РОДСТВЕННЫЕ ЗАЯВКИ
По настоящей заявке испрашивается приоритет предварительной заявки на патент США №63/130,153, поданной 23 декабря 2020 г., предварительной заявки на патент США №63/185,425, поданной 7 мая 2021 г., и предварительной заявки на патент США №63/282,244, поданной 23 ноября 2021 г. Полное содержание каждой из этих заявок полностью включено в настоящее описание посредством ссылки.
ССЫЛКА НА СПИСОК ПОСЛЕДОВАТЕЛЬНОСТЕЙ
Настоящая заявка подана в электронном виде через EFS-Web и включает представленный в электронном виде список последовательностей в формате.txt. Файл.txt размером 220 КБ содержит список последовательностей под названием «PC072713_ST25_17Nov2021.txt», созданный 17 ноября 2021 г. Список последовательностей, содержащийся в этом файле.txt, является частью описания и полностью включен в настоящее описание посредством ссылки.
ОБЛАСТЬ ТЕХНИКИ, К КОТОРОЙ ОТНОСИТСЯ ИЗОБРЕТЕНИЕ
Настоящее изобретение относится к мутантным полипептидам FimH Escherichia coli и способам их применения.
УРОВЕНЬ ТЕХНИКИ
Инфекции мочевыводящих путей (ИМП, UTI) хотя бы раз в жизни поражают каждую пятую женщину и являются причиной значительного уровня заболеваемости и смертности, что ложится тяжелым бременем на системы здравоохранения. Хотя вызывать ИМП могут несколько разных бактерий, наиболее распространенной причиной (90-95% случаев) являются грамотрицательные бактерии Escherichia coli (Е.coli). Большинство вызываемых Е.coli ИМП вызываются уропатогенными Е.coli (UPEC), которые колонизируют желудочно-кишечный тракт и мигрируют из фекальной флоры в урогенитальный тракт, где прикрепляются к уроэпителиальным клеткам хозяина, создавая таким образом резервуар для восходящих инфекций мочевыводящих путей. Адгезии способствуют фимбриальные адгезины, в том числе фимбрии типа 1, которые связываются с маннозилированными гликопротеинами в эпителиальном слое или секретируются в мочу. Фимбрии типа 1 являются высококонсервативными среди клинических изолятов UPEC и кодируются кластером генов, называемым fim, которые кодируют акцессорные белки (FimC, FimD), различные структурные субъединицы (FimE, FimF, FimG) и адгезии, называемый FimH. Все характеристики инфекции ИМП на мышиных моделях, которые имитируют аспекты инфекции мочевого пузыря человека, зависят от FimH (Hannan et al. PLoS Pathog. 2010 Aug 12; 6(8): e1001042; doi: 10.1371/journal.ppat.1001042; Schwartz et al. Infect Immun. 2011 Oct; 79(10): 4250-9. doi: 10.1128/IAI.05339-11).
Низкомолекулярные ингибиторы, воздействующие на FimH путем имитации маннозилированных рецепторов, являются дополнительным подтверждением роли FimH в ИМП и демонстрируют многообещающие терапевтические свойства на животных моделях (Cusumano С.K., et al. Sci Transl Med. 2011; 3(109):109ra115. doi: 10.1126/scitranslmed.3003021). Кроме того, FimH подвергается положительной селекции в изолятах Е.coli при цистите человека (Chen SL, et al. Proc Natl Acad Sci U.S.A. 2009 Dec 29; 106(52):22439-44. doi: 10.1073/pnas.0902179106), и остатки полученные в результате положительного отбора, могут влиять на вирулентность в мышиных моделях цистита (Schwartz, D.J. et al. Proc Natl Acad Sci U.S.A. 110, 15530-15537, doi:10.1073/pnas.1315203110 (2013)).
FimH состоит из двух доменов: лектин-связывающего домена (FimHLD), ответственного за связывание с маннозилированными гликопротеинами, и пилиновый домена. Пилиновый домен служит для связывания FimH с другими структурными субъединицами пилуса, такими как FimG, посредством механизма, называемого обменом донорной цепи (Le Trong, I et al., J. Struct Biol. 2010 Dec; 172(3):380-8. doi: 10.1016/j.jsb.2010.06.002). Пилиновый домен FimH формирует неполную укладку иммуноглобулина, в результате чего образуется бороздка, которая обеспечивает сайт связывания N-концевой β-цепи FimG, образуя сильную межмолекулярную связь между FimH и FimG. Хотя FimHLD может экспрессироваться в растворимой стабильной форме, полноразмерный FimH сам по себе является нестабильным (Vetsch, М., et al. J. Mol. Biol. 322:827-840 (2002); Barnhart MM, et al., Proc. Natl Acad Sci U.S.A. (2000) Jul 5; 97(14):7709-14), за исключением случаев, когда он находится в комплексе с шапероном FimC или дополнен пептидом донорной цепи FimG в пептидной форме или находится в виде слитого белка (Barnhart MM, et al., Proc Natl Acad Sci U.S.A. (2000) Jul 5; 97(14): 7709-14; Sauer MM et al. Nat Commun. (2016) Mar 7; 7:10738; Barnhart MM et al., J Bacteriol. 2003 May; 185(9):2723-30). Разработка и экспрессия полноразмерной молекулы FimH путем связывания донорного пептида FimG с полноразмерным FimH через глицин-сериновый линкер были описаны ранее (см. международную публикацию РСТ №WO2021/084429, опубликованную 6 мая 2021 г.), а сам продукт обозначен как FimH-DSG.
FimHLD считается плохим иммуногеном с точки зрения его способности стимулировать функциональную иммуногенность. Некоторые исследования предполагают, что, хотя титры связывающих антител могут быть инициированы FimHLD с адъювантом и без него, функциональные титры нейтрализующих антител наблюдались только в присутствии адъюванта (см. международную публикацию РСТ №WO2021/084429, опубликованную 6 мая 2021 г.). Исследования показывают, что фиксирование FimH в открытой конформации с пониженным сродством к маннозидным лигандам улучшает функциональную иммуногенность (Kisiela, D.I. et al., Proc Natl Acad Sci U.S.A. 110, 19089-19094 (2013)). Соответственно, в данной области техники существует потребность в новых мутантах FimH с пониженным сродством к маннозидным лигандам и улучшенными биохимическими свойствами, которые обеспечивают повышенную функциональную иммуногенность по сравнению с FimH дикого типа.
СУЩНОСТЬ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к разработке мутантных полипептидов FimH Е.coli, которые обладают улучшенными биохимическими свойствами и иммуногенностью, композициям, содержащим такие полипептиды, и их применению. Например, в одном из аспектов настоящее изобретение относится к мутантному полипептиду FimH, который содержит по меньшей мере одну аминокислотную мутацию относительно аминокислотной последовательности полипептида FimH дикого типа, где положение мутации выбирают из группы, состоящей из: F1, Р12, G14, G15, G16, А18, Р26, V27, V28, Q32, N33, L34, V35, R60, S62, Y64, G65, L68, F71, Т86, L107, Y108, L109, V112, S113, А115, G116, V118, А119, А127, L129, Q133, F144, V154, V155, V156, Р157, Т158, V163 и V185, где положения аминокислот пронумерованы в соответствии с SEQ ID NO:59.
В еще одном аспекте представлен мутантный полипептид FimH, содержащий по меньшей мере одну мутацию, выбранную из группы, состоящей из: F1I; F1L; F1V; F1M; F1Y; F1W; Р12С; G14C; G15A; G15P; G16A; G16P; А18С; Р26С; V27A; V27C; V28C; Q32C; N33C; L34C; L34N; L34S; L34T; L34D; L34E; L34K; L34R; V35C; R60P; S62C; Y64C; G65A; L68C; F71C; Т86С; L107C; Y108C; L109C; V112C; S113C; A115V; G116C; V118C; А119С; A119N; A119S; А119Т; A119D; А119Е; А119К; A119R; А127С; L129C; Q133K; F144C; V154C; V156C; Р157С; Т158С; V163I; и V185I или любую их комбинацию. Например, мутантный полипептид FimH содержит мутации G15A и G16A. Кроме того, мутантный полипептид FimH содержит мутации Р12С и А18С. Кроме того, мутантный полипептид FimH содержит мутации G14C и F144C. Кроме того, мутантный полипептид FimH содержит мутации Р26С и V35C. Кроме того, мутантный полипептид FimH содержит мутации Р26С и V154C. Кроме того, мутантный полипептид FimH содержит мутации Р26С и V156C. Кроме того, мутантный полипептид FimH содержит мутации V27C и L34C. Кроме того, мутантный полипептид FimH содержит мутации V28C и N33C. Кроме того, мутантный полипептид FimH содержит мутации V28C и Р157С.Кроме того, мутантный полипептид FimH содержит мутации Q32C и Y108C. Кроме того, мутантный полипептид FimH содержит мутации N33C и L109C. Кроме того, мутантный полипептид FimH содержит мутации N33C и Р157С. Кроме того, мутантный полипептид FimH содержит мутации V35C и L107C. Кроме того, мутантный полипептид FimH содержит мутации V35C и L109C. Кроме того, мутантный полипептид FimH содержит мутации S62C и Т86С. Кроме того, мутантный полипептид FimH содержит мутации S62C и L129C. Кроме того, мутантный полипептид FimH содержит мутации Y64C и L68C. Кроме того, мутантный полипептид FimH содержит мутации Y64C и А127С. Кроме того, мутантный полипептид FimH содержит мутации L68C и F71C. Кроме того, мутантный полипептид FimH содержит мутации V112C и Т158С. Кроме того, мутантный полипептид FimH содержит мутации S113C и G116C. Кроме того, мутантный полипептид FimH содержит мутации S113C и Т158С. Кроме того, мутантный полипептид FimH содержит мутации V118C и V156C. Кроме того, мутантный полипептид FimH содержит мутации А119С и V155C. Кроме того, мутантный полипептид FimH содержит мутации L34N и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34S и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34T и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34D и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34E и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34K и V27A. Кроме того, мутантный полипептид FimH содержит мутации L34R и V27A. Кроме того, мутантный полипептид FimH содержит мутации A119N и V27A. Кроме того, мутантный полипептид FimH содержит мутации A119S и V27A. Кроме того, мутантный полипептид FimH содержит мутации А119Т и V27A. Кроме того, мутантный полипептид FimH содержит мутации A119D и V27A. Кроме того, мутантный полипептид FimH содержит мутации А119Е и V27A. Кроме того, мутантный полипептид FimH содержит мутации A119K и V27A. Кроме того, мутантный полипептид FimH содержит мутации A119R и V27A. Кроме того, мутантный полипептид FimH содержит мутации G15A и V27A. Кроме того, мутантный полипептид FimH содержит мутации G16A и V27A. Кроме того, мутантный полипептид FimH содержит мутации G15P и V27A. Кроме того, мутантный полипептид FimH содержит мутации G16P и V27A. Кроме того, мутантный полипептид FimH содержит мутации G15A, G16A и V27A. Кроме того, мутантный полипептид FimH содержит мутации G65A и V27A. Кроме того, мутантный полипептид FimH содержит мутации V27A и Q133K. Кроме того, мутантный полипептид FimH содержит мутации G15A, G16A, V27A и Q133K. Кроме того, например, мутантный полипептид FimH содержит последовательность любой из SEQ ID NO: 2-58 и 60-64. Кроме того, например, мутантный полипептид FimH раскрыт в настоящем описании, где полипептид является выделенным.
В другом примере настоящее изобретение относится к фармацевтической композиции, содержащей (i) мутантный полипептид FimH, раскрытый в настоящем описании, и (ii) фармацевтически приемлемый носитель.
В другом примере настоящее изобретение относится к иммуногенной композиции, содержащей мутантный полипептид FimH, раскрытый в настоящем описании. Например, иммуногенная композиция дополнительно содержит по меньшей мере один дополнительный антиген, такой как полисахарид, или гликоконъюгат, или белок. Кроме того, например, иммуногенная композиция дополнительно содержит по меньшей мере один адъювант.
В другом примере настоящее изобретение относится к молекуле нуклеиновой кислоты, содержащей нуклеотидную последовательность, кодирующую аминокислотную последовательность мутантного полипептида FimH, раскрытого в настоящем описании.
В другом примере настоящее изобретение относится к мутантному полипептиду FimH, раскрытому в настоящем описании, где указанный полипептид является иммуногенным.
Настоящее изобретение также относится к рекомбинантной клетке млекопитающего, содержащей полинуклеотид, кодирующий мутантный полипептид FimH, раскрытый в настоящем описании.
Настоящее изобретение также относится к культуре, содержащей рекомбинантную клетку, раскрытую в настоящем описании, где объем указанной культуры составляет не менее 5 литров, не менее 10 литров, не менее 20 литров, не менее 50 литров, не менее 100 литров, не менее 200 литров, не менее 500 литров, не менее 1000 литров или не менее 2000 литров.
Настоящее изобретение также относится к способу получения мутантного полипептида FimH, раскрытого в настоящем описании, включающему культивирование рекомбинантной клетки млекопитающего, раскрытой в настоящем описании, в подходящих условиях, обеспечивая таким образом экспрессию полипептида; и сбор полипептида.
В настоящем описании также представлен способ (i) индукции иммунного ответа у субъекта против внекишечной патогенной Е.coli или (ii) индукции продуцирования опсонофагоцитарных и/или нейтрализующих антител у субъекта, специфичных к внекишечной патогенной Е.coli, где способ включает введение субъекту эффективного количества композиции, раскрытой в настоящем описании. В одном из примеров субъект подвержен риску развития инфекции мочевыводящих путей. В другом примере субъект подвержен риску развития бактериемии. В другом примере субъект подвержен риску развития сепсиса. В другом примере субъект подвержен риску развития болезни Крона.
Настоящее изобретение также относится к способу индукции иммунного ответа против Е.coli у млекопитающего, включающему введение млекопитающему эффективного количества композиции, раскрытой в настоящем описании. В одном из примеров иммунный ответ включает опсонофагоцитарные и/или нейтрализующие антитела к Е.coli. В другом примере иммунный ответ защищает млекопитающего от инфекции Е.coli.
Настоящее изобретение также относится к способу профилактики, лечения или облегчения бактериальной инфекции, заболевания или состояния у субъекта, включающему введение субъекту иммунологически эффективного количества композиции, раскрытой в настоящем описании.
КРАТКОЕ ОПИСАНИЕ РИСУНКОВ
На фиг.1А-1В показаны спектры кругового дихроизма мутантов FimHLD и FimH-DSG. На фиг.1А показаны спектры кругового дихроизма в ближней УФ области, и на фиг.1В показаны спектры кругового дихроизма в дальней УФ области.
На фиг.2 показана относительная иммуногенность мутантов FimHLD в анализе нейтрализации связывания дрожжевого маннана в присутствии PD3.
На фиг.3А-3В показана иммуногенность мутантов FimHLD и FimH-DSG в анализе нейтрализации связывания дрожжевого маннана в присутствии PD2 (фиг.3А) и PD3 (фиг.3В).
Фиг. 4 представляет собой диаграмму, показывающую основные этапы очистки, используемые для выделения His-меченных FimH-DsG дикого типа и мутантных форм.
На фиг.5А-5В показан профиль очистки FimH-DSG WT.
Фиг. 5А представляет собой профиль элюирования FimH-DSG WT на колонке с SP-сефароза, и фиг.5В представляет собой SDS-PAGE анализ элюированных фракций.
На фиг.6А-6В показан профиль очистки мутанта FimH-DSG G15A G16A V27A.
На фиг.6А показан профиль элюирования мутанта FimH-DSG G15A G16A V27A на колонке с SP-сефарозой, и на фиг.6В показан SDS-PAGE анализ элюированных фракций.
На фиг.7А-7В показана аналитическая SEC белков FimH-DSG. Аналитическая SEC FimH-DSG G15A G16A V27A показана на фиг.7А, а FimH-DSG WT дикого типа показана на фиг.7В.
На фиг.8 показано схематическое изображение механизма образования HMW комплекса FimH.
На фиг.9 показан график иммунизации нечеловекообразных приматов и последующее заражение, описанное в примере 21 настоящего описания.
На фиг.10 показано повышение уровня серотип-специфических антител к О-антигену после вакцинации NHP мутантом FimH-DSG G15A G16A V27A+4-валентными О-антигенами Е.coli (O1a, O2, О6 и O25b). Условные обозначения: Плацебо (круги); FimH-DSG G15A G16A V27A (квадраты); FimH-DSG G15A G16A V27A+4-валентный O-антиген (треугольники).
На фиг.11А-11В показано, что иммунизация FimH-DSG G15A G16A V27A+/-4plex О-антигенами вызывает сильные функциональные ответы анти-FimH антител.
На фиг.11А показаны результаты прямого анализа анти-FiinH IgG в системе Luminex, и на фиг.11В показаны результаты анализа ингибирования связывания Е.coli. В контексте настоящего описания термин «4plex» имеет такое же значение как «4-валентный», которые являются взаимозаменяемыми.
На фиг.12 показано, что бактериурия снижается у вакцинированных нечеловекообразных приматов (NHP) после заражения, как описано в примере 21. Условные обозначения: плацебо (круги); FimH-DSG G15A G16A V27A (квадраты); FimH-DSG G15A G16A V27A+4-валентный О-антиген (треугольники); *р≤0,007 относительно группы плацебо.
На фиг.13А-13С показано, что биомаркеры инфекции снижаются у вакцинированных NHP после заражения трех групп: плацебо, только FimH-DSG G15A G16A V27A и FimH-DSG G15A G16A V27A+4-валентные О-антигены, как описано в примере 21. На фиг.13А показано количественное определение МРО в моче, на фиг.13В показано количественное определение IL-8 в моче, а на фиг.13С показан процент животных с повышенным содержанием PMN в отложениях мочи. Условные обозначения: плацебо (круги); FimH-DSG G15A G16A V27A (квадраты); FimH-DSG G15A G16A V27A+4-валентный О-антиген (треугольники).
ИДЕНТИФИКАТОРЫ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
SEQ ID NO: 1 представляет аминокислотную последовательность FimH-LD Е.coli дикого типа (FimHLD_WT).
SEQ ID NO: 2 представляет аминокислотную последовательность мутантного FimHLD_G65A_V27A Е. coll.
SEQ ID NO: 3 представляет аминокислотную последовательность мутантного FimHLD_F1I Е.coli.
SEQ ID NO: 4 представляет аминокислотную последовательность мутантного FimHLD_F1L Е.coli.
SEQ ID NO: 5 представляет аминокислотную последовательность мутантного FimHLD_F1V Е.coli.
SEQ ID NO: 6 представляет аминокислотную последовательность мутантного FimHLD_F1M Е.coli.
SEQ ID NO: 7 представляет аминокислотную последовательность мутантного FimHLD_F1Y Е.coli.
SEQ ID NO: 8 представляет аминокислотную последовательность мутантного FimHLD_F1W Е.coli.
SEQ ID NO: 9 представляет аминокислотную последовательность мутантного FimHLD_Q133K Е.coli.
SEQ ID NO: 10 представляет аминокислотную последовательность мутантного FimHLD_G15A Е.coli.
SEQ ID NO: 11 представляет аминокислотную последовательность мутантного FimHLD_G15P Е.coli.
SEQ ID NO: 12 представляет аминокислотную последовательность мутантного FimHLD_G16A Е.coli.
SEQ ID NO: 13 представляет аминокислотную последовательность мутантного FimHLD_G16P Е.coli.
SEQ ID NO: 14 представляет аминокислотную последовательность мутантного FimHLD_G15A_G16A Е.coli.
SEQ ID NO: 15 представляет аминокислотную последовательность мутантного FimHLD_R60P Е.coli.
SEQ ID NO: 16 представляет аминокислотную последовательность мутантного FimHLD_G65A Е.coli.
SEQ ID NO: 17 представляет аминокислотную последовательность мутантного FimHLD_P12C_А18С Е.coli.
SEQ ID NO: 18 представляет аминокислотную последовательность мутантного FimHLD_G14C_F144C Е.coli.
SEQ ID NO: 19 представляет аминокислотную последовательность мутантного FimHLD_P26C_V35C Е.coli.
SEQ ID NO: 20 представляет аминокислотную последовательность мутантного FimHLD_P26C_V154C Е.coli.
SEQ ID NO: 21 представляет аминокислотную последовательность мутантного FimHLD_P26C_V156C Е.coli.
SEQ ID NO: 22 представляет аминокислотную последовательность мутантного FimHLD_V27C_L34C Е.coli.
SEQ ID NO: 23 представляет аминокислотную последовательность мутантного FimHLD_V28C_N33C Е.coli.
SEQ ID NO: 24 представляет аминокислотную последовательность мутантного FimHLD_V28C_P157C Е.coli.
SEQ ID NO: 25 представляет аминокислотную последовательность мутантного FimHLD_Q32C_Y108C Е.coli.
SEQ ID NO: 26 представляет аминокислотную последовательность мутантного FimHLD_N33C_L109C Е.coli.
SEQ ID NO: 27 представляет аминокислотную последовательность мутантного FimHLD_N33C_P157C Е.coli.
SEQ ID NO: 28 представляет аминокислотную последовательность мутантного FimHLD_V35C_L107C Е.coli.
SEQ ID NO: 29 представляет аминокислотную последовательность мутантного FimHLD_V35C_L109C Е.coli.
SEQ ID NO: 30 представляет аминокислотную последовательность мутантного FimHLD_S62C_T86C Е.coli.
SEQ ID NO: 31 представляет аминокислотную последовательность мутантного FimHLD_S62C_L129C Е.coli.
SEQ ID NO: 32 представляет аминокислотную последовательность мутантного FimHLD_Y64C_L68C Е.coli.
SEQ ID NO: 33 представляет аминокислотную последовательность мутантного FimHLD_Y64C_А127С Е.coli.
SEQ ID NO: 34 представляет аминокислотную последовательность мутантного FimHLD_L68C_F71C Е.coli.
SEQ ID NO: 35 представляет аминокислотную последовательность мутантного FimHLD_V112C_Т158С Е.coli.
SEQ ID NO: 36 представляет аминокислотную последовательность мутантного FimHLD_S113C_G116C Е.coli.
SEQ ID NO: 37 представляет аминокислотную последовательность мутантного FimHLD_S113C_Т158С Е.coli.
SEQ ID NO: 38 представляет аминокислотную последовательность мутантного FimHLD_V118C_V156C Е.coli.
SEQ ID NO: 39 представляет аминокислотную последовательность мутантного FimHLD_А119С_V155C Е.coli.
SEQ ID NO: 40 представляет аминокислотную последовательность мутантного FimHLD_L34N_V27A Е.coli.
SEQ ID NO: 41 представляет аминокислотную последовательность мутантного FimHLD_L34S_V27A Е.coli.
SEQ ID NO: 42 представляет аминокислотную последовательность мутантного FimHLD_L34T_V27A Е.coli.
SEQ ID NO: 43 представляет аминокислотную последовательность мутантного FimHLD_A119N_V27A Е.coli.
SEQ ID NO: 44 представляет аминокислотную последовательность мутантного FimHLD_A119S_V27A Е.coli.
SEQ ID NO: 45 представляет аминокислотную последовательность мутантного FimHLD_А119Т_V27A Е.coli.
SEQ ID NO: 46 представляет аминокислотную последовательность мутантного FimH-DSG_A115V Е.coli.
SEQ ID NO: 47 представляет аминокислотную последовательность мутантного FimH-DSG_V163I Е.coli.
SEQ ID NO: 48 представляет аминокислотную последовательность мутантного FimH-DSG_V185I Е.coli.
SEQ ID NO: 49 представляет аминокислотную последовательность мутантного FimH-DSG_DSG_V3I Е.coli.
SEQ ID NO: 50 представляет аминокислотную последовательность мутантного FimHLD_G15A_V27A Е.coli.
SEQ ID NO: 51 представляет аминокислотную последовательность мутантного FimHLD_G16A_V27A Е.coli.
SEQ ID NO: 52 представляет аминокислотную последовательность мутантного FimHLD_G15P_V27A Е.coli.
SEQ ID NO: 53 представляет аминокислотную последовательность мутантного FimHLD_G16P_V27A Е.coli.
SEQ ID NO: 54 представляет аминокислотную последовательность мутантного FimHLD_G15A_G16A_V27A Е.coli.
SEQ ID NO: 55 представляет аминокислотную последовательность мутантного FimHLD_V27A_R60P Е.coli.
SEQ ID NO: 56 представляет аминокислотную последовательность мутантного FimHLD_G65A_V27A Е.coli.
SEQ ID NO: 57 представляет аминокислотную последовательность мутантного FimHLD_V27A_Q133K Е.coli.
SEQ ID NO: 58 представляет аминокислотную последовательность мутантного FimHLD_G15A_G16A_V27A_Q133K Е.coli.
SEQ ID NO: 59 представляет аминокислотную последовательность полноразмерного FimH Е.coli дикого типа, включая донорную цепь пептида FimG соединенную через линкер (FimH-DSG_WT).
SEQ ID NO: 60 представляет аминокислотную последовательность мутантного FimH-DSG_V27A Е.coli.
SEQ ID NO: 61 представляет аминокислотную последовательность мутантного FimH-DSG_G15A_V27A Е.coli.
SEQ ID NO: 62 представляет аминокислотную последовательность DSG_G15A_G16A_V27A мутантного FimH Е.coli.
SEQ ID NO: 63 представляет аминокислотную последовательность DSG_V27A_Q133K мутантного FimH Е.coli.
SEQ ID NO: 64 представляет аминокислотную последовательность DSG_G15A_G16A_V27A_Q133K мутантного FimH Е.coli.
SEQ ID NO: 65 представляет аминокислотную последовательность сигнального пептида каппа-цепи Ig мыши.
SEQ ID NO: 66-108 представляют аминокислотные последовательности и последовательности нуклеиновых кислот для относящегося к наноструктуре полипептида или его фрагмента.
SEQ ID NO: 109 - праймер для ПЦР.
SEQ ID NO: 110 - праймер для ПЦР.
SEQ ID NO: 111 - зонд для ПЦР.
SEQ ID NO: 112 представляет аминокислотную последовательность O25b 2401 WzzB.
SEQ ID NO: 113 представляет аминокислотную последовательность 025а:K5:Н1 WzzB.
SEQ ID NO: 114 представляет аминокислотную последовательность 025а ETEC АТСС WzzB.
SEQ ID NO: 115 представляет аминокислотную последовательность K12 W3110 WzzB.
SEQ ID NO: 116 представляет аминокислотную последовательность LT2 WzzB Salmonella.
SEQ ID NO: 117 представляет аминокислотную последовательность O25b 2401 FepE.
SEQ ID NO: 118 представляет аминокислотную последовательность O25a:K5:H1 FepE.
SEQ ID NO: 119 представляет аминокислотную последовательность O25a ETEC АТСС FepE.
SEQ ID NO: 120 представляет аминокислотную последовательность O157 FepE.
SEQ ID NO: 121 представляет аминокислотную последовательность LT2 FepE Salmonella.
SEQ ID NO: 122 представляет последовательность праймеров для LT2wzzB_S.
SEQ ID NO: 123 представляет последовательность праймеров для LT2wzzB_AS.
SEQ ID NO: 124 представляет последовательность праймеров для 025bFepE_S.
SEQ ID NO: 125 представляет последовательность праймеров для 025bFepE_A.
SEQ ID NO: 126 представляет последовательность праймеров для P1_S wzzB.
SEQ ID NO: 127 представляет последовательность праймеров для P2_AS wzzB.
SEQ ID NO: 128 представляет последовательность праймеров для Р3_S wzzB.
SEQ ID NO: 129 представляет последовательность праймеров для P4_AS wzzB.
SEQ ID NO: 130 представляет последовательность праймеров для FepE_S 0157.
SEQ ID NO: 131 представляет последовательность праймеров для FepE_AS 0157.
SEQ ID NO: 132 представляет последовательность праймеров для pBAD33_adaptor_S.
SEQ ID NO: 133 представляет последовательность праймеров для pBAD33 adaptor AS.
SEQ ID NO: 134 представляет последовательность праймеров для JUMPSTART_r.
SEQ ID NO: 135 представляет последовательность праймеров для gnd_f.
SEQ ID NO: 136 представляет аминокислотную последовательность сигнального пептида большой субъединицы р51 FcRn рецептора человеческого IgG.
SEQ ID NO: 137 представляет аминокислотную последовательность человеческого сигнального пептида белка IL-10.
SEQ ID NO: 138 представляет аминокислотную последовательность сигнального пептида слияния гликопротеина F0 респираторно-синцитиального вируса А человека (штамм А2).
SEQ ID NO: 139 представляет аминокислотную последовательность сигнального пептида гемагглютинина вируса гриппа А.
SEQ ID NO: 140-147 представляют сигнальные последовательности Р4.1 (DTU Bioinformatics), полученные из различных видов, используемые для предсказания сигнальных пептидов.
ПОДРОБНОЕ ОПИСАНИЕ ИЗОБРЕТЕНИЯ
Настоящее изобретение относится к мутантным полипептидам Е. FimH coli (мутантам), композициям, содержащим мутанты FimH, способам получения и очистки мутантов FimH, нуклеиновым кислотам, которые кодируют мутанты FimH, клеткам-хозяевам, которые содержат такие нуклеиновые кислоты, и способам применения композиций, содержащих мутанты FimH.
В контексте настоящего описания указание диапазонов значений приводится только в качестве сокращенного способа ссылки на каждое отдельное значение, попадающее в указанный диапазон. Если в описании не указано иное, каждое отдельное значение включено в описание, как если бы оно было приведено отдельно. Все способы, раскрытые в настоящем описании, могут быть выполнены в любом подходящем порядке, если в настоящем описании не указано иное, или если иное в явном виде не противоречит контексту. Использование любого и всех примеров или иллюстративного языка (например, «такой как»), предусмотренное в настоящем описании, предназначено только для дополнительной иллюстрации изобретения и не налагает ограничения на объем формулы изобретения. Никакую формулировку в описании не следует рассматривать как указывающую на любой незаявленный элемент, существенный для осуществления изобретения.
По ходу всего изложения текста настоящего описания изобретения цитируются несколько документов. Каждый из документов, процитированных в настоящем описании (включая все патенты, заявки на получение патента, научные публикации, спецификации производителя, инструкции и т.д.), как выше, так и ниже, включены в настоящее описание во всей своей полноте посредством ссылки. Ничто в настоящем описании не следует рассматривать как допущение того, что подобные публикации составляют предшествующий уровень техники в отношении прилагаемой формулы изобретения.
Определения
В контексте настоящего описания термин «примерно» означает приблизительно или почти, а в контексте числового значения или диапазона, представленного в одном из приведенных в описании вариантов осуществления, означает ±20%, ±10%, ±5% или ±3% от указанного или заявленного числового значения или диапазона.
Термины «а», «an» и «the» и аналогичные ссылки, используемые в контексте описания изобретения (особенно в контексте формулы изобретения), следует рассматривать как охватывающие как единственное, так и множественное число, если иное не указано в настоящем описании или если иное в явном виде не противоречит контексту.
«Фрагмент» по отношению к аминокислотной последовательности (пептиду или белку) относится к части аминокислотной последовательности, т.е. последовательности, которая представляет собой аминокислотную последовательность, укороченную на N-конце и/или С-конце. Фрагмент, укороченный на С-конце (N-концевой фрагмент), можно получить, например, путем трансляции укороченной открытой рамки считывания, в которой отсутствует 3'-конец открытой рамки считывания. Фрагмент, укороченный на N-конце (С-концевой фрагмент), можно получить, например, путем трансляции укороченной открытой рамки считывания, в которой отсутствует 5'-конец открытой рамки считывания, при условии, что укороченная открытая рамка считывания содержит стартовый кодон, который служит для инициации трансляции. Фрагмент аминокислотной последовательности содержит, например, по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% аминокислотных остатков аминокислотной последовательности. Фрагмент аминокислотной последовательности предпочтительно содержит по меньшей мере 6, в частности, по меньшей мере 8, по меньшей мере 12, по меньшей мере 15, по меньшей мере 20, по меньшей мере 30, по меньшей мере 50 или по меньшей мере 100 последовательных аминокислот из аминокислотной последовательности.
В контексте настоящего описания термин «дикий тип» или «WT», или «нативный» относится к встречающейся в природе аминокислотной последовательности, включая аллельные варианты. Аминокислотная последовательность, пептид или белок дикого типа имеет аминокислотную последовательность, которая не была преднамеренно модифицирована.
В контексте настоящего описания термин «варианты» аминокислотной последовательности (пептида, белка или полипептида) или «мутанты», или ссылка на «мутантный» полипептид содержит варианты/мутанты со вставкой аминокислот, варианты/мутанты с добавлением аминокислот, варианты/мутанты с делецией аминокислот и/или варианты/мутанты с аминокислотной заменой. Термин «вариант» или «мутант» включает все мутанты, варианты сплайсинга, посттрансляционно модифицированные варианты, конформации, изоформы, аллельные варианты, варианты видов и гомологи видов, в частности такие, которые встречаются в природе. Термин «вариант» или «мутант» включает, в частности, фрагменты аминокислотной последовательности.
Варианты со вставкой аминокислот содержат вставки одной, двух или более аминокислот в конкретную аминокислотную последовательность. В случае вариантов аминокислотной последовательности, имеющих вставку, один или более аминокислотных остатков вводят в конкретный участок аминокислотной последовательности, хотя также возможна случайная вставка с соответствующим скринингом полученного продукта. Варианты с добавлением аминокислот содержат амино- и/или карбокси-концевые слияния одной или более аминокислот, например 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот. Варианты с делецией аминокислот характеризуются удалением одной или более аминокислот из последовательности, например удалением 1, 2, 3, 5, 10, 20, 30, 50 или более аминокислот. Делеции могут находиться в любом положении белка. Варианты с делецией аминокислот, которые содержат делецию на N-конце и/или С-конце белка, также называют укороченными вариантами по N-концу и/или С-концу. Варианты с аминокислотной заменой характеризуются удалением по меньшей мере одного остатка в последовательности и вставкой другого остатка на его место. Предпочтение отдается модификациям, находящимся в положениях аминокислотной последовательности, которые не являются консервативными, между гомологичными белками или пептидами и/или замене аминокислот другими, обладающими сходными свойствами. Предпочтительно аминокислотные замены в пептидных и белковых вариантах представляют собой консервативные аминокислотные замены, т.е. замены аналогично заряженных или незаряженных аминокислот. Консервативная аминокислотная замена включает замену одной аминокислоты из семейства аминокислот, родственных по своим боковым цепям. Встречающиеся в природе аминокислоты обычно подразделяют на четыре семейства: кислые (аспартат, глутамат), основные (лизин, аргинин, гистидин), неполярные (аланин, валин, лейцин, изолейцин, пролин, фенилаланин, метионин, триптофан) и незаряженные полярные (глицин, аспарагин, глутамин, цистеин, серии, треонин, тирозин) аминокислоты. Фенилаланин, триптофан и тирозин иногда вместе классифицируют как ароматические аминокислоты. В одном из вариантов осуществления консервативные аминокислотные замены включают замены в пределах следующих групп:
глицин, аланин;
валин, изолейцин, лейцин;
аспарагиновая кислота, глутаминовая кислота;
аспарагин, глутамин;
серин, треонин;
лизин, аргинин; и
фенилаланин, тирозин.
Предпочтительно степень сходства, предпочтительно идентичности, между данной аминокислотной последовательностью и аминокислотной последовательностью, которая является вариантом указанной данной аминокислотной последовательности, будет составлять по меньшей мере примерно 60%, 70%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%. Степень сходства или идентичности приводится предпочтительно для аминокислотной области, длина которой составляет по меньшей мере примерно 10%, по меньшей мере примерно 20%, по меньшей мере примерно 30%, по меньшей мере примерно 40%, по меньшей мере примерно 50%, по меньшей мере примерно 60%, по меньшей мере примерно 70%, по меньшей мере примерно 80%, по меньшей мере примерно 90% или примерно 100% длины эталонной аминокислотной последовательности. Например, если эталонная аминокислотная последовательность состоит из 200 аминокислот, то в некоторых вариантах осуществления непрерывных аминокислот степень сходства или идентичности приводится предпочтительно для по меньшей мере примерно 20, по меньшей мере примерно 40, по меньшей мере примерно 60, по меньшей мере примерно 80, по меньшей мере примерно 100, по меньшей мере примерно 120, по меньшей мере примерно 140, по меньшей мере примерно 160, по меньшей мере примерно 180 или примерно 200 аминокислот. В некоторых вариантах осуществления степень сходства или идентичности указана относительно полной длины эталонной аминокислотной последовательности. Выравнивание для определения сходства последовательностей, предпочтительно идентичности последовательностей, может быть выполнено с помощью известных в данной области инструментов, предпочтительно с помощью наилучшего способа выравнивания последовательностей, например Align, используя стандартные настройки, предпочтительно EMBOSS::needle, Matrix: Blosum62, штраф за открытие пробела 10,0, штраф за удлинение пробела 0,5.
В контексте настоящего описания термин «сходство последовательностей» указывает на процент аминокислот, которые либо являются идентичными, либо представляют собой консервативные аминокислотные замены. «Идентичность последовательностей» между двумя аминокислотными последовательностями указывает процент аминокислот, идентичных между последовательностями. «Идентичность последовательностей» между двумя последовательностями нуклеиновых кислот указывает процент нуклеотидов, идентичных между последовательностями.
Термины «идентичный на %», «% идентичности» или аналогичные термины предназначены для обозначения, в частности процента нуклеотидов или аминокислот, которые идентичны при оптимальном выравнивании сравниваемых последовательностей. Упомянутый процент является чисто статистическим, и различия между двумя последовательностями могут быть распределены, необязательно, случайным образом по всей длине сравниваемых последовательностей. Сравнение двух последовательностей обычно выполняют путем сравнения последовательностей, после оптимального выравнивания, относительно сегмента или «окна сравнения», чтобы идентифицировать локальные области соответствующих последовательностей. Оптимальное выравнивание для сравнения может быть выполнено вручную или с помощью алгоритма локальной гомологии Смита-Ватермана (Smith and Waterman), 1981, Ads App.Math. 2, 482, с помощью алгоритма локальной гомологии Нидлмана-Вунша (Neddleman and Wunsch), 1970, J. Mol. Biol. 48, 443, с помощью алгоритма поиска подобия Пирсона и Липмана (Pearson and Lipman), 1988, Proc. Natl Acad. Sci. USA 88, 2444, или с помощью компьютерных программ, в которых используются указанные алгоритмы (GAP, BESTFIT, FASTA, BLAST Р, BLAST N и TFASTA в Wisconsin Genetics Software Package, Genetics Computer Group, 575 Science Drive, Madison, WI). В некоторых вариантах осуществления процент идентичности двух последовательностей определяют с помощью алгоритма BLASTN или BLASTP, доступного на веб-сайте Национального центра биотехнологической информации США (NCBI) (например, по адресу blast.ncbi.nlm.nih.gov/Blast.cgi?PAGE_TYPE=BlastSearch&BLAST_SPEC=blast2seg&LINK_LOC=align2seg). В некоторых вариантах осуществления параметры, используемые для алгоритма BLASTN на веб-сайте NCBI, включают: (i) ожидаемый порог: 10; (ii) размер слова: 28; (iii) максимальное количество совпадений в диапазоне запроса: 0; (iv) оценка совпадения/несоответствия: 1, -2; (v) штрафы за пропуск: Линейный; и (vi) используется фильтр для областей низкой сложности. В некоторых вариантах осуществления параметры, используемые для алгоритма BLASTP на веб-сайте NCBI, включают: (i) ожидаемый порог: 10; (ii) размер слова: 3; (iii) максимальное количество совпадений в диапазоне запроса: 0; (iv) матрица: BLOSUM62; (v) штраф за удлинение пробела: 11, удлинение:11; и (vi) условная композиционная поправка матрицы замен.
Процент идентичности получают путем определения количества идентичных положений, в которых соответствуют сравниваемые последовательности, путем деления этого количества на количество сравниваемых положений (например, количество положений в эталонной последовательности) и умножения этого результата на 100.
В некоторых вариантах осуществления указана степень сходства или идентичности для области, длина которой составляет по меньшей мере примерно 50%, по меньшей мере примерно 60%, по меньшей мере примерно 70%, по меньшей мере примерно 80%, по меньшей мере примерно 90% или примерно 100% длины эталонной последовательности. Например, если эталонная последовательность нуклеиновой кислоты состоит из 200 нуклеотидов, то в некоторых вариантах осуществления непрерывных нуклеотидов степень идентичности приводится для по меньшей мере примерно 100, по меньшей мере примерно 120, по меньшей мере примерно 140, по меньшей мере примерно 160, по меньшей мере примерно 180 или примерно 200 нуклеотидов. В некоторых вариантах осуществления степень сходства или идентичности указана для полной длины эталонной последовательности.
Гомологичные аминокислотные последовательности согласно изобретению имеют по меньшей мере 40%, в частности по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, по меньшей мере 90% и предпочтительно по меньшей мере 95%, по меньшей мере 98 или по меньшей мере 99% идентичных аминокислотных остатков. Варианты/мутанты аминокислотной последовательности, представленные в настоящем описании, могут быть легко получены специалистом в данной области техники, например, путем манипуляций с рекомбинантной ДНК. Манипуляции с ДНК последовательностями для получения пептидов или белков, имеющих замены, добавления, вставки или делеции, подробно описаны, например, в Sambrook et al. (1989). Кроме того, пептиды и варианты аминокислот, представленные в описании, могут быть легко получены известными методами синтеза пептидов, такими как, например твердофазный синтез, и аналогичными методами.
В одном из аспектов фрагмент или вариант/мутант аминокислотной последовательности (пептид или белок) предпочтительно представляет собой «функциональный фрагмент» или «функциональный вариант». Термин «функциональный фрагмент» или «функциональный вариант/мутант» аминокислотной последовательности относится к любому фрагменту или варианту/мутанту, проявляющему одно или более функциональных свойств, идентичных или сходных со свойствами аминокислотной последовательности, из которой он получен, т.е. он является функционально эквивалентным. В отношении антигенов или антигенных последовательностей одной конкретной функцией является одна или более иммуногенных активностей, проявляемых аминокислотной последовательностью, из которой получен фрагмент или вариант. Термин «функциональный фрагмент» или «функциональный вариант/мутант», используемый в настоящем описании, в частности, относится к вариантной/мутантной молекуле или последовательности, которая содержит аминокислотную последовательность с измененной одной или несколькими аминокислотами по сравнению с аминокислотной последовательностью родительской молекулы или последовательности и которая все еще способна выполнять одну или более функций родительской молекулы или последовательности, например, индуцировать иммунный ответ. В одном из аспектов модификации в аминокислотной последовательности родительской молекулы или последовательности не оказывают существенного влияния на характеристики молекулы или последовательности и не изменяют их. В различных вариантах осуществления функция функционального фрагмента или функционального варианта может быть снижена, но все еще в значительной степени присутствовать, например, иммуногенность функционального варианта может составлять по меньшей мере 50%, по меньшей мере 60%, по меньшей мере 70%, по меньшей мере 80%, или по меньшей мере 90% родительской молекулы или последовательности. Однако в других вариантах осуществления иммуногенность функционального фрагмента или функционального варианта может быть увеличена по сравнению с родительской молекулой или последовательностью.
В контексте настоящего описания термин «выделенный» означает измененный или удаленный из естественного состояния. Например, нуклеиновая кислота или пептид, естественным образом присутствующие в живом организме, не являются «выделенными», а та же самая нуклеиновая кислота или пептид, частично или полностью отделенные от материалов, сосуществующих с ним в его естественном состоянии, являются "выделенными". Выделенная нуклеиновая кислота или белок могут существовать в практически очищенной форме или могут существовать в ненативной среде, такой как, например, клетка-хозяин.
I. Полипептиды FimH Е.coli
Фимбриальные адгезины, включая фимбрии типа 1, связываются с маннозилированными гликопротеинами в эпителиальном слое или секретируются в мочу. Фимбрии типа 1 являются высококонсервативными среди клинических изолятов UPEC и кодируются кластером генов, называемым fim, которые кодируют акцессорные белки (FimC, FimD), различные структурные субъединицы (FimE, FimF, FimG) и адгезии, называемый FimH. FimH состоит из двух доменов: лектинсвязывающего домена (FimHLD), ответственного за связывание с маннозилированными гликопротеинами, и пилинового домена. Пилиновый домен служит для связывания FimH с другими структурными субъединицами пилуса, такими как FimG, посредством механизма, называемого обменом донорной цепи. Пилиновый домен FimH образует неполную укладку иммуноглобулина, в результате чего образуется бороздка, которая обеспечивает сайт связывания N-концевой β-цепи FimG, образуя сильную межмолекулярную связь между FimH и FimG. Хотя FimHLD может экспрессироваться в растворимой стабильной форме, полноразмерный FimH сам по себе является нестабильным, если только он не находится в комплексе с шапероном FimC или не дополнен пептидом донорной цепи FimG в пептидной форме или не находится в виде слитого белка. Соответственно, экспрессия стабильной полноразмерной молекулы FimH возможна путем связывания донорного пептида FimG с С-концом полноразмерного FimH через глицин-сериновый линкер с получением продукта, обозначенного как FimH-DSG.
В одном из аспектов настоящее изобретение относится к мутантным полипептидам FimH, таким как показанные в таблице 1. Такие мутанты содержат мутации в аминокислотной последовательности относительно аминокислотной последовательности соответствующего полипептида FimH дикого типа (WT). В некоторых аспектах такие мутанты являются иммуногенными в отношении белка FimH дикого типа или бактерий, экспрессирующих полипептид FimH дикого типа. В некоторых аспектах мутанты FimH обладают определенными полезными характеристиками, такими как повышенные иммуногенные свойства по сравнению с соответствующим полипептидом FimH дикого типа.
В контексте настоящего описания термин «полипептид FimH» относится к любому домену полноразмерного полипептида FimH Е.coli дикого типа, любой комбинации доменов полноразмерного полипептида FimH Е.coli дикого типа или к полноразмерному полипептиду FimH Е.coli дикого типа или любому его фрагменту. Например, в одном из вариантов осуществления настоящее изобретение относится к мутантному полипептиду FimH, который представляет собой мутантный полипептид FimHLD или полипептид FimH-DSG. Настоящее изобретение относится к новым мутантам FimHLD и FimH-DSG со сниженным сродством к маннозидным лигандам (подтвержденным биохимическим и биофизическим анализами), что улучшает функциональную иммуногенность, и описывает оценку нейтрализующих ответов этих мутантов по сравнению с FimHLD дикого типа.
Введенные аминокислотные мутации в мутантных полипептидах FimH могут включать аминокислотные замены, делеции или добавления. В некоторых аспектах единственными мутациями в аминокислотной последовательности мутантов полипептида FimH являются аминокислотные замены относительно белка FimH дикого типа.









Аминокислотная последовательность полипептида FimH дикого типа хорошо известна в данной области. Например, аминокислотная последовательность домена FimHLD представлена в настоящем описании как SEQ ID NO:1. Полноразмерный полипептид FimH дикого типа, включая донорный пептид FimG, связанный с С-концом полноразмерного FimH посредством глицин-серинового линкера, представлен в настоящем описании как SEQ ID NO:59. Последовательности нуклеиновых кислот, кодирующие такие аминокислотные последовательности, также хорошо известны в данной области.
В одном из аспектов изобретения некоторые мутантные полипептиды FimH позволяют получить зафиксированную открытую конформацию, что приводит к снижению сродства к маннозидным лигандам и к улучшенной функциональной иммуногенности. Соответственно, такие мутанты FimH могут быть использованы в качестве антигенов в иммуногенной композиции, такой как вакцина, против инфекции Е.coli. Поскольку FimHLD дикого типа считается плохим иммуногеном с точки зрения его способности стимулировать функциональную иммуногенность, такие мутанты FimH позволяют получить улучшенные антигены, которые можно использовать в таких иммуногенных композициях.
В одном из аспектов, описанном в примере 1, создают мутанты FimH, позволяющие зафиксировать лектиновый домен FimH в открытой конформации, чтобы уменьшить сродство к маннозидным лигандам. Такие мутанты могут включать по меньшей мере 1, 2, 3, 4, 5 или более мутаций. Мутации могут включать: встречающиеся в природе аминокислотные замены, которые распространены среди изолятов инфекций мочевыводящих путей (такие как V27A); замены в лиганд-связывающем сайте FimHLD (например, в положениях F1 и Q133); мутации переключения глицина в FimHLD (например, в положениях G15, G16 и G65); введение цистеиновых пар для стабилизации дисульфидной связи в FimHLD (например, в парах положений Р12 -А18; G14 - F144; Р26 - V35; Р26 - V154; Р26 - V156; V27 - L34; V28 - N33; V28 - Р157; Q32 - Y108; N33 - L109; N33 - Р157; V35 -L107; V35 - L109; S62 - Т86; S62 - L129; Y64 - А127; L68 - F71; V112 - Т158; S113 - Т158; V118 - V156; и/или А119 - V155); неполярные-в-полярные мутации в FimHLD (например, в положениях V27, L34, А119 или в любой их комбинации); мутации заполнения полости на поверхности контакта пилин-лектин FimH-DSG (например, в положениях А115, V163, V185 или V3 в последовательности DSG); или любая комбинация этих типов мутаций и в указанных выше положениях аминокислот. В другом аспекте настоящее изобретение относится к мутантам FimH, представленным в SEQ ID NO: 2-58 и 60-64, или к любой комбинации мутантов, указанных в любой из таких последовательностей. В другом аспекте в настоящем изобретении предложен мутант FimH с любой из SEQ ID NO: 23, 50, 51, 52, 53, 54, 60 и 62. В другом аспекте в настоящем изобретении предложен мутант FimH с SEQ ID NO: 62.
В еще одном аспекте настоящее изобретение относится к любому из мутантов FimH с SEQ ID NO: 2-58 и 60-64, где указанные мутанты являются выделенными. Например, в одном из аспектов настоящего изобретения предложен мутант FimH с любой из SEQ ID NO: 23, 50, 51, 52, 53, 54, 60 и 62, где указанный мутант FimH является выделенным. В еще одном аспекте настоящего изобретения предложен мутант FimH в соответствии с SEQ ID NO: 62, где указанный мутант FimH является выделенным.
Соответственно, в некоторых конкретных аспектах настоящее изобретение относится к мутанту FimH, содержащему комбинацию введенных мутаций, где мутант содержит комбинацию мутаций, указанных в любом из мутантов, представленных в таблице 1 (т.е. в SEQ ID NO: 2-58 и 60-64). Любая комбинация аминокислотных замен, представленных в каждом из мутантов в таблице 1, может быть использована в последовательности полипептида FimH дикого типа с получением различных мутантов FimH. Мутанты FimH, основанные на нативной последовательности полипептида FimH любого другого подтипа или штамма, содержащие любую комбинацию мутаций, представленных в настоящем описании, также входят в объем настоящего изобретения.
Еще одним аспектом настоящего изобретения является полипептид, который является на по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичным любой из SEQ ID NO: 1-64. В предпочтительном аспекте полипептид является на по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентичным SEQ ID NO: 62. В другом аспекте настоящего изобретения речь идет о полипептиде, который является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 99,9% идентичным любой из SEQ ID NO: 1-64. В предпочтительном аспекте полипептид является на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 99,9% идентичным SEQ ID NO: 62.
Мутанты FimH, представленные в настоящем изобретении, могут быть получены обычными способами, известными в данной области, такими как экспрессия в рекомбинантной системе-хозяине с использованием подходящего вектора. Подходящие рекомбинантные клетки-хозяева включают, например, клетки насекомых, клетки млекопитающих, клетки птиц, бактерии и дрожжевые клетки. Примеры подходящих клеток насекомых включают, например, клетки Sf9, клетки Sf21, клетки Tn5, клетки Schneider S2 и клетки High Five (клональный изолят, полученный из родительской клеточной линии Trichoplusia ni BTI-TN-5B1-4 (Invitrogen)). Примеры подходящих клеток млекопитающих включают клетки яичника китайского хомячка (СНО), эмбриональные клетки почек человека (клетки HEK293 или Expi 293, обычно трансформированные ДНК аденовируса типа 5, фрагментированной в результате гидродинамического сдвига), клетки NIH-3T3, клетки 293-Т, клетки Vero и клетки HeLa. Подходящие птичьи клетки включают, например, стволовые клетки куриного эмбриона (например, клетки ЕВх®), фибробласты куриного эмбриона, зародышевые клетки куриного эмбриона, фибробласты перепела (например, ELL-О) и клетки утки. Подходящие системы экспрессии клеток насекомых, такие как системы на основе бакуловирусного вектора, известны специалистам в данной области и описаны, например, в Summers and Smith, Texas Agricultural Experiment Station Bulletin No. 1555 (1987). Материалы и способы для систем экспрессии бакуловирус/клетки насекомых коммерчески доступны в форме наборов, среди прочего, от Invitrogen, San Diego Calif. Системы экспрессии птичьих клеток также известны специалистам в данной области и описаны, например, в патенте США №№5,340,740; 5,656,479; 5,830,510; 6,114,168; и 6,500,668. Аналогично, системы экспрессии клеток бактерий и млекопитающих также известны в данной области и описаны, например, в Yeast Genetic Engineering (Barr et al., eds., 1989) Butterworths, London.
Множество подходящих векторов для экспрессии рекомбинантных белков в клетках насекомых или млекопитающих хорошо известно и традиционно используются в данной области. Подходящие векторы могут содержать ряд компонентов, включая, без ограничения, один или более из следующего: точку начала репликации; селектируемый маркерный ген; один или более элементов контроля экспрессии, таких как элемент контроля транскрипции (например, промотор, энхансер, терминатор) и/или один или более сигналов трансляции; и сигнальную последовательность или лидерную последовательность для нацеливания на секреторный путь в выбранной клетке-хозяине (например, происходящей от млекопитающего или от гетерологичных млекопитающих или видов, не относящихся к млекопитающим). Например, для экспрессии в клетках насекомых используют подходящий бакуловирусный вектор экспрессии, такой как pFastBac (Invitrogen), для получения рекомбинантных бакуловирусных частиц. Бакуловирусные частицы амплифицируют и используют для заражения клеток насекомых для экспрессии рекомбинантного белка. Для экспрессии в клетках млекопитающих используют вектор, который будет управлять экспрессией конструкции в желаемой клетке-хозяине млекопитающего (например, в клетках яичника китайского хомячка).
Мутантные полипептиды FimH могут быть выделены с помощью любых подходящих способов. Например, в данной области техники известны способы очистки мутантных полипептидов белка FimH с помощью иммуноаффинной хроматографии. Ruiz-Arguello et al., J. Gen. Virol., 85:3677-3687 (2004). Подходящие способы очистки желаемых белков, включая преципитацию и различные типы хроматографии, такие как хроматография гидрофобного взаимодействия, ионообменная, аффинная, хелатирующая и эксклюзионная хроматография, хорошо известны в данной области. Подходящие схемы очистки могут быть созданы с использованием двух или более из этих или других подходящих способов. При желании мутантные полипептиды FimH могут включать «метку», облегчающую очистку, такую как эпитопную метку или гистидиновую (His) метку. Такие меченые полипептиды удобно выделять, например, из кондиционированных сред с помощью хелатирующей хроматографии или аффинной хроматографии.
В контексте настоящего описания термин «антиген» относится к молекуле, которая может распознаваться антителом. Примеры антигенов включают полипептиды, пептиды, липиды, полисахариды и нуклеиновые кислоты, содержащие антигенные детерминанты, такие как детерминанты, распознаваемые иммунной клеткой.
II. Нуклеиновые кислоты, кодирующие мутанты FimH
В другом аспекте настоящее изобретение относится к молекулам нуклеиновой кислоты, которые кодируют мутант FimH, раскрытый в настоящем описании. Такие молекулы нуклеиновой кислоты включают последовательности ДНК, кДНК и РНК. В одном из вариантов осуществления молекула нуклеиновой кислоты может быть включена в вектор, такой как вектор экспрессии.
В одном из аспектов представлены нуклеиновые кислоты, кодирующие мутантные полипептиды FimH Е.coli или любой их фрагмент. Одну или более конструкций нуклеиновых кислот, кодирующих мутантные полипептиды FimH или их фрагменты, можно использовать для геномной интеграции и последующей экспрессии полипептида. Например, одна конструкция нуклеиновой кислоты, кодирующая мутантный полипептид FimH или его фрагмент, может быть введена в клетку-хозяина. Альтернативно кодирующие последовательности полипептида могут быть перенесены двумя или более конструкциями нуклеиновых кислот, которые затем вводят в клетку-хозяина одновременно или последовательно.
Например, в одном из иллюстративных вариантов осуществления одна конструкция нуклеиновой кислоты кодирует лектиновый домен и пилиновый домен FimH Е.coli. В другом типичном варианте осуществления одна конструкция нуклеиновой кислоты кодирует лектиновый домен, а вторая конструкция нуклеиновой кислоты кодирует пилиновый домен FimH Е.coli. В некоторых аспектах достигается геномная интеграция.
Конструкция нуклеиновой кислоты может содержать геномную ДНК, содержащую один или более интронов, или кДНК. Некоторые гены экспрессируются более эффективно, когда присутствуют интроны. В некоторых аспектах последовательность нуклеиновой кислоты пригодна для экспрессии экзогенных полипептидов в указанной клетке млекопитающего.
В некоторых аспектах нуклеиновая кислота, кодирующая полипептид или его фрагмент, оптимизирована по кодонам для повышения уровня экспрессии в любой конкретной клетке.
В некоторых аспектах конструкция нуклеиновой кислоты включает сигнальную последовательность, которая кодирует пептид, управляющий секрецией полипептида, полученного из Е.coli, или его фрагмент. В некоторых аспектах нуклеиновая кислота включает нативную сигнальную последовательность полипептида, полученного из FimH Е.coli. В некоторых аспектах, где полипептид, полученный из Е.coli, или его фрагмент включает эндогенную сигнальную последовательность, последовательность нуклеиновой кислоты, кодирующая сигнальную последовательность, может быть оптимизирована по кодонам для повышения уровня экспрессии белка в клетке-хозяине.
В некоторых аспектах сигнальная последовательность имеет любую длину из следующих: 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29 и 30 аминокислот в длину. В некоторых аспектах длина сигнальной последовательности составляет 20 аминокислот. В некоторых аспектах длина сигнальной последовательности составляет 21 аминокислоту.
В некоторых аспектах, где полипептид или его фрагмент включает сигнальную последовательность, для повышения уровня экспрессии полипептида или его фрагмента в культивируемых клетках эндогенная сигнальная последовательность, естественным образом связанная с полипептидом, может быть заменена сигнальной последовательностью, не связанной с полипептидом дикого типа. Соответственно, в некоторых аспектах нуклеиновая кислота не включает нативную сигнальную последовательность полипептида, полученного из Е.coli, или его фрагмента. В некоторых аспектах нуклеиновая кислота не включает нативную сигнальную последовательность полипептида, полученного из FimH Е.coli. В некоторых аспектах полипептид, полученный из Е.coli, или его фрагмент может быть экспрессирован гетерологичным пептидом, который предпочтительно представляет собой сигнальную последовательность, или другим пептидом, имеющим специфический сайт расщепления на N-конце зрелого белка или полипептида, полученного из Е.coli, или его фрагмента. Например, полипептид, полученный из FimH Е.coli, или его фрагмент может быть экспрессирован гетерологичным пептидом (например, сигнальной последовательностью IgK), который предпочтительно представляет собой сигнальную последовательность, или другим пептидом, имеющим специфический сайт расщепления на N-конце зрелого белка FimH Е.coli. В предпочтительных аспектах специфический сайт расщепления на N-конце зрелого белка FimH Е.coli находится непосредственно перед исходным фенилаланиновым остатком зрелого белка FimH Е.coli. Выбранная гетерологичная последовательность предпочтительно является последовательностью, которая распознается и процессируется (т.е. расщепляется сигнальной пептидазой) клеткой-хозяином.
В предпочтительных аспектах сигнальная последовательность представляет собой сигнальную последовательность IgK. В некоторых аспектах нуклеиновая кислота кодирует полипептид, имеющий аминокислотную последовательность, указанную в любой из SEQ ID NO: 1-64. В некоторых аспектах нуклеиновая кислота кодирует аминокислотную последовательность SEQ ID NO: 23, 50, 51, 52, 53, 54, 60, 61 или 62. В некоторых аспектах нуклеиновая кислота кодирует полипептид, имеющий аминокислотную последовательность, приведенную в SEQ ID NO: 62. В предпочтительных аспектах сигнальная последовательность представляет собой сигнальную последовательность мышиного IgK.
Подходящие векторы экспрессии млекопитающих для получения мутантных полипептидов FimH или их фрагментов известны в данной области и могут быть коммерчески доступными, например вектор экспрессии pSecTag2 от Invitrogen™. Пример последовательности сигнального пептида каппа-цепи Ig мыши включает последовательность ETDTLLLWVLLLWVPGSTG (SEQ ID NO: 65). В некоторых аспектах вектор включает вектор экспрессии млекопитающих pBudCE4.1 от Thermo Fisher. Дополнительные иллюстративные и подходящие векторы включают вектор экспрессии млекопитающих pcDNA™3.1 (Thermo Fisher).
В некоторых аспектах сигнальная последовательность не включает сигнальную последовательность гемагглютинина.
В некоторых аспектах нуклеиновая кислота включает нативную сигнальную последовательность полипептида FimH или его фрагмента. В некоторых аспектах сигнальная последовательность не является сигнальной последовательностью IgK. В некоторых аспектах сигнальная последовательность включает сигнальную последовательность гемагглютинина.
В одном из аспектов в настоящем описании раскрыты векторы, которые включают кодирующие последовательности мутантного полипептида FimH или его фрагмента. Примеры векторов включают плазмиды, которые способны реплицироваться автономно или реплицироваться в клетке млекопитающего. Типичные векторы экспрессии содержат подходящие промоторы, энхансеры и терминаторы, которые можно использовать для регуляции экспрессии кодирующей последовательности(ей) в экспрессионной конструкции. Векторы также могут включать маркеры селекции для обеспечения фенотипического признака для селекции трансформированных клеток-хозяев (например, путем придания устойчивости к антибиотикам, таким как ампициллин или неомицин).
Подходящие промоторы известны в данной области. Типичные промоторы включают, например, промотор CMV, аденовируса, EF1a, промотор металлотионина GAPDH, ранний промотор SV-40, поздний промотор SV-40, промотор вируса опухоли молочной железы мышей, промотор вируса саркомы Рауса, промотор полиэдрина и т.д. Промоторы могут быть конститутивными или индуцибельными. Можно использовать один или более векторов (например, один вектор, кодирующий все субъединицы или домены, или их фрагменты, или несколько векторов, вместе кодирующих субъединицы, или домены, или их фрагменты).
Также можно использовать внутренний сайт посадки рибосомы (IRES) и последовательности пептида 2А. IRES и пептид 2А обеспечивают альтернативные подходы к совместной экспрессии нескольких последовательностей. IRES представляет собой нуклеотидную последовательность, которая позволяет инициировать трансляцию в середине последовательности матричной РНК (мРНК) как часть более крупного процесса синтеза белка. Обычно у эукариот трансляция может инициироваться только на 5'-конце молекулы мРНК. Элементы IRES позволяют экспрессировать несколько генов в одном транскрипте. Полицистронные векторы на основе IRES, которые экспрессируют несколько белков из одного транскрипта, могут уменьшать ускользание неэкспрессирующих клонов от селекции. Пептид 2А позволяет транслировать несколько белков в одной открытой рамке считывания в полипротеин, который впоследствии расщепляется на отдельные белки с помощью механизма пропуска рибосом. Пептид 2А может обеспечивать более сбалансированную экспрессию множества белковых продуктов. Примеры последовательностей IRES включают, например, EV71 IRES, EMCV IRES, HCV IRES. Что касается геномной интеграции, то она может быть сайт-специфичной или случайной. Сайт-специфическая рекомбинация может быть достигнута путем введения гомологичной последовательности(ей) в конструкции нуклеиновых кислот, представленные в настоящем описании. Такая гомологичная последовательность по существу соответствует эндогенной последовательности в конкретном сайте-мишени в геноме хозяина. В качестве альтернативы можно использовать случайную интеграцию. Иногда уровень экспрессии белка может изменяться в зависимости от сайта интеграции. Следовательно, для идентификации клона, который достигает желаемого уровня экспрессии, можно отобрать несколько клонов в соответствии с уровнем экспрессии рекомбинантного белка.
Примеры конструкций нуклеиновых кислот дополнительно описаны на фигурах, например фиг.2А-2Т, международной публикации РСТ №WO2021/084429, опубликованной 6 мая 2021 г., которая включена в настоящее описание в качестве ссылки.
В одном из аспектов последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая на по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере, по меньшей мере 97%, по меньшей мере 98%, по меньшей мере 99% идентична любой из SEQ ID NO: 1-64. В предпочтительном аспекте последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая на по меньшей мере 90%, по меньшей мере 91%, по меньшей мере 92%, по меньшей мере 93%, по меньшей мере 94%, по меньшей мере 95%, по меньшей мере 96%, по меньшей мере 97%, по меньшей мере 98% или по меньшей мере 99% идентична SEQ ID NO: 62. В другом аспекте настоящего изобретения последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 99,9% идентична любой из SEQ ID NO: 1-64. В предпочтительном аспекте последовательность нуклеиновой кислоты кодирует аминокислотную последовательность, которая на по меньшей мере 70%, 71%, 72%, 73%, 74%, 75%, 76%, 77%, 78%, 79%, 80%, 81%, 82%, 83%, 84%, 85%, 86%, 87%, 88%, 89%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98%, 99% или 99,9% идентична SEQ ID NO: 62.
В некоторых аспектах настоящего изобретения РНК представляет собой информационную РНК (мРНК), которая относится к РНК-транскрипту, кодирующему пептид или белок. Как установлено в данной области, мРНК обычно содержит 5'-нетранслируемую область (5'-UTR), область, кодирующую пептид, и 3'-нетранслируемую область (3'-UTR). В некоторых вариантах осуществления РНК получают транскрипцией in vitro или химическим синтезом. В одном из вариантов осуществления мРНК получают путем транскрипции in vitro с использованием матрицы ДНК, где ДНК относится к нуклеиновой кислоте, содержащей дезоксирибонуклеотиды. В одном из аспектов представленная в настоящем описании РНК может содержать модифицированные нуклеозиды. В некоторых аспектах РНК содержит модифицированный нуклеозид вместо по меньшей мере одного (например, каждого) уридина.
В некоторых вариантах осуществления композиции или медицинские препараты, раскрытые в настоящем описании, содержат РНК, кодирующую аминокислотную последовательность, содержащую мутантный полипептид FimH. Аналогично, способы, раскрытые в настоящем описании, включают введение такой РНК. Одна из платформ, подходящих для использования в контексте изобретения, основана на вакцине на основе антиген-кодирующей РНК для индукции сильных нейтрализующих антител и сопровождающего/сопутствующего Т-клеточного ответа для достижения защитной иммунизации предпочтительно минимальными дозами вакцины. Вводимая РНК предпочтительно представляет собой РНК, транскрибированную in vitro. Особенно предпочтительными являются три различные платформы РНК, а именно немодифицированная уридин-содержащая мРНК (уРНК), нуклеозид-модифицированная мРНК (модРНК) и самоамплифицирующаяся РНК (саРНК). В одном особенно предпочтительном аспекте РНК представляет собой РНК, транскрибируемую in vitro.
III. Клетки-хозяева
В одном из аспектов изобретение относится к клеткам, в которых последовательности, кодирующие мутантный полипептид FimH или его фрагмент, экспрессируются в клетке-хозяине млекопитающего. В одном из вариантов осуществления полипептид временно экспрессируется в клетке-хозяине. В другом варианте осуществления полипептид стабильно интегрирован в геном клеток-хозяев и при культивировании в подходящих условиях экспрессирует полипептид или его фрагмент. В предпочтительном варианте осуществления полинуклеотидная последовательность экспрессируется с высокой эффективностью и является генетически стабильной.
Подходящие клетки-хозяева млекопитающих известны в данной области. Предпочтительно клетка-хозяин пригодна для продуцирования белка в промышленных масштабах. Примеры клеток-хозяев млекопитающих включают любые из следующих клеток и их производных: клетки яичника китайского хомяка (СНО), клетки COS (линия клеток, полученная из почки обезьяны, африканской зеленой мартышки), клетки Vero, клетки Hela, клетки почки детеныша хомячка (ВНК), клетки эмбриональных почек человека (HEK), клетки NSO (линия клеток мышиной миеломы) и клетки С127 (линия неопухолевых мышиных клеток). Дополнительные типичные клетки-хозяева млекопитающих включают мышиные клетки Sertoli (ТМ4), клетки печени крысы-буйвола (BRL 3А), клетки опухоли молочной железы мыши (ММТ), клетки гепатомы крысы (НТС), клетки мышиной миеломы (NSO), клетки мышиной гибридомы (Sp2/0), клетки мышиной тимомы (EL4), клетки яичника китайского хомячка (СНО) и производные клеток СНО, клетки эмбриона мыши (NIH/3T3, 3Т3 Li), клетки миокарда крысы (Н9с2), мышиные миобласты (С2С12) и клетки почки мыши (miMCD-3). Другие примеры клеточных линий млекопитающих включают NS0/1, Sp2/0, Hep G2, PER.C6, COS-7, ТМ4, CV1, VERO-76, MDCK, BRL 3А, W138, ММТ 060562, TR1, MRC5 и FS4.
В соответствии с настоящим изобретением может быть использована любая клетка, позволяющая получить клеточную культуру. В некоторых аспектах клетка представляет собой клетку млекопитающего. Неограничивающие примеры клеток млекопитающих, которые можно использовать в соответствии с настоящим изобретением, включают линию мышиной миеломы BALB/c (NSO/1, ЕСАСС №: 85110503); ретинобласты человека (PER.C6, CruCell, Leiden, The Netherlands); линию почки обезьяны CV1, трансформированную SV40 (COS-7, АТСС CRL 1651); линию эмбриональных почек человека (клетки 293 или 293, субклонированные для роста в суспензионной культуре, Graham et al., J. Gen Virol., 36:59, 1977); клетки почек детенышей хомячка (ВНК, АТСС CCL 10); клетки яичника китайского хомячка+/-DHFR (СНО, Urlaub and Chasin, Proc. Natl. Acad. Sci. USA, 77:4216, 1980); мышиные 28yophil клетки (TM4, Mather, Biol. Reprod., 23:243-251, 1980); клетки почки обезьяны (CV1 АТСС CCL 70); клетки почки африканской зеленой мартышки (VERO-76, АТСС CRL-1587); клетки карциномы шейки матки человека (HeLa, АТСС CCL 2); клетки почки собаки (MDCK, АТСС CCL 34); клетки печени крысы-буйвола (BRL 3А, АТСС CRL 1442); клетки легкого человека (W138, АТСС CCL 75); клетки печени человека (Hep G2, НВ 8065); клетки опухоли молочной железы мыши (ММТ 060562, АТСС CCL51); клетки TRI (Mather et al., Annals N.Y. Acad. Sci., 383:44-68, 1982); MRC 5 клетки; FS4 клетки; и линию гепатомы человека (Hep G2). В некоторых предпочтительных вариантах осуществления клетки представляют собой клетки СНО. В некоторых предпочтительных аспектах клетки представляют собой GS-клетки.
Кроме того, в соответствии с настоящим изобретением можно использовать любое количество коммерчески доступных и некоммерческих клеточных линий гибридомы. Термин «гибридома», используемый в настоящем описании, относится к клетке или потомству клетки, полученному в результате слияния иммортализованной клетки и клетки, продуцирующей антитело. Такая полученная гибридома представляет собой иммортализованную клетку, продуцирующую антитела. Отдельные клетки, используемые для создания гибридомы, могут быть получены из любого млекопитающего, включая, без ограничения, крысу, свинью, кролика, овцу, свинью, козу и человека. В некоторых аспектах гибридома представляет собой клеточную линию триомы, которая возникает, когда потомство слияний гетерогибридной миеломы, которые являются продуктом слияния между клетками человека и клеточной линией мышиной шеломы, впоследствии сливают с плазматической клеткой. В некоторых аспектах гибридома представляет собой любую иммортализованную линию гибридных клеток, которая продуцирует антитела, такие как, например, квадромы (см. например, Milstein et al., Nature 537:3053 (1983)). Специалисту в данной области будет понятно, что гибридные клеточные линии могут иметь разные требования к питанию и/или могут потребоваться различные условия культивирования для оптимального роста, которые специалист способен изменять по мере необходимости.
В некоторых аспектах клетка содержит первый представляющий интерес ген, при этом первый представляющий интерес ген является хромосомно-интегрированным. В некоторых аспектах первый представляющий интерес ген содержит репортерный ген, селективный ген, представляющий интерес ген (например, кодирующий полипептид, полученный из Е.coli, или его фрагмент), вспомогательный ген или их комбинацию. В некоторых аспектах ген, представляющий терапевтический интерес, содержит ген, кодирующий трудно экспрессируемый белок (DtE).
В некоторых аспектах первый представляющий интерес ген располагают между двумя отдельными целевыми сайтами рекомбинации (RTS) в клетке млекопитающего посредством сайт-специфической интеграции (SSI), где два RTS являются хромосомно-интегрированными в локус NL1 или локус NL2. См., например, публикацию заявки на патент США №20200002727 для описания локуса NL1, локуса NL2, локуса NL3, локуса NL4, локуса NL5 и локуса NL6. В некоторых аспектах первый представляющий интерес ген расположен в локусе NL1. В некоторых аспектах клетка содержит второй представляющий интерес ген, где второй представляющий интерес ген является хромосомно-интегрированным. В некоторых аспектах второй представляющий интерес ген содержит репортерный ген, селективный ген, представляющий терапевтический интерес ген (такой как мутантный полипептид FimH или его фрагмент), вспомогательный ген или их комбинацию. В некоторых аспектах представляющий терапевтический интерес ген содержит ген, кодирующий белок DtE. В некоторых аспектах второй представляющий интерес ген расположен между двумя RTS. В некоторых аспектах второй представляющий интерес ген расположен в локусе NL1 или локусе NL2. В некоторых аспектах первый представляющий интерес ген расположен в локусе NL1, а второй представляющий интерес ген расположен в локусе NL2. В некоторых аспектах клетка содержит третий представляющий интерес ген, где третий представляющий интерес ген является хромосомно-интегрированным. В некоторых аспектах третий представляющий интерес ген содержит репортерный ген, селективный ген, представляющий терапевтический интерес ген (такой как полипептид, полученный из Е.coli или его фрагмент), вспомогательный ген или их комбинацию. В некоторых аспектах представляющий терапевтический интерес ген содержит ген, кодирующий белок DtE. В некоторых аспектах третий представляющий интерес ген расположен между двумя RTS. В некоторых аспектах третий представляющий интерес ген расположен в локусе NL1 или локусе NL2. В некоторых аспектах третий представляющий интерес ген расположен в локусе, отличном от локуса NL1 и локуса NL2. В некоторых аспектах первый представляющий интерес ген, второй представляющий интерес ген и третий представляющий интерес ген находятся в трех отдельных локусах. В некоторых аспектах по меньшей мере один из первого представляющего интерес гена, второго представляющего интерес гена и третьего представляющего интерес гена находится в локусе NL1, и по меньшей мере один из первого представляющего интерес гена, второго представляющего интерес гена и третьего представляющего интерес гена находится в локусе NL2. В некоторых аспектах клетка содержит сайт-специфический ген рекомбиназы. В некоторых аспектах сайт специфический ген рекомбиназы является хромосомно-интегрированным.
В другом аспекте настоящее изобретение относится к клетке млекопитающего, содержащей по меньшей мере четыре разных RTS, при этом клетка содержит (а) по меньшей мере два разных RTS, хромосомно-интегрированных в локус NL1 или локус NL2; (b) первый представляющий интерес ген, встроенный между по меньшей мере двумя RTS из (а), где первый представляющий интерес ген содержит репортерный ген, ген, кодирующий белок DtE, вспомогательный ген или их комбинацию; (с) и второй представляющий интерес ген, хромосомно-интегрированный во второй локус, отличный от локуса (а), где второй представляющий интерес ген содержит репортерный ген, ген, кодирующий белок DtE (такой как полипептид, полученный из Е.coli или его фрагмент), вспомогательный ген или их комбинацию. В некоторых аспектах настоящее изобретение относится к клетке млекопитающего, содержащей по меньшей мере четыре разных RTS, при этом клетка содержит (а) по меньшей мере два разных RTS, хромосомно-интегрированных в локус Fer1L4; (b) по меньшей мере два разных RTS, хромосомно-интегрированных в локус NL1 или локус NL2; (с) первый представляющий интерес ген, хромосомно-интегрированный в локус Fer1L4, при этом первый представляющий интерес ген содержит репортерный ген, ген, кодирующий белок DtE, вспомогательный ген или их комбинацию; и (d) второй представляющий интерес ген, хромосомно-интегрированный в локус NL1 или локус NL2 из (b), где второй представляющий интерес ген содержит репортерный ген, ген, кодирующий белок DtE (такой как полученный из полипептида из Е.coli или его фрагмента), вспомогательный ген или их комбинацию.
В некоторых аспектах настоящее изобретение относится к клетке млекопитающего, содержащей по меньшей мере шесть разных RTS, где клетка содержит (а) по меньшей мере два разных RTS и первый представляющий интерес ген, хромосомно-интегрированные в локус Fer1L4; (b) по меньшей мере два разных RTS и второй представляющий интерес ген, хромосомно-интегрированные в локус NL1; и (с) по меньшей мере два разных RTS и третий представляющий интерес ген, хромосомно-интегрированные в локус NL2.
В контексте настоящего описания термины «в функциональной комбинации», «в функциональном порядке» и «функционально связанный» относятся к связыванию последовательностей нуклеиновых кислот таким образом, что продуцируется молекула нуклеиновой кислоты, способная направлять транскрипцию данного гена и/или синтез желаемой белковой молекулы. Этот термин также относится к соединению аминокислотных последовательностей таким образом, что образуется функциональный белок. В некоторых аспектах представляющий интерес ген функционально связан с промотором, причем представляющий интерес ген является хромосомно-интегрированным в клетке-хозяине. В некоторых аспектах представляющий интерес ген функционально связан с гетерологичным промотором; где представляющий интерес ген является хромосомно-интегрированным в клетке-хозяине. В некоторых аспектах вспомогательный ген функционально связан с промотором, где вспомогательный ген является хромосомно-интегрированным в геном клетки-хозяина. В некоторых аспектах вспомогательный ген функционально связан с гетерологичным промотором; где вспомогательный ген является хромосомно-интегрированным в геном клетки-хозяина. В некоторых аспектах ген, кодирующий белок DtE, функционально связан с промотором, при этом ген, кодирующий белок DtE, является хромосомно-интегрированным в геном клетки-хозяина. В некоторых аспектах ген, кодирующий белок DtE, функционально связан с гетерологичным промотором, где ген, кодирующий белок DtE, является хромосомно-интегрированным в геном клетки-хозяина. В некоторых аспектах ген рекомбиназы функционально связан с промотором, где ген рекомбиназы является хромосомно-интегрированным в клетке-хозяине. В некоторых аспектах ген рекомбиназы функционально связан с промотором, при этом ген рекомбиназы не интегрирован в геном клетки-хозяина. В некоторых аспектах ген рекомбиназы функционально связан с гетерологичным промотором, где ген рекомбиназы не является хромосомно-интегрированным в геном клетки-хозяина. В некоторых аспектах ген рекомбиназы функционально связан с гетерологичным промотором, где ген рекомбиназы не является хромосомно-интегрированным в геном клетки-хозяина.
В контексте настоящего описания термин «хромосомно-интегрированный» или «хромосомная интеграция» относится к стабильному включению последовательности нуклеиновой кислоты в хромосому клетки-хозяина, например клетку млекопитающего, т.е., последовательность нуклеиновой кислоты, которая является хромосомно-интегрированной в геномной ДНК (гДНК) клетки-хозяина, например клетки млекопитающего. В некоторых аспектах последовательность нуклеиновой кислоты, интегрированная в хромосому, является стабильной. В некоторых аспектах последовательность нуклеиновой кислоты, которая является хромосомно-интегрированной, не расположена на плазмиде или в векторе. В некоторых аспектах последовательность нуклеиновой кислоты, которая является хромосомно-интегрированной, не вырезается. В некоторых аспектах хромосомная интеграция опосредована сгруппированными короткими палиндромными повторами с регулярными промежутками (CRISPR) и системой редактирования генов CRISPR-ассоциированного белка (Cas) (CRISPR/CAS).
IV. Композиции и составы
В одном из аспектов настоящее изобретение относится к композиции, содержащей по меньшей мере один мутантный полипептид FimH или его фрагмент, представленный в настоящем описании. В некоторых аспектах композиция вызывает иммунный ответ, включая антитела, которые могут обеспечить иммунитет к патогенным видам Е.coli.
В некоторых аспектах композиция содержит мутантный полипептид FimH в качестве единственного антигена. В некоторых аспектах композиция не включает конъюгат.
В некоторых аспектах композиция содержит мутантный полипептид FimH и по меньшей мере один дополнительный антиген. В некоторых аспектах композиция содержит мутантный полипептид FimH и дополнительный антиген Е.coli. В некоторых аспектах композиция содержит мутантный полипептид FimH и гликоконъюгат Е.coli.
В некоторых аспектах композиция содержит мутантный полипептид FimH и полипептид, полученный из FimC Е.coli, или его фрагмент.
В одном из вариантов осуществления изобретение относится к композиции, содержащей мутантный полипептид FimH и сахарид, содержащий любую структуру, выбранную из структур сахаридов, раскрытых в международных публикациях РСТ №WO2021/084429, опубликованной 6 мая 2021 г., и WO2020/039359, опубликованной 27 февраля 2020 г., и в публикации США №US2020/0061177, опубликованной 27 февраля 2020 г., каждая из которых полностью включена в настоящее описание посредством ссылки. В одном из аспектов изобретение относится к композиции, содержащей мутантный полипептид FimH; и сахарид, содержащий структуру, выбранную из: формулы O1 (например, формулы O1A, формулы O1 В, и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы 05ab и формулы 05ас (штамм 180/С3)), формулы О6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B, и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы 023А), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D-1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187, где n представляет собой целое число от 1 до 100, предпочтительно от 31 до 90.
В некоторых вариантах осуществления композиция включает любой из раскрытых в настоящем описании сахаридов. В предпочтительных вариантах осуществления композиция включает любой из раскрытых в настоящем описании конъюгатов.
В некоторых вариантах осуществления композиция включает по меньшей мере один гликоконъюгат из Е.coli серотипа O25, предпочтительно серотипа O25b. В одном из вариантов осуществления композиция включает по меньшей мере один гликоконъюгат из Е.coli серотипа O1, предпочтительно серотипа O1a. В одном из вариантов осуществления композиция включает по меньшей мере один гликоконъюгат из Е.coli серотипа O2. В одном из вариантов осуществления композиция включает по меньшей мере один гликоконъюгат из Е.coli серотипа O6.
В одном из вариантов осуществления композиция содержит по меньшей мере один гликоконъюгат, выбранный из любого из следующих серотипов Е.coli: O25, O1, O2 и O6, предпочтительно из: O25b, O1a, O2 и O6. В одном из вариантов осуществления композиция содержит по меньшей мере два гликоконъюгата, выбранных из любого из следующих серотипов Е.coli: O25, O1, O2 и O6, предпочтительно из: O25b, O1a, O2 и O6. В другом варианте осуществления композиция содержит по меньшей мере три гликоконъюгата, выбранных из любого из следующих серотипов Е.coli: O25, O1, O2 и O6, предпочтительно из: O25b, O1a, O2 и O6. В другом варианте осуществления композиция содержит гликоконъюгат каждого из следующих серотипов Е.coli: O25, O1, O2 и O6, предпочтительно O25b, O1a, O2 и О6.
В предпочтительном варианте осуществления гликоконъюгат любой из вышеуказанных композиций индивидуально конъюгирован с CRM197. В другом предпочтительном варианте осуществления любой гликоконъюгат из вышеуказанных композиций индивидуально конъюгирован с SCP.
Соответственно, в некоторых вариантах осуществления композиция включает мутантный полипептид FimH и О-антиген по меньшей мере одного серотипа Е.coli. В предпочтительном варианте осуществления композиция включает мутантный полипептид FimH и О-антиген из более чем 1 серотипа Е.coli. Например, композиция может включать О-антиген от двух разных серотипов Е.coli (или «v», валентность) и вплоть до 12 разных серотипов (12v). В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 3 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 4 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 5 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 6 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 7 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 8 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 9 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 10 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 11 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 12 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 13 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 14 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 15 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 16 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 17 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 18 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-антиген из 19 разных серотипов. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 20 разных серотипов.
Предпочтительно количество сахаридов Е.coli может меняться от 1 серотипа (или «v», валентности) до 26 разных серотипов (26v). В одном из вариантов осуществления присутствует один серотип. В одном из вариантов осуществления присутствуют 2 разных серотипа. В одном из вариантов осуществления присутствуют 3 разных серотипа. В одном из вариантов осуществления присутствуют 4 разных серотипа. В одном из вариантов осуществления присутствуют 5 разных серотипов. В одном из вариантов осуществления присутствуют 6 разных серотипов. В одном из вариантов осуществления присутствуют 7 разных серотипов. В одном из вариантов осуществления присутствуют 8 разных серотипов. В одном из вариантов осуществления присутствуют 9 разных серотипов. В одном из вариантов осуществления присутствуют 10 разных серотипов. В одном из вариантов осуществления присутствуют 11 разных серотипов. В одном из вариантов осуществления присутствуют 12 разных серотипов. В одном из вариантов осуществления присутствуют 13 разных серотипов. В одном из вариантов осуществления присутствуют 14 разных серотипов. В одном из вариантов осуществления присутствуют 15 разных серотипов. В одном из вариантов осуществления присутствуют 16 разных серотипов. В одном из вариантов осуществления присутствуют 17 разных серотипов. В одном из вариантов осуществления присутствуют 18 разных серотипов. В одном из вариантов осуществления присутствуют 19 разных серотипов. В одном из вариантов осуществления присутствуют 20 разных серотипов. В одном из вариантов осуществления присутствует 21 разный серотип. В одном из вариантов осуществления присутствуют 22 разных серотипа. В одном из вариантов осуществления присутствуют 23 разных серотипа. В одном из вариантов осуществления присутствуют 24 разных серотипа. В одном из вариантов осуществления присутствуют 25 разных серотипов. В одном из вариантов осуществления присутствуют 26 разных серотипов. Сахариды конъюгированы с белком-носителем с образованием гликоконъюгатов, раскрытых в настоящем описании.
В одном из аспектов композиция включает мутантный полипептид FimH; и гликоконъюгат, который включает О-антиген из по меньшей мере одной серогруппы Е.coli, где О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH; и О-антиген из более чем 1 серотипа Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 2 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 3 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 4 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 5 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 6 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 7 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 8 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 9 разных серотипов Е.coli, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-антиген из мутантного полипептида FimH и 10 разных серотипов Е.coli, где каждый O-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-антиген из мутантного полипептида FimH и 11 разных серотипов Е.coli, где каждый O-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 12 разных серотипов, где каждый O-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 13 разных серотипов, где каждый O-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 14 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 15 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 16 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 17 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 18 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 19 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-антиген из 20 разных серотипов, где каждый О-антиген конъюгирован с белком-носителем.
В другом аспекте композиция включает О-полисахарид по меньшей мере одного серотипа Е.coli. В предпочтительном варианте осуществления композиция включает О-полисахарид из более чем 1 серотипа Е.coli. Например, композиция может включать О-полисахарид из двух разных серотипов Е.coli и вплоть до 12 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 3 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 4 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 5 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 6 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает O-полисахарид из 7 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 8 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 9 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 10 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает О-полисахарид из 11 разных серотипов Е.coli. В одном из вариантов осуществления композиция включает O-полисахарид из 12 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 13 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 14 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 15 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 16 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 17 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 18 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 19 разных серотипов. В одном из вариантов осуществления композиция включает О-полисахарид из 20 разных серотипов.
В предпочтительном варианте осуществления композиция включает О-полисахарид по меньшей мере одного серотипа Е.coli, где О-полисахарид конъюгирован с белком-носителем. В предпочтительном варианте осуществления композиция включает O-полисахарид из более чем 1 серотипа Е.coli, где каждый O-полисахарид конъюгирован с белком-носителем. Например, композиция может включать О-полисахарид из двух разных серотипов Е.coli и вплоть до из 12 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 3 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 4 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает O-полисахарид из 5 разных серотипов Е.coli, где каждый O-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 6 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 7 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает O-полисахарид из 8 разных серотипов Е.coli, где каждый O-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 9 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 10 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 11 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 12 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 13 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 14 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 15 разных серотипов, где каждый O-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 16 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 17 разных серотипов, где каждый O-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 18 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 19 разных серотипов, где каждый O-полисахарид конъюгирован с белком-носителем. В одном из вариантов осуществления композиция включает О-полисахарид из 20 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем.
В наиболее предпочтительном варианте осуществления композиция включает О-полисахарид по меньшей мере одного серотипа Е.coli, где О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает O-антиген и коровый (центральный) сахарид. В предпочтительном варианте осуществления композиция включает О-полисахарид из более чем 1 серотипа Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает O-антиген и коровый сахарид. Например, композиция может включать О-полисахарид из двух разных серотипов Е.coli и вплоть до из 12 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 3 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 4 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 5 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 6 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 7 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 8 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 9 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 10 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 11 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 12 разных серотипов, где каждый O-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 13 разных серотипов, где каждый О-полисахарид конъюгирован с белком носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 14 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает O-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 15 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 16 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 17 разных серотипов, где каждый O-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 18 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 19 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где О-полисахарид включает O-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает О-полисахарид из 20 разных серотипов, где каждый О-полисахарид конъюгирован с белком-носителем, и где O-полисахарид включает О-антиген и коровый сахарид. В предпочтительном варианте осуществления белок-носитель представляет собой CRM197.
В другом предпочтительном варианте осуществления композиция включает мутантный полипептид FimH и О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O25a, где n равно по меньшей мере 30, и коровый сахарид. В предпочтительном варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где O-полисахарид включает формулу O25b, где n равно по меньшей мере 40, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O1a, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O2, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O6, где n равно по меньшей мере 30, и коровый сахарид.
В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O17, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O15, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу 018А, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O75, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O4, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O16, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O13, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления композиция дополнительно включает O-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O7, где n равно по меньшей мере 30, и коровый сахарид.
В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где O-полисахарид включает формулу O8, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления О-полисахарид включает формулу O8, где n равно 1-20, предпочтительно 2-5, более предпочтительно 3. В другом варианте осуществления композиция дополнительно включает О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O9, где n равно по меньшей мере 30, и коровый сахарид. В другом варианте осуществления О-полисахарид включает формулу O9, где n равно 1-20, предпочтительно 4-8, более предпочтительно 5. В другом варианте осуществления О-полисахарид включает формулу O9a, где n равно 1-20, предпочтительно 4-8, более предпочтительно 5.
В некоторых вариантах осуществления О-полисахарид выбирают из формулы O20ab, формулы О20ас, формулы O52, формулы O97 и формулы О101, где n равно 1-20, предпочтительно 4-8, более предпочтительно 5.
Как описано выше, композиция может включать мутантный полипептид FimH и любую комбинацию конъюгированных O-полисахаридов (антигенов). В одном из иллюстративных вариантов осуществления композиция включает полисахарид, который включает формулу O25b, полисахарид, который включает формулу O1a, полисахарид, который включает формулу O2, и полисахарид, который включает формулу O6. Более конкретно, такая композиция включает: (i) О-полисахарид, конъюгированный с CRM197, где O-полисахарид включает формулу O25b, где n равно по меньшей мере 30, и коровый сахарид; (ii) O-полисахарид, конъюгированный с CRM197, где O-полисахарид включает формулу O1a, где n равно по меньшей мере 30, и коровый сахарид; (iii) O-полисахарид, конъюгированный с CRM197, где O-полисахарид включает формулу O2, где n равно по меньшей мере 30, и коровый сахарид; и (iv) О-полисахарид, конъюгированный с CRM197, где О-полисахарид включает формулу O6, где n равно по меньшей мере 30, и коровый сахарид.
В одном из вариантов осуществления композиция включает мутантный полипептид FimH и по меньшей мере один О-полисахарид, полученный из любого серотипа Е.coli, где серотип не является O25a. Например, в одном из вариантов осуществления композиция не включает сахарид, который включает формулу O25a. Такая композиция может включать, например, О-полисахарид, который включает формулу O25b, О-полисахарид, который включает формулу O1a, О-полисахарид, который включает формулу O2, и O-полисахарид, который включает формулу О6.
В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 2 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 3 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает O-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 4 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 5 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 6 различных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 7 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 8 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 9 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 10 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где O-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 11 разных серотипов Е.coli, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 12 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 13 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-полисахарид из 14 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 15 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 16 разных серотипов, где каждый O-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 17 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 18 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и O-полисахарид из 19 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид. В одном из вариантов осуществления композиция включает мутантный полипептид FimH и О-полисахарид из 20 разных серотипов, где каждый О-полисахарид конъюгирован с CRM197, и где О-полисахарид включает О-антиген и коровый сахарид.
В одном из аспектов изобретение относится к композиции, которая содержит мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O25b, где n равно 15±2. В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O25b, где n равно 17±2. В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O25b, где n равно 55±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O25b, где n равно 51±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O25b, где n представляет собой целое число больше 30, предпочтительно n представляет собой целое число от 31 до 100. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R1 корового сахарида Е.coli. В другом варианте осуществления сахарид дополнительно включает фрагмент K12 корового сахарида Е.coli. В другом варианте осуществления сахарид дополнительно включает фрагмент KDO. Предпочтительно белок-носитель представляет собой CRM197. В одном из вариантов осуществления конъюгат получают в результате одноконцевой конъюгации. В одном из вариантов осуществления конъюгат получают химическим методом восстановительного аминирования, предпочтительно в DMSO буфере. В одном из варианте осуществления сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер. Предпочтительно композиция дополнительно включает фармацевтически приемлемый разбавитель.
В одном из аспектов иммуногенная композиция индуцирует у людей образование антител IgG, причем указанные антитела способны связываться с полисахаридом Е.coli серотипа O25b в концентрации по меньшей мере 0,2 пг/мл, 0,3 пг/мл, 0,35 пг/мл, 0,4 пг/мл или 0,5 пг/мл, как определено с помощью анализа ELISA. Таким образом, можно выполнить сравнение активности ОРА сыворотки до и после иммунизации иммуногенной композицией по изобретению и сравнить ответы на серотип O25b, чтобы оценить потенциальное увеличение числа пациентов с ответом на лечение. В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны убивать Е.coli серотипа O25b, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция индуцирует образование у людей функциональных антител, причем указанные антитела способны убивать Е.coli серотипа O25b, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипа O25b (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция вызывает титр по меньшей мере 1:8 против Е.coli серотипа O25b по меньшей мере у 50% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению вызывает титр по меньшей мере 1:8 против Е.coli серотипа O25b по меньшей мере у 60%, 70%, 80% или по меньшей мере 90% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипа O25b (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает титры ОРА у людей против Е.coli серотипа O25b по сравнению с ранее иммунизированной популяцией.
В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O1a, где n представляет собой целое число больше 30, предпочтительно n представляет собой целое число от 31 до 100. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O1a, где n равно 39±2. В другом аспекте изобретение относится к композиции, которая включает полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O1a, где n равно 13±2. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R1 корового сахарида Е.coli. В одном из вариантов осуществления сахарид дополнительно включает фрагмент KDO. Предпочтительно белок-носитель представляет собой CRM197. В одном из вариантов осуществления конъюгат получают в результате одноконцевой конъюгации. В одном из вариантов осуществления конъюгат получают химическим методом восстановительного аминирования,
предпочтительно в DMSO буфере. В одном из вариантов осуществления сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер. Предпочтительно композиция дополнительно включает фармацевтически приемлемый разбавитель.
В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны связываться с полисахаридом Е.coli серотипа O1a в концентрации по меньшей мере 0,2 пг/мл, 0,3 пг/мл, 0,35 пг/мл, 0,4 пг/мл или 0,5 пг/мл, как определено с помощью анализа ELISA. Следовательно, можно выполнить сравнение активности ОРА сыворотки до и после иммунизации иммуногенной композицией по изобретению и сравнить ответы на серотип O1a, чтобы оценить потенциальное увеличение числа пациентов с ответом на лечение. В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны убивать Е.coli серотипа O1a, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция индуцирует функциональные антитела у людей, причем указанные антитела способны убивать Е.coli серотипа O1a, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипа O1a (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция вызывает титр по меньшей мере 1:8 против Е.coli серотипа O1a по меньшей мере у 50% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению вызывает титр по меньшей мере 1:8 против Е.coli серотипа O1a по меньшей мере у 60%, 70%, 80% или по меньшей мере 90% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает долю пациентов, с ответом на лечение инфекции Е.coli серотипа O1a (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает титры ОРА у людей против Е.coli серотипа O1a по сравнению с ранее иммунизированной популяцией.
В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O2, где n представляет собой целое число больше 30, предпочтительно n равно целое число от 31 до 100. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O2, где n равно 43±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O2, где n равно 47±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O2, где n равно 17±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O2, где n равно 18±2. В одном варианте осуществления сахарид дополнительно включает фрагмент R1 корового сахарида Е. coll. В другом варианте осуществления сахарид дополнительно включает фрагмент R4 корового сахарида Е. coll. В другом варианте осуществления сахарид дополнительно включает фрагмент KDO. Предпочтительно белок-носитель представляет собой CRM197. В одном из вариантов осуществления конъюгат получают в результате одноконцевой конъюгации. В одном из вариантов осуществления конъюгат получают химическим методом восстановительного аминирования, предпочтительно в DMSO буфере. В одном из вариантов осуществления сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер. Предпочтительно композиция дополнительно включает фармацевтически приемлемый разбавитель.
В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны связываться с полисахаридом Е.coli серотипа O2 в концентрации по меньшей мере 0,2 пг/мл, 0,3 пг/мл, 0,35 пг/мл, 0,4 пг/мл или 0,5 пг/мл, как определено с помощью анализа ELISA. Таким образом, можно выполнить сравнение активности ОРА сыворотки до и после иммунизации иммуногенной композицией по изобретению и сравнить ответы на серотип O2, чтобы оценить потенциальное увеличение пациентов с ответом на лечение. В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны убивать Е.coli серотипа O2, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция индуцирует образование у людей функциональных антител, причем указанные антитела способны убивать Е.coli серотипа O2, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипа O2 (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция вызывает титр по меньшей мере 1:8 против Е.coli серотипа O2 по меньшей мере у 50% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению вызывает титр по меньшей мере 1:8 против Е.coli серотипа O2 по меньшей мере у 60%, 70%, 80% или по меньшей мере 90% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипов O2 (т.е. индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает титры ОРА у людей против Е.coli серотипа O2 по сравнению с ранее иммунизированной популяцией.
В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу О6, где n представляет собой целое число больше 30, предпочтительно n представляет собой целое число от 31 до 100. В одном из аспектов изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу О6, где n равно 42±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O6, где n равно 50±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу O6, где n равно 17±2. В другом аспекте изобретение относится к композиции, которая включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает формулу О6, где n равно 18±2. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R1 корового сахарида Е.coli. В одном из вариантов осуществления сахарид дополнительно включает фрагмент KDO. Предпочтительно белок-носитель представляет собой CRM197. В одном из вариантов осуществления конъюгат получают в результате одноконцевой конъюгации. В одном из вариантов осуществления конъюгат получают химическим методом восстановительного аминирования, предпочтительно в DMSO буфере. В одном из вариантов осуществления сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер. Предпочтительно композиция дополнительно включает фармацевтически приемлемый разбавитель.
В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны связываться с полисахаридом Е.coli серотипа O6 в концентрации по меньшей мере 0,2 пг/мл, 0,3 пг/мл, 0,35 пг/мл, 0,4 пг/мл или 0,5 пг/мл, как определено с помощью анализа ELISA. Таким образом, можно выполнить сравнение активности ОРА сыворотки до и после иммунизации иммуногенной композицией по изобретению и сравнить ответы на серотип O6, чтобы оценить потенциальное увеличение числа пациентов с ответом на лечение. В одном из вариантов осуществления иммуногенная композиция индуцирует образование антител IgG у людей, причем указанные антитела способны убивать Е.coli серотипа О6, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция индуцирует образование у людей функциональных антител, причем указанные антитела способны убивать Е.coli серотипа О6, как определено с помощью опсонофагоцитарного анализа in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению увеличивает долю пациентов с ответом на лечение инфекции Е.coli серотипа О6 (т.е. индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция вызывает титр по меньшей мере 1:8 против Е.coli серотипа O6 по меньшей мере у 50% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению вызывает титр по меньшей мере 1:8 против Е.coli серотипа О6 по меньшей мере у 60%, 70%, 80% или по меньшей мере 90% субъектов, как определено с помощью анализа опсонофагоцитарного уничтожения in vitro. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает долю пациентов с ответом на лечение против Е.coli серотипа O6 (т.е., индивидуумов с сывороткой, имеющей титр по меньшей мере 1:8, определенный по ОРА in vitro) по сравнению с ранее иммунизированной популяцией. В одном из вариантов осуществления иммуногенная композиция по изобретению значимо увеличивает титры ОРА у людей против Е.coli серотипа O6 по сравнению с ранее иммунизированной популяцией.
В одном из аспектов композиция включает мутантный полипептид FimH и конъюгат, включающий сахарид, ковалентно связанный с белком-носителем, где сахарид включает структуру, выбранную из любой из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45 rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы О60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187, где n представляет собой целое число от 1 до 100, предпочтительно от 31 до 90. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R1 корового сахарида Е.coli. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R2 корового сахарида Е.coli. В одном из вариантов осуществления сахарид дополнительно включает фрагмент R3 корового сахарида Е.coli. В другом варианте осуществления сахарид дополнительно включает фрагмент R4 корового сахарида Е.coli. В одном из вариантов осуществления сахарид дополнительно включает фрагмент К12 корового сахарида Е.coli. В другом варианте осуществления сахарид дополнительно включает фрагмент KD0. Предпочтительно белок-носитель представляет собой CRM197. В одном из вариантов осуществления конъюгат получают в результате одноконцевой конъюгации. В одном из вариантов осуществления конъюгат получают химическим методом восстановительного аминирования, предпочтительно в DMSO буфере. В одном из вариантов осуществления сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер. Предпочтительно композиция дополнительно включает фармацевтически приемлемый разбавитель. В одном из вариантов осуществления композиция дополнительно включает по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28 или 29 дополнительных конъюгатов, максимально до 30 дополнительных конъюгатов, где каждый конъюгат включает сахарид, ковалентно связанный с белком-носителем, и где сахарид включает структуру, выбранную из любой из указанных формул.
А. Сахариды
1. Сахариды и О-полисахариды
В одном из вариантов осуществления сахарид получают путем экспрессии (не обязательно сверхэкспрессии) различных белков Wzz (например, WzzB) для контроля размера сахарида.
В контексте настоящего описания термин «сахарид» относится к одному сахарному фрагменту или моносахаридному звену, а также к комбинациям двух или более отдельных сахарных фрагментов или моносахаридных звеньев, ковалентно связанных с образованием дисахаридов, олигосахаридов и полисахаридов. Сахарид может быть линейным или разветвленным.
В одном из вариантов осуществления сахарид продуцируют в рекомбинантной грамотрицательной бактерии. В одном из вариантов осуществления сахарид продуцируют в рекомбинантной клетке Е.coli. В одном из вариантов осуществления сахарид продуцируют в рекомбинантной клетке сальмонеллы. Примеры бактерий включают Е.coli O25K5H1, Е.coli BD559, Е.coli GAR2831, Е.coli GAR865, Е.coli GAR868, Е.coli GAR869, Е.coli GAR872, Е.coli GAR878, Е.coli GAR896, Е.coli GAR1902, Е.coli O25a ETC NR-5, Е.coli O157:Н7:K-, штамм LT2 серовара Typhimurium Salmonella enterica, Е.coli GAR2401, CVD 1943 серотипа Enteritidis Salmonella enterica, CVD 1925 серотипа Typhimurium Salmonella enterica, CVD 1902 серотипа Paratyphi A Salmonella enterica и CVD 1208S Shigella flexneri. В одном из вариантов осуществления бактерия не является Е.coli GAR24 O1. Этот генетический подход к продуцированию сахаридов позволяет эффективно продуцировать О-полисахариды и молекулы О-антигена в качестве компонентов вакцины.
Термин «белок wzz», используемый в настоящем описании, относится к полипептиду, определяющему длину цепи, такому как, например, wzzB, wzz, wzzSF, wzzST, fepE, wzzfepE, wzzl и wzz2. Номера доступа в GenBank для иллюстративных последовательностей генов wzz являются следующими: AFO11910 для Е4991/76, AF011911 для F186, AF011912 для М70/1-1, AF011913 для 79/311, AF011914 для Bi750-41, AF01915 для С664-1992, AF01916 для С258-94, AF01917 для С722-89 и AF01919 для EDL933. Номера доступа в GenBank для последовательностей генов G7 и Bi316-41 wzz являются следующими: U3930 и U3930, соответственно. Дополнительные номера доступа в GenBank для типичных последовательностей генов wzz являются следующими: NP 459581 для Salmonella enterica подтип Enterica, серовар Typhimurium, штамм LT2 FepE; AIG66859 для штамма Е.coli O157:Н7 EDL933 FepE; NP_46104 для Salmonella enterica, подтип Enterica, серовар Typhimurium, штамм LT2 WzzB; NP_416531 для E.coli K-12, подштамм MG1655 WzzB; NP_415119 для E.coli K-12, подштамм MG1655 FepE. В предпочтительных аспектах белок семейства wzz представляет собой любой из белков wzzB, wzz, wzzSF, wzzST, fepE, wzzfepE, wzzl и wzz2, наиболее предпочтительно wzzB, более предпочтительно fepE.
Примеры последовательностей wzzB включают последовательности, представленные в SEQ ID No: 112-116. Примеры последовательностей FepE включают последовательности, представленные в SEQ ID No: 117-121.
В некоторых аспектах модифицированный сахарид (модифицированный относительно соответствующего сахарида дикого типа) может быть получен путем экспрессии (не обязательно сверхэкспрессии) белка семейства wzz (например, fepE) из грамотрицательной бактерии и/или путем выключения (т.е. репрессии, делеции, удаления) в грамотрицательной бактерии второго гена wzz (например, wzzB) с получением высокомолекулярных сахаридов, таких как липополисахариды, содержащие промежуточные или длинные цепи О-антигена, которые имеют повышенное количество повторяющихся звеньев относительно соответствующего О-полисахарида дикого типа. Например, модифицированные сахариды могут быть получены путем экспрессии (не обязательно сверхэкспрессии) wzz2 и выключения wzz1 Либо, в качестве альтернативы, модифицированные сахариды могут быть получены путем экспрессии (не обязательно сверхэкспрессии) wzz/fepE и выключения wzzB. В другом варианте осуществления модифицированные сахариды могут быть получены путем экспрессии (не обязательно сверхэкспрессии) wzzB, но с выключением wzz/fepE. В другом варианте осуществления модифицированные сахариды могут быть получены путем экспрессии fepE. Предпочтительно белок семейства wzz получают из штамма, гетерологичного клетке-хозяину. Способы определения длины сахаридов известны в данной области. Такие методы включают, без ограничения, ядерный магнитный резонанс, масс-спектроскопию и эксклюзионную хроматографию. Способы получения раскрытых в настоящем описании высокомолекулярных сахаридов, таких как липополисахариды, содержащих промежуточные или длинные цепи О-антигены, описаны в международной публикация РСТ WO 2020/039359 и соответствующей публикации США US 2020/0061177, каждая из которых полностью включена в настоящее описание посредством ссылки.
В некоторых вариантах осуществления сахарид получают путем экспрессии белка семейства wzz, имеющего аминокислотную последовательность, которая на по меньшей мере 30%, 50%, 70%, 75%, 80%, 85%, 90%, 95%, 98% 99% или 100% идентична последовательности любой из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121. В одном из вариантов осуществления белок семейства wzz включает последовательность, выбранную из любой из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121. Предпочтительно, белок семейства wzz имеет последовательность, которая на по меньшей мере 30%, 50%, 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентична последовательности из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116. В некоторых вариантах осуществления сахарид получают путем экспрессии белка, аминокислотная последовательность которого на по меньшей мере 30%, 50% 70%, 75%, 80%, 85%, 90%, 95%, 98%, 99% или 100% идентична последовательности белка fepE.
В одном из аспектов изобретение относится к сахаридам, полученным путем экспрессии белка семейства wzz, предпочтительно fepE, в грамотрицательной бактерии с образованием высокомолекулярных сахаридов, содержащих промежуточные или длинные цепи О-антигена, которые увеличены на по меньшей мере 1, 2, 3, 4 или 5 повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа. В одном из аспектов изобретение относится к сахаридам, продуцируемым грамотрицательной бактерией в культуре, которая экспрессирует (не обязательно сверхэкспрессирует) белок семейства wzz (например, wzzB) из грамотрицательной бактерии с образованием высокомолекулярных сахаридов, содержащих промежуточные или длинные цепи О-антигена, которые увеличены на по меньшей мере 1, 2, 3, 4 или 5 повторяющихся звеньев по сравнению с соответствующим О-антигеном дикого типа. Описание O-полисахаридов и О-антигенов для дополнительных иллюстративных сахаридов, имеющих увеличенное количество повторяющихся звеньев по сравнению с соответствующими сахаридами дикого типа, представлено ниже. Желаемой длиной цепи является длина цепи, обеспечивающая улучшенную или максимальную иммуногенность в контексте данной конструкции вакцины.
В другом варианте осуществления сахарид включает любую формулу, выбранную из Таблицы А, где количество повторяющихся звеньев n в сахариде больше, чем количество повторяющихся звеньев в соответствующем О-полисахариде дикого типа на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более повторяющихся звеньев. Предпочтительно сахарид увеличен на по меньшей мере 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа. Способы определения длины сахаридов известны в данной области. Такие способы включают ядерный магнитный резонанс, масс-спектроскопию и эксклюзионную хроматографию.
В предпочтительном варианте осуществления изобретение относится к сахариду, продуцируемому в рекомбинантной клетке-хозяине Е.coli, где ген эндогенного регулятора длины О-антигена wzz (например, wzzB) удален и заменен (вторым) геном wzz из грамотрицательной бактерии, гетерологичной рекомбинантной клетке-хозяину Е.coli (например, Salmonella fepE), с образованием высокомолекулярных сахаридов, таких как липополисахариды, содержащие промежуточные или длинные цепи O-антигена. В некоторых вариантах осуществления рекомбинантная клетка-хозяин Е.coli включает ген wzz из Salmonella, предпочтительно из Salmonella enterica.
В одном из вариантов осуществления клетка-хозяин включает гетерологичный ген белка семейства wzz в виде стабильно поддерживаемого плазмидного вектора. В другом варианте осуществления клетка-хозяин включает гетерологичный ген белка семейства wzz в виде гена, встроенного в хромосомную ДНК клетки-хозяина. Способы стабильной экспрессии плазмидного вектора в клетке-хозяине Е.coli и способы интеграции гетерологичного гена в хромосому клетки-хозяина Е.coli известны в данной области. В одном из вариантов осуществления клетка-хозяин включает гетерологичные гены О-антигена в виде стабильно поддерживаемого плазмидного вектора. В другом варианте осуществления клетка-хозяин включает гетерологичные гены О-антигена в виде гена, встроенного в хромосомную ДНК клетки-хозяина. Способы стабильной экспрессии плазмидного вектора в клетке-хозяине Е.coli и клетке-хозяине Salmonella известны в данной области. Способы интеграции гетерологичного гена в хромосому клетки-хозяина Е.coli и клетки-хозяина Salmonella известны в данной области.
В одном из аспектов рекомбинантную клетку-хозяин культивируют в среде, содержащей источник углерода. Источники углерода для культивирования Е.coli известны в данной области техники. Примеры источников углерода включают сахарные спирты, полиолы, альдольные сахара или кетосахара, включая арабинозу, целлобиозу, фруктозу, глюкозу, глицерин, инозитол, лактозу, мальтозу, маннит, маннозу, рамнозу, раффинозу, сорбит, сорбозу, сахарозу, трегалозу, пируват, сукцинат и метиламин. В предпочтительном варианте осуществления среда включает глюкозу. В некоторых вариантах осуществления среда включает в качестве источника углерода полиол или альдольный сахар, например, маннит, инозитол, сорбозу, глицерин, сорбит, лактозу и арабинозу. Все источники углерода могут быть добавлены в среду до начала культивирования, либо их можно добавлять поэтапно или непрерывно во время культивирования.
Типичная культуральная среда для рекомбинантной клетки-хозяина включает элемент, выбранный из любого из: KH2PO4, K2HPO4, (NH4)2SO4, цитрата натрия, Na2SO4, аспарагиновой кислоты, глюкозы, MgSO4, FeSO4-7H2O, Na2MoO4-2H2O, H3BO3, CoCl2-6Н2О, CuCl2-2H2O, MnCl2-4H2O, ZnCl2 и CaCl2-2Н2О. Предпочтительно среда включает KH2PO4, K2HPO4, (NH4)2SO4, цитрат натрия, Na2SO4, аспарагиновую кислоту, глюкозу, MgSO4, FeSO4-7H2O, Na2MoO4-2H2O, H3BO3, CoCl2-6Н2О, CuCl2-2H2O, MnCl2-4H2O, ZnCl2 и CaCl2-2H2O.
Используемая в контексте настоящего описания среда может быть твердой или жидкой, синтетической (т.е. искусственной) или природной и может включать достаточное количество питательных веществ для культивирования рекомбинантной клетки-хозяина. Предпочтительно среда представляет собой жидкую среду.
В некоторых вариантах осуществления среда может дополнительно включать подходящие неорганические соли. В некоторых вариантах осуществления среда может дополнительно включать микроэлементы питательных веществ. В некоторых вариантах осуществления среда может дополнительно включать факторы роста. В некоторых вариантах осуществления среда может дополнительно включать дополнительный источник углерода. В некоторых вариантах осуществления среда может дополнительно включать подходящие неорганические соли, микроэлементы, факторы роста и дополнительный источник углерода. Неорганические соли, следовые питательные вещества, факторы роста и дополнительные источники углерода, подходящие для культивирования Е.coli, известны в данной области.
В некоторых вариантах осуществления при необходимости среда может включать дополнительные компоненты, такие как пептон, N-Z амин, ферментативный гидрозилат сои, дополнительный дрожжевой экстракт, солодовый экстракт, дополнительные источники углерода и различные витамины. В некоторых вариантах осуществления среда не включает такие дополнительные компоненты, как пептон, N-Z амин, ферментативный гидрозилат сои, дополнительный дрожжевой экстракт, солодовый экстракт, дополнительные источники углерода и различные витамины.
Иллюстративные примеры подходящих дополнительных источников углерода включают, без ограничения, другие углеводы, такие как глюкоза, фруктоза, маннит, крахмал или гидролизат крахмала, гидролизат целлюлозы и меласса; органические кислоты, такие как уксусная кислота, пропионовая кислота, молочная кислота, муравьиная кислота, яблочная кислота, лимонная кислота и фумаровая кислота; и спирты, такие как глицерин, инозитол, маннит и сорбит.
В некоторых вариантах осуществления среда дополнительно включает источник азота. Источники азота, подходящие для культивирования Е.coli, известны в данной области. Иллюстративные примеры подходящих источников азота включают, без ограничения, аммиак, включая газообразный аммиак и водный раствор аммиака; аммониевые соли неорганических или органических кислот, такие как хлорид аммония, нитрат аммония, фосфат аммония, сульфат аммония и ацетат аммония; мочевину; нитратные или нитритные соли и другие азотсодержащие материалы, включая аминокислоты в чистом или неочищенном виде, мясной экстракт, пептон, рыбную муку, рыбный гидролизат, кукурузный экстракт, гидролизат казеина, гидролизат соевого жмыха, дрожжевой экстракт, сухие дрожжи, этанол-дрожжевой дистиллят, соевую муку, хлопковую муку и т.п.
В некоторых вариантах осуществления среда включает неорганическую соль. Иллюстративные примеры подходящих неорганических солей включают, без ограничения, соли калия, кальция, натрия, магния, марганца, железа, кобальта, цинка, меди, молибдена, вольфрама и других микроэлементов, а также фосфорную кислоту.
В некоторых вариантах осуществления среда включает соответствующие факторы роста. Иллюстративные примеры подходящих микроэлементов, факторов роста и т.п. включают, без ограничения, кофермент А, пантотеновую кислоту, пиридоксин-HCl, биотин, тиамин, рибофлавин, флавинмононуклеотид, флавинадениндинуклеотид, DL-6,8-тиоктовую кислоту, фолиевую кислоту, витамин В12, другие витамины, аминокислоты, такие как цистеин и гидроксипролин, основания, такие как аденин, урацил, гуанин, тимин и цитозин, тиосульфат натрия, н- или п-аминобензойную кислоту, ниацинамид, нитрилоацетат и т.п., либо в виде чистых или частично очищенных химических соединений, либо в виде природных материалов. Количества могут быть определены специалистом в данной области эмпирически в соответствии со способами и методиками, известными в данной области.
В другом варианте осуществления модифицированный сахарид (по сравнению с соответствующим сахаридом дикого типа), представленный в настоящем описании, получают синтетическим путем, например, in vitro. Синтетическое производство или синтез сахаридов может способствовать отказу от дорогостоящих и длительных производственных процессов. В одном из вариантов осуществления сахарид синтезируют, например, согласно стратегии последовательного гликозилирования или комбинации последовательного гликозилирования и стратегии [3+2]-блока синтеза из подходящим образом защищенных моносахаридных промежуточных соединений. Например, при гликозилировании в качестве доноров гликозила могут быть использованы тиогликозиды и производные гликозил-три-хлорацетимидата. В одном из вариантов осуществления сахарид, который синтетически синтезирован in vitro, имеет структуру, идентичную сахариду, полученному рекомбинантными способами, такими как манипулирование белком семейства wzz, описанным выше.
Сахарид, полученный (с помощью рекомбинантных или синтетических средств), включает структуру, полученную из любого серотипа Е.coli, включая, например, любой из следующих серотипов Е.coli: O1 (например, O1A, O1B, и O1C), O2, O3, O4 (например, O4:K52 и O4:K6), O5 (например, O5ab и O5ac (штамм 180/С3), O6 (например, O6:K2; К13; К15 и O6:K54), O7, O8, O9, O10, O11, O12, O13, O14, O15, O16, O17, O18 (например, O18A, O18ac, O18A1, O18B, и O18B1), O19, O20, O21, O22, O23 (например, O23A), O24, O25 (например, O25a и O25b), O26, O27, O28, O29, O30, O32, O33, O34, O35, O36, O37, O38, O39, O40, O41, O42, O43, O44, O45 (например, O45 и O45 rel), O46, O48, O49, O50, O51, O52, O53, O54, O55, O56, O57, O58, O59, O60, O61, O62, 62D1, O63, O64, O65, O66, O68, O69, O70, O71, O73 (например, O73 (штамм 73-1)), O74, O75, O76, O77, O78, O79, O80, O81, O82, O83, O84, O85, O86, O87, O88, O89, O90, O91, O92, O93, O95, O96, O97, O98, O99, O100, O101, O102, O103, O104, O105, O106, O107, O108, O109, О110, O111, O112, O113, O114, O115, O116, O117, O118, O119, O120, O121, O123, O124, O125, O126, O127, O128, O129, O130, O131, O132, O133, O134, O135, O136, O137, O138, O139, O140, O141, O142, O143, O144, O145, O146, O147, O148, O149, O150, O151, O152, O153, O154, O155, O156, O157, O158, O159, O160, O161, O162, O163, O164, O165, O166, O167, O168, O169, O170, O171, O172, O173, O174, O175, O176, O177, O178, O179, O180, O181, O182, O183, O184, O185, O186 и O187.
Индивидуальные полисахариды обычно очищают (обогащают относительно количества конъюгата полисахарид-белок) способами, известными в данной области техники, такими как, например, диализ, операции концентрирования, операции диафильтрации, фильтрация в тангенциальном потоке, осаждение, элюирование, центрифугирование, преципитация, ультрафильтрация, глубинная фильтрация и/или колоночная хроматография (ионообменная хроматография, мультимодальная ионообменная хроматография, DEAE и хроматография гидрофобного взаимодействия). Полисахариды предпочтительно очищают методом, который включает фильтрацию в тангенциальным потоке.
Очищенные полисахариды могут быть активированы (например, химически активированы), что наделяет их способностью реагировать (например, либо непосредственно с белком-носителем, либо через линкер, такой как спейсер еТЕС), а затем включены в гликоконъюгаты по изобретению, как дополнительно раскрыто в настоящем описании.
В одном из предпочтительных вариантов осуществления сахарид по изобретению получают из серотипа Е.coli, где серотип представляет собой O25a. В другом предпочтительном варианте осуществления серотип представляет собой O25b. В другом предпочтительном варианте осуществления серотип представляет собой O1A. В другом предпочтительном варианте осуществления серотип представляет собой O2. В другом предпочтительном варианте осуществления серотип представляет собой O6. В другом предпочтительном варианте осуществления серотип представляет собой O17. В другом предпочтительном варианте осуществления серотип представляет собой O15. В другом предпочтительном варианте осуществления серотип представляет собой O18A. В другом предпочтительном варианте осуществления серотип представляет собой O75. В другом предпочтительном варианте осуществления серотип представляет собой O4. В другом предпочтительном варианте осуществления серотип представляет собой O16. В другом предпочтительном варианте осуществления серотип представляет собой O13. В другом предпочтительном варианте осуществления серотип представляет собой O7. В другом предпочтительном варианте осуществления серотип представляет собой O8. В другом предпочтительном варианте осуществления серотип представляет собой O9.
Используемая в настоящем описании ссылка на любой из перечисленных выше серотипов относится к серотипу, который включает структуру повторяющихся звеньев (0-звена, как описано ниже), известную в данной области, и является уникальной для соответствующего серотипа. Например, термин серотип «O25a» (также известный в данной области как серотип «O25») относится к серотипу, который охватывает формулу O25, приведенную в таблице А. В качестве другого примера термин серотип «O25b» относится к серотипу, который охватывает формулу O25b, приведенную в Таблице А.
Используемые в настоящем описании серотипы упоминаются в общем, если не указано иное, например, термин «формулы «O18» в общем относится к формуле O18A, формуле O18ac, формуле 18А1, формуле O18B и формуле O18B1.
Используемый в настоящем описании термин «O1» в общем относится к разновидностям формулы, названия которых включают общий термин «O1» в соответствии с таблицей А, например к любой из: формулы O1A, формулы O1A1, формулы O1B и формулы O1C, каждая из которых показана в таблице А. Соответственно, «серотип O1» в общем относится к серотипу, который охватывает любую формулу из: формулы O1A, формулы O1A1, формулы O1B и формулы O1C.
В контексте настоящего описания термин «O6» в общем относится к разновидностям формулы, названия которых включают общий термин «O6» в соответствии с Таблицей А, например к любой формуле из: O6:K2; К13; К15; и O6:K54, каждая из которых показана в таблице А. Соответственно, «серотип O6» в общем относится к серотипу, который охватывает любую из формул: O6:K2; K13; K15; и O6:K54.
Другие примеры терминов, которые в общем относятся к разновидностям формулы, названия которых включают общий термин в соответствии с Таблицей А, включают: «O4», «O5», «O18» и «O45».
В контексте настоящего описания термин «O2» относится к формуле O2, показанной в таблице А. Термин «О-антиген O2» относится к сахариду, который охватывает формулу O2, показанную в таблице А.
В контексте настоящего описания ссылка на О-антиген указанного выше серотипа относится к сахариду, который охватывает формулу с указанием соответствующего названия серотипа. Например, термин «О-антиген O25 В» относится к сахариду, который охватывает формулу O25 В, показанную в Таблице А.
В качестве другого примера термин «О-антиген O1» в общем относится к сахариду, который охватывает формулу, включающую термин «O1», например формулы O1A, формулы O1A1, формулы O1B и формулы O1C, каждая из которых показана в Таблица А.
В качестве другого примера термин «О-антиген О6» в общем относится к сахариду, который охватывает формулу, включающую термин «О6», например формулы O6:K2; формулы 0б:K13; формулы O6:K15 и формулы О6:K54, каждая из которых показана в Таблице А.
В контексте настоящего описания термин «О-полисахарид» относится к любой структуре, которая включает О-антиген, при условии, что структура не включает цельную клетку или липид А. Например, в одном из вариантов осуществления О-полисахарид включает липополисахарид, где липид А не является связанным. Этап удаления липида А известен в данной области техники и включает, например, термическую обработку с добавлением кислоты. Иллюстративный процесс включает обработку 1% уксусной кислотой при 100°С в течение 90 минут. Этот процесс сочетается с процессом выделения липида А в качестве удаленного. Иллюстративный способ выделения липида А включает ультрацентрифугирование.
В одном из вариантов осуществления О-полисахарид относится к структуре, состоящей из О-антигена, и в этом случае O-полисахарид является синонимом термина О-антиген. В одном из предпочтительных вариантов осуществления О-полисахарид относится к структуре, которая включает повторяющиеся звенья О-антигена без корового сахарида. Соответственно, в одном из вариантов осуществления О-полисахарид не включает кор R1 Е.coli. В другом варианте осуществления О-полисахарид не включает кор R2 Е.coli. В другом варианте осуществления О-полисахарид не включает кор R3 Е.coli. В другом варианте осуществления О-полисахарид не включает кор R4 Е.coli. В другом варианте осуществления O-полисахарид не включает кор K12 Е.coli. В другом предпочтительном варианте осуществления О-полисахарид относится к структуре, которая включает О-антиген и коровый сахарид. В другом варианте осуществления О-полисахарид относится к структуре, которая включает О-антиген, коровый сахарид и фрагмент KD0.
Способы очистки О-полисахарида, который включает коровый олигосахарид, от LPS известны в данной области техники. Например, после очистки LPS очищенный LPS можно гидролизовать путем нагревания в 1% (об./об.) уксусной кислоте в течение 90 минут при 100 градусах Цельсия с последующим
ультрацентрифугированием при 142000 g в течение 5 часов при 4 градусах Цельсия. Супернатант, содержащий О-полисахарид, сушат путем лиофилизации и хранят при 4 градусах Цельсия. В некоторых вариантах осуществления описано удаление генов, необходимых для синтеза капсулы, для обеспечения простой очистки О-полисахарида.
О-полисахарид может быть выделен способами, включая, без ограничения, мягкий кислотный гидролиз для удаления липида А из LPS. Другие варианты осуществления могут включать использование гидразина в качестве агента для получения О-полисахарида. Получение LPS можно осуществить известными в данной области способами.
В некоторых вариантах осуществления в конъюгированных вакцинах используют О-полисахариды, очищенные от модифицированных или аттенуированных штаммов грамотрицательных бактерий дикого типа, которые экспрессируют (не обязательно сверхэкспрессируют) белок Wzz (например, wzzB). В предпочтительных вариантах осуществления О-полисахаридную цепь очищают от штамма грамотрицательных бактерий, экспрессирующего (не обязательно сверхэкспрессирующего) белок wzz, для использования в качестве вакцинного антигена либо в виде конъюгата, либо в виде комплексной вакцины.
В одном из вариантов осуществления О-полисахарид имеет молекулярную массу, которая увеличена примерно в 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 раз или более по сравнению с соответствующим О-полисахаридом дикого типа. В предпочтительном варианте осуществления О-полисахарид имеет молекулярную массу, увеличенную в по меньшей мере 1 раз, но не более чем в 5 раз по сравнению с соответствующим О-полисахаридом дикого типа. В другом варианте осуществления О-полисахарид имеет молекулярную массу, увеличенную в по меньшей мере 2 раза, но не более чем в 4 раза по сравнению с соответствующим О-полисахаридом дикого типа. Увеличение молекулярной массы О-полисахарида по сравнению с соответствующим О-полисахаридом дикого типа предпочтительно связано с увеличением количества повторяющихся звеньев О-антигена. В одном из вариантов осуществления увеличение молекулярной массы О-полисахарида обусловлено белком семейства wzz.
В одном из вариантов осуществления О-полисахарид имеет молекулярную массу, которая увеличена на примерно 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 кДа или более по сравнению с соответствующим О-полисахаридом дикого типа. В одном из вариантов осуществления О-полисахарид по изобретению имеет молекулярную массу, увеличенную на по меньшей мере 1, но не более чем на 200 кДа по сравнению с соответствующим О-полисахаридом дикого типа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 5, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 12, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 15, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличивается на по меньшей мере 18, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 21, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 22, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 30, но не более чем на 200 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 1, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 5, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 12, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 15, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 100 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 1, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 5, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 12, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 15, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 18, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 30, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 90 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 12, но не более чем на 85 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 75 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 70 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 60 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 50 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 49 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 48 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 47 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 10, но не более чем на 46 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 45 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 44 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 43 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 42 кДа. В одном из вариантов осуществления молекулярная масса увеличена на по меньшей мере 20, но не более чем на 41 кДа. Такое увеличение молекулярной массы О-полисахарида по сравнению с соответствующим О-полисахаридом дикого типа предпочтительно связано с увеличением количества повторяющихся звеньев О-антигена. В одном из вариантов осуществления увеличение молекулярной массы О-полисахарида обусловлено белком семейства wzz.
В другом варианте осуществления О-полисахарид включает любую формулу, выбранную из Таблицы А, где количество повторяющихся звеньев n в О-полисахариде больше, чем количество повторяющихся звеньев в соответствующем О-полисахариде дикого типа на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более повторяющихся звеньев. Предпочтительно сахарид увеличен на по меньшей мере 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа.
2. О-антиген
О-антиген входит в состав липополисахарида (LPS) внешней мембраны грамотрицательных бактерий. О-антиген находится на поверхности клетки и является вариабельным компонентом клетки. Вариабельность О-антигена служит основой для серотипирования грамотрицательных бактерий. Текущая схема серотипирования Е.coli включает О-полисахариды с 1 по 181.
О-антиген включает олигосахаридные повторяющиеся звенья (О-звенья), структура дикого типа которых обычно содержит от двух до восьми остатков Сахаров широкого спектра. О-единицы типичных О-антигенов Е.coli описаны в таблице А и в международной публикации РСТ № WO 2021/084429, опубликованной 6 мая 201 г., которая полностью включена в настоящее описание посредством ссылки. В некоторых вариантах осуществления настоящее изобретение включает композицию, содержащую по меньшей мере один мутантный полипептид FimH и по меньшей мере один из О-антигенов, описанных в Таблице Айв международной публикации РСТ № WO 2021/084429, опубликованной 6 мая 2021 г., которая полностью включена в настоящее описание посредством ссылки.
В одном из вариантов осуществления сахарид по изобретению может представлять собой одно олигосахаридное звено. В одном из вариантов осуществления сахарид по изобретению представляет собой одно повторяющееся олигосахаридное звено соответствующего серотипа. В таких вариантах осуществления сахарид может включать структуру, выбранную из формулы O1a, формулы O2, формулы O6, формулы O8, формулы O9a, формулы O9, формулы O20ab, формулы О20ас, формулы O25b, формулы O52, формулы O97 и формулы O101. В другом варианте осуществления сахарид может включать структуру, выбранную из формулы O1a, формулы O2, формулы O6 и формулы O25b.
В одном из вариантов осуществления сахарид по изобретению может представлять собой олигосахарид. Олигосахариды имеют небольшое количество повторяющихся звеньев (обычно 5-15 повторяющихся звеньев), и их обычно получают синтетическим путем или путем гидролиза полисахаридов. В таких вариантах осуществления сахарид может включать структуру, выбранную из формулы O1a, формулы O2, формулы O6, формулы O8, формулы O9a, формулы O9, формулы O20ab, формулы О20ас, формулы O25b, формулы O52, формулы O97 и формулы O101. В другом варианте осуществления сахарид может включать структуру, выбранную из формулы O1a, формулы O2, формулы O6 и формулы O25b.
Предпочтительно, чтобы все сахариды по настоящему изобретению, и в иммуногенных композициях по настоящему изобретению, были полисахаридами. Высокомолекулярные
полисахариды могут индуцировать определенные иммунные ответы антител, обусловленные эпитопами, присутствующими на антигенной поверхности. Использование в конъюгатах, композициях и способах по настоящему изобретению предпочтительно предусматривает выделение и очистку высокомолекулярных полисахаридов.
В некоторых вариантах осуществления количество повторяющихся O-звеньев в каждом отдельном полимере О-антигена (и, следовательно, длина и молекулярная масса полимерной цепи) зависит от регулятора длины цепи wzz, белка внутренней мембраны. Разные белки wzz обеспечивают разные диапазоны модальной длины (от 4 до >100 повторяющихся звеньев). Термин «модальная длина» относится к количеству повторяющихся О-звеньев. Грамотрицательные бактерии часто имеют два разных белка Wzz, которые обеспечивают две разные модальные длины цепей Oag: одну длиннее, а другую короче. Экспрессия (не обязательно сверхэкспрессия) белков семейства wzz (например, wzzB) в грамотрицательных бактериях позволяет манипулировать длиной О-антигена, чтобы сдвинуть или сместить бактериальное продуцирование О-антигенов, имеющих длину в определенном диапазоне, и чтобы увеличить продуцирование липополисахаридов с высоким выходом и высокой молекулярной массой. В одном из вариантов осуществления «короткая» модальная длина, используемая в настоящем описании, относится к небольшому количеству повторяющихся О-звеньев, например 1-20. В одном из вариантов осуществления «длинная» модальная длина, используемая в настоящем описании, относится к количеству повторяющихся O-звеньев, превышающему 20 с максимальным количеством 40. В одном из вариантов осуществления «очень длинная» модальная длина, используемая в настоящем описании, относится к более чем 40 повторяющимся O-звеньям.
В одном из вариантов осуществления полученный сахарид увеличен на по меньшей мере 10 повторяющихся звеньев, 15 повторяющихся звеньев, 20 повторяющихся звеньев, 25 повторяющихся звеньев, 30 повторяющихся звеньев, 35 повторяющихся звеньев, 40 повторяющихся звеньев, 45 повторяющихся звеньев, 50 повторяющихся звеньев, 55 повторяющиеся звенья, 60 повторяющихся звеньев, 65 повторяющихся звеньев, 70 повторяющихся звеньев, 75 повторяющихся звеньев, 80 повторяющихся звеньев, 85 повторяющихся звеньев, 90 повторяющихся звеньев, 95 повторяющихся звеньев или 100 повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа.
В другом варианте осуществления сахарид по изобретению увеличен на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа. Предпочтительно сахарид увеличен на по меньшей мере 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49 или 50 повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа. См., например, Таблицу 21. Способы определения длины сахаридов известны в данной области. Такие способы включают ядерный магнитный резонанс, масс-спектроскопию и эксклюзионную хроматографию, как описано в примере 13.
Способы определения количества повторяющихся звеньев в сахариде также известны в данной области. Например, количество повторяющихся звеньев (или «n» в формуле) можно рассчитать путем деления молекулярной массы полисахарида (без молекулярной массы корового сахарида или остатка KDO) на молекулярную массу повторяющегося звена (т.е. молекулярную массу структуры в соответствующей формуле, показанной, например, в Таблице 1, которая теоретически может быть рассчитана в виде суммы молекулярных масс каждого моносахарида в формуле). Молекулярная масса каждого моносахарида в формуле известна в данной области техники. Молекулярная масса повторяющегося звена формулы O25b, например, составляет примерно 862 Да. Молекулярная масса повторяющегося звена формулы O1a, например, составляет примерно 845 Да. Молекулярная масса повторяющегося звена формулы O2, например, составляет примерно 829 Да. Молекулярная масса повторяющегося звена формулы O6, например, составляет примерно 893 Да. При определении количества повторяющихся звеньев в конъюгате при вычислении учитываются молекулярная масса белка-носителя и соотношение белок:полисахарид. Как определено в настоящем описании, «n» относится к количеству повторяющихся звеньев (представленных в скобках в таблице 1) в молекуле полисахарида. Как известно в данной области, в биологических макромолекулах повторяющиеся структуры могут перемежаться областями несовершенных повторов, такими как, например, отсутствующие ответвления. Кроме того, в данной области техники известно, что полисахариды, выделенные и очищенные из природных источников, таких как бактерии, могут быть гетерогенными по размеру и разветвлению. В таком случае n может представлять собой среднее или медианное значение n для молекул в популяции.
В одном из вариантов осуществления О-полисахарид увеличен на по меньшей мере одно повторяющееся звено О-антигена по сравнению с соответствующим О-полисахаридом дикого типа. Повторяющиеся звенья О-антигенов показаны в таблице 1. В одном из вариантов осуществления О-полисахарид включает 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более повторяющихся звеньев. Предпочтительно сахарид имеет в общей сложности от по меньшей мере 3 до не более 80 повторяющихся звеньев. В другом варианте осуществления О-полисахарид увеличен на 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89, 90, 91, 92, 93, 94, 95, 96, 97, 98, 99, 100 или более повторяющихся звеньев по сравнению с соответствующим О-полисахаридом дикого типа.
В одном из вариантов осуществления сахарид включает O-антиген, где n в любой из формул О-антигена (такой как, например, формулы, показанные в таблице 1 (см. также фиг.9А-9С и фиг.10А-10 В)) является целым числом, равным по меньшей мере 1, 2, 3, 4, 5, 10, 20, 21, 22, 23, 24, 25, 26, 27, 28, 29, 30, 31, 32, 33, 34, 35, 36, 37, 38, 39, 40 и не более 200, 100, 99, 98, 97, 96, 95, 94, 93, 92, 91, 90, 89, 88, 87, 86, 81, 80, 79, 78, 77, 76, 75, 74, 73, 72, 71, 70, 69, 68, 67, 66, 65, 60, 59, 58, 57, 56, 55, 54, 53, 52, 51 или 50. Для определения диапазона могут быть объединены любое минимальное значение и любое максимальное значение. Иллюстративные диапазоны включают, например, от по меньшей мере 1 до не более 1000; от по меньшей мере 10 до не более 500; и от по меньшей мере 20 до не более 80, предпочтительно не более 90. В одном из предпочтительных вариантов осуществления n составляет от по меньшей мере 31 до не более 90. В предпочтительном варианте осуществления n составляет от 40 до 90, более предпочтительно от 60 до 85.
В одном из вариантов осуществления сахарид включает О-антиген, где n в любой из формул О-антигена составляет от по меньшей мере 1 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 5 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 10 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 25 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 50 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 75 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 100 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 125 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 150 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 175 до не более 200. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 1 до не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 5 до не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 10 до не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 25 до не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 50 до не более не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 75 до не более 100. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 1 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 5 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 10 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 20 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 25 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 30 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 40 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 50 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 30 до не более 90. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 85. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 75. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 70. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 60. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 50. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 50. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 49. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 48. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 47. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 35 до не более 46. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 36 до не более 45. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 37 до не более 44. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 38 до не более 43. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 39 до не более 42. В одном из вариантов осуществления n в любой из формул О-антигена составляет от по меньшей мере 39 до не более 41. Например, в одном из вариантов осуществления n в сахариде равно 31, 32, 33, 34, 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68, 69, 70, 71, 72, 73, 74, 75, 76, 77, 78, 79, 80, 81, 82, 83, 84, 85, 86, 87, 88, 89 или 90, наиболее предпочтительно 40. В другом варианте осуществления n составляет от не менее 35 до не более 60. Например, в одном из вариантов осуществления n равно любому числу из 35, 36, 37, 38, 39, 40, 41, 42, 43, 44, 45, 46, 47, 48, 49, 50, 51, 52, 53, 54, 55, 56, 57, 58, 59 и 60, предпочтительно 50. В другом предпочтительном варианте осуществления n составляет от по меньшей мере 55 до не более 75. Например, в одном из вариантов осуществления n равно 55, 56, 57, 58, 59, 60, 61, 62, 63, 64, 65, 66, 67, 68 или 69, наиболее предпочтительно 60.
Структура сахарида может быть определена способами и инструментами, известными в уровне техники, такими как, например, ЯМР, включая 1D, 1Н и/или 13С, 2D TOCSY, DQF-COSY, NOESY и/или HMQC.
В некоторых вариантах осуществления очищенный полисахарид перед конъюгацией имеет молекулярную массу от 5 кДа до 400 кДа. В других таких вариантах осуществления сахарид имеет молекулярную массу от 10 кДа до 400 кДа; от 5 кДа до 400 кДа; от 5 кДа до 300 кДа; от 5 кДа до 200 кДа; от 5 кДа до 150 кДа; от 10 кДа до 100 кДа; от 10 кДа до 75 кДа; от 10 кДа до 60 кДа; от 10 кДа до 40 кДа; от 10 кДа до 100 кДа; 10 кДа и 200 кДа; от 15 кДа до 150 кДа; от 12 кДа до 120 кДа; от 12 кДа до 75 кДа; от 12 кДа до 50 кДа; от 12 до 60 кДа; от 35 кДа до 75 кДа; от 40 кДа до 60 кДа; от 35 кДа до 60 кДа; от 20 кДа до 60 кДа; от 12 кДа до 20 кДа; или от 20 кДа до 50 кДа. В дополнительных вариантах осуществления полисахарид имеет молекулярную массу от 7 кДа до 15 кДа; от 8 кДа до 16 кДа; от 9 кДа до 25 кДа; от 10 кДа до 100; от 10 кДа до 60 кДа; от 10 кДа до 70 кДа; от 10 кДа до 160 кДа; от 15 кДа до 600 кДа; от 20 кДа до 1000 кДа; от 20 кДа до 600 кДа; от 20 кДа до 400 кДа; от 30 кДа до 1000 кДа; от 30 кДа до 60 кДа; от 30 кДа до 50 кДа или от 5 кДа до 60 кДа. Любое целое число в любом из вышеуказанных диапазонов рассматривается как вариант осуществления изобретения.
В контексте настоящего описания термин «молекулярная масса» полисахарида или конъюгата белок-носитель-полисахарид относится к молекулярной массе, рассчитанной с помощью эксклюзионной хроматографии (SEC) в комбинации с использованием детектора многоуглового лазерного светорассеяния (MALLS).
Во время обычных процедур очистки полисахарид может немного уменьшиться в размерах. Кроме того, как раскрыто в настоящем описании, перед конъюгацией полисахарид может быть подвергнут сортировке по размеру. Может быть использована механическая или химическая сортировке по размеру. Химический гидролиз можно выполнять с применением уксусной кислоты. Механическая сортировка по размеру может быть выполнена с помощью гомогенизации под высоким давлением с усилием сдвига. Упомянутые выше диапазоны молекулярной массы относятся к очищенным полисахаридам перед конъюгацией (например, перед активацией).
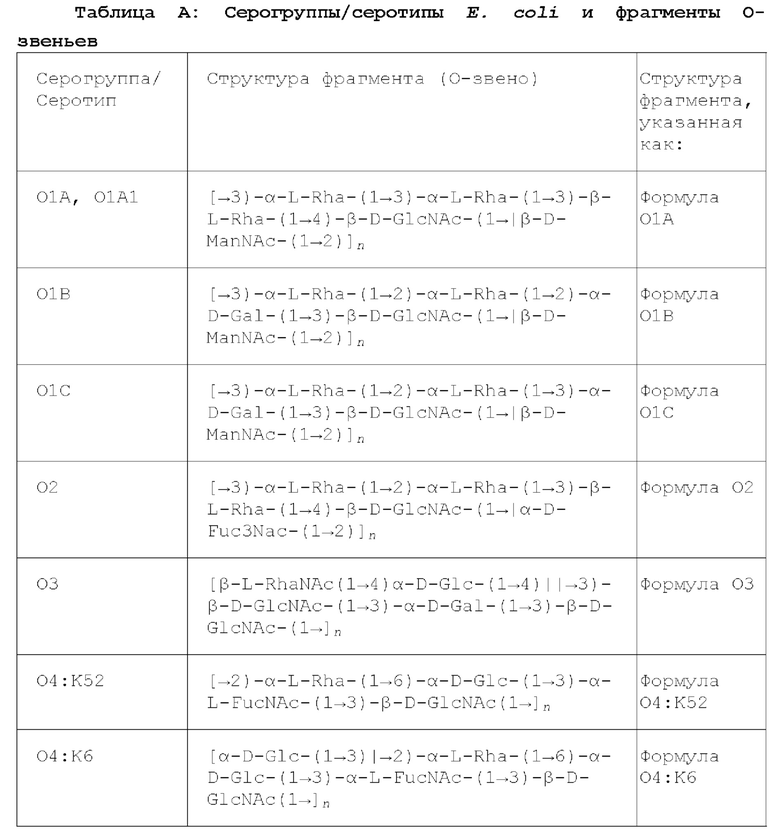
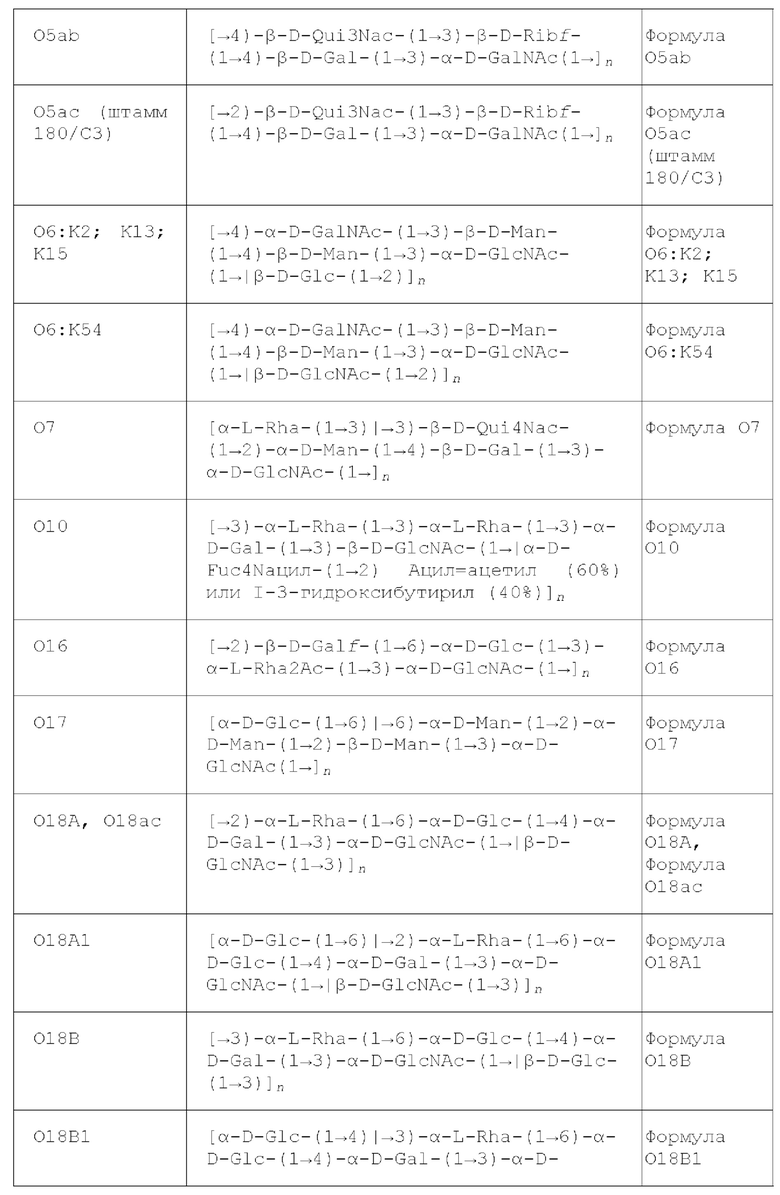
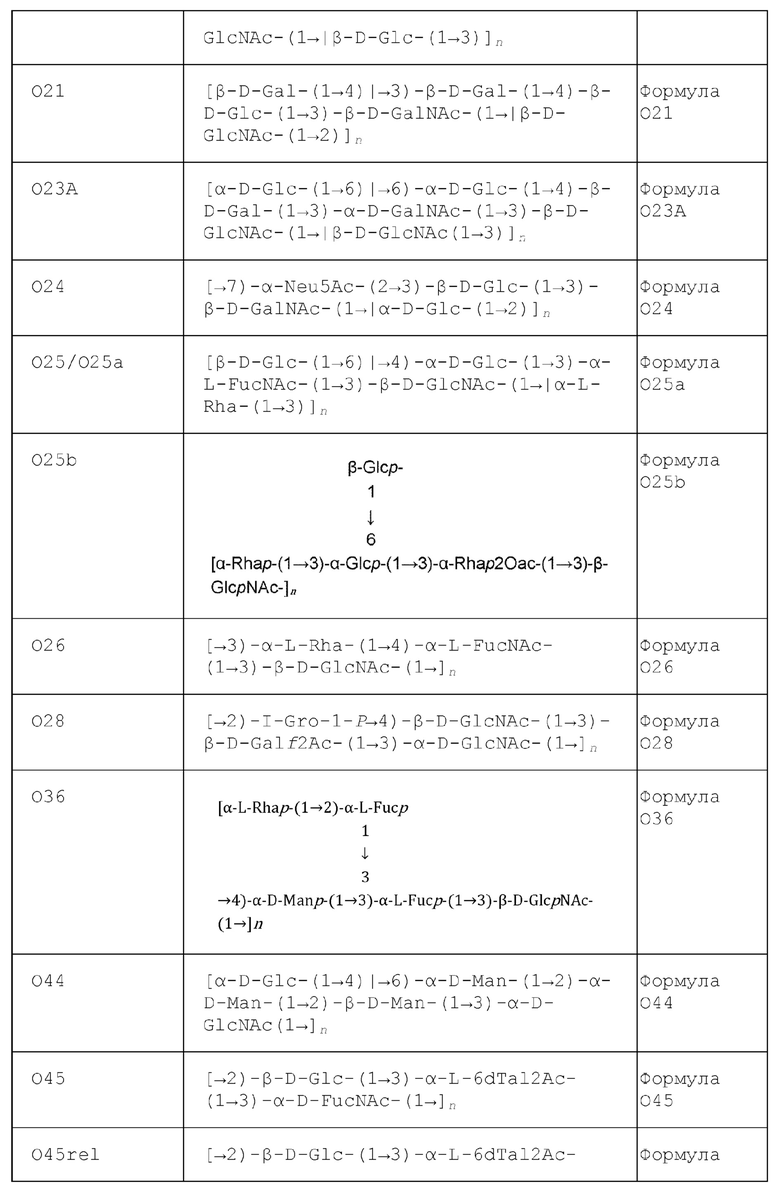

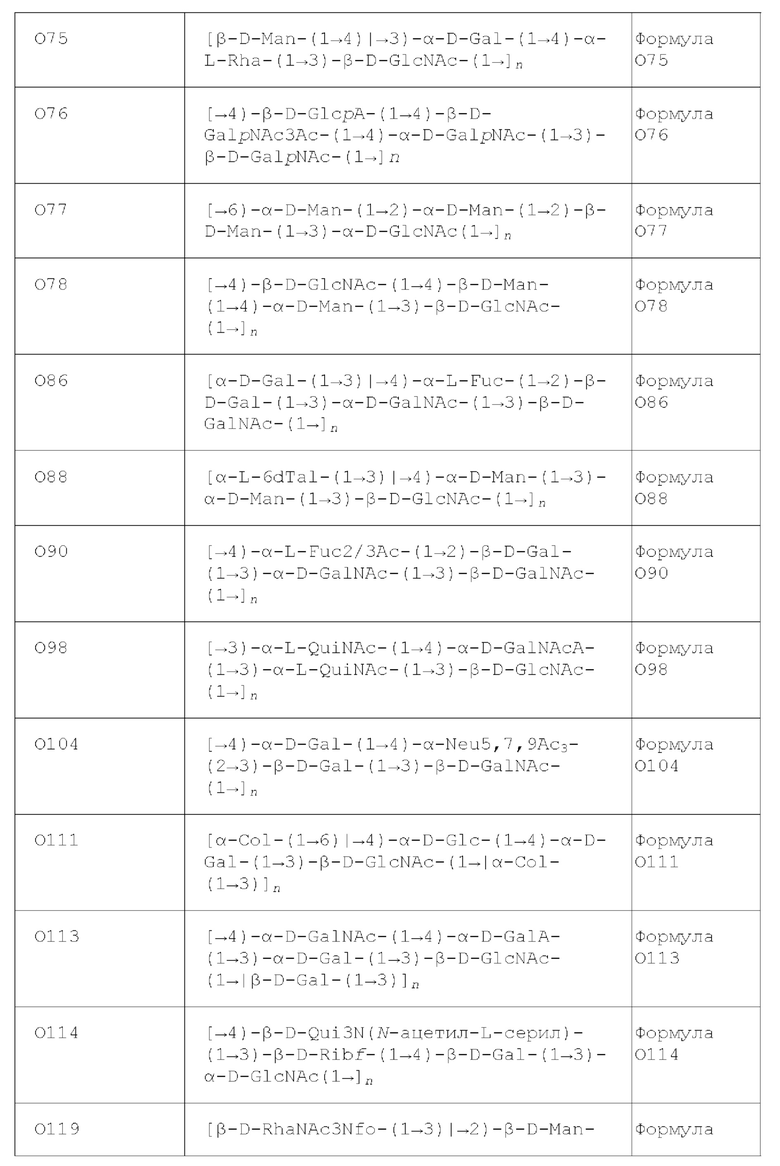

3. Коровый олигосахарид
Коровый олигосахарид расположен между липидом А и внешней областью О-антигена в LPS Е.coli дикого типа. Более конкретно, коровый олигосахарид представляет собой часть полисахарида, которая включает связь между О-антигеном и липидом А в Е.coli дикого типа. Эта связь включает кетозидную связь между полукетальной функциональной группой самого внутреннего остатка 3-дезокси-d-манно-окт-2-улозоновой кислоты (KDO)) и гидроксильной группой остатка GlcNAc липида А. Область корового олигосахарида демонстрирует высокую степень сходства среди штаммов Е.coli дикого типа. Обычно она включает ограниченное количество Сахаров. Коровый олигосахарид включает внутреннюю область кора и внешнюю область кора.
Более конкретно, внутренний кор состоит в основном из L-глицеро-D-манно-гептозы (гептозы) и остатков KDO. Внутренний кор очень консервативный. Остаток KDO включает следующую формулу KDO:
Внешняя область корового олигосахарида демонстрирует больше вариаций, чем внутренний кор, и эта область различается пятью хемотипами Е.coli: R1, R2, R3, R4 и К-12. Обобщенные структуры углеводного остова олигосахаридов внешнего кора пяти известных хемотипов хорошо известны в данной области. HepII является последним остатком олигосахарида внутреннего кора. Хотя все олигосахариды внешнего кора имеют общую структурную тему с (гексозой)3 углеводным остовом и двумя остатками боковой цепи, порядок гексоз в остове, а также природа, положение и связь остатков боковой цепи могут изменяться. Структуры олигосахаридов внешнего кора R1 и R4 очень похожи и различаются только одним β-связанным остатком.
Классификация коровых олигосахаридов Е.coli дикого типа в данной области основана на структуре дистального олигосахарида у пяти разных хемотипов: Е.coli R1, Е.coli R2, Е.coli R3, Е.coli R4 и Е.coli K12.
В предпочтительном варианте осуществления композиции, представленные в настоящем описании, включают гликоконъюгаты, у которых О-полисахарид включает коровый олигосахарид, связанный с О-антигеном. В одном из вариантов осуществления композиция индуцирует иммунный ответ против по меньшей мере любого из коровых хемотипов Е.coli: Е.coli R1, Е.coli R2, Е.coli R3, Е.coli RA и Е.coli K12. В другом варианте осуществления композиция индуцирует иммунный ответ против по меньшей мере двух коровых хемотипов Е.coli. В другом варианте осуществления композиция индуцирует иммунный ответ против по меньшей мере трех коровых хемотипов Е.coli. В другом варианте осуществления композиция индуцирует иммунный ответ против по меньшей мере четырех коровых хемотипов Е.coli. В другом варианте осуществления композиция индуцирует иммунный ответ против всех пяти коровых хемотипов Е.coli.
В другом предпочтительном варианте осуществления композиции, представленные в настоящем описании, включают гликоконъюгаты, в которых О-полисахарид не включает коровый олигосахарид, связанный с О-антигеном. В одном из вариантов осуществления такая композиция индуцирует иммунный ответ против по меньшей мере любого из коровых хемотипов Е.coli: Е.coli R1, Е.coli R2, Е.coli R3, Е.coli R4 и Е.coli К12, несмотря на гликоконъюгат, содержащий О-полисахарид, который не включает коровый олигосахарид.
Серотипы Е.coli можно охарактеризовать в соответствии с одним из пяти хемотипов. В таблице В перечислены иллюстративные серотипы, охарактеризованные в соответствии с хемотипом. Серотипы, выделенные жирным шрифтом, представляют собой серотипы, которые чаще всего связаны с указанным основным хемотипом. Соответственно, в предпочтительном варианте осуществления композиция индуцирует иммунный ответ против по меньшей мере любого из коровых хемотипов Е.coli: Е.coli R1, Е.coli R2, Е.coli R3, Е.coli R4 и Е.coli К12, который включает иммунный ответ против любого из соответствующих серотипов Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип R1, например, выбранный из сахарида, имеющего формулу O25a, формулу O6, формулу O2, формулу O1, формулу O75, формулу O4, формулу O16, формулу O8, формулу O18, формулу O9, формулу O13, формулу O20, формулу O21, формулу O91 и формулу O163, где n составляет от 1 до 100. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает коровый фрагмен R1 Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип R1, например, выбранный из сахарида, имеющего формулу O25a, формулу O6, формулу O2, формулу O1, формулу O75, формулу O4, формулу O16, формулу O18, формулу O13, формулу O20, формулу O21, формулу O91 и формулу O163, где n составляет от 1 до 100, предпочтительно от 31 до 100, предпочтительно от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает сахарид, включающий коровый фрагмент R1 Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип R2, например, выбранный из сахарида, имеющего формулу O21, формулу O44, формулу O11, формулу O89, формулу O162 и формулу O9, где n составляет от 1 до 100, предпочтительно от 31 до 100, предпочтительно от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает коровый фрагмент R2 Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип R3, например, выбранный из сахарида, имеющего формулу O25b, формулу O15, формулу O153, формулу O21, формулу O17, формулу O11, формулу O159, формулу O22, формулу O86 и формулу O93, где n составляет от 1 до 100, предпочтительно от 31 до 100, предпочтительно от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает коровый фрагмент R3 Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип R4, например, выбранный из сахарида, имеющего формулу O2, формулу O1, формулу O86, формулу O7, формулу O102, формулу O160 и формулу O166, где n составляет от 1 до 100, предпочтительно от 31 до 100, предпочтительно от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает коровый фрагмент R4 Е.coli.
В некоторых вариантах осуществления композиция включает сахарид, который включает структуру, полученную из серотипа, имеющего хемотип К-12 (например, выбранного из сахарида, имеющего формулу O25b, и сахарида, имеющего формулу O16), где n составляет от 1 до 1000, предпочтительно от 31 до 100, предпочтительно от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65. В некоторых вариантах осуществления сахарид в указанной композиции дополнительно включает коровый фрагмент К-12 Е.coli.
В некоторых вариантах осуществления сахарид включает коровый сахарид. Соответственно, в одном из вариантов осуществления О-полисахарид дополнительно включает коровый фрагмент R1 Е.coli. В другом варианте осуществления О-полисахарид дополнительно включает коровый фрагмент R2 Е.coli. В другом варианте осуществления О-полисахарид дополнительно включает коровый фрагмент R3 Е.coli. В другом варианте осуществления О-полисахарид дополнительно включает коровый фрагмент R4 Е.coli. В другом варианте осуществления O-полисахарид дополнительно включает коровый фрагмент К12 Е.coli.
В некоторых вариантах осуществления сахарид не включает коровый сахарид. Соответственно, в одном из вариантов осуществления О-полисахарид не включает коровый фрагмент R1 Е.coli. В другом варианте осуществления О-полисахарид не включает коровый фрагмент R2 Е.coli. В другом варианте осуществления O-полисахарид не включает коровый фрагмент R3 Е.coli. В другом варианте осуществления О-полисахарид не включает коровый фрагмент R4 Е.coli. В другом варианте осуществления O-полисахарид не включает коровый фрагмент К12 Е.coli.
4. Конгьюгиро ванные О-антигены
Химическая связь О-антигенов или предпочтительно О-полисахаридов с белковыми носителями может улучшить иммуногенность О-антигенов или О-полисахаридов. Однако изменчивость размера полимера на практике является производственной проблемой. При коммерческом использовании размер сахарида может влиять на совместимость с разными стратегиями синтеза при конъюгации, на однородность продукта и иммуногенность конъюгата. Контроль экспрессии регулятора длины цепи белка семейства Wzz путем манипуляции в ходе синтеза О-антигена позволяет получать желаемую длину цепей О-антигена в различных штаммах грамотрицательных бактерий, включая Е.coli.
В одном из вариантов осуществления очищенные сахариды химически активируют для получения активированных сахаридов, способных реагировать с белком-носителем. После активации каждый сахарид отдельно конъюгируют с белком-носителем с образованием конъюгата, а именно гликоконъюгата. В контексте настоящего описания термин «гликоконъюгат» относится к сахариду, ковалентно связанному с белком-носителем. В одном из вариантов осуществления сахарид связан непосредственно с белком-носителем. В другом варианте осуществления сахарид связан с белком через спейсер/линкер.
Конъюгаты могут быть получены по схемам, которые позволяют связывать носитель с О-антигеном в одном или более сайтах вдоль О-антигена, или по схемам, которые позволяют активировать по меньшей мере один остаток корового олигосахарида.
В одном из вариантов осуществления каждый сахарид конъюгирован с одним и тем же белком-носителем.
Если белок-носитель является одинаковым для 2 или более сахаридов в композиции, сахариды могут быть конъюгированы с одной и той же молекулой белка-носителя (например, молекулы-носители имеют конъюгированные с ней 2 или более разных сахаридов).
В предпочтительном варианте осуществления каждый из сахаридов индивидуально конъюгирован с разными молекулами белка-носителя (каждая молекула белка-носителя имеет конъюгированный с ней только один тип сахарида). В указанном варианте осуществления сахариды, как говорят, индивидуально конъюгированы с белком-носителем.
Химическая активация сахаридов и последующая конъюгация с белком-носителем могут быть достигнуты раскрытыми в настоящем описании способами активации и конъюгации. После конъюгации полисахарида с белком-носителем гликоконъюгаты очищают (обогащают относительно количества конъюгата полисахарид-белок) различными способами. Эти способы включают операции концентрации/диафильтрации, осаждения/элюирования, колоночной хроматографии и глубинной фильтрации. После очистки отдельных гликоконъюгатов их компаундируют с получением иммуногенной композиции по настоящему изобретению.
а. Активация. Настоящее изобретение дополнительно относится к активированным полисахаридам, полученным в соответствии с любым из описанных здесь вариантов осуществления настоящего изобретения, где полисахарид активируют химическим реагентом с образованием реакционноспособных групп для конъюгации с линкером или белком-носителем. В некоторых вариантах осуществления сахарид по изобретению активируют перед конъюгацией с белком-носителем. В некоторых вариантах осуществления степень активации не приводит к значительному снижению молекулярной массы полисахарида. Например, в некоторых вариантах осуществления степень активации не приводит к расщеплению полисахаридного остова. В некоторых вариантах осуществления степень активации не оказывает существенного влияния на степень конъюгации, измеряемую количеством остатков лизина, модифицированных в белке-носителе, таком как CRM197 (определенным с помощью аминокислотного анализа). Например, в некоторых вариантах осуществления степень активации не приводит к значительному увеличению количества модифицированных остатков лизина (определенному с помощью аминокислотного анализа) в белке-носителе в 3 раза по сравнению с количеством остатков лизина, модифицированных в белке-носителе конъюгата с эталонным полисахаридом при той же степени активации. В некоторых вариантах осуществления степень активации не увеличивает уровень неконъюгированного свободного сахарида. В некоторых вариантах осуществления степень активации не снижает оптимальное соотношение сахарид/белок.
В некоторых вариантах осуществления активированный сахарид имеет процент активации, при котором количество молей тиола на сахаридное повторяющееся звено активированного сахарида составляет от 1 до 100%, например, от 2 до 80%, от 2 до 50%, от 3-30% и от 4-25%. Степень активации составляет по меньшей мере 1%, 2%, 3%, 4%, 5%, 6%, 7%, 8%, 9%, 10%, 11%, 12%, 13%, 14%, 15%, 16%, 17%, 18%, 19%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%, или примерно 100%. Предпочтительно степень активации составляет не более 50%, более предпочтительно не более 25%. В одном из вариантов осуществления степень активации составляет не более 20%. Любое минимальное значение и любое максимальное значение могут быть объединены для определения диапазона.
В одном из вариантов осуществления полисахарид активируют тетрафторборатом 1-циано-4-диметиламинопиридиния (CDAP) с образованием цианатного эфира. Затем активированный полисахарид связывают непосредственно или через спейсерную (линкерную) группу с аминогруппой на белке-носителе (предпочтительно CRM197 или столбнячном анатоксине).
Например, спейсером может быть цистамин или цистеамин для получения тиолированного полисахарида, который может быть связан с носителем через тиоэфирную связь, полученную в результате взаимодействия с малеимид-активированным белком-носителем (например, с применением сложного эфира N-[Y-малеимидобутирилокси]сукцинимида (GMBS)) или галоацетилированным белком-носителем (например, с применением 76yophi1ized76e, N-сукцинимидилбромацетата (SBA; SIB), N-сукцинимидил(4-иодацетил)аминобензоата (SIAB), сульфосукцинимидил(4-иодацетил)аминобензоата (сульфо-SIAB), N-сукцинимидилиодацетата (SIA) или сукцинимидил-3-[бромацетамидо]пропионата (SBAP)). В одном из вариантов осуществления цианатный сложный эфир (необязательно полученный с помощью CDAP-химии) связывают с гександиамином или дигидразидом адипиновой кислоты (ADH), а аминопроизводный сахарид конъюгируют с белком-носителем (например, CRM197) посредством карбодиимидной (например, EDAC или EDC) химии через карбоксильную группу на белке-носителе.
В других подходящих методах конъюгации используют карбодиимиды, гидразиды, активные сложные эфиры, норборан, пара-нитробензойную кислоту, N-гидроксисукцинимид, S-NHS, EDC, TSTU. Конъюгация может включать карбонильный линкер, который может быть образован в результате взаимодействия свободной гидроксильной группы сахарида с CDI с последующим взаимодействием с белком с образованием карбаматной связи. Этот процесс может включать восстановление аномерного конца до первичной гидроксильной группы, необязательную защиту/удаление защиты первичной гидроксильной группы, взаимодействие первичной гидроксильной группы с CDI с образованием карбаматного промежуточного соединения CDI и сочетание карбаматного промежуточного соединения CDI с аминогруппой на белке (химия CDI).
b. Молекулярная масса. В некоторых вариантах осуществления гликоконъюгат содержит сахарид с молекулярной массой от 10 кДа до 2000 кДа. В других вариантах осуществления сахарид имеет молекулярную массу от 50 кДа до 1000 кДа. В других вариантах осуществления сахарид имеет молекулярную массу от 70 кДа до 900 кДа. В других вариантах осуществления сахарид имеет молекулярную массу от 100 кДа до 800 кДа. В других вариантах осуществления сахарид имеет молекулярную массу от 200 кДа до 600 кДа. В дополнительных вариантах осуществления сахарид имеет молекулярную массу от 100 кДа до 1000 кДа; от 100 кДа до 900 кДа; от 100 кДа до 800 кДа; от 100 кДа до 700 кДа; от 100 кДа до 600 кДа; от 100 кДа до 500 кДа; от 100 кДа до 400 кДа; от 100 кДа до 300 кДа; от 150 кДа до 1000 кДа; от 150 кДа до 900 кДа; от 150 кДа до 800 кДа; от 150 кДа до 700 кДа; от 150 кДа до 600 кДа; от 150 кДа до 500 кДа; от 150 кДа до 400 кДа; от 150 кДа до 300 кДа; от 200 кДа до 1000 кДа; от 200 кДа до 900 кДа; от 200 кДа до 800 кДа; от 200 кДа до 700 кДа; от 200 кДа до 600 кДа; от 200 кДа до 500 кДа; от 200 кДа до 400 кДа; от 200 кДа до 300; от 250 кДа до 1000 кДа; от 250 кДа до 900 кДа; от 250 кДа до 800 кДа; от 250 кДа до 700 кДа; от 250 кДа до 600 кДа; от 250 кДа до 500 кДа; от 250 кДа до 400 кДа; от 250 кДа до 350 кДа; от 300 кДа до 1000 кДа; от 300 кДа до 900 кДа; от 300 кДа до 800 кДа; от 300 кДа до 700 кДа; от 300 кДа до 600 кДа; от 300 кДа до 500 кДа; от 300 кДа до 400 кДа; от 400 кДа до 1000 кДа; от 400 кДа до 900 кДа; от 400 кДа до 800 кДа; от 400 кДа до 700 кДа; от 400 кДа до 600 кДа; от 500 кДа до 600 кДа. В одном из вариантов осуществления гликоконъюгат, имеющий такую молекулярную массу, получают одноконцевой конъюгацией. В другом варианте осуществления гликоконъюгат, имеющий такую молекулярную массу, получают с помощью химии восстановительного аминирования (RAC) в водном буфере. Любое целое число в любом из вышеуказанных диапазонов рассматривается как вариант осуществления изобретения.
В некоторых вариантах осуществления гликоконъюгат по изобретению имеет молекулярную массу от 400 кДа до 15000 кДа; от 500 кДа до 10000 кДа; от 2000 кДа до 10000 кДа; от 3000 кДа до 8000 кДа; или от 3000 кДа до 5000 кДа. В других вариантах осуществления гликоконъюгат имеет молекулярную массу от 500 кДа до 10000 кДа. В других вариантах осуществления гликоконъюгат имеет молекулярную массу от 1000 кДа до 8000 кДа. В других вариантах осуществления гликоконъюгат имеет молекулярную массу от 2000 кДа до 8000 кДа или от 3000 кДа до 7000 кДа. В дополнительных вариантах осуществления гликоконъюгат по изобретению имеет молекулярную массу от 200 кДа до 20000 кДа; от 200 кДа до 15000 кДа; от 200 кДа до 10000 кДа; от 200 кДа до 7500 кДа; от 200 кДа до 5000 кДа; от 200 кДа до 3000 кДа; от 200 кДа до 1000 кДа; от 500 кДа до 20000 кДа; от 500 кДа до 15000 кДа; от 500 кДа до 12500 кДа; от 500 кДа до 10000 кДа; от 500 кДа до 7500 кДа; от 500 кДа до 6000 кДа; от 500 кДа до 5000 кДа; от 500 кДа до 4000 кДа; от 500 кДа до 3000 кДа; от 500 кДа до 2000 кДа; от 500 кДа до 1500 кДа; от 500 кДа до 1000 кДа; от 750 кДа до 20000 кДа; от 750 кДа до 15000 кДа; от 750 кДа до 12500 кДа; от 750 кДа до 10000 кДа; от 750 кДа до 7500 кДа; от 750 кДа до 6000 кДа; от 750 кДа до 5000 кДа; от 750 кДа до 4000 кДа; от 750 кДа до 3000 кДа; от 750 кДа до 2000 кДа; от 750 кДа до 1500 кДа; от 1000 кДа до 15000 кДа; от 1000 кДа до 12500 кДа; от 1000 кДа до 10000 кДа; от 1000 кДа до 7500 кДа; от 1000 кДа до 6000 кДа; от 1000 кДа до 5000 кДа; от 1000 кДа до 4000 кДа; от 1000 кДа до 2500 кДа; от 2000 кДа до 15000 кДа; от 2000 кДа до 12500 кДа; от 2000 кДа до 10000 кДа; от 2000 кДа до 7500 кДа; от 2000 кДа до 6000 кДа; от 2000 кДа до 5000 кДа; от 2000 кДа до 4000 кДа; или между 2000 кДа и 3000 кДа. В одном из вариантов осуществления гликоконъюгат, имеющий такую молекулярную массу, получают конъюгацией через еТЕС, раскрытой в настоящем описании. В другом варианте осуществления гликоконъюгат, имеющий такую молекулярную массу, получают с помощью химии восстановительного аминирования (RAC). В другом варианте осуществления гликоконъюгат с такой молекулярной массой получают с помощью химии восстановительного аминирования (RAC) в DMSO.
В дополнительных вариантах осуществления гликоконъюгат по изобретению имеет молекулярную массу от 1000 кДа до 20000 кДа; от 1000 кДа до 15000 кДа; от 2000 кДа до 10000 кДа; от 2000 кДа до 7500 кДа; от 2000 кДа до 5000 кДа; от 3000 кДа до 20000 кДа; от 3000 кДа до 15000 кДа; от 3000 кДа до 12500 кДа; от 4000 кДа до 10000 кДа; от 4000 кДа до 7500 кДа; от 4000 кДа до 6000 кДа; или от 5000 кДа до 7000 кДа. В одном из вариантов осуществления гликоконъюгат, имеющий такую молекулярную массу, получают с помощью химии восстановительного аминирования (RAC). В другом варианте осуществления гликоконъюгат с такой молекулярной массой получают с помощью химии восстановительного аминирования (RAC) в DMSO. В другом варианте осуществления гликоконъюгат с такой молекулярной массой получают с помощью раскрытой в настоящем описании конъюгации через еТЕС.
В дополнительных вариантах осуществления гликоконъюгат по изобретению имеет молекулярную массу от 5000 кДа до 20000 кДа; от 5000 кДа до 15000 кДа; от 5000 кДа до 10000 кДа; от 5000 кДа до 7500 кДа; от 6000 кДа до 20000 кДа; от 6000 кДа до 15000 кДа; от 6000 кДа до 12500 кДа; от 6000 кДа до 10000 кДа или от 6000 кДа до 7500 кДа.
Молекулярную массу гликоконъюгата можно измерить с помощью SEC-MALLS. Любое целое число в любом из вышеуказанных диапазонов рассматривается как вариант осуществления изобретения. Гликоконъюгаты по изобретению также могут быть охарактеризованы отношением (масс./масс.) сахарида к белку-носителю. В некоторых вариантах осуществления отношение полисахарида к белку-носителю в гликоконъюгате (масс./масс.) составляет от 0,5 до 3 (например, примерно 0,5, примерно 0,6, примерно 0,7, примерно 0,8, примерно 0,9, примерно 1,0, примерно 1,1, примерно 1,2, примерно 1,3, примерно 1,4, примерно 1,5, примерно 1,6, примерно 1,7, примерно 1,8, примерно 1,9, примерно 2,0, примерно 2,1, примерно 2,2, примерно 2,3, примерно 2,4, примерно 2,5, примерно 2,6, примерно 2,7, примерно 2,8, примерно 2,9 или примерно 3,0). В других вариантах осуществления отношение сахарида к белку-носителю (масс./масс.) составляет от 0,5 до 2,0, от 0,5 до 1,5, от 0,8 до 1,2, от 0,5 до 1,0, от 1,0 до 1,5 или от 1,0 до 2,0. В дополнительных вариантах осуществления отношение сахарида к белку-носителю (масс./масс.) составляет от 0,8 до 1,2. В предпочтительном варианте отношение полисахарида к белку-носителю в конъюгате составляет от 0,9 до 1,1. В некоторых таких вариантах осуществления белок-носитель представляет собой CRM197.
Гликоконъюгаты также можно охарактеризовать по распределению их молекул по размерам (Kd). Среду эксклюзионной хроматографии (CL-4B) можно использовать для определения относительного распределения молекул конъюгата по размеру. Эксклюзионную хроматографию (SEC) выполняют, используя колонки с гравитационной подачей, для получения профиля распределения молекул конъюгата по размеру. Крупные молекулы, исключенные из пор среды, элюируют быстрее, чем мелкие. Для сбора элюата колонки используют коллекторы фракций. Фракции тестируют колориметрически с помощью анализа сахаридов. Для определения Kd колонки калибруют, чтобы установить долю, при которой молекулы полностью исключаются (V0), (Kd=0), и долю, представляющую максимальное удерживание (Vi), (Kd=1). Доля, при которой достигается указанный атрибут образца (Ve), связана с Kd следующим выражением: Kd=(Ve - Vo)/(Vi - V0).
с. Свободный сахарид. Гликоконъюгаты и иммуногенные композиции по изобретению могут включать свободный сахарид, который не является ковалентно конъюгировэнным с белком-носителем, но, тем не менее, присутствует в композиции гликоконъюгата. Свободный сахарид может быть связан нековалентными связями с гликоконъюгатом (т.е., быть нековалентно связанным, адсорбированным на нем или захваченным им или вместе с ним). В предпочтительном варианте осуществления гликоконъюгат содержит не более 50%, 45%, 40%, 35%, 30%, 25%, 20% или 15% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит менее примерно 25% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит не более примерно 20% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит не более примерно 15% свободного полисахарида относительно общего количества полисахарида. В другом предпочтительном варианте осуществления гликоконъюгат содержит не более примерно 20%, 19%, 18%, 17%, 16%, 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит менее примерно 8% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит не более примерно 6% свободного полисахарида относительно общего количества полисахарида. В предпочтительном варианте осуществления гликоконъюгат содержит не более примерно 5% свободного полисахарида относительно общего количества полисахарида.
d. Ковалентная связь. В других вариантах осуществления конъюгат содержит по меньшей мере одну ковалентную связь между белком-носителем и сахаридом на каждые 5-10 сахаридных повторяющихся звеньев; на каждые 2-7 сахаридных повторяющихся звеньев; на каждые 3-8 сахаридных повторяющихся звеньев; на каждые 4-9 сахаридных повторяющихся звеньев; на каждые 6-11 сахаридных повторяющихся звеньев; на каждые 7-12 сахаридных повторяющихся звеньев; на каждые 8-13 сахаридных повторяющихся звеньев; на каждые 9-14 сахаридных повторяющихся звеньев; на каждые 10-15 сахаридных повторяющихся звеньев; на каждые 2-6 сахаридных повторяющихся звеньев, на каждые 3-7 сахаридных повторяющихся звеньев; на каждые 4-8 сахаридных повторяющихся звеньев; на каждые 6-10 сахаридных повторяющихся звеньев; на каждые 7-11 сахаридных повторяющихся звеньев; на каждые 8-12 сахаридных повторяющихся звеньев; на каждые 9-13 сахаридных повторяющихся звеньев; на каждые 10-14 сахаридных повторяющихся звеньев; на каждые 10-20 сахаридных повторяющихся звеньев; на каждые 4-25 сахаридных повторяющихся звеньев или на каждые 2-25 сахаридных повторяющихся звеньев. В частых вариантах осуществления белок-носитель представляет собой CRM197. В другом варианте осуществления по меньшей мере одна связь между белком-носителем и сахаридом возникает на каждые 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 сахаридных повторяющихся звеньев полисахарида. В одном из вариантов осуществления белок-носитель представляет собой CRM197. Любое целое число в любом из вышеуказанных диапазонов рассматривается как вариант осуществления изобретения.
е. Остатки лизина. Другой способ, которым можно охарактеризовать гликоконъюгаты по изобретению, заключается в количестве остатков лизина в белке-носителе (например, CRM197), которые становятся конъюгированными с сахаридом, что может быть охарактеризовано как уровень конъюгированных лизинов (степень конъюгации). Доказательства модификации лизина белка-носителя вследствие ковалентных связей с полисахаридами могут быть получены с помощью аминокислотного анализа с применением обычных методов, известных специалистам в данной области. Конъюгация приводит к уменьшению количества извлеченных остатков лизина по сравнению с исходным материалом белка-носителя, используемым для получения материалов конъюгата. В предпочтительном варианте осуществления степень конъюгации гликоконъюгата по изобретению составляет от 2 до 15, от 2 до 13, от 2 до 10, от 2 до 8, от 2 до 6, от 2 до 5, от 2 до 4, от 3 до 15, от 3 до 13, от 3 до 10, от 3 до 8, от 3 до 6, от 3 до 5, от 3 до 4, от 5 до 15, от 5 до 10, от 8 до 15, от 8 до 12, от 10 до 15 или от 10 до 12. В одном из вариантов осуществления степень конъюгации гликоконъюгата по изобретению составляет примерно 2, примерно 3, примерно 4, примерно 5, примерно 6, примерно 7, примерно 8, примерно 9, примерно 10, примерно 11, примерно 12, примерно 13, примерно 14 или примерно 15. В предпочтительном варианте осуществления степень конъюгации гликоконъюгата по изобретению составляет от 4 до 7. В некоторых таких вариантах осуществления белком-носителем является CRM197.
Частота присоединения сахаридной цепи к лизину на белке-носителе является еще одним параметром, которым могут быть охарактеризованы гликоконъюгаты по изобретению. Например, в некоторых вариантах осуществления по меньшей мере одна ковалентная связь между белком-носителем и полисахаридом приходится на каждые 4 сахаридных повторяющихся звена полисахарида. В другом варианте осуществления между белком-носителем и полисахаридом возникает по меньшей мере одна ковалентная связь на каждые 10 сахаридных повторяющихся звеньев полисахарида. В другом варианте осуществления между белком-носителем и полисахаридом возникает по меньшей мере одна ковалентная связь на каждые 15 сахаридных повторяющихся звеньев полисахарида. В другом варианте осуществления между белком-носителем и полисахаридом возникает по меньшей мере одна ковалентная связь на каждые 25 сахаридных повторяющихся звеньев полисахарида.
f. О-ацетилирование. В некоторых вариантах осуществления сахариды по изобретению являются О-ацетилированными. В некоторых вариантах осуществления гликоконъюгат содержит сахарид со степенью О-ацетилирования от 10 до 100%, от 20 до 100%, от 30 до 100%, от 40 до 100%, от 50 до 100%, от 60 до 100%, от 70 до 100%, от 75 до 100%, от 80 до 100%, от 90 до 100%, от 50 до 90%, от 60 до 90%, от 70 до 90% или от 80 до 90%. В других вариантах осуществления степень О-ацетилирования составляет 10%, 20%, 30%, 40%, 50%, 60%, 70%, 80% или 90%, или примерно 100%. Под % О-ацетилирования подразумевается процентное содержание данного сахарида по отношению к 100% (где каждое повторяющееся звено полностью ацетилировано относительно его ацетилированной структуры).
В некоторых вариантах осуществления гликоконъюгат получают восстановительным аминированием. В некоторых вариантах осуществления гликоконъюгат представляет собой конъюгированный сахарид с одной концевой связью, где сахарид ковалентно связан непосредственно с белком-носителем. В некоторых вариантах осуществления гликоконъюгат ковалентно связан с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер.
g. Восстановительное аминирование. В одном из вариантов осуществления сахарид конъюгирован с белком-носителем путем восстановительного аминирования (например, как описано в публикациях заявок на патенты США №№2006/0228380, 2007/0231340, 2007/0184071 и 2007/0184072, WO 2006/110381, WO 2008/079653 и WO 2008/143709).
Восстановительное аминирование включает (1) окисление сахарида, (2) восстановление активированного сахарида и белка-носителя с образованием конъюгата. Перед окислением сахарид необязательно гидролизуют. Можно использовать механический или химический гидролиз. Химический гидролиз можно выполнять с применением уксусной кислоты.
Этап окисления может включать реакцию с периодатом. В контексте настоящего описания термин «периодат» относится как к периодату, так и к периодной кислоте. Этот термин также включает как метапериодат (IO4-), так и ортопериодат (IO65-), а также различные соли периодата (например, периодат натрия и периодат калия). В одном из вариантов осуществления полисахарид окисляют в присутствии метапериодата, предпочтительно в присутствии периодата натрия (NalO4). В другом варианте осуществления полисахарид окисляют в присутствии ортопериодата, предпочтительно в присутствии йодной кислоты.
В одном из вариантов осуществления окислитель представляет собой стабильное нитроксильное или нитроксильное радикальное соединение, такое как пиперидин-N-окси или пирролидин-N-окси соединение, в присутствии окислителя для селективного окисления первичных гидроксилов. В указанной реакции фактическим окислителем является соль N-оксоаммония в каталитическом цикле. В одном из аспектов указанное стабильное нитроксильное или нитроксильное радикальное соединение представляет собой пиперидин-N-окси или пирролидин-N-окси соединение. В одном из аспектов указанное стабильное нитроксильное или нитроксильное радикальное соединение несет фрагмент TEMPO (2,2,6,6-тетраметил-1-пиперидинилокси) или PROXYL (2,2,5,5-тетраметил-1-пирролидинилокси). В одном из аспектов указанное стабильное нитроксильное радикальное соединение представляет собой TEMPO или его производное. В одном из аспектов указанный окислитель представляет собой молекулу, несущую фрагмент N-галогена. В одном из аспектов указанный окислитель выбирают из любого из N-хлорсукцинимида, N-бромсукцинимида, N-иодосукцинимида, дихлоризоциануровой кислоты, 1,3,5-трихлор-1,3,5-триазинан-2,4,6-триона, дибромизоциануровой кислоты, 1,3,5-трибром-1,3,5-триазинан-2,4,6-триона, дийодизоциануровой кислоты и 1,3,5 трииод-1,3,5-триазинан-2,4,6-триона. Предпочтительно указанный окислитель представляет собой N-хлорсукцинимид.
После этапа окисления сахарида говорят, что сахарид активирован, и далее в настоящем описании он упоминается как «активированный». Активированный сахарид и белок-носитель могут быть лиофилизированы (высушены сублимацией) либо независимо (дискретная лиофилизация), либо вместе (совместная лиофилизация). В одном из вариантов осуществления активированный сахарид и белок-носитель лиофилизируют вместе. В другом варианте осуществления активированный полисахарид и белок-носитель лиофилизируют независимо.
В одном из вариантов осуществления лиофилизация происходит в присутствии невосстанавливающего сахара, возможные невосстанавливающие сахара включают сахарозу, трегалозу, раффинозу, стахиозу, мелецитозу, декстран, маннит, лактит и палатинит.
Следующим этапом процесса конъюгации является восстановление активированного сахарида и белка-носителя с образованием конъюгата (так называемое восстановительное аминирование) с применением восстанавливающего агента. Подходящие восстанавливающие агенты включают цианоборгидриды, такие как цианоборгидрид натрия, триацетоксиборгидрид натрия или боргидрид натрия или цинка (в присутствии кислот Бренстеда или Льюиса), аминобораны, такие как пиридинборан, 2-пиколинборан, 2,6-диборан-метанол, диметиламин-боран, t-BuMe'PrN-ВН3, бензиламин-ВН3 или 5-этил-2-метилпиридинборан (РЕМВ), боран-пиридин или боргидрид-обменная смола. В одном из вариантов осуществления восстановителем является цианоборгидрид натрия.
В одном из вариантов осуществления реакцию восстановления проводят в водном растворителе (например, выбранном из PBS, MES, НЕ PES, Bis-tris, ADA, PIPES, MOPSO, BES, MOPS, DIPSO, MOBS, HEPPSO, POPSO, TEA, EPPS, Bicine или НЕРВ, при pH от 6,0 до 8,5, от 7,0 до 8,0 или от 7,0 до 7,5), в другом варианте осуществления реакцию проводят в апротонном растворителе. В одном из вариантов осуществления реакцию восстановления проводят в растворителе DMSO (диметилсульфоксид) или в DMF (диметилформамид). Растворитель DMSO или DMF можно использовать для восстановления лиофилизированных активированного полисахарида и белка-носителя.
В конце реакции восстановления в конъюгатах могут остаться непрореагировавшие альдегидные группы, которые можно блокировать с помощью подходящего блокирующего агента. В одном из вариантов осуществления этот блокирующего агент представляет собой боргидрид натрия (NaBH4). После конъюгации (реакции восстановления и, необязательно, кэпирования) гликоконъюгаты могут быть очищены (обогащены по отношению к количеству конъюгата полисахарид-белок) различными методами, известными специалисту в данной области. Эти методы включают диализ, операции концентрирования/диафильтрации, осаждения/элюирования фильтрации в тангенциальном протоке, колоночную хроматографию (DEAE или хроматографию гидрофобного взаимодействия) и глубинную фильтрацию. Гликоконъюгаты могут быть очищены диафильтрацией, и/или ионообменной хроматографией, и/или эксклюзионной хроматографией. В одном из вариантов осуществления гликоконъюгаты очищают диафильтрацией или ионообменной хроматографией или эксклюзионной хроматографией. В одном из вариантов осуществления гликоконъюгаты подвергают стерильной фильтрации.
В предпочтительном варианте осуществления гликоконъюгат Е.coli выбирают из любого из серотипов: O25b, O1a, O2 и О6, полученного восстановительным аминированием. В предпочтительном варианте осуществления гликоконъюгаты Е.coli серотипов O25b, O1a, O2 и О6 получают восстановительным аминированием.
В одном из аспектов изобретение относится к конъюгату, который включает белок-носитель, например, CRM197, связанный с сахаридом формулы O25b, представленной:
где n представляет собой любое целое число, большее или равное 1. В предпочтительном варианте осуществления n представляет собой целое число, равное по меньшей мере 31, 32, 33, 34, 35, 36, 37, 38, 39, 40 и не более 200, 100, 99, 98, 97, 96, 95, 94, 93, 92, 91, 90, 89, 88, 87, 86, 81, 80, 79, 78, 77, 76, 75, 74, 73, 72, 71, 70, 69, 68, 67, 66, 65, 60, 59, 58, 57, 56, 55, 54, 53, 52, 51 или 50. Любое минимальное значение и любое максимальное значение могут быть объединены для определения диапазона. Иллюстративные диапазоны включают, например, от не менее 1 до не более 1000; от не менее 10 до не более 500; и от не менее 20 до не более 80. В одном из предпочтительных вариантов осуществления n составляет от не менее 31 до не более 90, более предпочтительно от 40 до 90, наиболее предпочтительно от 60 до 85.
В другом аспекте изобретение относится к конъюгату, который включает белок-носитель, например, CRM197, связанный с сахаридом, имеющим любую из структур, приведенных в таблице А, где n представляет собой целое число, большее или равное 1.
Не ограничиваясь какой-либо теорией или механизмом, в некоторых вариантах осуществления считается, что стабильный конъюгат требует такого уровня модификации сахаридного антигена, который не приводит к нарушению структурной целостности критических иммуногенных эпитопов антигена.
h. Активация и образование альдегида. В некоторых вариантах осуществления сахарид по изобретению активируют с образованием альдегида. В таких вариантах осуществления с активированным сахаридом процент (%) активации (или степень окисления (DO)) относится к молям сахаридного повторяющегося звена на моль альдегида активированного полисахарида. Например, в некоторых вариантах осуществления сахарид активируют через периодатное окисление вицинальных диолов на повторяющемся звене полисахарида с образованием альдегида. Изменение молярных эквивалентов (мэкв) периодата натрия относительно сахаридного повторяющегося звена и температуры во время окисления приводит к различным уровням степени окисления (DO).
Концентрации сахаридов и альдегидов обычно определяют с помощью колориметрических анализов. Альтернативным реагентом является комбинация TEMPO (2,2,6,6-тетраметилпиперидин-1-оксил (свободный радикал))-N-хлорсукцинимид (NCS), которая приводит к образованию альдегидов из первичных спиртовых групп.
В некоторых вариантах осуществления активированный сахарид имеет степень окисления, при которой количество молей сахаридного повторяющегося звена на моль альдегида активированного сахарида составляет от 1 до 100, например, от 2 до 80, от 2 до 50, от 3 до 30 и от 4 до 25. Степень активации составляет по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, ≥20, ≥30, ≥40, ≥50, ≥60, ≥70, ≥80 или>90, или примерно 100. Предпочтительно степень окисления (DO) составляет от не менее 5 до не более 50, более предпочтительно от не менее 10 до не более 25. В одном из вариантов осуществления степень активации составляет от не менее 10 до не более 25. Любое минимальное значение и любое максимальное значение могут быть объединены для определения диапазона. Степень окисления может быть представлена в процентах (%) активации. Например, в одном из вариантов осуществления значение DO, равное 10, относится к одной активированной сахаридной повторяющейся единице на 10 сахаридных повторяющихся звеньев в активированном сахариде, и в этом случае значение DO, равное 10, может быть представлено как 10% активации.
В некоторых вариантах осуществления конъюгат, полученный с помощью восстановительного аминирования, включает белок-носитель и сахарид, где сахарид имеет структуру, выбранную из: формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O6O, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы О104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187. В некоторых вариантах осуществления сахарид в конъюгате включает формулу, где n представляет собой целое число от 1 до 1000, от 5 до 1000, предпочтительно от 31 до 100 или от 31 до 90, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65.
i. Конъюгаты с одной концевой связью. В некоторых вариантах осуществления конъюгат представляет собой конъюгированный сахарид с одной концевой связью, где сахарид ковалентно связан на одном конце сахарида с белком-носителем. В некоторых вариантах осуществления конъюгированный полисахарид с одной концевой связью имеет концевой сахарид. Например, конъюгат имеет одну концевую связь, если один из концов (концевой сахаридный остаток) полисахарида ковалентно связан с белком-носителем. В некоторых вариантах осуществления конъюгат имеет одну концевую связь, если концевой сахаридный остаток полисахарида ковалентно связан с белком-носителем через линкер. Такие линкеры могут включать, например, цистаминовый линкер (А1), линкер на основе 3,3'-дитио-бис(пропановый дигидразидна) (А4) и линкер на основе 2,2'-дитио-N, N'-бис(этан-2,1-диил)бис(2-(аминоокси)ацетамидна) (А6).
В некоторых вариантах осуществления сахарид конъюгирован с белком-носителем через остаток 3-дезокси-с1-манно-окт-2-улозоновой кислоты (KDO) с образованием с конъюгата с одной концевой связью.
В некоторых вариантах осуществления конъюгат
предпочтительно не является биоконъюгатом. Термин «биоконъюгат» относится к конъюгату между белком (например, белком-носителем) и антигеном, например О-антигеном (например, O25B), полученным с помощью клетки-хозяина, где механизм клетки-хозяина связывает антиген с белком (например, N-связями). Гликоконъюгаты включают биоконъюгаты, а также конъюгаты сахарного антигена (например, олиго- и полисахарида) с белком, полученные способами, не требующими получения конъюгата в клетке-хозяине, например путем конъюгирования через химическое связывание белка с сахаридом.
j. Тиол-активированные сахариды. В некоторых вариантах осуществления сахарид по изобретению активируют тиолом. В таких вариантах осуществления с тиол-активированным сахаридом процент (%) активации относится к молям тиола на сахаридное повторяющееся звено активированного полисахарида. Концентрации сахаридов и тиолов обычно определяют с помощью анализа Эллмана для количественного определения сульфгидрилов. Например, в некоторых вариантах осуществления сахарид включает активацию 2-кето-3-дезоксиоктановой кислоты (KDO) дисульфид-аминовым линкером. В некоторых вариантах осуществления сахарид ковалентно связан с белком-носителем через двухвалентный гетеробифункциональный линкер (также называемый в настоящем описании «спейсером»). Линкер предпочтительно обеспечивает тиоэфирную связь между сахаридом и белком-носителем, что приводит к образованию гликоконъюгата, называемого в настоящем описании «тиоэфирным гликоконъюгатом». В некоторых вариантах осуществления линкер дополнительно обеспечивает карбаматные и амидные связи, такие как, например, (2-((2-оксоэтил)тио)этил)карбамат (еТЕС).
В некоторых вариантах осуществления конъюгат с одной концевой связью, включает белок-носитель и сахарид, где сахарид включает структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы 017, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы О60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187. В некоторых вариантах осуществления сахарид в конъюгате включает формулу, где n представляет собой целое число от 1 до 1000, от 5 до 1000, предпочтительно от 31 до 100, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65.
Например, в одном из вариантов осуществления конъюгат с одной концевой связью включает белок-носитель и сахарид, имеющий структуру, выбранную из формулы O8, формулы O9a, формулы O9, формулы O20ab, формулы О20ас, формулы O52, формулы O97 и формулы O101, где n представляет собой целое число от 1 до 10.
5. еТЕС-связанные конъюгаты
В одном из аспектов изобретение в целом относится к гликоконъюгатам, содержащим сахарид, полученный из описанной выше Е.coli, ковалентно конъюгированный с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер (как описано, например, в патенте США 9517274 и публикации международной патентной заявки WO 2014027302, которые полностью включены в настоящее описание посредством ссылки), включая иммуногенные композиции, содержащие такие гликоконъюгаты, и способы получения и применения таких гликоконъюгатов и иммуногенных композиций. Указанные гликоконъюгаты содержат сахарид, ковалентно конъюгированный со спейсером еТЕС через один или более спейсеров еТЕС, где сахарид ковалентно конъюгирован со спейсером еТЕС посредством карбаматной связи, и где белок-носитель ковалентно конъюгирован со спейсером еТЕС посредством амидной связи. Спейсер еТЕС включает семь линейных атомов (т.е. -С(О)NH(СН2)2SCH2C(О)-) и обеспечивает стабильные тиоэфирные и амидные связи между сахаридом и белком-носителем.
еТЕС-связанные гликоконъюгаты по изобретению могут быть представлены общей формулой (I):
где атомы, составляющие спейсер еТЕС, содержатся в центральном квадрате.
В указанных гликоконъюгатах по изобретению сахарид может представлять собой полисахарид или олигосахарид.
Белки-носители, включенные в гликоконъюгаты по изобретению, выбирают из группы белков-носителей, обычно подходящих для таких целей, как описано здесь далее или известно специалистам в данной области. В конкретных вариантах осуществления белок-носитель представляет собой CRM197.
В другом аспекте изобретение относится к способу получения гликоконъюгата, содержащего сахарид, раскрытый в настоящем описании, конъюгированный с белком-носителем через спейсер еТЕС, включающий этапы а) взаимодействия сахарида с производным угольной кислоты в органическом растворителе с получением активированного сахарид; b) взаимодействие активированного сахарида с цистамином или цистеамином или их солями с получением тиолированного сахарида; с) взаимодействие тиолированного сахарида с восстанавливающим агентом с получением активированного тиолированного сахарида, содержащего один или более свободных сульфгидрильных остатков; d) взаимодействие активированного тиолированного сахарида с активированным белком-носителем, содержащим одну или более α-галогенацетамидных групп, с получением конъюгата тиолированного сахарида с белком-носителем; и е) взаимодействие конъюгата тиолированного сахарида с белком-носителем с (i) первым кэпирующим реагентом, способным кэпировать неконъюгированные α-галогенацетамидные группы активированного белка-носителя; и/или (ii) вторым кэпирующим реагентом, способным кэпировать неконъюгированные свободные сульфгидрильные остатки активированного тиолированного сахарида; в результате чего образуется еТЕС-связанный гликоконъюгат.
В частых вариантах осуществления производное угольной кислоты представляет собой 1,1'-карбонил-ди-(1,2,4-триазол) (CDT) или 1,1'-карбонилдиимидазол (CDI). Предпочтительно производное угольной кислоты представляет собой CDT, а органический растворитель представляет собой полярный апротонный растворитель, такой как диметилсульфоксид (DMSO). В предпочтительных вариантах осуществления тиолированный сахарид получают реакцией активированного сахарида с бифункциональным симметричным тиоалкиламиновым реагентом, цистамином или их солями. Альтернативно, тиолированный сахарид может быть образован в результате взаимодействия активированного сахарида с цистеамином или его солью. еТЕС-связанные гликоконъюгаты, полученные способами по изобретению, могут быть представлены общей формулой (I).
В частых вариантах осуществления первым кэпирующим реагентом является N-ацетил-L-цистеин, который реагирует с неконъюгированными α-галогенацетамидными группами на остатках лизина белка-носителя с образованием остатка S-карбоксиметилцистеина (CMC), ковалентно связанного с активированным остатком лизина через тиоэфирная связь.
В других вариантах осуществления второй копирующий реагент представляет собой иодацетамид (IAA), который взаимодействует с неконъюгированными свободными сульфгидрильными группами активированного тиолированного сахарида с получением копированного тиоацетамида. Часто этап е) включает копирование как первым копирующим реагентом, так и вторым копирующим реагентом. В некоторых вариантах осуществления этап е) включает копирование N-ацетил-L-цистеином в качестве первого копирующего реагента и IAA в качестве второго копирующего реагента.
В некоторых вариантах осуществления этап е) копирования дополнительно включает взаимодействие с восстанавливающим агентом, например, DTT, ТСЕР или меркаптоэтанолом, после реакции с первым и/или вторым копирующим реагентом.
еТЕС-связанные гликоконъюгаты и иммуногенные композиции по изобретению могут включать свободные сульфгидрильные остатки. В некоторых случаях активированные тиолированные сахариды, образованные способами, представленными в настоящем описании, будут включать несколько свободных сульфгидрильных остатков, некоторые из которых могут не подвергаться ковалентной конъюгации с белком-носителем на этапе конъюгации. Такие остаточные свободные сульфгидрильные остатки блокируют через взаимодействие с блокирующим реагентом, не относящимся к тиол-реактивному блокирующему реагенту, например с иодацетамидом (IAA), для блокирования потенциально реакционноспособной функциональной группы. Также предусмотрено использование других копирующих реагентов, реагирующих с тиолами, например реагентов, содержащих малеимид и т.п.
Кроме того, еТЕС-связанные гликоконъюгаты и иммуногенные композиции по изобретению могут включать остаточный неконъюгированный белок-носитель, который может включать активированный белок-носитель, претерпевший модификацию во время отапов процесса копирования.
В некоторых вариантах осуществления этап d) дополнительно включает предоставление активированного белка-носителя, содержащего одну или более α-галогенацетамидных групп, перед взаимодействием активированного тиолированного сахарида с активированным белком-носителем. В частых вариантах осуществления активированный белок-носитель содержит одну или более α-бромацетамидных групп.
В другом аспекте изобретение относится к еТЕС-связанному гликоконъюгату, содержащему раскрытый в настоящем описании сахарид, конъюгированный с белком-носителем через спейсер еТЕС, полученный в соответствии с любым из описанных здесь способов.
В некоторых вариантах осуществления белок-носитель представляет собой CRM197, и между CRM197 и полисахаридом образуется по меньшей мере одна ковалентная связь через спейсер еТЕС на каждые 4, 10, 15 или 25 сахаридных повторяющихся звеньев полисахарида.
Для каждого из аспектов изобретения, в конкретных вариантах осуществления способов и композиций, представленных в настоящем описании, еТЕС-связанный гликоконъюгат содержит представленный в настоящем описании сахарид, такой как сахарид, полученный из Е.coli.
В другом аспекте изобретение относится к способу профилактики, лечения или облегчения бактериальной инфекции, заболевания или состояния у субъекта, включающий введение субъекту иммунологически эффективного количества иммуногенной композиции по изобретению, где указанная иммуногенная композиция содержит еТЕС-связанный гликоконъюгат, содержащий раскрытый в настоящем описании сахарид. В некоторых вариантах осуществления сахарид получают из Е.coli.
В некоторых вариантах осуществления еТЕС-связанный гликоконъюгат содержит белок-носитель и сахарид, в котором указанный сахарид имеет структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы 038, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O6O, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187. В некоторых вариантах осуществления сахарид в конъюгате включает формулу, где n представляет собой целое число от 1 до 1000, от 5 до 1000, предпочтительно от 31 до 100, более предпочтительно от 35 до 90, наиболее предпочтительно от 35 до 65.
Количество остатков лизина в белке-носителе, которые становятся конъюгированными с сахаридом, можно охарактеризовать как диапазон конъюгированных лизинов. Например, в некоторых вариантах осуществления иммуногенных композиций CRM197 может содержать от 4 до 16 остатков лизина из 39, ковалентно связанных с сахаридом. Другой способ выражения этого параметра состоит в том, что примерно от 10% до примерно 41% остатков лизина в CRM197 ковалентно связаны с сахаридом. В других вариантах осуществления CRM197 может содержать от 2 до 20 остатков лизина из 39, ковалентно связанных с сахаридом. Другой способ выражения этого параметра заключается в том, что примерно от 5% до примерно 50% остатков лизина в CRM197 ковалентно связаны с сахаридом.
В частых вариантах осуществления белок-носитель представляет собой CRM197, и между CRM197 и полисахаридом образуется по меньшей мере одна ковалентная связь через спейсер еТЕС на каждые 4, 10, 15 или 25 сахаридных повторяющихся звеньев полисахарида.
В других вариантах осуществления конъюгат содержит по меньшей мере одну ковалентную связь между белком-носителем и сахаридом на каждые 5-10 сахаридных повторяющихся звеньев; на каждые 2-7 сахаридных повторяющихся звеньев; на каждые 3-8 сахаридных повторяющихся звеньев; на каждые 4-9 сахаридных повторяющихся звеньев; на каждые 6-11 сахаридных повторяющихся звеньев; на каждые 7-12 сахаридных повторяющихся звеньев; на каждые 8-13 сахаридных повторяющихся звеньев; на каждые 9-14 сахаридных повторяющихся звеньев; на каждые 10-15 сахаридных повторяющихся звеньев; на каждые 2-6 сахаридных повторяющихся звеньев, на каждые 3-7 сахаридных повторяющихся звеньев; на каждые 4-8 сахаридных повторяющихся звеньев; на каждые 6-10 сахаридных повторяющихся звеньев; на каждые 7-11 сахаридных повторяющихся звеньев; на каждые 8-12 сахаридных повторяющихся звеньев; на каждые 9-13 сахаридных повторяющихся звеньев; на каждые 10-14 сахаридных повторяющихся звеньев; на каждые 10-20 сахаридных повторяющихся звеньев; или на каждые 4-25 сахаридных повторяющихся звеньев.
В другом варианте осуществления между белком-носителем и сахаридом образуется по меньшей мере одна связь на каждые 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20, 21, 22, 23, 24 или 25 сахаридных повторяющихся звеньев полисахарида.
6. Белки-носители
Компонент гликоконъюгата по изобретению представляет собой белок-носитель, с которым конъюгирован сахарид. Термины «белок носитель» или «носитель-белок» или «носитель» могут использоваться в настоящем описании взаимозаменяемо. Белки-носители должны поддаваться стандартным процедурам конъюгации.
Одним из компонентов конъюгата является белок-носитель, с которым конъюгирован О-полисахарид. В одном из вариантов осуществления конъюгат включает белок-носитель, конъюгированный с коровым олигосахаридом О-полисахарида. В одном из вариантов осуществления конъюгат включает белок-носитель, конъюгированный с О-антигеном О-полисахарида.
Термины «белок-носитель» или «носитель-белок» или «носитель» могут использоваться в настоящем описании взаимозаменяемо. Белки-носители должны поддаваться стандартным процедурам конъюгации.
В предпочтительном варианте осуществления белок-носитель конъюгатов независимо выбирают из любого одного из мутантов ТТ, DT, DT (таких как CRM197), белка D Н. influenzae, слитых белков PhtX, PhtD, PhtDE (в частности, описанных в WO O1/98334 и WO O3/54007), детоксифицированного пневмолизина, PorB, белка N19, PspA, ОМРС, токсина А или В С. Difficile и PsaA. В одном из вариантов осуществления белок-носитель конъюгатов по изобретению представляет собой DT (дифтерийный анатоксин). В другом варианте осуществления белок-носитель конъюгатов по изобретению представляет собой ТТ (столбнячный анатоксин). В другом варианте осуществления белок-носитель конъюгатов по изобретению представляет собой PD (белок D Haemophilus influenzae - см., например, ЕР O594610 В). В некоторых вариантах осуществления белок-носитель включает поли(L-лизин) (PLL).
В предпочтительном варианте осуществления сахариды конъюгированы с белком CRM197. Белок CRM197 представляет собой нетоксичную форму дифтерийного токсина, но иммунологически не отличимого от дифтерийного токсина. CRM197 продуцируется С. diphtheriae, инфицированным нетоксигенным фагом β197tox, созданным в результате нитрозогуанидинового мутагенеза токсигенного коринефага бета. Белок CRM197 имеет ту же молекулярную массу, что и дифтерийный токсин, но отличается от него одной заменой основания (гуанин на аденин) в структурном гене. Это единственное изменение основания вызывает аминокислотную замену глицина на глутаминовую кислоту в зрелом белке и устраняет токсические свойства дифтерийного токсина. Белок CRM197 является Т-клеточно-зависимым безопасным и эффективным носителем сахаридов.
Соответственно, в некоторых вариантах осуществления конъюгаты по изобретению включают CRM197 в качестве белка-носителя, где сахарид ковалентно связан с CRM197.
В предпочтительном варианте осуществления белок-носитель гликоконъюгатов выбирают из группы, состоящей из DT (дифтерийного токсина), ТТ (столбнячного анатоксина) или фрагмента С ТТ, CRM197 (нетоксичного, но антигенно идентичного варианта дифтерийного токсина), других мутантов DT (таких как CRM176, CRM228, CRM45 (Uchida et al. J. Biol. Chem. 218; 3838-3844, 1973), CRM9, CRM45, CRM102, CRM103 или CRM107; и другие мутации, описанные Nicholls and Youle в Genetically Engineered Toxins, Ed: Frankel, Maecel Dekker Inc, 1992; делеции или мутации Glu-148 в Asp, Gin или Ser и/или Ala 158 в Gly и другие мутации, раскрытые в US 4709017 или US 4950740; мутации по меньшей мере одного или более остатков из: Lys 516, Lys 526, Phe 530 и/или Lys 534, и другие мутации, раскрытые в US 5917017 или US 6455673; или фрагмент, раскрытый в US 5843711), пневмококкового пневмолизина (Kuo et al (1995) Infect lmmun 63; 2706-13), включая детоксифицированный каким-либо образом Ply, например dPLY-GMBS (WO O4081515, РСТ/ЕР2005/010258) или dPLY-формол, PhtX, включая PhtA, PhtB, PhtD, PhtE (последовательности PhtA, PhtB, PhtD или PhtE раскрыты в WO O0/37105 или WO O0/39299) и слитые белки Pht, например, слитые белки PhtDE, слитые белки PhtBE, Pht А-Е (WO O1/98334, WO O3/54007, WO 2009/000826), OMPC (менингококкового белка наружной мембраны, обычно экстрагируемого из JV. meningitidis серогруппы В ЕР O372501), PorB (из N. meningitidis), PD (белка D Haemophilus influenzae - см., например, ЕР O594610 В) или их иммунологически функциональных эквивалентов, синтетических пептидов (ЕР O378881, ЕР O427347), белков теплового шока (WO 93/17712, WO 94/03208), белков коклюша (WO 98/58668, ЕР0471177), цитокинов, лимфокинов, факторов или гормонов роста (WO 91/01146), искусственных белков, содержащих множественные CD4+ Т-клеточные эпитопы человека из антигенов, полученных из различных патогенов (Falugi et al (2001) Eur J Immunol 31; 3816-3824), например белка N19 (Baraldoi et al. (2004) Infect lmmun 72; 4884-7), пневмококкового поверхностного белка PspA (WO O2/091998), железосвязывающих белков (WO O1/72337), токсина А или В С. difficile (WO O0/61761), трансферрин-связывающих белков, пневмококкового белка адгезии (PsaA), рекомбинантного окзотоксина A Pseudomonas aeruginosa (в частности, их нетоксичных мутантов (таких как окзотоксин А с заменой глутаминовой кислоты 553 (Uchida Cameron DM, RJ Collier. 1987. J. Bacteriol. 169:4967-4971)). В качестве белков-носителей также могут быть использованы другие белки, такие как овальбумин, гемоцианин лимфы улитки (KLH), бычий сывороточный альбумин (BSA) или очищенное производное белка туберкулина (PPD). Другие подходящие белки-носители включают инактивированные бактериальные токсины, такие как холерный анатоксин (например, как описано в международной патентной заявке № WO 2004/083251), LT Е.colif ST Е.coli и окзотоксин А из Pseudomonas aeruginosa.
В некоторых вариантах осуществления белок-носитель выбирают из любого из, например, CRM197, фрагмента дифтерийного токсина В (DTFB), С8 DTFB, дифтерийного анатоксина (DT), столбнячного анатоксина (ТТ), фрагмента С ТТ, коклюшного анатоксина, холерного анатоксина или экзотоксина А из Pseudomonas aeruginosa.; детоксифицированного экзотоксина А P. aeruginosa (ЕРА), белка, связывающего мальтозу (МБР), флагеллина, детоксифицированного гемолизина A S. aureus, фактора слипания А, фактора слипания В, субъединицы холерного токсина В (СТВ), пневмолизина Streptococcus pneumoniae и их детоксифицированных вариантов, AcrA С.jejuni, природных гликопротеинов С. jejuni и стрептококковой пептидазы С5а (SCP). В одном из вариантов осуществления белок-носитель представляет собой детоксифицированный экзотоксин Pseudomonas (ЕРА). В другом варианте осуществления белок-носитель представляет собой недетоксифицированный экзотоксин Pseudomonas (ЕРА). В одном из вариантов осуществления белок-носитель представляет собой флагеллин. В другом варианте осуществления белок-носитель не является флагеллином.
В предпочтительном варианте осуществления белок-носитель гликоконъюгатов независимо выбирают из группы, состоящей из мутантов ТТ, DT, DT (таких как CRM197), белка D И. influenzae, гибридов PhtX, PhtD, PhtDE (в частности, описанных в WO O1/98334 и WO O3/54007), детоксифицированного пневмолизина, PorB, белка N19, PspA, ОМРС, токсина А или В С. Difficile и PsaA. В одном из вариантов осуществления белок-носитель гликоконъюгатов по изобретению представляет собой DT (дифтерийный анатоксин). В другом варианте осуществления белок-носитель гликоконъюгатов по изобретению представляет собой ТТ (столбнячный анатоксин). В другом варианте осуществления белок-носитель гликоконъюгатов по изобретению представляет собой PD (белок D Haemophilus influenzae - см., например, ЕР O594610 В).
В предпочтительном варианте осуществления капсульные сахариды по изобретению конъюгированы с белком CRM197. Белок CRM197 представляет собой нетоксичную форму дифтерийного токсина, но иммунологически не отличимую от дифтерийного токсина. CRM197 продуцируется С. diphtheriae, инфицированным нетоксигенным фагом β197tox, созданным в результате нитрозогуанидинового мутагенеза токсигенного коринефага бета (Uchida, Т. et al. 1971, Nature New Biology 233:8-11). Белок CRM197 имеет ту же молекулярную массу, что и дифтерийный токсин, но отличается от него одной заменой основания (гуанин на аденин) в структурном гене. Это единственное изменение основания вызывает аминокислотную замену глицина на глутаминовую кислоту в зрелом белке и устраняет токсические свойства дифтерийного токсина. Белок CRM197 является Т-клеточно-зависимым безопасным и эффективным носителем сахаридов. Более подробную информацию о CRM197 и его производстве можно найти, например, в США 5,614,382.
Соответственно, в частых вариантах осуществления гликоконъюгаты по изобретению содержат CRM197 в качестве белка-носителя, где капсулярный полисахарид ковалентно связан с CRM197.
В другом варианте осуществления белком-носителем гликоконъюгатов является SCP (стрептококковая пептидаза С5а). Все изоляты р-гемолитических стрептококков человека продуцируют высококонсервативный белок клеточной стенки SCP (стрептококковая пептидаза С5а), который специфически инактивирует С5а. Гены scp кодируют полипептид, содержащий от 1134 до 1181 аминокислоты (Brown et al., PNAS, 2005, vol. 102, no. 51 pp. 18391-18396). Первые 31 остаток представляют собой пре-последовательность сигнала экспорта и удаляются при прохождении через цитоплазматическую мембрану. Следующие 68 остатков служат про-последовательностью и должны быть удалены для получения активного SCP. Следующие 10 остатков могут быть удалены без потери протеазной активности. На другом конце, начиная с Lys-1034, находятся четыре последовательных мотива из 17 остатков, за которыми следует сигнал сортировки клеток и прикрепления к клеточной стенке. Этот комбинированный сигнал состоит из гидрофильной последовательности из 20 остатков, содержащей последовательность LPTTND, гидрофобную последовательности из 17 остатков и короткий основной карбоксильный конец.
SCP может быть разделен на домены (см. фиг. 1 В у Brown et al., PNAS, 2005, vol. 102, no. 51 pp.18391-18396). Эти домены представляют собой домен Pre/Pro (который включает пре-последовательность сигнала экспорта (обычно первые 31 остаток) и про-последовательность (обычно следующие 68 остатков)), домен протеазы (который разделен на две части (часть 1 домена протеазы, обычно остатки 89-333/334, и часть 2 домена протеазы, обычно остатки 467/468-583/584), ассоциированный с протеазой домен (домен РА) (обычно остатки 333/334-467/468), три домена фибронектина типа III (Fn) (Fn1, обычно остатки 583/584-712/713; Fn2, обычно остатки 712/713-928/929/930; Fn3, обычно остатки 929/930-1029/1030/1031) и якорный домен клеточной стенки (обычно остатки 1029/1030/1031 до С-конца).
В одном из вариантов осуществления белок-носитель гликоконъюгатов по изобретению представляет собой SCP из GBS (SCPB). Пример SCPB приведен в SEQ ID NO: 3 в WO 97/26008. См. также SEQ ID NO: 3 в WO O0/34487.
В другом варианте осуществления белок-носитель гликоконъюгата по изобретению представляет собой SCP из GAS (SCPA). Примеры SCPA можно найти в SEQ ID NO: 1 и SEQ ID NO: 2 в WO 97/26008. См. также SEQ ID No: 1, 2 и 23 в WO O0/34487.
В другом варианте осуществления белок-носитель гликоконъюгата по изобретению представляет собой SCP, как указано в SEQ ID NO: 150 или 151 в WO 2014/136064.
В. Адъюванты
В некоторых аспектах раскрытые в настоящем описании иммуногенные композиции могут дополнительно содержать по меньшей мере один, два или три адъюванта. Термин «адъювант» относится к соединению или смеси, которые усиливают иммунный ответ на антиген. Антигены могут действовать в первую очередь как система доставки, в первую очередь как иммуномодулятор, или могут иметь сильные признаки обоих. Подходящие адъюванты включают адъюванты, подходящие для применения у млекопитающих, включая человека.
Примеры известных подходящих адъювантов типа системы доставки, которые можно использовать у людей, включают, без ограничения, квасцы (например, фосфат алюминия, сульфат алюминия или гидроксид алюминия), фосфат кальция, липосомы, эмульсии масло-в-воде, такие как MF59 (4,3% масс./об. сквалена, O,5% масс./об. полисорбата 80 (Tween 80), O,5% масс./об. сорбитан-триолеат (Span 85)), эмульсии вода-в-масле, такие как Montanide, и поли(D, L-лактид-со-гликолид) (PLG) в виде микрочастицы или наночастицы.
В одном из аспектов раскрытые в настоящем описании иммуногенные композиции содержат соли алюминия (квасцы) в качестве адъюванта (например, фосфат алюминия, сульфат алюминия или гидроксид алюминия). В еще одном аспекте раскрытые в настоящем описании иммуногенные композиции содержат фосфат алюминия или гидроксид алюминия в качестве адъюванта. В одном из аспектов раскрытые в настоящем описании иммуногенные композиции содержат от O,1 мг/мл до 1 мг/мл или от O,2 мг/мл до O,3 мг/мл элементарного алюминия в форме фосфата алюминия. В одном из аспектов раскрытые в настоящем описании иммуногенные композиции содержат примерно O,25 мг/мл элементарного алюминия в форме фосфата алюминия.
Примеры известных подходящих адъювантов иммуномодулирующего типа, которые можно использовать у людей, включают, без ограничения, сапониновые экстракты из коры дерева Aquilla (QS21, Quil А), агонисты TLR4, такие как MPL (монофосфорил-липид А), 3DMPL (3-О-деацилированный MPL) или GLA-AQ, LT/CT мутанты, цитокины, такие как различные интерлейкины (например, IL-2, IL-12) или GM-CSF, AS01 и т.п.
Примеры известных подходящих адъювантов иммуномодулирующего типа как со свойствами доставки, так и с иммуномодулирующими характеристиками, которые можно использовать у людей, включают, без ограничения, ISCOMS (см., например, Sjolander et al. (1998) J. Leukocyte Biol. 64:713); WO 90/03184, WO 96/11711, WO O0/48630, WO 98/36772, WO O0/41720, WO 2006/134423 и WO 2007/026190) или GLA-EM, который представляет собой комбинацию агониста TLR4 и эмульсии масло-в-воде.
Для ветеринарных применений можно использовать, например, не ограничиваясь экспериментами на животных, полный адъювант Фрейнда (CFA), неполный адъювант Фрейнда (IFA), эмульсиген, N-ацетилмурамил-L-треонил-В-изоглутамин (thr-MDP), N-ацетил-нор-мурамил-L-аланил-В-изоглутамин (CGP 11637, обозначаемый как нор-МБР), N-ацетилмурамил-L-аланил-D-изоглутаминил-L-аланин-2-(1'-2'-дипальмитоил-зп-глицеро-3-гидроксифосфорилокси)-этиламин (CGP 19835А, обозначаемый как МТР-РЕ) и RIBI, который содержит три компонента, экстрагированные из бактерий, монофосфориловый липид А, димиколат трегалозы и скелет клеточной стенки (MPL+TDM+CWS) в 2% омульсии сквален/Tween 80.
Дополнительные иллюстративные адъюванты для повышения оффективности иммуногенных композиций, раскрытых в настоящем описании, включают, без ограничения, (1) составы омульсий типа «масло-в-воде» (с другими специфическими иммуностимулирующими агентами, такими как мурамиловые пептиды (см. ниже) или без них, или компоненты бактериальной клеточной стенки), такие как, например, (a) SAF, содержащий 10% сквалана, O,4% Tween 80, 5% блокированного плюроником полимера L121 и thr-MDP, либо микрофлюидизированный в субмикронную эмульсию, либо перемешанный на вортексе с получением омульсии большего размера, и (b) адъювантная система RIBI™ (RAS) (Ribi Immunochem, Hamilton, Mont.), содержащая 2% сквалена, O,2% Tween 80 и один или более компонентов бактериальной клеточной стенки, таких как монофосфорилипид A (MPL), трегалозодимиколат (TDM) и скелет клеточной стенки (CWS), предпочтительно MPL+CWS (DETOX™); (2) сапониновые адъюванты, такие как QS21, STIMULON™ (Cambridge Bioscience, WO rcester, Mass.), ABISCO® (Isconova, Sweden) или ISCOMATRIX® (Commonwealth Serum Laboratories, Australia), или полученные из них частицы, такие как ISC0M (иммуностимулирующие комплексы), причем ISC0M могут быть лишены дополнительного детергента (например, WO O0/07621); (3) полный адъювант Фрейнда (CFA) и неполный адъювант Фрейнда (IFA); (4) цитокины, такие как интерлейкины (например, IL-1, IL-2, IL-4, IL-5, IL-6, IL-7, IL-12 (например, WO 99/44636)), интерфероны (например, гамма-интерферон), макрофагальный колониестимулирующий фактор (M-CSF), фактор некроза опухоли (TNF) и т.д.; (5) монофосфориллипид А (MPL) или 3-О-деацилированный MPL (3dMPL) (см., например, GB2220211, ЕР0689454) (см., например, WO O0/56358); (6) комбинацию 3dMPL, например, с QS21 и/или омульсиями масло-в-воде (см., например, ЕР0835318, ЕР0735898, ЕР0761231); (7) полиоксиотиленовый простой офир или полиоксиэтиленовый сложный офир (см., например, WO 99/52549); (8) поверхностно-активное вещество на основе сложного эфира полиоксиотиленсорбитана в комбинации с октоксинолом (например, WO O1/21207) или поверхностно-активное вещество на основе алкилового или сложного офира полиоксиэтилена в комбинации с по меньшей мере одним дополнительным неионогенным поверхностно-активным веществом, таким как октоксинол (например, WO O1/21152); (9) сапонин и иммуностимулирующий олигонуклеотид (например, олигонуклеотид CpG) (например, WO O0/62800); (10) иммуностимулятор и частица соли металла (см., например, WO O0/23105); (11) сапонин и эмульсия масло-в-воде (например, WO 99/11241); (12) сапонин (например, QS21)+3dMPL+IM2 (необязательно+стерол) (например, WO 98/57659); (13) другие вещества, которые действуют как иммуностимулирующие агенты для повышения эффективности композиции. Мурамиловые пептиды включают N-ацетилмурамил-L-треонил-L-изоглутамин (тр-MDP), N-25-ацетил-нормурамил-L-аланил-D-изоглутамин (нор-MDP), И-ацетилмурамил-L-аланил-L-изоглутарнинил-L-аланин-2-(1'-2'-дипальмитоил-зп-глицеро-3-гидроксифосфорилокси)-этиламин МТР-РЕ) и т.д.
В другом варианте осуществления адъювант представляет собой липосомальный состав Quillaja Saponaria-21 (QS21), содержащий 5,1 мг/мл QS-21, 5 мМ сукцината, 60 мМ NaCl, O,1% PS80, рН 5,6. В другом варианте осуществления адъювант представляет собой композицию липосомального монофосфориллипида A (MPLA, Synthetic, PHAD®, Avanti), содержащую 15 мМ фосфатного буфера, рН 6,1, 4 мг/мл 1,2-диолеоил-зп-глицеро-3-фосфохолина (DOPC), 1 мг/мл холестерина, O,2 мг/мл MPLA (лот O0714551-0018-2XLipoMPL), с размером липосомальных частиц 71 нм, определенным методом динамического светорассеяния. В еще одном варианте осуществления адъювант представляет собой липосомальный состав MPLA/QS21, содержащий 15 мМ фосфатного буфера, рН 6,1, 4 мг/мл DOPC, 1 мг/мл холестерина, O,2 мг/мл MPLA и O,2 мг/мл QS-21 (лот O0714551-0018-2XlipoMQ), с размером MPLA-QS21 липосомальных частиц 75 нм, определенный методом динамического светорассеяния.
В еще одном аспекте настоящего изобретения иммуногенные композиции, раскрытые в настоящем описании, содержат олигонуклеотид CpG в качестве адъюванта. Олигонуклеотид CpG, используемый в настоящем описании, относится к иммуностимулирующему олигодезоксинуклеотиду CpG (CpG ODN), и, соответственно, эти термины используются взаимозаменяемо, если не указано иное. Иммуностимулирующие олигодезоксинуклеотиды CpG содержат один или более иммуностимулирующих мотивов CpG, которые представляют собой неметилированные цитозин-гуаниновые динуклеотиды, необязательно в определенных предпочтительных контекстах оснований. Статус метилирования иммуностимулирующего мотива CpG обычно относится к остатку цитозина в динуклеотиде. Иммуностимулирующий олигонуклеотид, содержащий по меньшей мере один неметилированный динуклеотид CpG, представляет собой олигонуклеотид, который содержит 5'-неметилированный цитозин, связанный фосфатной связью с 3'-гуанином и который активирует иммунную систему посредством связывания с Toll-подобным рецептором 9 (TLR-9). В другом варианте осуществления иммуностимулирующий олигонуклеотид может содержать один или более метилированных динуклеотидов CpG, которые будут активировать иммунную систему через TLR9, но не так сильно, как если бы мотив(ы) CpG был/были неметилированы.
Иммуностимулирующие олигонуклеотиды CpG могут содержать один или более палиндромов, которые, в свою очередь, могут охватывать динуклеотид CpG. Олигонуклеотиды CpG описаны в ряде выданных патентов, опубликованных патентных заявках и других публикациях, включая патенты США №№6,194,388; 6,207,646; 6,214,806; 6,218,371; 6,239,116; и 6,339,068.
Были идентифицированы различные классы иммуностимулирующих олигонуклеотидов CpG. Они обозначены как классы А, В, С и Р и более подробно описаны в WO 2010/125480 (стр. 3, строка 22; стр. 12, строка 36). Способы по настоящему изобретению включают использование этих различных классов иммуностимулирующих олигонуклеотидов CpG.
V. Методы очистки и производства
В одном из аспектов настоящее изобретение относится к способам получения мутантного полипептида FimH. Такие способы могут включать, например, культивирование клетки млекопитающего в подходящих условиях, тем самым обеспечивая экспрессию мутантного полипептида FimH. Способ может дополнительно включать сбор полипептида из культуры. Способ может дополнительно включать очистку полипептида.
В некоторых аспектах способ обеспечивает мутантный полипептид FimH с выходом примерно от O,1 г/л до O,5 г/л. В некоторых аспектах выход мутантного полипептида FimH составляет по меньшей мере примерно 1 мг/л, по меньшей мере примерно 2 мг/л, по меньшей мере примерно 3 мг/л, по меньшей мере примерно 4 мг/л, по меньшей мере примерно 5 мг/л, по меньшей мере примерно 6 мг/л, по меньшей мере примерно 7 мг/л, по меньшей мере примерно 8 мг/л, по меньшей мере примерно 9 мг/л, по меньшей мере примерно 10 мг/л, по меньшей мере примерно 11 мг/л, по меньшей мере примерно 12 мг/л, по меньшей мере примерно 13 мг/л, по меньшей мере примерно 14 мг/л, по меньшей мере примерно 15 мг/л, по меньшей мере примерно 16 мг/л, по меньшей мере примерно 17 мг/л, по меньшей мере примерно 18 мг/л, по меньшей мере примерно 19 мг/л, по меньшей мере примерно 20 мг/л, по меньшей мере примерно 25 мг/л, по меньшей мере примерно 30 мг/л, по меньшей мере примерно 35 мг/л, по меньшей мере примерно 40 мг/л, по меньшей мере примерно 45 мг/л, по меньшей мере примерно 50 мг/л, по меньшей мере примерно 55 мг/л, по меньшей мере примерно 60 мг/л, по меньшей мере примерно 65 мг/л, по меньшей мере примерно 70 мг/л, по меньшей мере примерно 75 мг/л, по меньшей мере примерно 80 мг/л, по меньшей мере примерно 85 мг/л, по меньшей мере примерно 90 мг/л, по меньшей мере примерно 95 мг/л или по меньшей мере примерно 100 мг/л.
В некоторых аспектах культура клеток, подходящая для настоящего изобретения, представляет собой периодически подпитываемую культуру. В контексте настоящего описания термин «подпитываемая культура» относится к способу культивирования клеток, при котором дополнительные компоненты добавляют в культуру во время или после начала процесса культивирования. Такие предоставляемые компоненты обычно содержат питательные компоненты для клеток, которые истощаются в процессе культивирования. Культивирование периодически подпитываемой культуры обычно останавливают в определенный момент времени, и клетки и/или компоненты среды собирают и, необязательно, выделяют.В некоторых аспектах периодически подпитываемая культура содержит базовую среду с добавлением подпиточной среды.
В некоторых аспектах клетки можно выращивать в периодических культурах или подпитываемых культурах, где культивирование останавливают после достижения достаточного уровня экспрессии полипептида, после чего экспрессированный полипептид собирают и необязательно выделяют.В некоторых аспектах клетки можно выращивать в перфузионных культурах, где культивирование не останавливают и к культуре периодически или непрерывно добавляются новые питательные вещества и другие компоненты, и периодически, во время подпитки, или непрерывно собирают экспрессированный полипептид.
В некоторых аспектах уровень экспрессии или активность мутантного полипептида FimH увеличивается в по меньшей мере 2 раза, в по меньшей мере 3 раза, в по меньшей мере 5 раз, в по меньшей мере 10 раз, в по меньшей мере 20 раз, в по меньшей мере 30 раз, в по меньшей мере 40 раз, в по меньшей мере 50 раз, в по меньшей мере 60 раз, в по меньшей мере 70 раз, в по меньшей мере 75 раз, в по меньшей мере 80 раз, в по меньшей мере 90 раз, в по меньшей мере 100 раз по сравнению с экспрессией мутантного полипептида FimH в бактериальной клетке, такой как, например, клетка-хозяин Е.coli.
В некоторых аспектах клетки можно выращивать в небольших реакционных сосудах с образованием клеточной культуры объемом от нескольких миллилитров до нескольких литров. В некоторых аспектах клетки можно выращивать в крупномасштабных коммерческих биореакторах с получением клеточной культуры, объем которой может изменяться от приблизительно по меньшей мере 1 литра до 10, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000 литров и более или любого промежуточного объема. В некоторых вариантах осуществления размер клеточной культуры может составлять от 10 л до 5000 л, от 10 л до 10000 л, от 10 л до 20000 л, от 10 л до 50000 л, от 40 л до 50000 л, от 100 л до 50000 л, от 500 л до 50000 л, от 1000 л до 50000 л, от 2000 л до 50000 л, от 3000 л до 50000 л, от 4000 л до 50000 л, от 4500 л до 50000 л, от 1000 л до 10000 л, от 1000 л до 20000 л, от 1000 л до 25000 л, от 1000 л до 30000 л, от 15 л до 2000 л, от 40 л до 1000 л, от 100 л до 500 л, от 200 л до 400 л, или может быть равно любому целому числу между ними.
Температуру клеточной культуры выбирают, прежде всего, с учетом диапазона температур, при которых клеточная культура остается жизнеспособной, диапазона температур, при которых продуцируется высокий уровень полипептида, диапазона температур, при которых образование или накопление продуктов метаболизма сводится к минимуму, и/или любой комбинации этих или других факторов, которые практикующий специалист считает важными. В качестве одного из неограничивающих примеров клетки СНО хорошо растут и продуцируют высокие уровни белка или полипептида примерно при 37°С.Обычно, большинство клеток млекопитающих хорошо растут и/или могут продуцировать высокие уровни белка или полипептида в диапазоне примерно от 25°С до 42°С, хотя способы, описанные в настоящем изобретении, не ограничиваются этими температурами. Некоторые клетки млекопитающих хорошо растут и/или могут продуцировать высокие уровни белка или полипептида в диапазоне примерно от 35°С до 40°С. В некоторых аспектах в процессе культивирования клеток культуру клеток выращивают один или несколько раз при температуре 20°С, 21°С, 22°С, 23°С, 24°С, 25°С, 26°С, 27°С, 28°С, 29°С, 30°С, 31°С, 32°С, 33°С, 34°С, 35°С, 36°С, 37°С, 38°С, 39°С, 40°С, 41°С, 42°С, 43°С, 44°С или 45°С.
Термины «культура» и «культура клеток», используемые в настоящем описании, относятся к клеточной популяции, которая суспендирована в среде в условиях, подходящих для выживания и/или роста клеточной популяции. Как будет понятно специалистам в данной области техники, в некоторых аспектах эти термины, используемые в настоящем описании, относятся к комбинации, включающей клеточную популяцию и среду, в которой суспендирована эта популяция. В некоторых аспектах клетки клеточной культуры включают клетки млекопитающих.
В некоторых аспектах клетки можно выращивать в одной из множества сред с определенным химическим составом, где компоненты среды известны и контролируются. В некоторых аспектах клетки можно выращивать в сложной среде, в которой не все компоненты среды известны и/или контролируются. В течение последних нескольких десятилетий были опубликованы разработки множества сред для выращивания культур клеток млекопитающих с определенным химическим составом. Все компоненты определенных сред хорошо охарактеризованы, поэтому определенные среды не содержат сложных добавок, таких как сыворотка или гидролизаты. Составы сред, разработанные для обеспечения роста клеток и поддержания их жизнеспособности в более ранние годы, создавались практически без учета продукции белка. Недавно разработанные составы сред специально создавали для поддержки высокопродуктивных клеточных культур, продуцирующих рекомбинантный белок. Такие среды предпочтительны для использования в способе изобретения. Такие среды обычно содержат большое количество питательных веществ и, в частности, аминокислот для поддержки роста и/или высокой плотности клеток. Для использования в способе по изобретению эти среды при необходимости могут быть модифицированы специалистом в данной области техники. Например, специалист в данной области техники может уменьшить количество фенилаланина, тирозина, триптофана и/или метионина в этих средах для их использования в качестве базовой среды или питательной среды в способе, представленном в настоящем описании.
Не все компоненты сложных сред хорошо охарактеризованы, поэтому сложные среды могут содержать, среди прочего, такие добавки, как простые и/или сложные источники углерода, простые и/или сложные источники азота и сыворотку. В некоторых аспектах комплексная среда, подходящая для настоящего изобретения, содержит добавки, такие как гидролизаты, в дополнение к другим компонентам определенной среды, раскрытой в настоящем описании. В некоторых аспектах среда с известным химическим составом обычно включает примерно пятьдесят химических соединений в известных концентрациях в воде. Большинство из них также содержат один или более хорошо охарактеризованных белков, таких как инсулин, IGF-1, трансферрин или BSA, но другие не требуют белковых компонентов и поотому называются средами с известным химическим составом, не содержащими белков. Типичные химические компоненты среды делятся на пять широких категорий: аминокислоты, витамины, неорганические соли, микроэлементы и другие категории, не поддающиеся четкой категоризации.
Среда для культивирования клеток необязательно может быть дополнена дополнительными компонентами. В контексте настоящего описания термин «дополнительные компоненты» относится к компонентам, которые усиливают рост и/или выживаемость выше минимального уровня, включая, без ограничения, гормоны и/или другие факторы роста, определенные ионы (такие как натрий, хлорид, кальций, магний и фосфат), буферы, витамины, нуклеозиды или нуклеотиды, микроэлементы (неорганические соединения обычно присутствуют в очень низких конечных концентрациях), аминокислоты, липиды и/или глюкозу или другой источник энергии. В некоторых аспектах к исходной культуре клеток могут быть добавлены дополнительные компоненты. В некоторых аспектах дополнительные компоненты могут быть добавлены после начала культивирования клеток. Как правило, микроэлементы относятся к различным неорганическим солям, содержащимся в микромолярных или более низких количествах. Например, часто включаемыми микроолементами являются цинк, селен, медь и другие. В некоторых аспектах для культивирования клеток в микромолярных концентрациях в качестве микроэлемента в исходную питательную среду для культивирования клеток может быть включено железо (двухвалентное железо или соли трехвалентного железа). В число микроолементов часто также включают марганец в виде двухвалентного катиона (MnCl2 или MnSO4) в диапазоне концентраций от наномолярных до микромолярных. Многочисленные менее распространенные микроолементы обычно добавляют в наномолярных концентрациях.
В некоторых аспектах среда, используемая в способе по изобретению, представляет собой среду, пригодную для поддержания высокой плотности клеток в культуре, такой как, например, 1×106 клеток/мл, 5×106 клеток/мл, 1×107 клеток/мл, 5×107 клеток/мл, 1×108 клеток/мл или 5×108 клеток/мл. В некоторых аспектах клеточная культура представляет собой периодическую культуру клеток млекопитающих, предпочтительно периодическую культуру клеток СНО.
В некоторых аспектах среда для культивирования клеток содержит фенилаланин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит тирозин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит триптофан в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит лейцин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит серии в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит треонин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит глицин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит два компонента из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин и тирозин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин и триптофан в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит тирозин и триптофан в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит тирозин и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит триптофан и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит три компонента из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин, тирозин и триптофан в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин, тирозин и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ.
В некоторых аспектах среда для культивирования клеток содержит фенилаланин, триптофан и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит тирозин, триптофан и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ.
В некоторых аспектах среда для культивирования клеток содержит четыре компонента из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ.
В некоторых аспектах среда для культивирования клеток содержит фенилаланин, тирозин, триптофан и метионин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит пять компонентов из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит шесть компонентов из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит семь компонентов из: фенилаланина, тирозина, триптофана, метионина, лейцина, серина, треонина и глицина, в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток содержит фенилаланин, тирозин, триптофан, метионин, лейцин, серии, треонин и глицин в концентрации ниже 2 мМ, ниже 1 мМ, от O,1 до 2 мМ, от O,1 до 1 мМ, от O,5 до 1,5 мМ или от O,5 до 1 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 13 компонентов из: глицина, валина, лейцина, изолейцина, пролина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата и аспарагина, в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит по меньшей мере 5 компонентов из: глицина, валина, лейцина, изолейцина, пролина, серина, треонина, лизина, аргинина, гистидина, аспартата, глутамата и аспарагина, в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит глицин, валин, лейцин, изолейцин, пролин, серии, треонин, лизин, аргинин, гистидин, аспартат, глутамат и аспарагин в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8 или 9 компонентов из: валина, изолейцина, пролина, лизина, аргинина, гистидина, аспартата, глутамата и аспарагина, в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит по меньшей мере 5 компонентов из: валина, изолейцина, пролина, лизина, аргинина, гистидина, аспартата, глутамата и аспарагина, в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток дополнительно содержит валин, изолейцин, пролин, лизин, аргинин, гистидин, аспартат, глутамат и аспарагин в концентрации выше 2 мМ, 3 мМ, 4 мМ, 5 мМ, 10 мМ, 15 мМ, предпочтительно 2 мМ. В некоторых аспектах среда для культивирования клеток содержит серии в концентрации выше 3 мМ, 5 мМ, 7 мМ, 10 мМ, 15 мМ или 20 мМ, предпочтительно 10 мМ. В некоторых аспектах среда для культивирования клеток содержит валин в концентрации выше 3 мМ, 5 мМ, 7 мМ, 10 мМ, 15 мМ или 20 мМ, предпочтительно 10 мМ. В некоторых аспектах среда для культивирования клеток содержит цистеин в концентрации выше 3 мМ, 5 мМ, 7 мМ, 10 мМ, 15 мМ или 20 мМ, предпочтительно 10 мМ. В некоторых аспектах среда для культивирования клеток содержит изолейцин в концентрации выше 3 мМ, 5 мМ, 7 мМ, 10 мМ, 15 мМ или 20 мМ, предпочтительно 10 мМ.
В некоторых аспектах среда для культивирования клеток содержит лейцин в концентрации выше 3 мМ, 5 мМ, 7 мМ, 10 мМ, 15 мМ или 20 мМ, предпочтительно 10 мМ. В некоторых аспектах указанная выше среда для культивирования клеток предназначена для применения в способе, раскрытом в настоящем описании. В некоторых аспектах вышеуказанная среда для культивирования клеток используется в раскрытом в настоящем описании способе в качестве базовой среды. В некоторых аспектах указанную выше среду для культивирования клеток используют в способе, раскрытом в настоящем описании, в качестве подпиточной среды.
Способы по настоящему изобретению можно использовать с любым способом культивирования клеток, который подходит для желаемого процесса (например, получения рекомбинантного белка). В качестве неограничивающего примера клетки можно выращивать в периодических культурах или подпиточных культурах, где культивирование останавливают после достижения достаточного уровня окспрессии рекомбинантного белка (например, антитела), после чего окспрессированный белок собирают.Альтернативно, в качестве еще одного неограничивающего примера, клетки можно выращивать в условиях периодической подпитки, когда культивирование не останавливают, а новые питательные вещества и другие компоненты периодически или непрерывно добавляют в культуру, и в процессе культивирования экспрессированный рекомбинантный белок периодически или непрерывно собирают. В данной области техники известны и другие подходящие способы (например, культивирование в центрифужных пробирках), которые могут быть использованы для практической реализации настоящего изобретения.
Клетки можно выращивать в любом удобном объеме, выбранном специалистом-практиком. Например, клетки можно выращивать в небольших реакционных сосудах объемом от нескольких миллилитров до нескольких литров. В качестве альтернативы клетки можно выращивать в крупномасштабных коммерческих биореакторах объемом примерно от 1 литра до 10, 50, 100, 250, 500, 1000, 2500, 5000, 8000, 10000, 12000, 15000, 20000 или 25000 литров или больше, или в любом объеме между указанными величинами.
Температуру клеточной культуры выбирают, прежде всего, исходя из диапазона температур, при котором клеточная культура остается жизнеспособной, и диапазона температур, при котором продуцируется высокий уровень желаемого продукта (например, рекомбинантного белка). О6ычно, большинство клеток млекопитающих хорошо растут и могут продуцировать желаемые продукты (например, рекомбинантные белки) в диапазоне температур примерно от 25°С до 42°С, хотя способы, изложенные в настоящем описании, не ограничены этими температурами. Некоторые клетки млекопитающих хорошо растут и могут продуцировать желаемые продукты (например, рекомбинантные белки или антитела) в диапазоне температур примерно от 35°С до 40°С. В некоторых аспектах в процессе культивирования клеток культуру клеток выращивают один или несколько раз при температуре 20°С, 21°С, 22°С, 23°С, 24°С, 25°С, 2б°С, 27°С, 28°С, 29°С, 30°С, 31°С, 32°С, 33°С, 34°С, 35°С, 36°С, 37°С, 38°С, 39°С, 40°С, 41°С, 42°С, 43°С, 44°С или 45°С.Специалисты в данной области смогут выбрать подходящую температуру или температуры для выращивания клеток в зависимости от конкретных потребностей клеток и конкретных производственных требований специалиста-практика. Клетки можно выращивать в течение любого периода времени, в зависимости от потребностей специалиста-практика и требований самих клеток. В некоторых вариантах осуществления клетки выращивают при 37°С. В некоторых аспектах клетки выращивают при 36,5°С.
В некоторых аспектах клетки можно выращивать во время начальной фазы роста (или фазы роста) в течение более или менее продолжительного периода времени, в зависимости от потребностей специалиста-практика и требований самих клеток. В некоторых аспектах клетки выращивают в течение периода времени, достаточного для достижения заданной плотности клеток. В некоторых аспектах клетки выращивают в течение периода времени, достаточного для достижения плотности клеток, которая представляет собой заданный процент от максимальной плотности клеток, которая была бы в конечном итоге достигнута клетками, если бы их рост не нарушался. Например, клетки можно выращивать в течение периода времени, достаточного для достижения желаемой плотности жизнеспособных клеток, составляющей 1, 5, 10, 15, 20, 25, 30, 35, 40, 45, 50, 55, 60, 65, 70, 75, 80, 85, 90, 95 или 99 процентов от максимальной плотности клеток. В некоторых аспектах клетки выращивают до тех пор, пока плотность клеток не увеличится более чем на 15%, 14%, 13%, 12%, 11%, 10%, 9%, 8%, 7%, 6%, 5%, 4%, 3%, 2% или 1% за 1 день культивирования. В некоторых аспектах клетки выращивают до тех пор, пока плотность клеток не увеличивается более чем на 5% за 1 день культивирования.
В некоторых аспектах клеткам позволяют расти в течение определенного периода времени. Например, в зависимости от исходной концентрации клеточной культуры, температуры, при которой выращивают клетки, и собственной скорости роста клеток клетки можно выращивать в течение O, 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12, 13, 14, 15, 16, 17, 18, 19, 20 или более дней, предпочтительно от 4 до 10 дней. В некоторых случаях клеткам дают возможность расти в течение месяца или более. Специалист по настоящему изобретению сможет выбрать продолжительность начальной фазы роста в зависимости от требований к продукции белка и потребностей самих клеток.
Во время начальной фазы культивирования культуру клеток можно перемешивать или встряхивать для увеличения оксигенации и улучшения доступа питательных веществ к клеткам. В соответствии с настоящим изобретением специалисту в данной области техники будет понятно, что на начальной стадии роста стоит контролировать или регулировать определенные внутренние условия биореактора, включая, помимо прочего, рН, температуру, оксигенацию и т.д.
В конце начальной фазы роста по меньшей мере одно из условий культивирования может быть изменено с получением второго набора условий культивирования, при котором в культуре происходит метаболический сдвиг. Метаболический сдвиг может быть достигнут, например, путем изменения температуры, рН, осмоляльности или уровня химического индуктивного вещества в клеточной культуре. В одном из неограничивающих вариантов осуществления условия культивирования изменяют путем изменения температуры культивирования. Однако, как известно в данной области техники, изменение температуры не является единственным механизмом, с помощью которого может быть достигнут соответствующий метаболический сдвиг. Например, такой метаболический сдвиг также может быть достигнут путем изменения других условий культивирования, включая, помимо прочего, рН, осмоляльность и уровни бутирата натрия. Время метаболического сдвига в культуре будет определять специалист, осуществляющий настоящее изобретение, исходя из требований к продуцированию белка или потребностей самих клеток.
При изменении температуры культуры, такое изменение можно выполнять относительно постепенно. Например, для полного изменения температуры может потребоваться несколько часов или дней. В качестве альтернативы изменение температуры может быть относительно резким. Например, изменение температуры может быть завершено менее чем за несколько часов. При наличии соответствующего производственного и контрольного оборудования, такого как стандартное оборудования для промышленного крупномасштабного производства полипептидов или белков, изменение температуры может быть завершено менее чем за час.
В некоторых аспектах после изменения условий культивирования клеток, как обсуждалось выше, клеточную культуру поддерживают в последующей фазе продуцирования при втором наборе условий культивирования, которые способствуют выживанию и жизнеспособности клеточной культуры и являются подходящими для экспрессии желаемого полипептида или белка на коммерчески приемлемом уровне.
Как обсуждалось выше, в культуре можно достичь сдвига путем изменения одного или более условий культивирования, включая, без ограничения, температуру, рН, осмоляльность и уровни бутирата натрия. В некоторых аспектах для сдвига культуры используют изменение температуры. В соответствии с этим вариантом осуществления во время последующей фазы продуцирования культуру поддерживают при температуре или в диапазоне температур, которые ниже, чем температура или диапазон температур на начальной фазе роста. Как обсуждалось выше, для увеличения плотности или жизнеспособности клеток или для увеличения экспрессии рекомбинантного белка можно использовать несколько дискретных изменений температурного режима.
В контексте настоящего описания термин «титр» относится, например, к общему количеству рекомбинантно экспрессированного белка, продуцированного клеточной культурой в заданном объеме среды. Титр обычно выражают в граммах белка на литр среды.
В некоторых аспектах рост клеток увеличивается на по меньшей мере 5%, 10%, 15%, 20% или 25% по сравнению с контрольной культурой. В некоторых аспектах рост клеток увеличивается на по меньшей мере 10% по сравнению с контрольной культурой. В некоторых аспектах рост клеток увеличивается на по меньшей мере 20% по сравнению с контрольной культурой.
В некоторых аспектах продуктивность определяют в титрах и/или в виде объемной продуктивности.
В некоторых аспектах продуктивность определяют в титрах. В некоторых аспектах продуктивность увеличивается на по меньшей мере 5%, 10%, 15%, 20% или 25% по сравнению с контрольной культурой. В некоторых аспектах продуктивность увеличивается на по меньшей мере 10% по сравнению с контрольной культурой. В некоторых аспектах продуктивность увеличивается на по меньшей мере 20% по сравнению с контрольной культурой.
Очистка
В некоторых аспектах способ получения мутантного полипептида FimH включает выделение и/или очистку полипептида. В некоторых аспектах экспрессированный полипептид секретируется в среду, и, таким образом, клетки и другие твердые вещества могут быть удалены центрифугированием и/или фильтрацией. В предпочтительном варианте осуществления полипептид или его фрагмент являются растворимыми.
Мутантный полипептид FimH, полученный в соответствии со способами, описанными в настоящем документе, может быть собран из клеток-хозяев и выделен любым подходящим способом, широко известным в данной области (например, Ruiz-Arguello et al., J. Gen. Virol., 85:3677-3687 (2004)). Подходящие способы очистки полипептида включают преципитацию и различные типы хроматографии, такие как хроматография гидрофобного взаимодействия, ионообменная, аффинная, хелатирующая и эксклюзионная хроматография, все из которых известны в данной области техники. Подходящие схемы очистки могут включать два или более из этих или других подходящих способов. В некоторых аспектах один или более полипептидов могут включать «метку», облегчающую очистку или детектирование. Примеры включают, например, His-метку (связывается с ионом металла, например, гексагистидином), антитело, мальтозо-связывающий белок (МВР) (связывается с амилозой), глутатион-Э-трансферазу (GST) (связывается с глутатионом), FLAG-метку (связывается с анти-FLAG антителом), стрептококковую метку (связывается со стрептавидином или его производным). Такие меченые полипептиды удобно выделять, например, из кондиционированных сред с помощью хелатирующей хроматографии или аффинной хроматографии. Необязательно последовательность метки может быть отщеплена после очистки. В одном из аспектов мутантный полипептид FimH не включает метку для очистки.
В одном из аспектов мутантные полипептиды FimH можно выделить путем получения сначала супернатанта клеточной культуры, а затем супернатант может быть подвергнут как ультрафильтрации, так и диафильтрации. Такие способы фильтрации известны специалистам в данной области. После ультрафильтрации и диафильтрации полученный бесклеточный раствор далее подвергают этапу хроматографии, такой как хроматография Ni-NTA с применением, например, никелевой аффинной смолы. Далее за этим этапом может следовать диализ, за которым может следовать катионообменная хроматография, например, на колонке SP. При определенных условиях, во время очистки на SP-сефарозе может быть желательным использование кислого рН (например, менее примерно 6,0, менее примерно 5,5, менее примерно 5,0, менее примерно 4,5, примерно 4,4, примерно 4,3, примерно 4,2, примерно 4,1 или примерно 4,0 или менее).
Хотя в настоящем описании могут быть указаны конкретные штаммы Е.colif следует понимать, что полипептид, полученный из Е.colif или его фрагмент не ограничены конкретными штаммами, если не указано иное.
VI. Применение композиций
В одном из аспектов настоящее изобретение относится к применению мутантного полипептида FimH, нуклеиновых кислот, кодирующих такой мутант, векторов для экспрессии такого мутанта, композиций, содержащих такой мутант или нуклеиновые кислоты, в качестве лекарственного средства или при производстве лекарственного средства для индукции иммунного ответа против инфекции Е.coli или для профилактики инфекции Е.coli у субъекта.
В других аспектах настоящее изобретение относится к способу индукции иммунного ответа против Е.coli у субъекта, такого как человек, включающему введение субъекту эффективного количества мутантного полипептида FimH, молекулы нуклеиновой кислоты, кодирующей мутантный полипептид FimH, или композиции, содержащей мутантный полипептид FimH или молекулу нуклеиновой кислоты. В настоящем изобретении также предложен способ профилактики инфекции Е.coli у субъекта, включающий введение субъекту эффективного количества фармацевтической композиции, такой как вакцина, содержащей мутантный полипептид FimH, нуклеиновую кислоту, кодирующую мутантный полипептид FimH, или вектор, экспрессирующий мутантный полипептид FimH. В некоторых конкретных аспектах фармацевтическая композиция содержит мутантный полипептид FimH, раскрытый в настоящем описании. В некоторых аспектах способов, представленных в настоящем описании выше, субъектом является человек.
В других аспектах настоящее изобретение относится к способу индукции иммунного ответа у субъекта против внекишечной патогенной Е.coli или индукции образования опсонофагоцитарных и/или нейтрализующих антител у субъекта, специфичных к внекишечной патогенной Е.coli, где способ включает введение субъекту эффективного количества любой из композиций, раскрытых в настоящем описании, таких как композиции, содержащие мутантный полипептид FimH, представленный в настоящем описании. В другом аспекте таких способов субъект подвержен риску развития инфекции мочевыводящих путей и/или риску развития бактериемии, и/или риску развития сепсиса.
В еще одном аспекте настоящее изобретение относится к способу индукции у млекопитающего иммунного ответа против Е.coli, включающему введение млекопитающему эффективного количества любой из раскрытых в описании композиций. Например, в одном из аспектов иммунный ответ включает образование опсонофагоцитарных и/или нейтрализующих антител к Е.coli. В еще одном аспекте иммунный ответ защищает млекопитающее от инфекции Е.coli.
В еще одном аспекте настоящего изобретения предложен способ профилактики, лечения или облегчения бактериальной инфекции, заболевания или состояния у субъекта, включающий введение субъекту иммунологически эффективного количества любой из описанных здесь композиций.
В способах по настоящему изобретению композицию можно вводить субъекту вместе с адъювантом или без него. Эффективное количество, вводимое субъекту, представляет собой количество, достаточное для индукции у субъекта иммунного ответа к антигену Е.coli, такому как белок FimH. Субъекты, которые могут быть выбраны для лечения, включают лиц с риском развития инфекции Е.coli, например, с риском развития инфекции мочевыводящих путей и/или с риском развития бактериемии, и/или с риском развития сепсиса, из-за воздействия или возможного воздействия Е.coli.
В контексте настоящего описания термин «субъект» означает млекопитающее, предпочтительно человека. В одном из вариантов осуществления субъект подвержен риску развития любого из состояний, выбранных из группы, состоящей из инфекции мочевыводящих путей, холецистита, холангита, диареи, гемолитико-уремического синдрома, неонатального менингита, уросепсиса, интраабдоминальной инфекции, менингита, осложненной пневмонии, раневой инфекции, инфекции, связанной с биопсией простаты, неонатального/младенческого сепсиса, нейтропенической лихорадки и других инфекций кровотока; пневмонии, бактериемии и сепсиса.
Введение композиций, представленных в настоящем описании, таких как фармацевтические композиции, можно осуществлять стандартными способами введения. Неограничивающие варианты осуществления включают парентеральное введение, такое как внутрикожное, внутримышечное, подкожное, чрескожное введение, введение через слизистую оболочку или пероральное введение.
Общая доза композиции, вводимой субъекту во время одного введения, может меняться, как известно специалистам-практикам.
Также возможно одно или более повторных введений одной или более иммуногенных композиций. В случае бустерной вакцинации, как правило, такую бустерную вакцинацию вводят тому же субъекту через промежуток времени от одной недели до 10 лет, предпочтительно от двух недель до шести месяцев, после первого введения композиции субъекту (которая в таких случаях называется «примирующей вакцинацией»). В альтернативных схемах бустерной вакцинации после примирующей вакцинации субъекту также можно вводить различные векторы, например, один или более аденовирусных или других векторов, таких как вектор модифицированного вируса осповакцины Анкара (MVA), или ДНК, или белок. Иммуногенные композиции, представленные в настоящем описании, можно использовать вместе с одной или более другими иммуногенными композициями.
Дозировки композиций
Режимы дозирования могут быть скорректированы для обеспечения оптимального желаемого ответа. Например, можно вводить одну дозу мутантного полипептида FimH, можно вводить несколько разделенных доз в течение времени или дозу можно пропорционально уменьшать или увеличивать в зависимости от остроты ситуации. Следует отметить, что величина дозировки может меняться в зависимости от типа и тяжести состояния, подлежащего облегчению, причем дозировка может включать однократные или многократные дозы. Кроме того, следует понимать, что для любого конкретного субъекта конкретные режимы дозирования должны корректироваться с течением времени в соответствии с индивидуальными потребностями и профессиональной оценкой лица, вводящего или контролирующего введение композиций, и что диапазоны дозировок, указанные в настоящем документе, являются исключительно иллюстративными и не предназначены для ограничения объема или применения заявленной композиции.
В некоторых аспектах количество мутантного полипептида FimH в композиции может составлять от примерно 10 мкг до примерно 300 мкг каждого белкового антигена. В некоторых аспектах количество мутантного полипептида FimH в композиции может составлять от примерно 20 мкг до примерно 200 мкг каждого белкового антигена.
Количество гликоконъюгата(ов) в каждой дозе подбирают таким образом, чтобы оно индуцировало иммунозащитный ответ без существенных неблагоприятных побочных эффектов в типичных вакцинах. Такое количество будет меняться в зависимости от того, какой конкретный иммуноген используется и как он представлен.
Количество конкретного гликоконъюгата в иммуногенной композиции можно рассчитать, исходя из общего количества полисахаридов для такого конъюгата (конъюгированного и неконъюгированного). Например, гликоконъюгат с 20% свободного полисахарида будет содержать примерно 80 г конъюгированного полисахарида и примерно 20 г неконъюгированного полисахарида в дозе 100 г полисахарида. Количество гликоконъюгата может меняться в зависимости от серотипа Е.coli. Концентрацию сахарида можно определить с помощью анализа уроновой кислоты.
«Иммуногенное количество» разных полисахаридных компонентов в иммуногенной композиции может отличаться, и каждое из них может составлять примерно 1,0 г, примерно 2,0 г, примерно 3,0 г, примерно 4,0 г, примерно 5,0 г, примерно 6,0 г, примерно 7,0 г, примерно 8,0 г, примерно 9,0 г, примерно 10,0 г, примерно 15,0 г, примерно 20,0 г, примерно 30,0 г, примерно 40,0 пг, примерно 50,0 пг, примерно 60,0 пг, примерно 70,0 пг, примерно 80,0 пг, примерно 90,0 пг или примерно 100,0 г любого конкретного полисахаридного антигена. Как правило, каждая доза будет содержать от O,1 г до 100 г полисахарида для данного серотипа, в частности от O,5 г до 20 г, более конкретно от 1 г до 10 г и еще более конкретно от 2 г до 5 г. Любое целое число в любом из вышеуказанных диапазонов рассматривается как вариант осуществления изобретения. В одном из вариантов осуществления каждая доза будет содержать 1 г, 2 г, 3 г, 4 г, 5 г, 6 г, 7 г, 8 г, 9 г, 10 г, 15 г или 20 г полисахарида для данного серотипа.
VII. КОМБИНАЦИЯ С САХАРИДОМ И/ИЛИ ПОЛИПЕПТИДОМ ИЛИ ИХ ФРАГМЕНТАМИ, ПРОИСХОДЯЩИМИ ОТ KLEBSIELLA PNEUMONIAE
Klebsiella pneumoniae (К. pneumoniae) представляет собой грамотрицательный патоген, который, как известно, вызывает инфекции мочевыводящих путей, бактериемию и сепсис. Инфекции K. pneumoniae с множественной лекарственной устойчивостью являются растущей причиной смертности среди уязвимых групп риска. Серотипы О-антигена широко распространены среди штаммов, вызывающих инвазивные заболевания во всем мире, и полученные гликоконъюгаты О-антигена привлекательны в качестве вакцинных антигенов.
В одном из аспектов любая из раскрытых здесь композиций может дополнительно содержать по меньшей мере один сахарид, который является производным по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12. В предпочтительном варианте осуществления любая из представленных в описании композиций может дополнительно содержать полипептид, происходящий от K. pneumoniae, выбранный из полипептида, полученного из фимбриального белка типа I K. pneumoniae, или его иммуногенного фрагмента; или полипептида, полученного из фимбриального белка типа III K. pneumoniae, или его иммуногенного фрагмента; или их комбинации.
Как известно в данной области техники, О-антигены K. pneumoniae O1 и O2 и соответствующие им подтипы v1 и v2 представляют собой полимерные галактаны, отличающиеся структурой своих повторяющихся звеньев. Антигены K. pneumoniae O1 и O2 содержат гомополимерные галактозные звенья (или галактаны). Каждый из антигенов O1 и O2 K. pneumoniae содержит звенья D-галактана I (иногда называемые повторяющимся звеном O2a), но антигены O1 отличаются тем, что они имеют кэп-структуру D-галактана II. D-галактан III (d-Gal-III) представляет собой вариант D-галактана I. Структуры основных галактанов I и III определяют два разных подтипа серотипа O2, O2v1 и O2v2; и структуры производных химер, полученных в результате копирования галактаном II, который дает подтипы O1v1 и O1v2, показаны в Kelly SD, et al. J Biol Chem 2019; 294:10863-76; and Clarke BR, et al. J Biol Chem 2018; 293:4666-79.
В некоторых вариантах осуществления сахарид, полученный из O1 K. pneumoniae, включает повторяющееся звено [→3)-β-D-Galf-(1→3)-α-D-Galp-(1→]. В некоторых вариантах осуществления сахарид, полученный из O1 K. pneumoniae, включает повторяющееся звено [→3)-α-D-Galp-(1→3)-β-D-Galp-(1→]. В некоторых вариантах осуществления сахарид, полученный из O1 K. pneumoniae, включает повторяющееся звено [→3)-β-D-Galf-(1→3)-α-D-Galp-(1→] и повторяющееся звено [→3)-α-D-Galp-(1→3)-β-D-Galp-(1→]. В некоторых вариантах осуществления сахарид, полученный из O1 K. pneumoniae, включает повторяющееся звено →3)-β-D-Galf-(1→3)-[α-D-Galp-(1→4)]-α-D-Galp-(1→] (упоминаемое как повторяющееся звено D-Gal-III) (Kol О., et al. (1992) Carbohydr. Res. 236, 339-344; Whitfield C, et al. (1991) J. Bacterid. 173, 1420-1431).
В некоторых вариантах осуществления сахарид, полученный из O2 K. pneumoniae, включает повторяющееся звено [→3)-α-D-Galp-(1→3)-β-D-Galf-(1→] (которое может быть элементом антигена серотипа O2a K. pneumoniae). В некоторых вариантах осуществления сахарид, полученный из O2 K. pneumoniae, включает повторяющееся звено [→3)-β-D-GlcpNAc-(1→5)-β-D-Galf-((которое может быть олементом антигена серотипа O2 с K. pneumoniae). В некоторых вариантах осуществления сахарид, полученный из O2 K. pneumoniae, включает модификацию повторяющегося звена O2a путем добавления в боковую цепь (1→4)-связанных остатков Galp (которые могут быть олементом антигена O2afg K. pneumoniae). В некоторых вариантах осуществления сахарид, полученный из O2 K. pneumoniae, включает модификацию повторяющегося звена O2a путем добавления в боковую цепь (1-2)-связанных остатков Galp (которые могут быть элементом антигена O2aeh K. pneumoniae). (Whitfield С, et al. (1992) J. Bacterid. 174, 4913-4919).
He ограничиваясь каким-либо механизмом или теорией, структура О-антигена полисахарида серотипов O3 и O5 K. pneumoniae, раскрытая в данной области, идентична таковой у серотипов O9a (формулы O9a) и O8 (формулы O8) Е.coli, соответственно.
В некоторых вариантах осуществления сахарид, полученный из O4 K. pneumoniae, включает повторяющееся звено [→4)-α-D-Galp-(1→2)-β-D-Ribf-(1→)]. В некоторых вариантах осуществления сахарид, полученный из O7 K. pneumoniae, включает повторяющееся звено [→2-a-L-Rhap-(1→2)-β-D-Ribf-(1→3)-α-L-Rhap-(1→3)-α-L-Rhap-(1→]. В некоторых вариантах осуществления сахарид, полученный из серотипа O8 K. pneumoniae, включает структуру повторяющихся звеньев, которая является такой же что и у O2a K. pneumoniae, но нестехиометрически О-ацетилированной. В некоторых вариантах осуществления сахарид, полученный из серотипа O12 K. pneumoniae, включает повторяющееся звено [α-Rhap-(1→3)-β-GlcpNAc]
дисахаридного повторяющегося звена.
В одном из аспектов изобретение относится к композиции, включающей полипептид, полученный из FimH Е.coli, или его фрагмент; и по меньшей мере один сахарид, который является или получен из по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12. В некоторых вариантах осуществления композиция включает сахариды из, или полученные из, одного или более серотипов O1, O2, O3 и O5 или их комбинации. В некоторых вариантах осуществления композиция включает сахариды из, или полученные из, каждого из серотипов O1, O2, O3 и O5.
В другом аспекте изобретение включает композицию, включающую по меньшей мере один сахарид из, или который получен из, по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12; и сахарид, полученный из О-антигена Е.coli, имеющего структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы О6 (например, формулы O6:K2; K13; K15 и формулы О6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы 0169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187, где n представляет собой целое число от 1 до 100. В некоторых вариантах осуществления композиция включает сахарид из, или полученный из, одного или более серотипов O1, O2, O3 и O5 K. pneumoniae или их комбинации. В некоторых вариантах осуществления композиция включает сахарид из, или полученный из, каждого из серотипов O1, O2, O3 и O5 K. pneumoniae. В некоторых вариантах осуществления композиция включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O9, и не включает сахарид, полученный из серотипа O3 K. pneumoniae. В некоторых вариантах осуществления композиция включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O8, и не включает сахарид, полученный из серотипа O5 K. pneumoniae.
В другом аспекте изобретение относится к композиции, включающей полипептид, полученный из FimH Е.coli, или его фрагмент; по меньшей мере один сахарид из, или полученный из, по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12; и сахарид, имеющий структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187, где n представляет собой целое число от 1 до 100, предпочтительно от 31 до 90. В некоторых вариантах осуществления композиция включает сахарид, полученный из О-антигена Е.coli, имеющий формулу O9, и не включает сахарид, полученный из серотипа O3 K. pneumoniae. В некоторых вариантах осуществления композиция включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O8, и не включает сахарид, полученный из серотипа O5 K. pneumoniae.
В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид, полученный из любого типа K. pneumoniae, выбранного из группы, состоящей из O1, O2, O3 и O5.
В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид, полученный из типа O1 K. pneumoniae. В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae выбирают из подтипа v1 (O1v1) или подтипа v2 (O1v2). В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae выбирают из подтипа v1 (O1v1) и подтипа v2 (O1v2). В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид, полученный из типа O2 K. pneumoniae. В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae выбирают из подтипа v1 (O2v1) или подтипа v2 (O2v2). В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae выбирают из подтипа v1 (O2v1) и подтипа v2 (O2v2). В другом аспекте О-антиген K. pneumoniae выбирают из группы, состоящей из: а) серотипа O1 подтипа v1 (O1v1), b) серотипа O1 подтипа v2 (O1v2), с) серотипа O2 подтипа v1 (O2v1) и d) серотипа O2 подтипа v2 (O2v2). В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae относится к подтипу v1 (O1v1). В одном из аспектов этого варианта осуществления O-антиген K. pneumoniae относится к подтипу v2 (O1v2). В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae относится к подтипу v1 (O2v1). В одном из аспектов этого варианта осуществления О-антиген K. pneumoniae относится к подтипу v2 (O2v2). В другом аспекте этого варианта осуществления композиция содержит один, два, три или четыре О-антигена K. pneumoniae, выбранных из группы, состоящей из: а) серотипа O1 подтипа v1 (O1v1), b) серотипа O1 подтипа v2 (O1v2), с) серотипа O2 подтипа v1 (O2v1), и d) серотипа O2 подтипа v2 (O2v2). В некоторых вариантах осуществления композиция включает комбинацию сахаридов, полученных из K. pneumoniae, где первый сахарид получен из любого одного типа K. pneumoniae, выбранного из группы, состоящей из O1, O2, O3 и O5; и второй сахарид получен из сахарида, полученного из любого одного типа K. pneumoniae, выбранного из группы, состоящей из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12. Например, в некоторых вариантах осуществления композиция включает по меньшей мере один сахарид, полученный из типа O1 K. pneumoniae, и по меньшей мере один сахарид, полученный из типа O2 K. pneumoniae. В предпочтительном варианте осуществления сахарид, полученный из K. pneumoniae, конъюгирован с белком-носителем; и сахарид, полученный из Е.coli, конъюгирован с белком-носителем.
В другом аспекте изобретение включает композицию, включающую полипептид, полученный из FimH Е.coli, или его фрагмент; и по меньшей мере один сахарид, полученный из любого типа K. pneumoniae, выбранного из группы, состоящей из O1, O2, O3 и O5.
В другом аспекте изобретение включает по меньшей мере один сахарид, полученный из любого типа K. pneumoniae, выбранного из группы, состоящей из O1, O2, O3 и O5; и по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы О6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы 0126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187. В некоторых вариантах осуществления композиция включает сахарид, полученный из O-антигена Е.coli, имеющего формулу O9, и не включает сахарид, полученный из серотипа O3 K. pneumoniae. В некоторых вариантах осуществления композиция включает сахарид, полученный из O-антигена Е.coli, имеющего формулу O8, и не включает сахарид, полученный из серотипа O5 K. pneumoniae.
В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид, полученный из типа O1 K. pneumoniae; и по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из группы, состоящей из формулы O8 и формулы O9. В другом варианте осуществления композиция включает по меньшей мере один сахарид, полученный из типа O2 K. pneumoniae; и по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из группы, состоящей из формулы O8 и формулы O9. В другом варианте осуществления композиция включает по меньшей мере один сахарид, полученный из типа O1 K. pneumoniae; по меньшей мере один сахарид, полученный из типа O2 K. pneumoniae; и по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из группы, состоящей из формулы O8 и формулы O9.
В одном из вариантов осуществления изобретение относится к способу индукции иммунного ответа на K. pneumoniae у субъекта, включающему введение субъекту иммунологически эффективного количества иммуногенной композиции, содержащей по меньшей мере один гликоконъюгат из серотипа O8 или O9 Е.coli, где указанный иммуногенная композиция не содержит гликоконъюгаты из серотипа 05 или O3 K. pneumoniae. В одном из аспектов композиция включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O8, и не включает сахарид, полученный из серотипа O5 K. pneumoniae. В другом аспекте композиция включает сахарид, полученный из O-антигена Е.coli, имеющего формулу O9, и не включает сахарид, полученный из серотипа O3 K. pneumoniae.
В другом варианте осуществления изобретение относится к способу индукции иммунного ответа на Е.coli у субъекта, включающему введение субъекту иммунологически эффективного количества иммуногенной композиции, содержащей по меньшей мере один гликоконъюгат из серотипа O5 или O3 K. pneumoniae или его вариант, где указанная иммуногенная композиция не содержит гликоконъюгаты из серотипа O8 или O9 Е.coli. В одном из аспектов композиция включает сахарид, полученный из серотипа O5 K. pneumoniae, и не включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O8. В другом аспекте композиция включает сахарид, полученный из серотипа O3 K. pneumoniae, и не включает сахарид, полученный из О-антигена Е.coli, имеющего формулу O9.
В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид из, или полученный из, по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12; по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из группы, состоящей из формулы O8 и формулы O9. В некоторых вариантах осуществления композиция включает по меньшей мере один сахарид из, или полученный из, по меньшей мере одного серотипа K. pneumoniae, выбранного из O1 (и вариантов d-Gal-III), O2 (и вариантов d-Gal-III), O2ac, O3, O4, O5, O7, O8 и O12; по меньшей мере один сахарид, полученный из Е.coli, имеющий структуру, выбранную из группы, состоящей из формулы O1A, формулы O1B, формулы O2, формулы О6 и формулы O25B.
В некоторых вариантах осуществления композиция дополнительно включает полипептид, полученный из K. pneumoniae, выбранный из полипептида, полученного из фимбриального белка типа I K. pneumoniae, или его иммуногенного фрагмента; или полипептида, полученного из фимбриального белка типа III K. pneumoniae, или его иммуногенного фрагмента, или их комбинации. Последовательности указанных полипептидов известны в данной области.
VIII. Наночастицы
В другом аспекте в настоящем описании раскрыт иммуногенный комплекс, включающий 1) наноструктуру; и 2) по меньшей мере один фимбриальный полипептидный антиген или его фрагмент.Предпочтительно фимбриальный полипептид или его фрагмент получают из фимбриального Н Е.coli (fimH). В предпочтительном варианте осуществления фимбриальный полипептид выбирают из любого из описанных выше фимбриальных полипептидов. Например, фимбриальный полипептид может содержать одну любую аминокислотную последовательность, выбранную из SEQ ID NO: 1-65.
В некоторых вариантах осуществления антиген слит или конъюгирован с внешней частью наноструктуры для стимуляции развития адаптивных иммунных ответов на экспонированные эпитопы. В некоторых вариантах осуществления иммуногенный комплекс дополнительно включает адъювант или другие иммуномодулирующие соединения, прикрепленные к внешней части и/или инкапсулированные внутри кейджа, что облегчает адаптацию типа иммунного ответа, генерируемого для каждого патогена.
В некоторых вариантах осуществления наноструктура включает одну сборку, включающую множество идентичных первых относящихся к наноструктуре полипептидов.
В альтернативных вариантах осуществления наноструктура включает множество сборок, включающих множество идентичных первых относящихся к наноструктуре полипептидов и множество вторых сборок, где каждая вторая сборка содержит множество идентичных вторых относящихся к наноструктуре полипептидов.
Для создания раскрытых в описании иммуногенных композиций можно использовать различные платформы наноструктур. В некоторых вариантах осуществления используемые наноструктуры образованы множеством копий одной субъединицы. В некоторых вариантах осуществления используемые наноструктуры образованы множеством копий множества разных субъединиц.
Наноструктуры обычно имеют шарообразную форму и/или обладают осевой симметрией (например, с осью 3-го и 5-го порядка), например, с икосаэдрической структурой, приведенной здесь в качестве примера.
В некоторых вариантах осуществления антиген презентирован на самособирающихся наночастицах, таких как самособирающиеся наноструктуры, полученные из ферритина (FR), Е2р, QB и 13-01. Е2р представляет собой переработанный вариант дигидролипоилацилтрансферазы из Bacillus stearothermophilus. 13-01 представляет собой сконструированный белок, обладающий способностью к самосборке в гиперстабильные наночастицы. Последовательности субъединиц этих белков известны в данной области. В первом аспекте в настоящем описании раскрыт относящийся к наноструктуре полипептид, содержащий аминокислотную последовательность, которая является идентичной аминокислотной последовательности относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105, по длине на по меньшей мере 75%, а также по меньшей мере в одном идентифицированном положении поверхности контакта. Относящиеся к наноструктуре полипептиды можно использовать, например, для получения наноструктур. Относящиеся к наноструктуре полипептиды разработаны со способностью к самосборке в пары с образованием наноструктур, таких как икосаэдрические наноструктуры.
В некоторых вариантах осуществления наноструктура включает (а) множество первых сборок, где каждая первая сборка содержит множество идентичных первых относящихся к наноструктуре полипептидов, причем первые относящиеся к наноструктуре полипептиды содержат аминокислотную последовательность относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105; и (b) множество вторых сборок, где каждая вторая сборка содержит множество идентичных вторых относящихся к наноструктуре полипептидов, где вторые относящиеся к наноструктуре полипептиды содержат аминокислотную последовательность относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105, и где второй относящийся к наноструктуре полипептид отличается от первого относящегося к наноструктуре полипептида; при этом множество первых сборок взаимодействуют без образования ковалентных связей со множеством вторых сборок с образованием наноструктуры.
Наноструктуры включают симметрично повторяющиеся, неприродные, не связанные ковалентно поверхности контакта полипептид-полипептид, которые ориентируют первую сборку и вторую сборку внутрь наноструктуры, такой как структура с икосаодрической симметрией.
SEQ ID NO: 66-105 обеспечивают аминокислотную последовательность иллюстративных относящихся к наноструктуре полипептидов. Количество остатков на поверхности контакта иллюстративных относящихся к наноструктуре полипептидов SEQ ID NO: 66-105 находится в диапазоне от 4 до 13 остатков. В различных вариантах осуществления относящиеся к наноструктуре полипептиды содержат аминокислотную последовательность, которая является идентичной аминокислотной последовательности относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105, по длине на по меньшей мере 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, а также в по меньшей мере 1, 2, 3, 4, 5, 6, 7, 8, 9, 10, 11, 12 или 13 идентифицированных положениях поверхности контакта (в зависимости от количества остатков на поверхности контакта данного относящегося к наноструктуре полипептида). В других вариантах осуществления относящиеся к наноструктуре полипептиды содержат аминокислотную последовательность, которая идентична аминокислотной последовательности относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105, по длине на по меньшей мере 75%, 80%, 85%, 90%, 91%, 92%, 93%, 94%, 95%, 96%, 97%, 98% или 99%, а также в по меньшей мере 20%, 25%, 33%, 40%, 50%, 60%, 70%, 75%, 80%, 90% или 100% идентифицированных положениях поверхности контакта. В дополнительных вариантах осуществления относящиеся к наноструктуре полипептиды включают относящийся к наноструктуре полипептид с аминокислотной последовательностью относящегося к наноструктуре полипептида, выбранного из группы, состоящей из SEQ ID NO: 66-105.
В одном из неограничивающих вариантов осуществления относящиеся к наноструктуре полипептиды могут быть модифицированы для облегчения образования ковалентной связи с представляющим интерес «грузом». В одном из неограничивающих примеров относящиеся к наноструктуре полипептиды могут быть модифицированы, например, путем введения различных остатков цистеина в определенные положения для связи с одним или более представляющими интерес антигенами с получением наноструктуры из относящихся к наноструктуре полипептидов, обеспечивающих каркас для доставки большого количества антигенов, в качестве вакцины для достижения улучшенного иммунного ответа.
В некоторых вариантах осуществления некоторые или все нативные остатки цистеина, которые присутствуют в относящихся к наноструктуре полипептидах, но не предназначены для конъюгации, могут быть заменены другими аминокислотами для облегчения конъюгации в определенных положениях. В другом неограничивающем варианте осуществления относящиеся к наноструктуре полипептиды могут быть модифицированы путем связывания (ковалентного или нековалентного) с фрагментом для облегчения «ускользания из эндосомы». Для применений, которые включают доставку представляющих интерес молекул в клетку-мишень, такую как адресная доставка, критическим этапом может быть ускользание из эндосомы, связанной с мембраной органеллы, которая является точкой входа средства доставки в клетку. Эндосомы превращаются в лизосомы, которые разлагают свое содержимое. Следовательно, если средство доставки каким-либо образом не «ускользнет» из эндосомы до того, как она станет лизосомой, оно будет разрушено и не сможет выполнить свою функцию. Существует множество липидов или органических полимеров, которые разрушают эндосомы и позволяют проникнуть в цитозоль. Таким образом, в этом варианте относящиеся к наноструктуре полипептиды могут быть модифицированы, например, путем введения остатков цистеина, которые обеспечат возможность осуществления химической конъюгации такого липида или органического полимера с мономером или полученной поверхностью сборки. В другом неограничивающем примере относящиеся к наноструктуре полипептиды могут быть модифицированы, например, путем введения остатков цистеина, которые обеспечат возможность осуществления химической конъюгации флуорофоров или других визуализирующих агентов, позволяющих визуализировать наноструктуры in vitro или in vivo.
Поверхностные аминокислотные остатки на относящихся к наноструктуре полипептидах могут быть мутированы для улучшения стабильности или растворимости белковых субъединиц или собранных наноструктур. Как известно специалистам в данной области, если относящийся к наноструктуре полипептид имеет высокую гомологию последовательности с существующим семейством белков, то можно использовать множественное выравнивание последовательностей других белков из этого семейства для выбора аминокислотных мутаций в неконсервативных положениях, которые могут повысить стабильность и/или растворимость белка; процесс, называемый консенсусным дизайном белка (9).
Аминокислотные остатки на поверхности относящихся к наноструктуре полипептидов могут быть мутированы в положительно заряженные (Arg, Lys) или отрицательно заряженные (Asp, Glu) аминокислоты, придавая поверхности белка общий положительный или общий отрицательный заряд. В одном из неограничивающих вариантов осуществления поверхностные аминокислотные остатки на относящихся к наноструктуре полипептидах могут быть мутированы для обеспечения высокого суммарного заряда на внутренней поверхности самособирающейся наноструктуры. Затем такую наноструктуру можно использовать для упаковки или инкапсуляции молекулы-груза с противоположным суммарным зарядом благодаря электростатическому взаимодействию между внутренней поверхностью наноструктуры и молекулой-грузом. В одном из неограничивающих вариантов осуществления поверхностные аминокислотные остатки на относящихся к наноструктуре полипептидах могут быть мутированы в первую очередь на остатки аргинина или лизина, придавая внутренней поверхности самособирающейся наноструктуры суммарный положительный заряд. Затем растворы, содержащие относящиеся к наноструктуре полипептиды, можно смешать в присутствии молекулы-носителя нуклеиновой кислоты, такой как двухцепочечная ДНК, одноцепочечная ДНК, двухцепочечная РНК, одноцепочечная РНК, кДНК, микроРНК, миРНК, кшРНК, пиРНК или другая нуклеиновая кислота, для инкапсуляции нуклеиновой кислоты внутри самособирающейся наноструктуры. Такую наноструктуру можно использовать, например, для защиты, доставки или концентрирования нуклеиновых кислот.
В одном из вариантов осуществления наноструктура имеет икосаэдрическую симметрию. В этом варианте осуществления наноструктура может содержать 60 копий первого относящегося к наноструктуре полипептида и 60 копий второго относящегося к наноструктуре полипептида. В одном из таких вариантов осуществления количество идентичных первых относящихся к наноструктуре полипептидов в каждой первой сборке отличается от количества идентичных вторых относящихся к наноструктуре полипептидов в каждой второй сборке. Например, в одном из вариантов осуществления наноструктура включает двенадцать первых сборок и двадцать вторых сборок; в этом варианте каждая первая сборка может, например, содержат пять копий идентичного первого относящегося к наноструктуре полипептида, и каждая вторая сборка может, например, содержать три копии идентичного второго относящегося к наноструктуре полипептида. В другом варианте осуществления наноструктура содержит двенадцать первых сборок и тридцать вторых сборок; в этом варианте осуществления каждая первая сборка может, например, содержать пять копий идентичного первого относящегося к наноструктуре полипептида, и каждая вторая сборка может, например, содержать две копии идентичного второго относящегося к наноструктуре полипептида.
В другом варианте осуществления наноструктура содержит двадцать первых сборок и тридцать вторых сборок; в этом варианте осуществления каждая первая сборка может, например, содержать три копии идентичного первого относящегося к наноструктуре полипептида, и каждая вторая сборка может, например, содержать две копии идентичного второго относящегося к наноструктуре полипептида. Все эти варианты осуществления способны формировать синтетические наноматериалы с правильной икосаэдрической симметрией.
ПРИМЕРЫ
Для лучшего понимания изобретения ниже приведены примеры. Эти примеры предназначены исключительно для целей иллюстрации и не должны рассматриваться как ограничивающие каким-либо образом объем изобретения.
ПРИМЕР 1: Создание антигена
Для того, чтобы зафиксировать лектиновый домен FimH в открытой конформации с целью улучшения функциональной иммуногенности, были разработаны мутации в FimHLD или FimH-DSG. Мутации относились к разным классам, описанным в таблицах 2-9 ниже. Аминокислотные последовательности для различных мутантных полипептидов FimH показаны в таблице 1.
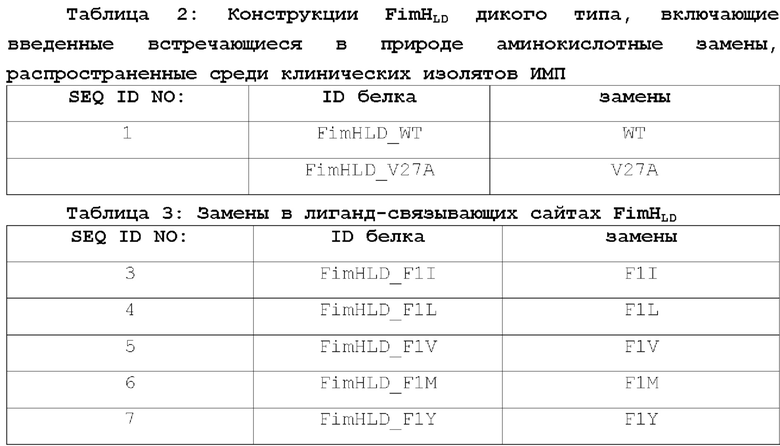
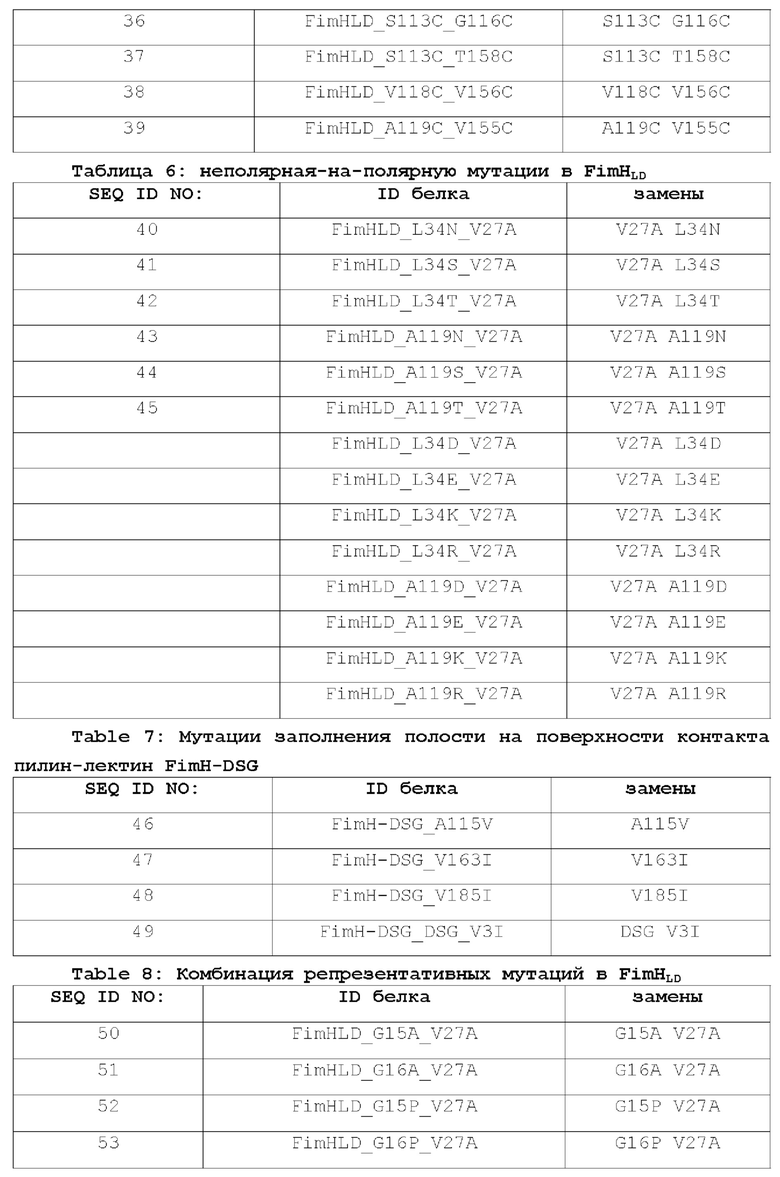
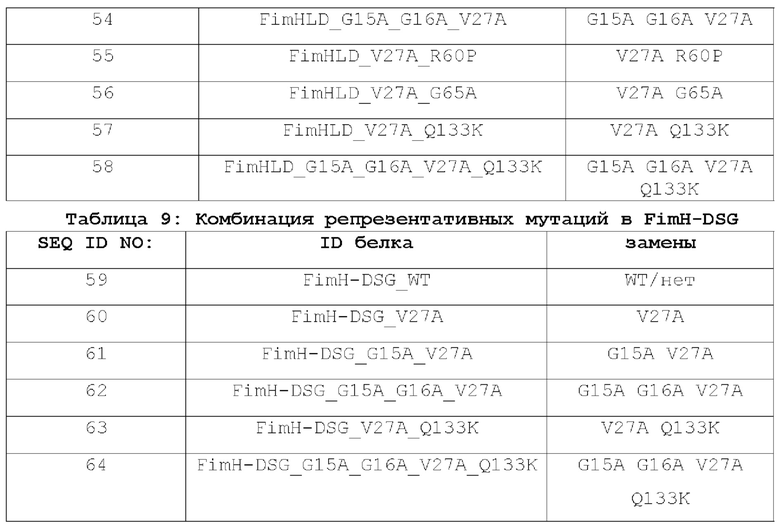
Пример 2: Экспрессия и очистка антигена
ДНК, кодирующую мутанты FimHLD и FimH-DSG, клонировали в pcDNA3.1, содержащую сигнальный пептид IgK мыши, и экспрессировали в клетках Expi293™, как описано ранее (международная публикация РСТ №WO 2021/084429, опубликованная 6 мая 2021 г.). Для характеристики белков и изучения иммуногенности белки выделяли с помощью никелевой аффинной смолы для и эксклюзионной хроматографии, как описано в международной публикация РСТ №WO 2021/084429, опубликованной 6 мая 2021 г.
Пример 3: Анализ поляризации флуоресценции
Для определения констант диссоциации мутантов FimH для маннозидных лигандов был разработан анализ поляризации флуоресценции на основе методов, описанных Rabbani et al. (J. Biol. Chem. 293:1835-1849 (2018)) с применением флуоресцеина, конъюгированного с маннозидными лигандами с высоким сродством к FimH. Белки FimH разбавляли в 20 мМ НЕ PES рН 7,4, 150 мМ NaCl, O,05 мг/мл плюс BSA O,05% при трехкратном титровании по 11 точкам в черном плоскодонном 96-луночном полипропиленовом планшете (Greiner) с конечным объемом 50 мкл. В каждую лунку добавляли 50 мкл флуоресцеин-октилбифенилманнопиранозидного лиганда в концентрации O,7 нМ в том же буфере. Планшеты инкубировали в течение ночи при комнатной температуре, встряхивая при 100 об/мин. Через 20-24 часа планшеты считывали с помощью планшетного ридера ClarioStar Plus при возбуждении флуоресцеина при 488 нм и испускании при 530 нм.
Пример 4: Анализ термической стабильности (анализ ThermoFluor)
Для определения температуры плавления выделенных белков в форме АРО (несвязанной) и в присутствии лиганда разрабатывали анализ термостабильности в 384-луночном планшете с использованием SYPRO оранжевого. Для анализа ассоциации с белком использовали соединения маннозида (метил α-D-маннопиранозид (Sigma М6882)), имитирующие природный лиганд FimH (маннозу). Исходные растворы белка FimH готовили путем разбавления белков в 40 мМ Трис, рН 8, 400 мМ NaCl (буфер для анализа) до 4 мкМ; оранжевый краситель SYPRO (Invitrogen S6650) разбавляли в отношении 1:10 в буфере для анализа. Мутанты FimH с концентрацией 4 мкМ (5 мкл) смешивали с оранжевым красителем SYPRO 1:10 (0,1 мкл) и буфером для анализа или лигандом, разведенным в буфере для анализа (5 мкл) до конечного реакционного объема 10 мкл в 384-луночном планшете MicroAmp EnduraPlate Optical (Applied Biosystems 4483285). Планшет анализировали по кривой плавления в системе ПЦР в реальном времени QuantStudio 5 (ThermoFisher) согласно протоколу диссоциации при температуре от 20°С до 98°С со скоростью 0,05°С/сек. TAMRA указывали в качестве мишени и репортера, ROX -в качестве пассивного эталона (однако в анализе не использовали). Данные наносили на график в виде распределения Максвелла-Больцмана с температурой (от 20°С до 98°С) по оси X и флуоресценцией из канала TAMRA по оси Y (каждой точке температуры, считываемой во время измерения кривой плавления, присваивали определенное значение возбуждения флуоресценции для репортера TAMRA). Для выравнивания интенсивности флуоресценции между лунками и образцами, для возможности сравнения значений флуоресценции оси Y между планшетами по шкале от 0 (отсутствие флуоресценции) до 1 (самая высокая зарегистрированная флуоресценция) разрабатывали алгоритм нормализации. Это уравнение показано ниже. С помощью функции поиска в Microsoft Excel (также ниже) было зарегистрировано относительное значение флуоресценции (после нормализации), равное 0,5 (указывающее, что диссоциировала приблизительно половина белка), что коррелировало с конкретной температурой. Эту температуру, измеренную температуру плавления (Tm) белка, вычисляли таким образом.
Изменение температуры плавления (ΔTm) определяли путем вычитания Tm белка+лиганда из состояния АРО. Сводные таблицы в Microsoft Excel использовали для организации Tin из макета планшета.
Уравнение для нормализации сигнала флуоресценции TAMRA:
Значение нормализации (от 0 до 1) = (необработанное значение флуоресценции - минимальное значение флуоресценции для всей лунки (от 20°С до 98°С)) / (максимальное значение флуоресценции для всей лунки минимальное значение флуоресценции для всей лунки)
Функция поиска в Excel для определения Tm (0,5 нормализованная флуоресценция или 50% плавления белка):
=LOOKUP (0,5, начало нормированных значений флуоресценции: конец значений, $ начало значений температуры: $ конец значений)
Пример 5: Подтверждение конформационного состояния мутантов FimH с помощью FimH-специфических нейтрализующих моноклональных антител
Нейтрализующие моноклональные антитела 299-3, 304-1 и 440-2 (собственной разработки) использовали для подтверждения конформационного состояния мутантов FimH; 229-3 и 304-1 связываются с такими же эпитопами, как MAb 475 и 926 (Kisiela, D.I. et al. Proc Natl Acad Sci USA 110, 19089-19094 (2013)), в то время как 440-2 распознает другой эпитоп и, по-видимому, предпочтительно связывается с FimHLD в состоянии открытой конформации. Ожидается, что варианты, сохраняющие структурную целостность, аналогичную дикому типу, будут связывать все антитела. Для измерения реактивности антител по отношению к каждому мутанту во всех экспериментах по кинетическому биомолекулярному взаимодействию в реальном времени использовали Octet НТХ от ForteBio. Эксперименты выполняли при 30°С при перемешивании со скоростью 1000 об/мин в 9б-луночных черных планшетах, содержащих по 240 мкл на лунку. До загрузки мутантных His-меченных белков FimH в концентрации 5 мкг/мл, которую осуществляли в течение 5 минут, биосенсоры Ni-NTA уравновешивали в буфере, содержащем 1х буфер PBS, дополненный 0,5% BSA и 0,05% Tween 20 (РВТ). До ассоциации с антителами из разных групп при 5 мкг/мл, которую выполняли в течение 5 минут, FimH-загруженным биосенсорам давали возможность восстановиться до исходного уровня в РВТ в течение 3 минут. Кинетический анализ стадии ассоциации и получения отклика в виде сдвига в нм (таблица) использовали программное обеспечение для анализа данных Octet.
Пример 6: Спектроскопия кругового дихроизма
Для мутантов FimHLD и FimH-DSG регистрировали спектры кругового дихроизма в дальней УФ области (320-250 нм) и ближней УФ (260-200 нм) области с помощью спектрополярометра JASCO J-810 (Jasco), оснащенного JASCO PTC-424S/15 (Jasco) и водяной баней Isotemp (Fisher Scientific). Для дальней УФ области использовали 1 мм кювету, а для ближней УФ области использовали 10 мм кювету. Белки разбавляли до 0,3 мг/мл в PBS, и спектры регистрировали при 20°С в кюветах с длиной оптического пути 1 мм (дальняя УФ область) или 10 мм (ближняя УФ область). Сканирование выполняли со скоростью 100 нм/мин, DIT устанавливали на 1 с, полосу пропускания - на 3 с, и шаг данных - на 0,1 нм. Чувствительность выставляли на стандарт. Накапливали и усредняли десять спектров для ближней УФ области и пять - для измерений в дальней УФ области, соответственно. Спектры корректировали относительно фона вручную с помощью спектров CD, полученных из пустых прогонов PBS, и преобразовывали в среднюю эллиптичность остатка с помощью уравнения 1, где θMRE представляет собой рассчитанную среднюю эллиптичность остатков, θEXP представляет собой экспериментально измеренный сигнал CD, MW представляет собой молекулярную массу белка, N является количеством аминокислотных остатков, С представляет собой концентрацию белка в мг/мл, 1 -длина оптического пути в см.
Уравнение 1:
θMRE =(θEXP ⋅ MW) / (10⋅N⋅C⋅1)
Пример 7: Исследование ЕС-1678 по изучению иммуногенности у животных
Мышей CD-1 в возрасте 6-8 недель получали из Charles River Laboratories. 20 животных каждой группы иммунизировали подкожно через 0, 4 и 8 недель 10 мкг белка FimH, смешанного с 20 мкг Quillaja Saponaria-21 (QS-21) из маточного раствора QS-21 с концентрацией 5,1 мг/мл, содержащего 5 мМ сукцината, 60 мМ NaCl, 0,1% PS80, рН 5,6.
Пример 8: Анализы нейтрализации FimH в цельных клетках Для оценки способности сыворотки вакцинированных животных ингибировать связывание фимбриированных Е.coli с маннозилированными субстратами использовали анализ нейтрализации в цельных клетках с использованием дрожжевого маннана, как описано в международной публикация РСТ №WO 2021/084429, опубликованной 6 мая 2021 г.
Пример 9: Очистка мутанта FimH-DSG WT и FimH-DSG G15A G16A V27A из клеток СНО
Белки экспрессировали в клетках СНО в виде секретируемых белков с С-концевыми His-метками. Собирали супернатант клеточной культуры и добавляли 1 М Трис, рН 7,4, и 5 М NaCl до конечных концентраций 20 мМ и 150 мМ, соответственно. Кассеты на 5 кДа для TFF промывали буфером и уравновешивали в 20 мМ Трис, рН 7,5, с 500 мМ NaCl и 40 мМ имидазолом. Супернатант концентрировали в 2 раза и подвергали диафильтрации против 6 объемов 20 мМ Трис, рН 7,5, 500 мМ NaCl, 40 мМ имидазола. Ретентат собирали и промывали 50-100 мл 20 мМ Трис, рН 7,5, 500 мМ NaCl, 40 мМ имидазола. Ретентат фильтровали и промывали 0,2 мкм бутылочным фильтром. Колонку XK26/20 наполняли смолой Ni-сефароза 6 Fast Flow (Cytiva Life Sciences) и уравновешивали 5 объемами колонки 20 мМ Tris, рН 7, 5, 500 мМ NaCl, 40 мМ имидазола. Ретентат наносили при половинной скорости потока и промывали до достижения стабильной исходной линии (примерно 55 объемов колонки). Связанный белок элюировали 20 мМ Трис, 500 мМ NaCl, 500 мМ имидазол, рН 7,5. Фракции, содержащие представляющий интерес белок, объединяли и подвергали диализу в диализной кассете на 2 кДа против 20 мМ ацетата натрия, рН 4,3, при 4°С с двумя заменами буфера. Белок наносили на катионообменную колонку SP-сефароза (Cytiva Life Sciences), уравновешенную тем же буфером. Материал, связанный с катионообменной смолой, элюировали линейным градиентом NaCl с использованием 20 мМ ацетата натрия, рН 4,3, 1 М буфера NaCl. Фракции объединяли и подвергали диализу против TBS, рН 7,4.
Пример 10: Аналитическая эксклюзионная хроматография (SEC) для определения FimH-DSG WT и мутанта FimH-DSG G15A G16A V27A
Аналитическую SEC выполняли с использованием колонки Waters X bridge Protein ВЕН SEC 125 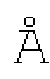 2,5 мкм, 4, 6×300 мм в буфере TBS, рН 7,4, содержащем 10 мМ EDTA, при 25°С. Объем инъекции составлял 10 мкл, скорость потока 0,5 мл/мин.
2,5 мкм, 4, 6×300 мм в буфере TBS, рН 7,4, содержащем 10 мМ EDTA, при 25°С. Объем инъекции составлял 10 мкл, скорость потока 0,5 мл/мин.
Пример 11: Анализ моносахаридов с помощью анионообменной хроматографии с высоким рН с импульсным амперометрическим детектированием (HPAEC-PAD)
Водные образцы FimH-DSG дикого типа и тройного мутанта FimH-DSG (G15A, G16A, V27A) расщепляли в течение 2 часов при 120°С в 2N трифторуксусной кислоте. По истечении этого времени образцы выпаривали досуха в вакууме при 45°С в течение 6 часов. Образцы восстанавливали в Milli-Q H2O и оценивали с помощью HPAEC-PAD в системе ионной хроматографии DIONEX ICS 3000. Использовали колонку Dionex CarboPac РА1 (4×250 мм) с изократическим элюированием смесью H2O и 200 мМ NaOH. Состав моносахаридов подтверждали путем сравнения времен удерживания пиков, обнаруженных в образцах FimH, с растворами известных стандартов моносахаридов.
Пример 12: Детектирование связывания сахарного фрагмента О-антигена с FimH-DSG WT и мутантом FimH-DSG G15A G16A V27A
Для измерения возможных взаимодействий О-антигена с FimH-DSG WT и мутантом FimH-DSG G15A G16A V27A во всех экспериментах по кинетическому биомолекулярному взаимодействию в реальном времени использовали октет НТХ от ForteBio. Эксперименты выполняли при 30°С при перемешивании со скоростью 1000 об/мин в 96-луночных черных планшетах, содержащих по 240 мкл на лунку. До загрузки His-меченого FimH-DSG WT или мутанта FimH-DSG G15A G16A V27A в биосенсор в концентрации 5 мкг/мл, которую выполняли в течение 5 мин, Ni-NTA биосенсоры уравновешивали в буфере, содержащем 1х буфер PBS с 0,5% BSA и 0,05% Tween 20 (РВТ). Биосенсорам, загруженным FimH-DSG WT или FimH-DSG G15A G16A V27A, давали восстановить исходный уровень в РВТ в течение 3 минут, а затем загружали с двукратным титрованием (200-3,125 мкг/мл) конъюгаты CRM с О-антигенным полисахаридом O9 или O25b, или O1a, или O2. В качестве эталона использовали загруженный антигеном биосенсор без какого-либо полисахарида. Биосенсоры, загруженные титрованием FimH и О-антигена, погружали в РВТ на 3 минуты для установления нового исходного уровня. Детектирование связывания О-антигена с мутантом тестировали на стадии ассоциации с 5 мкг/мл О-антиген-специфических mAb в течение 5 мин (MAb 601 для O9, MAb ЕСО-80-11 для O25b, MAb ЕСО-48-2 для O1a и MAb ЕСО-172-13 для O2). Для кинетического анализа стадии ассоциации с вычетом эталонного значения и получения отклика в виде сдвига в нм (в таблице) использовали программное обеспечение для анализа данных Octet.
Пример 13: Экспрессия и очистка белка
Мутанты FimHLD и FimH-DSG экспрессировали в клетках Expi293 и выделяли из супернатантов путем аффинного захвата никеля с последующей эксклюзионной хроматографией. Следует отметить, что некоторые мутанты имели низкий уровень экспрессии и не прошли биохимическую или биофизическую оценку (например, FimHLD Р26С V35C, N33C Р157С, N33C L109C, V35C L107C, V35C L109C, S113C Т158С). Мутанты, для которых можно было получить достаточный выход, оценивали в анализах термостабильности и связывания лиганда.
Пример 14: Идентификация мутантов FimHLD и FimH-DSG с улучшенной термической стабильностью и уменьшенным сдвигом температуры плавления в присутствии маннозидного лиганда
Температуры плавления мутантных белков FimH определяли с помощью дифференциальной сканирующей флуориметрии по тепловому сдвигу оранжевого красителя SYPRO, где Tm обозначает температуру, при которой разворачивается 50% белка. Нековалентные лиганды часто стабилизируют белковые мишени при специфическом связывании, что приводит к повышению температуры плавления белка. Поэтому температуры плавления определяли в присутствии метил-альфа-D-маннопиранозида, который является производным альфа-Б-маннозы и обладает микромолярным сродством к FimH (Bouckaert, J. et al. Mol Microbiol. 55, 441-455 (2005)), и вычисляли разницу температур плавления (ΔTm) белка в присутствии лиганда относительно АРО формы.
Белки FimH-BSG дикого типа (WT) демонстрировали значительно более высокие температуры плавления по сравнению с FimHLD WT и имели более низкую ΔTm в присутствии лиганда (таблица 10, таблица 11). FimH-BSG WT имел температуру плавления 71,66°С, в то время как температура плавления FimHLD WT была значительно ниже (61,54°С). В присутствии метил-альфа-D-маннопиранозида температура плавления FimHLD WT сдвинулась на 10,99°С, а температура плавления FimH-BSG в присутствии лиганда сдвинулась только на 2,13°С. Это предполагает, что FimHLD более эффективно стабилизируется лигандом по сравнению с FimH-DSG, что может отражать уменьшение связывания лиганда с FimH-DSG.
Мутации влияли на температуру плавления белков FimH в АРО состоянии и в присутствии лиганда. Зафиксированный мутант FimHLD V27C L34C, описанный ранее (Kisiela, D.I. et al. Proc Natl Acad Sci U.S.A. 110, 19089-19094 (2013); Rodriguez, V.B. et al. J Biol Chem 288, 24128-24139 (2013)) показал более низкую температуру плавления (51,42°С) по сравнению с FimHLD дикого типа (61,54°С), что согласуется с опубликованными данными (Kisiela, D.I. et al. Proc Natl Acad Sci U.S.A. 110, 19089-19094 (2013); Rodriguez, V.B. et al. J. Biol Chem 288, 24128-24139 (2013)). Инкубация FimHLD V27C L34C с метил-альфа-D-маннопиранозидом повышала температуру плавления на 7,27°С по сравнению с АРО формой, что позволяет предположить, что этот мутант частично стабилизирован лигандом и может иметь остаточную эффективность связывания лиганда. Что касается мутанта FimH-DSG V27C L34C, то он был менее термостабильным по сравнению с WT (Tm=63,29°С), а температурный сдвиг мутанта V27C L34C в присутствии лиганда уменьшился незначительно по сравнению с WT (ΔTm=1,29°С). Пять из шести мутантов Phel FimHLD имели пониженную температуру плавления по сравнению с FimHLD дикого типа, за исключением F1L, который имел аналогичную температуру плавления. Значение ΔTm у четырех из шести мутантов FimHLD Phel в присутствии лиганда было небольшим, что указывает на плохую стабилизацию лигандом. Напротив, наиболее консервативные аминокислотные замены F1W и F1Y показали промежуточные и сопоставимые значения ΔTm, соответственно, по сравнению с FimHLD Phel дикого типа. Что касается общей термической стабильности, эталонная мутация R60P (описанная ранее - Rabbani et al. J. Biol. Chem. 293:1835-1849 (2018); Rodriguez, V.B. et al. J Biol Chem 288, 24128-24139 (2013)) и несколько новых мутаций, разработанных собственными силами, значительно повысили температуру плавления по сравнению с V27C L34C. Интересно, что FimHLD V28C N33C (оба сайта смещены всего на один остаток от эталонного FimHLD V27C L34C) имел самую высокую температуру плавления среди всех мутантов FimHLD (Tm=65,77°C) и имел ΔTm 2,81°С в присутствии метил-альфа-D-маннопиранозида, что свидетельствует о сниженном сродстве к лиганду. Мутации FimHLD в области глициновой петли (G15A, G16A) значительно повысили термостабильность по сравнению с V27C L34C, и в присутствии лиганда наблюдался очень небольшой сдвиг температуры плавления. Мутации глициновой петли также слегка повышали термическую стабильность FimH-DSG, но в присутствии лиганда температурный сдвиг не наблюдался, что в совокупности позволяет предположить, что мутанты FimHLD и FimH-DSG не стабилизируются лигандом и, следовательно, могут иметь сниженную эффективность связывания по сравнению с диким типом.
Последовательность FimHLD WT получали из ассоциированного с ИМП изолята J96 Е.coli (Hull, R.A. et al., Infect Immun 33, 933-938 (1981)). V27A представляет собой природный вариант, ассоциированный с ИМП вирулентными изолятами, и изолятами, связанными с болезнью Крона (Schwartz, D.J. et al., Proc Natl Acad Sci USA 110, 15530-15537 (2013); Cespedes et al., Front Microbiol 8:639 (2017)). Включение V27A в FimHLD немного снижало температуру плавления FimHLD WT, и более слабый сдвиг наблюдали для V27A в присутствии метил-альфа-D-маннопиранозида по сравнению с WT. С другой стороны, V27A, по-видимому, оказывает стабилизирующее действие в контексте мутантов FimHLD G15A, G16A, G15P, G16P по глициновой петле, все из которых имели более высокую температуру плавления с V27A по сравнению с отсутствием этой мутации, и имели ΔTm<2°С в присутствии V27A по сравнению с 6,05°С (G16P) без этой мутации. Кроме того, мутанты FimH-DSG, содержащие V27A, имели слегка улучшенную термостабильность, и в присутствии лиганда не демонстрировали температурного сдвига. В совокупности это предполагает, что V27A оказывает стабилизирующее действие на температуру плавления FimH и снижает эффективность связывания лиганда.
Несколько мутантов FimHLD по дисульфиду и из неполярного на полярный остаток экспрессировались на низких уровнях и имели либо низкую термостабильность, либо демонстрировали значительные температурные сдвиги в присутствии соединения маннозида, что позволяет предположить, что они сохраняли эффективность связывания лиганда (таблица 12). Эти мутанты были протестированы в единичных повторах и исключены из дальнейшего анализа. Аналогичным образом была проанализирована термостабильность нескольких других мутантов FimH-DSG, два из которых показали улучшенную термостабильность и уменьшенный сдвиг в присутствии лиганда (FimH-DSG V27A Q133K и FimH-DSG G15A G16A V27A Q133K) (таблица 13). Мутация Q133K представляет собой мутацию в связывающем кармане FimH, которая устраняет связывание лиганда, о чем было описано ранее (Schwartz et al., Proc Natl Acad Sci USA 110:15530-15537 (2013)). Эти мутанты были исключены из последующего анализа.
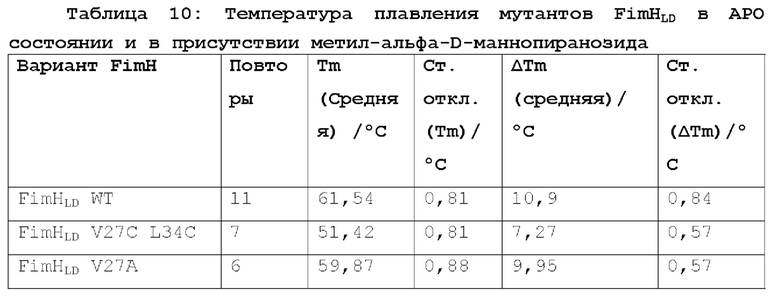
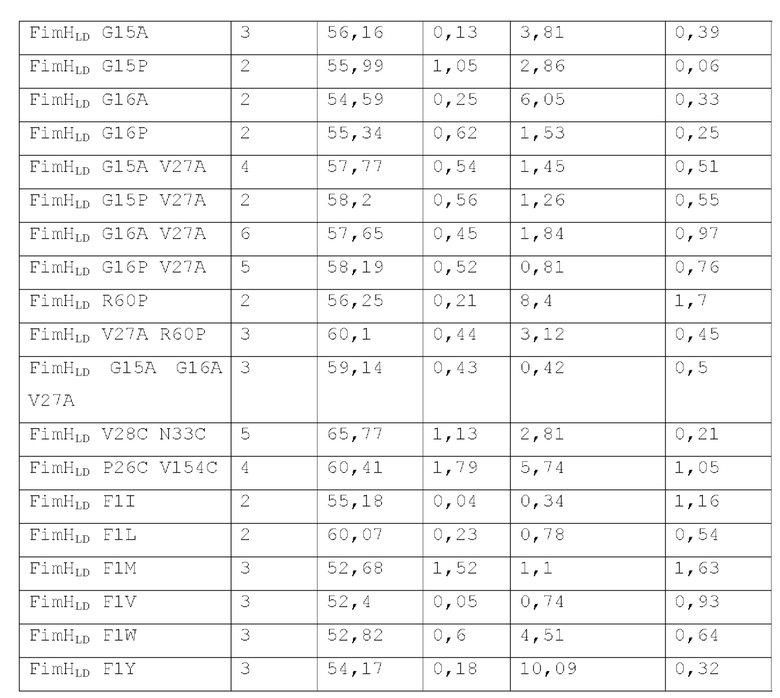
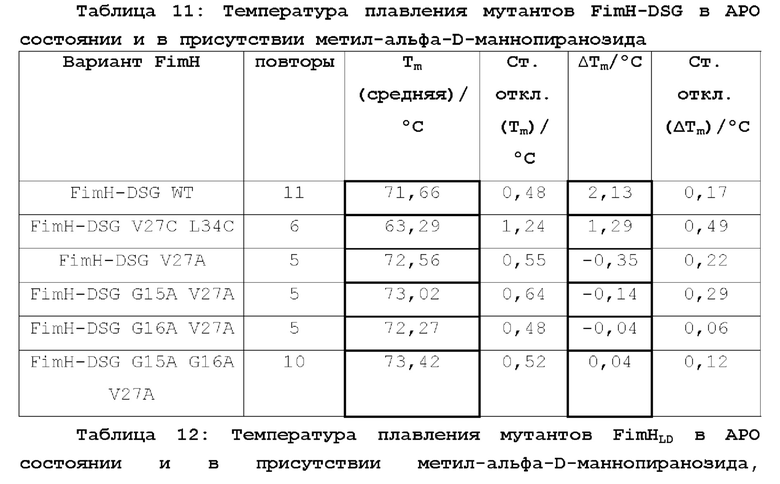
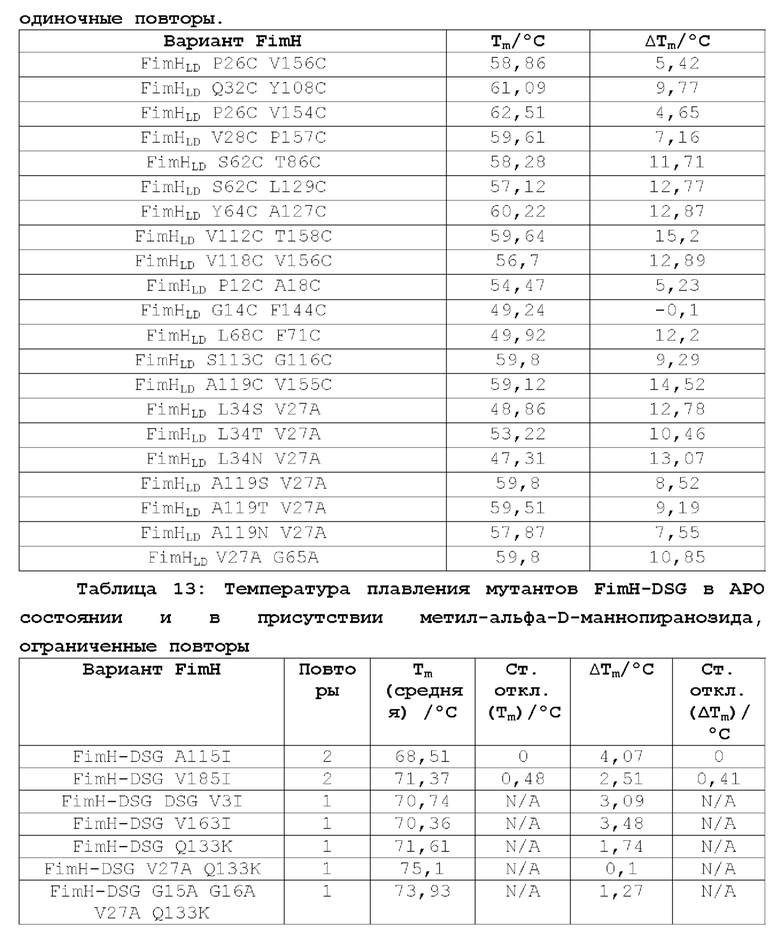
Пример 15: Идентификация мутантов FimH со сниженным сродством к маннозидному лиганду
Константы диссоциации (Kd) мутантов FimH для маннозидного лиганда определяли с помощью анализа поляризации флуоресценции при непосредственном связывании с флуоресцеин-конъюгированным октилбифенилманнопиранозидным (ВРМР) лигандом. Значения Kd мутантов FimHLD относительно WT показаны в таблице 14. FimHLD WT и V27A показали аналогичное высокое сродство к ВРМР. Эталонный зафиксированный мутант FimHLD V27C L34C (Kisiela, D.I. et al., Proc Natl Acad Sci USA 110, 19089-19094 (2013); Rodriguez, V.V. et al., J Biol Chem 288:24128-24139 (2013)) имел в 91 раз более низкое сродство к лиганду по сравнению с FimHLD WT, в то время как сродство FimHLD R60P V27A (Rabbani et al., J Biol Chem 293:1835-1849 (2018)) было ниже в 179 раз. Описанные здесь мутанты сравнивали с эталонными зафиксированными мутантами. Мутанты FimHLD по глициновой петле G15A V27A, G15P V27A, G16P V27A и G16A G16A V27A не показали связывание, в то время как Kd G16A V27A было в 156 раз выше, чем у WT. Все мутации глициновой петли в комбинации с V27A показали значительно более высокое значение Kd, чем мутанты только глициновой петли, что позволяет предположить, что V27A обладает статистически неопределимым стабилизирующим эффектом, хотя и с малым влиянием на Kd FimHLD WT. Включение V27A также дополнительно снижало сродство связывания FimHLD R60P.
В этом анализе были протестированы три новых мутанта с фиксирующими дисульфидными связями, все из которых имели умеренное уменьшение сродства связывания лиганда по сравнению с FimHLD WT (сродство ниже в 33-43 раза). Мутанты FimHLD, содержащие неполярные-на-полярные мутации (А119Т V27A, A119N V27A, L34T V27A, L34N V27A), обладали высоким сродством к ВРМР, аналогичным с таковым у FimHLD WT. Мутанты FimHLD F1 продемонстрировали плохое связывание, за исключением F1Y со сродством связывания, аналогичным сродству у FimHLD WT.
Сродство связывания лиганда у конструкций FimH-DSG показано в таблице 15. Значение Kd у FimH-DSG WT более чем в 100 раз выше, чем у FimHLD WT, что, вероятно, является отражением различных конформационных состояний двух форм FimH. FimH-DSG V27A также имел более низкое сродство по сравнению с FimH-DSG WT. Это согласуется с предыдущими данными, показывающими, что полноразмерный FimH с А27 в комплексе с FimC и FimG имеет пониженное сродство связывания с маннозидом по сравнению с FimH V27 (Schwartz et al., Proc Natl Acad Sci USA 110:15530-15537 (2013)). Введение фиксирующей мутации V27C L34C в FimH-DSG снижало сродство к ВРМР в 2,5 раза, в то время как мутант FimH-DSG по глициновой петле G15A G16A V27A имел сродство в 28 раз более низкое по сравнению с FimH-DSG WT. Для FimH-DSG G15A V27A и FimH-DSG G16A V27A не удалось рассчитать Kd, что свидетельствует о том, что эти мутанты не могут связываться с ВРМР. Мутации, предназначенные для стабилизации FimH-DSG в открытой конформации посредством изменения поверхности контакта пилин-лектинового домена (A115I, V185I), улучшали сродство связывания по сравнению с FimH-DSG WT, как следует из данных по термостабильности (пример 14).
Таким образом, были идентифицированы мутации глициновой петли в белке FimHLD или FimH-DSG, которые имели очень низкое сродство связывания с ВРМР. Для оценки в исследованиях функциональной иммуногенности на мышах глициновые мутанты отбирали на основании этих данных и данных по термостабильности.
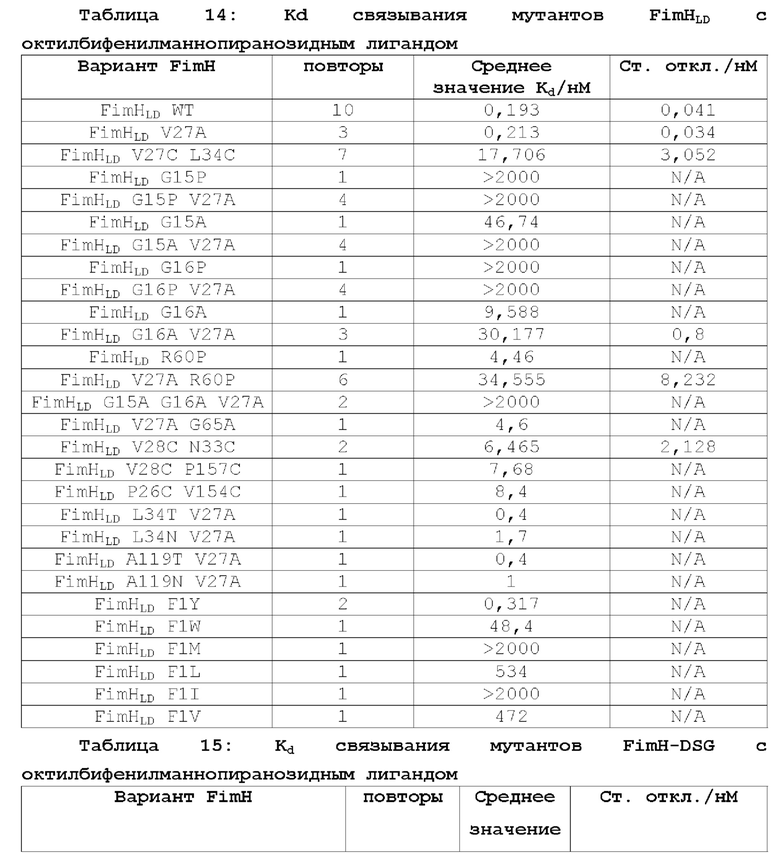
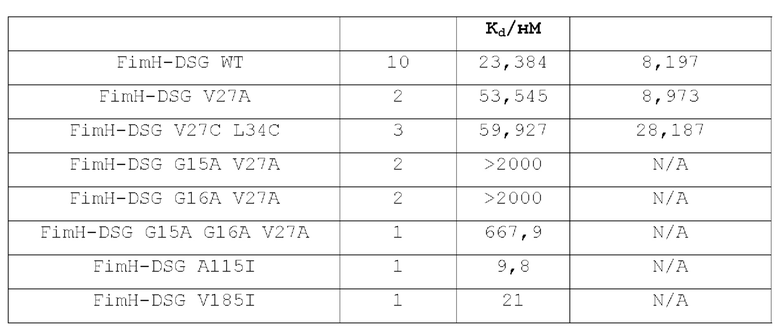
Пример 16: Подтверждение конформационного состояния мутантов FimH с помощью спектроскопии кругового дихроизма
Вторичную и третичную структуры мутантов FimHLD и FimH-DSG, демонстрирующие повышенную термическую стабильность и пониженное сродство связывания с маннозидным лигандом (примеры 14 и 15), анализировали методом кругового дихроизма (CD). FimHLD дикого типа и конформационно зафиксированные мутанты FimHLD имеют разные третичные профили CD (Rabbani et al., J Biol Chem 293:1835-1849 (2018)). Вторичные и третичные структуры выбранных мутантов FimHLD и FimH-DSG и дикого типа исследовали с помощью CD в дальней УФ области (вторичная структура) и CD в ближней УФ области (третичная структура) (см. фиг. 1). CD-спектр для FimHLD в дальней УФ области согласуется с ранее опубликованными данными (Rabbani et al. J Biol Chem 293:1835-1849 (2018)), и спектр для FimHLD и FimH-DSG в дальней УФ области характерен для белка с высоким содержанием бета-листов. Спектр для FimHLD V27C L34C в дальней УФ области немного отличался от спектра для FimHLD дикого типа, согласно наблюдениям других исследователей (Rabbani et al. J Biol Chem 293:1835-1849 (2018)), что отражает открытое конформационное состояние. Профиль вторичной структуры природного мутанта FimHLD V27A также несколько отличался. В целом, вторичные структуры мутантов FimHLD или FimH-DSG очень похожи на вторичные структуры белков дикого типа (фиг.1), что позволяет предположить, что общая вторичная структура у этих мутантов не изменилась. Профили третичной структуры мутантов FimHLD, полученные авторами изобретения, имели очень похожие спектры, которые также совпадали с опубликованными спектрами CD для FimHLD V27 L34C и V27A R60P, стабилизированными в открытом конформационном состоянии (Rabbani et al. J Biol Chem 293:1835-1849 (2018)). Профили описанных здесь мутантов также значимо отличались от профилей FimHLD дикого типа или FimHLD V27A. В совокупности эти данные позволяют предположить, что введенные мутации сдвигают конформацию FimHLD в сторону открытой конформации, при этом при введении проанализированных в настоящем описании стабилизирующих конформацию мутаций третичная структура FimH-DSG остается в значительной степени неизменной.
Пример 17: Характеристика мутантов FimH с помощью нейтрализующих моноклональных антител
Конформации нескольких выбранных антигенов FimH были охарактеризованы методом биослойной интерферометрии с использованием моноклональных антител, специфичных к лектиновым доменам 299-3, 304-1 и 440-2. Эксперименты с использованием конкурентного анализа (не показаны) продемонстрировали, что антитела 229-3 и 304-1 связываются с такими же эпитопами лиганд-связывающего сайта, что и MAb 475 и 926 (Kisiela et al., Proc Natl Acad Sci USA 110:19089-19094 (2013)). Моноклональное антитело 440-2 связывается с другим эпитопом и, по-видимому, предпочтительно связывается с FimHLD в открытой конформации. Антитела 229-3 и 304-1 оказались способными распознавать все варианты FimHLD (таблица 16) и FimH-DSG (таблица 17), хотя уровень связывания FimHLD V27A был более низким. Напротив, ответы на антитело 440-2 были выше у всех мутантов FimHLD или FimH-DSG по сравнению с WT или V27A. Это согласуется с профилями спектроскопии CD, показанными на фиг. 1, что позволяет предположить, что мутанты FimHLD находятся в открытой конформации. Реакция на 440-2 была также увеличена у FimH-DSG WT, что в комбинации с перекрывающимися профилями CD-спектроскопии мутантов FimH-DSG и WT (пример 16) позволяет предположить, что этот белок находится в состоянии открытой конформации независимо от присутствия или отсутствия стабилизирующих мутаций.
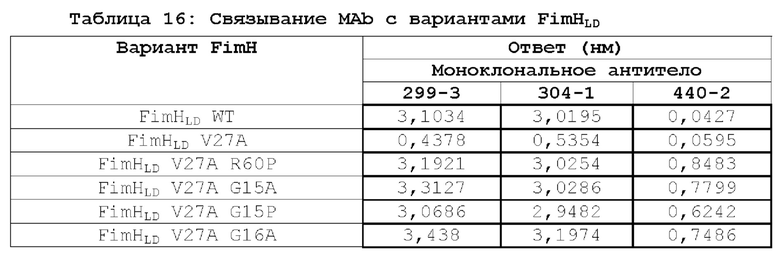
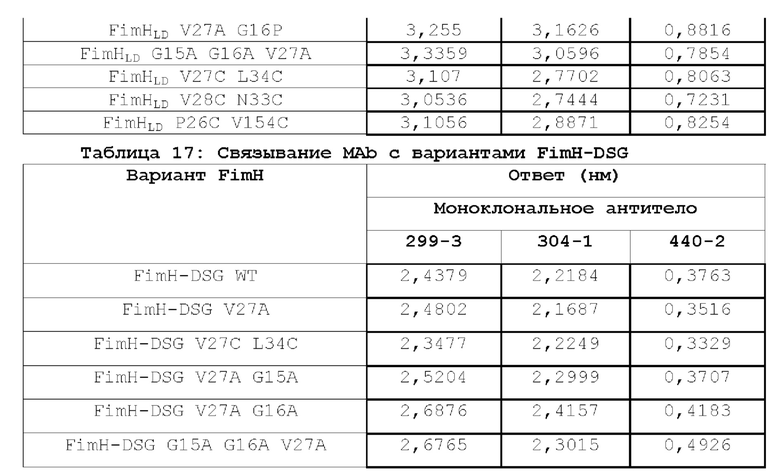
Пример 18: Данные по нейтрализации мутанта FimH
Для оценки относительной иммуногенности выбранных мутантов мышей вакцинировали мутантами FimH. Способность мутантов FimH вызывать титры функциональных антител количественно оценивали с помощью анализа нейтрализации маннана цельных дрожжей, описанного выше и ранее опубликованного (международная публикация РСТ №WO 2021/084429, опубликованная 6 мая 2021 г.). Вкратце, фимбриированные Е.coli инкубировали с сывороткой и давали возможность связаться с микротитрационным планшетом, покрытым дрожжевым маннаном. Планшет промывали и с помощью люминесцентного зонда определяли количество жизнеспособных Е.coli, связанных с планшетом. Титры нейтрализации сыворотки, которые ингибируют связывание фимбриированных бактерий с дрожжевым маннаном, определяли из серии двукратных разведений сыворотки вакцинированных мышей по восьми точкам. Титры представляют собой обратную величину разведения сыворотки, при которой 50% бактерий остаются связанными с планшетом. Суммарная информация по средним титрам и ответам показана в таблице 18. Графики индивидуальных ответов IC50 мышей после введения дозы 2 и 3 показаны на фиг. 2 и фиг. 3.

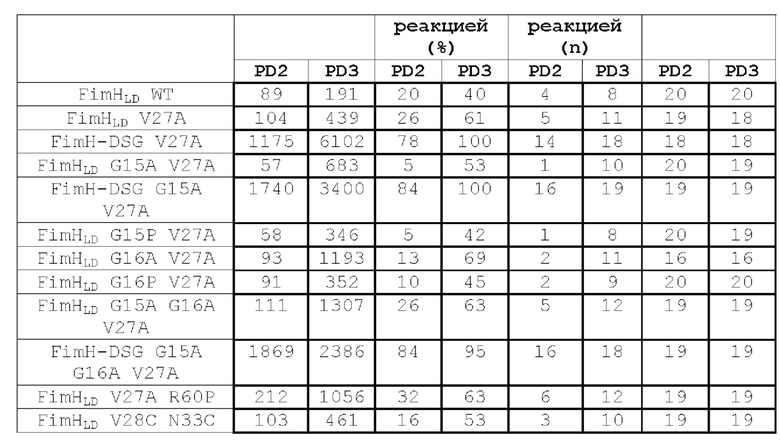
В предыдущей работе (международная публикация РСТ No. WO 2021/084429, опубликованная 6 мая 2021 г.) было показано, что ранее описанный мутантный FimHLD V27C L34C с фиксирующими дисульфидными связями (Kisiela et al., Proc Natl Acad Sci USA 110:19089-19094 (2013)) не улучшал функциональную иммуногенность по сравнению с FimHLD WT. Непосредственное сравнение функциональной иммуногенности новых мутантов FimHLD с иммуногенностью другого ранее описанного конформационно ограниченного мутанта FimHLD V27A R60P (Rabbani et al., J Biol Chem 293:1835-1849 (2018)) показано на фиг. 2. Мутанты FimHLD G16A V27A, FimHLD G15A G16A V27A и FimHLD V27A R60P обеспечили более высокое количество мышей с реакцией и более высокие титры (значение р<0,05), чем FimHLD WT. Другие мутации (G15A V27A, G16P V27A, V28C N33C) не приводили к значительному увеличению функциональной иммуногенности, и хотя высокие титры наблюдали у мышей, которые действительно имели ответ, количество мышей с ответом в этих группах было таким же, как и в группе FimHLD WT. Таким образом, несколько мутантов, созданных для повышения функциональной иммуногенности FimHLD путем фиксации FimHLD в открытой конформации, улучшали функциональную иммуногенность по сравнению с FimHLD WT. После вакцинации 2-мя дозами мутантов FimHLD и FimH-DSG в группах, вакцинированных FimH-DSG, нейтрализующие титры наблюдали у значительно более высокого количества животных по сравнению с группой FimHLD (фиг. 3). Эта тенденция сохранялась после введения 3-ей дозы, где 95-100% мышей продемонстрировали реакцию в группах, вакцинированных FimH-DSG V27A, FimH-DSG G15A V27A и FimH-DSG G15A G16A V27A. IC50 средних геометрических титров (GMT) также были значительно выше во всех группах, вакцинированных мутантами FimH-DSG после введения дозы 3. Аналогичные мутанты FimH-DSG (V27A, G15A, V27A, G15A, G16A, V27A) генерировали более высокие значения GMT по сравнению с мутантами FimHLD (значение р<0,05).
Пример 19: FimH-DSG G15A G16A V27A не связывается с гликанами хозяина и может быть выделен в гомогенном состоянии
Мутантные белки FimH-DSG WT и FimH-DSG G15A G16A V27A экспрессировали в клетках СНО в виде секретируемых белков, содержащих С-концевые His-метки. Очистку рекомбинантных His-меченых форм FimH-DSG выполняли, как показано на фиг. 4.
После ультрафильтрации и диафильтрации бесклеточную культуральную среду, содержащую FimH-DSG WT, выделяли на никелевой аффинной смоле и подвергали катионообменной хроматографии. Элюированный пик был довольно широким и имел несколько отчетливых плеч, что свидетельствует о возможной гетерогенности видов FimH-DSG WT (фиг. 5). Интересно, что когда элюированные фракции анализировали с помощью SDS-PAGE, в каждой фракции были обнаружены только отдельные полосы, соответствующие FimH-DSG WT.
В процессе очистки FimH-DSG проявлял свойства, указывающие на его плохую растворимость. В частности, Ni-сефарозные элюаты FimH-DSG WT всегда выглядели мутными при визуальном осмотре. Сдвиг в кислую сторону рН (4,3) для последующей очистки на SP-сефарозе приводил к осветлению раствора белка. Однако плохую растворимость препаратов FimH-DSG WT и склонность к агрегации вновь наблюдали после переноса (диализа) выделенного белка в TBS, рН 7,4. Это приводило к постепенной потере белка из-за агрегации и преципитации. Удаление осадка центрифугированием не останавливало и не замедляло процесс агрегации, хотя концентрация белка в этот момент обычно снижалась до 0,2-0,4 мг/мл. Потерю белка из-за агрегации можно контролировать в присутствии 10% глицерина, включенного в буфер для хранения (TBS, рН 7,4). Однако присутствие глицерина не препятствовало образованию растворимых HMW агрегатов, которые выявляли спектрофотометрически по рассеянию света при 350 нм.
Когда тот же процесс использовали для выделения мутанта FimH-DSG G15A G16A V27A, было выявлено несколько отличий. Во-первых, к ним относится отсутствие каких-либо признаков помутнения при элюировании белка на колонке с Ni-сефарозной смолой. Во-вторых, профиль пика, элюированного на колонки SP-сефароза, не был таким широким, как профиль, наблюдаемый для FimH-DSG WT на фиг. 6. И, наконец, мутант FimH-DSG G15A G16A V27A оставался полностью растворимым после переноса в буфер с физиологическим рН (TBS рН 7,4). В концентрациях до 5-6 мг/мл мутант FimH-DSG G15A G16A V27A не проявлял признаков агрегации или преципитации.
Анализ выделенного FimH-DSG WT и мутанта FimH-DSG G15A G16A V27A также выявил дополнительные различия между этими двумя вариантами FimH-DSG. Аналитическая эксклюзионная хроматография (SEC) продемонстрировала, что мутант FimH-DSG G15A G16A V27A элюируется в виде одного пика со временем удерживания, соответствующим его молекулярной массе. Напротив, профиль элюирования FimH-DSG дикого типа состоял из нескольких пиков, где время удерживания основного пика было меньше, чем у мутанта, показанного на фиг. 7. Эти данные ясно свидетельствуют о том, что FimH-DSG WT образует комплексы с высокой молекулярной массой, обнаруживаемые с помощью SEC. Наличие HMW комплексов, образованных FimH-DSG WT, и склонность к агрегации могут быть связаны с функциональной активностью его N-концевого лектин-связывающего домена. Авторами изобретения было выдвинуто предположение, что во время СНО ферментации и при секреции в культуральную среду FimH-DSG WT связывается с молекулой(ами) гликана, высвобождаемой(ыми) с поверхности клеток СНО-хозяина. Из-за разветвленной природы гликанов каждый гликан может содержать более одной копии молекулы FimH-DSG. Непрерывная «декорация» гликана увеличенным количеством FimH-DSG возможно и приводила к образованию различных HMW комплексов и, в конечном счете, к потере растворимости и преципитации (см. фиг. 8). Чтобы проверить эту гипотезу, выделенные виды дикого типа и мутантные виды FimH-DSG (и FimHLD) подвергали анализу с помощью анионообменной хроматографии при высоких значениях рН с импульсным амперометрическим (электрохимическим) детектированием (HPAEC-PAD). Этот метод позволяет идентифицировать олигосахариды или гликаны в образце белка, а также предоставляет информацию о составе этих олигосахаридов. Вкратце, для высвобождения моносахаридов проводят кислотный гидролиз с последующим анализом пиков относительно моносахаридного стандарта. Результаты анализа HPAEC-PAD показали, что выделенные препараты FimH-DSG WT (гликозилированные) и FimHLD (негликозилированные, данные не показаны) содержат значительные количества моносахаридов. Суммарная информация идентифицированных моносахаридов у FimH-DSG WT и мутанта FimH-DSG G15A G16A V27A показана в таблице 19. Содержание моносахаридов у мутанта FimH-DSG G15A G16A V27A было значительно ниже, чем у FimH-DSG WT. Кроме того, вполне возможно, что обнаруженное низкое содержание моносахаридов представляет собой сахарные фрагменты N-гликана, которые, как предполагается, модифицируют N235.
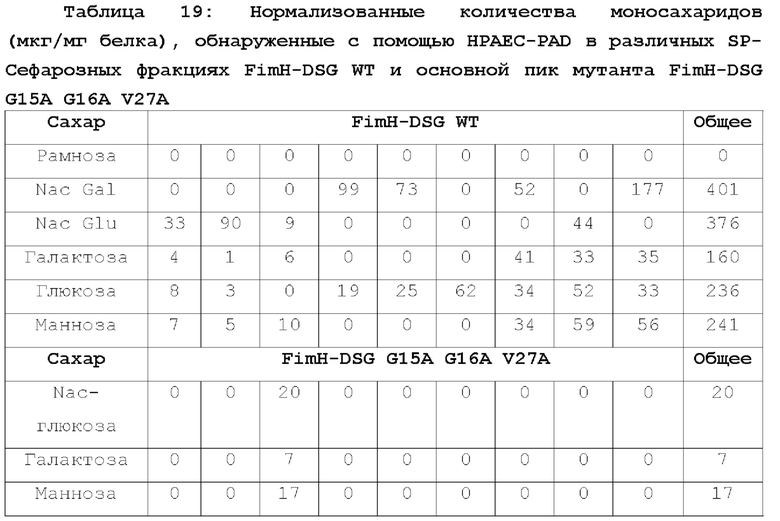
Пример 20: Мутанты FimHLD и FimH-DSG G15A G16A V27A не связываются с О-антигенами Е.coli
Повторяющиеся звенья О-антигенов O8 и O9 Е.coli состоят из остатков полиманнозы. В связи с этим возникает вопрос, может ли FimH связываться с О-антигенными конъюгатами в комбинированной вакцине конъюгата FimH-O-антиген. Для проверки способности FimHLD WT связываться либо со свободным полисахаридом O9, либо с CRM-конъюгированным полисахаридом O9, и такой же способности у мутанта FimHLD G15A G16A V27A, который в анализах связывания показал недетектируемое сродство к маннозидному лиганду, разрабатывали эксперименты по биослойной интерферометрии. Данные по связыванию О-антигена показаны в Таблице 20. При высоких концентрациях свободного полисахарида ответ наблюдали для FimHLD WT. Более высокие ответы наблюдали с полисахаридом, конъюгированным с CRM. Однако в случае мутантного белка FimHLD G15A G16A V27A детектируемое связывание отсутствовало. Аналогичные эксперименты выполняли для FimH-DSG, где была протестирована способность мутанта FimH-DSG G15A G16A V27A связываться со свободными или CRM-конъюгированными О-антигенами различных серотипов (O9, O25b, O1a и O2) (см. Таблицу 21). Как и ожидалось, исходя из свойств связывания маннозидного лиганда, общее значение сродства связывания у FimH-DSG WT было значительно ниже, чем у соответствующего варианта FimHLD. При самой высокой концентрации CRM-конъюгата некоторое титруемое связывание наблюдали у FimH-DSG WT, при этом уровни связывания лишь немного превышали фоновые уровни, наблюдаемые для свободного полисахарида. Как и в случае мутанта FimHLD G15A G16A V27A, белок FimH-DSG G15A G16A V27A не связывался ни с одним из свободных или CRM-конъюгированных полисахаридов. В заключение, в отличие от родительских антигенов FimHLD WT или FimH-DSG WT, производные мутанты G15A G16A V27A не могут связывать 0-антигены, предоставляя возможности для разработки комбиниоованной вакцины FimH и О-антигена.
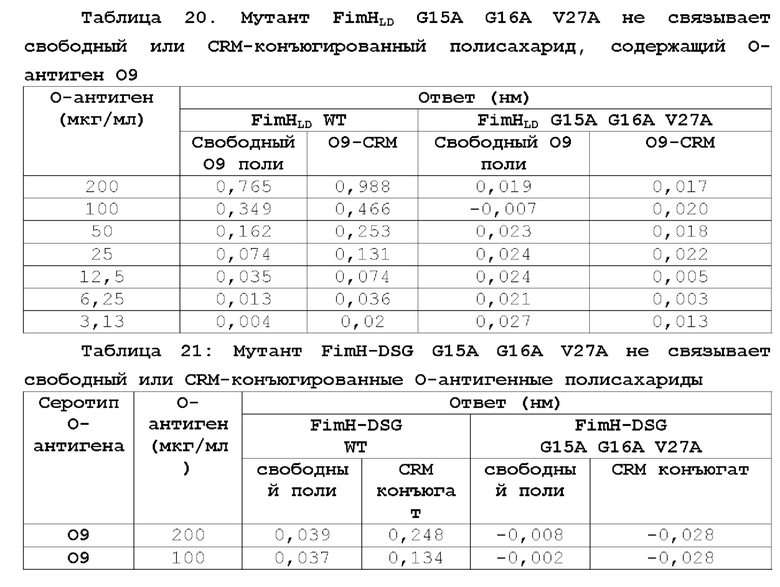
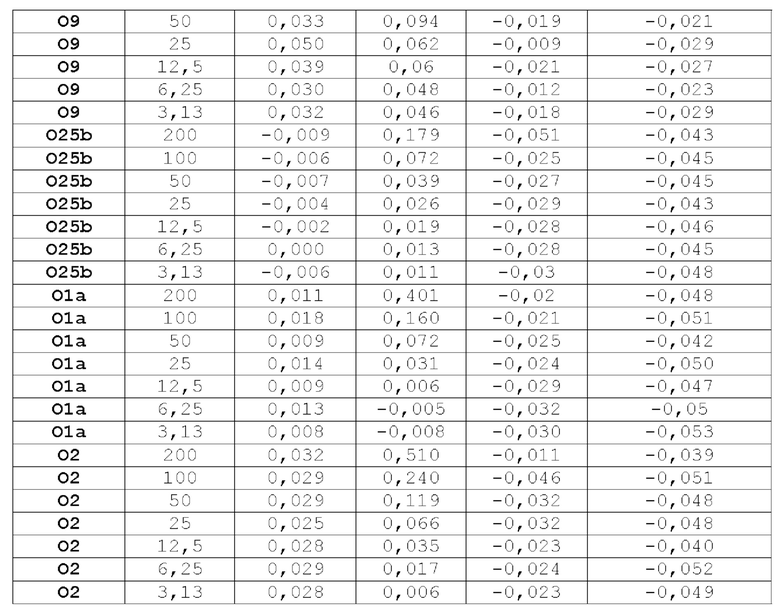
Пример 21. Нечеловекообразные приматы, вакцинированные мутантным FimH-DSG G15A G16A V27A с О-антигенами и без них А.
Методы
1. FimH IgG dLIA
Мутантный фимбриальный антиген FimH-DSG G15A Gl6A V27A Е.coli, связанный со спектрально отличающимися микрогранулами MagPlex-C (Luminex), разводили в блокирующем буфере до концентрации 50000 гранул/мл в течение 1-2 часов при комнатной температуре при встряхивании непосредственно перед первичной инкубацией до анализа. Разбавленную смесь микрогранул добавляли в планшеты для анализа, содержащие соответствующим образом разведенные образцы сыворотки нечеловекообразных приматов, контроли и эталонный стандарт, собственное гуманизированное моноклональное антитело (FimH Y202), которое связывается с пилиновым доменом FimH-DSG, для инкубации в течение ночи при 2-8°С при встряхивании. После промывки несвязавшихся компонентов к смеси микрогранул добавляли очищенное вторичное R-фикоэритрин-конъюгированное козье антитело к Fcγ-фрагменту человеческого IgG (Jackson ImmunoResearch Labortories, 109-116-170) и инкубировали в течение 90 минут при комнатной температуре при встряхивании. Величина флуоресцентного сигнала РЕ, измеренная считывателем Luminex FLEXMAP 3D, была прямо пропорциональна количеству анти-FimH-DSG IgG, связанного с микрогранулами, связанными с белком. Данные анализировали с помощью пользовательского приложения SAS, в котором использовали модель логарифмической линейной регрессии стандартной кривой для интерполяции концентраций антиген-специфических антител (мкг/мл) из средней интенсивности флуоресценции. Нижний предел количественного определения (LLOQ), равный 0,763 мкг/мл, вычисляли, исходя из смещения стандартной кривой.
2. 4-валентный O-Ag IgG dLIA
Полисахариды длинного О-антигена Е.coli серотипов O25b, O1a, O2 и O6 ковалентно конъюгировали с поли-L-лизином, и полученные конъюгаты связывали со спектрально различающимися микрогранулами MagPlex-C (Luminex) согласно стандартному EDC/NHS-опосредованному протоколу реакции сочетания.
Микрогранулы инкубировали в течение ночи при 2-8°С с серийно разбавленными образцами сыворотки нечеловекообразных приматов, контролями и поликлональным стандартом при встряхивании. После промывки связанный серотип-специфический IgG детектировали с помощью РЕ-конъюгированного вторичного козьего антитела, специфического к Fcγ-фрагменту человеческого IgG, (Jackson ImmunoResearch Labortories, 109-115-098), после инкубации в течение 90 минут при комнатной температуре при встряхивании. Флуоресценцию измеряли с помощью ридера Luminex 200 для каждой из четырех спектрально отличающихся областей и выражали в виде средней интенсивности флуоресценции. График стандартной кривой титрования поликлонального стандарта давал линейный график, выраженный в условных единицах, из которого сигналы можно было интерполировать в виде уровней серотип-специфических антител (Ед/мл).
3. Нечеловекообразные приматы (NHP)
Самок Cynomolgus Macagues (Масаса fasicularis) изначально получали из Charles River Laboratories (Хьюстон, Техас), а затем переводили в Pfizer, Pearl River, NY (возрастной диапазон: 4-5 лет, весовой диапазон: 3,1-5,9 кг). NHP содержали в стандартных четырехместных клетках со свободным доступом к воде и пище. Животных подкожно чипировали для контроля внутренней температуры. В экспериментальные группы включали только NHP без инфекции Е.coli на основании отрицательных результатов количественной ПЦР мочи (см. раздел 10 метода ниже).
4. Вакцинация и забор крови
Яванских макак иммунизировали внутримышечно (0,55 мл) на 0-ой, 4-ой и 14-ой неделях либо контрольным носителем (PBS, рН 6,2), мономерным фимбриальным антигеном FimH-DSG G15A G16A V27A (50 мкг/доза), либо смесью конъюгатов 4-валентных O25b, O1a, O2 и О6 O-антигенных полисахаридов (1 мкг/доза) в комбинации с мономерным фимбриальным антигеном FimH-DSG G15A G16A V27A (50 мкг/доза). Вакцинные антигены адъювировали AS01b (50 мкг MPL и 50 мкг QS-21 на дозу).
На 0-ой, 6-ой и 16-ой неделях собирали 10 мл крови через бедренную вену в 1 сепараторную пробирку для сыворотки (BD Vacutainer) с помощью безопасной 21g иглы/вакутейнера. Пробирки для сбора оставляли при комнатной температуре на 30 мин и центрифугировали при 3000 g в течение 10 мин. Сыворотку в супернатанте собирали, разделяли на аликвоты и хранили при 80°С.
5. Клинический изолят Е.coli
Один репрезентативный клинический изолят ST131 O25b выбирали на основе возраста пациента и происхождения коллекции образцов (PFEEC0578, мужчина, возраст 38 лет, получен из мочевого пузыря) из штаммов UPEC, собранных в рамках спонсируемой Pfizer базы данных Pfizer-sponsored Antimicrobial Testing Leadership and Surveillance (ATLAS), которая поддерживается клинической лабораторией International Health Management Associates (IHMA). Штамм несет гены, кодирующие продуцирование капсульных полисахаридов неизвестного типа.
6. Подготовка штамма UPEC
Маточный раствор Е.coli готовили путем инокуляции 12 мл бульона LB (Teknova, #L8198) с последующей инкубацией в течение ночи при 37°С при перемешивании со скоростью 275 об/мин. Через 18 часов 12 мл культуры разводили в 113 мл бульона LB в 250 мл колбе (Corning, #431407). Культуру инкубировали в течение 2-3 часов при 37°С при ~275 об/мин до значений OD600 между 2,1 и 2,7. К культуре примешивали 25 мл глицерина (80%, MP, #3055-044). Аликвоты по 5 мл замораживали при -80°С для длительного хранения. Концентрацию жизнеспособных бактерий во флаконе подтверждали посевом серийных разведений маточного раствора на чашки с TSA (BD, триптиказо-соевый агар BBL (соевый казеиновый расщепленный агар), № по каталогу В21283Х) и анализировали после 18-часовой инкубации при 30°С.
7. Модель цистита у нечеловекообразных приматов.
NHP анестезировали смесью кетамин/дексдомитор, вводимой внутримышечно. Для предотвращения контаминации мочевого пузыря при катетеризации уретры аногенитальную область протирали стерильной марлей, смоченной стерильным физиологическим раствором и/или антисептическими салфетками, содержащими бензалкония хлорид. Через мочеиспускательный канал в мочевой пузырь осторожно вводили стерильный красный резиновый катетер 5 French, предварительно покрытый Surgi Lube для предотвращения раздражения тканей. Затем мочевой пузырь опорожняли от любой мочи либо естественным потоком, либо путем аспирации с помощью шприца. Через катетер непосредственно в мочевой пузырь вводили 1 мл, содержащий 108 КОЕ UPEC штамма PFEEC0578.
8. Мониторинг животных после заражения
После контрольного заражения за животными наблюдали два раза в сутки в течение 1 недели и один раз в сутки в последующие недели. За NHP наблюдали в течение 30 дней после заражения. Мониторинг включал наблюдение за появлением диуреза, изменением поведения или аппетита, признаками боли/дискомфорта и измерением температуры тела.
9. Сбор мочи через установку катетера.
Для сбора чистых образцов мочи мочевой пузырь анестезированных NHP катетеризировали, как описано выше. После установки катетера мочевой пузырь опорожняли либо естественным потоком, либо аспирацией с помощью шприца. Когда мочевые пузыри не содержали мочи, вводили 10 мл физиологического раствора и повторяли аспирацию через катетер. Все собранные образцы сразу переносили на лед для хранения.
10. Экстракция ДНК
Экстракцию ДНК Е.coli из трех повторов образцов мочи NHP выполняли с помощью наборов для выделения ДНК Qiagen Minelute (Qiagen, номер по каталогу 51306, количество иногда ограничивалось объемом собранного образца). Протокол производителя Blood and Body Fluid Spin использовали со следующими изменениями: начальный объем образца был увеличен до 500 мкл (если позволял объем образца), объем буфера AL был увеличен до 500 мкл, объем протеиназы K был увеличен до 50 мкл, 50 мкл воды молекулярной чистоты (Corning Inc., Ref# 46-000-СМ) нагревали до 37°С и использовали вместо элюирующего буфера ЕВ. Наконец, после добавления воды молекулярной чистоты при температуре 37°С спин-колонки инкубировали в течение 5 минут при комнатной температуре перед окончательным вращением.
11. Количественная ПЦР в реальном времени (кПЦР)
Количественную ПЦР в реальном времени (кПЦР) использовали для оценки бактериальной нагрузки в образцах мочи NHP. ДНК, специфичную к серотипу O25b Е.coli, амплифицировали с помощью количественной ПЦР с использованием следующих праймеров: прямой, TTGAAAGTGATGGTTTGGTAAGAAAT (SEQ ID NO: 109); обратный, Т GCAGCACGTATGATAACTTCAAAG (SEQ ID NO: 110), и для количественной оценки репликации использовали зонд с флуоресцентным репортером Fain и последовательностью AGGATATTTTACCCAGCAGTGCCCCGT (SEQ ID NO: 111).
Ампликоны, специфические к серотипу O25b, соответствуют частям области orf10 серотипа O25b. Праймеры и зонд, разработанные по индивидуальному заказу, приобретали в лиофилизированном виде у Integrated DNA Technologies, и восстанавливали в буфере ТЕ (Corning Inc., номер по кат.46-009-СМ) до концентрации 100 нмоль/мл.
Образцы ДНК из описанной выше процедуры выделения ДНК анализировали в 96-луночных реакционных планшетах Applied Biosystems MicroAmp Optical (Applied Biosystems, номер no каталогу N8010560). Реакцию кПЦР выполняли в общем объеме 25 мкл с использованием 12,5 мкл смеси Applied Biosystems 2Х Taqman Fast Advanced Master Mix (Applied Biosystems, номер по каталогу 4444554), 0,125 мкл каждого восстановленного праймера, 0,5 мкл зонда, 1,75 мкл воды для молекулярной биологии (Corning Inc., номер по каталогу 46-000-СМ) и 10 мкл образца на лунку.
Условия реакции для амплификации ДНК были следующими: 50°С в течение 2 минут, затем 95°С в течение 2 минут и 40 циклов при 95°С в течение 3 секунд и при 60°С в течение 30 секунд, которые осуществляли либо в системе ПЦР в реальном времени Applied Biosystems 7500, либо в системе ПЦР в реальном времени Applied Biosystems QuantStudio 6 (Applied Biosystems).
Для количественного определения получали линейную стандартную кривую. Аликвоты того же замороженного маточного раствора Е.coli, который использовали в каждом эксперименте по заражению (подготовка описана выше), разбавляли до 1×109 КОЕ/мл в стерильном PBS (Corning, 21-040-СМ). Последующие серийные разведения выполняли для получения растворов, содержащих Е.coli в концентрациях 1×108, 1×107, 1×106, 1×105, 1×104, 1000, 100 и 10 КОЕ/мл. Серийные разведения готовили в стерильном PBS (Corning, 21-040-СМ) или объединяли дважды отфильтрованную мочу NHP, собранную у субъектов перед инокуляцией. Позже было обнаружено, что разведения в PBS и объединенной, дважды отфильтрованной моче NHP эквивалентны. Количество жизнеспособных бактерий, присутствующих в каждом разведении, подтверждали посевом на чашки с TSA (BD, BBL Trypticase Soy Agar (Soybean Casein Digest Agar), номер по каталогу B21283X). Экстракцию ДНК выполняли при каждом серийном разведении теми же методами, которые применяли для выделения ДНК из образцов, как описано выше. Эти стандарты количественной ПЦР запускали на каждом планшете для анализа количественной ПЦР в двух экземплярах.
Линейный регрессионный анализ выполняли с помощью программного обеспечения Applied Biosystems QuantStudio. Статистический анализ показал, что построенная стандартная кривая вела себя линейно на отрезке между 100 и 1×108 бактерий/мл. Следовательно, нижний предел количественного определения (LLOQ) был определен равным 100 бактерий/мл. В некоторых случаях образцы достигали порога флуоресценции, но в цикле, соответствующем количеству ниже LLOQ, в других случаях порог флуоресценции вообще не достигался (значение не определено). При возникновении любого из этих условий, такие значения указаны в настоящем описании как значение LLOQ (100 бактерий/мл).
12. ELISA для определения миелопероксидазы (МРО)
Наборы Invitrogen Myeloperoxidase Instant ELISA (Invitrogen, номер по каталогу BMS2038INST) использовали для количественного определения миелопероксидазы (МРО) в моче NHP. Буфер PIPES добавляли к чистым образцам мочи до конечной концентрации 5% 0,5 М буфера PIPES, рН 6,8 (Alfa Aesar, номер по каталогу J61786-AK). Образцы встряхивали в течение 15 секунд, а затем разбавляли в отношении 1:1 разбавителем для образцов, поставляемым производителем. Образцы анализировали в двух повторах, и в оставшейся части анализа следовали инструкциям производителя. В конечной точке интенсивность цвета при 450 нм измеряли на приборе Spectramax Plus (Molecular Devices). Стандарты, включенные в набор для анализа, использовали для построения стандартной кривой. Для анализа стандарт с низкой концентрацией (156,25 пг/мл) набора для анализа считали нижним пределом детектирования (LLOD). Сопутствующее программное обеспечение прибора Spectramax (Softmax, Molecular Devices) позволяет экстраполировать результаты за пределы низкого стандарта. Значение, соответствующее половине LLOD (78,125 пг/мл), или если результат анализа полностью был ниже предела обнаружения, заменяли любыми значениями, экстраполированными ниже этого значения.
13. Анализ IL-8 Luminex
Интерлейкин-8 (IL-8) измеряли с помощью пользовательского набора Bio-Rad IL-8 Human Cytokine Screening Panel Luminex Assay (BioRad Laboratories Inc., номер по каталогу 17005177). Буфер PIPES добавляли к беспримесным образцам мочи до конечной концентрации 5% 0,5 М буфера PIPES, рН 6,8 (Alfa Aesar, номер по каталогу J61786-AK). Образцы встряхивали в течение 15 секунд, а затем разбавляли в отношении 1:1 50% модифицированным буфером LXA-4 (PBS IX, 0,5% BSA, 0,025% азида натрия). Образцы анализировали в двух повторах, и в оставшейся части анализа следовали инструкциям производителя. Планшет для анализа считывали на приборе BioPlex 200 Luminex (BioRad Laboratories Inc.). Сопутствующее программное обеспечение прибора Bio-Plex 200 Luminex, «BioPlex Manager», и стандарты, включенные в набор для анализа, использовали для построения стандартной кривой и экстраполяции концентрации образца по интенсивности флуоресценции. Нижний предел количественного определения (LLOQ) определяли с помощью программного обеспечения BioPlex Manager.
14. Общий подсчет ядерных клеток и анализ осадка мочи под световым микроскопом.
В течение 1 часа после сбора образцы мочи (от 500 мкл до 1 мл) фиксировали формалином до конечной концентрации 1% и отправляли на ночь на лед в Pfizer Groton, СТ. При получении подсчитывали общее количество ядерных клеток (эпителиальные клетки и полиморфноядерные клетки) на гемоцитометре. В двойную цитоворонку Thermo Scientific Shandon EZ в целом загружали примерно 300 мкл: 100 мкл в одну воронку и 200 мкл в другую воронку. Образцы центрифугировали в течение 5 мин при 750 об/мин на предметных стеклах с покрытием Shandon Double Cytoslide Microscope на цитоцентрифуге Thermo Scientific CytoSpin 4. Для образцов с высоким количеством клеток перед цитоцентрифугированием мочу сначала разбавляли в отношении 1:10 0,9% физиологическим раствором. Затем с помощью прибора Sysmex SP-10 цитопрепарированные предметные стекла кратковременно фиксировали метанолом и окрашивали красителями Гимзы и Май-Грюнвальда. Для каждого образца мочи одно предметное стекло на образец мочи готовили с двумя объемами образца, указанными выше.
Слайды Cytospin, содержащие концентрированный и окрашенный осадок мочи, оценивали с помощью световой микроскопии на наличие или отсутствие повышенного количества полиморфноядерных клеток (PMN, т.е. гранулоцитов с сегментированными или почковидными ядрами, обычно состоящими из нейтрофилов, но также включая эозинофилы и базофилы). Случаи отсутствия наблюдаемых PMN или редко наблюдаемых PMN определяли как отсутствие увеличенных клеток PMN., Случаи наблюдения более чем редких PMN по сравнению с фоновой популяцией эпителиальных клеток определяли как наличие повышенного уровня PMN.
В. Результаты
1. Вакцинация мутантным FimH-DSG G15A G16A V27A с O-антигенами и без них вызывает образование эффективных суммарных и нейтрализующих антител у нечеловекообразных приматов.
Нечеловекообразных приматов вакцинировали мутантом FimH-DSG G15A G16A V27A с 4-валентным (O25b, 06, O1a, O2) O-антигенным конъюгатом или без него и адъювантом AS01b (фиг. 9). В группе, вакцинированной FimH-DSG G15A, G16A, V27A и 4-валентным O-антигеном, титры антител, специфичных к серотипу O-антигена, возросли примерно в 400 раз через 6 недель после вакцинации (фиг. 10 и таблица 22). Общие титры анти-FimH антител количественно определяли с помощью прямого иммуноанализа Luminex (dLIA) (фиг. 11А и таблица 23). Животные в группе плацебо имели титры ниже предела количественного определения. В обеих вакцинированных группах титры повышались после двух доз и могли быть усилены третьей дозой. В целом титры были немного ниже у животных, вакцинированных FimH-DSG G15A G16A V27A с O-антигенами, по сравнению с FimH-DSG G15A G16A V27A, введенным отдельно.
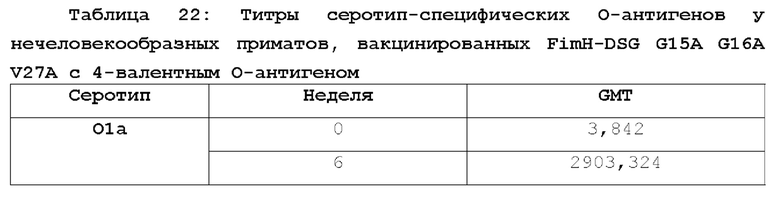
Сыворотки оценивали в анализе ингибирования связывания Е.coli для оценки способности анти-FimH антител блокировать связывание Е.coli с маннаном дрожжей (фиг. 11В и таблица 24). Среднее значение IC50 сыворотки животных, вакцинированных только FimH-DSG G15A G16A V27A, повысилось до 293,65 после 2-ой дозы и до 1698,39 после 3-ей дозы, в то время как среднее значение IC50 животных, вакцинированных FimH-DSG G15A G16A V27A в комбинации с О-антигенами, составило 480,12 после 2-ой дозы и 756,45 после 3-ей дозы.


В совокупности эти данные показывают, что FimH-DSG G15A G16A V27A вызывает сильный гуморальный ответ у нечеловекообразных приматов, а комбинация с О-антигенами приводит к высоким титрам, хотя и несколько ниже, чем при введении одного FimH.
2. Вакцинация мутантным FimH-DSG G15A G16A V27A с O-антигенами и без них снижает бактериурию и биомаркеры инфекции в модели нечеловекообразных приматов.
Через пять недель после последней бустерной вакцинации вакцинированным и обработанным плацебо NHP вводили 108 КОЕ изолята UPEC PFEEC0578 посредством внутрипузырной катетеризации. Бактериурию отслеживали в моче, собранной через катетер, в течение 28 дней. У всех животных, получавших плацебо, инстилляция живых бактерий приводила к высокому уровню бактериурии на 2-ые и 7-ые сутки после заражения (среднее геометрическое составило приблизительно 106 бактерий/мл мочи). По сравнению с группой плацебо, животные, вакцинированные FimH-DSG G15A G16A V27A или FimH-DSG G15A G16A V27A+4-валентный O-антиген, на 2-е и 7-е сутки после заражения демонстрировали соответственно 300-кратное или 1000-кратное снижение среднегеометрического значения бактериурии.
На 14-е сутки приблизительно у 50% животных, вакцинированных плацебо, по-прежнему наблюдали бактериурию, >105 бактерий/мл мочи. Наконец, большинство NHP группы плацебо избавились от инфекции на 21 и 28 сутки. Напротив, у большинства животных, вакцинированных FimH-DSG G15A G16A V27A или FimH-DSG G15A G16A V27A+4-валентный 0-антиген, инфекция исчезла к 14-м суткам (фиг. 12).
Затем в моче зараженных NHP отслеживали различные воспалительные биомаркеры. На 7-е сутки после заражения все животные, получавшие плацебо, демонстрировали повышенные уровни полиморфноядерных (PMN) клеток в осадке мочи, которые подтверждали с помощью цитологического анализа. Напротив, менее 25% NHP, вакцинированных FimH-DSG G15A G16A V27A, и ни одно животное, иммунизированное FimH-DSG G15A G16A V27A+4-валентным О-антигеном, не имели повышенных уровней клеток PMN в осадке мочи (фиг. 13С). Параллельно в образцах мочи измеряли уровни миелопероксидазы (МРО) и интерлейкина 8 (IL-8) в течение 7 дней. На 2-е сутки после контрольного заражения в обеих вакцинированных группах наблюдали 2-кратное снижение уровней МРО (среднее геометрическое ~200 пг/мл) по сравнению с группой плацебо (среднее геометрическое 470 пг/мл) (фиг. 13А).
Кроме того, на 2-е и 7-е сутки после заражения концентрация IL-8 в моче у животных, вакцинированных FimH-DSG G15A G16A V27A, снизилась примерно в 10 и 5 раз, соответственно (среднее геометрическое 5,9 пг/мл и 9,8 пг/мл), по сравнению с уровнями, измеренными в моче NHP, получавших плацебо (среднее геометрическое 54,2 пг/мл и 32,7 пг/мл). С аналогичной тенденцией уровни IL-8 в моче на 2-е и 7-е сутки у группы NHP, иммунизированной FimH-DSG G15A G16A V27A в комбинации с О-антигенами, снизились примерно в 5 и 3 раза, соответственно (среднее геометрическое 11,3 пг/мл), по сравнению с концентрациями IL-8, наблюдаемыми у животных, получавших плацебо (фиг. 13В).
С. Выводы
Мутант FimH-DSG G15A G16A V27A индуцирует высокие титры анти-FimH IgG у NHP, которые можно повысить путем введения 3-й дозы. Животные, вакцинированные комбинацией мутанта FimH-DSG G15A G16A V27A с 4-валентными О-антигенами, показали высокие титры IgG к О-антигену.
FimH-DSG G15A G16A V27A индуцирует образование эффективных нейтрализующих антител у нечеловекообразных приматов. Комбинация с 4-валентными O-антигенами также является иммуногенной.
Мутант FimH-DSG G15A G16A V27A с 4-валентными О-антигенами или без них снижает бактериурию и количество биомаркеров инфекции в модели инфекции мочевыводящих путей у яванских макак.
Приведенные ниже пункты описывают дополнительные аспекты изобретения:
С1. Мутантный полипептид FimH, содержащий по меньшей мере одну аминокислотную мутацию относительно аминокислотной последовательности полипептида FimH дикого типа, где положение мутации выбирают из группы, состоящей из: F1, Р12, G14, G15, G16, А18, Р26, V27, V28, Q32, N33, L34, V35, R60, S62, Y64, G65, L68, F71, Т86, L107, Y108, L109, V112, S113, А115, G116, V118, А119, А127, L129, Q133, F144, V154, V155, V156, Р157, Т158, V163 и V185, где положения аминокислот пронумерованы в соответствии с SEQ ID NO: 59.
С2. Мутантный полипептид FimH по п. С1, содержащий по меньшей мере одну мутацию, выбранную из группы, состоящей из: F1I; F1L; F1V; F1M; F1Y; F1W; Р12С; G14C; G15A; G15P; G16A; G16P; А18С; Р26С; V27A; V27C; V28C; Q32C; N33C; L34C; L34N; L34S; L34T; L34D; L34E; L34K; L34R; V35C; R60P; S62C; Y64C; G65A; L68C; F71C; Т86С; L107C; Y108C; L109C; V112C; S113C; A115V; G116C; V118C; А119С; A119N; A119S; А119Т; A119D; А119Е; A119K; A119R; А127С; L129C; Q133K; F144C; V154C; V156C; Р157С; Т158С; V163I; и V185I или любой их комбинации.
С3. Мутантный полипептид FimH по п. С2, содержащий мутации G15A и G16A.
С4. Мутантный полипептид FimH по п. С2, содержащий мутации Р12С и А18С.
С5. Мутантный полипептид FimH по п. С2, содержащий мутации G14C и F144C.
С6. Мутантный полипептид FimH по п. С2, содержащий мутации Р26С и V35C.
С7. Мутантный полипептид FimH по п. С2, содержащий мутации Р26С и V154C.
С8. Мутантный полипептид FimH по п. С2, содержащий мутации Р26С и V156C.
С9. Мутантный полипептид FimH по п. С2, содержащий мутации V27C и L34C.
С10. Мутантный полипептид FimH по п. С2, содержащий мутации V28C и N33C.
С11. Мутантный полипептид FimH по п. С2, содержащий мутации V28C и Р157С.
С12. Мутантный полипептид FimH по п. С2, содержащий мутации Q32C и Y108C.
С13. Мутантный полипептид FimH по п. С2, содержащий мутации N33C и L109C.
С14. Мутантный полипептид FimH по п. С2, содержащий мутации N33C и Р157С.
С15. Мутантный полипептид FimH по п. С2, содержащий мутации V35C и L107C.
С16. Мутантный полипептид FimH по п. С2, содержащий мутации V35C и L109C.
С17. Мутантный полипептид FimH по п. С2, содержащий мутации S62C и Т86С.
С18. Мутантный полипептид FimH по п. С2, содержащий мутации S62C и L129C.
C19. Мутантный полипептид FimH по п. С2, содержащий мутации Y64C и L68C.
С20. Мутантный полипептид FimH по п. С2, содержащий мутации Y64C и А127С.
С21. Мутантный полипептид FimH по п. С2, содержащий мутации L68C и F71C.
С22. Мутантный полипептид FimH по п. С2, содержащий мутации V112C и Т158С.
С23. Мутантный полипептид FimH по п. С2, содержащий мутации S113C и G116C.
С24. Мутантный полипептид FimH по п. С2, содержащий мутации S113C и Т158С.
С25. Мутантный полипептид FimH по п. С2, содержащий мутации V118C и V156C.
С26. Мутантный полипептид FimH по п. С2, содержащий мутации А119С и V155C.
С27. Мутантный полипептид FimH по п. С2, содержащий мутации L34N и V27A.
С28. Мутантный полипептид FimH по п. С2, содержащий мутации L34S и V27A.
С29. Мутантный полипептид FimH по п. С2, содержащий мутации L34T и V27A.
С30. Мутантный полипептид FimH по п. С2, содержащий мутации L34D и V27A.
С31. Мутантный полипептид FimH по п. С2, содержащий мутации L34E и V27A.
С32. Мутантный полипептид FimH по п. С2, содержащий мутации L34K и V27A.
С33. Мутантный полипептид FimH по п. С2, содержащий мутации L34R и V27A.
С34. Мутантный полипептид FimH по п. С2, содержащий мутации A119N и V27A.
С35. Мутантный полипептид FimH по п. С2, содержащий мутации A119S и V27A.
С36. Мутантный полипептид FimH по п. С2, содержащий мутации А119Т и V27A.
С37. Мутантный полипептид FimH по п. С2, содержащий мутации A119D и V27A.
С38. Мутантный полипептид FimH по п. С2, содержащий мутации А119Е и V27A.
С39. Мутантный полипептид FimH по п. С2, содержащий мутации А119К и V27A.
С40. Мутантный полипептид FimH по п. С2, содержащий мутации A119R и V27A.
С41. Мутантный полипептид FimH по п. С2, содержащий мутации G15A и V27A.
С42. Мутантный полипептид FimH по п. С2, содержащий мутации G16A и V27A.
С43. Мутантный полипептид FimH по п. С2, содержащий мутации G15P и V27A.
С44. Мутантный полипептид FimH по п. С2, содержащий мутации G16P и V27A.
С45. Мутантный полипептид FimH по п. С2, содержащий мутации G15A, G16A и V27A.
С46. Мутантный полипептид FimH по п. С2, содержащий мутации G65A и V27A.
С47. Мутантный полипептид FimH по п. С2, содержащий мутации V27A и Q133K.
С4 . Мутантный полипептид FimH по п. С2, содержащий мутации G15A, G16A, V27A и Q133K.
С49. Мутантный полипептид FimH по п. С2, содержащий последовательность любой из SEQ ID NO: 2-58 и 60-64.
С50. Мутантный полипептид FimH по любому из пп. С1-С49, где указанный полипептид является выделенным.
С51. Фармацевтическая композиция, содержащая (i) мутантный полипептид FimH по любому из пп. С1-С50 и (ii) фармацевтически приемлемый носитель.
С52. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С1-С50.
С53. Иммуногенная композиция по п. С52, дополнительно содержащая по меньшей мере один дополнительный антиген.
С54. Иммуногенная композиция по п. С53, где по меньшей мере один дополнительный антиген представляет собой сахарид или полисахарид, или гликоконъюгат, или белок.
С55. Иммуногенная композиция по п. С52, дополнительно содержащая по меньшей мере один адъювант.
С56. Молекула нуклеиновой кислоты, содержащая нуклеотидную последовательность, кодирующую аминокислотную последовательность мутантного полипептида FimH по любому из пп. С1-С49.
С57. Мутантный полипептид FimH по любому из пп. С1-С50, где указанный полипептид является иммуногенным.
С58. Рекомбинантная клетка млекопитающего, содержащая полинуклеотид, кодирующий мутантный полипептид FimH по любому из пп. С1-С50.
С59. Культура, содержащая рекомбинантную клетку по п. С58, где размер указанной культуры составляет не менее 5 литров.
С60. Способ получения мутантного полипептида FimH по любому из пп. С1-С50, включающий культивирование рекомбинантной клетки млекопитающего по п. С58 в подходящих условиях, тем самым обеспечивая экспрессию полипептида; и сбор полипептида.
С61. Способ (i) индукции иммунного ответа у субъекта к внекишечной патогенной Е.coli или (ii) индукции образования опсонофагоцитарных и/или нейтрализующих антител у субъекта, специфичных к внекишечной патогенной Е.coli, где способ включает введение субъекту эффективного количества композиции по любому из пп. С51-С55.
С62. Способ по п. С61, где субъект подвержен риску развития инфекции мочевыводящих путей.
С63. Способ по п. С61, где субъект подвержен риску развития бактериемии.
С64. Способ по п. С61, где субъект подвержен риску развития сепсиса.
С65. Способ индукции иммунного ответа против Е.coli у млекопитающего, включающий введение млекопитающему эффективного количества композиции по любому из пп. С51-С55.
С66. Способ по п. С65, где иммунный ответ включает опсонофагоцитарные и/или нейтрализующие антитела к Е.coli.
С67. Способ по п. С65, где иммунный ответ обеспечивает защиту млекопитающего от инфекции Е.coli.
С68. Способ профилактики, лечения или облегчения бактериальной инфекции, заболевания или состояния у субъекта, включающий введение субъекту иммунологически эффективного количества композиции по любому из пп. С51-С55.
С69. Иммуногенная композиция по п. С54, где дополнительный антиген представляет собой сахарид, содержащий структуру, выбранную из формулыО1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы О6 (например, формулы O6:K 2; K13; K15 и формулы О6:K54), формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O19, формулы O20, формулы O21, формулы O22, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45 (например, формулы O45 и формулы O45 rel), формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O74, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O109, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O17 О, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы О180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186 и формулы O187, где n представляет собой целое число от 1 до 100.
С70. Иммуногенная композиция по п. С69, где сахарид содержит структуру, выбранную из формулы O1(например, формулы O1A, формулы O1B, и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O10, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O21, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O28, формулы O44, формулы O45 (например, формулы O45 и формулы O45 rel), формулы O55, формулы O56, формулы O58, формулы O64, формулы O69, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O75, формулы O77, формулы O78, формулы O86, формулы O88, формулы O90, формулы O98, формулы O104, формулы O111, формулы O113, формулы O114, формулы O119, формулы O121, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O136, формулы O138, формулы O141, формулы O142, формулы O143, формулы O147, формулы O149, формулы O152, формулы O157, формулы O158, формулы O159, формулы O164, формулы O173, формулы 62D1, формулы O22, формулы O35, формулы O65, формулы O66, формулы O83, формулы O91, формулы O105, формулы O116, формулы O117, формулы O139, формулы O153, формулы O167 и формулы O172, где n представляет собой целое число от 31 до 100.
С71. Иммуногенная композиция по п. С70, где сахарид содержит структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B, и формулы O1C), формулы O2, формулы O3, формулы O4 (например, формулы O4:K52 и формулы O4:K6), формулы O5 (например, формулы O5ab и формулы O5ac (штамм 180/С3)), формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O7, формулы O10, формулы O16, формулы O17, формулы O18 (например, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B и формулы O18B1), формулы O21, формулы O23 (например, формулы O23A), формулы O24, формулы O25 (например, формулы O25a и формулы O25b), формулы O26, формулы O28, формулы O44, формулы O45 (например, формулы O45 и формулы O45 rel), формулы O55, формулы O56, формулы O58, формулы O64, формулы O69, формулы O73 (например, формулы O73 (штамм 73-1)), формулы O75, формулы O77, формулы O78, формулы O86, формулы O88, формулы O90, формулы O98, формулы O104, формулы O111, формулы O113, формулы O114, формулы O119, формулы O121, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O136, формулы O138, формулы O141, формулы O142, формулы O143, формулы O147, формулы O149, формулы O152, формулы O157, формулы O158, формулы O159, формулы O164, формулы O173 и формулы 62D1, где n равно целому числу от 31 до 100.
С72. Иммуногенная композиция по п. С70, содержащая структуру, выбранную из формулы O1 (например, формулы O1A, формулы O1B и формулы O1C), формулы O2, формулы O6 (например, формулы O6:K2; K13; K15 и формулы O6:K54), формулы O15, формулы O16, формулы O21, формулы O25 (например, формулы O25a и формулы O25b) и формулы O75.
С73. Иммуногенная композиция по п. С70, содержащая структуру, выбранную из формулы O4, формулы O11, формулы O21 и формулы O75.
С74. Иммуногенная композиция по п. С69, где сахарид не содержит структуру, выбранную из формулы O8, формулы O9a, формулы O9, формулы O20ab, формулы O2Oac, формулы O52, формулы 097 и формулы O101.
С75. Иммуногенная композиция по п. С69, где сахарид не содержит структуру, выбранную из формулы O12.
С76. Иммуногенная композиция по п. С72, где сахарид получают путем экспрессии белка семейства wzz в грамотрицательной бактерии с образованием указанного сахарида.
С77. Иммуногенная композиция по п. С7б, где белок семейства wzz выбирают из группы, состоящей из wzzB, wzz, wzzSF, wzzST, fepE, wzzfepE, wzz1 и wzz2.
C78. Иммуногенная композиция по п. С76, где белок семейства wzz представляет собой wzzB.
С79. Иммуногенная композиция по п. С76, где белок семейства wzz представляет собой fepE.
С80. Иммуногенная композиция по п. С76, где белок семейства wzz представляет собой wzzB и fepE.
С81. Иммуногенная композиция по п. С76, где белок семейства wzz получают из Salmonella enterica.
С82. Иммуногенная композиция по п. С76, где белок семейства wzz содержит последовательность, выбранную из любой из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121.
C83. Иммуногенная композиция по п. С76, где белок семейства wzz содержит последовательность, которая на по меньшей мере 90% идентична любой из последовательностей SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115 и SEQ ID NO: 116.
C84. Иммуногенная композиция по п. С76, где белок семейства wzz содержит последовательность, выбранную из любой из SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121.
C85. Иммуногенная композиция по п. С69, где сахарид синтезирован синтетическим путем.
С86. Иммуногенная композиция по любому из пп. С69-С85, где сахарид дополнительно содержит фрагмент R1 Е. coll.
С87. Иммуногенная композиция по любому из пп. С69-С85, где сахарид дополнительно содержит фрагмент R2 Е. coll.
С88. Иммуногенная композиция по любому из пп. С69-С85, где сахарид дополнительно содержит фрагмент R3 Е. coll.
С89. Иммуногенная композиция по любому из пп. С69-С85, где сахарид дополнительно содержит фрагмент R4 Е. coll.
С90. Иммуногенная композиция по любому из пп. С69-С85, где сахарид дополнительно содержит фрагмент K-12 Е. coll.
С91. Иммуногенная композиция по любому из пп. С69-С90, где сахарид дополнительно содержит фрагмент 3-дезокси-d-манно-окт-2-улозоновой кислоты (KDO).
С92. Иммуногенная композиция по любому из пп. С69-С85, где сахарид не содержит дополнительно фрагмент R1 Е.coli.
С93. Иммуногенная композиция по любому из пп. С69-С85, где сахарид не содержит дополнительно фрагмент R2 Е.coli.
С94. Иммуногенная композиция по любому из пп. С69-С85, где сахарид не содержит дополнительно фрагмент R3 Е.coli.
С95. Иммуногенная композиция по любому из пп. С69-С85, где сахарид не содержит дополнительно фрагмент R4 Е.coli.
С96. Иммуногенная композиция по любому из пп. С69-С85, где сахарид не содержит дополнительно фрагмент К-12 Е.coli.
С97. Иммуногенная композиция по любому из пп. С69-С90, где сахарид не содержит дополнительно фрагмент 3-дезокси-d-манно-окт-2-улозоновой кислоты (KDO).
С98. Иммуногенная композиция по любому из пп. С69-С91, где сахарид не содержит липид А.
С99. Иммуногенная композиция по любому из пп. С69-С98, где полисахарид имеет молекулярную массу от 10 кДа до 2000 кДа или от 50 кДа до 2000 кДа.
С100. Иммуногенная композиция по любому из пп. С69-С99, где сахарид имеет среднюю молекулярную массу 20-40 кДа.
С101. Иммуногенная композиция по любому из пп. С69-С100, где сахарид имеет среднюю молекулярную массу от 40000 до 60000 кДа.
С102. Иммуногенная композиция по любому из пп. С69-С101, где n представляет собой целое число от 31 до 90.
С103. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С69-С50, и конъюгат, содержащий сахарид, ковалентно связанный с белком-носителем, где сахарид получен из Е.coli.
С104. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С69-С50, и конъюгат, содержащий сахарид по любому из пп. С69-С102, ковалентно связанный с белком-носителем.
С105. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С69-С50 или его фрагмент; и конъюгат по любому из пп. С69-С102, где белок-носитель выбирают из любого из: поли (L-лизина), CRM197, фрагмента В дифтерийного токсина (DTFB), С8 DTFB, дифтерийного анатоксина (DT), столбнячного анатоксина (ТТ), фрагмента С ТТ, коклюшного анатоксина, холерного анатоксина или экзотоксина A Pseudomonas aeruginosa.; детоксифицированного экзотоксина А P. aeruginosa (ЕРА), белка, связывающего мальтозу (МВР), детоксифицированного гемолизина A S. aureus, фактора слипания А, фактора слипания В, субъединицы холерного токсина В (СТВ), пневмолизина Streptococcus pneumoniae и его детоксифицированных вариантов, АсгА С.jejuni, природных гликопротеинов С.jejuni и стрептококковой пептидазы С5а (SCP).
С106. Иммуногенная композиция по любому из пп. С103-С105, где белок-носитель представляет собой CRM197.
С107. Иммуногенная композиция по любому из пп. С103-С105, где белок-носитель представляет собой столбнячный анатоксин (ТТ).
С108. Иммуногенная композиция по любому из пп. С103-С105, где белок-носитель представляет собой поли(L-лизин).
С109. Иммуногенная композиция по любому из пп. С103-С107, где конъюгат получают восстановительным аминированием.
С110. Иммуногенная композиция по любому из пп. С103-С107, где конъюгат получают с помощью химии CDAP.
С111. Иммуногенная композиция по любому из пп. С103-С107, где конъюгат представляет собой конъюгированный сахарид с одной концевой связью.
С112. Иммуногенная композиция по любому из пп. С103-С107, где сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер.
С113. Иммуногенная композиция по п. С112, где сахарид конъюгирован с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер, где сахарид ковалентно связан со спейсером еТЕС карбаматной связью, и где белок-носитель ковалентно связан со спейсером еТЕС амидной связью.
С114. Иммуногенная композиция по любому из пп. С112-С113, где CRM197 содержит от 2 до 20 или от 4 до 16 остатков лизина, ковалентно связанных с полисахаридом через спейсер еТЕС.
С115. Иммуногенная композиция по любому из пп. С103-С114, где соотношение сахарид : белок-носитель (вес/вес) составляет от 0,2 до 4.
С116. Иммуногенная композиция по любому из пп. С103-С114, где отношение сахарида к белку составляет не менее 0,5 и не более 2.
С117. Иммуногенная композиция по любому из пп. С103-С114, где отношение сахарида к белку составляет от 0,4 до 1,7.
С118. Иммуногенная композиция по любому из пп. С111-С117, где сахарид конъюгирован с белком-носителем через остаток 3-дезокси-d-манно-окт-2-улозоновой кислоты (KDO).
С119. Иммуногенная композиция по п. С69, где конъюгат содержит сахарид, ковалентно связанный с белком-носителем, при этом сахарид содержит структуру, выбранную из формулы O8, формулы O9a, формулы O9, формулы O20ab, формулы О20ас, формулы O52, формулы O97 и формула O101, где n представляет собой целое число от 1 до 10.
С120. Иммуногенная композиция, содержащая мутантный полипептид FimH и сахарид по любому из пп. С69-С102, и фармацевтически приемлемый разбавитель.
С121. Иммуногенная композиция, содержащая мутантный полипептид FimH и гликоконъюгат по любому из пп. С103-С119, и фармацевтически приемлемый разбавитель.
С122. Иммуногенная композиция по п. С121, содержащая не более примерно 25% свободного сахарида относительно общего количества сахарида в композиции.
С123. Иммуногенная композиция по любому из пп. С120-С121, дополнительно содержащая адъювант.
С124. Иммуногенная композиция по любому из пп. С120-С121, дополнительно содержащая алюминий.
С125. Иммуногенная композиция по любому из пп. С120-С121, дополнительно содержащая QS-21.
С126. Иммуногенная композиция по любому из пп. С120-С121, дополнительно содержащая олигонуклеотид CpG.
С127. Иммуногенная композиция по любому из пп. С120-С121, где композиция не включает адъювант.
С128. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С69-С118 и сахарид, полученный из Е.coli, конъюгированный с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер, где полисахарид ковалентно связан со спейсером еТЕС карбаматной связью, и где белок-носитель ковалентно связан со спейсером еТЕС амидной связью.
С129. Иммуногенная композиция по п. С128, где сахарид представляет собой О-антиген, полученный из Е.coli.
С130. Иммуногенная композиция по п. С128, дополнительно содержащая фармацевтически приемлемый наполнитель, носитель или разбавитель.
С131. Иммуногенная композиция по п. С128, где сахарид представляет собой О-антиген, полученный из Е.coli.
С132. Иммуногенная композиция, содержащая мутантный полипептид FimH и сахарид по любому из пп. С69-С85, конъюгированный с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер, где полисахарид ковалентно связан со спейсером еТЕС карбаматной связью, и где белок-носитель ковалентно связан со спейсером еТЕС амидной связью.
С133. Иммуногенная композиция, содержащая мутантный полипептид FimH и (i) конъюгат антигена O25 В Е.coli, ковалентно связанный с белком-носителем, (ii) конъюгат антигена 01А Е. coli, ковалентно связанный с белком-носителем, (iii) конъюгат антигена O2 Е.coli, ковалентно связанный с белком-носителем, и (iv) конъюгат антигена 06, ковалентно связанный с белком-носителем, где антиген O25B Е.coli содержит структуру формулы O25B, где n является целым числом больше 30.
С134. Иммуногенная композиция по п. С133, где белок-носитель выбирают из любого из: поли (L-лизина), CRM197, фрагмента В дифтерийного токсина (DTFB), С8 DTFB, дифтерийного анатоксина (DT), столбнячного анатоксина (ТТ), фрагмента С ТТ, коклюшного анатоксина, холерного анатоксина или экзотоксина A Pseudomonas aeruginosa; детоксифицированного экзотоксина А P. aeruginosa (ЕРА), белка, связывающего мальтозу (МВР), детоксифицированного гемолизина A S. aureus, фактора слипания А, фактора слипания В, субъединицы холерного токсина В (СТВ), пневмолизина Streptococcus pneumoniae и его детоксифицированных вариантов, AcrA С.jejuni, природных гликопротеинов С. jejuni и стрептококковой пептидазы С5а (SCP).
С135. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С69-С118 и (i) конъюгат антигена O25B Е.coli, ковалентно связанный с белком-носителем, (ii) конъюгат антигена O4 Е.coli, ковалентно связанный с белком-носителем, (iii) конъюгат антигена O11 Е.coli, ковалентно связанный с белком-носителем, и (iv) конъюгат антигена O21, ковалентно связанный с белком-носителем, где антиген O25B Е.coli содержит структуру формулы O75, где n представляет собой целое число больше 30.
С136. Иммуногенная композиция по п. С135, где белок-носитель выбирают из любого из: поли (L-лизина), CRM197, фрагмента В дифтерийного токсина (DTFB), С8 DTFB, дифтерийного анатоксина (DT), столбнячного анатоксина (ТТ), фрагмента С ТТ, коклюшного анатоксина, холерного анатоксина или экзотоксина A Pseudomonas aeruginosa; детоксифицированного экзотоксина А P. aeruginosa (ЕРА), белка, связывающего мальтозу (МВР), детоксифицированного гемолизина A S. aureus, фактора слипания А, фактора слипания В, субъединицы холерного токсина В (СТВ), пневмолизина Streptococcus pneumoniae и его детоксифицированных вариантов, АсгА С.jejuni, природных гликопротеинов С. jejuni и стрептококковой пептидазы С5а (SCP).
С137. Способ получения иммуногенной композиции, содержащей мутантный полипептид FimH и конъюгат, содержащий сахарид, конъюгированный с белком-носителем через (2-((2-оксоэтил)тио)этил)карбаматный (еТЕС) спейсер, включающий этапы: а) взаимодействия сахарида с 1,1'-карбонил-ди-(1,2,4-триазолом) (CDT) или 1,1'-карбонилдиимидазолом (CDI) в органическом растворителе с получением активированного сахарида; b) взаимодействия активированного сахарида с цистамином или цистеамином или их солями с получением тиолированного сахарида; с) взаимодействия тиолированного сахарида с восстанавливающим агентом с получением активированного тиолированного сахарида, содержащего один или более свободных сульфгидрильных остатков; о!) взаимодействия активированного тиолированного сахарида с активированным белком-носителем, содержащим одну или более α-галогенацетамидных групп, с получением конъюгата тиолированного сахарида с белком-носителем; и е) взаимодействия конъюгата тиолированного сахарида с белком-носителем с (i) первым кэпирующим реагентом, способным кэпировать неконъюгированные α-галогенацетамидные группы активированного белка-носителя; и/или (ii) вторым кэпирующим реагентом, способным кэпировать неконъюгированные свободные сульфгидрильные остатки; посредством чего получают гликоконъюгат, связанный с еТЕС, где сахарид получают из Е.coli; дополнительно включающий экспрессию полинуклеотида, кодирующего полипептид, полученный из FimH, или его фрагмент в рекомбинантной клетке млекопитающего, и выделение указанного полипептида или его фрагмента.
С138. Способ по п. С137, включающий получение иммуногенной композиции по любому из пп. С69-С102.
С139. Способ по любому из пп. С137-С138, где этап кэпирования е) включает взаимодействие конъюгата тиолированного сахарида с белком-носителем с (i) N-ацетил-L-цистеином в качестве первого копирующего реагента, и/или (ii) иодацетамидом в качестве второго копирующего реагента.
С140. Способ по любому из пп. С137-С139, дополнительно включающий этап компаундирования сахарида путем взаимодействия с триазолом или имидазолом с получением компаундированного сахарида, где компаундированный сахарид замораживают, лиофилизируют и восстанавливают в органическом растворителе перед этапом а).
С141. Способ по любому из пп. С137-С140, дополнительно включающий очистку тиолированного полисахарида, полученного на этапе с), где этап очистки включает диафильтрацию.
С142. Способ по любому из пп. С137-С141, где способ дополнительно включает очистку гликоконъюгата, связанного с еТЕС, путем диафильтрации.
С143. Способ по любому из пп. С137-С142, где органический растворитель на этапе а) представляет собой полярный апротонный растворитель, выбранный из любого из следующего: диметилсульфоксида (DMSO), диметилформамида (DMF), диметилацетамида (DMA), N-метил-2-пирролидона (NMP), ацетонитрила, 1,3-диметил-3,4,5,6-тетрагидро-2(1Н)-пиримидинона (DMPU) и гексаметилфосфорамида (НМРА) или их смеси.
С144. Способ по любому из пп. С137-С142, где среда содержит элемент, выбранный из любого из: KH2PO4, K2HPO4, (NH4)2SO4, цитрата натрия, Na2SO4, аспарагиновой кислоты, глюкозы, MgSO4, FeSO4-7H2O, Na2MoO4-2H2O, H3BO3, CoCl2-6H2O, CuCl2-2H2O, MnCl2-4H2O, ZnCl2 и CaCl2-2H2O.
С145. Среда по п. С144, где указанную среду используют для культивирования Е. coll.
С146. Способ получения сахарида по любому из пп. С69-С102, включающий культивирование рекомбинантной Е.coli в среде; продуцирование указанного сахарида путем культивирования указанной клетки в указанной среде; при этом указанная клетка продуцирует указанный сахарид.
С147. Способ по п. С146, где среда содержит элемент, выбранный из любого из: KH2PO4, K2HPO4, (NH4)2SO4, цитрата натрия, Na2SO4, аспарагиновой кислоты, глюкозы, MgSO4, FeSO4-7H2O, Na2MoO4-2H2O, H3BO3, CoCl2-6H2O, CuCl2-2H2O, MnCl2-4H2O, ZnCl2 и CaCl2-2H2O.
С148. Способ по п. С146, где среда содержит гидролизат сои.
С149. Способ по п. С146, где среда содержит дрожжевой экстракт.
С150. Способ по п. С146, где среда не содержит дополнительно гидролизат сои и дрожжевой экстракт.
С151. Способ по п. С146, где клетка Е.coli содержит гетерологичный белок семейства wzz, выбранный из любого из: wzzB, wzz, wzzSF, wzzST, fepE, wzzfepE, wzzl и wzz2.
C152. Способ по п. C146, где клетка E.coli содержит белок семейства wzz Salmonella enterica, выбранный из любого из: wzzB, wzz, wzzSF, wzzST, fepE, wzzfepE, wzzl и wzz2.
C153. Способ по п. C152, где белок семейства wzz содержит последовательность, выбранную из любой из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115 и SEQ ID NO: 116.
С154. Способ по п. С146, где культивирование дает выход >120 OD600/мл.
С155. Способ по п. С146, дополнительно включающий очистку сахарида.
С156. Способ по п. С146, где этап очистки включает любое из следующего: диализ, операцию концентрирования, операцию диафильтрации, фильтрацию в тангенциальном потоке, осаждение, элюирование, центрифугирование, осаждение, ультрафильтрацию, глубинную фильтрацию и колоночную хроматографию (ионно обменную хроматографию, мультимодальную ионообменную хроматографию, DEAE и хроматографию гидрофобного взаимодействия).
С157. Способ индукции иммунного ответа у субъекта, включающий введение субъекту композиции по любому из пп. С69-С136.
С158. Способ по п. С157, где иммунный ответ включает индукцию образования антител, специфических к сывороточным O-полисахаридам Е coli.
С159. Способ по п. С157, где иммунный ответ включает индукцию образования анти-E. coli антитела IgG.
С160. Способ по п. С157, где иммунный ответ включает индукцию бактерицидной активности против Е.coli.
С161. Способ по п. С157, где иммунный ответ включает индукцию образования опсонофагоцитарных антител к Е.coli.
С162. Способ по п. С157, где иммунный ответ включает уровень среднего геометрического титра (GMT) по меньшей мере от 1000 до 200000 после первоначального введения дозы.
С163. Способ по п. С157, где композиция содержит сахарид, содержащий формулу 025, где n представляет собой целое число от 40 до 100, при этом иммунный ответ включает уровень среднего геометрического титра (GMT) по меньшей мере от 1000 до 200000 после введения начальной дозы.
С164. Способ по п. С157, где субъект подвержен риску развития любого из состояний, выбранных из: инфекции мочевыводящих путей, холецистита, холангита, диареи, гемолитико-уремического синдрома, неонатального менингита, уросепсиса, интраабдоминальной инфекции, менингита, осложненной пневмонии, раневой инфекции, инфекции, связанной с биопсией простаты, неонатального/младенческого сепсиса, нейтропенической лихорадки и других инфекций кровотока; пневмонии, бактериемии и сепсиса.
С165. Способ по п. С157, где субъект представляет собой млекопитающее.
С166. Способ (i) индукции иммунного ответа у субъекта против внекишечной патогенной Е.coli или (ii) индукции образования у субъекта опсонофагоцитарных антител, специфичных к внекишечной патогенной Е.coli, где способ включает введение субъекту эффективного количества композиции по любому из пп. С69-С136.
С167. Способ по п. С166, где субъект подвержен риску развития инфекции мочевыводящих путей.
С168. Способ по п. С166, где субъект подвержен риску развития бактериемии.
С169. Способ по п. С166, где субъект подвержен риску развития сепсиса.
С170. Способ индукции иммунного ответа у субъекта, включающий введение субъекту композиции по любому из пп. С69-С136.
С171. Способ по п. С170, где иммунный ответ включает индукцию образования антитела, специфического к сывороточным полисахаридам Е.coli.
С172. Способ по п. С170, где антитело, специфическое к сывороточным полисахаридам Е.coli, представляет собой антитело IgG.
С173. Способ по п. С170, где антитело, специфическое к сывороточным полисахаридам Е.coli, представляет собой антитело IgG, обладающее бактерицидной активностью по отношению к Е.coli.
С174. Иммуногенная композиция по п. С69, где n больше, чем количество повторяющихся звеньев в соответствующем полисахариде Е.coli дикого типа.
С175. Композиция по п. С174, где n представляет собой целое число от 31 до 100.
С176. Композиция по п. С174, где сахарид содержит структуру согласно любой из: формулы O1A, формулы O1B и формулы O1C, формулы O2, формулы O6 и формулы O25B.
С177. Композиция по п. С174, где сахарид продуцируют в рекомбинантной клетке-хозяине, которая экспрессирует белок семейства wzz, который на по меньшей мере 90% идентичен последовательности любой из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115, SEQ ID NO: 116, SEQ ID NO: 117, SEQ ID NO: 118, SEQ ID NO: 119, SEQ ID NO: 120 и SEQ ID NO: 121.
С178. Композиция по п. С177, где белок содержит любую из SEQ ID NO: 112, SEQ ID NO: 113, SEQ ID NO: 114, SEQ ID NO: 115 и SEQ ID NO: 116.
C179. Сахарид по п. C174, где сахарид синтезирован синтетическим путем.
С180. Иммуногенная композиция, содержащая мутантный полипептид FimH по любому из пп. С1-С55 и (а) конъюгат, содержащий белок-носитель, ковалентно связанный с сахаридом формулы O25b, где n представляет собой целое число от 31 до 90, (b) конъюгат, содержащий белок-носитель, ковалентно связанный с сахаридом формулы O1A, где n представляет собой целое число от 31 до 90, (с) конъюгат, содержащий белок-носитель, ковалентно связанный с сахаридом формулы O2, где n представляет собой целое число от 31 до 90, и (d) конъюгат, содержащий белок-носитель, ковалентно связанный с сахаридом, содержащим формулу O6, где n представляет собой целое число от 31 до 90.
С181. Иммуногенная композиция по п. С180, дополнительно содержащая конъюгат, содержащий белок-носитель, ковалентно связанный с сахаридом, имеющим структуру, выбранную из любой из: формулы O15, формулы O16, формулы O17, формулы O18 и формулы O75, где n представляет собой целое число от 31 до 90.
С182. Иммуногенная композиция по п. С180, содержащая не более 25% свободного сахарида относительно общего количества сахарида в композиции.
С183. Способ индукции иммунного ответа против Е.coli у млекопитающего, включающий введение млекопитающему эффективного количества композиции по любому из пп. С180-С182.
С184. Способ по п. С183, где иммунный ответ включает опсонофагоцитарные антитела к Е.coli.
С185. Способ по п. С183, где иммунный ответ защищает млекопитающее от инфекции Е.coli.
С186. Рекомбинантная клетка млекопитающего, содержащая (а) первый представляющий интерес ген, кодирующий мутантный полипептид FimH по любому из пп. С1-С55, где указанный ген интегрирован по меньшей мере между двумя сайтами-мишенями рекомбинации (RTS).
С187. Вариант осуществления по пп. С186, где два RTS являются хромосомно-интегрированными в локус NL1 или локус NL2.
С188. Вариант осуществления по п. С186, где первый представляющий интерес ген дополнительно содержит репортерный ген, ген, кодирующий трудно экспрессируемый белок, вспомогательный ген или их комбинацию.
С189. Вариант осуществления по п. С186, дополнительно содержащий второй представляющий интерес ген, хромосомно-интегрированный во второй локус, отличный от локуса (а), где второй представляющий интерес ген содержит репортерный ген, ген, кодирующий трудно экспрессируемый белок, вспомогательный ген или их комбинацию.
С190. Рекомбинантная клетка по п. С186, где полинуклеотидная последовательность интегрирована в геномную ДНК указанной клетки млекопитающего.
С191. Рекомбинантная клетка по п. С186, где полинуклеотидная последовательность оптимизирована по кодонам для экспрессии в клетке.
С192. Рекомбинантная клетка по п. С186, где клетка представляет собой клетку эмбриональной почки человека.
С193. Рекомбинантная клетка по п. С192, где клетка эмбриональной почки человека содержит клетку НЕК293.
С194. Рекомбинантная клетка по п. С193, где клетку HEK293 выбирают из любой из клеток HEK293T, клеток HEK293TS и клеток HEK293E.
С195. Рекомбинантная клетка по п. С186, где клетка представляет собой клетку СНО.
С196. Рекомбинантная клетка по п. С195, где указанная клетка СНО представляет собой клетку CHO-К1, клетку CHO-DUXB11, CHO-DG44 или клетку CHO-S.
С197. Рекомбинантная клетка по п. С186, где полипептид является растворимым.
С198. Рекомбинантная клетка по п. С186, где полипептид секретируется из клетки.
С199. Культура, содержащая рекомбинантную клетку по п. С186, где указанная культура имеет размер по меньшей мере 5 литров.
С200. Культура по п. С199, где выход полипептида или его фрагмента составляет по меньшей мере 0,05 г/л.
С201. Культура по п. С199, где выход полипептида или его фрагмента составляет по меньшей мере 0,10 г/л.
С202. Способ получения полипептида, полученного из Е.coli, или его фрагмента, включающий культивирование рекомбинантной клетки млекопитающего по п. С186 в подходящих условиях, тем самым обеспечивая экспрессию полипептида или его фрагмента; и сбор полипептида или его фрагмента.
С203. Способ по п. С202, дополнительно включающий очистку полипептида или его фрагмента.
С204. Иммуногенная композиция по п. С54, дополнительно содержащая по меньшей мере один сахарид, полученный из любого типа K. pneumoniae, выбранного из группы, состоящей из O1, O2, O3 и O5.
С205. Иммуногенная композиция по п. С204, дополнительно содержащая сахарид, полученный из KK. pneumoniae типа O1.
С206. Иммуногенная композиция по п. С204, дополнительно содержащая сахарид, полученный из K. pneumoniae типа O2.
С207. Композиция по п. С204, дополнительно содержащая сахарид, полученный из K. pneumoniae типа 03.
С208. Иммуногенная композиция по п. С204, дополнительно содержащая сахарид, полученный из K. pneumoniae типа O5.
С209. Иммуногенная композиция по п. С204, дополнительно содержащая сахарид, полученный из K. pneumoniae типа O1, и сахарид, полученный из K. pneumoniae типа O2.
С210. Иммуногенная композиция по любому из пп. С204-C2O9, дополнительно содержащая по меньшей мере один сахарид, содержащий структуру, выбранную из формулы O1, формулы O1A, формулы O1B, формулы O1C, формулы O2, формулы O3, формулы O4, формулы O4:K52, формулы O4:K6, формулы O5, формулы O5ab, формулы O5ac, формулы O6, формулы O6:K2; K13; K15, формулы O6:K54, формулы O7, формулы O8, формулы O9, формулы O10, формулы O11, формулы O12, формулы O13, формулы O14, формулы O15, формулы O16, формулы O17, формулы O18, формулы O18A, формулы O18ac, формулы O18A1, формулы O18B, формулы O18B1, формулы O19, формулы O20, формулы O21, формулы O22, формулы O23, формулы O23A, формулы O24, формулы O25, формулы O25a, формулы O25b, формулы O26, формулы O27, формулы O28, формулы O29, формулы O30, формулы O32, формулы O33, формулы O34, формулы O35, формулы O36, формулы O37, формулы O38, формулы O39, формулы O40, формулы O41, формулы O42, формулы O43, формулы O44, формулы O45, формулы O45, формулы O45 rel, формулы O46, формулы O48, формулы O49, формулы O50, формулы O51, формулы O52, формулы O53, формулы O54, формулы O55, формулы O56, формулы O57, формулы O58, формулы O59, формулы O60, формулы O61, формулы O62, формулы 62D1, формулы O63, формулы O64, формулы O65, формулы O66, формулы O68, формулы O69, формулы O70, формулы O71, формулы O73, формулы O73, формулы O 4, формулы O75, формулы O76, формулы O77, формулы O78, формулы O79, формулы O80, формулы O81, формулы O82, формулы O83, формулы O84, формулы O85, формулы O86, формулы O87, формулы O88, формулы O89, формулы O90, формулы O91, формулы O92, формулы O93, формулы O95, формулы O96, формулы O97, формулы O98, формулы O99, формулы O100, формулы O101, формулы O102, формулы O103, формулы O104, формулы O105, формулы O106, формулы O107, формулы O108, формулы O1O9, формулы О110, формулы O111, формулы O112, формулы O113, формулы O114, формулы O115, формулы O116, формулы O117, формулы O118, формулы O119, формулы O120, формулы O121, формулы O123, формулы O124, формулы O125, формулы O126, формулы O127, формулы O128, формулы O129, формулы O130, формулы O131, формулы O132, формулы O133, формулы O134, формулы O135, формулы O136, формулы O137, формулы O138, формулы O139, формулы O140, формулы O141, формулы O142, формулы O143, формулы O144, формулы O145, формулы O146, формулы O147, формулы O148, формулы O149, формулы O150, формулы O151, формулы O152, формулы O153, формулы O154, формулы O155, формулы O156, формулы O157, формулы O158, формулы O159, формулы O160, формулы O161, формулы O162, формулы O163, формулы O164, формулы O165, формулы O166, формулы O167, формулы O168, формулы O169, формулы O170, формулы O171, формулы O172, формулы O173, формулы O174, формулы O175, формулы O176, формулы O177, формулы O178, формулы O179, формулы O180, формулы O181, формулы O182, формулы O183, формулы O184, формулы O185, формулы O186, формулы O187, где n представляет собой целое число от 1 до 100, более предпочтительно от 31 до 90.
С211. Иммуногенная композиция по п. С210, где сахарид, полученный из K. pneumoniae, конъюгирован с белком-носителем; и сахарид, полученный из Е.coli, конъюгирован с белком-носителем.
С212. Способ индукции иммунного ответа против Е.coli у млекопитающего, включающий введение млекопитающему эффективного количества композиции по любому из пп. С204-С211.
С213. Способ по п. С212, где иммунный ответ включает опсонофагоцитарные антитела к Е.coli.
С214. Способ по п. С212, где иммунный ответ защищает млекопитающее от инфекции Е.coli.
С215. Способ индукции иммунного ответа против K. pneumoniae у млекопитающего, включающий введение млекопитающему эффективного количества композиции по любому из пп. С204-С211.
С216. Способ по п. С215, где иммунный ответ включает опсонофагоцитарные антитела к K. pneumoniae.
С217. Способ по п. С215, где иммунный ответ защищает млекопитающее от инфекции K. pneumoniae.
С218. Композиции и способы по любому из пп. С204-С217, где серотип O1 K. pneumoniae содержит вариант O1V1 или O1V2.
С219. Композиции и способы по любому из пп. С204-С217, где серотип O2 K. pneumoniae содержит вариант O2V1 или O2V2.
С220. Применение композиций по любому из пп. С1-С219, представленных в настоящем описании.
С221. Композиция по п. С211, где О-антиген K. pneumoniae выбирают из группы, состоящей из а) серотипа O1 подтипа v1 (O1v1), b) серотипа O1 подтипа v2 (O1v2), с) серотипа O2 подтипа v1 (O2v1) и d) серотипа O2 подтипа v2 (O2v2).
С222. Нуклеиновая кислота, содержащая нуклеотиды, кодирующие полипептиды по любому из пп. С1-С221.
С223. Нуклеиновая кислота по п. С222, где нуклеиновая кислота представляет собой РНК.
С224. Наночастица, содержащая нуклеиновую кислоту по п. С222 или п. С223.
С225. Иммуногенная композиция по настоящему изобретению, дополнительно содержащая один или более конъюгатов, содержащих сахарид, выбранный из группы, состоящей из: формулы O4, формулы O11, формулы O13, формулы O21 и формулы O86, где n представляет собой целое число от 1 до 100, предпочтительно от 31 до 90.
С226. Иммуногенная композиция по настоящему изобретению, дополнительно содержащая один или более конъюгатов, содержащих сахарид, выбранный из группы, состоящей из формулы O1a, формулы O2, формулы O6 и формулы O25b, где n представляет собой целое число от 1 до 100, предпочтительно от 31 до 90.
--->
СПИСКИ ПОСЛЕДОВАТЕЛЬНОСТЕЙ
<110> Pfizer Inc.
Che, Ye
Chorro, Laurent Oliver
Donald, Robert George Konrad
Griffor, Matthew Curtis
Silmon de Monerri, Natalie Clare
<120> Мутанты FimH E. coli и их применение
<130> PC072713
<160> 147
<170> PatentIn version 3.5
<210> 1
<211> 160
<212> Белок
<213> Escherichia coli
<400> 1
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 2
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 2
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Ala Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 3
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 3
Ile Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 4
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 4
Leu Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 5
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 5
Val Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 6
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 6
Met Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 7
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 7
Tyr Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 8
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 8
Trp Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 9
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 9
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Lys Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 10
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 10
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 11
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 11
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Pro Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 12
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 12
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 13
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 13
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Pro
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 14
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 14
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 15
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 15
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Pro Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 16
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 16
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Ala Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 17
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 17
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Cys Ile Gly Gly Gly
1 5 10 15
Ser Cys Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 18
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 18
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Cys Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Cys
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 19
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 19
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Cys Val Val Asn Val Gly Gln
20 25 30
Asn Leu Cys Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 20
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 20
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Cys Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Cys Val Val Pro Thr Gly Gly
145 150 155 160
<210> 21
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 21
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Cys Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Cys Pro Thr Gly Gly
145 150 155 160
<210> 22
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 22
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Cys Val Asn Val Gly Gln
20 25 30
Asn Cys Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 23
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 23
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Cys Asn Val Gly Gln
20 25 30
Cys Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 24
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 24
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Cys Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Cys Thr Gly Gly
145 150 155 160
<210> 25
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 25
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Cys
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Cys Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 26
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 26
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Cys Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Cys Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 27
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 27
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Cys Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Cys Thr Gly Gly
145 150 155 160
<210> 28
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 28
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Cys Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Cys Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 29
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 29
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Cys Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Cys Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 30
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 30
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Cys Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Cys Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 31
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 31
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Cys Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Cys Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 32
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 32
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Cys
50 55 60
Gly Gly Val Cys Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 33
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 33
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Cys
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Cys Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 34
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 34
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Cys Ser Ser Cys Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 35
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 35
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Cys
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Cys Gly Gly
145 150 155 160
<210> 36
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 36
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Cys Ser Ala Cys Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 37
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 37
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Cys Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Cys Gly Gly
145 150 155 160
<210> 38
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 38
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Cys Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Cys Pro Thr Gly Gly
145 150 155 160
<210> 39
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 39
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Cys Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Cys Val Pro Thr Gly Gly
145 150 155 160
<210> 40
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 40
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Asn Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 41
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 41
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Ser Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 42
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 42
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Thr Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 43
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 43
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Asn Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 44
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 44
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ser Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 45
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 45
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Thr Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 46
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 46
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Val Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 47
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 47
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Ile Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 48
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 48
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Ile Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 49
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 49
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Ile Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 50
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 50
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 51
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 51
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 52
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 52
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Pro Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 53
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 53
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Pro
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 54
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 54
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 55
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 55
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Pro Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 56
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 56
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Ala Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 57
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 57
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Lys Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 58
<211> 160
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 58
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Lys Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
<210> 59
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 59
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 60
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 60
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 61
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 61
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 62
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 62
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Gln Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 63
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 63
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Gly Gly
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Lys Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 64
<211> 300
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 64
Phe Ala Cys Lys Thr Ala Ser Gly Thr Ala Ile Pro Ile Gly Ala Ala
1 5 10 15
Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Ala Val Asn Val Gly Gln
20 25 30
Asn Leu Val Val Asp Leu Ser Thr Gln Ile Phe Cys His Asn Asp Tyr
35 40 45
Pro Glu Thr Ile Thr Asp Tyr Val Thr Leu Gln Arg Gly Ser Ala Tyr
50 55 60
Gly Gly Val Leu Ser Ser Phe Ser Gly Thr Val Lys Tyr Ser Gly Ser
65 70 75 80
Ser Tyr Pro Phe Pro Thr Thr Ser Glu Thr Pro Arg Val Val Tyr Asn
85 90 95
Ser Arg Thr Asp Lys Pro Trp Pro Val Ala Leu Tyr Leu Thr Pro Val
100 105 110
Ser Ser Ala Gly Gly Val Ala Ile Lys Ala Gly Ser Leu Ile Ala Val
115 120 125
Leu Ile Leu Arg Lys Thr Asn Asn Tyr Asn Ser Asp Asp Phe Gln Phe
130 135 140
Val Trp Asn Ile Tyr Ala Asn Asn Asp Val Val Val Pro Thr Gly Gly
145 150 155 160
Cys Asp Val Ser Ala Arg Asp Val Thr Val Thr Leu Pro Asp Tyr Pro
165 170 175
Gly Ser Val Pro Ile Pro Leu Thr Val Tyr Cys Ala Lys Ser Gln Asn
180 185 190
Leu Gly Tyr Tyr Leu Ser Gly Thr Thr Ala Asp Ala Gly Asn Ser Ile
195 200 205
Phe Thr Asn Thr Ala Ser Phe Ser Pro Ala Gln Gly Val Gly Val Gln
210 215 220
Leu Thr Arg Gln Gly Thr Ile Ile Pro Ala Asn Asn Thr Val Ser Leu
225 230 235 240
Gly Ala Val Gly Thr Ser Ala Val Ser Leu Gly Leu Thr Ala Asn Tyr
245 250 255
Ala Arg Thr Gly Gly Gln Val Thr Ala Gly Asn Val Gln Ser Ile Ile
260 265 270
Gly Val Thr Phe Val Tyr Gln Gly Gly Ser Ser Gly Gly Gly Ala Asp
275 280 285
Val Thr Ile Thr Val Asn Gly Lys Val Val Ala Lys
290 295 300
<210> 65
<211> 19
<212> Белок
<213> Mus musculus
<400> 65
Glu Thr Asp Thr Leu Leu Leu Trp Val Leu Leu Leu Trp Val Pro Gly
1 5 10 15
Ser Thr Gly
<210> 66
<211> 207
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 66
Met Glu Gly Met Asp Pro Leu Ala Val Leu Ala Glu Ser Arg Leu Leu
1 5 10 15
Pro Leu Leu Thr Val Arg Gly Gly Glu Asp Leu Ala Gly Leu Ala Thr
20 25 30
Val Leu Glu Leu Met Gly Val Gly Ala Leu Glu Ile Thr Leu Arg Thr
35 40 45
Glu Lys Gly Leu Glu Ala Leu Lys Ala Leu Arg Lys Ser Gly Leu Leu
50 55 60
Leu Gly Ala Gly Thr Val Arg Ser Pro Lys Glu Ala Glu Ala Ala Leu
65 70 75 80
Glu Ala Gly Ala Ala Phe Leu Val Ser Pro Gly Leu Leu Glu Glu Val
85 90 95
Ala Ala Leu Ala Gln Ala Arg Gly Val Pro Tyr Leu Pro Gly Val Leu
100 105 110
Thr Pro Thr Glu Val Glu Arg Ala Leu Ala Leu Gly Leu Ser Ala Leu
115 120 125
Lys Phe Phe Pro Ala Glu Pro Phe Gln Gly Val Arg Val Leu Arg Ala
130 135 140
Tyr Ala Glu Val Phe Pro Glu Val Arg Phe Leu Pro Thr Gly Gly Ile
145 150 155 160
Lys Glu Glu His Leu Pro His Tyr Ala Ala Leu Pro Asn Leu Leu Ala
165 170 175
Val Gly Gly Ser Trp Leu Leu Gln Gly Asp Leu Ala Ala Val Met Lys
180 185 190
Lys Val Lys Ala Ala Lys Ala Leu Leu Ser Pro Gln Ala Pro Gly
195 200 205
<210> 67
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 67
Met Thr Lys Lys Val Gly Ile Val Asp Thr Thr Phe Ala Arg Val Asp
1 5 10 15
Met Ala Glu Ala Ala Ile Arg Thr Leu Lys Ala Leu Ser Pro Asn Ile
20 25 30
Lys Ile Ile Arg Lys Thr Val Pro Gly Ile Lys Asp Leu Pro Val Ala
35 40 45
Cys Lys Lys Leu Leu Glu Glu Glu Gly Cys Asp Ile Val Met Ala Leu
50 55 60
Gly Met Pro Gly Lys Ala Glu Lys Asp Lys Val Cys Ala His Glu Ala
65 70 75 80
Ser Leu Gly Leu Met Leu Ala Gln Leu Met Thr Asn Lys His Ile Ile
85 90 95
Glu Val Phe Val His Glu Asp Glu Ala Lys Asp Asp Asp Glu Leu Asp
100 105 110
Ile Leu Ala Leu Val Arg Ala Ile Glu His Ala Ala Asn Val Tyr Tyr
115 120 125
Leu Leu Phe Lys Pro Glu Tyr Leu Thr Arg Met Ala Gly Lys Gly Leu
130 135 140
Arg Gln Gly Arg Glu Asp Ala Gly Pro Ala Arg Glu
145 150 155
<210> 68
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 68
Met Thr Lys Lys Val Gly Ile Val Asp Thr Thr Phe Ala Arg Val Asp
1 5 10 15
Met Ala Ser Ala Ala Ile Leu Thr Leu Lys Met Glu Ser Pro Asn Ile
20 25 30
Lys Ile Ile Arg Lys Thr Val Pro Gly Ile Lys Asp Leu Pro Val Ala
35 40 45
Cys Lys Lys Leu Leu Glu Glu Glu Gly Cys Asp Ile Val Met Ala Leu
50 55 60
Gly Met Pro Gly Lys Ala Glu Lys Asp Lys Val Cys Ala His Glu Ala
65 70 75 80
Ser Leu Gly Leu Met Leu Ala Gln Leu Met Thr Asn Lys His Ile Ile
85 90 95
Glu Val Phe Val His Glu Asp Glu Ala Lys Asp Asp Ala Glu Leu Lys
100 105 110
Ile Leu Ala Ala Arg Arg Ala Ile Glu His Ala Leu Asn Val Tyr Tyr
115 120 125
Leu Leu Phe Lys Pro Glu Tyr Leu Thr Arg Met Ala Gly Lys Gly Leu
130 135 140
Arg Gln Gly Phe Glu Asp Ala Gly Pro Ala Arg Glu
145 150 155
<210> 69
<211> 209
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 69
Met Ser Thr Ile Asn Asn Gln Leu Lys Ala Leu Lys Val Ile Pro Val
1 5 10 15
Ile Ala Ile Asp Asn Ala Glu Asp Ile Ile Pro Leu Gly Lys Val Leu
20 25 30
Ala Glu Asn Gly Leu Pro Ala Ala Glu Ile Thr Phe Arg Ser Ser Ala
35 40 45
Ala Val Lys Ala Ile Met Leu Leu Arg Ser Ala Gln Pro Glu Met Leu
50 55 60
Ile Gly Ala Gly Thr Ile Leu Asn Gly Val Gln Ala Leu Ala Ala Lys
65 70 75 80
Glu Ala Gly Ala Thr Phe Val Val Ser Pro Gly Phe Asn Pro Asn Thr
85 90 95
Val Arg Ala Cys Gln Ile Ile Gly Ile Asp Ile Val Pro Gly Val Asn
100 105 110
Asn Pro Ser Thr Val Glu Ala Ala Leu Glu Met Gly Leu Thr Thr Leu
115 120 125
Lys Phe Phe Pro Ala Glu Ala Ser Gly Gly Ile Ser Met Val Lys Ser
130 135 140
Leu Val Gly Pro Tyr Gly Asp Ile Arg Leu Met Pro Thr Gly Gly Ile
145 150 155 160
Thr Pro Ser Asn Ile Asp Asn Tyr Leu Ala Ile Pro Gln Val Leu Ala
165 170 175
Cys Gly Gly Thr Trp Met Val Asp Lys Lys Leu Val Thr Asn Gly Glu
180 185 190
Trp Asp Glu Ile Ala Arg Leu Thr Arg Glu Ile Val Glu Gln Val Asn
195 200 205
Pro
<210> 70
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 70
Met Pro Ile Phe Thr Leu Asn Thr Asn Ile Lys Ala Thr Asp Val Pro
1 5 10 15
Ser Asp Phe Leu Ser Leu Thr Ser Arg Leu Val Gly Leu Ile Leu Ser
20 25 30
Lys Pro Gly Ser Tyr Val Ala Val His Ile Asn Thr Asp Gln Gln Leu
35 40 45
Ser Phe Gly Gly Ser Thr Asn Pro Ala Ala Phe Gly Thr Leu Met Ser
50 55 60
Ile Gly Gly Ile Glu Pro Ser Lys Asn Arg Asp His Ser Ala Val Leu
65 70 75 80
Phe Asp His Leu Asn Ala Met Leu Gly Ile Pro Lys Asn Arg Met Tyr
85 90 95
Ile His Phe Val Asn Leu Asn Gly Asp Asp Val Gly Trp Asn Gly Thr
100 105 110
Thr Phe
<210> 71
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 71
Met Asn Gln His Ser His Lys Asp Tyr Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Asp Ile Val Asp Ala Cys Val Glu Ala
20 25 30
Phe Glu Ile Ala Met Ala Ala Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Ser Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Arg Tyr Arg Asp Ser Ala Glu His His Arg
115 120 125
Phe Phe Ala Ala His Phe Ala Val Lys Gly Val Glu Ala Ala Arg Ala
130 135 140
Cys Ile Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 72
<211> 205
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 72
Met Lys Met Glu Glu Leu Phe Lys Lys His Lys Ile Val Ala Val Leu
1 5 10 15
Arg Ala Asn Ser Val Glu Glu Ala Ile Glu Lys Ala Val Ala Val Phe
20 25 30
Ala Gly Gly Val His Leu Ile Glu Ile Thr Phe Thr Val Pro Asp Ala
35 40 45
Asp Thr Val Ile Lys Ala Leu Ser Val Leu Lys Glu Lys Gly Ala Ile
50 55 60
Ile Gly Ala Gly Thr Val Thr Ser Val Glu Gln Cys Arg Lys Ala Val
65 70 75 80
Glu Ser Gly Ala Glu Phe Ile Val Ser Pro His Leu Asp Glu Glu Ile
85 90 95
Ser Gln Phe Cys Lys Glu Lys Gly Val Phe Tyr Met Pro Gly Val Met
100 105 110
Thr Pro Thr Glu Leu Val Lys Ala Met Lys Leu Gly His Thr Ile Leu
115 120 125
Lys Leu Phe Pro Gly Glu Val Val Gly Pro Gln Phe Val Lys Ala Met
130 135 140
Lys Gly Pro Phe Pro Asn Val Lys Phe Val Pro Thr Gly Gly Val Asn
145 150 155 160
Leu Asp Asn Val Cys Glu Trp Phe Lys Ala Gly Val Leu Ala Val Gly
165 170 175
Val Gly Ser Ala Leu Val Lys Gly Thr Pro Asp Glu Val Arg Glu Lys
180 185 190
Ala Lys Ala Phe Val Glu Lys Ile Arg Gly Cys Thr Glu
195 200 205
<210> 73
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 73
Met Asn Gln His Ser His Lys Asp Tyr Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Glu Ile Val Asp Ala Cys Val Ser Ala
20 25 30
Phe Glu Ala Ala Met Ala Asp Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Ser Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Arg Tyr Arg Asp Ser Asp Ala His Thr Leu
115 120 125
Leu Phe Leu Ala Leu Phe Ala Val Lys Gly Met Glu Ala Ala Arg Ala
130 135 140
Cys Val Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 74
<211> 177
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 74
Met Phe Thr Lys Ser Gly Asp Asp Gly Asn Thr Asn Val Ile Asn Lys
1 5 10 15
Arg Val Gly Lys Asp Ser Pro Leu Val Asn Phe Leu Gly Asp Leu Asp
20 25 30
Glu Leu Asn Ser Phe Ile Gly Phe Ala Ile Ser Lys Ile Pro Trp Glu
35 40 45
Asp Met Lys Lys Asp Leu Glu Arg Val Gln Val Glu Leu Phe Glu Ile
50 55 60
Gly Glu Asp Leu Ser Thr Gln Ser Ser Lys Lys Lys Ile Asp Glu Ser
65 70 75 80
Tyr Val Leu Trp Leu Leu Ala Ala Thr Ala Ile Tyr Arg Ile Glu Ser
85 90 95
Gly Pro Val Lys Leu Phe Val Ile Pro Gly Gly Ser Glu Glu Ala Ser
100 105 110
Val Leu His Val Thr Arg Ser Val Ala Arg Arg Val Glu Arg Asn Ala
115 120 125
Val Lys Tyr Thr Lys Glu Leu Pro Glu Ile Asn Arg Met Ile Ile Val
130 135 140
Tyr Leu Asn Arg Leu Ser Ser Leu Leu Phe Ala Met Ala Leu Val Ala
145 150 155 160
Asn Lys Arg Arg Asn Gln Ser Glu Lys Ile Tyr Glu Ile Gly Lys Ser
165 170 175
Trp
<210> 75
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 75
Met Asn Gln His Ser His Lys Asp Tyr Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Asp Ile Val Asp Gln Cys Val Arg Ala
20 25 30
Phe Glu Glu Ala Met Ala Asp Ala Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Ser Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Arg Tyr Arg Ser Ser Arg Glu His His Glu
115 120 125
Phe Phe Arg Glu His Phe Met Val Lys Gly Val Glu Ala Ala Ala Ala
130 135 140
Cys Ile Thr Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 76
<211> 201
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 76
Met Gly His Thr Lys Gly Pro Thr Pro Gln Gln His Asp Gly Ser Ala
1 5 10 15
Leu Arg Ile Gly Ile Val His Ala Arg Trp Asn Lys Thr Ile Ile Met
20 25 30
Pro Leu Leu Ile Gly Thr Ile Ala Lys Leu Leu Glu Cys Gly Val Lys
35 40 45
Ala Ser Asn Ile Val Val Gln Ser Val Pro Gly Ser Trp Glu Leu Pro
50 55 60
Ile Ala Val Gln Arg Leu Tyr Ser Ala Ser Gln Leu Gln Thr Pro Ser
65 70 75 80
Ser Gly Pro Ser Leu Ser Ala Gly Asp Leu Leu Gly Ser Ser Thr Thr
85 90 95
Asp Leu Thr Ala Leu Pro Thr Thr Thr Ala Ser Ser Thr Gly Pro Phe
100 105 110
Asp Ala Leu Ile Ala Ile Gly Val Leu Ile Lys Gly Glu Thr Met His
115 120 125
Phe Glu Tyr Ile Ala Asp Ser Val Ser His Gly Leu Met Arg Val Gln
130 135 140
Leu Asp Thr Gly Val Pro Val Ile Phe Gly Val Leu Thr Val Leu Thr
145 150 155 160
Asp Asp Gln Ala Lys Ala Arg Ala Gly Val Ile Glu Gly Ser His Asn
165 170 175
His Gly Glu Asp Trp Gly Leu Ala Ala Val Glu Met Gly Val Arg Arg
180 185 190
Arg Asp Trp Ala Ala Gly Lys Thr Glu
195 200
<210> 77
<211> 237
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 77
Met Tyr Glu Val Asp His Ala Asp Val Tyr Asp Leu Phe Tyr Leu Gly
1 5 10 15
Arg Gly Lys Asp Tyr Ala Ala Glu Ala Ser Asp Ile Ala Asp Leu Val
20 25 30
Arg Ser Arg Thr Pro Glu Ala Ser Ser Leu Leu Asp Val Ala Cys Gly
35 40 45
Thr Gly Thr His Leu Glu His Phe Thr Lys Glu Phe Gly Asp Thr Ala
50 55 60
Gly Leu Glu Leu Ser Glu Asp Met Leu Thr His Ala Arg Lys Arg Leu
65 70 75 80
Pro Asp Ala Thr Leu His Gln Gly Asp Met Arg Asp Phe Gln Leu Gly
85 90 95
Arg Lys Phe Ser Ala Val Val Ser Met Phe Ser Ser Val Gly Tyr Leu
100 105 110
Lys Thr Val Ala Glu Leu Gly Ala Ala Val Ala Ser Phe Ala Glu His
115 120 125
Leu Glu Pro Gly Gly Val Val Val Val Glu Pro Trp Trp Phe Pro Glu
130 135 140
Thr Phe Ala Asp Gly Trp Val Ser Ala Asp Val Val Arg Arg Asp Gly
145 150 155 160
Arg Thr Val Ala Arg Val Ser His Ser Val Arg Glu Gly Asn Ala Thr
165 170 175
Arg Met Glu Val His Phe Thr Val Ala Asp Pro Gly Lys Gly Val Arg
180 185 190
His Phe Ser Asp Val His Leu Ile Thr Leu Phe His Gln Arg Glu Tyr
195 200 205
Glu Ala Ala Phe Met Ala Ala Gly Leu Arg Val Glu Tyr Leu Glu Gly
210 215 220
Gly Pro Ser Gly Arg Gly Leu Phe Val Gly Val Pro Ala
225 230 235
<210> 78
<211> 138
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 78
Met Gly Met Lys Glu Lys Phe Val Leu Ile Ile Thr His Gly Asp Phe
1 5 10 15
Gly Lys Gly Leu Leu Ser Gly Ala Glu Val Ile Ile Gly Lys Gln Glu
20 25 30
Asn Val His Thr Val Gly Leu Asn Leu Gly Asp Asn Ile Glu Lys Val
35 40 45
Ala Lys Glu Val Met Arg Ile Ile Ile Ala Lys Leu Ala Glu Asp Lys
50 55 60
Glu Ile Ile Ile Val Val Asp Leu Phe Gly Gly Ser Pro Phe Asn Ile
65 70 75 80
Ala Leu Glu Met Met Lys Thr Phe Asp Val Lys Val Ile Thr Gly Ile
85 90 95
Asn Met Pro Met Leu Val Glu Leu Leu Thr Ser Ile Asn Val Tyr Asp
100 105 110
Thr Thr Glu Leu Leu Glu Asn Ile Ser Lys Ile Gly Lys Asp Gly Ile
115 120 125
Lys Val Ile Glu Lys Ser Ser Leu Lys Met
130 135
<210> 79
<211> 154
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 79
Met Lys Tyr Asp Gly Ser Lys Leu Arg Ile Gly Ile Leu His Ala Arg
1 5 10 15
Trp Asn Leu Glu Ile Ile Ala Ala Leu Val Ala Gly Ala Ile Lys Arg
20 25 30
Leu Gln Glu Phe Gly Val Lys Ala Glu Asn Ile Ile Ile Glu Thr Val
35 40 45
Pro Gly Ser Phe Glu Leu Pro Tyr Gly Ser Lys Leu Phe Val Glu Lys
50 55 60
Gln Lys Arg Leu Gly Lys Pro Leu Asp Ala Ile Ile Pro Ile Gly Val
65 70 75 80
Leu Ile Lys Gly Ser Thr Met His Phe Glu Tyr Ile Cys Asp Ser Thr
85 90 95
Thr His Gln Leu Met Lys Leu Asn Phe Glu Leu Gly Ile Pro Val Ile
100 105 110
Phe Gly Val Leu Thr Cys Leu Thr Asp Glu Gln Ala Glu Ala Arg Ala
115 120 125
Gly Leu Ile Glu Gly Lys Met His Asn His Gly Glu Asp Trp Gly Ala
130 135 140
Ala Ala Val Glu Met Ala Thr Lys Phe Asn
145 150
<210> 80
<211> 164
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 80
Met Ala Val Lys Gly Leu Gly Glu Val Asp Gln Lys Tyr Asp Gly Ser
1 5 10 15
Lys Leu Arg Ile Gly Ile Leu His Ala Arg Trp Asn Arg Lys Ile Ile
20 25 30
Leu Ala Leu Val Ala Gly Ala Val Leu Arg Leu Leu Glu Phe Gly Val
35 40 45
Lys Ala Glu Asn Ile Ile Ile Glu Thr Val Pro Gly Ser Phe Glu Leu
50 55 60
Pro Tyr Gly Ser Lys Leu Phe Val Glu Lys Gln Lys Arg Leu Gly Lys
65 70 75 80
Pro Leu Asp Ala Ile Ile Pro Ile Gly Val Leu Ile Lys Gly Ser Thr
85 90 95
Met His Phe Glu Tyr Ile Cys Asp Ser Thr Thr His Gln Leu Met Lys
100 105 110
Leu Asn Phe Glu Leu Gly Ile Pro Val Ile Phe Gly Val Leu Thr Cys
115 120 125
Leu Thr Asp Glu Gln Ala Glu Ala Arg Ala Gly Leu Ile Glu Gly Lys
130 135 140
Met His Asn His Gly Glu Asp Trp Gly Ala Ala Ala Val Glu Met Ala
145 150 155 160
Thr Lys Phe Asn
<210> 81
<211> 175
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 81
Met Gly Ala Asn Trp Tyr Leu Asp Asn Glu Ser Ser Arg Leu Ser Phe
1 5 10 15
Thr Ser Thr Lys Asn Ala Asp Ile Ala Glu Val His Arg Phe Leu Val
20 25 30
Leu His Gly Lys Val Asp Pro Lys Gly Leu Ala Glu Val Glu Val Glu
35 40 45
Thr Glu Ser Ile Ser Thr Gly Ile Pro Leu Arg Asp Met Leu Leu Arg
50 55 60
Val Leu Val Phe Gln Val Ser Lys Phe Pro Val Ala Gln Ile Asn Ala
65 70 75 80
Gln Leu Asp Met Arg Pro Ile Asn Asn Leu Ala Pro Gly Ala Gln Leu
85 90 95
Glu Leu Arg Leu Pro Leu Thr Val Ser Leu Arg Gly Lys Ser His Ser
100 105 110
Tyr Asn Ala Glu Leu Leu Ala Thr Arg Leu Asp Glu Arg Arg Phe Gln
115 120 125
Val Val Thr Leu Glu Pro Leu Val Ile His Ala Gln Asp Phe Asp Met
130 135 140
Val Arg Ala Phe Asn Ala Leu Arg Leu Val Ala Gly Leu Ser Ala Val
145 150 155 160
Ser Leu Ser Val Pro Val Gly Ala Val Leu Ile Phe Thr Ala Arg
165 170 175
<210> 82
<211> 208
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 82
Met Thr Asp Tyr Ile Arg Asp Gly Ser Ala Ile Lys Ala Leu Ser Phe
1 5 10 15
Ala Ile Ile Leu Ala Glu Ala Asp Leu Arg His Ile Pro Gln Asp Leu
20 25 30
Gln Arg Leu Ala Val Arg Val Ile His Ala Cys Gly Met Val Asp Val
35 40 45
Ala Asn Asp Leu Ala Phe Ser Glu Gly Ala Gly Lys Ala Gly Arg Asn
50 55 60
Ala Leu Leu Ala Gly Ala Pro Ile Leu Cys Asp Ala Arg Met Val Ala
65 70 75 80
Glu Gly Ile Thr Arg Ser Arg Leu Pro Ala Asp Asn Arg Val Ile Tyr
85 90 95
Thr Leu Ser Asp Pro Ser Val Pro Glu Leu Ala Lys Lys Ile Gly Asn
100 105 110
Thr Arg Ser Ala Ala Ala Leu Asp Leu Trp Leu Pro His Ile Glu Gly
115 120 125
Ser Ile Val Ala Ile Gly Asn Ala Pro Thr Ala Leu Phe Arg Leu Phe
130 135 140
Glu Leu Leu Asp Ala Gly Ala Pro Lys Pro Ala Leu Ile Ile Gly Met
145 150 155 160
Pro Val Gly Phe Val Gly Ala Ala Glu Ser Lys Asp Glu Leu Ala Ala
165 170 175
Asn Ser Arg Gly Val Pro Tyr Val Ile Val Arg Gly Arg Arg Gly Gly
180 185 190
Ser Ala Met Thr Ala Ala Ala Val Asn Ala Leu Ala Ser Glu Arg Glu
195 200 205
<210> 83
<211> 128
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 83
Met Ile Thr Val Phe Gly Leu Lys Ser Lys Leu Ala Pro Arg Arg Glu
1 5 10 15
Lys Leu Ala Glu Val Ile Tyr Ser Ser Leu His Leu Gly Leu Asp Ile
20 25 30
Pro Lys Gly Lys His Ala Ile Arg Phe Leu Cys Leu Glu Lys Glu Asp
35 40 45
Phe Tyr Tyr Pro Phe Asp Arg Ser Asp Asp Tyr Thr Val Ile Glu Ile
50 55 60
Asn Leu Met Ala Gly Arg Ser Glu Glu Thr Lys Met Leu Leu Ile Phe
65 70 75 80
Leu Leu Phe Ile Ala Leu Glu Arg Lys Leu Gly Ile Arg Ala His Asp
85 90 95
Val Glu Ile Thr Ile Lys Glu Gln Pro Ala His Cys Trp Gly Phe Arg
100 105 110
Gly Arg Thr Gly Asp Ser Ala Arg Asp Leu Asp Tyr Asp Ile Tyr Val
115 120 125
<210> 84
<211> 235
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 84
Met Gly Ser Asp Leu Gln Lys Leu Gln Arg Phe Ser Thr Cys Asp Ile
1 5 10 15
Ser Asp Gly Leu Leu Asn Val Tyr Asn Ile Pro Thr Gly Gly Tyr Phe
20 25 30
Pro Asn Leu Thr Ala Ile Ser Pro Pro Gln Asn Ser Ser Ile Val Gly
35 40 45
Thr Ala Tyr Thr Val Leu Phe Ala Pro Ile Asp Asp Pro Arg Pro Ala
50 55 60
Val Asn Tyr Ile Asp Ser Val Pro Pro Asn Ser Ile Leu Val Leu Ala
65 70 75 80
Leu Glu Pro His Leu Gln Ser Gln Phe His Pro Phe Ile Lys Ile Thr
85 90 95
Gln Ala Met Tyr Gly Gly Leu Met Ser Thr Arg Ala Gln Tyr Leu Lys
100 105 110
Ser Asn Gly Thr Val Val Phe Gly Arg Ile Arg Asp Val Asp Glu His
115 120 125
Arg Thr Leu Asn His Pro Val Phe Ala Tyr Gly Val Gly Ser Cys Ala
130 135 140
Pro Lys Ala Val Val Lys Ala Val Gly Thr Asn Val Gln Leu Lys Ile
145 150 155 160
Leu Thr Ser Asp Gly Val Thr Gln Thr Ile Cys Pro Gly Asp Tyr Ile
165 170 175
Ala Gly Asp Asn Asn Gly Ile Val Arg Ile Pro Val Gln Glu Thr Asp
180 185 190
Ile Ser Lys Leu Val Thr Tyr Ile Glu Lys Ser Ile Glu Val Asp Arg
195 200 205
Leu Val Ser Glu Ala Ile Lys Asn Gly Leu Pro Ala Lys Ala Ala Gln
210 215 220
Thr Ala Arg Arg Met Val Leu Lys Asp Tyr Ile
225 230 235
<210> 85
<211> 162
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 85
Met Ser Gly Met Arg Val Tyr Leu Gly Ala Asp His Ala Gly Tyr Glu
1 5 10 15
Leu Lys Gln Ala Ile Ile Ala Phe Leu Lys Met Thr Gly His Glu Pro
20 25 30
Ile Asp Cys Gly Ala Leu Arg Tyr Asp Ala Asp Asp Asp Tyr Pro Ala
35 40 45
Phe Cys Ile Ala Ala Ala Thr Arg Thr Val Ala Asp Pro Gly Ser Leu
50 55 60
Gly Ile Val Leu Gly Gly Ser Gly Asn Gly Glu Gln Ile Ala Ala Asn
65 70 75 80
Lys Val Pro Gly Ala Arg Cys Ala Leu Ala Trp Ser Val Gln Thr Ala
85 90 95
Ala Leu Ala Arg Glu His Asn Asn Ala Gln Leu Ile Gly Ile Gly Gly
100 105 110
Arg Met His Thr Leu Glu Glu Ala Leu Arg Ile Val Lys Ala Phe Val
115 120 125
Thr Thr Pro Trp Ser Lys Ala Gln Arg His Gln Arg Arg Ile Asp Ile
130 135 140
Leu Ala Glu Tyr Glu Arg Thr His Glu Ala Pro Pro Val Pro Gly Ala
145 150 155 160
Pro Ala
<210> 86
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 86
Met Gly Asp Asp Ala Arg Ile Ala Ala Ile Gly Asp Val Asp Glu Leu
1 5 10 15
Asn Ser Gln Ile Gly Val Leu Leu Ala Glu Pro Leu Pro Asp Asp Val
20 25 30
Arg Ala Ala Leu Ser Ala Ile Gln His Asp Leu Phe Asp Leu Gly Gly
35 40 45
Glu Leu Cys Ile Pro Gly His Ala Ala Ile Thr Glu Asp His Leu Leu
50 55 60
Arg Leu Ala Leu Trp Leu Val His Tyr Asn Gly Gln Leu Pro Pro Leu
65 70 75 80
Glu Glu Phe Ile Leu Pro Gly Gly Ala Arg Gly Ala Ala Leu Ala His
85 90 95
Val Cys Arg Thr Val Cys Arg Arg Ala Glu Arg Ser Ile Lys Ala Leu
100 105 110
Gly Ala Ser Glu Pro Leu Asn Ile Ala Pro Ala Ala Tyr Val Asn Leu
115 120 125
Leu Ser Asp Leu Leu Phe Val Leu Ala Arg Val Leu Asn Arg Ala Ala
130 135 140
Gly Gly Ala Asp Val Leu Trp Asp Arg Thr Arg Ala His
145 150 155
<210> 87
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 87
Met Ile Leu Ser Ala Glu Gln Ser Phe Thr Leu Arg His Pro His Gly
1 5 10 15
Gln Ala Ala Ala Leu Ala Phe Val Arg Glu Pro Ala Ala Ala Leu Ala
20 25 30
Gly Val Gln Arg Leu Arg Gly Leu Asp Ser Asp Gly Glu Gln Val Trp
35 40 45
Gly Glu Leu Leu Val Arg Val Pro Leu Leu Gly Glu Val Asp Leu Pro
50 55 60
Phe Arg Ser Glu Ile Val Arg Thr Pro Gln Gly Ala Glu Leu Arg Pro
65 70 75 80
Leu Thr Leu Thr Gly Glu Arg Ala Trp Val Ala Val Ser Gly Gln Ala
85 90 95
Thr Ala Ala Glu Gly Gly Glu Met Ala Phe Ala Phe Gln Phe Gln Ala
100 105 110
His Leu Ala Thr Pro Glu Ala Glu Gly Glu Gly Gly Ala Ala Phe Glu
115 120 125
Val Met Val Gln Ala Ala Ala Gly Val Thr Leu Leu Leu Val Ala Met
130 135 140
Ala Leu Pro Gln Gly Leu Ala Ala Gly Leu Pro Pro Ala
145 150 155
<210> 88
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 88
Met Thr Lys Lys Val Gly Ile Val Asp Thr Thr Phe Ala Arg Val Asp
1 5 10 15
Met Ala Ser Ala Ala Ile Leu Thr Leu Lys Met Glu Ser Pro Asn Ile
20 25 30
Lys Ile Ile Arg Lys Thr Val Pro Gly Ile Lys Asp Leu Pro Val Ala
35 40 45
Cys Lys Lys Leu Leu Glu Glu Glu Gly Cys Asp Ile Val Met Ala Leu
50 55 60
Gly Met Pro Gly Lys Lys Glu Lys Asp Lys Val Cys Ala His Glu Ala
65 70 75 80
Ser Leu Gly Leu Met Leu Ala Gln Leu Met Thr Asn Lys His Ile Ile
85 90 95
Glu Val Phe Val His Glu Asp Glu Ala Lys Asp Asp Ala Glu Leu Lys
100 105 110
Ile Leu Ala Ala Arg Arg Ala Ile Glu His Ala Leu Asn Val Tyr Tyr
115 120 125
Leu Leu Phe Lys Pro Glu Tyr Leu Thr Arg Met Ala Gly Lys Gly Leu
130 135 140
Arg Gln Gly Phe Glu Asp Ala Gly Pro Ala Arg Glu
145 150 155
<210> 89
<211> 209
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 89
Met Asp Asp Ile Asn Asn Gln Leu Lys Arg Leu Lys Val Ile Pro Val
1 5 10 15
Ile Ala Ile Asp Asn Ala Glu Asp Ile Ile Pro Leu Gly Lys Val Leu
20 25 30
Ala Glu Asn Gly Leu Pro Ala Ala Glu Ile Thr Phe Arg Ser Ser Ala
35 40 45
Ala Val Lys Ala Ile Met Leu Leu Arg Ser Ala Gln Pro Glu Met Leu
50 55 60
Ile Gly Ala Gly Thr Ile Leu Asn Gly Val Gln Ala Leu Ala Ala Lys
65 70 75 80
Glu Ala Gly Ala Asp Phe Val Val Ser Pro Gly Phe Asn Pro Asn Thr
85 90 95
Val Arg Ala Cys Gln Ile Ile Gly Ile Asp Ile Val Pro Gly Val Asn
100 105 110
Asn Pro Ser Thr Val Glu Gln Ala Leu Glu Met Gly Leu Thr Thr Leu
115 120 125
Lys Phe Phe Pro Ala Glu Ala Ser Gly Gly Ile Ser Met Val Lys Ser
130 135 140
Leu Val Gly Pro Tyr Gly Asp Ile Arg Leu Met Pro Thr Gly Gly Ile
145 150 155 160
Thr Pro Asp Asn Ile Asp Asn Tyr Leu Ala Ile Pro Gln Val Leu Ala
165 170 175
Cys Gly Gly Thr Trp Met Val Asp Lys Lys Leu Val Arg Asn Gly Glu
180 185 190
Trp Asp Glu Ile Ala Arg Leu Thr Arg Glu Ile Val Glu Gln Val Asn
195 200 205
Pro
<210> 90
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 90
Met Pro Ile Phe Thr Leu Asn Thr Asn Ile Lys Ala Asp Asp Val Pro
1 5 10 15
Ser Asp Phe Leu Ser Leu Thr Ser Arg Leu Val Gly Leu Ile Leu Ser
20 25 30
Lys Pro Gly Ser Tyr Val Ala Val His Ile Asn Thr Asp Gln Gln Leu
35 40 45
Ser Phe Gly Gly Ser Thr Asn Pro Ala Ala Phe Gly Thr Leu Met Ser
50 55 60
Ile Gly Gly Ile Glu Pro Asp Lys Asn Arg Asp His Ser Ala Val Leu
65 70 75 80
Phe Asp His Leu Asn Ala Met Leu Gly Ile Pro Lys Asn Arg Met Tyr
85 90 95
Ile His Phe Val Asn Leu Asn Gly Asp Asp Val Gly Trp Asn Gly Thr
100 105 110
Thr Phe
<210> 91
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 91
Met Pro Ile Phe Thr Leu Asn Thr Asn Ile Lys Ala Asp Asp Val Pro
1 5 10 15
Ser Asp Phe Leu Ser Leu Thr Ser Arg Leu Val Gly Leu Ile Leu Ser
20 25 30
Glu Pro Gly Ser Tyr Val Ala Val His Ile Asn Thr Asp Gln Gln Leu
35 40 45
Ser Phe Gly Gly Ser Thr Asn Pro Ala Ala Phe Gly Thr Leu Met Ser
50 55 60
Ile Gly Gly Ile Glu Pro Asp Lys Asn Glu Asp His Ser Ala Val Leu
65 70 75 80
Phe Asp His Leu Asn Ala Met Leu Gly Ile Pro Lys Asn Arg Met Tyr
85 90 95
Ile His Phe Val Asp Leu Asp Gly Asp Asp Val Gly Trp Asn Gly Thr
100 105 110
Thr Phe
<210> 92
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 92
Met Asn Gln His Ser His Lys Asp His Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Asp Ile Val Asp Ala Cys Val Glu Ala
20 25 30
Phe Glu Ile Ala Met Ala Ala Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Asp Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Arg Tyr Arg Asp Ser Asp Glu His His Arg
115 120 125
Phe Phe Ala Ala His Phe Ala Val Lys Gly Val Glu Ala Ala Arg Ala
130 135 140
Cys Ile Glu Ile Leu Asn Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 93
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 93
Met Asn Gln His Ser His Lys Asp His Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Asp Ile Val Asp Ala Cys Val Glu Ala
20 25 30
Phe Glu Ile Ala Met Ala Ala Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asp Gly Gly Ile Tyr Asp His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Asp Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Glu Tyr Glu Asp Ser Asp Glu Asp His Glu
115 120 125
Phe Phe Ala Ala His Phe Ala Val Lys Gly Val Glu Ala Ala Arg Ala
130 135 140
Cys Ile Glu Ile Leu Asn Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 94
<211> 205
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 94
Met Lys Met Glu Glu Leu Phe Lys Lys His Lys Ile Val Ala Val Leu
1 5 10 15
Arg Ala Asn Ser Val Glu Glu Ala Ile Glu Lys Ala Val Ala Val Phe
20 25 30
Ala Gly Gly Val His Leu Ile Glu Ile Thr Phe Thr Val Pro Asp Ala
35 40 45
Asp Thr Val Ile Lys Ala Leu Ser Val Leu Lys Glu Lys Gly Ala Ile
50 55 60
Ile Gly Ala Gly Thr Val Thr Ser Val Glu Gln Cys Arg Lys Ala Val
65 70 75 80
Glu Ser Gly Ala Glu Phe Ile Val Ser Pro His Leu Asp Glu Glu Ile
85 90 95
Ser Gln Phe Cys Lys Glu Lys Gly Val Phe Tyr Met Pro Gly Val Met
100 105 110
Thr Pro Thr Glu Leu Val Lys Ala Met Lys Leu Gly His Asp Ile Leu
115 120 125
Lys Leu Phe Pro Gly Glu Val Val Gly Pro Gln Phe Val Lys Ala Met
130 135 140
Lys Gly Pro Phe Pro Asn Val Lys Phe Val Pro Thr Gly Gly Val Asn
145 150 155 160
Leu Asp Asn Val Cys Glu Trp Phe Lys Ala Gly Val Leu Ala Val Gly
165 170 175
Val Gly Asp Ala Leu Val Lys Gly Asp Pro Asp Glu Val Arg Glu Lys
180 185 190
Ala Lys Lys Phe Val Glu Lys Ile Arg Gly Cys Thr Glu
195 200 205
<210> 95
<211> 205
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 95
Met Lys Met Glu Glu Leu Phe Lys Lys His Lys Ile Val Ala Val Leu
1 5 10 15
Arg Ala Asn Ser Val Glu Glu Ala Ile Glu Lys Ala Val Ala Val Phe
20 25 30
Ala Gly Gly Val His Leu Ile Glu Ile Thr Phe Thr Val Pro Asp Ala
35 40 45
Asp Thr Val Ile Lys Ala Leu Ser Val Leu Lys Glu Lys Gly Ala Ile
50 55 60
Ile Gly Ala Gly Thr Val Thr Ser Val Glu Gln Cys Arg Lys Ala Val
65 70 75 80
Glu Ser Gly Ala Glu Phe Ile Val Ser Pro His Leu Asp Glu Glu Ile
85 90 95
Ser Gln Phe Cys Lys Glu Lys Gly Val Phe Tyr Met Pro Gly Val Met
100 105 110
Thr Pro Thr Glu Leu Val Lys Ala Met Lys Leu Gly His Asp Ile Leu
115 120 125
Lys Leu Phe Pro Gly Glu Val Val Gly Pro Glu Phe Val Glu Ala Met
130 135 140
Lys Gly Pro Phe Pro Asn Val Lys Phe Val Pro Thr Gly Gly Val Asp
145 150 155 160
Leu Asp Asp Val Cys Glu Trp Phe Asp Ala Gly Val Leu Ala Val Gly
165 170 175
Val Gly Asp Ala Leu Val Glu Gly Asp Pro Asp Glu Val Arg Glu Asp
180 185 190
Ala Lys Glu Phe Val Glu Glu Ile Arg Gly Cys Thr Glu
195 200 205
<210> 96
<211> 205
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 96
Met Lys Met Glu Glu Leu Phe Lys Lys His Lys Ile Val Ala Val Leu
1 5 10 15
Arg Ala Asn Ser Val Glu Glu Ala Ile Glu Lys Ala Val Ala Val Phe
20 25 30
Ala Gly Gly Val His Leu Ile Glu Ile Thr Phe Thr Val Pro Asp Ala
35 40 45
Asp Thr Val Ile Lys Ala Leu Ser Val Leu Lys Glu Lys Gly Ala Ile
50 55 60
Ile Gly Ala Gly Thr Val Thr Ser Val Glu Gln Cys Arg Lys Ala Val
65 70 75 80
Glu Ser Gly Ala Glu Phe Ile Val Ser Pro His Leu Asp Glu Glu Ile
85 90 95
Ser Gln Phe Cys Lys Glu Lys Gly Val Phe Tyr Met Pro Gly Val Met
100 105 110
Thr Pro Thr Glu Leu Val Lys Ala Met Lys Leu Gly His Asp Ile Leu
115 120 125
Lys Leu Phe Pro Gly Glu Val Val Gly Pro Gln Phe Val Lys Ala Met
130 135 140
Lys Gly Pro Phe Pro Asn Val Lys Phe Val Pro Thr Gly Gly Val Asn
145 150 155 160
Leu Asp Asn Val Cys Lys Trp Phe Lys Ala Gly Val Leu Ala Val Gly
165 170 175
Val Gly Lys Ala Leu Val Lys Gly Lys Pro Asp Glu Val Arg Glu Lys
180 185 190
Ala Lys Lys Phe Val Lys Lys Ile Arg Gly Cys Thr Glu
195 200 205
<210> 97
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 97
Met Asn Gln His Ser His Lys Asp His Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Glu Ile Val Asp Ala Cys Val Ser Ala
20 25 30
Phe Glu Ala Ala Met Arg Asp Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Asp Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Arg Tyr Arg Asp Ser Asp Ala His Thr Leu
115 120 125
Leu Phe Leu Ala Leu Phe Ala Val Lys Gly Met Glu Ala Ala Arg Ala
130 135 140
Cys Val Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 98
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 98
Met Asn Gln His Ser His Lys Asp His Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Glu Ile Val Asp Ala Cys Val Ser Ala
20 25 30
Phe Glu Ala Ala Met Arg Asp Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asp Gly Gly Ile Tyr Asp His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Asp Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Glu Tyr Glu Asp Ser Asp Ala Asp Thr Leu
115 120 125
Leu Phe Leu Ala Leu Phe Ala Val Lys Gly Met Glu Ala Ala Arg Ala
130 135 140
Cys Val Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 99
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 99
Met Asn Gln His Ser His Lys Asp His Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Glu Ile Val Asp Ala Cys Val Ser Ala
20 25 30
Phe Glu Ala Ala Met Arg Asp Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Asn Gly Gly Ile Tyr Arg His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asn Gly Met Met Asn Val Gln Leu Asn Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Asn Tyr Asp Lys Ser Lys Ala His Thr Leu
115 120 125
Leu Phe Leu Ala Leu Phe Ala Val Lys Gly Met Glu Ala Ala Arg Ala
130 135 140
Cys Val Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 100
<211> 156
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (70)..(70)
<223> Xaa представляет собой A или K
<400> 100
Met Thr Lys Lys Val Gly Ile Val Asp Thr Thr Phe Ala Arg Val Asp
1 5 10 15
Met Ala Ser Ala Ala Ile Leu Thr Leu Lys Met Glu Ser Pro Asn Ile
20 25 30
Lys Ile Ile Arg Lys Thr Val Pro Gly Ile Lys Asp Leu Pro Val Ala
35 40 45
Cys Lys Lys Leu Leu Glu Glu Glu Gly Cys Asp Ile Val Met Ala Leu
50 55 60
Gly Met Pro Gly Lys Xaa Glu Lys Asp Lys Val Cys Ala His Glu Ala
65 70 75 80
Ser Leu Gly Leu Met Leu Ala Gln Leu Met Thr Asn Lys His Ile Ile
85 90 95
Glu Val Phe Val His Glu Asp Glu Ala Lys Asp Asp Ala Glu Leu Lys
100 105 110
Ile Leu Ala Ala Arg Arg Ala Ile Glu His Ala Leu Asn Val Tyr Tyr
115 120 125
Leu Leu Phe Lys Pro Glu Tyr Leu Thr Arg Met Ala Gly Lys Gly Leu
130 135 140
Arg Gln Gly Phe Glu Asp Ala Gly Pro Ala Arg Glu
145 150 155
<210> 101
<211> 209
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (2)..(2)
<223> Xaa представляет собой S или D
<220>
<221> MISC_FEATURE
<222> (3)..(3)
<223> Xaa представляет собой T или D
<220>
<221> MISC_FEATURE
<222> (10)..(10)
<223> Xaa представляет собой A или R
<220>
<221> MISC_FEATURE
<222> (85)..(85)
<223> Xaa представляет собой T или D
<220>
<221> MISC_FEATURE
<222> (119)..(119)
<223> Xaa представляет собой A или Q
<220>
<221> MISC_FEATURE
<222> (163)..(163)
<223> Xaa представляет собой S или D
<220>
<221> MISC_FEATURE
<222> (189)..(189)
<223> Xaa представляет собой T или R
<400> 101
Met Xaa Xaa Ile Asn Asn Gln Leu Lys Xaa Leu Lys Val Ile Pro Val
1 5 10 15
Ile Ala Ile Asp Asn Ala Glu Asp Ile Ile Pro Leu Gly Lys Val Leu
20 25 30
Ala Glu Asn Gly Leu Pro Ala Ala Glu Ile Thr Phe Arg Ser Ser Ala
35 40 45
Ala Val Lys Ala Ile Met Leu Leu Arg Ser Ala Gln Pro Glu Met Leu
50 55 60
Ile Gly Ala Gly Thr Ile Leu Asn Gly Val Gln Ala Leu Ala Ala Lys
65 70 75 80
Glu Ala Gly Ala Xaa Phe Val Val Ser Pro Gly Phe Asn Pro Asn Thr
85 90 95
Val Arg Ala Cys Gln Ile Ile Gly Ile Asp Ile Val Pro Gly Val Asn
100 105 110
Asn Pro Ser Thr Val Glu Xaa Ala Leu Glu Met Gly Leu Thr Thr Leu
115 120 125
Lys Phe Phe Pro Ala Glu Ala Ser Gly Gly Ile Ser Met Val Lys Ser
130 135 140
Leu Val Gly Pro Tyr Gly Asp Ile Arg Leu Met Pro Thr Gly Gly Ile
145 150 155 160
Thr Pro Xaa Asn Ile Asp Asn Tyr Leu Ala Ile Pro Gln Val Leu Ala
165 170 175
Cys Gly Gly Thr Trp Met Val Asp Lys Lys Leu Val Xaa Asn Gly Glu
180 185 190
Trp Asp Glu Ile Ala Arg Leu Thr Arg Glu Ile Val Glu Gln Val Asn
195 200 205
Pro
<210> 102
<211> 114
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (13)..(13)
<223> Xaa представляет собой T или D
<220>
<221> MISC_FEATURE
<222> (33)..(33)
<223> Xaa представляет собой K или E
<220>
<221> MISC_FEATURE
<222> (71)..(71)
<223> Xaa представляет собой S или D
<220>
<221> MISC_FEATURE
<222> (74)..(74)
<223> Xaa представляет собой R или E
<220>
<221> MISC_FEATURE
<222> (101)..(101)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (103)..(103)
<223> Xaa представляет собой N или D
<400> 102
Met Pro Ile Phe Thr Leu Asn Thr Asn Ile Lys Ala Xaa Asp Val Pro
1 5 10 15
Ser Asp Phe Leu Ser Leu Thr Ser Arg Leu Val Gly Leu Ile Leu Ser
20 25 30
Xaa Pro Gly Ser Tyr Val Ala Val His Ile Asn Thr Asp Gln Gln Leu
35 40 45
Ser Phe Gly Gly Ser Thr Asn Pro Ala Ala Phe Gly Thr Leu Met Ser
50 55 60
Ile Gly Gly Ile Glu Pro Xaa Lys Asn Xaa Asp His Ser Ala Val Leu
65 70 75 80
Phe Asp His Leu Asn Ala Met Leu Gly Ile Pro Lys Asn Arg Met Tyr
85 90 95
Ile His Phe Val Xaa Leu Xaa Gly Asp Asp Val Gly Trp Asn Gly Thr
100 105 110
Thr Phe
<210> 103
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa представляет собой Y или H
<220>
<221> MISC_FEATURE
<222> (82)..(82)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (87)..(87)
<223> Xaa представляет собой R или D
<220>
<221> MISC_FEATURE
<222> (105)..(105)
<223> Xaa представляет собой S или D
<220>
<221> MISC_FEATURE
<222> (119)..(119)
<223> Xaa представляет собой R или E
<220>
<221> MISC_FEATURE
<222> (121)..(121)
<223> Xaa представляет собой R или E
<220>
<221> MISC_FEATURE
<222> (124)..(124)
<223> Xaa представляет собой A или D
<220>
<221> MISC_FEATURE
<222> (126)..(126)
<223> Xaa представляет собой H или D
<220>
<221> MISC_FEATURE
<222> (128)..(128)
<223> Xaa представляет собой R или E
<220>
<221> MISC_FEATURE
<222> (150)..(150)
<223> Xaa представляет собой A или N
<400> 103
Met Asn Gln His Ser His Lys Asp Xaa Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Asp Ile Val Asp Ala Cys Val Glu Ala
20 25 30
Phe Glu Ile Ala Met Ala Ala Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Xaa Gly Gly Ile Tyr Xaa His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Asp Gly Met Met Asn Val Gln Leu Xaa Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Xaa Tyr Xaa Asp Ser Xaa Glu Xaa His Xaa
115 120 125
Phe Phe Ala Ala His Phe Ala Val Lys Gly Val Glu Ala Ala Arg Ala
130 135 140
Cys Ile Glu Ile Leu Xaa Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 104
<211> 205
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (126)..(126)
<223> Xaa представляет собой T или D
<220>
<221> MISC_FEATURE
<222> (139)..(139)
<223> Xaa представляет собой Q или E
<220>
<221> MISC_FEATURE
<222> (142)..(142)
<223> Xaa представляет собой K или E
<220>
<221> MISC_FEATURE
<222> (160)..(160)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (163)..(163)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (166)..(166)
<223> Xaa представляет собой E или K
<220>
<221> MISC_FEATURE
<222> (169)..(169)
<223> Xaa представляет собой D или K
<220>
<221> MISC_FEATURE
<222> (179)..(179)
<223> Xaa представляет собой S, D или K
<220>
<221> MISC_FEATURE
<222> (183)..(183)
<223> Xaa представляет собой K или E
<220>
<221> MISC_FEATURE
<222> (185)..(185)
<223> Xaa представляет собой T, D или K
<220>
<221> MISC_FEATURE
<222> (192)..(192)
<223> Xaa представляет собой D или K
<220>
<221> MISC_FEATURE
<222> (195)..(195)
<223> Xaa представляет собой A, E или K
<220>
<221> MISC_FEATURE
<222> (198)..(198)
<223> Xaa представляет собой E или K
<220>
<221> MISC_FEATURE
<222> (199)..(199)
<223> Xaa представляет собой E или K
<400> 104
Met Lys Met Glu Glu Leu Phe Lys Lys His Lys Ile Val Ala Val Leu
1 5 10 15
Arg Ala Asn Ser Val Glu Glu Ala Ile Glu Lys Ala Val Ala Val Phe
20 25 30
Ala Gly Gly Val His Leu Ile Glu Ile Thr Phe Thr Val Pro Asp Ala
35 40 45
Asp Thr Val Ile Lys Ala Leu Ser Val Leu Lys Glu Lys Gly Ala Ile
50 55 60
Ile Gly Ala Gly Thr Val Thr Ser Val Glu Gln Cys Arg Lys Ala Val
65 70 75 80
Glu Ser Gly Ala Glu Phe Ile Val Ser Pro His Leu Asp Glu Glu Ile
85 90 95
Ser Gln Phe Cys Lys Glu Lys Gly Val Phe Tyr Met Pro Gly Val Met
100 105 110
Thr Pro Thr Glu Leu Val Lys Ala Met Lys Leu Gly His Xaa Ile Leu
115 120 125
Lys Leu Phe Pro Gly Glu Val Val Gly Pro Xaa Phe Val Xaa Ala Met
130 135 140
Lys Gly Pro Phe Pro Asn Val Lys Phe Val Pro Thr Gly Gly Val Xaa
145 150 155 160
Leu Asp Xaa Val Cys Xaa Trp Phe Xaa Ala Gly Val Leu Ala Val Gly
165 170 175
Val Gly Xaa Ala Leu Val Xaa Gly Xaa Pro Asp Glu Val Arg Glu Xaa
180 185 190
Ala Lys Xaa Phe Val Xaa Xaa Ile Arg Gly Cys Thr Glu
195 200 205
<210> 105
<211> 157
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<220>
<221> MISC_FEATURE
<222> (9)..(9)
<223> Xaa представляет собой Y или H
<220>
<221> MISC_FEATURE
<222> (38)..(38)
<223> Xaa представляет собой A или R
<220>
<221> MISC_FEATURE
<222> (82)..(82)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (87)..(87)
<223> Xaa представляет собой R или D
<220>
<221> MISC_FEATURE
<222> (97)..(97)
<223> Xaa представляет собой N или D
<220>
<221> MISC_FEATURE
<222> (105)..(105)
<223> Xaa представляет собой S, N или D
<220>
<221> MISC_FEATURE
<222> (119)..(119)
<223> Xaa представляет собой R, E или N
<220>
<221> MISC_FEATURE
<222> (121)..(121)
<223> Xaa представляет собой R, E или D
<220>
<221> MISC_FEATURE
<222> (122)..(122)
<223> Xaa представляет собой K или D
<220>
<221> MISC_FEATURE
<222> (124)..(124)
<223> Xaa представляет собой K или D
<220>
<221> MISC_FEATURE
<222> (126)..(126)
<223> Xaa представляет собой H или D
<400> 105
Met Asn Gln His Ser His Lys Asp Xaa Glu Thr Val Arg Ile Ala Val
1 5 10 15
Val Arg Ala Arg Trp His Ala Glu Ile Val Asp Ala Cys Val Ser Ala
20 25 30
Phe Glu Ala Ala Met Xaa Asp Ile Gly Gly Asp Arg Phe Ala Val Asp
35 40 45
Val Phe Asp Val Pro Gly Ala Tyr Glu Ile Pro Leu His Ala Arg Thr
50 55 60
Leu Ala Glu Thr Gly Arg Tyr Gly Ala Val Leu Gly Thr Ala Phe Val
65 70 75 80
Val Xaa Gly Gly Ile Tyr Xaa His Glu Phe Val Ala Ser Ala Val Ile
85 90 95
Xaa Gly Met Met Asn Val Gln Leu Xaa Thr Gly Val Pro Val Leu Ser
100 105 110
Ala Val Leu Thr Pro His Xaa Tyr Xaa Xaa Ser Xaa Ala Xaa Thr Leu
115 120 125
Leu Phe Leu Ala Leu Phe Ala Val Lys Gly Met Glu Ala Ala Arg Ala
130 135 140
Cys Val Glu Ile Leu Ala Ala Arg Glu Lys Ile Ala Ala
145 150 155
<210> 106
<211> 142
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 106
taatgcttaa gtcgaacaga aagtaatcgt attgtacacg gccgcataat cgaaattaat 60
acgactcact ataggggaat tgtgagcgga taacaattcc ccatcttagt atattagtta 120
agtataagaa ggagatatac tt 142
<210> 107
<211> 18
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 107
taaagaagga gatatcat 18
<210> 108
<211> 17
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 108
tgagaaggag atatcat 17
<210> 109
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 109
ttgaaagtga tggtttggta agaaat 26
<210> 110
<211> 25
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 110
tgcagcacgt atgataactt caaag 25
<210> 111
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 111
aggatatttt acccagcagt gccccgt 27
<210> 112
<211> 325
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 112
Met Arg Val Glu Asn Asn Asn Val Ser Gly Gln Asn His Asp Pro Glu
1 5 10 15
Gln Ile Asp Leu Ile Asp Leu Leu Val Gln Leu Trp Arg Gly Lys Met
20 25 30
Thr Ile Ile Ile Ser Val Ile Val Ala Ile Ala Leu Ala Ile Gly Tyr
35 40 45
Leu Ala Val Ala Lys Glu Lys Trp Thr Ser Thr Ala Ile Ile Thr Gln
50 55 60
Pro Asp Val Gly Gln Ile Ala Gly Tyr Asn Asn Ala Met Asn Val Ile
65 70 75 80
Tyr Gly Gln Ala Ala Pro Lys Val Ser Asp Leu Gln Glu Thr Leu Ile
85 90 95
Gly Arg Phe Ser Ser Ala Phe Ser Ala Leu Ala Glu Thr Leu Asp Asn
100 105 110
Gln Glu Glu Pro Glu Lys Leu Thr Ile Glu Pro Ser Val Lys Asn Gln
115 120 125
Gln Leu Pro Leu Thr Val Ser Tyr Val Gly Gln Thr Ala Glu Gly Ala
130 135 140
Gln Met Lys Leu Ala Gln Tyr Ile Gln Gln Val Asp Asp Lys Val Asn
145 150 155 160
Gln Glu Leu Glu Lys Asp Leu Lys Asp Asn Ile Ala Leu Gly Arg Lys
165 170 175
Asn Leu Gln Asp Ser Leu Arg Thr Gln Glu Val Val Ala Gln Glu Gln
180 185 190
Lys Asp Leu Arg Ile Arg Gln Ile Gln Glu Ala Leu Gln Tyr Ala Asn
195 200 205
Gln Glu Gln Val Thr Lys Pro Gln Val Gln Gln Thr Glu Asp Val Thr
210 215 220
Gln Asp Thr Leu Phe Leu Leu Gly Ser Glu Ala Leu Glu Ser Met Ile
225 230 235 240
Lys His Glu Ala Thr Arg Pro Leu Val Phe Ser Ser Asn Tyr Tyr Gln
245 250 255
Thr Arg Gln Asn Leu Leu Asp Ile Glu Ser Leu Lys Val Asp Asp Leu
260 265 270
Asp Ile His Ala Tyr Arg Tyr Val Met Lys Pro Thr Leu Pro Ile Arg
275 280 285
Arg Asp Ser Pro Lys Lys Ala Ile Thr Leu Ile Leu Ala Val Leu Leu
290 295 300
Gly Gly Met Val Gly Ala Gly Ile Val Leu Gly Arg Asn Ala Leu Arg
305 310 315 320
Asn Tyr Asn Ala Lys
325
<210> 113
<211> 326
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 113
Met Arg Val Glu Asn Asn Asn Val Ser Gly Gln Asn Asn Asp Pro Glu
1 5 10 15
Gln Ile Asp Leu Ile Asp Leu Leu Val Gln Leu Trp Arg Gly Lys Met
20 25 30
Thr Ile Ile Ile Ser Val Ile Val Ala Ile Ala Leu Ala Ile Gly Tyr
35 40 45
Leu Ala Val Ala Lys Glu Lys Trp Thr Ser Thr Ala Ile Ile Thr Gln
50 55 60
Pro Asp Val Gly Gln Ile Ala Gly Tyr Asn Asn Ala Met Asn Val Ile
65 70 75 80
Tyr Gly Gln Ala Ala Pro Lys Val Ser Asp Leu Gln Glu Thr Leu Ile
85 90 95
Gly Arg Phe Ser Ser Ala Phe Ser Ala Leu Ala Glu Thr Leu Asp Asn
100 105 110
Gln Asp Glu Pro Glu Lys Leu Thr Ile Glu Pro Ser Val Lys Asn Gln
115 120 125
Gln Leu Pro Leu Thr Val Ser Tyr Val Gly Gln Thr Ala Glu Gly Ala
130 135 140
Gln Met Lys Leu Ala Gln Tyr Ile Gln Gln Val Asp Asp Lys Val Asn
145 150 155 160
Gln Glu Leu Glu Lys Asp Leu Lys Asp Asn Ile Ala Leu Gly Arg Lys
165 170 175
Asn Leu Gln Asp Ser Leu Arg Thr Gln Glu Val Val Ala Gln Glu Gln
180 185 190
Lys Asp Leu Arg Ile Arg Gln Ile Gln Glu Ala Leu Gln Tyr Ala Asn
195 200 205
Gln Ala Gln Val Thr Lys Pro Gln Ile Gln Gln Thr Gly Glu Asp Ile
210 215 220
Thr Gln Asp Thr Leu Phe Leu Leu Gly Ser Glu Ala Leu Glu Ser Met
225 230 235 240
Ile Lys His Glu Ala Thr Arg Pro Leu Val Phe Ser Pro Asn Tyr Tyr
245 250 255
Gln Thr Arg Gln Asn Leu Leu Asp Ile Glu Ser Leu Lys Val Asp Asp
260 265 270
Leu Asp Ile His Ala Tyr Arg Tyr Val Met Lys Pro Thr Leu Pro Ile
275 280 285
Arg Arg Asp Ser Pro Lys Lys Ala Ile Thr Leu Ile Leu Ala Val Leu
290 295 300
Leu Gly Gly Met Val Gly Ala Gly Ile Val Leu Gly Arg Asn Ala Leu
305 310 315 320
Arg Asn Tyr Asn Ala Lys
325
<210> 114
<211> 326
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 114
Met Arg Val Glu Asn Asn Asn Val Ser Gly Gln Asn His Asp Pro Glu
1 5 10 15
Gln Ile Asp Leu Ile Asp Leu Leu Val Gln Leu Trp Arg Gly Lys Met
20 25 30
Thr Ile Ile Ile Ser Val Val Val Ala Ile Ala Leu Ala Ile Gly Tyr
35 40 45
Leu Ala Val Ala Lys Glu Lys Trp Thr Ser Thr Ala Ile Ile Thr Gln
50 55 60
Pro Asp Val Gly Gln Ile Ala Gly Tyr Asn Asn Ala Met Asn Val Ile
65 70 75 80
Tyr Gly Gln Ala Ala Pro Lys Val Ser Asp Leu Gln Glu Thr Leu Ile
85 90 95
Gly Arg Phe Ser Phe Ala Phe Ser Ala Leu Ala Glu Thr Leu Asp Asn
100 105 110
Gln Lys Glu Pro Glu Lys Leu Thr Ile Glu Pro Ser Val Lys Asn Gln
115 120 125
Gln Leu Pro Leu Thr Val Ser Tyr Val Gly Gln Thr Ala Glu Asp Ala
130 135 140
Gln Met Lys Leu Ala Gln Tyr Ile Gln Gln Val Asp Asp Lys Val Asn
145 150 155 160
Gln Glu Leu Glu Lys Asp Leu Lys Asp Asn Leu Ala Leu Gly Arg Lys
165 170 175
Asn Leu Gln Asp Ser Leu Arg Thr Gln Glu Val Val Ala Gln Glu Gln
180 185 190
Lys Asp Leu Arg Ile Arg Gln Ile Gln Glu Ala Leu Gln Tyr Ala Asn
195 200 205
Gln Ala Gln Val Thr Lys Pro Gln Ile Gln Gln Thr Gly Glu Asp Ile
210 215 220
Thr Gln Asp Thr Leu Phe Leu Leu Gly Ser Glu Ala Leu Glu Ser Met
225 230 235 240
Ile Lys His Glu Ala Thr Arg Pro Leu Val Phe Ser Pro Asn Tyr Tyr
245 250 255
Gln Thr Arg Gln Asn Leu Leu Asp Ile Glu Asn Leu Lys Val Asp Asp
260 265 270
Leu Asp Ile His Ala Tyr Arg Tyr Val Met Lys Pro Thr Leu Pro Ile
275 280 285
Arg Arg Asp Ser Pro Lys Lys Ala Ile Thr Leu Ile Leu Ala Val Leu
290 295 300
Leu Gly Gly Met Val Gly Ala Gly Ile Val Leu Gly Arg Asn Ala Leu
305 310 315 320
Arg Asn Tyr Asn Ser Lys
325
<210> 115
<211> 326
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 115
Met Arg Val Glu Asn Asn Asn Val Ser Gly Gln Asn His Asp Pro Glu
1 5 10 15
Gln Ile Asp Leu Ile Asp Leu Leu Val Gln Leu Trp Arg Gly Lys Met
20 25 30
Thr Ile Ile Ile Ser Val Ile Val Ala Ile Ala Leu Ala Ile Gly Tyr
35 40 45
Leu Ala Val Ala Lys Glu Lys Trp Thr Ser Thr Ala Ile Ile Thr Gln
50 55 60
Pro Asp Val Gly Gln Ile Ala Gly Tyr Asn Asn Ala Met Asn Val Ile
65 70 75 80
Tyr Gly Gln Ala Ala Pro Lys Val Ser Asp Leu Gln Glu Thr Leu Ile
85 90 95
Gly Arg Phe Ser Ser Ala Phe Ser Ala Leu Ala Glu Thr Leu Asp Asn
100 105 110
Gln Glu Glu Arg Glu Lys Leu Thr Ile Glu Pro Ser Val Lys Asn Gln
115 120 125
Gln Leu Pro Leu Thr Val Ser Tyr Val Gly Gln Thr Ala Glu Gly Ala
130 135 140
Gln Met Lys Leu Ala Gln Tyr Ile Gln Gln Val Asp Asp Lys Val Asn
145 150 155 160
Gln Glu Leu Glu Lys Asp Leu Lys Asp Asn Ile Ala Leu Gly Arg Lys
165 170 175
Asn Leu Gln Asp Ser Leu Arg Thr Gln Glu Val Val Ala Gln Glu Gln
180 185 190
Lys Asp Leu Arg Ile Arg Gln Ile Gln Glu Ala Leu Gln Tyr Ala Asn
195 200 205
Gln Ala Gln Val Thr Lys Pro Gln Ile Gln Gln Thr Gly Glu Asp Ile
210 215 220
Thr Gln Asp Thr Leu Phe Leu Leu Gly Ser Glu Ala Leu Glu Ser Met
225 230 235 240
Ile Lys His Glu Ala Thr Arg Pro Leu Val Phe Ser Pro Asn Tyr Tyr
245 250 255
Gln Thr Arg Gln Asn Leu Leu Asp Ile Glu Ser Leu Lys Val Asp Asp
260 265 270
Leu Asp Ile His Ala Tyr Arg Tyr Val Met Lys Pro Met Leu Pro Ile
275 280 285
Arg Arg Asp Ser Pro Lys Lys Ala Ile Thr Leu Ile Leu Ala Val Leu
290 295 300
Leu Gly Gly Met Val Gly Ala Gly Ile Val Leu Gly Arg Asn Ala Leu
305 310 315 320
Arg Asn Tyr Asn Ala Lys
325
<210> 116
<211> 327
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 116
Met Thr Val Asp Ser Asn Thr Ser Ser Gly Arg Gly Asn Asp Pro Glu
1 5 10 15
Gln Ile Asp Leu Ile Glu Leu Leu Leu Gln Leu Trp Arg Gly Lys Met
20 25 30
Thr Ile Ile Val Ala Val Ile Ile Ala Ile Leu Leu Ala Val Gly Tyr
35 40 45
Leu Met Ile Ala Lys Glu Lys Trp Thr Ser Thr Ala Ile Ile Thr Gln
50 55 60
Pro Asp Ala Ala Gln Val Ala Thr Tyr Thr Asn Ala Leu Asn Val Leu
65 70 75 80
Tyr Gly Gly Asn Ala Pro Lys Ile Ser Glu Val Gln Ala Asn Phe Ile
85 90 95
Ser Arg Phe Ser Ser Ala Phe Ser Ala Leu Ser Glu Val Leu Asp Asn
100 105 110
Gln Lys Glu Arg Glu Lys Leu Thr Ile Glu Gln Ser Val Lys Gly Gln
115 120 125
Ala Leu Pro Leu Ser Val Ser Tyr Val Ser Thr Thr Ala Glu Gly Ala
130 135 140
Gln Arg Arg Leu Ala Glu Tyr Ile Gln Gln Val Asp Glu Glu Val Ala
145 150 155 160
Lys Glu Leu Glu Val Asp Leu Lys Asp Asn Ile Thr Leu Gln Thr Lys
165 170 175
Thr Leu Gln Glu Ser Leu Glu Thr Gln Glu Val Val Ala Gln Glu Gln
180 185 190
Lys Asp Leu Arg Ile Lys Gln Ile Glu Glu Ala Leu Arg Tyr Ala Asp
195 200 205
Glu Ala Lys Ile Thr Gln Pro Gln Ile Gln Gln Thr Gln Asp Val Thr
210 215 220
Gln Asp Thr Met Phe Leu Leu Gly Ser Asp Ala Leu Lys Ser Met Ile
225 230 235 240
Gln Asn Glu Ala Thr Arg Pro Leu Val Phe Ser Pro Ala Tyr Tyr Gln
245 250 255
Thr Lys Gln Thr Leu Leu Asp Ile Lys Asn Leu Lys Val Thr Ala Asp
260 265 270
Thr Val His Val Tyr Arg Tyr Val Met Lys Pro Thr Leu Pro Val Arg
275 280 285
Arg Asp Ser Pro Lys Thr Ala Ile Thr Leu Val Leu Ala Val Leu Leu
290 295 300
Gly Gly Met Ile Gly Ala Gly Ile Val Leu Gly Arg Asn Ala Leu Arg
305 310 315 320
Ser Tyr Lys Pro Lys Ala Leu
325
<210> 117
<211> 377
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 117
Met Ser Ser Leu Asn Ile Lys Gln Gly Ser Asp Ala His Phe Pro Asp
1 5 10 15
Tyr Pro Leu Ala Ser Pro Ser Asn Asn Glu Ile Asp Leu Leu Asn Leu
20 25 30
Ile Ser Val Leu Trp Arg Ala Lys Lys Thr Val Met Ala Val Val Phe
35 40 45
Ala Phe Ala Cys Ala Gly Leu Leu Ile Ser Phe Ile Leu Pro Gln Lys
50 55 60
Trp Thr Ser Ala Ala Val Val Thr Pro Pro Glu Pro Val Gln Trp Gln
65 70 75 80
Glu Leu Glu Lys Ser Phe Thr Lys Leu Arg Val Leu Asp Leu Asp Ile
85 90 95
Lys Ile Asp Arg Thr Glu Ala Phe Asn Leu Phe Ile Lys Lys Phe Gln
100 105 110
Ser Val Ser Leu Leu Glu Glu Tyr Leu Arg Ser Ser Pro Tyr Val Met
115 120 125
Asp Gln Leu Lys Glu Ala Lys Ile Asp Glu Leu Asp Leu His Arg Ala
130 135 140
Ile Val Ala Leu Ser Glu Lys Met Lys Ala Val Asp Asp Asn Ala Ser
145 150 155 160
Lys Lys Lys Asp Glu Pro Ser Leu Tyr Thr Ser Trp Thr Leu Ser Phe
165 170 175
Thr Ala Pro Thr Ser Glu Glu Ala Gln Thr Val Leu Ser Gly Tyr Ile
180 185 190
Asp Tyr Ile Ser Thr Leu Val Val Lys Glu Ser Leu Glu Asn Val Arg
195 200 205
Asn Lys Leu Glu Ile Lys Thr Gln Phe Glu Lys Glu Lys Leu Ala Gln
210 215 220
Asp Arg Ile Lys Thr Lys Asn Gln Leu Asp Ala Asn Ile Gln Arg Leu
225 230 235 240
Asn Tyr Ser Leu Asp Ile Ala Asn Ala Ala Gly Ile Lys Lys Pro Val
245 250 255
Tyr Ser Asn Gly Gln Ala Val Lys Asp Asp Pro Asp Phe Ser Ile Ser
260 265 270
Leu Gly Ala Asp Gly Ile Glu Arg Lys Leu Glu Ile Glu Lys Ala Val
275 280 285
Thr Asp Val Ala Glu Leu Asn Gly Glu Leu Arg Asn Arg Gln Tyr Leu
290 295 300
Val Glu Gln Leu Thr Lys Ala His Val Asn Asp Val Asn Phe Thr Pro
305 310 315 320
Phe Lys Tyr Gln Leu Ser Pro Ser Leu Pro Val Lys Lys Asp Gly Pro
325 330 335
Gly Lys Ala Ile Ile Val Ile Leu Ser Ala Leu Ile Gly Gly Met Val
340 345 350
Ala Cys Gly Gly Val Leu Leu Arg Tyr Ala Met Ala Ser Arg Lys Gln
355 360 365
Asp Ala Met Met Ala Asp His Leu Val
370 375
<210> 118
<211> 377
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 118
Met Ser Ser Leu Asn Ile Lys Gln Gly Ser Glu Ala His Phe Pro Glu
1 5 10 15
Tyr Pro Leu Ala Ser Pro Ser Asn Asn Glu Ile Asp Leu Leu Asn Leu
20 25 30
Ile Glu Val Leu Trp Arg Ala Lys Lys Thr Val Met Ala Val Val Phe
35 40 45
Ala Phe Ala Cys Ala Gly Leu Leu Ile Ser Phe Ile Leu Pro Gln Lys
50 55 60
Trp Thr Ser Ala Ala Val Val Thr Pro Pro Glu Pro Val Gln Trp Gln
65 70 75 80
Glu Leu Glu Lys Thr Phe Thr Lys Leu Arg Val Leu Asp Leu Asp Ile
85 90 95
Lys Ile Asp Arg Thr Glu Ala Phe Asn Leu Phe Ile Lys Lys Phe Gln
100 105 110
Ser Val Ser Leu Leu Glu Glu Tyr Leu Arg Ser Ser Pro Tyr Val Met
115 120 125
Asp Gln Leu Lys Glu Ala Lys Ile Asp Pro Leu Asp Leu His Arg Ala
130 135 140
Ile Val Ala Leu Ser Glu Lys Met Lys Ala Val Asp Asp Asn Ala Ser
145 150 155 160
Lys Lys Lys Asp Glu Ser Ala Leu Tyr Thr Ser Trp Thr Leu Ser Phe
165 170 175
Thr Ala Pro Thr Ser Glu Glu Ala Gln Lys Val Leu Ala Gly Tyr Ile
180 185 190
Asp Tyr Ile Ser Ala Leu Val Val Lys Glu Ser Ile Glu Asn Val Arg
195 200 205
Asn Lys Leu Glu Ile Lys Thr Gln Phe Glu Lys Glu Lys Leu Ala Gln
210 215 220
Asp Arg Ile Lys Thr Lys Asn Gln Leu Asp Ala Asn Ile Gln Arg Leu
225 230 235 240
Asn Tyr Ser Leu Asp Ile Ala Asn Ala Ala Gly Ile Lys Lys Pro Val
245 250 255
Tyr Ser Asn Gly Gln Ala Val Lys Asp Asp Pro Asp Phe Ser Ile Ser
260 265 270
Leu Gly Ala Asp Gly Ile Glu Arg Lys Leu Glu Ile Glu Lys Ala Val
275 280 285
Thr Asp Val Ala Glu Leu Asn Gly Glu Leu Arg Asn Arg Gln Tyr Leu
290 295 300
Val Glu Gln Leu Thr Lys Thr Asn Ile Asn Asp Val Asn Phe Thr Pro
305 310 315 320
Phe Lys Tyr Gln Leu Arg Pro Ser Leu Pro Val Lys Lys Asp Gly Gln
325 330 335
Gly Lys Ala Ile Ile Val Ile Leu Ser Ala Leu Val Gly Gly Met Val
340 345 350
Ala Cys Gly Gly Val Leu Leu Arg His Ala Met Ala Ser Arg Lys Gln
355 360 365
Asp Ala Met Met Ala Asp His Leu Val
370 375
<210> 119
<211> 377
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 119
Met Ser Ser Leu Asn Ile Lys Gln Gly Ser Asp Ala His Phe Pro Asp
1 5 10 15
Tyr Pro Leu Ala Ser Pro Ser Asn Asn Glu Ile Asp Leu Leu Asn Leu
20 25 30
Ile Ser Val Leu Trp Arg Ala Lys Lys Thr Val Met Ala Val Val Phe
35 40 45
Ala Phe Ala Cys Ala Gly Leu Leu Ile Ser Phe Ile Leu Pro Gln Lys
50 55 60
Trp Thr Ser Ala Ala Val Val Thr Pro Pro Glu Pro Val Gln Trp Gln
65 70 75 80
Glu Leu Glu Lys Ser Phe Thr Lys Leu Arg Val Leu Asp Leu Asp Ile
85 90 95
Lys Ile Asp Arg Thr Glu Ala Phe Asn Leu Phe Ile Lys Lys Phe Gln
100 105 110
Ser Val Ser Leu Leu Glu Glu Tyr Leu Arg Ser Ser Pro Tyr Val Met
115 120 125
Asp Gln Leu Lys Glu Ala Lys Ile Asp Glu Leu Asp Leu His Arg Ala
130 135 140
Ile Val Ala Leu Ser Glu Lys Met Lys Ala Val Asp Asp Asn Ala Ser
145 150 155 160
Lys Lys Lys Asp Glu Pro Ser Leu Tyr Thr Ser Trp Thr Leu Ser Phe
165 170 175
Thr Ala Pro Thr Ser Glu Glu Ala Gln Thr Val Leu Ser Gly Tyr Ile
180 185 190
Asp Tyr Ile Ser Thr Leu Val Val Lys Glu Ser Leu Glu Asn Val Arg
195 200 205
Asn Lys Leu Glu Ile Lys Thr Gln Phe Glu Lys Glu Lys Leu Ala Gln
210 215 220
Asp Arg Ile Lys Thr Lys Asn Gln Leu Asp Ala Asn Ile Gln Arg Leu
225 230 235 240
Asn Tyr Ser Leu Asp Ile Ala Asn Ala Ala Gly Ile Lys Lys Pro Val
245 250 255
Tyr Ser Asn Gly Gln Ala Val Lys Asp Asp Pro Asp Phe Ser Ile Ser
260 265 270
Leu Gly Ala Asp Gly Ile Glu Arg Lys Leu Glu Ile Glu Lys Ala Val
275 280 285
Thr Asp Val Ala Glu Leu Asn Gly Glu Leu Arg Asn Arg Gln Tyr Leu
290 295 300
Val Glu Gln Leu Thr Lys Ala His Val Asn Asp Val Asn Phe Thr Pro
305 310 315 320
Phe Lys Tyr Gln Leu Ser Pro Ser Leu Pro Val Lys Lys Asp Gly Pro
325 330 335
Gly Lys Ala Ile Ile Val Ile Leu Ser Ala Leu Ile Gly Gly Met Val
340 345 350
Ala Cys Gly Gly Val Leu Leu Arg Tyr Ala Met Ala Ser Arg Lys Gln
355 360 365
Asp Ala Met Met Ala Asp His Leu Val
370 375
<210> 120
<211> 377
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 120
Met Ser Ser Leu Asn Ile Lys Gln Gly Ser Asp Ala His Phe Pro Asp
1 5 10 15
Tyr Pro Leu Ala Ser Pro Ser Asn Asn Glu Ile Asp Leu Leu Asn Leu
20 25 30
Ile Ser Val Leu Trp Arg Ala Lys Lys Thr Val Met Ala Val Val Phe
35 40 45
Ala Phe Ala Cys Ala Gly Leu Leu Ile Ser Phe Ile Leu Pro Gln Lys
50 55 60
Trp Thr Ser Ala Ala Val Val Thr Pro Pro Glu Pro Val Gln Trp Gln
65 70 75 80
Glu Leu Glu Lys Thr Phe Thr Lys Leu Arg Val Leu Asp Leu Asp Ile
85 90 95
Lys Ile Asp Arg Thr Glu Ala Phe Asn Leu Phe Ile Lys Lys Phe Gln
100 105 110
Ser Val Ser Leu Leu Glu Glu Tyr Leu Arg Ser Ser Pro Tyr Val Met
115 120 125
Asp Gln Leu Lys Glu Ala Lys Ile Asp Glu Leu Asp Leu His Arg Ala
130 135 140
Ile Val Ala Leu Ser Glu Lys Met Lys Ala Val Asp Asp Asn Ala Ser
145 150 155 160
Lys Lys Lys Asp Glu Pro Ser Leu Tyr Thr Ser Trp Thr Leu Ser Phe
165 170 175
Thr Ala Pro Thr Ser Glu Glu Ala Gln Thr Val Leu Ser Gly Tyr Ile
180 185 190
Asp Tyr Ile Ser Ala Leu Val Val Lys Glu Ser Ile Glu Asn Val Arg
195 200 205
Asn Lys Leu Glu Ile Lys Thr Gln Phe Glu Lys Glu Lys Leu Ala Gln
210 215 220
Asp Arg Ile Lys Met Lys Asn Gln Leu Asp Ala Asn Ile Gln Arg Leu
225 230 235 240
Asn Tyr Ser Leu Asp Ile Ala Asn Ala Ala Gly Ile Lys Lys Pro Val
245 250 255
Tyr Ser Asn Gly Gln Ala Val Lys Asp Asp Pro Asp Phe Ser Ile Ser
260 265 270
Leu Gly Ala Asp Gly Ile Glu Arg Lys Leu Glu Ile Glu Lys Ala Val
275 280 285
Thr Asp Val Ala Glu Leu Asn Gly Glu Leu Arg Asn Arg Gln Tyr Leu
290 295 300
Val Glu Gln Leu Thr Lys Ala Asn Ile Asn Asp Val Asn Phe Thr Pro
305 310 315 320
Phe Lys Tyr Gln Leu Ser Pro Ser Leu Pro Val Lys Lys Asp Gly Pro
325 330 335
Gly Lys Ala Ile Ile Val Ile Leu Ser Ala Leu Ile Gly Gly Met Val
340 345 350
Ala Cys Gly Ser Val Leu Leu Arg Tyr Ala Met Ala Ser Arg Lys Gln
355 360 365
Asp Ala Met Met Ala Asp His Leu Val
370 375
<210> 121
<211> 378
<212> Белок
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 121
Met Pro Ser Leu Asn Val Lys Gln Glu Lys Asn Gln Ser Phe Ala Gly
1 5 10 15
Tyr Ser Leu Pro Pro Ala Asn Ser His Glu Ile Asp Leu Phe Ser Leu
20 25 30
Ile Glu Val Leu Trp Gln Ala Lys Arg Arg Ile Leu Ala Thr Val Phe
35 40 45
Ala Phe Ala Cys Val Gly Leu Leu Leu Ser Phe Leu Leu Pro Gln Lys
50 55 60
Trp Thr Ser Gln Ala Ile Val Thr Pro Ala Glu Ser Val Gln Trp Gln
65 70 75 80
Gly Leu Glu Arg Thr Leu Thr Ala Leu Arg Val Leu Asp Met Glu Val
85 90 95
Ser Val Asp Arg Gly Ser Val Phe Asn Leu Phe Ile Lys Lys Phe Ser
100 105 110
Ser Pro Ser Leu Leu Glu Glu Tyr Leu Arg Ser Ser Pro Tyr Val Met
115 120 125
Asp Gln Leu Lys Gly Ala Gln Ile Asp Glu Gln Asp Leu His Arg Ala
130 135 140
Ile Val Leu Leu Ser Glu Lys Met Lys Ala Val Asp Ser Asn Val Gly
145 150 155 160
Lys Lys Asn Glu Thr Ser Leu Phe Thr Ser Trp Thr Leu Ser Phe Thr
165 170 175
Ala Pro Thr Arg Glu Glu Ala Gln Lys Val Leu Ala Gly Tyr Ile Gln
180 185 190
Tyr Ile Ser Asp Ile Val Val Lys Glu Thr Leu Glu Asn Ile Arg Asn
195 200 205
Gln Leu Glu Ile Lys Thr Arg Tyr Glu Gln Glu Lys Leu Ala Met Asp
210 215 220
Arg Val Arg Leu Lys Asn Gln Leu Asp Ala Asn Ile Gln Arg Leu His
225 230 235 240
Tyr Ser Leu Glu Ile Ala Asn Ala Ala Gly Ile Lys Arg Pro Val Tyr
245 250 255
Ser Asn Gly Gln Ala Val Lys Asp Asp Pro Asp Phe Ser Ile Ser Leu
260 265 270
Gly Ala Asp Gly Ile Ser Arg Lys Leu Glu Ile Glu Lys Gly Val Thr
275 280 285
Asp Val Ala Glu Ile Asp Gly Asp Leu Arg Asn Arg Gln Tyr His Val
290 295 300
Glu Gln Leu Ala Ala Met Asn Val Ser Asp Val Lys Phe Thr Pro Phe
305 310 315 320
Lys Tyr Gln Leu Ser Pro Ser Leu Pro Val Lys Lys Asp Gly Pro Gly
325 330 335
Lys Ala Ile Ile Ile Ile Leu Ala Ala Leu Ile Gly Gly Met Met Ala
340 345 350
Cys Gly Gly Val Leu Leu Arg His Ala Met Val Ser Arg Lys Met Glu
355 360 365
Asn Ala Leu Ala Ile Asp Glu Arg Leu Val
370 375
<210> 122
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 122
gaagcaaacc gtacgcgtaa ag 22
<210> 123
<211> 20
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 123
cgaccagctc ttacacggcg 20
<210> 124
<211> 36
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 124
gaaataggac cactaataaa tacacaaatt aataac 36
<210> 125
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 125
ataattgacg atccggttgc c 21
<210> 126
<211> 27
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 126
gctatttacg ccctgattgt cttttgt 27
<210> 127
<211> 22
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 127
attgagaacc tgcgtaaacg gc 22
<210> 128
<211> 24
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 128
tgaagagcgg ttcagataac ttcc 24
<210> 129
<211> 21
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 129
cgatccggaa acctcctaca c 21
<210> 130
<211> 26
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 130
gattattcgc gcaacgctaa acagat 26
<210> 131
<211> 23
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 131
tgatcattga cgatccggta gcc 23
<210> 132
<211> 70
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 132
cggtagctgt aaagccaggg gcggtagcgt ggtttaaacc caagcaacag atcggcgtcg 60
tcggtatgga 70
<210> 133
<211> 78
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 133
agcttccata ccgacgacgc cgatctgttg cttgggttta aaccacgcta ccgcccctgg 60
ctttacagct accgagct 78
<210> 134
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 134
ggtagctgta aagccagggg cggtagcgtg 30
<210> 135
<211> 30
<212> ДНК
<213> Искусственная последовательность
<220>
<223> Синтетический
<400> 135
ccataccgac gacgccgatc tgttgcttgg 30
<210> 136
<211> 23
<212> Белок
<213> Homo sapiens
<400> 136
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly
20
<210> 137
<211> 18
<212> Белок
<213> Homo sapiens
<400> 137
Met His Ser Ser Ala Leu Leu Cys Cys Leu Val Leu Leu Thr Gly Val
1 5 10 15
Arg Ala
<210> 138
<211> 25
<212> Белок
<213> Респираторно-синцитиальный вирус человека А (штамм A2)
<400> 138
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly
20 25
<210> 139
<211> 15
<212> Белок
<213> Респираторно-синцитиальный вирус человека А (штамм A2)
<400> 139
Met Ala Ile Ile Tyr Leu Ile Leu Leu Phe Thr Ala Val Arg Gly
1 5 10 15
<210> 140
<211> 365
<212> Белок
<213> Homo sapiens
<400> 140
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Ala Glu Ser His Leu Ser Leu Leu Tyr
20 25 30
His Leu Thr Ala Val Ser Ser Pro Ala Pro Gly Thr Pro Ala Phe Trp
35 40 45
Val Ser Gly Trp Leu Gly Pro Gln Gln Tyr Leu Ser Tyr Asn Ser Leu
50 55 60
Arg Gly Glu Ala Glu Pro Cys Gly Ala Trp Val Trp Glu Asn Gln Val
65 70 75 80
Ser Trp Tyr Trp Glu Lys Glu Thr Thr Asp Leu Arg Ile Lys Glu Lys
85 90 95
Leu Phe Leu Glu Ala Phe Lys Ala Leu Gly Gly Lys Gly Pro Tyr Thr
100 105 110
Leu Gln Gly Leu Leu Gly Cys Glu Leu Gly Pro Asp Asn Thr Ser Val
115 120 125
Pro Thr Ala Lys Phe Ala Leu Asn Gly Glu Glu Phe Met Asn Phe Asp
130 135 140
Leu Lys Gln Gly Thr Trp Gly Gly Asp Trp Pro Glu Ala Leu Ala Ile
145 150 155 160
Ser Gln Arg Trp Gln Gln Gln Asp Lys Ala Ala Asn Lys Glu Leu Thr
165 170 175
Phe Leu Leu Phe Ser Cys Pro His Arg Leu Arg Glu His Leu Glu Arg
180 185 190
Gly Arg Gly Asn Leu Glu Trp Lys Glu Pro Pro Ser Met Arg Leu Lys
195 200 205
Ala Arg Pro Ser Ser Pro Gly Phe Ser Val Leu Thr Cys Ser Ala Phe
210 215 220
Ser Phe Tyr Pro Pro Glu Leu Gln Leu Arg Phe Leu Arg Asn Gly Leu
225 230 235 240
Ala Ala Gly Thr Gly Gln Gly Asp Phe Gly Pro Asn Ser Asp Gly Ser
245 250 255
Phe His Ala Ser Ser Ser Leu Thr Val Lys Ser Gly Asp Glu His His
260 265 270
Tyr Cys Cys Ile Val Gln His Ala Gly Leu Ala Gln Pro Leu Arg Val
275 280 285
Glu Leu Glu Ser Pro Ala Lys Ser Ser Val Leu Val Val Gly Ile Val
290 295 300
Ile Gly Val Leu Leu Leu Thr Ala Ala Ala Val Gly Gly Ala Leu Leu
305 310 315 320
Trp Arg Arg Met Arg Ser Gly Leu Pro Ala Pro Trp Ile Ser Leu Arg
325 330 335
Gly Asp Asp Thr Gly Val Leu Leu Pro Thr Pro Gly Glu Ala Gln Asp
340 345 350
Ala Asp Leu Lys Asp Val Asn Val Ile Pro Ala Thr Ala
355 360 365
<210> 141
<211> 62
<212> Белок
<213> Homo sapiens
<400> 141
Met Gly Val Pro Arg Pro Gln Pro Trp Ala Leu Gly Leu Leu Leu Phe
1 5 10 15
Leu Leu Pro Gly Ser Leu Gly Phe Ala Cys Lys Thr Ala Asn Gly Thr
20 25 30
Ala Ile Pro Ile Gly Gly Gly Ser Ala Asn Val Tyr Val Asn Leu Ala
35 40 45
Pro Val Val Asn Val Gly Gln Asn Leu Val Val Asp Leu Ser
50 55 60
<210> 142
<211> 574
<212> Белок
<213> Респираторно-синцитиальный вирус человека А (штамм A2)
<400> 142
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Gln Asn Ile Thr Glu Glu Phe
20 25 30
Tyr Gln Ser Thr Cys Ser Ala Val Ser Lys Gly Tyr Leu Ser Ala Leu
35 40 45
Arg Thr Gly Trp Tyr Thr Ser Val Ile Thr Ile Glu Leu Ser Asn Ile
50 55 60
Lys Glu Asn Lys Cys Asn Gly Thr Asp Ala Lys Val Lys Leu Ile Lys
65 70 75 80
Gln Glu Leu Asp Lys Tyr Lys Asn Ala Val Thr Glu Leu Gln Leu Leu
85 90 95
Met Gln Ser Thr Pro Pro Thr Asn Asn Arg Ala Arg Arg Glu Leu Pro
100 105 110
Arg Phe Met Asn Tyr Thr Leu Asn Asn Ala Lys Lys Thr Asn Val Thr
115 120 125
Leu Ser Lys Lys Arg Lys Arg Arg Phe Leu Gly Phe Leu Leu Gly Val
130 135 140
Gly Ser Ala Ile Ala Ser Gly Val Ala Val Ser Lys Val Leu His Leu
145 150 155 160
Glu Gly Glu Val Asn Lys Ile Lys Ser Ala Leu Leu Ser Thr Asn Lys
165 170 175
Ala Val Val Ser Leu Ser Asn Gly Val Ser Val Leu Thr Ser Lys Val
180 185 190
Leu Asp Leu Lys Asn Tyr Ile Asp Lys Gln Leu Leu Pro Ile Val Asn
195 200 205
Lys Gln Ser Cys Ser Ile Ser Asn Ile Glu Thr Val Ile Glu Phe Gln
210 215 220
Gln Lys Asn Asn Arg Leu Leu Glu Ile Thr Arg Glu Phe Ser Val Asn
225 230 235 240
Ala Gly Val Thr Thr Pro Val Ser Thr Tyr Met Leu Thr Asn Ser Glu
245 250 255
Leu Leu Ser Leu Ile Asn Asp Met Pro Ile Thr Asn Asp Gln Lys Lys
260 265 270
Leu Met Ser Asn Asn Val Gln Ile Val Arg Gln Gln Ser Tyr Ser Ile
275 280 285
Met Ser Ile Ile Lys Glu Glu Val Leu Ala Tyr Val Val Gln Leu Pro
290 295 300
Leu Tyr Gly Val Ile Asp Thr Pro Cys Trp Lys Leu His Thr Ser Pro
305 310 315 320
Leu Cys Thr Thr Asn Thr Lys Glu Gly Ser Asn Ile Cys Leu Thr Arg
325 330 335
Thr Asp Arg Gly Trp Tyr Cys Asp Asn Ala Gly Ser Val Ser Phe Phe
340 345 350
Pro Gln Ala Glu Thr Cys Lys Val Gln Ser Asn Arg Val Phe Cys Asp
355 360 365
Thr Met Asn Ser Leu Thr Leu Pro Ser Glu Ile Asn Leu Cys Asn Val
370 375 380
Asp Ile Phe Asn Pro Lys Tyr Asp Cys Lys Ile Met Thr Ser Lys Thr
385 390 395 400
Asp Val Ser Ser Ser Val Ile Thr Ser Leu Gly Ala Ile Val Ser Cys
405 410 415
Tyr Gly Lys Thr Lys Cys Thr Ala Ser Asn Lys Asn Arg Gly Ile Ile
420 425 430
Lys Thr Phe Ser Asn Gly Cys Asp Tyr Val Ser Asn Lys Gly Met Asp
435 440 445
Thr Val Ser Val Gly Asn Thr Leu Tyr Tyr Val Asn Lys Gln Glu Gly
450 455 460
Lys Ser Leu Tyr Val Lys Gly Glu Pro Ile Ile Asn Phe Tyr Asp Pro
465 470 475 480
Leu Val Phe Pro Ser Asp Glu Phe Asp Ala Ser Ile Ser Gln Val Asn
485 490 495
Glu Lys Ile Asn Gln Ser Leu Ala Phe Ile Arg Lys Ser Asp Glu Leu
500 505 510
Leu His Asn Val Asn Ala Gly Lys Ser Thr Thr Asn Ile Met Ile Thr
515 520 525
Thr Ile Ile Ile Val Ile Ile Val Ile Leu Leu Ser Leu Ile Ala Val
530 535 540
Gly Leu Leu Leu Tyr Cys Lys Ala Arg Ser Thr Pro Val Thr Leu Ser
545 550 555 560
Lys Asp Gln Leu Ser Gly Ile Asn Asn Ile Ala Phe Ser Asn
565 570
<210> 143
<211> 64
<212> Белок
<213> Респираторно-синцитиальный вирус человека А (штамм A2)
<400> 143
Met Glu Leu Leu Ile Leu Lys Ala Asn Ala Ile Thr Thr Ile Leu Thr
1 5 10 15
Ala Val Thr Phe Cys Phe Ala Ser Gly Phe Ala Cys Lys Thr Ala Asn
20 25 30
Gly Thr Ala Ile Pro Ile Gly Gly Gly Ser Ala Asn Val Tyr Val Asn
35 40 45
Leu Ala Pro Val Val Asn Val Gly Gln Asn Leu Val Val Asp Leu Ser
50 55 60
<210> 144
<211> 178
<212> Белок
<213> Homo sapiens
<400> 144
Met His Ser Ser Ala Leu Leu Cys Cys Leu Val Leu Leu Thr Gly Val
1 5 10 15
Arg Ala Ser Pro Gly Gln Gly Thr Gln Ser Glu Asn Ser Cys Thr His
20 25 30
Phe Pro Gly Asn Leu Pro Asn Met Leu Arg Asp Leu Arg Asp Ala Phe
35 40 45
Ser Arg Val Lys Thr Phe Phe Gln Met Lys Asp Gln Leu Asp Asn Leu
50 55 60
Leu Leu Lys Glu Ser Leu Leu Glu Asp Phe Lys Gly Tyr Leu Gly Cys
65 70 75 80
Gln Ala Leu Ser Glu Met Ile Gln Phe Tyr Leu Glu Glu Val Met Pro
85 90 95
Gln Ala Glu Asn Gln Asp Pro Asp Ile Lys Ala His Val Asn Ser Leu
100 105 110
Gly Glu Asn Leu Lys Thr Leu Arg Leu Arg Leu Arg Arg Cys His Arg
115 120 125
Phe Leu Pro Cys Glu Asn Lys Ser Lys Ala Val Glu Gln Val Lys Asn
130 135 140
Ala Phe Asn Lys Leu Gln Glu Lys Gly Ile Tyr Lys Ala Met Ser Glu
145 150 155 160
Phe Asp Ile Phe Ile Asn Tyr Ile Glu Ala Tyr Met Thr Met Lys Ile
165 170 175
Arg Asn
<210> 145
<211> 57
<212> Белок
<213> Homo sapiens
<400> 145
Met His Ser Ser Ala Leu Leu Cys Cys Leu Val Leu Leu Thr Gly Val
1 5 10 15
Arg Ala Phe Ala Cys Lys Thr Ala Asn Gly Thr Ala Ile Pro Ile Gly
20 25 30
Gly Gly Ser Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val
35 40 45
Gly Gln Asn Leu Val Val Asp Leu Ser
50 55
<210> 146
<211> 562
<212> Белок
<213> Вирус гриппа А (штамм A/Japan/305/1957 H2N2)
<400> 146
Met Ala Ile Ile Tyr Leu Ile Leu Leu Phe Thr Ala Val Arg Gly Asp
1 5 10 15
Gln Ile Cys Ile Gly Tyr His Ala Asn Asn Ser Thr Glu Lys Val Asp
20 25 30
Thr Asn Leu Glu Arg Asn Val Thr Val Thr His Ala Lys Asp Ile Leu
35 40 45
Glu Lys Thr His Asn Gly Lys Leu Cys Lys Leu Asn Gly Ile Pro Pro
50 55 60
Leu Glu Leu Gly Asp Cys Ser Ile Ala Gly Trp Leu Leu Gly Asn Pro
65 70 75 80
Glu Cys Asp Arg Leu Leu Ser Val Pro Glu Trp Ser Tyr Ile Met Glu
85 90 95
Lys Glu Asn Pro Arg Asp Gly Leu Cys Tyr Pro Gly Ser Phe Asn Asp
100 105 110
Tyr Glu Glu Leu Lys His Leu Leu Ser Ser Val Lys His Phe Glu Lys
115 120 125
Val Lys Ile Leu Pro Lys Asp Arg Trp Thr Gln His Thr Thr Thr Gly
130 135 140
Gly Ser Arg Ala Cys Ala Val Ser Gly Asn Pro Ser Phe Phe Arg Asn
145 150 155 160
Met Val Trp Leu Thr Lys Glu Gly Ser Asp Tyr Pro Val Ala Lys Gly
165 170 175
Ser Tyr Asn Asn Thr Ser Gly Glu Gln Met Leu Ile Ile Trp Gly Val
180 185 190
His His Pro Ile Asp Glu Thr Glu Gln Arg Thr Leu Tyr Gln Asn Val
195 200 205
Gly Thr Tyr Val Ser Val Gly Thr Ser Thr Leu Asn Lys Arg Ser Thr
210 215 220
Pro Glu Ile Ala Thr Arg Pro Lys Val Asn Gly Gln Gly Gly Arg Met
225 230 235 240
Glu Phe Ser Trp Thr Leu Leu Asp Met Trp Asp Thr Ile Asn Phe Glu
245 250 255
Ser Thr Gly Asn Leu Ile Ala Pro Glu Tyr Gly Phe Lys Ile Ser Lys
260 265 270
Arg Gly Ser Ser Gly Ile Met Lys Thr Glu Gly Thr Leu Glu Asn Cys
275 280 285
Glu Thr Lys Cys Gln Thr Pro Leu Gly Ala Ile Asn Thr Thr Leu Pro
290 295 300
Phe His Asn Val His Pro Leu Thr Ile Gly Glu Cys Pro Lys Tyr Val
305 310 315 320
Lys Ser Glu Lys Leu Val Leu Ala Thr Gly Leu Arg Asn Val Pro Gln
325 330 335
Ile Glu Ser Arg Gly Leu Phe Gly Ala Ile Ala Gly Phe Ile Glu Gly
340 345 350
Gly Trp Gln Gly Met Val Asp Gly Trp Tyr Gly Tyr His His Ser Asn
355 360 365
Asp Gln Gly Ser Gly Tyr Ala Ala Asp Lys Glu Ser Thr Gln Lys Ala
370 375 380
Phe Asp Gly Ile Thr Asn Lys Val Asn Ser Val Ile Glu Lys Met Asn
385 390 395 400
Thr Gln Phe Glu Ala Val Gly Lys Glu Phe Gly Asn Leu Glu Arg Arg
405 410 415
Leu Glu Asn Leu Asn Lys Arg Met Glu Asp Gly Phe Leu Asp Val Trp
420 425 430
Thr Tyr Asn Ala Glu Leu Leu Val Leu Met Glu Asn Glu Arg Thr Leu
435 440 445
Asp Phe His Asp Ser Asn Val Lys Asn Leu Tyr Asp Lys Val Arg Met
450 455 460
Gln Leu Arg Asp Asn Val Lys Glu Leu Gly Asn Gly Cys Phe Glu Phe
465 470 475 480
Tyr His Lys Cys Asp Asp Glu Cys Met Asn Ser Val Lys Asn Gly Thr
485 490 495
Tyr Asp Tyr Pro Lys Tyr Glu Glu Glu Ser Lys Leu Asn Arg Asn Glu
500 505 510
Ile Lys Gly Val Lys Leu Ser Ser Met Gly Val Tyr Gln Ile Leu Ala
515 520 525
Ile Tyr Ala Thr Val Ala Gly Ser Leu Ser Leu Ala Ile Met Met Ala
530 535 540
Gly Ile Ser Phe Trp Met Cys Ser Asn Gly Ser Leu Gln Cys Arg Ile
545 550 555 560
Cys Ile
<210> 147
<211> 54
<212> Белок
<213> Вирус гриппа А (штамм A/Japan/305/1957 H2N2)
<400> 147
Met Ala Ile Ile Tyr Leu Ile Leu Leu Phe Thr Ala Val Arg Gly Phe
1 5 10 15
Ala Cys Lys Thr Ala Asn Gly Thr Ala Ile Pro Ile Gly Gly Gly Ser
20 25 30
Ala Asn Val Tyr Val Asn Leu Ala Pro Val Val Asn Val Gly Gln Asn
35 40 45
Leu Val Val Asp Leu Ser
50
<---
| название | год | авторы | номер документа |
|---|---|---|---|
| КОМПОЗИЦИИ ESCHERICHIA COLI И СПОСОБЫ НА ИХ ОСНОВЕ | 2021 |
|
RU2821929C1 |
| СПОСОБ ПОВЫШЕНИЯ ИММУНОГЕННОСТИ БЕЛКОВОГО/ПЕПТИДНОГО АНТИГЕНА | 2021 |
|
RU2830627C1 |
| Композиции с Neisseria meningitidis и способы их применения | 2020 |
|
RU2839906C1 |
| МУЛЬТИМЕРИЗУЮЩИЕСЯ ПОЛИПЕПТИДЫ, ПРОИСХОДЯЩИЕ ОТ ДОМЕНА С УКЛАДКОЙ ТИПА РУЛЕТ ОСНОВАНИЯ ПЕНТОНА АДЕНОВИРУСА | 2019 |
|
RU2820522C2 |
| СКОНСТРУИРОВАННЫЙ БОТУЛИНИЧЕСКИЙ НЕЙРОТОКСИН | 2017 |
|
RU2789302C2 |
| СКОНСТРУИРОВАННЫЙ БОТУЛИНИЧЕСКИЙ НЕЙРОТОКСИН | 2016 |
|
RU2746736C2 |
| АНТИГЕННЫЕ ПОЛИПЕПТИДЫ НА ОСНОВЕ ПОСЛЕДОВАТЕЛЬНОСТИ OspA | 2019 |
|
RU2816208C2 |
| СПОСОБЫ ПРОИЗВОДСТВА ПРОТЕОЛИТИЧЕСКИ ПРОЦЕССИРОВАННЫХ ПОЛИПЕПТИДОВ | 2018 |
|
RU2719164C1 |
| Вакцины против Chlamydia sp. | 2014 |
|
RU2723046C2 |
| АДЕНОВИРУСЫ И СПОСОБЫ ПРИМЕНЕНИЯ АДЕНОВИРУСОВ | 2019 |
|
RU2782528C1 |
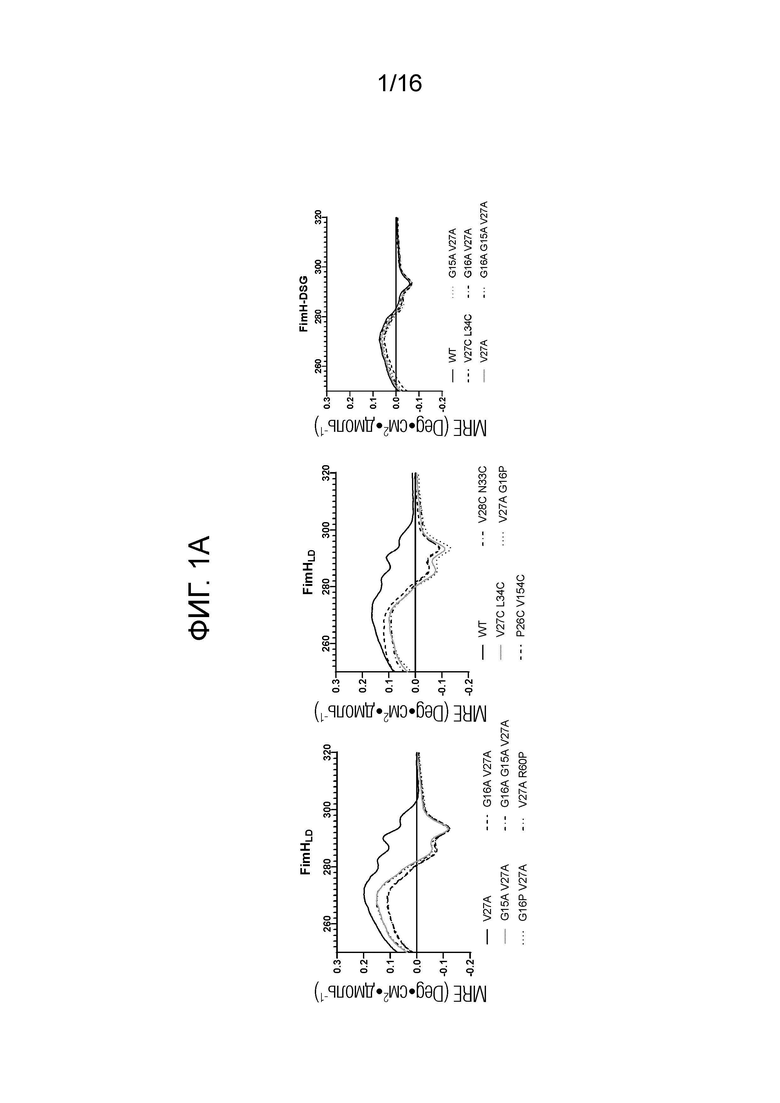
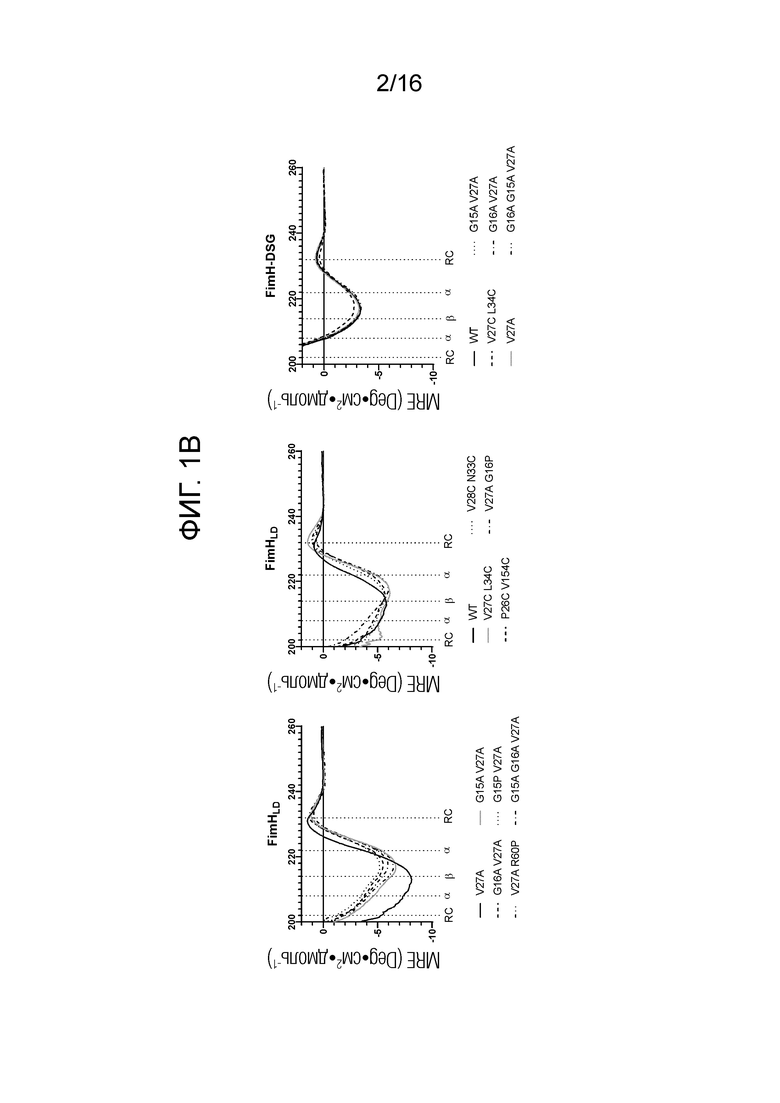
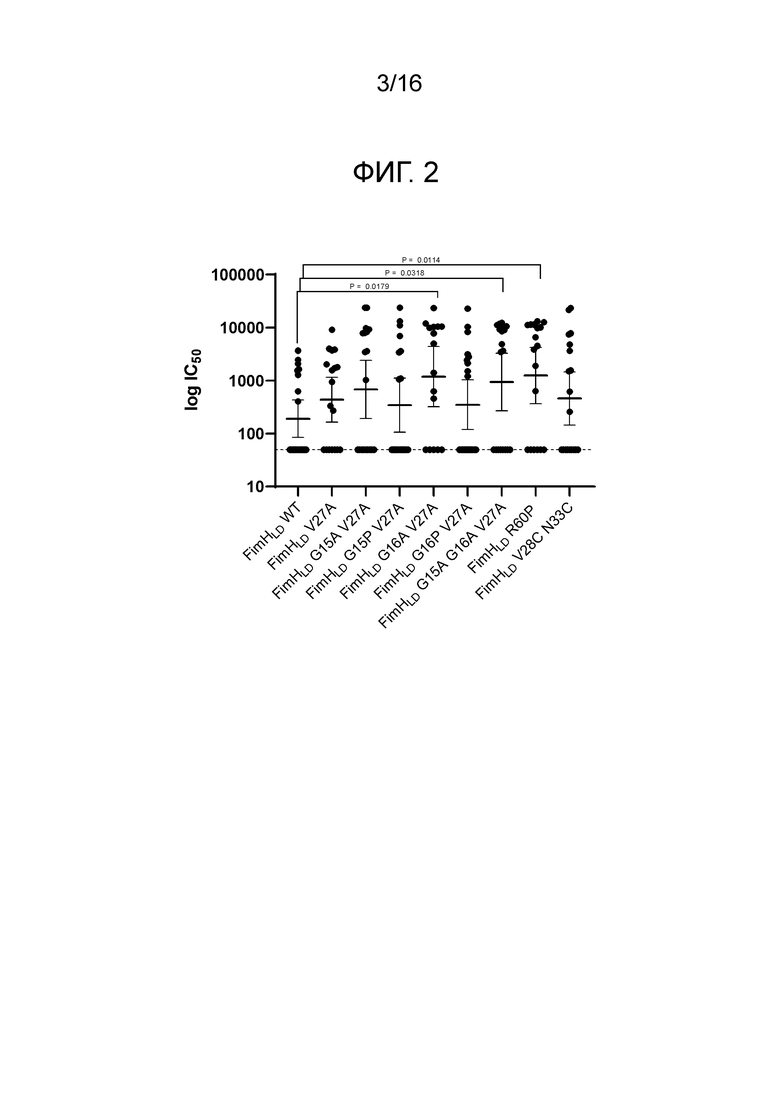
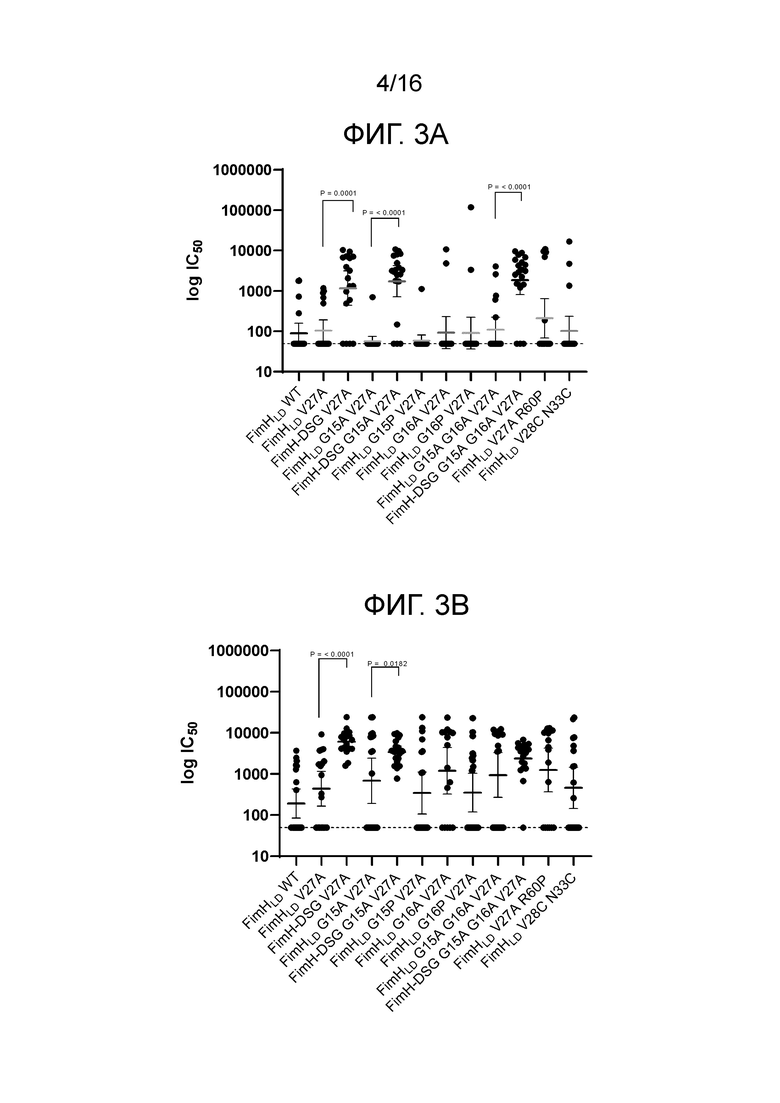
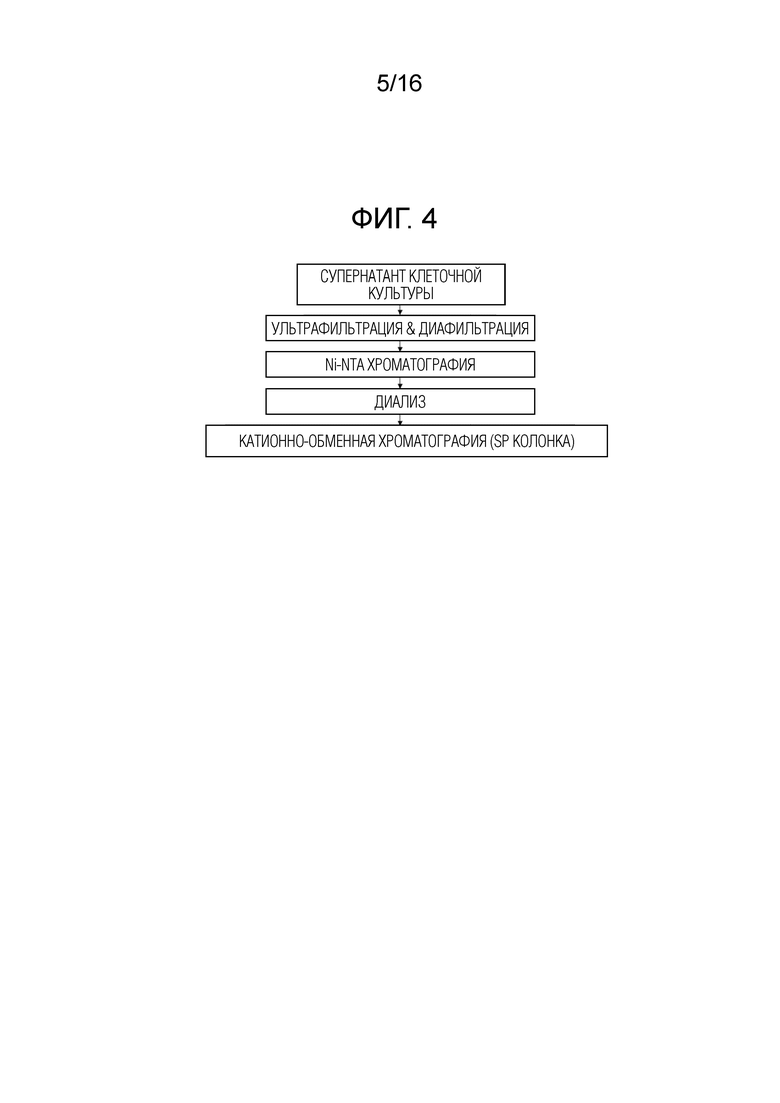
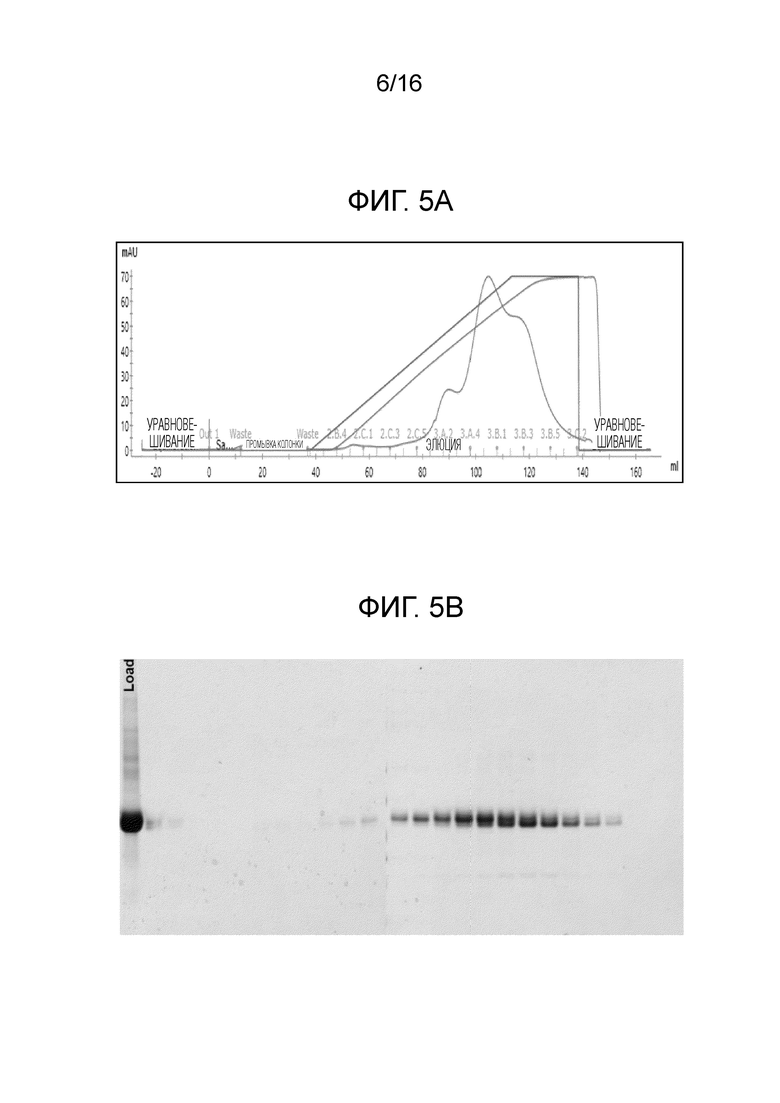
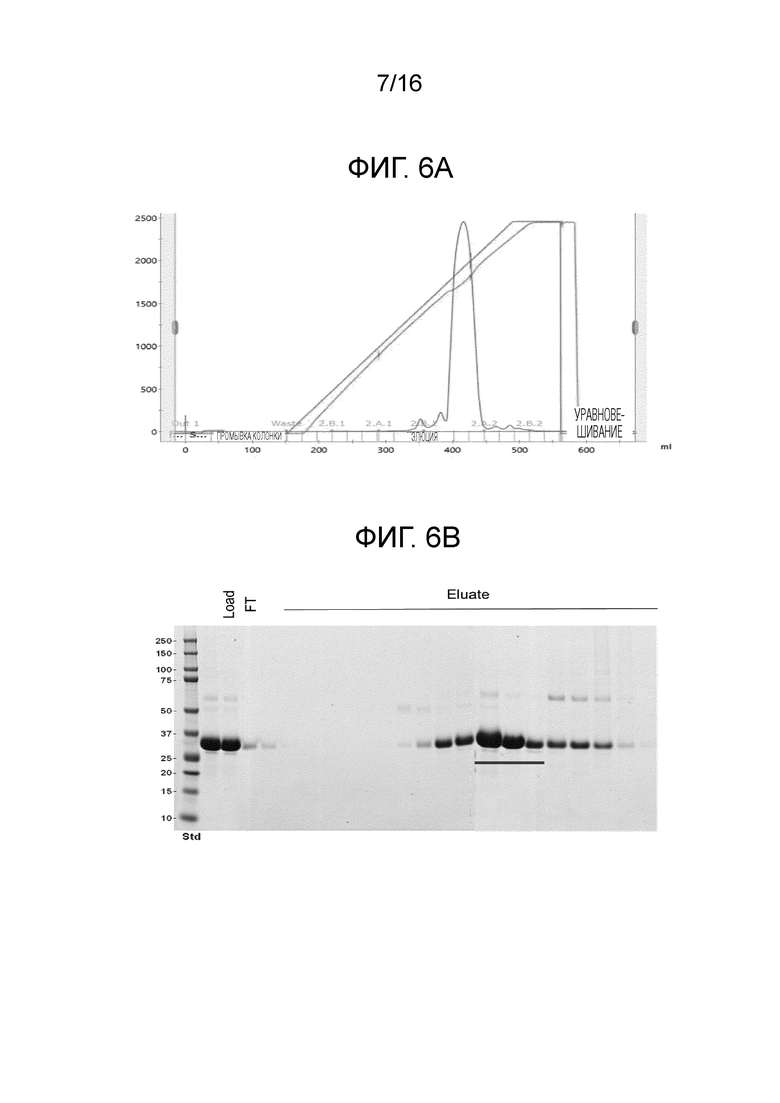
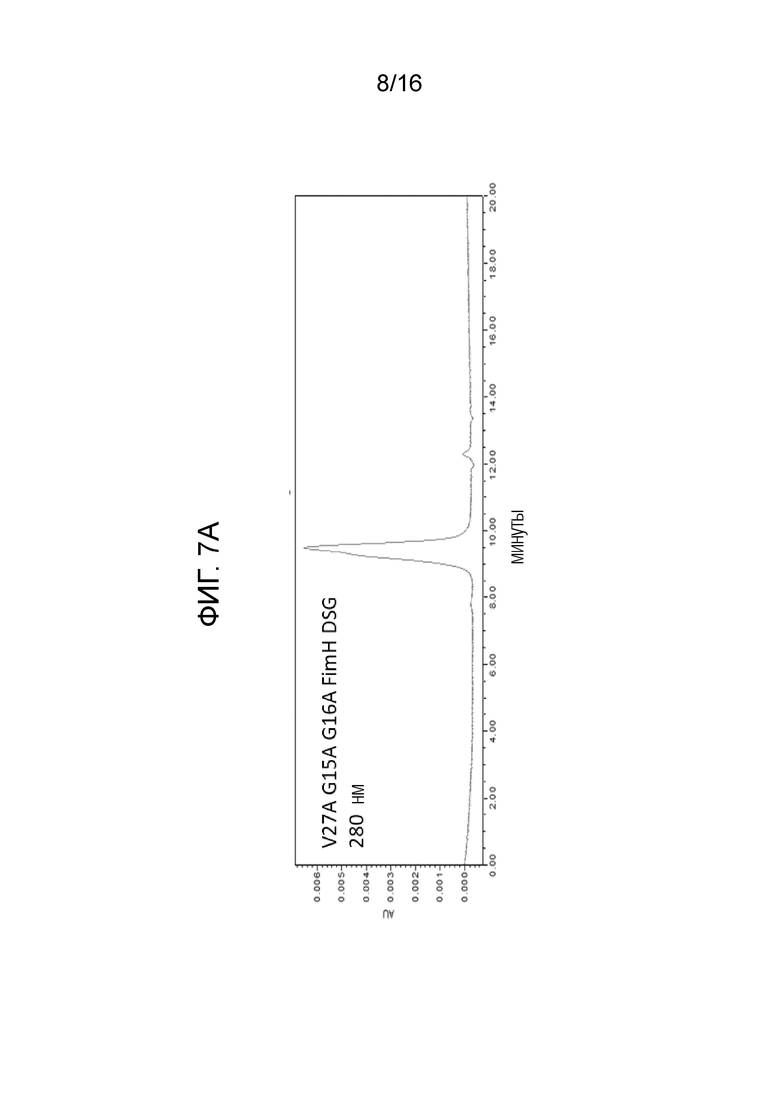
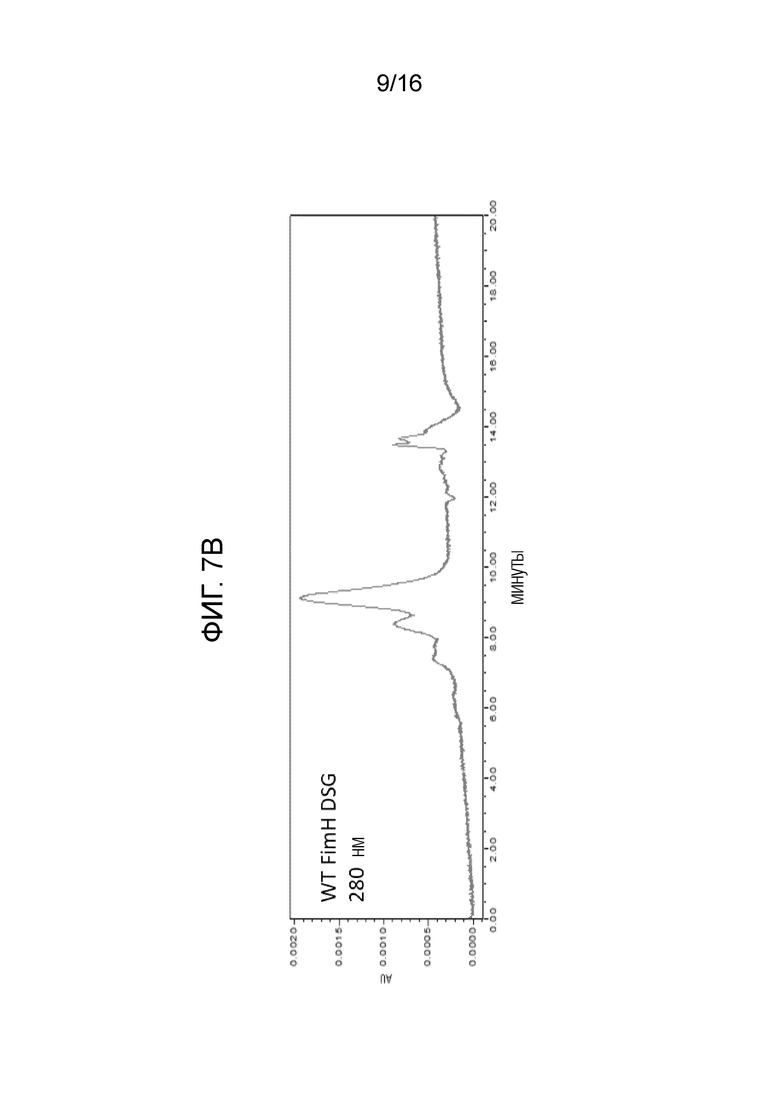
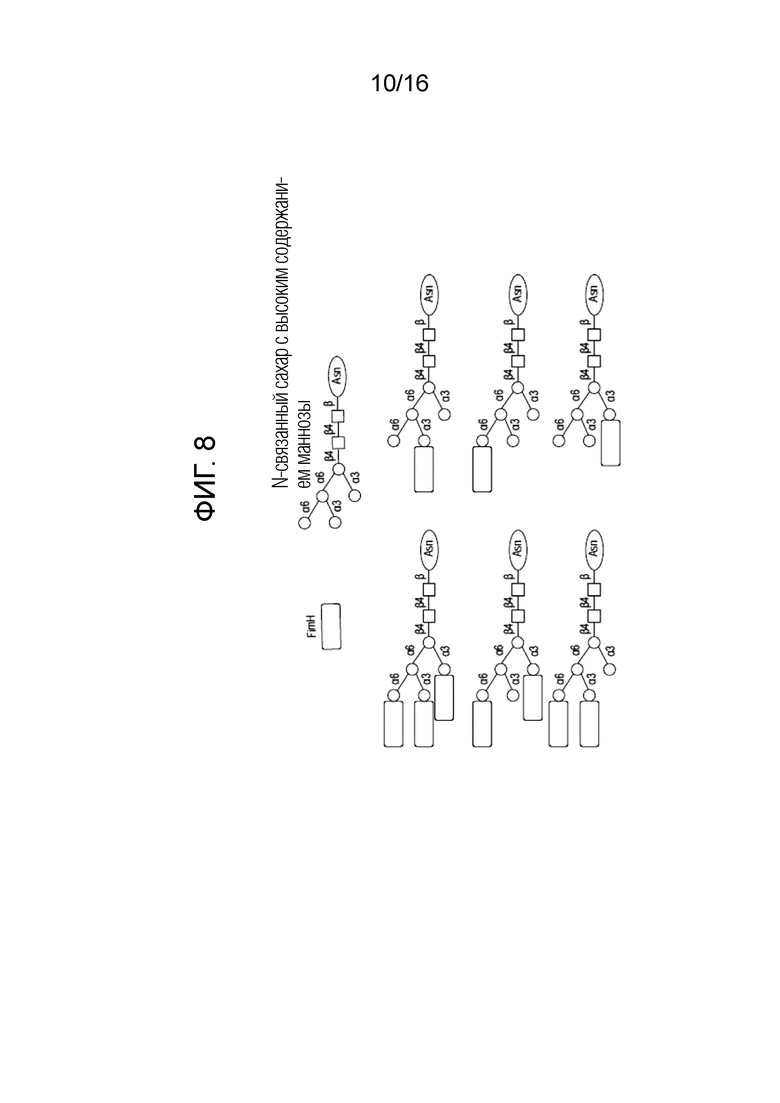
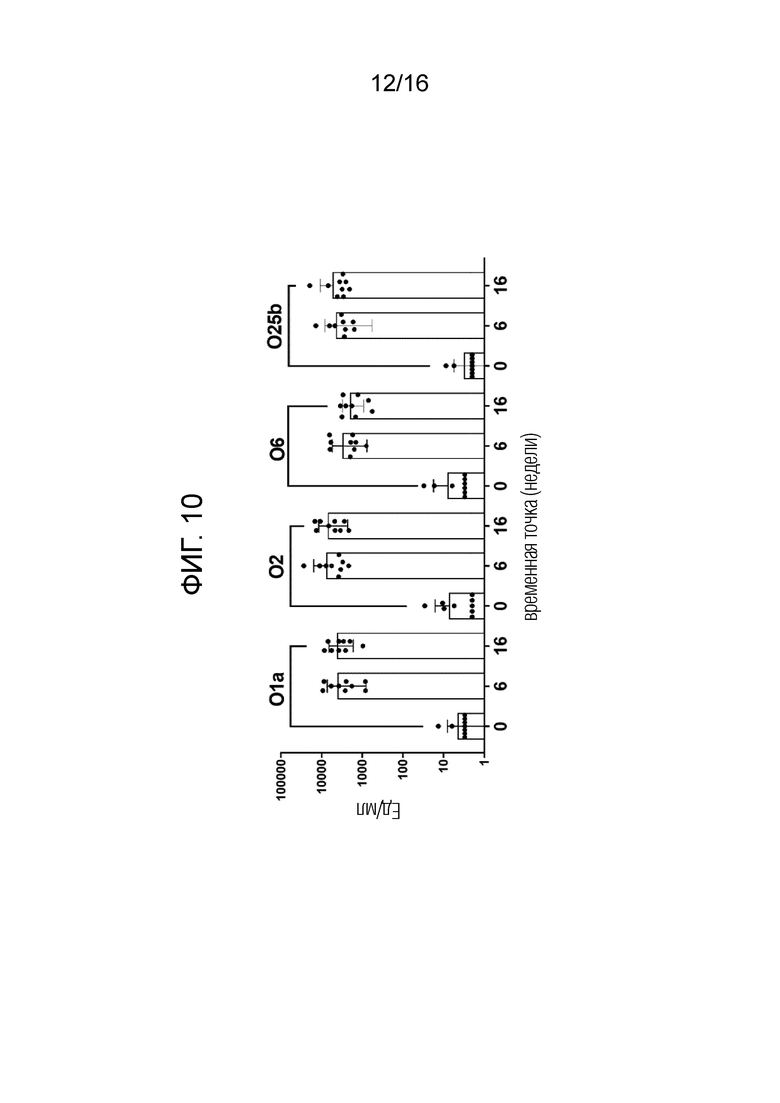
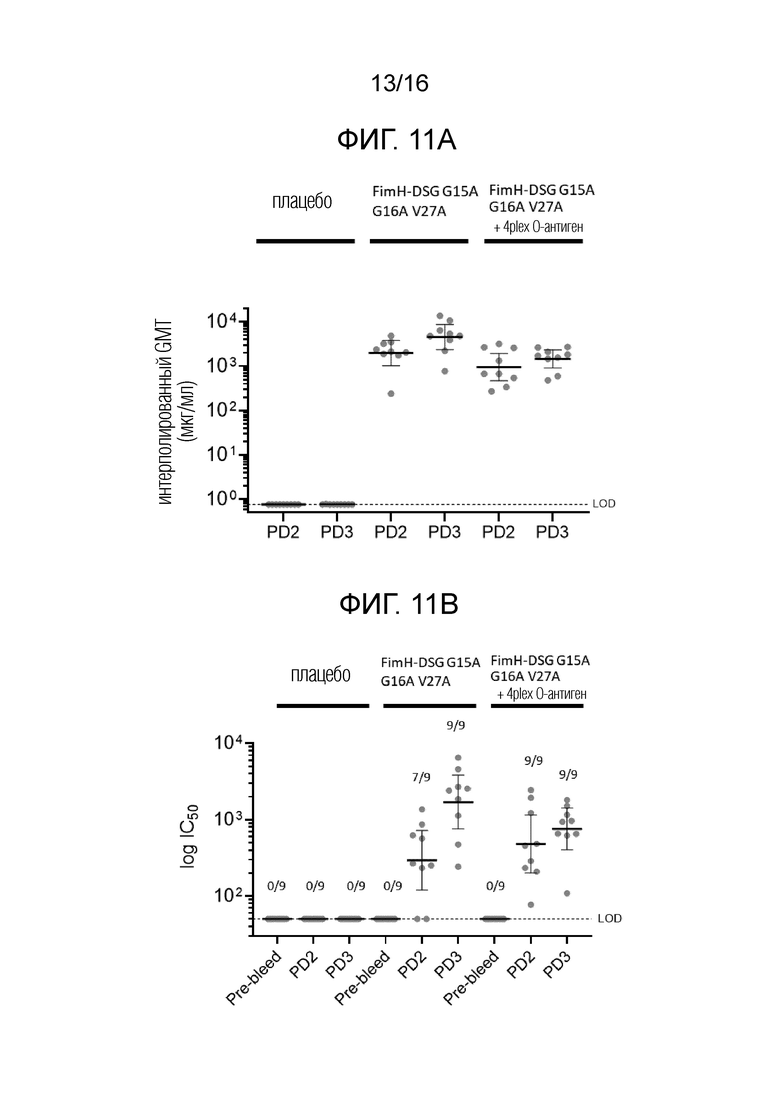
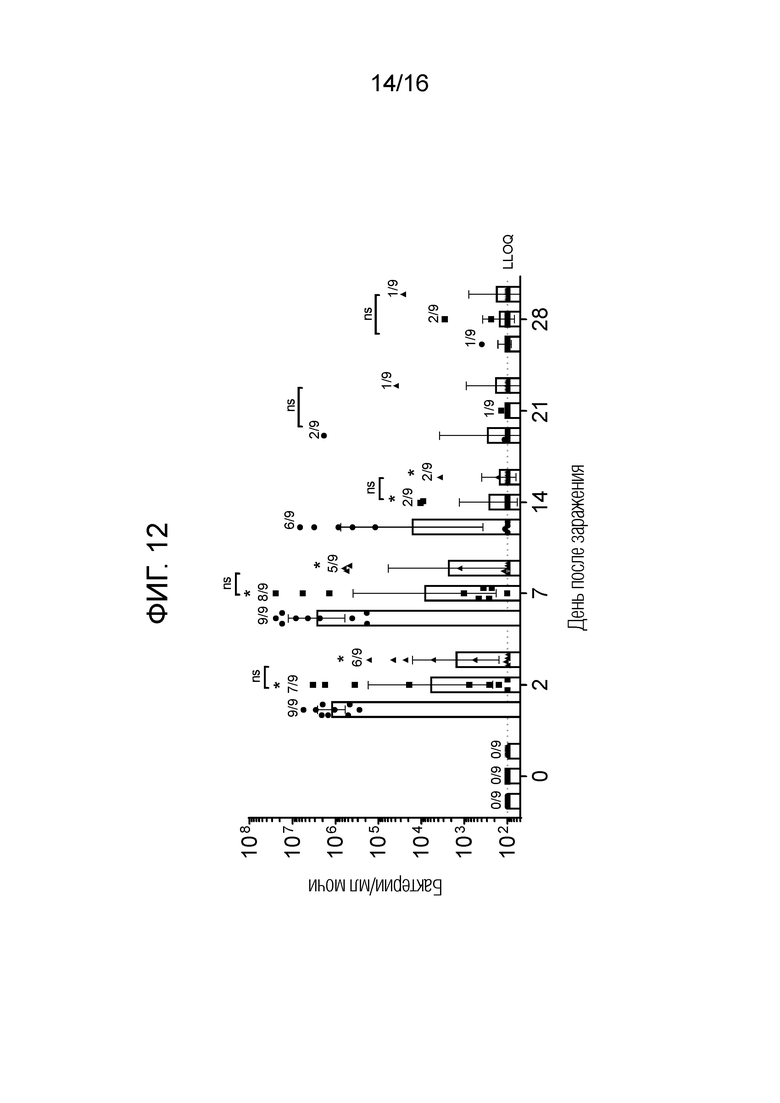
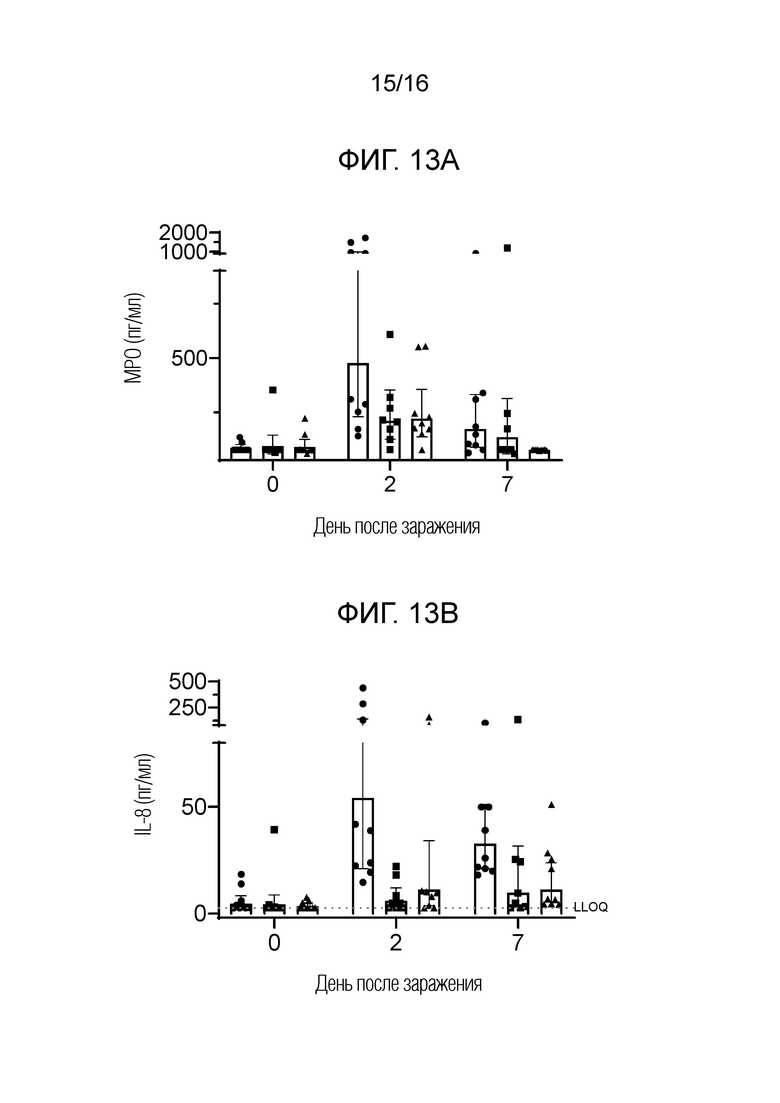
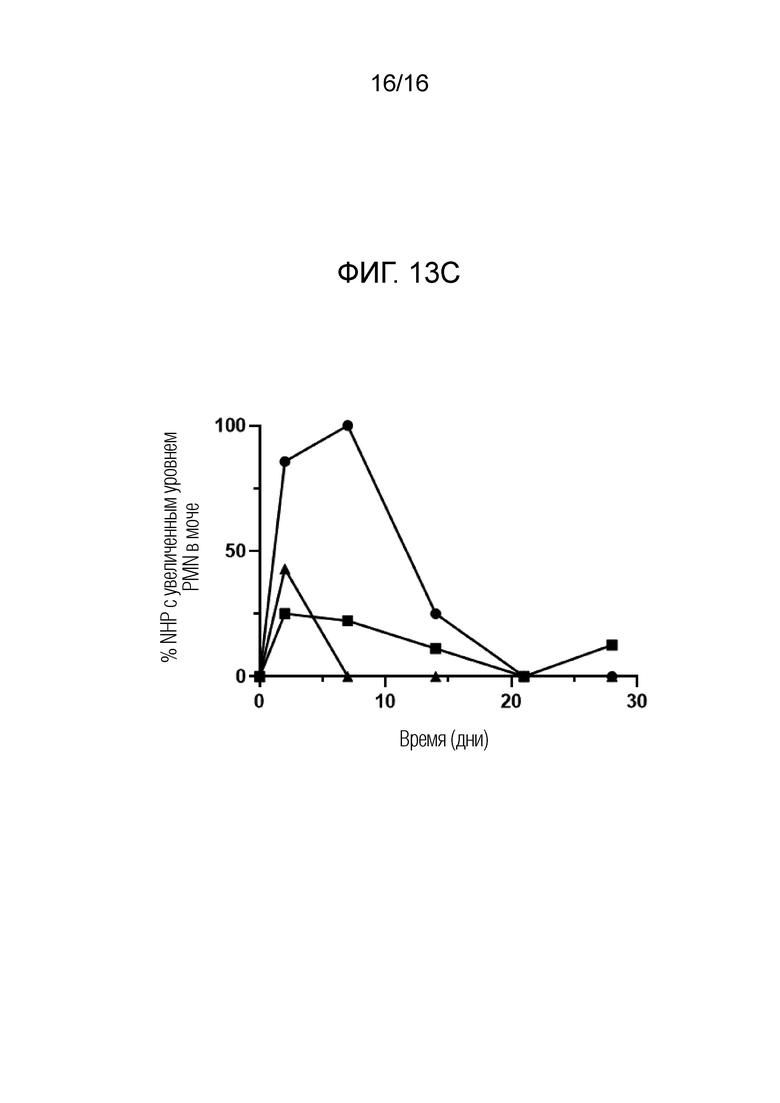
Изобретение относится к биотехнологии, в частности к мутантным полипептидам FimH Escherichia coli и их применению. Предложены варианты мутантного полипептида FimH для индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта. Также предложены молекула нуклеиновой кислоты, кодирующая указанный полипептид, фармацевтическая композиция, содержащая указанные полипептид или молекулу нуклеиновой кислоты, рекомбинантная клетка млекопитающего для продуцирования мутантного полипептида FimH, культура для продуцирования мутантного полипептида FimH, способ получения мутантного полипептида FimH, способ индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта, а также способ профилактики, лечения или облегчения инфекции уропатогенных E.coli у субъекта. Изобретение обеспечивает получение мутантных полипептидов FimH Е.coli с пониженным сродством к маннозидным лигандам и улучшенными биохимическими свойствами, которые обеспечивают повышенную функциональную иммуногенность по сравнению с FimH дикого типа. 10 н. и 11 з.п. ф-лы, 13 ил., 24 табл., 21 пр.
1. Мутантный полипептид FimH для индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта, который содержит аминокислотные мутации относительно аминокислотной последовательности полипептида FimH дикого типа, где мутации выбраны из группы, состоящей из:
a) мутаций G15A и V27A;
b) мутаций G16A и V27A; и
c) мутаций G15A, G16A и V27A, и где положения аминокислот пронумерованы в соответствии с SEQ ID NO: 59.
2. Мутантный полипептид FimH для индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта, содержащий последовательность, выбранную из группы, состоящей из SEQ ID NO: 50, 51, 54, 61 и 62.
3. Мутантный полипептид FimH для индукции продуцирования антител, специфичных к уропатогенным E.coli у субъекта, который включает аминокислотные мутации относительно аминокислотной последовательности полипептида FimH дикого типа, и где указанные мутации включают G15A, G16A и V27A.
4. Мутантный полипептид FimH по любому из пп.1-3, где полипептид дополнительно включает пептид FimG, связанный через глицин-сериновый линкер.
5. Мутантный полипептид FimH по любому из пп.1-4, где полипептид является иммуногенным.
6. Мутантный полипептид FimH по любому из пп.1-5, где полипептид является выделенным.
7. Молекула нуклеиновой кислоты, включающая нуклеотидную последовательность, которая кодирует аминокислотную последовательность мутантного полипептида FimH по любому из пп.1-5.
8. Молекула нуклеиновой кислоты по п.7, где нуклеиновая кислота представляет собой РНК.
9. Фармацевтическая композиция для индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта, содержащая (i) эффективное количество мутантного полипептида FimH по любому из пп.1-6 или молекулы нуклеиновой кислоты по п.7 или 8 и (ii) фармацевтически приемлемый носитель.
10. Фармацевтическая композиция по п.9, дополнительно содержащая по меньшей мере один дополнительный антиген.
11. Фармацевтическая композиция по п.9, где по меньшей мере один дополнительный антиген представляет собой полисахарид или белок.
12. Фармацевтическая композиция по п.9, дополнительно содержащая по меньшей мере один адъювант.
13. Рекомбинантная клетка млекопитающего для продуцирования мутантного полипептида FimH, содержащая полинуклеотид, кодирующий мутантный полипептид FimH по любому из пп.1-5.
14. Культура для продуцирования мутантного полипептида FimH, содержащая рекомбинантную клетку по п.13 и среду для культивирования клеток, где указанная культура имеет размер не менее 5 литров.
15. Способ получения мутантного полипептида FimH по любому из пп.1-5, включающий культивирование рекомбинантной клетки млекопитающего по п.13 в подходящих условиях, обеспечивая таким образом экспрессию полипептида; и сбор полипептида.
16. Способ индукции продуцирования антител, специфичных к уропатогенным E.coli, у субъекта, где способ включает введение субъекту эффективного количества композиции по любому из пп.9-12.
17. Способ по п.16, где субъект подвержен риску развития инфекции мочевыводящих путей.
18. Способ по п.16, где субъект подвержен риску развития бактериемии.
19. Способ по п.16, где субъект подвержен риску развития сепсиса.
20. Способ по п.16, где антитела защищают млекопитающее от инфекции уропатогенных E.coli.
21. Способ профилактики, лечения или облегчения инфекции уропатогенных E.coli у субъекта, включающий введение субъекту иммунологически эффективного количества композиции по любому из пп.9-12.
| KISIELA D | |||
| I | |||
| et al | |||
| Conformational inactivation induces immunogenicity of the receptor-binding pocket of a bacterial adhesion | |||
| PROCEEDINGS OF THE NATIONAL ACADEMY OF SCIENCES, vol | |||
| Прибор, автоматически записывающий пройденный путь | 1920 |
|
SU110A1 |
| Способ очищения сернокислого глинозема от железа | 1920 |
|
SU47A1 |
| Приспособление для обрезания края подошв | 1929 |
|
SU19089A1 |
| MUNERA D | |||
| et al | |||
| Печь для непрерывного получения сернистого натрия | 1921 |
|
SU1A1 |

