ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к антителу или его антигенсвязывающей части, которые могут связываться с белком 1 программируемой клеточной смерти (PD-1) человека. Антитело по настоящему изобретению, кроме того, используют в получении лекарственного средства для лечения заболеваний, при которых экспрессия PD-1 является вредной.
УРОВЕНЬ ТЕХНИКИ ДЛЯ ИЗОБРЕТЕНИЯ
PD-1 (CD279) представляет собой белок из 288 аминокислот, ингибирующий рецептор, экспрессированный на активированных T-клетках и B-клетках, клетках естественных киллерах и моноцитах. PD-1 является членом семейства T-клеточных коингибирующих рецепторов CD28/CTLA-4 (антигена цитотоксических T-лимфоцитов)/ICOS (индуцируемого костимулятора). Рецептор PD-1 имеет два лиганда, а именно, белок лиганд смерти 1 (PD-L1) и белок лиганд смерти 2 (PD-L2). PD-L1 (CD274, B7H 1) широко экспрессируется как на лимфоидных, так и на нелимфоидных тканях, таких как CD4 и CD8 T-клетки, клетки линий макрофагов, периферические ткани, так же как на клетках опухолей, инфицированных вирусом клетках и клетках аутоиммунной ткани. PD-L2 (CD273, B7-DC) имеет более ограниченную экспрессию, чем PD-L1, являясь экспрессированным на активированных дендритных клетках и макрофагах (1). PD-L1 экспрессируется в большинстве злокачественных опухолей человека, включая меланому, глиому, немелкоклеточный рак легкого, плоскоклеточную карциному головы и шеи, лейкоз, рак поджелудочной железы, почечноклеточную карциному и печеночноклеточную карциному, и может являться индуцируемым почти во всех типах злокачественных опухолей (2). Связывание PD-1 с его лигандами приводит к уменьшенной пролиферации T-клеток и секреции цитокинов, супрессируя гуморальный и клеточный иммунные ответы и ухудшая течение заболеваний, где активный иммунный ответ в ином случае облегчал бы состояние заболевания. Эту иммуносупрессию можно обращать посредством ингибирования локального взаимодействия PD-1 с PD-L1, и эффект является аддитивным также, когда взаимодействие PD-1 с PD-L2 блокировано (3, 4).
Блокирование PD-1 с использованием антагонистов, включая моноклональные антитела, исследовали при лечении злокачественных опухолей и хронических вирусных инфекций (5).
Моноклональные антитела против PD-1 известны в данной области и описаны, например, в Патентных документах WO2006121168, WO2009114335, WO2008156712, WO2012145493, WO2015036394, WO2015112800, WO2016015685 и WO2018128939.
Три антитела, нацеленные на PD-1 человека, для лечения различных злокачественных опухолей, в комбинации с общепринятыми лекарственными средствами, являются коммерчески доступными. Эти три антитела представляют собой ниволумаб, пембролизумаб и цемиплимаб.
Несмотря на клинический успех антител против PD-1, эти терапевтические антитела имеют несколько недостатков, включая высокую стоимость, ограниченное время полужизни и иммуногенность.
Соответственно, в данной области существует постоянная необходимость основанного на пути PD-1/PD-L1 лечения заболеваний с использованием антител, которые могут эффективно связывать PD-1 человека и блокировать его связывание с PD-L1 или PD-L2, а также обеспечивать улучшение, по отношению к некоторым из проблем существующих видов терапии. Настоящее изобретение относится к таким новым антителам.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым антителам против PD-1, которые имеют одну или несколько улучшенных характеристик, например, относительно известных антител против PD-1, используемых для терапевтических целей. Антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению связывает с высокой аффинностью PD-1 человека. Аминокислотная последовательность константной области антитела против PD-1 состоит из константной области IgGl, IgG2, IgG3, IgG4, IgA, IgE, IgM или IgD, предпочтительно, IgGl или IgG4. Кроме того, одно или несколько антител против PD-1 по настоящему изобретению имеют модифицированную или уменьшенную, или отсутствующую активность ADCC и/или CDC. В некоторых аспектах, настоящее изобретение относится к антителам против PD-1, имеющим высокую активность ADCC и/или CDC, которые могут приводить к лизису экспрессирующих PD-1 клеток. Антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет KD 10-10 M или менее, более предпочтительно, 10-11 M или менее и даже более предпочтительно, 10-12 M или менее для антигена PD-1. Значение KD представляет собой измерение аффинности связывания антитела по отношению к его антигену-мишени. Антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно использовать для лечения заболеваний, где взаимодействие PD-1 с PDL1 и/или PDL2 вовлечено в модификацию состояния заболевания, например, при инфекциях и различных злокачественных опухолях. В одном аспекте, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению блокирует взаимодействие рецептора PD-1 с его природным лигандом PD-L1 и/или PD-L2. В другом аспекте изобретения, антитело против PD-1 или его антигенсвязывающая часть уничтожает T-клетки, экспрессирующие PD-1. Настоящее изобретение относится также к способам получения новых антител против PD-1 и содержащих их фармацевтических композиций.
КРАТКОЕ ОПИСАНИЕ ЧЕРТЕЖЕЙ
На фиг. 1 изображена общая схема вектора для векторной конструкции pZRCIIhyg LC-HC против PD-1, которую используют для получения полноразмерного моноклонального антитела против PD-1 по настоящему изобретению.
На фиг. 2 изображена общая схема вектора pSEX83, используемого для получения библиотеки, для получения Fab против PD-1 по настоящему изобретению.
На фиг. 3 изображена общая схема вектора для векторной конструкции pZRCIIIhyg LC-HC против PD-1, которую используют для получения полноразмерного моноклонального антитела против PD-1 из Fab против PD-1 по настоящему изобретению.
На фиг. 4 изображены результаты эксперимента проточной цитометрии, показывающие, что антитела против PD-1 IP-H4.2L1, IP-H4L1 (IgG1) и IP-H4L1 (IgG4) по настоящему изобретению, направленные против PD-1 человека, связывают PD-1 на клеточной поверхности PBMC человека.
На фиг. 5 изображены результаты эксперимента проточной цитометрии с использованием линий клеток HEK 293T, показывающие, что антитела против PD-1 IP-H4.2L1, IP-H4.19L1, IP-H4.36L1 по настоящему изобретению, направленные против PD-1 человека, связывают PD-1 на клеточной поверхности.
На фиг. 6 изображены результаты эксперимента проточной цитометрии с использованием линий клеток HEK 293T, показывающие, что антитела против PD-1 N5NL, N6NL, N7NL, N9NL и N10NL по настоящему изобретению, направленные против PD-1 человека, связывают PD-1 на клеточной поверхности.
На фиг. 7 изображены результаты эксперимента, показывающие, что антитела против PD-1 IP-H4.2L1, IP-H4.19L1, IP-H4.36L1 по настоящему изобретению, направленные против PD-1 человека, стимулируют секрецию интерлейкина-2 (IL-2) в анализе реакции смешанной культуры лимфоцитов.
На фиг. 8 изображены результаты эксперимента, показывающие, что антитела против PD-1 IP-H4.2L1, IP-H4.19L1, IP-H4.36L1 по настоящему изобретению, направленные против PD-1 человека, стимулируют секрецию IFN-гамма (IFN-γ) в анализе реакции смешанной культуры лимфоцитов.
На фиг. 9 изображены результаты эксперимента, показывающие, что антитела против PD-1 N5NL, N6NL, N9NL и N10NL по настоящему изобретению, направленные против PD-1 человека, стимулируют секрецию IL-2 в анализе реакции смешанной культуры лимфоцитов.
На фиг. 10 изображены результаты эксперимента, показывающие, что антитела против PD-1 N5NL, N6NL, N9NL и N10NL по настоящему изобретению, направленные против PD-1 человека, стимулируют секрецию IFN-γ в анализе реакции смешанной культуры лимфоцитов.
Определения
Термин «антитело», как обозначено в настоящем описании, включает полноразмерные антитела и любой их антигенсвязывающий фрагмент (т.е., «антигенсвязывающую часть») или их одиночные цепи. «Антитело» относится к гликопротеину, содержащему по меньшей мере две тяжелые (H) цепи и две легкие (L) цепи, соединенные между собой дисульфидными связями, или его антигенсвязывающей части. Каждая тяжелая цепь состоит из вариабельной области тяжелой цепи (сокращенно обозначенной в настоящем описании как VH) и константной области тяжелой цепи (сокращенно обозначенной в настоящем описании как CH). Константная область тяжелой цепи состоит из трех доменов, CH1, CH2 и CH3. Каждая легкая цепь состоит из вариабельной области легкой цепи (сокращенно обозначенной в настоящем описании как VL) и константной области легкой цепи. Константная область легкой цепи состоит из одного домена, CL. Области VH и VL можно дополнительно подразделять на области гипервариабельности, названные определяющими комплементарность областями (CDR), перемежающиеся областями, которые являются более консервативными, названными каркасными областями (FR). Каждая VH и VL состоит из трех CDR и четырех FR, аранжированных от амино-конца к карбокси-концу в следующем порядке: FRl, CDRl, FR2, CDR2, FR3, CDR3, FR4. Вариабельные области тяжелых и легких цепей содержат связывающий домен, взаимодействующий с антигеном. Константные области антител могут опосредовать связывание иммуноглобулина с тканями или факторами хозяина, включая различные клетки иммунной системы (например, иммуноэффекторные клетки) и первый компонент (C1q) классической системы комплемента.
Термин «функционально связанный» предназначен для обозначения того, что ген антитела лигирован в вектор таким образом, что последовательности для контроля транскрипции и трансляции внутри вектора служат для предназначенной для них функции регуляции транскрипции и трансляции гена антитела.
Термины «Ka» и Kd хорошо известны специалисту в данной области, где «Ka» представляет собой скорость связывания для конкретного взаимодействия антитело-антиген, в то время как термин «Kd» представляет собой скорость диссоциации для конкретного взаимодействия антитело-антиген. Термин «KD» представляет собой константу аффинности, которую получают из соотношения Kd и Ka. Ее можно измерять с использованием способа поверхностного плазмонного резонанса, который хорошо известен в данной области. Значение KD является измерением аффинности связывания антитела по отношению к его антигену-мишени. Термин «KD» определен также в WO 2006121168. Содержание этого Патентного документа приведено в настоящем описании в качестве ссылки.
Термины «моноклональные антитело» или «композиция моноклональных антител», в рамках изобретения, относятся к препарату молекул антител одного молекулярного состава. Композиция моноклональных антител имеет одну специфичность связывания и аффинность для конкретного эпитопа.
Термин «биспецифическое антитело» относится к гомогенной популяции антител, вовлеченной в высоко специфическое узнавание и связывание двух различных антигенных детерминант или эпитопов.
Термин «рекомбинантное антитело» по настоящему изобретению, включает моноклональные антитела, которые получают рекомбинантным способом с использованием синтетических генов тяжелых и легких цепей. Рекомбинантные антитела по этому изобретению представляют собой моноклональные антитела (mAb), которые не получены с использованием традиционных технологий на основе гибридомы, и не нуждаются в гибридомах и животных в процессе получения.
Термин «иммуноэффекторная функция», в рамках изобретения, представляет собой биохимическое событие, возникающее в результате взаимодействия области Fc антитела с рецептором или лигандом Fc. Эффекторные функции включают, но без ограничения ADCC, ADCP и CDC. Термин также представляет физиологическое событие, такое как время полужизни в кровотоке лекарственного средства или нацеливание лекарственного средства на конкретный тип клетки или ткани.
Термин «ADCC» или «антителозависимая опосредованная клетками цитотоксичность», в рамках изобретения, представляет собой опосредованную клетками реакцию, где неспецифические цитотоксические клетки, которые экспрессируют FcγR, узнают связанное антитело на клетке-мишени и затем вызывают лизис клетки-мишени.
Термин «ADCP» или «антителозависимый опосредованный клетками фагоцитоз», в рамках изобретения, представляет собой опосредованную клетками реакцию, где неспецифические цитотоксические клетки, которые экспрессируют FcγR, узнают связанное антитело на клетке-мишени и впоследствии вызывают фагоцитоз клетки-мишени.
Термин «иммуноэффекторная клетка», в рамках изобретения, представляет собой клетку, которая экспрессирует один или несколько рецепторов Fc и опосредует одну или несколько эффекторных функций. Эффекторные клетки включают, но без ограничения, моноциты, макрофаги, нейтрофилы, дендритные клетки, эозинофилы, тучные клетки, тромбоциты, B-клетки, большие гранулярные лимфоциты, клетки Лангерганса, клетки естественные киллеры (NK), и γδ - T-клетки, и могут происходить из любого организма, включая, но без ограничения, человека, мышей, крыс, кроликов и обезьян.
Термин фрагмент «Fc», название которого отражает его способность легко кристаллизоваться. В молекулах IgG, область Fc получают посредством расщепления папаином на N-конце от Cys 226. Область Fc является центральной для иммуноэффекторных функций антител.
Термин «белок Fc», в рамках изобретения, относится к части одной тяжелой цепи иммуноглобулина, начинающейся в шарнирной области непосредственно выше участка расщепления папаином и заканчивающийся на C-конце антитела., соответственно, полный домен Fc содержит по меньшей мере часть шарнирного (например, верхней, средней и/или нижней шарнирной области) домена, домена CH2 и домена CH3.
Термин «фармацевтический состав» относится к препаратам, которые находятся в такой форме, чтобы обеспечивать однозначно эффективную биологическую активность активных ингредиентов. Термин «фармацевтический состав» или «фармацевтическая композиция», или «композиция» могут быть использованы в настоящем описании взаимозаменяемо.
Термин «наполнитель» относится к средству, которое можно добавлять к составу для стабилизации активной лекарственной субстанции в составленной форме для коррекции и поддержания осмоляльности и pH фармацевтических препаратов. Примеры общеупотребительных наполнителей включают, но без ограничения, сахара, полиолы, аминокислоты, поверхностно-активные вещества и полимеры. «Фармацевтически приемлемые» наполнители представляют собой наполнители, которые можно целесообразно вводить субъекту-млекопитающему для обеспечения эффективной дозы используемого активного ингредиента.
Термин «лечение» или «терапевтические средства», в рамках изобретения, относится к любому лечению заболевания у млекопитающего, в частности, у человека. Он включает: (a) предотвращение возникновения заболевания у субъекта, который может являться предрасположенным к заболеванию или подверженным риску получения заболевания, но еще не был диагностирован как имеющий его; (b) ингибирование заболевания, т.е., остановка его развития; и (c) облегчение заболевания, т.е., вызов регрессии заболевания.
Термины «пациент» и «субъект» использованы взаимозаменяемо и использованы в своем общепринятом смысле для обозначения живого организма, страдающего от состояния или подверженного состоянию, которое можно предотвращать или лечить посредством введения композиции по настоящему изобретению, и включают как человека, так и не относящихся к человеку животных. Примеры субъектов включают, но без ограничения, человека, шимпанзе и другие виды обезьян старого и нового света; сельскохозяйственных животных, таких как крупный рогатый скот, овцы, свиньи, козы и лошади; домашних млекопитающих, таких как собаки и кошки; лабораторных животных, включая грызунов, таких как мыши, крысы и морские свинки; птиц, включая домашних, диких и промысловых птиц, таких как куры, индюшки и другие куриные, утки, гуси и т.п. Термин не обозначает конкретный возраст. Таким образом, взрослые, молодые и новорожденные индивидуумы представляют интерес.
«Эффективное количество» антитела по изобретению, или его композиции представляет собой количество, которое доставляют субъекту-млекопитающему, либо в однократной дозе, либо в качестве части из серий, которое является эффективным для индукции иммунного ответа против антигена-мишени у указанного субъекта. Это количество меняется в зависимости от состояния здоровья и физического состояния индивидуума, подлежащего лечению, таксономической группы индивидуума, подлежащего лечению, и других важных факторов. Ожидают, что количество может попадать в относительно широкий диапазон, который можно определять посредством общепринятых исследований.
«Фармацевтически эффективная доза» или «терапевтически эффективная доза» представляет собой дозу, необходимую для лечения или предотвращения, или облегчения одного или нескольких связанных с PD-1 нарушений или симптомов у субъекта, предпочтительно, по настоящему изобретению, для злокачественной опухоли или инфекции, или аутоиммунного заболевания. Фармацевтически эффективная доза зависит, среди прочего, от конкретного соединения (в настоящем описании оно представляет собой антитело против PD-1 или его комбинацию или конъюгат, или биспецифическое антитело) для введения, тяжести симптомов, чувствительности субъекта к побочным эффектам, типа заболевания, используемой композиции, способа введения, типа млекопитающего, подвергаемого лечению, физических характеристик конкретного млекопитающего, принимаемых во внимание, таких как состояние здоровья и физическое состояние, сопутствующее лекарственное средство, желательная степень защиты и другие факторы, хорошо известные специалисту в данной области.
Сокращенные наименования аминокислот, как использовано в настоящей заявке, представлены в таблице ниже.
Другие сокращения, используемые в настоящей заявке:
ACT: Адоптивный перенос клеток
ADCC: Антителозависимая клеточная цитотоксичность
ADCP: Антителозависимый клеточный фагоцитоз
CDC: Комплементзависимая цитотоксичность
CDR: Определяющая комплементарность область
КОЕ: Колониеобразующая единица
CH: Константная область тяжелой цепи
cHL: Классическая лимфома Ходжкина
CL: Константная область легкой цепи
dNTP: Дезоксирибонуклеотидтрифосфат
ESCC: Плоскоклеточная карцинома пищевода
FACS: Активированная флуоресценцией сортировка клеток
HC: Тяжелая цепь
HCC: Печеночноклеточная карцинома
HCVR: Вариабельная область тяжелой цепи
HEK: Эмбриональная почка человека
hyg: гигромицин
IFN-γ: IFN-гамма
LC: Легкая цепь
LCVR: Вариабельная область легкой цепи
mAb: Моноклональное антитело
MCC: Карцинома из клеток Меркеля
MLR: Реакция смешанной культуры лимфоцитов
MOI: Множественность инфекции
NSCLC: Немелкоклеточный рак легкого
PBMC: Мононуклеарные клетки периферической крови
PD-1/PD 1: Рецептор 1 программируемой клеточной смерти
PD-L1/PD L1: Лиганд 1 программируемой смерти
PD-L2/PD L2: Лиганд 2 программируемой смерти
Pfx: ДНК-полимераза с корректирующей активностью, Pfx™ от Invitrogen.
PMBCL: Первичная медиастинальная крупноклеточная B-клеточная лимфома
sRCA: Избирательная амплификация по типу катящегося кольца
RCC: Почечноклеточная карцинома
об/мин: Оборотов в минуту
SCCHN: Плоскоклеточная карцинома головы и шеи
SCLC: Мелкоклеточный рак легкого
SEQ/seq: Последовательность
SPR: Поверхностный плазмонный резонанс
VH: Вариабельная область тяжелой цепи
VL: Вариабельная область легкой цепи.
ВАРИАНТЫ ОСУЩЕСТВЛЕНИЯ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к новым антителам против PD-1, которые можно использовать для терапевтических целей.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению связывает с высокой аффинностью PD-1 человека.
В одном варианте осуществления, аминокислотная последовательность константной области антитела против PD-1 состоит из константной области IgGl, IgG2, IgG3, IgG4, IgG2/G4, IgA, IgE, IgM или IgD, предпочтительно, IgGl или IgG4.
В другом варианте осуществления, одно или несколько антител против PD-1 по настоящему изобретению имеют модифицированную или уменьшенную, или отсутствующую активность ADCC и/или CDC. В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть имеет уменьшенный потенциал для вызова проблемы с безопасностью ADCC и CDC. В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть имеет активность ADCC и/или CDC.
В другом варианте осуществления, одно или несколько антител против PD-1 по настоящему изобретению имеет активность ADCP.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет KD 10-8 M или менее, более предпочтительно, 10-10 M или менее и даже более предпочтительно, 10-11 M или менее, даже более предпочтительно, 10-12 M или менее для антигена PD-1. Значение KD представляет собой измерение аффинности связывания антитела по отношению к его антигену-мишени.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению вступает в перекрестную реакцию с PD-1 из видов, отличных от человека.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет более высокую специфичность связывания для PD-1 человека.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет увеличенное время полужизни у субъекта.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению блокирует функцию PD-1.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению предотвращает связывание PD-1 с экспрессирующими PD-L1 клетками-мишенями.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению предотвращает связывание PD-1 с экспрессирующими PD-L2 клетками-мишенями.
В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению уничтожает T-клетки, экспрессирующие PD-1.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет улучшенное время полужизни в кровотоке.
В другом варианте осуществления, антитело против PD-1 или антигенсвязывающая часть по настоящему изобретению является способным связывать PD-1 обезьяны, обеспечивая простоту разработки лекарственного средства посредством предоставления соответствующей модели фармакологии и токсикологии на животных.
В одном варианте осуществления, настоящее изобретение относится к последовательности нуклеиновой кислоты, кодирующей антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению.
В одном варианте осуществления, настоящее изобретение относится к экспрессирующему вектору, содержащему последовательность нуклеиновой кислоты, кодирующую антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению.
В одном варианте осуществления, настоящее изобретение относится к клетке-хозяину, содержащей экспрессирующий вектор, где экспрессирующий вектор содержит последовательность нуклеиновой кислоты, кодирующую антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению.
В одном из вариантов осуществления, настоящее изобретение относится к композиции, содержащей антитело против PD-1 или его антигенсвязывающую часть, специфически связывающие PD-1 человека, и приемлемый носитель.
В одном из вариантов осуществления, настоящее изобретение относится к иммуноконъюгату, содержащему антитело против PD-1 или его антигенсвязывающую часть, связанные с лекарственным средством.
В одном из вариантов осуществления, настоящее изобретение относится к биспецифической молекуле, содержащей антитело против PD-1 или его антигенсвязывающую часть, связанные с второй функциональной группой, имеющей специфичность связывания, отличную от указанного антитела или его антигенсвязывающей части.
В одном из вариантов осуществления, настоящее изобретение относится к комбинации, содержащей по меньшей мере два или более антитела или их антигенсвязывающих частей, где по меньшей мере одно антитело или его антигенсвязывающая часть представляет собой антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению.
В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно использовать для лечения заболеваний, при которых активность PD-1 является вредной, таких как различные злокачественные опухоли, различные инфекции. В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно использовать для лечения аутоиммунных нарушений.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно использовать для лечения злокачественной опухоли, выбранной из NSCLC, SCLC, RCC, cHL, SCCHN, уротелиальной карциномы, колоректального рака, ESCC, первичной медиастинальной крупноклеточной B-клеточной лимфомы, злокачественной опухоли с высокой микросателлитной нестабильностью, рака желудка, рака шейки матки, карциномы из клеток Меркеля, эндометриальной карциномы и злокачественной опухоли с высокой мутационной нагрузкой опухоли (TMB-H), где активность PD-1 умножена.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению связывает с высокой аффинностью PD-1 человека.
В одном варианте осуществления, CDR1 тяжелой цепи (далее в настоящем описании обозначено как CDRH1) антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность общей формулы (I): X1a-A1a-X2a-A2a, где
X1a представляет собой аминокислоту, выбранную из аспарагина, глицина и треонина;
A1a представляет собой дипептид, выбранный из Tyr-Tyr и Ile-Thr;
X2a представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, фенилаланина, метионина и аланина;
A2a представляет собой одиночную аминокислоту или дипептид, или трипептид, или тетрапептид, выбранные из тирозина, аспарагина, серина, глицина, Asn-Ser, Asn-Ser-Gly и Ser-Asn-Ser-Gly, при условии, что аспарагин в качестве X1a и метионин в качестве X2a не присутствуют совместно.
В одном из предпочтительных вариантов осуществления, CDRH1 тяжелой цепи антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность формулы (Ia): X1a-Y-Y-X2a-Y, где
X1a представляет собой аминокислоту, выбранную из аспарагина и треонина;
X2a представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, метионина и аланина, при условии, что аспарагин в качестве X1a и метионин в качестве X2a не присутствуют совместно.
В более предпочтительном варианте осуществления, CDRH1 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой TYYIY.
В альтернативном предпочтительном варианте осуществления, CDRH1 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой GITFSNSG.
В одном варианте осуществления, CDR2 тяжелой цепи (далее в настоящем описании обозначено как CDRH2) антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность общей формулы (II): A1b-X1b-A2b-X2b-X3b-A3b-X4b-X5b-X6b-X7b-X8b-A4b, где
A1b необязательно присутствует, и когда присутствует, представляет собой глицин;
X1b представляет собой аминокислоту, выбранную из метионина, изолейцина, лейцина, глицина, валина и аланина;
A2b представляет собой трипептид или тетрапептид, выбранный из Asn-Pro-Ser-Asn или Trp-Tyr-Asp;
Каждый из X2b и X3b независимо представляет собой аминокислоту, выбранную из глицина и серина;
A3b представляет собой одиночную аминокислоту или дипептид, выбранные из лизина или Thr-Asn;
X4b представляет собой аминокислоту, выбранную из тирозина, аргинина и фенилаланина;
X5b представляет собой аминокислоту, выбранную из серина, тирозина и аспарагина;
X6b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из глутаминовой кислоты и глутамина;
X7b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из аспарагина и лизина;
X8b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из тирозина и фенилаланина;
A4b может необязательно присутствовать, и когда присутствует, представляет собой лизин.
В одном из предпочтительных вариантов осуществления, CDRH2 тяжелой цепи антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность формулы (IIa): G-X1b-N-P-S-N-X2b-X3b-T-N-X4b-X5b-X6b-X7b-X8b-K, где
X1b представляет собой аминокислоту, выбранную из метионина, изолейцина, лейцина, глицина, валина и аланина;
Каждый из X2b и X3b независимо представляет собой аминокислоту, выбранную из глицина и серина;
Каждый из X4b и X8b независимо представляет собой аминокислоту, выбранную из тирозина и фенилаланина;
X5b представляет собой аминокислоту, выбранную из серина и аспарагина;
X6b представляет собой аминокислоту, выбранную из глутаминовой кислоты и глутамина;
X7b представляет собой аминокислоту, выбранную из аспарагина и лизина.
В более предпочтительном варианте осуществления, CDRH2 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению выбрана из GVNPSNGGTNYNENYK, GVNPSNGGTNYNQNYK или GVNPSNSGTNYNENYK.
В альтернативном предпочтительном варианте осуществления, CDRH2 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой IWYDGSKRY.
В одном варианте осуществления, CDR3 тяжелой цепи (далее в настоящем описании обозначено как CDRH3) антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность общей формулы (III): A1c-X1c-A2c-X2c-A3c-X3c-A4c-X4c-X5c-A5c, где
A1c может необязательно присутствовать, и когда присутствует, представляет собой аргинин;
X1c представляет собой аминокислоту, выбранную из аспарагиновой кислоты, аспарагина и глутаминовой кислоты;
A2c представляет собой аминокислоту или дипептид, выбранные из аспарагина, серина и треонина или Tyr-Arg;
X2c представляет собой аминокислоту, выбранную из тирозина, гистидина, аспарагиновой кислоты, глутаминовой кислоты, глицина и фенилаланина;
A3c представляет собой аминокислоту, выбранную из аспарагиновой кислоты, тирозина, изолейцина, фенилаланина, гистидина и триптофана;
X3c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, глутамина и метионина;
A4c может необязательно присутствовать, и когда присутствует, представляет собой глицин;
X4c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из тирозина, гистидина и фенилаланина;
X5c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
A5c может необязательно присутствовать, и когда присутствует, представляет собой тирозин.
В одном из вариантов осуществления, CDRH3 тяжелой цепи антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность формулы (IIIa): R-X1c-Y-R-X2c-D-X3c-G-X4c-X5c-Y, где
Каждый из X1c и X5c независимо представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
Каждый из X2c и X4c независимо представляет собой аминокислоту, выбранную из тирозина, гистидина и фенилаланина;
X3c представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, глутамина и метионина.
В более предпочтительном варианте осуществления CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению выбран из RDYRYDMGFDY или RDYRYDMGYDY, или RDYRYDMGHDY.
В альтернативном предпочтительном варианте осуществления, CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой ESEY или NNDI, или NSDF, или NSDH, или NSDY, или NSGY, или NTDW, или NTDY.
В одном варианте осуществления, CDR1 легкой цепи (далее в настоящем описании обозначено как CDRL1) антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению имеет аминокислотную последовательность общей формулы (IV): A1d-X1d-A2d-X2d-X3d-A3d-X4d-A4d-X5d-A5d, где
A1d представляет собой дипептид или трипептид, выбранный из Gln-Ser или Arg-Ala-Ser;
X1d представляет собой аминокислоту, выбранную из валина, глутаминовой кислоты и лизина;
A2d представляет собой аминокислоту, выбранную из глицина и серина;
X2d представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, серина и метионина;
X3d представляет собой аминокислоту, выбранную из серина, тирозина и глутаминовой кислоты;
A3d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из треонина;
X4d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты.
A4d может необязательно присутствовать, и когда присутствует, представляет собой тетрапептид Gly-Tyr-Ser-Tyr;
X5d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина;
A5d может необязательно присутствовать, и когда присутствует, представляет собой гистидин.
В одном из предпочтительных вариантов осуществления, CDRL1 легкой цепи (антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению) имеет аминокислотную последовательность формулы (IVa): R-A-S-X1d-G-X2d-X3d-T-X4d-G-Y-S-Y-X5d-H, где
X1d представляет собой аминокислоту, выбранную из глутаминовой кислоты и лизина;
Каждый из X2d и X5d независимо представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина;
X3d представляет собой аминокислоту, выбранную из серина и глутаминовой кислоты;
X4d представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты.
В более предпочтительном варианте осуществления CDRL1 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой RASKGVSTSGYSYLH.
В альтернативном предпочтительном варианте осуществления CDRL1 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой QSVSSY.
В одном варианте осуществления, CDRL2 легкой цепи (далее в настоящем описании обозначено как CDRL2) антитела против PD-1 или его антигенсвязывающей части имеет аминокислотную последовательность общей формулы (V): A1e-X1e-A2e-X2e-A3e, где
A1e может необязательно присутствовать, и когда присутствует, представляет собой дипептид Leu-Ala;
X1e представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
A2e представляет собой аминокислоту, выбранную из тирозина или аланина;
X2e представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, серина и метионина;
A3e может необязательно присутствовать, и когда присутствует, представляет собой дипептид Glu-Ser.
В одном из предпочтительных вариантов осуществления, CDRL2 легкой цепи антитела против PD-1 или его антигенсвязывающей части имеет аминокислотную последовательность формулы (Va): L-A-X1e-Y-X2e-E-S, где
X1e представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
X2e представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина.
В более предпочтительном варианте осуществления CDRL2 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой LASYLES.
В альтернативном предпочтительном варианте осуществления CDRL2 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой DAS.
В одном варианте осуществления, CDR3 легкой цепи (далее в настоящем описании обозначено как CDRL3) антитела против PD-1 или его антигенсвязывающей части имеет аминокислотную последовательность общей формулы (VI): A1f-X1f-A2f-X2f-X3f-X4f-X5f-X6f, где
A1f представляет собой дипептид, выбранный из Gln-His и Gln-Gln;
X1f представляет собой аминокислотную последовательность, выбранную из серина, аргинина, аспарагиновой кислоты и глутаминовой кислоты;
A2f представляет собой аминокислоту, выбранную из аргинина и серина;
X2f представляет собой аминокислоту, выбранную из аспарагиновой кислоты, аспарагина и глутаминовой кислоты;
Каждый из X3f и X5f независимо представляет собой аминокислотную последовательность, выбранную из изолейцина, лейцина, валина, аланина, триптофана, аргинина и метионина;
X4f представляет собой пролин;
X6f представляет собой треонин.
В одном из предпочтительных вариантов осуществления, CDRL3 легкой цепи антитела против PD-1 или его антигенсвязывающей части имеет аминокислотную последовательность формулы (VIa): Q-H-X1f-R-X2f-X3f-P-X4f-T, где
X1f представляет собой аминокислотную последовательность, выбранную из серина, аргинина, аспарагиновой кислоты и глутаминовой кислоты;
X2f представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
Каждый из X3f и X4f независимо представляет собой аминокислотную последовательность, выбранную из изолейцина, лейцина, валина, аланина и метионина.
В более предпочтительном варианте осуществления CDRL3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой QHSRDLPLT.
В альтернативном предпочтительном варианте осуществления CDRL3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляет собой QQSSNWPRT.
В одном из предпочтительных вариантов осуществления, CDRH1, CDRH2, CDRH3, CDRL1, CDRL2 и CDRL3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению выбраны из аминокислотных последовательностей, как приведено ниже в таблице 1.
В одном из вариантов осуществления, настоящее изобретение относится к антителу против PD-1 или его антигенсвязывающей части, содержащим:
(a) CDRH1 общей формулы (I): X1a-A1a-X2a-A2a;
(b) CDRH2 общей формулы (II): A1b-X1b-A2b-X2b-X3b-A3b-X4b-X5b-X6b-X7b- X8b-A4b;
(c) CDRH3 общей формулы (III): A1c-X1c-A2c-X2c-A3c-X3c-A4c-X4c-X5c-A5c;
(d) CDRL1 общей формулы (IV): A1d-X1d-A2d-X2d-X3d-A3d-X4d-A4d-X5d-A5d;
(e) CDRL2 общей формулы (V): A1e-X1e-A2e-X2e-A3e и
(f) CDRL3 общей формулы (VI): A1f-X1f-A2f-X2f -X3f- X4f -X5f-X6f,
где
X1a представляет собой аминокислоту, выбранную из аспарагина, глицина и треонина;
A1a представляет собой дипептид, выбранный из Tyr-Tyr и Ile-Thr;
X2a представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, фенилаланина, метионина и аланина;
A2a представляет собой одиночную аминокислоту или дипептид, или трипептид, или тетрапептид, выбранные из тирозина, аспарагина, серина, глицина, Asn-Ser, Asn-Ser-Gly и Ser-Asn-Ser-Gly;
A1b необязательно присутствует, и когда присутствует, представляет собой глицин;
X1b представляет собой аминокислоту, выбранную из метионина, изолейцина, лейцина, глицина, валина и аланина;
A2b представляет собой трипептид или тетрапептид, выбранный из Asn-Pro-Ser-Asn или Trp-Tyr-Asp;
Каждый из X2b и X3b независимо представляет собой аминокислоту, выбранную из глицина и серина;
A3b представляет собой одиночную аминокислоту или дипептид, выбранные из лизина или Thr-Asn;
X4b представляет собой аминокислоту, выбранную из тирозина, аргинина и фенилаланина;
X5b представляет собой аминокислоту, выбранную из серина, тирозина и аспарагина;
X6b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из глутаминовой кислоты и глутамина;
X7b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из аспарагина и лизина;
X8b может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из тирозина и фенилаланина;
A4b может необязательно присутствовать, и когда присутствует, представляет собой лизин;
A1c может необязательно присутствовать, и когда присутствует, представляет собой аргинин;
X1c представляет собой аминокислоту, выбранную из аспарагиновой кислоты, аспарагина и глутаминовой кислоты;
A2c представляет собой аминокислоту или дипептид, выбранные из аспарагина, серина и треонина или Tyr-Arg;
X2c представляет собой аминокислоту, выбранную из тирозина, гистидина, аспарагиновой кислоты, глутаминовой кислоты, глицина и фенилаланина;
A3c представляет собой аминокислоту, выбранную из аспарагиновой кислоты, тирозина, изолейцина, фенилаланина, гистидина и триптофана;
X3c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, глутамина и метионина;
A4c может необязательно присутствовать, и когда присутствует, представляет собой глицин;
X4c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из тирозина, гистидина и фенилаланина;
X5c может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
A5c может необязательно присутствовать, и когда присутствует, представляет собой тирозин;
A1d представляет собой дипептид или трипептид, выбранный из Gln-Ser или Arg-Ala-Ser;
X1d представляет собой аминокислоту, выбранную из валина, глутаминовой кислоты и лизина;
A2d представляет собой аминокислоту, выбранную из глицина и серина;
X2d представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, серина и метионина;
X3d представляет собой аминокислоту, выбранную из серина, тирозина и глутаминовой кислоты;
A3d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из треонина;
X4d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
A4d может необязательно присутствовать, и когда присутствует, представляет собой тетрапептид Gly-Tyr-Ser-Tyr;
X5d может необязательно присутствовать, и когда присутствует, представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина;
A5d может необязательно присутствовать, и когда присутствует, представляет собой гистидин;
A1e может необязательно присутствовать, и когда присутствует, представляет собой дипептид Leu-Ala;
X1e представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
A2e представляет собой аминокислоту, выбранную из тирозина или аланина;
X2e представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, серина и метионина;
A3e может необязательно присутствовать, и когда присутствует, представляет собой дипептид Glu-Ser;
A1f представляет собой дипептид, выбранный из Gln-His и Gln-Gln;
X1f представляет собой аминокислотную последовательность, выбранную из серина, аргинина, аспарагиновой кислоты и глутаминовой кислоты;
A2f представляет собой аминокислоту, выбранную из аргинина и серина;
X2f представляет собой аминокислоту, выбранную из аспарагиновой кислоты, аспарагина и глутаминовой кислоты;
Каждый из X3f и X5f независимо представляет собой аминокислотную последовательность, выбранную из изолейцина, лейцина, валина, аланина, триптофана, аргинина и метионина;
X4f представляет собой пролин;
X6f представляет собой треонин,
при условии, что аспарагин в качестве X1a и метионин в качестве X2a не присутствуют совместно.
В одном из предпочтительных вариантов осуществления, настоящее изобретение относится к антителу против PD-1 или его антигенсвязывающей части, содержащим:
(a) CDRH1, содержащую аминокислотную последовательность формулы (Ia): X1a-Y-Y-X2a-Y;
(b) CDRH2, содержащую аминокислотную последовательность формулы (IIa):G-X1b-N-P-S-N-X2b-X3b-T-N-X4b-X5b-X6b-X7b-X8b-K, и
(c) CDRH3, содержащую аминокислотную последовательность формулы (IIIa): R-X1c-Y-R-X2c-D-X3c-G-X4c-X5c-Y;
(d) CDRL1, содержащую аминокислотную последовательность формулы (IVa): R-A-S-X1d-G-X2d-X3d-T-X4d-G-Y-S-Y-X5d-H;
(e) CDRL2, содержащую аминокислотную последовательность формулы (Va): L-A-X1e-Y-X2e-E-S;
(f) CDRL3, содержащую аминокислотную последовательность формулы (VIa): Q-H-X1f-R- X2f-X3f-P-X4f-T,
где
X1a представляет собой аминокислоту, выбранную из аспарагина и треонина;
X2a представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, метионина и аланина;
X1b представляет собой аминокислоту, выбранную из метионина, изолейцина, лейцина, глицина, валина и аланина;
Каждый из X2b и X3b независимо представляет собой аминокислоту, выбранную из глицина и серина;
Каждый из X4b и X8b независимо представляет собой аминокислоту, выбранную из тирозина и фенилаланина;
X5b представляет собой аминокислоту, выбранную из серина и аспарагина;
X6b представляет собой аминокислоту, выбранную из глутаминовой кислоты и глутамина;
X7b представляет собой аминокислоту, выбранную из аспарагина и лизина;
Каждый из X1c и X5c независимо представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
Каждый из X2c и X4c независимо представляет собой аминокислоту, выбранную из тирозина, гистидина и фенилаланина;
X3c представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина, глутамина и метионина;
X1d представляет собой аминокислоту, выбранную из глутаминовой кислоты и лизина
Каждый из X2d и X5d независимо представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина;
X3d представляет собой аминокислоту, выбранную из серина и глутаминовой кислоты;
X4d представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
X1e представляет собой аминокислоту, выбранную из серина, аспарагиновой кислоты и глутаминовой кислоты;
X2e представляет собой аминокислоту, выбранную из изолейцина, лейцина, валина, аланина и метионина;
X1f представляет собой аминокислотную последовательность, выбранную из серина, аргинина, аспарагиновой кислоты и глутаминовой кислоты;
X2f представляет собой аминокислоту, выбранную из аспарагиновой кислоты и глутаминовой кислоты;
Каждый из X3f и X4f независимо представляет собой аминокислотную последовательность, выбранную из изолейцина, лейцина, валина, аланина и метионина,
при условии, что аспарагин в качестве X1a и метионин в качестве X2a не присутствуют совместно.
В одном из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNENYK и RDYRYDMGFDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNENYK и RDYRYDMGYDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNENYK и RDYRYDMGHDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNSGTNYNENYK и RDYRYDMGFDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNSGTNYNENYK и RDYRYDMGYDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNSGTNYNENYK и RDYRYDMGHDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNQNYK и RDYRYDMGFDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNQNYK и RDYRYDMGYDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой TYYIY, GVNPSNGGTNYNQNYK и RDYRYDMGHDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой GITFSNSG, IWYDGSKRY и NSDF, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой GITFSNSG, IWYDGSKRY и NSDH, соответственно.
В одном из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой GITFSNSG, IWYDGSKRY и NSDY, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой GITFSNSG, IWYDGSKRY и NTDW, соответственно.
В одном из предпочтительных вариантов осуществления, аминокислотные последовательности CDRH1, CDRH2 и CDRH3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой GITFSNSG, IWYDGSKRY и NTDY, соответственно.
В одном из предпочтительных вариантов осуществления, аминокислотные последовательности CDRL1, CDRL2 и CDRL3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой RASKGVSTSGYSYLH, LASYLES и QHSRDLPLT, соответственно.
В другом из предпочтительных вариантов осуществления, аминокислотные последовательности CDRL1, CDRL2 и CDRL3 антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению представляют собой QSVSSY, DAS и QQSSNWPRT, соответственно.
В другом варианте осуществления, настоящее изобретение относится к антителу или его антигенсвязывающей части, содержащим:
(a) CDRH1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:1, 2, 3, 4, 5, 6 и 7;
(b) CDRH2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 и 30;
(c) CDRH3, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54 и 55;
(d) CDRL1, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79,80 и 81;
(e) CDRL2, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100, 101, 102, 103, 104, 105, 106, 107, 108, 109, 110, 111 и 112; и
(f) CDRL3, содержащую аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:113, 114, 115, 116, 117, 118, 119, 120, 121, 122, 123, 124, 125, 126, 127, 128, 129, 130, 131, 132, 133, 134, 135 и 136;
где антитело специфически связывает PD-1, предпочтительно, PD-1 человека.
В одном из вариантов осуществления, HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению выбраны из аминокислотных последовательностей, как приведено ниже в таблице 2.
No.
IP-H4
IP-H4.2
IP-H4.2
(Y.H)
IP-H4.19
IP-H4.19
(F.Y)
IP-H4.19
(Y.H)
IP-H4.36
IP-H4.36
(F.Y)
IP-H4.36
(Y.H)
N5
N6
N7
N9
N10
IP-L1
NL
В одном из вариантов осуществления, вариабельная область тяжелой цепи антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению состоит из CDRH1, CDRH2 и CDRH3, содержащих аминокислотные последовательности, выбранные из приведенной ниже таблицы 3.
В одном из вариантов осуществления, вариабельная область легкой цепи антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению состоит из CDRL1, CDRL2 и CDRL3, содержащих аминокислотные последовательности, выбранные из приведенной ниже таблицы 4.
Соответственно, настоящее изобретение относится к антителу против PD-1 или его антигенсвязывающей части, содержащим вариабельную область тяжелой цепи и вариабельную область легкой цепи, где:
(a) вариабельная область тяжелой цепи содержит аминокислотную последовательность, выбранную из группы, состоящей из SEQ ID NO:137, 138, 139, 140, 141, 142, 143, 144, 145146147, 148, 149 и 150;
(b) вариабельная область легкой цепи содержит аминокислотную последовательность из SEQ ID NO:151 и 152.
Предпочтительные комбинации CDR:
Предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:8;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:32;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:9;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:32;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:26;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:32;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Более предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:8;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:33;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Более предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:8;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:47;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:9;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:33;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:9;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:47;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:26;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:33;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:4;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:26;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:47;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:77;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:108; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:132.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:7;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:30;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:50;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:81;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:112; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:136.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:7;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:30;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:51;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:81;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:112; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:136.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:7;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:30;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:52;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:81;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:112; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:136.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:7;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:30;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:54;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:81;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:112; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:136.
Другая предпочтительная комбинация CDR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) CDRH1 вариабельной области тяжелой цепи, содержащую SEQ ID NO:7;
(b) CDRH2 вариабельной области тяжелой цепи, содержащую SEQ ID NO:30;
(c) CDRH3 вариабельной области тяжелой цепи, содержащую SEQ ID NO:55;
(d) CDRL1 вариабельной области легкой цепи, содержащую SEQ ID NO:81;
(e) CDRL2 вариабельной области легкой цепи, содержащую SEQ ID NO:112; и
(f) CDRL3 вариабельной области легкой цепи, содержащую SEQ ID NO:136.
Предпочтительные комбинации вариабельных областей:
Предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:137; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:138; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:139; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:140; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:141; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:142; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:143; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:144; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:145; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:151.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:146; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:152.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:147; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:152.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:148; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:152.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:149; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:152.
Другая предпочтительная комбинация HCVR и LCVR антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению содержит:
(a) вариабельную область тяжелой цепи, содержащую аминокислотную последовательность из SEQ ID NO:150; и
(b) вариабельную область легкой цепи, содержащую аминокислотную последовательность из SEQ ID NO:152.
В одном из вариантов осуществления, антитело или его антигенсвязывающая часть по настоящему изобретению имеет по меньшей мере одну из следующих характеристик:
(a) вступает в перекрестную реакцию с PD-1 из видов, отличных от человека;
(b) более высокая специфичность связывания для PD-1 человека;
(c) стимулирует секрецию цитокина IL-2;
(d) стимулирует более высокую секрецию IFN-гамма;
(e) блокирует взаимодействие рецептора PD-1 с его природным лигандом PD-L1 и/или PD-L2;
(f) стимулирует пролиферацию T-клеток и
(g) имеет улучшенное время полужизни в кровотоке.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть имеет KD 10-10 M или менее, более предпочтительно, 10-11 M или менее, для антигена PD-1. В предпочтительном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению связывает PD-1 человека с KD 10-10 M или менее, предпочтительно, 10-11 M или менее. Значение KD представляет собой измерение аффинности связывания антитела по отношению к его антигену-мишени. Такую аффинность связывания и кинетику связывания антител против PD-1 можно исследовать посредством анализа SPR. Указанный анализ является очень хорошо известным в данной области для измерения аффинности антитела или его антигенсвязывающей части по отношению к его антигену-мишени, и специалисту в данной области хорошо известны способы проведения такого анализа.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет высокую специфичность для PD-1. Антитело против PD-1 по настоящему изобретению имеет высокую аффинность для антигена/эпитопа PD-1 человека. В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет высокую специфичность связывания для PD-1 человека. Специфичность связывания исследуют способом SPR. «Высокая специфичность для PD-1 или для PD-1 человека» означает, что антитело или его антигенсвязывающая часть не связывает другие коингибирующие рецепторы T-клетки с такой же аффинностью.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть вступает в перекрестную реакцию с PD-1 из другого вида, такого как яванский макак и т.д. с достаточной аффинностью.
В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению блокирует взаимодействие PD-1 с его лигандом в анализе поверхностного плазмонного резонанса.
В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению уничтожает T-клетки, экспрессирующие PD-1.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению стимулирует секрецию цитокина IL-2. В таком варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению, когда являются доступными в ходе MLR, приводят к увеличенной продукции IL-2, как измерено посредством ELISA.
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению стимулирует более высокую секрецию IFN-гамма. Термин «более высокая» обозначен в настоящем описании по отношению к известному антителу против PD-1. В другом варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению, когда являются доступными в ходе MLR, приводят к увеличенной продукции IFN-гамма, как измерено посредством ELISA. В эксперименты по трансплантации костного мозга с использованием мышей IFN-γR-/- включали IFN-γ в качестве существенного звена для контроля опосредованной PD-1-инфильтрации опухоли T-клетками. Также оценивали и подтвердили, что другие индуцируемые IFN-γ хемокины также могут играть роль в синергическом эффекте антитела против PD-1 на ACT. (7) Кроме того, опубликовано, что IFN-γ является важным маркером для прогнозирования ответа на блокирование иммунной контрольной точки (8).
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению стимулирует пролиферацию T-клеток. В таком варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению, когда являются доступными в ходе MLR, приводят к увеличенной пролиферации T-клеток, как измерено посредством анализа измерения пролиферации. Такие анализы известны специалисту в данной области.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет улучшенные фармакокинетические свойства.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению имеет увеличенное время полужизни у человека.
Фармакокинетический профиль антитела против PD1 по настоящему изобретению определяют у яванского макака. Поскольку намеченным способом введения у пациентов является внутривенный, антитело против PD1 вводят внутривенно животным в исследованиях, в буферном растворе. Кратко, исследование включает 3 животных/пол/группу, которых подвергают введению на уровнях дозирования вплоть до 50 мг/кг. Исследуют PD эффекты антитела против PD1, такие как модуляция популяции периферических иммуноцитов, и изменение абсолютного количества лимфоцитов (ALC).
В одном варианте осуществления, антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению блокирует взаимодействие рецептора PD-1 с его природными лигандами PD-L1 и/или PD-L2. Антитела против PD-1 по настоящему изобретению тестируют по их способности блокировать связывание лигандов PD-L1 и PD-L2 с PD-1, экспрессированным на клетках, с использованием реакции смешанной культуры лимфоцитов (MLR). Антитела против PD-1 по настоящему изобретению тестируют по их способности блокировать связывание лигандов PD-L1 и PD-L2 с PD-1 посредством SPR.
В одном из вариантов осуществления, эффект связывания антитела против PD-1 или его антигенсвязывающей части по настоящему изобретению с PD-1 на активацию T-клеток анализируют посредством подходящих анализов, известных в данной области. Антитело против PD-1 или его антигенсвязывающая часть по настоящему изобретению связывает PD-1, экспрессированный на экспрессирующей PD-1 трансфицированной линии клеток. Кроме того, антитела против PD-1 или их антигенсвязывающая часть по настоящему изобретению связываются с мононуклеарными клетками периферической крови (PBMC).
Антитела по настоящему изобретению могут являться полноразмерными (например, антитело IgG1 или IgG4) или могут содержать только антигенсвязывающую часть (например, фрагмент Fab, F(ab')2 или scFv), и необязательно, являться модифицированными для влияния на функциональность, например, для уничтожения остаточных эффекторных функций, таких как активность ADCC и CDC.
В одном из вариантов осуществления, аминокислотные последовательности константной области антитела против PD-1 представляют собой константную область IgGl, IgG2, IgG3, IgG4, IgA, IgE, IgM, IgD или гибрид упомянутых константных областей, таких как IgG2/IgG4, предпочтительно, IgG1 или IgG4, более предпочтительно, IgG4.
В другом варианте осуществления, область Fc антител по настоящему изобретению изменяют посредством замены по меньшей мере одного аминокислотного остатка на другой аминокислотный остаток для изменения эффекторной функции (функций) антитела.
В одном из предпочтительных вариантов осуществления, аминокислотные последовательности константной области тяжелой цепи антитела против PD-1 по настоящему изобретению имеют мутацию в его шарнирной области, предпочтительно, мутацию серина до пролина в шарнирной области константной области антитела IgG4. Человеческие антитела IgG4 могут существовать в двух формах, которые ассоциированы с гетерогенностью шарнира. В одной форме, антитело содержит стабильную четырехцепочечную конструкцию приблизительно 150-160 кДа, в которой димеры удерживаются вместе дисульфидной связью между тяжелыми цепями. Во второй форме димеры не связаны межцепочечными дисульфидными связями, и образуется молекула приблизительно 75-80 кДа, состоящая из ковалентно связанных легкой и тяжелой цепей (полуантитело). Последние формы было чрезвычайно сложно отделять от полноразмерного антитела, даже после аффинной очистки. Частота образования второй формы в различных интактных изотипах IgG4 обусловлена, но без ограничения, структурными различиями, ассоциированными с шарнирной областью изотипа антитела. Одиночная аминокислотная замена в шарнирной области шарнира человеческого IgG4 может значительно уменьшать образование второй формы (6), до уровней, обычно наблюдаемых с использованием человеческого шарнира IgG1. Полноразмерные антитела, содержащие CDR или вариабельные области по настоящему изобретению дополнительно содержат указанную одиночную аминокислотную замену (т.е. S228P), когда оно разработано в форме IgG4.
В одном варианте осуществления, антитело против PD-1 по настоящему изобретению имеет активность ADCC и/или CDC. В предпочтительном варианте осуществления, антитело против PD-1 по настоящему изобретению с модифицированной или уменьшенной, или отсутствующей активностью ADCC и/или CDC представляет собой антитело против PD-1 с константной областью IgG1 или IgG4, или IgG2/IgG4. В другом варианте осуществления, область Fc антител по настоящему изобретению изменяют посредством замены по меньшей мере одного аминокислотного остатка на другой аминокислотный остаток для увеличения активность ADCC и/или CDC. В одном из предпочтительных вариантов осуществления, антитело против PD-1 по настоящему изобретению с увеличенной активностью ADCC и/или CDC представляет собой антитело против PD-1 с константной областью IgG1 или IgG1 с по меньшей мере одним измененным аминокислотным остатком. IgG1 с константной областью с по меньшей мере одним измененным аминокислотным остатком, как обозначено в настоящем описании, представляет собой модифицированную IgG1, которое может обеспечивать более высокую активность ADCC и/или CDC, по сравнению с IgG1 без такой модификации.
В другом предпочтительном варианте осуществления, антитело против PD-1 по настоящему изобретению с уменьшенной или отсутствующей активностью ADCC и/или CDC представляет собой антитело против PD-1 с константной областью IgG1 или IgG1 с по меньшей мере одним измененным аминокислотным остатком. IgG1 с константной областью с по меньшей мере одним измененным аминокислотным остатком, как обозначено в настоящем описании, представляет собой модифицированное IgG1, которое может обеспечивать уменьшенную или отсутствующую активность ADCC и/или CDC, по сравнению с IgG1 без такой модификации.
В другом предпочтительном варианте осуществления, антитело против PD-1 по настоящему изобретению с уменьшенной или отсутствующей активностью ADCC и/или CDC представляет собой антитело против PD-1 с IgG4 или IgG4 с константной областью с по меньшей мере одним измененным аминокислотным остатком. IgG4 с константной областью с по меньшей мере одним измененным аминокислотным остатком, как обозначено в настоящем описании, представляет собой модифицированное IgG4, которое может обеспечивать уменьшенную или отсутствующую активность ADCC и/или CDC, по сравнению с IgG4 без такой модификации.
В другом аспекте антитело против PD-1 по настоящему изобретению имеет увеличенное связывание FcRn и увеличенное время полужизни с модифицированной или уменьшенной, или отсутствующей активностью ADCC и/или CDC. В одном из вариантов осуществления, антитело против PD-1 по настоящему изобретению имеет аминокислотные последовательности константной области IgG1 или IgG4 с мутацией P329G и/или M428L и N434S. Константные области антитела против PD-1 с упомянутыми всеми тремя мутациями в константной области IgG1 и IgG4 обозначены в настоящем описании как IgG1(GLS) и IgG4(GLS), соответственно.
В предпочтительном варианте осуществления, антитело против PD-1 по настоящему изобретению содержит одиночную аминокислотную замену, выбранную из S228P, P329G, M428L, N434S и их подходящих комбинаций.
В одном из вариантов осуществления, антитело против PD-1 по настоящему изобретению представляет собой моноклональное антитело или биспецифическое антитело, или поликлональное антитело, предпочтительно, моноклональные антитело.
В одном из вариантов осуществления, антитело или его антигенсвязывающая часть, нацеленные на антиген PD-1, по настоящему изобретению, являются мышиными, химерными, рекомбинантными или гуманизированными по характеру, предпочтительно, рекомбинантными по характеру.
Иммуноконъюгаты и биспецифические антитела
В одном из вариантов осуществления, антитело против PD-1 по настоящему изобретению является конъюгированным с лекарственным средством, предпочтительно, цитотоксическим средством, необязательно, через линкер. Настоящее изобретение также относится к иммуноконъюгату, содержащему антитело по изобретению или его антигенсвязывающую часть, связанные с другим лекарственным средством, таким как цитотоксин или радиоактивный изотоп. Такие иммуноконъюгаты, полученные по настоящему изобретению, специфически связываются с экспрессирующими PD1 иммуноцитами, приводя к их лизису. Эти иммуноциты могут включать T-клетки, клетки NK, B-клетки, моноциты и т.п., экспрессирующие PD-1. Таким образом, иммуноконъюгаты, полученные по настоящему изобретению, помогают лечить заболевание, при котором активность экспрессирующих PD-1 иммуноцитов является вредной.
В одном из вариантов осуществления, антитело против PD-1 по настоящему изобретению используют для получения биспецифической молекулы. Биспецифическая молекула, содержащая антитело или его антигенсвязывающую часть, по настоящему изобретению, имеющие два уникальных антигенсвязывающих плеча или функциональных группы, таких, что одно связывает PD-1 и другое имеет отличную специфичность связывания. В одном из вариантов осуществления, вторая функциональная группа по настоящему изобретению может связывать антиген, выбранный из CTLA-4, PD-L1, LAG-3, TIGIT и HER2.
Молекулы нуклеиновой кислоты, кодирующие антитела против PD-1, векторы и клетки-хозяева
Настоящее изобретение относится к молекулам нуклеиновой кислоты или последовательностям нуклеиновой кислоты, кодирующим антитела или их антигенсвязывающие части по настоящему изобретению, так же как к экспрессирующим векторам, содержащим такие последовательности нуклеиновой кислоты, и к клеткам-хозяевам, содержащим такие экспрессирующие векторы. В настоящей заявке, векторы pZRCII, так же как pZRCIII, используют для клонирования и экспрессии антител против PD-1 по настоящему изобретению. Вектор pZRCII (pZRCIIhyg) и вектор pZRCIII описаны в Патентных документах WO 2007/017903 и WO 2012/046255A2, соответственно. Клетка-хозяин по настоящему изобретению представляет собой прокариотическую или эукариотическую клетку, предпочтительно, клетка-хозяин представляет собой клетку E. coli или клетку млекопитающего, такую как клетка CHO или клетка NS0, или клетка CHO-GS, или клетка CHO-S или клетка CHO-K1.
Получение антител
По настоящему изобретению, антитело против PD-1 или его антигенсвязывающую часть получают с использованием рекомбинантным способов. В рекомбинантном способе по настоящему изобретению, используют подходящие экспрессирующие векторы. Вектор для получения антител по настоящему изобретению может представлять собой вектор, хорошо известный специалисту в данной области. Экспрессирующий вектор и последовательности для контроля экспрессии выбирают, чтобы они являлись совместимыми с используемой экспрессирующей клеткой-хозяином. Ген легкой цепи антитела и ген тяжелой цепи антитела можно вставлять в отдельный вектор или, более типично, оба гена вставляют в один и тот же экспрессирующий вектор. Гены антитела вставляют в экспрессирующий вектор посредством стандартных способов (например, лигирования комплементарных участков рестрикции в фрагменте ген антитела и векторе, или лигирования тупых концов, если не присутствует участков рестрикции), известных специалистам в данной области. Например, для экспрессии антител, или фрагментов антител из них, ДНК, кодирующие частичные или полноразмерные легкие и тяжелые цепи, можно получать посредством стандартных способов молекулярной биологии (например, амплификации ПЦР или клонирования кДНК с использованием гибридомы, которая экспрессирует представляющее интерес антитело), и ДНК можно вставлять в экспрессирующие векторы таким образом, чтобы гены являлись функционально связанными с последовательностями для контроля транскрипции и трансляции. В дополнение к генам цепей антител и регуляторным последовательностям, рекомбинантные экспрессирующие векторы по изобретению могут нести дополнительные последовательности, такие как последовательности, регулирующие репликацию вектора в клетках-хозяевах (например, точки начала репликации) и гены селективных маркеров. Вариабельные области тяжелых цепей антител, описанных в настоящем описании, можно использовать для получения полноразмерных генов антител любого изотипа антитела посредством их вставки в экспрессирующие векторы, уже кодирующие константную область тяжелой цепи и полноразмерную легкую цепь желательного изотипа таким образом, чтобы фрагмент VH являлся функционально связанным с фрагментом(фрагментами) CH внутри вектора. Дополнительно или альтернативно, рекомбинантный экспрессирующий вектор может кодировать сигнальный пептид, облегчающий секрецию цепи антитела из клетки-хозяина. Для экспрессии легких и тяжелых цепей, экспрессирующий вектор(ы), кодирующие тяжелые и легкие цепи, трансфицируют в клетку-хозяина посредством стандартных способов, таких как опосредованная липидами трансфекция, электропорация, преципитация фосфатом кальция, трансфекция с использованием DEAE-декстрана и т.п. Предпочтительные клетки-хозяева млекопитающих для экспрессии рекомбинантных антител по изобретению включают клетки яичника китайского хомяка (клетки CHO) (включая клетки DHFR- CHO), клетки миеломы NSO, клетки COS и клетки SP2. В частности, для использования с клетками миеломы NSO, другой предпочтительной системой экспрессии является система экспрессии генов GS. Антитела, полученные по настоящему изобретению, можно, кроме того, получать посредством известных способов культивирования клеток для крупномасштабной продукции антител. Антитела можно выделять из культуральной среды с использованием стандартных способов очистки белка. При использованием рекомбинантных способов, антитело может быть продуцировано внутриклеточно, в периплазматическом пространстве, или непосредственно секретировано в среду. Если антитело продуцировано внутриклеточно, в качестве первой стадии, частицы дебриса, либо клеток-хозяев, либо лизированных фрагментов, удаляют, например, посредством центрифугирования или ультрафильтрации. Антитела по изобретению можно тестировать по связыванию с PD-1, посредством, например, стандартного ELISA.
Антитела против PD-1 по настоящему изобретению также получают с использованием способа библиотек фагового дисплея после получения библиотек Fab с использованием способа сайт-направленного мутагенеза, как описано в примерах в настоящем описании. В этом способе, с использованием способа библиотеки фагов, получают области Fab против PD-1, которые далее конвертируют в полноразмерные с использованием рекомбинантных способов на основе экспрессирующего вектора, как описано в настоящей заявке.
Комбинирование антитела против PD-1 по настоящему изобретению с другими лекарственными средствами
Настоящее изобретение относится к комбинации, содержащей по меньшей мере два или более антител или их антигенсвязывающих частей, где по меньшей мере одно антитело или его антигенсвязывающая часть представляет собой антитело против PD-1 по настоящему изобретению. Комбинация по настоящему изобретению может содержать второе антитело или его антигенсвязывающую часть, выбранные из антитела против LAG3, антитела против CTLA4, антитела против HER2, антитела против VEGF, антитела против PDL1, антитела против PD-1, в комбинации с антителом против PD-1 или его антигенсвязывающей частью по настоящему изобретению. В другом варианте осуществления, настоящее изобретение относится к комбинации, состоящей из антитела против PD-1 или его антигенсвязывающей части(частей) и пептида, или к комбинации, содержащей антитело против PD-1 или его антигенсвязывающую часть(части) и цитокин (предпочтительно, интерлейкин).
Фармацевтические композиции
Может быть разработана фармацевтическая композиция, содержащая одно или комбинацию из моноклональных антител или их антигенсвязывающей части(частей), или комбинацию из антитела против PD-1 или его антигенсвязывающей части(частей) и пептида, или комбинацию из антитела против PD-1 или его антигенсвязывающей части(частей) и цитокина (предпочтительно, интерлейкина), по настоящему изобретению, составленные вместе с фармацевтически приемлемым носителем. Настоящее изобретение относится также к композиции, например, фармацевтической композиции, содержащей одно или комбинацию из моноклональных антител или их антигенсвязывающей части(частей), или комбинацию из антитела против PD-1 или его антигенсвязывающей части(частей) и пептида, или комбинацию из антитела против PD-1 или его антигенсвязывающей части(частей) и цитокина (предпочтительно интерлейкина), по настоящему изобретению, составленные вместе с фармацевтически приемлемым носителем. Такие композиции могут включать одно или комбинацию из (например, двух или более различных) антител или иммуноконъюгатов, или биспецифических молекул по изобретению. Например, фармацевтическая композиция по изобретению может содержать комбинацию антител (или иммуноконъюгатов или биспецифических антител), которые связываются с различными эпитопами на антигене-мишени или с различными эпитопами на различных антигенах-мишенях, или которые имеют комплементирующую активность.
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающую часть можно комбинировать с другим подходящим лекарственным средством для лечения различных злокачественных опухолей, инфекций или аутоиммунных нарушений.
Антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно комбинировать или можно использовать в комбинации с другим подходящим химически синтезированным терапевтическим средством(средствами). Указанное химически синтезированное терапевтическое средство может представлять собой противораковые или противоинфекционные лекарственные средства, или комбинацию таких лекарственных средств.
Антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно комбинировать с химиотерапевтическим средством(средствами) в клинических условиях.
Антитело против PD-1 или его антигенсвязывающую часть по настоящему изобретению можно использовать в комбинации с вакциной(вакцинами) для индукции иммунных ответов T- и B-клеток, возможно, посредством продления индуцированной вакциной пролиферации T- или B-клеток, как в профилактических, так и в терапевтических условиях.
Терапевтические применения
В одном из вариантов осуществления, антитело против PD-1 или его антигенсвязывающую часть, или комбинацию, или конъюгат, или биспецифическое антитело по настоящему изобретению можно использовать для лечения заболеваний, при которых активность PD-1 или экспрессия PD-1 является вредной. Такие заболевания хорошо известны специалисту в данной области.
Антитело против PD-1 или его антигенсвязывающую часть, или комбинацию, или конъюгат, или биспецифическое антитело по настоящему изобретению можно использовать для лечения различных злокачественных опухолей, таких как NSCLC, SCLC, RCC, cHL, SCCHN, рак молочной железы, уротелиальная карцинома, колоректальный рак, ESCC, первичная медиастинальная крупноклеточная B-клеточная лимфома, злокачественная опухоль с высокой микросателлитной нестабильностью, рак желудка, рак яичника, рак предстательной железы, глиомы, глиобластома, астроцитомы, мультиформная глиобластома, синдром Баннаяна-Зонана, болезнь Каудена, болезнь Лермитта-Дюкло, воспалительный рак молочной железы, опухоль Вильмса, саркома Юинга, рабдомиосаркома, эпендимома, медуллобластома, рак почки, рак печени, меланома, рак поджелудочной железы, саркома, остеосаркома, гигантоклеточная опухоль кости, рак щитовидной железы, лимфобластный T-клеточный лейкоз, хронический миелогенный лейкоз, хронический лимфоцитарный лейкоз, волосатоклеточный лейкоз, острый лимфобластный лейкоз, острый миелогенный лейкоз, AML, хронический нейтрофильный лейкоз, острый лимфобластный T-клеточный лейкоз, плазмацитома, иммунобластный крупноклеточный лейкоз, лейкоз из клеток мантийной зоны, множественная миелома, мегакариобластный лейкоз, множественная миелома, острый мегакариоцитарный лейкоз, промиелоцитарный лейкоз, эритролейкоз, злокачественная лимфома, лимфома Ходжкина, неходжкинская лимфома, лимфобластная T-клеточная лимфома, лимфома Беркитта, фолликулярная лимфома, нейробластома, рак шейки матки, карцинома из клеток Меркеля, эндометриальная карцинома, злокачественная опухоль с высокой мутационной нагрузкой опухоли (TMB-H), рак женских наружных половых органов, рак слюнных желез, рак носоглотки, буккальный рак, злокачественная опухоль полости рта, GIST (стромальная опухоль желудочно-кишечного тракта) и рак яичка.
Антитело против PD-1 или его антигенсвязывающую часть, или комбинацию, или конъюгат, или биспецифическое антитело по настоящему изобретению можно использовать для лечения различных инфекций. Примерами таких инфекций могут являться инфекции, вызванные патогенами, выбранными из HIV, инфекции гепатита (A, B и C), гриппа, герпеса, жиардии, малярии, лейшмании, Staphylococcus aureus, Pseudomonas aeruginosa, SARS-CoV, MERS-CoV, SARS-CoV-2, Ebola и т.д.
Антитело против PD-1 или его антигенсвязывающую часть, или комбинацию или конъюгат, или биспецифическое антитело по настоящему изобретению можно использовать для лечения различных аутоиммунных нарушений. Примерами таких аутоиммунных нарушений являются ревматоидный артрит, псориаз, псориатический артрит, анкилозирующий спондилит, ювенильный ревматоидный артрит, болезнь Крона, язвенный колит, сепсис, атопическое заболевание, такое как астма, экзема, аллергия, рассеянный склероз, системная красная волчанка, васкулит, миастения, болезнь Грэйвса, тиреоидит Хашимото, сепсис, аутоиммунная гемолитическая анемия, пернициозная анемия, идиопатическая тромбоцитопеническая пурпура, синдром Гудпасчера, буллезный пемфигоид, обычный пемфигоид, тиреоидит Хашимото, инсулинзависимый сахарный диабет (IDDM), диабет CIDP, воздействие антианемических средств (против бета-талассемии), гематопоэтических средств, офтальмологических лекарственных средств, заболевания кости, неврологические генетические нарушения, связанная с возрастом дегенерация желтого пятна, диабетическая ретинопатия, макулярные заболевания, офтальмологические генетические нарушения, гемофилия B, воздействие средств против (недостаточности фактора свертывания IX (9)), сердечно-сосудистые заболевания, диабет типа 2, воздействие иммунодепрессантов, лечения отторжения трансплантата, терапии миелодиспластического синдрома, ретинопатия, апластическая анемия, воздействие анальгетических лекарственных средств, гематологические генетические нарушения, мультисистемные генетические нарушения, остеоартрит, склеродермия, воздействие лечения аутоиммунных заболеваний, лечения подагры, крапивница, воздействие терапии сердечной недостаточности, терапии лимфомы, нефрит, респираторные нарушения и воздействие терапии врожденных нарушений метаболизма.
В предпочтительном варианте осуществления, настоящее изобретение относится к эффективному количеству антитела против PD-1 или его антигенсвязывающей части(частей), как раскрыто по настоящему изобретению, которое может уменьшать опосредованную PD-1 иммуносупрессию. Эти антитела можно вводить общепринятыми способами и в общепринятых дозах, таких как «фармацевтически эффективная доза» или «терапевтически эффективная доза».
Примеры
Следующие примеры являются иллюстративными, и их не следует рассматривать как ограничивающие объем настоящего изобретения никаким образом, поскольку множество вариантов и эквивалентов станут очевидными специалисту в данной области после прочтения настоящего описания. Полное содержание всех патентов, патентных заявок и публикаций, процитированных в настоящем описании, приведено в настоящем описании в качестве ссылки.
Пример 1: Получение векторов pZRCII hyg для легкой и тяжелой цепи антитела против PD-1
Конструирование вектора pZRCII hygIgG4
Химически синтезированную константную область человеческого IgG4 (SEQ ID NO:156) амплифицировали для включения выступающих концов участков рестрикционных ферментов SalI, ApaI на 5’- и выступающих концов участка NotI на 3’-концах посредством 30 циклов ПЦР с использованием 0,2 пМ специфических олигонуклеотидных праймеров, содержащих вышеуказанные участки рестрикции, в объеме 50 мкл, содержащем 1× буфер для Pfx, 1,5 мМ MgSO4, 2,5 мкМ каждый из 4 dNTP и 1 единицу полимеразы Pfx. Каждый цикл амплификации ПЦР состоял из инкубаций при 95°C в течение 30 секунд (денатурация), 55°C в течение 45 секунд (гибридизация) и 72°C в течение 1 минуты (удлинение). Амплифицированный продукт реакции ПЦР разделяли в 1% агарозном геле. Желательный фрагмент размером приблизительно 980 пар оснований вырезали из геля и очищали с использованием набора для экстракции из геля Qiagen. Этот очищенный фрагмент продукта ПЦР и вектор pZRCII с геном устойчивости к гигромицину расщепляли с использованием рестрикционных ферментов SalI и NotI. Вектор pZRCII получали, как описано в патентной публикации WO 2007/017903. Фрагмент вектора приблизительно 7692 пар оснований очищали из геля с использованием набора для экстракции из геля Qiagen. Расщепленный фрагмент продукта ПЦР также очищали с использованием набора для экстракции из геля Qiagen. Расщепленный и очищенный продукт ПЦР и вектор pZRCII hyg лигировали, и продукт лигирования трансформировали в E. coli штамма Top 10F'. Трансформанты оценивали на основании устойчивости к канамицину. Плазмидную ДНК, выделенную из таких колоний, анализировали по присутствию фрагмента константной области IgG4 посредством рестрикционного расщепления с использованием рестрикционных ферментов SalI и NotI. Одна из плазмид, для которой показан ожидаемый профиль рестрикции, имеющая фрагмент константной области IgG4, интегрированный в вектор pZRCII, названа pZRC II IgG4 константная область. Таким же способом, химически синтезированный Fc IgG1 (SEQ ID NO:157) клонирован в вектор pZRCII и назван вектор pZRCIIhyg.IgG1.
Конструирование векторных конструкций pZRCIIhyg LC-HC против PD-1
Гены легкой цепи и тяжелой цепи антитела против PD1 химически синтезировали и клонировали в клонирующий вектор pMA (Geneart, Germany). Для клонирования гена легкой цепи в вектор pZRCIIhyg.IgG4 константная область, векторы pMA с легкой цепью и pZRCIIhyg.IgG4 расщепляли с использованием рестрикционных ферментов XhoI и kpnI. Легкую цепь размером приблизительно 737 п.о. (SEQ ID NO:163) и вектор pZRCIIhyg.IgG4 приблизительно 8672 пар оснований вырезали из геля и очищали с использованием набора для экстракции из геля Qiagen. Расщепленные и очищенные легкую цепь и вектор pZRCIIhyg.IgG4 лигировали и трансформировали в E. coli штамма Top 10F'. Трансформанты отбирали на основании устойчивости к канамицину. Плазмидную ДНК, выделенную из приблизительно 20 таких колоний, анализировали по присутствию гена легкой цепи посредством рестрикционного расщепления с использованием различных рестрикционных ферментов. Одна такая плазмида, имеющая ген легкой цепи, интегрированный в первый транскрипционный блок вектора pZRCIIhyg.IgG4, выбрана для получения векторов pZRCIIhyg. LC-HC. Для клонирования генов тяжелой цепи, химически синтезированные вариабельные фрагменты гена тяжелой цепи (SEQ ID No.:158, 159, 160, 161 и 162), все размером приблизительно 446 пар оснований, и вектор pZRCII hyg. LC-IgG4, имеющий легкую цепь из SEQ ID NO:163, расщепляли с использованием SalI и ApaI. Расщепленные и очищенные фрагменты гена вариабельной области тяжелой цепи (SEQ ID NO:158, 159, 160, 161 и 162) индивидуально лигировали в вектор pZRCIIhyg LC IgG4 против PD-1 и трансформировали в E. coli Top 10F'. Трансформанты отбирали на основании устойчивости к канамицину. Плазмидную ДНК, выделенную из трансформантов, анализировали по присутствию гена тяжелой цепи посредством рестрикционного расщепления с использованием SalI и NotI. Конечные векторы, содержащие легкую цепь из SEQ ID NO:163 и тяжелую цепь из SEQ ID No.:158, легкую цепь из SEQ ID NO:163 и тяжелую цепь из SEQ ID NO:159, легкую цепь из SEQ ID NO:163 и тяжелую цепь из SEQ ID NO:160, легкую цепь из SEQ ID NO:163 и тяжелую цепь из SEQ ID NO:161, легкую цепь из SEQ ID NO:163 и тяжелую цепь из SEQ ID NO:162, подтверждали посредством нуклеотидного секвенирования тяжелых и легких цепей. Конечные конструкции, содержащие различные комбинации тяжелой цепи и легкой цепи, названы как IP-H4L1, IP-H4.2L1, IP-H4.2(Y.H)L1, IP-H4.19L1, и IP-H4.36L1, соответственно. Указанные различные конструкции кодируют аминокислотные последовательности HCVR и LCVR антител против PD-1 по настоящему изобретению, как описано в таблице 5.
Общая схема вектора указанной конструкции представлена на фигуре 1. IP-H4L1 получали также с изотипом IgG1, как проиллюстрировано в настоящем описании, с использованием химически синтезированной константной области человеческого IgG1 (SEQ ID NO:157).
Аминокислотные последовательности вариабельных областей тяжелых цепей и легких цепей, представлены в таблице 2, и константные области IgG1, IgG4 и легкой цепи приведены в конце настоящего описания. Таким же образом, нуклеотидные последовательности вариабельных областей тяжелых цепей, константной области IgG1 и IgG4 и легкой цепи также приведены в конце настоящего описания.
Пример 2: Получение mAb против PD-1, синтезированных с использованием вектора pZRCII
Для получения материала для исследований для подтверждения концепции, различные конструкции антител против PD-1, как получено в примере 1 (IP-H4L1, IP-H4.2L1, IP-H4.2(Y.H)L1, IP-H4.19L1 и IP-H4.36L1) экспрессировали с использованием системы экспрессии ExpiCHO™ (Gibco). За одни сутки до трансфекции, ExpiCHO-S™ рассевали до конечной плотности 3 × 106 жизнеспособных клеток/мл в встряхиваемой колбе/пробирке для культивирования и позволяли расти в течение ночи. На следующие сутки, клетки подсчитывали и доводили до конечной плотности 6 × 106 жизнеспособных клеток/мл с использованием свежей среды для экспрессии ExpiCHO™, предварительно нагретой до 37°C. Подготавливали комплексы реагента для трансфекции CHO ExpiFectamine™ и плазмидной ДНК. Раствор медленно добавляли к суспензии клеток с осторожным перемешиванием. Клетки инкубировали в инкубаторе 37°C с увлажненной атмосферой и 5% CO2 в орбитальном встряхивателе. Температуру переводили на 32°C на сутки 1. Усилитель продукции белка ExpiFectamine™ CHO Enhancer и подпитку добавляли на сутки 1, с последующей второй подпиткой на сутки 5 после трансфекции. Культуры собирали (на сутки 8-12), и одностадийную очистку проводили с использованием матрикса агарозы-белка A (смола для очистки антител MAbselect SuRe, GE). Связанное антитело элюировали с использованием глицинового буфера, pH 2,5, и нейтрализовали до pH 7 с использованием Трис-буфера, pH 9. Концентрацию белка для очищенного антитела определяли способом HPLC с белком A.
Пример 3: Получение библиотеки фагового дисплея Fab
(a) Получение плазмиды с Fab
Для получения библиотеки, оптимизированную для E. coli область Fab с выступающими концами участков для EcoRI и BamHI химически синтезировали и клонировали в векторы pMA/pMK, полученные из Geneart, Germany.
Ген Fab (SEQ ID No. 164) выделяли из векторной конструкции pMA после расщепления с использованием EcoRI и BamHI. Аминокислотная последовательность клонированного гена Fab представлена в настоящем описании как SEQ ID NO:165 для VH и 166 для VL.
Модифицировали фагмидный вектор pSEX81 (кат. No: PR3005, Progen), в котором ген pIII фагмидного вектора укорачивали и расщепляли с использованием EcoRI и BamHI, и линеаризованный вектор лигировали с расщепленным геном Fab. Полученный модифицированный вектор обозначен в настоящем описании как pSEX83. Карта вектора pSEX83 приведена в настоящем описании как фиг. 2. Расщепленные фрагменты Fab и вектора pSEX83 анализировали в агарозном геле и очищали из геля с использованием набора для экстракции из геля QIAquick (Qiagen кат. no 28706). Продукты лигирования как Fab (вставки), так и pSEX83 (вектора) трансформировали в электрокомпетентные клетки E. coli TG1 (кат. No: 60502-1, Lucigen). Трансформированные клетки рассевали на чашки с агаром 2xYT, содержащие ампициллин (100 мкг/мл). Клоны анализировали посредством рестрикционного расщепления EcoRI и BamHI, и секвенирования ДНК с использованием способа Сэнгера.
(b) Получение библитотек
Следовали двум стадиям для получения библиотеки фагового дисплея Fab.
Стадия 1: Сайт-направленный мутагенез на основе праймеров для ПЦР в CDRH1
В данном случае нацеливались на область CDRH1 Fab для получения вариантов. Мутации в выбранные области вводили с использованием ПЦР с соответствующей парой праймеров (NI-CDRH1F и NI-CDRH1R), имеющих участок рестрикции SapI (Таблица 6). Каждую пару праймеров конструировали таким образом, чтобы они амплифицировали полную плазмиду в линейной форме, что после рестрикционного расщепления посредством SapI с последующим лигированием с использованием ДНК-лигазы T4 приводило к получению кольцевой плазмиды с мутациями, введенными посредством пары праймеров.
100 нг ДНК-матрицы гена Fab в векторе pSEX83 использовали для амплификации с использованием пар праймеров, упомянутых в таблице 6. После амплификации матрицы с использованием рандомизированных праймеров, продукт расщепляли с использованием SapI для получения липких концов. После очистки продукта ПЦР, расщепленный продукт лигировали в течение ночи при 16°C. Затем лигированной ДНК трансформировали свежеприготовленные электрокомпетентные клетки (TG1). Трансформированные клетки культивировали для получения фагов. Получение фагов выполняли отдельно для каждой библиотеки, как объяснено ранее (9).
Стадия 2: мутагенез по Кункелю и sRCA
Мутагенез по Кункелю проводили с использованием sRCA, следуя Huovinen et al 2012. Небольшое количество культуры клеток CJ236 инфицировали фагами, несущими плазмиду с подвергнутым мутагенезу в CDRH1 геном Fab. Инфицированные клетки выращивали в 2xYT, содержащей уридин (6 мкг/мл) и антибиотик карбенициллин (100 мкг/мл). Фаги продуцировали после суперинфекции фагом-помощником VCS M13 (Agilent technologies, USA) и очищали посредством преципитации с использованием PEG6000 (4%) и NaCl (500 мМ). Одноцепочечную уридилированную ДНК (оц(U)ДНК), выделяли из очищенных фагов с использованием набора для очистки M13 из E.Z.N.A.® M13 DNA mini kit (OMEGA bio-tek, USA) следуя инструкциям производителя.
Эту оц(U)ДНК использовали в качестве матрицы в мутагенезе по Кункелю, и различные праймеры со случайными мутациями использовали для получения библиотеки. Праймеры, нацеленные на различные CDR, т.е., CDRH2, CDRH3 и CDRL3 Fab ниволумаба (Таблица 6) гибридизовали отдельно с матрицей оц(U)ДНК и удлиняли посредством способа мутагенеза по Кункелю. Продукт обрабатывали UDG и избирательно амплифицировали посредством RCA (10). Продукт RCA расщепляли с использованием Hind III, циркуляризовали с использованием ДНК-лигазы T4 и трансформировали в SS320 клетки. Фаги (вторичную библиотеку) продуцировали отдельно для каждого праймера для мутагенеза посредством суперинфекции фагом-помощником, как описано ниже.
Пример 4: Продукция фага с мутантным Fab и его очистка
В колбы, содержащие среды 2xYT с карбенициллином или ампициллином (100 мкл/мл), инокулировали клетки из препарата для хранения в глицерине из вышеописанной библиотеки при начальной концентрации 0,06 OD600. Культуры выращивали при 37°C с встряхиванием при 250 об/мин, пока они не достигали OD600 вплоть до ~ 0,4-0,6. Затем фаги-помощники, либо VCSM13 (Agilent, кат. no. 200251), либо M13KO7 (GE healthcare, кат. no. 27152401), добавляли в культуру при множественности инфекции (MOI) 20, и инкубировали сначала без встряхивания при 37°C в течение 40 минут, с последующими другими 40 минут инкубации при 37°C с встряхиванием. Канамицин затем добавляли в среды, и культуру выращивали в течение ночи при 26°C при 150 об/мин. В течение ночи культуру фагов центрифугировали при 4000 g, и осадок клеток отбрасывали. Раствор PEG (20%)/NaCl (2,5 М) в соотношении 1:5 добавляли к супернатанту для преципитации фагов. Ресуспендированный раствор инкубировали на льду в течение 20 минут, с последующим центрифугированием при 14000 g в течение 15 минут при 4°C. Супернатант отбрасывали, и осадок ресуспендировали в 1 мл стерильного PBS с 0,01% азидом натрия. Фаги хранили при 4°C до дальнейшего использования.
Пример 5: Биопэннинг фагов, экспрессирующих высокоаффинные варианты Fab
Библиотеку (1×1012 бое), полученную в примере 4, подвергали скринингу по высокоаффинным Fab, связывающим рецептор PD1. Первый цикл пэннинга против антигена рекомбинантного рецептора PD1 (Sino biological, кат. No, 10377-H08H) при pH 7,4, проводили с использованием имеющих иммобилизованный антиген иммунопробирок (Quidel, USA), которые подготавливали посредством их инкубации с раствором 5 мкг/мл белка PD1 в карбонатном буфере (0,1 M, pH 9,6) в течение ночи при 4°C. Иммунопробирки промывали 3 раза с использованием PBS и затем инкубировали с фагами в PBS в течение 2 час при 25°C с постоянным вращением. Пробирки промывали десять раз с использованием 4 мл PBS с tween 20 (0,1%) и затем 10 раз с использованием PBS. Связанные фаги элюировали с использованием глицина-HCL pH 2,1 (0,1 M). Элюированные фаги спасали посредством инфекции клеток TG1 E. coli, рассевали, и фаги продуцировали, как описано в примере 4.
Второй и третий цикл пэннинга проводили в покрытых антигеном PD1 иммунопробирках таким образом, что использовали выходные фаги из первого цикла пэннинга (1×1011 КОЕ) и второго (1×1010 КОЕ) цикла пэннинга, в качестве входных фагов для второго и третьего цикла пэннинга, соответственно. Фагами после третьего цикла пэннинга инфицировали клетки TG1, как упомянуто ранее, и фагмидную ДНК выделяли для клонирования антитела против PD-1 в подходящем экспрессирующем векторе.
Пример 6: Скрининг индивидуальных клонов антител против PD-1 в форме растворимых белков Fab
Клонирование в экспрессирующий вектор и продукция белка
Гены антител против PD-1 из обогащенной библиотеки, полученной после 3 циклов пэннинга, клонировали в экспрессирующий вектор pOPE101 (устойчивый к карбенициллину) (Progen, Germany), таким образом, чтобы индивидуальные клоны антитела против PD-1 можно было получать в форме меченных His слитых продуктов. Как векторную ДНК (pOPE101), так и фагмидную ДНК, расщепляли с использованием рестрикционных ферментов (EcoRI и BamHI) для выделения вектора и генов антитела против PD-1, соответственно. Рестрикционно расщепленные как вектор, так и ген антитела против PD-1 лигировали и трансформировали в электрокомпетентные клетки TG1. Трансформированные клетки рассевали на чашки с агаром 2xYT, содержащие антибиотик карбенициллин. Индивидуальные клоны отбирали с чашек с агаром 2xYT и культивировали в 15 мл пробирках с 5 мл сред 2xYT, содержащих карбенициллин или ампициллин (100 мкл/мл), в течение ночи при 32°C при 200 об/мин. На следующие сутки, культуры повторно инокулировали в свежую среду 2xYT, содержащую карбенициллин (100 мкг/мл) и глюкозу (0,1%) в объемном соотношении 1:200. Культуры выращивали до достижения OD600 ~ 0,6-0,8. После этого 1 мМ изопропил-β-D-1-тиогалактопиранозид (IPTG) добавляли в культуру и выращивали в течение ночи при 30°C при 200 об/мин. Ночную культуру центрифугировали при 10800 g в течение 15 минут, и супернатант удаляли. Осадок клеток ресуспендировали в 1/20й объема исходной культуры 1x PBS буфера pH-7,4, содержащего 2 мг/мл лизоцима, 0,1% triton X и 1 ед./100 мл бензоназы, и инкубировали при 37°C в течение 1 час. Ресуспендированный осадок центрифугировали при 10800 g в течение 15 минут, и супернатант собирали в форме лизата клеток, содержащего растворимые меченные His варианты Fab. Эту фракцию лизата клеток использовали в иммуноанализах для исследования связывания антитела против PD-1 после количественной оценки растворимого Fab в периплазматической фракции.
Определение концентрации растворимых Fab против PD-1 в периплазматической фракции
Клоны культивировали индивидуально для продукции растворимых вариантов Fab, как описано выше. Тотальные Fab в лизате клеток или периплазматической фракции оценивали количественно посредством иммобилизации растворимых Fab на планшетах maxisorp, предварительно покрытых антителом козы против человеческого IgG (специфическим для Fab) (Sigma-Aldrich, кат. no. I5260). Покрытие осуществляли в течение ночи с использованием антитела козы против человеческого IgG (специфического для Fab) (Sigma-Aldrich, кат. no. I5260) при концентрациях 1 мкг/мл (100 мкл/лунку) в буфере для покрытия (0,1M NaHCO3, pH 9,6). После промывки планшета дважды, добавляли лизат клеток 100 мкл (1/20 об/об в 1x PBS) и инкубировали в течение 1 час при 25°C. Известные концентрации серийно разведенных человеческих Fab использовали в качестве стандартов для расчета количества белка Fab в лизате клеток. Планшет промывали 3 раза с использованием PBST (0,1% Tween20 в 1x PBS). 100 мкл (1:10000) конъюгированного с HRP антитела козы против человеческого IgG (специфического для Fab) (Sigma-Aldrich, кат. no. A0293) добавляли в каждую лунку планшета и инкубировали в течение 1 час. Планшет затем промывали 5 раз с использованием PBST, с последующим добавлением субстрата o-фенилендиаминдигидрохлорида (OPD) (100 мкл/лунку) на 15 минут при 37°C. Реакцию останавливали с использованием 1Н H2SO4 (100 мкл/лунку), и оптическую плотность (OD) измеряли при 450 нм с использованием TECAN INFINITE® M1000pro. Концентрацию Fab в лизате клеток затем рассчитывали с использованием эталонных стандартов.
Качественный анализ связывающих Fab против PD-1
Рекомбинантным антигеном PD1 покрывали полистирольный планшет (100 нг/100 мкл/лунку), в течение ночи при 4°C в буфере для покрытия (0,1 М NaHCO3, pH 9,6). После промывки планшета дважды, 100 нг связывающего Fab, разведенного в 100 мкл PBS, pH 6,0, добавляли в каждую лунку и инкубировали в течение 1 час при 25°C. Планшеты промывали 4 раза с использованием PBST (0,1% Tween20 в 1x PBS, pH 6,0) с последующим добавлением 100 мкл (1:10000) конъюгированного с HRP антитела козы против человеческого IgG (специфического для Fab) (Sigma-Aldrich, кат. no. A0293) и инкубации в течение 1 час при 25°C. Затем планшет промывали 5 раз с использованием PBST, с последующим добавлением субстрата o-фенилендиаминдигидрохлорида (OPD) (100 мкл/лунку) в течение 15 минут при 37°C. Реакцию останавливали с использованием 1 Н H2SO4 (100 мкл/лунку), и оптическую плотность измеряли при 450 нм с использованием TECAN INFINITE® M1000pro. Индивидуальные клоны с относительно высоким сигналом OD, по сравнению с исходным Fab, включали в укороченный список для дальнейшей характеризации.
Пример 7: Получение векторов pZRCIII hyg для легкой и тяжелой цепи антитела против PD-1
Пять клонов включены в укороченный список, на основании результатов, полученных из качественного анализа связывающих Fab против PD-1. Эти пять клонов обозначены в настоящем описании как N5, N6, N7, N9 и N10, на основании области HCVR связывающих Fab.
Гены, кодирующие вариабельные области тяжелой цепи из N5, N6, N7, N9 и N10 (аминокислотные последовательности SEQ ID No.:146-150 и нуклеотидные последовательности SEQ ID NO:169-173, соответственно), и ген легкой цепи (SEQ ID NO:167 и 174 представляют собой аминокислотную и нуклеотидную последовательность, соответственно.) химически синтезировали и клонировали в вектор pMA (GeneArt, Germany). Для клонирования легкой цепи в экспрессирующий вектор для млекопитающих, вектор pZRCIII hyg.IgG4 расщепляли с использованием рестрикционных ферментов XhoI и KpnI. Вставку легкой цепи, расщепленную с использованием таких же рестрикционных ферментов, лигировали в расщепленный с использованием XhoI и KpnI вектор pZRCIII hyg.IgG4 и трансформировали в E. coli штамма Top 10F'. Трансформанты отбирали на основании устойчивости к канамицину. Плазмидную ДНК, выделенную из таких колоний, анализировали по присутствию гена легкой цепи посредством рестрикционного расщепления с использованием рестрикционных ферментов XhoI и KpnI. Одну из плазмид, для которой показан ожидаемый профиль рестрикции, дополнительно расщепляли с использованием рестрикционных ферментов SalI и ApaI для вставки генов, кодирующих вариабельные области тяжелой цепи из N5, N6, N7, N9 и N10 (аминокислотные последовательности SEQ ID No.:146-150 и нуклеотидные последовательности SEQ ID NO:169-173, соответственно) в рамке считывания с константной областью IgG4 (SEQ ID NO:168 и 175 представляют собой аминокислотную и нуклеотидную последовательность, соответственно), из вектора pZRCIII hyg.IgG4. Вектор и вставку, расщепленные с использованием рестрикционных ферментов SalI и ApaI, лигировали и трансформировали в E. coli штамма Top 10F'. Трансформанты отбирали на основании устойчивости к канамицину. Плазмидную ДНК из трансформантов расщепляли с использованием рестрикционных ферментов SalI и ApaI для подтверждения присутствия соответствующей вариабельной области. Конечные векторы, содержащие легкую цепь из SEQ ID NO:174 и тяжелую цепь из SEQ ID No.:169; легкую цепь из SEQ ID NO:174 и тяжелую цепь из SEQ ID NO:170; легкую цепь из SEQ ID NO:174 и тяжелую цепь из SEQ ID NO:171; легкую цепь из SEQ ID NO:174 и тяжелую цепь из SEQ ID NO:172; и легкую цепь из SEQ ID NO:174 и тяжелую цепь из SEQ ID NO:173, подтверждали посредством нуклеотидного секвенирования тяжелых и легких цепей. Эти конечные векторы обозначены как N5NL, N6NL, N7NL, N8NL, N9NL и N10NL, соответственно.
Указанные различные конструкции кодируют аминокислотные последовательности HCVR и LCVR антител против PD-1 по настоящему изобретению, как описано в таблице 7.
Общая схема вектора для указанной конструкции представлена на фигуре 3.
Аминокислотные последовательности вариабельных областей тяжелых цепей и легких цепей представлены в таблице 2, и константные области IgG1, IgG4 и легкой цепи приведены в конце настоящего описания. Таким же образом, нуклеотидные последовательности вариабельных областей тяжелых цепей, константные области IgG1 и IgG4, и легкой цепи также приведены в конце настоящего описания.
Пример 8: Получение mAb против PD-1, синтезированных с использованием вектора pZRCIII
Для получения материала для исследований для подтверждения концепции, различные конструкции антител против PD-1, как получено в примере 7 (N5NL, N6NL, N7NL, N9NL и N10NL), экспрессировали в клетках CHO-S (Invitrogen). Клетки общепринятым образом культивировали в средах CDM от Lonza. Клетки рассевали за ~24 часа до трансфекции. Трансфекции проводили посредством электропорации с использованием системы для трансфекции Neon (Invitrogen) с использованием предварительно оптимизированных условий. После трансфекции, клетки рассевали в 24-луночный планшет, содержащий 1 мл предварительно нагретых бессывороточных сред. Клетки инкубировали в увлажненном инкубаторе при 37°C в присутствии 5% CO2. Отбор трансфицированного пула выполняли в присутствии 600 мкг/мл гигромицина в бессывороточной среде. Количество клеток из пула мониторировали регулярно в ходе процесса отбора в течение периода 2-3 недель и проводили регулярные замены сред. Экспрессирующие антитела против PD-1 пулы стабильных трансфектантов размножали, и антитела получали с использованием периодических культур с подпиткой. Клетки рассевали при плотности 0,3×106 клеток/мл в 125 мл встряхиваемых колбах Эрленмейера в химически определенных средах. Колбы инкубировали в увлажненном встряхивателе-инкубаторе Kuhner при температуре 37°C, уровне CO2 5%, со скоростью встряхивания 110 об/мин. Фиксированному режиму ежесуточной подпитки следовали на протяжении всего культивирования. Использовали химически определенные подпитки от Hyclone, GE. Подпитку начинали через 72 часа культивирования и продолжали до сбора культуры. Культуры собирали (на сутки 8-12), и одностадийную очистку проводили с использованием матрикса агарозы-белка A (смола для очистки антител MAbselect SuRe, GE). Связанное антитело элюировали с использованием глицинового буфера, pH 2,5, и нейтрализовали до pH 7 с использованием Трис-буфера, pH 9. Концентрацию белка для очищенного антитела определяли способом HPLC с белком A.
Пример 9: Анализ аффинности антитела против PD-1 человека способом поверхностного плазмонного резонанса (SPR)
Константы аффинности для связывания антител-кандидатов против PD-1 (IP-H4L1 (IgG4), IP-H4L1 (IgG1), IP-H4.2L1, IP-H4.19L1, IP-H4.36L1, N5NL, N6NL, N7NL, N9NL и N10NL) с PD-1 человека определяли с использованием Proteon XPR (Biorad).
PD-1 человека (AcroBiosystems) напрямую иммобилизовали на чипе GLC (ProteOn) с использованием способа связывания по аминогруппе для получения уровня иммобилизации лиганда приблизительно 500 RU. 10 мМ фосфатно-солевой буфер (PBS) (10 мМ фосфатный буфер, pH 7,4, 300 мМ NaCl, 0,005% поверхностно-активного вещества P20) использовали в качестве рабочего буфера для проведения кинетических измерений. Все образцы антител после одностадийной очистки разводили в рабочем буфере, описанном выше, при подходящей концентрации, в сериях концентраций от 10 нМ до 0,09 нМ. Эти образцы инъецировали поверх чипа при скорости потока 50 мкл/мин со временем связывания 300 с и временем диссоциации 1800 с. После анализа каждого образца, поверхность чипа регенерировали с использованием 10 мМ глицинового буфера pH 1,5 в течение 18 с при скорости потока 100 мкл/мин.
Значения KD (кинетические константы) определяли как отношение скорости диссоциации (kd) к скорости связывания (ka), т.е. KD=kd/ka, с использованием программного обеспечения ProteOn manager v3.1.0.6.
Результаты представлены в таблице 8. Данные анализировали с использованием модели связывания Ленгмюра.
Ka (1/Мс)
Kd (1/с)
KD (M)
(IgG4)
(IgG1)
Пример 10: Анализ антител против PD-1 с использованием проточного цитометрического анализа в клетках PBMC
Человеческие мононуклеарные клетки периферической крови (PBMC), истощенные по моноцитам, активировали с использованием планетов, покрытых антителом против CD3 (клон UCHT-1; BD Biosciences), в течение 3 суток. После промывки, клетки инкубировали с различными концентрациями антител против PD-1 IP- H4.2L1, IP-H4L1 (IgG1) и IP-H4L1 (IgG4). Связывание этих антител против PD-1 с PD1, экспрессированным на активированных клетках, оценивали с использованием конъюгированного с R-фикоэритрином фрагмента F(ab)2 козы против человеческого IgG, специфического для фрагмента Fcγ, в качестве вторичного реагента, и оценки образцов с использованием проточного цитометра (BD FACS Canto™ II). Данные связывания показаны на фигуре 4. Они показывают специфичность связывания IP-H4.2L, IP-H4L1 (IgG1) и IP-H4L1 (IgG4) для PD1 человека, экспрессированного на клетках.
Пример 11: Анализ антител против PD-1 с использованием проточного цитометрического анализа в клетках HEK-293T
Клетки HEK 293T, стабильно трансфицированные для экспрессии PD-1 человека, инкубировали с различными концентрациями антител против PD-1 IP-H4.19L1, IP-H4.2L1 и IP-H4.36L1. Связывание этих антител с PD1 на клетках HEK-293T оценивали с использованием конъюгированного с R-фикоэритрином фрагмента F(ab)2 козы против человеческого IgG, специфического для фрагмента Fcγ, в качестве вторичного реагента, оцененного с использованием проточного цитометра (BD FACS CantoTM II). Данные показаны на фигуре 5. Они показывают специфичность связывания IP- H4.19L1, IP-H4.2L1 и IP-H4.36L1 для PD1 человека, экспрессированного на клетках.
Сходным способом, другие антитела против PD-1 N5NL, N6NL, N7NL, N9NL и N10NL анализировали с использованием проточного цитометрического анализа в клетках HEK-293T. Данные показаны на фигуре 6.
Пример 12: Анализ антитела против PD-1 в реакции смешанной культуры лимфоцитов
Дендритные клетки (DC) получали посредством культивирования моноцитов, выделенных из PBMC, in vitro в течение 7 суток с 50 нг/мл интерлейкина-4 (IL-4) и 100 нг/мл GM-CSF (11). Истощенные по моноцитам PBMC (5×104) и аллогенные DC (1×104) совместно культивировали в присутствии или в отсутствие различных концентраций антител против PD1 IP-H4.2L1, IP-H4.36L1, IP- H4.19L1, IP-H4L1 (IgG4) и аналога ниволумаба, добавленных в начале анализа. Через 5 суток, культуральные супернатанты собирали и анализировали по секреции IL2 и IFN гамма посредством ELISA (BD Biosciences). Увеличенную секрецию либо IL-2, либо IFN гамма, обусловленную данным антителом против PD-1, рассчитывали после вычитания значений, полученных с использованием контроля изотипа IgG4 из соответствующей группы. Результаты IL2 и IFN-гамма показаны на фигурах 7 и 8, соответственно. Полученные результаты показывают, что антитела против PD1, антитела против PD-1 по настоящему изобретению являются способными стимулировать секрецию IL-2 и IFN-гамма T-клетками в MLR.
Подобным образом, другие антитела против PD-1 N5NL, N6NL, N9NL и N10NL анализировали в реакции смешанной культуры лимфоцитов для анализа увеличенной секреции IL2 и IFN гамма посредством ELISA (BD Biosciences). Результаты для IL2 и IFN гамма показаны на фигурах 9 и 10, соответственно.
Список аминокислотных последовательностей, используемых по настоящему изобретению:
SEQ ID NO:153 - Константная область IgG4
ASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
SEQ ID NO:154 - Константная область IgG1
ASTKGPSVFPLAPSSKSTSGGTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTQTYICNVNHKPSNTKVDKKVEPKSCDKTHTCPPCPAPELLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSHEDPEVKFNWYVDGVEVHNAKTKPREEQYNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKALPAPIEKTISKAKGQPREPQVYTLPPSREEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSKLTVDKSRWQQGNVFSCSVMHEALHNHYTQKSLSLSPGK
SEQ ID NO:155 - Константная область легкой цепи IP-L1
RTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:165 - Вариабельная область тяжелой цепи ниволумаба (полученная синтетически)
QVQLVESGGGVVQPGRSLRLDCKASGITFSNSGMHWVRQAPGKGLEWVAVIWYDGSKRYYADSVKGRFTISRDNSKNTLFLQMNSLRAEDTAVYYCATNDDYWGQGTLVTVSSASTKGPSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPC
SEQ ID NO:166 - Легкая цепь ниволумаба (полученная синтетически)
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQSSNWPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:167 - Легкая цепь NL
EIVLTQSPATLSLSPGERATLSCRASQSVSSYLAWYQQKPGQAPRLLIYDASNRATGIPARFSGSGSGTDFTLTISSLEPEDFAVYYCQQSSNWPRTFGQGTKVEIKRTVAAPSVFIFPPSDEQLKSGTASVVCLLNNFYPREAKVQWKVDNALQSGNSQESVTEQDSKDSTYSLSSTLTLSKADYEKHKVYACEVTHQGLSSPVTKSFNRGEC
SEQ ID NO:168 - Константная область IgG4
PSVFPLAPCSRSTSESTAALGCLVKDYFPEPVTVSWNSGALTSGVHTFPAVLQSSGLYSLSSVVTVPSSSLGTKTYTCNVDHKPSNTKVDKRVESKYGPPCPPCPAPEFLGGPSVFLFPPKPKDTLMISRTPEVTCVVVDVSQEDPEVQFNWYVDGVEVHNAKTKPREEQFNSTYRVVSVLTVLHQDWLNGKEYKCKVSNKGLPSSIEKTISKAKGQPREPQVYTLPPSQEEMTKNQVSLTCLVKGFYPSDIAVEWESNGQPENNYKTTPPVLDSDGSFFLYSRLTVDKSRWQEGNVFSCSVMHEALHNHYTQKSLSLSLGK
Список нуклеотидных последовательностей, используемых по настоящему изобретению:
SEQ ID NO:156 - Константная область IgG4
GCCTCCACCAAGGGCCCCTCCGTGTTCCCTCTGGCCCCTTGCTCCCGGTCCACCTCCGAGTCTACCGCCGCTCTGGGCTGCCTGGTGAAAGACTACTTCCCCGAGCCCGTGACCGTGTCCTGGAACTCTGGCGCCCTGACCAGCGGCGTGCACACCTTCCCTGCCGTGCTGCAGTCCTCCGGCCTGTACTCCCTGTCCTCCGTGGTGACCGTGCCCTCCTCCAGCCTGGGCACCAAGACCTACACCTGTAACGTGGACCACAAGCCCTCCAACACCAAGGTGGACAAGCGGGTGGAATCTAAGTACGGCCCTCCCTGCCCCCCCTGCCCTGCCCCTGAATTTCTGGGCGGACCTTCCGTGTTCCTGTTCCCCCCAAAGCCCAAGGACACCCTGATGATCTCCCGGACCCCCGAAGTGACCTGCGTGGTGGTGGACGTGTCCCAGGAAGATCCCGAGGTGCAGTTCAATTGGTACGTGGACGGCGTGGAAGTGCACAACGCCAAGACCAAGCCCAGAGAGGAACAGTTCAACTCCACCTACCGGGTGGTGTCTGTGCTGACCGTGCTGCACCAGGACTGGCTGAACGGCAAAGAGTACAAGTGCAAGGTGTCCAACAAGGGCCTGCCCTCCAGCATCGAAAAGACCATCTCCAAGGCCAAGGGCCAGCCCCGCGAGCCCCAGGTGTACACCCTGCCCCCTAGCCAGGAAGAGATGACCAAGAACCAGGTGTCCCTGACCTGTCTGGTGAAAGGCTTCTACCCCTCCGATATCGCCGTGGAATGGGAGTCCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACTCCGACGGCTCCTTCTTCCTGTACTCTCGGCTGACCGTGGACAAGTCCCGGTGGCAGGAAGGCAACGTGTTCTCCTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACCCAGAAGTCCCTGTCCCTGAGCCTGGGTAAGTGATGA
SEQ ID NO:157 - Константная область IgG1
GCTAGCACCAAGGGCCCATCGGTCTTCCCCCTGGCACCCTCCTCCAAGAGCACCTCTGGGGGCACAGCGGCCCTGGGCTGCCTGGTCAAGGACTACTTCCCCGAACCGGTGACGGTGTCGTGGAACTCAGGCGCCCTGACCAGCGGCGTGCACACCTTCCCGGCTGTCCTACAGTCCTCAGGACTCTACTCCCTCAGCAGCGTGGTGACCGTGCCCTCCAGCAGCTTGGGCACCCAGACCTACATCTGCAACGTGAATCACAAGCCCAGCAACACCAAGGTGGACAAGAAAGTTGAGCCCAAATCTTGTGACAAAACTCACACATGCCCACCGTGCCCAGCACCTGAACTCCTGGGGGGACCGTCAGTCTTCCTCTTCCCCCCAAAACCCAAGGACACCCTCATGATCTCCCGGACCCCTGAGGTCACATGCGTGGTGGTGGACGTGAGCCACGAAGACCCTGAGGTCAAGTTCAACTGGTACGTGGACGGCGTGGAGGTGCATAATGCCAAGACAAAGCCGCGGGAGGAGCAGTACAACAGCACGTACCGTGTGGTCAGCGTCCTCACCGTCCTGCACCAGGACTGGCTGAATGGCAAGGAGTACAAGTGCAAGGTCTCCAACAAAGCCCTCCCAGCCCCCATCGAGAAAACCATCTCCAAAGCCAAAGGGCAGCCCCGAGAACCACAGGTGTACACCCTGCCCCCATCCCGGGAGGAGATGACCAAGAACCAGGTCAGCCTGACCTGCCTGGTCAAAGGCTTCTATCCCAGCGACATCGCCGTGGAGTGGGAGAGCAATGGGCAGCCGGAGAACAACTACAAGACCACGCCTCCCGTGCTGGACTCCGACGGCTCCTTCTTCCTCTACAGCAAGCTCACCGTGGACAAGAGCAGGTGGCAGCAGGGGAACGTCTTCTCATGCTCCGTGATGCATGAGGCTCTGCACAACCACTACACGCAGAAGAGCCTCTCCCTGTCTCCGGGTAAATGA
SEQ ID NO:158 - Вариабельная область тяжелой цепи IP-H4
CAGGTGCAGCTGGTGCAGTCCGGCGTGGAAGTGAAGAAACCTGGCGCCTCCGTGAAGGTGTCCTGCAAGGCCTCCGGCTACACCTTCACCACCTACTACATCTACTGGGTCCGACAGGCCCCAGGCCAGGGCCTGGAATGGATGGGCGGCGTGAACCCTTCCAACGGCGGCACCAACTACAACGAGAACTACAAGAACAGAGTGACCCTGACCACCGACTCCTCCACCACCACCGCCTACATGGAACTGAAGTCCCTGCAGTTCGACGACACCGCCGTGTACTACTGCGCCAGACGGGACTACAGATACGACATGGGCTTCGACTACTGGGGCCAGGGCACCACCGTGACCGTGTCCTCT
SEQ ID NO:159 - Вариабельная область тяжелой цепи IP-H4.2
CAGGTTCAATTGGTGCAGTCCGGCGTGGAAGTGAAGAAACCTGGCGCTTCTGTGAAGGTGTCCTGCAAGGCCTCTGGCTACACCTTTACCACCTACTACATCTACTGGGTCCGACAGGCTCCTGGACAGGGACTTGAATGGATGGGCGGCGTGAACCCTTCTAACGGCGGCACCAACTACAACGAGAACTACAAGAACAGAGTGACCCTGACCACCGACTCCTCTACCACCACCGCCTACATGGAACTGAAGTCCCTGCAGTTCGACGACACCGCCGTGTACTACTGCGCCAGAAGAGACTACAGATACGACATGGGCTACGACTACTGGGGCCAGGGCACAACAGTGACCGTGTCCTCT
SEQ ID NO:160 - Вариабельная область тяжелой цепи IP-H4.2 (Y.H)
CAGGTGCAGCTGGTGCAGTCCGGCGTGGAAGTGAAGAAACCTGGCGCCTCCGTGAAGGTGTCCTGCAAGGCCTCCGGCTACACCTTCACCACCTACTACATCTACTGGGTCCGACAGGCCCCAGGCCAGGGCCTGGAATGGATGGGCGGCGTGAACCCTTCCAACGGCGGCACCAACTACAACGAGAACTACAAGAACAGAGTGACCCTGACCACCGACTCCTCCACCACCACCGCCTACATGGAACTGAAGTCCCTGCAGTTCGACGACACCGCCGTGTACTACTGCGCCAGACGGGACTACAGATACGACATGGGCCATGACTACTGGGGCCAGGGCACCACCGTGACCGTGTCCTCTGCCTCCACCAAGGGCCC
SEQ ID NO:161 - Вариабельная область тяжелой цепи IP- H4.19
CAGGTTCAATTGGTGCAGTCCGGCGTGGAAGTGAAGAAACCTGGCGCTTCTGTGAAGGTGTCCTGCAAGGCCTCTGGCTACACCTTTACCACCTACTACATCTACTGGGTCCGACAGGCTCCTGGACAGGGACTTGAATGGATGGGCGGCGTGAACCCTTCCAACTCCGGCACCAACTACAACGAGAACTACAAGAACAGAGTGACCCTGACCACCGACTCCTCTACCACCACCGCCTACATGGAACTGAAGTCCCTGCAGTTCGACGACACCGCCGTGTACTACTGCGCCAGAAGAGACTACAGATACGACATGGGCTTCGACTACTGGGGCCAGGGCACAACAGTGACCGTGTCCTCT
SEQ ID NO:162 - Вариабельная область тяжелой цепи IP-H4.36
CAGGTTCAATTGGTGCAGTCCGGCGTGGAAGTGAAGAAACCTGGCGCTTCTGTGAAGGTGTCCTGCAAGGCCTCTGGCTACACCTTTACCACCTACTACATCTACTGGGTCCGACAGGCTCCTGGACAGGGACTTGAATGGATGGGCGGCGTGAACCCTTCTAACGGCGGCACCAACTACAACCAGAACTACAAGAACAGAGTGACCCTGACCACCGACTCCTCTACCACCACCGCCTACATGGAACTGAAGTCCCTGCAGTTCGACGACACCGCCGTGTACTACTGCGCCAGAAGAGACTACAGATACGACATGGGCTTCGACTACTGGGGCCAGGGCACAACAGTGACCGTGTCCTCT
SEQ ID NO:163 - Легкая цепь IP-L1
GAGATCGTGCTGACCCAGTCCCCCGCCACCCTGTCTCTGTCTCCCGGCGAGAGAGCCACCCTGAGCTGCAGAGCCTCCAAGGGCGTGTCCACCTCCGGCTACTCCTACCTGCACTGGTATCAGCAGAAGCCCGGCCAGGCCCCTCGGCTGCTGATCTACCTGGCCTCCTACCTGGAATCCGGCGTGCCCGCCAGATTCTCCGGCTCTGGCTCTGGCACCGACTTCACCCTGACCATCTCCAGCCTGGAACCCGAGGACTTCGCCGTGTACTACTGCCAGCACTCCCGGGACCTGCCCCTGACCTTTGGCGGAGGCACCAAGGTGGAAATCAAGCGGACCGTGGCCGCTCCCTCCGTGTTCATCTTCCCACCCTCCGACGAGCAGCTGAAGTCCGGCACCGCCTCCGTCGTGTGCCTGCTGAACAACTTCTACCCCCGCGAGGCCAAGGTGCAGTGGAAGGTGGACAATGCCCTGCAGTCCGGCAACTCCCAGGAATCCGTCACCGAGCAGGACTCCAAGGACAGCACCTACTCCCTGTCCTCCACCCTGACCCTGTCCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGCGAAGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGTCCTTCAACCGGGGCGAGTGCTAATGAGGTACC
SEQ ID NO:164 - Ген Fab ниволумаба
GGATAACAATTTCACACAGAATTCATTAAAGAGGAGAAATTAACCATGAAAAAGAATATCGCATTTCTTCTTGCATCTATGTTCGTTTTTTCTATTGCTACAAATGCCTATGCATCCGAAATCGTGCTGACCCAGTCGCCCGCTACATTATCGTTGTCACCGGGGGAACGCGCGACATTGTCCTGTCGCGCATCACAGAGCGTATCTTCGTATTTGGCCTGGTATCAGCAAAAGCCAGGCCAAGCCCCCCGTCTTTTAATTTATGACGCCTCTAATCGCGCAACCGGCATTCCTGCACGCTTTTCCGGCTCAGGGAGTGGCACGGATTTCACTTTGACGATCAGTAGTCTTGAACCAGAAGACTTCGCGGTTTACTACTGCCAACAATCCTCGAACTGGCCTCGTACATTTGGTCAAGGTACCAAGGTGGAAATCAAACGCACCGTAGCCGCACCGTCAGTCTTCATCTTCCCTCCCTCTGACGAACAATTAAAATCTGGCACAGCTTCCGTTGTATGCCTTCTTAACAACTTTTATCCACGTGAAGCGAAAGTCCAATGGAAAGTTGATAATGCACTTCAGTCTGGGAACTCACAGGAGTCCGTAACGGAACAGGACTCTAAGGACTCAACCTATAGCTTATCTTCAACATTGACATTGTCAAAGGCAGACTATGAGAAGCACAAAGTGTACGCTTGTGAAGTCACCCATCAGGGCTTGTCTTCTCCAGTAACCAAAAGTTTCAATCGCGGGGAGTGTGGTGGTTCTGATTACAAAGATGACGATGACAAATAATTAACTCGAGGCTGAGCAAAGCAGACTACTAATAACATAAAGTCTACGCCGGACGCATCGTGGCCCTAGTACGCAAGTTCACGTAAAAAGGGTAACTAGAGGTTGAGGTGATTTTATGAAAAAGAATATCGCATTTCTTCTTGCATCTATGTTCGTTTTTTCTATTGCTACAAACGCGTACGCTGAGATCTCCCAAGTCCAGCTGGTTGAATCAGGTGGGGGAGTTGTTCAACCGGGGCGTTCCCTGCGTCTGGACTGCAAAGCCTCAGGAATTACTTTTTCAAACTCCGGGATGCACTGGGTTCGTCAAGCCCCCGGAAAAGGGTTGGAGTGGGTAGCAGTGATCTGGTATGATGGCTCAAAACGTTACTACGCGGACTCGGTTAAAGGTCGCTTTACTATTTCACGCGACAACTCGAAAAATACTTTGTTCTTACAGATGAACTCTCTTCGCGCCGAAGATACGGCAGTGTACTATTGCGCCACAAATGATGATTATTGGGGTCAAGGTACATTAGTGACCGTCAGTAGCGCGAGTACGAAGGGACCTAGCGTGTTTCCCTTAGCACCATGTAGTCGTTCGACAAGCGAATCCACAGCGGCCTTAGGGTGCTTAGTGAAGGATTATTTTCCCGAGCCAGTAACAGTATCGTGGAATAGCGGCGCATTAACTTCCGGCGTTCATACCTTCCCTGCCGTTTTACAGTCATCCGGTTTGTATTCTCTTTCGTCAGTAGTAACAGTGCCTTCCTCCTCGCTGGGGACCAAGACCTATACCTGCAATGTGGACCATAAGCCTTCAAACACAAAAGTAGACAAACGTGTTGAATCAAAGTACGGCCCGCCTTGTCCGCCCTGCGGATCCAAAGATATCAGATCTGGT
SEQ ID NO:169 - Вариабельная область тяжелой цепи N5
CAGGTTCAATTGGTTGAATCTGGCGGCGGAGTGGTGCAGCCTGGAAGAAGTCTGAGACTGGATTGCAAGGCCTCCGGCATCACCTTCTCCAACTCTGGCATGCACTGGGTCCGACAGGCCCCTGGAAAAGGACTGGAATGGGTCGCCGTGATTTGGTACGACGGCTCTAAGCGGTACTACGCCGACTCCGTGAAGGGCAGATTCACCATCTCTCGGGACAACTCCAAGAACACCCTGTTTCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCACCAACTCTGATTTTTGGGGCCAGGGCACACTGGTCACCGTGTCCTCTGCTTCTACAAAGGGCCC
SEQ ID NO:170 - Вариабельная область тяжелой цепи N6
CAGGTTCAATTGGTTGAATCTGGCGGCGGAGTGGTGCAGCCTGGAAGAAGTCTGAGACTGGATTGCAAGGCCTCCGGCATCACCTTCTCCAACTCTGGCATGCACTGGGTCCGACAGGCCCCTGGAAAAGGACTGGAATGGGTCGCCGTGATTTGGTACGACGGCTCTAAGCGGTACTACGCCGACTCCGTGAAGGGCAGATTCACCATCTCTCGGGACAACTCCAAGAACACCCTGTTTCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCACCAATTCTGATCATTGGGGCCAGGGCACACTGGTCACCGTGTCCTCTGCTTCTACAAAGGGCCC
SEQ ID NO:171 - Вариабельная область тяжелой цепи N7
CAGGTTCAATTGGTTGAATCTGGCGGCGGAGTGGTGCAGCCTGGAAGAAGTCTGAGACTGGATTGCAAGGCCTCCGGCATCACCTTCTCCAACTCTGGCATGCACTGGGTCCGACAGGCCCCTGGAAAAGGACTGGAATGGGTCGCCGTGATTTGGTACGACGGCTCTAAGCGGTACTACGCCGACTCCGTGAAGGGCAGATTCACCATCTCTCGGGACAACTCCAAGAACACCCTGTTTCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCACCAACTCTGATTATTGGGGCCAGGGCACACTGGTCACCGTGTCCTCTGCTTCTACAAAGGGCCC
SEQ ID NO:172 - Вариабельная область тяжелой цепи N9
CAGGTTCAATTGGTTGAATCTGGCGGCGGAGTGGTGCAGCCTGGAAGAAGTCTGAGACTGGATTGCAAGGCCTCCGGCATCACCTTCTCCAACTCTGGCATGCACTGGGTCCGACAGGCCCCTGGAAAAGGACTGGAATGGGTCGCCGTGATTTGGTACGACGGCTCTAAGCGGTACTACGCCGACTCCGTGAAGGGCAGATTCACCATCTCTCGGGACAACTCCAAGAACACCCTGTTTCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCACAAATACCGATTGGTGGGGCCAGGGCACACTGGTCACAGTGTCCTCTGCTTCCACAAAGGGCCC
SEQ ID NO:173 - Вариабельная область тяжелой цепи N10
CAGGTTCAATTGGTTGAATCTGGCGGCGGAGTGGTGCAGCCTGGAAGAAGTCTGAGACTGGATTGCAAGGCCTCCGGCATCACCTTCTCCAACTCTGGCATGCACTGGGTCCGACAGGCCCCTGGAAAAGGACTGGAATGGGTCGCCGTGATTTGGTACGACGGCTCTAAGCGGTACTACGCCGACTCCGTGAAGGGCAGATTCACCATCTCTCGGGACAACTCCAAGAACACCCTGTTTCTGCAGATGAACTCCCTGAGAGCCGAGGACACCGCCGTGTACTACTGTGCCACCAATACCGATTATTGGGGCCAGGGCACACTGGTCACCGTGTCCTCTGCTTCTACAAAGGGCCC
SEQ ID NO:174 - Легкая цепь NL
GAGATCGTGCTGACCCAGTCCCCCGCCACCCTGTCTCTGAGCCCTGGCGAGAGAGCCACCCTGAGCTGCAGAGCCTCCCAGTCCGTGTCCTCCTACCTGGCCTGGTATCAGCAGAAGCCCGGCCAGGCCCCTCGGCTGCTGATCTACGACGCCTCCAACCGGGCCACCGGCATCCCTGCCAGATTCTCCGGCTCTGGCTCCGGCACCGACTTCACCCTGACCATCTCCAGCCTGGAACCCGAGGACTTCGCCGTGTACTACTGCCAGCAGTCCTCCAACTGGCCCCGGACCTTCGGCCAGGGCACCAAGGTGGAAATCAAGCGGACCGTGGCCGCTCCCTCCGTGTTCATCTTCCCACCCTCCGACGAGCAGCTGAAGTCCGGCACCGCCTCCGTGGTCTGCCTGCTGAACAACTTCTACCCCCGCGAGGCCAAGGTGCAGTGGAAGGTGGACAACGCCCTGCAGTCCGGCAACTCCCAGGAATCCGTGACCGAGCAGGACTCCAAGGACAGCACCTACTCCCTGTCCTCCACCCTGACCCTGTCCAAGGCCGACTACGAGAAGCACAAGGTGTACGCCTGCGAAGTGACCCACCAGGGCCTGTCCAGCCCCGTGACCAAGTCCTTCAACCGGGGCGAGTGCTGATGA
SEQ ID NO:175 - Константная область IgG4
GGGCCCCTCCGTGTTCCCTCTGGCCCCTTGCTCCCGGTCCACCTCCGAGTCTACCGCCGCTCTGGGCTGCCTGGTGAAAGACTACTTCCCCGAGCCCGTGACCGTGTCCTGGAACTCTGGCGCCCTGACCAGCGGCGTGCACACCTTCCCTGCCGTGCTGCAGTCCTCCGGCCTGTACTCCCTGTCCTCCGTGGTGACCGTGCCCTCCTCCAGCCTGGGCACCAAGACCTACACCTGTAACGTGGACCACAAGCCCTCCAACACCAAGGTGGACAAGCGGGTGGAATCTAAGTACGGCCCTCCCTGCCCCCCCTGCCCTGCCCCTGAATTTCTGGGCGGACCTTCCGTGTTCCTGTTCCCCCCAAAGCCCAAGGACACCCTGATGATCTCCCGGACCCCCGAAGTGACCTGCGTGGTGGTGGACGTGTCCCAGGAAGATCCCGAGGTGCAGTTCAATTGGTACGTGGACGGCGTGGAAGTGCACAACGCCAAGACCAAGCCCAGAGAGGAACAGTTCAACTCCACCTACCGGGTGGTGTCTGTGCTGACCGTGCTGCACCAGGACTGGCTGAACGGCAAAGAGTACAAGTGCAAGGTGTCCAACAAGGGCCTGCCCTCCAGCATCGAAAAGACCATCTCCAAGGCCAAGGGCCAGCCCCGCGAGCCCCAGGTGTACACCCTGCCCCCTAGCCAGGAAGAGATGACCAAGAACCAGGTGTCCCTGACCTGTCTGGTGAAAGGCTTCTACCCCTCCGATATCGCCGTGGAATGGGAGTCCAACGGCCAGCCCGAGAACAACTACAAGACCACCCCCCCTGTGCTGGACTCCGACGGCTCCTTCTTCCTGTACTCTCGGCTGACCGTGGACAAGTCCCGGTGGCAGGAAGGCAACGTGTTCTCCTGCTCCGTGATGCACGAGGCCCTGCACAACCACTACACCCAGAAGTCCCTGTCCCTGAGCCTGGGCAAGTGATGA
Ссылки, включенные в настоящую патентную заявку:
1. Dong et al. B7-H1, a third member of the B7 family, co-stimulates T-cell proliferation and interleukin-10 secretion, Nature Med., 1999 Dec;5(12):1365-9, 1999.
2. Zou et al. Inhibitory B7-family molecules in the tumour microenvironment, Nature Review Immunol., Jun;8(6):467-77, 2008.
3. Iwai et al. Involvement of PD-L1 on Tumor Cells in the Escape From Host Immune System and Tumor Immunotherapy by PD-L1 Blockade, Proc. Nat 7. Acad. ScL USA 99: 12293-7, 2002.
4. Brown et al. Blockade of Programmed Death-1 Ligands on Dendritic Cells Enhances T Cell Activation and Cytokine Production, J. Immunol. 170:1257-66, 2003.
5. Sheridan et al. Cautious optimism surrounds early clinical data for PD-1 blocker, Nature Biotechnology 30: 729-730, 2012.
6. Angal S., King D.J., Bodmer M.W., Turner A., Lawson A.D., Roberts G., Pedley B., Adair J.R. A single amino acid substitution abolishes the heterogeneity of chimeric mouse/human (IgG4) antibody. Mol Immunol. 1993; 30(1): 105-8.
7. Weiyi Peng, Chengwen Liu et al. PD-1 blockade enhances T cell migration to tumors by elevating IFN-γ inducible chemokines Cancer Res. 2012 October 15; 72(20): 5209-5218.
8. Niki Karachaliou, Maria Gonzalez-Cao, Guillermo Crespo et al. Interferon gamma, an important marker of response to immune checkpoint blockade in non-small cell lung cancer and melanoma patients Therapeutic Advances in Medical Oncology, 2018, vol. 10: 1-23, 2018.
9. Brockmann et al. Synthetic single-framework antibody library integrated with rapid affinity maturation by VL shuffling, Protein Engineering, Design & Selection, vol. 24 no. 9 pp. 691-700, 2011.
10. Huovinen T., Brockmann E.-C., Akter S., Perez-Gamarra S., Ylä-Pelto J. et al. Primer Extension Mutagenesis Powered by Selective Rolling Circle Amplification, volume 7, issue 2, e31817, 2012.
11. Palucka et al. Dendritic Cells as the Terminal Stage of Monocyte Differentiation, J Immunology, 160:4587-4595, 1998.
Включение в качестве ссылки
Полное содержание каждого из Патентных документов и научных статей, на которые ссылаются в настоящем описании, приведено в качестве ссылки для всех целей.
Эквиваленты
Изобретение можно осуществлять в других конкретных формах без отклонения от содержания или его важных характеристик. Приведенные выше варианты осуществления, таким образом, следует считать во всех отношениях иллюстративными, а не ограничивающими изобретение, описанное в настоящем описании. Объем изобретения, таким образом, указан в прилагаемой формуле изобретения, а не в приведенном выше описании, и все изменения, которые попадают в рамки значения и диапазона эквивалентности пунктов формулы изобретения, предназначены для охвата ими.
--->
СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> КАДИЛА ХЕЛЗКЭР ЛИМИТЕД
<120> Антитела против рецептора программируемой смерти PD-1 человека
<130> CHL- PCT 0802
<150> 201921027443
<151> 2019-07-09
<160> 175
<170> PatentIn версии 3.5
<210> 1
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 1
Asn Tyr Tyr Leu Tyr
1 5
<210> 2
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 2
Asn Tyr Tyr Val Tyr
1 5
<210> 3
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 3
Asn Tyr Tyr Ile Tyr
1 5
<210> 4
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 4
Thr Tyr Tyr Ile Tyr
1 5
<210> 5
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 5
Thr Tyr Tyr Val Tyr
1 5
<210> 6
<211> 5
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 6
Thr Tyr Tyr Met Tyr
1 5
<210> 7
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 7
Gly Ile Thr Phe Ser Asn Ser Gly
1 5
<210> 8
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 8
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 9
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 9
Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 10
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 10
Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 11
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 11
Gly Val Asn Pro Ser Asn Gly Ser Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 12
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 12
Gly Met Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 13
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 13
Gly Met Asn Pro Ser Asn Gly Ser Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 14
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 14
Gly Val Asn Ser Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 15
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 15
Gly Ile Asn Ser Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 16
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 16
Gly Val Asn Ser Ser Asn Gly Ser Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 17
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 17
Gly Leu Asn Ser Ser Asn Gly Ser Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 18
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 18
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Ser Glu Lys Tyr Lys
1 5 10 15
<210> 19
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 19
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Ser Glu Lys Phe Lys
1 5 10 15
<210> 20
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 20
Gly Leu Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 21
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 21
Gly Leu Asn Pro Ser Asn Gly Ser Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 22
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 22
Gly Ile Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 23
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 23
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Phe Asn Glu Asn Tyr Lys
1 5 10 15
<210> 24
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 24
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Lys Tyr Lys
1 5 10 15
<210> 25
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 25
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Phe Lys
1 5 10 15
<210> 26
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 26
Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 27
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 27
Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 28
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 28
Gly Val Asn Pro Ser Asn Gly Ser Thr Asn Tyr Asn Gln Asn Tyr Lys
1 5 10 15
<210> 29
<211> 16
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 29
Gly Met Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr Lys
1 5 10 15
<210> 30
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 30
Ile Trp Tyr Asp Gly Ser Lys Arg Tyr
1 5
<210> 31
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 31
Arg Asp Tyr Arg Phe Asp Met Gly Tyr Asp Tyr
1 5 10
<210> 32
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 32
Arg Asp Tyr Arg Tyr Asp Met Gly Phe Asp Tyr
1 5 10
<210> 33
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 33
Arg Asp Tyr Arg Tyr Asp Met Gly Tyr Asp Tyr
1 5 10
<210> 34
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 34
Arg Glu Tyr Arg Phe Asp Met Gly Tyr Asp Tyr
1 5 10
<210> 35
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 35
Arg Glu Tyr Arg Phe Asp Met Gly Tyr Glu Tyr
1 5 10
<210> 36
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 36
Arg Glu Tyr Arg Tyr Asp Met Gly Tyr Glu Tyr
1 5 10
<210> 37
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 37
Arg Asp Tyr Arg Tyr Asp Leu Gly Tyr Asp Tyr
1 5 10
<210> 38
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 38
Arg Glu Tyr Arg Tyr Asp Met Gly Phe Asp Tyr
1 5 10
<210> 39
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 39
Arg Asp Tyr Arg Tyr Asp Met Gly Phe Glu Tyr
1 5 10
<210> 40
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 40
Arg Asp Tyr Arg Tyr Asp Met Gly Tyr Glu Tyr
1 5 10
<210> 41
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 41
Arg Glu Tyr Arg Tyr Asp Met Gly Tyr Glu Tyr
1 5 10
<210> 42
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 42
Arg Asp Tyr Arg Tyr Asp Ile Gly Phe Asp Tyr
1 5 10
<210> 43
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 43
Arg Glu Tyr Arg Tyr Asp Ile Gly Tyr Asp Tyr
1 5 10
<210> 44
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 44
Arg Glu Tyr Arg Tyr Asp Ile Gly Tyr Glu Tyr
1 5 10
<210> 45
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 45
Arg Asp Tyr Arg Tyr Asp Gln Gly Tyr Asp Tyr
1 5 10
<210> 46
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 46
Arg Glu Tyr Arg Tyr Asp Gln Gly Tyr Glu Tyr
1 5 10
<210> 47
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 47
Arg Asp Tyr Arg Tyr Asp Met Gly His Asp Tyr
1 5 10
<210> 48
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 48
Glu Ser Glu Tyr
1
<210> 49
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 49
Asn Asn Asp Ile
1
<210> 50
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 50
Asn Ser Asp Phe
1
<210> 51
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 51
Asn Ser Asp His
1
<210> 52
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 52
Asn Ser Asp Tyr
1
<210> 53
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 53
Asn Ser Gly Tyr
1
<210> 54
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 54
Asn Thr Asp Trp
1
<210> 55
<211> 4
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 55
Asn Thr Asp Tyr
1
<210> 56
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 56
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Phe Leu His
1 5 10 15
<210> 57
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 57
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Trp Leu His
1 5 10 15
<210> 58
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 58
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Tyr Leu Tyr
1 5 10 15
<210> 59
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 59
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Tyr Ile Tyr
1 5 10 15
<210> 60
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 60
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Phe Leu Tyr
1 5 10 15
<210> 61
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 61
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Thr Tyr Leu Tyr
1 5 10 15
<210> 62
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 62
Arg Ala Ser Lys Asn Val Ser Thr Ser Gly Tyr Ser Tyr Leu Tyr
1 5 10 15
<210> 63
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 63
Arg Ala Ser Glu Gly Val Ser Thr Ser Gly Tyr Ser Tyr Leu His
1 5 10 15
<210> 64
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 64
Arg Ala Ser Glu Gly Val Ser Thr Ser Gly Phe Ser Tyr Ile His
1 5 10 15
<210> 65
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 65
Arg Ala Ser Lys Asn Val Ser Thr Thr Gly Phe Ser Tyr Leu His
1 5 10 15
<210> 66
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 66
Arg Ala Ser Lys Asn Val Ser Ser Thr Ser Tyr Ser Tyr Leu His
1 5 10 15
<210> 67
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 67
Arg Ala Ser Lys Asn Val Ser Ser Thr Ser Phe Ser Tyr Leu Tyr
1 5 10 15
<210> 68
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 68
Arg Ala Ser Lys Gly Val Ser Ser Thr Ser Phe Ser Tyr Ile His
1 5 10 15
<210> 69
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 69
Arg Ala Ser Arg Gly Ile Ser Thr Ser Gly Tyr Ser Tyr Ile His
1 5 10 15
<210> 70
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 70
Arg Ala Ser Glu Gly Ile Ser Thr Ser Gly Tyr Ser Tyr Ile His
1 5 10 15
<210> 71
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 71
Arg Ala Ser Glu Gly Leu Asp Thr Ser Gly Tyr Ser Tyr Ile His
1 5 10 15
<210> 72
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 72
Arg Ala Ser Glu Gly Leu Glu Thr Ser Gly Tyr Ser Tyr Ile His
1 5 10 15
<210> 73
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 73
Arg Ala Ser Glu Gly Ile Ser Thr Ser Gly Tyr Ser Tyr Val His
1 5 10 15
<210> 74
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 74
Arg Ala Ser Lys Gly Ile Ser Thr Asp Gly Tyr Ser Tyr Met His
1 5 10 15
<210> 75
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 75
Arg Ala Ser Lys Gly Ile Ser Thr Glu Gly Tyr Ser Tyr Met His
1 5 10 15
<210> 76
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 76
Arg Ala Asp Lys Gly Val Ser Thr Ser Gly Tyr Ser Tyr Met His
1 5 10 15
<210> 77
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 77
Arg Ala Ser Lys Gly Val Ser Thr Ser Gly Tyr Ser Tyr Leu His
1 5 10 15
<210> 78
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 78
Arg Ala Ser Lys Ser Val Ser Thr Ser Gly Phe Ser Tyr Leu His
1 5 10 15
<210> 79
<211> 11
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 79
Arg Ala Ser Gln Ser Val Ser Ser Tyr Leu Ala
1 5 10
<210> 80
<211> 15
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 80
Arg Gly Ser Lys Gly Val Ser Ser Gly Ile Tyr Ser Tyr Leu His
1 5 10 15
<210> 81
<211> 6
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 81
Gln Ser Val Ser Ser Tyr
1 5
<210> 82
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 82
Ile Ala Ser Tyr Leu Glu Ser
1 5
<210> 83
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 83
Met Ala Ser Tyr Leu Glu Ser
1 5
<210> 84
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 84
Val Ala Ser Tyr Leu Glu Ser
1 5
<210> 85
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 85
Phe Ala Ser Tyr Leu Glu Ser
1 5
<210> 86
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 86
Ala Ala Ser Tyr Leu Glu Ser
1 5
<210> 87
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 87
Ile Ala Ser Phe Leu Glu Ser
1 5
<210> 88
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 88
Ile Ala Ser Trp Leu Glu Ser
1 5
<210> 89
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 89
Ile Ala Ser His Leu Glu Ser
1 5
<210> 90
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 90
Met Ala Ser Phe Leu Glu Ser
1 5
<210> 91
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 91
Met Ala Ser Trp Leu Glu Ser
1 5
<210> 92
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 92
Leu Ala Ser Tyr Leu Gln Ser
1 5
<210> 93
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 93
Ile Ala Ser Tyr Leu Gln Ser
1 5
<210> 94
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 94
Ile Ala Ser Phe Leu Gln Ser
1 5
<210> 95
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 95
Leu Ala Ser Phe Leu Gln Ser
1 5
<210> 96
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 96
Leu Ala Ser Trp Leu Gln Ser
1 5
<210> 97
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 97
Ile Ala Ser Trp Leu Gln Ser
1 5
<210> 98
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 98
Val Ala Ser Trp Leu Gln Ser
1 5
<210> 99
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 99
Ala Ala Ser Phe Leu Glu Ser
1 5
<210> 100
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 100
Leu Ala Asp Tyr Leu Glu Ser
1 5
<210> 101
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 101
Leu Ala Asp Tyr Ile Glu Ser
1 5
<210> 102
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 102
Leu Ala Asp Tyr Val Glu Ser
1 5
<210> 103
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 103
Leu Ala Glu Tyr Met Glu Ser
1 5
<210> 104
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 104
Leu Ala Glu Tyr Val Glu Ser
1 5
<210> 105
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 105
Leu Ala Asp Tyr Leu Glu Asp Tyr
1 5
<210> 106
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 106
Leu Ala Asp Tyr Ile Glu Asp Tyr
1 5
<210> 107
<211> 8
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 107
Leu Ala Glu Tyr Leu Glu Asp Tyr
1 5
<210> 108
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> LASYLES
<400> 108
Leu Ala Ser Tyr Leu Glu Ser
1 5
<210> 109
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 109
Leu Ala Ser Asn Leu Glu Ser
1 5
<210> 110
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 110
Asp Ala Ser Asn Arg Ala Thr
1 5
<210> 111
<211> 7
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 111
Lys Ala Ser Ser Leu Glu Ser
1 5
<210> 112
<211> 3
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 112
Asp Ala Ser
1
<210> 113
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 113
Gln His Ser Arg Glu Leu Pro Leu Thr
1 5
<210> 114
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 114
Gln His Ser Arg Asn Leu Pro Leu Thr
1 5
<210> 115
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 115
Gln His Ser Arg Glu Ile Pro Leu Thr
1 5
<210> 116
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 116
Gln His Ser Arg Asn Ile Pro Leu Thr
1 5
<210> 117
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 117
Gln His Ser Arg Asp Ile Pro Leu Thr
1 5
<210> 118
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 118
Gln His Ser Arg Asp Phe Pro Leu Thr
1 5
<210> 119
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 119
Gln His Ser Arg Asn Phe Pro Leu Thr
1 5
<210> 120
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 120
Gln His Ser Arg Asp Ile Pro Ile Thr
1 5
<210> 121
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 121
Gln His Ser Arg Asp Phe Pro Ile Thr
1 5
<210> 122
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 122
Gln Arg Ser Arg Asp Leu Pro Leu Thr
1 5
<210> 123
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 123
Gln Lys Ser Arg Asp Leu Pro Leu Thr
1 5
<210> 124
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 124
Gln Tyr Ser Arg Asp Leu Pro Ile Thr
1 5
<210> 125
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 125
Gln Arg Ser Arg Asp Ile Pro Leu Thr
1 5
<210> 126
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 126
Gln Lys Ser Arg Asp Ile Pro Leu Thr
1 5
<210> 127
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 127
Gln Tyr Ser Arg Asp Ile Pro Leu Thr
1 5
<210> 128
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 128
Gln His Glu Arg Asp Leu Pro Leu Thr
1 5
<210> 129
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 129
Gln His Glu Arg Asp Ile Pro Leu Thr
1 5
<210> 130
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 130
Gln His Arg Arg Asp Ile Pro Val Thr
1 5
<210> 131
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 131
Gln His Asp Arg Asp Leu Pro Met Thr
1 5
<210> 132
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 132
Gln His Ser Arg Asp Leu Pro Leu Thr
1 5
<210> 133
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 133
Gln His Ser Trp Glu Leu Pro Leu Thr
1 5
<210> 134
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 134
Gln His Tyr Ser Asn Tyr Pro Leu Thr
1 5
<210> 135
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 135
Gln His Ser Arg Asp Ala Pro Leu Thr
1 5
<210> 136
<211> 9
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 136
Gln Gln Ser Ser Asn Trp Pro Arg Thr
1 5
<210> 137
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 137
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 138
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 138
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Tyr Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 139
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 139
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly His Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 140
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 140
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 141
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 141
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Tyr Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 142
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 142
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Ser Gly Thr Asn Tyr Asn Glu Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly His Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 143
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 143
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Gln Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Phe Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 144
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 144
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Gln Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly Tyr Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 145
<211> 120
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 145
Gln Val Gln Leu Val Gln Ser Gly Val Glu Val Lys Lys Pro Gly Ala
1 5 10 15
Ser Val Lys Val Ser Cys Lys Ala Ser Gly Tyr Thr Phe Thr Thr Tyr
20 25 30
Tyr Ile Tyr Trp Val Arg Gln Ala Pro Gly Gln Gly Leu Glu Trp Met
35 40 45
Gly Gly Val Asn Pro Ser Asn Gly Gly Thr Asn Tyr Asn Gln Asn Tyr
50 55 60
Lys Asn Arg Val Thr Leu Thr Thr Asp Ser Ser Thr Thr Thr Ala Tyr
65 70 75 80
Met Glu Leu Lys Ser Leu Gln Phe Asp Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Arg Arg Asp Tyr Arg Tyr Asp Met Gly His Asp Tyr Trp Gly Gln
100 105 110
Gly Thr Thr Val Thr Val Ser Ser
115 120
<210> 146
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 146
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Ser Asp Phe Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly
115
<210> 147
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 147
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Ser Asp His Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly
115
<210> 148
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 148
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Ser Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly
115
<210> 149
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 149
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Thr Asp Trp Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly
115
<210> 150
<211> 118
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 150
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Thr Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly
115
<210> 151
<211> 111
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 151
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Lys Gly Val Ser Thr Ser
20 25 30
Gly Tyr Ser Tyr Leu His Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro
35 40 45
Arg Leu Leu Ile Tyr Leu Ala Ser Tyr Leu Glu Ser Gly Val Pro Ala
50 55 60
Arg Phe Ser Gly Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser
65 70 75 80
Ser Leu Glu Pro Glu Asp Phe Ala Val Tyr Tyr Cys Gln His Ser Arg
85 90 95
Asp Leu Pro Leu Thr Phe Gly Gly Gly Thr Lys Val Glu Ile Lys
100 105 110
<210> 152
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 152
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys
100 105
<210> 153
<211> 327
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 153
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg
1 5 10 15
Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr
65 70 75 80
Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro
100 105 110
Glu Phe Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys
115 120 125
Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys Val Val Val
130 135 140
Asp Val Ser Gln Glu Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp
145 150 155 160
Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe
165 170 175
Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu His Gln Asp
180 185 190
Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu
195 200 205
Pro Ser Ser Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg
210 215 220
Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys
225 230 235 240
Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp
245 250 255
Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys
260 265 270
Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser
275 280 285
Arg Leu Thr Val Asp Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser
290 295 300
Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser
305 310 315 320
Leu Ser Leu Ser Leu Gly Lys
325
<210> 154
<211> 330
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 154
Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Ser Ser Lys
1 5 10 15
Ser Thr Ser Gly Gly Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr
20 25 30
Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser
35 40 45
Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser
50 55 60
Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Gln Thr
65 70 75 80
Tyr Ile Cys Asn Val Asn His Lys Pro Ser Asn Thr Lys Val Asp Lys
85 90 95
Lys Val Glu Pro Lys Ser Cys Asp Lys Thr His Thr Cys Pro Pro Cys
100 105 110
Pro Ala Pro Glu Leu Leu Gly Gly Pro Ser Val Phe Leu Phe Pro Pro
115 120 125
Lys Pro Lys Asp Thr Leu Met Ile Ser Arg Thr Pro Glu Val Thr Cys
130 135 140
Val Val Val Asp Val Ser His Glu Asp Pro Glu Val Lys Phe Asn Trp
145 150 155 160
Tyr Val Asp Gly Val Glu Val His Asn Ala Lys Thr Lys Pro Arg Glu
165 170 175
Glu Gln Tyr Asn Ser Thr Tyr Arg Val Val Ser Val Leu Thr Val Leu
180 185 190
His Gln Asp Trp Leu Asn Gly Lys Glu Tyr Lys Cys Lys Val Ser Asn
195 200 205
Lys Ala Leu Pro Ala Pro Ile Glu Lys Thr Ile Ser Lys Ala Lys Gly
210 215 220
Gln Pro Arg Glu Pro Gln Val Tyr Thr Leu Pro Pro Ser Arg Glu Glu
225 230 235 240
Met Thr Lys Asn Gln Val Ser Leu Thr Cys Leu Val Lys Gly Phe Tyr
245 250 255
Pro Ser Asp Ile Ala Val Glu Trp Glu Ser Asn Gly Gln Pro Glu Asn
260 265 270
Asn Tyr Lys Thr Thr Pro Pro Val Leu Asp Ser Asp Gly Ser Phe Phe
275 280 285
Leu Tyr Ser Lys Leu Thr Val Asp Lys Ser Arg Trp Gln Gln Gly Asn
290 295 300
Val Phe Ser Cys Ser Val Met His Glu Ala Leu His Asn His Tyr Thr
305 310 315 320
Gln Lys Ser Leu Ser Leu Ser Pro Gly Lys
325 330
<210> 155
<211> 107
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 155
Arg Thr Val Ala Ala Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu
1 5 10 15
Gln Leu Lys Ser Gly Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe
20 25 30
Tyr Pro Arg Glu Ala Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln
35 40 45
Ser Gly Asn Ser Gln Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser
50 55 60
Thr Tyr Ser Leu Ser Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu
65 70 75 80
Lys His Lys Val Tyr Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser
85 90 95
Pro Val Thr Lys Ser Phe Asn Arg Gly Glu Cys
100 105
<210> 156
<211> 987
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 156
Gly Cys Cys Thr Cys Cys Ala Cys Cys Ala Ala Gly Gly Gly Cys Cys
1 5 10 15
Cys Cys Thr Cys Cys Gly Thr Gly Thr Thr Cys Cys Cys Thr Cys Thr
20 25 30
Gly Gly Cys Cys Cys Cys Thr Thr Gly Cys Thr Cys Cys Cys Gly Gly
35 40 45
Thr Cys Cys Ala Cys Cys Thr Cys Cys Gly Ala Gly Thr Cys Thr Ala
50 55 60
Cys Cys Gly Cys Cys Gly Cys Thr Cys Thr Gly Gly Gly Cys Thr Gly
65 70 75 80
Cys Cys Thr Gly Gly Thr Gly Ala Ala Ala Gly Ala Cys Thr Ala Cys
85 90 95
Thr Thr Cys Cys Cys Cys Gly Ala Gly Cys Cys Cys Gly Thr Gly Ala
100 105 110
Cys Cys Gly Thr Gly Thr Cys Cys Thr Gly Gly Ala Ala Cys Thr Cys
115 120 125
Thr Gly Gly Cys Gly Cys Cys Cys Thr Gly Ala Cys Cys Ala Gly Cys
130 135 140
Gly Gly Cys Gly Thr Gly Cys Ala Cys Ala Cys Cys Thr Thr Cys Cys
145 150 155 160
Cys Thr Gly Cys Cys Gly Thr Gly Cys Thr Gly Cys Ala Gly Thr Cys
165 170 175
Cys Thr Cys Cys Gly Gly Cys Cys Thr Gly Thr Ala Cys Thr Cys Cys
180 185 190
Cys Thr Gly Thr Cys Cys Thr Cys Cys Gly Thr Gly Gly Thr Gly Ala
195 200 205
Cys Cys Gly Thr Gly Cys Cys Cys Thr Cys Cys Thr Cys Cys Ala Gly
210 215 220
Cys Cys Thr Gly Gly Gly Cys Ala Cys Cys Ala Ala Gly Ala Cys Cys
225 230 235 240
Thr Ala Cys Ala Cys Cys Thr Gly Thr Ala Ala Cys Gly Thr Gly Gly
245 250 255
Ala Cys Cys Ala Cys Ala Ala Gly Cys Cys Cys Thr Cys Cys Ala Ala
260 265 270
Cys Ala Cys Cys Ala Ala Gly Gly Thr Gly Gly Ala Cys Ala Ala Gly
275 280 285
Cys Gly Gly Gly Thr Gly Gly Ala Ala Thr Cys Thr Ala Ala Gly Thr
290 295 300
Ala Cys Gly Gly Cys Cys Cys Thr Cys Cys Cys Thr Gly Cys Cys Cys
305 310 315 320
Cys Cys Cys Cys Thr Gly Cys Cys Cys Thr Gly Cys Cys Cys Cys Thr
325 330 335
Gly Ala Ala Thr Thr Thr Cys Thr Gly Gly Gly Cys Gly Gly Ala Cys
340 345 350
Cys Thr Thr Cys Cys Gly Thr Gly Thr Thr Cys Cys Thr Gly Thr Thr
355 360 365
Cys Cys Cys Cys Cys Cys Ala Ala Ala Gly Cys Cys Cys Ala Ala Gly
370 375 380
Gly Ala Cys Ala Cys Cys Cys Thr Gly Ala Thr Gly Ala Thr Cys Thr
385 390 395 400
Cys Cys Cys Gly Gly Ala Cys Cys Cys Cys Cys Gly Ala Ala Gly Thr
405 410 415
Gly Ala Cys Cys Thr Gly Cys Gly Thr Gly Gly Thr Gly Gly Thr Gly
420 425 430
Gly Ala Cys Gly Thr Gly Thr Cys Cys Cys Ala Gly Gly Ala Ala Gly
435 440 445
Ala Thr Cys Cys Cys Gly Ala Gly Gly Thr Gly Cys Ala Gly Thr Thr
450 455 460
Cys Ala Ala Thr Thr Gly Gly Thr Ala Cys Gly Thr Gly Gly Ala Cys
465 470 475 480
Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr Gly Cys Ala Cys Ala
485 490 495
Ala Cys Gly Cys Cys Ala Ala Gly Ala Cys Cys Ala Ala Gly Cys Cys
500 505 510
Cys Ala Gly Ala Gly Ala Gly Gly Ala Ala Cys Ala Gly Thr Thr Cys
515 520 525
Ala Ala Cys Thr Cys Cys Ala Cys Cys Thr Ala Cys Cys Gly Gly Gly
530 535 540
Thr Gly Gly Thr Gly Thr Cys Thr Gly Thr Gly Cys Thr Gly Ala Cys
545 550 555 560
Cys Gly Thr Gly Cys Thr Gly Cys Ala Cys Cys Ala Gly Gly Ala Cys
565 570 575
Thr Gly Gly Cys Thr Gly Ala Ala Cys Gly Gly Cys Ala Ala Ala Gly
580 585 590
Ala Gly Thr Ala Cys Ala Ala Gly Thr Gly Cys Ala Ala Gly Gly Thr
595 600 605
Gly Thr Cys Cys Ala Ala Cys Ala Ala Gly Gly Gly Cys Cys Thr Gly
610 615 620
Cys Cys Cys Thr Cys Cys Ala Gly Cys Ala Thr Cys Gly Ala Ala Ala
625 630 635 640
Ala Gly Ala Cys Cys Ala Thr Cys Thr Cys Cys Ala Ala Gly Gly Cys
645 650 655
Cys Ala Ala Gly Gly Gly Cys Cys Ala Gly Cys Cys Cys Cys Gly Cys
660 665 670
Gly Ala Gly Cys Cys Cys Cys Ala Gly Gly Thr Gly Thr Ala Cys Ala
675 680 685
Cys Cys Cys Thr Gly Cys Cys Cys Cys Cys Thr Ala Gly Cys Cys Ala
690 695 700
Gly Gly Ala Ala Gly Ala Gly Ala Thr Gly Ala Cys Cys Ala Ala Gly
705 710 715 720
Ala Ala Cys Cys Ala Gly Gly Thr Gly Thr Cys Cys Cys Thr Gly Ala
725 730 735
Cys Cys Thr Gly Thr Cys Thr Gly Gly Thr Gly Ala Ala Ala Gly Gly
740 745 750
Cys Thr Thr Cys Thr Ala Cys Cys Cys Cys Thr Cys Cys Gly Ala Thr
755 760 765
Ala Thr Cys Gly Cys Cys Gly Thr Gly Gly Ala Ala Thr Gly Gly Gly
770 775 780
Ala Gly Thr Cys Cys Ala Ala Cys Gly Gly Cys Cys Ala Gly Cys Cys
785 790 795 800
Cys Gly Ala Gly Ala Ala Cys Ala Ala Cys Thr Ala Cys Ala Ala Gly
805 810 815
Ala Cys Cys Ala Cys Cys Cys Cys Cys Cys Cys Thr Gly Thr Gly Cys
820 825 830
Thr Gly Gly Ala Cys Thr Cys Cys Gly Ala Cys Gly Gly Cys Thr Cys
835 840 845
Cys Thr Thr Cys Thr Thr Cys Cys Thr Gly Thr Ala Cys Thr Cys Thr
850 855 860
Cys Gly Gly Cys Thr Gly Ala Cys Cys Gly Thr Gly Gly Ala Cys Ala
865 870 875 880
Ala Gly Thr Cys Cys Cys Gly Gly Thr Gly Gly Cys Ala Gly Gly Ala
885 890 895
Ala Gly Gly Cys Ala Ala Cys Gly Thr Gly Thr Thr Cys Thr Cys Cys
900 905 910
Thr Gly Cys Thr Cys Cys Gly Thr Gly Ala Thr Gly Cys Ala Cys Gly
915 920 925
Ala Gly Gly Cys Cys Cys Thr Gly Cys Ala Cys Ala Ala Cys Cys Ala
930 935 940
Cys Thr Ala Cys Ala Cys Cys Cys Ala Gly Ala Ala Gly Thr Cys Cys
945 950 955 960
Cys Thr Gly Thr Cys Cys Cys Thr Gly Ala Gly Cys Cys Thr Gly Gly
965 970 975
Gly Thr Ala Ala Gly Thr Gly Ala Thr Gly Ala
980 985
<210> 157
<211> 993
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 157
Gly Cys Thr Ala Gly Cys Ala Cys Cys Ala Ala Gly Gly Gly Cys Cys
1 5 10 15
Cys Ala Thr Cys Gly Gly Thr Cys Thr Thr Cys Cys Cys Cys Cys Thr
20 25 30
Gly Gly Cys Ala Cys Cys Cys Thr Cys Cys Thr Cys Cys Ala Ala Gly
35 40 45
Ala Gly Cys Ala Cys Cys Thr Cys Thr Gly Gly Gly Gly Gly Cys Ala
50 55 60
Cys Ala Gly Cys Gly Gly Cys Cys Cys Thr Gly Gly Gly Cys Thr Gly
65 70 75 80
Cys Cys Thr Gly Gly Thr Cys Ala Ala Gly Gly Ala Cys Thr Ala Cys
85 90 95
Thr Thr Cys Cys Cys Cys Gly Ala Ala Cys Cys Gly Gly Thr Gly Ala
100 105 110
Cys Gly Gly Thr Gly Thr Cys Gly Thr Gly Gly Ala Ala Cys Thr Cys
115 120 125
Ala Gly Gly Cys Gly Cys Cys Cys Thr Gly Ala Cys Cys Ala Gly Cys
130 135 140
Gly Gly Cys Gly Thr Gly Cys Ala Cys Ala Cys Cys Thr Thr Cys Cys
145 150 155 160
Cys Gly Gly Cys Thr Gly Thr Cys Cys Thr Ala Cys Ala Gly Thr Cys
165 170 175
Cys Thr Cys Ala Gly Gly Ala Cys Thr Cys Thr Ala Cys Thr Cys Cys
180 185 190
Cys Thr Cys Ala Gly Cys Ala Gly Cys Gly Thr Gly Gly Thr Gly Ala
195 200 205
Cys Cys Gly Thr Gly Cys Cys Cys Thr Cys Cys Ala Gly Cys Ala Gly
210 215 220
Cys Thr Thr Gly Gly Gly Cys Ala Cys Cys Cys Ala Gly Ala Cys Cys
225 230 235 240
Thr Ala Cys Ala Thr Cys Thr Gly Cys Ala Ala Cys Gly Thr Gly Ala
245 250 255
Ala Thr Cys Ala Cys Ala Ala Gly Cys Cys Cys Ala Gly Cys Ala Ala
260 265 270
Cys Ala Cys Cys Ala Ala Gly Gly Thr Gly Gly Ala Cys Ala Ala Gly
275 280 285
Ala Ala Ala Gly Thr Thr Gly Ala Gly Cys Cys Cys Ala Ala Ala Thr
290 295 300
Cys Thr Thr Gly Thr Gly Ala Cys Ala Ala Ala Ala Cys Thr Cys Ala
305 310 315 320
Cys Ala Cys Ala Thr Gly Cys Cys Cys Ala Cys Cys Gly Thr Gly Cys
325 330 335
Cys Cys Ala Gly Cys Ala Cys Cys Thr Gly Ala Ala Cys Thr Cys Cys
340 345 350
Thr Gly Gly Gly Gly Gly Gly Ala Cys Cys Gly Thr Cys Ala Gly Thr
355 360 365
Cys Thr Thr Cys Cys Thr Cys Thr Thr Cys Cys Cys Cys Cys Cys Ala
370 375 380
Ala Ala Ala Cys Cys Cys Ala Ala Gly Gly Ala Cys Ala Cys Cys Cys
385 390 395 400
Thr Cys Ala Thr Gly Ala Thr Cys Thr Cys Cys Cys Gly Gly Ala Cys
405 410 415
Cys Cys Cys Thr Gly Ala Gly Gly Thr Cys Ala Cys Ala Thr Gly Cys
420 425 430
Gly Thr Gly Gly Thr Gly Gly Thr Gly Gly Ala Cys Gly Thr Gly Ala
435 440 445
Gly Cys Cys Ala Cys Gly Ala Ala Gly Ala Cys Cys Cys Thr Gly Ala
450 455 460
Gly Gly Thr Cys Ala Ala Gly Thr Thr Cys Ala Ala Cys Thr Gly Gly
465 470 475 480
Thr Ala Cys Gly Thr Gly Gly Ala Cys Gly Gly Cys Gly Thr Gly Gly
485 490 495
Ala Gly Gly Thr Gly Cys Ala Thr Ala Ala Thr Gly Cys Cys Ala Ala
500 505 510
Gly Ala Cys Ala Ala Ala Gly Cys Cys Gly Cys Gly Gly Gly Ala Gly
515 520 525
Gly Ala Gly Cys Ala Gly Thr Ala Cys Ala Ala Cys Ala Gly Cys Ala
530 535 540
Cys Gly Thr Ala Cys Cys Gly Thr Gly Thr Gly Gly Thr Cys Ala Gly
545 550 555 560
Cys Gly Thr Cys Cys Thr Cys Ala Cys Cys Gly Thr Cys Cys Thr Gly
565 570 575
Cys Ala Cys Cys Ala Gly Gly Ala Cys Thr Gly Gly Cys Thr Gly Ala
580 585 590
Ala Thr Gly Gly Cys Ala Ala Gly Gly Ala Gly Thr Ala Cys Ala Ala
595 600 605
Gly Thr Gly Cys Ala Ala Gly Gly Thr Cys Thr Cys Cys Ala Ala Cys
610 615 620
Ala Ala Ala Gly Cys Cys Cys Thr Cys Cys Cys Ala Gly Cys Cys Cys
625 630 635 640
Cys Cys Ala Thr Cys Gly Ala Gly Ala Ala Ala Ala Cys Cys Ala Thr
645 650 655
Cys Thr Cys Cys Ala Ala Ala Gly Cys Cys Ala Ala Ala Gly Gly Gly
660 665 670
Cys Ala Gly Cys Cys Cys Cys Gly Ala Gly Ala Ala Cys Cys Ala Cys
675 680 685
Ala Gly Gly Thr Gly Thr Ala Cys Ala Cys Cys Cys Thr Gly Cys Cys
690 695 700
Cys Cys Cys Ala Thr Cys Cys Cys Gly Gly Gly Ala Gly Gly Ala Gly
705 710 715 720
Ala Thr Gly Ala Cys Cys Ala Ala Gly Ala Ala Cys Cys Ala Gly Gly
725 730 735
Thr Cys Ala Gly Cys Cys Thr Gly Ala Cys Cys Thr Gly Cys Cys Thr
740 745 750
Gly Gly Thr Cys Ala Ala Ala Gly Gly Cys Thr Thr Cys Thr Ala Thr
755 760 765
Cys Cys Cys Ala Gly Cys Gly Ala Cys Ala Thr Cys Gly Cys Cys Gly
770 775 780
Thr Gly Gly Ala Gly Thr Gly Gly Gly Ala Gly Ala Gly Cys Ala Ala
785 790 795 800
Thr Gly Gly Gly Cys Ala Gly Cys Cys Gly Gly Ala Gly Ala Ala Cys
805 810 815
Ala Ala Cys Thr Ala Cys Ala Ala Gly Ala Cys Cys Ala Cys Gly Cys
820 825 830
Cys Thr Cys Cys Cys Gly Thr Gly Cys Thr Gly Gly Ala Cys Thr Cys
835 840 845
Cys Gly Ala Cys Gly Gly Cys Thr Cys Cys Thr Thr Cys Thr Thr Cys
850 855 860
Cys Thr Cys Thr Ala Cys Ala Gly Cys Ala Ala Gly Cys Thr Cys Ala
865 870 875 880
Cys Cys Gly Thr Gly Gly Ala Cys Ala Ala Gly Ala Gly Cys Ala Gly
885 890 895
Gly Thr Gly Gly Cys Ala Gly Cys Ala Gly Gly Gly Gly Ala Ala Cys
900 905 910
Gly Thr Cys Thr Thr Cys Thr Cys Ala Thr Gly Cys Thr Cys Cys Gly
915 920 925
Thr Gly Ala Thr Gly Cys Ala Thr Gly Ala Gly Gly Cys Thr Cys Thr
930 935 940
Gly Cys Ala Cys Ala Ala Cys Cys Ala Cys Thr Ala Cys Ala Cys Gly
945 950 955 960
Cys Ala Gly Ala Ala Gly Ala Gly Cys Cys Thr Cys Thr Cys Cys Cys
965 970 975
Thr Gly Thr Cys Thr Cys Cys Gly Gly Gly Thr Ala Ala Ala Thr Gly
980 985 990
Ala
<210> 158
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 158
Cys Ala Gly Gly Thr Gly Cys Ala Gly Cys Thr Gly Gly Thr Gly Cys
1 5 10 15
Ala Gly Thr Cys Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
20 25 30
Gly Ala Ala Gly Ala Ala Ala Cys Cys Thr Gly Gly Cys Gly Cys Cys
35 40 45
Thr Cys Cys Gly Thr Gly Ala Ala Gly Gly Thr Gly Thr Cys Cys Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Thr Ala
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Ala Cys Cys Ala Cys Cys Thr Ala Cys
85 90 95
Thr Ala Cys Ala Thr Cys Thr Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Ala Gly Gly Cys Cys Ala
115 120 125
Gly Gly Gly Cys Cys Thr Gly Gly Ala Ala Thr Gly Gly Ala Thr Gly
130 135 140
Gly Gly Cys Gly Gly Cys Gly Thr Gly Ala Ala Cys Cys Cys Thr Thr
145 150 155 160
Cys Cys Ala Ala Cys Gly Gly Cys Gly Gly Cys Ala Cys Cys Ala Ala
165 170 175
Cys Thr Ala Cys Ala Ala Cys Gly Ala Gly Ala Ala Cys Thr Ala Cys
180 185 190
Ala Ala Gly Ala Ala Cys Ala Gly Ala Gly Thr Gly Ala Cys Cys Cys
195 200 205
Thr Gly Ala Cys Cys Ala Cys Cys Gly Ala Cys Thr Cys Cys Thr Cys
210 215 220
Cys Ala Cys Cys Ala Cys Cys Ala Cys Cys Gly Cys Cys Thr Ala Cys
225 230 235 240
Ala Thr Gly Gly Ala Ala Cys Thr Gly Ala Ala Gly Thr Cys Cys Cys
245 250 255
Thr Gly Cys Ala Gly Thr Thr Cys Gly Ala Cys Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Cys
275 280 285
Gly Cys Cys Ala Gly Ala Cys Gly Gly Gly Ala Cys Thr Ala Cys Ala
290 295 300
Gly Ala Thr Ala Cys Gly Ala Cys Ala Thr Gly Gly Gly Cys Thr Thr
305 310 315 320
Cys Gly Ala Cys Thr Ala Cys Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Cys Ala Cys Cys Ala Cys Cys Gly Thr Gly Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Thr
355 360
<210> 159
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 159
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Gly Cys
1 5 10 15
Ala Gly Thr Cys Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
20 25 30
Gly Ala Ala Gly Ala Ala Ala Cys Cys Thr Gly Gly Cys Gly Cys Thr
35 40 45
Thr Cys Thr Gly Thr Gly Ala Ala Gly Gly Thr Gly Thr Cys Cys Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Thr Gly Gly Cys Thr Ala
65 70 75 80
Cys Ala Cys Cys Thr Thr Thr Ala Cys Cys Ala Cys Cys Thr Ala Cys
85 90 95
Thr Ala Cys Ala Thr Cys Thr Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Thr Cys Cys Thr Gly Gly Ala Cys Ala
115 120 125
Gly Gly Gly Ala Cys Thr Thr Gly Ala Ala Thr Gly Gly Ala Thr Gly
130 135 140
Gly Gly Cys Gly Gly Cys Gly Thr Gly Ala Ala Cys Cys Cys Thr Thr
145 150 155 160
Cys Thr Ala Ala Cys Gly Gly Cys Gly Gly Cys Ala Cys Cys Ala Ala
165 170 175
Cys Thr Ala Cys Ala Ala Cys Gly Ala Gly Ala Ala Cys Thr Ala Cys
180 185 190
Ala Ala Gly Ala Ala Cys Ala Gly Ala Gly Thr Gly Ala Cys Cys Cys
195 200 205
Thr Gly Ala Cys Cys Ala Cys Cys Gly Ala Cys Thr Cys Cys Thr Cys
210 215 220
Thr Ala Cys Cys Ala Cys Cys Ala Cys Cys Gly Cys Cys Thr Ala Cys
225 230 235 240
Ala Thr Gly Gly Ala Ala Cys Thr Gly Ala Ala Gly Thr Cys Cys Cys
245 250 255
Thr Gly Cys Ala Gly Thr Thr Cys Gly Ala Cys Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Cys
275 280 285
Gly Cys Cys Ala Gly Ala Ala Gly Ala Gly Ala Cys Thr Ala Cys Ala
290 295 300
Gly Ala Thr Ala Cys Gly Ala Cys Ala Thr Gly Gly Gly Cys Thr Ala
305 310 315 320
Cys Gly Ala Cys Thr Ala Cys Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Cys Ala Cys Ala Ala Cys Ala Gly Thr Gly Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Thr
355 360
<210> 160
<211> 377
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 160
Cys Ala Gly Gly Thr Gly Cys Ala Gly Cys Thr Gly Gly Thr Gly Cys
1 5 10 15
Ala Gly Thr Cys Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
20 25 30
Gly Ala Ala Gly Ala Ala Ala Cys Cys Thr Gly Gly Cys Gly Cys Cys
35 40 45
Thr Cys Cys Gly Thr Gly Ala Ala Gly Gly Thr Gly Thr Cys Cys Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Thr Ala
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Ala Cys Cys Ala Cys Cys Thr Ala Cys
85 90 95
Thr Ala Cys Ala Thr Cys Thr Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Ala Gly Gly Cys Cys Ala
115 120 125
Gly Gly Gly Cys Cys Thr Gly Gly Ala Ala Thr Gly Gly Ala Thr Gly
130 135 140
Gly Gly Cys Gly Gly Cys Gly Thr Gly Ala Ala Cys Cys Cys Thr Thr
145 150 155 160
Cys Cys Ala Ala Cys Gly Gly Cys Gly Gly Cys Ala Cys Cys Ala Ala
165 170 175
Cys Thr Ala Cys Ala Ala Cys Gly Ala Gly Ala Ala Cys Thr Ala Cys
180 185 190
Ala Ala Gly Ala Ala Cys Ala Gly Ala Gly Thr Gly Ala Cys Cys Cys
195 200 205
Thr Gly Ala Cys Cys Ala Cys Cys Gly Ala Cys Thr Cys Cys Thr Cys
210 215 220
Cys Ala Cys Cys Ala Cys Cys Ala Cys Cys Gly Cys Cys Thr Ala Cys
225 230 235 240
Ala Thr Gly Gly Ala Ala Cys Thr Gly Ala Ala Gly Thr Cys Cys Cys
245 250 255
Thr Gly Cys Ala Gly Thr Thr Cys Gly Ala Cys Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Cys
275 280 285
Gly Cys Cys Ala Gly Ala Cys Gly Gly Gly Ala Cys Thr Ala Cys Ala
290 295 300
Gly Ala Thr Ala Cys Gly Ala Cys Ala Thr Gly Gly Gly Cys Cys Ala
305 310 315 320
Thr Gly Ala Cys Thr Ala Cys Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Cys Ala Cys Cys Ala Cys Cys Gly Thr Gly Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Thr Gly Cys Cys Thr Cys Cys Ala Cys
355 360 365
Cys Ala Ala Gly Gly Gly Cys Cys Cys
370 375
<210> 161
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 161
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Gly Cys
1 5 10 15
Ala Gly Thr Cys Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
20 25 30
Gly Ala Ala Gly Ala Ala Ala Cys Cys Thr Gly Gly Cys Gly Cys Thr
35 40 45
Thr Cys Thr Gly Thr Gly Ala Ala Gly Gly Thr Gly Thr Cys Cys Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Thr Gly Gly Cys Thr Ala
65 70 75 80
Cys Ala Cys Cys Thr Thr Thr Ala Cys Cys Ala Cys Cys Thr Ala Cys
85 90 95
Thr Ala Cys Ala Thr Cys Thr Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Thr Cys Cys Thr Gly Gly Ala Cys Ala
115 120 125
Gly Gly Gly Ala Cys Thr Thr Gly Ala Ala Thr Gly Gly Ala Thr Gly
130 135 140
Gly Gly Cys Gly Gly Cys Gly Thr Gly Ala Ala Cys Cys Cys Thr Thr
145 150 155 160
Cys Cys Ala Ala Cys Thr Cys Cys Gly Gly Cys Ala Cys Cys Ala Ala
165 170 175
Cys Thr Ala Cys Ala Ala Cys Gly Ala Gly Ala Ala Cys Thr Ala Cys
180 185 190
Ala Ala Gly Ala Ala Cys Ala Gly Ala Gly Thr Gly Ala Cys Cys Cys
195 200 205
Thr Gly Ala Cys Cys Ala Cys Cys Gly Ala Cys Thr Cys Cys Thr Cys
210 215 220
Thr Ala Cys Cys Ala Cys Cys Ala Cys Cys Gly Cys Cys Thr Ala Cys
225 230 235 240
Ala Thr Gly Gly Ala Ala Cys Thr Gly Ala Ala Gly Thr Cys Cys Cys
245 250 255
Thr Gly Cys Ala Gly Thr Thr Cys Gly Ala Cys Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Cys
275 280 285
Gly Cys Cys Ala Gly Ala Ala Gly Ala Gly Ala Cys Thr Ala Cys Ala
290 295 300
Gly Ala Thr Ala Cys Gly Ala Cys Ala Thr Gly Gly Gly Cys Thr Thr
305 310 315 320
Cys Gly Ala Cys Thr Ala Cys Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Cys Ala Cys Ala Ala Cys Ala Gly Thr Gly Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Thr
355 360
<210> 162
<211> 360
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 162
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Gly Cys
1 5 10 15
Ala Gly Thr Cys Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
20 25 30
Gly Ala Ala Gly Ala Ala Ala Cys Cys Thr Gly Gly Cys Gly Cys Thr
35 40 45
Thr Cys Thr Gly Thr Gly Ala Ala Gly Gly Thr Gly Thr Cys Cys Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Thr Gly Gly Cys Thr Ala
65 70 75 80
Cys Ala Cys Cys Thr Thr Thr Ala Cys Cys Ala Cys Cys Thr Ala Cys
85 90 95
Thr Ala Cys Ala Thr Cys Thr Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Thr Cys Cys Thr Gly Gly Ala Cys Ala
115 120 125
Gly Gly Gly Ala Cys Thr Thr Gly Ala Ala Thr Gly Gly Ala Thr Gly
130 135 140
Gly Gly Cys Gly Gly Cys Gly Thr Gly Ala Ala Cys Cys Cys Thr Thr
145 150 155 160
Cys Thr Ala Ala Cys Gly Gly Cys Gly Gly Cys Ala Cys Cys Ala Ala
165 170 175
Cys Thr Ala Cys Ala Ala Cys Cys Ala Gly Ala Ala Cys Thr Ala Cys
180 185 190
Ala Ala Gly Ala Ala Cys Ala Gly Ala Gly Thr Gly Ala Cys Cys Cys
195 200 205
Thr Gly Ala Cys Cys Ala Cys Cys Gly Ala Cys Thr Cys Cys Thr Cys
210 215 220
Thr Ala Cys Cys Ala Cys Cys Ala Cys Cys Gly Cys Cys Thr Ala Cys
225 230 235 240
Ala Thr Gly Gly Ala Ala Cys Thr Gly Ala Ala Gly Thr Cys Cys Cys
245 250 255
Thr Gly Cys Ala Gly Thr Thr Cys Gly Ala Cys Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Cys
275 280 285
Gly Cys Cys Ala Gly Ala Ala Gly Ala Gly Ala Cys Thr Ala Cys Ala
290 295 300
Gly Ala Thr Ala Cys Gly Ala Cys Ala Thr Gly Gly Gly Cys Thr Thr
305 310 315 320
Cys Gly Ala Cys Thr Ala Cys Thr Gly Gly Gly Gly Cys Cys Ala Gly
325 330 335
Gly Gly Cys Ala Cys Ala Ala Cys Ala Gly Thr Gly Ala Cys Cys Gly
340 345 350
Thr Gly Thr Cys Cys Thr Cys Thr
355 360
<210> 163
<211> 666
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 163
Gly Ala Gly Ala Thr Cys Gly Thr Gly Cys Thr Gly Ala Cys Cys Cys
1 5 10 15
Ala Gly Thr Cys Cys Cys Cys Cys Gly Cys Cys Ala Cys Cys Cys Thr
20 25 30
Gly Thr Cys Thr Cys Thr Gly Thr Cys Thr Cys Cys Cys Gly Gly Cys
35 40 45
Gly Ala Gly Ala Gly Ala Gly Cys Cys Ala Cys Cys Cys Thr Gly Ala
50 55 60
Gly Cys Thr Gly Cys Ala Gly Ala Gly Cys Cys Thr Cys Cys Ala Ala
65 70 75 80
Gly Gly Gly Cys Gly Thr Gly Thr Cys Cys Ala Cys Cys Thr Cys Cys
85 90 95
Gly Gly Cys Thr Ala Cys Thr Cys Cys Thr Ala Cys Cys Thr Gly Cys
100 105 110
Ala Cys Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys Ala Gly Ala Ala
115 120 125
Gly Cys Cys Cys Gly Gly Cys Cys Ala Gly Gly Cys Cys Cys Cys Thr
130 135 140
Cys Gly Gly Cys Thr Gly Cys Thr Gly Ala Thr Cys Thr Ala Cys Cys
145 150 155 160
Thr Gly Gly Cys Cys Thr Cys Cys Thr Ala Cys Cys Thr Gly Gly Ala
165 170 175
Ala Thr Cys Cys Gly Gly Cys Gly Thr Gly Cys Cys Cys Gly Cys Cys
180 185 190
Ala Gly Ala Thr Thr Cys Thr Cys Cys Gly Gly Cys Thr Cys Thr Gly
195 200 205
Gly Cys Thr Cys Thr Gly Gly Cys Ala Cys Cys Gly Ala Cys Thr Thr
210 215 220
Cys Ala Cys Cys Cys Thr Gly Ala Cys Cys Ala Thr Cys Thr Cys Cys
225 230 235 240
Ala Gly Cys Cys Thr Gly Gly Ala Ala Cys Cys Cys Gly Ala Gly Gly
245 250 255
Ala Cys Thr Thr Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala
260 265 270
Cys Thr Gly Cys Cys Ala Gly Cys Ala Cys Thr Cys Cys Cys Gly Gly
275 280 285
Gly Ala Cys Cys Thr Gly Cys Cys Cys Cys Thr Gly Ala Cys Cys Thr
290 295 300
Thr Thr Gly Gly Cys Gly Gly Ala Gly Gly Cys Ala Cys Cys Ala Ala
305 310 315 320
Gly Gly Thr Gly Gly Ala Ala Ala Thr Cys Ala Ala Gly Cys Gly Gly
325 330 335
Ala Cys Cys Gly Thr Gly Gly Cys Cys Gly Cys Thr Cys Cys Cys Thr
340 345 350
Cys Cys Gly Thr Gly Thr Thr Cys Ala Thr Cys Thr Thr Cys Cys Cys
355 360 365
Ala Cys Cys Cys Thr Cys Cys Gly Ala Cys Gly Ala Gly Cys Ala Gly
370 375 380
Cys Thr Gly Ala Ala Gly Thr Cys Cys Gly Gly Cys Ala Cys Cys Gly
385 390 395 400
Cys Cys Thr Cys Cys Gly Thr Cys Gly Thr Gly Thr Gly Cys Cys Thr
405 410 415
Gly Cys Thr Gly Ala Ala Cys Ala Ala Cys Thr Thr Cys Thr Ala Cys
420 425 430
Cys Cys Cys Cys Gly Cys Gly Ala Gly Gly Cys Cys Ala Ala Gly Gly
435 440 445
Thr Gly Cys Ala Gly Thr Gly Gly Ala Ala Gly Gly Thr Gly Gly Ala
450 455 460
Cys Ala Ala Thr Gly Cys Cys Cys Thr Gly Cys Ala Gly Thr Cys Cys
465 470 475 480
Gly Gly Cys Ala Ala Cys Thr Cys Cys Cys Ala Gly Gly Ala Ala Thr
485 490 495
Cys Cys Gly Thr Cys Ala Cys Cys Gly Ala Gly Cys Ala Gly Gly Ala
500 505 510
Cys Thr Cys Cys Ala Ala Gly Gly Ala Cys Ala Gly Cys Ala Cys Cys
515 520 525
Thr Ala Cys Thr Cys Cys Cys Thr Gly Thr Cys Cys Thr Cys Cys Ala
530 535 540
Cys Cys Cys Thr Gly Ala Cys Cys Cys Thr Gly Thr Cys Cys Ala Ala
545 550 555 560
Gly Gly Cys Cys Gly Ala Cys Thr Ala Cys Gly Ala Gly Ala Ala Gly
565 570 575
Cys Ala Cys Ala Ala Gly Gly Thr Gly Thr Ala Cys Gly Cys Cys Thr
580 585 590
Gly Cys Gly Ala Ala Gly Thr Gly Ala Cys Cys Cys Ala Cys Cys Ala
595 600 605
Gly Gly Gly Cys Cys Thr Gly Thr Cys Cys Ala Gly Cys Cys Cys Cys
610 615 620
Gly Thr Gly Ala Cys Cys Ala Ala Gly Thr Cys Cys Thr Thr Cys Ala
625 630 635 640
Ala Cys Cys Gly Gly Gly Gly Cys Gly Ala Gly Thr Gly Cys Thr Ala
645 650 655
Ala Thr Gly Ala Gly Gly Thr Ala Cys Cys
660 665
<210> 164
<211> 1678
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 164
Gly Gly Ala Thr Ala Ala Cys Ala Ala Thr Thr Thr Cys Ala Cys Ala
1 5 10 15
Cys Ala Gly Ala Ala Thr Thr Cys Ala Thr Thr Ala Ala Ala Gly Ala
20 25 30
Gly Gly Ala Gly Ala Ala Ala Thr Thr Ala Ala Cys Cys Ala Thr Gly
35 40 45
Ala Ala Ala Ala Ala Gly Ala Ala Thr Ala Thr Cys Gly Cys Ala Thr
50 55 60
Thr Thr Cys Thr Thr Cys Thr Thr Gly Cys Ala Thr Cys Thr Ala Thr
65 70 75 80
Gly Thr Thr Cys Gly Thr Thr Thr Thr Thr Thr Cys Thr Ala Thr Thr
85 90 95
Gly Cys Thr Ala Cys Ala Ala Ala Thr Gly Cys Cys Thr Ala Thr Gly
100 105 110
Cys Ala Thr Cys Cys Gly Ala Ala Ala Thr Cys Gly Thr Gly Cys Thr
115 120 125
Gly Ala Cys Cys Cys Ala Gly Thr Cys Gly Cys Cys Cys Gly Cys Thr
130 135 140
Ala Cys Ala Thr Thr Ala Thr Cys Gly Thr Thr Gly Thr Cys Ala Cys
145 150 155 160
Cys Gly Gly Gly Gly Gly Ala Ala Cys Gly Cys Gly Cys Gly Ala Cys
165 170 175
Ala Thr Thr Gly Thr Cys Cys Thr Gly Thr Cys Gly Cys Gly Cys Ala
180 185 190
Thr Cys Ala Cys Ala Gly Ala Gly Cys Gly Thr Ala Thr Cys Thr Thr
195 200 205
Cys Gly Thr Ala Thr Thr Thr Gly Gly Cys Cys Thr Gly Gly Thr Ala
210 215 220
Thr Cys Ala Gly Cys Ala Ala Ala Ala Gly Cys Cys Ala Gly Gly Cys
225 230 235 240
Cys Ala Ala Gly Cys Cys Cys Cys Cys Cys Gly Thr Cys Thr Thr Thr
245 250 255
Thr Ala Ala Thr Thr Thr Ala Thr Gly Ala Cys Gly Cys Cys Thr Cys
260 265 270
Thr Ala Ala Thr Cys Gly Cys Gly Cys Ala Ala Cys Cys Gly Gly Cys
275 280 285
Ala Thr Thr Cys Cys Thr Gly Cys Ala Cys Gly Cys Thr Thr Thr Thr
290 295 300
Cys Cys Gly Gly Cys Thr Cys Ala Gly Gly Gly Ala Gly Thr Gly Gly
305 310 315 320
Cys Ala Cys Gly Gly Ala Thr Thr Thr Cys Ala Cys Thr Thr Thr Gly
325 330 335
Ala Cys Gly Ala Thr Cys Ala Gly Thr Ala Gly Thr Cys Thr Thr Gly
340 345 350
Ala Ala Cys Cys Ala Gly Ala Ala Gly Ala Cys Thr Thr Cys Gly Cys
355 360 365
Gly Gly Thr Thr Thr Ala Cys Thr Ala Cys Thr Gly Cys Cys Ala Ala
370 375 380
Cys Ala Ala Thr Cys Cys Thr Cys Gly Ala Ala Cys Thr Gly Gly Cys
385 390 395 400
Cys Thr Cys Gly Thr Ala Cys Ala Thr Thr Thr Gly Gly Thr Cys Ala
405 410 415
Ala Gly Gly Thr Ala Cys Cys Ala Ala Gly Gly Thr Gly Gly Ala Ala
420 425 430
Ala Thr Cys Ala Ala Ala Cys Gly Cys Ala Cys Cys Gly Thr Ala Gly
435 440 445
Cys Cys Gly Cys Ala Cys Cys Gly Thr Cys Ala Gly Thr Cys Thr Thr
450 455 460
Cys Ala Thr Cys Thr Thr Cys Cys Cys Thr Cys Cys Cys Thr Cys Thr
465 470 475 480
Gly Ala Cys Gly Ala Ala Cys Ala Ala Thr Thr Ala Ala Ala Ala Thr
485 490 495
Cys Thr Gly Gly Cys Ala Cys Ala Gly Cys Thr Thr Cys Cys Gly Thr
500 505 510
Thr Gly Thr Ala Thr Gly Cys Cys Thr Thr Cys Thr Thr Ala Ala Cys
515 520 525
Ala Ala Cys Thr Thr Thr Thr Ala Thr Cys Cys Ala Cys Gly Thr Gly
530 535 540
Ala Ala Gly Cys Gly Ala Ala Ala Gly Thr Cys Cys Ala Ala Thr Gly
545 550 555 560
Gly Ala Ala Ala Gly Thr Thr Gly Ala Thr Ala Ala Thr Gly Cys Ala
565 570 575
Cys Thr Thr Cys Ala Gly Thr Cys Thr Gly Gly Gly Ala Ala Cys Thr
580 585 590
Cys Ala Cys Ala Gly Gly Ala Gly Thr Cys Cys Gly Thr Ala Ala Cys
595 600 605
Gly Gly Ala Ala Cys Ala Gly Gly Ala Cys Thr Cys Thr Ala Ala Gly
610 615 620
Gly Ala Cys Thr Cys Ala Ala Cys Cys Thr Ala Thr Ala Gly Cys Thr
625 630 635 640
Thr Ala Thr Cys Thr Thr Cys Ala Ala Cys Ala Thr Thr Gly Ala Cys
645 650 655
Ala Thr Thr Gly Thr Cys Ala Ala Ala Gly Gly Cys Ala Gly Ala Cys
660 665 670
Thr Ala Thr Gly Ala Gly Ala Ala Gly Cys Ala Cys Ala Ala Ala Gly
675 680 685
Thr Gly Thr Ala Cys Gly Cys Thr Thr Gly Thr Gly Ala Ala Gly Thr
690 695 700
Cys Ala Cys Cys Cys Ala Thr Cys Ala Gly Gly Gly Cys Thr Thr Gly
705 710 715 720
Thr Cys Thr Thr Cys Thr Cys Cys Ala Gly Thr Ala Ala Cys Cys Ala
725 730 735
Ala Ala Ala Gly Thr Thr Thr Cys Ala Ala Thr Cys Gly Cys Gly Gly
740 745 750
Gly Gly Ala Gly Thr Gly Thr Gly Gly Thr Gly Gly Thr Thr Cys Thr
755 760 765
Gly Ala Thr Thr Ala Cys Ala Ala Ala Gly Ala Thr Gly Ala Cys Gly
770 775 780
Ala Thr Gly Ala Cys Ala Ala Ala Thr Ala Ala Thr Thr Ala Ala Cys
785 790 795 800
Thr Cys Gly Ala Gly Gly Cys Thr Gly Ala Gly Cys Ala Ala Ala Gly
805 810 815
Cys Ala Gly Ala Cys Thr Ala Cys Thr Ala Ala Thr Ala Ala Cys Ala
820 825 830
Thr Ala Ala Ala Gly Thr Cys Thr Ala Cys Gly Cys Cys Gly Gly Ala
835 840 845
Cys Gly Cys Ala Thr Cys Gly Thr Gly Gly Cys Cys Cys Thr Ala Gly
850 855 860
Thr Ala Cys Gly Cys Ala Ala Gly Thr Thr Cys Ala Cys Gly Thr Ala
865 870 875 880
Ala Ala Ala Ala Gly Gly Gly Thr Ala Ala Cys Thr Ala Gly Ala Gly
885 890 895
Gly Thr Thr Gly Ala Gly Gly Thr Gly Ala Thr Thr Thr Thr Ala Thr
900 905 910
Gly Ala Ala Ala Ala Ala Gly Ala Ala Thr Ala Thr Cys Gly Cys Ala
915 920 925
Thr Thr Thr Cys Thr Thr Cys Thr Thr Gly Cys Ala Thr Cys Thr Ala
930 935 940
Thr Gly Thr Thr Cys Gly Thr Thr Thr Thr Thr Thr Cys Thr Ala Thr
945 950 955 960
Thr Gly Cys Thr Ala Cys Ala Ala Ala Cys Gly Cys Gly Thr Ala Cys
965 970 975
Gly Cys Thr Gly Ala Gly Ala Thr Cys Thr Cys Cys Cys Ala Ala Gly
980 985 990
Thr Cys Cys Ala Gly Cys Thr Gly Gly Thr Thr Gly Ala Ala Thr Cys
995 1000 1005
Ala Gly Gly Thr Gly Gly Gly Gly Gly Ala Gly Thr Thr Gly Thr
1010 1015 1020
Thr Cys Ala Ala Cys Cys Gly Gly Gly Gly Cys Gly Thr Thr Cys
1025 1030 1035
Cys Cys Thr Gly Cys Gly Thr Cys Thr Gly Gly Ala Cys Thr Gly
1040 1045 1050
Cys Ala Ala Ala Gly Cys Cys Thr Cys Ala Gly Gly Ala Ala Thr
1055 1060 1065
Thr Ala Cys Thr Thr Thr Thr Thr Cys Ala Ala Ala Cys Thr Cys
1070 1075 1080
Cys Gly Gly Gly Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr
1085 1090 1095
Thr Cys Gly Thr Cys Ala Ala Gly Cys Cys Cys Cys Cys Gly Gly
1100 1105 1110
Ala Ala Ala Ala Gly Gly Gly Thr Thr Gly Gly Ala Gly Thr Gly
1115 1120 1125
Gly Gly Thr Ala Gly Cys Ala Gly Thr Gly Ala Thr Cys Thr Gly
1130 1135 1140
Gly Thr Ala Thr Gly Ala Thr Gly Gly Cys Thr Cys Ala Ala Ala
1145 1150 1155
Ala Cys Gly Thr Thr Ala Cys Thr Ala Cys Gly Cys Gly Gly Ala
1160 1165 1170
Cys Thr Cys Gly Gly Thr Thr Ala Ala Ala Gly Gly Thr Cys Gly
1175 1180 1185
Cys Thr Thr Thr Ala Cys Thr Ala Thr Thr Thr Cys Ala Cys Gly
1190 1195 1200
Cys Gly Ala Cys Ala Ala Cys Thr Cys Gly Ala Ala Ala Ala Ala
1205 1210 1215
Thr Ala Cys Thr Thr Thr Gly Thr Thr Cys Thr Thr Ala Cys Ala
1220 1225 1230
Gly Ala Thr Gly Ala Ala Cys Thr Cys Thr Cys Thr Thr Cys Gly
1235 1240 1245
Cys Gly Cys Cys Gly Ala Ala Gly Ala Thr Ala Cys Gly Gly Cys
1250 1255 1260
Ala Gly Thr Gly Thr Ala Cys Thr Ala Thr Thr Gly Cys Gly Cys
1265 1270 1275
Cys Ala Cys Ala Ala Ala Thr Gly Ala Thr Gly Ala Thr Thr Ala
1280 1285 1290
Thr Thr Gly Gly Gly Gly Thr Cys Ala Ala Gly Gly Thr Ala Cys
1295 1300 1305
Ala Thr Thr Ala Gly Thr Gly Ala Cys Cys Gly Thr Cys Ala Gly
1310 1315 1320
Thr Ala Gly Cys Gly Cys Gly Ala Gly Thr Ala Cys Gly Ala Ala
1325 1330 1335
Gly Gly Gly Ala Cys Cys Thr Ala Gly Cys Gly Thr Gly Thr Thr
1340 1345 1350
Thr Cys Cys Cys Thr Thr Ala Gly Cys Ala Cys Cys Ala Thr Gly
1355 1360 1365
Thr Ala Gly Thr Cys Gly Thr Thr Cys Gly Ala Cys Ala Ala Gly
1370 1375 1380
Cys Gly Ala Ala Thr Cys Cys Ala Cys Ala Gly Cys Gly Gly Cys
1385 1390 1395
Cys Thr Thr Ala Gly Gly Gly Thr Gly Cys Thr Thr Ala Gly Thr
1400 1405 1410
Gly Ala Ala Gly Gly Ala Thr Thr Ala Thr Thr Thr Thr Cys Cys
1415 1420 1425
Cys Gly Ala Gly Cys Cys Ala Gly Thr Ala Ala Cys Ala Gly Thr
1430 1435 1440
Ala Thr Cys Gly Thr Gly Gly Ala Ala Thr Ala Gly Cys Gly Gly
1445 1450 1455
Cys Gly Cys Ala Thr Thr Ala Ala Cys Thr Thr Cys Cys Gly Gly
1460 1465 1470
Cys Gly Thr Thr Cys Ala Thr Ala Cys Cys Thr Thr Cys Cys Cys
1475 1480 1485
Thr Gly Cys Cys Gly Thr Thr Thr Thr Ala Cys Ala Gly Thr Cys
1490 1495 1500
Ala Thr Cys Cys Gly Gly Thr Thr Thr Gly Thr Ala Thr Thr Cys
1505 1510 1515
Thr Cys Thr Thr Thr Cys Gly Thr Cys Ala Gly Thr Ala Gly Thr
1520 1525 1530
Ala Ala Cys Ala Gly Thr Gly Cys Cys Thr Thr Cys Cys Thr Cys
1535 1540 1545
Cys Thr Cys Gly Cys Thr Gly Gly Gly Gly Ala Cys Cys Ala Ala
1550 1555 1560
Gly Ala Cys Cys Thr Ala Thr Ala Cys Cys Thr Gly Cys Ala Ala
1565 1570 1575
Thr Gly Thr Gly Gly Ala Cys Cys Ala Thr Ala Ala Gly Cys Cys
1580 1585 1590
Thr Thr Cys Ala Ala Ala Cys Ala Cys Ala Ala Ala Ala Gly Thr
1595 1600 1605
Ala Gly Ala Cys Ala Ala Ala Cys Gly Thr Gly Thr Thr Gly Ala
1610 1615 1620
Ala Thr Cys Ala Ala Ala Gly Thr Ala Cys Gly Gly Cys Cys Cys
1625 1630 1635
Gly Cys Cys Thr Thr Gly Thr Cys Cys Gly Cys Cys Cys Thr Gly
1640 1645 1650
Cys Gly Gly Ala Thr Cys Cys Ala Ala Ala Gly Ala Thr Ala Thr
1655 1660 1665
Cys Ala Gly Ala Thr Cys Thr Gly Gly Thr
1670 1675
<210> 165
<211> 222
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 165
Gln Val Gln Leu Val Glu Ser Gly Gly Gly Val Val Gln Pro Gly Arg
1 5 10 15
Ser Leu Arg Leu Asp Cys Lys Ala Ser Gly Ile Thr Phe Ser Asn Ser
20 25 30
Gly Met His Trp Val Arg Gln Ala Pro Gly Lys Gly Leu Glu Trp Val
35 40 45
Ala Val Ile Trp Tyr Asp Gly Ser Lys Arg Tyr Tyr Ala Asp Ser Val
50 55 60
Lys Gly Arg Phe Thr Ile Ser Arg Asp Asn Ser Lys Asn Thr Leu Phe
65 70 75 80
Leu Gln Met Asn Ser Leu Arg Ala Glu Asp Thr Ala Val Tyr Tyr Cys
85 90 95
Ala Thr Asn Asp Asp Tyr Trp Gly Gln Gly Thr Leu Val Thr Val Ser
100 105 110
Ser Ala Ser Thr Lys Gly Pro Ser Val Phe Pro Leu Ala Pro Cys Ser
115 120 125
Arg Ser Thr Ser Glu Ser Thr Ala Ala Leu Gly Cys Leu Val Lys Asp
130 135 140
Tyr Phe Pro Glu Pro Val Thr Val Ser Trp Asn Ser Gly Ala Leu Thr
145 150 155 160
Ser Gly Val His Thr Phe Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr
165 170 175
Ser Leu Ser Ser Val Val Thr Val Pro Ser Ser Ser Leu Gly Thr Lys
180 185 190
Thr Tyr Thr Cys Asn Val Asp His Lys Pro Ser Asn Thr Lys Val Asp
195 200 205
Lys Arg Val Glu Ser Lys Tyr Gly Pro Pro Cys Pro Pro Cys
210 215 220
<210> 166
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 166
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 167
<211> 214
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 167
Glu Ile Val Leu Thr Gln Ser Pro Ala Thr Leu Ser Leu Ser Pro Gly
1 5 10 15
Glu Arg Ala Thr Leu Ser Cys Arg Ala Ser Gln Ser Val Ser Ser Tyr
20 25 30
Leu Ala Trp Tyr Gln Gln Lys Pro Gly Gln Ala Pro Arg Leu Leu Ile
35 40 45
Tyr Asp Ala Ser Asn Arg Ala Thr Gly Ile Pro Ala Arg Phe Ser Gly
50 55 60
Ser Gly Ser Gly Thr Asp Phe Thr Leu Thr Ile Ser Ser Leu Glu Pro
65 70 75 80
Glu Asp Phe Ala Val Tyr Tyr Cys Gln Gln Ser Ser Asn Trp Pro Arg
85 90 95
Thr Phe Gly Gln Gly Thr Lys Val Glu Ile Lys Arg Thr Val Ala Ala
100 105 110
Pro Ser Val Phe Ile Phe Pro Pro Ser Asp Glu Gln Leu Lys Ser Gly
115 120 125
Thr Ala Ser Val Val Cys Leu Leu Asn Asn Phe Tyr Pro Arg Glu Ala
130 135 140
Lys Val Gln Trp Lys Val Asp Asn Ala Leu Gln Ser Gly Asn Ser Gln
145 150 155 160
Glu Ser Val Thr Glu Gln Asp Ser Lys Asp Ser Thr Tyr Ser Leu Ser
165 170 175
Ser Thr Leu Thr Leu Ser Lys Ala Asp Tyr Glu Lys His Lys Val Tyr
180 185 190
Ala Cys Glu Val Thr His Gln Gly Leu Ser Ser Pro Val Thr Lys Ser
195 200 205
Phe Asn Arg Gly Glu Cys
210
<210> 168
<211> 322
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 168
Pro Ser Val Phe Pro Leu Ala Pro Cys Ser Arg Ser Thr Ser Glu Ser
1 5 10 15
Thr Ala Ala Leu Gly Cys Leu Val Lys Asp Tyr Phe Pro Glu Pro Val
20 25 30
Thr Val Ser Trp Asn Ser Gly Ala Leu Thr Ser Gly Val His Thr Phe
35 40 45
Pro Ala Val Leu Gln Ser Ser Gly Leu Tyr Ser Leu Ser Ser Val Val
50 55 60
Thr Val Pro Ser Ser Ser Leu Gly Thr Lys Thr Tyr Thr Cys Asn Val
65 70 75 80
Asp His Lys Pro Ser Asn Thr Lys Val Asp Lys Arg Val Glu Ser Lys
85 90 95
Tyr Gly Pro Pro Cys Pro Pro Cys Pro Ala Pro Glu Phe Leu Gly Gly
100 105 110
Pro Ser Val Phe Leu Phe Pro Pro Lys Pro Lys Asp Thr Leu Met Ile
115 120 125
Ser Arg Thr Pro Glu Val Thr Cys Val Val Val Asp Val Ser Gln Glu
130 135 140
Asp Pro Glu Val Gln Phe Asn Trp Tyr Val Asp Gly Val Glu Val His
145 150 155 160
Asn Ala Lys Thr Lys Pro Arg Glu Glu Gln Phe Asn Ser Thr Tyr Arg
165 170 175
Val Val Ser Val Leu Thr Val Leu His Gln Asp Trp Leu Asn Gly Lys
180 185 190
Glu Tyr Lys Cys Lys Val Ser Asn Lys Gly Leu Pro Ser Ser Ile Glu
195 200 205
Lys Thr Ile Ser Lys Ala Lys Gly Gln Pro Arg Glu Pro Gln Val Tyr
210 215 220
Thr Leu Pro Pro Ser Gln Glu Glu Met Thr Lys Asn Gln Val Ser Leu
225 230 235 240
Thr Cys Leu Val Lys Gly Phe Tyr Pro Ser Asp Ile Ala Val Glu Trp
245 250 255
Glu Ser Asn Gly Gln Pro Glu Asn Asn Tyr Lys Thr Thr Pro Pro Val
260 265 270
Leu Asp Ser Asp Gly Ser Phe Phe Leu Tyr Ser Arg Leu Thr Val Asp
275 280 285
Lys Ser Arg Trp Gln Glu Gly Asn Val Phe Ser Cys Ser Val Met His
290 295 300
Glu Ala Leu His Asn His Tyr Thr Gln Lys Ser Leu Ser Leu Ser Leu
305 310 315 320
Gly Lys
<210> 169
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 169
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Thr Gly
1 5 10 15
Ala Ala Thr Cys Thr Gly Gly Cys Gly Gly Cys Gly Gly Ala Gly Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Gly Ala
35 40 45
Ala Gly Thr Cys Thr Gly Ala Gly Ala Cys Thr Gly Gly Ala Thr Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Ala Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Ala Ala Cys Thr Cys Thr
85 90 95
Gly Gly Cys Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Thr Gly Gly Ala Ala Ala
115 120 125
Ala Gly Gly Ala Cys Thr Gly Gly Ala Ala Thr Gly Gly Gly Thr Cys
130 135 140
Gly Cys Cys Gly Thr Gly Ala Thr Thr Thr Gly Gly Thr Ala Cys Gly
145 150 155 160
Ala Cys Gly Gly Cys Thr Cys Thr Ala Ala Gly Cys Gly Gly Thr Ala
165 170 175
Cys Thr Ala Cys Gly Cys Cys Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Ala Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Thr Cys Gly Gly Gly Ala Cys Ala Ala Cys Thr Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Cys Cys Thr Gly Thr Thr Thr
225 230 235 240
Cys Thr Gly Cys Ala Gly Ala Thr Gly Ala Ala Cys Thr Cys Cys Cys
245 250 255
Thr Gly Ala Gly Ala Gly Cys Cys Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Cys Ala Cys Cys Ala Ala Cys Thr Cys Thr Gly Ala Thr Thr
290 295 300
Thr Thr Thr Gly Gly Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala Cys
305 310 315 320
Ala Cys Thr Gly Gly Thr Cys Ala Cys Cys Gly Thr Gly Thr Cys Cys
325 330 335
Thr Cys Thr Gly Cys Thr Thr Cys Thr Ala Cys Ala Ala Ala Gly Gly
340 345 350
Gly Cys Cys Cys
355
<210> 170
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 170
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Thr Gly
1 5 10 15
Ala Ala Thr Cys Thr Gly Gly Cys Gly Gly Cys Gly Gly Ala Gly Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Gly Ala
35 40 45
Ala Gly Thr Cys Thr Gly Ala Gly Ala Cys Thr Gly Gly Ala Thr Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Ala Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Ala Ala Cys Thr Cys Thr
85 90 95
Gly Gly Cys Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Thr Gly Gly Ala Ala Ala
115 120 125
Ala Gly Gly Ala Cys Thr Gly Gly Ala Ala Thr Gly Gly Gly Thr Cys
130 135 140
Gly Cys Cys Gly Thr Gly Ala Thr Thr Thr Gly Gly Thr Ala Cys Gly
145 150 155 160
Ala Cys Gly Gly Cys Thr Cys Thr Ala Ala Gly Cys Gly Gly Thr Ala
165 170 175
Cys Thr Ala Cys Gly Cys Cys Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Ala Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Thr Cys Gly Gly Gly Ala Cys Ala Ala Cys Thr Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Cys Cys Thr Gly Thr Thr Thr
225 230 235 240
Cys Thr Gly Cys Ala Gly Ala Thr Gly Ala Ala Cys Thr Cys Cys Cys
245 250 255
Thr Gly Ala Gly Ala Gly Cys Cys Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Cys Ala Cys Cys Ala Ala Thr Thr Cys Thr Gly Ala Thr Cys
290 295 300
Ala Thr Thr Gly Gly Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala Cys
305 310 315 320
Ala Cys Thr Gly Gly Thr Cys Ala Cys Cys Gly Thr Gly Thr Cys Cys
325 330 335
Thr Cys Thr Gly Cys Thr Thr Cys Thr Ala Cys Ala Ala Ala Gly Gly
340 345 350
Gly Cys Cys Cys
355
<210> 171
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 171
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Thr Gly
1 5 10 15
Ala Ala Thr Cys Thr Gly Gly Cys Gly Gly Cys Gly Gly Ala Gly Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Gly Ala
35 40 45
Ala Gly Thr Cys Thr Gly Ala Gly Ala Cys Thr Gly Gly Ala Thr Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Ala Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Ala Ala Cys Thr Cys Thr
85 90 95
Gly Gly Cys Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Thr Gly Gly Ala Ala Ala
115 120 125
Ala Gly Gly Ala Cys Thr Gly Gly Ala Ala Thr Gly Gly Gly Thr Cys
130 135 140
Gly Cys Cys Gly Thr Gly Ala Thr Thr Thr Gly Gly Thr Ala Cys Gly
145 150 155 160
Ala Cys Gly Gly Cys Thr Cys Thr Ala Ala Gly Cys Gly Gly Thr Ala
165 170 175
Cys Thr Ala Cys Gly Cys Cys Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Ala Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Thr Cys Gly Gly Gly Ala Cys Ala Ala Cys Thr Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Cys Cys Thr Gly Thr Thr Thr
225 230 235 240
Cys Thr Gly Cys Ala Gly Ala Thr Gly Ala Ala Cys Thr Cys Cys Cys
245 250 255
Thr Gly Ala Gly Ala Gly Cys Cys Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Cys Ala Cys Cys Ala Ala Cys Thr Cys Thr Gly Ala Thr Thr
290 295 300
Ala Thr Thr Gly Gly Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala Cys
305 310 315 320
Ala Cys Thr Gly Gly Thr Cys Ala Cys Cys Gly Thr Gly Thr Cys Cys
325 330 335
Thr Cys Thr Gly Cys Thr Thr Cys Thr Ala Cys Ala Ala Ala Gly Gly
340 345 350
Gly Cys Cys Cys
355
<210> 172
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 172
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Thr Gly
1 5 10 15
Ala Ala Thr Cys Thr Gly Gly Cys Gly Gly Cys Gly Gly Ala Gly Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Gly Ala
35 40 45
Ala Gly Thr Cys Thr Gly Ala Gly Ala Cys Thr Gly Gly Ala Thr Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Ala Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Ala Ala Cys Thr Cys Thr
85 90 95
Gly Gly Cys Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Thr Gly Gly Ala Ala Ala
115 120 125
Ala Gly Gly Ala Cys Thr Gly Gly Ala Ala Thr Gly Gly Gly Thr Cys
130 135 140
Gly Cys Cys Gly Thr Gly Ala Thr Thr Thr Gly Gly Thr Ala Cys Gly
145 150 155 160
Ala Cys Gly Gly Cys Thr Cys Thr Ala Ala Gly Cys Gly Gly Thr Ala
165 170 175
Cys Thr Ala Cys Gly Cys Cys Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Ala Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Thr Cys Gly Gly Gly Ala Cys Ala Ala Cys Thr Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Cys Cys Thr Gly Thr Thr Thr
225 230 235 240
Cys Thr Gly Cys Ala Gly Ala Thr Gly Ala Ala Cys Thr Cys Cys Cys
245 250 255
Thr Gly Ala Gly Ala Gly Cys Cys Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Cys Ala Cys Ala Ala Ala Thr Ala Cys Cys Gly Ala Thr Thr
290 295 300
Gly Gly Thr Gly Gly Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala Cys
305 310 315 320
Ala Cys Thr Gly Gly Thr Cys Ala Cys Ala Gly Thr Gly Thr Cys Cys
325 330 335
Thr Cys Thr Gly Cys Thr Thr Cys Cys Ala Cys Ala Ala Ala Gly Gly
340 345 350
Gly Cys Cys Cys
355
<210> 173
<211> 356
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 173
Cys Ala Gly Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Thr Gly
1 5 10 15
Ala Ala Thr Cys Thr Gly Gly Cys Gly Gly Cys Gly Gly Ala Gly Thr
20 25 30
Gly Gly Thr Gly Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Gly Ala
35 40 45
Ala Gly Thr Cys Thr Gly Ala Gly Ala Cys Thr Gly Gly Ala Thr Thr
50 55 60
Gly Cys Ala Ala Gly Gly Cys Cys Thr Cys Cys Gly Gly Cys Ala Thr
65 70 75 80
Cys Ala Cys Cys Thr Thr Cys Thr Cys Cys Ala Ala Cys Thr Cys Thr
85 90 95
Gly Gly Cys Ala Thr Gly Cys Ala Cys Thr Gly Gly Gly Thr Cys Cys
100 105 110
Gly Ala Cys Ala Gly Gly Cys Cys Cys Cys Thr Gly Gly Ala Ala Ala
115 120 125
Ala Gly Gly Ala Cys Thr Gly Gly Ala Ala Thr Gly Gly Gly Thr Cys
130 135 140
Gly Cys Cys Gly Thr Gly Ala Thr Thr Thr Gly Gly Thr Ala Cys Gly
145 150 155 160
Ala Cys Gly Gly Cys Thr Cys Thr Ala Ala Gly Cys Gly Gly Thr Ala
165 170 175
Cys Thr Ala Cys Gly Cys Cys Gly Ala Cys Thr Cys Cys Gly Thr Gly
180 185 190
Ala Ala Gly Gly Gly Cys Ala Gly Ala Thr Thr Cys Ala Cys Cys Ala
195 200 205
Thr Cys Thr Cys Thr Cys Gly Gly Gly Ala Cys Ala Ala Cys Thr Cys
210 215 220
Cys Ala Ala Gly Ala Ala Cys Ala Cys Cys Cys Thr Gly Thr Thr Thr
225 230 235 240
Cys Thr Gly Cys Ala Gly Ala Thr Gly Ala Ala Cys Thr Cys Cys Cys
245 250 255
Thr Gly Ala Gly Ala Gly Cys Cys Gly Ala Gly Gly Ala Cys Ala Cys
260 265 270
Cys Gly Cys Cys Gly Thr Gly Thr Ala Cys Thr Ala Cys Thr Gly Thr
275 280 285
Gly Cys Cys Ala Cys Cys Ala Ala Thr Ala Cys Cys Gly Ala Thr Thr
290 295 300
Ala Thr Thr Gly Gly Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala Cys
305 310 315 320
Ala Cys Thr Gly Gly Thr Cys Ala Cys Cys Gly Thr Gly Thr Cys Cys
325 330 335
Thr Cys Thr Gly Cys Thr Thr Cys Thr Ala Cys Ala Ala Ala Gly Gly
340 345 350
Gly Cys Cys Cys
355
<210> 174
<211> 648
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 174
Gly Ala Gly Ala Thr Cys Gly Thr Gly Cys Thr Gly Ala Cys Cys Cys
1 5 10 15
Ala Gly Thr Cys Cys Cys Cys Cys Gly Cys Cys Ala Cys Cys Cys Thr
20 25 30
Gly Thr Cys Thr Cys Thr Gly Ala Gly Cys Cys Cys Thr Gly Gly Cys
35 40 45
Gly Ala Gly Ala Gly Ala Gly Cys Cys Ala Cys Cys Cys Thr Gly Ala
50 55 60
Gly Cys Thr Gly Cys Ala Gly Ala Gly Cys Cys Thr Cys Cys Cys Ala
65 70 75 80
Gly Thr Cys Cys Gly Thr Gly Thr Cys Cys Thr Cys Cys Thr Ala Cys
85 90 95
Cys Thr Gly Gly Cys Cys Thr Gly Gly Thr Ala Thr Cys Ala Gly Cys
100 105 110
Ala Gly Ala Ala Gly Cys Cys Cys Gly Gly Cys Cys Ala Gly Gly Cys
115 120 125
Cys Cys Cys Thr Cys Gly Gly Cys Thr Gly Cys Thr Gly Ala Thr Cys
130 135 140
Thr Ala Cys Gly Ala Cys Gly Cys Cys Thr Cys Cys Ala Ala Cys Cys
145 150 155 160
Gly Gly Gly Cys Cys Ala Cys Cys Gly Gly Cys Ala Thr Cys Cys Cys
165 170 175
Thr Gly Cys Cys Ala Gly Ala Thr Thr Cys Thr Cys Cys Gly Gly Cys
180 185 190
Thr Cys Thr Gly Gly Cys Thr Cys Cys Gly Gly Cys Ala Cys Cys Gly
195 200 205
Ala Cys Thr Thr Cys Ala Cys Cys Cys Thr Gly Ala Cys Cys Ala Thr
210 215 220
Cys Thr Cys Cys Ala Gly Cys Cys Thr Gly Gly Ala Ala Cys Cys Cys
225 230 235 240
Gly Ala Gly Gly Ala Cys Thr Thr Cys Gly Cys Cys Gly Thr Gly Thr
245 250 255
Ala Cys Thr Ala Cys Thr Gly Cys Cys Ala Gly Cys Ala Gly Thr Cys
260 265 270
Cys Thr Cys Cys Ala Ala Cys Thr Gly Gly Cys Cys Cys Cys Gly Gly
275 280 285
Ala Cys Cys Thr Thr Cys Gly Gly Cys Cys Ala Gly Gly Gly Cys Ala
290 295 300
Cys Cys Ala Ala Gly Gly Thr Gly Gly Ala Ala Ala Thr Cys Ala Ala
305 310 315 320
Gly Cys Gly Gly Ala Cys Cys Gly Thr Gly Gly Cys Cys Gly Cys Thr
325 330 335
Cys Cys Cys Thr Cys Cys Gly Thr Gly Thr Thr Cys Ala Thr Cys Thr
340 345 350
Thr Cys Cys Cys Ala Cys Cys Cys Thr Cys Cys Gly Ala Cys Gly Ala
355 360 365
Gly Cys Ala Gly Cys Thr Gly Ala Ala Gly Thr Cys Cys Gly Gly Cys
370 375 380
Ala Cys Cys Gly Cys Cys Thr Cys Cys Gly Thr Gly Gly Thr Cys Thr
385 390 395 400
Gly Cys Cys Thr Gly Cys Thr Gly Ala Ala Cys Ala Ala Cys Thr Thr
405 410 415
Cys Thr Ala Cys Cys Cys Cys Cys Gly Cys Gly Ala Gly Gly Cys Cys
420 425 430
Ala Ala Gly Gly Thr Gly Cys Ala Gly Thr Gly Gly Ala Ala Gly Gly
435 440 445
Thr Gly Gly Ala Cys Ala Ala Cys Gly Cys Cys Cys Thr Gly Cys Ala
450 455 460
Gly Thr Cys Cys Gly Gly Cys Ala Ala Cys Thr Cys Cys Cys Ala Gly
465 470 475 480
Gly Ala Ala Thr Cys Cys Gly Thr Gly Ala Cys Cys Gly Ala Gly Cys
485 490 495
Ala Gly Gly Ala Cys Thr Cys Cys Ala Ala Gly Gly Ala Cys Ala Gly
500 505 510
Cys Ala Cys Cys Thr Ala Cys Thr Cys Cys Cys Thr Gly Thr Cys Cys
515 520 525
Thr Cys Cys Ala Cys Cys Cys Thr Gly Ala Cys Cys Cys Thr Gly Thr
530 535 540
Cys Cys Ala Ala Gly Gly Cys Cys Gly Ala Cys Thr Ala Cys Gly Ala
545 550 555 560
Gly Ala Ala Gly Cys Ala Cys Ala Ala Gly Gly Thr Gly Thr Ala Cys
565 570 575
Gly Cys Cys Thr Gly Cys Gly Ala Ala Gly Thr Gly Ala Cys Cys Cys
580 585 590
Ala Cys Cys Ala Gly Gly Gly Cys Cys Thr Gly Thr Cys Cys Ala Gly
595 600 605
Cys Cys Cys Cys Gly Thr Gly Ala Cys Cys Ala Ala Gly Thr Cys Cys
610 615 620
Thr Thr Cys Ala Ala Cys Cys Gly Gly Gly Gly Cys Gly Ala Gly Thr
625 630 635 640
Gly Cys Thr Gly Ala Thr Gly Ala
645
<210> 175
<211> 976
<212> БЕЛОК
<213> Искусственная последовательность
<220>
<223> Рекомбинантная последовательность
<400> 175
Gly Gly Gly Cys Cys Cys Cys Thr Cys Cys Gly Thr Gly Thr Thr Cys
1 5 10 15
Cys Cys Thr Cys Thr Gly Gly Cys Cys Cys Cys Thr Thr Gly Cys Thr
20 25 30
Cys Cys Cys Gly Gly Thr Cys Cys Ala Cys Cys Thr Cys Cys Gly Ala
35 40 45
Gly Thr Cys Thr Ala Cys Cys Gly Cys Cys Gly Cys Thr Cys Thr Gly
50 55 60
Gly Gly Cys Thr Gly Cys Cys Thr Gly Gly Thr Gly Ala Ala Ala Gly
65 70 75 80
Ala Cys Thr Ala Cys Thr Thr Cys Cys Cys Cys Gly Ala Gly Cys Cys
85 90 95
Cys Gly Thr Gly Ala Cys Cys Gly Thr Gly Thr Cys Cys Thr Gly Gly
100 105 110
Ala Ala Cys Thr Cys Thr Gly Gly Cys Gly Cys Cys Cys Thr Gly Ala
115 120 125
Cys Cys Ala Gly Cys Gly Gly Cys Gly Thr Gly Cys Ala Cys Ala Cys
130 135 140
Cys Thr Thr Cys Cys Cys Thr Gly Cys Cys Gly Thr Gly Cys Thr Gly
145 150 155 160
Cys Ala Gly Thr Cys Cys Thr Cys Cys Gly Gly Cys Cys Thr Gly Thr
165 170 175
Ala Cys Thr Cys Cys Cys Thr Gly Thr Cys Cys Thr Cys Cys Gly Thr
180 185 190
Gly Gly Thr Gly Ala Cys Cys Gly Thr Gly Cys Cys Cys Thr Cys Cys
195 200 205
Thr Cys Cys Ala Gly Cys Cys Thr Gly Gly Gly Cys Ala Cys Cys Ala
210 215 220
Ala Gly Ala Cys Cys Thr Ala Cys Ala Cys Cys Thr Gly Thr Ala Ala
225 230 235 240
Cys Gly Thr Gly Gly Ala Cys Cys Ala Cys Ala Ala Gly Cys Cys Cys
245 250 255
Thr Cys Cys Ala Ala Cys Ala Cys Cys Ala Ala Gly Gly Thr Gly Gly
260 265 270
Ala Cys Ala Ala Gly Cys Gly Gly Gly Thr Gly Gly Ala Ala Thr Cys
275 280 285
Thr Ala Ala Gly Thr Ala Cys Gly Gly Cys Cys Cys Thr Cys Cys Cys
290 295 300
Thr Gly Cys Cys Cys Cys Cys Cys Cys Thr Gly Cys Cys Cys Thr Gly
305 310 315 320
Cys Cys Cys Cys Thr Gly Ala Ala Thr Thr Thr Cys Thr Gly Gly Gly
325 330 335
Cys Gly Gly Ala Cys Cys Thr Thr Cys Cys Gly Thr Gly Thr Thr Cys
340 345 350
Cys Thr Gly Thr Thr Cys Cys Cys Cys Cys Cys Ala Ala Ala Gly Cys
355 360 365
Cys Cys Ala Ala Gly Gly Ala Cys Ala Cys Cys Cys Thr Gly Ala Thr
370 375 380
Gly Ala Thr Cys Thr Cys Cys Cys Gly Gly Ala Cys Cys Cys Cys Cys
385 390 395 400
Gly Ala Ala Gly Thr Gly Ala Cys Cys Thr Gly Cys Gly Thr Gly Gly
405 410 415
Thr Gly Gly Thr Gly Gly Ala Cys Gly Thr Gly Thr Cys Cys Cys Ala
420 425 430
Gly Gly Ala Ala Gly Ala Thr Cys Cys Cys Gly Ala Gly Gly Thr Gly
435 440 445
Cys Ala Gly Thr Thr Cys Ala Ala Thr Thr Gly Gly Thr Ala Cys Gly
450 455 460
Thr Gly Gly Ala Cys Gly Gly Cys Gly Thr Gly Gly Ala Ala Gly Thr
465 470 475 480
Gly Cys Ala Cys Ala Ala Cys Gly Cys Cys Ala Ala Gly Ala Cys Cys
485 490 495
Ala Ala Gly Cys Cys Cys Ala Gly Ala Gly Ala Gly Gly Ala Ala Cys
500 505 510
Ala Gly Thr Thr Cys Ala Ala Cys Thr Cys Cys Ala Cys Cys Thr Ala
515 520 525
Cys Cys Gly Gly Gly Thr Gly Gly Thr Gly Thr Cys Thr Gly Thr Gly
530 535 540
Cys Thr Gly Ala Cys Cys Gly Thr Gly Cys Thr Gly Cys Ala Cys Cys
545 550 555 560
Ala Gly Gly Ala Cys Thr Gly Gly Cys Thr Gly Ala Ala Cys Gly Gly
565 570 575
Cys Ala Ala Ala Gly Ala Gly Thr Ala Cys Ala Ala Gly Thr Gly Cys
580 585 590
Ala Ala Gly Gly Thr Gly Thr Cys Cys Ala Ala Cys Ala Ala Gly Gly
595 600 605
Gly Cys Cys Thr Gly Cys Cys Cys Thr Cys Cys Ala Gly Cys Ala Thr
610 615 620
Cys Gly Ala Ala Ala Ala Gly Ala Cys Cys Ala Thr Cys Thr Cys Cys
625 630 635 640
Ala Ala Gly Gly Cys Cys Ala Ala Gly Gly Gly Cys Cys Ala Gly Cys
645 650 655
Cys Cys Cys Gly Cys Gly Ala Gly Cys Cys Cys Cys Ala Gly Gly Thr
660 665 670
Gly Thr Ala Cys Ala Cys Cys Cys Thr Gly Cys Cys Cys Cys Cys Thr
675 680 685
Ala Gly Cys Cys Ala Gly Gly Ala Ala Gly Ala Gly Ala Thr Gly Ala
690 695 700
Cys Cys Ala Ala Gly Ala Ala Cys Cys Ala Gly Gly Thr Gly Thr Cys
705 710 715 720
Cys Cys Thr Gly Ala Cys Cys Thr Gly Thr Cys Thr Gly Gly Thr Gly
725 730 735
Ala Ala Ala Gly Gly Cys Thr Thr Cys Thr Ala Cys Cys Cys Cys Thr
740 745 750
Cys Cys Gly Ala Thr Ala Thr Cys Gly Cys Cys Gly Thr Gly Gly Ala
755 760 765
Ala Thr Gly Gly Gly Ala Gly Thr Cys Cys Ala Ala Cys Gly Gly Cys
770 775 780
Cys Ala Gly Cys Cys Cys Gly Ala Gly Ala Ala Cys Ala Ala Cys Thr
785 790 795 800
Ala Cys Ala Ala Gly Ala Cys Cys Ala Cys Cys Cys Cys Cys Cys Cys
805 810 815
Thr Gly Thr Gly Cys Thr Gly Gly Ala Cys Thr Cys Cys Gly Ala Cys
820 825 830
Gly Gly Cys Thr Cys Cys Thr Thr Cys Thr Thr Cys Cys Thr Gly Thr
835 840 845
Ala Cys Thr Cys Thr Cys Gly Gly Cys Thr Gly Ala Cys Cys Gly Thr
850 855 860
Gly Gly Ala Cys Ala Ala Gly Thr Cys Cys Cys Gly Gly Thr Gly Gly
865 870 875 880
Cys Ala Gly Gly Ala Ala Gly Gly Cys Ala Ala Cys Gly Thr Gly Thr
885 890 895
Thr Cys Thr Cys Cys Thr Gly Cys Thr Cys Cys Gly Thr Gly Ala Thr
900 905 910
Gly Cys Ala Cys Gly Ala Gly Gly Cys Cys Cys Thr Gly Cys Ala Cys
915 920 925
Ala Ala Cys Cys Ala Cys Thr Ala Cys Ala Cys Cys Cys Ala Gly Ala
930 935 940
Ala Gly Thr Cys Cys Cys Thr Gly Thr Cys Cys Cys Thr Gly Ala Gly
945 950 955 960
Cys Cys Thr Gly Gly Gly Cys Ala Ala Gly Thr Gly Ala Thr Gly Ala
965 970 975
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| Антитела против лиганда-1 запрограммированной смерти (PD-L1) и их применение | 2017 |
|
RU2721582C1 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2832918C2 |
| АНТИТЕЛА К PD-L1, СВЯЗЫВАЮЩИЕ PD-L1 СОБАКИ | 2015 |
|
RU2722562C2 |
| АНТИТЕЛА ПРОТИВ PD-L1 И ИХ ВАРИАНТЫ | 2017 |
|
RU2770590C2 |
| КОМБИНИРОВАННЫЕ ЛЕКАРСТВЕННЫЕ СРЕДСТВА, НАЦЕЛЕННЫЕ НА PD-1, TIM-3 И LAG-3 | 2018 |
|
RU2795232C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2815059C2 |
| СТАБИЛЬНЫЕ СОСТАВЫ АНТИТЕЛ ПРОТИВ TIGIT, ОТДЕЛЬНО И В КОМБИНАЦИИ С АНТИТЕЛАМИ ПРОТИВ РЕЦЕПТОРА 1 ПРОГРАММИРУЕМОЙ СМЕРТИ (PD-1), И СПОСОБЫ ИХ ПРИМЕНЕНИЯ | 2018 |
|
RU2820576C2 |
| НОВЫЕ КОМПОЗИЦИИ АНТИТЕЛ ДЛЯ ИММУНОТЕРАПИИ РАКА | 2019 |
|
RU2804490C2 |
| СОБАЧЬИ АНТИТЕЛА С МОДИФИЦИРОВАННЫМИ ПОСЛЕДОВАТЕЛЬНОСТЯМИ CH2-CH3 | 2014 |
|
RU2801209C2 |
| АНТИТЕЛА ПРОТИВ БЕЛКА-1 ЗАПРОГРАММИРОВАННОЙ КЛЕТОЧНОЙ СМЕРТИ (PD-1) И ИХ ПРИМЕНЕНИЕ | 2017 |
|
RU2725950C1 |
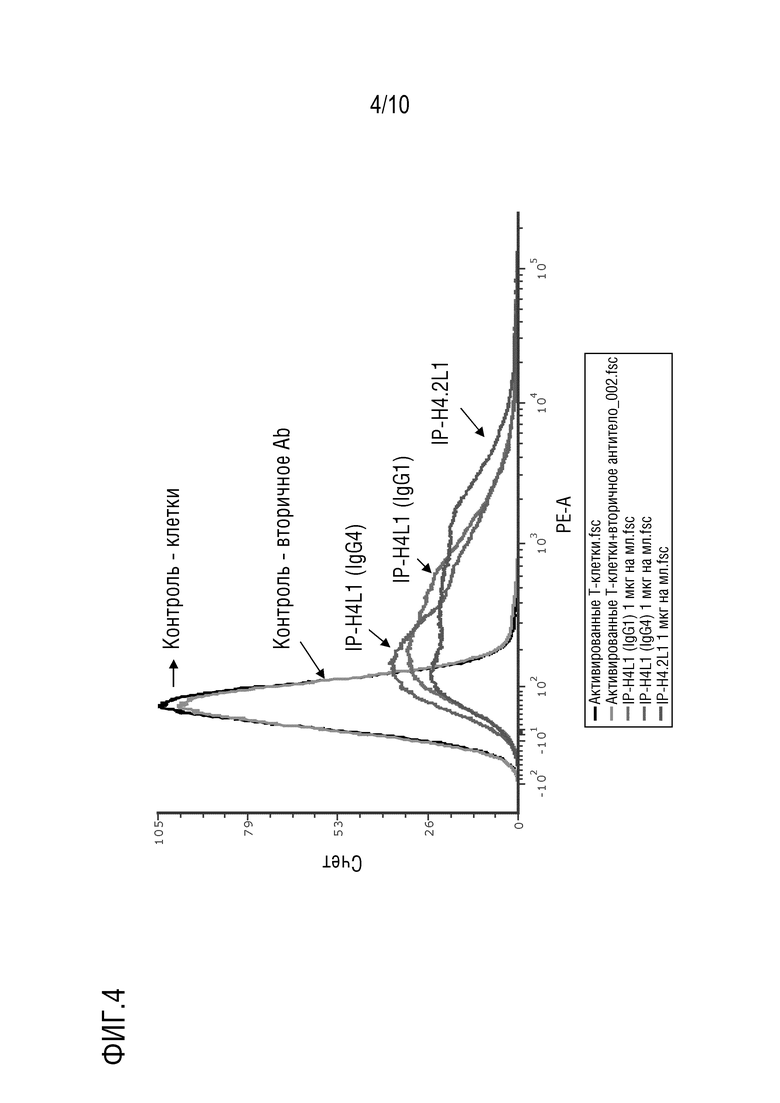
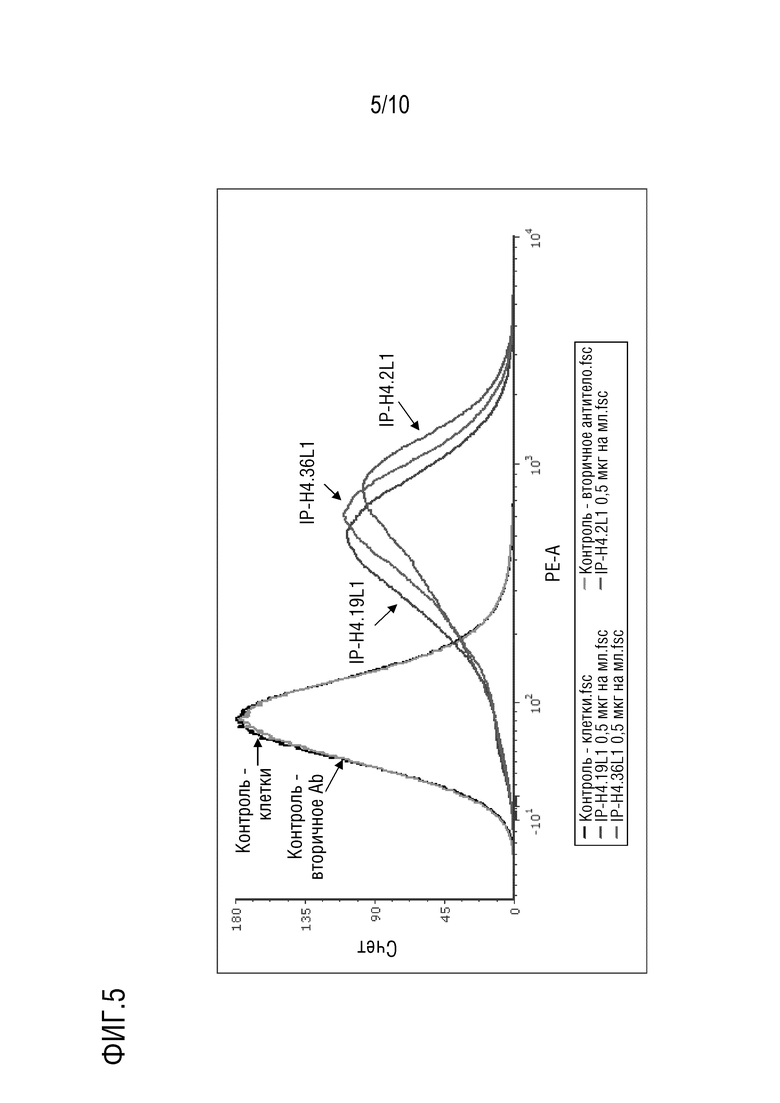
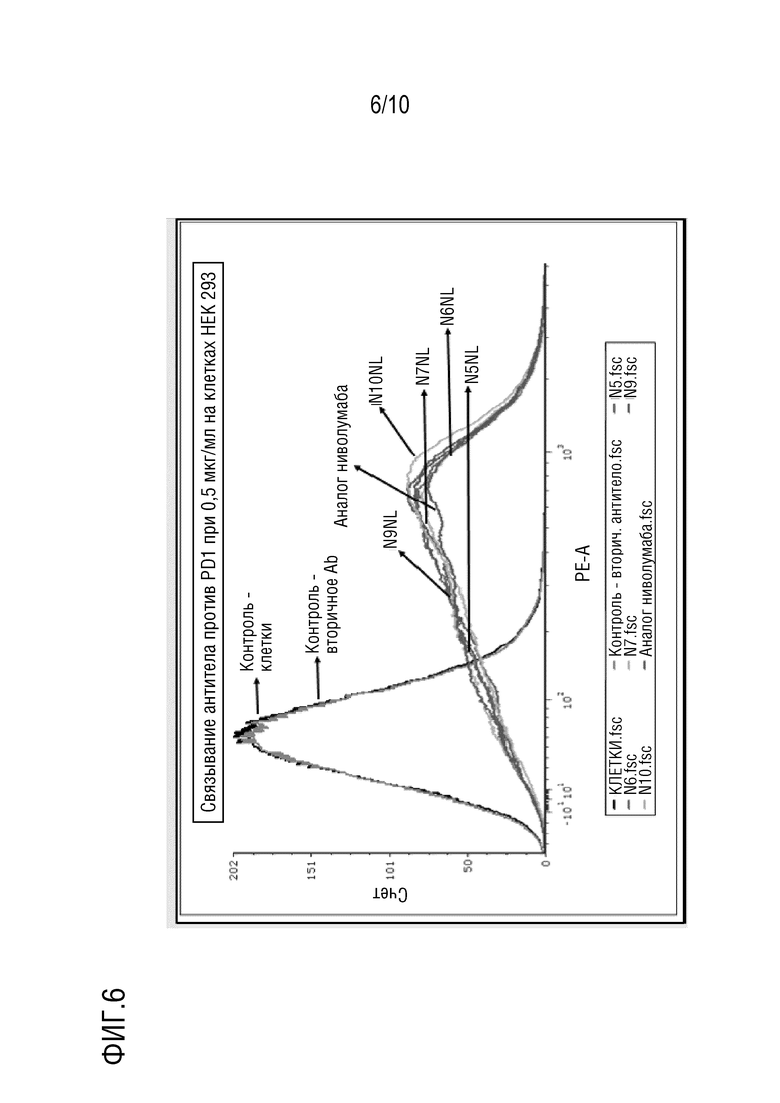
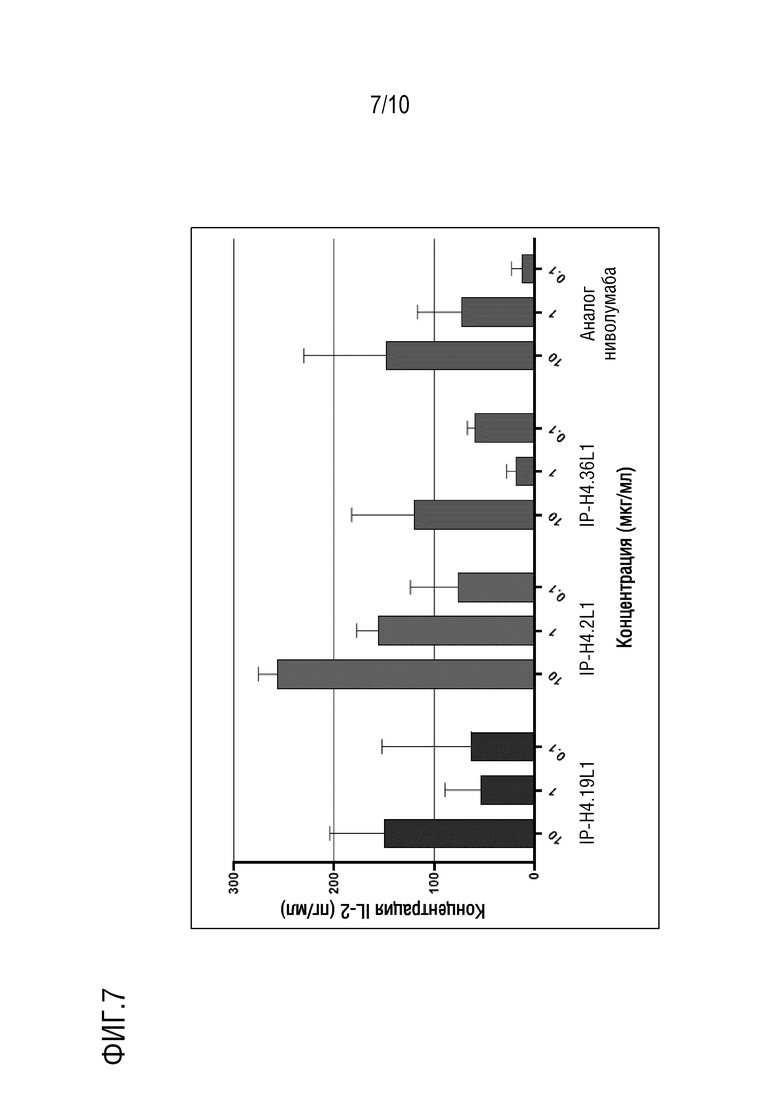
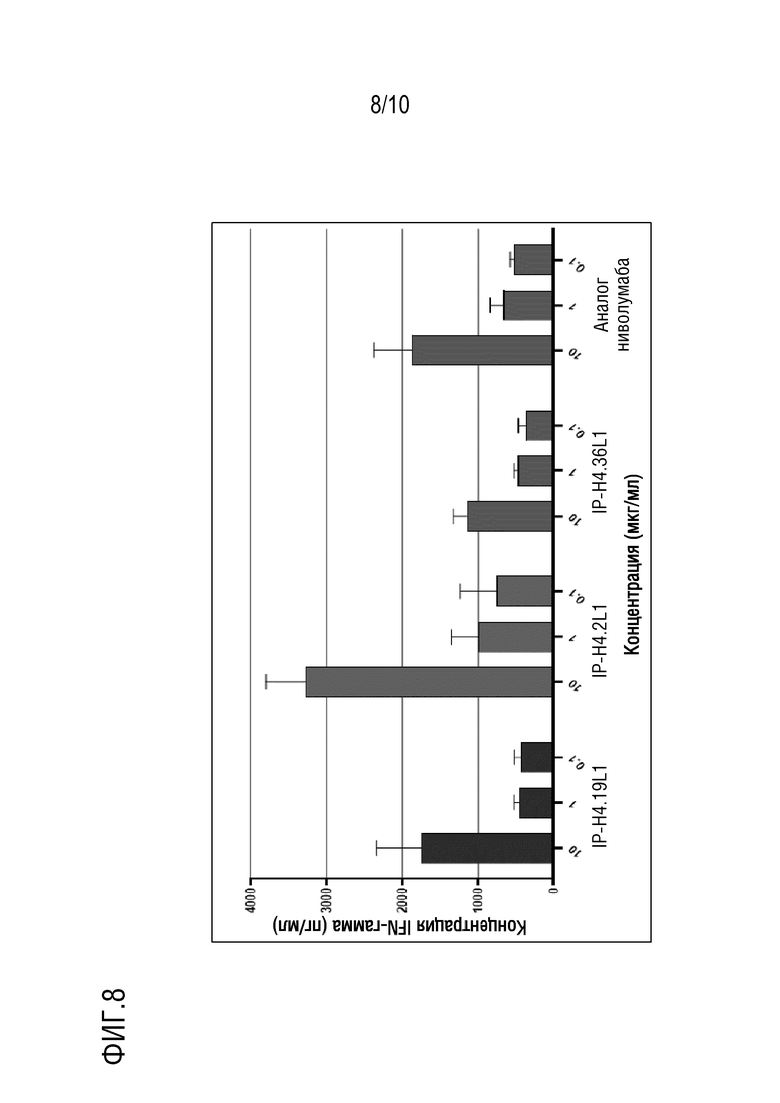
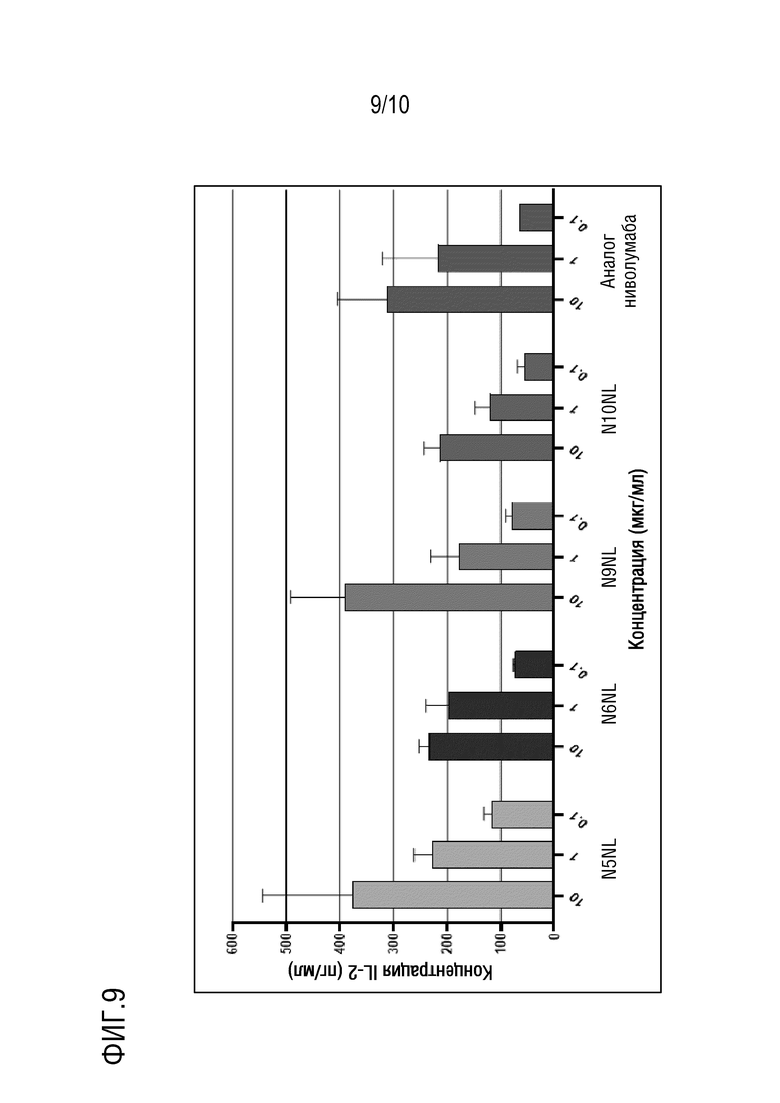
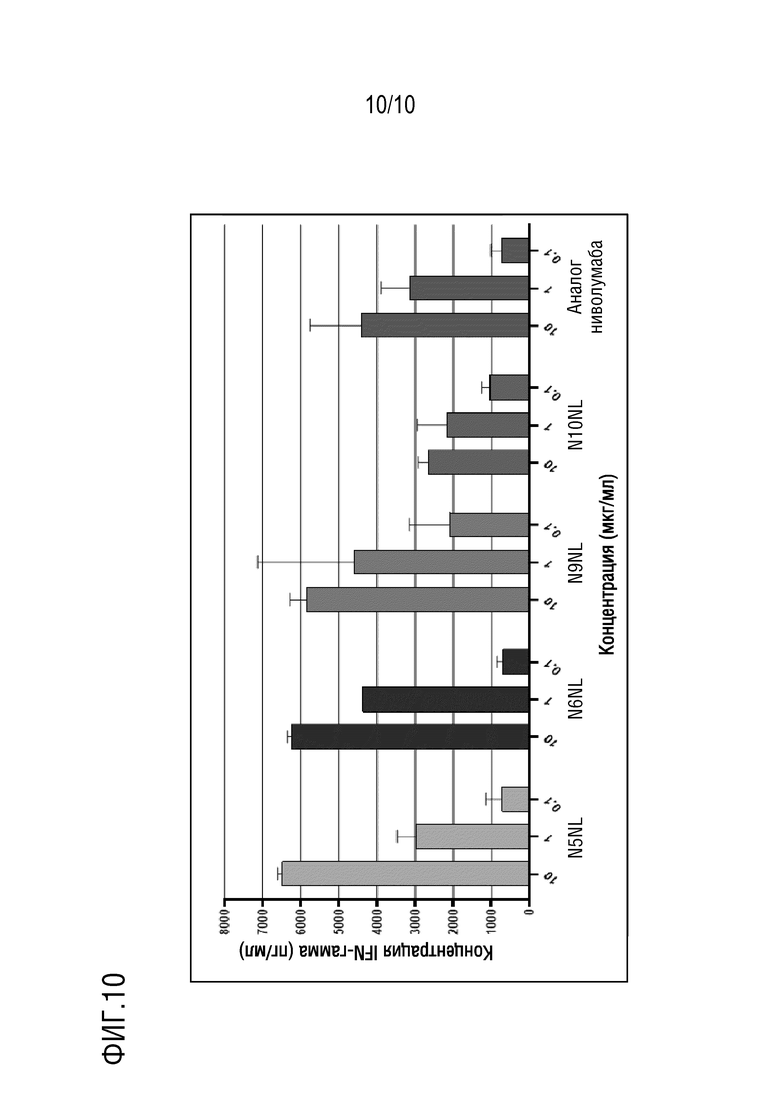
Изобретение относится к биотехнологии. Описано антитело против PD-1, или его антигенсвязывающая часть, содержащее аминокислотную последовательность SEQ ID NO: 149 и вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 152, где антитело против PD-1 содержит константную область IgG4 с аминокислотной последовательностью SEQ ID NO: 153, содержащей единичную аминокислотную замену S228P, P329G, M428L и N434S. Изобретение расширяет арсенал средств для лечения заболеваний, при которых экспрессия PD-1 является вредной. 4 з.п. ф-лы, 10 ил., 9 табл., 12 пр.
1. Антитело против PD-1, или его антигенсвязывающая часть, содержащее
вариабельную область тяжелой цепи, содержащую аминокислотную последовательность SEQ ID NO: 149, и
вариабельную область легкой цепи, содержащую аминокислотную последовательность SEQ ID NO: 152,
где антитело против PD-1 содержит константную область IgG4 с аминокислотной последовательностью SEQ ID NO: 153, содержащей единичную аминокислотную замену S228P, P329G, M428L и N434S.
2. Антитело, или его антигенсвязывающая часть, по п.1, которое связывает с PD-1 человека.
3. Антитело, или его антигенсвязывающая часть, по п.1, которое связывается с PD-1 человека со значением KD, равным 10-10 M или менее, предпочтительно, 10-11 M или менее.
4. Антитело, или его антигенсвязывающая часть, по п.1, обладающее активностью ADCC и/или CDC.
5. Антитело, или его антигенсвязывающая часть, по п.1, где антитело, или его антигенсвязывающая часть, имеет по меньшей мере одну из следующих характеристик:
(a) вступает в перекрестную реакцию с PD-1 видов, отличных от человека;
(b) имеет более высокую специфичность связывания в отношении PD-1 человека;
(c) стимулирует секрецию цитокина IL-2;
(d) стимулирует более высокую секрецию IFN-гамма;
(e) блокирует взаимодействие рецептора PD-1 с его природным лигандом PD-L1 и/или PD-L2;
(f) стимулирует пролиферацию T-клеток;
(g) имеет повышенное время полужизни у индивида.
| WO 2018087143 A2, 17.05.2018 | |||
| US 9492539 B2, 15.11.2016 | |||
| WO 2017011580 A2, 19.01.2017 | |||
| АНТИТЕЛО ПРОТИВ БЕЛКА ЗАПРОГРАММИРОВАННОЙ СМЕРТИ КЛЕТКИ 1 (PD-1) И ЕГО ПРИМЕНЕНИЕ | 2015 |
|
RU2661678C1 |

