115 120 125
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
130 135 140
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
145 150 155 160
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
165 170 175
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
180 185 190
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
195 200 205
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
210 215
<210> 44
<211> 440
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 44
Gln Val Gln Leu Lys Glu Ser Gly Pro Gly Leu Val Ala Pro Ser Glu
1. 5 10 15
Ser Leu Ser Ile Thr Cys Thr Val Ser Gly Phe Ser Leu Ile Asp Tyr
20 25 30
Gly Val Asn Trp Ile Arg Gln Pro Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Val Ile Trp Gly Asp Gly Thr Thr Tyr Tyr Asn Pro Ser Leu Lys
50 55 60
Ser Arg Leu Ser Ile Ser Lys Asp Asn Ser Lys Ser Gln Val Phe Leu
65 70 75 80
Lys Val Thr Ser Leu Thr Thr Asp Asp Thr Ala Met Tyr Tyr Cys Ala
85 90 95
Arg Ile Val Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ala Ala
100 105 110
Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys Ser
115 120 125
Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe
130 135 140
Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly
145 150 155 160
Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu
165 170 175
Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr Tyr
180 185 190
Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys Lys
195 200 205
Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys Pro
210 215 220
Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys
225 230 235 240
Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val
245 250 255
Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp Tyr
260 265 270
Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu
275 280 285
Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His
290 295 300
Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys
305 310 315 320
Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln
325 330 335
Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu Met
340 345 350
Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro
355 360 365
Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn
370 375 380
Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu
385 390 395 400
Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn Val
405 410 415
Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln
420 425 430
Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 45
<211> 216
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 45
Gln Pro Val Leu Thr Gln Pro Pro Ser Val Ser Lys Asp Leu Arg Gln
1. 5 10 15
Thr Ala Thr Leu Thr Cys Thr Gly Asn Ser Asn Asn Val Gly Asn Gln
20 25 30
Gly Ala Thr Trp Leu Gln Gln His Gln Gly His Pro Pro Lys Leu Leu
35 40 45
Ser Tyr Lys Asn Asn Asn Arg Pro Ser Gly Ile Ser Glu Arg Phe Ser
50 55 60
Ala Ser Arg Ser Gly Asn Thr Ala Ser Leu Thr Ile Thr Gly Leu Gln
65 70 75 80
Pro Glu Asp Glu Ala Asp Tyr Tyr Cys Ser Ala Trp Asp Ser Ser Leu
85 90 95
Ser Ala Trp Val Phe Gly Gly Gly Thr Gln Leu Thr Val Leu Gly Gln
100 105 110
Pro Lys Ala Ala Pro Ser Val Thr Leu Phe Pro Pro Ser Ser Glu Glu
115 120 125
Leu Gln Ala Asn Lys Ala Thr Leu Val Cys Leu Ile Ser Asp Phe Tyr
130 135 140
Pro Gly Ala Val Thr Val Ala Trp Lys Ala Asp Ser Ser Pro Val Lys
145 150 155 160
Ala Gly Val Glu Thr Thr Thr Pro Ser Lys Gln Ser Asn Asn Lys Tyr
165 170 175
Ala Ala Ser Ser Tyr Leu Ser Leu Thr Pro Glu Gln Trp Lys Ser His
180 185 190
Arg Ser Tyr Ser Cys Gln Val Thr His Glu Gly Ser Thr Val Glu Lys
195 200 205
Thr Val Ala Pro Thr Glu Cys Ser
210 215
<210> 46
<211> 441
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 46
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Val Ser Ser Asn
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ser Val Ile Tyr Ser Gly Gly Ser Thr Tyr Tyr Ala Asp Ser Val Lys
50 55 60
Gly Arg Phe Thr Ile Ser Arg His Asn Ser Lys Asn Thr Leu Tyr Leu
65 70 75 80
Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys Ala
85 90 95
Arg Gly Tyr Val Val Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
100 105 110
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
115 120 125
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
130 135 140
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
145 150 155 160
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
165 170 175
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
180 185 190
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
195 200 205
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
210 215 220
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
225 230 235 240
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
245 250 255
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
260 265 270
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
275 280 285
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
290 295 300
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
305 310 315 320
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
325 330 335
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
340 345 350
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
355 360 365
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
370 375 380
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
385 390 395 400
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
405 410 415
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
420 425 430
Gln Lys Ser Leu Ser Leu Ser Pro Gly
435 440
<210> 47
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 47
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 48
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 48
Asp Val Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 49
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 49
Asp Val Gln Val Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 50
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 50
Asp Val Gln Val Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Phe Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 51
<211> 111
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 51
Asp Ile Gln Val Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Glu Ser Val Glu Tyr His
20 25 30
Gly Thr Ser Leu Met His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro
35 40 45
Lys Leu Leu Ile Tyr Ala Ala Ser Asn Val Glu Ser Gly Val Pro Ser
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Gln Pro Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Arg
85 90 95
Lys Val Pro Trp Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 52
<211> 121
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 52
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Lys Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Pro Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asp Ser Lys Asn Thr
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Lys Thr Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser Ala Ser
115 120
<210> 53
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 53
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 54
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 54
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 55
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 55
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Thr Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 56
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 56
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Val Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 57
<211> 119
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 57
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Thr Asp Phe
20 25 30
Tyr Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Leu
35 40 45
Gly Phe Ile Arg Asn Lys Ala Asn Gly Tyr Thr Thr Glu Tyr Ser Ala
50 55 60
Ser Val Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser
65 70 75 80
Leu Tyr Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr
85 90 95
Tyr Cys Ala Arg Val Tyr Gly Ser Thr Leu His Tyr Trp Gly Gln Gly
100 105 110
Thr Leu Val Thr Val Ser Ser
115
<210> 58
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 58
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 59
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 59
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 60
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 60
Asp Ile Gln Leu Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Trp Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Tyr Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 61
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 61
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Arg Leu Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 62
<211> 106
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 62
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Ser Ala Ser Ser Ser Val Asn Tyr Ile
20 25 30
His Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile Tyr
35 40 45
Glu Thr Ser Arg Leu Ala Ser Gly Val Pro Ser Arg Phe Ser Gly Ser
50 55 60
Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro Glu
65 70 75 80
Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Trp Ser Ser Asn Pro Leu Thr
85 90 95
Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 63
<211> 122
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 63
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Arg Ser Tyr
20 25 30
Gly Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Thr Ile Ser Ser Gly Gly Thr Tyr Thr Phe Tyr Pro Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Gly Glu Asp Tyr Arg Gly Ala Leu Glu His Trp Gly Gln
100 105 110
Gly Thr Leu Val Thr Val Ser Ser Ala Ser
115 120
<210> 64
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 64
atcacgcggc cgcctcacca tgaagttgcc tgttaggctg ttggtgctg 49
<210> 65
<211> 48
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (26)..(26)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (41)..(41)
<223> n может быть c или t
<400> 65
atcacgcggc cgcctcacca tggagncaga cacactcctg ntatgggt 48
<210> 66
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (45)..(45)
<223> n может быть c или g
<400> 66
atcacgcggc cgcctcacca tgagtgtgct cactcaggtc ctggngttg 49
<210> 67
<211> 52
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (26)..(26)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (38)..(38)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (41)..(41)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (46)..(46)
<223> n может быть a или c
<220>
<221> прочий признак
<222> (47)..(47)
<223> n может быть a или t
<400> 67
atcacgcggc cgcctcacca tgaggncccc tgctcagntt nttggnntct tg 52
<210> 68
<211> 49
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (28)..(28)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (41)..(41)
<223> n может быть a или t
<400> 68
atcacgcggc cgcctcacca tggatttnca ggtgcagatt ntcagcttc 49
<210> 69
<211> 46
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (27)..(27)
<223> n может быть g или t
<220>
<221> прочий признак
<222> (29)..(29)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (30)..(30)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (33)..(33)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (35)..(35)
<223> n может быть g или c
<220>
<221> прочий признак
<222> (38)..(38)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (40)..(40)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (44)..(44)
<223> n может быть g или a
<400> 69
atcacgcggc cgcctcacca tgaggtncnn tgntnagntn ctgngg 46
<210> 70
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (26)..(26)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (43)..(43)
<223> n может быть g или t
<220>
<221> прочий признак
<222> (44)..(44)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (45)..(45)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (46)..(46)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (48)..(48)
<223> n может быть a или t
<400> 70
atcacgcggc cgcctcacca tgggcntcaa gatggagtca cannnncngg 50
<210> 71
<211> 50
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (29)..(29)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (32)..(32)
<223> n может быть g или t
<220>
<221> прочий признак
<222> (36)..(36)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (38)..(38)
<223> n может быть a или c
<220>
<221> прочий признак
<222> (39)..(39)
<223> n может быть a или c
<400> 71
atcacgcggc cgcctcacca tgtgggganc tntttncnnt ttttcaattg 50
<210> 72
<211> 44
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (25)..(25)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (29)..(29)
<223> n может быть a или t
<220>
<221> прочий признак
<222> (32)..(32)
<223> n может быть c или g
<400> 72
atcacgcggc cgcctcacca tggtntccnc anctcagttc cttg 44
<210> 73
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 73
atccacgcgg ccgcctcacc atgtatatat gtttgttgtc tatttct 47
<210> 74
<211> 47
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 74
atcacgcggc cgcctcacca tggaagcccc agctcagctt ctcttcc 47
<210> 75
<211> 34
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 75
cagttggtgc agcatccgta cgtttgattt ccag 34
<210> 76
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (33)..(33)
<223> n может быть c или g
<400> 76
actagcggcc gcatgaaatg cagctgggtc atnttcttc 39
<210> 77
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (26)..(26)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (33)..(33)
<223> n может быть c или g
<220>
<221> прочий признак
<222> (34)..(34)
<223> n может быть c или t
<400> 77
actagcggcc gcatgggatg gagctntatc atnntct 37
<210> 78
<211> 40
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (19)..(19)
<223> n может быть a или t
<400> 78
actagcggcc gcatgaagnt gtggttaaac tgggtttttt 40
<210> 79
<211> 37
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (16)..(16)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (25)..(25)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (34)..(34)
<223> n может быть a или g
<400> 79
actagcggcc gcatgnactt tgggntcagc ttgnttt 37
<210> 80
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 80
actagcggcc gcatggactc caggctcaat ttagttttcc tt 42
<210> 81
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (22)..(22)
<223> n может быть c или t
<220>
<221> прочий признак
<222> (24)..(24)
<223> n может быть g или a
<220>
<221> прочий признак
<222> (26)..(26)
<223> n может быть c или g
<220>
<221> прочий признак
<222> (30)..(30)
<223> n может быть g или a
<400> 81
actagcggcc gcatggctgt cntngngctn ctcttctgc 39
<210> 82
<211> 38
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (17)..(17)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (25)..(25)
<223> n может быть g или t
<220>
<221> прочий признак
<222> (28)..(28)
<223> n может быть a или g
<220>
<221> прочий признак
<222> (34)..(34)
<223> n может быть c или a
<400> 82
actagcggcc gcatggnatg gagcnggntc tttntctt 38
<210> 83
<211> 35
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 83
actagcggcc gcatgagagt gctgattctt ttgtg 35
<210> 84
<211> 42
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (17)..(17)
<223> n может быть a или c
<220>
<221> прочий признак
<222> (29)..(29)
<223> n может быть a или c
<400> 84
actagcggcc gcatggnttg ggtgtgganc ttgctattcc tg 42
<210> 85
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 85
actagcggcc gcatggattt tgggctgatt tttttattg 39
<210> 86
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 86
actagcggcc gcatgggcag acttacattc tcattcctg 39
<210> 87
<211> 39
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 87
actagcggcc gcatgatggt gttaagtctt ctgtacctg 39
<210> 88
<211> 31
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<220>
<221> прочий признак
<222> (20)..(20)
<223> n может быть a или c
<220>
<221> прочий признак
<222> (21)..(21)
<223> n может быть g или a
<220>
<221> прочий признак
<222> (27)..(27)
<223> n может быть g или a
<400> 88
gggggtgtcg tcgacgctgn ngagacngtg a 31
<210> 89
<211> 330
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 89
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1. 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 90
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 90
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1. 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 91
<211> 113
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 91
Glu Val Gln Leu Val Glu Ser Gly Gly Gly Leu Val Gln Pro Gly Gly
1. 5 10 15
Ser Leu Arg Leu Ser Cys Ala Ala Ser Gly Phe Thr Phe Ser Ser Tyr
20 25 30
Trp Met Ser Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Asn Ile Lys Gln Asp Gly Ser Glu Lys Tyr Tyr Val Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ala Lys Asn Ser Leu Tyr
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Tyr Phe Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser
<210> 92
<211> 11
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 92
Trp Gly Gln Gly Thr Leu Val Thr Val Ser Ser
1. 5 10
<210> 93
<211> 107
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 93
Asp Ile Gln Met Thr Gln Ser Pro Ser Ser Leu Ser Ala Ser Val Gly
1. 5 10 15
Asp Arg Val Thr Ile Thr Cys Arg Ala Ser Gln Ser Ile Ser Ser Tyr
20 25 30
Leu Asn Trp Tyr Gln Gln Lys Pro Gly Lys Ala Pro Lys Leu Leu Ile
35 40 45
Tyr Ala Ala Ser Ser Leu Gln Ser Gly Val Pro Ser Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Gln Pro
65 70 75 80
Glu Asp Phe Ala Thr Tyr Tyr Cys Gln Gln Ser Tyr Ser Thr Pro Leu
85 90 95
Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105
<210> 94
<211> 10
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 94
Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
1. 5 10
<210> 95
<211> 333
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 95
gacatccaga tgacccagag cccttccagc ctgagcgctt ccgtgggaga tagggtgacc 60
atcacctgca gggccagcga gtccgtggag taccacggca ccagcctgat gcactggtac 120
cagcagaagc ccggcaaggc ccccaagctg ctgatctacg ccgccagcaa cgtggagagc 180
ggcgtgccta gcagattcag cggcagcgga agcggcaccg acttcaccct gaccattagc 240
agcctgcagc ccgaggactt cgccacctac tactgtcagc agagcaggaa ggtgccctgg 300
accttcggcc agggcaccaa ggtcgagatc aag 333
<210> 96
<211> 357
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 96
gaggtgcagc tggtggagag cggaggagga ctggtgcagc ctggcggaag cctgagactg 60
agctgcgccg ccagcggctt cacctttacc gacttctaca tgagctgggt gaggcaggct 120
cccggcaaag gactggagtg ggtgggtttc atcaggaaca aggccaacgg ctacaccacc 180
gagtatagcg cctccgtgaa gggcaggttc accatcagca gggacaacgc caagaacagc 240
ctgtacctgc agatgaacag cctgagggcc gaggacaccg ccgtgtacta ctgcgccaga 300
gtgtacggca gcacactgca ctactggggc cagggcaccc tggtgaccgt gagcagc 357
<210> 97
<211> 657
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 97
gacatccaga tgacccagag cccttccagc ctgagcgctt ccgtgggaga tagggtgacc 60
atcacctgca gggccagcga gtccgtggag taccacggca ccagcctgat gcactggtac 120
cagcagaagc ccggcaaggc ccccaagctg ctgatctacg ccgccagcaa cgtggagagc 180
ggcgtgccta gcagattcag cggcagcgga agcggcaccg acttcaccct gaccattagc 240
agcctgcagc ccgaggactt cgccacctac tactgtcagc agagcaggaa ggtgccctgg 300
accttcggcc agggcaccaa ggtcgagatc aagcgtacgg tggctgcacc atctgtcttc 360
atcttcccgc catctgatga gcagttgaaa tctggaactg cctctgttgt gtgcctgctg 420
aataacttct atcccagaga ggccaaagta cagtggaagg tggataacgc cctccaatcg 480
ggtaactccc aggagagtgt cacagagcag gacagcaagg acagcaccta cagcctcagc 540
agcaccctga cgctgagcaa agcagactac gagaaacaca aagtctacgc ctgcgaagtc 600
acccatcagg gcctgagctc gcccgtcaca aagagcttca acaggggaga gtgttag 657
<210> 98
<211> 1347
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 98
gaggtgcagc tggtggagag cggaggagga ctggtgcagc ctggcggaag cctgagactg 60
agctgcgccg ccagcggctt cacctttacc gacttctaca tgagctgggt gaggcaggct 120
cccggcaaag gactggagtg ggtgggtttc atcaggaaca aggccaacgg ctacaccacc 180
gagtatagcg cctccgtgaa gggcaggttc accatcagca gggacaacgc caagaacagc 240
ctgtacctgc agatgaacag cctgagggcc gaggacaccg ccgtgtacta ctgcgccaga 300
gtgtacggca gcacactgca ctactggggc cagggcaccc tggtgaccgt gagcagcgcg 360
tcgaccaagg gcccatcggt cttccccctg gcaccctcct ccaagagcac ctctgggggc 420
acagcggccc tgggctgcct ggtcaaggac tacttccccg aaccggtgac ggtgtcgtgg 480
aactcaggcg ccctgaccag cggcgtgcac accttcccgg ctgtcctaca gtcctcagga 540
ctctactccc tcagcagcgt ggtgaccgtg ccctccagca gcttgggcac ccagacctac 600
atctgcaacg tgaatcacaa gcccagcaac accaaggtgg acaagaaagt tgagcccaaa 660
tcttgtgaca aaactcacac atgcccaccg tgcccagcac ctgaactcct ggggggaccg 720
tcagtcttcc tcttcccccc aaaacccaag gacaccctca tgatctcccg gacccctgag 780
gtcacatgcg tggtggtgga cgtgagccac gaagaccctg aggtcaagtt caactggtac 840
gtggacggcg tggaggtgca taatgccaag acaaagccgc gggaggagca gtacaacagc 900
acgtaccgtg tggtcagcgt cctcaccgtc ctgcaccagg actggctgaa tggcaaggag 960
tacaagtgca aggtctccaa caaagccctc ccagccccca tcgagaaaac catctccaaa 1020
gccaaagggc agccccgaga accacaggtg tacaccctgc ccccatcccg ggaggagatg 1080
accaagaacc aggtcagcct gacctgcctg gtcaaaggct tctatcccag cgacatcgcc 1140
gtggagtggg agagcaatgg gcagccggag aacaactaca agaccacgcc tcccgtgctg 1200
gactccgacg gctccttctt cctctatagc aagctcaccg tggacaagag caggtggcag 1260
caggggaacg tcttctcatg ctccgtgatg catgaggctc tgcacaacca ctacacgcag 1320
aagagcctct ccctgtcccc gggttga 1347
<210> 99
<211> 990
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 99
gcgtcgacca agggcccatc ggtcttcccc ctggcaccct cctccaagag cacctctggg 60
ggcacagcgg ccctgggctg cctggtcaag gactacttcc ccgaaccggt gacggtgtcg 120
tggaactcag gcgccctgac cagcggcgtg cacaccttcc cggctgtcct acagtcctca 180
ggactctact ccctcagcag cgtggtgacc gtgccctcca gcagcttggg cacccagacc 240
tacatctgca acgtgaatca caagcccagc aacaccaagg tggacaagaa agttgagccc 300
aaatcttgtg acaaaactca cacatgccca ccgtgcccag cacctgaact cctgggggga 360
ccgtcagtct tcctcttccc cccaaaaccc aaggacaccc tcatgatctc ccggacccct 420
gaggtcacat gcgtggtggt ggacgtgagc cacgaagacc ctgaggtcaa gttcaactgg 480
tacgtggacg gcgtggaggt gcataatgcc aagacaaagc cgcgggagga gcagtacaac 540
agcacgtacc gtgtggtcag cgtcctcacc gtcctgcacc aggactggct gaatggcaag 600
gagtacaagt gcaaggtctc caacaaagcc ctcccagccc ccatcgagaa aaccatctcc 660
aaagccaaag ggcagccccg agaaccacag gtgtacaccc tgcccccatc ccgggaggag 720
atgaccaaga accaggtcag cctgacctgc ctggtcaaag gcttctatcc cagcgacatc 780
gccgtggagt gggagagcaa tgggcagccg gagaacaact acaagaccac gcctcccgtg 840
ctggactccg acggctcctt cttcctctat agcaagctca ccgtggacaa gagcaggtgg 900
cagcagggga acgtcttctc atgctccgtg atgcatgagg ctctgcacaa ccactacacg 960
cagaagagcc tctccctgtc cccgggttga 990
<210> 100
<211> 324
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетическая конструкция
<400> 100
cgtacggtgg ctgcaccatc tgtcttcatc ttcccgccat ctgatgagca gttgaaatct 60
ggaactgcct ctgttgtgtg cctgctgaat aacttctatc ccagagaggc caaagtacag 120
tggaaggtgg ataacgccct ccaatcgggt aactcccagg agagtgtcac agagcaggac 180
agcaaggaca gcacctacag cctcagcagc accctgacgc tgagcaaagc agactacgag 240
aaacacaaag tctacgcctg cgaagtcacc catcagggcc tgagctcgcc cgtcacaaag 300
agcttcaaca ggggagagtg ttag 324
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| АНТИ-αvβ8 АНТИТЕЛА И КОМПОЗИЦИИ И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2812478C2 |
| АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2018 |
|
RU2774716C2 |
| ГУМАНИЗИРОВАННЫЕ АНТИ-C5 АНТИТЕЛА И ИХ ПРИМЕНЕНИЕ | 2019 |
|
RU2792237C2 |
| ТЕРАПЕВТИЧЕСКИЕ СРЕДСТВА И СПОСОБЫ ДЛЯ ЛЕЧЕНИЯ TLR2-ОПОСРЕДОВАННЫХ ЗАБОЛЕВАНИЙ И НАРУШЕНИЙ | 2019 |
|
RU2791022C2 |
| EGFL6-СПЕЦИФИЧЕСКИЕ МОНОКЛОНАЛЬНЫЕ АНТИТЕЛА И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2017 |
|
RU2779902C2 |
| КОНСТРУКЦИИ ДНК-АНТИТЕЛ ДЛЯ ПРИМЕНЕНИЯ ПРОТИВ БОЛЕЗНИ ЛАЙМА | 2017 |
|
RU2813829C2 |
| ХИМЕРНЫЕ АНТИГЕННЫЕ РЕЦЕПТОРЫ ДЛЯ ЛЕЧЕНИЯ РАКА | 2017 |
|
RU2826270C2 |
| ВИДЫ КОМБИНИРОВАННОЙ ТЕРАПИИ С ИСПОЛЬЗОВАНИЕМ ХИМЕРНЫХ АНТИГЕННЫХ РЕЦЕПТОРОВ И ИНГИБИТОРОВ PD-1 | 2017 |
|
RU2809160C2 |
| КОМБИНАЦИИ АНТИТЕЛ К STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2804815C2 |
| АНТИТЕЛА, НАПРАВЛЕННЫЕ ПРОТИВ ЛЕЙКОТОКСИНОВ STAPHYLOCOCCUS AUREUS | 2019 |
|
RU2805969C2 |
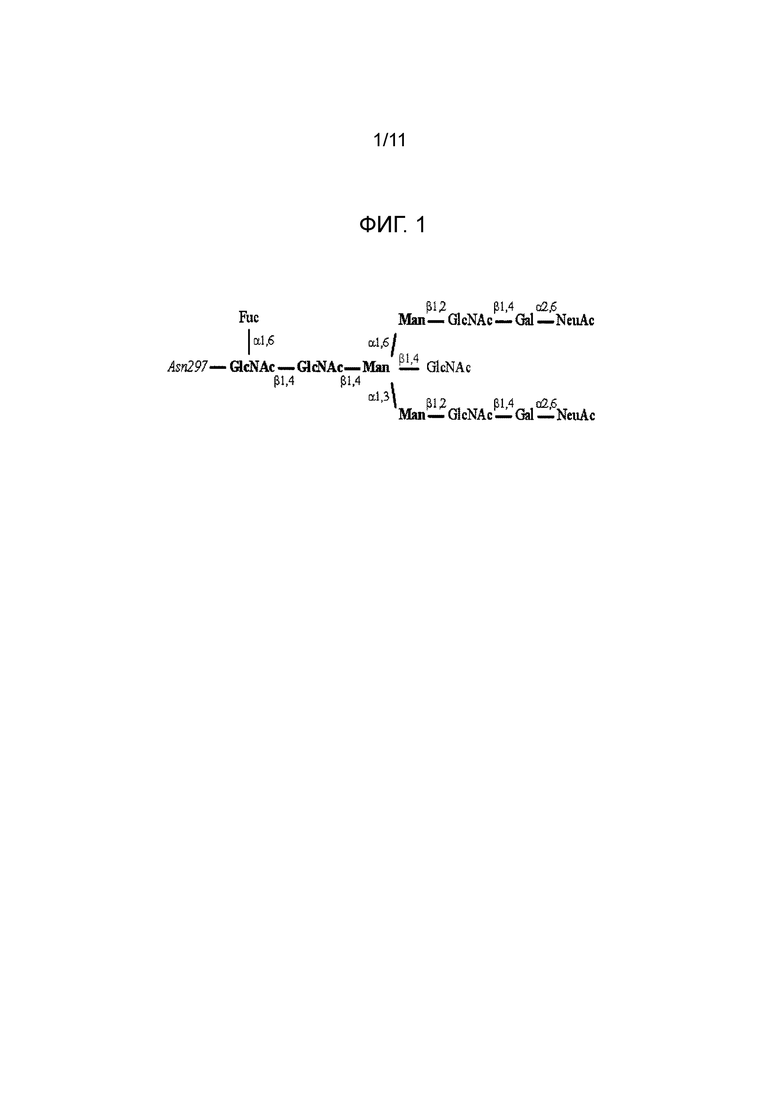
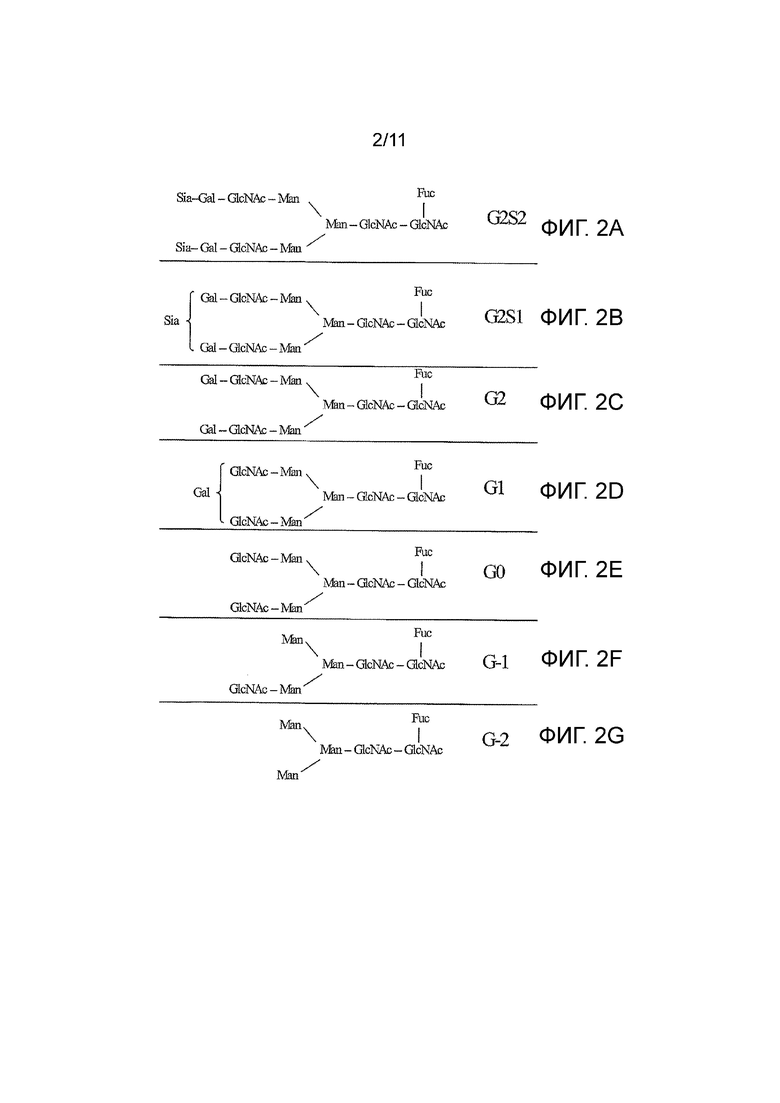
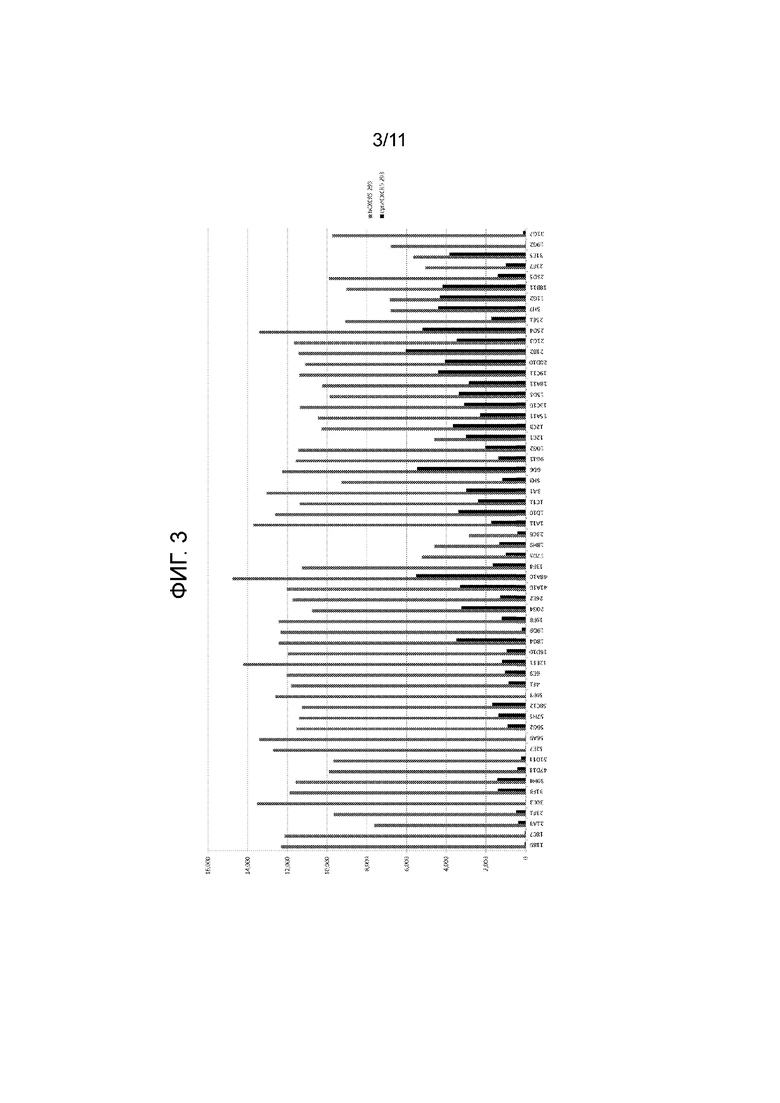
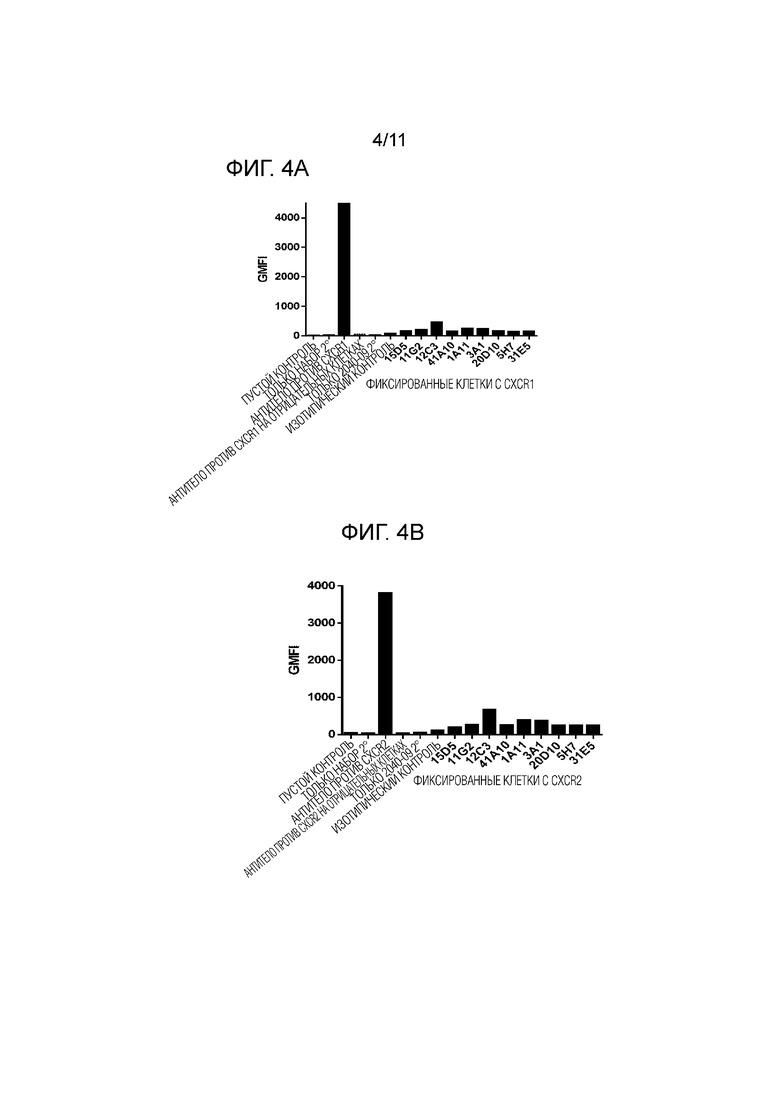
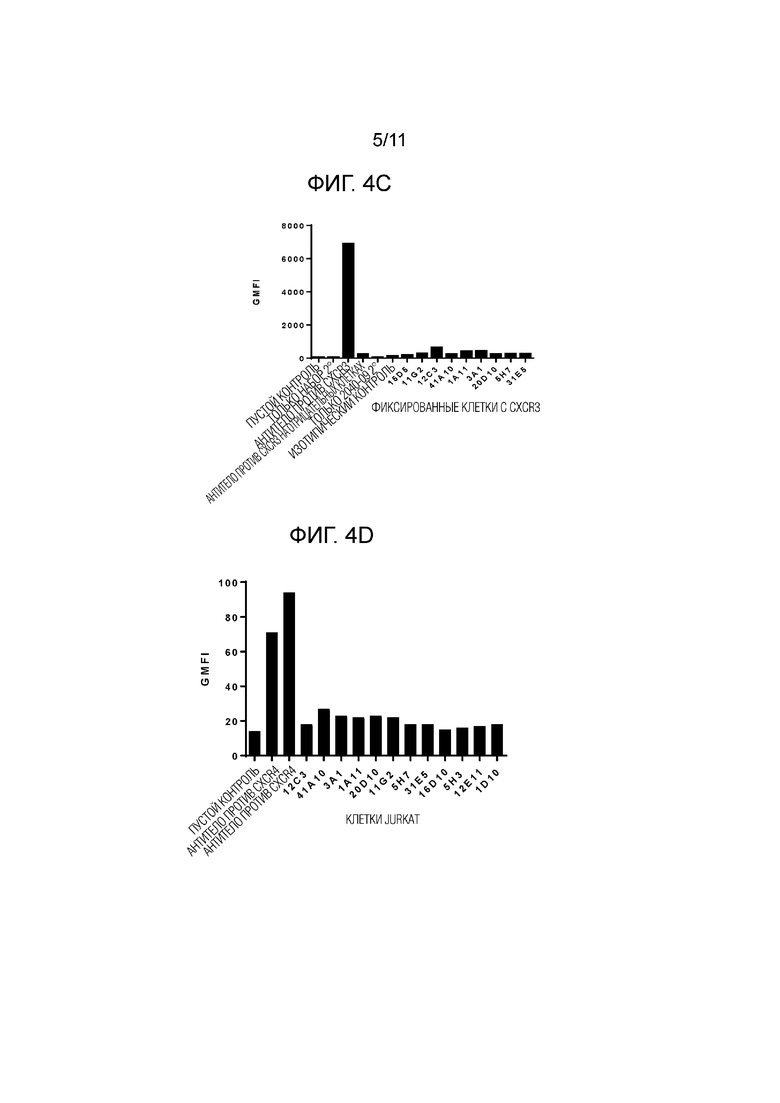
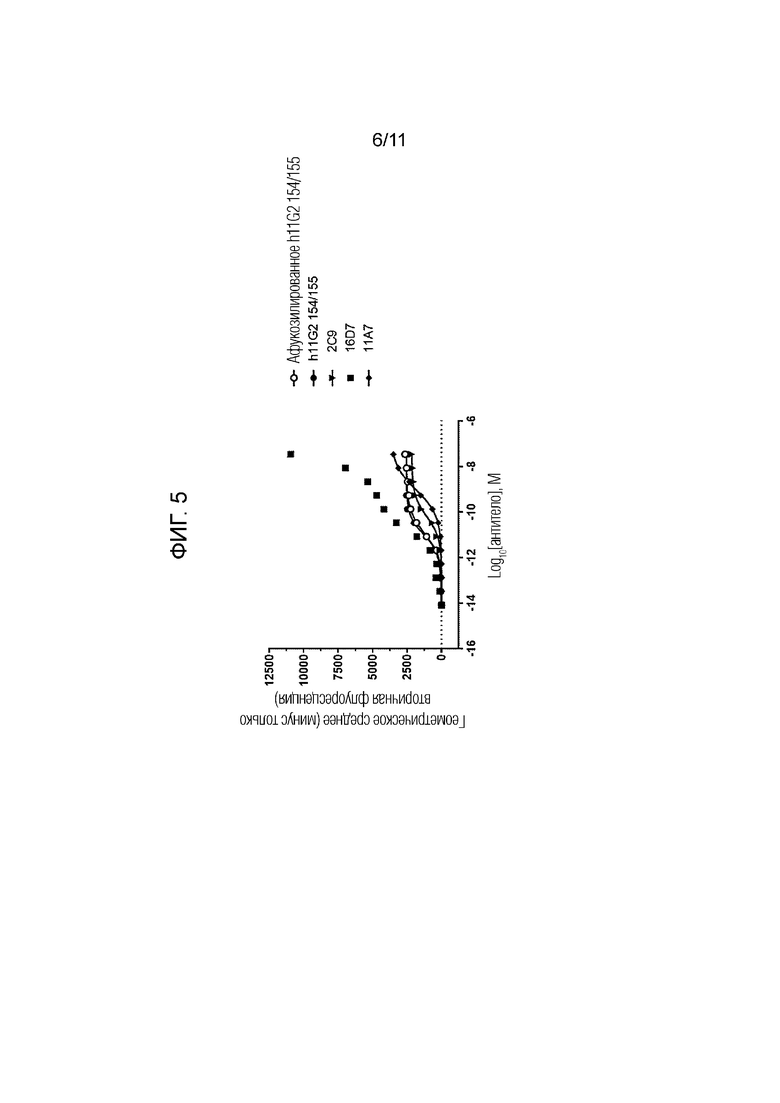

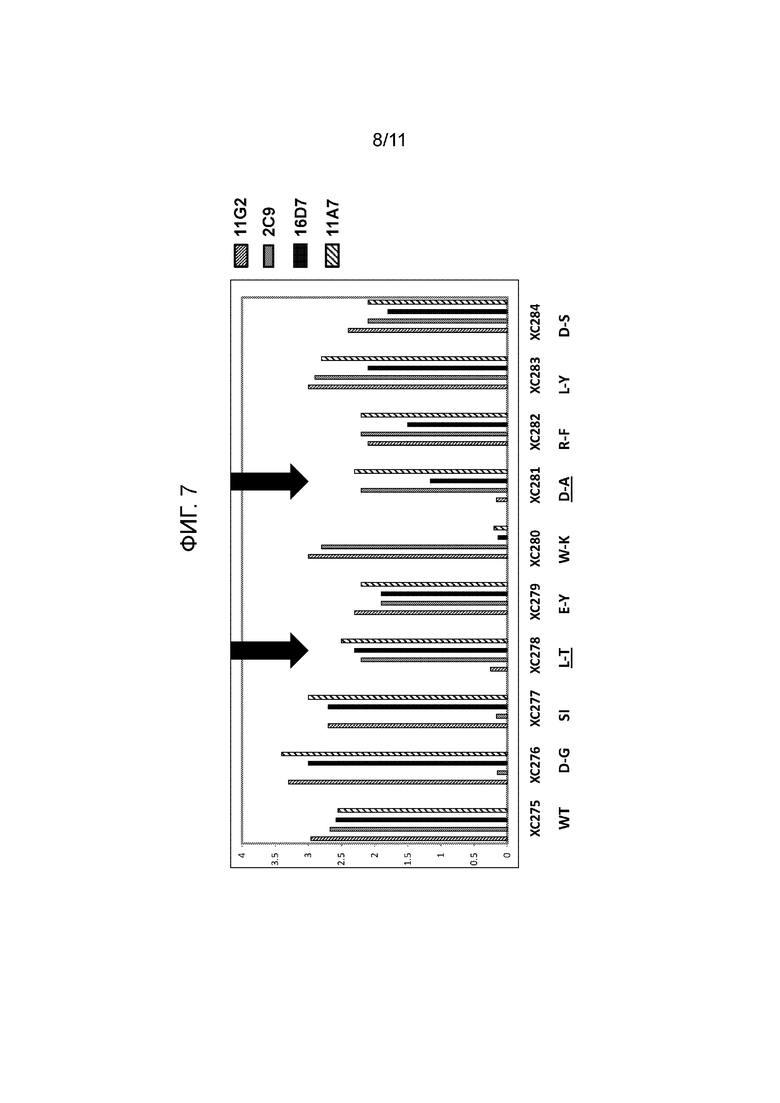
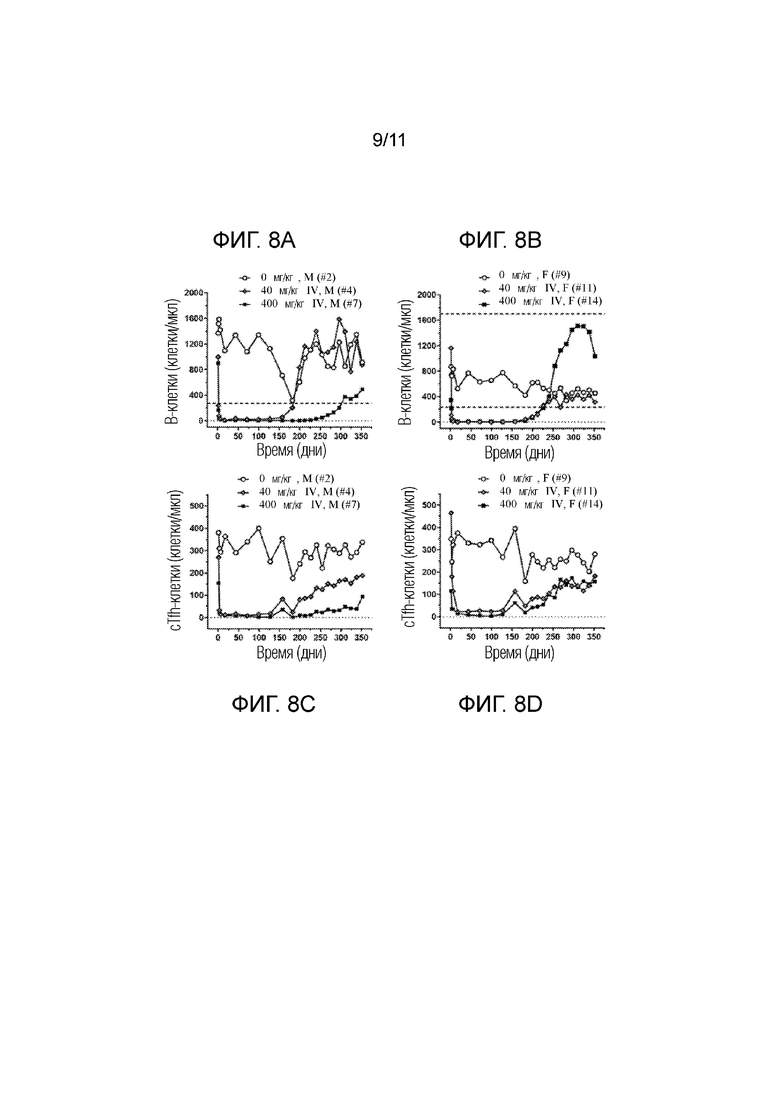
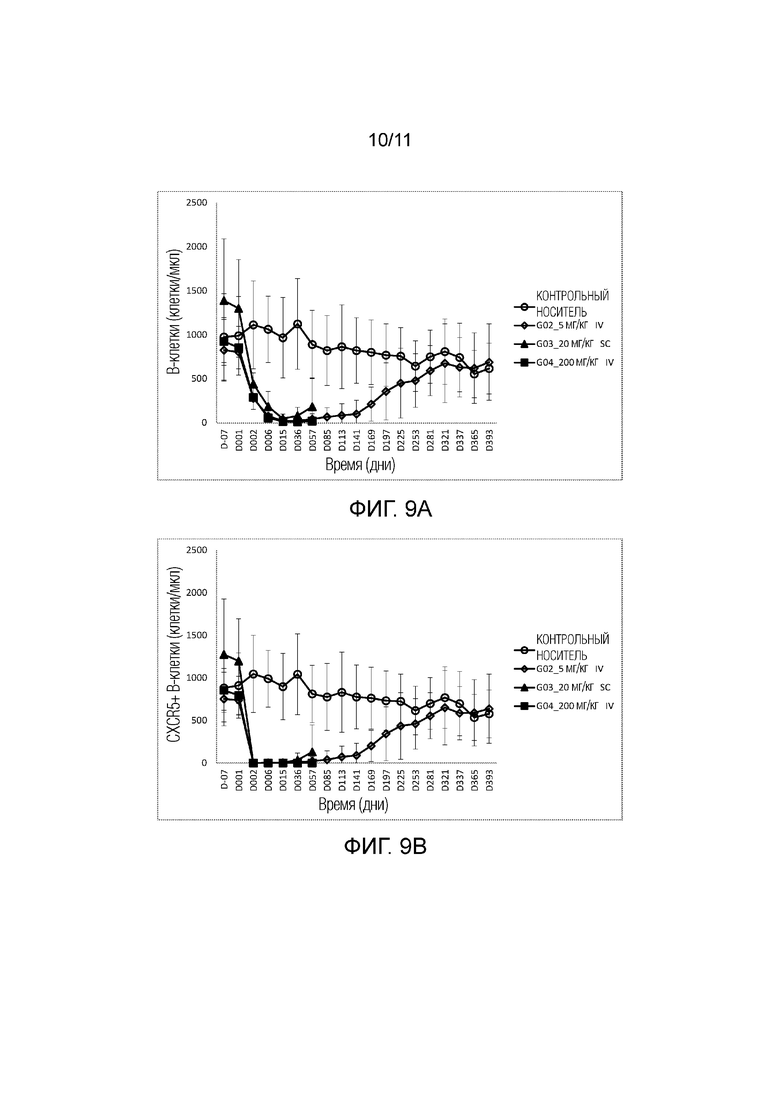
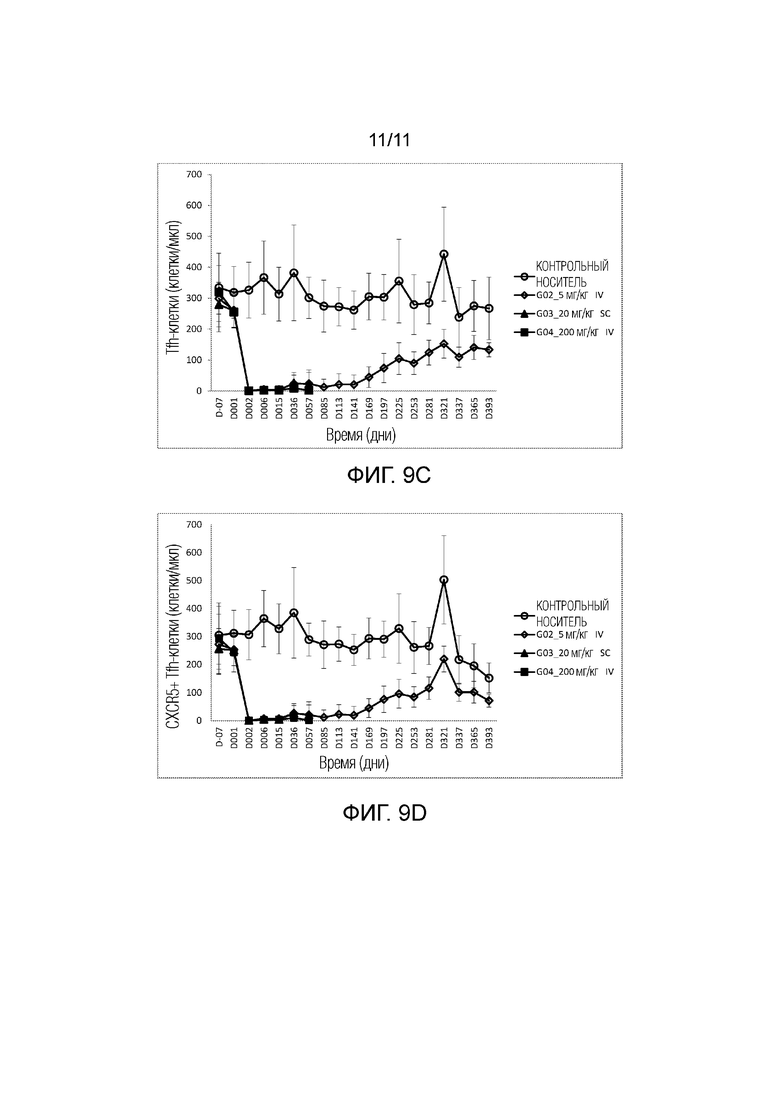
Изобретение относится к области биотехнологии, в частности к выделенному антителу или его антигенсвязывающему фрагменту, специфически связывающемуся с рецептором С-Х-С-хемокина 5 (CXCR5), к композиции, его содержащей, а также к способу его получения. Также раскрыты выделенная нуклеиновая кислота, кодирующая вышеуказанное антитело или его фрагмент, а также к вектору и клетке, ее содержащим. Изобретение также относится к способу детекции CXCR5 в образце, ткани или клетке, предусматривающему использование вышеуказанного антитела или его фрагмента. Изобретение эффективно для лечения или профилактики иммунологического заболевания, нарушения или состояния, опосредованного CXCR5. 18 н. и 29 з.п. ф-лы, 9 ил., 16 табл., 17 пр.
1. Выделенное антитело или его антигенсвязывающий фрагмент, специфически связывающееся с рецептором С-Х-С-хемокина 5 (CXCR5), где антитело является по меньшей мере одним из антител, выбранных из группы, состоящей из:
a) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9;
b) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 11;
c) антитела, содержащего CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 14; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 15; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 16; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 19; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 20; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 21;
d) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 1, и VH, содержащую аминокислотную последовательность SEQ ID NO: 6;
e) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 13, и VH, содержащую аминокислотную последовательность SEQ ID NO: 18;
f) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 47, и VH, содержащую аминокислотную последовательность SEQ ID NO: 52;
g) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 5, и VH, содержащую аминокислотную последовательность SEQ ID NO: 6;
h) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 5, и VH, содержащую аминокислотную последовательность SEQ ID NO: 10;
i) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 13, и VH, содержащую аминокислотную последовательность SEQ ID NO: 17;
j) антитела, содержащего VL, содержащую аминокислотную последовательность SEQ ID NO: 1, и VH, содержащую аминокислотную последовательность SEQ ID NO: 12;
k) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO: 22, и HC, содержащую аминокислотную последовательность SEQ ID NO: 23;
l) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO: 24, и HC, содержащую аминокислотную последовательность SEQ ID NO: 25;
m) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO: 26, и HC, содержащую аминокислотную последовательность SEQ ID NO: 27; и
n) антитела, содержащего LC, содержащую аминокислотную последовательность SEQ ID NO: 28, и HC, содержащую аминокислотную последовательность SEQ ID NO: 29.
2. Антитело или его антигенсвязывающий фрагмент по п.1, где указанное антитело или его антигенсвязывающий фрагмент проявляет по меньшей мере одну из следующих биологических активностей:
a) связывается с CXCR5+ B-клеток человека с кажущейся аффинностью EC50, равной менее приблизительно 26×10-12 молей (пМ), но не связывается с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика;
b) имеет активность антагониста в отношении ингибирования CXCL13 высвобождения цАМФ, запускаемого форсколином;
c) запускает ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и в TMC человека-донора;
d) связывается с CXCR5 человека, но не связывается с хемокиновыми рецепторами CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1 человека;
e) истощает В-клетки периферической крови;
f) истощает Tfh-подобные клетки в периферической крови;
g) истощает истинные клетки Tfh в селезенке; и
h) нарушает гуморальный ответ иммунной памяти.
3. Антитело или его антигенсвязывающий фрагмент по п.1 или 2, содержащее VL, содержащую аминокислотную последовательность SEQ ID NO: 1, и VH, содержащую аминокислотную последовательность SEQ ID NO: 6.
4. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-3, содержащий LC, содержащую аминокислотную последовательность SEQ ID NO: 28, и HC, содержащую аминокислотную последовательность SEQ ID NO: 29.
5. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-4, где антитело или его антигенсвязывающий фрагмент афукозилированы.
6. Выделенное антитело, которое специфически связывается с рецептором C-X-C-хемокина 5 (CXCR5), содержащее LC, состоящую из аминокислотной последовательности SEQ ID NO: 28, и HC, состоящую из аминокислотной последовательности SEQ ID NO: 29.
7. Выделенное антитело, которое специфически связывается с C-X-C-хемокиновым рецептором 5 (CXCR5), содержащее LC, содержащий аминокислотную последовательность SEQ ID NO: 28, и HC, содержащий аминокислотную последовательность SEQ ID NO: 29.
8. Выделенное антитело, которое специфически связывается с C-X-C-хемокиновым рецептором 5 (CXCR5), содержащее область VL антитела, содержащую аминокислотную последовательность SEQ ID NO: 1, и область VH антитела, содержащую аминокислотную последовательность SEQ ID NO: 6.
9. Выделенное антитело, которое специфически связывается с рецептором C-X-C-хемокина 5 (CXCR5), содержащее:
область VL антитела, содержащую первую, вторую и третью CDR из области VL, содержащую аминокислотную последовательность SEQ ID NO: 1; и
область VH антитела, содержащую первую, вторую и третью CDR из области VH, содержащую аминокислотную последовательность SEQ ID NO: 6.
10. Выделенное антитело, которое специфически связывается с рецептором C-X-C-хемокина 5 (CXCR5), содержащее:
а) область VL антитела, содержащую первую, вторую и третью CDR из области VL, содержащую аминокислотную последовательность SEQ ID NO: 1;
область VH антитела, содержащую первую, вторую и третью CDR из области VH, содержащую аминокислотную последовательность SEQ ID NO: 6;
константную область легкой цепи (CL) антитела, содержащую аминокислотную последовательность SEQ ID NO: 30; и
константную область тяжелой цепи (СН) антитела, содержащую аминокислотную последовательность SEQ ID NO: 31; или
b) область VL антитела, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 1 и содержащую СDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 4; и область VH антитела, которая по меньшей мере на 95% идентична аминокислотной последовательности SEQ ID NO: 6 и содержащую CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9.
11. Выделенное антитело, которое специфически связывается с рецептором C-X-C-хемокина 5 (CXCR5), где указанное антитело содержит CDR-L1, содержащую аминокислотную последовательность SEQ ID NO: 2; CDR-L2, содержащую аминокислотную последовательность SEQ ID NO: 3; CDR-L3, содержащую аминокислотную последовательность SEQ ID NO: 4; CDR-H1, содержащую аминокислотную последовательность SEQ ID NO: 7; CDR-H2, содержащую аминокислотную последовательность SEQ ID NO: 8; и CDR-H3, содержащую аминокислотную последовательность SEQ ID NO: 9.
12. Выделенное антитело, которое специфически связывается с CXCR5, по любому из пп.6-11, где антитело афукозилировано.
13. Выделенное антитело или его антигенсвязывающий фрагмент по п.11 или 12, которое специфически связывается с рецептором C-X-C-хемокина 5 (CXCR5),
где антитело или его антигенсвязывающий фрагмент представляет собой по меньшей мере одно антитело, выбранное из группы, состоящей из:
а) антитела или его антигенсвязывающего фрагмента, которое связывается с hCXCR5, экспрессируемым на В-клетках человека, с кажущейся аффинностью ЕС50, равной приблизительно 6,60×10-12 молей (пМ) со стандартным отклонением примерно плюс или минус 2,33 пМ;
b) антитела или его антигенсвязывающего фрагмента, которое связывается с hCXCR5, экспрессируемым на циркулирующих фолликулярных Т-хелперных клетках человека, с кажущейся аффинностью ЕС50, равной приблизительно 5,89 пМ со стандартным отклонением примерно плюс или минус 1,40 пМ;
c) антитела или его антигенсвязывающего фрагмента, которое связывается с hCXCR5, экспрессируемым на фолликулярных Т-хелперных (Tfh) клетках человека с кажущимся сродством EC50 около 10,6 пМ;
d) антитела или его антигенсвяющего фрагмента, которое связывается с cynoCXCR5, экспрессируемым на В-клетках яванского макака, с кажущейся аффинностью EC50, равной приблизительно 1,32 пМ;
e) антитела или его антигенсвязывающего фрагмента, которое связывается с cynoCXCR5, экспрессируемым на Tfh-подобных клетках яванского макака, с кажущейся аффинностью EC50, равной приблизительно 10,5 пМ;
f) антитела или его антигенсвязывающего фрагмента, которое обладает антагонистической активностью на передачу сигнала CXCR5-CXCL13 в репортерном анализе цАМФ с EC50, равной приблизительно 961 пМ;
g) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против B-клеток человека, экспрессирующих hCXCR5, с EC50, равной приблизительно 2,01 пМ со стандартным отклонением приблизительно плюс или минус 2,28 пМ;
h) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против Tfh-подобных клеток человека, экспрессирующих hCXCR5, с EC50, равной приблизительно 4,28 пМ со стандартным отклонением приблизительно плюс или минус 2,88 пМ;
i) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против Tfh-клеток человека, экспрессирующих hCXCR5, с EC50, равной приблизительно 0,11 пМ;
j) антитела или его антигенсвязывающего фрагмента, проявляющего активность ADCC против B-клеток яванского макака, экспрессирующих cynoCXCR5, с EC50, равной приблизительно 15,3 пМ со стандартным отклонением приблизительно плюс или минус 11,7 пМ;
k) антитела или его антигенсвязывающего фрагмента, связывающегося с hCXCR5, но не связывающегося детектируемо с хемокиновыми рецепторами CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 и XCR1 человека;
l) антитела или его антигенсвязывающего фрагмента, ингибирующего связывание CXCR5 с CXCL13;
m) антитела или его антигенсвязывающего фрагмента, связывающегося с CXCR5+ B-клетками человека с кажущейся аффинностью EC50, равной менее приблизительно 26 пМ, но не связывающегося с клетками, экспрессирующими ортологи CXCR5 мыши, крысы или кролика;
n) антитела или его антигенсвязывающего фрагмента, антагонистически действующего на ингибирование CXCL13 высвобождения цАМФ, запускаемого форсколином;
o) антитела или его антигенсвязывающего фрагмента, запускающего ADCC CXCR5-экспрессирующих клеток в PBMC человека-донора и яванского макака и TMC человека-донора;
p) антитела или его антигенсвязывающего фрагмента, связывающегося с CXCR5 человека, но не связывающегося с хемокиновыми рецепторами CCR1, CCR2, CCR3, CCR4, CCR5, CCR6, CCR7, CCR8, CCR9, CCR10, CMKLR1, CXCR3R1, CXCR1, CXCR2, CXCR3, CXCR4, CXCR6, CXCR7 или XCR1 человека;
q) антитела или его антигенсвязывающего фрагмента, истощающего B-клетки в периферической крови;
r) антитела или его антигенсвязывающего фрагмента, истощающего Tfh-подобные клетки в периферической крови;
s) антитела или его антигенсвязывающего фрагмента, истощающего истинные Tfh-клетки в селезенке; и
t) антитела или его антигенсвязывающего фрагмента, нарушающего гуморальный анамнестический иммунный ответ.
14. Выделенная нуклеиновая кислота, кодирующая антитело или его антигенсвязывающий фрагмент по любому из пп.1-13.
15. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 95.
16. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 96.
17. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 95 и SEQ ID NO: 96.
18. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 97.
19. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 98.
20. Выделенная нуклеиновая кислота по п.14, содержащая последовательность нуклеиновой кислоты SEQ ID NO: 97 и SEQ ID NO: 98.
21. Вектор экспрессии, содержащий нуклеиновую кислоту по любому из пп.14-20.
22. Клетка-хозяин для получения антитела или его антигенсвязывающего фрагмента по любому из пп.1-13, содержащая вектор экспрессии по п.21.
23. Клетка-хозяин по п.22, где указанная клетка-хозяин является клеткой млекопитающего, выбранной из группы, состоящей из клетки яичников китайского хомяка (CHO), клетки почек обезьяны (COS), линии 293 эмбриональных клеток почек человека (HEK-293), линии клеток миеломы мыши BALB/c NS0, клетки ретинобласты человека PER.C6® или клетки Sp2.0.
24. Клетка-хозяин по п.23, где в указанной клетке отсутствует функциональная альфа-1,6-фукозилтрансфераза (FUT8).
25. Клетка-хозяин по п.24, где указанная клетка является клеткой Potelligent® CHOK1SV или клеткой Lec13 CHO.
26. Способ получения антитела или его антигенсвязывающего фрагмента, включающий культивирование клетки Potelligent® CHOK1SV по п.25 в условиях, в которых указанное антитело или его антигенсвязывающий фрагмент экспрессируются указанной клеткой-хозяином и являются афукозилированными.
27. Способ по п.26, дополнительно включающий выделение указанного антитела или его антигенсвязывающего фрагмента.
28. Антитело или его антигенсвязывающий фрагмент по любому из пп.6-13, где антитело является афукозилированным и демонстрирует повышенную антитело-зависимую клеточную цитотоксичность (ADCC) по сравнению с в остальном идентичным антителом или его антигенсвязывающим фрагментом, являющимся фукозилированным.
29. Фармацевтическая композиция для лечения или профилактики иммунного заболевания, нарушения или состояния, опосредованного рецептором С-Х-С-хемокина 5 (CXCR5), содержащая антитело или его антигенсвязывающий фрагмент по любому из пп.1-13 и 28 и фармацевтически приемлемый носитель или эксципиент.
30. Антитело или его антигенсвязывающий фрагмент по любому из пп.1-13, 28 и фармацевтическая композиция по п.29 для применения в лечении иммунологического заболевания, нарушения или состояния.
31. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-13, 29 или фармацевтической композиции по п.29 для лечения иммунологического заболевания, нарушения или состояния.
32. Способ лечения или профилактики иммунологического заболевания, нарушения или состояния, опосредованного рецептором С-Х-С хемокина 5 (CXCR5), у больного, включающий введение индивидууму эффективного количества фармацевтической композиции по п.29, где указанное заболевание, нарушение или состояние выбрано из группы, состоящей из воспалительных ответов, таких как воспалительные заболевания кожи, включая псориаз и дерматит; дерматомиозита; системной склеродермии и склероза; ответов, ассоциированных с воспалительным заболеванием кишечника; респираторного дистресс-синдрома; дерматита; менингита; энцефалита; увеита; колита; гастрита; гломерулонефрита; аллергических состояний, таких как экзема и астма и другие состояния, включающие инфильтрацию T-клетками и хронические воспалительные ответы; атеросклероза; дефицита адгезии лейкоцитов; ревматоидного артрита (RA); системной красной волчанки (SLE); сахарного диабета; рассеянного склероза; синдрома Рейно; аутоиммунного тиреоидита; аллергического энцефаломиелита; синдрома Шегрена; ювенильного диабета; и иммунных ответов, ассоциированных с острой и замедленной реакцией гиперчувствительности, опосредованных цитокинами и T-лимфоцитами, как правило, обнаруживаемых при туберкулезе, саркоидозе, полимиозите, гранулематозе и васкулите; гранулематоза Вегенера; пернициозной анемии; заболеваний, включающих диапедез лейкоцитов; воспалительного нарушения центральной нервной системы (ЦНС); синдрома полиорганной недостаточности; гемолитической анемии; миастении; заболеваний, опосредованных комплексами антиген-антитело; болезни анти-БМК-антител; антифосфолипидного синдрома; аллергического неврита; болезни Грейвса; миастенического синдрома Ламберта-Итона; буллезного пемфигоида; пемфигуса; аутоиммунных полиэндокринопатий; витилиго; болезни Рейтера; синдрома мышечной скованности; болезни Бехчета; гигантоклеточного артериита; иммунокомплексного нефрита; IgA-нефропатии; IgM-полинейропатий; иммунной тромбоцитопенической пурпуры (ITP) или аутоиммунной тромбоцитопении и аутоиммунных гемолитических заболеваний; тиреоидита Хашимото; аутоиммунного гепатита; аутоиммунной гемофилии; аутоиммунного лимфопролиферативного синдрома (ALPS); аутоиммунного увеоретинита; синдрома Гийена-Барре; синдрома Гудпасчера; смешанного заболевания соединительной ткани; аутоиммунно-ассоциированного бесплодия; нодозного полиартериита; гнездной алопеции; идиопатической микседемы; реакции "трансплантат против хозяина"; мышечной дистрофии и злокачественных опухолей, экспрессирующих CXCR5.
33. Способ по п.32, где дерматит представляет собой атопический дерматит.
34. Способ по п.32, где воспалительное заболевание кишечника представляет собой болезнь Крона и язвенный колит.
35. Способ по п.32, где респираторный дистресс-синдром представляет собой респираторный дистресс-синдром взрослых (ARDS).
36. Способ по п.32, где сахарный диабет представляет собой сахарный диабет I типа или инсулинозависимый сахарный диабет.
37. Способ по п.32, где пернициозная анемия связана с болезнью Аддисона.
38. Способ по п.32, где гемолитическая анемия представляет собой криоглобулинемию или аутоиммунную гемолитическую анемию с неполными тепловыми агглютининами.
39. Способ по п.32, где мышечная дистрофия представляет собой дистрофию Дюшена, дистрофию Беккера, миотоническую дистрофию, конечностно-поясную дистрофию, плече-лопаточно-лицевую дистрофию, врожденную дистрофию, окулофарингеальную дистрофию, дистальную дистрофию, дистрофию Эмери-Дрейфуса.
40. Способ по п.32, где злокачественные клетки, экспрессирующие CXCR5, относятся к раку поджелудочной железы, раку толстого кишечника, раку мочевого пузыря, T-клеточному лейкозу и B-клеточному лейкозу.
41. Способ по п.32, где указанное заболевание является SLE или ревматоидным артритом.
42. Применение антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или 28 для получения лекарственного средства для лечения иммунологического заболевания, нарушения или состояния.
43. Способ детекции рецептора С-Х-С-хемокина 5 (CXCR5) в образце, ткани или клетке с использованием антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или 28, включающий приведение образца, ткани или клетки в контакт с антителом и детекцию CXCR5 в образце, ткани, клетке путем детекции антитела.
44. Способ снижения биологической активности рецептора С-Х-С-хемокина 5 (CXCR5) у больного, включающий введение терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или 28 или фармацевтической композиции по п.29.
45. Способ по п.44, где антитело опосредует истощение по меньшей мере одной клетки, экспрессирующей CXCR5, выбранной из группы, состоящей из Tfh-клетки в селезенке, B-клетки в периферической крови и Tfh-подобной клетки в периферической крови.
46. Способ ингибирования гуморального иммунного ответа у больного, включающий введение терапевтически эффективного количества антитела или его антигенсвязывающего фрагмента по любому из пп.1-13 или 28 или фармацевтической композиции по п.29.
47. Способ по п.46, где антитело опосредует истощение по меньшей мере одной клетки, экспрессирующей CXCR5, выбранной из группы, состоящей из фолликулярной Т-хелперной (Tfh)-клетки в селезенке, B-клетки в периферической крови и Tfh-подобной клетки в периферической крови.
| WO 2012010582 A1, 26.01.2012 | |||
| WO 2016028573 A1, 25.02.2016 | |||
| ГУМАНИЗИРОВАННЫЕ АНТИТЕЛА К CXCR5, ИХ ПРОИЗВОДНЫЕ И ИХ ПРИМЕНЕНИЕ | 2008 |
|
RU2571515C2 |
| NAOKO YAMANE-OHNUKI et al., Establishment of FUT8 Knockout Chinese Hamster Ovary Cells: An Ideal Host Cell Line for Producing Completely Defucosylated Antibodies With Enhanced Antibody-Dependent Cellular Cytotoxicity, Biotechnology and Bioengineering, 2004, | |||
