Группа изобретений относится к области биотехнологии и генетики, в частности, к способу очистки и выделения высокомолекулярных нуклеиновых кислот ДНК или РНК из биологических образцов для последующего секвенирования с длинным считыванием, к устройству и гелю для очистки и выделения нуклеиновых кислот.
ПРЕДПОСЫЛКИ ИЗОБРЕТЕНИЯ
В настоящее время очистка и выделения ДНК или РНК из многокомпонентных биологических образцов представляет собой первый шаг в широком спектре молекулярно-биологических протоколов, используемых в генетике, молекулярной медицине, судебной экспертизе и биотехнологии. Конечный успех многих из этих протоколов зависит от эффективного отделения целевых нуклеиновых кислот от различных примесей, таких как полипептиды и белки (например, нуклеазы), олигонуклеотиды, полисахариды, полифенолы, липиды, пигменты, вторичные метаболиты, гуминовые вещества, различные ингибиторы ферментов и т.д. Одним из конкретных способов использования, успех которых в большой степени зависит от чистоты ДНК, является определение их нуклеотидной последовательности (секвенирование). В эпоху секвенирования по Сэнгеру, азатем в эпоху массового параллельного секвенирования нового поколения (Next-generation sequencing, NGS)важной предпосылкой для получения хороших результатов являлось выделение высокоочищенной ДНК. В настоящее время выделение высокоочищенной ДНК остается важнейшим условием для успешного секвенирования с использованием платформ третьего поколения (third-generation sequencing, TGS), которые способны считывать десятки и даже сотни тысяч пар оснований. Однако, чтобы в полной мере воспользоваться таким длинным считыванием, ДНК должна быть не только высокого качества, но и максимально возможной длины. Таким образом, для любого метода секвенирования нового и последующего поколений, с длинным считыванием, необходимо сочетать высокомолекулярную ДНК и с высокой степенью её очистки. Для решения данной проблемы было исследовано несколько подходов.
УРОВЕНЬ ТЕХНИКИ
В настоящее время описаны многочисленные экспериментальные протоколы выделения и очистки ДНК (1-5). Эти протоколы обычно включают этап лизиса клеток с последующим отделением ДНК от примесей. Наиболее часто используемые стратегии разделения можно разделить на три категории: преципитация ДНК, жидкофазная и твердофазная экстракция.
Первая стратегия разделения основана на способности полярных растворителей (например, этанола) и солей (например, ацетата натрия) осаждать ДНК из водных буферов (6). Фактическому осаждению ДНК могут предшествовать дополнительные этапы очистки, такие как денатурация белков и перевод их в нерастворимую форму (7). Хотя эта стратегия подходит для очистки высокомолекулярной ДНК и даже используется в коммерческих наборах, выход и чистота высокомолекулярной ДНК могут быть субоптимальными, особенно для сложных образцов. При жидкофазной экстракции полярная ДНК остается в водной фазе, а неполярные загрязнители, такие как денатурированные белки и липиды, переходят в органическую фазу (обычно фенол-хлороформ) или концентрируются на границе раздела фаз (8).Чтобы получить ДНК приемлемой чистоты, процедуру органической экстракции рекомендуется проводить повторно, что приводит к снижению выхода. При твердофазной экстракции ДНК связывают с твердым сорбентом, а загрязнения удаляют промыванием. Хотя многие типы твердых сорбентов могут быть использованы для очистки ДНК, силикатные сорбенты (силикагели) стали особенно популярны в последние несколько десятилетий. Коммерческие наборы на основе твердофазной экстракции широко доступны, а их преимуществами являются сокращение времени выделения ДНК и параллельная обработка большого количества образцов. Тем не менее, эти наборы тоже имеют свои недостатки: выделение ДНК с их помощью происходит в несколько этапов и требует использования коммерческих спин-колонок, а также специализированного оборудования, такого как центрифуги или устройства для вакуумной фильтрации. Более того, обычные протоколы твердофазной экстракции и коммерческие наборы на их основе, как правило, не подходят для выделения высокомолекулярной и высокоочищенной ДНК. При использовании этих протоколов большая часть выделяемой ДНК проходит мимо колонки и не сорбируется, что в конечном итоге приводит к низкому выходу. Таким образом, для выделения и очистки высокомолекулярной ДНК доступен ряд способов, но все они имеют индивидуальные преимущества и ограничения, такие как низкий выход конечной ДНК, ограничения по молекулярному размеру (вследствие механической деградации высокомолекулярной ДНК), низкая степень очистки (так как полисахариды и другие высокомолекулярные соединения остаются связанными с ДНК), стоимость (при очистке на магнитных шариках), и возможное использование токсичных химических веществ (фенол, хлороформ, тиоцианаты). Скорее всего, на данный момент не существует универсального способа выделения высокомолекулярной ДНК, который бы не зависел от природы и химического состава образца. Отсутствие такого способа представляет особую проблему, если исходный материал является сложной смесью химически разных молекул, содержащих лишь небольшое количество целевой ДНК.
Одним из подходов к выделению высококачественной ДНК из сложных смесей является гель-электрофорез с последующей электроэлюцией из вырезанных фрагментов геля, содержащих целевую ДНК. Наиболее распространенным вариантом этого метода является электроэлюция в диализных мешках (9,10). Другие варианты, такие как электроэлюция в градиенте глицерина (11), также были описаны в литературе. Однако значительная трудоемкость этих традиционных методов ограничивает возможность их применения в повседневной лабораторной практике. Электрофорез, за которым непрерывно следует электроэлюция (12, 13), является интересным альтернативным решением, с помощью которого можно получить высокоочищенную ДНК из геля. Такая ДНК будет совместима с большинством приложений молекулярной биологии. Одним из примеров такого подхода является система Sage ELF (Sage Science, Inc., США), в которой ДНК загружается в кассету с агарозным гелем, и после электрофореза разделенные фрагменты ДНК электроэлюируются из геля с использованием боковых электродов в двенадцать отдельных лунок для сбора образцов. Полученные фракции ДНК можно использовать для секвенирования нуклеотидов и других приложений без дополнительной очистки. Хотя Sage ELF является хорошим инструментом для фракционирования и очистки ДНК, к его недостаткам относятся дорогостоящее оборудование и расходные материалы, верхнее ограничение по массе выделяемых фрагментов 40 т.п.н. и невозможность одновременной обработки большого количества образцов.
Из уровня техники известен патент на изобретение RU 2650865 от 17.04.2018 (45), в котором описан набор реактивов для выделения ДНК из биологических объектов. Набор включает лизирующий буферный раствор, промывочные буферные растворы №1 и №2, элюирующий буферный раствор. Лизирующий буферный раствор содержит: сильный хаотропный агент - гуанидинтиоцианат, неионогенный детергент Тритон Х-100,рН-буфер на основе трис-гидрохлорида, и сорбент в виде суспензии из магнитных микросфер в солевом растворе. Промывочный буфер №1 содержит гуанидинтиоцианат, трис-гидрохлорид и этиловый спирт. Промывочный буфер №2 содержит трис-гидрохлорид, хлорид натрия и этиловый спирт. Элюирующий раствор представляет собой деионизированную воду. Изобретение обеспечивает увеличение выхода ДНК.
Известен патент на полезную модель RU 84381 U1 от 10.07.2009 (46), в котором описано устройство для автоматизированного выделения нуклеиновых кислот из биологических образцов, состоящее из (а) картриджа, содержащего резервуары с реагентами для лизиса клеток, микроорганизмов и вирусных частиц, очистки и элюции нуклеиновых кислот, каналы, микроколонки и клапаны, и (б) управляющей установки-контроллера, осуществляющей подачу давления, нагрев, перемешивание реагентов в индивидуальных резервуарах картриджа и перемещение реакционных смесей и растворов в резервуарах картриджа. При этом картридж содержит приемную камеру для ввода биологического образца, резервуары с сухими реакционными смесями буферов для лизиса и буфера для промывки, микроколонку с твердофазным сорбентом для связывания нуклеиновых кислот, резервуары для этанола, смеси вода-этанол и воды для растворения сухих компонентов промывочного буфера, промывки микроколонки и элюции нуклеиновых кислот, связавшихся с твердофазным сорбентом в микроколонке, выходной порт, совместимый со стандартной микропробиркой, резервуар для сбора отходов реакций, а управляющая установка-контроллер содержит блок соленоидов для управления клапанами картриджа, блок нагревателей, блок электромагнитных мешалок, компрессор и блок электронного управления.
Недостатками вышеуказанных запатентованных технических решений является большая длительность и многостадийность процедуры очистки ДНК, низкий выход ДНК и низкая степень очистки для «сложных» образцов.
Известен источник информации - протокол Zarzosa -Alvarezetal. (19), где вырезанный фрагмент агарозного геля, содержащий целевую ДНК, помещают в прибор для электроэлюции (электроэлютер), а его V-образный канал заполняют буфером с повышенным содержанием соли (1 М хлорид натрия). Во время электроэлюции фрагмент ДНК мигрирует из агарозного геля в соляную ловушку. На заключительном этапе протокола буфер с повышенным содержанием соли, содержащий электроэлюированную ДНК, извлекают пипеткой и осаждают этанолом. Указанный источник информации выбран наиболее близким аналогом (прототипом).Недостатком прототипа является то, что этот способ является достаточно трудоемким, а именно требует вырезания полос ДНК из агарозного геля под УФ-А излучением, переноса полученного фрагмента агарозы в электроэлютор и отдельного проведения процесса электроэлюции. Таким образом, в данной подходе допускается только стадия очистки ДНК и требуется специальное оборудование (электроэлютер). Дополнительно, недостатком этого способа является то, что на выходе целевая ДНК растворена в высокосолевом буфере и непригодна для дальнейшего применения без дополнительной очистки. Осаждение ДНК из солевого раствора обязательна, что в свою очередь, может привести к снижению выхода ДНК при сильно разбавленном растворе.
В результате проведенного анализа уровня техники можно сделать вывод о том, что предлагаемая группа изобретений может быть признана соответствующей критериям патентоспособности «новизна» и «изобретательский уровень» до даты испрашиваемого приоритета.
РАСКРЫТИЕ СУЩНОСТИ ИЗОБРЕТЕНИЯ
Для решения технической проблемы, а также устранения недостатков имеющихся аналогов и прототипа, заявителем предложены способ и устройство для очистки и выделения высокомолекулярной ДНК или РНК из биологических образцов, а также гель для очистки и выделения высокомолекулярной ДНК или РНК и способ изготовления устройства. Группа изобретений объединена единым изобретательским замыслом.
Технологии секвенирования с длинным считыванием требуют ДНК не только высокой чистоты и целостности, но и высокой молекулярной массы, которую может быть трудно получить из сложных биологических образцов.
В этой связи авторами разработан способ очистки и выделения высокомолекулярной ДНК, который включает электрофорез с последующей электроэлюцией и основан на том факте, что электрофоретическая подвижность нуклеиновых кислот замедляется в гелях и растворах с высокой ионной силой. Образец высокомолекулярной ДНК сначала очищают электрофорезом в разделительном канале, заполненномгелем (например, агарозным). После достаточной очистки электрофорез приостанавливают и перед полосой ДНК (ниже по направлению ее движения) помещают блокирующий гель, содержащий повышенную концентрацию соли. Отбор очищенной высокомолекулярной нуклеиновой кислоты может осуществляться из резервуара(промежутка) для сбора проб, расположенного между основным гелем и гелем, содержащем повышенную концентрацию соли. После установки блокирующего солевого геля ток подается повторно до тех пор, пока ДНК не будет электроэлюирована из разделяющего геля в резервуар для сбора (зазор между гелями), где она постепенно замедляется и накапливается. Очищенную ДНК можно легко собрать из резервуара с помощью пипетки и использовать без дополнительного обессоливания в реакциях секвенирования с длинным считыванием. Преимущества этого способа заключаются в том, что он прост и недорог, а также позволяет получать высокомолекулярную ДНК или РНК высокой очистки и с высоким выходом даже из сложных биологических образцов. Другими преимуществами является то, что предложенный способ совместим с секвенированием с длинным считыванием и имеет потенциал для автоматизации в вертикальном или горизонтальном формате.
Целью предлагаемого технического решения является создание способа, устройства и составных частей для осуществления качественной очистки высокомолекулярных нуклеиновых кислот из многокомпонентных биологических образцов с целью их дальнейшего секвенирования методами третьего поколения. Существует потребность в разработке новых способов выделения и очистки ДНК из биологических образцов, и в новых, более эффективных устройствах и материалах для реализации таких способов.
Технический результат заключается в упрощении способа выделения и очистки высокомолекулярной ДНК или РНК, пригодной для секвенирования с длинным считыванием, в повышении эффективности выделения высокомолекулярной ДНК или РНК из различных сложных биологических образцов(например, растительных образцов, образцов почвы, фекалий и т.д.), в улучшении качества очистки высокомолекулярных нуклеиновых кислот для секвенирования с длинным считыванием, в ускорении выделения и очистки высокомолекулярных нуклеиновых кислот за счет отсутствия этапа переноса материала из прибора для электрофореза в электроэлютер, в отсутствии необходимости дополнительной очистки нуклеиновых кислот от соли, а также в обеспечении возможности одновременной обработки большого количества образцов в предлагаемом устройстве заявленным способом, что позволяет масштабировать и роботизировать предлагаемые технические решения. Благодаря этому становится возможным, в частности, получать ДНК (или РНК) с высокой степенью чистоты (от 50% до 95 %) и высоким выходом (от 50% до 90%).
ОБЪЕКТ ИЗОБРЕТЕНИЯ
В настоящей группе изобретений описан простой и эффективный способ очистки и выделения высокомолекулярной ДНК (или РНК), устройство и гель для очистки и выделения высокомолекулярной ДНК (или РНК), способ изготовления устройства.
Указанный технический результат достигается совокупностью существенных признаков, представленной в формуле изобретения.
В одной неограничивающей реализации изобретения предложен способ очистки и выделения высокомолекулярных нуклеиновых кислот из образца, включающий:
- заполнение зоны электрофорезного канала агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой;
- нанесение образца, содержащего целевую высокомолекулярную нуклеиновую кислоту, на агарозный разделяющий гель, содержащий буфер для электрофореза с низкой ионной силой;
- очистку высокомолекулярной нуклеиновой кислоты методом электрофореза;
- остановку электрофореза после отделения высокомолекулярной нуклеиновой кислоты от загрязнений до того, как очищенная нуклеиновая кислота выйдет из агарозного разделяющего геля, содержащего буфер для электрофореза с низкой ионной силой;
- установку агарозного блокирующего геля с содержанием соли в переделах 0,5 - 1 М хлорида натрия в электрофорезный канал в направлении движения нуклеиновых кислот, после агарозного разделяющего геля, содержащего буфер для электрофореза с низкой ионной силой, в котором находится очищенная нуклеиновая кислота, с зазором между гелями не менее 0,5 см;
- заполнение зазора между гелями буфером для электрофореза с низкой ионной силой;
- возобновление электрофореза до тех пор, пока целевая нуклеиновая кислота не мигрирует в зазор между гелями;
- отбор очищенной высокомолекулярной нуклеиновой кислоты из зазора между гелями.
Ещё в одном примере реализации изобретения отбор очищенной нуклеиновой кислоты из зазора между гелями проводится с помощью автоматической пипетки или лабораторного робота.
Ещё в одном примере реализации изобретения отбор очищенной нуклеиновой кислоты из зазора между гелями проводится в микропробирку для дальнейшего использования.
Ещё в одном примере реализации изобретения перед очисткой высокомолекулярной нуклеиновой кислоты методом электрофореза определяют продолжительность электрофореза.
Ещё в одном примере реализации изобретения зазор между основным и агарозным блокирующим гелями заполнен буфером для электрофореза с низкой ионной силой. Под низкой ионной силой понимается ионная сила, меньшая, чем у агарозного блокирующего геля.
Ещё в одном примере реализации изобретения буфер для электрофореза с низкой ионной силой представляет собой 20 мМТрис и 20 мМ HEPES.
Ещё в одном примере реализации изобретения образец дополнительно содержит смесь биологических молекул, включающую нецелевые нуклеиновые кислоты или короткие олигонуклеотиды, или белки, или пептиды, или полисахариды, или липиды, или вторичные метаболиты, или гуминовые вещества, или пигменты, или полифенолы.
Также технический результатдостигается за счет того, что устройство для очистки и выделения высокомолекулярных нуклеиновых кислот из образца включает:
- по меньшей мере один электрофорезный канал;
- зону электрофорезного канала для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой;
- зону электрофорезного канала для установки агарозного блокирующего геля с содержанием соли в переделах 0,5 - 1 М хлорида натрия,
- зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля с содержанием соли в переделах 0,5 - 1 М хлорида натрия;
- при этом зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля выполнен с возможностью заполнения буфером для электрофореза с низкой ионной силой и составляет не менее 0,5 см. Ещё в одном примере реализации изобретения электрофорезный канал выполнен горизонтальным или вертикальным.
Ещё в одном примере реализации изобретения электрофорезный канал выполнен горизонтальным или вертикальным.
Ещё в одном примере реализации изобретения электрофорезный канал выполнен открытым или закрытым.
Ещё в одном примере реализации изобретения буфер для электрофореза с низкой ионной силой представляет собой 20 мМ Трис и 20 мМ HEPES.
Ещё в одном примере реализации изобретения устройство изготовлено из пластика, прозрачного для видимого и УФ излучения, например,315-400 нм.
Ещё в одном примере реализации изобретения устройство дополнительно включает электроды для осуществления электрофореза в электрофорезном канале.
Ещё в одном примере реализации изобретения устройство дополнительно включает сенсор для сбора информации о выделенных высокомолекулярных нуклеиновых кислотах, соединенный с блоком обработки информации и блоком отображения.
КРАТКОЕ ОПИСАНИЕ ФИГУР, ТАБЛИЦ, ИНЫХ ГРАФИЧЕСКИХ МАТЕРИАЛОВ
Изобретение иллюстрируются следующими графическими материалами.
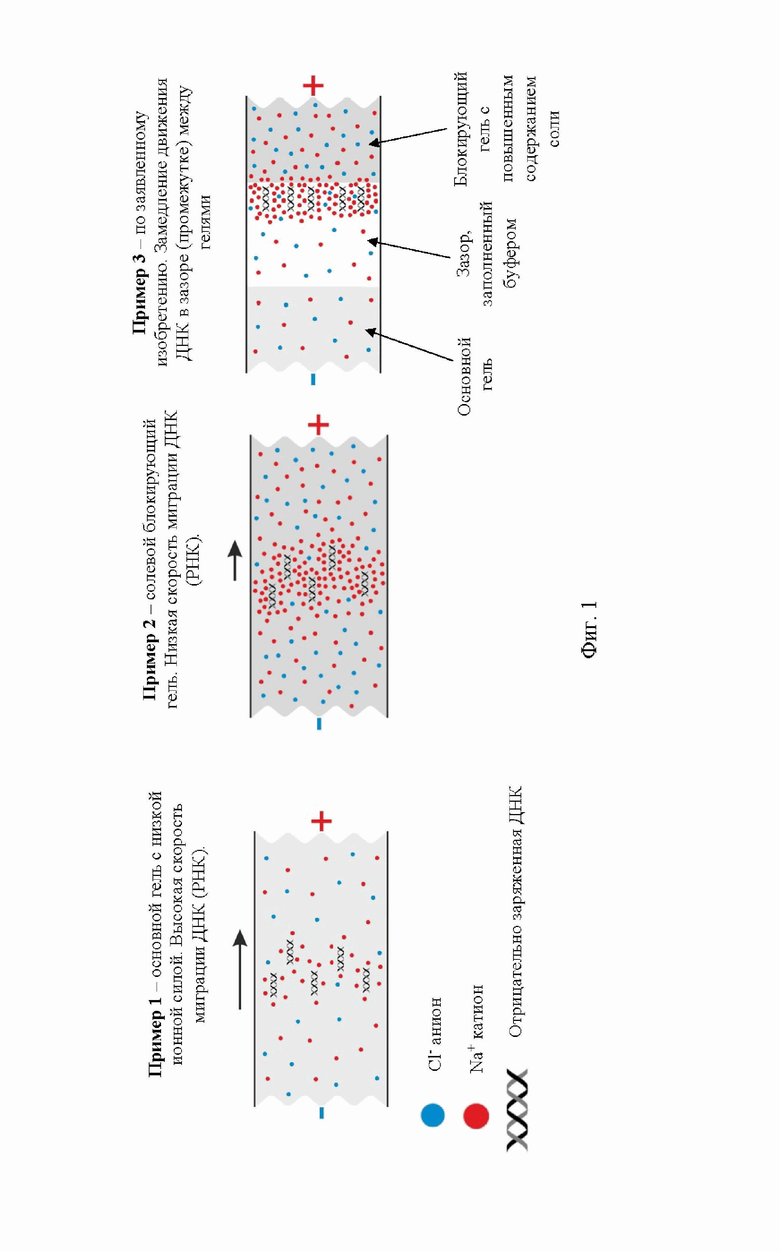
На фигуре 1 показан принцип захвата электроэлюированной ДНК в зазоре между гелями с низкой и высокой ионной силой.
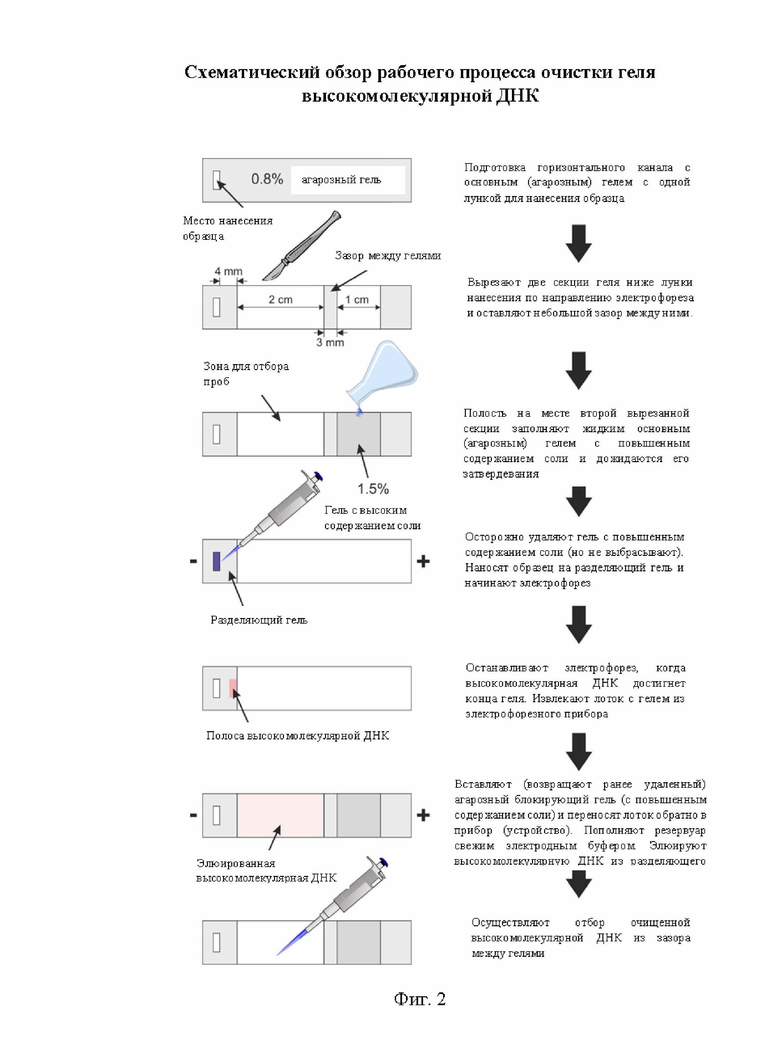
На фигуре 2 схематически показан способ (обзор рабочего процесса) очистки геля высокомолекулярной ДНК.
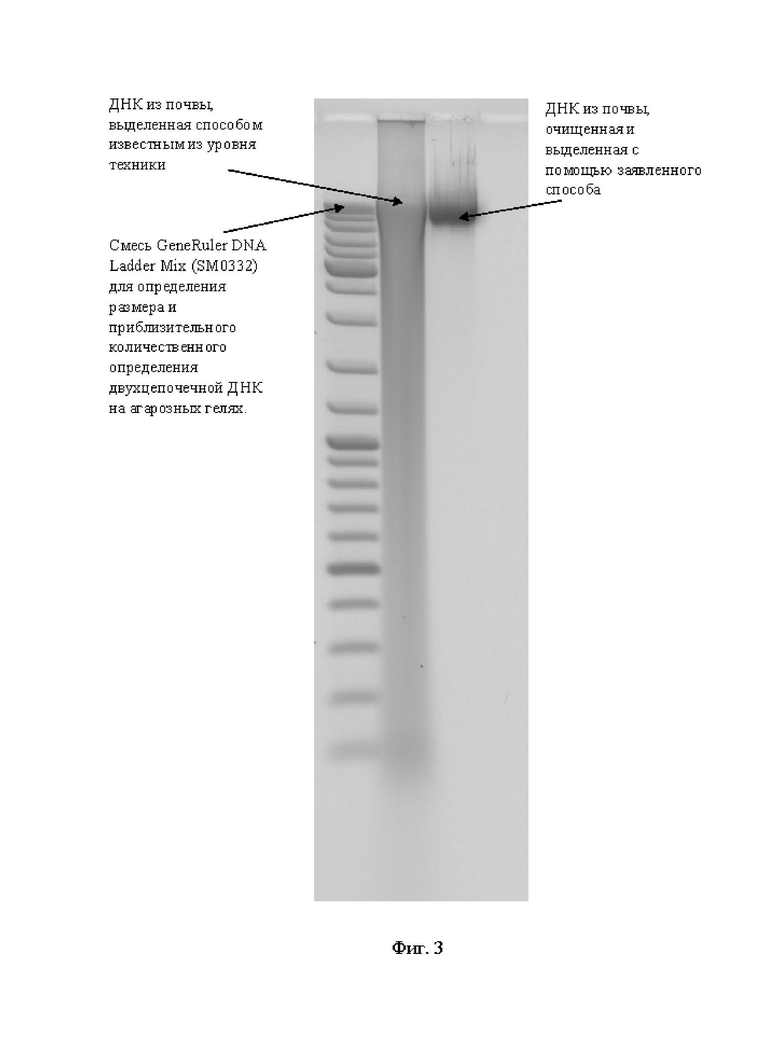
На фигуре 3 представлено фото смеси GeneRuler DNA LadderMix (ThermoFisherScientific, SM0332) для определения размера и приблизительного количественного определения двухцепочечной ДНК на агарозных гелях и сопоставлены результаты очистки ДНК способом известным из уровня техники (с помощью способа, описанного в прототипе) и ДНК, очищенного с помощью заявленного способа.
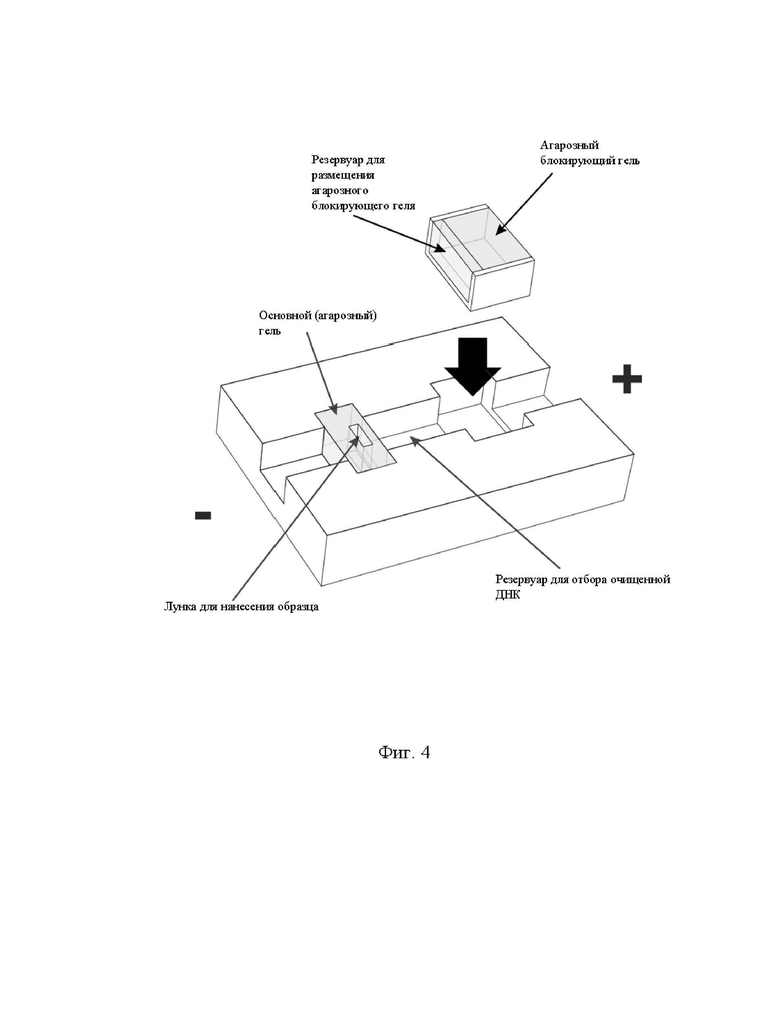
На фигуре 4 показана модель устройства для очистки и выделения высокомолекулярных нуклеиновых кислот из образца.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Определения и общая терминология.
Далее будут приведены ссылки на определенные варианты осуществления изобретения, примеры которых проиллюстрированы предлагаемым способом и устройством. Подразумевается, что изобретение охватывает все альтернативы, модификации и эквиваленты, которые могут быть включены в объем настоящего изобретения, как определено в формуле изобретения.
Специалисту в данной области известны многие способы и материалы, аналогичные или эквивалентные описанным в данном документе, которые могут быть использованы при практическом осуществлении настоящей группы изобретений.
Настоящее изобретение не ограничивается изложенными в данном документе способами и материалами. В случае, если один или несколько из включенных литературных ссылок, патентов и аналогичных материалов отличается от или противоречит данной заявке, включая, в частности, определенные термины, использование терминов, описанные приемы и т.п., преимущественную силу имеет настоящая заявка.
Далее следует принять во внимание, что определенные признаки изобретения, которые для ясности описаны в контексте отдельных вариантов осуществления изобретения, могут быть представлены также совместно в одном варианте. И наоборот, разнообразные признаки изобретения, которые для краткости описаны в контексте одного варианта осуществления изобретения, могут быть представлены отдельно или в любой подходящей подкомбинации.
Если представлен диапазон значений, то следует понимать, что каждое промежуточное значение, до десятых долей единицы нижнего предела, если из контекста явно не следует иное между верхней и нижней границами этого диапазона и любые другие указанные или промежуточные значения в этом установленном интервале охватываются настоящим изобретением. Верхние и нижние пределы этих меньших диапазонов могут быть независимо включены в меньшие диапазоны, что также входит в объем изобретения, с учетом любого конкретно исключенного предела в указанном диапазоне. Когда указано, что диапазон включает один или оба предела, диапазоны, исключающие любое из обоих включенных пределов, также входят в объем изобретения.
Все публикации, патенты и заявки на патенты включены в настоящий документ посредством ссылки. Хотя в вышеприведенном описании это изобретение было описано в отношении некоторых предпочтительных вариантов его осуществления, и многие детали были изложены в целях иллюстрации, для специалистов в данной области техники будет очевидно, что изобретение допускает дополнительные варианты осуществления и что некоторые детали, описанные в данном документе, могут значительно изменяться без отклонения от сущности изобретения.
Использование терминов в единственном числе в контексте описания изобретения должно толковаться как охватывающее как единственное, так и множественное число, если иное не указано в данном документе или явно не противоречит контексту.
Термины «состоящий из», «имеющий», «включающий» и «содержащий» следует толковать как неограничивающие термины, т.е. означающие «включая, но не ограничиваясь», если не указано иное.
Использование любых и всех примеров или иллюстративного языка (например, «такой как»), представленных в данном документе, предназначено просто для лучшего описания изобретения и не налагает ограничения на объем изобретения, если иное не заявлено.
Никакие формулировки в описании не следует истолковывать как указывающие на какой-либо не заявленный элемент как существенный для практического применения изобретения.
Здесь описаны варианты осуществления этого изобретения, включая лучший из известных изобретателям способа осуществления изобретения. Разновидности этих вариантов осуществления могут стать очевидными для специалистов в данной области техники после прочтения предшествующего описания. Авторы ожидают, что квалифицированные специалисты будут использовать такие варианты в зависимости от обстоятельств, и авторы предполагают, что изобретение будет реализовано на практике иначе, чем конкретно описано в данном документе. Соответственно, это изобретение включает в себя все модификации и эквиваленты признаков, изложенных в прилагаемой формуле изобретения, как это разрешено действующим законодательством. Более того, любая комбинация вышеописанных признаков во всех их возможных вариациях охватывается изобретением, если иное не указано в данном документе или иным образом явно не противоречит контексту.
Для описания настоящего изобретения используются следующие термины.
Все технические и научные термины, использованные в данном документе, имеют такое значение, какое обычно понимается специалистами в области, к которой относится изобретение, если не указано иное. Все патенты и публикации, на которые имеются ссылки в данном документе, полностью включены в изобретение посредством ссылки.
Сокращения, используемые в настоящем описании, включая приведенные в иллюстративных схемах и последующих примерах, хорошо известны среднему специалисту. Некоторые из сокращений используют как нижеследующие:
ДНК - дезоксирибонуклеиновая кислота.
РНК - рибонуклеиновая кислота.
Нецелевая нуклеиновая кислота –примесная нуклеиновая кислота, которая не является объектом очистки и выделения.
Электрофорезный канал – канал в устройстве с возможностью проведения электрофореза, который подвергается электрофорезу.
NGS - секвенирование нового поколения (англ. next generation sequencing, NGS).
HEPES -(4-(2-гидроксиэтил)-1-пиперазинэтансульфоновая кислота) представляет собой цвиттер-ионный органический буферный агент; один из двадцати в списке буферных растворов Гуда.
THE - электрофорезный буфер (Трис-HEPES-ЭДТА), а также для растворения и хранения ДНК, 1-кратный буфер содержит 20 мМ Трис и 20 мМ HEPES и 0.1 мМ ЭДТА.
TE - буфер для растворения и хранения ДНК, 1-кратный буфер содержит 10 мМ Трис-HCl (pH 8.0) и 0.1мМ ЭДТА.
TBE – электрофорезный буфер (Трис-борат-ЭДТА), 1-кратный буфер содержит 45 мМ Трис, 45 мМ борную кислоту и 0.5мМ ЭДТА, рН 8.3.
TAE - электрофорезный буфер (трис-ацетат-ЭДТА), 1-кратный буфер содержит 40 мМ Трис, 20 мМ ацетат и 0.5мМ ЭДТА, рН 8.0. Фасовка 5 л поставляется в кубическом контейнере.
ЭДТА - этилендиаминтетрауксусная кислота, (от англ. EDTA).
SDS - додецилсульфат натрия, анионоактивное поверхностно-активное вещество.
CTAB - цетилтриметиламмония бромид (англ. cetril trimethyl ammonium bromide), синтетическое поверхностно-активное вещество, обладает антисептическим и антистатическим действием.
TGS - платформа секвенирования третьего поколения (от англ. third-generation sequencing, TGS).
SYBR GreenI - асимметричный цианиновый краситель, используемый в молекулярной биологии для окрашивания нуклеиновых кислот.
EtBr - бромистый этидий (3,8-диамино-5-этил-6-фенилфенантридиум бромид), флуоресцентный краситель, как интеркалирующий агент в молекулярной биологии для выявления нуклеиновых кислот, при электрофорезе ДНК в агарозном геле.
XyleneCyanol FF - ксиленцианол, используется в качестве следящего красителя при электрофорезе агарозы или полиакриламидного геля. Он имеет небольшой отрицательный заряд и мигрирует в том же направлении, что и ДНК, позволяя пользователю следить за продвижением молекул через гель. Скорость миграции зависит от состава геля.
PFGE - гель-электрофорез в импульсном поле.
ЭКСПЕРИМЕНТАЛЬНЫЕ ПРОЦЕДУРЫ
Заявленный способ выделения и очистки высокомолекулярных нуклеиновых кислот из образца включает заполнение зоны электрофорезного канала основным гелем, затем нанесение образца в слот (лунку для нанесения образца).
Далее осуществляют очистку высокомолекулярной нуклеиновой кислоты методом электрофореза. Затем осуществляют остановку электрофореза после отделения высокомолекулярной нуклеиновой кислоты от загрязнений и до того, как очищенная нуклеиновая кислота выйдет из основного геля. Затем производят установку агарозного блокирующего геля с повышенным содержанием соли в электрофорезный канал после основного геля, в котором находится очищенная нуклеиновая кислота, с образованием зазора между гелями не менее 0,5 см. На следующем этапе заполняют зазор между гелями буфером для электрофореза и возобновляют электрофорез до тех пор, пока целевая нуклеиновая кислота не мигрирует в зазор между гелями. На последнем этапе реализации способа осуществляют отбор очищенной высокомолекулярной нуклеиновой кислоты из зазора между гелями.
Заявленный способ включает метод гель-электрофореза, за которым следует электроэлюция. Он основан на том факте, что электрофоретическая подвижность ДНК уменьшается с увеличением концентрации соли(ионной силы) в среде электрофореза. После достаточного отделения ДНК от различных загрязнений гелевый блок с повышенным содержанием соли помещают на пути дальнейшего электрофореза ДНК (или РНК), а резервуар для сбора очищенного образца располагают между разделяющим гелем и гелем с повышенным содержанием соли. Электроэлюция из основного (разделяющего) геля может продолжаться до тех пор, пока целевая ДНК не мигрирует в резервуар для сбора (зазор между гелями), где она постепенно замедляется и накапливается. Очищенную ДНК можно легко извлечь из резервуара с помощью пипетки и использовать либо сразу, либо после обессоливания, в зависимости от дальнейшего применения. Предлагаемый способ очистки ДНК особенно полезен, когда другие методы неэффективны или нецелесообразны, например, для обработки многокомпонентных образцов, содержащих сложные смеси биомолекул. Кроме того, заявленный способ экономически эффективен и не требует сложных реагентов или инструментов.
В ионных растворах отрицательно заряженная нуклеиновая кислота окружена положительно заряженными катионами (противоионами) и отрицательно заряженными анионами (коионами), которые образуют так называемую ионную атмосферу. Ионная атмосфера имеет более высокую концентрацию притягиваемых катионов (накопление противоионов) и более низкую концентрацию отталкиваемых анионов (истощение коионов) и сильно зависит от ионной силы раствора.
Известно, что увеличение ионной силы вызывает электростатическое экранирование отрицательного заряда ДНК противоионами (фиг. 1), тем самым уменьшая ее подвижность в растворе во время капиллярного электрофореза.
На фиг.1 показан принцип захвата электроэлюированной ДНК в зазоре между гелями с низкой и высокой ионной силой (фиг. 1 пример 3). Схематическое изображение ионной атмосферы, окружающей ДНК в гелях, содержащих различные концентрации соли. Красные и синие кружки представляют собой катионы Na+ и анионы Cl-. Более высокое накопление катионов вокруг ДНК и РНК в геле с повышенным содержанием соли (фиг. 1 пример 2) по сравнению с обычным гелем (фиг. 1 пример 1) вызывает более сильное электростатическое экранирование отрицательного заряда ДНК, тем самым уменьшая ее электрофоретическую подвижность. Это явление можно использовать для улавливания ДНК в заполненном буфером промежутке перед гелем с повышенным содержанием соли, где подвижность ДНК замедляется и происходит ее накопление (фиг. 1 пример 3).
Основываясь на принципе, лежащем в основе вышеуказанной методологии, что электрофоретическая подвижность нуклеиновых кислот зависит от концентрации солей и может сильно снижаться в присутствии высокой концентрации соли, когда солевой блок (блокирующий гель), содержащий повышенную концентрацию соли, располагается по направлению движения нуклеиновых кислот. Чтобы продемонстрировать это явление, был вырезан фрагмент геля перед мигрирующей ДНК и его заменили гелем, содержащим повышенную концентрацию соли. При приближении к высокосолевому гелю (агарозному блокирующему гелю) ДНК показала заметное снижение электрофоретической подвижности, свидетельствующее об электростатическом экранировании избытком противоионов (фиг.1). Основываясь на этой проверке концепции, разработан способ очистки высокомолекулярной ДНК (или РНК), который сочетает в себе гель-электрофорез и электроэлюцию и включает улавливание ДНК (РНК)с помощью гелевого блока (добавлением агарозного блокирующего геля) с повышенным содержанием соли.
Предлагаемый способ и устройство использует очистку высокомолекулярной ДНК электрофорезом, например, в горизонтальных каналах. Формовочный лоток для гелей можно легко изготовить самостоятельно с помощью 3D-печати или на фрезерном станке для обработки пластиковой заготовки, прозрачной для видимого и УФ излучения. Ширина и высота каждого канала может составлять примерно 1 см, что достаточно для эффективного разделения и очистки стандартных образцов ДНК и для размещения однолуночной гребенки для электрофореза.
На фиг. 2схематически показан процесс очистки геля высокомолекулярной ДНК, предложенныйв изобретении.
Две секции вырезали ниже по течению от места для нанесения образца, создавая резервуар для сбора очищенного образца(зазор) и формовочную полость для размещения агарозного блокирующего геля.
На следующем этапе полученную формовочную полость могут заполнять расплавленным раствором агарозного блокирующего геля, содержащим повышенную концентрацию соли. В одном из вариантов реализации способа после затвердевания агарозного блокирующего геля с повышенным содержанием соли, его осторожно удаляли из формовочного лотка и откладывали для дальнейшего использования во время электрофореза.
Лоток (резервуар для размещения агарозного блокирующего геля см. фиг. 4), который теперь содержал только основной гель, переносили в резервуар для электрофореза.
Образец, содержащий целевую высокомолекулярную ДНК, наносили в лунку на основном геле и подвергали электрофорезу.
Электрофорез продолжался до тех пор, пока высокомолекулярная ДНК не достигала конца основного геля, ток временно отключали и блокирующий гель вынимали из резервуара для электрофореза.
Резервуар для сбора образцов (зазор) заполняли свежим электрофорезным буфером с низкой ионной силой. Устанавливали агарозный блокирующий гель и снова включали ток для электроэлюции очищенной высокомолекулярной ДНК из разделяющего геля в резервуар.
Дальнейшая миграция ДНК через заполненный буфером зазор постепенно замедлялась за счет нейтрализации отрицательного заряда ДНК избытком катионов, двигающихся навстречу из агарозного блокирующего геля. Это приводило к накоплению очищенной высокомолекулярной ДНК в зазоре, из которого ее можно было легко извлечь с помощью пипетки.
Гель-электрофорез представляет собой способ выбора для очистки длинных олигонуклеотидов, поскольку он предлагает один из самых высоких уровней чистоты среди доступных методов очистки. То же самое верно для очистки полноразмерной ДНК.
Результат применения заявленного способа представлен на фиг. 3, на которой наблюдается эффективно очищенная и концентрированная ДНК.
Способ, предложенный в этом изобретении, не предполагает измельчения геля; вместо этого он основан на непрерывном процессе электрофореза и электроэлюции, который не нарушает структуры геля, тем самым устраняя загрязнения образца полисахаридами из агарозы.
В предлагаемом способе электроэлюированная ДНК накапливается в резервуаре для сбора образцов (зазоре между гелями), заполненном буфером для электрофореза. Предпочтительно, чтобы буфер был совместим с последующими ферментативными реакциями и длительным хранением образцов. Электрофорезный буфер трис-борат-ЭДТА (ТВЕ) является одним из наиболее распространенных рабочих буферов, используемых в электрофорезе коротких фрагментов нуклеиновых кислот. Однако борат-ионы образуют комплексы с ДНК (22, 23) и белками (24-26), а также ингибируют активность многих ферментов (27).Это делает TBE непригодным для данного протокола. Наиболее популярной альтернативой TBE является буфер трис-ацетат-ЭДТА (TAE). Буфер ТАЕ на основе уксусной кислоты и триса, может быть использован. Однако, высокое содержание уксусной кислоты в ТАЕ буфере, потенциально может ингибировать некоторые ферментативные реакции, такие как ПЦР (28) и данный буфер быстро теряет буферную емкость и нагревается при длительном использовании. Присутствие в буферахвысокой концентрации ЭДТА, может также ингибирует ПЦР (28) и ряд ферментов, модифицирующих ДНК, таких как щелочная фосфатаза (29) и нуклеаза S1 (30).
Определение локализации интересующей полосы целевой высокомолекулярной ДНК имеет значение для определения момента, когда необходимо использовать блокирующий гель с повышенным содержанием соли, останавливающий подвижность ДНК и РНК. Самый простой способ приблизительно оценить (без использования УФ-А излучения), когда высокомолекулярная ДНК приблизится к концу разделяющего геля, это использовать медленно мигрирующий лидирующий краситель, такой как ксиленцианол (XyleneCyanol FF). Поскольку ксиленцианол мигрирует в 0,8% агарозном геле примерно с той же скоростью, что и фрагмент двуцепочечной ДНК размером не ниже 10т.п.н., блокирующий гель с повышенным содержанием соли можно использовать после того, как краситель ксиленцианол начнет выходить из основного геля.
В качестве альтернативы, можно оценить миграцию высокомолекулярной ДНК на основе положения фрагментов, предварительно окрашенных (например, окрашенных интеркалирующими красителями (SYBR Green I, EvaGreen, SYBR Safe, GelRed или GelGreen),с использованием, например, ближнего УФ-А излучения (LED источник черного светав диапазоне 365-395нм), которое не будет разрушать нуклеиновые кислоты. Наиболее распространенным способом обнаружения ДНК в геле является использование флуоресцентного интеркалирующего красителя, небольшой молекулы, которая обратимо встраивается между соседними парами оснований двойной спирали. Бромистый этидий (EtBr) остается наиболее широко используемым флуоресцентным интеркалирующим красителем в электрофорезе ДНК несмотря на то, что он является потенциальным канцерогеном и предпочтительным использованием УФ-B (210 нм и 285 нм) излучения для визуализации, что приводит к повреждению ДНК. Использование, например, ближнего УФ-А излучения (источник черного света) допускается при окрашивании ДНК с помощью красителя EtBr. Альтернативы красителю EtBr включают, среди прочего, SYBR Green I, EvaGreen, SYBR Safe, GelRed и GelGreen, все из которых имеют лучшие профили безопасности, чем EtBr, и могут быть визуализированы с использованием, например, ближнего УФ-А излучения (источник черного свет, в диапазоне 365-395 нм) или синего света. Тем не менее, из-за своей ДНК-связывающей природы все интеркалирующие красители изменяют структуру и механические свойства ДНК (31-33) и потенциально будут мешать последующим ферментативным реакциям(34-36). Поэтому, если последующее применение чувствительно к присутствию интеркалирующего красителя, рекомендуется выполнить дополнительную стадию очистки, с помощью осаждения ДНК изопропанолом (см. абзац ниже). Однако самый простой способ избежать описанной выше проблемы - сначала определить время, когда флуоресцентно окрашенная высокомолекулярная ДНК достигнет конца разделяющего геля, а затем рутинно использовать полученные параметры (время электрофореза и используемое напряжение) для выделения аналогичных, но неокрашенных, ДНК. Этот подход может быть особенно ценным при параллельном выделении высокомолекулярных ДНК одинакового размера в крупномасштабных экспериментах.
При выборе концентрации соли в блокирующем геле необходимо соблюдать баланс. С одной стороны, концентрация соли должна быть достаточной для замедления миграции ДНК в зазоре между гелями. С другой стороны, слишком высокая концентрация соли загрязнит образец ДНК и может привести к остановке ДНК уже в разделяющем геле до того, как она попадет в резервуар для сбора образцов. После проведения ряда тестов определили, что оптимальная концентрация (т.е. дополнительно повышение достижения технического результата) соли составляет приблизительно в переделах 0,5 - 1 М хлорида натрия. Это значение согласуется с ранее опубликованными данными, показывающими, что электрофоретическая подвижность двухцепочечной ДНК в свободном растворе уменьшалась с увеличением ионной силы до тех пор, пока она не начала выравниваться примерно при 0,6 М ацетат натрия (16). При 0,5 М хлорида натрия в блокирующем геле концентрация ионов натрия в выделенном образце ДНК составляла от 30 до 40 мМ по результатам определения с помощью оптической эмиссионной спектроскопии с индуктивно-связанной плазмой (ICP-OES). Большинство ферментативных реакций, даже наиболее чувствительных к соли (например, лигирование ДНК-лигазой Т4), совместимы с этой концентрацией соли, особенно с учетом разбавления образца в реакционной смеси. Таким образом, полученная ДНК подходит без дополнительного обессоливания для различных последующих ферментативных реакций, включая лигирование адаптера для секвенирования с длинным считыванием. Однако для некоторых специфических применений, таких как подготовка образца для секвенирования с использованием платформ третьего поколения, для анализа ДНК с помощью масс-спектрометрии с лазерной десорбцией/ионизацией (MALDI) (37), может потребоваться дополнительная стадия обессоливания и концентрирование образца. Преципитация нуклеиновых кислот с помощью смешения сравным объемом солевой смеси с изопропанолом является первым методом выбора для обессоливания и концентрирования ДНК. Этот метод требует наличие настольной центрифуги с охлаждением и определённого времени для инкубации образца при -20°в морозилке, но может привести к неполному осаждению ДНК из разбавленных образцов. Другим подходом для одновременного обессоливания и концентрирования образцов является использование коммерческих центрифужных мембранных фильтрующих устройств (например, Amicon Ultra-2 CentrifugalFilterUnit от Millipore). Дополнительно, существуют альтернативные методы обессоливания, такие как гель-фильтрация на спин-колонках с последующей (при необходимости) экстракцией бутанолом (38) для концентрирования образца. Спин-колонка может быть как коммерческой, так и заполненной сорбентом для гель-фильтрации (39) в собственнойлаборатории. Таким образом, существует несколько методов эффективного обессоливания и концентрирования высокомолекулярной ДНК. Эти методы, помимо выполнения своих основных функций, также отделяют любые остаточные лидирующие и/или интеркалирующие красители от целевой высокомолекулярной ДНК.
Основное преимущество предлагаемого способа заключается в том, что предлагается простое и эффективное решение для выделения и очистки высокомолекулярной ДНК, пригодное для секвенированияс длинным считыванием. Современные технологии длинных считываний/TGS (40) были разработаны для преодоления основного ограничения секвенирования второго поколения, а именно коротких прочтений длиной менее 600 нуклеотидов. Более длинные прочтения (в диапазоне от 10 т.п.н. до более чем 400 т.п.н.), генерируемые TGS, существенно улучшают качество и полноту сборки генома и особенно полезны для характеристики высоко повторяющихся геномных областей. Текущая реакционная смесь, использующая при TGS чувствительна к присутствию загрязнителей в образцах ДНК. Поэтому важно получать ДНК как с высокой молекулярной массой, так и с высокой чистотой, чтобы воспользоваться возможностями, предлагаемыми TGS.
Несмотря на то, что верхний предел разрешения 0,8% агарозных гелей, использованных в настоящем исследовании, составляет около 50т.п.н., более крупные молекулы ДНК могут также быть извлечены из таких гелей, если ДНК входит в гель и мигрирует в нем в рамках процесса, называемого рептация (41). Таким образом, предложенный способ может быть успешно использован для отделения ДНК размером более 50т.п.н. от примесей меньшего размера. Однако, если есть необходимость отделить друг от друга отдельные молекулы ДНК размером более 50т.п.н., верхний предел разрешения гель-электрофореза должен быть расширен. Теоретически этого можно достичь, используя низкопроцентные жидкие агарозные гели (0,1–0,3%), которые способны разделять молекулы ДНК длиной до 750 т.п.н. Однако, такие гели в горизонтальном варианте являются хрупкими, требуют особого обращения и разделение в них требует много времени (42,43). Использование коротких вертикальных и закрытых каналов с жидким гелем, позволяет хорошо и быстро элюировать целую высокомолекулярную ДНК любого размера (вирусы, хромосомы). Другой способ расширить верхний предел разрешения агарозных гелей — это использование гель-электрофореза в импульсном поле (PFGE), который основан на приложении электрического поля к гелю с разных направлений. В отличие от обычного электрофореза в стационарном поле, PFGE способен разделять ДНК длиной до 5 миллионов пар оснований(44). Заявленный способ может использоваться для выделения высокомолекулярной ДНК, разделенной с помощью PFGE. С точки зрения теории электрофореза, нет никаких препятствий для использования ДНК-ловушки на основе блокирующего геля с повышенным содержанием соли в сочетании с PFGE. Таким образом, заявленный способ имеет потенциал для выделения высокомолекулярной ДНК размером до нескольких миллионов оснований с чистотой, подходящей для TGS и других требовательных медицинских и технологических применений.
Еще одним преимуществом предлагаемого способа является способность гель-электрофореза отделять интересующую ДНК от химически различных биомолекул за одну стадию. Эти молекулы могут включать, но не ограничиваться, следующими: нежелательные длинноцепочечные ДНК или РНК, короткие олигонуклеотиды ДНК и РНК, полисахариды, полифенолы, гуминовые вещества, белки, пептиды, липиды, пигменты и вторичные метаболиты. Обычные методы очистки ДНК часто требуют нескольких этапов при работе со сложными образцами. В отличие от этих методов, гель-электрофорез позволяет отделить целевую ДНК от различных химически родственных и неродственных молекул за одну стадию, основываясь на их различиях в заряде и размере. Кроме того, гель-электрофорез обычно не требует сложной предварительной обработки образца для достижения хорошего разделения. В наших экспериментах для обработки образца было достаточно простого и недорогого протокола, который позволяет сохранить первичный размер высокомолекулярной ДНК, основанного на использовании додецилсульфта натрия и протеиназы K. Еще одно преимущество очистки ДНК с помощью гель-электрофореза заключается в том, что для нее требуется меньше исходной ДНК, чем для других способов очистки. Это особенно ценно, когда очищенный образец содержит лишь небольшое количество целевой ДНК. Кроме того, нанесение несколько микрограмм ДНК для разделения в геле, вызывает размытие полос, и эта проблема усугубляется по мере увеличения размера ДНК. Для достижения наилучших результатов рекомендуется использовать меньше исходного материала, чем обычно требуется для других способов. Например, всего 10 мкг образца ткани с нормальным содержанием ДНК может быть достаточно для получения до 1 мкг очищенной высокомолекулярной ДНК. В соответствии с вышеизложенным предлагаемый способ является эффективным для одноэтапной обработки сложных образцов, содержащих химически разнообразные примеси и лишь небольшое количество целевой ДНК.
Важной особенностью предлагаемого способ является его потенциал масштабируемости и автоматизации. Существует возможность масштабирования системы для размещения 8, 12, 24, 48 или даже большего количества разделительных каналов в вертикальном или горизонтальном варианте, и оптимизации форм каналов для конкретных высокопроизводительных задач.
После первоначальной оптимизации с помощью 3D-печати подходящий прототип устройства может быть изготовлен на станке с ЧПУ из пластика, прозрачного для УФ-А излучения. На фиг. 4 представлена модель устройства для электрофореза/электроэлюции для крупномасштабной очистки высокомолекулярной ДНК, имеющее канал с двумя расширениями, первое из которых содержит предварительно залитый основной (разделяющий) агарозный гель. Образец, содержащий целевую ДНК, загружают в лунку для нанесения образца и проводят электрофорез до тех пор, пока высокомолекулярная ДНК не достигнет конца разделяющего геля, а все нежелательные молекулы выйдут из него. Электрофорез останавливают, резервуар для сбора очищенного образца наполняют свежим буфером и во второе расширение электрофорезного канала вставляют лоток с основным и с блокирующим гелем. Ток подается повторно до тех пор, пока высокомолекулярная ДНК не мигрирует из основного геля в заполненный буфером резервуар для сбора, где она замедляется и накапливается. Устройство можно масштабировать для размещения нескольких каналов разделения. Большая часть рабочего процесса может быть автоматизирована, включая загрузку образцов, добавление свежего буфера, введение геля с повышенным содержанием соли и отбор ДНК из резервуара для сбора очищенного образца.
Таким образом, предложенные устройство и способ для выделения и очистки ДНК являются эффективными инструментами для выделения высокомолекулярной ДНК для секвенирования с длинным считыванием из эукариотических и прокариотических клеток, митохондрий, хлоропластов, крупных вирусов и т.д. Устройство или способ можно использовать, когда другие способы неэффективны, например, для одноступенчатой очистки высокомолекулярной ДНК из сложных биологических образцов, содержащих большое количество химически разнообразных примесей и только небольшое количество целевой ДНК. Кроме того, способ экономически эффективен и имеет потенциал для масштабируемости и высокой степени автоматизации.
Измерения содержания натрия проводили с помощью оптической эмиссионной спектроскопии с индуктивно-связанной плазмой (ICP-OES) с использованием прибора Agilent 720 (AgilentTechnologies, Inc., Санта-Клара, Калифорния, США). Были выбраны следующие атомные линии натрия: 568,821 нм, 588,995 нм и 589,592 нм. Среднее содержание натрия рассчитывали по двум повторности. Перед анализом все образцы разбавляли сверхчистой водой в 20 раз. Точность оценивалась с использованием стандартов образцов с добавками в соответствии с рекомендациями ISO 5725-4 (2020).
Используемый 50-кратный THE буфер для электрофореза:1 М Трис, 1 М HEPES (121,14 г трис-основание, 238,3 г HEPES (свободная кислота); растворяют в сверхчистой воде и доводят конечный объем до 1 литр). pH не нуждается в корректировке и должен быть в пределах от 8,0 до 8,1.
Последовательность реализации заявленного способа может быть представлена следующим образом.
1. Приготовить двухлитровую бутыль с 1-кратный THE электрофорезным буфером, растворив 40 мл 50-кратный THE буфера в общем объеме 2 л дистиллированной воды.
2. Приготовить 5 М раствор NaCl, растворяют 292 г NaCl в сверхчистой воде и доводят конечный объем до 1 литра.
3. Чтобы приготовить 0,8%-й основной, разделяющий гель, добавить 0,8 г сверхчистой агарозы к 100 мл буфера 1-кратным THE в стеклянном бутыле на 250 мл с завинчивающейся крышкой. Объема раствора, обозначенного далее, как раствор агарозы 1 (AS1), более чем достаточно для заливки пяти гелевых каналов.
4. Нагреть колбу с AS1 в микроволновой печи при максимально мощности (например, 800 Вт), время от времени осторожно встряхивая, пока вся агароза полностью не растворится, образуя прозрачный раствор. Следить за тем, чтобы раствор не выкипел. Хранить расплавленную агарозу при 45-55°C для дальнейшего использования.
5. Чтобы приготовить 1%-ный блокирующий гель с повышенным содержанием соли, добавляют1 г сверхчистой агарозы к 90 мл буфера 1-кратным THE в стеклянном бутыле на 250 мл с завинчивающейся крышкой. Раствор далее обозначается как раствор агарозы 2 (AS2).
6. Нагревают колбу с AS2 в микроволновой печи при максимально мощности (например, 800 Вт), время от времени осторожно встряхивая, пока вся агароза полностью не растворится, образуя прозрачный раствор. Следить за тем, чтобы раствор не выкипел. После полного растворения агарозы, добавить 10 мл раствора 5М NaCl, хорошо перемешать. Хранить расплавленную агарозу при 45-55°C для дальнейшего использования.
7. Вставляют однолуночную гребенку для электрофореза в верхнюю часть канала для формования геля. Канал с обеих сторон закрыт липкой лентой, которую удаляют перед началом электрофореза. Наливают раствор AS1 в канал, заполнив всю полость канала, и следят за тем, чтобы пузырьки воздуха не попали под зубья гребенки. Дают гелю затвердеть при комнатной температуре, образуя горизонтальный разделительный канал из агарозы шириной и глубиной в 1 см. Гель станет слегка непрозрачным после полного затвердевания.
9. Срезают чистым скальпелем участок геля длиной 1 см на расстоянии ~ 4 мм следующим за гребенкой (это будет тот гель, который будет использоваться для электрофреза и очистки ДНК), создав резервуар для сбора образцов. Акцизируют еще один участок геля длиной 1 см на ~ 3 мм, следующий за резервуаром для сбора образца ДНК, создавая форму для блокирующего геля с повышенным содержанием соли. Заливают раствор AS2 в полость формы и позволяют блокирующему гелю затвердеть, при комнатной температуре.
10. Аккуратно выделяют шпателем высокосолевой блокирующий гель, в окружении с короткими фрагментами низкосолевым электрофорезным гелем, для дальнейшего использования. Удаляют низкосолевой агарозный гель, со стороны резервуара, окружающую блокирующий солевой гель, используемую при формировании этого блока.
11. Помещают блок с подготовленными каналами в емкость для электрофореза так, чтобы гребенка была направлена к катоду. Заполняют резервуар буфером 1X THE, пока гель не будет покрыт. Аккуратно снимают гребень. Заливка геля буфером перед снятием гребенки предотвращает деформацию стенок загрузочной лунки и их прилипание друг к другу.
12. Предварительно, смешивают образец ДНК с 10-кратный загрузочным буфером (20 % (вес/объем) Ficoll 400, 100 мМ Трис-HCl (pH 8,0), 5 мМ ЭДТА, 0,01% ксиленцианол FF) до конечной концентрации 1-2 кратностью.
13. Осторожно загружают образец в лунку и подключают электроды к источнику питания.
14. Включают электрофорез при постоянном напряжение 100 В, пока следящий краситель (ксиленцианол) начнет мигрировать из геля. В качестве альтернативы отслеживают миграцию ДНК во время электрофореза и останавливают прогон, когда интересующая ДНК достигает конца геля. Следят за тем, чтобы целевая ДНК не начала мигрировать из геля.
15. После отключения питания, удаляют и промывают оставшийся рабочий буфер в канале.
16. Осторожно возвращают агарозный блокирующий гель в исходное положение в канал.
17. Помещают блок с каналами обратно в камеру для электрофореза. Заполняют резервуар 1-кратным буфером THE, чтобы уровень буфера соответствовал уровню поверхности геля.
18. Продолжают электрофорез при постоянном напряжение 100 В, дополнительностью10 - 40 мин, чтобы элюировать гель-очищенную ДНК в резервуар для сбора образцов.
19. Отключают питание, и переносят буфер с элюированной ДНК из канала для сбора с помощью пипетки в чистую микропробирку.
Сущность и промышленная применимость заявленной группы изобретений поясняются следующими примерами, ни в коей мере не уменьшая притязания по формуле изобретения во всех частных формах воплощения признаков.
Пример 1
Выделение ДНК из растительной, животной или из тканей насекомых, грибов и микроорганизмов. Протокол для непосредственной очистки общей ДНК из смеси клеток (ткани) с помощью лизирующего раствора с SDS и протеиназой К.
1. В микропробирку на 1.5 мл помещается гомогенизированный кусочек ткани растительной, животного или другого происхождения не более 100 мкг, полученного через замораживание в жидком азоте или при -80°С, и измельчением в охлажденной ступке или мешалке. Альтернативный способ гомогенизации ткани, это измельчение ткани с помощью пестика в микропробирке при комнатной температуре в присутствии лизирующего раствора.
2. В пробирку с гомогенизированной тканью добавляется 500 мкллизирующего1% раствора SDS раствора (1% SDS, 10 мМ EDTA, 50 мМ Трис-HCl, pH 8.0) и добавить до конечной концентрации 100-200 мкг/мл протеиназы К (эндопептидаза K; КФ 3.4.21.64) и хорошо перемешивается на мешалке.
3. Эту смесь инкубируют при 55°C в течение, минимум одного часа.
4. Предварительная подготовка образца для нанесения в канал. Часть образца в лизирующем буфере, в объеме 100 мкл переносится в новую микропробирку, содержащую20 мкл10-кратного буферадля нанесения в гель с красителем XyleneCyanol FF, и хорошо перемешивается на мешалке.
5. На этом этапе, солевой блок (AS2)временно удаляют из канала. Таким образом, чтобы канал был заполнен только буфером.
6. Нанести в слот для нанесения, не более 50 мкл подготовленного образца, с буфером для нанесения. Начать электрофорез при постоянном напряжении 100 В, в течение которого можно наблюдать вхождение образца ДНК с красителем XyleneCyanol FF в агарозный блок.
7. После того, как краситель XyleneCyanol FF начал свободно двигаться по каналу, электрофорез останавливают. Сливают весь буфер из канала и возвращают в канал солевой блок (AS2). Заполняют электрофорезным буфером пространство для элюции (в объеме 1 мл, до полного покрытия геля) и продолжают электрофорез при постоянном напряжении 100 В,в течение 10-20 минут. Собирают весь элюат из канала, содержащий ДНК, в микропробирку.
8. Собранный элюат будет содержать остатки красителя XyleneCyanol FF, которые не мешают последующим ферментативным реакциям. Но который также можно удалить вместе с остатками соли из раствора ДНК с помощью ультрофильтрации на колонках Amicon Ultra-2 CentrifugalFilterUnit (Millipore) или стандартной процедурой осаждением ДНК в изопропаноле.
В результате получили 2 мкг ДНК из 10 мкг ткани, что соответствует 90% суммарной ДНК в растительной ткани к массе ткани молодых проростков. Чистоту ДНК измеряли спектрофотометрически на наличие полисахаридов и полипептидов. Спектрометрические данные показали полное отсутствие в образце полипептидов и наличие остатков полисахаридов, мигрирующие в образец из агарозы. Соотношение величин поглощения нуклеиновых кислот на А260 (абсорбция при 260 нм) к величине поглощения для полисахаридов на А230 (абсорбция при 230 нм) находится на величине выше 2. Что соответствует минимальному количеству в образце остатков агарозы попавших из геля, и тем самым количество чистой ДНК превышает величину 70-80%.
Пример 2
Выделение и очистка общей ДНК из почвы.
Протокол может использоваться также для других «сложных» образцов (например, фекалии) и тканей, содержащих высокое количество полимерных молекул, пигментов, отделить которые невозможно никаким другим способом. Масштаб в данном примере взят для получения ДНК из образцов весом 1-5 грамма, но может масштабироваться на микрообъем и массу образца.
Предварительная подготовка образца
1. Образцы почвы измельчаются при -80°C, полученного через замораживание в жидком азоте или при -80°C и измельчением в охлажденной до -80°C ступке или мешалке.
2. Образец гомогенизированной почвы массой не более 5 граммов помещают в 50 мл пробирку, в который добавляется CTAB раствор (2% CTAB, 1.5 М NaCl, 10 мМ Na3EDTA, 0.1 M HEPES-кислота, рН 5.3) до конечного объема в 40 мл. Очень интенсивно перемешивается на мешалке и инкубируется в течение ночи, при 65°C.
3. Пробирки с образцами почвы центрифугируется для осветления и осаждения осадка почвы, в центрифуге для больших пробирок, при охлаждении +4°C,в течение 10 минут при 10 тысячах оборотах в минуту.
4. Осветленный надосадочный раствор разделяют на две равные порции по 20 мл и переносится в новые пробирки на 50 мл, в которые уже добавлен равный объём 100% изопропанола (20 мл), охлаждённого до -20°C, и интенсивно перемешивается на мешалке.
5. Пробирки центрифугируются при +4°C в течение 10 минут при 10 тысячах оборотах в минуту, и вся надосадочная жидкость сливается. На дне пробирки образовался коричневый осадок ДНК, содержащий компоненты почвы.
6. В пробирки с осадком добавляется 10 мл 70% этанола для промывки осадка, хорошо перемешивают на мешалке. Пробирки центрифугируются при +4°C в течение 3 минут при 10 тысячах оборотах в минуту.
7. Надосадочный раствор спирта полностью сливается, не сушат, и сразу к осадку добавляется 1 мл 1-кратного раствора TE, хорошо перемешивают и инкубируют при 65°C до полного растворения осадка ДНК. Растворы ДНК из обоих пробирок объединяются и объеденный раствор ДНК может храниться в микропробирках на 2-5 мл.
Очистка ДНК из образца почвы после осаждения и растворения.
Дальнейшие шаги по очистки образцов почвы, начинаются с шага 4 (Предварительная подготовка образца для нанесения в канал) соответствующего, по очистки ДНК из примера 1.
В результате получили 5 мкг высокоочищенной и высокомолекулярной ДНК из суммарной массы 10 мкг почвенной ДНК, полученной при осаждении изопропанолом, без очистки. Что соответствует 50% от общей массы ДНК, так как большая часть почвенной ДНК составляли фрагменты разной длины, и ДНК размером ниже 10 тысяч пар нуклеотидов не были целью очистки и были отброшены при очистке. Чистоту ДНК измеряли спектрофотометрически на наличие полисахаридов и полипептидов. Спектрометрические данные показали полное отсутствие в образце полипептидов и наличие остатков полисахаридов, мигрирующие в образец из агарозы. Соотношение величин поглощения нуклеиновых кислот на А260 (абсорбция при 260 нм) к величине поглощения для полисахаридов на А230 (абсорбция при 230 нм) находится на величине выше 2. Что соответствует минимальному количеству в образце остатков агарозы попавших из геля, и тем самым количество чистой ДНК превышает величину 75%.
Источники литературы:
1. Green, M.R. and Sambrook, J. (2012) Molecular Cloning: A Laboratory Manual. Cold Spring Harbor Laboratory Press.
2. Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. and Struhl, K. (2003) Short Protocols in Molecular Biology. John Wiley & Sons Inc.
3. Ausubel, F.M., Brent, R., Kingston, R.E., Moore, D.D., Seidman, J.G., Smith, J.A. and Struhl, K. (eds.) (1988) Current Protocols in Molecular Biology. John Wiley & Sons, Inc., New York.
4. Tan, S.C. and Yiap, B.C. (2009) DNA, RNA, and protein extraction: the past and the present. J Biomed Biotechnol, 2009, 574398.
5. Ali, N., Rampazzo, R.C.P., Costa, A.D.T. and Krieger, M.A. (2017) Current Nucleic Acid Extraction Methods and Their Implications to Point-of-Care Diagnostics. Biomed Res Int, 2017, 9306564.
6. Shapiro, D.J. (1981) Quantitative ethanol precipitation of nanogram quantities of DNA and RNA. Anal Biochem, 110, 229-231.
7. Miller, S.A., Dykes, D.D. and Polesky, H.F. (1988) A simple salting out procedure for extracting DNA from human nucleated cells. Nucleic acids research, 16, 1215.
8. Sambrook, J. and Russell, D.W. (2006) Purification of nucleic acids by extraction with phenol:chloroform. CSH Protoc, 2006.
9. McDonell, M.W., Simon, M.N. and Studier, F.W. (1977) Analysis of restriction fragments of T7 DNA and determination of molecular weights by electrophoresis in neutral and alkaline gels. J Mol Biol, 110, 119-146.
10. Chouikh, Y., Volovitch, M. and Yot, P. (1979) A simple and fast electrophoretic method for elution of nucleic acids from gels. Mol Biol Rep, 5, 237-239.
11. Saha, B.K., Strelow, S. and Schlessinger, D. (1983) Electrophoretic elution of nucleic acids from acrylamide and agarose gels. J Biochem Biophys Methods, 7, 277-284.
12. Zhen, L. and Swank, R.T. (1993) A simple and high yield method for recovering DNA from agarose gels. Biotechniques, 14, 894-898.
13. Kormanec, J., Homerova, D., Sevcikova, B. and Rezuchova, B. (2001) A method for isolation of small DNA fragments from agarose and polyacrylamide gels. Anal Biochem, 293, 138-139.
14. Lipfert, J., Doniach, S., Das, R. and Herschlag, D. (2014) Understanding nucleic acid-ion interactions. Annual review of biochemistry, 83, 813-841.
15. Stellwagen, E. and Stellwagen, N.C. (2003) Probing the electrostatic shielding of DNA with capillary electrophoresis. Biophys J, 84, 1855-1866.
16. Stellwagen, E. and Stellwagen, N.C. (2020) Electrophoretic Mobility of DNA in Solutions of High Ionic Strength. Biophys J, 118, 2783-2789.
17. Yeon-Bo, C. and Woo-Jin, N. (1995) Electroelution into an Agarose-plugged Salt Channel. Mol. Cells, 5, 243-247.
18. Zassenhaus, H.P., Butow, R.A. and Hannon, Y.P. (1982) Rapid electroelution of nucleic acids from agarose and acrylamide gels. Anal Biochem, 125, 125-130.
19. Zarzosa-Alvarez, A.L., Sandoval-Cabrera, A., Torres-Huerta, A.L. and Bermudez-Cruz, R.M. (2010) Electroeluting DNA fragments. J Vis Exp. (Прототип).
20. Strongin, A.Y., Kozlov, Y.I., Debabov, V.G., Arsatians, R.A. and Zlochevsky, M.L. (1977) A reliable technique for large-scale DNA separation. Anal Biochem, 79, 1-10.
21. Wu, R. (1971) Inhibition of polynucleotide kinase by agar, dextran sulfate and other polysaccharide sulfates. Biochemical and biophysical research communications, 43, 927-934.
22. Stellwagen, N.C., Gelfi, C. and Righetti, P.G. (1997) The free solution mobility of DNA. Biopolymers, 42, 687-703.
23. Stellwagen, N.C., Gelfi, C. and Righetti, P.G. (2000) DNA and buffers: the hidden danger of complex formation. Biopolymers, 54, 137-142.
24. Parker, W.C. and Bearn, A.G. (1963) Boric Acid-Induced Heterogeneity of Conalbumin by Starch-Gel Electrophoresis. Nature, 199, 1184-1186.
25. Zittle, C.A. (1951) Reaction of borate with substances of biological interest. Adv Enzymol Relat Subj Biochem, 12, 493-527.
26. Cann, J.R. (1966) Multiple electrophoretic zones arising from protein-buffer interaction. Biochemistry, 5, 1108-1112.
27. Woods, W.G. (1994) An introduction to boron: history, sources, uses, and chemistry. Environ Health Perspect, 102 Suppl 7, 5-11.
28. Qiagen, n.d. (2016), Maximizing PCR and RT-PCR Success, Third Edition, pp. 15.
29. Whisnant, A.R. and Gilman, S.D. (2002) Studies of reversible inhibition, irreversible inhibition, and activation of alkaline phosphatase by capillary electrophoresis. Anal Biochem, 307, 226-234.
30. Rittie, L. and Perbal, B. (2008) Enzymes used in molecular biology: a useful guide. J Cell Commun Signal, 2, 25-45.
31. Lerman, L.S. (1961) Structural considerations in the interaction of DNA and acridines. J Mol Biol, 3, 18-30.
32. Vladescu, I.D., McCauley, M.J., Nunez, M.E., Rouzina, I. and Williams, M.C. (2007) Quantifying force-dependent and zero-force DNA intercalation by single-molecule stretching. Nature methods, 4, 517-522.
33. Smith, S.B., Finzi, L. and Bustamante, C. (1992) Direct mechanical measurements of the elasticity of single DNA molecules by using magnetic beads. Science, 258, 1122-1126.
34. Hurley, L.H. (2002) DNA and its associated processes as targets for cancer therapy. Nat Rev Cancer, 2, 188-200.
35. Koster, D.A., Palle, K., Bot, E.S., Bjornsti, M.A. and Dekker, N.H. (2007) Antitumour drugs impede DNA uncoiling by topoisomerase I. Nature, 448, 213-217.
36. Nath, K., Sarosy, J.W., Hahn, J. and Di Como, C.J. (2000) Effects of ethidium bromide and SYBR Green I on different polymerase chain reaction systems. J Biochem Biophys Methods, 42, 15-29.
37. Berkenkamp, S., Kirpekar, F. and Hillenkamp, F. (1998) Infrared MALDI mass spectrometry of large nucleic acids. Science, 281, 260-262.
38. Green, M.R. and Sambrook, J. (2017) Concentrating Nucleic Acids by Extraction with Butanol. Cold Spring Harb Protoc, 2017, pdb prot093401.
39. Miller, D.N., Bryant, J.E., Madsen, E.L. and Ghiorse, W.C. (1999) Evaluation and optimization of DNA extraction and purification procedures for soil and sediment samples. Appl Environ Microbiol, 65, 4715-4724.
40. Athanasopoulou, K., Boti, M.A., Adamopoulos, P.G., Skourou, P.C. and Scorilas, A. (2021) Third-Generation Sequencing: The Spearhead towards the Radical Transformation of Modern Genomics. Life (Basel), 12.
41. Slater, G.W. (2009) DNA gel electrophoresis: the reptation model(s). Electrophoresis, 30 Suppl 1, S181-187.
42. Fangman, W.L. (1978) Separation of very large DNA molecules by gel electrophoresis. Nucleic acids research, 5, 653-665.
43. Serwer, P. (1980) Electrophoresis of duplex deoxyribonucleic acid in multiple-concentration agarose gels: fractionation of molecules with molecular weights between 2 X 10(6) and 110 X 10(6). Biochemistry, 19, 3001-3004.
44. Lopez-Canovas, L., Martinez Benitez, M.B., Herrera Isidron, J.A. and Flores Soto, E. (2019) Pulsed Field Gel Electrophoresis: Past, present, and future. Anal Biochem, 573, 17-29.
45. Патент на изобретение RU 2650865 от 17.04.2018 (Общество с ограниченной ответственностью "Магнэтик" (RU))" Набор реактивов для выделения ДНК".
46. Патент на полезную модель RU 84381 U1 от 10.07.2009 (Учреждение Российской академии наук Институт молекулярной биологии им. В.А. Энгельгардта РАН (ИМБ РАН) (RU)) "Устройство для автоматизированного выделения нуклеиновых кислот из биологических образцов".
| название | год | авторы | номер документа |
|---|---|---|---|
| Способ определения числа тандемных макросателлитных повторяющихся последовательностей на каждом из аллелей в геноме | 2020 |
|
RU2747117C1 |
| СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ НУКЛЕИНОВЫХ КИСЛОТ | 1996 |
|
RU2119954C1 |
| СПОСОБ ДЕТЕКЦИИ ПРОДУКТА ПОЛИМЕРАЗНОЙ ЦЕПНОЙ РЕАКЦИИ | 2000 |
|
RU2169771C1 |
| Способ генотипирования изолятов и штаммов калицивируса кошек посредством филогенетического анализа с применением оргинальных олигонуклеотидных праймеров для высоковариабельного участка генов ORF2-ORF3 | 2023 |
|
RU2822162C1 |
| Способ устранения ложноположительных результатов, вызванных контаминацией ДНК-полимераз фрагментами чужеродной ДНК, при индентификации генов blaTEM | 2019 |
|
RU2728317C1 |
| СПОСОБ АВТОМАТИЗИРОВАННОГО ВЫДЕЛЕНИЯ С ОДНОВРЕМЕННОЙ ОЧИСТКОЙ НУКЛЕИНОВЫХ КИСЛОТ ИЗ НЕСКОЛЬКИХ БИОЛОГИЧЕСКИХ ОБРАЗЦОВ | 2014 |
|
RU2595374C2 |
| СМЕННЫЙ МИКРОФЛЮИДНЫЙ МОДУЛЬ ДЛЯ АВТОМАТИЗИРОВАННОГО ВЫДЕЛЕНИЯ И ОЧИСТКИ НУКЛЕИНОВЫХ КИСЛОТ ИЗ БИОЛОГИЧЕСКИХ ОБРАЗЦОВ И СПОСОБ ВЫДЕЛЕНИЯ И ОЧИСТКИ НУКЛЕИНОВЫХ КИСЛОТ С ЕГО ИСПОЛЬЗОВАНИЕМ | 2008 |
|
RU2380418C1 |
| СПОСОБ ОЦЕНКИ УСТОЙЧИВОСТИ МЯГКОЙ ПШЕНИЦЫ К НАНОЧАСТИЦАМ ТЯЖЕЛЫХ МЕТАЛЛОВ | 2018 |
|
RU2688478C1 |
| МАГНИТНЫЙ СОРБЕНТ, СПОСОБ ЕГО ПОЛУЧЕНИЯ И СПОСОБ ВЫДЕЛЕНИЯ МОЛЕКУЛ НУКЛЕИНОВЫХ КИСЛОТ | 2017 |
|
RU2653130C1 |
| ТЕРМОСТАБИЛЬНАЯ ДНК-ПОЛИМЕРАЗА, СПОСОБ ЕЕ ПОЛУЧЕНИЯ, ФРАГМЕНТ ВЫДЕЛЕННОЙ ДНК, СПОСОБ АМПЛИФИКАЦИИ ДНК, СПОСОБ ВВЕДЕНИЯ МЕТКИ В ДНК И СПОСОБ ОБРАТНОЙ ТРАНСКРИПЦИИ | 1997 |
|
RU2197524C2 |
Группа изобретений относится к области биотехнологии и генетики. Описан способ очистки и выделения высокомолекулярных нуклеиновых кислот из образца. Осуществляют заполнение зоны электрофорезного канала агарозным разделяющим гелем; нанесение образца, содержащего целевую высокомолекулярную нуклеиновую кислоту, на агарозный разделяющий гель; очистку высокомолекулярной нуклеиновой кислоты методом электрофореза; остановку электрофореза после отделения высокомолекулярной нуклеиновой кислоты от загрязнений; заполнение зазора между гелями буфером для электрофореза с низкой ионной силой; возобновление электрофореза до тех пор, пока целевая нуклеиновая кислота не мигрирует в зазор между гелями; отбор очищенной высокомолекулярной нуклеиновой кислоты из зазора между гелями. Также описано устройство для очистки и выделения высокомолекулярных нуклеиновых кислот из образца, включающее: по меньшей мере один электрофорезный канал; зону электрофорезного канала для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой; зону электрофорезного канала для установки агарозного блокирующего геля с содержанием соли в переделах 0,5-1 М хлорида натрия, зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля с содержанием соли в переделах 0,5-1 М хлорида натрия; при этом зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля выполнен с возможностью заполнения буфером для электрофореза с низкой ионной силой и составляет не менее 0,5 см. Технический результат заключается в упрощении способа выделения и очистки высокомолекулярной ДНК или РНК, пригодной для секвенирования с длинным считыванием, в повышении эффективности выделения высокомолекулярной ДНК или РНК из различных сложных биологических образцов, например, растительных образцов, образцов почвы, фекалий и т.д., в улучшении качества очистки высокомолекулярных нуклеиновых кислот для секвенирования с длинным считыванием, в ускорении выделения и очистки высокомолекулярных нуклеиновых кислот, в отсутствии необходимости дополнительной очистки нуклеиновых кислот от соли, а также в обеспечении возможности одновременной обработки большого количества образцов, что позволяет масштабировать и роботизировать предлагаемые технические решения. Благодаря этому становится возможным, в частности, получать ДНК или РНК с высокой степенью чистоты от 50% до 95 % и высоким выходом от 50% до 90%. 2 н. и 11 з.п. ф-лы, 4 ил., 2 пр.
1. Способ очистки и выделения высокомолекулярных нуклеиновых кислот из образца, включающий:
- заполнение зоны электрофорезного канала агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой;
- нанесение образца, содержащего целевую высокомолекулярную нуклеиновую кислоту, на агарозный разделяющий гель, содержащий буфер для электрофореза с низкой ионной силой;
- очистку высокомолекулярной нуклеиновой кислоты методом электрофореза;
- остановку электрофореза после отделения высокомолекулярной нуклеиновой кислоты от загрязнений до того, как очищенная нуклеиновая кислота выйдет из агарозного разделяющего геля, содержащего буфер для электрофореза с низкой ионной силой;
- установку агарозного блокирующего геля с содержанием соли в переделах 0,5-1 М хлорида натрия в электрофорезный канал в направлении движения нуклеиновых кислот, после агарозного разделяющего геля, содержащего буфер для электрофореза с низкой ионной силой, в котором находится очищенная нуклеиновая кислота, с зазором между гелями не менее 0,5 см;
- заполнение зазора между гелями буфером для электрофореза с низкой ионной силой;
- возобновление электрофореза до тех пор, пока целевая нуклеиновая кислота не мигрирует в зазор между гелями;
- отбор очищенной высокомолекулярной нуклеиновой кислоты из зазора между гелями.
2. Способ по п. 1, в котором отбор очищенной нуклеиновой кислоты из зазора между гелями проводится с помощью автоматической пипетки или лабораторного робота.
3. Способ по п. 1, в котором отбор очищенной нуклеиновой кислоты из зазора между гелями проводится в микропробирку для дальнейшего использования.
4. Способ по п.1, в котором перед очисткой высокомолекулярной нуклеиновой кислоты методом электрофореза определяют продолжительность электрофореза.
5. Способ по п.1, в котором буфер для электрофореза с низкой ионной силой представляет собой 20 мМ Трис и 20 мМ HEPES.
6. Способ по п.1, в котором образец дополнительно содержит смесь биологических молекул, включающую нецелевые нуклеиновые кислоты или короткие олигонуклеотиды, или белки, или пептиды, или полисахариды, или липиды, или вторичные метаболиты, или гуминовые вещества, или пигменты, или полифенолы.
7. Устройство для очистки и выделения высокомолекулярных нуклеиновых кислот из образца способом по п.1, включающее:
- по меньшей мере один электрофорезный канал;
- зону электрофорезного канала для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой;
- зону электрофорезного канала для установки агарозного блокирующего геля с содержанием соли в переделах 0,5-1 М хлорида натрия,
- зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля с содержанием соли в переделах 0,5-1 М хлорида натрия;
- при этом зазор между зоной для заполнения агарозным разделяющим гелем, содержащим буфер для электрофореза с низкой ионной силой, и зоной для установки агарозного блокирующего геля выполнен с возможностью заполнения буфером для электрофореза с низкой ионной силой и составляет не менее 0,5 см.
8. Устройство по п.7, в котором электрофорезный канал выполнен горизонтальным или вертикальным.
9. Устройство по п.7, в котором электрофорезный канал выполнен открытым или закрытым.
10. Устройство по п.7, в котором буфер для электрофореза с низкой ионной силой представляет собой 20 мМ Трис и 20 мМ HEPES.
11. Устройство по п.7, изготовленное из пластика, прозрачного для видимого и УФ излучения.
12. Устройство по п.7, которое дополнительно включает электроды для осуществления электрофореза в электрофорезном канале.
13. Устройство по п.7, которое дополнительно включает сенсор для сбора информации о выделенных высокомолекулярных нуклеиновых кислотах, соединенный с блоком обработки информации и блоком отображения.
| ZARZOSA-ALVAREZ A.L., SANDOVAL-CABRERA A., TORRES-HUERTA A.L | |||
| AND BERMUDEZ-CRUZ R.M | |||
| Electroeluting DNA fragments | |||
| J Vis Exp., 2010 | |||
| Способ получения огнеупорных материалов | 1949 |
|
SU84381A1 |
| СПОСОБ ОБНАРУЖЕНИЯ ЦЕЛЕВОЙ НУКЛЕИНОВОЙ КИСЛОТЫ, НАБОР (ВАРИАНТЫ), ДНК ДУПЛЕКС И КОМПОЗИЦИЯ ДЛЯ ОСУЩЕСТВЛЕНИЯ СПОСОБА | 1994 |
|
RU2158765C2 |
| EP 3445850 В1, 27.10.2021 | |||
| ОСТЕРМАН Л.А | |||
| Методы исследования белков и нуклеиновых кислот | |||
| М: Наука, 1981, стр.16-42. | |||

